
Volumen 2 / No. 2 / Julio-Diciembre 2008 - :::SCCH ... no.2/Revista_SCC… · Efecto de la...
124
Transcript of Volumen 2 / No. 2 / Julio-Diciembre 2008 - :::SCCH ... no.2/Revista_SCC… · Efecto de la...

Volumen 2 / N
o. 2 / Julio-Diciem
bre 2008Revista C
olombiana de C
iencias Hortícolas
Editorial 131
Seccióndefrutales
Efectodelaaplicaciónprecosechadegiberelinasycalcioenlaproducción,calidadyrajadodelfrutodeuchuva(Physalis peruvianaL.)Effect of preharvest application of gibberellins and calcium in production, fruit quality and cracking of cape gooseberry (Physalis peruviana L.)
NathalyAmézquita,HelberEnriqueBalaguera-LópezyJavierGiovanniÁlvarez-Herrera 133
Aspectosanatómicosdelasemilladepapaya(Carica papayaL.)Anatomical aspects of papaya (Carica papaya L.) seed
ArletteIvonneGilyDiegoMiranda 145
Comportamientode26cultivaresdenaranjaencondicionesdelpiedemontedelMeta,ColombiaPerformance of 26 orange cultivars in conditions of Piedmont of Meta, Colombia
JavierOrlandoOrduz-RodríguezyFranciscoAvella 157
EfectodelencalamientodesuelossulfatadosácidosenCórdoba(Colombia)sobreelfrutodepiña(Ananas comosus(L.)Merr.)Effect of liming of acid sulfate soils in Cordoba (Colombia) on pineapple (Ananas comosus (L.) Merr.) fruitEnriqueCombattC.,AlfredoJarmaO.yAlejandroClevesL. 175
Seccióndehortalizas
Acumulacióndemetalespesadosensueloyplantasdecuatrocultivoshortícolas,regadosconaguadelríoBogotáAccumulation of heavy metals in soil and plants of four vegetable crops irrigated with water of Bogota river DiegoMiranda,CarlosCarranza,CarlosAndrésRojas,CarlosMartínJerez,
GerhardFischeryJorgeZurita 180
Caracterizacióndelasdeficienciasdemacronutrientesenplantasdecebollín(Allium schoenoprasum L.)Characterization of macronutrient deficiencies in chives (Allium schoenoprasum L.) plantsDiegoAndrésBernal,LeidyCarolinaMorales,GerhardFischer,
JairoCuervoyStanislavMagnitskiy 192
EfectodelaaplicacióndecachazafrescaydedossistemasdeproducciónmaízymaízenasocioconfríjolsobrelaspropiedadesfísicasdeunInceptisolEffect of application of fresh filter cake and two production systems of corn and corn associated with beans on the physical properties of an Inceptisol
FabioEmilioForero,JaimeTorresyHelberEnriqueBalaguera-López. 205
Seccióndeornamentales
Determinacióndelmomentodecortedelaflorenlimonio(Limoniumsp.)apartirdeseisestadosfenológicosDetermination of timing of flower cut in sea lavender (Limonium sp.) starting from six phenological stagesFánorCasierra-PosadayJuanCarlosAbril 217
Efectodelsustratoytamañodelpropáguloenelenraizamientodelgingerrojo(Alpinia purpurata)Effect of substrate type and cutting size on red ginger (Alpinia purpurata) rootingIsidroE.Suárez,GledyLuzMarrugoyMaryorikPeña 225
Artículoderevisión
ConsideracionessobreladispersióndelabrocadelcaféHypothenemus hampeienrelaciónconelempleodetrampasconatrayente.UnarevisiónConsiderations about dispersion of the coffee berry borer Hypothenemus hampei related to the use of attractan-baited traps. A reviewDanielRodríguez,JoséR.CureH.yFernandoCantorR. 232
Políticaeditorialeinstruccionesalosautores 240


PRESIDENTEDiego Miranda
COMITÉ EDITORIAL
COMITÉ CIENTÍFICO
VICEPRESIDENTEMaría Soledad Hernández
EDITORFánor Casierra–Posada
COEDITORGerhard Fischer
Marco CabezasBernardo Chaves
Miguel EspitiaRebecca Lee
Nelson RodríguezEdison Valencia Kris Wyckhuys
SOCIEDAD COLOMBIANA DE CIENCIAS HORTÍCOLAS (SCCH)BOGOTÁ – COLOMBIA
Miguel A. AltieriUniversity of California, Berkely (Estados Unidos)
Galdino Andrade FilhoUniversidade Estadual de Londrina (Brasil)
Raúl CabreraTexas A+M University (Estados Unidos)
Ana María CastagninoUniversidad Nacional del Centro de la Provincia de Buenos Aires (Argentina)
Daniel H. DíazLaboratorios Agroenzimas (México)
Georg EbertKali + Salz Gruppe (Alemania)
José Miguel GuzmánUniversidad de Almería (España)
Miguel JordánUniversidad Mayor, Santiago (Chile)
Jairo Antonio OsorioCorporación Colombiana de Investigación Agropecuaria (Colombia)
Marcelo Francisco PompelliUniversidade Federal de Pernambuco (Brasil)
Ricardo RamírezUniversity of Guelph (Canadá)
Philip A. StanslyUniversity of Florida (Estados Unidos)

© 2008 Revista Colombiana de Ciencias HortícolasSociedad Colombiana de Ciencias HortícolasISSN: 2011-2173
INFORMACIÓN, CORRESPONDENCIA Y CANJE
Sociedad Colombiana de Ciencias HortícolasFacultad de Agronomía Universidad Nacional de ColombiaA.A. 14490, Bogotá, ColombiaTeléfono: 571 - 316 5000 Ext. 19041/19043Tel/fax: 571 - 316 [email protected], [email protected]
ASISTENTE EDITORIALJulián F. Cárdenas-Hernández
CORRECTOR DE ESTILO EN INGLÉSStanislav Magnitskiy
DISEÑO GRÁFICO Y ARMADA ELECTRÓNICAEquilibrio gráfico/editorial [email protected]
IMPRESIÓNOffset Gráfico Editores S.A.
Bogotá, ColombiaDiciembre de 2008
La presente publicación es de carácter científico (artículo 4, Resolución 1508, Ministerio de Cultura, octubre 3 de 2000).
A nivel internacional, la Revista Colombiana de Ciencias Hortícolas está integrada en las bases de datos de CAB Abstracts y AGRIS (FAO).

JUNTA DIRECTIVA DE LA SOCIEDAD COLOMBIANA DE CIENCIAS HORTÍCOLAS (SCCH)
Gerhard FischerUniversidad Nacional de Colombia, Bogotá
Wilson de Jesús Piedrahita Universidad Nacional de Colombia, Bogotá
VOCALES
Diego MirandaUniversidad Nacional de Colombia, Bogotá
María Soledad HernándezInstituto Sinchi, Bogotá
Marlon Hans Rodríguez Sociedad Colombiana de Ciencias Hortícolas
Alfredo JarmaUniversidad de Córdoba, Monterí[email protected]
Hernán PinzónAsesor Hortícola, Bogotá [email protected]
Stanislav MagnitskiyUniversidad Nacional de Colombia, Bogotá
PRESIDENTE VICEPRESIDENTE SECRETARIO EJECUTIVO TESORERO
Luis Enrique FlórezUniversidad de Bogotá
Jorge Tadeo [email protected]
FISCAL
Fánor Casierra–PosadaUniversidad Pedagógica y Tecnológica
de Colombia, [email protected]
EDITOR REVISTA


CONTENIDOVolumen 2 - No. 2 - 2008
REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS
PÁG.
Editorial 131
Sección de frutales
Efecto de la aplicación precosecha de giberelinas y calcio en la producción, calidad y rajado del fruto de uchuva (Physalis peruviana L.)Effect of preharvest application of gibberellins and calcium in production, fruit quality and cracking of cape gooseberry (Physalis peruviana L.)
Nathaly Amézquita, Helber Enrique Balaguera-López y Javier Giovanni Álvarez-Herrera ...................................................................... 133
Aspectos anatómicos de la semilla de papaya (Carica papaya L.)Anatomical aspects of papaya (Carica papaya L.) seed
Arlette Ivonne Gil y Diego Miranda ...................................................................................................................................................... 145
Comportamiento de 26 cultivares de naranja en condiciones del piedemonte del Meta, ColombiaPerformance of 26 orange cultivars in conditions of Piedmont of Meta, Colombia
Javier Orlando Orduz-Rodríguez y Francisco Avella .............................................................................................................................. 157
Efecto del encalamiento de suelos sulfatados ácidos en Córdoba (Colombia) sobre el fruto de piña (Ananas comosus (L.) Merr.)Effect of liming of acid sulfate soils in Cordoba (Colombia) on pineapple (Ananas comosus (L.) Merr.) fruit
Enrique Combatt C., Alfredo Jarma O. y Alejandro Cleves L. ................................................................................................................ 175
Sección de hortalizas
Acumulación de metales pesados en suelo y plantas de cuatro cultivos hortícolas, regados con agua del río Bogotá Accumulation of heavy metals in soil and plants of four vegetable crops irrigated with water of Bogota river
Diego Miranda, Carlos Carranza, Carlos Andrés Rojas, Carlos Martín Jerez, Gerhard Fischer y Jorge Zurita ....................................... 180
Caracterización de las deficiencias de macronutrientes en plantas de cebollín (Allium schoenoprasum L.) Characterization of macronutrient deficiencies in chives (Allium schoenoprasum L.) plants
Diego Andrés Bernal, Leidy Carolina Morales, Gerhard Fischer, Jairo Cuervo y Stanislav Magnitskiy ................................................. 192

REV. COLOMB. CIENC. HORTÍC.
130
Efecto de la aplicación de cachaza fresca y de dos sistemas de producción maíz y maíz en asocio con fríjol sobre las propiedades físicas de un InceptisolEffect of application of fresh filter cake and two production systems of corn and corn associated with beans on the physical properties of an Inceptisol
Fabio Emilio Forero, Jaime Torres y Helber Enrique Balaguera-López . ................................................................................................. 205
Sección de ornamentales
Determinación del momento de corte de la flor en limonio (Limonium sp.) a partir de seis estados fenológicosDetermination of timing of flower cut in sea lavender (Limonium sp.) starting from six phenological stages
Fánor Casierra-Posada y Juan Carlos Abril ........................................................................................................................................... 217
Efecto del sustrato y tamaño del propágulo en el enraizamiento del ginger rojo (Alpinia purpurata)Effect of substrate type and cutting size on red ginger (Alpinia purpurata) rooting
Isidro E. Suárez, Gledy Luz Marrugo y Maryorik Peña .......................................................................................................................... 225
Artículo de revisión
Consideraciones sobre la dispersión de la broca del café Hypothenemus hampei en relación con el empleo de trampas con atrayente. Una revisión Considerations about dispersion of the coffee berry borer Hypothenemus hampei related to the use of attractan-baited traps. A review
Daniel Rodríguez, José R. Cure H. y Fernando Cantor R. ...................................................................................................................... 232
Política editorial e instrucciones a los autores 240

EDITORIALFÁNOR CASIERRA–POSADADirector Comité Editorial
Revista Colombiana de Ciencias Hortícolas
La Revista de la Sociedad Colombiana de Ciencias Hortícolas se creó, entre otros, con el ánimo de constituir un espacio de referencia de la investigación científica en el campo de la horticultura tropical. En la actualidad, la distribución de esta publicación a los afiliados a nuestra sociedad y las bibliotecas, a nivel nacional e internacional, es un éxito para la horticultura y la comunidad científica. Es así, como la revista se ha consolidado como un espacio de divulgación y desarrollo de nuevas ideas e iniciativas para nuestra comunidad.
En la entrega actual ofrecemos a nuestros lectores cuatro secciones, dedicadas a fru-tales, hortalizas, ornamentales y a textos de revisión. En la primera sección se pro-ponen alternativas viables para contrarrestar el rajado de los frutos de uchuva, fruto de gran importancia para el mercado de exportación. Se hace también un estudio detallado de la anatomía de las semillas de papaya. Se presenta el reporte de un estudio realizado durante cerca de diez años, en el que se llevaron registros del com-portamiento de 26 cultivares de naranja. Para cerrar esta sección, se presentan los resultados de un trabajo en el que se evaluó la calidad del fruto en plantas de piña que crecían en suelos sulfatados ácidos que habían sido encalados.
En lo relacionado con hortalizas, publicamos un estudio de la presencia de metales pesados en el suelo y en las plantas, cuando se tiene al río Bogotá como fuente de agua para riego. Se presenta un cuadro de síntomas de deficiencias de macronutrien-tes en plantas de cebollín, lo cual se constituye en un método de diagnóstico muy práctico para los cultivadores y para los técnicos. También se pone a consideración de nuestros lectores, el resultado de un ensayo en el que se evaluaron los beneficios al suelo, de la utilización de la cachaza fresca.
Los expertos y cultivadores de limonio y ginger rojo se ven favorecidos con la presen-tación de sendos trabajos en la sección dedicada a ornamentales. En uno de ellos se dan orientaciones relacionadas con el punto de corte de la flor de limonio en relación con el estadio fenológico en el que se encuentran las plantas. En el otro artículo se presentan resultados relevantes conducentes a la selección del material de propaga-ción en ginger rojo.
Vol. 2 - No.2 - 2008

REV. COLOMB. CIENC. HORTÍC.
132
En vista de que nuestra sociedad y, por tanto, su órgano de divulgación son abiertos al intercambio científico con otras agremiaciones diferentes a aquellas que guardan estre-cha relación con la horticultura, se presenta, en la sección de artículos de revisión, un texto sobre la dispersión de Hypothenemus hampei, coleóptero conocido por ser la plaga que más daño causa a los cultivos de café en el mundo.
Finalmente, es importante destacar, como actividad de nuestra sociedad, la exitosa celebración del Simposio Internacional sobre Tomate en el Trópico, evento celebrado en Villa de Leyva, en septiembre del presente año. A la convocatoria acudieron alrededor de 200 participantes de diferentes países y fue un importante escenario de intercambio entre cultivadores, académicos y el sector comercial, que seguramente redundará en beneficios para todos los sectores que se aglutinan alrededor de este cultivo.

Nathaly amézquita1 helber eNrique balaguera-lópez1
Javier giovaNNi Álvarez-herrera1, 2
reSumeN
La uchuva actualmente es la fruta exótica de mayor exportación en Colombia. Uno de los principales proble-mas fisiológicos que afecta la calidad y la rentabilidad del cultivo de uchuva es el rajado del fruto, además, se hace necesario aumentar la productividad y la calidad del fruto para hacerlo más competitivo en el mercado. Por tanto, en un diseño completamente al azar se evaluó el efecto de la aplicación en precosecha vía foliar de siete tratamientos correspondientes a diferentes concentraciones de giberelinas (5, 10 ó 15 mg L-1) y de calcio (0,5; 1,0 ó 1,5 g L-1) y un testigo absoluto sobre la producción, calidad y rajado del fruto de uchuva. Las aplicaciones de los tratamientos se realizaron a los 3, 10, 17 y 24 días después de antesis. Las aplicaciones foliares de calcio y giberelinas aumentaron significativamente la producción total de uchuva, pero no hubo diferencias en la calidad extra y calidad I. Se presentó menor porcentaje de rajado de frutos con 10 mg L-1 de giberelinas, seguido del tratamiento de 0,5 g L-1 de calcio. La aplicación de 0,5 g L-1 de calcio generó mayor firmeza y contenido de sólidos solubles totales en los frutos, mientras que todos los tratamientos fueron estadísticamente superiores al testigo en la acidez total titulable e inferiores en la relación de madurez, esto indica que las giberelinas y el calcio retrazan la madurez de los frutos de uchuva y potencialmente pueden tener mayor vida en poscosecha.
1 Grupo de Investigaciones Agrícolas, Facultad de Ciencias Agropecuarias, Universidad Pedagógica y Tecnológica de Colombia, Tunja (Colombia).
2 Autor para correspondencia. [email protected]
Efecto de la aplicación precosecha de giberelinas y calcio en la producción, calidad y rajado del fruto de uchuva (Physalis peruviana L.)
Effect of preharvest application of gibberellins and calcium on production, fruit quality and cracking of cape gooseberry (Physalis peruviana L.)
REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 2 - No.2 - pp. 133-144, 2008
Fruto de uchuva en su punto óptimo de cosecha.Foto: G. Fischer
palabras clave adicionales: firmeza, sólidos solubles totales, acidez total titulable, relación de madurez.

REV. COLOMB. CIENC. HORTIC.
134
iNtroDuCCióN
additional key words: firmness, total soluble solids, total titratable acidity, maturity ratio.
Fecha de recepción: 21-10-2008 Aprobado para publicación: 02-12-2008
abStraCt
Cape gooseberry is currently the main exotic fruit of export in Colombia. Fruit cracking is one of the main physiological problems that affect the quality and rentability of cape gooseberry crop; also it is necessary to increase the productivity and fruit quality to make this plant more competitive in the market. Therefore, in a completely randomized design, the effect of preharvest foliar application of seven treatments corresponding to different concentrations of gibberellins (5, 10 or 15 mg L-1) and calcium (0.5, 1.0 or 1.5 g L-1) and a control was evaluated on the production, quality and fruit cracking of cape gooseberry. The applications of treatments were done at 3, 10, 17 and 24 days after anthesis. Foliar applications of calcium and gibberellins significantly increased the total production of cape gooseberry, but no differences in the quality extra and quality I were observed. It was presented a low percentage of fruit cracking with 10 mg L-1 gibberellins followed by treatment of 0.5 g L-1 calcium. The application of 0.5 g L-1 calcium generated higher firmness and higher soluble solids content in fruit, while all treatments were statistically higher than control in the total acidity and lower in terms of maturity ratio. This suggests that gibberellins and calcium delay the maturity of the fruits of cape gooseberry and potentially contribute longer post-harvest life.
AMézquITA/BALAGuERA-LópEz/ÁLVAREz-HERRERA
La uchuva (Physalis peruviana L.) es la fruta exótica de mayor exportación en Colombia. Para el 2008, el país contó con una producción de 15.463 t, 841 ha cosechadas y una tasa de crec-imiento anual de 42,6% y 33,7% para la produc-ción y el área cosechada respectivamente, donde los departamentos de mayor producción en or-den fueron: Boyacá, Cundinamarca y Antioquia (Agronet, 2008). Estas importantes cifras jus-tifican la necesidad de continuar mejorando el nivel tecnológico del cultivo para aumentar la producción, la productividad, la calidad del fruto y, por ende, la rentabilidad del cultivo, todo esto mediante la investigación y desarrollo de nuevas técnicas de manejo.
Uno de los principales problemas fisiológicos que afectan la calidad y la rentabilidad del cultivo de uchuva es el rajado de los frutos. Esta anomalía hace que en cosecha y en poscosecha se pierda más de 30% de la producción (Gordillo et al.,
2004) y hasta 50% de los frutos son rechazados por el exportador en la época de lluvia (Fischer, 2005), lo que arroja pérdidas tanto para los pro-ductores de fruta como para los comercializado-res y consumidores.
El rajado de los frutos, se debe a una serie de factores intrínsecos y extrínsecos. Aunque el mecanismo por el cual se genera el rajado no es completamente conocido, se ha sugerido que es un fenómeno osmótico (Sekse, 1995; Pratella, 1996), acompañado por pérdida de la integridad de las membranas celulares; por consiguiente, concentraciones de calcio que generen mayor firmeza en el fruto, contribuirán a disminuir el rajado. El rajado también puede darse debido a que el parénquima de los frutos de uchuva no es compacto y presenta numerosas “lagunas”, cuyo número y tamaño aumenta a medida que el fruto madura (Valencia, 1985); el grado de co-hesión celular está directamente relacionado con

Vol. 2 - No.2 - 2008
135EFECTO DE LA ApLICACIóN pRECOSECHA DE GIBERELINAS y CALCIO EN EL FRuTO DE uCHuVA
la capacidad de retención de líquidos dentro de la pulpa de los frutos y, por tanto, con las fuerzas de presión en el interior de los mismos (Opara, 1997). Del mismo modo, los rendimientos po-tenciales del cultivo y la calidad en la cosecha se ven disminuidas por dosis inadecuadas de Ca y boro (Cooman et al., 2005).
En tomate, se deben evitar cambios bruscos de humedad en el suelo durante la época de cosecha, ya que si esto ocurre, se presentan rajaduras en los frutos (Peet, 1992). Los incrementos súbitos en la humedad del suelo disminuyen la fuerza de ten-sión normal de la epidermis del fruto, reduciendo su fuerza de elasticidad y haciendo que ésta se agrande rápidamente y desarrolle el rajado (Gor-dillo et al., 2004). Sin embargo, mediante la apli-cación foliar de calcio se logra aumentar la rigidez celular y con giberelinas es posible incrementar la extensibilidad y elasticidad de la célula, así como expresar una modificación de la estructura de la pared celular (Podestá et al., 2001), factores que pueden amortiguar el efecto negativo de los cam-bios bruscos de humedad en el suelo y disminuir el rajado del fruto de uchuva.
El Ca está implicado en varias funciones impor-tantes dentro de la planta, principalmente en la estabilización y rigidez de membranas y paredes celulares (Marschner, 2002). Esto se da porque en la pared celular el calcio forma enlaces con los grupos carboxilos en cadenas de poligalac-turonanos adyacentes presentes en la lámina media de las paredes celulares, esto contribuye a la adhesión y cohesión célula-célula (Hernán-dez-Muñoz et al., 2008). A nivel de membranas, el calcio genera estabilidad al formar enlaces con los fosfolípidos y proteínas mediante los puentes fosfatos y carboxílicos, preferencialmente en la superficie de membranas (Marschner, 2002).
El calcio, por su capacidad para disminuir la per-meabilidad hidráulica de las membranas celula-res, reducir la absorción de agua y aumentar la firmeza de la pulpa en algunas especies (Brown et al., 1996), es el factor nutricional más estudia-
do respecto al rajado de los frutos (Huang et al., 2005). Brown et al. (1996) proponen que la de-ficiencia de calcio está asociada con el rajado en cerezas, sin embargo, Podestá et al. (2001) encon-traron que su aplicación precosecha no aumentó la resistencia al rajado. Respecto a la aplicación foliar de calcio, fue reportado que reduce el ra-jado en manzanas, duraznos, naranjas, peras, ciruelas e higos, mientras que hay reportes que no muestran efecto significativo en melón, uva y cerezas (Xu-Ming et al., 2008).
Las giberelinas están asociadas, entre otros, pro-cesos, con la división y elongación celular (Taiz y Zeiger, 2006), la iniciación floral, desarrollo de los órganos florales, desarrollo de la fructifica-ción e inducción de algunas enzimas hidrolíticas (Matsuoka, 2003). Según Taiz y Zeiger (2006), las giberelinas pueden favorecer el cuajado y crecimiento de algunos frutos como manzanas, además promueven el crecimiento celular debido a que incrementan la hidrólisis de almidón, fruc-tosa y sacarosa, con lo que se originan moléculas de fructosa y glucosa, que contribuyen a la for-mación de la pared celular. La información sobre la acción de las giberelinas en el rajado de frutos no es consistente, algunos resultados indican que disminuyó el rajado (Looney y Lidster, 1980); sin embargo, Cline (1995, citado por Sansavini y Lu-gli, 1997) señala que no tuvo acción eficaz sobre este desorden fisiológico.
Para aumentar el tamaño de los frutos se han utilizado comercialmente aplicaciones foliares de ácido giberélico (Facteau et al., 1992). Tam-bién se ha encontrado que esta hormona retrasa la maduración, aumenta la firmeza de la pulpa (Looney y Lidster, 1980; Facteau et al., 1992) e incrementa del contenido de sólidos solubles (Facteau et al., 1992).
Peet (1992) constata que todas las prácticas cul-turales que producen uniformidad y crecimiento relativamente lento ofrecen alguna protección contra el rajado del fruto. Esto significa que se de-ben evitar prácticas culturales que aumenten en

REV. COLOMB. CIENC. HORTIC.
136
exceso el crecimiento del fruto, sea un riego o una fertilización demasiado abundante o una aplica-ción de hormonas de crecimiento no adecuada.
Por tanto, el objetivo de esta investigación fue evaluar el efecto de la aplicación foliar en preco-secha de diferentes concentraciones de gibereli-nas y calcio sobre la producción, calidad y rajado de frutos de uchuva.
materialeS y métoDoS
El experimento se realizó en una plantación co-mercial de uchuva en la finca El Saque, localiza-da en la Vereda San Antonio, en el municipio de Tuta, Boyacá, que se encuentra a 2.600 msnm, 5°41’36’’ N y 73°13’51’’ W. Este municipio cuenta con una temperatura promedio de 14°C, hume-dad relativa del 80% y una precipitación prome-dio anual de 935 mm.
Plantas de la Physalis peruviana, ecotipo Colom-bia, se sembraron con distancias de 2 x 2 m, en sistema de espaldera, conducidas en vaso abierto. A los 80 d después de la siembra, una vez las flo-res entraron en antesis, se seleccionaron las plan-tas a utilizar, las cuales presentaron condiciones homogéneas en función del porte, diámetro de tronco y altura de planta. A estas plantas se les realizaron las labores culturales típicas para la zona, el riego se suministró mediante goteo y la fertilización edáfica se hizo con base en el resul-tado del análisis de suelos (tabla 1). Al momento de la siembra se aplicó 200 g/planta del la fórmu-la 10-20-20 + 30 g/planta de Nutrimins (N 8%; P2O5 5%; CaO 18%; MgO 6%; S 1,6%; Cu 0,14%; B 1%; Mo 0,005% y Zn 2,5%) + 50 g/planta de micorrizas + 2 kg/planta de materia orgánica (Abimgra). A los 6 meses se fertilizó en corona con 100 g/planta de 15-15-15 + 50 g/planta de Nitrabor (N 15,4%; CaO 25,6%; B 0,3%) y 50 g/planta de Nutrimins.
Se utilizó un diseño completamente al azar con siete tratamientos correspondientes a la aplica-
ción de cuatro concentraciones de giberelinas (GA3) (5, 10 ó 15 mg L-1) y cuatro concentracio-nes de calcio (0,5; 1,0 ó 1,5 g L-1) y un testigo absoluto sin aplicación. Cada tratamiento tuvo tres repeticiones, para un total de 21 unidades experimentales (UE). Cada UE estuvo compues-ta por tres plantas de uchuva. Los tratamientos fueron aplicados vía foliar con fumigadora de es-palda de 20 L a los 3, 10, 17 y 24 d después de an-tesis. La fuente de calcio utilizada fue Ca(NO3)2, mientras que para las giberelinas se utilizó ácido giberélico al 10% (Progibb, Bayer CropScience, Bogotá).
La cosecha de los frutos de uchuva se llevó a cabo en el punto de madurez recomendado por la Norma Técnica Colombiana NTC 4580 (Icon-tec, 1999): color 5 y 6 (frutos de color anaranja-do y anaranjado intenso). Una vez realizada la cosecha se midieron las siguientes variables: (1) Producción por calidades comerciales (kg ha-1) de c/u de las calidades comerciales según norma NTC 4580, calidad extra, calidad I y calidad II; (2) Producción (kg ha-1) como sumatoria de la producción obtenida en c/u de las calidades co-merciales; (3) Frutos rajados (%): se calculó el porcentaje con base en el peso de todos los frutos
propiedad Valor unidadespH 5,62
Ca 6,22 cmol kg-1 de suelo
Mg 0,24 cmol kg-1 de suelo
K 0,46 cmol kg-1 de suelo
Na 0,26 cmol kg-1 de suelo
P 141,90 mg kg-1 de suelo
Fe 431,63 mg kg-1 de suelo
Cu 0,88 mg kg-1 de suelo
Zn 2,35 mg kg-1 de suelo
Mn 2,96 mg kg-1 de suelo
S 19,40 mg kg-1 de suelo
B 0,56 mg kg-1 de suelo
CIC 7,27 cmol kg-1 de suelo
CE 0,25 dS m-1
tabla 1. propiedades fisicoquímicas del suelo antes de la siembra.
AMézquITA/BALAGuERA-LópEz/ÁLVAREz-HERRERA
Vol. 2 - No.2 - 2008
137
que presentaron los síntomas del rajado durante la cosecha por cada calidad comercial.
Se realizó la evaluación de las variables de ca-lidad de fruto, estas fueron: (1) Acidez total ti-tulable (ATT) mediante cálculos con datos de volumen de NaOH, incorporado en 5 g de jugo de frutos agregando tres gotas de fenoftaleína en una titulación potenciométrica hasta pH de 8,2; (2) Sólidos solubles totales (SST) a través de mediciones de grados Brix con un refractómetro manual marca Atago; (3) Relación de madurez (RM) se determinó mediante la relación entre los SST y la ATT; (4) Firmeza del fruto (N) me-diante la utilización de un penetrómetro digital PCE-PTR200.
Con los datos obtenidos se realizó un análisis de varianza y se hizo una prueba de comparación de promedios de Tukey al 5%. Para el análisis de los datos se utilizó el programa SAS v. 8e.
reSultaDoS y DiSCuSióN
producción
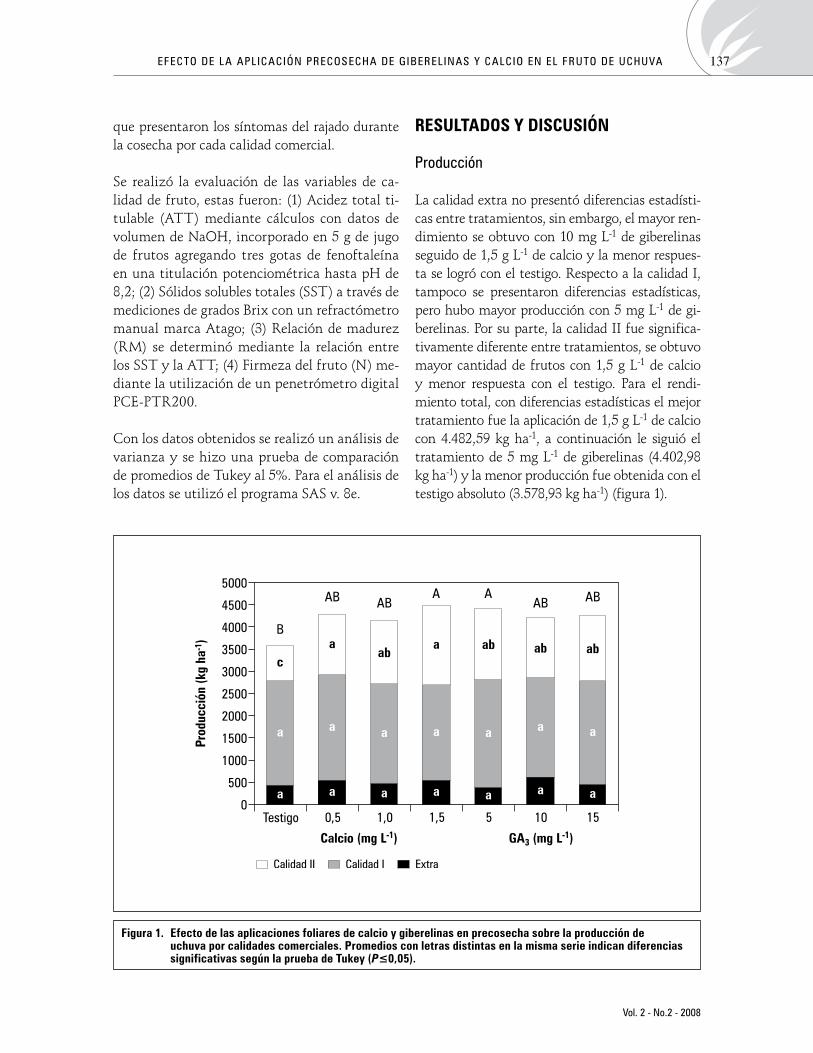
La calidad extra no presentó diferencias estadísti-cas entre tratamientos, sin embargo, el mayor ren-dimiento se obtuvo con 10 mg L-1 de giberelinas seguido de 1,5 g L-1 de calcio y la menor respues-ta se logró con el testigo. Respecto a la calidad I, tampoco se presentaron diferencias estadísticas, pero hubo mayor producción con 5 mg L-1 de gi-berelinas. Por su parte, la calidad II fue significa-tivamente diferente entre tratamientos, se obtuvo mayor cantidad de frutos con 1,5 g L-1 de calcio y menor respuesta con el testigo. Para el rendi-miento total, con diferencias estadísticas el mejor tratamiento fue la aplicación de 1,5 g L-1 de calcio con 4.482,59 kg ha-1, a continuación le siguió el tratamiento de 5 mg L-1 de giberelinas (4.402,98 kg ha-1) y la menor producción fue obtenida con el testigo absoluto (3.578,93 kg ha-1) (figura 1).
EFECTO DE LA ApLICACIóN pRECOSECHA DE GIBERELINAS y CALCIO EN EL FRuTO DE uCHuVA
Figura 1. efecto de las aplicaciones foliares de calcio y giberelinas en precosecha sobre la producción de uchuva por calidades comerciales. promedios con letras distintas en la misma serie indican diferencias significativas según la prueba de tukey (P≤0,05).

REV. COLOMB. CIENC. HORTIC.
138
Los resultados indican que el calcio y las gibere-linas influyen sobre la producción de frutos de uchuva. El calcio aumentó el rendimiento debido posiblemente a que es un elemento esencial para las plantas, indispensable para la formación del tubo polínico (Barker y Pilbeam, 2007), consti-tuye la lámina media, las paredes y membranas de la célula, participa en la división y extensión celulares, influye en la compartimentalización celular, modula la acción de hormonas y seña-les, y contribuye al equilibrio iónico de la célula (Marschner, 2002); además, participa en la dis-minución de la respiración del fruto y en el au-mento de la fotosíntesis de la planta, procesos que en conjunto pueden aumentar la productivi-dad de los cultivos. Resultados similares fueron obtenidos con la aplicación de nitrato de calcio en melón, pues se aumentó el número y el peso de los frutos por planta y el diámetro polar y ecuatorial del fruto (Román y Gutiérrez, 1998).
La producción favorable que fue obtenida con la aplicación de giberelinas, principalmente en calidad extra, se explica probablemente porque esta hormona aumenta la divisón y elongación celular y la entrada de agua a las células (Taiz y Zeiger, 2006), procesos que se traducen en aumento del tamaño final del fruto (Grange, 1996) y mayor acumulación de masa fresca, lo cual concuerda con los resultados obtenidos por Usenik et al. (2005), quienes demostraron que la aplicación de ácido giberélico en frutos de cereza aumentó el rendimiento y la calidad de estos. En uva también existen reportes de que las gibe-relinas aumentaron los rendimientos (Botelho et al., 2004; Vieira et al., 2008). Por el contrario, Paroussi et al. (2002) encontraron que al apli-car diferentes concentraciones de giberelinas en fresa, no se presentó un efecto positivo sobre el rendimiento.
Rajado
No se observaron diferencias estadísticas en el porcentaje de rajado para la calidad extra, sin em-
bargo, todos los frutos con aplicaciones de calcio y giberelinas mostraron un menor rajado que el testigo, principalmente con el tratamiento de 10 mg L-1 de giberelinas. En la calidad I el rajado fue estadísticamente diferente, en este caso, en los frutos del testigo absoluto se presentó el menor porcentaje de rajado, mientras que en la calidad II la respuesta fue contraria pero sin diferencias estadísticas. El porcentaje de rajado en frutos de uchuva es más alto a medida que los frutos son más grandes (Fischer, 2005), razón por la cual, se presentó mayor rajado en la calidad extra, segui-do de la calidad I y por último la calidad II. Para el porcentaje total de rajado, no hubo diferencias estadísticas, no obstante, con 10 mg L-1 de gi-berelinas el rajado de frutos fue menor pero se presentó mayor rajado con 1,5 g L-1 de calcio aún con más rajado que el testigo (figura 2).
Este último resultado no se esperaba, pues se ha mencionado que el calcio incrementa la firmeza del fruto al darle mayor estabilidad y rigidez a pa-redes y membranas celulares (Marschner, 2002) y esto aumenta la resistencia al rajado (Brown et al., 1996); sin embargo, la propiedad del calcio de reducir la extensibilidad de la pared en las di-cotiledóneas (Taiz y Zeiger, 2006) podría ser un factor crítico relacionado con la sensibilidad al rajado, lo cual también fue encontrado en cere-zas (Podestá et al., 2001). Es importante aclarar que este resultado desfavorable se debió a un alto porcentaje de rajado en las calidades I y II, y que las demás dosis de calcio sí lograron disminuir el rajado con relación al testigo.
Existe evidencia de que en frutos de Litchi chi-nensis el contenido de calcio está implicado en la resistencia al rajado, pues el pericarpio de frutos rajados tuvo significativamente más bajas con-centraciones de calcio que frutos normales, esta situación se observó en el mismo árbol y aun en la misma panícula (Lin, 2001), lo cual se debe posiblemente a la diferencia en la actividad me-tabólica entre frutos y a las auxinas exportadas basipetalamente (Xu-Ming et al., 2008).
AMézquITA/BALAGuERA-LópEz/ÁLVAREz-HERRERA

Vol. 2 - No.2 - 2008
139
De acuerdo con Marschner (2002), el Ca permi-tiría una menor disgregación de los tejidos debi-do a que inhibe la síntesis de poligalacturonasa, responsable de la degradación de los pectatos de calcio. Según resultados experimentales obteni-dos por Meheriuk et al. (1991) la aplicación fo-liar de Ca redujo hasta en un 75% el rajado en cerezas. En concordancia, compuestos de calcio pulverizado aplicados durante la precosecha en cerezas pueden favorecer la firmeza de los frutos y su resistencia al rajado (Podestá et al., 1998).
Ben-Airie et al. (1986) afirman que la aplicación de giberelinas reduce el metabolismo respirato-rio y la producción de etileno, lo que disminuye la síntesis de enzimas hidrolíticas. Al respecto, Andrews y Shulin (1995) encontraron que las giberelinas disminuyen la actividad de la poliga-lacturonasa y pectinmetilesterasa, enzimas hi-drolíticas que se encuentran en la pared celular y que aumentan el rajado del fruto, lo cual podría
explicar la disminución del rajado en los frutos de uchuva con la aplicación de giberelinas. Una alta firmeza producida por la aplicación de 10 mg L-1 de giberelinas podría expresar una modificación de la estructura de la pared celular que aumente la resistencia al rajado. Se ha sugerido que el au-mento en la firmeza producida por las aplicacio-nes de giberelinas podría deberse a algún efecto del regulador de crecimiento sobre la estructura de la pared celular (Andrews y Shulin, 1995).
Sólidos solubles totales
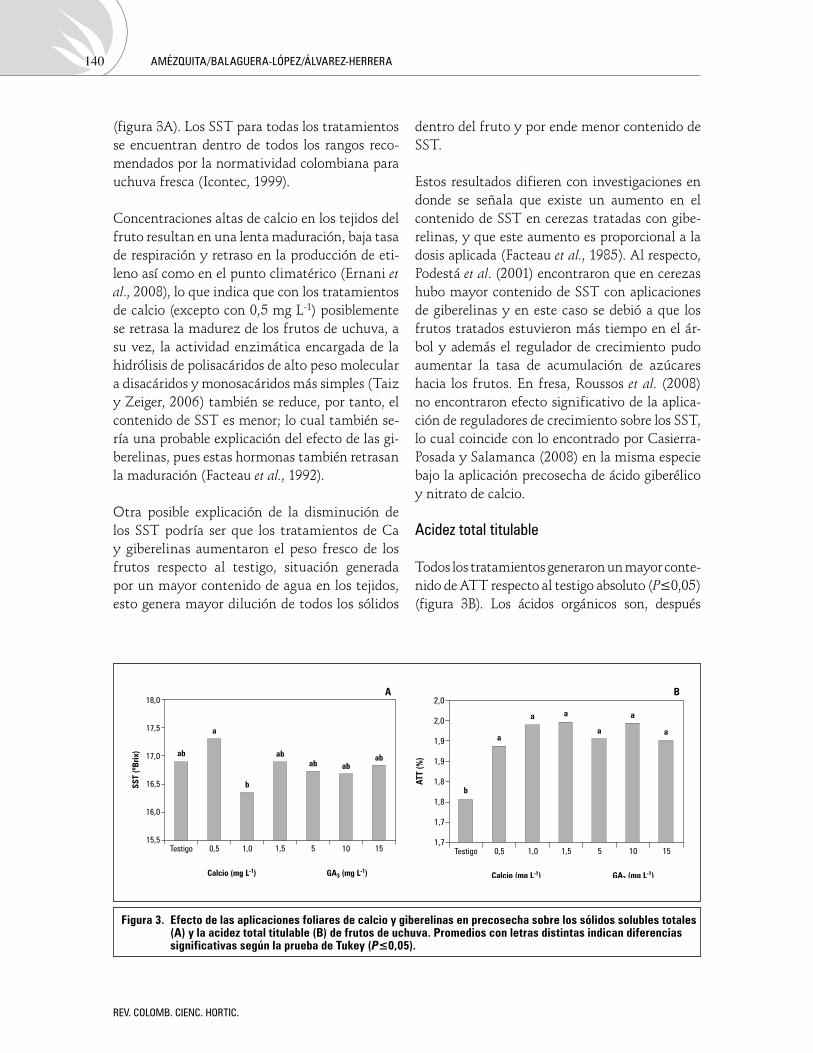
La aplicación de los tratamientos presentó di-ferencias estadísticamente significativas en el contenido de SST, la dosis de 1 mg L-1 de calcio presentó valores de 16,2 y fue el tratamiento de menor contenido de azúcares, mientras que con 0,5 mg L-1 de calcio se obtuvo el valor más alto de SST, todos los valores presentados con la apli-cación de giberelinas fueron inferiores al testigo
EFECTO DE LA ApLICACIóN pRECOSECHA DE GIBERELINAS y CALCIO EN EL FRuTO DE uCHuVA
Figura 2. efecto de las aplicaciones foliares de calcio y giberelinas en precosecha sobre el rajado de frutos de uchuva por calidades comerciales. promedios con letras distintas en la misma serie indican diferencias significativas según la prueba de tukey (P≤0,05).
REV. COLOMB. CIENC. HORTIC.
140
(figura 3A). Los SST para todas los tratamientos se encuentran dentro de todos los rangos reco-mendados por la normatividad colombiana para uchuva fresca (Icontec, 1999).
Concentraciones altas de calcio en los tejidos del fruto resultan en una lenta maduración, baja tasa de respiración y retraso en la producción de eti-leno así como en el punto climatérico (Ernani et al., 2008), lo que indica que con los tratamientos de calcio (excepto con 0,5 mg L-1) posiblemente se retrasa la madurez de los frutos de uchuva, a su vez, la actividad enzimática encargada de la hidrólisis de polisacáridos de alto peso molecular a disacáridos y monosacáridos más simples (Taiz y Zeiger, 2006) también se reduce, por tanto, el contenido de SST es menor; lo cual también se-ría una probable explicación del efecto de las gi-berelinas, pues estas hormonas también retrasan la maduración (Facteau et al., 1992).
Otra posible explicación de la disminución de los SST podría ser que los tratamientos de Ca y giberelinas aumentaron el peso fresco de los frutos respecto al testigo, situación generada por un mayor contenido de agua en los tejidos, esto genera mayor dilución de todos los sólidos
dentro del fruto y por ende menor contenido de SST.
Estos resultados difieren con investigaciones en donde se señala que existe un aumento en el contenido de SST en cerezas tratadas con gibe-relinas, y que este aumento es proporcional a la dosis aplicada (Facteau et al., 1985). Al respecto, Podestá et al. (2001) encontraron que en cerezas hubo mayor contenido de SST con aplicaciones de giberelinas y en este caso se debió a que los frutos tratados estuvieron más tiempo en el ár-bol y además el regulador de crecimiento pudo aumentar la tasa de acumulación de azúcares hacia los frutos. En fresa, Roussos et al. (2008) no encontraron efecto significativo de la aplica-ción de reguladores de crecimiento sobre los SST, lo cual coincide con lo encontrado por Casierra-Posada y Salamanca (2008) en la misma especie bajo la aplicación precosecha de ácido giberélico y nitrato de calcio.
Acidez total titulable
Todos los tratamientos generaron un mayor conte-nido de ATT respecto al testigo absoluto (P≤0,05) (figura 3B). Los ácidos orgánicos son, después
AMézquITA/BALAGuERA-LópEz/ÁLVAREz-HERRERA
Figura 3. efecto de las aplicaciones foliares de calcio y giberelinas en precosecha sobre los sólidos solubles totales (A) y la acidez total titulable (B) de frutos de uchuva. Promedios con letras distintas indican diferencias significativas según la prueba de tukey (P≤0,05).

Vol. 2 - No.2 - 2008
141
de los carbohidratos, el depósito energético más importante para el fruto (Osterloh et al., 1996). Durante la respiración del fruto son utilizados los ácidos orgánicos y convertidos a azucares, y va-rios de estos ácidos son componentes esenciales en el ciclo respiratorio de los ácidos tricarboxílicos (Kays, 2004). Esto indica que un mayor conteni-do de ácidos en la cosecha garantizaría una mayor vida útil en anaquel de los frutos de uchuva, por tanto, la aplicación de calcio y giberelinas favo-recerían la duración poscosecha en uchuva. Una mayor vida útil en poscosecha con aplicaciones de ácido giberélico y nitrato de calcio fue encontrado en fresa, aunque sin diferencias estadísticas en la ATT (Casierra-Posada y Salamanca, 2008).
Aplicaciones de cloruro de calcio al 0,5% en dife-rentes frecuencias (0, 4, 8, 12 veces/año) a árboles de manzano no afectaron significativamente la ca-lidad de los frutos respecto a la firmeza, SST, ATT, contenido de almidón y color (Ernani et al., 2008). Mientras que en cereza, un suministro foliar de calcio en precosecha aumentó la ATT, los SST y la relación de madurez (Podestá et al., 2001)
Relación de madurez
Para la RM se presentaron diferencias estadísti-cas entre los tratamientos, de tal forma que el testigo presentó el mayor valor de RM (figura 4A). Para el caso de la curuba Tellez et al. (2007), manifestaron que frutos con una alta RM tien-den a ser mas agradables, pues una mayor RM indica un contenido predominante de azúcares sobre ácidos, lo que indica que los frutos del tra-tamiento testigo presentarían un mejor sabor, no obstante, una RM más baja con la aplicación de calcio y giberelinas en los frutos de uchuva posiblemente indica un retraso en la madurez que favorecería una mayor vida en poscosecha. Sin embargo, estos resultados fueron contrarios a los encontrados por Podestá et al. (2001) con aplicaciones precosecha de calcio y giberelinas en cereza, donde la RM fue mayor con estos tra-tamientos respecto al testigo.
Firmeza
La firmeza mostró diferencias significativas en-tre tratamientos, la mayor firmeza fue obteni-da con la aplicación de 0,5 g L-1 de Ca. En gene-ral se encontró que tanto la aplicación de calcio como la de giberelinas presentó mayor firmeza que el testigo (excepto 5 mg L-1 de giberelinas) (figura 4B).
Las aplicaciones de calcio exógenas son efecti-vas para reducir el ablandamiento en todos los frutos (Saftner et al., 2003). Estas aplicaciones permiten que el calcio suministrado se una a las cargas negativas de los residuos de ácidos uró-nicos desesterificados que son generados por la enzima pectinmetilesterasa durante la madura-ción, aumentando la resistencia mecánica de los tejidos (Magee et al., 2003).
La aplicación de 3 g kg-1 de Ca en precosecha a frutos de Citrus sinenisis incrementó significati-vamente el contenido de Ca en la pulpa y corte-za respecto a dosis menores (1 ó 2 g kg-1 de Ca), sin embargo, con cualquiera de las aplicaciones se disminuyó la actividad de poligalacturonasa, pectinesterasa y B-galactosidasa, mediante la influencia de la expresión de genes, y por tan-to, la disolución e hidrólisis de pectinas fueron restringidas. También modificaron la estructura de los tejidos de la membrana, siendo estos más compactos y homogéneos con mayor presencia de Ca (Dong et al., 2009).
El aumento de la firmeza por aplicaciones de gi-berelinas podría deberse a algún efecto del regula-dor de crecimiento sobre la estructura de la pared celular (Andrews y Shulin, 1995), o simplemente a un retraso en la maduración de los frutos, que permitiría mantener la integridad de las membra-nas y paredes celulares por más tiempo al inhibir la acción de enzimas hidrolíticas que actúan sobre la pectina. Una mayor firmeza y un retraso en la maduración por acción de las giberelinas fue en-contrado por Facteau et al. (1992).
EFECTO DE LA ApLICACIóN pRECOSECHA DE GIBERELINAS y CALCIO EN EL FRuTO DE uCHuVA

REV. COLOMB. CIENC. HORTIC.
142
agraDeCimieNtoS
CoNCluSioNeS
Bajo las condiciones del experimento, la aplica-ción de calcio y giberelinas favoreció la produc-ción de frutos de uchuva y disminuyó el porcen-taje de rajado de los mismos, especialmente en
concentraciones de 0,5 y 1,0 mg L-1 de calcio y 10 y 15 mg L-1 de GA3. Además, estos tratamientos presentaron un incremento en la firmeza y ATT y una disminución en los SST y la RM, lo cual indica que retrasan la madurez y pueden conser-var la calidad de los frutos por más tiempo.
Los autores agradecemos al ingeniero agrónomo Alfonso Molano, propietario de la finca El Saque, por su valiosa colaboración para que la presente investigación llegara a buen término.
AMézquITA/BALAGuERA-LópEz/ÁLVAREz-HERRERA
Figura 4. Efecto de las aplicaciones foliares de calcio y giberelinas en precosecha sobre la relación de madurez (RM) (A) y la firmeza (B) de frutos de uchuva. Promedios con letras distintas indican diferencias significativas según la prueba de tukey (P≤0,05).

