
Toll like receptors
66
Toll Like Receptors TLR
-
Upload
yomi-s-mtz -
Category
Health & Medicine
-
view
30 -
download
0
Transcript of Toll like receptors

Toll Like Receptor
sTLR
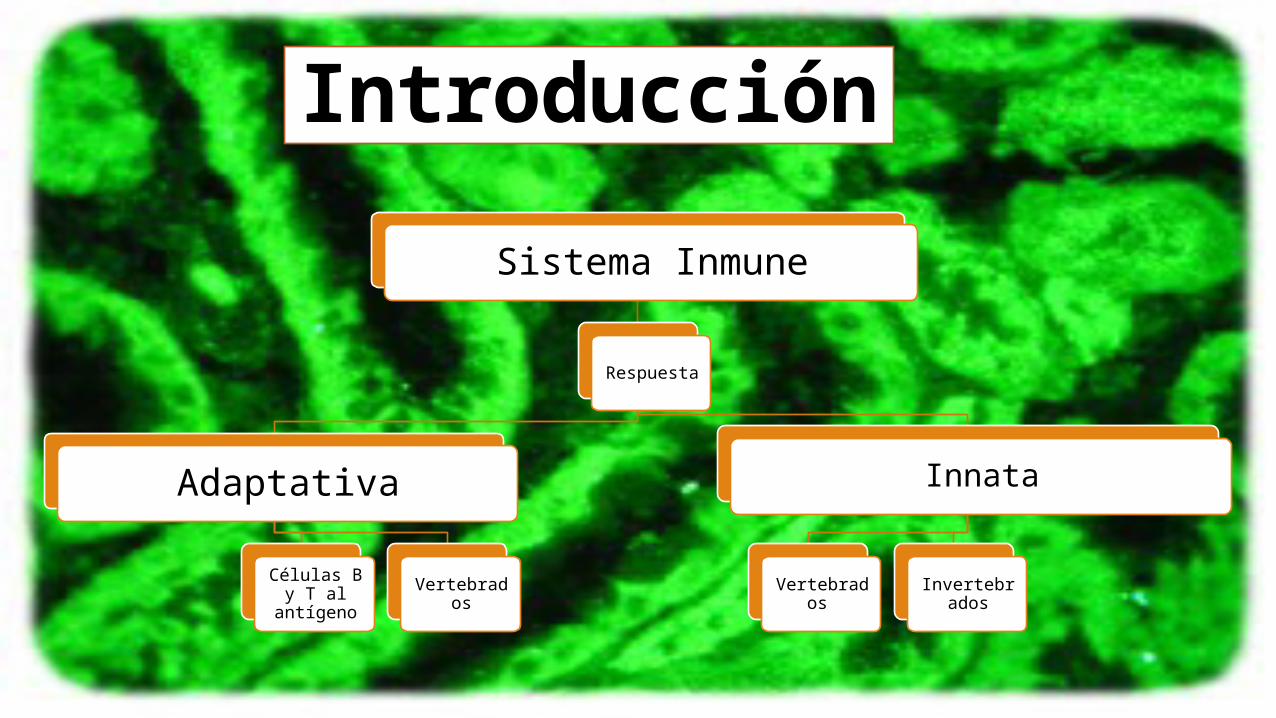
Introducción
Sistema Inmune
Respuesta
Adaptativa
Células B y T al antígeno Vertebrados
Innata
Vertebrados Invertebrados

Elementos codificados por el
ADN
Primer Línea de Defensa
Determina Magnitud y Localización del
Desafío
Facilita la Adaptación de la Respuesta Inmune• Mediada por los genes• Mantienen en la línea Germinal• Codifican Proteínas que no reconocen patrones estructurales conservados en microorganismos
Inmunidad Innata

Receptores de Reconocimiento de Patrones (PRR’s)
son proteínas .
presentes en las células del sistema inmunitario como fagocitos
Identifican moléculas asociadas con patógenos microbianos o
estres celulares
Se clasifican
PRR de señalamiento
Toll Membrana
NOD Citoplasma
PRR endocíticos

Definición
Los receptores tipo Toll (TLR’s) son receptores transmembrana tipo 1

Los TLR’s son proteínas transmembrana altamente conservadas a través de la evolución presente en animales invertebrados hasta los vertebrados más evolucionados como el hombre.
Los TLR’s son expresados en diferentes tipos celulares, Principalmente células del sistema inmune tales como células:
DendríticasMacrófagosneutrófilos Linfocitosendoteliales epitelialesentre otras

El primer miembro de la familia de TLR identificado fue la proteína Toll,
que se caracterizó por primera vez en la mosca del vinagre Drosophila melanogaster
Los TLR’s no parecen tener una función de desarrollo, pero han demostrado que juegan un papel central en el sistema inmune.

