
vyo182formacion la vision del movimiento - … Formación Continuada VER Y OÍR - ENERO - FEBRERO...
16
20 Formación Continuada VER Y OÍR - ENERO - FEBRERO 2004 L PERCEPCIÓN VISUAL Análisis del movimiento por el sistema visual El análisis del movimiento de los objetos de nuestro entorno es una de las tareas más importantes y complejas que realiza nuestro sistema visual. Para los animales y para nosotros, los cambios en el mundo exterior son mucho más importantes que las condiciones estáticas; pensemos tanto en la supervivencia del depredador como en la de la presa. La interacción con nuestro entorno va a depender del conocimiento y del análisis de la información La visión del movimiento (I) M.D. de Fez* y P. Capilla ** * E.U. de Óptica y Optometría. Universitat de Alicante ** Departament. d’Òptica. Universitat de Valencia Correspondencia M.D. de Fez Escuela Universitaria de Óptica y Optometría Apartado 99 03080 Alicante [email protected] a sensibilidad al movimiento es un aspecto fundamental de la visión. Por un lado analizamos el movimiento de los objetos de nuestro campo visual para poder interaccionar con ellos. Por otro, al movernos dentro de nuestro entorno, se producen también cambios en la imagen retiniana de los objetos que nos rodean aunque éstos permanezcan estáticos. En ambos casos se producen cambios espaciotemporales de luminancia en la imagen percibida y estos cambios son la fuente de información sobre la disposición del entorno y el movimiento del observador con respecto a ese entorno. El análisis de esta información permite establecer la existencia de los límites de detección del movimiento y las características diferenciales entre los movimientos reales y los movimientos aparentes. obtenida a partir de los cambios que se produzcan en las imágenes proyectadas en nuestra retina. La finalidad principal de obtener la información sobre el movimiento es, sin lugar a dudas, la interacción de la persona con los objetos del entorno. Se pueden contemplar dos casos diferentes, un observador estacionario o un observador que se mueve en un entorno estacionario. Para un observador estacionario Si el observador es estacionario, la información que se deduce de los cambios espaciotemporales que ocurren en la retina nos sirve para saber cómo se mueven los objetos del entorno y poder interaccionar con ellos. Obviamente estos cambios van a depender de la distancia al objeto (fig. 1). Hay razones por las que resulta importante conocer la velocidad de los movimientos, por ejemplo, para poder realizar movimientos oculares de fijación o de
Transcript of vyo182formacion la vision del movimiento - … Formación Continuada VER Y OÍR - ENERO - FEBRERO...

20
Formación Continuada
VER Y OÍR - ENERO - FEBRERO 2004
L
PERCEPCIÓN VISUAL
Análisis del movimientopor el sistema visual
El análisis del movimiento de los objetos denuestro entorno es una de las tareas másimportantes y complejas que realiza nuestrosistema visual. Para los animales y para nosotros,los cambios en el mundo exterior son muchomás importantes que las condiciones estáticas;pensemos tanto en la supervivencia deldepredador como en la de la presa. Lainteracción con nuestro entorno va a dependerdel conocimiento y del análisis de la información
La visión del movimiento (I)
M.D. de Fez* y P. Capilla**
*E.U. de Óptica y Optometría. Universitat de Alicante**Departament. d’Òptica. Universitat de Valencia
CorrespondenciaM.D. de FezEscuela Universitaria de Óptica y OptometríaApartado 9903080 [email protected]
a sensibilidad al movimiento es un aspecto fundamental de la visión. Por un lado analizamos elmovimiento de los objetos de nuestro campo visual para poder interaccionar con ellos. Por otro, almovernos dentro de nuestro entorno, se producen también cambios en la imagen retiniana de losobjetos que nos rodean aunque éstos permanezcan estáticos. En ambos casos se producen cambiosespaciotemporales de luminancia en la imagen percibida y estos cambios son la fuente de informaciónsobre la disposición del entorno y el movimiento del observador con respecto a ese entorno. Elanálisis de esta información permite establecer la existencia de los límites de detección del movimientoy las características diferenciales entre los movimientos reales y los movimientos aparentes.
obtenida a partir de los cambios que seproduzcan en las imágenes proyectadas ennuestra retina. La finalidad principal de obtenerla información sobre el movimiento es, sin lugara dudas, la interacción de la persona con losobjetos del entorno. Se pueden contemplar doscasos diferentes, un observador estacionario oun observador que se mueve en un entornoestacionario.
Para un observador estacionario
Si el observador es estacionario, lainformación que se deduce de los cambiosespaciotemporales que ocurren en la retina nossirve para saber cómo se mueven los objetosdel entorno y poder interaccionar con ellos.Obviamente estos cambios van a depender dela distancia al objeto (fig. 1). Hay razones porlas que resulta importante conocer la velocidadde los movimientos, por ejemplo, para poderrealizar movimientos oculares de fijación o de

21
Formación Continuada
VER Y OÍR - ENERO - FEBRERO 2004
rastreo y no precisar de movimientos bruscos(sacádicos), para evitar un choque con un objetoque se nos viene encima, para conducir, etc. Elobjetivo es mantener la imagen en la fóvea porrazones obvias: es aquí donde disponemos delas mejores prestaciones de nuestro sistemavisual (máxima agudeza visual, máximasensibilidad al contraste, mejor detectabilidaden general de patrones espaciotemporales). Enla extrafóvea estas características decaen demanera importante.
Para un observador moviéndoseen un entorno estacionario
Si el entorno es estacionario y es el observadorel que se mueve, el mismo análisis nos servirápara posicionar los objetos en el entorno y poderinteraccionar con ellos. Las velocidades de lasdistintas partes de la imagen retiniana estánrelacionadas con la distancia de los objetoscorrespondientes y con la velocidad del propioobservador (en módulo y dirección). Haciendouso pues de tal información, el observadorpuede controlar la trayectoria de su propiomovimiento (en módulo y dirección) en aquellastareas que lo requieran, por ejemplo, paramodificar el tiempo de impacto contra unasuperficie. Se puede interpretar que elobservador móvil genera un patrón demovimientos (o de velocidades) de los objetosque están en su campo visual y ese patrón es loque utiliza de forma dinámica para ajustar sutrayectoria. En las figuras 2 y 3 se muestran dosejemplos en los que un sujeto se estádesplazando horizontalmente hacia el horizonte(típicamente un desplazamiento en coche) yverticalmente hacia el suelo (un helicópteroaterrizando).
Sobre la complejidad de la tarea:interacciones del movimiento conotras dimensiones de la percepción
La percepción del movimiento es una tareacompleja. En la figura 4 se muestran algunosejemplos. Si fijamos la mirada en el pájaro y elcoche atraviesa nuestro campo visual, la imagendel coche proyectada en la retina sufre undesplazamiento, marcado por la flecha (fig. 4a).El movimiento de la imagen proyectada en retinanos permite decir que el coche se mueve dederecha a izquierda. Supongamos ahora quefijamos la mirada en el coche, ¿qué es lo queocurre con la imagen en la retina? (fig. 4b).Ahora el ojo debe girar para seguir elmovimiento y mantener la imagen en la mismazona de la retina, pero es ésta la que ha cambiado
Fig. 1. Para un observador estacionario, la información delos cambios espaciotemporales en la retina sirve para sabercómo se mueven los objetos del entorno y poderinteraccionar con ellos.
Fig. 2. Observador moviéndose en un entorno estaciona-rio, desplazándose horizontalmente hacia el horizonte(típicamente un desplazamiento en coche).
Fig. 3. Observador moviéndose en un entorno estaciona-rio, desplazándose verticalmente hacia el suelo (porejemplo, un helicóptero aterrizando).

