
UNIVERSIDAD NACIONAL MAYOR DE SAN...
104
UNIVERSIDAD NACIONAL MAYOR DE SAN MARCOS (Universidad del Perú, Decana de América) ESCUELA DE POSGRADO FACULTAD DE CIENCIAS BIOLÓGICAS UNIDAD DE POSGRADO PRODUCCIÓN DE mRNA PARA CITOQUINAS HEMATOPOYÉTICAS (IL-3, GM-CSF e IL-7) EN RATONES INMUNOSUPRIMIDOS TRATADOS CON EXTRACTO ACUOSO DE Lepidium meyenii WALPERS (MACA) Tesis para optar al Grado Académico de MAGÍSTER EN BIOLOGÍA MOLECULAR Bachiller Evelyn Katy Alvarez Salazar LIMA – PERÚ 2013
Transcript of UNIVERSIDAD NACIONAL MAYOR DE SAN...

UNIVERSIDAD NACIONAL MAYOR DE SAN MARCOS
(Universidad del Perú, Decana de América)
ESCUELA DE POSGRADO
FACULTAD DE CIENCIAS BIOLÓGICAS
UNIDAD DE POSGRADO
PRODUCCIÓN DE mRNA PARA CITOQUINAS
HEMATOPOYÉTICAS (IL-3, GM-CSF e IL-7) EN RATONES
INMUNOSUPRIMIDOS TRATADOS CON EXTRACTO ACUOSO
DE Lepidium meyenii WALPERS (MACA)
Tesis para optar al Grado Académico de
MAGÍSTER EN BIOLOGÍA MOLECULAR
Bachiller Evelyn Katy Alvarez Salazar
LIMA – PERÚ
2013

Mira que te mando que te esfuerces y seas valiente; no temas ni desmayes,
porque Jehová tu Dios estará contigo en dondequiera que vayas
(Josué 1: 9)

AGRADECIMIENTOS
A Dios, por acompañarme y guiarme a lo largo de mi vida, por ser mi fortaleza
en los momentos de debilidad y por brindarme una vida llena de aprendizajes,
experiencias y sobre todo felicidad.
A mis padres, Uldarico y Dionicia, por su ejemplo de lucha y honestidad,
porque creyeron en mí y me dieron la oportunidad de tener una profesión.
A mis hermanas Elizabeth y Rosa, por llenar mi vida de alegrías y amor
cuando más lo he necesitado.
A mi asesora Libertad Alzamora por aceptarme en su laboratorio, por haber
compartido conmigo sus conocimientos y sobre todo su amistad.
A todas aquellas personas que de una u otra forma, colaboraron o participaron
en la realización de esta investigación, hago extensivo mi más sincero
agradecimiento.
Al Consejo Nacional de Ciencia, Tecnología e Innovación Tecnológica por el
apoyo económico para la culminación del grado y el desarrollo de la Tesis.

CONTENIDO
PAG
RESUMEN I
ABSTRACT II
LISTA DE TABLAS III
LISTA DE FIGURAS IV
LISTA DE ABREVIATURAS V
1. INTRODUCCIÓN 1
2. ANTECEDENTES
2.1 Lepidium meyenii WALPERS (MACA) 3
2.2 HEMATOPOYESIS Y CITOQUINAS 6
2.3 CICLOFOSFAMIDA 9
2.4 EMPLEO DE PLANTAS COMO PROTECTORES FRENTE A DROGAS
CITOTÓXICAS
11
3. HIPÓTESIS Y OBJETIVOS 13
4. MATERIALES Y MÉTODOS
4.1 ELABORACIÓN DEL EXTRACTO ACUOSO (EAc) DE MACA E
IDENTIFICACIÓN FITOQUÍMICA DE SUS PRINCIPALES COMPONENTES
14
4.2 INMUNOSUPRESIÓN EXPERIMENTAL CON CICLOFOSFAMIDA 18
4.3 EVALUACIÓN DEL EFECTO MODULADOR DEL EAc DE MACA SOBRE
LA PRODUCCIÓN DE mRNA PARA IL-3, GM-SCF e IL-7 EN EL BAZO Y
LA MÉDULA ÓSEA DE RATONES INMUNOSUPRIMIDOS
22
4.4 EVALUACIÓN DEL EFECTO MODULADOR DEL EAc DE MACA SOBRE
LA PROLIFERACIÓN CELULAR Y PRODUCCIÓN DE mRNA PARA IL-3,
GM-CSF E IL-7 EN CULTIVOS DE CÉLULAS MONONUCLEARES DE LA
MÉDULA ÓSEA
35

4.5 EVALUACIÓN DE LA INFLUENCIA DEL TRATAMIENTO CON EL EAc DE
MACA SOBRE LA PROLIFERACIÓN DE LAS CÉLULAS
HEMATOPOYÉTICAS DE RATONES INMUNOSUPRIMIDOS
39
4.6 ANÁLISIS ESTADÍSTICO 43
5. RESULTADOS
5.1 IDENTIFICACIÓN DE LOS PRINCIPALES COMPONENTES PRESENTES
EN EL EXTRACTO ACUOSO (EAc) DE MACA
44
5.2 INMUNOSUPRESIÓN EXPERIMENTAL CON CICLOFOSFAMIDA Y
TRATAMIENTO CON EL EAc DE MACA EN RATONES
45
5.3 EFECTO DEL EAc DE MACA SOBRE LA PRODUCCIÓN DE mRNA PARA
IL-3, GM-SCF e IL-7 EN CÉLULAS DEL BAZO Y LA MÉDULA ÓSEA DE
RATONES INMUNOSUPRIMIDOS
48
5.4 EFECTO MODULADOR DEL EAc SOBRE LA PROLIFERACIÓN
CELULAR Y PRODUCCIÓN DE mRNA PARA IL-3, GM-CSF E IL-7 EN
CULTIVOS DE CÉLULAS MONONUCLEARES DE LA MÉDULA ÓSEA
56
5.5 INFLUENCIA DEL TRATAMIENTO CON EL EAc DE MACA SOBRE LA
PROLIFERACIÓN DE LAS CÉLULAS HEMATOPOYÉTICAS DE RATONES
INMUNOSUPRIMIDOS
58
6. DISCUSIÓN 67
7. CONCLUSIONES 78
8. RECOMENDACIONES 79
9. REFERENCIAS BIBLIOGRÁFICAS 80
10. ANEXOS 92

RESUMEN
Lepidium meyenni (maca) es un cultivo tradicional de los Andes Centrales
del Perú. Se han demostrado sus propiedades antitumorales e
inmunomoduladoras, que lo convierten en un excelente candidato para
investigar su actividad hematopoyética.
En el presente estudio, se trataron ratones Balb/c a dosis de 200 mg/kg de
extracto acuoso (EAc) de maca amarilla por vía oral durante 2 meses previo a
la inmunosupresión (IS) con ciclofosfamida (CF), y se evaluó la producción de
mRNA para las citoquinas hematopoyeticas: interleuquina 3 (IL-3), factor
estimulador de colonias de granulocitos y monocitos (GM-CSF) e interleuquina
7 (IL-7) en el bazo y la médula ósea, tanto en los ratones tratados como en
sus controles. Se realizaron cultivos de células mononucleares de médula
ósea y se evaluó el efecto del EAc sobre la proliferación celular y producción
de mRNA para las 3 citoquinas hematopoyéticas.
La administración de EAc en ratones IS, incrementó (p<0.05) la producción
de mRNA para las tres citoquinas en el bazo y de IL-7 en la médula ósea, dos
días después de la IS; e IL-3 y GM-CSF en la médula ósea 5 días post-IS, al
compararlos con los grupos no tratados con el extracto. En cultivos de médula
ósea, el EAc en la dosis de 100 µg/ml, estimuló (p<0.05) la proliferación celular
y la producción de mRNA para IL-7. Además, los ratones IS y tratados con
EAc mostraron mayor recuento de células de médula ósea, sangre periférica,
unidades formadoras de colonias endógenas en el bazo (CFU-S) y respuesta
proliferativa a mitógenos de los linfocitos, cinco días después de la IS.
El EAc de maca estimula la producción de mRNA para las tres citoquinas
hematopoyéticas. La administración de EAc a ratones inmunocomprometidos
puede revertir los efectos supresores de la ciclofosfamida.
Palabras clave: Lepidium meyenii, maca amarilla, extracto acuoso,
inmunomodulación, citoquinas hematopoyéticas.

ABSTRACT
Lepidium meyenni (maca) is a traditional crop of the Central Andes of Peru.
Studies demonstrated their antitumor and immunomodulatory properties, which
makes it an excellent candidate to investigate its hematopoietic activity.
In the present study, Balb/c mice were orally treated with aqueous extract
(EAc) of maca at the doses of 200 mg/kg for two months prior to
immunosuppression (IS) with a single doses of cyclophosphamide (CF) (130
mg/kg) to evaluate the mRNA production of hematopoietic cytokines:
interleukin 3 (IL-3), granulocyte-monocyte colony stimulating factor (GM-CSF)
and interleukin-7 (IL-7) in spleen and bone marrow, both in treated and controls
mice. Murine bone marrow cells were isolated and co-incubated with aqueous
extract (EAc) of yellow maca to evaluate its effects on the cells proliferation
and mRNA production of the three hematopoietic cytokines.
The administration of EAc in immunosuppressed mice, increased mRNA
production for the three cytokines by spleen and IL-7 mRNA by bone marrow
on day 2 after immunosuppression, and IL-3 and GM-CSF mRNA production
by bone marrow on days 5 after IS, compared with the untreated group. EAc of
maca at 100 μg/ml stimulated the proliferation of mononuclear cells and IL-7
mRNA production of cultured bone marrow cells. The extract also induced in
EAc-treated mice and IS, an increase in bone marrow and peripheral blood cell
counts, endogenous colony forming units-spleen (CFU-S) and the proliferative
response of lymphocytes on days 5 after IS.
The aqueous extract of maca stimulates the mRNA production for three
hematopoietic cytokines. The administration of EAc to immunocompromised
mice can reverse the suppressive effects of cyclophosphamide.
Keywords: Lepidium meyenii, yellow maca, aqueous extract,
immunomodulation, hematopoietic cytokines.

LISTA DE TABLAS
Tabla 1. Protocolo empleado para el tratamiento con el extracto acuoso de
maca en ratones hembras balb/c.
Tabla 2. Secuencias de los primers empleados para el RT-PCR en tiempo
real.
Tabla 3. Resultados del tamizaje fitoquímico del extracto acuoso de Lepidium
meyenii (maca).
Tabla 4. Efecto del extracto acuoso de maca sobre los niveles séricos de la
enzima transaminasa glutámico pirúvica en los ratones inmunosuprimidos con
ciclofosfamida.
Tabla 5. Efecto del extracto acuoso de maca sobre la expresión de mRNA de
IL-3, GM-SCF e IL-7 en los cultivos de células mononucleares de la médula
ósea.
Tabla 6. Efecto del extracto acuoso de maca sobre el recuento diferencial de
sangre circulante en ratones inmunosuprimidos con ciclofosfamida.

LISTA DE FIGURAS
Figura 1. Hipocótilos del Lepidium meyenii Walpers (maca), ecotipo amarillo.
Figura 2. Elaboración del EAc de maca.
Figura 3. Administración del extracto acuoso de maca a los ratones.
Figura 4. Electroforesis de los RNA totales de las muestras analizadas.
Figura 5. Resultado del análisis de los primers para IL-3 mediante el Software
Primer-Blast.
Figura 6. Amplificación por PCR convencional con los primers específicos
para los genes evaluados.
Figura 7. Programa de análisis de imagen TotalLab Quant.
Figura 8. Curva de disociación correspondiente a los fragmentos amplificados
del transcrito del gen IL-7, se observa un solo pico con un Tm aproximado de
75,6°C en todas las muestras analizadas para IL-7.
Figura 9. Gráfico de la segunda derivada de la fluorescencia emitido por cada
reacción, mediante el análisis del Método de Cuantificación Comparativa del
software Rotor Gene (versión 1.7).
Figura 10. Gráfico de la segunda derivada generado por el software Rotor
Gene, para los fragmentos amplificados del transcrito del gen IL-7.
Figura 11. Obtención de la médula ósea de los fémures de los ratones.
Figura 12. Ensayo del MTT. El formazán formado es disuelto con
solubilizantes (DMSO, alcohol isopropílico, etc.) y la intensidad de la
coloración es proporcional a la cantidad de células vivas.
Figura 13. Colonias macroscópicas formadas en la superficie del bazo de
ratones inmunosuprimidos con ciclofosfamida.
Figura 14. Efecto de diferentes dosis de ciclofosfamida sobre el porcentaje de
células nucleadas de la médula ósea.
Figura 15. Efecto del extracto acuoso de maca sobre la expresión de mRNA
de IL-3 por esplenocitos de ratones inmunosuprimidos con ciclofosfamida.
Figura 16. Efecto del extracto acuoso de maca sobre la expresión de mRNA
de GM-CSF por esplenocitos de ratones inmunosuprimidos con ciclofosfamida.

Figura 17. Efecto del extracto acuoso de maca sobre la expresión de mRNA
de IL-7 por esplenocitos de ratones inmunosuprimidos con ciclofosfamida.
Figura 18. Efecto del extracto acuoso de maca sobre la expresión de mRNA
de IL-3 en la médula ósea de ratones inmunosuprimidos con ciclofosfamida
Figura 19. Efecto del extracto acuoso de maca sobre la expresión de mRNA
de GM-CSF en la médula ósea de ratones inmunosuprimidos con
ciclofosfamida.
Figura 20. Efecto del extracto acuoso de maca sobre la expresión de mRNA
de IL-7 en la médula ósea de ratones inmunosuprimidos con ciclofosfamida.
Figura 21. Efecto del extracto acuoso de maca sobre la proliferación de las
células mononucleares de la médula ósea.
Figura 22. Efecto del extracto acuoso de maca sobre el recuento de células
nucleadas de médula ósea en ratones inmunosuprimidos con ciclofosfamida.
Figura 23. Efecto del extracto acuoso de maca sobre el recuento de células
nucleadas de sangre circulante en ratones inmunosuprimidos con
ciclofosfamida.
Figura 24. Macrocolonias en el bazo de ratones inmunosuprimidos con
ciclofosfamida y evaluadas 5 días después de la inmunosupresión.
Figura 25. Efecto del extracto de maca sobre el recuento de las unidades
formadoras de colonia esplénica (CFU-S) de ratones inmunosuprimidos con
ciclofosfamida.
Figura 26. Efecto del extracto acuoso de maca sobre la respuesta proliferativa
a concanavalina A de los linfocitos del bazo de ratones inmunosuprimidos con
ciclofosfamida.
Figura 27. Efecto del extracto acuoso de maca sobre la respuesta proliferativa
a Lipopolisacárido de los linfocitos del bazo de ratones inmunosuprimidos con
ciclofosfamida.
Figura 28. Posible mecanismo de acción del extracto acuoso de maca sobre
las células hematopoyéticas.

LISTA DE ABREVIATURAS
EAc: Extracto acuoso
CF: Ciclofosfamida
IS: Inmunosuprimido
p.c.: Peso corporal
IL-3: Interleuquina 3
IL-7: Interleuquina 7
GM-CSF: Factor estimulador de colonias de granulocitos y macrófagos
CFU-S: Unidades formadoras de colonias endógenas en el bazo
CMH: Célula madre hematopoyética
CPH: Célula progenitora hematopoyética
RT-PCR: Reacción en cadena de la polimerasa con transcriptasa inversa
mRNA: Ácido ribonucleico mensajero

1
1. INTRODUCCIÓN
La hematopoyesis es el proceso de formación, desarrollo y maduración de
eritrocitos, leucocitos y plaquetas a partir de un precursor celular común e
indiferenciado conocido como célula madre que en el adulto se encuentra en
la médula ósea. Estas células requieren de un microambiente específico
conocido como estroma medular y de factores de crecimiento hematopoyético
(FCH) para su normal evolución. Los FCH son un conjunto de proteínas
llamadas citoquinas que actúan sobre las poblaciones inmaduras potenciando
su maduración y proliferación. Entre las más importantes están: Factor
Estimulador de Colonias Granulocito-Macrófago (GM-CSF), Factor Estimulador
de Células Precursoras (SCF), IL-3, IL-7 y Eritropoyetina (Epo) (Abbas, 2008;
Mayani et al., 2007).
La médula ósea es un lugar de continua proliferación y renovación de
células sanguíneas, por lo tanto es el órgano más afectado durante cualquier
terapia de inmunosupresión con fármacos citotóxicos como la ciclofosfamida.
La pérdida de células madre y la incapacidad de regeneración de nuevas
células sanguíneas de la médula ósea se traducirán en anemia, leucopenia y
trombocitopenia (Anderson et al., 1995; Haubitz, 2007). Actualmente, se están
aplicando estas drogas antineoplásicas en combinación con varios agentes
detoxificantes e inmunomoduladores con el fin de reducir o eliminar sus
efectos tóxicos adversos (Jena et al., 2010).
En los últimos años se está produciendo una tendencia de rápido
crecimiento mundial hacia la revalorización de los conocimientos sobre el uso
de plantas tradicionales, por sus posibles efectos medicinales y/o
nutricionales. Muchas de las plantas utilizadas en la medicina tradicional han
demostrado poseer actividades moduladoras de la médula ósea,
contrarrestando los efectos tóxicos de las radiaciones y drogas
antineoplásicas, además de mejorar los mecanismos inmunológicos
encargados de combatir las células malignas y proteger al organismo de las
infecciones (Bhushan y Manish, 2007).

2
Lepidium meyenii Walpers (maca) es una especie nativa adaptada a las
condiciones extremas existentes en los pisos ecológicos más altos de la
cordillera de los Andes Centrales del Perú. Es apreciada por su alto valor
nutritivo y energético, por lo que su importancia económica está vinculada
principalmente a sus propiedades revitalizantes, vigorizantes y estimulantes de
la reproducción (Obregón, 1998).
Es así como en los últimos años se han evaluado científicamente muchas
de sus propiedades, confirmando las mencionadas y aportando otras (Wanga
et al., 2007; Gonzales, 2012). Se comprobaron sus propiedades antitumorales
e inmunomoduladoras, al estimular tanto la respuesta inmune humoral como
celular (Alzamora, 2003; Alzamora et al., 2004). Sobre los estudios de la maca
en la hematopoyesis se reportaron que el extracto acuoso de maca ecotipo
amarillo favoreció significativamente el incremento de glóbulos blancos,
hemoglobina y recuento de células de médula ósea (Torres, 2008), en
animales inmunosuprimidos con 50 mg/kg de ciclofosfamida, señalando un
posible efecto estimulatorio en las células madre/progenitoras
hematopoyéticas, que la convierte en un recurso con potencial
quimioprotector.
Dado que estas células inmaduras responden a una serie de citoquinas
hematopoyéticas del estroma medular, importantes en la regulación de la
proliferación y diferenciación a células maduras, el estudio de estas citoquinas
es importante para dilucidar el mecanismo de acción de la maca a nivel
hematopoyético. Por lo que en este estudio se evaluará el efecto de la maca
sobre la expresión génica de tres citoquinas hematopoyéticas (IL-3, IL-7 y GM-
CSF) en un modelo de mielosupresión murina, y de esta manera se
proporcionará el soporte básico-molecular, indispensable para la explicación
adecuada de sus propiedades, en este caso moduladora de la hematopoyesis.

3
2. ANTECEDENTES
2.1 Lepidium meyenii WALPERS (MACA)
Es un cultivo tradicional de los Andes Centrales Peruanos, que crece y se
desarrolla en los ecosistemas Suni y Puna de los departamentos de Junín y
Pasco, en altitudes que oscilan entre los 3700 hasta los 4450 m.s.n.m. Es una
planta que reúne calidad alimenticia, alta productividad y adaptación a
condiciones ecológicas extremas donde otro cultivo no podría prosperar,
debido a las bajas temperaturas, heladas, granizadas y sequías; por lo que es
considerada la única especie del género Lepidium domesticada en los Andes
(Obregón, 1998). La maca fue un producto valioso para los incas, no solo por
su alto valor nutricional sino por su uso medicinal, especialmente como
revitalizante, afrodisiaco y potenciador de la fertilidad, razón por la cual esta
planta era considerada en la categoría de las plantas mágicas en los ritos que
los Incas y sus descendientes realizaban (Tello et al., 1992).
Los cultivares de maca que existen en la actualidad se diferencian
principalmente por el color externo del hipocótilo (ecotipos), que es la porción
comestible y pueden ser blancos, amarillos, negros, rojos y morados, siendo
los amarillos los más consumidos y de mayor producción. La diversidad de
esta coloración se debe principalmente a la presencia de antocianinas y
probablemente a xantofilas (Yllescas, 1994; Obregón, 1998). La población
nativa peruana de los Andes Centrales consume los hipocótilos de la maca
después de hacerlas secar naturalmente y en cantidades de 20 - 50 g/diario.
Los hipocótilos secos son consumidos preferentemente hervidos en agua o
leche, en forma de jugos o cocteles (Vílchez, 2001).
El nombre científico Lepidium meyenii Walpers fue designado por el doctor
Gerhard Walpers en 1843, en base a la observación de un espécimen silvestre
de la planicie de Pisacoma en Puno; sin embargo en 1989 la doctora Gloria
Chacón señaló que no había correspondencia entre los caracteres botánicos
del espécimen cultivado tradicionalmente en los Andes Centrales Peruanos y

4
las características de Lepidium meyenii Walpers, razón por la cual planteó la
necesidad del cambio del nombre a Lepidium peruvianum Chacón (Chacón,
1990), por lo que el nombre científico de la maca fue cuestionado a partir de
ese momento. Sin embargo en el 2011, el Doctor Destéfano y colaboradores,
determinaron mediante el análisis de código de barras de DNA del cloroplasto,
que ambos "especímenes" cultivado y silvestre, eran prácticamente lo mismo,
por lo que señalaron que no habría necesidad de cambiar el nombre
designado por el doctor Walpers (Destéfano, 2011).
Los análisis bromatológicos realizados a la maca comprobaron su alto valor
nutricional por su alto contenido de macro y micronutrientes, por lo que es
utilizada como suplemento alimenticio. Se concluye que su valor nutricional es
comparable al de cereales como maíz, arroz y trigo, superando en contenido
calórico, proteínas y carbohidratos a otros vegetales, además presenta un
contenido proteico superior a otros hipocótilos y tubérculos, y altos valores de
calcio y hierro. Entre otros nutrientes encontrados están los lípidos, fibras,
vitaminas y aminoácidos esenciales (Fairlie et al., 1999; Canales et al., 2000,
Bianchi, 2003).
En 1961 se reportó por primera vez la presencia de alcaloides, glucósidos,
taninos y saponinas (Castaño, 2008). Las investigaciones fitoquímicas
mencionan principalmente:
Alcaloides, hasta 4 fracciones macaína 1, 2 3 y 4 (Dini et al., 1994)
actualmente identificados como: (1R,3S)-1 methyltetrahidro-b-carboline-
3-oic-acid, un derivado de la dihidropiridina (macaridina) y dos de tipo
imidazólico (lepidilina A y lepidilina B) y (Cui et al., 2003; Muhammad et
al., 2002).
Glucosinolatos: Benzylglucosinolato (Glucotropaeolin) y m-
methoxybenzylglucosinolato (Dini et al., 2002). Otros investigadores
afirman la presencia de 8 glucosinolatos (Valentová y Ulrichová, 2003).
Isotiocianatos, producto de la hidrólisis de los glucosinolatos, por acción
de la mirosinasa. Son responsables del fuerte olor y sabor picante de la
maca (Piacente et al., 2002).

5
Macaeno y Macamidas (alkamidas benziladas), son ácidos grasos poli-
insaturados (Zhao et al., 2005).
Además de esteroles (Gutiérrez et al., 2009), antocianinas, flavonoides
(Sandoval et al; 2002), taninos y saponinas (Yllescas, 1994).
Algunas de las propiedades atribuidas a la maca han sido comprobadas
científicamente en base a experimentos con animales y humanos, la mayoría
realizados con maca ecotipo amarillo, por ser el más abundante en las
cosechas (Gonzales, 2012): En ratas se demostró que la maca revierte
parcialmente el efecto deletéreo de acetato de plomo y de la altura sobre la
espermatogénesis (Rubio et al., 2006; Gonzales et al., 2004). En humanos, se
reportó que la maca gelatinizada mejora el recuento, movilidad de
espermatozoides y el deseo sexual en varones adultos, pero los efectos
reproductivos son independientes de cambios hormonales, puesto que estos
no se modifican (Gonzales et al., 2003), en mujeres se reportó que reduce los
síntomas psicológicos y la disfunción sexual de la post-menopausia, pero
independiente de la actividad estrogénica y androgénica (Brooks et al., 2008).
También se reportó que la maca favorece la tasa de crecimiento y
supervivencia de peces juveniles (Lee et al., 2004) y que presenta un efecto
favorable para el tratamiento de la osteoporosis, al mejorar la densidad ósea y
restaurar la red trabecular en un modelo de ratas ovariectomizadas tratadas
con extracto etanólico de maca (Zhang et al., 2006). Otros estudios
demostraron el potencial antioxidante del extracto acuoso de maca, por su
capacidad de atrapar los radicales libres y proteger a las células contra el
estrés oxidativo (Sandoval et al., 2002), no inducir hemólisis de eritrocitos
(Rosas y Pino, 2005), no ejercer efectos citotóxicos en cultivos de hepatocitos
con dosis crecientes de maca (Valentová et al., 2006) y revertir los parámetros
lipídicos, de glucosa y el nivel de enzimas anti-oxidantes en ratas con
hipertrigliceridemia hereditaria, alimentadas con maca como suplemento
dietario (Vecera et al., 2007). Además, que el extracto acuoso favorece la
respuesta del organismo en una situación estresante y físicamente extenuante
(Suárez et al., 2009); y mejora el aprendizaje, memoria y defensas

6
antioxidantes del cerebro en ratas recién destetadas, proponiendo a la maca
como un adaptógeno (Oré et al., 2011).
Esta planta también ha demostrado poseer actividades
inmunomoduladoras, estimulando tanto la respuesta inmune humoral y celular.
Se ha demostrado que la administración oral del extracto clorofórmico de
maca ecotipo amarillo en ratones Balb/c, conduce a un incremento del título de
anticuerpos y de la capacidad fagocítica, favorece la recuperación de la
respuesta celular natural en animales inmunosuprimidos con metilprednisona y
estimula la producción de óxido nítrico en cultivos de macrófagos peritoneales.
Resultados similares se obtuvieron con el extracto acuoso (Alzamora, 2003;
Alzamora et al., 2004; 2007).
2.2 HEMATOPOYESIS Y CITOQUINAS
Es el proceso a través del cual se generan las células de la sangre y ocurre
bajo condiciones muy específicas en la llamada médula ósea, donde las
células hematopoyéticas se desarrollan en un ambiente específico
denominado microambiente hematopoyético, que consiste en una estructura
tridimensional altamente organizada de células del estroma (macrófagos,
fibroblastos, adipocitos, osteoblastos, células endoteliales, entre otras), células
accesorias (linfocitos) y sus productos (matriz extracelular, citoquinas,
quimiocinas entre otras) que regulan la sobrevida, autorenovación,
proliferación, maduración y migración de las células hematopoyéticas y resulta
en una progresión ordenada del sistema hematopoyético. Dicho
microambiente es crucial para la regulación de la hematopoyesis y la
alteración de algunos de sus componentes puede contribuir al desarrollo de
enfermedades hematológicas (Mayani et al., 2007; Ruiz, 2009).
El sistema hematopoyético está formado por diferentes tipos celulares
organizados jerárquicamente, y se pueden agrupar en:
Células madre hematopoyéticas (CMH) (0.01%), son las más primitivas
y se distinguen por su capacidad de autorenovación y

7
multipotencialidad. Solo una pequeña parte de su población se
encuentra en ciclo celular (1 al 25%) en consonancia con las
necesidades hematopoyéticas del momento, el resto se encuentra en
estado de quiescencia.
Células progenitoras hematopoyéticas (CPH) (0.5%), han perdido su
capacidad de autorenovación pero conservan su potencial proliferativo,
pueden ser multi, bi o mono-potenciales. La mayoría de ellas (55 al
75%) están en ciclo celular, por lo tanto son las más afectadas por las
drogas citotóxicas.
Células precursoras (>90%) son inmaduras, tienen escasa actividad
proliferativa, pueden ser identificadas en los frotis de médula ósea
Células maduras, son diferenciadas, tienen una identidad y función
definitiva.
Las células progenitoras son las que proliferan a gran escala, proveyendo
billones de células sanguíneas por día que son necesarias para mantener la
homeostasis en el individuo; pero cuando el número de estas células
disminuyen, por ejemplo a causa de una terapia mieloablativa, las células
madre hematopoyéticas son liberadas de su inhibición y comienzan a dividirse
y diferenciarse según los requerimientos del organismo (Mera et al., 2007;
Welsch, 2010).
La hematopoyesis no solo puede ocurrir en la médula ósea sino también en
órganos fuera de ella, siempre que el lugar contenga células estromales donde
se puedan acomodar las CMH/CPH y una producción local de citoquinas
hematopoyéticos que mantengan e induzcan la diferenciación de CMH/CPH, a
esto se lo conoce como hematopoyesis extramedular y en el humano puede
ocurrir en condiciones patológicas. A diferencia del humano, la hematopoyesis
extramedular persiste en la vida adulta del ratón y el bazo constituye el
principal sitio de hematopoyesis extramedular, donde las CMH/CPH presentes
en la pulpa roja, no solo circulan sino también residen, aunque su número es
inferior a los de la médula ósea, ellas contribuyen a la hematopoyesis en
condiciones de estrés, formando colonias macroscópicas que son visibles
como discretos nódulos en la superficie del bazo, por ejemplo como una

8
reacción compensatoria a la mielosupresión inducida por drogas citotóxicas,
ya que la médula ósea se vuelve inadecuada para mantener la hematopoyesis
normal (Oziemlak, 2005; Herbert et al., 2008; Massberg y Von Adrian 2009;
Kim, 2010).
Las citoquinas son piezas claves en la regulación de la hematopoyesis,
porque son capaces de inducir la sobrevivencia y proliferación de células
progenitoras, conduciéndolas hacia linajes específicos; además de ser
responsables de las acciones fisiológicas de las células maduras de la sangre.
Son glicoproteínas, en general de bajo peso molecular, liberadas por las
células del estroma y las células accesorias y actúan como mediadores
celulares, a través de receptores específicos que se encuentran en la
superficie de la célula diana hematopoyética. La unión de la citoquina a su
receptor inicia una cascada de fosforilación con la activación de varias vías de
señalización como las JAK/STAT cinasa y RAS/MAP cinasa. Mientras algunas
de ellas tienen un amplio rango de acción sobre muchas progenitoras
primitivas conduciéndolas a un aumento de todas las líneas celulares y a su
diferenciación; otras actúan de una manera más restringida, en líneas
celulares específicas comprometidas o diferenciadas (Soto et al., 1999; Roitt et
al., 2008; Abbas et al., 2008).
La Interleuquina 3 (IL-3) también conocida como factor estimulador de
colonias de multi-linajes, es producida principalmente por los linfocitos T
activados y actúa sobre las células progenitoras muy primitivas, expandiendo y
preparando esta población para la exposición posterior al GM-CSF y otros
factores de crecimiento, estimulando su diferenciación hacia la línea mieloide
(Córdova, 2003). Además, participa en la supervivencia de las células
hematopoyéticas y regulación de las funciones efectoras de las células
mieloides diferenciadas en su estado terminal. En el ratón, el gen de la IL-3
esta codificada en el cromosoma 11, tiene un tamaño de 2.2 Kb y posee 5
exones. Su mRNA tiene un tamaño de 849 bp, su cadena polipeptídica
contiene 166 residuos de aminoácidos (Nimer y Uchida, 1995; Hara y
Miyajima, 1996; Reddy et al., 2000).

