







Idiomas
Páginas
Jurídico


1
DIVERSIDAD DE ARAÑAS (ARACHNIDA: ARANEAE) EN UN PAISAJE RURAL
CAFETERO DEL DEPARTAMENTO DE RISARALDA, COLOMBIA
CARLOS ALBERTO GUZMÁN RUIZ
Trabajo de grado presentado como requisito parcial para optar al título de
Biólogo
Director
ALEXANDER SABOGAL-GONZÁLEZ
Magister en Ciencias Agrarias - Entomología
UNIVERSIDAD DEL TOLIMA
FACULTAD DE CIENCIAS
PROGRAMA DE BIOLOGÍA
IBAGUÉ - TOLIMA
2015

2

3
DEDICATORIA
A mis padres Stella y Alberto
Amo la libertad del ser y del pensamiento... Amo la naturaleza que me renueva las
esperanzas... Amo la música que me renueva las energías y las ganas de vivir...
C. Guzmán-Ruiz
"Me convertí en un loco con largos intervalos de horrible cordura"
E.Poe
“Creo que las cosas imposibles se pueden conseguir, que los besos con los ojos
cerrados son los únicos que cuentan, que las heridas no siempre cierran, y que
todo el mundo se enamora alguna vez. Creo que quien te quiere se queda, que
tiene más cuenta buscar a alguien que limitarse a esperarle, y que si no luchas
por esa persona luego no puedes ir llorando por los rincones. Creo que nos
empeñamos en hacer las cosas más difíciles de lo que son. Creo que un mundo
mejor es posible, y sobre todo creo en las personas que luchan para conseguirlo.
Creo que después de llorar siempre hay quién te saca una sonrisa. Creo que hay
que disfrutar del ahora. Creo en el destino, y creo que nosotros mismos lo
elegimos.”
J. Cortazar

4
AGRADECIMIENTOS
A Wildlife Conservation Society Programa Colombia (WCS), MacArthur Foundation y
Fondo de Investigaciones y Desarrollo Científico de la Universidad del Tolima por la
financiación y apoyo logístico para la ejecución de este proyecto.
A la profe Gladys Reinoso Flórez, por su paciencia y apoyo incondicional.
A Alexander Sabogal-González, por su orientación y buena energía.
A las familias campesinas que me abrieron cálidamente las puertas sus hogares para
hospedarme y lograr llevar a buen término la fase de campo de este proyecto.
A los integrantes del Grupo de Investigación en Zoología, y a todas las personas que de
una u otra manera participaron he hicieron posible la realización de éste trabajo.

5
CONTENIDO
INTRODUCCIÓN .......................................................................................................... 15
1. OBJETIVOS........................................................................................................... 18
1.1 OBJETIVO GENERAL 18
1.2 OBJETIVOS ESPECÍFICOS 18
2. MARCO REFERENCIAL ....................................................................................... 19
2.1 ANTECEDENTES Y ESTADO ACTUAL DEL ESTUDIO DE ARAÑAS EN COLOMBIA
19
2.2 BREVE INTRODUCCIÓN AL ORDEN ARANEAE 21
2.3 INTERRELACIONES ECOLÓGICAS DE LAS ARAÑAS 23
2.4 ESTRUCTURA GREMIAL EN ARAÑAS (GRUPOS FUNCIONALES) 24
2.5 EL PAISAJE RURAL 25
3. METODOLOGÍA .................................................................................................... 27
3.1 ÁREA DE ESTUDIO 27
3.1.1 Aspectos físicos. 28
3.1.2 Aspectos bióticos. 28
3.1.3 Coberturas de muestreo. 29
3.1.3.1 Bosque 29
3.1.3.2 Cafetales 30

6
3.2 MÉTODOS DE CAMPO 30
3.2.1 Unidad de muestreo. 31
3.2.2 Métodos de captura 33
3.2.2.1 Colecta manual 33
3.2.2.2 Agitación de vegetación (Beating) 34
3.3 MÉTODOS DE LABORATORIO 36
3.3.1 Organización de las morfoespecies (mfsp) 36
3.3.2 Determinación taxonómica y disposición final de los organismos 37
3.4 ANÁLISIS DE DATOS 37
3.4.1 Abundancia. 37
3.4.2 Evaluación de la diversidad alfa (α) 38
3.4.2.1 Riqueza 38
3.4.2.2 Curvas de acumulación de especies 38
3.4.2.3 Índices de abundancia proporcional 38
3.4.2.4 Modelos de abundancia 39
3.4.3 Evaluación de la diversidad beta (β): 40
3.4.3.1 Índice de similitud de Chao-Jaccard 40
3.4.4 Grupos funcionales de arañas 40
3.4.5 Preferencia de microhábitat 40
3.4.6 Densidad de arañas por unidad de muestreo 41
4. RESULTADOS ...................................................................................................... 42
4.1 COMPOSICIÓN Y ABUNDANCIA DE LAS ARAÑAS EN UN PAISAJE RURAL
CAFETERO 42
4.1.1 Composición 42

7
4.1.2 Abundancia 43
4.2 EVALUACIÓN DE LA DIVERSIDAD ALFA (α) 48
4.2.1 Riqueza específica 48
4.2.2 Estimaciones de riqueza y representatividad del muestreo 49
4.2.3 Índices de abundancia proporcional 52
4.2.4 Modelos de abundancia. 53
4.3 EVALUACIÓN DE LA DIVERSIDAD BETA (β) 54
4.3.1 Índice de similitud de Chao-Jaccard. 54
4.4 GRUPOS FUNCIONALES DE ARAÑAS 54
4.5 PREFERENCIA DE MICROHÁBITAT 57
4.5.1 Bosque 57
4.5.2 Cafetal. 61
4.6 DENSIDAD DE ARAÑAS POR UNIDAD DE MUESTREO 65
5. DISCUSIÓN ........................................................................................................... 66
5.1 COMPOSICIÓN Y ABUNDANCIA DE LAS ARAÑAS EN UN PAISAJE RURAL
CAFETERO 66
5.2 DIVERSIDAD DE LA COMUNIDAD DE ARAÑAS EN UN PAISAJE RURAL
CAFETERO 68
5.3 GRUPOS FUNCIONALES DE LA COMUNIDAD DE ARAÑAS 72
5.4 PREFERENCIA DE MICROHÁBITAT 74
5.5 DENSIDAD DE ARAÑAS POR UNIDAD DE MUESTREO 74
6. CONCLUSIONES .................................................................................................. 76

8
RECOMENDACIONES ................................................................................................. 78
REFERENCIAS ............................................................................................................ 80
ANEXOS ....................................................................................................................... 94

9
LISTA DE TABLAS
Tabla 1 Área de las coberturas presentes en la ventana de trabajo: paisaje rural cafetero
de El Brillante, Risaralda. .............................................................................................. 28
Tabla 2 Coordenadas de las estaciones de muestreo definidas para la caracterización
de la comunidad de arañas en un paisaje rural cafetero de El Brillante, Risaralda. ...... 31
Tabla 3 Tipos de estratos definidos para la caracterización de preferencia por
microhábitat de la comunidad de arañas en las coberturas bosque y cafetal de la ventana
de trabajo: paisaje rural cafetero de El Brillante, Risaralda. .......................................... 41
Tabla 4 Familias colectadas en un paisaje rural cafetero de Risaralda, Colombia. ...... 42
Tabla 5 Morfoespecies presentes en sólo una de las dos coberturas evaluadas en un
paisaje rural cafetero de Risaralda, Colombia. .............................................................. 47
Tabla 6 Estimaciones de riqueza calculadas para las coberturas bosque y cafetal en un
paisaje rural cafetero de Risaralda, Colombia. .............................................................. 51
Tabla 7 Índices de diversidad α para las arañas de un paisaje rural cafetero de Risaralda,
Colombia. ...................................................................................................................... 52
Tabla 8 Índice de similitud de Chao-Jaccard calculado para bosque y cafetal de un
paisaje rural cafetero de Risaralda, Colombia. .............................................................. 54
Tabla 9 Gremios de arañas encontrados en un paisaje rural cafetero de Risaralda,
Colombia. ...................................................................................................................... 55
Tabla 10 Familias y morfoespecies presentes en sólo uno de los tres estratos evaluadas
en el bosque de un paisaje rural cafetero de Risaralda, Colombia. .............................. 59
Tabla 11 Familias y morfoespecies presentes en sólo uno de los dos estratos evaluados
en el cultivo de café de un paisaje rural cafetero de Risaralda, Colombia. ................... 63
Tabla 12 Área total muestreada en un paisaje rural cafetero de Risaralda, Colombia.
...................................................................................................................................... 65
Tabla 13 Número total de arañas colectadas en un paisaje rural cafetero de Risaralda,
Colombia. ...................................................................................................................... 65

10
LISTA DE FIGURAS
Figura 1 Ubicación del área de estudio: paisaje rural cafetero en la vereda El Brillante,
Risaralda. ...................................................................................................................... 27
Figura 2 Coberturas evaluadas: remanente de bosque y cafetales de sol, en un paisaje
rural en la vereda El Brillante, Risaralda. ...................................................................... 31
Figura 3 Esquema del cuadrante de muestreo y del recorrido efectuado. ................... 32
Figura 4 Métodos de colecta manual de suelo y aéreo. ............................................... 33
Figura 5 Polvera artesanal con fécula de maíz y telaraña antes y después de revelar. 34
Figura 6 Trampa de agitación (Paraguas japonés). ..................................................... 35
Figura 7 Aspirador entomológico, recipientes y elementos para la captura de arañas. 36
Figura 8 Abundancia relativa de las familias de arañas registradas en un paisaje rural
cafetero de Risaralda, Colombia. .................................................................................. 43
Figura 9 Abundancia relativa de las familias de arañas encontradas en bosque y cafetal
en un paisaje rural cafetero de Risaralda, Colombia. .................................................... 44
Figura 10 Abundancia relativa de las morfoespecies de arañas registradas en un paisaje
rural cafetero de Risaralda, Colombia. .......................................................................... 45
Figura 11 Abundancia relativa de las morfoespecies de arañas encontradas en bosque
y cafetal de un paisaje rural cafetero de Risaralda, Colombia. ..................................... 46
Figura 12 Riqueza específica de las familias de arañas registradas en un paisaje rural
cafetero de Risaralda, Colombia. .................................................................................. 48
Figura 13 Riqueza específica de las familias de arañas registradas en bosque y cafetal
de un paisaje rural cafetero de Risaralda, Colombia. .................................................... 49
Figura 14 Curva de acumulación de especies para un paisaje rural cafetero de Risaralda,
Colombia. ...................................................................................................................... 50
Figura 15 Modelos de abundancia (log-normal) para bosque y cafetal de un paisaje rural
cafetero de Risaralda, Colombia. .................................................................................. 53
Figura 16 Proporción de los gremios encontrados en un paisaje rural cafetero de
Risaralda, Colombia. ..................................................................................................... 56

11
Figura 17 Discriminación de los gremios en cada una de las coberturas evaluadas de un
paisaje rural cafetero de Risaralda, Colombia. .............................................................. 56
Figura 18 Número de las familias y morfoespecies de arañas registradas en los estratos
arbustivo, herbáceo y rasante del bosque en un paisaje rural cafetero de Risaralda,
Colombia. ...................................................................................................................... 57
Figura 19 Abundancia relativa de las familias y morfoespecies de arañas registradas en
los estratos arbustivo, herbáceo y rasante del bosque en un paisaje rural cafetero de
Risaralda, Colombia. ..................................................................................................... 58
Figura 20 Distribución de los gremios de arañas en los estratos arbustivo, herbáceo y
rasante del bosque en un paisaje rural cafetero de Risaralda, Colombia. .................... 60
Figura 21 Número de las familias y morfoespecies de arañas registradas en los estratos
vegetativo y rasante del cultivo de café en un paisaje rural cafetero de Risaralda,
Colombia. ...................................................................................................................... 61
Figura 22 Abundancia relativa de las familias y morfoespecies de arañas registradas en
los estratos vegetativo y rasante del cultivo de café en un paisaje rural cafetero de
Risaralda, Colombia. ..................................................................................................... 62
Figura 23 Distribución de los gremios de arañas en los estratos vegetativo y rasante del
cultivo de café en un paisaje rural cafetero de Risaralda, Colombia. ............................ 64

12
LISTA DE ANEXOS
Anexo A. Anatomía externa del cuerpo de una araña: (a) vista lateral, (b) vista ventral.
E= epigino (en una hembra adulta). .............................................................................. 95
Anexo B. Tipos de quelíceros y uñas en el Orden Araneae. ........................................ 96
Anexo C. Esquema de clasificación taxonómico del orden Araneae. ........................... 98
Anexo D. Coordenadas de las estaciones y puntos de muestreo definidas en la cobertura
bosque secundario para la caracterización de la comunidad de arañas en un paisaje rural
cafetero de El Brillante, Risaralda. ................................................................................ 99
Anexo E. Coordenadas de las estaciones y puntos de muestreo definidas en la cobertura
cafetal de sol para la caracterización de la comunidad de arañas en un paisaje rural
cafetero de El Brillante, Risaralda. .............................................................................. 100
Anexo F. Formato para la consignación de datos en campo de los muestreos de arañas
(Arachnida: Araneae) en un paisaje rural de Risaralda, Colombia. ............................. 101
Anexo G. Listado de morfoespecies colectadas en un paisaje rural cafetero del
departamento de Risaralda. ........................................................................................ 102
Anexo H. Registro climatológico de la zona de estudio: Paisaje Rural Cafetero de
Risaralda, Colombia. ................................................................................................... 105

13
RESUMEN
Se evaluó la diversidad de arañas en un paisaje rural cafetero ubicado en El Brillante,
municipio de La Celia, Risaralda en diferentes coberturas vegetales. Se realizaron
muestreos diurnos en tres cafetales de sol y tres bosques remanentes (siete puntos de
muestreo por cobertura), mediante colecta manual de suelo (0 – 0,5 m), colecta manual
aérea (0,5 – 2 m) y agitación de follaje (2 – 5 m). Se colectaron 1425 individuos adultos
pertenecientes a 16 familias, agrupados en 87 morfoespecies (mfsp) de las cuales 28
fueron determinadas a nivel de especie y 29 de ellas a género. Las familias más
abundantes fueron Linyphiidae (473 indv.) y Pholcidae (266), y las de mayor riqueza
Theridiidae (27 mfsp) y Araneidae (18). Los cafetales fueron la cobertura que presentó
mayor abundancia, riqueza y diversidad con 848 individuos y 62 mfsp (H’= 2.83, 1-D=
0.12), respecto de los bosques que presentaron 577 individuos y 47 mfsp (H’= 2.60, 1-
D= 0.18). Las coberturas comparadas con Chao-Jacard presentaron una baja similitud
(0.45). El gremio dominante fue el de las arañas constructoras de telas tridimensionales.
A pesar de tener una menor extensión, los remanentes de bosque albergan cerca del
50% de la composición de arañas; sin embargo, es interesante que el resto de la riqueza
está asociada a una cobertura vegetal altamente homogénea (Cafetal), lo que indica el
potencial de este elemento del paisaje rural cafetero para la conservación y estudio de
la biodiversidad de arácnidos en Colombia.
Palabras Clave: café, agroecosistemas, bosque secundario, conservación.

14
ABSTRACT
Spider diversity in coffee countryside located in El Brillante, municipality of La Celia,
Risaralda in different coverages was evaluated. Diurnal samples were taken in three sun
coffee plantations and three remaining forests (seven sampling points for each coverage)
using looking down (0 - 0.5 m), looking up (0.5 - 2 m) and beating (2-5 m). 1425 adult
individuals distributed in 16 families and 87 morphospecies (mpsp), of which 28 were
determinanted until species and 29 until genre, were collected. Linyphidae (473 indv.)
and Pholcidae (266) were the most abundant families while Theridiidae (27 mpsp) and
Araneidae (18) were the most richest. Coffe plantations were the coverage with the
highest abundance, richness and diversity with 848 individuals and 62 mpsp (H '= 2.83,
1-D = 0.12), with respect to forests that had 577 individuals and 47 mpsp (H' = 2.60, 1-D
= 0.18). According to Chao-Jacard index the coverages had low similarity value between
them (0.45). Space web builders spiders were the most dominant guild for this study.
Despite having a lesser extent, the remnants of forest are home to nearly 50% of the
composition of spiders; however, it is interesting that the rest of wealth is associated with
a highly homogeneous vegetation cover (Cafetal), indicating the potential of this element
of the coffee countryside for conservation and study of biodiversity of arachnids in
Colombia.
Keywords: coffee plantations, agroecosystem, secondary forest, conservation.

15
INTRODUCCIÓN
Los bosques montanos y premontanos de los Andes colombianos han sido catalogados
como uno de los ecosistemas con mayor biodiversidad y paradójicamente como uno de
los más amenazados del mundo por la intensa presión antrópica, debido a que gran parte
de la población del país (70%) está concentrada en esta zona (Gómez, Anaya & Álvarez,
2005; Kattan, Franco, Rojas & Morales, 2004; Marín, Toro & Uribe, 2008; Wildlife
Conservation Society, 2009). No obstante, estos bosques conservan una parte
importante de la biodiversidad original y proveen diferentes tipos de valores asociados:
albergan alta diversidad biológica, alta proporción de endemismos, suministro de agua,
recursos, interacciones bióticas y alta diversidad de cultivos (Mendoza, Jiménez, Lozano,
Caycedo y Renjifo, 2006).
Si bien, parte de dichos bosques se encuentran dentro del Sistema Nacional de Áreas
Protegidas – SINAP, en Colombia los bosques montanos están representados por
33.288.000 Ha, de las cuales 4.770.000 Ha son áreas protegidas (Mendoza et al., 2006),
buena parte de la biodiversidad de estos, también reside en zonas intervenidas por el
hombre, especialmente en los denominados paisajes rurales agrícolas o ganaderos que
de acuerdo a Lozano, Jiménez y Caycedo (2006) han originado mosaicos de hábitats
donde los procesos ecológicos para el mantenimiento de la diversidad, están altamente
influenciados por los procesos culturales.
Aunque las áreas protegidas son y se mantendrán como una estrategia central de
conservación, es probable que solo puedan asegurar la sobrevivencia a largo plazo, de
una pequeña fracción de la biodiversidad. Éstas áreas son demasiado pequeñas, pocas,
aisladas y también sujetas a cambios. Además, muchos de los beneficios que su
biodiversidad proporciona a la humanidad son obtenidos a escala local y regional; ello
significa que su suministro dependerá de la capacidad de la biota de los paisajes rurales
para generarlos (Harvey y Sáenz, 2008). En Colombia, una buena proporción de la
superficie nacional se encuentra actualmente en áreas de paisajes rurales dominadas

16
por agroecosistemas. El grado de modificación de las coberturas vegetales nativas en
regiones como la Caribe (82,3%), Andina (61,8%) y Orinoquía (59,9%) revelan cifras
dramáticas que implican una reducción neta de éstas áreas naturales disponibles para
la fauna y la flora nativa Lozano, 2009.
Una razón de peso para incorporar los paisajes rurales, más exactamente paisajes
rurales cafeteros del departamento de Risaralda en estudios para la caracterización de
la biodiversidad se debe a su posición estratégica, al estar inmerso en los corredores
ambientales de la cordillera Occidental (PNN del Tatamá), Central (PNN de los Nevados)
y del río Cauca, y al presentar relaciones de vecindad, conectividad y continuidad con el
Chocó Biogeográfico en el occidente, el valle del Magdalena en el oriente, Antioquia en
el norte y Valle del Cauca y Macizo Colombiano en el sur (Corporación Autónoma
Regional de Risaralda, 2008).
No obstante, y debido a su particular posición geográfica, los paisajes naturales de
Risaralda se ven afectados por la fuerte presión antrópica que se ejerce sobre estos, al
ser uno de los principales departamentos productores de café para el país Bustillo, 2007.
De esta manera, se enmarca al paisaje cafetero de Risaralda como un área crítica. No
solo para el futuro de la biodiversidad y de los servicios ecológicos que presta a nivel
local y regional, sino también para los beneficios sociales derivados de sus ecosistemas
y la biodiversidad que contienen Harvey y Sáenz, 2008. Es así como se destaca la
importancia de incrementar el conocimiento de la composición y estructura de las
comunidades bióticas, en este tipo de paisajes transformados.
Como modelo biológico para analizar parte de la dinámica de los paisajes cafeteros, se
seleccionó a las arañas como grupo de estudio, ya que cumplen con los criterios del
sistema de evaluación para la selección de indicadores de diversidad propuesto por
Villarreal et al., (2006). Asimismo, debido al papel ecológico que desempeñan las arañas
en los ecosistemas terrestres como agentes estabilizadores y/o reguladores de
poblaciones de insectos, de otros artrópodos e incluso de pequeños vertebrados (Flórez,
1996). Adicionalmente, se han empleado como indicadores de la calidad ambiental

17
(Ambrecht & Perfecto, 2001; Cardoso, Silva, De Oliveira & Serrano, 2004) pues sus
comunidades son fuertemente influenciadas por el tipo de hábitat y uso del suelo (Weeks
y Holtzser, 2000 citado por Sabogal, 2011).
El presente estudio fue orientado a evaluar la estructura y composición de la comunidad
de arañas en un paisaje rural cafetero de Risaralda, además, se comparó si existen
diferencias significativas de esta diversidad según la composición de los elementos del
paisaje, especialmente en relación a la proporción de hábitats naturales (bosques
remanentes) versus sistemas agrícolas (cafetales de sol), como una medida de la
intensidad de uso de suelo; que permita dar una contribución inicial al conocimiento de
la diversidad biológica de este grupo taxonómico y su distribución dentro de este tipo de
paisaje rural en Colombia.

