
1. INTRODUCCIÓNucv.altavoz.net/prontus_unidacad/site/artic/20061205/asocfile/... · cepas...
64
1. INTRODUCCIÓN El tomate (Lycopersicon esculentum Mill) es una de las hortalizas de mayor importancia a nivel mundial debido a su gran difusión comercial. Sus frutos se consumen frescos y también son materia prima para la agroindustria. Es una de las plantas que ha sido más investigada por los estudiosos en todos sus aspectos básicos y agrícolas (GIACONI Y ESCAFF, 1993; CHAMARRO, 1995). En Chile, se estima que su superficie cubre unas 18.800 ha, de las cuales 11.200 ha corresponden a tomate industrial, 6.300 ha de tomate al aire libre y 1.300 ha bajo invernadero. Es la hortaliza con mayor superficie a nivel nacional (ODEPA, 2002). El cultivo bajo invernadero frío, se concentra en la V región, principalmente en la provincia de Quillota (INDAP, 1998). En la zona central de Chile, el principal objetivo de la producción es cosechar tomates fuera de la época estival, para así entrar al mercado cuando la oferta de tomates para consumo fresco es baja, alcanzando de este modo los mejores precios. Es importante destacar la diferencia que existe entre la incidencia de enfermedades en el mismo cultivo al aire libre o bajo plástico, debido a que en este último se modifican las condiciones ambientales y aumentan el desarrollo de enfermedades, especialmente las causadas por hongos y bacterias (BESOAIN, 1989).
-
Upload
trinhhuong -
Category
Documents
-
view
221 -
download
0
Transcript of 1. INTRODUCCIÓNucv.altavoz.net/prontus_unidacad/site/artic/20061205/asocfile/... · cepas...

1. INTRODUCCIÓN
El tomate (Lycopersicon esculentum Mill) es una de las hortalizas de mayor
importancia a nivel mundial debido a su gran difusión comercial. Sus frutos
se consumen frescos y también son materia prima para la agroindustria. Es
una de las plantas que ha sido más investigada por los estudiosos en todos
sus aspectos básicos y agrícolas (GIACONI Y ESCAFF, 1993; CHAMARRO,
1995).
En Chile, se estima que su superficie cubre unas 18.800 ha, de las cuales
11.200 ha corresponden a tomate industrial, 6.300 ha de tomate al aire libre y
1.300 ha bajo invernadero. Es la hortaliza con mayor superficie a nivel
nacional (ODEPA, 2002).
El cultivo bajo invernadero frío, se concentra en la V región, principalmente
en la provincia de Quillota (INDAP, 1998).
En la zona central de Chile, el principal objetivo de la producción es cosechar
tomates fuera de la época estival, para así entrar al mercado cuando la oferta
de tomates para consumo fresco es baja, alcanzando de este modo los
mejores precios.
Es importante destacar la diferencia que existe entre la incidencia de
enfermedades en el mismo cultivo al aire libre o bajo plástico, debido a que
en este último se modifican las condiciones ambientales y aumentan el
desarrollo de enfermedades, especialmente las causadas por hongos y
bacterias (BESOAIN, 1989).

2
El medio ambiente tiene gran influencia en la multiplicación de las bacterias,
especialmente con alta humedad relativa y temperatura entre 20-22 °C se
beneficia su difusión (BLANCARD, 1990; RECHE, 1991; APABLAZA, 1999).
Esta combinación de factores en cultivos bajo plásticos se ve favorecida
sobre todo por la humedad relativa, al permitir que existan condiciones de
sobresaturación o agua libre por varias horas (BESOAIN, 1989).
Durante la temporada pasada (2003) se diagnosticó una patología asociada
a la presencia de Pseudomonas syringae en tomate, afectando
principalmente al cultivar Fortaleza en invernaderos de la V región. Esta
enfermedad se caracteriza por la presencia de manchas necróticas rodeadas
de halo clorótico en los folíolos, y por sobre todo un severo atizonamiento de
los tallos, caracterizado por la presencia de lesiones corticales necróticas sin
comprometer el tejido vascular. Las plantas severamente atacadas detienen
su crecimiento, fructifican pobremente y eventualmente mueren (BESOAIN,
McLEAN y LATORRE, 2004). Estos síntomas difieren de la enfermedad
conocida como peca bacteriana, la que se caracteriza por producir sólo
manchas necróticas en hojas y frutos.
A partir de un total de 12 muestras de cv. Fortaleza, recolectadas en cinco
diferentes predios ubicados en Quillota, Limache y La Cruz, se efectuaron
aislamientos en medio B de King, obteniéndose en forma consistente la
presencia de colonias bacterianas fluorescentes, a las cuales se les realizó el
test de LOPAT y se probó patogenicidad en limones y cerezas. Las cepas
aisladas se identificaron tentativamente como Pseudomonas syringae. Tres
de estos aislamientos fueron patogénicos en tomate luego de asperjar y
frotar levemente una suspensión bacteriana. Los síntomas aparecieron luego
de siete días de la inoculación y a los 35 días las plantas presentaron un
atizonamiento similar al inicialmente descrito. Se reaisló Pseudomonas

3
syringae cumpliéndose de este modo con los postulados de Koch (BESOAIN,
McLEAN y LATORRE, 2004).
Debido a la gran importancia que tiene la V región como zona productora de
tomates bajo plástico y a la magnitud de los daños observados, surge la
necesidad de realizar nuevos estudios que permitan incrementar el
conocimiento de esta nueva sintomatología. De acuerdo a ello, esta
investigación tuvo como objetivos:
• Determinar a qué patovar de Pseudomonas syringae, corresponden las
cepas asociadas a una nueva sintomatología que afecta a plantas de
tomate cv. Fortaleza, en la V región.
• Comparar daño y comportamiento de las cepas bacterianas obtenidas, en
comparación con Pseudomonas syringae pv. syringae.
• Evaluar la susceptibilidad de cuatro variedades de tomate con la cepa
más virulenta causante de la nueva sintomatología.

4
2. REVISIÓN BIBLIOGRÁFICA
2.1. Antecedentes generales de Pseudomonas syringae:
El género Pseudomonas Migula 1894 (EUZÉBY, 2005), pertenece a la
familia Pseudomonadacea Winslow et al. 1917 (EUZÉBY, 2005), del Reino
Bacteria. Corresponde a un organismo ampliamente distribuido en cultivos
hospederos y malezas, esto significa que en ausencia de síntomas aún es
posible detectar poblaciones bacterianas en diversos órganos de la planta
(BRUNA, 1993).
Pseudomonas syringae van Hall 1902 (DYE et al., 1980) es una de las
especies fitopatógenas que se denominan fluorescentes, debido a que al
crecer en un medio nutritivo con bajo contenido de hierro, produce pigmentos
fluorescentes, de color verde amarillo, con capacidad de difundirse (SMITH et
al., 1992). Se trata de un organismo estrictamente aerobio, gram-negativo,
que posee forma de bastón y se moviliza utilizando un flagelo polar. Esta
bacteria, responde en forma negativa a la producción de citocromo c-
oxidasa, a la de arginina dihidrolasa y que aún siendo muy patogénica para
muchas plantas, no produce pudrición en papa. Además, causa una reacción
de hipersensibilidad cuando se infiltra en hojas de tabaco, esta reacción es
utilizada para el diagnóstico entre pseudomonas fitopatógenas de las no
fitopatógenas (HIRANO y UPPER, 1990). BRADBURY (1986); BRAUN-
KIEWNICK y SANDS (2001) y GONZÁLEZ, RODICIO y MENDOZA (2003)
concuerdan con lo citado anteriormente y añaden que Pseudomonas
syringae origina colonias convexas en un medio hipersacarosado dando una
respuesta positiva a la producción de levano.

5
De acuerdo a la clasificación recién mencionada, Pseudomonas syringae
puede ser diferenciada de las otras Pseudomonas fluorescentes, y esto se
resume en el denominado perfil LOPAT que está compuesto por las
siguientes pruebas: Levano (+), Oxidasa (-), Papa (-), Arginina (-) e
hipersensibilidad en Tabaco (+) (LELLIOTT, BILLING y HAYWARD, 1966).
2.2. Sistema de agrupación de Pseudomonas syringae:
Entre las pseudomonadas fluorescentes la más representativa es
Pseudomonas syringae, que comprende 45 diferentes patovares, los que se
basan en la patogenicidad específica en relación con la planta hospedera.
Sin embargo, de acuerdo a las características fenotípicas y a la
patogenicidad cruzada que presenta un patovar, confrontado a hospederos
específicos de otros patovares, se ha evidenciado que muchos de éstos
presentan notable parecido y pueden inducir síntomas en plantas que
normalmente se clasifican como no hospederas (SCORTICHINI, 1995).
2.3. Peca bacteriana, mancha del halo o “bacterial speck”:
2.3.1. Agente causal El agente causal de esta enfermedad corresponde a Pseudomonas syringae
pv. tomato (Okabe 1933) Young, Dye y Wilkie 1978 (DYE et al., 1980). Se
han reportado dos razas, denominadas “0” y “1” (GABOR y WIEBE, 1997).
Pseudomonas syringae pv. tomato raza 0 corresponde al primer registro de
la peca bacteriana a nivel mundial, esta raza se logró controlar mediante la
utilización de cultivares resistentes, estos poseían el gen PTO de resistencia
a Pseudomonas syringae pv. tomato. En 1986 aparece el primer reporte de

6
Pseudomonas syringae pv. tomato raza 1 (LAWNTON y MAcNEILL, 1986).
Posteriormente en 1998, la enfermedad fue encontrada nuevamente en
numerosos cultivos resistentes a raza 0 en campos a través del valle de
Sacramento, California, Estados Unidos. En varios de estos predios causó
defoliación severa de los plantines de tomate, los síntomas incluían hojas
café oscuras hasta negras y lesiones en los tallos rodeadas por halos
amarrillos, por lo tanto corresponden a los mismos síntomas descritos para la
raza 0 (ARREDONDO y DAVIS, 2000).
El gen PTO de resistencia a la raza 0 fue introducido en numerosas
variedades de tomates orientadas principalmente al mercado fresco. Sin
embargo, este gen no confiere resistencia a la raza 1 de Pseudomonas
syringae pv. tomato (WILSON et al., 2002).
2.3.2. Hospederos
Pseudomonas syringae pv. tomato ataca exclusivamente a las solanáceas
pimentón (Capsicum annuum L) y tomate (Lycopersicon esculentum Mill), sin
embargo, existen reportes que también puede ser patogénica al realizar
inoculaciones en berenjena (Solanum melongena L) (BRADBURY, 1986).
2.3.3. Sintomatología
La sintomatología se manifiesta sobre hojas, tallos, pedúnculos y sépalos, en
forma de pústulas negras de 2 a 3 mm de diámetro que adquieren una
morfología redondeada, pudiendo o no estar rodeadas por un halo amarillo
(RECHE, 1991; LATORRE, 1992; MESSIAEN et al., 1995; FLOYD, 2004).
Otros autores señalan que las lesiones en tallos y pecíolos se pueden

7
desarrollar con formas ovaladas a elongadas de color café oscuro (JONES,
1993; POHRONEZNY y VOLIN, 2002; DAVIS et al., 2003).
Las lesiones negras con bordes amarillos también pueden ocurrir en los
márgenes de las hojas donde se acumulan las gotas por efecto del proceso
de gutación; cuando estas lesiones se unen, grandes áreas de tejido foliar
pueden morir. Generalmente las lesiones en el fruto son pequeñas (1 mm),
con forma de lunar y superficiales, sin embargo, también pueden ser más
grandes y hendidas, y en frutos inmaduros están rodeadas por un halo verde
(GABOR y WIEBE, 1997).
En los frutos las manchas se desarrollan superficialmente y no afectan la
pulpa (RECHE, 1991). JONES (1993) señala que también en los frutos se
desarrollan lesiones o punteados minúsculos de coloración oscura y rara vez
con diámetro mayor a un mm. El tejido alrededor de cada punto puede ser de
un verde más intenso que en las áreas no afectadas. Las lesiones pueden
ser levemente elevadas o inicialmente planas. En algunas instancias los
puntos son hendidos y un halo verde oscuro puede estar asociado. Además,
VENETTE, LAMEY y SMITH (1996) indican que al ser los tejidos inmaduros
son más susceptibles, al infectarse la fruta temprano, las lesiones con forma
de punto pueden causar orificios, debido a que el tejido fino infectado crece
más lento que tejido fino sano. Las frutas maduras son resistentes como
resultado de su alta acidez.
La multiplicación de esta enfermedad puede generar un amarillamiento
generalizado, seguido de desecación foliar (MESSIAEN et al., 1995). En
casos severos, las plantas infectadas presentan retraso en la madurez de su
fruta y reducen la producción (DAVIS et al., 2003).

