Trabajo de Mitocondrias
34
Facultad de ciencias medicas ALUMNA : DANA ALMONTE RIVERA TEMA : ORGANELO CELULAR “EL MITOCONDRIAS” CURSO : BIOLOGÍA MOLECULAR Y CELULAR PROFESOR: GUILLERMO BORNAZ ZCOSTA AÑO : PRIMERO TACNA - PERU
-
Upload
danacatacora -
Category
Documents
-
view
211 -
download
2
Transcript of Trabajo de Mitocondrias

Facultad de ciencias medicas
ALUMNA : DANA ALMONTE RIVERA
TEMA : ORGANELO CELULAR “EL MITOCONDRIAS”
CURSO : BIOLOGÍA MOLECULAR Y CELULAR
PROFESOR: GUILLERMO BORNAZ ZCOSTA
AÑO : PRIMERO
TACNA - PERU
2010

LA CELULALa célula es la forma más sencilla de organización biológica. Es considerada la unidad estructural, funcional y hereditaria mas pequeña de materia capaz de realizar todas las actividades inherentes a lo seres vivos. La célula posee distintos organelos .
Ahora estudiaremos el organelo del mitocondrias :

Dedicatoria:
Este trabajo esta dedicado con el fin de conocer un poco más sobre la
estructura y composición del organelo de la célula como lo es la mitocondrias.
Agradecimiento:
Un agradecimiento en especial a mi profesor por permitirme realizar este
trabajo y así podérselo presentárselo .
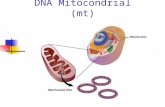
INTRODUCCIONMitocondria del griego: mitocondrias, thread + chondros, gránulo es el sitio donde se lleva a cabo la respiración celular, i.e. el metabolismo aerobio, en la mayoría de los organismos eucariontes (con núcleo verdadero). Lo anterior fue demostrado por A. Lenhinger y E.

Kennedy en 1948. Este organelo descubierto en el siglo XIX, varía en tamaño y forma dependiendo de la fuente y el estado metabólico, pero a menudo son elipsoides de aproximadamente 5 µm de diámetro y 1 µm de largo, del tamaño de una bacteria típica. Una célula eucarionte típica contiene mas de 2000 mitocondrias, lo que ocupa alrededor de la quinta parte del volumen celular; esta cantidad es necesaria porque este organelo es la central energética de la célula. Las células de mamíferos que contienen más mitocondrias son las células cardiacas y los espermatozoides..
Las microscopias electrónicas de las mitocondrias, revelan la presencia de dos membranas, una de ellas lisa, en la parte externa del organelo y otra muy plegada, a cada pliegue se le denomina cresta. El número de crestas varía con la actividad respiratoria del tipo particular de célula en estudio. Lo anterior se debe a que las enzimas que llevan a cabo el transporte de electrones y las fosforilación oxidativa, están unidas a esta membrana. En los hepatocitos, las mitocondrias tienen pocas crestas, en las células cardiacas, hay muchas. Debido a lo anterior, en este organelo existen dos espacios, el intermembranal y la matriz. Las enzimas respiratorias, forman parte tanto de la membrana interna, como de la matriz gelatinosa (50 % H2O). Estas enzimas acoplan la energía producida por la oxidación de los nutrientes a la energía necesaria para sintetizar adenosín trifosfato (ATP), que después de transportarse a los diversos organelos, es utilizado como combustible en diversos procesos.Las mitocondrias tienen más similitudes con las bacterias que únicamente tamaño y forma. La matriz mitocondrial, contiene DNA, RNA y ribosomas que participan en la síntesis de algunos componentes mitocondrias (el resto es importado del citoplasma después de su expresión desde el núcleo). A pesar de esto, se reproducen por fisión binaria, además el proceso respiratorio que llevan a cabo, es muy semejante al que se observa en bacterias aerobias actuales.. Los nutrientes abastecidos por el eucarionte y consumidos por la bacteria, fueron presumiblemente reembolsados con creces, debido a la alta eficiencia metabólica oxidativa que el procarionte (sin núcleo), le dio al eucarionte. La corroboración de la teoría endosimbiótica se da en la amiba Pelomyxa palustris, uno de los pocos eucariontes que carece de mitocondrias. Esta amiba, permanentemente hace endosimbiosis con bacterias aerobias. El origen de diversos organelos también se puede explicar a partir de la teoría endosimbiótica.
A la mitocondria, se le considera la central energética de las células, ahí se lleva a cabo el metabolismo oxidativo de los eucariontes: actividades de piruvato deshidrogenasa, enzimas del ciclo del ácido cítrico, oxidación de los ácidos grasos y las enzimas y proteínas redox que llevan a cabo el transporte de electrones y la fosforilación oxidativa.
SUMARIO
Dedicatoria

Agradecimiento Introducción
1 Historia 2 Características 3Estructura y composición
o 3.1 Membrana externa o 3.2 Membrana interna o 3.3 Espacio intermembranoso o 3.4 Matriz mitocondrial
4 Función 5 Origen 6 Enfermedades mitocondriales 7 Conclusiones 8 Anexos 9 Bibliografía
.
1.-HISTORIAEl descubrimiento de la mitocondria podría decirse que es un hecho colectivo como ninguno. Prueba de ello está en el gran número de términos acuñados que se refieren al

