
UNIVEUNIVERSIDAD VERACRUZANA ERACRUZANA FACULTAD DE ... · ... del proceso de enraizamiento a...
85
1 ESTABLECIMIENTO DE UNA METODOLOGÍA PARA LA MICROPROPAGACIÓN DE PATRONES TOLERANTES AL VIRUS DE LA TRISTEZA DE LOS CÍTRICOS (VTC) TESIS QUE PARA OBTENER EL GRADO DE MAESTRO EN CIENCIAS ESPECIALIDAD: BIOTECNOLOGIA DE PLANTAS PRESENTA: PAULA ROJAS MENCIO CORDOBA, VER. JUNIO DEL 2001 UNIVEUNIVERSIDAD VERACRUZANA ERACRUZANA FACULTAD DE CIENCIAS BIOLOGICAS Y AGROPECUARIAS
Transcript of UNIVEUNIVERSIDAD VERACRUZANA ERACRUZANA FACULTAD DE ... · ... del proceso de enraizamiento a...

1
ESTABLECIMIENTO DE UNA
METODOLOGÍA PARA LA
MICROPROPAGACIÓN DE PATRONES
TOLERANTES AL VIRUS DE LA
TRISTEZA DE LOS CÍTRICOS (VTC)
TESIS
QUE PARA OBTENER EL GRADO DE MAESTRO EN CIENCIAS
ESPECIALIDAD: BIOTECNOLOGIA DE PLANTAS
PRESENTA:
PAULA ROJAS MENCIO
CORDOBA, VER. JUNIO DEL 2001
UNIVEUNIVERSIDAD VERACRUZANA ERACRUZANA
FACULTAD DE CIENCIAS BIOLOGICAS Y AGROPECUARIAS

1
INTRODUCCIÓN
Los cítricos constituyen el cultivo de mayor importancia en la fruticultura
mexicana, tanto por el área que se destina a su cultivo en 17 estados de la
República, como por los empleos que genera en la producción, la industria y la
comercialización nacional e internacional (De la Osa, 1998; Gómez et al., 1998).
Actualmente, México ocupa el cuarto lugar como productor de cítricos en
el ámbito mundial de naranjas y primer lugar en la producción de limas ácidas
solamente superado por Brasil, Estados Unidos y China (De la Osa, 1998). La
superficie cultivada con cítricos en México es de 485,000 ha, que producen en
promedio anual 5.6 millones de toneladas de fruta, con una valor estimado de
4,659 millones de pesos (Villarreal, 2001). De la superficie total cultivada, la
naranja ocupa el 69 %, el limón mexicano el 21 %, el limón persa el 3 % y la
mandarina, toronja, otros limones y pomelo ocupan el 7 % (Chagolla, 1990;
Villarreal 2001).
Veracruz con 176,000 ha ocupa el primer lugar en la producción de
cítricos, le sigue San Luis Potosí con 44,000 ha, Tamaulipas con 38,000 ha,
Michoacán con 35,000 ha, Nuevo León y Colima, con 32,000 ha cada uno; en
conjunto estas entidades concentran el 83 % de la superficie cultivada en el país
(Villarreal, 2001). De la superficie cultivada en el estado de Veracruz, alrededor del
85.0 % corresponde a naranjas, el 10.2 % a mandarinas, el 4.5 % a limón persa y
el 0.3 % a toronja (Gómez et al., 1998).
La propagación de este grupo de especies se lleva a cabo casi
exclusivamente mediante injertos sobre patrones de semillas, que en muchos
casos tienen un origen apomíctico. Este tipo de propagación, que en general
requiere una gran superficie y resulta algo más tardado, se encuentra limitado por
la época del año y la obtención de semilla adecuada, además de que puede

2
promover la transmisión de enfermedades, especialmente sistémicas (Chagolla,
1990).
El Virus de la Tristeza de los Cítricos (VTC) constituye hoy en día la
enfermedad viral, de carácter letal, que más daño ha causado a la citricultura
mundial (Orozco, 1996). Desde fines de la década del 30 hasta los últimos años,
esta enfermedad ha ocasionado la muerte de 50 millones de árboles en diversos
países (Orozco, 1996). En particular, el VTC ha destruido la citricultura de
Sudamérica (especialmente en países como Argentina, Brasil y Venezuela) y ha
dañado sensiblemente la citricultura de Estados Unidos, España, e Israel
(Roistacher et al., 1994; RIAC, 1995). En gran parte, estos daños pueden ser
atribuidos al hecho de que más del 90 % de los árboles de los 6 países de mayor
producción y exportación de cítricos en la región de América Latina y el Caribe, se
encuentran injertados sobre patrones de naranjo agrio (Citrus aurantium L.) (RIAC,
1995) el cual, además de ser altamente susceptible al virus de la tristeza de los
cítricos, resulta también muy sensible a la psorosis y a los nemátodos (Curti-Díaz
et al., 1996).
México, por su parte, posee el 95 % del total de sus plantaciones
susceptibles al VTC, ya que están injertados sobre patrones de naranjo agrio o
son de "pie franco" (sin injertar) como los limones "Colima" o "Criollo" (C.
aurantifolia L.), todos ellos extremadamente susceptibles al VTC (Orozco, 1996).
El primer registro de VTC en México fue en plantaciones comerciales en
Tamaulipas en 1983, y posteriormente en viveros del estado de Veracruz en 1986-
87. Durante 1993-95, el Centro Nacional de Referencia en un diagnóstico
fitosanitario de la Dirección General de Sanidad Vegetal de la SAGAR, detectó
plantaciones infectadas con VTC en Nuevo León, Quintana Roo y Veracruz. En
1997 se reporta en Baja California, en 1998 en Colima y Michoacán, en 1999 y
2000 en Quintana Roo, Campeche y Yucatán (Villarreal, 2001).

3
El vector más eficiente de la enfermedad, es el áfido Toxoptera citricidus
Kirkardly (Brunt et al., 1997), elemento que desencadena con su diseminación la
alta mortalidad de plantas. Ha sido reportado en varios países de Centro América,
en migración hacia el Norte, así como en las islas del Caribe incluyendo a Cuba.
Desde noviembre de 1995 se le ha detectado también en el estado de la Florida y
en octubre de 1996 llegó a Belice (De la Osa, 1998).
Como medida de prevención, ante esta seria enfermedad, desde hace
años la mayoría de países citricultores afectados por el VTC ha ido sustituyendo el
naranjo agrio (Citrus aurantium L.), por patrones tolerantes al VTC como son: los
Citranges "troyer y "carrizo" (Citrus sinensis L. Osbeck x Poncirus trifoliata L. Raf.),
la mandarina "Cleopatra" (Citrus reticulata Blanco), la lima "Rangpur" (Citrus
aurantifolia L.) y el limón Volkameriano (Citrus volkameriana Pasq.) (RIAC, 1995;
Medina, 1995; Orozco, 1996). Algunos de estos patrones como los Citranges
"troyer" y "carrizo" (Citrus sinensis L. Osbeck x Poncirus trifoliata L. Raf.) además
de tolerantes al VTC, pertenecen al grupo de patrones cuyo efecto sobre la calidad
del fruto es alto (Padrón, 1991).
Aunque en México se han desarrollado diversas acciones preventivas
contra el VTC (López, 1994; Colli y Cárdenas, 1995; INEGI, 1996), no se cuenta,
en la magnitud de lo necesario, con un programa de certificación de plantas de
cítricos, que permita la producción de patrones tolerantes al VTC, ni de yemas
certificadas de las principales variedades comerciales, de manera que los
productores puedan contar con plantas certificadas como medio de defensa ante
esta enfermedad (Orozco, 1996). Es por ello que la mayoría de los productores de
la región del Golfo siguen empleando el naranjo agrio (Citrus aurantium L.) como
patron, debido fundamentalmente a la poca disponibilidad de semilla y el gran
desconocimiento que tienen sobre el comportamiento de otros patrones en sus
plantaciones; por estas razones, algunos citricultores han optado por plantar
alternadamente un patrón agrio y uno tolerante al VTC, dentro de cada hilera; de
este modo esperan asegurar una supervivencia de al menos un 50 % de sus

4
árboles en caso de que se presenten algunas cepas severas de este virus (Curti-
Díaz et al., 1996).
De acuerdo con López (1994), en la zona norte de Veracruz, donde se
encuentran diferencias notables en suelos y clima, no se puede recomendar el uso
de un solo patrón como sustituto del naranjo agrio, por lo que afirma que la nueva
tecnología exige precisión en la selección de un grupo numeroso de patrones, ya
que cada uno de ellos presenta limitantes a las propiedades extremas físicas y
química del suelo y otras enfermedades.
Cabe resaltar que ante esta situación, desde hace algunos años, se han
estado desarrollando algunas acciones, para lograr un adecuado control y
prevención del VTC, por parte del Comité Estatal de Sanidad Vegetal de Veracruz
(CESVVER) en Coordinación con el Instituto Tecnológico Agropecuario No. 18
(ITA-18) de Ciudad Cardel, Ver., lo que ha conllevado, entre otras, al
establecimiento de un vivero de patrones de cítricos tolerantes al VTC, que se ha
venido caracterizando desde un punto de vista morfoagronómico (Coto y
Rodríguez, 1998; Gómez et al., 1998).
Las investigaciones dirigidas en particular al estudio, selección y
propagación de patrones, que proporcionen las mejores características hortícolas,
con los mejores requerimientos de control de plagas y enfermedades, constituye
uno de los fundamentos necesarios, para cualquier industria citrícola y por ello
cada país, con una citricultura viable, debe, con alta prioridad, adoptar un
programa adecuado de producción de material de propagación (RIAC, 1995).
Por todo lo anterior, la aplicación de las técnicas biotecnológicas con este
propósito puede constituir una vía muy útil para apoyar los programas de
producción y certificación de patrones y yemas tolerantes al VTC.

5
Sobre la base de todo lo antes expuesto, se propuso el desarrollo del
presente trabajo con el objetivo general de establecer las condiciones para la
inducción del proceso morfogenético en segmentos nodales de dos patrones de
cítricos C. troyer y C. carrizo, tolerantes al Virus de la Tristeza.
Como parte de este objetivo general, se propone el desarrollo de diversos
experimentos con vistas a establecer; la técnica de desinfección y termoterapia
más adecuada para la obtención de cultivos asépticos, la técnica para la inducción
de brotes múltiples a partir de segmentos nodales, la combinación de reguladores
de crecimiento más apropiada para la inducción del proceso de enraizamiento a
partir de brotes y el tipo de sustrato más apropiado para la aclimatación de las
vitroplántulas obtenidas a partir de la inducción del proceso morfogenético.

6
REVISIÓN DE LA LITERATURA
2.1. Origen y distribución de los cítricos
Los cítricos son originarios de una vasta región comprendida por la
Conchinchina, el Archipiélago Malayo y partes adyacentes de Asia (Palacios,
1978; Cameron y Soost, 1987). El conocimiento sobre la utilización de sus frutos,
así como el cultivo de los árboles, se extendió desde China e India pasando a
través de Persia y Palestina, hasta conocerse en Africa del Norte y Europa, en las
áreas adyacentes a la cuenca del Mediterráneo (Palacios, 1978; Morín, 1985).
La cidra, el naranjo agrio y el limonero, estuvieron entre las primeras
especies conocidas, y fueron introducidas en Europa alrededor del año 1200
(Loussert, 1992). El naranjo dulce es originario de China y estuvo entre las
primeras especies citrícolas más difundidas al resto del mundo (Palacios, 1978).
Cuando Cristóbal Colón realizó los primeros viajes hacia América, llevó consigo
semillas de naranjo dulce. En esa época los cítricos ya se habían distribuido en los
países de la cuenca del mediterráneo, sobre todo en España, Italia y Grecia
(Loussert, 1992). Los portugueses fueron quiénes introdujeron el naranjo dulce en
Europa desde la India y China, durante los viajes que realizaban a través del Cabo
de Buena Esperanza (Palacios, 1978).
Se presume que el mandarino es originario de China y Conchinchina y
pasó al Japón que es actualmente uno de los países de mayor producción
(Palacios, 1978). En el año 1493, realizando Colón su segundo viaje, introdujo
semillas de agrios a la Isla "La Española" (Santo Domingo) y "La Isabela" (Islas
Bahamas) (Palacios, 1978; Loussert, 1992). Posteriormente se difundió a México y
Cuba en 1517 (Palacios, 1978). Los portugueses en el año 1530, introdujeron el
naranjo en Brasil cuando se lanzan a colonizar el Amazonas (Palacios, 1978; Coto
y Rodríguez, 1998). En el año 1565, se introdujeron los primeros agrios a la
Península de Florida, los Jesuitas llevaron los primeros cítricos a California años

7
después, distribuyéndolos en las zonas adyacentes del Pacífico (Palacios, 1978;
Loussert, 1992). Los Jesuitas difundieron los cultivos de naranjos y limoneros con
material proveniente de Paraguay. Por los años 1750 existían importantes huertos
citrícolas en Paraguay, Brasil, Perú y Argentina (Palacios, 1978; Morín 1985).
2.2. Clasificación botánica
De acuerdo con Swingle y Reece (1967), los patrones de cítricos C. troyer
y C. carrizo se ubican taxonómicamente dentro de la familia Rutaceae y la
subfamilia Aurantioideae y pertenecen a la subtribu Citrinae. De acuerdo con estos
autores existen dentro de esta subtribu 3 géneros que son: Citrus, Fortunella y
Poncirus y dentro del género Citrus existen 10 especies, las que se relacionan en
la Tabla 1.
Tabla 1. Relación de especies del género Citrus de acuerdo con Swingle y Reece
(1967)
No. ESPECIES DE Citrus
1. Citrus sinensis (L.) Osbeck.- Naranjo dulce 2. C. aurantium L.- Naranjo agrio
C. aurantium var. Myrtifolia Ker-Gawl.- Naranja mirtifoliado 3. C. reticulata Blanco.- Mandarinero
C. reticulata var. Austera Swing.- Mandarina agria 4. C. grandis (L.) Osbeck.- Shaddock 5. C. paradisi Macf.- Toronjo 6. C. medica L.- Citrón
C. medica var. Sarcodactylid (Noot.) Swing-Citrón de dedos C. medica var. Ethrog Engl.- Citrón Etrog
7. C. limon (L.) Burn.- Limonero Real 8. C. aurantifolia (Christm.) Swing.- Lima ácida o limonero sutil 9. C. indica Tan.- Naranjo silvestre hindú 10. C. tachibana (Mak.) Tan.- Naranjo Tachibana

8
2.3. Importancia del uso de patrones en cítricos
Uno de los aspectos de mayor importancia para el éxito de una huerta
citrícola, es la correcta selección de los patrones (Curti-Díaz et al., 1996).
Wutscher (1979), ha indicado que los patrones han contribuido quizá más que
ningún otro factor al éxito o fracaso de la industria citrícola en el mundo.
De acuerdo con Castle et al. (1993), el uso de patrones tiene tres
propósitos generales:
Reducir el período juvenil o período improductivo del árbol, mediante el
injerto de yemas maduras provenientes de árboles en producción. Tolerar las condiciones adversas de suelo, clima y agentes patógenos, para
ampliar el área citrícola hacia regiones donde los cítricos normalmente no se desarrollan adecuadamente.
Mejorar el comportamiento frutícola del árbol.
2.3.1. Patrones tolerantes al virus de la tristeza de los cítricos (VTC)
2.3.1.1. Citrange troyer (Poncirus trifoliata (L.) Raf. X Citrus sinensis (L.) Osbeck)
Este Citrange fue obtenido polinizando flores Washington navel (Citrus
sinensis L. Osbeck) con polen de Poncirus trifoliata L. Raf. (Forner-Valero, 1985).
Este patrón tiene muy buena aceptación en casi todas las regiones del mundo
(Padrón, 1991). En lo que a virosis se refiere, es tolerante a la tristeza, psoriasis y
cachexia-xyloporosis, pero es sensible a la exocortis (Padrón, 2001). El patrón es
considerado como resistente a Phytophthora sp. pero sensible a exocortis. Tiene
buena afinidad con variedades de naranjo dulce, mandarina y pomelo, pero es
incompatible con el limón Eureka (Padrón, 1991; 2001). Este patrón induce buena

9
productividad y vigor, precocidad en la producción, frutos de alta calidad, es
sensible a los altos contenidos de caliza activa en el suelo y a la salinidad (Coto y
Rodríguez, 1998; Padrón, 1991; 2001).
2.3.1.2. Citrange carrizo (Poncirus trifoliata (L.) Raf. X Citrus sinensis (L.) Osbeck)
Este Citrange procede vegetativamente del mismo híbrido que originó a
Citrange troyer, por lo que son prácticamente iguales desde el punto de vista
morfológico, solo con pequeñas diferencias respecto a su comportamiento
agronómico (Coto y Rodríguez, 1998; Padrón, 2001). Este híbrido variedad
muestra una mayor tolerancia a la gomosis, al virus de la tristeza, la exocortis, la
cachexia y la agalla de la madera. Resulta por otra parte, tolerante a la pudrición
de la raíz, pero es sensible a Armillaria y a los nemátodos de los cítricos, y
muestra resistencia a Phytophthora. Las variedades injertadas sobre este patrón
son más productivas que sobre C. troyer (Coto y Rodríguez, 1998; Padrón, 2001).
Este patrón presenta una respuesta aceptable al Boro y a los suelos arenosos,
limosos y arcillosos, no así para los cloruros y suelos calcáreos (Ferguson et al.,
1990). En cuanto al clima, es resistente al frío, pero la respuesta a la asfixia
radicular es insatisfactoria (Coto y Rodríguez, 1998).
En relación a sus características de vigor, tamaño y tolerancia a la sequía
posee un comportamiento medio. Agronómicamente tiene un comportamiento
medio para producción y tamaño del fruto y posee un alto porcentaje de zumo,
acidez y sólidos solubles totales (Ferguson et al., 1990; Padrón, 1991).

10
2.4. Virus de la tristeza de los cítricos (VTC)
La tristeza de los cítricos (VTC) es una enfermedad que afecta
prácticamente a todas las especies citrícolas, pero principalmente al naranjo,
toronjo y lima (Agrios, 1998).
2.4.1. Etiología
El virus de la tristeza de los cítricos, es un miembro del género
Closterovirus, de la familia Closteroviridae (Orozco, 1995; Agrios, 1998; Isidrón et
al., 2001). Microscópicamente se ha determinado que son partículas en forma de
"varilla flexible" que mide 2,000 nm. (Orozco,1996; Agrios, 1998). Se han aislado,
de preparaciones purificadas, algunos tipos de ácidos ribonucléicos de cadena
simple con pesos moleculares de 5.4 a 6.5 por 10 y proteínas de cápside, con
pesos de 28,000, 23,000 y 21,000 daltones (Medina, 1995; Agrios, 1998). El virus
se encuentra por lo regular limitado al floema de los árboles afectados aunque
también se ha detectado en la corteza de los tejidos jóvenes (Orozco, 1996; Brunt
et al., 1997).
2.4.2. Epidemiología y transmisión por vectores
El VTC se transmite por áfidos, principalmente Toxoptera citricidus
Kirkaldy (RIAC,1995; Rocha-Peña et al.,1995; Agrios, 1998; Brunt et al., 1997;
SAGAR, 1993; Francis, 2001) y Aphis gossypii Glover (Roistacher y Bar-Joseph,
1987; Yokomi y Damgsteet, 1991). También por injerto de yema, transmisión
mecánica en tallos de hospedantes susceptibles (toronja y limón mexicano)
(Garnsey y Muller, 1988). La transmisión por semilla no ha sido demostrada (Bar-
Joseph y Lee, 1989).

11
El grado de eficiencia en el proceso de la diseminación de la enfermedad
varía con las especies de pulgones, razas del virus y especies hospederas (Isidrón
et al., 2001). La especie Cuscuta sp. puede transmitir la enfermedad (Garnsey y
Lee, 1988; Orozco, 1996).
En 1930 se introdujeron a Brasil y Argentina cítricos provenientes de Africa
del Sur y Asia, los cuales venían infectados por el VTC junto con el T. citricidus,
diseminándose el virus (Roistacher y Moreno, 1991; Roistacher et al., 1991). En
1920-1940, El VTC, se introdujo sin el vector, de los países asiáticos a California,
Florida, Israel y España por importaciones masivas de cítricos. Treinta a cincuenta
años después de efectuadas las importaciones por los países antes mencionados,
se descubrió un cambio en la transmisibilidad de muchos aislados del VTC
severos, eficientemente diseminados por A. gossypii (Wallace, 1978; Moreno et
al., 1983; Roistacher y Bar-Joseph, 1987; Yokomi y Garnsey, 1987; Garnsey y
Lee, 1988; Yokomi y Damgsteet, 1991; Orozco, 1996).
Los áfidos: A. gossypii, A. spiracoela y T. aurantii, son vectores poco
eficientes en la transmisión del VTC (Roistacher y Bar-Joseph, 1987; Roistacher et
al., 1991).
2.4.3. Variabilidad del virus de la tristeza de los cítricos
El VTC ocurre en la naturaleza en aislados o razas, las cuales pueden
variar en gran medida, y la reacción y sintomatología en diversos hospedantes
(Lee y Peña-Rocha, 1992; Isidrón et al., 2001).
Las razas existentes en América del Sur, Sudáfrica, Australia y Asia
producen daños severos en toronjas (C. paradisi L.) y naranja dulce (C. sinensis

12
L.). En Africa y Brasil, existen razas muy virulentas que además de afectar a las
toronjas, afectan a todas las variedades del naranjo dulce y algunos patrones
tolerantes, naranjo trifoliado, lima Rangpur, limón rugoso y mandarina Cleopatra
(Moreno et al., 1983; Garnsey y Lee,1988; Lee et al., 1992; Orozco, 1996).
2.4.4. Sintomatología de la enfermedad
Los síntomas de VTC en árboles afectados dependen de si se presenta en
árboles injertados sobre patrones susceptibles o tolerantes a la enfermedad
(Orozco, 1996).
2.4.4.1. Síntomas producidos en árboles injertados sobre naranjo agrio (C. aurantium L.)
Los síntomas en árboles sobre patrones de naranjo agrio (C. aurantium L.)
producen decaimiento de todas las variedades comerciales de naranja dulce (C.
sinensis), toronja (C. paradisi), mandarina (C. reticulata Blanco). El limon
mexicano (C. aurantium), el italiano (C. limon) y el eureka se han observado que
se infectan menos (INIFAP, 1996; Padrón, 2001). La muerte de los árboles se
debe a la obstrucción de los vasos conductores de la savia elaborada que sirve de
alimento a las raíces (Wallace, 1978; Moreno et al., 1983; Garnsey y Lee, 1988;
Lee y Rocha, 1992; Orozco, 1996). El colapso puede ser rápido con
marchitamiento foliar de color cenizo y muerte repentina en 1 o 2 semanas,
quedando secos por completo y solo persistiendo los frutos (INIFAP, 1996; Reyes
de Acevedo y Palmieri, 1997).
Declinamiento lento. En árboles jóvenes se presenta una abundante
floración y amarre del fruto, el cual puede colorear prematuramente las nervaduras

13
de las hojas; éstas se aclaran y después se amarillan y caen, causando la muerte
de las ramas y el acortamiento de los brotes vegetativos, reduciéndose el tamaño
de la copa (Isidrón et al., 2001). Se produce además, lenta y gradualmente, la
muerte de las raicillas, presentándose un descortezamiento y pudrición en todo el
sistema radical en la unión patrón-injerto; al levantar la corteza (2 a 3 cm de
ancho) con una navaja, se observan abundantes punteaduras en la cara interna
de la corteza de naranjo agrio (C. aurantium L.) conocido como panal de abeja
(Brunt et al., 1997; Isidrón et al., 2001), y en el tronco se presentan prolongaciones
agudas, parecidas a las cerdas cortas y fibrosas de un cepillo (INIFAP,1996;
Reyes de Acevedo y Palmieri, 1997). Los frutos producidos son de tamaño
pequeño que se amarillean prematuramente y muestran un color más pálido que
lo normal.
Árboles sin declinamiento. Árboles que no muestran síntomas de la
enfermedad; algunas veces presentan menor altura, hinchazón arriba de la línea
de injerto y punteaduras en la corteza del patrón. Eso indica que está infectado
con una raza débil del virus; lo anterior es muy importante, primero porque son
focos de infección y segundo porque el comportamiento de estos árboles suele ser
inestable, pudiendo manifestar declinamiento y muerte, transcurrido algún tiempo.
Algunas de estas razas han sido utilizadas para la protección cruzada con el
objeto de reducir la diseminación de razas severas de VTC (Orozco, 1995;
INIFAP, 1996; Reyes de Acevedo y Palmieri, 1997; Isidrón et al., 2001).
2.4.4.2. Síntomas en árboles sobre patrones tolerantes
El síntoma más característico es la presencia de acanaladuras en la
madera ocasionando que las ramas sean muy quebradizas y se rompan con
facilidad con el viento; el follaje puede presentar síntomas de decaimiento y
deficiencias nutricionales, lo que conduce a una disminución en la producción y a
una baja calidad de los frutos (Orozco, 1996).