9
El Factor estimulador de colonias de granulocitos y macrófagos (GM-
CSF) induce la diferenciación y proliferación de las células progenitoras
hematopoyéticas de granulocitos/macrófagos. Además, regula la
supervivencia y función de varias líneas celulares maduras (granulocitos,
neutrófilos y células dendríticas) y funciona como un mediador inflamatorio,
actuando sobre un número de diferentes tipos celulares y modulando su
función celular como presentadora de antígeno. Un amplio rango de tipos
celulares puede producir GM-CSF, pero a menudo requieren de estímulos.
Células T, macrófagos, células endoteliales, fibroblastos y mastocitos son
ejemplos de células que secretan GM-CSF luego de una estimulación. En el
ratón, el gen para GM-CSF está ubicado en el cromosoma 11, con 4 exones y
su mRNA posee un tamaño de 1,033 bp. Pertenece a la misma familia de la
IL-3 (Guthridge et al., 1998; Shi et al., 2006; Hercus et al., 2009).
La Interleuquina 7 (IL-7) es una glicoproteína de 20-28 KD, producida por
las células estromales del timo, medula ósea, y los órganos linfoides
secundarios. Es una citoquina pleitrópica pero no redundante, es decir
absolutamente necesario para el desarrollo de los linfocitos T y B en el ratón, y
T en el humano. Además, regula la homeostasis periférica de las células T
vírgenes y de memoria. El gen de la IL-7 murino consiste aproximadamente de
42 kb y está localizado en el cromosoma 3, posee 5 exones y su mRNA tiene
un tamaño de 2,475 bp (Fry y Mackall, 2005; Huang y Luther, 2012; Ceredig y
Rolink, 2012).
2.3 CICLOFOSFAMIDA
La ciclofosfamida (CF) es una de las drogas alquilantes más utilizada en
los protocolos quimioterapeúticos debido a su amplio espectro antitumoral, en
la prevención de rechazo de injertos y el tratamiento de algunas enfermedades
autoinmunes; y puede ser empleada sola o en combinación con otros
productos. Debe ser metabolizada por las enzimas microsomales del hígado
para ejercer sus efectos citostáticos, a través de sus dos metabolitos
alquilantes: Fosforamida y Acroleína. Gran parte de sus efectos se debe a la
inhibición de replicación de DNA, por lo que la CF no solo está restringida a las

10
células cancerosas sino también a otras células con proliferación activa como
los del sistema hematopoyético (CMH/CPH), gastrointestinal, epitelial, folículos
pilosos y las glándulas genitales. Por lo que la quimioterapia conlleva a varios
efectos tóxicos e implicaciones clínicamente significantes como la leucopenia,
anemia y daño intestinal, causales de la discontinuación del tratamiento
(Anderson et al., 1995; Liberman et al., 2008; Llopis, 2009).
La supresión de las funciones de la médula ósea o mielosupresión es uno
de los efectos adversos más frecuentes y limitantes de la administración de
ciclofosfamida, aunque es reversible implica la interrupción del tratamiento
hasta que los valores leucocitarios estén dentro del rango normal permitido
para la aplicación de otra dosis. La supresión de la médula ósea se produce
debido a la constante renovación de las células hematopoyéticas, que las hace
muy vulnerables a los citostáticos, y se refleja en anemia, leucopenia y
trombocitopenia en los recuentos de sangre periférica, que pueden llevar a
una inadecuada función inmunológica, infecciones severas e incluso la muerte
(Haubitz, 2007).
Salem et al. (2012) reportaron que la administración de 4 mg ciclofosfamida
(aproximadamente 150 mg/kg p.c.) a ratones C57BL/6, en dosis única, induce
una profunda leucopenia en la periferia entre los días 3 a 15, y en bazo y
médula ósea entre los días 3 a 6 (al 9no día hay un rebote, al 12avo día se
evidencia completa recuperación). En la fase de recuperación, el retorno a los
valores normales conlleva primero a un incremento significativo de células
mieloides en el bazo y médula ósea en el 9no día (efecto rebote), retornando a
los valores normales el 12avo día, los linfocitos B demoran 3 semanas en
recuperarse, de ahí la efectividad de la CF en el tratamiento enfermedades
autoinmunes.
Sefc et al. (2003) en un trabajo realizado con ratones C57B1/10SnPh
inoculados con 135 mg/kg de CF, dosis única, reportaron que la celularidad de
la médula ósea retornó a los valores normales absolutos después de 6 a 7
días, aunque la composición celular se mantuvo alterada hasta el día 14. El
mismo patrón se observa en el bazo, donde los eritrocitos y linfocitos son casi

11
eliminados, y el incremento de celularidad del bazo 5 días post-CF se debe
principalmente a la expansión de las células mieloides, que está relacionada
con el incremento significativo de las células progenitoras en el bazo (que
pasa del 4% al 69%), acompañado con una reducción de ellas en la médula.
Actualmente, se están aplicando estas drogas antineoplásicas en
combinación con varios agentes detoxificantes e inmunomoduladores, como
son los factores de crecimiento hematopoyético sintéticos (G-CSF, GM-CSF,
eritropoyetina, etc) y los agentes citoprotectores (Amifostina) con el fin de
acelerar la recuperación hematopoyética y/o evitar daños citotóxicos, y así
permitir un tratamiento más drástico y por ende más eficaz (Krawczenko et al.,
2005; Meenu, 2008; Jena et al., 2010).
2.4 EMPLEO DE PLANTAS COMO PROTECTORES FRENTE A DROGAS
CITOTÓXICAS
Muchas de las plantas utilizadas en la medicina tradicional poseen efecto
protector demostrado frente a la toxicidad de las radiaciones y drogas,
sugiriendo sus potenciales usos como suplemento alimenticio en pacientes
cuyo sistema hematopoyético se ve alterado por la quimio/radioterapia
(Bhushan y Manish, 2007).
La administración de los constituyentes de 4 plantas chinas (Fórmula Si-
Wu-Tang) en ratones irradiados, resultó en un incremento del recuento
leucocitario y de todos los progenitores hematopoyéticos de la médula ósea,
que son afectados por la irradiación (Liang et al., 2006). Por otro lado, se
demostró la eficacia de Myelophil que es una mezcla de extractos de Astragali
radix y Salviae radix, en incrementar los parámetros hematológicos, el
recuento de progenitores hematopoyéticos en ensayos clonogénicos y sobre-
regular la expresión de IL-3 en el bazo de ratones inmunosuprimidos con una
dosis de 0.3 g/kg de 5-Fluorouracilo (Shin et al., 2008).
Patra et al. (2012) reportaron que la administración previa de los
componentes fenólicos de la planta oriental Cinnamomum cassia en ratones

12
inmunosuprimidos con 50 mg/kg de CF redujo la hipocelularidad inducida por
CF en el bazo, médula ósea y sangre periférica, lo que se correlaciona con la
reducción de células hiploides e incremento de los niveles de enzimas anti-
oxidantes, reduciendo el estrés oxidativo hepático y medular producto del
metabolismo de la CF. Otro estudio señaló que el extracto acuoso de Polygoni
multiflori rico en polisacáridos, es capaz de incrementar significativamente los
niveles de IL-2, parámetros hematológicos, perfil anti-oxidante y promover la
hematopoyesis esplénica mediante la sobreexpresión del receptor de la
eritropoyetina y el factor de transcripción GATA-1 en animales
inmunosuprimidos por aplicación de 40 mg/kg/día de CF durante 5 días
consecutivos (Chen et al., 2012).
Con respecto Lepidium meyenii, se reportó que el extracto acuoso de maca
ecotipo amarillo en una dosis de 300 mg/kg p.c., favorece significativamente el
incremento de glóbulos blancos, hemoglobina y recuento de células de médula
ósea) en animales inmunosuprimidos con 50 mg/kg de ciclofosfamida (Torres,
2008), los mismos resultados se observaron con el extracto metanólico de
maca ecotipo morado, señalando por primera vez un posible efecto
estimulador de la maca sobre la hematopoyesis (Alvarez, 2008).
Aunque la administración de dosis subletales de CF conlleva a una
profunda supresión de la respuesta humoral, celular y de la médula ósea; el
tiempo y duración de la mielosupresión depende no sólo de la dosis empleada,
sino también del organismo (edad, estado nutricional, funcionamiento de la
médula), una recuperación rápida implica una menor probabilidad de contraer
infecciones severas y continuar con otro ciclo de inmunosupresión.

13
3. HIPÓTESIS Y OBJETIVOS
3.1 Hipótesis
El extracto acuoso de Lepidium meyenii (maca) estimula la producción de
mRNA para tres citoquinas hematopoyéticas: Interleuquina 3, Factor
estimulador de colonias de granulocitos / macrófagos e Interleuquina 7 en
ratones inmunosuprimidos con ciclofosfamida.
3.2 Objetivo General
Determinar la producción de mRNA para citoquinas hematopoyéticas (IL-3,
GM-CSF e IL-7) en ratones inmunosuprimidos con ciclofosfamida y
tratados con extracto acuoso de Lepidium meyenii Walpers (maca).
3.3 Objetivos específicos
3.3.1 Elaborar el extracto acuoso de maca y realizar la identificación
fitoquímica de sus principales componentes.
3.3.2 Determinar la dosis óptima de inmunosupresión con ciclofosfamida en
ratones.
3.3.3 Demostrar el efecto del extracto acuoso sobre la producción de mRNA
para las tres citoquinas hematopoyéticas en el bazo y la médula ósea
de ratones inmunosuprimidos.
3.3.4 Demostrar el efecto modulador del extracto acuoso sobre la
proliferación celular y producción de mRNA para las tres citoquinas
hematopoyéticas en cultivos de células mononucleares de la médula
ósea.
3.3.5 Determinar la influencia del tratamiento con el extracto sobre la
proliferación de las células hematopoyéticas de ratones
inmunosuprimidos.

14
4. MATERIALES Y MÉTODOS
4.1 ELABORACIÓN DEL EXTRACTO ACUOSO (EAc) DE MACA E
IDENTIFICACIÓN FITOQUÍMICA DE SUS PRINCIPALES
COMPONENTES
4.1.1 Elaboración del extracto acuoso de maca
Se trabajó con hipocótilos de maca, ecotipo amarillo, procedentes del Valle
de Junín, Departamento de Junín. Los hipocótilos de maca fueron
seleccionados según su estado de conservación y luego se procedió a
limpiarlos, cortarlos en pequeños trozos, secarlos a 45°C por 48 horas y
pulverizarlos con la ayuda de un molino (Figura 1). La harina de maca
obtenida se mezcló con agua destilada (1:10 P/V respectivamente) y se
sometió a hervido durante 15 minutos. Luego de enfriar, la mezcla se
centrifugó a 2000 rpm x 10 minutos y se recolectó el sobrenadante en placas
petri de plástico, en total esterilidad, que fueron puestas dentro de un
deshidratador hasta total evaporación del agua. El residuo seco, que es el
extracto acuoso deshidratado, se recolectó mediante raspado en un frasco y
se almacenó a 4°C (Figura 2).
Figura 1. Hipocótilos del Lepidium meyenii Walpers (maca), ecotipo
amarillo. Los hipocótilos fueron cortados en pequeños trozos empleando un
procesador de alimentos.

15
Figura 2. Elaboración del EAc de maca.
A. Sobrenadante proveniente del hervido de la harina de maca.
B. Residuo seco de maca, después de la evaporación total del agua.
4.1.2 Identificación fitoquímica de los principales componentes del EAc
Para la identificación de algunos de los componentes del EAc de maca, se
preparó una solución de trabajo, para ello se disolvió el extracto acuoso en
agua destilada a la dosis de 30 mg/ml, que corresponde a la dosis de 200
mg/kg de peso corporal promedio de un ratón. Se utilizaron los procedimientos
descritos en algunos libros de investigación fitoquímica (Lock, 1994; Sharapin,
2000; Marcano y Hasegawa, 2002).
Carbohidratos. A 2ml de la solución de trabajo se le adicionó 2 gotas del
reactivo de Molish (alfa-naftol al 1% en etanol 95%), y por las paredes del tubo
se depositó cuidadosamente 2 ml de H2SO4 concentrado, la prueba es positiva
para carbohidratos totales si se forma un anillo violeta en la interface. La
identificación de almidones, se realizó adicionando 2 gotas del reactivo de
Lugol (KI al 0,06% e I al 0,04% en H20dd) a 2 ml de la solución, la aparición de
color violeta indica reacción positiva. Se procedió a determinar la
concentración de carbohidratos totales presentes en la solución de trabajo,

16
mediante el método colorimétrico fenol/H2SO4. Para ello se diluyó 10 µl de la
solución de trabajo con 990 µl de H2Odd, a la dilución se le adicionó 500 µl del
reactivo fenol (fenol al 5% en H20dd) y 2,5 ml de H2SO4, después de mezclar se
dejó reposar por 10 minutos en oscuridad, para luego llevar los tubos a baño
maría por 30 minutos a 30°C. Cumplido el tiempo se realizó la lectura a 492
nm. La concentración de carbohidratos en mg/ml se determinó usando como
estándar una curva de glucosa.
Proteínas. Se mezclaron 250 µl de la solución de trabajo con 2,5 ml del
reactivo Bradford, se dejó reposar por 2 minutos y se realizó la lectura a 595
nm en espectrofotómetro. Se determinó la concentración de proteínas en
mg/ml mediante una curva estándar para seroalbúmina bovina.
Flavonoides. A 2ml de la solución de trabajo se le adicionó 0,5 ml de NaOH al
30%, el cambio de color de la solución a amarillo intenso es indicativo de la
presencia de flavonoides. Luego se determinó la concentración de flavonoides
(flavonoles y antocianinas) mediante el método de Lees y Francis (1972): A
0,5 gramos del extracto acuoso deshidratado, se le adicionó 9,5 ml de alcohol
acidificado (HCl 1,5 N en etanol 95%, en proporción 85:15 v/v). Se
homogenizó bien y se dejó macerar en refrigeración durante toda una noche.
Al cabo del tiempo, se filtró el macerado con papel Whatman N°1 y se
completó el volumen a 100 ml. Después de incubar la muestra en oscuridad
por 2 horas, se midió la absorbancia del filtrado a 374 nm (flavonoles) y 535
nm (antocianinas) en un espectrofotómeto. Se utilizaron las siguientes
fórmulas:
Flavonoles (mg/g extracto) = (A x V) / (98,2 x W)
Antocianinas (mg/g extracto) = (A x V) / (76,5 x W)
Donde:
A = Absorbancia
V = Volumen total del extracto en ml
W = Peso de la muestra en gramos

17
Alcaloides. Se disolvieron 0,5 gramos del extracto acuoso deshidratado en 10
ml de HCl 10% y se calentó la mezcla por 10 minutos a 60 °C. Después de
dejar enfriar, se filtró la mezcla con papel Whatman N°1 y se procedió a lavar
el papel filtro con 10 ml de HCl 10%. El filtrado resultante se utilizó para las
pruebas de alcaloides, en donde a cada 2 ml del filtrado se añadió 3 gotas del
reactivo de Hager (ácido pícrico saturado) o Mayer (MgCl2 al 1.4% y KI al 5%
en H2Odd). Se consideran como positivas, las pruebas en las que aparece una
turbidez definida (Mayer) o precipitado blanquecino (Hager y Mayer).
Triterpenos / esteroles. Se disolvieron 0,5 gramos del extracto en 3 ml de
H2SO4 y se dejó macerar durante 10 minutos. Se tomó 1 ml de la suspensión y
se mezcló con 1ml de anhidrido acético, se formaron 2 fases. La presencia de
un anillo guinda en la interfase es indicativo de triterpenos y de una fase
superior verdosa es indicativo de esteroles (López-Casamayor, 2007).
Taninos. A 1 ml de la solución de trabajo se le adicionó 250 µl de solución
FeCl3 (FeCl3 al 5% en HCl 0,5N), el viraje de color a azul-negruzco o pardo-
verdoso es indicativo de la presencia de taninos.
Saponinas. Se tomaron 10 ml de la solución de trabajo, se agitó
vigorosamente por 5 minutos. La prueba se considera positiva si aparece
espuma en la superficie del líquido de más de 2 mm de altura y persiste por
más de 2 minutos.
Todas las lecturas de absorbancia se hicieron empleando el espectrofotómetro
UNICO 1100.

18
4.2 INMUNOSUPRESIÓN EXPERIMENTAL CON CICLOFOSFAMIDA
4.2.1 Animales de experimentación
Se utilizaron ratones hembras Balb/c, de 4 semanas de edad, con pesos
promedios de 25 gramos, procedentes del Instituto Nacional de Salud y se
observaron durante una semana para confirmar su buen estado de salud.
Todos los ratones fueron mantenidos en condiciones estándar de temperatura,
humedad y luz y recibieron ad libitum agua y alimento balanceado para
roedores (UNALM). El manejo de los animales se hizo en base al código de
ética sobre experimentación animal y en los principios éticos internacionales
que guían la investigación biomédica con animales.
4.2.2 Determinación de la dosis óptima de inmunosupresión con
ciclofosfamida
El inmunosupresor empleado fue la ciclofosfamida en polvo estéril
(NEOPHOS 200). Se trata de una droga antineoplásica sintética, del tipo de
las mostazas nitrogenadas; se presenta como un polvo blanco cristalino,
soluble en agua, solución fisiológica y etanol. Su nombre químico es 2-[bis (2-
cloroetil) amino] tetrahidro-2H-13,2-oxazafosforina 2-oxido monohidrato; su
fórmula molecular es C7H5C12N2O2PH2O y su peso molecular es 279,1. Este
profármaco requiere su bioactivación en los microsomas hepáticos para
ejercer su mecanismo de acción, que se basa en la alquilación de las bases
nitrogenadas del DNA. La alquilación provoca la pérdida de la configuración
espacial de las cadenas de DNA e imposibilita su síntesis durante la mitosis y
de esta forma ocasiona la muerte celular.
Con la finalidad de escoger la dosis adecuada para inducir supresión de la
función de la médula ósea sin poner en riesgo la vida del animal, y evaluar los
resultados en la fase inicial de recuperación post-inmunosupresión con
ciclofosfamida (post-IS), se realizó un estudio piloto con tres ratones cada
grupo, probando tres dosis diferentes de ciclofosfamida: 65, 130 y 200 mg/kg
de peso corporal (pc), en un volumen de 0,2 ml y por vía intraperitoneal. La

19
evaluación se realizó a los 2, 5 y 7 días después de la inmunosupresión, se
obtuvieron suspensiones celulares de la médula ósea de los fémures de cada
ratón y se realizó el recuento al microscopio, utilizando una cámara de
Neubauer de las suspensiones medulares diluidas en solución de TURK, que
hemolisa a los eritrocitos. Se eligió la dosis de 130 mg/ kg pc de ciclofosfamida
y realizar la evaluación a los 2 y 5 días después de la inmunosupresión.
4.2.3 Tratamiento con el extracto acuoso de maca
Se formaron 6 grupos de 12 ratones cada uno, de acuerdo a la siguiente
relación:
Grupo IS2d – EAc : Inmunosuprimidos, evaluados 2 días
después de la inmunosupresión, recibieron tratamiento con EAc de maca
Grupo IS2d – Sin EAc : Inmunosuprimidos, evaluados 2 días
después de la inmunosupresión, no recibieron tratamiento con EAc de
maca
Grupo IS5d – EAc : Inmunosuprimidos, evaluados 5 días
después de la inmunosupresión, recibieron tratamiento con EAc de maca
Grupo IS5d – Sin EAc : Inmunosuprimidos, evaluados 5 días
después de la inmunosupresión, no recibieron tratamiento con EAc de
maca
Grupo EAc : No inmunosuprimidos, recibieron
tratamiento con EAc de maca
Grupo Sin EAc : No inmunosuprimidos, no recibieron
tratamiento con EAc de maca
Se preparó una dilución del extracto acuoso (EAc) deshidratado, en agua
bidestilada estéril a una dosis de 200 mg/kg de peso corporal (p.c). La dosis
de EAc contenida en un volumen de 0,2 ml, se aplicó diariamente por vía oral,
en una solo dosis, a los ratones de los grupos IS2d-EAc, IS5d-EAc y EAc
durante todo el experimento, que tuvo una duración de 2 meses (Figura 3).
Los otros 3 grupos (IS2d-Sin EAc, IS5d-Sin EAc, Sin EAc) sólo recibieron
agua. La inmunosupresión se realizó en la última semana del experimento; los

20
grupos IS2d-EAc, IS2d-Sin EAc, IS5d-EAc e IS5d-Sin EAc recibieron una
dosis de 130 mg/kg de peso corporal de ciclofosfamida (NEOPHOS 200)
contenida en 0,2 ml, por vía intraperitoneal. Los ratones de los grupos IS2d-
EAc e IS2d-Sin EAc fueron sacrificados por dislocación cervical, 2 días
después de la inmunosupresión. A diferencia de los ratones de los grupos
IS5d-EAc e IS5d-Sin EAc, que fueron sacrificados 5 días después de la
inmunosupresión. El protocolo de tratamiento con el extracto acuoso para los
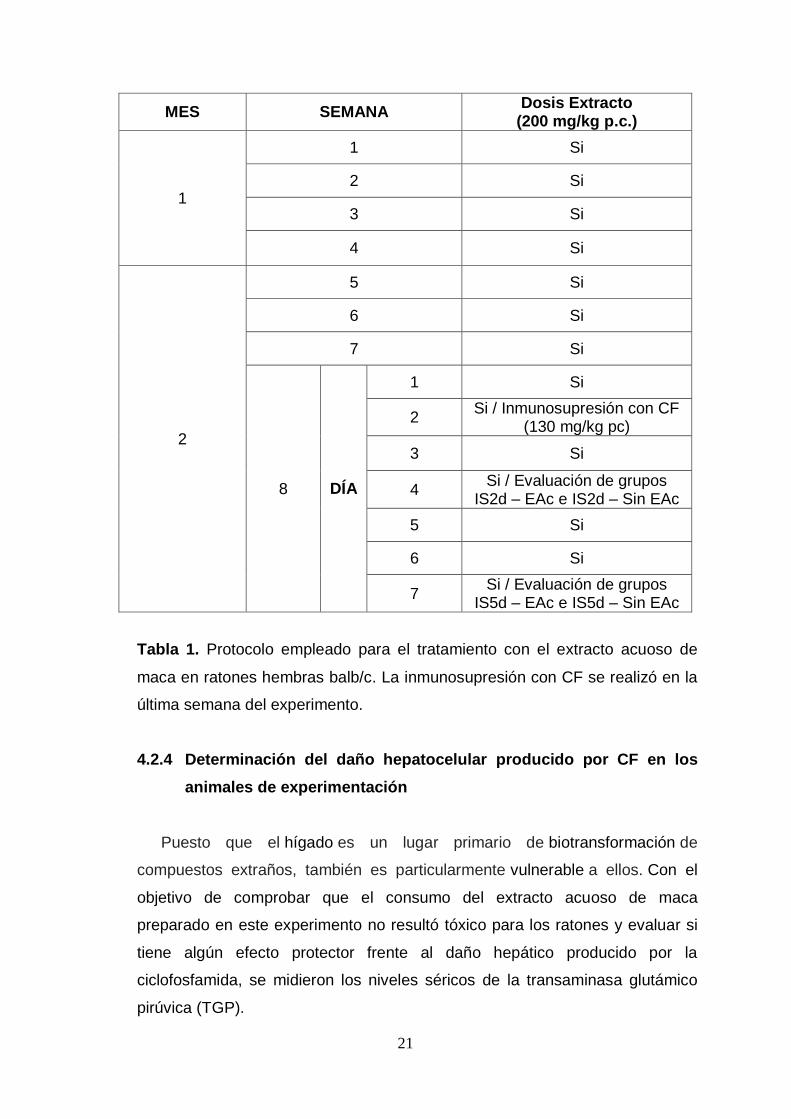
ratones inmunosuprimidos y no inmunosuprimidos se presenta en la Tabla 1.
Figura 3. Administración del extracto acuoso de maca a los ratones.
21
MES SEMANA Dosis Extracto
(200 mg/kg p.c.)
1
1 Si
2 Si
3 Si
4 Si
2
5 Si
6 Si
7 Si
8 DÍA
1 Si
2 Si / Inmunosupresión con CF
(130 mg/kg pc)
3 Si
4 Si / Evaluación de grupos
IS2d – EAc e IS2d – Sin EAc
5 Si
6 Si
7 Si / Evaluación de grupos
IS5d – EAc e IS5d – Sin EAc
Tabla 1. Protocolo empleado para el tratamiento con el extracto acuoso de
maca en ratones hembras balb/c. La inmunosupresión con CF se realizó en la
última semana del experimento.
4.2.4 Determinación del daño hepatocelular producido por CF en los
animales de experimentación
Puesto que el hígado es un lugar primario de biotransformación de
compuestos extraños, también es particularmente vulnerable a ellos. Con el
objetivo de comprobar que el consumo del extracto acuoso de maca
preparado en este experimento no resultó tóxico para los ratones y evaluar si
tiene algún efecto protector frente al daño hepático producido por la
ciclofosfamida, se midieron los niveles séricos de la transaminasa glutámico
pirúvica (TGP).