mismo orgánulo: Blefaroplasto, condrioconto, condriómitos, condrioplastos, condriosomas, condriosferas, fila, gránulos fucsinofílicos, Korner, Fadenkörper, mitogel, cuerpos parabasales, vermículas, sarcosomas, cuerpos intersticiales, plasmosomas, plastocondrios, bioblastos. Cowdry y Lehninger intentaron en 1918 sistematizar y unificar todos los términos.13
Probablemente las primeras observaciones se deben al botánico suizo Kolliker, quien en 1880-1888 anotó la presencia de unos gránulos en células musculares de insectos a los que denominó sarcosomas. Llegó incluso a la conclusión de que presentaban membrana.14 En 1882, el alemán Walther Flemming descubrió una serie de inclusiones a las que denominó fila.15 En 1884 también fueron observados por Richard Altmann, quien más tarde en su obra publicada en Leipzig Die Elementarorganismen describe una serie de corpúsculos que observa mediante una tinción especial que incluye fucsina. Especula que se trata de una suerte de parásitos independientes, con su propio metabolismo y los denomina bioblastos. El hallazgo fue rechazado como un artefacto de la preparación, y sólo más tarde fue reconocido como mitocondrias por N.H. Cowdry (1916).16 También los "plastídulos" del protozoólogo italiano Leopoldo Maggi podrían tratarse de observaciones tempranas de mitocondrias.17
Sin embargo, el nombre de "mitocondria", que es el que alcanzó mayor fortuna, se debe a Carl Benda, quien en 1889 denominó así a unos gránulos que aparecían con gran brillo en tinciones de violeta cristal y alizarina, y que anteriormente habían sido denominados "citomicrosomas" por Velette St. George.16 14 En 1904 F. Meves confirma su presencia en una planta, concretamente en células del tapete de la antera de Nymphaea, y en 1913 Otto Heinrich Warburg descubre la asociación con enzimas de la cadena respiratoria, aunque ya Kingsbury, en 1912 había relacionado estos orgánulos con la respiración celular. En 1934 fueron aisladas por primera vez a partir de homogeneizados de hígado y en 1948 Hogeboon, Schneider y Palade establecen definitivamente la mitocondria como el lugar donde se produce la respiración celular.18
La presencia del ADN mitocondrial fue descubierta por Margit M. K. Nass y Sylvan Nass en 1963.14 19
2.-CARACTERISTICAUna característica peculiar de las mitocondrias es que son de origen maternoo, ya que sólo el óvulo aporta las mitocondrias a la célula original, y cómo la mitocondria posee ADN , podemos decir que esta información va pasando a las generaciones exclusivamente a través de las mujeres.
3.-ESTRUCTURA Y COMPOSICIÓN

Estructura de una mitocondria
Sin las mitocondrias las células dependerían de la glucólisis anaeróbica para formar ATP. Pero este proceso solo es capaz de liberar una pequeña cantidad de la energía disponible en la glucosa. En las mitocondrias el metabolismo de los azúcares está integrado: el piruvato (glucóilisis) es importado dentro de la mitocondria y oxidado por el O2 a CO2 y H2O. La energía liberada es almacenada de una manera tan eficiente que por cada glucosa oxidada se producen aprox. 30 ATP.La morfología de la mitocondria es difícil de describir puesto que son estructuras muy plásticas que se deforman, se dividen y fusionan. Normalmente se las representa en forma alargada. Su tamaño oscila entre 0,5 y 1 μm de diámetro y hasta 7 μ de longitud.20 Su número depende de las necesidades energéticas de la célula. Al conjunto de las mitocondrias de la célula se le denomina condrioma celular.Las mitocondrias están rodeadas de dos membranas claramente diferentes en sus funciones y actividades enzimáticas, que separan tres espacios: el citosol, el espacio intermembrana y la matriz mitocondrial. Cada mitocondria esta limitada por dos membranas muy especializadas. Definen dos compartimientos: Matriz y el espacio intermembranoso.

2.1 Membrana externa
Es una bicapa lipídica exterior permeable a iones, metabolitos y muchos polipéptidos. Eso es debido a que contiene proteínas que forman poros, llamadas porinas o VDAC (de canal aniónico dependiente de voltaje), que permiten el paso de grandes moléculas de hasta 10.000 dalton y un diámetro aproximado de 20 Å. La membrana externa realiza relativamente pocas funciones enzimáticas o de transporte. Contiene entre un 60 y un 70% de proteínas. Contiene numerosas proteínas que regulan los intercambios de sustancias con el citosol. Destacan las proteínas de canal, las cuales forman grandes poros que la hacen muy permeable, es la membrana que cubre a todas las demás.
2.2 Membrana interna
La membrana interna contiene más proteínas, carece de poros y es altamente selectiva; contiene muchos complejos enzimáticos y sistemas de transporte transmembrana, que están implicados en la translocación de moléculas. Esta membrana forma invaginaciones o pliegues llamadas crestas mitocondriales, que aumentan mucho la superficie para el asentamiento de dichas enzimas. En la mayoría de los eucariontes, las crestas forman tabiques aplanados perpendiculares al eje de la mitocondria, pero en algunos protistas tienen forma tubular o discoidal. En la composición de la membrana interna hay una gran abundancia de proteínas (un 80%), que son además exclusivas de este orgánulo:
1. La cadena de transporte de electrones, compuesta por cuatro complejos enzimáticos fijos y dos transportadores de electrones móviles:
1. Complejo I o NADH deshidrogenasa que contiene flavina mononucleótido (FMN)
2. Complejo II o succinato deshidrogenasa; ambos ceden electrones al coenzima Q o ubiquinona;
3. Complejo III o citocromo bc1 que cede electrones al citocromo c 4. Complejo IV o citocromo c oxidasa que cede electrones al O2 para producir
dos moléculas de agua. 2. Un complejo enzimático, el canal de H+ ATP-sintasa que cataliza la síntesis de
ATP (fosforilación oxidativa). 3. Proteínas transportadoras que permiten el paso de iones y moléculas a su través,
como ácidos grasos, ácido pirúvico, ADP, ATP, O2 y agua; pueden destacarse: 1. Nucleótido de adenina translocasa. Se encarga de transportar a la matriz
mitocondrial el ADP citosólico formado durante las reacciones que consumen energía y, paralelamente transloca hacia el citosol el ATP recién sintetizado durante la fosforilación oxidativa.
2. Fosfato translocasa. Transloca fosfato citosólico junto con un protón a la matriz; el fosfato es esencial para fosforilar el ADP durante la fosforilación oxidativa.
2.3 Espacio intermembranoso
Entre ambas membranas queda delimitado un espacio intermembranoso que está compuesto de un líquido similar al hialoplasma; tienen una alta concentración de protones como resultado del bombeo de los mismos por los complejos enzimáticos de la cadena