14
2.4.5. Métodos de detección del VTC
Los métodos de diagnóstico de la enfermedad se han incrementado en las
últimas décadas y va desde la utilización de plantas indicadoras, serología,
técnicas basadas en el análisis de ácidos nucléicos, la utilización de sondas
marcadas, la amplificación por cadena de polimerasa y la combinación de
diferentes técnicas (Francis, 2001).
El VTC está constituido de proteínas y doble cadena de ARN (Guerri et al.,
1990; Brunt et al., 1997). La detección se realiza por métodos electroforéticos,
conociéndose 7 tipos de virus que causan la enfermedad en base a la posición de
bandas de ARN (Moreno et al., 1983). Entre los métodos de detección del VTC
que más se han empleado se encuentra la prueba de ELISA, que detecta la
presencia de la enfermedad en plantas, aún cuando no se manifiestan sus
síntomas (SAGAR, 1997b; Garnsey et al., 1993). Otro método consiste en el uso
de los anticuerpos monoclonales (ACM), y se emplea como herramienta para
conocer la diversidad de epítopes existentes en la cubierta proteica de los aislados
de VTC. Estos estudios permiten establecer serogrupos de aislados teniendo en
cuenta su reacción frente a diferentes anticuerpos monoclonales (Gutiérrez, 1996).
2.4.6. Control legal
a) . Norma Oficial Mexicana. La operatividad de la campaña fitosanitaria se
establece en el proyecto de Norma oficial mexicana NOM-031-FITO-1995, que
tiene por objeto establecer las medidas fitosanitarias que deben aplicarse para
prevenir, combatir, controlar y/o erradicar al VTC en las diferentes zonas citrícolas
(SAGAR, 1997a).

15
b) . Dispositivo Nacional de Emergencia. Con fundamento en el artículo 46
de la Ley Federal de Sanidad Vegetal se establecerá el dispositivo nacional de
emergencia cuando se detecte la presencia de plagas que pongan en situación de
emergencia a una o varias especies vegetales en todo o en parte del territorio
nacional; en caso de requerirse es factible el dispositivo nacional de emergencia
para la destrucción de brotes del virus de la tristeza de los cítricos (SAGAR,
1997a).
c) . Regulación cuarentenaria
El objetivo de esta medida es evitar la entrada ilegal de material vegetativo
procedente de áreas donde el VTC está presente (Garnsey y Lee, 1988; Moreno
et. al., 1983; Orozco, 1996).
c.1. Nacional. Debido a la presencia de VTC en el estado de Veracruz,
catalogado como el principal estado proveedor de material propagativo, y
considerando la potencial existencia de VTC y su posible diseminación a otras
entidades, se establece como requisito fitosanitario que la movilización de material
propagativo (plantas) debe hacerse únicamente mediante el certificado
fitosanitario para la movilización nacional, previo diagnóstico fitosanitario negativo
por un laboratorio aprobado (SAGAR, 1997a).
c.2. Internacional. Un alto riesgo de introducción del VTC y su vector,
representa la importación directa de germoplasma de cítricos procedentes de
áreas donde el VTC es endémico, o de aquellos países que han sufrido la
introducción del patógeno a su territorio (SAGAR, 1997a). Se regula la importación
de plantas, yemas o cualquier material propagativo de cítricos de los países
afectados por el virus de la tristeza, a excepción de aquellos donde se compruebe
mediante verificación, en origen, por personal aprobado o acreditado de la
Secretaria de Agricultura, que los productos proceden de áreas libres del patógeno

16
(SAGAR, 1997a). Se permite la introducción de material propagativo de plántulas
in vitro, previo cumplimiento de los requisitos fitosanitarios establecidos por la
Secretaria y certificado por el país de origen (SAGAR, 1997a).
2.4.6.1. Programa de certificación del material libre de virus
La amenaza que representa para toda la citricultura de México el VTC y su
vector, hace a la campaña contra el VTC como una prioridad dentro de la
fitosanidad nacional, por lo que es imperativo establecer medidas fitosanitarias
extremas para prevenir una situación de daño casi irreversible para la citricultura
nacional, siendo el eje principal el establecimiento de centros de producción de
material propagativo de cítricos certificados o viveros certificados, que
proporcionen material propagativo tolerante certificado y de calidad varietal para la
sustitución y establecimiento de nuevas plantaciones (SAGAR, 1997a).
2.4.6.2. Erradicación
Esta medida conlleva la eliminación total de los árboles enfermos en un
área determinada, que pongan en riesgo la citricultura nacional (SAGAR, 1997a).
2.4.6.3. Uso de patrones tolerantes a VTC
La utilización de patrones tolerantes como medida de combate es una
práctica necesaria, sobre todo en países donde existen razas altamente virulentas,
capaces de causar daños directos en variedades infectadas sobre los patrones
(Orozco ,1996). La necesidad de utilizar patrones tolerantes al VTC, con
cualidades deseables, ha propiciado la evaluación de cultivares, híbridos y
parientes de los cítricos, originando con ello información sobre el comportamiento

17
de los cítricos injertados sobre muchos patrones diferentes. Los principales
criterios de selección son: tolerancia a enfermedades y nemátodos, tolerancia a la
salinidad y tipo de suelo, y efecto del patrón sobre el árbol (compatibilidad)
(Padrón, 2001).
Actualmente, en México se utilizan los siguientes patrones tolerantes a
VTC: mandarina Cleopatra, mandarina Sunki, mandarina Sun chu sha, naranja
trifoliada, citranges (troyer y carrizo), citrumelos swingle, limón rugoso, C.
volkameriana y lima Rangpur (Padrón, 2001).
2.4.6.4. Protección cruzada
El método de protección cruzada consiste en infectar la planta con una
variante atenuada de un virus, con vistas a desarrollar una sintomatología menos
severa de la enfermedad, al ser infectadas por una variante más severa del mismo
virus (Gonsalves y Garnsey, 1989; Gutiérrez, 1996). Este método, sin embargo,
puede generar algunas dificultades, dado entre otras, a que la variante atenuada
puede mutar y convertirse en una variante severa; por otra parte, puede existir
sinergismo entre la variante protectora (atenuada) y la variante severa, o con otro
virus, ocasionando una enfermedad más severa. Otro problema que reviste este
método es la dificultad que conlleva la identificación y selección de una variante
atenuada adecuada, ya que puede resultar un proceso muy largo. Incluso la
variante atenuada seleccionada en una área o región determinada puede
comportarse como variante severa en otra área. La resistencia observada durante
la protección cruzada puede deberse a la interacción especifica entre uno o más
genes virales de la variante protectora y uno o más genes de la variante atacante,
resultando en la reducción y/o inhibición de la multiplicación de la variante
atacante (Fitchen y Beachy, 1993; Gutiérrez, 1996). Esta medida ha sido
implementada en Brasil (Muller et al., 1988), Africa del Sur (Garnsey y Lee, 1988)
y Australia (Broadbent et al., 1991). En estos casos, la medida más efectiva para
combatir el VTC es la protección de las diferentes especies de cítricos con cepas

18
benignas. En Brasil este método se ha utilizado para proteger los cítricos de razas
de virus que ocasionan el picado del tallo (Orozco, 1996).
2.5. Embrionía sexual y nucelar
La mayoría de las especies cítricas presentan poliembrionía, o sea, que
una sola semilla puede tener dos o más embriones; sin embargo, sólo uno de
éstos proviene de la fusión de los gametos femenino y masculino dentro del óvulo;
es decir, en una semilla poliembriónica solo un embrión es de origen sexual y los
restantes se originan de las células somáticas del nucelo de la planta madre y se
extienden fuera del saco embrionario (Palacios, 1978). Salvo la posibilidad de
mutaciones, las plántulas provenientes de embriones nucelares, mantienen una
constitución genética idéntica a la planta madre y se caracterizan por su vigor y
uniformidad (Palacios, 1978).
Las plantas originadas por embriones sexuales o gaméticos, se distinguen
fácilmente de las nucelares por débil crecimiento, diferencias morfológicas y de
desarrollo, y es frecuente que el embrión nucelar domine al sexual y no lo deje
desarrollar (Palacios, 1978). Por eso en algunas especies y variedades, las
semillas producen casi exclusivamente plantas nucelares, lo cual indica que el
embrión sexual estuvo obstaculizado o fue demasiado débil para vivir (Cameron y
Soost, 1987).
Existen algunas especies cítricas monoembriónicas entre las que se
encuentra la toronja (Citrus grandis Osbeck), los mandarinos (Citrus reticulata
Blanco) y las cidras (C. medica L.) (Palacios, 1978).

19
2.6. Métodos de propagación de los cítricos 2.6.1. Métodos convencionales de propagación 2.6.1.1. Propagación por semilla
Los cítricos se pueden reproducir por semilla, sin embargo este método de
propagación presenta diversas desventajas, entre las que se pueden incluir:
juvenilidad, producción errática, gran tamaño y susceptibilidad a la pudrición de pie
por Phytophthora.
Esta enfermedad fue el factor que influyó en gran medida para que se
optara por los árboles injertados, en vez de árboles por semilla (Castle, 1987), por
lo que actualmente los cultivares comerciales de cítricos se propagan por injerto
de yema, sobre patrones, siendo las fuentes preferidas para estos últimos los de
origen nucelar (Cameron y Soost, 1987). Es por ello que la poliembrionía en los
cítricos y sus géneros afines Poncirus y Fortunella, es de gran importancia en los
trabajos de micropropagación de microinjertos, pues aún multiplicando por semilla,
la embrionía nucelar permite obtener plantas muy uniformes con todas las
características de la planta madre (Coto y Rodríguez, 1998).
2.6.1.2. Propagación por injerto
El injerto es sin duda alguna el procedimiento tradicional más utilizado en
la propagación de los cítricos, pues es el que ofrece las mayores ventajas sobre
los otros métodos de reproducción (Padrón, 2001). La técnica consiste en
conseguir la unión íntima de dos especies diferentes, de tal manera que ambas se
sueldan, permanezcan unidas y continúen su vida de esa manera, formando una
especie de simbiosis. Una de las especies, utilizada como patrón contribuye con
su sistema radical. Este debe provenir de plantas nucelares; la otra especie, el
cultivar o variedad, dará lugar a la parte aérea, pudiendo haberse derivado de una
yema simple o de una vareta o púa, que provenga de fuentes sanas y certificadas

20
(Padrón, 2001). La unión íntima de ambas partes solo se puede llevar a cabo
cuando el contacto se realice entre el cambium de una con el cambium de la otra.
Es decir, para que puedan soldarse las dos especies es necesario poner en
contacto estrecho sus meristemos secundarios. Estas dos partes llegan a
constituir una completa unidad en su fisiología y en su funcionamiento total, como
si se tratara de un solo individuo, pero permaneciendo invariable su constitución
genética de cada una de ellas. Por ello, la parte aérea producirá frutos
correspondientes a un tipo de variedad o clon especial al que pertenezca y toda su
morfología y características intrínsecas permanecerán invariables, salvo algunas
influencias que sobre ella determine el patrón (Calderón, 1993).
En los cítricos se pueden aplicar varias técnicas de injerto, siendo el injerto
de yema o escudete el de uso más frecuente porque se puede obtener un número
máximo de planta a partir de una cantidad limitada de varetas por yemas
(Palacios, 1978). Los injertos de púa se pueden emplear también como los del
enchapado lateral, en aquellos casos en los que las varetas y los patrones son
demasiado pequeños para utilizar yemas (Cameron y Soost, 1987).
Los patrones de cítricos se deben injertar cuando alcancen una edad de
12 a 15 meses, o alcancen una altura de cuando menos 45 cm. Cuando presenten
cualquiera de estas condiciones deben prepararse para el injerto; éste se realiza
removiendo las hojas y espinas a una altura de 45 cm una semana antes de ser
injertados (Opeke, 1982). Por otra parte, las yemas que se empleen en el injerto
de los patrones, se deben colectar en la mañana del mismo día y deben ser
utilizados lo más pronto posible (Opeke, 1982).
Actualmente existe un gran número de patrones y variedades comerciales
para la propagación de los cítricos, pero la elección de las combinaciones para los
injertos deben efectuarse con sumo cuidado pues es una decisión clave para el
éxito de cualquier explotación citrícola, ya que algunas combinaciones patrón-

21
injerto, no son compatibles y las plantas injertadas pueden declinar e inclusive
morir (Castle, 1988). A los injertos se les atribuyen algunas desventajas, tales
como menor vida comercial y un menor desarrollo de las plantas, comparadas a
las obtenidas por semilla. Sin embargo, varios especialistas, entre ellos Padrón
(1991), coinciden en que en la propagación por injerto los problemas que se
presentan deben ser atribuidos a una mala selección del patrón, el cual puede ser
poco vigoroso, mal adaptado al suelo o tener una compatibilidad deficiente.
2.6.2. Métodos biotecnológicos de propagación
2.6.2.1. Microinjertación
Esta técnica desarrollada primeramente por Murashige et al. (1972) y
puesta en práctica por Navarro et al. (1975) y Francis (2001), permite la obtención
de plantas libres de virus, mediante la inducción al desarrollo de un explante de
meristemo apical, sobre el epicotilo decapitado de una plántula originada a partir
de una semilla, en un medio apropiado y bajo condiciones asépticas. De acuerdo
con Navarro et al. (1975) es posible lograr entre un 30% - 50% de éxito en la
aplicación de esta técnica y en algunos casos se ha reportado hasta un 90%
(Chen et al., 1992). La técnica de microinjerto posee además la ventaja de que
permite la eliminación no solo del virus de la tristeza sino también de otros virus
como la psorosis y la exocortis (Monteverde et al., 1999; González et al., 1980),
como indicaran Gravina y Piestum (1991), quienes mencionan que la no detección
de virus en 400 plantas de distintas variedades de Citrus obtenidas por
microinjerto y cuya fuente original se encontraba afectada por uno o más de los
virus de psorosis, amarillamiento, concavidad gomosa, hoja rasgada, infección
jaspeada, xyloporosis, exocortis y Spiroplasma citri, entre otros, demuestra la alta
eficiencia de esta técnica para la eliminación de distintas razas de estos virus. Por
otra parte, se ha indicado (Navarro et al., 1975) que los microinjertos obtenidos no
muestran características de juvenilidad, lo que les permite incluso florecer y

22
fructificar en menos de un año. Es por ello que esta técnica se ha utilizado en
muchos países cítrícolas del mundo para obtener yemas libres de virus, con vistas
a su uso en la propagación comercial de cítricos (González et al.,1977;
Starrantino, 1992; Monteverde et al., 1999; Gravina y Piestum, 1991; Paiva et al.,
1993; Zarei y Rahimian, 1997; Mukhopadhyay et al., 1997; Parthasarathy et al.,
1997).
La frecuencia de éxito de los microinjertos se incrementa con el tamaño
del explante meristemático, pero disminuye el porcentaje de plantas libres de virus
(Navarro et al., 1975), aunque muchas veces esta relación depende del patógeno
(Litz et al.,1985). Por ello, se considera óptimo, para lograr un buen equilibrio entre
el porcentaje de microinjertos prendidos y la eliminación de virus , el empleo de
explantes meristemáticos cuyos tamaños oscilan entre 0.12-0.18 mm y que
posean de 2 a 4 primordios foliares (Navarro et al.,1975; Gravina y Piestum,1991).
En México, como apuntara Chagolla (1990), se han logrado obtener plantas de
cítricos libres de virus, siguiendo la metodología propuesta por Navarro et al.
(1975) gracias entre otros, a los trabajos desarrollados en el Campo Experimental
de "General Terán" del INIFAP.
2.6.2.2. Embriogénesis somática
De igual forma, se ha logrado inducir en cítricos el proceso de
embriogénesis somática especialmente en aquellas especies poliembriónicas
(Kunitake et al., 1991; Gill et al., 1994; Gill et al., 1995; Belkoura et al., 1995). Sin
embargo, los callos embriogénicos producidos, pueden presentar cierta
inestabilidad genética, que no los hacen muy apropiados para su uso como
método de propagación (Pierik, 1990).

23
2.6.2.2.1. Inducción de embriones nucelares
Los primeros trabajos de propagación in vitro en cítricos se realizaron
desde la década de los sesenta con la inducción de embriones nucelares en tres
especies monoembriónicas de cítricos (Rangan et al., 1968). Desde entonces se
han venido desarrollando numerosos trabajos para la propagación de cítricos a
partir de tejidos nucelares de óvulos fertilizados (Rangan et al., 1969; Chaturvedi y
Mitra, 1974) y no fertilizados (Button y Borman, 1971).
Sin embargo, los métodos antes mencionados son tediosos y poseen
entre otros inconvenientes el que solamente se puedan disponer de óvulos que se
encuentren en un estadio apropiado de desarrollo en un corto período del año,
además de que las plántulas obtenidas por esta vía muestran características de
juvenilidad (Chaturvedi y Mitra, 1974). Estas características se manifiestan en
forma de árboles muy altos y espinosos, más lentos para entrar en fructificación,
producción alternante más marcada, un mayor grosor de la corteza del fruto y de
la pulpa (Monteverde et al., 1999).
2.6.2.3. Organogénesis directa
También ha sido muy empleada la regeneración de plantas de cítricos,
principalmente por la vía de la organogénesis directa (Litz et al.,1985). Esta
técnica ha sido descrita en diversos tejidos como raíces (Sauton et al., 1982;
Starrantino y Caruso, 1986; Raman et al., 1992) secciones de hojas (Chaturvedi y
Mitra,1974) y secciones de tallo (Moore,1986; Raman et al., 1992 ; Mas et al.,
1994; Harada y Murai, 1996; Thirumalai y Thamburaj, 1996; Pérez Molphe-Balch y
Ochoa- Alejo, 1997). Dentro de éstas se han logrado muy buenos resultados con
el empleo en particular de explantes a partir de segmentos de tallo (Can et al.,
1992; Pérez-Molphe- Balch y Ochoa-Alejo,1997).