18
1. OBJETIVOS
1.1 OBJETIVO GENERAL
Evaluar la diversidad de la comunidad de arañas (Arachnida: Araneae) encontradas en
cafetales de sol y remanente de bosque en un paisaje rural del municipio de La Celia,
departamento de Risaralda, Colombia.
1.2 OBJETIVOS ESPECÍFICOS
Determinar taxonómicamente las arañas encontradas en cafetales de sol y remanente
de bosque en la vereda El Brillante (La Celia, Risaralda).
Establecer la estructura y composición de las comunidades de arañas presentes en
cafetales de sol y remanente de bosque evaluados.
Estimar la diversidad, riqueza y abundancia de las comunidades de arañas presentes en
las coberturas evaluadas.
Comparar los valores de diversidad y riqueza de la comunidad de arañas de cada
cobertura evaluada.

19
2. MARCO REFERENCIAL
2.1 ANTECEDENTES Y ESTADO ACTUAL DEL ESTUDIO DE ARAÑAS EN COLOMBIA
El gran número de arañas y su notable presencia en los ecosistemas ha motivado a
varios investigadores, quienes durante décadas han desarrollado valiosos estudios en
América Latina y por supuesto Colombia Alape, 2008. Los primeros trabajos con arañas
en el país se remontan al siglo XIX, los cuales consistían en las descripciones de
especies colectadas por expediciones científicas procedentes principalmente de Europa.
A comienzos del siglo XX y debido a la presencia de los Hermanos La Sallistas,
interesados en el reconocimiento de la fauna y flora del país, se establecieron las
primeras colecciones biológicas nacionales. Sin embargo, sólo fue hasta finales de la
década de 1960 que se generan los primeros aportes nacionales Flórez, 1996.
En la década de 1970 William Eberhard realiza los primeros estudios detallados sobre
arañas en el departamento del Valle del Cauca, quien abordó en sus investigaciones dos
líneas base que fueron: la ecología y la etología de varias especies de arañas de la
región. De otra parte, promovió numerosas descripciones de nuevas especies, o
ampliaciones de los rangos de distribución de diversas arañas vallecaucanas. Asimismo,
impulsó trabajos con estudiantes de biología de la Universidad del Valle (Flórez, 1996).
Otros aportes significativos de Eberhard durante su permanencia en la región, tuvieron
que ver con los diseños de trampas pegajosas imitadoras de las telas naturales, y el
“Espolvoreador” o “revelador” de harina, para facilitar la visualización de detalles
estructurales de las telarañas Flórez, 2004.
Tal vez el único aracnólogo colombiano que trabajó paralelamente a Eberhard durante
esa época fue Nicolás Paz, quien realizó un gran aporte con sus trabajos faunísticos
regionales en el departamento de Antioquia. Desde finales de los setenta, hasta la
actualidad, el interés por el estudio de las arañas en Colombia ha ido en aumento,

20
particularmente gracias a los trabajos realizados o dirigidos por Eduardo Flórez,
considerado el aracnólogo con mayor trayectoria en el país Sabogal, 2010.
El número de trabajos con arañas en Colombia en los últimos 30 años (1980 – 2010),
asciende a más de 100, la mayoría de ellos han sido enfocados al reconocimiento de la
biodiversidad de este grupo faunístico en ecosistemas naturales, principalmente en áreas
boscosas, alcanzando el 50% de las investigaciones. Los proyectos en agroecosistemas
ocupan el segundo lugar con un 25%. Pocos se han enfocado en aspectos taxonómicos
y con menor participación los que involucran estudios comportamentales, esto
corresponde al 25% restante Sabogal, 2010.
Con respecto al paisaje en zonas cafeteras se encuentran los trabajos realizados por
Alape 2008 y Cepeda 2009. Alape 2008, realizó una comparación entre cultivo de café,
el bosque adyacente y el borde del bosque en una reserva forestal del municipio de
Ibagué, encontrando diferencias en términos de composición de la araneofauna en los
tres hábitats, siendo el cafetal el que presentó el mayor número de individuos (767) y el
borde de bosque el que presentó el mayor número de especies (40 mfsp), no obstante,
aunque la riqueza y abundancia en el bosque no fueron las más elevadas se mantuvieron
constantes en las muestras; la cobertura más diversa fue el bosque (H’=2.69) y la menos
diversa fue el Cafetal (H’=2.24), debido a que las coberturas de interior de bosque tienen
una mayor disponibilidad de sustratos para la permanencia de las arañas tejedoras, lo
que se traduce en un valor de diversidad alto para éste grupo. Concluyendo, que la
complejidad estructural de la vegetación influye en la riqueza y abundancia de especies
de arañas tejedoras.
Por su parte, Cepeda 2009, realizó una comparación de la diversidad de arañas y
coleópteros en dos fincas bajo diferente sistema producción (convencional y orgánica)
en el departamento de Cundinamarca, observando que el manejo convencional
disminuye la biodiversidad y afecta la calidad ecosistémica, mientras el orgánico
promueve la biodiversidad funcional dentro del cultivo. También concluyó que los
coleópteros son más sensibles al tipo de manejo, pues este afectó aspectos de la

21
diversidad en estos organismos a excepción de la equidad. Por otra parte las arañas
parecen más resistentes, ya que el manejo convencional sólo afectó su abundancia, tal
vez esto sea un indicativo de que las prácticas propias del cultivo del café, aunque exista
aplicación de insecticidas, ayudan a mantener la riqueza de arañas.
2.2 BREVE INTRODUCCIÓN AL ORDEN ARANEAE
Las arañas conforman uno de los grupos de artrópodos con mayor diversidad global en
términos de especies descritas o estimadas dentro del Reino Animal, ubicándose en el
séptimo lugar después de los coleópteros, himenópteros, lepidópteros, dípteros,
hemípteros y ácaros (Barnes y Ruppert, 1996; Coddington y Levy, 1991). Son
organismos de distribución cosmopolita, y han logrado colonizar una amplia variedad de
ecosistemas terrestres desde las islas árticas, hasta las regiones desérticas y están muy
bien representadas en los ecosistemas tropicales Foelix, 2011. No es exagerado decir
que las arañas han conquistado todos los posibles nichos ecológicos en la tierra Turnbull,
1973.
La mayoría de las arañas son relativamente pequeñas, en un rango entre 2-10 mm de
longitud corporal (sin tener en cuenta la longitud de las patas), sin embargo, algunas
grandes tarántulas pueden alcanzar un tamaño de 80-90 mm. Las arañas macho, son
casi siempre más pequeñas y tienen una vida más corta que las hembras Flórez, 1996;
Foelix, 2011.
Todas las arañas son de hábitos exclusivamente depredadores de presas vivas y
móviles. Muchas se han especializado como constructoras de redes o telas que actúan
a manera de trampa (arañas tejedoras o sedentarias), mientras que otras cazan de
manera activa buscando y persiguiendo a sus presas sobre el suelo o entre la vegetación
(arañas errantes). También, hay un grupo que vive cerca o sobre las telas de otras
arañas, alimentándose de los residuos de las presas de las arañas residentes (arañas
comensales). Los insectos constituyen la principal fuente de alimento para las arañas;

22
sin embargo, pueden consumir a menudo algunos otros grupos de invertebrados
terrestres y pequeños vertebrados (presas inusuales) (Flórez, 1996; 1997).
El cuerpo de una araña se compone de dos partes principales: una parte anterior, el
prosoma (o cefalotórax), y una parte posterior, el opistosoma (o abdomen). Estos están
conectados por un tallo estrecho, el pedicelo (Anexo A).
El prosoma está cubierto por una placa dorsal y otra ventral, el caparazón y el esternón,
respectivamente. Sirve de lugar de unión para seis pares de apéndices articulados: un
par de quelíceros, un par de pedipalpos y cuatro pares de patas; en las arañas macho
adultas, los pedipalpos son modificados en órganos copuladores. En el prosoma también
se alojan los órganos de la visión, el aparato bucal, las glándulas de veneno y el sistema
nervioso central. El opistosoma es generalmente no segmentado, excepto en algunas
arañas, que se considera que han evolucionado a partir de especies antiguas
(Mesothelae). En contraste con el prosoma, el opistosoma es un poco más blando y
globoso; en su parte ventral se encuentra el epigino, que es un placa esclerotizada que
cubre las aperturas de los poros genitales; en la parte más distal se encuentran las hileras
que corresponden a dos o tres pares de apéndices cilíndricos o cónicos que guardan en
su interior los conductos excretores de las glándulas de la seda. Además, en el
opistosoma se encuentran los sistemas digestivo, circulatorio, respiratorio y excretor
(Flórez, 1996; Dippenaar-Schoeman y Jocqué, 1997; Jocqué & Dippenaar-Schoeman,
2006; Foelix, 2011).
En cuanto a la sistemática y clasificación de las arañas, en la actualidad los taxónomos
reconocen alrededor de 44.906 especies válidamente descritas, que se agrupan en 3.935
géneros y 114 familias (World Spider Catalog, 2014), para Colombia se hallan registradas
cerca de 1244 especies, 554 géneros y 66 familias (Perafán et al., 2013). El orden de las
arañas (Araneae), por lo general se divide en tres subórdenes: Mesothelae,
Mygalomorphae y Araneomorphae. Hasta hace poco los migalomorfos se conocían como
ortognatos (Orthognatha) debido a la alineación casi paralela de su quelíceros, mientras
que los araneomorfos correspondían a los labidognatos (Labidognatha), que poseen

23
quelíceros verticales opuestos entre sí (Anexo B). El suborden Mesothelae representa a
las arañas filogenéticamente más antiguas debido a que presentan un abdomen
claramente segmentado, así como varios otros caracteres "primitivos". Los
Mygalomorphae comprenden a todas las tarántulas, y se caracterizan por presentar
quelíceros casi paralelos entre sí e hileras a menudo reducidas. Más del 90% de todas
las arañas pertenecen a los Araneomorphae, los cuales se clasifican en dos grupos:
Cribellatae y Ecribellatae, basado en la presencia o ausencia de una placa quitinosa
(cribelo) situada en la región anterior de las hileras; la presencia de cribelo es distintivo
del grupo de las arañas Cribellatae. Entre el grupo de las arañas Ecribellatae, algunas
familias presentan estructuras genitales simples (Haplogynae) y estructuras genitales
complejas (Entelegynae); a su vez el grupo de las arañas Entelegynae se divide en
Dionycha y Trionycha, dependiendo de si las patas tienen dos o tres uñas tarsales
respectivamente (Anexo C) (Dippenaar-Schoeman y Jocqué, 1997; Jocqué y Dippenaar-
Schoeman, 2006; Foelix, 2011).
2.3 INTERRELACIONES ECOLÓGICAS DE LAS ARAÑAS
Las arañas son conocidas como los cazadores del micromundo y desempeñan un papel
importante como integrantes intermedios dentro de las redes tróficas, ya que su actividad
depredadora impacta muchas especies, permitiendo que se conserve un equilibrio entre
las distintas poblaciones (Blanco y Salas, 2007). Asimismo, son activamente depredadas
por especies como aves, lagartos, avispas u otras arañas (Flórez, 1997). Wise citado por
Calixto (2004) argumenta que las arañas en su mayoría son depredadores generalistas,
cuya base alimenticia consiste principalmente de insectos y otros invertebrados de
pequeña y media talla. No obstante, se ha logrado establecer que este espectro
alimenticio es aún más amplio e incluye el consumo de huevos de artrópodos, de polen
de plantas y de animales muertos (presas inusuales) como ratones, aves murciélagos,
serpientes, sapos, lagartos, peces, escorpiones y cangrejos (Flórez, 1997).
Además, el hábito depredador de las arañas, ha llevado a considerar la utilización
potencial de algunas especies como posibles reguladoras de plagas en diversos cultivos.

24
Pese a que no exhiben una marcada especificidad por sus presas, si ofrecen un efecto
“amortiguador” generalizado sobre las poblaciones de insectos presentes en los cultivos,
contribuyendo de manera efectiva a reducir las explosiones demográficas de especies
fitófagas Flórez, 1997. Según Nentwig citado por Calixto (2004), solo unas pocas
especies exhiben hábitos especialistas alimentándose de un número limitado de
especies presa.
2.4 ESTRUCTURA GREMIAL EN ARAÑAS (GRUPOS FUNCIONALES)
El concepto de “gremio” o grupo funcional lo definió Root (1967) como “un grupo de
especies que explotan la misma clase de recursos del medio ambiente de una manera
similar”. Basado en este concepto Uetz, Halaj y Cady (1999) caracterizaron dos grandes
gremios de arañas: cazadoras y constructoras de telas, conforme a la manera como
realizan la cacería de sus presas. Las cazadoras buscan activamente sus presas por la
superficie del suelo o la vegetación, mientras que las constructoras de telas emplean su
seda para fabricar trampas (telarañas) donde quedan atrapadas las presas. Asimismo,
dividieron estos dos gremios en siete subgremios (cazadoras errantes en vegetación,
cazadoras errantes en suelo, cazadoras al asecho, cazadoras por emboscada,
constructoras de telas tridimensionales, constructoras de telas en sábana o maraña y
constructoras de telas orbiculares).
Existen dos aspectos importantes que deben considerarse al proponer gremios para
agrupar las arañas: 1) Las arañas son depredadores generalistas de artrópodos; pese a
la diversidad de niveles tróficos que sus presas representan (herbívoros, detritívoros,
parasitoides, depredadores). 2) A pesar de la condición generalista, existen diferentes
estrategias de forrajeo que pueden afectar la utilización del recurso, haciendo que el
mismo recurso se subdivida y a su vez sea explotado como un recurso diferente (Uetz et
al., 1999). Por ejemplo, las arañas tejedoras son casi estrictamente insectívoras, y sus
telas están adaptadas para capturar en su mayoría insectos voladores, mientras que las
errantes se alimentan de insectos y otros artrópodos, y sus presas son generalmente
cursoriales Nyffeler, 1999.

25
Si se parte de la premisa de que cada uno de los gremios de arañas está utilizando un
recurso diferente, quiere decir que cada gremio está actuando como depredador de un
grupo de presas diferente. Esto puede ser utilizado en estudios para realizar
aproximaciones sobre el papel que desempeñan las arañas frente a poblaciones de
insectos, particularmente en lo referente a insectos plaga en agroecosistemas Uetz et
al., 1999. Además, los gremios pueden ser útiles en el estudio comparativo de
comunidades; el mismo gremio puede ser representado en varias comunidades
diferentes y así servir de base para comparar la diversidad de especies, el grado de
diferencias, biomasa, etc. en diferentes biotopos Root, 1967.
2.5 EL PAISAJE RURAL
El término de paisaje se define en su sentido más simple como un área espacial
heterogénea Halffter, Moreno y Pineda, 2001; Turner, Gardner y O’Neill, 2001. Los
paisajes rurales (Countryside) son áreas extensas de territorio que han sido modificadas
por la intervención humana y como resultado se transformaron en un mosaico de
remanentes de hábitats naturales y sistemas productivos en donde estos últimos
constituyen la matriz del paisaje (Harvey y Sáenz, 2008; Lozano, Jiménez y Caycedo,
2006). El concepto de paisaje no se reduce o circunscribe a las unidades biofísicas y
silvestres sino que además incluye todas aquellas fracciones antrópicas o humanizadas
del territorio, es decir considera también las porciones de naturaleza bajo manejo
humano Toledo, 2005. Por esta razón, el paisaje tiene que ver con los resultados de la
interrelación entre actividad humana y naturaleza que responde en primera instancia a
las necesidades económicas, sociales y políticas de la comunidad que los habita Zorrilla
2005.
Por otro lado, es importante destacar los servicios ecológicos que brinda este paisaje,
tales como alimento (plantas y animales), fármacos, madera, energía (combustibles y
agua), productos industriales (ceras, tintas, aceites, etc.), biodiversidad (recursos
genéticos), procesos de regeneración (reciclaje, filtración, desintoxicación,
descomposición, etc.), procesos de traslado (dispersión de semillas y polinización),

26
procesos de estabilización (mitigación de sequías e inundaciones, estabilización parcial
del clima, etc.) y funciones de realización (provisión cultural, intelectual e inspiración
espiritual) que como lo afirman Harvey y Sáenz, 2008, apoyan y sustentan la vida del ser
humano y garantizan las condiciones de vida y las opciones para el futuro.

27
3. METODOLOGÍA
3.1 ÁREA DE ESTUDIO
El área de estudio corresponde a una muestra del paisaje de 500 ha (denominada
ventana). Se encuentra localizada en la vertiente oriental de la Cordillera Occidental, en
jurisdicción de la vereda El Brillante, 6 km al norte del caso urbano del municipio de La
Celia, subregión II del departamento de Risaralda (Figura 1).
Figura 1 Ubicación del área de estudio: paisaje rural cafetero en la vereda El Brillante,
Risaralda.
Fuente: el autor
Espacialmente la ventana se ubica en el sistema de coordenadas geográficas con origen
Bogotá a los 5° 1’ 47,81” de latitud norte y 76° 0’ 18,15” de longitud oeste (norte 388591
m y oeste 556035 m en coordenadas UTM) en su vértice inferior izquierdo dirección sur-
occidente y a los 5° 3’ 0,17” de latitud norte y 75° 58’ 56,64” de longitud oeste (norte

28
391119 m y oeste 558273 m) en su vértice superior derecho dirección norte-oriente; en
un rango altitudinal que va desde los 1471 m a los 2051 m.
3.1.1 Aspectos físicos. De acuerdo con la clasificación de Holdridge (1967) el área de
estudio se ubica en la zona de transición entre las formaciones vegetales “bosque muy
húmedo premontano” (69%) y “bosque muy húmedo montano bajo” (31%) (p. 20). El
clima está influenciado por la circulación de vientos cálidos provenientes del valle del río
Risaralda que al encontrarse con masa de aire frío que provienen de las partes más altas
de los cerros Tatamá, Tamaná y Caramanta, conforman un área de alta precipitación que
alcanza niveles del orden de los 2240 mm anuales. Las temperaturas medias anules
oscilan entre 10 y 24 °C. Se caracteriza por presentar montañas de crestas altas con
cañones y arroyos profundos y colinas en los valles. Es atravesada por las quebradas
Boriana, Pabellon, Quebraditas y Las Mellizas. Entre los diferentes elementos del paisaje
que componen la ventana de estudio encontramos fragmentos de bosque secundario,
bosques ribereños, cultivos de café y agroecosistemas ganaderos (Tabla 1) (Vásquez &
Serrano, 2009; Ríos, 2010).
Tabla 1 Área de las coberturas presentes en la ventana de trabajo: paisaje rural cafetero
de El Brillante, Risaralda.
Cobertura Área (Ha) Área (m2) %Área
Agroecosistema de caña 6,12 61,200 1
Agroecosistema ganadero 31,01 310,100 6
Agroecosistemas cafeteros 315,03 3’150,300 63
Bosque 147,84 1’478,400 30
Fuente: Ríos, 2010.
3.1.2 Aspectos bióticos. Hasta hace unos 60 años, la región del área de trabajo se
encontraba cubierta en su totalidad por bosque andinos, los cuales según Vásquez &
Serrano (2009) fueron virtualmente eliminados para extraer maderas de alto valor
comercial y establecer potreros y cultivos. En la actualidad los ecosistemas naturales de

29
la zona, se limitan a remanentes o relictos de bosque secundario hacia las partes altas
de las montañas, nacimientos y causes de quebradas.
En la composición florística del área se resalta la presencia de especies de valor
económico como comino (Aniba perutilis), flor morado (Tabebuia rosea), nogal cafetero
(Cordia alliodora), mediacara (Pouteria lucuma), roble (Quercus humboldtii), encenillo
(Weinmannia sp.) amarillos (Nectandra spp., Ocotea spp.), quina (Cinchona pubescens)
y nogal (Juglans neotropica).
A pesar de las actividades de cacería adelantadas en la zona desde tiempo atrás, aún
se encuentran especies de mamíferos de porte mediano entre las que se destacan
perezoso (Choloepus hoffmanni), venado (Odocoileus virginianus y Mazama americana)
cusumbo (Nasua nasua), comadreja (Mustela sp.), armadillo (Dasypus novemcinctus) y
zorro (Cerdocyon thous).
Los registros de aves muestran la presencia de especies endémicas, tales como tángara
(Habia cristata), candelita (Myioborus ornatus), toche (Ramphocelus flammigerus) y
clorocrisa esmeralda (Chlorochrysa phoenicotis) (Vásquez y Serrano, 2009).
3.1.3 Coberturas de muestreo. Al interior de la ventana de estudio se evaluaron dos tipos
de coberturas:
3.1.3.1 Bosque. Parche de bosque nativo en diferentes estados de sucesión, ubicado
hacia la parte alta de cerro “Pelahuevos” o “El Brillante”. Cuenta con una extensión
aproximada de 250 ha, siendo uno de los bosque más grandes que se conservan en el
paisaje de la ventana de estudio. El bosque de “Pelahuevos” se destaca como un área
de importancia estratégica a nivel local dado el papel que cumple en la protección de
centenares de nacimientos de agua que nutren directamente a los ríos Peñas Blancas y
Los Monos, que de acuerdo a Vásquez & Serrano (2009) abastecen al acueducto
municipal de La Celia y surten el agua para el consumo humano de pobladores y la
producción agropecuaria.