respiratoria. En él se localizan diversos enzimas que intervienen en la transferencia del enlace de alta energía del ATP, como la adenilato quinasa o la creatina quinasa. También se localiza la carnitina, una molécula implicada en el transporte de ácidos grasos desde el citosol hasta la matriz mitocondrial, donde serán oxidados (beta-oxidación).
2.4 Matriz mitocondrial
La matriz mitocondrial o mitosol contiene menos moléculas que el citosol, aunque contiene iones, metabolitos a oxidar, ADN circular bicatenario muy parecido al de las bacterias, ribosomas tipo 70S similares a los de bacterias, llamados mitorribosomas, que realizan la síntesis de algunas proteínas mitocondriales, y contiene ARN mitocondrial; es decir, tienen los orgánulos que tendría una célula procariota de vida libre. En la matriz mitocondrial tienen lugar diversas rutas metabólicas clave para la vida, como el ciclo de Krebs y la beta-oxidación de los ácidos grasos; también se oxidan los aminoácidos y se localizan algunas reacciones de la síntesis de urea y grupos hemo.
3.-FUNCIÓNDel apartado anterior se deduce que la principal función de las mitocondrias es la oxidación de metabolitos (ciclo de Krebs, beta-oxidación de ácidos grasos) y la obtención de ATP mediante la fosforilación oxidativa, que es dependiente de la cadena transportadora de electrones; el ATP producido en la mitocondria supone un porcentaje muy alto del ATP sintetizado por la célula. También sirve de almacén de sustancias como iones, agua y algunas partículas como restos de virus y proteínas. La cadena de transporte de electrones separa los electrones y los protones de cada uno de los diez átomos de hidrógeno. Los diez electrones se envían a lo largo de la cadena y acaban por combinarse con oxígeno y los protones para formar agua. La energía se libera a medida que los electrones pasan desde las coenzimas a los átomos de oxígeno y se almacena en compuestos de la cadena de transporte de electrones. A medida que éstos pasan de uno a otro, los componentes de la cadena bombean aleatoriamente protones desde la matriz hacia el espacio comprendido entre las membranas interna y externa. Los protones sólo pueden volver a la matriz por una vía compleja de proteínas integradas en la membrana interior. Este complejo de proteínas de membrana permite a los protones volver a la matriz sólo si se añade un grupo fosfato al compuesto difosfato de adenosina (ADP) para formar ATP en un proceso llamado fosforilación

Ciclo de Krebs:
Es la ruta final de la oxidación del piruvato, ácidos grasos y cadenas de carbono de los aminoácidos.
Paso 1:El enlace energético se rompe y con otro grupo acetilo forman una molécula de citrato.
Paso 2:Los átomos del citrato se reacomodan cuando se elimina una molécula de agua y se agrega otra molécula de agua ,por estas reacciones se convierte en un isocitrato.
Paso 3:El isocitrato es deshidrogenizado y descarbolixado para formar un compuesto de 5 carbonos el ððcetoglutarato y dioxido de carbono.
Paso 4:El ððcetoglutarato se somete a una descarboxilacion oxidativa para formar un compuesto de 4 carbonos la succinil coenzima A Y CO2

Paso 5:En esta fase ocurre la fosforilacion a nivel de sustrato. El enlace que une la coenzima A con el succinato, como el de acetil CoA,es energetico y se señala
como (~S).En esta etapa la succinil CoA se convierte en succinato
Paso 6:El succinato es oxidado y 2 de sus atomos de hidrogeno se trasfieren al FADH2.El compuesto formado por el succinato es el fumarato.
Paso 7:Con la adicion del agua el fumarato se convierte en malato.
Ciclo de Krebs
Ocurre en la matriz mitocondrial.
Resultado: CO2 y electrones ricos en energía, que pasan vía NADH y FADH2 a la cadena respiratoria.
El CO2 se elimina como producto de deshecho, mientras que los electrones de alta energía se desplazan por la cadena respiratoria y finalmente se combinan con O2 y forman H2O

EL CICLO DE KREBS
4.-ORIGENLa científica estadounidense Lynn Margulis, junto con otros científicos, recuperó en torno a 1980 una antigua hipótesis, reformulándola como teoría endosimbiótica. Según esta versión actualizada, hace unos 1.500 millones de años, una célula procariota capaz de obtener energía de los nutrientes orgánicos empleando el oxígeno molecular como oxidante, se fusionó en un momento de la evolución con otra célula procariota o eucariota primitiva al ser fagocitada sin ser inmediatamente digerida, un fenómeno frecuentemente observado. De esta manera se produjo una simbiosis permanente entre ambos tipos de seres: la procariota fagocitada proporcionaba energía, especialmente en forma de ATP y la célula hospedadora ofrecía un medio estable y rico en nutrientes a la otra. Este mutuo beneficio hizo que la célula invasora llegara a formar parte del organismo mayor, acabando por convertirse en parte de ella: la mitocondria. Otro factor que apoya esta teoría es que las bacterias y las mitocondrias tienen mucho en común, tales como el

