







Idiomas
Páginas
Jurídico


Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
ALEFSI DAVID SANCHEZ REINOSO
Universidad Nacional de Colombia
Facultad de Ciencias Agrarias, Departamento de Agronomía
Bogotá D.C., Colombia
2016


Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
ALEFSI DAVID SANCHEZ REINOSO
Tesis o trabajo de investigación presentada(o) como requisito parcial para optar al título
de: Magister en Ciencias Agrarias – Línea Fisiología de Cultivos
Director:
Profesor Hermann Restrepo Díaz
Codirector:
Profesor. Gustavo Adolfo Ligarreto Moreno
Línea de Investigación:
Fisiología de Cultivos
Universidad Nacional de Colombia
Facultad de Ciencias Agrarias, Departamento de Agronomía
Bogotá D.C., Colombia
2016


Dedicatoria
A Dios por darme la salud, perseverancia e
inteligencia para lograr todas mis metas. A mis padres
Fanny Reinoso Mosquera y José Crispín Sánchez
Rincón, por todo su apoyo incondicional en cada
decisión que he tomado. A mis hermanos y amigos
que me acompañaron en cada una de las etapas de
mi vida.


Agradecimientos
Quiero agradecer sinceramente a todas aquellas personas que compartieron
conmigo diferentes experiencias y conocimientos en el desarrollo de esta tesis. A
la Universidad Nacional de Colombia, mi alma máter. A los profesores Hermann
Restrepo, Gustavo Ligarreto y Augusto Ramírez por su apoyo y consejos durante
mi trabajo en la maestría. A mis compañeros Nixon Flórez Velasco y Cristian
Camilo Chávez Arias por su apoyo en los diferentes muestreos realizados en las
etapas del desarrollo de la tesis. A los laboratoristas Rafael Cruz y Wadith de León
Camelo por su generoso apoyo y asesorías prestadas en múltiples protocolos. Al
programa de Jóvenes Investigadores de Colciencias y al programa Estímulos
Thomas Van der Hammen del Jardín Botánico José Celestino Mutis, por la
financiación de gran parte del proyecto. A la beca exención de derechos
académicos de la Facultad de Ciencias Agrarias.


Resumen y Abstract IX
Resumen
La disponibilidad del recurso hídrico es uno de los principales factores ambientales
que determina el potencial desarrollo y rendimiento de una especie cultivada. En
los últimos años las precipitaciones han variado su frecuencia y volumen, donde
Colombia no ha sido la excepción incrementando las áreas expuestas a sequías.
Este aumento de la temperatura perjudica el crecimiento, desarrollo y producción
de los cultivos, dentro de ellos el fríjol. Esta especie es una de las principales
fuentes de proteína y minerales en la dieta humana, por lo que cualquier reducción
en el rendimiento afecta la seguridad alimentaria del mundo. En el presente estudio
se caracterizó la respuesta de cinco cultivares de fríjol arbustivo a estrés por déficit
hídrico de 15 días en estado de formación de la tercera hoja verdadera y durante
la apertura de las flores (30% – 40%). Para esto se evaluaron variables como
fotosíntesis, parámetros relacionados con estrés oxidativo, y algunos parámetros
de rendimiento. Los resultados indicaron que: i) el déficit hídrico afecta los procesos
fisiológicos (en estado vegetativo) desde los 10 días después de tratamiento e
exposición y que estos varían con el tiempo ii) el déficit hídrico disminuye la
acumulación de materia seca en los diferentes órganos debido a una menor
asimilación de CO2; iii) el déficit hídrico reduce los diferentes parámetros de
rendimiento en todos los cultivares evaluados, pero el más afectado fue Bacatá en
la etapa vegetativa y reproductiva; iv) Los índices de tolerancia y/o susceptibilidad
al estrés permitieron categorizar los diferentes cultivares; v) Se sugieren las
siguientes variables como posibles marcadores de estrés déficit hídrico en fríjol
arbustivo: asimilación de CO2, conductancia estomática, uso eficiente del agua
acumulación de masa en la raíz, MDA y contenido de prolina, índices de tolerancia
y/o susceptibilidad al estrés por déficit hídrico.
Palabras clave: limitación estomática, estrés oxidativo, peroxidación lipídica, déficit
hídrico, tolerancia al estrés.

X Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
Abstract
Water deficit is one of the main environmental factors that determines the potential
development and performance in crops. In recent years the rains have changed their
frequency and volume, where Colombia has been no exception increasing drought prone
areas. This temperature increase impairs growth, development and production of crops,
beans within them. This specie is one of the main source of proteins and minerals in the
human diet, so some reduction in yield affects food security in the world. In the present
study the response five cultivars of bush beans stress was characterized by water deficit
of 15 days in the third true leaf formation and during the opening of flowers (30% - 40%).
For this reason, variables like photosynthesis, parameters related to oxidative stress, and
some performance parameters were evaluated. The results indicated that: i) water deficit
affects physiological processes (vegetative state) from the 10 days of exposure and these
change over time ii) water deficit decreases dry matter accumulation due to a lower
assimilation CO2; iii) water deficit reduces different performance parameters in all cultivars
evaluated, but Bacatá was the most affected in the vegetative and reproductive stage; iv)
tolerance and / or susceptibility index to stress allowed categorize the different cultivars; v)
the following variables as potential markers of water deficit stress in bush beans are
suggested: CO2 assimilation, stomatal conductance, water use efficiency, mass
accumulation in the root, MDA and proline content, indexes of tolerance and / or
susceptibility to stress water deficit.
Keywords: stomatal limitation, oxidative stress, lipid peroxidation, water deficit,
stress tolerance.

Contenido XI
Contenido
Pág.
Resumen IX
Lista de figuras ............................................................................................................. XV
Lista de tablas ........................................................................................................... XVIII
Lista de Símbolos y abreviaturas ................................................................................ XX
Introducción .................................................................................................................... 1
1. Capítulo 1. Marco Teórico .......................................................................................... 5
1.1 El estrés por déficit hídrico y condiciones que lo generan. ............................... 5
1.2 La importancia del agua en las plantas. ........................................................... 5
1.3 Efectos a nivel fisiológico del déficit hídrico en plantas .................................... 6
1.4 Las respuestas de las plantas a condiciones de déficit hídrico ........................ 8
1.5 Fenología y el efecto del déficit hídrico sobre la fenología del fríjol (P. vulgaris
L.) 9
1.6 Antecedentes en el cultivo de fríjol en condiciones de déficit hídrico. ............ 12
1.7 La utilidad de los Índices de Tolerancia en la caracterización de genotipos de
plantas cultivadas. .................................................................................................... 14
2. Capítulo. Identificación del periodo crítico para estrés hídrico sobre variables
fisiológicas de un cultivar de fríjol arbustivo (Phaseolus vulgaris L.). ..................... 16
2.1 Resumen ....................................................................................................... 16
2.2 Introducción ................................................................................................... 17
2.3 Materiales y métodos ..................................................................................... 19
2.3.1 Material vegetal y condiciones de crecimiento .................................... 19
2.3.2 Tratamientos de déficit hídrico y toma de datos en los muestreos....... 20
2.3.3 Diseño experimental y análisis de los datos ........................................ 21

XII Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico
2.3.4 Variables fisiológicas .......................................................................... 21
2.3.5 Variables bioquímicas ......................................................................... 23
2.4 Resultados .................................................................................................... 24
2.4.1 Intercambio gaseoso y relaciones hídricas. ........................................ 24
2.4.2 Parámetros de fluorescencia del fotosistema II, contenido de clorofilas
y carotenoides totales ....................................................................................... 29
2.4.3 Daño de la membrana por estrés oxidativo y la producción de
osmolitos. ......................................................................................................... 31
2.5 Discusión ....................................................................................................... 34
2.6 Conclusiones ................................................................................................. 36
3. Capítulo 3. Evaluación fisiológica y bioquímica de cinco cultivares de fríjol
arbustivo (Phaseolus vulgaris L.) bajo condición de estrés por déficit hídrico. ...... 37
3.1 Resumen ....................................................................................................... 37
3.2 Introducción ................................................................................................... 38
3.3 Materiales y métodos .................................................................................... 41
3.3.1 Material vegetal y condiciones de crecimiento .................................... 41
3.3.2 Tratamiento de déficit hídrico y determinación de las variables .......... 42
3.3.3 Diseño experimental y análisis de los datos ........................................ 42
3.3.4 Variables fisiológicas .......................................................................... 43
3.3.5 Variables bioquímicas ......................................................................... 45
3.4 Resultados .................................................................................................... 46
3.4.1 Relaciones hídricas e intercambio gaseoso de la hoja ........................ 46
3.4.2 Acumulación de materia seca y relación parte aérea/raíz ................... 53
3.4.3 Daño de la membrana por estrés oxidativo y la producción de
osmolitos. ......................................................................................................... 56
3.4.4 Índice Relativo de Tolerancia (RTI) ..................................................... 59
3.5 Discusión ....................................................................................................... 60
3.6 Conclusiones ................................................................................................. 62
4. Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) a condiciones de estrés por déficit hídrico en dos etapas
fenológicas. ................................................................................................................... 63
4.1 Resumen ....................................................................................................... 63

Contenido XIII
4.2 Introducción ................................................................................................... 64
4.3 Materiales y métodos ..................................................................................... 66
4.3.1 Material vegetal y condiciones de crecimiento .................................... 66
4.3.2 Tratamiento de déficit hídrico y determinación de las variables ........... 67
4.3.3 Diseño experimental y análisis de los datos ........................................ 68
4.3.4 Variables fisiológicas ........................................................................... 68
4.3.5 Variables bioquímicas ......................................................................... 70
4.4 Resultados ..................................................................................................... 71
4.4.1 Humedad del suelo, Relaciones hídricas e intercambio gaseoso de la
hoja ............................................................................................................ 71
4.4.2 Contenido de clorofilas, carotenoides y parámetros de fluorescencia, 78
4.4.3 Concentración de Malondialdehido y Prolina ....................................... 81
4.5 Conclusiones ................................................................................................. 82
5. Capítulo 5. Evaluación de índices de tolerancia a la sequía para la selección de
cultivares de fríjol arbustivo (Phaseolus vulgaris L.) por inducción de estrés por
déficit hídrico. ................................................................................................................ 83
5.1 Resumen ....................................................................................................... 83
5.2 Introducción ................................................................................................... 84
5.3 Materiales y métodos ..................................................................................... 85
5.3.1 Material vegetal y condiciones de crecimiento .................................... 85
5.3.2 Tratamiento de déficit hídrico .............................................................. 86
5.3.3 Diseño experimental y análisis de los datos ........................................ 87
5.3.4 Contenido Relativo de Agua ................................................................ 88
5.3.5 Parámetros de rendimiento ................................................................. 88
5.3.6 Índice de tolerancia y susceptibilidad al estrés por sequía .................. 88
5.4 Resultados y discusión .................................................................................. 92
5.4.1 Humedad del suelo y Contenido Relativo de Agua .............................. 92
5.4.2 Parámetros de rendimiento ................................................................. 94
5.4.3 Comparación de cultivares con base a los índices de
resistencia/tolerancia ......................................................................................... 97
5.4.4 Análisis de correlación ...................................................................... 100
5.5 Conclusiones ............................................................................................... 102
6. Discusión final ........................................................................................................ 103

XIV Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico
7. Conclusiones y recomendaciones ........................................................................ 105
7.1 Conclusiones ............................................................................................... 105
7.2 Recomendaciones ....................................................................................... 106
8. Modelo conceptual ................................................................................................. 108
Bibliografía .................................................................................................................. 111

Contenido XV
Lista de figuras
.................................................................................................................... Pág.
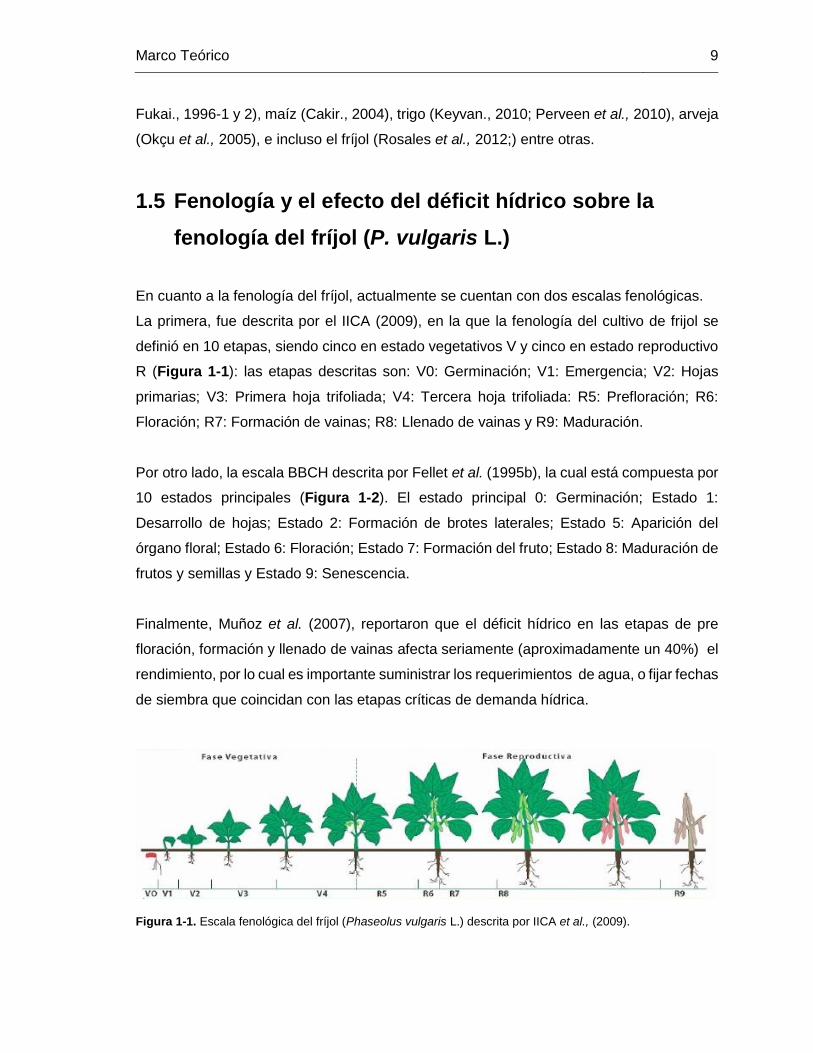
Figura 1-1. Escala fenológica del fríjol (Phaseolus vulgaris L.) descrita por IICA et al.,
(2009). .............................................................................................................................. 9
Figura 1-2. Escala fenológica BBCH del fríjol (Phaseolus vulgaris L.) descrita por Feller et
al., (1995b). .................................................................................................................... 11
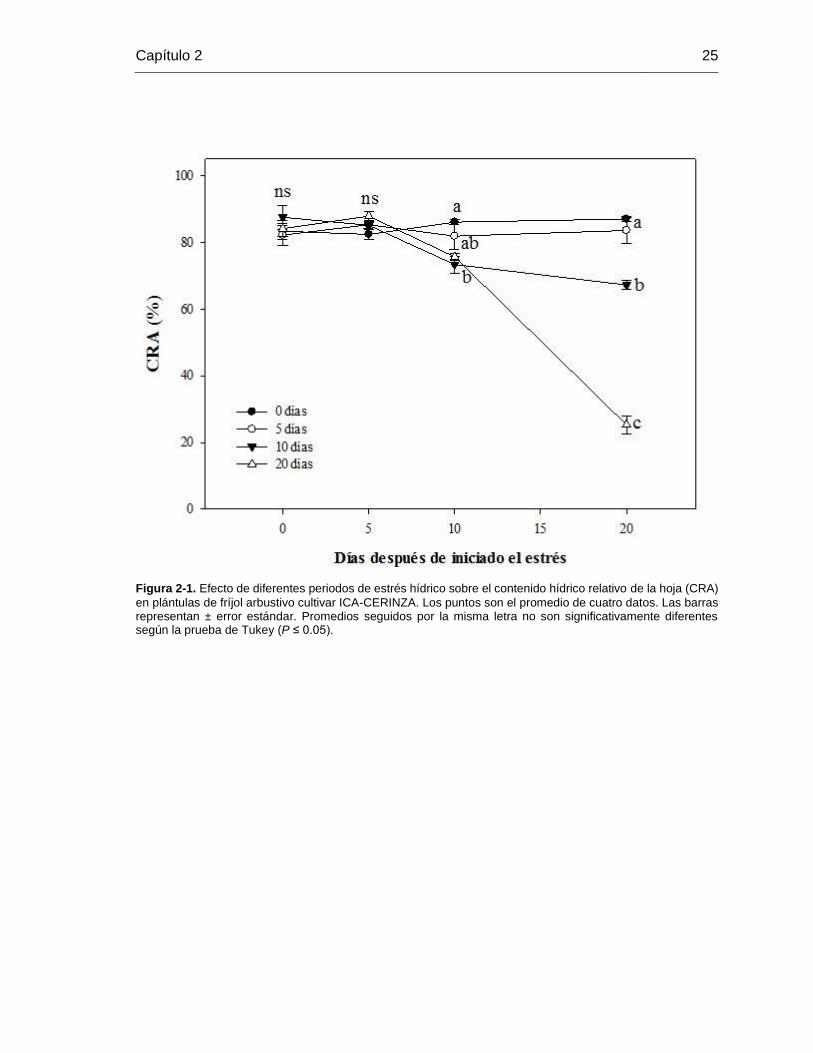
Figura 2-1. Efecto de diferentes periodos de estrés hídrico sobre el contenido hídrico
relativo de la hoja (CRA) en plántulas de fríjol arbustivo cultivar ICA-CERINZA. Los puntos
son el promedio de cuatro datos. Las barras representan ± error estándar. Promedios
seguidos por la misma letra no son significativamente diferentes según la prueba de Tukey
(P ≤ 0.05). ...................................................................................................................... 25
Figura 2-2. Efecto de diferentes periodos de estrés hídrico sobre parámetros de
intercambio gaseoso de la hoja en plántulas de fríjol arbustivo cultivar ICA-CERINZA: A).
Fotosíntesis neta (Pn); B). Conductancia estomática (gs) y C). Transpiración (E). Las barras
son el promedio de cuatro datos ± error estándar. Promedios seguidos por la misma letra
no son significativamente diferentes según la prueba de Tukey (P ≤ 0.05). .................... 27
Figura 2-3. Efecto de diferentes periodos de estrés hídrico sobre parámetros de
fluorescencia del fotosistema II plántulas de fríjol arbustivo cultivar ICA-CERINZA: A).
Eficiencia máxima del quantum en el fotosistema II (Fv/Fm); B). Quenching fotoquímico
(qP); C). La eficiencia real del fotosistema II (Y(II)) y D). Quenching no fotoquímico (QNP)
en plántulas de fríjol arbustivo. Las barras son el promedio de cuatro datos ± error estándar.
Promedios seguidos por la misma letra no son significativamente diferentes según la
prueba de Tukey (P ≤ 0.05). ........................................................................................... 30
Figura 2-4. Efecto de diferentes periodos de estrés hídrico sobre contenido de clorofilas y
carotenoides totales en plántulas de fríjol arbustivo cultivar ICA-CERINZA: A). Contenido
de clorofila total; B). Contenido de clorofila a; C). Contenido de clorofila b y D). Contenido
de carotenoides totales en plántulas de fríjol arbustivo. Las barras son el promedio de
cuatro datos ± error estándar. Promedios seguidos por la misma letra no son
significativamente diferentes según la prueba de Tukey (P ≤ 0.05). ............................... 31

XVI Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico
Figura 2-5. Efecto de diferentes periodos de estrés hídrico sobre el daño de la membrana
por estrés oxidativo y la producción de osmolitos: A). Fuga de electrolitos; B). Producción
de malondialdehido (MDA) y C). Producción de prolina en plántulas de fríjol arbustivo. Las
barras son el promedio de cuatro datos ± error estándar. Promedios seguidos por la misma
letra no son significativamente diferentes según la prueba de Tukey (P ≤ 0.05). ............ 33
Figura 3-1. Efecto del estrés hídrico sobre el contenido hídrico relativo de la hoja (CRA)
en plántulas de cinco cultivares de fríjol arbustivo. Las barras son el promedio de cuatro
datos ± error estándar. .................................................................................................... 50
Figura 3-2. Efecto del estrés hídrico sobre contenido de clorofilas y carotenoides totales
en plántulas de cinco cultivares de fríjol arbustivo: A). Fotosíntesis neta (Pn); B).
Conductancia estomática (gs); C). Transpiración (E) es en plántulas de fríjol arbustivo. .
Las barras son el promedio de cuatro datos ± error estándar. ........................................ 52
Figura 3-3. Efecto del estrés hídrico sobre contenido de clorofilas y carotenoides totales
en plántulas de cinco cultivares de fríjol arbustivo: A). Contenido de clorofila total; B).
Contenido de clorofila a; C). Contenido de clorofila b y D). Contenido de carotenoides
totales en plántulas de fríjol arbustivo. . Las barras son el promedio de cuatro datos ± error
estándar. ......................................................................................................................... 56
Figura 3-4. Efecto del estrés hídrico sobre la eficiencia máxima del quantum en el
fotosistema II (Fv/Fm) en plántulas de cinco cultivares de fríjol arbustivo: Las barras son el
promedio de cuatro datos ± error estándar. .................................................................... 56
Figura 3-5. Efecto del estrés hídrico sobre el daño de la membrana por estrés oxidativo y
la producción de osmolitos: A). Fuga de electrolitos; B). Producción de malondialdehido
(MDA) y C). Producción de prolina en plántulas de cinco cultivares de fríjol arbustivo. Las
barras son el promedio de cuatro datos ± error estándar. ............................................... 58
Figura 3-6. Efecto del estrés hídrico sobre el índice relativo de tolerancia (RTI) en plántulas
de cinco cultivares de fríjol arbustivo: Las barras son el promedio de cuatro datos ± error
estándar. ......................................................................................................................... 59
Figura 4-1. Efecto del estrés por déficit hídrico en dos etapas fenológicas diferentes sobre
la humedad del suelo (A) en porcentaje de la capacidad de campo entre los factores de
las diferentes etapas fenológicas y el contenido relativo de agua de la hoja de cinco
cultivares de fríjol arbustivo (B corresponden al muestreo después de finalizar el periodo
de estrés en etapa vegetativa; C corresponden al muestreo después de finalizar el periodo
de estrés en etapa reproductiva. Las barras representan ± error estándar. .................... 72

Contenido XVII
Figura 4-2. Efecto del estrés por déficit hídrico en dos etapas fenológicas diferentes sobre
la fotosíntesis (A y B), conductancia estomática (C y D) y limitación estomática (E y F) de
cinco cultivares de fríjol arbustivo (A, C y E corresponden al muestreo después de finalizar
el periodo de estrés en etapa vegetativa; B, D y F corresponden al muestreo después de
finalizar el periodo de estrés en etapa reproductiva. Las barras representan ± error
estándar. ........................................................................................................................ 75
Figura 4-3. Efecto del estrés por déficit hídrico en dos etapas fenológicas diferentes sobre
la clorofila total (A y B) y carotenoides totales (C y D) de cinco cultivares de fríjol arbustivo
(A y C corresponden al muestreo después de finalizar el periodo de estrés en etapa
vegetativa; B y D corresponden al muestreo después de finalizar el periodo de estrés en
etapa reproductiva. Las barras representan ± error estándar. ........................................ 79
Figura 4-4. Efecto del estrés por déficit hídrico en dos etapas fenológicas diferentes sobre
la producción de malondialdehido (A y B) y producción de prolina (C y D) de cinco cultivares
de fríjol arbustivo (A y C corresponden al muestreo después de finalizar el periodo de estrés
en etapa vegetativa; B y D corresponden al muestreo después de finalizar el periodo de
estrés en etapa reproductiva. Las barras representan ± error estándar. ......................... 82
Figura 5-1. Efecto del estrés por déficit hídrico en dos etapas fenológicas diferentes sobre
la humedad del suelo (A) en porcentaje de la capacidad de campo entre los factores de
las diferentes etapas fenológicas y el contenido relativo de agua de la hoja de cinco
cultivares de fríjol arbustivo (B corresponden al muestreo después de finalizar el periodo
de estrés en etapa vegetativa; C corresponden al muestreo después de finalizar el periodo
de estrés en etapa reproductiva. Las barras representan ± error estándar. .................... 93
Figura 5-2. Biplot de Análisis de Componente Principal (PCA) de diferentes variables de
rendimiento en plantas de cinco genotipos de fríjol arbustivo bajo condiciones de estrés en
dos etapas fenológicas diferentes. Gpod: Número de granos por vaina; Pod: Número de
vainas por planta; Pcg: Peso de 100 granos; RtoP: Rendimiento por planta; RtoHa:
Rendimiento por hectárea; GRP: Número de granos por planta. C: Control; E1: Estrés en
etapa Vegetativa; E2: Estrés en etapa Reproductiva; Cz: plantas del cultivar Cerinza; Bch:
plantas del cultivar Bachue; NUA: plantas del cultivar NUA35; Ba: plantas del cultivar
Bacatá y Bl: plantas del cultivar Bianca. ......................................................................... 97
Figura 8-1. Modelo conceptual de las respuestas de las plantas de fríjol arbustivo
(Phaseolus vulgaris L.) a condiciones de estrés por déficit hídrico. ...............................109

Contenido XVIII
Lista de tablas
Pág. Tabla 1-1. Resumen de los estudios en caracterización de las respuestas fisiológicas de
fríjol bajo condiciones de sequía. Adaptado de Beebe et al. (2013). ............................... 12
Tabla 2-1. Efecto de diferentes periodos de estrés hídrico sobre el Uso Eficiente del Agua
(UEA), Carbono intercelular (Ci) y Relación Carbono intercelular sobre el Carbono
atmosférico de la hoja (Ci/Ca) en plántulas de fríjol arbustivo cultivar ICA-CERINZA. ..... 28
Tabla 3-1. Resumen del análisis de varianza de los efectos del estrés hídrico sobre el
comportamiento fisiológico de las plántulas de cinco cultivares de fríjol arbustivo. ......... 49
Tabla 3-2. Efecto del estrés hídrico sobre el Uso Eficiente del Agua (UEA) y la relación
Carbono intercelular sobre el Carbono atmosférico (Ci/Ca) en plántulas de cinco cultivares
de fríjol arbustivoa. .......................................................................................................... 51
Tabla 3-3. Efecto del estrés hídrico sobre el Peso Seco de la Parte Aérea (PSA), Peso
Seco de la Raíz (PSR), Peso Seco Total (PST) y Relación Parte Aérea : Raíz (PA/R) en
plántulas de cinco cultivares de fríjol arbustivoa. ............................................................. 54
Tabla 4-1. Resumen del análisis de varianza de los efectos del estrés hídrico sobre el
comportamiento fisiológico y parámetros de rendimiento de cinco genotipos de fríjol
arbustivo. *, **, y *** significativamente diferente en los niveles de probabilidad a 0.05, 0.01
y 0.001, respectivamente. NS, no significativo con a = 0.05. ........................................... 73
Tabla 4-2. Efecto del estrés por déficit hídrico en dos etapas fenológicas diferentes sobre
parámetros de intercambio gaseoso de cinco genotipos de fríjol arbustivo (55 DDE y 65
DDE). .............................................................................................................................. 77
Tabla 4-3. Efecto del estrés por déficit hídrico en dos etapas fenológicas diferentes sobre
parámetros de fluorescencia de cinco genotipos de fríjol arbustivo (55 DDE y 65 DDE). 80
Tabla 5-1. Resumen del análisis de varianza de los efectos del estrés hídrico sobre el
comportamiento fisiológico y parámetros de rendimiento de cinco genotipos de fríjol
arbustivo. ........................................................................................................................ 91
Tabla 5-2. Efecto del estrés por déficit hídrico en dos etapas fenológicas diferentes sobre
los parámetros de rendimiento de cinco genotipos de fríjol arbustivo. ............................. 96

Contenido XIX
Tabla 5-3. Índice de tolerancia y susceptibilidad de cinco genotipos de fríjol arbustivo bajo
condiciones de estrés por sequía. .................................................................................. 99
Tabla 5-4. Coeficiente de correlación entre los diferentes índices de tolerancia y
susceptibilidad al estrés por déficit hídrico en cinco genotipos de fríjol arbustivo. .........101

Contenido XX
Lista de Símbolos y abreviaturas
Símbolos con letras latinas Símbolo Término Unidad SI Definición
A Área m2 ∬ 𝑑𝑥 𝑑𝑦
D Diámetro m L Longitud m DF T Temperatura C DF T Tiempo s DF
V Volumen L ∫ 𝑑𝑟3
Superíndices Superíndice Término
N Exponente, potencia
Abreviaturas Abreviatura Término
CRA Contenido Relativo de Agua DDE Días después de Emergencia Pn Fotosíntesis neta gs Conductancia estomática E Transpiración UEA Uso Eficiente del Agua UEAi Uso Eficiente del Agua intrínseco Ci/Ca Relación Carbono intercelular sobre Carbono atmosférico Ls Limitación estomática Pn/Ci Eficiencia de la carboxilación Chl total Clorofila total Chl a Clorofila a Chl b Clorofila b Cx + c Carotenoides totales Fv/Fm Eficiencia máxima potencial del fotosistema II Y(II) Eficiencia real del fotosistema II qP Quenching fotoquímico

Contenido XXI
Abreviatura Término NPQ Quenching no fotoquímico MDA Malondialdehido PS Peso Seco PF Peso Fresco PT Peso Turgente RTI Índice Relativo de Tolerancia CHS Contenido de Humedad del Suelo Ys Promedio del rendimiento bajo condición de estrés Yp Promedio del rendimiento sin estrés
�̅�𝑆 Rendimiento promedio de todos los genotipos sin estrés
�̅�𝑃 Rendimiento promedio de todos los genotipos con estrés SSI Índice de Susceptibilidad al Estrés TOL Tolerancia MP Productividad Media GMP Productividad Media Geométrica STI Índice de Tolerancia al Estrés YSI Índice de Estabilidad del Rendimiento YI Índice de Rendimiento HM Media Armónica SDI Índice de Sensibilidad a la Sequía


Introducción
Una de las leguminosas de grano más importante en el mundo es el frijol común
(Phaseolus vulgaris L.), debido a que sus semillas constituyen una de las principales
fuentes de proteína y minerales de origen vegetal en la dieta de las comunidades de bajos
recursos (Broughton et al., 2003; Beebe, 2012). Es una especie nativa de Meso y
Suramérica, perteneciente a la familia Fabaceae (Kalavacharla et al., 2011). En los países
de América Latina, el consumo nacional per de los granos es típicamente entre 12 y 18 kg
por año, pero estas cifras no discriminan las zonas urbanas de las rurales (Broughton et
al., 2003).
Aunque la producción mundial es difícil de calcular con precisión debido al desconcierto
con otras leguminosas, se estima que sea entre 11 y 12 millones de toneladas (FAO, 2007).
América Latina es la región de mayor producción de frijol, abarcando alrededor del 50%
de la cosecha mundial y en segundo lugar se encuentra África con un 25%. Los sistemas
productivos de frijol son tradicionalmente pequeñas parcelas campesinas, que
tradicionalmente se siembran en sistemas agrícolas complejos en asociación o rotación
con las especies como el sorgo, plátano, café en etapas tempranas, maíz, entre otros
(Broughton et al., 2003). Existen diferentes fríjoles con hábitos de crecimiento de tipos
arbustivos determinados a escaladores vigorosos de crecimiento indeterminado, como
también la gama de los ciclos de crecimiento de 2 a 10 meses de duración, los cuales
producen granos en cultivos que se adaptan muchos nichos de producción (Beebe et al.,
2013).
Por otro lado, la estructura genética del frijol ha sido estudiada ampliamente (Gepts y
Debouck, 1991; Singh et al., 1991; Broughton et al, 2003; Beebe et al., 2013). Los frijoles
cultivados presentan una estructura genética bien definida que se origina en el ancestro
de frijol silvestre (Beebe et al., 2013). Según Toro et al. (1990), éste ancestro crece como

2 Introducción
una planta herbácea anual en regiones de bosque pre-montano sub-húmedo que van
desde el norte de México hasta el norte de Argentina. Los principales bancos de
germoplasma han clasificado los fríjoles en razas basados en la morfología de la planta,
rango de adaptación y las características agronómicas (Beebe et al., 2013). Finalmente,
las colecciones genéticas de origen mesoamericano se han dividido en tres razas: i)
Jalisco: frijol con crecimiento voluble de las tierras altas húmedas del centro de México; ii)
Mesoamérica: fríjoles con crecimiento determinado de semilla pequeña, encontrado en
tierras bajas de América central y México; y iii) Durango: fríjoles de tipo postrado con
crecimiento determinado con semillas de tamaño medio, encontrado en las tierras altas y
secas de México (Singh et al., 1991).
La variabilidad climática es uno de los fenómenos ambientales de mayor impacto en los
últimos años, incidiendo en las superficies afectadas por sequías que ocasionan
condiciones de estrés hídrico. Asimismo, el Niño/ Oscilación del Sur (ENOS) es uno de los
fenómenos que genera mayores anomalías en precipitación pluvial y temperaturas
(Monasterio et al., 2011). Por otro lado, Pabón y Torres (2007) evaluaron la variabilidad
climática asociada al ciclo ENOS para la Sabana de Bogotá, encontrando una relación
inversa entre las anomalías de la temperatura de la superficie del mar y la precipitación,
de tal forma que condiciones cálidas (anomalías asociadas al Niño) reducen la
precipitación regional. Igualmente, la producción agrícola es afectada fuertemente por las
condiciones ambientales desfavorables (exceso o déficit de precipitaciones),
especialmente, bajo condiciones de ciclo ENOS (Fenómeno del Niño o la Niña) que causa
climas extremos como inundaciones, altas temperaturas o sequías en varias regiones del
mundo (Holmgren et al., 2001; Ministerio de Agricultura y Desarrollo Rural, 2011).
Adicionalmente, el cambio climático ha tenido un gran impacto en las zonas agrícolas,
incrementando las áreas afectadas por condiciones de baja disponibilidad de agua,
propiciando más episodios de estrés hídrico (Monasterio et al., 2011), y para el caso de
Colombia, gran parte de esta área se encuentra en la región andina donde el fríjol es un
cultivo predominante (Comunidad Andina, 2009).
Asimismo, se ha reportado que diferentes genotipos de frijol presentan distintas respuestas
a las condiciones de estrés (Beebe et al., 2013). Además, identificar y priorizar parámetros
fisiológicos y bioquímicos son de gran utilidad para los programas de fitomejoramiento
(Yousfi et al., 2012), ya que nos permite determinar posibles respuestas de aclimatación y

Introducción 3
sobrevivencia a la condición ambiental adversa estudiada. Finalmente, la implementación
de índices de tolerancia a sequía en los métodos de selección de genotipos de fríjol
tolerantes y/o susceptibilidad a la sequía no son muy usados.
Bajo dicho contexto, el presente estudio tuvo como objetivo evaluar las respuestas
fisiológicas y bioquímicas de cinco cultivares de fríjol arbustivo bajo condiciones de estrés
por déficit hídrico. Para ello, se plantearon tres objetivos específicos los cuales fueron: i)
Identificar la duración de un periodo de estrés moderado por déficit hídrico para diseñar
ensayo al que se someterán los cinco genotipos de fríjol arbustivo (Phaseolus vulgaris L.);
ii) Medir y comparar las respuestas fisiológicas, relaciones hídricas, respuesta oxidativa y
anatomía de los cinco cultivares de fríjol arbustivo bajo condiciones de estrés hídrico en
variables bioquímicas como producción de prolina y malondialdehido de cinco genotipos
de fríjol arbustivo; iii) Evaluar los efectos del estrés por déficit hídrico en una etapa
vegetativa y en una etapa reproductiva sobre parámetros de rendimiento e índices de
tolerancia de cinco cultivares de fríjol arbustivo.
Adicionalmente, es importante reconocer y aprovechar el potencial de los genotipos
colombianos como fuente de genes de tolerancia a estrés por déficit hídrico, considerando
que para los cultivares de fríjol arbustivo de Colombia se desconoce dicha información.
Igualmente, este trabajo aporta información fundamental acerca de la caracterización de
la respuesta de plantas de cinco cultivares de fríjol arbustivo ante la condición de déficit
hídrico.
En este documento está presentado diferentes capítulos, en el que el primero reúne el
marco teórico como estado del arte del déficit hídrico y unos antecedentes para el caso del
fríjol. Posteriormente se presentan cuatro capítulos adicionales los cuales reúnen los
resultados de las diferentes etapas de investigación para el alcance de los objetivos de la
tesis. La primera etapa corresponde al primer objetivo específico, en el que se buscó
identificar el periodo de estrés moderado por déficit hídrico para un cultivar comercial de
fríjol arbustivo. Seguidamente, al identificar el período de estrés, se realizó la
caracterización de los cinco cultivares de fríjol arbustivo en una etapa fenológica de la fase
vegetativa. Finalmente, se dividen los resultados de la última etapa de investigación en dos
capítulos; uno muestra los parámetros fisiológicos y bioquímicos evaluados; y el otro los
diferentes parámetros de rendimiento e índices de tolerancia de los cinco cultivares de fríjol

4 Introducción
arbustivo en dos etapas fenológicas: una en la fase vegetativa y una en la fase
reproductiva.

1. Capítulo 1. Marco Teórico
1.1 El estrés por déficit hídrico y condiciones que lo generan.
El estrés por déficit hídrico está definido como aquella condición en el que la tasa
transpiratoria de la planta supera la tasa de absorción de agua por parte de las raíces
(Lacape et al., 1998). Asimismo, se considera que un estrés moderado por déficit hídrico
es aquel que genera limitaciones en la fotosíntesis de tipo estomático, mientras que un
estrés severo por déficit hídrico cuenta con limitaciones de tipo estomática y no estomática
de la fotosíntesis, generalmente por daños en el fotosistema II (Hsiao., 1973). Son múltiples
los agentes del entorno que actúan como fuentes de estrés.
1.2 La importancia del agua en las plantas.
El agua es uno de los recursos renovables del planeta, aunque su disponibilidad es limitada
y variable (Pimentel et al., 1997). Según proyecciones de la FAO (2016), para el año 2025,
cerca de 3200 millones de personas vivirán en regiones del mundo en las que se tendrán
influencias de déficit hídrico severo. Según Oki y Kanae (2006), la agricultura con
suministro de riego comprende el 20% de las tierras con vocación agrícola e incluye la
mitad de la producción mundial.
Igualmente, la importancia del agua para los cultivos hortícolas es particularmente amplia
ya que la mayoría de la producción hortícola se vende por peso fresco, siendo un alto
porcentaje de agua el componente principal, por lo que a menudo se prioriza en asegurar
el suministro de agua a través de un control preciso de riego (Jones and Tardieu., 1998).