22
Las transaminasas son enzimas que catalizan la transferencia reversible de
un grupo amino entre un aminoácido y un cetoácido. La transaminasa
glutámico pirúvica (TGP) es una enzima citosólica que se encuentra en altas
concentraciones en el hígado. La lesión hepatocelular desencadena la
liberación de esta enzima en la circulación, por lo que constituye un excelente
marcador de la lesión hepatocelular (Alvarez, 2005).
Al finalizar el experimento, se tomaron muestras de sangre por punción
cardiaca en tubos sin anticoagulante de cada animal. Los tubos se
centrifugaron a 3500 rpm por 10 minutos para la obtención del suero. Los
niveles séricos de la TGP se determinaron según el procedimiento señalado
en el inserto del Kit diagnóstico Transaminasas color (Valtek diagnostic). El
fundamento del método es el siguiente: Las transaminasas del suero
reaccionan con el substrato, formando el producto piruvato que reacciona con
2,4 dinitrofenilhidrazina, generando en medio alcalino una hidrazona coloreada
que se lee a 505 nm. La cantidad de transaminasas en el suero se determinó
por interpolación de las absorbancias en la curva de calibración del reactivo
estándar. Se descartaron las muestras con hemolisis visible, ya que se pueden
obtener valores falsamente elevados.
4.3 EVALUACIÓN DEL EFECTO MODULADOR DEL EAc DE MACA SOBRE
LA PRODUCCIÓN DE mRNA PARA IL-3, GM-SCF e IL-7 EN EL BAZO Y
LA MÉDULA ÓSEA DE RATONES INMUNOSUPRIMIDOS
4.3.1 Análisis semicuantitativo de la expresión de mRNA de las
citoquinas IL-3, GM-SCF e IL-7 del bazo
Las suspensiones del bazo fueron obtenidos por presión del órgano con
ayuda de un émbolo para disgregar el tejido y liberar las células, dentro de una
placa petri con 5 ml de medio RPMI. Se juntaron las suspensiones celulares
de 3 ratones por grupo (se tomaron 500 µl de suspensión del bazo de cada
ratón), para proceder a extraer el RNA con Trizol, determinar su integridad por

23
electroforesis, su pureza y concentración por espectrofotometría y realizar la
transcripción reversa para la síntesis de DNA complementario.
4.3.1.1 Extracción de RNA
La extracción de RNA se realizó mediante el método de fenol-cloroformo
usando el reactivo Trizol (InvitrogenTM, Life Technologies) según el protocolo
establecido por Invitrogen. Todos los materiales de plástico utilizado en la
extracción, eran libres de RNasas, tratados con NaOH 0,5 M y autoclavados,
para evitar la degradación del RNA. Las centrifugaciones se realizaron a 4°C
en un microcentrífuga refrigerada.
Al finalizar el experimento, se obtuvieron suspensiones celulares del bazo
que fueron recolectadas en crioviales estériles que se centrifugaron a 1500
rpm por 3 minutos. Después se eliminó el sobrenadante y se adicionó 500 µl
de Trizol, luego de 5 minutos de incubación se añadió 100 µl de cloroformo y
se centrifugaron los crioviales por 15 minutos a 15000 rpm, obteniéndose tres
fases. La fase superior acuosa que contiene el RNA se retiró a otro criovial,
sobre el cual se adicionó 250 µl de alcohol isopropílico para precipitar el RNA,
formándose un pellet. El RNA precipitado se lavó dos veces con etanol al 70%,
y se dejó secar por 10 minutos, luego de resupenderlo en 50 µl de agua para
PCR, se separaron en alicuotas y almacenaron a -20°C.
4.3.1.2 Determinación de la integridad, pureza y concentración del RNA
extraído
La integridad de las muestras del RNA extraído fue verificada a través de
electroforesis en gel de agarosa al 1,5 %. Se mezclaron 6 µl de RNA con 4 µl
de formamida y 2 µl de buffer de carga (0,25 % de azul de bromofenol, 50% de
glicerol, EDTA 1 mM) dentro de crioviales, que se incubaron en baño maría a
60°C por 3 minutos y se pasó rápidamente al hielo. Se tomó 10 µl de cada
mezcla para cargar los pocillos del gel de agarosa en buffer TAE 1X (40 mM
tris, 20 mM ácido acético, 1 mM EDTA, ph 7,7). La electroforesis se llevó a
cabo a 100 voltios por 60 minutos, al finalizar los geles fueron teñidos con

24
bromuro de etidio (0,5 µg/ml) por 5 minutos. El RNA se observó en un
transiluminador con radiación UV (Biometra TI 1). Debido a que el mRNA
comprende el 1-3% del RNA total, es difícil detectarlo incluso con los métodos
más sensible; por otra parte, el RNA ribosomal comprende más del 80% y la
mayoría son las subunidades 18s y 28s, con un peso molecular aproximado de
2 kb y 5 kb respectivamente. La presencia de dos bandas definidas (18s y 28s
de los RNA ribosomales) es indicativa de la integridad del RNA (Figura 4).
Figura 4. Electroforesis de los RNA totales de las muestras analizadas.
Las fechas señalan la presencia de dos bandas de RNA ribosomales, en
los carriles 3, 4, 6 y 7. El primer carril corresponde al marcador de 1kb – 10
kb.
La pureza y concentración del RNA extraído fue determinado mediante
espectrofotometría UV (UNICO modelo 2100 UV). Se preparó una dilución
1:200 en buffer TE (10 mM tris, 1 mM EDTA, ph 7,5) estéril y se midió la
absorbancia en el espectrofotómetro a 260 nm y 280 nm. Con los valores de
absorbancia se calcularon la pureza y concentración de RNA, a partir de las
siguientes ecuaciones:

25
Pureza de RNA = A260 / A280
Valores entre 1,8 - 2 considera que el RNA extraído está libre de
contaminantes (proteína, sales)
Concentración RNA (µg/µl) = (A260 x 40 x Factor de dilución) / 1000
Dado que 1 unidad de Absorbancia (A) a 260 nm corresponde a 40 µg/ml
de RNA
Se consideraron las muestras de RNA que alcanzaron buenos estándares
de integridad, calidad y cantidad (Rapley y Manning, 1998).
4.3.1.3 Síntesis de DNA complementario (cDNA)
El RNA extraído fue sometido a una transcripción reversa para la síntesis
de cDNA, para lo cual se empleó el Kit High capacity RNA-to-cDNA (Applied
Biosystems), siguiendo las instrucciones del protocolo. Este Kit permite la
síntesis de DNA en solo dos pasos, ya que entre sus componentes se tiene al
Buffer mix RT 2X (contiene los dNTPs, random primers) y la Enzima mix RT
20X (contiene la enzima transcriptasa reversa Multiscribe™MuLV, proteína
inhibidora de RNAsas). Brevemente, a 40 µl de la mezcla RT (25 µl de Buffer
mix RT 2X, 2,5 ul Enzima mix RT 20X y 12,5 µl de H20) se le añadió 10 µl de
RNA (0,2 µg/µl) de manera que la concentración final de RNA sea 0,04 µg/µl.
Esta mezcla se sometió a un ciclo de síntesis de cDNA a 37°C por 60 minutos
y a un ciclo final de desnaturalización de la enzima a 95°C por 5 minutos en el
termociclador (Bioer). Una vez que terminaron los ciclos, el cDNA se almacenó
a - 20°C.
4.3.1.4 Elección de los Primers
Después de hacer una selección exhaustiva considerando el tipo de tejido
analizado y la metodología empleada, se escogieron aquellos primers que se
ajusten a los parámetros sugeridos para el diseño de primers (Apéndice IV del
protocolo Power SYBR® Green Cells-to-CT™ Kit). Se realizó un búsqueda
bibliográfica de secuencias de primers utilizados para cada uno de los

26
transcritos de los genes murinos de IL-3, IL-7, GM-CSF y Gliceraldehído 3
fosfato deshidrogenasa (GAPDH), donde el último fue considerado como
control de expresión constitutiva (housekeeping) para normalizar los
resultados del PCR en tiempo real (Ullmannovi y Haakovec, 2003; Willems et
al., 2006).
Las eficiencias de los primers se probaron virtualmente mediante Primer
BLAST disponible en http://www.ncbi.nlm.nih.gov/tools/primer-blast., el cual
emplea el software Primer 3 para diseñar los oligonocleótidos y después los
analiza en BLAST para evitar la formación de dímeros así como hibridaciones
inespecíficas (Figura 5) (Ye et al., 2012). Alternativamente, los primers fueron
analizados mediante la herramienta bioinformática Oligoanalyzer 3,1 para
verificar el contenido de G-C, formación de hetero - y homo - dímeros,
formación de horquillas y la determinación de su Tm, con el fin de obtener
unos primers óptimos (Cano et al., 2010). Finalmente, los primers se enviaron
a sintetizar a la casa comercial Invitrogen Life Technologies ®. La secuencia
de los primers, el tamaño esperado de los productos amplificados para cada
transcrito y el número de acceso en el GenBank, se enlistan en la Tabla 2.

27
Figura 5. Resultado del análisis de los primers para IL-3 mediante el
Software Primer-Blast.
Nombre del Gen
Secuencias de los primers (5´ - 3´) Tamaño del
amplicón esperado (pares de bases)
N° de acceso Gen Bank
IL-3 FW: TACATCTGCGAATGACTCTGC RV: GGCTGAGGTGGTCTAGAGGTT
132 NM_010556.4
GM-CSF FW: ACCACCTATGCGGATTTCAT RW: TCATTACGCAGGCACAAAAG
146 NM_009969-4
IL-7 FW: GTGCTGCTCGCAAGTTGAAG RW: AGTTCACCAGTGTTTGTGTGC
102 NM_008371.4
GAPDH FW: TGCACCACCAACTGCTTAGC RV: GGCATGGACTGTGGTCATGAG
258 NM_008084
Tabla 2. Secuencias de los primers empleados para el RT-PCR en tiempo real

28
Con el fin de evaluar la especificidad de los primers sintetizados, se
realizaron amplificaciones por PCR convencional de los genes evaluados, esto
además permitió corroborar la calidad de los RNA totales obtenidos, así como
analizar la eficiencia de la transcripción reversa. Las amplificaciones
obtenidas se evaluaron mediante electroforesis en gel de agarosa 2%, teñido
con bromuro de etidio. Las condiciones de amplificación por PCR convencional
se explican más adelante.
Por comparación con el marcador de peso molecular incluido en el gel, se
pudo establecer que los productos amplificados correspondieron al tamaño
esperado en todas las reacciones. El tamaño de las bandas fueron
determinadas con el Programa de análisis de imagen TotalLab Quant (Figura
6).
Figura 6. Amplificación por PCR convencional con los primers específicos
para los genes evaluados. El primer carril corresponde al marcador de
peso molecular de 100 – 1000 pares de bases.

29
4.3.1.5 Reacción en cadena de la polimerasa (PCR)
El cDNA sintetizado fue posteriormente amplificado mediante PCR
convencional. La reacción del PCR se preparó en un volumen final de 25 µl, en
una mezcla de reacción que contenía: 1 µl de cDNA, 2,5 µl de Buffer PCR 10X
(Appied Biosystems), 0,5 µl de MIX dNTP 10 mM, 1,5 µl de MgCl 15 mM
(Applied Biosystems), 1 µl de cada uno de los primers (forward y reverse, 10
µl), 0,25 µl de Taq-polimerasa (5U/µl) (Invitrogen TM Lifes Technologies) y
17,25 µl de H20 para PCR.
Las amplificación del cDNA se realizó en un termociclador (Bioer) y las
condiciones fueron: un paso de 94°C por 5 minutos (desnaturalización inicial) y
35 ciclos de 94°C por 30 s (desnaturalización), 55°C por 30 s (alineamiento) y
72°C por 45 s (extensión). De los 25 µl del volumen total de reacción de PCR
se tomaron 4 µl y se mezclaron con 1 µl de buffer de carga (0,25 % de azul de
bromofenol, 50% de glicerol, EDTA 1 mM).
Los productos obtenidos se separaron en geles de agarosa horizontal al
2% por electroforesis en buffer TAE 1X (40 mM Tris, 20 mM ácido acético, 1
mM EDTA, ph 7,7) a 100 voltios durante 50 minutos. Después de la corrida,
los geles fueron teñidos con bromuro de etidio (0,5 µg/ml) y las bandas
resultantes se visualizaron en un transiluminador de luz UV (Biometra TI 1)
que fueron fotografiadas con una cámara digital. Las imágenes digitales
obtenidas en color real fueron seleccionadas y transformadas en una imagen
en escala de grises, para estimar la intensidad de las bandas por
densitometría de una manera indirecta, midiendo el nivel de gris medio de las
bandas (Arrebola, 1999). Todo esto se realizó mediante el Programa de
análisis de imagen TotalLab Quant (www.totallab.com/products/totallabquant/)
(Figura 7). Todos los valores, expresados en niveles de gris, fueron
normalizados a su respectivo gen de referencia (GAPDH) como sigue:
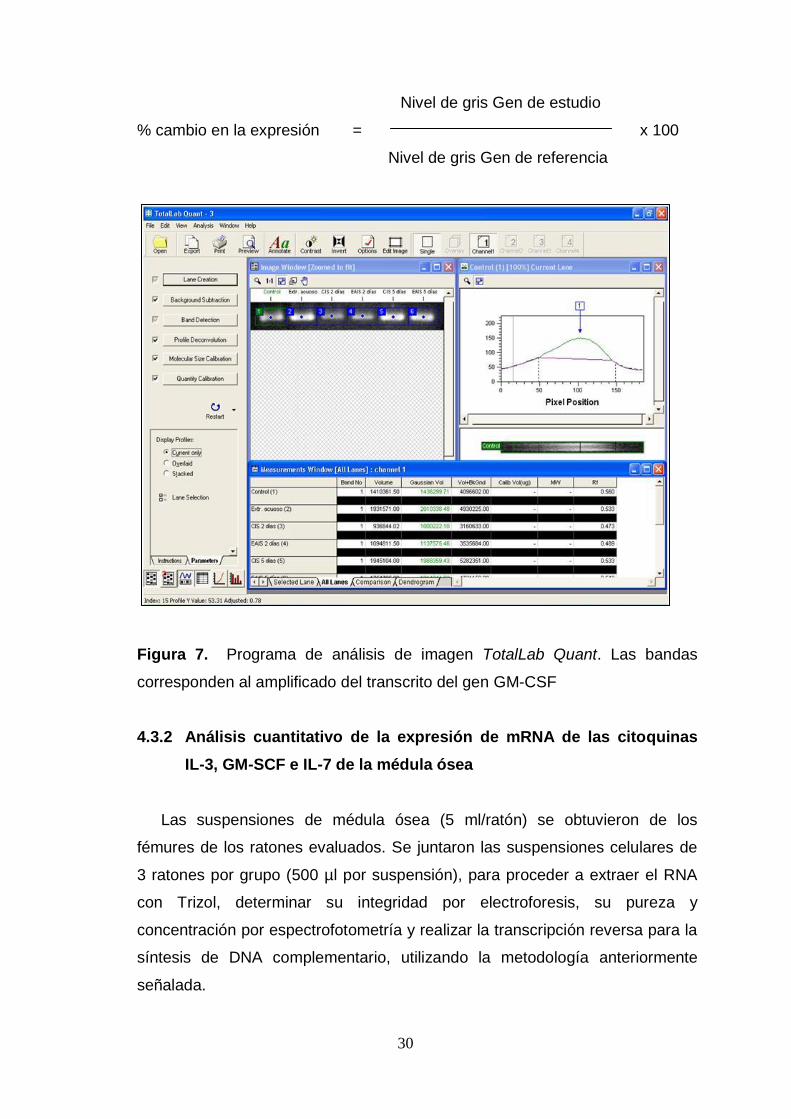
30
Nivel de gris Gen de estudio
% cambio en la expresión = x 100
Nivel de gris Gen de referencia
Figura 7. Programa de análisis de imagen TotalLab Quant. Las bandas
corresponden al amplificado del transcrito del gen GM-CSF
4.3.2 Análisis cuantitativo de la expresión de mRNA de las citoquinas
IL-3, GM-SCF e IL-7 de la médula ósea
Las suspensiones de médula ósea (5 ml/ratón) se obtuvieron de los
fémures de los ratones evaluados. Se juntaron las suspensiones celulares de
3 ratones por grupo (500 µl por suspensión), para proceder a extraer el RNA
con Trizol, determinar su integridad por electroforesis, su pureza y
concentración por espectrofotometría y realizar la transcripción reversa para la
síntesis de DNA complementario, utilizando la metodología anteriormente
señalada.

31
4.3.2.1 Reacción en cadena de la polimerasa (PCR) en tiempo real
La expresión génica de IL-3, GM-SCF, IL-7 y GAPDH fue evaluada por
PCR en tiempo real utilizando un equipo ROTOR GENE 3000 (Corbett
Research, Mortlake, Australia) Para la prueba se emplearon los primers
específicos para cada gen, el cDNA sintetizado y el reactivo Master Mix Power
SyBr®Green PCR (Applied Byosistem) que contiene todos los componentes
necesarios para la amplificación (AmpliTaq Gold DNA polimerase LD,
SybrGreen I, dNTPs, referencia interna pasiva). Se siguieron las instrucciones
del protocolo establecido para el kit Power SYBR® Green Cells-to-CT™.
Se realizó la siguiente mezcla de reacción en tubos ópticos: 10 µl de
Master Mix, 1 µl de cada primer (Forward y Reverse, 10 µM) y 4 µl de Agua
para PCR. A cada mezcla se añadió 4 µl de cDNA (diluido 1:3 con agua para
PCR). El volumen final fue de 20 µl, y las concentraciones finales de cada
primer y cDNA fueron de 500 nM y 2.7 ng/µl (si se considera una
retrotranscripción 100% eficiente), respectivamente; los controles fueron los
tubos con la mezcla de reacción y agua pura en vez de cDNA. Las muestras
fueron corridas por triplicado y en dos ocasiones separadas, las condiciones
de PCR utilizadas fueron las mismas para los cuatro genes: Activación de la
enzima (95° por 10 minutos), 40 ciclos de PCR (95°C por 15 segundos y 60°C
por 1 minuto) y por último la curva de disociación (melting), que incluye un
calentamiento gradual cada 0,1°C, desde 50 a 99°C.
El SYBR Green es un flourocromo que tiene la propiedad de unirse a toda
molécula de DNA de doble cadena, de manera inespecífica, por lo que el
incremento de la señal de fluorescencia es proporcional a la cantidad de DNA
de doble cadena presente en la reacción. Sin embargo esta inespecificidad
puede resultar en falsos positivos porque se puede unir a dímeros de primers.
Una forma de asegurar la especificidad de la reacción es analizando las
curvas de disociación o de melting. La curva de disociación se basa en una
gradiente de temperaturas crecientes, que permite monitorizar la cinética de
disociación de los fragmentos amplificados a partir de su temperatura de
disociación (Tm) que es específica para cada fragmento amplificado, ya que
32
depende del tamaño de los amplicones y secuencia de los mismos (contenido
de G y C). La presencia de dos o más picos sugiere que se ha obtenido más
de un amplificado y que el proceso de amplificación no fue específico para el
DNA blanco (Erali y Wittwer, 2010).
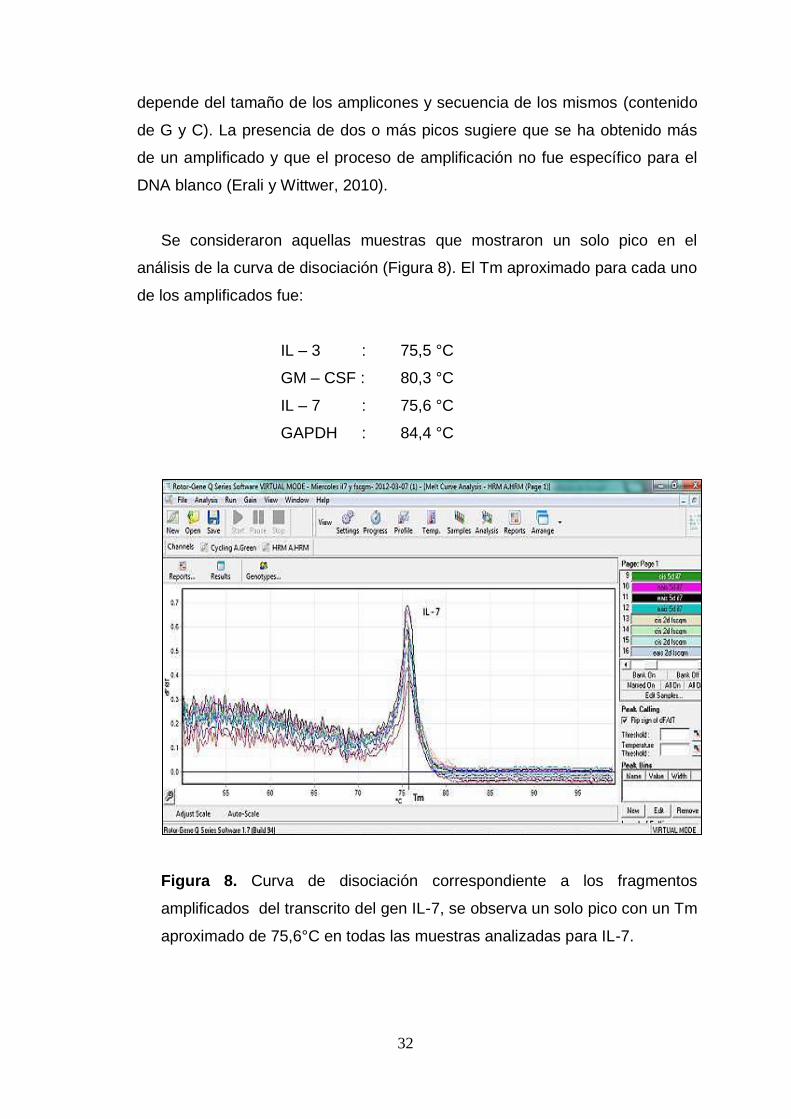
Se consideraron aquellas muestras que mostraron un solo pico en el
análisis de la curva de disociación (Figura 8). El Tm aproximado para cada uno
de los amplificados fue:
IL – 3 : 75,5 °C
GM – CSF : 80,3 °C
IL – 7 : 75,6 °C
GAPDH : 84,4 °C
Figura 8. Curva de disociación correspondiente a los fragmentos
amplificados del transcrito del gen IL-7, se observa un solo pico con un Tm
aproximado de 75,6°C en todas las muestras analizadas para IL-7.
33
La recolección y análisis de los datos generados fueron realizados
mediante el Rotor Gene Software (versión 1.7) y se utilizó el Método de
Cuantificación Comparativa incluido en el software. Utilizando éste método ya
no es necesario escoger arbitrariamente el nivel del umbral para generar los Ct
(valor umbral de ciclo) y tampoco realizar una curva estándar de eficiencia por
cada gen analizado, requeridos para calcular la expresión génica. Este
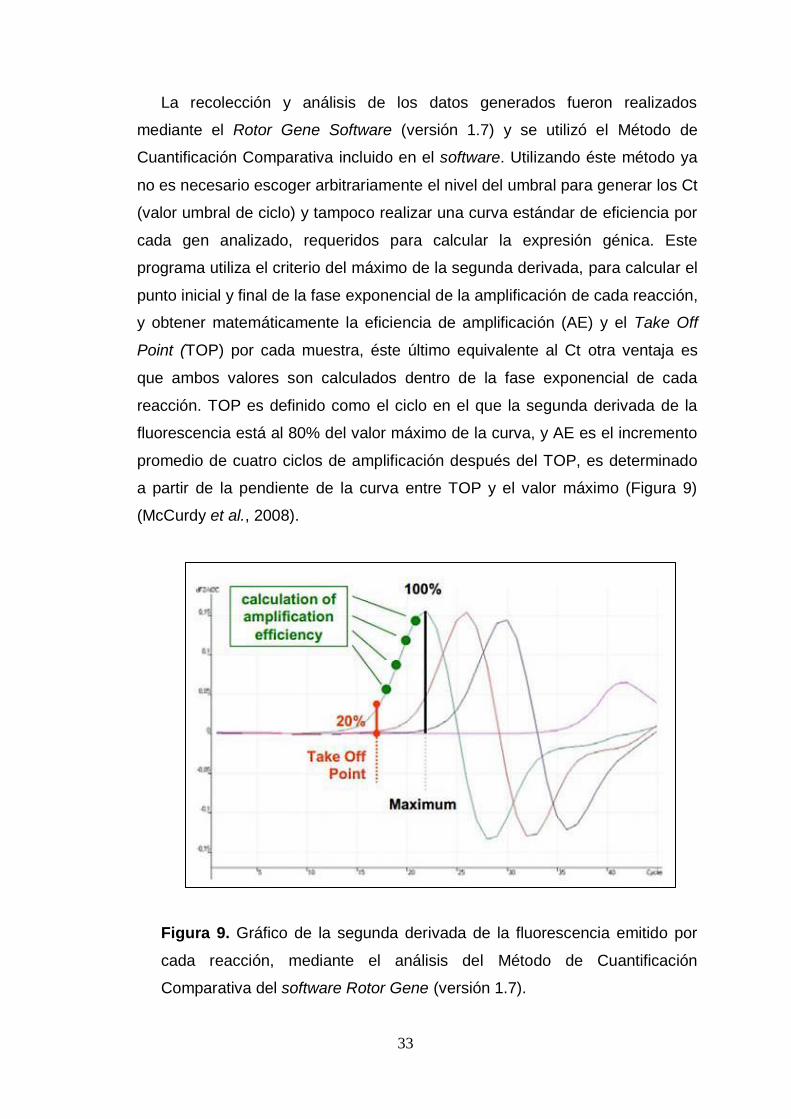
programa utiliza el criterio del máximo de la segunda derivada, para calcular el
punto inicial y final de la fase exponencial de la amplificación de cada reacción,
y obtener matemáticamente la eficiencia de amplificación (AE) y el Take Off
Point (TOP) por cada muestra, éste último equivalente al Ct otra ventaja es
que ambos valores son calculados dentro de la fase exponencial de cada
reacción. TOP es definido como el ciclo en el que la segunda derivada de la
fluorescencia está al 80% del valor máximo de la curva, y AE es el incremento
promedio de cuatro ciclos de amplificación después del TOP, es determinado
a partir de la pendiente de la curva entre TOP y el valor máximo (Figura 9)
(McCurdy et al., 2008).
Figura 9. Gráfico de la segunda derivada de la fluorescencia emitido por
cada reacción, mediante el análisis del Método de Cuantificación
Comparativa del software Rotor Gene (versión 1.7).

34
Los valores AE y TOP generados, fueron usados en el modelo matemático
descrito por Pfaffl, M. (2001) para calcular la cuantificación relativa de la
expresión génica, según la fórmula:
(EA blanco) ΔTOP blanco
Cuantificación relativa =
(EA referencia) ΔTOP referencia
Donde:
EA blanco : Es la eficiencia de amplificación del gen en estudio (IL-
3, GM-CSF, IL-7)
EA referencia : Es la eficiencia de amplificación del gen de referencia
(GAPDH)
ΔTOP blanco : TOP control - TOP tratamiento, del gen en estudio
ΔTOP referencia : TOP control - TOP tratamiento, del gen de referencia
Los cálculos se realizaron con los promedios de los AE y TOP generados
por muestra, ya que fueron corridos por triplicado. Aquellas muestras con
valores de amplificación (AE) menores que 1,5 o mayores de 2 fueron
descartadas (Figura 10).
35
Figura 10. Gráfico de la segunda derivada generado por el software Rotor
Gene, para los fragmentos amplificados del transcrito del gen IL-7.
4.4 EVALUACIÓN DEL EFECTO MODULADOR DEL EAc DE MACA SOBRE
LA PROLIFERACIÓN CELULAR Y PRODUCCIÓN DE mRNA PARA IL-3,
GM-SCF e IL-7 EN CULTIVOS DE CÉLULAS MONONUCLEARES DE LA
MÉDULA ÓSEA
4.4.1 Determinación de la dosis óptima del EAc sobre la proliferación
celular
4.4.1.1 Preparación de la dosis
Se pesó el extracto obtenido y se diluyó en medio de cultivo DMEM
suplementado con L-glutamina 2mM, HEPES 10 mM, bicarbonato de sodio
2g/L, gentamicina 50 µg/ml y estreptomicina 100 µg/ml. El preparado se
esterilizó empleando un filtro milipore 0.2 µm (CA-membrane). Se preparó una
dosis patrón de 400 µg/ml, a partir de la cual se hicieron las diluciones
respectivas para usarlas en los cultivos. Se evaluaron las dosis de 50, 100 y
200 µg/ml.

36
4.4.1.2 Obtención de células mononucleares de médula ósea
Los ratones fueron sacrificados por dislocación cervical y se extrajeron los
fémures de cada ratón. Se cortaron ambos extremos de cada hueso con un
bisturí y se utilizaron agujas N° 25 con jeringa de 10 ml llenos de medio RPMI-
1640 para obtener la médula ósea, al pasar la jeringuilla por medio del canal
del fémur (por desplazamiento de volúmenes con el medio) (Figura 11). El
contenido se recolectó en tubos estériles y se disgregó por pipeteo continuo.
La suspensión celular (10 ml) se depositó sobre 4 ml de Ficoll-Hypaque en
tubos cónicos y se centrifugaron a 2500 rpm por 25 minutos. Se extrajo la
capa de células mononucleares, succionando cuidadosamente con una pipeta,
y seguidamente se lavaron tres veces mediante centrifugación a 1000 rpm por
5 minutos. Después de la última centrifugación se resuspendió el taco celular
en medio de cultivo completo (DMEM suplementado con suero bovino fetal
10%, L-glutamina 2mM, HEPES 10 mM, bicarbonato de sodio 2g/L,
gentamicina 50 µg/ml y estreptomicina 100 µg/ml), se realizó el recuento de
células vivas de la suspensión obtenida mediante el colorante vital Azul de
Tripán en una cámara de Neubauer y se diluyó la suspensión celular a la
concentración de 1 x 106 células/ml. Todo el proceso se realizó en completa
esterilidad.
Figura 11. Obtención de la médula ósea de los fémures de los ratones.

37
4.4.1.3 Cultivo de células mononucleares de médula ósea
Se realizaron en placas de 24 pocillos, por triplicado y en un volumen final
de 0,5 ml. En cada pocillo se mezclaron 250 µl de la suspensión celular (1 x
106 células/ml) y 250 µl del medio de cultivo completo conteniendo el extracto
acuoso preparado, en las dosis finales de 50, 100 y 200 µg/ml (tratadas). Se
consideró como control los cultivos sin EAc. Las placas se colocaron dentro de
una incubadora con 5% de CO2, a una temperatura de 37°C y en un ambiente
húmedo, durante 48 horas. Se realizó un recuento diario de todos los cultivos
con azul de Tripán, para comprobar la viabilidad celular.
4.4.1.4 Medición de la proliferación celular
Se realizó mediante el ensayo colorimétrio del MTT, que se basa en la
reducción metabólica del Bromuro de 3-(4,5-dimetiltiazol-2-ilo)-2,5-
difeniltetrazol (MTT) a un compuesto insoluble de color violeta (formazán),
realizada por la enzima mitocondrial succinato-deshidrogenasa. Este método
es muy utilizado para medir supervivencia y proliferación celular. La cantidad
de células vivas es proporcional a la cantidad de formazán producido (Figura
12) (Mosmann, 1983).
Figura 12. Ensayo del MTT. El formazán formado es disuelto con
solubilizantes (DMSO, alcohol isopropílico, etc.) y la intensidad de la
coloración es proporcional a la cantidad de células vivas.