22
Formación Continuada
VER Y OÍR - ENERO - FEBRERO 2004
de posición. La traslación de la retina nospermite decir que el coche se mueve de derechaa izquierda. Pero aún nos queda una terceraforma de seguir el movimiento del coche (fig.4c). Supongamos que los ojos permanecenquietos y es la cabeza la que gira para seguir elmovimiento. De nuevo, el desplazamientoexperimentado, en este caso por la cabeza, nospermite decir que el coche se desplaza dederecha a izquierda. Otro aspecto que el sistemavisual tiene en cuenta es la distancia a la que seencuentra el objeto del ojo (fig. 4d).Supongamos el caso en el que fijamos la miradaen el pájaro y pensemos en dos coches adiferentes distancias. Si los coches se mueven ala misma velocidad, el ángulo subtendido porel desplazamiento entre dos instantesdeterminados es diferente en cada caso, es decir,la información sobre la retina es diferente. Sinembargo, el observador percibe la mismavelocidad para ambos. Lo que está ocurriendoes que la información de la profundidad tambiénesta de alguna manera actuando. Y es que lainformación sobre el movimiento interaccionacon la información relativa, no sólo a laprofundidad, sino a cualquier otra dimensiónde la percepción visual, incluida la forma o elcolor. Más adelante veremos algunos ejemplos.
Los límites del movimiento
La detección de un movimiento depende deuna gran cantidad de factores. Consideremosun objeto sencillo, por ejemplo, un circulito ouna barrita, que se desplaza delante de los ojosde un observador, desde el instante t=0,durante un tiempo Dt. Para saber cual ha deser la velocidad a la que, como mínimo, ha dedesplazarse el objeto para que el movimientosea percibido, le pediremos a nuestroobservador que nos diga hacia que lado, derechao izquierda, se ha desplazado el objeto.Repetiremos el ejercicio N veces con diferentesvalores de la velocidad y en cada presentacióncambiaremos aleatoriamente el sentido delmovimiento. Aquella velocidad para la que elobservador acierte, por ejemplo, un 75% delas veces, cual ha sido el sentido del movimiento,lo consideraremos como el “umbral delmovimiento”. Sin embargo, se pueden definirhasta cuatro tipos distintos de límites demovimiento, a saber:
• Límite inferior de movimiento: la velocidadmínima a partir de la cual percibimos elmovimiento.
• Límite superior de movimiento coherente:velocidad máxima a la que todavía percibimosun movimiento coherente del objeto, esto es,el objeto se percibe sin distorsiones espaciales.
• Límite superior de movimiento conservandoel patrón espacial: velocidad máxima a la quepercibimos movimiento, aunque ya sininformación sobre la forma del objeto.
• Límite de visibilidad absoluta delmovimiento: velocidad a partir de la cual ya nosomos capaces de percibir el movimiento.
Aunque el estudio de estos cuatro límitespermite analizar diferentes aspectos de lanaturaleza de los mecanismos de visión demovimientos, centraremos nuestra atención enel límite inferior. Las medidas realizadas conredes sinusoidales que se muestran en la figura5 indican que para un amplio rango defrecuencias espaciales y contrastes, empezamosa ver movimiento cuando la velocidad alcanzaun cierto valor mínimo y constante (Harris,1984,Johnston y Wright, 1985). Este resultado sugierela existencia de un mecanismo limitado por lavelocidad. Como veremos más adelante, estecomportamiento es sorprendente ya que
Fig. 4. Complejidad de la tarea. a) Punto de fijación en el pájaro,el coche atraviesa el campo visual y la información se recogedirectamente en la retina. b) Punto de fijación en el coche, lainformación del movimiento la proporciona el giro del ojo. c)Punto de fijación en el coche, la información del movimiento laproporciona el giro de la cabeza. d) La información de laprofundidad también está de alguna manera actuando.

23
Formación Continuada
VER Y OÍR - ENERO - FEBRERO 2004
diferentes experimentos psicofísicos (medidas desensibilidad al contraste, postefectos de movimiento)parecen, por el contrario indicar que es la frecuenciatemporal y no la velocidad el parámetro realmenterelevante. Por otra parte, existen asimismo evidenciasque apuntan hacia un mecanismo diferente queoperaría en visión central y especialmente paraestímulos breves (hasta 0,5 segundos) y que no estaríalimitado ni por la velocidad, ni por la frecuenciatemporal, sino por el desplazamiento (Boulton,1987).Presumiblemente estos dos mecanismos operaríanen paralelo y el límite estaría determinado por elmás sensible de los dos. El mecanismo limitado porla velocidad podría ser el dominante cuando elestímulo es visto durante un tiempo prolongado y elmecanismo limitado por el desplazamiento sería mássensible cuando el movimiento percibido esespecialmente breve. Esta posibilidad es consistentecon los resultados que se muestran en la figura 6(Leibowitz, 1955). Alternativamente, se ha sugeridouna segunda hipótesis según la cual, el mecanismolimitado por el desplazamiento sería más sensiblecon frecuencias espaciales altas y frecuenciastemporales bajas, o lo que es lo mismo, paramovimientos lentos, mientras el mecanismo limitado
por la velocidad lo sería con frecuencias espacialesbajas y frecuencias temporales altas, o lo que es lomismo, con movimientos de velocidades medias yaltas (Bonnet, 1977). Evidencias en favor de estahipótesis proceden de distintas medidas del umbralde contraste realizadas con un amplio conjunto decombinaciones de las frecuencias espacial y temporalen las que se pedía al observador que detectara elmovimiento del patrón espaciotemporal. Téngaseen cuenta que en el umbral de detección de unpatrón espaciotemporal, puede detectarse elmovimiento o el patrón espacial, por lo que esimportante que el observador entienda cual es latarea que ha de realizar. Cuando las medidas deBurr y Ross se representan en función de la frecuenciatemporal para cada valor constante de la velocidad(fig. 7) se encuentra que todas las velocidadesproducen una misma función de sintonizado salvojustamente cuando la velocidad es especialmentebaja (Burr y Ross, 1982). Por otra parte, los umbralesque se obtienen cuando la velocidad es baja se desvíande manera significativa de las predicciones que sederivarían de un modelo tradicional para la deteccióndel movimiento (Murray et al., 1983).
Fig. 6. Velocidad límite en función del tiempo de exposición( Leibowitz, 1955).
Fig. 5. Límite inferior de movimiento (Harris, 1984).
Fig. 7. Medida del contraste umbral en función de la frecuenciatemporal para diferentes velocidades (en grados/seg). Sólo paravelocidades bajas la función de sintonizado es sensiblementediferente al resto (Burr y Ross, 1982).
Movimiento real y movimientoaparente
Existen dos tipos de movimiento. Por unaparte, un movimiento real que es percibidocomo tal: cuando el coche, (esto es, un estímulofísico), se mueve, cambia su posición realmenteen el espacio objeto. Pero también hemos detener en cuenta que muchos de los movimientosque experimentamos pueden ser clasificadoscomo aparentes, es decir, que es posible percibir

24
Formación Continuada
VER Y OÍR - ENERO - FEBRERO 2004
movimiento cuando el estímulo no se estámoviendo, cuando no hay cambio de posiciónen el espacio objeto. Veamos a continuaciónalgunos ejemplos.