8
2.3.4. Diseminación y sobrevivencia
Es una enfermedad que puede penetrar por heridas, estomas y que se
puede transmitir por semilla contaminada y a través del suelo (RECHE,
1991). Además puede sobrevivir en la rizosfera de las raíces y en restos
vegetales, en forma epífita en tomate y malezas, pudiendo también sobrevivir
en las semillas (RISTA, 2005).
GABOR y WIEBE (1997), concuerdan con lo citado en el párrafo anterior ya
que argumentan que el organismo puede sobrevivir en el suelo y en hojas de
muchos cultivos y malezas, además de semillas que también pueden
infectarse. Sin embargo, señalan que la transmisión por semilla es
usualmente de menor importancia, JONES (1993) añade que algunos
autores no reconocen a dicho órgano como una vía de transmisión, aun
cuando sería una forma de explicar el incremento de los reportes mundiales
de esta enfermedad. El mismo autor indica que entre plantas de un mismo
cultivo la bacteria es diseminada por las salpicaduras de gotas de lluvia y por
los utensilios usados en el proceso de transplante de almácigos.
En condiciones de verano la bacteria puede sobrevivir en la superficie de
plantas voluntarias de tomate, pero en una cantidad muy baja
(POHRONEZNY y VOLIN, 2002).
2.3.5. Factores predisponentes
La enfermedad se favorece por un clima frío (13-20°C), y condiciones de alta
humedad y lluvias. Usualmente se requiere de un día de elevada humedad
constante en la hoja para que la enfermedad se desarrolle (BRUNA, 1993;

9
JONES, 1993; GABOR y WIEBE, 1997; RISTA, 2005). Los síntomas de la
enfermedad se hacen presentes 8 a 10 días después de la inoculación
(RISTA, 2005)
MESSIAEN et al. (1995) señalan que esta enfermedad es muy dañina en
primaveras lluviosas o bajo plástico, por generarse condiciones demasiado
húmedas. Además POHRONEZNY y VOLIN (2002), mencionan que en el
año 1977-1978 en la localidad de Homestead, Florida del Sur, Estados
Unidos, el invierno se desarrolló en condiciones inusuales de altas
precipitaciones y de bajas temperaturas, favoreciendo un fuerte brote de
Pseudomonas syringae pv. tomato.
Esta bacteria a menudo aparece en zonas con agroclimas secos y donde se
usa riego por aspersión, pero muy pocas veces ocurre cuando en dichos
lugares se usa goteo o riego por surco (GABOR y WIEBE, 1997).
2.3.6. Incidencia e importancia económica
La peca bacteriana ha llegado a ser económicamente importante en todo el
mundo, desde mediados de la década de 1970 (AGRIOS, 1996). JONES
(1993) señala un incremento en los reportes de esta enfermedad en esta
última década.
2.3.7. Estrategias de control
Según RECHE (1991), el control se basa en eliminar restos vegetales y de
plantas enfermas, utilizar semilla sana y certificada, favorecer la aireación del
invernadero para disminuir la humedad y aplicar productos bactericidas.

10
JONES (1993) concuerda y sugiere además evitar plantar en el mismo
terreno por dos años consecutivos, producir plantines libre de la enfermedad
en lugares donde no se haya producido tomate anteriormente, y el
tratamiento a las semillas debe ser un procedimiento de rutina. Por otro lado,
se deben mantener todos los terrenos de producción libres de malezas y
plantas voluntarias, y no amontonar desechos vegetales en o cerca de las
zonas de producción.
Referente al tratamiento a las semillas FLOYD (1990) y VENETTE, LAMEY y
SMITH (1996) indican que se deben utilizar sólo semillas tratadas. Una
opción consiste en sumergir las semillas por 25 minutos en agua caliente a
50 °C, con este proceso se logra controlar las bacterias al interior de las
semillas, pero reduce la germinación. Otra alternativa consiste en bañar las
semillas por un minuto en hipoclorito de sodio al 1 %, en este caso se
eliminan sólo las bacterias superficiales de la semilla. Por otra parte SIKORA,
GAZAWAY y MULLEN (1998) sugieren tratar las semillas con una solución
de ácido acético al 0,6 %, para asegurar que las semillas estén libres de la
enfermedad.
GABOR y WIEBE (1997) señalan que el uso de variedades resistentes es
una forma efectiva de controlar la enfermedad. Los autores también
mencionan que la aspersión temprana de cobre puede reducir su ocurrencia.
El riego por aspersión aumenta la incidencia cuando la bacteria está
presente, y por lo tanto se debe usar en lo posible riego por goteo o surcos.
Referente al uso de variedades resistentes VENETTE, LAMEY y SMITH
(1996) señalan que no existen cultivares de tomate totalmente resistentes a
la peca bacteriana y que sólo algunas líneas soportan la enfermedad mejor
que otras.

11
POHRONEZNY y VOLIN (2002) y ZITTER (2005), indican que aun cuando
no se conocen productos químicos específicos para el control de la peca
bacteriana, el uso de mancozeb, cobre y antibióticos como la estreptomicina
pueden utilizarse como estrategias de control. JETT (2003) y RISTA (2005)
agregan que aplicaciones de cobre en forma preventiva como caldo bordelés
o como oxicloruro de cobre, pueden disminuir la incidencia y la dispersión del
organismo patógeno, y concuerda con la utilización de mancozeb, ya que
trabajos recientes han demostrado que el cobre asociado con este fungicida
de la familia de los ditiocarbamatos aumenta la eficacia del cobre. Del mismo
modo, WICK (1991); SIKORA, GAZAWAY y MULLEN (1998) y DAVIS et al.
(2003) señalan que al adicionar hidróxido de cobre y mancozeb se aumenta
la efectividad del cobre y lo sugieren como una medida de control efectiva.
En otro ámbito, recientes experiencias indican que Actigard 50WG
(Acibenzolar-S-metil) tiene la habilidad de inducir la resistencia sistémica
adquirida SAR para un gran número de patógenos en diferentes cultivos.
Esto se evaluó en el manejo de la peca bacteriana en experimentos
realizados en tomate en distintas localidades de Estados Unidos,
concluyendo después de un periodo de cuatro años de estudio que Actigard
50WG puede ser integrado como una alternativa viable junto a bactericidas a
base de cobre para el manejo de esta enfermedad, particularmente donde
predominen las poblaciones resistentes al cobre (LOUWS et al., 2001). En
relación con esto, JETT (2003) añade que actuales estudios de la
Universidad de Kentucky, indican que programas de aspersiones de 14 días
con Actigard 50WG pueden proporcionar un aceptable control de la
enfermedad, al igual que MAcNAB (2004a) que también lo sugiere como una
herramienta de control.

12
RIEDEL (2000) señala que Actigard 50WG tiene un modo de la acción único,
distinto al de los fungicidas y bactericidas actualmente disponibles, puesto
que no tiene ninguna actividad directa sobre los patógenos de la planta, por
lo tanto la probabilidad de desarrollar resistencias o insensibilidad en el
patógeno es escasa.
Es importante destacar que el cobre proporciona un control parcial de la
enfermedad, por lo cual se debe aplicar al aparecer los primeros síntomas y
repetir a intervalos de 10 o 14 días si las condiciones frescas y húmedas
prevalecen. Se utiliza idealmente como producto preventivo, por lo cual debe
ser aplicado antes de que ocurra el periodo de la infección. Una o dos
aplicaciones serán suficientes para proteger a las plantas de tomates durante
las etapas más susceptibles del crecimiento (DAVIS et al., 2003).
Por otra parte, el uso de la estreptomicina debe ser restringida
exclusivamente a aplicaciones en plántulas de tomate previo al transplante,
idealmente cuando la primera hoja verdadera aparezca, continuando las
aplicaciones cada cuatro o cinco días hasta transplantar (WICK, 1991;
VENETTE, LAMEY y SMITH, 1996; MAcNAB, 2004b; ZITTER, 2005).
Finalmente DAVIS et al. (2003) se refieren a los controles culturales. Los
autores mencionan el retraso de las plantaciones para escapar de las
condiciones frescas y húmedas que favorecen el desarrollo de la
enfermedad, y las aspersiones de cobre tempranas en la temporada, ambos
como métodos de control aceptados para la producción orgánica certificada.

13
Además RISTA (2005), añade que para complementar un manejo integrado
en invernaderos es conveniente el control de la humedad, evitando la
presencia de agua libre en las plantas, ventilando en forma constante.
2.4. Mancha foliar de syringae o “syringae leaf spot”:
2.4.1. Agente causal
El agente causal de esta enfermedad es Pseudomonas syringae pv. syringae
van Hall 1902 (DYE et al., 1980).
2.4.2. Hospederos
Pseudomonas syringae pv. syringae, es sin lugar a dudas la bacteria
fitopatógena más polífaga y ubicua, capaz de causar infección en alrededor
de 170 especies diferentes y además puede ser encontrada fácilmente en
forma epífita (SCORTICHINI, 1995).
2.4.3. Sintomatología
En tomate los síntomas en las hojas pueden variar desde manchas café que
no tienen halo, hasta manchas negras con aureolas amarillo fuerte, que se
asemejan bastante a aquellas causadas por el patovar tomato, pero de
mayor tamaño. Sin embargo, es necesario aislar la bacteria y realizar
pruebas de laboratorio para determinar qué patógeno está involucrado
(GABOR y WIEBE, 1997).
GITAITIS (1993) indica que los síntomas varían desde pequeñas lesiones
color café sin halos a lesiones casi negras con halos color amarillo fuerte,

14
igual a aquellos causados por P. syringae pv. tomato. En ocasiones el
patógeno puede originar necrosis marginal aislada, extensas áreas marchitas
y tejidos que aparentemente han sufrido heladas, siendo difícil reproducir los
síntomas en un invernadero.
2.4.4. Diseminación y sobrevivencia
P. syringae pv. syringae en plantas de tomate es considerado un patógeno
débil y oportunista, que se encuentra frecuentemente en heridas y como un
organismo secundario, presente en infecciones mixtas con otros patógenos
que causan manchas en hoja (GITAITIS, 1993; GABOR y WIEBE, 1997). La
bacteria sobrevive tanto en plantas hospederas como no hospederas en un
estado no parasitario, y puede diseminarse desde esas plantas cuando las
condiciones medio ambientales frías y húmedas favorecen el desarrollo de la
enfermedad (GABOR y WIEBE, 1997).
En frutales de carozo la infección se produce a través de estomas, la bacteria
se propaga intercelularmente produciendo el colapso y muerte de las células,
dando origen a pequeñas manchas de forma irregular. Cuando el clima es
húmedo, las bacterias exudan de las manchas y se propagan hacia otras
hojas por contacto directo, a través de insectos, lluvia y otros factores
(AGRIOS, 1996)
2.4.5. Factores predisponentes
El patógeno requiere de heridas para infectar el tejido vegetal y puede invadir
lesiones ya existentes causadas por otra enfermedad (GABOR y WIEBE,
1997).