38
Al cabo del periodo de incubación, se añadió a cada pocillo 50 µl de MTT
(Sigma Chemical Co., St Luis, Mo) a la concentración de 5 µg/ml, y las placas
volvieron a la incubadora toda la noche en las mismas condiciones de cultivo.
Al día siguiente se retiró cuidadosamente todo el medio de cultivo de cada
pocillo, se añadió 1 ml de alcohol isopropílico y se mezcló hasta disolver los
precipitados formados (formazán), producto del metabolismo de MTT.
Después de 4 horas de incubación a oscuridad, se realizaron las lecturas a
540 nm para determinar sus absorbancias (Abs). El porcentaje de proliferación
se obtuvo con la siguiente fórmula:
(Abs Tratadas - Abs Control)
Porcentaje de proliferación = x 100
Abs Control
Abs = Absorbancia
4.4.2 Análisis de la expresión del mRNA de las citoquinas
hematopoyéticas IL-3, GM-CSF e IL-7
Se volvieron a realizar los cultivos con la dosis de EAc que mejor estimuló
la proliferación y en las mismas condiciones señaladas, para la obtención de
los RNA totales en tres periodos de tiempo: 2 horas, 18 horas y 48 horas de
cultivo. Se consideraron estos tiempos con el fin de observar el curso del nivel
de expresión génica: 0 horas es el nivel basal donde aún no se registra una
expresión significativa, 18 horas es donde se da el mayor porcentaje de
expresión génica (70%) y 48 horas donde no se registraría una expresión
significativa, porque los mRNA fueron traducidos a proteínas (Denzler et al.,
2010).
Al cumplir el tiempo de cultivo establecido, los sedimentos celulares de
cada pocillo de la placa de cultivo, se resuspendieron con su mismo medio y
se recolectaron en crioviales estériles para proceder a extraer el RNA con
Trizol, determinar su integridad por electroforesis, su pureza y concentración

39
por espectrofotometría y realizar la transcripción reversa para la síntesis de
DNA complementario, utilizando la metodología anteriormente señalada.
La medición de la expresión génica a nivel de mRNA de IL-3, IL-7, GM-
SCF y GAPDH se realizó por PCR en tiempo real utilizando el equipo ROTOR
GENE 3000 (Corbett Research, Mortlake, Australia). Se evaluó la especificidad
de la amplificación para cada uno de los fragmentos de los genes investigados
mediante el análisis de la curva de disociación.
La recolección y análisis de los datos generados se hicieron mediante el
software Rotor Gene (versión 1.7) y se utilizó el Método de Cuantificación
Comparativa incluido en el software.
4.5 EVALUACIÓN DE LA INFLUENCIA DEL TRATAMIENTO CON EL EAc
DE MACA SOBRE LA PROLIFERACIÓN DE LAS CÉLULAS
HEMATOPOYÉTICAS DE RATONES INMUNOSUPRIMIDOS
4.5.1 Recuento de células nucleadas de la médula ósea y sangre
circulante
Al final del tratamiento con el extracto acuoso de maca se obtuvieron
suspensiones celulares de la médula ósea de los fémures de cada ratón, al
cortar los extremos de cada hueso con un bisturí y pasar una jeringuilla con 5
ml de medio RPMI por el medio del canal del fémur. El contenido (5 ml/ratón)
se recolectó en tubos estériles y se disgregó mecánicamente por pipeteo
continuo. Las muestras de sangre se extrajeron por punción cardiaca y se
colectaron de forma individual en tubos con EDTA. Para facilitar el recuento
total, se realizaron diluciones de las suspensiones medulares (1/10) y
muestras sanguíneas (1/20) en solución de TURK, que hemolisa los
eritrocitos. La celularidad se determinó por recuento al microscopio utilizando
una cámara de Neubauer.
Se realizó un frotís de una gota de sangre sobre un portaobjeto, que
posteriormente fue teñida con el colorante Wright. La identificación de las

40
diferentes poblaciones leucocitarias (blastos, granulocitos, linfocitos y
monocitos) se realizó a través del microscopio con objetivo 100X, los valores
expresados en porcentajes se multiplicaron por el recuento total, para obtener
las poblaciones leucocitarias en número absoluto. Alteraciones importantes en
el número de leucocitos, conlleva a conocer no sólo fórmula porcentual, sino,
también los valores absolutos de cada tipo celular, para así evitar falsas
interpretaciones.
4.5.2 Evaluación de la hematopoyesis en el bazo
A lo largo de la vida adulta del ratón y otros mamíferos, el bazo contribuye
a la hematopoyesis en condiciones de estrés (hematopoyesis extramedular)
por ejemplo, como una reacción compensatoria a la mielosupresión inducida
por drogas, ya que la médula ósea se vuelve inadecuada para mantener una
hematopoyesis normal. Las células progenitoras hematopoyéticas presentes
en el bazo, proliferan formando colonias macroscópicas que son visibles como
discretos nódulos en su superficie (Soto et al., 1999).
La evaluación de la hematopoyesis esplénica, se realizó mediante el
ensayo de las unidades formadoras de colonias endógenas en la superficie del
bazo (CFU-S). Al término del tratamiento, se fijaron los bazos de los ratones
en solución Bouin por 24 horas a 4°C, luego se lavaron con agua corriente
hasta que el agua deje de ser amarilla. Tras el lavado, se contó el número de
colonias macroscópicas formadas en su superficie identificadas como nódulos
(Figura 13), con ayuda de un microscopio estereoscópico LEICA Zoom
2000™.

41
Figura 13. Colonias macroscópicas formadas en la superficie del bazo de
ratones inmunosuprimidos con ciclofosfamida.
4.5.3 Respuesta proliferativa de los linfocitos del bazo a mitógenos
policlonales
Se realizó mediante la prueba de proliferación de linfocitos inducidos por
mitógenos, in vitro. Este ensayo permite evaluar la capacidad de los linfocitos
para llevar a cabo la proliferación clonal en respuesta a un estímulo no
específico y se usa para medir la función inmunológica después de la
administración de algún fármaco o en pacientes bajo sospecha de
inmunodeficiencias (Janeway et al., 2003). Las suspensiones celulares del
bazo, obtenidas en los pasos anteriores se usaron para realizar los cultivos
celulares. La suspensión celular (10 ml) procedente de 3 ratones, se depositó
sobre 4 ml de Ficoll-Hypaque en tubos cónicos y se centrifugó a 2500 rpm por
25 minutos. Se extrajo la capa de células mononucleadas, succionando
cuidadosamente con una pipeta, y seguidamente se lavaron tres veces
mediante centrifugación a 1000 rpm por 5 minutos. Después de la última
centrifugación se resuspendió el taco celular en medio de cultivo completo
RPMI-1640 (suplementado con suero bovino fetal 10%, L-glutamina 2mM,
HEPES 10 mM, bicarbonato de sodio 2g/L, gentamicina 50 µg/ml y

42
estreptomicina 100 µg/ml), se realizó el recuento de células vivas de la
suspensión obtenida mediante el colorante vital azul de Tripán en una cámara
de Neubauer y se diluyó la suspensión celular a la concentración de 1 x 106
células /ml.
Los mitógenos, concanavalina A (Con A, lote C5275) y Lipopolisacárido
(LPS, lote L4391) fueron adquiridos en Sigma Chemical Co (St. Luis, Mo) y se
reconstituyeron en medio de cultivo RPMI-1640, siguiendo las instrucciones
del inserto, se prepararon soluciones en concentraciones de 500 µg/ml para
Con A y 100 µg/ml para LPS, que fueron alicuotadas y congeladas a -20 °C
hasta su uso.
Los cultivos de células mononucleares se realizaron en placas de 24
pocillos, por triplicado y en un volumen final de 0.5 ml. En cada pocillo se
mezclaron 250 µl de la suspensión celular (1 x 106 células/ml) y 250 µl del
medio de cultivo completo sólo (control) o conteniendo los mitógenos a las
concentraciones finales de 5 µg/ml de Con A y 0,5 µg/ml para los cultivos de
esplenocitos (dosis óptimas determinadas en pruebas anteriores). Las placas
se colocaron dentro de una incubadora en atmósfera de CO2 (5%), a 37°C y
en ambiente húmedo, durante 48 horas.
Al cabo del periodo de incubación, se añadió a cada pocillo 50 µl de 3-(4,5-
dimetiltiazol-2-ilo)-2,5-difeniltetrazol (MTT) a la concentración de 5 µg/ml se
incubaron toda la noche. Al día siguiente se retiró cuidadosamente todo el
medio de cultivo de cada pocillo, se añadió 1 ml de alcohol isopropílico y se
incubó 4 horas en oscuridad, se realizaron las lecturas de la mezcla a 540 nm
en un espectrofotómetro. El porcentaje de proliferación se obtuvo con la
siguiente fórmula:
(Abs tratadas con mitógeno - Abs no tratadas con mitógeno)
% Proliferación = x 100
Abs no tratadas con mitógeno

43
4.6 ANÁLISIS ESTADÍSTICO
Los resultados fueron analizados usando el paquete estadístico SPSS
(versión 15). Los resultados se expresaron como la media ± desviación
estándar y las diferencias entre dos grupos se determinaron mediante el
análisis de varianza Levene seguido de la prueba t de Student. Las
comparaciones múltiples se realizaron mediante el análisis de varianza
ANOVA seguido del test de comparación múltiple HSD de Tukey (varianzas
homogéneas) o T3 de Dunnett (varianzas no homogéneas). Los valores
p<0.05 fueron considerados estadísticamente significativos.
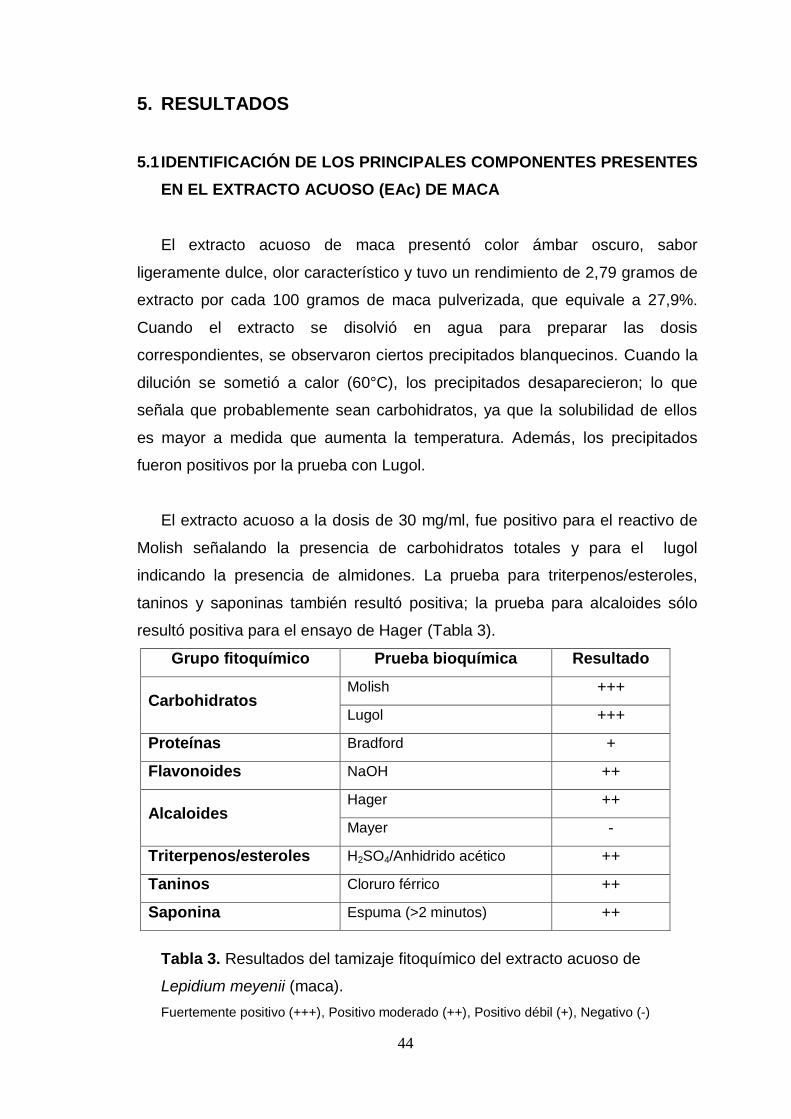
44
5. RESULTADOS
5.1 IDENTIFICACIÓN DE LOS PRINCIPALES COMPONENTES PRESENTES
EN EL EXTRACTO ACUOSO (EAc) DE MACA
El extracto acuoso de maca presentó color ámbar oscuro, sabor
ligeramente dulce, olor característico y tuvo un rendimiento de 2,79 gramos de
extracto por cada 100 gramos de maca pulverizada, que equivale a 27,9%.
Cuando el extracto se disolvió en agua para preparar las dosis
correspondientes, se observaron ciertos precipitados blanquecinos. Cuando la
dilución se sometió a calor (60°C), los precipitados desaparecieron; lo que
señala que probablemente sean carbohidratos, ya que la solubilidad de ellos
es mayor a medida que aumenta la temperatura. Además, los precipitados
fueron positivos por la prueba con Lugol.
El extracto acuoso a la dosis de 30 mg/ml, fue positivo para el reactivo de
Molish señalando la presencia de carbohidratos totales y para el lugol
indicando la presencia de almidones. La prueba para triterpenos/esteroles,
taninos y saponinas también resultó positiva; la prueba para alcaloides sólo
resultó positiva para el ensayo de Hager (Tabla 3).
Grupo fitoquímico Prueba bioquímica Resultado
Carbohidratos Molish +++
Lugol +++
Proteínas Bradford +
Flavonoides NaOH ++
Alcaloides Hager ++
Mayer -
Triterpenos/esteroles H2SO4/Anhidrido acético ++
Taninos Cloruro férrico ++
Saponina Espuma (>2 minutos) ++
Tabla 3. Resultados del tamizaje fitoquímico del extracto acuoso de
Lepidium meyenii (maca).
Fuertemente positivo (+++), Positivo moderado (++), Positivo débil (+), Negativo (-)

45
La cantidad de carbohidratos totales y proteínas por gramo de extracto
acuoso, resultó ser 636.7 mg y 8.4 mg respectivamente. Además se
registraron 0,108 mg de antocianinas y 1,47 mg de flavonoles por cada gramo
del extracto acuoso de maca.
5.2 INMUNOSUPRESIÓN EXPERIMENTAL CON CICLOFOSFAMIDA Y
TRATAMIENTO CON EL EAc DE MACA EN RATONES
5.2.1 Dosis óptima de ciclofosfamida (CF) para inmunosupresión (IS) de
ratones
Los resultados se presentan en la Figura 14 y están expresados en
porcentajes con respecto al día 0 (100%). La dosis de 65 mg/kg de CF indujo
una mielosupresión de 62,8%, 2 días post-IS; pero 5 días después de la IS ya
se observaba una recuperación casi total de células nucleadas (89,9%). Con la
dosis de 130 mg/kg, la mielosupresión fue más severa 2 días después de la IS
(26,9%) y los recuentos aún se mantenían bajos 5 días después de la IS
(64,7%). A los 7 días hay una recuperación total con ambas dosis. La dosis de
200 mg/kg resultó ser muy tóxica para los animales, uno de ellos murió al día
siguiente y los otros 2 se mostraron inapetentes y presentaron caída de pelo
en todo el cuerpo, poniendo en riesgo la vida del animal y además el éxito del
experimento. Después de comparar los datos y de acuerdo al objetivo del
estudio se optó por la dosis de 130 mg/kg peso corporal y evaluarlas 2 y 5 días
después de la inmunosupresión.
Los ratones consumieron la dosis de extracto acuoso con facilidad, además
los animales tratados con el extracto se mostraron muy activos en
comparación a los no tratados. La CF ocasionó que los animales de todos los
grupos se mostraran inapetentes, con somnolencia y con muy poca actividad.
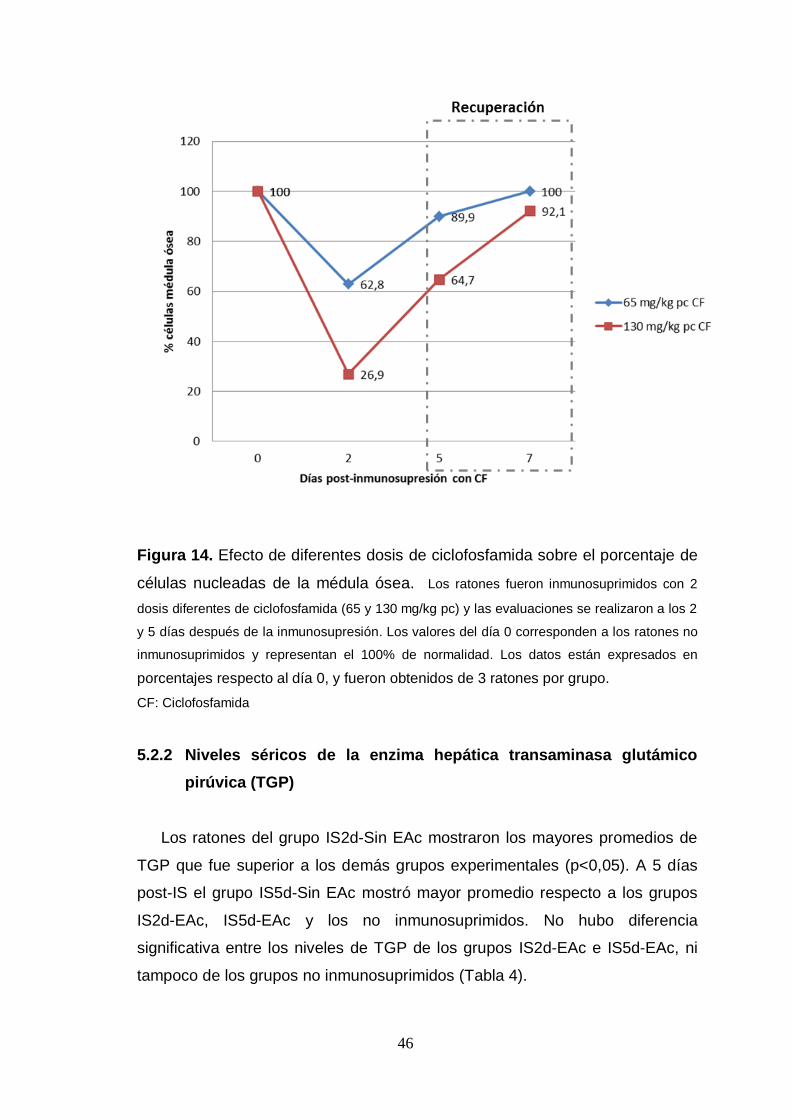
46
Figura 14. Efecto de diferentes dosis de ciclofosfamida sobre el porcentaje de
células nucleadas de la médula ósea. Los ratones fueron inmunosuprimidos con 2
dosis diferentes de ciclofosfamida (65 y 130 mg/kg pc) y las evaluaciones se realizaron a los 2
y 5 días después de la inmunosupresión. Los valores del día 0 corresponden a los ratones no
inmunosuprimidos y representan el 100% de normalidad. Los datos están expresados en
porcentajes respecto al día 0, y fueron obtenidos de 3 ratones por grupo.
CF: Ciclofosfamida
5.2.2 Niveles séricos de la enzima hepática transaminasa glutámico
pirúvica (TGP)
Los ratones del grupo IS2d-Sin EAc mostraron los mayores promedios de
TGP que fue superior a los demás grupos experimentales (p<0,05). A 5 días
post-IS el grupo IS5d-Sin EAc mostró mayor promedio respecto a los grupos
IS2d-EAc, IS5d-EAc y los no inmunosuprimidos. No hubo diferencia
significativa entre los niveles de TGP de los grupos IS2d-EAc e IS5d-EAc, ni
tampoco de los grupos no inmunosuprimidos (Tabla 4).
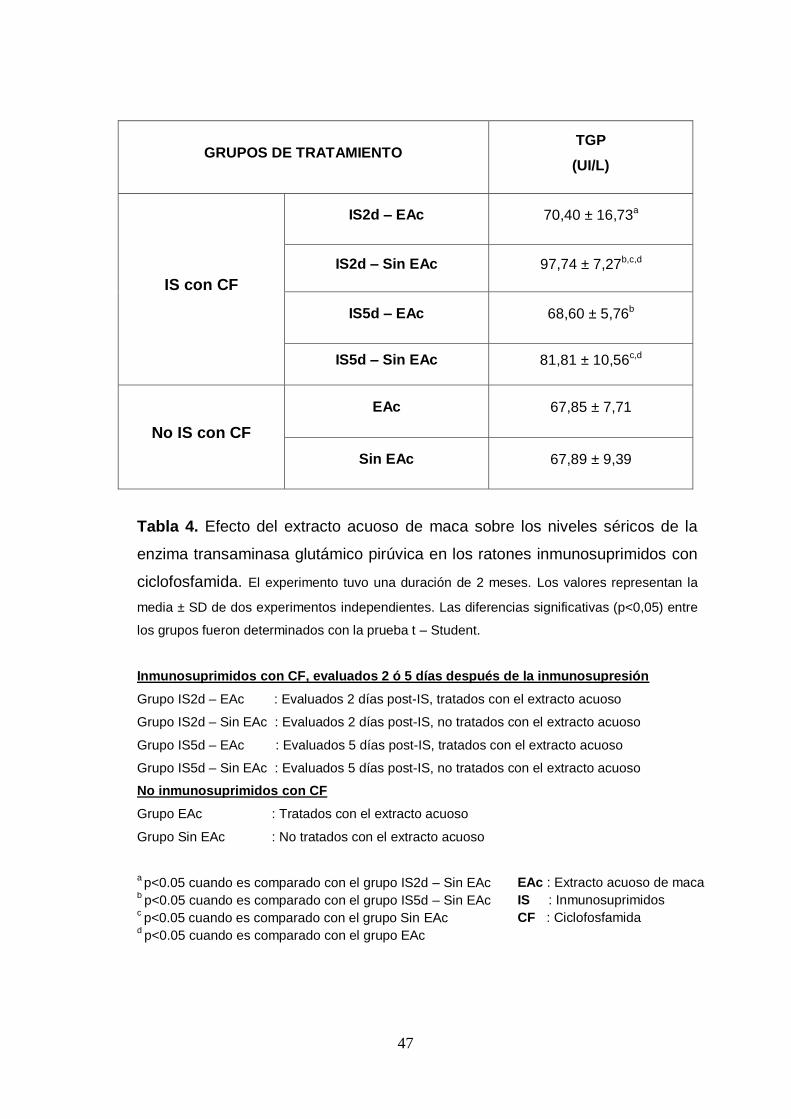
47
GRUPOS DE TRATAMIENTO TGP
(UI/L)
IS con CF
IS2d – EAc 70,40 ± 16,73a
IS2d – Sin EAc 97,74 ± 7,27b,c,d
IS5d – EAc 68,60 ± 5,76b
IS5d – Sin EAc 81,81 ± 10,56c,d
No IS con CF
EAc 67,85 ± 7,71
Sin EAc 67,89 ± 9,39
Tabla 4. Efecto del extracto acuoso de maca sobre los niveles séricos de la
enzima transaminasa glutámico pirúvica en los ratones inmunosuprimidos con
ciclofosfamida. El experimento tuvo una duración de 2 meses. Los valores representan la
media ± SD de dos experimentos independientes. Las diferencias significativas (p<0,05) entre
los grupos fueron determinados con la prueba t – Student.
Inmunosuprimidos con CF, evaluados 2 ó 5 días después de la inmunosupresión
Grupo IS2d – EAc : Evaluados 2 días post-IS, tratados con el extracto acuoso
Grupo IS2d – Sin EAc : Evaluados 2 días post-IS, no tratados con el extracto acuoso
Grupo IS5d – EAc : Evaluados 5 días post-IS, tratados con el extracto acuoso
Grupo IS5d – Sin EAc : Evaluados 5 días post-IS, no tratados con el extracto acuoso
No inmunosuprimidos con CF
Grupo EAc : Tratados con el extracto acuoso
Grupo Sin EAc : No tratados con el extracto acuoso
a p<0.05 cuando es comparado con el grupo IS2d – Sin EAc EAc : Extracto acuoso de maca
b p<0.05 cuando es comparado con el grupo IS5d – Sin EAc IS : Inmunosuprimidos
c p<0.05 cuando es comparado con el grupo Sin EAc CF : Ciclofosfamida
d p<0.05 cuando es comparado con el grupo EAc

48
5.3 EFECTO DEL EAc DE MACA SOBRE LA PRODUCCIÓN DE mRNA
PARA IL-3, GM-SCF e IL-7 EN CÉLULAS DEL BAZO Y LA MÉDULA
ÓSEA DE RATONES INMUNOSUPRIMIDOS
5.3.1 Niveles de expresión de los genes IL-3, GM-SCF e IL-7 por
esplenocitos
La inmunosupresión redujo drásticamente los niveles de expresión de IL-3,
GM-CSF e IL-7 en el bazo, a dos días post-IS (Grupo IS2d-Sin EAc) al
compararlos con los no inmunosuprimidos (p<0.05), pero el tratamiento con el
extracto acuoso amortiguo estos efectos. A 5 días post-IS se observó el
incremento significativo en el nivel de expresión de las tres citoquinas
evaluadas, siendo superior incluso a los valores obtenidos en los ratones no
inmunosuprimidos (EAc y Sin EAc).
En la Figura 15 se observa que el grupo IS2d-EAc presentó mayor
expresión génica de IL-3 con respecto al grupo IS2d-Sin EAc (10 veces más),
pero fue menor al presentado por los grupos evaluados 5 días post-IS, donde
es notorio un incremento en los niveles de expresión para esta citoquina en
ambos grupos inmunosuprimidos, aunque el grupo IS5d-Sin EAc mostró
mayor producción que el grupo IS5d-EAc (pero solo fue 1,2 veces más). Entre
los grupos no inmunosuprimidos, los esplenocitos de los ratones tratados con
el extracto (grupo EAc) mostraron mayor expresión génica para IL-3 con
respecto a grupo Sin EAc (2,5 veces más).
La Figura 16 muestra que el tratamiento con el EAc provocó un aumento
en los niveles de mRNA de GM-CSF en el grupo IS2d-EAc respecto al grupo
IS2d-Sin EAc (1,3 veces más). No se encontraron diferencias de expresión
entre los grupos evaluados 5 días post-IS, ni entre los grupos no
inmunosuprimidos. Los mismos resultados se observaron para los niveles de
expresión de IL-7, sólo se observaron diferencias significativas entre los
ratones evaluados 2 días post-IS, donde el grupo EAcIS-2d mostró mayor
expresión génica para IL-7 (1,5 veces más) respecto al grupo IS2d-Sin EAc
(Figura 17).
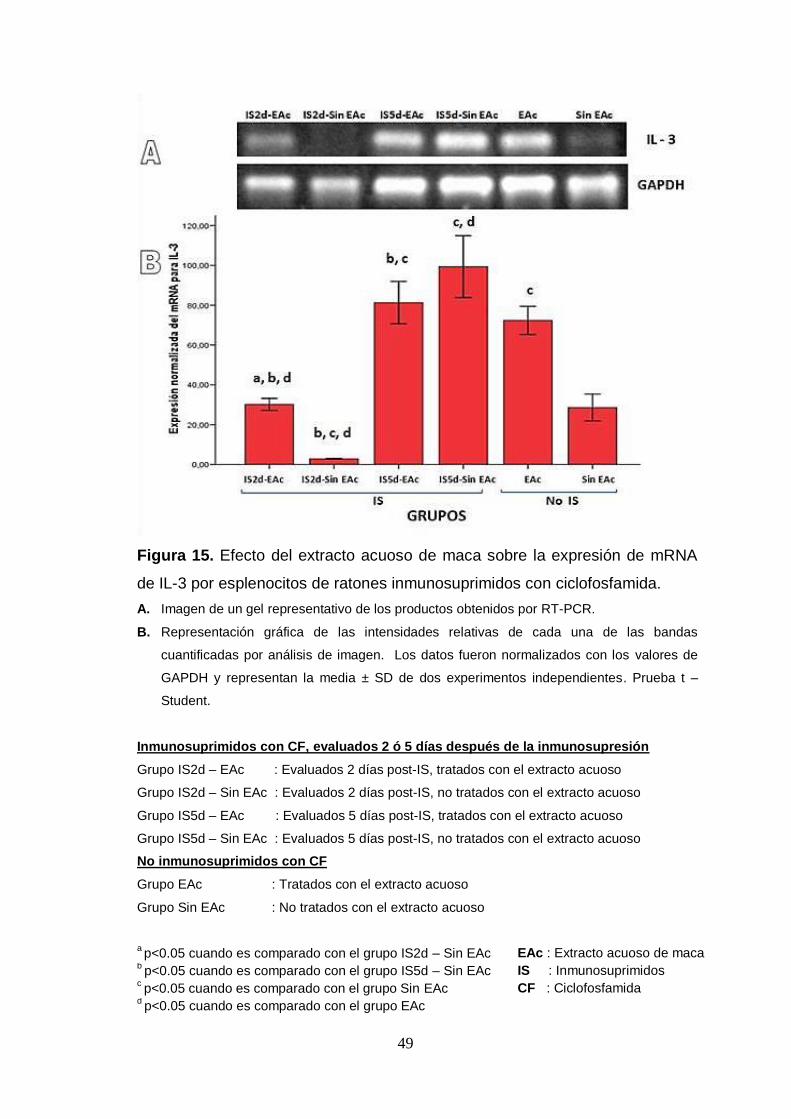
49
Figura 15. Efecto del extracto acuoso de maca sobre la expresión de mRNA
de IL-3 por esplenocitos de ratones inmunosuprimidos con ciclofosfamida.
A. Imagen de un gel representativo de los productos obtenidos por RT-PCR.
B. Representación gráfica de las intensidades relativas de cada una de las bandas
cuantificadas por análisis de imagen. Los datos fueron normalizados con los valores de
GAPDH y representan la media ± SD de dos experimentos independientes. Prueba t –
Student.
Inmunosuprimidos con CF, evaluados 2 ó 5 días después de la inmunosupresión
Grupo IS2d – EAc : Evaluados 2 días post-IS, tratados con el extracto acuoso
Grupo IS2d – Sin EAc : Evaluados 2 días post-IS, no tratados con el extracto acuoso
Grupo IS5d – EAc : Evaluados 5 días post-IS, tratados con el extracto acuoso
Grupo IS5d – Sin EAc : Evaluados 5 días post-IS, no tratados con el extracto acuoso
No inmunosuprimidos con CF
Grupo EAc : Tratados con el extracto acuoso
Grupo Sin EAc : No tratados con el extracto acuoso
a p<0.05 cuando es comparado con el grupo IS2d – Sin EAc EAc : Extracto acuoso de maca
b p<0.05 cuando es comparado con el grupo IS5d – Sin EAc IS : Inmunosuprimidos
c p<0.05 cuando es comparado con el grupo Sin EAc CF : Ciclofosfamida
d p<0.05 cuando es comparado con el grupo EAc
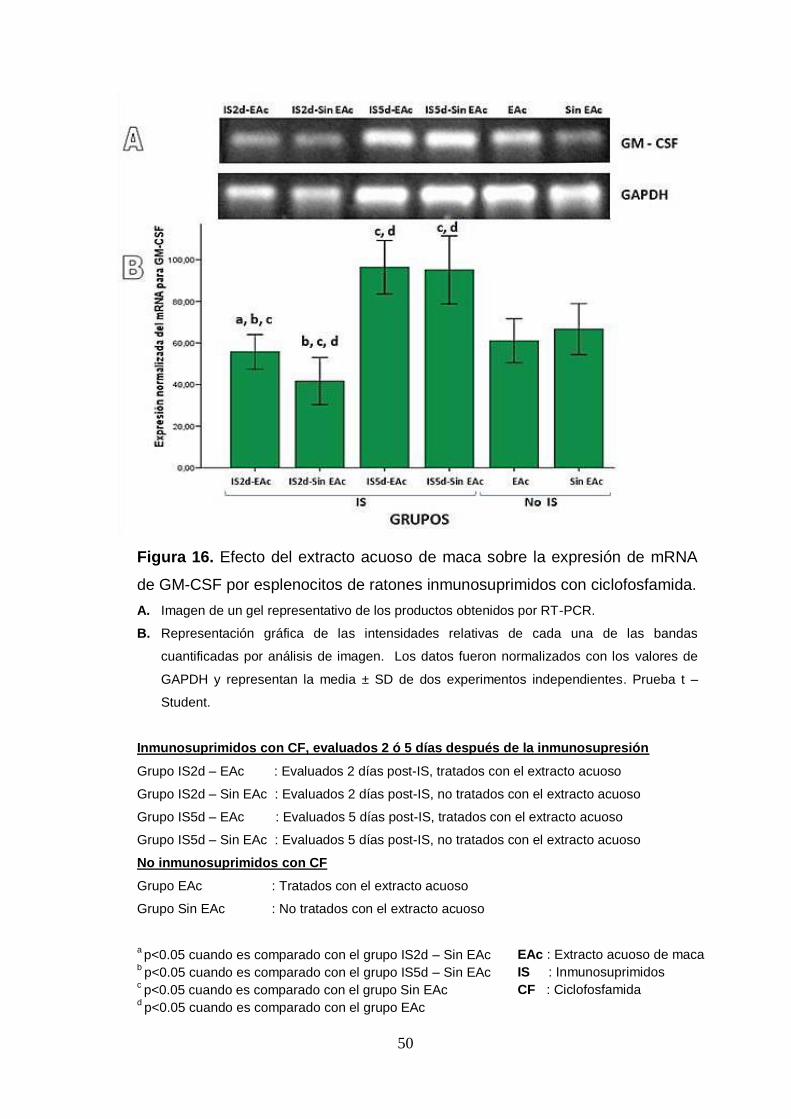
50
Figura 16. Efecto del extracto acuoso de maca sobre la expresión de mRNA
de GM-CSF por esplenocitos de ratones inmunosuprimidos con ciclofosfamida.
A. Imagen de un gel representativo de los productos obtenidos por RT-PCR.
B. Representación gráfica de las intensidades relativas de cada una de las bandas
cuantificadas por análisis de imagen. Los datos fueron normalizados con los valores de
GAPDH y representan la media ± SD de dos experimentos independientes. Prueba t –
Student.
Inmunosuprimidos con CF, evaluados 2 ó 5 días después de la inmunosupresión
Grupo IS2d – EAc : Evaluados 2 días post-IS, tratados con el extracto acuoso
Grupo IS2d – Sin EAc : Evaluados 2 días post-IS, no tratados con el extracto acuoso
Grupo IS5d – EAc : Evaluados 5 días post-IS, tratados con el extracto acuoso
Grupo IS5d – Sin EAc : Evaluados 5 días post-IS, no tratados con el extracto acuoso
No inmunosuprimidos con CF
Grupo EAc : Tratados con el extracto acuoso
Grupo Sin EAc : No tratados con el extracto acuoso
a p<0.05 cuando es comparado con el grupo IS2d – Sin EAc EAc : Extracto acuoso de maca
b p<0.05 cuando es comparado con el grupo IS5d – Sin EAc IS : Inmunosuprimidos
c p<0.05 cuando es comparado con el grupo Sin EAc CF : Ciclofosfamida
d p<0.05 cuando es comparado con el grupo EAc
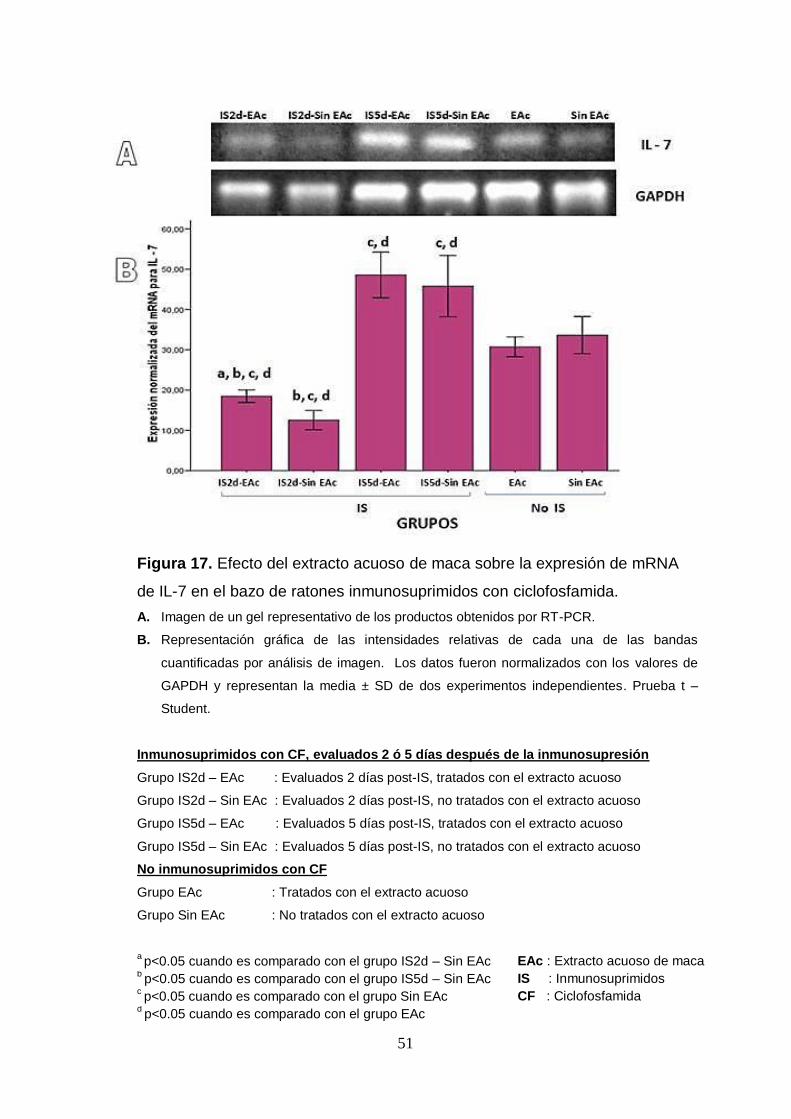
51
Figura 17. Efecto del extracto acuoso de maca sobre la expresión de mRNA
de IL-7 en el bazo de ratones inmunosuprimidos con ciclofosfamida.
A. Imagen de un gel representativo de los productos obtenidos por RT-PCR.
B. Representación gráfica de las intensidades relativas de cada una de las bandas
cuantificadas por análisis de imagen. Los datos fueron normalizados con los valores de
GAPDH y representan la media ± SD de dos experimentos independientes. Prueba t –
Student.
Inmunosuprimidos con CF, evaluados 2 ó 5 días después de la inmunosupresión
Grupo IS2d – EAc : Evaluados 2 días post-IS, tratados con el extracto acuoso
Grupo IS2d – Sin EAc : Evaluados 2 días post-IS, no tratados con el extracto acuoso
Grupo IS5d – EAc : Evaluados 5 días post-IS, tratados con el extracto acuoso
Grupo IS5d – Sin EAc : Evaluados 5 días post-IS, no tratados con el extracto acuoso
No inmunosuprimidos con CF
Grupo EAc : Tratados con el extracto acuoso
Grupo Sin EAc : No tratados con el extracto acuoso
a p<0.05 cuando es comparado con el grupo IS2d – Sin EAc EAc : Extracto acuoso de maca
b p<0.05 cuando es comparado con el grupo IS5d – Sin EAc IS : Inmunosuprimidos
c p<0.05 cuando es comparado con el grupo Sin EAc CF : Ciclofosfamida
d p<0.05 cuando es comparado con el grupo EAc