Vol. 2 - No.2 - 2008
143
reFereNCiaS bibliogrÁFiCaS
Agronet. 2008. Producción y área cosechada de uchuva en Colombia en 2008. Ministerio de Agricultura y Desarrollo Rural. En: http://www.agronet.gov.co/; consulta: diciembre de 2008.
Andrews, P.K. y L. Shulin. 1995. Cell wall hydrolytic en-zyme activity during development of nonclimac-teric sweet cherry (Prunus avium L.) fruit. J. Hort. Sci. 70 (4), 561-567.
Barker, A. y D. Pilbeam. 2007. Handbook of plant nu-trition. CRC Press Taylor & Francis Group, New York, NY.
Ben-Airie, R.; H. Bazak y A. Blumenfeld. 1986. Gibberel-lin delays harvest and prolongs life of persimmon fruits. Acta Hort. 179, 807-813.
Botelho, R.V.; E.J.P. Pires y M.M. Terra. 2004. Efeitos de reguladores vegetais na qualidade de uvas ‘Niagara Rosada’ na região noroeste do Estado de São Paulo. Rev. Bras. Frutic. 26(1), 74-77.
Brown, G.S.; S. Wilson; W. Boucher; B. Graham y B. McGlasson. 1996. Effects of copper-calcium sprays on fruit cracking in sweet cherry (Prunus avium). Scientia Hort. 62, 75-80.
Casierra-Posada, F. y R. Salamanca. 2008. Influencia del ácido giberélico y del nitrato de calcio sobre la du-ración poscosecha de frutos de fresa (Fragaria sp.). Rev. Colomb. Cienc. Hortic. 2(1), 33-42.
Cooman, A.; C. Torres y G. Fischer. 2005. Determina-ción de las causas del rajado del fruto de uchuva (Physalis peruviana L.) bajo cubierta: II. Efecto de la oferta de calcio, boro y cobre. Agron. Colomb. 23(1), 74-82.
Dong, T.; R. Xia; Z. Xiao; P. Wanga y W. Song. 2009. Effect of pre-harvest application of calcium and boron on dietary fibre, hydrolases and ultrastruc-ture in ‘Cara Cara’ navel orange (Citrus sinensis L. Osbeck) fruit. Scientia Hort. 121 (en imprenta).
Ernani, P.R.; J. Dias; C.V.T. Do Amarante; D.C. Ribeiro y D.A. Rogeri. 2008. Preharvest calcium sprays were not always needed to improve fruit quality of ‘Gala’ apples in Brazil. Rev. Bras. Frutic. 30(4), 892-896.
Facteau, T.J.; N.E. Chestnut; K.E. Rowe y C. Payne. 1992. Brine quality of gibberellic acid-treated «Napoleon» sweet cherries. HortScience 27(2), 118-122.
Facteau, T.J.; K.E. Rowe y N.E. Chestnut. 1985. Re-sponse patterns of gibberellic acid-treated sweet cherry fruit at different soluble solids levels and leaf/fruit ratios. Scientia Hort. 27, 257-262.
Gordillo, O.; G. Fischer y R. Guerrero. 2004. Efecto del riego y de la fertilización sobre la incidencia del rajado en frutos de uchuva (Physalis peruviana L.) en la zona de Silvania (Cundinamarca). Agron. Co-lomb. 22 (1), 53 – 62.
Grange, R. 1996. Crecimiento del fruto. pp. 449-462. En: Azcón-Bieto, J. y M.Talón (eds.). Fisiología y bioquímica vegetal. Interamericana-McGraw-Hill, Madrid.
Fischer, G. 2005. El problema del rajado del fruto de uchuva y su posible control. pp. 55-82. En: Fischer, G., D. Miranda, W. Piedrahita y J. Romero (eds.) Avances en cultivo, poscosecha y exportacion de la uchuva (Physalis peruviana L.) en Colombia. Fa-cultad de Agronomia, Universidad Nacional de Co-lombia; Unibiblos, Bogotá.
Hernández-Muñoz, P.; E. Almenar; V. Del Valle; D. Ve-lez y R. Gavara. 2008. Effect of chitosan coating combined with postharvest calcium treatment on strawberry (Fragaria x ananassa) quality during re-frigerated storage. Food Chem. 110, 428-435.
Huang, X.M.; H.C. Wang; J.G. Li; J.H. Yin; W.Q. Yuan; J.M. Lu y H.B. Huang. 2005. An overview of calci-um’s role in lychee fruit cracking. Acta Hort. 665, 231-240.
Icontec. 1999. Frutas frescas. Uchuva. Especificaciones. Norma Técnica Colombiana NTC 4580. Instituto Colombiano de Normas Técnicas y Certificación, Bogotá.
Kays, S. 2004. Postharvest biology. Exon Press, Athens.
Lin, L.W. 2001. Effect of mineral nutrient on fruit crack-ing rate of Litchi chinensis Sonn. Soil Environ. Sci. 10, 55-56.
Looney, N.E. y P.D. Lidster. 1980. Some growth regula-tor effects on fruit quality, mesocarp composition, and susceptibility to postharvest surface marking of sweet cherries. J. Amer. Soc. Hort. Sci. 105(1), 130-134.
Magee, R.L.; F. Caporaso y A. Prakash. 2003. Effects of exogenous calcium salt treatments on inhibiting
EFECTO DE LA ApLICACIóN pRECOSECHA DE GIBERELINAS y CALCIO EN EL FRuTO DE uCHuVA

REV. COLOMB. CIENC. HORTIC.
144
irradiation-induced softening in diced roma toma-toes. J. Food Sci. 68, 2430-2435.
Marschner, H. 2002. Mineral nutrition of higher plants. 2a ed. Academic Press, London.
Matsuoka, M. 2003. Gibberellins signaling: how do plant cells respond to GA signals? Plant Growth Regul. 22, 123-125.
Meheriuk, M.; G.H. Neilsen y D.L. Mckenzie. 1991. In-cidence of rain splitting in sweet cherries treated with calcium or coating materials. Can. J. Plant Sci. 71, 23 l-234.
Opara, L. 1997. Fruit skin splitting and cracking. Hort. Rev. 19, 217-262.
Osterloh, A.; G. Ebert; W. Held; H. Schulz y E. Urban. 1996. Lagerung von Obst und Südfrüchten. Verlag Ulmer, Stuttgart, Alemania.
Paroussi, G.; D.G. Voyiatzis; E. Paroussi y P.D. Drogour. 2002. Growth, flowering and yield responses to GA3 of strawberry grown under different environ-mental conditions. Scientia Hort. 9,103-113.
Peet, M.M. 1992. Fruit cracking in tomato. Hort Tech-nol. 2, 216-223.
Podestá, L.; F. Gil; M. Rodríguez y C. Arjona. 2001. Efec-to del ácido giberélico y del calcio sobre el tamaño, agrietamiento y otros parámetros de calidad en frutos de cerezo (Prunus avium L.) cv. Bing. Invest. Agr. Prod. Prot. Veg. 16 (1), 37-48.
Podestá, L.; F. Gil; M. Rodríguez y C. Arjona. 1998. Efec-to del calcio y el ácido giberélico sobre el tamaño, el cracking y otros parámetros de calidad en frutos de cerezo (Prunus avium L.) cv. Bing. XXI Congreso Ar-gentino de Horticultura. Departamento de Produc-ción Agropecuaria, Universidad Nacional de Cuyo, Mendoza, Argentina.
Pratella G.C. 1996. La screpolatura delle ciliege. Rivista di Frutticoltura e di Ortofloricoltura 10, 71-73.
Román, L.F. y M.A. Gutiérrez. 1998. Evaluación de áci-dos carboxílicos y nitrato de calcio para incremen-
tar la calidad, cantidad y vida en anaquel en tres tipos de melón. Terra 16(1), 49-54.
Roussos, P.A.; N.K. Denaza y T. Damvakaris. 2008. Strawberry fruit quality attributes alter applica-tion of plant growth stimulating compounds. Scientia Hort. 119(2), 138-146.
Saftner, R.A.; J. Bai; J.A. Abbott y Y.S. Lee. 2003. Sani-tary dips with calcium propionate, calcium chlo-ride, or calcium amino acid chelates maintain quality and shelf stability of fresh-cut honeydew. Postharv. Biol. Technol. 29, 257-269.
Sansavini, S. y S. Lugli. 1997. Prospettive e innovazioni tecnico-scientifiche per la cerasicoltura italiana. Rivista di Frutticoltura e di Ortofloricoltura 6, 9-18.
Sekse, L. 1995. Cuticular fracturing in fruit cherry (Prunus avium L.) resulting from changing soil wa-ter contents. J. Hort. Sci. 70(4), 631-635.
Taiz L. y E. Zeiger. 2006. Plant physiology. 4a ed. Sinauer Associates Publishers, Sunderland, MA.
Téllez, C.P.; G. Fischer y O. Quintero. 2007. Compor-tamiento fisiológico y fisicoquímico de frutos de curuba (Passiflora mollissima Bailey) encerados y almacenados a dos temperaturas. Rev. Colomb. Cienc. Hort. 1(1), 67-80.
Usenik, V.; D. Kastelec y F. Tampar. 2005. Physicochemi-cal changes of sweet cherry fruits related to applica-tion of gibberellic acid. Food Chem. 90(4), 663-671.
Valencia, M. 1985. Anatomía del fruto de la uchuva. Acta Biol. Colomb. 1(2), 63-89.
Vieira, C.R.Y.; E.J. Pires; M.M. Terra; M.A. Tecchio y R.V. Botelho. 2008. Efeitos do ácido giberélico e do thidiazuron sobre as características dos frutos e do mosto da uva ‘Niagara Rosada’. Rev. Bras. Frutic. 30(1), 12-19.
Xu-Ming, H.; W. Hui-Cong; Z. Wei-Liang; Y. Wei-Qun; L. Jie-Mei y L. Jian-Guo. 2008. Spraying calcium is not an effective way to increase structural calcium in litchi pericarp. Scientia Hort. 117, 39-44.
AMézquITA/BALAGuERA-LópEz/ÁLVAREz-HERRERA

arlette ivoNNe gil1,3 Diego miraNDa2
Aspectos anatómicos de la semilla de papaya (Carica papaya L.)
Anatomical aspects of papaya (Carica papaya L.) seed
Frutos de papaya con semillas. Foto: D. Miranda L.
1 Facultad de Ciencias Agropecuarias, Universidad de Cundinamarca, Fusagasugá (Colombia).2 Departamento de Agronomía, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá (Colombia).3 Autor para correspondencia. [email protected]
REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 2 - No.2 - pp. 145-156, 2008
reSumeN
Debido a la falta de estudios anatómicos en semillas de especies de interés económico, y el poco conocimiento de este tópico para los ingenieros agrónomos, se procedió a la realización de una investigación base para su entendimiento y posterior ampliación en futuros estudios. Con este fin se analizaron diversos aspectos anató-micos de la semilla de papaya, para lo cual se tomaron frutos inmaduros de 60 días y frutos maduros entre 120 y 150 días, entre madurez fisiológica y comercial, en una plantación sembrada con el híbrido Tainung-1. Las semillas extraídas de los frutos mencionados fueron llevadas al laboratorio y adecuadas mediante la técnica de parafina, para posteriormente realizar los cortes en micrótomo de rotación con el propósito de describir ana-tómicamente los tejidos y estructuras que las componen, bajo microscopía óptica. La semilla de papaya está compuesta por la cubierta seminal, el endospermo y el embrión. Por tratarse de una semilla bitegumentada, se observaron la testa y el tegmen con sus correspondientes componentes. El endospermo está compuesto prin-cipalmente de lípidos y proteínas agrupados en granos de aleurona. Los cotiledones están compuestos además de la epidermis, por cuatro capas celulares internas y en la zona radicular se pudo apreciar el esbozo de raíces secundarias. Este estudio tuvo como objetivo generar conocimiento acerca de la anatomía de la semilla de papaya, para el entendimiento de aspectos básicos de la fisiología de semillas como vigor, viabilidad y latencia en esta especie, previo para futuras investigaciones.
palabras clave adicionales: híbrido Tainung-1, testa, endospermo, embrión, granos de aleurona.

REV. COLOMB. CIENC. HORTIC.
146
abStraCt
Due to the lack of the anatomical studies in seeds of the species of economical interest and a few knowledge of this matter by the agronomists, the investigation as a base for understanding of this problem and further development in future studies was done. With this purpose, several anatomical aspects of papaya seed were analyzed from immature fruits of 60 days age and mature fruits of 120 and 150 days between physiological and commercial maturity were taken from Tainung-1 hybrid plantation. The seeds extracted from fruits were transported to the laboratory and adequated with paraffin technique to make the microtome sections in order to describe anatomically the tissues and structures under optical microscopy. Papaya seed is composed of the seed coat, endosperm, and embryo. As a bitegumental seed, the testa and tegmen were observed with their own parts. The endosperm is composed mainly of lipids and proteins grouped in aleurone grains. The cotyledons are composed by the epidermis and four-layer inner cells and, in the radicle zone, initiation of the secondary roots was observed. The objective of this study was to generate knowledge about the anatomy of papaya seed for understanding the basic aspects of seed physiology, such as vigour, viability, and dormancy in this species, prior to future researches.
La papaya (Carica papaya L.) es considerada de ori-gen Americano, específicamente de Centroaméri-ca, entre México y Costa Rica (León, 1987). La identificación de las distintas características mor-fológicas y fisiológicas es un paso inicial impor-tante en el estudio del desarrollo de las semillas en las diversas especies (Ren y Bewley, 1998). El me-gasporangio (óvulo) es la estructura a partir de la cual se forman las semillas. En las angiospermas está formado por uno o dos tegumentos, ocasio-nalmente por tres, y muy raramente por cuatro. En la semilla madura, los tegumentos que rodean al óvulo se transforman en la cubierta seminal (Niembro, 1988; Reiser y Fischer, 1993).
Las semillas de las angiospermas pueden ser divi-didas en tres partes, de origen genéticamente di-ferente: el embrión, el endospermo y la cubierta seminal. El cigoto, a partir del cual se desarrolla el embrión, combina los genotipos del haploide masculino y el gameto femenino. Después de la
iNtroDuCCióN
additional key words: Tainung-1 hybrid, testa, endosperm, embryo, aleurone grains.
Fecha de recepción: 30-07-2008 Aprobado para publicación: 02-12-2008
fertilización, durante el desarrollo del embrión y la subsiguiente maduración de la semilla, los integumentos experimentan cambios morfológi-cos y se convertirán en la testa, la cual posee el genotipo materno (Fosket, 1994; León-Klooster-ziel et al., 1994). Debido a que el embrión está encerrado por el endospermo, el cual a su vez está dentro del integumento del óvulo, estas tres estructuras deben coordinar su desarrollo para producir una semilla madura de tamaño adecua-do (García et al., 2003).
Cuando la semilla proviene de un primordio se-minal con un solo tegumento, es unitegumen-tada y su cubierta seminal se denomina testa. Igualmente si el primordio posee dos tegumen-tos, los tejidos derivados del tegumento externo constituyen la testa y los derivados del tegu-mento interno conforman el tegmen, cuando la semilla es bitegumentada (Becerra y Chaparro, 1999).
GIL/MIRANDA

Vol. 2 - No.2 - 2008
147
El endospermo es un tejido que se origina a par-tir de la fusión de uno de los núcleos espermáti-cos del grano de polen con dos núcleos polares del saco embrionario, dando como resultado un tejido triploide (Becerra y Chaparro, 1999). El endospermo acumula la mayor parte de las reser-vas nutritivas en la mayoría de los casos cuando los cotiledones no lo hacen (Esau, 1985; Besnier, 1988). Algunas semillas pueden presentar un en-dospermo muy delgado, limitado a una o varias células de grosor, o bien pueden estar provistas de uno masivo, parenquimatoso (Fahn, 1985). El endospermo es único en diversos aspectos de su desarrollo, biología celular y bioquímica. Su aparente simplicidad reflejada en la presencia de unos cuantos tipos de células diferenciadas, ha permitido estudios detallados en histodiferen-ciación y biosíntesis de ciertos metabolitos al-macenados (Lopes y Larkins, 1993).
El encerramiento del embrión y del endospermo en integumentos de origen estrictamente mater-nos, ha llevado a interacciones entre los tejidos de diferentes contribuciones parentales en la pro-tección del embrión y del endospermo, y en el control de la protrusión radicular de las semillas. Tales interacciones influencian la germinación, latencia, calidad de la semilla, en particular vigor y longevidad (Debeaujon et al., 2000).
El embrión de las angiospermas consta de un breve eje que lleva uno o dos cotiledones u hojas embrionales. El punto de fijación de los cotile-dones al eje, designado como nudo cotiledonar, divide al eje en dos regiones: basal o hipocotilo que termina en una raíz embrionaria o radícula, y la región superior o epicotilo que puede presen-tar una yema denominada plúmula (Becerra y Chaparro, 1999).
El objetivo de esta investigación consistió en generar un conocimiento básico acerca de la es-tructura de la semilla de papaya, con el fin de servir como base para futuros estudios en el área de anatomía y fisiología vegetal.
materialeS y métoDoS
En la finca Santa Helena propiedad del grupo Grajales S.A., se realizó el muestreo de frutos de papaya del híbrido Tainung-1 cuyo origen es taiwanés, con el fin de extraer sus semillas. La plantación se encuentra ubicada en el municipio de Roldanillo (Valle del Cauca, Colombia), a una altura de 935 msnm, temperatura de 24,2°C, precipitación anual de 1.015 mm y humedad re-lativa de 71,1%.
Se tomaron frutos inmaduros, de 60 d de edad, que contenían rudimentos de semillas (primor-dios seminales) y frutos maduros (madurez fisio-lógica y comercial) entre 120 y 150 d. Las semi-llas (figura 1) se extrajeron de los frutos y fueron acondicionadas según la metodología de Johan-sen (1940), en la cual el material vegetal (semilla) fue incluido en bloques de parafina para poste-riormente proceder a los cortes en micrótomo de rotación, con el fin de estudiar su anatomía bajo microscopio óptico. Para el teñido del material se utilizó Fast Green y safranina.
reSultaDoS y DiSCuSióN
primordios seminales
En el corte transversal medio realizado a un fru-to inmaduro de 60 d, se observó que las semillas en formación (primordios seminales) se encon-traban adheridas a las paredes internas del fruto por medio del funículo, lo que se conoce como placentación parietal (figura 2). Esta caracterís-tica ayuda a clasificar a la papaya dentro del or-den de las parietales, lo que ratifica lo afirmado por Font Quer (1965). Al interior del funículo se observaron de seis a siete capas de células muy alargadas, correspondientes al haz conductor, con unas pocas diferenciadas en xilema, de en-grosamientos anulares. En este estado, las célu-las del rudimento presentaban la apariencia de células embrionarias, con paredes delgadas y nú-
ASpECTOS ANATóMICOS DE LA SEMILLA DE pApAyA

REV. COLOMB. CIENC. HORTIC.
148
cleos prominentes, como evidencia de la división activa a la que se encontraban sometidas.
Stadler et al. (2005) mostraron en semillas de Arabidopsis thaliana que el integumento exter-no es una extensión simplástica del floema del funículo, sugiriendo que los nutrientes al final de este pueden ser descargados a través de este tejido. El transporte de sacarosa en el funículo indica que esta es una vía común para la des-carga de nutrientes a las semillas en desarrollo (Skinner et al., 2004). La asimilación de carbono es suplida hacia el embrión en desarrollo. Los ha-ces vasculares están conectados al óvulo a través del funículo al final del tejido nucelar. Los tejidos vasculares no se extienden más allá del funículo dentro del óvulo (Imlau et al., 1999).
La epidermis que recubría el funículo y la parte externa de la semilla correspondía a la continua-ción de la epidermis interna del fruto, consti-tuida por células de forma rectangular, un poco más altas que anchas, con la pared externa algo arqueada, a modo de las células epidermales típi-cas. Al analizar la parte interna del rudimento, se observaron células un poco más anchas, muy juntas, que se unían sin dejar espacios entre sí. La cubierta seminal, que es la estructura de protec-ción de la semilla contra los agentes externos, se deriva de estos tegumentos dispuestos de forma concéntrica (figura 3). Las seis capas subyacen-tes a la epidermis, constituyentes del tegumento externo, midieron 7 μm de espesor, presentando forma aplanada y disposición regular. Hacia la parte interna se observaron unas 10 capas celula-res, correspondientes al tegumento interno, con paredes angulares de formas y tamaños diversos, cuyo espesor fue de 7,5 μm.
Los óvulos son el sitio de los procesos esenciales para la reproducción sexual de las plantas, inclui-das la formación del megagametofito, fertiliza-ción, embriogénesis y, finalmente, la formación de la semilla. En Arabidopsis, los óvulos se inician en los primordios con una forma alargada a par-tir de la placenta de la superficie interna de los
carpelos (Skinner et al., 2004). Los integumen-tos interno y externo surgen de cada primordio del óvulo, con su región de origen definiendo la chalaza, la cual separa la nucela apical del funí-culo. Los dos integumentos crecen para cubrir y encapsular la nucela, teniendo una pequeña abertura, el micrópilo. El funículo provee la con-ducción de nutrientes al óvulo en desarrollo y al embrión, y determina parcialmente la posición del micrópilo. Los integumentos son requeridos para encerrar el saco embrionario, contribuyen-do al posicionamiento del óvulo para posterior-mente formar la cubierta seminal. La nucela provee el inicio celular para la diferenciación del megasporocito, el cual, por procesos de mitosis y meiosis, produce un megagametofito llamado saco embrionario (Webb y Gunning, 1990).
Semilla madura
Al observar una vista parcial del corte transme-diano de estas semillas, se diferenciaron clara-mente tres zonas de tejidos: la cubierta seminal, el endospermo y el embrión (figura 4).
La elongación celular en la cubierta seminal está dirigida por el crecimiento del endospermo y es el mayor responsable en el control del tamaño de la semilla. La proliferación celular ocurre en la cubierta seminal después de la fertilización y también puede cumplir un papel importante en el tamaño de la semilla (Garcia et al., 2005; Ingouff et al., 2006; Schruff et al., 2006). Sin em-bargo, es desconocido si la proliferación celular es regulada en la cubierta seminal en respuesta a la fertilización. Además las interacciones invo-lucran la proliferación celular y la elongación en la cubierta seminal, y la evidencia ha sido obte-nida por la interacción de la diferenciación entre el endospermo y la cubierta seminal en cebada (Hordeum vulgare L.) y maíz (Zea mays L.) (Chau-dhury y Berger, 2001).
La cubierta seminal estuvo constituida por cuatro capas de coloración diferencial, denominadas exo-testa, mesotesta, endotesta y tegmen (figura 5).
GIL/MIRANDA

Vol. 2 - No.2 - 2008
149
La zona más externa consistió en varias capas de tejido de paredes delgadas y transparentes, con forma rectangular y dispuestas en empalizada, elongadas radialmente y organizadas sin dejar espacios entre sí, recordando la organización tí-pica de las capas epidermales. Estas capas consti-tuyeron la sarcotesta, que corresponde a la exo-testa jugosa que recubre a las semillas de papaya. Hacia el interior se encontraron hasta 15 capas de células escleróticas cuboides o elongadas tan-gencialmente, que constituyeron la parte dura y oscura de la cubierta seminal. Estas células po-seían lúmenes grandes que al unirse dejaban pe-queños espacios entre sí, dando la apariencia de una malla con unidades de forma estrellada. Al final de las aristas formadas por las paredes, en la conjunción de tres células, se formaba un es-pacio en forma de triángulo. Esta capa formaba la parte rugosa de la semilla, que es posible ob-servar una vez se ha removido la exotesta. El nú-mero de capas celulares que la constituía deter-minaba los valles y los picos en las rugosidades, y conformaban la mesotesta externa de la semilla. Inmediatamente por debajo de esta capa, se en-contró otra de unas 25 filas de células también esclerenquimáticas, de entre 0,5 a 1 μm de alto por 1 μm de ancho, con un grosor de 16 μm. Las células fueron de forma ovoide, más anchas que altas y se unían entre sí sin dejar espacios. En conjunto esta capa se denomina mesotesta inter-na (figura 6).
La capa subyacente o endotesta estuvo constitui-da por una fila de células esclerenquimáticas re-dondeadas, con la pared tangencial interna ligera-mente engrosada y lignificada, con una altura de 3 μm y ancho de 1,5 μm. Debajo de esta capa fue posible observar el exotegmen y el mesotegmen, que estaban formados por células entrecruzadas. La capa más interna de la cubierta seminal, co-rrespondiente al endotegmen, estuvo constitui-da por una fila de células rectangulares, de 0,7 μm de alto por 2 μm de ancho, cuyo contenido celular estaba constituido por taninos. Seguida-mente se observó la cutícula nucelar, en contacto con el endospermo de la semilla (figura 7).
Durante el desarrollo de la semilla, cada integu-mento se encamina hacia un dramático proceso de diferenciación para formar la cubierta semi-nal madura: algunos de los tejidos que son ge-nerados en los estados tempranos e intermedios del desarrollo de la semilla desaparecen en los es-tados tardíos en semillas de Brassicaceae (Beeck-man et al., 2000; Western et al., 2000; Windsor et al., 2000). Durante el proceso de diferencia-ción, el tegumento interno llega a comprimirse e impregnarse con pigmentos que le imparten la característica de color marrón, sin embargo, los mecanismos moleculares bioquímicos base de este proceso no son del todo entendibles (Nakaune et al., 2005).
El endospermo se observó de consistencia carno-sa y suave, externo al embrión. Cada lado de este tuvo cerca de 16 capas de células parenquimáti-cas. Las células de las seis más externas fueron de forma rectangular, mientras que las capas internas estuvieron constituidas por células iso-diamétricas de forma poliédrica. Los lípidos y las proteínas son las sustancias de reserva almace-nadas en el endospermo de la semilla de papaya. Los lípidos se pudieron observar como oleoso-mas, mientras que las proteínas se encontraron almacenadas en pequeñas estructuras llamadas cuerpos proteínicos o granos de aleurona, las cua-les presentan gran similitud en tamaño y forma con los granos de almidón (figura 8). Las paredes celulares en este tejido de reserva son delgadas, de acuerdo a lo reportado por Niembro (1988).
El endospermo es un tejido único. Debido a su rol central en la nutrición y protección del em-brión, el desarrollo del endospermo está sujeto a los procesos adaptativos, los cuales conllevan a la evolución del origen parental dependiente de la regulación genética (Baroux et al., 2007). El endospermo es un tejido efímero restringido al periodo de producción de semilla y germinación. Es mayormente consumido tanto por el embrión en desarrollo o después de la germinación de la semilla. La inusual constitución genética del en-dospermo puede ser vista desde la contribución
ASpECTOS ANATóMICOS DE LA SEMILLA DE pApAyA

REV. COLOMB. CIENC. HORTIC.
150
parental como una fuente de heterocigosis prove-yendo vigor poliploide -un estado genético bene-ficioso para la alta tasa de proliferación y metabo-lismo del endospermo (Lopes y Larkins, 1993)-. Además de su función como tejido de reserva, el endospermo ha demostrado ejercer control sobre la germinación, al secretar enzimas que aflojan la pared celular debilitando la resistencia mecánica para facilitar la protrusión radicular (Penfield et al., 2006).
Haig y Westoby (1989) proponen que la triplo-día y el origen biparental del endospermo puede ser el resultado de conflictos genéticos parenta-les sobre la localización de la fuente a partir del esporofito materno a su progenie. La doble ferti-lización pudo primero evolucionar para transmi-tir los intereses del padre en promocionar su tasa de crecimiento en la progenie, mientras que el doble genoma materno puede ser visto como una respuesta evolutiva para reforzar los intereses de la madre en la repartición equitativa de recursos entre hermanos.
Las semillas acumulan lípidos con el fin de suplir los requerimientos energéticos para el crecimien-to de la plántula después de la germinación. Tales lípidos son generalmente acumulados como tria-cilgliceroles (TAG) en compartimientos esféricos conocidos como esferosomas, oleosomas o más comúnmente, cuerpos lipídicos. Esos organelos emergen del retículo endoplasmático, el cual es responsable de la síntesis de TAG (Siloto et al., 2006). La glicólisis es una vía ubicua esencial para la producción de aceite en semillas en desarro-llo de Arabidopsis thaliana y cultivos oleaginosos. Una importante función metabólica del desarro-llo de la semilla de esta especie, es la deposición de sustancias de reserva -aceite en la forma de triacilgliceroles, pero también proteínas y oligo y polisacáridos (Andre et al., 2007)-. En semillas no endospérmicas, como Brassica spp, los cuerpos lipídicos se encuentran en los cotiledones y el eje embrionario. En Ricinus communis y otras semillas
endospérmicas, los cuerpos lipídicos pueden ser encontrados en el endospermo (Anil et al., 2003).
El embrión estuvo limitado por la protodermis, la cual estaba formada por células entre 0,7 a 1,2 μm de alto por 3,0 a 3,5 μm de ancho. Las células pro-todermales fueron homogéneas y se unían entre sí, sin dejar espacios. Por debajo de la protodermis, en el plano medio del embrión, se encontraron de ocho a nueve capas de células, que constituían el parénquima fundamental cortical. Las células se observaron de forma cuadrangular (de 0,8 a 1,2 μm de alto por 1,2 a 1,4 μm de ancho), agrupadas sin dejar espacios intercelulares y formando co-lumnas. Al interior se encontraron de siete a ocho capas de procambium a cada lado del embrión. Las células midieron de 2,0 a 2,8 μm de alto por 0,6 a 1,0 μm de ancho. La parte interna estuvo constituida por cinco capas de células, que midie-ron entre 1,7 y 2,0 μm de alto por 2,0 a 2,4 μm de ancho, las cuales se denominan parénquima fun-damental medular (figura 9).
Durante la embriogénesis, se establece un plan para el establecimiento del cuerpo de la planta que consiste en el meristemo aéreo, cotiledones, hipocotilo, raíz y meristemo radicular a lo largo del eje apical-basal y un arreglo concéntrico de la epidermis, el tejido subepidermal y el cilindro central vascular a lo largo del eje radial -para establecer esta organización, las células del em-brión necesitan especializarse y deben diferen-ciarse (Laux et al., 2004)-.
En el ápice del embrión se encontró la túnica, que es el promeristemo que origina primero a la pro-todermis y posteriormente a la epidermis de la planta, de acuerdo con Barton y Poethig (1993). Estas células se dividieron de forma anticlinal y en las semillas de papaya estuvo constituida por una sola capa. Debajo de la túnica se encontró el cuerpo o corpus, que es el promeristemo del cual se originan los demás órganos de la planta. Las células que lo constituyeron fueron en su mayo-
GIL/MIRANDA

Vol. 2 - No.2 - 2008
151
ría de forma redondeada, con cuatro a seis capas de células (figura 10).
El crecimiento del ápice del embrión puede ser visto en términos de tres parámetros celulares. El primero se manifiesta como una polaridad re-gional que puede distinguirse por los planos de división celular en diferentes capas en el meris-temo apical. En las capas superficiales que cons-tituyen la túnica, las divisiones celulares son predominantemente anticlinales (nuevas pare-des celulares se forman perpendicularmente a la superficie del eje). En contraste, las células más internas constituyen el cuerpo o corpus y expe-rimentan divisiones en una variedad de planos (Szymkowiak y Sussex, 1996).
Los cotiledones fueron ovados y de posición recta, planos y delgados. Además de la epidermis abaxial y adaxial, poseían cuatro capas celulares internas, cuyo grosor fue de 14 μm, en donde se observó cierta organización de las capas parenquimáticas internas: la primera de ellas en empalizada, la subyacente estuvo constituida por células ovoides de gran tamaño alternando con grupos de tres células de forma similar pero de menor tamaño, y las dos últimas se constituían de células más o menos cuadrangulares, por debajo de las cuales se encontró la epidermis abaxial (figura 11).
En dicotiledóneas, los cotiledones son órganos laterales y el meristemo apical está en una posi-ción central, mientras que en monocotiledóneas el cotiledón es terminal y el meristemo apical está en una posición lateral. Los cotiledones se forman durante la embriogénesis y también están defini-dos como epígeos si emergen por encima del suelo, se expanden después de la germinación y llegan a ser fotosintéticos, o hipógeos, si ellos permanecen por debajo del suelo, no se expanden y permane-cen no fotosintéticos. Los cotiledones pueden ser estructuras efímeras, persistiendo solamente por unos pocos días después de la emergencia, o pue-den ser retenidos por la planta a través de su ciclo de vida (Chandler, 2008).
La polaridad de los cotiledones y las hojas es establecida para ambos órganos con respecto al meristemo apical, siendo la superficie adaxial la más cercana al eje de la planta o al meristemo apical, y derivándose a partir del dominio cen-tral apical del embrión, y el lado abaxial, opues-to y generado por el dominio periférico del embrión. Las superficies adaxiales y abaxiales de los cotiledones son morfológicamente distin-tas con respecto al tamaño celular y densidad estomática: la epidermis adaxial posee células de tamaño uniforme y tiene una superficie apla-nada con una baja densidad estomática, mien-tras que la superficie abaxial tiene entremezcla-das células grandes y pequeñas, una superficie áspera y alta densidad estomática (Siegfried et al., 1999).
En la zona radicular fue posible observar el esbo-zo de raíces secundarias de forma semicircular (figura 12). Las raíces laterales se originan por divisiones en el periciclo, dando como resultado una masa pequeña de células, que crecen atra-vesando la endodermis, el córtex, la epidermis, para brotar como una raíz joven. La epidermis (rizodermis) estuvo conformada por un estrato de células estrechamente unidas. En el córtex, las células fueron de forma alargada, de paredes delgadas, como las células parenquimáticas. En la endodermis, las células se observaron muy unidas, sin dejar espacios. El periciclo constó de una sola capa de células de paredes delgadas.
En A. thaliana, las raíces laterales se forman a partir de las células del periciclo adyacentes a los polos del xilema. El desarrollo de las raíces late-rales está regulado antagónicamente por auxinas y citoquininas. Mientras que es ampliamente conocido cómo las auxinas promueven el desa-rrollo de raíces laterales, el mecanismo represor de las citoquininas aún no es claro. Elevados ni-veles de citoquininas desorganizan la iniciación de raíces laterales y el patrón de división regular que caracteriza el desarrollo lateral de raíces en Arabidopsis (Laplaze et al., 2007).
ASpECTOS ANATóMICOS DE LA SEMILLA DE pApAyA

REV. COLOMB. CIENC. HORTIC.
152
Figura 1. Tipo de semillas utilizadas para los cortes en micrótomo. A, primordios seminales de 60 días; B, semilla de fruto de 120 días; C, 150 días.
a b C
Figura 2. Panorámica de los primordios seminales (ps) en papaya. Funículo (f), mesocarpo del fruto (mf). aumento de 4X.
mf ps f
Figura 3. primordio seminal con capas celulares concéntricas correspondientes a los tegumentos. Epidermis (e), funículo (f), nucela (n), tegumento externo (te), tegumento interno (ti). Aumento de 10X.
f e n te ti
Figura 4. Vista parcial de semilla de papaya de 120 días. Cubierta seminal (C), endospermo (En), embrión (Em). Aumento de 10X.
C em en
Figura 5. Componentes de la cubierta seminal de la semilla de papaya. Exotesta (ex), mesotesta (mt), endotesta (et) y tegmen (tg). Aumento de 10X.
tg et mt et
GIL/MIRANDA

Vol. 2 - No.2 - 2008
153
Las raíces laterales se originan a partir de un pe-queño número de células diferenciadas situadas en la periferia de los tejidos vasculares. En Ara-bidopsis, las raíces laterales son derivadas a par-tir de las células del periciclo adyacentes a los polos xilemáticos, llamadas células fundadoras del periciclo (Casimiro et al., 2001). Dichas célu-las experimentan un programa definido de divi-siones y expansiones para formar un primordio lateral radicular. El primer paso en el desarrollo lateral de raíces ocurre en las tres filas de células adyacentes al periciclo. Una división anticlinal polarizada tiene lugar en dos células fundadoras por fila conllevando a la formación de dos pe-queñas células hijas encerradas por dos células más grandes (estado I). Divisiones anticlinales, expansión celular y divisiones periclinales dan lugar a un primordio lateral radicular de cuatro capas (estado IV). Más divisiones y expansión resultan en la formación de un estado complejo (estado VI) cuya organización es similar al me-ristemo de la raíz primaria. La expansión celular conlleva a la emergencia de primordios laterales radiculares a partir de la raíz primaria (estado VIII) (Laplaze et al., 2007).
CoNCluSioNeS
La semilla de papaya está constituida por tres partes principales: la cubierta seminal, el endos-permo y el embrión.
Por ser bitegumentada, la cubierta seminal de la semilla de papaya posee testa y tegmen. La tes-ta se compone de la sarcotesta o exotesta jugosa, la cual recubre la mesotesta, compuesta a su vez por la mesotesta externa e interna, y la endotesta, que corresponde a la parte más interna. El tegmen está compuesto por tres partes denominadas exo-tegmen, mesotegmen y endotegmen, en estrecho contacto con el endospermo de la semilla.
El endospermo de la semilla de papaya está cons-tituido principalmente por lípidos y proteínas, almacenados en estructuras denominadas gra-nos de aleurona.
En el embrión se encontró que los cotiledones están compuestos por la epidermis adaxial y abaxial, además de cuatro capas celulares inter-nas ordenadas, y en la zona radicular se observó el esbozo de raíces secundarias laterales.
agraDeCimieNtoS
A William López, laboratorista de la sección de Histotecnia, Departamento de Biología, Universidad Nacional de Colombia, por su colaboración para la realización de los cortes en micrótomo y el teñido del material vegetal presentado en este artículo.
A la profesora Martha de Valencia, Departamento de Biología, Universidad Nacional de Colombia, por su ayuda en la identificación de algunas estructuras anatómicas del material vegetal.
ASpECTOS ANATóMICOS DE LA SEMILLA DE pApAyA

REV. COLOMB. CIENC. HORTIC.
154
Figura 6. Detalle de la exotesta (ex), mesotesta externa (mte) y mesotesta interna (mti) en la semilla de papaya. Aumento de 45X.
mti mte ex
Figura 8. Granos de aleurona (ga) en el endospermo de semilla de papaya. Aumento de 45X.
ga
Figura 9. vista parcial del embrión de la semilla de papaya. Protodermis (pr), parénquima cortical (pc), parénquima medular (pm), procambium (p), cotiledón (co). Aumento de 10X.
co pr pm pc
Figura 10. Detalle del ápice del embrión mostrando la túnica (tun) y el cuerpo (cuer). Aumento de 45X.
tun
Figura 11. vista parcial de las capas celulares de los cotiledones (co). Aumento de 45X.
co
Figura 7. Endotesta (et), exotegmen (extg), mesotegmen (mtg), endotegmen (etg) y cutícula nucelar (cn) en la semilla de papaya. Endospermo (En). Aumento de 45X.
en cn mtg extgetget
p
cuer
Figura 12. Esbozo de raíces secundarias (rs). Endodermis (ed), córtex (cr), periciclo (pr), cilindro central (cc), epidermis (e). Aumento de 10X.
ed
rs
cr pr cc e
GIL/MIRANDA

Vol. 2 - No.2 - 2008
155
Andre, C.; J.R. Froehlich; M.R. Moll y C. Benning. 2007. A heteromeric plastidic pyruvate kinase complex involved in seed oil biosynthesis in Arabidopsis. Plant Cell 19, 2006-2022.
Anil, V.S.; A.C. Harmon y K.S. Rao. 2003. Temporal as-sociation of Ca2+ -dependent protein kinase with oilbodies during seed development in Santalum al-bum L.: Its biochemical characterization and signi-ficance. Plant Cell Physiol.44, 367-376.
Baroux, C.; A. Pecinka; J. Fuchs; I. Schubert y S. Gross-niklau. 2007. The triploid endosperm genome of Arabidopsis adopts a peculiar, parental-dosage-de-pendent chromatin organization. Plant Cell 19, 1782-1794.
Barton, M.K. y R.S. Poethig. 1993. Formation of the shoot apical meristem in Arabidopsis thaliana: an analysis of development in the wild type and in the shoot meristemless mutant. Development 119, 823-831.
Becerra, N. y M. Chaparro. 1999. Morfología y anato-mía vegetal. Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia, Bogotá.
Beeckman, T.; R. de Rycke; R. Viane y R. Inze. 2000. Histological study of seed coat development in Arabidopsis thaliana. J. Plant Res. 113, 139-148.
Besnier, F. 1988. Semillas: biología y tecnología. Edicio-nes Mundi-Prensa, Madrid.
Casimiro, I.; A. Marchant; R. Bhalerao; T. Beeckman; S. Dhoge; R. Swarup; N. Graham; D. Inze; G. Sand-berg; P. Casero y M. Bennet. 2001. Auxin transport promotes Arabidopsis lateral root initiation. Plant Cell 13, 843-852.
Chandler, J.W. 2008. Cotyledon organogénesis. J. Exp. Bot. 59(11), 2917-2931.
Chaudhury, A.M. y F. Berger. 2001. Maternal control of seed development. Cell Dev. Biol. 12, 381-386.
Debeaujon, I.; K.M. León-Kloosterziel y M. Koornneef. 2000. Influence of the testa on seed dormancy, ger-mination, and longevity in Arabidopsis. Plant Phy-siol. 122, 403-413.
Esau, K. 1985. Anatomía vegetal. Ediciones Omega, Bar-celona, España. pp. 641-659.
Fahn, A. 1985. Anatomía vegetal. Ediciones Pirámide, Madrid. pp. 528-554.
reFereNCiaS bibliogrÁFiCaS
Font Quer, P. 1965. Diccionario de botánica. Editorial Labor, Barcelona, España.
Fosket, D.E. 1994. Plant growth and development: a mo-lecular approach. Academic Press, San Diego, CA.
Garcia. D.; J. Fitzgerald y F. Berger. 2005. Maternal con-trol of integument cell elongation and zygotic con-trol of endosperm growth are coordinated to deter-mine seed size in Arabidopsis. Plant Cell 17, 52-60.
García, D.; V, Saingery; P, Chambrier; U, Mayer; G, Jür-gens y F. Berger. 2003. Arabidopsis haiku mutants reveal new controls of seed size by endosperm. Plant Physiol. 131, 1661-1670.
Haig, D. y M. Westoby. 1989. Parent-specific gene ex-pression and the triploid endosperm. Amer. Nat. 134, 147-155.
Imlau, A.; E. Truernit y N. Sauer. 1999. Cell-to-cell and long-distance trafficking of the green fluorescent protein in the phloem and symplastic unloading of the protein into sink tissues. Plant Cell 11, 309-322.
Ingouff, M; P.E. Jullien y F. Berger. 2006. The female ga-metophyte and the endosperm control cell prolife-ration and differentiation of the seed coat in Arabi-dopsis. Plant Cell 18, 3491-3501.
Johansen, D.A. 1940. Plant microtechnique. Mc Graw-Hill Book Company, New York, NY.
Laplaze, L.; E. Benkova; I. Casimiro; L. Maes; S. Van-neste; R. Swarup; D. Weijers; V. Calvo; B. Parizot; M.B. Herrera-Rodríguez; R. Offringa; N. Graham, P. Doumas; J. Friml; D. Bogusz; T. Beeckman y M. Bennet. 2007. Cytokinins act directly on lateral root founder cells to inhibit root initiation. Plant Cell 19, 3889-3900.
Laux, T.; T. Würschum y H. Breuninger. 2004. Genetic regulation of embryonic pattern formation. Plant Cell 16, S190-S202.
León, J. 1987. Botánica de los cultivos tropicales. Segun-da edición. Instituto Interamericano de Coopera-ción para la Agricultura (IICA), San José. pp. 375-379.
Léon-Kloosterziel, K.M.; C.J. Keijzer y M. Koornneef. 1994. A seed shape mutant of Arabidopsis that is affected in integument development. Plant Cell 6, 385-392.
Lopes, M.A. y B.A. Larkins. 1993. Endosperm origin, de-velopment and function. Plant Cell 5, 1383-1399.
ASpECTOS ANATóMICOS DE LA SEMILLA DE pApAyA

