







Idiomas
Páginas
Jurídico


UNIVERSIDAD AUTONOMA GABRIEL RENE MORENO FACULTAD DE CIENCIAS AGRICOLAS CARRERA DE INGENIERIA FORESTAL
2014
DOCUMENTO CIENTIFICO N° 5-2014
Determinación del crecimiento de la Cacha
(Aspidosperma quebracho blanco) mediante el
método de dendrocronología
Richard Chaca
Pedro Saravia

Determinación del crecimiento de la Cacha (Aspidosperma quebracho blanco) mediante
el método de dendrocronología
Richard Chaca
Pedro Saravia
Referencia
Chaca, Richard; Saravia, Pedro. 2014. Determinación del crecimiento de la cacha (Aspidosperma quebracho blanco) mediante el método de dendrocronología. Documento Científico N° 5-2014. Universidad Gabriel René Moreno, Carrera de Ingeniería Forestal. Santa Cruz, Bolivia.
Editor:
Eduardo Sandoval H., Ph.D. Carrera de Ingeniería Forestal Facultad de Ciencias Agrícolas –UAGRM -, Km 8,5 Carretera al Norte [email protected] Santa Cruz, Bolivia
Reservados todos los derechos. Ninguna parte de esta publicación se puede reproducir,
almacenar en sistema de recuperación ni transmitir en forma alguna por medios electrónicos,
mecanismos, fotocopia o cualquier otro medio, sin una adecuada referencia a la fuente.
Santa Cruz, Bolivia
2014

CONTENIDO
1. INTRODUCCION ....................................................................................................................... 1
2. OBJETIVOS ................................................................................................................................. 2
2.1 Objetivo general .............................................................................................................................. 2 2.2 Objetivos específicos ..................................................................................................................... 2 2.3 Hipótesis ............................................................................................................................................ 2
3. REVISION DE LITERATURA 2 3.1 Dendrocronología en los trópicos ............................................................................................ 2 3.2 Estructura anatómica de los anillos de crecimiento ......................................................... 3 3.3 Potencialidades de la dendrocronología en el manejo forestal .................................... 4 3.4 Estudios dendrocronológicos de especies en Bolivia ....................................................... 5 3.5 Incremento en diámetro .............................................................................................................. 6 3.6 Índice de espacio vital................................................................................................................... 7 3.7 Descripción de Aspidosperma quebracho-blanco .............................................................. 7
4. MATERIALES Y METODOS .................................................................................................... 9
4.1 Descripción de área de estudio ................................................................................................. 9 4.2 Materiales y equipo .................................................................................................................... 11 4.3 METODOS ....................................................................................................................................... 11 4.3.1 Obtención de la muestra 11 4.3.2 Preparación de las rodajas 12 4.3.3 Conteo y datación 13 4.3.4 Medición de los anillos de crecimiento 13 4.3.5 Análisis estadístico 14
5. RESULTADOS Y DISCUSION ................................................................................................ 15
5.1 Caracterización de los anillos de crecimiento .................................................................. 15 5.2 Crecimiento de la especie en bosque natural y cortinas rompe vientos ................ 16 5.3 Ancho de anillos de crecimiento en los dos sitios de estudio..................................... 18 5.4 Incremento en diámetro de la especie en estudio. ........................................................ 20 5.5 Índice de espaciamiento vital entre árboles .................................................................... 23
6. CONCLUSIONES ...................................................................................................................... 23
8. LITERATURA CITADA .......................................................................................................... 24

RESUMEN
El conocimiento de la edad de los árboles tropicales es de importancia para la comprensión
sobre la dinámica del crecimiento de los bosques y para obtener información sobre las tasas de
crecimiento a largo plazo para el manejo forestal. El presente estudio se realizó en la
Propiedad de Yabaré, perteneciente a la Universidad Autónoma Gabriel René Moreno,
ubicada en los Municipios de Pailón y Concepción del Departamento de Santa Cruz. El
objetivo fue analizar el crecimiento de la Cacha (Aspidosperma quebracho-blanco), para
establecer el incremento diamétrico en dos sitios de la propiedad de Yabaré. Se seleccionaron
al azar 15 árboles (5 árboles del bosque y 10 árboles de las cortinas), se tomaron sus datos
dendrologicos y luego fueron derribados para obtener rodajas de 5 a10 cm de espesor a la
altura de 30 cm del suelo. Las rodajas se lijaron y pulieron siguiendo las técnicas
dendrocronológicas hasta visualizar los anillos de crecimiento para realizar el conteo,
datación y la medición del ancho de los anillos de crecimiento para su análisis. Los resultados
mostraron que los anillos de A. quebracho-blanco están delimitados por fibras comprimidas
con paredes gruesas. Las tasas de crecimiento presentaron una gran variación analizando los
dos sitios, alcanzando el diámetro mínimo de corta (DMC) de 40 cm entre 110 y 220 años.
Realizando un análisis del ancho de anillos se observó periodos de bajo crecimiento en ambos
sitios. El incremento acumulado en diámetro también mostró una variación entre sitos,
alcanzando el DMC de 40 cm a los 190 años, presentando un crecimiento positivo en el
diámetro lo que sugiere un ciclo de corta mayor a 20 años al establecido por la ley forestal.
Los incrementos también presentaron variación entre sitios, presentando un incremento
corriente máximo de 1,33 mm a la edad de 204 años y un incremento anual medio máximo de
1,10 mm a la edad de 233 años, prolongándose así la culminación del crecimiento el cual en
esta especie es muy lento en los bosques transicionales, lo que sugiere tomarse con cautela
para su aprovechamiento en los planes de manejo forestal. El índice de espaciamiento vital
mostró que los árboles competidores del bosque están más cerca al árbol objeto. Asimismo, no
se encontraron diferencias significativas entre ambos sitios.
Palabras claves:Cacha, Aspidosperma quebracho blanco, anillos de crecimiento, dendro-cronología

1
1. INTRODUCCION
La realización de un manejo forestal adecuado en un ecosistema forestal exige el
conocimiento de la renta que produce el capital monte, representado por el crecimiento del
bosque (Ríos et al. 2001). Este crecimiento se puede estimar a partir de datos obtenidos de
parcelas permanentes de muestreo y de anillos de crecimiento de los árboles (Bowman et al.
2013). Sin embargo los datos de crecimiento obtenidos de parcelas permanentes de muestreo
solo cubren periodos cortos de la vida de un árbol, a comparación del análisis de los anillos de
crecimiento que proporcionan un medio para reconstruir las tasas de crecimiento de toda la
vida de un árbol, que fácilmente se extiende por más de un siglo (Zuidema et al. 2012).
La dendrocronología es la ciencia que estudia los anillos de crecimiento de árboles para
reconstruir el crecimiento de un árbol en una determinada región. A través de los anillos de
crecimiento se obtiene información, acerca del clima, de las condiciones ambientales pasadas
e información para el manejo forestal de los bosques (Szejner 2012). Las aplicaciones de esta
ciencia son múltiples y abarcan la climatología, ecología, geología, arqueología, criminología
y el estudio del cambio climático (Llorente 2012).
La mayor parte de los estudios dendrocronológicos en los trópicos no consideraban viable
medir anillos de crecimiento anuales sucesivos. Uno de los principales motivos es que muchos
árboles de regiones tropicales no forman anillos claramente distinguibles o anuales, debido a
la aparente falta de una estacionalidad climática, que introduzca la latencia y detenga el
crecimiento de los árboles (Mariaux 1983). No obstante, la marcada estacionalidad de la
precipitación que se observa en diversos climas tropicales, interrumpe temporalmente el
crecimiento de las plantas, produciendo la formación de anillos distinguibles (Roig 2000).
En las últimas décadas, el desarrollo de cronologías de anillos en árboles tropicales tuvo
importantes beneficios potenciales para la climatología, la ecología de los bosques y la gestión
forestal (Stahle 1999). Demostrado el potencial de los anillos de crecimiento para determinar
las edad de los árboles tropicales (Ecksteins et al. 1983, Worbes 1999), para la evaluación de
variables climáticas (Paredes 2012), restauración de caudales de ríos (Urritia et al. 2005) y la
reconstrucción de incendios forestales. Así también su aplicación en el manejo forestal, para
obtener información sobre las tasas de crecimiento en diámetro de los árboles (López et al.
2011, Paredes 2012), como apoyo para determinar los Diámetros Mínimos de Corta (Brienen
y Zuidema 2006, Rozendaal 2010), evaluación de ciclos de corta (López 2011), estimación de
cortas permisibles (Rozendaal et al. 2010) y estado en sucesión del bosque (Brienen et al.
2009) entre otras aplicaciones.
En Bolivia, en los últimos años se han realizado estudios dendrocronológicos en Bosques de la
Amazonia, Bosques del Cerrado y Bosques de los Andes (Killen et al. 1993), con especies que
presentan anillos de crecimiento claramente visibles y distinguibles con algunas dificultades,
que han permitido conocer el crecimiento anual de los árboles en diferentes ambientes
naturales. Sin embargo en los Bosques del Gran Chaco, la mayor parte de las especies poseen
maderas duras, en las cuales es posible distinguir los anillos de crecimiento en algunas
especies, aunque con dificultades (Roig 2000).

2
En Bolivia los planes de manejo forestal están basados en ciclos de corta de 20 años y en
diámetros mínimos de corta (DMC), que pueden estar ocasionando sobre explotación del
bosque. En este sentido la dendrocronología se torna en una herramienta importante para el
entendimiento del crecimiento de los árboles y la dinámica del bosque.
El presente estudio permite estimar el crecimiento e incremento en diámetro de Aspidosperma
quebracho-blanco a partir de anillos de crecimiento como contribución para su manejo
forestal en un bosque transicional chaqueño-chiquitano.
2. OBJETIVOS
2.1 Objetivo general
Analizar el crecimiento de la Cacha (Aspidosperma quebracho-blanco), para establecer el
incremento diamétrico en la chiquitanía.
2.2 Objetivos específicos
Evaluar el crecimiento de la especie en bosque natural y cortinas rompe vientos.
Comparar el ancho de anillos de crecimiento, en los dos sitios en estudio.
Establecer el incremento diamétrico promedio de la especie en estudio.
Determinar un índice de espaciamiento vital entre árboles vecinos.
2.3 Hipótesis
El crecimiento de la especie es igual en las dos áreas (bosque natural y cortinas rompe viento).
3. REVISION DE LITERATURA
3.1 Dendrocronología en los trópicos
En los climas templados el crecimiento se produce en la estación vegetativa (primavera y
verano) cuando las condiciones climáticas son favorables. El cambium se divide en dirección
radial provocando un ensanchamiento del xilema y generando nueva corteza. Cuando llegan
las condiciones desfavorables en otoño e invierno, el cambium cambia de estado de
crecimiento activo a inactivo produciendo una marca oscura y de alta densidad en la madera
llamada anillo de crecimiento (Botosso 2009).
La dendrocronología del griego dendros, árbol; cronos, tiempo y logos, conocimiento. Es una
disciplina científica dotada de un conjunto de principios, técnicas y métodos que permiten
datar los anillos de crecimiento anuales de los árboles, extraer, separar e interpretar la
información que contienen de los diferentes factores que han influido en su crecimiento
(Gutiérrez 2008). En la actualidad se reconoce como padre de la dendrocronología al
astrónomo Andrew E. Douglas, quien durante la primera mitad del siglo XX fundó el