15
GITAITIS (1993) coincide señalando que el desarrollo de la enfermedad en
tomate se favorece por alta humedad y heridas. Además, las únicas fuentes
conocidas de inóculo son otros hospederos, siendo los más comunes: el
centeno, que se usa como cubierta verde de invierno y árboles de cerezo
silvestre que bordean las plantaciones. Áreas con gran número de cerezos
tiene una alta incidencia de P. syringae pv. syringae cada año. En el estado
de Georgia, Estados Unidos, esta enfermedad se concentra en abril durante
la época fría, en cambio, P. syringae pv. tomato y Xanthomonas campestris
pv. versicatoria se desarrollan en mayo, cuando las temperaturas promedio
durante el día han aumentado.
2.4.6. Incidencia e importancia económica
Esta enfermedad usualmente no se encuentra en tomate y en caso de
presentarse no es común que se produzca daño económico (GABOR y
WIEBE, 1997).
GITAITIS (1993) señala que el diagnóstico correcto es crítico, ya que P.
syringae pv. syringae es relativamente inocua. Consecuentemente, en el
programa de certificación para la producción de plantines de tomate del
Estado de Georgia (Estados Unidos), sólo las plántulas que tienen P.
syringae pv. syringae se les permite mantener su certificado libre de
enfermedades, y las plantas que tienen P. syringae pv. tomato o
Xanthomonas campestris pv. versicatoria son puestas en cuarentena.
El mismo autor indica que en la zona del sureste de Estados Unidos se
presentó una mayor incidencia de P. syringae pv. syringae, y por lo tanto, fue
necesario un análisis de laboratorio más específico y costoso en el programa
de certificación de esta área.

16
2.4.7. Estrategias de control
GABOR y WIEBE (1997) recomiendan hacer aspersiones de cobre, pero al
no ser comunes los daños económicos esta práctica generalmente no es
necesaria. Si se sospecha de esta enfermedad se debe verificar que los
síntomas son causados por P. syringae pv. syringae y no por otra
enfermedad bacterial que requeriría un control más estricto.
GITAITIS (1993) coincide con los autores mencionados en el párrafo anterior
y además recomienda aplicaciones semanales de bactericidas para controlar
esta enfermedad adecuadamente. Asimismo, advierte la importancia de
comprobar que efectivamente P. syringae pv. syringae es el causante del
problema, para lo cual se debe realizar una caracterización acuciosa y
rigurosa del agente causal, especialmente en aquellos casos en que la
enfermedad requiera tratamientos de control más duros.
2.5. Caracterización de los aislados:
La correcta identificación de enfermedades bacterianas que causan manchas
pequeñas sobre hojas de plantines de tomate, es muy difícil si el diagnóstico
está basado exclusivamente en los síntomas. Sin embargo, existen pruebas
útiles para distinguir entre P. syringae pv. syringae y P. syringae pv. tomato,
patógenos que probablemente pueden ser confundidos (GITAITIS, 1993).
Similares antecedentes entrega SCORTICCHINI (1995), quien indica que la
gran versatilidad bioquímica de las bacterias, permite discriminar entre las
especies y patovares. Ello se realiza en base a la capacidad del aislado de
utilizar compuestos orgánicos particulares, producir pigmentos, crecer en

17
condiciones de temperatura específicas y provocar reacciones características
de hipersensibilidad, entre otras pruebas específicas.
Otra característica fenotípica útil para caracterizar un aislado en etapa de
estudio, es la producción de proteínas que desarrollan funciones
catalizadoras en la formación de núcleos de hielo. Este es uno de los
factores de virulencia más desarrollados entre las bacterias fitopatógenas y
que permite discriminar entre ellas (BRAUN-KIEWNICK y SANDS, 2001).
Por su parte YOUNG y TRIGGS (1994) indican que existen ensayos claves
para la caracterización de los patovares syringae y tomato de Pseudomonas
syringae, estos son el estudio de la capacidad para formar núcleos de hielo
“ice nucleation” y la utilización de los sustratos orgánicos eritritol y DL-lactato
como únicas fuentes de carbono.
Los dos organismos mencionados en el párrafo anterior también se pueden
diferenciar por sus perfiles de ácidos grasos. Aunque los dos perfiles son
casi idénticos, el patovar syringae contiene una cantidad significativa de
ácido δ-cis-9 hexadecanoico, 10-metileno, compuesto que está ausente en el
patovar tomato. Por lo tanto, las pruebas más importantes para discriminar
entre estos patovares son la utilización de eritritol, y DL-lactato por parte de P.
syringae pv. syringae, que lo diferencia de P. syringae pv. tomato el cual no
utiliza estos compuestos. Además, las cepas de P. syringae pv. syringae son
activas a la nucleación de hielo, lo cual también puede diferenciar éste
organismo de P. syringae pv. tomato (Cuadro 1).
Es importante destacar que las cepas de P. syringae pv. syringae inducen
una reacción de patogenicidad marcada en el cultivar caupí (Vigna
unguiculata (L.)), variedad California Blackeye N°3, lo cual también puede

18
diferenciar éste organismo de P. syringae pv. tomato. (GITAITIS, 1993).
Referente a esto, OTTA y ENGLISH (1971) junto a LAY y HASS (1973),
coinciden en señalar al test de patogenicidad en frejol caupí como una
instancia adecuada para el reconocimiento de P. syringae pv. syringae, por
ser una planta indicadora específica para este patovar y no así para patovar
tomato.
CUADRO 1. Identificación de cepas de Pseudomonas syringae pv. syringae y Pseudomonas syringae pv tomato
Reacción Test P. syringae pv.
syringae P. syringae pv
tomato Reacción gram - -
Arginina dihidrolasa - - Reacción citocromo oxidasa - - Hipersensibilidad en tabaco + +
Fluorescencia en MB de King + + Utilización de eritritol + - Utilización DL-lactato + -
Formación de núcleos de hielo + - Fuente: BRADBURY (1986)
Esta información aún está vigente y ha sido citada por SCHAAD (1980),
JONES (1993), GITAITIS (1993), YOUNG y TRIGGS (1994), SCORTICCHINI
(1995), LUDWIG y KLENK (2001) y BRAUN-KIEWNICK y SANDS (2001).
El sistema de clasificación en patovares se adapta a las necesidades
prácticas de los patólogos vegetales, pero no refleja necesariamente las
relaciones genéticas reales entre las cepas. Sin embargo, la llegada de la
biología molecular ha causado un cambio significativo en el tipo de
herramienta utilizada, permitiendo una caracterización y clasificación mucho
más rápida y fiable. Los protocolos que analizan ácidos nucleicos reducen el

19
tiempo utilizado, parecen producir resultados más estables y repetitivos, con
frecuencia reflejan más fielmente las relaciones filogenéticos y son útiles para
ordenar las cepas en grupos (ROSALES, 2005).
CINTAS, BULL y KOIKE (2002) mencionan que una alternativa actualmente
muy utilizada corresponde a la caracterización genotípica mediante rep-PCR,
donde una reacción en cadena de la polimerasa reconoce cadenas repetidas
de ADN bacteriano. Con esta reacción de la polimerasa de ADN en cadena
usando iniciadores de óligonucleótidos dirigidos a distintas secuencias, se
pueden generar patrones de fragmentos de ADN, dichos patrones se llaman
huellas digitales del ADN. Estos métodos se usan tanto para investigar la
diversidad genética dentro de las poblaciones de cepas como para agrupar
las cepas bacterianas, y se basan en la variación genética en las secuencias
blanco localizadas en los genomas que sirven para anclar a los iniciadores.
Los productos sintetizados por la reacción de PCR se separan por
electroforesis en base a su tamaño molecular y su secuencia de nucleótidos.
Los iniciadores comunes usados en la caracterización de bacterias son los
de las secuencias “BOX” (BOX-PCR) y los que sirven para las secuencias
repetidas extragénicas y palindrómicas (rep-PCR) entre otros (WANG y
MARTÍNEZ-ROMERO, 2000).
2.6. Principales variedades de tomate utilizadas en la provincia de Quillota, V
región:
Fortaleza. Híbrido indeterminado precoz, para producciones de ciclo corto en
condiciones de frió y baja luminosidad. Variedad multilocular estructural, de
vigor moderado, con hojas pequeñas que permite una alta densidad de
plantación, una buena ventilación, penetración de luz y productos
fitosanitarios, estas características hacen que sea una variedad ampliamente

20
utilizada en el valle de Quillota para el tomate de cultivo primor
(BIOAMERICA, 2004).
Naomi. Híbrido indeterminado precoz y de cosecha concentrada. Variedad
multilocular estructural, caracterizada por un excelente vigor, abierta con
buena ventilación, se utiliza como tomate de otoño en la zona central de
Chile, por el mayor rendimiento en comparación con los multiloculares
tradicionales (BIOAMERICA, 2004).
FA-593. Variedad larga vida indeterminada, caracterizada por un buen vigor,
tendiente al crecimiento vegetativo. Se utiliza como cultivo trastomate o
tomate de otoño en el valle central de Chile (DEVIA, 2004)*.
Yonit. Al igual que FA-593, es una variedad larga vida. Tomate hibrido
indeterminado, que se caracteriza principalmente por su escaso vigor, su
gran precocidad y rusticidad. Ideal para el uso en invernadero
(TATTERSALL, 2003).
* DEVIA, J. Ing.Agr. 2004. Profesor cátedra cultivos forzados. Universidad
Católica de Valparaíso. Facultad Agronomía. Comunicación personal.

21
3. MATERIALES Y MÉTODOS
Los ensayos fueron llevados a cabo en el Laboratorio e Invernadero del área
de Fitopatología de la Facultad de Agronomía de la Pontificia Universidad
Católica de Valparaíso, ubicada en la comuna de Quillota, V región
(coordenadas 32º Lat Sur y 71º Long Oeste).
Se trabajó en un invernadero de madera tipo capilla con cubierta de
polietileno transparente, de dimensiones: 9 m de ancho, 15 m de longitud y
una altura de 3,10 m en el punto más alto. En el Anexo 1, se presenta el
comportamiento térmico y de humedad relativa, registrados en un
invernadero similar en la localidad de La Palma.
Los aislados de Pseudomonas se obtuvieron a partir de 12 muestras de
tomate cv. Fortaleza de cinco diferentes predios, las cuales fueron
identificadas tentativamente como Pseudomonas syringae. La sintomatología
que presentaron las plantas naturalmente infectadas (Figura 3 a), a partir de
las cuales se aislaron las cepas bacterianas, se caracterizó por la presencia
de manchas necróticas rodeadas de halo clorótico en los folíolos y un severo
atizonamiento de pecíolos y tallos, esta presencia de lesiones corticales
necróticas no comprometía el tejido vascular y su movimiento a través del
tallo se iniciaba desde la zona basal de las plantas hasta la parte alta, donde
se detiene el atizonamiento sin comprometer frutos.
Los aislados 627.1, 627.2, 627.3, 593.1, 593.2 593.3, 593.4, 588.1 588.2,
290, Sta. Rosa y McLean, fueron recuperadas a partir de conservados
liofilizados, los que habían sido aislados desde plantas enfermas en la
primavera del año 2003. Para su reactivación las cepas se sembraron sobre

22
un medio B de KING (MBK) (LELLIOTT y STEAD, 1987) y tras cinco días de
incubación a 23ºC se repicaron a partir de una colonia sobre placas del
mismo medio de cultivo, para purificar y obtener nuevo inóculo.
3.1. Caracterización de aislados de Pseudomonas syringae (in vitro):
3.1.1. Utilización de sustratos orgánicos
A todos los aislados mencionadas en el párrafo anterior, junto con la cepa
112 correspondiente a P. syringae pv. syringae, se les realizó pruebas de
crecimiento de colonias sobre dos distintas fuentes de carbono: eritritol y DL-
lactato. Se utilizó 1 g de la fuente de carbono en 1000 ml del medio de sales
minerales Ayers, el cual para un litro de medio requiere: 1 g de NH4H2PO4,
0,2 g de KCl, 0,2 g de MgSO4x7H2O, 1 ml de bromotimol azul 1,6% p/v en
95% de etanol y 12 g de agar. Antes de autoclavar el medio se ajustó el pH a
7,2. Luego de 20 minutos en autoclave se dejó enfriar, se traspasó a placas
Petri, luego de dos días se sembró las bacterias mediante un estriado y se
incubó a 27°C por 3, 7 y 14 días. El crecimiento se comparó con placas que
no contenían una fuente de carbono (LELLIOTT y STEAD, 1987;
SCORTICHINI, 1995).
3.1.2. Capacidad de formar núcleos de hielo
Inicialmente los aislados 290, 588, 593, Sta Rosa y 112 se hicieron crecer
por dos días a 25ºC sobre el medio LPGA, el cual está compuesto por 5 g de
extracto de levadura, 5 g de peptona, 5 g de glucosa y 15 g de agar. Luego
de dos días de crecimiento se preparó una suspensión de 1x109 u.f.c./ml que
se introdujo en tubos estériles, además se prepararon tubos control que
contenían sólo agua destilada estéril. Posteriormente los tubos se