Están involucrados en las enfermedades infecciosas a través del reconocimiento de diversos productos microbianos y mediante la activación de cascadas de señalización.
La activación inadecuada puede dar lugar a fallos sépticos multiorgánicos, incluyendo el daño pulmonar, insuficiencia cardiaca y daño cerebral

Además, los TLR’s son capaces de reaccionar con antígenos del medio ambiente e incluso antígenos propios y por lo tanto pueden contribuir a
la generación de varias enfermedades no infecciosas, como alergias o enfermedades autoinmunes.

Los TLR´s también son considerados receptores de "vigilancia", lo que indica que son capaces de vigilar los
tejidos para los estados de la enfermedad
compuestos
Espacio extracelular
Regi
ones
rica
s en
Leu
cina
Región
transmembranal
Dominio TIR
Citoplasma

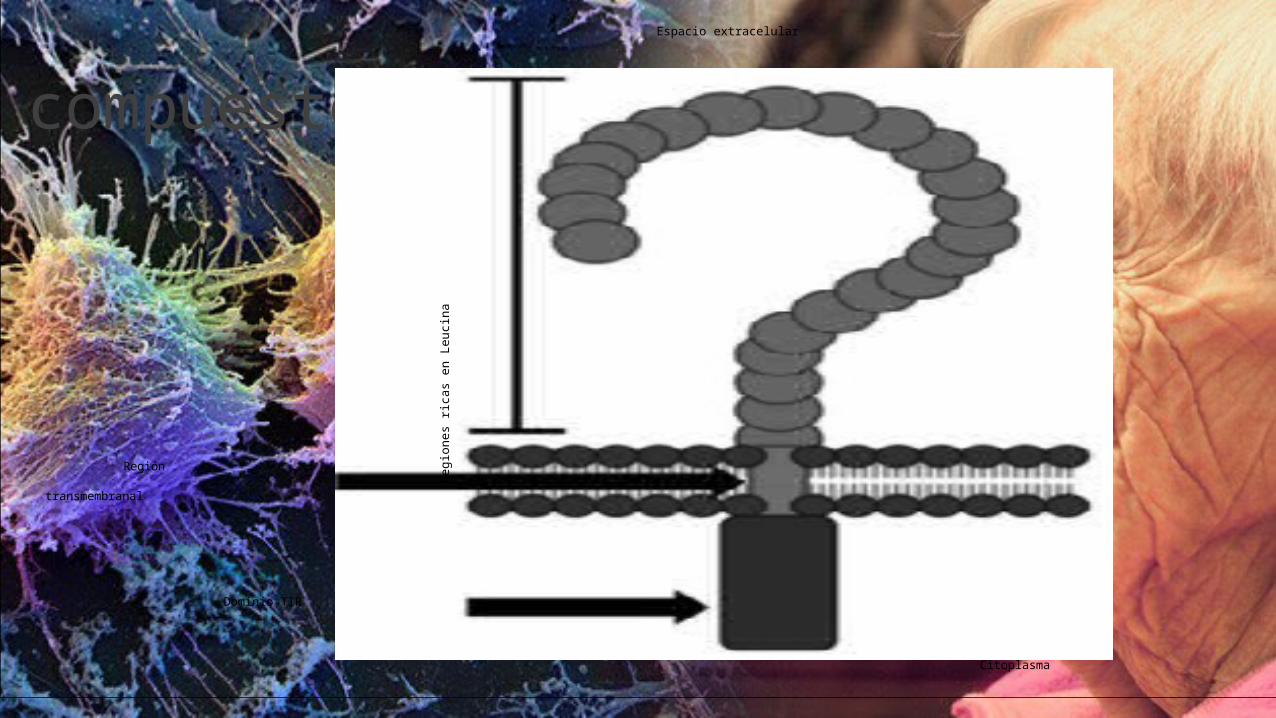
Estructura
Todos los TLR’s comparten la misma estructura: ◦ un gran dominio extracelular (550 a 980 aminoácidos) que consiste en repeticiones ricas en leucina◦ un dominio transmembrana ◦ una porción citoplasmática similar al receptor de IL-1 llamado TIR (como Toll/IL-1R-) de unos 200 aminoácidos de longitud.
Espacio extracelular
Regi
ones
rica
s en
Leu
cina
Regióntransmembranal
Dominio TIR
Citoplasma

El dominio extracelular tiene la capacidad de unión al ligando
mediante las Repeticiones Ricas en Leucina (LRR’s)
siendo las responsables del reconocimiento de los diferentes Patrones moleculares asociados a patógenos (PAMP’s),
y el dominio TIR media la señal intracelular
Espacio extracelular
Regi
ones
rica
s en
Leu
cina
Regióntransmembranal
Dominio TIR
Citoplasma

Función
-Detectar la presencia y lograrla diferenciar acorde al inmunógeno estimulador el tipo de patógeno
-Generar de manera rápida una respuesta frente al patógeno
-Estimulando el desarrollo de una respuesta adaptativa duradera y más eficaz.
-Inducen como resultado de su estimulación la producción de citosinas proinflamatorias y la expresión de moléculas coestimuladoras en células maduras que resultan en señales de alerta inmunológica.