30
La historia de actividades agrícolas y de explotación maderera de la región explica el alto
grado de intervención del bosque de “Pelahuevos” donde, la cobertura vegetal
predomínate corresponde a bosque secundario localizado en su mayor parte hacia el
borde del bosque, mientras que, en la porción más alta del cerro se encuentran
sucesiones hasta de 15-20 años, así como parches de bosque primario. El área del
bosque mejor conservado presenta alturas hasta de 25 m y un alto grado de epifitismo,
resultado del nivel de humedad allí imperante, así como abundante regeneración natural
y presencia de lianas y bejucos. Los rastrojos adyacentes a la periferia del bosque, son
coberturas que también fueron sustancialmente intervenidas pero están en la etapa
inicial de la sucesión natural. Presentan una estructura de matorral denso con dominio
de especies de las familias Melastomataceae y Asteraceae (Vásquez & Serrano, 2009).
3.1.3.2 Cafetales. Corresponden a sistemas de producción tecnificado de lotes de Coffea
arabica variedad Caturra, los cuales han sido trazados y establecidos al sol, en una
población mayor a 2.500 plantas por hectárea. Se seleccionaron cultivos en producción
con más de dos años de edad, los cuales presentaban características similares como:
control de malezas dos veces por año, uno químico (herbicidas como glifosato) y otro
mecánico (machete); y abono del cultivo mediante el uso de urea y/o DAP.
3.2 MÉTODOS DE CAMPO
El muestreo se realizó entre los meses de septiembre y octubre de 2010, en jornadas
diurnas entre las 07:00 y 17:00 horas con esfuerzo de un único colector. Se trabajó en
dos coberturas diferentes: bosque remanente y cafetal de sol, estableciéndose tres
estaciones de muestreo por cobertura (Tabla 2; Figura 2). Cada estación constaba de
siete puntos o cuadrantes de muestreo (Anexos D y E) escogidos al azar, distanciados
aproximadamente 10 m uno del otro.
31
Tabla 2 Coordenadas de las estaciones de muestreo definidas para la caracterización
de la comunidad de arañas en un paisaje rural cafetero de El Brillante, Risaralda.
Estaciones de
Muestreo Coordenadas UTM Coordenadas Geográficas
Altitud (m)
Tipo de Cobertura
Bos1 390379 X 557230 Y 5° 2' 26,3" N 75° 59' 20,0" W 1948 Bosque
Bos2 390317 X 557169 Y 5° 2' 24,6" N 75° 59' 22,5" W 1844 Bosque
Bos3 390255 X 557108 Y 5° 2' 22,2" N 75° 59' 24,7" W 1906 Bosque
Cf1 389300 X 556679 Y 5° 2' 08,0" N 75° 59' 55,6" W 1645 Cafetal
Cf2 389916 X 556586 Y 5° 2' 05,0" N 75° 59' 35,5" W 1839 Cafetal
Cf3 389670 X 557170 Y 5° 2' 24,0" N 75° 59' 43,1" W 1774 Cafetal
Fuente: el autor
Figura 2 Coberturas evaluadas: remanente de bosque y cafetales de sol, en un paisaje
rural en la vereda El Brillante, Risaralda.
Fuente: el autor
3.2.1 Unidad de muestreo. Como unidad de muestreo se empleó la metodología de
cuadrantes propuesta por Sabogal (2011), puesto que permite: 1) medir el área de

32
muestreo, 2) cuantificar la densidad de arañas en cada cuadrante y 3) asegurar un área
característica de la cobertura evaluada, evitando zonas de transición como bordes, el
paso de un cuerpo de agua o cualquier otro tipo de discontinuidad que pudiera afectar la
distribución de las arañas.
Como ya se mencionó, se establecieron siete puntos o cuadrantes de 5 x 5 m,
correspondiente a un área de 25 m2 en cada estación de muestreo. Una vez delimitado
el cuadrante (no se realizó la demarcación con cuerda, ya que este procedimiento puede
destruir las telas y alterar el muestreo), se procedió a realizar dentro del mismo,
recorridos en forma de “S” de aproximadamente un metro de ancho, lo que equivale a
recorrer cinco transectos de 5 m2 cada uno, solo que para el caso particular se
encontraban uno al lado del otro (Figura 3).
En las ocasiones donde fue necesario revisar un área mayor a los 25 m2, se continuó
con el desplazamiento en “S” y al terminar el tiempo de muestreo, se cuantificó el
recorrido exacto.
Figura 3 Esquema del cuadrante de muestreo y del recorrido efectuado.
Fuente: el autor; modificado de Sabogal, 2011.
5 m
5 m
1 m
1
2
3
4
5

33
3.2.2 Métodos de captura. Se emplearon tres métodos de captura en cada punto o
cuadrante de muestreo: colecta manual de suelo (CMS), colecta manual aérea (CMA) y
agitación de vegetación (A); los cuales se describen a continuación.
3.2.2.1 Colecta manual. Es un método de captura directa propuesto por Coddington,
Griswold, Silva, Peñaranda, & Larcher, 1991. Consiste en la búsqueda activa de las
arañas realizando un examen cuidadoso sobre diferentes sustratos como: hojarasca,
depósitos de detritus, debajo de rocas, troncos en descomposición, madrigueras, frutos
caídos, corteza de árboles y arbustos, hojas, ramas huecas y partes de flores, epifitas,
telas de araña, entre otros. Este método se implementa por estratos y dependiendo de
este, recibe diferentes nombres. La Colecta Manual Rasante o de Suelo (Looking down)
se implementa sobre los sustratos encontrados entre los 0 y 0,5 m y la Colecta Manual
Aérea (Looking up) se realiza sobre la vegetación desde los 0,5 m hasta la máxima
extensión del brazo del colector (2 m aproximadamente), la primera debe hacerse
arrastrándose sobre el suelo, la segunda se hace caminando (Figura 4).
Figura 3 Métodos de colecta manual de suelo y aéreo.
Fuente: el autor

34
Para una mejor visualización de las telas y ubicación de las arañas en los métodos de
colecta manual se aplicó la técnica del revelado propuesta por Eberhard 1976, que
consiste en espolvorear fécula de maíz, sobre los posibles sitios donde pueden ser
encontrados los hilos de las telarañas (Figura 5). Una vez adheridas las partículas de la
harina a las telas, éstas son revisadas hasta encontrar el sitio de refugio de las arañas.
Las arañas fueron recolectadas con ayuda de una pinza o pincel y depositadas en
recipientes plásticos con alcohol etílico al 70%, adicionalmente se anotaron datos de
colecta que se consignaron en la ficha de campo (Anexo F).
Figura 4 Polvera artesanal con fécula de maíz y telaraña antes y después de revelar.
Fuente: el autor
El esfuerzo de muestreo para los métodos de colecta manual corresponde a una (1) hora
de muestreo ininterrumpido. Se contabilizó el tiempo mediante el empleo de un
cronómetro digital, el cual se detuvo en casos particulares para no afectar el esfuerzo de
muestreo. Una vez se consiguieron las condiciones para continuar la colecta se dio
nuevamente inicio al conteo del tiempo (Coddington et al. 1991).
3.2.2.2 Agitación de vegetación (Beating). Es un método de captura indirecto y se empleó
para colectar las arañas que se encuentran entre los 2 y 5 m. Consistió en golpear o

35
sacudir vigorosamente una unidad vegetal (rama, arbusto o árbol joven) por periodos
cortos (10 segundos aproximadamente). La unidad vegetal era escogida al azar, y su
agitación se daba por terminada cuando después de tres sacudidas consecutivas no caía
ningún individuo sobre la trampa de agitación o paraguas japonés (Figura 6). Los
ejemplares fueron colectados de la trampa mediante el uso de un aspirador entomológico
y depositados en recipientes plásticos con alcohol etílico al 70% (Figura 7); para este
método el esfuerzo de muestreo fue la agitación de vegetación durante una (1) hora. Al
finalizar el tiempo de agitación se registró en la ficha de campo el número de unidades
vegetales sacudidas (Anexo F). La medición del tiempo se hizo siguiendo las
recomendaciones de Coddington et al. (1991).
Figura 5 Trampa de agitación (Paraguas japonés).
Fuente: el autor

36
Figura 6 Aspirador entomológico, recipientes y elementos para la captura de arañas.
Fuente: el autor
3.3 MÉTODOS DE LABORATORIO
Los organismos colectados se trasladaron al Laboratorio de Zoología del Grupo de
Investigación en Zoología (GIZ) de la Universidad del Tolima, con el propósito de realizar
una determinación taxonómica preliminar de los individuos, la cual fue corroborada en el
laboratorio de Artrópodos del Centro Internacional de Física en Bogotá, adscrito a la
Universidad Nacional de Colombia, Sede Bogotá.
3.3.1 Organización de las morfoespecies (mfsp). Inicialmente, el material colectado fue
determinado a nivel de familia mediante el uso de claves especializadas y, separado de
acuerdo a su etapa de desarrollo (inmaduros, subadulto y adultos) y sexo (hembras y
machos). A continuación se organizaron y agruparon los organismos con base en
características fenotípicas particulares de cada uno de ellos (forma, tamaño y coloración
general del cuerpo, así como presencia de espinas, tubérculos, manchas, puntos o
bandas en alguna parte del abdomen, caparazón, patas, esternón, etc. de la araña).
Finalmente se definieron las respectivas morfoespecies comparando las estructuras
reproductivas tanto de machos como de hembras, utilizando las revisiones disponibles.
Los individuos adultos y subadultos fueron determinados hasta la categoría taxonómica
de familia, género o especie cuando fue posible. La asignación de los individuos

37
inmaduros a las diferentes morfoespecies no siempre fue posible, muchos no presentan
los patrones morfológicos de los adultos, por esta razón para los análisis estadísticos
sólo se incluyeron ejemplares adultos y subadultos, por ser estos los que pueden ser
asociados confiablemente.
3.3.2 Determinación taxonómica y disposición final de los organismos. Para la
determinación taxonómica de los organismos se empleó un estereomicroscopio Motic
SMZ-168 y las claves dicotómicas de Kaston (1978), Roth (1993), Crespo, Iglesias y
Valverde (1999), Florez (1996), Dippenaar-Schoeman y Jocqué (1997), Ubick, Paquin,
Cushing y Roth (2005) y Jocqué y Dippenaar-Schoeman (2006) a nivel de familia. Para
géneros y especies las claves fueron las de Agnarsson (2006), Glueck (1993), Levi (1955,
1959, 1963, 1964a, 1964b, 1978, 1985, 1986, 1988, 1991a, 1991b, 1992, 1995, 1999,
2002, 2004, 2007), Levi y Levi (1962), Millidge (1985, 1991) y Opell (1979). Finalizado el
proceso de determinación se procedió a depositar el material colectado en la Colección
Zoológica de la Universidad del Tolima (CZUT). De igual forma se dejará una copia de
estos especímenes en la Colección Aracnológica del Instituto de Ciencias Naturales de
la Universidad Nacional de Colombia (ICN).
3.4 ANÁLISIS DE DATOS
3.4.1 Abundancia. Con los datos obtenidos a nivel de paisaje y por cobertura, se calculó
y graficó la abundancia relativa (%AR) por familias y morfoespecies. La abundancia
relativa se define como:
% AR = (n1 / N) x 100
Dónde:
AR= Abundancia relativa del taxón
n1= El número de individuos capturados u observados de la familia o especie
N= El número total de individuos capturados u observados (Lasso, Señoris, Lasso &
Castroviejo, 1995 citado por Alape, 2008)

38
3.4.2 Evaluación de la diversidad alfa (α):
3.4.2.1 Riqueza. Se obtuvo la riqueza específica (S) para la ventana y coberturas
muestreadas. La riqueza específica se basa únicamente en el número de especies
(morfoespecies) presentes, sin tomar en cuenta el valor de importancia de las mismas
(Moreno, 2001). Es la forma más sencilla de evaluar la diversidad puntual, aunque se ve
afectada por el tamaño de la muestra (Halffter, Moreno y Pineda, 2001).
3.4.2.2 Curvas de acumulación de especies. Este método se emplea para estimar el
número de especies esperadas a partir de un muestreo (Villareal et al., 2006), por tanto,
permite evaluar que tan completos han sido los muestreos en registrar todas las especies
esperadas (Halffter et al., 2001) y así, dar fiabilidad a los inventarios biológicos y
posibilitar su comparación (Jiménez-Valverde y Hortal, 2003). Se utilizó el programa
estadístico EstimateS versión 9.1.0 Colwell, 2013 para calcular los valores de los
estimadores de riqueza (índices basados en la abundancia: ACE y Chao1 y basados en
presencia/ausencia: ICE, Chao2, Jacknife1, Jacknife2 y Bootstrap) (Colwell y
Coddington, 1994; Moreno, 2001) esgrimidos en la elaboración de las curvas de
acumulación de especies, empleándose 100 aleatorizaciones en todos los cálculos.
Se graficó el promedio de los siete estimadores haciendo uso del programa Microsoft
Office Excel 2007 ®; este promedio se realizó con los valores por muestra de cada uno
de los estimadores de riqueza y permite realizar una curva de acumulación de especies
consolidada que reúne el comportamiento de los estimadores calculados.
Adicionalmente a la curva de acumulación de especies esperada se incluyen en la
gráfica, la curva de acumulación de las especies observadas y las curvas de las especies
raras: Singletons (Singulares: especies con un solo individuo), Doubletons (Dobles:
especies con dos individuos), Uniques (Únicas: especies registradas en una muestra) y
Duplicates (Duplicadas: especies registradas en dos muestras) (Sabogal, 2011).
3.4.2.3 Índices de abundancia proporcional. Se estimó la diversidad alfa para cada
cobertura muestreada con los índices de Shannon-Wiener y Simpson; calculados con el

39
programa PAST (PAleontological STatistics) versión 2.17c (Hammer, Ø., Harper, D. A.
T., & Ryan, P. D. 2001.). Este programa, realiza procedimientos estadísticos
(bootstraping y permutación) que permiten evaluar la existencia de diferencias entre los
valores de estos índices, para cada una de las coberturas vegetales estudiadas Sabogal,
2011.
Índice de equidad de Shannon-Wiener (H’): Asume que todas las especies están
representadas en las muestras; indica qué tan uniformes están representadas las
especies (en abundancia) teniendo en cuenta todas las especies muestreadas
(Villareal et al., 2006). El valor suele hallarse entre 1,5 (diversidad baja) y 3,5
(diversidad alta) (Coddington et al., 1991; Magurran, 2004).
Índice de dominancia de Simpson (D): Muestra la probabilidad de que dos individuos
sacados al azar de una muestra correspondan a la misma especie (Villareal et al.,
2006). El valor máximo que puede tomar este índice es uno, si la dominancia es alta
la diversidad será baja por eso al restarle a uno la dominancia debería resultar el valor
de la diversidad (Fernández y Leiva 2002 citado por Sabogal, 2011).
3.4.2.4 Modelos de abundancia. Se graficó la distribución de abundancias de las
especies (morfoespecies) por tipo de cobertura con el programa PAST versión 2.17c
(Hammer et al., 2001) con el fin de evaluar el modelo (Serie geométrica, logarítmica,
distribución log-normal o vara quebrada) que se ajusta mejor a los datos de abundancia.
Estos modelos matemáticos describen de forma gráfica la relación entre la abundancia
y las especies ordenadas en categorías de la más a la menos abundante (Villareal et al.,
2006); de esta forma, permiten comparar los diferentes ensambles de mfsp y explicar el
comportamiento de su abundancia relativa.
Para verificar el ajuste de los ensambles de morfoespecies a los modelos de abundancia
preestablecidos, el programa realiza una prueba de bondad de ajuste de Chi-cuadrado
(χ2), donde la Ho (Los datos de abundancia se ajustan al modelo) es rechazada si la
probabilidad de la prueba es mayor al nivel de significancia (α= 0,05) (Magurran, 1988).

40
3.4.3 Evaluación de la diversidad beta (β):
3.4.3.1 Índice de similitud de Chao-Jaccard. Para evaluar el grado de recambio de
especies o diversidad beta con el programa estadístico EstimateS versión 9.1.0 (Colwell,
2013) se calculó el índice de similitud Chao-Jaccard; este índice además de tener en
cuenta el apareamiento de la abundancia de las especies (como lo hacen los índices de
Morisita-Horn o Bray-Curtis) en los ensambles, también tiene en cuenta la similitud de
las abundancias relativas, siendo más apropiado que los índices clásicos
correspondientes para la evaluación de la similitud en la composición entre muestras de
diferentes tamaños, así como en situaciones reales o sospechadas de submuestreo, y
cuando es probable que los ensambles tengan numerosas especies raras (Chao,
Chazdon, Colwell y Tsung-Jen, 2005).
3.4.4 Grupos funcionales de arañas. Para entender mejor la comunidad de arañas,
analizar la estructura de los grupos funcionales en cada cobertura y establecer como las
diferencias en la estructura de los hábitats pueden reflejarse en las estrategias de
forrajeo de estos organismos, las arañas fueron grupadas en gremios (grupos) con base
en la estrategia de forrajeo (Root, 1967).
El diseño de los gremios se basó en las categorías descritas por Cardoso, Pekár, Jocqué
y Coddington, 2011. Y teniendo en cuenta las características ecológicas de las familias
colectadas (modo de forrajeo, tipo de tela, uso de microhábitat), y para las morfoespecies
encontradas el método de colecta con el cual fueron capturadas. Se graficó el número
de morfoespecies por gremio en cada cobertura.
3.4.5 Preferencia de microhábitat. Adicional a los análisis planteados para este estudio,
y con el propósito de detallar un poco más la comunidad de arañas presente en la
ventana de trabajo, se calculó y graficó la riqueza y abundancia relativa de familias,
morfoespecies y gremios a nivel de los estratos de cada una de las coberturas.

41
Se definieron los estratos de acuerdo a los métodos de captura y teniendo en cuenta las
descripciones realizadas por Rangel & Lozano (1986) y Villarreal et al., (2006) para la
caracterización de la distribución vertical de la vegetación (Tabla 3).
Tabla 3 Tipos de estratos definidos para la caracterización de preferencia por
microhábitat de la comunidad de arañas en las coberturas bosque y cafetal de la ventana
de trabajo: paisaje rural cafetero de El Brillante, Risaralda.
Cobertura Estrato Rango
Bosque
Rasante 0 - 0,5 m
Herbáceo 0,5 - 2 m
Arbustivo 2 - 5 m
cafetal Rasante 0 - 0,5 m
Vegetativo 0,5 - 2 m
Fuente: el autor; modificado de Rangel & Lozano, 1986 y Villarreal et al., 2006.
3.4.6 Densidad de arañas por unidad de muestreo. De forma complementaria para este
trabajo se tuvo en cuenta la densidad de arañas por unidad de muestreo y estrato
vegetal, calculada a partir del número total de individuos colectados y el área revisada
por muestra para cada método de captura. Particularmente, para el método de agitación
de vegetación no se midió de forma directa el área muestreada, no obstante, se tuvo en
cuenta el número de unidades vegetales (arbustos) sacudidos sobre el paragua japonés,
el cual presenta un área de muestreo de 1 m2.