REV. COLOMB. CIENC. HORTIC.
156
Nakaune, S.; K. Yamada; M. Nondo; T. Kato; T. Satoshi; M. Nishimura y I. Hara-Nishimura. 2005. A va-cuolar processing enzyme, VPE, is involved in seed coat formation at the early stage of seed develop-ment. Plant Cell 17, 876-887.
Niembro, A. 1988. Semillas de árboles y arbustos: onto-genia y estructura. Limusa, México.
Penfield, S.; Y. Li; A. Gilday; S. Graham e I. Graham. 2006. Arabidopsis ABA INSENSITIVE4 regulates li-pid mobilization in the embryo and reveals repres-sion of seed germination by the endosperm. Plant Cell 18, 1887-1899.
Reiser, L. y R. Fischer. 1993. The ovule and the embryo sac. Plant Cell 5, 1291-1301.
Ren, C. y J.D. Bewley. 1998. Seed development, testa structure and precocious germination of Chinese cabbage (Brassica rapa subsp. pekinensis). Seed Sci. Res. 8, 385-397.
Schruff, M.; M. Spielman; S. Tiwari; S. Adams; N. Fenby y R. Scott. 2006. The AUXIN RESPONSE FACTOR 2 gene of Arabidopsis links auxin signalling, cell division, and the size of seeds and other organs. Development 133, 251-261.
Siegfried, K.R.; Y. Ehed; S.F. Baum; D. Otsuga; G. Drews y J. Bowman. 1999. Members of the YABBY gene family specify abaxial cell fate in Arabidopsis. De-velopment 126, 4117-4128.
Siloto, R.; K. Findlay; A. Lopez-Villalobos; E. Yeung; C. Nykiforuk y M. Moloney. 2006. The accumulation of oleosins determines the size of seed oilbodies in Arabidopsis. Plant Cell 18, 1961-1974.
Skinner, D.; T. Hill y C. Gasser. 2004. Regulation of Ovule Development. Plant Cell 16, S32-S45.
Stadler, R.; C. Lauterbach y N. Sauer. 2005. Cell-to-cell movement of green fluorescent protein reveals post-phloem transport in the outer integument and identifies symplastic domains in Arabidopsis seeds and embryos. Plant Physiol. 139, 701-712.
Szymkowiak, E.J. e I.M. Sussex. 1996. What chimeras can tell us about plant development. Annu. Rev. Plant Physiol. Plant Mol. Biol. 47, 351-376.
Webb, M. y B. Gunning. 1990. Embryo sac development in Arabidopsis thaliana. 1. Megasporogenesis, in-cluding the microtubular cytoskeleton. Sex. Plant Reprod. 3, 244-256.
Western, T.L.; D.J. Skinner y G.W. Haughn. 2000. Diffe-rentiation of mucilage secretory cells of the Arabi-dopsis seed coat. Plant Physiol. 122, 345-355.
Windsor, J.B.; V. Symonds; J. Mendenhall y A.M. Lloyd. 2000. Arabidopsis seed coat development: Morpho-logical differentiation of the outer integument. Plant J. 22, 483-493.
GIL/MIRANDA

reSumeN
Con el propósito de seleccionar nuevos materiales de cítricos para las condiciones ambientales del trópico bajo en el piedemonte del Meta, se evaluó el comportamiento en crecimiento, producción y calidad de 26 geno-tipos entre variedades y cultivares de naranja. Se establecieron seis plantas por cada cultivar en el año 1999 en el Centro de Investigación La Libertad de Corpoica, Villavicencio (Meta, Colombia). Se llevaron registros anuales de variables vegetativas, productivas y de calidad del fruto durante cinco cosechas (2004 a 2008). La clasificación de las variedades en grupos homogéneos se realizó utilizando un análisis de conglomerados que agrupa los objetos que tengan características similares de las variables seleccionadas. Las variables utilizadas fueron: producción, peso fresco del fruto, porcentaje de jugo, sólidos solubles totales (SST o grados Brix), acidez total titulable (ATT) e índice de madurez (SST/ATT). Los genotipos se clasificaron en cinco grupos así: (1) grupo de genotipos de alta producción y calidad de los frutos sobresalientes: Cuban Queen, Pera del río, Enterprise, Pineapple II, Hamlin, Madame Vinois, Palmira Ruby, Naranjal excelente y Salerma; (2) cultivares que presentan una menor producción y calidad de fruto aceptable: García Valencia, Rico, Salustiana, VN7, Lerma, Nativa naranjal, VN8 y Pineapple; (3) cultivares de producción y calidad de los frutos bajas: Galicia, Parson Brown y Naranjal ombligona; (4) frutos con los valores más bajos de producción y calidad (Atwood Navel, Valle Washington y Valle Washington II); (5) genotipos sin producción en las cinco cosechas: Jaffa, Moro Blood y Lane Late.
Javier orlaNDo orDuz-roDríguez1, 2 FraNCiSCo avella1
Comportamiento de 26 cultivares de naranja en condiciones del piedemonte del Meta, Colombia
Performance of 26 orange cultivars in conditions of Piedmont of Meta, Colombia
Naranja Valencia en madurez de cosecha, C.i. la libertad, villavicencio.Foto: H. Velásquez
1 Centro de Investigación La Libertad, Corporación Colombiana de Investigación Agropecuaria (Corpoica), Villavicencio (Meta, Colombia).
2 Autor para correspondencia. [email protected]
REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 2 - No.2 - pp. 157-172, 2008

REV. COLOMB. CIENC. HORTIC.
158
iNtroDuCCióN
additional key words: production, fruit quality, Citrus sinensis, tropical low land, Eastern piedmont.
Fecha de recepción: 04-10-2008 Aprobado para publicación: 02-12-2008
Según la FAO (2004), los cítricos son el prin-cipal cultivo de frutas en el mundo. En los úl-timos 30 años se ha duplicado su producción, pasando de 46 a 96 millones de toneladas de la década de 1970 al año 2005. El área cultivada con cítricos en Colombia se calcula en 43.000 ha para el año 2005 (IICA-MADR, 2005). De acuerdo con información del Dane (2007), en el año 2015 Colombia tendrá una población de 53 millones de habitantes y teniendo en cuenta un consumo per capita de 18 kg de fruta cítrica por año, se estima que para ese año se va a ne-cesitar un millón de toneladas de fruta para el mercado en fresco y para el consumo interno. El país puede necesitar entre 60.000 y 70.000 ha en producción en el año 2020. Lo anterior señala que es necesario aumentar el área de cí-tricos en 15.000 ha y remplazar por lo menos 20.000 ha que terminan su ciclo productivo.
Los llanos orientales poseen cerca de 6.500 ha plantadas con cítricos, que producen entre 100.000 y 120.000 t año-1 y generan alrededor de 1.800 empleos de forma directa. Se calcula que el 60% del área plantada son naranjas. El área de cultivo de cítricos se va a duplicar en los próximos 10 años. La principal debilidad de la citricultura llanera es la excesiva dependencia de una sola variedad por cada especie cultivada siendo para la naranja la variedad Valencia que logra la madurez de consumo 9 meses después de la antesis. Lo anterior señala la necesidad urgente de desarrollar variedades o cultivares de naranja precoces, de producción y calidad sobresaliente en estas condiciones ambienta-les y preferiblemente de madurez temprana o intermedia que permitan ampliar la época de cosecha con el consiguiente beneficio para el productor, al mejorar el precio de venta de
ORDuz-RODRÍGuEz/AVELLA
abStraCt
In order to select new materials of citric crops suitable for the environmental conditions of tropical lowlands in the Piedmont of Meta, were evaluated the growth, production and quality performance of cultivars and varieties of 26 orange genotypes. Six plants were established for each cultivar in 1999 at the Research Center La Libertad of Corpoica, Villavicencio (Meta). Annual records were carried out for vegetative and production variables and fruit quality for five harvests from 2004 to 2008. The classification of varieties in homogeneous groups was performed using a cluster analysis that groups together objects with similar characteristics of selected variables. The variables used were production, fruit fresh weight, percentage of juice, total soluble solids (TSS or Brix grades), total titratable acids (TAA), and maturity index (TSS/TAA). The genotypes were classified into five groups: (1) group of high production genotypes and fruit quality: Cuban Queen, Pera del río, Enterprise, Pineapple II, Hamlin, Madame Vinois, Palmira Ruby, Naranjal excellent, and Salerma, which are premature, give more production and improved fruit quality; (2) cultivars with lower fruit production and acceptable fruit quality: García Valencia, Rico, Salustiana, VN7, Lerma, Native naranjal VN8, and Pineapple; (3) cultivars of low production and fruit quality: Galicia, Parson Brown, and Naranjal ombligona; (4) fruits with the lowest levels of production and quality: Atwood Navel, , Valle Washington, and Valle Washington II; (5) genotypes without fruit production during five harvests: Jaffa, Moro Blood and Lane Late.
palabras clave adicionales: producción, calidad del fruto, Citrus sinensis, trópico bajo, piedemonte llanero.

Vol. 2 - No.2 - 2008
159
la fruta y para el consumidor por tener más oferta de fruta en los meses de septiembre a noviembre, en el cual la oferta de naranja es escasa en el país (Orduz, 2003).
El piedemonte llanero presenta 4.930 unidades de calor acumuladas para el cultivo de los cí-tricos (horas diarias por encima de 12,5ºC que se considera el cero biológico de los cítricos) y se clasifica dentro de las regiones del trópico bajo (Orduz, 2007a) y su cultivo se realiza en suelos ácidos con altos contenidos de aluminio toxico y bajos contenidos de bases (Ca, Mg y K); y bajo contenido de materia orgánica y de elementos menores.
El cultivo comercial de las especies cítricas se basa en la exaltación de los caracteres ge-néticos y en el conocimiento de la influencia que el medio ambiente (factores climáticos y edáficos) ejerce sobre su desarrollo. Las res-puestas fisiológicas de la planta provienen de la interacción del genotipo (compuesto por la variedad y el patrón) con el ambiente y se constituyen en los mecanismos endógenos que controlan el crecimiento y desarrollo de las plantas cítricas (Agustí, 2003). Las varie-dades de naranja para consumo en fresco se han desarrollado para las condiciones de los cinturones citrícolas entre los 22 y 40º de lati-tud N y S, y con la mitad o tercera parte de las unidades de calor acumuladas en condiciones del trópico bajo. Por otro lado, casi toda la in-formación científica y las prácticas de manejo del cultivo de cítricos se han generado para las regiones productoras del subtrópico (la región mediterránea, el sur de América, Australia y California para fruta fresca; y Florida y Sao Paulo para concentrado de jugo de naranja) (Davies y Albrigo, 1994); mientras que el de-sarrollo científico y tecnológico es escaso en las regiones tropicales (Reuther, 1977).
Con el presente trabajo se busca determinar los cultivares que presentan mejor comporta-miento productivo y de calidad de la fruta a las condiciones climáticas del trópico bajo y
representa la primera fase para la selección de nuevos cultivares o variedades recomendados para su cultivo en el piedemonte llanero. El de-sarrollo de nuevos cultivares permitirá forta-lecer la citricultura llanera al tener una mayor opción de recursos genéticos disponibles para ampliar la época de cosecha, buscar la segmen-tación de los mercados y mejorar el retorno de la inversión de los productores de cítricos en las condiciones de evaluación.
materialeS y métoDoS
Los cultivares se recolectaron de lotes experi-mentales de Corpoica en la zona cafetera y de colecciones de clones en la finca El Naranjal de Villavicencio. Los materiales se injertaron en el año 1998 sobre mandarina Cleopatra (Citrus reti-culata Blanco). Este patrón es utilizado tradicio-nalmente en la región debido a su tolerancia a Phytophthora y a las principales enfermedades vi-rales de cítricos transmitidas por injerto (Castle, 1987). En la tabla 1 se presenta la información de los cultivares evaluados, su identificación y pro-cedencia de los materiales evaluados.
Los árboles fueron plantados en abril de 1999 en un lote del C.I. La Libertad de Corpoica en Villavicencio (Meta) (4º03’ N; 73º29’ W, y 336 msnm), con una distancia de plantación de 8 x 6 m (204 plantas/ha). El suelo se clasifica como Typic Haplustox de textura franco arcillosa; es-tos suelos se conocen como suelos clase IV (te-rrazas altas) en la clasificación regional y son los recomendados para cítricos en la región (Orduz y Baquero, 2003); se caracterizan por poseer buen drenaje interno y externo, y adecuada pro-fundidad efectiva. En la tabla 2 se presenta el re-sultado del análisis de químico del suelo del lote utilizado en la evaluación.
Para corregir la acidez y aumentar la saturación de bases al 60% (Ca, Mg) antes del transplan-te se aplicó e incorporó cal dolomita (2 t ha-1) y Escorias Thomas (1 t ha-1) como correctivos, posteriormente se incorporaron con rastra y so-
COMpORTAMIENTO DE 26 CuLTIVARES DE NARANjA EN CONDICIONES DEL p IEDEMONTE

REV. COLOMB. CIENC. HORTIC.
160
tabla 1. identificación y origen de los cultivares de naranja de la investigación.
Textura pH MO P Al Ca Mg K Na Fe B Cu Mn Zn
FArA % mg kg-1 cmol(+) kg-1 mg kg-1
5,0 3,3 17 1,2 0,96 0,34 0,14 0,13 26 0,32 1,8 4,0 1,8
Tabla 2. Análisis químico del suelo del lote experimental.
ORDuz-RODRÍGuEz/AVELLA
No. IdentificaciónCultivares de naranja
(Cirus sinensis)procedencia del material
evaluado
1 A Atwood Navel Zona Cafetera
2 C Cuban Queen Zona Cafetera
3 E Enterprise Zona Cafetera
4 G Galicia Zona Cafetera
5 Gv Garcia Valencia Zona Cafetera
6 H Hamlin Zona Cafetera
7 J Jaffa Zona Cafetera
8 L Lane Late Zona Cafetera
9 Le Lerma Zona Cafetera
10 M Madame Vinois Zona Cafetera
11 m Moro Blood Zona Cafetera
12 VNEx Naranjal excelente Villavicencio
13 N5 Nativa naranjal Zona Cafetera
14 N4 Naranjal ombligona Villavicencio
15 Pa Palmira Ruby Zona Cafetera
16 Pbn Parson Brown Villavicencio
17 Pr Pera del río Zona Cafetera
18 P Pinneapple Zona Cafetera
19 PII Pinneapple II Zona Cafetera
20 R Rico Zona Cafetera
21 Sn Salerma naranjal Villavicencio
22 S Salustiana Zona Cafetera
23 VN7 Valencia naranjal 1 Villavicencio
24 VN8 Valencia naranjal 2 Villavicencio
25 V Valle Washington Zona Cafetera
26 VII Valle Washington II Zona Cafetera

Vol. 2 - No.2 - 2008
161
bre la superficie del suelo se aplicó 1,51 t ha-1 de yeso agrícola. Las dosis fueron calculadas usan-do la fórmula de Malavolta (1995). El área entre las filas fue sembrada con maní forrajero como cobertura viva en el año 2000. Anualmente reci-bieron dos aplicaciones de glifosato en el área de plateo y dos pases de guadaña de tractor entre las filas. La fertilización se aplicó tres veces al año utilizando el fertilizante compuesto 17-6-18-2. Toda la fertilización fue edáfica y la dosis fue creciente siendo 0,5 kg por árbol en el primer año, aumentando la dosis hasta 3 kg por árbol a partir del quinto año. Cada 3 años se aplicó el 30% de los correctivos aplicados al inició, en las calles del cultivo. Solo se aplicó riego el primer verano después del establecimiento del experi-mento. La fertilización, manejo agronómico y sanitario se realizó siguiendo las recomendacio-nes para el cultivo de Orduz y Baquero (2003) para el piedemonte llanero.
El piedemonte del Meta presenta un régimen pluviométrico monomodal en el cual la tem-porada húmeda se inicia en marzo y termina en noviembre. La precipitación promedio anual oscila entre 2.500 y 3.800 mm (en promedio 2.918 mm) y con una temperatura media de
26ºC (Ideam, 1997). La clasificación climática de la región corresponde a bosque húmedo tro-pical (IGAC, 2004). En la figura 1 se presenta el balance hídrico para el lugar de evaluación con información climática de 20 años (Orduz y Fischer, 2007). El estrés hídrico se termina después del inicio de la precipitación que nor-malmente se presenta a mediados del mes de marzo y 15 d después se presenta la antesis de la principal floración anual de la naranja.
Las seis plantas se ubicaron en la misma fila y de forma aleatoria entre cultivares. Anual-mente se evaluaron en cada árbol las variables de crecimiento vegetativo (altura de planta, ancho de copa N-S y E-W, grosor del tronco 10 cm arriba y debajo de la zona de injerto, datos no presentados); estas se tomaron anualmente en la época seca (dic.-feb.) al finalizar la tem-porada de crecimiento. El volumen de la copa fue calculado usando la formula de Turrel (1946), V = 0,5236·H·D2, en el que H = al-tura de la copa y D = diámetro de la copa. La producción se inició al cuarto año, se llevaron registros de producción por árbol (peso de fru-tos, número de frutos y producción por árbol) durante cinco cosechas anuales. Los frutos se
Figura 1. balance hídrico entre la precipitación promedio anual y la evapotranspiración del cultivo de cítricos en el C.I. La Libertad, Villavicencio (Meta).
COMpORTAMIENTO DE 26 CuLTIVARES DE NARANjA EN CONDICIONES DEL p IEDEMONTE
mm
PrecipitaciónEvapotranspiración
050
100150200250300350400450500
Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic Ene Feb Mar

REV. COLOMB. CIENC. HORTIC.
162
cosecharon a partir de noviembre y hasta ene-ro dependiendo de la época de floración. Des-pués de la cosecha se tomó una muestra de 10 frutos por cada parcela, a los cuales se les realizó los análisis de calidad de fruta. Los fru-tos fueron pesados y se les extrajo el jugo con un exprimidor eléctrico. El contenido de jugo fue medido con probeta graduada; los sólidos solubles totales (SST) fueron determinados usando un refractómetro manual y los ácidos totales titulables (ATT) (como equivalente del ácido cítrico) por titulación con NaOH 0,1 N.
Para las variables de rendimiento y volumen de la copa se utilizó la estadística descriptiva. A las variables de calidad del fruto se les cal-culó el promedio con la desviación estándar de la información de seis plantas. Las varia-bles principales de mayor peso en el análisis son: producción (kg/árbol) y calidad del fruto (peso promedio del fruto, porcentaje de jugo, grados Brix e índice de madurez -SST/ATT-. Para la clasificación de las variedades en gru-pos homogéneos, se realizó un análisis de con-glomerados que agrupa los objetos que tenían características similares de las variables selec-cionadas (Díaz, 2002). Este análisis se reali-zó mediante el procedimiento CLUSTER del paquete estadístico Statistic Analysis System (SAS versión 9.1).
reSultaDoS y DiSCuSióN
Comportamiento vegetativo
Volumen de copa
El volumen de la copa es un indicador de la in-fluencia de las condiciones climáticas sobre el cre-cimiento vegetativo de la planta. El volumen de todos los cultivares fue en aumento con relación a los años, de manera que su crecimiento se dio de forma ascendente sin excepción (tabla 3).
Teniendo en cuenta los valores del año 2008, los cultivares García Valencia y Valle Washington, al igual que en la variable altura de la planta, son las que presentan los valores más bajos con volúmenes de 9,88 y 10,67 m3. Los cultivares que indujeron mayor volumen de copa fueron Hamlin, Salustiana, Parson Brown y Madame Vinois con volúmenes de copa entre 37,25 y 40,67 m3. El volumen de copa de estos cultiva-res duplican, triplican y en ocasiones cuadru-plican, el volumen de copa de cultivares como García Valencia (9,88 m3), Valle Washington (10,67 m3), Jaffa (12,27 m3), Moro Blood (12,61 m3), Pineapple (16,68 m3), VN7 (16,98 m3), Naranjal excelente (20,32 m3). Los demás cul-tivares se encuentran en un rango de 20,58 a 35,81 m3. Los cultivares Madame Vinois, Par-son Brown y Salustiana, han desarrollado ár-boles de tamaño mediano a grande en regiones subtropicales (Agustí, 2003), lo que sugiere una amplia capacidad de estos cultivares para adap-tarse a las dos condiciones climáticas.
Los materiales que presentan un mayor de-sarrollo vegetativo estarían presentando una mejor adaptación a la alta acumulación de uni-dades de calor anual prevalentes en las condi-ciones del piedemonte del Meta que es de 4.927 unidades de calor al año (Orduz, 2007b).
Comportamiento productivo
Producción anual y acumulada
La producción promedio anual y la producción acumulada por árbol, permitió agrupar los cul-tivares según el comportamiento productivo de cada una de ellas, de manera que se identifican los cultivares de temprana, media y tardía pro-ducción. El patrón Cleopatra presenta un com-portamiento tardío que disminuye los rendi-mientos en las primeras cosechas (Castle, 1987). En la tabla 4 se presenta el comportamiento me-dio anual de los cultivares evaluados.
ORDuz-RODRÍGuEz/AVELLA

Vol. 2 - No.2 - 2008
163
Variedad 2000 2003 2005 2006 2007 2008atwood Navel 0,4 1.2 6,6 11,4 17,6 20,6
Cuban queen 1,0 6,4 20,9 22,1 25,4 31,1
enterprise 0,9 3,5 20,2 24,3 30,3 35,3
galicia 0,5 1,5 11,2 14,1 16,3 23,5
garcía valencia 0,3 0,3 4,6 6,7 8,2 9,9
hamlin 0,6 3,3 16,3 20,9 28,7 37,2
Jaffa 0,1 1,4 5,5 9,5 9,9 12,3
lane late 0,5 2,3 11,7 17,1 26,0 30,6
lerma 0,5 2,7 11,6 15,8 21,3 25,8
madame vinois 0,7 3,9 22,3 28,1 37,3 40,7
moro blood 0,2 1,9 4,6 7,7 9,3 12,6
Naranjal excelente 0,4 2,9 9,0 13,3 24,6 20,3
Naranjal ombligona 0,5 1,8 20,8 27,3 38,2 39,2
Nativa naranjal 0,4 2,2 9,9 15,4 19,5 27,8
palmira ruby 0,4 3,1 18,9 19,2 21,5 26,9
parson brown 0,4 2,6 14,9 25,3 35,6 40,0
pera del río 0,5 2,5 11,6 16,1 22,2 26,3
pinneaple 0,2 0,9 4,5 8,5 11,8 16,7
pinneaple ii 0,4 3,5 13,0 18,2 27,7 35,8
rico 0,5 2,7 9,5 16,8 20,3 27,6
Salerma 0,2 1,8 9,6 14,7 20,8 29,4
Salustiana 0,7 3,8 16,5 22,0 24,8 38,1
v. Washington 0,4 0,9 3,4 5,5 9,4 10,7
v. Washington ii 0,5 2,5 10,5 14,9 22,7 29,0
vN7 0,2 0,7 5,1 9,6 13,9 17,0
vN8 0,4 3,0 11,1 15,1 19,7 32,2
tabla 3. volumen de copa (m3/planta) de 26 cultivares de naranja. C.I. La Libertad, Villavicencio. Árboles plantados en 1999.
Los cultivares se clasificaron según su compor-tamiento productivo acumulado en cultivares de producción alta: Cuban Queen (145,3 kg/árbol), Pera del río (131,7 kg/árbol), Pineapple II (114,6 kg/árbol), Enterprise (114,1 kg/árbol) y Salerma (95,3 kg/árbol). Estos cultivares no solo presen-taron las producciones más altas sino que se pueden considerar como cultivares precoces. Los cultivares de producciones intermedia fueron: Naranjal excelente (91,9 kg/árbol), Madame Vi-nois (89,1 kg/árbol), Galicia (81,8 kg/árbol), Ha-mlin (78,7 kg/árbol), Naranjal ombligona (77,5 kg/árbol), Palmira Ruby (77,0 kg/árbol), Parson Brown (64,4 kg/árbol) y VN8 (62,5 kg/árbol). Los cultivares que presentaron las produccio-
nes más bajas fueron: Lerma (49,4 kg/árbol), Nativa naranjal (42,9 kg/árbol), Pineapple (42,9 kg/árbol), Rico (34,6 kg/árbol), Salustiana (29,8 kg/árbol), VN7 (29,0 kg/árbol), García Valencia (28,9 kg/árbol), Valle Washington II (22,5 kg/ár-bol), Valle Washington (9,2 kg/árbol) y Atwood Navel (3,5 kg/árbol). Tres de los 26 cultivares (Moro Blood, Lane Late y Jaffa) no presentaron ninguna producción de fruto en los años de eva-luación, descartándolas como cultivares promi-sorios para las condiciones del trópico bajo. La causa de este fenómeno posiblemente sea que los cultivares por ser seleccionados en latitudes altas tienen una baja fotosíntesis neta y, por tanto, no presentan una buena adaptación a las con-
COMpORTAMIENTO DE 26 CuLTIVARES DE NARANjA EN CONDICIONES DEL p IEDEMONTE

REV. COLOMB. CIENC. HORTIC.
164
Variedad 2004 2005 2006 2007 2008 producción acumulada
atwood Navel 0 0 1,6 0 1,9 3,5
Cuban queen 39,2 55,7 20,2 3,6 26,6 145,3
enterprise 12,6 29,1 15,9 4,4 52,2 114,1
galicia 8,6 28,0 19,5 1,8 24,0 81,8
garcía valencia 13,6 0,0 7,4 3,6 4,3 28,9
hamlin 3,7 20,3 28,2 5,4 21,1 78,7
Jaffa 0 0 0 0 0 0
lane late 0 0 0 0 0 0
lerma 6,1 7,4 18,4 0,0 17,4 49,4
madame vinois 10,1 36,8 28,3 2,6 11,3 89,1
moro blood 0 0 0 0 0 0
Naranjal excelente 36,1 0 19,8 6,2 29,8 91,9
Naranjal ombligona 6,4 33,5 16,1 4,6 16,8 77,5
Nativa naranjal 5,6 3,0 11,1 4,1 19,1 42,9
palmira ruby 10,5 44,0 8,4 4,1 10,0 77,0
parson brown 2,2 11,0 12,8 5,8 32,6 64,4
pera del río 4,7 42,1 53,2 8,1 23,5 131,7
pinneaple 0,6 10,5 6,2 2,6 23,0 42,9
pinneaple ii 16,6 25,6 27,0 6,7 38,7 114,6
rico 0,5 1,9 16,5 3,6 12,2 34,6
Salerma 21,6 48,6 14,2 2,6 8,4 95,3
Salustiana 1,3 2,5 22,7 0 3,3 29,8
v. Washington 0 0 3,6 3,4 2,2 9,2
v. Washington ii 3,5 2,1 6,7 1,4 8,7 22,5
vN7 15,1 0 8,0 0,9 5,0 29,0
vN8 8,9 0 38,9 4,3 10,4 62,5
Tabla 4. Producción anual y acumulada por árbol (kg). Durante cinco cosechas de 2004 a 2008 de 26 cultivares de naranja. C.i. la libertad, villavicencio. Árboles plantados en 1999.
diciones ambientales del piedemonte del Meta. Los cultivares del grupo de naranjas de ombligo como Valle Washington y Atwood Navel pre-sentan una baja producción y por consiguiente deficiente adaptación. Estas variedades son tem-pranas y tienen un buen comportamiento en condiciones del mediterráneo con 1.600 a 1.800 unidades anuales de calor acumuladas (Davies y Albrigo, 1994).
Pineapple y Enterprise son mencionadas por Davies y Albrigo (1994) como variedades de
media estación cultivadas principalmente en Florida; presentan producciones altas en con-diciones de trópico bajo; al igual que la Pera del río que es la más cultivada en el estado de Sao Paulo en Brasil para concentrado de jugo de na-ranja, durante el periodo de evaluación.
Calidad de los frutos
Por ser un fruto no climatérico, este pueden lle-gar a presentar disminución de la tasa de res-piración, ocasionando cambios poco notorios
ORDuz-RODRÍGuEz/AVELLA

Vol. 2 - No.2 - 2008
165
principalmente en los contenidos de azucares y ácidos; por lo que es necesario cosecharlo con las características apropiadas para su consumo (Agustí, 2003).
Peso fresco del fruto
Los cultivares Lerma (173,9 g), García Valencia (174,5 g), Pinneaple (177,5 g), Hamlin (180,4 g), Pinneaple II (184,0 g) desarrollaron los pesos promedio más bajos, mientras que los cultivares Nativa naranjal (234,8 g), Naranjal ombligona (264,4 g), Valle Washington (295,1 g), Valle Was-hington II (313,4 g) y Atwood Navel (339,5 g) presentaron los pesos promedios más altos den-tro del estudio; las demás variedades presentaron pesos promedios de frutos desde 192,4 g (Pera del río) hasta 228,9 g (Naranjal excelente) (figura 2).
Los cultivares que presentan el mayor tamaño de naranja son los que tienen una menor produc-ción de los cultivares evaluados y, por tanto, no presentan ningún interés desde el punto de vis-ta comercial. Los cultivares de naranja que pro-duzcan frutos de menos de 200 g presentan un menor interés en el mercado de fruta para con-
sumo en fresco; dentro de estas se encuentran los cultivares Pineapple y la Hamlin. Figueiredo (1991) reporta que para las condiciones de Sao Paulo en Brasil, ‘Hamlin’, ‘Ruby’, ‘Pera del río’ y las Valencias presentan unos pesos de fruto inferiores a los registrados bajo las condiciones de evaluación; aspecto de menor interés para las frutas destinadas a la agroindustria como es el caso de ese estado de Brasil.
Contenido de jugo
Las variedades que presentaron los mejores cin-co contenidos promedios de de jugo en cinco cosechas fueron; Salustiana (45,6%), Naranjal Excelente (47,6%), Nativa naranjal (48,3%), VN8 (48,29%) y García Valencia (49,13%) (figura 3).
El contenido de jugo de algunas variedades está por debajo del 40%, porcentaje mínimo de jugo para la aceptación comercial, por lo que esta va-riable permite descartar cultivares que no pre-senten una calidad de frutos deseable. Este es el caso de las Navel, ‘Pinneaple’, ‘Parson Brown’, ‘Hamlin’ y ‘Cuban Queen’. Aunque en el caso de estas tres últimas por ser variedades tempranas
Figura 2. peso fresco promedio de los frutos de cinco cosechas de 23 cultivares de naranja, nueve meses después de antesis, en villavicencio. las barras sobre las columnas indican la desviación estándar.
COMpORTAMIENTO DE 26 CuLTIVARES DE NARANjA EN CONDICIONES DEL p IEDEMONTE
0
50
100
150
200
250
300
350
400
P H PII Pr R Pbn Pa C G S E M Sn VN7 VN8 VNEx VN5 VN4 VII V A
Cultivar
Peso
fresc
ode
fruto
(g)

REV. COLOMB. CIENC. HORTIC.
166
o intermedias, al evaluar la calidad de la fruta en el noveno mes después de antesis es posible que estén afectadas por este hecho.
Sólidos solubles totales (SST)
La variedad Valle Washington es la única que presenta valores por debajo de 8,0, con 7,8 ºBrix (figura 4). Las variedades VN8 (9,5 ºBrix), Na-ranjal excelente (9,7 ºBrix), Naranjal ombligo-na (9,9 ºBrix), Lerma (10,1 ºBrix) y VN7 (10,8 ºBrix), son los cultivares con mayor contenido de SST. Cuatro de los cinco materiales pertene-cen a los cultivares regionales, lo que señala un aspecto de gran interés con estos cultivares si se mejorara la precocidad y la producción total al ser evaluados en patrones que transmitan esas características a las copas.
Davies y Albrigo (1994) caracterizan las varieda-des Valencia y las ombligonas como las de altos niveles de SST. Estos autores reportan que el cul-tivar Hamlin presenta los SST más bajos de todos
los cultivares importantes de naranja dulce; sin embargo bajo condiciones de piedemonte llanero, desarrolla un buen contenido de SST, superando incluso a cultivares de gran importancia comercial como Pera del río. El cultivar Pineapple se carac-teriza por presentar altos contenidos de SST en condiciones de Florida (Davies y Albrigo, 1994). Figueiredo (1991) reporta que bajo condiciones de Sao Paulo los cultivares del grupo de las blancas presentan contenidos de SST superiores a los pre-sentados bajo condiciones de piedemonte llanero. El promedio de los cultivares evaluados muestra concentraciones de SST cercanos o superiores a 8 y el promedio no supera a los 10 ºBrix; es probable que los días cortos de las condiciones tropicales y las altas temperaturas medias impidan una ma-yor diferencia entre cultivares de diferente origen; también, la cantidad total de grados Brix por fru-to varía con el contenido de jugo. Se deben reali-zar estudios específicos sobre estos aspectos para determinar cuál o cuáles cultivares pueden ser entregados a los productores teniendo en cuenta el mercado objetivo.
ORDuz-RODRÍGuEz/AVELLA
Figura 3. Contenido de jugo por fruto promedio de cinco cosechas de 23 cultivares de naranja nueve meses después de antesis, en villavicencio. las barras sobre las columnas indican la desviación estándar.

Vol. 2 - No.2 - 2008
167
Figura 4. Concentración de sólidos solubles totales (ºBrix) promedio de cinco cosechas de 23 cultivares de naranja nueve meses después de antesis, en villavicencio. las barras sobre las columnas indican la desviación estándar.
Acidez total titulable (ATT)
Las variedades regionales VN7 (0,9%), Naranjal excelente (1,0%), VN8 (1,0%) y Nativa naranjal (1,0%), al igual que en los valores de grados Brix, presentan los porcentajes de ácido cítrico más al-tos; lo que permite deducir que estas variedades presentan buenas características de calidad de las frutas. Las demás variedades se encuentran en un rango entre 0,3% (Parson Brown) y 0,9% (García Valencia) (figura 5).
La variedad Galicia se considera como de alto porcentaje de acidez bajo condiciones de Pal-mira, Colombia (Sinclair, 1961), presentando similares características bajo las condiciones del trópico bajo. Figueiredo (1991) reporta que bajo condiciones de Sao Paulo, los culti-vares del grupo de las blancas, presentan un porcentaje de ácido cítrico superior a los cul-tivares bajo condiciones de piedemonte llane-ro. La disminución de la ATT está influida en primer orden por la temperatura (acumulación
de unidades de calor) (Davies y Albrigo, 1994) y por el patrón. Algunos autores (Reuther y Ríos-Castaño, 1969; Davies y Albrigo, 1994) han identificado que cuanto más alto es el ré-gimen térmico día/noche, más baja es la con-centración de ácidos. El descenso en la acidez se ha atribuido a la rápida respiración de áci-dos orgánicos que se incrementa al aumentar las temperaturas medias. Por este motivo las variedades nativas se destacan de las demás presentando una mayor acidez, lo que permite una cosecha tardía sin perder las característi-cas organolépticas para su consumo. Los culti-vares de baja o media acidez acompañados con adecuados rendimientos pueden tener interés para su desarrollo como variedades tempranas o intermedias en el trópico bajo como es el caso de Parson Brown, Enterprise, Pineapple, Madame Vinois y Pera del río. Se confirma la mayor acidez de los cultivares del grupo Va-lencia lo que les permite ser de cosecha tardía realizándose su recolección entre 9 a 12 meses en los cultivos de la región.
COMpORTAMIENTO DE 26 CuLTIVARES DE NARANjA EN CONDICIONES DEL p IEDEMONTE
Cultivar
0
2
4
6
8
10
12
14
Le Gv P H PII Pr R Pbn Pa C G S E M Sn VN7 VN8 VNEx VN5 VN4 VII V A
°Brix

REV. COLOMB. CIENC. HORTIC.
168
Figura 5. Acidez total titulable (% ácido cítrico) promedio de cinco cosechas de 23 cultivares de naranja nueve meses después de antesis, en villavicencio. las barras sobre las columnas indican la desviación estándar.
Índice de madurez (SST/ATT)
Las variedades que presentaron los valores más bajos en esta variable fueron Salerma (8,5), Na-tiva naranjal (8,9), VN8 (9,0), Naranjal excelente (9,6), Cuban Queen (10,4), VN7 (10,5), García Valencia (10,7) y Pera del río (10,7). Las varie-dades con valores más altos fueron: Atwood Navel (18,7), Naranjal ombligona (23,3), Galicia (25,5), Valle Washington (25,7) y Parson Brown (27,6).
La alta acumulación de unidades de calor influye sobre el mejor momento de consumo de la fruta, por la influencia que tiene la acidez sobre el ba-lance de los SST/ATT, que determina el momen-to de madurez (Reuther y Ríos-Castaño, 1969). Los mejores valores para el consumo de la fruta están entre un índice de madurez de 10 hasta 12 ó 13. Los cultivares que presentan un bajo índice de madurez a los 9 meses después de la antesis pueden tener interés para ser evaluados como va-riedades tardías ya que podrían almacenarse en el árbol un mayor periodo.
Figueiredo (1991) reporta que bajo condiciones de Sao Paulo, el cultivar Hamlin presenta un índice de madurez menor al que presentó bajo condiciones de trópico bajo. Además reporta que cultivares como Ruby, Pera del río y las Valencia presentan un índice de madurez superior en con-diciones de Sao Paulo, que los presentados por estos cultivares en el piedemonte llanero.
Análisis de componentes principales de los cultivares evaluados
El cluster permitió agrupar las variedades de ma-nera que se identificaron los cultivares que pre-sentaron características similares de las variables más importantes en producción y calidad, como son producción acumulada, peso promedio del fruto, contenido de jugo, SST e índice de madu-rez; teniendo en cuenta que estas representan interés tanto para el productor como para el con-sumidor.
El dendrograma permite agrupar los cultivares de acuerdo al comportamiento similar que presen-
ORDuz-RODRÍGuEz/AVELLA
Le Gv P H PII Pr R Pbn Pa C G S E M Sn VN7 VN8 VNEx VN5 VN4 VII V A0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
CultivarCultivar
Ácid
ocí
trico
(%)

Vol. 2 - No.2 - 2008
169
Figura 6. Índice de madurez (SST/ATT) promedio de cinco cosechas de 23 cultivares de naranja nueve meses después de antesis, en villavicencio. las barras sobre las columnas indican la desviación estándar.
tan bajo condiciones de piedemonte, de manera que se distinguen cinco grandes grupos hasta la quinta cosecha (figura 7):
1. Cultivares con sobresaliente calidad de los frutos y producción por planta. Estos cultivares se caracterizan por ser precoces o de producciones tempranas, presentan las mayores producciones y buena calidad de fruto, señalando que son cultivares que de-ben continuar en evaluación para llegar a ser recomendados a los citricultores del piede-monte llanero. Estos cultivares son: Cuban Queen, Pera del río, Enterprise, Pinneaple II, Hamlin, Madame Vinois, Palmira Ruby, Na-ranjal excelente y Salerma. De este grupo tie-nen menor interés los que producen naranjas pequeñas como el caso de la ‘Hamlin’.
2. Cultivares de buena producción y ca-lidad de frutos. En este grupo se ubican los cultivares regionales considerados como tardíos que presentan una calidad de fruto
aceptable y otros introducidos de buen com-portamiento. Estos cultivares son: García Valencia, Rico, Salustiana, VN7, Lerma, Na-tiva naranjal, VN8 y Pineapple; este último presenta valores muy buenos en la calidad de los frutos pero su producción es muy baja y debería ser evaluada en otros patrones que mejoren estas características.
3. Cultivares de producción y calidad de frutos regular. Este grupo presenta una baja producción acumulada y deficiente ca-lidad de los frutos. Estos cultivares son: Ga-licia, Parson Brown y Naranjal ombligona. Esta colección muestra las mejores caracte-rísticas de interés frutícola y debería ser eva-luada en otro patrón que acentúe precocidad y producción.
4. Cultivares de producción y calidad de frutos no aceptable. A pesar de que son cultivares que presentaron producción, fue-ron los rendimientos más bajos y su cali-
COMpORTAMIENTO DE 26 CuLTIVARES DE NARANjA EN CONDICIONES DEL p IEDEMONTE

REV. COLOMB. CIENC. HORTIC.
170 ORDuz-RODRÍGuEz/AVELLA
Figu
ra 7
. D
endr
ogra
ma
de 2
6 cu
ltiva
res
de N
aran
ja. C
.I. L
a Li
bert
ad, V
illav
icen
cio.
Árb
oles
pla
ntad
os e
n 19
99.

Vol. 2 - No.2 - 2008
171
dad fue muy regular, demostrando su poca adaptación a las condiciones del piedemonte llanero. Estos cultivares son: Atwood Navel, Valle Washington y Valle Washington II. Estos genotipos pueden presentar su mejor comportamiento en regiones tropicales de mayor altitud, como la zona cafetera de Co-lombia.
5. Cultivares sin producción. En este grupo se encuentran las variedades que no presen-taron producción y adaptación a las condicio-nes del trópico bajo del piedemonte llanero, que son ‘Jaffa’, ‘Moro Blood’ y ‘Lane Late’.
CoNCluSioNeS
•Las variedades Cuban Queen, Madame Vinois, Salerma, Palmira Ruby, Enterprise, Hamlin y Pinneaple II presentaron los resultados más favorables para las primeras cinco cosechas, re-ferente a las variables de producción y calidad de los frutos; siendo el grupo de cultivares más
precoces de los evaluados y muestran su adap-tación a las condiciones del trópico bajo.
•Las variedades Jaffa, Moro Blood y Lana Late, Atwood Navel confirman su escasa adapta-ción en condiciones de alta acumulación de unidades de calor.
•Los cultivares Valle Washington, Valle Was-hington II y García Valencia, recomendados para su cultivo en las condiciones de Palmira (Valle), bajo condiciones de piedemonte llane-ro, presentan valores bajos en producción.
•El uso del patrón Cleopatra (tolerante a enfer-medades) retrasa la entrada a producción de los materiales evaluados.
•La acumulación de unidades de calor puede ser el principal aspecto involucrado en las diferentes respuestas obtenidas por los genotipos evaluados en las condiciones del trópico bajo y debe tenerse en cuenta en los programas de desarrollo y selec-ción de variedades en la citricultura tropical.
agraDeCimieNtoS
Los autores expresan sus agradecimientos a los directivos y personal administrativo de la Cor-poración Colombiana de Investigación Agropecuaria (Corpoica), C.I. La Libertad. A los señores Heberth Velázquez, Capitolino Ciprian, Alfredo Pardo, David López y la compañera Melba Mora, por el acompañamiento y apoyo durante el desarrollo del proyecto; y al agrónomo Daniel Acosta por la colaboración en el documento final.
COMpORTAMIENTO DE 26 CuLTIVARES DE NARANjA EN CONDICIONES DEL p IEDEMONTE

REV. COLOMB. CIENC. HORTIC.
172
Agustí, M. 2003. Citricultura. Ediciones Mundi-Prensa, Madrid.
Castle, W.S. 1987. Citrus rootstocks. pp. 361-369. En: Rom, R.C. y Carlson, R.C. (eds.). Rootstocks for fruit crops. John Wiley and Sons, New York, NY.
Dane, 2007. Indicadores de la dinámica demográfica. Bogotá.
Davies, F.S. y L.G. Albrigo. 1994. Citrus. CAB Interna-tional, Wallingford, UK.
Díaz, L.G. 2002. Estadística multivariada: inferencia y métodos. Departamento de Estadística, Facultad de Ciencias, Universidad Nacional de Colombia, Bogotá.
FAO. 2004. Citrus fresh and processing. Annual statistics. FAO, Roma.
Figueiredo, J.O. 1991 Variedades copas. pp. 228-264. En: Rodriguez, O. (ed.). Citricultura brasileira. Vol. 1. 2a ed. Fundacao Cargill, Campinas, Brasil.
IDEAM. 1997. Balance hídrico climático de las estacio-nes Aeropuerto Vanguardia, Barranca de Upia, Ca-rimagua, Lejanías y Villavicencio, Colombia.
IGAC. 2004. Estudio general de suelos y zonificación de tierras, Departamento de Meta. Bogotá.
IICA-Ministerio de Agricultura y Desarrollo Rural. 2005. Cadena de cítricos. pp. 613-670. En: La competitividad de las cadenas agroproductivas en Colombia. Análisis de su estructura y dinámica (1991-2004). Observatorio de Agrocadenas, Anua-rio 2004. Bogotá.
Malavolta, E. 1995. Nutrición y fertilización de los cí-tricos. pp. 130-170. En: Guerrero, R. (ed.). Fertili-zación de cultivos de clima medio. Monómeros Colombo Venezolanos, Bogotá.
reFereNCiaS bibliogrÁFiCaS
Orduz, J.O. 2003. Enfoque de la investigación de cítri-cos en los llanos orientales de Colombia. Revista Achagua 7, 3-6.
Orduz, J.O. 2007a. Estudios ecofisiológicos y caracteri-zación morfológica y molecular de la mandarina ‘Arrayana’ (Citrus reticulata Blanco) en el piedemon-te llanero de Colombia. Tesis doctoral. Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá.
Orduz, J.O. 2007b. Ecofisología de los cítricos en el trópi-co: revisión y perspectivas. pp. 67-76. Memorias Se-gundo Congreso Colombiano de Horticultura, So-ciedad Colombiana de Ciencias Hortícolas, Bogotá.
Orduz, J.O. y J. Baquero. 2003. Aspectos básicos para el cultivo de los cítricos en el piedemonte llanero. Revista Achagua 7, 7-20.
Orduz, J.O. y G. Fischer. 2007. Balance hídrico e influencia del estrés hídrico en la inducción y desarrollo floral de la mandarina ‘Arrayana’ en el piedemonte llanero de Colombia. Agron. Colomb. 25(2), 255-263.
Reuther, W. 1977. Citrus. pp. 409-439. En: Alvim, P. y T. Kozlowski (eds.). Ecophisiology of tropical crops. Academic Press, New York, NY.
Reuther, W. y D. Ríos-Castaño. 1969. Comparison of growth, maduration and composition of citrus fruit in subtropical California and tropical Colom-bia. Proc. First Intl. Citrus Symp. 3, 277-300.
Ríos-Castaño, D. 1967. Naranjas criollas en Colombia. Revista ICA 2(4), 37-51.
Sinclair, W.B. 1961. The orange: its biochemistry and physiology. Univ. California. Div. Agric. Science, Riverside, CA.
Turrel, F.M. 1946. Tables of surfaces and volumes of spheres and of prolates and oblates spheroides and spheroidal coefficients. University of California Press, Berkeley, CA.
ORDuz-RODRÍGuEz/AVELLA

eNrique miguel Combatt C.1, 3 alFreDo De JeSúS Jarma o.1 aleJaNDro CleveS l.2
Efecto del encalamiento de suelos sulfatados ácidos en Córdoba (Colombia) sobre el fruto de piña (Ananas comosus (L.) Merr.)
Effect of liming of acid sulfate soils in Cordoba (Colombia) on pineapple (Ananas comosus (L.) Merr.) fruit
piña Criolla iniciando fructificación. Foto: A. jarma O.
reSumeN
En el departamento de Córdoba (Colombia) existen entre 20.000 y 30.000 ha de suelos sulfatados ácidos (SSA), los cuales causan toxicidad principalmente por la elevada solubilidad del aluminio, poca disponibilidad de fósforo, baja saturación de las bases y salinidad, entre otros. Lo anterior es más grave aún si se considera que existe un gran vacío en el conocimiento del comportamiento de especies como la piña en este tipo de suelos. Por lo anterior, el presente trabajo determinó el efecto de tres niveles de encalamiento en SSA del municipio de Ciénaga de Oro (Córdoba), sobre algunos aspectos de la calidad del fruto de dos variedades de piña Cayena lisa y Reina de Zarandelo o Criolla. Se realizó un diseño de parcelas divididas bajo bloques completamente al azar en donde las parcelas principales fueron los genotipos y las subparcelas las diferentes dosis de cal agrícola (3, 6 ó 9 t ha-1 de CaCO3). Los resultados indicaron que tanto el diámetro ecuatorial como la firmeza del fruto fueron dependientes solamente de la variedad; el diámetro del fruto de Cayena lisa fue superior en 10% y la firmeza de ‘Criolla’ fue superior en 26%. Los valores de pH del fruto indicaron que esta variable es afectada por la interacción del genotipo con la dosis de encalamiento, registrando un comportamiento menos estable la variedad ‘Cayena lisa’ cuando se varían los niveles de cal en el suelo.
1 Facultad de Ciencias Agrícolas, Universidad de Córdoba, Montería (Colombia). 2 Facultad Seccional Duitama, Universidad Pedagógica y Tecnológica de Colombia, Duitama (Colombia). 3 Autor para correspondencia. [email protected]
REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 2 - No.2 - pp. 173-179, 2008
palabras clave adicionales: enmiendas, SSA, recuperación de suelos, fruticultura.