Movimientos aparentes clásicos
Quizás el movimiento aparente porantonomasia es el denominado efecto f,descubierto por Wertheimer en 1912(Wertheimer, 1912). Por ejemplo, cuando dosluces separadas una cierta distancia sonencendidas secuencialmente con determinadointervalo, se tiene la sensación de movimientoaunque ningún objeto se está desplazando.Este efecto se denomina también movimientoestroboscópico y, comúnmente, es usado paracrear esa ilusión de movimiento medianteluces en la televisión, películas y monitoresde ordenador. También lo hemos visto en lailuminación festiva de calles o en la navideñade algunos establecimientos comerciales quetodos tenemos en mente. Supongamos uncírculo de luz (aunque no es relevante laforma) con luminancia L1 y longitud de ondal1, que se enciende durante un instante t1. Untiempo Dt después de que éste se apague, seenciende otro círculo de luz, con luminanciaL2 y longitud de onda l 2 durante un tiempo t2.Los dos círculos están separados espacialmenteuna distancia Dx. Tenemos un montón deparámetros con los que jugar, así es quesimplifiquemos el asunto. Supongamos quelas luminancias y los colores son iguales.Supongamos que los tiempos de encendidoson también iguales y además muy cortos encomparación con Dt. Si la distancia Dx esconstante, sólo nos queda como variable elintervalo de tiempo Dt. Si Dt es mayor que,por ejemplo, 200 ms, los estímulos se percibensucesivamente, o sea, uno después del otro.Si por el contrario, Dt es menor que porejemplo, 30 ms, los estímulos se percibensimultáneamente. Entre ambas situaciones, entorno a un Dt de 60 ms parece que el estímulose desplaza realmente de un lado al otro. Es,naturalmente, un movimiento aparente: elmovimiento f. En 1915, Korte estableció unconjunto de reglas empíricas que gobernaríaneste efecto, en función de todos y cada unode los parámetros comentados anteriormente(Korte, 1915). Estas reglas se conocen hoycomo leyes de Korte.
Un ejemplo especialmente ilustrativo de estetipo de movimiento es la técnica utilizada en
los dibujos animados: dibujos ligeramentediferentes presentados con una cadencia rápidaaparentan un movimiento continuo.
Cinematogramas
Consideremos la experiencia conocida como elcuadrado de puntos que se mueven: consiste enun conjunto de imágenes de puntos aleatorios quese presentan secuencialmente, con una zonacuadrada que se repite, desplazada espacialmente,una pequeña cantidad respecto de la imagenanterior, (del orden de 15’ de arco). En ningúnfotograma puede verse contorno alguno. Perocuando el sistema visual compara imágenesconsecutivas detecta un movimiento aparente:parece que el cuadrado se ha movido. Así pues,la percepción del movimiento (comparación entreimágenes) precede a la percepción de la formadentro de cada imagen (véase la web).
Consideremos ahora una variante de laexperiencia anterior: una secuencia de imágenesde puntos aleatorios pero ahora con la zonacuadrada compuesta de pequeños puntos condiferente textura o color que los del fondo (fig.8). Lo que ocurre ahora es que el sistema visualidentifica en primer lugar la forma dentro de laimagen y luego la localiza en la imagen siguiente:la percepción de forma precede en este caso ala percepción del movimiento. El sistema visualpercibe movimiento por agrupamiento deimágenes idénticas, lo que nos indica tambiénque tiene cierta tolerancia a la deformación, yaque los bordes del cuadrado no tienen por quéser los mismos en todas las imágenes (veáse laweb).
Algunas características de los movimientosaparentes con estímulos clásicos (por ejemplo,el efecto phi) son sin embargo distintas a las delos movimientos aparentes producidos concinematogramas de puntos aleatorios. Porejemplo, la distancia máxima entre imágenesque puede haber en un cinematograma,denominada Dmax, es del orden de 15 minutosde arco, lo que significa que el límite espacialpara percibir esta clase de movimiento aparentees mucho menor que el que se requiere parapercibir, por ejemplo, efecto phi. Basándose enestas y otras características diferenciales,Braddick propuso la existencia de dos tipos demecanismos o de procesos para el análisis delmovimiento, denominados de corto alcance yde largo alcance (Braddick, 1974). Sin embargo,el punto de vista moderno es que, en realidad,estas diferencias se pueden analizar en términos

25
Formación Continuada
VER Y OÍR - ENERO - FEBRERO 2004
de los distintos espectros espaciotemporales decada escena que deben ser analizados por unaorganización única de mecanismos espacio-temporales (Cavanagh y Mather, 1989).
Postefectos
Otro efecto de percepción de movimiento sinque realmente se mueva un objeto, son lospostefectos de movimiento. Podemos encontrarinfinidad de ejemplos que se han estudiadodesde hace mucho tiempo. Por ejemplo, elefecto que se conoce como ilusión de la cascada(fig. 9), que se produce cuando miramos a una
imagen estática después de habernos adaptadomirando a la cascada: “parece que el agua subehacia arriba” (véase la web). La interpretaciónde este efecto es sencilla. Basta con asumir queexisten detectores de movimiento “hacia arriba”y “hacia abajo”, es decir, selectivos a laorientación del movimiento. La adaptaciónmirando el movimiento del agua de la cascadahacia abajo, reduce la sensibilidad de losdetectores de movimiento en ese sentido (saturalos detectores). El desequilibrio que se producese manifiesta al mirar la imagen estática. Comolos detectores hacia abajo están saturados, sólodarán señal los detectores hacia arriba y, porconsiguiente, se percibe la cascada subiendohacia arriba.
Movimientos inducidos
Cuando un estímulo estacionario estásuperpuesto sobre un objeto grande enmovimiento, parece que es el estímulo el quese mueve en sentido contrario; esto es lo queocurre cuando miramos a la luna cubiertaparcialmente por nubes que se desplazan enun sentido: parece que es la luna la que sedesplaza en sentido contrario.
Los enigmáticos
Por último un apartado especial para una seriede efectos que no pueden ser clasificados en lascategorías anteriores, ya que se percibe unmovimiento al mirar una imagen totalmenteestática. Estos efectos son comúnmente llamadosilusiones enigma (Zeki et al, 1993). Existennumerosos ejemplos, de los que presentamosdos en la figura 10. En ambos casos, al fijar lamirada en cualquier punto de la imagen, sepercibe un giro de los objetos distribuidos enlas regiones extrafoveales.
Fig. 8. Secuencia de cinematogramas de puntos aleatorios en los que aparece un cuadrado con diferente color al fondo, desplazadoligeramente en cada fotograma respecto al anterior. La presentación secuencial de los fotogramas produce la sensación de movimiento.
Fig. 9. Ilusión de la cascada. Si nos adaptamos al movi-miento del agua de la cascada hacia abajo y miramos luegola imagen estática, se produce un postefecto de movimien-to y nos parece que el agua sube hacia arriba.

26
Formación Continuada
VER Y OÍR - ENERO - FEBRERO 2004
Una última reflexión
Podríamos plantearnos si tenemos en nuestrosistema visual un único mecanismo con el quevemos el movimiento real y el aparente, o unmecanismo distinto para cada tipo demovimiento. Veremos que esta posibilidad sepuede descartar. De lo dicho hasta el momento,parece que el sistema visual trata exactamenteigual los movimientos reales que losmovimientos aparentes. Por ejemplo, unmovimiento aparente puede generar postefectosde movimiento. Sin embargo, cabría preguntarseentonces por qué en ciertas condiciones unmovimiento aparente no se percibe suave ocontinuo como uno real, lo que ofrecería todavíaalgunas dudas sobre si existe en el sistema visualun mecanismo específico para los movimientosaparentes. En la próxima lección analizaremos
Fig. 10: Ilusiones enigma. Al mirar una imagen totalmenteestática, se percibe un movimiento de giro en regionesextrafoveales.
cuales son estas condiciones en el contexto delos fundamentos fisiológicos para el análisis delmovimiento.
