
Ad 2009501
293
Mt ~4? UNIVERSIDAD COMPLUTENSE DE MADRID Departamento de Nutrición y Bromatología II I (Higiene y Tecnología de los Alimentos) FACULTAD DE VETERINARIA CARACTERIZACION PARCIAL, BIOQUíMICA E INMUNOLOGICA, DE UNA SUSTANCIA ANTIMICROBIANA PRODUCIDA POR Lactobacillus sake 14 8 Memoria que, para optar al grado de Doctor en Veterinaria, presenta el Licenciado: ODON JULIAN SOBRINO ABUJA
-
Upload
jairo-garcia -
Category
Documents
-
view
102 -
download
1
Transcript of Ad 2009501

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 1/293
Mt ~4?
UNIVERSIDAD COMPLUTENSE DE MADRIDDepartamento de Nutrición y Bromatología III
(Higiene y Tecnología de los Alimentos)
FACULTAD DE VETERINARIA
CARACTERIZACION PARCIAL,BIOQUíMICA E INMUNOLOGICA,
DE UNA SUSTANCIAANTIMICROBIANA PRODUCIDA
POR Lactobacillus sake 148
Memoria que, para optar al gradode Doctor en Veterinaria,
presenta el Licenciado:
ODON JULIAN SOBRINO ABUJA

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 2/293
PABLO ELPIDIO HERNANDEZ CRUZA, CATEDRATICO DE NUTRICION Y
BROMATOLOGíA DE LA FACULTAD DE VETERINARIA DE LA UNIVERSIDAD
COMPLUTENSE DE MADRID,
CERTIFICA:
Que la tesis doctoral titulada ‘CARACTERIZACION PARCIAL, BIOQUIMICA E
INMUNOLOGICA. DE UNA SUSTANCIA ANTIMICROBIANA PRODUCIDA POR
LACTOBACILLUS SAKE 148, de la que es autor D. Odón Julián Sobrino Abuja, ha sido
realizada en el Departamento de Nutrición y Bronntología III (Higiene y Tecnología de los
Alimentos), bajo la dirección conjunta del catedrático D. Bernabé Sanz Pérez, de la profesora
titular Dña. María E. Fernández Alvarez y del que suscribe y cumple las condiciones exigidas
para optar al título de Doctor en Veterinaria.
Madrid, 1 9 de enero de 1993
cnAQNcttx~c {OwkJ) __
.—rd~EÉ~rnabé Sanz Pérez Edo: Maria FiFétrá~d~~ Alvarez Edo: Pablo E. Hernández Cruza

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 3/293
AGRADECIMIENTOS
En primer lugar, quiero agradecer al Prof D. Bernabé Sanz Pérez la confianza
depositada en mi al acogerme en el Departamento, que tan acertadamente ha dirigido durante
años y, en el que se ha desarrollado este trabajo. Deseo agradecerle también su paciencia y
comprensión durante mis primeros pasos, torpes y vacilantes, porel laboratorio así como sus
enriquecedoras orientaciones, que han contribuido sobremanera a mi completa formacion.
De manera especial quiero mostrar mi agradecimiento al Prof. D. Pablo E. Hernández
Cmza, verdadera piedra angular y soporte científico de este trabajo. Agradecerle, sobre tQdo.
su ejemplo, modelo de trabajo, dedicación, abnegación y sacrificio. Su acertado
asesoramiento, apoyo, comprensión y constante estímulo han conseguido, además de hacerme
permanecer cientos de horas (incluidos fines de semana y festivos) en el laboratorio,
mculcanne una enorme pasión porel trabajo de investigación desarrollado.
No puedo olvidar, por supuesto, expresar mi agradecimiento a los restantes miembros
del “Hogar del Lactobacilo”; a Juan Miguel, ‘socio cofundador” y compañero de las
situaciones más inverosímiles que en un laboratorio presentarse puedan (Rodríguez, 1991); a
Wagner Luiz, por su amistad y por compartir conmigo peripecias, desaguisados y muchas
horas de trabajo, jalonadas en algunas ocasiones, de pequeños éxitos; al resto de lo s miembros
del “Hogar”, a Luis por su entusiasmo y, en especial a Fernanda por su peculiar sentido del
humor y por la ayuda, comprensión y cariño demostrados en lo s momentos más dificiles de
este trabajo.
También quiero agradecer al resto de los componentes del Departamento de Nutrición
y Bromatología III su contribución al clima de amistad y trabajo que han alimentado el
desarrollo de esta tesis. A Almudena y Manuela por las agradables y terapéuticas tertulias, a

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 4/293
Olga por su colaboración en las técnicas de HPLC, esencial en la obtención de algunos de los
resultados de este trabajo. A todos, en general, agradecerles su ilusión por mantener viva la
titilante llama de la investigación y el compañerismo.
A Industrias Cabo, S.A. por su generosacolaboración al proporcionamos las muestras
de embutidos cuando fueron requeridas.
Al personal del hospital militar ‘Gómez Ulla” por su contribución al desarrollo de laspruebas inmunológicas.
A la Comunidad de Madrid por la concesión de una Becade Formación de Personal
Investigador, con la que se ha realizado parte de esta tesis. Asimismo, este trabajo ha sido
parcialmente subvencionado por el BRIDGE (Biotechnology Research for Innovation,
Development aud Growth in Europe) T-Provect on Lactic Acid Bacteria (Contract BIOT-0263)y por la Comisión Interministerial de Ciencia y Tecnología (CICYT ALI91-0255).
Por último, mi más especial agradecimiento a Cristina, por haber renunciado a más de
un fin de semana y fiestas de guardar, y por la comprensión y cariño demostrado durante todos
los años que ha durado la ejecución de este trabajo.

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 5/293
INDICE
CAPITULO 1 : EXPOSICION GENERAL DEL PROBLEMA
A INVESTIGAR
CAPITULO II: REVISION BIBLIOGRÁFICA 6
II. 1.- Utilización de agentes antimicrobianos en los alimentos;
presente y futuro 71.- Temperatura 7
2.- Actividad del agua 8
3.- pH 9
4.- Gases y atmósferas modificadas o controladas 9
5.- Ácidos orgánicos 1 1
6.- Sales de curado 1 1
7.- Antibióticos 12
8.- Radiaciones 129.- Envasado 1 3
10.- Interacción de factores 13
II.- Las bacterias lácticas y su posible empleo como conservadores de los
alimentos 14
II. 2.- Bacterias lácticas; ¡ntroducción histórica 14
1.- Características generales 17
2.- Taxonomia 20
II. 3.- Género Lactobacillus 23
3. 1.- Morfología 23
1.- Motilidad 24
2.- Tinción 24
3. 2.- Pared celular y ultraestructura 25
3. 3.- Desarrollo y características de las colonias 25
3. 4.- Necesidades nutritivas 26
¡

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 6/293
1.- Medios de cultivo 27
3. 5.- Metabolismo 28
3. 6.- Consideraciones taxonómicas; Clasificación 316. 1.- Grupo 1.- Homofermentativos obligados 32
6. 2.- Grupo II.- Heterofermentativos facultativos 33
2. 1.- Estreptobacterias atípicas 33
1.- Lactobacillus sake 33
6. 3.- Grupo III.- Heterofermentativos obligados 34
3. 7.- Ecología, habitats y biotecnología 34
II. 4.- Actividad antimicrobiana de la s bacterias lácticas 354. 1.- Actividad antimicrobiana de los ácidos orgánicos 36
4. 2.- Actividad antimicrobiana de componentes inorgánicos 37
1.-Dióxido de carbono 37
2.- Peróxido de hidrógeno 38
3.- Diacetilo 39
4.- Reuterina 39
4. 3.- Efecto antimicrobiano de las bacteriocinas y sustancias afines 40
II. 5.- Bacteriocinas; Definición 40
5. 1.- Detección de bacterias productoras de bactenocinas 41
1.- Detección de las cepas productoras 41
2.- Prueba del antagonismo directo 41
3.- Prueba del antagonismo diferido 42
5. 2.- La acción de las bacteriocinas y su diferenciación de otras posibles
causas de inhibición 42
5. 3.- Producción óptima de bacteriocinas 445. 4.- Purificación 44
5. 5.- Propiedades de las bacteriocinas de las bacterias lácticas 45
1.- Composición química 45
2.- Propiedades físicas 46
3.- Espectro de acción 46
4.- Mecanismo de acción 47
5. 6.- Regulación genética 48
II

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 7/293
5. 7.- Bacteriocinas de las bacterias lácticas 50
1.- Bacteriocinas producidas por especies del género Loctobacillus 50
2.- Bacteriocinas producidas por especies del género Lactococcus 513.- Bacteriocinas producidas por especies del género Pediococcus 51
4.- Bacteriocinas producidas por especies del género Carnobacteriwn. 52
5.- Bacteriocinas producidas por especies del género Leuconostoc 52
CAPITULO III: MATERIALES Y METODOS 53
III. 1.- Materiales 54
1. 1.- Material biológico 54
- Microorganismos empleados 54
2.- Obtención de los inmunosueros 54
1. 2.- Productos y reactivos 55
1. 3.- Material de laboratorio 55
III. 2.- Métodos 60
2. 1.- Medios de cultivo empleados en el crecimiento de lo s
microorganismos 60
1.- Medios de cultivo para el crecimiento de las bacterias lácticas 60
2.- Medios de cultivo para el crecimiento de microorganismos
indicadores distintos de las bacterias lácticas 62
2. 2.- Aislamiento y selección de las bacterias lácticas de lo s embutidos
crudos madurados 63
2. 3.- Actividad antimicrobiana de las bacterias lácticas seleccionadas 65
3. 1.- Soluciones tampón empleadas 653. 2.- Pruebas directas de antagonismo 65
1.- Prueba directa de antagonismo en pocillos 65
3. 3.- Actividad inhibidora dc lo s sobrenadantes concentradoslibres de células 66
1.- Preparación de lo s sobrenadantes 662.- Detección de la actividad inhibidorade lo s sobrenadantes
concentrados 20 veces 67
3. 4.- Efecto de la catalasa en la actividad inhibidora de las bacterias
III

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 8/293
lácticas seleccionadas 67
2. 4.- Identificación y caracterización bioquímica de algunas bacterias
lácticas con actividad antimicrobiana 681.- Morfología y tinción por el método de Gram 68
2.- Prueba de la catalasa 68
3.- Producción de gas a partir de glucosa 68
4.-Hidrólisis de la arginina 70
5.- Producción de ácido sulfhídrico 71
6.- Prueba de Voges-Proskauer 72
7.- Fermentación de carbohidratos 73
8.- Tolerancia al cloruro sádico 759.- Tolerancia al pH de 3,9 75
10.- Desarrollo a diversas temperaturas 75
2. 5.- Espectro antlinicrobiano de las sustancias inhibidoras producidas
por las cepas de L. sake números 77, 90, 148 y 180 76
2. 6.- Efecto de lacomposición del medio de cultivo en la actividad
antimicrobiana de Lactobacillus .sake 148 76
2. 7.- Crecimiento de L. sake 148, cinética del desarrollo y producción de
sustancia antimicrobiana a diversas temperaturas 777. 1.- Crecimiento de los cultivos 77
7. 2.- Parámetros cinéticos del desarrollo microbiano 78
1.- Velocidad específica de crecimiento (g) 78
2.- Tiempo de duplicación (td) 79
3.- Número de generaciones por hora (g/h) 79
4.- Determinación del peso celular seco 80
2. 8.- Purificación parcial de la actividad antimicrobiana exocelular de
L. sake 148 82
8. 1.- Cromatografía de filtración en geles 82
1.- Soluciones tampón empleadas 82
2.-Geles 82
3.- Condiciones de trabajo 83
8. 2.- Determinación de la proteina 84
2. 9.- Caracterización parcial de la actividad antimicrobiana exocelular
parcialmente purificada de L. sake 148 86
Iv

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 9/293
9. 1.- Preparación de la solución patrón de la sustancia antimicrobiana
parcialmente purificada 86
9.2.- Efecto de diversos enzimas en la actividad inhibidora de L. sake 148 87
9. 3.- Efecto de diferentes pHs en la actividad inhibidora de
L. sake 148 87
- Preparación de las soluciones tampón de distintos pHs 87
2.- Condiciones de trabajo 88
9. 4.- Cinética de tennodestrucción de la actividad inhibidora de
L. sake 148 89
1.- Tratamiento térmico 892.- Parámetros cinéticos de termodestruccion 89
9. 5.- Determinación del peso molecular de la sustancia inhibidora de
L. sakc 148 por cromatografía de filtración en Sephadex 0-50... 94
9 . 6- Electroforesis en geles de poliacrilarnida con dodecil sulfato sódico
(SDS-PAGE) 94
6. 1.- Técnica de Swank y Munkres (1971) 96
1.- Tampones, geles y soluciones empleadas 96
2.- Preparación de las muestras 983.- Preparación de los geles 98
4.- Electroforesis 99
5.- Tinción de los geles 99
6.- Determinación del peso molecular 99
6. 2.- Técnica de Laemli (1970) 100
1.- Tampones, geles y soluciones empleadas 100
2.- Preparación de las muestras 103
3.- Preparación de los geles y electroforesis 103
4.- Tinción de los geles 103
5.- Determinación del peso molecular 104
9. 7.- Determinación de la composición aminoacídica de la sustancia
antimicrobiana de L. saLe 148 porcromatografía líquida de alta
resolución (HPLC) 104
1.-Tampones y reactivos 104
y

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 10/293
2.- Digestión ácida de la muestra problema 106
3.- Preparación de los patrones 106
4.- Derivatización de los aminoácidos 1065.- Técnica de la cromatografía en HPLC 107
2. 10.- Mecanismo de acción de la sustancia inhibidora deL sake 148 108
2. 11.- Concentración inhibidora mínima de la sustancia antimicrobiana
parcialmente purificada en diversos microorganismos indicadores. 109
2. 12.-Métodos inmunológícos 109
12. 1.- Obtención de los inmunosueros 109
1.- Pauta de inmunización 109
2.- Sangría final 1103.- Obtención y conservación del suero 112
12. 2.- Inmunodifusión en geles de agarosa 113
1.- Preparación del gel 113
2.- Preparación de las placas de inmunodifusión 114
3.- Llenado de los pocillos e incubación de las placas 114
4.- Lavado y secado de los geles 114
5.- Tinción 115
12. 3.- Técnicas inmunoenzimáticas 115
1.- Técnicas del ELISA indirecto 115
2.- Técnica del ELISA indirecto clásico 115
3.- Técnica del ELISA indirecto utilizando el sistema de amplificación
biotina-avidina 119
CAPITULO IV: RESULTADOS 123
IV.1.-Evolución de las bacterias lacticas durante la maduración
de un lote de embutidos crudos curados 124
IV. 2.- Actividad antimicrobiana de las bacterias lácticas
seleccionadas 124
2. 1.- Actividad inhibidora directa 124
2. 2.- Actividad inhibidora de los sobrenadantes de lo s cultivos libres de
células 152
VI

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 11/293
2. 3.- Efecto de lacatalasa en la actividad inhibidora de las bacterias
lácticas seleccionadas 152
IV. 3.- Identificación y caracterización bioquímica parcial de las
bacterias lácticas seleccionadas por su actividad
antimicrobiana 154
1.- Morfología y tinción por el método de Gram 154
2.- Prueba de la catalasa 154
3.- Producción de CO2 a partir de glucosa 154
4.- Hidrólisis de la arginina 154
5.- Producción de ácido sulfhídrico 155
6.- Prueba de Voges-Proskañer 155
7.- Fermentación de carbohidratos 155
8.- Tolerancia al cloruro sódico 156
9.- Tolerancia al pH de 3,9 157
10.- Crecimiento a diversas temperaturas 157
IV. 4.- Espectro antimicrobiano de la sustancia inhibidoraexocelular d e l a s cepas de L. sake 77 , 90 , 148 y 180 162
IV. 5.-Efecto de la composición del medio de cultivo en la
actividad antirnicrobiana de Lactobacillus sake 148 165
IV. 6.- Parámetros cinéticos y actividad inhibidora de L. sake 148a distintas temperaturas 167
1.- Crecimiento y actividad antimicrobiana de L. sake 148 1672.- Parámetros cinéticos del crecimiento microbiano 173
IV. 7.- Purificación parcial de la actividad antimicrobiana
exocelular de L. sake 148 175
1.- Cromatografía de filtración en Sephadex G-150, G-75 y G-50 175
VII

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 12/293
IV. 8.- Caracterización parcial de la actividad antimicrobiana de
Lactobacillus sake148 182
Efecto de diversos enzimas en la actividad inhibidora. 1822.- Efecto del pH en la actividad inhibidora 184
3.- Cinética de termodestrucción de la actividad antimicrobiana
exocelular de Lactobaciltus sake 148 184
4.- Determinación del peso molecular por cromatografía de filtración en
Sephadex G-50 188
5.- Determinación del peso molecular por electroforésis en geles de
poliacrilamida con dodecil sulfato sódico 188
1.- Electroforesis según la técnica de Swank y Munkxes 1902.- Electroforesis en geles del 20% poliacrilarnida con SDS
(Laemly, 1979) 190
6.- Determinación de la composición aminoacídica de la sustancia
antimicrobiana parcialmente purificada 191
IV. 9.- Mecanismo de acción de la sustancía antimicrobiana
exocelular de Lactobacillus sake 148 194
IV. 10.- Concentración inhibidora mínima (CIM) de la sustancia
antimicrobiana exocelular de L. sake 148 194
IV. 11.- Caracterización inmunológica parcial de la sustancia
antimicrohiana de L. sake 148 19911. 1.- Análisis por inmunodifusión en geles de agarosa, de lo s inmuno-
sueros de conejos cuando se emplea como antígeno la sustancia
anfimicrobiana exocelular de L. sake 148 199
11. 2.- Detección de la sustancia antimicrobiana parcialmente purificada
de L. sake 148, mediante técnicas inmunoenzimáticas (ELISA).... 200
2. 1.- Detección por la técnica del ELISA indirecto clásico 200
Evaluación de la concentración idónea de lo s reactivos que
intervienen en el ensayo 200
2.- Determinación de la respuesta inmunológica de los conejos
frente al extracto antigénico inoculado 201
VIII

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 13/293
2. 2.- Detección de la sustancia antimicrobiana deL. sake 148 en el
medio de cultivo MM-triptosa 203
1.- Evaluación de la concentración idóneade lo s reactivos queintervienen en el ensayo 203
11. 3.- Detección de la sustancia antimicrobiana deL. sake 148 mediante
la técnica del ELISA indirecto con el sistema de amplificación
biotina-avidina 207
CAPITULO V: DISCUSION 209
V. 1.- Aislamiento de bacterias lácticas de embutidos crudos
curados 210
V. 2.- Actividad antimicrobiana de las bacterias lácticas
seleccionadas 211
2. 1.- Elección de las pruebas de antagonismo microbiano 211
1.- Pruebas directas de antagonismo microbiano 211
2.- Antagonismo de lo s sobrenadantes libres de células213
2. 2.- Actividad inhibidora de los cultivos de las bacterias lácticas
seleccionadas 215
2. 3.- Actividad inhibidora de los sobrenadantes libres de células 217
2. 4.- Efecto de lacatalasa en la actividad inhibidora de las bacterias
lácticas seleccionadas 218
y. 3.- Identificación y caracterización bioquímica parcial de las
bacterias lácticas co n actividad antimicrobiana 219
V. 4.- Espectro antimicrobiano de los sobrenadantes libres de
células de las cepas de L. sake seleccionadas, una vez
neutralizados y concentrados 225
V. 5.- Efecto de la composición del medio d e cultivo e n l a
producción de actividad inhibidora 229
Ix

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 14/293
V. 6.- Parámetros cinéticos y producción de la actividad inhibidora
de L. sake 148 a diversas temperaturas 231
V. 7.- Caracterización bioquímica parcial de la actividad
antimicrobiana de L. sake 148 232
V. &- Mecanismo de acción de la bacteriocina producida por
L. sake 148 241
V. 9.- Propiedades antigénicas de la bacteriocina de L. sake 148.. 248
CAPITULO VI: CONCLUSIONES 251
CAPITULO VII: TRABAJO FUTURo.... 254
CAPITULO VIII: BIBLIOGRAFíA 257
x

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 15/293
EXP. GRAL. DEL PROBLEMA A IN VES? lOAR
CAPITULO 1
EXPOSICION GENERAL
DEL
PROBLEMA A INVESTIGAR

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 16/293
EXP. ORAL. DEL PROBLEMA A INVESTIGAR
La masificación y complejidad de los medios de producción y distribución de
alimentos y el empleo de la refrigeración como principal método de conservación de los
mismos, obliga a pensar en el desarrollo de nuevas técnicas que, unidas a las ya existentes,
permitan, no sólo garantizar la calidad higiénica, sino prolongar la vida útil de los alimentos.
Para alcanzar estos objetivos en los últimos años se han aplicado en la carne y productos
cárnicos otras metodologías de conservación. Algunas podrían mejorarse empleando técnicas
basadas en el antagonismo biológico existente entre algunos géneros microbianos saprofitos y
los posibles microorganismos patógenos o alterantes cuya presencia se quiere eliminar.
Si se dispusiese de métodos que destruyeran selectivamente la flora patógena
potencialmente presente en la carne o si se inhibiese el desarrollo de su flora psicotrofa
alterante, la calidad higiénica y la vida útil de este alimento mejo¡-ai-ían considerablemente
(Molin y Ternstom, 1982; Shaw y Latty, 1982, 1984; Dainty, 1986). Si bien éstos objetivos
pueden cumplirse con la utilización de aditivos, antibióticos o radiaciones ionizantes, talesmétodos presentan muchos inconvenientes. Asimismo, es conveniente disponer de otras
alternativas que aumenten la calidad higiénica y la vida útil de las carnes envasadas al vacío o
de las que, a pesar de someterse a la acción del calor, sufren .u n proceso tecnológico previo que
puede permitir un desarrollo microbiano excesivo que determina una pérdida de su calidad
tecnológica cuando no un peligro sanitario para el consumidor.
Las bacterias lácticas constituyen un grupo de microorganismos que, además de no Ser
peligrosos para el consumidor, inhiben el crecimiento de los microorganismos patógenos
potencialmente presentes en la carne yen sus productos. Aunque el mecanismo íntimo de la
acción antimicrobiana de éstas bacterias no seconoce completamente, se sabe que sintetizan
metabolitos como ácidos orgánicos, peróxido de hidrógeno, etc. que desempeñan un papel
importante en la inhibición del desarrollo de otros microorganismos. Además también
sintetizan otras sustancias inhibidoras, que cuando son de naturaleza proteica reciben el
2

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 17/293
EXP. ORAL. DEL PROBLEMA A INVESTIGAR
nombre genérico de bacteriocinas.
La posible utilización de las bacterias lácticas o de las bacteriocinas que producen como
conservadores o factores de “seguridad” en un amplio número de productos alimenticios,
constituye una línea de trabajo desarrollada recientemente por algunos investigadores (Barefoot
y Klaenhamrner, 1984; Pucci y col, 1988; Daeschel, 1989: Bhunia y col.. 1991) que han
logrado purificar y, en algunos casos, caracterizar algunas bacteriocinas.
El objetivo de éste trabajo consiste en la búsqueda. identificación, purificación, y
caracterización bioquímica e inmunológica parcial de sustancias antintcrobianas elaboradas por
las bacterias lácticas procedentes de embutidos crudos madurados. Una vez purificadas y
establecidas sus características bioquímicas se estudiará su posible utilización como factores de
seguridad que incrementan la calidad higiénica y la vida útil de la carne y de los derivados
cárnicos. Presumimos que, como factores de “seguridad”, podrían utilizarse tanto las bacteriaslácticas inhibidoras, como las sustancias antimicrobianas que producen, una vez que hayan
sido purificadas.
Para lograr los objetivos propuestos se necesita desarrollar el programa de trabajo que
se describe a continuación:
1v).- De lo s embutidos crudos madurados, que son teóricamente los sustratos cárnicos
más adecuados pal-a el desarrollo de bacterias lácticas productoras de sustancias inhibidoras, se
seleccionará al azar un número de colonias microbianas cuyo potencial inhibidor del desarrollo
de otros microorganismos será convenientemente evaluado.
29.- El antagonismo bacteriano se estudiará tanto en las colonias aisladas como en el
medio extracelular.
3

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 18/293
EXP. CHAL. DEL PROBLEMA A INVESTICAR
39).~ Los grupos microbianos frente a los que se evaluará el potencial inhibidor de las
bacterias lácticas escogidas serán: bacterias lácticas de diversos origenes (leche, carne,
hortalizas), micrococáceas, enterococos, Pseudonionas sp. psicotrofas de la carne refrigerada,
microorganismos de las carnes envasadas al vacío (Brochothrix therínosphacta y
enterobacteriáceas) y microorganismos patógenos diversos como E. coli enteropatógeno,
Staphylococcus orn-cus, Lisreria tnonocyíogenes, Sairnonella spp., Clostridium boutulinum y
Clostridiuni peifringens.
4$.- Mediante técnicas microbiológicas adecuadas se identificarán el género y la
especie de las bacterias lácticas con mayor poder antagonista.
5Q).~ Se evaluará el efecto de diversos parámetros como composición del medio de
cultivo, pH, temperatura, tiempo de crecimiento, etc, en la síntesis de las sustancias
inhibidoras.
6$.- Mediante técnicas de separación de filtración en geles se intentará purificar la
actividad inhibidora extracelular de la que resulte más interesante.
79.- Se procederá a la caracterización bioquímica parcial de L a sustancia inhibidora
seleccionada, determinando su peso molecular y su sensibilidad al pH, al calor, y a los
enzimas proteolíticos. lipolíticos y amilolítícos.
8~).- A partir de conejos se obtendrán anticuerpos policlonales frente a la bacteriocina
parcialmente purificada.
9$.- Finalmente se establecerá, mediante técnicas inmunoenzimáticas (ELISA). la
capacidad de lo s anticuemos obtenidos para caractenzar inmunológicamente dicha bacteriocina
4

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 19/293
EXP. GRAL. DEL PROBLEMA A INVESTIGAR
que se comparará con las producidas por otras bacterias lácticas. También se estudiará la
utilidad de los anticuerpos en la posible detección y cuantificación de la bacteriocina en medios
de cultivo liquidos y en sustratos cárnicos.
5

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 20/293
REVISION BIALIOGRAFIC’A
CAPITULO J I
REVISION
BIBLIOGRAFICA
6

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 21/293
REVISION il/BLIOGRA PICA
II. 1.- Utilización de agentes antimicrobianos en lo s alimentos: presente y futuro
Uno de los mayores problemas con lo s que el hombre se ha enfrentado a lo largo de su
historia ha sido la necesidad de evitar, de una forma u otra, la alteración de los alimentos,
incrementando su vida útil. Para solventarlo ha utilizado los materiales y técnicas a su alcance
como miel, sal, especias, ahumado, desecación y otros sistemas sencillos de conservación. La
mayor parte de los mismos se siguen empleando actualmente, en algunos casos sin variación
tecnológica alguna y, en otros, con las modificaciones necesarias introducidas por losadelantos técnicos. A medida que crecen los conocimientos sobre los alimentos y la causa de
su alteración, aumentan también las metodologías necesarias para evitar ésta última. Los
métodos empleados para evitar la alteración de lo s alimentos e incrementar su vida útil se basan
en la utilización de:
II. 1. 1.- Temperatura
La temperatura desempeña un papel muy importante en el desarrollo bacteriano y al
modificarla se retrasa e incluso se elimina el desarrollo de los microorganismos patógenos y
alterantes (ICMSF, 1980). Las temperaturas de refrigeración sólamente retrasan el crecimiento
de los microorganismos mesófilos o termófilos, permitiendo el desarrollo de los psicrófilos,
entre los que se encuentran muchos alterantes de los alimentos y otros patógenos, por lo que la
refrigeración no se puede considerar, por sí sola, como un método que permita asegurar unos
alimentos sanos, en periodos de tiempo que superen una o, en el mejor de los casos, dos
semanas.
Si la temperatura es tan baja que tiene lugar la congelación del agua, incluso se inhibe
el desarrollo de lo s microorganismos psicrófilos, permitiendo la conservación de lo s alimentos
durante largos periodos de tiempo (Speck y Ray, 1977). Pero la utilización de temperaturas de
7

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 22/293
REVISION BIBLIOGRA PICA
congelación no implica la eliminación de los microorganismos patógenos o alterantes que
pueden permanecer viables (Obafemi y Davies, 1986). Además la congelación provoca daños
celuLares en las bacterias aunque. con el tiempo, también ocasiona modificaciones perjudiciales
en los alimentos.
El empleo de temperaturas altas permite la destrucción de la mayor parte de los
microorganismos (pasterización) asi como de sus formas de resistencia (esterilización), sin
embargo no pueden aplicarse a todos los alimentos por que modifican su estructura y/o suscaracterísticas organolépticas.
La temperatura ambiental puede emplearse como factor de seguridad si favorece el
desarrollo de ciertos microorganismos que, además de modificar los caracteres organolépticos
de los alimentos, mejoran su calidad microbiológica. Los procesos más representativos de la
industria alimentaria que tienen lugar a temperatura ambiente son los que se basan en lafermentación láctica y/o alcohólica desarrollada por microorganismos distintos (Gibbs, 1987).
II. 1 . 2.- Actividad del agua
El desarrollo de los microorganismos depende, invariablemente, de la presencia de
agua libre (ICMSF, 1980). La actividad del agua (a~) de un alimento orienta, objetivamente,
de la cantidad de agua de que disponen los microorganismos para su desarrollo. La
determinación de la a~ limitante del desarrollo de un microorganismo dado, es importante para
pronosticar su viabilidad en un alimento concreto. La aw disminuye al aumentar la
concentración de solutos (azúcares o sales) y al eliminar el agua disponible para el desarrollo
microbiano por calentamiento o liofilización.
8

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 23/293
REI’ISION BIBLIOGRA PICA
IL 1.3.- p1 1
Al igual que ocurre co n la temperatura, los microorganismos poseen un pH mínimo,
óptimo y máximo para su crecimiento. El pH de lo s alimentos es uno de lo s factores que
determinan los microorganismos que se desarrollan en ellos, originando su alteración, una
fermentación deseable o un riesgo para la salud del consumidor.
La disminución natural o artificial del pH se ha utilizado, durante cientos de años, paragarantizar la estabilidad microbiológica de los alimentos. Como ejemplos de disminución
natural del pH en conservación de alimentos citaremos las fermentaciones controladas de la
leche, carne y hortalizas, que originan productos tan estables y seguros como el queso, yogur,
embutidos y encurtidos. La acidificación artificial de los alimentos se consigue añadiéndoles
ácidos orgánicos.
Los alimentos ácidos (pH<4,6). aunque pueden alterarse, es muy dificil que permitan
la proliferación de lo s microorganismos patógenos o la germinación de esporas, en cambio
esto si ocurre en los alimentos débilmente ácidos (pH>4,6) cuyo pH no garantiza, por sí
mismo, su estabilidad microbiológica (Tanaka y col., 1986).
II. 1. 4. - Gases y atmósferas modificadas o controladas
Durante los últimos años se ha generalizado el empleo de ciertos gases como
conservadores de lo s alimentos. En la acción inhibidora de estos gases influyen, además de la
naturaleza del gas, un gran número de factores relacionados con el alimento, el proceso
tecnológico utilizado y el tipo de microorganismo implicado. Así porejemplo. cl CO2 inhibe el
desarrollo de un número variable de microorganismos, dependiendo de la concentración del
gas, temperatura de almacenamiento del alimento y actividad de agua del medio. En general, se
9

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 24/293
REVISION BII)LJOORAFICA
considera que las mezclas gaseosas con un 10% de CO2 inhiben el desarrollo del 50% de lo s
microorganismos (Ledward y col., 1971). Este gas se emplea para inhibir el crecimiento de
microorganismos en productos envasados al vacío y en atmósferas modificadas; sin embargo,
la evidencia de que el CO2 puede favorecer el crecimiento de algunos esporulados, como lo s
del género Clostr¡diurn, hace necesario un estudio previo de las características del alimento en
el que se va a utilizar
Otros gases experimentados han sido; el monóxido de carbono (CO) (Davidson y col.,
1983), que se ha comprobado que inhibe el desarrollo de psicotrofos, hongos y levaduras
(Clark y Takacks, 1980) pero cuya toxicidad para quienes lo manipulan desaconseja su
empleo, y el N2 que inhibe el desarrollo de lo s mohos. El dióxido de azufre (SO2) es efectivo
frente a bacterias, hongos y levaduras (Dziezak, 1986), por formar sulfitos con los
compuestos orgánicos de las soluciones acuosas que inhiben el desarrollo microbiano. Sinembargo, solamente es activa la forma iónica que se produce a pH=4.de otro lado originaen
las personas sensibles reacciones de hipersensibilidad, por lo que no es aconsejable su uso en
los alimentos. El óxido de etileno también se ha empleado para reducir el número de
microorganismos de lo s alimentos, pero su elevada toxicidad limita su utilización. El óxido de
propileno, menos estudiado que el anterior, también se ha empleado para inhibir el desarrollo
microbiano (Skinner y Hugo, 1976). Y, por último, citaremos el ozono, cuyo empleo selimita a la esterilización de líquidos (Torricelíl, 1959).
Las atmósferas modificadas o controladas se consiguen mediante la mezcla de varios
gases que se introducen en el interior del envase que protege al alimento, pudiéndose mantener
constante la mezcla gaseosa inicial (atmósferas controladas) o no (atmósferas modificadas). En
lo s alimentos envasados al vacío lo que se hace es extraer el aire del interior del alimento
empaquetado. El objetivo de lo s métodos descritos es limitar el desarrollode la flora alterante
10

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 25/293
REVISION BIBLIOGRA PICA
de los alimentos. El problema es que con ello aumenta la posibilidad del desarroLlo de
anaerobios como C. botulinum.
II. 1 . 5.- Acidos or2ánicos
A los ácidos orgánicos se les considera los antimicrobianos naturales más difundidos
en la naturaleza. Los más empleados en la industria alimentaria son el acético, benzoico,
propiónico y sórbico. El ácido acético y sus sales ejercen su mayor acción inhibidora a pH
próximo a 4,5. El ácido benzoico es más eficaz en el control de mohos y levaduras que en el de
bacterias (Chichester y Tanner, 1972). Generalmente en los alimentos se emplea la sal sódica.
El ácido propiónico y sus sales inhiben a los mohos, pero muy poco a bacterias y levaduras. El
ácido sórbico es activo en alimentos cuyo p l - ] está próximo a 6,0. Generalmente los sorbatos
son más efectivos en las levaduras y mohos que en las bacterias (Ldck. 1976).
II. 1 . 6.- Sales de curado
El curado es uno de los métodos más antiguos de conservación de los alimentos;
inicialmente se realizaba de forma empfrica co n sa l común contaminada con nitro o salitre, más
tarde, a la sal común se le añadía nitrato sódico; las impurezas o contaminantes del cloruro
sódico, es decir el salitre, eran lo s responsables de la aparición de la tonalidad rosa-rojiza típicade los alimentos curados. Los nitritos, procedentes de la reducción de lo s nitratos, originan
NO que reaccionando con la mioglobina dom a lo s alimentos curados de un color atractivo para
el consumidor e impide el desarrollo bacteriano pero, a su vez, participa en la fonnación de
nitrosaminas cancerígenas por lo que el empleo de nitratos y nitritos está regulado por la
legislación alimentaria.
1 1

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 26/293
REVISION BIBLIOGRA PICA
II. 1 . 7.- Antibióticos
Los antibióticos, producidos por microorganismos o por síntesis química, inhiben el
desarrollo de muchos microorganismos incluso a concentraciones pequeñas. Sin embargo, su
presencia en los alimentos y lo s riesgos que ello conlíeva para el consumidor, limitan su
empleo. La nisina, un polipéptido producido por Lacrococcus facÉis, ha sido aceptado como
aditivo para su empleo en los alimentos por la comunidad científica internacional desde 1956. y
sus condiciones de empleo sc encuentran reguladas por la ED A (Food and Drug
Administration) (CFR, 1988).
En los últimos años se ha descrito la síntesis por las bacterias lácticas, de sustancias
antimicrobianas de naturaleza proteica. denominadas bacteriocinas, cuyas propiedades han
permitido concebir grandes esperanzas sobre su potencialidad como conservadores de lo s
alimentos.
U. 1 . 8.- Radiaciones
Las radiaciones ionizantes se caracterizan por su gran poder de penetración y su
letalidad a nivel celular (ICMSF. 1980). La aplicación práctica de las radiaciones ionizantes
para destruir microorganismos en productos alimenticios se ha desarrollado en las últimasdécadas. Durante los últimos 30 años, el empleo de las radiaciones ha sufrido constantes
cambios. Así, mientras a principios de los 60 se permitía su empleo en algunos alimentos, un
poco más tarde la administración americana las prohibió, basándose en que quizá éste método
de conservación pudiera inducir la formación de compuestos potencialmente mutagénicos,
cancerígenos o teratógenos. Por último, lo s resultados de múltiples estudios han demostrado la
seguridad de éste método: de ahí que la Comisión de expertos de la FAO/WHO haya
recomendado su empleo como procedimiento de conservación de los alimentos
12

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 27/293
REVISION BIRLIOORA PICA
(FAO/IAEA/WHO Expert Committee. 1977).
La utilización de las radiaciones ionizantes presenta una serie de ventajas sobre otros
métodos higienizantes de conservación de los alimentos ya que, a dosis pequeñas (<0,5 Mrad)
no se producen cambios organolépticos apreciables en los alimentos e incluso a dosis más
elevadas (>1 Mrad), los cambios químicos son mínimos. Además, la penetración de la
radiación es instantánea, homogénea y profunda, permitiendo un control exacto del proceso, lo
que no ocurre en lo s tratamientos térmicOs.
II. 1 . 9.- Envasado
El envase que envuelve a lo s alimentos durante su almacenamiento, transporte y
manipulaciones posteriores, les proporciona una protección; química, impidiendo el paso de
agua, oxigeno y otros gases: física, al proteger al alimento de la luz, polvo o suciedad, de lapérdida de peso y de las alteraciones de origen mecánico y aú n biológico, al prevenir la entrada
de microorganismos e insectos. Los factores de lo s que depende el desarrollo microbiano en
los alimentos envasados son, aparte de la naturaleza del alimento y del tipo de
microorganismo, la permeabilidad del material del envase al 02, CO2 y vapor de agua, sobre
todo cuando en el envasado se utilizan el vacío o las atmósferas modificadas (Calvet, 1968).
II. 1 . 10.- Interacción de factores
Ninguno de lo s métodos descritos pueden, aisladamente, garantizar la calidad higiénica
de los alimentos, lo que lleva a eniplearlos conjuntamente para conseguir una actividad aditiva
o sinérgica.
1 3

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 28/293
REVISION BIBLIOGRA PICA
II. 1. II.- Las bacterias lácticas y su posible empleo como conservadores de los
alimentos
Los mecanismos mediante los que las bacterias lácticas ejercen su acción aniinicrobiana
se describirán en la sección II. 4. Las bacterias lácticas reunen una gran parte de las actividades
antimicrobianas utilizadas actualmente para la conservación de los alimentos en la industria
agroalimentaria; además poseen otros mecanismos que, si bien aisladamente no tienen gran
importancia, conjuntamente contribuyen a mejorar la efectividad antimicrobiana.
La síntesis por las bacterias lácticas de productos metabólicos finales como el CO2,
H2 02 y los ácidos orgánicos, el descenso del pH que ello conlíeva y la producción de
sustancias antimicrobianas como la reuterina y las bacteriocinas, hacen que, cada vez más, se
profundice en el estudio de las bacterias lácticas que han sido seleccionadas por su actividad
antimicrobiana, para su empleo como conservantes de los alimentos. (Friend y col.. 1983;
Raccach. 1981; Rubin y col., 1982)
II .2.- Bacterias lácticas: introducción histórica
Bajo el nombre genérico de “bacterias lácticas” se agrupan un número de
microorganismos cuya principal característica es la producción de ácido láctico a partir de
hidratos de carbono fermentables. El término “Bacterium acidi lactici” se debe a WeiEmamn
que lo propuso en 1899, aunque las primeras referencias a este tipo de bacterias se deban a
Hueppe (1884), quien describió una parte de la flora microbiana responsable de la acidificación
de la leche y productos lácteos, a la que se denominó “Milchsauerbacillus”.
Las bacterias lácticas se encuentran en los más variados habitats, entre los que se
1 4

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 29/293
REI’LSVON BIBLICORA PICA
incluyen la leche y productos lácteos, carne y productos cárnicos, pescado, productos
vegetales, tracto gastrointestinal y urinario de personas y animales e, incluso, aguas residuales
y estiercol. Esta diversidad ecológica también origina una diversidad morfológica y fisiológica
de las especies de este grupo, lo que entraña una gran dificultad para establecer los rasgos de
semejanza que permiten reunir a todas las especies en un grupo homogéneo desde el punto de
vista taxonómico.
Orla-Jensen (1919) fué responsable de lo s primeros intentos de demarcación de éstegrupo microbiano, y son esos mismos criterios lo s que constituyen, todavía en la actualidad,
las bases de su clasificación:
1).- Criterios morfológicos: El grupo estada constituido por cocos y bacilos Gram-
positivos, no esporulados e inmóviles.
2).- Criterios fisiológicos: Microorganismos fermentativos con producción final de
ácido láctico, catalasa-negativos, microaerófilos o anaerobios, mesófilos, y con complejos
requerimientos nutritivos.
A pesar de que las características esenciales no se han alterado desde su
establecimiento por Orla-Jensen, s i se ha modificado la nomenclatura del grupo. Así por
ejemplo, de los seis géneros establecidos inicialmente (Bembaceiun;, Streptobacter¡un¡,
Thermobaceriuni, Betacoccus, So-eptococcus y Teti-acoccus) (Orla-Jensen. 1919), sólamente
se mantiene el género Sffepwcoccus. Los otros 5 han sido sustituidos por lo s géneros
Lac~robaciIIus (englobaría a Betabacterium, Sueptobacu’riuni y Thcrmobacteriu,n ),
Leuconoswc y Pediococcus. Estos cambios en la asignación de los nombres genéricos no son
más que un reflejo del dinamismo y fluidez de lo s criterios de clasificación, ya que, a medida
que se producen avances en las metodologías analíticas y en la filosoña de su clasificación,
15

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 30/293
REVISION BIBLIOORAFICA
también se producen cambios en la denominación de géneros y especies, trasvase de especies
de unos géneros a otros y aumento del número total de especies. Como mejor se aprecia esta
evolución es observando las especies que se describen en las tres últimas ediciones del manual
de Bergey (Tabla I I . 1) (Breed y col., 1957; Buchanan y Gibsons, 1974; Kandler y Weiss,
1986).
Tabla II. 1.- Modificaciones en el número de especies de las bacterias lácticas
N ~ d e e s p e c i e s l i s t a d a s e n e l m a n u a l d e Be rg e v e n
l a s e d i c i o n e s d e
1 9 5 7 1 9 7 4 1 9 8 6Bacterias
B a c i l o s
Gén. Lactobacillus
Cocos
Gén.
Gén.
Gén.
1 5
Streptococcus
Pediococcus
Leuconostoc
1 9
2
3
N 9 t o t a l d e e s p e c i e s 3 9
2 7
2 1
5
6
59
45
29
8
4
86
1 6

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 31/293
REVISION BIBLIOGRA PICA
II . 2. 1.- Características generales
L o s m i c r o o r g a n i s m o s d e es t e gr u p o s e c a r a c t e r i z a n pri n c i pa l m e n t e p o r s e r b a c t e r i a s
Gram-positivas, catalasa-negativas, no esporuladas, inmóviles, anaerobias o microaerófilas,
ácido-tolerantes, no reductoras del nitrito y p r o d u c t o r a s d e á c i d o l á c t i c o como p ro d u ct o f i n a l d e
l a f e rm e n t ac ió n d e l o s h i d r a t o s d e c a r b o n o .
S in e m b a r g o l a t i n c i ó n d e Gram e s l a ú n i c a c a r a c t e r í s t i c a d e f i n i t o r i a , e xcl u yé n d o se d e l
gr u p o t od o m i c r o o r g a n i s m o Gra m- n e gat ivo. L a s o t r a s , n i s o n e x c l u s i v a s d e l a s b a c t e r i a s
l á c t i c a s , n i s o n con st ant es de nt ro d e l g r u p o . A s í p or e j e mp l o, a p e s a r d e con sid e ra rse
c a t a l a s a - n e g a t i v o s , s e c o n o c e d e sd e h ac e t ie m p o l a s r e a c c i o n e s c a t a l a s a - p o s i t i v a s d e l o s
Pediococcus ( F e l t o n y c o l . , 1 9 5 3 ) e incluso de otros géneros, incluido el Lactobacillus
( W h it t e n b u r y , 1 9 6 4 ) .
Las bacterias lácticas son generalmente inmóviles, aunque se han descrito un gran
número de cepas móviles con flagelos peritricos, especialmente cuando proceden de medios
diferentes de la leche y sus derivados; así se han descrito en la carne (Deibel y Niven, 1958),
en zumos de frutas (Hays y Reister, 1952) y en diversos productos fermentados (Vankova,
1957). El carácter microaerófilo es muy variable (Whittenbury, 1963), considerándose que es
la tensión de CO2. más que la anaerobiosis. la que favorece el desarrollo de las bacterias
lácticas.
La reducción de lo s nitratos a nitritos no se detecta en lo s medios que se emplean en el
desarrollo de éstas bacterias (Rogosa, 1961). La concentración de glucosa (2%) de estos
medios origina una elevada producción de ácido láctico y, consecuentemente, un gran
descenso de pH que inhibe la reducción. La capacidad reductora sólamente se aprecia
empleando medios de cultivo con un 0,1-1% de glucosa (Rogosa. 1961). De esta manera se ha
1 7

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 32/293
REVISlON I3IBLIOGRA PICA
visto que la mayor parte de las bacterias lácticas aisladas de carnes curadas reducen los
nitratos.
La producción de ácido láctico o, más exactamente, la síntesis de este metabolito como
producto final de la fermentación de lo s hidratos de carbono, es uno de lo s principales criterios
para la diferenciación fisiológica de lo s distintos componentes del grupo. Las bacterias lácticas
se dividen en tres grupos fisiológicos distintos en virtud de sus diferencias enzimáticas (Buyze
y col., 1957):
1$.- Heterofermentativas obligadas: poseen los enzimas glucosa- 6-fosfato
deshidrogenasa y 6-fosfogluconato deshidrogenasa, pero carecen de la fructosa difosfato
aldolasa, originando como productos finales de la fermentación de las hexosas CO2 y ácido
láctico. Como fuente de hidratos de carbono sólamente pueden utilizar las hexosas.
2v).- Homofermentativas obligadas: poseen fructosa difosfato aldolasa, pero carecen
de las dos deshidrogenasas. Al igual que las anteriores son incapaces de utilizar las pentosas
como fuente de hidratos de carbono.
3$.- Homofermentativas facultativas: poseen las dos deshidrogenasas pero
metabolizan la glucosa por la vía de Embden-Meyerhof (glicolisis); utilizan las pentosas y se
corresponden con las Streptobacteria de Orla-Jensen.
Los esterolsómeros (D, L, DL) del ácido láctico resultante de la utilización de los
hidratos de carbono sirven como criterios de clasificación del grupo. Las especies del género
Leuconosíoc producen el isómero D(-) exclusivamente, mientras que lo s lactobacilos
heteiofermentativos producen la fonna racémica (DL). Esta diferenciación bioquímica se refleja
en la Tabla II. 2.
18

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 33/293
REVISION BIBLIOGRA PiCA
Tabla II. 2.- Diferenciación bioquímica de las bacterias lácticas
Tipo de Producto final Configuración
Género fermentación del metabolismo del ácido láctico
Streptococcus Homofermentativa ácido láctico L(+)
Pediococcus Homofermentativa ác. láctico DL, L(+)
Lactobacillus:
Thermobac’terium Homofermentativa ác. láctico D(-), L(+), DL
Streptobacteriwn H om of e rm e n t at iv a á c . l á c t i c o D (- ) , L ( ± ) ,DL
Heterofermentativa (1) ~ láctico: ác . acético
(1:1) D(-), L(+), DL
Retabacterl ¡¡n i Heterofermentativa ác. láctico:ác. acético:C02
(1:1:1) DL
Leuconostoc Heterofermentativa ác. láctico:ác. acético:C02
(1:1:1) D (— )
(1 ) Utilización d e pentosas.
Fuente: Kandler, 1983.
19

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 34/293
REVISION BIBLIOGRAFICA
IL 2. 2.- Taxonomia
La clasificación de las bacterias lácticas según la última edición del manual de Bergey
Kandler y Weiss, 1986) se muestra en la Tabla II . 3. Los Grupos 1 , 1 1 y 1 1 1 sustituyen a las
denominaciones de Therrnobacreriurn, Streptobacreriuní y Betabacíeriurn establecidas por
Orla-Jensen. Sin embargo, desde la publicación de ésta edición en 1986, y gracias a la
flexibilidad taxonómica y modificaciones de lo s criterios de clasificación anteriormente citados,
ya se han producido nuevos cambios en la taxonomía de este grapo.
Recientemente (Schillinger y Holzapfel, 1990), se ha propuesto la creación de un
nuevo género, denominado Carnobacteriurn, integrado por especies relacionadas con los
lactobacilos pero con peculiaridades diferenciadoras, como son el no poder desaiTollarse en
agar acetato, y hacerlo en medios con un pH alto (pH 8,5-8,9) y el sintetizar ácido oleico en
lugar de ácido cis-vaccénico. Actualmente Lactobacillus divergens y L. piscicola sedenominan, respectivamente, Carnobacteriuní divergens y C. piscicola, mientras C. mobile y
C. gauinarurn son los restantes componentes del nuevo género.
La sugerencia de separar a los estreptococos lácticos (tipo N de Lancefleld) del resto de
los estreptococos, creando con ellos un nuevo género denominado Lactococcus (Schleifer y
col., 1985), cristalizó en 1986 con la validación del nuevo género por la “International Unionof Microbiological Societies”. En éste caso, los criterios empleados para establecer la nueva
clasificación resultaron de tecnologías basadas en la hibridación de ácidos nucleicos, afinidad
ininunológica, composición lipídica y características químicas de la pared bacteriana (Sandine,
1988).
20

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 35/293
REVíSlON RIIJLIOORA PICA
Tabla II. 3.- Taxonornia de las bacterias lácticas(a)
Género N9 de especies
Lactobacillus
Grupo 1 : Homofermentativos obligados 16
Grupo II : Heterofermentativos facultativos 1 1
Grupo III: Heterofermentativos obligados 18
Strepíococcus
Estreptococos piógenos 5
Estreptococos orales 9
Enterococos (tipo D de Lancefield) 4
Estreptococos lácticos (tipo N de Lanceñeld) 2
Estreptococos anaeróbicos 4
Otros Estreptococos 5
Pediococcus 8
Leuconostoc 4
N 2 total de especies 86
(a) Fuente: Kandlery Weiss, 1986; Hardie. 1986 yGarvie, 1986a, ]986b.
2 1

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 36/293
REVISION BIBLIOGRA PICA
Estas modificaciones taxonómicas se muestran en la Tabla 11 -4 : actualmente el grnpo
de las bacterias lácticas está constituido por microorganismos pertenecientes a lo s géneros
Lactobacillus, Lactococcus, Carnobaceriun¡, Pediococcus y &U¿co,,ostoc
Tabla II. 4.- Modificaciones recientes en la taxonomia de las bacterias lácticas
Nueva denominación
Género Loctococcus
Lactococcus Itictis subsp.
Lactococcus ¡cicÉ is subsp.
Lactococcus ladis subsp.
Lactococcus lactis subsp.
Lactococcus. garv¡ae
Lactococcus plantaruní (1)
Loctococcus raifinolacús
Género Carnobacíeriuní
Carnobacteri uní divergens
Carnobacterium piscicola
Car,¡obacterium niobile
Carnobacteriun, gaulinrn-un2
It,ctis
lacas
cremoris
hordinae
Antigua denominación
Streptococcus lactis subsp. lactis
Streptococcu.s ¡o c fis subsp. diacetvlactis
Streptococcus lacris subsp. crernoris
Lactobacillus hordince
Streptococcus garx’¡ae
Streptococcus planíau-u¡n
Streptococ-cus raifinolactis
Lociobacillus divergens
Lactobacillus piscicola
(1) No confundir con Lactobacillus plan taruní
Fuente: Sandine, 1988 y Schillinger y Holzapfel, 1990
22

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 37/293
REVISION BIBLIOGRA PICA
I I . 3.- Género Locrobacillus
Realizar una descripción profunda de los componentes de las bacterias lácticas
resultaría larga, tediosa e incluso improcedente, dado el tema central de este trabajo. Sin
embargo, si es interesante conocer algunas de las características morfológicas, fisiológicas y
bioquímicas más relevantes del género Lactobacillus.
La última edición del “Bergev’s Manual of Svstematic Bacteriologv” (1986) clasifica algénero Lactobacillus en el volumen 2. sección 14 como “bacilos Gram-positivos no
esporulados”. La sección comprende 7 géneros (Lacrobacillus, ErysipelothrLr. Brochothrix,
Listeria, Kurthia, Ca¡yophanon y Renibacrerium) que poseen en común ser bacilos
Gram-positivos, no esporulados, mesófilos, y que requieren medios complejos para su
crecimiento.
II. 3. 1.- Morfología
La morfología de los lactobacilos es variable, desde bacilos largos, rectos o
ligeramente curvados, hasta cocobacilos. Su longitud y curvatura depende de la edad del
cultivo, la composición del medio y la tensión de oxígeno. Como ejemplo de variabilidad en su
morfología podría citarse a L. delbrueckii subsp. buí garicus que en medios de cultivo líquidos
o en agar MRS crece en formaciones espirales debido a que las células permanecen unidas
después de la división, mientras que en la leche origina cadenas largas y filamentosas.
Según Bottazzi (1988), los distintos lactobacilos podrían distinguirse por sus
diferencias morfológicas. Así por ejemplo, lo s lactobacilos mesófilos son bacilos cortos,
gruesos, regulares y unidos, formando filamentos (Srreptobacteria ). Las especies wrmóñlas
23

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 38/293
REVISION BiBLIOGRA PICA
son bacilos largos y rectos, de células separadas, en parejas o en empalizada. En las especies
heteroferínentativas obligadas, la morfología típica es la de bacilos cortos, rectos y separados.
Algunas especies de lactobacilos productores de gas (L. ferrnentum o L. brevis) pueden
presentar una morfología mezcla de bacilos largos y cortos, en el mismo cultivo e, incluso, en
una misma colonia (Kandler y Weiss. 1986).
Los cocobacilos pueden ser tan extremadamente cortos que pueden llegar a
confundirse con los Leuconostoc o estreptococos. De otra parte, algunos estreptococosligeramente alargados se han confundido durante mucho tiempo con lactobacilos (Garvie y
col., 1981). La división celular de lo s lactobacilos tiene lugaren un solo plano, mientras que la
tendencia a formar cadenas varía entre las distintas especies e, incluso, entre las distintas
cepas, dependiendo de la fase de desarrollo y del pH (Rhee y Pack, 1980).
1 1 . 3. 1 . 1.- Motilidad
El género Lactobacillus comprende bacterias inmóviles, aunque algunas especies se ha
visto que son móviles por poseer flagelos peritricos. En cualquier caso su motilidad depende
del medio y de la edad del cultivo, y sólo se observa en el momento del aislamiento,
perdiéndose al resembrarías en diversos medios de cultivo.
1 1 . 3. 1 . 2.- Tinción
Quizá la característica común de éste género (al igual que en las bacterias lácticas en
general) sea la reacción positiva a la tinción de Gram. Sólo las bacterias de cultivos viejos y las
deterioradas muestran resultados variables. Si se emplea la tinción de Gram o la del Azul de
Metileno pueden observarse granulaciones internas, sobre todo cuando se trata de lactobacilos
homofermentativos que forman bacilos Largos. Observados con microscopia electrónica
24

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 39/293
RE VLSION BIBLIOORA PICA
En los medios de cuhivo líquidos las bacterias sediiíientan rápidamente cuando cesa su
desarrollo. El sedimento es liso y homogéneo, siendo muy rara la presencia de precipitados
granulares o mucilaginosos. Los lactobacilos no forman películas superficiales ni originan
olores específicos durante su crecimiento en lo s medios de cultivo ordinarios, a pesar de
contribuir al desarrollo del aroma de lo s alimentos fermentados. Este aroma es consecuencia de
la producción de compuestos volátiles, como diacetilo y derivados. 1 - 1 2 5 y aminas, que se
liberan al medio durante el desarrollo microbiano (Sharpe y Franklin, 1962).
II. 3. 4.- Necesidades nutritivas
El género Lactobacillus, al igual que el resto de las bacterias lácticas, tiene unas
necesidades nutritivas muy complejos ya que necesitan como fuente de energia y de carbono,
además de hidratos de carbono. nucleótidos, aminoácidos y vitaminas. Todas las especies del
género, (salvo algunas cepas) requieren ácido pantoténico y nicotínico, mientras que la tiamina
sólo es necesaria para el desarrollo de los lactobacilos heterofermentativos. Las necesidades de
vitaminas son variables dependiendo de la especie, pero la mayoríanecesita riboflavina y, muy
pocas, biotina o vitamina B2.
A pesar de necesitar medios muy complejos para su desarrollo, la mayor parte de lo scomponentes del género (sino todos) tienen en su genoma la información necesaria para
sintetizar todos los nutrientes citados. Además, se ha visto que los mutantes de L. casei
sintetizan determinados aminoácidos en medios carentes de los mismos (aunque crecen mucho
más lentamente), dicha capacidad la pierden cuando crecen en medios más complejos, lo que
acelera su velocidad de crecimiento (Morishita y col., 1974). Estos complejos requerimientos
nutritivos de los Lactobacilos actuales reflejan la selección natural de inutantes deficientes endeterminados rutas biosintéticas. lo que les permite crecer más rápidamente que las cepas
26

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 40/293
REVISION BII3LIOCRA PICA
originarias en medios de cultivo con muchos nutrientes
II. 3. 4. 1.- Medios de cultivo
Los medios empleados para el crecimiento de lo s lactobacilos cubren sus necesidades
nutritivas y están constituidos por hidratos de carbono fermentables, extractos de carne y
extracto de levadura. Además, la mayor parte de las especies necesitan otros compuestos
específicos que favorezcan su crecimiento; en algunos casos son esenciales para el desarrollo,tal como ocurre con el jugo de tomate, manganeso, acetato y ésteres del ácido oleico
(especialmente Tween 80). Los compuestos citados, excepto el jugo de tomate, forman parte
del medio de cultivo MRS (De Man y col., 1960) que es. sin duda, el más utilizado para el
crecimiento de los lactobacilos.
Por otra parte, algunos lactobacilos adaptados a sustratos muy particulares necesitan
factores de crecimiento especiales. Algunas especies aisladas del vino de arroz, necesitan ácido
D-mevalónico, mientras que L. sanfranciscus necesita un péptido de bajo peso molecular
aislado del extracto de levadura (Berg y col., 1981).
El pH del medio también influye en el crecimiento que es mayor en medios ligérarnente
ácidos, cuyo pH inicial varía de 6,4 a 4,5. El crecimiento se detiene cuando se alcanza un pH
de 4,0 a 3,6 dependiendo de la especie y cepa. La aireación también es importante ya que,
aunque la mayor parte de las cepas se desarrollan en condiciones normales de aerobiosis, el
crecimiento será óptimo bajo condiciones de anaerobiosis o microaerofilia. Pero, más que la
anaerobiosis, lo que influye en el crecimiento es la tensión de CO2 (más incluso que la tensión
de 02 ) , considerándose óptima la presencia de un 5-10% de CO2 (Kandler y Weiss. 1986).
27

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 41/293
R IP VISION BII3LIOGRA PICA
La temperatura de incubación óptima, que fué uno de los criterios iniciales de
clasificación de las bacterias lácticas (Orla-Jensen, 1919), también varía con la especie. La
mayor parte de las especies (mesófilas) tienen una temperatura óptima de crecimiento de 25-30
con un límite superior de 40 0C. Algunas crecen a temperaturas menores de 1 5 0C y, muy
raramente, a menos de 5 0C siendo el limite inferior lo s 4 0 C , temperatura a la que crecen
algunas estreptobacterias atípicas de indudable interés bromatológico (Schillinger y Locke.
1987a).
1 1 . 3. 5.- Metabolismo
Desde el punto de vista metabólico se considera que lo s lactobacilos se encuentran en
el límite entre el metabolismo anaeróbico y aeróbico. No poseen quinonas isoprenoides
(excepto L. vanrnnwshiensis y L casel subsp. rhwnnosus ) (Collins y iones, 1981) , ni
sistemas de citocromos con los que realizar la fosforilación oxidativa típica del metabolismoaeróbico. Sin embargo, poseen oxidasas y peroxidasas que oxidan el NADH
2 utilizando al 02
como aceptor final de electrones. A pesar de no tener catalasas ni peróxido~dismutasas también
metabolizan lo s peróxidos mediante un sistema enzimático catalizado por el Mg++ (Gótz y
col., 1980).
los lactobacilos son de metabolismo esencialmente fenuentativo; degradan las hexosas
utilizando dos vías enzimáticas (fig. 2.1):
lo).- Via de Em~en-Meverhof (glucolisis), por la que convierten un niol de hexosa en
dos moles de ácido láctico (fermentación homoláctica).
28