6 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
Asimismo, la sequía es una limitación importante para el rendimiento del cultivo en aquellos
sistemas agrícolas que son de secano predominante, pero asociadas con bajas pérdidas
de rendimiento (muy por debajo del 10% de todos reportó pérdidas) para los cultivos en
los sistemas agrícolas con buen sistema de irrigación (Li et al., 2011). Adicionalmente, la
duración del cultivo de frijol para cosechar grano seco oscila entre 90 a 100 días, con una
demanda hídrica de aproximadamente 350 a 500 mm de agua por ciclo la cual varía con
respecto al cultivar, la profundidad del suelo y el clima (Beebe et al. 2010).
1.3 Efectos a nivel fisiológico del déficit hídrico en plantas
Las condiciones de estrés por déficit hídrico afectan drásticamente el rendimiento del
cultivo de fríjol (Rainey y Griffiths, 2005, Beebe et al., 2013). Igualmente, White e Izquierdo
(1991), reportaron que en periodos prolongados de ausencia de lluvias o reducción en la
humedad del suelo afectan más del 60% de la producción de frijol en todo el mundo.
Asimismo, las plantas presentan diferentes respuestas a nivel anatómico, fisiológico,
bioquímico y genético a las condiciones de estrés. Yousfi et al. (2012) encontraron que
materiales de alfalfa tolerantes a estrés hídrico presentan un menor número de estomas
en la hoja, mayor presencia de tricomas en la lámina foliar, mayor eficiencia en el uso del
agua, menor tasa de transpiración, los parámetros de rendimiento no se reducen
significativamente, siendo estas características de gran importancia para identificar
materiales tolerantes a estrés hídrico. Según Farooq et al. (2009), las plantas responden
al estrés hídrico con la reducción en la transpiración debido al cierre estomático inducido
por el incremento de las concentraciones de ácido abscísico (ABA), el crecimiento de la
células se ve restringido por la reducción en el turgor celular debido al descenso en el
potencial hídrico. Dado lo anterior, es importante enfocar investigaciones en conocer el
comportamiento de diferentes genotipos de fríjol bajo estas condiciones que puedan
contribuir en programas de fitomejoramiento.
La fotosíntesis es el proceso fisiológico más susceptible a cualquier condición de estrés
abiótico (Restrepo-Díaz y Garcés-Varón, 2013). Asimismo, ambientes contrastantes

Marco Teórico 7
también pueden afectar el contenido de clorofila en las hojas, la eficiencia máxima del PSII
fotoquímica (Fv/Fm), quenching no fotoquímico y el estado de activación de la enzima
Rubisco (Cao et al., 2008). En cuanto a las relaciones hídricas, Miyashita et al. (2005)
reportan que cuando plantas de fríjol se encuentran sometidas a condiciones de estrés
hídrico alteran las relaciones hídricas, con la finalidad de disminuir la pérdida de agua por
procesos de transpiración.
Se han estudiado ampliamente factores involucrados en la tolerancia a estrés ambiental,
que podrían ser usados como criterios de selección (Blumm y Jordan, 1989; Morgan y
Condon, 1986). La prolina, un aminoácido que aumenta en plantas bajo estrés, se ha
sugerido como un parámetro para la selección de materiales resistentes al estrés hídrico
(Bates et al., 1973). Sun et al., (2013), encontraron que condiciones de estrés hídrico
estimulan la producción de prolina, involucrada en funciones de las plantas tales como la
regulación osmótica, el mantenimiento de la estabilidad de la membrana, ser fuente de
carbono, entre otras. Adicionalmente, Ahmed y Hassan (2011) encontraron que valores
altos de prolina están asociados a genotipos con mejor adaptación a estrés ambiental.
Asimismo, mantener el balance osmótico es una importante adaptación fisiológica para
minimizar el efecto del estrés; algunas moléculas orgánicas específicas se acumulan en el
citoplasma celular, entre ellas se pueden mencionar azúcares solubles, betainas, polioles
y prolina (Sánchez et al., 1998). En efecto, variables como la producción de
malondialdehido y prolina se consideran como indicador de la integridad estructural de las
membranas sometidas a condiciones de estrés por déficit hídrico (Cao et al., 2008).
Sun et al. (2013) reportan que plantas susceptibles y tolerantes sometidas a condiciones
de estrés hídrico presentan actividad enzimática antioxidante con el fin de mitigar los
efectos provocados por los radicales libres de oxígeno. Los incrementos en las
concentraciones de enzimas como el superóxido dismutasa (SOD), Peroxidasa (POD) y
Catalasas (CAT) indican que las plantas han activado mecanismos de respuestas a
condiciones estresantes (Rosales et al., 2012).

8 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
1.4 Las respuestas de las plantas a condiciones de déficit
hídrico
Según Levitt. (1972) y Farooq et al. (2009) se reconocen tres tipos de mecanismos de a la
sequía: escape; evitación y tolerancia, las cuales se explican a continuación:
1. El escape a la sequía es definido como la capacidad de un genotipo vegetal para
completar su ciclo de vida antes de un grave déficit de agua del suelo y los cultivos
se desarrollan. Este mecanismo se caracteriza por un rápido desarrollo fenológico
reduciendo el tiempo para alcanzar la primera floración como una madurez
temprana, la plasticidad del desarrollo (variación en la duración del período de
crecimiento en función del grado de déficit de agua), y la removilización de
fotoasimilados al grano.
2. La evitación a la sequía es entendida como la capacidad del cultivo para mantener
alto el potencial de agua de los diferentes tejidos relativamente, a pesar de la baja
disponibilidad de humedad del suelo. Para ello, las plantas producen raíces más
largas para alcanzar una mayor profundidad en el suelo, un sistema de raíces
eficiente y el aumento de la conductancia hidráulica, y una menor pérdida de agua
a través del cierre de estomas en la hoja, la reducción de la absorción de la
radiación por el movimiento de las hojas, al igual que una reducción de la superficie
foliar para disminuir la evaporación.
3. La tolerancia a la sequía se define como la capacidad de una especie vegetal para
soportar el déficit hídrico a un bajo potencial de agua de los tejidos. Esto se logra a
través del mantenimiento de la turgencia mediante un ajuste osmótico, aumento de
la elasticidad de células y tolerancia a la desecación por la resistencia
protoplasmática.
El estudio de los diferentes mecanismos respuestas al déficit hídrico ha sido ampliamente
abordado en diferentes especies de importancia agronómica como el arroz (Boonjung y
Marco Teórico 9
Fukai., 1996-1 y 2), maíz (Cakir., 2004), trigo (Keyvan., 2010; Perveen et al., 2010), arveja
(Okçu et al., 2005), e incluso el fríjol (Rosales et al., 2012;) entre otras.
1.5 Fenología y el efecto del déficit hídrico sobre la
fenología del fríjol (P. vulgaris L.)
En cuanto a la fenología del fríjol, actualmente se cuentan con dos escalas fenológicas.
La primera, fue descrita por el IICA (2009), en la que la fenología del cultivo de frijol se
definió en 10 etapas, siendo cinco en estado vegetativos V y cinco en estado reproductivo
R (Figura 1-1): las etapas descritas son: V0: Germinación; V1: Emergencia; V2: Hojas
primarias; V3: Primera hoja trifoliada; V4: Tercera hoja trifoliada: R5: Prefloración; R6:
Floración; R7: Formación de vainas; R8: Llenado de vainas y R9: Maduración.
Por otro lado, la escala BBCH descrita por Fellet et al. (1995b), la cual está compuesta por
10 estados principales (Figura 1-2). El estado principal 0: Germinación; Estado 1:
Desarrollo de hojas; Estado 2: Formación de brotes laterales; Estado 5: Aparición del
órgano floral; Estado 6: Floración; Estado 7: Formación del fruto; Estado 8: Maduración de
frutos y semillas y Estado 9: Senescencia.
Finalmente, Muñoz et al. (2007), reportaron que el déficit hídrico en las etapas de pre
floración, formación y llenado de vainas afecta seriamente (aproximadamente un 40%) el
rendimiento, por lo cual es importante suministrar los requerimientos de agua, o fijar fechas
de siembra que coincidan con las etapas críticas de demanda hídrica.
Figura 1-1. Escala fenológica del fríjol (Phaseolus vulgaris L.) descrita por IICA et al., (2009).

1
0
Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.

Figura 1-2. Escala fenológica BBCH del fríjol (Phaseolus vulgaris L.) descrita por Feller et al., (1995b).

1.6 Antecedentes en el cultivo de fríjol en condiciones de
déficit hídrico.
A través de la historia se han realizado diferentes investigaciones para caracterizar las
respuestas fisiológicas de genotipos de las plantas de fríjol a condiciones de sequía. Beebe
et al. (2013), resumió las principales publicaciones realizadas en fríjol en condiciones de
estrés por sequía, en las cuales se evalúan las diferentes estrategias de las plantas a la
condición adversa las cuales incluyen el escape, la evitación y la tolerancia, las cuales se
relacionan en la Tabla 1-1.
Tabla 1-1. Resumen de los estudios en caracterización de las respuestas fisiológicas de fríjol bajo condiciones de sequía. Adaptado de Beebe et al. (2013).
Artículo Título Autores
1
Physiological analysis of common bean (Phaseolus
vulgaris L.) cultivars uncovers characteristics related to
terminal drought resistance.
Rosales et al. (2012)
2
Water use efficiency among dry bean landraces and
cultivars in drought-stressed and non-stressed
environments
Muñoz-Perea et al. (2007)
3 Differential adaptation of two varieties of common bean to
abiotic stress - II. Acclimation of photosynthesis Wentworth et al. (2006)
4
Differential adaptation of two varieties of common bean to
abiotic stress: I. Effects of drought on yield and
photosynthesis
Lizana et al. (2006)
5
Physiological Response to Drought Stress of Common
Bean (Phaseolus vulgaris L) Genotypes Differing in
Drought Resistance
Gebeyehu (2006)
6
Comparative effects of partial root drying (PRD) and
regulated deficit irrigation (RDI) on water relations and
water use efficiency in common bean (Phaseolus vulgaris
L).
Wakrim et al. (2005)
7 Comparative drought postponing and tolerance potentials
of two tepary bean lines in relation to seed yield Mohamed et al. (2005)
8 Leaf movements and photoinhibition in relation to water
stress in field-grown beans. Pastenes et al. (2005)
9 Paraheliotropism can protect water-stressed bean
(Phaseolus vulgaris L) plants against photoinhibition. Pastenes et al. (2004)
10 Breeding beans for resistance to terminal drought in the
lowland tropics Frahm et al. (2004)
11 Traits associated with dry edible bean (Phaseolus vulgaris
L) productivity under diverse soil moisture environments Shenkut y Brick (2003)
12 Mechanisms of drought resistance in grain legumes I:
osmotic adjustment Amede et al. (2003)

Marco Teórico 13
Artículo Título Autores
13 Mechanisms of drought resistance in grain legumes II:
stomatal regulation and root growth Amede y Schubert (2003)
14 Dehydration-avoidance responses of tepary bean lines
differing in drought resistance Mohamed et al. (2002)
15 Stomatal responses to leaf water status in common bean
(Phaseolus vulgaris L) is a function of time of day Mencuccini et al. (2000)
16 Indirect selection for drought resistance: a pilot study Dowkiw et al. (2000)
17
Acumulação de biomassa, características fisiológicas e
rendimento de grãos em cultivares de feijoeiro irrigado e
sob sequeiro
Gomes et al. (2000)
18
Differences in growth and water relations among
Phaseolus vulgaris cultivars in response to induced drought
stress
Costa et al. (2000)
19 Intrinsic water use efficiency at the pollination stage as a
parameter for drought tolerance in Phaseolus vulgaris Pimentel et al. (1999)
20 N2 fixation response to drought in common bean
(Phaseolus vulgaris L) Serraj y Sinclair (1998)
21 Traits related to drought resistance in common bean Ramirez-Vallejo y Kelly
(1998)
22 Adaptação do feijoeiro (Phaseolus vulgaris L) a seca. I.
Densidade e efficiência radicular Guimarães et al. (1996)
23 Moisture stress impact on N partitioning, N remobilization
and N-use efficiency in beans (Phaseolus vulgaris L) Foster et al. (1995)
24 Phenological plasticity as an adaptation by common bean
to rainfed environments
Acosta-Gallegos y White
(1995)
25
Relations of carbon isotope discrimination and other
physiological traits to yield in common bean (Phaseolus
vulgaris) under rainfed conditions
White et al. (1994)
26 Variation in leaf movements among common bean
cultivars Kao et al. (1994)
27
Stomatal response to humidity in common bean (Phaseolus
vulgaris): implications for maximum transpiration rate,
water-use efficiency and productivity
Comstock y Ehleringer
(1993)
28 Evaluation of diverse shoot genotypes on selected root
genotypes of common bean under soil water deficits
Jeffrey White y Castillo
(1992)
29 Saturated rates of photosynthesis in water-stressed leaves
of common bean and tepary bean
Castonguay y Markhart
(1991)
30 Carbon isotope discrimination and transpiration efficiency
in common bean Ehleringer et al. (1991)
31
Associations between productivity, root growth and carbon
isotope discrimination in Phaseolus vulgaris under water
deficit
White et al. (1990)
32 Relative effect of root and shoot genotypes and yield on
common bean under drought stress White y Castillo (1989)
33
Root growth of four common bean cultivars in relation to
drought tolerance in environments with contrasting soil
types
Sponchiado et al. (1989)

1
4
Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
Artículo Título Autores
34 Comparative water relations of Phaseolus vulgaris L and
Phaseolus acutifolius Gray Markhart (1985)
35 Effects of water stress on the water relations of Phaseolus
vulgaris and the drought resistant Phaseolus acutifolius Parsons y Howe (1984)
36 Photosynthetic response to water stress in Phaseolus
vulgaris O'Toole et al. (1977)
La mayoría de los estudios están enfocados en las diferentes variables fisiológicas
determinadas a nivel foliar, y pocos en las respuestas a nivel de raíz. Adicionalmente, los
estudios que se reportan anteriormente desarrollados en Colombia fueron realizados por
investigadores de CIAT, donde se han evaluado parámetros a nivel morfológico a nivel de
raíz (longitud, formación de raíces secundarias, acumulación de materia seca, etc.) y
fisiológico teniendo en cuenta variables como la eficiencia en el uso del agua el uso
eficiente del agua, transpiración, fotosíntesis, entre otros. Sin embargo, el uso de variables
bioquímicas como la producción de prolina y la acumulación de malondialdehido no ha sido
fuertemente estudiado en el país, e incluso el uso de tecnologías como el polietilenglicol
en sistemas de hidroponía, para estudiar detalladamente las raíces es escaso.
1.7 La utilidad de los Índices de Tolerancia en la caracterización de genotipos de plantas cultivadas.
La selección de genotipos con tolerancia a la sequía no es sencilla, debido principalmente
a las fuertes interacciones entre genotipos y el medio ambiente, además del escaso
conocimiento de la función y el papel de los diferentes mecanismos de tolerancia (Naghavi
et al., 2013). En este sentido, la implementación de índices de tolerancia a sequía en los
métodos de selección de genotipos de fríjol tolerantes y/o susceptibilidad al déficit hídrico
no son muy estudiados.
Actualmente, investigadores como Khakwani et al. (2011); Farshadfar et al. (2013) y
Naghavi et al. (2013), han trabajado en los diferentes métodos de selección de genotipos
con tolerancia a la sequía en cultivos de maíz y trigo mediante el uso de índices de
tolerancia y/o susceptibilidad. Igualmente, Fernández (1992), sugiere que el Índice de

Marco Teórico 15
tolerancia al estrés (STI) está definido como un marcador útil para determinar el potencial
de alta tolerancia y rendimiento el estrés de los genotipos evaluados. No obstante, se han
propuesto novedades de índices alternos útiles para la identificación de genotipos tales
como el Índice de Susceptibilidad al Estrés (SSI) por Fischer y Maurer (1978); la Tolerancia
al Estrés (TOL) y la Productividad Media (MP) por Rosielle y Hamblin (1981); la
Productividad Media Geométrica (GMP) por Fernández (1992), entre otros.

2. Capítulo. Identificación del periodo crítico para estrés hídrico sobre variables fisiológicas de un cultivar de fríjol arbustivo (Phaseolus vulgaris L.).
2.1 Resumen
El cambio climático es uno de los fenómenos ambientales de mayor impacto en los últimos
años, incidiendo en las superficies afectadas por sequías, e incrementando el área de las
mismas en zonas que eran influenciadas por la baja disponibilidad de agua, propiciando
condiciones de estrés hídrico. Por tal motivo, el objetivo de esta investigación fue
determinar la duración de un periodo crítico de estrés hídrico en fríjol arbustivo evaluando
las respuestas fisiológicas y bioquímicas bajo condiciones controladas. Sembrando
semillas de fríjol ICA-Cerinza, se realizó un diseño completamente al azar con cuatro
tratamientos (0, 5, 10 y 20 días de duración de estrés hídrico). Para cuantificar los
desórdenes fisiológicos, se realizaron medidas de intercambio gaseoso, fluorescencia del
PSII, pigmentos y pruebas bioquímicas (prolina y malondialdehido). La tasa de fotosíntesis,
conductancia estomática, clorofilas, y relaciones de fluorescencia se reducen en todos los
tratamientos, pero el efecto es prolongado en plantas sometidas a 10 y 20 días de estrés.
Asimismo, se evidencia una mayor fuga de electrolitos, acumulación de carotenoides
totales, MDA y prolina en las plantas que fueron afectadas por un periodo prolongado de
estrés hídrico. En conclusión, un estrés hídrico de 20 días genera daños fisiológicos
irreversibles en las plantas de fríjol arbustivo. Periodos de 5 y 10 días de estrés hídrico no
genera efectos severos sobre las variables fisiológicas de la especie estudiada. Un periodo
de estrés de 15 días permite generar un efecto moderado de estrés hídrico en las plantas
de fríjol sin ocasionar su muerte.
Palabras clave: Fotosíntesis, uso eficiente del agua, peroxidación lipídica,
regulación osmótica.

Capítulo 2 17
2.2 Introducción
El fríjol (Phaseolus vulgaris L.) es una especie de leguminosa nativa de Meso y
Suramérica, perteneciente a la familia Fabaceae (Kalavacharla et al., 2011). Esta
leguminosa es consumida ampliamente en Colombia como grano seco por su contenido
de proteínas, carbohidratos, vitaminas y minerales (FAO, 2007). Asimismo, esta especie
ocupó una producción de 62,974 toneladas durante el primer semestre del año 2015
(Fenalce, 2016), causando un impacto positivo sobre la generación de empleo e ingresos
para la economía campesina (FAO, 2007).
Los cultivos hortícolas se han visto sometidos a condiciones climáticas adversas
acentuadas por condiciones de cambio climático durante los últimos años en Colombia
(Miranda, 2009). Este sentido, Monasterio et al. (2011) afirman que el cambio climático ha
incidido en el incremento de áreas afectadas por condiciones de baja disponibilidad de
agua, propiciando más episodios de estrés hídrico. Asimismo, fenómenos de variabilidad
climática como el Niño/ Oscilación del Sur (ENOS) también causan mayores reducciones
de las precipitaciones e incremento de las temperaturas (Li et al., 2011; Ruiz y Pabón,
2013). En este aspecto, se ha reportado que Fenómenos ENOS pueden causar una
disminución en la oferta hídrica del 50% en más de 180 municipios del país (UNGRD,
2014). Igualmente, en el país existe un área alrededor de los 59.000 km2 expuestas a
sequía, encontrando gran parte de esta área en la región andina donde el fríjol es
comúnmente cultivado (Comunidad Andina, 2009).
Las plantas presentan diferentes tipos de respuestas a déficit hídrico como cambios
anatómicos, fisiológicos y bioquímicos a periodos transitorios o prolongados de baja
disponibilidad de agua, ocasionando afectaciones negativas sobre el crecimiento,
desarrollo y rendimiento (Segura-Monroy et al., 2015). Türkan et al. (2005) reportan en
plantas de Phaseolus vulgaris L. y Phaseolus acutifolius L. expuestas a diferentes días de
estrés hídrico se ven afectadas negativamente en parámetros como la acumulación de
materia fresca y seca, longitud de raíz y parte aérea de la planta entre otros, pero dicha
afectación es más significativa cuando el periodo de estrés es más prolongado (14 días de

1
8
Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
duración). Con respecto a las variables fisiológicas, las propiedades de intercambio
gaseoso de la hoja (fotosíntesis, transpiración y conductancia estomática), se ha reportado
ampliamente que bajo condiciones de estrés por déficit hídrico la tasa fotosintética y la
transpiración tienden a disminuir como consecuencia del cierre estomático (Yousfi et al.,
2012; Rosales et al., 2012). En cuanto a las relaciones hídricas, Miyashita et al. (2005)
reportan que cuando plantas de fríjol se encuentran sometidas a condiciones de estrés
hídrico afectan las relaciones hídricas, con la finalidad de disminuir la pérdida de agua por
procesos de transpiración. Adicionalmente, varios estudios han reportado que periodos de
déficit hídrico provocan alteraciones de los componentes de la fase lumínica tales como
una disminución sobre el contenido de clorofila en las hojas, la eficiencia máxima del PSII
fotoquímica (Fv/Fm) y el quenching fotoquímico disminuyen; mientras un incremento en el
quenching no fotoquímico como una estrategia de disipación de energía en forma de calor
(Aranjuelo et al., 2011).
Por otro lado, el estrés por sequía también induce una serie de respuestas bioquímicas
que son indicadores de tolerancia o susceptibilidad de plantas (Ahmed y Hassan, 2001;
Aranjuelo et al, 2011; Mohamed y Ibrahim, 2011). Las respuestas fisiológicas al igual que
las bioquímicas de las plantas a estrés por sequía están condicionadas por la duración del
periodo de estrés (Ashraf y Harris 2013). En este contexto, periodos cortos o prolongados
de estrés hídrico pueden afectar la estabilidad de la membrana debido a la peroxidación
lipídica, causando una producción de iones de peróxido y malondialdehido (MDA)
(Sánchez-Rodríguez et al., 2009; Sánchez-Reinoso et al., 2013). Del mismo modo,
cambios en la concentración de MDA han sido considerado como otro indicador de la
integridad estructural de las membranas sometidas a condiciones de estrés por estrés
hídrico (Cao et al., 2008). Adicionalmente, Sun et al. (2013) encontraron que condiciones
de estrés hídrico estimulan la producción de metabolitos secundarios tales como prolina,
que cumplen diversas funciones en las plantas en procesos como la regulación osmótica,
el mantenimiento de la estabilidad de la membrana, ser fuente de carbono y entre otras.
Igualmente, Ahmed y Hassan (2011) reportaron que una alta producción de prolina está
asociada a genotipos con mejor aclimatación a estreses ambientales. Por otro lado, el
estudio de la duración de diferentes periodos de estrés hídrico ha permitido conocer las
posibles estrategias de las plantas que les ayudan a sobrellevar esta condición abiótica
adversa y asimismo, tener una comprensión de los mecanismos fisiológicos involucrados

Capítulo 2 19
en las respuestas de aclimatación que repercuten en el funcionamiento de la planta y
posteriormente en el rendimiento del cultivo (Chaves et al., 2002; Farooq, et al., 2009, Xu
et al., 2010). Asimismo, la literatura disponible sobre el comportamiento de materiales
arbustivos comerciales de frijol a la duración de periodos de estrés hídrico en Colombia es
aún escasa, ya que la información disponible se ha centrado principalmente en la
caracterización de genotipos en un periodo puntual de sequía (Polanía et al, 2009;
Cardona-Ayala et al., 2014). En este sentido, evaluar las respuestas fisiológicas y
bioquímicas en plántulas de fríjol a diferentes periodos de duración del estrés por sequía
es de gran importancia, ya que nos permite determinar posibles respuestas de aclimatación
y sobrevivencia a la condición ambiental adversa estudiada. Por tal motivo, el objetivo de
esta investigación fue estudiar el efecto de diferentes periodos de estrés hídrico sobre las
respuestas fisiológicas y bioquímicas de plántulas de un cultivar de fríjol arbustivo
(Phaseolus vulgaris L.) determinando variables tales como la fotosíntesis, conductancia
estomática, transpiración, carbono interno, contenido relativo hídrico de la hoja, pérdida de
electrolitos, producción de malondialdehido y prolina
2.3 Materiales y métodos
2.3.1 Material vegetal y condiciones de crecimiento
El ensayo fue llevado a cabo en los invernaderos y laboratorios de la Facultad de Ciencias
Agrarias de la Universidad Nacional de Colombia ubicada en la ciudad de Bogotá a una
altura de 2556 msnm, (4° 35’56’’LN y 74°04’51’’LO) en el periodo comprendido entre los
meses de septiembre a noviembre del 2014. En el invernadero durante el periodo de la
evaluación, la temperatura promedio osciló entre los 15 y 22°C, la humedad relativa estuvo
entre el 60 a 90% y se tuvo un fotoperiodo natural de 12 h, con radiación fotosintéticamente
activa al medio día en promedio de 1000 µmol m−2 s−1). Para el establecimiento del
experimento, semillas del cultivar de fríjol arbustivo ICA-CERINZA fueron sembradas en
macetas de 1,5 litros de capacidad conteniendo arena cuarzítica, con el fin de mantener
un sustrato de baja retención de humedad y garantizar una condición de estrés por déficit
hídrico. Después de la germinación de semillas, las plantas fueron fertirrigidas con una

2
0
Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
solución nutritiva conteniendo la siguiente composición: 2.08 mM Ca(NO3)2 4H2O, 1.99 mM
Mg SO4 7 H2O, 2.00 mM NH4H2PO4, 10.09 mM KNO3, 46.26 nM H3BO3, 0.45 nM Na2MoO4
2H2O, 0.32 nM CuSO4 5H2O, 9.19 nM MnCl2 4H2O, 0.76 nM ZnSO4 7H2O, 19.75 nM FeSO4
H2O (Flórez-Velasco et al., 2015). El volumen de fertirriego fue calculado de acuerdo a la
capacidad de matera y los requerimientos hídricos de las plantas, realizando aplicaciones
cada dos días de 100 mL en las etapas iniciales (1-32 días después de emergencia (DDE))
y de 200 mL en etapas finales del experimento (33-65 DDE).
2.3.2 Tratamientos de déficit hídrico y toma de datos en los muestreos
El inicio de aplicación de todos los tratamientos fue cuando las plantas alcanzaron el
estado fenológico según escala BBCH 13-14 (formación de 3 a 4 hojas trifoliadas
completamente expandidas) aproximadamente a los 45 DDE según la metodología
reportada por Ghanbari et al. (2013). Los tratamientos consistieron en establecer cuatro
diferentes periodos de exposición a un déficit hídrico. Adicionalmente, plantas de los
tratamientos de 5 y 10 días contaron con un periodo de rehidratación hasta el muestreo a
los 20 días de inicio del estrés. El déficit hídrico fue establecido mediante el procedimiento
descrito por Moftha y Al Humaid (2005) y Segura-Monroy et al. (2015) donde se cubrían el
50% de las necesidades de evapotranspiración, las cuales eran determinadas mediante la
pérdida de peso de las materas. Asimismo, el criterio de tomar un 50% de humedad en el
suelo se fundamentó porque las plantas presentan reducción alteraciones en sus
relaciones hídricas en humedades del sustrato entre el 40 y 60 % (Costa-França et al.,
2000; Segura-Monroy et al., 2015).
Se desarrollaron cuatro muestreos de variables fisiológicas y bioquímicas a los 0, 5, 10 y
20 días de iniciado los diferentes tratamientos de estrés (DDT), donde las plantas con
periodo de estrés de 5 y 10 días contaron con 15 y 10 días de recuperación al final del
experimento, respectivamente. En general, las medidas fisiológicas fueron realizadas en
la segunda hoja trifoliada completamente expandida según la metodología utilizada por
Ghambari et al. (2013). Con respecto a las variables bioquímicas, se colectó material
vegetal, el cual fue homogenizado con nitrógeno líquido y posteriormente almacenado a -

Capítulo 2 21
80°C. Finalmente, el experimento duró aproximadamente 65 días a partir de la emergencia
de la totalidad de las plántulas
2.3.3 Diseño experimental y análisis de los datos
Se aplicó un diseño completamente al azar en el que se evaluaron cuatro tratamientos de
duración del estrés por déficit hídrico (cero, cinco, diez y veinte días). Cada tratamiento
consistió de cuatro repeticiones por tratamiento-muestreo (16 plántulas por tratamiento; 64
plántulas por todo el experimento). Posteriormente, cuando se presentaron diferencias
significativas en el ANAVA, se empleó la prueba comparativa de medias de Tukey a p ≤
0.05. Los datos se analizaron usando el programa Statistix v 9.0 (software analítico,
Tallahassee, FL, US).
2.3.4 Variables fisiológicas
Relaciones hídricas e intercambio gaseoso de la hoja
El contenido relativo de agua (CRA) fue determinado en hojas totalmente expandidas de
la parte media del follaje. Las hojas fueron recolectadas a los cinco, diez y veinte días
después del inicio del periodo de estrés hídrico para las plantas de cada uno de los
tratamientos. El CRA fue calculado mediante la siguiente ecuación
𝐶𝑅𝐴 = [𝑃𝐹 − 𝑃𝑆
𝑃𝑇 − 𝑃𝑆] 𝑋100 (𝐸𝑐𝑢𝑎𝑐𝑖ó𝑛 1)
Siendo PF el peso fresco, PT el peso en turgencia medido después de 24 horas de
saturación en agua destilada a 4 °C en la oscuridad y PS es el peso seco determinado
después de 48 horas en un horno a 70°C.

2
2
Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
La fotosíntesis fue estimada usando un medidor de fotosíntesis portable (LI-COR 6200,
Lincoln, NE, USA), a los 45, 50, 55 y 65 DDE (0, 5, 10 y 20 DDT) entre los periodos
comprendidos de las 930 a 1100 h y 1330 a 1500 h durante tres días seguidos a partir del
penúltimo día antes de finalizar el periodo de estrés de cada tratamiento. Asimismo, este
equipo estimó la concentración interna de dióxido de carbono. Adicionalmente, se calculó
la relación Carbono intercelular entre el Carbono atmosférico (Ci/Ca). La conductancia
estomática y la traspiración se midió en la segunda hoja trifoliada completamente
expandida con la ayuda de un porómetro portable (LI-COR 1600, Lincoln, NE, USA). Del
mismo modo, se calculó la eficiencia en el uso del agua como producto de la relación entre
la fotosíntesis y la transpiración. Las mediciones se realizaron entre los periodos
comprendidos de las 930 a 1100 h y 1330 a 1500 h. Durante las mediciones las condiciones
fueron las siguientes: Radiación fotosintéticamente activa 800 μmol m-2 s-1, temperatura de
la hoja 27 ± 5 °C, diferencia de presión de vapor del aire de la hoja 1.8 ± 0.5 kPa.
Parámetros de fluorescencia, contenido de clorofilas y carotenoides
Para la determinación de los parámetros de fluorescencia de PSII, se empleó un
fluorómetro modulado (MINI-PAM, Walz, Effeltrich, GE). Las medidas fueron tomadas
entre las 700 y 1000 h en la segunda hoja trifoliada completamente expandida desde la
parte superior del dosel a los 45, 50, 55 y 65 DDE (0, 5, 10 y 20 DDT). Las relaciones de
fluorescencia fueron determinadas con un pulso máximo de intensidad de luz de hasta
2.600 µmol m-2 s-1 en la superficie de la muestra de la hoja, adaptando previamente a la
oscuridad durante 10 minutos. Para las mediciones de clorofilas y carotenoides, las
muestras de tejidos foliar (0.03 g aprox. de la segunda hoja trifoliada completamente
expandida) fueron homogenizadas en 2 mL de acetona (80%), centrifugada para eliminar
partículas y el sobrenadante diluido a un volumen final de 6 mL adicionando acetona (Sims
y Gamon, 2002). El contenido de clorofila fue determinado a 663 y 646 nm y carotenoides
a 470 nm utilizando un espectrofotómetro (Spectronic BioMate 3 UV–vis Thermo, Madi-
son, WI, USA). Para obtener el contenido de pigmentos fotosintéticos en hoja, se
emplearon las ecuaciones para acetona descritas por Wellburn (1994).

Capítulo 2 23
2.3.5 Variables bioquímicas
MDA y daño en la permeabilidad de la membrana en fuga de electrolitos
Para la determinación de la oxidación lipídica (Malondialdehido-MDA), se empleó el
método del ácido tiobarbitúrico (TBA) descrito por Hodges et al. (1999) tomando aprox. 0.3
g de material vegetal homogenizado y almacenado con nitrógeno líquido. Las muestras
luego de ser centrifugadas se les estimó las absorbancias a 440, 532 y 600 nm con un
espectrofotómetro (Spectronic BioMate 3 UV - Vis, Thermo, Madison, WI, USA). Por último,
se utilizó un coeficiente de extinción (157 M mL-1) para la obtención de la concentración de
MDA.
El daño de la membrana plasmática estimada en el porcentaje de fuga de electrolitos se
determinó por el método descrito por Jiang and Zhang (2001) modificado. Extrayendo 5
discos de 0.5 dm de diámetro de la segunda hoja trifoliada completamente expandida. Se
utilizó 25 mL de agua desionizada como medio en tubos falcon de 50 mL de capacidad, a
las cuales se les registró la conductividad eléctrica inicial (CE1) luego de dos horas en baño
de agua a 30°C y la conductividad eléctrica final (CE2) del medio después de 20 min en
baño de agua a 90°C. El porcentaje de electrolitos se calculó aplicando la siguiente
ecuación:
𝐹𝑢𝑔𝑎 𝑑𝑒 𝑒𝑙𝑒𝑐𝑡𝑟𝑜𝑙𝑖𝑡𝑜𝑠 (%) = (𝐶𝐸1
𝐶𝐸2) 𝑥 100 (𝐸𝑐𝑢𝑎𝑐𝑖ó𝑛 2)
Concentración de Prolina
Para la determinación del contenido de prolina, utilizamos el método descrito por Bates et
al. (1973), extrayendo de la segunda hoja trifoliada completamente expandida
aproximadamente 0.3 g de material vegetal homogenizado y almacenado con nitrógeno
líquido. La absorbancia se medió a 520 nm usando un espectrofotómetro (Spectronic

2
4
Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
BioMate 3 UV - Vis, Thermo, Madison, WI, USA). El contenido de prolina se determinó
mediante una curva estándar y calculada en peso fresco basado en la ecuación 3:
𝜇𝑚𝑜𝑙 𝑃𝑟𝑜𝑙𝑖𝑛𝑎
𝑔 𝑚𝑎𝑡𝑒𝑟𝑖𝑎𝑙 𝑣𝑒𝑔𝑒𝑡𝑎𝑙 𝑓𝑟𝑒𝑠𝑐𝑜=
[(
𝜇𝑔 𝑃𝑟𝑜𝑙𝑖𝑛𝑎𝑚𝐿
𝑥 𝑚𝐿 𝑇𝑜𝑙𝑢𝑒𝑛𝑜)
115.5 𝜇𝑔𝜇𝑚𝑜𝑙
]
[𝑔 𝑚𝑢𝑒𝑠𝑡𝑟𝑎
5 ] (𝐸𝑐𝑢𝑎𝑐𝑖ó𝑛 3)
2.4 Resultados
2.4.1 Intercambio gaseoso y relaciones hídricas.
Resultados del Contenido Relativo de Agua (CRA) en la hoja se muestran en la Figura
2-1. En general, plantas de los tratamientos control y cinco días de déficit hídrico mostraron
un CRA aproximadamente del 90% a lo largo de los diferentes periodos de exposición al
estrés. Asimismo, diferencias significativas se observaron a los 10 días de iniciado los
tratamientos de estrés hídrico (DIT). En este punto, plántulas de frijol de los tratamientos
de 10 y 20 días de duración de estrés empezaron a registrar un menor CRA (73.3 y 75.7%
respectivamente). Adicionalmente, un CRA del 30% fue obtenido cuando las plántulas
estuvieron bajo una condición estresante de 20 días, situando a este grupo de plantas
cerca del punto de marchitez permanente, en el que se han reportado que valores
inferiores al 40% en el CRA están relacionados con condiciones de punto de marchitez
permanente (Lacape et al., 1998; Korir et al., 2006). Finalmente, estos resultados permiten
indicar que se poseían diferentes estatus hídricos durante los diferentes periodos de
exposición.
Capítulo 2 25
Figura 2-1. Efecto de diferentes periodos de estrés hídrico sobre el contenido hídrico relativo de la hoja (CRA)
en plántulas de fríjol arbustivo cultivar ICA-CERINZA. Los puntos son el promedio de cuatro datos. Las barras representan ± error estándar. Promedios seguidos por la misma letra no son significativamente diferentes según la prueba de Tukey (P ≤ 0.05).

2
6
Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
Los resultados de las variables de intercambio gaseoso de la hoja (fotosíntesis (Pn),
conductancia estomática (gs) y transpiración (E)), los resultados son resumidos en la
Figura 2-2 (A, B y C, respectivamente). La Figura 2-2A muestra las diferencias
significativas a partir de los primeros 5 días de iniciado el estrés hídrico, donde la tasa de
fotosíntesis se redujo en más del 50% en las plantas de los tratamientos de cinco, diez y
veinte días de estrés. Asimismo, esta tendencia se mantuvo entre tratamientos a los 10
DDT. Al final del experimento (20 DDT), la Pn siguió en los mismos bajos valores en las
plantas que fueron sometidas durante 10 y 20 días a estrés. Mientras los valores de Pn en
el tratamiento de 5 días de estrés mostraron una recuperación y alcanzaron niveles
cercanos a las plantas control. Por otro lado, la gs también se vio afectada negativamente
en los diferentes tratamientos a partir de los 10 DDT con un descenso en los valores de
aproximadamente el 45% en todos los tratamientos de estrés hídrico con respecto al
control. Asimismo, esta variable de intercambio gaseoso mostro una mayor reducción
(alrededor del 95%) en el tratamiento de veinte días de estrés al final del ensayo. Mientras,
las plantas expuestas cinco y diez días 5 y 10 días de duración del periodo de estrés
mostraron una recuperación en la gs con respecto a las plantas control (Figura 2-2B). En
este sentido, la reducción de la conductancia estomática está relacionada con los bajos
valores de fotosíntesis y las mismas tendencias para todos los Figura 1-1tratamientos
fueron observadas en los resultados de la tasa de transpiración, donde los valores más
bajos se pudieron apreciar en el tratamiento de 20 días de estrés, con una disminución
mayor al 70% al final del ensayo (Figura 2-2C). Sin embargo, el uso eficiente del agua
(UEA) mostró diferencias significativas entre los tratamientos a partir de los 5 días de
iniciado el periodo de estrés, aunque los cambios más marcados se evidenciaron en el
último muestreo (20 DDT). En este punto el UEA en plantas que estuvieron bajo 20 días
de periodo de estrés fue diez veces mayor en comparación con los demás tratamientos
(Tabla 2-1).