Bibliografía
- Bonnet C. Visual motion detection models: feature andfrequency filters. Perception 1977;6:491-500.- Boulton JC. Two mechanisms for the detection of slowmotion. J Opt Soc Am A 1987;4:1634-1642.- Braddick O. A short- range process in apparent motion.Vision Research 1974;14:519-527.- Burr DC, Ross J. Contrast sensitivity at high velocities.Vision Rersearch 1982;22:479-484.- Cavangh P, Mather G. Motion: the long and short of it.Spatial Vision 1989;4:103-129.- Harris MG. The role of pattern and flicker mechanisms indetermining the spatiotemporal limits of velocity perception 2.The lower movement threshholds. Perception 1984;13:409-415.- Johnston A, Wright MJ. Lower thresholds of motion forgratings as a function of eccentricity and contrast. VisionResearch 1985;25:179-185.- Korte A. Kinematoskopische untersuchungen. Z Psychol1915;72:193-206.- Leibowitz HW. The relation between the rate thresholdfor the perception of movement and luminance for variousdurations of exposure. J Exp Psy 1955;49:209-214.- Murray I, MacCana F, Kulikowski JJ. Contribution of twomovement detecting mechanisms to central and peripheralvision. Vision Research 1983;2:151-159.- Wertheimer M. Experimentelle studien uber das sehenvon bewegung. Zeitschrift F. Psychol 1912;61:161-265.Experimental studies on the seeing of motion. Reprinted inT. Shipley (1961). Classics in Psychology. New York:Philosophical Library.- Zeki S, Watson JPG, Frackowiak RSJ. Going beyond theinformation given: the relation of illusory visual motion to brainactivity. Proc Royal Soc London Series B 1993;252:215-222.
currículumPascual Capilla Perea, es licenciado en Ciencias Físicas por la Universitat de València en 1984, Doctor en Ciencias
Físicas por la Universitat de València en 1990, ayudante de la Escuela Universitaria de la Universidad de Alicantedesde el 1 de diciembre de 1986. Titular de Universidad de la Universitat de València en 1995.
Ha sido en distintas ocasiones profesor en las asignaturas de Óptica Fisiológica, Psicofísica de la Visión, PercepciónVisual y Colorimetría y Visión del color, todas ellas de la diplomatura de Óptica y Optometría, así como de laasignatura Teoría del color, de la Licenciatura en Física.
Es coautor de los libros Problemas de Óptica Fisiológica, Óptica Fisiológica, Psicofísica de la Visión, Fundamentosde Colorimetría y Tecnología del color.
Como investigador, ha dirigido 4 Tesis Doctorales, 2 Tesinas de Licenciatura y 4 de Diplomatura. Es autor de 28publicaciones en revistas de investigación nacionales e internacionales y de 9 publicaciones en revistas de divulgacióncientífica. Ha presentado 53 comunicaciones en congresos nacionales e internacionales.
M. Dolores de Fez Saiz, Doctora en Ciéncias Físicas por la Universidad de Valencia (orientación óptica). Profesoratitular de la Escuela de Óptica y Optometría de la Universidad de Alicante, donde imparte docencia de las asignaturasdel área de Óptica Fisiológica. Miembro del equipo de investigación GVC: Grupo de Visión y Color de la Universidadde Alicante.

134
Formación Continuada
VER Y OÍR - ABRIL 2004
E
PERCEPCIÓN VISUAL
La visión del movimiento (II)Fundamentos fisiológicos y modelos
M.D. de Fez* y P. Capilla**
*E.U. de Óptica y Optometría. Universitat de Alicante**Departament. d’Òptica. Universitat de Valencia
CorrespondenciaM.D. de FezEscuela Universitaria de Óptica y OptometríaApartado 9903080 [email protected]
Problema del muestreo temporal
Recordemos que un movimiento aparente esun conjunto de imágenes diferentes que alpresentarse secuencialmente nos dan lapercepción de movimiento continuo. Nodisponemos pues de toda la información, comoen un movimiento real, sólo de parte de ella.Pensemos en una imagen con una distribuciónúnicamente espacial: cuando dicha imagen seforma sobre un mosaico de detectores discretoseparados una distancia ∆d, se está muestreandoespacialmente porque toda la información quecae entre dos detectores se pierde (fig. 1). Lafrecuencia de muestreo espacial se define como:
l concepto de movimiento aparente, introducido en la primera lección sobre la visión demovimientos, sienta las bases para entender porque el sistema visual puede interpretar movimientosdebido al muestreo temporal que realiza de la información. El problema del muestreo surge en elprocesamiento debido a la comprensión de la información del objeto. En la segunda aparte de estalección se introduce el estudio de los fundamentos fisiológicos de la visión de movimientos, un pasonecesario para establecer los modelos para el análisis de movimientos.
Fig. 1. Ejemplo de muestreo espacial con una red defotorreceptores separados ∆d (frecuencia de muestreo 1/∆d).
135
Formación Continuada
VER Y OÍR - ABRIL 2004
Un problema análogo tiene lugar en el dominiotemporal cuando “tomamos fotografías” de unobjeto en movimiento a intervalos de tiempo ∆t.La frecuencia de muestreo es análogamente:
Un ejemplo sencillo se ilustra en la figura 2.Un círculo negro se mueve en el plano (x,y)partiendo de una cierta posición inicial (x0,y0)en el instante t0. Podemos representar esemovimiento como el resultado de undeslizamiento continuo. A partir del instanteinicial se toman fotografías cada ∆t. El resultadode cada fotografía mostraría el círculo en unaposición (x, y) diferente que se iría desplazandoen la dirección del movimiento. Al exponer lasdiferentes fotografías secuencialmente, elmovimiento aparece como cambios entre unaimagen y la siguiente, aunque falte informaciónintermedia. Pero si las diferencias de una imagena la siguiente son muy grandes, entonces lasensación de movimiento continuo se rompe.
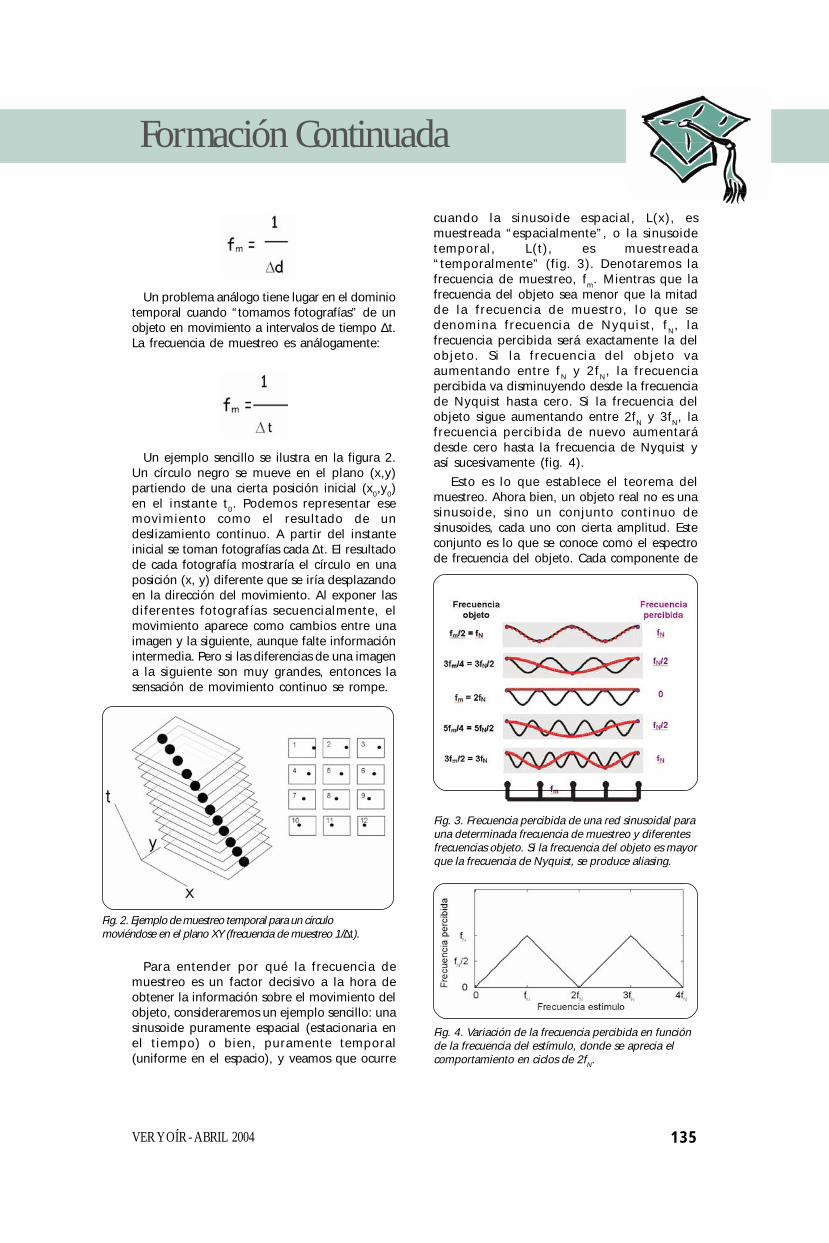
cuando la sinusoide espacial, L(x), esmuestreada “espacialmente”, o la sinusoidetemporal, L(t), es muestreada“temporalmente” (fig. 3). Denotaremos lafrecuencia de muestreo, fm. Mientras que lafrecuencia del objeto sea menor que la mitadde la frecuencia de muestro, lo que sedenomina frecuencia de Nyquist, fN, lafrecuencia percibida será exactamente la delobjeto. Si la frecuencia del objeto vaaumentando entre fN y 2fN, la frecuenciapercibida va disminuyendo desde la frecuenciade Nyquist hasta cero. Si la frecuencia delobjeto sigue aumentando entre 2fN y 3fN, lafrecuencia percibida de nuevo aumentarádesde cero hasta la frecuencia de Nyquist yasí sucesivamente (fig. 4).