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 42/293
REVISION BIBLIOGRA PICA
ciio
« 1- ‘ -1
m
ti)
ti)*4 -4os .lti )4 -)eJ
Tríasa - 32
ADP
P
rFiruvato
-6P
62 - Gluconato
Xiluiosa 5 2 + 00 2
4—
Triosa - 3P + Acetíl - 2
AD 2
AT 2
Piruvato
l f rLactato Lactato
r1
Acetato ( E t a n o ] >
VIA EMBDEN-MEYERHOF VTA 6 — FOSFOGLUCOPJATO
(Otucotisis)
(Homofermentativas) (Heterofermentat i v a s >
Figui-a 2. 1.- Repi-esentación esquemática de las rutas metabólicas del género Lacwbacillus.
( A d a p t a d o d e K a n d l e r , 1 9 8 3 ) .
Fructosa — 1,6 F Glucosa
o
r
E n
o
tiIt ji‘-1
uIt
14 - 1
1
Ho4Jti )- uo41
o4 - 1
m
29

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 43/293
REVIS ION IIIBLIOGRA PICA
2v).- Vía del 64osfogluconato, por la que producen un mol de CO2, un mol de etanol
( o d e á cid o a c é t i c o ) y u n ml d e á c i d o l á c t i c o ( f e r m e n t a c i ó n h e t c r o l á c t i c a ) . E n co n d ici on e s d e
a e ¡ o b i o s i s , l a m a y o r pa r t e d e l a s c e p a s u t i l i z a n a l 0 2 como a c e p t o r f i n a l d e e l e c t r o n e s , mien t ras
no reducen el Acetil-CoA (Kandler, 1983).
El ácido pirúvico es un metabolito intermediario dc las dos rutas metabólicas
f e r m e n t a t i v a s , s i e n d o l a c a n t i d a d d e h e x o s a d e l me d io l a q u e r e g u l a e l t ip o d e f e r m e n t a c i ó n , d e
forma que si escasea éste azucar, la fennentación homoláctica se conviene en heteroláctica con,
p r o d u c i ó n f i n a l d e á c i d o s a c é t i c o . f ó r m i c o y d e e t a n o l ( T h o m a s y c o l . . 1 9 7 9 ) . E l á cid o láctico
formado, puede oxidarse originando ácido acético y fórmico mediante mecanismos no bien
conocidos (Kandler, 1983).
L ad i f e r e n c i a e n t r e
l a c t o b a c i l o s homoy h e t e ro f e rm e n t at iv os s e e n c u e n t r a e n l a
p r e s e n ci a o au se n ci a d e lo s enzimas difosfofructo-aldolasa y fosfocetolasa. Así, mientras los
lactobacilos heterofermentativos obligados sintetizan fosfocetolasa pero no Mdolasa, los
homofermentativos sintetizan aldolasa pero no fosfocetolasa.
Los lactobacilos heterofermentativos hidrolizan las pentosas gracias al enzima
fosfocetolasa, originando cantidades equimolares de lo s ácidos acético y láctico. Sin embargo,algunos lactobacilos liomofermentativos (Su-epíobacíeriun¡ de Oila-Jensen) poseen un aldolasa
y un fosfocetolasa que se activan únicamente en presencia de pentosas, de manera que
hidrolizan las hexosas siguiendo la vía Embden-Meyerhoff, originando ácido láctico, mientras
que las pentosas por la v í a d e l 6 - f o s f o g l u c o n a t o d an l u g a r a c a n t i d a d e s e qu im ol ar e s d e á cid os
láctico y acético, pero no originan CO2, convirtiéndose así en los denominados
heteroferníentarivos facultativos (Kandler. 1983).
30

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 44/293
REVISION BIBLIOGRA PICA
El ácido láctico procedente de la fermentación de los hidratos de carbono, posee una
configuración D- o L- dependiendo de la estercoespecificidad del lactato deshidrogenasacelular. En presencia de O- y L-lactato deshidrogenasas, el ácido láctico se produce en su
forma racémica (DL). Lo mismo que ocurre cuando coexisten L-lactato deshidrogenasa y
lactato racemasa inducible. Esta última circunstancia es la que se produce en L. casel, L.
cuivatus y L. sa/ce (De Vries y c o l . , 1 9 7 0 ) . L a m a y o r p a r t e d e l o s e n z i m a s n o s o n a l o s t é r i c o s ,
pero algunas especies (L. cusei, L. cur’.’atus, L. sa/ce y L- bavaricus), poseen L-lactato
deshidrogenasas alostéricos cuyos activadores son la fructosa difosfato y el Mn±±(Hensel y
c o l . , 1 9 7 7 ) .
L o s l a c t o b a c i l o s t am b ié n e m p l e a n l o s h i d r a t o s d e ca rb on o c o n o t r o s f in es a d e m á s d e l a
obtención de energía. La sacarosa se utiliza en la síntesis de mucopolisacáridos y la fructosa
puede utilizarse como aceptor de electrones dando lugar a la formación de manitol. La mayoria
de lo s monosacáridos y oligosacáridos que utilizan las células como fuente de energía proceden
del medio extracelular, penetrando en su interior con ayuda de permeasas específicos, no
obstante, en algunas espeéies la lactosa y la galactosa se transportan mediante un sistema
enzimático fosfoenol piruvato fosfotransferasa (PEP-PTS) (Chassy y Thompson, 1983).
T a m b i é n s e c o n o c e n o t r o s s i s t e m a s d e t ra n sp on e a c t i v o d e a m i n a s y a m i n o á c i d o s ( L a w y
Kolstad, 1983).
I I . 3 . 6 . - C o n s i d e r a c i o n e s t a x o n ó m i c a s ~ C l a s i f i c a c i ó n
P a r a l a c l a s i f i c a c i ó n d e l a s d is t in t as e s p e c i e s d e l gé n e ro Lactobacillus se han
co n sid e ra d o n u m e ro s o s p a r á m e t r o s , e n t r e l o s q u e s c e n cu e n t ra n l a d e t e rmi n ac i ó n d e l o s t i p o s
d e p e p t id ogl ican o d e l a p a r e d c e l u l a r ( W i l l i a n s , 1 9 7 5 ) , l a f e rm e n t ac ió n d e l o s az ú ca re s y l a
temperatura de crecimiento (Schillinger y L o c k e . 19 8 7b ) , l a m o vi l i d a d e l e c t r o f o r é t i c a d e
d i v e r s o s e n z i m a s ( Lo n d o n , 1 9 7 6 ) , e l an á li si s i n mu n o l ó gi co d e l a s l a c t a t o deshidrogenasas
3 1

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 45/293
REi’I~SION fi IBLIOGRA PICA
homólogas (London, 1976), el grupo antigénico (Sharpe, 1970), la determinación de los
isómeros de ácido láctico y, por último, la de hibridación del DNA, que es la más emplea da en
la actualidad para determinar L a proximidad filogenética entre dos especies (Simonds y col.,
1971).
El primer intento de clasificación de lo s lactobacilos lo realizó Orla-Jensen (1919),
quién los dividió en tres géneros d istintos según las temperaturas óptimas de crecimiento y los
productos finales de su metabolismo fermentativo. Estos tres géneros, (“Thermobacteriurn“Stre¡nobacíe¡-iurn “ y “Betabacíe,-iurn” ) , ya no aparecen como tales en la última edición del
“Bergey’s Manual of Systematic Bacteriology” (Kandler y Weiss., 1986) al considerarse que
la división de Orla-Jensen no origina grupos homogéneos desde el punto de vista filogenético.
Así, y a pesar de que la mayoi-ía de las especies de cada uno de lo s grupos actuales
(denominados 1 , II y III ) coincide con las características d e las “termobacterias”,
“estreptobaterias” y “betabacterias” respectivamente, algunas de las especies descubiertas
recientemente no cumplen dichas características. La clasificación en tres grupos se realiza en
virtud de su metabolismo fermentativo.
II. 3 . 6. 1.- Gruno 1 .- Homofermentativos obligados
Este grupo se encuentra formado por dos subgrupos de especies relacionadas entre si:
en el primero se encuentran L. delbrucck-ii, L. bulgaricus, L. lactis y L. leichrnanníi, mientras
en el segundo aparecen L. acidophilus, L. gasseri y L. crispatus. Dicho grupo comprende la
mayor parte de las termobacterias de Orla-Jensen.
3 2

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 46/293
REt’íSION BíBLIOCRAFICA
II. 3. 6. 2.- Grupo II.- Heterofermentativos facultativos
El grupo lo integran tres subgrupos: Uno primero, representado mayoritariamente por
L. plantaruni, un segundo con distintas subespecies de L. casei y un tercero que comprende a
L. sa/ce, L. curvatus y L. bavaricus . Los dos primeros subgrupos de esta división se
corresponderían con las antiguas “estreptobacterias”, mientras el tercero comprendería las
denominadas “estreptobacterias atípicas” (Thornley y Sharpe, 1959).
II . 3 . 6 . 2. 1.- Estreytobacterias atínicas
Las dos primeras especies de éste grupo, L. sa/ce y L. curva/as, muestran un alto
porcentaje de homología según las técnicas de hibridación del DNA. Además, las dos poseen
una racernasa inducible por ácido láctico, que convierte el ácido L(±)-lácticoen ácido láctico
racémico (Kandler y Weiss, 1986). Esta circunstancia, y otras características fenotípicas
diferencian éstas dos especies del tercer componente del grupo, L. bava¡-icus. La diferenciación
entre L. sa/ce y L. curvatus se fundamenta en su capacidad de utilizar diversos azúcares
(Schillinger y Liicke, 1987b). Este subgrupo también se considera como los “lactobacilos
psicrotofos verdaderos” (Reuter, 1981), al ser lo s que se desarrollan a temperatura más baja de
todos los lactobacilos (2-4 0C) . Otra característica que las diferencia de las estrepto- bacterias
típicas es su aspecto cocoide cuando se siembran en medios sólidos (Reuter, 1981).
II. 3. 6. 2. 1 . 1.- Lactobacillus saLe
Lactobacillus saL-e recibe esta denominación (Katagiri y col.. 1934) por haberse
a i s l a d o p or p ri me ra v e z d e e s t e p ro d u ct o j a p o n é s, s i e n d o s u s p r i n c i p a l e s c a r a c t e r í s t i c a s
(Champomier y col., 1987) las de poseer una morfología cocobacilar ligeramente curvada y
regular, sobre todo en la fase estacionaria, con un tamaño de 0,8-1 p.m de ancho por 2-3 gm
33

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 47/293
REVISION BIBLIOGRA PICA
de largo y por desarrollarse aislado o en cadenas co¡-tas Heterofermentativo facultativo,
produce lo s isómeros D- y L- del ácido láctico, sedesaiTolla a 2-4 0C y 1 5 0C, pero no a 45
crece a pH 3,9 y en medios con un 8% de NaCí, pero no cuando tienen un 10% de NaCí y
utiliza los siguientes hidratos de carbono: D-ribosa, D-galactosa, D-glucosa, D-fructosa,
D-manosa, N-acetilglucosamina, D-melibiosa, 0-trealosa, sacarosa y gluconato, pero no estos
otros: dulcitol, inulina, D-melicitosa, D-rafinosa, glucógeno, D-wgatosa, L-fucosa, D-arabitol,
D-arabinosa, D-xilosa, L-sorbosa, ramnosa, D-manitol y sorbitol. L. sa/ce también se ha
aislado de ensilados, carne, embutidos crudos madurados (Schillinger y Lúcke, 1987b. 1989)
y d e p rod u ct os c á r n i c o s e n vasa d os a l v a c í o .
II. 2. 6. 3.- Grupo ¡IL- l-ieterofennentativos obligados
E st e g ru p o l o f o r m a n c a s i i n t e g r a m e n t e l a s “ b e t a b a c t e r i a s ” d e O r l a - J e n s e n , a l a s q u e s e
han unido otras especies de reciente descubrimiento y comprende 18 especies entre las que
d e s t a c a n : L. bife,-mentans, L. vi¡-idescens, L. safranciscus L. ferrnenturn, L. reuteri, L.
confusus, L. brevis, L. halotolerans, L. kefir y L. fructivoraus.. En la última edición del
Ber2ev’s Manual of Svstematic Bacteriolotzv (Kandler y Weiss,1986) también se incluye en
e s t e gr u p o a L. diveí-gens, a u n q u e act ual men t e s e admite que esta especie pertenece al género
Carnobacteriun;.
1 1 . 3. T- Ecolo2ía. habitats y biotecnología
E l g é n e ro Lactobacillus, como el resto de las bacterias lácticas, se encuentra
ampliamente distribuido en la naturaleza debido a que es capaz de desarrollarse en las
co n d ici on e s am b ie n t al e s m a s d i v e r s a s , s i e mp r e y c u a n d o e x i s t a n e n e l m e d i o l o s complejos
nutrientes que necesita (hidratos de carbono, péptidos y aminoácidos, ácidos nucleicos yv i t a m i n a s ) . E l d e s a r r o l l o e n a n a e r o b i o s i s y e n t r e u n am p l io i n t e r v a l o d e t e mp e ra t u r as (2 - 4 5
3 4

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 48/293
REVISIÓN BIBLIOCRA PICA
0 C ) . a s í como s u t o l e r a n c i a a u n a t e n s i ó n d e 0 2 b a j a , pe rmit e d e t e c t a r l o s e n l o s m á s v a r i a d o s
habitats, como en los ensilados y productos fermentados vegetales, zumos de frutas, cerveza,
vino (Whitting, 1975; Sharpe, 1981: Kandler, 1984), queso (Botazzi y col., 1973), yogur
(Davis, 1975) y otras leches fermentadas (Kandler y Kunath, 1983), productos cárnicos
refrigerados (Reuter. 1975). embutidos crudos curados y productos cárnicos envasados al
vacío (Holzapfel y Gerber, 1983). También se han aislados de pescados (Cone, 1982) del
tracto gastrointestinal y urinario de personas y an im al e s ( S h a r p e y c o l . , 1 9 7 3 ; S h a r p e , 1 9 8 1 ;
Dent y Willians, 1982), asi como del estiércol y de las aguas residuales (Okada y col., 1979;Weiss y col., 1981).
El efecto beneficioso de éstos microorganismos al impedir de diversas maneras el
desarrollo de otros microorganismos patógenoso alterantes, así como los cambios
organolépticos y nutritivos favorables que originan al multiplicarse en diversos sustratos.
hacen que algunas especies del género Lacrobacillus se hayan utilizado desde hace tiempo enl a e l a b o r a c i ó n d e al ime nt os f e r n i e n t a d o s ( y o g u r , k e f i r , l e c h e s acidificadas, embutidos crudos
curados) y ensilados. Son microorganismos tecnológicos indispensables y seguirán siéndolo
en la futura tecnología de lo s alimentos y, sin duda, aumentará su interés en salud humana y
animal.
1 1 . 4.- Actividad antimicrobiana de l a s b a c t e r i a s l á c t i c a s
La actividad antimicrobiana de las bacterias lácticas se conoce desde hace tiempo, s i
bien de una forma empfrica. Hasta hace muy poco tiempo no se inició el estudio científico del
mecanismo, o los mecanismos mediante los que las bacterias lácticas manifiestan su actividad
a n t i m i c r o b i an a.
A c t u a l m e n t e s e p i e n s a q u e e l p ri me r m e c a n i s m o an t im ic ro b ia n o e j e i c i d o p or l a s
3 5

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 49/293
RE VISíaN BIBLIOGRA PICA
bacterias lácticas pod¡ia serel de la simple competencia por lo s nuti-ientes, como consecuencia
de las distintas velocidades de crecimiento de lo s microorganismos, es muy posible que una
determinada especie bacteriana de crecimiento rápido agote lo s nutrientes c¡íticos presentes en
el sustrato, perjudicando la existencia de otros microorganismos. Este sistema no es exclusivo
de las bacterias lácticas, aunque s i lo s so n los que a continuación se detallan.
II. 4. 1.- Actividad antimicrobiana de lo s ácidos orgánicos
Como consecuencia de su metabolismo fermentativo, las bacterias lácticas sintetizan
ácidos orgánicos, lo que se acompaña de una notable caida del pH del medio.
L a acción antimicrobiana de los ácidos orgánicos puede deberse a tres factores
( Be u c h a t y G o l d e n , 1 9 8 9 ) :
i Q ) . ~ A l e f e c t o a n t a g o n i s t a d e l a c a i d a d e l p H , qu e i n h i b e e l d e s a r r o l l o d e o t r o s
microorganismos ácido-sensibles.
2Q)< A la actividad antagonista de la forma no disociada del ácido que penetra en el
interior de las células e inhibe algunas reacciones enzimáticas metabolicamente importantes.
39.- A l a a c c i ó n e s p e c í f i c a d e l o s á c i d o s o r g á n i c o s , y a q u e l o s d e m á s d e 4 á t o m o s d e
c a rbo n o a u m e n t a n s u a c t i v i d a d a n t i m i c ro bi a n a a m e d i d a q u e l o h ac e l a l o n g i t u d d e l a c a d e n a .
No obs t ant e, l a e sca sa l i p o s o l u b i l i d a d d e l o s á c i d o s d e m á s d e 8 á t o m o s d e ca rb on o,
d i s m i n u y e s u a c t i v i d a d e n l a s b ac t er ia s G r am- n e gat ivas ( B a i r d P a r k e r , 1 9 8 0 ) .
L o s á c i d o s o rgá n i c o s d e ca d e n a c o r t a , como e l l á c t i c o y a c é t i c o , s o n l i p o f í l i c o s c u a n d o
no están disociados, por lo que pueden penetrar al interior de lo s microorganismos
36

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 50/293
REVÍSION BíBLIOGRA PICA
disminuyendo el pH intracelular, alterando sustratos enzimáticos e inhibiendo funciones
metabólicas esenciales corno la fosforilación oxidativa (Haird Parker. 1980). E L queel ácido
acético sea más inhibidor en mohos y levaduras (Baird Parker, 1980) que el láctico podría
deberse a que a pH 4,0-4,6 (e l natural del entorno donde se desarrollan las bacterias lácticas)
este ácido está entre 2 y 4 veces menos disociado que el láctico (Lindgren y Dobrogosz,
1 9 9 0 ) . L o s á cido s a c é t i c o y l á c t i c o p u e d e n a c t u a r s i n é rg i c a m e n t e i n h i b i e n d o e l cr e cim ie n t o d e
salmonelas (Rubin, 1978) y levaduras (Moon, 1983) ya que el bajo pH que proporciona el
ácido láctico aumenta la concentración del ácido acético no disociado. Otros ácidos producidospor las bacterias lácticas en diversos sustratos, como fórmico, cítrico y málico, tienen una
actividad antimicrobiana menor que lo s citados.
II. 4. 2.- Actividad antimicrobiana de componentes inorgánicos
Las bacterias lácticas también producen otros compuestos antimicrobianos como CO2,
H202 y el diacetilo.
I I . 4 . 2 . 1.- Dióxido de carbono
El CO2 producido como consecuencia del metabolismo heterofermentativo obligado
e j e r c e s u a c c i ó n a n t i b a c t e r i a n a c r e a n d o un ambiente anaeróbico al reemplazar el 02, impidiendo
así el desarrollo de los microorganismos aeróbios obligados, o bien mediante una actividad
inhibidora intrinseca y variable según el microorganismo de que se trate (Clark y Takacs,
1980).
A u n q u e n o s e c o n o c e c o n e x a c t i t u d e l mecanismo íntimo por el que el CO2 ejerce su
a c c i ó n a n t i m i c r o b i a n a , q u i z á s e d e b a a l e f e c t o d e d ich o c o m p u e s t o e n l a i n h i b i c i ó n d e
37

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 51/293
REl’ISION BIBLIOGRA PICA
d e sca rb ox il acio n e s e n z i m á t i c a s ( Ki n g y N a g e l , 1 9 7 5 ) o p o r a c ú m u l o d e ¡ CO2 e n l a b i c a p a
lipidica de la membrana celular, alterando su permeabilidad (Eklund, 1984).
1 1 . 4. 2. 2.- Peróxido de hidró2eno
A l g u n a s b a c t e r i a s lá ct ic as s i n t e t i z a n H202 e n p r e s e n c i a d e o x í ge n o g r a c i a s a l a
actividad de determinadas flavoprotein-oxidasas o NADH-peroxidasas. Este H202 se
acumula en el medio exocelular ya que las bacterias lácticas carecen, de actividad catalasa
(Kandler y Weiss, 1986). La producción de H202 por las bacterias lácticas depende de la
c e p a y d e l a c a n t i d a d d e ox ig e n o p r e s e n t e ( C o l l i n s y A r a m a k i , 1 9 8 0 ) . D a h i y a y S p e c k (1 96 8 )
y P r i c e y L e e ( 1 9 7 0 ) . d e mo st rar on e l e f e c t o i n h i b i d o r d e l H202 p rod u ci d o p o r l a s b a c t e r i a s
l á c t i c a s , e n d i v e r s a s c e p a s d e Sraphylococcus al/leus y Pseudornonas spp. El efecto bactericida
d e l H202 s e a t r i b u y e a s u p o t e n t e a c c i ó n o x i d a n t e e n l a c é l u l a b a c t e r i a n a ( F o o s t e r y c o l . , 1 9 5 7 )
y a la destrucción de estructuras moleculares básicas de las proteinas celulares (Sykes, 1965).
Asimismo, algunas bacterias lácticas presentes en la leche, activan el sistema
a n t i b a c t e r i a n o d e l a l a c t o p e r o x i d a s a ( B j ó r c k , 1 9 8 5 ) , d e b i d o a l a s í n t e s i s e n a e r o b i o s i s d e H202
(Carlsson y col., 1983). La actividad antimicrobiana del H202 es variable, no afectando
prácticamente a las bacterias Gram-positivas entre las que se incluyen las bacterias lácticas,
mientras que las Gram-negativas, como Escherichia coIi, Salmon ella s p p . y Pseudomonas
spp. son muy sensibles a su actividad (Bjórck, 1985). Se cree que el efecto antimicrobiano del
sistema lactoperoxidasa se debe a la oxidación de lo s grupos -SH de algunos enzimas
metabólicos vitales, como la hexoquinasa, aldolasa y gliceraldehido-3P deshidrogenasa
(Bjórck, 1985).
3 8

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 52/293
REVISIÓN I3IULIOGRAFICA
II. 4. 2. 3.- Diacetilo
Algunas especies de bacterias lácticas producen, a partir del citrato, díacetílo
(2.3-butanodiona) (Jay,1982), siguiendo la vía del piruvato que es el metabolito intermediario
( Ka n d l e r , 1 9 8 3 ) . A p e s a r d e q u e e l me t ab ol is mo d e l a s h e x o sa s i n h i b e l a p ro d u cc ió n d e
diacetilo, éste se acumula en el medio y, además de generar el aroma que caracteriza a ciertos
productos como la mantequilla, ejerce una actividad inhibidora sobre un gran número de
microorganismos, tanto Gram-positivos como Gram-negativos (Jay, 1982). Esta actividad sefavorece cuando el pH del medio es bajo (Jay, 1982). El diacetilo inhibe el desaiTollo de los
microorganismos Gram-negativos al interaccionar con el metabolismo de la arginina,
impidiendo la utilización de éste aminoácido (Jay. 1986).
II. 4. 2. 4.- Reuterina
Lactobacillus reuteril es el único lactobacilo y q u i z a s l a ú n i c a b a c t e r i a , q u e p ro d u ce
reuterina y la libera en el medio exocelular como consecuencia del metabolismo anaeróbico del
glicerol.
La reuterina (Axelsson y col., 1989), es una sustancia antimicrobiana activa frente a
bacterias Gram-positivas, Gram-negativas, levaduras, hongos y protozoos. Químicamente es
una mezcla homogénea de monómeros, monómeros hidratados y dímeros cíclicos del aldehido
3-hidroxipropiónico (Talaricco y Dobrogosz, 1990) que proceden del glicerol mediante una
reacción catalizada por una coenzima glicerol-deshidratasa dependiente de la vitamina 812
(Talaricco y Dobrogosz, 1990; Talaricco y col., 1990). De ésta formaL. reuter¡i es la única
bacteria conocida capaz de utilizar el glicerol como aceptor alternativo de hidrógeno durante la
fermentación de los hidratos de carbono, produciendo mayor cantidad de aldehido
3-hidroxipropiónico del que es capaz de reducir, por lo que éste se libera en el medio
3 9

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 53/293
REVISION BIBLiOGRA PICA
extracelular donde manifiesta su actividad antimicrobiana. Entre los microorganismos sensibles
a esta sustancia se encuentran Salmondlla sp., Shigella sp., clostridios, estafilococos,
listerias, candidas y tripanosomas.
II. 4. 3.- Efecto antimicrobiano de las bacteriocinas y s u s t a n c i a s a f i n e s
La primera noticia sobre la producción por las bacterias lácticas de sustancias
antagonistas, distintas de los productos finales del metabolismo microbiano, se atribuye a
Rogers (1928), quien describió la actividad antimicrobiana de &reptococcus lactis
( ac t ual me n t e Lacococcus ladús) sobre L. ac¡dophilus . D e s d e e n t o n c e s s e h an i d e n t i f i c a d o u n
c i e r t o n ú m e r o d e s us t a n c i a s p r o t e i c a s , c o n act ivid ad an t im ic ro b ia n a sob re ot ros
microorganismos. Este grupo está constituido por una serie de sustancias de composición
q u í m i c a , m e c a n i s m o d e a c c i ó n y e s p e c t r o a n t i m i c ro bi a n o va r ia b le .
Por la importancia que tiene este grupo de sustancias, al que pertenece el agente
an t imi cro bi an o a i s l a d o y c a r a c t e r i z a d o par cial ment e e n es t e t r ab aj o , s e á n o s p e r m i t i d o d e s c r i b i r
y r e v i s a r c o n c i e r t o d e t a l l e s u s a s p e c t o s m á s so b r es al ie n t es .
I I . 5.- Bacteriocinas: Definición
L a s b a c t e r i o c i n a s s e d e f i n e n como p r o t e i n a s o c o m p l e j o s d e n a t u r a l e z a p r o t e i c a c o n
acción bactericida en microorganismos filogenéticamente muy próximos a la bacteria
productora (Tagg y col., 1976). Este grupo heterogéneo de sustancias, generados tanto por
m i c r o o r g a n i s m o s G ra m- p os it ivo s como G ra m- n e ga t iv os , v a r í a mucho e n p ro p ie d a d e s
b i o q u í m i c a s , e s p e c t r o a n t i m i c ro bi a n o y m e c a n i s m o d e a c c i ó n .
4 0

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 54/293
REI’ISION BIBLIOGRA PICA
1 1 . 5. 1 .- Detección de bacterias nroductoras de bacteriocinas
La detección de bacterias con actividad antirnicrobiana, a t r i b u i b l e a l a s b a c t e i i o c i n a s , s e
realiza en dos fases o etapas: 1).- detección de la cepa productora de la actividad antagonista, y
2).- diferenciación de la actividad antimicrobiana debida a las bacteriocinas de otras posibles
causas de inhibición.
I I . 5. 1 . 1 . - Detección de las ceDas uroductoras
Silo que se pretende es detectarla posible actividad antimicrobiana de un gran número
de bacterias lácticas aisladas de diversos sustratos alimentarios, el método de elección consiste
en utilizar la denominada “prueba general de antagonismo”, que se realiza en un medio de
cultivo sólido en el que se observan fácilmente los halos resultantes de la inhibición del
desarrollo de un microorganismo indicador producida al crecer en la misma placa cada una delas bacterias lácticas aisladas. Los métodos experimentales más utilizados son el “antagonismo
directo” y el “antagonismo diferido” (Tagg y col.,1976). En cualquier caso, se basan en la
siembra de colonias aisladas de la cepa problema en un medio sólido apropiado, sobre el que
se deposita una capa de medio sólido o semisólido que lleva el microorganismo indicador.
II. 5. 1 . 2.- Prueba del anta2onismo directo
La prueba más sencilla y utilizada se basa en el método desarrollado por Orada (1946).
En este caso, la cepa problema y el cultivo indicador se desarrollan simultáneamente y la
v i s u a l i z a c i ó n de la actividad antimicrobiana sólo es posible s i se produce durante las primeras
f a s e s d e cr e ci mi e n t o d e l a c e p a p r o b l e m a ( Ta gg y c o l . , 1 9 7 6 ) . T a m b i é n e s i m p o l - t a n t e l a
densidad del cultivo indicador utilizado (Kuttner, 1966). El cultivo indicador se deposita en las
placas inmediatamente después de inoculadas co n la cepa problema.
4 1

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 55/293
REVISION BI/?LIOGRA PICA
Una variante de éste método consiste en cortar pocillos en las placas de medio sólido y
rellenarlos con una gota del mismo medio conteniendo la cepa problema. Unavez solidificado
el agardel pocillo se siembra sobre la placa el microorganismo indicador (Tagg y col., 1976).
Pero quizás el método de elección sea el empleado por De Klerk (1967). En esta
variante la cepa problema se siembra en profundidad, con un asa de platino o un palillo, en el
m e d i o d e c u l t i v o s ó l i d o y d e sp u és s e i n o c u l a s u su per f ici e c o n e l m i c r o o rg a n i s m o i n d i c a d o r .
I I . 5 . 1 . 3 . - P r u e b a d e l an t ag on ismo d i f e r i d o
En e s t a p ru e b a l a b a c t e r i a p ro b l e ma s e s i e m b r a y d e s a r r o l l a e n p l a c a s d e m e d i o s ó l i d o ,
d u r a n t e u n p e ri od o d e t ie m p o d a d o , i n a c t i v á d o l a a c o n t i n u a c i ó n p o r e xp osi ci ó n a l c a l o r o a l
cl orof ormo. Más t a r d e , e l c u l t i v o i n d i c a d o r m e z c l ad o c o n a g a r s e m i s ó l i d o , s e si e mb ra e n l a s
p l a c a s i n a c t i v a d a s como q u e d a d i c h o ( F r e d e r i c q , 1 9 4 8 ; B a r e f o o t y K l a e n h a m m e r , 1 9 8 4 ) . E st e
si st e ma t i e n e l a v e n t a j a d e p e n n i t i r e l c r e c i m i e n t o d e l a b a c t e r i a p r o b l e m a y d e l i n d i c a d o r e n
co n d ic io n e s ó p t im a s d e t e m p e r a t u r a y a q u e l a i n c u b a c i ó n n o e s s i m u l t á n e a . No o b s t a n t e , p u e d e
ocurrir que el cultivo indicador sea sensible a los métodos que inactivan el cultivo problema
(Tagg y col., 1976).
I I . 5 . 2 . - L a a c c i ó n d e l a s b a c t e r i o c i n a s y su diferenciación d e o t r a s p o s i b l e s cau sas d e
i n h i b i c i ó n
L o s m é t o d o s a n t e s d e s c r i t o s n o g a r a n t i z a n q u e l a a c t i vi d a d an t ím íc ro b ia n a d e l
m i c r o o r g a n i s m o e s t u d i a d o s e d e b a a l a s b a c t e r i o c i n a s , e n c on se cu e n c ia d e b e e va l u ar se l a
a c t i v i d a d ant imicr obia na d e l o s m i c r o o r g a n i s m o s d e i n t e r é s u t i l i z a n d o e n sa y o s e xp e ri m e n t al e s
co mp l e me n t ari os . En e s t o s e n sa yo s s e t r a t a d e e l i m i n a r l a p r e s e n c i a e n e l m e d i o d e c u l t i v o d e
4 2

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 56/293
REl’LSION BIBLIOCRAPICA
otros microorganimos o productos metabólicos con actividad antimicrobiana.
Consecuentemente, la bacteria problema que interesa se siembra en un medio líquido que se
cultiva en condiciones óptimas para la producción máxima de actividad antimicrobiana Tales
condiciones se determinarán experimentalmente ya que no tienen porqué coincidir con las
ó p t i ma s d e c r e c i m i e n t o ( T a g g y c o l . , 1 9 7 6 ) . a c o n t i n u a c i ó n s e e s t e r i l i z a e l m e d i o d e c u l t i v o p o r
filtración para eliminar la posible presencia de bacteriófagos (Tagg y col., 1976), que podrían
ser los responsables de los halos de inhibición observados al cultivar las bacterias problema
con el microorganismo indicador
L a a c t i vi d a d a n t a g o n i s t a d e l H20 2 q u e p u d i e r a p r o d u c i r l a c e p a d e i n t e r é s s e el imin a
incubandoel microorganismo en anaerobiosis estricta (Schillinger y Liicke, 1989), o también
añadiendo al medio de cultivo catalasa (Su, 1949) o peroxidasa (Holmberg y Hallander,
1 9 7 3 ) , a n t e s d e d e t e rm i n a r s u a c t i v i d a d a n t i m i c r o b i a n a f r e n t e a l m i c r o o r g a n i s m o i n d i c a d o r .
F in a l m e n t e , l a a c i d e z d e l m e d i o d e c u l t i v o s e n e u t r a l i z a t r a t á n d o l o co n u n a s o l u c i ó n
básica fuerte (Tramer, 1966>.
De esta manera, una vez eliminadas otras causas de inhibición distintas de las
b a c t e r i o c i n a s , s e eval úa l a a c t i v i d a d a n t i m i c ro bi a n a d e l o s m e d i o s d e c u l t i v o l i b r e s d e cé lu la s.
Esto se realiza de tres formas:
1).- Depositando una gota del medio líquido problema neutralizado, en una placa de
Petri con agar semisólido en la que se desarrolla el microorganismo indicador (Daeschel y
Klaenhammer, 1985).
2).- Practicados unos pocillos en placas de medio sólido, se recubre su superficie con
a g a r y s e d e p os it a u n a go t a d e l m e d i o l í q u i d o p r o b l e m a p re vi a m e n t e n e u t r a l i z a d o , s e d e j a
4 3

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 57/293
RPA’ISION BIBLIOCRA PICA
transcurrir un tiempo suficiente para que difunda y, finalmente, se rellenan los pocillos con
agar y se deposita en ellos el microorganismo indicador (Daeschel y col., 1990; Ahn y Stiles,
1 9 9 0 a) ; t am b ié n s e p u e d e s e p a r a r e l aga r d e l a p l a c a d e P e t r i q u e l o c o n t i e n e y , después de
i n v e r t i d o , s e si e mb ra e l m i c r o o r g a n i s m o i n d i c a d o r e n l a o t r a c a r a d e l a ca p a d e agar ( T a g g y
McGiven, 1971; Schillingery Lúcke, 1989).
3).- Saturando un disco de papel de filtro con una cantidad suficiente del medio líquido
neutralizado. Una vez seco el disco, se coloca sobre el microorganismo indicador sembrado enagar semisólido (Bhunia y col., 1987; Bhunia y col., 1988).
II. 5. 3.- Producción óptima de bacteriocinas
Tanto l a c o m p o s i c i ó n d e l o s m e d i o s d e cu lt iv o , como l a s co n d ic io n e s d e d e s a r r o l l o
n e c e s a r i a s p a r a u n a p rod u cc ió n ó p t im a d e b a c t e r i o c i n a s v a r i a n d e u n a s b a c t e r i a s pro d uct oras a
o t r a s ( Ta gg y c o l . , 1 9 7 6 ) . A s í p o r e j e m p l o , a l g u n a s b a c t e r i o c i n a s s ó l o s e p ro d u c e n c u a n d o e l
m i c r o o r g a n s i m o s e c u l t i v a e n m e d i o s s ó l i d o s o s e m i s ó l i d o s ( G e i s y c o l . , 1 9 8 3 ) , mi e n t ras q u e
o t r a s ba c t e ri a s pr od uct oras n e c e s i t a n s u p l e m e n t o s e s p e c í f i c o s como e x t r a c t o d e levadura
( N i e l s e n y c o l . , 1 9 9 0 ) o gl u co sa ( Bhu n i a y c o l . , 1 9 8 7 ) . L o s m e d i o s s e m i s i n t é t i c o s , d e
composición más sencilla, se emplean para favorecer las labores de purificación de las
bacteriocinas (Bhunia y col., 1987). La temperatura, el tiempo de incubación el cultivo conagitación o estacionario también influyen en la producción de bacteriocinas (Biswas y col.,
1991).
II. 5. 4.- Purificación
En la caracterización bioquímica parcial de las bacteriocinas se necesita su purificación
previa a partir del medio de cultivo libre de células en el que se encuentra la sustancia
44

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 58/293
REVISION BIBLIOGRA PICA
inhibidora. Con esta purificación se pretende separar la sustancla activa del resto de
componentes del medio de cultivo y de las células que en él crecieron. Debido a [a
heterogeneidad de estas sustancias es muy dificil establecer un protocolo modelo de
purificación, cada tipo de bacteriocina requiere una técnica específica que debe determinarse
empíricamente, al igual que las condiciones de cultivo, (Tagg y col., 1976).
En general, el primer paso de la purificación de las bacteriocinas consiste en
concentrarías en el medio de cultivo neutralizado mediante precipitación con sales, ácidos oetanol, diálisis o liofilización. Una vez obtenido el concentrado, se procede a la separación de
los distintos compuestos por su tamaño molecular (cromatografía de filtración, ultrafiltración,
centrifugación, etc.) o por su distinto potencial eléctrico (cromatografía de intercambio iónico,
electroforesis, etc.)
El único punto en el que coinciden todos lo s métodos de purificación es en la pérdidade actividad, a veces masiva, durante la purificación de las bacteriocinas (Tagg y col., 1976).
Por eso es importante determinar, en cada paso del proceso de purificación, [a cantidad de
proteina y su actividad específica (actividad inhibidora/mg de proteina) a fin de evitar aquellos
pasos que impliquen una gran pérdida de actividad (Tagg y col., 1976).
II. 5. 5.- Propiedades de las bacteriocinas de las bacterias lácticas
U. 5 . 5 . 1.- Composición química
Las bacteriocinas son, según la definición de Tagg y col. (1976), sustancias
antagonistas de naturaleza proteica. Por tanto, la inhibición o desaparición de la actividad
antagonista mediante la utilización de enzimas proteoliticos es definitiva para catalogar a una
sustancia inhibidora como bacteriocina. No obstante, aunque las bacteriocinas suelen ser
45

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 59/293
REVISION IJIBLIOGRAPICA
moléculas sencillas (agregadas o no), algunas presentan en su composición al menos durante
los estadios previos a su purificación, además de proteina, componentes [ipídicos y/o
hidrocarbonados (De Klerck y Smith, 1967; Upreti y Hinsdill, 1973).
II. 5. 5. 2.- Propiedades físicas
El peso molecular de estas sustancias varía desde los 2.700 daltons de la pediocina
AcH (Bhunia y col., 1988), hasta lo s 37.000 daltons de la helveticina J que puede llegar aformar agregados de hasta 300000 daltons; agregados de éste tamaño sólo se separan
mediante electroforesis en geles de poliacrilamida con dodecil sulfato sódico (SDS-PAGE)
(Joerger y Klaenhammer, 1986). El peso molecular suele determinarse por cromatografía de
filtración en geles empleando agentes disociantes para evitar la formación de agregados
(Pulusani y col., 1979; Joerger y Klaenhammer, 1986) también puede hacerse por
electroforesis en SDS-PAGE (Bhunia y col., 1987).
Una característica que prácticamente la presentan todas las bacteriocinas es su
termoestabilidad (West y Warner, 1988; Daeschel y col., 1990; Schillinger y Lúcke, 1989);
sin embargo, esta característica es díficil de cuantificar, ya que depende de su grado de
purificación, del pH, de la fuerza iónica y de la presencia de moléculas termoprotectoras (Tagg
y c o l . , 1 9 7 6 ) .
I I . 5. 5 . 3 .- Espectro de acción
En cuanto a su espectro de acción, las bacteriocinas se han dividido arbitrariamente en
dos tipos (Klaenhammer, 1988):
19.- Bacteriocinas con un espectro de acción muy restringido, limitado a especies
46

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 60/293
REVISION BIBLIOGRAPICA
bacterianas muy relacionadas filogenéticamente con la cepa productora. lo que concuerda con
la definición clásica de bacteriocinas (Tagg y col., 1976) y entre las que se incluyen muchas
bacteriocinas caracterizadas hasta ahora, como la diplococina, lactacinas B y F y lactocina 27.
2v).- Bacteriocinas con un espectro más amplio activas, además de frente a las especies
con ellas relacionadas, frente a otras bacterias Gram-positivas. Como ejemplo de este grupo se
cita la nisina, que impide la esporulación y crecimiento de especies bacterianas de los géneros
Clostridiurn y Racillus. No obstante, todavía no se han descrito bacceriocinas activas frente abacterias Gram- negativas (Klaenhammer, 1988).
II. 5. 5. 4.- Mecanismo de acción
El mecanismo de acción suele determinarse adicionando la sustancia, más o menos
purificada, a un cultivo líquido en crecimiento inoculado con una población determinada delmicroorganismo sensible. Durante su incubación (microoganismo±bacteriocina), se toman
muestras a intervalos de tiempo regulares y se determinan las bacterias viables mediante
recuentos en placa. La sustancia será bactericida si disminuye el número de microorganismos
viables y bacteriostática silos recuentos se mantienen constantes.
Según la definición clásica de Tagg y col. (1976), las bacteriocinas son bactericidas
para los microorganismos sensibles. El mecanismo mediante el que ejercen esta acción no es
bien conocido ya que, aunque algunos autores (Tagg y col., 1976) describen esta actividad en
dos etapas: a).- adsorción a la pared bacteriana y b).- destrucción de la pared bacteriana
seguida de lisis celular, otros (Barefoot y Klaenhammer, 1983; Harding y Shaw, 1990) han
puesto de manifiesto la adsorción de la bacteriocina incluso en las paredes de bacterias
resistentes; por tanw, la actividad antimicrobiana no estaría ligada a la presencia de receptores
específicos en la pared celular.
4 7

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 61/293
R IE l ISIOiV BII?LIOGRA PICA
También se han descrito sustancias con características idénticas a las bacteriocinas,
pero cuyo mecanismo de acción es bacteriostático. Entre ellas se encuentran la lactocina 27
(U pr et i y H i n s d i l l , 1 9 7 5 ) y o t r a s t o d a v í a n o c a r a c t e r i z a d a s b io qu imi ca me n t e ( R a c c - a c h y c o l . ,
1989). El mecanismo por el que la lactocina 27 ejerce su acción bacteriostática se debe a su
adsorción a la pared celular de las bacterias sensibles y a la posterior inhibición de la síntesis
proteica celular sin afectar a la del ADN, ARN ni ATP.
1 1 .
5. 6.- Regulación senética
Una de las cuestiones sobre las que lo s investigadores han puesto mayor énfasis en los
ú l t im os a ñ o s e s l a d e c a r a c t e r i z a c i ó n g e n é t i c a , a n i v e l mol ecul ar, d e l a p ro d u cc ió n d e
bacteriocinas. De hecho, lo s ensayos destinados al estudio de lo s determinantes genéticos que
re g u l an s u p ro d u cc ió n y l a r e s i s t e n c i a a s u a c c i ó n , a s í c o m o e l e s t u d i o d e l o s m e c a n i s m o s d e
transferencia genética de éstos determinantes han contribuido significativamente al desarrollo y
aplicación de las modernas técnicas genéticas (Klaenhammer, 1988).
Se ha demostrado con ello que la producción y la resistencia a las bacteriocinas suele
encontrarse codificada en plásmidos, generalmente en un mismo plásmido (Klaenhammer,
1988). Pero la producción de bacteriocinas no está siempre codificada en estos elementos
genéticos. Así por ejemplo, la lactacina B producida por L. acidophilus N 2 n o s e e n c u e n t r a
a s o c i a d a a l a p r e s e n c i a d e p l á s mi d o s y l o m i s m o o c u r r e c o n l a l a c t a c i n a F ( Bar e f oot y
Klaenhammer, 1984). Tampoco se ha determinado la existencia de plásmidos ni de otros
elementos genéticos extracromosomales en la producción de la lactostrepeina Las 5 de L. lacás
subsp. c’ernoiis (Dobrzanski y col., 1982).
Pero como ya se ha dicho, son numerosos lo s investigadores que han comprobado que
la producción de bacteriocinas se encuentra ligada a la presencia de plásmidos. De esta forma.
48

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 62/293
R íE VISION BIBLIOCRA PICA
la diplococina de L. lactis subsp. crernoris 346 se halla ligada a un plásmido de 54
Megadaltons (Davey, 1984), mientras la pediococina A de P. pentosaceus FBB6I estácodificada en un plásmido de 13.6 Megadaltons (Daeschel y Klaenhamrner, 1985). Asimismo
la producciónde nisina por diversas cepas de L. ¡actis se encuentra codificada en un plásmido
d e 2 8 - 3 0 M e g ad al t on s, e n e l q u e t am b ié n s e c o d i f i c a l a r e s i s t e n c i a a d i c h a b a c t e r i o c i n a y l a
fermentación de la sacarosa (Gonzalez y Kunka, 1987).
Además, no sólo se han identificado los plásmidos que codifican la síntesis debacteriocinas, sino que se ha llegado, incluso, a donar los genes responsables de su
producción y de la resistencia a la acción de dicha bacteriocina en otras bacterias receptoras
( V a n B e l k u m y c o l . , 1 9 8 9 ) , d e t e r m i n á n d o s e l a se cu e n ci a d e n u c l e ó t i d o s d e l o s op e ro n e s q u e
l o s regu l an ( Va n B e l k u m y c o l . , 1 9 9 1 ) .
El rápido desarrollo de las tecnologías genéticas en los últimos años puede permitir el
d e s a r r o l l o d e b a c t e r i a s l á c t i c a s d e i n t e r é s p o r t a n d o p l á sm id os d e p ro d u cc ió n y r e s i s t e n c i a a l a s
bacteriocinas, facilitando su supervivencia al crecer con otras bacterias carentes de éstos
plásmidos (Klaenhammer, 1988). Estas técnicas también permitirían expresar en una misma
bacteria los determinantes de varias bacteriocinas, pero no ya sólo de las bacteriocinas de otras
bacterias lácticas, sino de otras especies bacterianas distintas, con lo que el espectro
antimicrobiano de las bacterias lácticas se vería notablemente aumentado. Además se les podría
p ro p or ci on ar r e s i s t e n c i a f r e n t e a t o d o s l o s c o m pu e s t o s a n t i m i c r o b i a n o s l o q u e su p on d rí a u n a
ventaja adicional al poder crecer en un medio hostil (Klaenhammer, 1988). El empleo de estas
metodologías genéticas en la producción y ‘mejora” de las bacterias lácticas productoras de
bacteriocinas (sobre todo en las cepas de mayor interés industrial), abre unas perspectivas
prometedoras a la industria de fermentación y conservación de alimentos (Klaenhammer,
1988).
49

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 63/293
REVISIÓN RIBLIOGRAPIC’A
1 1 . 5. 7.- Bacteriocinas de las bacterias lácticas
Prácticamente todos lo s géneros de las bacterias lácticas presentan especies o cepas
productoras de bactenocinas:
II. 5. 7. 1.- Bacteriocinas producidas porespecies del 2énero Lactobacillus
La primera especie de este género en la que se identificó una actividad antimicrobianaasociada a una bacteriocina fué en L. ferment¡i (De Klerk y Goetzze, 1961). A partir de
e n t o n ce s s e h an i d e n t i f i c a d o y , e n al gun os c a s o s c a r a c t e r i z a d o , s u s t a n c i a s d e é s t e t ip o e n o t r a s
especies del género
a) L. helveticus .- Se conocen dos bacteriocinas producidas por esta especie, la
lactocina 27 (Upretti y Hinsdill, 1973), y la helveticina J (Joerger y Klaenhamer, 1986).
b) L. acidophilus .- A la primera sustancia con actividad bacteriocina identificada en
esta especie se le denominó acidofilina (Vicent y col., 1959), aunque más tarde se identificaron
la lactacina B (Barefoot y Klaenhammer, 1984) y la lactacina F (Muriana y Klaenhammer,
1991).
c) L. plantarurn .- En esta especie se h a n descrito tres bacteriocinas, la lactolina
(Kodama, 1952), la plantaricina SIK-83 (Andersson y col., 1988) y la plantaricina A
(Daeschel y col., 1990).
d) L.sake .- En los últimos años y de forma casi simultánea se han identificado dos
bacteilocinas producidas por microorganismos de esta especie, la sakacina A (Schillinger y
Lticke, 1989), y la lactocina 5 (M0rtvedt y Nes, 1990).
50

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 64/293
REVISION RIBLJOGRA PICA
También se ha citado la producción de bacteriocinas en L. delb,-ueckii subsp. ladis
(Toba y col., 1991). L. bulgaricus (Abdel-Bar y col., 1987) y en cepas de lactobacillosaislados a partir de intestino humano (Silva y col, 1987) y de la flora vaginal saprofita
(McGroaríy y Reid, 1988).
II. 5. 7. 2.- Bacteriocinas producidas por especies del género Lactococcus
La nisina producida por L. lactis subsp. lactis es, quizá, la bacteriocina mejoridentificada y caracterizada de todas las producidas por las bacterias lácticas (Hurst, 19813. No
obstante, además de la nisina se han descrito otras bacteriocinas, como la diplococina
producida por L. lactis subsp. cA-enioris (Davey y Richardson, 1981) y las lactostrepcinas,
distintas a la nisina y producidas por diversas cepas de L. lacris subsp. lactis y L. lactis
subsp. crernoris (Kózak y col, 1978; Dobrzanski y col, 1982). También se han identificado,
en otras especies de este género, 8 tipos diferentes de bacteriocinas que son claramente
distintas de la nisina y las lactostrepcinas, (Geis y col., 1983).
II. 5. 7. 3.- Bacteriocinas producidas por especies del género Pediococcus
Los primeros indicios de producción de bacteriocinas por parte de este género datande
mediados de lo s años 70, cuando se identificó una actividad inhibidora distinta del H202 en
una cepa de P. cerevisae (Fleming y col., 1975). Posteriormente se ha caracterizado la
pediococina A de?. pentosaceus (Daeschel y Klaenhammer, 1985) y las pediococina PA-1
(Gonzalez y Kunka, 1987) y pediococina AcH (Ehunia y col., 1988) producidas por 1’.
acidilacrici.
5 1

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 65/293
REVISiÓN BIBLIOGRAPICA
II. 5. 7. 4.- Bacteriocinas producidas por especies del género Carnobacwrium
Aunque es un género de nueva creación (Collins y col., 1987), ya se ha descrito [a
producción de bacteriocinas por C . piscicola (Ahn y Stiles, 1990) y C . d¡veigens (Schillinger
y Holzapfe¡, ¡990).
II. 5. 7. 5.- Bacteriocinas producidas porespecies del género Leuconostoc
A este género solamente se le atribuía actividad inhibidora debida a la producción de
ácidos orgánicos y diacetilo, hasta que se ha caracterizado la producción una bacteriocina por
una cepade Leu. gelidun¡ (Harding y Shaw, 1990; Hasting y Síiles, 1991).
5 2

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 66/293
MATERIALES Y METODOS
CAPITULO III
MATERIALES
Y
METODOS
5 3

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 67/293
MATERIALES Y MEFODOS
¡U . 1 .- MATERIALES
III. 1 . 1.- Material b i o l ó 2 i c o
Iii 1 . 1 . 1.- Microoreanismos empleados
Las bacterias lácticas utilizadas en la realización de este trabajo se han aislado de
embutidos crudos madurados, elaborados sin cultivo iniciador industrial y donados
generosamente por la empresa cárnica Industrias Cabo S.A., de Madrid. Las cepas se aislaron
siguiendo un método aleatorio y la mayor pai-te de ellas se identificaron tentativamente como
Lactobacillus sake s i g u i e n d o e l e s q u e m a d e i d e n t i f i c a c i ó n d e S c h i l l i n g e r y L i i c k e ( 1 9 8 7 b ) .
En la tabla 1 1 1 .1 se muestran lo s microorganismos utilizados como indicadores para
evaluar la actividad antimicrobiana de las bacterias lácticas procedentes de los embutidos
crudos madurados, así como su origen. Todos los microorganismos se liofilizaron en un
medio de leche desnatada al 10% para garantizar su conservación. Los “stocks” de trabajo se
mantuvieron en congelación a -20 oc en caldo nutritivo con un 15% de glicerol,
revitalizándolos cadados meses mediante dos o tres pases en el mismo caldo.
II. 1. 1. 2. - Obtención de lo s inmunosueros
Los inmunosueros se obtuvieron por inoculación de la sustancia antirnicrobiana
parcialmente purificada en conejos machos de raza “New Zealand”, de 2,5-3 Kg de peso,
cedidos por el animalario del Departamento de Cirugía Experimental de Hospital Militar Central
“Gómez Ulla” de Madrid.
5 4

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 68/293
MAIIERIALES Y MIETDDOS
III. 1 . 2. - Productos y reactivos
Los productos químicos utilizados en las experiencias descritas en este trabajo fueron
de calidad reactivo y suministrados por las siguientes firmas: “Merck”, “Probus”, “Sigma”,
“FLuka” o “Panreac”.
Los productos utilizados en las experiencias microbiológicas procedían de las firmas
“Difco” y “Oxoid”, excepto los componentes minerales, azúcares y colorantes que fueronsuministrados por “Merck”, “Panreac” y “BDH”.
En las técnicas cromatográficas y electroforéticas se emplearon productos
suministrados por “Pharmacia” y “Bio-Rad” y los utilizados en los ensayos enzimáticos,
inmunológicos e inmunoenziniáticos lo fueron por “Sigma”, “Difco”, “Pharmacia” y
“Nordik”.
II. 1 . 3. - Material de laboratorio
En la preparación de lo s medios de cultivo y soluciones acuosas se ha empleado agua
destilada, obtenida de un destilador “Afora”(1) y desmineralizada en un intercambiador iónico
“Seta” mod. R600.
Las pesadas ordinarias se realizaron en balanzas monoplato “Sauter” mod. S-1000 y
las de precisión en balanzas analíticas “Sartorius” mod. 2443 y “Microwa” mod. 6620. Para
equilibrar las muestras a centrifugar se utilizó una balanza biplaso “Cobos” mod. 28
( 1) La cita de marcas comerciales no significa que el autor las recomiende
con preferencia a otras del mercado.
5 5

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 69/293
MATERIALES YMIPTODOS
T ab l a I I I . 1 . - M i c r o o r g a n i s m o s in d ic ad or e s e m p l e a d o s p ar a e val u ar l a ac t ivi d ad
a n t i m i c ro b i a n a d e l a s b ac t er ia s l á c t i c a s aisl a d a s d e e m b u t i d o s cr u d os m a d u r a d o s
Microorganismo indicador Cepa n ~ Origena
Bacterias Gram-positivas
Lactobacillus acidophilus 289 CEOE
Lactobacillus brevis L b 2 2 6 IMTH Lactobacillus carnis L v 6 l FRIB LaCtobaCillus casel 4 7 5 CEOE Lactobacillus curvatus L b 7 2 6 IMTH Lactobacillus farcinzinis L b 3 4 I M T I - I Lactobacillusfermentunz 2 8 5 CEOE Lactobacillus n)eseItteiiCus 3 9 4 CEOE Lactobocillus plantarwn 2 2 1 CEOE
L b S ? ? IMTH Lactobacillus sake L b 6 8 4 IMTH Lactobacillus mesentericus MR 3 6 4 FRIB
Carnobacteriunz divergens Lvi 3 FRIBCainoba Cte/lun? piscicola MR37 1 FRIB
Micrococcus varians 230 CEOE
Enteí-ococcusfaecalis 481 CEOE Enterococcusfaecalis (var. liquefaciens) 1 84 CEOEEnterococcusfaeciunz 410 CEOE
Bacillus stearoterrnophilus 49 CEOE
Bacillusce,~eus
148 CEOE Brochotrív thermosphacua 10018 NCIB
Staphylococc’us xylosus 237 CEOEStaphylococcus aureus 59 CEOEStaphylococcus aureus 1 37 FR 1Staphylococcus aureus 361 FRI
56
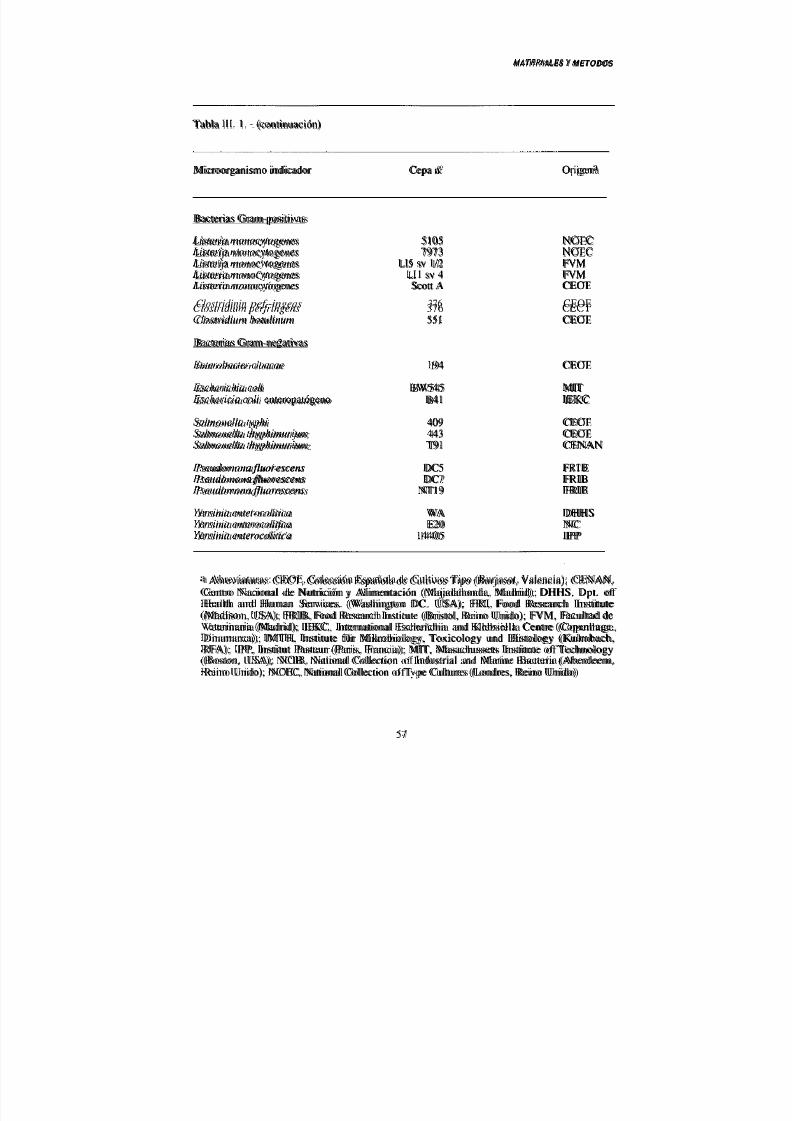
7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 70/293
MATI?í?IALES Y METODOS
Tabla 1 1 1 . 1 . - (continuación)
Microorganismo indicador Cepa & O¡.igena
Bacterias Gram-positivas
Listeria rnonocytogenesListeija nhonocytogenes
Listeija rnonoc’vtogenesListeria rnonoCytogenesListeria nzonocyíogenes
51057973
LIS sv 1/2LI 1 sv 4Scott A
NOECNOEC
FVMFVMCEOE
Clostridinin pef,-ingeas
Clostridiurn botulinurn
376
551
CEOE
CEOE
Bacterias Gram-ne2ativas
194 CEOE Enterobacter cloacae
Escherichia coli
Escheí-icia Coli enteropatógeno
BW545
B4 1
MIT
IEKC
Salnzonella typhiSalmonella thyphimurjan;Salmonella thyphimuriunz
409443T9 1
CEOECEOECENAN
Pseudornonafluoí-escensPseudomonafluorescensPseudornonafluorescens
DC5DC?
NT1 9
FRT EFR IBFRIE
Yersinia enteí~ocoliricaYersinia enterocolificaYersinia enterocoliric’a
WAE20
14405
DHHSNCIPP
a Abreviaturas: CEOE, Colección Española de Cultivos Tipo (Burjasot, Valencia); CENAN,Centro Nacional de Nutrición y Alimentación (Majadahonda, Madrid); DHHS. Dpt. of Health and Human Services. (Washington DC. USA); FRI, Food Research Institute(Madison, USA); FRIB, Food Research Insticute (Bristol, Reino Unido); FVM, Facultad deVeterinaria (Madrid); IEKC, International Escheñchia and Klebsiella Centre (Copenhage,Dinamarca); IMTH, Institute flir Mikrobiology, Toxicology und Histology (Kulmbach,RFA); IPP, Institut Pasteur (Paris, Francia); MIT, Masachussets Institute of Technology(Boston, USA); NCIB, National Collection of Industrial an d Marine Bacteria (Aberdeem,Reino Unido); NOEC, National Collection of T~pe Cultures (Londres, Reino Unido)
5 - 7