Capítulo 2 27
Figura 2-2. Efecto de diferentes periodos de estrés hídrico sobre parámetros de intercambio gaseoso de la hoja en plántulas de fríjol arbustivo cultivar ICA-CERINZA: A). Fotosíntesis neta (Pn); B). Conductancia estomática (gs) y C). Transpiración (E). Las barras son el promedio de cuatro datos ± error estándar. Promedios seguidos por la misma letra no son significativamente diferentes según la prueba de Tukey (P ≤ 0.05).
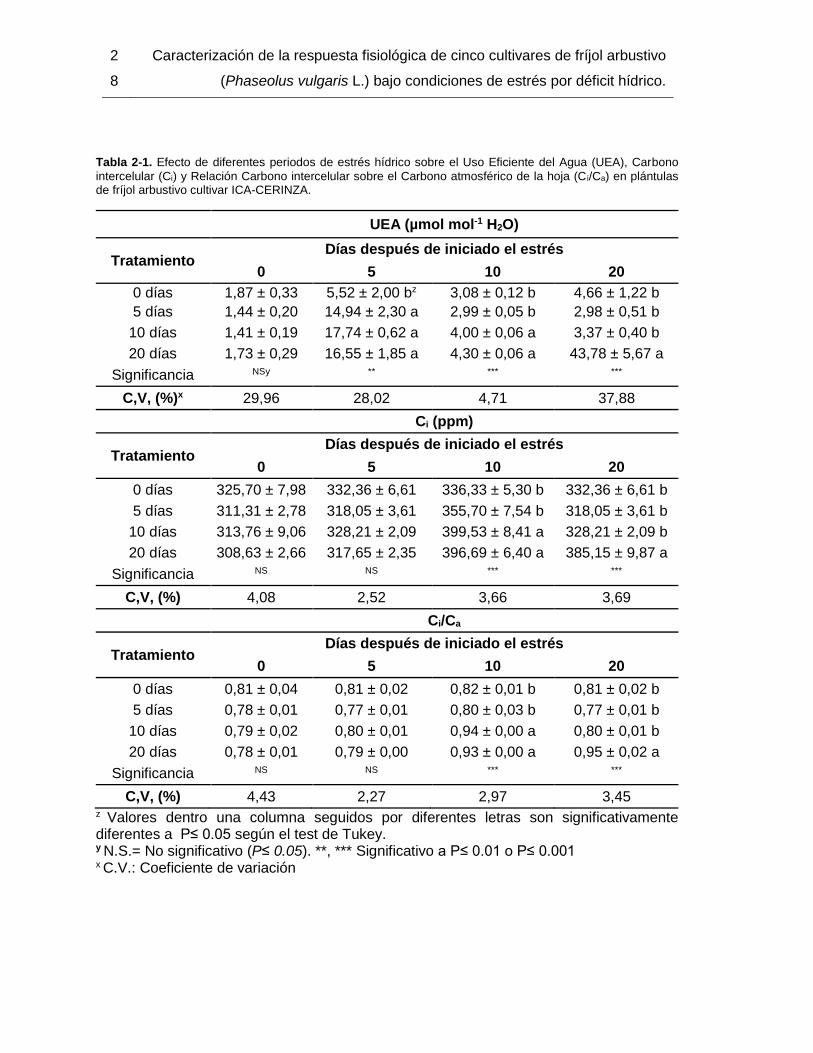
2
8
Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
Tabla 2-1. Efecto de diferentes periodos de estrés hídrico sobre el Uso Eficiente del Agua (UEA), Carbono
intercelular (Ci) y Relación Carbono intercelular sobre el Carbono atmosférico de la hoja (C i/Ca) en plántulas de fríjol arbustivo cultivar ICA-CERINZA.
UEA (µmol mol-1 H2O)
Tratamiento Días después de iniciado el estrés
0 5 10 20
0 días 1,87 ± 0,33 5,52 ± 2,00 bz 3,08 ± 0,12 b 4,66 ± 1,22 b
5 días 1,44 ± 0,20 14,94 ± 2,30 a 2,99 ± 0,05 b 2,98 ± 0,51 b
10 días 1,41 ± 0,19 17,74 ± 0,62 a 4,00 ± 0,06 a 3,37 ± 0,40 b
20 días 1,73 ± 0,29 16,55 ± 1,85 a 4,30 ± 0,06 a 43,78 ± 5,67 a
Significancia NSy ** *** ***
C,V, (%)x 29,96 28,02 4,71 37,88
Ci (ppm)
Tratamiento Días después de iniciado el estrés
0 5 10 20
0 días 325,70 ± 7,98 332,36 ± 6,61 336,33 ± 5,30 b 332,36 ± 6,61 b
5 días 311,31 ± 2,78 318,05 ± 3,61 355,70 ± 7,54 b 318,05 ± 3,61 b
10 días 313,76 ± 9,06 328,21 ± 2,09 399,53 ± 8,41 a 328,21 ± 2,09 b
20 días 308,63 ± 2,66 317,65 ± 2,35 396,69 ± 6,40 a 385,15 ± 9,87 a
Significancia NS NS *** ***
C,V, (%) 4,08 2,52 3,66 3,69
Ci/Ca
Tratamiento Días después de iniciado el estrés
0 5 10 20
0 días 0,81 ± 0,04 0,81 ± 0,02 0,82 ± 0,01 b 0,81 ± 0,02 b
5 días 0,78 ± 0,01 0,77 ± 0,01 0,80 ± 0,03 b 0,77 ± 0,01 b
10 días 0,79 ± 0,02 0,80 ± 0,01 0,94 ± 0,00 a 0,80 ± 0,01 b
20 días 0,78 ± 0,01 0,79 ± 0,00 0,93 ± 0,00 a 0,95 ± 0,02 a
Significancia NS NS *** ***
C,V, (%) 4,43 2,27 2,97 3,45 z Valores dentro una columna seguidos por diferentes letras son significativamente diferentes a P≤ 0.05 según el test de Tukey. y N.S.= No significativo (P≤ 0.05). **, *** Significativo a P≤ 0.01 o P≤ 0.001 x C.V.: Coeficiente de variación

Capítulo 2 29
Adicionalmente, en el contenido de carbono intercelular (Ci) se encontraron diferencias
significativas a partir de los 10 días de estrés, evidenciándose un incremento
aproximadamente del 15% en las plantas sometidas a tres diferentes periodos de duración
de déficit de agua en el suelo comparadas con plantas control. Al final del experimento,
las plantas de frijol que estuvieron durante 20 días de estrés continuaron mostrando una
mayor Ci Mientras, las plantas con cinco y diez días de estrés sus niveles internos CO2
estuvieron cercanos a los observados en las plantas control. Finalmente, la relación
Carbono intercelular sobre el Carbono atmosférico presentó diferencias significativas a los
10 y 20 DDT. Se pudo observar que las mayores valores fueron obtenido para los
tratamientos de mayor duración de sequía (10 y 20 días de sequía respectivamente) con
respecto al tratamiento control y cinco días de duración de estrés con respecto al tercer
muestreo (10 DDE). A los 20 DDE, el único tratamiento que mostro diferencias con
respecto a los otros tres tratamiento fueron aquellas plántulas que tuvieron el periodo de
estrés más prolongado (20 días). En este punto de muestro, la relación Ci/Ca fue 18%
mayor comparado al control.
2.4.2 Parámetros de fluorescencia del fotosistema II, contenido de clorofilas y carotenoides totales
En cuanto a los parámetros de fluorescencia, resultados de la eficiencia máxima del
quantum en el fotosistema II (Fv/Fm), Eficiencia real del fotosistema II (YII), quenching no
fotoquímico (QNP) y quenching fotoquímico (qP) se muestran en la Figura 2-3(A, B, C y D
respectivamente). Diferencias significativas fueron encontradas en las plantas de todos los
tratamientos a los 20 días después de haber iniciado el estrés; donde las mayores
reducciones en la Fv/Fm, Y(II) y qP se evidencian en las plantas que fueron sometidas a un
periodo de 20 días de estrés. Sin embargo, una tendencia opuesta fue observada sobre el
QPN donde este índice presentó un incremento, indicando que una estrategia de
aclimatación es la disipación de energía en el fotosistema II a través de la producción de
calor.

3
0
Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
Por otro lado, la Figura 2-4 se muestra los resultados obtenidos en la cuantificación del
contenido de clorofila a, b, total y contenido de carotenoides totales. Se puede observar
que para los tres primeros muestreos (a los cero, cinco y diez DDE) no se obtuvieron
diferencias significativas en la concentración de los pigmentos fotosintéticos evaluados. En
el último muestro (20 DDT) se observó una reducción en el contenido de clorofila a, b y
total del ~25%, ~7% y ~17% respectivamente en las plantas de frijol que estuvieron
sometidas al periodo de estrés de mayor duración. Sin embargo, la producción de
carotenoides totales presento un incremento en general para los tratamientos que
contaron con algún periodo de estrés. Asimismo, las plántulas que estuvieron bajo 20 días
de sequía mostraron un incremento aproximadamente del doble de la concentración de los
carotenoides totales obtenida en el tratamiento control.
Figura 2-3. Efecto de diferentes periodos de estrés hídrico sobre parámetros de fluorescencia del fotosistema II plántulas de fríjol arbustivo cultivar ICA-CERINZA: A). Eficiencia máxima del quantum en el fotosistema II (Fv/Fm); B). Quenching fotoquímico (qP); C). La eficiencia real del fotosistema II (Y(II)) y D). Quenching no
fotoquímico (QNP) en plántulas de fríjol arbustivo. Las barras son el promedio de cuatro datos ± error estándar. Promedios seguidos por la misma letra no son significativamente diferentes según la prueba de Tukey (P ≤ 0.05).
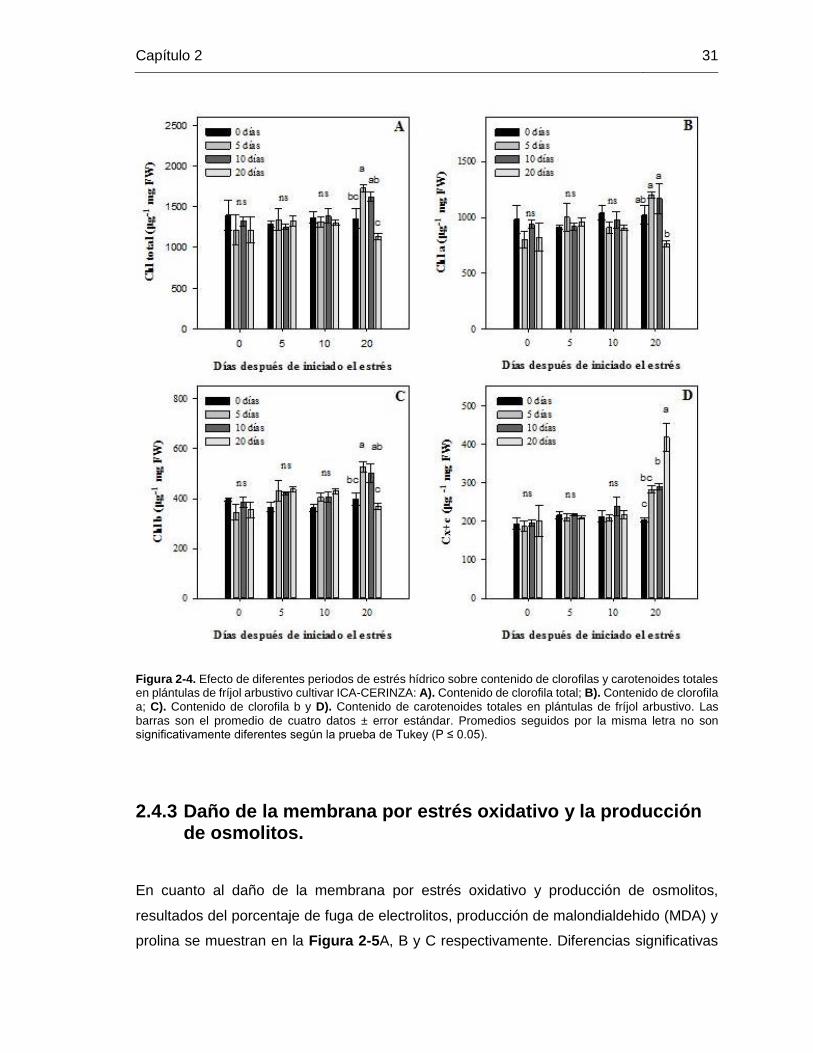
Capítulo 2 31
Figura 2-4. Efecto de diferentes periodos de estrés hídrico sobre contenido de clorofilas y carotenoides totales en plántulas de fríjol arbustivo cultivar ICA-CERINZA: A). Contenido de clorofila total; B). Contenido de clorofila a; C). Contenido de clorofila b y D). Contenido de carotenoides totales en plántulas de fríjol arbustivo. Las
barras son el promedio de cuatro datos ± error estándar. Promedios seguidos por la misma letra no son significativamente diferentes según la prueba de Tukey (P ≤ 0.05).
2.4.3 Daño de la membrana por estrés oxidativo y la producción de osmolitos.
En cuanto al daño de la membrana por estrés oxidativo y producción de osmolitos,
resultados del porcentaje de fuga de electrolitos, producción de malondialdehido (MDA) y
prolina se muestran en la Figura 2-5A, B y C respectivamente. Diferencias significativas

3
2
Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
fueron observadas entre los diferentes tratamiento en las variables anteriormente
mencionada a partir de los 10 DDT, sin embargo, estas diferencias fueron más notorias al
final del experimento (20 DDT). En este punto de muestreo, la mayor pérdida de electrolitos
se evidencia en el tratamiento de 20 días de duración del periodo de estrés, alcanzando
valores cercanos al 50%; mientras que los tratamientos de 5 y 10 días no superaron niveles
del 20% y 18%, respectivamente; siendo los niveles del tratamiento control no mayores al
16% (Figura 2-5A). Lo anterior se puede relacionar con el incremento en la producción de
MDA donde los mayores valores también se obtuvieron en el tratamiento de 20 días de
estrés, En este grupo de plantas fue encontrada una producción de MDA cercana al doble
a los valores presentados en el tratamiento control (Figura 2-4B). Asimismo, la producción
de prolina fue favorecida en todos los tratamientos que tuvieron estrés hídrico a los cinco
y diez DDT. Sin embargo, las plantas expuestas a 20 días de déficit de agua se presentaron
los mayores valores de 350 µmol g-1 PF comparado con los 25 µmol g-1 FW encontrados
en hojas de plantas frijol control y sometidas a cinco y diez días de sequía.
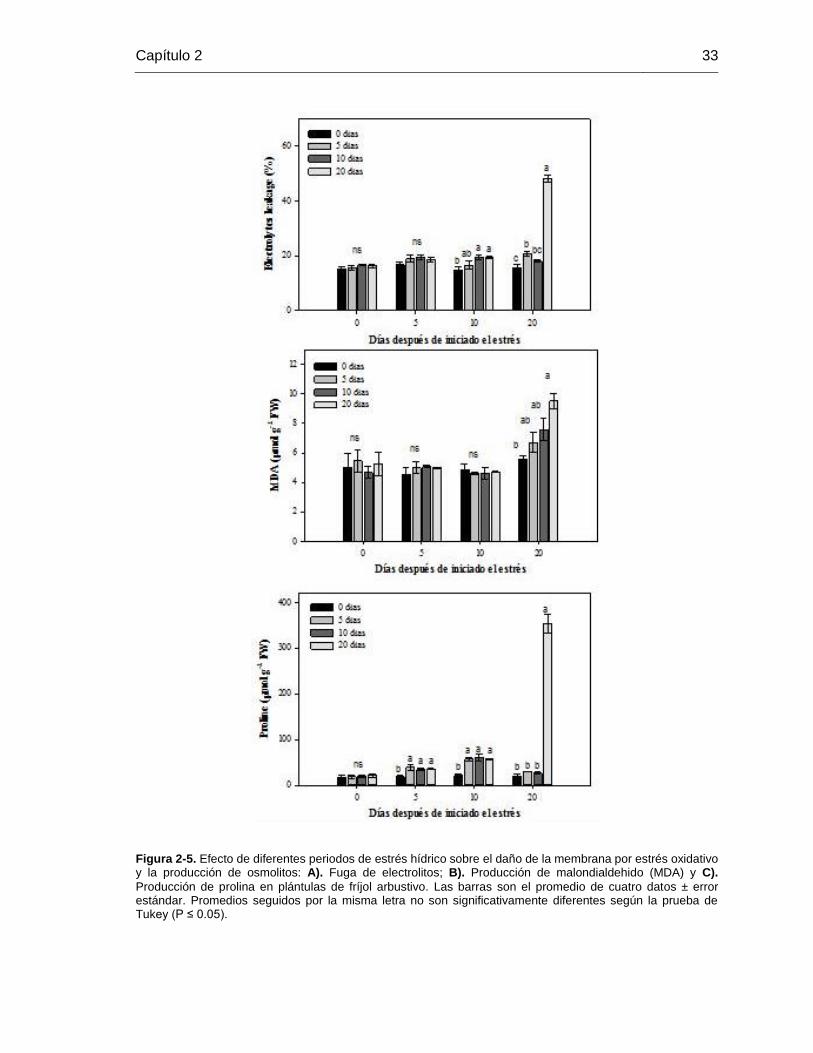
Capítulo 2 33
Figura 2-5. Efecto de diferentes periodos de estrés hídrico sobre el daño de la membrana por estrés oxidativo y la producción de osmolitos: A). Fuga de electrolitos; B). Producción de malondialdehido (MDA) y C).
Producción de prolina en plántulas de fríjol arbustivo. Las barras son el promedio de cuatro datos ± error estándar. Promedios seguidos por la misma letra no son significativamente diferentes según la prueba de Tukey (P ≤ 0.05).

3
4
Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
2.5 Discusión
El contenido relativo de agua de la hoja (CRA) es una de las variables de gran importancia
fisiológica que ayuda a cuantificar el estado hídrico de una planta tanto en condiciones
normales como de sequía (Keyvan., 2010). Cuando el CRA presenta una reducción en los
valores ayuda a inferir que las relaciones hídricas pueden estar siendo afectadas por una
condición abiótica adversa, lo cual puede repercutir en algunos procesos fisiológicos como
una reducción de la tasa fotosintética (Costa-França et al., 2000). Los resultados
encontrados en el presente estudio muestran que a medida que el estrés por déficit hídrico
era más prolongado, se podía evidenciar una reducción de los valores del CRA en las
plantas de los diferentes tratamientos, especialmente en las plantas que estuvieron
sometidas durante 20 días de estrés. Costa-França et al., 2000 y De Laat, 2014
argumentan que un menor CRA está relacionado con una baja tasa de fotosíntesis
principalmente debido a una limitación por cierre de estomas, la cual se refleja en los
valores bajos de la conductancia estomática y transpiración. Adicionalmente, Huang et al.,
(2011) señalan que un ajuste estomático es una estrategia común para evitar una
condición de sequía en plantas y que una menor Pn puede ser ocasionada por: i) una
limitación estomática que reduce simultáneamente la gs y Ci o ii) una limitación no
estomática como el aumento de la Ci. En este estudio, la Pn mostro una reducción
alrededor de un 50% y la Cí aumento alrededor de un 398 ppm y 385 ppm cuando las
plantas estuvieron entre 10 y 20 días de exposición a un déficit hídrico, respectivamente.
Lo anterior indica que las plantas de frijol presentaron una limitación no estomática, lo cual
se reflejó en una menor Pn y a su vez en un incremento de la Ci (Farquhar y Sharkey,
1982). Por otro lado, el UEA fue mayor cuando el periodo de estrés hídrico aumentaba en
este trabajo. En este aspecto, Huang et al. (2011) señalan que un alto UEA provee una
evidencia del papel de los estomas en mantener un alto contenido de agua en los tejidos
de la hoja bajo condiciones de déficit hídrico. Asimismo, un alto UEA puede conducir a una
alta actividad fotosintética, sin embargo, ellos señalan que estos morfogenéticos y
metabólicos son requeridos con el propósito de un mejor entendimiento de la relación entre
cierre estomático y UEA. Por otro lado, el efecto negativo que puede generar una
condición de déficit hídrico sobre la eficiencia máxima del PSII fotoquímica (Fv/Fm), la
eficiencia real del fotosistema II (Y(II)), quenching fotoquímico y no fotoquímico (qP y QNP)

Capítulo 2 35
depende de la duración y la severidad del estrés (Aranjuelo et al, 2011). En este contexto,
los resultados obtenidos indican que un periodo de 20 días de duración de estrés hídrico
redujo significativamente los parámetros del fotosistema II (Fv/Fm, Y(II) y qP).
Adicionalmente, un déficit hídrico generó un estrés moderado ya que solo presentó
limitaciones de la fotosíntesis de tipo estomático, mientras que un periodo de 20 días de
déficit hídrico generó un estrés severo, ya que se evidenciaron limitaciones de la
fotosíntesis te tipo estomático y no estomático. Similares observaciones fueron obtenidas
por Dias y Brüggemann. (2010) en Phaseolus vulgaris donde encontraron que plantas bajo
condición de estrés hídrico reducen el qP considerablemente comparadas con las plantas
control. De igual manera, se evidencia un incremento en el NPQ como medida principal de
disipación de energía en el PSII bajo condiciones de estrés abiótico (Perveen et al., 2010;
Aranjuelo et al, 2011; Gorbe y Catalayud., 2012).En el presente trabajo, se observó
también un incremento del NPQ, especialmente, en las plantas con 20 días de déficit
hídrico. Cousins et al. 2002 señalan que un mayor NPQ puede ser un mecanismo de
protección en contra de la fotoinhibición y fotodaño bajo estrés hídrico. Con respecto a la
producción de pigmentos fotosintéticos, se observó que el contenido de clorofilas totales
mantuvo valores cercanos a los obtenidos en las plantas control a los 0, 5 y 10 DDT (~1400
µg mg-1 PF). Similares observaciones fueron obtenidas por Aranjuelo et al., (2011) donde
la clorofila total no varió significativamente entre las plantas con estrés hídrico vs las
plantas control, debido a que el estrés hídrico de 5 y 10 días de duración en fríjol no
degrada los pigmentos fotosintéticos. Sin embargo, las plántulas de frijol con un periodo
de estrés de 20 días si mostraron un descenso en el contenido de clorofila total con
respecto al control. La disminución del contenido clorofila bajo condiciones de déficit
hídrico ha sido también reportada en otras especies, dependiendo la severidad y la
duración del estrés (Mafakheri et al., 2010). En este aspecto, una reducción del contenido
total de clorofila bajo estrés hídrico señala una baja capacidad de los centros de reacción
de la luz con el propósito evitar daños de especies reactivas de oxígeno, ya que la
producción de estas es impulsado principalmente por el exceso de absorción de energía
en el aparato fotosintético (Herbinger et al., 2002). Por otro lado, la concentración de
carotenoides totales se incrementó en las plantas a medida que el periodo de estrés era
mayor (Fig. 4). Los resultados sugieren que la producción de carotenoides obedecen a una
estrategia de aclimatación a la condición estresante, la cual está involucrada en las
funciones antioxidantes de dichos pigmentos (Efeoğlu et al., 2009).

3
6
Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
El uso de marcadores bioquímicos como MDA y prolina también han sido ampliamente
evaluados para evaluar el grado de aclimatación de las plantas a condiciones de estrés
por déficit hídrico (Keyvan., 2010; Türkan et al., 2005). En el presente estudio, la
concentración de prolina era mayor comparada en las plantas control, especialmente
cuando las plantas del cultivar Cerinza tenían un estrés de 20 días de déficit hídrico (353.6
µmol g-1 PF en plantas estresadas vs 20.04 µmol g-1 PF en plantas control). En general, la
literatura menciona que la relación entre la acumulación de prolina y estrés ambiental
sugiere que este aminoácido puede tener una función de tipo de protección que ayuda a
la planta en su proceso de aclimatación a esa condición de déficit hídrico (Farooq et al.,
2009). Adicionalmente, Cao et al. (2008) reporta que la producción y acumulación de MDA
en plantas expuestas a condiciones de estrés es considerado como un indicador de
peroxidación lipídica, es decir que a mayor valor mayor daño a la membrana de la
integridad estructural de las membranas sometidas a condiciones de estrés hídrico.
Similares observaciones han sido reportadas por Türkan et al. (2005) donde encontraron
una mayor acumulación de MDA (50% mayor en plantas estresadas), lo cual les indico que
las plantas sufrieron un daño severo causado por la condición estresante (Türkan et al.,
2005; Farooq et al., 2009; Keyvan., 2010).
2.6 Conclusiones
• En el presente estudio se identificó que un periodo de déficit hídrico (50% humedad
del suelo) superior a quince días causó una afectación negativa sobre la fisiología
de plantas de frijol arbustivo (ICA-CERINZA) debido a un mayor estrés oxidativo y
una menor fotosíntesis.

3. Capítulo 3. Evaluación fisiológica y bioquímica de cinco cultivares de fríjol arbustivo (Phaseolus vulgaris L.) bajo condición de estrés por déficit hídrico.
3.1 Resumen
La agricultura mundial se ha visto afectada negativamente por la baja disponibilidad de
agua en épocas de sequía, como consecuencia de la variabilidad climática y el cambio
climático, generando condiciones de estrés en las plantas por déficit hídrico, afectando
cultivos de importancia en la dieta humana como el fríjol. Por tal motivo, el objetivo de esta
investigación fue evaluar las respuesta fisiológicas y bioquímicas de cinco cultivares de
fríjol arbustivo (Phaseolus vulgaris L.) bajo una condición de estrés por déficit hídrico de
15 días en una etapa vegetativa. Sembrando semillas de los cultivares de fríjol ICA-
Cerinza, ICA-Bachue, NUA35, Bacatá y Bianca se realizó un diseño completamente al azar
en arreglo factorial (factor principal el estatus hídrico y factor secundario los cultivares)
para un total de 10 tratamientos con cuatro repeticiones. Para cuantificar las respuestas
fisiológicas, se realizaron medidas de fotosíntesis, fluorescencia del PSII, acumulación de
materia seca, pigmentos, prolina y malondialdehido. La tasa de fotosíntesis, conductancia
estomática, clorofilas, y relaciones de fluorescencia se reducen en todos cultivares que
tuvieron condición de estrés. Asimismo, se evidencia una mayor fuga de electrolitos,
acumulación de carotenoides totales, MDA y prolina en las plantas que fueron afectadas
por un periodo de 15 días de déficit hídrico. En conclusión, los cinco cultivares de fríjol
arbustivo evaluados en la etapa vegetativa se vieron afectados negativamente en variables
fisiológicas evaluadas a causa de un déficit hídrico de 15 días, aunque los cultivares más
afectados fueron Bacatá y Bianca.
Palabras clave: Polietilenglicol, estrés oxidativo, partición de asimilados.

38 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
3.2 Introducción
La producción agrícola está siendo afectada negativamente por condiciones climáticas
adversas acentuadas en los escenarios de cambio climático durante los últimos años en
Colombia (Miranda, 2009). Este sentido, el cambio climático ha incidido en el incremento
de áreas afectadas por condiciones de baja disponibilidad de agua, propiciando
condiciones de estrés hídrico (Monasterio et al., 2011). Asimismo, fenómenos de
variabilidad climática tales como El Niño/ Oscilación del Sur (ENOS) han generado un
incremento de las temperaturas y menor volumen y frecuencia de las precipitaciones
durante ciertas épocas del año (Li et al., 2011; Ruiz y Pabón, 2013). En este aspecto, se
ha reportado que Fenómenos del Niño pueden causar una disminución en la oferta hídrica
alrededor del 50% en más de 180 municipios del país (UNGRD, 2014). Igualmente, según
reportes de la Comunidad Andina (2009), Colombia posee un área cercana de los 59.000
km2 expuesta a sequía, encontrando gran parte de esta área en la región andina donde el
fríjol es comúnmente cultivado y esta puede incrementar por el escenario de cambio
climático tienden a incrementar.
Asimismo, el fríjol (Phaseolus vulgaris L.) es una especie de leguminosa perteneciente a
la familia Fabaceae cuyo centro de domesticación es meso y Suramérica (Kalavacharla et
al., 2011; Kumar et al., 2015), de gran importancia en la dieta familiar en América Latina
por su contenido de proteínas, carbohidratos, vitaminas y minerales (FAO, 2007). En
Colombia, esta especie alcanzó una producción de 62,974 toneladas durante el primer
semestre del año 2015 con un rendimiento promedio de 1.3 ton ha-1 (Fenalce, 2016).
Por otro lado, las plantas presentan diferentes tipos de respuestas como cambios
anatómicos, fisiológicos y bioquímicos a condiciones de baja disponibilidad de agua,
ocasionando afectaciones negativas sobre el crecimiento, desarrollo y rendimiento (Farooq
et al., 2009; Sánchez-Reinoso et al., 2014; Segura-Monroy et al., 2015). Con respecto a
las variables fisiológicas, las propiedades de intercambio gaseoso de la hoja (fotosíntesis,
conductancia estomática y transpiración), se ha reportado que bajo condiciones de estrés
por déficit hídrico la tasa fotosintética y la transpiración tienden a disminuir como
consecuencia a un cierre de estomático (Rosales et al., 2012; Yousfi et al., 2012). En

Capítulo 39
cuanto al estatus hídrico de la hoja, las plantas de fríjol sometidas a condiciones de estrés
por déficit hídrico presentan cambios en el estatus hídrico de la hoja tales como incremento
del potencial hídrico, reducción del Contenido Relativo de Agua y un incremento en el Uso
Eficiente del Agua tanto extrínseco como intrínseco, con la finalidad de disminuir la pérdida
de agua por procesos de transpiración (Miyashita et al., 2005; Farooq et al., 2009; Keyvan.,
2010; De Laat, 2014). Adicionalmente, varios estudios han reportado que periodos de
déficit hídrico provocan cambios en los componentes de la fase lumínica de la fotosíntesis
tales como una disminución sobre el contenido de clorofila en las hojas, la eficiencia
máxima del PSII fotoquímica (Fv/Fm) y el quenching fotoquímico (qP); de lo contrario, se
ha observado un incremento en el quenching no fotoquímico (NPQ) como una estrategia
de disipación de energía en forma de calor (Perveen et al., 2010; Aranjuelo et al, 2011;
Gorbe y Catalayud., 2012).
El estrés por déficit hídrico también genera una serie de respuestas bioquímicas que son
indicadores de tolerancia o susceptibilidad de plantas (Ahmed y Hassan, 2001; Aranjuelo
et al, 2011; Mohamed y Ibrahim, 2011;). En este contexto, las plantas de tomate expuestas
a un periodo de 22 días de estrés por déficit hídrico presentaron afectaciones a nivel de la
estabilidad de la membrana debido a la peroxidación lipídica, causando una producción de
iones de peróxido y malondialdehido (MDA) (Sánchez-Rodríguez et al., 2010). Del mismo
modo, cambios en la concentración de MDA pueden ser considerado otro indicador de
estrés oxidativo de las membranas sometidas a condiciones de estrés hídrico (Cao et al.,
2008). Adicionalmente, Sun et al. (2013) encontraron que condiciones de estrés hídrico
estimulan la producción de metabolitos secundarios tales como prolina, que cumplen
diversas funciones en las plantas en procesos como la regulación osmótica, el
mantenimiento de la estabilidad de la membrana, ser fuente de carbono y entre otras.
Igualmente, Ahmed y Hassan (2011) reportaron que valores altos de prolina están
asociados a genotipos con mejor aclimatación a estreses ambientales. Por otro lado, el
estudio de las respuestas fisiológicas de diferentes cultivares bajo estrés por déficit hídrico
ha permitido conocer las posibles estrategias de las plantas que les ayudan a sobrellevar
esta condición abiótica adversa y asimismo, tener una comprensión de los mecanismos
fisiológicos involucrados en sus respuestas de aclimatación que repercuten en el
funcionamiento de la planta y posteriormente en el rendimiento del cultivo (Chaves et al.,
2002; Farooq, et al., 2009, Xu et al., 2010).

40 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
Asimismo, se han desarrollado estudios en diferentes genotipos de frijol a condiciones de
déficit hídrico y han observado diferencias en sus comportamiento fisiológico y bioquímico
bajo esta condición adversa (Beebe et al., 2013). Muchos estudios reportan los efectos en
el rendimiento de genotipos con rasgos de tolerancia a condiciones están más
influenciados por una mayor longitud de raíz (White y Castillo., 1992). Lo anterior, también
relaciona el tamaño de las raíces con el estatus hídrico de la hoja (Sanders y Markhart.,
1992). También, Rao et al. (2006a) concluyeron que una de las características de dos
genotipos de fríjol con rasgos de tolerancia a estrés por sequía es la capacidad de movilizar
fotosintatos al grano en desarrollo con el fin de utilizar el nitrógeno adquirido de forma más
eficiente para la producción de granos. Por lo tanto, identificar y priorizar parámetros
fisiológicos y bioquímicos tales como menor reducción de la tasa fotosintética, menor tasa
de transpiración, mayor eficiencia en el uso del agua, producción de metabolitos
secundarios como prolina, menor daño a nivel de membrana han sido características de
gran importancia para clasificar genotipos susceptibles y/o tolerantes a estrés hídrico
(Farooq, et al., 2009; Mohamed y Ibrahim, 2011; Yousfi et al., 2012).
Dado lo anterior, es importante desarrollar investigaciones con el propósito conocer el
comportamiento de diferentes genotipos de fríjol bajo estas condiciones que puedan
contribuir en programas de fitomejoramiento y/o zonificación agrícola (Polanía et al, 2009;
Restrepo-Díaz et al., 2010; Yousfi et al., 2012; Cardona-Ayala et al., 2014). Hong-Bo et al.
(2006) encontraron que en 10 diferentes genotipos de trigo sometidos a condición de estrés
por sequía producían prolina en diferentes concentraciones, y aquellos con mayor nivel de
prolina tenían un mayor ajuste osmótico. Del mismo modo, Sánchez-Rodríguez et al.
(2010) reportaron que en plantas de cinco diferentes genotipos de tomate tenían diferente
grado de respuestas en parámetros como el contenido relativo de agua de la hoja (con
genotipos susceptibles perdían mayor cantidad de agua en la hoja), acumulaban menor
cantidad de materia seca, la producción de MDA y peróxido de hidrógeno incrementa
(cuyos niveles eran más altos en genotipos susceptibles), al igual que una alta producción
de sustancias antioxidantes tales como peroxidasas, catalasas, glutatión reductasa, entre
otros en los genotipos tolerantes. Igualmente, otro beneficio de evaluar las respuestas
fisiológicas y bioquímicas en plántulas de diferentes cultivares de fríjol expuestas a estrés
por déficit hídrico es caracterizar posible respuestas de aclimatación y sobrevivencia a la
condición ambiental adversa estudiada. En consecuencia, el objetivo de esta investigación

Capítulo 41
fue evaluar las respuesta fisiológicas como la fotosíntesis, conductancia estomática,
transpiración, carbono interno, contenido relativo hídrico de la hoja y pérdida de electrolitos;
y bioquímicas como la producción de malondialdehido y prolina de cinco cultivares de fríjol
arbustivo (Phaseolus vulgaris L.) bajo una condición de estrés por déficit hídrico.
3.3 Materiales y métodos
3.3.1 Material vegetal y condiciones de crecimiento
Semillas de cinco cultivares de fríjol arbustivo fueron usadas en el presente estudios. Los
cultivares estudiados fueron los siguientes: i) ICA-Cerinza e ICA-Bachue son dos
materiales con 20 años de siembra en la agricultura tradicional colombiana, ii) NUA35 es
un genotipo con siete años de comercialización desde su liberación y iii) Bianca y Bacatá
son dos cultivares de reciente liberación (menos de dos años). Las semillas fueron
pregerminadas en una cámara de crecimiento durante siete días bajo condiciones de
oscuridad con 75% HR y 25 °C de temperatura ambiental antes de ser trasplantadas.
Posteriormente, las plántulas de cada genotipo fueron trasplantadas a un sistema de
hidroponía flotante con recipientes de vidrio de 80 L de capacidad. El ensayo fue llevado a
cabo en el laboratorio de Fisiología de Cultivos de la Facultad de Ciencias Agrarias de la
Universidad Nacional de Colombia ubicado en la ciudad de Bogotá a una altura de 2556
msnm, (4°35’56’’ y 74°04’51’’) durante 55 días (septiembre a noviembre del 2014). Las
condiciones de crecimiento del laboratorio durante la evaluación fueron temperatura
promedio de 22°C, humedad relativa de 60 a 80% y un fotoperiodo artificial con lámparas
de luz incandescente de 12 h con una PAR de 800 µmol m-2 s-1.
Las plantas fueron establecidas en una solución de hidroponía (Nutri-Ponic, Walco S.A.,
Bogotá D.C., Colombia) a una dosis de 2.5 mL L-1. La composición de la solución nutritiva
fue: N nítrico, 40.3 g L-1, N amoniacal, 4 g L-1; P asimilable (P2O5), 20.4 g L-1; K soluble
(K2O), 50.6 g L-1; 28.8 g L-1 Ca; 11.4 g L-1 Mg; 1.0 g L-1 S; 1120 mg L-1 Fe; 112 mg L-1 Mn;
12.0 mg L-1 Cu; 26.4 mg L-1 Zn; 106 mg L-1 B; 1.2 mg L-1 Mo y 0.36 mg L-1 Co.

42 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
Adicionalmente, la solución tenía una conductividad eléctrica (C.E) de 2.0 mmhos cm-3 -
pH en una solución 1:200 (v/v) 5.8. El sustrato líquido se cambiaba cada 15 días
suministrando la concentración inicial, manteniendo un volumen aproximado de solución
nutritiva en todos los recipientes de 42 L.
3.3.2 Tratamiento de déficit hídrico y determinación de las variables
El inicio del periodo de estrés hídrico fue establecido cuando las plantas alcanzaron el
estado fenológico BBCH (Feller et al., 1995b) 13-14 de (formación de 3 a 4 hojas trifoliadas
completamente expandidas) aproximadamente a los 40 días después del trasplante (DDT)
según la metodología reportada por Ghanbari et al. (2013). El tratamiento de déficit hídrico
en las plantas de frijol fue inducido mediante la adición de polietilenglicol (PEG 6000,
PanReac, Barcelona, España) a la solución nutritiva, a una dosis modificada de 20 g L-1
(DeLaat et al. 2014); la cual permite generar un estrés hídrico a partir de las 24 horas
después de aplicación del PEG 6000. El periodo exposición de las plantas a la solución
con polietilenglicol fue de quince días. Posteriormente, las diferentes las variables
fisiológicas y bioquímicas se estimaron al final del periodo de estrés (entre los 52 y 55
DDE). En general, las medidas fisiológicas fueron desarrollas en la segunda hoja trifoliada
completamente expandida entre los 52 y 55 días después de la emergencia (DDE). Con
respecto a las variables bioquímicas, se colectó 1.5 g de material vegetal, el cual fue
homogenizado con nitrógeno líquido y posteriormente almacenado a -80°C. Finalmente, el
experimento duró aproximadamente 55 DDE.
3.3.3 Diseño experimental y análisis de los datos
Se realizó un diseño completamente al azar en arreglo factorial donde el primer factor fue
el estado hídrico y el segundo factor fueron los cinco cultivares evaluados para un total de
10 tratamientos con cuatro repeticiones para un total de 40 plántulas. Antes del análisis,
los valores de porcentaje fueron transformados con la fórmula del arcoseno.