Esto es lo que establece el teorema delmuestreo. Ahora bien, un objeto real no es unasinusoide, sino un conjunto continuo desinusoides, cada uno con cierta amplitud. Esteconjunto es lo que se conoce como el espectrode frecuencia del objeto. Cada componente de
Fig. 2. Ejemplo de muestreo temporal para un círculomoviéndose en el plano XY (frecuencia de muestreo 1/∆t).
Para entender por qué la frecuencia demuestreo es un factor decisivo a la hora deobtener la información sobre el movimiento delobjeto, consideraremos un ejemplo sencillo: unasinusoide puramente espacial (estacionaria enel tiempo) o bien, puramente temporal(uniforme en el espacio), y veamos que ocurre
Fig. 4. Variación de la frecuencia percibida en funciónde la frecuencia del estímulo, donde se aprecia elcomportamiento en ciclos de 2fN.
Fig. 3. Frecuencia percibida de una red sinusoidal parauna determinada frecuencia de muestreo y diferentesfrecuencias objeto. Si la frecuencia del objeto es mayorque la frecuencia de Nyquist, se produce aliasing.

136
Formación Continuada
VER Y OÍR - ABRIL 2004
un objeto cuya frecuencia sea mayor que lafrecuencia de muestreo, será mal muestreada.Por ejemplo, una frecuencia de 3/2 de fN sepercibirá como 1/2 de fN. Si en el objeto yaexiste una componente de esta frecuencia,entonces la amplitud resultante será la suma delas contribuciones debidas a la componenteexistente y al muestreo de la componente 3/2de fN. El efecto resultante se denomina aliasing.Si el espectro del objeto fuera discreto, (lo quesólo ocurre si el objeto es periódico), y nocontuviera una componente de frecuencia 1/2de fN, entonces ésta aparecería como unafrecuencia espúrea. Aunque esto es también unaliasing, no es lo que suele ocurrir, porque losespectros de los objetos son, en general,continuos.
Para evitar los problemas de aliasing queacabamos de describir se deberá realizar elmuestreo con una frecuencia de Nyquist queno sea menor que la frecuencia de corte (fc) dela CSF temporal. Si fuese menor, entre la fN y lafc tendríamos problemas de aliasing. Si fuesemayor que la fc estaríamos muestreandofrecuencias que no son visibles para el sistemavisual, lo que sería un gasto inútil. Teniendo encuenta que aumentar la frecuencia de muestreoes, en términos económicos enormemente caro,la solución entonces más eficiente es hacer quefN y fc simplemente coincidan.
Pero recuperemos ahora nuestro problemaoriginal. Muestrear un objeto cualquiera enmovimiento es un problema complejo, porqueno se trata de hacer un muestreo espacial deL(x), en general de L(x, y), o un muestreotemporal de L(t), sino de hacer un muestreotemporal de un objeto que es de la forma L(x,y, t). Hacer un muestreo temporal de este objetoa una cierta frecuencia de muestro, esto es, aintervalos ∆t, significa multiplicar la función L(x,y, t) por una función de muestreo temporal. Siasumimos que la duración de las “fotos” esdespreciable con respecto a ∆t, la función demuestreo será simplemente un peine de deltasen el tiempo. Multiplicar dos funciones en eldominio de posiciones y tiempos es equivalente,en el dominio de frecuencias espaciales ytemporales, a la convolución entre suscorrespondientes espectros, esto es, el espectrode frecuencias de L(x, y, t) y el espectro defrecuencias del peine de deltas. Puesto que elespectro de un peine de deltas en el tiempo esun peine de deltas en la frecuencia temporal, elresultado de la convolución es que el espectrode la imagen se repite a intervalos ∆ft. En la
figura 5a se muestra el espectro de una líneaque se mueve hacia la derecha con velocidadv, muestreado a dos frecuencias distintas. En5b el muestreo se realiza a una frecuencia baja,esto es, con un ∆t grande, en 5c el muestreo serealiza a una frecuencia alta, esto es, con un ∆tpequeño. Nótese que la separación entre lasdeltas del peine en frecuencias esta inversamenterelacionada con la separación entre las deltasdel peine en el tiempo. Recordemos que, de lasCSFs espacial y temporal del sistema visualsabemos que existen unas ciertas frecuenciasde corte, tanto en el dominio espacial (fce) comoen el dominio temporal (fct). Para las frecuenciasque son más altas que las de corte, la sensi-bilidad al contraste es cero, o en otras palabras,no se ven. Se denomina ventana de visibilidada la frontera que delimita la región defrecuencias espaciales y temporales que sonvistas por el sistema visual. Si representamosesta frontera poniendo, por ejemplo, lafrecuencia temporal en el eje x y la frecuenciaespacial en el eje y, los límites en los propiosejes serán las frecuencias de corte correspon-dientes. Asumiremos que la forma de la fronteraes circular. Esta ventana corresponde a la zonasombreada en la figura 5. Si las replicas delespectro de la imagen introducidas por elmuestreo caen fuera de la ventana de visibilidadno se verán y por consiguiente es como si noestuvieran: la imagen muestreada, querecordemos era un objeto en movimiento, escompletamente indistinguible de la imagen real.Si por el contrario, una o varias replicas caendentro de la ventana de visibilidad, entonces
Fig. 5. a) Espectro de una línea que se mueve hacia laderecha con velocidad v. b) Muestreo a una frecuen-cia baja (intervalo temporal grande). c) Muestreo auna frecuencia alta (intervalo temporal pequeño).
137
Formación Continuada
VER Y OÍR - ABRIL 2004
tendremos distorsiones de la imagen (aliasing)como consecuencia del muestreo y lo queveremos será cualquier cosa, pero desde luego,algo que no tiene por qué parecerse en absolutoal movimiento real del objeto. Como habíamosrazonado previamente, un muestreo eficientedebería realizarse de manera que las frecuenciasde Nyquist espacial y temporal coincidan conlas correspondientes frecuencias de corte, fce yfct, respectivamente.
Ahora que hemos analizado el muestreo conmás detenimiento, podemos entender porqueun movimiento aparente, que resulta demuestrear un movimiento real es, en ocasiones,igual que el real y en otras no, sin necesidadalguna de admitir que existan en el sistemavisual mecanismos independientes para ver cadaclase de movimiento.