3
Laboratorio de investigación de anillos de crecimiento (Tree-Ring Research Laboratory) en la
Universidad de Arizona, Estados Unidos.
Según Worbes (1995), la formación de anillos de crecimiento en especies de árboles, esta
normalmente relacionada con las variaciones de las estaciones de crecimiento, como la
temperatura de invierno en las regiones templadas, la precipitación y las inundaciones
temporales (bosques inundables) en las regiones tropicales y altitudes elevadas, induciendo la
reducción y/o dormancia cambial y consecuentemente la formación de capas de anillos de
crecimiento. Lisi et al. (2008) analizaron el crecimiento de 24 especies en un Bosque
Estacional Semi-decidual en la región sudeste de Brasil y relacionaron estas con las variables
meteorologías y actividades fenológicas. Los mayores incrementos fueron encontrados durante
la estación lluviosa y los menores en la estación seca, siendo que las especies pierden sus
hojas en el inicio de la estación seca y se renuevan al final de la misma temporada.
Sin embargo, el descubrimiento de anillos anuales en varias especies bajo un clima ecuatorial
con precipitaciones superiores a 4000 mm anuales y promedios mensuales superiores a 100
mm de la Estación Biológica La Selva, Costa Rica, ha hecho afirmar que la anualidad de los
anillos en árboles tropicales se debe extender a toda las condiciones en que pueden crecer los
árboles en el trópico (Fichtler et al. 2003). También se han encontrado anillos anuales en la
vegetación arbórea de las Zonas Andinas de Bolivia, donde se han desarrollado las cronologías
a mayor altitud del mundo con especies de Polylepis pepei y Polylepis tarapacana que están
ligados a un ciclo estacional de crecimiento (Roig et al 2001, Argollo et al. 2004).
De acuerdo con Fahn et al. (1983), existen dos tipos de árboles tropicales, aquellos que
muestran distintas zonas de crecimiento y aquellos en los cuales las zonas de crecimiento son
indistinguibles. En árboles con distintas zonas de crecimiento, son posibles al menos tres
explicaciones: zonas que representan incrementos anuales, zonas que reflejan algunos cambios
periódicos externos o internos ocurridos durante el año y las zonas debidas a ciclos no anuales.
Algunas de las zonas pueden ser anillos falsos incompletos. Las causas de los anillos falsos
pueden incluir inundaciones, sequias, heladas, fuego, defoliación, extenso crecimiento de una
o algunas ramas durante el periodo de baja actividad del cambium (Fahn et al. 1983). Como
resultado de este último puede desarrollarse en estas ramas el completo crecimiento de los
anillos, y anillos incompletos en el tallo principal.
Los métodos empleados para investigación de la periodicidad con que se forman los anillos de
crecimiento, pueden ser realizados mediante las observaciones de los eventos fenológicos,
evaluando el incremento a través de fajas dendrometricas y realizando heridas cambiales
(ventanas de Mariaux) con posterior análisis de las cicatrices a través de la colecta destructiva
(disco) o no destructiva (barreno de Pressler) del tronco de los árboles (Roig 2000).
3.2 Estructura anatómica de los anillos de crecimiento
Roig (2000) menciona tres tipos de arreglos estructurales importantes que definen el borde de
un anillo de crecimiento: i) diferencias notables en el diámetro de los vasos en el leño
temprano respecto de los del tardío en dos anillos contiguos, ii) presencia de fibras o
traqueidas del leño tardío cuyas paredes son aplastadas radialmente y más gruesas respecto a

4
las fibras o traqueidas del leño temprano y iii) presencia de una banda marginal continua de
parénquima axial. Boninsegna et al. (1989) realizaron estudios anatómicos de la madera de 13
especies de la Selva Misionera en Argentina encontrando diferencias de las estructuras
anatómicas en los anillos de crecimiento: anillos porosos o semi-porosos en Cedrela fissilis y
Cordia trichotoma; parénquima marginal uni o multiseriado en Parapiptadenia rigida,
Peltophorum dubium, Myrocarpus frondosus; zonas de crecimiento constituida por varias
hileras de fibras con pequeño diámetro radial y paredes gruesas en Chorisia speciosa,
Aspidosperma polyneuron, Ocotea puberula, Nectandra saligna; de parénquima a lo largo de
las capas de crecimiento en Cabrelea oblongifolia.
Según Beltrán y Valencia (2013), se estudiaron la anatomía de anillos de crecimiento de 80
especies en la Selva Central del Perú. Los resultados muestran que 24 de las 80 especies
mostraron un potencial para estudios dendrocronológicos, 25 tienen problemas para el análisis
de los anillos de crecimiento y 31 no tienen potencial. Asimismo, 12 especies Lauráceas
presentaron anillos de crecimiento delimitados por una banda de tejido acortado radialmente y
de las 13 especies fabáceas, 4 presentaron anillos de crecimiento delimitados por una banda de
parénquima marginal.
López (2011) estudió los anillos de crecimiento de 11 especies en bosques tropicales secos del
Cerrado Boliviano. Los resultados muestran que todas las especies presentaron anillos de
crecimiento anuales. En Amburana cearensis, Cariniana ianeirensis y Ficus boliviana los
anillos de crecimiento están delimitados por fibras comprimidas; en Cedrela fissilis y Zeyheria
tuberculosa por elementos porosos y parénquima terminal/inicial y en Anadenanthera
colubrina, Centrolobium microchaete, Copaifera chodatiana, Hymenaea courbaril,
Machaerium scleroxylon y Platymiscium ulei por parénquima terminal/inicial. En algunas
especies se observaron la presencia de lentes de crecimiento y anillos falsos.
3.3 Potencialidades de la dendrocronología en el manejo forestal
Villalba et al. (2006) afirman que el empleo de los anillos de crecimiento permite cuantificar
en forma precisa las velocidades de crecimiento radial de los árboles en distintos ambientes,
así como determinar diferencias en las estructuras poblacionales del bosque en relación a
factores tales como el clima, el suelo, los disturbios naturales y la historia de su uso.
Asimismo, todo estudio que requiera información sobre la estructura de edad y la dinámica de
los bosques, la determinación de sus turnos de aprovechamiento, el establecimiento de las
relaciones entre el crecimiento leñoso, fructificación y/o variaciones climáticas o simplemente
la relación del bosque a un tratamiento silvícola determinado, necesita del fechado y la
evaluación de los ritmos pasados y presentes del crecimiento (Villalba et al. 2000, Villalba et
al. 2006).
En 1856, Sir Dietrich Brandis reporto la existencia de anillos de crecimiento anuales en los
árboles de Teca (Tectona grandis) y los empleó para estimar las tasas de crecimiento en
bosques tropicales de la India y Burma; y en base a los anillos estableció la corta anual
permisible para un sistema silvicultural sostenible (Mariaux 1983, Worbes 2002). Boninsegna
et al. (1989) analizaron los anillos de crecimiento de 15 árboles de Cedrela fissilis, para
determinar las tasas de crecimiento radial y en área basal en Selva Misionera de Argentina.

5
Los resultados muestran que la culminación del incremento radial se produce a los 63 años y
el incremento medio anual (IMA) en área basal a los 153 años con una variación de 61 a 180
años, independiente del diámetro de los árboles. Constituyéndose así una importante
herramienta para el manejo forestal de los bosques tropicales.
3.4 Estudios dendrocronológicos de especies en Bolivia
Los estudios dendrocronológicos en Bolivia se iniciaron en las últimas décadas. Roig (2000),
presentó una tabla de 39 especies que crecen en Bolivia, indicando el potencial
dendrocronológico según las formaciones forestales, fenología de las especies y anillos de
crecimiento de los árboles. Iniciándose así los primeros estudios dendrocronológicos y
construyéndose la primera cronología de ancho de anillos de 135 años a partir de muestras de
madera de Polylepis pepei en la zona andina (Roig et al 2001). Asimismo se ha podido
demostrar que Polylepis tarapacana registró las condiciones ambientales del altiplano
boliviano de los últimos 500-700 años (Argollo y Miranda 2002, Argollo et al. 2004).
De acuerdo con Miranda et al. (2002), se estudiaron la formación de anillos de árboles de 44
especies en bosques tropicales de Bolivia, mediante la colecta de muestra de madera y
muestras obtenidas con barreno. Los autores encontraron la formación de anillos en 11
especies delimitados por bandas de parénquima, densidad de vasos, diferencias de vasos por
tamaños y fibras terminales; 15 no presentaron anillos diferenciables y 18 especies no
presentaron anillos visibles. De las 11 especies Cedrela odorata y Swietenia macrophylla
forman anillos anuales y en el caso de Tabebuia heptaphylla, Hymenaea coubarril,
Cyphomandra oblongifolia, Terminalia oblonga, Junglans soratensis, Junglans boliviana,
Vismia glabra, Aspidosperma ramiflorum y H. parviflora no se sabe si la formación de los
anillos es anual. Asimismo para la misma zona, Torrico y Roig (2002) mencionan a las
siguientes especies como formadoras de anillos de crecimiento claramente distinguibles:
Cedro (Cedrela odorata), Mara (Swietenia macrophylla), Goma (Hevea brasiliensis),
Amarillo (Aspidosperma vargassi), Taruma (Vitex sp), Tumi (Amburana cearensis),
Paquiocillo (Hymenaea parvifolia), y Paquio (H. coubaril).
Miranda y Fernández (2002) afirman que en las sabanas húmedas de Moxos, el género
Tabebuia, es un excelente grupo para la realización de análisis dendrocronológicos por la
evidencia y claridad de los anillos de crecimiento, que pueden ser relacionadas a efectos
climatológicos (precipitación y temperatura) y a influencias antropogéneticas, como el fuego.
Brienen y Zuidema (2003), realizaron una guía metodología para el uso de los anillos de
crecimiento en el manejo forestal. Asimismo realizaron una revisión de 32 especies
maderables y especies maderables bolivianas como Vochsia vismiifolia, Dipteryx odorata,
Moronobea sp., Cariniana micrantha, Pithecellobium corymbosum, Ceiba pentandra,
Manilkara bidentata, Peltogyne heterophylla, Tachigali cf. paniculata, Hymenaea sp.,
Schizolobium sp., Parkia pendula, C. odorata, Astronium lecointei, Tetragastris altissima,
Cedrelinga catenaeformis, Clarisia racemosa y A. cearensis que son potenciales para su
aplicación de análisis de anillos de crecimiento para el manejo forestal. Asimismo realizaron
simulaciones de la recuperación del volumen maderable bajo diferentes ciclos de corta para
algunas especies.