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 71/293
MATERIALES Y MIITODOS
Los ajustes y determinaciones de pH se llevaron a cabo co n pH-metros “Crison” mod.
Digit 501 y “Radiometer” mod. 28.
Las esterilizaciones de los medios de cultivo y de las soluciones cuya naturaleza así lo
permitía se realizaton en autoclaves “Averly’ y “Selecta” mod. 437-G. La esterilización del
material de vidrio se efectuó por calor seco en una estufa de aire forzado “I-Ieraeus” mod.
KFTU-K. La esterilización de otras soluciones tuvo lugar por filtración con filtros “Millipore”
de 0,22 y 0,45 gm de diámetro de poro.
Las homogenizaciones se llevaron a cabo en un homogenizador-Stomacher
‘Colworth” mod. 400.
Las centrifugaciones se efectuaron en una centrífuga refrigerada “Sorvalí” mod.
RC-5B, equipada con rotores SS-34 y GSA y en una microcentrífuga “Heraeus” Christ, mod.Biofuge, equipada con un rotor tipo 1220.
Las siembras microbianas se realizaron en una cámara de flujo laminar “Telstar” mod.
CE-A. Las incubaciones se efectuaron en estufas “Heraeus”, mods. KB-500 y B6200 y
“Selecta”. mod. Termotronic 338, termostatadas a la temperatura deseada. Los tratamientos
térmicos e incubaciones que requerían un control de la temperatura más preciso se realizaron en
baños de agua o de glicerina provistos de termostatos “Selecta”, mod. Tectron.
El crecimiento de lo s cultivos microbianos se estimó porturbidometría. empleando un
colorímetro “Klett-Summerson” mod. 800-3. Los recuentos microbianos se hicieron en un
contador de colonias “WTW” mod. BZG-24. Las observaciones microscópicas se realizaron
en un microscopio óptico “Nikon”, ruod. L-ke, equipado con un dispositivo de contraste de
fases.
58

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 72/293
MATERIALíIS Y MEPODOS
Las determinaciones espectrofotométricas se llevaron a cabo en espectrofotómetros
“Kontron”, mod. Uvikon 820, e “Hitachi”, mod. U-2000, registrándose lo s resultados en una
impresoi-a térmica “Kontron”, mod. Uvikon LS Thennoprinter 4B .
Las muestras se conservaron en arcones congeladores “Kelvinator”, mods. AC-550 y
ACK-55 y “Liebherr”, mod. GT6102, así como en frigoríficos “Aspes”. “Kelvinator” mod.
AKR y “Liebherr” y en un armario frigorífico termostatado a 4 0C±1 0C construido por una
firma local.
Las liofilizaciones se efectuaron en una aparato “Terruzzi Melvisa”, mod. TP-3.
Las cromatografias de filtración en geles se realizaron utilizando columnas de
diferentes dimensiones de las marcas “Phai-macia” y ‘Wright”. Las fracciones cromatográficas
se recogieron en colectores de fracciones “LKB-Bromma” mod. 7000 Ultrarac y 2212 Helirac.
Las electroforesis en geles de poliacrilamida co n dodecil sulfato sódico se practicaron
en una cubeta de electroforesis “Bio-Rad’. mod. Protean II, empleando como fuente de
alimentación un aparato “Shandon”, mod. SAE 2761.
En los ensayos inmunoenzimáticos (ELISA), se emplearon placas de ELISA marca“Costar” mod. 3590 de 96 pocillos y la lectura de las mismas se efectuó en un lector de placas
ELISA “Titerek Multiskan” mod. Plus.
En la determinación de la composición aminoacídica de la susíancia antimicrobiana
p arc ia l me n t e p u r i f i c a d a m e d i a n t e t é n i c a s d e c r o m a t o g r a f í a l í q u i d a d e a l t a r e s o l u c i ó n ( H P L C ) .
se empleó un cromatógrafo Beckman mod. 332. dotado de una columna (25 cm x 4,6 mm) de
fase reversa Spherisorb ODS-2 (Supelco) y un detector UV. Beckman mod. 160. El
59

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 73/293
MATER/ALES Y MÉTODOS
cromatograma se obtuvo en un integrador Hewlett Packard mod. HP-3394A.
Como material general de laboratorio se han utilizado pipetas, micropipetas
automáticas “Gilson”, mods. P-20, P-200. P-l000, agitadores, mecheros de gas,
termómetros, etc. El material de vidrio empleado fué siempre del tipo “Pirex” -
I I I . 2. - METODOS
III. 2. 1 . - Medios de cultivo empleados en el crecimiento de lo s mlcroor2antsmos
1 1 1 . 2. 1 . 1 . - Medios de cultivo para el crecimiento de las bacterias lácticas
1. - Medio de Man. Rogosa y Sharpe <MRS. de “Oxoid”
Contiene:
Peptona 10,0
Extracto de carne 8,0
Extracto de levadura 4,0
Dextrosa 20,0
A c e t a t o s ó d i c o 5 , 0
Fosfato dipotásico 2,0
Citrato triamónico 2,0
5 ulfato magnésico 0,2
Sulfato de manganeso 0,05
Tween 80 í ml
pH 6,2
60

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 74/293
MATERIALES Y MÉTODOS
Preparación
Se suspenden 52 g del medio de M RS en 11500 ml de agua destilada calentando la
solución hasta su disolución y esterilizándola en un autoclave a 121 0C durante 15 minutos.
Cuando el medio se depositaba en placas de Peni, a lo s componentes citados se les añadía 10,0
g/l de agar bacteriológico (DIFCO) y 7,5 gil cuando se elaboraba el agar de MRS semisólido
para los experimentos de inhibición.
2. - Medio base
Contiene: g/l
Dextrosa 10,0
Extracto de levadura 5,0
Cloruro sódico 2,0
Citrato amónico 2,0Fosfato potásico dihidrogenado 1 ,0
Fosfato dipotásico 1 ,0
Sulfato magnésico 0,20
Sulfato de manganeso 0,05
Sulfato feaoso 0,01
Tween 80 1 mlpH final llevado a 6,2 co n HCI iN
Pieparación.
Para estudiar la influencia de la fuente de nitrógeno en la producción de las sustancias
antibacterianas, este medio base se suplementó con 1 5 g/l de uno de lo s siguientes extractos:
triptona, triptosa. proteosa-peptona, casitona, peptona (suministrados por Difeo) o extracto de
61

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 75/293
MA TER/ALE S Y MÉTODOS
carne (Oxoid>. Los componentes citados se disolvieron en 1.000 ml de agua destilada. La
preparación ulterior de cada uno de lo s medios semisintéticos así obten idos es idéntica a laseñalada para el medio de MRS (sección III. 2. 1 . 1) El medio base suplementado con 1 5 gil
con triptosa (MM-Triptosa) se empleó para la purificación de las sustancias antimicrobianas
producidas por las bacterias lácticas.
III. 2. 1 . 2.- Medio de cultivo para el crecimiento de microorganismos indicadores
distintos de las bacterias lácticas
1 . Caldo infusión de cerebro y corazón (BHI, “Oxoid”
Contiene: g/ l
Extracto de ce jebro 12,5
Extracto de corazón 5,0
Proteosa-peptona 5,0
Cloruro sódico 5.0
Dextrosa 2,0
Fosfato disódico 2,5
pH 7,4
Preparación
Se suspenden 37 g del producto deshidratado en polvo en un litro de agua destilada.
La preparación ulterior del medio así obtenido es idéntica a la señalada para el medio de MRS
( s e c c i ó n I I I . 2 . 1 . 1 )
62

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 76/293
MATERIALES Y MI7FODOS
2. Caldo con Tween para todos lo s uropósitos (APT. “Difco”~
Contiene:
Triptona
Dextrosa
Extracto de levadura
Citrato sódico
Cloruro sódico
Fosfato dipotásicoSulfato magnésico
Cloruro de manganeso
Sulfato ferroso
Clorhidrato de tiamina
Tween 80
pH 6,7
g/ l
12,5
10,1
7,5
5,0
5,0
5,00,8
0,14
0,04
0,001
0,2
Preparación
Se suspenden 46,2 g del polvo deshidratado en un litro de agua destilada. La
preparación ulterior del medio así obtenido es idéntica a la señalada para el medio de MRS
(sección III. 2. 1 . 1).
III. 2. 2. - Aislamiento y selección de las bacterias lácticas de lo s embutidos crudos
madurados
Las bacterias lácticas (cepas LAB) utilizadas en este trabajo se aislaron durante la
maduración de un lote de embutidos comerciales a lo s que no se les había añadido ningún
63

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 77/293
MATERIALES Y MEXODOS
cultivo iniciador. Las muestras se tomaron de la masa cárnica durante lo s siguientes dias de
maduración: 0, 2, 6, 10, 21 y 35. Para el aislamiento de las bacterias lácticas se recogieron
asépticamente 20 g de la porción central de los embutidos que se homogeneizaron durante 10
minutos en 180 mIde un medio que contenía peptona (1 gIl) y cloruro sódico (8,5 gIl) en agua
destilada.
El recuento total de microorganismos presentes en el homogeneizado se realizó en
placas de PCA suplementado con cloruro sódico (1%). El agar de recuentos en placa (PCA,“Oxoid”) contiene (por litro); 5,0 g de triptona, 2,5 g de extracto de levadura, 1 ,0 g de
dextrosa y 9,0 g de agar. Su pH final es de 7,0 y se prepara suspendiendo 17,5 g de polvo del
medio deshidratado en un litro de agua destilada. El resto de las operaciones de preparación
del medio son idénticas a las indicadas para el medio de MRS (sección III. 2. 1 . 1 . 1). El
recuento de bacterias lácticas se efectuó en agar de MRS. Las placas de PCA y de MRS se
incubaron a 320C durante 3 días.
De las colonias crecidas en agar de MRS se seleccionaron 180, en grupos de 30,
durante lo s dias 0, 2, 6, 10 , 21 y 35 de maduración siguiendo el método aleatorio descrito por
Ordóñez (1979). Las colonias seleccionadas se recogieron con una asa de platino y se
sembraron en tubos que contenían caldo de MRS, manteniendo los tubos durante 16 h a 32
0C. A los cultivos obtenidos se les adicionó un 15% de glicerol estéril y se almacenaron encongelación a -20 0C hasta su posterior utilización. Alícuotas de cada una de las cepas se
almacenaron en liofilización con leche desnatada al 10 %.
64

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 78/293
MATERIALES Y MEFODOS
III. 2. 3. - Actividad antimicrobianade las bacterias lácticas seleccionadas
1 1 1 . 2. 3. 1 . - Soluciones tampón empleadas
1.) Tampón de fosfato 4 mM, pH 7,0.
a. Solución de Na2HPO4, 4m iN 4.
Contiene 0,76 g de Na2HPO4 en 1 litro de agua destilada.
b. Solución de NaH2PO4, 4 mM.
Contiene 0,55 g de NaH2PO4 en 1 litro de agua destilada.
Preparación
Se añade la solución (b) a la (a) hasta que su pH alcanza el valor de 7,0.
III. 2. 3 . 2 . - Pruebas directas de antagonismo
111.2.3.2. 1.- Prueba directa de antaEonismo en pocillos
Para detectar la actividad inhibidora de las bacterias lácticas (LAB), aisladas de lo s
embutidos crudos madurados, frente a lo s microorganismos indicadores, se ideé un sistema
consistente en utilizar placas de Petri con medio MPS sólido en las que, con un sacabocados
de? mm de diámetro, se hicieron pocillos, cuyo fondo se tapizó con 1 0 ~fldel mismo medio.
A continuación se depositaron en cada pocillo 20 pl de los cultivos de las cepas LAB
escogidas y cultivadas en caldo MRS, después de laxadas y resuspendidas en el mismo medio.
65

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 79/293
MATERIALES Y MEIVDOS
Las placas se prepararon por duplicado y cada pocillo se rellenó con agar MRS.
Posteriormente, los microorganismos indicadores a una concentí-ación ap¡-oxiinada de 1 x ío~ufc/ml, se sembraron en una de las placas del par, que contenía agar MRS, cuando se trataba
de bacterias lácticas y agar BHI en el resto de los casos. Tanto las placas a las que se les había
añadido el indicador como sus compañeras del par carentes del mismo, se incubaron a 32 0C
durante 24 horas. Transcurrido éste tiempo se añadió indicador a las placas del par que
carecían del mismo y cuyas cepas LAB habían obtenido una ventaja de crecimiento de 24
horas, siguiendo la pauta citada (prueba de actividad inhibidora diferida). Estas placas seincubaron durante a 32 0C durante otras 24 horas.
Con esta técnica la inhibición ejercida por las bacterias lácticas en el microorganismo
indicador se manifiesta porla aparición de halos en la superficie de las placas. Dicha inhibición
se cuantifica determinando el área de la corona circular del halo de inhibición (mm2) por
unidad de densidad óptica de los cultivos en unidades Klett y por 0,020 ml de medio de
cultivo.
III. 2. 3 . 3 . - Actividad inhibidora de los sobrenadantes concentrados libres de células
¡U. 2. 3 . 3 . 1 - Preparación de los sobrenadantes
Las 24 bacterias lácticas seleccionadas (20 con gran actividad inhibidora directa y 4
con una actividad manifiestamente menor), se cultivaron en caldo MRS durante 16-18 h a 32
y los sobrenadantes libres de células se obtuvieron centrifugando lo s cultivos a 12.000 g
durante 10 minutos. El pH de los sobrenadantes se ajustó a 6,2 con NaOH IN y, a
continuación, se filtraron por un filtro de 0,22 gm de diámetro de poro. Los filtrados se
liofilizaron y antes de su utilización se concentraron 20 veces, resuspendiéndolos en tampón de
fosfato 4 mM, de pl] 7,0.
66

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 80/293
MATERIALES Y MÉTODOS
III. 2. 3. 3. 2. - Detección de la actividad inhibidora de los sobrenadantes
concentrados 20 veces
Para detectar y cuantificar la actividad inhibidora de dichos sobrenadantes se utilizaron
discos de papel de filtro Whatman n 9 3. de 7 mm de diámetro, que contenían 0,030 ml de los
sobrenadantes concentrados 20 veces. Estos discos se colocaron en placas de Petri que tenían
su fondo tapizado con una fina capa de agar base y en las que se depositaron aproximadamente
3 x io~ células de los microorganismos indicadores de interés señalados en la tabla III 1,
resuspendidos en 6 ml de agar semisólido MRS o BHI (0,75 % de agar en todos los casos).
Las placas se incubaron a 32 0C durante 24 h y la actividad inhibidora de los sobrenadantes se
determinó midiendo lo s halos de inhibición formados alrededor de lo s discos de papel de filtro.
Para ello se utilizó la expresión: Diámetro del halo de inhibición (Radio de la zona de
inhibición x 2) + d i á m e t r o del papel de filtro (7 mm).
III. 2. 3. 4. - Efecto de la catalasa en la actividad inhibidora de las bacterias lácticas
seleccionadas
La influencia del H202 en la actividad inhibidora de las bacterias lácticas se evaluó
adicionando catalasa (130 UI/mí) a lo s cultivos libres de células neutralizados y filtrados. La
actividad inhibidora se determinó empleando el sistema de discos en placas descrito en la
sección III. 2. 3. 3. 2. La actividad inhibidora debida al H202 se determinó cuantificando la
diferencia de diámetro entre los halos de inhibición de las muestras problema y las de los
controles a las que no se les había adicionado catalasa.
67

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 81/293
MATERIALES Y MI7iODOS
IIL 2 . 4 . - Identificación y carac¡erización bi~uímica de al2unas bacterias lácticas con
actividad antimicrobiana
III. 2. 4. 1 . - Morfolo2ía y tinción por el método de Gram
Los cultivos de las 6 bacterias lácticas seleccionadas por la mayor actividad inhibidora
de sus cultivos libres de células, se tiñeron por el método Gram y se observaron al
microscopio para determinar su morfología y sus afinidades tintoriales.
TU. 2 . 4 . 2 . - Prueba de la catalasa
La producción de catalasa se realizó añadiendo una gota de peróxido de hidrógeno
(110 vol) a una alícuota del cultivo a analizar. Como control positivo se empleó un cultivo de
S. aureus. La prueba se basa en la descomposición del peróxido de hidrógeno por la catalasadando agua y oxígeno, manifestándose este último por la presencia de burbujas.
1 1 1 . 2. 4. 3. - Producción de gas a partir de glucosa
La producción de C O2 a partir de L a glucosa se determinó de dos mane;-as diferentes:
1.) —
composición
cultivos, éste
Introduciendo campanas de Durham en tubos con caldo MRS de cuya
se suprime el citrato. Si las cepas producen gas durante la incubación de los
se manifiesta por la presencia de burbujas en el interior de la campana.
2.) - Colocando en lo s tubos de medio CS-T modificado tapones de agar (2 % p/v) y
observando si durante la incubación de aquellos se produce un desplazamiento de lo s tapones
debido a la presencia de gas.
68

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 82/293
MA ¡‘ERIALES Y METODOS
El medio CS-T modificado contiene:
g/l
Casaminoácidos~1> 5,0
Extracto de levadura 5,0
Fosfato monopotásico 0,6
Fosfato dipotásico 0,6
Sulfato magnésico (7 H20) 0,2
Sulfato de manganeso (1 H20) 0,05
Sulfato ferroso 0,01
Clomro sódico 0,01
Bromotimol azul 0,08
Agar 2,5
Tween80
1,0 mlpH 6,2
Preparación
A un litro de medio de cultivo esterilizado a 1210C durante 1 5 minutos, se le añade
una solución de glucosa al 10 % en una proporción del 10% (y/y). La solución de glucosa se
esteriliza por filtración con filtros Millipore de 0,22 ~.tmde tamaño de poro. Una vez enfriado
el medio, se le adicionan 150 jil del cultivo a investigar realizándose a continuación picaduras
en el medio semisólido. Se le añaden 2,5 ml del agar al 2 %. La formación de gas se detecta
porel desplazamiento del tapón hacia arriba después de 3 días de incubados los tubos a 32 0C.
Una mezcla comercial de aminoácidos obtenidos por hidrólisis de caseína
69

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 83/293
MATERIALES Y MCI ODOS
III. 2. 4. 4. - Hidrólisis de la ar2inina
L) Caldo Arginina
Contiene: g / 1
Peptona 10,0
Extracto de levadura 4,0
Fosfato dipotásico 2,0
Acetato sódico 5,0Citrato sódico 2.0
Sulfato magnésico 0,2
Sulfato de manganeso 0,02
L-Arginina 3,0
Tween 80 1,0
Preparación.
Una vez disueltos los componentes del medio, se distribuye en tubos que se esterilizan
en autoclave a 121 0Cdurante 1 5 minutos.
2.) Reactivo de Nessler
a.) Solución 1 .
Contiene:
Yoduro potásico 7 g
Yoduro mercúrico lO g
Agua destilada 40 mi
70

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 84/293
MATERIALES Y M171ODOS
b.) Solución 2.
Contiene 1 0 g de NaOH en 50 ml de agua destilada.
Preparación.
Una vez preparadas las dos soluciones (y dejada enfriar la solución 2), se mezclan en
un matraz aforado de 100 ml y se añade agua destilada hasta enrasar. Se deja que sedimente el
precipitado, se decanta el líquido sobrenadante claro, que se guarda, y se desecha elprecipitado.
Sembradas las bacterias lácticas de interés en caldo de Arginina, se incuban a 32 0C
durante 2 días; a 1 mIde éstos cultivos se le añade 1 ml del reactivo de Nessler. La hidrólisis
de la arginina, con formación de amoníaco, produce una alcalinidad que se manifiesta por el
desarrollo deun
colorentre anaranjadoy
marrón.
¡U . 2. 4. 5. - Producción de ácido sulfhídrico
III. 2.4. 5. 1. - Técnica de Shav y E~an (l981~
1 . Agar base de acetato de plomo (Difco).Contiene: g/l
Peptona 15,0
Pioteosa 5,0
Dextrosa 1 ,0
Acetato de plomo 0,2
Tiosulfato sódico 0,08
Agar 15,0
71

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 85/293
MA ¡ERIALES Y MÉTODOS
pH 6,6
Pre pw-ocíón.
Se suspenden 36 g del medio deshidratado, 1 ml de Tween 80 y 0,05 g de sulfato de
manganeso en 1 litro de agua destilada La suspension se calienta hasta ebullición y se
esteriliza en autoclave a 121 0C durante 1 5 minutos. El medio esterilizado sedeja enfriar hasta
45 0C para distribuirlo finalmente en placas.
Si las colonias desarrolladas en las placas incubadas a 32 0C durante 3 días producen
ácido sulfhídrico, se observa un ennegrecimiento del medio.
III. 2 . 4 . 5 . 2 . - Asar hierro de Klieser
Los cultivos seleccionados se inocularon en tubos inclinados de agar hierro de Klieger,
tanto en picadura como en estría. Los tubos se incubaron durante 7 días a 32 0C ,
manifestándose la producción de ácido sulfhídrico por un ennegrecimiento del medio.
H l. 2. 4 . 6 . - Prueba de Voses-Proskauer
a.) Caldo MR-VP.
Contiene: g/l
Peptona 7.0
Fosfato dipotásico 5.0
Dexuosa 5,0
pl] 7,5
72

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 86/293
MATERIALES Y MElaDOS
a.) Solución de a-naftol en etanol
Contiene:
a-naftol
Etanol
c.) Solución de hidróxido potásico.
Contiene:K O I- I
Agua destilada
Prepún -ación.
Una vez disuelto el medio a.) se distribuye en tubos que se esterilizan en autoclave a
121 0C durante 1 5 minutos Sembrado el inóculo de la cepa problema e incubados los tubos a
32 0C, a 5 ml de los cultivos se añaden 0,5 ml de la solución b.) y 0,5 ml de la cj y se observa
el cambio de coloración del medio. El desarrollo de un color rojo intenso se debe a la
producción de acetil metil carbinol a partir de la dextrosa que, en presencia de ICOR, reacciona
con el a-naftol.
III. 2. 4 . 7. - Fermentación de carbohidratos
1.) Medio API Cí-IL (BioMérieux)
Contiene:
Peptona
Extracto de levadura
Tween 80
gil
10,0
5,0
1 ,0
6
100
40
100
g
ml
g
ml
73

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 87/293
MATERIALES Y MÉTODOS
Fosfato dipotásico 2,0
Acetato sódico 5,0
Citrato diamónico 2,0
Sulfato magnésico 0,20
Sulfato de manganeso 0.05
Bromocresol púrpura 0,17
pH 6,9-7,0
2.) Tiras API 5 0 C H (BioMérieux).
Cada tira contiene los siguientes carbohidratos: 0. control; 1 . glicerol; 2. eritritol; 3.
D-arabinosa; 4. L-arabinosa; 5. ribosa; 6. D-xilosa; 7. L-xilosa; 8. adonitol; 9. ~3-metil
D-xilósido; 10. galactosa; 11 . glucosa; 12 . fructosa; 13 . manosa; 14 . sorbosa; 15. ramnosa;
16. dulcitol; 17. inositol; 18. manitol; 19 . sorbitol; 20. a-metil D-manósido; 21. a-metil
D-glucósido; 22. N-acetil glucosamina; 23. amigdalina; 24. arbutina; 25. esculina; 26. salicina;
27. celobiosa; 28. maltosa; 29. lactosa; 30. melibiosa; 31. sacarosa; 32. trealosa; 33. inulina;
34. melicitosa; 35. rafinosa; 36. almidón; 37. glucógeno; 38. xilitol; 39. gentibiosa; 40.
D-turanosa; 41. D-lixosa; 42. D-tagatosa; 43. D-fucosa; 44. L-fucosa; 45. D-arabitol 46.
L-arabitol; 47. gluconato; 48. 2-cetogluconato y 49. 5-cetogluconato.
Procedimiento.
Los cultivos seleccionados, desarrollados previamente en caldo MRS, se centrifugaron
a 12.000 g durante 1 0 minutos y el sedimento (las células) obtenido se resuspendió en el
medio API CHL. El sedimento resuspendido se depositó en microtubos en lo s que se
encuentran los carbohidratos La zona aeróbica de los microtubos (superficie) se cubrió con
aceite de parafina para crear las condiciones de anaerobiosis requeridas. Las tiras con lo s
74

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 88/293
MATERIALES Y MÉTODOS
distintos microtubos se incubaron a 32 0C, realizando dos lecturas de las mismas a las 24 y 48
h, respectivamente. La fermentación de lo s carbohidratos se manifiesta por un cambio de colordel microtubo, debido a la producción anaeróbica de ácido, que es detectado por el indicador
de pH incluido en el medio API CHL.
III. 2 . 4 . 8 . - Tolerancia al cloruro sódico
La tolerancia de las bacterias seleccionadas a la sal se determinó en caldo MRSsuplementado con un? y un 1 0 % de NaCí, respectivamente. El medio, una vez inoculado, se
incubó durante 6 días a 32 0C.
III. 2. 4. 9. - Tolerancia al pH de 3.9
El crecimiento de los cultivos a pH 3,9 se evaluó en caldo MRS cuyo pH se había
ajustado a 3,9 con HCI IN. El medio se incubó durante 5 días a 32 0C.
III. 2. 4. 10 . - Desarrollo a diversas temperaturas
El desarrollo de lo s cultivos a diversas temperaturas se analizó depositando alícuotas
de cada uno de ellos en tubos de ensayo que contenían caldo MRS. Una vez inoculados los
tubos, se procedió a su incubación a las diversas temperaturas durante periodos variables de
tiempo. El desarrollo de lo s cultivos se determinó a intervalos de tiempo regulares midiendo su
absorbancia en Unidades Klett.
‘7 5

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 89/293
MATERíALES Y MEraDOS
III. 2. 5.- Esnectro antimicrobiano de las sustancias inhibidoras producidas por las
cepas deL. sate números 77. 90. 148 y 180
El espectro antimicrobiano de las 4 cepas de U sake que mostraron una mayor
actividad antagonista, se detenninó viendo la actividad inhibidora frente a lo s microorganismos
indicadores citados en la tabla III. 1 del sobrenadante concentrado libre de células, e laborado
según se describe en la sección Hl. 2. 3. 3. La actividad inhibidora se determinó midiendo lo s
halos de inhibición alrededor de lo s circulos de papel de filtro y se expresó como: Diámetro de
inhibición — (Radio de la zona de inhibición x 2) + d i á m e t r o del papel de filtro (7 mm).
III. 2. 6. - Efecto de la composición del medio de cultivo en la actividad
antimicrobiana de Locwbacillus s a k e 148
Para estudiar el efecto del medio de cultivo en la actividad antimicrobiana deL. saLe
148, este se sembró en diversos medios de cultivo que se incubaron durante 1 6 h a 32 ‘C . Los
medios empleados fueron los siguientes:
1.) Medio de APT
2.) Medio de APT suplementado:
a) con 1 0 g de triptosa/l (APT-Triptosa)
b ) con 1 0 g de proteosa-peptona/l (APT-Proteosa)
c) con 1 0 g de peptonail (APT-Peptona)
d) con 1 0 g de extracto de came/l (APT-Extracto de carne)
3.) Medio EHI
4.) Medio BHI con las supletnentaciones descritas:
a) BHI-Triptosa
b) BHI-Proteosa
c) Rl-II-Peptona
76

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 90/293
MATERIALES Y METOI)OS
d) EHI-Extiacto de carne
5.)
Medio base suplementado:a) con 15 g de triptosall (MM-Triptosa)
b) con 15 g de proteosa/l (MM-Proteosa)
c) con 15 g de peptona/l (MM-Peptona)
d) con 15 g de extracto de carne/l (MM-Extracto de carne)
e) con 15 g de caseina/1 (MM-Caseina)
fl co n 1 5 g de triptonall (MM-Triptona)
Terminada la incubación se prepararon lo s sobrenadantes concentrados libres de células
y se determinó su actividad antimicrobiana de la manera descrita en la sección III. 2. 3. 3.,
utilizando a L.fermcntunz CEGI’285 como el microorganismo indicador
III. 2. 7. - Crecimiento de L. saLe 148. cinética del desarrollo y producción de
sustancia antimicrobiana a diversas temperaturas
III. 2.7. 1. - Crecimiento de los cultivos
Alícuotas de 100 ~.dde un cultivo de L. sake 148 se inocularon en tubos con caldo
MRS y MM-Triptosa y se incubaron a 4, 8,15,20 y 32 0C durante? días, 5 días, 30 h, 1 6 h y12 h, respectivamente. El desarrollo de lo s cultivos se estimó por turbidometría a determinados
intervalos de tiempo; con los datos obtenidos se calcularon los parámetros cinéticos del
desarrollo microbiano que se describen en la sección III. 2. 6 . 2 . En lo s mismos intervalos se
determiné el valor del pH del cultivo.
La actividad antimicrobiana de los sobrenadantes obtenidos en cada intervalo, se
determinó como se indica en la sección III. 2 . 3 . 3., empleando a Lferrnentum CECT285
77

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 91/293
MATERIALES Y MITODOS
como el microoraanismo indicador.
1 1 1 . 2. 7. 2. - Parámetros cinéticos del desarrollo microbiano
111.2.7.2. 1.- Velocidad específica de crecimiento (u
Se define como el aumento de la masa celular de lo s cultivos respecto del tiempo de
incubación y se calcula a partir de la ecuación de la recta de regresión de una gráfica que
representa la masa celular de lo s cultivos en función del tiempo de incubación (figura 3.1)
xc
t
Figura 3. 1.- Representación gráfica teórica del incremento de la masa celular de lo s
cultivos en función del tiempo de incubacion.
1 - ’
78

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 92/293
MATERIALES Y MÉTODOS
De la figura 31 se deduce que:
dx uxdt
siendo “x ” la densidad óptica de lo s cultivos en unidades Klett, “ t ” e l tiempo de i n c u b a c i ó n e n
horas y “ 1 ” l a v e l o c i d a d e sp e cíf ic a d e c r e c i m i e n t o .
Asimismo,
X 2 dx
5 5 x o
gdt;
X2
In
X 2
In = jit ; p o r t ant o,
t
III. 2. 7. 2. 2. - Tiempo de dunlicación ( t1
Se define como el tiempo que tardan lo s cultivos en duplicar su masa celular a una
temperatura de incubación determinada. Su valor se calcula de la expresión:
x2In ____ r ¡ i t ; cuando X2 = 2X1;
xl
2X1In =jlt<j: ln
2=Jitd;0,693
td =
xl
III. 2. 7. 2 3. - Número de generaciones por hora (st/h
Este término indica el número de generaciones microbianas de lo s cultivos en una hora
a una temperatura detenninada y su valor se corresponde con el inverso del tiempo de
79

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 93/293
MATERIALES Y MÉTODOS
duplicación «a).
I I I . 2. 7. 2 . 4 . - Determinación del peso celular seco
El peso celular seco se determinó por interpolación en una gráfica (figura 3. 2) en la
que se representa el peso celular seco con respecto a la absorbancia en Unidades Klett
III. 2. 7. 2. 4 . 1 . - Elaboración de la recta patrón para la determinación del yeso celular
seco
Para determinar la recta patrón se prepararon una serie de tubos con medios de cultivo
estériles de MRS y MM-triptosa, que se inocularon con J i. saLe 148 y se incubaron a 32 0C.
La absorbancia de los cultivos se determinó cada ciertos intervalos de tiempo al
progresar el tiempo de incubación; para ello se tomaban alícuotas de 5 ml de lo s medios de
cultivo a intervalos regulares en lo s que se determinaron las Unidades Klett hasta que éstas se
estacionaron (ya no aumentaban más).
Las alícuotas se filtraron por filtros Millipore, de 0,45 ~m de diámetro de poro,
previamente tarados. Los filtros, en los que habían quedado retenidas las células, se
introdujeron en placas de Petri de vidrio y se desecaron durante 10 h a 80 0C. El peso celular
seco (n Jml), correspondiente a cada absorbancia se estableció determinando la diferencia en
peso del filtro con y sin microorganismos dividida por 5. La absorbancia en Unidades Klett se
relacionó con el peso celular seco mediante una recta de regresión (fig 3. 2).
80
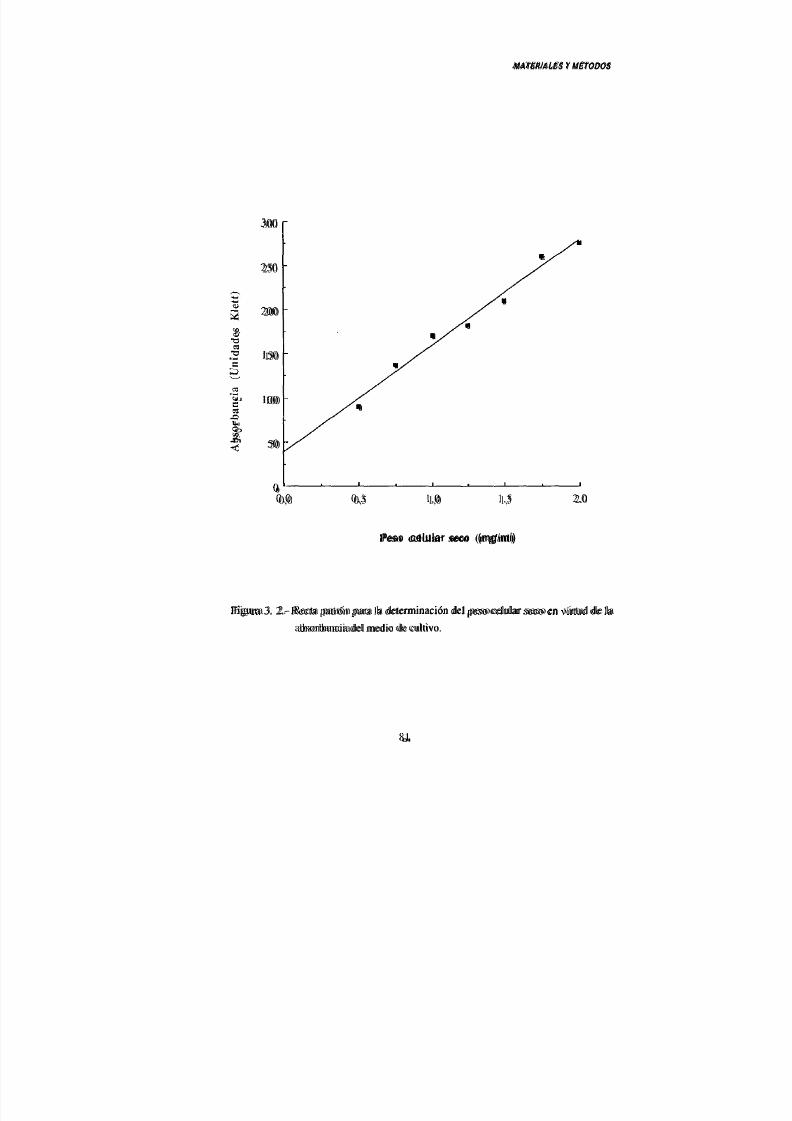
7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 94/293
MATER/A LE S Y MÉTODOS
300
250
y,
u
2- aoy,n
200
150
100
50
o0,0 2.0
Peso celular seco (mg/mí)
Figura 3. 2.- Recta patrón para la determinación del peso celular seco en virtud de la
absorbancia del medio de cultivo.
E
u
uu
u
u
0,5 1,0 1 ,5
81

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 95/293
MATERIALES Y MÉTODOS
III. 2. 8 . — Purificación parcial de la actividad antimicrobiana exocelular deL. saLe 148
III. 2. 8. 1 . - Cromatografía de filtración en celes
Esta técnica cromatográfica se basa en la separación de las moléculas por su tamaño
molecular.
III. 2 . 8 . 1 . 1 . - Soluciones tampón emoleadas
1.) Tampón de ácido cítrico-fosfato, de pH 5,6.
(a.) Solución de ácido cítrico, 0,1 M.
Contiene 21,01 gde ácido cítrico (1 H20) por litro de agua destilada.
(b.) Solución de Na,HPO4, 0,2 M.
Contiene 28,4 g de Na2HPO4 por litro de agua destilada.
Preparación.
Se mezclan 42 ml de ácido cítrico, 0 ,1 M con 58 ml de Na9HPO4. 0,2
M.
2.) Tampón de ácido cítrico-fosfato, de pH 5,6, con urea 0 ,1 M, y 1 M
A la solución tampón base (sección III. 2. 8. 1 . 1 . 1), se l e a ñ a d e n , re sp e c t iv am e n t e , 6
g ó 60 g de urea p or l i t r o d e s o l u c t o n .
I I I . 2. 8. 1 . 2. - Geles
El Sephadex es un polímero resultante de la formación de enlaces ciuzados entre las
moléculas de dextrano y de epiclorhidrina. Debido al gran número dc grupos hidroxilo de su
molécula este polímero es muy hidrofílico y se hincha fácilmente en el agua y en soluciones
82

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 96/293
MA TiRRiALE S Y MÉTODOS
electrolíticas. Los tipos de Sephadex difieren en su grado de entrecruzamiento, por lo que se
utilizan par-a alcanzar fraccionamientos co n diversos intervalos de tamaño molecular y que en elcaso de los Sephadex 0-150 fino, G-75 y 0-50 son, respectivamente, de 5.000 a 300.000
daltons, de 3.000 a 80.000 daltons y de 1.500 a 30.000 daltons.
Los geles se prepararon y activaron según las normas de la casa suministradora
(“Pharmacia Fine Chemicals). Los geles hidratados se conservaron en refrigeración hasta su
utilización.
III. 2 . 8 . 1 . 3. - Condiciones de trabaio
La cromatografía se realizó en una cámara termostatada a 4±10C y el eluato
cromatografiado se recogió en un colector de fracciones.
El sobrenadante, libre de células y liofilizado, de cultivos de Ji . saLe 148
desarrollados en MM-triptosa durante 1 6 h a 32 0C , se resuspendió en el tampón de ácido
cítrico-fosfato, de pH 5,6 y urea 1 M, hasta una concentración 20 veces mayor que la inicial. A
continuación 20 ml de esta solución se depositaron en una columna (3,2 x 4 4 3 cm) de Sephadex
G-150, previamente equilibrada con el tampón de ácido cítrico-fosfato, de pH 5,6 y urea 0 ,1
M. El eluato, en fracciones de 5 mí, se sometió a una lectura espectrofotométrica a 280 nmpara determinar su contenido proteico, mientras que su actividad inhibidora se estableció de la
manera descrita en la sección III. 2 . 3 . 3 . 2 . utilizando L. fermentum CECT285 como el
microorganismo indicador. Las fracciones cromatográficas con actividad antimicrobiana se
juntaron, liofilizaron y resuspendieron en el tampón de elución para depositarias de nuevo en
una columna (2,5 x 90 cm) de Sephadex 0-75, previamente equilibrada con cl tampón de
clución. Las fracciones eluidas dotadas de actividad antimicrobiana se manipularon corno se ha
descrito antes y se depositaron de nuevo en una tercera columna (1,6 x 90 cm) de Sephadex
83

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 97/293
MATERIALES Y MÉTODOS
G-50. Las fracciones cromatográficas con actividad antimicrobiana se juntaron, liofilizaron y
se mantuvieron en un desecador a 4 0C hasta su utilización posterior.
III. 2. 8. 2. - Determinación de la proteína
Se realizó por la técnica de Lowry, según la modificación de Markwell y col. (1978).
La técnica se basa en el desarrollo de color al poner en contacto las proteínas con los reactivos
que posteriormente se detallan. El desarrollo del color se debe a una combinación de
reacciones:
a.) Formación de un complejo entre lo s enlaces peptídicos de las proteínas con el cobre
en tin medio alcalino (reacción tipo Biuret).
b.) Reducción del reactivo fosfomolíbdico-fosfotúngstico por la tirosinay
eltriptófano.
Esta técnica pone de manifiesto lo s grupos fenoles presentes en las proteínas; por ello
es necesario extrapolar los resultados a una curva patrón construida con anterioridad. Como
proteína estándar para construir la curva patrón se empleó la fracción V de la seroalbúmina
bovina (figura 3. 3).
Reactivos:
- Solución A: Na,C03 al 2 %, NaOH a l 0.4 % y tartrato sódico
p ot á sic o (NaKC4H4O6) (4 H90) al 0,16 % en agua destilada.
84

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 98/293
MATERIALES Y MÉTODOS
0,4
0,3
0,2
01
0,0120
Proteina (~ag/mI)
Figura 3.3.- Recta patrón para la determinación de proteína por el método de Lowry
modificado.
u
uoso‘o
u
‘O
o‘di
.0
o 20 40 60 80 100
85

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 99/293
MATERIALES Y M5IODOS
- Solución B: CuSO4 (5 H-,O) al 4 % en agua destilada.
- Solución C: Se obtiene mezclando 100 v o l ú m e n e s d e l a so l u ci ó n A c o n
1 volumen de la solución R
- Solución D: Reactivo de Folin-Ciocalteau diluido en agua destilada en la
proporción 1:1 (y/y).
P,ocedimie,ito.
A 1 ml de una muestra que contenga entre 1 0 y 100 ¡ig de proteína se le añaden 3 ml
de la solución C; la mezcla se deja en reposo a temperatura ambiente durante 1 0 minutos. A
c o n t i n u a c i ó n s e a d i c i o n a n 0,3 ml de la solución D , a gi t á n d ol a i n m e d i a t a m e n t e y d e j á n d o l a
reaccionardurante 45 minutos, al término de los cuales se mide el aumento de la absorbancia a
660 nm con referencia a un blanco preparado de la misma manera pe¡o con agua destilada.
III. 2. 9. - Caracterización parcial de la actividad antimicrobiana exocelular
parcialmente purificada d e J i. saLe 148
III. 2. 9. 1. - P r e p a r a c i ó n d e l a s o l u c i ó n p a t ró n d e l a su st an ci a a n t i m i c r o b i a n a
parcialmente purificada
Para realizar las pruebas de estabilidad enzimática y térmica, se resuspendieron 200 mg
de la sustancia antimicrobianaparcialmente purificada, obtenida de la columna de Sephadex
G-50 (sección III. 2 . 8 . 2), en 1 ml de una solución t a m p ó n d c c i t r a t o - f o s f a t o 4mM, d e p H ? .
Esta solución se preparaba justo antes de su empleo.
86

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 100/293
MATERIALES Y MÉTODOS
I I I . 2 . 9. 2 . E f e c t o d e d iv e rso s e n z i m a s e n l a a c t i v i d a d i n h i b i d o i - a d e J i saLe 148
Para determinar la sensibilidad de la actividad inhibidora a los enzimas proteolíticos
(tripsina, pepsina, papaína, proteasa II y proteasa XIV), lipolíticos (lipasa 1 y lipasa VII) y
amilolíticos (a-amilasa), se preparó una solución de la sustancia inhibidora parcialmente
purificada (sección III. 2 . 9 . 1). A lo s pocillos de placas de ELISA en los que se habían
depositado 60 gí de las dos soluciones (2 y 1 0 mg/mí) de lo s distintos enzimas en tampón de
citrato-fosfato, se añadieron 60 ¡il de la solución conteniendo la sustancia inhibidora, de talmanera que la concentración final del enzima fué de 1 y 5 mg/ml respectivamente en cada dos
pocillos. Como control positivo en un pocillo se colocó la sustancia antimicrobiana diluida al
50% con el tampón citado. Para determinar la posible acción inhibidora o estimulante del
crecimiento del microorganismo indicador, ejercida por los enzimas empleados, se determinó
l a p osi b l e act ivid ad an t imi cr ob ian a a l a s co n ce n t rac io n e s d e s c r i t a s . L a act ivid ad a n t i m i c r o b i a n a
re si d u al d e l o s po ci l l os s e d e t e r m i n ó t r a s 2 . 6 , y 1 2 h o r a s d e i n c u b a c i ó n d e l a s m e z c l a s a 3 2
0C , segúnel método descrito en la sección [It. 2.3. 3. 2; como microorganismo indicador se
u t i l i z ó L.ferrnentwn CECT285.
III. 2 . 9 . 3 . - Efecto de diferentes vHs en la actividad inhibidora de J i. saLe 148
1 1 1 . 2. 9. 3 . 1.-
Preparación de las soluciones tampón de distintos vHs
E) Tampón universal (Dawson y col., 1969).
a) Solución A.
Contiene: gp
A c i d o c í t r i c o 6 , 0 0 8
Fosfato monopotásico 3,893
87

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 101/293
MATERIALES Y MErODOS
Acido bórico 1,769
Acido dietilbarbitújico 5,266Agua destilada hasta 1000 ml
b) Solución B.
Contiene 0,8 g de NaOH en 1.000 ml de agua destilada.
Procedimiento:
A 100 ml de la solución A se le añaden lo s ml de la solución B necesarios para obtener
el valor de pH deseado. Se realizaron soluciones con valores de pH de 2,6, 3,6, 4,6, 5,6, 6 , 6 . .
7 , 6 , 8 , 6 , 9 , 6 , 1 0 , 6 , 11 , 6 y 1 2 , 0 .
III. 2 . 9 . 3 . 2 . - Condiciones de trabaio
Para establecer la estabilidad de la actividad antimicrobiana parcialmente purificada
(sección III. 2. 8. 2.) a lo s diferentes pHs, se distribuyeron en tubos alícuotas de 1 ml de la
solución tampón. Después de incubar los tubos 24 h a 2 40 C , se determinó su actividad
antimicrobiana por el sistema de los discos (sección III. 2. 3. 3. 2), utilizando a L.fermentunz
CECT285 como microorganismo indicador. Para comprobar que lo s tampones de diferente pHno ejercían acción inhibidora en el microorganismo indicador, se realizó la misma prueba de
inhibición con soluciones de la sustancia con actividad antimicrobiana.
88

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 102/293
MATERIALES Y AtETaDOS
I I I . 2 . 9 . 4. - Cinética de tennodestrucción de la actividad inhibidora deL, saLe 148
III. 2.9.4. 1.-Tratamiento térmico
En viales de vidrio (10 x 30 mm) herméticamente cenados se colocaron 0,1 ml de la
solución de sustancia antimicrobiana parcialmente purificada de Ji . saLe 148 que se calentaron
a 80, 100, 121, 135 y 150 0C en un baño de glicerina termostatado, durante intervalos de
tiempo variables en función de la temperatura de calentamiento. Finalizado el tratamiento
ténnico L as muestras se enfriaron rápidamente en un baño de hielo picado y se determinó su
actividad antimicrobiana residual de la manera descrita en la sección III. 2. 3. 3. 2. empleando
como indicador a L.fermentunz CECT285.
III. 2. 9. 4. 2. - Parámetros cinéticos de termodestrucción
A) Valor “D ’ o tiempo de reducción decimal
Se define como el tiempo necesario para disminuir en un 90 % a u n a t e m p e ra t u ra
determinada, la actividad inhibidora inicial de la sustancia antimicrobiana parcialmente
purificada de Ji . saLe 148, y se corresponde con el tiempo en el que la curva de supervivencia
a t r a v i e s a u n c i c l o l o g a r í t m i c o . S e c a l c u l a a p a r t i r d e l a e c u a c i ó n d e l a r ect a d e r e g r e s i ó n d e u n a
g r á f i c a d e t e rm od e st ruc ci ó n , q u e r e p r e s e n t a e l l oga rit mo d e l po rc e n t a je d e l a a c t i vi d a d
inhibidora inicial en función del tiempo de calentamiento (figura 3. 4).
De la figura 3. 4, se deduce que:
log x=
-kt + C
89

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 103/293
MATERIALES Y MÉTODOS
o
t
F igu ra 3 . 4 .- R e p re se n t ac ió n g r á f i c a t e ó r i c a d e l a a c t i v i d a d i n h i b i d o r a e n f u n ci ó n d e l
t ie mp o d e c a i e n t a m i e n t o .
siendo X” el % dc actividad inhibidora residual; ‘1” la constante de inactivación en m m - y
‘t’ el tiempo de calentamiento de las muestras
A s i m i s m o :
( l o g x ~ - log xí) = -k (t2- t1>
log x~ - l o g x i 1 logx,-logx~
l o g X2 - l o g x 1
o
k
7
tI - D
ti - t 2
90

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 104/293
MATERIALES Y METODOS
B) Tiempo medio It l/2~
S e d e f i n e c o m o e l t ie mp o e n e l q u e l a a c t i v i d a d i n h i b i d o r a i n i c i a l s e r e d u c e e n u n 5 0 %
a u n a t e m p e ra t u ra d e t e rm i n ad a. E l t ér mi n o p u e d e c a l c u l a r s e gr á f ic am e n t e ( f i g u r a 3 . 5 ) ,
representando la variación de la actividad inhibidora inicial en función del tiempo de
calentamiento, o bien matemáticamente, asumiendo que el cambio de la actividad
inhibidora con respecto al tiempo es función de la actividad inhibidora inicial (ecuaciones 1 y
2 ) ,
dX
- kX(l)
dt
dX
— k d t ( 2 ) ;
x
d o n d e x representa l a a c t i v i d a d i n h i b i d o r a , ‘t’ el tiempo de calentamiento a una determinada
temperatura y ‘k ” l a constante de inactivación d e l a a c t i v i d a d inhibidora en min1.
Integrando la expresión (2) entre dos valores de x diferentes (x0 y x) a (t0 y t) y siendo
x 0 l a a c t i v i d a d i n h i b i d o r a inicial en el tiempo t0, resulta:
t
dr In _x0 —
to x
k(t-t0) ó 2 ,3 logxo=k(t~t0)
x
Si x =50, 2,3 log100 2,3 log 2
____ =ktíp; t1¡2=
50 kti!2 ZQSAi23
k
x
- s dx k 5
x
91

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 105/293
MATERIALES 1 ’ MÉTODOS
~100
y
y,4 >i- 50
-‘-a
u
t 1/2
F ig u ra 3 . 5 . - Representación gi-áfica teórica de l a a c t i v i d a d i n h i b i d o r a e n f u n c i ó n d e l
tiempo de calentamiento.
C) Valor ‘7
S e d e f i n e como l a t emp erat ura n e c e s a r i a p a r a d i s m i n u i r e l v a l o r D e n u n 9 0 %, y s e
calcula a partir de la ecuación de la recta de regresión, obtenida al representar gráficamente el
l oga rit mo d e l o s v a l o r e s “ D e n f u n c i ó n d e l a t e m p e r a t u r a a l a q u e f u e r o n o b t e n i d o s ( f i g u r a 3 .
6).
l)e la figura 3. 6 se deduce que:
l o g D a T + C
donde “D es el tiempo de reducción decimal a una determinada temperatura y ‘Y’ es la
t
9 2

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 106/293
MATERIALES Y MÉTODOS
T
Figura 3 . 6 . - Representación gráfica teórica de lo s valores D en función de la
temperatura.
1 — lSipordefinición, a=- — , logD=- T±C
z z
1
l o g D2 - log D1 = - (T2 - l o g D2 - l o g D 1 = ____
z z
lo g D2 - lo g D¡
temperatura del tratamiento térmico en0C.
o o >o
z
a
(T1 - T2)
93

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 107/293
MATERIALES Y MEFODOS
III. 2. 9. 5. - Determinación del peso molecular de la sustancia inhibidora deL. sake
148 por cromatoiirafía de filtración en Sephadex 0-50
La cromatografía de filtración en geles de Sephadex 0-50 para determinar el peso
molecular de la sustancia antimicrobiana parcialmente purificada, se realizó de la manera
descrita en la sección III. 2. 8 . 1 . 3. Para ello 20 mg de la sustancia parcialmente purificada,
disuelta en 3 ml de tampón de ácido cítrico-fosfato, de pH 5 ,6 y urea 1 M se depositaron en la
columna (1,6 x 90 cm) que contenía el Sephadex 0-50, determinándose posteriormente laabsorbancia a 280 nm y la actividad antimicrobiana de las fracciones eluidas resultantes.
A continuación, en la misma columna se depositaron 3 ml de una solución de 5 mg de
dextrano azul, 6 mg de a-quimotripsinógeno A (25.000 daltons), 8 mg de RNasa pancreática
bovina (13.700 daltons) y 1 mg de vitamina B12 (1.300 daltons), disueltos en el mismo
tampón en el que se disolvió la sustancia antimicrobiana parcialmente purificada de L. sake
148. La absorbancia de tas fracciones etuidas se determiné a 280 nrn, mientras el peso
molecular de la sustancia problema se determinó por interpolación en una gráfica (figura 3. 7),
en la que se representaba el logaritmo de los pesos moleculares de las proteínas estándar en
función de la fracción cromatográfica en la que se encontraban.
III. 2. 9. 6. - Electroforesis en 2eles de poliacrilamida con dodecil sulfato sódico
(SOS-PACE
Esta técnica permite separar mezclas complejas de polipéptidos en función de su
tamaño molecular. La electroforesis en geles de poliacrilamida con dodecil sulfato sádico
(SDS-PAGE). se realizó según las técnicas de Swank y Munkres (1971) yde Laemli (1970).
El dodecil sulfato sódico es un detergente que con otros agentes, como el mercaptoetanol y el
calor, ínte¡-vicne en la desnaturalización de las proteínas a subunidades y. además, proporciona
94
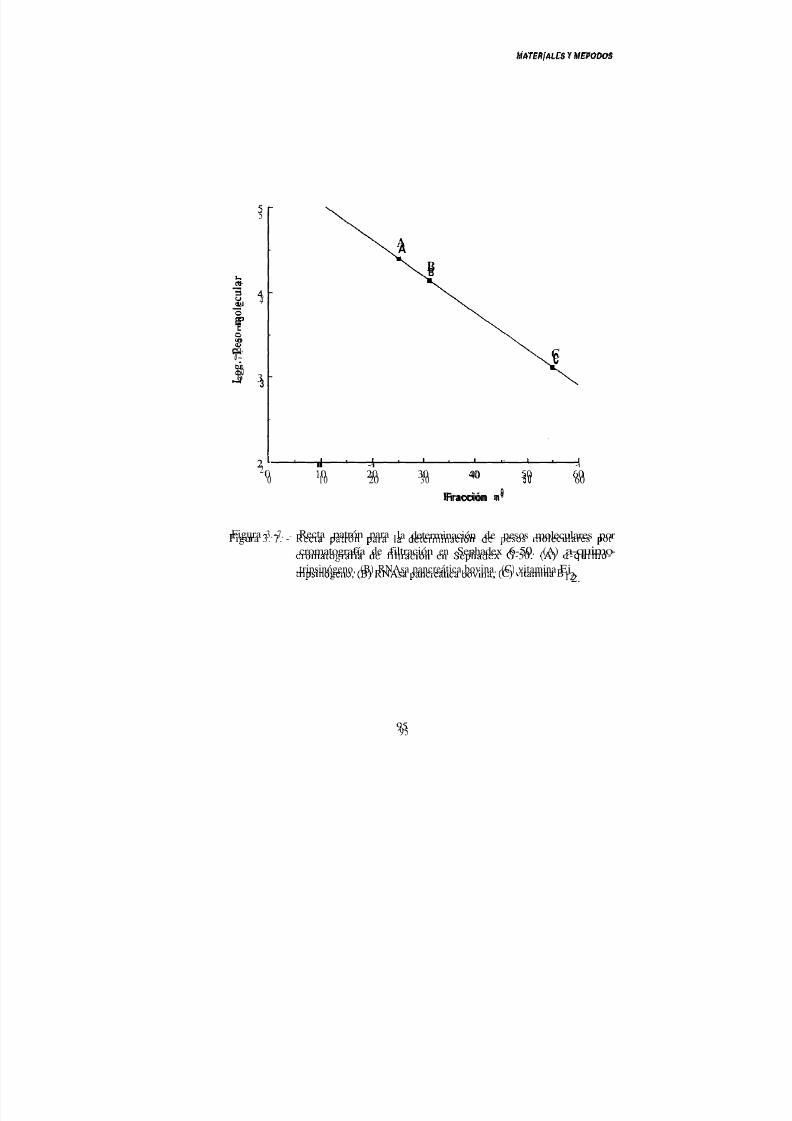
7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 108/293
MATER¡ALLS Y MEPODOS
1 • ¡
40
Fracción n 9
Figura 3. 7. - Recta patrón para la determinación de pesos moleculares porcromatografía de filtración en Sephadex 6-50. (A) a-quimo-
tripsinógeno, (B) RNAsa pancreática bovina. (C) vitamina Ej2.
5
A
B1~
u
o Eoo,
0 -~
b iJ o
4
3
c
20
II
10
-1
20 30 5 0
-1
60
95

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 109/293
A lA TIERIALES Y MUFODOS
a las cadenas polipeptídicas una densidad decarga similar. De esta forma cuando el complejo
SDS-proteína se somete a electroforesis en un gel que contiene SDS, su velocidad de
migración viene deterirtinada principalmente por la masa de la partícula SDS-polipéptido según
el pricipio de exclusión molecular. El campo eléctrico, en este caso, sólo suministra la fuerza
impulsora.
III. 2. 9. 6 . 1 . - Técnica de Swank y Munkres (1971
III. 2. 9. 6. 1 . 1 . - Tampones. geles y soluciones emuleadas
1.) Tampón para solubilizar las muestras.
Se compone de:
Urea
3-mercaptoetanol
SDS
Acido ortofosfórico
2.) Tampón de ácido fosfórico 1
Condene:
4,8
0,5
0.25
10,00,01 M hasta
g
in I
g
ml
M-SDS, pH 6.8.
Acido ortofosfórico (85 %)SDS
Agua destilada, hasta
El pH se ajuna a 6,8 con Tris.
3.) Solución de acrilamida-bisacrilamida.
Está formado por:Acri lamida
33,75,0
500,0
mlg
ml
18,75 g
96

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 110/293
MATERIALES Y MIETODOS
N,N’-metilén-bisacrilamida 1,87 g
Agua destilada hasta 50,0 ml
4.) Gel de separación.
Consta de:
Solución de acrilamida-bisacrilarnida 9,99 ml
Tampón de ácido fosfórico 1 M-SDS 3,0 ml
Urea 14,4 g
Agua destilada 29,0 ml
TEMED (N,N,N’.N’, tetrametilén-etilen-diamina) 9,0 gí
Persulfato amónico (6 %) 0,3 ml
5.) Gel de concentración.
Le forman:
Solución de acrilamida-bisacrilamida 1,65 ml
Tampón de ácido fosfórico 1 M-SDS 0,5 ml
Urea 2,45 g
Agua destilada 10,0 ml
TEMED 1,5 gí
Persulfato amónico (6%) 0 ,1 ml
6.) Tampón de electroforesis.
Contiene:
Tampón de ácido fosfórico 1 M-SDS 0,225 ml
Agua destilada 2,225 ml
9 7

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 111/293
MATERiALES YMETODOS
7.) Solución de fijación.
Consta de isopropanol: ácido acético: agua destilada (2,5:10:6,5 y/y).
8.) Solución de tinción.
Consiste en una solución de azul brillante de Coomassie al 2 % en ácido acético al
7 %.
9.) Solución de lavado.
Es una solución al 7 % de ácido acético en agua destilada.
III. 2 . 9 . 6 . 1 . 2. - Preparación de las muestras
Las muestras solubilizadas en tampón de solubilización, con 50, 100 y 200 gg de la
sustancia antimicrobiana, parcialmente purificada, se mantuvieron durante 5 minutos en bañode agua hirviendo antes de depositar 30 gí de cada solución en el gel de concentracion.
III. 2. 9. 5. 1 . 3. - Preparación de lo s Leles
Los geles se prepararon de la manera descrita por Swank y Munkres (1971). El gel
consta de dos porciones: fase inferior (gel de separación) y fase superior (gel deconcentración). Los geles se prepararon como se describe en las secciones III. 2. 9 . 6 . 1 . 4 y
III. 2. 9. 6. 1 . 5. Para evitar la presencia de burbujas de aire en los geles, la mezcla se
desgasificó a temperatura ambiente por sonicación en un baño, durante 5 minutos, antes de
añadirle el TEMED y el persulfato amónico. Los receptáculos de formación de lo s geles se
llenaron primero con la solución del gel de separación hasta unos 3 cmde su extremo superior,
mientras en la superficie de la mezcla, y para que no se formaran meniscos, se depositó un
pequeño volumen de una solución saturada de butanol. La mezcla se mantuvo durante 1 h a 37
98