tamaño, la estructura, componentes de su membrana y la forma en que producen energía, etc.
Esta hipótesis tiene entre sus fundamentos la evidencia de que las mitocondrias poseen su propio ADN y está recubierta por su propia membrana. Otra evidencia que sostiene esta hipótesis es que el código genético del ADN mitocondrial no suele ser el mismo que el código genético del ADN nuclear.21 A lo largo de la historia común la mayor parte de los genes mitocondriales han sido transferidos al núcleo, de tal manera que la mitocondria no es viable fuera de la célula huésped y ésta no suele serlo sin mitocondrias.
5.-ENFERMEDADES MITOCONDRIALES
El ADN mitocondrial humano contiene información genética para 13 proteínas mitocondriales y algunos ARN;no obstante, la mayoría de las proteínas de las mitocondrias proceden de genes localizados en el ADN del núcleo celular y que son sintetizadas por ribosomas libres del citosol y luego importadas por el orgánulo. Se han descrito más de 150 enfermedades mitocondriales, como la enfermedad de Luft o la neuropatía óptica hereditaria de Leber. Tanto las mutaciones del ADN mitocondrial, como del ADN nuclear dan lugar a enfermedades genéticas mitocondriales, que originan un mal funcionamiento de procesos que se desarrollan en las mitocondrias, como alteraciones de enzimas, ARN, componentes de la cadena de transporte de electrones y sistemas de transporte de la membrana interna; muchas de ellas afectan al músculo esquelético y al sistema nervioso central.
El ADN mitocondrial puede dañarse con los radicales libres formados en la mitocondria; así, enfermedades degenerativas relacionadas con el envejecimiento, como la enfermedad de Parkinson, la enfermedad de Alzheimer y las cardiopatías pueden tener relaciones con lesiones mitocondriales
Patología mitocondrial en las enfermedades del miocardio
Las mutaciones en el genoma mitocondrial pueden ocurrir en cualquier órgano o sistema. La función cardíaca, al igual que la de otros órganos como el músculo esquelético y el cerebro, depende de la energía generada en las mitocondrias por ß-oxidación principalmente de los ácidos grasos y de los carbohidratos mediante el mecanismo de fosforilación oxidativa. Todas las proteínas codificadas por el genoma mitocondrial son subunidades del complejo mecanismo de fosforilación oxidativa, de tal manera que las mutaciones del ácido desoxiribonucleico mitocondrial conllevan a alteraciones de este crucial proceso que se manifiestan en el corazón con enfermedades miocárdicas (miocardiopatías), trastornos de conducción y ocasionalmente, arritmias letales.
Todas las enfermedades del músculo cardíaco cursan con cambios estructurales y funcionales de las mitocondrias en grado variable e igualmente, las mitocondrias pueden por si solas, ser el origen de una serie de alteraciones específicas del músculo cardíaco (Miocardiopatías mitocondriales) y de otros órganos. La importancia de esto radica en que pueden surgir terapéuticas adecuadas a raíz del conocimiento del papel que las mismas desempeñan en el funcionamiento cardíaco normal y anormal. Es importante, por lo tanto, advertir sobre la existencia "no tan rara" de dichos desórdenes.

Clasificación de las miocardiopatías mitocondriales
La clasificación de las enfermedades mitocondriales aún no está definida completamente y aparentemente no existe uniformidad en los criterios para agrupar este tipo de afecciones del miocardio. La Organización Mundial de la Salud (OMS), en su última clasificación, hace más de un quinquenio, no las identifica separadamente (31). Obviamente, estas enzimopatías mitocondriales entran en el grupo de las miocardiopatías específicas del miocardio en el renglón de las miocardiopatías metabólicas.
Desde las primeras descripciones de las miocardiopatías idiopáticas con mitocondrias gigantes en 1970, han sido descritas numerosas alteraciones metabólicas mitocondriales que cursan con miocardiopatías, miopatías, sordera, diabetes, encefalopatías y se han reconocido síndromes clínicos definidos (32). Actualmente, la mayoría de las miocardiopatías mitocondriales han sido identificadas con defectos genéticos de la cadena de enzimas respiratorias mitocondriales especialmente de los complejos I, III y IV, y /o a defectos del transporte de moléculas desde el citosol a la mitocondria.
La clasificación genética (33) incluye: Formas esporádicas (síndrome de Kearns-Sayre y de Pearson); Por herencia mendeliana (miocardiopatías excesivamente raras) ocasionadas por mutaciones en el ADN nuclear, el cual codifica polipéptidos para las mitocondrias, o a defectos en la comunicación intergenómica como la miocardiopatía y oftalmoplejía externa progresiva autosómica recesiva y las formas maternalmente heredadas. Estas últimas son las miocardiopatías más frecuentes y representan una gran proporción de todas las miocardiopatías familiares. Son la consecuencia de mutaciones puntuales del ADNmit y se pueden asociar con otros signos de alteración en la OXPHOS tales como la encefalomiopatía o acidemia láctica o pueden ser la única característica clínica de la enfermedad. Ejemplos de esta entidad lo constituyen el síndrome de MELAS (Mitochondrial Encephalomyopathy, Lactic Acidosis and Stroke-likes episodes) y el MERRF (Myoclonic Epilepsy with Ragged-Red Fibers).
La clasificación de Gilbert Barness y col. (34) comprende todas las enfermedades ocasionadas por desórdenes mitocondriales (con o sin mitocondrias anormales), que incluyen los causados por: Alteraciones de la cadena respiratoria; la miocardiopatía hipertrófica con miopatía mitocondrial del músculo esquelético y cataratas; la enfermedad de Leigh y la deficiencia sistémica de carnitina (Cuadro 1).
Cuadro 1
Clasificación de las miocardiopatías mitocondriales
• Por deficiencia de las enzimas de la cadena respiratoria.
Deficiencia del complejo I (coenzima citocromo c reductasa).
Deficiencia del complejo II.
Deficiencia del complejo III (citocromo b).