Capítulo 43
Posteriormente, cuando se presentaron diferencias significativas en el ANAVA, se empleó
la prueba comparativa de medias de Tukey a P ≤ 0.05. Los datos se analizaron usando el
programa Statistix v 9.0 (software analítico, Tallahassee, FL, US). Los resultados del
análisis de varianza son resumidos en la Tabla 3-1.
3.3.4 Variables fisiológicas
Relaciones hídricas e intercambio gaseoso de la hoja
El contenido relativo de agua (CRA) fue determinado en hojas totalmente expandidas de
la parte media. Las hojas fueron recolectadas al final del periodo de estrés hídrico para
cada uno de los cultivares de fríjol evaluados. El CRA fue calculado mediante la siguiente
ecuación:
𝐶𝑅𝐴 = [𝑃𝐹 − 𝑃𝑆
𝑃𝑇 − 𝑃𝑆] 𝑋100 (𝐸𝑐𝑢𝑎𝑐𝑖ó𝑛. 1)
Siendo PF el peso fresco, PT el peso en turgencia medido después de 24 horas de
saturación en agua destilada a 4 °C en la oscuridad y PS es el peso seco determinado
después de 48 horas en un horno a 70°C.
La fotosíntesis fue estimada en la segunda hoja trifoliada completamente expandida del
porción media superior de la planta usando un medidor de fotosíntesis portable (LI-COR
6200, Lincoln, NE, USA), entre los periodos comprendidos de las 930 a 1100 h y 1330 a
1500 h durante tres días seguidos a partir del penúltimo día antes de finalizar el periodo
de estrés. Asimismo, este equipo estimó la concentración interna de CO2. Adicionalmente,
se calculó la relación Carbono intercelular entre el Carbono atmosférico (Ci/Ca). La
conductancia estomática y la traspiración se midió en la segunda hoja trifoliada
completamente expandida con la ayuda de un porómetro portable (LI-COR 1600, Lincoln,
NE, USA). Del mismo modo, se calculó el uso eficiente del agua como producto de la
relación entre la fotosíntesis y la transpiración. La toma de los datos se realizó entre las

44 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
1000 y las 1500 horas entre los 52 y 55 DDE. Durante las mediciones las condiciones
fueron las siguientes: Radiación fotosintéticamente activa 800 μmol m-2 s-1, temperatura
de la hoja 27 ± 5 °C, diferencia de presión de vapor del aire de la hoja 1.8 ± 0.5 kPa.
Pesos secos y relación parte aérea/raíz
Las plantas fueron recolectadas los 55 DDT. Luego, las plantas se secaron en una estufa
de aire seco comprimido (ThelcoMod 27, Chicago, USA) a 70 °C durante 48 horas con el
propósito de obtener los pesos secos de hojas, tallos, raíces y total de la planta.
Igualmente, la relación parte aérea / raíz (PA/R) se calculó mediante la relación entre el
peso seco de la parte aérea sobre el peso seco de la raíz.
Parámetros de fluorescencia, contenido de clorofilas y carotenoides
Para la determinación de los parámetros de fluorescencia de PSII, se utilizó un fluorómetro
modulado (MINI-PAM, Walz, Effeltrich, GE). Las medidas fueron tomadas entre las 700 y
1000 h en la segunda hoja trifoliada completamente expandida desde la parte superior del
dosel. Las relaciones de fluorescencia fueron determinadas con un pulso máximo de
intensidad de luz de hasta 2.600 µmol m-2 s-1 en la superficie de la muestra de la hoja,
adaptándola previamente a la oscuridad durante 10 minutos. Para las mediciones de
clorofilas y carotenoides, las muestras de tejidos (30 mg aprox. de la segunda hoja trifoliada
completamente expandida) fueron homogenizadas en 4 mL de acetona (80%),
posteriormente se llevaron las muestras a una centrífuga (Model 420101, Becton Dickinson
Primary Care Diagnostics, Maryland, USA) a 5000 rpm durante 10 min para eliminar
partículas y el sobrenadante diluido a un volumen final de 6 ml adicionando acetona (Sims
y Gamon, 2002). El contenido de clorofila fue determinado a 663 y 646 nm y carotenoides
a 470 nm utilizando un espectrofotómetro (Spectronic BioMate 3 UV–vis Thermo, Madison,
WI, USA). Para obtener el contenido de pigmentos fotosintéticos en hoja, se emplearon las
ecuaciones para acetona descritas por Wellburn (1994).

Capítulo 45
Índice Relativo de Tolerancia (RTI)
El cálculo del Índice Relativo de Tolerancia (RTI) fue obtenido mediante la metodología
empleada por Dutta Gupta et al. (1995), basada en el uso de la acumulación de material
seca total de la planta en los diferentes tratamientos y se utilizó la siguiente fórmula:
𝑅𝑇𝐼 = (𝑇𝑜𝑡𝑎𝑙 𝑑𝑒 𝑙𝑎 𝑚𝑎𝑠𝑎 𝑏𝑎𝑗𝑜 𝑠𝑒𝑞𝑢í𝑎
𝑇𝑜𝑡𝑎𝑙 𝑑𝑒 𝑙𝑎 𝑚𝑎𝑠𝑎 𝑐𝑜𝑛𝑡𝑟𝑜𝑙 𝑖𝑟𝑟𝑖𝑔𝑎𝑑𝑜) 𝑥 100 𝐸𝑐𝑢𝑎𝑐𝑖ó𝑛 2
3.3.5 Variables bioquímicas
MDA e incremento de la permeabilidad de la membrana en fuga de electrolitos
Para la determinación de la oxidación lipídica (Malondialdehido-MDA), se empleó el
método del ácido tiobarbitúrico (TBA) descrito por Hodges et al. (1999), donde se tomaron
0.3 g aprox. de material vegetal homogenizado y almacenado con nitrógeno líquido. Las
muestras luego de ser centrifugadas se les estimó las absorbancias a 440, 532 y 600 nm
con un espectrofotómetro (Spectronic BioMate 3 UV - Vis, Thermo, Madison, WI, USA).
Por último, se utilizó un coeficiente de extinción (157 M mL-1) para la obtención de la
concentración de MDA.
Por otro lado, el daño de la membrana plasmática estimada en el porcentaje de fuga de
electrolitos se determinó por el método descrito por Jiang and Zhang (2001) modificado.
Extrayendo 5 discos de 0.5 dm de diámetro de la segunda hoja trifoliada completamente
expandida. Se utilizó 25 mL de agua desionizada como medio en tubos falcon de 50 mL
de capacidad, a las cuales se les registró la conductividad eléctrica inicial (CE1) luego de
dos horas en baño de agua a 30°C y la conductividad eléctrica final (CE2) del medio
después de 20 min en baño de agua a 90°C. El porcentaje de electrolitos se calculó por el
uso de la siguiente ecuación:

46 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
𝐹𝑢𝑔𝑎 𝑑𝑒 𝑒𝑙𝑒𝑐𝑡𝑟𝑜𝑙𝑖𝑡𝑜𝑠 (%) = (𝐶𝐸1
𝐶𝐸2) 𝑥 100 (𝐸𝑐𝑢𝑎𝑐𝑖ó𝑛 3)
Concentración de Prolina
Para la determinación del contenido de prolina, se utilizó el método descrito por Bates et
al. (1973) donde se recolectaron de la segunda hoja trifoliada completamente expandida
aproximadamente 0.3 g de material vegetal homogenizado y almacenado con nitrógeno
líquido. La absorbancia se medió a 520 nm empleando el espectrofotómetro que se utilizó
en la cuantificación de pigmentos fotosintéticos (Spectronic BioMate 3 UV - Vis, Thermo,
Madison, WI, USA). El contenido de prolina se determinó mediante una curva estándar y
calculada en peso fresco basado en la ecuación 3:
𝜇𝑚𝑜𝑙 𝑃𝑟𝑜𝑙𝑖𝑛𝑎
𝑔 𝑚𝑎𝑡𝑒𝑟𝑖𝑎𝑙 𝑣𝑒𝑔𝑒𝑡𝑎𝑙 𝑓𝑟𝑒𝑠𝑐𝑜=
[(
𝜇𝑔 𝑃𝑟𝑜𝑙𝑖𝑛𝑎𝑚𝐿
𝑥 𝑚𝐿 𝑇𝑜𝑙𝑢𝑒𝑛𝑜)
115.5 𝜇𝑔𝜇𝑚𝑜𝑙
]
[𝑔 𝑚𝑢𝑒𝑠𝑡𝑟𝑎
5 ] (𝐸𝑐𝑢𝑎𝑐𝑖ó𝑛 4)
3.4 Resultados
3.4.1 Relaciones hídricas e intercambio gaseoso de la hoja
Diferencias significativas fueron solamente observadas el factor de estatus hídrico sobre
el Contenido Relativo de Agua (CRA) con P≤0.001 (Tabla 3-1).En general, las plantas bajo
condiciones de déficit hídrico de todos los cultivares evaluados presentaron valores

Capítulo 47
cercanos al 60% del CRA, mientras que las plantas control presentaron valores del 85%
(Figura 3-1). Esta variación en el CRA indica la existencia de un grupo de plantas se
encontraba con dos estados hídricos diferentes debido a los dos tratamientos de riego. En
cuanto a las propiedades de intercambio gaseoso de la hoja (fotosíntesis (Pn),
conductancia estomática (gs) y transpiración (E)) se encuentran resumidas en la Figura
3-2 (A, B y C, respectivamente). Se encontraron diferencias significativas solamente a nivel
de los tres factores: estatus hídrico, cultivar e interacción (Tabla 3-1); estatus hídrico en la
Pn, mientras que en la conductancia estomática y transpiración se evidenciaron diferencias
significativas a nivel de interacción estatus hídrico × cultivar con P≤0.05 (Tabla 1). Con
respecto a la Pn, plántulas no estresadas en todos los cultivares, en general, presentaron
valores alrededor 10 µmol m-2 s-1. Sin embargo, bajo condiciones de estrés por déficit
hídrico una reducción de aproximadamente el 60% (3 µmol m-2 s-1) se observó en esta
variable (Figura 3-2A).
Por otro lado, la conductancia estomática mostró que los cultivares no sometidos a
condiciones de déficit hídrico tuvieron una gs alrededor de 0.5 µmol m-2 s-1. Bajo estrés, la
gs mostró una reducción, siendo esta reducción mayor especialmente en los genotipos
NUA35 (0.21 µmol m-2 s-1) y Bianca (0.18 µmol m-2 s-1) en comparación a las plantas de
Bacatá (0.31 µmol m-2 s-1) y Bachue (0.29 µmol m-2 s-1) que tuvieron unas reducciones
aproximadamente de 30% y 40%, respectivamente Figura 3-2B). Finalmente, se
obtuvieron diferencias también entre cultivares y déficit hídrico sobre la tasa de
transpiración (E) donde Bacatá, Bianca y NUA35 fueron los cultivares con una mayor tasa
de transpiración en condiciones normales (6.65 mol m-2 s-1). Asimismo, un periodo de déficit
hídrico de 15 días también causo una reducción en la transpiración donde los cultivares
Bachue, Bianca y NUA35 tuvieron una mayor caída (1.04, 1.13 y 1.12 mol m-2 s-1,
respectivamente) en comparación a Cerinza y Bacatá (1.15 y 1.37 mol m-2 s-1,
respectivamente) (Figura 3-2C). Lo anterior concuerda con o reportado por Miyashita et
al. (2005); Keyvan. (2010); De Laat. (2014), donde el estrés por déficit hídrico reduce la
tasa fotosintética y la transpiración como consecuencia a un cierre de estomático.
Por otro lado, el uso eficiente del agua (UEA) y la relación carbono intercelular sobre el
carbono atmosférico (Ci/Ca) mostraron diferencias significativas a nivel de interacción
estatus hídrico × cultivar (P≤0.001) (Tabla 3-1). En general, los valores más altos de UEA
fueron obtenidos en las plantas que estuvieron sometidas a condiciones de estrés por

48 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
déficit hídrico. En este contexto, las plantas del cultivar Bianca tuvieron los mayores valores
(28.08 µmol CO2 mol-1 H2O), seguidos por los cultivares NUA35, Bachue y Cerinza (20.46,
20.11 y 18.21 µmol CO2 mol-1 H2O, respectivamente). Con respecto a la relación Ci/Ca,
ésta también mostró un incremento en las plantas que estuvieron expuestas a condiciones
de estrés donde plantas cultivares Cerinza y Bachue en condiciones normales tuvieron los
siguientes valores 0.82 y 0.85 y por la condiciones esta relación sufrió un aumento (0.93 y
0.93 respectivamente). Mientras, los genotipos NUA35, Bacatá y Bianca no presentaron
variaciones con respecto a las plantas en condiciones normales, ya que presentaron
valores de Ci/Ca cercanos a 0.90 en ambas condiciones (Tabla 3-2). Mismas tendencias
fueron encontradas por Rosales et al. (2012) y Yousfi et al. (2012).
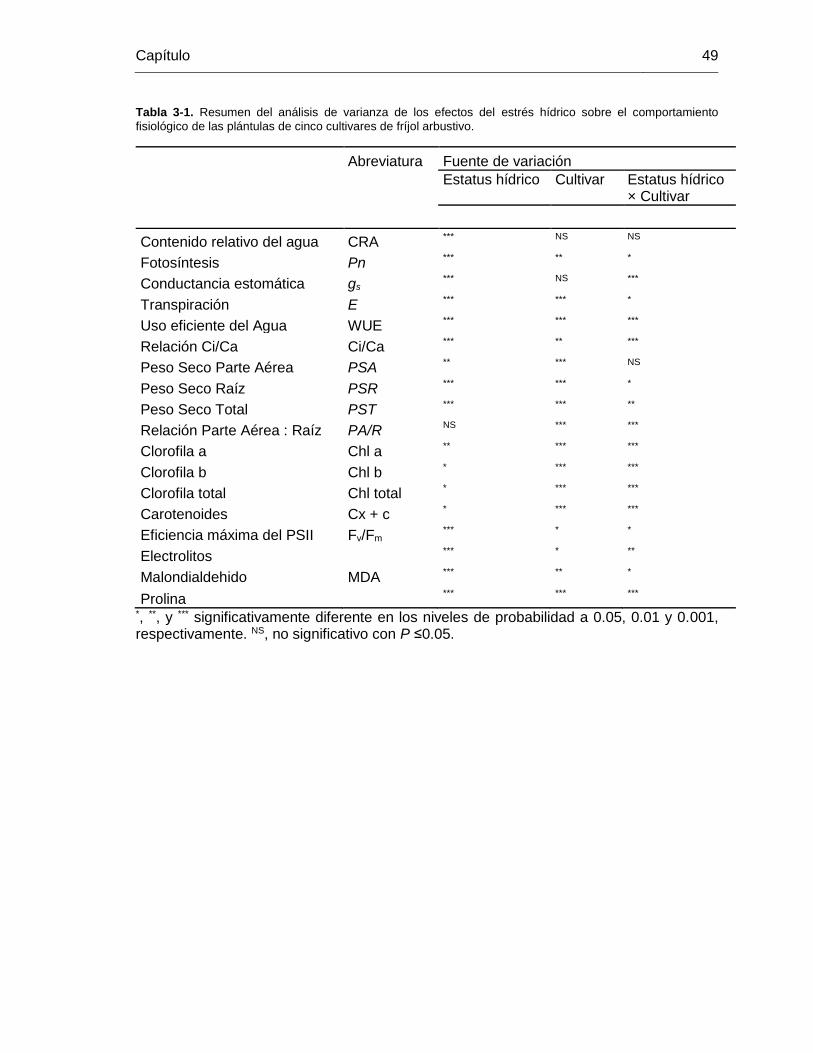
Capítulo 49
Tabla 3-1. Resumen del análisis de varianza de los efectos del estrés hídrico sobre el comportamiento
fisiológico de las plántulas de cinco cultivares de fríjol arbustivo.
Abreviatura Fuente de variación
Estatus hídrico Cultivar Estatus hídrico × Cultivar
Contenido relativo del agua CRA *** NS NS
Fotosíntesis Pn *** ** *
Conductancia estomática gs *** NS ***
Transpiración E *** *** *
Uso eficiente del Agua WUE *** *** ***
Relación Ci/Ca Ci/Ca *** ** ***
Peso Seco Parte Aérea PSA ** *** NS
Peso Seco Raíz PSR *** *** *
Peso Seco Total PST *** *** **
Relación Parte Aérea : Raíz PA/R NS *** ***
Clorofila a Chl a ** *** ***
Clorofila b Chl b * *** ***
Clorofila total Chl total * *** ***
Carotenoides Cx + c * *** ***
Eficiencia máxima del PSII Fv/Fm *** * *
Electrolitos *** * **
Malondialdehido MDA *** ** *
Prolina *** *** ***
*, **, y *** significativamente diferente en los niveles de probabilidad a 0.05, 0.01 y 0.001, respectivamente. NS, no significativo con P ≤0.05.

50 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
Figura 3-1. Efecto del estrés hídrico sobre el contenido hídrico relativo de la hoja (CRA) en plántulas de cinco
cultivares de fríjol arbustivo. Las barras son el promedio de cuatro datos ± error estándar.
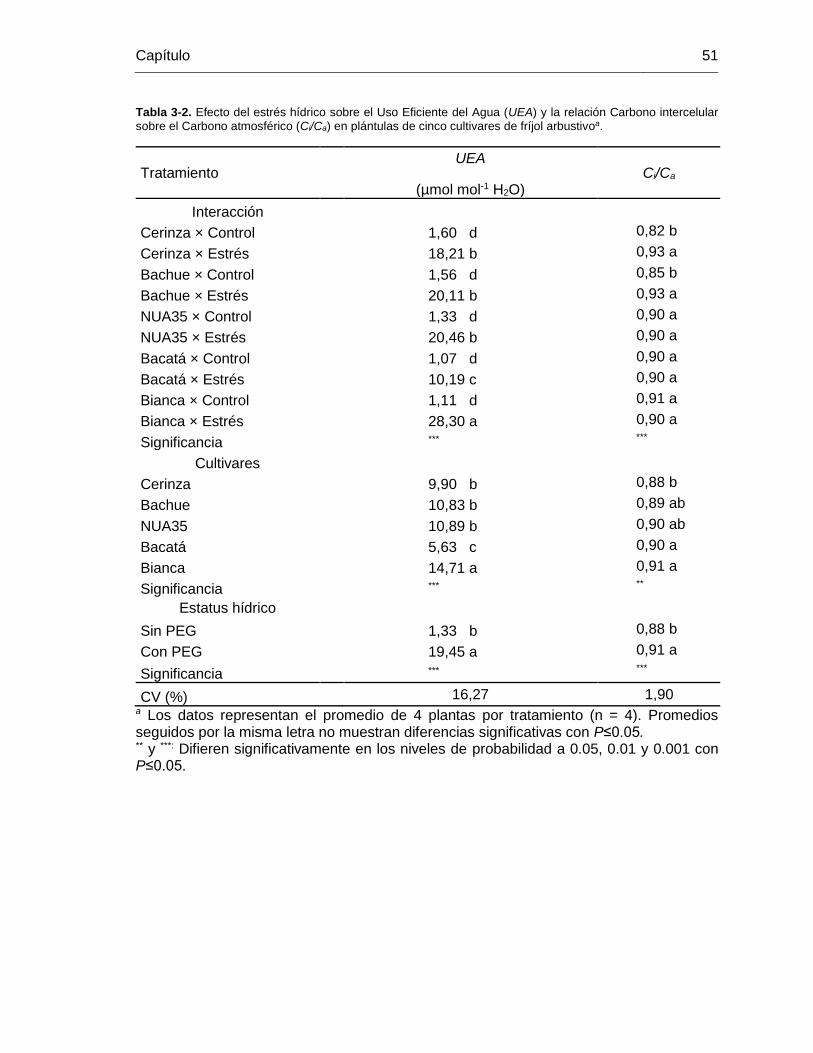
Capítulo 51
Tabla 3-2. Efecto del estrés hídrico sobre el Uso Eficiente del Agua (UEA) y la relación Carbono intercelular sobre el Carbono atmosférico (Ci/Ca) en plántulas de cinco cultivares de fríjol arbustivoa.
Tratamiento UEA
Ci/Ca (µmol mol-1 H2O)
Interacción
Cerinza × Control 1,60 d 0,82 b
Cerinza × Estrés 18,21 b 0,93 a
Bachue × Control 1,56 d 0,85 b
Bachue × Estrés 20,11 b 0,93 a
NUA35 × Control 1,33 d 0,90 a
NUA35 × Estrés 20,46 b 0,90 a
Bacatá × Control 1,07 d 0,90 a
Bacatá × Estrés 10,19 c 0,90 a
Bianca × Control 1,11 d 0,91 a
Bianca × Estrés 28,30 a 0,90 a
Significancia *** ***
Cultivares
Cerinza 9,90 b 0,88 b
Bachue 10,83 b 0,89 ab
NUA35 10,89 b 0,90 ab
Bacatá 5,63 c 0,90 a
Bianca 14,71 a 0,91 a
Significancia *** **
Estatus hídrico
Sin PEG 1,33 b 0,88 b
Con PEG 19,45 a 0,91 a
Significancia *** ***
CV (%) 16,27 1,90 a Los datos representan el promedio de 4 plantas por tratamiento (n = 4). Promedios seguidos por la misma letra no muestran diferencias significativas con P≤0.05. ** y ***: Difieren significativamente en los niveles de probabilidad a 0.05, 0.01 y 0.001 con P≤0.05.
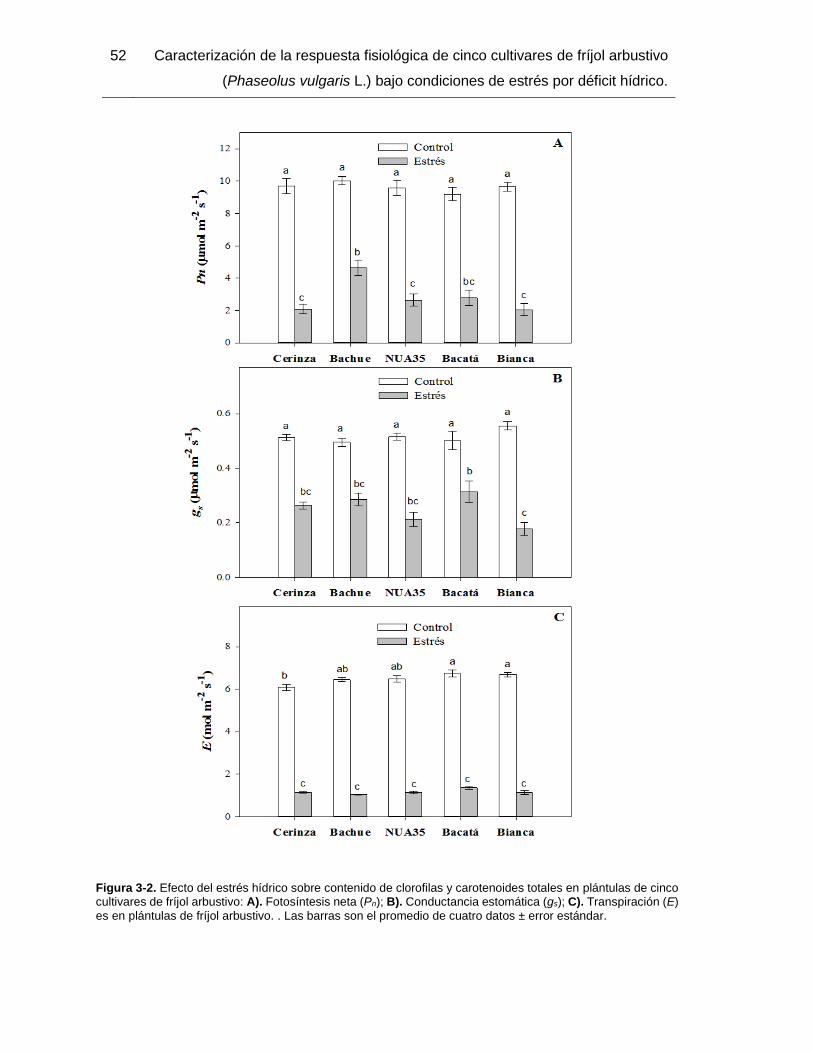
52 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
Figura 3-2. Efecto del estrés hídrico sobre contenido de clorofilas y carotenoides totales en plántulas de cinco cultivares de fríjol arbustivo: A). Fotosíntesis neta (Pn); B). Conductancia estomática (gs); C). Transpiración (E)
es en plántulas de fríjol arbustivo. . Las barras son el promedio de cuatro datos ± error estándar.

Capítulo 53
3.4.2 Acumulación de materia seca y relación parte aérea/raíz
Resultados de la acumulación de materia seca en la parte aérea, raíz, total y relación parte
aérea sobre la raíz son resumidos en la Tabla 3-3. Diferencias fueron encontradas en la
interacción déficit hídrico vs cultivares sobre la materia seca de la parte aérea (PSA
P≤0.05), raíz (P≤0.01), total y la relación Aérea/Raíz (PA/R P≤0.001). En general, las
plantas del cultivar Cerinza bajo condiciones de estrés hídrico presentaron la mayor
reducción en la acumulación de materia seca en la parte aérea en comparación a las
plantas de los demás cultivares, alcanzando valores cercanos a 0.82 g. Por otro lado, la
acumulación de materia seca en la raíz se vio más afectada en las plántulas de los
cultivares Bianca y Bacatá bajo condiciones de estrés hídrico (0.68 y 0.83 g
respectivamente). Asimismo, la materia seca total tuvo una menor acumulación cuando las
plantas de todos los cultivares estuvieron sometidas a un déficit hídrico de 15 días, ya que
este grupo de plantas mostraron valores alrededor de 3.42 g mientras las plantas sin
periodo de déficit presentaron valores de 4.86 g. Igualmente, es importante señalar que las
plantas del cultivar Bachue bajo condiciones de estrés hídrico presentaron la mayor
acumulación de materia seca total con respecto a los demás cultivares evaluados (5.36 g).
En contraste, las plantas de los cultivares Cerinza y Bianca con estrés hídrico fueron las
más afectadas y tuvieron los menores valores (2.50 y 1.71 g respectivamente). Finalmente,
la relación (PA/R) también fue condicionada por las condiciones de baja disponibilidad de
agua. Plantas con estrés por déficit hídrico de los cultivares Cerinza, Bianca y Bachue
presentaron los menores valores en la PA/R (aproximadamente entre 1.45 y 1.65) en
comparación a los cultivares Bacatá y NUA35 que mostraron una mayor relación (alrededor
3.65). Similares resultados fueron reportados por Huang et al. (2011), donde la mayor
acumulación de materia seca se presenta en las plantas con características de tolerancia
al estrés por déficit hídrico.
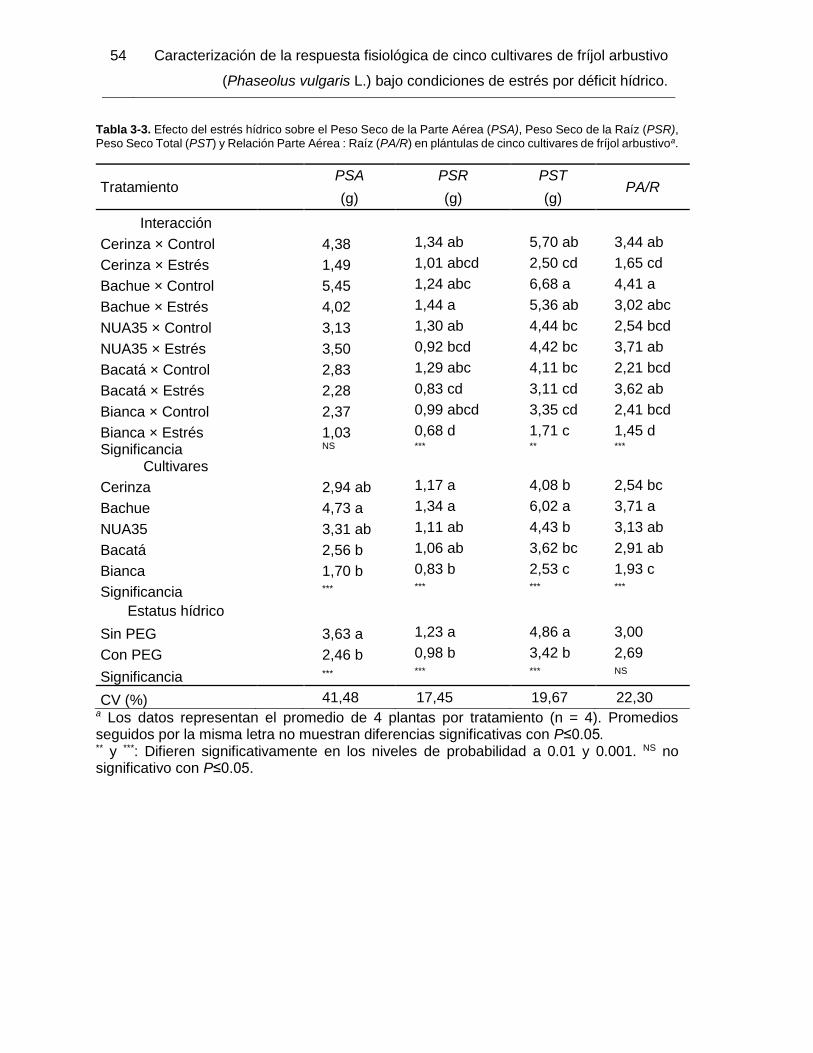
54 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
Tabla 3-3. Efecto del estrés hídrico sobre el Peso Seco de la Parte Aérea (PSA), Peso Seco de la Raíz (PSR), Peso Seco Total (PST) y Relación Parte Aérea : Raíz (PA/R) en plántulas de cinco cultivares de fríjol arbustivoa.
Tratamiento PSA PSR PST
PA/R (g) (g) (g)
Interacción
Cerinza × Control 4,38 1,34 ab 5,70 ab 3,44 ab
Cerinza × Estrés 1,49 1,01 abcd 2,50 cd 1,65 cd
Bachue × Control 5,45 1,24 abc 6,68 a 4,41 a
Bachue × Estrés 4,02 1,44 a 5,36 ab 3,02 abc
NUA35 × Control 3,13 1,30 ab 4,44 bc 2,54 bcd
NUA35 × Estrés 3,50 0,92 bcd 4,42 bc 3,71 ab
Bacatá × Control 2,83 1,29 abc 4,11 bc 2,21 bcd
Bacatá × Estrés 2,28 0,83 cd 3,11 cd 3,62 ab
Bianca × Control 2,37 0,99 abcd 3,35 cd 2,41 bcd
Bianca × Estrés 1,03 0,68 d 1,71 c 1,45 d
Significancia NS *** ** ***
Cultivares
Cerinza 2,94 ab 1,17 a 4,08 b 2,54 bc
Bachue 4,73 a 1,34 a 6,02 a 3,71 a
NUA35 3,31 ab 1,11 ab 4,43 b 3,13 ab
Bacatá 2,56 b 1,06 ab 3,62 bc 2,91 ab
Bianca 1,70 b 0,83 b 2,53 c 1,93 c
Significancia *** *** *** ***
Estatus hídrico
Sin PEG 3,63 a 1,23 a 4,86 a 3,00
Con PEG 2,46 b 0,98 b 3,42 b 2,69
Significancia *** *** *** NS
CV (%) 41,48 17,45 19,67 22,30 a Los datos representan el promedio de 4 plantas por tratamiento (n = 4). Promedios seguidos por la misma letra no muestran diferencias significativas con P≤0.05. ** y ***: Difieren significativamente en los niveles de probabilidad a 0.01 y 0.001. NS no significativo con P≤0.05.

Capítulo 55
- Parámetros de fluorescencia, contenido de clorofilas y carotenoides
Diferencias significativas fueron encontradas en la concentración de los pigmentos
fotosintéticos evaluados a nivel de estatus hídrico vs cultivar (Figura 3-3). En general, el
contenido de clorofila a, b y total de la mayoría de los cultivares bajo condiciones de estrés
por déficit hídrico estresados no sufrieron mayores variaciones con respecto al control a
excepción del cultivar Bianca. En este material, se observó el doble de la concentración de
dichos pigmentos comparados con los niveles encontrados en las plantas control (Chl a de
3384.39 µg mg-1 PF; Chl b de 1358.29 µg mg-1 PF, Chl total de 4566.73 µg mg-1 PF. ). Una
similar tendencia también fue observada para la concentración carotenoides totales, cuyo
incremento alcanzó valores cercanos de ~617.64 µg mg-1 PF. Por otro lado, la Figura 3-4
resume los resultados obtenidos en la eficiencia máxima del quantum en el fotosistema II
(Fv/Fm) donde las plántulas del cultivar Bianca mostraron la mayor reducción de la Fv/Fm
con valores cercanos a 0.50, mientras que la Fv/Fm de los cultivares Cerinza, Bachue,
NUA35 y Bacatá bajo estrés por déficit hídrico estuvo cerca del 0.71. Esto concuerda con
lo reportado por Perveen et al, (2010); Aranjuelo et al. (2011); Gorbe y Catalayud. (2012).
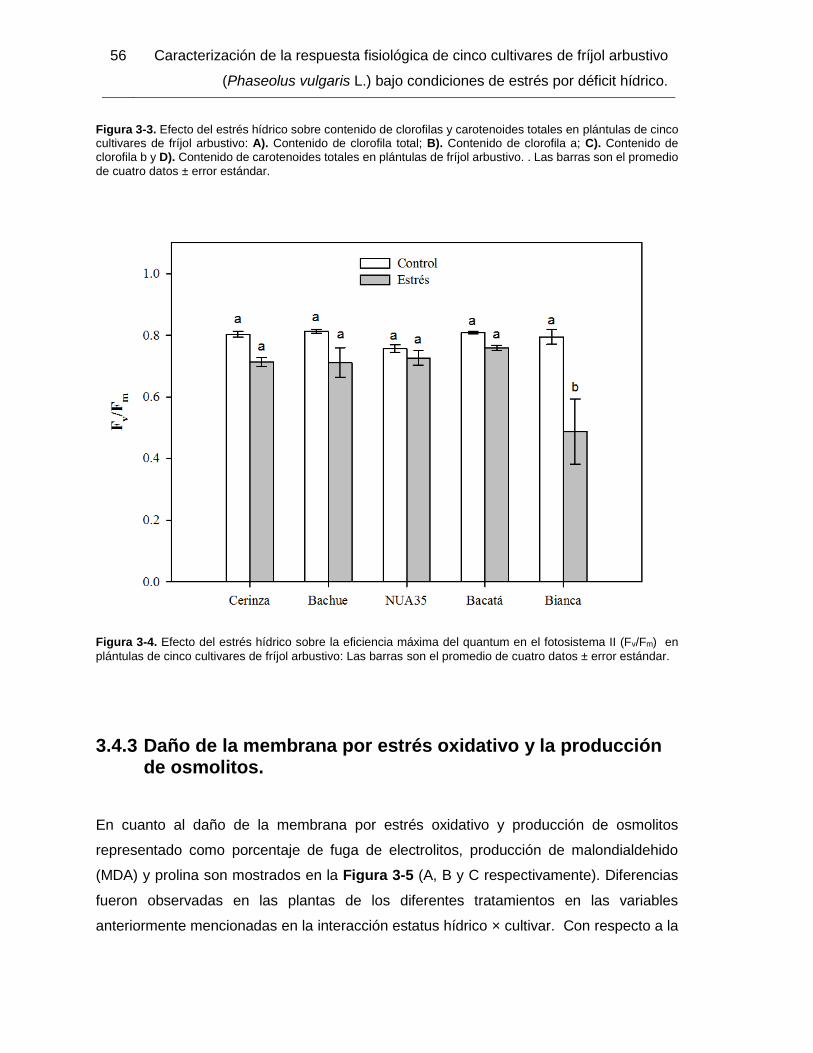
56 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
Figura 3-3. Efecto del estrés hídrico sobre contenido de clorofilas y carotenoides totales en plántulas de cinco cultivares de fríjol arbustivo: A). Contenido de clorofila total; B). Contenido de clorofila a; C). Contenido de clorofila b y D). Contenido de carotenoides totales en plántulas de fríjol arbustivo. . Las barras son el promedio
de cuatro datos ± error estándar.
Figura 3-4. Efecto del estrés hídrico sobre la eficiencia máxima del quantum en el fotosistema II (Fv/Fm) en
plántulas de cinco cultivares de fríjol arbustivo: Las barras son el promedio de cuatro datos ± error estándar.
3.4.3 Daño de la membrana por estrés oxidativo y la producción de osmolitos.
En cuanto al daño de la membrana por estrés oxidativo y producción de osmolitos
representado como porcentaje de fuga de electrolitos, producción de malondialdehido
(MDA) y prolina son mostrados en la Figura 3-5 (A, B y C respectivamente). Diferencias
fueron observadas en las plantas de los diferentes tratamientos en las variables
anteriormente mencionadas en la interacción estatus hídrico × cultivar. Con respecto a la

Capítulo 57
fuga de electrolitos, los cultivares Cerinza, NUA35, Bacatá y Bianca pasaron de un
17.75% a un 27.70% de perdida cuando las plántulas se encontraron en condiciones de
estrés a excepción del cultivar Bachue que mantuvo los niveles cercanos a los encontrados
en las plantas control (17.28%) (Figura 3-5A). Por otro lado, la producción de
malondialdehido (MDA) fue mayor en todos los cultivares evaluados cuando estuvieron
sometidas a un estrés por déficit hídrico, siendo los genotipos Bianca y Bacatá los de mayor
producción (20 y 16 µmol g-1 PF, respectivamente). Asimismo, la producción de prolina fue
favorecida bajo estrés donde Cerinza y Bianca fueron los cultivares con mayor producción
de este osmolito (92 y 86 µmol g-1 PF respectivamente) y seguidos por Bacatá, NUA35 y
Bachue ( ̴50 µmol g-1 PF) (Figura 3-5B). Ahmed y Hassan (2011), reportan que plantas
con alta producción de prolina son relacionadas como genotipos con tolerancia al déficit
hídrico.

58 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
Figura 3-5. Efecto del estrés hídrico sobre el daño de la membrana por estrés oxidativo y la producción de osmolitos: A). Fuga de electrolitos; B). Producción de malondialdehido (MDA) y C). Producción de prolina en
plántulas de cinco cultivares de fríjol arbustivo. Las barras son el promedio de cuatro datos ± error estándar.
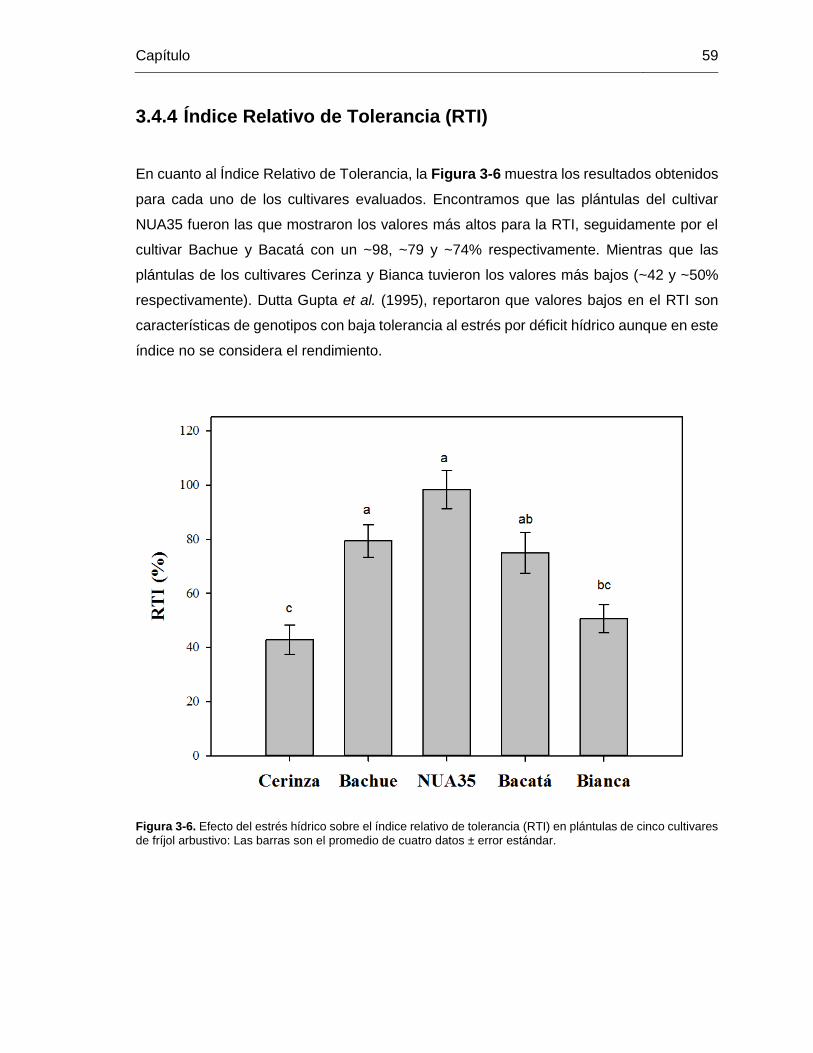
Capítulo 59
3.4.4 Índice Relativo de Tolerancia (RTI)
En cuanto al Índice Relativo de Tolerancia, la Figura 3-6 muestra los resultados obtenidos
para cada uno de los cultivares evaluados. Encontramos que las plántulas del cultivar
NUA35 fueron las que mostraron los valores más altos para la RTI, seguidamente por el
cultivar Bachue y Bacatá con un ~98, ~79 y ~74% respectivamente. Mientras que las
plántulas de los cultivares Cerinza y Bianca tuvieron los valores más bajos (~42 y ~50%
respectivamente). Dutta Gupta et al. (1995), reportaron que valores bajos en el RTI son
características de genotipos con baja tolerancia al estrés por déficit hídrico aunque en este
índice no se considera el rendimiento.
Figura 3-6. Efecto del estrés hídrico sobre el índice relativo de tolerancia (RTI) en plántulas de cinco cultivares
de fríjol arbustivo: Las barras son el promedio de cuatro datos ± error estándar.