23
sumergieron por cinco minutos en un recipiente de plástico con una solución
anticongelante (NaCl 2 M) a una temperatura estable de -4°C, que se logró
utilizando un congelador horizontal con termostato ajustable, el cual se
instaló dentro de una cámara frigorífica a temperatura constante de -1°C
perteneciente al Laboratorio de Poscosecha. Se registró un resultado
positivo, cuando el líquido contenido en el tubo donde fue puesta la
suspensión bacteriana, se congeló claramente. Se verificó que en los tubos
control no se presentara congelación. Esta prueba se repitió dos veces
(SCORTICHINI, 1995; CINTAS, BULL y KOIKE, 2002).
3.2. Ensayos de virulencia, susceptibilidad varietal y patogenicidad (in vivo):
Se realizaron tres ensayos bajo condiciones de invernadero, el primero
consistió en evaluar en un cultivo de tomate cv. Fortaleza, la virulencia de
cinco cepas de las cuales, una era P. syringae pv. syringae y las cuatro
restantes correspondieron a aislados causantes de la nueva sintomatología
en plantas de tomate. En el segundo ensayo se utilizó uno de los cuatro
aislados anteriormente analizados causantes de la nueva sintomatología,
para evaluar susceptibilidad en cuatro variedades de tomate. En el tercer
ensayo se realizó la prueba de patogenicidad en frejol caupí, utilizando las
mismas cepas del Ensayo 1.
3.2.1. Ensayo 1: Evaluación de virulencia de cuatro aislados de P. syringae
causantes de la nueva sintomatología.
Este primer ensayo constó con seis tratamientos, con cinco cepas
bacterianas y un testigo en plantas de tomate con cinco hojas verdaderas. Se
empleó una cepa de Pseudomonas syringae pv. syringae contrastada con

24
cuatro aislados de Pseudomonas syringae causantes de la nueva
sintomatología.
Se trabajó con plántulas de tomate variedad Fortaleza donadas por Quintil
S.A., las cuales fueron sembradas el 20 de abril en bandejas almacigueras
de 286 alvéolos. El día 28 de mayo fueron transplantadas en bolsas
individuales de polietileno negro con un volumen de sustrato de siete litros y
llevadas a invernadero frío, donde se mantuvieron todo el ensayo. El sustrato
consistió en una mezcla de 60% de tierra de hoja, 20% de suelo franco
arcilloso y 20 % de arena, el cual fue vaporizado y esterilizado el día 26 de
mayo.
Una semana después del transplante se inoculó con las cepas
correspondientes a cada uno de los tratamientos (ver sección 3.2.1.2.),
conservadas en forma liofilizada y estriadas a MBK para su purificación y
multiplicación.
El cultivo presentó durante el periodo del ensayo ataques de mosquita blanca
(Trialeurodes vaporariorum) y de polilla del tomate (Tuta absoluta). Para su
control se utilizó productos tales como Confidor 350 SC (i.a. imidacloprid) y
Evisect-S (i.a. thiocyclam) en dosis comerciales para mantener la sanidad
vegetal de la parte aérea mediante aspersiones foliares. Durante el ensayo
no se empleó ningún bactericida o fungicida.
En cuanto al manejo de riego y fertilización, se realizaron riegos semanales y
para esto se utilizó un sistema de riego presurizado, formado por una matriz
a la cual estaban conectadas cuatro líneas de riego de 16 mm y a estas los
microtubos con gasto de 4 l/h (un emisor por planta). La fertilización consistió

25
en la aplicación semanal vía riego de tres fertilizantes, las dosis utilizadas se
presentan en el Anexo 2.
3.2.1.1. Método de obtención y aislamiento del inóculo
Las cinco cepas se obtuvieron a partir de la recuperación de conservados
liofilizados. Para reactivarlas se sembraron sobre un MBK. Tras cuatro días
de incubación a 23°C se repicaron nuevamente sobre otras placas del mismo
medio para aumentar la cantidad del inóculo.
Una vez recuperadas las cinco cepas a inocular, se procedió a repicar
nuevamente cada una de ellas, a partir de una colonia en una nueva placa
de MBK, con el fin de obtener un cultivo puro para todas las cepas. Una vez
transcurridas 48 horas (máximo) desde el repique se realizó el test de
LOPAT (LELLIOTT, BILLING y HAYWARD, 1966) a todas las cepas.
3.2.1.2. Forma de inoculación
Para la obtención del inóculo de las cinco cepas, se procedió a sembrar en
placas con cada una de ellas. Luego de dos días de incubación con un
crecimiento bacteriano abundante, se diluyó el contenido de cada placa en
40 ml de agua destilada estéril, midiendo posteriormente la transmitancia de
la suspensión en un espectrofotómetro a 580 nm, aceptándose una lectura
de 50% para asegurar una adecuada concentración del patógeno (en este
caso corresponderá a 1 x 108 ufc/ml, cantidad suficiente para lograr un
adecuado y homogéneo nivel de desarrollo de Pseudomonas syringae).
La solución bacteriana se aplicó sobre las hojas de las plantas de tomate una
semana luego del transplante (4 de junio). Previo a la inoculación, las hojas

26
de las plantas de cada tratamiento se asperjaron con carborundum, y luego
la superficie se rozó para generar microheridas en la epidermis de las plantas
y facilitar el ingreso del patógeno. Luego se utilizó un asperjador manual
estéril, hasta dejar la epidermis notoriamente húmeda con la suspensión
bacteriana. Es importante destacar que las plantas testigo también fueron
frotadas con carborundum y posteriormente rociadas con agua destilada
estéril.
Inmediatamente post inoculación de cada planta, se cubrió cada una de ellas
con una bolsa de polietileno transparente, cerrándola con un elástico para
evitar el contagio entre tratamientos y lograr un efecto de cámara húmeda. El
plástico se retiró luego de tres días.
Para evitar el contagio entre tratamientos se inició la inoculación por los
testigos (T1) y siguiendo uno a uno en orden con los otros tratamientos. En la
medida que se avanzó en la inoculación, luego de cada tratamiento por
bloque (10 plantas) se realizó una minuciosa desinfección de manos con
hipoclorito de sodio (0,1%).
3.2.1.3. Tratamientos
Con el fin de evaluar y comparar el comportamiento y daño ocasionado por
los aislados causantes de la nueva sintomatología versus P. syringae pv.
syringae, se realizaron los siguientes tratamientos:
Tratamiento 1: Testigo, sin inoculación
Tratamiento 2: Cepa 112 de P. syringae pv. syringae
Tratamiento 3: Aislado Cepa 290
Tratamiento 4: Aislado 588

27
Tratamiento 5: Aislado 593
Tratamiento 6: Aislado Sta. Rosa
3.2.1.4. Diseño estadístico
El experimento se condujo mediante un diseño de bloques completos al azar.
La unidad experimental correspondió a 10 plantas, las que fueron
observadas y evaluadas. Se realizaron seis tratamientos incluyendo un
testigo, los cuales se distribuyeron aleatoriamente sobre cada una de las
cuatro repeticiones (cuatro bloques) en el invernadero del ensayo. Los
factores de bloqueo fueron riego y luminosidad dentro del invernadero. La
distribución de este ensayo se representa en el Anexo 3.
Para la evaluación del experimento, los datos se sometieron a un análisis de
varianza mediante la Prueba de significancia de Fisher (prueba F) y cuando
el valor P asociado al estadístico F fue ≤ 0,05, se realizó una comparación de
medias con el test de Tukey (α=0,05).
3.2.1.5. Variables evaluadas
Las variables se eligieron de acuerdo al tipo de daño ocasionado por el
patógeno, por tratarse de una enfermedad que se caracteriza por producir
lesiones a nivel foliar y de tallo, se evaluó el efecto del patógeno en la planta
considerando daños en el follaje, pecíolos y tallo, a través del estudio del
porcentaje de incidencia de la enfermedad (usando como criterio la
observación de presencia y ausencia de síntomas en las plantas de cada
tratamiento mediante un seguimiento de éstos), y estimaciones del índice de
daño de la enfermedad (utilizando una escala de daño o lesiones causadas
por las distintas cepas de Pseudomonas syringae).

28
La fórmula utilizada es la siguiente:
% de Incidencia (I) = Nº de plantas enfermas por unidad x 100
Total de plantas observadas (sanas más enfermas)
(COMMONWEALTH MYCOLOGICAL INSTITUTE, 1985).
Para efectos del índice de daño, se utilizó la siguiente escala:
0 Ausencia de síntomas, planta sana.
1 Manchas necróticas puntiformes en escaso número sobre las
hojas.
2 Manchas necróticas puntiformes sobre las hojas y atizonamiento
de pecíolos.
3 Manchas necróticas puntiformes esparcidas sobre las hojas y
atizonamiento del tallo.
A través de la observación visual se estableció una nota dentro de la escala
de 0 a 3 para cada una de las plantas y posteriormente se obtuvieron los
promedios para cada tratamiento.
La evaluación fue realizada un mes después del transplante, cuando se
evidenciaron los síntomas en las plantas (2 de julio).
3.2.2. Ensayo 2: Evaluación de la susceptibilidad de cuatro variedades de
tomate al aislado 593.
En este ensayo se trabajó con plántulas de tomate de diferentes variedades:
Fortaleza, Naomi, FA 593 y Yonit. Estas fueron producidas por Euro Plant

29
Chile S.A., con siembra el 15 de junio en bandejas almacigueras de 286
alvéolos para la obtención del almácigo. El día 26 de julio de 2004 fueron
transplantadas en bolsas individuales de polietileno negro con un volumen de
sustrato de siete litros y llevadas a invernadero frío. El sustrato fue formulado
de igual manera que en el Ensayo 1.
El día 5 de agosto se procedió con la inoculación de los tratamientos. El
testigo con agua destilada estéril y el resto con el aislado 593 recuperado
desde frascos conservados en forma liofilizada.
En relación a los manejos fitosanitarios, de riego y fertilización de cultivo,
éstos se desarrollaron del mismo modo que los descritos en el Ensayo 1.
3.2.2.1. Método de obtención y aislamiento del inóculo
Para obtener el aislado 593 se procedió con la recuperación de conservados
liofilizados, se reactivó la cepa sembrándose sobre un medio B de King y tras
cuatro días de incubación se repicó nuevamente sobre otras placas de este
mismo medio para aumentar la cantidad del inóculo. Luego se volvió a
repicar sobre 20 placas, cada una de ellas a partir de colonias individuales.
3.2.2.2. Forma de inoculación
Luego de dos días de incubación con un crecimiento bacteriano abundante,
se diluyeron las 20 placas en 800 ml de agua destilada estéril y al igual que
el Ensayo 1, se midió su transmitancia a 580 nm en un espectrofotómetro,
ajustada a una lectura de 50%.

30
El proceso de inoculación fue el mismo realizado en el Ensayo 1 (también los
tratamientos testigos).
La solución bacteriana fue aplicada sobre hojas de las plantas de tomate el
día 5 de agosto de 2004, 10 días después de la plantación utilizando un
asperjador manual estéril, hasta dejar la epidermis a punto de escurrimiento.
Luego de la inoculación, al igual que el Ensayo 1, se procedió a cubrir cada
planta con una bolsa de polietileno transparente. El plástico se retiró el 7 de
agosto.
3.2.2.3. Tratamientos
El ensayo constó de los siguientes tratamientos:
Tratamiento 1: Variedad Fortaleza sin inoculación
Tratamiento 2: Variedad Fortaleza inoculada con aislado 593
Tratamiento 3: Variedad Naomi sin inoculación
Tratamiento 4: Variedad Naomi inoculada con aislado 593
Tratamiento 5: Variedad FA 593 sin inoculación
Tratamiento 6: Variedad FA 593 inoculada con aislado 593
Tratamiento 7: Variedad Yonit sin inoculación
Tratamiento 8: Variedad Yonit inoculada con aislado 593
3.2.2.4. Diseño estadístico
Los experimentos fueron conducidos mediante un diseño de bloques
completos al azar. La unidad experimental correspondió a 10 plantas, las que
fueron observadas y evaluadas. Se realizaron ocho tratamientos incluyendo

31
un testigo los cuales se distribuyeron aleatoriamente sobre cada una de los
cuatro bloques en la nave de invernadero del ensayo. Se procedió a bloquear
los tratamientos debido a las variables riego y luminosidad dentro del
invernadero. La distribución de los tratamientos se observa en el Anexo 4.
El análisis de datos fue igual al Ensayo 1.
3.2.2.5. Variables evaluadas
Al igual que en el Ensayo 1, el daño fue estudiado mediante estimaciones del
índice de daño y el porcentaje de incidencia de la enfermedad. Se
consideraron además las variables altura, peso fresco y peso seco, con la
intención de establecer diferencias en la agresividad del ataque de acuerdo a
la variedad.
La evaluación de la escala de daño fue realizada un mes después de la
inoculación, el día 2 de septiembre de 2004, cuando se evidenciaron los
síntomas en la parte aérea de las plantas.
Para la evaluación de las variables altura de plantas, peso fresco y peso
seco, se cosecharon las plantas extrayéndolas con el sistema radical lo más
íntegro posible. La altura de la planta se obtuvo midiendo la distancia desde
el cuello hasta el ápice de las plantas, utilizándose una huincha graduada en
centímetros. Consecutivamente se procedió a lavar las raíces, las cuales
fueron secadas con papel absorbente y debidamente pesadas junto con el
follaje. Posteriormente cada planta se secó a 62ºC hasta lograr un peso
constante, para luego medir el peso seco.