52
5.3.2 Niveles de expresión de los genes IL-3, GM-SCF e IL-7 de la médula
ósea
El patrón de expresión de las 3 citoquinas evaluadas no fue el mismo en la
médula ósea que en el bazo, la mayor expresión génica en la médula ósea se
observó a los 2 días post-IS (IS2d-EAc e IS2d-Sin EAc) y fue superior
respecto a los demás grupos (p<0,05), sin embargo se reduce drásticamente a
los 5 días post-IS hasta llegar a niveles similares a los grupos no
inmunosuprimidos.
No se observaron diferencias de niveles de expresión de IL-3, entre los
grupos IS2d-EAc e IS2d-Sin EAc. Sin embargo a 5 días post-IS el nivel de
expresión de IL-3 en el grupo IS5d-EAc fue superior al grupo IS5d-Sin EAc
(1,8 veces más) igualándose al grupo EAc. Entre los ratones no
inmunosuprimidos el grupo tratado con el extracto (EAc) mostró mayor nivel de
expresión con respecto al grupo Sin EAc (1,4 veces más) (Figura 18).
La Figura 19 muestra que los ratones del grupo IS5d-EAc presentan mayor
expresión de GM-CSF con respecto al grupo IS5d-Sin EAc (aunque solo fue
1,2 veces más). No se encontraron diferencias de expresión entre los grupos
IS2d-EAc e IS2d-Sin EAc, ni tampoco entre los grupos no inmunosuprimidos
(EAc y control).
Los ratones tratados con el extracto y evaluados dos días post-IS (grupo
IS2d-EAc) tuvieron mayor expresión de IL-7 con respecto al grupo IS2d-Sin
EAc (aunque solo fue 1,2 veces más). No se observaron diferencias de
expresión para IL-7 entre los grupos IS5d-EAc e IS5d-Sin EAc. Entre los
grupos no inmunosuprimidos, los ratones tratados con EAc mostraron mayor
expresión génica de IL-7 (2,5 veces más), al compararlo con el grupo Sin EAc.
El grupo EAc también registró mayor expresión de IL-7 que los ratones
evaluados 5 días post-IS (grupos IS5d-EAc e IS5d-Sin EAc) (Figura 20).
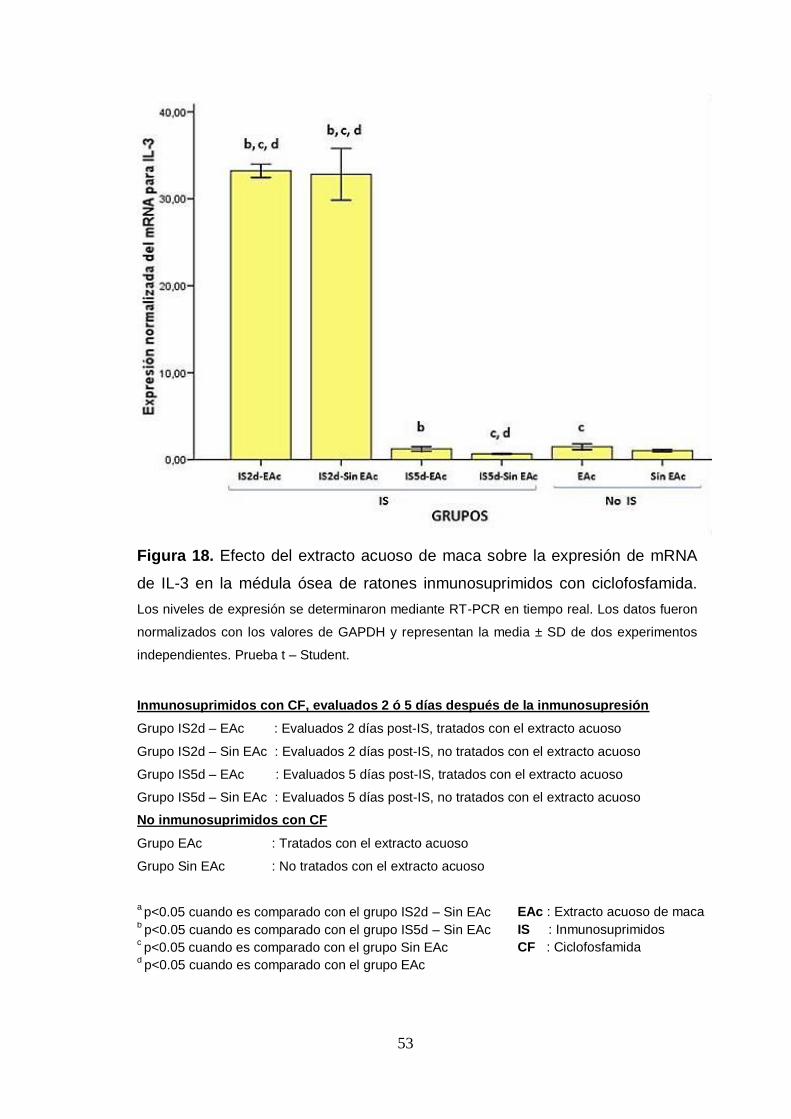
53
Figura 18. Efecto del extracto acuoso de maca sobre la expresión de mRNA
de IL-3 en la médula ósea de ratones inmunosuprimidos con ciclofosfamida.
Los niveles de expresión se determinaron mediante RT-PCR en tiempo real. Los datos fueron
normalizados con los valores de GAPDH y representan la media ± SD de dos experimentos
independientes. Prueba t – Student.
Inmunosuprimidos con CF, evaluados 2 ó 5 días después de la inmunosupresión
Grupo IS2d – EAc : Evaluados 2 días post-IS, tratados con el extracto acuoso
Grupo IS2d – Sin EAc : Evaluados 2 días post-IS, no tratados con el extracto acuoso
Grupo IS5d – EAc : Evaluados 5 días post-IS, tratados con el extracto acuoso
Grupo IS5d – Sin EAc : Evaluados 5 días post-IS, no tratados con el extracto acuoso
No inmunosuprimidos con CF
Grupo EAc : Tratados con el extracto acuoso
Grupo Sin EAc : No tratados con el extracto acuoso
a p<0.05 cuando es comparado con el grupo IS2d – Sin EAc EAc : Extracto acuoso de maca
b p<0.05 cuando es comparado con el grupo IS5d – Sin EAc IS : Inmunosuprimidos
c p<0.05 cuando es comparado con el grupo Sin EAc CF : Ciclofosfamida
d p<0.05 cuando es comparado con el grupo EAc
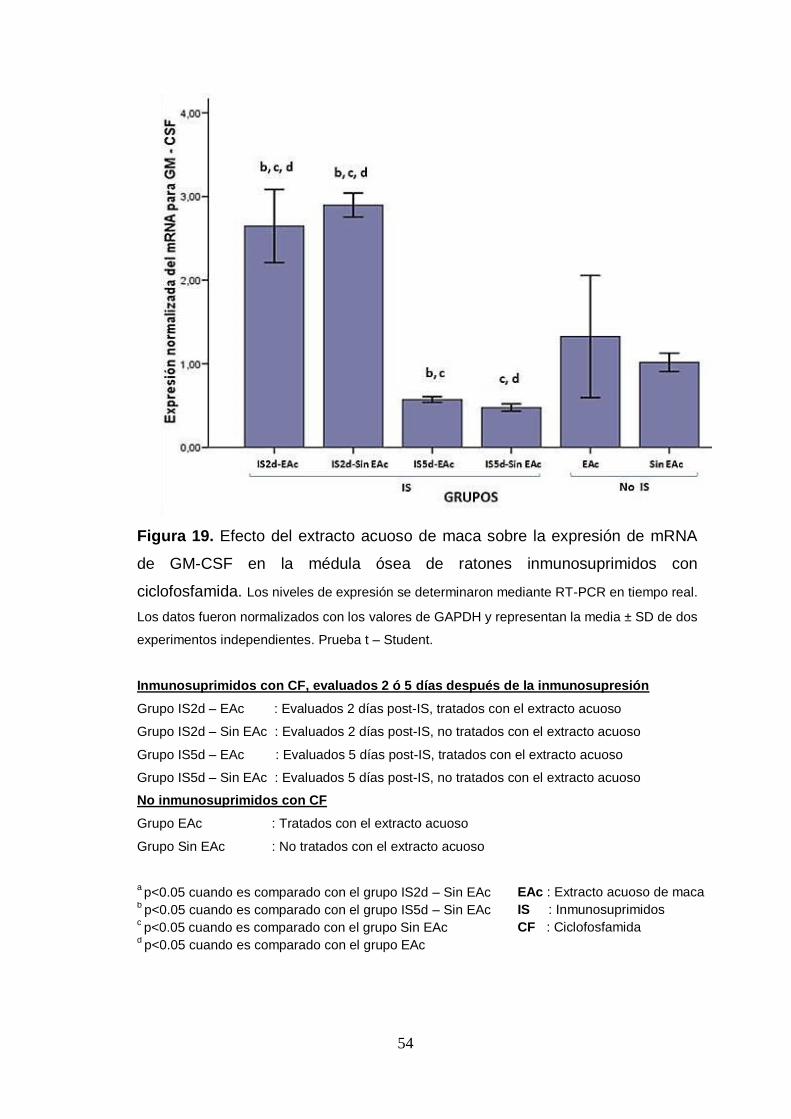
54
Figura 19. Efecto del extracto acuoso de maca sobre la expresión de mRNA
de GM-CSF en la médula ósea de ratones inmunosuprimidos con
ciclofosfamida. Los niveles de expresión se determinaron mediante RT-PCR en tiempo real.
Los datos fueron normalizados con los valores de GAPDH y representan la media ± SD de dos
experimentos independientes. Prueba t – Student.
Inmunosuprimidos con CF, evaluados 2 ó 5 días después de la inmunosupresión
Grupo IS2d – EAc : Evaluados 2 días post-IS, tratados con el extracto acuoso
Grupo IS2d – Sin EAc : Evaluados 2 días post-IS, no tratados con el extracto acuoso
Grupo IS5d – EAc : Evaluados 5 días post-IS, tratados con el extracto acuoso
Grupo IS5d – Sin EAc : Evaluados 5 días post-IS, no tratados con el extracto acuoso
No inmunosuprimidos con CF
Grupo EAc : Tratados con el extracto acuoso
Grupo Sin EAc : No tratados con el extracto acuoso
a p<0.05 cuando es comparado con el grupo IS2d – Sin EAc EAc : Extracto acuoso de maca
b p<0.05 cuando es comparado con el grupo IS5d – Sin EAc IS : Inmunosuprimidos
c p<0.05 cuando es comparado con el grupo Sin EAc CF : Ciclofosfamida
d p<0.05 cuando es comparado con el grupo EAc
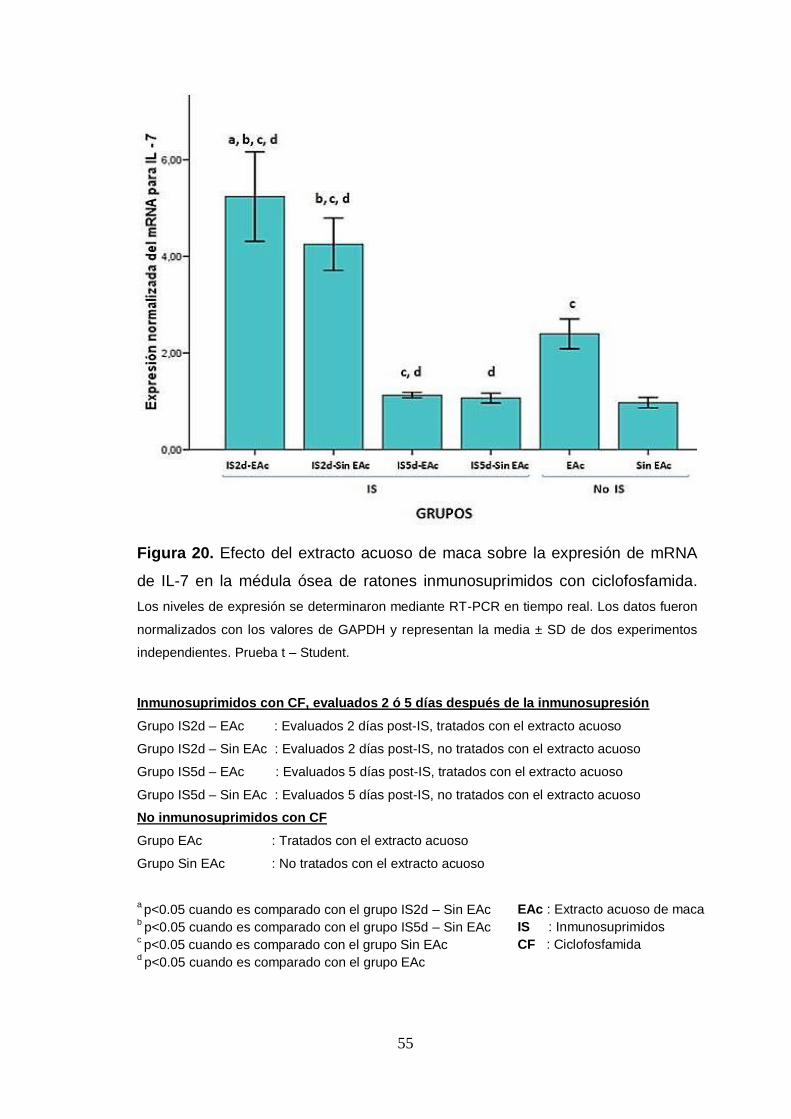
55
Figura 20. Efecto del extracto acuoso de maca sobre la expresión de mRNA
de IL-7 en la médula ósea de ratones inmunosuprimidos con ciclofosfamida.
Los niveles de expresión se determinaron mediante RT-PCR en tiempo real. Los datos fueron
normalizados con los valores de GAPDH y representan la media ± SD de dos experimentos
independientes. Prueba t – Student.
Inmunosuprimidos con CF, evaluados 2 ó 5 días después de la inmunosupresión
Grupo IS2d – EAc : Evaluados 2 días post-IS, tratados con el extracto acuoso
Grupo IS2d – Sin EAc : Evaluados 2 días post-IS, no tratados con el extracto acuoso
Grupo IS5d – EAc : Evaluados 5 días post-IS, tratados con el extracto acuoso
Grupo IS5d – Sin EAc : Evaluados 5 días post-IS, no tratados con el extracto acuoso
No inmunosuprimidos con CF
Grupo EAc : Tratados con el extracto acuoso
Grupo Sin EAc : No tratados con el extracto acuoso
a p<0.05 cuando es comparado con el grupo IS2d – Sin EAc EAc : Extracto acuoso de maca
b p<0.05 cuando es comparado con el grupo IS5d – Sin EAc IS : Inmunosuprimidos
c p<0.05 cuando es comparado con el grupo Sin EAc CF : Ciclofosfamida
d p<0.05 cuando es comparado con el grupo EAc
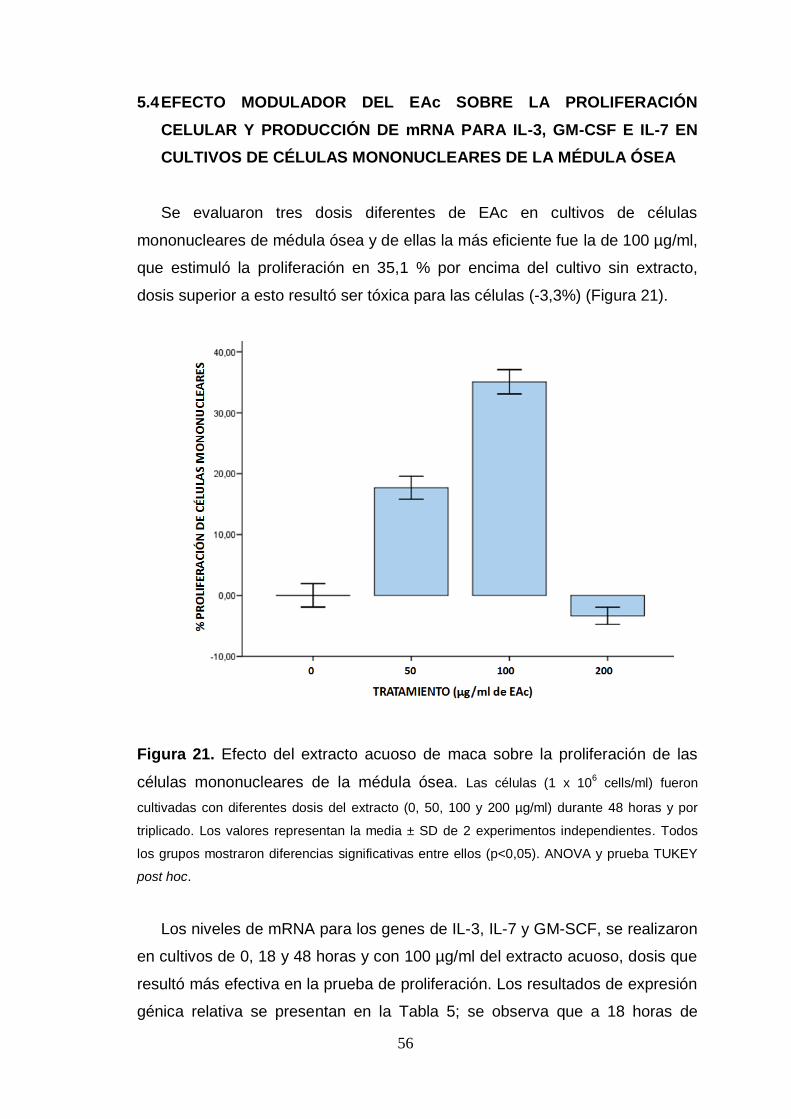
56
5.4 EFECTO MODULADOR DEL EAc SOBRE LA PROLIFERACIÓN
CELULAR Y PRODUCCIÓN DE mRNA PARA IL-3, GM-CSF E IL-7 EN
CULTIVOS DE CÉLULAS MONONUCLEARES DE LA MÉDULA ÓSEA
Se evaluaron tres dosis diferentes de EAc en cultivos de células
mononucleares de médula ósea y de ellas la más eficiente fue la de 100 µg/ml,
que estimuló la proliferación en 35,1 % por encima del cultivo sin extracto,
dosis superior a esto resultó ser tóxica para las células (-3,3%) (Figura 21).
Figura 21. Efecto del extracto acuoso de maca sobre la proliferación de las
células mononucleares de la médula ósea. Las células (1 x 106 cells/ml) fueron
cultivadas con diferentes dosis del extracto (0, 50, 100 y 200 µg/ml) durante 48 horas y por
triplicado. Los valores representan la media ± SD de 2 experimentos independientes. Todos
los grupos mostraron diferencias significativas entre ellos (p<0,05). ANOVA y prueba TUKEY
post hoc.
Los niveles de mRNA para los genes de IL-3, IL-7 y GM-SCF, se realizaron
en cultivos de 0, 18 y 48 horas y con 100 µg/ml del extracto acuoso, dosis que
resultó más efectiva en la prueba de proliferación. Los resultados de expresión
génica relativa se presentan en la Tabla 5; se observa que a 18 horas de
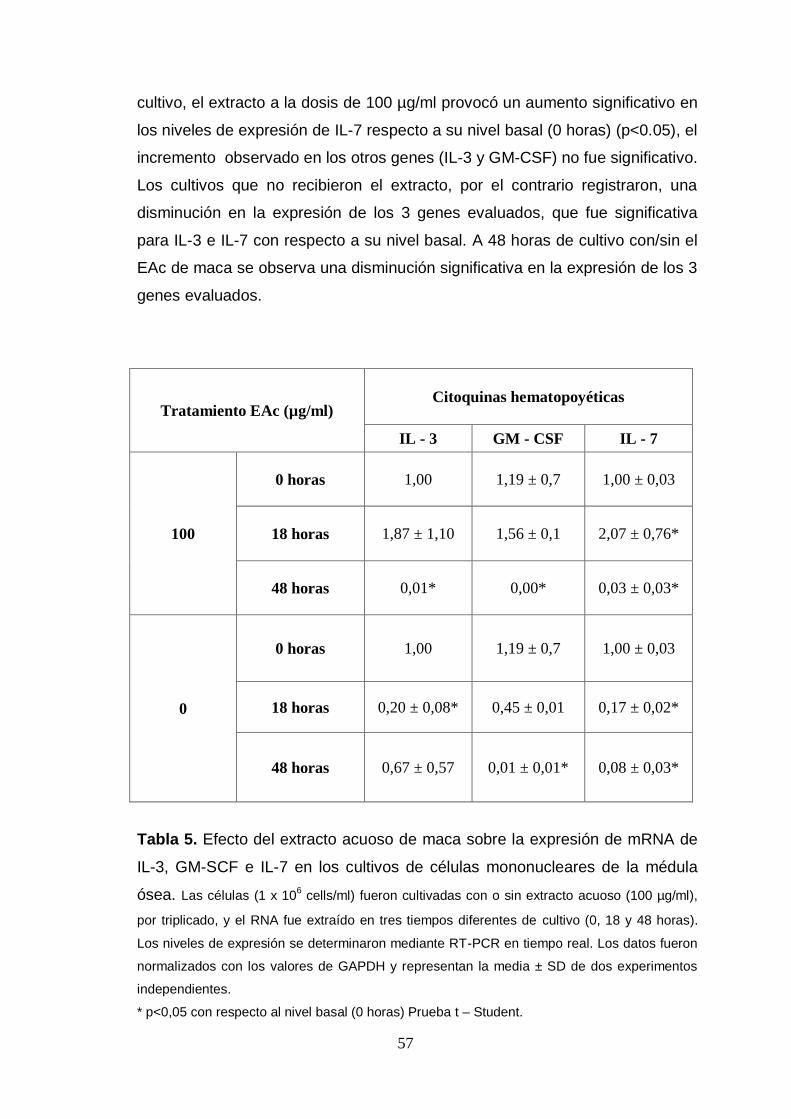
57
cultivo, el extracto a la dosis de 100 µg/ml provocó un aumento significativo en
los niveles de expresión de IL-7 respecto a su nivel basal (0 horas) (p<0.05), el
incremento observado en los otros genes (IL-3 y GM-CSF) no fue significativo.
Los cultivos que no recibieron el extracto, por el contrario registraron, una
disminución en la expresión de los 3 genes evaluados, que fue significativa
para IL-3 e IL-7 con respecto a su nivel basal. A 48 horas de cultivo con/sin el
EAc de maca se observa una disminución significativa en la expresión de los 3
genes evaluados.
Tratamiento EAc (µg/ml)
Citoquinas hematopoyéticas
IL - 3 GM - CSF IL - 7
100
0 horas 1,00 1,19 ± 0,7 1,00 ± 0,03
18 horas 1,87 ± 1,10 1,56 ± 0,1 2,07 ± 0,76*
48 horas 0,01* 0,00* 0,03 ± 0,03*
0
0 horas 1,00 1,19 ± 0,7 1,00 ± 0,03
18 horas 0,20 ± 0,08* 0,45 ± 0,01 0,17 ± 0,02*
48 horas 0,67 ± 0,57 0,01 ± 0,01* 0,08 ± 0,03*
Tabla 5. Efecto del extracto acuoso de maca sobre la expresión de mRNA de
IL-3, GM-SCF e IL-7 en los cultivos de células mononucleares de la médula
ósea. Las células (1 x 106 cells/ml) fueron cultivadas con o sin extracto acuoso (100 µg/ml),
por triplicado, y el RNA fue extraído en tres tiempos diferentes de cultivo (0, 18 y 48 horas).
Los niveles de expresión se determinaron mediante RT-PCR en tiempo real. Los datos fueron
normalizados con los valores de GAPDH y representan la media ± SD de dos experimentos
independientes.
* p<0,05 con respecto al nivel basal (0 horas) Prueba t – Student.