42
4. RESULTADOS
4.1 COMPOSICIÓN Y ABUNDANCIA DE LAS ARAÑAS EN UN PAISAJE RURAL
CAFETERO
4.1.1 Composición. Se colectó un total de 2491 individuos en 124 muestras, de los cuales
fueron seleccionados 1425 individuos (920 adultos y 505 subadultos) para la realización
de los diferentes análisis estadísticos; los 1066 individuos restantes no fueron tenidos en
cuenta por ser organismos inmaduros, lo que dificulta la asignación confiable de cada
uno de ellos a las morfoespecies definidas con las características morfológicas de los
individuos adultos.
Entre los individuos seleccionados se determinaron 16 familias que agrupan 87
morfoespecies (mfsp), 29 de las cuales se identificaron a género y 28 a especie (Tabla
4; Anexo G); estas 16 familias de arañas encontradas representan el 24% y 14% de las
familias reportadas para Colombia y el mundo respectivamente (Perafán et al., 2013;
World Spider Catalog, 2014).
Tabla 4 Familias colectadas en un paisaje rural cafetero de Risaralda, Colombia.
Familia Subadultos Adultos Total Familia Subadultos Adultos Total
Linyphiidae 154 319 473 Ochyroceratidae 3 6 9
Pholcidae 48 218 266 Uloboridae 1 7 8
Theridiidae 114 147 261 Oxyopidae 1 3 4
Tetragnathidae 90 88 178 Thomisidae 0 2 2
Araneidae 47 61 108 Amaurobiidae 0 1 1
Lycosidae 33 17 50 Liocranidae 0 1 1
Salticidae 8 38 46 Oonopidae 0 1 1
Anyphaenidae 6 10 16 Theridiosomatidae 0 1 1
Total 16 505 920 1425
Fuente: el autor

43
Con respecto a las coberturas muestreadas, en el bosque remanente se colectó un total
de 1057 individuos (42,43%), de los que se seleccionaron 577 para los análisis
estadísticos; se determinaron 12 familias que agrupan 47 mfsp; 27 mfsp se lograron
identificar a género y 13 a especies. En el cafetal se colectaron 1434 individuos (57,57%).
De estos, se eligieron 848 para los respectivos análisis y se determinó un total de 13
familias que agrupan 62 mfsp, de las cuales 21 se identificaron a género y 23 a especies
(Anexo G).
4.1.2 Abundancia. De las familias colectadas Linyphiidae (473 indv.), Pholcidae (266),
Theridiidae (261), Tetragnathidae (178) y Araneidae (108) presentaron las mayores
abundancias. Lycosidae (50), Salticidae (46) y Anyphaenidae (16) mostraron una
abundancia media. Las demás familias exhibieron una abundancia igual o menor a 9
individuos y corresponden al 1,89% del total de las familias colectadas (Figura 8).
Figura 8 Abundancia relativa de las familias de arañas registradas en un paisaje rural
cafetero de Risaralda, Colombia.
Fuente: el autor
En algunas familias solo se colectó uno o dos individuos, considerándose estas como
familias con morfoespecies raras, debido a su mínima abundancia; así, las familias
0,070,070,070,070,140,280,560,631,12
3,233,51
7,5812,49
18,3218,67
33,19
0 5 10 15 20 25 30 35
Amaurobiidae
Liocranidae
Oonopidae
Theridiosomatidae
Thomisidae
Oxyopidae
Uloboridae
Ochyroceratidae
Anyphaenidae
Salticidae
Lycosidae
Araneidae
Tetragnathidae
Theridiidae
Pholcidae
Linyphiidae
% Abundancia
Fam
ilia
s

44
Amaurobiidae, Liocranidae, Oonopidae y Theridiosomatidae registraron un (1) solo
individuo y Thomisidae dos (2) individuos (Tabla 4; Anexo G).
La familia más abundante en el bosque fue Pholcidae (238 indv.), seguida de Linyphiidae
(99), Theridiidae (83) y Araneidae (59). Las familias Oonopidae, Theridiosomatidae y
Thomisidae presentaron la menor abundancia para esta cobertura con el registro cada
una de un sólo individuo. En el cafetal las familias Linyphiidae (374), Theridiidae (178),
Tetragnathidae (141) y Lycosidae (50) presentaron la mayor abundancia; en contraste, y
con la presencia de un solo organismo para cada una, las familias Amaurobiidae,
Liocranidae y Thomisidae presentaron la menor abundancia (Figura 9).
Figura 9 Abundancia relativa de las familias de arañas encontradas en bosque y cafetal
en un paisaje rural cafetero de Risaralda, Colombia.
Fuente: el autor
Es de mencionar que familias como Ochyroceratidae, Oonopidae y Theridiosomatidae
sólo se registraron en el bosque, y por su parte Amaurobiidae, Liocranidae, Lycosidae y
Oxyopidae sólo fueron registradas en el cafetal (Anexo G).
0,17
0,17
0,17
0,69
1,56
2,08
5,72
6,41
10,23
14,38
17,16
41,25
0 20 40 60
Oonopidae
Theridiosomatidae
Thomisidae
Uloboridae
Ochyroceratidae
Anyphaenidae
Salticidae
Tetragnathidae
Araneidae
Theridiidae
Linyphiidae
Pholcidae
% Abundancia Bosque
Fam
ilia
s
0,12
0,12
0,12
0,47
0,47
0,47
1,53
3,30
5,78
5,90
16,63
20,99
44,10
0 10 20 30 40 50
Amaurobiidae
Liocranidae
Thomisidae
Anyphaenidae
Oxyopidae
Uloboridae
Salticidae
Pholcidae
Araneidae
Lycosidae
Tetragnathidae
Theridiidae
Linyphiidae
% Abundancia Cafetal

45
Entre las morfoespecies (mfsp) más abundantes se destacaron Linyphiidae sp1 (294
indv.), Pholcidae sp1 (247) y Dubiaranea sp (101), las cuales agrupan el 45,05% del total
de individuos colectados. Las morfoespecies restantes (84) se organizan de la siguiente
forma: 19 de las morfoespecies colectadas presentaron una abundancia de 70 a 10
individuos (39,91%), 35 mfsp mostraron una abundancia menor a 10 individuos (12,32%),
8 mfsp con registro de 2 individuos (0,98%) y 22 mfsp con registro de 1 individuo (1,54%)
(Figura 10; Anexo G).
Figura 10 Abundancia relativa de las morfoespecies de arañas registradas en un paisaje
rural cafetero de Risaralda, Colombia.
Mfsp Un indv. (Agrupa Mfsp con un solo individuo registrado); Mfsp Dos indv. (Agrupa
Mfsp con dos individuos registrados); Otras Mfsp -10 indv. (Agrupa Mfsp con menos de
diez individuos registrados).
Fuente: el autor
En cuanto a las coberturas, para el bosque la morfoespecie Pholcidae sp1 (228 indv.)
fue la más abundante, seguida de Erigoninae sp3 (51) y Linyphiidae sp1 (41). En el
0,700,700,700,770,840,910,981,051,191,541,541,821,89
2,882,883,023,373,513,653,72
4,777,09
12,3217,33
20,63
0 5 10 15 20 25
Pholcidae sp2Chrysso sp
Thwaitesia sp1Cyclosa rubronigra
Theridiidae sp17Theridiidae sp24
Mfsp Dos indv.Chrysometa calima
Salticidae sp4Erigoninae sp2
Mfsp Un indv.Mangora eberhardi
Micrathena lucasiChrysometa alboguttata
Theridion frizzellorumTetragnathidae sp1Anelosimus baeza
Lycosidae sp1Erigoninae sp3
Theridion nigricepsLeucauge argentea
Dubiaranea spOtras Mfsp -10 indv.
Pholcidae sp1Linyphiidae sp1
%Abundancia
Mo
rfo
esp
ecie
s (
mfs
p)

46
cafetal Linyphiidae sp1 (253), Dubiaranea sp (101) y Lycosidae sp1 (50) presentaron las
mayores abundancias (Figura 11).
Figura 11 Abundancia relativa de las morfoespecies de arañas encontradas en bosque
y cafetal de un paisaje rural cafetero de Risaralda, Colombia.
Mfsp Un indv. (Agrupa Mfsp con un solo individuo registrado); Mfsp Dos indv. (Agrupa
Mfsp con dos individuos registrados); Otras Mfsp -10 indv. (Agrupa Mfsp con menos de
diez individuos registrados).
Fuente: el autor
Referente a las morfoespecies raras para cada cobertura, en el bosque se registraron 5
mfsp con una abundancia de dos individuos cada una y 15 mfsp con abundancia de un
solo individuo; en el cafetal 6 de las mfsp colectadas presentaron una abundancia de dos
0,520,520,690,690,690,870,871,211,211,211,391,391,391,561,561,731,731,731,731,731,912,602,953,474,514,687,118,84
39,51
0 20 40 60
Anyphaeninae sp3
Parawixia rimosa
Achaearanea trapezoidalis
Philoponella sp1
Theridiidae sp5
Alpaida queremal
Phoroncidia studo
Chrysometa carmelo
Erigoninae sp2
Pholcidae sp3
Salticidae sp9
Tetragnathidae sp1
Theridiidae sp2
Anyphaeninae sp2
Ochyroceratidae sp1
Msp Dos indv.
Chrysso sp
Cyclosa rubronigra
Theridiidae sp24
Thwaitesia sp1
Micrathena lucasi
Msp Un indv.
Salticidae sp4
Leucauge argentea
Mangora eberhardi
Theridion nigriceps
Linyphiidae sp1
Erigoninae sp3
Pholcidae sp1
% Abundancia Bosque
Mo
rfo
esp
ecie
s (
mfs
p)
1,42
1,42
1,77
1,77
1,89
2,24
2,48
3,07
4,13
4,83
4,83
5,66
5,66
5,90
11,20
11,91
29,83
0 10 20 30 40
Theridiidae sp17
Msp Dos indv.
Chrysometa calima
Erigoninae sp2
Micrathena lucasi
Pholcidae sp1
Msp Un indv.
Theridion nigriceps
Tetragnathidae sp1
Chrysometa alboguttata
Theridion frizzellorum
Anelosimus baeza
Leucauge argentea
Lycosidae sp1
Otras Msp Menos de 10 indv.
Dubiaranea sp
Linyphiidae sp1
% Abundancia Cafetal

47
individuos y 21 mfsp mostraron una abundancia de un individuo (Figura 11). Las especies
raras se concentraron en las familias Theridiidae, Araneidae y Salticidae (Anexo G).
Asimismo se destaca el gran número de morfoespecies que se registraron sólo en una
de las coberturas, es así como el bosque presentó un total de 24 mfsp y el cafetal 38
mfsp (Tabla 5). Nuevamente Theridiidae, Araneidae y Salticidae agruparon el mayor
número de este tipo de morfoespecies.
Tabla 5 Morfoespecies presentes en sólo una de las dos coberturas evaluadas en un
paisaje rural cafetero de Risaralda, Colombia.
Mfsp en Bosque Mfsp en Cafetal
1 Alpaida queremal 1 Acacesia hamata 25 Salticidae sp3
2 Anyphaeninae sp2 2 Amaurobiidae sp1 26 Spintharus sp
3 Chrysometa carmelo 3 Anelosimus baeza 27 Theridiidae sp10
4 Chrysometa serachui 4 Anelosimus cf tungurahua 28 Theridiidae sp13
5 Chrysometa sp 1 5 Anyphaeninae sp1 29 Theridiidae sp16
6 Chrysso sp 6 Argiope argentata 30 Theridiidae sp17
7 Episinus sp1 7 Chrysometa alboguttata 31 Theridiidae sp18
8 Lyssomanes sp1 8 Chrysometa calima 32 Theridiidae sp19
9 Mangora eberhardi 9 Chrysometa yotoco 33 Theridiidae sp22
10 Ochyroceratidae sp1 10 Coleosoma blandum 34 Theridiidae sp33
11 Ocrepeira sp 11 Cyclosa triquetra 35 Theridiidae sp9
12 Oonopidae sp1 12 Dubiaranea sp 36 Uloborus sp2
13 Pholcidae sp4 13 Erigoninae sp1 37 Wagneriana tauricornis
14 Phoroncidia studo 14 Eriophora sp 38 Wagneriana undecimtuberculata
15 Salticidae sp16 15 Eustala scutigera
16 Salticidae sp4 16 Gasteracantha cancriformis
17 Salticidae sp5 17 Liocranidae sp1
18 Salticidae sp8 18 Lycosidae sp1
19 Salticidae sp9 19 Metazygia keyserlingi
20 Theridiidae sp2 20 Neoscona sp
21 Theridiidae sp34 21 Peucetia sp1
22 Theridiidae sp37 22 Salticidae sp10
23 Theridiosomatidae sp1 23 Salticidae sp17
24 Thwaitesia sp1 24 Salticidae sp2
Fuente: el autor

48
4.2 EVALUACIÓN DE LA DIVERSIDAD ALFA (α)
4.2.1 Riqueza específica. La mayor riqueza de morfoespecies estuvo representada por
la familia Theridiidae, con 27 mfsp, correspondientes al 31% del total de mfsp
encontradas. Le siguen en importancia de riqueza dentro del área de estudio las familias
Araneidae con 18 mfsp, Salticidae con 12 mfsp y Tetragnathidae con 8 mfsp (Figura 12).
Este comportamiento se reflejó de manera similar si se observan las coberturas
evaluadas por separado, no obstante, si registran diferencias a nivel de las familias de
menor riqueza. Para el bosque se encuentran Ochyroceratidae, Oonopidae,
Theridiosomatidae, Thomisidae y Uloboridae, y en cafetales Amaurobiidae, Liocranidae,
Lycosidae, Oxyopidae y Thomisidae como las familias de menor número de
morfoespecies (Figura 13).
Figura 12 Riqueza específica de las familias de arañas registradas en un paisaje rural
cafetero de Risaralda, Colombia.
Fuente: el autor
1
1
1
1
1
1
1
1
2
3
4
5
8
12
18
27
1,15
1,15
1,15
1,15
1,15
1,15
1,15
1,15
2,30
3,45
4,60
5,75
9,20
13,79
20,69
31,03
0 10 20 30 40 50 60 70
Amaurobiidae
Liocranidae
Lycosidae
Ochyroceratidae
Oonopidae
Oxyopidae
Theridiosomatidae
Thomisidae
Uloboridae
Anyphaenidae
Pholcidae
Linyphiidae
Tetragnathidae
Salticidae
Araneidae
Theridiidae
Fam
ilia
s
No. de Mfsp. por familia % Mfsp. por familia

49
Figura 13 Riqueza específica de las familias de arañas registradas en bosque y cafetal
de un paisaje rural cafetero de Risaralda, Colombia.
Fuente: el autor
4.2.2 Estimaciones de riqueza y representatividad del muestreo. Con el propósito de
evaluar la eficiencia del muestreo se elaboró una curva de acumulación de especies con
los datos del muestreo general, empleando el promedio de los valores de riqueza
calculados por el programa EstimateS versión 9.1.0 (Colwell, 2013) para los 7
estimadores no paramétricos utilizados (Chao 1, Chao 2, Jacknife de primer y segundo
orden, ACE, ICE y Bootstrap). En los muestreos por cobertura, solamente se describen
los valores estimados de morfoespecies y el respectivo porcentaje de eficiencia.
De acuerdo a la curva de acumulación de especies general se observó una eficiencia del
muestreo del 81% al comparar el número de morfoespecies observado (87 mfsp)
respecto al número promedio de morfoespecies esperado (108 mfsp) (Figura 14; Tabla
6). La curva de acumulación no alcanza la asíntota lo que evidencia que aún existe un
porcentaje de especies en el sistema que no han sido colectadas por este estudio,
demostrando que un mayor esfuerzo de muestreo permitirá conocer el número “real” de
especies. No obstante, y dado que el valor de la curva de acumulación se encuentra
1
1
1
1
1
2
2
3
5
7
8
13
2,22
2,22
2,22
2,22
2,22
4,44
4,44
6,67
11,11
15,56
17,78
28,89
0 10 20 30 40 50
Ochyroceratidae
Oonopidae
Theridiosomatidae
Thomisidae
Uloboridae
Anyphaenidae
Pholcidae
Linyphiidae
Tetragnathidae
Salticidae
Araneidae
Theridiidae
Fam
ilia
s
Bosque
No. de Mfsp. por familia
1
11
1
12
2
25
5
615
20
1,61
1,611,61
1,61
1,613,23
3,23
3,238,06
8,06
9,6824,19
32,26
0 20 40 60
Amaurobiidae
LiocranidaeLycosidae
Oxyopidae
ThomisidaeAnyphaenidae
PholcidaeUloboridae
Linyphiidae
TetragnathidaeSalticidae
Araneidae
Theridiidae
Cafetal
% Mfsp. por familia

50
cercano al valor teórico de eficiencia del 85% (Villareal et al., 2006), es posible asegurar
que se tiene una buena representación de la araneofauna presente en el paisaje rural
cafetero de estudio.
Figura 14 Curva de acumulación de especies para un paisaje rural cafetero de Risaralda,
Colombia.
Sobs (Morfoespecies observadas); PromEstRiqTot (Promedio del total de los
estimadores de riqueza).
Fuente: el autor
Adicionalmente se observa que las curvas de acumulación de especies raras, tanto las
basadas en la abundancia (Singletons y doubletons), como las basadas en la
presencia/ausencia (Uniques y duplicates) tienden a decrecer (Figura 14), lo que indica
que al aumentar el número de estaciones muestreadas se fueron adicionando ejemplares
a las especies menos comunes. El número total de singletons y doubletons fue de 30
mfsp (34%) y de uniques y duplicates fue de 35 mfsp (40%); es de señalar que tales
porcentajes de especies raras se encuentran por encima del rango de aceptación de
especies raras, el cual no debe superar el 30% del total de especies colectadas
(Coddington, 2008 citado por Sabogal, 2011).
0
20
40
60
80
100
120
0 20 40 60 80 100 120 140
No
. M
orf
oesp
ecie
s
No. Muestras
Sobs Singletons Doubletons Uniques Duplicates PromEstRiqTot
% Eficiencia= 81%

51
Igualmente se estableció para cada una de las coberturas evaluadas la eficiencia del
muestreo (Tabla 6), encontrando que aún faltan por determinar un número importante de
especies. El bosque corresponde a la cobertura que estuvo más cerca de alcanzar el
número “real” de especies, en contraste con el cafetal. Al calcular la eficiencia con cada
estimador, se encontró que algunos presentan una proporción alta de riqueza conocida.
El estimador Bootstrap fue el más conservador en todos los tratamientos mostrando
valores cercanos al número de especies observadas (Sobs); Jacknife 1 y Chao 2
mostraron estimaciones intermedias para el bosque y el cafetal respectivamente, y el
estimador de riqueza de especies Jacknife 2 mostró siempre los valores estimados más
elevados (Tabla 6).
Tabla 6 Estimaciones de riqueza calculadas para las coberturas bosque y cafetal en un
paisaje rural cafetero de Risaralda, Colombia.
*Morfoespecies observadas; **Promedio del total de los estimadores de riqueza.
Variables de Diversidad Bosque Cafetal General
Muestras 61 63 124
Sobs* 47 62 87
Singletons 15 21 22
Doubletons 5 6 8
Uniques 16 23 23
Duplicates 8 9 12
ACE 61.09 82.86 104
ICE 63.55 88.68 104.99
Chao 1 64.47 91.96 112.65
Chao 2 60.11 86.9 106.3
Jack 1 62.74 84.63 109.81
Jack 2 70.6 98.33 120.73
Bootstrap 54.06 72 97.78
PromEstRiqTot** 62 86 108
% Eficiencia 76 72 81
Fuente: el autor

52
4.2.3 Índices de abundancia proporcional. De acuerdo con los resultados arrojados por
el programa PAST versión 2.17c (Hammer et al., 2001), se observa que el índice de
diversidad de Shannon-Wiener presenta diferencias significativas entre las coberturas
evaluadas, como era de esperarse. El cafetal presentó el mayor valor (2.83) lo que es
congruente con su mayor riqueza (62 mfsp); en comparación, el bosque presentó un
menor valor del índice (2.60) que se relaciona a su menor riqueza (47 mfsp) (Tabla 7).
Acorde a esta información se puede mencionar que la comunidad de arañas en la
ventana evaluada correspondiente a una muestra del paisaje rural cafetero del
departamento de Risaralda presenta una diversidad media, conforme con Magurran
(2004), quien afirma que el rango de la diversidad de dicho índice oscila entre 1.5 y 3.5
sobrepasando raramente un valor de 4.5.
Tabla 7 Índices de diversidad α para las arañas de un paisaje rural cafetero de Risaralda,
Colombia.
Fuente: el autor
Los valores de dominancia calculados con el índice de Simpson (D) asimismo
presentaron diferencias significativas con respecto de las coberturas y, permitieron
corroborar los valores de diversidad anteriormente tratados: el cafetal con mayor
diversidad obtuvo la menor dominancia (0.12) presentando así una distribución de
abundancias de las mfsp mucho más homogénea; el bosque por su parte, con menor
diversidad y mayor dominancia (0.18) coincide con el menor número de mfsp
encontradas (Tabla 7).
Índices Bosque Cafetal Boot p(eq) Perm p(eq)
Especies 47 62 0.046 0.129
Individuos 577 848 0 0
Shannon H 2.60 2.83 0.004 0.012
Simpson D 0.18 0.12 0.001 0.001

53
4.2.4 Modelos de abundancia. Se evaluaron los modelos de abundancia de las
morfoespecies entre coberturas, las cuales se ajustaron al modelo log-normal. La
probabilidad (p) para cada una de las pruebas de χ2 fue mayor al nivel de significancia
utilizado (0.05); para bosque fue de p= 0.3737 y para cafetal de p= 0.2143.
La representación gráfica del ajuste al modelo log-normal en ambas coberturas no
muestra una línea velo, por lo que para ambos casos la primera octava es la dominante
respecto a las demás frecuencias de morfoespecies (Figura 15).
Figura 15 Modelos de abundancia (log-normal) para bosque y cafetal de un paisaje rural
cafetero de Risaralda, Colombia.
Fuente: el autor
0 2.5 5 7.5 10 12.5 15
Octave
0
2
4
6
8
10
12
14
16
18
Spec
ies p
er o
ctav
e
0 2.5 5 7.5 10 12.5 15
Octave
0
3
6
9
12
15
18
21
24
27
Spec
ies p
er o
ctav
e
CAFETAL
BOSQUE

54
4.3 EVALUACIÓN DE LA DIVERSIDAD BETA (β)
4.3.1 Índice de similitud de Chao-Jaccard. El análisis de Chao-Jaccard exhibe una
similitud de las morfoespecies presentes en las coberturas evaluadas del orden del 45%
(0,452) (Tabla 8), indicando que existe un recambio amplio de morfoespecies, las cuales
no se encuentran para ambas coberturas, coincidiendo con lo expresado en el numeral
4.1.2, tabla 5.
Tabla 8 Índice de similitud de Chao-Jaccard calculado para bosque y cafetal de un
paisaje rural cafetero de Risaralda, Colombia.
Sobs
Bosque
Sobs
Cafetal
Sobs
Compartidas Chao-Jaccard Recambio
47 62 22 0,452 0,548
Fuente: el autor
4.4 GRUPOS FUNCIONALES DE ARAÑAS
Siguiendo los criterios de Cardoso et al. (2011), las 16 familias de arañas se agruparon
en seis gremios (Tabla 9). El gremio más representativo dentro de la araneofauna
asociada a la muestra de paisaje rural cafetero evaluado fue el de las constructoras de
telas tridimensionales, con un 36.98% del total de individuos capturados, seguido por las
constructoras de telas en sábana o maraña con el 21.33% (Figura 16). Se destaca a las
arañas constructoras de telas como el grupo de arañas de mayor representatividad para
el área de estudio con el 79% de los individuos recolectados frente a de las arañas
cazadoras con el 21% (Figura 16). De acuerdo al muestreo por coberturas se encontró
para el bosque que las arañas constructoras de telas tridimensionales (56%), presentan
la mayor representatividad seguidas por los gremios de otras cazadoras y las
constructoras de telas orbiculares (18% c/u). En el cafetal la mayor representatividad de
los gremios fue para las arañas constructoras de telas en sábana o maraña (30%),
seguidas por las constructoras de telas tridimensionales (24%) y las constructoras de