6
Brienen y Zuidema (2005), determinaron en la Amazonia Boliviana la ocurrencia de anillos
anuales, correlacionando el ancho de anillos con datos de precipitación mensual para A.
cearensis, C. odorata, C. catenaeformis y Tachigali vasquezii, contando anillos en árboles de
edad conocida y por radiocarbono para P. heterophylla y Bertholletia excelsa. Lo que permitió determinar sus edades y reconstruir tasas de crecimiento históricas toda su vida (Brienen y
Zuidema 2006a). Usando los mismos datos determinaron su diámetro mínimo de corta y la
estimación del volumen maderable para el segundo ciclo de corta para 4 especies (Brienen y
Zuidema 2006b, Brienen y Zuidema 2007).
López (2003) y López et al. (2012b), determinaron anillos de crecimiento de 11 especies en el
municipio de Guarayos, Santa Cruz. Los resultados muestran que la mayoría de las especies
estudiadas presentan bandas de crecimiento anuales, delimitados por parénquima terminal en
Ficus boliviana, Ampelocera ruizii, Albizia niopoides, Sweetia fruticosa, Pseudolmedia laevis,
Terminalia oblonga; fibras comprimidas en Pouteria nemorosa, Aspidosperma
cylindrocarpon, Hura crepitans, Centrolobium microchaete, y Cariniana ianeirensis.
Destacando la presencia de lentes y falsos anillos en algunas especies lo que dificultó su
fechado.
López (2011) y López et al. (2011,2012a y 2013), proponen para los bosques tropicales secos
del Cerrado Boliviano, como alternativa el empleo de los anillos de crecimiento para la
determinación de tasas de crecimiento. Donde se determinó los incrementos corrientes anuales
(ICA), los incrementos medios anules (IMA) y los turnos biológicos de corte (TBC) para A.
cearensis, Anadenanthera colubrina, C. ianeirensis, Cedrela fissilis, C. microchaete,
Copaifera chodatiana, Ficus boliviana, H. courbaril, Machaerium scleroxylon, Platymiscium
ulei y Zeyheria tuberculosa. En base a estos parámetros también se evaluó los ciclos de corte y
los diámetros mínimos de corta establecidos en las pautas actuales del manejo forestal en
Bolivia. Paredes (2012) y Paredes-Villanueva et al. (2013), determinaron en los bosques
semideciduos chiquitanos de Bolivia, el potencial dendrocronológico de M. scleroxylon y su
respuesta a las variables climáticas como apoyo en la toma de decisiones en el manejo forestal.
3.5 Incremento en diámetro
El crecimiento de los árboles puede definirse como el resultado del aumento de sus
dimensiones en un periodo de tiempo, es un fenómeno muy complejo en el que intervienen
numerosos factores internos y externos determinantes (Araujo et al. 2007). También depende
de las reservas acumuladas por el árbol, pero depende más del medio ambiente, en ciertos
límites el incremento es mayor cuando cuenta con más espacio, lo mismo sucede con la luz. El
incremento en diámetro se manifiesta en los anillos de crecimiento, el ancho de los anillos no
es igual a lo largo de todo el fuste. Los anillos de crecimiento son más anchos en la parte
superior del fuste inmediatamente debajo de las primeras ramas; más abajo el ancho de los
anillos comienza a decrecer y en la base del árbol ensancha otra vez, lo que le da a la porción
inferior una forma especial (Klepac 1983).
Según Imaña y Encinas (2008), el incremento puede ser definido como el crecimiento del
árbol o de un rodal forestal en un determinado tiempo. Este periodo puede ser expresado en
días, meses, años o décadas. Mencionando así los siguientes incrementos:

7
El incremento corriente anual: Es el crecimiento que logra un árbol o una masa en el curso
de un año. Este crecimiento también es conocido como crecimiento acumulado, incremento
corriente anual (ICA) o simplemente como incremento anual (IA), correspondiendo a lo que el
árbol creció en el periodo de un año.
El incremento medio anual: Es el promedio anual del crecimiento total a cierta edad del
árbol, el IMA se obtiene dividiendo las dimensiones de un árbol o una masa entre su edad.
3.6 Índice de espacio vital
El crecimiento de los árboles es el resultado de su tendencia al crecimiento limitado
(dependiente de su potencial abiótico y actividad fotosintética) y de las restricciones impuestas
por el medio (competencia con otros individuos y proceso de envejecimiento). Si bien se trata
de factores cuyo análisis es complejo, se acepta que la interacción espacial con los árboles
cercanos (denominada competencia) tiene el mayor peso en el proceso.
Según Ford y Sorrensen (1992) citado por Álvarez et al. (2003), la relación entre el
crecimiento y la competencia se puede sintetizar en cinco principios básicos:
Las plantas modifican su entorno al crecer, reduciendo los recursos disponibles para
otras plantas con las que compiten.
El principal mecanismo de competición es la interacción espacial.
Como consecuencia de la disminución de los recursos, las plantas ven reducido su
crecimiento y el efecto final de la competencia es la muerte.
Las plantas de adaptan a los cambios de su entorno, responden a la competencia y
modifican la naturaleza de la competición.
En los procesos de competición hay diferencias entre especies.
Según Prodan et al. (1997), los índices de competencia se agrupan en dos categorías, los que
representan la competencia a nivel rodal y a árboles individuales. Este último proporciona
información más detallada que otras metodologías de modelización y generalmente funciona
mejor que los índices a nivel rodal para proyecciones a corto plazo (García 1994).
Spiecker (1981), indica que la competencia en el árbol individual puede ser definida por la
distancia y la dimensión de los árboles vecinos.
Por lo tanto, la situación de la competencia de cada árbol es cuantificada en términos de
índices de competencia, el cual está en función del espacio vital del árbol y la distancia de sus
vecinos (Prodan et al. 1997).
3.7 Descripción de Aspidosperma quebracho-blanco
Distribución: Abarca el Gran Chaco que se extiende desde Santa Cruz (Bolivia) a Matto
Grosso (Brasil), el oeste de Paraguay y la parte central septentrional de Argentina. Su
distribución natural en Bolivia es el Chaco seco, selva en transición, provincia Luis Calvo en
Tarija, provincia Cordillera y Chiquitos en Santa Cruz.

8
Características dendrológicas: Es un árbol de la familia Apocinaceae, mediano hasta alto,
que llega hasta los 25 m de altura y alrededor de 60 cm de diámetro, fuste recto y cilíndrico
Corteza amarilla grisácea muy rugosa, con hendiduras longitudinales muy pronunciadas de
color ocre hacia la parte más profunda; con exudados blancos escasos, ligeramente viscoso.
Hojas simples, coriáceas, en verticilos, lanceoladas marcadamente acuminadas, ápice
mucronado. Flores pequeñas, tubulosas y amarillentas dispuestas en racimos axilares. Fruto
apocárpico, con dos folículos, elípticos, gris-verdoso y péndulo con semillas aladas
membranosas más o menos circulares, blancas y numerosas con consistencia papirácea (Lara
1988, Justiniano 2006, Mostacedo et al. 2003).
Regeneración natural: En todas partes es buena y abundante, aunque una parte de las
numerosas semillas producidas no es viable. La capacidad germinativa es de corta duración;
por ello las semillas destinadas a la siembra solamente deberían ser almacenadas poco tiempo
y en lugar seco.
Ecología: Especie siempre verde, heliófita, emergente típica del Bosque Chaqueño, menos
abundante en el Bosque Chiquitano. Se desarrolla en suelos pesados. Florece en abril y
fructifican de julio a diciembre (Justiniano 2006).
Descripción de la madera: No tiene mucha diferencia de color entre albura y duramen,
ambos son de color amarillo ocre a rosado, presenta un veteado suave, textura fina y
homogénea y brillo escaso. El grano es oblicuo a entrecruzado. Produce madera juvenil hasta
un DAP de 18 a 30 cm, lo que indica que árboles aprovechados con un diámetro de 30 cm
todavía presentan un alto porcentaje de madera juvenil. Las características anatómicas de la
madera juvenil son responsables de la menor densidad básica y la mayor contracción
longitudinal, transversal y menor resistencia que la madera dura, lo que explicaría la cierta
forma de comportamiento inestable y su tendencia a sufrir alabeos. Madera muy pesada de
densidad 0.85 gr/cm3 fácil de trabajar y se emplea como madera de construcción (vigas),
tornería, postes, durmientes y para la confección de morteros y otros objetos. (Moglia et al.
2009)
Descripción macroscópica y microscópica: Los anillos de crecimiento son visibles a simple
vista, como líneas oscuras y a nivel microscópico por 2 a 5 estratos de fibras comprimidas de
paredes engrosadas con un rango de ancho de anillos de 1-4 mm. Presenta porosidad difusa,
dispuestos en patrón no uniforme, exclusivamente solitarios. Fibras con paredes de espesor
medio a paredes gruesas. Parénquima axial apotraqueal, difuso y difuso en agregados. Los
radios con la parte multiseriada de igual ancho como la parte uniseriada. Presenta cristales
prismáticos, localizados en células de los radios. Presenta un xilema altamente especializado
en tolerar el estrés hídrico, desarrollando una estrategia adaptativa diferente a la mayoría de
las especies de la Región Chaqueña. Los vasos solitarios proporcionan eficiencia en la
conducción, mientras las traqueidas vasicéntricas y fibrotraqueidas intervienen en la
conducción y forman un sistema de subsidiario e intervienen también proporcionando
seguridad en la conducción. El xilema de esta especie funciona a manera de una red
interconectada, que le ha permitido tener una amplia zona de distribución y extenderse más
allá de los límites de la región chaqueña, donde las condiciones ambientales son más severas
(Moglia et al. 2009, Giménez y Moglia 1993, Richter y Dallwitz 2000).