58
5.5 INFLUENCIA DEL TRATAMIENTO CON EL EAc DE MACA SOBRE LA
PROLIFERACIÓN DE LAS CÉLULAS HEMATOPOYÉTICAS DE
RATONES INMUNOSUPRIMIDOS
5.5.1 Recuento de células nucleadas de la médula ósea y sangre
circulante
La inmunosupresión provocó un descenso significativo en la celularidad de
la médula ósea con respecto a los ratones no inmunosuprimidos (p<0,05). No
se registraron diferencias de celularidad de médula ósea entre los grupos
IS2d-EAc e IS2d-Sin EAc. A 5 días post-IS se evidenció una recuperación de
la celularidad, los ratones del grupo IS5d-EAc mostraron mayor recuento con
respecto al grupo IS5d-Sin EAc (p<0,05), pero fueron inferiores respecto a los
no inmunosuprimidos. Entre los grupos no inmunosuprimidos, los ratones
tratados con el extracto (EAc) mostraron mayor recuento de células de médula
ósea con respecto a los no tratados (Sin EAc) (Figura 22).
Todos los grupos inmunosuprimidos mostraron menor recuento de sangre
periférica con respecto a los ratones no inmunosuprimidos. No se encontró
diferencia de recuento entre los grupos IS2d-EAc e IS2d-Sin EAc. Los ratones
del grupo IS5d-EAc mostraron mayor recuento promedio de sangre circulante
con respecto al grupo IS5d-Sin EAc (p<0,05), pero fue inferior con relación a
los ratones no inmunosuprimidos. No hubo diferencia entre los promedios
celulares de los grupos EAc y Sin EAc (Figura 23).
En cuanto al recuento diferencial, los ratones del grupo IS2d-EAc
mostraron mayores promedios de blastos, neutrófilos y monocitos que el grupo
IS2d-Sin EAc (p<0,05). A 5 días post-IS, el grupo IS5d-EAc mostró mayor
recuento de neutrófilos y monocitos que el grupo IS5d-Sin EAC; no se
encontraron diferencias de recuento de linfocitos, aunque el grupo IS5d-EAc
mostró mayor promedio (p= 0,065). No se observan diferencias significativas
entre los grupos no inmunosuprimidos (EAc y Sin EAc), pero ellos fueron
superiores a los grupos inmunosuprimidos (Tabla 5).
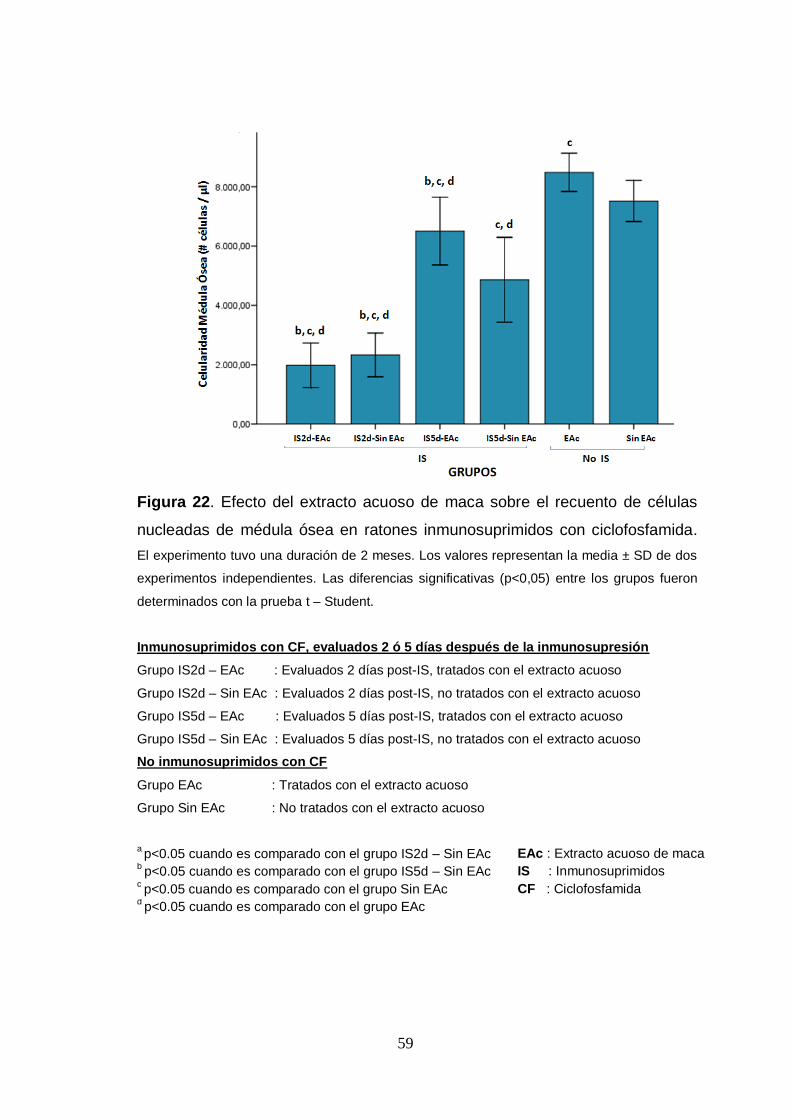
59
Figura 22. Efecto del extracto acuoso de maca sobre el recuento de células
nucleadas de médula ósea en ratones inmunosuprimidos con ciclofosfamida.
El experimento tuvo una duración de 2 meses. Los valores representan la media ± SD de dos
experimentos independientes. Las diferencias significativas (p<0,05) entre los grupos fueron
determinados con la prueba t – Student.
Inmunosuprimidos con CF, evaluados 2 ó 5 días después de la inmunosupresión
Grupo IS2d – EAc : Evaluados 2 días post-IS, tratados con el extracto acuoso
Grupo IS2d – Sin EAc : Evaluados 2 días post-IS, no tratados con el extracto acuoso
Grupo IS5d – EAc : Evaluados 5 días post-IS, tratados con el extracto acuoso
Grupo IS5d – Sin EAc : Evaluados 5 días post-IS, no tratados con el extracto acuoso
No inmunosuprimidos con CF
Grupo EAc : Tratados con el extracto acuoso
Grupo Sin EAc : No tratados con el extracto acuoso
a p<0.05 cuando es comparado con el grupo IS2d – Sin EAc EAc : Extracto acuoso de maca
b p<0.05 cuando es comparado con el grupo IS5d – Sin EAc IS : Inmunosuprimidos
c p<0.05 cuando es comparado con el grupo Sin EAc CF : Ciclofosfamida
d p<0.05 cuando es comparado con el grupo EAc
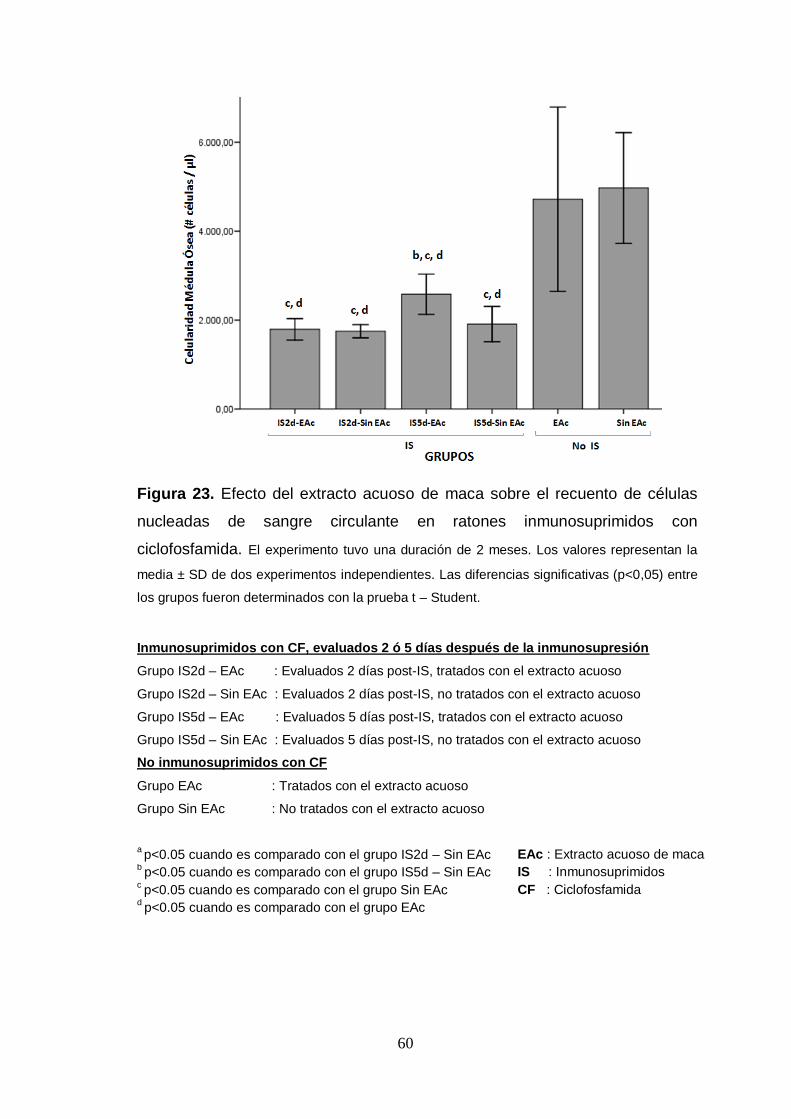
60
Figura 23. Efecto del extracto acuoso de maca sobre el recuento de células
nucleadas de sangre circulante en ratones inmunosuprimidos con
ciclofosfamida. El experimento tuvo una duración de 2 meses. Los valores representan la
media ± SD de dos experimentos independientes. Las diferencias significativas (p<0,05) entre
los grupos fueron determinados con la prueba t – Student.
Inmunosuprimidos con CF, evaluados 2 ó 5 días después de la inmunosupresión
Grupo IS2d – EAc : Evaluados 2 días post-IS, tratados con el extracto acuoso
Grupo IS2d – Sin EAc : Evaluados 2 días post-IS, no tratados con el extracto acuoso
Grupo IS5d – EAc : Evaluados 5 días post-IS, tratados con el extracto acuoso
Grupo IS5d – Sin EAc : Evaluados 5 días post-IS, no tratados con el extracto acuoso
No inmunosuprimidos con CF
Grupo EAc : Tratados con el extracto acuoso
Grupo Sin EAc : No tratados con el extracto acuoso
a p<0.05 cuando es comparado con el grupo IS2d – Sin EAc EAc : Extracto acuoso de maca
b p<0.05 cuando es comparado con el grupo IS5d – Sin EAc IS : Inmunosuprimidos
c p<0.05 cuando es comparado con el grupo Sin EAc CF : Ciclofosfamida
d p<0.05 cuando es comparado con el grupo EAc
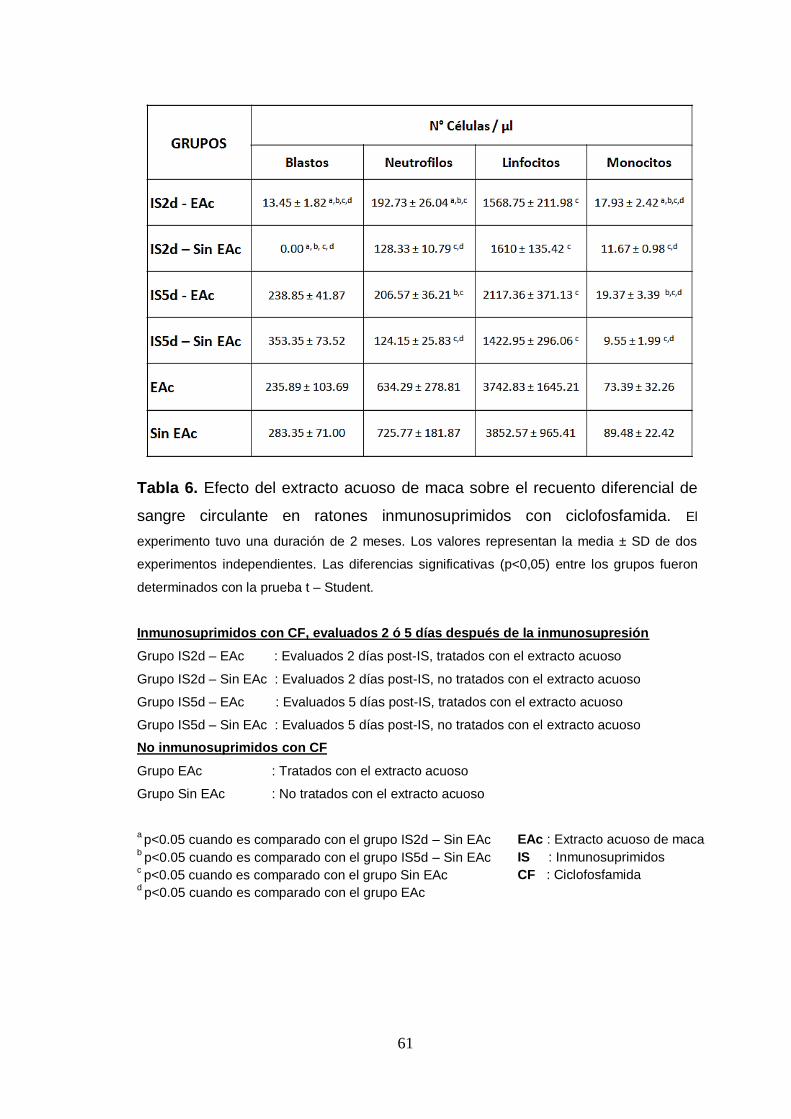
61
Tabla 6. Efecto del extracto acuoso de maca sobre el recuento diferencial de
sangre circulante en ratones inmunosuprimidos con ciclofosfamida. El
experimento tuvo una duración de 2 meses. Los valores representan la media ± SD de dos
experimentos independientes. Las diferencias significativas (p<0,05) entre los grupos fueron
determinados con la prueba t – Student.
Inmunosuprimidos con CF, evaluados 2 ó 5 días después de la inmunosupresión
Grupo IS2d – EAc : Evaluados 2 días post-IS, tratados con el extracto acuoso
Grupo IS2d – Sin EAc : Evaluados 2 días post-IS, no tratados con el extracto acuoso
Grupo IS5d – EAc : Evaluados 5 días post-IS, tratados con el extracto acuoso
Grupo IS5d – Sin EAc : Evaluados 5 días post-IS, no tratados con el extracto acuoso
No inmunosuprimidos con CF
Grupo EAc : Tratados con el extracto acuoso
Grupo Sin EAc : No tratados con el extracto acuoso
a p<0.05 cuando es comparado con el grupo IS2d – Sin EAc EAc : Extracto acuoso de maca
b p<0.05 cuando es comparado con el grupo IS5d – Sin EAc IS : Inmunosuprimidos
c p<0.05 cuando es comparado con el grupo Sin EAc CF : Ciclofosfamida
d p<0.05 cuando es comparado con el grupo EAc
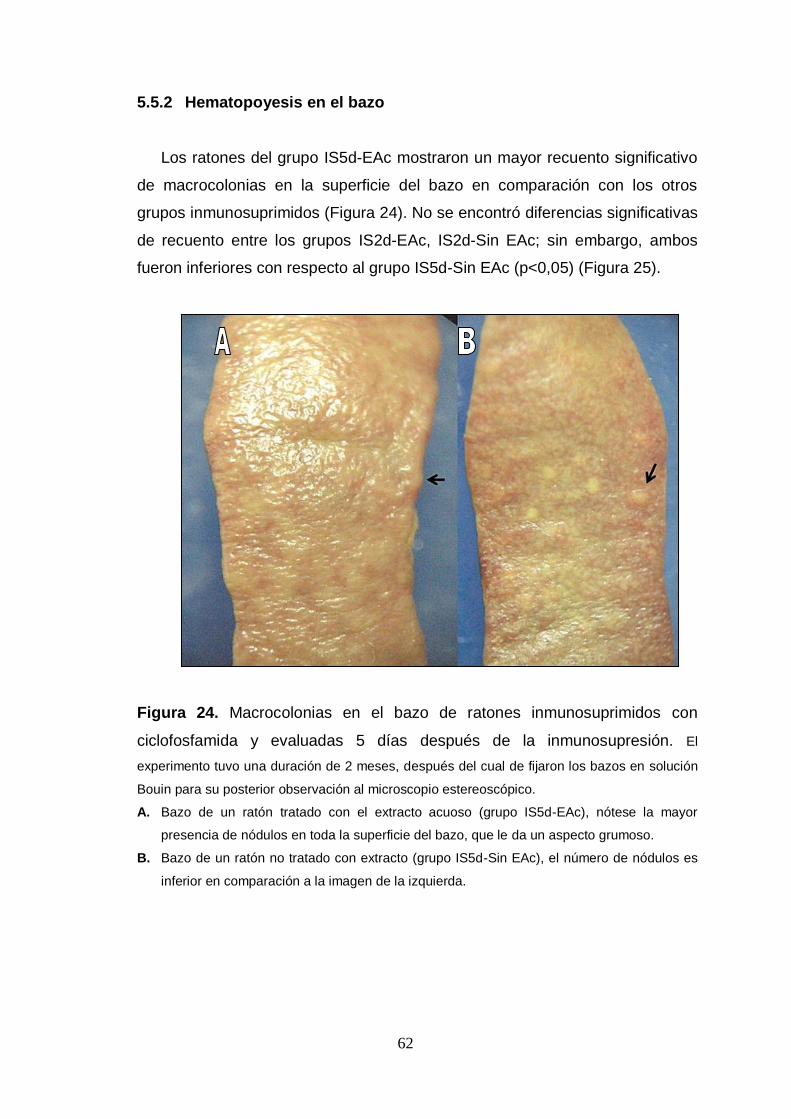
62
5.5.2 Hematopoyesis en el bazo
Los ratones del grupo IS5d-EAc mostraron un mayor recuento significativo
de macrocolonias en la superficie del bazo en comparación con los otros
grupos inmunosuprimidos (Figura 24). No se encontró diferencias significativas
de recuento entre los grupos IS2d-EAc, IS2d-Sin EAc; sin embargo, ambos
fueron inferiores con respecto al grupo IS5d-Sin EAc (p<0,05) (Figura 25).
Figura 24. Macrocolonias en el bazo de ratones inmunosuprimidos con
ciclofosfamida y evaluadas 5 días después de la inmunosupresión. El
experimento tuvo una duración de 2 meses, después del cual de fijaron los bazos en solución
Bouin para su posterior observación al microscopio estereoscópico.
A. Bazo de un ratón tratado con el extracto acuoso (grupo IS5d-EAc), nótese la mayor
presencia de nódulos en toda la superficie del bazo, que le da un aspecto grumoso.
B. Bazo de un ratón no tratado con extracto (grupo IS5d-Sin EAc), el número de nódulos es
inferior en comparación a la imagen de la izquierda.
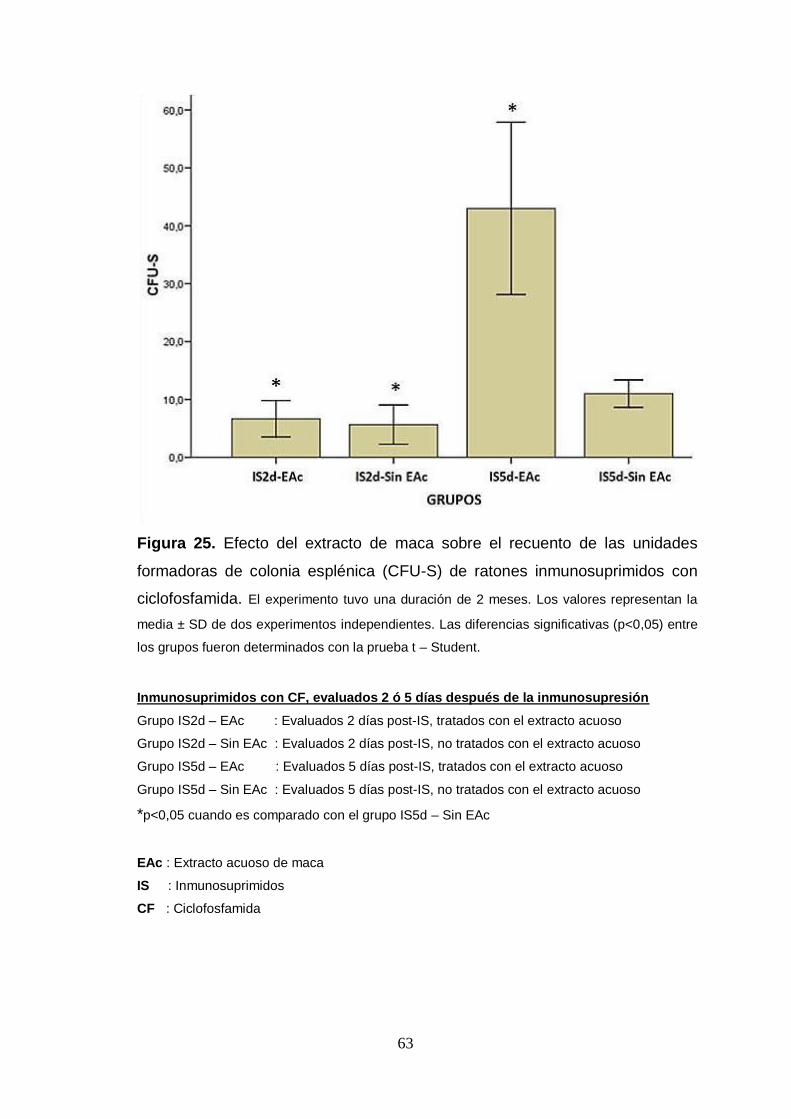
63
Figura 25. Efecto del extracto de maca sobre el recuento de las unidades
formadoras de colonia esplénica (CFU-S) de ratones inmunosuprimidos con
ciclofosfamida. El experimento tuvo una duración de 2 meses. Los valores representan la
media ± SD de dos experimentos independientes. Las diferencias significativas (p<0,05) entre
los grupos fueron determinados con la prueba t – Student.
Inmunosuprimidos con CF, evaluados 2 ó 5 días después de la inmunosupresión
Grupo IS2d – EAc : Evaluados 2 días post-IS, tratados con el extracto acuoso
Grupo IS2d – Sin EAc : Evaluados 2 días post-IS, no tratados con el extracto acuoso
Grupo IS5d – EAc : Evaluados 5 días post-IS, tratados con el extracto acuoso
Grupo IS5d – Sin EAc : Evaluados 5 días post-IS, no tratados con el extracto acuoso
*p<0,05 cuando es comparado con el grupo IS5d – Sin EAc
EAc : Extracto acuoso de maca
IS : Inmunosuprimidos
CF : Ciclofosfamida

64
5.5.3 Respuesta proliferativa de los linfocitos del bazo a mitógenos
policlonales
La inmunosupresión redujo drásticamente la respuesta proliferativa de los
linfocitos del bazo a concanavalina A y LPS a 48 horas de cultivo. Los
linfocitos procedentes del bazo de ratones inmunosuprimidos tratados con el
extracto acuoso (grupos IS2d-EAc e IS5d-EAc) mostraron mayor respuesta
proliferativa a concanavalina, respecto a los grupos no tratados (grupos IS2d-
Sin EAc e IS5d-Sin EAc), aunque fue inferior en comparación a los ratones del
grupo EAc. Entre los grupos no inmunosuprimidos, el grupo EAc mostró mayor
promedio, que además fue significativo respecto a todos los grupos
experimentales. No se encontraron diferencias entre los grupos EAc e IS5d-
EAc, pero ambos fueron superiores al grupo IS5d-Sin EAc (Figura 26).
No hubo diferencia entre los promedios de proliferación de los linfocitos del
bazo a LPS de los grupos EAc y Sin EAc, pero ambos fueron significativos
(p<0.05) con respecto a los grupos inmunosuprimidos. Los linfocitos de los
ratones inmunosuprimidos y tratados con el extracto (grupos IS2d-EAc e IS5d-
EAc) mostraron mayor capacidad de respuesta a LPS que los grupos IS2d-Sin
EAc e IS5d-Sin EAc. No se encontró diferencia de respuesta entre los grupos
IS2d-Sin EAc e IS5d-Sin EAc. El grupo IS5d-EAc mostró mayor promedio que
el grupo IS2d-EAc (Figura 27).
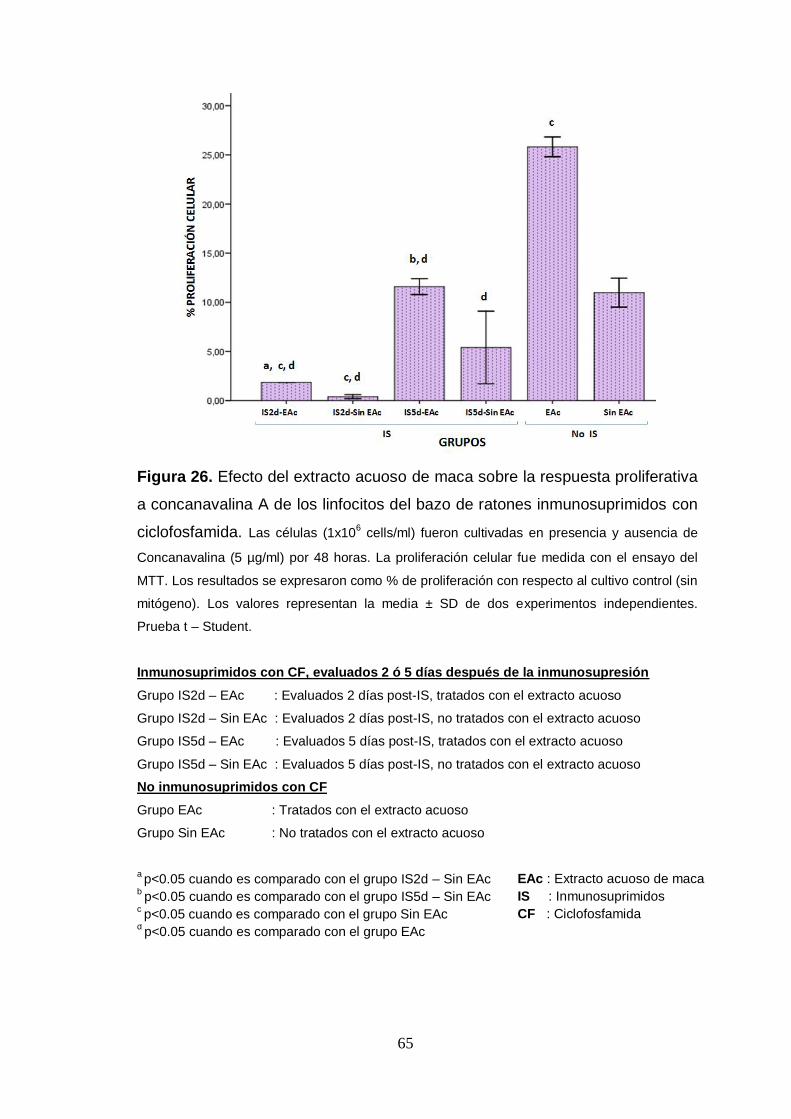
65
Figura 26. Efecto del extracto acuoso de maca sobre la respuesta proliferativa
a concanavalina A de los linfocitos del bazo de ratones inmunosuprimidos con
ciclofosfamida. Las células (1x106 cells/ml) fueron cultivadas en presencia y ausencia de
Concanavalina (5 µg/ml) por 48 horas. La proliferación celular fue medida con el ensayo del
MTT. Los resultados se expresaron como % de proliferación con respecto al cultivo control (sin
mitógeno). Los valores representan la media ± SD de dos experimentos independientes.
Prueba t – Student.
Inmunosuprimidos con CF, evaluados 2 ó 5 días después de la inmunosupresión
Grupo IS2d – EAc : Evaluados 2 días post-IS, tratados con el extracto acuoso
Grupo IS2d – Sin EAc : Evaluados 2 días post-IS, no tratados con el extracto acuoso
Grupo IS5d – EAc : Evaluados 5 días post-IS, tratados con el extracto acuoso
Grupo IS5d – Sin EAc : Evaluados 5 días post-IS, no tratados con el extracto acuoso
No inmunosuprimidos con CF
Grupo EAc : Tratados con el extracto acuoso
Grupo Sin EAc : No tratados con el extracto acuoso
a p<0.05 cuando es comparado con el grupo IS2d – Sin EAc EAc : Extracto acuoso de maca
b p<0.05 cuando es comparado con el grupo IS5d – Sin EAc IS : Inmunosuprimidos
c p<0.05 cuando es comparado con el grupo Sin EAc CF : Ciclofosfamida
d p<0.05 cuando es comparado con el grupo EAc
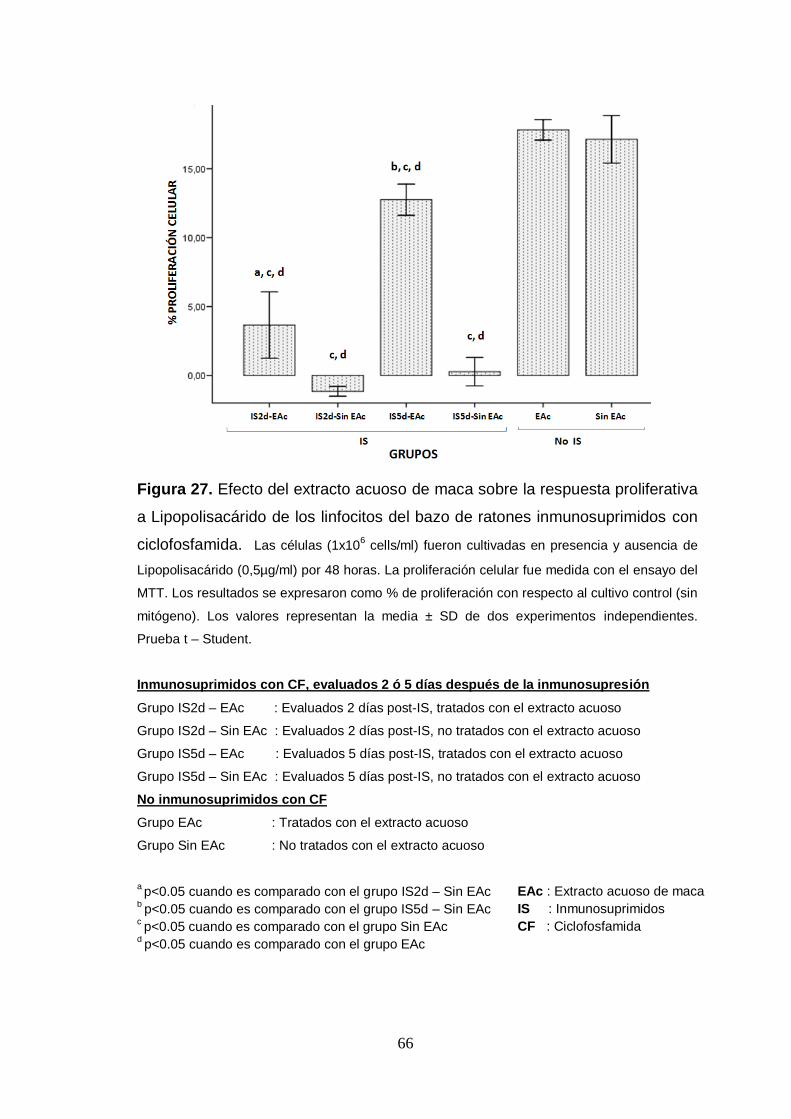
66
Figura 27. Efecto del extracto acuoso de maca sobre la respuesta proliferativa
a Lipopolisacárido de los linfocitos del bazo de ratones inmunosuprimidos con
ciclofosfamida. Las células (1x106 cells/ml) fueron cultivadas en presencia y ausencia de
Lipopolisacárido (0,5µg/ml) por 48 horas. La proliferación celular fue medida con el ensayo del
MTT. Los resultados se expresaron como % de proliferación con respecto al cultivo control (sin
mitógeno). Los valores representan la media ± SD de dos experimentos independientes.
Prueba t – Student.
Inmunosuprimidos con CF, evaluados 2 ó 5 días después de la inmunosupresión
Grupo IS2d – EAc : Evaluados 2 días post-IS, tratados con el extracto acuoso
Grupo IS2d – Sin EAc : Evaluados 2 días post-IS, no tratados con el extracto acuoso
Grupo IS5d – EAc : Evaluados 5 días post-IS, tratados con el extracto acuoso
Grupo IS5d – Sin EAc : Evaluados 5 días post-IS, no tratados con el extracto acuoso
No inmunosuprimidos con CF
Grupo EAc : Tratados con el extracto acuoso
Grupo Sin EAc : No tratados con el extracto acuoso
a p<0.05 cuando es comparado con el grupo IS2d – Sin EAc EAc : Extracto acuoso de maca
b p<0.05 cuando es comparado con el grupo IS5d – Sin EAc IS : Inmunosuprimidos
c p<0.05 cuando es comparado con el grupo Sin EAc CF : Ciclofosfamida
d p<0.05 cuando es comparado con el grupo EAc