55
telas orbiculares (23%). Para ambas coberturas, el gremio de arañas cazadoras por
emboscada presentó la menor representatividad al capturar un (1) sólo individuo de este
gremio en cada una de ellas (Figura 17).
Tabla 9 Gremios de arañas encontrados en un paisaje rural cafetero de Risaralda,
Colombia.
Gremios
Según Cardoso et al., 2011 Familia
No. de
Mfsp. No. Indv.
Porcent.
%
Constructoras de telas
tridimensionales
(CTT)
Pholcidae
31 527 36.98 Theridiidae
Constructoras de telas en
sábanas
(CTS)
Amaurobiidae
3 304 21.33 Linyphiidae
(Linyphiidae sp1)
Ochyroceratidae
Constructoras de telas
orbiculares
(CTO)
Araneidae
29 295 20.70 Tetragnathidae
Theridiosomatidae
Uloboridae
Otras cazadoras
(OC)
Oxyopidae
20 245 17.19
Salticidae
Anyphaenidae
Linyphiidae
(Dubiaranea;
Erigoninae)
Cazadoras errantes en suelo
(CES)
Liocranidae
3 52 3.65 Lycosidae
Oonopidae
Cazadoras por emboscada
(CE) Thomisidae 1 2 0.14
Fuente: el autor
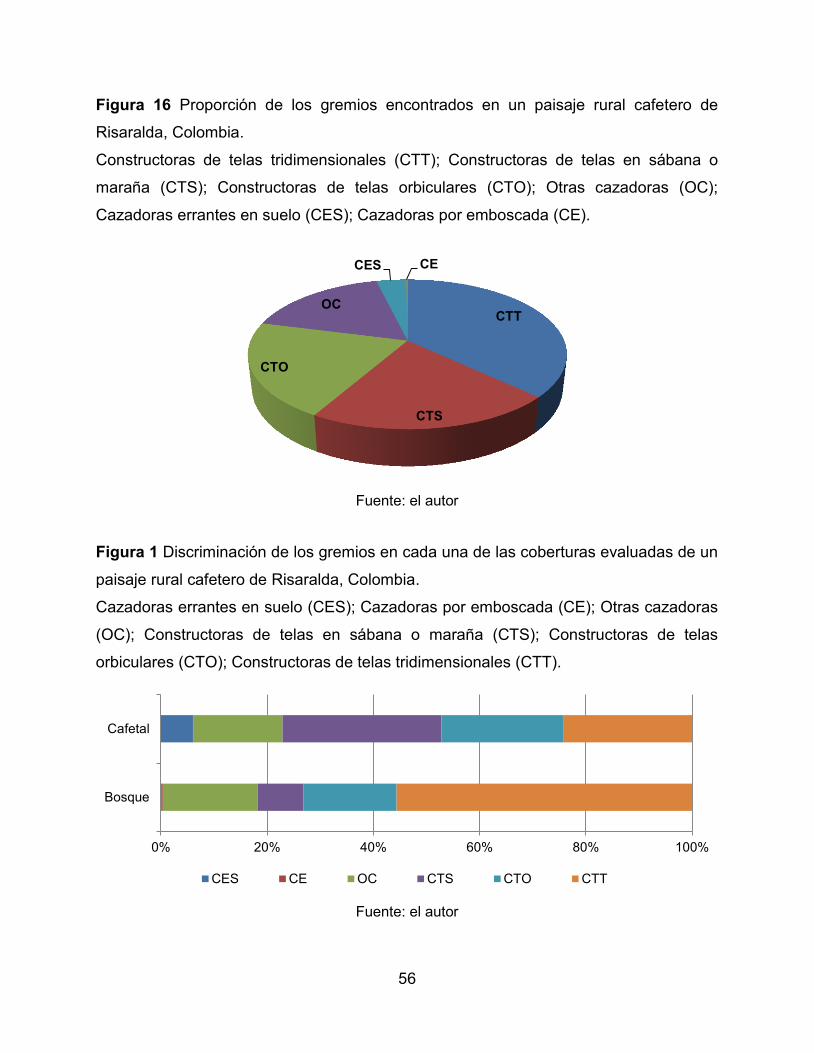
56
Figura 16 Proporción de los gremios encontrados en un paisaje rural cafetero de
Risaralda, Colombia.
Constructoras de telas tridimensionales (CTT); Constructoras de telas en sábana o
maraña (CTS); Constructoras de telas orbiculares (CTO); Otras cazadoras (OC);
Cazadoras errantes en suelo (CES); Cazadoras por emboscada (CE).
Fuente: el autor
Figura 1 Discriminación de los gremios en cada una de las coberturas evaluadas de un
paisaje rural cafetero de Risaralda, Colombia.
Cazadoras errantes en suelo (CES); Cazadoras por emboscada (CE); Otras cazadoras
(OC); Constructoras de telas en sábana o maraña (CTS); Constructoras de telas
orbiculares (CTO); Constructoras de telas tridimensionales (CTT).
Fuente: el autor
CTT
CTS
CTO
OC
CES CE
0% 20% 40% 60% 80% 100%
Bosque
Cafetal
CES CE OC CTS CTO CTT

57
4.5 PREFERENCIA DE MICROHÁBITAT
4.5.1 Bosque. El número de familias de la araneofauna presente en los estratos
evaluados fue muy similar, presentándose 9 familias para los estratos arbustivo y
rasante, y 8 para el estrato herbáceo. El estrato arbustivo exhibió la mayor riqueza de
morfoespecies (30), seguido de los estratos herbáceo (25) y rasante (20) (Figura 18).
Respecto de la abundancia, el estrato herbáceo aportó el 49% de los individuos
colectados dentro del bosque. La familia con mayor número de individuos fue Pholcidae
(Pholcidae sp1) para los estratos arbustivo y herbáceo, y Linyphiidae (Erigoninae sp3)
para el estrato rasante (Figura 19). Se encontraron familias que sólo se registraron para
uno de los estratos, así Ochyroceratidae y Thomisidae se encontraron sólo en el estrato
arbustivo, Uloboridae en el estrato herbáceo y Oonopidae y Theridiosomatidae en el
estrato rasante. Del mismo modo se registraron morfoespecies para uno solo de los
estratos evaluados, es así como el estrato arbustivo presentó un total de 11 mfsp, y los
estratos herbáceo y rasante 7 mfsp cada uno (Tabla 10).
Figura 2 Número de las familias y morfoespecies de arañas registradas en los estratos
arbustivo, herbáceo y rasante del bosque en un paisaje rural cafetero de Risaralda,
Colombia.
Fuente: el autor
0
5
10
15
20
25
30
35
E. Arbustivo E. Herbáceo E. Rasante
Riq
ueza
No. Familias No. Morfoespecies

58
Figura 3 Abundancia relativa de las familias y morfoespecies de arañas registradas en
los estratos arbustivo, herbáceo y rasante del bosque en un paisaje rural cafetero de
Risaralda, Colombia.
Mfsp Un indv. (Agrupa Mfsp con un solo individuo registrado); Mfsp Dos indv. (Agrupa
Mfsp con dos individuos registrados).
Fuente: el autor

59
Tabla 10 Familias y morfoespecies presentes en sólo uno de los tres estratos evaluadas
en el bosque de un paisaje rural cafetero de Risaralda, Colombia.
Familia Morfoespecie E. Arbustivo E. Herbáceo E. Rasante
Anyphaenidae Anyphaeninae sp3 X
Araneidae
Araneus bogotensis X
Cyclosa caroli X
Ocrepeira sp X
Liocranidae Liocranidae sp1 X
Ochyroceratidae Ochyroceratidae sp1 X
Oonopidae Oonopidae sp1 X
Pholcidae
Pholcidae sp2 X
Pholcidae sp3 X
Pholcidae sp4 X
Salticidae Salticidae sp16 X
Salticidae sp5 X
Tetragnathidae
Chrysometa carmelo X
Chrysometa serachui X
Chrysometa sp 1 ns X
Theridiidae
Episinus sp1 X
Theridiidae sp24 X
Theridiidae sp26 X
Theridiidae sp27 X
Theridiidae sp34 X
Theridiidae sp37 X
Theridiidae sp5 X
Theridiosomatidae Theridiosomatidae sp1 X
Thomisidae Thomisidae sp1 X
Uloboridae Philoponella sp1 X
Fuente: el autor

60
En cuanto a la distribución de los gremios encontrados para cada uno de los estratos del
bosque, se registró la siguiente información: las familias de arañas se agruparon en cinco
gremios para el estrato arbustivo y cuatro gremios para cada uno de los estratos
herbáceo y rasante. En el estrato arbustivo las arañas constructoras de telas
tridimensionales (56%) presentan la mayor representatividad seguidas por los gremios
de otras cazadoras (20%) y las constructoras de telas orbiculares (17%). En el estrato
herbáceo la mayor representatividad de los gremios fue para las arañas constructoras
de telas tridimensionales (64%), seguidas por las constructoras de telas orbiculares
(18%) y las constructoras de telas en sábana o maraña (13%). Finalmente, para el estrato
rasante los gremios de otras cazadoras (47%) y constructoras de telas tridimensionales
(35%) mostraron la mayor representatividad (Figura 20).
Figura 20 Distribución de los gremios de arañas en los estratos arbustivo, herbáceo y
rasante del bosque en un paisaje rural cafetero de Risaralda, Colombia.
Constructoras de telas tridimensionales (CTT); Constructoras de telas orbiculares
(CTO); Constructoras de telas en sábana o maraña (CTS); Otras cazadoras (OC);
Cazadoras por emboscada (CE); Cazadoras errantes en suelo (CES).
Fuente: el autor
0,0
0,6
19,6
7,3
16,8
55,9
0,0
0,0
4,6
13,2
18,1
64,1
0,9
0,0
47,0
0,0
17,1
35,0
0% 10% 20% 30% 40% 50% 60% 70% 80% 90% 100%
CES
CE
OC
CTS
CTO
CTT
% AR - E. Arbustivo %AR - E. Herbáceo %AR - E. Rasante

61
4.5.2 Cafetal. El número de familias presentes en los estratos evaluados para esta
cobertura fue muy similar, registrándose 9 familias para el estrato rasante, y 8 para el
estrato vegetativo. El estrato vegetativo exhibió la mayor riqueza de morfoespecies (35)
respecto del estrato rasante (25) (Figura 21). En relación a la abundancia, el estrato
vegetativo aportó el 60% de los individuos colectados dentro del cafetal. La familia más
abundante fue Linyphiidae para ambos estratos; esta misma familia presentó las
morfoespecies con mayor número de individuos, Linyphiidae sp1 y Dubiaranea sp para
el estrato vegetativo y Linyphiidae sp1 para el estrato rasante. Lycosidae sp1 presentó
la segunda mayor abundante dentro de las morfoespecies del estrato rasante (Figura
22).
Se encontraron familias que sólo se registraron para uno de los estratos; en
consecuencia Uloboridae se encontró sólo en el estrato vegetativo y Amaurobiidae y
Lycosidae en el estrato rasante. Del mismo modo se registraron morfoespecies para uno
solo de los estratos evaluados, así que el estrato vegetativo presentó un total de 18 mfsp
y el rasante 8 mfsp (Tabla 11).
Figura 24 Número de las familias y morfoespecies de arañas registradas en los estratos
vegetativo y rasante del cultivo de café en un paisaje rural cafetero de Risaralda,
Colombia.
Fuente: el autor
0
5
10
15
20
25
30
35
40
E. Vegetativo E. Rasante
Riq
ueza
No. Familias No. Morfoespecies

62
Figura 22 Abundancia relativa de las familias y morfoespecies de arañas registradas en
los estratos vegetativo y rasante del cultivo de café en un paisaje rural cafetero de
Risaralda, Colombia.
Mfsp Un indv. (Agrupa Mfsp con un solo individuo registrado); Mfsp Dos indv. (Agrupa
Mfsp con dos individuos registrados).
Fuente: el autor
0,3
0,6
1,2
1,2
7,2
15,3
22,0
52,3
0 20 40 60
Uloboridae
Oxyopidae
Pholcidae
Salticidae
Araneidae
Tetragnathidae
Theridiidae
Linyphiidae
%Abundancia relativa
0,90,90,91,21,21,41,42,02,32,62,92,93,54,04,3
7,28,1
22,529,8
0 10 20 30 40
Chrysometa alboguttata
Gasteracantha cancriformis
Theridiidae sp24
Pholcidae sp1
Theridiidae sp22
Cyclosa triquetra
Theridiidae sp19
Theridiidae sp17
Mfsp Dos indv.
Micrathena lucasi
Chrysometa calima
Theridion nigriceps
Theridion frizzellorum
Mfsp Un indv.
Leucauge argentea
Tetragnathidae sp1
Anelosimus baeza
Dubiaranea sp
Linyphiidae sp1
%Abundancia relativa
0,3
0,6
0,6
2,9
3,5
10,5
14,6
19,0
48,0
0 20 40 60
Amaurobiidae
Oxyopidae
Salticidae
Araneidae
Pholcidae
Theridiidae
Lycosidae
Tetragnathidae
Linyphiidae
%Abundancia relativa
2,6
2,3
0,9
0,9
1,2
2,3
2,9
3,2
4,1
5,3
7,6
8,2
14,6
43,9
0 10 20 30 40 50
Mfsp Un indv.
Mfsp Dos indv.
Argiope argentata
Theridiidae sp26
Micrathena lucasi
Anelosimus baeza
Tetragnathidae sp1
Pholcidae sp1
Erigoninae sp2
Theridion frizzellorum
Chrysometa alboguttata
Leucauge argentea
Lycosidae sp1
Linyphiidae sp1
%Abundancia relativa
Estrato Vegetativo
Estrato Rasante

63
Tabla 11 Familias y morfoespecies presentes en sólo uno de los dos estratos evaluados
en el cultivo de café de un paisaje rural cafetero de Risaralda, Colombia.
Familia Morfoespecies E. Vegetativo E. Rasante
Amaurobiidae Amaurobiidae sp1 X
Araneidae
Acacesia hamata X
Araneus bogotensis X
Cyclosa rubronigra X
Cyclosa triquetra X
Eriophora sp X
Gasteracantha cancriformis X
Metazygia keyserlingi X
Neoscona sp X
Parawixia rimosa X
Wagneriana tauricornis X
Linyphiidae Dubiaranea sp X
Erigoninae sp2 X
Lycosidae Lycosidae sp1 X
Pholcidae Pholcidae sp2 X
Salticidae Salticidae sp17 X
Salticidae sp7 X
Theridiidae
Spintharus sp X
Theridiidae sp17 X
Theridiidae sp18 X
Theridiidae sp19 X
Theridiidae sp24 X
Theridiidae sp27 X
Theridiidae sp33 X
Theridiidae sp5 X
Uloboridae Uloborus sp2 X
Fuente: el autor

64
En la distribución de los gremios en cada uno de los estratos del cafetal se registró que
las familias de arañas encontradas para el estrato vegetativo se agruparon en seis
gremios y las familias del estrato rasante se concentraron en cinco gremios. Para el
estrato vegetativo las arañas constructoras de telas en sábano o maraña exhiben la
mayor representatividad con casi el 30% de las abundancias, le siguen los gremios de
otras cazadoras, las constructoras de telas tridimensionales y las constructoras de telas
orbiculares con un rango entre el 22 – 24% de las abundancias registradas para este
estrato. En el estrato rasante la mayor representatividad de los gremios fue para las
arañas constructoras de telas en sábana o maraña con el 40% de las abundancias. Los
demás gremios registrados para este estrato mostraron una representatividad menor al
22% (Figura 23).
Figura 23 Distribución de los gremios de arañas en los estratos vegetativo y rasante del
cultivo de café en un paisaje rural cafetero de Risaralda, Colombia.
Constructoras de telas tridimensionales (CTT); Constructoras de telas orbiculares (CTO);
Constructoras de telas en sábana o maraña (CTS); Otras cazadoras (OC); Cazadoras
por emboscada (CE); Cazadoras errantes en suelo (CES).
Fuente: el autor
0,3
0,3
24,3
29,8
22,8
23,1
14,6
0,0
5,3
44,2
21,9
14,0
0% 10% 20% 30% 40% 50% 60% 70% 80% 90% 100%
CES
CE
OC
CTS
CTO
CTT
%AR - E. Vegetativo %AR - E. Rasante

65
4.6 DENSIDAD DE ARAÑAS POR UNIDAD DE MUESTREO
En el área de estudió que comprendió una muestra del paisaje rural cafetero se muestreó
un área total efectiva de 1044 m2 (Tabla 12) donde se colectó un total de 2491 individuos
(Tabla 13) que corresponde a una densidad de 2.4 arañas por metro cuadrado. En
relación a las coberturas evaluadas, para el bosque se muestreó un área total de 571 m2
donde se colectaron 1057 individuos mostrando una densidad de 1.9 arañas por metro
cuadrado; el cafetal con un área total de 474 m2 muestreados y 1434 individuos
colectados presentó una densidad de 3.0 arañas por metro cuadrado. En cuanto a los
estratos evaluados por cobertura, en el bosque, el estrato arbustivo presentó el mayor
valor de densidad con 2.6 arañas/m2. Los estratos herbáceo y rasante presentaron las
menores densidades con 1.6 y 1.5 arañas/m2 respectivamente. En el cafetal el estrato
rasante mostró la mayor densidad (5.4 arañas/m2) respecto del estrato vegetativo (2.4
arañas/m2).
Tabla 12 Área total muestreada en un paisaje rural cafetero de Risaralda, Colombia.
Cobertura
No. Unidades de Muestreo
(1UM= 25 m2)
Área Total UM (m2)
Área Muestreada por Técnica de captura
(m2)
Área Total Muestreada
(m2)
Porcentaje (%)
CMS CMA A
Bosque 21 525 96.5 339 135 571 55
Cafetal 21 525 125.5 243 105 474 45
Total 42 1050 222 582 240 1044 100
Fuente: el autor
Tabla 1 Número total de arañas colectadas en un paisaje rural cafetero de Risaralda,
Colombia.
Métodos de Captura Bosque Cafetal Total
Agitación 355 171 526
C. Manual Aérea 555 580 1135
C. Manual Suelo 147 683 830
Total 1057 1434 2491
Fuente: el autor

66
5. DISCUSIÓN
5.1 COMPOSICIÓN Y ABUNDANCIA DE LAS ARAÑAS EN UN PAISAJE RURAL
CAFETERO
En el presente estudio se registraron 87 morfoespecies de arañas agrupadas en 16
familias que componen el 24 % del total de familias reportadas para Colombia (Sabogal,
2011), evidenciando la importancia del paisaje rural cafetero como fuente de
araneofauna a la diversidad de artrópodos del país.
Se observa además, que la comunidad de arañas en el área de estudio está
representada por familias consideradas de amplia distribución como los son Araneidae,
Linyphiidae, Pholcidae, Salticidae, Tetragnathidae y Theridiidae (Dippenaar y Jocque,
1997), información que coincide con los resultados obtenidos en estudios de diversidad
realizados en distintas comunidades de arañas del país (Flórez, 1999; Blanco-Vargas,
Amat-García y Flórez, 2003; Rico-G., Beltrán, Álvarez & Flórez, 2005; Cepeda y Flórez,
2007; Alape, 2008; Cabra-García, 2009; Cepeda, 2009; Cabra-García, Montealegre y
Arce, 2010; Guasca y Reina, 2010; Reyna, 2010; Sabogal, 2011). Estas familias son
unas de las más diversas a nivel mundial, y se caracterizan por presentar especies con
capacidad de explotar los diversos recursos disponibles.
La abundancia de las familias y morfoespecies varía entre las coberturas evaluadas,
indicando que las arañas se ven influenciadas quizás por factores como disponibilidad
de sustratos, topografía, factores ambientales, entre otros. Los resultados demuestran
que el cafetal presentó los valores más altos de abundancia en comparación con el
bosque. Al respecto, Cortés citado por Alape (2008) señala el bosque como un hábitat
donde la riqueza y abundancia de especies no es considerablemente alta, pero sí es
constante, circunstancia que pueden ser contemplada en los resultados del presente
estudio.

67
De acuerdo con lo anterior se resalta a Pholcidae como la familia de mayor abundancia
en los bosques evaluados, pese a que no es una familia frecuentemente reportada como
predominante en ecosistemas del trópico. Su alta abundancia en esta zona puede
explicarse en relación con la diversa de microhábitat disponibles en esta cobertura
boscosa, ya que los fólcidos pueden encontrarse en casi todos los hábitat viables para
las arañas, desde la hojarasca y la parte inferior de las piedras hasta la copa de los
árboles (Huber, 2000), además, la menor luminosidad de los bosques en contraste con
los cafetales de sol, es otra característica que hace estos ecosistemas propicios para
estos organismos.
Linyphiidae fue la familia más abundante en el cafetal; se caracteriza porque muchas de
sus especies (68% de los linífidos encontrados en el cafetal) tejen telas en sábana o
maraña en estrato rasante sobre vegetación herbácea (Ubick, et al. 2005), por tanto, este
hábitat les ofrece las condiciones de sustrato adecuadas, en contraposición a lo percibido
en los bosques donde no es tan abundante. Esto sugiere la preferencia de Linyphiidae
por áreas abiertas en donde la vegetación herbácea u hojarasca es abundante, la
iluminación solar es constante y por tanto la temperatura es más elevada, características
que no se cumplen al interior del bosque, al menos para la mayoría de las especies, pues
algunas pertenecientes a la subfamilia Erigoninae fueron encontradas en esta cobertura,
aunque en menor proporción a las encontradas para el cafetal.
Las familias Theridiidae, Tetragnathidae y Araneidae mostraron para ambas coberturas
una abundancia media y las riquezas más altas, sugiriendo un carácter generalista o de
amplia distribución. Estos resultados son comparables con lo encontrado por Gómez
(2005), Rico-G. et al. (2005), Alape (2008), Guasca y Reina (2010) y Reyna (2010),
donde el comportamiento de estas familias con respecto a sus abundancias fue muy
similar a lo encontrado para este estudio. La menor abundancia y riqueza de familias
como Thomisidae, Theridiosomatidae, Oonopidae, Liocranidae y Amaurobiidae pudo
deberse a los hábitos crípticos y tamaños de pequeño a mediano que presentan los
individuos de estas familias, que desfavorecen la detección de este tipo de ejemplares.