9
4. MATERIALES Y METODOS
4.1 Descripción de área de estudio
El área de estudio está ubicada en los Municipios de Pailón y Concepción, de las Provincias
Chiquitos y Ñuflo de Chávez a una distancia de 130 km al este del Departamento de Santa
Cruz. Específicamente se encuentra en la propiedad de Yabaré, perteneciente a la Facultad de
Ciencias Agrícolas de la UAGRM, ubicada a 24 km al norte del Cantón “Tres Cruces” (Fig.
1). Se tiene acceso a través de la red caminera de tipo vecinal conectada con la carretera
bioceánica que une Santa Cruz – Puerto Suárez y la vía férrea Santa Cruz – Puerto Suárez
(PDMP 2001-2005). Actualmente, esta propiedad cuenta con una superficie total de 5194 ha,
de las cuales 1710 ha están desmontadas para uso agrícola bajo normas y una distribución
consecuente con las necesidades de defensa de la misma. Su ubicación geográfica es el
paralelo 62°11'53'' latitud oeste y el paralelo 17°26'24'' latitud sur y una elevación de 272
msnm.
Por las características de precipitación y temperatura, la propiedad de Yabaré se encuentra
clasificada como clima subhúmedo seco megatermal con nulo o exceso de agua
(CORDECRUZ 1988). De acuerdo a la estación meteorológica de Pailón la precipitación
promedio anual es de 1086 mm y el periodo de lluvias comienza en noviembre y se prolonga
hasta abril y una temperatura promedio de 24,9 °C con extremos de 37 °C y mínimas de 3°C.
En la estación de “Tres Cruces” la precipitación promedio anual es de 1045 mm y el periodo
de lluvias empieza en noviembre y extiende hasta abril (Fig. 2), y una temperatura promedio
de 24,2 °C con extremos de 38 °C y mínimas de 3°C que ocasionalmente puede existir heladas
y presenta vientos predominantes del noroeste a una velocidad de 10-25 km/h.
La geología pertenece a la segunda fase de deposición cuaternaria, donde el sistema de drenaje
actual está asociado con depósitos de arena, limo y arcilla (Aguilera 2001), que corresponden
al aluvión reciente que se encuentra en la mayor parte del Departamento de Santa Cruz. Estas
zonas, constituyen la matriz del paisaje vegetal, ocupando suelos medianamente drenados a
algo imperfectamente drenados, con texturas dominantes arcillo-limosas, franco-limosas o
franco-arcillosas y desarrollo más o menos marcado de microrelieves gilgai (sartenejal)
(Navarro y Ferreira 2007).

10
Fig. 1. Ubicación del área de estudio
Fuente: SENAMHI, 2013
Fig. 2. Clima diagrama base estaciones meteorológicas de Pailón y Tres Cruces
El bosque presenta, dosel y estrato medio que están, de cierta manera, fusionados, aunque con
especies emergentes bien contrastantes. El dosel promedio posee una altura entre 5 y 7 m; las

11
especies más comunes son el Alcaparra (Capparis speciosa), Sao (Trithrinax schizophylla),
Choroquete (Ruprechtia triflora); los árboles emergentes se presentan de manera aislada y
miden hasta 12 m, y son en su mayoría individuos de Cacha (A. quebracho-blanco) y Cala
(Diplokeleba floribunda). Entre otras especies como Guayacán morado (Bulnesia sarmientoi),
Cuchi (Astronium urundeuva), Cuta (Phyllostylom rhamnoides), Curupaú (A. colubrina),
Jichituriqui (Aspidosperma spp.), Algarrobillo (Caesalpinia poraguariensis) además de cactus
arbóreos como Carapari (Stetsonia corine) y Oreja de perro (Quiabentia verticillata).
4.2 Materiales y equipo
Los materiales utilizados fueron arboles de cacha (Aspidosperma quebracho blanco),
motosierra para obtener las rodajas, marcadores indelebles, cinta métrica, cinta diamétrica,
brújula y GPS. El análisis tecnológico fue realizado en el Laboratorio de Tecnología de la
Madera de la Carrera de Ingeniería Forestal de la Universidad Autónoma Gabriel Rene
Moreno. Los materiales utilizados para la preparación de las rodajas fueron amoladora y
lijadora de bandas con lijas de diferente granulometría (60, 120, 240, 600 y 1200) y en el
Laboratorio de Dendrocronología los materiales, equipos y programas utilizados fueron
estéreomicroscopio Leica, sistema de medición LINTAB 6 acoplado al software TSAP-Win,
programa COFECHA (Holmes 1983), Programa AGE del Dendrochronology Program Library
(DPL) y Microsoft Excel 2010.
4.3 METODOS
4.3.1 Obtención de la muestra
La muestra estuvo conformada por 15 árboles seleccionados al azar (5 árboles del bosque y 10
árboles de las cortinas rompe vientos) de diferentes diámetros (29-59 cm). Se registraron
datos de diámetro a la altura del pecho (DAP), altura del fuste, altura total, calidad de fuste,
inclinación del fuste, posición sociológica, estado fitosanitario, forma de copa, iluminación de
la copa y dirección de los vientos. También se tomaron datos de DAP y altura total del árbol
competidor 1 más cercano, árbol competidor 2 más cercano y sus distancias en relación al
árbol en estudio, para el cálculo de índice espaciamiento entre árboles vecinos.
Debido a la dureza de la madera y al crecimiento irregular en árboles tropicales (Brienen y
Zuidema 2003), se decidió trabajar con rodajas en lugar de tarugos extraído con barreno. Para
extraer las rodajas se derribaron los árboles con una motosierra y se tomaron rodajas de 5 a
10 cm de grosor a la altura de 30 cm del suelo, todas las rodajas se codificaron previamente en
campo y fueron colectadas en octubre del año 2011 (Fig. 3).

12
Fig. 3. Obtención de las rodajas y codificado de las mismas
4.3.2 Preparación de las rodajas
Las rodajas fueron transportadas al Laboratorio de Tecnología de la Madera, donde se siguió
los procedimientos descritos por Stokes y Smiley (1968). Para eliminar el contenido de
humedad de las rodajas estas fueron depositadas bajo sombra a temperatura ambiente. Una vez
secas, las rodajas fueron lijadas con lija N° 60 para uniformizar grietas dejadas por la
motosierra. Luego se las pulió con una lijadora de banda, con lijas de granulometría mediana a
muy fina hasta obtener una superficie lisa que permita visualizar los anillos de crecimiento
(Fig. 4).
a) Lijado de las rodajas b) Rodaja lijada, anillos de crecimiento visibles
Fig. 4: Lijado y pulido de las rodajas de A. Quebracho blanco
a
b

13
4.3.3 Conteo y datación
Para realizar el conteo de anillos de crecimiento las rodajas fueron transportadas al laboratorio
de dendrocronología. Para el conteo de los anillos se trazaron 3 radios en cada rodaja desde la
corteza hasta la medula, de tal manera que el promedio de los 3 radios represente el radio
promedio del árbol (Fig. 5a).
A partir del año 2010, con la ayuda de un estéreo microscopio Leica, se fueron marcando los
anillos con lápiz. Para identificar los anillos sobrepuestos que no se observaban, se humedeció
con agua permitiendo así, mejor visibilidad de los anillos de crecimiento durante el conteo en
cuenta regresiva desde la parte externa del leño hasta llegar al primer año de formación en la
medula (Fig. 5b). La datación se realizó según la convención de Schulman (1956), a cada
anillo se le asignó el año que se inició el crecimiento, por ejemplo el anillo asignado al 2010
corresponde al periodo de crecimiento que incluye la primavera del 2010 y el verano del 2011.
Cada década fue marcado con un punto, medio siglo con dos puntos y cada siglo con tres
puntos para facilitar el proceso de medición.
a) Trazado de los radios en la rodaja b) Conteo de anillos de crecimiento
Fig. 5: Conteo de anillos en las rodjas de A. quebracho blanco
4.3.4 Medición de los anillos de crecimiento
La medición del ancho de los anillos de crecimiento se realizó con la ayuda del
estéreomicroscopio Leica que ajusta el ángulo de medición perpendicular de los anillos de
crecimiento y el sistema de medición LINTAB 6 acoplado al software TSAP-Win con una
precisión de 0,01 mm, quedando así registrado la medida en archivo, el cual es fuente para
otros programas. También este programa realiza el control del fechado y permite hacer la
identificación de los segmentos de las series de anillos que tienen baja correlación con la serie
maestra y puede representar los posibles errores en las mediciones (Fig. 6).

14
a) Rodaja datada b) Medición de ancho de anillos
Fig. 6: Datación y medición de los anillos
4.3.5 Análisis estadístico
Para la identificación de posibles errores tales como la presencia de anillos ausentes o falsos se
utilizó COFECHA (Holmes 1983), que permite calcular la correlación entre las series
individuales de ancho de anillos (Grissino-Mayer 2001). La misma permitió realizar las
correcciones necesarias y sincronizar las series de anchos de anillos de todos los árboles. En
base a los datos obtenidos de la medición del ancho de los anillos de crecimiento, se
promediaron los crecimientos de los 3 radios de las rodajas, para obtener el crecimiento
acumulado en diámetro que resulta de la sumatoria del ancho de los anillos de crecimiento.
Por tanto el crecimiento acumulado en diámetro (CAD) se expresa de la siguiente manera:
CAD = ∑ A1+A2+……+An
Donde: CAD=Crecimiento acumulado en diámetro
A1 es el ancho del primer anillo de crecimiento,
A2 el del segundo anillo y
An el ancho del anillo formado al año n.
Luego los crecimientos acumulados en diámetro fueron ordenados por la edad biológica, en el
entendido que la edad biológica representa el asignar al anillo central cerca a la medula el año
t= 1 de cada serie y no por su fecha calendario. El análisis de ancho de anillos se realizó en 30
series para las cortinas y 15 series para el bosque por año calendario, de las cuales se
obtuvieron promedios para cada sitio para detectar diferencias en el ancho de anillos de
crecimiento. También se tomaron promedios de los últimos 18 años, que son años que han
transcurrido desde el establecimiento de las cortinas en el año 1992 hasta la corta de los
árboles en el año 2011.
El Programa AGE de la biblioteca de programas de Dendrocronología (DPL), permite evaluar
el crecimiento por la edad biológica de los individuos, asignándole el año 1 al primer anillo en
cada muestra (Holmes 1999). Este programa se empleó para calcular el crecimiento
acumulado en diámetro (CA), el incremento corriente anual (ICA) e incremento medio anual
(IMA) para cada sitio y un promedio para la zona de estudio. El ICA resulta de restar el

15
crecimiento acumulado (CA) correspondiente al año menos el tamaño que tenía el árbol en el
año anterior t-1 y este corresponde al incremento anual para un determinado año. Mientras que
el IMA resulta de dividir el incremento acumulado (CA) para el año t por t años. Como se
muestran en las siguientes ecuaciones:
ICA=CAt -CAt-1 IMA=CAt /t
Con el propósito de reducir en las series de ancho de anillos su variabilidad debido a las
variaciones climáticas o la dinámica del rodal (variaciones de periodos cortos o de alta
frecuencia) en las estimaciones del IMA, las series de ancho de anillos fueron suavizados
mediante el uso de un filtro digital spline de 32 años. Con el fin de maximizar la tendencia
biológica del crecimiento (variaciones de periodos largos o de baja frecuencia).
Para conocer el espacio vital del árbol objeto y entre árboles vecinos competidores en las
cortinas y el bosque, primero se analizó para cada árbol objeto y su competidor 1 y 2
aplicando el índice de Pelz. Luego se obtuvo un promedio de los individuos analizados para
cada uno de los sitios.
Ko= 0.5x∑So_i x [Do2/ (Do2+Di2)] Pelz, R. (1978)
Dónde: Ko= Distancia de espacio vital del árbol objeto So_i=Distancia del árbol objeto con el competidor Do=Diámetro del árbol objeto o referencia Di=Diámetro del árbol competidor según su proximidad al árbol objeto (1,2)
5. RESULTADOS Y DISCUSION
5.1 Caracterización de los anillos de crecimiento
La estructura de los anillos de crecimiento de A. quebracho-blanco, observados y analizados
en las rodajas, se caracterizan por presentar un leño inicial y tardío difícilmente diferenciado.
Presentando una porosidad difusa visibles a simpe vista (con lupa de 10X). El límite de los
anillos se observan como líneas oscuras macroscópicamente y a nivel microscópico se
caracterizó por la concentración de estratos de fibras comprimidas (Fig. 7a, b y c).
La estructura anatómica de A. quebracho-blanco fue descrita por (Richter y Dallwitz 2000,
Moglia et al. 2009) quienes la caracterizaron por tener una porosidad difusa no uniforme y los
anillos de crecimiento delimitados por 2 a 5 estratos de fibras comprimidas con paredes
gruesas. Las muestras del leño de esta especie presentan una gran variabilidad en los anillos
de crecimiento, siendo estos más estrechos próximos a la medula y la corteza (Fig. 7c). Esta
variabilidad en el incremento demuestra la sensibilidad de los árboles a las condiciones de
factores climáticos y de sitio que influyen en el crecimiento. Por otra parte presentaron
cicatrices causado por disturbios pasados en el límite de los anillos en algunas rodajas,
estableciendo que los anillos de crecimiento de esta especie se forman anualmente (Fig. 7c).