Sobre los fundamentos fisiológicosde la visión del movimiento
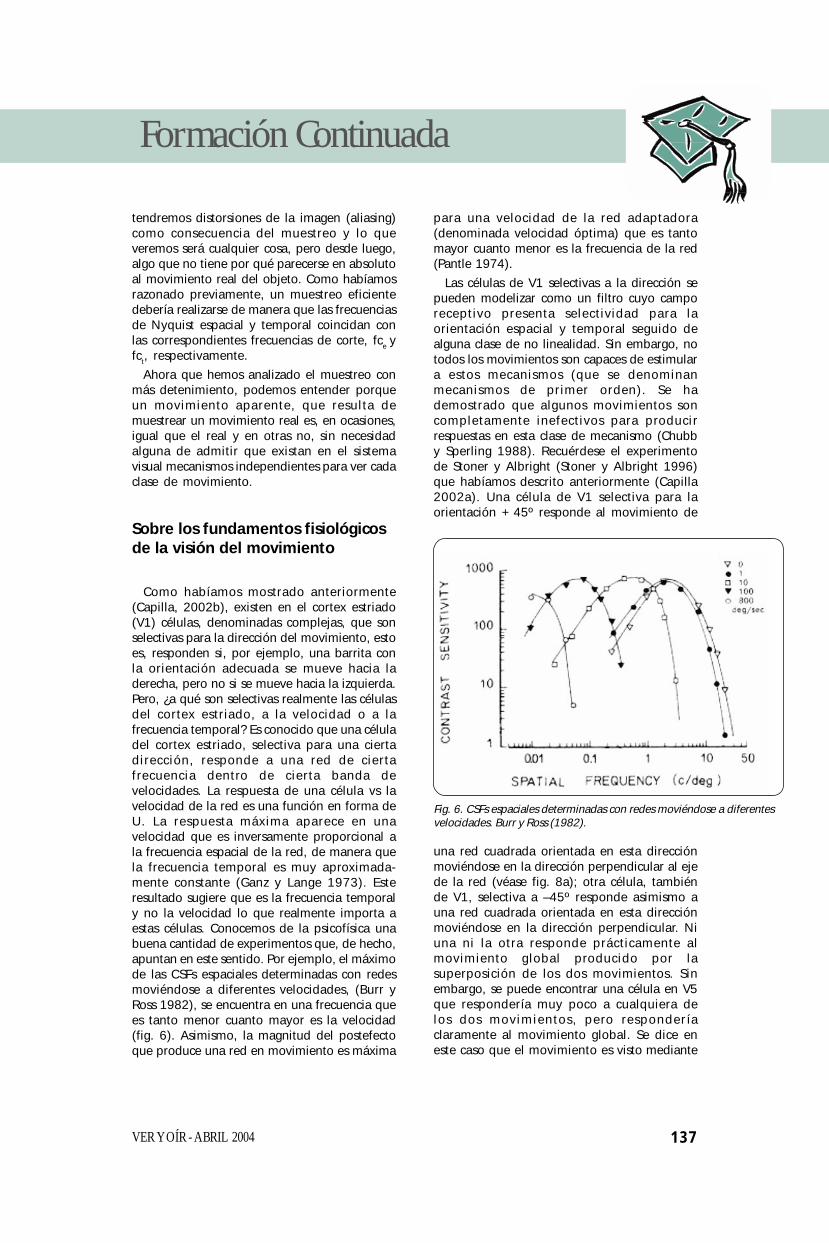
Como habíamos mostrado anteriormente(Capilla, 2002b), existen en el cortex estriado(V1) células, denominadas complejas, que sonselectivas para la dirección del movimiento, estoes, responden si, por ejemplo, una barrita conla orientación adecuada se mueve hacia laderecha, pero no si se mueve hacia la izquierda.Pero, ¿a qué son selectivas realmente las célulasdel cortex estriado, a la velocidad o a lafrecuencia temporal? Es conocido que una céluladel cortex estriado, selectiva para una ciertadirección, responde a una red de ciertafrecuencia dentro de cierta banda develocidades. La respuesta de una célula vs lavelocidad de la red es una función en forma deU. La respuesta máxima aparece en unavelocidad que es inversamente proporcional ala frecuencia espacial de la red, de manera quela frecuencia temporal es muy aproximada-mente constante (Ganz y Lange 1973). Esteresultado sugiere que es la frecuencia temporaly no la velocidad lo que realmente importa aestas células. Conocemos de la psicofísica unabuena cantidad de experimentos que, de hecho,apuntan en este sentido. Por ejemplo, el máximode las CSFs espaciales determinadas con redesmoviéndose a diferentes velocidades, (Burr yRoss 1982), se encuentra en una frecuencia quees tanto menor cuanto mayor es la velocidad(fig. 6). Asimismo, la magnitud del postefectoque produce una red en movimiento es máxima
para una velocidad de la red adaptadora(denominada velocidad óptima) que es tantomayor cuanto menor es la frecuencia de la red(Pantle 1974).
Las células de V1 selectivas a la dirección sepueden modelizar como un filtro cuyo camporeceptivo presenta selectividad para laorientación espacial y temporal seguido dealguna clase de no linealidad. Sin embargo, notodos los movimientos son capaces de estimulara estos mecanismos (que se denominanmecanismos de primer orden). Se hademostrado que algunos movimientos soncompletamente inefectivos para producirrespuestas en esta clase de mecanismo (Chubby Sperling 1988). Recuérdese el experimentode Stoner y Albright (Stoner y Albright 1996)que habíamos descrito anteriormente (Capilla2002a). Una célula de V1 selectiva para laorientación +45º responde al movimiento de
Fig. 6. CSFs espaciales determinadas con redes moviéndose a diferentesvelocidades. Burr y Ross (1982).
una red cuadrada orientada en esta direcciónmoviéndose en la dirección perpendicular al ejede la red (véase fig. 8a); otra célula, tambiénde V1, selectiva a –45º responde asimismo auna red cuadrada orientada en esta direcciónmoviéndose en la dirección perpendicular. Niuna ni la otra responde prácticamente almovimiento global producido por lasuperposición de los dos movimientos. Sinembargo, se puede encontrar una célula en V5que respondería muy poco a cualquiera delos dos movimientos, pero responderíaclaramente al movimiento global. Se dice eneste caso que el movimiento es visto mediante

138
Formación Continuada
VER Y OÍR - ABRIL 2004
un mecanismo de segundo orden. Movshon ycolaboradores se refieren a los mecanismos deprimer y de segundo orden como mecanismosselectivos para el componente y para el patrón,respectivamente (Movshon et al. 1985). Elsoporte fisiológico para ambos mecanismos seencontraría en V1 y V5, respectivamente. Laparticipación de V5 en la percepción delmovimiento es hoy día incuestionable, toda vezque se conoce que personas con lesiones enesta área realizan con gran dificultad, o no soncapaces de realizar, tareas relacionadas con lavisión de movimientos, mientras que no tienenproblema alguna para realizar cualquier tareaque involucre la agudeza visual, la visión delcolor o la estereopsis.
Modelos para el análisisdel movimiento
El movimiento de un objeto dentro de unaimagen 3D se caracteriza por el vector velocidad:la dirección nos indica hacia dónde se mueve elobjeto y el módulo la rapidez con la que lohace. El sistema visual debe ser capaz decomputar ambas cosas y un modelo para lavisión del movimiento debe imitar estacapacidad.