24
Algunos autores (Rangaswamy, 1961; Murashige y Tucker, 1969),
consideran que las condiciones de cultivo más adecuadas para la inducción de la
organogénesis en cítricos, implican mantener los cultivos a una intensidad
lumínica de 3,000 luxes por un período diario de 12 a 16 horas y a una
temperatura de 25 a 30° C. Los mismos autores coinciden en señalar que la
sacarosa satisface los requisitos en la mayoría de los tejidos de cítricos si se
emplea en una concentración de 5% (p/v).
De forma general, el análisis de los resultados sobre cultivo in vitro en
cítricos muestran diferencias en las respuestas de los explantes aún usando
técnicas similares, dada la influencia entre otros de factores como son: tipo de
tejido, preparación de los mismos, composición química del medio de cultivo y
condiciones de cultivo (Espinoza,1994).
2.6.2.3.1. Cultivo de meristemos
El cultivo de meristemos, que ha resultado exitoso en varias especies
vegetales para limpiar materiales infectados por virus, no siempre ha dado buenos
resultados en cítricos (Litz et al., 1985 ; Gravina y Piestum, 1991). Esta técnica es
la que constituye la ruta más fácil y la que menos cambios genéticos provoca ya
que no requiere de la iniciación y diferenciación de órganos adventicios, sino
solamente desarrollar los puntos de crecimiento presentes en las yemas. Esta vía
ha sido empleada, entre otros, por Mosella y Ascui (1985), Lukman et al. (1990),
Omura y Hidaka (1992) y Baruah et al. (1996). Hoy en día, como indicara Baldwin
(1993), el cultivo de meristemos se ha adaptado muy bien para la producción
comercial de yemas de cítricos libres de virus, fundamentalmente en Florida.
Lukman et al. (1990) obtuvieron mejores resultados en el cultivo de
meristemos de Citrange troyer empleando primeramente medio MS líquido
(Murashige y Skoog, 1962) suplementado con 1 mg.l-l de ácido giberélico (AG3) y

25
0.8 mg .l-1 de benciladenina (BA) y luego medio MS sólido suplementado con 0.01
mg.l-1 de ácido naftalenacético (ANA), 0.5 mg.l-1 de BA y 1.0 mg.l-1 de AG3.
Singh et al. (1994) lograron inducir un mayor número de brotes (alrededor
de 6 brotes por yema) a partir de cultivo de ápices de 5 a 6 mm de longitud
provenientes de plantas maduras de Citrus reticulata Blanco cv. mandarina "Khasi"
y C. limon Burm f. cv. "limón Asam" en medio MS, suplementado con BAP (6-
bencilaminopurina), ANA y cinetina (6- furfurilaminopurina). Estos autores
constataron la importancia del empleo de una combinación de ANA, AIB (ácido
indolbutírico) y BAP para lograr un mayor éxito en la formación de brotes múltiples
y durante el proceso de enraizamiento, y recomendaron el empleo del medio MS
suplementado con 1 mg.l-1 de BA, 0.5 mg.l-1 de cinetina y 0.5 mg.l-1 de ANA para
lograr una tasa más elevada de proliferación de brotes en ambas especies.
2.6.2.3.2. Cultivo de segmentos nodales
Con este nombre se le conoce al aislamiento de una yema junto con una
porción de tallo, para obtener un vástago o brote a partir de la yema. Este es el
método más natural de propagación vegetativa de las plantas in vitro, ya que
también puede aplicarse in vivo. Cada una de las yemas que se encuentran en las
axilas de las hojas idénticas a las del ápice del tallo, pueden ser cultivadas en un
medio nutritivo, intentándose así su desarrollo in vitro, realizándose los repicados
cuando son necesarios (López y Avitia, 1991). Cuando se obtiene un número
suficientemente grande de vástagos o brotes, éstos son enraizados y finalmente
se realiza la transferencia al suelo (López y Avitia, 1991). El aislamiento de yemas
y ápices del vástago, es una técnica donde en principio no se añaden citocininas
para romper la dominancia apical (Pierik, 1990). En algunos casos, las giberelinas
y las citocininas pueden estimular la ruptura de la dormancia de las yemas. La
etiolación (causada por la oscuridad o la baja irradiancia) puede producir la ruptura
de la dormancia de las yemas (Pierik, 1990). Esta técnica no es recomendable

26
cuando existen infecciones internas. La velocidad de propagación depende del
número de yemas disponibles y de la tasa de multiplicación in vitro (Pierik, 1990).
2.7. Importancia del cultivo in vitro en cítricos
El empleo de las técnicas biotecnológicas como la micropropagación, ya
sea a través del cultivo de meristemos y de microinjertos, en combinación o no con
las técnicas de termoterapia, así como la inducción del proceso de embriogénesis
somática, constituyen en la actualidad alternativas muy prometedoras para
obtener plantas de cítricos libres de virus (Duran-Vila y Marín, 1991; Iglesias et al.,
2000; Francis, 2001). Es por ello que en los últimos años se ha puesto un mayor
énfasis en el empleo de las técnicas de cultivo de células somáticas y otras
técnicas moleculares en el mejoramiento genético de los cítricos (Gill et al.,1995).
En particular en cítricos, los métodos de multiplicación aséptica presentan
numerosas ventajas con respecto a los métodos tradicionales de propagación,
debido entre otras, a que permiten obtener altas tasas de multiplicación en corto
tiempo, empleando espacios reducidos y sin las limitaciones impuestas por la
época del año, además de que posibilitan la eliminación de patógenos no
obligados y promueven la liberación de patógenos sistémicos (Chagolla,1990).
Asimismo, como indicaran Mas et al. (1991), mediante esta vía se facilita el
transporte e introducción de material vegetativo, sin el riesgo de transmisión de
enfermedades.

27
MATERIALES Y MÉTODOS
3.1. Localización
La presente investigación se realizó en el Laboratorio de Cultivo de
Tejidos del Instituto de Genética Forestal de la Universidad Veracruzana, ubicado
en el Parque Ecológico "El Haya", en el kilómetro 1.5 carretera vieja Xalapa-
Coatepec, Ver., durante el período comprendido entre Septiembre de 1998 al mes
de Marzo del 2001.
3.2. Material Vegetal
Para el desarrollo del presente trabajo se emplearon semillas importadas
de Citrange troyer (C. troyer) y Citrange carrizo (C. carrizo) (Citrus sinensis
Osbeck X Poncirus trifoliata Raf.) provenientes de Willits & New Comb, Inc. Arvin
California, Estados Unidos (ANEXO 1).
3.2.1. Medios de Cultivo
El medio base de cultivo utilizado para la presente investigación fue el de
Murashige y Skoog (1962) y las vitaminas de Gamborg (ANEXO 2) conteniendo
6 g.l-1 de agar y 30 g.l-1 de sacarosa. El medio se ajustó a un pH de 5.7 ± 0.1 con
una solución de NaOH 0.1 N. En todos los experimentos in vitro se utilizaron tubos
de ensaye de 120 X 25 mm; a cada tubo se le añadió una alícuota de 25 ml de
medio. Los tubos fueron sellados con papel aluminio y Kleenpack*. La
esterilización se efectuó en una autoclave a 121° C y a una presión de vapor de

28
1.05 Kg.cm-2 durante 15 minutos. Los medios esterilizados y gelificados a
temperatura ambiente fueron mantenidos en área aséptica hasta su siembra.
3.2.2. Condiciones de incubación
Las semillas se cultivaron en completa oscuridad, a 27 ± 1° C. Para la
inducción de brotes y enraizamiento de las partes aéreas, se mantuvieron a 25 ±
2° C con una intensidad lumínica de 1,200 lux, utilizando lámparas Osram*
Daylight. Los tubos de ensaye fueron sellados con papel aluminio y Kleenpack * a
fin de evitar la contaminación exógena, y posteriormente se colocaron en gradillas
sobre anaqueles, a una distancia de 50 cm de las lámparas de luz blanca.
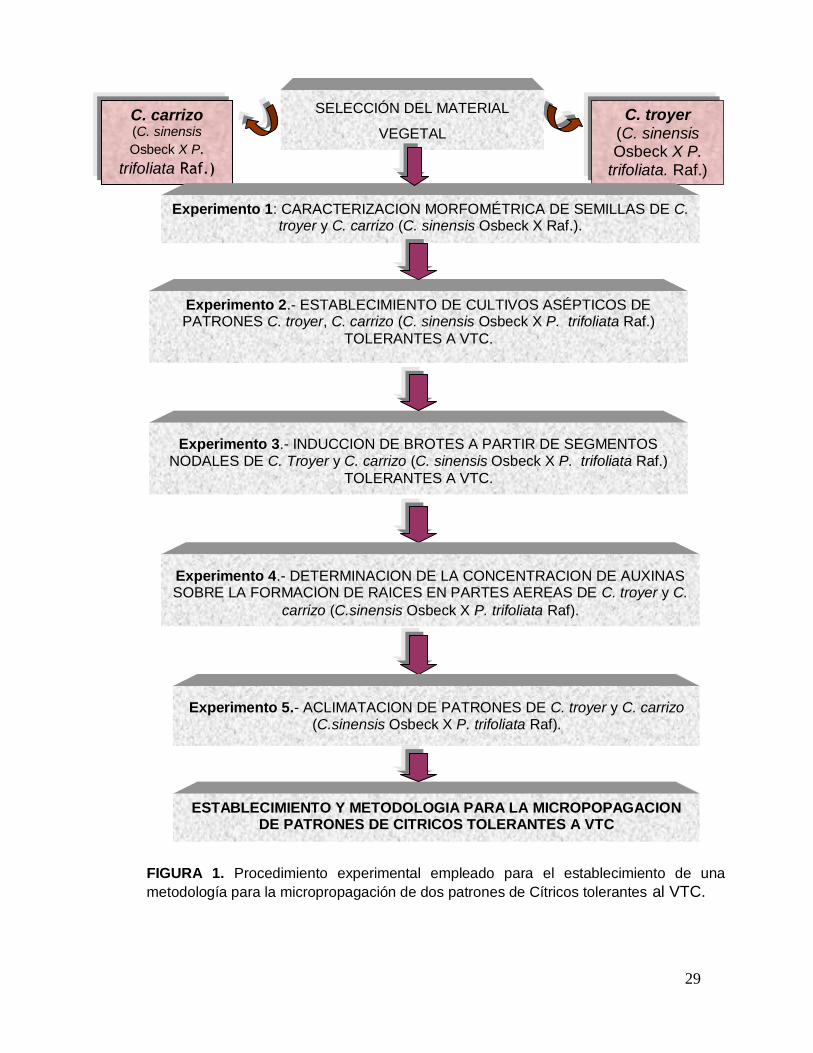
3.3. Procedimiento experimental
Para el desarrollo del presente estudio fue seguido el esquema de trabajo
que se muestra en la Figura 1.
29
C. carrizo (C. sinensis
Osbeck X P.
trifoliata Raf.)
C. troyer (C. sinensis Osbeck X P.
trifoliata. Raf.)
Experimento 2.- ESTABLECIMIENTO DE CULTIVOS ASÉPTICOS DE PATRONES C. troyer, C. carrizo (C. sinensis Osbeck X P. trifoliata Raf.)
TOLERANTES A VTC.
Experimento 3.- INDUCCION DE BROTES A PARTIR DE SEGMENTOS NODALES DE C. Troyer y C. carrizo (C. sinensis Osbeck X P. trifoliata Raf.)
TOLERANTES A VTC.
ESTABLECIMIENTO Y METODOLOGIA PARA LA MICROPOPAGACION DE PATRONES DE CITRICOS TOLERANTES A VTC
Experimento 1: CARACTERIZACION MORFOMÉTRICA DE SEMILLAS DE C. troyer y C. carrizo (C. sinensis Osbeck X Raf.).
SELECCIÓN DEL MATERIAL
VEGETAL
Experimento 4.- DETERMINACION DE LA CONCENTRACION DE AUXINAS SOBRE LA FORMACION DE RAICES EN PARTES AEREAS DE C. troyer y C.
carrizo (C.sinensis Osbeck X P. trifoliata Raf).
Experimento 5.- ACLIMATACION DE PATRONES DE C. troyer y C. carrizo (C.sinensis Osbeck X P. trifoliata Raf).
FIGURA 1. Procedimiento experimental empleado para el establecimiento de una
metodología para la micropropagación de dos patrones de Cítricos tolerantes al VTC.

30
3.3.1. Experimento 1. Caracterización morfométrica de semillas de C. troyer y C. carrizo (C. sinensis Osbeck X P. trifoliata Raf.)
Para la caracterización morfométrica de la semilla de los dos patrones en
estudio se tomó una muestra aleatoria de 30 semillas, a fin de evaluar los
caracteres: largo, ancho, grosor, relación largo/ancho y peso de la semilla. Las
mediciones morfométricas fueron realizadas con una regla vernier. Se utilizó
película Kodak 35 mm, ASA 200 para la fotografía de las semillas de los dos
patrones en estudio.
Los datos se procesaron estadísticamente por medio de un análisis de
varianza (ANOVA) de clasificación simple de efectos fijos, mediante el programa
STATISTICA (Versión 4.5). Se calcularon los valores de Coeficiente de Variación
(CV) y Factor de Variación (FV). Este último se determinó mediante la metodología
propuesta por el CIAT (1985). Se empleó el Programa Excel (versión 97) para la
presentación gráfica de los resultados obtenidos.
3.3.2. Experimento 2. Establecimiento de cultivos asépticos de los patrones C. troyer y C. carrizo (C. sinensis Osbeck x P. trifoliata Raf.) tolerantes al VTC
3.3.2.1. Desinfección y termoterapia de la semilla
La predesinfección de la semilla se llevó a cabo con abundante agua y
jabón; enjuagándose por tres veces con agua destilada. Para la desinfección de
las semillas de los patrones fue empleada la metodología propuesta por Navarro
et al. (1975) en donde se aplicaron diferentes tratamientos con Benomyl (Metil 1-
butil carbamoil) -2- bencimidazol carbamato) (González, 1988), conocido
comercialmente como Benlate* (1 g.l-1) y Clorox* (Tabla 2), y tres enjuagues
posteriores con agua destilada estéril. Por último, fueron sometidas a diferentes

31
tratamientos de temperatura, según lo recomendado por Gravina y Piestum
(1991), Calavan et al. (1972), y Roistacher y Calavan (1972).
Tabla 2. Tratamientos de desinfección y termoterapia de las semillas de patrones de C. troyer y C. carrizo
TRATAMIENTOS BENLATE*
(1g.l-1)/TIEMPO (min)
HIPOCLORITO DE SODIO (%)/TIEMPO
(min)
TEMPERATURA (° C)/TIEMPO (min)
1 20 min. 25 / 10 52-55 ° C / 10 2 20 min. 25 / 10. 52-55 ° C / 05 3 20 min. 30 / 05. 52-55 ° C / 10 4 20 min. 30 / 05 52-55 ° C / 05 5 25 min. 25 / 10 52-55 ° C / 10 6 25 min. 25 / 10 52-55 ° C / 05 7 25 min. 30 / 05 52-55 ° C / 10 8 25 min. 30 / 05 52-55 ° C / 05
3.3.2.2. Siembra de semilla
Las semillas se manejaron bajo condiciones asépticas, en cámara de flujo
laminar. Realizada la desinfección y la termoterapia, se extirpó el tegumento a las
semillas con una pinza estéril y bisturí, bajo el microscopio estereoscópico con
aumento de 10X. Estas se sembraron bajo un diseño completamente aleatorizado,
con 5 repeticiones por tratamiento, en medio MS al 50%, adicionando 6 g.l -1 de
Agar. A cada tubo de ensaye, de 120 X 25 mm, se le añadió una alícuota de 25 ml
de medio, ajustado a un pH de 5.7 ±. 0.1 con una solución de NaOH 0.1N .
Finalmente los cultivos se incubaron bajo completa oscuridad, a 27 ± 1° C,
por 15 días, según lo recomendado por Gravina y Piestum (1991).
A las 72 horas de incubación, se evaluó visualmente la contaminación
bacteriana y fúngica, manifestadas por turbidez y/o presencia de hifas en el medio

32
de cultivo. Se contabilizó el número de semillas contaminadas por tratamiento, así
como el número de semillas germinadas, a los 15 días posteriores a la siembra.
Para efectuar la prueba serológica de ELISA, para el diagnóstico de VTC, se
enviaron 5 muestras de plantas de ambos patrones al Laboratorio de Alta
Tecnología de Xalapa, A.C. (LATEX).
Se empleó un diseño experimental con arreglo factorial de 2 X 8 (2
patrones X 8 tratamientos) y gráficas de barras del programa Excel (versión 97)
para la presentación de los resultados. Se utilizó película Kodak* 35 mm, ASA
200, para la fotografía de los mismos.
3.3.3. Experimento 3. Inducción de brotes a partir de segmentos nodales de C. troyer y C. carrizo (C. sinensis Osbeck X P. trifoliata Raf.) tolerantes al VTC.
3.3.3.1. Efecto de diferentes combinaciones hormonales en la inducción de brotes a partir de segmentos nodales utilizando BAP, ANA y AG3
A fin de conocer el efecto de diferentes reguladores de crecimiento sobre
el proceso morfogenético en cítricos, se sembraron bajo condiciones asépticas, en
campana de flujo laminar, secciones de segmentos nodales de aproximadamente
2 cm de longitud de los 2 patrones en estudio (Pierik, 1990). Estas se sembraron
bajo un diseño completamente aleatorizado con tres repeticiones por tratamiento
hormonal aplicados en tubos de 120 X 25 mm, con alícuotas de 20 ml de medios
de MS, conteniendo las combinaciones hormonales que se indican en la Tabla 3,
así como las vitaminas de Gamborg, 6 g.l-1 de agar, 30 g.l-1 de sacarosa. En todos
los casos se ajustó el medio a un pH de 5.7 ± 0.1 con una solución de NaOH 0.1
N.
33
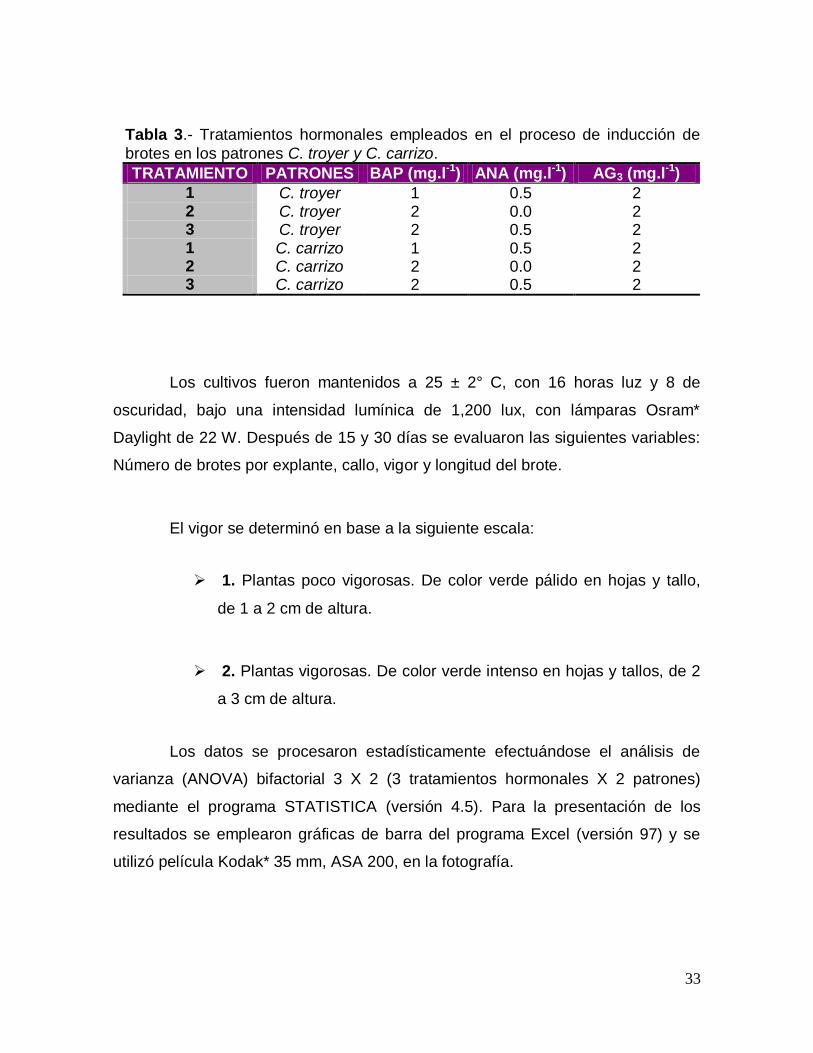
Tabla 3.- Tratamientos hormonales empleados en el proceso de inducción de
brotes en los patrones C. troyer y C. carrizo.
TRATAMIENTO PATRONES BAP (mg.l-1) ANA (mg.l-1) AG3 (mg.l-1)
1 C. troyer 1 0.5 2 2 C. troyer 2 0.0 2 3 C. troyer 2 0.5 2 1 C. carrizo 1 0.5 2 2 C. carrizo 2 0.0 2 3 C. carrizo 2 0.5 2
Los cultivos fueron mantenidos a 25 ± 2° C, con 16 horas luz y 8 de
oscuridad, bajo una intensidad lumínica de 1,200 lux, con lámparas Osram*
Daylight de 22 W. Después de 15 y 30 días se evaluaron las siguientes variables:
Número de brotes por explante, callo, vigor y longitud del brote.
El vigor se determinó en base a la siguiente escala:
1. Plantas poco vigorosas. De color verde pálido en hojas y tallo,
de 1 a 2 cm de altura.
2. Plantas vigorosas. De color verde intenso en hojas y tallos, de 2
a 3 cm de altura.
Los datos se procesaron estadísticamente efectuándose el análisis de
varianza (ANOVA) bifactorial 3 X 2 (3 tratamientos hormonales X 2 patrones)
mediante el programa STATISTICA (versión 4.5). Para la presentación de los
resultados se emplearon gráficas de barra del programa Excel (versión 97) y se
utilizó película Kodak* 35 mm, ASA 200, en la fotografía.

34
3.3.3.2. Evaluación de diferentes concentraciones de BAP en los procesos de inducción de brotes en dos patrones de cítricos: C. troyer y C. carrizo
Las plántulas de los dos patrones provenientes del experimento número 2,
se manejaron bajo condiciones asépticas en campana de flujo laminar. Estas se
seccionaron con un bisturí en varios segmentos nodales de aproximadamente 2
cm de longitud (Pierik, 1990). Los segmentos nodales obtenidos se sembraron
bajo un diseño completamente aleatorizado, con tres repeticiones por cada
tratamiento aplicado, a cada patrón en estudio (Tabla 4). La siembra se efectúo en
tubos de 120 X 25 mm con alícuotas de 20 ml de medio MS, adicionado con
vitaminas de Gamborg, 6 g.l-1 de agar y 30 g.l-1 de sacarosa; el medio se ajustó a
pH de 5.7 ± 0.1 con una solución de NaOH 0.1N.
Tabla 4.- Tratamientos hormonales empleados en el proceso de inducción de
brotes de C. troyer y C. carrizo
PATRONES TRATAMIENTOS DE BAP
C. troyer 2 mg.l-1 4 mg.l-1 6 mg.l-1 C. carrizo 2 mg.l-1 4 mg.l-1 6 mg.l-1
Los cultivos se mantuvieron a una temperatura de 25 ± 2° C, con 16 horas
luz y 8 de oscuridad, bajo una intensidad lumínica de 1,200 lux, con lámparas
Osram* Daylight de 22 W. A los 30 días se evaluaron las siguientes variables:
Número de brotes por explante, longitud del brote y vigor (de acuerdo a la escala
definida con anterioridad).
Los datos se procesaron estadísticamente efectuándose el análisis de
varianza (ANOVA) bifactorial 3 X 2 (3 concentraciones X 2 patrones) mediante el
programa STATISTICA (versión 4.5).