Cascada de Señalización◦ La activación de la cascada de señalización de los TLR’s se origina en el dominio TIR, el cual posee cuatro
adaptadores,
◦ MyD88◦ TIRAP/MAL◦ TRIF◦ TRAM
◦ Estos adaptadores están asociados con las interacciones del dominio TIR, siendo diferentes los adaptadores que se activan según el tipo de TLR, existiendo diferentes combinaciones de estos adaptadores
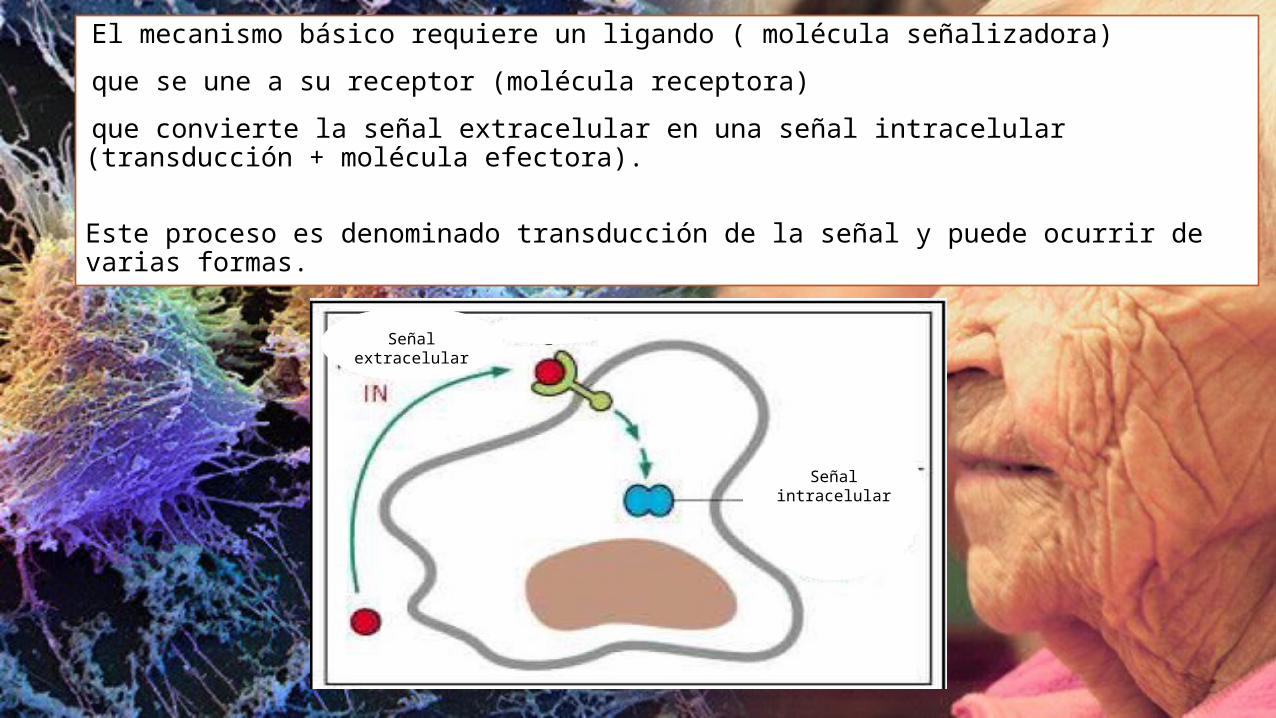
El mecanismo básico requiere un ligando ( molécula señalizadora)
que se une a su receptor (molécula receptora)
que convierte la señal extracelular en una señal intracelular (transducción + molécula efectora).
Este proceso es denominado transducción de la señal y puede ocurrir de varias formas.
Señal extracelular
Señal intracelular

Características de la cascada de señalización:
◦ 1. Transferencia física de la señal (RECEPTORA) del sitio de recepción – membrana plasmática o citosol – hacia la maquinaria celular donde ocurrirá la respuesta.
◦ 2. Transformación de la señal (TRANSDUCTORA) en forma molecular con propiedad de provocar una respuesta.
◦ 3. Amplificación de la señal (EFECTORA) de una forma suficiente para provocar la respuesta, de modo que muy poca cantidad de moléculas es suficiente para provocar una respuesta en la célula.
◦ 4. Distribución de la señal (INTEGRADORA) hacia varios sitios en el interior de la célula con el fin de influenciar diversos procesos simultáneamente.
◦ 5. Cada etapa del proceso de señalización podrá ser modificada o alterada (MODULADORA) por otros factores mientras se produzca la señal.