Deficiencia del complejo III con miocardiopatía histiocitoide.
Deficiencia del complejo IV (citocromo c oxidasa o COX).
Deficiencia del complejo V (ATPsintetasa).
Defectos múltiples de la cadena respiratoria o síndromes de superposición.
Síndrome de Kearns Sayre.
Síndrome de MELAS.
Síndrome de MERRF.
Síndrome de Barth (X-unido a la miopatía mitocondrial y granulocitopenia).
Síndrome de MELAS/MERRF.
• Miocardiopatía hipertrófica hereditaria con miopatía esquelética y cataratas.
• Miocardiopatía hipertrófica con enfermedad de Leigh (encefalomiopatía aguda necrotizante).
• Deficiencia de carnitina.
BASES DE LA ENFERMEDAD
Las enfermedades mitocondriales son resultado de la falla de las mitocondrias, los compartimentos especializados presentes en cada célula del cuerpo, con excepción de los glóbulos rojos de la sangre. Las mitocondrias son las responsables de la creación de más del 90% de la energía que el cuerpo necesita para mantener la vida y apoyar el crecimiento. Cuando fallan, se genera cada vez menos energía al interior de la célula. Puede entonces presentarse lesión celular o incluso la muerte de la célula. Si este proceso se repite en todo el cuerpo, los sistemas completos comienzan a fallar y la vida de la persona que lo sufre, está en grave riesgo. Esta enfermedad afecta principalmente a los niños, pero los brotes en adultos se están volviendo más y más comunes.
Las enfermedades de las mitocondrias parecen ocasionar el mayor daño a las células del cerebro, del corazón, del hígado, músculo esqueléticas, del riñón así como a los sistemas endocrino y respiratorio.

Dependiendo de qué células resulten afectadas, los síntomas pueden incluir pérdida de control motor, debilidad muscular y dolor; desórdenes gastrointestinales y dificultades para deglutir; crecimiento deficiente, enfermedades cardiacas, del hígado, diabetes, complicaciones respiratorias, convulsiones, problemas visuales y auditivos, acidosis láctica, retrasos en el desarrollo y susceptibilidad a contraer infecciones.
CUÁNDO SOSPECHAR QUE HAY UNA DISFUNCIÓN MITOCONDRIAL
No hay una característica única para identificar una enfermedad mitocondrial. Los pacientes presentan varios problemas que pueden surgir desde el nacimiento hasta en la edad adulta madura. Piense en las mitocondrias cuando:
Una “enfermedad común” presente características atípicas que la distingan del resto.
Haya tres o más órganos involucrados Se presenten recaídas recurrentes, o cuando ocurran brotes de infección en una
enfermedad crónica.
Las enfermedades mitocondriales o citopatías, deberán considerarse como posibles en diagnósticos diferenciales, cuando aparezcan estas características inexplicables, especialmente cuando ocurran en combinación con:

Síntomas
Encefalopatía o Convulciones o Retraso en el Desarrollo o Regresión (incluyendo demencia temprana o
episodios tardíos) o Mioclono o Desórdenes de Movimiento (distonia, disquinesias, corea, etc.) o Migraña Complicada o Infartos
Neuropatía Defectos en los Conductos Cardiacos
o Cardiomiopatías Deficiencias en la Audición Estatura corta Desórdenes de Músculos Extraoculares
o tosis o estrabismo adquirido o oftalmoplegia
Diabetes Enfermedad Renal Tubular Pérdida de la Visión
o retinitis pigmentosa o atrofia óptica
Acidosis láctica (puede ser leve)
Problemas Asociados a las Citopatías Mitocondriales
SISTEMA DE ÓRGANOS
PROBLEMAS POSIBLES
Cerebro
Retraso en el desarrollo, retardo mental, demencia, convulsiones, desórdenes neuro-psiquiátricos, paralsis cerebral atípica, migrañas, infartos

Nervios
Debilidad (que puede ser intermitente), dolor neuropático, ausencia de reflejos, problemas gastrointestinales (reflujo gastroesofágeo, vaciado gástrico retrasado, constipación, pseudo obstrucción), desmayos, ausencia o exceso de sudor relacionados con problemas de regulación de la temperatura
Músculos Debilidad, hipotonia, calambres, dolor muscular
Riñones
Desgaste proximal renal tubular que provoca pérdida de proteínas, magnesio, fósforo, calcio y otros electrolitos
Corazón
Defectos en los conductos cardiacos (bloqueos del corazón), cardiomiopatía
Hígado Hipoglicemia (niveles de azúcar bajos en la sangre), falla del hígado
Ojos Pérdida de visión y ceguera
Oídos Pérdida auditiva y sordera
Páncreas
Diabetes y falla pancreatítica exocrina (incapacidad para generar encimas digestivas)
Sistémico
Incapacidad para subir de peso, corta estatura, fatiga, problemas respiratorios incluyendo sofocamientos intermitentes
Enfermedades mitocondriales con pérdidas visuales y auditivas con sus descripciones
Hay gran número de enfermedades mitocondriales listadas en el sitio web de la United Mitochondrial Disease Foundation. Algunas de éstas tienen varios efectos visuales y de pérdida auditiva asociadas. Algunas de las que se nombran incluyen:
Enfermedad de Alpers
Nombre completo: Poliodistrofia Infantil Progresiva
Síntomas: convulsiones, demencia, espaticidad, ceguera, disfunción del hígado y degeneración cerebral.