REV. COLOMB. CIENC. HORTIC.
174
abStraCt
In the province of Cordoba (Colombia), exist between 20,000 and 30,000 ha of acid sulphate soils (ASS). The chemical problems that present ASS for crop production include toxicity, principally due to the high solubility of aluminum, low availability of phosphorus, low saturation of bases, salinization, among others. There is a lack of knowledge about the behavior of species, such as pineapple in this type of soils, for which was studied the effect of three liming levels of ASS located in the municipality Cienaga de Oro on yield and quality of fruits of two pineapple varieties Cayena Lisa and Reyna de Zarandelo or Criolla. The experiment was done in a completely randomized block design, where the principal plots were genotypes and the subplots were the different doses of agricultural lime (3, 6 or 9 t ha-1 CaCO3). The results indicated that both the equatorial diameter and the fruit firmness were dependent only on variety; the fruit diameter of ‘Cayena Lisa’ was 10% wider than that of fruits of ‘Criolla’ and firmness values of ‘Criolla’ was 26 % higher than that values of Cayena. The pH values indicated that this variable is affected by the interaction genotype-dose of liming registering a less stable behavior of the variety Cayena Lisa when the levels of lime are changed in soil.
additional key words: amendments, ASS, soils recovery, pomology.
Fecha de recepción: 03-09-2008 Aprobado para publicación: 02-12-2008
La toxicidad por aluminio limita en 30% la pro-ducción de los cultivos en los suelos arables del mundo que incluyen muchas áreas del trópico y subtrópico, donde predomina el aluminio en forma de Al3+, lo cual afecta el crecimiento de las raíces y aumenta la deficiencia de nutrientes (Nowak y Friend, 2005; Pérez y Martínez, 2006).
Los suelos sulfatados ácidos (SSA) reducen con-siderablemente la productividad agrícola en el trópico; el ácido soluble disminuye el pH a tal punto que las altas concentraciones de iones de hidrógeno en la solución del suelo promueven el ataque a la estructura cristalina, lo que permite que se liberen iones de aluminio que luego pasan a ocupar lugares en los sitios de cambio (Bello y Gómez, 2001). Los procesos de oxidación tam-
iNtroDuCCióN
bién contribuyen en estos suelos a la composi-ción de sales ácidas influyendo en conductivi-dades eléctricas hasta de 10 dS m-1, además de afectar su estructura, constituyendo suelos ma-sivos e impermeables por efectos cementantes y floculantes de los compuestos de oxihidróxido de hierro y aluminio (NSWDLWC, 1999).
Algunas especies han creado mecanismos natu-rales de adaptación a este tipo de suelos. Diver-sos autores indican procesos eficientes como la exudación del aluminio a través de ácidos orgá-nicos por las raíces, aunque esto no está comple-tamente identificado (Watanabe y Osaki, 2002; Taylor et al., 2000). Otros mecanismos de resis-tencia al aluminio también han sido reportados, como el incremento del pH en la rizosfera y la
COMBATT C./jARMA O./CLEVES L.

Vol. 2 - No.2 - 2008
175
formación de complejos de aluminio con ácidos orgánicos o quelatos, que secuestran esos com-plejos en las vacuolas.
Para contrarrestar los efectos de los SSA, ge-neralmente se ha usado el encalamiento con el propósito de neutralizar la acidez ocasionada por el hidrógeno, aluminio, manganeso, hierro y ácidos de la materia orgánica, con el fin de permitir el crecimiento normal de las plantas (Welch et al., 2009). Sin embargo, algunos auto-res dicen que esta práctica es a menudo cara y poco eficiente en muchas partes de los trópicos (Sierra et al., 2003), y otros recomiendan realizar lavados antes de encalar y posteriormente ferti-lizar (Mathew et al., 2001).
En Colombia, la piña (Ananas comosus (L.) Merr.) se cultiva en una amplia gama de suelos, sin que en muchos casos se hagan las aplicaciones o co-rrectivos necesarios, y peor aún, no existe infor-mación sobre el comportamiento de esta en SSA. Se usan suelos con pH desde 3,7 en Lebrija (San-tander), hasta 7,5, en Yotoco (Valle del Cauca) y contenidos nutricionales muy diferentes (Sala-zar, 1994). La piña es una planta exigente en nu-trientes; extrae aproximadamente 350, 30 y 500 kg ha-1 de N, P y K respectivamente en cultivos con una densidad de población de 50.000 plantas/ha (Teixeira et al., 2002). El peso y la calidad del fruto de la piña dependen, entre otros factores, especialmente del estado nutricional y el estado de crecimiento alcanzado por la planta cuando se da la diferenciación floral (Cunha y Cabral, 1999). Según Paula et al. (1998), los nutrientes más exigi-dos por piña son K, N y Ca.
El fruto de la piña es una infrutescencia relativa-mente comprimida formada por “frutillos” que maduran en orden creciente de abajo hacia arriba (Coppens y Leal, 2003), de modo que los de la parte inferior tienen edades fisiológicas mayores que los de las partes medianas y superior, lo que puede resultar en variaciones muy significati-vas en atributos de calidad de la pulpa del fruto (Reinhardt, 2004).
materialeS y métoDoS
El trabajo se realizó en un SSA del municipio de San Carlos, Vereda La Coroza (departamento de Córdoba, Colombia), ubicada a 08º55’47,5” N y 075º42’16,5” W a una altura de 15 msnm, hume-dad relativa promedio de 85%, precipitación de 1.250 mm año-1 y temperatura media anual de 28ºC. Se usó un diseño experimental de bloques completos al azar con estructura de parcelas divididas, donde las parcelas principales fueron los genotipos de piña ‘Cayena lisa’ y ‘Reina de Zarandelo’ o Criolla y las subparcelas fueron las dosis de cal (3, 6 y 9 t ha-1). La siembra se realizó 30 d después del encalamiento (dde). La fertili-zación se realizó con base en los resultados del análisis de suelo en cantidades por hectárea de 69 kg de N (urea), 46 kg de fosfato diamónico en presiembra (P2O5) y 60 kg de K como K2O.
Al observar un término medio de maduración de los frutos (alrededor de 150 d después de la for-mación de la inflorescencia), sobre cinco frutos escogidos al azar por tratamiento se midieron las variables resistencia a la penetración, medida en kg cm-2 y diámetro ecuatorial, medido en cm y pH. Para el análisis estadístico se usó el pa-quete Statistical Analysis System (SAS), versión 6.0 de la Universidad de Córdoba; la compara-ción de los efectos simples se realizó mediante la prueba de comparación de medias DMS a un nivel del 5% de probabilidad y las interacciones que resultaron se descompusieron al mismo ni-vel de significancia con tablas de doble entrada, midiendo los efectos de los niveles del factor ge-notipo dentro de los niveles del factor dosis y viceversa.
reSultaDoS y DiSCuSioN
El análisis de los cuadrados medios de las varia-bles dependientes indicó que la resistencia y el diámetro ecuatorial del fruto fueron afectados significativamente de manera exclusiva por el genotipo (P≤0,05), pero el pH registró un com-
EFECTO DEL ENCALAMIENTO DE SuELOS SuLFATADOS ÁCIDOS SOBRE EL FRuTO DE p IñA

REV. COLOMB. CIENC. HORTIC.
176
portamiento que es afectado por la interacción del genotipo y la dosis de cal utilizada (tabla 1).
Resistencia y diámetro ecuatorial del fruto
Estas variables solo fueron afectadas por el efec-to del genotipo indicando que no son influencia-das por el nivel de encalamiento y que sus valo-res son explicados por el grado de madurez del fruto de cada variedad, por lo que solo se discu-tirá este aspecto. Los resultados indicaron que la variedad Criolla registró mayores promedios de resistencia a la penetración (P≤0,05) respecto a Cayena lisa (8,47 y 6,29 kg cm-2 respectivamen-te) (tabla 2). También se observan los valores de diámetro ecuatorial del fruto, registrándose un comportamiento similar y consecuente con el de resistencia, en el sentido que los frutos con menor grado de madurez en este caso de la va-riedad criolla, registraron 0,9 cm menos de lon-gitud (P≤0,05), respecto a la variedad Cayena lisa que mostró valores promedio de 9,19 cm. Esta última variedad por tanto, registraría fru-tos de piña de mayor diámetro ecuatorial y con un mayor grado de madurez respecto a los de criolla en esta región del país, cuando es cultiva-da en SSA de manera independiente al nivel de encalamiento.
A pesar de que algunos estudios confirman que a los 150 d después de la aparición de la inflo-rescencia se presenta un estado de madurez in-termedio en términos generales para la especie (Arias y Toledo, 2000), estos resultados serían los primeros reportes sobre la fenología y la re-sistencia del fruto de estas variedades para las condiciones de Córdoba, por lo que es difícil re-ferenciarlos con otros trabajos.
Aunque el estado de madurez en piña depende en gran medida del genotipo (Carvalho et al., 2005), es importante considerar que las condi-ciones ambientales y algunos factores de estrés como la salinidad del suelo, pueden inducir ma-yor síntesis de etileno endógeno que probable-mente derivan en una mayor precocidad a flora-ción (Bernal, 2006; Bartholomew y Malézieux, 1994; Bartholomew y Kadzimin, 1997).
Trabajos que describan el desarrollo del fruto y de sus características en esta región del país son escasos. Algunos valores de resistencia en piña (Ananas comosus cv. MD-2) han sido reportados por De la Cruz et al. (2007), en trabajos de pos-cosecha con rangos que van desde 12 (protegidas del daño por frío) a 2 kg cm-2 (sin proteger). Por su parte, en un trabajo publicado por la Univer-sidad de Antioquia (1993), se reportan valores de 10,43 cm en el diámetro central de la variedad Cayena lisa para las condiciones del Valle del Cauca.
tabla 1. Cuadrados medios y significancia estadística del diámetro ecuatorial, resistencia a la penetración y ph del fruto de dos variedades de piña cultivadas en suelos sulfatados ácidos de Ciénaga de oro (Córdoba, Colombia).
F de v resistencia Diámetro ecuatorial ph
Variedad (V) 38,06 * 13,08 * 0,2363 **
Dosis (D) 4,68 ns 2,97 ns 0,0104 **
V x D 0,91 ns 1,51 ns 0,0136 **
CV (%) 13,10 11,70 11,4
ns = no significativo; * y ** significativos al 10 y 5% respectivamente según el test de DMS.
tabla 2. promedios de las mediciones del diámetro ecuatorial y resistencia a la penetración del fruto de dos variedades de piña cultivadas en suelos sulfatados ácidos de Ciénaga de oro (Córdoba, Colombia).
*Promedios con letras distintas en la misma columna indican diferencia significativa según el test protegido de DMS (P≤0,05).
Resistencia (kg cm-2)
Diámetro ecuatorial (cm)
Cayena lisa 6,29 a (± 0,727) 9,19 a (± 0,769)
Criolla 8,47 b (± 2,187) 8,31 b (± 1,361)
CV (%) 23 11,7
COMBATT C./jARMA O./CLEVES L.

Vol. 2 - No.2 - 2008
177
pH del fruto
Tal como se indicó anteriormente, los resulta-dos del pH del fruto fueron diferenciales con la variedad al interactuar con el nivel de enca-lamiento. La tabla 3 registra la descomposición de la interacción, la cual se analizó inicialmente en función de la respuesta de la variedad dentro de cada nivel de encalamiento (tabla 3a) y pos-teriormente en función de la respuesta de cada nivel de encalamiento dentro del factor variedad (tabla 3b).
Al considerar el efecto de la variedad dentro de cada dosis de encalamiento, los resultados indi-caron que las variedades se comportaron de ma-nera similar en cualquier nivel de encalamiento siendo siempre el valor del pH del fruto menor en ‘Criolla’ respecto a ‘Cayena lisa’. Estos resul-tados no explicarían los efectos significativos registrados en el análisis de la interacción, por lo que se estudiaron los efectos de las dosis den-tro de las variedades (tabla 3b). En este caso, sí se detectaron diferencias (P≤0,05), que indican que para los frutos de la variedad Cayena lisa el pH presenta un comportamiento errático en función del nivel de cal aunque con tendencia a ser menor en la medida que se aumenta el en-calamiento. A diferencia de Cayena lisa, para la variedad Criolla, el pH es significativamente mayor cuando se aplica cal en cualquiera de las tres dosis estudiadas, sugiriendo una respuesta directa del pH del fruto al encalamiento.
Lo anterior indicaría que la eficiencia de la prácti-ca de encalamiento podría ser explicada en parte porque la cal influyó en la neutralización de la acidez generada por el Al3+ permitiendo la dispo-nibilidad de otros nutrientes como el nitrógeno, lo cual tendría consecuencias directas sobre el pH del fruto (Watanabe y Osaki, 2002; Taylor et al., 2000). Este efecto podría estar explicado además porque la variedad Criolla, por presentar mayores niveles de adaptación a los suelos y condiciones
de la región, podría presentar mecanismos de to-lerancia al Al3+ más eficientes que los de Cayena lisa, explicados probablemente por los cambios en los ácidos orgánicos de los exudados de la raíz y a la proteína soluble de los ápices de esta, tal como lo proponen Le Van y Masuda (2004).
En los sistemas de producción de pequeños pro-ductores como los agricultores de piña en Cór-doba, no es raro observar que al escoger la semi-lla de su próxima siembra, seleccionen material vegetal de manera natural a partir de plantas sanas y vigorosas, lo que indirectamente se tra-duce en una selección de ecotipos adaptados a las condiciones de los SSA típicos de la región; esto pudiera ser una explicación adicional al comportamiento más estable de ‘Criolla’ respec-to a ‘Cayena lisa’.
Dosis de cal (t ha-1)
Variedad
Cayena lisa Criolla
0 4,12 ab 3,82 a
3 4,12 a 3,97 b
6 4,08 ab 3,97 b
9 4,06 b 3,92 b
Tabla 3. Análisis de la interacción variedad x dosis del ph del fruto de dos variedades de piña cultivadas en suelos sulfatados ácidos de Ciénaga de Oro (Córdoba-Colombia).
a. análisis del efecto de la variedad dentro del nivel de encalamiento
variedadDosis de cal (t ha-1)
0 3 6 9
Cayena lisa 4,12 a 4,12 a 4,08 a 4,06 a
Criolla 3,82 b 3,97 b 3,97 b 3,92 b
b. análisis del efecto de la dosis de encalamiento dentro de la variedad
Promedios con letras distintas en la misma columna indican diferencia significativa según el test protegido de DMS (P≤0,05).
EFECTO DEL ENCALAMIENTO DE SuELOS SuLFATADOS ÁCIDOS SOBRE EL FRuTO DE p IñA

REV. COLOMB. CIENC. HORTIC.
178
Arias, C. y J. Toledo. 2000. Manual de manejo postcose-cha de frutas tropicales. Papaya, piña, plátano, cí-tricos. En: http://www.fao.org/inpho/content/do-cuments/vlibrary/ac304s/ac304s00.htm; consulta: junio de 2007.
Bartholomew, D. y S. Kadzimin. 1997. Pineapple. pp. 113-156. En: Alvim, P. de T. y T.T. Kozlowski (eds.). Eco-physiology of tropical crops. Academic Press, NY.
Bartholomew, D.P. y E. Malézieux. 1994. Pineapple. pp. 243-291. En: Schaffer, B. y P.C. Anderson (eds.). Handbook on environmental physiology of fruit crops, Vol. II. Subtropical and tropical crops. CRC Press, Boca Raton, Florida, FL.
Bello, F. y M. Gómez. 2001. Recuperación de suelos sulfa-tados ácidos en el área del distrito de riego del Alto Chicamocha, Tibasosa. Trabajo de grado. Facultad de Ciencias Agropecuarias, Universidad Pedagógica y Tecnológica de Colombia, Tunja.
Bernal, M. 2006. Madurez prematura en piña afecta la cosecha. En: El Universo.http://archivo.eluniverso.com/2006/01/21/0001/71/953BE7C7FD69406B80382F2320F6F5FD.aspx; consulta: enero de 2006.
Carvalho, S.; C. Neves; R. Bürkle y C.J. Marur. 2005. Flo-ral induction period and thermal time requirements from the flowering to the harvest period for Smooth Cayenne pineapple. Rev. Bras. Frutic. 27(3), 430-433.
Coppens, G. y F. Leal. 2003. Morphology, anatomy and taxonomy. pp. 13-32 En: Bartholomew, D.; R. Paull y K. Rohrbach (eds.). The pineapple: botany, production and uses. CAB International, New York, NY.
CoNCluSioNeS
Los resultados de la investigación permiten con-cluir que el encalamiento de los SSA de Ciénaga de Oro en el departamento de Córdoba (Colom-bia) no tuvo un efecto importante sobre la resis-tencia y el diámetro del fruto de la piña en las
variedades Cayena lisa y Criolla. Sin embargo, el pH del fruto mostró un comportamiento dife-rencial dentro de las variedades al cambiar el ni-vel de encalamiento. En este sentido, la variedad Criolla presentó una respuesta más directa al en-calamiento probablemente como consecuencia de su adaptación a los SSA de la región.
reFereNCiaS bibliogrÁFiCaS
Cunha, G. y J. Cabral. 1999. Taxonomia, espécies, culti-vares e morfologia. pp. 17-51. En: Cunha, G.; J. Ca-bral y L. Souza, L. (eds.). O abacaxizeiro: cultivo, agroindústria e economia. Embrapa Comunicação para Transferência de Tecnologia, Brasília.
De la Cruz, J.; P. Hernández; A. Rebolledo y H. García. 2007. Efecto de la aplicación de metil jasmonato sobre la fisiología poscosecha de piña (Ananas co-mosus cv. MD-2). pp. 82-91. En: Memorias V Con-greso Iberoamericano de Tecnología Postcosecha y Agroexportaciones. Murcia, España.
Le Van, H. y T. Masuda. 2004. Physiological and bio-chemical studies on aluminum tolerance in pine-apple. Aust. J. Soil Res. 42(5-6), 699-707.
Mathew, E.; R. Panda y M. Nair. 2001. Influence of subsurface drainage on crop production and soil quality in a low-lying acid sulphate soil. Agr. Water Manage. 47, 191-209.
New South Wales Department of Land and Water Con-servation (NSWDLWC). 1999. Acid sulfate soil survey in NSW. Sidney.
Nowak, J. y A. Friend. 2005. Aluminum fractions in root tips of slash pine and loblolly pine families differ-ing in Al resistance. Tree Physiol. 25, 245-250.
Paula, M.; H. Mesquita y F. Nogueira. 1998. Nutrição e Adubação do Abacaxizeiro. Informe Agropecuário, (Belo Horizonte) 19(195), 33-39.
Pérez F. y J. Martínez 2006. Aluminio. En: http://pdf.rin-condelvago.com/aluminio_1.html; consulta: abril de 2006.
COMBATT C./jARMA O./CLEVES L.

Vol. 2 - No.2 - 2008
179
Reinhardt, D. 2004. Quality gradients in ‘Pérola’ pineap-ple in function of fruit size and maturation stage. Rev. Bras. Frutic. 26(3), 544-546.
Salazar, R. 1994. Situación del cultivo de la piña en Co-lombia. pp. 151-159. En: Frutas tropicales. ICA, Bogotá.
Sierra, J.; C. Noel; L. Dufour; H. Ozier-Lafontaine; C. Welcker y L. Desfontaines. 2003. Mineral nutrition and growth of tropical maize as affected by soil acidity. Plant Soil 252, 215-226.
Taylor, G.; J. McDonald-Stephens; D. Hunter; P. Bertsch; D. Elmore; Z. Rengel y R. Reid. 2000. Direct mea-surement of aluminum uptake and distribution in single cells of Chara corallina. Plant Physiol. 123, 987-996.
Teixeira, L.; A. Spironello y P. Furlani. 2002. Split appli-cation of NPK fertilizers on pineapple. Rev. Bras. Frutic. 24(1), 219-224.
Universidad de Antioquia. 1993. Producción, transfor-mación y comercialización. Pulpas de frutas tropi-cales. En: http://huitoto.udea.edu.co/FrutasTropi-cales/pina.html; consulta: junio de 2009.
Watanabe, T. y M. Osaki. 2002. Mechanisms of adapta-tion to high aluminum condition in native plant species growing in acid soils: a review. Comm. Soil Sci. Plant Anal. 33(7-8), 1247-1260.
Welch, S.; A. Christy; L. Isaacson y D. Kirste. 2009. Mine-ralogical control of rare earth elements in acid sul-fate soils. Geochimica et Cosmochimica Acta 73, 44-64.
EFECTO DEL ENCALAMIENTO DE SuELOS SuLFATADOS ÁCIDOS SOBRE EL FRuTO DE p IñA

Diego miraNDa1,2 CarloS CarraNza1 CarloS aNDréS roJaS1 CarloS martíN Jerez1 gerharD FiSCher1 Jorge zurita1
plantulas de tomate. Foto: Álvarez-Herrera.
Acumulación de metales pesados en suelo y plantas de cuatro cultivos hortícolas, regados con agua del río Bogotá
Accumulation of heavy metals in soil and plants of four vegetable crops irrigated with water of Bogota river
planta de lechuga, cultivada en la cuenca media del río bogotá. Foto: C. Carranza
REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 2 - No.2 - pp. 180-191, 2008
reSumeN
En los cultivos hortícolas establecidos en la Sabana de Bogotá los productores desarrollan actividades de riego utilizando las aguas del río Bogotá a través del distrito de riego La Ramada, que cubre unas 6.400 ha. Su uso ha generado una problemática de contaminación en las especies hortícolas causando deterioro de la calidad de los productos que en su mayoría son consumidos en fresco. El presente estudio se desarrolló en plantaciones de lechuga (‘Coolguard’), apio (‘Tall Utah’), repollo (híbrido Delus) y brócoli (híbrido Legacy) en una finca en Soacha (vereda Canoas), Cundinamarca, ubicada en la cuenca media del río Bogotá, analizando los niveles de metales pesados plomo (Pb), cadmio (Cd), arsénico (As) y mercurio (Hg) en agua, suelo y en la parte comestible de las plantas. Se encontró que las concentraciones de Cd en el agua del riego estuvieron cercanas a los límites establecidos por las normas vigentes nacionales e internacionales, mientras que los niveles en el suelo de las dos plantaciones estuvieron dentro del rango normal. La lechuga y el apio, a los 74 días después transplante, con 0,40 y 0,43 mg kg-1 peso fresco, respectivamente, presentaron contaminación con Cd superando el límite de la norma de la Unión Europea. Se discuten las posibles razones agroecológicas, fisiológicas y de muestreo para este comportamiento. En las cuatro hortalizas, la concentración de Pb superó la concentración máxima permitida en alimentos para lactantes y niños de corta edad establecida por la Unión Europea. En general, la lechuga acumuló niveles más altos de metales pesados que las otras tres especies.
1 Departamento de Agronomía, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá (Colombia).2 Autor para correspondencia. [email protected]
palabras clave adicionales: lechuga, apio, repollo, brócoli, contaminación, cadmio, plomo, arsénico, mercurio.

Vol. 2 - No.2 - 2008
181
abStraCt
In horticultural crops established in the Colombian Bogota Plateau, farmers develop activities of irrigation utilizing water of the Bogota river through the Irrigation District “La Ramada” which is covering 6,400 ha of land. This water use generated a problem of contamination in these crops causing quality deterioration of the produce, which in mainly consumed freshly. The present study was carried out in plantations of lettuce (‘Coolguard’) celery (‘Tall Utah’), cabbage (Delus hybrid) and broccoli (Legacy hybrid) in a farm in Soacha (Canoas vereda), Cundinamarca province. The farm is located in the middle part of the Bogota river and the heavy metals lead (Pb), cadmium (Cd), arsenic (As) and mercury (Hg) in water, soil and edible part of plants were analyzed. It was found that concentrations of Cd in the irrigation water were near the limits established by national and international legal standards. Heavy metal levels of the soil in the two plantations were in the normal range. Lettuce and celery plants, at 74 days after transplanting, with 0.40 and 0.43 mg kg-1 fresh weight, respectively, were contaminated with Cd exceeding the limits of the European Union norms. Possible agroecological, physiological and sampling reasons for this behavior are discussed. In the four vegetables lead contents exceeded the maximum allowed concentration in foods for children still on milk-feeded and young children, established by the European Union. En general, lettuce plants accumulated higher levels of heavy metals than the other three vegetable species.
additional key words: lettuce, celery, cabbage, broccoli, contamination, heavy metals, cadmium, lead, arsenic, mercury.
Fecha de recepción: 26-06-2008 Aprobado para publicación: 02-12-2008
iNtroDuCCióN
La cuenca del río Bogotá tiene una de las estruc-turas económicas más diversificadas en el país. Está estrechamente relacionada con la utiliza-ción de los recursos naturales por medio de ac-tividades como agricultura, minería y transfor-mación industrial (Secretaría de Agricultura de Cundinamarca, 2004). La cuenca del río Bogotá se divide en tres tramos, el primero, la cuenca alta que se encuentra en la zona norte de la ciu-dad de Bogotá, la cuenca media, y la cuenca baja que es la que recibe las aguas residuales de la misma ciudad y las de algunos municipios de la zona sur hasta su desembocadura en el río Mag-dalena (EAAB, 2002). La cuenca media del río Bogotá es la principal fuente de agua para la pro-ducción agrícola en los municipios aledaños del noroccidente y sur de la ciudad de Bogotá, que comprende 650 ha de cultivos hortícolas.
Los vertimientos a los que se ve sometido el río Bogotá a lo largo de su recorrido generan proble-
mas de salud en la población, tanto en la mar-gen oriental como occidental de su cauce, pero el problema más grave se genera sobre la población que habita la margen oriental del río, principal-mente por la producción de hortalizas desarro-llada en la localidad de Bosa y el municipio de Soacha (EAAB, 2002).
La contaminación del río Bogotá por los verti-mientos de aguas residuales de los municipios de la cuenca ha sido un problema a resolver desde la primera mitad del siglo anterior. En su reco-rrido drena las aguas de 6.000 km2, donde ha-bitan alrededor de 8,04 millones de habitantes en 42 municipios, incluyendo el Distrito Capital (CONPES, 2004). Han sido muchos los estudios realizados por diferentes investigadores y entida-des (por ejemplo: CAR, 1991; González y Mejía, 1995; Vargas y Mejía, 1996; Montenegro, 2000) en los que se ha demostrado el alto grado de con-taminación del río, con elementos traza como el
ACuMuLACIóN DE METALES pESADOS EN SuELO y pLANTAS DE CuATRO CuLTIVOS HORTÍCOLAS

REV. COLOMB. CIENC. HORTIC.
182
arsénico (As), cadmio (Cd), cobalto (Co), cromo (Cr), cobre (Cu), hierro (Fe), mercurio (Hg), man-ganeso (Mn), níquel (Ni), plomo (Pb), estaño (Sn), cinc (Zn), y una amplia variedad de sustancias orgánicas e inorgánicas potencialmente tóxicas.
En el distrito de riego La Ramada que cubre un área bruta de 6.400 ha, los suelos están desarro-llados principalmente sobre sedimentos aluviales finos del valle del río Bogotá en un 75% del área aproximadamente. En comparación con el tipo de suelo que predomina en la Sabana de Bogotá, estos suelos aluviales poseen texturas arcillosas pesadas, nivel freático fluctuante y drenaje natu-ral pobre (CPT, 1985).
La producción de hortalizas en Colombia, en tér-minos generales, es una actividad riesgosa que se caracteriza por los siguientes aspectos: culti-vos de tamaño pequeño (0,5-10 ha), dispersos, uso intensivo de mano de obra, altos costos de producción, mercado inestable y variable, caren-cia de tecnología apropiada, manejo poscosecha deficiente y debilidad empresarial y gremial (Va-llejo, 2004). La producción de hortalizas en esta zona se caracteriza porque vincula en su ma-yoría a pequeños y medianos productores que cumplen un papel importante como proveedores de las principales centrales de abasto, no solo de la región, sino de otras zonas del país.
La lechuga es tal vez la más importante hortaliza cultivada en Cundinamarca con un área cultiva-da de cerca de 636 ha con una producción anual de 9.276 t (Asohofrucol, 2007). En repollo, las zonas más productoras del país son la Sabana de Bogotá, con una área sembrada de 500 ha y una producción anual de 14.475 t anuales, también se incluyen, parte de Cundinamarca y Boyacá, la región del oriente de Antioquia, los alrededores de Pasto (Nariño), la región de Anaime en To-lima, y los alrededores de Manizales en Caldas (Semillas Arroyave, 2006; Asohofrucol, 2007).
Los departamentos más productores de brócoli son Cundinamarca (861 ha), seguido por Boyacá
(146 ha), Nariño (116 ha) y Antioquia (80 ha), con productividades promedios de 11, 21, 6 y 17,4 t ha-1, respectivamente, con diferentes materia-les híbridos entre los cuales se destacan Legacy, Marathon, Centenario F1 y Centauro F1, entre otras (Asohofrucol, 2007). En apio se reporta un área plantada de 90 ha con un volumen de pro-ducción de 1.708 t, distribuidas en los departa-mentos de Cundinamarca, Antioquia y Boyacá principalmente, siendo uno de los cultivos con mayores tasas de crecimiento entre 2000 y 2006 en área cosechada (23%) (Asohofrucol, 2007).
Los metales pesados están presentes natural-mente en los suelos, pero en los últimos años se ha presentado una acumulación antropogénica por las actividades industriales, agrícolas y la dis-posición de residuos de todo tipo (Giuffré et al., 2005). Se considera metal pesado aquel elemento que tiene una densidad igual o superior a 5 g cm3 cuando está en forma elemental, o cuyo número atómico es superior a 20 (excluyendo los metales alcalinos y alcalino-térreos) (García y Dorron-soro, 2005), aunque en esta definición, encajan también elementos esenciales para las plantas como Fe, Cu, Mn, Zn u otros esenciales para los animales como Co, Cr o Ni (Mendoza, 2006). Los elementos As y Se, se consideran como me-taloides pero por su efecto contaminante se in-cluyen dentro de los metales pesados.
Los metales pesados más comunes y ampliamen-te distribuidos como contaminantes ambienta-les incluyen plomo (Pb), cadmio (Cd), mercurio (Hg) y el metaloide arsénico (As) (Reilly, 2002). Sin embargo, los metales pesados en las plantas, como en otros seres vivos, son esenciales, ya que son componentes estructurales o catalizadores de los procesos bioquímicos de los organismos, las actividades humanas vierten sobre los recur-sos de suelo y agua grandes cantidades de esos elementos, generando excesos por acumulación de Cd, Hg, Ni y Pb, entre otros, afectando así las relaciones de las plantas y otros organismos, lo cual origina toxicidades en los ecosistemas (Montenegro, 2002). En general, estos metales
MIRANDA/CARRANzA/ROjAS/jEREz/FISCHER/zuRITA

Vol. 2 - No.2 - 2008
183
no son biodegradables y, por consiguiente, pue-den acumularse en órganos vitales del cuerpo humano, produciendo efectos tóxicos progre-sivos (Queirolo et al., 2000). La mayoría de los metales pesados entran en la cadena alimentaria principalmente a través de los cultivos que ab-sorben el agua de riego contaminada.
Las plantas han desarrollado mecanismos alta-mente específicos para absorber, translocar y acumular sustancias (Lasat, 2000); sin embar-go, algunos metales y metaloides no esenciales para los vegetales son absorbidos, translocados y acumulados en la planta debido a que presentan un comportamiento electroquímico similar a los elementos nutritivos requeridos. La fitotoxicidad de los metales pesados se manifiesta particular-mente en los suelos ácidos y afecta al crecimien-to y la formación de raíces laterales y secunda-rias (Tadeo y Gómez-Cadenas, 2008).
El objetivo de este estudio fue determinar las concentraciones de metales pesados contami-nantes Pb, Cd, As y Hg, presentes en los suelos y el tejido foliar de los cultivos de lechuga, apio, repollo y brócoli, regados con agua proveniente del río Bogotá en el distrito de riego La Ramada.
materialeS y métoDoS
El material vegetal estuvo compuesto por la le-chuga ‘Coolguard’, el repollo híbrido Delus, el brócoli híbrido Legacy (los tres de marca Semi-nis de Semillas Arroyave, Bogotá) y el apio ‘Tall Utah’, marca Seminis de Semillas Saenz, Bogotá.
El experimento, que se llevó a cabo entre los me-ses de julio y noviembre de 2006, se estableció en lotes comerciales en una finca del municipio de Mosquera, Cundinamarca, vereda San Fran-cisco (4°42’N y 74°13’W) y en una del munici-pio de Soacha, Cundinamarca, vereda Canoas (4°30’N y 74°13’W). El clima en esta zona está caracterizado por una temperatura media anual de 12,6°C, una precipitación de 670 mm año-1 y unas 4,2 h de brillo solar por día.
Para cada uno de los cultivos se demarcó un área experimental consistente en tres franjas de 30 m de largo por 1,4 m de ancho, en un diseño de bloques completos al azar, con tres repeticiones. Estas parcelas se ubicaron hacia el centro de los cultivos para asegurar un riego más uniforme y para disminuir efectos de borde.
En cada muestreo se tomaron muestras de suelo, colectado según el modelo de muestreo plantea-do por Osorio y Ruíz (2005), entre 0 y 20 cm de profundidad, de las parcelas correspondientes a cada uno de los cultivos y sus repeticiones. De acuerdo con la topografía de los lotes, se realizó un método de muestreo en equis sobre las áreas de los lotes de cada cultivo (Gómez, 2006). Las muestras de agua, se colectaron directamente de los aspersores de riego, 1 L por muestra, y fueron llevadas al Laboratorio de Suelos y Aguas de la Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá, para las determinaciones correspondientes.
El muestreo del suelo en los ensayos correspon-dió: pH 5,59; CO 4,08%; N 0,35%; Ca 10,5, K 0,46, Mg 3,51, Na 4,94 y CIC 27,8 meq/100 g, respectivamente; P 48,1, Cu 1,04, Fe 4,26, Mn 1,49, Zn 21,8 y B 0,94 mg kg-1, respectivamente; Ar 17%, L 47% y A 36%, con una textura Fr.
Los días de muestreo de material vegetal se rea-lizaron, dependiendo del crecimiento de cada es-pecie, para la lechuga y el apio a los 30, 68 y 103 d después del transplante (ddt), y para el repollo y brócoli a los 85, 105 y 120 ddt. El muestreo con-sistió en tomar 15 plantas al azar, en las etapas iniciales del desarrollo del cultivo, disminuyendo la cantidad a medida que estas iban creciendo, esto con el fin de reunir el peso requerido de la muestra para cada uno de los análisis realizados. Las muestras (sin raíz) se secaron en estufas con aire forzado por 48 h a 70°C. Aproximadamen-te 150 g por muestra se pasaron por un molino eléctrico con un tamiz de 2 mm y fueron alma-cenadas para su posterior análisis. Las muestras procesadas de material vegetal, del agua fresca y del suelo fueron enviadas al laboratorio de In-
ACuMuLACIóN DE METALES pESADOS EN SuELO y pLANTAS DE CuATRO CuLTIVOS HORTÍCOLAS

REV. COLOMB. CIENC. HORTIC.
184
geniería Ambiental de la Universidad Nacional de Colombia, Bogotá, para el análisis de metales pesados.
La determinación de los metales pesados corres-pondió a lo recomendado por la Association of Official Analytical Chemists (AOAC), usando la espectrofotometría de absorción atómica: para el Cd el método 973.34; para el Pb el método 972.25 (absorción de llama, análisis tipo III) (FAO, 2007); para el As el método 952.13 (co-lorimetría, dietilditiocarbamato, análisis tipo II) y para el Hg el método 972.15 (empleando el espectrofotómetro de absorción atómica marca Varian Spectra AA 220 FS, equipado con gene-rador de hidruros) (FAO, 2007). Para el análisis estadístico de los datos, en cada muestreo se to-maron tres repeticiones de cada muestra.
reSultaDoS y DiSCuSióN
Metales pesados en aguas de riego
No se presentaron diferencias en el nivel de los metales pesados en el agua según los días del muestreo (tabla 1). Siendo los niveles para el uso agrícola en Colombia admisibles hasta 5 mg L-1 para Pb; 0,01 mg L-1 para Cd y 0,1 mg L-1 para As (Ministerio de Agricultura, 1984), el Cd se encuentra en el límite permitido para el riego de los cultivos con el agua del distrito La Ramada. Al comparar este resultado (tabla 1) con lo re-portado por la CAR (2006) sobre los niveles de Pb y Cd en la cuenca media del río Bogotá, estos resultados superaron los valores de 2006, cuan-do se determinaron niveles de 0,032 mg L-1 Pb y 0,003 mg L-1 Cd.
El resultado del presente estudio coincide con lo reportado por Montenegro (2002) sobre la pre-sencia de importantes concentraciones de ele-mentos como Pb, Cd, Cr, Cu, Fe, Ni y Zn en los ríos Juan Amarillo, Tunjuelo y Fucha que son los principales afluentes del río Bogotá.
El agua puede ser contaminada por metales de manera natural o a través de diferentes proce-sos de captación, tratamiento, almacenamiento y distribución, afectando su calidad en el punto final de consumo. La situación actual del Dis-trito con el uso de estas aguas correspondería al tramo de contaminación creciente a alta del río, que va desde el Salitre hasta Tocaima.
Montenegro (2002) informa que el río Bogotá vertía diariamente en su desembocadura al Mag-dalena 79 kg de Pb, 70 kg de Cr, 20 t de Fe, 5 t de detergentes y 1.473 t de sólidos en suspensión causando un proceso de contaminación difu-sa. A pesar de que en los muestreos realizados los valores no superaron valores referenciales, sí existe presencia de metales pesados que inci-den en la baja calidad del agua del río Bogotá, situación que genera conflictos con los usuarios actuales y los potenciales, en especial para riego en el sur y en el suroccidente de la sabana, donde se encuentra el más importante distrito de riego La Ramada, y los acueductos en la cuenca media y baja (Pérez, 2000).
Metales pesados en suelo
Los resultados muestran para Pb, Cd y As con-centraciones cercanas a los rangos normales, de-terminados por Bowie y Thornton (1985). Estos autores establecieron valores de concentración de
tabla 1. Concentraciones de metales pesados en agua para riego. río bogotá, distrito la ramada.
Días de muestreoplomo
(mg L-1)Cadmio(mg L-1)
Arsénico (mg L-1)
Mercurio (mg L-1)
30 <0,045 <0,01 <0,003 <0,000368 <0,045 <0,01 <0,003 <0,0003
103 <0,045 <0,01 <0,003 <0,0003
MIRANDA/CARRANzA/ROjAS/jEREz/FISCHER/zuRITA

Vol. 2 - No.2 - 2008
185
metales pesados, para Pb entre 10 y 150 mg kg-1; para Cd entre 1 y 2 mg kg-1 y para As entre 5 y 40 mg kg-1, válidos para suelos considerados “norma-les”. Por otro lado, las concentraciones encontra-das no superan los máximos aceptables en suelos agrícolas de los países de la Unión Europea, repor-tados por García-Navarro (2005) con 300 mg kg-1 suelo seco para Pb, 3 mg kg-1 para Cd, 20-50 mg kg-1 para As y 1,5 mg kg-1 para Hg. Sin embargo, los resultados mostraron incrementos considera-bles con respecto a los determinados por la CAR (2006) en diversos sectores de la cuenca del río Bogotá. Para Pb, nuestros valores estarían sepa-rados estos valores de los propuestos por Davies (1977) que fueron de 70 mg kg-1 para un suelo normal. La acidez creciente del suelo puede au-mentar la solubilidad del Pb, pero su movilización en el caso del suelo empleado en nuestro trabajo pudo deberse a que la materia orgánica del suelo favorece su acumulación, como lo mencionan Ka-bata y Pendias (2000). La concentración media de Hg para los suelos del mundo, raramente excede los 4 μg kg-1 asociándose principalmente a suelos orgánicos Kabata-Pendias (2000).
González y Mejía (1995) encontraron en seis zo-nas hortícolas diferentes en la Sabana de Bogotá, regadas con el agua del río Bogotá, niveles de Cd entre 0,95 y 2,00 mg kg-1, en una profundidad entre 0 y 30 cm del suelo, que son muy similares al presente estudio, mientras entre 30 y 60 cm de profundidad las concentraciones de Cd fueron solamente entre 0,65 y 1,40 mg kg-1, lo que mues-tra la mayor acumulación en la parte superficial del suelo y el peligro para las plantas que enraí-zan solamente en esta capa. Esta misma tenden-cia la encontraron González y Mejía (1995) para
As con valores entre 6,8 y 12,5 mg kg-1 para la zona entre 0 y 30 cm del suelo y entre 5,8 y 9,0 mg kg-1 As en 30 a 60 cm. Bergmann (1993) re-porta para el As una concentración máxima 20 mg kg-1 como tolerable en suelos agrícolas y que una buena aireación del suelo disminuye la ad-sorción de este elemento por la planta.
Las condiciones de texturas francas de los suelos de la zona de estudio, posiblemente favorecieron la acumulación de estos metales hasta este grado. En forma general, los metales pesados que ingre-san en pequeñas cantidades a los suelos se acu-mulan en lugares específicos de adsorción donde son retenidos fuertemente en los coloides orgáni-cos e inorgánicos del suelo (Soto, 2006). Los me-tales pesados cuando se acumulan en concentra-ciones excesivas son tóxicos para la mayoría de los organismos y frecuentemente se han repor-tado efectos tóxicos en los microorganismos del suelo (Giller et al., 1998), especialmente en suelos poco estructurados y con baja materia orgánica (Acevedo et al., 2005), como fue encontrada en la zona del estudio con 4,08% carbono orgánico.
García-Navarro (2005) comenta que solo se pue-de hablar de contaminación por metales pesados cuando el contenido en los mismos excede consi-derablemente de los valores habituales en el tipo de suelo que se está considerando, situación no observada en el presente trabajo. Los materiales constitutivos de los suelos como limos y arcillas fueron determinantes en la acumulación y dis-ponibilidad de estos elementos (Soto, 2006).
De acuerdo con Vargas y Mejía (1996) el Hg y el Pb pueden presentarse en una o más de las siguientes
Tabla 2. Concentración de metales pesados determinados en suelo cultivado con las cuatro hortalizas en Soacha, Cundinamarca.
Muestreoplomo
(mg kg-1)Cadmio
(mg kg-1)Arsénico (mg kg-1)
Mercurio (mg kg-1)
1 (30 ddt) 11,08 a 0,86 b 1,42 b <0,03 (nd)2 (68 ddt) 9,53 b 0,68 c 1,35 c <0,03 (nd)3 (103 ddt) 11,83 a 1,73 a 1,75 a <0,03 (nd)
Promedios con letras distintas indican diferencia significativa según la prueba de Tukey (P≤0,05); ddt, días después de transplante; nd, niveles no detectables.
ACuMuLACIóN DE METALES pESADOS EN SuELO y pLANTAS DE CuATRO CuLTIVOS HORTÍCOLAS
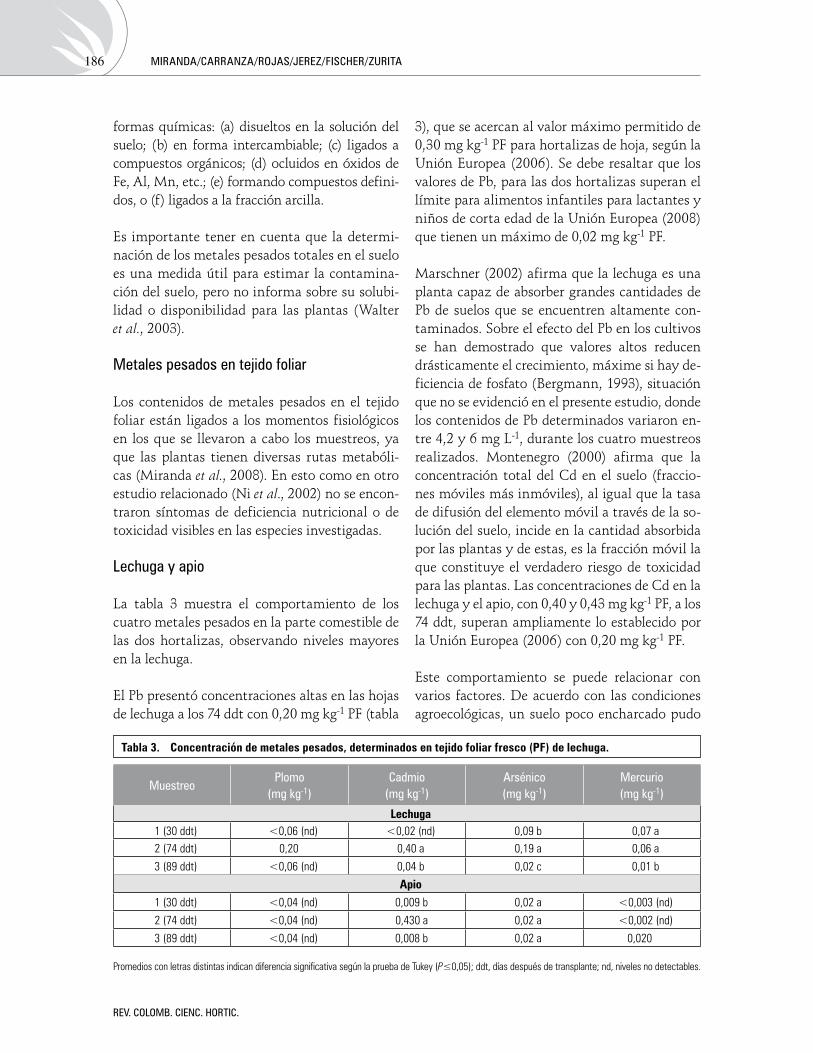
REV. COLOMB. CIENC. HORTIC.
186
formas químicas: (a) disueltos en la solución del suelo; (b) en forma intercambiable; (c) ligados a compuestos orgánicos; (d) ocluidos en óxidos de Fe, Al, Mn, etc.; (e) formando compuestos defini-dos, o (f) ligados a la fracción arcilla.
Es importante tener en cuenta que la determi-nación de los metales pesados totales en el suelo es una medida útil para estimar la contamina-ción del suelo, pero no informa sobre su solubi-lidad o disponibilidad para las plantas (Walter et al., 2003).
Metales pesados en tejido foliar
Los contenidos de metales pesados en el tejido foliar están ligados a los momentos fisiológicos en los que se llevaron a cabo los muestreos, ya que las plantas tienen diversas rutas metabóli-cas (Miranda et al., 2008). En esto como en otro estudio relacionado (Ni et al., 2002) no se encon-traron síntomas de deficiencia nutricional o de toxicidad visibles en las especies investigadas.
Lechuga y apio
La tabla 3 muestra el comportamiento de los cuatro metales pesados en la parte comestible de las dos hortalizas, observando niveles mayores en la lechuga.
El Pb presentó concentraciones altas en las hojas de lechuga a los 74 ddt con 0,20 mg kg-1 PF (tabla
3), que se acercan al valor máximo permitido de 0,30 mg kg-1 PF para hortalizas de hoja, según la Unión Europea (2006). Se debe resaltar que los valores de Pb, para las dos hortalizas superan el límite para alimentos infantiles para lactantes y niños de corta edad de la Unión Europea (2008) que tienen un máximo de 0,02 mg kg-1 PF.
Marschner (2002) afirma que la lechuga es una planta capaz de absorber grandes cantidades de Pb de suelos que se encuentren altamente con-taminados. Sobre el efecto del Pb en los cultivos se han demostrado que valores altos reducen drásticamente el crecimiento, máxime si hay de-ficiencia de fosfato (Bergmann, 1993), situación que no se evidenció en el presente estudio, donde los contenidos de Pb determinados variaron en-tre 4,2 y 6 mg L-1, durante los cuatro muestreos realizados. Montenegro (2000) afirma que la concentración total del Cd en el suelo (fraccio-nes móviles más inmóviles), al igual que la tasa de difusión del elemento móvil a través de la so-lución del suelo, incide en la cantidad absorbida por las plantas y de estas, es la fracción móvil la que constituye el verdadero riesgo de toxicidad para las plantas. Las concentraciones de Cd en la lechuga y el apio, con 0,40 y 0,43 mg kg-1 PF, a los 74 ddt, superan ampliamente lo establecido por la Unión Europea (2006) con 0,20 mg kg-1 PF.
Este comportamiento se puede relacionar con varios factores. De acuerdo con las condiciones agroecológicas, un suelo poco encharcado pudo
Tabla 3. Concentración de metales pesados, determinados en tejido foliar fresco (PF) de lechuga.
Muestreoplomo
(mg kg-1)Cadmio
(mg kg-1)Arsénico (mg kg-1)
Mercurio (mg kg-1)
lechuga 1 (30 ddt) <0,06 (nd) <0,02 (nd) 0,09 b 0,07 a2 (74 ddt) 0,20 ( 0,40 a 0,19 a 0,06 a
3 (89 ddt) <0,06 (nd) 0,04 b 0,02 c 0,01 b
apio
1 (30 ddt) <0,04 (nd) 0,009 b 0,02 a <0,003 (nd)
2 (74 ddt) <0,04 (nd) 0,430 a 0,02 a <0,002 (nd)
3 (89 ddt) <0,04 (nd) 0,008 b 0,02 a 0,020(nd)
Promedios con letras distintas indican diferencia significativa según la prueba de Tukey (P≤0,05); ddt, días después de transplante; nd, niveles no detectables.
MIRANDA/CARRANzA/ROjAS/jEREz/FISCHER/zuRITA