Molécula señalizadora
Las moléculas señalizadoras son Hidrofílicas y no tienen la habilidad de difundir a través de la membrana. Necesitan de un receptor de superficie celular que genera una señal intracelular en la célula diana.
Algunas moléculas señalizadoras Hidrofóbica (hormonas) pueden difundir a través de la membrana y unirse a receptores intracelulares localizados en el núcleo o en el citoplásma de la célula diana.

Receptores tienen dos funciones:
• Unir el ligando (hormona u otra molécula usada para transmitir la señal)
• Transformar el mensaje del ligando en una respuesta celular
Receptores
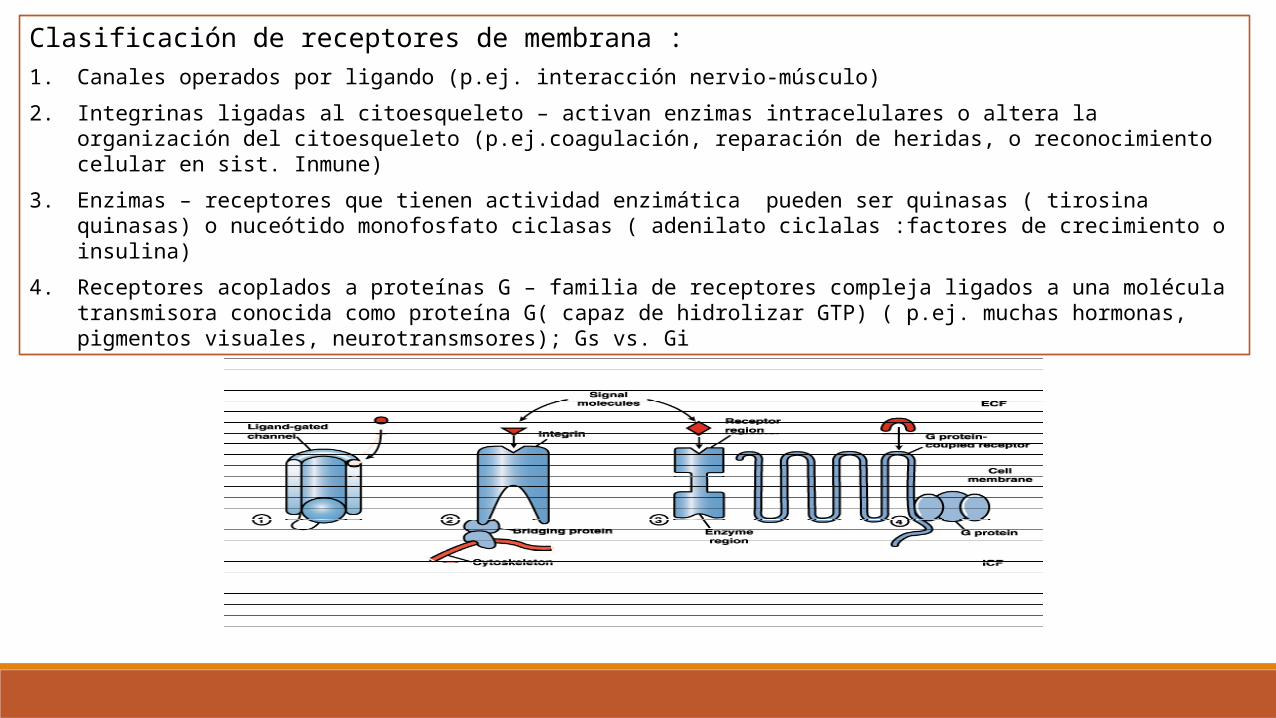
Clasificación de receptores de membrana :1. Canales operados por ligando (p.ej. interacción nervio-músculo)
2. Integrinas ligadas al citoesqueleto – activan enzimas intracelulares o altera la organización del citoesqueleto (p.ej.coagulación, reparación de heridas, o reconocimiento celular en sist. Inmune)
3. Enzimas – receptores que tienen actividad enzimática pueden ser quinasas ( tirosina quinasas) o nuceótido monofosfato ciclasas ( adenilato ciclalas :factores de crecimiento o insulina)
4. Receptores acoplados a proteínas G – familia de receptores compleja ligados a una molécula transmisora conocida como proteína G( capaz de hidrolizar GTP) ( p.ej. muchas hormonas, pigmentos visuales, neurotransmsores); Gs vs. Gi