60 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
3.5 Discusión
El déficit hídrico es uno de los estreses abióticos que más limitan la productividad de los
cultivos. Asimismo, las diferentes respuestas de la planta a condiciones de baja humedad
en el suelo pueden ser usadas para identificar características que ayuden a identificar
diferentes niveles de tolerancia (Celular, bioquímico, fisiológico y molecular) que ayudan a
proveer información de las diferentes estrategias de aclimatación que posteriormente
puedan ser usadas en programas de mejoramiento (Larbi, 2004; Farooq et al., 2009). En
este contexto, el contenido relativo de agua de la hoja (CRA) es una de las variables de
gran importancia fisiológica que ayuda a cuantificar el estado hídrico de una planta tanto
en condiciones normales como de sequía (Keyvan., 2010). Cuando el CRA presenta una
reducción en los valores ayuda a inferir que las relaciones hídricas pueden estar siendo
afectadas por una condición abiótica adversa, lo cual puede repercutir en algunos procesos
fisiológicos como una reducción de la tasa fotosintética (Costa-França et al., 2000). En
este sentido, el RWC ha sido usado como una variable fisiológica para caracterizar
genotipos tolerantes al déficit hídrico (Larbi, 2004). Sin embargo, los resultados
encontrados en el presente estudio mostraron una reducción de los valores del CRA
solamente en el factor sequía y esta variable no arrojo diferencias entre genotipos debido
a las condiciones estresantes (Tabla 1). Por otro lado, la habilidad de mantener el
contenido de clorofilas bajo condiciones de estrés abiótico ha sido usada como herramienta
en la selección de genotipos tolerantes en programas de mejoramiento (Kiani-Pouya y
Rasouli., 2014). Asimismo, el contenido de otro pigmento fotosintético como los
carotenoides también juega un papel importante en la tolerancia a condiciones de sequía
(Farooq et al., 2009). En este contexto, los contenidos de Clorofila y carotenoides fueron
favorecidos bajos condiciones de estrés hídrico, especialmente, en Bianca. Por lo tanto,
un incremento del contenido de estos pigmentos puede ser considerado como un
mecanismo de respuesta para reducir los efectos negativos del estrés hídrico tales como
la fotoinhibición (Silva et al., 2007). Sin embargo, el genotipo Bianca fue el que también
presentó una mayor reducción sobre la relación Fv/Fm, obteniéndose el menor valor. En
este sentido, se ha reportado ampliamente que el estrés por déficit hídrico causa
alteraciones a la eficiencia del PSII debido un daño a las membranas tilacoidales en parte
por un proceso de fotoinhibición (Farooq et al., 2009). En este aspecto, estudios

Capítulo 61
desarrollados por Rahbarian et al., (2011) encontraron que los genotipos tolerantes de
garbanzo presentaban una menor variación en la relación Fv/Fm y contenido de clorofila,
tal como ocurrió en nuestro ensayo. Continuando con variables del tipo bioquímico, la
prolina y MDA también han sido usados como indicadores para estimar la susceptibilidad
o tolerancia de genotipos a condiciones de estreses abióticos (Sánchez-Reinoso et al.,
2014). En este estudio, todos los genotipos estudiados mostraron un incremento de la
concentración de prolina bajo condiciones de déficit hídrico, siendo el incremento más
notorio en los genotipos ICA-Cerinza y Bianca. Un incremento del contenido de prolina
bajo condiciones de estrés hídrico tiene el propósito de mantener la hidratación en la célula
y prevenir daño por las condiciones estresantes. Igualmente, este aminoácido actúa como
osmolito y fuente de almacenamiento de carbono y nitrógeno que son posteriormente
utilizados para estabilizar macromoléculas, proteínas y membranas celulares en los tejidos
de las plantas (George et al., 2015). Por lo anterior, la determinación de la concentración
de prolina en los tejidos de la hoja se ha recomendado como un marcador bioquímico para
mirar la tolerancia de genotipos al estrés hídrico (Naser et al., 2010). Con respecto al MDA,
el cultivar Bianca presentó una mayor acumulación de este producto bajo condiciones de
estrés. En consecuencia, una mayor acumulación de MDA está indicando un mayor daño
a nivel celular debido a un incremento de la peroxidación lipídica de las membranas
(Sánchez-Reinoso et al., 2014). En general, se ha reportado que plantas que muestren
características fisiológicas tales como un alto CRA, contenido de prolina y una baja
producción de MDA bajo déficit hídrico son indicadores de tolerancia y que estas
características pueden ser usadas para la selección de genotipos tolerantes (Liu et al.,
2013).
El índice de tolerancia relativo del genotipo NUA35 fue el mayor (Fig. 6), dando a entender
que fue el material que presentó una mayor producción de biomasa (Tabla 3) y permitiendo
considerar como un genotipo con tolerancia a estrés hídrico. Dichas observaciones están
en concordancia por lo reportado por Kausar et al. (2012) donde genotipos de sorgo
considerados tolerantes a salinidad presentaron la mayor acumulación de biomasa que a
su vez se reflejaban en un mayor índice. Es ampliamente citado que una mayor
acumulación de biomasa bajo condiciones de estrés hídrico es una característica
importante a la ahora de caracterizar genotipos y este incremento en la biomasa puede
deberse a un mejor uso eficiente del agua, ya que se está optimizando la producción de

62 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
biomasa por unidad de agua transpirada (Farooq et al., 2009). En este aspecto, todos los
genotipos seleccionados para este estudio mostraron un mayor UEA cuando fueron
sometidas a un periodo de estrés de déficit hídrico por 15 días (Tabla 2). Finalmente, el
déficit hídrico afecta la fotosíntesis de las plantas por limitaciones estomáticas y no
estomáticas. En este sentido, la fotosíntesis fue seriamente afectada por el estrés hídrico.
La reducción de la tasa fotosintética en todos los genotipos de frijol puede deberse al hecho
que la plantas de frijol
3.6 Conclusiones
En condiciones ambientales asociadas a las condiciones de estrés en estudio,
debería tenerse en cuenta que cultivares como las anteriormente mencionadas
exhibirían una mayor afectación por estrés, resultados soportados en las
respuestas evaluadas en el trabajo.
El uso de PEG6000 en el estudio del estrés por déficit hídrico en cultivares de fríjol
arbustivo permite evaluar las características de la raíz sin perder material vegetal,
por lo cual es más preciso la toma de datos.
En el estado vegetativo de tercera hoja verdadera, se observó que el cultivar Bianca
fue el de mayor susceptibilidad evidenciando un mayor daño oxidativo y un menor
producción de biomasa e índice de tolerancia relativo.

4. Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo (Phaseolus vulgaris L.) a condiciones de estrés por déficit hídrico en dos etapas fenológicas.
Efecto sobre las propiedades de Intercambio Gaseoso de
la hoja, eficiencia de la fluorescencia de la clorofila a,
malondialdehido y prolina.
4.1 Resumen
El objetivo de esta investigación fue evaluar las respuestas fisiológicas y bioquímicas de cinco cultivares de fríjol arbustivo (Phaseolus vulgaris L.) bajo condición de estrés por déficit hídrico en dos etapas fenológicas diferentes (una vegetativa y una reproductiva) en variables como la fotosíntesis, conductancia estomática, transpiración, carbono interno, contenido relativo hídrico de la hoja y pérdida de electrolitos producción de malondialdehido y prolina. Se sembraron semillas de los cultivares de fríjol ICA-Cerinza, ICA-Bachue, NUA35, Bacatá y Bianca. Se realizó un diseño en bloques completamente al azar en arreglo factorial (factor principal la etapa fenológica y factor secundario los cultivares) para un total de 15 tratamientos con cuatro repeticiones. Se cuantificaron variables como el Contenido Relativo de Agua (CRA), la fotosíntesis, parámetros de fluorescencia del PSII, pigmentos, prolina y malondialdehido. La tasa de fotosíntesis, conductancia estomática y relaciones de fluorescencia disminuyen en todos cultivares que tuvieron condición de estrés en las dos etapas evaluadas, aunque en la etapa vegetativa fue mayor, especialmente en el cultivar Bacatá. Asimismo, se evidenció una mayor producción MDA, carotenoides totales y prolina en las plantas que fueron afectadas por un periodo de 15 días de déficit hídrico, principalmente en la etapa vegetativa. En conclusión, los cinco cultivares de fríjol arbustivo evaluados en las etapa vegetativa se vieron afectados negativamente en variables fisiológicas evaluadas a causa de un déficit hídrico de 15 días, aunque los cultivares más afectados fueron Bacatá y Bianca en las dos etapas fenológicas evaluadas.
Palabras clave: Uso eficiente del agua, fenología, peroxidación lipídica, antioxidante.

64 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
4.2 Introducción
Las plantas presentan una gran variedad de respuestas a condiciones de déficit hídrico
que pueden influir negativamente sobre el rendimiento de los cultivos (Segura-Monroy et
al., 2015). Asimismo, los efectos nocivos de la sequía sobre el rendimiento también
dependen de la etapa de crecimiento del cultivo y de la intensidad del estrés (Farooq et al.,
2012).En este contexto, un estrés hídrico en etapas vegetativas (33 a 67 días después de
siembra) reduce significativamente variables tales como el contenido relativo de agua de
la hoja, uso eficiente del agua, acumulación de materia seca, y afecta drásticamente los
diferentes componentes de rendimiento en arroz (Boonjung and Fukai., 1996-1 y 2).
Igualmente, Okçu et al. (2005) ha reportado también que un estrés hídrico en etapas
vegetativas tempranas puede reducir significativamente la acumulación de materia seca
en genotipos de Pisum sativum L. susceptibles a sequía. La caída en los rendimiento
puede deberse en parte que déficit hídrico causa una reducción de la tasa de asimilación
de carbono y transpiración como consecuencia del cierre de estomas en plantas (Rosales
et al., 2012; Yousfi et al., 2012). Adicionalmente, se ha mencionado que el tipo de
respuesta de un cultivo puede variar según la etapa fenológica en la que inicia el estrés
por déficit hídrico (Cakir, 2004). Por otro lado, los procesos involucrados en la fase lumínica
de la fotosíntesis (concentración de pigmentos fotosintéticos, eficiencias del fotosistema II
y mecanismos de disipación de energía fotoquímica y no fotoquímica) han sido utilizados
para entender los mecanismos de respuesta y aclimatación de los cultivos a estrés por
déficit hídrico (Keyvan., 2010; Perveen et al., 2010; Aranjuelo et al, 2011). Sin embargo,
estudios donde se relacionen los parámetros de la fase lumínica de la fotosíntesis bajo
condiciones de estrés por déficit hídrico en etapas fenológicas diferentes siguen siendo
escasos (Chaves et al., 2002; Moradi and Ismail., 2007;).
Dentro del proceso de caracterización de respuestas fisiológicas de cultivares a
condiciones de déficit hídrico, el estudio de variables bioquímicas ha sido de gran utilidad
para ser marcadores de susceptibilidad y/o tolerancia de genotipos a la condición de estrés
(Sánchez-Rodríguez et al., 2010; Aranjuelo et al, 2011; Mohamed y Ibrahim, 2011). En
este sentido, se ha reportado que los genotipos susceptibles acumulaban menor cantidad
de materia seca y producían prolina en diferentes concentraciones, es decir, aquellos

Capítulo 4 65
materiales con mayor contenido de prolina tenían un mejor ajuste osmótico (Sánchez-
Rodríguez et al., 2010). También, un déficit hídrico favorece la producción de MDA y
peróxido de hidrógeno incrementa, cuyos niveles eran más altos en genotipos
susceptibles. Por otro lado, el estudio de las respuestas fisiológicas de diversos genotipos
bajo estrés por déficit hídrico en etapas fenológicas diferentes ha permitido conocer las
posibles estrategias de las plantas para sobrellevar esta condición abiótica adversa, como
también tener una comprensión de los mecanismos fisiológicos involucrados en la
aclimatación que repercuten en el funcionamiento de la planta y posteriormente en el
rendimiento del cultivo (Winterhalter et al., 2011; Beebe et al., 2013; Osman., 2015). Por
lo tanto, identificar y priorizar la etapa fenológica más sensible al estrés por déficit hídrico,
al igual que los parámetros fisiológicos y bioquímicos tales como menor reducción de la
tasa fotosintética, menor tasa de transpiración, mayor eficiencia en el uso del agua,
producción de prolina, menor producción de malondialdehido son características de gran
importancia para identificar materiales tolerantes a estrés hídrico (Farooq, et al., 2009;
Mohamed y Ibrahim, 2011; Yousfi et al., 2012). Dado lo anterior, es importante enfocar
investigaciones en conocer el comportamiento de diferentes genotipos de fríjol bajo estas
condiciones en diferentes etapas fenológicas que puedan contribuir en programas de
fitomejoramiento y procesos de evaluación de tierras (Polanía et al, 2009; Yousfi et al.,
2012; Cardona-Ayala et al., 2014). En este aspecto, evaluar las respuestas fisiológicas y
bioquímicas en plántulas de diversos cultivares fríjol en etapas fenológicas diferentes bajo
un periodo de 15 días de estrés por déficit hídrico es de gran importancia, ya que nos
permite caracterizar posible respuestas de aclimatación y sobrevivencia a la condición
ambiental adversa estudiada, ya que la producción agrícola ha venido enfrentando
escenarios de cambio climático durante los últimos años en Colombia donde se han
afectado negativamente en el rendimiento e ingresos de los productores (Miranda, 2009).
Igualmente, el cultivo del frijol es sembrado principalmente en la región andina en Colombia
como una fuente ingresos para pequeños y mediados campesinos (Fenalce, 2016).
Asimismo, esta zona del país ha venido siendo afectada por el efecto de cambio climático
y/o variabilidad climática (Fenómenos ENOS), ya que causan incrementos en la
temperatura y alteraciones de los patrones normales de precipitación (Pabón y Torres.,
2007; Li et al., 2011; Monasterio et al., 2011). Por tal motivo, el objetivo de esta
investigación fue evaluar las respuesta fisiológicas y bioquímicas de cinco cultivares de
fríjol arbustivo (Phaseolus vulgaris L.) bajo un periodo moderado de déficit hídrico en dos
diferentes etapas fenológicas (vegetativa vs reproductiva), determinando el

66 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
comportamiento de variables como la fotosíntesis, conductancia estomática, transpiración,
carbono interno, contenido relativo hídrico de la hoja, pérdida de electrolitos, producción
de malondialdehido y prolina.
4.3 Materiales y métodos
4.3.1 Material vegetal y condiciones de crecimiento
Semillas de cinco cultivares de fríjol arbustivo fueron usadas en el presente estudio. Los
cultivares estudiados fueron los siguientes: i) ICA-Cerinza e ICA-Bachue son dos
materiales con 20 años de siembra en la agricultura tradicional colombiana, ii) NUA35 es
un genotipo con siete años de comercialización desde su liberación y iii) Bianca y Bacatá
son dos cultivares de reciente liberación (menos de dos años). Las semillas fueron
sembradas en el invernadero de la Facultad de Ciencias Agrarias de la Universidad
Nacional de Colombia ubicado en la ciudad de Bogotá a una altura de 2556 msnm,
(4°35’56’’ y 74°04’51’’) durante los meses de septiembre del 2015 a enero del 2016 en
parcelas de 2.1 m2 (3 surcos de 1 m lineal de largo). La parcela experimental contó con un
marco de plantación era 16 × 70 cm entre planta y entre surcos, respectivamente (20
semillas por parcela = 85.000 plantas ha-1). Las características físico-químicas del suelo
en el invernadero eran las siguientes: i) una textura Franco Arcilloso (26% arena, 42% de
limo y 32% de Arcilla), ii) características químicas: Nitrógeno total: 0.36%, Ca: 10.6, K:
0.98, Mg: 1.75, Na: 0.24 meq 100 g-1, Cu: 1.67, Fe: 310, Mn: 3.21, Zn: 15.5, B: 0.48 y P:
>116 mg kg-1, iii) pH 5.4 y iv) Capacidad de Intercambio Catiónico Efectiva (CICE) 13.8
meq/100g. Las condiciones de crecimiento del invernadero durante la evaluación fueron
temperatura promedio de 27°C, humedad relativa de 60 a 80% y un fotoperiodo natural de
12 h con una PAR de 1000 µmol m-2 s-1. Después de la emergencia, las plantas fueron
fertilizadas con dos fertilizantes edáficos: i) 4 g planta-1 (350 Kg ha-1) de un fertilizante
compuesto de grado 15-15-15 (TRIAN 15® Yara, Colombia) como fuente de nitrógeno,
fósforo y potasio; ii) y 2 g planta-1 (170 Kg Ha-1) de microelementos (Agrimins granulado,
Colinagro®, Colombia) respectivamente, repartidas en dos aplicaciones (los fertilizantes

Capítulo 4 67
edáficos fueron suministrados en dos aplicaciones de la mitad de la dosis
anteriormente mencionada a los 15 y 40 DDE)). Finalmente, la composición del
fertilizante Agrimins era: Nitrogeno total 8% (Úrea N 7% y Amonio N 1%), Fósforo
asimilable (P2O5) 5.0%, Calcio (CaO) 18%, Magnesio (MgO) 6%, Azufre Total 1.6%,
Cobre 0.75%, Boro 1%, Molibdeno 0.005% y Zinc 2.5%.
4.3.2 Tratamiento de déficit hídrico y determinación de las variables
Plantas de frijol por cada cultivar fueron separadas en dos grupos para ser sometidas a
dos diferentes periodos de déficit hídrico. Un primer grupo, las plantas sufrieron un periodo
de estrés de 15 días en etapa vegetativa cuando las plantas alcanzaron el estado
fenológico 13-14 según escala BBCH (Feller et al., 1995b) (formación de 3 a 4 hojas
trifoliadas completamente expandidas), el cual se alcanzó aproximadamente a los 40 días
después de la emergencia de la totalidad de las plantas (DDE). En el segundo grupo, el
estrés fue desarrollado durante la etapa reproductiva en el estado 63-64 (30 a 40% de las
flores abiertas) aproximadamente a los 50 días después de la emergencia de la totalidad
de las plantas según la metodología reportada por Ghanbari et al. (2013). El déficit de agua
fue establecido mediante la suspensión del agua de riego durante 15 días según estudios
realizados previamente. Posteriormente, se monitoreo el contenido de humedad del suelo
mediante una sonda de humedad (Kelway HB-2 Soil Acidity and Moisture Tester, Kel
Instruments Co., Inc. NJ, USA), buscando tener una humedad entre el 45% y 55% de la
capacidad de campo durante el periodo de estrés, teniendo en cuenta que el 100% de la
capacidad de campo corresponde a un 29.87% de humedad del suelo en peso.
Adicionalmente, el criterio de tomar un 50% de humedad en el suelo se fundamentó porque
las plantas presentan reducción alteraciones en sus relaciones hídricas en humedades del
sustrato entre el 40 y 60 % (Costa-França et al., 2000; Segura-Monroy et al., 2015).
Se realizaron muestreos de todas las variables fisiológicas y bioquímicas al final del
periodo de estrés (15 días después del inicio del déficit hídrico, es decir a los 55 DDT para
la etapa vegetativa y 65 DDT para la etapa reproductiva). En general, las variables
fisiológicas fueron registradas en la segunda hoja trifoliada completamente expandida de

68 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
la parte superior del dosel. Con respecto a las variables bioquímicas, se colectó
aproximadamente 2 g material vegetal, fue homogenizado con nitrógeno líquido y
posteriormente almacenado a -80°C hasta sus respectivos análisis. El experimento duró
aproximadamente 115 días a partir de la emergencia de la totalidad de las plántulas.
4.3.3 Diseño experimental y análisis de los datos
Se realizó un diseño en bloques al azar con arreglo factorial donde el primer factor fue la
etapa fenológica en la que se inició el estrés por sequía y el segundo factor fueron los cinco
genotipos evaluados para un total de 15 tratamientos con cuatro repeticiones para un total
de 60 parcelas de 2.1 m2 (3 surcos de 1 m lineal de largo). Posteriormente, cuando se
presentaron diferencias significativas en el ANAVA, se empleó la prueba comparativa de
medias de Tukey a P ≤ 0.05. Los datos se analizaron usando el programa Statistix v 9.0
(software analítico, Tallahassee, FL, US). Los resultados del análisis de varianza son
resumidos en la Tabla 4-1.
4.3.4 Variables fisiológicas
Relaciones hídricas e intercambio gaseoso de la hoja
El contenido relativo de agua (CRA) fue determinado en hojas totalmente expandidas de
la parte media. Las hojas fueron recolectadas al final del periodo de estrés hídrico para
cada uno de los genotipos de fríjol evaluados. El CRA fue calculado mediante la siguiente
ecuación:
𝐶𝑅𝐴 = [𝑃𝐹 − 𝑃𝑆
𝑃𝑇 − 𝑃𝑆] × 100 (𝐸𝑐𝑢𝑎𝑐𝑖ó𝑛 1)

Capítulo 4 69
Siendo PF el peso fresco, PT el peso en turgencia medido después de 24 horas de
saturación en agua destilada a 4 °C en la oscuridad y PS es el peso seco determinado
después de 48 horas en un horno a 70°C.
La fotosíntesis fue estimada en la segunda hoja trifoliada completamente expandida del
porción media superior de la planta usando un medidor de fotosíntesis portable (LI-COR
6200, Lincoln, NE, USA), entre los periodos comprendidos de las 930 a 1100 h y 1330 a
1500 h durante tres días seguidos a partir del penúltimo día antes de finalizar el periodo
de estrés. Asimismo, este equipo estimó la concentración interna de CO2. Adicionalmente,
se calculó la relación Carbono intercelular entre el Carbono atmosférico (Ci/Ca) y las
limitaciones estomáticas se obtuvo usando la ecuación Ls = 1 - (Ci/Ca) (Huang et al. 2011).
La conductancia se midió en la segunda hoja trifoliada completamente expandida con la
ayuda de un porómetro portable (LI-COR 1600, Lincoln, NE, USA). Del mismo modo, se
calculó el uso eficiente del agua como producto de la relación entre la fotosíntesis y la
transpiración. Las medidas de intercambio gaseoso se desarrollaron entre las 1000 y 1500
horas los 55 y 65 DDT. Durante las mediciones las condiciones fueron las siguientes:
Radiación fotosintéticamente activa 1000 μmol m-2 s-1, temperatura de la hoja 27 ± 5 °C,
diferencia de presión de vapor del aire de la hoja 1.8 ± 0.5 kPa.
Parámetros de fluorescencia, contenido de clorofilas y carotenoides
Para la determinación de los parámetros de fluorescencia de PSII, se utilizó un fluorómetro
modulado (MINI-PAM, Walz, Effeltrich, GE). Las medidas fueron tomadas entre las 700 y
1000 h en la segunda hoja trifoliada completamente expandida desde la parte superior del
dosel. Las relaciones de fluorescencia fueron determinadas con un pulso máximo de
intensidad de luz de hasta 2.600 µmol m-2 s-1 en la superficie de la muestra de la hoja,
adaptándola previamente a la oscuridad durante 10 minutos. Para las mediciones de
clorofilas y carotenoides, las muestras de tejidos (30 mg aprox. de la segunda hoja trifoliada
completamente expandida) fueron homogenizadas en 4 ml de acetona (80%),
posteriormente se llevaron las muestras a una centrífuga (Model 420101, Becton Dickinson
Primary Care Diagnostics, Maryland, USA) a 5000 rpm durante 10 min para eliminar
partículas y el sobrenadante diluido a un volumen final de 6 ml adicionando acetona (Sims

70 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
y Gamon., 2002). El contenido de clorofila fue determinado a 663 y 646 nm y carotenoides
a 470 nm utilizando un espectrofotómetro (Spectronic BioMate 3 UV–vis Thermo, Madi-
son, WI, USA). Para obtener el contenido de pigmentos fotosintéticos en hoja, se
emplearon las ecuaciones para acetona descritas por Wellburn (1994).
4.3.5 Variables bioquímicas
Malondialdehido
Para la determinación de la oxidación lipídica (Malondialdehido-MDA), se empleó el
método del ácido tiobarbitúrico (TBA) descrito por Hodges et al. (1999), tomando 0.3 g
aprox. de material vegetal homogenizado y almacenado con nitrógeno líquido. Las
muestras luego de ser centrifugadas se les estimó las absorbancias a 440, 532 y 600 nm
con un espectrofotómetro (Spectronic BioMate 3 UV - Vis, Thermo, Madison, WI, USA).
Por último, se utilizó un coeficiente de extinción (157 M mL-1) para la obtención de la
concentración de MDA.
Concentración de Prolina
Para la determinación del contenido de prolina, se usó el método descrito por Bates et al.
(1973), extrayendo de la segunda hoja trifoliada completamente expandida
aproximadamente 0.3 g de material vegetal homogenizado y almacenado con nitrógeno
líquido. La absorbancia se medió a 520 nm usando un espectrofotómetro (Spectronic
BioMate 3 UV - Vis, Thermo, Madison, WI, USA). El contenido de prolina se determinó
mediante una curva estándar y calculada en peso fresco basado en la ecuación 2:
𝜇𝑚𝑜𝑙 𝑃𝑟𝑜𝑙𝑖𝑛𝑎
𝑔 𝑚𝑎𝑡𝑒𝑟𝑖𝑎𝑙 𝑣𝑒𝑔𝑒𝑡𝑎𝑙 𝑓𝑟𝑒𝑠𝑐𝑜=
[(
𝜇𝑔 𝑃𝑟𝑜𝑙𝑖𝑛𝑎𝑚𝐿 𝑥 𝑚𝐿 𝑇𝑜𝑙𝑢𝑒𝑛𝑜)
115.5 𝜇𝑔𝜇𝑚𝑜𝑙
]
[𝑔 𝑚𝑢𝑒𝑠𝑡𝑟𝑎
5 ] (𝐸𝑐𝑢𝑎𝑐𝑖ó𝑛 2)

Capítulo 4 71
4.4 Resultados
4.4.1 Humedad del suelo, Relaciones hídricas e intercambio gaseoso de la hoja
En general, la humedad de suelo (CHS) en el tratamiento control estuvo alrededor 90% de
capacidad de campo durante el ensayo. Con respecto al CHS de los otros tratamientos, se
observó que este estuvo cercano al 55% al final de los 15 días de déficit hídrico iniciado
en la etapa vegetativa (55DDE). Asimismo, el CHS del tratamiento de estrés durante la
etapa reproductiva presentaba una humedad alrededor del 53.5% a los 65 DDE (Figura
4-1A). Los resultados obtenidos en la cuantificación de la humedad del suelo señalan que
un grupo de plantas estuvo con una menor disponibilidad de agua en algún momento de
su desarrollo. Igualmente, los resultados del Contenido Relativo de Agua (CRA) en la hoja
son resumidos en la Figura 4-1(B y C). Diferencias significativas solamente fueron
observadas para el factor de estatus hídrico (Tabla 1). En el muestreo de los 55 DDE,
plantas que iniciaron su periodo de estrés en la etapa vegetativa presentaron un
CRA ̴70% en comparación a las plantas control que presentaron valores cercanos al 85%
a los 55 DDE. Asimismo, las plantas que iniciaron a sufrir un déficit hídrico en la etapa
reproductiva mostraron un valor ̴ 77.43% en ese punto (Figura 4-1B). Cuando finalizó el
periodo de estrés de la fase reproductiva (65 DDE), las plantas control continuaron con un
CRA del 82.57% mientras las plantas con condiciones de estrés en la etapa vegetativa
fue aproximadamente del 75.22% y un 73.34% en fase reproductiva (Figura 4-1C).
Nuestros resultados concuerdan con lo reportado por DeLat et al. (2014) y Huang et al.
(2011), donde un estrés por déficit hídrico redujo significativamente el CRA en las plantas,
y ésta reducción está dada principalmente por la baja disponibilidad de agua en el suelo.
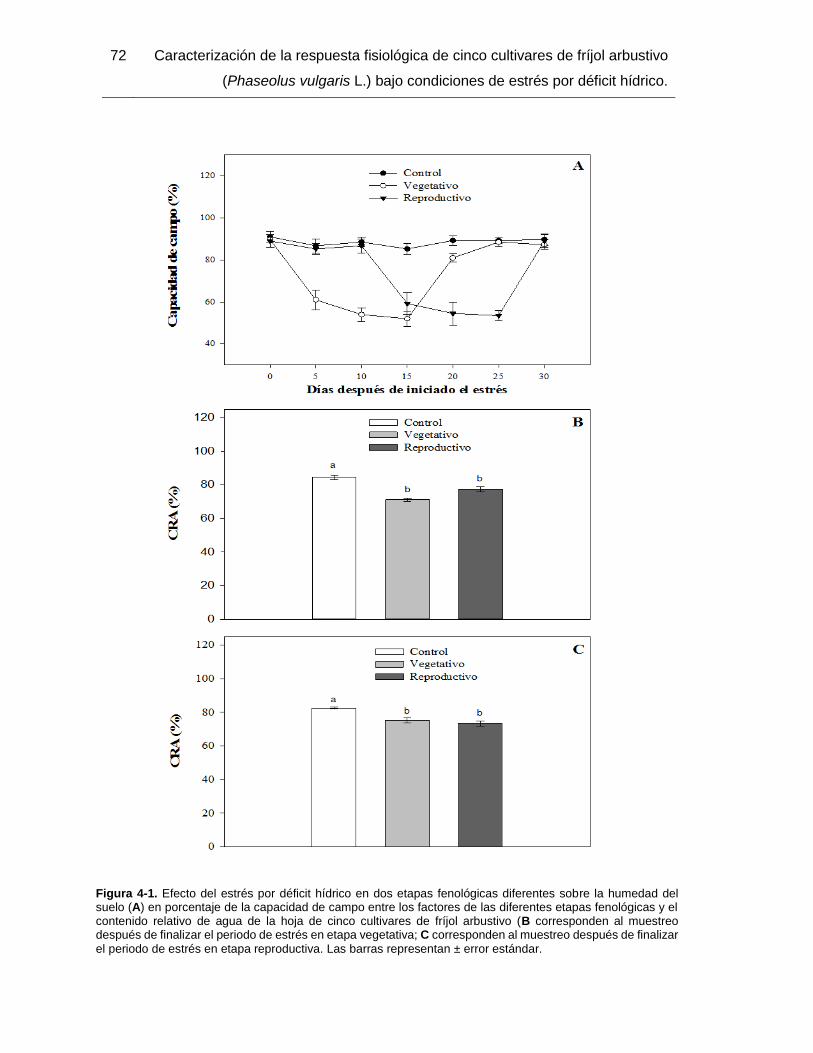
72 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
Figura 4-1. Efecto del estrés por déficit hídrico en dos etapas fenológicas diferentes sobre la humedad del suelo (A) en porcentaje de la capacidad de campo entre los factores de las diferentes etapas fenológicas y el contenido relativo de agua de la hoja de cinco cultivares de fríjol arbustivo (B corresponden al muestreo después de finalizar el periodo de estrés en etapa vegetativa; C corresponden al muestreo después de finalizar
el periodo de estrés en etapa reproductiva. Las barras representan ± error estándar.
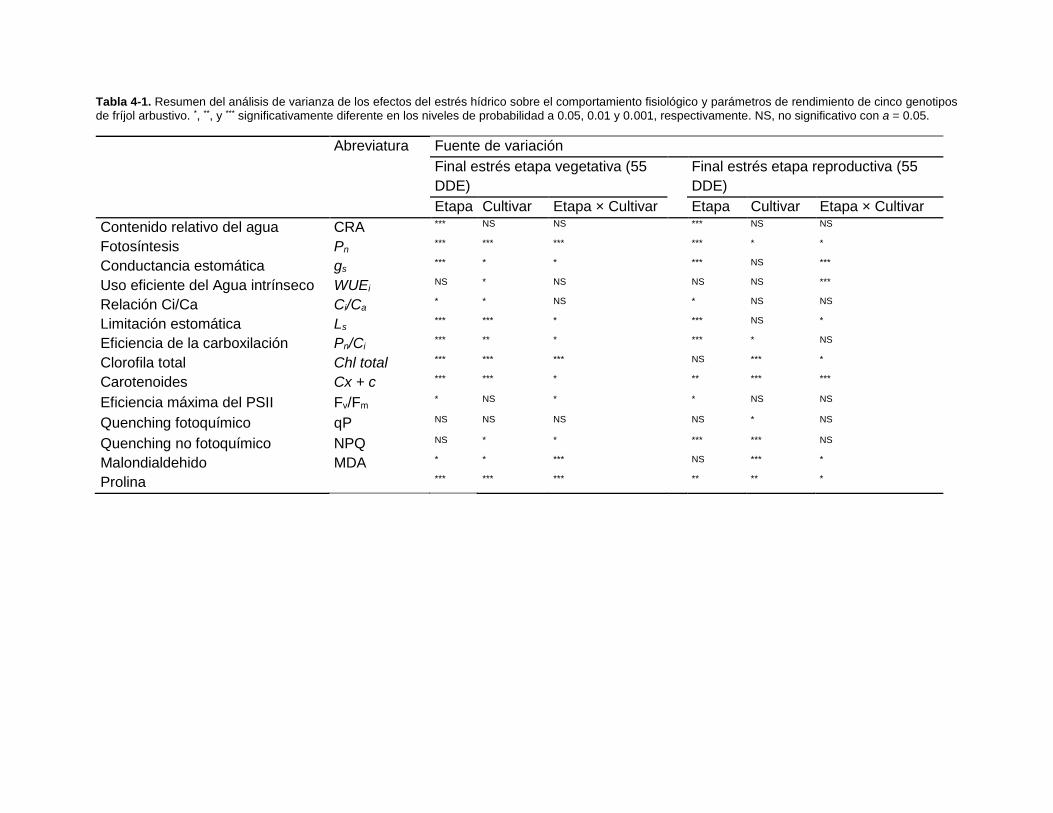
Tabla 4-1. Resumen del análisis de varianza de los efectos del estrés hídrico sobre el comportamiento fisiológico y parámetros de rendimiento de cinco genotipos de fríjol arbustivo. *, **, y *** significativamente diferente en los niveles de probabilidad a 0.05, 0.01 y 0.001, respectivamente. NS, no significativo con a = 0.05.
Abreviatura Fuente de variación
Final estrés etapa vegetativa (55
DDE)
Final estrés etapa reproductiva (55
DDE)
Etapa Cultivar Etapa × Cultivar Etapa Cultivar Etapa × Cultivar
Contenido relativo del agua CRA *** NS NS *** NS NS
Fotosíntesis Pn *** *** *** *** * *
Conductancia estomática gs *** * * *** NS ***
Uso eficiente del Agua intrínseco WUEi NS * NS NS NS ***
Relación Ci/Ca Ci/Ca * * NS * NS NS
Limitación estomática Ls *** *** * *** NS *
Eficiencia de la carboxilación Pn/Ci *** ** * *** * NS
Clorofila total Chl total *** *** *** NS *** *
Carotenoides Cx + c *** *** * ** *** ***
Eficiencia máxima del PSII Fv/Fm * NS * * NS NS
Quenching fotoquímico qP NS NS NS NS * NS
Quenching no fotoquímico NPQ NS * * *** *** NS
Malondialdehido MDA * * *** NS *** *
Prolina *** *** *** ** ** *

En cuanto a las propiedades de intercambio gaseoso de la hoja (fotosíntesis (Pn),
conductancia estomática (gs) y limitaciones estomáticas (Ls)), los resultados son
presentados en la Figura 4-2. Se encontraron diferencias significativas la interacción etapa
de desarrollo × cultivar en la Pn, gs y Ls al finalizar cada periodo de estrés (55 y 65 DDE).
A los 55 DDE, la Pn, en general, era mayor (18.20 µmol m-2 s-1) en todos los cultivares
estudiados sin déficit hídrico. Asimismo, la tasa de intercambio de CO2 fue un ~45 % (9.07
µmol m-2 s-1) menor en las plantas de los cultivares Bacatá y Bianca bajo condiciones de
estrés por estrés hídrico tanto en la fase vegetativa como en la reproductiva a excepción
de las plantas del cultivar Cerinza que presentaron una reducción solamente del ~37%
(12.10 µmol m-2 s-1) en la etapa vegetativa y ~17% (15.80 µmol m-2 s-1) en la reproductiva.
Finalmente, las plantas del cultivar NUA35 tuvieron reducciones en la Pn mayor en las
plantas con estrés en la etapa reproductiva cercanos al 42% (19.13 µmol m-2 s-1) (Figura
4-2A). Con respecto a las mediciones desarrolladas a los 65 DDE, se obtuvieron
tendencias similares a las observadas en el muestreo anterior. En este aspecto, las plantas
control de todos los cultivares presentaron valores iguales de fotosíntesis entre ellas (20.81
µmol m-2 s-1). Mientras, un periodo de déficit hídrico, en general, redujo la Pn cerca del 55%
(12.01 µmol m-2 s-1) tanto en etapa vegetativa o reproductiva en todos los casos (Figura
4-2B). Con respecto a la gs, ésta también fue mayor en el tratamiento control donde se
registraron lecturas promedios alrededor de 0.33 µmol m-2 s-1 y 0.51 µmol m-2 s-1 a los 55
y 65 DDE, respectivamente. Asimismo, un periodo de 15 días déficit hídrico tanto en la
fase vegetativa o reproductiva también causó una reducción significativa en los valores de
esta variable de intercambio gaseoso. En general, todos los cultivares mostraron una gs un
50% menor (~0.28 µmol m-2 s-1) en ambas etapas muestreos (Figura 4-2C y D).
Finalmente, la limitación estomática (Figura 4-2E y F) se mantuvo generalmente constante
en la mayoría de cultivares (Ls ~0.34) en ambos tratamientos a los 55 DDE, excepto las
plantas del genotipo NUA35 que mostraron una mayor valor en los dos periodos de déficit
hídrico (0.33) (Figura 4-2E). Asimismo, Ls fue mayor específicamente en las plantas de
frijol Cerinza sometidas a un periodo de 15 días de déficit hídrico durante la etapa
reproductiva a los 65 DDE (Ls bajo estrés: 0.27 y Ls Control: 0.21). No obstante, los
cultivares Bachue, NUA35, Bacatá y Bianca (0.24) mantuvieron los valores cercanos a los
encontrados en las plantas control (0.25) (Figura 4-2F).