Vol. 2 - No.2 - 2008
187
ocasionar una mayor absorción de Cd (Acevedo et al., 2005); sumado a esto, la cercanía de los lotes de muestreo a las factorías que se ubican alrededor del sitio experimental, que favorecen la contaminación aérea, la cual puede influir la concentración en la planta (Bergmann, 1993) en esta fase del cultivo y, posiblemente, la dificultad para retirar la capa contaminante adherida a las hojas muestreadas.
Otros factores que pudieron influenciar el incre-mento de Cd se relacionan con las diferencias en la morfología de las raíces, teniendo la lechuga numerosas raíces delgadas que pueden acumular más metales que otras plantas con raíces más gruesas y suculentas (Schierup y Larsen, 1981). Además, se ha comprobado que raíces de la le-chuga translocan hacía los brotes la mayor canti-dad del Cd absorbido comparado con otras espe-cies (Jarvis et al., 1976). De igual manera, factores de muestreo que pudieron haber influenciado la respuesta tienen que ver con el uso de hojas más viejas (externas) de la planta que se caracterizan por su mayor capacidad de acumular metales pe-sados que las hojas jóvenes (internas) (Das et al., 1997), debido a la presencia de péptidos que ligan estos metales (Vogeli-Lange y Wagner, 1990).
El Cd es uno de los metales traza del suelo más solubles (Jansson, 2002), peligroso debido a su alta movilidad inhibe drásticamente el crecimien-to celular y de toda la planta en concentraciones tóxicas (Prasad, 1995). Mengel et al. (2001) repor-tan que la tasa de absorción del Cd por la lechu-ga es particularmente alta, comparado con otras especies. Estudios realizados por Insuasty et al. (2008), evaluando la incorporación de dosis cre-cientes de Cd al cultivo de papa, mostraron que la acumulación de este metal en la planta es mayor en las concentraciones altas de este elemento en el suelo y que se acumula en mayor porcentaje en los primeros 15 cm de este, lo que manifies-ta el potencial de toxicidad para los cultivos de porte bajo con un sistema de raíces superficial.
Los síntomas más generales de toxicidad por Cd son atrofia y clorosis, esta puede aparecer debido a una interacción directa o indirecta con el Fe, el Zn, el P y el Mn. Das et al. (1998) reportan que aun cuando los efectos del Cd varían a ni-vel de especie, e incluso varietal, en general el Cd interfiere en la absorción y transporte de varios elementos (Ca, Mg, P y K) y del agua.
González y Mejía (1995) regaron lechugas en bol-sas de PE (2 L) con agua contaminada por Cd en 0,1 mg L-1 (que corresponde a una concentración 10 veces mayor que en el estudio presente) lo que aumentó el nivel de Cd en el tejido foliar a 6,12 mg kg-1 PS , concentración muy superior compa-rada con el límite internacional permitido.
Por otro lado, McLaughlin et al. (1999) reportan que la acumulación del Cd en las hortalizas pue-de ser más alta en suelos salinos, que es el caso de los terrenos en la zona de influencia del río Bogo-tá (Carranza et al., 2009), debido a que la forma-ción de complejos entre el Cl y el Cd aumenta la movilidad y la absorción de este por las plantas.
Para As y Hg la Unión Europea (2006) no esta-blece niveles tóxicos en hortalizas, solamente para el Hg los ha fijado en 0,5-1,0 mg kg-1 en di-ferentes tipos de pescado.
La absorción de Hg por la lechuga fue menor que la de los otros metales pesados estudiados, debi-do, muy posiblemente, por la menor concentra-ción en agua (tabla 1) y suelo (tabla 2) y la fuerte adsorción de los iones Hg en la sustancia orgánica del suelo (Bergmann, 1993). También, es de con-siderar que la absorción máxima de Hg2+ ocurre en rangos de pH entre 4,0 y 5,0 (McLaughlin et al., 1999), que fue mayor en el presente estudio y, además, la absorción de Hg se disminuye signifi-cativamente con la presencia de Cl (Hogg et al., 1978), situación que está dada por la salinidad del agua y del suelo en la zona de influencia del río Bogotá (Carranza et al., 2009).
ACuMuLACIóN DE METALES pESADOS EN SuELO y pLANTAS DE CuATRO CuLTIVOS HORTÍCOLAS

REV. COLOMB. CIENC. HORTIC.
188
Repollo y brócoli
En general, la acumulación de los metales Pb y Cd en el tejido comestible de repollo y brócoli (tabla 4) fue menor que en el de la lechuga (ta-bla 3), teniendo en cuenta que esta última es un bioacumulador de dichos metales (Mengel et al., 2001). También Bergmann (1993) clasifica la acumulación relativa del repollo para metales pesados como baja a mediana comparada con la lechuga que es alta. No obstante, para el caso del Hg el repollo pasó en la última medición (120 ddt) los niveles encontrados en lechuga.
Para el Pb los valores detectados en el tejido foliar fueron <0,05 mg kg-1 e inferiores a los niveles de detección exacta del equipo (tabla 4) y así por debajo de la norma de la Unión Europea (2007) para hortalizas del género Brassica con 0,3 mg kg-
1 PF. Teniendo en cuenta que para la elaboración de alimentos infantiles para lactantes y niños de corta edad rigen normas más estrictas, este grupo de población no debería consumir repollo ni brócoli contaminados porque pasan los 0,02 mg kg-1 de Pb en el alimento preparado (Unión Europea, 2008). En Colombia, la NTC 1474 es-tablece los límites máximos de contaminantes para lactantes y niños de corta edad para el Pb (límite máximo 0,02 mg kg-1 PF), determinado de acuerdo con lo indicado en la norma AOAC 999.11 (Icontec, 2009).
Promedios con letras distintas indican diferencia significativa según la prueba de Tukey (P≤0,05); ddt, días después transplante; nd, niveles no detectables.
Tabla 4. Concentración de metales pesados determinados en tejido foliar fresco (PF) de repollo y brócoli.
Muestreoplomo
(mg kg-1)Cadmio
(mg kg-1)Arsénico (mg kg-1)
Mercurio (mg kg-1)
repollo 1 (85 ddt) <0,05 (nd) <0,010 (nd) 0,06 a <0,003 (nd)2 (105 ddt) <0,04 (nd) <0,008 (nd) 0,04 a <0,002 (nd)
3 (120 ddt) <0,04 (nd) <0,008 (nd) 0,05 a 0,150
brócoli
1 (85 ddt) <0,05 (nd) <0,01 (nd) 0,05 a <0,003 (nd)
2 (105 ddt) <0,06 (nd) <0,01 (nd) 0,05 a <0,004 (nd)
3 (120 ddt) <0,05 (nd) <0,01 (nd) 0,05 a 0,004 (nd)
El Cd en la parte comestible de repollo y brócoli no pasó los niveles máximos establecidos por la Unión Europea (2006) con 0,20 mg kg-1 PF (ta-bla 4). Verma et al. (2007) modelaron la toma de Cd por la planta de repollo y encontraron un nivel notablemente menor que en zanahoria (máxima absorción) y espinaca (mediana absor-ción), mientras la toma de Cd por el rábano fue mínimo.
Los niveles de As en la localidad de Soacha fueron mayores que los contenidos medios reportados por Kabata-Pendias (2000) para repollo con 0,02-0,05 mg kg-1 peso seco (PS), los cuales estuvieron entre 0,48 y 0,57 mg kg-1 (PS) (que correspondió a 0,04-0,06 mg kg-1 PF en la tabla 4). Resultado que muestra las concentraciones altas de este meta-loide en el repollo en la zona de estudio compara-do con el nivel promedio internacional.
El Hg encontrado en el tejido comestible de repo-llo y brócoli (tabla 4) fue menor que en lechuga (tabla 3), solamente el tercer muestreo en repollo (tabla 4) superó a las demás mediciones de este metal, que se podría ver como un efecto de acu-mulación de Hg en la planta que también se pre-sentó en apio, pero no en brócoli y lechuga (tabla 3). Como se mencionó para el caso de la lechuga, por un exceso de Cl- la absorción de Hg+ en las partículas minerales del suelo y la materia orgá-nica disminuye debido a la formación de com-
MIRANDA/CARRANzA/ROjAS/jEREz/FISCHER/zuRITA

Vol. 2 - No.2 - 2008
189
plejos de Hg-Cl altamente estables que son poco adsorbidos (Acevedo et al., 2005). Por lo general, la absorción de Hg desde el suelo a las plantas es baja, constituyéndose en una barrera para la tras-locación desde las raíces a las partes aéreas de las plantas (Patra y Sharma, 2000), en consecuencia la cadena alimenticia no se ve tan afectada como en el caso del Cd (Bergmann, 1993).
CoNCluSioNeS
• Se observó en el agua de riego, proveniente del río Bogotá, que las concentraciones de metales pesados, especialmente el cadmio, están cerca-nas a los límites establecidos por las normas vigentes nacionales e internacionales. Los re-sultados mostraron incrementos considerables con respecto a los determinados anteriormen-te por la CAR en diversos sectores de la cuenca del río Bogotá.
• En el suelo de la plantación hortícola no se en-contraron niveles tóxicos de metales pesados estudiados.
• A los 74 ddt, se presentaron niveles tóxicos del cadmio en la lechuga y el apio que sobrepasan el nivel máximo permitido por la Unión Eu-ropea.
• No se encontró contaminación excesiva por plomo, arsénico y mercurio en las hortalizas investigadas.
• Sin embargo, las concentraciones de plomo su-peraron los niveles permitidos en las cuatro hor-talizas, para ser usadas en alimentos infantiles.
• Los suelos hortícolas urbanos y periurbanos deben ser cuidadosamente monitoreados con-siderando el riesgo de entrada de metales en la cadena alimentaria.
agraDeCimieNtoS
Los autores agradecen a los expertos de los laboratorios de suelos y aguas de la Facultad de Agrono-mía y al personal técnico del Instituto de Ensayos e Investigaciones (IEI) de la Universidad Nacional de Colombia por su apoyo en la ejecución de esta investigación.
Acevedo, E.; M.A. Carrasco; O. León; E. Martínez; P. Sil-va; G. Castillo; I. Ahumada; G. Borie y S. González. 2005. Criterios de calidad de suelo agrícola. Gobier-no de Chile, Ministerio de Agricultura (SAG), San-tiago.
Asohofrucol. 2007. Apio, brócoli, lechuga y repollo. En: http://frutasyhortalizas.com.co; consulta: diciem-bre de 2007.
Bergmann, W. 1993. Ernährungsstörungen bei Kulturp-flanzen. Gustav Fischer Verlag, Jena, Alemania.
Bowie, S.H.U. e I. Thornton (eds.). 1985. Environmen-tal geochemistry and health. D. Reidel Publishing Company, Lancaster, PA.
reFereNCiaS bibliogrÁFiCaS
CAR. 1991. Programa de aforo y muestreo de los ríos Bogotá, Ubaté y Suárez y sus afluentes; muestreo a embalses, lagos y lagunas, preservación y trans-porte de muestras. Informe final. Ingeseries Ltda., Bogotá.
CAR. 2006. Objetivos de calidad del agua para la cuenca del río Bogotá a lograr en el año 2020. Acuerdo No. 43 del 17 de octubre de 2006. Corporación Autóno-ma Regional de Cundinamarca, Bogotá.
Carranza, C.; O. Lanchero; D. Miranda y B. Chaves. 2009. Análisis de crecimiento de lechuga (Lactuca sativa L.) ‘Batavia’ cultivada en un suelo salino de la Sabana de Bogotá. Agron. Colomb. 27(1), 41-48.
ACuMuLACIóN DE METALES pESADOS EN SuELO y pLANTAS DE CuATRO CuLTIVOS HORTÍCOLAS

REV. COLOMB. CIENC. HORTIC.
190
CONPES. 2004. Estrategia para el manejo ambiental del Río Bogotá. Documento Conpes 3320. Consejo Nacional de Política Económica y Social. Departa-mento Nacional de Planeación. Ministerio de Am-biente, Vivienda y Desarrollo Territorial, Bogotá.
CPT. 1985. Estudio para la ampliación del Distrito de Rie-go La Ramada. Anexo 2-Agrología. Corporación Au-tónoma Regional de Cundinamarca (CAR), Bogotá.
Das, P.; S. Smarantay y G.R. Rout. 1998. Studies in Cd tox-icity in plants: A review. Environ. Pollution 98, 29-36.
EAAB. 2002. Estudio sobre la calidad del agua del río Bogotá. Empresa de Acueducto y Alcantarillado de Bogotá, Bogotá.
FAO. 2007. Código de prácticas de higiene para las fru-tas y hortalizas frescas. pp. 162-192. En: Codex alimentarius. Organización Mundial de la Salud. Organización de las Naciones Unidas para la Agri-cultura y la Alimentación, Roma.
García, I. y C. Dorronsoro. 2005. Contaminación por metales pesados. En: Tecnología de suelos. Depar-tamento de Edafología y Química Agrícola, Uni-versidad de Granada, Granada, España.
García-Navarro, A. 2003. Gestión y conservación del suelo. Lección 4. Contaminación del suelo. Con-taminantes específicos. Metales pesados. Departa-mento de Biología y Producción de los Vegetales, Universidad de Extremadura, Badajoz, España.
Giller, K.E.; E. Witter y S.P. McGrath. 1998. Toxicity of heavy metals to microorganism and microbial processes in agricultural soils: a review. Soil Biol. Biochem. 30(10-11), 1389-1414.
Giuffré, L.; S. Ratto; L. Marbán; J. Schonwald y R. Ro-maniuk 2005. Riesgo por metales pesados en horti-cultura urbana. Ciencia del Suelo 23(1), 101-106.
Gómez, M.I. 2006. Manual técnico de fertilización de cultivos. Microfertisa S.A., Produmedios, Bogotá.
González, S. y L. Mejía. 1995. Contaminación con cad-mio y arsénico en suelos y hortalizas de un sector de la cuenca del río Bogotá. Suelos Ecuatoriales 25, 51-56.
Hogg, T.J.; K.W.B. Stewart y J.R. Bettany. 1978. Influ-ence of the chemical form of mercury on its ad-sorption and ability to leach through soils. J. Envi-ron. Qual. 7, 440-445.
Icontec. 2009. Alimentos complementarios para niños lactantes y de corta edad. NTC 1474. Segunda ac-tualización. Instituto Colombiano de Normas Téc-nicas y Certificación, Bogotá.
Insuasty, L.; H. Burbano y J. Menjivar. 2008. Dinámica del cadmio en suelos cultivados con papa en Nari-ño, Colombia. Acta Agron. 57(1), 51-54.
Jansson, G. 2002. Cadmium in arable crops. The influ-ence of soil factors and liming. Tesis doctoral. De-partment of Soil Sciences, Swedish University of Agricultural Sciences, Uppsala, Suecia.
Jarvis, S.C.; L.H.P. Jones y M.J. Hopper. 1976. Cadmium uptake from solution by plants and its transport from roots do shoots. Panta Soil 44, 179-191.
Kabata-Pendias, A. 2000. Trace elements in soils and plants. 3a. ed. CRC Press, Boca Raton, FL.
Lasat, M.M. 2000. The use of plants for the removal of toxic metals from contaminated soil. American As-sociation for the Advancement of Sciencie, Environ-mental Protection Agency (EPA), Washington DC.
Marschner, H. 2002. Mineral nutrition of higher plants. Academic Press, London.
McLaughlin, M.J., D.R. Parker y J.M. Clarke. 1999. Me-tals and micronutrients – food safety issues. Field Crops Res. 60, 143-163.
Mendoza. P.M. 2006. Estudio de metales pesados en suelos bajo cultivos hortícolas de la provincia de Castellón. Publicaciones Server. Universidad de Va-lencia, Valencia, España.
Mengel, K.; E.A. Kirkby; H. Kosegarten y T. Appel. 2001. Principles of plant nutrition. Kluwer Academic Pu-blishers, Dordrecht, The Netherlands.
Ministerio de Agricultura. 1984. Decreto 1594 del 26 de junio de 1984 en cuanto a usos del agua y residuos líquidos. Bogotá.
Miranda L., D., C. Carranza y G. Fischer. 2008. Calidad del agua de riego en la Sabana de Bogotá. Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá.
Montenegro, R.O. 2002. Contaminación química de suelos y cultivos. Estrategias para la productividad de los suelos agrícolas. Sociedad Colombiana de la Ciencia del Suelo, Capitulo Tolima, Bogotá.
Ni, W.Z.; X.X. Long y X.E. Wang. 2002. Studies o the criteria of cadmium pollution in growth media of vegetable crops based on the hygienic limit of cad-mium in food. J. Plant Nutr. 25(5), 957-968.
Osorio, N. y O. Ruíz. 2005. Guía para el muestreo de suelos. Escuela de Geociencias, Facultad de Cien-cias, Universidad Nacional de Colombia, Medellín, Colombia.
MIRANDA/CARRANzA/ROjAS/jEREz/FISCHER/zuRITA

Vol. 2 - No.2 - 2008
191
Patra, M. y A. Sharma. 2000. Mercury toxicity in plants. Bot. Rev. 66, 379-422.
Pérez, A. 2000. La estructura ecológica especial de la Sa-bana de Bogotá. Sociedad Geográfica de Colombia, Academia de Ciencias Geográficas, Bogotá.
Prasad, M.N.V. 1995. Cadmium toxicity and tolerance in vascular plants. Environ. Exp. Bot. 35(4), 525-545.
Queirolo, F.; S. Stegen; M. Restovic; M. Paz; P. Ostapc-zuk; M.J. Schwuger y L. Muñoz. 2000. Total arse-nic, lead, and cadmium levels in vegetables culti-vated at the Andean villages of northern Chile. Sci. Total Environ. 255, 75-84.
Reilly, C. 2002. Metal contamination of food. Its signifi-cance for food quality and human health. Black-well Science Ltd., Oxford, UK.
Schierup, H. y V.J. Larsen. 1981. Macrophyte cycling of Zn, Cu, Pb and Cd in the littoral zone of a polluted and non-polluted lake. I. Availability, uptake and translocation of heavy metals in Phragmites aus-tralis (Cav.). Trin. Aquatic Bot. 11, 179-210.
Semillas Arroyave. 2006. Repollo Delus. En: http://www.semillasarroyave.com. 2 p.; consulta: di-ciembre de 2007.
Soto. J.S.H. 2006. Contaminación del río Rimac por me-tales pesados y efecto en la agricultura en el cono este de Lima metropolitana. Trabajo de grado. Uni-versidad Nacional Agraria La Molina, Lima.
Tadeo, F.R. y A. Gómez-Cadenas. 2008. Fisiología de las plantas y el estrés. pp. 577-597. En: Azcón-Bieto, J. y M. Talón. Fundamentos de fisiología vegetal. 2a ed. McGraw-Hill Interamericana, Madrid.
Unión Europea. 2006. Contenido máximo de determi-nantes contaminantes en los productos alimenti-cios. Reglamento CE 1881/2006 de la Comisión. Diario Oficial de la Unión Europea 20.12.2006, L 364/5-L364/24.
Unión Europea. 2008. Contenidos máximos de deter-minados contaminantes. En: http://europa.eu/le-gislation_summaries/food_safety/contamination_environmental_factors/l21290_es.htm; consulta: noviembre de 2008.
Vallejo, F.A. 2004. Hortalizas en Colombia. Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia, Palmira.
Vargas, O. y L. Mejía. 1996. Evaluación de la contamina-ción por Hg y Pb en suelos hortícolas de la cuenca alta del río Bogotá y del efecto del Hg y Pb de las aguas de riego y de suelos en varias hortalizas. II Simposio Internacional de Geoquímica Ambiental. Ingeominas, Universidad Nacional de Colombia, Cartagena, Colombia.
Verma, P., K.V. George; H.V. Singh y R.N. Singh. 2007. Modeling cadmium accumulation in radish, car-rot, spinach and cabbage. Appl. Math. Model. 31, 1652-1661.
Vogeli-Lange, R. y G.J. Wagner. 1990. Subcellular local-ization of cadmium-binding peptides in toabacco leaves. Plant Physiol. 92, 1086-1093.
Walter, I.; F. Martínez y G. Cuevas. 2003. Dinámica de los metales pesados en un suelo degradado enmen-dado con residuos orgánicos urbanos. pp. 367-371. En: Álvarez-Benedí, J. y P. Marinero (eds.). Estudios de la Zona No Saturada del Suelo Vol. VI. Departa-
ACuMuLACIóN DE METALES pESADOS EN SuELO y pLANTAS DE CuATRO CuLTIVOS HORTÍCOLAS

Diego aNDréS berNal1 leiDy CaroliNa moraleS1 gerharD FiSCher1, 2 Jairo Cuervo1 StaNiSlav magNitSkiy1
plantulas de tomate. Foto: Álvarez-Herrera.
Caracterización de las deficiencias de macronutrientes en plantas de cebollín (Allium schoenoprasum L.)
Characterization of macronutrient deficiencies in chives (Allium schoenoprasum L.) plants
plantación de cebollín en invernadero.Foto: j. Cuervo
reSumeN
El cebollín se encuentra dentro de la gama de plantas aromáticas exportadas desde Colombia al mercado inter-nacional el cual es muy exigente en términos de volumen y calidad. Para caracterizar el efecto de deficiencias nutricionales de macroelementos en el crecimiento y desarrollo de las plantas, éstas fueron cultivadas bajo in-vernadero en Bogotá en materas de plástico (2 L) con sustrato arena cuarcítica durante dos ciclos de producción (primer corte a los 69 días después de la siembra, dds, y segundo a los 110 dds). Se evaluaron seis tratamientos; comparándolos con el testigo (T0) con fertilización completa, generados a partir de una solución de Hoagland y Arnon modificada: (T1) fertilización completa y N al 25%; (T2) fertilización completa –P; (T3) fertilización completa –K; (T4) fertilización completa -Ca; (T5) fertilización completa –Mg y (T6) fertilización completa –S. Se evaluaron las variables número, diámetro y longitud de hojas y masa seca (MS) de hojas y raíces. La nutrición deficiente afectó especialmente el crecimiento y desarrollo del cebollín en el segundo ciclo de cultivo, en el cual la carencia de P redujo drásticamente el diámetro y la longitud foliar, la acumulación de MS en hoja y raíz, y el número de hojas en el primer ciclo. La concentración de N al 25% disminuyó significativamente el diámetro y la longitud foliar durante el segundo ciclo de cultivo. La carencia de Mg aumentó el número y la biomasa de hojas en el segundo corte por el incremento de N, P, Ca y S en la masa seca foliar de este tratamiento. Los síntomas más comunes fueron la clorosis de las hojas con puntas amarillas hasta necróticas.
1 Departamento de Agronomía, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá (Colombia).2 Autor para correspondencia. [email protected]
REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 2 - No.2 - pp. 192-204, 2008
palabras clave adicionales: solución nutritiva, crecimiento, hoja, raíz, masa seca.

Vol. 2 - No.2 - 2008
193
additional key words: nutrient solution, growth, leaf, root, dry mass.
Fecha de recepción: 28-08-2008 Aprobado para publicación: 02-12-2008
El cebollín (Allium schoenoprasum L.) es una es-pecie de hierba aromática, perteneciente a la familia Alliaceae. La planta posee un hábito de crecimiento perenne y presenta muchos ciclos de autorrenovación de sus estructuras vegetati-vas a través de los bulbillos (Delahaut y Newen-house, 2003; Abello et al., 2006). Las hojas son el órgano de interés económico; estas son cilín-dricas, tubulares y delgadas, con una longitud superior a 17 cm, poseen un delicado aroma que las hace ser preferidas por los gourmet para des-tacar el sabor de diferentes platos, su consumo es generalmente en fresco. El cebollín se produ-ce en climas fríos de Suramérica tropical, entre 2.000 y 2.800 msnm y bajo invernadero, aunque también se puede establecer a libre exposición, adaptándose a diferentes tipos de suelo (Barre-ño, 2006).
abStraCt
Chives plants belong to the group of aromatic plants exported from Colombia to the international market that requires products of high quality. In order to characterize the effect of deficiencies of macronutrients on the growth and development of chives plants, these were cultivated in greenhouse in Bogotá in 2 L plastic pots filled with quartz sand during two cycles of production (first harvest at 69 days and a second one at 110 days after planting). Six treatments were evaluated compared with the control (T0), which consisted in a complete fertilization using a modified solution of Hoagland and Arnon: (T1) complete fertilization with N at 25%, (T2) complete fertilization –P, (T3) complete fertilization –K, (T4) complete fertilization –Ca, (T5) complete fertilization –Mg, and (T6) complete fertilization –S. Number, diameter and length of leaves and dry matter (DM) of leaves and roots were evaluated. The deficient nutrition affected, especially, the growth and development of chives during the second crop cycle, in which the lack of P drastically reduced diameter and length of leaves, accumulation of DM in leaves and roots, and also leaf number during the first cycle. The concentration of 25% N decreased significantly diameter and length of leaves during the second crop cycle. Deficiency of Mg increased the number and biomass of leaves in the second cycle by increasing concentrations of N, P, Ca, and S in the leaf DM of this treatment. The most common visual symptoms of macronutrient deficiencies were leaf chlorosis and yellow to necrotic spots in leaf apices.
iNtroDuCCióN
Investigaciones con especies aliáceas mostraron una asociación protectiva de estas contra el cán-cer (Steinmetz y Potter, 1996), especialmente cáncer de estómago, debido a su contenido en compuestos como el dialil sulfido y alil metil trisulfido que inducen sistemas enzimáticos de detoxificación (You et al., 1989).
La ubicación geográfica en el trópico y su produc-ción durante todo el año le han permitido a esta especie cobrar gran fuerza en las exportaciones colombianas con una participación del 8% con respecto a las demás hierbas aromáticas culina-rias vendidas en fresco. Los países principales a los cuales se exporta cebollín son Estados Uni-dos, Canadá, Inglaterra, Alemania y Holanda (Barreño y Clavijo, 2006). La gran exigencia de los mercados internacionales requiere de plantas
CARACTERIzACIóN DE LAS DEFICIENCIAS DE MACRONuTRIENTES EN pLANTAS DE CEBOLLÍN

REV. COLOMB. CIENC. HORTIC.
194
de calidad, para lo cual es indispensable que la planta esté bien nutrida, esto permitirá el cre-cimiento y desarrollo adecuado de la raíz y de tejido foliar (Castro, 1998; Clavijo, 2006).
Los macronutrientes son elementos constituyen-tes de las biomoléculas estructurales (N, P, S), tales como proteínas, lípidos o carbohidratos, o pueden actuar como osmolitos (K), sus con-centraciones en los tejidos de las plantas pueden variar considerablemente dependiendo de la es-pecie, la edad de la planta y el nivel de otros ele-mentos (Bonilla, 2008). En cantidades óptimas, los nutrientes esenciales garantizan una buena producción y calidad de los cultivos siempre y cuando no se encuentren afectadas por otros fac-tores; las plantas pueden afectarse por la falta de algún elemento, aun cuando los demás nutrien-tes estén presentes en cantidades adecuadas y disponibles para las plantas (Sabino et al., 2007). Las plantas responden a un aporte insuficiente de un elemento esencial, presentando síntomas característicos a esta deficiencia, muchos de es-tos son visuales, tal como la disminución en el crecimiento, clorosis, necrosis, entre otros (Mar-tínez et al., 2009).
El nitrógeno forma parte de un gran número de compuestos orgánicos, incluidas hormonas de crecimiento (Wild y Jones, 1992); participa en la estructura de todas las proteínas, de los ácidos nucléicos, y además se encuentra como consti-tuyente de las clorofilas y enzimas del grupo de los citrocromos y en varias coenzimas (Navarro y Navarro, 2000).
El fósforo forma enlaces anhídridos altos en energía, como es el caso del ATP y ADP, desem-peñando, por tanto, un papel clave en la foto-síntesis, respiración y el metabolismo energético (Bonilla, 2008). Los fosfatos son, también, cons-tituyentes de los ácidos nucléicos y fosfolípidos (Wild y Jones, 1992), los cuales son fundamenta-les en las membranas (Bonilla, 2008).
El potasio activa muchas enzimas que son esen-ciales en la fotosíntesis y la respiración (Salisbury y Ross, 2000) y actúa como osmorregulador, controlando en las células guarda la apertura de los estomas (Bennet, 1993). También participa en la producción de ATP, la síntesis de almidón y proteínas, y en el proceso de la fotosíntesis y el metabolismo de los carbohidratos (Navarro y Navarro, 2000).
Una de la principales funciones del Ca en la plan-ta es formar parte de la estructura de la proto-pectina, como agente cementante para mantener las células unidas, estando localizado en la lámi-na media y en la pared primaria celular, además, participa en la regulación de la absorción de nitró-geno, la traslocación de los carbohidratos y proteí-nas, la neutralización de los ácidos orgánicos y la activación de algunas enzimas, tales como amila-sa y fosfolipasa (Navarro y Navarro, 2000),
El Mg es un componente específico de la cloro-fila (porfirina magnésica) en la que un átomo de magnesio está ligado a cuatro anillos pirrólicos (Wild y Jones, 1992). Cumple un rol específico como activador de enzimas imvolucradas en la respiración, fotosíntesis y síntesis de ADN y ARN (Taiz y Zeiger, 2006). Actúa como cofac-tor de la mayor parte de las enzimas que inter-vienen en la fosforilación, y su importancia es grande en la transferencia de la energía (Wild y Jones, 1992).
Los compuestos orgánicos azufrados participan en la biosíntesis de lípidos, clorofilas, carotenos y ácidos orgánicos, aminoácidos sulfurados como cisteína, cistina y metionina y además, en la for-mación del acetilcoenzima A, eslabón básico de conexión entre glucólisis y el ciclo de Krebs (Na-varro y Navarro, 2000), y la interacción del ni-trógeno y azufre influye en la pungencia (aroma) de las plantas Allium (Liu et al., 2009), compues-tos que pueden ser importantes como repelentes contra herbívoros (Öpik y Rolfe, 2005).
BERNAL/MORALES/FISCHER/CuERVO/MAGNITSkIy

Vol. 2 - No.2 - 2008
195
El conocimiento de los síntomas de deficiencias minerales le permite determinar qué tipo de de-ficiencia nutricional puede tener el cultivo, esta-blecer la mejor forma de aplicación y el momento más adecuado para fertilizar (Salisbury y Ross, 2000), permitiendo con ello generar modelos de nutrición mineral eficiente del cultivo (Le Bot et al., 1998). Si la concentración de un elemento nu-triente esencial en el tejido vegetal está por debajo del nivel necesario para un óptimo crecimiento, indica que la planta es deficiente en ese elemento, y se produce así una alteración en la ruta metabó-lica en la que participa dicho elemento, afectando además otros procesos inmediatamente involu-crados (Epstein y Bloom, 2005).
En Colombia no se registra un estudio enfoca-do hacia la caracterización de las deficiencias nutricionales en el cultivo de cebollín. Por tanto este trabajo tiene como objetivo caracterizar las deficiencias visuales de N, P, K, Ca, Mg y S a ni-vel foliar en plantas de cebollín estableciendo el efecto sobre diferentes variables de crecimiento, bajo una fertilización deficiente y, de esta mane-ra, proporcionar herramientas de diagnóstico.
materialeS y métoDoS
El trabajo se llevó a cabo en un invernadero de plástico de la Facultad de Agronomía de la Uni-versidad Nacional de Colombia, Bogotá (2.556 msnm), que presentó en el interior una tempera-tura promedio de 23ºC y una humedad relativa promedio de 61%. Durante el estudio incidieron 4,5 h brillo solar/día.
El trabajo inició con la siembra de las semillas de cebollín en materas de plástico, las cuales se lle-naron con 2 kg de arena cuarcítica con un tama-ño de grano de 0,5 mm. Cada semilla fue sem-brada a una profundidad de 0,5 cm con distancia de 3 cm (15 plantas/matera) de forma circular, las materas fueron ubicadas en una superficie plana de los bancos de propagación distanciadas
entre sí a 30 cm. Posterior a la siembra se cubrió con una lámina plástica de Vinipel (policloruro de vinilo) que permite un intercambio gaseoso entre las plantas y el ambiente, mantiene la hu-medad, la temperatura estable para acelerar la germinación de las semillas y bajar la pérdida de agua (Castro et al., 2003).
Se implementó un diseño de fertilización com-pletamente al azar con seis tratamientos y cuatro repeticiones, comparados con el testigo (T0) fer-tilización completa: (T1) Fertilización completa y nitrógeno al 25% (N 25%), (T2): fertilización completa menos fósforo (-P), (T3): fertilización completa menos potasio (-K), (T4): fertilización completa menos calcio (-Ca), (T5): fertilización completa menos magnesio (-Mg) y (T6): fertili-zación completa menos azufre (-S).
Para evaluar el efecto de los tratamientos, se utilizó la matriz de fertilización de Hoagland y Arnon modificada por H. Bolhuis (comunica-ción personal, 2008) (tabla 1), la cual fue mez-clada para generar las soluciones en canecas de 20 L, estas fueron aplicadas vía fertirriego 1, 2 ó 3 veces por día con regadera dependiendo de las condiciones ambientales que se presentaran. Las semillas y plántulas se regaron con agua desti-lada durante 21 días después de siembra (dds) y para el día 22 se inició con la solución nutritiva que contenía todos los elementos hasta el día 35; con esta solución se buscó que las plantas tuvieran un buen desarrollo y fueran homogé-neas al inicio de los tratamientos. Durante los tres días siguientes se regó solo con agua destila-da con el fin de lavar el sustrato de los minerales que pudieran estar disponibles posteriormente para las plantas. Para el riego y preparación de las soluciones nutritivas se utilizó agua destila-da; las fuentes de fertilizantes, suministrados por Brenntag Colombia S.A., Mosquera, fueron: Agrofeed Potasio Líquido, Potasa Cáustica, Áci-do Fosfórico, MAP, Nitrato de Potasio, Nitrato de Amonio, Sulfato de Potasio y los micronu-trientes.
CARACTERIzACIóN DE LAS DEFICIENCIAS DE MACRONuTRIENTES EN pLANTAS DE CEBOLLÍN

REV. COLOMB. CIENC. HORTIC.
196
Las evaluaciones se realizaron con una frecuen-cia de 7 d a partir del día 40 después de la siem-bra, durante dos ciclos de cultivo. Se evaluaron variables morfológicas, tal como longitud de hojas y de raíz (cm), diámetro medio de la hoja (mm), número de las hojas, masa seca de hojas y raíz (g) y para cada uno de los tratamientos se realizó un análisis foliar a los 124 dds. El primer corte (cosecha) se realizó a los 69 dds y el segun-do a los 110 dds.
Para el análisis de los datos se utilizó el paquete estadístico SAS® 9.1 y en el análisis de varianza se compararon promedios mediante la prueba de Duncan (P≤0,05) y contrastes ortogonales.
Los materiales de laboratorio utilizados fueron un pie de rey digital (marca Minutoyo-Vernier) para los diámetros de las hojas, balanza de pre-cisión (0,001 g) marca Chyo Jupiter SDT-200 y estufa marca WTB binder para el secado del material vegetal a una temperatura de 70ºC por 24 h.
NutrienteT0 T1 T2 T3 T4 T5 T6
Testigo N 25% -p -k -Ca -Mg -S
N 224,00 56,00 224,00 224,00 224,00 224,00 224,00
p 31,00 31,00 - 31,00 31,00 31,00 31,00
k 235,00 235,00 235,00 - 235,00 235,00 235,00
Ca 160,00 160,00 160,00 160,00 - 160,00 160,00
Mg 24,00 24,00 24,00 24,00 24,00 - 24,00
S 32,00 32,00 32,00 32,00 32,00 32,00 -
Mn 0,11 0,11 0,11 0,11 0,11 0,11 0,11
zn 0,13 0,13 0,13 0,13 0,13 0,13 0,13
Cu 0,03 0,03 0,03 0,03 0,03 0,03 0,03
Fe 1,50 1,50 1,50 1,50 1,50 1,50 1,50
B 0,27 0,27 0,27 0,27 0,27 0,27 0,27
Mo 0,05 0,05 0,05 0,05 0,05 0,05 0,05
Cl 140,00 140,00 - - - - -
tabla 1. Niveles de nutrientes (mg l-1) en solución nutritiva de Hoagland y Arnon, modificada según H. Bolhuis (comunicación personal, 2008) y aplicada diariamente.
reSultaDoS y DiSCuSióN
Contenido de los nutrientes en las hojas
La concentración foliar de los diferentes elemen-tos en las plantas testigo (tabla 2) mostró en orden decreciente K>N>Ca>Mg>P>S lo que coincide, en los primeros tres elementos, con lo encontra-do por Pôrto et al. (2006) en plantas de cebolla (hoja + bulbo) con K>N>Ca>S>Mg>P.
En las hojas de plantas deficientes se encontraron diferencias significativas (P≤0,05) para la con-centración de los seis elementos, especialmente para K, N, P, Ca y Mg (tabla 2). En el tratamien-to N25%, la concentración de N foliar se redujo 29% comparado con el testigo. La tabla 2 indica que la reducción de N está relacionada con el in-cremento del nivel de S, debido a que el S cum-ple funciones metabólicas similares en las cuales es importante el nitrógeno, de esta forma busca equilibrar el bajo contenido de este elemento (Sa-bino et al., 2007). Esto refleja la relación N/S que
BERNAL/MORALES/FISCHER/CuERVO/MAGNITSkIy

Vol. 2 - No.2 - 2008
197
TratamientoN p k Ca mg S
porcentaje en masa seca
Testigo 3,98 c 0,29 b 6,13 a 0,51 cd 0,32 a 0,16 b
N 25% 2,82 d 0,18 c 4,98 b 1,19 b 0,18 c 0,50 a
-p 4,15 c 0,13 c 3,88 c 1,10 bc 0,13 cd 0,56 a
-k 5,09 a 0,43 a 1,60 d 1,90 a 0,28 ab 0,53 a
-Ca 4,86 ab 0,52 a 3,68 c 0,44 d 0,25 b 0,51 a
-Mg 4,86 ab 0,41 a 3,87 c 1,73 ab 0,10 d 0,55 a
-S 4,30 bc 0,44 a 4,04 bc 1,72 ab 0,16 cd 0,13 b
tabla 2. efecto de la deficiencia de N, p, k, Ca, mg y S sobre la concentración de nutrientes en hojas de cebollín, a los 124 dds.
Promedios con letras distintas en la misma columna indican diferencia significativa según la prueba de Duncan (P≤0,05).
en el testigo es 24,9, mientras en plantas N25% alcanza solamente 5,6.
En el tratamiento -P se observó 55% menos de la concentración de este elemento comparado con el testigo (tabla 2). La concentración óptima en la cual se debe encontrar el P en el tejido foliar del cebollín es de 0,3 a 0,5% (Ramírez, 2006), muy similar a la óptima en hojas de cebolla (0,25 a 0,40%) reportada por Bender (1993), lo que muestra el bajo contenido de este elemento en las hojas analizadas (0,13%).
La concentración de K en las hojas del tratamien-to –K fue 73,8% menor que el testigo (tabla 2), lo que confirma que los Allium son sumamente susceptibles a deficiencias de este elemento (Ben-der, 1993). En este tratamiento, las concentra-ciones de N, P, Ca y S muestran un incremento significativo, que puede ser debido a una serie de cambios químicos que generan una disminución en el contenido de almidón o la acumulación de compuestos solubles de nitrógeno. Marschner (2002) reporta que la falta de K suprime la sínte-sis de enzimas como la nitrato reductasa, lo que lleva a una acumulación de N en forma de nitra-to en las hojas
La omisión de Ca en la solución nutritiva generó solamente una reducción del 13,7% de la concen-tración de este elemento en la hoja que no fue
significativa comparado con el testigo y que fue compensado por el aumento de otros nutrientes como el N, P y S en este órgano (tabla 2).
La deficiencia absoluta de Mg disminuyó la con-centración de este elemento en las hojas en un 68,7% comparado con el testigo, pero se generó un aumento de los elementos N, P, Ca y S en este tratamiento (tabla 2), dentro de los cuales, el Ca puede parcialmente reemplazar el Mg en varias funciones (Bergmann, 1993).
La concentración de azufre muestra una dismi-nución no significativa (P≤0,05) de 15,0% en las hojas comparada con el testigo (tabla 2), gene-rando incrementos en el N y P y reducción en el K. Alteraciones similares encontraron Alves et al. (2007) en plantas de Dimorphandra mollis.
Síntomas foliares de la planta
Las plantas de cebollín con deficiencia de N fue-ron más pequeñas (figura 1, tabla 4), con hojas más delgadas, cortas (tabla 4) y un color verde pálido (Bergmann, 1993), comparadas con el tes-tigo. Se formó una clorosis basal en las hojas del primer ciclo, mientras que las maduras mostra-ron un mayor número de puntas (ápices) ama-rillas las cuales se necrosaron a medida que se acercó su senescencia. Los síntomas iniciaron en la parte inferior de las hojas maduras (Martínez
CARACTERIzACIóN DE LAS DEFICIENCIAS DE MACRONuTRIENTES EN pLANTAS DE CEBOLLÍN

REV. COLOMB. CIENC. HORTIC.
198
et al., 2009) donde se generó progresivamente una coloración amarilla en la medida que la defi-ciencia se acentuaba (Bender, 1993).
Las plantas de cebollín, carentes de P, mostraron bajo crecimiento (Bender, 1993) y menor núme-ro de hojas (figura 1, tablas 3 y 4) que se agudi-zó, especialmente a partir de 113 dds, con hojas completamente delgadas, pequeñas y puntiagu-das y en un 60% de estas se observaron puntas amarillas.
Las plantas con deficiencia de K expresaron hojas entorchadas hacía el ápice y un 50% de puntas amarillas, produciéndose necrosis del tejido foliar en dirección basipétala especialmente en las ho-jas más viejas, lo cual coincide con lo descrito por Bergmann (1993). Las hojas afectadas presentaron una coloración más oscura, como reporta Bender (1993) para cebolla (figura 1).
Las plantas con deficiencia de Ca no presenta-ron alteraciones en su crecimiento en el primer ciclo, sin embargo mostraron puntas amarillas en 80% de sus hojas. A los 92 dds se observaban hojas flácidas, débiles, con pérdida de turgencia y partidas. En el segundo ciclo empezaron a to-mar colores más oscuras, con hojas quebradas, clorosis desde la base hacía el ápice, similar a los síntomas observados en plantas de uchuva (Mar-tínez et al., 2009).
La deficiencia de Mg se inició a partir de 58 dds con la aparición de una línea blanca-clorótica en dirección vertical; a partir de 68 dds 30% de las hojas presentaron puntas amarillas que se trans-formaron después en necrósis conforme con la cebolla (Bender, 1993). Cuando 70% de las hojas mostraron puntas amarillas perdieron su turgen-cia progresivamente y algunas se entorcharon (figura 1).
Figura 1. Desarrollo de plantas de cebollín con una nutrición ausente en P, K, Ca, Mg y S, así como N al 25%, comparado con el testigo (Total) en el primer corte (69 dds).
N 25% -p -k -Ca -mg -S total
BERNAL/MORALES/FISCHER/CuERVO/MAGNITSkIy