1.ligando + receptor intracelular = transcripción génica / síntesis proteica
2. ligando + canales iónicos = potencial membrana x citosol / muchas
3. ligando + proteína G (enzimas ± canales iónico) = 2º mensajeros / muchas
4. ligando + receptor enzimático = fosforilación / muchas
Transducción de la Señal – Mecanismos generales
ligando + receptor = transducción / respuesta
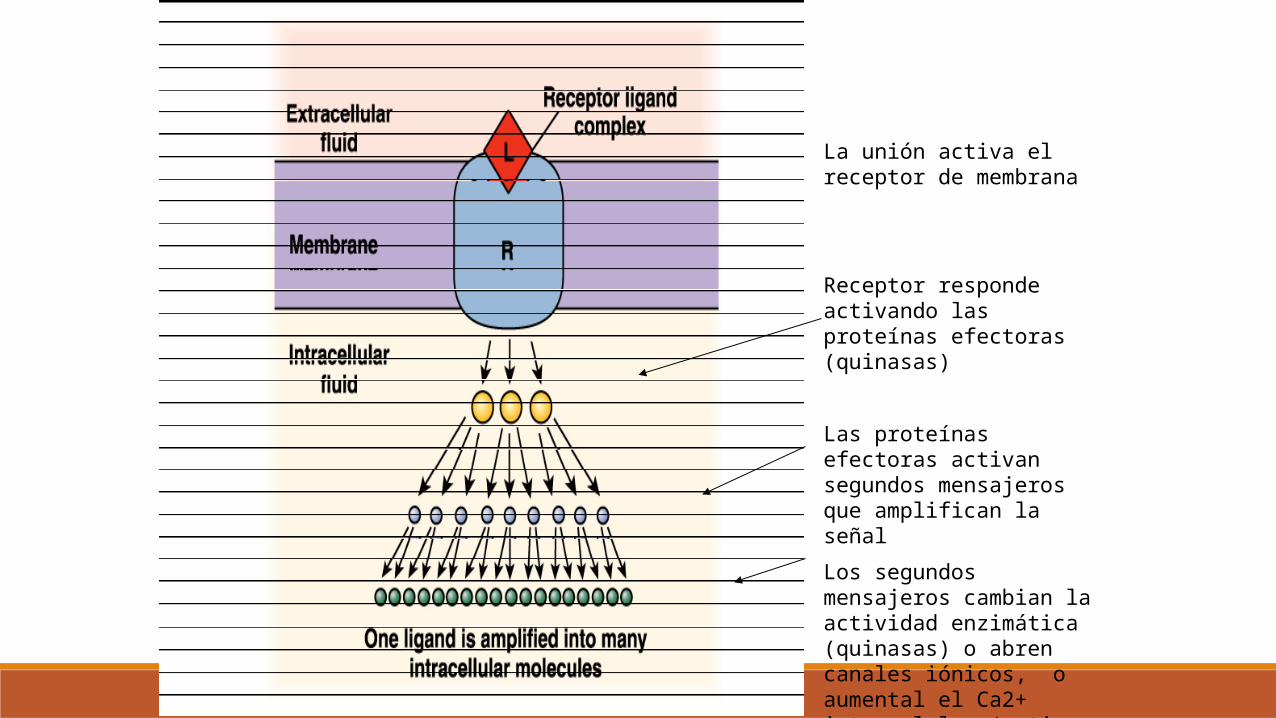
La unión activa el receptor de membrana
Receptor responde activando las proteínas efectoras (quinasas)
Las proteínas efectoras activan segundos mensajeros que amplifican la señal
Los segundos mensajeros cambian la actividad enzimática (quinasas) o abren canales iónicos, o aumental el Ca2+ intracelular (activa otras quinasas)