68
5.2 DIVERSIDAD DE LA COMUNIDAD DE ARAÑAS EN UN PAISAJE RURAL
CAFETERO
Una medida de amplio uso para evaluar la diversidad es la riqueza de especies
(Magurran, 2004). Trabajar con la riqueza de especies es más adecuado que la sola
abundancia para describir la comunidad o la diversidad regional o local (Gotelli & Colwell,
2001). Al analizar la diversidad en términos de la riqueza (número de morfoespecies)
obtenida en cada cobertura, se observa diferencias significativas entre estas, indicada
por los procedimientos estadísticos de bootstraping y permutación realizados a los
índices de abundancia proporcional de equidad y dominancia. El cafetal de sol concentró
una mayor diversidad (H’= 2.83, 1-D= 0.12) en comparación al remanente de bosque
(H’= 2.60, 1-D= 0.18) muestreado, lo que sugiere una distribución de abundancias mucho
más homogénea entre las morfoespecies del cafetal, con valores relativamente bajos de
dominancia y una riqueza superior a la del bosque. Estos resultados difieren de lo
esperado, debido a que la diversidad de las especies animales, en especial las arañas,
se correlaciona con la heterogeneidad estructural de la vegetación, que les proporcionan
una mayor cantidad de sustratos y recursos, por lo que debería haber una más alta
diversidad en el bosque que en cualquier monocultivo de café que presenta una menor
complejidad en su estructura (Pianka, 1966; Uetz, 1979; McNett y Rypstra, 2000; Tews
et al., 2004; Loyola y Martins, 2008; Cava, 2013).
Los resultados obtenidos en el presente estudio podrían estar sujetos al efecto de la
actividad humana en la zona evaluada, ya que produce perturbaciones importantes en
los sistemas naturales (Callicott, Crowder y Mumford, 1999). La diversidad de especies
en una comunidad está determinada por la frecuencia de perturbaciones ambientales
(bióticas y abióticas) y las causadas por los seres humanos. Sousa (1984) reportó que la
diversidad alfa puede ser más alta en ensambles fuertemente perturbados y no
equilibrados que en ensambles no perturbados. Cuando las perturbaciones son muy
frecuentes o muy poco frecuentes, la diversidad de especies es baja. Por el contrario,
cuando la frecuencia de las perturbaciones es intermedia, la diversidad de especies es
alta.

69
Connell, 1978 sugiere que las perturbaciones de magnitud y frecuencia intermedias
mantienen los mayores niveles de diversidad, al no dejar que transcurra tiempo suficiente
para que tengan lugar exclusiones competitivas interespecíficas ni que las
perturbaciones de gran intensidad reduzcan la diversidad. Extrapolando esta información
al presente estudio, se propone que: las perturbaciones ejercidas por el hombre sobre
las áreas boscosas en la zona de estudio y que atañen a la explotación de madera o tala
para sembradíos de café principalmente, se presentan con una frecuencia mucho menor
en comparación a la constante intervención que sufren los cafetales por cuestiones de
su manejo y administración (Establecimiento, podas, zoqueo, manejo cultural de
malezas, uso de plaguicidas y herbicidas, entre otros), hecho que lleva a propiciar la
mayor diversidad de morfoespecies para el cafetal de sol.
Se ha propuesto que la perturbación intermedia fomenta el aumento de la diversidad y la
abundancia de algunas especies (Paine y Levin, 1981), lo cual se ha documentado en
cafetales en México y Costa Rica (Moguel y Toledo, 1996; 1999; Perfecto, Rice,
Greenberg y Van der Voort, 1996), y se acerca a lo observado en este estudio. Esta
información ha sido útil para considerar a los cafetales como ambientes que fomentan la
conservación (Macip, López y Muñoz, 2013).
El esfuerzo y la cobertura del muestreo respecto al hábitat y al organismo estudiado, así
como la heterogeneidad ambiental son algunos factores que influyen en la medición y
estimación de la riqueza (Palmer, 1990). Es importante relevar que los resultados de los
estimadores de riqueza, permiten considerar el muestreo como representativo, debido a
que se registró el 81% de las especies estimadas para el área de estudio.
Es preciso indicar que los estimadores no paramétricos empleados valoran la riqueza de
especies “puntual” del área de estudio. Por lo que este valor de riqueza es una
subestimación de la riqueza “real” del área (Coddington, Young y Coyle, 1996), es decir,
que solo calculan la riqueza que fue asequible a los métodos de muestreo empleados, al
tiempo de muestreo y a los individuos adultos y subadultos analizados. Por tanto, la

70
riqueza local de especies de la ventana evaluada está condicionada a la escala espacio-
temporal que se definió para el estudio.
Las especies raras juegan un papel importante en el cálculo de la mayoría de los
estimadores al garantizar la confiabilidad de los resultados obtenidos, dado que los
estimadores de riqueza están basados en su cuantificación (Rico et al., 2005). El número
de especies raras fue alto, por lo que es relevante señalar que en los inventarios de
arañas es común que éste dato acostumbre a ser elevado (New, 1999) y, generalmente
las curvas de acumulación suelen quedar lejos de alcanzar la asíntota (Jiménez-Valverde
y Hortal, 2003). Este gran porcentaje de especies raras se puede atribuir a factores como:
submuestreo, efecto de borde, individuos adultos fuera de la temporada normal de
apareamiento, individuos que ocupan un microhábitat que no es muestreado
adecuadamente, individuos que prefieren un microhábitat no incluido en el protocolo de
muestreo (Cabra-García et al., 2010).
Eventualmente, el aumento del esfuerzo de colecta y/o la intensidad del muestreo podría
registrar algunas de las especies faltantes y reducir proporcionalmente las especies
raras. Sin embargo, llegar a realizar un inventario completo de las especies que
componen la araneofauna de un área particular durante un muestreo puntual es una
tarea muy compleja, mucho más cuando se trabaja con uno de los llamados grupos
hiperdiversos, aquellos ricos en especies pero del que se posee un desconocimiento
mayor (Jiménez-Valverde y Hortal, 2003). Las arañas son muy abundantes y diversas en
los ecosistemas terrestres; no obstante, el conocimiento taxonómico a nivel de especie
es muy pobre en la mayoría de los grupos y los datos sobre distribución son muy escasos
(New, 1999).
La justificación ecológica para emplear los modelos de distribución de abundancia como
la descripción matemática más completa de los datos de una comunidad se debe a que
estos modelos paramétricos utilizan toda la información recopilada para analizar la
diversidad en relación a las fluctuaciones de los recursos y sus usos relativos por las
especies (Magurran, 1988). Además, excluyen los sesgos de los estimadores no

71
paramétricos (Brose, Martínez & Williams, 2003) y pueden compararse con los patrones
arrojados por las curvas de acumulación de especies (Gray, Bjørgesæter y Ugland,
2005).
Para este estudio, los ensambles para cada cobertura en la ventana de trabajo, se
ajustaron mejor a la distribución Log-Normal, basado en la inspección del modelo de
ajuste de bondad. De acuerdo con Krebs (1985), cuando la distribución de la abundancia
de especies se ajusta al modelo Log-Normal, es posible estimar el número total de
especies de la comunidad, incluyendo a las especies raras aún no registradas. Esto se
hace explorando la curva acampanada por debajo de la clase de mínima abundancia, y
midiendo el área. Esta puede ser una propiedad útil para las comunidades en las que no
todas las especies pueden verse y tabularse eficazmente.
La estimación del número de especies para el bosque remanente (59,44 mfsp) y el cafetal
de sol (83,22 mfsp), calculas a partir del modelo de distribución Log-Normal, coincide con
los resultados arrojados por los índices de estimación no paramétricos. Asimismo, se
observa que la representación gráfica de la distribución Log-Normal está truncada en el
punto en que las especies están representadas por un solo individuo. Por lo que, un
muestreo más intensivo haría que la distribución se desplace hacia la derecha y descubra
el sector oculto de las especies raras (Preston, 1948 citado por Krebs, 1985). Este
modelo Log-Normal truncado a la izquierda es más probable que describa muestras de
comunidades pequeñas o en desarrollo donde la competencia interespecífica puede ser
menos importante en restringir la abundancia de las especies raras, y donde las especies
de abundancias intermedias se vuelven más comunes (Hughes, 1986; Magurran, 1988),
esto último como resultado de la presencia de diversas asociaciones estables, que
suceden en ambientes con bajo estrés y elevada oferta de recursos, así como una amplia
variedad de hábitats a nivel local (May, 1981 citado por Sabogal, 2011).
Se ha demostrado que las arañas son sensibles a los cambios ambientales y antrópicos
(Avalos, Rubio, Bar y González, 2007), por lo que la sustitución de ambientes naturales
por agroecosistemas genera un recambio de especies en la comunidad local de arañas

72
(Simó, Laborda, Jorge y Castro, 2001). Lo anterior se puede evidenciar en el presente
estudio, donde el análisis de similitud permite inferir que el bosque remanente y el cafetal
de sol se definen de acuerdo a los diferentes arreglos de las abundancias de las
morfoespecies registradas para cada una de estas coberturas. Los resultados obtenidos
sugieren que se presentan características particulares (probablemente hayan incidido
factores microclimáticos como humedad relativa, temperatura, intensidad lumínica u
otros vinculados con la estructura vegetal) para el establecimiento de los ensambles de
morfoespecies reportados en cada una de las coberturas evaluadas.
Un ejemplo del recambio de especies, se puede apreciar en la araña lobo Lycosidae sp1,
que vive en madrigueras excavadas en la tierra o entre grietas (Flórez, 1996), y que sólo
se registró en los cafetales de sol. Su ausencia en el remanente de bosque podría
deberse al uso de suelo, ya que es común observarla en pastizales y agroecosistemas
(Dippenaar-Schoeman y Jocqué, 1997). Debido a ello esta morfoespecie podría ser
considerada como indicador del impacto antrópico. Si bien el cafetal comparte un
porcentaje de morfoespecies con el bosque remanente, se apreció que el
agroecosistema permite el establecimiento de un gran número de nuevas morfoespecies
(38 mfsp) como respuesta al cambio ambiental.
5.3 GRUPOS FUNCIONALES DE LA COMUNIDAD DE ARAÑAS
Con el propósito de comprender como se relaciona la comunidad de arañas con su
entorno y específicamente al interior del paisaje rural cafetero estudiado, se realizó la
clasificación gremial basada en lo propuesto por Cardoso et al. (2011). Los grupos
funcionales o gremios representan diferentes ensambles de arañas que disponen de
determinados mecanismos para la obtención de los recursos. En los estudios de la
araneofauna de agroecosistemas, la identificación de los gremios predominantes permite
conocer las principales estrategias de captura de la comunidad y en particular de aquella
que resulta importante para establecer el control biológico (Simó et al., 2001), asimismo,
permite reconocer las preferencias de hábitat y sustrato de estos organismos.

73
Para el presente estudio, los gremios dominantes registrados para los dos tipos de
cobertura fueron: el gremio de las arañas constructores de telas tridimensionales (de
mayor abundancia en los bosques), constituido por las familias Pholcidae y Theridiidae,
y el gremio de las arañas constructores de telas en sábana o maraña (de mayor
abundancia en los cafetales), representado por las familias Amaurobiidae, Linyphiidae
(Linyphiidae sp1) y Ochyroceratidae. Estos resultados son similares a lo reportado por
Gómez (2005), Guasca y Reina (2010) y Reyna (2010) donde predominan los gremios
de las arañas constructoras de telas.
La mayor abundancia de gremios de arañas tejedoras en el bosque, como el de las
constructoras de telas tridimensionales y orbiculares, concuerda con lo expuesto por
Reyna (2010), en cuya revisión de literatura encontró que la riqueza de la vegetación es
un factor importante en la distribución de estos grupos de arañas; por tanto, se espera
una mayor abundancia de estos gremios en los bosques comparados con los cafetales,
los cuales representan un ambiente más simple en cuanto a estructura. Sin embargo,
hay que indicar que las familias que conforman este grupo de arañas tejedoras son de
una amplia distribución y logran colonizar una extensa variedad de sustratos y
microambientes. Aunque no es un gremio de gran abundancia en este estudio, las arañas
cazadoras errantes en suelo fueron representativas en el cafetal de sol. Este suceso se
puede atribuir a la estructura de los cafetales, pues muchas de las especies de arañas
cazadoras errantes en suelo están adaptadas al microclima de la estructura de suelos
desnudos o con poca vegetación (Gibson, Hamber y Brown, 1992), que puede
presentarse en los cultivos de café.
Es de resaltar, que la predominancia del gremio de arañas constructoras de telas
tridimensionales, relega al de las constructoras de telas orbiculares a un segundo plano,
es un resultado que se diferencia de la tendencia general exhibida en otros tipos de
bosque tropical en los que por lo general las arañas orbiculares son dominantes.

74
5.4 PREFERENCIA DE MICROHÁBITAT
El comportamiento de la comunidad de arañas por microhábitat o estratos, no arrojó
información muy diferente a la ya analizada en los resultados obtenidos a nivel general
del estudio; sin embargo, se presentaron algunas particularidades. En el bosque se
destaca la preferencia de las arañas constructoras de telas tridimensionales de la familia
Pholcidae por los estratos aéreos (arbustivo y herbáceo), lo que posiblemente está ligado
a que construir sus telas en esta parte del bosque les brinda una mayor amplitud de
captura de presas, abarcando gran variedad de insectos voladores (Huber, 2000). Se
registró a las constructoras de telas en sábana de la familia Linyphiidae (Linyphiidae sp1)
como las más abundantes para ambos estratos del cafetal de sol. Se observaron
empleando la base de las plantas de café, en la interface con el suelo para la
construcción de sus telas; y aunque se conoce poco sobre su biología y depredación
(Breene, Dean, Nyffeler y Edwards, 1993 citado por Gómez, 2005), se asume que
consumen insectos y artrópodos pequeños que podrían ser plagas potenciales de los
cultivos de café, como son los áfidos.
La dominancia de los gremios de arañas tejedoras evidencia la preferencia de estos
organismos por los microhábitat ubicados entre la vegetación arbustiva y arbórea que
brinda el bosque y la parte vegetativa de los cultivos de café, y que corresponde a los
ambientes que ofrecen una mayor cantidad de sustratos. Uezt (1975) citado por Guasca
& Reina (2010) encontró que hábitats con más estratos verticales presentan valores de
abundancia de araneofauna más altos; así, la heterogeneidad espacial del sustrato
puede ser el factor más importante que afecta la diversidad animal en una comunidad,
encontrando que la arquitectura del hábitat y microhábitat juegan un papel importante en
la diversidad de los depredadores invertebrados.
5.5 DENSIDAD DE ARAÑAS POR UNIDAD DE MUESTREO
El protocolo de muestreo empleado en este estudio permitió realizar una aproximación a
la densidad de las arañas en la muestra de paisaje rural cafetero evaluado. No obstante,

75
resulta una densidad casi cincuenta veces menor que la reportada por Turnbull (1973)
quien sintetiza la información procedente de 37 trabajos realizados sobre densidad
poblacional de arañas en diferentes partes del mundo, donde haya un promedio de 130.8
arañas por metro cuadrado (citado por Flórez, 1997), o por Bristowe (1971) que estimó
poblaciones cercanas a un millón de individuos por hectárea (citado por Calixto, 2004).
Habitualmente se ha considerado que las densidades poblacionales de la fauna tropical
exhiben poca variabilidad en virtud a la noción de que las comunidades complejas y
diversas son más estables, y a la relativa estabilidad de las condiciones climáticas
(Flórez, 1999), sin embargo, cambios drásticos en las condiciones medio ambientales
(Temperatura, humedad, luz solar, corrientes de aire, entre otros) pueden influir sobre la
distribución de la comunidad de arañas (Ávalos, 2007). En el año 2010, se presentó un
episodio de variabilidad climática denominado fenómeno de “La Niña”, el cual se
caracteriza por un aumento considerable de la precipitación y una disminución en la
temperatura, y que además, se consideró como un componente atmosférico importante
correspondiente a uno de los eventos más intensos del último siglo, con un fuerte
acoplamiento océano-atmósfera (Euscátegui y Hurtado, 2011). Quizá este evento con la
consecuente fluctuación de la temperatura y el aumento de las lluvias (Anexo H)
afectaron los valores de densidad de las arañas acá encontrados.

76
6. CONCLUSIONES
La ventana de paisaje rural cafetero evaluada es un centro importante de conservación
ya que presenta una buena porción en cuanto a la abundancia y riqueza de especies de
arañas; no obstante, los resultados obtenidos son una estimación preliminar de la riqueza
real que puede albergar este paisaje.
Se encontró que la comunidad de arañas de la muestra de paisaje rural cafetero evaluado
está compuesta por 16 familias que agruparon 87 morfoespecies. En general el muestreo
es representativo, ya que se observó el 81% de las especies estimadas.
El carácter cosmopolita, la alta diversidad tanto a nivel mundial como en Colombia y
posiblemente la capacidad de explotar diversos recursos, hicieron que familias como
Araneidae, Linyphiidae, Pholcidae, Salticidae, Tetragnathidae y Theridiidae fueran las
más diversas, ricas y abundantes para la muestra de paisaje rural cafetero evaluado.
La cobertura cafetal de sol presentó la mayor diversidad en comparación con el bosque
remanente. Se cree que la heterogeneidad estructural de la vegetación no es tan
influyente en la distribución y diversidad de las arañas encontradas, como sí lo sería el
grado y frecuencia de perturbación que se ejerce sobre cada una de las coberturas
evaluadas en la venta de estudio. A pesar de tener una menor extensión, los remanentes
de bosque albergan cerca del 50% de la composición de arañas para el paisaje evaluado.
La estructura de las comunidades de arañas en cada una de las coberturas evaluadas
se ajustó al modelo de abundancia Log-normal, indicando que estos ensambles se
encuentran estables y habitan en ambientes con una elevada oferta de recursos y
hábitats a nivel local.
El análisis de la diversidad beta permitió evidenciar que el bosque remanente y el cafetal
de sol se definen de acuerdo a los diferentes arreglos de las abundancias de las

77
morfoespecies registradas para cada una de estas coberturas. Sugiriendo que cada uno
de ellas presenta características bióticas y abióticas particulares para el establecimiento
de los ensambles de morfoespecies registradas.
En términos generales la estructura gremial de la comunidad de arañas de las coberturas
evaluadas en la ventana de estudio es similar, siendo los gremios de arañas tejedoras
los más representativos.
Finalmente, se considera que la fragmentación de los bosques puede afectar a la
araneofauna, en el sentido de que se disminuye la oferta de sustratos y por tanto la
estructura de la comunidad de arañas puede afectarse negativamente, al verse reducido
el número de individuos y de especies que pueden habitar una cobertura. Sin embargo,
es interesante observar que buena parte de la riqueza de especies encontradas está
asociada a una cobertura vegetal altamente homogénea (Cafetal), lo que indica el
potencial de este elemento del paisaje rural cafetero para la conservación y estudio de
la biodiversidad de arácnidos en Colombia.

78
RECOMENDACIONES
El empleo de 42 cuadrantes de muestreo permitió la colecta del 81% de las especies en
las dos coberturas muestreadas. Desde el punto de vista matemático el empleo de 5
cuadrantes más por cada cobertura (52 cuadrantes en total) permitirían la recolección de
un porcentaje más cercano a la “totalidad” de especies presentes en el área de estudio.
Para un muestreo como el que se planteó en este trabajo aconsejo aumentar el número
de colectores en campo, con el fin de reducir el tiempo de muestreo. Cabe aclarar que
es pertinente que ambos investigadores presenten una experiencia similar en lo que
respecta a la colecta de arañas para reducir un poco el sesgo o error inherente al
colector.
Realizar muestreos nocturnos ya que existen comunidades de arañas cuya actividad es
más alta en horas de la noche.
Debido al alto número de juveniles colectados, es recomendable un previo entrenamiento
para la diferenciación entre adultos y juveniles en campo, ya que para los análisis de
datos es muy importante contar con ejemplares adultos.
Las comunidades animales no son estáticas en el espacio y en el tiempo, por lo que es
necesario realizar estudios fenológicos de la comunidad de arañas de este paisaje rural
cafetero, para dilucidar el patrón de distribución y dinámica de las especies en los en los
diferentes periodos climáticos (altas y bajas lluvias) que se presentan en el país.
Debido a los vacíos de información, el uso de unidades taxonómicas reconocibles
(morfoespecies) ha demostrado ser útil para una rápida estimación de la biodiversidad;
no obstante, se requiere realizar una determinación taxonómica más fina, para obtener
una información más acorde con la realidad de las comunidades de arañas en los
diversos ecosistemas que podemos encontrar en Colombia.