Deficiencia del Complejo I
Nombre completo: Deficiencia NADH dehidrogenasa (NADH-Co! Reductasa)
Síntomas: Tres formas principales:
1. Desorden fatal infantil multisistémico, caracterizado por un retraso en el desarrollo, debilidad muscular, enfermedad cardiaca, acidosis láctica congénita y paro respiratorio.
2. Miopatía que inicia en la infancia o en la vida adulta, manifestándose como intolerancia al ejercicio o debilidad. Es común la elevación del ácido láctico.
3. Encefalomiopatía mitocondrial (incluyendo MELAS), que puede comenzar en la infancia o en la edad adulta y que consiste en combinaciones variables de síntomas y signos, incluyendo oftalmoplegia, convulsiones, demencia, ataxia, pérdida auditiva, retinopatía pigmentosa, neuropatía sensorial y movimientos incontrolables. Puede provocar el Síndrome de Leigh.
Deficiencia del Complejo III
Nombre completo: Deficiencia ubiquinona-citocroma c óxidoreductasa
Síntomas: Cuatro formas principales:
1. Encefalomiopatía mortal infantil, acidosis láctica congénita, hipotonia, postura distrófica, convulsiones y coma. Es común la fibrosis roja.
2. Encefalomiopatía de brotes posteriores (desde la infancia hasta la edad adulta): varias combinaciones de debilidad, estatura corta, ataxia, demencia, pérdida auditiva, neuropatía sensorial, retinopatía pigmentosa y síntomas piramidales. Es común la fibrosis roja. Posibilidad de acidosis láctica.
3. Miopatía, con intolerancia al ejercicio que deviene en debilidad continua. Es común la fibrosis roja. Posibilidad de acidosis láctica.
4. Cardiomiopatía histiocitoide infantil.
Deficiencia del Complejo IV / Deficiencia COX
Nombre completo: La deficiencia citocroma c óxidoreductasa es ocasionada por un defecto en el Complejo IV de la cadena respiratoria.
Síntomas: Dos formas principales:
1. Encefalomiopatía: normal durante los primeros 6 a 12 meses de vida; después presenta regresión en el desarrollo, ataxia, acidosis láctica, atrofia óptica, oftalmoplegia, nistagmus, distonia, signos piramidales y problemas respiratorios. Frecuentes convulsiones Puede provocar el Síndrome de Leigh.
2. Miopatía: dos variantes principales: a. Miopatía infantil mortal: puede empezar pronto después del nacimiento
acompañada de hipotonia, debilidad, acidosis láctica, fibrosis, paro respiratorio y problemas de riñón.

b. Miopatía infantil benigna: puede empezar pronto después del nacimiento acompañada de hipotonia, debilidad, acidosis láctica, fibrosis, falla respiratoria, pero (si el niño sobrevive) le sigue un mejoramiento espontáneo.
ANEXOS : enfermedad mitocondrial progresiva resultando de una mutación sin sentido nueva en el gene mitocondrial de la DNA ND3.

Describimos a un hombre de 42 años que presentó con una historia progresiva de la epilepsia, movimiento-como episodios, atrofia óptica bilateral, y la declinación cognoscitiva. La investigación de su biopsia del músculo reveló un defecto específico en actividad del complejo I. El análisis subsecuente del genoma mitocondrial identificó una mutación heteroplasmic nueva de T10191C en el gene ND3. La mutación estaba presente en los niveles inferiores en sangre del paciente y de los parientes maternales inafectados y es la primera mutación mitocondrial patógena de la DNA en el gene ND3 que se describirá.
Los intermedios de la oxidación no saturada del ácido graso se incorporan en triglicéridos pero no en fosfolípidos en tejidos de pacientes con defectos mitocondriales de la beta-oxidación.
La composición de ácido graso era resuelta del hígado, del músculo esquelético y del corazón obtenidos post mortem de pacientes con la deficiencia de la deshidrogenasa del acil-CoA de la medio-cadena (MCADD), la deficiencia múltiple de la deshidrogenasa del acil-CoA (MADD) y la deficiencia muy de larga cadena de la deshidrogenasa del acil-CoA (VLCADD). Cantidades crecientes de 4 5 del 12:1 ácido dodecenoic (n-7), 5 del 14:1 ácido tetradecenoic (n-9), 5.8 del 14:2 ácido tetradecadienoic (n-6) y 7.10 ácidos 16:2 hexadecadienoic del 10:1 ácido decenoic (n-6), (n-6)--intermedios de la oxidación no saturada del ácido graso--fueron encontrados. El fraccionamiento en diversas clases del lípido demostró que estos ácidos grasos estaban exclusivamente presentes en la fracción del triglicérido. No podían ser detectados en la fracción del ácido graso libre o en la fracción del fosfolípido. Nuestros resultados sugieren que los intermedios de la oxidación no saturada del ácido graso que acumulan como consecuencia de MCADD, de MADD y de VLCADD estén transportados al retículo endoplásmico para la esterificación en glycerolipids neutrales. El patrón de la acumulación es característico para cada enfermedad, que hace análisis del ácido graso del lípido total de tejidos post mortem una herramienta útil en la detección de defectos mitocondriales de la oxidación del ácido graso en los pacientes que murieron inesperado, por ejemplo con síndrome de la muerte infantil repentina
Tubulopathy mitocondrial: las muchas caras de desordenes mitocondriales.
Divulgamos una presentación rara del desorden mitocondrial en un niño con los espasmos carpopedales recurrentes debido al hypocalcemia y al hypomagnesemia, secundaria al hypoparathyroidism tubulopathy y posible próximo renal. Por lo menos dos especies mitocondriales de la DNA del mutante fueron identificadas, y las mitocondrias anormales fueron encontradas en el músculo y los especímenes renales de la biopsia. El caso ilustra el espectro y la diversidad de presentaciones mitocondriales, presentándose debido a heteroplasmy de las mutaciones y del tipo de órganos afectados.

Las muestras clínicas y del laboratorio de la disfunción mitocondrial secundarias a la terapia antiretroviral análoga del nucleósido son reversibles.
Durante 27 meses de tratamiento con el diario del stavudine del magnesio 400 didanosine del magnesio y 80 pero ninguna terapia del inhibidor de proteasa, una mujer seropositiva y-vieja 50 perdió gradualmente 13 kilogramos en peso, sus brazos, las piernas y las nalgas disminuidas en volumen y ella experimentaron fatiga y polineuropatía. Los pruebas de laboratorio demostraron aumentos leves en niveles de la enzima del lactato y del hígado del plasma. Dieciocho meses después del retiro de la droga antiretroviral, el paciente estaba libre de fatiga y de polineuropatía y había recuperado 7 kilogramos en peso así como la mayor parte de el volumen de sus brazos, piernas y nalgas.
El ácido de Valproic acciona rhabdomyolysis agudo en un paciente con el tipo deficiencia del palmitoyltransferase de la carnitina de II.
Un hombre de 47 años que sufría de un desorden bipolar y de un myoglobinuria intermitente presentó con rhabdomyolysis agudo con falta renal después de comenzar terapia con el ácido valproic. En la examinación morfológica, el músculo esquelético reveló almacenaje creciente del lípido. Bioquímico, la actividad enzimática disminuida del tipo II del palmitoyltransferase de la carnitina (CPT) con los niveles de la carnitina en el límite más bajo fue encontrada. El análisis genético detectó la substitución común de Ser113Leu en un alelo del gene CPT2. Concluimos que el ácido valproic se debe evitar en pacientes con el tipo deficiencia de CPT de II.
Caso de la autopsia de la paraplegia espástica hereditaria recesiva de un autosoma referente a la patología muscular.