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 112/293
MATERIALES YMFTODOS
0C y, una vez polimerizada, se retiró el butanol y se lavó su superficie con agua destilada. A
continuación se depositó el gel de concentración y se introdujo el peine que forma los pocillos
donde se depositarán L a s muestras. El gel se polimeriza durante 30 minutos a 37 0C . quedando
listo para realizar la electroforesis.
III. 2 . 9 . 6 . 1 . 4. - Electroforesis
La electroforesis tuvo lugar pasando por el gel una corriente de 18-20 mA y evitando
las temperaturas inferiores a 1 6 0C, para minimizar el riesgo de precipitación de la urea
Finalizada la electroforesis, el gel se extrajo de lo s vidrios del soporte.
III. 2. 9 . 6 . 1 . 5. - Tinción de los geles
Terminada la electroforesis, los geles se sumergieron en la solución de fijación y se
mantuvieron fijándose durante, aproximadamente 8 h, cambiando la solución de fijación 2 ó 3
veces. A continuación, se introdujeron en la solución de tinción donde se mantuvieron durante
2 h a temperatura ambiente. La eliminación del colorante no fijado se realizó con la solución de
lavado.
III. 2. 9. 6. 1 . 6-- Determinación del peso molecular
El peso molecular se determiné por interpolación en una gráfica en la que se representa
el logaritmo del peso molecular de las proteínas estándar frente a su distancia de migración en
el gel (figura 3. 8). Las proteínas utilizadas en la confección de la recta patrón procedían de un
“kit” comercial que contenía las siguientes proteínas estándar de bajo peso molecular:
mioglobina 1 1 1 (2,5 KDa), mioglobina 11(6,2 KDa). mioglobina 1(8,1 KDa). mioglobina 1 y
11(14 KDa) y mioglobina (16,9 KDa).
99

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 113/293
MATERIALES Y MÉTODOS
111.2.9.6.2. - Técnicade Laemli (1970
III. 2. 9.6. 2. 1 . - Tampones. Eeles y soluciones empleadas
1.) Tampón para solubilizar las muestras.
Se compone de:
Tris HCI 0,5 M, pH 6,8
GlicerolSDS (10%)
j3-rnercaptoetanol
Azul de bromofenol (0,05 %)
Agua destilada
2.) Solución de acrilamida-bisacrilamida.Está formada por:
Acrilamida
N,N-metilén-bisacrilamida
Agua destilada hasta
3.) Gel de separación.
Contiene:
Solución de acrilarnida-bisacrilamida
Tris HCI 1,5 M, pH 8,8
Agua destilada
SDS (10%)
Persulfato amónico (10 %)
TEMED
1,0 ml
0,8 ml
1 ,6 ml
0,4 ml
0.2 ml
4,0 ml
14,6 g
0,4 g
50,0 ml
20,0 ml
7,5 ml
1,9 ml
0,3 ml
0,15 ml
15,0 1 . 1 1
100

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 114/293
MATERIALES Y MEFODOS
4.) Gel de concentración.
Contiene:
Solución de acrilamida-bisacrilamida 1,3 ml
Tris HCI 0,5 M. pH 6,8 2,5 ml
Agua destilada 6,1 ml
SDS (10%) 0,1 ml
Persulfato amónico (10 %) 50,0 gí
TEMED 10,0 gí
5.) Tampón de electroforesís.
Consta de:
Tris base 6,6 g
Glicina 28,8 g
SDS 2,0 gAgua destilada 2,2 1
6.) Solución de tinción.
Está formado por:
Ácido acético 10,0 ml
Alcohol metflico 40,0 ml
Agua destilada 50,0 ml
Azul de coomasie 0,25 g
7.) Solución desteñidora
Se componede:
Acido acético 10.0 ml
Alcohol metílico 40,0 ml
101
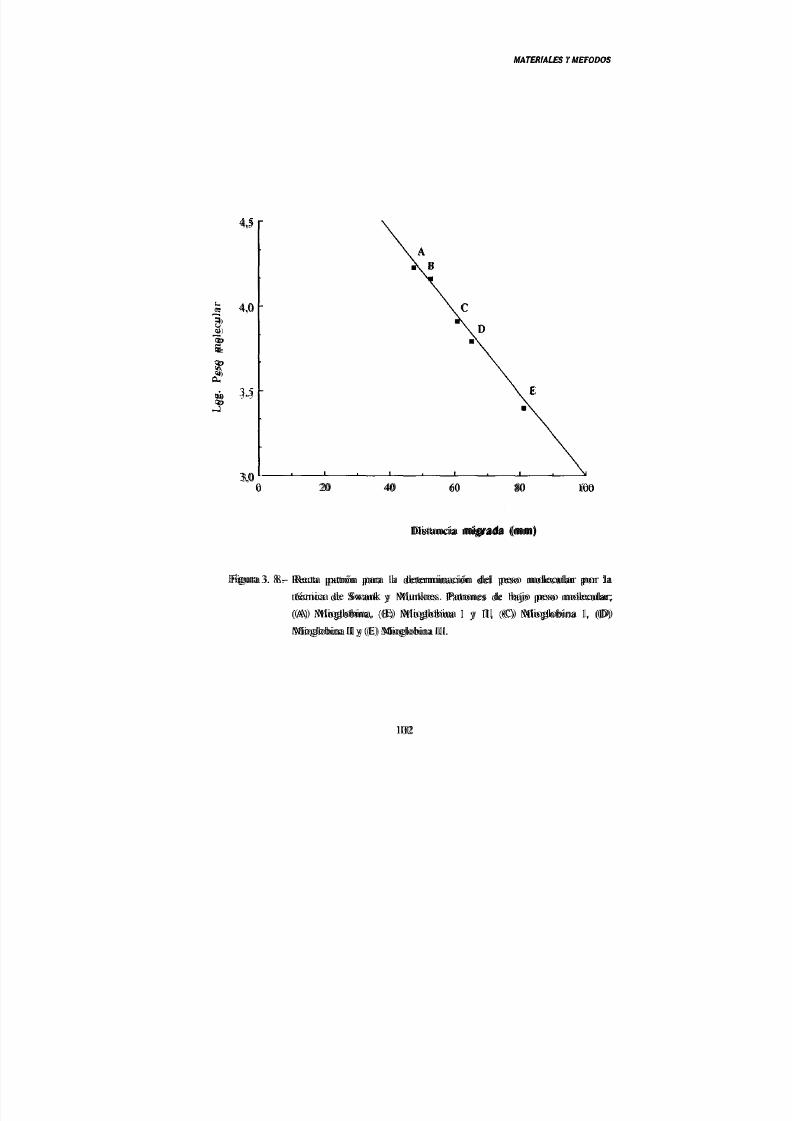
7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 115/293
MATERIALES Y MEFODOS
4,5
~ 4,04)
4)
o E
o o,4)
b) 3~5o
3,0o íoo
Distancia migrada (mm)
Figura 3. 8.- Recta patrón para la determinación del peso molecular por la
técnica de Swank y Munkres. Patrones de bajo peso molecular;
(A) Mioglobina, (E) Mioglobina 1 y T I, (C) Mioglobina 1 , (D)
Mioglobina II y (E ) Mioglobina 1 1 1 .
A
B
C
D
E
u
20 40 60 80
102

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 116/293
MATERIALES Y MÉTODOS
Agua destilada 50,0 ml
8 .) Solución de fijación:
Etanol:ácido ac¿tico:agua destilada (10:5:85 y/y).
Hl. 2. 9. 6. 2. 2. - Preparación de las muestras
Cinco, 10 y 15 gg de la sustancia antimicrobiana parcialmente purificada se
disolvieron y se mantuvieron durante 5 minutos en un baño de agua hirviendo, antes de
depositar lO jil decada solución en el gel de concentración.
III. 2. 9 . 6 . 2. 3. - Preparación de lo s eeles yelectroforesis
Tanto la preparación de lo s geles como la electroforesis correspondiente se realizó de [amanera descrita en las secciones III. 2 . 9 . 6 . 1 . 3 . y III. 2. 9.6. 1.4.
III. 2. 9. 6. 2. 4. - Tinción de lo s 2eles
III. 2. 9. 6. 2.4. 1. - Técnica del nitrato de plata
Los geles se tiñeron con el reactivo de nitrato de plata distribuido comercialmente por
la casa Bio-Rad. Este reactivo, elaborado según el método de Merril y col. (1981), es unas
10-50 veces más sensible que el azul brillante de Coomassie para visualizar las proteínas en
los geles de poliacrilamida con SDS. La tinción se realizó siguiendo estrictamente las
instrucciones recomendadas por la casa suministradora.
103

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 117/293
MATERIALES Y MÉTODOS
III. 2. 9. 6. 2 . 5 . - Determinación del peso molecular
El peso molecular se deterruinó interpolando en una gráfica los pesos moleculares de
las proteínas estándar frente a la distancia recorrida en su migración en el gel. Las proteínas
estándar utilizadas en la confección de la recta patrón procedían de un “kit” comercial que
contenía las proteínas de bajo peso molecular descritas en la sección 111.2.9.6. 1.6.
III. 2. 9. ~7.- la composición aminoacídica de la sustancia
- L. sake 148 por cromatografía líquida de alta
Determinación de
antimicrobiana de
resolución (HPLC
La determinación de lacomposición aminoacídica se realizó segúnla técnicadescrita
por Beaver y col (1986).
III. 2. 9. 7. 1 . - Tampones y reactivos
1.) Solución A: tampón acetato-¡rietilan,ina. de pH 6.8
a) Solución de acetato sódico 0,03M.
Disolver 2,46 g de acetato sádico en 1 litro de agua destilada.
b) Solución de trietilamina al 0,05% y/y.
Diluir 0,5 mIde trietilamina en 1 litro de agua destilada.
104

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 118/293
MATERIALES Y MÉTODOS
Procedimiento:
Mezclar ambas soluciones hasta obtener un pH de 6,8.
2.) Solución 1 3 : acetonitrilo al 90%
A 900 ml de acetonitrilo de calidad HPLC, añadirle agua Milli-Q hasta 1 litro.
3.) Solución de acetonitrilo en tampón fosfato 0.5M. de pH 7.4
a) Solución de fosfato sódico dibásico. 0.5M
Disolver 7 g de fosfato sódico dibásico en 100 mIde agua destilada.
b) Solución de fosfato sódico monobásico, 0.5M
Disolver 6,8 g de fosfato sádico monobásico en 100 ml de agua destilada.
Procedimiento
Añadir solución b) a la a) hasta obtener un pH de 7,4. La solución así obtenida se filtra
por un filtro de 0,45 ¡linde diámetro de poro y se añade acetonitrilo calidad HPLC hasta una
proporción del 5% (y/y).
4.) Solución de fenilisotiocianato
Contiene: ¡tI
Alcohol etílico 500
Trietilarnina 350
105

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 119/293
MATERIALES Y MÉTODOS
Fenilisotiocianato 100
Agua destilada 50
III. 2. 9. 7. 2 - DiLestión ácida de la muestra problema
En viales de vidrio (10 x 30 mm) que se cerraron herméticamente al vacío, se
colocaron previamente 10 mg de proteina de la sustancia antimicrobiana (sección 111.2. 8. 1.),
disuelta en 1 ml de HCI6 N ;
a continuación se calentaron a120
oc durante 24 horas pararealizar la hidrólisis ácida de la muestra. Una vez hidrolizada se liofiliza y el liofilizado
obtenido después se resuspende en 750 ¡ t I de HCI 0,2N y se le añaden 100 ¡ f i de una solución
acuosa de norleucina a l 1% (plv) en agua Milli-Q. Este aminoácido, que no se encuentra
normalmente en las muestras, actúacomo patrón interno.
III. 2. 9. 7 . 3 .-
Preparación de los patrones
De cada uno de los aminoácidos patrón y de la norleucina se pesaron 1 0 mg, que se
disolvieron en 10 ml de una solución de 0,2N HCI. Les aminoácidos patrón empleados fueron
los contenidos en un kit” comercial de la casa Sigmt
III. 2. 9. 7. 4.-
Derivatización de los aminoácidos
Tanto la solución de aminoácidos procedente de la hidrólisis de la sustancia
antimicrobiana como la de los aminoácidos patrón, se derivatizaron haciéndolas reaccionar
durante lO minutos en el doble de volumen de la solución de derivatización de
Etanol:Trietilamina: Fenilisotiocinato (7:2:1). Una vez terminada la reacción, el medio líquido
se evaporó en corriente de N 9. El material desecado resultante se resuspendió en una solución
de acetonitrílo en tampón fosfato 0,5M, de pH 7,4.
106

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 120/293
MATERIALES Y MÉTODOS
III. 2. 9. 7. 5. - Técnica de la cromatoLrafía en HPLC
En un cromatógrafo HPLC dotado de una columna de fase reversa (Supelco lSc)
termostatada a 35 0C se inyectaron alícuotas de 20 ¡tI de la solución problema y de lo s
aminoácidos patrón derivatizados. La fase móvil estaba constituida por un gradiente de la
Solución B en la A (tabla III. 2) y la detección de lo s aminoácidos derivatizados se realizó
determinando su absorbancia a 254 nm. La composición aminoacídica, cualitativa y
cuantitativa, de la sustancia antimicrobiana parcialmente purificada se determinó comparando el
cromatograma obtenido con elde lo s aminoácidos patrón.
Tabla III. 2.- Condiciones de trabajo para la determinación, mediante cromatografía líquida
de alta resolución (HPLC), de la composición aminoacídica de la sustancia
antimicrobiana parcialmente purificada de Ii sake 148
Tiempo
(mm)
Flujo
(ml/niin) % de E en A
Duración
(mm)
0 1 4 0
0,5 1 6 5
5,5 1 15,5 9,5
15 1 35 7
22 1 38,5 10
32 1,5 99 5
107

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 121/293
MATERIALES Y MÉTODOS
1 1 1 . 2. 10 . - Mecanismo de acción de la sustancia inhibidora deL. sake 148
Su determinación se realizó esencialmente de la forma descrita por Schillinger y Lucke
(1989). Se procedió como sigue: El sobrenadante de un cultivo de L. saLe 148, crecido en
medio MRS, se concentró unas 20 veces.
De este concentrado se tomó una alícuota de 0,5 ml que se vertió en 5 ml de medio
MRS o 1381 (dependiendo del microorganismo indicador utilizado) recién inoculado con un
microorganismo indicador; la concentración del inóculo fué en todos los casos de unas 5 x
u fc/ml.
Como microorganismos indicadores se emplearon los siguientes: L. fern¡entunz
CECT285, L. curvarus CECT726, Car. diuergens LVI3, Leu. mesenteroides CECT394, L.
monocytogenes NCTC5 105, L. monocwogenes LIS sv 1/2, L. nzonocyw genes Scott A, S.
typhiínuriun¡ T91, Y . enterocolihca E20. S. aureus FRII3Y, S. aureus FR1361 y C.
botulinun¡ CECT5Sí. C. pezfrin4’elzs CECT3J6. Los cultivos se incubaron, a 32 0C o37 oc y
en aerobiosis o anaerobiosis según el microorganismo indicador, durante 5 h y, a
continuación, se hicieron lo s correspondientes recuentos microbianos depositando alícuotas de
100 ¡ t I de dicho cultivo en placas de MRS o EHI. Como control se realizó una experiencia
similar en la que al microoraanismo indicador se le añadieron 0,5 ml del sobrenadante
concentrado de un cultivo deL. saLe 23. microorganismo cuyo sobrenadante concentrado no
mostraba ninguna actividad inhibidora detectable.
El mecanismo de acción de la suslancia antimicrobiana de L.sake 148 sería bactericida
SI SC observase una disminución de la viabilidad del microorganismo indicador y
bacteriostática, s i las ufc/ml del microor2anismo indicador permanecieran constantes respectode su valor inicial.
108

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 122/293
MATERIALES Y MÉTODOS
Hl. 2. 11.- Concentración inhibidora mfnima de la sustancía antimicrobiana en
diversos microorganismos indicadores
Para determinar la concentración inhibidora mínima (CIM), se preparó una solución de
la proteína purificada en tampón de fosfato 4 mM, de pH 7,0 a una concentración de 8 mg/mi;
a continuación se diluyó 2, 4, 8 y 1 6 veces. De cada dilución (1, 1/2, 1/4, 1/8, 1/16) se
tomaron alícuotas de 30 ¡ tI cuya actividad antimicrobiana se evaluó según la técnica descrita en
¡a seción III. 2. 3. 3. 2. Los microorganismos indicadores empleados fueron L. ferrnentumCECT285, C. divergens LV 13 , L. nbonocywgenes 7973, LIS sv 1/2 y Scott A, S. aureus
FRI 137, FRI 349 y FRI 362, C. botulinuin 551 y C.pe;fringcns 376.
La concentración inhibidora mínima (dM), se define como la concentración mínima
de proteína que produce un halo de inhibición en el medio sólido de crecimiento del
microorganismo indicador empleado. En este ensayo, únicamente se consideraron como halosdc inhibición aquéllos cuya corona circular tenía un radio mayor de 1 mm.
III. 2. 12 . - Métodos inmunológicos
III. 2. 12 . 1 . - Obtención de lo s inmunosueros
La stístancia responsable de la actividad inhibidora de L. saLe 148 (sección hL 2. 8.
1), se empleó para la inmunización de conejos con el fin de obtener lo s correspondientes
inmunosueros portadores de anticuemos.
III. 2. 1 2 . 1 . 1 - - Pauta (le i nífluni zacióil
(Domo animales de experi¡nenlación se empIcaron 2 conejos machos dc la raza ‘New
109

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 123/293
MAI’ERIALES Y MÉTODOS
Zealand” de 2,5-3 Kg de peso a los que se ¡noculó la sustancia inhibidora parcialmente
purificada. Depilada y desinfectada (con alcohol) la zona de inoculación se inyectaron
subcutáneamente a los conejos, en ambos lados de la espina dorsal, comenzando por la zona
más proxima a la región cervical. Como inóculo se emplearon 1-4 mg del antígeno (sustancia
inhibidora parcialmente purificada), emulsionados en una mezcla de 0,5 ml del Adyuvante
Completo o Incompleto de Freund (Difco) y 1 ml de aguadestilada estéril; las inyecciones se
llevaron a cabo durante un periodo de 91 días a intervalos de? días (tabla III. 3).
Antes de la primera inoculación se realizó una sangría parcial inicial (S0), para
comprobar la ausencia de reactividad de los animales frente a los antígenos utilizados.
Asimismo a los 21, 42, 63, 84 y a lo s 91 días del proceso de inmunización y, con el fin de
verificar la efectividad de dicho proceso, se realizaron sangrías parciales (S¡, 52, 53 y 54).
En las sangrias parciales la sangre se obtuvo de la vena marginal de la oreja, según el
siguiente procedimiento: El animal se introducía en una caja, de la que sobresalía la cabeza,
para inmovilizarlo; la zona elegida de la oreja se frotaba con algodón empapado en xilol para
producir una vasodilatación que favoreciera la sangría y, a continuación, se efectuaba una
punción con aguja o bien se seccionaba el vaso cuidadosamente con bisturí, recogiéndose por
goteo dc 5 a 10 ml desangre.
lii. 2. ¡2. 1 . 2. - SanEría final
Después de anestesiado el animal por vía intramuscu[ar con Keto!ar (clorhidrato de
ketamina) a una dosis de 1 0 mg/kg de peso, se colocaba en una mesa en posición de decúbito
supino y se inmovilizaba por las 4 extremidades.
Ho

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 124/293
MATERIALES Y MÉTODOS
Tabla III. 3. - Patita de inmunización de lo s conejos por inoculación subcutánea de la
sustancia con actividad antimicrobiana deL. sake 148
Adyuvante Adyuvante
completo incompleto
Días
Exuacto antigénico
(mg)
de Freund
(mí)
de Freund
(mí) Sangría
1
1
14
21
28
35
42
49
56
63
70
77
84
91
4
4
4
4
4
4
4
4
4
4
4
4
4
0,5
0,5
0,5
0,5
0,5
0,5
0,5
0,5
0,5
0,5
0,5
0,5
0,5
0,5 Sf
= sangría inicial
S~= sangría parcial día 2 1
= sangría parcial día 42
53 =sangría parcial día 63
=sangría parcial día 84Sf = sangría final día 91
111

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 125/293
MATERIALES YMETODOS
Una vez depilada y desinfectada perfectamente la zona cervical inferior, con el material
quirúrgico apropiado se practicaban las incisiones cutáneas siguientes:
a) longitudinal a lo largo de la línea media ventral
b) transversal a nivel de la segunda vertebra cervical
c) transversal a nivel de la sexta vertebra cervical
A continuación se disecan los músculos ventrales del cuello visibles en este área: Ms.
cleidomastoideus (parte del Ms. cleidocephalicus y éste a su vez del Ms. brac/iiocephalicus),
Ms. sternornastoideus (parte del Ms. sternocephalicus), Ms. sternohyoideus y Ms.
síernothyroideus.
Por el borde lateral del Ms. síernothyoideus se diseca en profundidad hasta llegar al
paquete vásculo-nervioso situado a uno y otro lado de la tráquea (A . carotis cornunis, V.
jugularis y Troncus vagosimpathicus). Se diseca la A. carotis comunis y con una pinza de tipo
“mosquito” se fija el N. vagus, que continúa unido al tronco vascular. De este modo al efectuar
la sección de la arteria se puede dirigir el flujo sanguíneo a un tubo o recipiente donde se
recoge la sangre.
Cuando disminuye el flujo de sangre, se recomienda aplicar un masaje cardiaco con el
fin de conseguir el mayor volumen de sangre posible.De cada animal se recogieron de 120 a150 ml de sangre cuando la sangría final se realizaba según las normas señaladas.
III. 2. 12 . 1 . 3. - Obtención y conservación del suero
La sangre obtenida se trasvasa lentamente a un tubo de centrífuga, a fin de evitar su
hemólisis, y se centrifuga a 2.000 g durante 1 0 minutos. A continuación, el coagulo formadose desprende de las paredes laterales con una espátula o aguja y el sobrenadante se recoge con
112

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 126/293
MATERIALES Y M E F O D O S
una jeringa. En caso de requerirlo, el sobrenadante se centrifuga de nuevo. Una vez obtenido
el suero se distribuye en fracciones dc 2 ml en viales, añadiéndoles una o dos gotas de una
solución de azida de sodio al 0,01 % que actúa como agente conservador. El suero se mantiene
en congelación a -20 0C hasta su utilización posterior.
Iii. 2. 12 . 2. - Inmunodifusión en Leles de agarosa
Esta técnica, que permite visualizar la interacción entre antígenos y anticuerpos, se
basa en su difusión por un medio semisólido (agarosa). Si ambos se corresponden, se forma
una línea de precipitación cuya posición dependerá del coeficiente de difusión de lo s antígenos
y anticuerpos, de sus respectivas concentraciones y del tiempo de reacción.
En este trabajo se ha empleado la técnica de inmunodifusión doble descrita por
Ouchterlony (1949), que permite verificar el proceso de inmunización de los animales de
experimentación y la correspondencia inmunológica entre lo s inmunosueros obtenidos y lo s
extractos antigénicos de interés.
Los extractos antigénicos utilizados fueron el sobrenadante neutralizado y concentrado
deL. sake 148 (sección III. 2. 3 . 3 . 1.) y la sustancia responsable de su actividad inhibidora
obtenida tal y como se describe en la sección III. 2. 8 .
III. 2. 12 . 2. 1. - Preparación del gel
El gel se preparaba inmediatamente antes de usarlo, constaba de agarosa al 1% en
solución de cloruro sódico al 0,85% (Suero fisiológico salino): se le adicionaba como agente
conservador un 0,0 1% de azida de sodio. Para prepararlo, la solución de agarosa en el suerosalino se calentaba con agitación hasta ebullición, añadiendo a continuación la solución de
113

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 127/293
MATERIALES Y MÉTODOS
azida de sodio.
III. 2. 12 . 2. 2. - Preparación de la ulacas de inmunodifusión
Se utilizaron portaobjetos de vidrio de 7.5 x 5 cm que se situaban sobre una mesa
niveladora depositando sobre cada uno de ellos 6 ml de la solución de agarosa caliente. Una
vez solidificada la agarosa, con la ayuda de moldes metálicos de sección circular, se realizaron
excavaciones: una central, de 1 2 mm de diámetro y 6 periféricas, dispuestas en forma de roseta
de 6 mm de diámetro que deistaban de la central 7 mm. Seguidamente, y con ayuda de una
aguja, se retiraron cuidadosamente lo s discos de agarosa añadiendo a continuación un par de
gotas de agarosa fundida en el fondo de lo s pocillos formados y evitar así posibles anomalías
en la difusión de los anticuemos.
III. 2. 12 . 2. 3. - Llenado de lo s pocillos e incubación de las placas
Preparadas las placas de inmunodifusión se depositaron 150 ¡tI del inmunosuero en el
pocillo central y 50 ¡tI de lo s extractos antigénicos en lo s pocillos periféricos. A continuación
se colocaron en unas bandejas que se introdujeron en una cubeta que contenía de 5 a 10 ml de
una solución de azida de sodio al 1%: de esta forma, además de inhibir el crecimiento
microbiano, se mantenía la humedad de la cubeta que, por último, se tapaba y se manteníadurante 18-24 horas a 37 0C.
III. 2. 12 . 2. 4. - Lavado y secado de los Leles
Una vez incubadas, las placas se lavaron en solución salina al 0,85% durante 48 horas
a temperatura ambiente; la solución de lavado se cambió tres veces cada día y el último lavadose realizó con agua destilada; la superficie de las placas se secó con una tira de papel de filtro
114

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 128/293
MATERIALES Y MÉTODOS
Whatman n 9 1 procurando eliminar las burbujas de aire que se sitúan entre el gel y el papel.
Unavez seco el gel, el papel se desprende con facilidad de la superficie de la placa.
III. 2. 12 . 2 . 5 . - Tinción
Tuvo lugar sumergiendo los portaobjetos durante 2 horas en una solución formada por
90 partes de metanol, 10 de ácido acético y un gramo de negro amida 1 0 1 3 (Merck).
Posteriormente se lavan 30 minutos con una solución de ácido acético al 5%, tras lo cual el
último se arratra con lavados repetidos de agua destilada.
III. 2. 12. 3. - Técnicas inmunoenzimáticas
III. 2. 12 . 3. 1 . - Técnica del ELISA indirecto
En este trabajo se han utilizado dosprocedimientos diferentes del ELISA indirecto. El
primero fué un ELISA indirecto clásico empleando un conjugado comercial de anticuerpos
anti-IgG de conejo conjugados al enzima peroxidasade rábano. En el segundo se utilizó un
sistema de amplificación biotina-avidina.
III. 2. 12 . 3. 2. - Técnica del ELISA indirecto clásico
En esta técnica los antígenos se inmovilizan por adsorción pasiva en una superficie
inerte; los anticuerpos específicos reconocen a sus correspondientes antígenos y el complejo
formado lo detecta un segundo anticuerpo marcado con un enzima, que reconoce como
antígenos a los anticuerpos anteriores. La reacción es visible porque al actuar cl enzima sobre
el sustrato se ¡ibera un compuesto coloreado.
115

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 129/293
MATERIALES YMETODOS
a) Antígenos
Como antígenos se emplearon la actividad antimicrobiana deL. sake 148 parcialmente
purificada (obtenida tal y como se describe en la sección III. 2. 8. 1.) así como los
sobrenadantes libres de células de L. sake 148 y de una cepa (L. sake 23), que no manifiesta
ninguna actividad antimicrobiana detectable; en ambos su desarrollo se llevó a cabo en
MM-Triptosa y en MRS.
b) Anticuerpos
Se utilizó el inmunosuero total obtenido al inmunizar los conejos con la sustancia
antimicrobiana parcialmente purificada.
c) Coniugado
El conjugado utilizado fué uno comercial de anti-inmunoglobulinas de conejo,
obtenidas en macho cabrio, y marcadas con el enzima peroxidasa de rábano (Nordic).
d) Tamuones y reactivos
1 . Tampón PES. de oH 7.2
NaCí 8,0 g
KH-,P04 0.2
Na2HPO4 (12 H,O)
KO
2,9
0,2
g
g
g
116

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 130/293
.LfATERIALES Y MÉTODOS
Agua destilada hasta 1000 ml
2. Tamnón PBST
Se prepara como el PES y se le añade un 5% de Tween 20.
3. Tampón de ácido cítrico-fosfato de pH 3.9
a) Preparar una solución de ácido cítrico monohidratado 0dM disolviendo 21,01
g en 1.000 ml de aguadestilada.
b) Preparar una solución de N a9HPO4. O,2M, disolviendo 28.4 gen 1.000 mIde
agua destilada.
c) Mezclar las dos soluciones hasta obtener un pH de 3,9.
4. Sustrato
Preparar una solución de ácido 2-2’-azino-bis-3-etil benzotiazolina sulfónico
(ABTS) (Sigma) en agua destilada a una concentración de 1 5 mg/ml y después preparar una
mezclade la siguiente composición:
- Solución de AETS (15 mg/mí) 0.4 ml
- Tampón cítrico-fosfato, pH 3,9
- H202 (30% en peso)
10.0 ml
20.0 i - ’ ~
117

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 131/293
MATERIALES Y M E F O D O S
5. Solución de frenado
- Fluoruro sódico (NaF) al 2% en agua destilada.
6. Solución de tapizado
- Gelatina a l 1% en tampón PES
e) Metodolo2ía del ELISA indirecto
Los pocillos dc las placas de ELISA (Costar 3590) se llenaron con 100 ¡tI de una
solución de la sustancia inhibidora en tampón PBS, de pH 7,2 o con 100 ¡tI de los
sobrenadantes de lo s cultivos de interés. Las placas se incubaron durante una hora a 37 0C, tras
lo cual se lavaron 5 veces con PEST para eliminarel exceso de antígeno no adsorbido en los
pocillos.
Unavez secas las placas y. para tapizar las zonas del pocillo en las que no se hubiese
adsorbido el antígeno, se añadieron a cada pocillo 200 ¡tI de gelatina a l 1% en PES,
incubándose las placas durante una hora a 37 0C. Después de lavadas otras cinco veces con
PBST, se añadieron a lo s pocillos 100 ¡tI de suero de conejo diluido en PBST; las placas se
incubaron en un agitador de placas de ELISA durante una hora a temperatura ambiente.
Terminada la incubación, se lavaron de nuevo cinco veces con PBST para eliminar los
complejos antígeno-anticuerpo formados en la neutralización y los restos de anticuerpo que no
hubieran reaccionado. Seguidamente se adicionaron a los pocillos 100 ¡tI del conjugado
anti-conejo diluido en PBST y, de nuevo, se incubaron las placas durante una hora atemperatura ambiente en el agitador.
lIS

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 132/293
MATERIALES Y MÉTODOS
Tras lavar las placas cinco veces con agua destilada para eliminar los restos de
conjugado libre, se añadieron a cada pocillo 150 ¡fl del sustrato y las placas se incubaron en el
agitador durante 45 minutos a temperatura ambiente; inmediatamente se para la reacción
adicionando a cada pocillo 50 ¡tI de NaF al 2%. El color verde de los pocillos se cuantificó
midiendo su absorbancia a 405 nm en un lector espectrofotométrico de placas de ELISA.
Además, en la prueba descrita se realizaban siempre lo s siguientes controles:
- Control del anticuerpo: Antígeno + Gelatina + Conjugado + Sustrato.
- Control del antígeno: Gelatina + Anticuerpo + Conjugado + Sustrato.
- Control del conjugado: Conjugado + Sustrato.
Si en alguno de lo s controles se alcanzaba una A~5 mayor de 0,150, el experimento se
consideraba nulo.
III. 2. 12 . 3 . 3 . - Técnica del ELISA indirecto utilizando el sistema de amplificación
biotina-avidina
Se diferencia de la técnica anterior en que los anticuerpos procedentes de•la
inmunización de lo s conejos se conjugan con la biotina, y en que la detección del complejo
antígeno-anticuerpo/biotinase realiza con un conjugado de avidina o de estreptavidina marcada
con peroxidasa de rábano.
En los últimos años, se ha visto que el complejo avidina-biotina es un mediador muy
útil y versátil en una gran variedad de aplicaciones analíticas, incluidas las técnicas
119

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 133/293
MATERIALES YMEFODOS
inmunoenzimáticas. El hecho de que en poco tiempo se haya generalizado su uso en campos
muy diversos, se debe a la elevadísima afinidad (1015 M-1) de la avidina por la biotina y a la
gran estabilidad deesta interacción no covalente.
Utilizando las propiedades del complejo biotina-avidina, se pueden amplificar las
reacciones inmunoenzimáticas (ELISA). En este caso, lo s anticuerpos específicos de la
reacción se conjugan con la biotina (biotinización) y se detectan por un conjugado de
avidina-enzima, en lugar de por las antí-inmunoglobulinas de la especie de la que procede el
anticuerpo unido al enzima.
III. 2. 12 . 3. 3. 1 . - Biotinización de lo s anticuemos
Los inmunosueros procedentes de la inmunización de los conejos, deben sufrir un
tratamiento previo antes de conjugarse a la biotina.
a.) Obtención de lo s anticuerpos a partir del inmunosuero
1) Tampones y reactivos
a) Solución saturada de Sulfato amónico
Se obtiene disolviendo 76,1 g de (NH4)2504 en 100 ml de agua destilada, hasta que la
solución precipUa. Momentos antes de su empleo, esta solución se filtra a través de un papel de
filtro Whatman n91 y su pH se ajusta a 7,4 con NaOH IN.
120

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 134/293
MATERIALES Y MÉTODOS
b) Tampón de PES. de nH 7.4
Obtenido tal y como se describe en la sección 111.2. 12. 3. 2. d.) 1.)
Procedimiento
25 ml del suero procedente de lo s conejos inmunizados se centrifugaron a 3.000 g
durante 30 minutos. Al sobrenadante obtenido se le añadió un volumen igual de solución
saturada de sulfato amónico y se dejó reposar a 4 0C durante 12 h. Pasado este tiempo, se
realizó una segunda centrifugación a 3.000 g durante 30 minutos y el precipitado obtenido tras
la eliminación del sobrenadante se diluyó en 12,5 ml de tampón PBS. Los anticuerpos asi
obtenidos se dializaron frente a PES y se liofilizaron antes de su conjugación con la biotina.
b.) Conjugación de los anticuerpos purificados con la biotina
Los anticuerpos precipitados se conjugaron con el éster biotin amidocaproato-
N-hidroxi-succinimida (Sigma), siguiendo la técnica de Bonnard y col. (1984), pero con
ligeras modificaciones. Se procedió como sigue:
1 . Preparar una solución del reactivo de biotina (1 mg/ml de dimetilsulfóxido).
2. En el tampón PES de pH 7,2, preparar una solución del anticuerpo precipitado (con
una concentración de, al menos, 1-3 mg/mí).
3. Añadir la solución de biotina a la del anticuerpo, en una relación molar
biotina/anticuerpo de 50/1.
1 2 1

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 135/293
MATERIALES YMETODOS
4. Incubar la mezcla durante 2 horas a temperatura ambiente.
5. Eliminar la biotina no reaccionante mediante diálisis de la mezcla frente al tampón
PBS (16ha40C).
6. Los anticuerpos conjugados se conservan en fracciones de 0,1 ml a -20 0C hasta el
momento de su utilización.
III. 2. 12 . 3 . 3 . 2 - MetodoloEía del ELISA indirecto utilizando el sistema de
amplificación biotina-avidina
La técnica sigue lo s mismos pasos y tiempos de incubación que los descritos en el
ELISA indirecto clásico (sección III. 2. 12 . 3. 2.), salvo que el conjugado empleado es uno
comercial de Estreptavidina-peroxidasa de rábano (Sigma).
122

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 136/293
RESULTADOS
CAPITULO IV
RESULTADOS
123

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 137/293
RESULTADOS
IV . 1 . - Evolución de las bacterias lácticas durante la maduración de un
lote de embutidos crudos curados
Cuando se analiza la relación entre carga microbiana total y carga de bacterias lácticas,
durante la maduración de un lote de embutidos se observa (Fig. 4.1) un rápido aumento del
número de bacterias lácticas que alcanza su punto álgido hacia el sexto día de maduración
igualándose con el recuento microbiano total, lo que sugiere que casi la totalidad de los
microorganismos de los embutidos crudos madurados pertenecen a este grupo. Los altos
recuentos de bacterias lácticas se mantienen, prácticamente sin sufrir modificaciones, a lo largo
de la maduración de los embutidos.
IV. 2. -Actividad antimicrobiana de las bacterias lácticas seleccionadas
IV.2.1.- Actividad inhibidora directa
Como ya se ha descrito en la sección 111.2.3.1 de este trabajo, durante la maduración de
un lote de embutidos crudos se aislaron 30 bacterias lácticas en los días 0, 2.6. 10 , 21 y 35 de
maduración, de forma que al final del periodo madurativo se reunieron 180 bacterias cuya
actividad antimicrobiana directa se estudió frente a diversos microorganismos indicadores. Les
resultados de la prueba directa de antagonismo se reflejan en las figuras 4. 2 a 4. 5. De lasmismas se deduce que la actividad inhibidora es distinta en cada cepa analizada, siendo
significativamente mayor cuando las bacterias se incubaron en las placas 24 h antes de la
incorporación de lo s microorganismos indicadores.
124

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 138/293
RESULTADOS
1 0
9
8
7
6
5
4
3
2
10 5 10 15 20 25 30 35 40
Días de maduración
Figura 4. 1.- Evolución de la flora microbiana total (O), y láctica (U ) durante la
maduración de lo s embutidos crudos curados.
12 5
o
.0
E
u4-
o
tÉo

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 139/293
RESULTADOS
La actividad antimicrobiana de las 180 bacterias lácticas seleccionadas se determinó
empleando como microorganismos indicadores a Lactobacillusfern;entunz CECT285 (fig. 4.
2), L. planrarum CECT22l (fig. 4. 3), Staphylococcus aureus CECTS9 (fig. 4. 4) y
Salmonella ¡yphimuriunz CECT443 (fig. 4. 5). Los resultados obtenidos indican una mayor
actividad inhibidora cuando las cepas seleccionadas se enfrentaron a otras bacterias lácticas,
tanto cuando crecían simultáneamente como cuando lo hacían 24 h antes de incorporales el
indicador. Frente a las bacterias patógenas como 5. aureus y S. typhimuriunz no se aprecia una
actividad inhibidora definida cuando se desarrollan simultáneamente con ellas, lo que contrastacon lo que ocurre cuando las bacterias lácticas seleccionadas se desarrollan en las placas 24 h
antes de la incorporación del indicador.
Es de destacar la presencia de halos de inhibición de bordes nitidos y definidos en
algunas de las cepas evaluadas frente a L. fermenturn y L. plantarum. Estas cepas, que
parecían poseer un mecanismo de inhibición diferenciado de las demás, representan un 25 %de las analizadas y en las figuras se señalan con un asterisco.
De las 180 cepas estudiadas, se seleccionaron 20 que estaban dotadas de una
manifiesta actividad antimicrobiana (halo de bordes nítidos) y 4 cuya actividad era menor. En
las figuras 4. 6 a 4. 29 se muestra la actividad antimicrobiana de las 24 cepas frente a L. brevis
LB826, L. carnis, L. curvalus LE726, L. dii’ergens MR375, L.farciminis LB34, L. sake
LE684, Leu. mesenteroides MR364, M. i’arians CECT23O, B. thermosphacta NCIB 10018,
S.faecium CECT4lO, E. cok LE841, B. stearothermophylus CECf49, E. cloacae CECT149,
8. cereus CECTI48, J’s.fluorescens DC7. Y . eníerocolitica NC E20, S.flexnerii CECT58S,
5. faecalis CECT48I, L. monocytogenes Lllsv4, L.fermeníum, L. plantarun> CECT22I. 5 .
typ/zimurium CECT443 y 5. ~weus CECT59. Las actividades inhibidoras observadas son
similares a las reseñadas antes y notablemente superiores cuando se emplean como indicadores
otras bacterias lácticas.
126

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 140/293
RESULTADOS
La mayor parte de las 24 cepas analizadas poseen una actividad antimicrobiana directa
claramentecuantificable frente a Y . enterocolláca E20 y 5. íyphiinuriunz CECT443 y. sobre
todo, frente a 5. aureus CECT59 y L. monocyíogenes LI lsv4, originando en los dos
últimos indicadores halos de inhibición definidos y de un diámetro apreciable, similares a los
observados frente a las bacterias lácticas. No obstante, frente a otros indicadores, como 8.
cereus CECTl48 y S.flexnerii CECT585, son pocas las cepas que ejercen acción inhibidora
definida.
127

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 141/293
RESULTADOS
1 1 1 u 1 — rl ~ -t .r,’.0 t— 20 ~ — rl en -t •n ‘c r— CC 0~’ ~— — rl en — ti rl rl rl u .ta ‘.0 20 0’ 0
rl rl rl rl ~l rl en
Ce ¡ n ~ n”
Figura 4. 2(1).- Actividad inhibidora de las 180 bacterias lácticas seleccionadas frente aL.
fern¡entunz CECT2S5.
m Actividad Inhibidora directa. ~ Actividad Inhibidora diferida.
12<)
1 1 < )
1 < > 0
90
$0
-e 70
-D ~
— — 50
rl 40
.30> rl
Q — 20 —j
¡0
o f i u
• 1
‘ I I H i l l ¡ I i i ~.1I L B I í i ~ í i u i t — rl ~ •~f. ~r, ‘.C r~ 20 0’ — rl rfl fl ‘Ci .0 r— 20 0~ — rl en - 1 • ‘Ci ‘O r— 20 0’ 0 en en en en en en en en en -t —t ‘-t —t t-t-.t. -t Ir. IC. •r,.r, .C. sc, SC, SC. St, SC, ‘.C
A *
Ce1ni u”Figura 4. 2(11).- Actividad inhibidora de las 180 bacterias lácticas seleccionadas frente a
L.fe,-menwm CECT2SS. Las columnas poseen el mismo significadoque en la figura 4. 2(1).
160
¡40
¡ 20
loo
.0 ~
— rl
o
> rl
Q
21
80
6<)
- 4 < )
20
o uII] I I
12 8

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 142/293
RESULTADOS
9 < )
so;
70
<dI
50
40
3 < )
20
10
< 1 1 ] 1 1 I i i f i t i I i F i l h F I l ~¡1~rl en ‘tIC, sc t— ~ Ci: 0’ — rs¡ en ? t SCj ‘.t t~ — CC Os —rl en tIt. sC U— CC 0’ t‘.0.0 ‘0 ‘5 ‘0 ‘SSO sS r— U-- U- U- U- U- U- U- U- U- 202020202020202020200’
(~ ? e ¡ni n’
Figura 4. 2(111).- Actividad inhibidora de las 180 bacterias lácticas seleccionadas frente a
L.fermentum CECT2S5. Las columnas poseen el mismo significado
que en la figura 4. 2(1).
— r~ en —t < t~ ~ ‘.5 U- 200 0—4 r-¡en’t lCa 5’ 0’ 050’ 050’ 0’ 050’
‘SU- C C Os — rl en Ú .4 ‘5 5— CC 0’ rl
(e¡:I II”
Figura 4. 2(1V).- Actividad inhibidora de las 180 bacterias lácticas seleccionadas frente a
Lfermentum CECT2S5. Las columnas poseen el mismo significado
que en la figura 4. 2(I).
¡ 2 <>
1 1 4 >
¡ < > 4 )
.0 ~.)
z _
-e ~~ C
-t o rl
u ~
o
¡2<>
110
10<)
90
80o
.0 ~
—
<‘1 j ‘ - t~e o
rl
u — — e
7< )
6 < >
50
40
3 < >
20
¡0
4 >
129
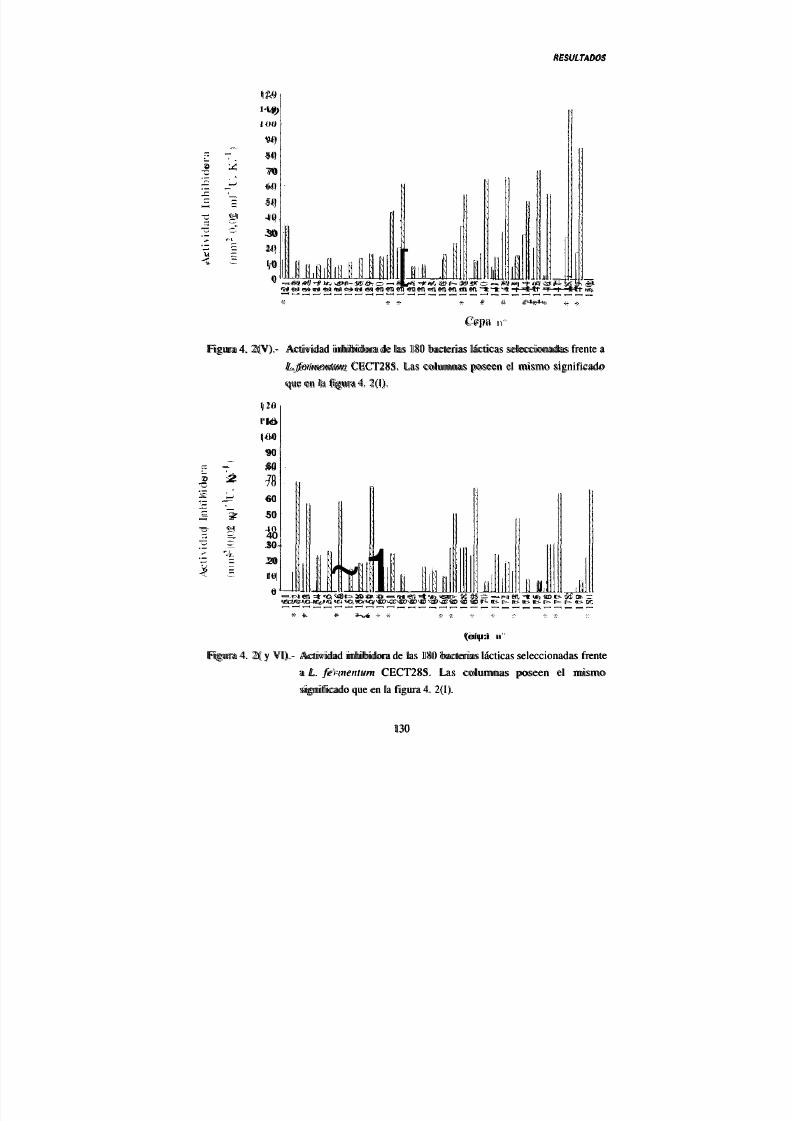
7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 143/293
RESULTADOS
1 2 < >
‘‘4)
0< )
$4)
70
6<)
5<)
-4 <)
30
2< )
¡0o
‘ f i [1 1 1 . 1 H Ii i
f iF
. 1
~....4 $9 efl ? bt.. .0 fi— CC O~ $9 eS t >r. SO 5-— 20 0’ .— — $9 eq — 1 - ‘Ci ‘.t U- Cc 0’arl $9 $9 rl $9 ti <9 rl $9 en en en en en <fl en en ffl en t t -t —-t-t ——-t-t mr.
- - * 4 $ *4+4+
Cepa u
Figura 4. 2(V).- Actividad inhibidora de las 180 bacterias lácticas seleccionadas frente a
L.ferinentunz CECT28S. Las columnas poseen el mismo significado
que en la figura 4. 2(1).
¡ 2 < >
‘lo
¡<>0
90
— $0
o 2 70
; ~ -~ 60
— z 50
— e-]
— 40 — 30~
•1
— 20u —
1 < )
o f i ~1u 1 1 L i I i U f i 1 ] L I u : t i — r~ - f i en -t ‘C, ‘-0 5-— 20 0’ 0 — rl en -t Ir, ‘SU- C C 05 — e — u en t Ir, SO U- C C OsÉCJ’C~.C~IC¿I C, •C~ •C, ICí •Cj SO ~.C SO SO SO ‘.0 ~C SO SO SO U- U- U— U- U- U- 5 — — 5-— 5 - -— U- CC
9- * **wt
(eíu:i it
Figura 4. 2( y VI).- Actividad inhibidora de las 180 bacterias lácticas seleccionadas frente
a L. fe’-,nenturn CECT28S. Las columnas poseen el mismo
significado que en la figura 4. 2(1).
o
u4-
e-]
130

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 144/293
RESULTADOS
‘nlL I t i — rl ‘5 -t Ifl SO U- C C Os O — rl rS -t ‘r, SO U- 20
4+
. 1
I t0’ 0 — 4+1 ‘5 -t In SO U- CC Cx O — tI ti $9 $9 <9 rl el $9 $1 $9 rS
4 +
(Ú¡h~ 11<’
Figura 4. 3<!).- Actividad inhibidora de las 180 bacterias lácticas seleccionadas frente a L.
plantarun; CECI’221. Las columnas poseen el mismo significado que en
la figura 4. 2(1).
[tiU 1 1 1 1 1 1 I I I I I j i 1 1 U 1 4 1 1 1 1 , ’ d i f l i n W ~ a<9’t ‘t It, SO U- CC Cx O — rl rfl -t OC . sC U- 20 0 — ti’t t •C4 SO U- 20 Cx O
rs en ‘senen ~. en < -e -. <n — 1 ~ -t —t -~t —t —t —1’ —, — r ~n ~r. ‘e, ~r> ti-, sr, ti-, ti-, ‘osr,xc
U <pa ti’’
Figura 4. 3(11).- Actividad inhibidora de las 180 bacterias lácticas seleccionadas frente a
L. plantarunr CECT221. Las columnas poseen el mismo significado que
en la figura 4. 2(1).
¡24)
¡ ¡ O
¡ < > 0
90
$4)
70
6< )
50
.0
.0 _e —
-t ti
‘.4
el
ej
40
30
2 < >1 . < )
< 1
¡20
III)
¡<>0
90
o
~2
— ti
— rs
-r
s < )
70
60
50
44)
3<)
20
¡4)
4 )
131

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 145/293
RESULTADOS
¡24)
1 1 < )
¡4)4)9< )
—. 84 )o1~ i — . 70.0 ~ _ 60
— — 50-e
c~ S 40-e o
30
e .> —— 20
O 10
o a b t u i i i 1 I I I f l t i f i M w f l ¡II-fi— $9 <-n <Ci SO U- CE QN O— $9 en fl ‘OSO U- 20050~ rl en-tsr, ‘o 5-~- 20050SOSO SO SO SO SO SO SO ‘O U- U- U- U- U- U- U- U- U- U- 20202020202020202020 Cx
4 +
( Ciflí u
Figura 4. 3(111).- Actividad inhibidora de las 180 bacterias lácticas seleccionadas frente a
L. pIa~aarurn CECT22I. Las columnas poseen el mismo significado
que en la figura 4. 2(1).
f i~ii— ú-~ tfl t < C ~ ‘.0U-20 QN O — e—] ‘t <0’.0 5 -— Sr 0’ = QNOsOsOsOsQNZNCN —
w ~id ii i t • i— Cx] r~, •1- «4 SO t- 20 Os O
$9
(epa <fi”
Figura 4. 3(1V).- Actividad inhibidora de las 180 bacterias lácticas seleccionadas frente a
L. plantarurn CECT22I. Las columnas poseen el mismo significado
que en la figura 4. 2(1).
¡20
- - fi O
1 < 1 < )
o :4-e
-J
‘.4 ci
0 -
9 < )
80
70
64 )
5< )
40
30
20
¡ 0
132

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 146/293
RESULTADOS
¡20
¡ 1 0
¡ 009 < >
$ < )
7< )
~(>0
— rl 40
34)
20: Ir1 1.
it’ ½ JWhibi1Li ji 1— rl ‘ e - , t•t. C U- C C 5 N — e - - f i en fl ~ ir, SOU- C i ? S N 0 — r~ en ~t ~tí ‘ - .0 U- 20 05 0<9<9 el C x l rlrj e - ~ f i r—ir—]enenenenenenenen’nen rr-r--r-r-r -t-t-t In
Figura 4. 3(V).- Actividad inhibidora de las 180 bacterias lácticas seleccionadas frente a
L. plantarun¡ CECT22L. Las columnas poseen el mismo significado
que en la figura 4. 2(1).
120
110
100
94 >
8 < >
:4 7<)
óo
5 < >
rl 44 )
34>
2 < >
1 < )
o
t cina u ’’
Figura 4. 3(y VI).- Actividad inhibidora de las 180 bacterias lácticas seleccionadas frentea L. planíarurn CECT22L Las columnas poseen el mismo significado
que en la figura 4. 2(1).
133
I I — e — f i en •t. SO U- 20 SN — e-.> en Ú‘ti SO U- ~t SN = — e--fi e5t If;j SO 1-- 20 SN 0It, SC, It. St, It. St, It. It, Ir. SO SO SOSO ‘SSO SO SO S O S O U- U- t~ U-U—U-U-U-U-U-CC

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 147/293
RESULTADOS
124)’
84 )
7<)
6 < )
5< )
40
.30
20
-o ”1 < )— e-—)en t ~ta SO U- 20 QN O — ti en -t Ita SO U- 20 QN
M U u t i II
O — Cx]’t ‘t •t, SO U- 20 050Cx] e--] e--] e-] r—t e-—, e-] e-] e--] r—i en
t’eí>:i I I
Figura 4. 4(1).- Actividad inhibidora de las 180 bacterias lácticas seleccionadas frente a 5.
aureus CECT59. Las columnas poseen el mismo significado que en la
figura 4. 2(I).
¡20
¡ 10
C d —
o :4
— e-
r t3-j
— < --5
-5
9 < )
$0
7< )
6< )
5<)
4<)
3 < )
2< )
¡4)’
4 >t i ’ u L i u r l 1 - 1 t i 1— C x ] en -, ‘C, SO U 20 QN O — Cx] ‘t -T •C, SO U- 20 0. o — e--] ‘t -t ~t,en ‘ten re-, en en en en en -t-t—— -T ~ -t -1- — l~ ‘ti íd-. IC~ St, I<~j S d ~ 4
sC U- 20 QN =•ta Ita •ti ta SO
(‘epa u”
Figura 4. 4(11).- Actividad inhibidora de las 180 bacterias lácticas seleccionadas frente a
5. ant-cus CECT59. Las columnas poseen el mismo significado que en
la figura 4. 2(1).
1 1 4 )
1 0< )
9 < >
.0
-1 -
~ rlOO
e-]- — e -
-
u —
o
134

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 148/293
RESULTADOS
- , ~1 ] Ñ u f i — e-’ ‘fl tfl ‘o U- CE QN O— e-] tS. It
4 SC U— 20 OsSOSO SOSO ‘SSO SO \C SO U-U-U- U- U- U- U- U- U- U-
0—rl C C 2020
H u5 t ‘ti ‘Ct~ 20 05
20fl20cC2020cS
C t— íní u”
Figura 4. 4(111).- Actividad inhibidora de las 180 bacterias lácticas seleccionadas frente a
S. aureus CECTS9. Las columnas poseen el mismo significado que en
la figura 4. 2(1).
1 2< )
1 1 < )
100
90
80
7<)
(>4)’
50
-lO
30
20’
lo
4 ) I I ¡ 1 . f i t i U I I t i 1 t i— e-fi en t ‘C~ SO U-CE Os O—rl en’? Ita SC U- C C 050 — r-i en’? ‘tí SO 5-— ~ 05 005Z’Z’05QNCN05ON —— Cx>
e p :m Ji ’
Figura 4.4(1V).- Actividad inhibidora de las 180 bacterias lácticas seleccionadas frente a
5. aureus CECTS9. Las columnas poseen el iiiismo significado que en
la figura 4. 2(1).
1 2 4 >
11<)
1 04 )
o :4
.0
E —
-e ~C d ‘r o— e-,
e . > —
-~ -t
90
$4 >
70
6 < )
5<)
40
30
20
1 < )
4 )
C d —
.0
-z _
cici‘.4 —
— 1 ~ ~ I
-4< =
135

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 149/293
RESULTADOS
3 2 < >
‘‘4)
1 4 1 0
9 4 >
— - 80o :4
70.2 ~
— 1 ‘ 5 < )
— — 50-o _
4 < )~0 0
3 < >
- 20
1 < )
( ) 1 ~ I - 1 u ¡ 1 [1 — ti rfl t It. SO U- 20 0’0 — ~en Ir> SO U- 20 Os O — $9 en’? Ir, SO U- 20 Os 0’e-) rl rl e-] e-] e-] rl ti e--fi fe-, e.t ‘t te-, te-, r5 te-, ‘t re-, ‘t ~? t t ~t 1 —t T —t It>
(.?cpzi II’’
Figura 4. 4(V).- Actividad inhibidora de las 180 bacterias lácticas seleccionadas frente a
5. aureus CECT59. Las columnas poseen el mismo significado que en
120
¡¡0
10 0
9 < )
8<)
74 )
6< )
54)
40
3 < >
2<)
1 < )
4 )
la figura 4. 2(1).
t i f i k u a 1 j i ’ t i — < --fi en It. S O U- 20 QN 0 — e-] en t It, SO U- 20 05 = — ~ re-. Id-a SO 5-- 20 050•Cí Ita íd-, Ita It, tCa MC, It. It. SO SO SO SO SO SO SO SO SO SO U- U- U- U- U- U- U- U- U- U- 20
Figura 4. 4(y VI).- Actividad inhibidora de las 180 bacterias lácticas seleccionadas frente a
5. aurcus CECT59. Las columnas poseen el mismo significado que enla figura 4. 2(l).
C d —
o
e- -
t
— .4
136

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 150/293
RESULTADOS
5.~ rl I I — e-] rs -t tCi SO U- CE 05 0 — e--fi en -t ~r¿ SO t— CC 05 =
rl
F i F f i i; L I I I — <—fi en ‘ti SO U- 20 05 0e-fi < - — f i Cx] Ci e -~ f i e--fi Cl e--fi < - - f i en
t’e ~
Figura 4. 5(I).- Actividad inhibidora de las 180 bacterias lácticas seleccionadas frente a S.
typhimuriurn CECT443. Las columnas poseen el mismo significado que
en la figura 4. 2(1).
‘ $ 1 I I—$9 en Ira SO U-en en en en en en en
ji’. j i H [1 1~ F - iC iD 05 0 — $9 ‘5 t ‘C, SO e- — 20 05 0 — e~fi ~e- •C, SO U- Y? 050en en t ‘t ~t ~t —t -t Ití tCj ICj ~t’a Ifl •Ca MC¿ I d -a SC, Ca SO
te1n, n’ ’
Figura 4. 5(11).- Actividad inhibidora dc las 180 bacterias lácticas seleccionadas frente a
S. ¡yphimurium CECT443. Las columnas poseen el mismo significadoque en la figura 4. 2(1).
¡24)
¡ ¡ 4 )
¡4)4)
o :4
.0
.0 -~
-e rlC d
-~ o ;,- rl
Q) —
21
9<)
8< )
70
6 < )
50
40
30
2010~
120
1 1 < )
14)0
9 < )
so70
6 < )
50
44 )
30
2<)
“-a —
o :4
z 2.2
— Cx]
‘ - - a‘0
— e - f i
u -
=
1 4 )
4 )
137

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 151/293
RESULTADOS
¡ 2 4 >
¡ [4)
[<‘4’
9’)‘< 4 )
74 )
64 )
5<)
40
3< )
24)
1 < ~ )O rl 1 ] j i ~lN I I f l I I F i F i
— $9 rfl Id-a’..0 U- C C 050 — r—fi’5 ,t ‘Ca SO U- C C 05 0 — < — 5 en íd-> SO U- 20 QN OSO SO ‘.0 SO SO SO SO SO SO U- U- U- U- U- U- U- U- U- U- 2020202020202020202005
(?eí>a i)<
Figura 4. 5(111).- Actividad inhibidora de las 180 bacterias lácticas se leccionadas frente a
S. typhimuriuni CECT443. Las columnas poseen el mismo significado
que en la figura 4. 2(1).
¡20 t en
A t ~
100
90’
8<)
70
60
50
4< )
3<)
20
¡ < )
I I t i 1 ’ 1 ~I-1 — Cx] ‘t t. SO U- 20 0 0 <-‘k en — t ‘ti —0 5 -— 20 0— 0 — rl en t tti SO U- CC 050
NQNSNONSN050’0’05 e-’
(-t.InI II
Figura 4. 5(1V).- Actividad inhibidora de las 180 bacterias lácticas seleccionadas frente a
S. iyp/ñmuriwn CEGI’443. Las columnas poseen el mismo significadoque en la figura 4. 2(l).
o
fl ~
.0
e-fi- C d O
~r o ~- el
21
o :4e- - -
— -4
-4 -
~ Cl
ti‘.4 —
— e-,
e . > —
138

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 152/293
RESULTADOS
¡24)
3 ¡ 4 >
1 04 )90
8< )o :4
2
—
— ti‘.4 —C j xi
-~ O>~ e-’
Q —
-5
7< )
6 < )
50 -40
30
20
1 < )
1 ) H u f l I I U l l . ~ . u r ¡ — rl en ‘? It) SO U- 20 OSO — <~1 en ‘? Ita SO U- 20 050 — $9 en -t ~CaSO U- 20 QN O
ti $9 rl rl <-‘fi rl $9 $9 <9enenenenenrsenenen’?’?tr’?-tÚ’o
(‘ epa u’
Figura 4. 5(V).- Actividad inhibidora de las 180 bacterias lácticas seleccionadas frentea
S. typhinturiutn CECT443. Las columnas poseen el mismo significado
que en la figura 4. 2(1).
~~11~
2 < >
1 ¡ < )
loo
90C d — y 80
o :470
— . 6< )
— — 50 — < - - f i
— — 4< )
— e-’ 30Q
— a 2< )
I4 )
4 > 1 4 Ix ’ ¡ 1 f I f i I I— $9 en’? It) SO U- 20 QN O — $9 en ‘t Ita -0 U- 20 0’. 0 — rl en -t It) ‘.0 U- 20 O~0
Ita fiCa It) 4 C ~ It4 It. It) ‘Ca fiCa SOSO ‘.0 SO SO SC SO SO SO SO U- U- U- U- U- U- U- U- U- U- 20
(tl>:fi II’
Figura 4. 5(y VI).- Actividad inhibidora de las 180 bacterias lácticas seleccionadas frente a
5. ryphimuriunz CECT443. Las columnas poseen el mismo
significado que en la figura 4. 2(1).
139

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 153/293
RESULTADOS
¡2<)
[1<)
¡ 4 ) 4 )
9 4 ’
$ 4 >O 4
7< ).0 -4
— . 60
54)-o <9
-~
-o 1~J 3 4 )
2< )
1 < )< 1
n
Figura 4. 6.- Actividad inhibidora de 24 bacterias lácticas seleccionadas frente a L. brevis
L8826. Las columnas poseen el mismo significado que en la figura 4. 2(I).
1 2 4 )
¡ ¡ o
¡ < ) < )
90
8< )o 4
74 )
( >4 )
5< ) — rl
-¿ 4 )
— e-fi
3020
1 4)
< 1
( ‘..¡>.~ ti
Figura 4. 7.- Actividad inhibidora de 24 bacterias lácticas seleccionadas frente a L. carnis
MR3Y 1 . Las columnas poseen el mismo significado que en ¡a figura 4 2(1).
140
‘t en SN — el U- — ‘e-. — e-i ir, Ci ? O- en < -n U- SO U- C C O— rl $9 <--1 < -n ‘? ‘? U- U- SN ~ $9 en ‘? -t t~ Ira SO SO U- U- U- C C
ji
trfltQN~-~tSOri5-—0—-en~efl&t.CiZ0’.nenr—CU-CCZ$9 $9 <-—1 T t~— U- 05 rl e-> t, ‘4+ ~t t t. S O SO U- U- 1 — C i?