Vol. 2 - No.2 - 2008
199
A final del primer ciclo, plantas deficientes de S presentaron algunos síntomas como clorosis, empezando en hojas nuevas (Bergmann, 1993) así como entorchamiento foliar, mientras en el segundo ciclo mostraron una clorosis basal que se agudizó a partir de 105 dds con un amarilla-miento muy marcado y homogéneo y puntas amarillas, síntomas comunes por falta de S (Sa-bino et al., 2007).
Número de hojas
En el primer corte, la deficiencia de P disminuyó significativamente el número de hojas, compara-do con el testigo (tabla 3), mientras la carencia de los demás elementos no afectó estadísticamente la emisión de hojas en cebollín.
El fosfato se redistribuye fácilmente de un ór-gano a otro acumulándose en las hojas jóvenes (Bonilla, 2008), por lo cual en el presente estudio se supone que la planta no contaba con una can-tidad suficiente de este elemento lo que dismi-nuyó la formación de hojas. Asimismo, la falta de ATP, ADP, AMP, los cuales son esenciales en el metabolismo energético, pudo haber causado una reducción en la emisión foliar (Salisbury y Ross, 2000; Jones, 1998).
Bender (1993) reporta que cebollas deficientes en azufre producen menos hojas; en cebollín este efecto no fue observado, debido, probablemente, a que la carencia de este elemento no fue total (tabla 2).
En el segundo corte se presentó un aumento del número de hojas en plantas deficientes del Mg, comparado con las -P en la solución nutritiva (ta-bla 3). Un efecto que pudo haber sido causado por el aumento de N, P, Ca y S en concentracio-nes significativas en el tratamiento –Mg, compa-rado con el testigo (tabla 2), teniendo en cuenta, por un lado, que elementos como el Ca, además del Mn y Zn (tabla 1) pueden reemplazar el Mg en varias de sus funciones (Bergmann, 1993) y,
por otro lado, la muy deficiente condición ener-gética de plantas faltantes en P.
Diámetro y longitud de hojas
Una de las características de las plantas de ce-bollín son sus hojas de forma cónica aguda, las cuales disminuyeron su diámetro significativa-mente (P≤0,05) con respecto a su diámetro nor-mal, lo que se observó en el primer corte en el tratamiento –Mg y al final del segundo ciclo en las plantas deficientes en P y K (tabla 4).
Se considera que al inicio del estudio las hojas de cebollín crecieron más lento en el tratamiento –Mg como reporta Bender (1993) para la cebolla, lo que podría haber afectado su aumento en diá-metro. Hasta el segundo corte, las deficiencia de P y N se agudizaron afectando marcadamente el desarrollo foliar (tabla 4), siendo el P esencial en el metabolismo energético, condición indispen-sable para el crecimiento normal de la planta. Marschner (2002) describe que en las plantas deficientes en fósforo hay un retardo en el creci-miento de la parte aérea y se limita el proceso de fotosíntesis debido a alteraciones producidas en la concentración de clorofila. También en cebo-llas, Bender (1993) observó hojas más angostas por falta de nitrógeno.
TratamientoNúmero de hojas
primer corte Segundo corte
Testigo 3,9 a 6,3 ab
N 25% 3,3 ab 6,1 ab
-p 3,1 b 4,7 b
-k 3,3 ab 6,0 ab
-Ca 3,4 ab 6,3 ab
-Mg 3,0 ab 8,1 a
-S 3,5 ab 6,9 ab
tabla 3. efecto de la deficiencia de N, p, k, Ca, mg y S sobre el número de hojas de cebollín en el primer y segundo corte.
Promedios con letras distintas en la misma columna indican diferencia significativa según la prueba de Duncan (P≤0,05).
CARACTERIzACIóN DE LAS DEFICIENCIAS DE MACRONuTRIENTES EN pLANTAS DE CEBOLLÍN

REV. COLOMB. CIENC. HORTIC.
200
La concentración de 0,13% de S en la masa seca foliar de cebollín, deficiente de este elemento (ta-bla 2), está muy cerca al valor mínimo de 0,10% que deben contener a hortalizas para evitar sín-tomas de deficiencia visibles (Bergmann, 1993). Este autor describe que en el caso de una defi-ciencia de azufre, el sulfato de las hojas maduras no es suficiente para un crecimiento normal de la planta, generando hojas jóvenes más pequeñas debido a una carencia de proteínas y clorofilas.
Para la variable longitud de hoja no se encontraron diferencias significativas entre los tratamientos en el primer corte. En el segundo corte se puede observar los valores más bajos (P≤0,05) en los tra-tamientos deficientes en P y N, debido a que estos elementos interfieren en los procesos fotosintéticos y de elongación celular (Salisbury y Ross, 1994). La falta del N debilita la planta, se desarrolla poco y las hojas permanecen pequeñas (Navarro y Nava-rro, 2000), por lo cual, Bonilla (2008) describe este síntoma como enanismo (figura 1).
El P se manifestó de nuevo como uno de los ele-mentos más importantes para el desarrollo de la planta de cebollín, especialmente en el segundo corte, donde redujo drásticamente el desarrollo foliar, lo que explica Bender (1993) como un cre-cimiento muy lento de la planta y un retardo en su madurez (Bonilla, 2008).
Masa seca de raíz y hojas
Entre el primer y segundo corte la masa seca, acumulada en raíz y hojas, aumentó considera-blemente (tabla 5).
Especialmente la ausencia del P en la solución nu-tritiva provocó una baja producción de biomasa radical que también perjudicó la acumulación de masa seca foliar (tabla 5) y de la planta en total (figura 2). Este resultado refleja la importancia del fósforo para la formación del sistema radi-cal (Gómez, 2006), siendo los fosfatos esenciales para la división celular y para el desarrollo de los tejidos meristemáticos (Wild y Jones, 1992) y además subraya el papel clave del P en el meta-bolismo energético de la planta (Bonilla, 2008) que se manifiesta en una formación deficiente de los órganos del cebollín. Las otras deficiencias no afectaron significativamente la acumulación de MS en la raíz del cebollín.
También, la deficiencia de P mostró los valores más bajos en MS foliar (tabla 5), que se redujo en un 67,87% con respecto al testigo, esto demues-tra nuevamente que el P es esencial para la trans-ferencia energética en la planta y afecta, según Bender (1993), todos los aspectos del crecimiento de las aliáceas y concuerda con lo que se encon-tró para cebollín (figura 2).
TratamientoDiámetro de hoja (cm) Longitud de hoja (cm)
primer corte Segundo corte primer corte Segundo corte
Testigo 1,67 ab 1,80 a 20,64 a 20,12 a
N 25% 1,82 a 1,34 b 19,97 a 17,84 b
-p 1,54 ab 1,14 b 18,46 a 14,29 b
-k 1,65 ab 1,50 ab 19,95 a 22,05 a
-Ca 1,59 ab 1,53 ab 19,53 a 23,89 a
-Mg 1,45 b 1,65 ab 18,57 a 22,62 a
-S 1,52 ab 1,53 ab 18,96 a 21,28 a
tabla 4. efecto de la deficiencia de N, p, k, Ca, mg y S sobre el diámetro y la longitud de hojas de cebollín en el primer y segundo corte.
Promedios con letras distintas en la misma columna indican diferencia significativa según la prueba de Duncan (P≤0,05).
BERNAL/MORALES/FISCHER/CuERVO/MAGNITSkIy

Vol. 2 - No.2 - 2008
201
En el segundo corte, la segunda producción más baja de la MS foliar fue mostrada por la aplica-ción del N al 25%, sin mostrar diferencias signi-ficativas con el testigo (tabla 5). Resende y Souza (2001) encontraron en ajo que la deficiencia de N afecta la producción de masa seca y Bender (1993) afirma que una carencia de N en cebolla ocasiona plantas pequeñas, lo que se confirma en lo presentado en la figura 1. Lemaire et al. (2008) constatan que la reducción del contenido de N por unidad de área foliar (tabla 2) reduce la eficiencia en el uso de la radiación que, en conse-cuencia, afecta la tasa fotosintética.
La ausencia de K en la fertilización no afectó mu-cho la acumulación de MS en los cebollines de este tratamiento (figura 2), debido a que en el te-jido foliar de estas plantas se aumentaron las con-centraciones de N, P y S comparado con el testigo (tabla 2). El N optimiza la tasa de expansión de la hoja y así la intercepción de la radiación fotosinté-ticamente activa (Lemaire et al., 2008).
Similar a la deficiencia de K, la de Ca no afec-tó significativamente la producción de MS de la planta, esto se debe a que la ausencia de este ele-mento en el agua de riego fue compensada por una mayor acumulación de N, P y S en el tejido foliar del cebollín. Tal vez, la poca reducción de
este elemento en la hoja (13,7%, tabla 2) generó condiciones que no fueron suficientemente ex-tremas para que se mostrara una reducción de la biomasa (Bergmann, 1993). En especies del ge-nero Aliaceae como la cebolla, la deficiencia de Ca es poco común, sin embargo, en el caso de una deficiencia de Ca2+ extrema las plantas no son capaces de sobrevivir y solamente en pocos casos pasan el estado de planta germinada (Ben-der, 1993).
La deficiencia absoluta de Mg en la solución nu-tritiva en el primer corte no tuvo diferencias con los demás tratamientos, mientras que en el se-gundo corte hubo un aumento significativo de la MS foliar, debido seguramente a las concentra-ciones incrementadas de N, P, Ca y S en este ór-gano, comparadas con el testigo (tabla 2), lo que se refleja en la tabla 5. Es conocido que el Mn, Zn y Ca pueden reemplazar el Mg en varias de sus funciones (Bergmann, 1993).
La deficiencia de azufre no produjo acumulación diferente (P≤0,05) de MS en la planta compara-da con el testigo (tabla 5), debido a la no reduc-ción del contenido de este elemento en plantas sin S (tabla 2). Así mismo, Bender (1993) repor-ta que esta deficiencia no es común en aliáceas, como por ejemplo en la cebolla.
TratamientoMasa seca raíz (g) Masa seca hoja (g)
primer corte Segundo corte primer corte Segundo corte
Testigo 0,034 ab 0,149 a 0,077 a 0,194 b
N 25% 0,039 a 0,148 a 0,061 a 0,161 bc
-p 0,016 b 0,069 b 0,036 b 0,063 c
-k 0,031 ab 0,116 ab 0,069 ab 0,194 bc
-Ca 0,023 ab 0,100 ab 0,058 ab 0,232 b
-Mg 0,018 b 0,132 ab 0,045 ab 0,324 a
-S 0,022 ab 0,113 ab 0,051 ab 0,209 b
Tabla 5. Efecto de la deficiencia de N, P, K, Ca, Mg y S sobre la masa seca en hojas y raíz en la planta de cebollín en el primer (69 dds) y segundo corte (110 dds).
Promedios con letras distintas en la misma columna indican diferencia significativa según la prueba de Duncan (P≤0,05).
CARACTERIzACIóN DE LAS DEFICIENCIAS DE MACRONuTRIENTES EN pLANTAS DE CEBOLLÍN

REV. COLOMB. CIENC. HORTIC.
202
CoNCluSioNeS
• El estudio determinó que una nutrición defi-ciente en P y N afecta el crecimiento y desa-rrollo del cebollín, especialmente durante el segundo ciclo del cultivo
• La deficiencia de P, que tiene funciones claves en el metabolismo energético de la planta, cau-só disminución notable en el área foliar y la acumulación de masa seca en hoja y raíz.
• Dentro de los seis elementos estudiados los más limitantes para el aumento de la bioma-
Figura 2. efecto de la deficiencia de N, p, k y S sobre la acumulación de la masa seca total en cebollín en el primer corte (69 dds). Promedios con letras distintas indican diferencia significativa a los 69 días del primer ciclo según la prueba de Duncan (P≤0,05).
sa seca de la planta, bajo las condiciones del experimento, fueron en un orden decreciente, en el primer corte: P>Mg>S>Ca>N>K y en el segundo corte: P>N>K>S>Ca>Mg.
• Las deficiencias de K y Ca no afectaron signifi-cativamente el crecimiento de las plantas.
• La carencia del Mg aumentó el número y la biomasa de las hojas en el segundo corte del cebollín por el incremento de N, P, Ca y S en la masa seca foliar de este tratamiento.
agraDeCimieNtoS
Los autores agradecen el apoyo para realizar este trabajo al Departamento de Agro de Brenntag Colombia S.A., Mosquera, especialmente a Hugo Bolhuis y Milton Galán.
BERNAL/MORALES/FISCHER/CuERVO/MAGNITSkIy

Vol. 2 - No.2 - 2008
203
reFereNCiaS bibliogrÁFiCaS
Abello, J.; J. Clavijo y P. Barreno. 2006. Estudio pre-liminar de algunos descriptores fisiológicos en cinco hierbas aromáticas. pp. 13-15. En: Memorias Curso de Extensión: Últimas tendencias en hierbas aromáticas culinarias para exportación en fresco. Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá.
Alves, C.; D. Alves; L.A. Fernandes; E.R. Martins; I.G. Souza; R.A. Sampaio y P.S. Lopes. 2007. Nutrição mineral da fava d’anta. Hortic. Brás. 25(1), 24-28.
Barreño, P. 2006. Hierbas aromáticas culinarias para ex-portación en fresco manejo agronómico, produc-ción y costos. pp. 65-72. En: Memorias Curso de Extensión: Últimas tendencias en hierbas aromáti-cas culinarias para exportación en fresco. Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá.
Barreño, P. y J. Clavijo. 2006. Hierbas aromáticas culina-rias para exportación en fresco. pp. 7-9. Memorias Curso de Extensión: Últimas tendencias en hierbas aromáticas culinarias para exportación en fresco. Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá.
Bender, D.A. 1993. Onions. pp. 131-135. En: Bennett, W.F. (ed.). Nutrient deficiencies and toxicities in crop plants. APS Press, St. Paul, MN.
Bergmann, W. 1993. Ernährungsstörungen bei Kulturpflan-zen. 3a ed. Gustav Fischer Verlag, Jena, Alemania.
Bonilla, I. 2008. Introducción a la nutrición mineral de las plantas. Los elementos minerales. pp. 102-121. En: Azcón-Bieto, A. y M. Talón. (eds.). Fundamen-tos de la fisiología vegetal. McGraw-Hill Interame-ricana, Madrid.
Castro, H.E. 1998. Capítulo 4: Hortalizas, producción y fertilización de hortalizas en Colombia. pp., 170-194. En: Guerrero, R. (ed.). Fertilización en cultivos de clima frío. Monómeros Colombo Venezolanos S.A., Colombia.
Castro, J.V.; L.B. Pfaffenbach; C.R.L Carvalho y C.J. Rossetto. 2003. Efecto del empaque plástico sobre la vida de anaquel del mango ‘Keitt’. Rev. Iberoa-mer. Tecnol. Poscos. 5(1), 33-37.
Clavijo, J. 2006. Fisiología de la producción de hierbas aromáticas. pp. 10-12. En: Memorias Curso de Ex-
tensión: Últimas tendencias en hierbas aromáticas culinarias para exportación en fresco. Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá.
Delahaut, K.A. y A.C. Newenhouse. 2003. Growing on-ions, garlic, leeks, and other Allium in Wisconsin: A guide for fresh-market growers. Cooperative Ex-tension, University of Wisconsin, Madison, WI.
Epstein, E. y A.J. Bloom. 2005. Mineral nutrition of plants: principles and perspectivas. 2a ed. Sinauer Associates Publishers, Sunderland, MA.
Gómez, M.I. 2006. Manual técnico de fertilización de cultivos. Microfertisa, Produmedios, Bogotá.
Jones, J.B. 1998. Manual of plant nutrition. CRC Press, Boca Raton, FL.
Le Bot, J.; S. Adamowicz y P. Robín. 1998. Modeling plant nutrition of horticultural crops: a review. Scientia Hort. 74(1-2), 47-82.
Lemaire, G.; E. Oosterom; M.H. Jeuffroy; F. Gastal y A. Massignam. 2008. Crop species present diffe-rent qualitative types of response to N deficiency during their vegetative growth. Field Crops Res. 105(1), 253-265.
Liu, S.; H. He; G. Feng y Q. Chen. 2009. Effect of nitro-gen and sulfur interaction on growth and pungen-cy of different pseudostem types of Chinese spring onion (Allium fistulosum L.). Scientia Hort. 121, en imprenta.
Marschner, H. 2002. Mineral nutrition of higher plants. Academic Press, Londres.
Martínez, F.E.; J. Sarmiento; G. Fischer y F. Jiménez. 2009. Síntomas de deficiencia de macronutrientes y boro en plantas de uchuva (Physalis peruviana L.). Agron. Colomb. 27(2), en imprenta.
Navarro, S. y G. Navarro. 2000. Química agrícola. Edi-ciones Mundi-Prensa, Madrid.
Öpik, H. y S. Rolfe. 2005. The physiology of flowering plants. Cambridge University Press, Cambridge, UK.
Pôrto, D,R.; A.B. Cecílio; A. May y J.C. Barbosa. 2006. Acúmulo de macronutrientes pela cebola ‘Optima’
CARACTERIzACIóN DE LAS DEFICIENCIAS DE MACRONuTRIENTES EN pLANTAS DE CEBOLLÍN

REV. COLOMB. CIENC. HORTIC.
204
estabelecida por semeadura directa. Hortic. Bras. 24(4), 470-475.
Ramírez, C. 2006. Nutrición mineral y producción vegetal. pp. 22-30. En: Memorias Curso de Exten-sión: Últimas tendencias en hierbas aromáticas culinarias para exportación en fresco. Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá.
Resende, G.M y R.J. Souza. 2001. Doses e épocas de apli-cação de nitrogênio sobre a productividade e carac-terísticas comerciais de alho. Hortic. Brás. 19(2), 126-129.
Sabino, H.; J. Lavres y M. Ferreira. 2007. Azufre como nutriente y agente de defensa contra plagas y en-fermedades. Informaciones Agronómicas. Interna-tional Plant Nutrition Institute 65(1), 1-4.
BERNAL/MORALES/FISCHER/CuERVO/MAGNITSkIy
Salisbury, F.B. y C.W. Ross. 2000. Fisiología de las plantas. Editorial Paraninfo Thomson Learning, Madrid.
Steinmetz, K.A. y J.D. Potter. 1996. Vegetables, fruit, and cancer prevention: A review. J. Amer. Diet. As-soc. 96(1), 1027-1039.
Taiz, L. y E. Zeiger. 2006. Plant physiology, 4a ed. Sinau-er Associates, Sunderland, MA.
Wild, A. y L.H.P. Jones. 1992. Nutrición mineral de las plantas cultivadas. pp. 73-119. En: Wild, A. (ed.). Condiciones del suelo y desarrollo de las plantas según Russel. Ediciones Mundi-Prensa, Madrid.
You, W.C.; W.J. Blot; Y.S. Chang; A. Ershow; Z.T. Yang; Q. An; B.E. Henderson; J.F. Fraumeni y T.-G. Wang. 1989. Allium vegetables and the reduced risk of stomach cancer. J. Natl. Cancer Inst. 81, 162-164.

resumen
La cachaza fresca de trapiche panelero es un subproducto obtenido del proceso de elaboración de panela, que tiene alto potencial como enmienda orgánica. Sin embargo, este residuo que equivale al 3% de la caña molida es arrojado a las fuentes hídricas o cerca de los trapiches generando problemas graves de contaminación am-biental. En la zona de la Hoya del Río Suárez (Boyacá, Colombia) el principal cultivo es la caña panelera, pero es apta para cultivos de pancoger como el maíz y el fríjol. Con el objetivo de evaluar el efecto de la cachaza fresca de trapiche panelero y dos sistemas de producción (maíz y maíz asocio fríjol) sobre las propiedades físicas de un Inceptisol, se aplicaron cuatro dosis de cachaza (0, 5, 10 ó 15 t ha-1) a cada uno de los sistemas de producción. Después de la cosecha se encontró que el sistema maíz asocio fríjol presentó el mayor diáme-tro ponderado medio en seco y húmedo y mayor porosidad, estas propiedades físicas junto con la densidad aparente fueron favorecidas con el incremento de la aplicación de cachaza. Con la aplicación de 15 t ha-1 de cachaza en el sistema maíz asocio fríjol se garantizó mayor efecto sobre las propiedades físicas del suelo.
Fabio emilio Forero u.1, 3 Jaime torreS b.2 helber eNrique balaguera-lópez1
Efecto de la aplicación de cachaza fresca y de dos sistemas de producción maíz y maíz con asocio fríjol sobre las propiedades físicas de un Inceptisol
Effect of application of fresh filter cake and two production systems of corn and corn associated with beans on the physical properties of an Inceptisol
Plantas de maíz fertilizadas con cachaza en Chitaraque, Boyacá.Foto: F.E. Forero
1 Grupo de Investigaciones Agrícolas, Facultad de Ciencias Agropecuarias, Universidad Pedagógica y Tecnológica de Colombia, Tunja (Colombia).
2 Departamento de Agronomía, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá (Colombia). 3 Autor para correspondencia. [email protected]
palabras clave adicionales: materia orgánica, suelo, sistema radical, porosidad del suelo.
REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 2 - No.2 - pp. 205-216, 2008

REV. COLOMB. CIENC. HORTIC.
206
abstract
The fresh filter cake of sugar cane mill is a byproduct of the process from the manufacturing of brown sugarloaf which has high potential as organic amendment. However, this waste is equivalent to 3% cane crushed that is discharged to water sources or thrown near the sugar mills creating serious problems of environmental contamination. In the “Hoya del Rio Suarez” zone (Boyaca, Colombia), the main crop is the sugar cane, but this land is suitable for other crops, such as corn and bean. In order to evaluate the effect of fresh filter cake of sugar cane mill and that of two production systems (maize and maize associated with bean) on the physical properties of the Inceptisol, four doses of filter cake (0, 5, 10 or 15 t ha-1) were applied to each of the production systems. After the harvest it was found that corn associated with beans had the greatest average weighted diameter dried and humid and the porosity; these properties together with bulk density were favored with the increase in the application of filter cake. With the application of 15 t ha-1 the filter cake in the system maize associated with bean guaranteed the major effect on soil physical properties.
additional key words: organic matter, soil, root system, soil porosity.
Fecha de recepción: 11-08-2008 Aprobado para publicación: 02-12-2008
iNtroDuCCióN
La cachaza es un material marrón oscuro, cons-tituido por una mezcla de fibra de caña, saca-rosa, coloides, coagulados, incluyendo la cera, fosfato de calcio y partículas de suelo (Zurro, 2005). Es considerada como el subproducto más importante de los ingenios azucareros y en la agroindustria artesanal de la caña panelera en la producción de panela, con cierto potencial como fertilizante y mejorador de las propiedades físicas del suelo (Zérega, 1993), producida a una tasa de 3 t húmedas por cada 100 t de caña molida. La cachaza tiene alto contenido de materia orgáni-ca y presenta potencial como acondicionador de suelos, ya que aporta cantidades altas de humus necesarias para mantener la fertilidad de suelos en zonas tropicales y subtropicales (Mohee y Panray, 1999).
En el año 2006, de la cachaza producida en la región de la Hoya del Río Suárez y Cundinamar-ca, se estaba utilizando 47% como alimento para animales domésticos y el 53% restante que co-rresponde aproximadamente a 21.812 t año-1 se
abandonaba al aire libre (Rodríguez y Gottret, 2006), incluyendo los cursos de agua, causando graves problemas de contaminación ambiental, ya que presenta una elevada demanda bioquími-ca de oxígeno, genera malos olores y atrae insec-tos, roedores y otras plagas.
En la región de la Hoya del Río Suárez, se des-conoce el uso que puede tener la cachaza en la producción agrícola como aporte de materia or-gánica (Arrieche y Mora, 2005), masa microbial (Khan et al., 2008) y nutrientes (Paturau, 1969), también se ha encontrado que afecta la salini-dad (Rasul et al., 2006), corrige acidez del suelo (Zérega, 1993), y principalmente, presenta alto potencial como mejorador de las propiedades fí-sicas del suelo (Painuli y Abrol, 1986).
Además, la costumbre ancestral es la siembra del monocultivo de la caña panelera, sin tener en cuenta que existen otros cultivos como el maíz (Zea mayz L.) y maíz en asocio con especies como el fríjol (Phaseolus vulgaris L.), los cuales son una
FORERO/TORRES/BALAGuERA-LópEz

Vol. 2 - No.2 - 2008
207
importante fuente de ingresos para muchos pro-ductores, y pueden ser una alternativa adecuada para la rotación de cultivos que permitan mante-ner o mejorar las propiedades físicas de los suelos de la zona.
Por lo anterior, el objetivo de esta investigación fue evaluar el efecto de la aplicación de cachaza fresca de trapiche panelero y de dos sistemas de producción maíz y maíz asocio fríjol sobre las propiedades físicas de un Inceptisol presente en la zona panelera de la Hoya del Río Suárez, con el propósito de darle una alternativa de uso a la cachaza producida en la zona.
materialeS y métoDoS
El trabajo de campo fue desarrollado en la Fin-ca La Florida del municipio de Chitaraque (Bo-yacá), que se encuentra ubicada a 5º59’87” N y 73º27’78” W, con una altura de 1.729 msnm. Las condiciones climáticas de la zona durante el periodo del experimento (4 meses) fueron las siguientes: precipitación 660 mm, temperatura promedio de 18,6ºC, temperatura máxima pro-medio 25,9°C, temperatura mínima promedio 13°C y humedad relativa promedio del 80%. Los análisis físicos de suelos se realizaron en el labo-ratorio de docencia de suelos de la Facultad de Ciencias Agropecuarias de la Universidad Peda-gógica y Tecnológica de Colombia, Tunja.
Se utilizó un diseño experimental en bloques completamente al azar, donde los bloques corres-pondieron a dos sistemas de producción (maíz y maíz asocio fríjol), cada bloque fue repetido tres veces, dentro de cada bloque se distribuyeron al azar cuatro dosis de cachaza fresca de trapiche panelero (0, 5, 10 ó 15 t ha-1), en total fueron 24 unidades experimentales (UE). Cada UE corres-pondió a una parcela de 6 x 4 m, separadas entre si a 0,5 m.
Para el sistema maíz se sembraron tres semillas de maíz por sitio con distancias de siembra de
0,9 m x 0,45 m. Después de la emergencia se dejaron solamente dos plántulas por sitio. En el sistema asocio maíz fríjol se utilizaron las mis-mas distancias de siembra para el maíz, y 15 días después de la siembra del maíz, en cada sitio se sembraron dos semillas de fríjol para finalmente dejar una planta. Se utilizó maíz ICA V-305 y fríjol cargamanto rojo.
La preparación del suelo se realizó de acuerdo a lo sugerido por los agricultores de la zona, me-diante arado con bueyes y dos cruzas. Luego, se realizó el trazo según el diseño experimental y se procedió a aplicar la cachaza fresca de trapi-che panelero, la cual fue aplicada manualmente al voleo y se incorporó con azadón, la siembra se realizó 20 d después de esta actividad, pues es necesario que la cachaza reaccione en el suelo y pierda su actividad fitotóxica, generada por el alto contenido de CO2 y CH4 que este material orgánico presenta (Zérega, 1993). La composi-ción química de la cachaza fresca de trapiche panelero se presenta en la tabla 2.
propiedad Valor unidadesArena 42,88 %
Arcilla 29,12 %
Limo 28,00 %
pH 4,60
Carbono orgánico 4,12 %
Ca 3,41 cmol kg-1 de suelo
Mg 0,56 cmol kg-1 de suelo
K 0,41 cmol kg-1 de suelo
Na 0,46 cmol kg-1 de suelo
P 8,50 mg kg-1 de suelo
Fe 97,70 mg kg-1 de suelo
Cu 2,43 mg kg-1 de suelo
Zn 3,00 mg kg-1 de suelo
Mn 1,93 mg kg-1 de suelo
Al+3 5,10 cmol kg-1 de suelo
H+ 0,10 cmol kg-1 de suelo
CICe 10,04 cmol kg-1 de suelo
CE 0,70 dS m-1
Taxonomía: Typic Dystrudepts.
tabla 1. propiedades fisicoquímicas del suelo antes de la siembra.
EFECTO DE LA ApLICACIóN DE CACHAzA FRESCA y DE DOS SISTEMAS DE pRODuCCIóN MAÍz

REV. COLOMB. CIENC. HORTIC.
208
Se realizó un muestreo de suelos previo a la pre-paración del mismo, con el fin de determinar las condiciones iniciales del experimento, para lo cual, se tomaron en zigzag seis submuestras a 20 cm de profundidad en toda el área expe-rimental con barreno holandés, las submuestras se mezclaron y se obtuvo una muestra de 1 kg para los análisis fisicoquímicos, cuyos resultados se reportan en la tabla 1. Este procedimiento fue repetido después de la cosecha (4 meses después de la siembra), no obstante, el muestreo se rea-lizó para cada UE y se cuantificó el diámetro ponderado medio seco (dpms) y húmedo (dpmh) siguiendo la metodología propuesta por el IGAC (2006). Además, se determinó la densidad apa-rente con el muestreador Bulman (0200, Soil Care Sampler), densidad real mediante método del picnómetro y porosidad total del suelo.
Los datos obtenidos fueron sometidos a un análisis de varianza (Anava) y se les realizó la prueba de comparación múltiple de Tukey con una confiabilidad del 95%. Para el análisis de los datos se utilizó el paquete estadístico SAS v.8e (Cary, NC.).
reSultaDoS y DiSCuSióN
El dpms fue afectado significativamente por el sistema de cultivo (P≤0,05), el sistema asocio maíz fríjol se caracterizó por aumentar en ma-yor grado los agregados, en este mismo sistema se observó que el dpms se incrementó lineal-mente en función de la dosis de cachaza aplicada (P≤0,05), mientras que en el sistema de maíz en monocultivo no se presentaron diferencias esta-dísticas, no obstante, hubo mayor respuesta con 10 t ha-1 (figura 1).
El sistema asocio maíz frijol fue significativa-mente (P≤0,05) más efectivo que el sistema maíz al generar mayor aumento del dpmh. Entre dosis de cachaza no hubo diferencias estadísticas, pero en los dos sistemas de cultivo se observó mayor respuesta con la dosis de 15 t ha-1 cachaza fresca de trapiche panelero (figura 2).
El sistema radical de las plantas de maíz y fríjol cumplen una función muy importante en la for-mación de agregados, pues, la influencia de la raíz puede ser mecánica, por el acomodo radicu-lar que permite la unión temporal de partículas y fragmentos minerales, y también bioquímica debido a la producción de mucílagos, humus, go-mas y otros compuestos orgánicos que actúan como enlaces transitorios y permanentes entre la fracción mineral, o bien, como cubiertas que protegen a los agregados incrementando su es-tabilidad (Velázquez-Rodríguez et al., 2001). No obstante, el aumento de la agregación del suelo puede ser optimizado con el aporte de materia orgánica de la cachaza y las raíces y a la solubili-zación de cementantes químicos.
atributo valor
% ceniza 8,925
% de humedad 49,6255
% de materia seca 50,3755
% de MO 21,1255
% CO 12,2355
% N 1,1555
% Ca 6,5255
% Mg 0,6655
% K 0,7355
% Na 0,3555
% P 0,0745
% Cu 0,0555
% Zn 0,0755
% Mn 0,0575
% Fe 0,0465
CE 3,7555
pH 4,0555
C/N 10,4455
Tabla 2. Composición química de la cachaza fresca de trapiche panelero.
FORERO/TORRES/BALAGuERA-LópEz

Vol. 2 - No.2 - 2008
209
Figura 1. Efecto de la aplicación de cachaza fresca de trapiche panelero y de dos sistemas de cultivo sobre el diámetro ponderado medio seco (dpms) de un Inceptisol. Promedios con letras distintas en la misma serie indican diferencia significativa según la prueba de tukey (P≤0,05).
Figura 2. Efecto de la aplicación de cachaza fresca de trapiche panelero y de dos sistemas de cultivo sobre el diámetro ponderado medio húmedo (dpmh) de un Inceptisol. Promedios con letras distintas en la misma serie indican diferencia significativa según la prueba de tukey (P≤0,05).
EFECTO DE LA ApLICACIóN DE CACHAzA FRESCA y DE DOS SISTEMAS DE pRODuCCIóN MAÍz

REV. COLOMB. CIENC. HORTIC.
210
Al respecto, Carpenedo y Mielniczuk (1990) afirman que el tipo de vegetación interfiere en la estructura del suelo, por ejemplo, las gramí-neas son más eficientes en aumentar y mantener la estabilidad de agregados que las leguminosas, por presentar un sistema radical más extenso y renovado constantemente (Harris et al., 1966). Sin embargo, el resultado que se encontró en el presente estudio indicó que el asocio maíz frí-jol es más eficiente que la siembra de solo maíz, porque genera mayor densidad radicular y aporte de nitrógeno que aumentan la tasa de descom-posición de los residuos como consecuencia de la disminución de la relación C/N (Carpenedo y Mielniczuk, 1990).
El efecto de la cachaza fresca de trapiche pane-lero como material orgánico junto con la mate-ria orgánica que generan las raíces muertas de las plantas de maíz y frijol, en la formación de agregados corresponde a lo afirmado por Tisdall y Oades (1982), quienes demostraron que la ma-teria orgánica ejerce un papel importante en la formación y estabilización de agregados del sue-lo debido a puentes entre polímeros orgánicos y una superficie orgánica mediada por cationes polivalentes. La materia orgánica actúa como agregante físico del suelo mediante su estructura compleja de largas cadenas de carbono que agre-gan las partículas minerales, además se encarga de proteger las partículas minerales de la acción del agua para evitar su disgregación y permitir así agregados más estables y persistentes en el suelo (Tisdall y Oades, 1982).
Cunha et al. (2007) determinaron que la mate-ria orgánica es el principal agente agregante del suelo, ya que a medida que aumentó el aporte de materia seca de pasturas como material orgáni-co, también lo hizo el diámetro medio pondera-do. Sánchez-Hernández et al. (2006) encontraron que el incremento en la dosis de vermicompost de cachaza aumentó la proporción de macroa-gregados estables en húmedo, específicamente los de tamaño mayor a 0,5 mm. Los agregados
menores a ese tamaño fueron más frecuentes en los suelos que recibieron menos cantidad de la enmienda. Solamente, los agregados mayores a 3,36 mm y entre 0,5-1,0 mm se diferenciaron estadísticamente, particularmente en los suelos que recibieron las dosis más altas.
Estos mismos autores encontraron que el dpms au-mentó en relación directa a las dosis de la enmien-da orgánica. De esta forma, el tratamiento de 60 t ha-1 presentó el mayor efecto, mientras que en el testigo la respuesta fue menor (Sánchez-Hernán-dez et al., 2006), lo cual coincide con lo encontra-do en este estudio mediante la adición de cachaza. Campos et al. (1995) afirman que los compuestos orgánicos y la acción microbiana favorecen la es-tabilidad de agregados por la alta correlación en-contrada entre el contenido de carbono orgánico (CO) y la actividad microbiana con el diámetro medio geométrico de los agregados. Similar res-puesta obtuvieron en dpmh. Esta situación puede también ser una posible explicación del incremen-to del tamaño del agregado en función de la dosis de cachaza suministrada al suelo.
También es posible que la cachaza panelera pro-mueva la liberación de gran cantidad de CO2 en su transformación formando H2CO3 y disuel-va cationes como lo reportan algunos autores (Van Rooyen y Webber, 1977; Zérega, 1993), y que sumado el efecto de los ácidos liberados por las raíces de maíz y fríjol coadyuden a la libe-ración de calcio, que también tiene alto efecto agregante (Fassbender, 1986). Así, de acuerdo con un modelo propuesto por Edwards y Bremer (1967), los cationes polivalentes, principalmente el calcio son cruciales para unir la fracción orgá-nica y los minerales de arcilla, sin esa unión, la fracción arcilla y la orgánica se dispersan porque las dos tiene cargas negativas permanentes. La predominancia del calcio en el complejo de cam-bio, además de promover la floculación adecuada de las arcillas contribuye con el aumento de la actividad biológica que favorece la formación de agregados (Fassbender, 1986).
FORERO/TORRES/BALAGuERA-LópEz

Vol. 2 - No.2 - 2008
211
Por otro lado, la acción conjunta de la cachaza y el efecto de los sistemas radiculares del maíz y el frí-jol parece ser que crean un ambiente propicio para aumentar las poblaciones de hongos y bacterias. Estos microorganismos son capaces de unir la frac-ción mineral del suelo en agregados estables, aun-que cada uno tiene una eficiencia específica para cumplir con este propósito (Harris et al., 1966).
No existe un consenso generalizado acerca del mecanismo general involucrado en la estabili-zación de los agregados, ya que en ello conflu-ye una serie de interacciones físicas, químicas y biológicas (Harris et al., 1966). El crecimiento y la liberación de exudados radicales de diferente tipo, el incremento de policationes en la rizós-fera, la producción de agentes cementantes por parte de la raíz y microorganismos, la naturaleza de la planta y condiciones de manejo agronómi-
co, actividad de lombrices y mesofauna, hongos filamentosos y actinomicetos, entre otros, son algunos de los factores más incidentes en la for-mación de agregados estables (Velázquez-Rodrí-guez et al., 2001), lo que explica un mayor au-mento en la formación de agregados cuando se da el efecto conjunto del sistema radical del maíz y el fríjol y la adición de cachaza.
La densidad aparente del suelo no presentó dife-rencias significativas en el sistema asocio. En el sistema maíz, una función cuadrática describió el comportamiento de la adición de cachaza so-bre la densidad aparente, donde la aplicación de 10 t ha-1 de cachaza presentó la menor densidad aparente (P≤0,05). Entre sistemas de cultivo no se presentaron diferencias estadísticas y los va-lores promedio fueron 1,08 g cm-3 para maíz y 1,09 g cm-3 para el sistema asocio (figura 3).
Figura 3. Efecto de la aplicación de cachaza fresca de trapiche panelero y de dos sistemas de cultivo sobre la densidad aparente de un inceptisol. promedios con letras distintas en la misma serie indican diferencia significativa según la prueba de tukey (P≤0,05).
EFECTO DE LA ApLICACIóN DE CACHAzA FRESCA y DE DOS SISTEMAS DE pRODuCCIóN MAÍz

REV. COLOMB. CIENC. HORTIC.
212
Las especies vegetales contribuyen decisivamen-te en la recuperación de suelos, en la formación de agregados y en la modificación de los que tie-nen una estructura deficiente, lo que se relaciona directamente con el efecto del sistema radical en la fragmentación, reorganización y estabili-zación del material y aumento del volumen del suelo (Tisdall, 1994).
Con la adición de cachaza al suelo es posible me-jorar la densidad aparente por su alto contenido de materia orgánica. Soane et al. (1981) determi-naron que la materia orgánica tiene una correla-ción elevada y negativa con la densidad aparente, debido a que esta, por su menor densidad, pro-duce dilución de la matriz del suelo y aumenta la estabilidad de agregados (MacRae y Mehuys, 1985). De esta manera, el agregado de materia orgánica aumenta la estabilidad estructural (Tis-dall y Oades, 1982), disminuye la densidad apa-rente, mejora las tasas de infiltración de agua y la conductividad hidráulica (Felton et al., 1995).
La densidad aparente es una de las propiedades más determinantes de las características de per-meabilidad y aireación del suelo, y su persisten-cia en el tiempo es reflejo de la estabilidad de los agregados. Así, diversos autores coinciden en señalar que aplicaciones de MO al suelo provo-can una disminución de la densidad, como con-secuencia de un aumento de la macroporosidad (Kay et al., 1997). Además, se obtienen incre-mentos notables en la infiltración del agua (Le Bissonnais y Arrouays, 1997) y un mayor con-tenido de agua aprovechable del suelo (Hudson, 1994). Por tanto, de la cachaza también se obten-drían efectos benéficos.
El aumento en la cantidad de raíces del sistema asocio incrementa la porosidad por acción mecá-nica, donde el crecimiento de raíces descompac-ta el suelo y aumenta su volumen, al igual que por el aporte de materia orgánica proveniente de raíces muertas, ya que según Hernández et al. (1995), la MO aportada por el sistema radical del cultivo de caña al suelo es de aproximadamente
2,5 t ha-1 año-1, que se convierte en una cantidad considerable con acción favorable sobre la ferti-lidad del suelo. La menor densidad aparente es el resultado de la mayor macroporosidad, ya que se generan espacios dentro de la matriz del suelo por efecto del material orgánico adicionado. Ma-yores cantidades de materia orgánica adicionados generan menores valores de densidad aparente y aumentos en los valores de porosidad total.
Unger y Jones (1998) afirman que la adición de materia orgánica al suelo puede reducir la densi-dad aparente; no afectarla (Acosta et al., 1999), y en algunas situaciones aumentarla (Bottenberg et al., 1999). El efecto puede variar por el tipo de suelo, antecedentes de las propiedades del suelo, tipo de materia orgánica, clima, uso del suelo y cantidades aplicadas (Mulumba y Lal, 2008). Por tanto, para el caso de la adición de cachaza fresca es evidente que disminuye la densidad aparente, y su efecto es mayor al aumentar las dosis.
Por otro lado, Sánchez-Hernández et al. (2006) demostraron que el aporte de materiales orgáni-cos en suelos vertisoles durante dos años, dismi-nuyó la densidad de 1,34 a 1,16 g cm-3, resultado que concuerda con lo reportado para cachaza en esta investigación.
Respecto a la porosidad total del suelo, no hubo diferencias estadísticas entre sistemas de cultivo, no obstante, el sistema asocio generó mayor poro-sidad (57,1%) que el sistema maíz (55,81%). En los dos sistemas de cultivo se aumentó la porosidad de forma lineal a medida que fue mayor el sumi-nistro de cachaza fresca (P≤0,05) (figura 4).
La aireación es uno de los factores físicos más im-portantes que puede limitar el desarrollo de los sistemas radiculares de las plantas, el crecimiento y producción de los cultivos (Boone y Veen, 1994). La porosidad del suelo interfiere en la aireación, conducción y retención del agua, resistencia a la penetración y a la ramificación de raíces en el suelo, y por ende en el aprovechamiento de agua y nutrientes disponibles. Un aumento de la poro-
FORERO/TORRES/BALAGuERA-LópEz

Vol. 2 - No.2 - 2008
213
sidad es también el resultado de la descomposi-ción de raíces y la liberación de exudados radicu-lares que generan una mayor actividad biológica del suelo, proporcionando mayor estabilidad de agregados (Tisdall y Oades, 1982).
Por lo anterior, las gramíneas incluyendo el maíz, por su sistema radical promueven un efec-to positivo en la aireación del suelo y estabilidad de agregados (Silva y Mielniczuk, 1998), aunque es posible que el asocio con leguminosas contri-buya a acelerar la descomposición de la materia orgánica, generando mayor aporte de nitrógeno y aumentando la porosidad del suelo (Cunha et al., 2007), lo constatado en la siembra de maíz asocio fríjol. Donde el mayor sistema radical lo-grado por el sistema asocio se reflejó en el mayor aumento en la porosidad del suelo estudiado. Lo que concuerda con el efecto del sistema radical
Figura 4. Efecto de la aplicación de cachaza fresca de trapiche panelero y de dos sistemas de cultivo sobre la porosidad total de un inceptisol. promedios con letras distintas en la misma serie indican diferencia significativa según la prueba de tukey (P≤0,05).
de abonos verdes, que lograron la recuperación de un suelo degradado, mejorando la porosidad y revirtiendo la compactación subsuperficial (Píc-colo, 1995).
El aumento de la porosidad del suelo observado en este experimento también puede ser debido al aporte de materia orgánica proveniente de la ca-chaza y a la descomposición de raíces de las plan-tas de maíz y fríjol, que son un aporte importan-te de materia orgánica y concuerda con Herrick y Lal (1995) quienes afirman que los aportes de materiales orgánicos disminuyen la densidad por mejoramiento en la estructura del suelo. Pues la variación de la porosidad depende del tamaño de las partículas y del grado de agregación, así, al favorecerse la formación de agregados hay un consecuente aumento de la porosidad. Además, la aplicación de materiales orgánicos mejora la
EFECTO DE LA ApLICACIóN DE CACHAzA FRESCA y DE DOS SISTEMAS DE pRODuCCIóN MAÍz

REV. COLOMB. CIENC. HORTIC.
214
estructura del suelo, incrementa la capacidad de aireación, lo que causa que las raíces se ex-tiendan y exploren un mayor volumen de suelo, esto a su vez aumenta la retención de humedad a través del perfil de suelo (Pernes-Debuyser y Tessier, 2004).
Mulumba y Lal (2008) determinaron que el apor-te de materia orgánica incrementó la porosidad siguiendo una curva descrita por un polinomio de segundo grado, donde la mayor porosidad se logró con 4 t ha-1 de materia orgánica. Emerson y McGarry (2003) encontraron que al aumentar el contenido de carbono orgánico, el volumen del suelo fue mayor debido a un aumento en el núme-ro de poros. Así, Cuevas et al. (2006) afirman que mayores cantidades de materia orgánica adiciona-das generan menores valores de densidad aparente y aumentos en los valores de porosidad total, fa-voreciendo el movimiento del aire y del agua.
Painuli y Abrol (1986) y Zérega (1993) recomien-dan que para lograr efectos prolongados en la porosidad del suelo superiores a 3 años se debe
hacer proporcionalmente mezclas de cachaza y enmiendas químicas como yeso o fosfoyeso; además, la mezcla de cachaza y materiales bajos en descomposición como bagazo por su alta rela-ción C/N son potencialmente importantes en el estudio para evaluar el mejoramiento de las pro-piedades físicas de los suelos.
CoNCluSioNeS
Con la aplicación de 15 t ha-1 de cachaza fres-ca de trapiche panelero en el sistema maíz aso-cio fríjol, se logró un efecto favorable sobre las propiedades físicas del suelo, pues se aumentó el diámetro ponderado medio en seco y en hú-medo y la porosidad, y se disminuyó la densidad aparente, convirtiéndose en una alternativa de manejo para los suelos de la zona cañera de la Hoya del Río Suárez, y también es una forma de darle un uso adecuado a la cachaza fresca y de esta manera disminuir la contaminación am-biental que produce.
reFereNCiaS bibliogrÁFiCaS
Acosta, M.V.; Z. Reicher; M. Bischoff y R.F. Turco. 1999. The role of tree leaf mulch and nitrogen fertilizer on turfgrass soil quality. Biol. Fertil. Soils 29(1), 55-61.
Arrieche, I. y O. Mora. 2005. Efecto de la aplicación de residuos orgánicos sobre el cultivo del maíz en sue-los agrícolas del estado de Yaracuy, Venezuela. Bio-agro 17(3), 155-159.
Boone, F.R. y B.W. Veen. 1994. Mechanisms of crop res-ponses to soil compaction. Developments in Ag-ricultural Engineering 11: Soil Compaction and Crop Production. Elsevier Science, Amsterdam, The Netherlands.
Bottenberg, H.; J. Masiunas y C. Eastman. 1999. Strip tillage reduces yield loss of snapbean planted in rye mulch. HortTechnol. 9(2), 235-240.
Campos, B.C.; D.J. Reinert y R. Nicolodi. 1995. Estabili-dade estrutural de um Latossolo Vermelho-Escuro distrófico após sete anos de rotação de culturas e sistemas de manejo de solo. Rev. Bras. Ci. Solo 19, 121-125.
Carpenedo, V. y J. Mielniczuk. 1990. Estado de agrega-ção e qualidade dos agregados de latossolos roxos, submetidos a diferentes sistemas de manejo. Rev. Bras. Ci. Solo 14, 99-105.
FORERO/TORRES/BALAGuERA-LópEz