67
6. DISCUSIÓN
La maca es ampliamente consumida por los pobladores andinos, debido a
sus propiedades nutricionales y medicinales. La divulgación de sus
propiedades atrajo el interés de investigadores que identificaron varios de sus
compuestos químicos a los que se les atribuyen las propiedades
farmacológicas. Como consecuencia se han incrementado los cultivos de la
especie, siendo el ecotipo amarillo el más común y comercial (47,8%), y se
han desarrollado a nivel industrial diferentes preparaciones como harina,
cápsulas y tabletas, que tienen gran acogida. La población nativa peruana
consume un promedio diario de 50 a 100 gramos de maca y la manera en que
la consumen es hirviéndola por varios minutos, y en forman de mermelada o
bebida (Gonzales, 2012). En este trabajo se utilizó el extracto acuoso de
maca, que es la forma que se encuentra directamente relacionado con su
consumo tradicional. Se obtuvieron 27,9% de extracto acuoso por cada 100
gramos de maca pulverizada y la dosis empleada fue de 200 mg/kg de peso
corporal (p.c.), por lo que a una persona adulta de peso promedio de 70 kilos,
le correspondería un consumo diario de 14 gramos del extracto acuoso, o de
50 gramos de harina de maca, que debe ser hervida antes de su consumo; por
lo que la dosis empleada está dentro del consumo tradicional de la población.
El análisis fitoquímico permitió identificar algunos de los componentes
presentes en el extracto acuoso utilizado en este estudio, que excluye la
posibilidad de pérdida de nutrientes en el extracto al pasar por un proceso de
ebullición; se identificaron los carbohidratos y proteínas, principales fuentes de
energía de los organismos, que participan en múltiples procesos biológicos. El
estudio cuantitativo determinó que el porcentaje de carbohidratos y proteínas
presentes en el extracto es del 63,7% y 0,84% respectivamente. La cantidad
de carbohidratos corresponde a lo reportado ampliamente en el hipocótilo seco
de maca, de un 50 - 70%. Sin embargo, el porcentaje obtenido de proteínas
fue muy inferior, ya que la literatura señala que el hipocótilo seco presenta de
10 - 16% de proteínas totales (Obregón, 1998), es probable que con la
extracción acuosa no se logre obtener todas las proteínas presentes y que la

68
mayoría de ellas se quedaron en el pellet no utilizado después de la
centrifugación. Los flavonoides son compuestos fenólicos con demostrada
acción antioxidante y eliminadora de radicales libres, entre ellos destacan los
flavonoles y las antocianinas; el extracto acuoso presentó antocianinas y
flavonoles en concentraciones reportadas por otros investigadores (Sandoval
et al., 2002). También resultó positiva para triterpenos/esteroles, taninos y
saponinas, metabolitos que estarían relacionados al efecto antioxidante de la
maca (Castaño, 2008). Es importante mencionar que algunos investigadores
recomiendan para la detección de saponinas, esperar la persistencia de la
espuma por 30 minutos; como en este trabajo sólo se consideró 3 minutos, es
necesario repetir el ensayo para confirmar su presencia en el extracto acuoso.
No se pudo confirmar la presencia de alcaloides con las dos reacciones
empleadas, ya que sólo reaccionó frente al reactivo de Hager, pero no sucedió
con el reactivo de Mayer. La detección de alcaloides mediante reacciones de
precipitación pueden dar falsos positivos y negativos, por lo que se conviene
realizar varias reacciones de detección para confirmar los resultados (López-
Casamayor, 2007).
Además de sus propiedades sobre la reproducción, la maca también ha
mostrado capacidad para estimular la respuesta inmune humoral y celular. Un
trabajo anterior señaló que ratones inmunosuprimidos con 50 mg/kg de
ciclofosfamida y tratados con el extracto acuoso de maca, presentaron un
mayor recuento de células sanguíneas, lo que indica un posible efecto sobre la
hematopoyesis (Torres, 2008).
Muchos pacientes que reciben quimio/radioterapia desarrollan una
supresión inmune y hematopoyética, que es el principal factor limitante para su
uso continuo en la clínica. En este sentido, el desarrollo de estrategias que
eviten las complicaciones provocadas por la quimioterapia, podrían disminuir
considerablemente el riesgo de padecer infecciones y reducir el número de
días en el hospital, además de mejorar la calidad de vida de los pacientes.
Como los efectos estimulatorios de un compuesto o suplemento nutricional
son difíciles de evaluar en personas o animales saludables, se usó la

69
ciclofosfamida en un modelo murino, como una aproximación para los estudios
de inmunomodulación en individuos inmunocomprometidos. La ciclofosfamida
(CF) es un agente alquilante muy usado en los regímenes quimioterapeúticos
debido a su amplio espectro antitumoral, y como inmunosupresor en las
enfermedades autoinmunes.
Esta droga por sí misma es inactiva y requiere su activación metabólica en
el hígado por las enzimas del citocromo P450 para formar mostaza de
fosforamida y acroleína, sustancias alquilantes responsables de su acción
citotóxica (Penel et al., 2012). El principal mecanismo de la CF como agente
antitumoral, es la inducción de entrecruzamiento de las cadenas de DNA/RNA
mediante reacciones de alquilación en células que se dividen activamente, por
lo que no solo está restringido a las células cancerosas sino también a otras
células con proliferación activa como las células madre y progenitoras de la
médula ósea. Es más tóxico para los tejidos altamente proliferantes como el
sistema hematopoyético, gastrointestinal, epitelial, folículos pilosos, las
glándulas genitales, además de ejercer una profunda depresión de la
respuesta inmune humoral y celular (Dolgova et al., 2012). Sin embargo la CF
tiene roles antineoplásicos e inmunomoduladores que son dosis-dependiente;
significando que dosis mínimas pueden reforzar la respuesta inmune. Por lo
tanto, una selección razonable de la dosis, tiempo y frecuencia, es primordial
para el establecimiento exitoso de inmunosupresión en modelos de animales
(Huyan et al., 2011).
La dosis empleada de CF para inducir una inmunosupresión varía de un
estudio a otro. Sefc et al. (2003) emplearon 135 mg/kg de CF en dosis única y
observaron que la celularidad de la médula ósea retornó a los valores
normales después de 6-7 días, aunque la composición celular se mantuvo
alterada hasta el día 14. Wang et al. (2011) emplearon la dosis 80 mg/kg de
CF durante tres días y evaluaron los resultados después de 4 días de la última
dosis, donde reportaron que los valores de leucocitos todavía se mantenían
bajos. En este estudio, se probaron tres dosis diferentes de CF y se evaluó el
efecto inmunosupresor mediante el recuento de células nucleares de la
médula ósea a los 2 y 5 días después de la inmunosupresión. Se consideraron

70
estos días con la finalidad de obtener datos consistentes sobre el efecto
modulador del extracto en la recuperación hematopoyética murina después de
un daño agudo por la CF. Se decidió trabajar con la dosis de 130 mg/kg de
peso corporal, ya que se obtuvieron bajos recuentos celulares a los 2 días
después de la inmunosupresión, que se mantuvieron bajos hasta 5 días post-
IS, a diferencia de la dosis de 65 mg/kg p.c., en el que se observó que 5 días
post-IS se produjo la recuperación casi total de la cantidad de células
nucleadas (89.9%) (Figura 14).
En el estudio realizado se utilizó el extracto acuoso de maca en la dosis de
200 mg/kg. Respecto a la dosis empleada, Alzamora L. (2003) administró la
dosis oral de 75 mg/kg del extracto clorofórmico y obtuvo resultados favorables
en la respuesta inmune humoral y celular. Torres D. (2008) empleó la dosis de
300 mg/kg del extracto acuoso, metanólico y clorofórmico, demostrando el
efecto modulador sobre la celularidad, hemograma y producción de
anticuerpos contra glóbulos rojos. Muchos estudios realizados con otras
plantas, señalaron que el incremento en las dosis produce una superación del
estado de inmunosupresión en menor tiempo (Bafna y Mishrash, 2005; Wang
et al, 2011; Patra et al., 2012). El tiempo de tratamiento fue de 2 meses, se
consideró este tiempo para confirmar el efecto hematopoyético del extracto de
maca en el ratón, es decir, garantizar que todos los efectos biológicos
observados están siendo influenciados por el extracto y que su consumo en un
tiempo prolongado no es hepatotóxico para el organismo.
El hígado es el lugar primario de biotransformación de compuestos
extraños, pero también es particularmente vulnerable a ellos. La
hepatotoxicidad del extracto acuoso de maca se evaluó midiendo los niveles
séricos de TGP, que es una enzima hepática muy utilizada como marcador de
daño hepático (Alvarez, 2005); los resultados confirmaron que el consumo por
2 meses del extracto acuoso de maca a la dosis de 200 mg/kg no resultó
dañino para los animales, ya que sus niveles de TGP fueron similares a los no
tratados con el extracto (67.65 vs 67.89). Reportes anteriores también indican
la ausencia de toxicidad de la maca en el desarrollo embrionario, la

71
descendencia, el hemograma y los niveles de enzimas hepáticas de los
roedores (Canales et al., 2000; D'Arriago et al., 2004; Castañeda et al., 2007).
Otros estudios además señalan el potencial antioxidante de la maca,
demostrado por su capacidad de atrapar los radicales libres y proteger a las
células contra el estrés oxidativo (Sandoval et al., 2002), revertir los
parámetros lipídicos, de glucosa y el nivel de enzimas anti-oxidantes en ratas
con hipertrigliceridemia hereditaria (Vecera et al., 2007) y aumentar la relación
glutatión reducido/glutatión oxidado (GSH/GSSG) a nivel plasmático,
contribuyendo con la mejora del estado redox en un modelo de diabetes
experimental (Cisneros et al., 2011). El tratamiento con CF produce niveles
altos de daño oxidativo en el tejido hepático, que es evidenciado por la
elevación de la concentración de malondialdehído (MDA) que indica un
incremento de la peroxidación lipídica en el hígado y por la inactivación de
funciones antioxidantes, causando un aumento de radicales libres de oxígeno
(ROS). El ROS incrementa la peroxidación lipídica, que a su vez altera la
integridad de las membranas, causa la pérdida de permeabilidad, disminución
de fluidez y por ende la fuga de enzimas como la TGP (Shathish et al., 2012;
Kim et al., 2012). En el presente trabajo, la administración de CF indujo un
aumento significativo en los niveles de TGP de todos los animales, sin
embargo los grupos tratados previamente con el extracto de maca no
modificaron sus niveles basales de TGP y fueron similares a los no
inmunosuprimidos con CF (Tabla 4), estos resultados confirman el efecto
hepatoprotector de la maca frente a un agente tóxico como la ciclofosfamida.
La inmunosupresión redujo drásticamente los niveles de expresión de IL-3,
GM-CSF e IL-7 en el bazo a 2 días post-IS (Grupo IS2d-Sin EAc), al
compararlos con los no inmunosuprimidos (p<0,05), pero el tratamiento con el
extracto acuoso amortiguó estos efectos (Grupo IS2d-EAc). El incremento en
la producción de las 3 citoquinas está asociada a una hematopoyesis creciente
en el bazo en ambos linajes: mieloides (IL-3, GM-CSF) y linfoide (IL-7);
además, de una mayor supervivencia y función efectora de las células
diferenciadas (Reddy et al., 2000). Como la producción de estas citoquinas es
realizada por las células estromales y accesorias, es probable que estas

72
células provenientes de ratones tratados con el extracto, respondan en menor
tiempo (2 post-IS) ante una emergencia y secreten estas citoquinas para
restaurar la homeostasis hematopoyética (Figuras 15, 16 y 17).
El patrón de expresión de las 3 citoquinas evaluadas no fue el mismo en la
médula ósea que en el bazo; la mayor expresión génica en la médula ósea se
observó a los 2 días post-IS (IS2d-EAc e IS2d-Sin EAc) y fue superior respecto
a los demás grupos (p<0.05); sin embargo, se reducen drásticamente a 5 días
post-IS hasta llegar a niveles similares a los grupos no inmunosuprimidos
(Figuras 18, 19 y 20), que es cuando se registra la mayor expresión en el bazo
(IS5d-EAc e IS5d-Sin EAc) indicando que la médula ósea responde primero
ante una emergencia hematopoyética y detiene su producción en el momento
en que aumenta la respuesta hematopoyética en el bazo, esto estaría
relacionado con la movilización de las células madre/ progenitoras, ya que
ellas están donde hay citoquinas para poder proliferar y diferenciarse. Sefc et
al. (2003) reportaron que al 5to día de la inmunosupresión con CF, hay un
incremento significativo de células progenitoras en el bazo que está
acompañado con una reducción de ellas en la médula ósea, debido a la
migración de los progenitores de la médula al bazo, porque el número de estas
células en el bazo pasa de 4% a 69%. Los ratones tratados con el extracto
mostraron mayor expresión de IL-7 a 2 días post-IS que indicaría mayor
proliferación y diferenciación de los progenitores hacia el linaje linfoide,
además de sus efectos sobre los linfocitos maduros (Meenu, W., 2008).
Aunque a 5 días post-IS hay disminución en la expresión de las 3 citoquinas,
los animales tratados con el extracto presentan mayor producción de IL-3 y
GM-CSF en la médula ósea (p<0.05). Todos estos resultados demuestran que
el extracto acuoso de maca puede atenuar la mielosupresión inducida por
ciclofosfamida, incrementando la producción de las 3 citoquinas evaluadas en
el bazo y médula ósea.
Para confirmar los efectos del extracto acuoso de maca sobre la
producción de citoquinas hematopoyéticas y su relación con las células
progenitoras de la médula ósea, se procedió a evaluar su acción en cultivos de
células mononucleares de médula ósea. El extracto acuoso de maca a la

73
dosis de 100 µg/ml estimuló la proliferación de células mononucleares de la
médula ósea a las 48 horas de cultivo (Figura 21) y esto acompañado de una
mayor producción de mRNA para las citoquinas hematopoyética IL-3, GM-CSF
e IL-7 a las 18 horas de cultivo, aunque solo fue significativo para IL-7 (Tabla
5). La IL-3 y el GM-CSF son secretados principalmente por las células T
activadas y están implicados en la supervivencia, proliferación y diferenciación
de las células progenitoras hacia el linaje mieloide, aunque a diferencia del
GM-CSF, la IL-3 actúa en los estados más primitivos de la célula
hematopoyética. La IL-7 es liberada por las células del estroma (fibroblastos,
células reticulares) y son absolutamente necesarios para el desarrollo de los
linfocitos T y B del ratón. Esto implica que las células mononucleares
obtenidas por gradiente en Ficoll Hypaque, constituyen una mezcla de
poblaciones celulares hematopoyéticas y estromales (Serrano et al., 2005).
Aunque en condiciones normales la hematopoyesis es controlada por
diversas citoquinas, pueden existir biomoléculas de origen y naturaleza distinta
a éstas que también pueden llegar a modular la proliferación y diferenciación
de las células hematopoyéticas. La estimulación de la proliferación de las
células de la médula ósea es un indicador que el extracto de maca es capaz
de promover la hematopoyesis medular, en este caso estimulando a las
células del entorno (linfocitos T y células del estroma) a producir las citoquinas
necesarias que regulan la proliferación y diferenciación de las células
hematopoyéticas; no se puede descartar un efecto directo de algunos
metabolitos del extracto sobre las mismas células madre/progenitoras.
Santiago et al. (2010) reportaron que la caseína es capaz de inducir la
proliferación de células mononucleares de ratón en más del 80%, además
señalaron que encontraron receptores para la caseína en las células
mieloides, que probablemente puedan incidir en su proliferación. Chen et al.
(2006) reportaron que el extracto crudo de la toda la planta de Angelica
sinensis estimula la proliferación de las células mononucleares
hematopoyéticas murinas y la producción de factores hematopoyéticos, a
través de las vías MAPK ERK1/2 y P38, que son importantes reguladores del
crecimiento, diferenciación o apoptosis celular. Los resultados obtenidos en los

74
cultivos in vitro de médula ósea, demuestran que la maca estimula la actividad
hematopoyética.
El recuento de células nucleadas de la médula ósea y sangre periférica es
el principal parámetro clínico que refleja el daño de las drogas citotóxicas. Los
ratones inmunosuprimidos presentaron los menores recuentos celulares, y
esto es debido a que el metabolismo de la CF por la enzimas hepáticas,
generan productos alquilantes que interfieren con las síntesis de DNA,
afectando a las células de división activa como las células madre/progenitoras
de la médula ósea, provocando la muerte celular (Patra et al., 2012); además
de los radicales libres que inducen estrés oxidativo en las células (Tripathi y
Jena, 2009). Sin embargo los grupos tratados con el extracto acuoso de maca
presentan un mayor recuento de células nucleadas en médula ósea y sangre
periférica a 5 días post-IS (Figuras 22 y 23); la estimulación de la proliferación
celular en la médula ósea pone en evidencia que el extracto es capaz de
promover la hematopoyesis medular, que se refleja en el mayor recuento total
y diferencial en la periferia. Es probable que la mayor producción de citoquinas
registradas en estos grupos, cumplan un rol importante en la repoblación de
las células de la médula ósea y sangre periférica, y según los resultados la IL-
3 estaría involucrada en el mayor recuento celular de la médula ósea, por ser
una de las citoquinas involucradas en los primeros estadíos de la
hematopoyesis. Además, su mayor producción no sólo se observó a los 5 días
post-IS sino también en el grupo EAc (no IS); tampoco se puede descartar al
GM-CSF e IL-7, que también se incrementaron en la médula ósea, por lo tanto
hay estimulación de ambos linajes. La presencia de blastos en la sangre
periférica del grupo IS2d-EAc en relación al grupo IS2d-Sin EAc, confirma el
efecto protector del extracto acuoso en las células inmaduras; este mismo
grupo también presentó mayores recuentos de granulocitos y monocitos
(p<0,05) (Tabla 6), probablemente relacionado con la mayor producción del
mRNA para GM-CSF, que está involucrado en la diferenciación de los
progenitores hacia granulocitos y macrófagos; sería interesante medir las
concentraciones sérica de esta citoquina, junto con la GM-CSF y M-CSF para
confirmar la estimulación del extracto al linaje granulocito/macrófago.

75
Existen muchos reportes que señalan que las células de sangre periférica
siguen un ritmo circadiano. En los ratones, animales nocturnos por naturaleza,
los leucocitos alcanzan su punto máximo en las primeras horas de la mañana
y van decreciendo a lo largo del día (Pelegri et al., 2003). Esto podría explicar
la gran magnitud de desviación estándar observada en el recuento total de
leucocitos (Figura 26, grupos No IS) y que se vio reflejada en el recuento
diferencial de sangre periférica (Tabla 6), por lo que se midió su coeficiente de
variación y resultó menor del 50% (datos no mostrados). Una forma de medir
el error muestral es a través del coeficiente de variación, que permite evaluar
la calidad estadística de las estimaciones, un coeficiente menor del 50% indica
que la media aritmética es representativa (Mitacc, 1996).
En el ratón, la hematopoyesis no solo se produce en la médula ósea sino
también en órganos distintos como el hígado y el bazo, en este trabajo se ha
comprobado la producción esplénica de mRNA para las 3 citoquinas
evaluadas; la hematopoyesis esplénica contribuye a la hematopoyesis medular
en condiciones de estrés, como una reacción compensatoria a la
inmunosupresión inducida por CF, ya que la médula ósea se vuelve
inadecuada para mantener una hematopoyesis normal (Kim, 2010). El
recuento de CFU-S es una medida indirecta de la frecuencia de células
madre/progenitoras con alta capacidad proliferativa que derivan de la médula
ósea formando discretos nódulos en la superficie del bazo (Parekkadan B y
Yarmush M, 2009). Los ratones del grupo IS5d-EAc mostraron incremento
significativo en el recuento de colonias (Figuras 24 y 25), lo que demuestra
que hay una mayor migración de células pluripotentes al bazo, donde
encuentran las citoquinas hematopoyéticas que propician condiciones
necesarias para proliferar y diferenciarse; esto concuerda con la mayor
producción de mRNA para la 3 citoquinas reportadas en los ratones del grupo
IS2d-EAc, cuyo resultado fue evidente a los 5 días post-IS en forma de
nódulos esplénicos y coincidiendo con investigadores como Shin et al. (2008)
quienes reportaron que Myelophil, que es una mezcla de extractos de Astragali
radix y Salviae radix, restauró la mielosupresión inducida por 5-Fluorouracilo
mediante la sobre-regulación de IL-3 en el bazo, además del incremento de los

76
parámetros hematológicos y el contaje de progenitores hematopoyéticos en
ensayos clonogénicos.
La CF también puede afectar la función (activación, proliferación y
diferenciación) de las células T y B, produciendo inmunosupresión. El ensayo
de la proliferación de linfocitos inducidos por mitógenos permite evaluar la
capacidad de los linfocitos para llevar a cabo una proliferación clonal en
respuesta a un estímulo no específico (Janeway et al., 2003). La interacción
de las células con el mitógeno, inicia una cascada de eventos bioquímicos y
expresión de genes que inducen a las células en reposo, a entrar al ciclo
celular para luego proliferar y diferenciarse, liberando citoquinas, factores de
crecimiento y anticuerpos (linfocitos B) que regulan otras funciones celulares
(Stoecker et al., 1978; Wang et al., 2005). En el presente estudio, la respuesta
proliferativa de los linfocitos T y B a sus respectivos mitógenos fue
drásticamente reducida en los grupos tratados con CF (IS2d-Sin EAc e IS5d-
Sin EAc). Sin embargo el tratamiento con el extracto acuoso de maca
promovió la recuperación de la respuesta proliferativa de las células T y B. La
mayor respuesta proliferativa de los linfocitos del bazo frente a Con A refleja la
mayor presencia de linfocitos T en los ratones tratados con el extracto de
maca, esto se relaciona con la mayor producción de mRNA de IL-3 y GM-CSF
observada en los grupos tratados con el extracto (Figuras 15 y 16), porque los
linfocitos T son las principales fuentes productoras de estas citoquinas, esta
afirmación se ve reforzada con la mayor expresión de IL-7 reportada en estos
grupos (Figura 17), esta citoquina participa en la regulación de las funciones
efectoras de los linfocitos T. La mayor respuesta proliferativa de los linfocitos
ante LPS, refleja la mejora en inmunidad humoral relacionada con los linfocitos
B, que explicaría el mayor título de anticuerpos reportados en trabajos
anteriores con ratones tratados con extractos de maca (Alzamora et al., 2007;
Torres, 2008).
El efecto tóxico de la CF sobre las funciones inmunes y hematopoyéticas
no solo es por el daño directo al DNA sino también por el incremento de la
producción de ROS, que puede llevar a la supresión de la médula ósea a
través de la inducción de apoptosis y senescencia en las células madre y

77
progenitoras (Chen et al., 2012), por lo que la presencia de metabolitos con
acción anti-oxidante e inmunoestimuladora en el extracto de maca serían
responsables de los efectos observados en la hematopoyesis. Estudios
realizados por Sandoval et al. (2002) empleando el extracto acuoso de maca
hervida (ecotipo amarillo) demostraron su capacidad de atrapar radicales libres
y proteger a las células contra el daño oxidativo. Oré (2008) reportó que el
extracto acuoso de maca amarilla fue capaz de ejercer efecto protector a nivel
hepático y aórtico en ratas hipercolesterolémicas y atribuye esta capacidad a
la presencia de fenoles y flavonoides. Son muchos los metabolitos de la maca
a los que se les puede atribuir estos efectos; los compuesto fenólicos (taninos
y flavonoides) y sus comprobadas propiedades quimio-protectoras (Huang et
al., 2010; Sandhar et al., 2011); los isotiocianatos, por su capacidad de reducir
la activación de los carcinógenos e incrementar su detoxificación (Wu et al.,
2009); los alcaloides β-carboline (reportado en la Maca por Piacente et al. en
2002) y su actividad anti-genotóxica y anti-mutagénica relacionado a sus
propiedades anti-oxidantes (Moura et al., 2007); las saponinas y sus
propiedades inmunoestimulantes (Rajput et al., 2007). Por otra parte, existen
muchos trabajos que señalan que también es fundamental el aporte energético
en los mecanismos inmunológicos (Calder y Kew, 2002; Wintergerst et al.,
2007; Marimuthu et al., 2012), por lo que el mayor contenido de nutrientes
presentes en el extracto acuoso de maca también contribuirían a los
resultados observados.

78
7. CONCLUSIONES
1. La dosis de 130 mg/kg de ciclofosfamida (CF) induce inmunosupresión
experimental que se mantuvo hasta 5 días después de su aplicación.
2. La administración del extracto acuoso de maca por dos meses no resultó
hepatotóxico para los animales de experimentación y resultó
hepatoprotector frente a la toxicidad inducida por ciclofosfamida.
3. El extracto acuoso de maca estimula la producción de mRNA para las
citoquinas IL-3, GM-CSF e IL-7 en el bazo de los ratones evaluados dos
días después de la inmunosupresión con ciclofosfamida.
4. En los ratones tratados con el extracto acuoso y evaluados dos días
después de la inmunosupresión muestran una mayor expresión de IL-7 en
la médula ósea. A 5 días post-IS, el extracto estimula la producción de
mRNA para IL-3 y GM-CSF.
5. El extracto acuoso de maca a la dosis de 100 µg/ml estimula la
proliferación de células mononucleares de médula ósea in vitro y provoca
el incremento significativo en los niveles de mRNA para IL-7.
6. El extracto acuoso de maca induce el incremento del recuento de células
nucleares de médula ósea, sangre periférica y colonias macroscópicas
endógenas en el bazo, además de una mayor respuesta proliferativa a
mitógenos de los linfocitos T y B en los ratones inmunosuprimidos con CF.

79
8. RECOMENDACIONES
Es importante profundizar los estudios a nivel celular y molecular del
hipocótilo de maca sobre la hematopoyesis, para determinar la población
celular sobre la que actúa y las vías intracelulares implicadas. Se puede
realizar cultivos de células de médula ósea en ensayos clonogénicos o de
formadoras de colonias en medio semisólido, que permitiría conocer la
población progenitora hematopoyética sobre la que tiene efecto el extracto
de maca.
Sería interesante realizar estudios del efecto de la maca a nivel de la
eritropoyesis. Se puede partir de animales con anemia inducida y evaluar si
la administración del extracto acuoso puede revertir la anemia, incrementar
la producción de eritropoyetina y la expresión de genes relacionados al
linaje eritroide.
Evaluar diferentes fracciones del extracto acuoso de maca sobre la
proliferación, producción de citoquinas hematopoyéticas y formación de
colonias de las células de médula ósea, e identificar el metabolito
responsable del efecto observado.

80
9. REFERENCIAS BIBLIOGRÁFICAS
Abbas A, Lichtman A, Pillai S. 2008. Inmunología celular y molecular. 6ta
edición. Elsevier Saunders, España.
Alvarez E. 2008. Estudio comparativo de la actividad moduladora del
extracto metanólico de cuatro ecotipos de Lepidium peruvianum Chacón
(maca) sobre la respuesta inmune humoral y celular en ratones. Tesis para
Titulación, Facultad de Ciencias Biológicas, Universidad Nacional Mayor de
San Marcos, Lima, Perú.
Alvarez H. 2005. El paciente con hipertransaminemia. Rev Fac Med UNAM
48: 58 - 65.
Alzamora L. 2003. Estudio del efecto antitumoral e inmunomodulador del
extracto clorofórmico de raíces de Lepidium peruvianum G. Chacón “Maca”
(Brassicaceae) en ratones. Tesis doctoral, Facultad de Ciencias Biológicas,
Universidad Nacional Mayor de San Marcos, Lima, Perú.
Alzamora L, Ávila G, Colona E, García J, Olivera J, Alzamora D. 2004.
Immunostimulation by aqueous extract from Lepidium peruvianum (Maca)
on cyclophosphamide-induced suppression mice. XIII Scientific Reunion of
the ICBAR. Biological Sciences Faculty, San Marcos University, Lima, Perú.
Alzamora L, Galván P, Alvarez E, Torres D, Colona E, Aliaga M, Marcelo A.
2007. Producción de IFN-γ en cultivos de linfocitos humanos por efecto de
los extractos metanólicos de cuatro ecotipos de Lepidium peruvianum
Chacón (Brassicaceae) Rev Peru biol, 13(3): 207 – 209.
Anderson D, Bishop J, Colin R, Ostrosky P, Selby P. 1995.
Cyclophosphamide: Review of its mutagenicity for an assessment of
potential germ cell risks. Mutation Research, 330: 115 - 181.
Arrebola F. 1999. Cuantificación de ácidos nucleicos desarrollados en gel
de agarosa. Diseño y validación de un método simple de análisis digital de
imagen. II Congreso virtual hispanoamericano de anatomía patológica.
Comunicación n° 046, II Congreso virtual hispanoamericano de anatomía
patológica.