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 154/293
RESULTADOS
¡ 2 4 >
114)
1 04>90
80o -~
70
— 60
-54 ) — < - — 1
40
30
20lo
Figura 4. 8.- Actividad inhibidora de 24 bacterias lácticas seleccionadas frente a L.
curvatus L8726. Las columnas poseen el mismo significado que en la
figura 4. 2(1).
12<)
¡ lO
4 00
9 4 )
8< )4
70
— . 6< )
50
4< )
-.3 0
20
[0
4 )
Ce ¡ni a’’
Figura 4. 9.- Actividad inhibidora de 24 bacterias lácticas seleccionadas frente a L.
divergens MR3’75. La s columnas poseen el mismo significado que en la
figura 4 . 2(1).
141
(?cpa u”
efl QN — SC < - — f i 5-’— ‘0 — en — <—1 ICí 20 Os e n e n ,—— ‘.0 U- 20 ‘0 — e-fi Cxl rl re-, t fl~ U- U- 05 Cxi Cxl -n -t ~t t Ita ‘.0 SO U- U- U- C C

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 155/293
RESULTADOS
12<>
¡ 1 (>
¡ 4 ) 0
90
o :474 )
.0 ~_ 60
— — 54)
40o
ej— 30’
— ze . > —
20~
¡ 4 )
1 ’ N
Y “5’
kS N
5 4
j i
‘ 5
1
j i
Fi
I t
‘ 1
ti
II
- n 1SS
‘5
I iitrn’?05tSOrflroen.Cx¡hnCC-~...e-U-’0U-
20— < ‘ f i < ‘ f i rl en U- U- 0’ e-fi e-fi en fl ‘? ‘t ? It. SO SO Cx— U- U- 20
(c¡ní u
Figura 4. 10.- Actividad inhibidora de 24 bacterias lácticas seleccionadas frente a L.
farcirninis L1334. Las columnas poseen el mismo significado que en la
figura 4. 2(I).
r i ‘ ; ~ .‘~ u U f i r , ¡ [it”’, -I . I’J-1~ rs -t 05 — -t SO e-fi I V — = — te-. — e-~ IF. C i? 0‘~‘ e-fi <‘fi e-fi < - - n t t~— t— 05 Cx] Cx] ‘~ —t $
1 í ~ i i
‘1’- C SO r - — - U- U- 20
( cpa it
Figura 4. II.- Actividad inhibidora de 24 bacterias lácticas seleccionadas frente aL. sakeL8684. Las columnas poseen el mismo significado que en la figura 4.
2(1).
¡ 2<)
¡ ¡ 0
1 0<)
90:
84 )
70 -o :4Cx
— -4
= -;
c rl
o.
:4- 1-fi
— el
e . > e.
-r =
< > 4 )
504 4 >
30
2<)
j 4):
4 )
142

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 156/293
RESULTADOS
¡24)
¡¡4)
1 009 4 )
— . 80o :42 . . 7< ) fl -~
— 60
— — 50 — rl
C d ‘0 — — 40
.3<)
E ‘ 24>
1 < )
4 )
Figura 4. 12.- Actividad inhibidora de 24 bacterias lácticas seleccionadas frente a Leu.
rnesenteroides MR364. Las columnas poseen el mismo significado que en
la figura 4. 2(1).
¡2< >
¡ ¡ o
14)0
90
— . 80o :4‘.4 7< )
.2 -4e-. . 60
— — 50— Cxi‘.4 —
-~ . 40
30t9
-A- = 24)
1 < )
4 ) U [II~“t el. t O ’. .—‘ ‘.1. SC f~-] I— ‘0 — efl —
— Cxl $9 <-fi rn t -t U- U- 05 $9 <‘fi fe-,
I i ~ 1 * ~—i
e-l Ita 20 0’ en en í- ‘o U- 20 ‘0‘? t Ir, SO SO U- U- U- C i?
Figura 4. 13.- Actividad inhibidora de 24 bacterias lácticas seleccionadas frente a M.
varians CECT23O. Las columnas poseen el mismo significado que en la
figura 4. 2(1).
Ce¡~í II”
1 4 3

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 157/293
RESULTADOS
F I ~ ~ ~
INi
N M k ’ 5 IN .N 1 1 ifil
r en -t QN — ‘.0 e—] U- ‘0 ‘ ‘n — $9 t, 20 05 — <‘fi <‘fi <‘f i en ‘? — l ~ U- U- QN <Ni e-fi en ‘T t t It,
‘SU- SO U- 200— O SO U- U- U- 20
Figura 4. 14.- Actividad inhibidora de 24 bacterias lácticas seleccionadas frente a B.
tern2osp harto NCIB 10018. Las columnas poseen el mismo significado que
en la figura 4. 2(I).
I ’ - 1ji ¡ - 1 i ~ ) [Jf ~
< - — f i e - f i rs t -t 5-—- U- 0’ rl e-> -5 ~t r — t- -t >r . en C x — SO U- 200SO SO U- U- U- 20
(epa u
Figura 4. 15< Actividad inhibidora de 24 bacterias Jácticas seleccionadas frente a S.
xdosus CECT237. Las columnas poseen el mismo significado que en la
figura 4. 2(1).
[24)
114)
¡(>4>
9 4 >
80
7 < >
6< )
5<)
o
- -4.2 _
— =
$9:1 ‘0
o — rl
— e.
e .? e-’=
44)
34 )
Lo
1<).
< 1
¡ 2<>
110
1 00
90
80 -C d —
E
— -4
a ’el
4- -
-r
74)
60
54)
- 4 < )
30
2 4 >
1 4 )
4 > ] — e-fi
144

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 158/293
RESULTADOS
H Ñ 1 ~ Ñ Ñ H ~ N Ñ N H‘5 k
k“ 5 -‘5
‘ 5 l i i i f i L l F i [1en~tOs.~,rSOrlU-0-4entiIC,20SN~n~nU-SOU-CC0
— $9 $9 < - — - f i ‘n — 1 - -t U- U- SN $9 $9 en t ‘t ‘~t Ir. -~ SO 5 - - — 5 — — 5 - - — 20
Úcíni ti
Figura 4. 16.- Actividad inhibidora de 24 bacterias lácticas seleccionadas frente a S.
faecium CECT4IO. Las columnas poseen el mismo siEnificado que en la
figura 4. 2(I).
¡20
¡ < >
¡ < ) O
9<)
t —. 8<)
‘.4 7<)
:~ <— — 5< ) — rl‘.4
C d o 40-e o— < - - f i 3< )-a -
e .) —- 20
1 < >
(1 I ~ 1 S i W ú ~ - 1 ¡ 1 U ~=, rs’ tu
en 05 — SO $9 U- ‘0 — en — $9 It, 20 0’ ~. U- SO 5--— C C =
— $9 rl < ~ — t en t ‘t U- U- Os e-] e-fi t t Ii, SO SO C x — U — 5 - ’ — - C C
(.e¡:¡ u’
hl ¡S\ ‘‘-‘u , - . rl
Figura 4. 17.- Actividad inhibidora de 24 bacterias lácticas seleccionadas frente a E. ccliLB841. Las columnas poseen el mismo significado que en la figura 4.
2(1).
I20
1 ¡ < )
¡ < ) 4 )
9<)
841
7<)
60
5< )
4<)
-e
e-
~e £ L ’d
-e rl
e .) ~-~ -z
3<)
2<)
E O
o
145

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 159/293
RESULTADOS
1 2 < >
1 1 4 >
¡ (> 4 >
9<)SI>
74)
t, 4)
5< )
-¡O
34.)
24 ’ )
¡ 4 )
<>11 ’’! M H r~iHt en tt Os — t ‘Ce—] U- ‘0 — en — ti It) ~Q 05 rs en U- SO U- 20 0
— Cxl $9 rl en t U- U- SN rl <9 en ? tt ¶t Ú Ita SO SO U- U- 1> 20
(..~el)a n’
Figura 4. 18.- Actividad inhibidora de 24 bacterias lácticas seleccionadas frente a B.
stearotermophylus CECT49. Las columnas poseen el mismo significado
que en la figura 4. 2(I).
¡2<)
1 1<)
¡00
90’
SO
74)
fil)
50
-fO
30
2 4 )
¡ < ) -
e—fi e-fi
L I‘ S I
‘ 5
‘ 5
‘ 5
SN — ‘t SO e - - ] Cx - 0 — en — Cx] Ifl 20 Os <n en U- SO U- C C ‘0e-fi en — t U- 5 -— - SN $9 <‘1 e n t -t ICa SO SO U- U- U- C C
(?~~>~ n’ ’
Figura 4. 19.- Actividad inhibidora de 24 bacterias lácticas seleccionadas frente a E.
clcacae CECTI49. Las columnas poseen el mismo significado que en la
figura 4. 2(1).
o
- -4
= —
CxlC d
‘0
— I’~fl
e.) e -
.5
o
- -4
.2
— e”
e .? —
« 1 ’ =
146

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 160/293
RESULTADOS
¡2<)
¡ ¡ 4 )
1 0<)9 < )
8<)
7 0
C d —
-e
-o ~ .2
-z
-e ~
C d ~-e o > ti
u —
-s
60
5 0
40
30 - -20
10
O H U M N l ¡ 1 l . N? ‘5 t QN — t ‘.0 e-fi U- O — e n — <‘~ Ir, YO SN efl ‘5 5-- ‘.t U- 20 ‘0
— $9 <‘-fi $9 en -t U- U- QN e-1 $9 <e-, C d ~ It. SO ‘O t~- U- U- 20
C — ¡> a u’
Figura 4. 20.- Actividad inhibidora de 24 bacterias lácticas seleccionadas frente a B.
cereus CECTL48. Las columnas poseen el mismo significado que en la
figura 4. 2(1).
ui ¡III Ñ F I L i Ñ L I I I fiL [1HÑ~IIt < e - , -t 0% — t ~C rl U- O — —. — $9 •t. 20 Os U- SO U- 00 0~
—~ < ‘ f i Cx ] $9 ‘S t U- U- SN e - f i e — f i ‘5 ‘t ~t Ú t It, ‘O SO U- U- U- 20
U~ ~>3 1 1 1 1
F ig u ra 4 . 2 1 . - Act ivid ad i n h i b i d o r a d e 2 4 b a c t e r i a s lácticas seleccionadas frente a Rs.
fluorcscens D C 7 . L a s c o l u m n a s p os e e n e l m i s m o s i g n i f i c a d o q u e e n l a
f i g u r a 4 . 2 ( 1 ) .
12<)
¡1<)
1 <)0
90C d —
-e
.2 _
— el
C dO’
e-fiel
u —
80
7 0
64 )
5< )
4<)
30
2 4 >
10
4 )
147

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 161/293
RESULTADOS
¡2<)
110
1 4)1 )
90
80’ 4
7< )
— 6< )
5 0<9
-z ‘‘
‘ .4 0
. ~ e - f i3<>
20
NÑN iliJLqI ‘3
en ‘t 0\ M SO <‘fi U- 0 en ‘~~‘ Cx) Ira 20 QN en en U- SO U- 20 ‘0
— <‘fi 4 +1 < NI en ~t U- U- 0\ e-fi <‘fi rs 7 T ~ It> ‘.0 SO U- C x — r’— CC
(‘epa n
Figura 4. 22.- Actividad inhibidora de 24 bacterias lácticas seleccionadas frente a Y.
enterocolitica E20. Las columnas poseen el mismo significado que en la
figura 4. 2(I).
¡2<)
‘¡O
LOO
941)
— . soO :4
74)
.0
e. . 6<)
— — 50
‘.4 rl
el 30
-1’ - 20
¡ O
4)
U cp:t ~>lI
Figura 4. 23.— Actividad inhibidora de 24 bacterias lácticas seleccionadas frente a S.
flexnerii CECT585. Las columnas poseen el mismo significado que en La
figura 4.2(l).
148
‘t 05 — SO ú-~ U- 0 — re - — r- ] ~ti YO SN efl rs U- SO Cx — 20 ‘0 — <‘fi e-fi <‘fi ‘t -t U- U- Q N < ‘~ fl e-fi ‘t t t -t ‘t It) 5- 0 St U- U- U- 20
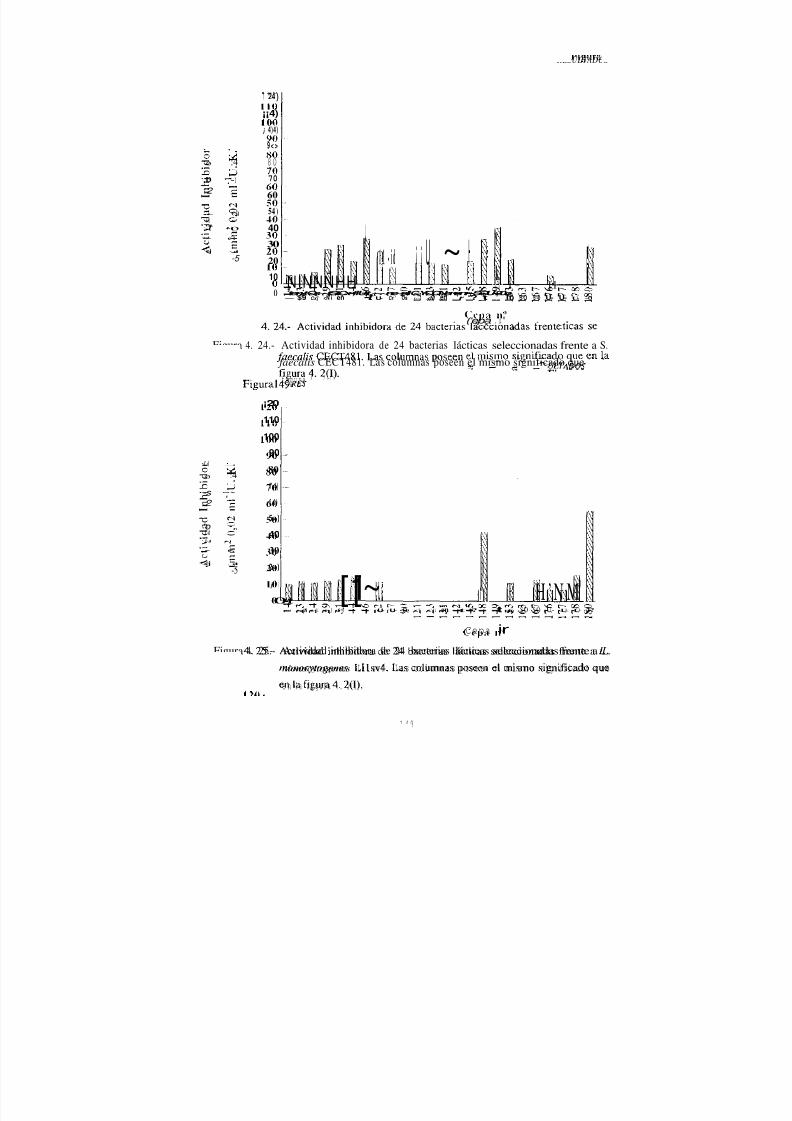
7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 162/293
RESULTADOS
1 24 )
¡¡4)
¡ 4 )4 )9 < >
8 0
70
60
54 )
40
30
20
10
O
F i
h1 1 - i d
¡ ¡ U ~H
NINNÑHH
Y
k1 ’ 5 u [1 H‘ten-tOs—tStCx-flU-t.—~ne~elIC,YOQNenenr--SOU-20o
— $9 Cx] <N i en -t U- Cx ’ Os < - ‘ - 3 <NI en r ‘? t It) S O 5 -0 U- U- U- ~Q
(‘epa u ”
Figura 4. 24.- Actividad inhibidora de 24 bacterias lácticas seleccionadas frente a S.
faecalis CECT481. Las columnas poseen el mismo significado que en la
f i g u r a 4 . 2 ( I ) .
80 -
74) --
6 4 )
5<) -
. ~ L I h U I I I [1[~uk1 ki ’ l . H H ’N M
<‘fi e - f i e- ] < -n U- U- Os < ‘ f i < ‘ f i en t ‘? t It) S O S O U- U- U- 2?
Cepa ir
Figura 4. 25.- Actividad inhibidora de 24 bacterias lácticas seleccionadas frente a L.
nionocytogenes Ll l sv4 . L a s c o l u m n a s p os e e n e l m i s m o s i g n i f i c a d o q u e
e n l a f i g u r a 4 . 2 ( 1 ) .
o :4
.D ~)
.2 _
— Cxl
C d ~“t o
— e-]
ej el
-5
¡20
110 -
100
90 -—
C d —
o :4
-s ~
.2 _
‘o C dt-.a
— e-,
ej el
-5
40 -
30
2<)
¡ < )
O-t
1 4 9
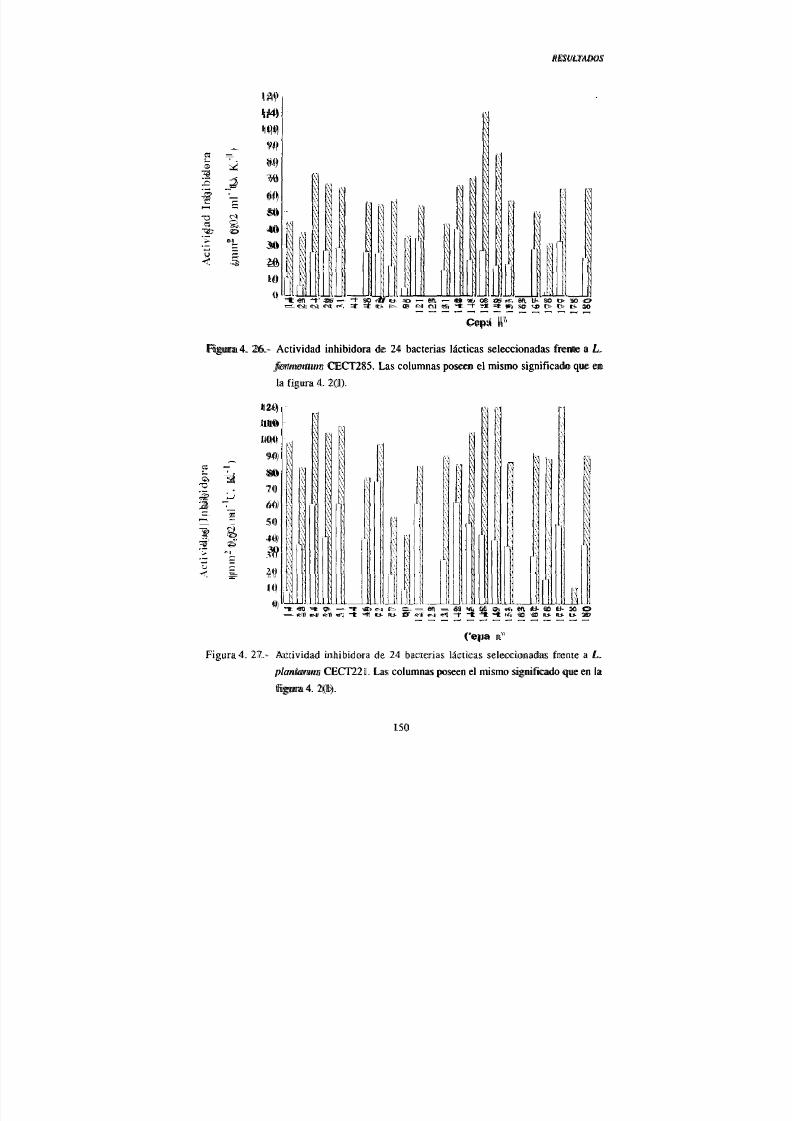
7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 163/293
RESULTADOS
1 2 4 )
C d —
~ -e
2.2 _
5-;
c o — e-]
e .? —
-s
¡¡4)
14 ) 4 )9 4 )
8<)
70
64 )
50
40
30
20
‘O
4 1 )
Ccp:í II ”
Figura 4. 26.- Actividad inhibidora de 24 bacterias lácticas seleccionadas frente a L.
fertnentunz CECT285. Las columnas poseen el mismo significado que en
l a f i g u r a 4 . 2 ( 1 ) .
12<)
110
‘41 )4 )
94 )
—. 80o :4
7 0.2 .2 _ 64 )
— e.
— — 5 0
o < -1— o -J e- 4<)-e O
30
= 20
1 < )
4 )
(‘e¡:a u ’ ’
F ig u ra 4 . 2 7 . - Act ivid ad i n h i b i d o r a d e 24 b a c t e r i a s lá ct ic as s e l e c c i o n a d a s f r e n t e a L.
planíarunz CECT22 1 . Las columnas poseen el mismo significado que en lafigura 4. 2 (1> .
t en ‘1’ 05 — SO ti U- ‘0 — en — ti It, CE 05 e - e - ~ en U- SO U- YO O — Cx] Cx-] <‘fi e . ~ . -t -t U- IV— 05 e--fi Cx] en t ..t -t Ir, SO ‘.0 U- U- U- 20
t en -t QN — t SO <‘~ 1-—- — — en — $9 It, C E QN en en U- SO U- Y O O — <‘fi e-fi <‘fi e”, t — t U- U- 0’ <‘fi e-~fi < -n t ‘t -t 1 <-a SO SO U- U- U- 20
1 5 0

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 164/293
RESULTADOS
120
110
14)4)
90,
80
74)
6< ) -
50
40 -
34)
20
1 4 1 )
4 )
HIv’
K
k K
t - i h u
‘ 5 ‘ 5 ‘ 5 ‘ 5 ‘ 5 ‘ 5
‘ 5 ‘ 5 ‘ 5 ‘ 5
‘ 5 ‘ 5en O S — ‘O $9 U- O — en - rl tt) 20 QN e n e n U- SO U- 20 o‘Ni ti e-] en ‘Ú ‘tt C x - — U- 0” <‘1 <‘fi en Ú ‘t “t ‘t ~t) S O SC U- Cx’- C x — 20
1~
Cepa n < ’
F ig u ra 4 . 2 8 . - Act ivid ad i n h i b i d o r a d e 2 4 - b a c t e r i a s l á c t i c a s seleccionadas frente a S.
typhirnurium C E C T 4 4 3 . L a s c o l u m n a s p os e e n e l m i s m o s i g n i f i c a d o q u e
e n l a f i g u r a 4 . 2 ( I ) .
120 -
110 —
100 -
90.—
80
7 0 -
60 - ‘5 0 —40 -
30
20
10
o u R l H u
I i i ”
F IN
rs QN —. ‘~t SO 4+-] U- O — — 4+-fi Ita 20 Q\ en en U- SO U- CC O— <‘fi t~fl <‘fi en ‘t U- U- 05 $9 rl en t t ‘t Ita SO SO U- U- U- 20
(Sepa i-i”
F ig u ra 4 . 2 9 . - Act ivid ad i n h i b i d o r a d e 24 bacterias lácticas seleccionadas frente a S.
aureus C E C T 5 9 . L a s c o l u m n a s p os e e n e l m i s m o s i g n i f i c a d o q u e e n laf i g u r a 4 . 2 ( 1 ) .
.2 _
~t rlC d
o
> <‘fi
et ~
C d —
-t
.2 _ — e .
‘o rl
C d ‘0)-‘o o
>- e-A — el
e .? e.- - A , o
151

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 165/293
RESULTADOS
IV. 2. 2. - Act ivid ad i n h i b i d o r a d e l o s s o b r e n a d a n t e s d e c u l t i v o s l i b r e s d e c é l u l a s
En l a t a b l a I V . 1 s e mu e st ra l a a c t i v i d a d an t im ic ro b ia n a d e l o s s o b re n ad an t e s d e
c u l t i v o s l i b r e s d e c é l u l a s d e l a s 2 4 ce p as p r e v i a m e n t e d e s c r i t a s d e s p u é s d e f i l t r a d o s ,
neutralizados y concentrados siguiendo el procedimiento descrito en la sección 1 1 1 . 2. 3. 3.
Como se observa, algunos presentan una actividad notable frente a ciertos microorganismos
i n d i c a d o r e s , sob re t od o f r e n t e a o t r a s b a c t e r i a s l á c t i c a s . Es d ign o d e m e n c i ó n q u e l o s
sobrenadantes de las cepas 148 y 180 presenten una notable actividad frente a L.
nz’onocytogenes. No obstante, frente a otros indicadores carecen de actividad inhibidora
detectable, lo que significa que, o bien éstos son insensibles a dicha actividad, o bien que la
concentración de la sustancia inhibidora es demasiado pequeña para detectarse con esta técnica.
IV. 2. 3. - Efecto de la catalasa en la actividad inhibidora de las bacterias lácticas
seleccionadas
Ya se ha descrito que el peróxido de hidrógeno (H,O,), producido por algunas
bacterias lácticas, inhibe el crecimiento de otras bacterias. Sin embargo, el tratamiento con 130
UY/ml de catalasa de los sobrenadantes de lo s cultivos libres de células con actividad inhibidora
no disminuye dicha actividad (no se muestran los resultados). Así pues, una vez comprobado
que, ni la acidez, ni la síntesis de H,02, ni la presencia de bacteriófagos estaban involucrados
en la actividad antimicrobiana de las cepas estudiadas, se pensó que éstas quizás sinteticen
o t r o s c o m pu e s t o s a c t i v o s d e b a j o pe so mo l e cu l ar y / o b a c t e r i o c i n a s . P or e l l o s c p r o c e d i ó a l a
identificación y caracterización bioquímica parcial de aquellas cepas cuyos sobrenadarnes libres
d e cé lu la s, u n a ve z co n ce n n ad os, ma n if e st ab an l a máxima actividad antimicrobiana
152

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 166/293
RESULTADOS
liii Os
5 -O1 1 ‘2
¡ 1 i 1 1 i 1 1 1 1 1 i 1 1
1 1 1 1 1 i 1 ¡ 1 i 1 1
1 1 1 i 1 i i 1 1 i 1
1 ¡ 1 i 1 1 ¡ i 1 i 1 1 i 1 ¡
¡ 1 ¡ 1 1 ¡ 1 1 1 1 ¡ i 1 1 ¡
a ¡ 1 1 ¡ 1 1 ¡ i 1 ¡ 1 1 ¡
1 1 ¡ i 1 i 1 1 1 1 1 ¡ 1
¡ r i 1 1 1 1 1 1 1 1 1 1 ¡ ¡ ¡
¡ r 1 i 1 1 1 1 1 i 1 ¡ 1
II i 1 1 i i i i i
i 1 1 ¡ i i 1 i 1 1 1 1 ¡ 1 ¡
1 1 1 1 1 1 ¡ i 1 1 i i
1 ¡ 1 1 1 1 1 1 1 1 i ¡
-e ‘Ou ¡a~§2 <9’o~’~’————
o-le-]00~o~nen en
¡——a:
¡i¡íZ~íuí¡Zí2
1 1 ¡ 1 1 1 i 1 i 1
¡ooc22 ztzfl ¡Cfl”-Cx40U-Cfl QV’DAC ¡2
a 1 i i 1 i i i
O’O.sflrn ¡ .-t’-O>U-
II
¡OSsOCCxt -~0ZZ
U- U-
~‘cn~O—n—Cx4Q’)20o\rncnr-—5-or—,Qo
E 12 & 5 > 4 -~
~--‘ U- .2
e::
u~
tnlri ~oc
u r2~ )
E ~
t~>O>
E .~ ~ u4 U ~
&# ‘ . 7 - )“-4 .. 3 ~0
~0
.2
cmU g.2~c~
e ~ :~ _
e . . . oQ)
o
C d ‘.0~~ ~
tHw—. ~‘
oUCJ e~ua .c,
‘Cd.’~’—. O
-a -. o ‘n
~ CJj
E ”-~ ~twC d
Cd2 2u:~-e
~ É~~-am ~
~.2 ..-g4¿~- C,~
1220 e..
-~
I5Z.2.2 ‘
,—~ L I~ ~ —-a
‘E-4
4,
½
4)SI
O
O
*
1-.
O-e Cd
O-e e
O
ECC
eC dtÉOOO
4)
e 0>‘--a
‘4-
‘ti
-e C deo OO4,
4)‘oCC
C dO
O‘C d
CC
C d
4,
OC d
-n
o-]4,
-e ‘ti O-c
C d
e4,
Oe o e)CC
C d
‘4)
O4)
-o CC
0)‘--a
-O
CC
4,— CC Co)Cd~
-e OC d t4,0
o CC
CC CCC
VCC
dCde- ÓL
o” -00‘e--O..O ,-,
.E E
t.-6
>2
Cd
C d
E.2
2’C d
¼-,
-4
O
5-4
C dO
‘- 4
.0
01
z C da
4)
u
1 . 5 3

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 167/293
RESULTADOS
I V . 3 . - I d e n t i f i c a c i ó n y c a r a c t e r i z a c i ó n b i o q u í m i c a p a r c i a l d e l a s b a c t e r i a s lá ct ica s
s e l e c c i o n a d a s n or s u a c t i v i d a d a n t i m i c ro bi a n a
L a s s e i s b a c t e r i a s l á c t i c a s c u y os s o b re n a d an t e s co n ce n t ra d o s l i b r e s d e c é l u l a s
ma n if e st aro n l a m á x i m a a c t i v i d a d a n t i m i c r o b i a n a , s e som e t ie ron a l a m a y o ría d e l a s p ru e b a s
r e c o m e n d a d a s p o r S c h i l l i n g e r y L ñ c k e ( 1 9 8 7 b ) , re se ñad as e n l a s e c c i ó n I I I . 2 . 4 . , p a r a l a
i d e n t i f i c a c i ó n r á p i d a d e l a s b a c t e r i a s l á c t i c a s p r o c e d e n t e s d e l a c a r n e y p ro d u ct os c á r n i c o s . L a s
ce p as sel ecci onad as f u e r o n l a s c o r r e s p o n d i e n t e s al o s n ú m e r o s 3 1 ,
7 7 ,9 0 ,
1 4 8 . 1 7 7 y 1 8 0 .
I V . 3. 1 . - Mor f ol ogía y t i n c i ó n p o r e l m é t o d o d e Gram
L a o b s e i ’ v a c i ó n mic ros có p ica d e l a s c e p a s seleccionadas, teñidas con el método de
G r a m , e v i d e n c i a u n a moifología de bacilos cortos Grarn positivos (tabla IV . 2).
I V . 3. 2 . - P r u e b a d e l a c a t a l a s a
To d a s l a s c e p a s a n a l i z a d a s f u e r o n c a t a l a s a n e g a t i v a s ( t a b l a I V . 2 >
I V . 3 . 3. - Pr od u cc ió n d e CO , a p a r t i r d e gl ucos a
L a ca p ac id ad d e l a s c e p a s a n a l i z a d a s d e p r o d u c i r C O , , s e investigó como se describe
e n L a s e c c i ó n 1 1 1 . 2 . 4 . 3 . N i n g u n a d e l a s c e p a s s e l e c c i o n a d a s p r o d u j o d i c h o g a s ( t a b l a I V . 2 ) .
I V . 3. 4. — H i d r ó l i s i s d e l a a r g i n i n a
L o s r e s u l t a d o s d e l a p r u e b a d e hidrólisis de la arginina se muestran en la tabla IV . 2,
154

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 168/293
RESULTADOS
n o o b s e rv á n d os e u n a re s pu e s t a u n if or me e n l a s d i f e r e n t e s ce p as se l e cci on ad as. D i c h a
re s p ue s t a v a r í a d e sd e l a r e a c c i ó n n e g a t i v a d e l a s c e p a s 3 1 , 7 7 , 9 0 yl S O , h a s t a l a p o s i t i v a d e l a s
ce p as 1 4 8 y 1 7 7 .
I V . 3 . 5 . - Producción de ácido sulfhídrico
L a r e a l i z a c i ó n d e l a s t é c n i c a s d e s c r i t a s e n l a s e c c i ó n I I I . 2 4 . 5.. n o e v i d e n c i é , e n
n ig ú n c a s o , l a p ro d u cc ió n d e á c i d o s u l f h í d r i c o p o r n in g u n a d e l a s c e p a s a n a l i z a d a s ( t a b l a I V .
2 ) .
I V . 3 . 6 . - P r u e b a d e Voges-ProskaUer
L a pr od uc ció n d e a c e t o í n a p o r l a s c e p a s seleccionadas seevaluó mediante la prueba de
Vo ge s- P ro sk aú e r ( s e c c i ó n I I I . 2 . 4 . 6 . ) . L o s r e s u l t a d o s o b t e n i d o s s e m u e st ra n e n l a t ab la I V .
2 ; d e ell o s s e d e d u c e q u e t o d a s l a s c e p a s p ro d u c e n a c e t o i n a e n c a n t i d a d e s m a y o re s ( c e p a s 7 7 y
90 ) o m e n o r e s ( c e p a s 3 1 , 1 7 7 y 1 8 0 ) . S i n e m b a r g o , l a c e p a 1 4 8 f u é n e g a t i v a e n e s t a p r u e b a .
I V . 3 . 7 . - F e rm e n t ac ió n d e c a r b o h i d r a t o s
L a cap acid ad fermentativa de carbohidratos de las cepas seleccionadas se evaluós i gu ie n d o e l procedimiento descrito en la sección III. 2. 4. 7 . L o s r e s u l t a d o s o b t e n i d os s e
mu e st ra n e n l a t a b l a I V . 3 . D e e l l o s s e d e d u c e q u e :
1 . ) To d a s l a c e p a s a n a l i z a d a s f e r m e n t an l a r i b o s a , g a l a c t o s a . D-gl uc o s a , D - f r u c t o s a ,
D - m a n o s a , N -ac et il gl ucos amin a. e s c u a l i n a , m e l i b i o s a , s a c a r o s a , t r ea lo sa y g l u c o n a t o .
2 . ) L a f e rm e n t ac ió n d e l o s a z ú c a r e s L—arabinosa, a-metil D-Qlucósido, D—turanosa,
155

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 169/293
RESULTADOS
s a l i c i l i n a , ma l t os a, lac t o sa , D - r a f i n o s a , c e l o bi o s a y 2 cet o-gl ucon at o e s v a r i a b l e d e p e n d i e n d o
d e l a s ce p as a n a l i z a d a s . A s í , mien t ras l a c e p a 1 7 7 e s l a ú n i c a q u e f e rm e n t a l a L - a r a b i n o s a ,
a- me t il D - g l u c ó si d o y D - t u r a n o s a , t o d a s , c o n e x c e p c i ó n d e l a 7 7 , u t ili z an l a s a l i c i l i n a . L a
ma l t os a e s f e r m e n t ad a p o r l a s c e p a s 3 1 , 7 7 y 1 7 7 y l a l a c t o s a p o r l a s c e p as 9 0 , 1 4 8 . 1 7 7 y
1 8 0 . L a s c e p as 3 1 , 7 7 y 1 7 7 u t i l i z a n l a D - r a f r n o s a , mie nt ras q u e l a 1 4 8 y 1 7 7 f e r m e n t an e l
2 - c e t o g l u c o n a t o . L a c e l o bi o s a s ó l o e s dé bil ment e f e r m e n t a d a p o r l a c e p a 1 8 0 .
3 . ) N i n g u n a d e l a s ce p as a n a l i z a d a s f e r m e n t a e l g li ce r o l, e r i t r i t o l , D - a r a b i n o s a ,
D-x i l o s a , L - x i l o s a , a d o n i t o l , ¡ 3 - m e t i l xi ló si d o , L - s o r b o s a , r a m n o s a , d u lci t o l, i n o s i t o l , m a n i t o l ,
s o r b it o l, a - m e t í m a n ó s i d o , a m i g d a l i n a . a r b u t i n a , in u li n a, m e l i z i t o s a , a l mi d ó n , gl u c ó ge n o.
x i l i t o l , ¡ 3 - g e n t i b i o s a , D - l i x o s a , D - t a g a t o s a , D- f u cos a, L - f u c o s a , D-arabitol, L-arabitol y 5
c e t o - g l u c o n a t o .
L a f e rm e n t ac i ó n d e l o s a z ú c a r e s m a n i t o l , r i b o s a , m e l i bi o s a , m a n o s a , s a c a r o s a y
t r eh al o sa , e s d e g r a n i m p o r t a n c i a e n l a c l a s i f i c a c i ó n d e l o s lactobacillos de origen cárnico. Con
l o s r e s u l t a d o s ob t e n id os y sigu ien do e l e s q u e m a d e i d e n t i f i c a c i ó n p ro p u e st o p or Sehillinger y
L ñ c k e ( 19 8 7b ) , t oda s l a s b a c t e r i a s lá ct ic as a n a l i z a d a s s e c l a s i f i c a r o n tentativamente como
Lactobacillus sake.
IV. 3 . 8 . -
Tolerancia al clomro sódico
En l a t a b l a I V. 2 s e mu e st ra e l d e s a r r o l l o d e l a s bacterias lácticas seleccionadas en
m e d i o MRS s u p l e m e n t a d o c o n u n 7 o u n 10% d e N aC í , i n cu b ad o a 3 2 0 C d u r a n t e u n t ie m p o
m á x i m o d e 6 d i a s . To d a s l a s c e p a s s e l e c c i o n a d a s , s a l v o l a 7 7 , c r e c e n a u n a co n ce n t rac ió n d e
N a C í d e l 7 % . s i n e mb argo , n in g u n a m a n i f i e s t a d e s a r r o l l o mensurable cuando la concentración
d e N a C í s e el eva a l 10%.
156

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 170/293
RESULTADOS
I V . 3 . 9 . - To l e ra n c i a a l pH d e 3 . 9
N i n g u n a c e p a s e d e s a r r o l l a e n e l m e d i o MRS a pH d e 3 , 9 ( a j u s t a d o c o n H C I , I N )
( t a b l a I V . 2 )
I V . 3 . 1 0 . - C re ci mi e n t o a d i v e r s a s t e m p e r a t u r a s
L a s t e m p e ra t ura s a l a s q u e l a sb ac t er ia s
l á c t i c a s s ed e s a r r o l l a n
s o n t am b ié n i m p o r t a n t e s
e n e l e s q u e m a d e i d e n t i f i c a c i ó n d e S c h i l l i n g e r y L i i c k e ( 1 9 8 7 b ) . En l a s e c c i ó n I I I . 2 . 4 . 1 0 s e
d e s c r i b e l a me t od ol ogí a s e g u i d a p a r a es t u d ia r el crecimiento bacteriano a l a s t e m pe ra t ura s d e 4 ,
1 5 y 45 0C, cuyos resultados se recogen en la tabla IV . 2. Todas las cepas seleccionadas se
d e s a r r o l l a n a 4 y 1 5 0 C , pe ro a 4 5 “ C ú n ic am e n t e l o h a c e n l a s c e p a s 1 4 8 y 1 7 7 .
1 5 7

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 171/293
RESULTADOS
T ab l a I V . 2 . - C a r a c t e r í s t i c a s m o rf o l ó gi c a s y b i o q u í m i c a s d e l a s b a c t e r i a s l á c t i c a s s e l e c c i o n a d a s
C e p a N ~
C a r a c t e r í s t i c a
3 1 77 9 0 1 4 8 1 7 7 1 8 0
M o r f o l o g í a
Ti n ció n Grambacilo
+
b a c i l o b a c i l o b a c i l o b a c i l o b a c i l o
+
C a t a l a s a
Pr od u cc ió n d e CO ,
H i d r ó l i s i s d e l a a r g i n i n a
Pr od u cc ió n SR2
Voges-ProskaUer
C re c i mi e n t o e n
7% N a C í
10% N a C í
Crecimiento a
pH 3 , 9
40C
¡ 5 0C
+ +
+
+
+
+
+
4 5 o c
+ + + +
+ +
+ + +
+ + + +
‘4 -
+
+
+
+
+ +
+ +
+
1 5 8

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 172/293
RESULTADOS
Ta b l a I V . 3 . - U t i l i z a c i ó n d e l o s c a r b o h i d r a t o s p o r l a s b a c t e r i a s l á c t i c a s s e l e c c i o n a d a s
C e p a N 9
Carbohidrato
3 1 7 7 9 0 1 4 8 1 7 7 1 8 0
G l i c e r o l - - - - -
E r i t r i t o l - - - - - -
D— arab in osa - - - - -
L - a r a b i n o s a - - - - +
R i b o s a + + + + + +
D- xi l osa - - - - - -
L - x i l o s a - - - - - -
Adon it ol
¡ 3 -m et il- xi lo sid o - - - - - -
G al act osa + + + + + +
D - G l u c o s a + + + + + +
D- F ru ct os a + + + + + +
D — M a n o s a - - --
- -
L -So rbo s a
R a m n o s a
D u l c i t o l
I n o s i t o l
1 5 9

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 173/293
RESULTADOS
T ab l a I V . 3 . - ( c o n t i n u a c i ó n )
C e p a N 9
C ar b oh id ra t o
3 1 7 7 9 0 1 4 8 1 7 7 1 8 0
Mani t ol - - - - -
S o r b i t o l - - - - -
a - m e t i l D — m a n o s i d o - - - - -
a-metil D-glucosido - - - - +
N-acetil glucosarnina + + + + + +
Amigdalina - - - - - -
Ar b u t in a - - - - - -
E s c u a l i n a + + + + + +
Salicilina + - + + + +
C e l o b i o s a - - - - - +
Ma l t os a + + - - + -
Lact osa - - + + + +
Me l ib ios a + + + + + +
S a c a r o s a + + + + + +
T rea l osa + + + + + +
I n u l i n a — — — — — —
D - r a f i n o s a + + - - + -
A l m i d ó n — — - — — —
1 6 0

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 174/293
RESULTADOS
T ab l a I V . 3 . - ( c o n t i n u a c i ó n . . y 2 )
C e p a N 2
C ar bo h id rat o
3 1 77 90 148 177 1 8 0
G l u c ó g e n o - - - - - -
X i l i t o l - - - - - -
¡ 3 - g e n t o b i o s a - - - - - -
D- t uran osa - . - - + -
D - l i x o s a - - -. - - -
D - t a g a t o s a - - - - - -
D- f u co sa - - - - - -
L- f u co sa - - - - - -
0 - a r a b i t o l - - - - - -
L - a r a b i t o l
G l u c o n a r o + + + + + +
2 ceto-gluconato - - - + + -
5 ceto-gluconato - - - - - -
1 6 1

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 175/293
RESULTADOS
I V . 4 . - Esoe ct ro an t imic rob ian o d e l a s u s t a n c i a i n h i b i d o r a e x o c e l u l a r d e l a s ce oas d e
L . s a L e 7 7 . 9 0 . 1 4 8 y 1 8 0
Una v e z c a r a c t e r i z a d a s b io qu imi ca me n e l a s s e i s b ac t er ia s l á c t i c a s d e i n t e r é s , e n c u a t r o
d e e l l a s s e e va l u ó d e n u e v o l a a c t i v i d a d i n h i b i d o r a d e s u s s o b r e n a d a n t e s co n ce n t ra d os l i b r e s d e
c é l u l a s f r e n t e a u n n u m e r o m a y o r y m á s d i v e r s o d e b a c t e r i a s al t er an t es y p a t ó g e n a s . Co n l a
m e t o d o l o gí a se ña l ad a e n l a s e c c i ó n 1 1 1 . 2 . 5 ( t a b l a I V . 4 ) s e ob se rvó q u e l o s s o b re n ad an t e s
c on ce n t rad os d e l a s 4 ce p as d e L. saLe a n a l i z a d a s e r a n a c t i v o s f r e n t e a c e p a s d e L. brevis, L.
curvatus, Carnobacteriurn di~’ergens, Leu. ¡nesenreroides, S. aui’eus, C’ botulinun,, C.
pe,f,’ingens y L. rnonocvtogenes.
L a a c t i v i d a d i n h i b i d o r a d e l a s c e p a s e s t u d i a d a s v a r i a b a n o s o l o d e s d e e l p u n t o d e v i s t a
c u a n t i t a t i v o s i n o t amb ié n c u a l i t a t i v o . A s í p o r e j e m p l o , e l sobrenadante d e L . saLe 1 4 8 f u é e l
ú n ic oq u e
s e m o s t ró a c t i v o f r e n t e a t o d a s l a s c e p a s d e L. monocytogenes e s t u d i a d a s ,a d e m á s
f u é t am b i é n e l ú n i c o q u e i n h i b í a e l d e s a r r o l l o d e l a s c e p a s CECT5 10 5 y S c o t t - A .
L a m a gn i t u d d e l a a c t i v i d a d i n h i b i d o r a , c u a n d o é st a e x i s t í a , t am b ié n f u é d is t in t a e n
c a d a c e p a e n s a y a d a s i e n d o , e n g e n e r a l , e l so b re n ad an t e d e L. saLe 1 4 8 e l m á s p o t e n t e , s i s e
e x c e p t ú a aL. saLe 1 8 0 c u y a capacidad de inhibir a S . aureus FR1137 o L. nzonocytogenes
L I S s v 1 /2 e r a m a y o r . S i n e m b a r g o , n o s e i n h i b i ó n in g u n a d e l a s b a c t e r i a s G r a r n - n e g a t i v a s
ensayadas, entre las que se incluían microorganismos potencialmente productores de
t o x i i n f e c c i o n e s a l i m e n t a r i a s como Salmonella £yphirnur¡wn y Ye,’sinia enterocolítica.
162

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 176/293
RESULTADOS
T ab l a I V . 4 . - E s p e c t r o a n t i m i c r o b i a n o d e l a s c e p a s d e Lactobadillus saLe s e l e c c i o n a d a s
Inhibición por el sobrenadante
concentrado de la cepa de L. sa/ce
Microoganismo Origena 77 90 148 180
&
B a c t er i a s G r a m - p o s i ti v a s
Lactobacillus ac¡dophilus 2 8 9 CECÍ O O 0 0
Loctobacillusbrei’is L b 2 8 9 IMTH 16 ,8 1 5 2 0 , 4 1 2
Loctobacillus ccn-n¡s L v 6 l F R I B O O O O
Lactobacillus casei 4 7 5 CECÍ O O O O
Lactobaci/lus curvatus L b 7 2 6 IMTH ¡ 4 , 2 15,6 17,2 13,8
Ca,-nobacteriun; divergeus L v 1 3 ERIE 1 7 1 7 2 1, 6 1 3
Lactobacillusfernientuni 2 8 5 CECT 1 9 , 4 1 7 , 4 2 3 , 8 1 4
Lactobacillus mesentericus 3 9 4 CECÍ O O O O
Lactobacilius planta¡’uni 2 2 1 CECÍ O O O O
Loctobacillus saLe L b 6 8 IMTH O O O O
Leuconostoc mescnw,’o¡des 3 9 4 CECÍ O O O O
Micrococcus i’arians 23 0 CECÍ O O O O
Staphy/ococcus riJosas 2 3 7 CECÍ O O O O
Sireptococcusfaecalis 4 8 1 CECÍ O O O O
Strepbococcusfaecalis
( v a r . Iicuejhc¡ens) 1 8 4 CECÍ O O O 1 )
Streptococcusfaúciuni 4 1 0 CECT 0 0 0 0
Bacillus stea¡’oíe¡’mophilus 4 9 CECÍ O O O O
BacH/as cen~us 1 4 8 CECÍ O O O O
B¡’oc’ho¡h,’Lv the¡-nmospl¡acu-¡ 1001 8 NCIB 0 0 ( 1 0
Swp/o’Iw’oc’c’us ameus 1 3 7 F R 1 O 8 . 4 9 1 0
Staphylococcus am-cus 3 6 1 FRI 0 8 . 8 9 , 2 8
1 6 3

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 177/293
RESULTADOS
T ab l a I V . 4 . - ( c o n t i n u a c i ó n )
I n h i b i c i ó n p o r e l so b re n ad an t e
co n ce n t ra d o d e l a c e p a d e L . s a k e
M i c r o o g a n i s m o O rige na 7 7 9 0 1 4 8
n 9
1 8 0
Listeria nzonocyrogenes
Listeria rnonocytogenes
Listeria rnonocytogenes
Listei’ia rnonocytogenes
Listei’ia nhonocytogenes
Clostridium botulinum
Clostridium pe,fringens
5 1 0 5
7 9 7 3
L I S s v 1 / 2
L I í s v 4
Scott A
551
376
Bacterias Gram-negativas
Ente,’obacter cloacac
Escherichia cok
Escherichia colie n t e ro pa t o ge n i c a
Salmonella ¡‘yphi
Salmon ella iyphinwrium
Pseudornonas fluorescens
Pseudonzonas fluorescens
Pseudomonas fluorescens
Ye,’sinia enterocolitica
Yersinia enterocolitica
194
BW545
B4 1
4 0 9
T91
DC5
DC?
NTI9
E20
1 4 4 0 5
CECÍ
MI T
IEKC
CECÍCENAN
FR 1 1 3
FRIB
FR I B
Nuestía
IPP
oo
0
o
o
o
0
0
colección O
4 1 )
a A b r e v i a t u r a s como e n t ab la I I I . 1
b Act ivi d ad i n h i b i d o r a e x p r e s a d a c o r n o di á me t ro d e h a l o d e i n h i b i c i ó n e n mm.
NCÍC
NCÍC
FVM
FVM
FVM
CECÍ
CECÍ
o
9
17,2
9
0
0
11,4
o
9 , 6
9 , 6
o
o
9,41 0
14 ,2
17
17
17
15,2
9 , 8
1 5
o
l o
1 9 , 4
9
0
10,2
10 ,6
o
o
o
o
o
( 3
o
O
o
o
oo
o
o
o
O
o
o
o
o
oo
ooo
o
o
0
o
o
164

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 178/293
RESULTADOS
De l o s r e s u l t a d o s o bt e n i d o s s e d e s p r e n d e q u e :
1). Las sustancias inhibidoras producidas por las cepas deL.sake seleccionadas son
d is t in t as , y a q u e v a r í a n t a n t o e n s u e s p e c t r o an t im ic ro b ia n o como e n l a m a g n i t u d d e s u
i n h i b i c i ó n .
2). La sensibilidad de lo s distintos microorganismos indicadores frente a las sustancias
i n h i b i d o r a s v a r í a d e a c u e r d o c o n la especie y cepa utilizada, de donde se desprende que la c e p a
p rod u ct or a n o s ó l o o r i g i n a u n a sustancia inhibidora específica, sino que la sensibilidad frente a
dicha actividad del microorganismo indicador empleado depende además de la especie también
d e l a c e p a .
3 ) . L a a c t i v i d a d i n h i b i d o r a d e L. sa/ce 1 4 8 f u é s u p e r i o r a l a d e l r e s t o d e l a s ce p as
e s t u d i a d a s , p or l o q u e s e s e l e c c i o n é d i c h o m i c r o o rg a n i s m o p a r a l a r e a l i z a c i ó nd e
e s t u d i o s
p o s t e r i o r e s .
I V . 5 . - E f e c t o d e l a comnosición del medio de cultivo en la actividad antimicrobiana
d e Loctobacillus sa/ce 148
El medio MRS, que se emplea generalernente p a r a e l d e s a r z ’ o l l o d e l a s b a c t e r i a s l á c t i c a s
es muy rico en sustancias proteicas por lo que dificultaría la purificación de las sustancias
proteicas antimicrobianas. Consecuentemente, la actividad inhibidora de L. saLe 148 se
detei’minó en otros medios de cultivo, unos complejos y otros más definidos, de acuerdo con
l a me t od ol og ía descrita en la sección III. 2 . 6. La actividad inhibidora de los sobrenadantes
co n ce n t rad os d e l o s c u l t i v o s l i b r e s d e c é l u l a s s e c o m p a r ó c o n l a cor re sp on d ie n t e d e L s a L e
1 4 S crec ido e n m e d i o MRS ( t a b l a I V . 5 ) .
1 6 5

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 179/293
RESULTADOS
T ab l a I V . 5 . - Act ivid ad an t imi cr ob ian a d e L. saLe 1 4 8 d e s a r r o l l a d o a 3 2 0 C e n d i v e r s o s
me d io s d e c u l t i v o
M e d i o d e c u l t i v o Actividad inhibidora a
MRS ++
ART
ART + Triptosa ++
APT + P r o t e o s a p e p t o n a ++
API + E x t r a c t o d e c a r n e ++
API’ + Peptona ++
BHI
BHI + Triptosa
BHI + P r o t e o s a p e p t o n a
BHI + E x t r a c t o d e c a r n e
BHI + Peptona
MM + Triptosa ++
MM + P r o t e o s a p e p t o n a +
MM + E x t r a c t o d e c a r n e ++
MM + P e p t on a +
MM + Tr’íptona
MM + C a s i t o n a
a Símbolos para el ensayo de difusión en agar: ++, Diámetro de inhibición grande (15-21
mm); +, Di á me t ro d e i n h i b i c i ó n pequeño (<15 mm); -.Sin inhibición.
1 6 6

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 180/293
RESULTADOS
Los resultados de la tabla IV . 5. indican que los medios de cultivo APT y EHI
(suplementado o no) no son adecuados para que se exprese la actividad antiínicrobiana de L.
saLe 148. Lo mismo ocurre con el medio base (MM), suplementado con triptona o casitona,
(hidrolizados proteicos derivados de la caseina). Sin embargo, la actividad inhibidora es
cuantificable cuando el microorganismo se cultiva en medio complejo de APT o en medio base
MM, suplementados con tripbosa, proteosa-peptona, peptona o extracto de carne (todos ellos
son hidrolizados proteicos de origen cárnico>. La actividad inhibidora de en el medio base
suplementado con triptosa (MM-triptosa), es similar a la obtenida en el medio MRS. Dado que
la triptosa es, como se ha dicho, un hidrolizado proteico cárnico que contiene,
fundamentalmente, aminoácidos y péptidos pequeños lo que facilita la purificación de la
sustancia antimicrobiana deL, s a L e 148.
IV. 6.- Parámetros cinéticos y actividad inhibidora de L saLe 148 a distintas
temperaturas
IV. 6. 1.- Crecimiento y actividad antimicrobianadeL. s a L e 148
En las figuras 4,30 a 4,34 se muestra la relación entre crecimiento y actividad
antimicrobiana de L. saLe 148, al desarrollarse en lo s medios M RS y MM-triptosa a diversas
temperaturas. De lo s resultados obtenidos se deduce que:
1 . ) L. saLe 1 4 8 manifiesta un crecimiento cuantificable a 4, 8, 16 , 25 y 32 0 C . s i b i e n
el desanollo es mayor a 25 y 3 2 oc.
167

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 181/293
RESULTADOS
3 00 -
2 5 0
200
1 5 0
loo
5 0
o
A
o l o 1 5 2 0 2 55 3 0
6 00
5 00C d1~o
4 00
300 ~
C d ‘o
200 ~zu
<‘fi
E
loo
O
Tiempo (días)
3 00
o
B
.2.
o 5 l o 1 5 20 2 5
6 00
5 0 0C d 1.,o
4 0 0
300 ~
C d ‘o
200 ~Zu
loo
03 0
< ‘ f i
EE
Tiempo (días)
Figura 4. 30.- Crecimiento ~o)y a c t i v i d a d an t imi cr ob ian a (u) de Lacm oba cil l us s a L e
1 4 8 c u l t i v a d o e n l o s m e d i o s MRS (A) y MM-triptosa (B) a 4 o c .
1 6 8
-5-.. 4 - - a
CC
‘o C d‘o
C d
u
C d.0 LI
oCC
.0
250
200
1 5 0
loo
5 0
CC
4 >e C d
‘o
C du
C d .0 1~
OCC
.0

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 182/293
RESULTADOS
Tiempo (días)
.1. E A
4 6 8
Tiempo (días)
Figura 4. 31.- C re ci m i e n t o ~o)y a c t i v i d a d a n t i m i c r o b i a n a ( M ~ d e Locwbacillus s a L e
1 4 8 cul t ivad o e n l o s me d io s MRS (A) y M M - t r i p t o s a (8) a 8 0 C .
169
3 00
4>
CC
4>e C d ‘o
C d
ue C d.0 1~
o u,.0
2 5 0
200
1 5 0
loo
5 0
6 00
5 0 0C d1-O
400
e
e 300 -o
C d
‘o
200 ~zu
E
fi-fi
Ee
o
3 00
o 2 4 6 8 1 0
loo
o12
B6 00
5 00- 5 - 1
~1
e>
CC
4>‘o C d ‘o
e
C d
ue C d .0
1-OCC
.0
250 -
200 -
1 5 0
loo
5 0 -
o
C d 1-O
400
e
e
3 0 0 - oC d
‘o
; 1.
200 zu
e-fi
e E
o.o 2
~1 loo
olo 12

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 183/293
RESULTADOS
3 00
25 0
200
150
loo
5 0
o
3 00 -
25 0
B
E
200 -
1 5 0 -
loo
5 0
o
6 0 < )
5 0 0C d 1-.o‘o
4 00 ~
e e
3 00 - oC d
200 zu
4+ 1
EA
loo
o
6 00
5 0 0 C d
o4 00
e
e
3 0 0 ~C d
2 0 0 • zu
4+.
EE
-
loo
1 1
O l O 2 0 3 0 4 0 5 0 6 0 7 0 8 0o
Tiempo (horas)
Figura 4. 32.- Crecimiento (O) y actividad antimicrobiana (U) de L saLe 148
c u l t i v a d o e n l o s f i f l e d i o s MRS (A) y MM - t ri p t os a (II) a 1 6 0 C .
1 7 0
4 - - a-4-a 4>
CC
e>‘o C d‘o
e
C d
u e C d
.0 oCC
.0
0 10 20 30 40 5 0 6 0 7 0 8 0
Tiempo (horas)
-4-4-ae>
u,a)‘o C d‘o
e
C dc ae C d.0 fi-o‘ti
.0

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 184/293
RESULTADOS
3 0 0
250
200
1 5 0
loo
5 0
o
300
~50
200
150
loo
5 0
o
6 00
5 00C d1-.o
4 00
e
e
3 0 4 1 ) oC d
‘o
200 ~-zc a
e-fi
EE
loo
o
6 00
5 00C d1-.O
400
e e
300 ~
C d
2 00 1=U
4+-fi
EE
1 0 < )
o
Tiempo (horas)
Figura 4. 33.- Crecimiento (o) y actividad antimicrobiana (u ) d e Lacrob acil l us s a L e
1 4 8 c u l t i v a d o e n los medios MRS (A) y MM—triptosa (II) a
250C.
e>
C Ce>
o C d
‘o e
C d
c ae C d .0
1~
OC C.0
o 6 1 2 1 8 24 3 0 3 6
Tiempo (horas)
- 5 - 14>
C Ce>
0 C d
e
C d
c ae C d.0
1-OC C
.0
o 6 1 2 1 8 24 3 0 3 6
171