35
Se emplearon gráficas de barra de acuerdo con el programa Excel
(Versión 97) y se tomaron fotografías con película Kodak* 35 mm, ASA 200 para la
presentación de los resultados.
3.3.4. Experimento 4. Efecto de tratamientos de auxinas sobre la formación de raíces en C. troyer y C. carrizo (C. sinensis Osbeck x P. trifoliata Raf)
3.3.4.1. Enraizamiento
Se seleccionaron los brotes mayores de 2 cm de longitud, se seccionaron
con un bisturí y se sembraron para estimular la formación de raíces en el medio de
MS, adicionado con las combinaciones hormonales (Tabla 5), bajo un diseño
completamente aleatorizado con arreglo factorial y tres repeticiones por
tratamiento. Además, se adicionaron vitaminas de Gamborg, 6 g.l -1 de agar y 30
g.l-1 de sacarosa. A cada tubo de 120 X 25 mm se le adicionó una alícuota de 20
ml de medio ajustado a un pH de 5.7 ± 0.1 con una solución de NaOH 0.1N.
Tabla 5.- Combinaciones hormonales empleadas en la inducción del proceso de rizogénesis en brotes de los patrones C. troyer y C. carrizo
TRATAMIENTO PATRONES ANA (mg.l-1) AIB (mg.l-1)
1 C. troyer 1.0 0.0 2 C. troyer 0.0 1.0 3 C. troyer 0.5 0.5 1 C. carrizo 1.0 0.0 2 C. carrizo 0.0 1.0 3 C. carrizo 0.5 0.5

36
Los cultivos se mantuvieron a una temperatura de 25 ± 2° C, con 16 horas
luz y 8 de oscuridad, bajo una intensidad lumínica de 1,200 lux, con lámparas
Osram* Dayliht de 22 W. La evaluación del porcentaje de brotes enraizados y
longitud de la raíz, se realizó a los 15 y 30 días. Los datos se procesaron por
análisis de varianza (ANOVA) bifactorial 3 X 2 (3 tratamientos hormonales X 2
patrones). Se emplearon gráficas de barra del programa Excel (Versión 97) y se
fotografiaron con película Kodak* 35 mm, ASA 200, para presentar los resultados.
3.3.5. Experimento 5. Aclimatación de patrones de C. troyer y C. carrizo C. sinensis Osbeck X P. trifoliata Raf).
Las vitroplántulas provenientes del experimento 4, que mostraron de 2 a 3
hojas expandidas y una longitud del tallo oscilando entre 4-6 cm, fueron extraídas
del tubo de ensaye y se lavaron perfectamente con agua corriente. Estas se
transfirieron, de acuerdo con lo recomendado por Hurtado y Merino (1994), a
bolsas de polietileno de color negro (10 X 20 cm), conteniendo: Tierra negra
(testigo) y lombricomposta al 10 y 20 %. Se empleó un diseño completamente
aleatorizado con arreglo factorial y tres repeticiones para cada combinación en
estudio (combinación de dos patrones y dos concentraciones del sustrato) (Tabla
6).
Para efectuar la prueba serológica de ELISA para el diagnóstico de VTC,
se enviaron 5 muestras de plantas de ambos patrones al Laboratorio de Alta
Tecnología de Xalapa, S.C. (LATEX).

37
Tabla 6.- Tratamientos empleados para la aclimatación de dos patrones de
cítricos.
PATRON TRATAMIENTOS LOMBRICOMPOSTA (%)
C. troyer 1 10
2 20
C. carrizo 1 10
2 20
Una vez transplantadas, se colocó a cada bolsa un soporte de madera en
forma de cruz, para cubrir adecuadamente cada vitroplántula con una bolsa de
polietileno transparente (30 X 40 cm). Al momento del transplante y cada tercer
día, por un período de 10 días, se adicionó a cada bolsa, aproximadamente 40 ml
de agua corriente. Las bolsas se cerraron con una liga y se colocaron en un área
sombreada a 26 ± 2° C con una humedad relativa de 80 a 90 %. A los 11 días de
efectuado el transplante, se retiraron las bolsas de polietileno y a partir de ese
momento se efectúo un riego semanal, hasta terminar el experimento. A los 30 y
45 días (6 semanas) se evaluaron los siguientes caracteres en todas las plantas:
largo del tallo, número de hojas, grosor del tallo y vigor.
Este último se determinó en base a la siguiente escala:
1. Plantas poco vigorosas. De color verde pálido en hojas y tallo, de
menos de 4 cm de altura.
2. Plantas vigorosas. De color verde intenso en hojas y tallos, de 4
a 6 cm de altura.

38
Los datos obtenidos se procesaron estadísticamente efectuando el
análisis de varianza (ANOVA), bifactorial 2 X 2 (2 patrones X 2 sustratos).
Posteriormente se realizó la prueba de Dunnet (Steel y Torrie, 1988) para
comparar las medias de los tratamientos con respecto a un grupo testigo.
39
RESULTADOS Y DISCUSION
4.1. Experimento 1. Caracterización morfométrica de semillas de C. troyer y
C. carrizo (C. sinensis Osbeck X Poncirus trifoliata Raf.)
Los resultados obtenidos de los análisis de varianza (ANOVA),
efectuados, no arrojaron diferencias significativas para las variables morfométricas
evaluadas en las semillas de los 2 patrones en estudio (Tabla 7).
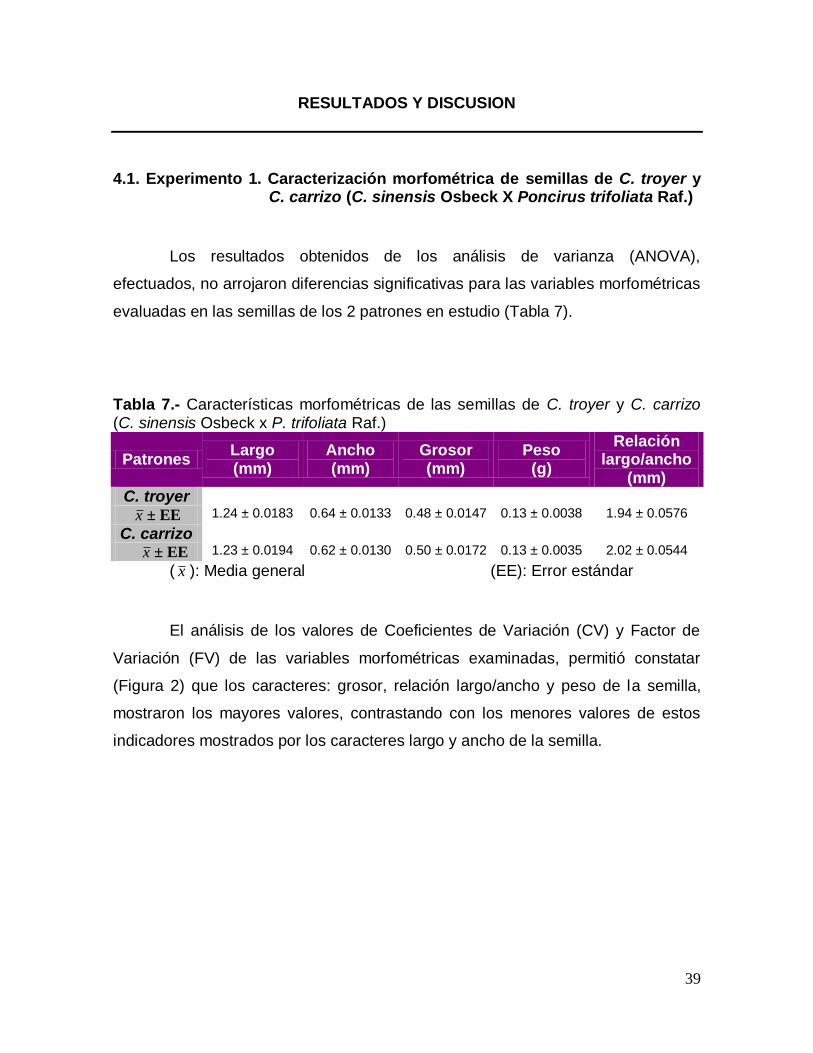
Tabla 7.- Características morfométricas de las semillas de C. troyer y C. carrizo (C. sinensis Osbeck x P. trifoliata Raf.)
Patrones Largo (mm)
Ancho (mm)
Grosor (mm)
Peso (g)
Relación largo/ancho
(mm) C. troyer
x ± EE 1.24 ± 0.0183 0.64 ± 0.0133 0.48 ± 0.0147 0.13 ± 0.0038 1.94 ± 0.0576
C. carrizo
x ± EE 1.23 ± 0.0194 0.62 ± 0.0130 0.50 ± 0.0172 0.13 ± 0.0035 2.02 ± 0.0544
( x ): Media general (EE): Error estándar
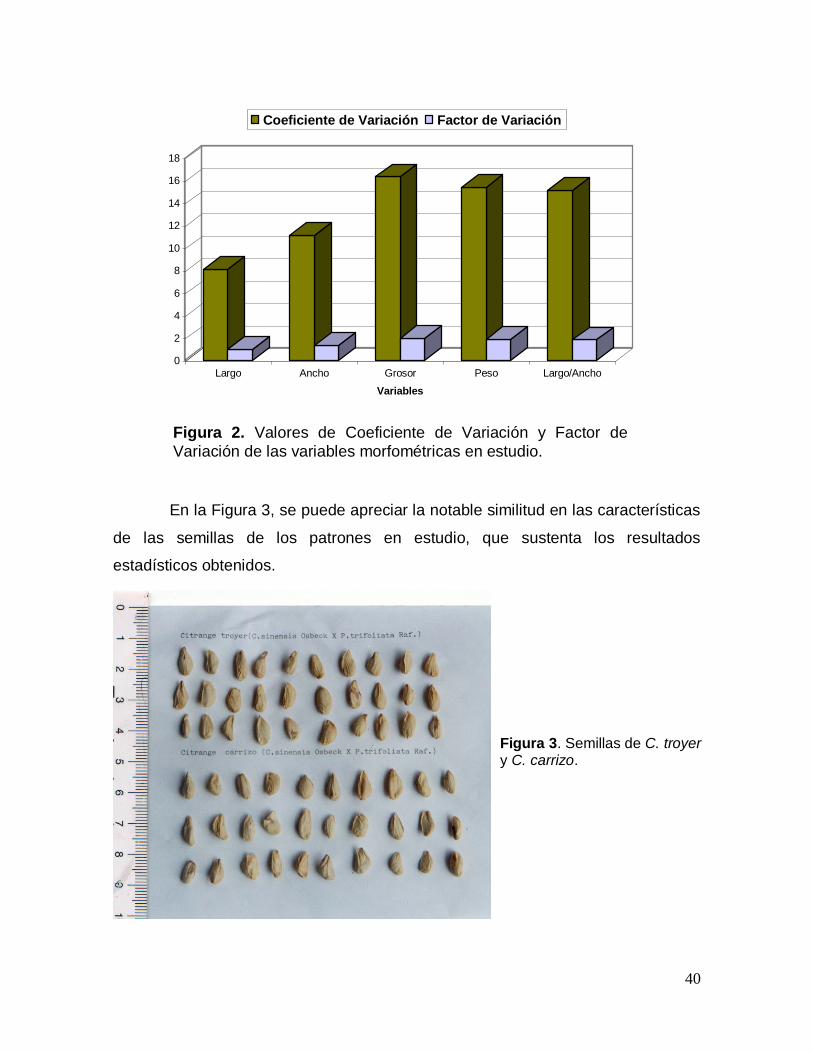
El análisis de los valores de Coeficientes de Variación (CV) y Factor de
Variación (FV) de las variables morfométricas examinadas, permitió constatar
(Figura 2) que los caracteres: grosor, relación largo/ancho y peso de la semilla,
mostraron los mayores valores, contrastando con los menores valores de estos
indicadores mostrados por los caracteres largo y ancho de la semilla.
40
0
2
4
6
8
10
12
14
16
18
Largo Ancho Grosor Peso Largo/Ancho
Variables
Coeficiente de Variación Factor de Variación
Figura 2. Valores de Coeficiente de Variación y Factor de
Variación de las variables morfométricas en estudio.
En la Figura 3, se puede apreciar la notable similitud en las características
de las semillas de los patrones en estudio, que sustenta los resultados
estadísticos obtenidos.
Figura 3. Semillas de C. troyer y C. carrizo.
41
Estos resultados sustentan la existencia de una estrecha relación genética
entre C. troyer y C. carrizo, dada la similitud en cuanto al origen genético de los
mismos (Palacios, 1978; Morín et al., 1985). Diversos autores (Gravina, 1989;
Kent y Levi, 1990; Chávez y González, 1999), han detectado de igual forma
similitudes en las características morfogenéticas de frutos y semillas de ambos
patrones.
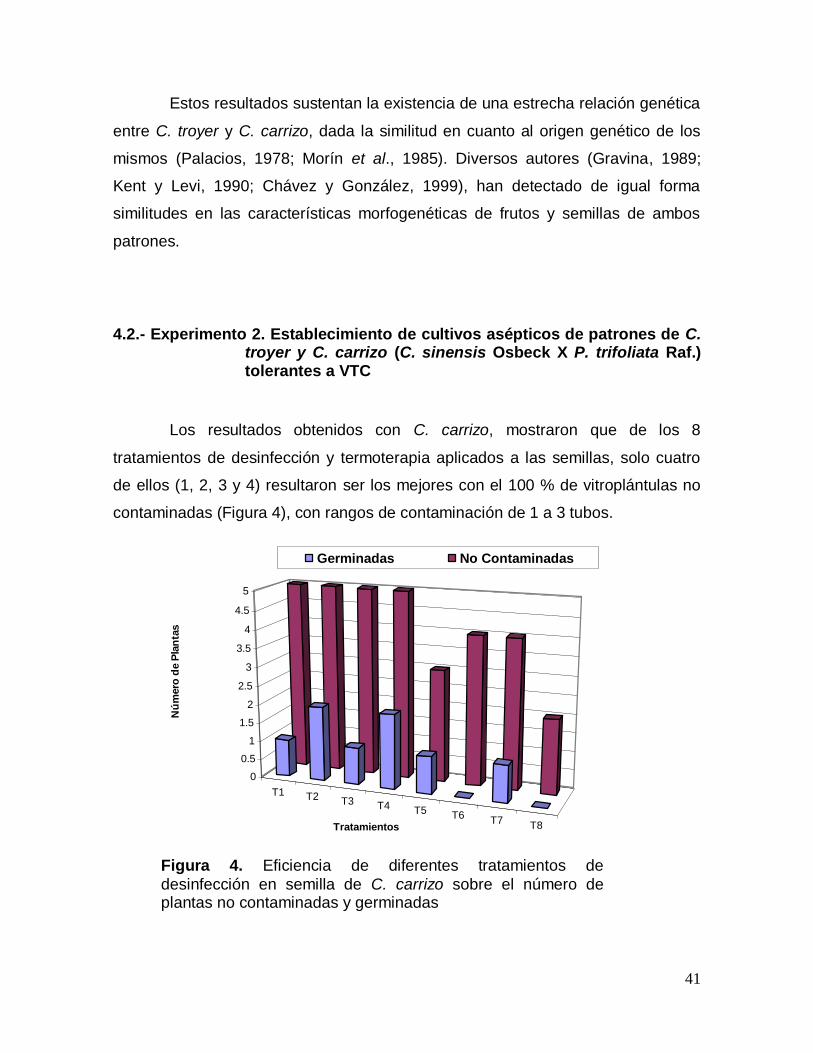
4.2.- Experimento 2. Establecimiento de cultivos asépticos de patrones de C. troyer y C. carrizo (C. sinensis Osbeck X P. trifoliata Raf.) tolerantes a VTC
Los resultados obtenidos con C. carrizo, mostraron que de los 8
tratamientos de desinfección y termoterapia aplicados a las semillas, solo cuatro
de ellos (1, 2, 3 y 4) resultaron ser los mejores con el 100 % de vitroplántulas no
contaminadas (Figura 4), con rangos de contaminación de 1 a 3 tubos.
T1 T2 T3 T4 T5 T6T7
T8
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
5
Tratamientos
Germinadas No Contaminadas
Nú
mero
de P
lan
tas
Figura 4. Eficiencia de diferentes tratamientos de
desinfección en semilla de C. carrizo sobre el número de plantas no contaminadas y germinadas

42
Los tratamientos 2 (Benlate*/20 minutos; Clorox* 25%/10 minutos a 52-55°
C/5 minutos) y 4 (Benlate*/20 minutos; Clorox* 30%/5 minutos a 52-55° C/5
minutos), aplicados a la semilla de C. carrizo permitieron obtener un mayor
porcentaje de germinaron in vitro (Figura 4) en comparación con el menor
porcentaje de germinación que se obtuvo en los tratamientos 1 (Benlate*/20
minutos; Clorox* 25%/10 minutos a 52-55° C/10 minutos) y 3 (Benlate*/20 minutos;
Clorox* 30%/5 minutos a 52-55° C/10 minutos), en los que se observaron también
un mayor control de la contaminación.
En la Figura 5, se muestran semillas de C. carrizo germinadas in vitro,
luego de aplicarse los tratamientos 2 y 4 de desinfección y termoterapia.
Figura 5. Germinación de
semillas de C. carrizo, empleando
los tratamientos 2 y 4.
Por otra parte, en C. troyer se detectaron dos tubos contaminados en el
tratamiento 6 (Benlate*/25 minutos; Clorox* 25%/10 minutos a 52-55° C/5 minutos)
(Figura 6). En los tratamientos restantes, se pudo lograr un adecuado control de la
contaminación, siendo ésta del 100 %.
43
T1 T2 T3 T4T5
T6T7
T8
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
5
Tratamientos
Germinadas No Contaminadas
Nú
mero
de P
lan
tas
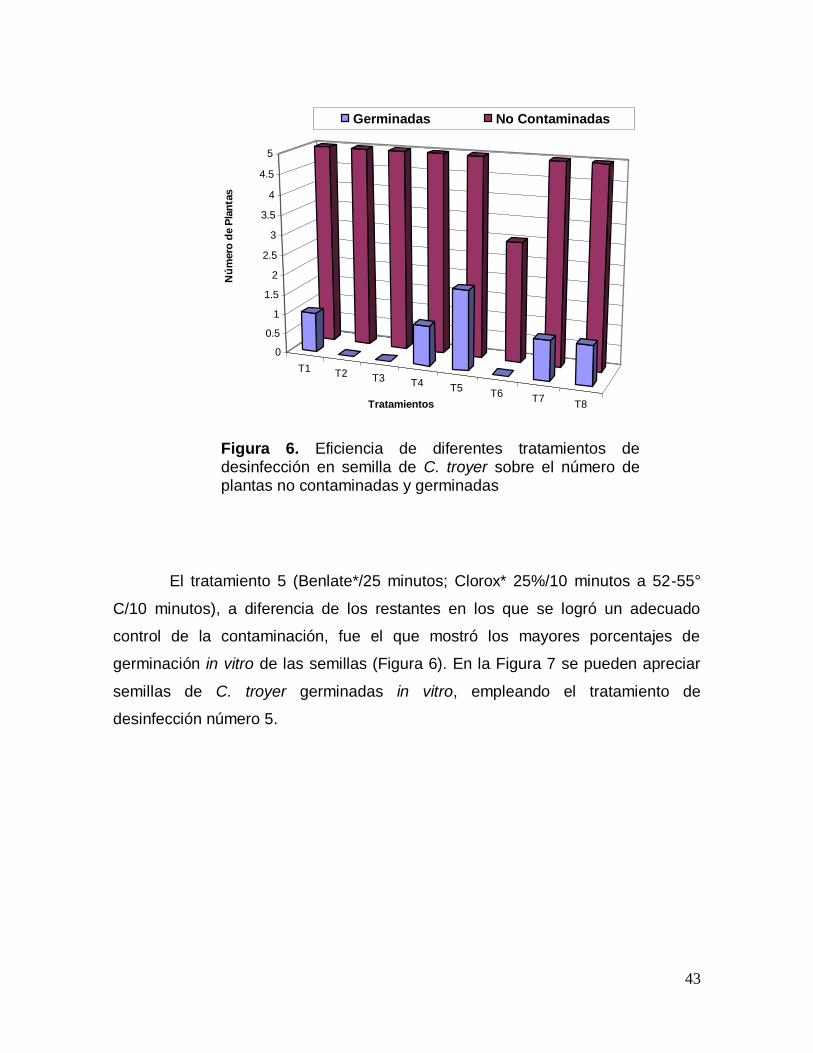
Figura 6. Eficiencia de diferentes tratamientos de desinfección en semilla de C. troyer sobre el número de plantas no contaminadas y germinadas
El tratamiento 5 (Benlate*/25 minutos; Clorox* 25%/10 minutos a 52-55°
C/10 minutos), a diferencia de los restantes en los que se logró un adecuado
control de la contaminación, fue el que mostró los mayores porcentajes de
germinación in vitro de las semillas (Figura 6). En la Figura 7 se pueden apreciar
semillas de C. troyer germinadas in vitro, empleando el tratamiento de
desinfección número 5.