Figura 4-2. Efecto del estrés por déficit hídrico en dos etapas fenológicas diferentes sobre la fotosíntesis (A y B), conductancia estomática (C y D) y limitación estomática (E y F) de cinco cultivares de fríjol arbustivo (A, C y E corresponden al muestreo después de finalizar el periodo de estrés en etapa vegetativa; B, D y F corresponden al muestreo después de finalizar el periodo de estrés en etapa reproductiva. Las barras representan ± error estándar.

76 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
Por otro lado, la Tabla 4-2 resume los resultados del uso eficiente del agua intrínseco
(UEAi), relación carbono intercelular sobre carbono atmosférico (Ci/Ca), y la Eficiencia de la
Carboxilación (Pn/Ci), respectivamente. Para el UEAi, se resaltan solamente las diferencias
significativas obtenidas en la interacción etapa × cultivar al finalizar el periodo de estrés por
déficit hídrico en la etapa reproductiva (65DDE). En este sentido, las plantas de los
cultivares Bachue, Bacatá y Bianca (45.21; 44.16 y 41.24 μmol CO2 mmol -1 H2O,
respectivamente) sometidas al periodo de estrés por déficit hídrico durante la etapa
reproductiva mostraron un mayor UEAi. Mientras, los cultivares NUA35 y Cerinza
expuestas a estrés durante un periodo de la fase reproductiva mostraron las menores
relaciones (21.37 y 39.79 μmol CO2 mmol-1 H2O, respectivamente). En cuanto a la relación
Ci/Ca, diferencias fueron obtenidas en el factor déficit hídrico tanto a los 55 y 66 DDE. En
este sentido, las plantas bajo estrés en etapa vegetativa presentaron los valores más bajos
en la Ci/Ca (aproximadamente de 0.69), mientras que las plantas de los cultivares bajo
condiciones control y estrés en la etapa reproductiva tuvieron valores de aproximadamente
0.72 a los 55 DDE (Tabla 4-2). Además, las plantas que estuvieron en estrés por déficit
hídrico en la etapa reproductiva tuvieron valores similares a las plantas en condiciones
control (Ci/Ca de 0.76) a los 65 DDE (Tabla 4-2).
Por otra parte, diferencias en la interacción etapa × cultivar (P≤0.05) fueron encontradas
sobre la eficiencia de carboxilación (Pn/Ci) a los 55 DDE. En este punto de muestreo,
plantas de todos los cultivares bajo alguna condiciones de estrés tuvieron una Pn/Ci en
promedio de 0.04 mol m-2 s-1 comparado con las plantas control (0.06 mol m-2 s-1). Del mismo
modo, para la Pn/Ci se encontraron diferencias significativas en el factor Etapa a los, DDE
donde las plantas de los cultivares bajo condición de estrés en la etapa vegetativa tuvieron
las relaciones más bajos en la Pn/Ci (aproximadamente de 0.03 mol m-2 s-1), mientras que
las plantas con estrés en la etapa reproductiva los valores de alrededor de 0.04 mol m-2 s-
1y las plantas en condiciones sin estrés por déficit hídrico mantuvieron valores cercanos a
0.07 mol m-2 s-1 (Tabla 4-2). Similares tendencias fueron reportadas por Boonjung y Fukai
(1996-1 y 2); Huang et al. (2011) Rosales et al. (2012); Yousfi et al. (2012), donde las
principales reducciones en los parámetros como la fotosíntesis neta, conductancia
estomática, e incrementos en las limitaciones estomáticas se evidencias cuando las plantas
se encuentran en déficit hídrico, especialmente en las etapas vegetativas tempranas.
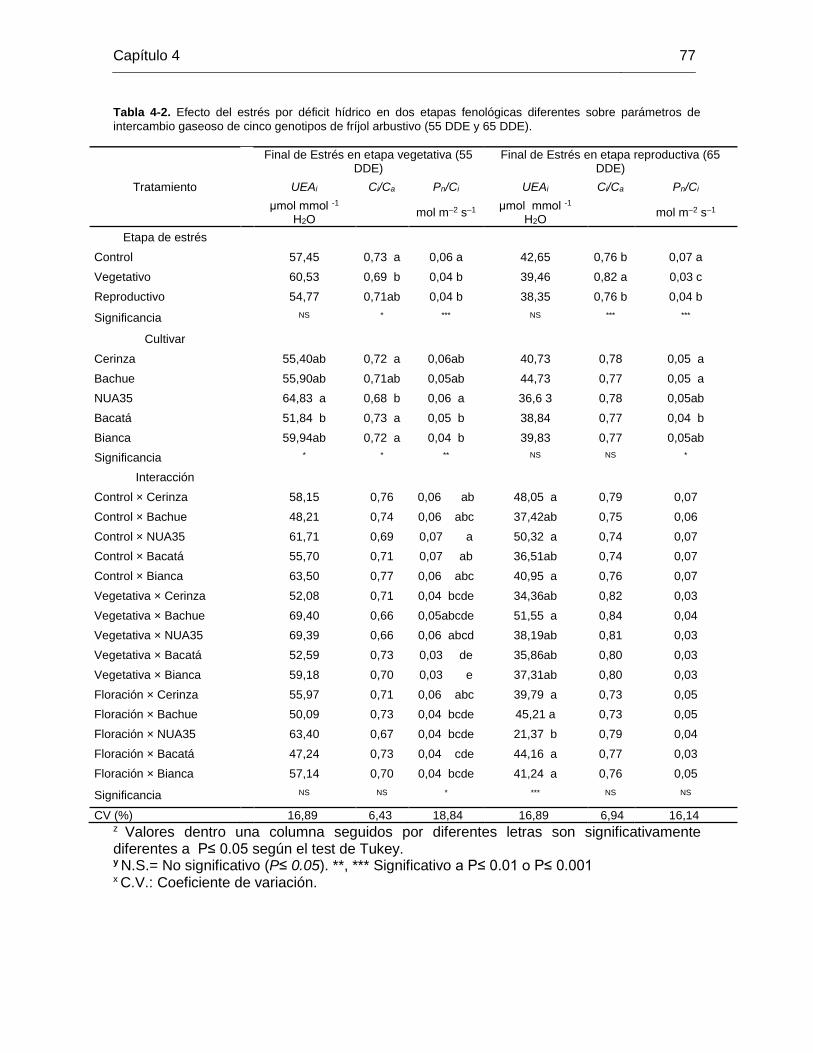
Capítulo 4 77
Tabla 4-2. Efecto del estrés por déficit hídrico en dos etapas fenológicas diferentes sobre parámetros de
intercambio gaseoso de cinco genotipos de fríjol arbustivo (55 DDE y 65 DDE).
Tratamiento
Final de Estrés en etapa vegetativa (55
DDE) Final de Estrés en etapa reproductiva (65
DDE)
UEAi Ci/Ca Pn/Ci UEAi Ci/Ca Pn/Ci
μmol mmol -1
H2O mol m–2 s–1
μmol mmol -1 H2O
mol m–2 s–1
Etapa de estrés
Control 57,45 0,73 a 0,06 a 42,65 0,76 b 0,07 a
Vegetativo 60,53 0,69 b 0,04 b 39,46 0,82 a 0,03 c
Reproductivo 54,77 0,71ab 0,04 b 38,35 0,76 b 0,04 b
Significancia NS * *** NS *** ***
Cultivar
Cerinza 55,40ab 0,72 a 0,06ab 40,73 0,78 0,05 a
Bachue 55,90ab 0,71ab 0,05ab 44,73 0,77 0,05 a
NUA35 64,83 a 0,68 b 0,06 a 36,6 3 0,78 0,05ab
Bacatá 51,84 b 0,73 a 0,05 b 38,84 0,77 0,04 b
Bianca 59,94ab 0,72 a 0,04 b 39,83 0,77 0,05ab
Significancia * * ** NS NS *
Interacción
Control × Cerinza 58,15 0,76 0,06 ab 48,05 a 0,79 0,07
Control × Bachue 48,21 0,74 0,06 abc 37,42ab 0,75 0,06
Control × NUA35 61,71 0,69 0,07 a 50,32 a 0,74 0,07
Control × Bacatá 55,70 0,71 0,07 ab 36,51ab 0,74 0,07
Control × Bianca 63,50 0,77 0,06 abc 40,95 a 0,76 0,07
Vegetativa × Cerinza 52,08 0,71 0,04 bcde 34,36ab 0,82 0,03
Vegetativa × Bachue 69,40 0,66 0,05abcde 51,55 a 0,84 0,04
Vegetativa × NUA35 69,39 0,66 0,06 abcd 38,19ab 0,81 0,03
Vegetativa × Bacatá 52,59 0,73 0,03 de 35,86ab 0,80 0,03
Vegetativa × Bianca 59,18 0,70 0,03 e 37,31ab 0,80 0,03
Floración × Cerinza 55,97 0,71 0,06 abc 39,79 a 0,73 0,05
Floración × Bachue 50,09 0,73 0,04 bcde 45,21 a 0,73 0,05
Floración × NUA35 63,40 0,67 0,04 bcde 21,37 b 0,79 0,04
Floración × Bacatá 47,24 0,73 0,04 cde 44,16 a 0,77 0,03
Floración × Bianca 57,14 0,70 0,04 bcde 41,24 a 0,76 0,05
Significancia NS NS * *** NS NS
CV (%) 16,89 6,43 18,84 16,89 6,94 16,14 z Valores dentro una columna seguidos por diferentes letras son significativamente diferentes a P≤ 0.05 según el test de Tukey. y N.S.= No significativo (P≤ 0.05). **, *** Significativo a P≤ 0.01 o P≤ 0.001 x C.V.: Coeficiente de variación.

78 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
4.4.2 Contenido de clorofilas, carotenoides y parámetros de fluorescencia,
Por otro lado, la Figura 4-3 se muestran los resultados obtenidos en la cuantificación del
contenido de clorofila total y el contenido de carotenoides totales. En general, se puede
observar que para los dos puntos de muestreo se obtuvieron diferencias significativas en
la interacción Etapa × Cultivar sobre la concentración de los pigmentos fotosintéticos
evaluados. En el primer muestro (55 DDT), se observó que la mayoría de plantas control
mostro un contenido de clorofila en hoja alrededor de 2166.26 µg mg-1 PF. Asimismo, es
importante señalar que se presentó una mayor reducción en el contenido de clorofila total
en las plantas del cultivar Bacatá que estuvieron sometidas condiciones de estrés por déficit
hídrico tanto en etapa vegetativa (1517.65 µg mg-1 PF) y reproductiva (1538.90 µg mg-1 PF).
Sin embargo, a los 65 DDE (final del estrés en la etapa reproductiva), las plantas del cultivar
Bachue con estrés en la etapa reproductiva presentaron los mayores contenidos de clorofila
total (~1709.29 µg mg-1 PF). Con respecto al contenido de carotenoides, se observó que el
éste pigmento disminuyo por el estrés hídrico tanto en fase vegetativo como reproductiva,
especialmente, en Bianca, Bacatá y NUA35 a los 55 DDE (Figura 4-3C). Por otro lado, la
concentración de estos carotenoides fue favorecida por una condición de estrés hídrico en
la etapa vegetativa en los cultivares Bacatá y Bianca tuvieron una concentración de estos
pigmento con un incremento cercano al 44% y 32% (279.46 y 356.13 µg mg-1 PF,
respectivamente) a los 65 DDE. Igualmente, las plantas del cultivar Bachue con estrés por
déficit hídrico en la etapa reproductiva tuvieron una mayor concentración (320.50 µg mg-1
PF) de carotenoides totales con relación a su control (230.39 µg mg-1 PF) en este punto de
muestreo (Figura 4-3D). Aranjuelo et al. (2011), reportaron similares resultados, donde la
clorofilas no se vieron significativamente afectadas por condiciones de estrés hídrico,
aunque una mayor producción de carotenoides totales puede ser considerada un
mecanismo de regulación del estrés como actividad antioxidante.
En cuanto a los parámetros de fluorescencia, la Tabla 4-3 presenta los resultados
encontrados en la eficiencia máxima del fotosistema II (Fv/Fm), quenching fotoquímico (qP)
y quenching no fotoquímico (NPQ). Diferencias significativas fueron encontradas en la
interacción Etapa × Cultivar para la relación Fv/Fm y el NPQ a los 55 DDE. Donde las
mayores reducciones en la Fv/Fm (0.67), y NPQ (0.71) se evidenciaron en las plantas del

Capítulo 4 79
cultivar NUA35 con un periodo de quince días de estrés en la etapa vegetativa en
comparación a las plantas control en el mismo cultivar (Fv/Fm de 0.78 y NPQ de 1.07).
Asimismo, no se encontraron diferencias significativas sobre qP en dicho punto de
muestreo. Por otro lado, al final del periodo de estrés por déficit hídrico en la etapa
reproductiva (65 DDE), se encontraron diferencias significativas a nivel de estrés entre
etapas sobre la relación Fv/Fm, donde los menores valores se encontraron en las plantas de
los cultivares que estuvieron bajo condiciones de estrés en la etapa vegetativa (Fv/Fm de
aproximadamente 0.72). Con respecto a los otros parámetros de la fluorescencia de la
clorofila a (qP y el NPQ), se obtuvieron diferencias principalmente en el factor cultivar. En
este aspecto, los valores más bajos en el qP fueron observados en las plantas del cultivar
Cerinza (0.56) y los más altos en las plantas del cultivar NUA35 (0.66); mientras que para
el NPQ los valores más bajos fueron encontrados en las plantas del cultivar NUA35 (0.97)
y los más altos en las plantas del cultivar Bacatá (1.60). Esto concuerda con lo reportado
con Keyvan. (2010) y Perveen et al. (2010).
Figura 4-3. Efecto del estrés por déficit hídrico en dos etapas fenológicas diferentes sobre la clorofila total (A y B) y carotenoides totales (C y D) de cinco cultivares de fríjol arbustivo (A y C corresponden al muestreo después de finalizar el periodo de estrés en etapa vegetativa; B y D corresponden al muestreo después de finalizar el
periodo de estrés en etapa reproductiva. Las barras representan ± error estándar.
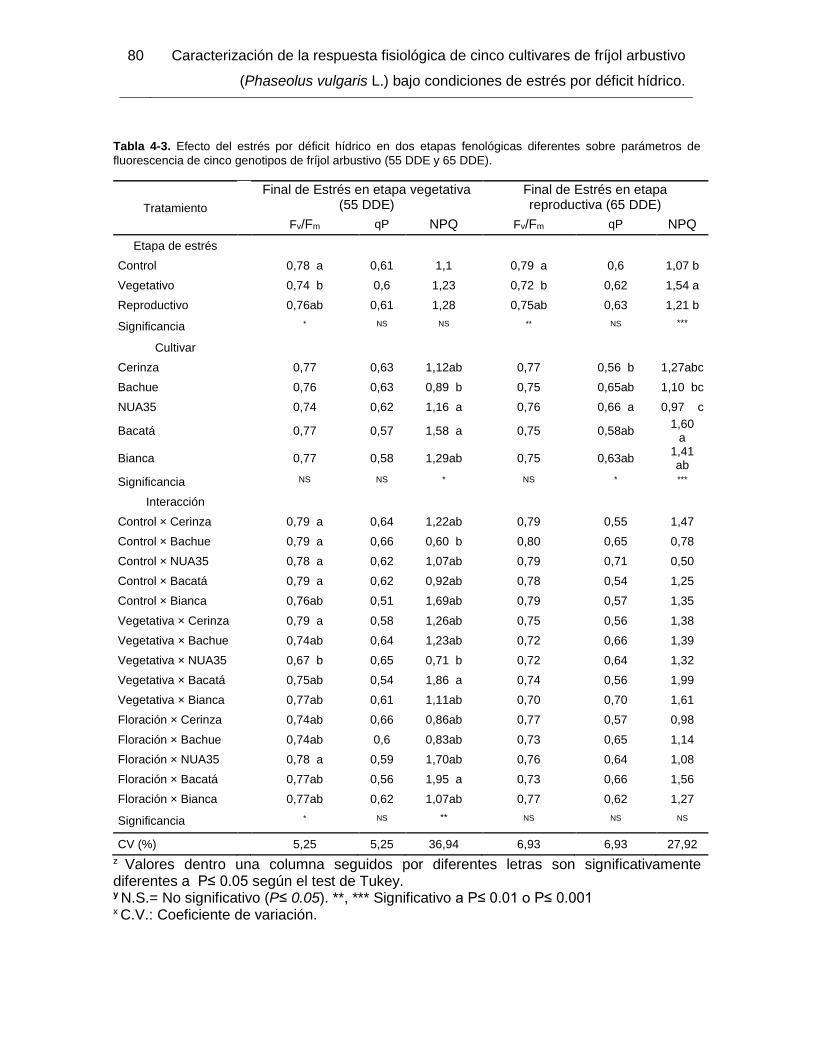
80 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
Tabla 4-3. Efecto del estrés por déficit hídrico en dos etapas fenológicas diferentes sobre parámetros de
fluorescencia de cinco genotipos de fríjol arbustivo (55 DDE y 65 DDE).
Tratamiento
Final de Estrés en etapa vegetativa (55 DDE)
Final de Estrés en etapa reproductiva (65 DDE)
Fv/Fm qP NPQ Fv/Fm qP NPQ
Etapa de estrés
Control 0,78 a 0,61 1,1 0,79 a 0,6 1,07 b
Vegetativo 0,74 b 0,6 1,23 0,72 b 0,62 1,54 a
Reproductivo 0,76ab 0,61 1,28 0,75ab 0,63 1,21 b
Significancia * NS NS ** NS ***
Cultivar
Cerinza 0,77 0,63 1,12ab 0,77 0,56 b 1,27abc
Bachue 0,76 0,63 0,89 b 0,75 0,65ab 1,10 bc
NUA35 0,74 0,62 1,16 a 0,76 0,66 a 0,97 c
Bacatá
0,77 0,57 1,58 a 0,75 0,58ab 1,60
a
Bianca
0,77 0,58 1,29ab 0,75 0,63ab 1,41 ab
Significancia NS NS * NS * ***
Interacción
Control × Cerinza 0,79 a 0,64 1,22ab 0,79 0,55 1,47
Control × Bachue 0,79 a 0,66 0,60 b 0,80 0,65 0,78
Control × NUA35 0,78 a 0,62 1,07ab 0,79 0,71 0,50
Control × Bacatá 0,79 a 0,62 0,92ab 0,78 0,54 1,25
Control × Bianca 0,76ab 0,51 1,69ab 0,79 0,57 1,35
Vegetativa × Cerinza 0,79 a 0,58 1,26ab 0,75 0,56 1,38
Vegetativa × Bachue 0,74ab 0,64 1,23ab 0,72 0,66 1,39
Vegetativa × NUA35 0,67 b 0,65 0,71 b 0,72 0,64 1,32
Vegetativa × Bacatá 0,75ab 0,54 1,86 a 0,74 0,56 1,99
Vegetativa × Bianca 0,77ab 0,61 1,11ab 0,70 0,70 1,61
Floración × Cerinza 0,74ab 0,66 0,86ab 0,77 0,57 0,98
Floración × Bachue 0,74ab 0,6 0,83ab 0,73 0,65 1,14
Floración × NUA35 0,78 a 0,59 1,70ab 0,76 0,64 1,08
Floración × Bacatá 0,77ab 0,56 1,95 a 0,73 0,66 1,56
Floración × Bianca 0,77ab 0,62 1,07ab 0,77 0,62 1,27
Significancia * NS ** NS NS NS
CV (%) 5,25 5,25 36,94 6,93 6,93 27,92
z Valores dentro una columna seguidos por diferentes letras son significativamente diferentes a P≤ 0.05 según el test de Tukey. y N.S.= No significativo (P≤ 0.05). **, *** Significativo a P≤ 0.01 o P≤ 0.001 x C.V.: Coeficiente de variación.

4.4.3 Concentración de Malondialdehido y Prolina
En cuanto al daño de la membrana por estrés oxidativo y producción de osmolitos,
resultados de la producción de malondialdehido (MDA) y prolina se muestran en la Figura
4-4A, B, C y D respectivamente). Diferencias significativas fueron también observadas
entre los diferentes tratamientos en las variables anteriormente mencionadas sobre la
interacción etapa de Estrés × Cultivar (P≤0.001 a los 55 DDE y P≤0.05 a los 65 DDE). Las
plantas del cultivar Cerinza con estrés hídrico durante la fase fenológica vegetativa
presentaron la mayor producción de MDA a los 55 DDE (con valores cercanos a 14 µmol
g-1 PF). Luego, los cultivares Bacatá (8.90 µmol g-1 PF) y Bianca (6.54 µmol g-1 PF) también
tuvieron un aumento en la producción de MDA cercano al 30% comparado a sus controles
(Figura 4-4A). A los 65 DDE, la producción de MDA fue especialmente mayor en el
genotipo Bacatá bajo condiciones de estrés en la etapa reproductiva (6.37 µmol g-1 PF),
seguido por el cultivar Cerinza que fue sometido a déficit en etapa vegetativa (5.96 µmol
g-1 PF) (Figura 4-4B).
Asimismo, la producción de prolina fue favorecida bajo las condiciones de estrés en las
etapas vegetativa y reproductiva en los dos puntos de muestreo (55 y 65 DDE,
respectivamente), donde Bachue fue el cultivar con mayor producción de este osmolito
(aproximadamente 76 y 58 µmol g-1 PF para la etapa vegetativa y reproductiva
respectivamente a los 55 DDE). Finalmente, los contenidos de prolina en general fueron
menores a los 65 DDE en relación a los observados a los 55 DDE. En este sentido, un
menor contenido de prolina fue registrado en los cultivares durante déficit hídrico en el
segundo punto de muestreo (65DDE). Es decir, los cultivares Cerinza, NUA35, Bacatá y
Bianca pasaron de un contenido promedio de prolina en plantas control de 40 µmol g-1 PF
a valores de 51 µmol g-1 PF bajo condiciones de sequía durante la eta reproductiva. (Figura
4-4 C y D). Similares resultados fueron reportados por Hong-Bo et al. (2006); Sánchez-
Rodríguez et al. (2010); Sun et al. (2013) y Osman. (2015), siendo los genotipos tolerantes
aquellos cuya producción de sustancias antioxidantes tales como la prolina y carotenoides
es mayor bajo condiciones de estrés.
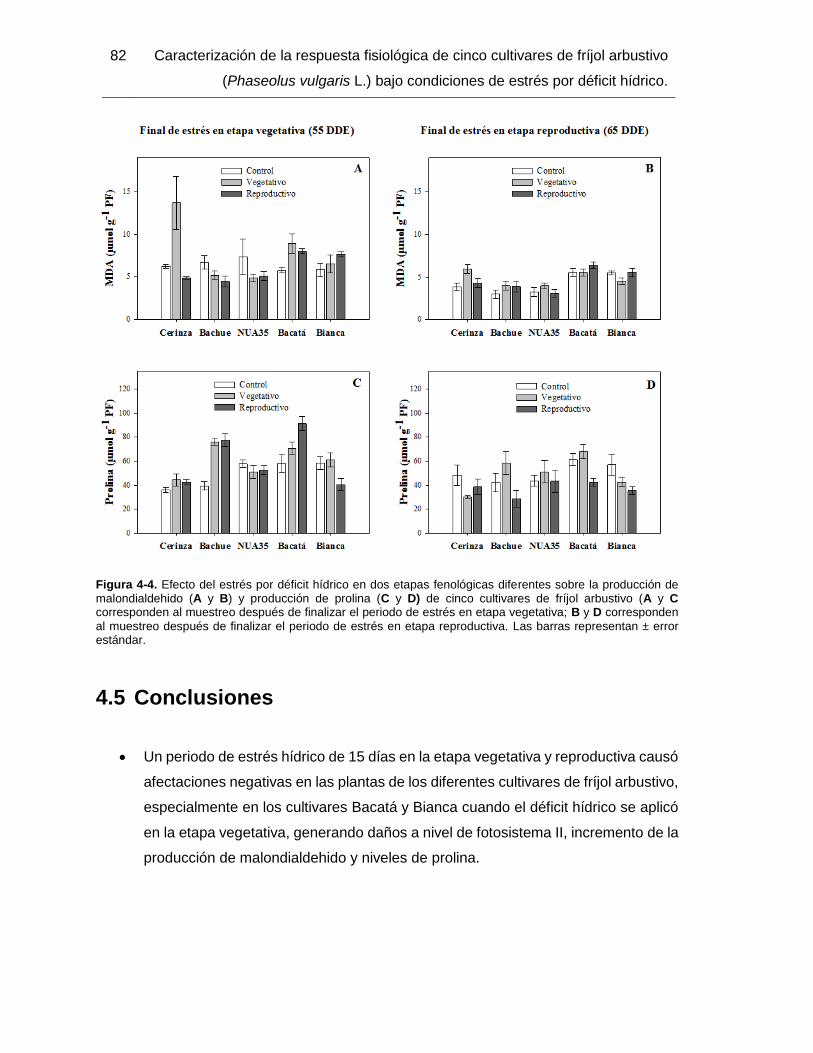
82 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
Figura 4-4. Efecto del estrés por déficit hídrico en dos etapas fenológicas diferentes sobre la producción de malondialdehido (A y B) y producción de prolina (C y D) de cinco cultivares de fríjol arbustivo (A y C corresponden al muestreo después de finalizar el periodo de estrés en etapa vegetativa; B y D corresponden
al muestreo después de finalizar el periodo de estrés en etapa reproductiva. Las barras representan ± error estándar.
4.5 Conclusiones
Un periodo de estrés hídrico de 15 días en la etapa vegetativa y reproductiva causó
afectaciones negativas en las plantas de los diferentes cultivares de fríjol arbustivo,
especialmente en los cultivares Bacatá y Bianca cuando el déficit hídrico se aplicó
en la etapa vegetativa, generando daños a nivel de fotosistema II, incremento de la
producción de malondialdehido y niveles de prolina.

5. Capítulo 5. Evaluación de índices de tolerancia a la sequía para la selección de cultivares de fríjol arbustivo (Phaseolus vulgaris L.) por inducción de estrés por déficit hídrico.
5.1 Resumen
El rendimiento es uno de los factores más afectados por condiciones de déficit hídrico.
Asimismo, se ha encontrado que existen etapas fenológicas más sensibles a condiciones
estresantes. Por tal motivo, el objetivo de esta investigación fue evaluar las respuestas en
parámetros de rendimiento de cinco cultivares de fríjol arbustivo (Phaseolus vulgaris L.)
bajo condición de estrés por déficit hídrico en dos etapas fenológicas diferentes (una
vegetativa y una reproductiva). Para ello se implementó un diseño en bloques
completamente al azar en arreglo factorial (factor principal la etapa fenológica y factor
secundario los cultivares) para un total de 15 tratamientos con cuatro repeticiones. Se
sembraron semillas de los cultivares de fríjol ICA-Cerinza, ICA-Bachue, NUA35, Bacatá y
Bianca se realizó un diseño. Adicionalmente se cuantificaron parámetros de rendimiento e
índices de susceptibilidad y/o tolerancia al estrés por déficit hídrico. Se encontró que el
cultivar Bacatá es el más sensible al déficit hídrico, especialmente en la etapa vegetativa,
lo cual se evidencia en la reducción de los diferentes parámetros de rendimiento.
Asimismo, el cultivar NUA35 fue considerado un cultivar tolerante al déficit hídrico en las
dos etapas evaluadas. Se recomienda el uso de índices para caracterizar genotipos a
condiciones de estrés por déficit hídrico.
Palabras clave: Susceptibilidad, tolerancia, sequía, rendimiento, fenología.

84 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
5.2 Introducción
El fríjol común (Phaseolus vulgaris L.) es el grano de legumbre más importante para la
alimentación humana (Beebe., 2012; Beebe et al., 2013). El consumo per cápita de fríjoles
en los países latinoamericanos oscila entre 12 Kg y 18 Kg por año, aunque éstas cifras no
discriminan el consumo urbano del rural (Broughton et al., 2003; Beebe et al., 2013). Es
consumido principalmente por su alto contenido de proteínas y minerales (FAO, 2007), y
América Latina es la región mundial con mayor producción de fríjol, cubriendo el 50% de
la producción mundial (Beebe et al., 2013), y Colombia en el primer semestre del 2015
alcanzó una producción de 62,974 toneladas (Fenalce., 2016).
Todos los estreses bióticos y abióticos causan en algún grado disminución sobre el
rendimiento de los cultivos; sin embargo, el estrés por déficit hídrico es el factor más
limitante de la producción agrícola del mundo (Mollasadeghi et al., 2011; Naghavi et al.,
2013). En este aspecto, la Unidad Nacional para la Gestión del Riesgo de Desastres en
Colombia (2014), ha reportado que Fenómenos del Niño pueden afectar a más de 180
municipios del país, causando una reducción de la oferta hídrica cercana al 50%.
Igualmente, según estudios de la Comunidad Andina (2009), en Colombia existe un área
de ~59.000 km2 expuestas a condiciones de sequía, en la que gran parte de esta área se
pertenece a la región andina, donde el fríjol es comúnmente cultivado.
Por otro lado, entender las respuestas de las plantas al estrés por sequía es de gran
importancia para la selección de genotipos tolerantes a condiciones ambientales adversas
(Zhao et al., 2008; Farshadfar et al., 2013). Asimismo, el comportamiento del rendimiento
relativo de los genotipos en condiciones tanto favorables como aquellos afectados por el
déficit hídrico parece ser un punto de partida importante en la identificación de genotipos
deseables para las regiones afectadas por la baja disponibilidad de agua (Nouri et al.,
2011; Farshadfar et al., 2013). La selección de genotipos con tolerancia a la sequía no es
fácil, debido a la ocurrencia de fuertes interacciones entre genotipos y el medio ambiente,

Capítulo 5 85
además del escaso conocimiento de la función y el papel de los diferentes mecanismos
de tolerancia (Naghavi et al., 2013).
Actualmente, numerosos investigadores han trabajado en los diferentes métodos de
selección de genotipos con tolerancia a la sequía mediante el uso de índices de tolerancia
y/o susceptibilidad (Yang et al., 2002; Khakwani et al., 2011; Farshadfar et al., 2013;
Naghavi et al., 2013). Fernández (1992), sugieren que el Índice de tolerancia al estrés (STI)
está definido como un marcador útil para determinar el potencial de alta tolerancia y
rendimiento el estrés de los genotipos evaluados. No obstante, se han propuesto
novedades de índices alternos útiles para la identificación de genotipos tales como el Índice
de Susceptibilidad al Estrés (SSI) por Fischer y Maurer (1978); la Tolerancia al Estrés
(TOL) y la Productividad Media (MP) por Rosielle y Hamblin (1981); la Productividad Media
Geométrica (GMP) por Fernández (1992), entre otros.
Sin embargo, la implementación de índices de tolerancia a sequía en los métodos de
selección de genotipos de fríjol tolerantes y/o susceptibilidad a la sequía no son muy
usados. Por tal motivo, el objetivo de esta investigación fue evaluar las respuestas en
parámetros de rendimiento de cinco cultivares de fríjol arbustivo (Phaseolus vulgaris L.)
bajo condición de estrés por déficit hídrico en dos etapas fenológicas diferentes (una
vegetativa y una reproductiva). Además, se realizó este estudio con el fin de evaluar los
criterios de selección para la identificación de tolerancia al déficit hídrico en cultivares de
fríjol, de modo que los cultivares adecuados puedan ser recomendados para el cultivo en
zonas propensas a la sequía en Colombia y áreas similares.
5.3 Materiales y métodos
5.3.1 Material vegetal y condiciones de crecimiento

86 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
Semillas de cinco cultivares de fríjol arbustivo fueron usadas en el presente estudio. Los
genotipos estudiados eran los siguientes: i) ICA-Cerinza e ICA-Bachue son dos materiales
con 20 años de siembra en la agricultura tradicional colombiana, ii) NUA35 es un genotipo
con siete años de comercialización desde su liberación y iii) Bianca y Bacatá son dos
cultivares de reciente liberación (menos de dos años). Las semillas fueron sembradas en
el invernadero de la Facultad de Ciencias Agrarias de la Universidad Nacional de Colombia
ubicado en la ciudad de Bogotá a una altura de 2556 msnm, (4°35’56’’ y 74°04’51’’) durante
los meses de septiembre del 2015 a enero del 2016 en parcelas de 2.1 m2 (3 surcos de 1
m lineal de largo). El marco de plantación era 16 × 70 cm entre planta y entre surcos,
respectivamente (20 semillas por parcela = 85.000 plantas ha-1). Las características físico-
químicas del suelo en el invernadero eran las siguientes: i) una textura Franco Arcilloso
(26% arena, 42% de limo y 32% de Arcilla), ii) características químicas: Nitrógeno total:
0.36%, Ca: 10.6, K: 0.98, Mg: 1.75, Na: 0.24 meq 100 g-1, Cu: 1.67, Fe: 310, Mn: 3.21, Zn:
15.5, B: 0.48 y P: >116 mg kg-1, iii) pH 5.4 y iv) Capacidad de Intercambio Catiónico
Efectiva (CICE) 13.8 meq/100g. Las condiciones de crecimiento del invernadero durante
la evaluación fueron temperatura promedio de 27°C, humedad relativa de 60 a 80% y un
fotoperiodo natural de 12 h con una PAR de 1000 µmol m-2 s-1. Después de la emergencia,
las plantas fueron fertilizadas con dos fertilizantes edáficos: i) 4 g planta-1 (350 Kg ha-1) de
un fertilizante compuesto de grado 15-15-15 (TRIAN 15® Yara, Colombia) como fuente de
nitrógeno, fósforo y potasio; ii) y 2 g planta-1 (170 Kg Ha-1) de microelementos (Agrimins
granulado, Colinagro®, Colombia) respectivamente, repartidas en dos aplicaciones (Los
fertilizantes edáficos fueron aplicados a los 15 y 40 DDE)). Finalmente, la composición
de Agrimins era: Nitrogeno total 8% (Úrea N 7% y Amonio N 1%), Fósforo asimilable
(P2O5) 5.0%, Calcio (CaO) 18%, Magnesio (MgO) 6%, Azufre Total 1.6%, Cobre
0.75%, Boro 1%, Molibdeno 0.005% y Zinc 2.5%.
5.3.2 Tratamiento de déficit hídrico
Plantas de frijol por cada cultivar fueron separadas en dos grupos para ser sometidas a
dos diferentes periodos de déficit hídrico. Un primer grupo, las plantas fueron sometidas a
un periodo de estrés en etapa vegetativa cuando las plantas alcanzaron el estado

Capítulo 5 87
fenológico 13-14 según escala BBCH (Feller et al., 1995b) (formación de 3 a 4 hojas
trifoliadas completamente expandidas), el cual se alcanzó aproximadamente a los 40 días
después de la emergencia de la totalidad de las plantas (DDE). En el segundo grupo, el
estrés fue aplicado desarrollado durante la etapa reproductiva en el estado 63-64 (30 a
40% de las flores abiertas) aproximadamente a los 50 días después de la emergencia de
la totalidad de las plantas según la metodología reportada por Ghanbari et al. (2013). El
déficit de agua fue establecido mediante la suspensión del agua de riego durante 15 días.
Este periodo de tiempo fue tomado según estudios desarrollados previamente donde
variables tales como fotosíntesis, perdida de electrolitos y producción de prolina tuvieron
afectaciones negativas. Posteriormente, se estuvo monitoreando el contenido de humedad
del suelo mediante una sonda de humedad (Kelway HB-2 Soil Acidity and Moisture Tester,
Kel Instruments Co., Inc. NJ, USA), buscando tener una humedad entre el 45% y 55% de
la capacidad de campo durante el periodo de estrés, teniendo en cuenta que el 100% de
la capacidad de campo corresponde a un 29.87% de humedad del suelo en peso.
Adicionalmente, el criterio de tomar un 50% de humedad en el suelo se fundamentó porque
las plantas presentan reducción y alteraciones en sus relaciones hídricas con humedades
del suelo entre el 40 y 60 % (Costa-França et al., 2000; Segura-Monroy et al., 2015).
5.3.3 Diseño experimental y análisis de los datos
Se aplicó un diseño en bloques al azar con arreglo factorial donde el primer factor fue la
etapa fenológica en la que se inició el estrés por sequía y el segundo factor fueron los cinco
cultivares evaluados para un total de 15 tratamientos con cuatro repeticiones para un total
de 60 parcelas. Posteriormente, cuando se presentaron diferencias significativas en el
ANAVA, se empleó la prueba comparativa de medias de Tukey a P ≤ 0.05. Los datos se
analizaron usando el programa Statistix v 9.0 (software analítico, Tallahassee, FL, US).
Adicionalmente, se realizó un análisis de componentes principales utilizando el programa
InfoStat 2016 (Di Rienzo et al., 2016). Los resultados del análisis de varianza son
resumidos en la Tabla 5-1.