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 185/293
RESULTADOS
3 00
250
200
1 5 0
loo
5 0
o
3 00
250
2 00
1 5 0
loo
5 0
o
Ligura 4. 34.-
6 00
500C d 1-o
4 00 ~e
e 3 00 c
C d
‘o
2 00 - zca
Cx.
EE
loo
o
6 00
5 00C d 1-o
4 00.0
e
3 00 ~C d ‘o
200 ca
rl
EE
loo
o
C re c im i e n t o (o) y a c t i v i d a d a n t i m i c ro bi a n a ( u > d e Lactobacillus s a / c e
1 4 8 c u l t i v a d o e n l o s me d io s MRS (A) y MM-triptosa (B) a 3 20 C .
172
. 5 - - a
a)
CC
e>‘o C d ‘o e
C d
c ae C d
.0
o u,
.0
~«
0 6 12 1 8 2 4 3 0 3 6 4 2 4 8
Tiempo (horas)
4>
u,a)
‘o C d
‘o
e
C d
c ae C d
.0 1-
ou,
.0
0 6 1 2 1 8 2 4 3 0 3 6 4 2 4 8
Tiempo (horas)

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 186/293
RESULTADOS
2 . ) D e n t ro d e u n a m i s m a t e m p e r a t u r a , e l d e s a r r o l l o ó p t i m o s e al c an z a e n e l m e d i o
M RS.
3 . ) A l a s t e m p e ra t ura s d e i n c u b a c i ó n e mpl e ad as , l a a c t i v i d a d antiinicrobiana se detectó
p o c o d e s pu é s d e i n i c i a r s e l a f a s e l o g a r í t m i c a d e c r e c i m i e n t o . P or t a n t o , l a act ivid ad
antimicrobiana de esta cepa es una propiedad ligada al desarrollo celular, siendo detectable y
cuantificable durante su crecimiento en un medio base o mínimo, suplementado con triptosa.
4 . ) T an t o e n e l m e d i o MRS como e n e l MM- t rip t osa , l a a c t i v i d a d an t im ic ro b ia n a f u é
máxima cuando L. saLe 148 se cultivaba a 32 0C, no obstante, la actividad fué siempre mayor
e n e l niedio complelo MRS. La actividad máxima a esta temperatura se detecta a las 16 horas
d e c r e c i m i e n t o ; po s t e ri o rm e n t e s e ob se rva b a u n l i g e r o d e s c e n s o h a s t a el final de la incubación.
IV. 6. 2. - Parámetros cinéticos del crecimiento microbiano
Los parámetros cinéticos estudiados en L. saLe 148 cultivado en lo s medios MRS y
MM-triptosa a 4, 8, 16 , 25 y 32 0 C , fueron los siguientes; velocidad específica de crecimiento
(gt), tiempo de duplicación celular (td), número de generaciones por llora (g/h) y peso celular
seco final (mg/mí).
Los parámetros citados, así como el pH y la actividad antimicrobiana final se muestran
en la tabla IV . 6. De ella se deduce que:
1.) La velocidad específica de crecimiento y el número de generaciones/tora fué mayor
y, consiguientemente, el tiempo de duplicación menor, cuando L . s a k e ¡48 se desarrollaba a
2 5 y 3 2 0 C . El crecimiento fué ligeramente inferior cuando el medio empleado era elMM-triptosa.
173

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 187/293
RESULTADOS
Ta bl a I V . 6 . - Pa rá me t ro s c i n é t i c o s d e c r e c i m i e n t o , pH final y actividad inhibidora de
Loctobacillus saLe 148, cultivado en lo s medios MRS y MM-triptosa ad i f e r e n t e s t e m p e ra t ura s
Pa rá me t ro ( a ) M e d i o T e mp e ra t u ra d e i n c u b a c i ó n ( 0 C )
4
MRSMM -t rip t osa
0,0010 , 0 0 1 .
8
0 , 0 0 60 , 0 0 5
1 6
0 , 0 3 50,035
25 32
0,0750,073
0,0930,087
MRS
MM -t rip t osa
MRS
MM -t rip t os a
MRS
MM-triptosa
MRS
MM- rri p t osa
MRS
MM- t rip t osa
MRS
MM -t r i p t o s a
5,4 5,2
5,2 4,8
137 160
101 101
0,8
0,8
0 .9
0 ,6
( a ) ~, velocidad específica de crecimiento (h~ 1);
generaciones/hora; pcs, peso celular seco (ng/mí):
td, tiempo d e d up l ica ció n (h), g/h,
pH, pH final: Al y AlE, actividad
inhibidora del sobrenadante (inín2/ml) y actividad inhibidora específica del mismo
(min2/rnl/Unidad K lem t)
td
pcs
563
700
0.002
0,00 1
0,82
0,83
108
137
0,009
0,007
0,89
0,87
19,2
19,8
0,050
0,050
1,14
1,15
9,2
9,9
0.108
0,100
1,25
1,20
7,4
7,9
0,130
0,120
1,34
1,28
pH
Al
AlE
4,9
4,4
170
108
0,7
0,5
4,5
4.3
541)3
502
1 ,9
2.2
4,5
4,3
513
485
1,9
2,2
174

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 188/293
RESULTADOS
2.) El peso celular seco final fué mayor a 32 0C. Con el medio de MM-triptosa se
o b t e n í a u n p e s o c e l u l a r l i g é r a m e n t e i n f e r i o r .
3 . ) E l pH final fué más bajo a las temperaturas óptimas de crecimiento, siendo menor
que el obtenido cuando el medio de cultivo empleado era el de MM-triptosa.
4.) La actividad inhibidora máxima se alcanzó a lo s 32 0C en el medio MRS, sin
embargo. la actividad inhibidora específica, es decir, el cociente de dividir la actividad
inhibidora máxima por la absorbancia del cultivo (Unidades Klett), fué igual a 25 y 32 0C y
ligeramente stiperior en el medio de MM-triptosa.
IV. 7. - Purificación parcial de la actividad antimicrobiana exocelular deL. sa/ce 1 4 8
Conocidas las condiciones óptimas de producción de la sustancia antimicrobiana
e xo ce l u l ar d e L. scA-e 148 en medio semisintético de MM-triptosa, se procedió a su
p u r i f i c a c i ó n por cromat ograf ía d e f i l t r a c i ó n e n geles.
IV. 7. 1 . - Cromatoizrafia de filtración en Sephadex G- 150. 0-75 y G-50
El sobrenadante concentrado libre de células de Lsake 148, obtenido como sedescribe en la sección 111.2. 3. 3. 1 . se purificó parcialmente en columnas de Sephadex 0-150
fino, 0-75 y G-50 de la manera descrita en la sección 111 .2 . 8. de este trabajo. El resultado de
ésta purificación se refleja en las figuras 4.35, 4. 36 y 4. 37 , en las que se muestra la actividad
antirnicrobiana y la absorbancia de las fracciones eluidas.
Al pasar el sobrenadante concentrado por la columna de Sephadex 0-150 fino (figura
4. 35), la actividad antiíuicrobiana eluye tardiarnente aunque antes de que aparezca el ~mayor de la absorbancia. Dado que este pico coincide con el volumen de eiución de Ja columna
175

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 189/293
RESULTADOS
se suponeque el peso molecular de la sustancia antimicrobiana se encuentra muy próximo al
límite inferior de exclusión del Sephadex G-150 fino, que es de unos 5.000 daltons. Para
obtener una mejor separación, las fracciones con mayor actividad antimicrobiana se juntaron y
liofilizaron, volviéndolas a pasar por una columna de Sephadex G-75.
El resultado de la segunda filtración se observa en la figura 4. 36 , apreciándose 2 picos
de absorbancia máxima no bien definidos. El primero de ellos, más pequeño, coincide con el
de la actividad inhibidora, mientras que el segundo coincide con el volumen de elución de lacolumna. La actividad inhibidora queda, por tanto, retenida en la columna. Sin embargo. Ja
capacidad de separación de ésta columna es muy amplia (3.000 - 80.000 Da) por lo que, para
obtener la mayor separación posible por cromatografía de filtración, se recurrió a una tercera
columna con un intervalo de separación entre 1.500 y 30.000 Da. De nuevo, las fracciones con
mayor actividad inhibidora procedentes de la segunda columna se juntaron y liofilizaron,
pasándolas a continuación por una columna de Sephadex G-50.
Los resultados obtenidos con la columna de Sephadex G-50 se reflejan en la figura 4.
37, en la que se observa que el pico de actividad se corresponde con el de absorbancia
máxima.
176

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 190/293
oo o
CI e e‘ o c
a
c
a
o c
C d
1 -
o‘ o o~1 -
o e - f i
o
4
w u O S Z
E
EI 3
U C t J J O S q~
o 0 0~-1
‘ -- a ,
4 , t
C
d
C
d
O
O
- e
O
. 0
u
C
d
O ( - 3
—
e :
o o
t
o c
—
el
C
d
C
d
e - )
~ O ,
~ O
O
C
d
‘ O
. 0
—
‘ -- a
o
o C
d
‘ o
o
. 0
t f l C
d
- e • 0 en
o
o
o
o
o
o
o c
S O
‘ t
O —
]
1
7
7

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 191/293
o c
O O
— ( - . 3
0 4
e - )
‘ o ; ~. <
4 ,- o C
d
O
o
. c
t . s t e-~
e C d
>
- a- ,
5-’
o
4 ,
~
—
el
C d
C d
C d
a 0 e
O
C
d
o
‘ 0 .
0 4
~ 5
C
d
‘ o
o . 0
t e
C d
0~.-’
en
o
O S O
4 2 )
WL i O
S Z
U
E i 3 uE q J o s q~
2 u - ’
1
7
8
O
O
O
O
O
L f l
en
el

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 192/293
O S
O
c
i
‘ o c
a
c
a
C
d
S-4
O- Ú O el O
t
t
i
u
I i
4
>
o c‘ =4LI ,
L
I
,
( .4 O )
~
C d
C
d
O
c e
- e
~ .- 5 f l
t c e
5
- e
t é C d
O
4 - >
0—
el
C d
—
C d
e - >
~ O ,
U O C d
‘ O
. 0
o~
i .f l
C d
o” - ,
U-
en-f -I t 2 )
u - .
e n
el
o
1
7
9

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 193/293
RESULTADOS
La purificación de la actividad antimicrobiana exocelular de L. saLe 148 se muestra en
la tabla IV. 7, en la que se detalla el grado de purificación y porcentaje de recuperación para
cada una de las etapas. La purificación final fué de 6,7, con una recuperación de la actividad
antimicrobiana del 8 %.
1 8 0

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 194/293
RESULTADOS
Tabla IV.?. - Purificación pai-cial de laactividad antimicrobiana de Lactobaciflus sa/ce 148
Actividad Actividad
Actividad Inhibidora Proteína Inhibidora Actividad Grado de
Volumen Inhibidora Total Total Específica Recuperad-a Purificación
(mí) (cm2/nil) (cm2) (mg/mi) (cin2/ml) (%) (veces)
Sobrenadantedel cultivo librede células 500 6,5 3.267 10 0,6 100 1
SephadexG-150 1 7 148,5 2.525 50 2,9 77 4,5
Sephadex4 1 3 - 7 5 6 111,6 670 25,9 2,2 21 3,2
SephadexG-50 3 88,3 265 30 4,4 8 6,7
La purificación se calcula corno la Actividad inhibidora específica de la fracción X/Actividad
inhibidora específica fracción 1 .
La recuperación se calcula como la Actividad inhibidora de la fracción X/Actividad inhibidora
de la fracción 1 multiplicado por 100.
181

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 195/293
RESULTADOS
IV. 8. - Caracterización parcial de la actividad antimicrobiana de Lactobacillus saLe
148
IV. 8 . 1 . - Efecto de diversos enzimas en la actividad inhibidora
Para determinar la sensibilidad de la actividad antimicrobiana parcialmente purificada a
los enzimas proteolíticos, lipolíticos y amiloliticos, se realizó el ensayo descrito en la sección
III. 2. 9. 2. Los resultados obtenidos se observan en la Tabla IV . 8, de la que se deduce que:
1.) - La actividad antimicrobiana de L. saLe 148 es destruida por los enzimas
proteolfticos inespecíficos papaina, proteasa XIV y proteasa II. Otros enzimas proteoliticos,
como la tripsina y pepsina, reducen la actividad antimicrobiana en un 92 % tras 12 horas de
incubación a 32 0 C .
2.) - Los enzimas lipolíticos lipasa 1 y lipasa VII, no modifican la actividad
antimicrobiana de Lactobacillus saL -e 148 y lo mismo ocurre cuando se emplean enzimas
amilolíticos.
Los resultados obtenidos indican que la actividad antimicrobiana de L. saL -e 148 es de
naturaleza proteica, sin que existan en su estructura componentes lipídicos ni hidrocarbonados
que intervengan en dicha actividad.
1 8 2

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 196/293
RESULTADOS
Tabla IV . 8. - Efecto de diversos enzimas en l a actividad antimicrobiana
purificada de Lactobacil/us saLe 148
parcialmente
Actividad residual (%)
EnzimaConcentraciónFinal Tiempo de incubación (h)
(mg/mí) 2 6 12
T r i p s i n a 1 46
5 31
P e p s in a 24
5 31
67Pa p ai n a
Proteasa XIV
P rot eas a I I
Lip asa 1
Lipasa VII
a-ami lasa
2 1
.7 ?1
5 37
1
5
loo
1 0 < )
loo5 loo
1 < 1 < )
5 loo
25 8
8 8
8
8
8
8
o5 5
16
26
1 6
100
1 0 0
1 0 0
1 0 0
O
16
o
1 0 0
100
100
1 0 0
1 0 01 0 0
10 < ) loo
183

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 197/293
RESULTADOS
IV. 8. 2. - Efecto del oH en la actividad inhibidora
El ensayo se realizó como se describe en la sección 1 1 1 . 2. 9. 3., variando el pH del
tampón utilizado en la disolución de la actividad antimicrobiana entre 2,6 y 12 y utilizando aL.
fernzentunz CECT285 como microorganismo indicador.
Los resultados obtenidos se muestran en la figura 4. 38, de laque se deduce que la
sustancia antimicrobiana es activa en el intervalo de pH analizado, aunque la mayor actividad
se observa a valores de pH entre 5.6 y 6,6. La actividad inhibidora es ligeramente inferior
cuando lo s valores de pH son extremos, tanto ácidos como básicos.
IV. 8. 3. - Cinética de termodestmcción de la actividad antimicrobiana exocelular de
Lactobacillus saLe 148
La cinética de termodestrucción de la actividad antimicrobiana de L sake 148, se
determinó depositando en viales de vidrio 20 mg de la sustancia parcialmente purificada en 100
gí de tampón citrato-fosfato 4 mM de pH 7, a continuación se cierran herméticamente, y se
calientan durante intervalos de tiempo variables, dependiendo de la temperatura de
calentamiento (sección III. 2. 9. 4).
La figura 4. 39 muestra la inactivación térmica de la sustancia antimicrobiana de L.
saLe 148 a80, 100,121, 135 y 150 0C ; deella se deduce:
La actividad antimicrobiana deL. saLe 148 es resistente a temperaturas de 80 y 100
0C. perdiendo únicamente un 40 % de su actividad inhibidora inicial cuando se calienta a 100
0Cdurante 30 mm.
184

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 198/293
RESULTADOS
1 0 0
8 0
6 0
40 -
2
20
o4 6 8 1 0 1 2
pH
Figura 4. 38. - Efecto del pH en la actividad de la sustancia antimicrobiana parcialmente
purificada deL. saLe 148.
185
Cd1~
O‘o
.0
.0
‘o C d
‘o
c a

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 199/293
RESULTADOS
Esta termorresistencia también se manifiesté cuando las temperaturas de calentamiento’
fueron de 121, 135 y iso 0C.
2.- A las temperaturas empleadas, la inactivación de la sustancia antimicrobiana se
ajustó, con ligerasvariaciones, a ecuaciones cinéticas de primer orden, mientras lo s parámetros
de termodestrucción “D”, ‘t
14’ y Z se calcularon, como se describe en la sección 1 1 1 . 2. 9.
5. 3 . 2 . , a partir de los resultados de termodestrucción (figura 4. 39)-
Los valores “D ’ a 80, 100, 121. 135 y 1500C, calculados asumiendo una cinética de
¡nactivación de primer orden, fueron de 285,7, 119, 23,8, 17,4 y 15,2 minutos
respectivamente. Los valores “t1~”, calculados par a las mismas temperaturas, fueron de 198,
82,5, 16,5, 12,05 y 10,6 minutos respectivamente. Finalmente, de la ecuación de la recta de
regresión obtenida al representar gráficamente el logaritmo de lo s valores ‘D” en función de la
temperatura a la que fueron obtenidos, se calculó el valor ‘7; dicho valor, procedente de la
recta comprendida entre 121 y 150 ‘=C,fué de 153,40C.
1 8 6

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 200/293
RESULTADOS
C d1~o
‘o
.0
.0
‘oC d
‘o
-ao
1 0 0
8 0
6 0
40
20
o6 0
Tiempo (mm)
Figura 4. 39.- Resistencia térmica de la actividad antimicrobiana parcialmente purificadade Lactobacillus saL -e 148 tras su calentamiento a 80 0C (m Il, 100 0C (o>,
121 0C(Á), 135 0C(A)y 1500C(•).
o 1 0 2 0 3 0 4 0 5 0
1 8 7

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 201/293
RESULTADOS
I V . 8 . 4 . - Determinación del ceso molecular por cromato2rafía de filtración en
Seohadex G-50
El peso molecular aparente de la sustancia antimicrobiana parcialmente purificadadeL.
saL-e 148, se determinó sometiéndolo a una nueva cromatografía de filtración en un gel de
Sephadex G-50. Como se describe en la sección III. 2. 9. 5, la representación gráfica de lo s
eluatos cromatográficos de marcadores de peso molecular estándar, en función del logarítmo
de sus pesos moleculares, permite determinar el peso molecular de una proteína problema por
interpolación en la gráfica de la fracción cromatográfica que la contiene.
La cromatografía se realizó en las condiciones descritas en la sección III. 2. 9 . 5 , y su
resultado se refleja en la tabla IV. 9. E n dicha tabla se muestran el peso molecular en daltons,
su logaritmo y las fracciones de elución de lo s compuestos estándar y de la actividad
antimicrobiana de L. saL-e 148. El peso molecular de la sustancia inhibidora, obtenido porinterpolación gráfica de lo s resultados obtenidos (fig. 3 . 7), fué de 4.640 daltons.
IV. 8. 5. - Determinación del Deso molecular por electroforesis en aeles de
poliactilamida co n dodecil sulfato sódico
Para confirmar el peso molecular de la sustancia antimicrobiana parcialmente purificadade L. saL-e 148. determinado por cromatografía de filtración, se utilizaron las técnicas de
electroforesis en geles de poliacril- amida con dodecil sulfato sódico y urea según la técnica de
Swank y Munkres (1971) y en geles de poliacrilamida con dodecil sulfato sódico
(SDS-PAGE) según la técnica de Laemli (1970).
188

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 202/293
RESULTADOS
Ta bl a IV. 9. - De t e rmi n ac ió n d e l p e s o mo l e c u l ar d e l a a c t i v i d a d an t im ic ro b ia n a p ar ci al me n t e
p u r i f i c a d a d e Lactobacillus saLe
S e p h a d e x G - 5 0
Compuesto
F r a c c i ó n d e
e l u c i ó n n 2
L o g a r i t m o d e l
pe so mol ec ul ar
Pe so mo l e c u l ar
( D a l t o n s )
a-quimotripsinógeno 27 4.40 25.000
RNAsa pancreático 32 4,14 13.700
Actividad
antimicrobiana 43 3,67 4.640
Vitamina 55 =13 1.355
1 8 9
1 4 8 , p o r c r o m a t o g r a f í a d e f il t r ac ió n e n

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 203/293
RESULTADOS
IV. 8. 5 . 1 . - E l e c t r o f o r e s i s s e 2 ú n la t é c n i c a d e S w a n k y M u n k r e s ( 1 9 7 1
E st a t é c n i c a p e r m i t e s e p a r a r ca d e n as p e p t í d i c a s d e p e q u e ñ o t a m a ñ o mo l e c u l ar g r a c i a s a
l a in corp orac ió n d e ur e a a l o s g e l e s d e p o l i a c r i l a m i d a . Co n t a l f i n s e p a r a d o r , 5 0 , 10 0 y 200 g g
d e l a a c t i vi d a d i n h i b i d o r a pa rc ia l me n t e p u r i f i c a d a d e L , saL -e 1 4 8 s e som e t ie ron a u n an á li si s
e l e c t r o f o r é t i c o e n g e l e s d e S D S - p o l i a c r i l a m i d a - u r e a , se gú n l a t é c n i c a d es cr it a e n l a s e c c i ó n I I I .
2 . 9 . 6 . 1 .
No o b s t a n t e , u n a v e z t e ñ i d o s l o s g e l e s n o s e a p r e c i ó n in g u n a b a n d a p r o t e i c a e n l o s
l u ga re s e n l o s q u e s e h ab ía n d e p os it ad o l a s sol uc ion e s q u e co n t e n ía n l a sust ancr a
an t im ic ro b ia n a parc ial ment e p u r i f i c a d a . C on si d e ra n d o q u e l a f a l t a d e bandas proteicas nítidas
en los geles podía ser consecuencia de una falta de poder separador del gel con urea o de un
fallo del método de tinción de los geles, se recurrió a la electroforesis en geles de
poliacrilamida con SDS según la técnica de La e ml i ( 1 9 7 0 ) .
I V . 8 . 5 . 2 . - E l e c t r o f o r e s i s e n g el es d e 20 % p o l i a c r i l a m i d a c o n SDS ( L a e m l i . 1 9 7 0
A u n q u e e s t a t é c n i c a e l e c t r o f o r é t i c a t ie n e u n m e n o r p o d e r d e r e s o l u c i ó n e n l o s péptidos
d e b a j o p e s o m o l e c u l ar q u e l a d e S w a n k y Mu n kr e s, a l u t i l i z a r e l método de tinción de Merril y
c o l . ( 1 9 8 ] ) , a b a s e d e n i t r a t o d e p l a t a , f a c i l i t a l a t i n c i ó n d e l a s pr o t ei n as . E s t a t i n c i ó n a p l i c a d a a
l a s pro t e í n a s d e l o s g e l e s d e p o l i a c r i l a m i d a c o n S D S , e s 1 0 - 5 0 v e c e s m á s s e n s i b l e q u e l a d e l
a z u l b r i l l a n t e d e C o o í n a s s i e .
S ig u ie n d o l a t é c n i c a d e s c r i t a e n l a s e c c i ó n I I I . 2 . 9 . 6 . 2 . s e a p l i c a r o n e n e l e c t r o f o r e s i s
5 , 1 0 y 1 5 ~igd e l a s u s t a n c i a a n t i m i c r o b i a n a p ar ci al me n t e p u r i f i c a d a d e LsaL-c 1 4 8 . En l a
figura 4 . 4 0 s e i n d i c a n l o s r e s u l t a d o s o b t e n i d o s . Como p u e d e observarse, no se detectan
bandas proteicas definidas en los lugares de lo s geles en lo s que se depositó la sustancia
1 9 0

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 204/293
RESULTADOS
a n t i í n i c r o b i a n a p a r c i a l m e n t e pu r if ic ad a. S ó l o e n el frente electroforético se puede distinguir una
b a n d a d e t i n c i ó n d i f u s a . L a d e n s i d ad d e e s t a banda aumenta a medida que crece la
co n ce n t ra ci ó n d e sust anci a a n t i í n i c r o b i a n a p ar c ia l me n t e purificada depositada en el
c o r r e s p o n d i e n t e p o c i l l o .
IV. 8. 6.- D e te r m in a c i ón d e l a c o m p o s i c i ó n a m i no a c í di c a d e l a su st an ci a
antimicrobiana narcialmente nurificada
L a u t i l i z a c i ó n d e l a t é c n i c a d e d e t e c c i ó n d e am in oá ci d os p o r cro mat ograf ía líquida de
alta resolución (HPLC). descrita en la sección III. 2. 9. 7 . a s í como e l e s t u d i o c o m p a r a t i v o d e
l o s c r o m a t o g i - a m a s d e l hidrolizado ácido de la sustancia antimicrobiana parcialmente purificada
y d e lo s aminoácidos patrón, dió lugar a lo s resultados que se reflejan en la tabla 1W 10 . De
ella se deduce que:
1 . ) L a s us t a n c i a an t imic rob ian a p ose e 1 4 am in oá ci d os d i s t i n t o s , d e l o s q u e 1 3 s e
corresponden a los siguientes: ácido aspártico. ácido glutámico, glicina, histidina, treonina,
a l a n i n a , pr o lin a, valina, tirosina, metionina, leucina, fenilalanina y lisina; el decimotercero, que
elufa entre el ácido glutámico y la glutamina, no pudo identificarse al no corresponder a
n in g u n o d e l o s am in oá cid os e m p l e a d o s c o r n o p a t r ó n .
191

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 205/293
RESULTADOS
16,9 Kd—14,4 Kd—
8,1 Kd—
6,2 Kd-»
2,5
F r e n t ee l e ct ro f o r é t i c o
F igu ra 4 . 4 0 . -
-h
1 234
Electroforesis por SDS-PAGE de la actividad antimicrobiana
parcialmente purificada de L. s a L -- e 148. Lineas: 1, marcadores de
peso molecular standard (de arriba a abajo, 16.949, 14.404, 8.159,
6.214 y 2.512 daltons); 2, 3 y 4 corresponden, respectivamente a
1 5 , ¡ 0 y 5 gg de la sustancia antimicrobiana parcialmente
purificada.
192

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 206/293
RESULTADOS
T ab l a I V . 1 0 . - C o m p o s i c i ó n a m i n o a c í d i c a d e l a sustancia antimicrobiana parcialmente
p u r i f i c a d a d e L. saL -e 1 4 8
Aminoácido Contenido
(nmol)
Residuos por molécula
(probable)
Acido aspártico 7,7 4
Ac id o g l u t ¿ i m i c o 1 0 , 9 6
No Identificado’~ 7,1 3(4)
G l i c i n a 8 , 6 4 ( 5 )
Histidina 1,9 ¡
Tr e on in a 4 , 3 2
Alanina 8,4 4
P r o l i n a 1 1 , 6 6
T i r o s i n a 2 1, 9 1
V a i n a 5 , 9 3
Metionina 2,1 1
Leucina 4,5 2Fenilalanina 1,5 1
Lisina 4,8 2 (3)
Pe so mo l e c u l ar asignado igual a l a me d ia d e l d e l o s am in oá ci d os e m p l e a d o s como p a t r ó n .
‘ 93

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 207/293
RESULTADOS
2.) La sustancia antimicrobiana es muy rica en aminoácidos de carácter hidrofóbico;
(alanina, leucina, valina, fenil alanina, metionina y prolina) que constituyen el 50% de la
molécula. Los más frecuentes son prolina (6 residuos/molécuLa), ácido glutámico (6) y glicina
(4-5), y los menos histidina (1), treonina (2), metionina (1). leucina (2) y fenil-alanina (1). El
aminoácido que no pudo identificarse eluye en una posición muy próxima de la glicina, y en
cada molécula hay unos 3-4 residuos (asumiendo un peso molecular medio de 130) .
3.) La sustancia antimicrobiana de L.sake 148 posee unos 40 a 43 aminoácidos por
molécula, lo que da idea de su pequeño tamaño molecular (5047); ello se corresponde bastante
bien con el peso molecular obtenido por cromatografía de filtración en Sephadex 0-50 (4640).
pero no con su comportamiento en la electroforesis en geles de poliacrilamida con dodecil
sulfato sódico (2500).
IV. 9 .- Mecanismo de acción de la sustancia antimicrobiana exocelular de Lactobaclihís
sa/ce 148
Cuando a los medios MRS, APT y BHI incubados con unas 5 x í0~ ufc/ml de diversos
microorganismos indicadores (sección III. 2. 10) se les adicionan 0,5 ml de sobrenadantes de
L. saLe 148 libres de células, debidamente concentrados, se detiene el Crecimiento de las
bacterias sensibles (tabla IV . 11), pero no disminuye el número de ufc/ml. Sin embargo, laadición de 0,5 ml de un cultivo de L saL -e 23, carente de actividad inhibidora, no afecta al
desarrollo de estos microorganismos indicadores. Los resultados obtenidos indican que la
actividad antimicrobiana deL. saL -e 148 posee un mecanismo de acción bacteriostático.
194

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 208/293
RESULTADOS
IV. 10. - Concentración inhibidora mínima <ClM~ de la sustancia antimicrobiana de L
su/cc 148
La concentración inhibidora mínima (CIM) de la sustancia antimicrobiana parcialmente
purificada deL. saL -e 148 frente a diversos microorganismos indicadores, se determinó de la
manera descrita en la sección 1 1 1 . 2. 11. Los resultados se muestran en la tabla IV. 12. Como
se ve, la CIM varía entre 1 y 1 8 mglml, dependiendo de los microorganismos indicadores
utilizados.
195

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 209/293
RESULTADOS
Tabla IV. 11.- Efecto de lo s sobrenadantes concentrados de Lactobacdlus sake 148 y 23,antagonista (1) y no antagonista (C) respectivamente, en el desarrollo a 32 oc
de algunos microorganismos indicadores.
Microorganismoa log ufc/m]
Oh 4h 8h 16h
Loctobacillusfernzentuni
Lactobacillus cwvatus
Carnobacterium diveigens
Leuconostoc mesenteroides
Listei-ic¡ nio¡zocytogenes
Listeria rnonocytogenes
285
726
LV 1 3
395
5105
7973
TC
Tc
TC
T
C TC
TC
5,35,5
5,65.6
5,75,6
5,7
5.75,85,7
5,35,3
5,3 5,46,8 8.7
5,6 5,76,9 8,5
5,7 5,36.9 8,7
5,8 5,9
6,9 8,95,8 5,76,9 8,9
5,2 5,76,8 8,7
Listeria rnonocytogenes LIS sv 1/2 1C
5,85,8
5,7 5,96,9 8.9
Listeria n¡onocytogenes
Salinon ella tvph¡niu’v¡nn
Yersinia enterocolitica
5/cpbviococcus am-cus
Scott A
T9 1
E20
T
CTC
1
13 7
5,8
5,8
5,55,4
5,75.8
5,35.2
C
1C
5,7 5,9
6,9 8,86,9 8,86.8 8,7
6,1 6,96,3 7,1
5,1 5,66.9 8,9
5,59,8
6,19,8
5,49,5
6,4
9,55,79,3
5,79,1
6,29,5
6,1
9,5
9,59,4
8.28.9
6,39,2
196

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 210/293
RESULTADOS
Tabla IV. 11. (continuación)
Microorganismoa log ufc/ml
Oh 4h 8h 16h
Staphylococcus aureus 371 CC
5,25,3
5,36,7
5.38,7
6,59,8
Clostridiun; botulinun; 551 C
C
4,9
4,9
4.8
5,0
4.9
5,7
4,9
6.3Clostridiun¡ pe4’ringens 376 C
C5,1
5,1
5,25,3
5,37.7
6,49,3
a Para conocer la procedencia de los microorganismos ver tabla III. 1
197

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 211/293
RESULTADOS
Tabla IV. 1%- Concentración Inhibidora Mínima (CIM) de la susíancia antimicrobiana
exocelular de L.sake 148, en diversos microorganismos indicadores.
Concentración
Microorganismo Inhibidora Mínima
indicador (¡ng/mi)
Loctobacillusferinentun; CECT28S
Carnobacter¡uin dit’e.’-gens LV 1 3 2
Listeria nzonocytogenes NCTC5 105 9
Listeria nhonocytogenes LIS sv 1/2 9
Listeria rnonocytogenes Scott A 9Staphvlococcus aureus FR1137 9
Staphvlococcus am-cus FR1349 1 8
Staphylococcus aureus FR136 1 1 8
C/osuldiurn botulinu.rn 551 4
Clostridiun; peífringens 376 4
198

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 212/293
RESULTADOS
IV. 11.- Caracterización inmunológica parcial de la susrancia antimicrobiana deL, sa/ce
148
IV. 11. 1.- Análisis por inmunodifusión en Leles de aaarosa. de los inmunosueros
de conejos cuando se emolea como antízeno la sustancia antimicrobiana
exocelulardeL. sa/ce 148
La sustancia antimicrobiana exocelular parcialmente purificada de L. sa/ce 148, se
obtuvo como se describe en la sección III. 2. 1 2 de este trabajo. Antes de inocularía a los
conejos se comprobó por inmunodifusión en geles de agarosa que lo s sueros de lo s conejos no
originaban líneas de precipitación visibles al enfrentarse con la sustancia.
Durante la inmunización de lo s animales se realizaron 4 sangrías parciales para
comprobar la produción de anticuerpos frente al extracto antigénico, así como para determinar
la presencia o ausencia de reacciones cruzadas con otros componentes procedentes del medio
MM-triptosa (concentrado 20 veces por liofilización); lo mismo se comprobó con el
sobrenadante libre de células de un cultivo de L sa L -e 148 en medio MM-triptosa y con el
mismo sobrenadante concentrado 20 veces por liofilización. No obstante, en ningún caso se
detectaron 1 incas de precipitación.
199

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 213/293
RESULTADOS
IV. 11 . 2.- Detección de la sustancia antimicrobiana parcialmente purificada deL, sa/ce
148 por técnicas inmuno-enzimáticas (ELISA
IV. 11 . 2. 1.- Detección por la técnica del ELISA indirecto clásico
Antes de la inoculación de los conejos con el extracto antigénico de interés, se
comprobó que ninguno de ellos presentaba reactividad inmunológica al someter una muestrade
su suero al ELISA indirecto que se describe a continuación.
IV. 11. 2 . 1 . 1.- Evaluación de la concentración idónea de los reactivos que
intervienen en el ensayo
Para garantizar que la sustancia antimicrobiana exocelular deL. sake 148 saturaba el
fondo de los pocillos de las placas de ELISA, se diluyeron 100 ~xgdel extracto antigénicocitado en 1 ml de tampón PBS. Con idénticos fines, y como extracto antigénico control, se
empleó el medio MM-triptosa liofilizado a la concentración dc 1 mg de proteina por ml de
tampón PBS.
Como conjugado se utilizó uno comercial, anti-IgG de conejo, obtenido de cabras y
marcado con peroxidasa de rábano (Nordic). Para establecer la dilución de trabajo de esteconjugado, se siguieron las indicaciones de l fabricante y el criterio aplicado pal-a la elección del
antígeno, es decir, garantizar la presencia de conjugado en exceso. La dilución elegida fué la
1/1000.
200

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 214/293
RESULTADOS
IV. 11 . 2. 1. 2.- Determinación de la respuesta inrnunoló~ica de lo s conejos frente al
extracto antigénico inoculado
Durante la inmunización de los conejos se realizaron varias sangrías parciales para
comprobar que producían anticuerpos frente al extracto antigénico inoculado, y que no se
originaban reacciones cruzadas frente al extracto antigénico control. En la figura 4. 41 se
muestran lo s resultados procedentes de estadeterminación. Dela figura se desprende que:
1.) En las sangrías parciales efectuadas a lo s 2 1 y 42 días de la primera inoculación
antigénica no aumenta significativamente la respuesta inmunológica de lo s conejos frente a lo s
antígenos de interés.
2.) En las sangrías realizadas a lo s 63 y 84 días de la primera inoculación la respuesta
inmunológica de los conejos aumentó significativamente, haciéndolo también pero ligeramenteen la sangría final que tuvo lugar a lo s 9 1 días de la inoculación iniciaL
201

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 215/293
RESULTADOS
3,0
2,5
2,0
1,5 -
1,0
0,5 -
0,01,0 1,5 2,5 3,0 3,5 4,0
Log. dilución del anticuerpo
Figura 4. 41.- Detección inmunológica por un ELISA indirecto, de la sustancla
antirnicrobiana parcialmente purificada de L.saLe 148 a lo s O (A),
21 (A). 42 (0), 56 (•), 84 (o) y 9 1 (U) días de inoculación en
conejo Concentración del andgeno 10 < ) gtg/ml. Dilución del
conjugado 1/1000.
E
U)
o
u
1-
oTJ~
.0
1—- —1
2,0
202

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 216/293
RESULTADOS
LV . 11 . 2. 2.- Detección de la sustancia antimicrobiana deL. sa/ce 148 en el medio de
cultivo MM-triptosa
La misma técnica del ELISA indirecto empleada para determinar la respuesta
inmunológica de los conejos inmunizados con la sustancia antimicrobiana deL, sa/ce 148, se
utilizó para evaluar la presencia de dicha sustancia en el medio de cultivo MM-triptosa.
IV. 11.2. 2. 1.- Determinación de la concentración idónea de los reactivos queintervienen en el ensayo
Las indicaciones del fabricante, junto con lo s resultados de experiencias preliminares,
permitieron establecer como dilución de trabajo del conjugado la de 1/1000.
Una vez ajustada la concentración de trabajo del conjugado, se procedió a establecer lamás idónea para el inmunosuero (cuya concentración inicial de proteína era de 20 mg/mI). Para
ello, en un ensayo previo, se determinó la dilución del inmunosuero más apta para diferenciar
la sustancia antimicrobiana parcialmente purificada disuelta en tampón PBS, de la disuelta en
medio MM-triptosa (figura 4 42). De L o s resultados obtenidos se desprende que los
componentes del medio de cultivo interfieren en la detección de la sustancia antimicrobiana de
L. sa/ce 148, s i bien el inmunosuero disuelto 1 6 veces en tampón PBSTes el que ¡nejordetectadicha sustancia en el medio de cultivo de MM-triptosa.
203

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 217/293
RESULTADOS
• • 1 • 1 • ¡ • ¡
2,0 2,5 3,0 3,5 4,0
Log. dilución de l anticuerpo
Figura 4. 42 .- ELISA indirecto dc
148 en tampón P135
parámetros como en
la sustancia antimicmbiana exocelular de LsaLc
( m ~ y en medio de MM-¡rip¡osa (ny El resto de lo s
la figura 4. 41.
3,0 -
2 ,5
E
U)oxl.
u
.0 1-o
.0
2,0
1,5
1,0
0,5
0,0
1,0 1,5
204

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 218/293
RESULTADOS
Posteriormente, se realizó un ensayo para detectar la sustancia antimicrobiana
exocelular de L. sa/ce 148 cultivado en medio de MM-triptosa. Los resultados del ELISAindirecto de la sustancia antimicrobiana de L. saLe 148 en tampón PBS, en medio de
MM-triptosa y en el sobrenadante libre dc células de un cultivo de este microorganismo
realizado en medio de MM-triptosa se muestran en la figura 4.43. De lo s resultados obtenidos
se deduce que:
1.- Cuando la bacteriocina parcialmente purificada de L. saLe 148 se disuelve entampón PBS se puede detectar la presencia de hasta 6 ~tgde proteina por cada pocillo de la
placa de ELISA.
2.- Sin embargo cuando se emplea el ELISA indirecto para detectar la sustancia
antimicrobiana de L. sa/ce 14 8 crecido en medio de cultivo MM-triptosa, los inmunosueros
empleados no detectan dicha sustancia.
Dada la imposibilidad de detectar la sustancia antimicrobiana en medio de MM-tríptosa
y con la finalidad de aumentar la sensibilidad del ELISA indirecto, se optó por mejorar la
sensibilidad del ensayo mediante la utilización de anticuemos conjugados a la biotina.
205

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 219/293
RESULTADOS
¿ tu
2,5
E~ 2,0
U)
o
t 1, 5
.0 1~oCC
.0 <1,0
0,5
0,0
0,0Log. dilución del antígeno
Figura 4. 43.- ELISA indirecto de la sustancia antimicrobiana exocelular de L. saL -e
148 en tampón PES (u), en medio de MM-triptosa (c) y en el
sobrenadante de un cultivo de t s a L - e 148 en MM-triptosa (•).
Dilución del anticuerpo 1/160. Dilución del conjugado l/10(XY
0,5 1,0 1,5 2,0 2.5 3,0 3,5
206

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 220/293
RESULTADOS
IV. 1 1 . 3.- Detección de la sustancia antimicrobíana de L. sa/ce 148 mediante un
ELISA indirecto con el sistema de amDlificación biotina-avidina
El empleo de esta técnica con los mismos extractos antigénicos y en las mismas
condiciones no mejoró los resultados ya que, paralelamente a la mejora observada en la
detección de la sustancía antimicrobiana parcialmente purificada, se aprecia también una mayor
interacción de los anticuerpos biotinizados con lo s extractos antigénicos empleados como
control (figura 4. 44). Se concluyó por lo tanto, que en el medio de MM-triptosa. a pesar detener una composición muy sencilla, existen compuestos que, de una forma u otra, interfieren
con la detección inmunológica de la actividad antimicrobiana.
207

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 221/293
RESULTADOS
3,0
2,5
2,0E
U)
o
c ~ 1, 5
u
.0 ~ 1, 0C C
.0
0,5
0,00,0
Figura 4. 44.- ELISA indirecto de la sustancia antimicrobiana exocelular de L. soke
148 en tampón PBS (u), en medio de MM-triptosa (o) y en el
sobrenadante libre de células de un cultivo deL sa L -e 148 desarrollado
en el medio de MM-triptosa (4). Dilución del anticuerpo biotinizado1/160. DiLución del conjugado 1/3000.
0,5 1,0 1,5 2.0 2,5 3.0 3,5
Log. dilución de l antígeno
208

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 222/293
DISCUSION
CAPITULO V
DISCUSION
209

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 223/293
DISCUSION
y. i. - Aislamiento de bacterias lácticas de embutidos crudos curados
Para la realización de este trabajo, que pretende el aislamiento y caracterización parcial
de bacterias lácticas utilizables para mejorar la calidad higiénica de la carne y sus productos,
primerose procedió al aislamiento de estas bacterias de embutidos cnidos curados. La elección
de estos productos como fuente de bacterias lácticas se debe a que las que contienen están
mejor adaptadas a la ecología bacteriana de la carne y lo s productos cárnicos que las aisladas de
otras fuentes, como leche, productos lácteos y hortalizas (Schillinger y LOcke, 1989).
Además las bacterias lácticas son, invariablemente, el grupo microbiano predominante
en la mayoría de los productos cárnicos yen las carnes envasados a vacio, constituyendo entre
el 75 y el 95 % de su población bacteriana total (Mol y col., 1971: Kitchell y Shaw, 1975;
Reuter, 1975; l-litchener y col., 1982; Shaw y Harding, 1984; Korkeala y Mátehl, 1989).
De otra parte, los recuentos de bacterias lácticas respecto del microbiano total
obtenidos por otros investigadores (LOcke, 1986; Holley y col., 1988> durante la maduración
de los embutidos crudos madurados, coinciden con lo s nuestros (Tabla 1W 1 ); en ambos casos
se ha visto que la mayor parte de la microflora cárnica está constituida por bacterias lácticas.
El número de bacterias lácticas al final de la maduración de las muestras estudiadas
(aproximadamente 8-9 x 108 ufc/g de embutido) concuerda con lo s resultados señalados por
otros investigadores para el bacon (Cavett, 1963), los embutidos crudos madurados (Egan,
1983) y las carnes envasadas al vacio (Hitchener y col.. 1982; Egan. 1983). En muchos
productos cárnicos, estas bacterias constituyen el principal componente de su microflora
debido a la supresión del desarrollo de otros grupos microbianos por mecanismos de
antagonismo microbiano no bien evaluados (Reuter, 1981).
210

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 224/293
EJISCUSION
V. 2. - Actividad antimicrobiana de las bacterias lácticas seleccionadas
y. 2. 1 . - Elección de las pruebas de anta2onismo microbiano
V. 2. 1 . 1 . - Pruebas directas de antaLonismo microbiano
El antagonismo microbiano puede evidenciarse de diversas maneras. En los estudios
preliminares, las pruebas de antagonismo se realizaron utilizando generalmente medios decultivo sólidos e implicaron la detección de la inhibición del crecimiento que ejercía el
microorganismo analizado (activo) en el microorganismo indicador (pasivo) (Tagg y col.,
1976).
Las dos pruebas enpleadas con este propósito son la directa o simultánea y la indirecta
o diferida. La concentración del microorganismo indicador es importante en la sensibilidad del
método (Kuttner, 1966).
En la prueba directa, el microorganismo a evaluar y la cepa indicadora se desarrollan al
mismo tiempo. Una de las pruebas directas más sencillas es la “prueba de la gota”, utilizada
originalmente por Gratia (1946), y cuyo empleo está ampliamente difundido (Geis y col.,
1983; Daeschel y Klaenhammer, 1985; Gonzalez y Kunka, 1987; Raccach y col.. 1989;
Spelhaug y Harlander, 1989; Daeschel y col., 1990; Ahn y Stiles, 1990a). En ella, una gota
del cutivo del microorganismo a evaluar se deposita en un medio de cultivo sólido,
previamente inoculado con el microorganismo indicador. Entre las variantes de este
procedimiento se incluye el empleo de pocillos excavados en el medio sólido que se rellenan
con una cantidad conocida del cultivo del microorganismo a evaluar (Sabine. 1963; Tagg y
McGiven. 1971; Tagg y col., 1976; Silva y col.. 1987; Harris y col.. 1989; Rodriguez y col..
1989).
211

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 225/293
DISCUSION
Otra modificación de gran utilidad es la siembra por picadura de los microorganismos a
estudiar en placas cuya superficie se ha sembrado con el microorganismo indicador (DeKlerk,1967; Gagliano y Hinsdill, 1970; Davey y Richardson, 1981; Barefoot y Klaenhammer,
1983).
Fredericq (1948) fué el primero en utilizar las pruebas diferidas de antagonismo. En
este caso, el microorganismo a evaluar se siembra e incuba en un medio sólido durante un
cierto tiempo, transcurrido el cual se cubre con otra capa de medio que contiene elmicroorganismo indicador. Las pruebas diferidas son a menudo más sensibles que las directas
(Taggycol., 1976).
Como en la prueba directa, una de las variantes de las pruebas diferidas más empleada
es la de la “gota (Andersson y col, 1988; Harris y col., 1989; Schillinger y LOcke, 1989;
Spelhaug y Harlander, 1989; Ahn y Stiles, 1990a). Con esta prueba y para resaltar más laacción .d e las sustancias inhibidoras difusibles de los cutivos, se inactivan las células del
microorganismo productor con calor (Geis y col, 1983; Ahn y Stiles, 1990a) o, mejor aún,
con vapores de cloroformo (Kekéssy y Piguet. 1970; Davey y Richardson, 1981; Geis y col.,
1983; Ahn y Stiles, 1990a). No obstante, esto conlíeva algunos inconvenientes como el que
algunos agentes inhibidores se inactivan al exponerse a los vapores de cloroformo. Brock y
Davie (1963) demostraron la sensibilidad al cloroformo de una sustancia inhibidora producidapor Enrerococcusfaecalis subsp. zvrnogenes. Los vapores de cloroformo también impiden el
empleo de placas de Petri de plástico; además el cloroformo residual de los medios de cultivo
puede originar resultados erróneos (Kekéssy y Piguet, 1970).
Otra alternativa, cuando se sospecha la existencia de sustancias antagonistas difusibles,
consiste en sembrar el cultivo indicador en una cara del medio de cultivo sólido y el cultivo a
evaluar en la opuesta, ya sea empleando el sistema de gotas (Kekéssy y Piguet. 1970) o el de
212

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 226/293
DISCUSION
pocillos (Tagg y McGiven, 1971; Barefoot y Klaenhammer, 1983; Spelhaug y Harlander,
1983). Una ventaja de este método es que así se excluye la actividad inhibidora debida abacteriófagos (Tagg y col., 1976).
Tanto en las pruebas de antagonismo directo, como diferido, es recomendable colocar
lo s cultivos de los microorganismos a analizar en pocillos abiertos en los medios sólidos, lo
que permite que si se deposita siempre el mismo número de microorganismos, pueden
establecerse comparaciones entre los diámetros de las zonas de inhibición obtenidas. Otraventaja de este método esel aumento de la sensibilidad, puesto que entre el microorganismo
estudiado y las cepas indicadoras únicamente se interpone una capa fina de agar; esto, además,
excluye la inhibición debida a bacteriófagos, que son microorganismos no difusibles (Tagg y
McGiven, 1971). Por todo ello, se eligió este método para la realización de las pruebas de
antagonismo, tanto directo como diferido, de las 180 bacterias lácticas aisladas y seleccionadas
de los embutidos crudos curados (sección IV. 1 ) .
V. 2. 1 . 2. - Antagonismo de los sobrenadantes libres de células
Cuando en las pruebas de antagonismo microbiano se emplean cultivos no es fácil
identificar la causa del mismo (Tagg y col., 1976). En la identificación de bacterias lácticas
productoras de sustancias antagonistas difusibles es importante excluir de los ensayos no sóloa los microorganismos productores de tales sustancias, sino también a otros metabolitos con
actividad inhibidora.
Las sustancias que en las pruebas de antagonismo bacteriano poseen una actividad
inhibidora similar a las bacteriocinas. pueden neutralizarse o eliminarse utilizando
sobrenadantes libres de células neutralizados o dializados. Los sobrenadantes se obtienen por
centrifugación de los cultivos y por filtración por filtros de 0.22 o 0.45 um de diámetro de
213

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 227/293
DISCUSlON
poro.
La neutralización de la actividad antagonista de los ácidos orgánicos puede realizarse
ajustando el pH de lo s sobrenadantes a un valor próximo a la neutralidad (pH 6,9-7,0) (Geis y
col., 1983: Barefoot y Klaenhammer, 1984; Rodriguez y col., 1989; Nielsen y col., 1990;
M0rtvedt y Nes, 1990), o dializando los sobrenadántes por membranas que permiten la
difusión de sustancias de un tamaño molecular menor de 6000 daltons. La diálisis puede ir
precedida de una precipitación de lo s sobrenadantes con sulfato amónico (Bhunia y col, 1987;González y Kunka. 1987; Ray y col., 1989). El precipitado, constituido fundamentalmente por
proteínas, se reconstituye y dializa. N o obstante, la mayoría de lo s autores prescinden de esta
precipitación (Andersson, 1986; Andersson y col., 1988; Harris y col., 1989; Schillinger y
LOcke, 1989; Daeschel y col., 1990; Ahn y Stiles, 1990b).
El efecto del peróxido de hidrógeno de lo s sobrenadantes puede evitarse incubando lo scutivos en anaerobiosis (Schillinger y LOcke, 1989), o tratando con catalasa los sobrenadantes
libres de células (Barefoot y Klaenhammer, 1983; Muriana y Klaenbammer, 1987; Ferreira y
Gilliland, 1988; Schillinger y LOcke, 1989; Rodriguez y col., 1989; ~arminati y col., 1989;
Ahn y Stiles, 1990a). Geis y col. (1983), añaden la catalasa directamente al medio de cultivo
del microorganismo indicador.
Los sobrenadantes libres de células también pueden concentrarse con el fin de
aumentar su actividad inhibidora. Entre lo s sistemas de concentracién más empleados destacan
la evaporación a vacío en rotavapores (Geis y col., 1983; Silva y col., 1987) y la liofilización
(Rodriguez y col., 1989).
La actividad inhibidora de lo s sobrenadantes puede detectarse con cualquiera de lasvariantes de antagonismo directo e indirecto citadas, estando muy difundidos lo s métodos “de
214

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 228/293
DISCUSION
la gota” (Geis y col., 1983; Ray y col.. 1989; Carminati y col., 1989: Ahn y Stiles, 1990a) y
la disposición de lo s sobrenadantes en pocillos abiertos en las placas (Joerger y Kiaenhammer,
1986; Ahn y Stiles, 1990a). Otro método muy empleado cuando se investigan bacteijas lácticas
bacteriocinogénicas, consiste en colocar lo s sobrenadantes en papeles de filtro estériles. Los
discos de papel de filtro, impregnados con el sobrenadante, se depositan en placas de agar
sembradas con el microorganismo indicador (Shahani y col, 1976; Ferreira y Gilliland. 1988)
o bien se deposita en lo s filtros una cantidad determinada de sobrenadante (Abdel-Bar y col.,
1987;Bhunia
ycol.,
1987;Bhunia
ycol.,
1988;Rodriguez y col., 1989). La segunda
opción
ha sido la utilizada en este trabajo, lo que permite comparar la actividad inhibidora de los
diversos sobrenadantes analizados.
V. 2. 2 . - Actividad inhibidora de lo s cultivos de las bacterias lácticas seleccionadas
Tanto las 180 bacterias lácticas examinadas en primera instancia, como las 24seleccionadas con posterioridad de entre las primeras, manifestaron actividad inhibidora frente
a alguno de los microorganismos indicadores empleados (figuras 4 6 a 4. 29). La actividad
inhibidora generalmente es mayorcuando lo s microorganismos a evaluar se siembran 24 horas
antes que lo s microorganismos indicadores, algo en lo que nuestros resultados coinciden con
los de Ahn y Stiles (1990a).
La sensibilidad de lo s microorganismos indicadores a la actividad inhibidora de las
bacterias lácticas analizadas es variable, lo que demuestra que dicha actividad es característica
de cadacepa y no de la especie bacteriana (Schillinger y LOcke, 1989: Daeschely col., 1990).
Los resultados obtenidos por otros investigadores que han evaluado la actividad
antimicrobiana de las bacterias lácticas varían bastante. Al igual que nosotros, Kozak y col.(1978) observaron que las 47 cepas deL. lacis subsp. lacús que estudiaron, eran activas
215

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 229/293
DISCUSION
frente a otras dos cepas de L. lacás subsp. ladis y frente a una de L. lacris subsp. Cremoris.
El mismo resultado obtuvieron Ahn y Stiles (1990a) al examinar la actividad inhibidora de 1 0
cepas de diversos géneros de bacterias lácticas frente a 1 0 indicadores que también eran
bacterias lácticas.
El 63 % de las 52 cepas de L acidophilus analizadas por Barefoot y Klaenhammer
(1983), inhibieron el desarrollo de diversas cepas de L. ¡eichn,anni, L. bulgaricus, L.
heIvetíCLis y L. lacds, porcentaje idéntico al obtenido por Raccach y coL (1989) con 1 1 cepas
de Lactobacillus y de Sireptococcus sp., empleando como indicadores diversas cepas de 6
especies distintas de bacterias lácticas y 4 cepas de L. monocytogenes. De otra parte,
únicamente el 15 % de las 79 cepas de Lacwbacillus sp. examinadas por Rammelsberg y
Radíer (1990) inhibieron una, al menos, de las 9 cepas indicadoras deL. brevis, P. daninosus
y Leuc. oenus ensayadas. El porcentaje de cepas inhibidoras registrado por Davey y
Richardson (1981) fue aún menor, ya que sólo el 7 % de las 150 cepas deL. ladis susbsp.
crenzork analizadas, inhibieron el desarrollo de variascepas de L. lactis subsp. crernoris y L.
lactis subsp. lactis.
Schillinger y LOcke (1989), al evaluar 142 cepas deL, saL-e, 4 deL. plantarwn y 75
de L. curvatus, observaron que el 13, el 75 y el 1 %, respectivamente, eran activas frente, al
menos, uno de los 31 microorganismos indicadores empleados; estos resultados contrastancon los de Ahn y Stiles (1990a) y con lo s nuestros, ya que en nuestro caso prácticamente todas
las cepas aisladas manifiestan actividad inhibidora sobre varios indicadores, contraste que
llama poderosamente la atención si se tiene en cuenta que en los tres trabajos las bacterias
lácticas proceden de una fuente común, lo s embutidos crudos madurados.
Lo expuesto anteriormente ofrece una idea de la dificultad que presenta el establecer laactividad inhibidora de las bacterias lácticas, lo que se agrava por la escasa uniformidad en las
216

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 230/293
DISCUSION
condiciones de trabajo y por la diversidad de microorganismos indicadores empleados. No
obstante, las bacterias lácticas analizadas en los trabajos citados son especialmente activas
frente a otras bacterias lácticas y nunca, o de forma excepcional. frente a las bacterias
Gram-negativas.
V. 2. 3. -Actividad inhibidora de los sobrenadantes libres de células
La actividad inhibidora de los sobrenadantes libres de células se estableció sólamente
en los aislados más activos. Los sobrenadantes libres de células de lo s cultivos de nuestras 24
bacterias lácticas seleccionadas, una vez concentrados, mostraron una actividad antimicrobiana
variable dependiendo de la cepay del indicador empleado (tabla IV. 1 ) ; fueron especialmente
sensibles las bacterias lácticas empleadas como indicadores.
Quizá sea conveniente recordar que las bacterias lácticas activas seleccionadas, se
aislaron durante la maduración de un lote de embutidos; en cada muestreo se seleccionaron 30
colonias. Esto significa, por ejemplo, que las cepas 24 y 29 procedían de la masa cárnica
original, la 31 y 46 del embutido en segundo día de maduración y, así sucesivamente, hasta la
cepa 180 que se alsió el último día (35) de maduración. De la observación de la tabla IV. 1, se
concluye que es posible aislar bacterias lácticas productoras de sustancias antimicrobianas,
distintas de los ácidos orgánicos o del agua oxigenada, en cualquier momento de la maduración
e, incluso, a partir de la masa cárnica inicial. Algunas de las cepas evaluadas mostraron
actividad frente a varias cepas de L. nionocytogenes y S. aureus, microorganismos
productores de toxiinfecciones alimentadas.
La actividad inhibidora de las bacterias lácticas frente a otras de su mismo grupo y a
menudo estrechamente relacionadas entre si, es un hecho ampliamente documentado (Geis y
col., 1983; Barefoot y Klaenhammer, 1984; Joerger y Klaenhaínmer, 1986; Andersson, 1986;
217

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 231/293
DISCUS ION
Gonzalez y Kunka, 1987; Bhunia y col., 1987; Schillinger y LOcke, 1989; Harris y col.,
1989; M0rtvedt y Nes, 1990; Ahn y Stiles, 1990a; Ahn y Stiles, 1990b). Sin embargo, son
mucho más escasos lo s estudios que demuestran que las bacterias lácticas inhiben el desarrollo
de cienos microorganismos patógenos Gram-positivos, como 5. aureus (Andersson, 1986;
Andersson y col., 1988; Bhunia y col., 1988) y L. inonocytogenes (Ehunia y col., 1988;
Carminati y col., 1989; Schillinger y LOcke, 1989; Harris y col., 1989; Spelhaug y Harlander,
1989; Ahn y Stiles, 1990b). La mayor parte de lo s autores citados coinciden en señalar que las
bacterias Gram-negativas son insensibles a las sustancias antibacterianas producidas por las
bacterias lácticas estudiadas. Quizá la excepción mejor conocida a este respecto la constituyan
las cepas de L. laCté productoras de nisina (Hurst, 1981). En general, la actividad
antimicrobiana es muy heterogénea y depende de cada cepa en particular (Harris y col., 1989).
V. 2. 4. - Efecto de la catalasa en la actividad inhibidora de las bacterias lácticas
seleccionadas
Los resultados obtenidos indican que la actividad antimicrobiana de los sobrenadantes
concentrados no puede ser, en ningún caso, atribuible al peróxido de hidrógeno. La capacidad
de algunas bacterias lácticas de producir peróxido de hidrógeno es una propiedad indeseable,
especialmente si en las emulsiones cárnicas existe un alto contenido de oxígeno, si se emplea
mucha grasa en la formulación del embutido o si la actividad de los enzimas que degradan el
peróxido de hidrógeno es pequeña (Schillinger y LOcke, 1987a). En estos casos, la presencia
en la carne y productos cárnicos de microorganismos productores de peróxido de hidrógeno
produce alteraciones del color o un enranciamiento precoz.
Las bacterias lácticas difieren en su capacidad de producir y degradar el peróxido de
hidrógeno. L-lastings y Hólzaptel (1987) observaron que el 50 % de las cepas de L. sa L -e de
origen cárnico producían peróxido de hidrógeno y, de ellas, la mayoría lo hacía muy
218

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 232/293
DISCUSION
débilmente. La falta de una buena selección de las bacterias lácticas autóctonas explica, en
parte, el que t. plantaruni, P. acidilacticí y P. pentosaceus sean lo s microorganismos más
empleados como cultivos iniciadores en la elaboración de productos cárnicos y no L. sa/ce o
L. curvatus, que son especies mucho más competitivas (LOcke y Hechelmann, 1987).
y. 3. - Identificación y caracterización bioquímica parcial de las bacterias lácticas con
actividad antimicrobiana
Cuando se aislan bacterias lácticas y en panicular lactobacilos, de la carne y productos
cárnicos, no es posible clasificarlos adecuadamente siguiendo lo s sistemas de identificación
empleados clásicamente, ya que éstos se basan en las propiedades de microorganismos
procedentes de otras fuentes (OrIa Jensen, 1919; Rogosa, 1970; Sharpe, 1979). Hasta muy
recientemente, las cepas difíciles de identificar se englobaron bajo la denominación de
estreptobacterias o lactobacilos atípicos” (Thornley y Sharpe, 1959; Reuter, 1975; Sharpe,
1979). No obstante, como sugirió Ingram (1975), el término de “estreptobacterias atípicas” es
poco afortunado ya que las bacterias lácticas de origen cárnico no son menos típicas que las
demás, en todo caso lo serán los criterios de identificación y clasificación, debido a la
ignorancia de las propiedades de las especies de origen cárnico.
Con el transcurso del tiempo se han realizado diversos intentos de identificar y
clasificar las estreptobacterias de origen cárnico (Reuter. 1975; Kitchell y Shaw, 1975;
Hitchener y col., 1982). Todos ellos se basan en reacciones bioquímicas variables que, al no
emplear métodos numéricos en el establecimiento de lo s grupos, han llevado a la creación de
grupos innecesariamente complicados y arbitrarios (Shaw y Harding. 1984). No obstante, en
1987, Schillinger y LOcke elaboraron un esquema de identificación rápidade lactobacillos de
origen cárnico, basado en sus características fisiológicas, la mayor parte de las cuales sonfácilmente determinables. Este esquema, que se muestra en la figura 5. 1 , incluye especies
219