44
Figura 7. Germinación de semillas de C. troyer, empleando el Tratamiento 5
De manera general, los resultados obtenidos mostraron un 100 % de
efectividad en los tratamientos de desinfección 2 y 4, en semillas de C. carrizo y el
tratamiento 5, en semillas de C. troyer. Es posible que la mayor efectividad
obtenida en el control de la contaminación de este trabajo se deba, entre otras, a
la mayor concentración de hipoclorito de sodio empleada, en relación con la
empleada por otros autores como Normah et al. (1997) y García et al. (1999).
Cabe destacar, sin embargo, que los bajos porcentajes de contaminación
obtenidos en este trabajo resultaron ser menores a los obtenidos en cítricos por
Rodríguez (1986), quien obtuvo el 90 % de tubos sin contaminación.
Con respecto al porcentaje de germinación, se apreciaron diferencias en la
respuesta a los diferentes tratamientos en ambos patrones (Figuras 4 y 6). En
general los valores de germinación detectados en ambos patrones no
sobrepasaron al 55%. Es posible que los tratamientos de desinfección y

45
termoterapia empleados hayan afectado el proceso germinativo tal como ha sido
reportado, entre otros, por Gravina (1989) y Boxus (1998).
Resultó interesante constatar, las diferencias mostradas por las semillas
de los dos patrones ante los tratamientos de desinfección y termoterapia
empleados. De hecho, a diferencia de lo observado en C. carrizo, el tratamiento 5
resultó ser el más adecuado para lograr un mejor control de la desinfección y un
porcentaje más elevado de germinación en C. troyer. Otro apecto a destacar de
estos resultados es que las pruebas de diagnóstico al VTC empleados en esta
fase, resultaron ser negativos (Anexo 3). Estos resultados concuerdan con lo
esperado si se tiene en cuenta los indicado por diversos autores (Gravina y
Piestun, 1991; Monteverde, 1999), en relación a que los virus y viroides de cítricos
se transmiten en bajo porcentaje a través de la semilla. Por otra parte, el uso de
tratamientos de temperatura, como el empleado en este trabajo han resultado
efectivos para la eliminación de algunos patógenos como la tristeza, psorosis,
concavidad gomosa e impietratura (Calavan et al., 1972; Roistacher y Calavan,
1972).
4.3. Experimento 3. Inducción de brotes a partir de segmentos nodales de C. troyer y C. carrizo (C. sinensis Osbeck X P. trifoliata Raf.) tolerantes al VTC
4.3.1.- Inducción de brotes a partir de segmentos nodales con BAP, ANA y AG3
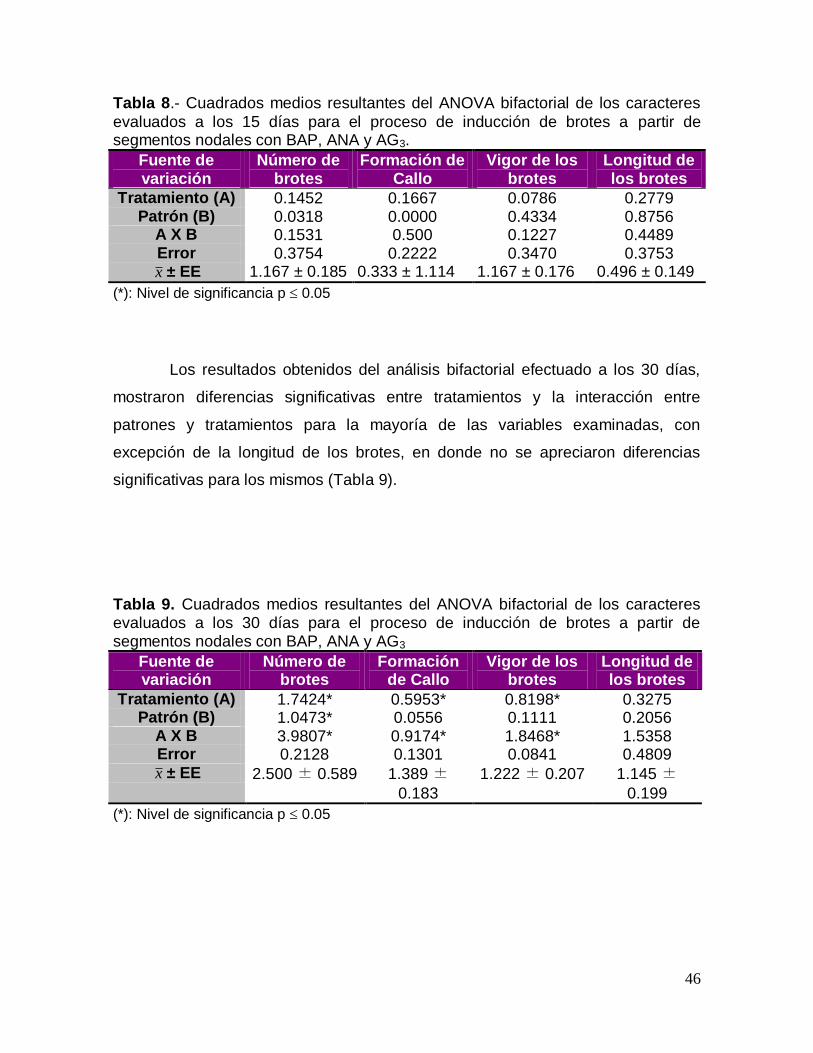
Los resultados obtenidos del análisis bifactorial efectuado en 15 días,
mostraron que no existieron diferencias significativas entre los tratamientos, entre
los patrones, ni entre la interacción para las variables número de brotes, callo,
vigor y longitud del brote (Tabla 8).
46
Tabla 8.- Cuadrados medios resultantes del ANOVA bifactorial de los caracteres
evaluados a los 15 días para el proceso de inducción de brotes a partir de segmentos nodales con BAP, ANA y AG3.
Fuente de variación
Número de brotes
Formación de Callo
Vigor de los brotes
Longitud de los brotes
Tratamiento (A) 0.1452 0.1667 0.0786 0.2779 Patrón (B) 0.0318 0.0000 0.4334 0.8756
A X B 0.1531 0.500 0.1227 0.4489 Error 0.3754 0.2222 0.3470 0.3753 x ± EE 1.167 ± 0.185 0.333 ± 1.114 1.167 ± 0.176 0.496 ± 0.149
(*): Nivel de significancia p 0.05
Los resultados obtenidos del análisis bifactorial efectuado a los 30 días,
mostraron diferencias significativas entre tratamientos y la interacción entre
patrones y tratamientos para la mayoría de las variables examinadas, con
excepción de la longitud de los brotes, en donde no se apreciaron diferencias
significativas para los mismos (Tabla 9).
Tabla 9. Cuadrados medios resultantes del ANOVA bifactorial de los caracteres evaluados a los 30 días para el proceso de inducción de brotes a partir de segmentos nodales con BAP, ANA y AG3
Fuente de variación
Número de brotes
Formación de Callo
Vigor de los brotes
Longitud de los brotes
Tratamiento (A) 1.7424* 0.5953* 0.8198* 0.3275 Patrón (B) 1.0473* 0.0556 0.1111 0.2056
A X B 3.9807* 0.9174* 1.8468* 1.5358 Error 0.2128 0.1301 0.0841 0.4809 x ± EE 2.500 ± 0.589 1.389 ±
0.183
1.222 ± 0.207 1.145 ±
0.199
(*): Nivel de significancia p 0.05

47
Para la variable número de brotes, se constató la existencia de diferencias
significativas para los tratamientos, patrones e interacción entre ambos factores
(Tabla 9). En la Tabla 10 se puede apreciar la respuesta promedio en el número
de brotes en función de la interacción entre los patrones y tratamientos.
Tabla 10.- Comparación de medias para la variable número de brotes en la
interacción entre patrones y tratamientos
Tratamiento Patrón Medias
2 Carrizo 2.1921 a 1 Carrizo 2.0680 a 2 Troyer 1.6261 a 3 Troyer 1.5201 ab 3 Carrizo 0.3333 bc 1 Troyer 0.0000 c
(*):Medias con letras diferentes son significativamente distintas según décima de
Tukey con p 0.05.
Para la variable longitud del brote, no se detectaron diferencias
significativas entre los tratamientos, ni entre los patrones, ni en la interacción entre
ambos (Tabla 9).
De acuerdo con estos resultados el tratamiento 2 (BAP 2 mg.l -1, ANA 0.0
mg.l-1 y AG3 2 mg.l-1), resultó en general como el más adecuado para lograr una
mayor inducción de brotes en ambos patrones (Tabla 10). Se observó una
diferencia en la respuesta morfogenética en la inducción de brotes en ambos
patrones ya que el tratamiento 1 (BAP 1 mg.l-1, ANA 0.5 mg.l-1 y AG3 2 mg.l-1) fue
igualmente satisfactorio que el tratamiento 2 en C. carrizo, no así para C. troyer.
48
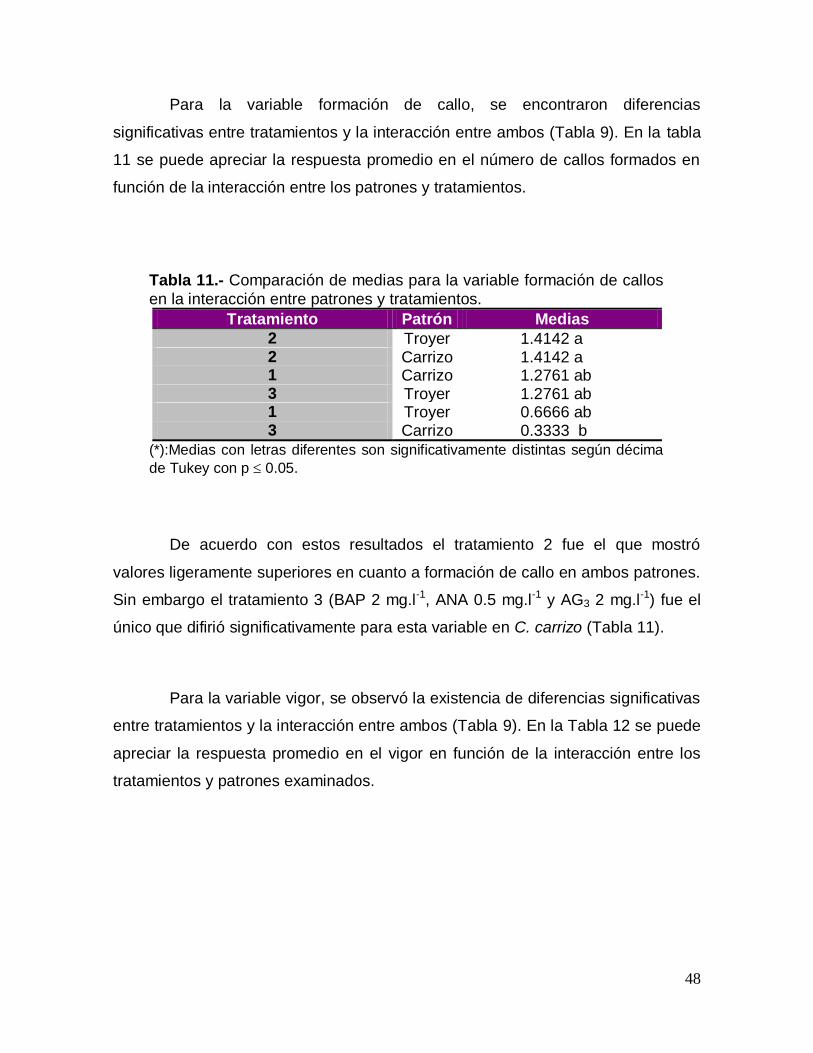
Para la variable formación de callo, se encontraron diferencias
significativas entre tratamientos y la interacción entre ambos (Tabla 9). En la tabla
11 se puede apreciar la respuesta promedio en el número de callos formados en
función de la interacción entre los patrones y tratamientos.
Tabla 11.- Comparación de medias para la variable formación de callos
en la interacción entre patrones y tratamientos.
Tratamiento Patrón Medias
2 Troyer 1.4142 a 2 Carrizo 1.4142 a 1 Carrizo 1.2761 ab 3 Troyer 1.2761 ab 1 Troyer 0.6666 ab 3 Carrizo 0.3333 b
(*):Medias con letras diferentes son significativamente distintas según décima
de Tukey con p 0.05.
De acuerdo con estos resultados el tratamiento 2 fue el que mostró
valores ligeramente superiores en cuanto a formación de callo en ambos patrones.
Sin embargo el tratamiento 3 (BAP 2 mg.l-1, ANA 0.5 mg.l-1 y AG3 2 mg.l-1) fue el
único que difirió significativamente para esta variable en C. carrizo (Tabla 11).
Para la variable vigor, se observó la existencia de diferencias significativas
entre tratamientos y la interacción entre ambos (Tabla 9). En la Tabla 12 se puede
apreciar la respuesta promedio en el vigor en función de la interacción entre los
tratamientos y patrones examinados.
49
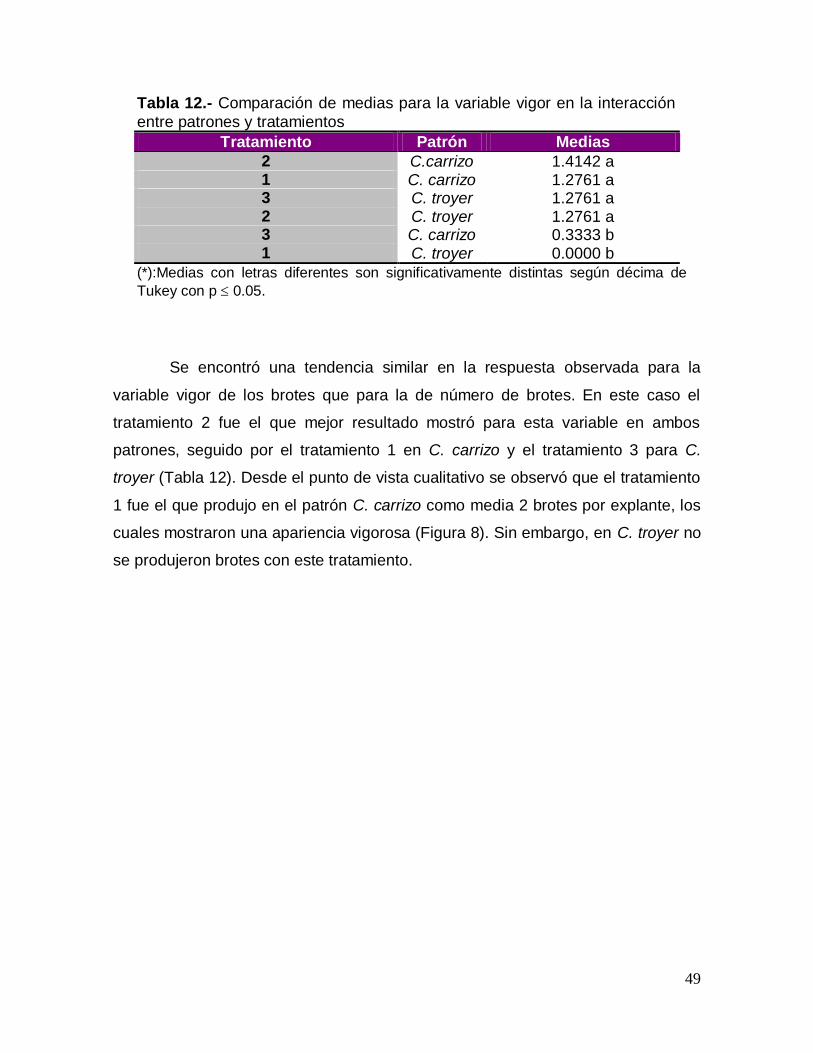
Tabla 12.- Comparación de medias para la variable vigor en la interacción
entre patrones y tratamientos
Tratamiento Patrón Medias
2 C.carrizo 1.4142 a 1 C. carrizo 1.2761 a 3 C. troyer 1.2761 a 2 C. troyer 1.2761 a 3 C. carrizo 0.3333 b 1 C. troyer 0.0000 b
(*):Medias con letras diferentes son significativamente distintas según décima de
Tukey con p 0.05.
Se encontró una tendencia similar en la respuesta observada para la
variable vigor de los brotes que para la de número de brotes. En este caso el
tratamiento 2 fue el que mejor resultado mostró para esta variable en ambos
patrones, seguido por el tratamiento 1 en C. carrizo y el tratamiento 3 para C.
troyer (Tabla 12). Desde el punto de vista cualitativo se observó que el tratamiento
1 fue el que produjo en el patrón C. carrizo como media 2 brotes por explante, los
cuales mostraron una apariencia vigorosa (Figura 8). Sin embargo, en C. troyer no
se produjeron brotes con este tratamiento.

50
Figura 8. Inducción de brotes a partir de segmentos nodales en C. carrizo, sembrados en el tratamiento 1.
De manera similar, se observó que el tratamiento 2 (Figura 9) produjo en
promedio 2 brotes por explante en C. carrizo y 1 brote por explante en C. troyer.
Cabe mencionar que en este tratamiento se formó un callo mucho más distintivo a
partir de los cuales se regeneraron los brotes en ambos patrones.

51
Figura 9.- Inducción de brotes a
partir de segmentos nodales en C. troyer, sembrados en el tratamiento 2.
En el tratamiento 3 (Figura 10) se observó, por otra parte, la formación de
pocos brotes por explante en ambos patrones. En este tratamiento, al igual que los
anteriores, se detectó la presencia de callo fundamentalmente en C. troyer así
como la presencia en la mayoría de los casos de cierta anormalidad en los brotes
formados (deformación de los tallos), probablemente asociada con la mayor
concentración de BAP aplicada.

52
Figura 10.- Inducción de brotes a partir
de segmentos nodales en C. troyer en el tratamiento 3.
En resumen, de acuerdo con estos resultados el tratamiento 1 fue el mejor
para la inducción de brotes en C. carrizo y el tratamiento 2 para C. troyer.
Diversos autores han logrado obtener un número de brotes similares a lo
obtenido en este estudio empleando segmentos nodales en diversas especies
vegetales (Pérez-Molphe y Ochoa-Alejo, 1997; Normah et al., 1997; Hernández y
Mejía, 1994; García et al., 1999). La diferente respuesta morfogenética observada
en este estudio entre los dos patrones puede deberse a la influencia del genotipo
en el proceso de inducción de brotes.

53
En C. carrizo Moore et al. (1992), reportó la inducción de brotes por
organogénesis directa sin formación de callo, sin embargo en otros genotipos de
cítricos se ha reportado que este proceso transcurre por organogénesis indirecta
(Raj y Arya, 1978; Durán-Vila et al., 1989).
Estudios al respecto efectuados en C. troyer por García et al. (1999)
mostraron que la vía morfogenética depende de la exposición del explante y que
de los explantes sembrados en posición vertical se originaban brotes del cambium
vascular por un proceso de morfogénesis directa, mientras que aquellos explantes,
en los que su extremo apical se encontraba en contacto con el medio, se producia
un proceso de regeneración indirecta. Es posible que lo anterior pueda también
haber influido en la variación de las respuestas morfogenéticas detectadas en este
estudio, sin descartar la influencia como indicara Espinoza (1994), de otros
factores relacionados con la composición química del medio de cultivo y
condiciones de cultivo.
4.3.2. Efecto de diferentes concentraciones de BAP (2, 4 y 6 mg.l-1) en el proceso de inducción de brotes
Los resultados obtenidos del análisis bifactorial, efectuados a los 30 días
para el efecto de diferentes concentraciones de BAP en el proceso de inducción
de brotes en dos patrones de cítricos en estudio, se muestran en la Tabla 13. Se
pudo observar que existieron diferencias significativas entre tratamientos para las
variables número de brotes (Figura 11), vigor (Figura 12) y longitud de los brotes
(Figura 13).
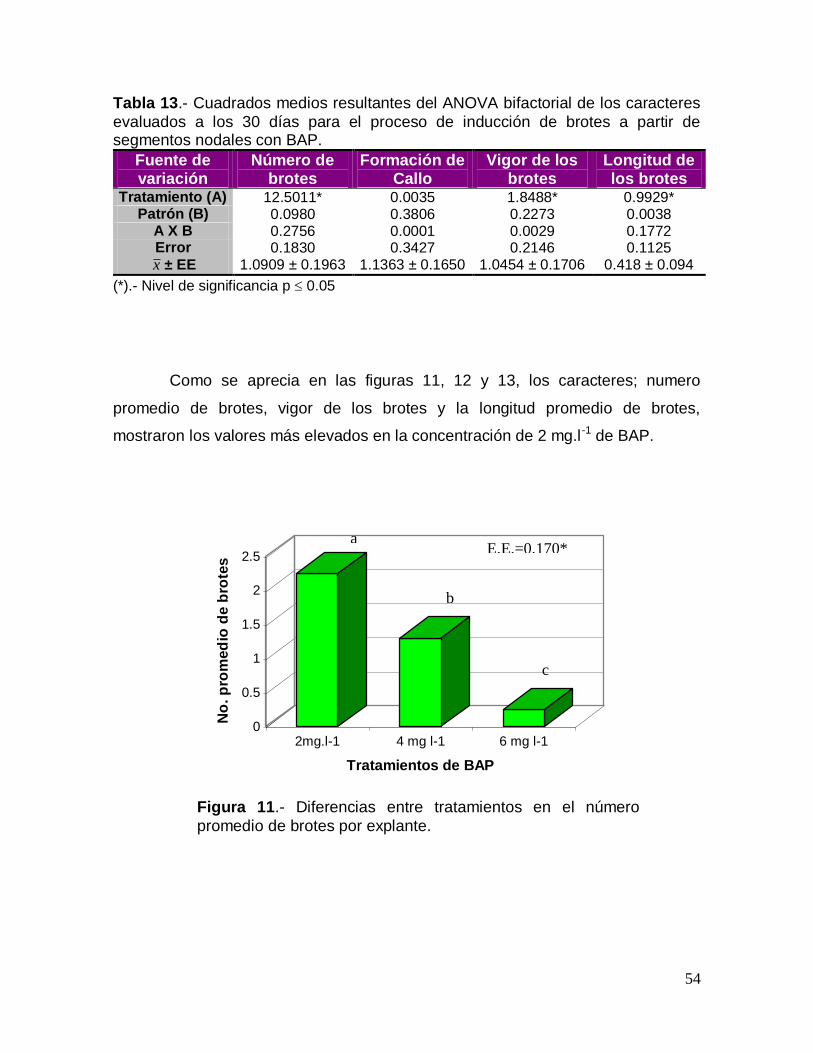
54
Tabla 13.- Cuadrados medios resultantes del ANOVA bifactorial de los caracteres
evaluados a los 30 días para el proceso de inducción de brotes a partir de segmentos nodales con BAP.
Fuente de variación
Número de brotes
Formación de Callo
Vigor de los brotes
Longitud de los brotes
Tratamiento (A) 12.5011* 0.0035 1.8488* 0.9929* Patrón (B) 0.0980 0.3806 0.2273 0.0038
A X B 0.2756 0.0001 0.0029 0.1772 Error 0.1830 0.3427 0.2146 0.1125 x ± EE 1.0909 ± 0.1963 1.1363 ± 0.1650 1.0454 ± 0.1706 0.418 ± 0.094
(*).- Nivel de significancia p 0.05
Como se aprecia en las figuras 11, 12 y 13, los caracteres; numero
promedio de brotes, vigor de los brotes y la longitud promedio de brotes,
mostraron los valores más elevados en la concentración de 2 mg.l-1 de BAP.
0
0.5
1
1.5
2
2.5
No
. p
rom
ed
io d
e b
rote
s
2mg.l-1 4 mg l-1 6 mg l-1
Tratamientos de BAP
Figura 11.- Diferencias entre tratamientos en el número
promedio de brotes por explante.
E.E.=0.170* a
b
c
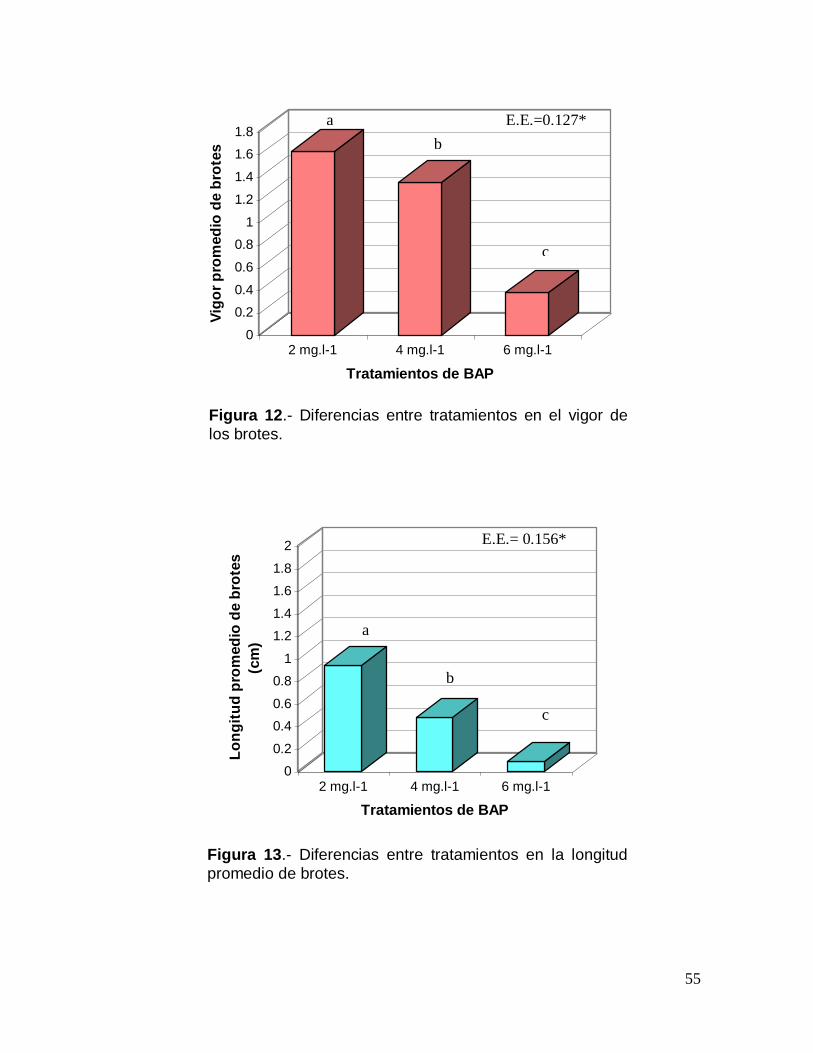
55
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
Vig
or
pro
med
io d
e b
rote
s
2 mg.l-1 4 mg.l-1 6 mg.l-1
Tratamientos de BAP
Figura 12.- Diferencias entre tratamientos en el vigor de
los brotes.
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
Lo
ng
itu
d p
rom
ed
io d
e b
rote
s
(cm
)
2 mg.l-1 4 mg.l-1 6 mg.l-1
Tratamientos de BAP
Figura 13.- Diferencias entre tratamientos en la longitud
promedio de brotes.
E.E.=0.127*
*****
a
b
c
E.E.= 0.156*
a
b
c

56
En el tratamiento de 2 mg.l-1 de BAP, el número de brotes por explante en
C. carrizo fue de 0 a 2 (Figura 14) y para C. troyer fue de 1 a 3 (Figura 15).
Figura 14.- Inducción de brotes a partir de segmentos nodales en C. carrizo, empleando 2 mg.l-1 de BAP
Figura 15.- Inducción de brotes a partir de segmentos nodales en C. troyer, empleando 2 mg.l-1 de BAP.