16
a) Microscópica b) Macroscópica
c) Cicatrices y variabilidad de anillos de crecimiento
Fig. 7: Estructura anatómica de anillos de crecimiento de A. quebracho-blanco
También se encontraron anillos falsos, discontinuos, lentes de crecimiento (López, 2011,
López y Villalba 2011, Paredes, 2012), que tienen características similares a las bandas
anuales y pueden ser fácilmente confundidas como anillos anuales si se analiza un solo radio
trazado. También se encontró sobre posición de los anillos, lo que demoró su identificación y
medición donde el crecimiento es excéntrico. Este crecimiento excéntrico de la especie se
debió probablemente a la necesidad de mantener el fuste recto y cilíndrico (Moglia et al.
2009).
5.2 Crecimiento de la especie en bosque natural y cortinas rompe
vientos
En la Fig. 8 se muestra el crecimiento acumulado en diámetro de 10 árboles de las cortinas
(líneas negras) y 5 árboles del bosque (líneas verdes). El ejemplar de mayor edad alcanzó 234
años y todos los árboles alcanzaron la edad bilógica de 100 años (flecha de puntos vertical),
donde se observaron una gran variabilidad en los ritmos de crecimiento. Si observamos los dos
sitios los ritmos de crecimiento varían entre 10 y 35 cm para una de 100 años. Los árboles de
rápido crecimiento podrían alcanzar diámetros de 40 cm entre 110 y 160 años. Sin embargo
los árboles de lento crecimiento demoran una edad mayor a 180 años (Fig. 8).
c

17
Fig. 8: Crecimiento acumulado en diámetro y edad
En la Fig. 8, la línea horizontal indica el Diámetro Mínimo de Corte (DMC) de A. quebracho-
blanco en Bolivia de 40 cm.
Así mismo se presenta el ancho de anillos de crecimiento promedio para cada árbol y el
diámetro alcanzado (Cuadro 1). Para los árboles de las cortinas y del bosque, la variación en el
crecimiento acumulado en diámetro en árboles individuales es muy grande. Si aplicamos el
diámetro mínimo de corta de 40 cm establecido por la norma técnica 248/98 de la Ley
Forestal de Bolivia (MDSP 1998) para esta especie, su crecimiento es muy lento, alcanzando
el diámetro mínimo de corta a edades comprendidas entre 110 y 220 años. Resultados
similares fueron encontrados por Brienen y Zuidema (2003) en la Amazonia boliviana donde
C. odorata necesita entre 80 a 140 años para alcanzar un DMC de 60 cm. Asimismo Brienen
et al. (2003) encontraron esta variación en Tetragastris altissima, Astronium lecointei, y C.
racemosa que demoran 140 años o más para llegar a su DMC. También P. heterophylla
necesita de 114 a 198 años para un DMC de 50 cm, C. racemosa de 112 a 241 años para un
DMC de 45cm mientras que C. catenaeformis alcanza un DMC de 60 cm en aproximadamente
30 a 107 años (Rozendaal et al. 2010).

18
Cuadro 1: Ancho de anillos de crecimiento promedio de árboles individuales de las
cortinas y el bosque
ArbolesAncho anillos
Promedio (mm)
Desviación estandar
ancho anillos
Numero de
anillos DAP (cm)
AQCSC1 0,95 0,523 234 44,74
AQCSC2 1,3 0,925 192 49,98
AQCSC3 1,84 1,291 146 53,95
AQCSC4 0,84 0,828 218 36,71
AQCSC5 1,07 0,984 199 42,77
AQCSC6 0,96 0,798 157 30,17
AQCSC7 1,22 0,888 187 45,74
AQCSC8 0,88 0,722 177 31,18
AQCSC9 1,33 1,249 147 39,18
AQCSC10 0,94 0,919 100 18,90
AQBSC1 0,96 0,674 169 32,58
AQBSC2 1,03 0,757 148 30,43
AQBSC3 0,99 0,839 178 35,19
AQBSC4 0,85 0,506 169 28,83
AQBSC5 0,77 0,525 204 31,22
Para los distritos biogeográficos del cerrado boliviano López (2011), observó en el distrito
biogeográfico Chiquitano que A. cearensis y Cedrela fissilis, alcanzan los DMC de 40 cm en
60–80 años. En cambio, especies con bajas tasas de crecimiento como C. chodatiana y Z.
tuberculosa, requieren entre 80–120 y 80–130 años de edad respectivamente, para alcanzar los
DMC establecidos para este distrito. En el distrito biogeográfico de Guarayos la mayoría de
las especies alcanzan los DMC a edades superiores a los 80 años. Sin embargo, C.
microchaete necesita edades superiores a 80 y 100 años para alcanzar DMC de 40 y 50 cm,
variando entre sitios y distritos biográficos del cerrado (López et al. 2011).
Villagra et al. (2002) afirman que la variabilidad se atribuye al incremento corriente que es
muy variable entre distintos bosques y entre individuos del mismo bosque. Es evidente que el
tiempo requerido para alcanzar el DMC coincide con la mayoría de los estudios reportados.
Aunque los resultados observados en A. quebracho-blanco concuerda más con los reportados
por Rozendaal et al. (2010) en C. racemosa. Demostrando así, que el crecimiento de esta
especie es más lento que lo establecido en los planes de manejo forestal.
5.3 Ancho de anillos de crecimiento en los dos sitios de estudio.
El control de calidad del ancho de anillos de crecimiento de 45 series de árboles de las cortinas
y el bosque no presentó una señal común, observándose pocos árboles de mayor edad (Fig.
9a). Estas mismas series promediadas para cada sitio, mostraron un periodo relativamente bajo
en el crecimiento del bosque en los años 1810 a 1835 seguido por un aumento hasta el año
1885 (Fig. 9b). Sin embargo, durante los últimos 18 años, se observó un bajo crecimiento en el
año 2003 en las cortinas, obteniendo un promedio de 1,50 y 1,17 mm para las cortinas y el
bosque respectivamente (Fig. 10).
Las variaciones en el ancho de anillos de las series analizadas son muy altas (Fig. 8), las
cuales fueron causadas por el crecimiento excéntrico observado en algunas rodajas en A.
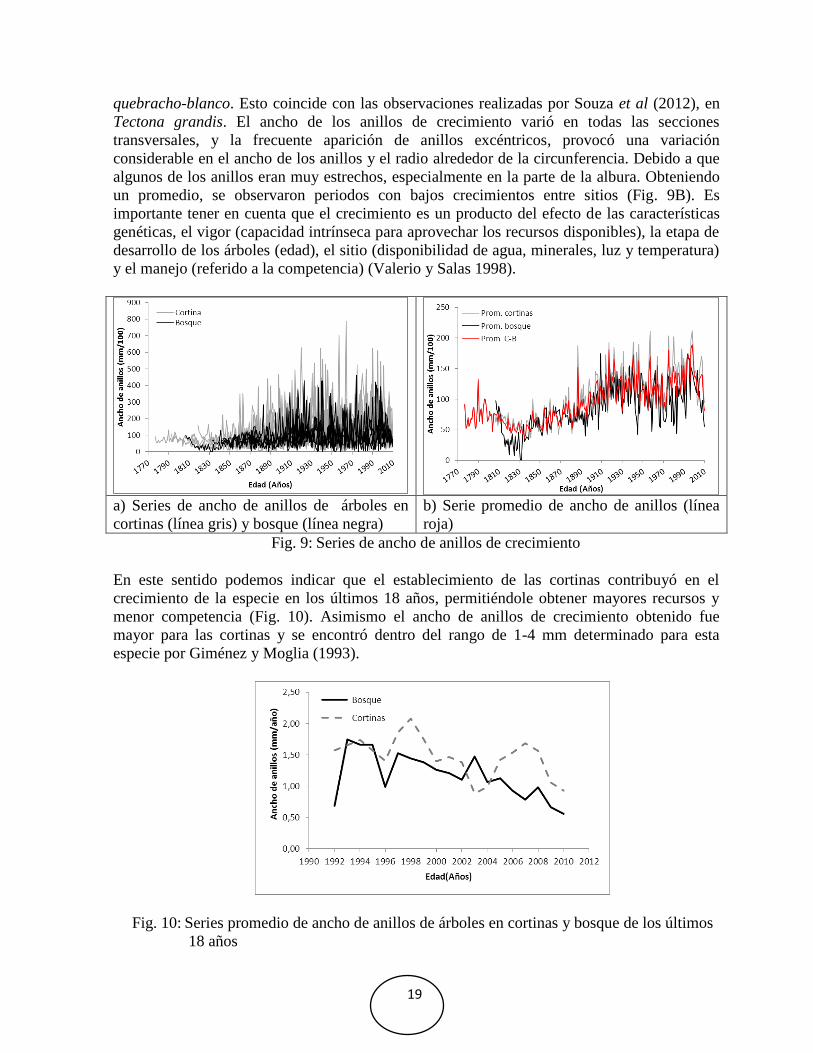
19
quebracho-blanco. Esto coincide con las observaciones realizadas por Souza et al (2012), en
Tectona grandis. El ancho de los anillos de crecimiento varió en todas las secciones
transversales, y la frecuente aparición de anillos excéntricos, provocó una variación
considerable en el ancho de los anillos y el radio alrededor de la circunferencia. Debido a que
algunos de los anillos eran muy estrechos, especialmente en la parte de la albura. Obteniendo
un promedio, se observaron periodos con bajos crecimientos entre sitios (Fig. 9B). Es
importante tener en cuenta que el crecimiento es un producto del efecto de las características
genéticas, el vigor (capacidad intrínseca para aprovechar los recursos disponibles), la etapa de
desarrollo de los árboles (edad), el sitio (disponibilidad de agua, minerales, luz y temperatura)
y el manejo (referido a la competencia) (Valerio y Salas 1998).
a) Series de ancho de anillos de árboles en
cortinas (línea gris) y bosque (línea negra)
b) Serie promedio de ancho de anillos (línea
roja)
Fig. 9: Series de ancho de anillos de crecimiento
En este sentido podemos indicar que el establecimiento de las cortinas contribuyó en el
crecimiento de la especie en los últimos 18 años, permitiéndole obtener mayores recursos y
menor competencia (Fig. 10). Asimismo el ancho de anillos de crecimiento obtenido fue
mayor para las cortinas y se encontró dentro del rango de 1-4 mm determinado para esta
especie por Giménez y Moglia (1993).
Fig. 10: Series promedio de ancho de anillos de árboles en cortinas y bosque de los últimos
18 años