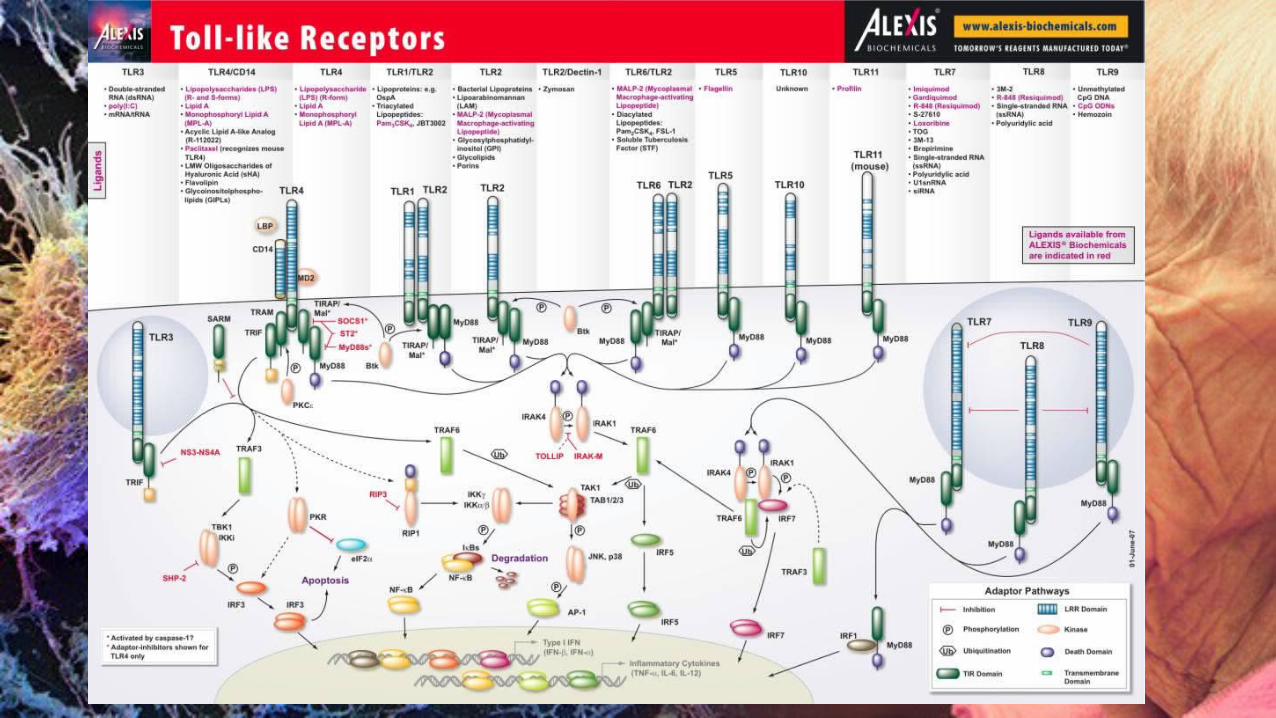
RUTAS DE SEÑALIZACIÓN DE LOS TLR
Los TLR reclutan y activan moléculas adaptadoras y quinasas para la transducción de señales. Existen dos rutas de señalización principales,
◦ la ruta dependiente de MyD88 ◦ la ruta independiente de MyD88.

Señalización Hay dos vías distintas de señalización asociadas con los TLRs:
la que requiere la proteína adaptora MyD88,
la MyD88-independiente.
Todos los TLRs menos TLR-3 señalan por la vía dependiente de MyD88.

Vía MyD88 Dependiente Cuando la activación de la vía es dependiente de MyD88, esta proteína se recluta y asocia con los TLR´s a través de los dominios TIR de ambas moléculas, este evento permite que MyD88 se una a IRAK-4 (cinasa 4 asociada al receptor de IL-1) a través de su dominio intermedio (DI) y a IRAK-1 mediante su dominio de muerte (DD); la proximidad entre ambas cinasas provoca que IRAK-4 fosforile a IRAK-1. IRAK-1 fosforilado se une a la proteína TRAF-6 (Factor asociado al receptor de TNF), ambos se disocian del complejo del receptor e interactúan con otro grupo proteico formado por TAK1 (cinasa activada de TGF- ) y TAB1 y 2 (proteínas de unión a TAK1).