32
Para el análisis estadístico de estas tres variables se procedió a realizar un
test de Tukey a las diferencias obtenidas entre los tratamientos testigos y los
tratamientos inoculados para cada variedad. El objetivo fue obtener el
porcentaje de disminución de cada una de las variables, para las cuatro
variedades.
3.2.2.6. Recuperación de Pseudomonas syringae desde lesiones y tejido
sano de plantas de tomate con nueva sintomatología.
Se realizó un seguimiento de la bacteria a lo largo de la planta, para lo cual
se eligieron al azar dos plantas enfermas por variedad y se efectuaron
aislamientos a partir de tejido de tres puntos de las plantas enfermas: desde
la zona de avance considerada como zona basal, a 5 cm del punto anterior
hacia el ápice o zona media y a 10 cm de la zona de avance o zona alta. Los
trozos de tallo se cortaron con un bisturí estéril. Los tejidos fueron
desinfectados bajo cámara de flujo laminar del laboratorio, con hipoclorito de
sodio al 1 % por 15 segundos; luego, se lavaron tres veces con agua
destilada estéril, se maceraron, y el extracto fue sembrado con un asa en
placas Petri en MBK. Estas placas se identificaron, se sellaron con parafilm y
luego se colocaron en estufa a 23ºC por dos días, para posteriormente
verificar la fluorescencia de las placas bajo luz cercana a ultravioleta (320
nm). Se evaluó desde que zona existía crecimiento bacteriano, para estudiar
si la enfermedad se desarrolla en forma sistémica.
3.2.3. Ensayo 3: Test de patogenicidad en frejol caupí.
Para efectuar el test de patogenicidad se emplearon las semillas de frejol
caupí variedad Blackeye, las cuales fueron facilitadas por el Ingeniero
Agrónomo, M. Sc. Gabriel Bascur B. del Departamento de Horticultura y

33
Cultivos del Instituto Nacional de Investigación Agropecuaria, CRI-INIA La
Platina.
El día 11 de noviembre de 2004, se inició el proceso de imbibición de las
semillas en agua destilada estéril por 24 horas; posteriormente se
introdujeron en placas Petri con papel absorbente levemente húmedo. Las
placas se incubaron a 23°C en estufa durante dos días para inducir el
proceso de germinación.
El día 15 de noviembre de 2004, se sembraron semillas previamente
germinadas y a las tres semanas de crecimiento se procedió con la
inoculación de las plantas. Al igual que en el primer y segundo ensayo se
utilizó carborundum, para generar microheridas en los folíolos de las plantas
y facilitar el ingreso del patógeno.
La recuperación de los aislados 588, 593, Sta Rosa, Mclean y 112
correspondió al mismo proceso realizado en los ensayos anteriores, y la
inoculación de las soluciones bacterianas se realizó mediante un asperjador
manual estéril, hasta dejar las hojas de las plantas de caupí notoriamente
húmedas. Las plantas testigo también fueron rociadas con carborundum y
posteriormente fueron asperjadas con agua estéril.
Post inoculación todas las plantas fueron cubiertas en bolsas transparentes
de polietileno por 36 hrs para proveer de alta humedad. Las plantas fueron
ubicadas en un invernadero climatizado en un rango de temperatura entre 15
y 25ºC. Los síntomas fueron registrados 15 días después de la inoculación.

34
3.3. Análisis realizados por IMI, Inglaterra:
El día 28 de diciembre 2004 fue enviado el aislado 593 al CAB International
Mycological Institute, Inglaterra, para corroborar la identificación del agente
causal. En este instituto se identificó la especie y patovar mediante análisis
de ácidos grasos y descripción molecular utilizando la técnica rep-PCR.

35
4. PRESENTACIÓN Y DISCUSIÓN DE RESULTADOS
4.1. Caracterización de los aislados de Pseudomonas syringae (in vitro):
4.1.1. Utilización de sustratos orgánicos
A través del cambio en la coloración en el medio de cultivo Ayers y a la
fluorescencia bajo luz ultra violeta, que indica crecimiento de la cepa
bacteriana a causa de la utilización de la fuente de carbono adicionada, se
evaluó el uso tanto de eritritol como DL-lactato, por parte de los aislados
causantes de la nueva sintomatología y de P. syringae pv. syringae (cepa
112), cuyos resultados se presentan en el Cuadro 2.
CUADRO 2. Análisis de aislados de Pseudomonas syringae enfrentándose a diferentes fuentes de carbono.
Medio Cepa 112 (pv. syringae) Aislados 290 - 593 - 588 - McLean - Fdo. Sta Rosa
Ayers Medio base de color celeste.
Sin fluorescencia en UV.
Medio base de color celeste .
Sin fluorescencia en UV.
Ayers con
Eritritol
Medio de color celeste amarillo.
Con fluorescencia
Cepa 112 eritritol positivo
Medio de color celeste claro.
No presentan fluorescencia
Respuesta negativa
Ayers con
DL-lactato
Medio vira a azul.
Con fluorescencia
Cepa 112 DL-lactato positivo
Medio de color celeste claro.
No presentan fluorescencia
Respuesta negativa

36
Se debe considerar que el medio base de sales minerales Ayers se utiliza
como control.
El análisis muestra en el Cuadro 2, que sólo la cepa 112 correspondiente a
P. syringae pv. syringae presenta un viraje en la coloración del medio, lo cual
corrobora lo descrito por SCORTICCHINI (1995), que indica un cambio de
coloración en el medio de cultivo como la respuesta positiva, debido al
crecimiento bacteriano.
Además SCHAAD (1980), GITAITIS (1993), YOUNG y TRIGGS (1994),
SCORTICCHINI (1995) y BRAUN-KIEWNICK y SANDS (2001), señalan que
P. syringae pv. syringae tiene respuesta positiva a la utilización de eritritol y
DL-lactato y no así P. syringae pv. tomato.
Asimismo, JONES, McCARTER y GITAITIS (1986), obtuvieron crecimiento y
fluorescencia, al analizar la utilización de DL-lactato como fuente de carbono
para las cepas de P. syringae pv. syringae; la misma respuesta obtuvieron al
utilizar como sustrato orgánico de crecimiento a eritritol. Por otro lado, las
pruebas realizadas a las cepas de P. syringae pv. tomato no mostraron
crecimiento ni fluorescencia para ambas fuente de carbono.
4.1.2. Capacidad de formar núcleos de hielo
La capacidad de formar núcleos de hielo por las bacterias en estudio, se
evaluó mediante la congelación de las suspensiones bacterianas a -4°C. Los
resultados se visualizan en la Figura 1 a, donde se evidencia claramente que
sólo una suspensión bacteriana se congela y es la correspondiente a la cepa
112 de P. syringae pv. syringae. Ninguno de los aislados causantes de la
nueva sintomatología fue capaz de congelarse.

37
Estos resultados coinciden con lo señalado por GITAITIS (1993) y LUDWIG y
KLENK (2001) quienes indican que las cepas de P. syringae pv. syringae son
activas a la nucleación de hielo, lo cual diferencia este organismo de P.
syringae pv. tomato.
Además los resultados concuerdan con estudios anteriores (JONES,
McCARTER y GITAITIS, 1981; BONN y GITAITIS, 1987; SCORTICHINI,
1995; CINTAS, KOIKE y BULL, 2002) que ratifican la capacidad de formar
núcleos de hielo en diferentes cepas de Pseudomonas syringae y
concuerdan con la utilidad de esta técnica para discriminar entre los
patovares en estudio.

38
FIGURA 1. (a) Respuesta a la capacidad de formar núcleos de hielo de
diferentes aislados bacterianos causantes de la nueva sintomatología, una cepa de P. syringae pv. syringae (2ª de izq. a derech.) y un tratamiento testigo. (b) Recuperación del aislado 593 desde tres puntos de la planta en placas con medio B de King observadas bajo luz ultravioleta. Las placas desde izquierda a derecha corresponden a la zona de avance, zona media (a 5 cm de la zona de avance hacia el ápice) y zona alta (a 5 cm de la zona anterior).
a
b

39
4.2. Ensayos de virulencia, susceptibilidad varietal y patogenicidad (in vivo):
4.2.1. Ensayo 1: Evaluación de virulencia de cuatro aislados de P. syringae
causantes de la nueva sintomatología.
De acuerdo a los resultados obtenidos en el Cuadro 3, se puede afirmar que
se observaron diferencias significativas en cuanto a incidencia de la
enfermedad, ya que se visualiza en forma clara que sólo los aislados
causantes de la nueva sintomatología tienen el máximo porcentaje de
incidencia, mientras que los tratamientos testigo (T1), como los inoculados
con la cepa 112 (T2) no presentan síntomas de enfermedad.
CUADRO 3. Efecto de diferentes aislados bacterianos causantes de la nueva sintomatología, una cepa de P. syringae pv. syringae y un tratamiento testigo sobre la incidencia de la nueva sintomatología en plantas de tomate cv. Fortaleza.
Tratamiento Incidencia de la Enfermedad (%)
T1: Testigo (planta sin inocular) 0 a
T2: Cepa P. syringae pv. syringae 0 a
T3: Aislado 290 100 b
T4: Aislado 588 100 b
T5: Aislado 593 100 b
T6: Aislado Sta. Rosa 100 b
Promedios con letras iguales dentro de una columna no presentan diferencias significativas (α=0,05), según test de Tukey.
Por otro lado, el máximo índice de daño es alcanzado por todos los
tratamientos inoculados con los aislados causantes de la nueva

40
sintomatología, los cuales difieren significativamente de los tratamientos
testigo (T1) y los inoculados con la cepa 112 (T2) (Figura 2).
Tanto el Cuadro 3 como la Figura 2, revelan que todas las cepas causantes
de la nueva sintomatología son igualmente virulentas, con una incidencia del
100% y con el grado de daño mayor de la escala. Lo anterior deja en
evidencia que las cepas en estudio no corresponden a P. syringae pv.
syringae y que todas ellas poseen igual capacidad para desarrollar la
enfermedad.
Estos resultados concuerdan con lo experimentado por LAWNTON y
MAcNEILL (1986) y CINTAS, KOIKE y BULL (2002), quienes señalan que no
hay desarrollo de la enfermedad en plantas de tomate al inocular con cepas
de P. syringae pv. syringae y si hay desarrollo de la enfermedad en las
plantas inoculadas con P. syringae pv. tomato. Lo observado concuerda con
lo descrito por GITAITIS (1993), quien advierte que es sumamente difícil
reproducir los síntomas de P. syringae pv. syringae bajo condiciones de
invernadero. Una posible explicación a esto es que P. syringae pv. syringae
en plantas de tomate actúa como un patógeno débil y oportunista, que se
encuentra frecuentemente en heridas y como un organismo secundario en
infecciones mixtas, sin desarrollar sintomatología, pero presente en forma
epífita en el vegetal (GITAITIS, 1993; GABOR y WIEBE, 1997).
Referente a la sintomatología observada, esta se presentó igual a la descrita
en plantas naturalmente infectadas (Figura 3 a). Post-inoculación los
síntomas se iniciaron en los folíolos en forma de pecas o pequeñas manchas
necróticas rodeadas de halo clorótico, posteriormente se ennegrecieron los
pecíolos y luego avanzó en forma contagiosa hacia la base del tallo de la
planta, iniciando un movimiento ascendente (Figura 3 b), el cual se detuvo en