Vol. 2 - No.2 - 2008
215
Cuevas J.B.; S.O. Seguel; A. Ellies y J. Dörner. 2006. Efectos de las enmiendas orgánicas sobre las pro-piedades físicas del suelo con especial referencias a la adición de lodos urbanos. J. Soil Sci. Plant Nutr. 6(2), 1-12.
Cunha, E.; L.C. Balbino; L.F. Stone; W.M. Leandro y G.C. De Oliveira. 2007. Influência de rotações de cultu-ras nas propriedades físico-hídricas de um Latosso-lo Vermelho em plantio direto. Eng. Agríc. 27(3), 665-674.
Edwards, A.P. y J.M. Bremer. 1967. Microagregates in soil. J. Soil Sci. 18, 64-73.
Emerson, W.W. y D. Mcgarry. 2003. Organic carbon and soil porosity. Aust. J. Soil Res. 41, 107-118.
Fassbender, W.H. 1986. Química de suelos. IICA, San José.
Felton, G.K. 1995. Temporal variation of soil hydraulic properties on municipal solid waste amended mine soils. Trans. ASAE 38, 775-782.
Harris, R.F.; G. Chesters y O.N. Allen. 1966. Dynamics of soil aggregation. Adv. Agron. 18, 107-169.
Hernández, I.; E. Medina y H.D. López. 1995. Respira-ción edáfica y aportes de materia orgánica por las raíces y la hojarasca en un cultivo de caña de azú-car. Agron. Trop. 45, 12-142.
Herrick, J.E. y R. Lal. 1995. Soil physical property chang-es during dung cescomposition in a tropical pas-ture. Soil Sci. Soc. Amer. J. 59, 908-912.
Hudson, B. 1994. Soil organic matter and available wa-ter capacity. J. Soil Water Conserv. 49, 189-194.
IGAC. 2006. Métodos analíticos de laboratorio de sue-los. 6a ed. Instituto Geográfico Agustín Codazzi, Bogotá.
Kay, B.D.; A.P. Da Silva y J.A. Baldock. 1997. Sensitivity of soil structure to changes in organic C content: predictions using pedotransfer functions. Can. J. Soil Sci. 77, 655-667.
Khan, K.; A. Gattinger; F. Buegger; M. Schloter y R.G. Joergensen. 2008. Microbial use of organic amend-ments in saline soils monitored by changes in the 13C/12C ratio. Soil Biol. Biochem. 40, 1217-1224.
Le Bissonnais, Y. y D. Arrouays. 1997. Aggregate stabi-lity and assessment of soil crustability and ero-dability: II. Application to humic loamy soils with
various organic carbon contents. Eur. J. Soil Sci. 48, 39-48.
MacRae, A. y G.R. Mehuys. 1985. The effect green ma-nuring on the physical properties of températe area soils. Adv. Soil Sci. 3, 71-94.
Mohee, R.R. y B. Panray. 1999. Life cycle analysis of compost incorporated sugarcane bioenergy sys-tems in Mauritius. Biomass Bioenergy 17, 73-83.
Mulumba, L. y N.R. Lal. 2008. Mulching effects on se-lected soil physical properties. Soil Tillage Res. 98, 106–111.
Painuli, D.K. y L.P. Abrol. 1986. Role of some organics in improving sodic soils. lndian J. Agríe. Sel. 56(4), 267-76.
Paturau, J.M. 1969. By-products of sugar cane industry. An introduction to their industrial utilization in América. Elservier Publ. Comp., London.
Pernes-Debuyser, A. y D. Tessier. 2004. Soil physical properties affected by long-term fertilization. Eur. J. Soil Sci. 55, 505-512.
Píccolo, G.A. 1995. Efecto de diferentes cultivos utiliza-dos como abonos verdes sobre un Rodudalf típico (Misiones, Argentina). Ciencia del Suelo 13, 101-103.
Rasul, G.; A. Appuhn; T. Muller y R.G. Joergensen. 2006. Salinity induced changes in the microbial use of sugar cane fillter cake material under acidic soil conditions. Appl. Soil Ecol. 31, 1-10.
Rodriguez, G. y M. Gottret. 2006. Aprendiendo del pasa-do para proyectarnos hacia el futuro e impacto de la tecnología de la panela en la Hoya del Río Suárez y Cundinamarca. Corpoica; CIAT, Cali, Colombia.
Sánchez-Hernández R.; V.M. Ordaz-Chaparro; G.S. Benedicto-Valdés; C.I. Hidalgo-Moreno y D.J. Pal-ma-López. 2006. Regeneración estructural de un suelo arcilloso por aportes de vermicompost en la chontalpa, tabasco, México. Universidad y Ciencia 22(1), 13-26.
Silva, I.R. y J. Mielniczuk. 1998. Sistemas de cultivo e características do solo afetando a estabilidade de agregados. Rev. Brasil. Ci. Solo 22(2), 311-17.
Soane, B.D.; P.S. Blackwell; J.W. Dickson y D.J. Painter. 1981. Compaction by agricultural vehicles: a re-view. 1. Soil and wheel characteristics. Soil Tillage Res. 1, 207-237.
EFECTO DE LA ApLICACIóN DE CACHAzA FRESCA y DE DOS SISTEMAS DE pRODuCCIóN MAÍz

REV. COLOMB. CIENC. HORTIC.
216
Tisdall J.M. 1994. Possible role of soil microorganisms in aggregation in soil. Plant Soil 159, 115-121.
Tisdall, J.M. y J.M. Oades. 1982. Organic matter and wa-ter-stable aggregates in soils. J. Soil Sci. 33, 141-163.
Unger, P.W. y O.R. Jones. 1998. Long-term tillage and cropping systems affect bulk density and penetra-tion resistance of soil cropped to dryland wheat and grain sorghum. Soil Till. Res. 45, 39-57.
Van Rooyen, P. C. y H. W. Webber. 1977. Long-term effects of five ameliorantson a saline-sodiesoil of South Africa. Geoderma 19, 213-225.
Velázquez-Rodríguez, A.; D. Flores-Román y O.A. Ace-vedo-Sandoval. 2001. Formación de agregados en tepetate por influencia de especies vegetales. Agro-ciencia 35, 311-320.
Zérega, L. 1993. Manejo y uso agronómico de la cacha-za en suelos cañameleros. Rev. Caña Azúcar 11(2), 71-92.
Zurro, P. 2005. Producción de abonos y fertilizantes a partir de subproductos de la industria azucarera or-gánica. En: http://www.elparanaense.com.ar/pdf/MemoriaabonosIngSanJavier MNES.pdf; consulta: noviembre de 2008.
FORERO/TORRES/BALAGuERA-LópEz

FÁNor CaSierra-poSaDa1, 3 JuaN CarloS abril2
Determinación del momento de corte de la flor en limonio (Limonium sp.) a partir de seis estados fenológicos
Determination of timing of flower cut in sea lavender (Limonium sp.) starting from six phenological stages
Cultivo de limonio en invernadero en Ubaté (Cundinamarca).Foto: j.C. Abril
reSumeN
La fenología es el estudio de los tiempos de los fenómenos naturales recurrentes en organismos vivos. Con-cierne principalmente a las épocas de ocurrencia, por primera vez, de los eventos naturales en el ciclo de las plantas. Las observaciones fenológicas las usan los agricultores desde hace siglos para maximizar la produc-ción agrícola, mediante la predicción de la época estacional de la cosecha. Es muy poca la disponibilidad de información relacionada con la planificación de la cosecha de la flor usando los estados fenológicos como referencia, en la producción de flores de corte. Se realizó un trabajo en invernadero en Ubaté (Cundinamarca, Colombia), para una mejor comprensión del crecimiento de los tallos a partir de seis estadios fenológicos dife-rentes hasta el momento de corte en cinco variedades de limonio ‘Supreme white light’, ‘Supreme blue night’, ‘Safora dark blue’, ‘Safora blue’ y ‘Beltlaard’. Se identificaron seis estadios fenológicos, y a partir de ellos se midió semanalmente la altura del tallo floral. La altura final de los tallos fue de 132,3; 130,1; 102,1; 85,6 y 113,2 cm para ‘Supreme white light’, ‘Supreme blue night’, ‘Safora dark blue’, ‘Safora blue’ y ‘Beltlaard’, res-pectivamente. Con la información colectada se calcularon las ecuaciones de regresión para el crecimiento. Para todas las variedades, a partir de cada estadio fenológico, se encontró un modelo cúbico con alto coeficiente de determinación (R2= 0,99 a 1,0). Las ecuaciones se pueden usar para la predicción del momento de corte para las variedades evaluadas, con propósitos prácticos.
1 Grupo de Investigación Ecofisiología Vegetal, Facultad de Ciencias Agropecuarias, Universidad Pedagógica y Tecnológi-ca de Colombia, Tunja (Colombia).
2 Facultad de Ciencias Agropecuarias, Universidad Pedagógica y Tecnológica de Colombia, Tunja (Colombia). 3 Autor para correspondencia. [email protected]
REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 2 - No.2 - pp. 217-224, 2008

REV. COLOMB. CIENC. HORTIC.
218
palabras clave adicionales: flores de corte, fenología, curvas de crecimiento, altura del tallo.
additional key words: cut flowers, phenology, growth curves, stem height.
Fecha de recepción: 08-07-2008 Aprobado para publicación: 02-12-2008
abstract
Phenology is the study of timing of natural phenomena recurring in living organisms. It has been principally concerned with the dates of first occurrence of natural events in the plant life cycle. Phenological observations have been used for centuries by farmers to maximize crop production by predicting the seasonal harvest time. Very little information is available on the planning of harvest of cut flowers using phenological stages as reference in cut flower production. In order to understand better the process of stem growth of five sea lavender varieties starting from six different phenological stages up to the cutting time, a study was carried out in greenhouse in Ubaté (Cundinamarca, Colombia). Plant material were sea lavender varieties ‘Supreme white light’, ‘Supreme blue night’, ‘Safora dark blue’, ‘Safora blue’ and ‘Beltlaard’. Six phenological stages were identified and starting from these stages, the growth of plant flower stem was weekly measured. The final stem height were 132.3, 130.1, 102.1, 85.6 and 113.2 cm for ‘Supreme white light’, ‘Supreme blue night’, ‘Safora dark blue’, ‘Safora blue’, and ‘Beltlaard’, respectively. With information collected, the regression equations for growth were calculated. For all varieties from each phenological stage a cubic model with high coefficients of determination (R2= 0.99 to 1.0) was found. The equations could be used for predicting the cut time for evaluated varieties for practical purposes.
iNtroDuCCióN
Limonium es un tipo de planta que produce una flor de corte utilizada en la elaboración de bouquets, en los que se combinan especies de colores variados, lo que otorga al ramo un valor agregado, razón por la cual, las compañías pro-ductoras de flores cultivan diferentes especies de este género con miras a lograr un mayor contras-te en el colorido del bouquet (Chamorro et al., 2007). El área destinada en Colombia, al cultivo de las flores, alcanzó 7.266 ha en 2007, en donde 79% de la producción tuvo lugar en la Sabana de Bogotá, 17% en Antoquia y en la región central y occidental del país se cultivó el 4% del área. De los productos cultivados, los bouquets están representados con un 32,48% de la producción nacional de flores de corte (Asocolflores, 2007).
En especies de Limonium se han desarrollado in-vestigaciones orientadas a determinar la respues-ta de las plantas a la temperatura y al fotope-riodo, encontrándose en L. sinuatum (statice) que esta especie no se ve afectada por el fotoperiodo, pero sí por la temperatura (Shillo, 1976; Escher, 1996). De igual manera, se ha encontrado en plantas de limonio (L. gmelinii) que la exposición de las plantas a bajas temperaturas en un estado adecuado del crecimiento, son necesarias para la floración; posteriormente, condiciones de día lar-go incrementan la tasa de floración, luego de que las plantas han evacuado su requerimiento de baja temperatura (Farina et al., 2000). En otros temas relacionados, se estudiaron el impacto de la cosecha de flores de Limonium carolinianum
CASIERRA-pOSADA/ABRIL

Vol. 2 - No.2 - 2008
219
sobre las posibilidades de supervivencia de las plantas (Baltzer et al., 2002) y Doi y Reid (1995) mejoraron la vida poscosecha en híbridos inte-respecíficos de Limonium
Los trabajos mencionados, desarrollados en es-pecies de Limonium tienen gran relevancia, sin embargo, se hace prioritaria la generación de in-formación que contribuya al desarrollo de la flo-ricultura nacional, y en especial, de las plantas de limonio cultivadas bajo las condiciones colom-bianas, dado que los trabajos realizados en esta área son bastante escasos. Al respecto, Casierra-Posada y Moreno (2007) realizaron un estudio en el que evaluaron el comportamiento de plantas de limonio expuestas a diferentes niveles de som-brío. Los autores hacen referencia a la alta sensi-bilidad de las plantas a la reducción de la luz.
A pesar de que el crecimiento y desarrollo de las plantas es un proceso continuo en el tiempo, re-sulta posible discernir en él, una serie de etapas, que si bien, pueden tener diferencias aún en indi-viduos de la misma especie, posee una regularidad que permite sacar conclusiones, cuando estas fa-ses se registran por periodos prolongados de tiem-po. El registro fenológico de una especie durante un tiempo más o menos considerable permite conocer las fechas probables en que se produci-rán las fases de crecimiento y desarrollo, así como también, las fechas límites (tempranas o tardías) y la frecuencia de las mismas, lo que posibilita el cálculo de la probabilidad porcentual de ocu-rrencias y la elaboración de mapas isócronos de la ocurrencia de las diferentes fases, lo que resulta de gran interés práctico en biología, agricultura y agrometeorología (Fuentes, 1992a, 1992b).
Por otro lado, para efectuar la comparación en-tre tipos de plantas o para realizar estudios de predicción, se debe elegir un estado de desarrollo o fenológico, y a partir de él, se deben realizar las mediciones requeridas para el logro del ob-jetivo propuesto. Por lo regular, no se parte de
una fecha calendario fija, puesto que se podría incurrir en errores, dado que a pesar de que la sucesión de los estados fenológicos es segura, puede variar en la duración de los intervalos en relación con el ambiente en el que se desarrollan las plantas (Sitte et al., 2004). Por tanto, cuando se aplica la fenología en plantas, se registra la dinámica del desarrollo, haciendo especial hin-capié en la sucesión de los diferentes estadios, más que en la influencia de los factores que los producen (Ghersa y Holt, 1995).
En cuanto a la aplicación de los estudios fenoló-gicos a la producción de flores de corte, se puede tomar en consideración los realizados en Anthu-rium andreanum en donde se determinaron los componentes del rendimiento para la producción de la flor. Las plantas presentan un crecimiento monopodial en la fase vegetativa, seguido de una fase simpodial, con producción de una flor por hoja (Dufour y Guérin, 2002). Con mayor exac-titud que en el caso de anturio, los estadios fe-nológicos se han determinado y se han usado en cultivos de rosas y claveles de corte. Además de los estadios fenológicos, se determinaron algu-nos índices de crecimiento como la tasa de creci-miento del cultivo, el índice de área foliar, la tasa relativa de crecimiento y la tasa de asimilación neta, los cuales son una herramienta importante para la toma de decisiones en este tipo de culti-vos (Chaparro et al., 2006; Cárdenas et al., 2006; Farías et al., 2006). A pesar de la importancia de la aplicación de estudios de fenología correlacio-nados con el rendimiento, orientados a la predic-ción y ajuste de épocas de corte de la flor para cumplir con las temporadas de máxima deman-da por parte del mercado internacional, son muy pocos los reportes al respecto en floricultura.
El propósito del presente estudio fue la determi-nación del tiempo en que se realizaría la cosecha en cinco cultivares de limonio (Limonium sp.), cuando se parte de seis estados fenológicos dife-rentes, bajo condiciones de invernadero.
DETERMINACIóN DEL MOMENTO DE CORTE DE LA FLOR EN L IMONIO

REV. COLOMB. CIENC. HORTIC.
220
materialeS y métoDoS
La investigación se realizó en un cultivo estable-cido en la empresa Flores Ubaté Ltda., ubicada a 5º18’24” N, 73º48’52” W y 2.560 msnm. Las plantas tenían cerca de 19 semanas de plantadas y fueron manejadas bajo condiciones de hidro-ponía con cascarilla de arroz tostada (80% de quemado en horno). Los fertilizantes para la pre-siembra se tomaron a partir de fuentes simples y se aplicó por cama (30 x 1 m) la siguiente mezcla: sulfato de calcio 3,5 kg; carbonato de calcio 2,5 kg; óxido de magnesio 0,9 kg; sulfato diamónico 1 kg y Rafos (Abocol®) 1,5 kg. La mezcla para la solución aplicada por el riego estuvo compues-ta por (mg L-1): N 145; Ca 100; P 20; K 120; Mg 30; Mn 0,5; Fe 2,5; Cu 0,5; Zn 0,5 y B 1,5. La solución se suministraba en dos pulsos por día y cada pulso estaba compuesto por 40 L por cama. Durante el ensayo se registró en promedio, una temperatura de 16,5ºC, con mínima de 8ºC y máxima de 25ºC. El material vegetal correspon-dió a variedades de Limonium sp., utilizadas co-múnmente en la producción de bouquets, como son: ‘Supreme white light’, ‘Supreme blue night’, ‘Safora dark blue’, ‘Safora blue’ y ‘Beltlaard’. Las cuatro primeras variedades fueron suministradas por la compañía Danziger-“Dan” Flower Farm, la última por Ball SB L.C.
Se tomó una cama por variedad y de esta, seis brotes en cada uno de los estadios fenológicos se-leccionados. Cada estadio fenológico tuvo cinco réplicas dentro de la misma cama y se les llevó un seguimiento de crecimiento cada semana desde el punto de partida de cada brote hasta el momento que fue cortado para su respectivo proceso de poscosecha.
A los diferentes estados fenológicos de brotes se les asignó un nombre, de la siguiente mane-ra: estado 01 ó punto mini, estado 02 ó punto pequeño, estado 03 ó punto medio, estado 04 ó punto espiga, estado 05 ó rayando color, y esta-do 06 ó punto marrón (figura 1). El estado 01 ó punto mini correspondió a la morfología del
brote recién emergido y a su vez, el estado seis o punto marrón fue el más cercano al corte. Se marcó cada uno de ellos mediante la utilización de etiquetas de marcación, las cuales contenían información adicional como la ubicación de la planta y fecha de inicio de las mediciones. Para medir la longitud de los tallos o brotes se utili-zó un flexómetro colocado de manera paralela a cada brote, colocando un extremo del mismo en la base de la planta, y el otro extremo hacia el ápice del brote, con lo cual se registró la altura de los tallos. Se monitoreó el tiempo necesario para alcanzar el punto de corte a partir de cada esta-dio fenológico, así como la duración de cada uno de los estados fenológicos de cada variedad.
punto mini punto pequeño
punto medio punto espiga
rayando color punto marrón
Figura 1. estadios fenológicos identificados en diferentes variedades de limonio (Limonium sp.)
CASIERRA-pOSADA/ABRIL

Vol. 2 - No.2 - 2008
221
El experimento consistió en un ensayo bifacto-rial, con cinco variedades como primer factor y seis estadios fenológicos como factor adicional. Se utilizó un diseño completamente al azar y cinco replicaciones, tomando una planta como unidad experimental. La información, se anali-zó mediante un análisis de varianza (Anava) y la prueba de separación de promedios de Tukey (P≤0,01). Adicionalmente se realizó un análisis de regresión para las curvas de crecimiento, con el programa SPSS versión 11.5.1.
reSultaDoS y DiSCuSióN
Acorde con el análisis de varianza, se presenta-ron diferencias altamente significativas (P≤0,01) para los factores variedad y estado fenológico, así como para todas las combinaciones de estos factores. Entre las variedades estudiadas se en-contró que ‘Supreme white light’ fue el material que presentó tallos florales más largos, seguido por ‘Supreme blue night’, ‘Beltlaard’, ‘Safora
dark blue’, y por último ‘Safora blue’ (figura 2). Esta información es de interés para los cultiva-dores de flores, con miras a determinar el desti-no de la flor cosechada, dado que la elaboración de bouquets requiere de flores con diferentes longitudes, acorde con el tamaño de las flores acompañantes. Adicionalmente se requiere el contraste de las flores complementarias con las de limonio, por tanto se hace necesario conside-rar la longitud de sus tallos.
De acuerdo con el estado fenológico de partida para la toma de la información, en los dos prime-ros estadios, los brotes crecen de manera acelera-da entre 36 y 43 d después de la aparición del es-tado fenológico, posteriormente, la velocidad del crecimiento disminuye de manera considerable (figura 3A y 3B). Este modelo de crecimiento sig-moide o hiperbólico representa el incremento en altura en plantas cuyas ramas tienen un tipo de crecimiento definido (Taiz y Zeiger, 2006). Las ramas presentan un crecimiento lento, poste-riormente el crecimiento en altura es logarítmico
Figura 2. Longitud máxima del tallo en cinco variedades de limonio (Limonium sp.) cultivadas bajo invernadero. La barra sobre la columna indica la desviación estándar. promedios con letras distintas indican diferencia significativa según la prueba de tukey (P≤0,01).
DETERMINACIóN DEL MOMENTO DE CORTE DE LA FLOR EN L IMONIO

REV. COLOMB. CIENC. HORTIC.
222
y por último se presenta una fase estacionaria. Esta última fase corresponde a la aparición de las flores en los meristemos apicales, por tanto, las ramas no pueden crecer más debido a su creci-miento determinado. En la figura 3A se observa este modelo de crecimiento en los cinco materia-les evaluados, sin embargo, la velocidad del creci-miento en cada una de ellas difiere en la medida en que el material es precoz como ‘Safora blue‘ y ‘Beltlaard’ o tardío como ‘Safora dark blue’, ‘Su-preme dark light’ y ‘Supreme blue night’.
Tomando en consideración los estadios fenológi-cos avanzados (figuras 3B a 3F), en la medida en que se tomó la información a partir de estadios más avanzados, empieza a desaparecer de la figura la fase de crecimiento logarítmica, quedando sólo la fase estacionaria (figuras 3E a 3F). La aplicación práctica de esta información radica en el hecho que una vez identificados los diferentes estadios en las variedades evaluadas se debe esperar un comportamiento de las variedades acorde con la ruta que describen las figuras y así, es posible ha-cer la predicción del corte de la flor con base en la información consignada en la tabla 1, en la cual aparecen las ecuaciones de regresión para cada una de las curvas presentadas en la figura 3, así como el tiempo restante de corte para cada variedad, cuan-do se parte de cada uno de los estadios evaluados.
Las dos fechas importantes para la venta de flor cortada corresponden al día de las madres y el día de San Valentín. Mientras que en Europa el día de las madres se celebra el primer domingo de mayo, en Estados Unidos se celebra el segundo domingo del mismo mes (Escuela Internacional, 2007). En el día de San Valentín se celebra el día de los enamorados, el 14 de febrero. Por tanto, la proyección de los puntos de corte para dar cum-plimiento a la exportación de la flor en estas dos fechas es motivo de una planificación minuciosa en las compañías colombianas productoras de flores para el mercado exterior.
El punto de corte es de gran relevancia para la ca-lidad de bouquet, dado que en las plantas de limo-
nio, las flores no continúan su apertura luego de cortadas. Adicionalmente se debe hacer énfasis en la importancia del color de la flor para lograr con-traste con las flores accesorias del bouquet. Los colores de los materiales utilizados en este estudio correspondieron a blanco aterciopelado para ‘Su-preme white light’, azul lavanda para ‘Supreme blue night’, púrpura profundo para ‘Safora dark blue’, púrpura-azul oscuro para ‘Safora blue’ y por último violeta para ‘Beltlaard’.
Se debe resaltar que de los materiales evaluados, solo en ‘Beltlaard’ es posible el cambio de color de la flor a través de su tinción, para lo cual, la flor se corta con una semana de anticipación a la fecha de envío, con miras a tener el tiempo sufi-ciente para teñir las flores. El procedimiento para tinte es reserva comercial de las empresas. Por otro lado, las flores que no se tinturan se cortan el mismo día de despacho. Al momento de corte, las flores de la variedad Beltlaard no han adqui-rido su color violeta característico, así que en ese punto presentan una tonalidad beige intenso, lo cual facilita el cambio artificial de color.
Con base en esta información y en la metodo-logía empleada en el estudio, los cultivadores de las variedades evaluadas podrán realizar la planificación del momento de corte de la flor, en la medida que la flor pueda o no tinturarse. Además, la utilización de los estadios fenológi-cos como indicador morfológico para predecir el momento de corte representa una herramienta práctica en las labores de planificación. Se debe hacer hincapié en la determinación del estadio fenológico con base en la escala mostrada en la metodología, con el fin de partir de un estadio determinado y no de una fecha calendario, dado que el desarrollo de los brotes es altamente de-pendiente de factores del entorno, como la tem-peratura (Sitte et al., 2004). Es así, como tempe-raturas altas pueden promover una mayor tasa de crecimiento como consecuencia de un meta-bolismo acelerado y por tanto, pueden acortar el lapso de tiempo entre la aparición de un estadio fenológico y el momento de corte.
CASIERRA-pOSADA/ABRIL

Vol. 2 - No.2 - 2008
223
Figura 3. Curvas de crecimiento en cinco variedades de limonio (Limonium sp.) a partir de seis estadios fenológicos. A) A partir de punto mini; B) A partir de punto pequeño; C) A partir de punto medio D) A partir de punto espiga; E) A partir de rayando color; F) A partir de punto marrón.
DETERMINACIóN DEL MOMENTO DE CORTE DE LA FLOR EN L IMONIO
Este estudio se desarrolló con el apoyo de la Dirección de Investigaciones (DIN) de la Universidad Pedagógica y Tecnológica de Colombia, en el marco del plan de trabajo del Grupo de Investigación Ecofisiología Vegetal, adscrito al programa de Ingeniería Agronómica de la Facultad de Ciencias Agropecuarias, Tunja.
agraDeCimieNtoS

REV. COLOMB. CIENC. HORTIC.
224
Asocolflores. 2007. Floricultura colombiana. Información general. En: http://www.asocolflores.org/asocolflo-res/servlet/Download?idExternalFile=236&name=HOJA-DATOS-2008-7.pdf; consulta: octubre de 2008.
Baltzer, J.L.; E.G. Reekie; H.L. Hewlin; P.D. Taylor y T.S. Boates. 2002. Impact of flower harvesting on the salt marsh plant Limonium carolinianum. Can. J. Bot. 80, 841-851.
Cárdenas, C.A.; I.F. Rivera; V.J. Flores R.; W. Piedrahita C. y B. Chaves C. 2006. Análisis de crecimiento en clavel estándar variedad ‘Nelson’ cultivado en sus-tratos. pp. 111-127. En: Florez R., V.J.; A.D.L.C. Fer-nández; D. Miranda L.; B. Chaves C. y J.M. Guzmán P. (eds.). Avances sobre fertirriego en la floricultura colombiana. Facultad de Agronomía, Universidad Nacional de Colombia; Unibiblos, Bogotá.
Casierra-Posada, F. y D.L. Moreno. 2007. Efecto del es-trés por sombra sobre la producción en plantas de limonio (Limonium sp. cv. Bluestream). Rev. Colomb. Cienc. Hortic. 1(2), 236-245.
Chamorro, A.H.; S.L. Martínez; J.C. Fernández; T. Mos-quera. 2007. Evaluación de diferentes concentra-ciones de algunos reguladores de crecimiento en la multiplicación y enraizamiento in vitro de Limonium var. Misty blue. Agron. Colomb. 25(1), 47-53.
Chaparro, L.A.; A. Farías A.; B. Chaves C.; D. Miranda L. y V.J. Flores R. 2006. Análisis de crecimiento en rosa variedad ‘Charlotte’ en los sistemas de cultivo en sustrato y en suelo. pp. 77-90. En: Florez R., V.J.; A.D.L.C. Fernández; D. Miranda L.; B. Chaves C. y J.M. Guzmán P. (eds.). Avances sobre ferti-rriego en la floricultura colombiana. Facultad de Agronomía, Universidad Nacional de Colombia; Unibiblos, Bogotá.
Doi, M. y M.S. Reid. 1995. Sucrose improves the post-harvest life of cut flowers of a hybrid Limonium. HortScience 30(5), 1058-1060.
Dufour, L. y V. Guérin. 2002. Growth, developmental features and flower production of Anthurium an-dreanum Lind. in tropical conditions. Scientia Hort. 98, 25-35.
reFereNCiaS bibliogrÁFiCaS
Escher, F. 1996. Kulturbeschreibungen. pp. 414-420. En: Escher, F.; H.-C. Gudehus; L. Mayr; W. Mezinger; P. Rehrmann y H. Thomale (eds.). Schnittblumen-kulturen. Ulmer Verlag, Stuttgart, Alemania.
Escuela Internacional. 2007. Día de la madre: Primer domingo de mayo. En: http://www.escuelai.com/spanish_culture/fiestas_espanolas/diadelamadre.html; consulta: octubre de 2008.
Farías, A.; L.A. Chaparro; A. Campos; B. Chaves y V.J. Flores. 2006. Curvas de crecimiento de rosa en siste-ma de cultivo sin suelo en la Sabana de Bogotá. pp. 91-110. En: Florez R., V.J.; A.D.L.C. Fernández; D. Miranda L.; B. Chaves C. y J.M. Guzmán P. (eds.). Avances sobre fertirriego en la floricultura colombia-na. Facultad de Agronomía, Universidad Nacional de Colombia; Unibiblos, Bogotá.
Farina, E.; C. Dalla Guda y E. Scordo. 2000. Effects of low temperatures and photoperiod on flowering of Limonium gmelinii. Acta Hort. 541, 193-199.
Fuentes, V. 1992a. Influencia de los factores climáticos y la fase fenológica en el contenido de metabolitos secundarios en las plantas. Experiencias en Cuba. p. 7. En: Resúmenes del seminario-taller provincial “La agrometeorología al servicio de la agricultura”. La Habana.
Fuentes, V. 1992b. Los estudios fenológicos en plantas medicinales en cuba. Experiencias en Cuba. p. 6. En: Resúmenes del seminario-taller provincial “La agrometeorología al servicio de la agricultura”. La Habana.
Ghersa, C.M. y J.S. Holt. 1995. Using phenology predic-tion in weed management: a review. Weed Res. 35, 461-470.
Shillo, R. 1976. Control of flower initiation and devel-opment of statice (Limonium sinuatum) by tempera-ture and daylength. Acta Hort. 64, 197-203.
Sitte, P.; E.W. Weiler; J.W. Kadereit; A. Bresinsky y C. Körner. 2004. Strassburger: Tratado de botánica. 35a ed. Ediciones Omega, Barcelona, España.
Taiz, L. y E. Zeiger. 2006. Plant physiology. Sinauer As-sociates, Inc., Publishers, Sunderland, MA.
CASIERRA-pOSADA/ABRIL

Efecto del sustrato y tamaño del propágulo en el enraizamiento de ginger rojo (Alpinia purpurata)
Effect of substrate type and cutting size on red ginger (Alpinia purpurata) rooting
iSiDro e. SuÁrez1, 2
gleDy luz marrugo1
maryorik peña1
Flor de ginger rojo en una plantación cerca de Santa marta, Colombia. Foto: G. Fischer
reSumeN
El efecto de diferentes tamaños de propágulo (2,0-4,0; 4,1-6,0 ó 6,1-8,0 cm) y diferentes tipos de sustrato (sue-lo, agua o aire) sobre el enraizamiento de Alpinia purpurata fue evaluado. Los tratamientos se establecieron bajo un cubrimiento con una polisombra del 30% de penetración de luz natural y suministro permanente de agua durante 60 días. Los datos demostraron que el tipo de sustrato que se utilice tiene un efecto sobre las variables número de raíces por propágulo, tamaño promedio de raíz, incremento de tamaño del propágulo con referencia al tamaño inicial e incremento en número de hojas del propágulo con respecto al número de hojas presentes al momento del establecimiento. El sustrato agua indujo la producción de un mayor número de raíces, mayor longitud radical, brotes con mayor tamaño e incremento en el número de hojas al compararse con la mezcla de suelo y con el enraizamiento al aire.
1 Departamento de Ingeniería Agronómica y Desarrollo Rural, Universidad de Córdoba, Montería (Colombia). 2 Autor para correspondencia: [email protected]
palabras clave adicionales: propagación, raíz, zingiberaceae.
REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 2 - No.2 - pp. 225-231, 2008

REV. COLOMB. CIENC. HORTIC.
226
additional key words: propagation, root, zingiberaceae.
Fecha de recepción: 31-07-2008 Aprobado para publicación: 02-12-2008
iNtroDuCCióN
abStraCt
The effect of cutting size (2.0-4.0, 4.1-6.0, or 6.1-8.0 cm) and substrate type (soil, water, or air) on rooting of Alpinia purpurata was evaluated. The treatments were placed under a mesh providing 30% light penetration and water supply was provided during 60 days. The data showed that substrate type significantly affected root number, rooth length, propagule size increase and number of new leaves. Water as a substrate induced a significant increase in the number of roots, root length, propagule size and leaf number compared with rooting in soil mixture and air.
El ginger rojo (Alpinia purpurata, Zingiberaceae) es una planta de gran importancia económica por el uso de sus productos tanto a nivel na-cional como en el mercado internacional, como flor de corte, follaje para arreglos y adecuación paisajística de ambientes. Sus inflorescencias tienen tamaños comprendidos entre 20 y 90 cm de largo, son erguidas cuando son jóvenes y curvadas o colgantes al acercarse a la senescen-cia. Su atractivo se debe a las brácteas de color rojo intenso y tamaño relativamente grande, en la base de las cuales emergen las verdaderas flo-res que son pequeñas, de color blanco o crema y poco abundantes (Neal, 1965; Criley, 1989; Hoyos, 1999; USDA-ARS, 1999). Aunque su in-troducción al mercado de las flores de corte, el de mayor competencia a nivel internacional, es relativamente reciente, su comercio ha tomado una gran atención por el enorme potencial que representa no solo por la belleza de sus flores, sino por la duración de la misma después del cor-te, la posibilidad de cosecha durante todo el año y los bajos costos de producción si se compara con otras flores (Builes, 2003).
Las plantas de ginger se propagan naturalmente mediante la formación de brotes caulinares que se originan en la estructura floral. Cuando la in-
florescencia comienza a marchitarse, de las axi-las de las brácteas se inicia el crecimiento de bro-tes que se desarrollan rápidamente. Estos brotes se separan fácilmente y, al ponerse en contacto con el sustrato, emiten raíces adventicias que le permiten crecer y formar una nueva planta. Alternativamente, la planta produce rizomas a partir de los cuales se desarrollan nuevos brotes que crecen y forman nuevos tallos unidos (Han-sen, 1993; Hoyos, 1999).
No obstante el gran potencial como cultivo comercial y las condiciones agroecológicas pro-picias para el crecimiento de ginger, no solo en Córdoba sino en todo el Caribe húmedo, el co-nocimiento técnico y científico fundamentales para la explotación de esta especie son escasos y las técnicas utilizadas tienen poco desarrollo tecnológico. Un ejemplo de esta situación es el uso convencional de rizomas como material para la propagación de nuevas plantas, lo cual además de tener una baja eficiencia en número de plantas obtenidas, se limita por la poca disponibilidad de material vegetativo en las condiciones y can-tidades necesarias para una producción masiva de acuerdo con los requerimientos y las expec-tativas del mercado, y ocasiona daños irreversi-bles en las plantas madres. Con el fin de abrir
SuÁREz/MARRuGO/pEñA

Vol. 2 - No.2 - 2008
227EFECTO DEL SuSTRATO y TAMAñO DEL pROpÁGuLO EN EL ENRAIzAMIENTO DE GINGER ROjO
las posibilidades para la producción comercial de ginger con miras al comercio en el mercado de las flores de corte, se ha planteado el presente trabajo que tuvo como objetivo evaluar el efecto de diferentes sustratos y tamaños de brote en la producción de nuevas plantas
materialeS y métoDoS
El presente estudio se realizó en el vivero de la de la Universidad de Córdoba (Montería, Colom-bia) a 13 msnm, con una temperatura promedio anual de 28°C, 12 h longitud del día y 83% de humedad relativa, con 1.200 mm de precipita-ción promedio anual en un régimen unimodal de lluvias. El material vegetal fue obtenido de plantas adultas establecidas en el Pasaje La Ron-da del Sinú (Montería, Córdoba), y consistió en brotes caulinares de ginger rojo (Alpinia purpu-rata) originados en la inflorescencia. Los brotes fueron separados de las inflorescencias y sumer-gidos en agua destilada por un periodo de 1 h, aproximadamente 1 cm a partir de la base para evitar la deshidratación. Las hojas más adultas fueron removidas y los brotes clasificados con respecto al tamaño en tres grupos diferentes a partir de la longitud de la base a la inserción de la hoja más joven: grupo 1: 2-4 cm, grupo 2: 4,1-6 cm, grupo 3: 6,1-8 cm. Los brotes de cada grupo fueron establecidos en tres sustratos diferentes. El primer sustrato consistió en una mezcla 1:1 v:v de material aluvial depositado por el río Sinú y cascarilla proveniente de la industria molinera del arroz. La mezcla fue homogenizada y des-infectada con formaldehido al 40% aplicando 1 L m-2 de mezcla, posteriormente se cubrió con un plástico por un periodo de 8 d, al final de los cuales la cubierta plástica fue removida, el suelo mezclado nuevamente y dejada sin cubrimiento por un espacio de 5 d cuando se procedieron a establecer los propágulos. Para evitar la deshidra-tación, se aplicaron tres riegos diarios con agua corriente potable utilizando regaderas manuales durante el desarrollo del experimento. El segun-do sustrato utilizado fue el agua destilada, la
cual fue envasada en vasos desechables de 200 mL, y a cada uno de ellos se le colocó una tapa de icopor ajustada a la superficie y perforada en el centro donde se procedió a ubicar el propágulo en cada vaso permitiendo que la parte basal del propágulo (aproximadamente 1/3 de la longi-tud total) quedara en permanente contacto con el agua. El tercer método utilizado consistió en ausencia completa de sustratos, para lo cual los propágulos fueron ubicados en los orificios de una bandeja de propagación plástica de 30 x 40 cm colocada en forma invertida, de forma que los orificios del fondo quedaron hacia arriba. Las bandejas fueron colocadas sobre un recipiente con una determinada cantidad de agua destilada con el fin de mantener una alta humedad relativa en la base del propágulo. Los tratamientos fue-ron distribuidos con un diseño de parcelas dividi-das con nueve tratamientos (tres sustratos x tres longitudes de propágulo); cada tratamiento fue repetido 10 veces para un total de 90 unidades experimentales. Transcurridos 60 d, se registró para cada tratamiento el número de plantas so-brevivientes, el número de raíces por propágulo, la longitud promedio de raíces (cm), incremento en longitud de propágulo (cm) con respecto al tamaño inicial del propágulo e incremento en número de hojas con respecto al número inicial del propágulo. Los datos obtenidos fueron anali-zados mediante un análisis de varianza de sepa-ración de promedios de Tukey (P≤0,05).
reSultaDoS y DiSCuSióN
Enraizamiento
La evaluación a los 60 d después de establecido el experimento demostró que en todos los trata-mientos la supervivencia de los propágulos fue del 100%. Igualmente, en todas las unidades ex-perimentales se observó la formación de raíces adventicias (tabla 1).
De acuerdo con la facilidad de formación de raíces, las plantas se clasifican en fácil de enraizar, me-

REV. COLOMB. CIENC. HORTIC.
228
dianamente fáciles de enraizar y recalcitrantes. Las dos primeras categorías se refieren a plantas cuyas estacas enraízan sin suplemento exógeno de auxinas o este incrementa significativamente el enraizamiento, respectivamente; mientras que las últimas forman muy pocas o ningunas raíces aun en presencia de auxinas (Hartmann et al., 2002). Los resultados del presente estudio permi-ten inferir que Alpinia purpurata es posiblemente una planta fácil de enraizar debido a que todos los propágulos desarrollaron raíces en todos los tratamientos, los cuales estaban completamente desprovistos de auxinas.
Número de raíces por propágulo
El análisis de varianza efectuado no permitió detectar efectos significativos (P≤0,05) con respecto a la interacción sustrato vs. tamaño de propágulo (tabla 1). Igualmente no se detec-taron diferencias significativas (P≤0,05) como resultado del efecto del tamaño de brote sobre el número promedio de raíces por brote. Inversa-mente, el tipo de sustrato influenció de manera significativa (P≤0,05) esta misma variable, ob-servándose que el mayor promedio de raíces fue observado cuando los brotes se establecieron en el sustrato agua (tabla 1). El uso del agua como sustrato de enraizamiento no ha sido reportado para Alpinia purpurata; sin embargo, sí ha sido previamente reportado en un estudio realizado en enraizamiento de Ixora coccinea L., observán-dose que las estacas enraizadas en agua desarro-llaron un mayor número de raíces en compara-
ción con aquellas establecidas en una mezcla de suelo (Méndez et al., 2004). El efecto favorable del agua en la producción de raíces adventicias puede ser el resultado de su acción como acti-vador de procesos fisiológicos a nivel de los me-ristemos presentes en la base de las estacas o a la acción disolvente sobre sustancias inhibidoras que limitan la respuesta de células competentes a estímulos inductores de enraizamiento (Bautis-ta et al., 1981). No obstante, es necesario aclarar que aunque para este caso la formación de raíces tuvo una respuesta favorable en un medio con ausencia total de oxígeno libre, para la mayoría de las especies la disponibilidad de oxígeno en el medio de enraizamiento es indispensable para la producción de raíces e incluso en algunas espe-cies leñosas se ha observado que el estrés hídrico hasta cierto nivel no afecta significativamente el enraizamiento (Murthy y Goldfarb, 2001; Hart-mann et al., 2002).
Longitud de raíz
El análisis de varianza permitió detectar que tan-to el tamaño de los brotes como el tipo de sus-trato de forma independiente tuvieron un efecto significativo (P≤0,05) sobre la longitud prome-dio de las raíces producidas (tabla 1). La mayor longitud promedio de raíces ocurrió cuando los brotes de mayor tamaño inicial (6,1-8,0 cm) fue-ron establecidos en agua (15,2 cm), seguidos por aquellas desarrolladas en ausencia de sustrato a partir de los brotes con tamaño intermedio (4,1-6,9 cm) (12,3 cm), mientras que las de menor
Tabla 1. Efecto del sustrato en el enraizamiento de brotes de Alpinia purpurata.
Promedios con letras distintas indican diferencia significativa según la prueba de Tukey (P≤0,05).
VariableSustrato
Suelo Agua Aire (sin sustrato)
Número de raíces por propágulo 3,3 b 7,5 a 2,5 b
Longitud de raíz (cm) 12,3 b 15,2 a 9,1 b
Longitud de propágulo (cm) 3,1 b 4,2 a 2,0 b
Número de hojas 1,5 b 2,0 a 1,5 b
SuÁREz/MARRuGO/pEñA

Vol. 2 - No.2 - 2008
229EFECTO DEL SuSTRATO y TAMAñO DEL pROpÁGuLO EN EL ENRAIzAMIENTO DE GINGER ROjO
longitud (9,1 cm) crecieron a partir de los brotes con menor tamaño (2,0-4,0 cm) establecidos en la mezcla de suelo.
Una mayor longitud de raíces adventicias produ-cidas en estacas de mayor tamaño puede expli-carse por el mayor contenido de materiales de reserva a disposición para el crecimiento de las raíces (Loach y Whalley, 1978; Hernández et al., 1995; Hamilton et al., 2002). Álvarez et al. (2007) reportaron el mayor tamaño de raíces en estacas de Rosmarium officinalis cuando estas tuvieron el mayor tamaño entre las estacas evaluadas (10 cm), respuesta que según los autores es el resulta-do de la mayor acumulación de fotoasimilados y mejor transporte de solutos para un crecimiento radical más acelerado. Alternativamente, el con-tenido hídrico a lo largo del perfil del suelo tiene un efecto en la distribución de raíces, así como también su morfología y su funcionamiento, ob-servándose que en presencia de un mayor con-tenido de agua en el suelo se produce un mayor crecimiento radical, entre otros aspectos por una menor resistencia del sustrato a la elongación de las raíces (Arias y Almanza, 2007). Este hallazgo puede explicar los resultados del presente estudio, donde se observó que cuando las raíces crecieron a partir de explantes establecidos en sustratos no sólidos (agua y aire) tuvieron una mayor longi-tud comparado con aquellas raíces que debieron interactuar con elementos que le ofrecieron una resistencia mecánica para su crecimiento
Incremento de longitud del propágulo
El análisis de varianza permitió constatar que la interacción tamaño del propágulo vs. sustrato y el tamaño del propágulo de forma individual no tuvieron efecto significativo (P≤0,05) sobre el incremento de la longitud del propágulo (tabla 1). Inversamente, esta variable sí fue afectada sig-nificativamente (P≤0,05), de forma individual, por el tipo de sustrato utilizado. La separación de promedios detectó que cuando los propágulos estuvieron en presencia de agua tuvieron un in-cremento del tamaño bastante mayor (4,2 cm),
que los establecidos en suelo (3,1 cm), y estos a su vez incrementaron significativamente su ta-maño sobre los que se establecieron en ausencia de sustrato (2 cm).
El agua es un factor ambiental crítico en el creci-miento vegetal y, en general, para todo el funcio-namiento fisiológico de la planta es afectado por el suministro de agua en células y tejidos metabó-licamente activos. Las plantas con déficit hídrico detienen la fotosíntesis, la respiración y los pro-cesos enzimáticos; el crecimiento, como resultado de la división y alargamiento celular, depende de la condición hídrica para responder al abastecimiento de compuestos orgánicos e inorgánicos necesarios para la síntesis de un nuevo citoplasma y una nue-va pared celular, que se expresan en el aumento de tamaño de los órganos (Kramer, 1983; Pérez-Mol-phe-Balch et al., 1996). Los resultados del presente trabajo, sugieren que la utilización de agua como sustrato básico de propagación, además de dismi-nuir los costos por manipulación de materiales só-lidos y reducir las implicaciones ambientales por uso de agentes químicos en la desinfección de las mezclas, contribuye a un mayor crecimiento de los propágulos en la etapa de vivero.
Incremento del número de hojas
Los resultados del análisis de varianza demostra-ron que ni la interacción tamaño del propágulo vs. sustrato, como tampoco el tamaño de propá-gulo por sí solo afectaron de manera significativa (P≤0,05) el número de nuevas hojas producidas (tabla 1); sin embargo, el mismo análisis permi-tió detectar diferencias importantes (P≤0,05) en los promedios de esta variable como resultado del efecto del sustrato. Cuando los propágulos fueron establecidos en agua el incremento del número de hojas (dos hojas) fue significativamente superior comparado con aquellos propágulos establecidos en suelo y en ausencia de sustrato (1,5 hojas). La relación entre un incremento en el número de hojas y la humedad disponible en el sustrato ha sido asociada con la utilización del agua como un insumo para el adecuado funcionamiento fisio-

REV. COLOMB. CIENC. HORTIC.
230
lógico de la planta (Calderón, 2001). Además, la producción de nuevas hojas durante la propaga-ción por estacas debe tomarse como un indicati-vo inequívoco de un proceso funcional, ya que la síntesis de factores necesarios para la formación de nuevas raíces ocurre en las hojas en forma-ción. Dentro de estos morfógenos radicales, las más importantes son las auxinas, cuyo efecto en la formación de raíces adventicias ha sido amplia-mente documentada (Davies, 1995; Castrillón et al., 2008).
Los autores dedican el presente trabajo a la memoria del profesor Ulises Caraballo Bagget.
CoNCluSioNeS• Alpinia purpurata es una especie fácil de enrai-
zar en mezclas de suelo, agua o al aire.
• El tamaño de los brotes no tiene efecto alguno sobre el enraizamiento de Alpinia purpurata.
• El sustrato agua incrementó significativamen-te el número de raíces por propágulo, la longi-tud de las raíces producidas, el tamaño de los brotes y el número de hojas de los brotes.
agraDeCimieNtoS
reFereNCiaS bibliogrÁFiCaS
Álvarez, J.; S. Lusardo y E. Chacon. 2007. Efecto de dife-rentes tamaños de esqueje y sustratos en la propa-gación del romero (Rosmarinus officinalis L.). Agron. Colomb. 25(2), 17-18.
Arias, I. y P. Almanza. 2007. Efecto de diferentes láminas de riego y sustratos sobre la propagación de esta-cas de vid (Vitis vinifera L.). p. 109. En: Memorias II Congreso Colombiano de Ciencias Hortícolas, Bogotá.
Bautista, D.; G. Vargas; J. Colmenares y Y. De Freitez. 1981. Efecto de algunos factores en el enraizamien-to y brotación de la vid ‘Criolla Negra’. Agron. Tropic. 31, 106-110.
Builes, M. 2003. Diagnóstico del mercado de los plata-nillos (Zingiberales) en el Departamento de Antio-quia. Tesis Administradora de Empresas Agropecua-rias. Corporación Universitaria Lasallista, Medellín, Colombia.
Calderón, F. 2001. Qué son los cultivos hidropónicos y el porqué la hidroponía. p. 20. En: Calderón (ed.). Me-morias Primer Curso de Hidroponía para la Floricul-tura. 31 de mayo al 2 de junio de 2001. Bogotá.
Castrillón, J.; E. Carvajal; G. Ligarreto y S. Magnitskiy. 2008. El efecto de auxinas sobre el enraizamiento de las estacas de agraz (Vaccinium meridionale Swartz) en diferentes sustratos. Agron. Colomb. 26(1), 6-22.
Criley, R. 1989. Development of Heliconia and Alpinia in Hawaii: Cultivar selection and culture. Acta Hort. 246, 247-258.
Davies, P. 1995. The plant hormones: Their nature, oc-currence and functions. pp. 1-12. En: Davies, P. (ed.). Plant hormones. Physiology, biochemistry and molecular biology. Kluwer Academic Publish-ers, The Netherlands.
Hamilton, C.; E. Emino y C. Bartuska. 2002. The ef-fect of cutting size, leaf area and shipping on Co-leus cutting quality parameters including rootings. Proc. Fla. State Hort. Soc. 115, 134-136.
Hansen, J. 1993. Field phenology of red ginger, Alpinia purpurata. Proc. Fla. State Hort. Soc. 106, 290-292.
Hartmann, H.; D. Kester; F. Davies y R. Geneve. 2002. Plant propagation. Principles and practices. Pren-tice Hall, Upper Saddle River, NJ.
SuÁREz/MARRuGO/pEñA

Vol. 2 - No.2 - 2008
231EFECTO DEL SuSTRATO y TAMAñO DEL pROpÁGuLO EN EL ENRAIzAMIENTO DE GINGER ROjO
Hernández, J.; H. Aramendiz y C. Cardona. 2005. In-fluencia del ácido indolbutírico y ácido naftale-noacético sobre el enraizamiento de esquejes de caña flecha (Gynerium sagittatum Aubl.). Rev. Te-mas Agrar. 10(1), 5-13.
Hoyos, J. 1999. Plantas tropicales ornamentales de tallo herbáceo. Sociedad de Ciencias Naturales, La Salle, Caracas.
Kramer, J. 1983. Water relations of plants. Academic Press, Orlando, FL.
Loach, K. y D. Whalley. 1978. Water and carbohydrate relationships during the rooting of cuttings. Acta Hort. 79, 161-168.
Méndez, J.; R. Salazar; M. Dautant; N. Alcorcés y J. Laynez. 2004. Efecto del medio de enraizamiento, número de hojas por estaca y lesionado de las esta-cas de (Ixora coccinea L.) con hormojardín N˚4. Rev. Cient. UDO Agríc. 4(1), 31-35.
Murthy, R. y B. Goldfarb. 2001. Effect of handling and water stress on water status and rooting of lob-lolly pine stem cuttings. New Forests 21(3), 217-230.
Neal, M. 1965. Gardens of Hawaii. Bishop Museum Press, Honolulu, HI.
Pérez-Molphe-Balch, E.; M. Gidekel; M. Segura-Nieto; L. Herrera-Estrella y N. Ochoa-Alejo. 1996. Effects of water stress on plant growth and root proteins in three cultivars of rice (Oryza sativa) with different levels of drought tolerance. Physiol. Plant. 96(2), 284-290.
USDA, ARS. 1999. National genetic resources program. Germplasm Resources Information Network (GRIN). En: National Germplasm Resources Labo-ratory, Beltsville, http://www. ars-grin.gov/cgibin/npgs/html/taxon.pl?2674; consulta: septiembre de 2008.