57
Los resultados obtenidos en este estudio con el empleo de BAP se
encuentran en concordancia con lo indicado por varios autores en relación con la
inducción de brotes múltiples a partir de diversos explantes de especies vegetales,
incluyendo al género Citrus (Durán-Vila et al., 1989;Beloualy, 1991 y Normah et
al., 1997).
Los valores más bajos detectados en todas las variables examinadas al
elevarse la concentración de BAP en el medio, pueden estar dados por la
presencia de inestabilidad genetica, que como apuntarán Evans y Sharp (1986) y
Christopher y Rajam (1994), acompañan por lo regular al empleo de dosis
elevadas de esta citocinina.
4.4. Experimento 4. Efecto de la concentración de auxinas sobre la formación de raíces en C. troyer y C. carrizo (C. sinensis Osbeck X P. trifoliata Raf)
Los resultados obtenidos del análisis bifactorial, efectuado a los 15 días,
revelaron la no existencia de diferencias significativas en los tratamientos
empleados para la inducción del proceso de rizogénesis en ambos patrones; es
decir, no existieron diferencias significativas ni entre tratamientos, patrones e
interacción (Tabla 14).
Tabla 14.- Valores de los cuadrados medios resultantes del ANOVA bifactorial de
los caracteres evaluados a los 15 días en el proceso de rizogénesis.
Fuente de variación Presencia de Raíz Longitud de Raíz Callo
Tratamiento (A) 0.3228 0.4050 0.1667 Patrón (B) 0.0095 0.0050 0.2222
A X B 0.0095 0.0050 0.3889 Error 0.1667 0.2050 0.2222 x ± EE 0.167 ± 0.121 0.150 ± 0.104 0.333 ± 0.114
(*): Nivel de significancia p 0.05
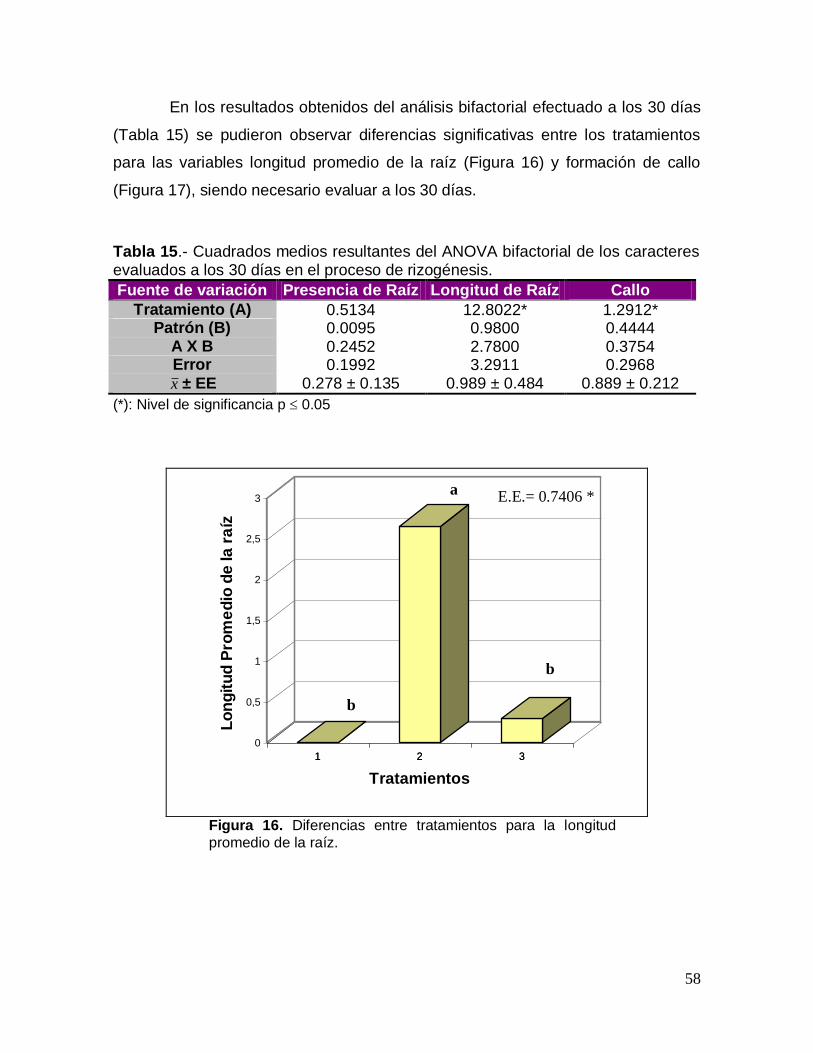
58
En los resultados obtenidos del análisis bifactorial efectuado a los 30 días
(Tabla 15) se pudieron observar diferencias significativas entre los tratamientos
para las variables longitud promedio de la raíz (Figura 16) y formación de callo
(Figura 17), siendo necesario evaluar a los 30 días.
Tabla 15.- Cuadrados medios resultantes del ANOVA bifactorial de los caracteres evaluados a los 30 días en el proceso de rizogénesis.
Fuente de variación Presencia de Raíz Longitud de Raíz Callo
Tratamiento (A) 0.5134 12.8022* 1.2912* Patrón (B) 0.0095 0.9800 0.4444
A X B 0.2452 2.7800 0.3754 Error 0.1992 3.2911 0.2968
x ± EE 0.278 ± 0.135 0.989 ± 0.484 0.889 ± 0.212
(*): Nivel de significancia p 0.05
1 2 3
0
0,5
1
1,5
2
2,5
3
Lo
ng
itu
d P
rom
ed
io d
e la
ra
íz
1 2 3
Tratamientos
Figura 16. Diferencias entre tratamientos para la longitud
promedio de la raíz.
b
a
b
b
E.E.= 0.7406 *
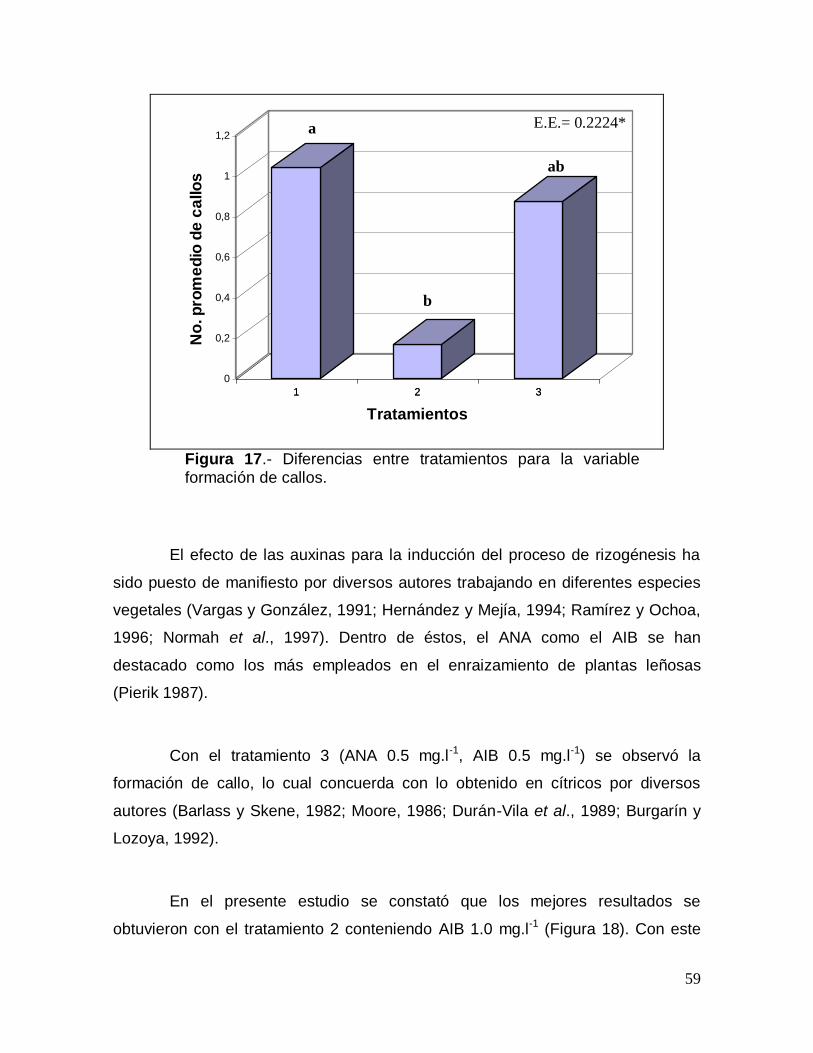
59
1 2 3
0
0,2
0,4
0,6
0,8
1
1,2
No
. p
rom
ed
io d
e c
allo
s
1 2 3
Tratamientos
Figura 17.- Diferencias entre tratamientos para la variable formación de callos.
El efecto de las auxinas para la inducción del proceso de rizogénesis ha
sido puesto de manifiesto por diversos autores trabajando en diferentes especies
vegetales (Vargas y González, 1991; Hernández y Mejía, 1994; Ramírez y Ochoa,
1996; Normah et al., 1997). Dentro de éstos, el ANA como el AIB se han
destacado como los más empleados en el enraizamiento de plantas leñosas
(Pierik 1987).
Con el tratamiento 3 (ANA 0.5 mg.l-1, AIB 0.5 mg.l-1) se observó la
formación de callo, lo cual concuerda con lo obtenido en cítricos por diversos
autores (Barlass y Skene, 1982; Moore, 1986; Durán-Vila et al., 1989; Burgarín y
Lozoya, 1992).
En el presente estudio se constató que los mejores resultados se
obtuvieron con el tratamiento 2 conteniendo AIB 1.0 mg.l-1 (Figura 18). Con este
a
ab
b
E.E.= 0.2224*

60
tratamiento no se presenta la formación de callo y la raíz llega a alcanzar una
longitud media de 5 cm en C. troyer y en C. carrizo hasta de 6 cm. Estos
resultados son similares a los obtenidos por Moore (1992) y por Pérez-Molphe-
Balch y Ochoa-Alejo (1997); sin embargo, difieren de lo obtenido por Kitto y Young
(1981) y Normah et al. (1997), quienes encontraron que el ANA inducía mejor
enraizamiento que el AIB en C. carrizo.
Figura 18. Inducción de rizogénesis en C. troyer, a partir de segmentos nodales en el tratamiento 2.
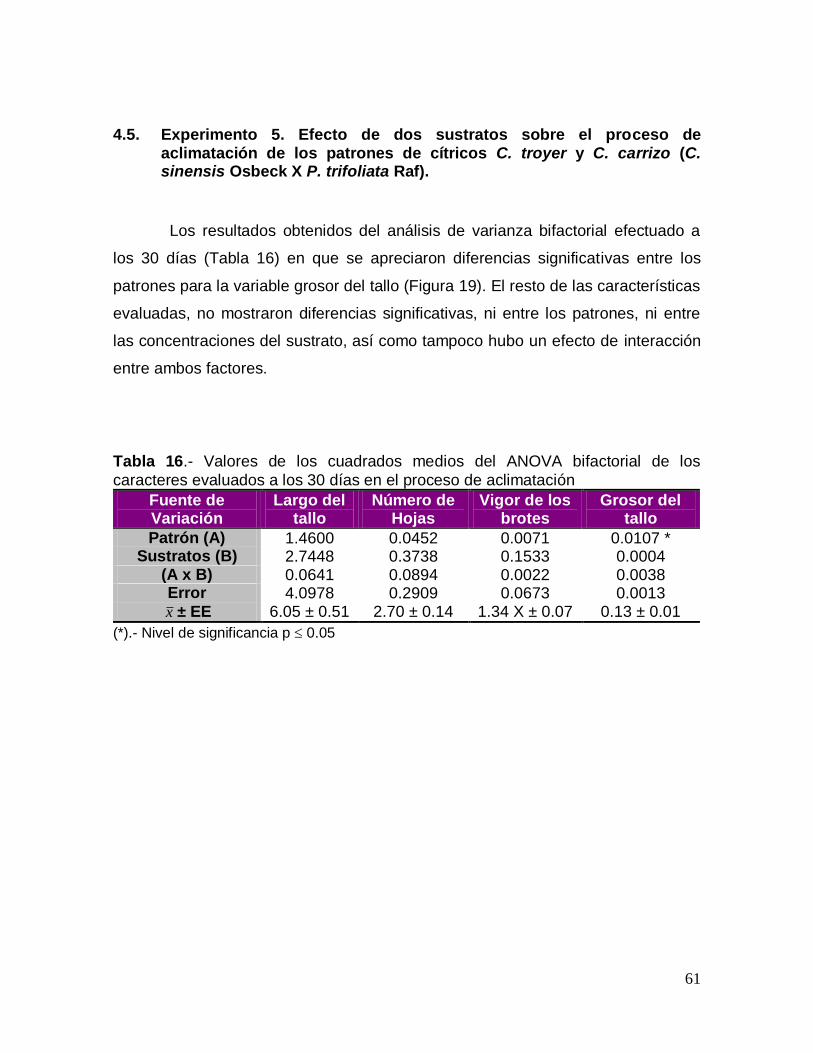
61
4.5. Experimento 5. Efecto de dos sustratos sobre el proceso de aclimatación de los patrones de cítricos C. troyer y C. carrizo (C. sinensis Osbeck X P. trifoliata Raf).
Los resultados obtenidos del análisis de varianza bifactorial efectuado a
los 30 días (Tabla 16) en que se apreciaron diferencias significativas entre los
patrones para la variable grosor del tallo (Figura 19). El resto de las características
evaluadas, no mostraron diferencias significativas, ni entre los patrones, ni entre
las concentraciones del sustrato, así como tampoco hubo un efecto de interacción
entre ambos factores.
Tabla 16.- Valores de los cuadrados medios del ANOVA bifactorial de los
caracteres evaluados a los 30 días en el proceso de aclimatación
Fuente de Variación
Largo del tallo
Número de Hojas
Vigor de los brotes
Grosor del tallo
Patrón (A) 1.4600 0.0452 0.0071 0.0107 * Sustratos (B) 2.7448 0.3738 0.1533 0.0004
(A x B) 0.0641 0.0894 0.0022 0.0038 Error 4.0978 0.2909 0.0673 0.0013 x ± EE 6.05 ± 0.51 2.70 ± 0.14 1.34 X ± 0.07 0.13 ± 0.01
(*).- Nivel de significancia p 0.05
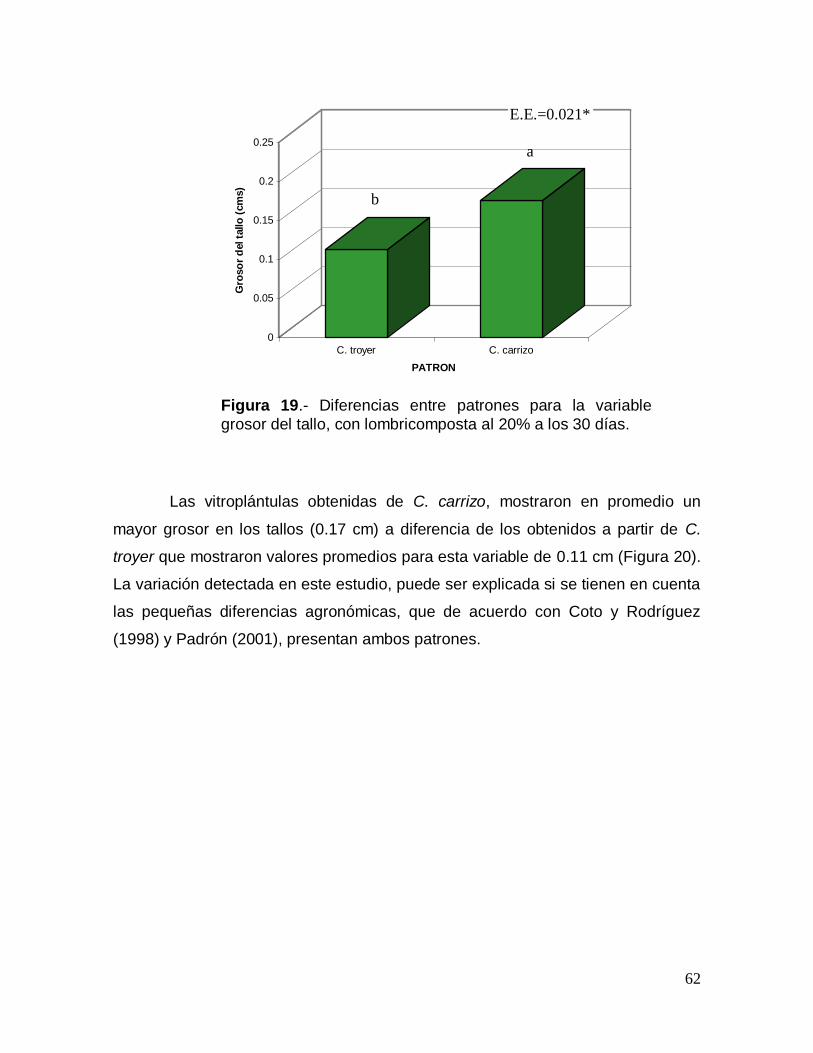
62
0
0.05
0.1
0.15
0.2
0.25
Gro
so
r d
el ta
llo
(cm
s)
C. troyer C. carrizo
PATRON
Figura 19.- Diferencias entre patrones para la variable
grosor del tallo, con lombricomposta al 20% a los 30 días.
Las vitroplántulas obtenidas de C. carrizo, mostraron en promedio un
mayor grosor en los tallos (0.17 cm) a diferencia de los obtenidos a partir de C.
troyer que mostraron valores promedios para esta variable de 0.11 cm (Figura 20).
La variación detectada en este estudio, puede ser explicada si se tienen en cuenta
las pequeñas diferencias agronómicas, que de acuerdo con Coto y Rodríguez
(1998) y Padrón (2001), presentan ambos patrones.
E.E.=0.021*
a
b
63
Figura 20.-
Plántulas de 30 días en fase de aclimatación de C. carrizo y C. troyer.
Por otro lado, los resultados del análisis efectuado a los datos de la
primera evaluación (30 días) para las variables largo del tallo, número de hojas,
vigor, grosor del tallo, comparados con los grupos testigo (prueba de Dunnet) se
presentan en la Tabla 17.
Tabla 17.- Comparación de Dunnet sobre las respuestas promedio obtenidas a los 30 días, en las combinaciones patrón-concentración, respecto al testigo, en el proceso de aclimatación
Patrones
Conc.
sustrato
(%)
Largo
tallo
(cm)
Testigo
Número
de
hojas
Testigo Vigor Testigo Grosor
tallo Testigo
C. troyer 10 5.88 5.38 2.57 2.194 1.22 1.31 0.10 0.14
C. troyer 20 6.72 5.38 2.76 2.194 1.49 1.31 0.12 0.14
C. carrizo 10 5.00 2.8 2.52 1.41 1.20 1.00 0.20* 0.32*
C. carrizo 20 6.15 2.8 3.07* 1.41* 1.41 1.00 0.15* 0.32*
(*).- Nivel de significancia p 0.05