1. Mecanismos de análisis del movimientopara objetos identificados y localizados
Consideremos el proceso más simple paraanalizar el movimiento del objeto: primero,identificar un objeto y localizarlo espacialmenteen el instante t1; segundo, localizarlo en elinstante t2>t1, y por último, medir la distanciarecorrida, Dx, y dividir por Dt, o sea, calcularla velocidad. Una vez que se ha identificado elobjeto, la clave es analizar posiciones y medirla velocidad. Pues bien, el sistema visual nodebe trabajar así, aunque este método tendríala ventaja de evitar el problema de los objetosque se ocultan en un momento dado detrásde un obstáculo. Pero si el movimiento es muyrápido, no daría tiempo para realizar tantasoperaciones porque necesitamos invertir unacantidad no despreciable en la tarea deidentificación que no es de bajo nivel, sino todolo contrario. De hecho, el sistema visual realizaya un análisis del movimiento a bajo nivel.Veamos algunos ejemplos en los que este hechoes evidente:
a) En el cuadrado de puntos que se mueven,(véase la lección anterior), es el movimiento loque ayuda a reconocer que los puntos que semueven forman un cuadrado y no al revés. Portanto, estamos detectando un movimiento sinidentificación previa del objeto y, obviamente,sin localización espacial alguna de ningún objeto.
b) En la ilusión de la cascada, (véase la lecciónanterior), tenemos sensación de movimiento sincambio alguno de posición.
c) Más aún: el efecto φ invertido, (variantedel efecto φ en el que el movimiento aparentees en el sentido opuesto al del cambio deposición del objeto), prueba que la sensaciónde movimiento no se deriva necesariamente delos cambios de posición del objeto en la imagenretiniana, sino de los cambios espaciotemporalesque se producen en la distribución de luminanciasde la imagen retiniana.
Veamos pues como podría funcionar unmodelo sin la necesidad de exigir la identificaciónprevia del objeto.
2. Mecanismos de análisis del movimientosin necesidad de identificación
Acabamos de decir que la sensación demovimiento se deriva de cambios espacio-temporales de la distribución de luminancias enla retina. Sin embargo, estos cambios puedenanalizarse directamente en el dominio espacio-tiempo, o alternativamente, en el dominio defrecuencias espaciales y temporales.
Modelos en el dominio espacio-tiempo
El modelo de Reichardt (Reichardt 1961) ilustrade forma sencilla lo que es un modelo en eldominio de espacio y tiempo (fig. 7a).Supongamos un punto moviéndose hacia laizquierda que pasa por encima del detector Aen el instante t=0. Un rato después pasará porencima del detector B. Supongamos que lavelocidad de conducción de la señal desde losdetectores A y B al multiplicador es infinita, perointroducimos un delay (parada) en el caminode A. Si el movimiento es muy rápido llegaráantes la señal de B, si es muy lento llegará antesla señal de A. Claro que lo de rápido o lentoestará en función de lo que dure el delay quefrena la señal en el cable que viene desde A. Sila distancia entre los detectores es ∆x y el retardoestá ajustado para que las dos señales lleguenal mismo tiempo, el multiplicador dará una

139
Formación Continuada
VER Y OÍR - ABRIL 2004
respuesta máxima. Esta maquinaria, que seconoce como unidad de Reichardt, constituyepues un detector de movimiento sintonizadopara una cierta velocidad y para un sentido delmovimiento particular (hacia la izquierda).Podemos pensar en una unidad simétrica paramovimiento hacia la derecha. Con un par deunidades de Reichardt se puede construir unaunidad más sofisticada que permitiría decidirsobre el sentido del movimiento (fig. 7b) (Poggioy Reichardt 1973). Si los multiplicadores de lasdos unidades restan sus respuestas seinterpretará que el movimiento es hacia laderecha o hacia la izquierda según si la diferenciaes positiva o negativa.
¿Tiene este tipo de modelos algún sentidodesde el punto de vista fisiológico? Ciertamentesí, ya que sabemos que existen células corticalesque justamente tienen la propiedad de serselectivas para la dirección del movimiento: lascélulas complejas.
En la primera edición del libro de Hubel(Hubel 1995), adaptando las ideas originales deBarlow y Levick [Barlow y Levick (1965) propuso
un posible circuito neural que funcionaríabásicamente como una unidad de Reichardt(fig. 8). La idea es relativamente sencilla.Supongamos que entre las células simples ylas células complejas existen unas célulasintermedias (blancas) que reciben inputsexcitatorios de una célula simple e inhibitoriosde otra célula simple vía una segunda célulaintermedia (grises). Los campos receptivos delas dos células intermedias son adyacentes.La célula intermedia gris actúa como un delay.Finalmente, varias células intermedias blancasestán conectadas con una célula master,compleja. Supongamos que el objeto semueve de derecha a izquierda. Si la velocidaddel movimiento es la adecuada, la célulaintermedia blanca recibirá una excitación justoen el momento en el que está tambiénrecibiendo una inhibición de la célulaintermedia gris por cuyo campo receptivoacababa de cruzar el objeto. Ambas señalesse cancelan y la célula compleja no recibiráseñal alguna. Pero si, por el contrario, elmovimiento es de izquierda a derecha, lainhibición llegará demasiado tarde para evitarque la célula blanca haya ya enviado señal ala célula compleja. La célula compleja tienepor tanto las propiedades de una célulaselectiva a la orientación, está sintonizada aun movimiento y a una velocidad. Sinembargo, nuevas evidencias experimentalessugieren organizaciones corticales mássofisticadas (Murray et al. 1983).
Fig. 7. Unidad de Reichardt para la detección delmovimiento con una determinada velocidad en unsentido (a) o en ambos sentidos (b).
Fig. 8. Modelos en el domino espacio-tiempo: circuitoneural propuesto por Hubel (1995).

140
Formación Continuada
VER Y OÍR - ABRIL 2004
Modelos en el dominio de frecuencias
Los modelos en el dominio de frecuenciasespaciales y temporales son matemáticamentemucho mas complicados, por lo que noscontentaremos con una idea cualitativa sobrecomo funcionaría un modelo de este tipo paraextraer la información de la velocidad. Nosapoyaremos para ello en el modelo de Watsony Ahumada (Watson y Ahumada 1985).
Para facilitar el estudio más sencillo de losdiferentes mecanismos que entran en juegovamos a plantearnos primero cómo realizar la
Fig. 9. Representación en el dominio espacio-temporal de una red unidimensional moviéndose con velocidad v=1.5grados/segundo hacia la izquierda.
Fig. 10. Representación en el dominio de frecuenciasespacio-temporales de una red unidimensionalmoviéndose con velocidad v=1.5 grados/segundohacia la izquierda y de un sólido rígido moviéndosecon v=3 grados/segundo hacia la derecha.
Fig. 11. Modelos en el domino de frecuenciasespaciales y temporales: modelo propuesto porWatson y Ahumada (1985).
representación gráfica de objetos en movimiento.Por ejemplo, consideremos para empezar unobjeto elemental, la red sinusoidal. El movimientode una sinusoide unidimensional con velocidadv hacia la izquierda se puede representargráficamente en un plano (x, t) como se ilustraen la figura 9. Una red estacionaria serepresentaría vertical, esto es, en el eje de tiemposno habría periodicidad alguna. El hecho de quela red se desplace hace que aparezca unafrecuencia temporal. En este ejemplo la frecuenciaespacial es 3 ciclos/grado, la frecuencia temporales 4,5 ciclos/segundo y la velocidad es 1,5 º/seg(v=ft/fe).
En el dominio de frecuencias espacio-temporales, el movimiento de esta sinusoide se

141
Formación Continuada
VER Y OÍR - ABRIL 2004
representaría como un par de puntos (+fe, +ft),(-fe, -ft) en cuadrantes opuestos. Esto es así yaque un valor de velocidad positivo implica quelos signos de ambas frecuencias son idénticospero pueden corresponder a dos valores positivoso a dos valores negativos (fig. 10) Si por elcontrario, el movimiento fuera con el mismomódulo pero hacia la derecha, el par de puntoscorrespondientes estaría situado en el segundoy el cuarto cuadrante (cada frecuencia con unsigno).