20
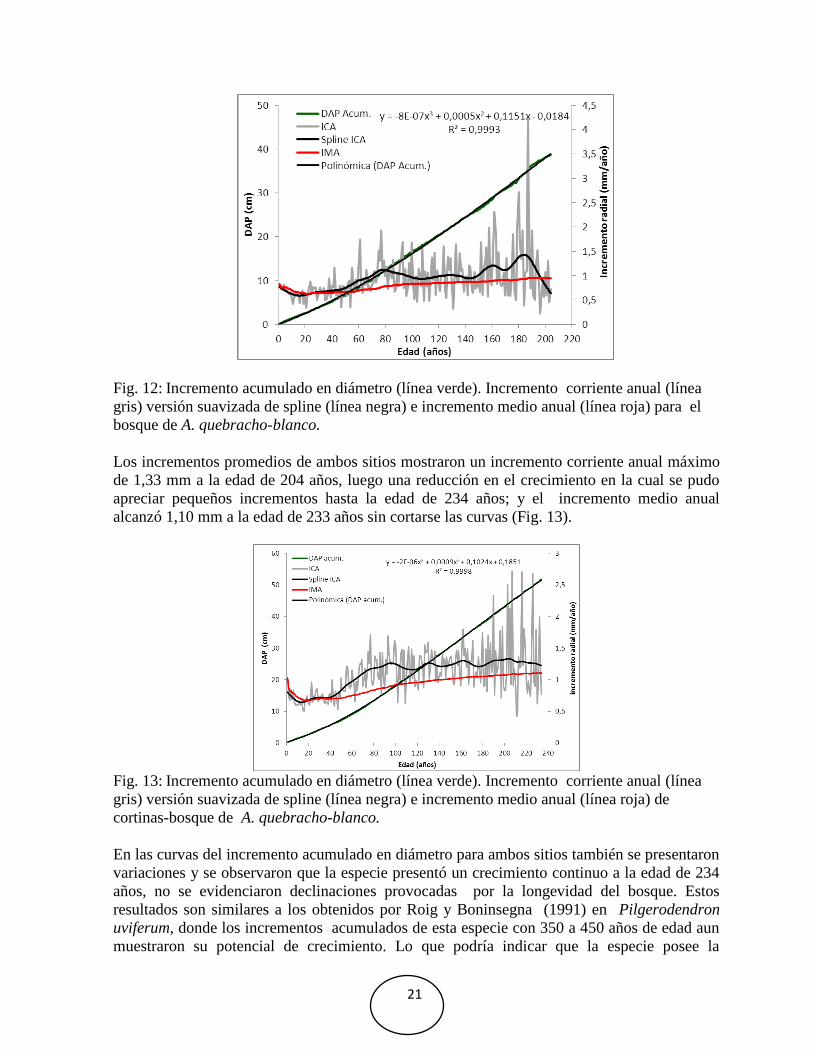
5.4 Incremento en diámetro de la especie en estudio.
El incremento acumulado en diámetro de las series analizadas presentó variación entre sitios.
Para el diámetro de 40 cm establecido por la norma técnica 248/98 para esta especie, las
edades oscilaron entre 180 años en las cortinas (Fig. 11) y una edad de 200 años para el
bosque (Fig. 12) y el promedio de los dos sitios a una edad de 190 años (Fig. 13). Estas
curvas se ajustan a una ecuación polinómica de tercer orden, donde el coeficiente de
determinación en cada sitio y promedio es cercano a 1 (Fig. 11, 12 y 13), lo que nos indica el
buen ajuste del modelo a los datos, que al 99 % de la variación del diámetro está en función
de la edad de los árboles.
El incremento corriente anual en las cortinas alcanzó un máximo de 1,37 mm a la edad de 98
años, observándose una reducción en el crecimiento en la cual se pudo apreciar pequeños
incrementos; y el incremento medio anual alcanzó 1,13 mm a la edad de 233 años sin cortarse
las curvas (Fig. 11). Sin embargo, en el bosque el incremento corriente anual alcanzó un
máximo de 1,43 mm a la edad de 185 años y alcanzando un incremento medio anual de 0,95
mm, cortándose las curvas a la edad de 195 años (Fig. 12).
Fig. 11: Incremento acumulado en diámetro (línea verde). Incremento corriente anual (línea
gris) versión suavizada de spline (línea negra) e incremento medio anual (línea roja) para
cortinas de A. quebracho-blanco.
21
Fig. 12: Incremento acumulado en diámetro (línea verde). Incremento corriente anual (línea
gris) versión suavizada de spline (línea negra) e incremento medio anual (línea roja) para el
bosque de A. quebracho-blanco.
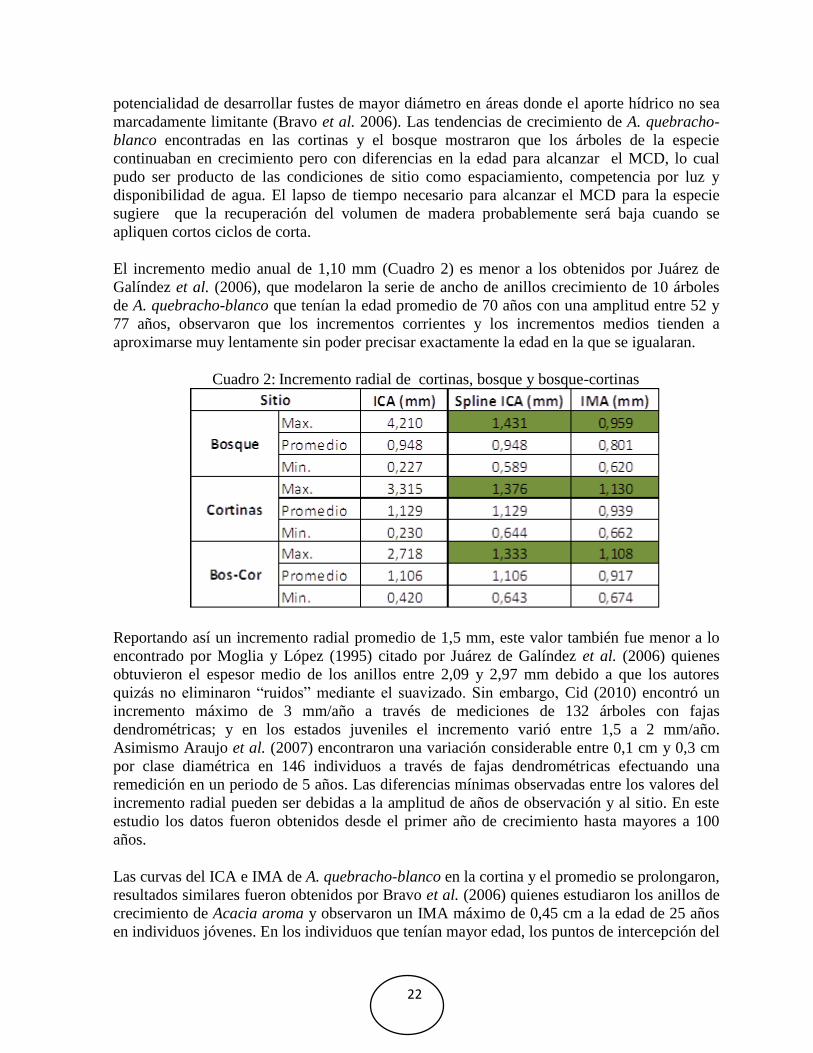
Los incrementos promedios de ambos sitios mostraron un incremento corriente anual máximo
de 1,33 mm a la edad de 204 años, luego una reducción en el crecimiento en la cual se pudo
apreciar pequeños incrementos hasta la edad de 234 años; y el incremento medio anual
alcanzó 1,10 mm a la edad de 233 años sin cortarse las curvas (Fig. 13).
Fig. 13: Incremento acumulado en diámetro (línea verde). Incremento corriente anual (línea
gris) versión suavizada de spline (línea negra) e incremento medio anual (línea roja) de
cortinas-bosque de A. quebracho-blanco.
En las curvas del incremento acumulado en diámetro para ambos sitios también se presentaron
variaciones y se observaron que la especie presentó un crecimiento continuo a la edad de 234
años, no se evidenciaron declinaciones provocadas por la longevidad del bosque. Estos
resultados son similares a los obtenidos por Roig y Boninsegna (1991) en Pilgerodendron
uviferum, donde los incrementos acumulados de esta especie con 350 a 450 años de edad aun
muestraron su potencial de crecimiento. Lo que podría indicar que la especie posee la
22
potencialidad de desarrollar fustes de mayor diámetro en áreas donde el aporte hídrico no sea
marcadamente limitante (Bravo et al. 2006). Las tendencias de crecimiento de A. quebracho-
blanco encontradas en las cortinas y el bosque mostraron que los árboles de la especie
continuaban en crecimiento pero con diferencias en la edad para alcanzar el MCD, lo cual
pudo ser producto de las condiciones de sitio como espaciamiento, competencia por luz y
disponibilidad de agua. El lapso de tiempo necesario para alcanzar el MCD para la especie
sugiere que la recuperación del volumen de madera probablemente será baja cuando se
apliquen cortos ciclos de corta.
El incremento medio anual de 1,10 mm (Cuadro 2) es menor a los obtenidos por Juárez de
Galíndez et al. (2006), que modelaron la serie de ancho de anillos crecimiento de 10 árboles
de A. quebracho-blanco que tenían la edad promedio de 70 años con una amplitud entre 52 y
77 años, observaron que los incrementos corrientes y los incrementos medios tienden a
aproximarse muy lentamente sin poder precisar exactamente la edad en la que se igualaran.
Cuadro 2: Incremento radial de cortinas, bosque y bosque-cortinas
Reportando así un incremento radial promedio de 1,5 mm, este valor también fue menor a lo
encontrado por Moglia y López (1995) citado por Juárez de Galíndez et al. (2006) quienes
obtuvieron el espesor medio de los anillos entre 2,09 y 2,97 mm debido a que los autores
quizás no eliminaron “ruidos” mediante el suavizado. Sin embargo, Cid (2010) encontró un
incremento máximo de 3 mm/año a través de mediciones de 132 árboles con fajas
dendrométricas; y en los estados juveniles el incremento varió entre 1,5 a 2 mm/año.
Asimismo Araujo et al. (2007) encontraron una variación considerable entre 0,1 cm y 0,3 cm
por clase diamétrica en 146 individuos a través de fajas dendrométricas efectuando una
remedición en un periodo de 5 años. Las diferencias mínimas observadas entre los valores del
incremento radial pueden ser debidas a la amplitud de años de observación y al sitio. En este
estudio los datos fueron obtenidos desde el primer año de crecimiento hasta mayores a 100
años.
Las curvas del ICA e IMA de A. quebracho-blanco en la cortina y el promedio se prolongaron,
resultados similares fueron obtenidos por Bravo et al. (2006) quienes estudiaron los anillos de
crecimiento de Acacia aroma y observaron un IMA máximo de 0,45 cm a la edad de 25 años
en individuos jóvenes. En los individuos que tenían mayor edad, los puntos de intercepción del
23
ICA e IMA ocurrieron a edad más avanzada sugiriendo que la edad de culminación de
crecimiento en individuos de mayor edad puede prolongarse. Además los cambios bruscos en
las curvas y su finalización en el bosque podrían atribuirse posiblemente al bajo número de
muestras promediadas (Roig y Boninsegna 1991). Sin embargo, Villagra et al. (2002) afirma
que esto puede atribuirse a la variabilidad propia de los individuos de la misma población.
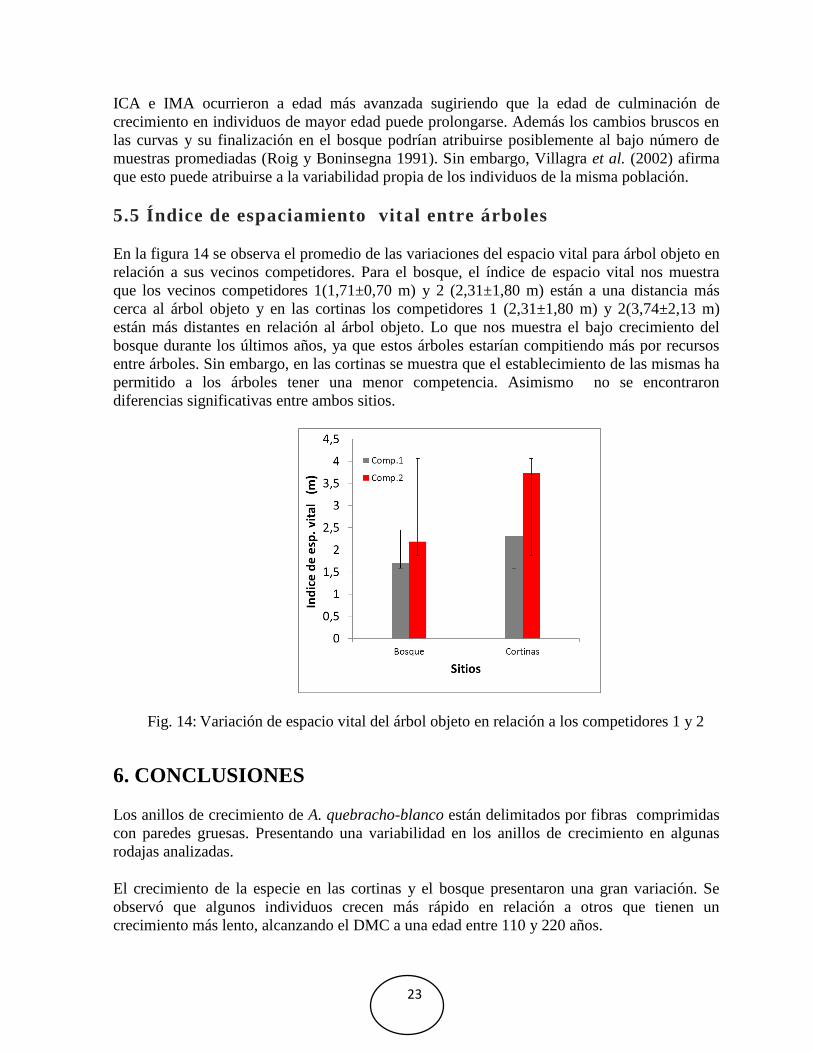
5.5 Índice de espaciamiento vital entre árboles
En la figura 14 se observa el promedio de las variaciones del espacio vital para árbol objeto en
relación a sus vecinos competidores. Para el bosque, el índice de espacio vital nos muestra
que los vecinos competidores 1(1,71±0,70 m) y 2 (2,31±1,80 m) están a una distancia más
cerca al árbol objeto y en las cortinas los competidores 1 (2,31±1,80 m) y 2(3,74±2,13 m)
están más distantes en relación al árbol objeto. Lo que nos muestra el bajo crecimiento del
bosque durante los últimos años, ya que estos árboles estarían compitiendo más por recursos
entre árboles. Sin embargo, en las cortinas se muestra que el establecimiento de las mismas ha
permitido a los árboles tener una menor competencia. Asimismo no se encontraron
diferencias significativas entre ambos sitios.
Fig. 14: Variación de espacio vital del árbol objeto en relación a los competidores 1 y 2
6. CONCLUSIONES
Los anillos de crecimiento de A. quebracho-blanco están delimitados por fibras comprimidas
con paredes gruesas. Presentando una variabilidad en los anillos de crecimiento en algunas
rodajas analizadas.
El crecimiento de la especie en las cortinas y el bosque presentaron una gran variación. Se
observó que algunos individuos crecen más rápido en relación a otros que tienen un
crecimiento más lento, alcanzando el DMC a una edad entre 110 y 220 años.