Un caso autopsied de la paraplegia espástica hereditaria recesiva de un autosoma con atrofia muscular neurogénica severa se describe adjunto. Este paciente, mujer de 16 años, presentada con disturbio del paso. Ella desarrolló la parálisis espástica progresiva de los miembros superiores y más bajos y de la deterioración mental. Ella hizo postrada en cama en aproximadamente 40 años de edad. La disartria empeoró en 45 años de edad. Ella murió de pulmonía en 50 años de edad. Su hermana más joven ha demostrado síntomas clínicos similares e hizo postrada en cama en 37 años de edad. Sus padres eran segundos primos. La autopsia reveló un cerebro seriamente atrófico, pesando 720 G. La corteza cerebral era fina, y la materia blanca fue reducida extremadamente en volumen. Microscópico, la pérdida neuronal y el astrogliosis variable con los cambios esponjosos difusos eran evidentes en la corteza cerebral, los núcleos talámicos, los ganglios básicos y el hipocampo. Las neuronas restantes fueron atrofiadas con la deposición pesada del lipofuscin. En la médula espinal, las zonas piramidales así como las zonas espinocerebelosas dorsales fueron degeneradas. Además, la pérdida marcada de las células de cuerno anterior fue considerada. La pérdida neuronal severa del núcleo gracilis también fue detectada. En cambio, solamente la degeneración suave de las zonas espinocerebelosas ventrales y el cuneatus del fasciulus en la médula espinal fueron observados. En las secciones congeladas del músculo esquelético, la atrofia neurogénica severa y la infiltración grasa eran evidentes. Además, varias vacuolas bordeadas fueron observadas en las fibras atróficas, y las fibras coxidase-deficientes del citocromo estaban presentes en el dinucleótido de adenina de niconamida reducido partición (NADH) - acumulación anormal reveladora reacción de la reductasa del tetrazolium de mitocondrias alrededor del centro de las fibras de músculo no-atróficas. Se sugiere que un análisis de la función mitocondrial del hemiplegia espástico hereditario recesivo de un autosoma japonés puede proporcionar la información adicional para aclarar la patogenesia.
enfermedad y movimiento mitocondriales. FONDO Y PROPÓSITO: Es bien sabido que algunos desordenes mitocondriales son responsables del infarto cerebral isquémico en pacientes jovenes. Nuestro propósito era determinar, en este estudio en curso anticipado, el movimiento isquémico es la única manifestación de un desorden mitocondrial en pacientes jovenes. métodos: Los pacientes envejecieron (+info)
síndrome hydranencephalic-hydrocephalic congénito con vasculopathy proliferativo: una relación posible con la disfunción mitocondrial.
Divulgamos el caso de un feto abortado en la semana 20 de la gestación debido a síndrome hydranencephalic-hydrocephalic. El feto era el tercer embarazo de un par nonconsanguineous cuyo primer niño exhibió el síndrome hydranencephalic-hydrocephalic congénito asociado a los resultados de la histología del músculo constantes con cytopathy mitocondrial y de la deficiencia de complejos III e IV de la cadena respiratoria y cuyo segundo embarazo había terminado en un aborto electivo en la detección de hidrocefalia progresiva en la semana 19 de la gestación.

El tercer embarazo tenía un curso normal según las examinaciones obstétricas y de la sonografía realizadas en las semanas 5, 10, y 15 de la gestación, y los resultados negativos fueron obtenidos en serológico estándar y la reacción en cadena de polimerasa (polimerización en cadena) prueba para las infecciones prenatales de la madre. Sin embargo, la examinación de la sonografía en la semana 18 de la gestación reveló la hidrocefalia, en respuesta a la cual los padres pedidos un aborto, que fue realizado en la semana 20 de la gestación; el feto era masculino y sin malformaciones externas evidentes. Los estudios histopatológicos del oblongata del cerebro y de la médula revelaron inclusiones vasculopathy (los recipientes del glomeruloid, las inclusiones intracitoplásmicas, y las microcalcificaciones) e intracitoplásmicas proliferativas en el músculo voluntario. Las pruebas microbiológicas y de la polimerización en cadena del tejido hepático y del bazo eran negativas para las infecciones prenatales. Debido al precedente de una hermana con la disfunción mitocondrial, estos resultados levantan el sibility de la posición que por lo menos algunos casos del síndrome familiar del síndrome hydranencephalic-hydrocephalic congénito con vasculopathy proliferativo se pueden atribuir a las alteraciones en la cadena respiratoria mitocondrial. (+info)
9/102. Parkinson' la enfermedad de s se asoció a la fosforilación oxidativa deteriorada. Parkinson' la enfermedad de s puede ser debido a los defectos primarios o secundarios de la fosforilación oxidativa (OXPHOS). En un hombre de 76 años con Parkinson' la enfermedad de s desde 1992, la fosfocinasa de creatina levemente pero recurrentemente elevada, la glucosa de sangre recurrentemente elevada, espesamiento del miocardio ventricular izquierdo, bloque y hypacusis bifascicular fueron encontradas. MRI cerebral demostró atrofia, demyelination periventricular, múltiplo, las lagunas diseminadas, supra e infratentoriales, y los depósitos de la hemosiderina en ambos cuernos posteriores. La biopsia del músculo demostró características típicas de un defecto de OXPHOS. Si la asociación de Parkinson' la enfermedad de s y OXPHOS deteriorado eran causativos o coincidente sigue siendo desconocido.
Posiblemente, el defecto mitocondrial actuaba como factor de riesgo adicional para Parkinson' la enfermedad de s o el defecto de OXPHOS empeoró las debilitaciones neurológicas preexistentes por un mecanismo acumulativo o sinérgico. En conclusión, este caso demuestra que Parkinson' la enfermedad de s se puede asociar a mitochondrially o nuclearly codificar defecto de OXPHOS, manifestando como hypacusis, polineuropatía myopathy, axonal, cardiomiopatía y los movimientos y las hemorragias isquémicos subclínicos recurrentes.
Muertes neonatales múltiples debido a una mutación mitocondrial homoplasmic de la DNA.