Supongamos un objeto con un cierto espectroespacial que se mueve con velocidad v hacia laderecha. Este movimiento se representaría en elplano de frecuencias como una línea recta dependiente v negativa (véase de nuevo la figura10), ya que, obviamente, todos los componentesdel espectro del objeto se moverán en tándemcon la misma velocidad y en el mismo sentido.Ahora tenemos que asumir algo muy importante:que en el sistema visual existe un cierto númerode filtros, cada cual sintonizado para una bandade frecuencias espaciales y temporales. Aunqueson posibles distintas hipótesis sobre los filtros, enel modelo que seguimos como ejemplo se suponeque la función de sintonizado espacio-temporales una gaussiana bidimensional, y porconsiguiente, la anchura de banda de cada filtroes la misma en frecuencia espacial que enfrecuencia temporal, o en otras palabras, los filtrosson circulares en el plano (fe, ft). Además,asumiremos que todos los filtros tienen la mismaanchura. Con estas ideas, el movimientoconsiderado se representaría como se ilustra en lafigura 11.
Ya sólo nos queda por admitir que existenpatrones de filtros cada cual sintonizado parauna determinada velocidad. Veamos.Supongamos que tenemos 9 filtros numeradosdel 1 al 9. Por ejemplo, un conjunto derespuestas, r1.....r9, sintonizado para lavelocidad v3, es un patrón en el que los filtros1, 2, 4, 6, 8 y 9 no dan ninguna respuesta. Elfiltro 5 es insensible a cualquier cambio develocidad dado que la sensibilidad espacial delfiltro habíamos quedado en que tiene simetríacircular. Un pequeño cambio de velocidad haceque los filtros 3 y 7 respondan un poco menos,y si seguimos cambiando la velocidad, los filtros3 y 7 responderán progresivamente menos yen un momento dado comenzarían a responderlos filtros adyacentes. De esta forma, tenemosun continuo de patrones de velocidad y, endefinitiva, aquel patrón que dé la máximacorrelación con nuestro movimiento será el quedetermine cual es la velocidad.
A modo de conclusión
Los filtros de los que se hace uso en el modelodescrito encuentran soporte fisiológico en loscampos receptivos orientados espacial ytemporalmente característicos de las células deV1. Este tipo de campo receptivo ha sido muyrecientemente evidenciado de maneraexperimental (De Angelis et al. 1993). La manerade implementar un campo receptivo con estascaracterísticas no es evidente. Sin embargo, debequedar claro que un mecanismo sintonizadopara la orientación espaciotemporal no es lomismo que un mecanismo sintonizado para lavelocidad. Un modelo para el análisis delmovimiento debería hoy incluir dos etapas deprocesado. La primera etapa tendría lugarciertamente en V1, la segunda en MT. Unpromediado local de las respuestas de unconjunto de células simples con una mismaorientación espaciotemporal serviría de inputpara una célula compleja. En la segunda etapa,un mecanismo con selectividad para ciertadirección y módulo de la velocidad resultaríacombinando los outputs de un conjunto decélulas complejas con los campos receptivossintonizados para una orientación espacio-temporal consistente con la velocidad deseada.(Simoncelli y Heeger 1998).
Bibliografía
- Barlow HB, Levick WR. The mechanism of directionally sensitiveunits in rabbit´s retina. J Physiol (Lond) 1965;178: 477-504.- Burr DC, Ross J. Contrast sensitivity at high velocities.Vision Rersearch 1982;22:479-484.- Capilla P. Caminos visuales paralelos I. Especializaciónfuncional. Ver y Oír 2002ª;19:44-54.- Capilla P. Caminos visuales paralelos II. Correlacionesentre la psicofísica y la fisiología. Ver y Oír 2002b;19:597-609.- Chubb C, Sperling G. Drift-balanced random stimuli: ageneral basis for studying( non-Fourier motion perception.J Opt Soc Am A 1988;5:1986-2006.- DeAngelis GC, Ohzawa I, Freeman RD. Spatiotemporalorganization of simple-cell receptive fields in the cat’sstriate cortex I: General characteristics and postnataldevelopment. J Neuro 1993;69:1091-1117.- DeAngelis GC, Ohzawa I, Freeman RD. Spatiotemporalorganization of simple-cell receptive fields in the cat’sstriate cortex II: Linearity of temporal and spatialsummation. J Neuro 1993;69:1118-1135.- Ganz L, Lange A. Changes in motion sensitivity of catvisual cortex neurons during the course of dark adaptation.

142
Formación Continuada
VER Y OÍR - ABRIL 2004
Meeting of the Association for Research in Vision andOphthalmology, Sarasota, Florida 1973.- Hubel DH. Ojo, cerebro y visión. Scientific AmericanLibrary (1995). Traducción del Servicio de Publicacionesde la Universidad de Murcia, 2000.- Movshon JA, Adelson EH, Gizzi MS, Newsome WT. Theanalysis of moving visual patterns. En Chagas C, GattassR, Gross C, (eds.), Pattern recognition mechanisms. NewYork:Springer-Verlag, 1985;117-151.- Murray I, MacCana F, Kulikowski JJ. Contribution oftwo movement detecting mechanisms to central andperipheral vision. Vision Research 1983;2:151-159.- Pantle A. Motion after-effect magnitude as a measureof the spatio-temporal response properties of direction-sensitive analyzers. Vision Research 1974;14:1229-1236.
- Poggio T, Reichardt W. Considerations on modelsof movements detection. Kybernetik 1973;13:223-227.
- Reichardt W. Autocorrelation, a principle for the evaluationof sensory information by the central nervous system. InSensory Communication, de. Rosenblith WA. New Yoek:Wiley 1961.
- Simoncelli EP, Heeger DJ. A model of neuronal responsesin visual area MT. Vision Res 1998;38:743-761.
- Stoner GR, Albright TD. The interpretation of visualmotion: evidence for surface segmentation mechanisms.Vision Research 1996;36:1291-1310.
- Watson AB, Ahumada AJ. Model of human visual-motionsensing. J Opticl Soc Am A 1985;2:322-341.
currículum
Dolores de Fez Saiz, Doctora en Ciencias Físicas por la Universidad de Valencia (orientación óptica). Profesoratitular de la Escuela de Óptica y Optometría de la Universidad de Alicante, donde imparte docencia de lasasignaturas del área de Óptica Fisiológica. Miembro del equipo de investigación GVC: Grupo de Visión y Colorde la Universidad de Alicante.
Pascual Capilla Perea, es licenciado en Ciencias Físicas por la Universitat de València en 1984, Doctor enCiencias Físicas por la Universitat de València en 1990, ayudante de la Escuela Universitaria de la Universidadde Alicante desde el 1 de diciembre de 1986. Titular de Universidad de la Universitat de València en 1995.
Ha sido en distintas ocasiones profesor en las asignaturas de Óptica Fisiológica, Psicofísica de la Visión,Percepción Visual y Colorimetría y Visión del color, todas ellas de la diplomatura de Óptica y Optometría, asícomo de la asignatura Teoría del color, de la Licenciatura en Física.
Es co-autor de los libros Problemas de Óptica Fisiológica, Óptica Fisiológica, Psicofísica de la Visión, Fundamentosde Colorimetría y Tecnología del color.
Como investigador, ha dirigido 4 Tesis Doctorales, 2 Tesinas de Licenciatura y 4 de Diplomatura. Es autor de28 publicaciones en revistas de investigación nacionales e internacionales y de 9 publicaciones en revistas dedivulgación científica. Ha presentado 53 comunicaciones en congresos nacionales e internacionales.
Es miembro de la Sociedad Española de Óptica


