24
Se observaron periodos de bajo crecimiento en el bosque, pero en los últimos 18 años las
cortinas también presentaron bajo crecimiento siendo el más visible el año 2003. Obteniendo
un promedio de 1,50 y 1,17 mm para las cortinas y el bosque.
El incremento acumulado en diámetro para ambos sitios presentó variación alcanzando el
diámetro de 40 cm a diferentes edades. El promedio de ambos sitios alcanzó el diámetro de
40 cm a la edad de 190 años, observándose que la especie aun muestra un crecimiento
continuo con la potencialidad de desarrollar fustes de mayor diámetro. Sugiriendo así, un ciclo
de corta para la especie mayor a los 20 años a los establecidos en la Ley forestal.
Los incrementos también presentaron variación entre sitios. Sin embargo, analizando ambos
sitios se observó que las curvas no tienden a cortarse, presentando un incremento corriente
máximo de 1,33 mm a la edad de 204 años y un incremento anual medio máximo de 1,10 mm
a la edad de 233 años, prolongándose así la culminación del crecimiento.
El índice de espaciamiento vital mostró que los arboles competidores del bosque están más
cerca del árbol objeto en relación a las cortinas y no se encontró diferencias significativas
entre ambos sitios.
8. LITERATURA CITADA
Araujo, P., Juárez de Galindez, M. y Iturre, M. 2007. Crecimiento de las especies principales
de un bosque en regeneración del Chaco Santiagueño. Quebracho 14:36-46.
Argollo, J. y Miranda, G. 2002. Dendrocronología como instrumento de reconstrucción de
paleoambientes. En: BOLFOR (Eds.). Avances y perspectivas para la investigación
forestal en Bolivia. BOLFOR, Santa cruz, Bolivia.
Argollo. J., Solíz, C., y Villalba, R. 2004. Potencialidad dendrocronologica de Polylepis
tarapacana en los andes centrales de Bolivia. Ecología en Bolivia, 39(1):5-24.
Beltrán, L. y Valencia, G. 2013. Anatomía de anillos de crecimiento de 80 especies arbóreas
potenciales para estudios dendrocronológicos en la Selva Central, Perú. Biología
Tropical 61(3): 1025-1037.
Boninsegna, J.A., Villalba,R., Amarilla, P. y Ocampo, J. 1989. Studies on tree rings, growth
rates and age-size relationships of tropical tree species in Misiones, Argentina. IAWA
Bulletin 10(2): 161-169.
Botosso, P. 2009. Identificacao macroscópica de madeiras: guía prático e nocoes básicas para
o seu reconocimiento. Documentos 194, Colombo: EMBRAPA, Brasil. 25 pag.
Bowman, D., Brienen, R.J.W., Gloor, E. Phillips, O. y Prior, L. 2013. Detecting trends in tree
growth: not so simple. Trends in Plant Science 18(1):11-17.
Bravo, S., Gimenez, A. y Moglia, J. 2006. Caracterizacion antomica del leño y evolución del
crecimiento en ejemplares de Acacia aroma y Acacia furcatispina en la región
chaqueña, Argentina. Bosque 27(2): 146-154.
Brienen, R.J.W. and Zuidema, P. 2006a. Lifetime growth patterns and ages of Bolivian rain
forest trees obtained by tree ring analysis. Journal of Ecology 94: 481-493.