Una vez formado este complejo proteico surgen dos vías independientes de señalización:
una que lleva a la activación de las MAP cinasas
otra que conduce a la activación del sistema NF-
En la primera ruta, la activación de TAK1 induce la fosforilación de las MAPK´s cinasas (ERK, JNK y p38) promoviendo la translocación nuclear del factor AP1
en la segunda ruta, TAK 1 fosforila el complejo de cinasas de I (IKK’s) que a su vez fosforilan a I marcándolo para su ubiquitinación y subsecuente destrucción por el proteasoma.
El dímero NF- B (p50, p65) se transloca al núcleo cuando la secuencia de localización nuclear queda expuesta, ya en el núcleo, el factor transcripcional se une a sus elementos de respuesta en los promotores de sus genes blanco
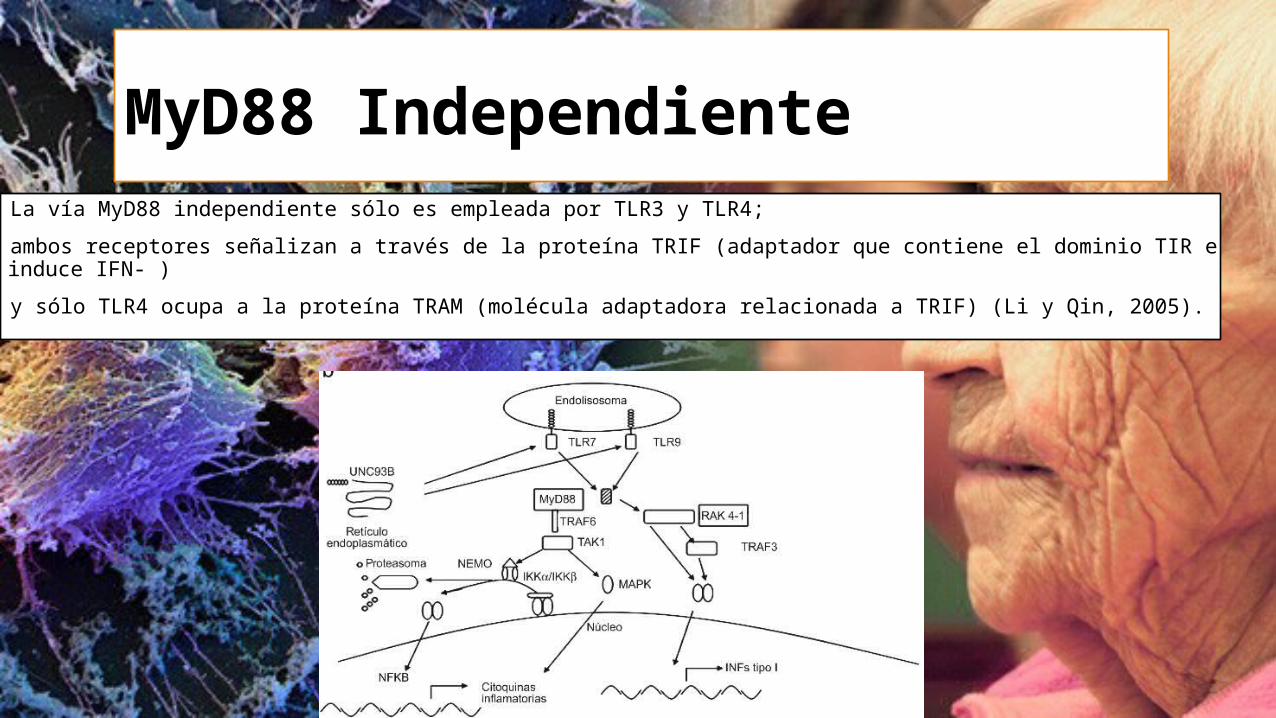
MyD88 Independiente La vía MyD88 independiente sólo es empleada por TLR3 y TLR4;
ambos receptores señalizan a través de la proteína TRIF (adaptador que contiene el dominio TIR e induce IFN- )
y sólo TLR4 ocupa a la proteína TRAM (molécula adaptadora relacionada a TRIF) (Li y Qin, 2005).

La señalización de la vía independiente a MyD88 abarca la siguiente secuencia:
La proteína acopladora TRIF recluta al complejo proteico TRAF6- TAK1- TAB2 que activa a las IKK’s permitiendo la liberación de NF- . A través de otra ruta, la molécula TRIF interactúa con el dímero TBK1/IKK -i ocasionando la translocación del factor nuclear IRF- 3 (Factor regulador de IFN-3), provocando la síntesis de interferón tipo I (IFN / ).

MOLÉCULAS SINTETIZADAS POR LA ESTIMULACIÓN DE LOS TLR
Gracias a los factores de transcripción activados por las vías de señalización mediadas por los TLR, se favorece la transcripción de moléculas blanco, importantes para la generación y mantenimiento de la respuesta inmune
La activación de NF-KB, es una parte clave en la regulación inmunológica e inflamatoria, ya que regula la transcripción de genes como el de
IL-1, IL-2, IL-6, IL-12, TNFA , LTA, LTB Y GM-CSF, algunas quimiocinas, moléculas de adhesión (ICAM, VACAM, ELAM)) proteínas de fase aguda, moléculas del MHC y Péptidos Naturales Antimicrobianos (PNA).

SÍNTESIS DE PÉPTIDOS NATURALES ANTIMICROBIANOS
Los PNA son un grupo diverso de moléculas producidas en gran variedad de tejidos y células en invertebrados, plantas y animales.
Su composición de aminoácidos, su anfipatía, su carga catiónica y su tamaño, les permiten unirse e insertarse en las membranas de los microorganismos para formar poros y matarlos.
Hasta el momento, se han aislado cerca de 850 péptidos diferentes, que se clasifican en 5 grupos según su composición de aminoácidos y su estructura.

Actualmente se ha comprobado la participación de los TLR en la producción de PNA.
Durante los últimos años ha aumentado la cantidad de PNA aislados y ahora se sabe el tejido o célula que los producen, ya sea de manera constitutiva o inducible, y en algunos casos el elemento regulador involucrado.