Las mutaciones de la DNA mitocondrial (mtDNA) son una causa importante de la enfermedad genética. Describimos a una familia con una mutación homoplasmic inusual que dio lugar a seis muertes neonatales y a un niño superviviente con el síndrome de Leigh. La madre es clínico normal, pero un defecto genético bioquímico y molecular severo estaba presente en un niño fatal afectado y la madre. Esta familia destaca el papel de las mutaciones homoplasmic mt-tRNA en enfermedad genética. (+info)
Investigación reciente sobre la mitocondria. Las mitocondrias se utilizan para buscar los ancestros de organismos que contienen células eucarióticas. Entre los mamíferos, las mitocondrias tienden a seguir una pauta de herencia materna. Cuando una célula se divide, las mitocondrias se reproducen con independencia del núcleo. Las dos células hijas formadas después de la división reciben cada una la mitad de las mitocondrias. Cuando el espermatozoide fecunda al óvulo, sus mitocondrias quedan fuera del huevo. El cigoto fecundado hereda sólo las mitocondrias de la madre. Esta herencia materna crea un árbol familiar que no se ve afectado por la recombinación de genes que tiene lugar entre el padre y la madre.
Una comparación reciente de muestras de mDNA humano sugiere que la humanidad desciende de una mujer que vivió en África hace entre 140.000 y 290.000 años. Muestras genéticas tomadas de grupos étnicos africanos, asiáticos, australianos, europeos y de Nueva Guinea han revelado un número específico de tipos de mDNA. La comparación de estos tipos ha permitido a los científicos
construir un árbol genealógico que sugiere que los distintos grupos empezaron probablemente a evolucionar por separado. En este árbol, el mDNA africano ocupa la rama más larga y antigua y de ella brotan los demás grupos étnicos. Probablemente había muchas otras mujeres vivas en la época de la llamada Eva mitocondrial, pero sus líneas de herencia materna se han extinguido. Esto ocurre habitualmente cuando una generación de una familia no produce ninguna hija. El análisis de mDNA se aplica también en investigación forense. Recientemente se ha establecido la identidad de unos esqueletos atribuidos a Nicolás II, último zar de Rusia, y a su familia utilizando mDNA. El obtenido de un pariente vivo de la familia del zar resultó ser idéntico al encontrado en los restos de Alejandra de Rusia, esposa de Nicolás, y en tres de sus hijos. Como el mDNA se hereda por línea materna, el del esqueleto del zar no coincidía con el hallado en los restos de la zarina y de sus hijos. Según investigaciones recientes, unas pocas enfermedades heredadas por línea materna son imputables a defectos del mDNA, entre ellas algunas patologías neuromusculares y ciertas formas de diabetes mellitus.

CONCLUSIONESEl estudio de este trabajo nos permite arriba a las siguientes conclusiones:
Por esta función que desempeñan, llamada respiración, se dice que las mitocondrias son el motor de la célula.
Se encuentran mitocondrias en las células eucarióticas (células con el núcleo delimitado por membrana). El número de mitocondrias de una célula depende de la función de ésta.
Una mitocondria está rodeada por una membrana mitocondrial externa, dentro de la cual hay otra estructura membranosa, la membrana mitocondrial interna, que emite pliegues hacia el interior para formar las llamadas crestas mitocondriales. Las mitocondrias reciben del núcleo un estimulo para su intensa multiplicación. En la célula se hallan en continuo movimiento.
Las mitocondrias se utilizan para buscar los ancestros de organismos que contienen células eucarióticas.
Cuando una célula se divide, las mitocondrias se reproducen con independencia del núcleo. En las células eucariotas (con núcleo verdadero), éste se encuentra separado del citoplasma por la membrana nuclear, que lo delimita.
El núcleo en reposo tiene estructuras y dimensiones características. La estructura del núcleo eucariótico varía considerablemente a lo largo de la vida de una célula. Los cambios de la estructura del núcleo son regulares y constantes, y están relacionados con la división celular.
El núcleo de las células eucarióticas es una estructura discreta que contiene los cromosomas, recipientes de la dotación genética de la célula. Está separado del resto de la célula por una membrana nuclear de doble capa y contiene un material llamado nucleoplasma. La membrana nuclear está perforada por poros que permiten el intercambio de material celular entre nucleoplasma y citoplasma.
El núcleo suele ocupar una posición central. Son particularmente frecuente en las células dotadas de gran actividad metabólica, en las que, naturalmente, los intercambios entre el núcleo y en citoplasma son intensos.
BIBLIOGRAFIA Carlos Gispert, Josè Gay, José A. Vidal, Mentor. ENCICLOPEDIA TEMATICA
ESTUDIANTIL OCEANO. Barcelona- España, Océano Grupo Editorial, 1999. Pág. 578,579,580.
J.H. Otto A. Towle. BIOLOGÌA MODERNA. Mèxico. Mc Graw-Hill, 1995. Pàg 621. Microsoft Coporation 1993-2001. ENCICLOPEDIA MICROSOFT ENCARTA 2002. HTTP://WWW.MITOCONDRIAL.COM HTTP://WWW.SCIENCEMAGAZINE.COM