DaNiel roDríguez1,2
JoSé r. Cure h.1
FerNaNDo CaNtor r.1
plantulas de tomate. Foto: Álvarez-Herrera.
Consideraciones sobre la dispersión de la broca del café Hypothenemus hampei en relación con el empleo de trampas con atrayente. una revisión
Considerations about dispersion of the coffee berry borer Hypothenemus hampei related to the use of attractan-baited traps. A review
Fruto de café brocado. Foto: D. Rodríguez
resumen
En este trabajo se presenta una revisión acerca de la investigación realizada en torno a los aspectos biológicos de la migración y establecimiento de la broca del café, como un aspecto relevante para el desarrollo de métodos de captura en trampas, útiles para fines de monitoreo o control de la plaga. La discusión en relación con la broca del café es complementada con información disponible para otros Scolytinae de zonas con estaciones, que presentan características de comportamiento similiares. A partir del diagnóstico realizado se proponen temas de investigación relevantes para próximos trabajos en esta temática.
1 Facultad de Ciencias, Universidad Militar Nueva Granada, Bogotá (Colombia).2 Autor para correspondencia. [email protected]
palabras clave adicionales: migración, trampas con semioquímico, Scolytinae.
REVISTA COLOMBIANA DE CIENCIAS HORTÍCOLAS - Vol. 2 - No.2 - pp. 232-239, 2008

Vol. 2 - No.2 - 2008
233
abstract
A review about the research carried out on the biological topics of migration and establishment of the coffee berry borer is presented in this work, as an important topic to know in order to develop catch methods based on the use of traps, which can be useful for monitoring or control of the pest. The discussion about the coffee berry borer is complemented with information of Scolytinae from zones with seasonal climate, which presents a similar behavior. According to the diagnostics obtained, some research topics are proposed for future studies.
additional key words: migration, traps lured with semiochemical, Scolytinae.
Fecha de recepción: 14-08-2008 Aprobado para publicación: 02-12-2008
Una estrategia para el control de la broca del café que ha venido implementándose con resultados variables, es el empleo de trampas cebadas con una mezcla de alcoholes como atrayente. La idea de emplear trampas para la captura de broca nace del uso de esta estrategia para el manejo de otros Scolytinae de cultivos forestales (Moeck, 1970; Moeck, 1981; Lindgren, 1983), con base en principios de comportamiento similares. Des-de entonces se han realizado estudios de com-portamiento para identificar los estímulos que determinan la atracción de la broca hacia los frutos (Ticheler, 1963; Prates, 1969; Mendoza, 1991; Mathieu et al., 2001; Ortíz et al., 2004), y otros estudios con el propósito de definir las con-diciones óptimas para el uso de la estrategia en campo (Mendoza, 1991; Mathieu, 1997b; Gon-zález y Dufour, 2000; Borbón et al., 2000; Cár-denas, 2000; Villacorta et al., 2001; Barrera et al., 2004; Barrera et al., 2005; Fernández y Cordero, 2005; Cardona y Bustillo, 2006; Cardona y Bus-tillo, 2007; Dufour y Frérot, 2008), consideran-do factores como el atrayente empleado, la tasa de liberación del mismo, el color de la trampa, su diseño, la altura a la cual deben colocarse las trampas y el espaciamiento de estas en el cultivo. Sin embargo, existen discrepancias importantes entre los hallazgos de diferentes autores en cuan-to a las características óptimas del muestreo en
campo, y en cuanto a la eficiencia de las trampas como estrategia de control. Un aspecto que re-sulta de gran relevancia para obtener una mejor comprensión de la interacción entre la broca y las trampas con atrayente, es el conocimiento de los aspectos biológicos asociados a la migración y detección de nuevos hospederos, tema que se desarrolla en la presente revisión. El empleo de las trampas con atrayente se basa en utilizar los mecanismos propios de la plaga para realizar el proceso de selección del hospedero. Esta selec-ción puede ser vista como parte de un proceso más amplio, que en el contexto de los Scolyti-nae plaga, de especies forestales, ha sido definido por Wood (1982), como la estrategia de coloni-zación del hospedero. Debido a las importantes similitudes en algunos aspectos de la biología de estos insectos plaga de los forestales y la broca, en la presente revisión se ha empleado el modelo propuesto por este autor como herramienta de aproximación al caso de la broca. La estrategia de colonización consta de las etapas que se des-criben detalladamente a continuación.
DiSperSióN
Inicia con el abandono del fruto de origen por parte de la hembra y termina con la respuesta
CONSIDERACIONES SOBRE LA DISpERSIóN DE LA BROCA DEL CAFé
iNtroDuCCióN

REV. COLOMB. CIENC. HORTIC.
234
a los estímulos del hospedero. Se ha sugerido que algunos Scolytinae vuelan aleatoriamente, sin ser atraídos hacia sus hospederos por nin-gún tipo de estímulo (Raffa y Berriman, 1983; Moeck et al., 1981; Byers, 1996). En términos evolutivos, se plantea que la etapa de vuelo y dispersión representa una alta vulnerabilidad para los individuos, que están expuestos a con-diciones ambientales adversas, predadores y alto gasto energético, por lo que la búsqueda aleato-ria de hospederos tendría un alto costo para la viabilidad de las especies. Por el contrario, los mecanismos que permiten reducir este periodo de exposición y con ello la mortalidad y el gasto energético, incrementan la viabilidad de la espe-cie. Uno de tales mecanismos es la identificación del hospedero a través del reconocimiento de sus compuestos volátiles y la orientación del vuelo en función de dichos compuestos. Este tipo de atracción ha sido reportado ampliamente en especies de Scolytinae (Pearson, 1931; Bedard, 1966; Byers et al., 1979; Chapman, 1962; Cobb et al., 1968; Coster y Gara, 1968; Johnson y Cos-ter, 1978; Moeck, 1970; Moeck, 1981; Renwick y Vité, 1969; Stark et al., 1968; Visser, 1986; Vité y Pitman, 1969; Wood, 1972). En la broca del café ha sido comprobada la atracción de las hembras en respuesta a alcoholes y mezclas de alcoholes (Benassi, 1990; Mendoza, 1991; Mathieu et al., 1997b, González y Dufour, 2000; Borbón et al., 2000; Cárdenas, 2000; Villacorta et al., 2001). Es oportuno recordar que en el caso de la broca del café, solamente las hembras migran y son res-ponsables del establecimiento de la especie en nuevas áreas; los machos son ápteros y perma-necen toda su vida dentro del fruto de origen.
El conjunto de características comportamentales que muestran las hembras con capacidad migra-toria ha sido denominado comportamiento de abandono (Mathieu et al., 2001). Estas hembras han sido fecundadas antes de abandonar el fruto donde emergieron, poseen alas funcionales, y sa-len del fruto por efecto de las lluvias (Baker et al., 1984) o la alta insolación (Mathieu et al., 1997a). Durante el periodo de crecimiento del fruto, las
hembras emergen de forma continua en busca de nuevos frutos. Al finalizar la cosecha, particular-mente en Centroamérica, se dan migraciones ma-sivas de la broca, que abandona los cultivos debido a la mínima oferta de frutos durante este periodo. Sin embargo, una parte de ellas permanece en los frutos sobremaduros que no fueron cosechados. En estos frutos la emergencia y la oviposición se detienen por completo, y se encuentra una gran concentración de adultos. Este fenómeno, conoci-do como “diapausa reproductiva” no ha sido estu-diado a profundidad en el caso de la broca, pero se acentúa en las regiones donde el periodo entre cosechas es largo y se presenta marcada escasez de frutos durante el mismo (Mansingh, 1991). Las lluvias fuertes que inducen la nueva floración del cafeto promueven la ruptura de esta diapausa, con lo que se da la emergencia masiva de hembras de los frutos secos (Barrera et al., 2004b). El térmi-no “brocas en tránsito” se emplea para referirse a estos individuos que provienen de los frutos secos del año anterior, que tras el rompimiento de la diapausa reproductiva inician la búsqueda de nue-vos frutos. Los individuos tienen una gran capaci-dad de vuelo, como se ha podido comprobar bajo condiciones de laboratorio, donde se han registra-do tiempos de 90 min de vuelo continuo y 180 min de vuelos intermitentes (Bustillo y Villalba, 2004). Sin embargo, no todas las hembras migran, pues algunas nunca abandonan el fruto de origen, y solo aquellas que lo abandonan presentan una respuesta marcada a los estímulos olfativos. Aun-que es una condición indispensable que la hembra haya sido fecundada para que abandone el fruto, hay una proporción de hembras que pese a haber sido fecundadas, no abandonan el fruto (Da Sil-va et al., 2006; Mathieu et al., 2001). Esto puede deberse a múltiples factores como la falta de de-sarrollo alar, carencia de depósitos de grasa, que constituyen la reserva de energía para el vuelo, o a una baja “propensión al vuelo”, factores que a su vez pueden estar sujetos a control genético (Roff y Fairbairn, 2007). Es necesario realizar investiga-ciones que permitan una mejor comprensión de este fenómeno en la broca del café. Las hembras que no abandonan los frutos tampoco responden
RODRÍGuEz/CuRE H./CANTOR R.

Vol. 2 - No.2 - 2008
235
a los estímulos olfativos y su progenie se desarro-lla en el mismo fruto de origen (Mathieu et al., 2001). En algunos Scolytinae ha sido reportada la necesidad de alguna cantidad de vuelo como re-quisito para la percepción de estímulos olfativos (Borden, 1967; Wood y Bushing, 1963). Se requie-ren trabajos de investigación que permitan una mejor comprensión de la relación entre capacidad de vuelo y capacidad de respuesta a estímulos ol-fativos en la broca.
SeleCCióN
Comprende el periodo entre la percepción de los estímulos olfativos provenientes del hospedero hasta el inicio de la alimentación de la hembra sobre el sustrato (Wood, 1982). En cuanto al ate-rrizaje, el proceso podría ser aleatorio (Wood, 1982; Franklin et al., 2000; Franklin y Grégoire, 1999; Raffa y Berriman, 1983; Moeck et al., 1981; Byers, 1996; Gara et al., 1984), en el cual los in-dividuos posarían indiscriminadamente sobre hospederos y no hospederos, sin embargo esto puede ser interpretado como una primera fase de aproximación a la zona apropiada y no necesa-riamente al fruto específico. Los modelos de se-lección de hospedero que involucran integración de estímulos visuales y químicos han adquirido relevancia en años recientes (Campbell y Borden, 2006a; Campbell y Borden, 2006b). Dicha inte-gración también ha sido propuesta para la broca del café por Mathieu et al. (2001), quienes plan-tean que una vezla hembra entra en la etapa de selección, los estímulos visuales, constituidos por la percepción del color y, posiblemente, la forma y el tamaño, podrían integrarse con los estímulos olfativos, para permitir a la hembra su llegada a una baya en el estado de madurez óptimo y con-tinuar con las etapas de concentración y estable-cimiento. También Bustillo y Villalba (2004) pro-ponen que la broca es inicialmente atraída por el olor, y luego por el color y la forma del fruto. Este planteamiento concuerda con lo encontrado por Mendoza (1991), quien ha verificado la respuesta diferencial de las hembras tanto a estímulos ol-
fativos de cuatro estados de desarrollo del fruto, como a sus respectivos colores.
CoNCeNtraCióN
En los Scolytinae de los cultivos forestales, esta etapa se da una vez que los primeros individuos colonizadores, previa evaluación de la aptitud del sustrato, comienzan a liberar feromonas de agregación que pueden tener un efecto sinérgico con volátiles de la planta, para orientar al resto de la población hacia su lugar de ubicación y así generar una concentración alta de la población en el hospedero (Wood, 1982). A este proceso se le ha denominado “atracción secundaria”, en contraposición a la “atracción primaria”, que se da cuando los primeros individuos coloniza-dores deben por sí mismos ubicar un sustrato adecuado para la alimentación y reproducción, únicamente mediante señales químicas del hos-pedero, o al azar. El tiempo transcurrido entre la detección del hospedero adecuado y la produc-ción de feromona de agregación, es un aspecto importante, pues un lapso de tiempo pequeño implica una mayor sobrevivencia de la población de adultos buscando hospedero, que reducen su gasto energético y su exposición a predadores y condiciones ambientales adversas. En varios tra-bajos (Mendoza, 1991; Bustillo y Villalba, 2004; Bustillo, 2006) se plantea la posibilidad que en Hypothenemus hampei existan los dos tipos de atracción, pues ocurre una atracción primaria mediada por los estímulos químicos y visuales antes mencionados, y posteriormente podría darse una etapa de atracción secundaria, me-diante sustancias producidas por las brocas co-lonizadoras, que podrían actuar en forma sinér-gica con los volátiles del café. Mendoza (1991) reporta que tanto frutos verdes como maduros ejercen una mayor atracción sobre las hembras cuando ya han sido atacados, y la agregación de las poblaciones en campo reportada inicialmente por Baker (1984), es consistente con la presencia de una feromona que genere dicha agregación. Las evidencias indican que estas feromonas de
CONSIDERACIONES SOBRE LA DISpERSIóN DE LA BROCA DEL CAFé

REV. COLOMB. CIENC. HORTIC.
236
agregación podrían encontrarse en las heces de las hembras (Bustillo y Villalba, 2004; Bustillo, 2006). Sin embargo hasta ahora no existen re-portes de compuestos aislados de las heces, o de cualquier otra parte de H. hampei, como po-sibles feromonas de agregación, lo que a su vez impide estudiar aspectos más específicos como el tiempo transcurrido entre la llegada a los fru-tos y la producción de la feromona y la medida en que este factor puede reducir la mortalidad de hembras buscando nuevos granos. Esto pone de manifiesto la necesidad de realizar trabajos de investigación sobre los procesos involucrados en la etapa de concentración en H. hampei.
eStableCimieNto
Esta etapa inicia con la perforación de las gale-rías, previa evaluación de la calidad del recurso y concluye con la oviposición (Wood, 1982). En H. hampei se han reportado diferencias en el tiempo de perforación de los frutos, según su estado de desarrollo: se emplea un tiempo de perforación de 5 h y 36 min en los frutos verdes, 5 h y 54 min en frutos pintones, 4 h y 50 min en frutos maduros, y 11 h y 21 min para penetrar frutos sobremaduros (Bustillo y Villalba, 2004). Es evi-dente la penetración más rápida en los frutos maduros. En el desarrollo temprano de los frutos (<120 d) el porcentaje de materia seca inferior al 20% en los frutos, hace que su consistencia sea inapropiada para la perforación de la galería. Si esto ocurre, la hembra permanece en un peque-ño canal de penetración cercano a la superficie, esperando a que el fruto alcance la consistencia adecuada. Durante este periodo el individuo es muy vulnerable pues se encuentra muy cerca
del exterior del fruto. El inicio de la oviposición puede tardar desde 63 días después de la llegada de la hembra, si el fruto aún es demasiado verde (89% de humedad) hasta cuatro días en frutos maduros (66% de humedad). En la broca del café, al igual que en Trypodendron lineatum Oliv. (Co-leoptera: Curculionidae: Scolytinae), las alas se degeneran después de que ha ocurrido perfora-ción u oviposición (Mathieu et al., 2001). Debi-do a esto la hembra no abandona el fruto una vez que ha iniciado la perforación de la galería, y generalmente se presenta una sola hembra por fruto (Barrera, 2004; Borbón, 2004). De acuerdo con Cantor et al. (2000) esto podría deberse a al-gún mecanismo de marcación química, pues no se observan marcas físicas evidentes, o también a un posible comportamiento territorial, dado que estos autores reportan la defensa del grano por parte de la hembra residente cuando una se-gunda hembra intenta establecerse en el mismo fruto. Sin embargo, recientemente se ha obser-vado en cultivos de la zona cafetera colombiana la presencia de perforaciones de varias hembras fundadoras en un mismo fruto, principalmen-te en las ramas del tercio inferior de la planta, donde el porcentaje de frutos con perforaciones múltiples oscila entre 2,0 y 16,5% (Benavides et al., 1997).
Finalmente es importante resaltar que los me-canismos de atracción primaria y secundaria, mediante compuestos producidos por la planta y una posible feromona respectivamente, son temas de estudio promisorios con miras a la ob-tención de compuestos atrayentes más eficien-tes que permitan obtener niveles significativos de control de la plaga bajo las condiciones co-lombianas.
RODRÍGuEz/CuRE H./CANTOR R.

Vol. 2 - No.2 - 2008
237
Baker, P.S. 1984. Some aspects of the behaviour of the coffee berry borer (Coleoptera:Scolytidae) in rela-tion to its control in southern México. Folia Ento-mol. Mexic. 62, 9-24.
Barrera, J.F.; J. Herrera y J. Valle. 2005. Efecto de la altura de la trampa en la captura de la “broca del café”: Implicaciones en dispersión y muestreo. Ento-mología Mexicana 4, 542-546.
Barrera, J.F.; A. Villacorta; J. Herrera; H. García y L. Cruz. 2004. Aplicación de trampas para el moni-toreo de la broca del café. pp. 95-113. En: Néri A. (ed.). Manejo da broca do café. Workshop Interna-cional, Londrina, Brasil.
Bedard, W.D. 1966. Variation in capacity of Ips confusus to reach attractive hosts. pp. 137-142. En: Gerhold, H.D.; E.J. Schreiner; R.E. McDermott y J.A. Winies-ki (eds.). Breeding pest resistant trees. Pergamon, Oxford, U.K.
Benassi, V.L.R.M. 1990. Resultados preliminares da flu-tuação populacional da broca-do-café Hypothene-mus hampei (Ferrari, 1867) (Coleoptera:Scolytidae) na região norte do Espírito Santo. p. 83. En: XVI Congressso Brasileiro de Pesquisas Cafeeiras. Rio de Janeiro, Brasil.
Benavides, P.; L.M. Constantino; L. Navarro y F.E. Ace-vedo. 1997. Herencia de caracteres genéticos y determinación de endogamia en la broca del café (Hypothenemus hampei). p. 46. Resúmenes XXXIV Congreso de la Sociedad Colombiana de Entomolo-gía (Socolen). Cartagena, Colombia.
Borbón, M.O.; O.M. Alfaro; A.M. Oelschlager y L.M. González. 2000. Proyecto de trampas, atrayentes y repelentes para el control de la broca del fruto del cafeto, Hypothenemus hampei L. (Coleoptera: Scolytidae). pp. 331-348. En: XIX Simposio Latino-americano de Caficultura. San José.
Borden, J.H. 1967. Factors influencing the response of Ips confusus to male attractant. Can. Entomol. 99,1164-1193.
Bustillo, A.E. 2006. Una revisión sobre la broca, Hy-pothenemus hampei (Coleoptera: Curculionidae: Scolytinae), en Colombia. Rev. Colomb. Entomol. 32(2), 101-116.
Bustillo, A.E. y D.A. Villalba. 2004. Efecto del clima y condiciones de cultivo del café en la biología y
comportamiento de la broca del café, Hypothenemus hampei (Ferrari) (Coleoptera: Curculionidae: Scoly-tinae). pp. En: Manejo da broca do café. Workshop Internacional. Londrina, Brasil.
Byers, J.A. 1996. An encounter rate model of bark beetle populations searching at random for susceptible host trees. J. Ecol. Modelling 91, 57-66.
Byers, J.A.; D.L. Wood; L.E. Browne; R.H. Fish; B. Piatek y L.B. Hendry. 1979. Relationship between a host plant compound, myrcene, and pheromone pro-duction in the bark beetle Ips paraconfusus. J. Insect Physiol. 25,477-482.
Campbell, S.A. y J. Borden. 2006a. Close-range, in flight integration of olfactory and visual information by a host-seeking bark beetle. Entomol. Exp. Appl. 120,91-98.
Campbell, S.A. y J. Borden. 2006b. Integration of visual and olfactory cues of hosts and non-hosts by three bark beetles. (Coleoptera: Scolytidae). Ecol. Ento-mol. 31, 437-449.
Cantor, F.; O. De Souza; E.F. Vilela; C.A. Faria y F. Gumier-Costa. 2000. Local deterrence of multiple attacks by the coffee Berry borer. p. 164. En: Libro de resúmenes I: XXI International Congress of En-tomology. Foz do Iguaçu, Brasil.
Cárdenas, R.M. 2000. Trampas y atrayentes para moni-toreo de poblaciones de broca del café Hypothen-emus hampei (Ferrari) (Col., Scolytidae). pp. 369-379. En: Memorias XIX Simposio Latinoamericano de Caficultura. San José.
Cardona, G.E. y A.E. Bustillo. 2006. Captura de broca del café Hypothenemus hampei (Ferrari) usando trampas en tres densidades diferentes. p. 133. En: Resúmenes XXXIII Congreso de la Sociedad Co-lombiana de Entomología (Socolen). Manizales, Colombia.
Cardona, G.E. y A.E. Bustillo. 2007. Evaluación de trampas cebadas con alcoholes para la captura de la broca del café, Hypothenemus hampei (Ferrari) (Coleoptera: Scolytidae). p. 75. Resúmenes XXX-IV Congreso de la Sociedad Colombiana de Ento-mología (Socolen). Cartagena, Colombia.
Chapman, J.A. 1962. Field studies on attack flight and log selection by the ambrosia beetle Trypodendron linea-tum (Olivier). Can. Entomol. 94, 74-92.
reFereNCiaS bibliogrÁFiCaS
CONSIDERACIONES SOBRE LA DISpERSIóN DE LA BROCA DEL CAFé

REV. COLOMB. CIENC. HORTIC.
238
Cobb, F.W.; D.L. Wood; R.W. Stark y P.R. Miller. 1968. Photochemical oxidant injury and bark beetle in-festation of ponderosa pine II. Effect of injury upon physical properties of oleoresin, moisture content and phloem thickness. Hilgardia 59, 127-134.
Coster, J.E. y R.I. Gara. 1968. Studies on the attack be-haviour of the southern pine beetle II. Response to attractive material. Contrib. Boyce Thompson Inst. 24, 69-76.
Da Silva, F.C.; M.U. Ventura y L. Morales. 2006. Capture of Hypothenemus hampei Ferrari (Coleoptera: Sco-lytidae) in response to trap characteristics. Scientia Agric. 63(6), 567-571.
Dufour, B.P. y B. Frérot. 2008. Optimization of coffe Berry borer, Hypothenemus hampei Ferrari (Col., Scolytidae) mass trapping with an attractant mix-ture. J. Appl. Entomol. 132, 591-600.
Fernández, S. y Cordero J. 2005. Evaluación de atrayen-tes alcohólicos en trampas artesanales para el mo-nitoreo y control de la broca del café, Hypothenemus hampei (Ferrari). Bioagro 17(3), 143-148.
Franklin, A.J.; C. Debruyne. y J.C. Grégoire. 2000. Re-capture of Ips typographus L. (Col.,Scolytidae) with attractants of low release rates: localized dis-persión and environmental influences. Agr. Forest Entomol. 2, 259-270.
Franklin, A. y J.C. Grégoire. 1999. Flight behaviour of Ips tipographus L. (Col., Scolytidae) in an environ-ment without pheromones. Ann. Forest Sci. 56(7), 591-598.
Gara, R.I.; D.R. Geizler y W.R. Littke. 1984. Primary at-traction of the mountain pine beetle Dendroctonus ponderosae to lodgepole pine, Pinus contorta in Or-egon. Ann. Entomol. Soc. Amer. 77, 333-334.
González, M.O. y B.P. Dufour. 2000. Diseño, desarrollo y evaluación del trampeo en el manejo integrado de la broca del café Hypothenemus hampei. pp. 381-396. En: XIX Simposio Latinoamericano de Caficultura. Costa Rica.
Johnson, P.C. y J.E. Coster. 1978. Probability of attack by the southern pine beetle in relation to distance from an attractive host tree. Forestry Sci. 24, 574-580.
Lindgren, B.S. 1983. A multifunnel trap for scolytid beteles (Coleoptera). Can. Entomol. 115(3), 299-302
Mansingh, A. 1991. Limitations of insecticides in the management of the coffe berry borer Hypothen-emus hampei Ferrari. J. Coffee Res.
Mathieu, F.; L.O. Brun y B. Frérot. 1997a. Factors rela-ted with native host abandon by the coffee berry borer. J. Appl. Entomol. 121, 181-186
Mathieu, F.; L.O. Brun; C. Marchillaud y B. Frérot. 1997b. Trapping of the coffee berry borer Hypothenemus hampei Ferr. (Col., Scolytidae) within a mesh-en-closed environment: Interaction of olfactory and visual stimuli. J. Appl. Entomol. 121(3), 181-186.
Mathieu, F. ; V. Gaudichon; L.O. Brun y B. Frérot. 2001. Effect of physiological status on olfactory and visual responses of female Hypothenemus hampei during host plant colonization. Physiol. Entomol. 26, 189-193.
Mendoza, M.J.R. 1991. Resposta da broca-do-café, Hy-pothenemus hampei, a estímulos visuais e semioquí-micos. Dissertaçao de Mestrado. Universidad Fede-ral de Viçosa, Brasil.
Moeck, H.A. 1970. Ethanol as the primary attrctant for the ambrosia beetle Trypodendrum lineatum (Cole-optera: Scolytidae). Can. Entomol. 102, 792-796.
Moeck, H.A. 1981. Ethanol induces attack on tres by spruce beteles, Dendroctonus rufipennis (Coleoptera: Scolytidae). Can. Entomol. 113(10), 939-942.
Ortíz, A.; F.E. Vega y F.J. Posada. 2004. Volatile composi-tion of coffee berries at different stages of ripennes, and their possible attraction to the coffee berry borer Hypothenemus hampei (Coleoptera: Curculio-nidae). J. Agr. Food Chem. 52, 5914-5918.
Pearson, H.L. 1931. Theory in explanation of the selec-tion of certain trees by the western pine beetle. J. Forestry 29, 696-699.
Prates, H.S. 1969. Observações preliminares de atração da broca-do-café Hypothenemus hampei (Ferrari, 1867) a extratos de frutos de cafeeiro (cereja e ver-de). O Solo 61(2), 13-14
Raffa, K.F. y A.A. Berryman. 1983. The role of host plant resistance in the colonization behaviour and ecol-ogy of bark beetles (Coleoptera: Scolytidae). Ecol. Monogr. 53, 27-49.
Renwick, J.A. y J.P. Vité. 1969. Bark Beetle Attractants: Mechanisms of colonization by Dendroctonus fron-talis. Nature 224, 1222-1223.
Roff, D.A. y D.J. Fairbairn. 2007. The evolution and genetics of migration in insects. BioScience 57(2), 155-164.
Stark, R.W.; P.R. Miller; F.W. Cobb; D.L. Wood y J.R. Parmeter. 1968. Photochemical oxidant injury and bark beetle infestation of ponderosa pine I. Inci-
RODRÍGuEz/CuRE H./CANTOR R.

Vol. 2 - No.2 - 2008
239
dence of bark beetle infestation of ponderosa pine. Hilgardia 39, 121-126.
Ticheler, J. H. G. 1963. Estudio analítico de la epidemio-logía del escolitídeo de los granos del café Stepha-noderes hampei Ferr. en Costa de Marfil. Revista Cenicafé 14(4), 223-294.
Villacorta, A.; A.F. Possagnolo; R.Z. Silva y P.S. Rodri-gues. 2001. Um modelo de armadilha com semio-químicos para o manejo integrado da broca do café Hypothenemus hampei (Ferrari) no Paraná. pp. 2093-2098. En: Simpósio de Pesquisa dos Cafés do Brasil. Vitória, Brasil.
Visser, J.H. 1986. Host odour perception in phytopha-gous insects. Annu. Rev. Entomol. 31, 121-144
Vité, J.P. y G.B. Pitman. 1969. Insect and host odours in the aggregation of the western pine beetle. Can. Entomol. 101, 113-117.
Wood, D.L. 1972. Selection and colonization of pon-derosa pine by bark beetles. pp. 101-117. En: Van Emden, H.F. (ed.). Plant relationships. Blackwell Science, Oxford, UK.
Wood, D.L. 1982. The role of pheromones, kairomones and allomones in the host selection and coloniza-tion behaviour of bark beetles. Annu. Rev. Ento-mol. 27, 411-446.
Wood, D.L. y R.W. Bushing. 1963. The olfactory response of Ips confusus (Lec.) to the secondary attraction in the laboratory. Can. Entomol. 95, 1066-1078.
CONSIDERACIONES SOBRE LA DISpERSIóN DE LA BROCA DEL CAFé

REV. COLOMB. CIENC. HORTIC.
240
POLÍTICA EDITORIAL
La Revista Colombiana de Ciencias Hortícolas es el órgano de divulgación de la Sociedad Colombiana de Ciencias Hortícolas (SCCH – www.soccolhort.com), adscrita a la ISHS (Internacional Society for Horticultural Science – www.ishs.org), con frecuencia semestral en el territorio nacional. Constituye una publicación abierta a la dis-cusión y difusión de trabajos técnico-científicos en el área de las ciencias agrícolas, con énfasis en horticultura (frutales, hortalizas, ornamentales, hierbas aromáticas, viveros) y disciplinas afines, propuestos por autores nacionales e internacionales. Busca divulgar trabajos inéditos desarrollados por investigadores de diversas uni-versidades y centros de investigación del país, y difundir y someter a discusión los avances científicos que se producen, con el fin de contribuir a la consolidación de una comunidad académica congregada en torno a las disciplinas afines a la horticultura.
Los artículos deben presentarse según las normas establecidas en las instrucciones para los autores. El Comité Editorial se reserva el derecho de aceptar o no las contribuciones que se hagan a la revista. Los manuscritos se enviarán a dos pares evaluadores expertos en el área específica del conocimiento, quienes decidirán si el artí-culo tiene el nivel científico necesario para publicarse. Si se llega a presentar alguna diferencia de conceptos en cuanto a la aprobación para publicación, el artículo será enviado a un tercer par, quien entregará la decisión final. Si el artículo se acepta para publicación, el autor debe realizar las correcciones sugeridas en el tiempo indicado por el editor.
La Revista Colombiana de Ciencias Hortícolas autoriza la reproducción y citación del contenido de la publi-cación, siempre y cuando se registre de forma explícita el nombre de la revista, nombre del autor(es), año, volumen, número y página del artículo consultado. Los artículos publicados en la Revista serán enviados en medio magnético a las diferentes bases de datos y sistemas de indexación (Publindex), para la divulgación de su contenido y se podrán consultar mediante Internet.
Las ideas y afirmaciones emitidas por los autores son responsabilidad de éstos y no expresan necesariamente las opiniones y políticas del Comité Editorial de la SCCH, ni de las instituciones a las cuales están vinculados quienes las proponen.
INSTRUCCIONES PARA LOS AUTORES
Los artículos puestos a consideración del Comité Editorial de la Revista Colombiana de Ciencias Hortícolas deben ser inéditos; en consecuencia, aquellos manuscritos que hayan sido publicados en otras revistas o publicacio-nes técnico-científicas no serán aceptados.
La Revista Colombiana de Ciencias Hortícolas acepta artículos originales de los siguientes tipos, en concordancia con lo requerido por el programa Publindex de Colciencias para la categoría B:
• Artículo de investigación científica y tecnológica: documento que presenta de manera detallada los resultados ori-ginales de proyectos de investigación. La estructura generalmente utilizada tiene cuatro partes esenciales: introducción, metodología (materiales y métodos), resultados y discusión, y conclusiones.
• Artículo de reflexión: documento que presenta resultados de investigación desde una perspectiva analítica, interpretativa y crítica del autor, sobre un tema específico y recurriendo a fuentes originales.

Vol. 3 - No.1 - 2009
241
• Artículo de revisión: documento resultado de una investigación en la que se analizan, sistematizan e integran los resultados de investigaciones, publicados o no, sobre un campo en ciencia o tecnología, con el fin de dar cuenta de los avances y tendencia de desarrollo; se caracteriza por presentar un soporte bibliográfico cuida-doso no menor a 50 referencias.
Extensión y remisión del artículo y de la ilustraciónLa extensión del artículo de investigación no debe exceder las 5.200 palabras (incluido literatura y las tablas), excepto el artículo de revisión, que puede contener hasta 6.500 palabras; tamaño carta, escritas a doble espa-ciado, letra o fuente Times New Roman con tamaño de 12 puntos, márgenes de 3 cm en la parte superior, 2 cm en la inferior y 2,5 cm en las márgenes laterales derecha e izquierda. Las tablas y figuras (es decir, los grá-ficos, dibujos, esquemas, diagramas de flujo, fotos y mapas) deben presentarse en hojas independientes y con numeración consecutiva (Tabla 1 ... Tabla n. Figura 1 ... Figura n; etc.). Los textos y tablas deben presentarse en el procesador de palabra MS-Word®; las tablas y los diagramas de frecuencia (barras y tortas) originales deben suministrarse en el archivo del manuscrito y también en su original de MS-Excel®; otras figuras, como fotografías sobre papel y dibujos, se pueden enviar en originales o escanearlas y remitirlas en el formato digital de compresión JPG (o JPEG), preferiblemente con una resolución de 600 x 600 dpi (mínimo 300 dpi). Como norma general, las tablas y figuras solo se aceptan en blanco y negro. El artículo con sus archivos en Word, Excel y JPG debe ser enviado al correo electrónico: [email protected], al editor profesor Dr. Fánor Casierra, [email protected] o coeditor profesor Dr. Gerhard Fischer, [email protected].
Título y autoresEl título es en minúscula y no debe exceder las 15 palabras. Es obligatoria su respectiva traducción al idioma inglés. Los nombres científicos de vegetales o animales se deben escribir con letra cursiva (itálica) y en minús-culas, y sólo con mayúsculas la primera letra del género y el clasificador. Debajo del título en inglés se escribe el(los) nombre(s) y apellido(s) del(de los) autor(es), sin sus respectivos títulos académicos ni cargos laborales, en una línea horizontal y de acuerdo con su contribución en la investigación o preparación del artículo. En la parte inferior de la primera página, es decir, en aquella que contiene el título del artículo, el nombre y la ciudad de ubicación de la entidad a la cual prestan sus servicios o del patrocinador para la realización del trabajo y el autor para correspondencia con su correo electrónico (e-mail).
Resumen, abstract y palabras clave adicionalesEl resumen debe escribirse en español, portugués o inglés, correspondiente al idioma del artículo. Los artículos en español o portugués deben contener un abstract en inglés; artículos en inglés, un resumen en español. Am-bos textos deben describir en forma breve el problema, los métodos utilizados, su justificación y los resultados obtenidos más relevantes, y no deben exceder de 250 palabras escritas en un único párrafo. Es obligatorio acompañar el resumen con máximo seis palabras clave, que no hayan sido usadas en el título, y de su traduc-ción a inglés (additional key words), las que tampoco deben figurar en el título en inglés.
IntroducciónEl texto que debe contener la situación actual del problema, su definición y la revisión de los trabajos previos relacionados con él; además, los objetivos y la justificación de la investigación. Es obligatorio acompañar los nombres vulgares con el(los) nombre(s) científico(s) y la(s) abreviatura(s) del clasificador en la primera men-ción en el artículo.
Materiales y métodosEn este apartado se deben describir de forma clara, concisa y secuencial los materiales (vegetales, animales, implementos agrícolas o de laboratorio) utilizados en desarrollo del trabajo, además de los procedimientos o protocolos seguidos y el diseño escogido para el tratamiento estadístico de los datos.
Resultados y discusiónLos resultados deben presentarse de manera lógica, objetiva y secuencial mediante textos, tablas y figuras; estos dos últimos apoyos deben ser fáciles de leer y se deben poder interpretar de manera autónoma, aunque deben citarse siempre en el texto. Las gráficas serán bidimensionales y a una sola tinta, con porcentajes de negro para las variaciones de las columnas; las líneas de las curvas deben ser de color negro, punteadas o con-tinuas (- - - - ó --------), usando convenciones como: l, , s, ∆, etc.

REV. COLOMB. CIENC. HORTIC.
242
Las tablas se deben elaborar con pocas columnas y renglones. La discusión de resultados debe ser completa y exhaustiva, contrastando los resultados obtenidos con la literatura más actual sobre el tema. En esta sección se relacionan los hallazgos más concluyentes de la investigación.
ConclusionesEn este apartado se relacionan los hallazgos más concluyentes de la investigación, es decir, aquellos que cons-tituyan un aporte significativo para el avance del campo temático explorado, además de un direccionamiento sobre futuras investigaciones.
AgradecimientosSi se considera necesario, se agradecen aquellas contribuciones importantes en la concepción, financiación o realización de la investigación: especialistas, firmas comerciales, entidades oficiales o privadas, asociaciones de profesionales y operarios.
Referencias bibliográficasPara las citas bibliográficas que sustentan las afirmaciones en el texto, se utilizará el sistema [autor(es), año] de forma uniforme; cuando la publicación citada tenga tres o más autores, se debe mencionar el apellido del primer autor acompañado de la expresión latina et al., equivalente a “y otros”, en cursivas y con el año (ej: García et al., 2003). La lista completa con las referencias bibliográficas mencionadas se debe incluir al final del artículo. Los apellidos y nombres de todos los autores deben escribirse en redondas, en el orden alfabético de sus apellidos; cuando se citan varias publicaciones del(de los) mismo(s) autor(es) deben listarse en orden cronológico. Se prevén algunos casos:
• Para libros: Autor(es), año. Título del libro, edición, casa editora y ciudad de su sede, páginas consultadas (pp. # - #). Ejemplo: Agrios, G. 1996. Fitopatología. Segunda edición. Editorial Limusa, México D.F.
• Para capítulos de libros: Autor(es), año. Título del capítulo, páginas consultadas (pp. # - #). En: Apellidos y nombres de los compiladores o editores (eds.)., título del libro, edición, casa editora y ciudad de su sede. Ejemplo: Bernal, H. 1996. Capítulo 6: Evapotranspiración. pp. 112-125. En: Agrios, G. (ed.). Fitopatología. 2a ed. Editorial Limusa, México D.F.
• Para revistas: Autor(es), año. Título del artículo, nombre abreviado de la revista volumen(número), página-página. Ejemplo: García, S.; W. Clinton; L. Arreaza y R. Thibaud. 2004. Inhibitory effect of flowering and early fruit growth on leaf photosynthesis in mango. Tree Physiol. 24(3), 387-399.
• Para citas de Internet: Autor(es), año. Título del artículo. En: Nombre de la publicación electrónica del web-site, portal o página y su URL, fecha de consulta. Ejemplo: Arafat, Y. 1996. Siembra de olivos en el desierto palestino. En: Agricultura Tropical, http://agrotropical.edunet.es; consulta: noviembre de 2003.

Forma de pago suscriptores nacionalesPago en efectivo o en cheque a nombre de la Sociedad Colombiana de Ciencias Hortícolas o por consignación nacional en el Banco de Bogotá, cuenta corriente 033811761, y envío posterior de copia de recibo y formato de suscripción diligenciado al fax (571) 316 5498.
Valor para el año 2008 (dos números, incluyendo costos postales para el envío): Colombia: $40.000 (pesos colombianos); América Latina y el Caribe: US$ 30; USA y Canadá: US$ 50; otras regiones y países: US$ 65.
SuScripción
revista colombiana de ciencias HortícolasSociedad Colombiana de Ciencias Hortícolas - Facultad de Agronomía Universidad Nacional de ColombiaA.A. 14490, Bogotá-ColombiaTeléfono: (571) 316 5000 ext. 19041/19043Tel/fax: (571) 316 [email protected], [email protected]
Nombre / Name
Documento de identidad / Identification document
Dirección / Address Ciudad / City
Teléfono / Phone
Departamento / Department Zona postal / Zip code
País / Country E-mail
Profesión / Profession
Institución / Employer
Dirección de envío / Mailing address
Último ejemplar recibido / Last issue mailed.
Año / Year Volumen/Volume Número/Number Fecha de envío / Mailing date



Volumen 2 / N
o. 2 / Julio-Diciem
bre 2008Revista C
olombiana de C
iencias Hortícolas
Editorial 131
Seccióndefrutales
Efectodelaaplicaciónprecosechadegiberelinasycalcioenlaproducción,calidadyrajadodelfrutodeuchuva(Physalis peruvianaL.)Effect of preharvest application of gibberellins and calcium in production, fruit quality and cracking of cape gooseberry (Physalis peruviana L.)
NathalyAmézquita,HelberEnriqueBalaguera-LópezyJavierGiovanniÁlvarez-Herrera 133
Aspectosanatómicosdelasemilladepapaya(Carica papayaL.)Anatomical aspects of papaya (Carica papaya L.) seed
ArletteIvonneGilyDiegoMiranda 145
Comportamientode26cultivaresdenaranjaencondicionesdelpiedemontedelMeta,ColombiaPerformance of 26 orange cultivars in conditions of Piedmont of Meta, Colombia
JavierOrlandoOrduz-RodríguezyFranciscoAvella 157
EfectodelencalamientodesuelossulfatadosácidosenCórdoba(Colombia)sobreelfrutodepiña(Ananas comosus(L.)Merr.)Effect of liming of acid sulfate soils in Cordoba (Colombia) on pineapple (Ananas comosus (L.) Merr.) fruitEnriqueCombattC.,AlfredoJarmaO.yAlejandroClevesL. 175
Seccióndehortalizas
Acumulacióndemetalespesadosensueloyplantasdecuatrocultivoshortícolas,regadosconaguadelríoBogotáAccumulation of heavy metals in soil and plants of four vegetable crops irrigated with water of Bogota river DiegoMiranda,CarlosCarranza,CarlosAndrésRojas,CarlosMartínJerez,
GerhardFischeryJorgeZurita 180
Caracterizacióndelasdeficienciasdemacronutrientesenplantasdecebollín(Allium schoenoprasum L.)Characterization of macronutrient deficiencies in chives (Allium schoenoprasum L.) plantsDiegoAndrésBernal,LeidyCarolinaMorales,GerhardFischer,
JairoCuervoyStanislavMagnitskiy 192
EfectodelaaplicacióndecachazafrescaydedossistemasdeproducciónmaízymaízenasocioconfríjolsobrelaspropiedadesfísicasdeunInceptisolEffect of application of fresh filter cake and two production systems of corn and corn associated with beans on the physical properties of an Inceptisol
FabioEmilioForero,JaimeTorresyHelberEnriqueBalaguera-López. 205
Seccióndeornamentales
Determinacióndelmomentodecortedelaflorenlimonio(Limoniumsp.)apartirdeseisestadosfenológicosDetermination of timing of flower cut in sea lavender (Limonium sp.) starting from six phenological stagesFánorCasierra-PosadayJuanCarlosAbril 217
Efectodelsustratoytamañodelpropáguloenelenraizamientodelgingerrojo(Alpinia purpurata)Effect of substrate type and cutting size on red ginger (Alpinia purpurata) rootingIsidroE.Suárez,GledyLuzMarrugoyMaryorikPeña 225
Artículoderevisión
ConsideracionessobreladispersióndelabrocadelcaféHypothenemus hampeienrelaciónconelempleodetrampasconatrayente.UnarevisiónConsiderations about dispersion of the coffee berry borer Hypothenemus hampei related to the use of attractan-baited traps. A reviewDanielRodríguez,JoséR.CureH.yFernandoCantorR. 232
Políticaeditorialeinstruccionesalosautores 240


