64
Para las variables largo del tallo y vigor, no se encontraron diferencias
estadísticamente significativas con respecto a los grupos de testigo para ambos
patrones y concentración de sustratos (Tabla 17). Para la variable número de
hojas, se encontraron diferencias significativas en la respuesta del experimento
con el 20 % para C. carrizo, en relación con el grupo testigo. Para C. troyer no se
encontraron diferencias significativas con respecto al grupo testigo (Tabla 17).
Para la variable grosor del tallo se encontraron diferencias significativas en
las respuestas del experimento con ambas concentraciones para C. carrizo en
relación con el grupo testigo. Para C. troyer no se encontraron diferencias
significativas con respecto al grupo testigo (Tabla 17).
Los resultados obtenidos del análisis de varianza bifactorial efectuado en
la segunda evaluación (45 días) (Tabla 18), muestra que para las variables largo
del tallo, número de hojas y vigor, no mostraron diferencias significativas entre los
patrones, ni entre las concentraciones, así como tampoco hubo un efecto de
interacción entre ambos factores.
Tabla 18.- Cuadrados medios resultantes del ANOVA bifactorial de los caracteres evaluados a 45 días en el proceso de aclimatación.
Fuente de Variación
Largo del tallo
No. de Hojas Vigor Grosor del
tallo
Patrón (A) 1.4300 0.1002 0.0174 0.0211 * Sustrato (B) 7.3293 0.2336 0.0174 0.0004
(A x B) 0.3776 0.1713 0.0174 0.0004 Error 3.8380 0.1823 0.0224 0.0008 x ± EE 6.90 ± 0.53 2.99 ± 0.11 1.51 ± 0.04 0.24 ± 0.01
(*).- Significancia p 0.05
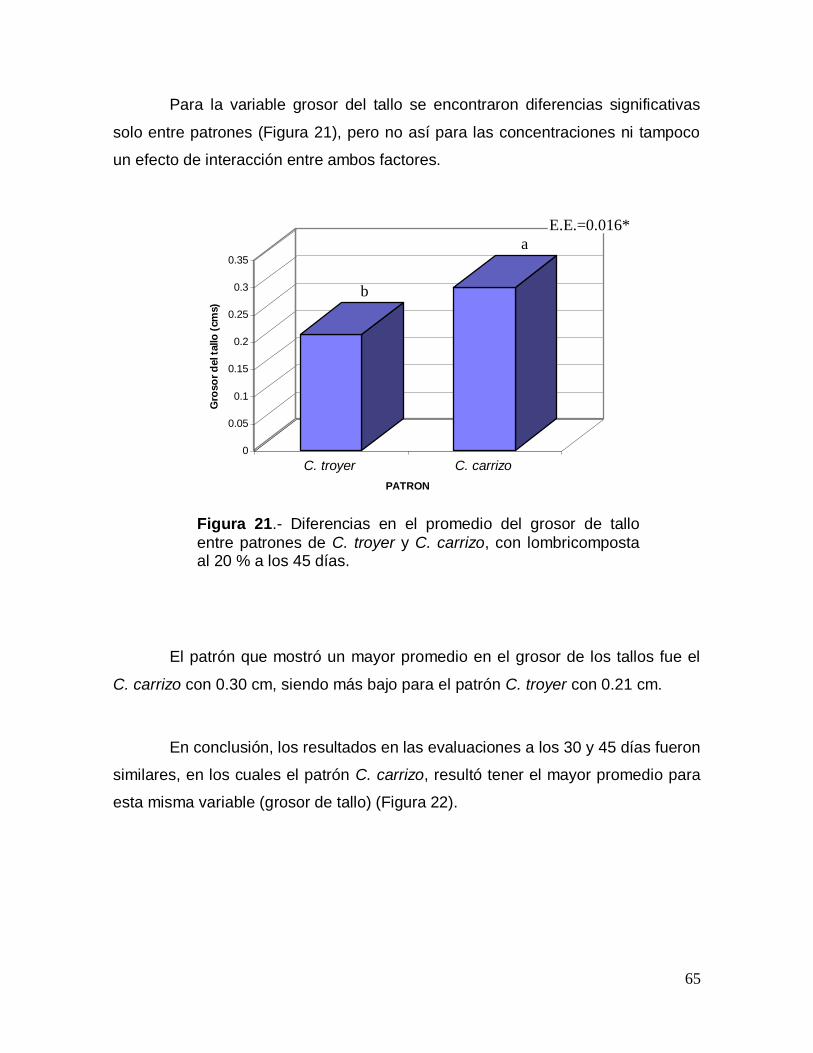
65
Para la variable grosor del tallo se encontraron diferencias significativas
solo entre patrones (Figura 21), pero no así para las concentraciones ni tampoco
un efecto de interacción entre ambos factores.
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
Gro
so
r d
el ta
llo
(cm
s)
C. troyer C. carrizo
PATRON
Figura 21.- Diferencias en el promedio del grosor de tallo
entre patrones de C. troyer y C. carrizo, con lombricomposta al 20 % a los 45 días.
El patrón que mostró un mayor promedio en el grosor de los tallos fue el
C. carrizo con 0.30 cm, siendo más bajo para el patrón C. troyer con 0.21 cm.
En conclusión, los resultados en las evaluaciones a los 30 y 45 días fueron
similares, en los cuales el patrón C. carrizo, resultó tener el mayor promedio para
esta misma variable (grosor de tallo) (Figura 22).
E.E.=0.016*
a
b

66
Figura 22.- Plantas de C. carrizo y C. troyer a los 45 días del proceso de aclimatación.
Los resultados del análisis estadístico de la segunda evaluación (45 días)
para las variables largo del tallo, número de hojas, vigor y grosor del tallo,
comparados con los grupos testigo (prueba de Dunnet), utilizando promedios de
cada variable analizada, se muestran en la Tabla 19.
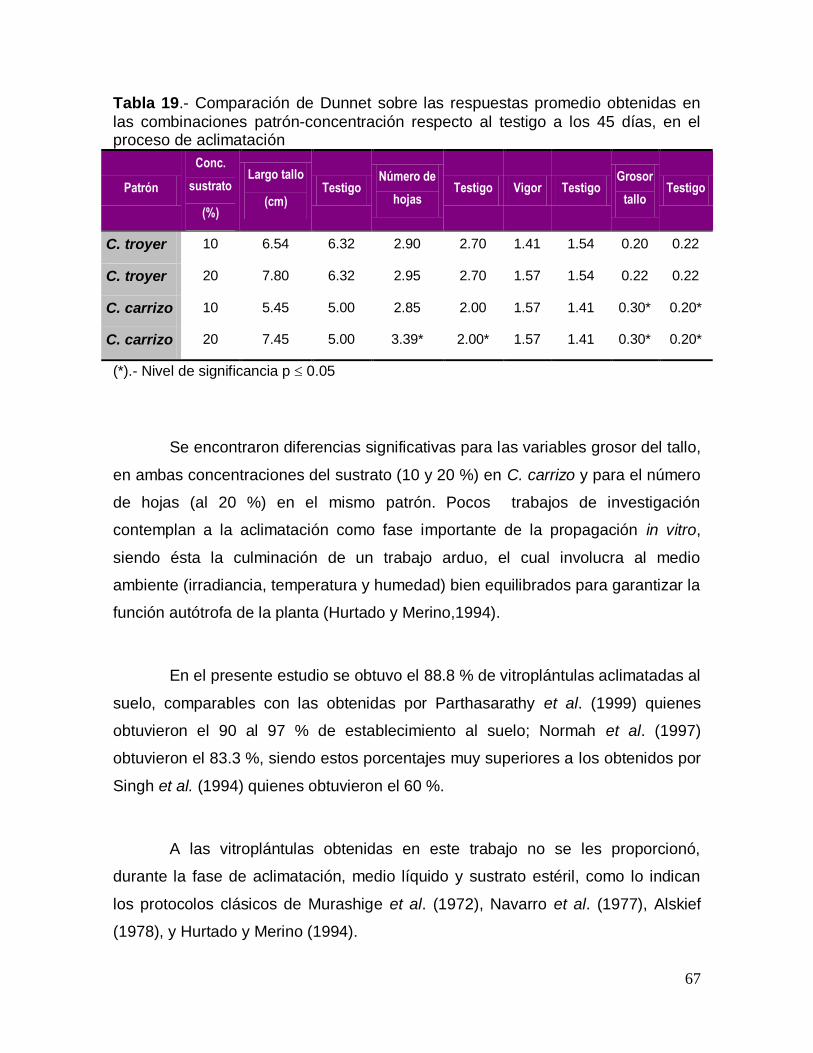
67
Tabla 19.- Comparación de Dunnet sobre las respuestas promedio obtenidas en
las combinaciones patrón-concentración respecto al testigo a los 45 días, en el proceso de aclimatación
Patrón
Conc.
sustrato
(%)
Largo tallo
(cm) Testigo
Número de
hojas Testigo Vigor Testigo
Grosor
tallo Testigo
C. troyer 10 6.54 6.32 2.90 2.70 1.41 1.54 0.20 0.22
C. troyer 20 7.80 6.32 2.95 2.70 1.57 1.54 0.22 0.22
C. carrizo 10 5.45 5.00 2.85 2.00 1.57 1.41 0.30* 0.20*
C. carrizo 20 7.45 5.00 3.39* 2.00* 1.57 1.41 0.30* 0.20*
(*).- Nivel de significancia p 0.05
Se encontraron diferencias significativas para las variables grosor del tallo,
en ambas concentraciones del sustrato (10 y 20 %) en C. carrizo y para el número
de hojas (al 20 %) en el mismo patrón. Pocos trabajos de investigación
contemplan a la aclimatación como fase importante de la propagación in vitro,
siendo ésta la culminación de un trabajo arduo, el cual involucra al medio
ambiente (irradiancia, temperatura y humedad) bien equilibrados para garantizar la
función autótrofa de la planta (Hurtado y Merino,1994).
En el presente estudio se obtuvo el 88.8 % de vitroplántulas aclimatadas al
suelo, comparables con las obtenidas por Parthasarathy et al. (1999) quienes
obtuvieron el 90 al 97 % de establecimiento al suelo; Normah et al. (1997)
obtuvieron el 83.3 %, siendo estos porcentajes muy superiores a los obtenidos por
Singh et al. (1994) quienes obtuvieron el 60 %.
A las vitroplántulas obtenidas en este trabajo no se les proporcionó,
durante la fase de aclimatación, medio líquido y sustrato estéril, como lo indican
los protocolos clásicos de Murashige et al. (1972), Navarro et al. (1977), Alskief
(1978), y Hurtado y Merino (1994).

68
Las vitroplántulas se lavaron con agua corriente, aún así el porcentaje de
sobreviviencia fue alto; esto demuestra que las plantas adquieren rápidamente la
capacidad de ser autótrofas y pueden sobrevivir a transplantes bruscos con un
tiempo de aclimatación mínimo de 45 días.
Después de la aclimatación de las plantas se llevó a cabo un segundo
diagnóstico de VTC con la prueba de ELISA, resultando plantas libres de virus
(ANEXO 4)

69
CONCLUSIONES
En base al estudio realizado sobre el establecimiento de una metodología
para la micropropagación de patrones tolerantes al virus de la tristeza de los
cítricos (VTC) se pudo arribar a las siguientes conclusiones:
1. Se comprobó que el mejor tratamiento para lograr desinfección y
termoterapia adecuadas en las semillas de ambos patrones, consistió en el
empleo de Benlate* por 25 minutos, hipoclorito de sodio 25% (v/v) de la
concentración comercial 10 minutos a 52-55° C por 10 minutos.
2. Se comprobó que la combinación hormonal más efectiva para la
inducción de brotes en C. carrizo y C. troyer fue la que contenía BAP 2.0 mg.l-1, y
AG3 2.0 mg.l-1.
3. El factor de crecimiento AIB en 1.0 mg.l-1, favoreció notablemente el
desarrollo armónico de la raíz en las plantas en un período de 30 días.
4. El tratamiento de lombricomposta al 20 % resultó el más adecuado para
lograr un elevado porcentaje de aclimatación (88.8 %) en ambos patrones.
5. Se constató que no existieron diferencias significativas para las
variables morfométricas evaluadas en las semillas de los 2 patrones en estudio, lo
que reafirma la notable similitud genética de las mismas.
6. La metodología propuesta permite la producción de vitroplántulas libres
de VTC con una tasa de multiplicación de 0.076, en un período de 105 días.

70
RECOMENDACIONES
Utilizar la metodología propuesta para la micropropagación (Figura
23) de genotipos valiosos de Citranges: C. troyer y C. carrizo, como variante
alternativa de la propagación vegetativa con vistas a:
1.- La multiplicación y regionalización a corto plazo de genotipos
promisorios; mantener la colección in vitro de variedades y establecer las bases
para los trabajos futuros relacionados con la producción masiva de estos patrones.
2.- Desarrollar estudios futuros encaminados a evaluar la homogeneidad
genética del material micropropagado.
3.- Emplear el esquema propuesto como base metodológica para el
establecimiento de la misma en otras especies cítricas de interés.
4.- Continuar profundizando en los estudios genético-fisiológicos y de
aclimatación para elevar la precisión de pruebas de progenie en estudios de
interacción genotipo-ambiente y en la selección temprana de caracteres
deseables.

71
5.- Continuar evaluando genéticamente las poblaciones obtenidas por esta
vía de propagación in vitro, con vistas a obtener información valiosa para su mejor
explotación en la práctica productiva.
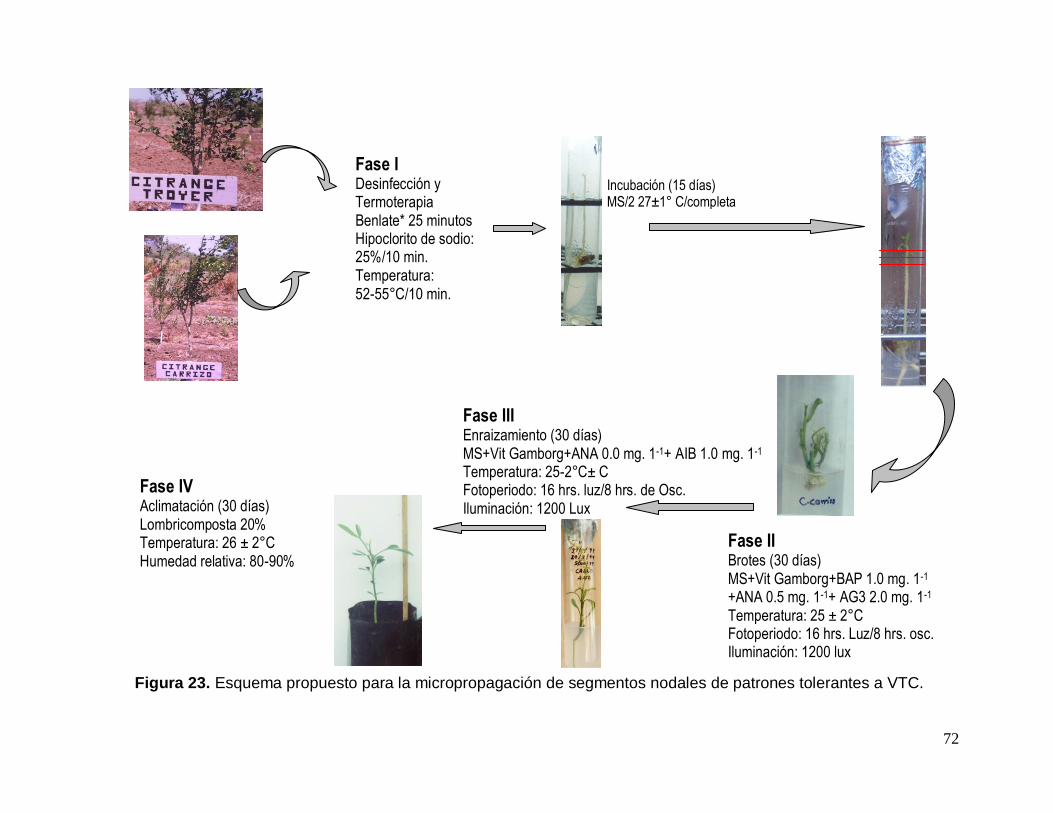
72
Fase I Desinfección y Termoterapia Benlate* 25 minutos Hipoclorito de sodio: 25%/10 min. Temperatura: 52-55°C/10 min.
Incubación (15 días) MS/2 27±1° C/completa osc.
Fase II Brotes (30 días) MS+Vit Gamborg+BAP 1.0 mg. 1-1 +ANA 0.5 mg. 1-1+ AG3 2.0 mg. 1-1
Temperatura: 25 ± 2°C Fotoperiodo: 16 hrs. Luz/8 hrs. osc. Iluminación: 1200 lux
Fase III Enraizamiento (30 días) MS+Vit Gamborg+ANA 0.0 mg. 1-1+ AIB 1.0 mg. 1-1
Temperatura: 25-2°C± C Fotoperiodo: 16 hrs. luz/8 hrs. de Osc. Iluminación: 1200 Lux
Fase IV Aclimatación (30 días) Lombricomposta 20%
Temperatura: 26 ± 2°C Humedad relativa: 80-90%
Figura 23. Esquema propuesto para la micropropagación de segmentos nodales de patrones tolerantes a VTC.

73
LITERATURA CITADA
AGRIOS, G.N. 1998. Fitopatología. UTEHA. Noriega Editores. Cuarta Impresión
de la segunda edición. México. ALSKIEF, J. 1978. La micropropagación in vitro de quelques arbres fruitiers. Tesis
(Doc) Université le sciences et techniques du Languedoc (USTL) Montpellier, Francia. 136 p.
BALDWIN, E.A. 1993. Citrus fruit. In: Biochemistry of Fruit Ripening. G. Seymour,
J. Taylor y G. Tucker (eds.). Chapman & Hall, London. Pp. 110-140. BAR-JOSEPH, M. AND LEE R. F. 1989. Citrus tristeza virus. Description of plant
viruses No. 353. Commonwealth Mycological Instituto/Association of applied Biologists. Kew Surrey, England. 7 p.
BAR-JOSEPH, M.; MARCUS R. AND LEE R. F.. 1989. The continuous challenge
of citrus tristeza virus control. Ann. Rev. Phytopathol. 27: 292-316. BARLASS, M. AND SKENE, K.G. 1982. In vitro plantlet formation from citrus
species and hybrids. Sci. Hort. 17: 333-341. BARUAH, A.; NAGARAJU, V. AND PARTHASARATHY, V.A. 1996.
Micropropagation of three endangered Citrus species. 1. Shoot proliferation in vitro. Ann. Plant Physiol. 10:124-128.
BELKOURA, L.; ISMAILI, M.; VASSEUR, J.; DUBOIS, J. AND DUBOIS, T. 1995.
Secondary somatic embryogenesis and plant regeneration of Troyer citrange from hypocotyl segments of nucellar embryos. Fruits, Paris. 50: 353-358.
BELOUALY, N. 1991. Plant regeneration from callus culture of three Citrus
roostock. Plant Cell Tiss. Org. Cult. 24:29-34. BOXUS, P. 1998. Plant biotechnology applied to horticultural crops. Hort. Science. BROADBENT, P.; BEVINGTON, K. B.., AND COOTE, B. G. 1991. Control of stem
pitting of grapefruit in Australia by mild strain protection. In: Brlansky, R. H., Lee, R. F. and Timmer, L. W. (Eds.). Proc. 11th Conf. Inter. Organ. Citrus Virol. Riverside, California. pp. 86-92.
BRUNT, A.A.; CRABTREE, K.; DALIWITZ, M.J.; GIBBS, A.J.; WATSON, L. AND
ZURCHER, E.J. (EDS). 1997. "Plant Viruses Online: Descriptions and Lists from the VIDE DATABASE”. Version 16th January 1997. URL http://bioloqy.anu.edu.au/Groups/MES/vide

74
BURGARIN, M.R.; LOZOYA, S.H. 1992. Propagación in vitro del portainjerto Rosa X Noisettiana.cv “Manettii” a partir de yemas axilares. Rev. Chapingo. Año XVI Num 78. Abr. Jun.
BUTTON, J. AND BORMAN, C.H. 1971. Develompment of nucellar plants from
unpollinated and unfertilized ovules of de Washington navel orange in vitro. South Afr. Bot. 37:127-134.
CALAVAN, E.; ROISTACHER, C.; NAUER E. 1972. Thermotherapy of citrus for
inactivation of certain viruses. Plant. Dis. Rep., 56: 976-980. CALDERON, A. I. 1993. Fruticultura General. 3ª. Edición. Ed. Limusa. México,
D.F. 763 p. CAMERON, J.W. AND SOOST, R.K. 1987. Citrus University of California,
Riverside. USA. Pag. 261-265. CAN, C.; KOC, N.K. AND CINAR, A. 1992. In vitro clonal propagation of sour
orange (Citrus aurantium var. Brezilia) by using epicotyls segments. Doga, Turk Tarim ve Ormancilik Dergisi. 16: 132-139.
CASTLE, W.S. 1987. In: Rootstocks for fruit crops, Rom, C.R. and Carlson, R.F.
(eds.). John Wiley and Sons, Inc. Ch 11. CASTLE, W.S. 1988. Patrones y Variedades. Memoria del seminario de
citricultura. Banco de México-FIRA. Merdida, Yuc. P 45-66. CASTLE, W.S.; TUCKER, D.P.H.; KREZDORN, A. H. AND YOUTSEY, C.O. 1993.
Rootstocks for Florida citrus. IFAS. Univ. Fla. CHAGOLLA, J.G. 1990. Cultivo de cítricos in vítro. FIRA. Boletín informativo. Vol.
XXII, No. 217. 12p CHATURVEDI, H.C.; Y MITRA, G.C. 1974. Clonal Propagation in vitro from
somatic callus cultures. HortScience, 9: 118-120. CHÁVEZ, J. L. Y GONZÁLEZ, C.F. 1999. Rev. Fitotec. Méx. 22: 27-41. CHEN, R.Z.; LI, G.G. AND ZHANG, L.Y. 1992. Study on in vitro grafting and
micropropagation of triploid Citrus plantlets. China Citrus. 21: 10-12. CHRISTOPHER, T. AND RAJAM, M.V. 1994. In vitro clonal propagation of
Capsicum spp. Plant Cell. Tiss. Org. Cult. 38:25-29. CIAT. 1983. Programa de Frijol. Informe Anual 1982. 278 p

75
COLLI, F.I.; Y CÁRDENAS, M.J.A. 1995. Campaña preventiva contra el VTC en México. In: Proceedings of the Third International Workshop. Lake Alfred, Florida, May 15-18. Lee, R. et al., University of Florida. USA. Pp. 211-214.
COTO, M.A.; Y RODRÍGUEZ, D.N. 1998. Evaluación de 4 patrones tolerantes al
virus de la tristeza de los cítricos en la región de Ursulo Galván, Veracruz. Memoria de Residencia Profesional. Dirección General de Educación Tecnológica Agropecuaria. Instituto Tecnológico Agropecuario No. 18. Ursulo Galván, Ver., 43p.
CURTI-DIAZ, S.A.; LOREDO, R.X.; DIAZ-ZORRILLA, U.; SANDOVAL, J.A. Y
HERNÁNDEZ, J. 1996. Manual de Producción de Limón Persa. (Tecnología de productores líderes para productores). INIFAP. Folleto No. 14.
DE LA OSA, F.J. 1998. Anteproyecto para el desarrollo de la fruticultura en el
CIICA. Documento Interno. Tapachula, Chiapas. 10p. DURAN-VILA, N.; ORTEGA, V. AND NAVARRO, L. 1989. Morphogenesis And
tissue cultures of three citrus species. Plant. Cell. Tiss. Org. Cult. 16:123-133. DURAN-VILA, N. AND MARIN, M.L. 1991. Conservation of citrus germoplasm in
vitro. J. Amer. Soc. Hort. Sci. 116: 740-746. ESPINOZA, G. 1994. Efectos de nutrición in vitro en los portainjertos Carrizo y
Yuma. Tesis para optar por el grado de Maestro en Ciencias. Colegio de Postgraduados. Montecillo, México. 103p.
EVANS, D.A. AND SHARP, W.R. 1986. Somaclonal and gametoclonal variation.
In: Evans, D.A. Sharp, W.R. and Ammirato, P.V. (Eds) Handbook of Plant Cell Culture. Vol. 4 Techniques and applications. Macmillan, New York. pp 97-132.
FERGUSON, L.; SOKOVICH, N.; AND ROOSE, M. 1990. California citrus
roostocks. Publication 21477. University of California. FITCHEN, J.H. AND BEACHY, R.N. 1993. Genetically engineered protection
against viruses in transgenic plants. Annu. Rev. Microbiol. 47:739-763. FORNER-VALERO, J.B. 1985. Combinaciones injerto-patrón en cítricos. Jornadas
Citrícolas Andaluzas. Junta de Andalucía, Consejería de Agricultura y Pesca. Dirección General de Estructuras y Producción Agraria. Sevilla España. pp. 41-56
FRANCIS, M. 2001. Principales métodos de detección del V.T.C. En: Symposium
Internacional “Virus tristeza de los cítricos”. Ciudad Victoria, Tamaulipas, México. 26 de Feb. Al 1 de Marzo. pp. 23.