25
Brienen, R.J.W. and Zuidema, P. 2006b. The use of tree rings in tropical forest management:
projecting timber yields of four Bolivian tree species. Forest Ecology and Management
226: 256–267.
Brienen, R.J.W. y Zudiema, P. 2003. Anillos de crecimiento de árboles maderables en Bolivia:
su potencial para el manejo de bosques y una guía metodológica. PROMAB/IGEMA,
informe técnico Nro. 7, Riberalta, Bolivia. 33 pp.
Brienen, R.J.W. y Zuidema, P. 2005. Relating tree growth to rainfall in Bolivian rain forests: a
test for six species using tree ring analysis. Ecología 146: 1-12.
Brienen, R.W. Zuidema, P. y Leigue, J. 2003. Recuperación del volumen madera bajo
diferentes ciclos de corta: resultados de simulaciones para seis especies maderables en el
norte de Bolivia. PROMAB, Informe Técnico No. 9, Riberalta, Bolivia. 32 pág.
Brienen, R.J.W., Lebrija-Trejos, E., Breugel, M., Pérez-García, E., Bongers, F., Meave, J. and
Martínez-Ramos, M. 2009. The potential of tree rings for the study of forest succession
in Southern Mexico. BIOTROPICA 41(2):186-195.
Eckstein, D., Ogden, J., Jacoby, G. y Ash, J. 1983. Determinación de la edad y ritmo de
crecimiento de árboles tropicales: La aplicación de métodos dendrocronológicos Pp. 86-
109. En: Bormann, F. H. y Berlin, G. (Eds.). Edad y tasa de crecimiento de los árboles
tropicales. Ed. CONTINENTAL. México, pp. 110-124
Fahn, A., Burley, J., Allan, K., Mariaux, A. y Barry, P. 1983. Posibles contribuciones de la
anatomía de la madera a la determinación de la edad de los árboles tropicales. En:
Bormann, F. H. y Berlin, G. (Eds.). Edad y tasa de crecimiento de los árboles tropicales.
Ed. CONTINENTAL. México, pp. 31-55.
Fichtler, E., Clark, D. y Worbes, M. 2003. Age and long-term growth of trees in a old-growth
tropical rain forest, based on analyses of tree rings and 14
C1. Biotropica 35: 306-317.
Giménez, A.M. y Moglia J. G. 1993. Determinación de patrones de crecimiento de especies
leñosas arbóreas de la región chaqueña seca. Yvyrareta: 4(4) 46-60.
Gutiérrez, E. 2008. La dendrocronología: métodos y aplicaciones. En: Nieto, X. y Cau, M.
(Eds.). Arqueología náutica mediterránea. UB, Barcelona. pp. 303-315.
Imaña, J. y Encinas, O. 2008. Epidometria forestal. Universidade de Brasilia, Departamento de
Engenharia Florestal, Brasilia. 68 pág.
Juárez de Galíndez, M., Moglia, J., Gimenez, A. M. y Pece, M. 2006. Comparación de dos
modelos de crecimiento de efectos fijos y errores independientes en quebracho blanco.
Revista Forestal Venezolana 50(1):65-73.
Killen, J., García, E. y Beck, G.1993. Guía de árboles de Bolivia. Publicado por Hervario
Nacional de Bolivia patrocinado por LIDEMA, La Paz, Bolivia. 92 pág.
Klepac, D. 1983. Crecimiento e incremento de árboles y masas forestales. Universidad
Autónoma de Chapingo, Chapingo, México. 365 pág.
Lisi, C., Tomazello, M., Botosso, P., Roig, F., Maria, V., Ferreira-Fedele, L. y Voigt, A. 2008.
Tree-rings formation, radial increment periodicity,and phenology of tree species from a
Seasonal Semi-Deciduos Forest in southeast Brazil. IAWA 29(2):198-207.
López, L. 2003. Estudio de anillos de crecimiento en once especies forestales. Tesis de Grado.
UAGRM. Santa Cruz, Bolivia. 85 p.
López, L. 2011. Una aproximación dendrocronología a la ecología y el manejo de los bosques
tropicales secos del cerrado boliviano. Tesis de doctorado, Centro Regional
Universitario Bariloche, Universidad Nacional del Comahue. Bariloche-Argentina. 280
p.

26
López, L. Villalba, R. y Peña- Claros, M. 2011. Los anillos de crecimiento de Centrolobium
microchaete (Fabaceae, Papilionoideae), una herramienta para evaluar el manejo forestal
de los bosques secos tropicales del cerrado boliviano. Ecología en Bolivia 46(2): 77-94.
López, L., Villalba, R. y Bravo, F. 2013. Cumulative diameter growth and biological rotation
age for seven tree species in the Cerrado biogeographical province of Bolivia. Forest
Ecology and Management 292: 49–55.
López, L., Villalba, R. and Peña-Claros, M. 2012b. Determining the annual periodicity of
growth rings in seven tree species of a tropical moist forest in Santa Cruz, Bolivia.
Forest Systems 21(3): 508-514.
López, L., Villalba, R. y Peña-Claros, M. 2012a. Ritmos de crecimiento diamétrico en los
bosques secos tropicales: aportes al manejo sostenible de los bosques de la provincia
biogeográfica del Cerrado Boliviano. BOSQUE 33(2): 211-219.
Llorente, C. 2012. La historia que nos explican los árboles. Biol. On-line 1(2).
Mariaux, A. 1983. Esfuerzos anteriores en la medida de la edad y el crecimiento anual en los
árboles tropicales. En: Bormann, F. H. y Berlin, G. (Eds.). Edad y tasa de crecimiento de
los árboles tropicales. Ed. CONTINENTAL. México, pp. 20-30.
Miranda, G. y Fernández, M. 2002. Tabebuia para la evidencia de cambios climáticos e
influencias antropogenéticas en islas de bosques en las sabanas húmedas de Moxos. En:
BOLFOR (Eds.). Avances y perspectivas para la investigación forestal en Bolivia.
BOLFOR, Santa cruz, Bolivia.
Miranda, G., Argollo, J., Sanjinés, N. y Muriel, M. 2002. Identificación y estudio de anillos
de árboles que crecen en bosques tropicales de Bolivia. En: BOLFOR (Eds.). Avances y
perspectivas para la investigación forestal en Bolivia. BOLFOR, Santa cruz, Bolivia.
Moglia, J.G., Bravo, S., Giménez, A.M. y López, C. 2009. ¿Son los caracteres estructurales de
la madera de Aspidosperma quebracho-blanco Schelkt causantes de su inestabilidad?.
Quebracho: 17 (1,2) 58-63.
Paredes, K. 2012. Potencial dendrocronológico de Machaerium scleroxylon Tul. (Morado) y
los anillos de crecimiento como indicadores de edad y fluctuaciones climáticas en Santa
Cruz, Bolivia. Tesis de maestría. UAGRM. Santa Cruz. Bolivia.
Paredes-Villanueva, K., Sanchez-Salguero, R., Manzanedo, R.D., Quevedo, R., Palacios, G. y
Navarro-Cerrillo, R.M. 2013. Growth rate and climatic response of Machaerium
scleroxylon in a dry tropical forest in southeasterrn Santa Cruz, Bolívia. TREE-RING
RESEARCH 69(2) 63-79.
Prodan, M.; Peters, R.; Cox, F. y Real, P., 1997. Mensura Forestal. Serie Investigación y
Educación en Desarrollo Sostenible. IICA/BMZ/GTZ. Litografía e Imprenta LIL S.A.
San José, Costa Rica. 561 p.
Richter, H.G., y Dallwitz, M.J. 2000. Commercial timbers: descriptions, illustrations,
identification, and information retrieval. In English, French, German, Portuguese, and
Spanish. Version: 25th June 2009. Consultado el 20 de agosto 2012. Disponible en
http://delta-intkey.com
Ríos, N., Giménez, A.M. y Moglia G. 2001. Crecimiento del Itín (Prosopis kuntsei Harms) en
la región chaqueña Argentina. Madera y Bosques 7(1): 47-56.
Roig, F. 2000. Dendrocronología en los bosques del trópico: revisión y prospección futura.
En: Roig, F. A. (Eds.). Dendrocronología en América Latina. Ed. de la Universidad
Nacional de Cuyo, Mendoza, Argentina, pp. 307-355.

27
Roig, F. y Boninsegna, J. 1991. Estudios sobre el crecimiento radial, basal y en altura de las
condiciones climáticas que afectan el desarrollo de Pilgerodendron uviferum. Revista
Chilena y Historia Natural, 64: 53-63.
Roig, F., Fernández, M., Gareca, E., Altamirano, S. y Monge, S. 2001. Estudios
dendrodronologicos en los ambientes húmedos de la puna Boliviana. Revista Boliviana
de Ecología y Conservación Ambiental, 9: 3-13.
Rozendaal, D.M.A. 2010. Looking backwards: using tree rings to evaluate long-term growth
patterns of Bolivian forest trees. PROMAB Scientific Series 12. Riberalta, Bolivia.
Rozendaal. DMA, Soliz-Gamboa, C. y Zuidema, P.2010. Timber yield projections for tropical
tree species: the influence of fast juvenile growth on timber volume recovery. Forest
Ecology and Management 259: 2292-2300.
Souza, V., Cardoso, S., Quilhó, T. y Pereira H. 2012. Growth rate and ring width variability of
teak, Tectona grandis (Verbenaceae) in an unmanaged forest in East Timor. Revista
Biologia Tropical 60(1): 483-494.
Stahle, D. 1999. Useful strategies for the development of tropical tree-ring crhonologies.
AIWA Journal: 20(3) 249-253.
Szejner, P. 2012. Aplicaciones de la dendrocronología para el manejo sustentable de los
recursos forestales. Curso on-line: Dendrocronología básica aplicada al manejo forestal
tropical. Universidad de Córdova. Córdova, España.
Urritia, R., Villalba, R. y Lara, A. 2005. Cómo ha variado la disponibilidad de agua en la
ecorregión de los bosques valdivianos en los últimos siglos. Revista Ambiente y
Desarrollo 21(3): 48-57.
Valerio, J. y Salas C.1998. Selección de prácticas silviculturales para bosques tropicales.
Manual técnico, Proyecto BOLFOR, Santa Cruz, Bolívia.83 pp.
Villagra, P, Morales, M., Villalba, R. y Boninsegna, J. 2002. Dendroecología de los
algarrobales de la zona árida argentina. Instituto Argentino de Nivología, Glaciología y
Ciencias Ambientales IANIGLA 1973-2002. p. 53-57.
Villalba, R., Delgado, S., Membiela, M. y Mendoza, D. 2006. Variabilidad interanual de los
caracteres anatómicos en el leño de Cedrela lilloi en el noroeste de Argentina. En
Pacheco, S. y Brown, A. (Eds.). Ecologia y producción de cedro (genero cedrela ) en las
yungas australes. LIEY-ProYungas,Argentina. pp. 59-82.
Worbes, M. 1995. How to measure growth dynamics in tropical trees a review. IAWA
Journal 16 (4): 337-351
Worbes, M. 1999. Annual growth rings, rainfall-dependent growth and long-term growth
patterns of tropical trees from the Caparo Forest Reserve in Venezuela. Journal of
Ecology 87: 391–403.
Worbes, M. 2002. One hundred years of tree-rings research in the tropics – a brief history and
an outlook to future challenges. Dendrocronologia 20(1-2): 217-231.
Zuidema, P., Brienen, R.J.W. y Schongart, J. 2012. Tropical forest warning: looking
backwards for more insights. Trends in Ecology and Evolution 27(4): 193-194.

28
s e
El “Documento Científico” es una publicación de la Carrera de Ingeniería Forestal de la Universidad Autónoma Gabriel René Moreno
(UAGRM) de Santa Cruz, Bolivia
Este documento tiene el fin de promover y fortalecer la investigación en ciencias forestales, difundir resultados de investigaciones realizadas por docentes investigadores, estudiantes e investigadores invitados por
la Carrera Forestal
Las opiniones y juicios expresados en este documento son de exclusiva responsabilidad del (los) autor (es) y no reflejan necesariamente la
opinión o política de la UAGRM.
Contactos
Carrera de Ingeniería Forestal
Facultad de Ciencias Agrícolas
Universidad Autónoma “Gabriel René Moreno” (UAGRM)
“Vallecito”, Carretera al Norte Km. 8.5, Santa Cruz de la Sierra, Bolivia
Telf./fax: +591 3 3436363
E-mail: [email protected]
Top Related