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 233/293
DISCUSJOJkI
recientemente caracterizadas, como L. sa/ce, L. cun’atus, C. piscicola y C. divergens, por le ’
que la hemos utilizado en la identificación y clasificación de nuestros aislados.
En las tablas IV . 2 y IV. 3 se muestran las características morfológicas y bioquímicas:
más significativas de nuestros aislados. De ellas se deduce que se trata de bacilos
homofermentativos, que crecen a 1 5 oc, no fennentan el manitol, pero s i la ribosa, sacarosa y
trealosa por lo que, siguiendo el esquema de la figura 5. 1 , los clasificamos tentativamente
como L. sa/ce; las características descritas coinciden con las señaladas por Kandler y Weiss
(1986) y por Champomier y col., (1987) para esta misma especie.
En nuestro caso, ninguna de las cepas aisladas crece en medio MRS de pH 3,9.
Schillinger y LOcke (1987b) han señalado que aproximadamente el 11% de las cepas de E.
sa/ce no crecen a dicho pR. A este respecto conviene destacar que lo s lactobacilos incluidos en
las “estreptobacterias atípicas” son menos acidificantes y acidotolerantes que los demás
(Reuter, 1975; Reuter, 1981).
Todas las cepas analizadas crecieron a una concentración de cloruro sódico del 7%, a
excepción de la número 77. lo que coincide con lo s resultados de Champomier y col. (1987),
Schillinger y LOcke (1987b), M0rtvedt y Nes (1990) y Rodríguez (1991). Sin embargo,
ninguna de las cepas estudiadas se desarrolló cuando la concentración de cloruro sódico fué del10%. Existen discrepancias en cuanto a la capacidad de E. sa/ce de desarrollarse a esta
concentración de cloruro sódico, ya que mientras Schillinger y LOcke (1987b) indican que el
93 % de 131 cepas de E. sa/ce crecen a esta concentración, Champomier y col. (1987)
mantienen que se trata de una característica excepcional.
220

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 234/293
DISCUSION
heremfermentativos
casei pseudo- ptontoruni
Figura 5. 1 . - Esquema de identificación rápida de bacterias de origen cárnico.
Fuente: Schillinger y LÍicke (1987b).
Lactococcus homofermentativos
LeuconosrocLacrobacillí
L.
subsp. subsp.
221

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 235/293
DISCUSION
Las cepas de L. sa/ce analizadas muestran una capacidad variable de hidrolizar la
arginina, pues mientras las cepas 90 y 148 producen amoniaco a partir de dicho aminoácido,
las cepas 31, 77, 1 7 7 y 1 8 0 n o lo producen (tabla IV. 2). Schillinger y LOcke (1987b),
observaron que la mayoría de las cepas de L. sa/ce poseían esta capacidad, lo que concuerda
con los resultados de Champomier y col. (1987). Sin embargo, resulta curioso que Kandler y
Weiss (1986) describan aL. sa/ce como incapaz de hidrolizar la arginina, ya que la cepa que
consideraron tipo, la L. sa/ce ATCC15521. carece de esta capacidad. Reuter (1981) y
Rodríguez(1991), concuerdan con nuestros resultados
enque la capacidad de hidrolizar la
arginina varía de unas cepas de L. sa/ce y de L. curvatus a otras.
La capacidad de algunos lactobacilos de producir ácido sulfhídrico es una característica
tecnológica perjudicial, ya que su proliferación en lo s alimentos acarrea su deterioro (Egan.
1983). Ninguna de las cepas que hemos ensayado producía ácido sulfhídrico en ninguno de
los medios estudiados (tabla IV. 2). No obstante, Schillinger y Locke (1987b), observaron enel 50 % de las cepas de L. sa/ce analizadas la producción de ácido sulfhídrico.
Las cepas de L. sa/ce comparten características básicas que las diferencian de otras
especies de lactobacilos. Sin embargo, poseen otras propiedades en las que la variabilidad
entre cepas es la nota predominanante. Las discrepancias entre la cepa tipo y las propiedades de
las de origen cárnico explican las dificultades de muchos investigadores para identificar susaislamientos (Reuter, 1981). En este sentido, la fermentación de la arabinosa, amigdalina.
esculina, arbutina, salicina, celobiosa, maltosa y lactosa, que en la descripción deL. sa/ce en
el Berizev’s Manual of Svstematic Bacterioloav (Kandler y Weiss, 1986) se admiten como
pruebas de identificación, variaron en las 13cepas de esta especie que analizaron Champomier
y col. (1987>. Quizás esto justifique la variabilidad de nuestras cepas en lo que concierne a
fermentación de azúcares.
222

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 236/293
DISCUSlON
Que todos nuestros aislados se hayan identificado como L. sa/ce no es sorprendente.
ya que algunos investigadores han observado que esta especie puede convertirse en la
predominante a las temperaturas de maduración normalmente empleadas en Europa en la
elaboración de los embutidos crudos (LOcke. 1986; Schillinger y LOcke, 1987a). LOcke y
Hechelmann (1987) apoyan esta observación al confirmar el predominio natural deL. sa/ce y
L. curvatus en embutidos europeos madurados tradicionalmente.
Holy y Holzapfel (1988) también han observado que entre las bacterias lácticasaisladas de carne de vacuno, envasada a vacío y almacenada a temperaturas de refrigeración,
hay un fuerte predominio de L. sa/ce, constituyendo L. sa/ce, L. curvatus y E. bavaricus casi
el 80 % del total de bacterias lácticas identificadas. Conviene recordar que L. bavaricus se
diferencia deL. sa/ce en que carece de racemasa (Kandler y Weiss, 1986a). Asimismo, Monte!
y col. (1989) consideran a L saL- -e como la bacteria láctica más abundante en la carne de
vacuno. Korkeala y Mákelá (1989), utilizando el esquema de clasificación de Sehillinger yLOcke (1987b), han establecido que L sa/ce es la bacteria láctica mayoritaria de lo s productos
cárnicos envasados a vacío. Hastings y col. (1986) también han observado que E. sa/ce es la
especie predominante en la carne picada de vacuno irradiada (5 KGy), adjudicando a esta
especie 100 de las 113 cepas aisladas.
Globalmente consideradas, y de acuerdo con numerosos estudios anteriores, las“estreptobacterias atípicas” también han constituido el grupo bacteriano más numeroso del
bacon (Cavett, 1963), de una gran variedad de productos cárnicos (Reuter, 1975) y de las
carnes envasadas a vacío (Mol y col., 1971; Reuter. 1975; Kitchell y Shaw, 1975; Hitchener y
col., 1982; Holzapfel y Gerber, 1986). Su presencia se ha considerado deseable, ya que
contribuyen al aumento de la vida útil de lo s productos citados (Egan. 1983). Recientemente, y
como contribución a una mejor comprensión de la clasificación de las bacterias lácticas deorigen cárnico, Champomier y col. (1987) han comprobado que lo s 6 grupos de lactobacilos
223

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 237/293
LIVSCUSION
aislados de productos cárnicos por Laban y col. (1978) y la mayoría de los integrantes del
grupo 1 1 de Shaw y Harding (1984). encajan perfectamente en la descripción tipo de U saL-e.
Actualmente L. saL-e y L. curvanis se consideran microorganismos psicrotrofos,
siendo de entre las bacterias lácticas lo s que crecen a las temperaturas más bajas (Reuter, 1981;
Kandler y Weiss, 1986 9 . Esta característica se refleja en todas las cepas analizadas, ya que
todas ellas se desarrollan a 4 oc (tabla IV. 2). Estos resultados coinciden con los obtenidos por
Schillinger y LOcke (1987b) y Champomier y col. (1987), dado que el 99 % de las cepas deL.
sa/ce estudiadas por los primeros, y todas las ensayadas por los segundos, crecieron a 4 0C.
Por el contrario, sólo el 50 % de las cepas de L. planrarwn se desarrollan, y débilmente, a esta
temperatura (Reuter, 1981; Schillinger y LOcke, 1987b). Hasting y Holzapfel (1987) han
observado que ninguna de la 100 cepas de L. sa/ce aisladas de carne radurizada se
desarrollaban ni a 4 oc ni a 8 0C . Quizás esta anomalía se deba a alteraciones de la fisiología
celularde lo s lactobacilos provocadas por la irradación.
De nuestras cepas de U sa/ce únicamente las 148 y 177 se desarrollan a 45 0C. La
mayoría de los estudios realizados hasta la fecha, coinciden en señalar que la mayor parte de
las cepas de L. sa/ce no se desarrollan a 45 oc (Reuter. 1975; Champomier y col., 1987;
Hasting y Holzapfel, 1987; Korkeala y Mákelá, 1989) o lo hacen muy débilmente (Reuter,
1981). Sin embargo, Schillinger y LOcke (1987b) han observado que el 87 % de las cepas de
L. sa/ce y el 77 % de las de L. cur”atus, crecen a dicha temperatura. La mayoría de las cepas
deL. plantarun¡ sise desarrollan a 45 0C (Reuter, 1981: Schillinger y LOcke, 1 987b).
La temperatura óptima de crecimiento de las bacterias lácticas constituye un parámetro
de gran importancia tecnológica en la elección de cultivos iniciadores cárnicos. Así, mientras
1 > . ac¡d¡lactic¡ se desarrolla bien a temperaturas superiores a 40 0C, las temperaturas óptimas de
crecimiento de It pentosaceus y L. planta¡-un¿ se sitúan entre 30 y 35 0C. siendo las deL.
224

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 238/293
DISCUSION
sa/ce y L. Curvatus algo menores (Locke y Hechelmann, 1987). En un estudio comparativo de
las bacterias lácticas normalmente incluidas en lo s cultivos iniciadores cárnicos (L. sa/ce, L
curvatus, L. planrarurn, P. acidflactici y 1 ’ . penzosaceus ), Schillinger y Locke (1989)
comprobaron que L. sa L -e era la especie que se desarrolla más rápidamente y con mayores
recuentos en una mezcla cárnica mantenida a 20 0C; su mayor rapidez de desarrollo fué
especialmente significativa respecto de lo s lactobacilos “nativos” de un lote control a] que no se
le había adicionado ningún cultivo iniciador.
V. 4.- Esoectro antimicrobiano de lo s sobrenadantes libres de células de las cepas de
L. sa/ce seleccionadas, una vez neutralizados y concentrados
El conocimiento exacto del espectro antimicrobiano de las bacteriocinas producidas por
las bacterias lácticas es muy importante, ya que su empleo podría inhibir el desarrollo de las
bacterias lácticas que se utilizan como cultivos iniciadores en la elaboración de productos
cárnicos. En este sentido. Hammes y col. (1990) observaron que cepas de E. curvatus y E.
sa/ce, productoras de sustancias inhibidoras no inhibian únicamente a las bacterias
contaminantes, como S. aureus, sino que también afectaban a microorganismos que se
emplean corrientemente como cultivos iniciadores como E. plantarunz o Micrococcus varians.
Por tanto, el conocimiento de las propiedades antimicrobianas de las bacterias lácticas,
facilitará la selección de las cepas que formen parte de un cultivo iniciador, de forma que se
minimicen lo s riesgos sanitarios provenientes lo s productos cárnicos.
Los sobrenadantes libres de células, neutralizados y concentrados, de 4 cepas de E.
saL-e (77, 90, 1 4 8 y 180), ejercieron una actividad antagonista diversa en distintos
microorganismos indicadores (tabla IV . 1). El espectro antimicrobiano de cada una de las
cepas es significativamente distinto, lo que induce a suponer que las sustancias antimicrobianas
son distintas entre sí . Este aspecto ya había sido observado por Bhunia y col. (1988), quienes
225

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 239/293
DISCUSION
comprobaron que algunas cepas de pediococos eran más activas que otras frente a diversos
microorganismos patógenos y alterantes de lo s alimentos.
La actividad antimicrobiana de las cepas de L. sa/ce estudiadas no se limita a especies
bacterianas filogenéticamente próximas, como parece ser la situación predominantante en las
sustancias inhibidoras producidas por el género Lactobacillus (Andersson y col., 1988;
Schillinger y Holzapfel, 1990; Toba y col., 1991), sino que también afecta a microorganismos
Gram-positivos, patógenosy
alterantesde
lo s alimentosde
graninterés en
la industriacárnica
como los pertenecientes a lo s géneros Lisreria, Swphylococcus y Closrridiunz. También es
importante destacar que las cepas ensayadas no inhiben el desarrollo de 5. carnosus ni de M.
varians, microorganismos cuya presencia en lo s alimentos no sólo es beneficiosa sino
deseable, ya que son lo s responsables de reacciones químicas que influyen beneficiosamente
en la calidad higiénica y en las características organolépticas de lo s alimentos.
Algunos investigadores han observado la reducción de los nitritos y el enrojecimiento
de los embutidos se suprimen cuando se utiliza como iniciador un cultivo compuesto de cepas
de L. sa/ce antagonistas de M. varians (Hammes y col., 1990). La resistencia de 5. carnosus y
M. varians, microorganismos utilizados como cultivos iniciadores comerciales, frente a la
acción antimicrobiana de las cepas de L. sa/ce que hemos aislado en este trabajo es muy
importante, ya que permite su empleo en los cultivos iniciadores al garantizar la calidadhigiénica de lo s alimentos sin inhibir reacciones bacterianas deseables.
Por otro lado, la posibilidad de inhibir el desarrollo de L. monocvtogenes es muy
interesante, sobre todo s i se tiene en cuenta que este microorganismo crece en un intervalo de
temperaturas muy amplio (1-45 0C) y en presencia de un lO % de NaCí (Seelinger y iones.
1986); además permanece viable durante el procesado y almnacenamniento de algunos alimentos
fermentados (Juntilla y col., 1989), proliferando en sustratos cuyo pH es ligéramente menor
226

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 240/293
DISCUSION
de 5,6 (Conner y col., 1986); no obstante, es relativamente sensible a los medios más ácidos
(Rodríguez, 1991).
Los alimentos pueden contaminarse durante su elaboración y manipulación con 5.
aureus que, posteriormente, puede desarrollarse y, si son cepas enterotoxigénicas, producir
toxinas (Smith y Palumbo, 1981), que pueden originar brotes de intoxicaciones alimentarias.
La actividad antimicrobiana de las cepas de L. sa/ce analizadas frente a 5. aureus
enterotoxigénico permite suponer que su empleo, junto a otros factores de seguridad (Hammes
y.col.. 1990), podría aumentar mucho la calidad higiénica de los alimentos y disminuir la
incidencia de estas toxiinfecciones alimentarias.
El crecimiento y producción de toxinas por C. boíulinurn se previene mediante la
adición a los alimentos de nitratos y nitritos; sin embargo, su incorporación a lo s alimentos
está limitado por su posible conversión en nitrosaminas, compuestos que son cancerígenos.Smith y Palumnbo (1981) comprobaron que las bacterias lácticas previenen el crecimiento y
producción de toxinas de C. borulinun;, únicamente cuando el sustrato contiene glucosa y
nitritos. Recientemente (CFR, 1988), la “Food and Drues Administration” ha permitido la
utilización de la nisina como antagonista de C. botulinum en algunos alimentos. Por ello, la
capacidad de nuestras cepas de L. sa/ce de inhibir el desarrollo de C. botulinunz, resulta
esperanzadora en cuanto a su posible utilización como inhibidor del desarrollo de este patógenoen los alimentos. Asimismo, la utilización de las cepas aisladas en este trabajo o las sustancias
inhibidoras elaboradas por ellas, permitirían disminuir los niveles de nitritos habitualmente
empleados en algunos derivados carnmcos.
Se sabe poco sobre el efecto de las bacterias lácticas en C. pe,fringens. Baran y
Stevenson (1975) demnostraron que este microorganismno supervivía en lo s embutidos crudosmadurados de pavo, elaborados con P. cerevisiae comno cultivo iniciador. Aunque no se
227

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 241/293
DISCUSION
conoce bien la persistencia o destrucción de C. perfringens durante el procesado,
almacenamiento y distribución de muchos productos cárnicos, es posible hipotetizar que las
sustancias antimicrobianas producidas por las cepas aisladas en nuestro trabajo dificultarían el
desarrollo de este patógeno.
La sensibilidad de las listerias a sustancias antagonistas de especies del género
Lactobacillus no es sorprendente, ya que estos dos géneros están estrechamente relacionados
(Wilkinson y Jones, 1977; Seelinger y Jones, 1986; Rhuland y Fiedíer, 1987). Además, sehan publicado numerosos trabajos que confirman la actividad antimicrobiana de diversos
lactobacilos sobre L. rnonocyrogenes (Harris y col., 1989; Schillinger y Holzapfel, 1990;
Harding y Shaw, 1990). Sin embargo, en lo s trabajos publicados, la actividad antagonista se
limita a otros lactobacilos y listerias. siendo muy rara su actividad frente a otros patógenos
Gram-positivos como 5. aureus (Andersson. 1986; Talarico y col., 1988). Hasta ahora, no
tenemos constancia de que existan cepas de L. sa/ce cuyo espectro antimicrobiano sea tanamplio corno el de las cepas que se han aislado en este trabajo.
El espectro antimicrobiano de estas cepas es similar al mostrado por cepas del género
Pediococcus. Sin embargo, la síntesis de sustancias antimicrobianas por especies de origen
cárnico pertenecientes a este género se limita únicamente a P. ac¡dilactici H con actividad
antimicrobiana frente a microorganismos Gram-positivos como C. perfringens, Listeria
nzonocyto genes y Staphylococcus aureus y dos especies Gram-negativas, Acromonas
hydrophila y Pseudornonas pulida (Bhunia y col.. ~988).
Por lo que respecta a lo s lactococos la nisina producida porLactococcus lactis inhibe el
desarrollo de L. monocvtogenes (Ehunia y col, 1988) y especies del género Clostridiurn
dificultando, además, la germinación de las esporas de C. botulinum (Daeschel. 1989). Sinembargo, es inactiva frente a microorganismos Gram-negativos. hongos o levaduras.
228

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 242/293
DiSCUSION
El amplio espectro de acción de las cepas de L. sa L -e estudiadas en este trabajo y, de la
cepa 148 en particular, permite concebir esperanzas sobre el futuro empleo de estosmicroorganismos como agentes conservadores de ciertas carnes y productos cárnicos, ya que
es precisamente la especificidad mostrada por las bacterias lácticas la que ha limitado el empleo
de estos microorganismos como cultivos iniciadores o protectores (Hastings y Stiles, 1991).
No obstante, antes de estimular su utilización en la industria cárnica. se necesita conocer silos
resultados de inhibición obtenidos “in vitro” son reproducibles en un sistema cárnico
experimental.
y. 5.- Efecto de la composición del medio de cultivo en la producción de
actividad inhibidora
Una vez seleccionado el microorganismo con mayor actividad antimícrobiana y con
unas características de crecimiento más favorables, se necesita establecer las condicionesóptimas de producción de su actividad para acometer su purificación con las mejores garantias
de éxito. Para ello se seleccionó la cepa L. sa/ce 148, cuyo intervalo de temperaturas de
crecimiento es muy amplio (4-45 oc) y cuyo sobrenadante cultural presenta una actividad
antimicrobiana comparable, sino superior, al de las otras cepas de L. sa/ce aisladas.
Tagg y col. (1976) ya señalaron que ciertos componentes del medio de cultivo podríanser críticos en la producción de ciertas bacteriocinas. Sin embargo. existen pocos trabajos
sobre el efecto de los distintos nutrientes en la producción de compuestos antagonistas por las
bacterias lácticas (Reddy y Ranganathan. 1983; Batish y col.. 1990; Biswas y col., 1991); en
la mayor parte de lo s casos no se ha llegado, a resultados concluyentes. Algunas cepas
productoras de sustancias antimicrobianas pierden esta capacidad cuando crecen en medios
líquidos y la recuperan cuando lo hacen en medios sólidos o semisólidos, tal es el caso de lalactacina B (Barefoot y Klaenhammer. 1983) y de la propionicina PLG-l (Lyon y Glatz,
229

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 243/293
DISCUSION
1991). Contrariamente a los resultados obtenidos por éstos investigadores, L. sa/ce 148
mostraba actividad inhibidora tanto en medios sólidos como líquidos.
De los resultados obtenidos en este trabajo (tabla IV . 5). debe destacarse la ausencia de
actividad antagonista cuando L. sa/ce 148 se desarrollaba en medios de cultivo semidefinidos
suplementados con hidrolizados de proteinas lácteas (caseinas). No sucede así cuando el
suplemento es un hidrolizado de proteinas cárnicas (peptona o extracto de carne), a pesar de
que la actividad antagonista es fácilmente mensurable y cuantificable. La producción óptimaacaece en el medio complejo MRS. Esta último observación concuerda con la de McGroarty y
Reid (1988) acerca de la producción de bacteriocinas por L. casei sp. rharnnosus y L.
acidophilus 76. Los resultados obtenidos refuerzan la teoría de Schillinger y LOcke (1989)
según la cual las bacterias lácticas aisladas de productos cárnicos, serían más adecuadas para
asegurar la calidad higiénica de la carne y productos cárnicos que las procedentes de otros
productos.
Tampoco se detectó la actividad antimicrobiana en L. sa/ce 148 cuando éste se
desarrollaba en caldo de APT sin suplementar. Sin embargo, cepas Carnobacterí un ; piscícola
(Ahn y Stiles, 1990a) y Leuconosíoc gelidunz (Hastings y Stiles, 1991), aisladas de productos
cárnicos envasados al vacío, sintetizan bacteriocinas en caldo de APT. Lo mismo ocurrió en el
medio BRI, que es un caldo de cultivo compejo de cuya composición forman partehidrolizados de cerebro y corazón bovino. En este caso resulta muy dificil explicar la ausencia
de actividad antagonista, tanto en el medio sin modificar como en el suplementado con
diversos extractos cárnicos, dada la extraordinaria riqueza nutritiva de este medio.
Las diferencias observadas en la producción de actividad antimicrobiana dependiendo
de la fuente proteica, se justificarian por la variación en algún componente específico(posiblemente aminoacídico) o por la existencia de mecanismos reguladores de origen
230

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 244/293
DISCUSION
genético. estimnulados o reprimidos por componentes del extracto proteico. Esto último
explicaría la falta de actividad inhibidora de L. saL -e 148 al crecer en el caldo BHI, tantosuplementado como sin suplemento. Morishita y col. (1974), demostraron que algunos
mutantes de L. cosed sintetizaban detenninados aminoácidos en medios carentes de lo s mismos;
dicha capacidad la perdían cuando crecían en medios complejos. De los resultados obtenidos se
deduce que sería conveniente estudiar más profundamente estos posibles mecanismos
reguladores, con el fin de predecir la posible inhibición o represión de la actividad antagonista
de L. sa/ce 148, cuando éste se desarrolle en la carne o en sus productos.
V. 6.- Parámetros cinéticos y producción de la actividad inhibidora de
L. sa/ce 148 a diversas temperaturas
L. sa/ce 148 se desarrolló a 4, 8. 16, 25 y 32 0C, tanto en medio complejo MRS como
en medio mínimo MM con triptosa (tabla IV. 6), obteniéndose unos parámetros de crecimiento
similares a lo s encontrados por otros investigadores con cepas de L. sa/ce y otras bacterias
lácticas de origen cárnico (Rodríguez. 1991).
Según Daeschel y col. (1990) la producción de bacteriocinas por las bacterias lácticas
puede iniciarse en cua[quier etapa del crecimiento, tanto en su comienzo (fase logarítmica)
como al final (fase estacionaria). La actividad antagonista de L. sa/ce 148 es detectable y
cuantificable al poco tiempo de iniciarse su crecimiento en todas las temperaturas ensayadas
(fig. 4. 30 a 4. 34). Estos resultados coinciden con los de la mayor parte de los trabajos
publicados (Gálvez y col., 1986; Ahn y Stiles, 1990b; Hastings y Stiles. 1991) en lo s que la
produción de sustancias inhibidoras parece ser una propiedad asociada al crecimiento.
Corno ya señalaron Ahny
Stiles (199Gb). la producción de bacrnriocinas en la fasemnicial del crecimniento bacteriano podría favorecer el predominio de la cepa productora en una
231

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 245/293
DISCUSlON
población bacteriana mixta con bacterias sensibles. Asimismo, la detección de la actividad
antagonista al comienzo del crecimiento indica que no corresponde a un metabolito secundario.
La disminución de la actividad durante la fase estacionaria (fm g. 4. 34), podría deberse a la
acción de enzimas proteoliticos endo o exocelulares, a que la sustancia responsable de la
actividad se convierte en otros metabolitos o a su inestabilidad ante el aumento de la acidez del
medio de crecimiento.
El incremento de la actividad antibacteriana al hacerlo la temperatura se debe, alparecer, al mayor desarrollo celular, ya que la actividad aumenta a medida que la temperatura
se aproxima a la óptima de crecimiento deL. sa/ce 148. Hasting y Stiles (1991) comprobaron
que la producción de bacteriocinas por Leuconosíoc gelidun; es directamente proporcional a la
masa celular del cultivo microbiano. Es de destacar que L. sa/ce 148 producía actividad
antimicrobiana a todas las temperaturas ensayadas.
La producción de la actividad antagonista a temperaturas de refrigeración, contribuida
a dificultar el desarrollo de los microorganismos alterantes y patógenos en lo s alimentos
refrigerados.
V. 7.- Caracterización biocuimica narcial de la actividad antimicrobiana deL, sa/ce 148
Una vez establecidas las condiciones óptimas de crecimiento y de producción de la
actividad antimicrobiana deL. sa/ce 148, el siguiente paso consistió en intentar su purificación,
con el fin de separar la entidad activa de otros productos o restos celulares y de las proteínas
del medio de cultivo. Hay que reconocer que el medio complejo M RS no es el más indicado
para iniciar la purificación de la actividad antagonista de L. sa/ce 148, ya que los
sobrenadantes procedentes de este medio contienen una concentración elevada de proteínas y
péptidos (Barefoot y Klaenhamnmer, 1984), lo que no sucede con el medio mínimno con triptosa
232

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 246/293
DISCUSION
que, siendo un medio semidefmnido sencillo, permite la síntesis de la actividad antimicrobiana
deL. saL- -e 148 (tabla IV . 6).
Debido a la heterogeneidad de las bacteriocinas, sus protocolos de purificación se han
diseñado empíricamente para cada una de ellas.
Barefoot y Klaenhammer (1984), tras emplear sin éxito la cromatografía de
intercambio iónico, purificaron la lactacina E creando condiciones disociantes (urea 8M) antes
de someterla a ultracentrifugación y a cromatografía de filtración en geles. Los resultados
obtenidos indicaron que el peso molecular de la bacteriocina parcialmente purificada era de
6.500 daltons. Sin embargo, cuando la lactacina B parcialmente purificada se sometió a
electroforesis en poliacrilamida con dodecil sulfato sódico (SDS-PAGE) no se apreciaron
bandas en los geles, a pesar de teñirlos con reactivos de plata, que es un proceder de gran
sensibilidad para la detección de las bandas de proteinas en los geles. Dos años más tarde,Joerger y Klaenhammer (1986) también purificaron la helveticina . 1 por cromatografía de
filtración en geles que llevaban dodecil sulfato sódico (SOS) como agente disociante. El peso
molecular de la bacteriocina purificada fué de unos 37.000 daltons.
De otra parte, Rammelsberg y Radíer (1990) han encontrado dificultades al intentar
purificar la brevicina 37 y la caseicina 80, utilizando lo s procedimientos de purificaciónproteica tradicionalmente empleados. Dichos investigadores han observado, asimismo. que la
concentración de los sobrenadantes por liofilización origina una pérdida importante de la
actividad antimicrobiana.
En contraste con la utilización de las técnicas cromatográficas de filtración en geles,
empleadas en la purificación de bacteriocinas producidas por lactobacilos, la purificación de lapediocina PA-l (Gonzalez y Kunka, 1987) y de la pediocina AcH (Bhunia y col.. 1988),
233

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 247/293
DISCUSION
producidas por pediococos, se ha realizado por cromatografía de intercambio iónico. El peso
molecular de la pediocina PA-I parcialmente purificada (16.500 daltons) se determinó por
cromatografía de filtración en geles, mientras que el de la pediocina Acl-i (2.700 daltons) se
estimó por SDS-PAGE.
En nuestro trabajo, la purificación de la actividad antimicrobiana exocelular deL. sa/ce
148, se realizó concentrando los sobrenadantes por liofilización y separando las proteínas por
cromatografía de filtración en geles. Utilizamos la urea (a una concentración 1 M) como agente
disociante de los posibles agregados proteicos, formados por la sustancia antimicrobiana
consigo misma o con otros componentes del medio del cultivo (figuras 4. 35 a 4. 37). El
proceso de purificación se tradujo en un aumento poco apreciable de la actividad inhibidora
específica (7 veces) y en una baja recuperación de la actividad inhibidora original (8 %) (tabla
IV. 7). Los resultados obtenidos confirman que, según se ha observado en otras ocasiones, las
bacteriocinas son, a veces, bastante resistentes a su purificación por las técnicas standar de
separación de proteinas (Ranimelsberg y Radíer, 1990). La concentración de las muestras por
liofilización antes de someterlas a cromatografía de filtración, posiblemente afecta
negativamente a la actividad antimicrobiana.
La pérdida de actividad, que a menudo es grande, producida durante la purificación
constituye un problemna corriente en las bacteriocinas originadas por las bacterias lácticas (Taggy col., 1976).
Barefoot y Klaenharnrner (1984) y Joerger y Klaenhammer (1986), únicamente
lograron recuperar el 0,4 % de la actividad inhibidora inicial de la lactacina E y el 4 % de la
helveticina J. Sin emnbargo, mientras la purificación final de la lactacina B fué de 3.222 veces y
1 - a de la helveticina 3 de 837 veces, la purificación final de la sustancia antimicrobiana de L.
sa/ce 148 es mucho menor. De ello se deducequeel método de purificación empleado es poco
234

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 248/293
DISCUSION
eficaz, por lo que conviene desarrollar otros más potentes.
La sustancia antimicrobiana de L. sa/ce 148 parece que se sintetiza en forma de
polipétido de pequeño tamaño molecular como lo demuestra que, en los experimentos de
filtración en geles de Sephadex G-150 fino, G-75 y G-50 (figuras 4. 35 a 4. 37), se eluye
lejos del volumen de vacio, volumen en el que se encuentran las moléculas de un tamaño
molecular mayor que el diámetro de poro del gel. En este aspecto, la sustancia antimicrobiana
de L. sa/ce 148 difiere de la lactocina 25 (Upreti y Hinsdill, 1973), lactacina B (Barefoot y
Klaenhammer, 1984), helveticina J (Joerger y Klaenhammer, 1986), lactacina F (Muriana y
Klaenhammer, 1991), propionicina PLG-1 (Lyon y Glatz, 1991) y la bacteriocina deL. sa/ce
449 (Rodríguez, 1991), que se aislaron primero, bien como agregados proteico-polisacáridos
complejos (partículas globulares parecidas a las micelas) o bien como grandes agregados
protéicos, que se separaron más tarde utilizando diversas técnicas analíticas.
Una vez realizada la purificación parcial de la actividad antimicrobiana conviene
realizar su caracterización físico-química. Uno de los criterios básicos para que una sustancia
antimicrobiana se considere una bacteriocina es que sea un componente proteico
biológicamente activo (‘Tagg y col.. 1976), lo que se determina por su sensibilidad a diversos
enzimas proteoliticos. En la tabla IV. 8 se m rm u estra el efecto de diversos enzimas proteolíticos,
lipolíticos y amilolíticos, en la actividad antimicrobiana exocelular parcialmente purificada de
L. saL- -e 148. En ella se observa que lo s enzimas inespecíficos, corno la papaína, proteasa II y
proteasa XIV, anulan la actividad inhibidora, mientras que otros enzimas más específicos la
disminuyen mucho; de ahí se deduce que la sustancia antimicrobiana deL. sa/ce 148 posee una
estructura proteica biológicamente activa.
Además, su resistenciaa
la acción de los enzimas lipoliticosy
amiloliticos indica queen su composición no entran a formar parte elementos lipídicos ni hidrocarbonados y, silo
235

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 249/293
DISCUSION
hacen, no influyen en su actividad antimicrobiana.
La sustancia antimicrobiana parcialmente purificada deL. sa/ce 148 es muy estable en
un amplio intervalo de valores de pH (fig. 4. 38), mostrando su mayor actividad a valores de
pH comprendidos entre 5,6 y 6,6; cuando el pH se situaba entre valores extremos (2,6-4,6;
10-12) la actividad disminuía ligeramente (sin superar nunca el 20%).
Gonzalez y Kunka (1987), comprobaron que la pediocina PA-1 producida por 1’.acidilacticí era estable entre valores de pH de 4 y 7, si bien mostraban ligeras pérdidas de
actividad a pH de 2.3 y 10. Bhunia y col. (1988) establecieron el intervalo de pH al que la
pediocina AcH era activa entre pH 2,5 y 1 0 y observaron una pérdida total de la actividad
cuando el pH era mayor de 10.
La nisina, producida por L. lactis es inactiva cuando el pH es superior a 7 (Hurst,1981; Liu y Hansen, 1990). La propionicina PLG-1 permanece estable a valores de pH entre 3
y 9, con ligeras pérdidas de actividad a pH de 3-4 (Lyon y Glatz, 1991).
La gran estabilidad frente al pH mostrada por la bacteriocina de L sa/ce 148 es muy
imnportante de cara a su posible utilización como agente antimicrobiano, tanto en lo s alimentos
fermentados como en otros productos alimenticios.
La resistencia al tratamiento térmico es una característica muy extendida entre las
bacteriocinas, no obstante, el criterio seguido para determinar la estabilidad de estas sustancias
varía bastante, ya que esta propiedad depende del grado de purificación de las bacteriocinas y
de otros factores como pH, fuerza iónica y presencia de moléculas termoprotectoras (Tagg y
col., 1976). La existencia de bacteriocinas termosensibles es menos corriente, aunque entre
éstas se encuentran la helveticina J (Joerger y Klaenhammer, 1986), la caseicina 80
236

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 250/293
DISCUSION
(Rammelsberg y Radíer, 1990), la bacteriocina sintetizada por L. delbruec/cii sp lactis (Toba y
col., 1991) y la propionicina PLG-í (Lyon y Glatz, 1991). La bacteriocina producida porL.
sa/ce 148 es muy termoestable (fig. 4. 39), pareciéndose en esto a las bacteriocinas de L.
acidophilus, (Barefoot y Klaenhammer, 1983; Muriana y Klaenhammer, 1987), L. helvericus
(Upreti y Hinsdill, 1973; Joerger y Klaenhammer, 1986), L. plantarurn (West y Warmer,
1988; Daeschel y col., 1990), L. sa/ce (Schillinger y LOcke, 1989; M0rvedt y Ness, 1990) y
L. brevis (Rammnelsberg y Raedler, 1990).
La gran termoestabilidad de la bacteriocina de L. sa L -e 148 le confiere ventajas en
cuanto a su posible utilización en alimentos higienizados por el calor, ya que permanece
biológicamente activa tras los tratamientos de pasterización y esterilización a los que se
somenten éstos productos.
La determinación del peso molecular aparente de la bacteriocina de L. sa/ce 148
mediante filtración en geles de Sephadex G-50, dió un peso molecular de 4640 daltons (tabla
IV. 9). La elución de esta bacteriocina en un solo pico de bajo peso molecular no es corriente,
dada la baja concentración del agente disociante empleado (urea 1M en el tampón de disolución
y urea 0,1M en el de elución). Lo corriente es que las bacteriocinas producidas por las
bacterias lácticas formen agregados de alto peso molecular (Muriana y Klaenhammer, 1991;
Rodríguez, 1991).
El peso molecular de las bacteriocinas de las bacterias lácticas es muy variable, con
moléculas grandes, como la helveticina J cuyo peso molecular es de 37.000 daltons (Joerger y
Klaenhammer, 1986), la caseicina 80 de un peso molecular entre 40.000 y 42.000 daltons
(Rammnelsberg y Raedíer, 1990) y la bacteriocina de L. sa L -e 449 de unos 30.000 daltons
(Rodríguez, 1991). No obstante, ta l y como ocurre con la bacteriocina de L. sa/ce 148, lamayor parte presentan un peso molecular relativamente pequeño, menor de 10.000 daltons. En
237

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 251/293
DISCUSION
este grupo se incluyen: nisina (Hurst. 1981), diplococina (Davey y Richardson, 1981),
lactacina B (Barefoot y Klaenhammer, 1984). pediocina AcH (Bhunia y col., ¡988). ¡actacinaF (Muriana y Klaenhammer, 1991), lactocina 5 (M0rtvedt y col., 1991) y carnocina U149
(Stoffels y col., 1992). Cuando las bacteriocinas forman agregados moleculares es difícil
determinar su peso molecular mediante técnicas de filtración en geles, por lo que se recurre a
la electroforesis en geles de poliacrilamida con dodecil sulfato sódico (SDS), con o sin urea.
Sin embargo, la utilización de la electroforesis en geles de poliacrilamida con SDS, cono sin urea, para determinarel peso molecular de Ja bacteriocina deL. sa/ce 148 dió resultados
negativos cuando la tinción se realizó con Azul Brillante de Coomasie (Laemmli, 1970). Esto
no es extraño, ya que Muriana y Klaenhammer (1991) obtuvieron el mismo resultado al
determinar el peso molecular de la lactacina F con este colorante. Por otra parte, Davey y
Richardson (1981) y Stoffels y col. (1992), observaron la existencia de una banda difusa
asociada al frente electroforético al determinar el peso molecular de la diplococina y lacarnocina U149 respectivamente. Cuando Barefoot y Klaenhammer (1984) utilizaron nitrato de
plata como agente tintorial para determinar el peso de la lactocina B obtuvieron, igual que
nosotros (fig. 4. 40), una banda de tinción difusa asociada al frente electroforético.
La dificultad de determinar el peso molecular de las bacteriocinas por técnicas
electroforéticas se atribuye a que las condiciones creadas durante la electiroforesis originan lahidrólisis espontánea de las moléculas proteicas (Muriana y Klaenhammer, 1991); de otra
parte, la cantidad de proteína depositada en lo s geles es lo suficientemente grande para poder
detectarse con el nitrato de plata, cuyo límite de detección es menor de 1 nanogramo de
proteína (Merril y col., 1981). Además, la falta de bandas proteicas en lo s geles indica que la
muestra se encuentra libre de otras proteinas contaminantes, lo que facilita la determinación de
la composición aminoacídica de las bacteriocinas parcialmente purificadas.
238

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 252/293
DISCUSION
Al comparar la composición aminoacídica de la bacteriocina de Ib . sa/ce 148, obtenida
por cromatografía líquida de alta resolución (HPLC) con la de otras bacteriocinas de peso
mnolecular semejante, se obtiene la tabla V.1. De esta tabla se desprende que, al igual que la
lactocina 5 de I b . sa/ce L45 (M0rtvedt y col., 1991), la bacteriocina de I b . sa/ce 148 tiene en su
composición un 50% de aminoácidos apolares, presentando ambas, además, una cierta
similitud en su composición cualitativa pero no cuantitativa, de aminoácidos. De la observación
de la tabla también se desprende que la bacteriocina de L. sa/ce 148 difiere de otras como la
nisina. subtilina y diplococina (Davey y Richardson, 1981), lactacina E (Muriana y
Klaenhammer, 1991) olacarnocina U149 (Stoffels y col., 1992)
La mejor conocida de las bacteriocinas producidas por las bacterias lácticas es la
nisina, de la que no sólo se conoce su composición de aminóacidos sino, además, la
disposición de los mismos en la cadena peptídica. Una característica fundamental que
diferencia a la nisina, subtilina, epidermina (Ailgaier y col., 1986), galliderinina (Kellner y
col., 1988), PepS (Kellner y col., 1989), ¡actocina 5 (M0rtvedt y col., 1991) y carnocina
U149 (Stoffels y col., 1992) de otras bacteriocinas como la diplococina y la lactacina E es la
presencia, en las tres primeras, de aminoácidos modificados postransíacionalmente como la
dehidroalanina, dehidrobutimna, lantionina y beta-metil lantionina. Esto determina que se haya
propuesto el nombre de “lantibioticos” para aquellas bacteriocinas que, como la nisina, posean
tales aminoácidos (Muriana y Klaenhammer, 1991).
En el cromatograma obtenido al análizar por HPLC el hidrolizado de la bacteriocina de
Ib . sa/ce 148 (resultados no mostrados), también se observa la presencia de un pico
cromatográfico que no se corresponde con ninguno de lo s patrones empleados.
239