81
Bafna AR, Mishra SH. 2005. Immunomodulatory activity of methanol extract
of roots of Cissampelos pareira Linn. Ars Pharm, 46 (3): 253 - 262.
Bhushan P, Manish G. 2007. Ethnopharmacology Approaches for Botanical
Immunomodulators and Chemoprotectants in Cancer Therapy. Alternative
Treatment For Cancer, 1: 255 - 284.
Bianchi A. 2003. MACA LEPIDIUM MEYENII. Boletín Latinoamericano y del
Caribe de Plantas Medicinales y Aromáticas, 2(003): 30 - 36.
Brooks NA, Wilcox G, Walker KZ, Ashton JF, Cox MB, Stojanovska L. 2008.
Beneficial effects of Lepidium meyenii (Maca) on psychological symptoms
and measures of sexual dysfunction in postmenopausal women are not
related to estrogen or androgen content. Menopause, 15(6): 1157 - 1162.
Calder PC, Kew S. 2002. The immune system: a target for functional
foods? Br J Nutr, 88(2): 165 - 177.
Canales M, Aguilar J, Prada A, Marcelo A, Huamán C, Carbajal L. 2000.
Evaluación nutricional de Lepidium meyenii (MACA) en ratones albinos y su
descendencia. Arch Latinoam Nutr, 50(2): 126 - 133.
Cano J, Flores A, Martínez J, Aguilar C, Rodríguez R. 2010. Diseño de
iniciadores para la amplificación del gen de penicilina G acilasa de Mucor
griseocyanus. Revista Científica de la Universidad Autónoma de Coahuila,
Volumen 2, No. 4.
Castañeda B, Castro de la Mata R, Manrique R, Ibañez L. 2007. Efectos
metabólicos de Lepidium meyenii Walpers “MACA” y Lupinus mutabilis
Sweet, “CHOCHO” en ratas. Revista Horizonte Médico, 7(1): 32 - 38.
Castaño MP. 2008. Maca (Lepidium peruvianum CHACÓN): composición
química y propiedades farmacológicas. Revista de Fitoterapia, 8(1): 21 -
28.
Ceredig R, Rolink A. 2012. The key role of IL-7 in lymphopoiesis. Review.
Seminars in immunology, 24: 159 - 164.
Chacón G. 1990. La Maca (Lepidium peruvianum Chacón sp.nov.) y su
hábitat. Rev Per Biol, 3(2): 169 - 272.
Chen Q, Zhang S, Ying H, Dai X, Li X, Yu C, Ye H. 2012. Chemical
characterization and immunostimulatory effects of a polysaccharide from

82
Polygoni multiflori radix praeparata in cyclophosphamide-induced anemic
mice. Carbohydrate Polymers, 88(4): 1476 - 1482.
Chen X, Chen J, Zhang P, Du J. 2006. Angelica stimulates proliferation of
murine bone marrow mononuclear cells by the MAPK pathway. Blood Cells
Mol Dis, 36(3):402 - 405.
Cisnero R, Oré R, Arnao I, Suárez S. 2011. Relación de glutatión
reducido/oxidado (GSH/GSSG) en ratas diabéticas tratadas con maca
(Lepidium meyenii walp). An Fac med, 72(2): 107 - 111.
Córdova A. 2003. Fisiología Dinámica. MASSON S.A. Barcelona, España.
Cui B, Zheng BL, He K, Zheng QY. 2003 Imidazole alkaloids from Lepidium
meyenii. J Nat Prod, 66: 1101 - 1103.
D´Arrigo G, Benavides V, Pino J. 2004. Preliminary Evaluation Effect of
Lepidium meyenii Walp on the embryonic development of mouse. Rev peru
biol, 11(1): 103 - 106.
Denzler K, Waters R, Jacobs B, Rochon Y, Langland J. 2010. Regulation of
inflammatory gene expression in PBMC by immunostimulatory Botanicals.
PloS One, 5(9): 1 - 15.
Destéfano L. 2011. Determinación del Código de barras de ADN para la
diferenciación taxonómica entre Lepidium meyenii y Lepidium peruvianum.
Congreso Peruano de Medicina tradicional alternativa y complementaria.
Colegio Médico del Perú, Lima, Perú.
Dini A, Migliuolo G, Rastrelli L, Saturnino P, Schettino O. 1994. Chemical
composition of Lepidium meyenii. Food Chem 49, 347 - 349.
Dini I, Tenore GC, Dini A. 2002. Glucosinolates from Maca (Lepidium
meyenii). Biochemical Systematics and Ecology, 30: 1087 - 1090.
Dolgova EV, Proskurina AS, Nikolin VP, Popova NA, Alyamkina EA,
Orishchenko KE, Rogachev VA, Efremov YR, Dubatolova TD, Prokopenko
AV, Chernykh ER, Ostanin AA, Taranov OS, Omigov VV, Zagrebelniy SN,
Bogachev SS, Shurdov MA. 2012. "Delayed death" phenomenon: a
synergistic action of cyclophosphamide and exogenous DNA. Gene, 10;
495(2): 134 - 145.
Erali M, Wittwer C. 2010. High resolution melting analysis for gene
scanning. Methods, 50: 250 - 261.

83
Fairlie T, Morales M, Holle M. 1999. Raíces y Tubérculos Andinos Avances
de Investigación. Centro Internacional de la Papa, Lima, Perú.
Fry T, Mackall C. 2005. The many faces of IL-7: From lymphopoiesis to
peripheral T cell maintenance. The journal of immunology, 174: 6571 -
6576.
Gonzalez GF, Córdova A, Vega K, Chung A, Villena A, Góñez C. 2003.
Effect of Lepidium meyenii (Maca), a root with aphrodisiac and fertility-
enhancing properties, on serum reproductive hormone levels in adult
healthy men. Journal of Endocrinology, 176(1): 163–168.
Gonzales GF, Gasco M, Córdova A, Chung A, Rubio J, Villegas L. 2004.
Effect of Lepidium meyenii (Maca) on spermatogenesis in male rats acutely
exposed to high altitude (4340 m). Journal of Endocrinology, 180(1): 87–95.
Gonzales GF. 2012. Ethnobiology and Ethnopharmacology of Lepidium
meyenii (Maca), a Plant from the Peruvian Highlands. Review Article,
Evidence-Based Complementary and Alternative Medicine, 1 - 10.
Guthridge MA, Stomski FC, Thomas D, Woodcock JM, Bagley CJ, Berndt
MC, Lopez AF. 1998. Mechanism of activation of the GM-CSF, IL-3, and IL-
5 family of receptors. Stem Cells, 16(5): 301 - 313.
Gutiérrez J, Montaño K, Bracho J, Rodríguez C, Chang A. 2009.
Caracterización de esteroles en la fracción lipídica de la maca (Walp.)
mediante técnicas cromatográficas. Rev Rev Soc Quím Perú, 75 (2): 254 -
265.
Hara T, Miyajima A. 1996. Function and signal transduction mediated by
the interleukin 3 receptor system in hematopoiesis. Stem cell, 14(6): 605 -
618.
Haubitz M. 2007. Acute and Long-term Toxicity of Cyclophosphamide.
Transplantationsmedizin, 19: 26 - 31.
Herbert K, Levesque JP, Haylock DN, Prince M. 2008. The use of
experimental murine models to assess novel agents of hematopoietic stem
and progenitor cell mobilization. Biology of blood and marrow
transplantation, 14: 603 - 621.
Hercus TR, Thomas D, Guthridge MA, Ekert PG, King-Scott J, Parker MW,
Lopez AF. 2009. The granulocyte-macrophage colony-stimulating factor

84
receptor: linking its structure to cell signaling and its role in disease. Blood,
114(7): 1289 - 1298.
Huang HY, Luther SA. 2012. Expression and function of interleukin-7 in
secondary and tertiary lymphoid organs. Review. Seminars in immunology,
24: 175 - 189.
Huang WY, Cai YZ, Zhang Y. 2010. Natural phenolic compounds from
medicinal herbs and dietary plants: potential use for cancer prevention. Nutr
Cancer, 62 (1):1 - 20.
Huyan XH, Lin YP, Gao T, Chen RY, Fan YM. 2011. Immunosuppressive
effect of cyclophosphamide on white blood cells and lymphocyte
subpopulations from peripheral blood of Balb/c mice. Int Immunopharmacol,
11(9):1293 - 1297.
Janeway C, Travers P, Walport M, Shlomchik. 2003. Inmunobiología. El
sistema inmunitario en condiciones de salud y enfermedad. MASSON S.A.
2da Edición, Barcelona, España.
Jena G, Vikram A, Tripathi DN, Ramarao P. 2010. Use of chemoprotectants
in chemotherapy and radiation therapy: the challenges of selecting an
appropriate agent. Integr Cancer Ther, 9(3): 253 - 258.
Kim CH. 2010. Homeostatic and pathogenic extramedullary hematopoiesis.
Review. Journal of blood medicine, 1: 13 - 19.
Kim SH, Lee IC, Lim JH, Moon C, Bae CS, Kim SH, Shin DH, Park SC, Kim
HC, Kim JC. 2011. Protective effects of pine bark extract on developmental
toxicity of cyclophosphamide in rats. Food Chem Toxicol, 50(2):109-115.
Krawczenko A, Kieda C, Dus D. 2005. The biological role and potential
therapeutic application of interleukin 7. Arch Immunol ther Exp, 53(6): 518 -
525.
Lee KJ, Dabrowski K, Rinchard J, Gomez C, Guz L, Vilchez C. 2004.
Supplementation of maca (Lepidium meyenii) tuber meal in diets improves
growth rate and survival of rainbow trout Oncorhynchus mykiss (Walbaum)
alevins and juveniles. Aquaculture Research, 35(3): 215 - 223.
Lees, D. H. y Francis, F. J. 1972. Standardization of pigment analysis in
cranberries. HortScience, 7: 83 – 84.

85
Liang QD, Gao Y, Tan HL, Guo P, Li YF, Zhou Z, Tan W, Ma ZC, Ma BP,
Wang SQ. 2006. Effects of four Si-Wu-Tang's constituents and their
combination on irradiated mice. Biol Pharm Bull, 29(7):1378 - 1382.
Liberman AC, Druker J, Refojo D, Arzt E. 2008. Mecanismos moleculares
de acción de algunas drogas inmunosupresoras. MEDICINA, 68: 455 - 464.
Llopis C. 2009. Modelado farmacocinético de ciclofosfamida en pacientes
con cáncer de mama. Tesis. Departamento de farmacia y tecnología
farmacéutica. Universidad de Barcelona, Barcelona, España.
Lock O. 1994. Investigación fitoquímica. Métodos en el estudio de
productos naturales. 2da edición, Fondo editorial de la Pontificia
Universidad Católica del Perú, Lima, Perú.
López-Casamayor E. 2007. Estudio fitoquímico y aproximación genética en
especies de la sección Plinthine del género Arenaria (Caryophyllaceae).
Tesis doctoral. Departamento de Botánica. Universidad de Granada,
Granada, España.
Marcano D, Hasegawa M. 2002. Fitoquímica orgánica. 2da edición,
Editorial Torino, Venezuela.
Marimuthu K, Varadhan KK, Ljungqvist O, Lobo DN. 2012. A meta-analysis
of the effect of combinations of immune modulating nutrients on outcome in
patients undergoing major open gastrointestinal surgery. Ann Surg,
255(6):1060 - 1068.
Massbereg S, Von Andrian UH. 2009. Novel trafficking for hematopoietic
stem and progenitor cells. Hematopoietic stem cell VII, 1176: 87 - 93.
Mayani H, Flores E, Pelayo R, Montesinos J, Flores P, Chávez A. 2007.
Hematopoyesis. Cancerología, 2: 95 – 107.
McCurdy, R, McGrath, J, Mackay, A. 2008. Validation of the comparative
quantification method of real-time PCR analysis and a cautionary tale of
housekeeping gene selection. Gene Ther Mol Biol, Vol 12: 15 - 24.
Meenu RT. 2008. Haematopoietic growth factors and their therapeutic use.
Thromb Haemost, 99: 863 - 873.
Mera C, Roa A, Ramirez S. 2007. Células madres hematopoyéticas,
generalidades y vías implicadas en su mecanismo de auto-renovación. Rev
Cienc Salud, 5(1): 67 – 89.

86
Mitacc M. 1996. Tópicos de estadística descriptiva y probabilidad. 1era
edición. Editorial Thales S.R.Ltda. Lima, Perú.
Mosmann T. 1983. Rapid colorimetric assay for cellular growth and survival:
application to proliferation and cytotoxicity assays. J Immunol Methods, 65:
55 - 63.
Moura DJ, Richter MF, Boeira JM, Pêgas Henriques JA, Saffi J. 2007.
Antioxidant properties of beta-carboline alkaloids are related to their
antimutagenic and antigenotoxic activities. Mutagenesis, 22(4):293 - 302.
Muhammad I, Zhao J, Chuck D, Khan I. 2002. Constituents of Lepidium
meyenii (maca). Phytochemistry, 59:105 – 110.
Nimer S, Uchida H. 1995. Regulation of granulocyte - macrophage colony -
stimulation factor and interleukin 3 expression. Stem cells, 13(4): 324 - 335.
Obregón L. 1998. “Maca” Planta medicinal y nutritiva del Perú. 1era
edición, Instituto de Fitoterapia Americano, Lima, Perú.
Oré R, Suárez S, Rojas L, Valdivieso R, Oriondo R. 2011. Efecto del
extracto acuoso de maca sobre la función cognitiva en ratas recién
destetadas. An Fac med, 72(1): 13 - 16.
Oré R. 2008. Efectos hipolipémico y antioxidante de Lepidium meyenii
Walp en ratas. Tesis de doctorado. Facultad de Ciencias Biológicas,
Universidad Nacional Mayor de San Marcos, Lima, Perú.
Oziemlak AM. 2005. Development origins of the murine hematopoietic
system. Tesis de doctorado, Departamento de Biología Celular.
Universidad de Erasmus, Rotterdam, Paises Bajos.
Parekkadan B y Yarmush M. 2009. Methods in Bioengineering: Stem Cell
Bioengineering. Artech House, United States.
Patra K, Bose S, Sarkar S, Rakshit J, Jana S, Mukherjee A, Roy A, Mandal
DP, Bhattacharjee S. 2012. Amelioration of cyclophosphamide induced
myelosuppression and oxidative stress by cinnamic acid. Chem Biol
Interact, 195(3): 231 - 239.
Penel N, Adenis A, Bocci G. 2012. Cyclophosphamide-based metronomic
chemotherapy: after 10 years of experience, where do we stand and where
are we going? Crit Rev Oncol Hematol, 82(1):40 - 50.

87
Pelegrí C, Vilaplana J, Castellote C, Rabanal M, Franch A, Castell M. 2003.
Circadian rhythms in surface molecules of rat blood lymphocytes. Am J
Physiol Cell Physiol. 284 (1):C67-76.
Pfaffl, M W. 2001. A new mathematical model for relative quantification in
real-time RT-PCR. Nucleic Acids Res, 29: 2002 - 2007
Piacente S, Carbone V, Plaza A, Zampelli A, Pizza C. 2002. Investigation of
the tuber constituents of maca (Lepidium meyenii Walp.). J Agric Food
Chem, 50: 5621 - 5625.
Rajput ZI, Hu SH, Xiao CW, Arijo AG. 2007. Adjuvant effects of saponins on
animal immune responses. J Zhejiang Univ Sci B, 8(3):153 - 161.
Rapley R, Manning D. 1998. RNA isolation and characterization protocols.
Editorial Hummana Press, USA.
Reddy EP, Korapati A, Chaturvedi P, Rane S. 2000. IL3 signaling and the
role of Src, JAKs and STATs: a covert liaison unveiled. Oncogene, 19: 2532
- 2547.
Roitt I, Delves P, Martin S, Burton D. 2008. Inmunología de Roitt:
Fundamentos. 11ava edición, Editorial médica panamericana, Buenos
aires, argentina.
Rosas J, Pino A. 2005. Efecto antioxidante de cuatro ecotipos y dos tipos
de harina del Lepidium peruvianum CHACÓN (Maca) in vitro. Encuentro
científico internacional ECIPERU, 2(1): 1 - 3.
Rubio J, Riqueros MI, Gasco M, Yucra S, Miranda S, Gonzales GF. 2006.
Lepidium meyenii (Maca) reversed the lead acetate induced-Damage on
reproductive function in male rats. Food and Chemical Toxicology, 44(7):
1114 - 1122.
Ruiz GJ. 2009. Fundamentos de Hematología. Editorial Médica
Panamericana. 4ta Edición. México.
Salem ML, Al-Khami AA, El-Nagaar SA, Zidan AA, Al-Sharkawi IM, Marcela
Díaz-Montero C, Cole DJ. 2012. Kinetics of rebounding of lymphoid and
myeloid cells in mouse peripheral blood, spleen and bone marrow after
treatment with cyclophosphamide. Cell Immunol, 276(1-2): 67 - 74.

88
Sandhar HK, Prasher S, Tiwari P, Salhan M, Sharma P. 2011. A review of
phytochemistry and pharmacology of flavonoids. Internationale
Pharmaceutica Sciencia 1: 25 - 41.
Sandoval M, Okuhama N, Angeles F, Melchor V, Conddezo L, Lao J, Miller
M. 2002. Antioxidant activity of the cruciferous vegetable Maca (Lepidium
meyenii). Food Chemistry, 79: 207 - 213.
Santiago E, Ledesma E, Weiss B, Muñoz L, Domínguez V, Aguiñiga I,
Córdova Y, Moreno L, Tiburcio R. 2010. Avances en la regulación de la
proliferación y diferenciación de células hematopoyéticas, tanto normales
como leucémicas, por la caseína y sus componentes. Hematología,
11(4):193 – 198.
Sefc L, Psenák O, Sýkora V, Sulc K, Necas E. 2003. Response of
hematopoiesis to cyclophosphamide follows highly specific patterns in bone
marrow and spleen. J Hematother Stem Cell Res, 12(1):47-61.
Serrano T, Alberti E, Lorigados L, Díaz I, Blanco L, Vallejo A. 2005.
Caracterización inmunofenotípica de las células de la médula ósea de
ratas. Biotecnología Aplicada, Vol.22, No.3. 234 - 236.
Sharapin N. 2000. Fundamento de tecnología de productos
fitoterapeúticos. 1era edición, Convenio Andrés Bello, Colombia.
Shathish K, Reena D, Guruvayoorappan C. 2012. Chemoprotective effect
of Decalepis hamiltonii against cyclophosphamide induced toxicity. J Exp
Ther Oncol, 9(4):291 - 301.
Shi Y, Liu CH, Roberts AI, Das J, Xu G, Ren G, Zhang Y, Zhang L, Yuan
ZR, Tan HS, Das G, Devadas S. 2006. Granulocyte-macrophage colony-
stimulating factor (GM-CSF) and T-cell responses: what we do and don't
know. Cell Res, 16(2): 126 - 133.
Shin JW, Lee MM, Son JY, Lee NH, Cho CK, Chung WK, Cho JH, Son CG.
2008. Myelophil, a mixture of Astragali radix and Salviae radix extract,
moderates toxic side effects of fluorouracil in mice. World J Gastroenterol,
14(15):2323 - 2328.
Soto I, Cáceres J, Mendoza J, Weiss B. 1999. Citoquinas en la
hematopoyesis y el sistema inmunológico: mecanismos celulares y
moleculares. 1era edición, Plaza y Valdéz editores, México.

89
Stoecker CA, Rickard BM, Abel CA. 1978. Effect of T- and B-lymphocyte
mitogens on interactions between lymphocytes and macrophages. Cell
Immunol, 35(2):362 - 377.
Suárez S, Oré R, Arnao I, Rojas L, Trabucco J. 2009. Extracto acuoso de
Lepidium meyenii Walp (maca) y su papel como adaptógeno, en un modelo
animal de resistencia física. An Fac med, 70(3): 181 - 185
Tello J, Hermann M, Calderón A. 1992. La maca (Lepidium meyenii Walp.)
cultivo alimenticio potencial para las zonas altoandinas. Bol Lima, vol 14:
59 - 66.
Torres D. 2008. Efecto modulador de la respuesta inmune humoral de
extractos de Lepidium peruvianum Chacón (Maca) en ratones
inmunosuprimidos con ciclosfosfamida. Tesis para Titulación, Facultad de
Ciencias Biológicas, Universidad Nacional Mayor de San Marcos, Lima,
Perú.
Tripathi DN, Jena GB. 2009. Intervention of astaxanthin against
cyclophosphamide - induced oxidative stress and DNA damage: a study in
mice. Chem Biol Interact, 180(3): 398 - 406.
Ullmannovi V, Haakovec C. 2003. The Use of Housekeeping Genes (HKG)
as an Internal Control for the Detection of Gene Expression by Quantitative
Real-Time RT-PCR. Folia Biologica (Praha), 49(6): 211 - 216.
Valentová K, Buckiová D, Kren V, Peknicová J, Ulrichová J, Šimánek V.
2006. The in vitro biological activity of Lepidium meyenii extracts. Cell
Biology and Toxicology, 22(2): 91 - 99.
Valentová K, Ulrichová J. 2003. Smallanthus Sonchifolius and Lepidium
meyenii-Prospective andean crops for the prevention of chronic diseases.
Biomed Papers, 147(2): 119 – 130.
Vecera R, Orolin J, Skottova N, Kazdova L, Oliyarnik O, Ulrichov J,
Simanek V. 2007. The Influence of Maca (Lepidium meyenii) on Antioxidant
Status, Lipid and Glucose Metabolism in Rat. Plant Foods for Human
Nutrition, 62(2): 59 – 63.
Vilchez JP. 2001. El cultivo de la maca y su consumo. CONCYTEC. Lima,
Perú.

90
Wang Y, Gao B, Tsan MF. 2005. Induction of cytokines by heat shock
proteins and concanavalin A in murine splenocytes. Cytokine, 32(3-4): 149 -
154.
Wang H, Wang M, Chen J, Tang Y, Dou J, Yu J, Xi T, Zhou C. 2011. A
polysaccharide from Strongylocentrotus nudus eggs protects against
myelosuppression and immunosuppression in cyclophosphamide-treated
mice. Int Immunopharmacol, 11(11): 1946 - 1953.
Wanga Y, Wanga Y, McNeilc B, Harveyc L. 2007. Maca: An Andean crop
with multi-pharmacological functions. Review. Food Research International,
40(7): 783 - 792.
Wu X, Zhou QH, Xu K. 2009. Are isothiocyanates potential anti-cancer
drugs? Acta Pharmacologica Sinica, 30: 501 - 512.
Welsch U. 2010. Histología. 2da Edición, Editorial Médica Panamericana,
México.
Willems E, Mateizel I, Kemp C, Cauffman G, Sermon K, Leyns L. 2006.
Selection of reference genes in mouse embryos and in differentiating
human and mouse ES cells. Int J Dev Biol, 50: 627 - 635.
Wintergerst ES, Maggini S, Hornig DH. 2007. Contribution of selected
vitamins and trace elements to immune function. Ann Nutr Metab, 51(4):
301 - 323.
Xiang Wu, Qing-hua Zhou, Ke Xu. 2009. Are isothiocyanates potential anti-
cancer drugs? Acta Pharmacologica Sinica, 30: 501 - 512.
Ye J, Coulouris G, Zaretskaya I, Cutcutache I, Rozen S, Madden T. 2012.
Primer-BLAST: A tool to design target-specific primers for polymerase chain
reaction. BMC Bioinformatics, 18; 13(1): 134.
Yllescas MG. 1994. Estudio Químico y Fitoquímico comparativo de 3
ecotipos de Lepidium meyenii Walp. “Maca” procedentes de Carhuamayo
(Junín). Tesis para Titulación, Facultad de Farmacia y Bioquímica,
Universidad Nacional Mayor de San Marcos, Lima, Perú.
Zhang Y, Yu L, Ao M, Jin W. 2006. Effect of ethanol extract of Lepidium
meyenii Walp. on osteoporosis in ovariectomized rat. Journal of
Ethnopharmacology, 105(1-2): 274 - 279.

91
Zhao J, Muhammad I, Chuck D, Mustafa J, Khan I. 2005. New Alkamides
from Maca (Lepidium meyenii). J Agric Food Chem, 53 (3): 690 - 693.

92
10. ANEXOS
Posible mecanismo modulador de la hematopoyesis producido por el
extracto acuoso de maca
A partir de los resultados obtenidos, se intentará representar gráficamente
las posibles relaciones entre el extracto acuoso empleado en las pruebas y
sus células blanco para la producción de las citoquinas estudiadas.
El extracto acuoso de maca podría actuar reduciendo la formación de
radicales libres producidos por la ciclofosfamida en las células con alta
actividad proliferativa, como las células madre/progenitoras hematopoyéticas,
esto podría estar mediado por los flavonoides u otros metabolitos presentes en
el extracto, también podría actuar inhibiendo directamente la acción alquilante
de CF sobre el DNA de las células proliferantes; aunque no se puede
descartar un efecto a nivel de membrana, con una acción similar a citoquinas
y/o activando vías de señalización intracelulares implicadas en la producción
de citoquinas hematopoyéticas. Esto daría como resultado una mejor
respuesta hematopoyética a la mielosupresión, como una mayor producción
de IL-3 y GM-CSF por parte de los linfocitos T y de IL-7 por parte de las
células estromales, principales productoras de estas citoquinas, demostrado
por la mayor expresión de mRNA para estas tres citoquinas. La IL-7 además
de estimular una mayor proliferación y diferenciación de los progenitores hacia
el linaje linfoide, tiene efecto sobre los linfocitos maduros, como los linfocitos
T. La sobreexpresión de estas tres citoquinas atenúa la mielosupresión
inducida por ciclofosfamida, reflejado en un mayor recuento celular de médula
ósea, sangre periférica y esplénico de células progenitoras y maduras (Figura
28).
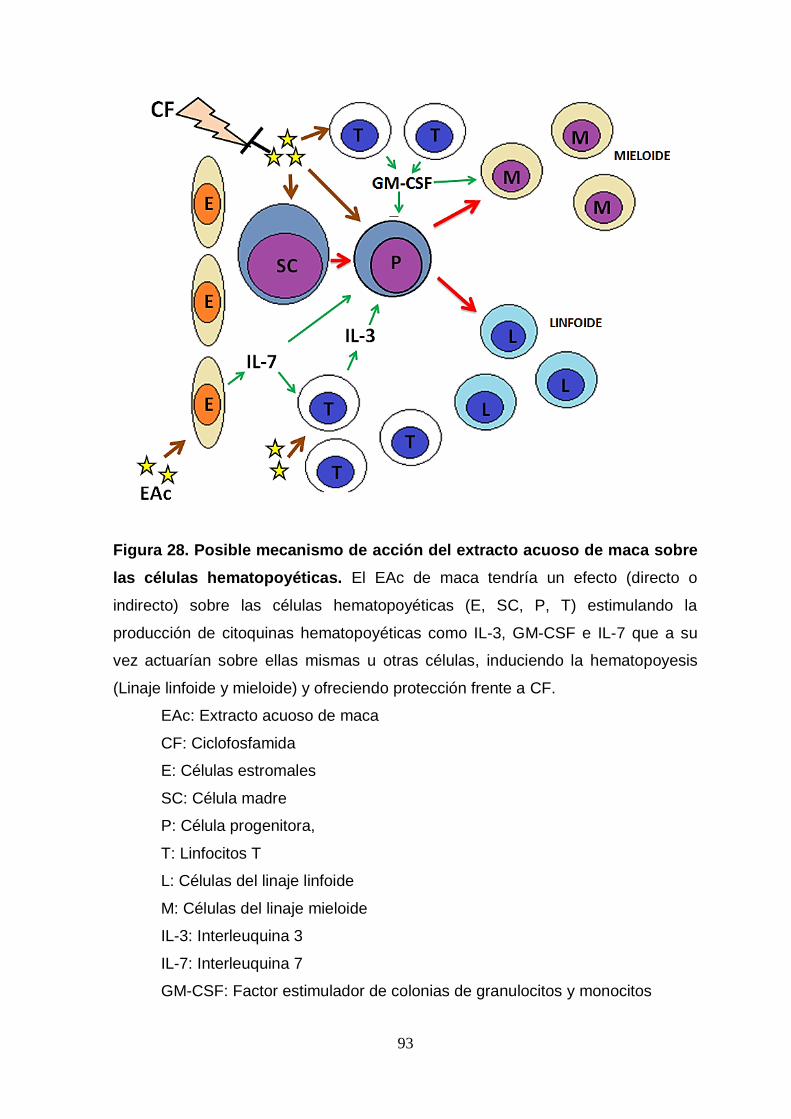
93
Figura 28. Posible mecanismo de acción del extracto acuoso de maca sobre
las células hematopoyéticas. El EAc de maca tendría un efecto (directo o
indirecto) sobre las células hematopoyéticas (E, SC, P, T) estimulando la
producción de citoquinas hematopoyéticas como IL-3, GM-CSF e IL-7 que a su
vez actuarían sobre ellas mismas u otras células, induciendo la hematopoyesis
(Linaje linfoide y mieloide) y ofreciendo protección frente a CF.
EAc: Extracto acuoso de maca
CF: Ciclofosfamida
E: Células estromales
SC: Célula madre
P: Célula progenitora,
T: Linfocitos T
L: Células del linaje linfoide
M: Células del linaje mieloide
IL-3: Interleuquina 3
IL-7: Interleuquina 7
GM-CSF: Factor estimulador de colonias de granulocitos y monocitos


