41
la parte superior, produciendo un severo atizonamiento a lo largo de los tallos
el cual no se prolongó a las hojas superiores ni a los frutos. El daño se
traduce en un menor desarrollo de las plantas (Figura 3 c). Cabe destacar
que estas lesiones corticales necróticas son superficiales y no comprometen
el tejido interno, lo que se visualiza al hacer cortes transversales. Esto
permite confirmar que la bacteria posee un movimiento periférico vía células
epidermicas o subepidérmicas.
Llama la atención la forma autónoma de avance de la lesión dentro de la
planta. Esto puede ser explicado por la falta de medios que propicien la
difusión de la bacteria como son la lluvia, el riego por aspersión, la
manipulación, el roce entre plantas por viento, maquinarias, etc. Factores
que ayudan a la dispersión y movimiento de la bacteria tanto dentro de las
plantas como entre ellas. Estas condiciones no se ven favorecidas bajo
invernadero, lo cual podría aclarar lo ocurrido tanto en los ensayos realizados
como en plantas naturalmente infectadas, donde se detiene el movimiento de
la bacteria y no hay presencia de síntomas en frutos y hojas superiores.
Por otra parte esta particular forma de avance de la lesión y el severo
atizonamiento en los tallos no corresponde a la tradicional descripción de la
sintomatología que ocasiona P. syringae pv. tomato, donde destaca el daño
en los frutos. En Chile ha sido descrita únicamente en tomate cultivado al
aire libre en la VI y VII región, específicamente en las zonas de Talca,
Rengo, Pelequen y Requinoa. En estos lugares la sintomatología origina
manchas negras de pequeño tamaño rodeadas de halo amarillo en las hojas
que pueden confluir llegando a secar foliolos. Tallos, pecíolos y bordes de los
sépalos, también pueden presentar manchas negras de contorno irregular.
Los frutos se afectan en estado verde, donde se observan pequeñas
manchas deprimidas (APABLAZA, 2000).

42
bbbb
aa0
1
2
3
Fortalezas/inocular
Fortaleza + cepapv. syringae
Fortaleza +aislado290
Fortaleza +aislado588
Fortaleza +aislado593
Fortaleza +aisladoSta.Rosa
Tratamientos
Indi
ce d
e D
año
FIGURA 2. Efecto de diferentes aislados bacterianos causantes de la nueva
sintomatología, una cepa de P. syringae pv. syringae y un tratamiento testigo, sobre el índice de daño de la nueva sintomatología en plantas de tomate cv. Fortaleza. Tratamientos seguidos por letras iguales no presentan diferencias significativas (α=0,05), según test de Tukey.

43
FIGURA 3. Plantas de tomate con nueva sintomatología provocada por
aislados de Pseudomonas syringae (a) Planta de tomate naturalmente infectada. (b) Planta de tomate con manchas necróticas puntiformes sobre las hojas y atizonamiento de tallo (nivel de daño 3). (c) Plantas de tomate variedad Fortaleza inoculadas con aislado 593 en comparación con sus testigos.
a
b
c
Plantas testigo Plantas
inoculadas

44
4.2.2. Ensayo 2: Evaluación de la susceptibilidad de cuatro variedades de
tomate al aislado 593.
De acuerdo a los resultados obtenidos en la evaluación de la susceptibilidad
de cuatro variedades de tomate (Cuadro 4), se puede afirmar que hay
diferencias significativas en cuanto a incidencia de la enfermedad, ya que se
observa que todas la variedades inoculadas con el aislado 593 tienen el
máximo porcentaje de incidencia y cada uno de sus respectivos tratamientos
testigo (sin inoculación) no desarrollan la enfermedad en las plantas.
CUADRO 4. Incidencia del aislado 593 causantes de la nueva sintomatología inoculado sobre diferentes variedades de tomate y sus respectivos testigos.
Tratamiento Incidencia de la Enfermedad (%)
T1: Variedad Fortaleza sin inocular 0 a
T2: Variedad Fortaleza inoculada con aislado 593 100 b
T3: Variedad Naomi sin inocular 0 a
T4: Variedad Naomi inoculada con aislado 593 100 b
T5: Variedad FA 593 sin inocular 0 a
T6: Variedad FA 593 inoculada con aislado 593 100 b
T7: Variedad Yonit sin inocular 0 a
T8: Variedad Yonit inoculada con aislado 593 100 b
Promedios con letras iguales dentro de una columna no presentan diferencias significativas (α=0,05), según test de Tukey.
Como se puede observar en la Figura 4, el máximo índice de daño es
alcanzado en todas las variedades en estudio por los tratamientos inoculados
con el aislado 593, causante de la nueva sintomatología, los cuales difieren
significativamente de sus respectivos tratamientos testigo.

45
FIGURA 4. Índice de daño de la aislado 593 causantes de la nueva
sintomatología inoculada sobre diferentes variedades de tomate y sus respectivos testigos. Tratamientos seguidos por letras iguales no presentan diferencias significativas (α=0,05), según test de Tukey.
a
b
a
bb
a
b
a
0
1
2
3
Fortaleza Naomi FA 593 Yonit
Tratamientos
Indi
ce d
e D
año

46
4.2.2.1. Altura, peso fresco y peso seco
En relación con la altura de planta, el peso fresco y el peso seco, se buscó
medir la diferencia existente entre las plantas testigos y las inoculadas, con la
intención de estudiar el comportamiento de las variables evaluadas para
cada una de las variedades.
CUADRO 5. Porcentaje de variación entre plantas testigos e inoculadas con aislado 593 en las variedades de tomate Fortaleza, FA 593, Yonit y Naomi, para las variables altura, peso fresco y peso seco.
Promedios con letras iguales dentro de una columna no presentan diferencias significativas (α=0,05), según test de Tukey.
Al analizar el Cuadro 5, la variable altura, se observa que la variedad FA 593
puede ser catalogada como la variedad que se ve significativamente menos
disminuida en tamaño en comparación con Fortaleza y Yonit.
Lo anterior deja en evidencia que si bien en el presente ensayo se obtuvo
diferencia en el tamaño de la parte aérea, estos resultados no expresan que
la variedad FA 593 sea resistente o tolerante, ya que en los ensayos
Variedad Disminución de altura (%)
Disminución de peso fresco (%)
Disminución de peso seco (%)
Fortaleza 35,02 a 66,88 a 67,08 a
FA 593
22,42 b 47,07 b 49,34 b
Yonit 34,82 a 54,63 a b 46,42 b
Naomi
29,28 a b
57,39 a b
50,03 b

47
realizados previamente se comprobó que todas las variedades obtienen el
máximo índice de daño y 100 % de incidencia de la enfermedad.
En relación con la disminución del peso fresco de las plantas inoculadas y
sus respectivos testigos, Yonit y Naomi mostraron estadísticamente el mismo
comportamiento, el cual se encuentra en un sitial intermedio entre FA 593 y
Fortaleza. FA 593 al igual que en la variable altura se ve menos afectada en
la disminución del peso fresco y se diferencia significativamente de Fortaleza,
variedad que se ve mayormente afectada por el aislado bacteriano.
Esto se acentúa aun más en la disminución de peso seco donde Fortaleza
marca una diferencia notable por un altísimo porcentaje de disminución, el
cual es significativamente mayor en comparación con las otras variedades.
Estos estudios concuerdan con lo observado en la Figura 3 c, puesto que en
lo referente al peso fresco, y altura de las plantas inoculadas en comparación
con sus respectivos testigos presentan diferencias evidentes.
Finalmente, se puede afirmar que la bacteria tuvo una influencia detrimental
en el desarrollo de las plantas inoculadas, y aun cuando todos los
tratamientos se ven severamente afectados en comparación con sus
respectivos testigos, en general es la variedad Fortaleza quien tiene la
diferencia mayor en todas las variables, por lo tanto, se convierte a la más
susceptible al ataque en comparación con las otras tres variedades incluidas
en este ensayo.
Esto concuerda con lo observado a nivel de campo, donde todos los predios
que presentaron este problema utilizaban la variedad Fortaleza.
48
4.2.2.2. Recuperación de Pseudomonas syringae desde lesiones y tejido
sano de plantas de tomate con nueva sintomatología
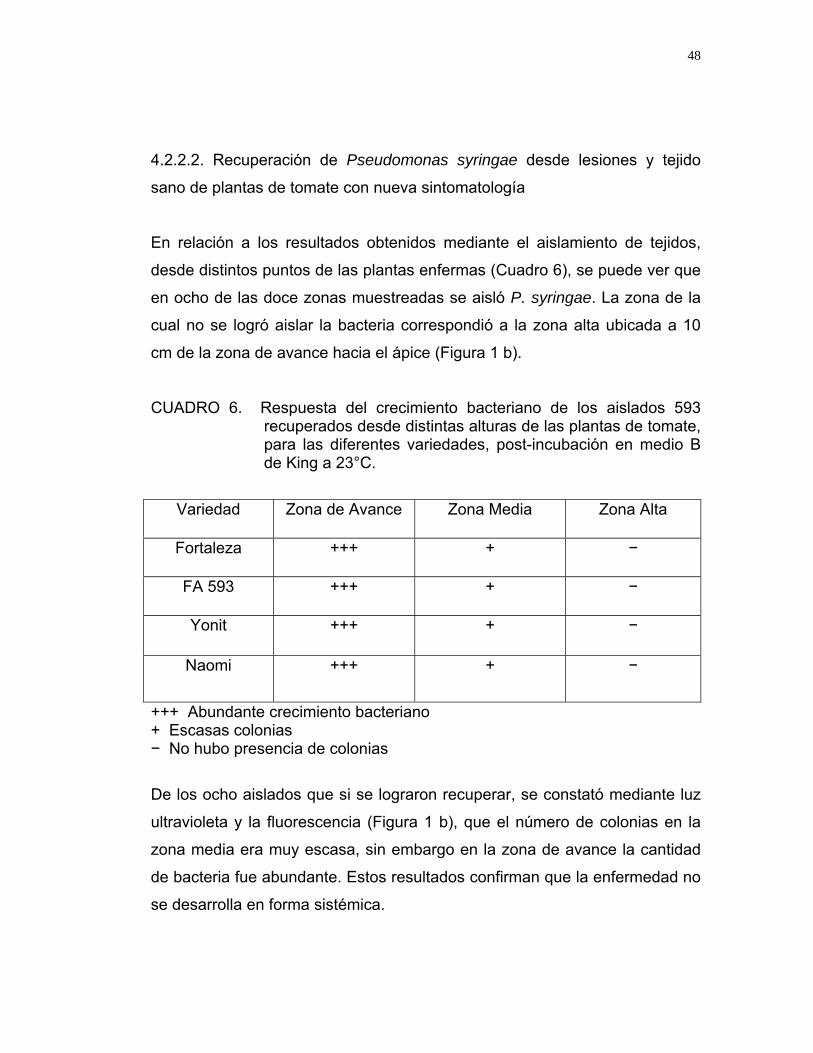
En relación a los resultados obtenidos mediante el aislamiento de tejidos,
desde distintos puntos de las plantas enfermas (Cuadro 6), se puede ver que
en ocho de las doce zonas muestreadas se aisló P. syringae. La zona de la
cual no se logró aislar la bacteria correspondió a la zona alta ubicada a 10
cm de la zona de avance hacia el ápice (Figura 1 b).
CUADRO 6. Respuesta del crecimiento bacteriano de los aislados 593 recuperados desde distintas alturas de las plantas de tomate, para las diferentes variedades, post-incubación en medio B de King a 23°C.
Variedad Zona de Avance Zona Media Zona Alta
Fortaleza +++ + −
FA 593 +++ + −
Yonit +++ + −
Naomi +++ + −
+++ Abundante crecimiento bacteriano + Escasas colonias − No hubo presencia de colonias
De los ocho aislados que si se lograron recuperar, se constató mediante luz
ultravioleta y la fluorescencia (Figura 1 b), que el número de colonias en la
zona media era muy escasa, sin embargo en la zona de avance la cantidad
de bacteria fue abundante. Estos resultados confirman que la enfermedad no
se desarrolla en forma sistémica.