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 253/293
t f l f l N C1f f l — s O O <f l ~-~
Cn N~’ C1 N
1
•
vi
vi —
—
o
s t 01~~’ f l mn C N- Ú
r~ N
—
———--~ en c1--’ —
1
y
111
111r <N O‘ n o
mi l l — o‘ n o e
n ‘ o
—I l mn—~vi —I —I l —l —i en t —I — t —vi ur---
vi ovi e n
e n
—I —I — en cÑl — cNl enl
dm1
s t vi
‘ o ¼D t n- ÚI cf l —’ c1 1 1 —— en
s t
0 1
o c i ~ a
. .E o
<1-’
o o
c e e
c e- a
c e c e
-— c e
c e
e
c e
u
Z
e
— s t — vi
O N O
d 1 u- ,
¡l i i i
s t cn o
c
o c e e
c e
~ c e . g
~
o t eE
~~~
ci a ) oE o ( Ji 4 )
c i ‘ 0 oE o u-
o c e -n o o
- e‘ 0 o u , oE o u
-D e on o c
e
- .2
c e
r e e ci o u o o , o c
e c e e
-n - o e
en
o ) - o c
e
O
C
a- s t
c i
—
o o
-~
c e
- o- o
1
u , c e e o o4
- )
o c e - o u , c
e u , a
—
,
o
- >
o - ,
- e c
e o o c
e
o c e e‘ o o u , o o
,E o u c
e - o c
e
1 -
I O N
Q N EN c
e
. C O N
e —
c e u ,
¡ e—
c e e ) t ~-4
¼ > u t - e
1 u , t t
0 0
‘ r er e
n o1 — Q
N
2 —’
‘ ——
u ,—
t u ¡ - e
~ 0 C )
¡ u , u ,
0 0
Qn
¡ c e
o
2 4 0

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 254/293
DiSCUSION
En nuestro caso fué imposible determinar con exactitud la identidad de este
aminoácido, sin embargo, su elución en la columna cromatográfica, en una posición muypróxima a la glicina, coincide con los resultados obtenidos al analizar, por el mismo sistema de
HPLC, la lactocina 5 (M0rtvedt y col.. 1991) y la carnocina U149 (Stoffels y col., 1992).
Estos autores demostraron que la lantionina eluye precisamente en esta posición, por lo que no
es descabellado plantear la hipótesis de que el aminoácido no identificado de la bacteriocina de
L. sa/ce 148 fuera la lantionina, lo que la convetiría, por definición, en un lantibiótico.
Si bien conviene conocer los aminoácidos que conforman la sustancia inhibidora,
también tiene un gran interés saber la localización o secuencia de dichos aminoácidos en la
estructura proteica. El conocimiento de la secuencia facilitaría la elucidación de los
determinantes antigénicos de esta bacteriocina, así como su donación, mediante técnicas de
amplificación por reacción en cadena de la polimerasa (PCR) en vectores genéticos de interés.
Tanto la secuenciación como el estudio de lo s determinantes, forman parte del esquema detrabajo futuro a desarrollar como continuación del actualmente en curso. La caracterización
completa de la bacteriocina de L. sa/ce 1 4 8 y el estudio genético de los mecanismos
responsables de su síntesis, permitirán conocer la relación existente entre su estructura y su
mecanismo de acción, sus relaciones estructurales con las bacteriocinas producidas por otras
bacterias lácticas y, por último, la biología molecular de la producción de bacteriocinas en el
género Lactobacillus.
V. 8. - Mecanismo de acción de la bactem-iocina producida por I b . sa/ce 148
La sustancia inhibidora elaborada por Ib . sa/ce 148 se aparta ligeramnente de la
definición que Tagg y col. (1976) dieron de las bacteriocinas, al considera¡-las como proteinas
o complejos proteicos con acción bactericida en especies bacterianas filogenéticamenterelacionadas con la cepa productora; esto no se cumple en la bacteriocina de I b . sa/ce 148 que,
241

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 255/293
DiSCUSION
como hemos visto, posee acción bacteriostática en lodos los microorganismos indicadores
estudiados (Tabla IV. 11).
La primera característica, su naturaleza proteica, se cumple en todas las bacteriocinas
de las bacterias lácticas descritas hasta la fecha. La segunda, su limitado espectro de acción, ya
fué cuestionado por Klaenhammer (1988), quién propuso la división de las bacteriocinas de las
bacterias lácticas en dos tipos: el primero constituido por sustancias con un espectro de acción
limitadoa otras
bacterias lácticas relacionadascon la
cepa productora; aeste grupo, que es
muy
numeroso, pertenecen entre otras, la lactacina E (Barefoot y Klaenhammer, 1983) y F
(Muriana y Klaenhammer, 1991), la lactocina 27 (Upreti y Hinsdill, 1975), la helveticina J
(Joerger y Klaenhammer, 1985), la sakacina A (Sehillinger y LOcke, 1989), y la lactocina 5
(M0rtved y Ness, 1990). El segundo grupo estaría formado por sustancias con un espectro de
acción más amplio en el que. además de las bacterias lácticas, se incluirían otros
microorganismos Gram-positivos como 5. aureus, L. monocytogenes, C. botulinunz y C. pe,f¡-ingens. Pertenecen a este grupo la nisina, la pediocina AcH y la sustancia antimicrobiana
objeto de este trabajo, producida por Ib . sa/ce 148.
En la tercera característica citada por Tagg y col. (1976), el mecanismo de acción
bactericida, es donde mayor es la discrepancia con los resultados de este trabajo.
Efectivamente, tai y como se aprecia en los recuentos microbianos realizados después de tratarvarios cultivos de diversos microorganismos sensibles con un sobrenadante neutralizado y
concentrado deL. sa/ce 148 (tabla IV. 11). el mecanismo de acción de esta bacteriocina es
bacteriostático.
La acción bactericida descrita para las bacteriocinas se produce en dos etapas (Tagg y
col.. 1976). En la primera, la bacteriocina se une a receptores específicos situados en lasuperficie de los microorganismos sensibles. Esta unión altera la permneabilidad de su
242

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 256/293
DISCUSION
membrana; en la segunda etapa de su mecanismo de acción se provoca la ruptura de la
membrana y la muerte celular. Esta acción bifásica se ha comprobado mediante examenmicroscópico, al apreciarse cambios morfológicos y posterior lisis celular en las bacterias
sensibles a la nisina (l-Iurst, 1981), plantaricina SIK-83 (Andersson y coL,1988) y lactacina F
(Muriana y Klaenhammer, 1991).
La acción de la nisina se atribuye a su elevada hidrofobicidad, lo que le permite
interaccionar con la membrana bacteriana y originar poros en la misma aumentando supermeabilidad y provocando la muerte celular por estallido (Hurst, 1981). Esta acción, similar
al efecto surfactante de ciertas moléculas hidrofóbicas, es también la causante de la acción
bactericida de la lactacina E (Muriana y Klaenhammer, 1991).
En ocasiones se puede apreciar un efecto bactericida sin que se produzca la lisis
celular, lo que ocurre en el caso de la pediocina AcH, cuyo mecanismo de acción ha sidoestudiadoen profundidad por Bhunia y col. (1991). E sta sustancia ejerce su actividad en varias
etapas: (1) inicialmente se une a receptores inespecificos de la pared celular, posiblemente
ácido lipoteicoico; (2 ) cuando se saturan estos receptores se une a receptores específicos,
originando alteraciones funcionales en la membrana que (3), provocan la pérdida de iones
potasio y la entrada en la célula de moléculas de pequeño tamaño; además, la bacteria pierde su
capacidad de multiplicarse. (4) Solo en algunas cepas especialmente sensibles la pared celularpierde su integridad estructural y el microorganismo se lisa. (5) La resistencia mostrada por
otras bacterias Gram-positivas se debería a ¡a falta o inaccesibilidad de los receptores
específicos y (6) la resistencia de las bacterias Gram-negativas estaría motivada por la carencia
tanto de receptores específicos, como inespecíficos. La lactacina B, que tiene acción
bactericida, tampoco origina alteraciones en la permeabilidad de la membrana ni lisis celular
(Earefoot y Klaenhammer. 1984).
243

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 257/293
DISCUSION
Las lactostrepcinas (Dobrzanski y col., 1982), lactacina E (Barefoot y Klaenhammer,
1983> , helveticina J (Joergery Klaenhammer, 1985), sakacina A (Schiljingery LOcke, 1989),
bacteriocina de Leuconostoc gelicluni (Harding y Shaw, 1990), plantaricina A (Daeschel y
col., 1991) y carnocina U149 (Stoffels y col., 1992), junto con las citadas nisina, lactacina F,
plantaricina SIK-83 y pediocina AcH son algunas de las bacteriocinas que, como la mayor
parte de las producidas por las bacterias lácticas, poseen un mecanismo de acción bactericida
independientemente del mecanismo intimo que conduce a la muerte bacteriana.
E] efecto bacteriostático de la bacteriocina de I b . sa/ce 148 contrasta con el bactericida
de la sakacina A, producida por otra cepa de I b . s a L -- e (Sehillinger y LOcke, 1989). Este
resultado refuerza la hipótesis, apuntada al analizar el espectro de acción, de que las
bacteriocinas producidas por distintas cepas de una misma especie no son idénticas entre sí.
Sin embargo esta acción bacteriostática no es única dentro de las bacterias lácticas, yaque este carácter se ha descrito en la lactocina 27 de I b . acidophilus (Upreti y Hinsdill, 1975),
aunque en algunos casos se ha observado un efecto bactericida sobre cepas de Ib .
ínonocytogenes (Raccach y col., 1989); el efecto bacteriostático también se ha descrito en
lactobacilos de origen cárnico identificados como Ib . sa/ce (Ahn y Stiles, 1990a) y en
Leuconostoc gelidurn (Hastings y Stiles, 1991).
Incluso el efecto bactericida atribuido a la pediocina AcH podría no ser tan evidente a
tenor de la observación de los resultados de Ehunia y col. (1988). Según dichos
mnvestigadores, la adición de pediocina AcH a un cultivo de Ib . plantarurn WSO-30 en
crecimiento (con una población inicial de 3 x io7 ufc/ml) originó una disminución de,
únicamnente, 0.8 unidades logarítmicas en el recuento de microorganismos viables efectuado
una hora después de dicha adición. En lo s recuentos realizados a las 3. 5, 7 y 1 1 horas
posteriores, se observa una recuperación, mientras posteriormente se mantienen los valores
244

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 258/293
DISCUSION
obtenidos al inicio del experimento (3 x io~ ufc/ml). Mientras tanto, el cultivo control continúa
su crecimiento normal hasta alcanzar valores de 2 x ío ~ ufc/ml. Hasting y Stiles (1991)
obtuvieron resultados similares a lo s anteriormente señalados y. a pesar de todo, adjudicaron
un mecanismo de acción bacteriostático a la bacteriocina producida por Leuconostoc gelidunz
UALl87.
Los resultados de Bhunia y col. (1988) contrastan con los claramente bactericidas de
Schillinger y LOcke (1989) quienes,empleando
un procedimiento idénticoal
nuestro y. muy
similar al de Bhunia y col. (1988), comprobaron que lo s recuentos del microorganismo
indicador (cuya población inicial era deS x i05 ufc/ml), disminuía 3,5 unidades logarítmicas
dos horas después de añadir el sobrenadante libre de células, neutralizado y concentrado de I b
sa/ce Lb706; en recuentos posteriores fué imposible detectarlo. Por otra parte, Ehunia y col.
(1991) señalan, que la bacteria indicadora es incapaz de multiplicarse; la ruptura de la pared
celular y la muerte bacteriana sería un destino reservado únicamente a cepas de
microorganismos especialmente sensibles.
Realmente, es difícil aceptar que ninguna de las cepas que emplemos como
microorganismos indicadores fuera sensible y que la bacteriocina de I b . sa/ce 148 ejerciera una
acción bactericida en otras cepas sensibles; por ello y, a diferencia de Bhunia y col. (1988),
consideramos que la sustancia inhibidora ejerce una acción bacteriostática ya que detiene el
crecimiento de los microorganismos sensibles, disminuyendo su vitalidad y no su viabilidad,
algo que no ocurre en las experiencias de Schillinger y LOcke con L. sa/ce LB706 .
La única bacteriocina de las bacterias lácticas de la que se conoce con exactitud la
formna en que ejerce su acción bacteriostática es L a lactocina 27, que detiene la síntesis proteica
sin afectar la del ADN o ARN, y sin modificar lo s niveles de APT (Upreti y Hinsdill, 1975).
Este mecanismo de acción, diferente del atribuido a otras bacteriocinas. es uno de lo s motivos
245

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 259/293
DISCUSION
que hace que cada vez se emplee más el término de “sustancias similares a bacteriocinas” para
referirse a las sustancias antimicrobianas que, como la lactocina 27 o la producida por Ib . sa /ce -
148, difieren en alguna de las propiedades atribuidas a las bacteriocinas clásicas
(Klaenhammer, 1988). En la tabla y. í se señalan algunas propiedades de las bacteriocinas
mejor conocidas, lo que permite comparar las coincidencias y diferencias de nuestra
bacteriocina parcialmente caracterizada y las de aquellas caracterizadas por otros
investigadores.
Nuestros resultados, junto a otros que ponen de manifiesto el efecto bacteriostático de
algunas bacterias lácticas (Upreti y Hinsdill, 1975; Raccach y col., 1989; Ahn y Stiles, 1990a;
Hastings y Stiles, 1991), explicarían porqué persiste pero no se desarrolla Ibisteria
monocytogenes en carne fresca de bovinos y en la carne y productos cárnicos de poílo en
presencia de una flora láctica competitiva Buchanan y col., (1989), Johnson y coL, (1988);
también explicaría el que Shelef (1989) haya observado que I b . monocytogenes no sedesarroua durante la maduración de embutidos elaborados con carne de bovinos.
La concentración inhibidora mínima de la sustancia antagonista parcialmente purificada
de Ib . sa/ce 148 en los diversos microorganismos ensayados (tabla IV. 12), oscila entre 1 y 18
i~gml, siendo las cepas de S. aureus FR1349 y FR1361 las más resistentes a su acción
inhibidora. Los resultados obtenidos sugieren, una vez más, la posible utilidad de las bacteriaslácticas y de sus bacteriocinas como factores de seguridad de la carne y de los productos
cárnicos.
246

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 260/293
+‘
C Ñ r—’ D C-4
— en
c e
e o
- >
‘ -~ c e e n
o~ en
o , cm
—
04~
0 C )
o )
o‘ -~—
o- t e 0 0 t —
c e~
( ‘ - 4
e c e
—‘ > 5
0~
O
N O
vi O
C
4. ,
4 —
c j ~
-~-z
~
~z ~
u o
n o
O C
e n
A
—A
‘ o
S n
c e
c e
c c c
0 0 0
r — 0 0 0 0 0
xl 0 0 0 0 0
‘ 0 0 0‘ o en o
xl 0 0 en‘ o O
t n & r - ,
—
AA
A
‘ o
en4E 2 . en
--- u--4 ) -~
o )
4 . .i
4 -~
o o , o
~.
c e
< cn
c e---
c e
e O
~
- 5
0 0
0 o‘ 0-~ 0
g s t
e
en
c e
—‘ —4
c e
‘ c e
e 0 o c
--‘ - u- u-- ,
5 t e t e
~
o o , o , o
( f l O C
c e c e c e 0
0 5
:
,- c em u
—-
4 5 -4 ,- ,4- ,
.z ¿
.4 .~
.± g t i
u , c e
e c e o
. 5
0 .
c e o~ e c o
u , 0 , 0
-—-— c e
4. j a--
~E
zE
t 4—~--~
c ~ Z
Z snr —‘ D vi
c~ c e c e c e
O t e e o ) 4 ) O C )
0 0 0 0 <
0
< 04 < c e c e c e
c c c
o- o- o-
0 0 0 o c en 0 0 0N O C
‘ f l en
A ¿A o S
n x t o4
- > ‘ 0 -z -L i e c e u- , ( nr .’
e n’ —
c e
c e
c a e--- t i .-
4 ) .—.4 >
o O - , o
0 0
0 N- s . s 0 0
o t i t i ci u
t E
c j ~ cz
U~
-~
~
~ U
u—- .
t i
-~
t z U
0 4 >
t i
- o
¡ 0 0
a ) ’ - ,
E —
t c e
‘ -4-’ e‘ 0 o u , o o , 5 o u4 ) a
-
-
,
-n 5 0 2
; c e
a
- ,
o o c e~
v a-- ,
( J o ,
z eE o e n
& o‘ o Q N u , o e o ) e
- o ) - o u , o e o
) c e
c e
c e
o e o
- >
u , u , o )
- c u , o~ 0 7
3
o ) - o o1-~
‘ L i
e u , c a o
- >
e‘ 4 . >
o -
4 ) a
-
-
.
e c e
‘ Ji c e
o ci ‘ c e
u , c e
- e o
) o c
e - o u , c
e o
-
>
- t i u , c e e o o- e o
) o c e
- o u , c
e eL i
t i ) c e
o
- >
- t u , c e
o u ,
a
- ,
o
- >
u c e
z c e
E —
2 4 7

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 261/293
DiSCUSION
V. 9.- Propiedades antigénicas de las bacteriocina de I b . sa/ce 148
El conococimiento de las propiedades inmunológicas de las bacteriocinas es interesante
por varios motivos. Primero, paradeterminar su seguridad para el consumidor, esto es,que no
le provoque reacciones alérgicas o indeseables. De otra parte, la obtención de anticuerpos
frente a las bacteriocinas permitiría su detección y cuantificación en las carnes y productos
cárnicos. Asimismo, lo s anticuerpos específicos facilitarían la purificación a homogeneidad de
la sustancia antimicrobiana por técnicas de cromatografía de afinidad, lo que permitiría supurificación en un solo paso, evitando la utilización de otros procedimientos de separación
proteica menos eficaces.
El primer aspecto, la salud del consumidor, parece garantizado ya que el carácter
proteico de la sustancia estudiada, su pequeño tamaño y su sensibilidad a lo s enzimas
proteoliticos hacen pensar que su uso per os, con lo s alimentos, carecerá de peligros para lasalud humana. Posiblemente esta característica es común a todas las bacteriocinas de las
bacterias lácticas, prueba de ello es que una de éstas sustancias cuyas propiedades antigénicas
no son del todo conocidas, la nisina, es la única bacteriocina autorizada en muchos paises para
su uso en alimentos dé consumo humano, ya que se ha demostrado que no origina reacciones
tóxicas en el consumidor (Hurst. 1981). Igualmente la bacteriocina deL. sa/ce 148 carece de
propiedades alergénicas, ya que su inoculación a lo s animales de experimentación no lesproduce reacciones alérgicas ni tóxicas.
En cuanto al segundo aspecto ennumerado, las características antigénicas de las
bacteriocinas, no está suficientemente estudiado ni siquiera en las sustancias mas ensayadas;
no obstante, se sabe que ciertas bacteriocinas de algunas bacterias Gram-negativas poseen
propiedades antigénicas (Gagliano y Hinsdill, 1970; Tubylewicz, 1970; Tagg y col., 1976).
248

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 262/293
DISCUSION
Unicamente Ehunia y col. (1990), han evaluado la antigenicidad de la pediocina AcH
de P. acidilacfici, llegando a la conclusión de que la inoculación de esta sustancia a ratones yconejos no genera la síntesis de anticuerpos neutralizantes. La explicación de este fracaso
inmunógeno habría que buscarla, según dichos autores, en el pequeño tamaño molecular de
ésta sustancia (2700 Da) que la haría comportarse como un hapteno; en que las técnicas de
mnmunización o lo s adyuvantes empleados no fueron lo s adecuados o en que el número de
animales de experimentación utilizados fué pequeño.
Además, la calidad del antígeno no era la mas indicada para estas experiencias, ya que
emplearon una bacteriocina parcialmente purificada cuya fracción activa (de 2700 daltons) iba
acompañada de, al menos, 2 proteinas de alto peso molecular visibles por electroforesis en
geles de poliacrilamida con dodecil sulfato sódico (SDS-PAGE).
Coincidiendo con nuestros resultados, Bhunia y col. (1990), mediante un sistemainmunoenzimático (ELISA) indirecto, observaron una respuesta inmunológica frente a la
pediocina AcH parcialmente purificada; tal respuesta la atribuyeron a las proteinas de alto peso
molecular, pero sin actividad inhibidora, presentes en la muestra inoculada. En nuestro caso, la
respuesta inmunológica no puede atribuirse a compuestos proteicos de elevado peso molecular,
ya que la electroforesis en geles de poliacrilamida (fig. 4. 40) demuestra claramente que la
preparación estaba libre de sustancias de este tipo.
De los resultados obtenidos, parece deducirse que la sustancia antimicrobiana
parcialmente purificada de I b . sa/ce 148 presenta cierta acción inmunógena, ya que el título de
anticuerpos obtenido en cada una de la sangrías parciales es superior a la anterior (fig. 4. 41).
Además, dicha sustancia puede identificarse y diferenciarse del antígeno empleado corno
control, cuando ambos se disuelven en tampón de PBS (ñg 4 42 ) Sin embargo, fué
imposible la detección de dicha sustancia cuando se añadió al medio de cultivo MM-triptosa
249

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 263/293
DISCUSION
debido, posiblemente, a la interferencia provocada por alguno de sus componentes. De los
resultados obtenidos puede concluirse, al igual que hicieron Ehunia y col. (1990), que labacteriocina deL. sa/ce 148 no es detectable mediante el ELISA indirecto cuando se encuentra
disuelta en un medio de cultivo, por lo que quizás sería necesario desarrollar una técnica más
sensible que evitara las interferencias citadas.
En cualquier caso, la respuesta inmunológica de lo s animales de experimentación fué
muy pequeña lo que pudo serdebido, aparte del tamaño de la partícula, a otros condicionantesya citados por Bhunia y col. (1990), como: a) que el número de animales inmunizados no
fuera suficiente para permitir la generalización de las conclusiones; b) que lo s adyuvantes, vías
y protocolos de inmunización no fueran lo s más adecuados o c) que el tamaño de la sustancia,
sea incapaz de generar una respuesta en el organismo receptor. En este caso sería necesario
incrementar la potencia inmunógena conjugándola con sustancias que le otorguen esta
capacidad como, por ejemplo, con la seroalbúmina bovina (BSA).
Las interferencias con otros componentes del medio podrían evitarse empleando otras
variantes del método ELISA. Pero quizá fuera más efectiva la obtención de anticuerpos
monoclonales que, al ser más específicos que los policlonales empleados en este trabajo,
evitarían reacciones indeseables con otros compuestos.
250

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 264/293
CONCLUSiONES
CAPITULO VI
CONCLUSIONES
251

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 265/293
CONCLUSIONES
1).- Las 1 8 0 bacterias lácticas seleccionadas durante la maduración de u n lote d e
embutidos crudos presentaron actividad inhibidora directa, variable y cuantificable, frente a
diversos microorganismos indicadores. De ellas, 20 manifestaron actividad antimicrobíana
exocelular; dichas cepas s e aislaron e n todas las etapas del proceso d e maduración, desde la
masa cárnica inicial hasta el producto final listo para s u consumo.
2).- Las 6 cepas con mayor actividad inhibidora s e identificaron como Lacrobacillus
sake. Todas s e desarrollaron a 4 y 1 5 oc, pero sólo las cepas 1 4 8 y IT 7 lo hicieron a 45 < ‘C . E l
espectro antimicrobiano d e las 4 cepas con mayor actividad inhibidora sugiere que las
sustancias inhibidoras son diferentes, ya que la sensibilidad d e los distintos microorganismos
indicadores e s variable dependiendo d e la cepa d e L. sake productora.
3).- L. sakc 148 e s capaz d e desarrollarse y producir actividad antimicrobiana
cuantificable a 4, 8 , 1 6 , 2 5 y 3 2 “C. La produción de es ta sustancia depende d e la composiciónde l medio de cultivo, s e detecta al poco t iempo d e iniciarse el crecimiento microbiano y alcanza
sus niveles óptimos a las 1 6 horas d e incubación a 3 2 0C .
4).- Cuando la sustancia antimicrobiana exocelular d e L. sake 148, s e purifica por
liofilización y cromatografía d e filtración e n geles, aumenta muy poco s u actividad inhibidora
específica (7 veces), siendo muy pequeña la recuperación d e la actividad inhibidora original
(8%).
5Y - La sustancia antimicrobiana parcialmente purificada de L saLe 148 t iene
naturaleza proteica, e s estable a valores d e pH comprendidos entre 2,6 y 1 2 , resistente al calor
y bacteriostática frente a los microorganismos sensibles, características que comparte con otras
bacteriocinas de las bacterias lácticas. La fracción activa d e la sustancia antimicrobianaparcialmente purificada por cromatografía d e filtración e n geles, reveló que t iene u n peso
252

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 266/293
CONCLUSIONES
molecular de 4.640 daltons. La sustancia parcialmente purificada s e compone de 1 3
aminoácidos; además, y por cromatografía HPLC, s e aprecia un pico que pertenece a un
aminoácido distinto d e los utilizados corno patrones. E s ta compuesto desconocido eluye junto
a la glicina, lo mismo que el aminoácido lanuonina.
6).- Consideramos que la sustancia antimicrobiana d e L. sake 1 4 8 e s bacteriostática,
ya que detiene el crecimiento d e los microorganismos sensibles, pero n o s u viabilidad. Su
concentración inhibidora mínima varía entre 1 y 1 8 mg/mI, dependiendo de losmicroorganismos ensayados.
7).- La inoculación d e la sustancia antimicrobiana a los animales d e experimentación
(conejos) originó una respuesta inmunógena débil. El desarrollo y puesta a punto d e u n ELISA
indirecto no permitió la detección d e la bacteriocina parcialmente purificada e n u n medio d e
cultivo semidefinido (medio d e MM-triptosa).
8).- Las características fisico-quimicas d e la bacteriocina producida por L i. saLe 1 4 8 y
su acción antimicrobiana frente a bacterias patógenas Gram-positivas d e gran interés e n la
industria alimentaria, hacen concebir esperanzas sobre la posible utilidad de las bacterias
lácticas o d e sus bacteriocinas, como factores d e seguridad tanto e n la carne y productos
cárnicos como e n otros sustratos alimentarios diversos.
253

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 267/293
TRAUIiIO FUTURO
CAPITULO VII
TRABAJO FUTURO
254

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 268/293
TRABAJO FUTURO
Las cepas de L. saLe con actividad antimicrobiana exocelular aisladas y caracterizadas
parcialmente, pueden ser d e gran utilidad e n la inhibición de l desarrollo d e microorganismos
alterantes y patógenos d e la carne y productos cárnicos, a s í como d e otros alimentos. S in
embargo, las condiciones d e u n sustrato alimentario sólido como la carne, son distintos d e las
que s e dan en los medios de cultivo líquidos. Por ello, la síntesis y la actividad de la s
sustancias antimicrobianas d e las bacterias lácticas descritas, deben examinarse e n sistemas
alimentarios reales, máxime ten iendo e n cuenta la incapacidad d e L. saLe 1 4 8 de producir la
actividad antimicrobiana cuando s e desarrolla e n diversos medios d e cultivo. En consecuencia,
parece conveniente acometer estudios tendentes a aclarar los mecanismos responsables de la
estimulación o represión d e la síntesis o actividad d e e s t a s sustancias antimicrobianas.
El empleo de cepas bacteriocinogénicas de L. sake, como cultivos iniciadores en la
elaboración de productos cárnicos, presupone tanto la prevención del desarrollo de
microorganismos indeseables, como un a mayor homogenidad de l proceso fermentativo. Porello, el desarrollo y puesta a punto d e sistemas novedosos d e transformación genética de las
bacterias lácticas, como la electroporación, permitiría introducirles genes que codifiquen
propiedades de importancia tecnológica. Gracias a la tecnología de l ADN recombinante, los
determinantes genéticos d e la p roducción d e bacteriocinas y d e la resistencia a las mismas
podrían utilizarse como marcadores genéticos o como genes d e interés para la producción d e
bacteriocinas como aditivos alimentarios. De aquí que constituya u n objetivo prioritario d e
nuestro trabajo futuro el estudiai-, co n la máxima amplitud posible, las características genéticas
d e los determinantes que codifican la produción d e bacteriocinas e n las cepas d e L i. sake d e
interes.
Otra 1 inca d e nvesti gac ión n tic sería importante desaíro ¡¿ ir ca el futuro e s la
determinación (le la secuencia aruinoacídica ( le la bacteriocina ( le L saLe 148, ya que elconocimiento de s u estructura proteica pemiitiría elucidar el papel que ésta desempeña e n s u
255

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 269/293
TRABAJO FUTURO
mecanismo de acción, favorecería el estudio d e lo s determinantes genéticos responsables de s u
síntesis y regulación y, por último, permitiría comparar las relaciones existentes entre
estructura-actividad e n la s distintas bacteriocinas producidas por la s bacterias lácticas.
De otra parte, el bajo poder inmunógeno mostrado por la bacteriocina de L i. sake 148
dificulta s u detección e n los alimentos por métodos inmunoenzimáticos (ELISA), este aspecto
e s fundamental, ya que uno de los requisitos que e s más importante cumplir para que s e
autorize la adición de e s t a bacteriocina a los alimentos e s , prec isamente, que s e pueda detectar
y cuantificar después d e incorporarse a los mismos. Por este motivo, s e pretende mejorar la
capacidad antigénica de e s t a sustancia empleando métodos d e inmunización alternativos, entre
los que s e incluiría s u conjugación a otras moléculas e , incluso, la obtención de anticuerpos
monoclonales específicos. Es necesario desarrollar y poner a punto otras variantes de la s
técnicas inmunoenzimáticas (ELISA) empleadas e n el presente trabajo para permitir la
detección y cuantificación de la sustancia antimicrobiana d e interés e n diversos sustratosalimentarios. Asimismo, los anticuerpos específicos podrían permitir la purificación a
homogeneidad de la sustancia antimicrobiana, por cromatografía d e afinidad, lo que permitida
s u purificación e n u n solo paso y evitada la utilización d e otros procedimientos de separación
proteica menos eficaces.
256

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 270/293
BIBLIOCRA E/A
CAPITULO VIII
BIBLIOGRAFíA
257

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 271/293
BIBLIOCRAFIA
ABDEL-BAR, N., HARRIS, N. D. y RILL, R . L. (1987)) . Purification an d properties of a n
antimicrobial substance produced by Lactobacillus bulgaricus. J. Food Sci.,2:
411-415.
AHN, C . y STILES, M. E . (1990a). Antibacterial activity of lactic acid bac teria isolated from
vacuum-packaged meats. J. AppI. Bacteriol., ~9s302-3 1 0 .
AHN, C . y STILES, M. E . (1990b). Plasmid-associated bacteriocin production by a strain of
Carnobacteriumpiscicola. Appl. Environ. Microbiol. 5fr 8: 2503-2510.
ALLGAIER, H., JUNG, G., WERNER, R. G., SCHNEIDER, U. y ZANHER, H. (1985).Elucidation of the structure of epidermin, a ribosomally synthesized tetracyclic
heterodetic polypeptide antibiotic. Angew. Chem. 24: 1 0 5 1-1053.
ANDERSSON, R. (1986). Inhibition of Staphylococcus aureus and spheroplasts of
Gram-negative bacteria by a n antagonistic compound produced by a strain of
Lacto bacillus plwztarum. mt. Y Food Microbiol., 3:149-160
ANDERSSON. R . E ., DAESCHEL, M. A. y HASSAN, H . M. (1988). Antibacterial activityof plantaricin SIK-83, a bacteriocin produced by Lacwbacillus plantarían.
Biochemie, 7 Q : 381-390.
ANONIMO (1986). mt. Journal System. Bacteriol. 36: 354-356.
AXELSSON, L. T., CHUNG, T. C.. DOBROGOSZ. W . J. y LINDGREN, 5. (1989).
Production of a broad spectrum antimicrobial substance by Lactobacillus renten.
Microb. Ecol. Hcalth Dis., 2: 1 3 1-136.
BAIRD-PARKER, A. C. (1980). Organic acids. En : “Mícrobíal Ecology of Foods’t,
vol. lpp.126-1135. I.C.M.S.F. (ed.). Academic, New York.
BARAN. W.L. y STEVENSON, K.E. (1975). Survival of selected pathogens during
processing of a fermerned turkey susage. Y Food Sci. 40: 6 18-620.
BAREFOOT. S. E. y KLAENHAMMER, T. R. (1983). Detection an d activity of lactacin B, abacteriocin produced by Lactobacillus acidophilus. Appl. Environ. Microbiol.,
258

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 272/293
BIBLIOCRA filA
45: 1808-1815.
BAREFOOT, 5. F. y KLAENHAMMER, T. R. (1984). Purification and characterization of tIte Lactobacillus acidophilus bacteriocin, ¡actacin B. Ant¡mocrob. Agents
Chemother.,~, 3: 328-334.
BATISH, y. K., LAL, R. y GROVER, 5. (1990). Studies on environmental andantinutritional factors on production of antifungal substance by Lactobacillus
acidophilus. R. Food Microbiol., 1:199-206.
BEAVER, R. W., WILSON, D. M., JONES, H. M. y HAYDON, K. D. (1986). Aminoacidanalysis in feeds an d feedstuffs using precolumn phenyl- isothiocyanate derivatization
and liquid chromatography - preliminary study. Y Assoc. 0ff. Anal. Chem., JjQ ,
3: 425-428.
BERO, R. W., SANDINE, W. E. y ANDERSON, A. W . (1981). Identification of a growth
stimulant for Lactobacillus sanfrancisco. Appl. Environ. MicrobioL. 42: 786-788.
BEUCHAT, L. R. Y GOLDEN, D.A. (1989). Antimicrobial ocurring naturally in foods.Food Teclinol., fl:134-142.
EHUNIA, A. K., JOHNSON, M. C . y RAY. B. (1987). Direct detection of a n antimicrobial
peptide of Pediococcus acidilactici in sodium dodecyl sulfate-polyacrylamida gel
electrophoresis. J. Industrial Microbiol., 2: 319-322.
BHUNIA, A. K., JOHNSON, M. C. y RAY. B. (1988). Purifícation, characterization an d
antimicrobial spectrum of a bacteriocin produced by Pediococcus acidilactici. J.Appl. Bacteriol., 65: 261-268.
I3HUNIA. A. fC., JOHNSON, M. C ., RAY. B. y BELDEN, E. L. (1990). Antigenic
property of pediocin AcH produced by Pediococcus acidilacticí H . J. Appl.
BacterioL, 69: 211-215.
BHUNIA, A. K., JOHNSON. M. C ., RAY, B. y KALCHAYANAND, N . (1991). Mode of
action of pediocin AcH form Pediococcus acidilachci H o n sensitive bacterial strains.J. App¡. Bacteriol., 70: 25-33.
259

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 273/293
BL’3LIOGRAFJA
BISWAS, 5. R., RAY, P., JOHNSON, M. C. y RAY, B. (1991). Influence of growth
conditions on the production of a bacteriocin, pediocin AcH. by Pediococcus
acídilactici H. AppI. Environ. Microbiol., 57 , 4:1265-1267.
EJÓRCK, L. (1985). The lactoperoxidase systern. En: “Natural Antimicrobial
Systems”, Pp. 18-30. IDF., 41 Square Vergote. 1040, Brussels.
BONNARD, C., PAPERMASTER, D. 5. y KRAEHENBUHL, J. PL (1984). The
streptavidin-biotin bridge technique: Application in ligth and electron microscope
immunocytochemistry. En: “Immunolabelling for Electron Microscopv”, P P .
95-109. J. M. Polak e 1 . M. Vamdell (eds.). Elsevier Science Publishers. London.
BOTAZZI, V. (1988). An introduction te rod-shaped lactic acid bacteria. Biochemie, 70
303-315.
BOTAZZI, V., VESCOVO, M. y DELLAGLIO, E. (1973). Microbiology of Grana cheese.
IX. Characteristics and distribution of Lactobacillus helveticus biotypes in natural
whey cheese starter. Sci. Tech. Latt.-casear_ 24: 23-39.
BREED, R. 5., MURRAY, E. G. D. y SMITH, N. R. (1975). Bergey’s Manual of
Determinative Bacteriology, 7 th e d . The Willians an d Wilkins Co., Baltimore.
BROCK, T. O. y DAVIE, J. M. (1963). Probable identity of a group D hemolysin with a
bacteriocin. J. Bacteriol., ~: 708-7 1 2 .
BUCHANAN, R. L. y GIBSONS, N. E. (1974). Bergey’s Manual of Determinative
Bacteriology, S th ed. The Williams an d Wilkins Co., Baltimore.
BUYZE. O., van DEN HAMER, 3. A. y DE HAAN, P. G. (1957). Correlation between
hexose-monophosphate shunt, glycolytic system an d fermentation type in lactobacilli.
Antonie van Lecuwenhoek, 23 : 34$350.
CALVET. 3 . J. (1968). The effects of newer fornis of packaging e n the microbiolo~y andstorage life of rneats, poultry a n d fish. Prog. md. Microbiol., 7: 77-85
CARLSSON, 3., IWAMI, Y. y YAMADA, T. (1983). Hydrogen peroxide excretion ny oral
260

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 274/293
BiBLIOGRA filA
streptococci an d effects o n lactoperoxidase-thiocyanate-hydrogen peroxide. Infect.
Immun.. 4 Q : 70-80.
CARMINATI, D. GIRAFFA, G. y BOSSI, lvi. (1989). Bacteriocin-Iike inhibitors of
Streptococcus lacús against Lisíeria rnonocyto genes. .1 . Food Protcct., 52. 9:
614-617.
CAVETT, J. J. (1963). A diagnostic key for identifying the Iactic acid bacteria of vacuum
packed bacon. 3. Appl. Bacteriol., ~: 453-470.
CFR. (1988). Nisin preparation; Affirmation of GRAS status as a direct human foodingredient. Code of Federal Regulations, TitIe 21, Section 184. U. 5.
Govt. Print. Office, Washington, D. C .
CHASSY, B. M. y THOMPSON, J. (1983). Regulation an d characterization of the
galactose-phosphoenolpyruvate-dependent phosphotransferase system in Lnciobacillus
cascí. J. Bacteriol. 154: 1204-1214.
CHAMPOMIER, M. C?, MONTEL. M. C. GRIMONT, F. y GRIMONT, 1 ’ . A. D. (1987).Genomic identification of meat lactobacilli a s Lactobacillus saLe . Ann. Inst.Pasteur/Microbiol. 13fl: 7 5 1-758.
CHICHESTER, D. F. y TANNER, .F . W. (1972). Antimícrobial food additives. En:
“Handbook of Food aditives” 2nd. al., P P . 115-150. T. E . Furia (cd.). Chem.Rubber Pub. Co. , Cleveland, Ohio.
CLARK, D. 5. y TAKACS, J . (1980). Gases a s preservatives. En: ‘Microbial Ecology
of Foods”, P p . 170-180. Silliker, J. H. (ed3. Academic Press, London.
COLLINS, E . B. y ARAMAKI, K. (1980). Production of hydrogen peroxide by
Lacrobacillus acidophilus. 3. Dairy ScL, 63 : 353-357.
COLLINS, M. D. y JONES, D. (1981). The distribution of isoprenoid quinone structural
types in bacteria an d their taxonomic ímplications. Microbiol. Rey. 45: 316-354.
261

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 275/293
8JULIO GRA filA
COLLINS, M. D., FARROW. J. A. E., PHILLIPS, 1 3 . A., FERUSU, 5. y JONES, D.
(1987). Classification of Loctobacillus divergens, lactobacillus piscicola, an d some
catalase-negative, asporogenous, rod-shaped bacteria from poultry in a new genus,Carnobacreriuni. m t. j. Systcm. Bacteriol., 3 4 : 310-316.
CONNER, D. E.. BRACKEIT, R. E. y BEUCHAT, L. R. (1986). Effect of temperature,
sodium chioride and pH on growth of Listeria tnonocytogenes in cabbage juice.
Appl. Environ. Microbiol. 52: 59-63.
CONE, D. K. (1982). A Lactobacillus s p . from diseased female rainbow trout, Salmo
gairdneri Richardson, in Newfounland, Canada. 3. F¡sb. Dis. ~: 479-485.
DAESCHEL, M. A. (1989). Antimicrobial substances from lactic acid bacteria for u s e a s food
preservatives. Food Technol_ 43 : 164-167.
DAESCHEL, M. A. y KLAENHAMMER, T. R . (1985). Association of a 13.6-megadalton
plasmid in Pediococcus pentosaceus with bacteriocin activity. AppI. Environ.
Microbiol., 50, 6:1538-1541.
DAESCHEL, M. A.. McKENNEY, M. C. y McDONALD, L. C. (1990). Bacteriocidal
activity of Lactobacillus planíarurn C-1 1. Food Microbiol., j: 9 1-98.
DAHIYA, R. 5. y SPECK, M. L. (1968). Hydrogen peroxide formation by lactobacilli and its
effects on Staphylococcus aureus. 3. Dairy Sci . , 51: 1568-1572.
DAINTY, R. H. (1986). Chemical changes associated with microbial growth on meat stored at
chilí temperatures. 32nd. Europ. Meet. Meat Res. Workers. Ghent, Belgium,1:179-186.
DAVEY, G. P . (1984). Pasmid associated with diplococcin production in Srreptococcus
cremoris. AppI. Environ. Microbiol. 45: 205-211.
DAVEY, G. P . y RICHARDSON, B . C . (1981). Purification and some properties of
diplococcin in Streptococcus crenioris 346. AppI. Environ. MicrobioL, 41
84-89.
262

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 276/293
BIBLIOGRA FíA
DAVIOSON, P. M., POST, L. 5., ERANEN, A. L. y McCLJRDY, A. R. (1983). Naturally
occurring an d miscellaneous food antimicroblais. En: “Antimicrobials in Foods’,
Pp . 3 7 1-390. A. 1 . Branen an d P . M. Davidson (eds.). Marcel Dekker, Inc., NewYork.
DAVIS, J. G. (1975). The microbiology of yoghurt. En: “Lactic Acid Bacteria in
Beverages and Foods”. pp. 245-263. J. 6. Carr, C . V. Cutting and G. C .
Whiting (eds.). Academic P r e s s , London.
DAWSON, R . M. C ., ELLIO’IT, D. C ., ELLIOfl. W. H. y JONES, K. M. (1969). E n
“Data br biochemical research’ 2nd cd., P P . 484-485. Oxford UniversityPress, Ely House, London.
DEIBEL, R. H. y NIVEN, C. F. Jr. (1958). Micmbiology of meat curing. 1 . The occurrence
an d significance of a motile microorganism of the genus Lacrobacillus in ham curing
brines. Appl. Microbiol. fr 323-327.
DE MAN, J. C ., ROGOSA. M. y SHARPE, M. E. (1960). A medium for the cultivation of
lactobacilli. 3 . Appl. Bact., 23: 130-135.
DENT, V. E . y WILLIAMS, R . A. D. (1982). Lactobacillus animalis s p . nov., a new species
of Lacrobacillus from the alimentary canal of animais. Zentralbí. Bakteriol.
Mikrobiol. Hyg. 1 Abt. Orig. C 3: 377-386.
DE KLERK, H . C . (1967). Bacteriocinogeny in Lactobadillus fermenti. Nature. London
214: 609.
DE KLERK, H . C . y GOETZEE. (1961). Antibiosis among lactobacilli. Nature. London
192: 340-341.
DE KLERK, H. C. y SMITH, . 1 . A. (1967). Properries of a Lacto/mci//ns ferrnenti
bacteriocin. 3. Gen. Microbiol. 48: 309-3 1 6 .
DE VRIES, W ., KAPTEIJN. W. M. C ., van DER BEEK, E. G. y STOUTHAMER. A. H.
(1970) Molar growth yields a u d fermentation balances of Lacrobacillus casel 1 3 inbatch cultures an d in continuous cultures. 3. Gen. Microbiol. 63 : 333-345.
263

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 277/293
BII3LIOCRAfilA
DOBRZANSKI, W. T., BARDOWSKI, J., KOZAK, W. y ZAJDEL, J. (1982).
Lactostrepcins: bacteriocins of lactic Síreprococci. En “Microbiology”, PP .
225-229. D. Schlessinger (ed.). A. 5. M., Washington, D. C.
DZIEZAK, J. D. (1986). Preservatives: Antimicrobial agents. Food Technol., 4fi, 9:
104- 125.
EGAN, A. F. (1983). Lactic acid bacteria of meat an d meat products. Antonie van
Leeuwenhoek 49 327-336.
EKLUND, T. (1984). The effect of carbon dioxide o n bacteria growth an d on uptakeprocesses iii the bacterial membrane vesicles. lid. 3. Food Microbiol_ 1
179-185.
FAO/IAEAIWHO Expert Committee. (1977). Wholesomeness of irradiatiated foods. Food an d
Agric. Org.¡Int. Atomic Energy Agency/World Health Org., Tech. Rep. Ser. 604.
World Health Org., Geneva Switzerland.
FELTON, E. A., EVANS, J. E. y NIVEN, C. E R. Jr. (1953). Production of catatase by thepediococci. 3 . Bact. ~5:48 1-482.
FERREIRA, C. L. y GILLILAND, 5. E. (1988). Bacteriocin involved in premature death of
Lactobacillus acidophilus NCFM during growth at pH 6. 3. Dairy Sci., 71:
306-315 .
FLEMING, H. P., ETCHELLS, J. L. y COSTILOW, R. N . (1975). Microbial inhibition by
a n isolate of Pediococcus from cucumber brines. Appl. Microbiol., 30: 1040-1042.
FOOSTER, E . M., NELSON, E E ., SPECK, M. L., DOLTSCH. R . N . y OLSON, J.-C.
(1957). En: ‘Da¡ry Microbiology’, P p . 106-107. Prentice-Hall, Englewood Cliffs,
NY.
FREDERICQ, P . (1948). Actions antibiot iques réciproques chez les Enterobacteriaceae. Rey.
BeIge. Pathol. Med. Expt. 1 9 (suppi. 4)1-107.
264

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 278/293
BU3LIOGRA filA
FRIEND, B. A., FIEDLIER. J. M. y SHAHANI, K. M. (1983). Influence of culture
selection on the flavor, antimicrobial activity, ¡3-galactosidase an d B-vitamins of
yogurt. Milchwissenschaft 38: 133-136.
GAGLIANO, y. J. y HINSDILL, R. D. (1970). Characterization of a Staphylococcus aureusbacteriocin. 3. Bacteriol., 104: 117-125.
GALVEZ, A., MAQUEDA, M., VALDIVIA, E., QUESADA, A. Y MONTOYA, E. (1986).
Characterization an d partial purification of a broad spectrum antibiotic AS-48 produced
by Streptococcus faecalis. Can. .1 . Food Microbio!., 32: 765-77 1.
GARVIE, E. 1 . (1986a). Genus Leuconostoc. En:
Bacter¡ology”, vol. 2., pp . 107 1-1075. P . H .
an d J . G. Holt (eds.). Williams & Wilkins Co.,
GARVIE, E . 1 . (1986b). Genus Pediococcus. En:
Bacteriology”, vol. 2., P P . 1075-1079. P . H .
an d J . G. Holt (eds.). Williams & Wilkins Co.,
“Bergey’s
A. Sneath,
Baltimore.
“Bergeys
A. Sneath,
Baltimore.
Manual of Systematic
N. 5. Mair, M. E. Sharpe
Manual of Systemat¡c
N. 5. Mair, M. E. Sharpe
GARVIE, E. 1 ., FARROW, J. A. E. y PHILLIPS, 8. A. (1981). A taxonomic study of some
strains of streptococci which grow at 1 0 0C but not at 45 0C including Streptococcus
lactis and Streptococcus crernoris. Zentralbí. Bakteriol. Mikrobiol. Hyg. 1
Abt. Orig. C 2:151-165
GEIS, A,, SINGH, J. y TEUBER, M. (1983) Potential of lactic Streptococci
bacteriocin. Appl. Environ. Microbio!. 45 k 205-211.
to produce
GIBBS, P . A. (1987). Novel u s e s for lactie acid fermentation. 3. AppI. Bacteriol. (Symp.
Suplem.): 515-585.
GONZALEZ, C . F. y KUNKA, B . 5. (1987). P]asmid-associated bacteriocin production an d
sucrose fermentation in Pediococcus aci/ilactici. Appl. Environ. Microbiol., 53
10 : 2534-2538.
GÓTZ, E., ELSTENER. E. E., SEDEWITZ, 1 3 . y LENGEELDER, E. (1980). Oxygenutilization by Lactobacillus planraruni . 1 1 . Superoxide an d superoxide dismutation.
265

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 279/293
BIBLIOGRA filA
Arcb. Microbiol. 125: 215-220.
GRATIA, A. (1946). Techniques sélect ives pour la recherche systématique de s germesantibiotiques. C . R . Sciences Soc. Biol. Paris 140: 1053-1055.
HAMMES, W. P., BANTLEON, A. y MIN, 5. (1990). Lactic acid bacteria in meat
ferrnentaúon. FEMS Microbiol. Rey., B Z : 165-174.
HARDIE, J. M . (1986). Genus Síreptococcus. En: “Bergey’s Manual of Systemat¡cBacteriology, vol. 2 . , Pp . 1043-1071. P . 1 - 1 . A. Sneath, N. 5. Mair, M. E . Sharpe
and J . G. Holt (eds.). Williams & Wilkins Co., Baltimore.
HARDING, C . D . y SHAW, 1 3 . G. (1990). Antimicrobial activity of Leuconostoc gelidum
against closely related species an d Listeria nionocytogenes. 3. Appl. BacterioL,
~i9:648-654.
HARRIS, L. J., DAESCREL, M. E ., STILES, M. A y KLAENHAMMER, T. R. (1989).
Antimicrobial activity of lactie acid bacteria against Listenc monocytogenes. . 1 . Food
Protee_ 52, 6: 384-387 .
HASTINGS, J. W. y HOLZAPFEL, W . H . (1987). Numerical taxonomy of lactobacilli
surviving radurization of meat. m t. 3. Food Microbiol,, 4: 33-49.
HASTINOS, J. W. y STILES, M. E . (1991). Antibiosis of Leuconostocgelidurn isolated
from meat. 3. Appl. Bacteriol,, 70: 127-134.
HASTINGS, J . W., HOLZAPFEL, W . H . y NIEMAND. J . G. (1986). Radiation resistanceof Lactobacillí isolated from radurized meat relative to growth an d environment.
Appl. Environ. Microbiol., 52 4 ~ 898-901.
HAYS, G. L. y REISTER, D. W . (1952). The control of “off-odour” spoilage in frozen
concentrated orange juice. Fd. Technol. Champaign., 6: 386-389.
HENSEL, R . U., MAYR, U.. STETTER. K. O. y KANDLER, 0. (1977). Comparative
studies of lactic acid dehydrogenases in lactie acid bacteria. 1 . Purification an d kinetics
of the allosteric L-lactic acid dehydrogenase from L. ccxci s s p . ccxci an d
266

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 280/293
BIBLIOCRA FíA
Lcuobccillus curvatus. Arch. Microbiol. 112: 8 1-93.
HITCHENER, 1 3 . J., EGAN, A. F. y ROGERS, P. J. (1982). Characteristies of lactie acidbacteria isolated from vacuum-packaged beef. 3. Appl. Bacteriol.. 52: 3 1-37.
HOLMBERG, K. y HALLANDER, U. 0. (1973). Production of bactericidal concentrations
of hydrogen peroxide by Síreptococcus sanguis. Arch. Oral Biol. fl: 423-434.
HOLY, A. V. y HOLZAPFEL, W. H . (1988). The influence of extrinsie factors on themicrobiological spoilage pattern of ground beef. m t. J. Food Microbiol., ~
269-280.
HOLLEY. R. A., JUL, P. A., WITI’MANN, M. y KWAN, P. (1988). Survival of 5. aureus
and 5. typhinzuniurn in raw r ipened dr>’ sausages formulated with mechanicalí>’
separated chiken meat. Fleischwirstch., 5 ~ , 2:194-201.
HOLZAPFEL, W. U. y GERBER, E. 5. (1983). Lactobacillus divergens s p . nov., a new
heterofermentative Lacrobacil¡us especies producing L(+)-lactate. Syst. Appl.
Microbiol. 4: 522-534.
HOLZAPFEL, W . U. y GERBER, E . 5. (1986). Predominance of Lactobecillus curx’atus
and Lactobecillus sa/ce in the spoilage association of vacuum packaged meat products.Abstr. 32nd Eur. Meet. Meat Research, Pp. 26 Ghent.
HUEPPE, F. (1884). Milchsáurebazillus syn. Bactenium acidi lactici. Zopf. Mitt. Kaiserl.
Gesundheitsamt., 2: 337-340.
HURST, A. (1981). Nisin. Adv. AppI. Microbio!., 27 : 85-123.
ICMSF. (1980). En: “Microbial Eco¡ogy of Foods. Vol. 1. Factors Affecting Lite
and Death of Microorgnanisms.” Intí. Comission on Microbiological
Specifications for Eoods. Academic P r e s s , New York.
LNGRAM, M. (1975). The lactic acid bacteria. A broad view. En “Lactie Acid Bacteria in
Beverages ami Foods’, P p . 1-13. J . O. Carr , C . y. Cutting an d O. C . Whiting(eds.). Academic Press, London.
267

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 281/293
81/H L lOGRAE/A
JAY, 1 . M. (1982). Antimicrobial propierties of diacetyl. Appl. Environ. Microbio!., 44525-531
JAY, J. M. (1986). En: “Modern food Microb¡ology” (3rd cd.), Pp . 275. Van
Nostrand Reinhoid, New York.
JOERGER, M. C . y KLAENHAMMER, T. R. (1986). Characterization an d purification of
helveticin J an d evidence for a chromosomally determined bacteriocin produced by
Lacrobacillus helveticu.s 481.3. Bacteriol., 167: 439-446.
JOHNSON, J. L., DOYLE, M. P . AN D CASSENS, R . G. (1988). Survival of Listeniarnonocytogenes in ground beef. ini 3. Food Microbiol., á : 243-247.
JUNTILLA, J. , HIRN, J . , HILL, P . y NURMI, E. (1989). Effect of different leveis of nitrite
an d nitrate on the survival of Listenia rnonocytogenes during the manufacture of
fermented sausage. 3. Food Protee. 52:158-161.
KANDLER, 0. (1983). Carbohydrate metabolism in lactic acid bacteria. Antonie van
Leeuwenhoek. 3. Microbiol. Serol. 42: 209-224.
KANDLER, 0. (1984). Current taxonomy of lactobacilli. md. Microbio!. 25: 109-123.
KANDLER, O. y KUNATH, P . (1983). En: Validation of the publication of new names an d
new combinaúons previonsí>’ effectively published outside the IJSB. List. n2 1 1 , m t.3. Syst. Bacteriol., 33 : 672-674.
KANDLER, O. y WEISS, N. (1986). Genus Lactobacil/us. En: “Bcrgey’s Manual of Systemat¡c Bacteriology”, vol. 2 . , pp. 1208-1234. 1 ? . H . A. Sneath, N. 5. Mair,
M. E . Sharpe an d J . G. Holt (eds.). Williams & Wilkins Co. , Baltimore.
KATAGIRI, H.. KITAHARA, K. y FUKAMI, K. 1934. The caracteristies of the Iactic acid
bacteria isolated from moto, yeast maslies of saké manufacture. IV. Classif¡cation of
the lactic acid bacteria. Bulletin of the Agricultural Society (Japan). 10
156- 157.
268

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 282/293
BIBLIOGRAFIA
KELLNER, R., JUNG, G., HÓRNER, T., ZAHNER, H., SCHNELL, N.. ENTIAN,
K.-D. y GÓTZ, F. (1988). Gallidermin: a new lanthionine-containing polypeptide
antibiotic. Eur. 3. Biochem. 117:53-59.
KELLNER, R ., JUNG, G., JOSTEN, M., KALE’TTA, C., ENTIAN, K.-D. y SAHL,
H.-G. (1989). PepS: stiructure elucidation of a large lantibiotic. Angw. Chem. m t.Ed. EngI. 2a:616-619.
KEKESSY, D. A. y PIGUET, J. D. (1970). New method for detecting bacteriocin
production. Appl. Microbio!., 20: 282-283.
KING, A. D. Jr. y NAGEL, C. W . (1975). Influence of carbon dioxide upon the metabolism
of Pseudomonas aeruginosa - 3. Food Sc¡., 40: 362-366.
KITCHELL, A. G. y SHAW, B. G. (1975). Lactic acid bacteria in fresh an d cured meat. En:
“Lactic Acid Bacteria in Beverages and Food”, Pp. 209-220. J. G. Carr, C.
V. Cutting and G. C . Whiting (eds.). Academic Press, London.
KLAENHAMMER, T. R . (1988). I3acter iocins of lactic acid bacteria. Biochemie, 7 .Q :337-349.
KNOX, K. W. y HALL, E. A. (1964). The relationship between the capsular an d cel wal
polysaccharides of strains of Lccrobacillus casel var. rhamnosus. 3. Gen.
Microbiol. 3 2 1 : 433-438.
KNOX, K. W. y WICKEN, A. J . (1973). Inmunological properties of teichoic acids.
Bacteriol. Rey., 37: 215-257.
KODAMA, R. (1952). Studies o n lactie acid bacteria. Lactolin, a new antibiotic substance
produced by lactic acid bacteria. . 1 . Antibiot_ 5 ~ 72
KORKEALA, H. y MAKELA, P. (1989). Characterization of lactic acid bacteria isolated from
vacuum-packed cooked ring sausages. lot. 3. Food Microbiol., 9: 33-43.
KOZAK, W., BARDOWSKI, J . y DOBRZANSKI. W. T. (1978). Lactostrepcins, acid
bacteriocins produced by lactic streptococci. 3. Dairv Res., 45: 247-254.
269

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 283/293
BIBLIOGRAFÍA
KUTTNER, A. G. (1966). Production of bacteriocins by group A streptococci with specialreference to the nephritogenic types. 3. Exp . Mcd. 124: 279-291.
LABAN, P., FAURE, C ., RAMET, F. Y LARPENT, J . P . (1978). Lactobacilli isolated from
french saucisson (taxonomic study). ZbI. Bakt., 1:105-111.
LAEMLI, U. K. (1970). Cleavage of structural proteins during the assembly of the head of
bacteriophage T4 . Nature, 2flj: 680-68 5.
LAW, 1 3 . A. y KOLSTAD, J. (1983). Proteolytic systems in lactic acid bacteria. Antonie
van Leeuwenboek, 4% 225-245.
LEOWARD, D. A.. NICOL, D. J. y SHAW, M. K. (1971). Microbiological an d colour
changes during aging of beef. Food Techno!. Austral., 7 5 : 30-35.
LINDGREN, 5. E . y DOBROGOSZ, W . J. (1990). Antagonistic activies of Iactic acid
bacteria in food an d feed fermentat ions. F.E.M.S. Microbiol. Rey., B Z : 149-164.
LIU, W. y HANSEN, J . N. (1990). Sorne chemical an d physical properties of nisin, asmall-protein antibiotic produced by Lactococcus lactis. Appl. Environ.
Microbio!. 5 6 8 ~ 2551-2558.
LONDON, J . (1976). The ecology an d taxonomic status of the lactobacilli. Ann. Rey.
Microbiol., 3Q: 279-301.
LUCK. E . (1976). Sorbic acid a s food preservative. IntI. Flavors Food Add., 7, 3 :
122-123.
LUCKE, F. K. (1986). Microbiological processes e n the manufacture of d r > ’ sausages and
raw ham. F!eischwírstch., 66 : 1505-1509.
LUCKE, F. K. y HELCHELMANN, H . (1987). Starter cultures for dry sausages and raw
ham. Fleiscbwírtsch_ 67 307-3 1 4 .
LYON. W. J . y GLATZ, B. A. (1991). Pardal purification an d characterizarion of a
bacteriocin produced by Propionobacíeriwn ¡hoe,zii. App!. Environ. Microbio!.,
270

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 284/293
BIBLIOGRA Fí A
51: 701-706.
MARKWELL, M. A. K., HAAS, 5. M., BIEBER, L. L . y TOLBERT, N. E. (1978). Amodification of the Lowry prodedure to simplify protein determination in membrane
and lipoprotein samples. Anal. B¡ochem., B Y : 206-210.
MERRIL, C. R., GOLDMAN, D., SEDMAN, 5. A. Y EBERTH, M.
Ultrasensitive stain for proteins in poíyacrilamide gels shows regionalcerebrospinal Huid proteins. Sc¡ence,2.li: 1437-1438.
H . (1981).
variation in
McGROARTY, J. A. y REID, G. (1988). Detection of a lactobacillus substance that inhibits Eschenichia coli. Can. 3. Microbio!.. 3 .4 : 974-978.
MOL, J. H. H., HIETBRING, J. A. E., MOLLEN, H . W . M. y
(1971). Observations on the microflora of vacuum-packed
products. 3. AppI. Bacteriol., 34: 377-379.
MOLIN, G. y TERNSTROM, A. (1982). Numerical taxonomy
pseudomonads. J. Gen. Microbio!., : 1249-1264.
VAN TINTERIN. J.
sliced cooked meat
of psychophilic
MONTEL, M. C., TALON, R . y CHAMPOMIER, M. C . (1989). Identifícation of some lactic
acid bacteria from meat. Proc. 3Mb mt. Cong. of Meat Sc¡ence and
Technology, vol. 1 . Pp . 299-301. Conpenhagen.
MOON, N. J . (1983). Inhibition of the growth of acid tolerant yeasts by acetate, lactate an d
propionate aud their synergistic mixtures. 3. App!. Bacteriol., 55 : 453-460.
MORISHITA, T., FUDADA, T., SHIROTA, M. y YURA, T. (1974). Genet ic basis.of
nutritional requeriments in Lactobacillus ccxci. 3. Bacteriol. 120: 1078-1084.
M0RTVEDT, C . 1 . y NES, 1 . E . (1990). Plasmid-associated bacteriocin production Lactobecillus sc/ce strain. 3. Gen. Microbiol., 136: 1601-1607.
by a
MORTVEDT, C . 1 ., NISSEN-MEYER, J., SLETTER, K. y NES, 1 . E. (1991). Purification
and aminoacid sequence of lactocin S , a bacteriocin produced by Lactobacillus sc/ceL45. AppI. Environ. Microbio! _ 57~ 1829-1834.
271

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 285/293
BÍBLIOCRAFÍA
MURIANA, P. M. y KLAENHAMMER. T. R . (1987). Conjugal transfer of plasmid encoded
determinants for bacteriocin production a n inmunity in Loctobacillus acidophilus 88.
App!. Environ. Microbiol., 53: 553-560.
MURIANA, P . M. y KLAENHAMMER, T. R. (1991). Purification and partial
characterization of lactacin F, a bacteriocin produced by Lacrobacillus acidophilus.
AppI. Environ. Microbio!., 57: 114-121.
NIELSEN, J. W., DICKSON, J. 5. y CROUSE. J. D. (1990). Use of a bacteriocin produced
by Pediococcus acidilactici to inhibit Listenia monocyrogenes associated with fresh
meat. AppI. Environ. Microbio!., S: 2142-2145.
OBAFEMI, A. y DAVIES, R. (1986). The destruction of Salmonella typhinzurium in chicken
exudate by different freeze-thaw treatments. J. AppI. Bacteriol., ~Q: 3 8 1-384.
OKADA, 5., SUZUKI, Y. y KOZAKI, M. (1979). A new heterofermentative Lacrobecillus
specie with rneso-diaminopimelic acid in peptidoglycan, Lactobacillus vaccinoswrcus,
Kozaki and Okada s p . nov. 3 . Gen. Appl. Microbio!. 25: 215-221.
ORDOÑEZ, J. A. (1979). Random number sampling method for estimation of lactic acid
bacteria. 3. Appl. Bacteriol., 46: 351-353.
ORLA-JENSEN, 5. (1919). “TIte Lactic Acid Bacteria”. Andr.
Copenhagen.
OUCHTERLONY, 0. (1949). Antigen-antibody reacttons in
Microbio!. Scand.,26: 507.
PRICE, R . J . y LEE, J. 5. (1970). Inhibition of Pseudomonas species
producing lactobacilli. J. M¡Ik Food Technol., 33:13-18.
Fred. Host and Son.,
gels. Acta PatIto!.
by hydrogen peroxide
PUCCI. M. J., VEDAMUTHU, E. R., KUNKA, 8. 5. y VANDENBERGH, P . A. (1988).Inhibition of Lis/ala rnonocywgenes by using bacteriocin PA- 1 produced by
Pediococcus acidilccrici PAC 1.0. AppI. Environ. Microbio!., 54 : 2349-2353.
272

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 286/293
BIBLIOGRAFM
PULUSANI, 5. R., RAO, D. R. y SUNKI, O. R. (1979). Ancimicrobia> activity of ]achic
cultures: partial purification an d characterization of antimicrobial compound(s)
produced by Su-epwcoccus rhermophilus. .1 . Food Sel., 44, 2: 575-578.
RACCACH, M. (1981). Control of Síaphylococcus aureus in dry sausage by a newly
developed meat starter culture an d phenolic-type antioxidants. 3. Food Protect., 44:
665-668.
RACCACH, M., McGRATH, R . y DAFTARIAN, H . (1989). Antibiosis of some lactic acid
bacteria including Lactobacillus acidophilus toward Listenia monocytogenes. m t. 3.
Food Microbiol., 2: 25-32.
RAMMELSBERG. M. y RADLER, F. (1990). Antibacterial polypeptides of Lactobacillus
species. 3. AppI. Bacterio!., ~% 177-184.
RAY, 5. K.. JOHNSON, M. C. y RAY, 1 3 . (1989). Bacteriocin plasmids of Pediococcus
acidilcctici. J. md. Microbio!., 4:163-171.
REDDY, 5. G. y RANAGANATHAN, B. (1983). Nutritional factors affecting growth an dproduction of antimicrobíal substance by Síreptococcus tactis subsp. dicce¡ylactis S1-67-C. 3. Food Protec., 4fr 514-517.
REUTER, G. (1975). Classification problems. ecology a n d some biochemical activities of
lactobacilli of meat products. En: “Lactic Acid Bacteria in Beverages and
Food”. pp. 221-229. J. G. Can , C . V. Cutting a n d O. C . Whiting (eds.). Academic
Press, London.
REUTER, 0. (1981). Psychrotrophic lactobacilli in meat products. En: “Psychrotopbic
M¡croorganisms in Spoilage and Pathogenicity”, Pp . 253-258. T. A.
Roberts, O. Hobbs, i. H . B . Christian an d N. Skovgaard (eds.). Academic Press,
London.
RHEE, 8. K. y PACK, M. Y. (1980). Effect of environmental pl-i on chain length of
Lccwbccillus bulganicus .3. Bacteriol. 144: 11254 136.
2 7 3

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 287/293
BIBLIOGRA FÍA
RODRíGUEZ, J. M. (1991). Aislamiento y caracterización parcial d e bacterias lácticas deorigen cárnico antagonistas del desarrollo de otros microorganismos. Tesis
Doctoral. Universidad Complutense. Madrid.
RODRíGUEZ, J . M., SOBRINO, O. J. , FERNANDEZ, M. F ., HERNANDEZ, P . E . and
SANZ, 1 3 . (1989). Antimicrobial activity of lactic acid bacteria isolated from Span ish
dry fermented sausages. Proc. 35tb mt. Cong. of Meat Sc¡ence and
Technology, vol. 2., P p . 308-312, Copenhagen, Denmark.
ROGERS, L. A. (1928). The inhibiting effect of Streptococcus lacris on Lactobacillus
bu/ge nicus. 3. Bacterio!., Th: 321.
ROGOSA, M. (1961). Experimental conditions for nitrate reduction by certain stirains of the
genus Lacrobacillus . 3. Gen. Microbiol. 24:401-408
ROGOSA, M. (1970). Characters used in the classif¡cation of lactobacilli. m t. 3. Syst.
Bacteriol., 2Q: 5 19-534.
RUBIN, H. E. (1978). Toxicological model for a two-acid system. Appl. Envíron.Microbiol., 36: 623-624.
RUBíN, H. E., NERAD, T. y VAUGRAN, F. (1982). Lactic acid inhibition of Sa/monet/a
ryphimuniuni in yoghurt. J. Dairy Sci., ~5L:197-199.
RHULAND, G. J. y FIEDLER, E . (1987). Ocurrence an d biochemistry of lipoteichoic acidsin the genus Listeria. Syst . AppI. Microbiol_ 9 ~ 40-46.
SABINE, D. 1 3 . (1963). An antibiotic-like effect of Lactobacil/us acidophi/us. Nature, 199
811-816.
SANDINE, W. E. (1988>. New nomenclature of the non-rod-shaped lactic acid bacteria.
Biochem. 70: 5 19-522.
SCH[LLINGER, U. y LOCKE, F . K. (1987a). Starter cultures for drv sausages an d raw
l iam. Fleischwirtsch_ 6 7 307-314.
274

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 288/293
BíBLIOGRA Fí A
SCHILLINGER, U. y LUCKE, F. K. (1987b). Identification of lactobaci¡li from meat aud
meat products. Food. Microbio!.. 4: 199-208.
SCHILLINGER, U. y LUCKE, F. K. (1989). Antibacterial activity of Lacwbacillus sa/ce
isolated from meat. Appl. Environ. MicrobioL, 55 : 1901-1906
SCHILLINGER. U. y HOLZAPFEL, W. H. (1990). Antibacterial activity of carnobacteria.
Food Microbiol., 7: 305-310.
SCHLEIFER. K. H. y KANDLER, 0. (1972). Peptidoglycan types of bacterial cdl walls an d
their taxonomic implications. Bacteriol. Rey. R: 407-477.
SCHLEIFER. K. H., KRAUS, J., DVORAK, C ., KILPPER-BALZ, R., COLLINS, M. D.y FISCHER, W. (1985). Transfer of Sireptococus lactis an d related streptococci to
the genus Lactococcus gen. nov. System. Appl. Microbiol., 6: 183-195.
SEELINGER, H. P. R. y JONES, 0. (1986). Genus Listenia. En: “Bergey’s Manual of Systematic Bacteriology”, vol. 2 . , Pp . 1235-1245. P . H . A. Sneath, N. 5. Mair,
M. E . Sharpe and J . 0. Holt (eds.). Williams & Wilkins Co. , Baltimore.
SHAHANI, K. M., VAKIL, J. R . y KILARA, A. (1976). Natural antibiotic activity of
Lacrobacillus acidophilus am i bulgaí-icus. 1 . Cultural conditions for the production of
antibiosis. Cu!t. Dairy Prod. J., II: 14-17.
SHARPE, M. E . (1970). Ceil wall as i d celí membrane ant igens us ed in the classification of
lactobacilli. m t. J. Syst . Bacteriol. 20: 509-518
SHARPE, M. E. (1979). Identification of the lactic acid bacteria. En: “Identification
Metliods for Microbiologists”, pp. 233-259. F. A. Skinner an d D. W. Lovelock
(eds.). Academic Press, London.
SHARPE, M. E. (1981). The genus Lactobaci//us. E n “The Prokaryotcs. A Handbook
on Habitats, Isolation and Identification of Bacteria”, pp. 1653-1679.
Starr, Stolp, Irtiper, Balows an d Schelegel (eds.). Springer. New York.
275

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 289/293
BIBLIOGRAFÍA
SHARPE, M. E . y FRANKLIN, J. 0. (1962). Production of hydrogen sulphide bylactobacilli isolated from Cheddar cheese. Proc. XVI Intern. Dairy Congr.,
Copenhagen,~.
SHARPE, M. E . GARVIE, E . 1 . y TILBURY, R. H . (1972). Some sume forming
heterofermentative species of the genus Lactobacillus . App!. Microbio!. 23 :3 89-397.
SHARPE, M. E., LATHAM, M. J., BARVIES, E. 1 ., ZIRNGIBL, J. y KANDLER, O.
(1973). Two new species of Lactobacil/us isolated from the bovine rumen,
Lactobacillus ruminis sp. nov. and Lacrobacillus vítu/inus sp. nov. J. Gen.Microbiol. 77 : 37-49.
SHAW, 1 3 . 0. y HARDING, C. D. (1984). A numer ical taxonomic study of lactic acidbacteria from vacuum-packed beef, pork, lamb an d bacon. 3. AppI. Bacteriol., 5~ :
25-40.
SHAW. 1 3 . J . y LA1TY, J . B. (1982). A n umer ical taxonomic study of Pseudon¡onas strains
from spoiled meat. 3 . Appl. Bacteriol., 52 : 219-228.
SHAW, B. J. y LATI’Y, J. 1 3 . (1984). A study of the relative incidence of different
Pseudomonas groups on meat us ing a computer assisted identification technique
employing only carbon source tests. 3. Appl. Bacteriol., 52: 59-67.
SHAY, 8. J. y EGAN, A. F. (1981). Hydrogen sulphide production an d spoilage of
vacuum-packed beef by a Lactobacillus. En: “Psychotroph¡c Microorganisms in
Spo¡Iage and Patliogenicity”, P p . 241-252. T. A. Roberts, G. Hobbs, J. H. B.Christian an d N . Skovgaard (eds.). Academic Press, London.
SHELEF, L. A. (1989). Survival of ListeRo rnonocyrogenes in ground beef an d liver afterstorage at 4 an d 2 5 0C . 3. Food Protect., 52:379-383
SILVA, M., JACOBUS, N. V., DENEKE, C . y GORBACH. S.L. (1987>. Antimicrobial
substance from a human Lacíobaclllus strain. Ant¡microb. Agenís Uhernotber.,
3 1 : 1 2 3 1 - 1 2 3 3 .
2 7 6

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 290/293
BIBLIOGRAFÍA
SIMONDS, J. , HANSE, P . A. y LAKSHAMANAN, 5. (1971). Deoxyribonicleic acid
hybridation among strains of lactobacilli. .1 . Bacteriol. 107: 382-384.
SKINNER, F. A. y HUGO, W . 1 3 . (1976). En: “Inhibition and Inactivation of
Vegetative Microbes”. Symp. Series 5, Soc. for Appl. Bacteriol. Academic Press,
New York.
SMITH, J. L. y PALUMBO, 5. A. (1981). Microorganisms as food aditives. 3. Food
Protect., 4 .4 : 936-955.
SPECK, M. L. y RAY, 1 3 . (1977). Effects of freezing an d storage o n microorganisms infrozen foods: A review. 3. Food Protect. 4W 333-MO.
SPELHAUG, 5. R. y HARLANDER, 5. K. (1989). Inhibition of foodborne bacterialpathogens by bacteriocins from Lacrococcus lactis an d Pediococcus penrosaceus. 3.
Food Protect., 52: 856-862.
SRIRANGANATHAN, N., SEILDER, R. J., SANDINE, W . E . y ELLIKER, P . R . (1973).
Cytological an d deoxyribonucleic acid-deoxyribonucleic acid hybridization studies on Lactobacillus isolated from San Francisco sour dough. AppI. Microbiol. 2 5 K :
46 1-470.
STTOFFELS, 0., NISSEN-MEYER, J., GUDMONDSDOTTIR, A., SLETTEN, K.,
1-JOLO, H . Y NES , I.F. (1992). Purification an d characrerization of a new bacteriocin
islolated from Carnobacrniunz s p . App!. Envirn. Microbiol., 58 : 1 4 17-1422
SU, T. L. (1949) Antibiotic producing organisms in faeces an d sewage. Br. 3. Exp.Pathol. 30: 466-473 .
SWANK, R . T. y MUNKRES, K. D . (1971). Molecular weighr analysis of oligopeptides byelectrophoresis in polyacrylamide gel with sodium dodecyl sulfate. Anal.
Biochcm., 39: 462-477.
SYKES, G. (1965). En:”Disinfection and stcrilization”, 2nd cd., p . 27 . J. B .
Lippincott (Ed.), Philadelphia.
2 7 7

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 291/293
BIBLIOGRAFíA
TAGG, J. R . y McGIVEN, A. R. (1971). Assay system for bacteriocins. AppI.Microbiol., 21: 943
TAGG, J . R., DAJANI, A. 5. y WANNAMAKER, L. W. (1976). Bacteriocins or
Gram-positive bacteria. Bacteriol. Rey. 4 Q : 722-7 5 6 .
TALARICCO, T. L. y DOBROOOSZ, W . J. (1990). Purification an d characterization of
glycerol dehidratase from Lactobacil/us reuteni. (1990). Appl. Environ.
Microbio!., 5 á : 1195-1197.
TALARICCO, T. L.. CASAS, 1 . A., CHUNG, T. C . y DOBROGOSZ, W. J. (1988).Production an d isolation of reuterin, a growth inhibitor produced by Lactobací/lus
reuteri. Antimicrob. Agents Chemother., 3 7 1 : 1854-1858.
TANAKA, N.. TRAISMAN, E., PLATINGA, P., FINN, L., FLOM, W., MESKE, L. y
GUGGISBERG, J . (1986). Evaluation of factors involved in antibotulinal properties
of pasteurized process cheese spread. J. Foad Protect., 49: 526-5 30.
THOMAS, T. D., ELLWOOD, D . C . y LONGYEAR, V. M. C . (1979). Change from homo-to heterolactic ferrnentation by Streptococcus /cctis resulting from glucose limitation in
anaerobie chemostat cultures. 3. Bacteriol. 138: 109-117.
THORNLEY, M. J . y S HA R PE , M. E . (1959). Microorganisms from chicken meat related to
both lactobacilli an d aerobic sporeformers. J. Appl. Bacteriol., 22 : 368-376.
TOBA, T., YOSHIOKA, E . y ITOR, T. (1981). Lacticin, a bacteriocin produced by
Lactobcci//us delbrueckii s ub s p . /cctis. Letters Appl. Microbio!_ 1 2 43-45.
TORRICELLI, A. (1959). Sterilization of empty containers for food industry. En: “Ozone
Chemistry and Technology”, P p . 375-376 . H . A. Leedy (Ed.). Adv. n Chem.
Series 21, Am. Chem. Soc., Washington, D . C
TRAMER, J . (1966) Inhibitory effect of Lacíobcci/Ius acidophilus. Nature. London. 211
204-205.
2 7 8

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 292/293
BIBLIOGRAFÍA
TUBYLEWICZ, F I. (1970). Antigenie properties of the bacteriocin preparations obtained from
Type A Clostridium perfringens strains. Buil. Polish Academy Sel. (BioIj,
fl: 253-257.
UPRETI, 0. C . y HINSDILL, R. D. (1973). Isolation an d characterization of a bacteriocin
from a homoferrnentative Lactobacillus. Antim¡crob. Agents Chemother. 4:
487-494.
UPRETI, 0. C . y HINSDILL, R. D. (1975). Production and mode of action of Laetocin 27 :
bacteriocin from a homofermentative Lactobacillus. Antimicrob. Agents
Chemother. 7:139-145
VAN BELKUM. M. J., HAYEMA, 1 3 . J., GEIS, A., KOK, J. y VENEMA, 0. (1989).
Cloning of two bacteriocin genes from a lactococcal bacteriocin plasmid. App.
Environ. Microbio!, vol 55.5: 1187-1191
VAN BELKUM, NI J., HAYEMA, 1 3 . J., JEENINGA, R. E., KOK, J. y VENEMA, 0.
(1991). Organization an d nucleot id sequences of two lactococcal b acter iocin operons.
AppI. Environ. Microbiol., 5 2 1 , 2: 492-498.
VANKOVA, J. (1957). Motile catalase producing strains of Lactobacíl/us delbrueckii.
Nature, London, 179: 204.
VICENT, J. 0., VEOMETT, R . C . y RILEY, R . F. (1959). Antibacterial activity associated
with Lactobacil/us acidophi/us . 3. Bacteriol. 78 : 477-484.
WEST, C . A. y WARNER, P . 3. (1988). Plantaricin 8, a bacteriocin produced by Lactobacillus p¡antarurn NCDO 1193. FEMS Microbiol. Lett_ 49• 163-165.
WEISS, N., SCHILLINGER, U., LATERNSER, M. y KANDLER, 0. (1981).
Lactobacil/us sharpeae s p . nov. an d Lactobacil/us agilis s p . nov., two new species of
homofermentative, meso-diaminopiinelic acid-containing lactobacilli isolated fromsewage. Zentralbí. Bakteriol. Mikrobiol. Hyg. 1 AM. Orig. Q7: 242-253.
WHITING, 0. C . (1975). Sonie biochemical an d flavour aspects of lactic acid bacteria inciders an d other alcoholic beverages. En: “Lactic Acid Bactcr¡a in Beverages
2 7 9

7/10/2019 Ad 2009501
http://slidepdf.com/reader/full/ad-2009501 293/293
BIBUOGRAFÍA


