88 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
5.3.4 Contenido Relativo de Agua
Se realizaron dos muestreos del contenido relativo del agua (CRA) de la hoja al final de
cada periodo de estrés (15 días después del inicio del déficit hídrico, es decir a los 55 DDT
para la etapa vegetativa y 65 DDT para la etapa reproductiva). El contenido relativo de
agua fue determinado en hojas totalmente expandidas de la parte media del dosel. Las
hojas solamente fueron recolectadas al final del periodo de estrés hídrico para cada uno
de los genotipos de fríjol evaluados. El CRA fue calculado mediante la siguiente ecuación:
𝐶𝑅𝐴 = [𝑃𝐹 − 𝑃𝑆
𝑃𝑇 − 𝑃𝑆] × 100 (𝐸𝑐𝑢𝑎𝑐𝑖ó𝑛 1)
Siendo PF el peso fresco, PT el peso en turgencia medido después de 24 horas de
saturación en agua destilada a 4 °C en la oscuridad y PS es el peso seco determinado
después de 48 horas en un horno a 70°C.
5.3.5 Parámetros de rendimiento
La cosecha de los granos se realizó al final del ciclo del cultivo (155 DDE) de los genotipos
(granos con aproximadamente el 14% de humedad). Al momento de la cosecha, se
seleccionaron al azar cinco plantas individuales para la evaluación de los parámetros de
rendimiento, teniendo en cuenta el número de granos por vaina, el número de semillas por
vaina, número de vainas por planta, número de semillas por planta, rendimiento por planta
y rendimiento por hectárea.
5.3.6 Índice de tolerancia y susceptibilidad al estrés por sequía
Para la estimación de la susceptibilidad y tolerancia de los genotipos al estrés por sequía
se aplicaron las ecuaciones propuestas por Fischer y Maurer (1978) para la determinación
del Índice de Susceptibilidad al Estrés (SSI); Rosielle y Hamblin (1981) para la Tolerancia
(TOL), Productividad Media (MP) y Productividad Media Geométrica (GMP); Fernández

Capítulo 5 89
(1992) para el Índice de Tolerancia al Estrés (STI); Bouslama y Schapaugh, (1984) para el
Índice de estabilidad del rendimiento (YSI); Gavuzzi et al. (1997) para el Índice de
rendimiento (YI), Farshadfar et al. (2013) para la Media armónica (HM), Farshadfar y
Javadinia, (2011) para el Índice de Sensibilidad a la Sequía (SDI).
𝑆𝑆𝐼 = 1 − (
𝑌𝑆𝑌𝑃
)
1 − (�̅�𝑆
�̅�𝑃)
(𝐸𝑐𝑢𝑎𝑐𝑖ó𝑛 3)
𝑇𝑂𝐿 = 𝑌𝑃 − 𝑌𝑆 (𝐸𝑐𝑢𝑎𝑐𝑖ó𝑛 4)
𝑀𝑃 = (𝑌𝑃 + 𝑌𝑆)
2 (𝐸𝑐𝑢𝑎𝑐𝑖ó𝑛 5)
𝐺𝑀𝑃 = √(𝑌𝑆 × 𝑌𝑃) (𝐸𝑐𝑢𝑎𝑐𝑖ó𝑛 6)
𝑆𝑇𝐼 = (𝑌𝑃 × 𝑌𝑆)
(�̅�𝑃)2 (𝐸𝑐𝑢𝑎𝑐𝑖ó𝑛 7)
𝑌𝑆𝐼 = 𝑌𝑆
𝑌𝑃 (𝐸𝑐𝑢𝑎𝑐𝑖ó𝑛 8)
𝑌𝐼 = 𝑌𝑆
�̅�𝑃
(𝐸𝑐𝑢𝑎𝑐𝑖ó𝑛 9)
𝐻𝑀 = 2 ( 𝑌𝑆)( 𝑌𝑃)
( 𝑌𝑆 + 𝑌𝑃) (𝐸𝑐𝑢𝑎𝑐𝑖ó𝑛 10)
𝑆𝐷𝐼 = 𝑌𝑆 − 𝑌𝑃
𝑌𝑃 (𝐸𝑐𝑢𝑎𝑐𝑖ó𝑛 11)
Donde YP es el promedio del rendimiento del genotipo sin condición de estrés, YS es el
promedio del rendimiento del genotipo bajo condición de estrés �̅�𝑃 es el rendimiento
promedio de todos los genotipos sin condición de estrés y �̅�𝑠 es el promedio de rendimiento
de todos los genotipos bajo condiciones de estrés.

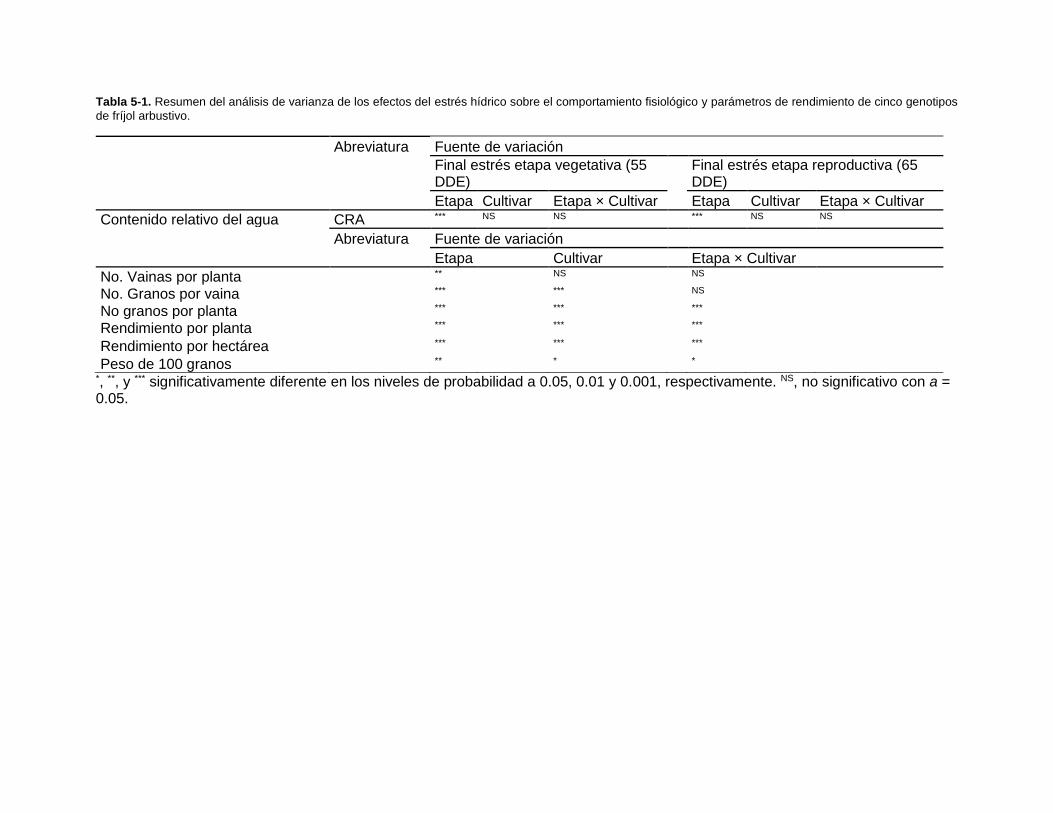
Tabla 5-1. Resumen del análisis de varianza de los efectos del estrés hídrico sobre el comportamiento fisiológico y parámetros de rendimiento de cinco genotipos
de fríjol arbustivo.
Abreviatura Fuente de variación
Final estrés etapa vegetativa (55 DDE)
Final estrés etapa reproductiva (65 DDE)
Etapa Cultivar Etapa × Cultivar Etapa Cultivar Etapa × Cultivar
Contenido relativo del agua CRA *** NS NS *** NS NS
Abreviatura Fuente de variación
Etapa Cultivar Etapa × Cultivar
No. Vainas por planta ** NS NS No. Granos por vaina *** *** NS
No granos por planta *** *** *** Rendimiento por planta *** *** ***
Rendimiento por hectárea *** *** ***
Peso de 100 granos ** * * *, **, y *** significativamente diferente en los niveles de probabilidad a 0.05, 0.01 y 0.001, respectivamente. NS, no significativo con a = 0.05.

92 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
5.4 Resultados y discusión
5.4.1 Humedad del suelo y Contenido Relativo de Agua
Resultados del contenido de humedad del suelo (CHS) y contenido relativo del agua CRA
se muestran en la Figura 5-1. En las plantas del tratamiento control estuvo alrededor 90%
de capacidad de campo durante el ensayo. Con respecto al CHS de los otros tratamientos,
se observó que este estuvo cercano al 55% al final de los 15 días de déficit hídrico iniciado
en la etapa vegetativa (55DDE). Asimismo, el CHS del tratamiento de estrés durante la
etapa reproductiva presentaba una humedad alrededor del 53.5% a los 65 DDE (Figura
5-1A). Los resultados obtenidos en la cuantificación de la humedad del suelo señalan que
un grupo de plantas estuvo con una menor disponibilidad de agua en algún momento de su
desarrollo. Igualmente, los resultados del Contenido Relativo de Agua (CRA) en la hoja son
resumidos en la Figura 5-1(B y C). Diferencias significativas solamente fueron observadas
para el factor de estatus hídrico (Tabla 5-1). En el muestreo de los 55 DDE, plantas que
iniciaron su periodo de estrés en la etapa vegetativa presentaron un CRA ̴70% en
comparación a las plantas control que presentaron valores cercanos al 85% a los 55 DDE.
Asimismo, las plantas que iniciaron a sufrir un déficit hídrico en la etapa reproductiva
mostraron un valor ̴ 77.43% en ese punto (Figura 5-1B). Cuando finalizó el periodo de
estrés de la fase reproductiva (65 DDE), las plantas control continuaron con un CRA del
82.57% mientras las plantas con condiciones de estrés en la etapa vegetativa fue
aproximadamente del 75.22% y un 73.34% en fase reproductiva (Figura 5-1C).
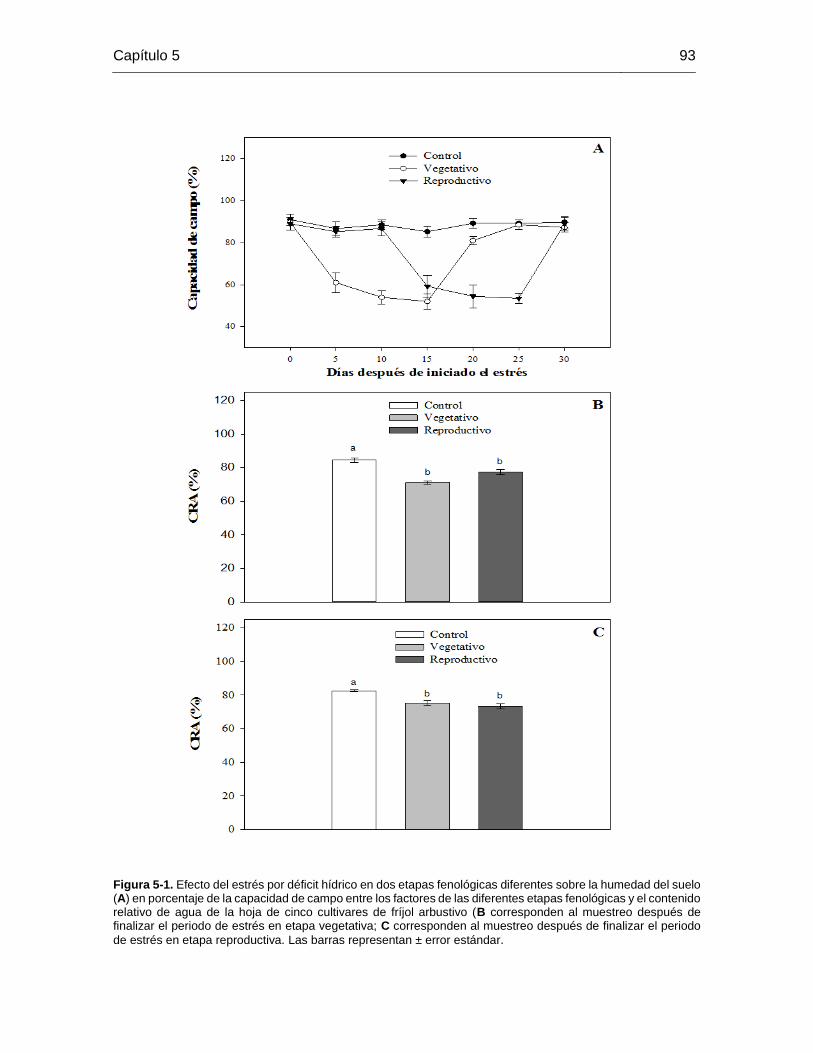
Capítulo 5 93
Figura 5-1. Efecto del estrés por déficit hídrico en dos etapas fenológicas diferentes sobre la humedad del suelo (A) en porcentaje de la capacidad de campo entre los factores de las diferentes etapas fenológicas y el contenido relativo de agua de la hoja de cinco cultivares de fríjol arbustivo (B corresponden al muestreo después de finalizar el periodo de estrés en etapa vegetativa; C corresponden al muestreo después de finalizar el periodo
de estrés en etapa reproductiva. Las barras representan ± error estándar.

94 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
5.4.2 Parámetros de rendimiento
La Tabla 5-2 resume las diferencias significativas en los parámetros de rendimiento para
cada uno de los cultivares bajo las condiciones de estrés evaluados. Una significancia de
P≤0.001 fue encontrada en la interacción Etapa × Cultivar para el número de granos por
planta, rendimiento por planta y rendimiento por hectárea, y una significancia de P≤0.05
para el peso de 100 granos. Sin embargo, para el número de granos por vaina se encontró
una significancia de P≤0.001 para el factor Cultivar, mientras que para el número de vainas
por planta solo se encontró diferencias significativas en el factor Etapa con P≤0.01 (Tabla
5-1). En general, un estrés hídrico de 15 días en la etapa vegetativa y reproductiva
disminuyen significativamente el número de granos por vaina (~3.80 en plantas control vs
~3.40 en plantas estresadas) y el número de vainas por planta (~7.30 en plantas control vs
~4.68 en plantas estresadas). Por otro lado, el número de granos por planta se vio afectado
negativamente en las dos etapas fenológicas evaluadas (~26.5 granos en plantas control
vs ~15.85 en plantas estresadas), aunque el cultivar Bacatá bajo condición de estrés por
déficit hídrico en la etapa vegetativa tuyo la mayor reducción en el número de granos,
pasando de ~25.50 a ~7.00 granos. Igualmente, fue el cultivar con la más baja producción
de granos cuando las plantas de los diferentes cultivares estuvieron bajo condición de
estrés por déficit hídrico en la etapa reproductiva (~12.33 granos por planta).
Adicionalmente, los que se afectaron en menor medida cuando el estrés inició en la etapa
vegetativa fueron Bachue, NUA35 y Cerinza, con una reducción del 30% (aproximadamente
18.42 granos por planta). Esta tendencia se mantuvo en el cultivar NUA35 cuando el déficit
hídrico inició en la etapa reproductiva, siendo el cultivar con mayor producción de granos
(cerca de 19.67 granos por planta), seguidamente del cultivar Cerinza, Bachue y Bianca
con 19.25, 17.50 y 13.75 granos por planta respectivamente (Tabla 5-2). En cuanto al peso
de 100 granos, del cultivar Bianca presentó el mayor rendimiento entre los cultivares que
se encontraron en condiciones control (72.03 g); y el cultivar Cerinza mostró el menor peso
(59.87g). No obstante, el cultivar Cerinza bajo condiciones de estrés por déficit hídrico en
la etapa vegetativa presentó un incremento del 20% en dicha variable comparado con las
plantas control (69.69 g), y mantuvo los valores cercanos a los encontrados en plantas

Capítulo 5 95
control cuando el estrés inició en la etapa reproductiva (59.48 g). Adicionalmente, los
cultivares Bacatá y Bianca presentaron las mayores reducciones en el peso de 100 granos
en las dos etapas evaluadas (46.54 g y 50.77 g respectivamente). Por otro parte, el cultivar
NUA35 no vario significativamente en el peso de 100 granos cuando las plantas tuvieron
influencia por déficit hídrico en la etapa vegetativa y reproductiva (Tabla 5-2).
En cuanto al rendimiento por hectárea, se observa que en general los cultivares bajo
condiciones control tienen un rendimiento cercano a 1427.75 Kg. Adicionalmente, el
rendimiento de los cultivares Cerinza, Bachue y NUA35 cuando estuvieron en condición de
déficit hídrico en la etapa vegetativa tuvieron la menor reducción, alcanzando una
producción de 1139.83 Kg, 901.04 Kg y 877.67 Kg, respectivamente y el rendimiento del
cultivar Bacatá en condición de déficit hídrico en la etapa vegetativa fue el más afectado,
con una reducción cercana al 80% (281.27 Kg). Esta tendencia se mantuvo cuando los
cultivares se encontraron en condiciones de estrés por déficit hídrico en la etapa
reproductiva, donde las plantas de NUA35, Cerinza y Bachue tuvieron el mayor rendimiento
(1074.88 Kg, 943.13 Kg y 780.62 Kg, respectivamente) y los rendimientos más bajos se
encontraron en los cultivares Bacatá y Bianca (472.78 Kg y 626.50 Kg, respectivamente).
El rendimiento por planta para todos los cultivares tuvo un comportamiento similar al
rendimiento por hectárea (Tabla 5-2).
En la Figura 5-2 se presenta el biplot para los primeros dos componentes principales, los
cuales representan el 94,1 % del total de la varianza. Las variables de rendimiento
evaluadas se representan mediante vectores, mientras que las variedades sometidas a
estrés en diferentes estados fisiológicos se identifican mediante puntos. Se puede apreciar
una clara separación de las muestras en dos grupos. El primer grupo correspondiente a las
muestras control, ubicadas hacia la parte positiva del componente 1; el segundo grupo,
ubicado en la parte negativa del primer componente principal, formado por las muestras
sometidas a estrés en las dos etapas vegetativas, con la excepción de Cz-E1 y NUA-E2.
Las variables de RtoP, RtoHa y GRP presentan gran similitud y tiene mayores valores en
las variedades Bl-C, NUA y Cerinza. Por su parte, las variedades de Ba-C y Bch-C
presentan menor RtoP, RtoHa y GRP que las otras tres variedades, pero tienen un Pcg
mayor.
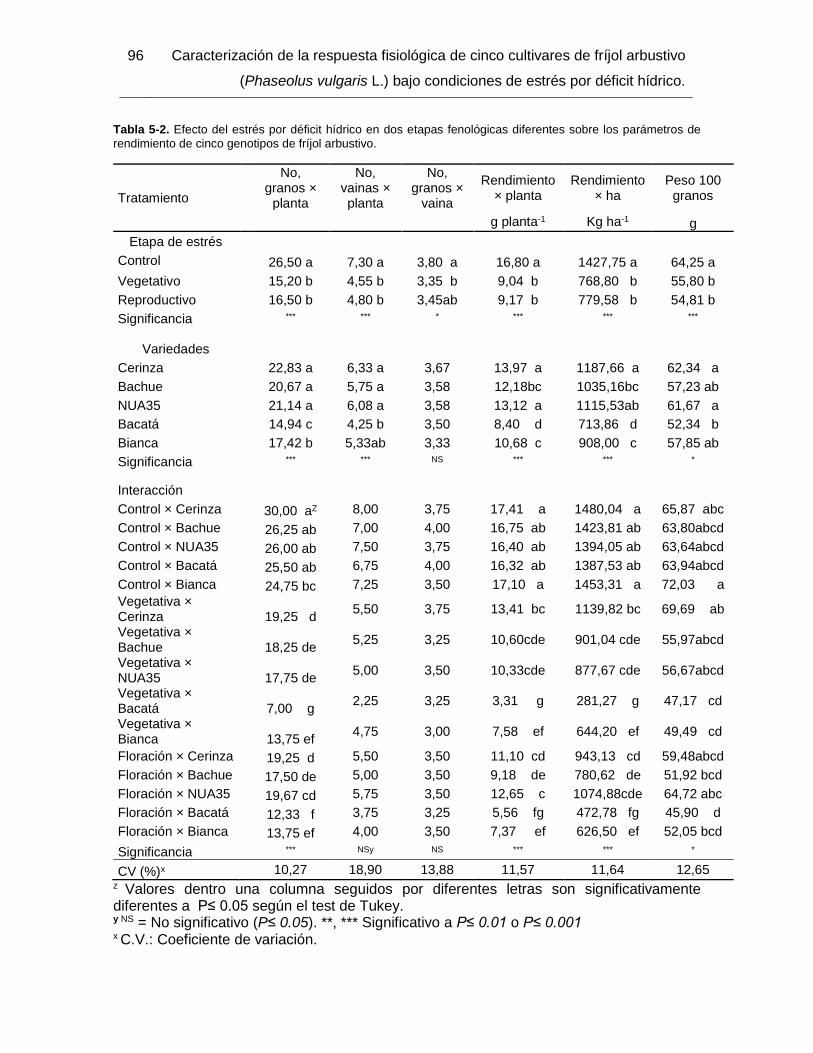
96 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
Tabla 5-2. Efecto del estrés por déficit hídrico en dos etapas fenológicas diferentes sobre los parámetros de
rendimiento de cinco genotipos de fríjol arbustivo.
Tratamiento
No, granos ×
planta
No, vainas × planta
No, granos ×
vaina
Rendimiento × planta
Rendimiento × ha
Peso 100 granos
g planta-1 Kg ha-1 g
Etapa de estrés
Control 26,50 a 7,30 a 3,80 a 16,80 a 1427,75 a 64,25 a
Vegetativo 15,20 b 4,55 b 3,35 b 9,04 b 768,80 b 55,80 b
Reproductivo 16,50 b 4,80 b 3,45ab 9,17 b 779,58 b 54,81 b
Significancia *** *** * *** *** ***
Variedades
Cerinza 22,83 a 6,33 a 3,67 13,97 a 1187,66 a 62,34 a
Bachue 20,67 a 5,75 a 3,58 12,18bc 1035,16bc 57,23 ab
NUA35 21,14 a 6,08 a 3,58 13,12 a 1115,53ab 61,67 a
Bacatá 14,94 c 4,25 b 3,50 8,40 d 713,86 d 52,34 b
Bianca 17,42 b 5,33ab 3,33 10,68 c 908,00 c 57,85 ab
Significancia *** *** NS *** *** *
Interacción
Control × Cerinza 30,00 aZ 8,00 3,75 17,41 a 1480,04 a 65,87 abc
Control × Bachue 26,25 ab 7,00 4,00 16,75 ab 1423,81 ab 63,80abcd
Control × NUA35 26,00 ab 7,50 3,75 16,40 ab 1394,05 ab 63,64abcd
Control × Bacatá 25,50 ab 6,75 4,00 16,32 ab 1387,53 ab 63,94abcd
Control × Bianca 24,75 bc 7,25 3,50 17,10 a 1453,31 a 72,03 a
Vegetativa × Cerinza 19,25 d 5,50 3,75 13,41 bc 1139,82 bc 69,69 ab
Vegetativa × Bachue 18,25 de 5,25 3,25 10,60cde 901,04 cde 55,97abcd
Vegetativa × NUA35 17,75 de 5,00 3,50 10,33cde 877,67 cde 56,67abcd
Vegetativa × Bacatá 7,00 g 2,25 3,25 3,31 g 281,27 g 47,17 cd
Vegetativa × Bianca 13,75 ef
4,75 3,00 7,58 ef 644,20 ef 49,49 cd
Floración × Cerinza 19,25 d 5,50 3,50 11,10 cd 943,13 cd 59,48abcd
Floración × Bachue 17,50 de 5,00 3,50 9,18 de 780,62 de 51,92 bcd
Floración × NUA35 19,67 cd 5,75 3,50 12,65 c 1074,88cde 64,72 abc
Floración × Bacatá 12,33 f 3,75 3,25 5,56 fg 472,78 fg 45,90 d
Floración × Bianca 13,75 ef 4,00 3,50 7,37 ef 626,50 ef 52,05 bcd
Significancia *** NSy NS *** *** *
CV (%)x 10,27 18,90 13,88 11,57 11,64 12,65 z Valores dentro una columna seguidos por diferentes letras son significativamente diferentes a P≤ 0.05 según el test de Tukey. y NS = No significativo (P≤ 0.05). **, *** Significativo a P≤ 0.01 o P≤ 0.001 x C.V.: Coeficiente de variación.
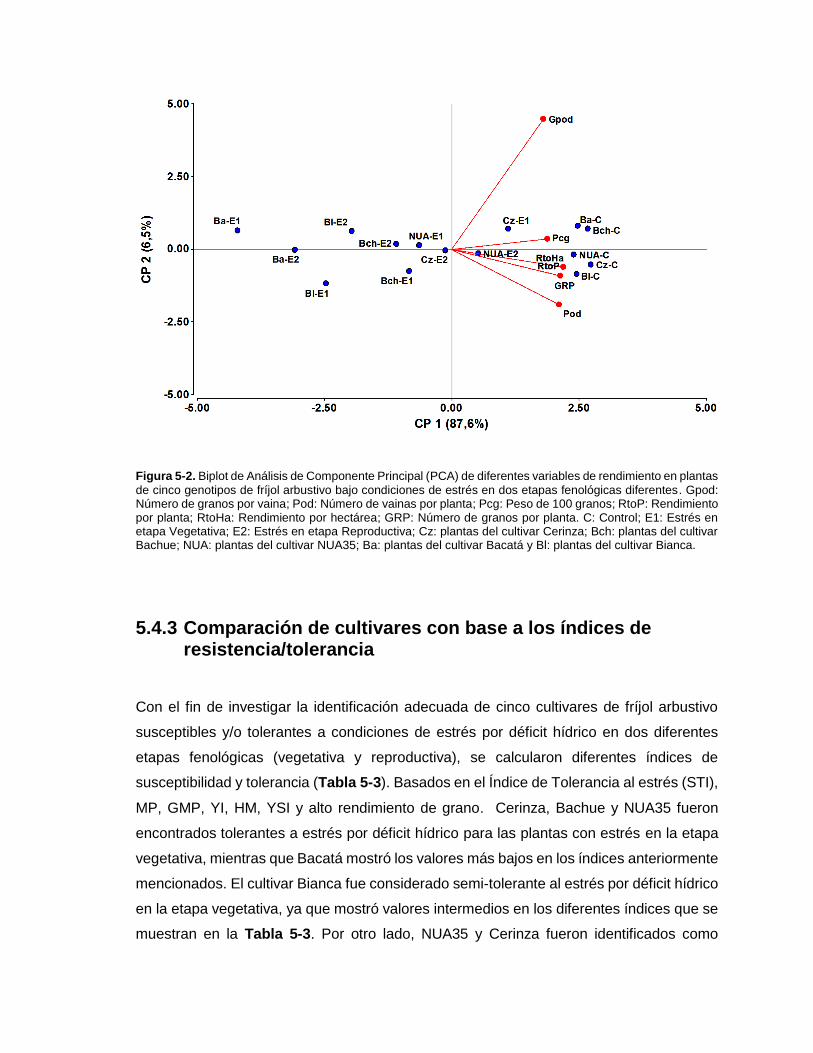
Figura 5-2. Biplot de Análisis de Componente Principal (PCA) de diferentes variables de rendimiento en plantas
de cinco genotipos de fríjol arbustivo bajo condiciones de estrés en dos etapas fenológicas diferentes. Gpod: Número de granos por vaina; Pod: Número de vainas por planta; Pcg: Peso de 100 granos; RtoP: Rendimiento por planta; RtoHa: Rendimiento por hectárea; GRP: Número de granos por planta. C: Control; E1: Estrés en etapa Vegetativa; E2: Estrés en etapa Reproductiva; Cz: plantas del cultivar Cerinza; Bch: plantas del cultivar Bachue; NUA: plantas del cultivar NUA35; Ba: plantas del cultivar Bacatá y Bl: plantas del cultivar Bianca.
5.4.3 Comparación de cultivares con base a los índices de resistencia/tolerancia
Con el fin de investigar la identificación adecuada de cinco cultivares de fríjol arbustivo
susceptibles y/o tolerantes a condiciones de estrés por déficit hídrico en dos diferentes
etapas fenológicas (vegetativa y reproductiva), se calcularon diferentes índices de
susceptibilidad y tolerancia (Tabla 5-3). Basados en el Índice de Tolerancia al estrés (STI),
MP, GMP, YI, HM, YSI y alto rendimiento de grano. Cerinza, Bachue y NUA35 fueron
encontrados tolerantes a estrés por déficit hídrico para las plantas con estrés en la etapa
vegetativa, mientras que Bacatá mostró los valores más bajos en los índices anteriormente
mencionados. El cultivar Bianca fue considerado semi-tolerante al estrés por déficit hídrico
en la etapa vegetativa, ya que mostró valores intermedios en los diferentes índices que se
muestran en la Tabla 5-3. Por otro lado, NUA35 y Cerinza fueron identificados como

98 Caracterización de la respuesta fisiológica de cinco cultivares de fríjol arbustivo
(Phaseolus vulgaris L.) bajo condiciones de estrés por déficit hídrico.
materiales tolerantes al estrés por déficit hídrico inició en la etapa reproductiva según el
STI, MP, GMP, YI, HM, YSI y el rendimiento de grano obtenido, mientras que los cultivares
Bachue y Bianca se categorizaron como materiales semi-tolerantes y el cultivar Bacatá
como susceptible (Tabla 5-3). Mismas tendencias fueron encontradas en genotipos de
trigo y maíz, identificando los genotipos más tolerantes por índices como el STI, MP, GMP
(Yang et al., 2002; Farshadfar et al., 2013; Naghavi et al., 2013).

Tabla 5-3. Índice de tolerancia y susceptibilidad de cinco genotipos de fríjol arbustivo bajo condiciones de estrés por sequía.
Genotipos YS YP SSI TOL MP GMP STI YSI YI HM SDI
Índice de tolerancia y susceptibilidad a estrés hídrico en etapa vegetativa
Cerinza 1,14 1,48 0,50 0,34 1,31 1,30 0,83 0,77 1,48 1,29 0,23
Bachue 0,90 1,42 0,80 0,52 1,16 1,13 0,63 0,63 1,17 1,10 0,37
NUA35 0,88 1,39 0,80 0,52 1,14 1,11 0,60 0,63 1,14 1,08 0,37
Bacatá 0,28 1,39 1,73 1,11 0,83 0,62 0,19 0,20 0,37 0,47 0,80
Bianca 0,64 1,45 1,21 0,81 1,05 0,97 0,46 0,44 0,84 0,89 0,56
Índice de tolerancia y susceptibilidad a estrés hídrico en etapa reproductiva
Cerinza 0,94 1,48 0,80 0,54 1,21 1,18 0,68 0,64 1,21 1,15 0,36
Bachue 0,78 1,42 1,00 0,64 1,10 1,05 0,55 0,55 1,00 1,01 0,45
NUA35 1,07 1,39 0,50 0,32 1,23 1,22 0,74 0,77 1,38 1,21 0,23
Bacatá 0,47 1,39 1,45 0,91 0,93 0,81 0,32 0,34 0,61 0,71 0,66
Bianca 0,63 1,45 1,25 0,83 1,04 0,95 0,45 0,43 0,80 0,88 0,57
Para SSI, TOL y SDI, los valores más bajos son deseables mientras que para YS, YP, MP, HM, GMP, YSI, YI y STI, los valores más altos son deseables.

5.4.4 Análisis de correlación
Con el fin de determinar los criterios de tolerancia al estrés por déficit hídrico más
convenientes, se calcularon los coeficientes de correlación entre YS, YP y otros índices
cuantitativos de la tolerancia a la sequía. En otras palabras, el análisis de correlación entre
los índices de tolerancia a la sequía y el rendimiento de grano pueden ser un buen criterio
para la selección de los mejores cultivares e índices utilizados (Tabla 5-4). La más alta
correlación positiva para las plantas con estrés en la etapa vegetativa fue observada entre
YS y YI al igual que entre el GMP y SDI (r = 1.00 con P≤0.001), mientras que la correlación
altamente negativa fue registrada entre SSI y YSI (r = -1.00 con P≤0.001). Sin embargo, la
más alta correlación positiva para las plantas con estrés en la etapa reproductiva se
registró entre YS y MP, MP y YI, GMP y YI, STI y YSI (r = 0.99 con P≤0.01), YS y GMP, YS
y STI, YS y YSI, YS y YI, YS y HM, TOL y SSI, MP y GMP, MP y STI, MP y YI, MP y HP,
GMP y STI, GMP y YI, GMP y HM, STI y YSI, STI y YI, STI y HM, YSI y YI, YI y HM (r =
0.99 con P≤0.001); entretanto que la correlación altamente negativa fue registrada entre
YS y SSI, TOL y YSI, TOL y YI, SSI y YSI, SSI y YI (r = -0.99 con P≤0.001). Esto concuerda
con lo reportado por Naghavi et al. (2013) y Farshadfar et al. (2013).

Capítulo 5 101
Tabla 5-4. Coeficiente de correlación entre los diferentes índices de tolerancia y susceptibilidad al estrés por déficit hídrico en cinco genotipos de fríjol arbustivo.
Índice de tolerancia y susceptibilidad al estrés por sequía en etapa vegetativa
YS TOL MP GMP SSI STI YSI YI HM SDI
YP 0,59 NS -0,51 NS 0,65 NS 0,61 NS -0,53 NS 0,64 NS 0,53NS 0,59 NS 0,58 NS 0,61 NS
YS -0,99*** 0,99*** 0,99 *** -0,99 *** 0,99*** 0,99*** 1,00*** 0,99*** 0,99 ***
TOL -0,98* -0,99 ** 0,99*** -0,99** -0,99*** -0,99*** -0,99** -0,99**
MP 0,99 *** -0,99 ** 0,99*** 0,99** 0,99*** 0,99*** 0,99***
GMP -0,99*** 0,99*** 0,99*** 0,99*** 0,99*** 1,00***
SSI -0,99*** -1,00*** -0,99*** -0,99*** -0,99***
STI 0,99*** 0,99*** 0,99*** 0,99***
YSI 0,99*** 0,99*** 0,99***
YI 0,99*** 0,99***
HM 0,99***
Índice de tolerancia y susceptibilidad al estrés por sequía en etapa reproductiva
YS TOL MP GMP SSI STI YSI YI HM SDI
YP 0,15 NS 0,26 NS 0,31 NS 0,27 NS -0,52 NS 0,23 NS 0,06NS 0,14NS 0,25NS 0,61NS
YS -0,98 * 0,99 ** 0,99 *** -0,99*** 0,99*** 0,99*** 0,99*** 0,99*** 0,83NS
TOL -0,94 * -0,95 * 0,99*** -0,97 ** -0,99*** -0,99*** -0,96* -0,73NS
MP 0,99*** -0,97 * 0,99*** 0,97** 0,99** 0,99*** 0,90*
GMP -0,97*** 0,99*** 0,98** 0,99** 0,99*** 0,89*
SSI -0,98 * -0,99*** -0,99*** -0,98** -0,77NS
STI 0,99** 0,99*** 0,99*** 0,87NS
YSI 0,99*** 0,98** 0,78NS
YI 0,99*** 0,82NS
HM 0,89*
NS = No significativo (P≤ 0.05). *, **, *** Significativo a P≤ 0.05, P≤ 0.01 o P≤ 0.001.

5.5 Conclusiones
• Los cultivares Bacatá y Bianca fueron los más susceptibles a condiciones de estrés
por déficit hídrico tantos en estado vegetativo como reproductivo, ya que
presentaron una mayor disminución de los componentes de rendimiento (granos
por planta, peso de 100 granos, rendimiento por planta y por hectárea), lo cual se
asoció con los menores índices de tolerancia (SSI, TOL, STI, YSI y SDI)

6. Discusión final
El fríjol (Phaseolus vulgaris L.) es una especie ampliamente cultivada en regiones
principalmente del África y América Latina, con gran importancia en la seguridad
alimentaria debido a sus altos contenidos de proteína y minerales (Beebe et al., 2010); y
presenta alta sensibilidad a la sequía debido a que posee un sistema radical poco profundo
(Beebe et al., 2013). Por consiguiente, la producción de los cultivos de fríjol en épocas
secas está limitada por la condición de déficit hídrico (White e Izquierdo., 1991). Esta
investigación tuvo como objetivo evaluar las respuestas fisiológicas y bioquímicas de cinco
cultivares de fríjol arbustivo bajo condiciones de estrés por déficit hídrico con el motivo de
conocer posibles mecanismos potenciales de respuesta de estos cultivares frente al estrés.
Los resultados presentados en los capítulos 2, 3, 4 y 5 del estudio permiten afirmar que de
los cinco cultivares evaluados. Los resultados son consistentes con el concepto de
tolerancia que plantea que un genotipo es tolerante cuando tiene la capacidad para crecer,
reproducirse y producir óptimo rendimiento bajo condiciones de restricción hídrica (Farooq
et al., 2009; Farooq et al., 2013).
Por otro lado, los resultados expuestos en este trabajo aportan información valiosa que
permite explicar los mecanismos utilizados por los cultivares de fríjol para aumentar su
tolerancia al déficit hídrico, mitigar el daño ocasionado por el estrés y proteger los
diferentes procesos fisiológicos sin disminuir el rendimiento. Adicionalmente, brinda
elementos importantes para la comprensión de cómo las variedades regulan y mitigan el
efecto negativo del déficit hídrico por 15 días; mostrando las diferencias fisiológicas,
bioquímicas y de rendimiento entre los cultivares evaluados y permiten indicar la existencia
diferencias genotípicas en la capacidad para generar diferentes respuestas para
sobrellevar un estrés.

10
4
Título de la tesis o trabajo de investigación
Para tolerar el período de déficit hídrico los cultivares de P. vulgaris L. respondieron
mediante la inducción de respuestas comunes relacionadas con los mecanismos
morfológicos de evitación, respuestas relacionadas con los mecanismos fisiológicos de
estabilidad de la membrana celular, ajuste osmótico y respuesta oxidativa. Además, el
mecanismo de evitación se evidenció a partir de respuestas como la reducción de la
conductancia estomática como estrategia de mitigación de la pérdida de agua
disminuyendo la transpiración (Xu et al., 2010; Farooq et al., 2009); permitiendo conservar
el contenido de agua a nivel celular sin interrumpir la actividad fisiológica durante el déficit
hídrico. Lo anterior fue acompañado de un incremento en el uso eficiente del agua
intrínseco, siendo más alta en aquellos cultivares con rasgos de tolerancia como Cerinza
en la etapa vegetativa y NUA35 en la etapa reproductiva, y ésta se relaciona con los altos
valores de rendimiento bajo condiciones de estrés (Boonjung and Fukai., 1996-1; Boutra
et al., 2010)
En resumen, la mayor capacidad de respuesta de los cultivares Cerinza y NUA35 para
tolerar un déficit hídrico de 15 días con respecto a los cultivares Bacatá, Bianca y Bachue
se relacionan con aquellas respuestas fisiológicas tales como: menor reducción en la
fotosíntesis neta, incremento en la concentración de clorofila total, mayor incremento del
Uso Eficiente del Agua intrínseco, menor transpiración, altos niveles de Carotenoides,
menor fuga de electrolitos, menor producción de malondialdehido y mayor acumulación de
prolina. La menor capacidad de respuesta del genotipo Bacatá y Bianca para tolerar un
déficit hídrico de 15 días está influenciada por una mayor reducción de la tasa fotosintética,
disminución en el WUEi, bajos niveles en el contenido de carotenoides y prolina, mayor
incremento MDA.
La mayor capacidad de respuesta a déficit hídrico presentada por Cerinza en la etapa
vegetativa y NUA35 en la etapa reproductiva se puede asociar con los índices de tolerancia
más altos encontrados en un mayor número de parámetros tanto fisiológicos como
bioquímicos y de componentes de rendimiento como la producción de granos por planta,
rendimiento por planta y por hectárea y el peso de cien granos con respecto a Bacatá y
Bianca (Tabla 5-3).

Anexo B. Nombrar el anexo B de acuerdo con su contenido 105
La información presentada en este trabajo aumenta el conocimiento acerca de los
mecanismos implicados en la tolerancia a la sequía de las variedades colombianas de
fríjol, considerando que para producir genotipos resistentes a sequía es necesario
comprender las respuestas de las plantas a estrés. También puede aportar para desarrollar
nuevas estrategias metodológicas que contribuyen para diferenciar los cultivares
tolerantes y/o susceptibles a estrés, utilizando parámetros fisiológicos y de rendimiento.
Finalmente, en la Figura 8-1se muestra un esquema que explica a manera general las
respuestas de las plantas de fríjol arbustivo evaluadas a condiciones de déficit hídrico, las
cuales incluyen la evitación, escape y tolerancia.
7. Conclusiones y recomendaciones
7.1 Conclusiones
1. El uso de variables relacionadas a la fase lumínica de la Pn como el contenido de
pigmentos fotosintéticos y parámetros de la fluorescencia de clorofila a mostraron
una utilidad en la selección y/o caracterización de genotipos de frijol arbustivo déficit
hídrico.
2. Un periodo de estrés hídrico de 20 días causo afectaciones negativas en las plantas
de fríjol arbustivo cultivar Cerinza. Lo anterior se evidencia en la caída de la tasa
de fotosíntesis, reducción del Fv/Fm, Y(II) y qP en el fotosistema II, incremento del
NPQ, fuga de electrolitos, MDA y niveles de prolina. Asimismo, periodos de 5 y 10
días de duración del estrés hídrico no genera efectos severos sobre las variables
fisiológicas de la especie estudiada.
3. Un periodo de estrés hídrico de 15 días con PEG 6000 en la etapa vegetativa causo
afectaciones negativas en las plantas de los diferentes cultivares de fríjol arbustivo,
especialmente en los cultivares Bacatá y Bianca. Lo anterior se evidencia en la
caída de la tasa de fotosíntesis, reducción de la masa seca, del Fv/Fm en el
fotosistema II, incremento de la fuga de electrolitos, MDA y niveles de prolina.