49
4.2.3. Ensayo 3: Test de patogenicidad en frejol caupí
Como se observa en el Cuadro 7, los aislados de P. syringae pv. syringae
produjeron lesiones necróticas rodeadas por halos en todas las repeticiones
de frejol caupí a los 15 días después de ser inoculadas. Ningún testigo ni
aislado de P. syringae produjo la reacción típica de síntomas.
CUADRO 7. Respuesta de las inoculaciones realizadas en frejol caupi con diferentes aislados bacterianos causantes de la nueva sintomatología, una cepa de P. syringae pv. syringae (cepa 112) y un tratamiento testigo. Se inocularon un total de cuatro plantas por aislado.
Aislado Plantas sanas Plantas con síntomas
593 4 0
Sta. Rosa 4 0
588 2 2*
McLean 4 0
112 0 4
Testigo 4 0
* Sintomatología diferente: áreas de foliolos con síntomas de escaldado.
Las dos plantas con síntomas correspondientes al aislado 588 no
presentaron los mismos síntomas que la cepa 112, sino que produjeron
aureolas blancas (escaldado). Se pueden considerar como una reacción
posible ya que ha sido descrita por JONES, McCARTER y GITAITS (1981),
quienes además señalan que la reacción típica de patogenicidad
corresponde a lesiones necróticas rodeadas por halos cloróticos, como los
manifestados en este ensayo, por lo que podemos decir que el test con frejol

50
caupí resultó de acuerdo a lo esperado, es decir, positivo sólo para P.
syringae pv. syringae.
Estos resultados concuerdan con lo descrito por GITAITIS (1993), quien
señala que la mayoría de las cepas de P. syringae pv. syringae recuperadas
desde tomate, inducen una reacción en frejol caupí cultivar California
Blackeye Nº3.
4.3. Análisis realizados por IMI, Inglaterra:
Los resultados de los análisis realizados por el Dr. L. Offord del IMI, registro
número 392758 (Anexo 5), revelaron que el agente causal corresponde a
Pseudomonas syringae y la respuesta al rep-PCR realizado reveló un perfil
idéntico al registrado tanto para P. syringae pv. tomato, como para P.
syringae pv. maculicola. Al ser el aislado 593 inicialmente recuperado desde
tomate, el IMI afirma con un alto grado de confianza que esta cepa
corresponde a P. syringae pv. tomato.
Lamentablemente la caracterización de P. syringae pv. tomato y P. syringae
pv. maculicola es idéntica, por lo cual tanto las pruebas bioquímicas como
biológicas arrojan la misma respuesta (LUDWIG y KLENK, 2001; BRAUN-
KIEWNICK y SANDS, 2001).
De acuerdo a esto, ZHAO et al. (2000) señalan en su estudio que al analizar
los perfiles oxidativos de la utilización de sustratos orgánicos entre cepas de
estos patovares, el porcentaje de semejanza corresponde a un 79,5%. Por
su parte WIEBE y CAMPBELL (1993) se refieren a la estrecha relación
existente entre P. syringae pv. maculicola y P. syringae pv. tomato y

51
aseveran que dentro de la especie P. syringae, estos son los patovares más
cercanos, debido a que poseen más del 75% de homología en su ADN.
Es importante destacar que P. syringae pv. maculicola en la caracterización
del rep-PCR posee dos perfiles definidos posibles, A o B. Esto indica que
puede haber convergido de al menos dos líneas evolucionarias. Por lo tanto,
al analizar los patrones de banda entre este patovar y P. syringae pv. tomato
puede ocurrir que este último revele el patrón de banda correspondiente al
grupo A de P. syringae pv. maculicola (ZHAO et al., 2001).
Por su parte CINTAS et al. (2001) y BRADBURY (1986) indican que todos los
hospederos de P. syringae pv. maculicola corresponden a crucíferas y la
mayor parte pertenece al género Brassica. Además LOPEZ y MONTESINOS
(2000) especifican como hospederos naturales a la col y la coliflor.
Sin embargo WIEBE y CAMPBELL (1993) prueban que al inocular en tomate
y coliflor cepas de los patovares maculicola y tomato, ambos fueron capaces
de infectar en tomate. No obstante la respuesta obtenida, aseveran que
estos patovares corresponden a entidades diferentes, ya que aún cuando las
cepas de P. syringae pv. maculicola causaron lesiones en hojas de tomate,
estas fueron diferentes a las naturalmente originadas por patovar tomato, el
cual no logró ser patogénico en coliflor. Esto concuerda con los estudios
realizados por CINTAS, BULL y KOIKE (2002) quienes indican que P.
syringae pv. tomato no fue patogénica en las crucíferas por ellos utilizadas
(sí en tomate), mientras que todas las cepas de P. syringae pv. maculicola si
reprodujeron enfermedad en crucíferas, y en tomate se describe una
reacción variable.

52
Esta información del rango de hospederos apoya la hipótesis de que el
patovar en estudio no se considera como una bacteria capaz de ocasionar
enfermedad en tomate y además no ha sido descrita en Chile (BRADBURY,
1986).
A partir de lo anterior, y considerando que no hay reportes que incluyan al
tomate como hospedero de P. syringae pv. maculicola, se puede afirmar que
el perfil descrito por el IMI corresponde a P. syringae pv. tomato.
4.4. Resumen de pruebas biológicas, bioquímicas y patogénicas:
De acuerdo a las diferentes pruebas biológicas, bioquímicas y de
patogenicidad realizadas (Cuadro 8), la nueva sintomatología originada por
los aislados en estudio, se ajusta a la especie y patovar P. syringae pv.
tomato, lo que concuerda con los resultados obtenidos por, las pruebas
bioquímicas conseguidas por LAWNTON y MAcNEILL (1986); CINTAS,
KOIKE y BULL (2002) y MALAVOLTA et al. (2002) y a la respuesta negativa
observada por JONES, McCARTER y GITAITIS (1986) en la formación de
núcleos de hielo.
Lo anterior, más la respuesta del IMI que corrobora que el aislado 593
corresponde a P. syringae pv. tomato permite afirmar que esta bacteria
causante de una sintomatología nunca antes reportada, corresponde a una
nueva variante o raza, sobre todo al considerar la magnitud del daño
ocasionado en las plantas afectadas a nivel de campo.

53
CUADRO 8. Caracterización de cepas de Pseudomonas syringae pv. syringae y Pseudomonas syringae causantes de la nueva sintomatología.
Reacción
Test P. syringae pv. syringae
P. syringae causante de nueva
sintomatología Levano + + Oxidasa - -
Pudrición en Papa - - Arginina dihidrolasa - -
Hipersensibilidad en Tabaco + + Fluorescencia en MB de King + +
Utilización de Eritritol + - Utilización DL-Lactato + -
Formación de núcleos de hielo + - Patogenicidad en caupí + -

54
5. CONCLUSIONES
En los tratamientos in vivo, se puede concluir que los aislados 290, 593, 588
y Sta. Rosa atacan con igual nivel de daño a la variedad Fortaleza, actuando
todas con igual grado de virulencia.
Las cuatro variedades evaluadas resultaron ser altamente susceptibles al
aislado bacteriano en estudio. Sin embargo FA-593 resultó ser más tolerante
y Fortaleza fue la variedad más susceptible.
De acuerdo a los resultados obtenidos en los ensayos in vitro, junto a la
respuesta del análisis molecular realizado por el CAB International
Mycological Institute, se puede concluir que los aislados causantes de la
nueva sintomatología corresponden a Pseudomonas syringae pv. tomato. De
acuerdo a la sintomatología observada, correspondería a una nueva variante
o raza de este patovar.

55
6. RESUMEN
Durante el año 2003, se observó una nueva sintomatología asociada a Pseudomonas syringae afectando plantas de tomate en invernadero frío. Se detectaron manchas necróticas rodeadas de halo clorótico en los folíolos y por sobre todo un severo atizonamiento en pecíolos y tallos. En un estudio preliminar se determinó que la especie correspondía a P. syringae [Fitopatología 39(1):54-55 (2004)]. En la presente investigación se realizaron pruebas bioquímicas para diferenciar patovares. Las pruebas de eritritol y DL-lactato indicaron que se trataría de P. syringae pv. tomato. Igual resultado se obtuvo al evaluar la capacidad de formar núcleos de hielo a -4ºC. Posteriormente se evaluó la virulencia de cuatro aislados en estudio (290, 588, 593 y Sta. Rosa) en comparación a P. syringae. pv. syringae (cepa 112, aislada de cerezo) y los respectivos testigos. Los cuatro aislados fueron capaces de producir síntomas iniciales de manchas necróticas circulares y halo clorótico en hojas, y al cabo de 10 días un atizonamiento de los tallos y pecíolos, aspectos que no fueron observados en las plantas tratadas con P. syringae pv. syringae, ni en las plantas testigo. Luego se diseñó un segundo ensayo, inoculándose el aislado 593 utilizado en el Ensayo 1, en los cvs. Fortaleza, FA 593, Naomi y Yonit. Todas las variedades presentaron una reducción en la altura, peso fresco y peso seco, en comparación con sus respectivos testigos. Sin embargo, se afectó en mayor grado el cv. Fortaleza. Paralelamente fue enviado el aislado 593 al CAB International Mycological Institute, Inglaterra, para la identificación del agente causal mediante rep-PCR. La respuesta del análisis molecular coincide con la caracterización de este aislado como patovar tomato. En base a estos resultados, y de acuerdo con los síntomas observados, que difieren a los descritos para P. syringae pv. tomato, se concluye la presencia de una nueva variante o raza de este patovar afectando a plantas de tomate bajo invernadero en la zona de Quillota y Limache, V región, Chile, problema denominado atizonamiento bacteriano del tomate.

56
7. ABSTRACT
During the year 2003, new symptoms related to Pseudomonas syringae were observed, affecting unheated greenhouse tomato plants. Severe necrotic spots surrounded by chlorotic halos and an overall blight in petioles and stems were detected. A preliminary study determined that this species corresponded to P. syringae [Fitopatología 39(1):54-55 (2004)]. In this study biochemical tests were done in order to differentiate pathovars. The erythritol and DL-lactate tests suggested that it could be P. syringae pv. tomato. The same result was obtained from evaluating the ice nucleation activity at -4ºC. Subsequently, the virulence of four isolates being studied (290, 588, 593 and Sta. Rosa) was evaluated and compared to P. syringae. pv. syringae (strain 112, isolated from cherry tree) and the corresponding negative controls. The four isolates produced the initial symptoms of circular necrotic spots and chlorotic halos on leaves, and after 10 days, blighting of stems and petioles, none of which were observed on plants treated with P. syringae. pv. syringae, nor in the negative control plants. A second test was then designed, in which the tomato cultivars Fortaleza, FA 593, Naomi and Yonit, were inoculated with isolate 593, used in Test 1. All of the varieties showed a reduction in height, fresh weight and dry weight, compared with their respective negative controls. Nevertheless, cv Fortaleza was the most strongly affected. At the same time, isolate 593 was sent to the CAB International Mycological Institute, England, for an identification of the causal agent by rep-PCR. The result of the molecular analysis coincided with the characterization of this strain as pathovar tomato. Based on these results, and on the symptoms observed, which differ from those described for P. syringae pv. tomato, the presence of a new variant of this pathovar is deduced, which affects greenhouse tomato plants in the area of Quillota and Limache, V region, Chile, and has been designated bacterial blight of tomatoes.

57
8. LITERATURA CITADA
AGRIOS, G. 1996. Fitopatología. Segunda Edición. México D.F., Editorial Limusa. 838 p. APABLAZA, G. 2000. Patología de cultivos. Epidemiología y control
holístico. Santiago, Ediciones Universidad Católica de Chile. 392 p. ARREDONDO, C.R., DAVIS, R.M. 2000. First report of Pseudomonas
syringae pv. tomato Race 1 on tomato in California. Plant Disease 84(11):371.
BIOAMERICA. 2004. Tomates indeterminados, Fortaleza. Santiago,
Bioamerica. 1 p. ________. 2004. Tomates indeterminados, Naomi. Santiago, Bioamerica
S.A. 1 p. BESOAIN, X. 1989. Manejo de enfermedades bajo plástico. Universidad
Católica de Valparaíso. Facultad de Agronomía. Curso Internacional. Uso de plástico en cultivos forzados de hortalizas y flores. Quillota. 2,3,4,5 de Octubre 1989. pp. L1-L9.
________. McLEAN, C., LATORRE, B. 2004. Presencia de un aparente
nuevo patovar o variante de Pseudomonas syringae encontrado en tomates de la V región. Fitopatología 39(1):54-55.
BLANCARD, D. 1990. Enfermedades del tomate. Madrid, Mundi Prensa.
212 p.