79
En la actualidad los inventarios de biodiversidad demandan protocolos sencillos, rápidos
y económicos. Sin embargo, teniendo como experiencia este trabajo, la afirmación
anterior no se cumple al pie de la letra para un taxón como las arañas. Si nos remitimos
al protocolo de muestreo aplicado en este estudio, en el que se evaluaron 2 coberturas,
21 cuadrantes de muestreo por cobertura y 3 métodos de captura por cuadrante (donde
la colecta manual requiere de 1 hora efectiva de muestreo por cada estrato rasante y
aéreo) obtenemos un total mínimo de 42 horas de muestreo efectivo, teniendo sólo en
cuenta el tiempo de los métodos de colecta manual. Si además, se muestrea como
mínimo 3 cuadrantes al día (teniendo acá en cuenta factores como la lluvia, distancia de
desplazamiento, entre otros, que podrían impedir el muestreo de un mayor número
cuadrantes diarios) con esfuerzo de un único colector, se tardaría un total de mínimo 7
días para el muestreo de los 21 cuadrantes de una solo cobertura vegetal, 14 días
efectivos de muestreo para las dos coberturas. Si se quisiera aumentar el número de
coberturas a estudiar, obviamente, aumentaría de manera proporcional los días de
muestreo. Considero pertinente revisar este tipo de metodología para el estudio de la
araneofauna del país. Y que por cuestiones metodológicas este grupo faunístico no sea
excluido de los estudios de biodiversidad, sino por el contrario sea cada vez más
empleado como un indicador del estado de los ecosistemas.

80
REFERENCIAS
Agnarsson, I. (2006). A revision of the New World eximius lineage of Anelosimus
(Araneae, Theridiidae) and a phylogenetic analysis using worldwide exemplars.
Zoological Journal of the Linnean Society, 146(4), 453–593.
Alape, D. E. (2008). Estructura de arañas tejedoras (Araneae: Araneomorpha) en la
Reserva Forestal Protectora Bellavista (Ibagué-Tolima). (Trabajo de grado). Facultad de
Ciencias: Universidad del Tolima, Ibagué, Colombia.
Armbrecht, L. & Perfecto, I. (2001). Diversidad de artrópodos en los agroecosistemas
cafeteros. Revista Protección Vegetal, Actas del Simposio Café y Biodiversidad, 2, 11-
16.
Avalos, G., Rubio, G., Bar, M. & González, A. (2007). Arañas (Arachnida: Araneae)
asociadas a dos bosques degradados del Chaco húmedo en Corrientes, Argentina.
Revista Biología Tropical, 55(3-4), 899-909.
Barnes, R. D., & Ruppert, E. (1996). Zoología de los Invertebrados. (6ª ed.). México:
McGraw-Hill interamericana S.A.
Blanco-Vargas E. & Salas G. (2007). Arácnidos, Guía de campo. (1ª ed.). Bogotá:
Bichoscolombia.
Blanco-Vargas, E., Amat-García, G. & Flórez, E. (2003). Araneofauna orbitelar (Araneae:
Orbiculariae) de los andes en Colombia: Comunidades en hábitats bajo regeneración.
Revista Ibérica de Aracnología, 7, 189 -203.
Brose, U., Martínez, N. & Williams, R. (2003). Estimating species richness: sensitivity to
sample coverage and insensitivity to spatial patterns. Ecology, 84(9), 2364–2377.

81
Bustillo, A. E. (2007). El manejo de cafetales y su relación con el control de la broca del
café en Colombia. (2ª ed.). Chinchiná, Caldas: Federación Nacional de Cafeteros de
Colombia - Centro Nacional De Investigaciones de Café CENICAFÉ.
Cabra-García, J. (2009). Estimación de la diversidad alfa, beta y gamma de arañas en
un ambiente heterogéneo: Parque Natural Regional El Vinculo (Valle, Colombia).
(Trabajo de grado). Facultad de Ciencias: Universidad del Valle, Cali, Colombia.
Cabra-García, J., Montealegre, L. & Arce, M. (2010). Evaluación rápida de la riqueza de
arañas en un bosque húmedo tropical del departamento del Cauca (Colombia). Boletín
del Museo de Entomología de la Universidad del Valle. 11(1), 1-9.
Callicott, J., Crowder, L. & Mumford, K. (1999). Current Normative Concepts in
Conservation. Conservation Biology, 13(1), 22–35.
Calixto, A. (2004). Diversidad y Conservación de las arañas en Agroecosistemas; el caso
del Nogal en Texas. En Calixto / XXXI Congreso de la Sociedad Colombiana de
Entomología - SOCOLEN (pp. 67-72). Bogotá, Colombia.
Cardoso, P., Silva, I., De Oliveira, N. G. & Serrano, A. (2004). Indicator taxa of spiders
(Araneae) diversity and their efficiency in conservation. Biological Conservation, 120,
517-524.
Cardoso P, Pekár S, Jocqué R & Coddington JA (2011) Global patterns of guild
composition and functional diversity of spiders. PLoS ONE 6(6): e21710.
doi:10.1371/journal.pone.0021710
Cava, M. B. (2013). Efecto de la Heterogeneidad del Hábitat sobre las Comunidades de
Artrópodos en Bosques de Chaco, Selva Paranaense y de la Isla Apipé Grande en la
provincia de Corrientes, Argentina. (Trabajo de grado). Facultad de Ciencias Naturales y
Museo: Universidad Nacional de La Plata, La Plata, Argentina.

82
Cepeda V., J. & Flórez D., E. (2007). Arañas tejedoras: uso de diferentes microhábitats
en un bosque andino de Colombia. Revista Ibérica de Aracnología. 14, 39-48.
Cepeda V., J. (2009). Comparación ecológica de comunidades de arañas y coleópteros
y análisis del impacto del manejo orgánico y convencional, en cultivos de café. (Trabajo
de grado). Facultad de Economía, Instituto de Estudios Ambientales: Universidad
Nacional de Colombia, Bogotá, Colombia.
Chao A, Chazdon R, Colwell R, Tsung-Jen S. (2005). Un nuevo método estadístico para
la evaluación de la similitud en la composición de especies con datos de incidencia y
abundancia. En Halffter G, Soberón J, Koleff P, Melic A, editores. Sobre Diversidad
Biológica: El significado de las Diversidades Alfa, Beta y Gamma. Zaragoza, España:
Grupo Diversitas-México - Sociedad Entomológica Aragonesa (SEA).
Coddington, J. A. & Levy. H. W. (1991). Systematics and evolution of spiders (Araneae).
Annual Review of Ecology and Systematics, 22, 565-592.
Coddington, J. A., Griswold, C. E., Silva D., D., Peñaranda, E. & Larcher, S. F. (1991).
Designing and testing sampling protocols to estimate biodiversity in tropical ecosystems.
En Dudley E. C. (ed.) The unity of evolutionary biology: Proceedings of the fourth
international congress of systematic and evolutionary biology. Portland, Oregon:
Dioscorides Press.
Coddington, J. A., Young, L. H. & Coyle, F. A. (1996). Estimating spider species richness
in a southern appalachian cove hardwood forest. The Journal of Arachnology, 24, 111–
128.
Colwell, R. K. & Coddington J. A. (1994). Estimating terrestrial biodiversity through
extrapolation. Philosophical Transactions of the Royal Society of London, 345, 101-118.

83
Colwell, R. K. (2013). EstimateS: Statistical estimation of species richness and shared
species from samples. Version 9. Recuperado de: http://purl.oclc.org/estimates
Connell, J. H. (1978). Diversity in tropical rain forests and coral reefs. Science, 199, 1302–
1309
Corporación Autónoma Regional de Risaralda. (2008). Risaralda bosque modelo para el
mundo. Pereira: CARDER.
Crespo, F. A., Iglesias, M. S. & Valverde, A. C. (1999). El ABC en la determinación de
artrópodos I. Argentina: Editorial CCC Educando.
Dippenaar-Schoeman, A. S. & Jocqué, R. (1997). African spiders: An identification
manual. South Africa: Agricultural research council.
Eberhard, W. G. 1976. Photography of orb webs in the field. Bull. Br. Arachnol. Soc., 3(7),
200-204.
Euscátegui & Hurtado. (2011). Análisis del impacto del fenómeno “La Niña” 2010-2011
en la hidroclimatología del país. Instituto de Hidrología, Meteorología y Estudios
Ambientales de Colombia - IDEAM.
Federación Nacional de Cafeteros de Colombia - CENICAFÉ (2011). Anuario
meteorológico cafetero 2010. Chinchiná, Caldas, Colombia.
Flórez D., E. (1996). Las Arañas del departamento del Valle del Cauca: Un manual
introductorio a su diversidad y clasificación. Santiago de Cali, Colombia: INCIVA-
COLCIENCIAS.
Flórez D., E. (1997). Las Arañas y la Naturaleza. En E. Flórez / XXIV Congreso de la
Sociedad Colombiana de Entomología - SOCOLEN (pp. 1-12). Pereira, Colombia.

84
Flórez D., E. (1999). Estructura y composición de una comunidad de arañas (Araneae)
en un bosque muy seco tropical de Colombia. Boletín de Entomología Venezuela, 14(1),
37–51.
Flórez D., E. (2004). Las arañas en Colombia: reseña histórica, presente y perspectivas
futuras. En E. Flórez / XXXI Congreso de la Sociedad Colombiana de Entomología -
SOCOLEN (pp. 73-76). Bogotá, Colombia.
Foelix, R. F. (2011). Biology of spiders. (3rd ed.). New York: Oxford University Press, Inc.
Gibson, C.W.; Hambler, C. & Brown, V. K. (1992). Changes in spider (Araneae)
assemblages in relation to succession and grazing management. Journal of Applied
Ecology. 29(1), 132-142.
Glueck, S. (1993). A taxonomic revision of the orb waver geneus Acacesia (Araneidae,
Araneidae). Psyche: A Journal of Entomology, 101, 59–84.
Gómez, A. M., Anaya, J. & Álvarez, E. (2005). Análisis de fragmentación de los
ecosistemas boscosos en una región de la cordillera central de los Andes colombianos.
Revista de Ingenierías Universidad de Medellín, 4(7), 13-27.
Gómez, L. A. (2005). Estudio comparativo de las comunidades de arañas en cultivos de
algodón convencional y transgénico en el departamento del Tolima, Colombia. Facultad
de Ciencias: Universidad Nacional de Colombia, Bogotá, Colombia.
Gotelli, N. J. & Colwell, R. K. (2001). Quantifying biodiversity: procedures and pitfalls in
the measurement and comparison of species richness. Ecology Letters, 4: 379-391.
Gray, J. S., Bjørgesæter, A. & Ugland, K. I. (2005). The impact of rare species on natural
assemblages. Journal of Animal Ecology, 74, 1131–1139.

85
Guasca D. & Reina R. (2010). Caracterización de la comunidad de arañas en el Cerro
Los Comuneros (Tenjo, Cundinamarca). (Trabajo de grado). Facultad de Ciencias
Básicas: Universidad El Bosque, Bogotá, Colombia.
Halffter, G., Moreno, C.E. & Pineda, E.O. (2001). Manual para la evaluación de la
biodiversidad en las Reservas de la Biosfera. Zaragoza, España: M&T- Manuales y Tesis
SEA.
Hammer, Ø., Harper, D. A. T., & Ryan, P. D. (2001). PAST: Paleontological Statistics
Software Package for Education and Data Analysis. Palaeontologia Electronica. 4(1), 1-
9.
Harvey, C. A. & Sáenz, J. C. (2008). Evaluación y conservación de biodiversidad en
paisajes fragmentados de Mesoamérica. Santo Domingo de Heredia, Costa Rica:
Instituto Nacional de Biodiversidad, INBios.
Holdridge, L. R. (1967). Life zone ecology. San José, Costa Rica: Tropical Science
Center.
Huber, B. (2000). New world pholcid spiders (Araneae: Pholcidae): a revision at generic
level. Bulletin of the American Museum of Natural History. 254, 1-348.
Hughes, R. G. (1986). Theories and models of species abundance. The American
Naturalist, 128(6), 879-899.
Jiménez-Valverde A & Hortal J. (2003). Las curvas de acumulación de especies y la
necesidad de evaluar la calidad de los inventarios biológicos. Revista Ibérica de
Aracnología, 8, 151-161.
Jocqué, R., & Dippenaar-Shoeman, A. S. (2006). Spider families of the world. Tervuren,
Belgium: Royal Museum for Central Africa.

86
Kaston, B. J. (1978). How to know the spiders. (3rd ed.). Dubuque, Iowa: The Pictured
Key Nature Series.
Kattan, G. H., Franco, P., Rojas, V. & Morales, G. (2004). Biological diversification in a
complex region: a spatial analysis of faunistic diversity and biogeography of the Andes of
Colombia. Journal of Biogeography, 31, 1829–1839.
Krebs, C. J. (1985). Ecology: The experimental analysis of distribution. (3rd ed.). Harper
and Row, Publishers, Inc.
Levi, H. W. (1955). The spider genera Coressa and Achaearanea in america North of
Mexico (Araneae, Theridiidae). Novitates, 1718, 1–33.
Levi, H. W. (1959). The spider genus Coleosoma (Araneae, Theridiidae). Breviora, 110,
1-10.
Levi, H. W. (1963). American spiders of the genus Theridion (Araneae, Theridiidae).
Bulletin of the American Museum of Natural History, 129(10), 481–589.
Levi, H. W. (1964a). American spiders of genus Phoroncidia (Araneae: Theridiidae).
Bulletin of the Museum of Comparative Zoology, 131(3), 65–86.
Levi, H. W. (1964b). American spider of the genus Episinus (Araneae: Theridiidae).
Bulletin of the Museum of Comparative Zoology, 131(1), 1–25.
Levi, H. W. (1978). The american orb weaver genera Colphepeira, Micrathena and
Gasteracantha North of Mexico (Araneae, Araneidae). Bulletin of the American Museum
of Natural History, 148(9), 417-442.
Levi, H. W. (1985). The spiny orb-weaver genera Micrathena and Chaetacis (Araneae,
Araneidae). Bulletin of the Museum of Comparative Zoology, 150(8), 429–618.

87
Levi, H. W. (1986). The neotropical orb-waever genus Chrysometa and Homalometa
(Araneae, Tetragnathidae). Bulletin of the American Museum of Natural History, 154(2),
6-53.
Levi, H. W. (1988). The neotropical orb-weaving spiders of the genus Alpaida (Araneae:
Araneidae). Bulletin of the Museum of Comparative Zoology, 151(7), 365–487.
Levi, H. W. (1991a). The neotropical and mexican species of the orb-weaver genera
Araneus, Dubiepeira and Aculepeira (Araneae: Araneidae). Bulletin of the American
Museum of Natural History, 152(6), 167–315.
Levi, H. W. (1991b). The neotropical org-weaver genera Edricus and Wagneriana
(Araneae: Araneidae). Bulletin of the American Museum of Natural History, 152(6), 363–
415.
Levi, H. W. (1992). Spider of the orb-waever genus Parawixia in America (Araneae,
Araneidae). Bulletin of the American Museum of Natural History, 151(3), 91-215.
Levi, H. W. (1995). The neotropical orb-waever genus Metazygia (Araneae, Araneidae).
Bulletin of the American Museum of Natural History, 154(2), 6-53.
Levi, H. W. (1999). The neotropical and Mexican orb weavers of the genera Cyclosa and
Allocyclosa (Araneae, Araneidae). Bulletin of The Museum of Comparative Zoology, 155,
299 – 379.
Levi, H. W. (2002). Keys to the genera of araneid orbweavers (Araneae, Araneidae) of
the Americas. The Journal of Arachnology, 30(3), 527–562.
Levi, H. W. (2004). Comments and new recoeds for the american genera Gea and
Argiope with description of new species (Araneae, Araneidae). Bulletin of the American
Museum of Natural History, 158(2), 47-66.

88
Levi, H. W. (2007). The orb weaver genus Mangora in South America (Araneae,
Araneidae). Bulletin of the American Museum of Natural History, 159(1), 1–144.
Levi, H. W., & Levi, L. R. (1962). The genera of spider family Theridiidae. Bulletin of the
Museum of Comparative Zoology, 127(1), 3–99.
Loyola, R. D. & Martins, R. P. (2008). Habitat structure components are effective
predictors of trap-nesting Hymenoptera diversity. Basic and Applied Ecology, 9, 735–742.
Lozano, F. H. (2009). Herramientas de manejo para la conservación de biodiversidad en
paisajes rurales. Bogotá: Instituto de Investigación de Recursos Biológicos Alexander
von Humboldt y Corporación Autónoma Regional de Cundinamarca (CAR).
Lozano, F. H., Jiménez, E. & Caycedo, P. (2006). Simposio - Conservación de
biodiversidad en paisajes rurales: aprendizajes en la cuenca media del río Barbas
(Filandia, Quindío). En F. H. Lozano et al. / Segundo Congreso Colombiano de Zoología
(pp. 43-59). Santa Marta, Magdalena.
Macip R., R., López A., S. & Muñoz A., A. (2013). Abundancia, uso de hábitat,
microhábitat y hora de actividad de Ameiva undulata (Squamata: Teidae) en un paisaje
fragmentado del Soconusco chiapaneco. Revista Mexicana de Biodiversidad, 84, 622-
629.
Magurran, A. (1988). Ecological diversity and Its measurement. (1ra ed.) United Kingdom:
Princenton University Press.
Magurran, A. (2004). Measuring biological diversity. (3ra ed.) United Kingdom: Blackwell
publishing company.

89
Marín, A., Toro, L. & Uribe, S. (2008). Conectividad estructural del paisaje cafetero en la
cuenca alta del río San Juan, suroeste antioqueño, Colombia. Boletín de ciencias de la
Tierra. No. 23, 43-54.
Mcnett, B. J. & Rypstra, A. L. (2000). Habitat selection in a large orb-weaving spider:
vegetational complexity determines site selection and distribution. Ecological
Entomology, 25(4), 423–432.
Mendoza S., J. E., Jiménez, E., Lozano Z., F. H., Caycedo R., P & Renjifo, L. M. (2006).
Identificación de elementos del paisaje prioritarios para la conservación de biodiversidad
en paisajes rurales de los Andes Centrales de Colombia. En J. E. Mendoza et al. /
Segundo Congreso Colombiano de Zoología (pp. 51-53). Santa Marta, Magdalena.
Millidge, A. F. (1985). Some linyphiid spiders from South America. American Museum
Novitates, 2836, 1–78.
Millidge, A. F. (1991). Further Linyphiid spiders (Araneae) from South America. Bulletin
of the American Museum of Natural History, 205, 1–199.
Moguel, P. & Toledo, V. M. (1996). El café en México: ecología, cultura indígena y
sustentabilidad. Ciencias, 43, 40-51.
Moguel, P. & Toledo, V. M. (1999). Biodiversity conservation in traditional coffee systems
of Mexico. Conservation Biology, 13(1), 5-12.
Moreno, C. E. (2001). Métodos para medir la Biodiversidad. Zaragoza, España: M&T-
Manuales y Tesis SEA.
New, T. R. (1999). Untangling the web: spiders and the challenges of invertebrate
conservation. Journal of Insect Conservation, 3, 251–256.

90
Nyffeler, M. (1999). Prey selection of spiders in the field. The Journal of Arachnology, 27,
317–324.
Opell, B. D. (1979). Revision of genera and tropical american species of the spider family
Uloboridae. Bulletin of the American Museum of Comparative Zoology, 148(10), 548–
549.
Paine, R. T. & Levin, S. A. (1981). Intertidal Landscapes: Disturbance and the Dynamics
of Pattern. Ecological Monographs, 51(2), 145-178.
Palmer, E. W. (1990). The estimation of species richness by extrapolation. Ecology,
71(3), 1195-1198.
Perafán, C. Sabogal-González, A., Moreno-González, J.A., García-Rincón, A., Luna-
Sarmiento, D. Romero-Ortíz, C. & Flórez D, E. (2013). Diagnóstico del estado actual de
la fauna de arácnidos y de su gestión en Colombia En C. Perafán et al. / 40° Congreso
de la Sociedad Colombiana de Entomología - SOCOLEN (pp. 308-335). Bogotá,
Colombia.
Perfecto, I., Rice, R. A., Greenberg, R. & Van der Voort, M. E. (1996). Shade Coffee: A
Disappearing Refuge for Biodiversity. BioScience, 46(8), 598-608.
Pianka, E. R. (1966). Latitudinal gradients in species diversity: a review of concepts. The
American Naturalist, 100(910), 33-46.
Rangel, J. O. & G. Lozano. (1986). Un perfil de vegetación entre La Plata (Huila) y el
volcán Puracé. Caldasia, 14(68-70), 53-547.
Reyna, J. (2010). Aproximación a la estructura y composición de la araneofauna del Valle
de Sogamoso (Boyacá, Colombia). Facultad de Ingeniería, Administración y Ciencias
Básicas: Universidad INCCA de Colombia, Bogotá, Colombia.

91
Rico-G., A., Beltrán-A., J. P. & Álvarez-D., A. (2005). Diversidad de arañas (Arachnida:
Araneae) en el Parque Nacional Natural isla Gorgona, pacífico colombiano. Biota
Neotropical. Número especial. 5(1a), 1-12.
Ríos, C. A. (2010). Respuesta al cambio climático: Planificación adaptativa para la
conservación en los andes colombianos: Caracterización de Ventanas de Trabajo. Cali:
Wildlife Conservation Society – Programa Colombia.
Root, R. (1967). The niche exploitation pattern of the Blue-Gray Gnatcatcher. Ecological
Monographs, 37(4), 317-350.
Roth, V. D. (1993). Spider genera of North America with keys to families and genera and
a guide to literature. (3rd ed.). Portal, Arizona.
Sabogal G., A. (2010). Estado actual del conocimiento de arañas (Araneae) en Colombia.
En A. Sabogal / XXXVII Congreso de la Sociedad Colombiana de Entomología -
SOCOLEN (pp. 123-136). Bogotá, Colombia.
Sabogal G., A. (2011). Estudio comparativo de las comunidades de arañas asociadas a
bosques conservados y áreas intervenidas en el Santuario de Flora y Fauna Otún
Quimbaya (Risaralda, Colombia). (Tesis de Maestría). Facultad de Agronomía:
Universidad Nacional de Colombia, Bogotá.
Simó, M., Laborda, A., Jorge, C., Castro, M. (2001). Las arañas en agroecosistemas:
bioindicadores terrestres de calidad ambiental. Revista del Laboratorio Tecnológico del
Uruguay, 6, 51-55.
Sousa, W. P. (1984). The Role of Disturbance in Natural Communities. Annual Review of
Ecology and Systematics, 15, 353-391.