10
6
Título de la tesis o trabajo de investigación
4. Un periodo de estrés hídrico de 15 días en la etapa vegetativa y reproductiva causo
afectaciones negativas en las plantas de los diferentes cultivares de fríjol arbustivo,
especialmente en los cultivares Bacatá y Bianca en la etapa vegetativa. Lo anterior
se evidencia en la caída de la tasa de fotosíntesis y conductancia estomática, del
Fv/Fm y qP en el fotosistema II, incremento del NPQ, MDA y niveles de prolina.
5. Los cultivares NUA35, Bachue y Cerinza durante el periodo de déficit hídrico
presentaron un aumento en el contenido de carotenoides, la eficiencia en el uso del
agua intrínseca y prolina temprana, indicando su alto potencial de respuesta a
estrés.
6. En los cinco cultivares el rendimiento potencial fue modificado por el déficit hídrico,
posiblemente porque hubo daño al aparato fotosintético; lo cual podría ser un
indicador de la baja capacidad de las plantas de fríjol para mantener el fotosistema
II funcional bajo estrés hídrico, siendo considerado un importante rasgo fisiológico
en la tolerancia a la sequía.
7. Asimismo, se identificó que el cultivar Bacatá es uno de los cultivares más
susceptibles a condiciones de estrés por déficit hídrico, lo cual se evidencia en los
diferentes índices de tolerancia y/o susceptibilidad.
8. Los índices de tolerancia (SSI, TOL, STI, YSI y SDI) fueron una herramienta
interesante en la caracterización del rendimiento de diferentes cultivares bajo
condiciones de déficit hídrico.
9. Este trabajo permitió caracterizar en general un grupo respuestas fisiológicas de
cultivares de frijol arbustivo a déficit hídrico a lo largo de su desarrollo.
7.2 Recomendaciones
1. A pesar de que varios de los mecanismos fisiológicos de tolerancia a un período de
15 días de déficit hídrico fueron explicados en este documento para los cultivares
Cerinza, Bachue, NUA35, Bacatá y Bianca, se recomienda hacer nuevos estudios
en los que se evalúe el efecto de diferentes períodos de déficit hídrico en etapas
fenológicas distintas tanto en invernadero como en campo para complementar los
resultados obtenidos en este estudio.
2. Es importante realizar estudios de las respuestas fisiológicas y bioquímicas en
plantas de diversos cultivares de fríjol arbustivo en diferentes etapas fenológicas.

Anexo B. Nombrar el anexo B de acuerdo con su contenido 107
3. Es necesario incluir nuevas variables bioquímicas como la producción de prolina y
MDA en tejidos de la raíz, como posibles marcadores del estrés, ya que estudios
en este nivel no se han realizado en fríjol.
4. El uso de los índices de tolerancia y/o susceptibilidad al estrés por déficit hídrico en
plantas de diversos cultivares de fríjol arbustivo en diferentes etapas fenológicas
pueden ser considerados marcadores de clasificación de susceptibilidad a la
sequía.
5. Realizar curvas de respuesta a la luz y CO2 con el fin de determinar puntos de
compensación de la fotosíntesis.
6. Caracterizar la actividad antioxidante enzimática y no enzimática como respuesta
al déficit hídrico.
7. Estudiar el comportamiento del uso eficiente de la radiación bajo condiciones de
déficit hídrico.

10
8
Título de la tesis o trabajo de investigación
8. Modelo conceptual
Este trabajo permitió caracterizar en general un grupo respuestas fisiológicas de cultivares
de frijol arbustivo a déficit hídrico a lo largo de su desarrollo. Estas respuestas son
resumidas en el modelo conceptual que se muestra en la Figura 8-1
En éste modelo se puede apreciar que dentro de las primeras respuestas de las plantas
de fríjol ante una condición de déficit hídrico se da a nivel de raíz, donde se han reportado
la recepción de señales a la sequía y se generan la producción de la hormona ácido
abscísico (ABA), la cual está involucrada en el mecanismo de apertura y cierre de estomas.
Seguidamente se da una reducción de la conductancia estomática, y por ende una
reducción de la transpiración y de la fotosíntesis neta, con el fin de incrementar la eficiencia
en el uso del agua. Dichas respuestas están relacionadas con la estrategia de la evitación
a la sequía, la cual ha sido reportada ampliamente en especies como el arroz, trigo, maíz
entre otras. Adicionalmente, se evidencia pérdida del turgor de la planta reflejada en una
reducción del contenido relativo de agua en la hoja, y estrés oxidativo por la producción de
especies reactivas de oxígeno (ROS), que generan daños a nivel de permeabilidad de
membranas. Ante ésta situación, dos de las respuestas de las plantas de fríjol arbustivo
que se encontraron en nuestro estudio relacionadas con el mecanismo de tolerancia fue la
producción de prolina como una estrategia de ajuste osmótico y la producción de
carotenoides, la cual ha sido reportada como una respuesta antioxidante de tipo no
enzimática. Por otro lado, debido a la reducción de la tasa de fotosíntesis, lo cual se
evidencia en una reducción de la acumulación de materia seca total, especialmente en los
órganos de la parte aérea. Igualmente, se evidencia una removilización de asimilados, en
donde cultivares con características de tolerancia al déficit hídrico mantienen los
parámetros de rendimiento en niveles cercanos a los encontrados en plastas bajo
condiciones control, lo cual se evidencia en los diferentes índices de tolerancia y/o
susceptibilidad al estrés por sequía.

Figura 8-1. Modelo conceptual de las respuestas de las plantas de fríjol arbustivo (Phaseolus vulgaris L.) a condiciones de estrés por déficit hídrico.


Bibliografía
Acosta-Gallegos, J. y J.W. White. 1995. Phenological Plasticity as an Adaptation by
Common Bean to Rainfed Environments. Crop Science, 35(1).
Ahmed, J. and M. Hassan. 2011. Evaluation of seedling proline content of wheat genotypes
in relation to heat tolerance. Bangladesh Journal of Botany 40, 17-22.
Amede, T., S. Schubert and K. Stahr. 2003. Mechanisms of drought resistance in grain
legumes I: Osmotic adjustment. SINET: Ethiopian Journal of Science, 26(1), 37-46.
Amede, T. and S. Schubert. 2003. Mechanisms of drought resistance in grain legumes II:
Stomatal regulation and root growth. Ethiopian Journal of Science, 26, 137-144.
Aranjuelo, I., G. Molero, G. Erice, J.C. Avice and S. Nogués. 2011. Plant physiology and
proteomics reveals the leaf response to drought in alfalfa (Medicago sativa L.).
Journal of Experimental Botany, 62(1), 111-123.
Ashraf M. and P.J.C. Harris. 2013. Photosynthesis under stressful environments: An
overview. Photosynthetica 51:163–190.
Bates, L., R. Waldren and I. Teare. 1973. Rapid determination of free proline for water-
stress studies. Plant and Soil 39, 205-207.
Beebe, S. E., I. M. Rao, M. W. Blair and J. A. Acosta-Gallegos. 2010. Phenotyping common
beans for adaptation to drought. In: J. M. Ribaut and P. Monneveux (eds.) Drought
phenotyping in crops: from theory to practice. Generation Challenge Program Special
Issue on Phenotyping, pp. 311-334
Beebe, S. E. 2012. Common bean breeding in the tropics. Plant Breed. Rev. 36, 357–426.
Beebe, S.E., I.M. Rao, M.W. Blair and J.A. Acosta-Gallegos. 2013. Phenotyping common
beans for adaptation to drought. Frontiers in physiology 4.
Blum, A. and W.R. Jordan. 1985. Breeding crop varieties for stress environments. Critical
Reviews in Plant Sciences 2, 199-238.

11
2
Título de la tesis o trabajo de investigación
Boonjung, H. and S. Fukai. 1996. Effects of soil water deficit at different growth stages on
rice growth and yield under upland conditions. 1. Growth during drought. Field crops
research, 48(1), 37-45.
Boonjung, H. and S. Fukai. 1996. Effects of soil water deficit at different growth stages on
rice growth and yield under upland conditions. 2. Phenology, biomass production and
yield. Field Crops Research, 48(1), 47-55.
Bouslama M and W.T. Schapaugh. 1984. Stress tolerance in soybean. Part 1: evaluation
of three screening techniques for heat and drought tolerance. Crop Sci J 24: 933-937
Broughton, W.J., G. Hernández, M. Blair, S. Beebe, P. Gepts and J. Vanderleyden. 2003.
Beans (Phaseolus spp.) – model food legumes. Plant Soil 252, 55–128.
Boutra T, A. Akhkha, A. Al-Shoaibi and A.M. Alhejeli. 2010. Effect of water stress on growth
and water use efficiency (WUE) of some wheat cultivars (Triticum durum) grown in
Saudi Arabia. J Taibah Univ Sci 3: 39-48
Cakir, R. 2004. Effect of water stress at different development stages on vegetative and
reproductive growth of corn. Field Crops Research, 89(1), 1-16.
Cao, Y.-Y., Duan, H., Yang, L.-N., Wang, Z.-Q., Zhou, S.-C., Yang, J.-C., 2008. Effect of
heat stress during meiosis on grain yield of rice cultivars differing in heat tolerance
and its physiological mechanism. Acta Agronomica Sinica 34, 2134-2142.
Cardona-Ayala, C., A. Jarma-Orozco y H. Araméndiz-Tatis. 2013. Mecanismos de
adaptación a sequía en caupí (Vigna unguiculata (L.) Walp.). Una revisión. Revista
Colombiana de Ciencias Hortícolas, 7(2), 277-288.
Castonguay, Y. and A.H. Markhart. 1991. Saturated Rates of Photosysnthesis in Water-
Stressed Leaves of Common Bean and Tepary Bean. Crop Science, 31(6).
Chaves, M. M., J.S. Pereira, J. Maroco, M.L. Rodrigues, C.P.P. Ricardo, M.L. Osório, I.
Carvalho, T. Faria and C. Pinheiro. 2002. How plants cope with water stress in the
field? Photosynthesis and growth. Annals of botany, 89(7), 907-916.
Comstock, J. and J. Ehleringer. 1993. Stomatal Response to Humidity in Common Bean
(Phaseolus vulgaris): Implications for Maximum Transpiration Rate, Water-Use
Efficiency and Productivity. Functional Plant Biology, 20(6), 669-691.
Comunidad Andina. 2009. Atlas de las dinámicas del territorio andino: población y bienes
expuestos a amenazas naturales. En: Por encima de la tierra - y cuando deja de

Bibliografía 113
llover. Feriva S.A. Cali Colombia. 185 pp. Consultado en diciembre de 2015:
http://www.comunidadandina.org/public/Atlas_12_Cuando_deja_de_llover.pdf
Comunidad Andina. 2009. Atlas de las dinámicas del territorio andino: población y bienes
expuestos a amenazas naturales. En: Por encima de la tierra - y cuando deja de
llover. Feriva S.A. Cali Colombia. 185 pp. Consultado en marzo de 2016:
http://www.comunidadandina.org/public/Atlas_12_Cuando_deja_de_llover.pdf
Costa França, M. G., A.T. Pham Thi, C. Pimentel, R.O. Pereyra Rossiello, Y. Zuily-Fodil
and D. Laffray. 2000. Differences in growth and water relations among Phaseolus
vulgaris cultivars in response to induced drought stress. Environmental and
Experimental Botany, 43(3), 227-237.
Cousins, A. B., N.R, Adam, G.W. Wall, B.A. Kimball, P.J. Pinter, M.J. Ottman, S.W. Leavitt
and A.N. Webber. 2002. Photosystem II energy use, non‐photochemical quenching
and the xanthophyll cycle in Sorghumbicolor grown under drought and free‐air CO2
enrichment (FACE) conditions. Plant, Cell & Environment, 25(11), 1551-1559.
Crafts, A. S., and C.E. Crisp. 1971. Phloem transport in plants. San Francisco, USA, WH
Freeman & Co.
DeLaat, D., C. Colombo, A. Chiorato and S. Carbonell. 2014. Induction of ferritin synthesis
by water deficit and iron excess in common bean (Phaseolus vulgaris L.). Molecular
Biology Reports, 41(3), 1427-1435.
Di Rienzo J.A., F. Casanoves, M.G. Balzarini, L. Gonzalez, M. Tablada y C.W. Robledo.
InfoStat versión 2016. Grupo InfoStat, FCA, Universidad Nacional de Córdoba,
Argentina. URL http://www.infostat.com.ar
Dias, M. C. and W. Brüggemann. 2010. Limitations of photosynthesis in Phaseolus vulgaris
under drought stress: gas exchange, chlorophyll fluorescence and Calvin cycle
enzymes. Photosynthetica, 48(1), 96-102.
Dowkiw, A., G. Wright, A. Cruickshank and R. Redden. 2000. Indirect selection for drought
resistance: a pilot study. ACIAR Food Legume Newslett, 31, 4-10.
Dutta Gupta, S., R.M. Auge, P.D. Denchev and B.V. Conger. 1995. Growth, proline
accumulation and water relations in NaCl-selected and non-selected callus lines of
Dactylis glomerata. Environ. Exp. Bot. 35, 83–92.

11
4
Título de la tesis o trabajo de investigación
Efeoğlu, B., Y. Ekmekci and N. Cicek. 2009. Physiological responses of three maize
cultivars to drought stress and recovery. South African Journal of Botany, 75(1), 34-
42.
Ehleringer, J. R., S. Klasen, C. Clayton, D. Sherrill, M. Fuller-Holbrook, F. Qing-nong and
T.A. Cooper. 1991. Carbon Isotope Discrimination and Transpiration Efficiency in
Common Bean. Crop Science, 31(6).
FAO, 2007. Manual Técnico de Buenas Prácticas Agrícolas en la producción de fríjol
voluble. 166 pp. En: http://www.fao.org.co/manualfrijol.pdf, Consultado en noviembre
de 2015.
FAOSTAT (2016) Production 2012. Food and Agriculture Organization, Roma. Web site at
http://www.fao.org., consultado en marzo de 2016
Farooq, M., A. Wahid, N. Kobayashi, D., Fujita and S.M.A. Basra. 2009. Plant drought
stress: effects, mechanisms and management. In Sustainable Agriculture (pp. 153-
188). Springer Netherlands.
Farooq, M., M. Hussain, A. Wahid, K.H.M Siddique. 2012. Drought stress in plants: an
overview. In: Aroca, R. (Ed.), Plant Responses to Drought Stress. Springer, Berlin
Heidelberg, pp. 1–33.
Farshadfar E and J. Javadinia. 2011. Evaluation of Chickpea (Cicer arietinum L.) genotypes
for drought tolerance. Seed and Plant Improv J 27(4): 517-537 (in Persian)
Farshadfar, E., R. Mohammadi, M. Farshadfar and S. Dabiri. 2013. Relationships and
repeatability of drought tolerance indices in wheat-rye disomic addition lines.
Australian Journal of Crop Science, 7(1), 130.
Feller, C., H. Bleiholder, L. Buhr, H. Hack, M. Hess, R. Klose, U. Meier, R. Stauss, T. Van
den Boom and E. Weber. 1995b: Phänologische Entwicklungsstadien von
Gemüsepflanzen: II. Fruchtgemüse und Hülsenfrüchte. Nachrichtenbl. Deut.
Pflanzenschutzd. 47, 217-232
Fenalce, 2012. Fondo Nacional de Leguminosas; Informe de Gestión año 2012. En:
www.fenalce.org_arc/i/os/e/nl2012.pdf, consulta: Julio de 2013.
Fenalce, 2016. Fondo Nacional de Leguminosas; Informe de Gestión año 2015. En:
http://www.fenalce.org/nueva/plantillas/arch_web/Salida_de_cosecha_leguminosas.
pdf, consulta: Marzo de 2016.

Bibliografía 115
Fernandez, G.C.J. 1992. Effective selection criteria for assessing stress tolerance. In: Kuo
CG (ed) Proceedings of the International Symposium on Adaptation of Vegetables
and Other Food Crops in Temperature and Water Stress, Publication, Tainan, Taiwan
Fischer R.A. and R. Maurer. 1978. Drought resistance in spring wheat cultivars. Australian
Journal of Agricultural Research. 29, 897-912.
Foster, E. F., A. Pajarito and J. Acosta-Gallegos. 1995. Moisture stress impact on N
partitioning, N remobilization and N-use efficiency in beans (Phaseolus vulgaris).
The Journal of Agricultural Science, 124(01), 27-37.
Frahm, M. A., J.C. Rosas, N. Mayek-Pérez, E. López-Salinas, J.A. Acosta-Gallegos and
J.D. Kelly. 2004. Breeding beans for resistance to terminal drought in the Lowland
tropics. Euphytica, 136(2), 223-232.
Gavuzzi P, F. Rizza, M. Palumbo, R.G. Campaline, G.L. Ricciardi, B. Borghi. 1997.
Evaluation of field and laboratory predictors of drought and heat tolerance in winter
cereals. Plant Sci 77:523-531.
Gebeyehu, S. 2006. Physiological response to drought stress of common bean (Phaseolus
vulgaris L.) genotypes differing in drought resistance. (Ph.D), Justus-Liebig-
Universität Giessen, Giessen, Germany. Retrieved from http://geb.uni-
giessen.de/geb/volltexte/2006/3726
Gepts, P., and D.G. Debouck. 1991. “Origin, domestication, and evolution of the common
bean (Phaseolus vulgaris L),” in Common Beans: Research for Crop Improvement,
eds A. van Schoonhoven and O. Voysest (Wallingford, UK: CAB International), 7–53.
Ghanbari, A. A., S.H. Mousavi, A.M. Gorji and I. Rao. 2013. Effects of water stress on
leaves and seeds of bean (Phaseolus vulgaris L.). Turkish Journal Of Field Crops,
18(1), 73-77.
Gomes, A. A., A.P. Araújo, R.O. Rossiello and C. Pimentel. 2000. Acumulação de
biomassa, características fisiológicas e rendimento de grãos em cultivares de
feijoeiro irrigado e sob sequeiro. Pesquisa Agropecuária Brasileira, 35, 1927-1937.
Gorbe, E., and A. Calatayud. 2012. Applications of chlorophyll fluorescence imaging
technique in horticultural research: A review. Scientia Horticulturae, 138, 24-35.
Guimarães, C.M., O. Brunini and L.F. Stone. 1996. Adaptação do feijoeiro (Phaseolus
vulgaris L.) à seca. I. Densidade e eficiência radicular. Pesquisa Agropecuária
Brasileira, 31(6), 393-399.

11
6
Título de la tesis o trabajo de investigación
Herbinger K, M. Tausz, A. Wonisch, G. Soja, A. Sorger and D. Grill. 2002 Complex
interactive effects of drought and ozone stress on the antioxidant defence systems of
two wheat cultivars. Plant Physiol. Biochem. 40: 691–696
Hoagland, D. R., and D.I. Arnon. 1950. The water-culture method for growing plants without
soil. Circular. California Agricultural Experiment Station, 347(2nd edit).
Hodges, D.M., DeLong, J.M., Forney, C.F., Prange, R.K., 1999. Improving the thiobarbituric
acid-reactive-substances assay for estimating lipid peroxidation in plant tissues
containing anthocyanin and other interfering compounds. Planta 207, 604-611.
Holmgren, M., M. Scheffer, E. Ezcurra, J.R. Gutiérrez, and G.M.J. Mohren. 2001. El Niño
effects on the dynamics of terrestrial ecosystems. Trends in Ecology and Evolution
16: 89-94.
Hong-Bo, S., C. Xiao-Yan, C. Li-Ye, Z. Xi-Ning, W. Gang, Y. Yong-Bing, Z. Chang-Xing and
H. Zan-Min. 2006. Investigation on the relationship of proline with wheat anti-drought
under soil water deficits. Colloids and Surfaces B: Biointerfaces, 53(1), 113-119.
Hsiao, T. C. 1973. Plant responses to water stress. Annual review of plant physiology.
24(1), 519-570.
Huang, Z., Z. Zou, C. He, Z. He, Z. Zhang and J. Li. 2011. Physiological and photosynthetic
responses of melon (Cucumis melo L.) seedlings to three Glomus species under
water deficit. Plant and soil, 339(1-2), 391-399.
IICA-RED SICTA-COSUDE. 2009, Guía técnica para el cultivo de frijol en los municipios
de Santa Lucía, Teustepe y San Lorenzo del Departamento de Boaco, Nicaragua
Irfan, M., Hayat, S., Hayat, Q., Afroz, S., Ahmad, A., 2010. Physiological and biochemical
changes in plants under waterlogging. Protoplasma 241, 3-17.
Jiang, M., Zhang, J., 2001. Effect of abscisic acid on active oxygen species, antioxidative
defence system and oxidative damage in leaves of maize seedlings. Plant and Cell
Physiology 42, 1265-1273.
Jones, H. G., and F. Tardieu. 1998. Modelling water relations of horticultural crops: a
review. Scientia Horticulturae, 74(1), 21-46.
Jones, H.G. 1985. Partitioning stomatal and non-stomatal limitations to photosynthesis.
Plant Cell Environ 8:95–104

Bibliografía 117
Kahn, B., Stoffella, P., Sandsted, R., Zobel, R., 1985. Influence of flooding on root
morphological components of young black beans. Journal of the American Society
for Horticultural Science 110.
Kalavacharla, V., Z. Liu, B.C. Meyers, J. Thimmapuram and K. Melmaiee. 2011.
Identification and analysis of common bean (Phaseolus vulgaris L.) transcriptomes
by massively parallel pyrosequencing. BMC plant biology, 11(1), 1
Kao, W.Y., J.R. Ehleringer and J.P. Comstock. 1994. Variation in leaf movements among
common bean cultivars. Crop Science, 34(5), 1273-1278.
Kaplan, L. 1981. What is the origin of the common bean? Economic Botany 35: 240-254.
Keyvan, S. 2010. The effects of drought stress on yield, relative water content, proline,
soluble carbohydrates and chlorophyll of bread wheat cultivars. J. Anim. Plant Sci,
8(3), 1051-1060.
Khakwani, A. A., M.D. Dennett and M. Munir. 2011. Drought tolerance screening of wheat
varieties by inducing water stress conditions. Songklanakarin J. Sci. Technol, 33(2),
135-142.
Kocheva, K. and G. Georgiev. 2003. Evaluation of the reaction of two contrasting barley
(Hordeum vulgare L.) cultivars in response to osmotic stress with PEG 6000.
Bulgarian Journal of Plant Physiology. 49, 290-294.
Korir, P.C., J.O. Nyabundi and P.K. Kimurto. 2006. Genotypic response of common bean
(Phaseolus vulgaris L.) to moisture stress conditions in Kenya. Asian Journal of Plant
Sciences.
Kumar, M.S., S.S. Chakravarthy, P.R. Babu, K.V. Rao and V.D. Reddy. 2015. Classification
of cytochrome P450s in common bean (Phaseolus vulgaris L.). Plant Systematics
and Evolution, 301(1), 211-216.
Lacape, M.J., J. Wery and D.J.M Annerose. 1998. Relationships between plant and soil
water status in five field-grown cotton (Gossypium hirsutum L.) cultivars. Field crops
research, 57(1), 29-43.
Li, R., Q., Min and Y. Fu. 2011. 1997/98 El Niño-induced changes in rainfall vertical
structure in the east Pacific. Journal of Climate, 24(24), 6373-6391.
Li, X., S.R. Waddington, J. Dixon, A.K. Joshi and M.C. De Vicente. 2011. The relative
importance of drought and other water-related constraints for major food crops in
South Asian farming systems. Food Security, 3(1), 19-33.

11
8
Título de la tesis o trabajo de investigación
Lizana, C., M. Wentworth, J.P. Martinez, D. Villegas, R. Meneses, E.H. Murchie and M.
Pinto. 2006. Differential adaptation of two varieties of common bean to abiotic
stress: I. Effects of drought on yield and photosynthesis. Journal of Experimental
Botany, 57(3), 685-697
Markhart, A. H. 1985. Comparative Water Relations of Phaseolus vulgaris L. and Phaseolus
acutifolius Gray. Plant Physiology, 77(1), 113-117.
Mencuccini, M., S. Mambelli and J. Comstock. 2000. Stomatal responsiveness to leaf water
status in common bean (Phaseolus vulgaris L.) is a function of time of day. Plant,
Cell & Environment, 23(10), 1109-1118.
Ministerio de Agricultura y Desarrollo Rural BC. 2011. Anuario Estadístico del Sector
Agropecuario Resultados Evaluaciones Agropecuarias Municipales 2011. In:
Documentación e información, datos estadísticos y sistemas de información.
Noviembre, 2012 ed. Bogotá, Colombia.
Miyashita, K., S. Tanakamaru, T. Maitani and K. Kimura. 2005. Recovery responses of
photosynthesis, transpiration, and stomatal conductance in kidney bean following
drought stress. Environmental and Experimental Botany, 53(2), 205-214.
Moftah, A. E. and A.R.I. Al-Humaid. 2005. Effects of antitranspirants on water relations and
photosynthetic rate of cultivated tropical plant (Polianthes tuberosa L.). Polish journal
of ecology, 53(2), 165-175.
Mohamed, M.F., N. Keutgen, A.A. Tawfika and G. Noga. 2002. Dehydration-avoidance
responses of tepary bean lines differing in drought resistance. Journal of Plant
Physiology, 159(1), 31-38.
Mohamed, M.F., M. Mohamed, N. Schmitz-Eiberger and G. Noga. 2005. Comparative
drought postponing and tolerance potentials of two tepary bean lines in relation to
seed yield. African Crop Science Journal, 13(1), 49-60.
Mohamed, S. A. and H. Ibrahim. 2011. Alleviation of adverse effects of drought stress on
common bean (Phaseolus vulgaris L.) by exogenous application of hydrogen
peroxide. Bangladesh Journal of Botany, 40(1), 75-83.
Mollasadeghi, V., M. Valizadeh, R.A. Shahryariand and A. Imani. 2011. Evaluation of end
drought tolerance of 12 wheat genotypes by stress in dices. MEJSR 7(2):241-247.
Monasterio, P. P., F. Pierre, T. Barreto, C. Marin, O. Mora, J. Tablante, C. Mendoza. 2011.
Influencia del fenómeno el niño/oscilación del sur sobre la precipitación y rendimiento

Bibliografía 119
del cultivo de maíz en el municipio peña, estado Yaracuy, Venezuela el
niño/southern. Agronomía Tropical, 61, 59-72.
Morgan, J., A. Condon. 1986. Water use, grain yield, and osmoregulation in wheat.
Functional plant biology 13, 523-532.
Muñoz-Perea, C.G., R.G. Allen, D.T. Westermann, J.L. Wright and S.P. Singh. 2007. Water
use efficiency among dry bean landraces and cultivars in drought-stressed and non-
stressed environments. Euphytica, 155(3), 393-402.
Naghavi, M.R., A.P. Aboughadareh and M. Khalili. 2013. Evaluation of drought tolerance
indices for screening some of corn (Zea mays L.) cultivars under environmental
conditions. Notulae Scientia Biologicae, 5(3), 388.
Nouri, A., A. Etminan, J.A. Teixeira da Silva and R. Mohammadi. 2011. Assessment of
yield, yield-related traits and drought tolerance of durum wheat genotypes (Triticum
turjidum var. durum Desf.). Aust J Crop Sci 5 (1): 8-16
Okçu, G., M.D. Kaya and M. Atak. 2005. Effects of salt and drought stresses on germination
and seedling growth of pea (Pisum sativum L.). Turkish journal of agriculture and
forestry, 29(4), 237-242.
Oki, T y S. Kanae. 2006. Global hydrological cycles and world resources. Science 313,
1068-1072
Osman, H. S. 2015. Enhancing antioxidant–yield relationship of pea plant under drought at
different growth stages by exogenously applied glycine betaine and proline. Annals
of Agricultural Sciences.
O'Toole, J. C., Ozbun, J. L. y Wallace, D. H. (1977). Photosynthetic Response to Water
Stress in Phaseolus vulgaris. Physiologia Plantarum, 40(2), 111-114.
Pabón, J. D., & Torres, G. 2007. Impacto socioeconómico de los fenómenos El Niño y La
Niña en la Sabana de Bogotá durante el siglo XX. Cuadernos de Geografía-Revista
Colombiana de Geografía. (16), 81-94.
Parsons, L.R. and T.K. Howe. 1984. Effects of water stress on the water relations of
Phaseolus vulgaris and the drought resistant Phaseolus acutifolius. Physiologia
Plantarum, 60(2), 197-202.
Pastenes, C., P. Pimentel and J. Lillo. 2005. Leaf movements and photoinhibition in relation
to water stress in field-grown beans. Journal of Experimental Botany, 56(411), 425-
433.

12
0
Título de la tesis o trabajo de investigación
Pastenes, C., V. Porter, C. Baginsky, P. Horton and J. González. 2004. Paraheliotropism
can protect water-stressed bean (Phaseolus vulgaris L.) plants against
photoinhibition. Journal of Plant Physiology, 161(12), 1315-1323.
Perveen, S., M. Shahbaz and M. Ashraf. 2010. Regulation in gas exchange and quantum
yield of photosystem II (PSII) in salt-stressed and non-stressed wheat plants raised
from seed treated with triacontanol. Pak J Bot, 42, 3073-81.
Pimentel, C., D. Laffray and P. Louguet. 1999. Intrinsic water use efficiency at the
pollination stage as a parameter for drought tolerance selection in Phaseolus
vulgaris. Physiologia Plantarum, 106(2), 184-189.
Pimentel, D., J. Houser, E. Preiss, O. White, H. Fang, L. Mesnick, T. Barsky, S. Tariche, J.
Schreck and S. Alpert. 1997. Water resources: agriculture, the environment, and
society. BioScience, 47(2), 97-106.
Polanía, J. A., I.M Rao, S. Beebe and R. García. 2009. Desarrollo y distribución de raíces
bajo estrés por sequía en frijol común (Phaseolus vulgaris L.) en un sistema de tubos
con suelo. Agronomía Colombiana, 27(1), 25.
Rainey, K.M., Griffiths, P.D., 2005. Evaluation of Phaseolus acutifolius A. Gray plant
introductions under high temperatures in a controlled environment. Genet Resour
Crop Evol 52, 117-120.
Ramirez-Vallejo, P. and J.D. Kelly. 1998. Traits related to drought resistance in common
bean. Euphytica, 99(2), 127-136.
Rao, I. M., S. Beebe, J. Polania, M.A. Grajales and R. Garcia. 2006a. “Differences in
drought resistance of advanced lines developed for the last 3 decades,” in Annual
Report 2006. Project IP-1: Bean Improvement for the Tropics (Cali, Colombia: CIAT),
2–6.
Restrepo H. and G. Garcés. 2013. Evaluation of low light intensity at three phonological
stages in the agronomic and physiological responses of two rice (Oryza sativa L.)
cultivars. Agronomia Colombiana 31(2): 195-200.
Restrepo-Díaz, H., J.C. Melgar and L. Lombardini. 2010. Ecophysiology of horticultural
crops: an overview. Agronomía Colombiana, 28(1), 71-79.
Rosales, M. A., E. Ocampo, R. Rodríguez-Valentín, Y. Olvera-Carrillo, J. Acosta-Gallegos
and A.A. Covarrubias. 2012. Physiological analysis of common bean (Phaseolus

Bibliografía 121
vulgaris L.) cultivars uncovers characteristics related to terminal drought resistance.
Plant Physiology and Biochemistry, 56, 24-34.
Rosielle, A.A. and J. Hamblin. 1981. Theoretical aspects of selection for yield in stress and
non-stress environments. Crop Science. 21, 943- 946.
Ruiz, A. D. C. y J. D. Pabón. 2013. Efecto de los fenómenos de El Niño y La Niña en la
precipitación y su impacto en la producción agrícola del departamento del Atlántico
(Colombia). Cuadernos de Geografía: Revista Colombiana de Geografía, 22(2), 35-
54.
Sánchez, F.J., M.A. Manzanares, E.F. de Andres, J.L. Tenorio and L. Ayerbe. 1998. Turgor
maintenance, osmotic adjustment and soluble sugar and proline accumulation in 49
pea cultivars in response to water stress. Field Crops Research 59, 225-235.
Sánchez-Reinoso, A. D., G. Garcés-Varón and H. Restrepo-Díaz. 2014. Biochemical and
physiological characterization of three rice cultivars under different daytime
temperature conditions. Chilean journal of agricultural research, 74(4), 373-379.
Sánchez-Rodríguez, E., M. Rubio-Wilhelmi, L.M., Cervilla, J.J. Blasco, J.J. Rios, M.A.
Rosales, L. Romero and J.M. Ruiz. 2010. Genotypic differences in some
physiological parameters symptomatic for oxidative stress under moderate drought
in tomato plants. Plant Science, 178(1), 30-40.
Sanders, P. L., and III. A.H. Markhart. 1992. Interspecific grafts demonstrate root system
control of leaf water status in water stressed Phaseolus. J. Exp. Bot. 43, 1563–1567.
Segura-Monroy, S. D., A. Uribe-Vallejo, A. Ramírez-Godoy y H. Restrepo-D. Effect of kaolin
application on growth, water use efficiency, and leaf epidermis characteristics of
Physalis peruviana L. seedlings under two irrigation regimes. Journal of agricultural
science and technology. 17(6): 1585-1596 pp.
Serraj, R. and T.R. Sinclair. 1998. N2Fixation Response to Drought in Common Bean
(Phaseolus vulgarisL.). Annals of Botany, 82(2), 229-234.
Shenkut, A.A. and M.A. Brick. 2003. Traits associated with dry edible bean (Phaseolus
vulgaris L.) productivity under diverse soil moisture environments. Euphytica,
133(3), 339-347.
Sims, D. A. and J.A. Gamon. 2002. Relationships between leaf pigment content and
spectral reflectance across a wide range of species, leaf structures and
developmental stages. Remote sensing of environment. 81(2), 337-354.

12
2
Título de la tesis o trabajo de investigación
Singh, S. P., P. Gepts and D.G. Debouck. 1991. Races of common bean (Phaseolus
vulgaris, Fabaceae). Econ. Bot. 45, 379–396.
Souza, R. P., E.C. Machado, J.A.B. Silva, A.M.M.A. Lagôa and J.A.G. Silveira. 2004.
Photosynthetic gas exchange, chlorophyll fluorescence and some associated
metabolic changes in cowpea (Vigna unguiculata) during water stress and recovery.
Environmental and experimental botany, 51(1), 45-56.
Sponchiado, B.N., J.W. White, J.A. Castillo and P.G. Jones. 1989. Root Growth of Four
Common Bean Cultivars in Relation to Drought Tolerance in Environments with
Contrasting Soil Types. Experimental Agriculture, 25(02), 249-257.
Sun, J., J. Gu, J. Zeng, S. Han, A. Song, F. Chen, S. Chen. 2013. Changes in leaf
morphology, antioxidant activity and photosynthesis capacity in two different drought-
tolerant cultivars of chrysanthemum during and after water stress. Scientia
Horticulturae, 161, 249-258.
Toro, O., J. Tohme and D.G. Debouck. 1990. Wild Bean (Phaseolus vulgaris L): Description
and Distribution. (Cali, Colombia: CIAT), 106.
Türkan, İ., M. Bor, F. Özdemir and H. Koca. 2005. Differential responses of lipid
peroxidation and antioxidants in the leaves of drought-tolerant P. acutifolius Gray and
drought-sensitive P. vulgaris L. subjected to polyethylene glycol mediated water
stress. Plant Science. 168(1), 223-231.
UNGRD. 2014. Plan Nacional de Contingencia ante el fenómeno del Niño 2014-2015. En:
http://portal.gestiondelriesgo.gov.co/Paginas/Noticias/2014/Activacion-planes-de-
contingencia-fenomeno-del-nino.aspx. Consultado en marzo de 2016.
Wahid, A., Gelani, S., Ashraf, M., Foolad, M., 2007. Heat tolerance in plants: an overview.
Environmental and Experimental Botany 61, 199-223.
Wakrim, R., S. Wahbi, H. Tahi, B. Aganchich and R. Serraj. 2005. Comparative effects of
partial root drying (PRD) and regulated deficit irrigation (RDI) on water relations and
water use efficiency in common bean (Phaseolus vulgaris L.). Agriculture,
Ecosystems & Environment, 106(2–3), 275-287.
Wellburn, A. R. 1994. The spectral determination of chlorophylls a and b, as well as total
carotenoids, using various solvents with spectrophotometers of different resolution.
Journal of plant physiology. 144(3), 307-313.

Bibliografía 123
Wentworth, M., E.H. Murchie, J.E. Gray, D. Villegas, C. Pastenes, M. Pinto and P. Horton.
2006. Differential adaptation of two varieties of common bean to abiotic stress - II.
Acclimation of photosynthesis. Journal of Experimental Botany, 57(3), 699-709.
White, J.W. y J.A, Castillo. 1989. Relative Effect of Root and Shoot Genotypes on Yield of
Common Bean under Drought Stress. Crop Science, 29(2).
White, J.W., and J.A. Castillo. 1992. Evaluation of diverse shoot genotypes on selected
root genotypes of common bean under soil water deficits. Crop Sci. 32, 762–765
White, J.W., J.A. Castillo, J.R. Ehleringer, J.A.C. Garcia and S.P. Singh. 1994. Relations of
carbon isotope discrimination and other physiological traits to yield in common bean
(Phaseolus vulgaris) under rainfed conditions. The Journal of Agricultural Science,
122(02), 275-284.
White, J., J. Castillo and J. Ehleringer. 1990. Associations Between Productivity, Root
Growth and Carbon Isotope Discrimination in Phaseolus vulgaris Under Water
Deficit. Functional Plant Biology, 17(2), 189-198.
White, J.W., Izquierdo, J., 1991. Physiology of yield potential and stress tolerance.
Common beans: Research for crop improvement. CAB International., Wallingford,
UK, 287-383.
Winterhalter, L., B. Mistele, S. Jampatong and U. Schmidhalter. 2011. High throughput
phenotyping of canopy water mass and canopy temperature in well-watered and
drought stressed tropical maize hybrids in the vegetative stage. European Journal of
Agronomy, 35(1), 22-32.
Xu, Z., G. Zhou and H. Shimizu. 2010. Plant responses to drought and rewatering. Plant
signaling & behavior, 5(6), 649-654.
Yang, J., R.G. Sears, B.S. Gill and G.M. Paulsen. 2002. Genotypic differences in utilization
of assimilate sources during maturation of wheat under chronic heat and heat shock
stresses. Euphytica, 125(2), 179-188.
Yousfi, N., I. Slama and C. Abdelly. 2012. Phenology, leaf gas exchange, growth, and seed
yield in contrasting Medicago truncatula and Medicago laciniata populations during
prolonged water deficit and recovery. Botany. 90(2), 79-91.