58
BONN, W., GITAITIS, R. 1987. Ice nucleation and disease caused by Pseudomonas syringae pv. syringae in tomato transplants. Phytopathology 72(12):1729.
BRADBURY, J.F. 1986 Pseudomonas. In: Guide to plant pathogenic
bacteria. England, CAB International Mycological Institute. pp 110-185. BRAUN-KIEWNICK, A., SANDS, D. 2001. Pseudomonas. In: Shaad, N.,
Jones, J., Chun, W. eds. Laboratory guide for identification of plant pathogenic bacteria. Third edition. Minnesota, American Phythopathological Society Press. pp.84-120.
BRUNA, A. 1993. Enfermedades del tomate y pepino de ensalada bajo
plástico y su control. Instituto de Investigación Agropecuaria. Curso internacional, producción de hortalizas protegidas bajo plástico. Santiago, noviembre de 2003. pp.10-20.
CHAMARRO, J. 1995. Anatomía y fisiología de la planta. In: Nuez, F. ed.
El cultivo del tomate. Madrid, Mundi-Prensa. pp.43-91. CINTAS, N.A., BULL, C.T., KOIKE, S.K. 2002. A new pathovar,
Pseudomonas syringae pv. alisalensis pv. nov.; proposed for the causal agent of bacterial blight of Broccoli and Broccoli Raab. Plant Disease 86(9):992-998.
CINTAS, N.A., BULL, C.T., KOIKE, S.K., BOUZAR, H. A. 2001. New
bacterial leaf spot disease of Broccolini, caused by Pseudomonas syringae pv. maculicola, in California. Plant Disease 85:1207.
COMMONWEALTH MYCOLOGICAL INSTITUTE. 1985. Manual para
patólogos vegetales. Surrey, Organización de las Naciones Unidas para la Agricultura y la Alimentación. Oficina regional FAO para América Latina y El Caribe. 438 p.

59
DAVIS, R.M., MIYAO, G., SUBBARAO, K.V., STAPLETON, J.J., FALK, B.W. 2003. UC Pest management guidelines. Tomato. bacterial speck, (on line). http://www.ipm.ucdavis.edu/PMG/r783101611.html.
DYE, D.W., BRADBURY, J.F., GOTO, M., HAYWARD, A. C., LELLIOT, R.
A., SCHROTH, M.N. 1980. International standards for naming pathovars of phytopathogenic bacteria and a list of pathovar names and pathogenic strains. Review of Plant Pathology 59:153-168.
EUZÉBY, J.P. 2005. List of bacterial names with standing in nomenclature,
(on line). http://www.bacterio.cict.fr/mr.html. FLOYD, R. 2004 Vegetable seed treatments. Farmnote 90/1990, (on line).
http://agspsrv34.agric.wa.gov.au/agency/pubns/farmnote/1990/F09090.HTM.
FLOYD, R. 1990. Bacterial and fungal diseases of tomatoes. Farmnote
93/91, (on line). http://agspsrv34.agric.wa.gov.au/agency/pubns/farmnote/1991/F09391.htm.
GABOR, B., WIEBE, W. 1997. Tomato Diseases. Saticoy. California,
Seminis Vegetable Seed, INC. 62 p. GIACONI, M., ESCAFF, M. 1993. Cultivo de hortalizas. Santiago, Editorial
Universitaria. 332 p. GITAITIS. R.D. 1993. Syringae Leaf Spot. In: Jones, J.B., Jones, J.P., Stall,
R.E., Zitter, T.A. eds. Compendium of tomato diseases. Minnesota, A.P.S. pp. 29-30.
GONZÁLEZ, A.J., RODICIO, M.R., MENDOZA, M.C. 2003. Pseudomonas
viridiflava, un patógeno emergente en la judía (Phaseolus vulgaris L.) en el principado de Asturias, (on line). http://phaselieu.cesga.es/Gonzalez.pdf.

60
HIRANO, S.S., UPPER, C.D. 1990. Population Biology and Epidemiology of Pseudomonas syringae. Annu. Rev. Phytopathol. 28:155-177.
INSTITUTO DE DESARROLLO AGROPECUARIO. 1998. Tomate de
Invernadero. Precios y Mercados para rubros de la pequeña agricultura. 30 p. (Boletín nº 11).
JONES, J.B 1993. Bacterial Speck In: Jones, J.B., Jones, J.P., Stall, R.E.,
Zitter, T.A. eds. Compendium of tomato diseases. Minnesota, A.P.S. pp. 29-30.
________. McCARTER, S.M., and GITAITIS. R.D. 1986. Fluorescence on
Single-Carbon Sources for Separation of Pseudomonas syringae pv. syringae, Pseudomonas syringae pv. tomato and P. viridiflava on Tomato Transplants. Plant Disease 70(2):151-153.
________. McCARTER, S.M., GITAITIS. R.D. 1981. Association of
Pseudomonas syringae pv. syringae with a leaf spot disease of tomato transplants in southern Georgia. Phythopathology 71(12):1281-1285.
JETT, L. 2003. What to do About Bacterial Speck, (on line).
http://agebb.missouri.edu/hort/meg/archives/v9n11/meg3.htm. LATORRE, B. 1992. Enfermedades de las plantas cultivadas. Tercera
edición. Santiago, Ediciones Universidad Católica de Chile. 928 p. LAWNTON, M.B., MAcNEILL, B.H. 1986. Ocurence of race 1 of
Pseudomonas syringae pv. tomato on field tomato in southwestern Ontario. Canadian Journal of Plant Pathology 8:85-88.
LAY, M., HASS, B. 1973. Reaction of cowpea seedlings to phytopathogenic
bacteria. Phythopathology 63:1099-1103.

61
LELLIOTT, R.A., STEAD, D.E 1987. Methods for the diagnosis of bacterial diseases of plant. Vol.2. British Society for Plant Pathology. 216 p.
LELLIOTT, R.A., BILLING, E., HAYWARD, A.C. 1966. A determinative
scheme for the fluorescent plant pathogenic pseudomonads. Journal of Applied Bacteriology 29:470-480.
LOPEZ, M.M., MONTESINOS, E. 2000. Enfermedades causadas por
bacterias fitopatógenas. In: Bello, A., Trapero, A., Lopez., Liacer, G. eds. Patologia Vegetal. Valencia, Sociedad Española de Fitopatología. pp.515-558.
LOUWS, F.J., WILSON, M., CAMPBELL, H.L., CUPPELS, D.A., JONES,
J.B., SHOEMAKER, P.B., SAHIN, F., MILLER, S.A. 2001. Field control of bacterial spot and bacterial speck of tomato using a plant activator. Plant Disease 85:481-488.
LUDWIG, W., KLENK, H.P. 2001. Overview: a phylogenetic backbone and
taxonomic framework for prokaryotic systematics. In: Boone, D.R. and Castenholz, R.W. eds. Bergey's manual of systematic bacteriology. New York, Springer-Verlag. pp.49-65.
MACNAB, A. 2004a. Tomatoes. Seed treatment. Disease control, (on line).
http://vegdis.cas.psu.edu/03Diseases/D429_Toma.html. MACNAB, A. 2004b. July disease reminders for commercial producers in
PA, (on line). http://vegdis.cas.psu.edu/03Diseases/D109.html. MALAVOLTA JÚNIOR, V.A., ALMEIDA, I.M.G., ROGRIGUES NETO,J.
BERIAM, L.O.S., DE MELO, P.C.T. 2002. Caracterização de Pseudomonas syringae pv. syringae em tomateiro no Brasil e reação de cultivares/genótipos de tomateiro a esse patovar e ao patovar tomato. Arquivos do Instituto Biológico 69(1):63-66.

62
MESSIAEN, C.M., BLANCARD, D., ROUXEL, F., LAFON, R. 1995. Enfermedades de las hortalizas. Madrid, Editorial Mundi-Prensa. 576 p.
OFICINA DE ESTUDIOS Y POLÍTICAS AGRARIAS. 2002. Superficie de
cultivos, (on line). http://www.odepa.cl. OTTA, J.D., ENGLISH, H. 1971. Serology and pathology Pseudomonas
syringae. Phythopathology. 61:443-452. POHRONEZNY, K., VOLIN, R.B. 2002. Bacterial speck of tomato, (on line).
http://plantpath.ifas.ufl.edu/takextpub/FactSheets/pp0010.pdf. RECHE, J. 1991. Enfermedades de hortalizas en invernadero. Madrid,
Ministerio de Agricultura, Pesca y Alimentación. 189 p. RIEDEL R.M. 2000. Actigard (tm) 50WG plant activator for protection
against certain diseases of leafy vegetables, tomatoes, and tobacco, (on line). http://www.ag.ohio-state.edu/~vegnet/news/vnew2700.htm.
RISTA, L.M. 2005. Enfermedades de cultivos en invernaderos, (on line).
http://www.fca.unl.edu.ar/intensivos/extension11.html. ROSALES, J. 2005. Encuentros en la biología, (on line).
http://www.encuentros.uma.es/encuentros84/84.pdf. SHAAD, N.W. 1980. Laboratory guide for the identification of plant
pathogenic bacteria. St. Paul. Minnesota, American Phythopathological Society. 72 p.
SIKORA, E.J., GAZAWAY, G., MULLEN, J. 1998. Foliar diseases of
tomatoes. Control measures for bacterial spot and bacterial speck, (on line). http://www.aces.edu/pubs/docs/A/ANR-0071/.

63
SCORTICHINI, M. 1995. Le malattie batteriche delle colture agrarie e delle specie forestali. Bologna, Edagricole. 436 p.
SMITH, I.M., DUNEZ, J., LELLIOT, R.A., PHILLIPS, D.H., ARCHER, S.A.
1992. Manual de enfermedades de plantas. Madrid, Mundi Prensa. 671 p.
TATTERSALL. 2003. Yonit. Santiago, Tattersall commercial S.A. 2 p. VENETTE, J.R., LAMEY, H.A., SMITH, R.C. 1996. Bacterial spot and
bacterial speck of tomato, (on line). http://www.ext.nodak.edu/extpubs/plantsci/hortcrop/pp736w.htm.
WANG, T., MARTÍNEZ-ROMERO, J. 2000. Taxonomía de Rhizobium, (on
line). http://www.microbiologia.org.mx/microbiosenlinea/CAPITULO_12/Capitulo12.pdf.
WICK, R. 1991. Bacterial spot and speck of tomato and pepper, (on line).
http://www.umassvegetable.org/soil_crop_pest_mgt/specific_crops/tomatoes/bac_spot_speck/.
WIEBE, W.J., CAMPBELL, R.N. 1993. Characterization of Pseudomonas
syringae pv. maculicola and comparison with P. s. tomato. Plant Disease 77(4):414-419.
WILSON, M., CAMPBELL,H.L,. JI, P., JONES, J.B., CUPPELS, D.A. 2002.
Biological control of bacterial speck of tomato under field conditions at several locations in North America. Phytopathology 92(12):1284-1292.
YOUNG, J.M., TRIGGS, C.M. 1994. Evaluation of determinative tests for
pathovars of Pseudomonas syringae van Hall 1902. Journal of Applied Bacteriology 77:195-207.

64
ZHAO, Y., DAMICONE, J.P., DEMEZAS, D.H., RANGASWAMY, V., BENDER, C.L. 2000 Bacterial leaf spot of leafy Crucifers in Oklahoma caused by Pseudomonas syringae pv. maculicola. Plant Disease 84(9):1015-1020.
ZITTER, T. 2005. Bacterial diseases of tomato. Bacterial speck, (on line).
http://vegetablemdonline.ppath.cornell.edu/factsheets/Tomato_Bacterial.htm.


