92
Tews, J., Brose, U., Grimm, V., Tielbörger, K., Wichmann, M. C., Schwager, M. & Jeltsch,
F. (2004). Animal species diversity driven by hábitat heterogeneity/diversity: the
importance of keystone structures. Journal of Biogeography, 31, 79–92
Toledo, V. M. (2005). Repensar la conservación: ¿áreas naturales protegidas o estrategia
bioregional?. Gaceta ecológica. 77; 67-83.
Turbull, A. L. (1973). Ecology of the true spiders. Annual Review of Entomol. 18, 305-
348.
Turner, M., Gardner, R & O’Neill, R. (2001). Landscape ecology in theory and practice.
United States of America: Springer.
Ubick D, Paquin P, Cushing P.E., & Roth V. (2005). Spiders of North America: an
identification manual. American Arachnological Society.
Uetz, G. (1979). The influence of variation in litter habitats on spider communities.
Oecologia, 40, 29-42.
Uetz, G., Halaj, J. & Cady, A. (1999). Guild structure of spiders in major crops. The
Journal of Arachnology, 27, 270-280.
Vásquez V., V. H. & Serrano G., M. A. (2009). Las áreas naturales protegidas de
Colombia. Bogotá: Conservación Internacional – Colombia & Fundación Biocolombia.
Villarreal H., M. Álvarez, S. Córdoba, F. Escobar, G. Fagua, F. Gast, H. Mendoza, M.
Ospina & A. M. Umaña. (2006). Manual de métodos para el desarrollo de inventarios de
biodiversidad. (2ª ed.). Bogotá, Colombia: Instituto de Investigación de Recursos
Biológicos Alexander von Humboldt.

93
Wildlife Conservation Society. (2009) Respuesta al cambio climático: planificación
adaptativa para la conservación en los andes colombianos. Cali: WCS – Programa
Colombia.
World Spider Catalog (2014). World Spider Catalog, version 15.5. Natural History
Museum Bern. Recuperado de: http://wsc.nmbe.ch
Zorrilla, M. (2005). La influencia de los aspectos sociales sobre la alteración ambiental y
la restauración ecológica. En: Sánchez, Peters, Márquez–Huitzil, Vega, Portales, Valdez
& Azuara (eds). Temas sobre restauración ecológica. (1ª ed.). Secretaría de Medio
Ambiente y Recursos Naturales, Instituto Nacional de Ecología, U.S. Fish & Wildlife
Service, Unidos para la Conservación, A.C.

94
ANEXOS

95
Anexo A Anatomía externa del cuerpo de una araña: (a) vista lateral, (b) vista ventral.
E= epigino (en una hembra adulta).
Fuente: Foelix, 2011.

96
Anexo B Tipos de quelíceros y uñas en el Orden Araneae.
a b
(a) Quelíceros paraxiales de una araña Mygalomorphae con una alineación más o menos
paralela. (b) Quelícero diaxiales de una araña Araneomorphae los cuales se observan
opuestos entre sí.
c d
(c) Diagrama del movimiento hacia adelante y hacia atrás de los quelíceros paraxiales.
(d) Movimiento lateral -perpendicular al eje del cuerpo- presentado por los quelíceros
diaxiales.

97
e f
(e) Las arañas antiguas probablemente tenían tres uñas tarsales en cada pata, como se
observa aquí en la araña "primitiva" Liphistius. (f) En algunas familias de arañas, las
Dionycha, la uña del medio se ha perdido supuestamente, sin embargo, una uña media
reducida (flecha) se observa todavía presente, como en este tarso de una araña saltarina
juvenil. Las dos garras principales (1, 2) son dentadas como un peine.
Fuente: Foelix, 2011.

98
Anexo C Esquema de clasificación taxonómico del orden Araneae.
Fuente: Flórez, 1996.
(6)
OR
DE
N
AR
AN
EA
E
Mygalomorpha
Barychelidae Ctenizidae
Dipluridae
Paratropididae Theraphosidae
Araneomorphae
Sección Cribellatae
(1)
Amaurobiidae Dictynidae
Uloboridae
Dinopidae
Sección
Ecribellatae (2)
Haplogynae (3)
Ochyroceratidae Oonopidae
Scytodidae
Segestriidae
Entelogynae (4)
Dionycha (5)
Anyphaenidae
Clubionidae Thomisidae Salticidae
Ctenidae
Trionycha I (7)
Pisauridae Lycosidae
Oxyopidae
Palpimanidae Trechaleidae
Mimetidae
Trionycha II (8)
Araneidae
Tetragnathidae Pholcidae
Linyphiidae
Pholcidae Theridiidae
Theridiosomatidae
(1): Arañas con cribelo y calamistro (2): Arañas sin cribelo, ni calamistro
(3): Arañas con epiginio simple
(4): Arañas con epiginio complejo
(5): Arañas con dos uñas en cada tarso (Dioniycha) (6): Arañas con tres uñas en cada tarso (Tryinycha) (7): Arañas tryonichas cazadoras (sin tela) (8): Arañas tryonichas tejedoras

99
Anexo D Coordenadas de las estaciones y puntos de muestreo definidas en la cobertura
bosque secundario para la caracterización de la comunidad de arañas en un paisaje rural
cafetero de El Brillante, Risaralda.
Estaciones de
Muestreo
Puntos de
Muestreo
Coordenadas UTM Coordenadas Geográficas Altitud (m)
X Y Latitud Longitud
Bos1
Bos1-P1 390379 557230 5° 2' 26.3" N 75° 59' 20.0" W 1948
Bos1-P2 390379 557230 5° 2' 26.3" N 75° 59' 20.0" W 1948
Bos1-P3 390379 557230 5° 2' 26.3" N 75° 59' 20.0" W 1948
Bos1-P4 390440 557261 5° 2' 27.0" N 75° 59' 18.9" W 1844
Bos1-P5 390410 557261 5° 2' 27.5" N 75° 59' 19.5" W 1844
Bos1-P6 390410 557261 5° 2' 27.5" N 75° 59' 19.5" W 1844
Bos1-P7 390317 557169 5° 2' 24.6" N 75° 59' 22.5" W 1844
Bos2
Bos2-P1 390317 557169 5° 2' 24.6" N 75° 59' 22.5" W 1844
Bos2-P2 390286 557138 5° 2' 23.7" N 75° 59' 23.0" W 1883
Bos2-P3 390286 557138 5° 2' 23.7" N 75° 59' 23.0" W 1883
Bos2-P4 390286 557108 5° 2' 22.8" N 75° 59' 23.2" W 1900
Bos2-P5 390286 557108 5° 2' 22.8" N 75° 59' 23.2" W 1900
Bos2-P6 390317 556954 5° 2' 17.5" N 75° 59' 22.5" W 1742
Bos2-P7 390317 556954 5° 2' 17.5" N 75° 59' 22.5" W 1742
Bos3
Bos3-P1 390255 557108 5° 2' 22.2" N 75° 59' 24.7" W 1906
Bos3-P2 390255 557108 5° 2' 22.2" N 75° 59' 24.7" W 1906
Bos3-P3 390255 557108 5° 2' 22.2" N 75° 59' 24.7" W 1906
Bos3-P4 390255 557108 5° 2' 22.2" N 75° 59' 24.7" W 1906
Bos3-P5 390255 557108 5° 2' 22.2" N 75° 59' 24.7" W 1906
Bos3-P6 390255 557108 5° 2' 22.2" N 75° 59' 24.7" W 1906
Bos3-P7 390255 557108 5° 2' 22.2" N 75° 59' 24.7" W 1906
Fuente: Autor

100
Anexo E Coordenadas de las estaciones y puntos de muestreo definidas en la cobertura
cafetal de sol para la caracterización de la comunidad de arañas en un paisaje rural
cafetero de El Brillante, Risaralda.
Estaciones de
Muestreo
Puntos de
Muestreo
Coordenadas UTM Coordenadas Geográficas Altitud (m)
X Y Latitud Longitud
Cf1
Cf1-P1 389300 556679 5° 2' 08.0" N 75° 59' 55.6" W 1645
Cf1-P2 389300 556710 5° 2' 09.4" N 75° 59' 55.8" W 1669
Cf1-P3 389269 556741 5° 2' 10.2" N 75° 59' 56.0" W 1673
Cf1-P4 389300 556741 5° 2' 10.0" N 75° 59' 55.2" W 1677
Cf1-P5 389300 556741 5° 2' 10.5" N 75° 59' 55.0" W 1688
Cf1-P6 389331 556741 5° 2' 10.2" N 75° 59' 54.1" W 1686
Cf1-P7 389331 556710 5° 2' 09.0" N 75° 59' 54.2" W 1683
Cf2
Cf2-P1 389916 556586 5° 2' 05.0" N 75° 59' 35.5" W 1839
Cf2-P2 389885 556586 5° 2' 05.0" N 75° 59' 36.5" W 1831
Cf2-P3 389885 556556 5° 2' 04.4" N 75° 59' 36.8" W 1828
Cf2-P4 389854 556525 5° 2' 03.8" N 75° 59' 37.3" W 1823
Cf2-P5 389854 556525 5° 2' 03.0" N 75° 59' 37.9" W 1816
Cf2-P6 389823 556525 5° 2' 03.0" N 75° 59' 38.6" W 1810
Cf2-P7 389793 556494 5° 2' 02.7" N 75° 59' 39.4" W 1808
Cf3
Cf3-P1 389670 557170 5° 2' 24.0" N 75° 59' 43.1" W 1774
Cf3-P2 389670 557170 5° 2' 24.6" N 75° 59' 43.1" W 1758
Cf3-P3 389701 557139 5° 2' 23.3" N 75° 59' 42.9" W 1765
Cf3-P4 389701 557139 5° 2' 23.8" N 75° 59' 42.5" W 1759
Cf3-P5 389701 557109 5° 2' 22.9" N 75° 59' 42.5" W 1768
Cf3-P6 389701 557139 5° 2' 23.2" N 75° 59' 42.2" W 1762
Cf3-P7 389701 557109 5° 2' 22.5" N 75° 59' 42.3" W 1768
Fuente: Autor

101
Anexo F Formato para la consignación de datos en campo de los muestreos de arañas
(Arachnida: Araneae) en un paisaje rural de Risaralda, Colombia.
Fuente: Autor

102
Anexo G Listado de morfoespecies colectadas en un paisaje rural cafetero del
departamento de Risaralda.
Familia Género/Especie/Mfsp No. Indv.
Bosque
No. Indv.
Cafetal
No. Total
Indv.
Amaurobiidae Amaurobiidae sp1 - 1 1
Anyphaenidae
Anyphaeninae sp1 - 3 3
Anyphaeninae sp2 9 - 9
Anyphaeninae sp3 3 1 4
Araneidae
Acacesia hamata - 5 5
Alpaida queremal 5 - 5
Araneus bogotensis 2 1 3
Argiope argentata - 4 4
Cyclosa caroli 1 2 3
Cyclosa rubronigra 10 1 11
Cyclosa triquetra - 5 5
Eriophora sp - 1 1
Eustala scutigera - 1 1
Gasteracantha cancriformis - 3 3
Mangora eberhardi 26 - 26
Metazygia keyserlingi - 1 1
Micrathena lucasi 11 16 27
Neoscona sp - 1 1
Ocrepeira sp 1 - 1
Parawixia rimosa 3 1 4
Wagneriana tauricornis - 3 3
Wagneriana
undecimtuberculata - 4 4
Linyphiidae
Dubiaranea sp - 101 101
Erigoninae sp1 - 4 4
Erigoninae sp2 7 15 22
Erigoninae sp3 51 1 52
Linyphiidae sp1 41 253 294
Liocranidae Liocranidae sp1 - 1 1

103
Anexo G. Continuación
Familia Género/Especie/Mfsp No. Indv.
Bosque
No. Indv.
Cafetal
No. Total
Indv.
Lycosidae Lycosidae sp1 - 50 50
Ochyroceratidae Ochyroceratidae sp1 9 - 9
Oonopidae Oonopidae sp1 1 - 1
Oxyopidae Peucetia sp1 - 4 4
Pholcidae
Pholcidae sp1 228 19 247
Pholcidae sp2 1 9 10
Pholcidae sp3 7 - 7
Pholcidae sp4 2 - 2
Salticidae
Lyssomanes sp1 1 - 1
Salticidae sp1 - 3 3
Salticidae sp10 - 1 1
Salticidae
Salticidae sp16 1 - 1
Salticidae sp17 - 2 2
Salticidae sp2 - 5 5
Salticidae sp3 - 1 1
Salticidae sp4 17 - 17
Salticidae sp5 2 - 2
Salticidae sp7 2 1 3
Salticidae sp8 2 - 2
Salticidae sp9 8 - 8
Tetragnathidae
Chrysometa alboguttata - 41 41
Chrysometa calima - 15 15
Chrysometa carmelo 7 - 7
Chrysometa serachui 1 - 1
Chrysometa sp 1 1 - 1
Chrysometa yotoco - 2 2
Leucauge argentea 20 48 68
Tetragnathidae sp1 8 35 43

104
Anexo G. Continuación
Familia Género/Especie/Mfsp No. Indv.
Bosque
No. Indv.
Cafetal
No. Total
Indv.
Theridiidae
Achaearanea trapezoidalis 4 4 8
Anelosimus baeza - 48 48
Anelosimus cf tungurahua - 1 1
Chrysso sp 10 - 10
Coleosoma blandum - 1 1
Episinus sp1 1 - 1
Phoroncidia studo 5 - 5
Spintharus sp - 5 5
Theridiidae sp10 - 1 1
Theridiidae sp13 - 3 3
Theridiidae sp16 - 2 2
Theridiidae sp17 - 12 12
Theridiidae sp18 - 4 4
Theridiidae sp19 - 7 7
Theridiidae sp2 8 - 8
Theridiidae sp22 - 9 9
Theridiidae sp24 10 3 13
Theridiidae sp26 1 5 6
Theridiidae sp27 1 2 3
Theridiidae sp33 - 1 1
Theridiidae sp34 1 - 1
Theridiidae sp37 1 - 1
Theridiidae sp5 4 1 5
Theridiidae sp9 - 2 2
Theridion frizzellorum - 41 41
Theridion nigriceps 27 26 53
Thwaitesia sp1 10 - 10
Theridiosomatida
e Theridiosomatidae sp1 1 - 1
Thomisidae Thomisidae sp1 1 1 2
Uloboridae Philoponella sp1 4 1 5
Uloborus sp2 - 3 3
Total (16) 87 577 848 1425
Fuente: Autor

105
Anexo H Registro climatológico de la zona de estudio: Paisaje Rural Cafetero de
Risaralda, Colombia.
Los registros climáticos reunidos en este anexo corresponden a las observaciones
meteorológicas obtenidas durante los días 20 a 30 de septiembre y 1 a 15 de octubre del
año 2010. Los registros de precipitación (mm) se obtuvieron de la red de estaciones que
posee la Federación Nacional de Cafeteros en el departamento de Risaralda (Tabla 1)
empleando un pluviómetro con boca colectora de 200 cm2 y reglilla de medida (FNC -
Cenicafé, 2011). La temperatura (°C) y la humedad relativa (%HR) fueron medidas en
cada punto de muestreo mediante el uso de un termohigrómetro digital TFA.
Tabla 1 Estaciones pluviométricas de la Federación Nacional de Cafeteros ubicadas en
la cordillera occidental vertiente oriental en el departamento de Risaralda.
Estación Municipio Altura Coordenadas
Los Naranjos Santuario 1580 5° 3' N - 75° 0' W
La Tribuna Balboa 1580 4° 57' N - 75° 58' W
Los Cambulos Belén de Umbría 1400 5° 9' N - 75° 53' W
Buenos Aires Guática 1814 5° 19' N - 75° 48' W
El Barranco Mistrató 1580 5° 20' N - 75° 53' W
La Oriental Quinchía 1730 5° 22' N - 75° 43' W
El Diamante Quinchía 1550 5° 19' N - 75° 42' W
Fuente: Autor
Los datos de precipitación se presentan a continuación en la tabla 2 y figura 1. La
temperatura y humedad relativa se muestran en la tabla 3 y figuras 2 y 3.

106
Tabla 2 Registro de la precipitación en los meses de septiembre y octubre de 2010.
Estaciones LN LT LC BA EB LO ED PROM
Mes Día
S O S O S O S O S O S O S O S O
1 21 0 12 0 22 0 2 0 3 0 10 4 21.1 0 13.0 0.6
2 0 12 0 3 0 5 1 0 0 3 3 0 0 0 0.6 3.3
3 5 27 10 0 3 5 10 11 2 4 0 10 50.2 0 11.5 8.1
4 4 11 50 0 0 0 6 2 3 0 0 3 0 0 9.0 2.3
5 0 37 0 0 2 10 0 7 0 13 0 5 46.7 0 7.0 10.3
6 35 0 0 0 7 0 14 1 10 0 15 0 0 0 11.6 0.1
7 5 2 7 40 21 1 13 9 14 1 10 0 38.5 0 15.5 7.6
8 13 0 62 0 48 1 8 1 3 0 18 0 0 9.8 21.7 1.7
9 0 0 0 0 0 0 0 0 0 0 0 0 8 0 1.1 0.0
10 4 0 7 0 0 0 0 0 4 0 20 0 0 0 5.0 0.0
11 0 0 0 0 0 0 0 0 7 0 5 0 0 0 1.7 0.0
12 0 0 8 0 0 0 0 0 3 0 0 0 0 0 1.6 0.0
13 15 0 0 0 0 0 10 16 0 3 3 0 3.4 0 4.5 2.7
14 3 0 0 0 12 0 0 0 2 2 2 22 4.3 18.2 3.3 6.0
15 17 37 5 10 10 0 0 20 7 2 8 25 0 9 6.7 14.7
16 0 7 0 18 0 10 10 22 10 2 20 20 28.7 0 9.8 11.3
17 2 10 0 0 0 10 0 8 5 3 14 18 20 17.9 5.9 9.6
18 9 30 0 6 2 0 5 0 7 0 0 22 0 0 3.3 8.3
19 11 0 74 0 18 5 10 20 12 3 5 15 15.2 0 20.7 6.1
20 16 8 8 0 11 0 20 9 3 0 10 0 7.1 0 10.7 2.4
21 12 29 0 20 0 1 8 8 0 0 4 10 0 0 3.4 9.7
22 9 10 24 19 15 10 3 8 4 2 0 5 31.1 36.8 12.3 13.0
23 21 36 0 30 10 12 10 10 4 3 4 22 1.2 0 7.2 16.1
24 0 10 0 20 25 6 8 1 0 3 20 30 0 0 7.6 10.0
25 0 11 0 30 12 5 1 8 2 2 1 0 0 24.6 2.3 11.5
26 5 5 17 40 0 4 20 5 4 4 40 4 0 0 12.3 8.9
27 2 8 0 50 0 5 2 12 2 2 20 18 0 0 3.7 13.6
28 7 15 0 10 3 8 1 2 3 2 1 5 0 10.4 2.1 7.5
29 25 30 0 18 1 40 0 13 0 3 5 6 30.9 28.5 8.8 19.8
30 0 2 0 6 9 0 9 2 2 3 12 22 0 0 4.6 5.0
31 10 10 1 15 2 0 0 5.4
Total 241 347 284 330 231 139 162 210 116 62 250 266 306.6 155.2 227.2 215.6
Días de lluvia 21 21 12 16 18 18 21 23 23 20 23 19 14 8 19 18
Estaciones: Los Naranjos (LN); La Tribuna (LT); Los Cambulos (LC); Buenos Aires (BA);
El Barranco (EB); La Oriental (LO); El Diamante (ED). Mes: Septiembre (S); Octubre (O).
Fuente: Autor

107
Figura 1 Fluctuación de la precipitación del 20 de septiembre al 15 de octubre de 2010,
en la región cafetera de la cordillera occidental vertiente oriental, Risaralda.
Fuente: Autor
Tabla 3 Precipitación total y promedio, y días de lluvia registrados durante el periodo del
20 de septiembre al 15 de octubre de 2010, en la región cafetera de la cordillera
occidental vertiente oriental, Risaralda.
Lluvia Total (mm) Lluvia Promedio (mm) Días de Lluvia
132.5 5.1 22
Fuente: Autor
108
Tabla 4 Registro de la temperatura y la humedad relativa en los meses de septiembre y
octubre de 2010.
Mes Día Temperatura (°C) Humedad Relativa (%HR)
Septiembre
20 20.8 88
21 24.5 76
22 21.3 92
23 20.9 93
24 19.6 94
27 20.6 89
28 22.2 88
29 21.8 87
Octubre
1 22.3 83
2 21.0 88
4 18.9 88
5 18.7 90
7 11.9 64
8 17.3 94
9 18.9 96
10 18.5 95
11 19.4 93
12 19.0 94
13 10.7 95
14 18.0 96
15 17.5 96
Promedio 19.2 89
Prom. Bosque 21 88
Prom. Cafetal 18.8 91
Fuente: Autor

109
Figura 2 Fluctuación de la temperatura del 20 de septiembre al 15 de octubre de 2010,
en los puntos de muestreo evaluados.
Fuente: Autor
Figura 3 Fluctuación de la humedad relativa del 20 de septiembre al 15 de octubre de
2010, en los puntos de muestreo evaluados.
Fuente: Autor
Top Related