76
GARCIA, L.A. Y BORDÓN, Y.; MOREIRA-DIAS, J.M.; MOLINA, R.V. AND GUARDIOLA, J.L. 1999. Explant orientation and polarity determine the morphogenic response of epicotyl segments of troyer citrange. Ann. Bot. 84: 715-723.
GARNSEY, S.M. AND MULLER G.W. 1988. Efficiency of mechanical transmission of citrus tristeza virus. In: Timmer, L. W., Garnsey, S. M. and Navarro, L. (Eds.). Proc. 10th conf. Inter. Organ. Citrus Virol. Riverside. Calif. Pag.
GARNSEY, S.M. AND LEE, R. F. 1988. Tristeza. In: Whiteside, J. O. Garnsey, S. M. and Timmer, L. W. (Eds.). Proc. 10th Conf. Intern. Organ. Citrus Virol. Riverside, Calif. pp. 48-50.
GARNSEY, S.M.; PERMAR, T. A.; CAMBRA, M. AND HENDERSON, C. T. 1993. Direct tissue blot Immunoassay (DTBIA) for detection of Citrus Tristeza Virus (CTV). In: Moreno, P. J. V. Da Graca, L. W. Timmer (Eds.). Proc. 12th Conf. Inter. Organ. Citrus Virol. Riverside. Calif. Pag. 39-50.
GILL, M.I.S.; SINGH, Z.; DHILLON, B.S. AND GOSAL, S.S. 1994. Somatic embryogenesis and plantlet regeneration on calluses derived from seedling explants of "Kinow' mandarin (Citrus nobilis Lour. X Citrus deliciosa Tenora). J. Amer. Hort. Sci. 69: 231-236.
GILL, M.I.S.; SINGH, Z.; DHILLON, B.S. AND GOSAL, S.S. 1995. Somatic embryogenesis and plantlet regeneration in mandarin (Citrus reticulata Blanco). Sci. Hort. 63: 167-174.
GONSALVES, D. AND GARNSEY, S.M. 1989. Cross-protection techniques for control of plant virus diseases in the tropics. Plant Dis. 75:592-597.
GONZÁLEZ, M.; PEÑA, L.; GONZÁLEZ, J.; ZAMORA, V. Y RODRIGUEZ, I. 1977. Introducción en Cuba del injerto in vitro de ápices de brotes en el género Citrus y géneros afines, como una forma de obtener plantas libres de virus. Agrotécnia Cuba. 9: 61-70.
GONZÁLEZ, M.; PEÑA, I. Y RODRÍGUEZ, I. 1980. Influencia de patrones y medios nutritivos sobre el prendimiento y desarrollo in vitro de injerto de ápices para la obtención de plantas libres del viroide de la exocortis a partir de un clon de Citrus limon infestado. Agrotécnia Cuba 12 : 67-76.
GONZALEZ, M.M.A. 1988. Diccionario De Especialidades Agroquímicas. 2da. ed. Ediciones P.L.M., S.A. de C.V. México, D.F.
GÓMEZ, S.; HERRERA, J. Y DIAZ, E. B. 1998. Comportamiento fonológico de variedades comerciales de cítricos sobre portainjertos tolerantes al virus de la tristeza en la región de Villa Ursulo Galván, Ver. Memoria de Residencia Profesional. Dirección General de Educación Tecnológica Agropecuaria. Instituto Tecno!ógico Agropecuarío No. 18. Ursulo Galván, Ver., 42p.

77
GRAVINA, T.A. 1989. Características viverísticas en 9 portainjertos de cítricos. Revista Chapingo (62-63):133-135.
GRAVINA, T.A. Y PIESTUN, D. 1991. Formación de un Banco de variedades de
citrus libres de virus: I. Obtención de plantas mediante la técnica de microinjerto. Boletín de Investigación No. 28. Facultad de Agronomía, Uruguay. 15P.
GUERRI, J.; MORENO, P. AND LEE, R, F. 1990. Identification of citrus tristeza
virus strains peptide maps of virion coat protein. Phyt. Int. J. 80:692. GUTIERREZ, E.M. A. 1996. Uso de la ingeniería genética en el control del virus de
la tristeza de los cítricos. En: Memorias del IV Simposium Internacional Sobre Sistemas de Producción en Cítricos. Vol. 1:95-99. E-mail:[email protected]
HARADA, H. ANS MURAI, Y. 1996. Clonal propagation of Poncirus trifoliata
through culture of shoots primordia. J. Hort. Sci. 71: 887-892. HERNÁNDEZ, P.F. Y MEJIA, M.J.M. 1994. Micropropagación de la Dalia (Dalia
variabilis Cav.). Revista Chapingo. Serie Horticultura Num 1:64-66. HURTADO, M..D.V. Y MERINO, M.M.E. 1994. Cultivo de Tejidos Vegetales. Ed.
Trillas. 3ra. Reimpresión. México, D.F. pp 154-161. IGLESIAS, L.; ROJAS, M. Y ENRIQUEZ, J. 2000. Utilización de las técnicas
biotecnológicas en los programas de prevención del virus de la tristeza de los cítricos en el estado de Veracruz. Cuaderno de Biodiversidad No. 6. 12 p
INEGI. 1996. Virus de la Tristeza de los Cítricos. Folleto Técnico No. 5 (2da.
Edición). División Agrícola. 37p. INIFAP. 1996. Manual de Producción de Limón Persa. Instituto Nacional de
Investigaciones Forestales, Agrícolas y Pecuarias. Centro de Investigación Regional Golfo Centro. Camp Experimental Ixtacuaco. Folleto Técnico Núm. 14. División Agrícola. Agosto 1996. Tlapacoyan, Ver. México. 145 p.
ISIDRON, H.L.; JUÁREZ, A.J.C.; VILLARREAL, G.L.A.; LLAUGUER, R.; BATISTA,
L.; PEÑA, I.; YABOR, L.; NIBLETT, CH.; PEREZ, M.E.; RIVERA, B.R. Y MARTINEZ, S.J.P. 2001. Caracterización de aislados mexicanos y cubanos del virus de la tristeza de los cítricos En: Symposium Internacional “Virus tristeza de los cítricos”. Ciudad Victoria, Tamaulipas, México. 26 de Feb. Al 1 de Marzo. pp. 7-22.

78
KENT, E.S. AND LEVIN, D.A. 1990. Quantitative genetics of seed size variation in Phlox. Evolutionary Ecology (4): 143-148.
KITTO, S.L. AND YOUNG, M.J. 1981. In vitro propagation of Carrizo Citrange.
HortScience 16: 305- 306. KUNITAKE, H., KAGAMI, H. AND MII, M. 1991. Somatic embryogenesis and plant
regeneration from protoplast of "Satsuma" mandarin (Citrus unshiu Marc.). Sci. Hort. 47:27-33.
LEE, R.F. AND ROCHA-PEÑA, M.A. 1992. Citrus tristeza virus. In: Plant deseases
of international importance III. A. N. Mukhapadhyay; H. S. Chaube; J. Kumar and U.S. Singh (Eds.). Prentice Hall. New Jersey.
LEE, R.F.; ROISTACHER, C.N.; NIBLETT, C.; LASTRA, R.; ROCHA, M.;
GARNSEY S.; YOKOMI, R.; GUMPF, D. AND DOBBS, J. 1992. Presence of Toxoptera citricidus in Central America. Citrus Ind. Mag. 73: 13-63.
LITZ, R.E.; MOORE, G.A. AND SRINIVASAN, CH. 1985. In vitro systems for
propagation and improvement of tropical fruits and palms. Hort. Rev. 7:157-197.
LOPEZ, B.J; AVITIA, G.E. 1991. Propagación in vitro de cuatro cultivares de
frambuesa (Rubus spp). Rev. Chapingo. Año XV: 73-74. Enero-Jun. LOPEZ, S. 1994. Problemática de la citricultura en la zona norte del estado de
Veracruz. Ponencia presentada al Programa Interdepartamental de Investigación y Servicio en Citricultura (PIISCI), en el IV Coloquio: "La citricultura en la cuenca del Golfo de México. Jalapa, Ver. Pp.60-63.
LOUSSERT, R. 1992. Los Agrios. Ed. Mundiprensa. Madrid, España. Pag. 19-42. LUKMAN, D.R.; JONARD, R. ANS VILLEMUR, P. 1990. Shoot apices culture of
Troyer citrange. lndonesian J. Tropical Agric. 1:72-74. MAS, O., VALLE, N.; RAMOS, M. AND DEL VALLE, N. 1994. In vítro propagation
of citremon. Proceedings of the Internacional Society of Citriculture, Volume 1. Taxonomy, breeding and varieties, rootstocks and propagation, plant physiology and ecology: 7th Internacional Cítrus Congress, Acireale, ltaly, 8-13 March: 318-320.
MEDINA, C.A. 1995. Propagación in vitro de tres portainjertos de cítricos
tolerantes a tristeza. Tesis para optar por el titulo de Maestro en Ciencias. Colegio de Postgraduados. Montecillo, México. 93 p.

79
MONTEVERDE, E.E.; GARCIA, M. L. Y BRICEÑO, M. 1999. Obtención de plantas cítricas libres de Psorosis y Exocortis en árboles infectados a través de la microinjertación de ápices in vitro. Agron. Tropical 36: 5-14.
MOORE, G.A. 1986. In vitro propagation of citrus rootstocks. HortScience 21:300-
301. MOORE, G.A.; JACONO, C.C.; NEIDIGH, J.L.; LAWRENCE, S.D. AND CLINE, K.
1992. Agrobacterium mediated transformation of citrus stem segments and regeneration of transgenic plants. Plant Cell Rep 11:238-242.
MORENO, P.; NAVARRO, L.; FUENTES, C.; PIÑA, J.A.; BALLESTER, J.F.;
HERMOSO DE MENDOZA, A.; JUÁREZ, J. Y CAMBRA, M. 1983. La tristeza de los agrios. Problemática en España. Hoja Técnica INIA No. 47. 28 pp
MORENO, P.; GUERRI, J. AND MUÑOZ, P. 1990. Identification of spanih of citrus
tristeza virus by análisis of double-stranded RNA . Phytopathology. 80:477. MORIN, C.H. 1985. Cultivo de Cítricos. IICA. San José de costa Rica. P. 203-228. MOSELLA, L. Y ASCUI, M. L. 1985. El cultivo in vitro como herramienta en la
investigación y multiplicación de plantas. I. Utilización de ápices meristemáticos en la micropropagación de cítricos. SIMIENTE. 55:54-60.
MULLER, G.W.; COSTA, A.S.; CASTRO, J.L. AND GUARADO, N. 1988. Results
from premunized tests to control the Capao Bonito strain of tristeza. In: Timmer, L.W., Garnsey, S.M., and Navarro, L. (Eds.). Proc. 10th Conf. Inter. Organ. Citrus Virol. Riverside, Calif. Pp. 82-86.
MUKOPADHYAY, S., JAISHREE, R., SHARMA, B. C. ANITA, G. SENGUPTA, R.
K. NATCH, P. S., RI, J. AND GURUNG, A. 1997. Micropropagation of Darjeeling orange (Citrus reticulata Blanco) by shoot tip grafting. J. Hort. Sci. 72(3):493-499.
MURASHIGE, T. AND SKOOG, F. 1962. A revised medium for rapid growth and
bioassay with tobacco tissue culture. Physiol. Plant. 15:473-497. MURASHIGE, T. AND TUCKER, 1969. Growth factor requeriments of Citrus tissue
culture. Proc. 1st. Itl. Citrus Symp. 3: 1155-1161. MURASHIGE, T.; BITTERS, W.P.; RANGAN, T.S.; NAUER, M.E.; ROISTACHER,
C.N. AND HOLLIDAY, P.B. 1972. A technique of shoot apex grafting and its utilization towards recovering virus free Citrus clones. HortScience 7:118-119.
NAVARRO, L.; ROISTACHER, C.N. AND MURASHIGE, T. 1975. lmprovement of
shoot-tip grafting in vitro for virus-free Citrus. J.Amer.Soc.Hort.Sci. 100:471- 479.

80
NAVARRO, L. AND JUAREZ, J. 1977. Elimination of citrus pathogens in
propagative budwood; II: In vitro propagation. Proc. Int. soc. Citriculture 3: 973-978.
NORMAH, N.N.; HAMIDAH, S. AND GHANI, F.D. 1997. Micropropagation of Citrus
halimii and endangered species of South-East Asia. Plant Cell Tiss. Org. Cult. 50: 225-227.
OMURA, M. AND HIDAKA, T. 1992. Shoot tip culture of Citrus. I. Culture
conditions. Bulletin of the Fruit Tree Research Station. No. 22:23-36. OPEKE, L.K. 1982. Tropical tree crops. John Wiley and Sons. USA. PP 202-214. OROZCO, S.M. 1995. Enfermedades presentes y potenciales de los cítricos en
México. UACH. 150p OROZCO, S.M. 1996. Enfermedades de los cítricos en México. Memorias sobre
sistemas de producción de cítricos. Universidad Autónoma Chapingo, PIISCI, pp. 115-128.
PADRÓN, J.E. 1991. Uso de patrones alternativos al naranjo agrio para naranja y
limón. En: Memorias del Primer Curso Internacional de Citricultura. PIISCI. Martínez de la Torre, Ver. , Marzo. Pp.33-36.
PADRÓN, J.E. 2001. Patrones tolerantes al VTC en México. En: Simposium
Internacional “Virus tristeza de los cítricos” Ciudad Victoria Tamaulipas, México. 26 de Feb. Al 1 de Marzo. pp. 27-28.
PAIVA, L.V.; CARVALHO, S. A.; SOUZA, M.; DE CARVALHO, S.A. AND DE
SOUZA, M. 1993. Obtaining a virus- free "Seleta Folha Mucha" through micrografting ín vítro. Pesq. Agrop. Brasileira 28: 1341-1344.
PALACIOS, J. 1978. Citricultura Moderna. Edit. Hemisferio Sur. Argentina. Pag. 1-
2. PARTHASARATHY, V.A.; NAGARAJU, V. AND RAHMAN, S.A.S. 1997. In vítro
grafting of Citrus reticulata Blanco. Folia Hort. 9: 87-90. PÉREZ-MOLPHE-BALCH, E. Y OCHOA-ALEJO, N. 1997. In vitro plant
regeneration of mexican lime and mandarin by direct organogenesis. HortScience 32:931-934.
PIERIK, R.L.M. 1987. In Vitro Culture of Higher Plants. Maritimus Nijhoft
Publishers, Dordrecht.

81
PIERIK, R.L.M. 1990. Cultivo in vitro de las plantas superiores. Ediciones Mundi-Prensa. Madrid, España. Pag. 69-87.
RAJ, B. R. AND ARYA H.C. 1978. Tissue culture propagation of citrus trees. Proc.
Int. Soc. Cit.: 135-140. RAMAN, H.; GOSAL, S.S. AND BRAR, D.S. 1992. Plant regeneration from callus
cultures of Citrus limon and C. jambhiri. Crop lmprov. 9:100-103. RAMIREZ, M.R. AND NEFTALI, O.A. 1996. An improved and reliable chili pepper
(Capsicum annuum L.) plant regeneration method. Plant Cell Rep. 16:226-231. RANGAN, T.S.; MURASHIGE, T. AND BITTERS, W.P. 1968. In vitro ínitiation of
nucellar embryos in monoembryonic Citrus. HortScience 3:226-227. RANGASWAMY, N.S. 1961. Experimental studies of females reproductive
structures of citrus microcarpa Bunge. Phytomorphology 11:109- 127. REYES, DE A. K. Y PALMIERI, M. 1997. El virus de la tristeza de los cítricos
(VTC) en Guatemala. Universidad del Valle de Guatemala. Rev. Ciencia en Acción Num. 2 pp 1-6.
RIAC. 1995. Declaración del taller de Lake Alfred sobre CTV. En: Aspectos
Técnicos y Prioridades para Mejorar la Viabilidad Financiera de la Producción y el Comercio de los Cítricos. Preparado por el Grupo de Trabajo sobre Manejo Integrado de Plagas de la Red Interamericana de Cítricos, RIAC. Mayo, 1995, Pp.11.
ROCHA-PEÑA, M.A.; LEE, R.F.; LASTRA, R.; NIBLETT, C.L.; OCHOA-CORONA,
F.M.; GARSNEY, S.M. AND YOKOMI, R.K. 1995. Citrus tristeza virus and its aphid vector Toxoptera citricida threats to Citrus production in the Caribbean and Central and North America. Plant Dis. 79(5):437-445.
RODRÍGUEZ, S.E.E. 1986. Propagación in vitro de limón mexicano (Citrus
aurantifolia). Tesis de Maestría en Ciencias. Centro de Fruticultura. Colegio de Postgraduados. Chapingo, México.
ROISTACHER, C.N. AND CALAVAN, E. 1972. Heat tolerance of preconditioned
citrus budwood for virus inactivation. Proc. 5th. Conf. Int. Org. Citrus Vir.: 256-261. U. Fla. Press, Gainesville.
ROISTACHER, C.N. AND BAR-JOSEPH, M. 1987. Aphid transmission of citrus
tristeza virus: A review. Phytophylactica 19: 163-167. ROISTACHER, C.N.; GUMPF, D.J.; DODDS, J.A. AND LEE, R.F. 1991. The threat
of “The citrus killer”. California Citograph 76: 4-12.

82
ROISTACHER, C.N. AND MORENO P. 1991. The worldwide theat from
destructive isolates of citrus tristeza virus – A review. In: Proc. 11th Inter. Organ. Citrus Virol. Riverside Calif. Pp. 7-19.
ROISTACHER, C.N.; GUMPF, J.A. Y LEE, R.F. 1994. Cada vez más cerca de
México el mayor enemigo de los cítricos. Resumen de Avances Citrícolas Pp.4-5.
SAGAR. 1993. Norma Oficial de Emergencia 001 SARH 1-1993 por la que se
establece con carácter obligatorio el Plan de Emergencia contra el virus de la Tristeza de los cítricos. Diario Oficial de la Federación 2 de Junio de 1993.
SAGAR. 1997a. Campaña Nacional Contra el Virus Tristeza de los Cítricos. En:
Taller “Virus Tristeza de los Cítricos en México” 10-12 de septiembre. CONASA. Dirección General de Sandiad Vegetal. México, D. F. Pag. 1-11.
SAGAR. 1997b. Guía ilustrada de la prueba de inmunoadsorción con enzimas
ligadas para la detección de fitopatógenos. CONASA. Dirección General de Sanidad Vegetal. Centro Nacional de Referencia de Diagnóstico Fitosanitario. México D.F. 23 p
SAUTON, A.; MOURAS, A. AND LITZ, A. 1982. Plant regeneration from Citrus root
meristerms. HortSience 57:227-231. SINGH, S.; RAY, B. K.; BHATTACHARYYA, S. AND DEKA, P.C. 1994. In vitro
propagation of Citrus reticulata Blanco and Citrus limon Burm. f. HortScience 29: 214- 216.
STARRANTINO, A. AND CARUSO, A. 1987. Experiences on the ín vitro
propagation of some Cítrus rootstocks. Acta Horticult. 112:471-478. STARRANTINO, A. 1992. In vitro micrografting of citrus; Petria 2: suplemento 1,
27-35; 13 ref. STEEL AND TORRIE. 1988. Bioestadística. Principios y Procedimientos. 2ª.
Edición. Ed. McGraw Hill. Pag. 182-183. SWINGLE, W.T. AND REECE, P.C. 1967. The botany of citrus and its wild
relatives. In: W. Reuther, H. J. Weber and L. D. Batchelar (eds). The citrus industry, Vol. 1 History World distribution, botany and varieties 190-430. University of California.
THIRUMALAI, S. AND THAMBURAJ, S. 1996. Plantlet regeneration in sweet
orange (Citrus sinensis Osbeck) var. Sathgudí and acid line (Citrus aurantifolia Swingle) var.PKM.1. South lndian Hort. 44:62-64.

83
VARGAS, H.J.J. Y GONZALEZ, R.H. 1991. Morfogénesis in vitro de Pinus patula Schl. et cham. Rev. Chapingo. Año XV. Núm. 76. Jul-Sep.
VILLARREAL, G.L.A. 2001. antecedents y situación del virus de la tristeza de los
cítricos en México. Dirección de Protección Fitosanitaria, DGSV-CONASAG. En: Memoria del Simposium Internacional “Virus Tristeza de los Cítricos”. Ciudad Victoria Tamaulipas, México. Pag. 1-6.
WALLACE, J.M. 1978. Virus and viruslike diseases. In: Reuther, W.; E. C. Calavan
and G. E. M. Carman (eds). The Citrus Industry IV: 89-109. Div. Agric. Sci. University of California, Riverside.
WUTSHCHER, H.K. 1979. Citrus rootstocks. In: J. Janik (Ed.) Hort. Rev. 1:237-
269. YOKOMI, R.K. AND GARNSEY, S.M. 1987. Transmission of citrus tristeza virus by
Aphis gossypii and Aphis citricola in Florida. Phytophylactica 19: 169-172. YOKOMI, R.K. AND DAMGSTEET, V.D. 1991. Comparison of citrus tristeza virus
isolates transmission efficiency between Toxoptera ctricidus and Aphis gossypii. In: Proc. Aphid-Plant Interactions: Populations and Molecules. Oklahoma State University. Stillwater, Oklahoma. 319 p.
ZAREI, A. AND RAHIMIAN, H. 1997. Elimination of Citrus tristeza virus from two
cultivars of Satsuma mandarin through shoot-tip grafting. lranian J. Plant Pathol. 33: 84-89.

84
ANEXOS
1. Certificado fitosanitario de la semilla.
2. Medio de Murashige y Skoog (1962) y vitaminas de Gamborg.
3. Primer diagnóstico de VTC, mediante la prueba de ELISA.
4. Segundo diagnóstico de VTC, mediante la prueba de ELISA.


