El modelo de peligro Desde hace más de 50 años los inmunólogos han basado sus pensamientos, experimentos y tratamientos clínicos, sobre la idea de que la respuesta inmune funciona haciendo una distinción entre lo propio y lo no propio.
El descubrimiento de las células presentadoras de antígeno (APC) permitió resolver problemas inmunológicos del antiguo modelo. Ahora se sabe que las APC discriminan lo “propio” y lo “no propio”, además de patógenos evolutivamente distantes, mediante los PRRs.

Recientemente, sobre las bases del modelo “propio no-propio”,
Polly Matzinger ha propuesto un nuevo modelo inmunológico llamado “Modelo del Peligro”,
sugiere que las APC además de ser activadas por los PAMPs de microorganismos patógenos, también son activadas por señales de alarma
producidas por células infectadas,
dañadas por toxinas o por daño mecánico entre otras causas
estas señales también son capaces de activar la Respuesta Inmune.

El Modelo del Peligro se basa en la idea de quela fuerza que dirige al sistema inmune es la necesidad de reconocer el
peligro
considerando “peligro” como cualquier cosa que causa estrés o destrucción en los tejidos.

En este modelo las señales de alarma no son provocadas por células sanas o por células apoptóticas
Por lo que un microorganismo no es la característica importante para responder en contra de lo no propio, y en
contraste, lo propio no es garantía de tolerancia.

Este modelo se sustenta en el descubrimiento de señales de alarma endógenas,
DNA,
RNA,
proteínas de choque térmico,
interferon-A,
interleucina 1B,
CD40-L
derivados del hialurón.

La lista de moléculas inmunoestimuladoras derivadas del huésped crece cada vez más, estás moléculas que no son expuesta normalmente a los PRRs puede ser nombradas Patrones Moleculares Asociados a Peligro (DAMPs), ya que sólo son expuestos debido a daños en células, órganos y tejidos y en respuesta a señales de peligro.
Por lo tanto, los TLR pueden reconocer tanto moléculas endógenas y exógenas (PAMPs)

Generalmente, nos acercamos al sistema inmune desde un punto de vista propio-no propio, sin embargo esta visión queda obsoleta desde la perspectiva de que el sistema inmune está más enfocado en detectar el peligro y la destrucción potencial, que en la distinción propio-no propio.
Esto quiere decir que las células del sistema inmune innato, pueden reconocer señales derivadas de tejidos en ausencia de patógenos, al igual que reconocer patógenos en ausencia de señales derivadas de tejidos.

Patologías que involucran a los TLR
El mecanismo de la señalización mediada por los TLR y su implicación en algunas enfermedades humanas es muy complejo debido a los factores ambientales y a
las diferencias genéticas entre los seres humanos.
Sin embargo, algunos estudios de polimorfismos en los genes que codifican los TLR o sus moléculas de señalización han ayudado a elucidar el papel de estos
receptores en las enfermedades humanas.

Sepsis
La sepsis representa un síndrome asociado a la infección sistémica principalmente por bacterias gram-negativas, aunque pueden presentarse casos causados por bacterias gram-positivas, por hongos o casos mixtos.
En humanos, el polimorfismo D299G en el gen del TLR4 es considerado un factor de riesgo para el desarrollo de sepsis causado por bacterias gram-negativas-
Tuberculosis
El polimorfismo R677W en el gen de TLR2 se ha asociado a lepra en población coreana y con tuberculosis en población Tunecina.
Además el polimorfismo R753Q en el mismo gen se considera protector durante la infección por Borrelia burgdorferi y Treponema pallidum, ya que disminuye la señalización de los heterodímeros TLR2/TLR1.
ATEROSCLEROSIS
La ateroesclerosis es un término genérico que engloba a tres clases de enfermedades vasculares que tienen en común el engrosamiento y la pérdida de elasticidad de las paredes vasculares.
La forma más frecuente es la ateroesclerosis, caracterizada por la formación de placasfibrosas en la íntima.
El polimorfismo D299G en el gen del TLR4 esta asociado con una reducción en el riesgo en la arterosclerosis de la arteria carótida y en eventos coronarios agudos.
Este polimorfismo se asocia incluso con una disminución de IL-6, fibrinógeno y VCAM-1, que son moléculas asociadas con el desarrollo de la inflamación, con la progresión de la arterosclerosis, la ruptura de plaquetas y la consecuente oclusión de vasos.

AUTOINMUNIDAD
El Lupus Eritematoso Sistémico (SLE) es una enfermedad autoinmune que se caracteriza por presentar anticuerpos en contra de antígenos propios, principalmente contra DNA.
El mecanismo involucrado en esta enfermedad podría estar relacionado con la ruta de señalización del TLR9, ya que el DNA de los mamíferos puede estimular este receptor cuando se presenta en forma de inmunocomplejos, los cuales son muy comunes en los pacientes con SLE

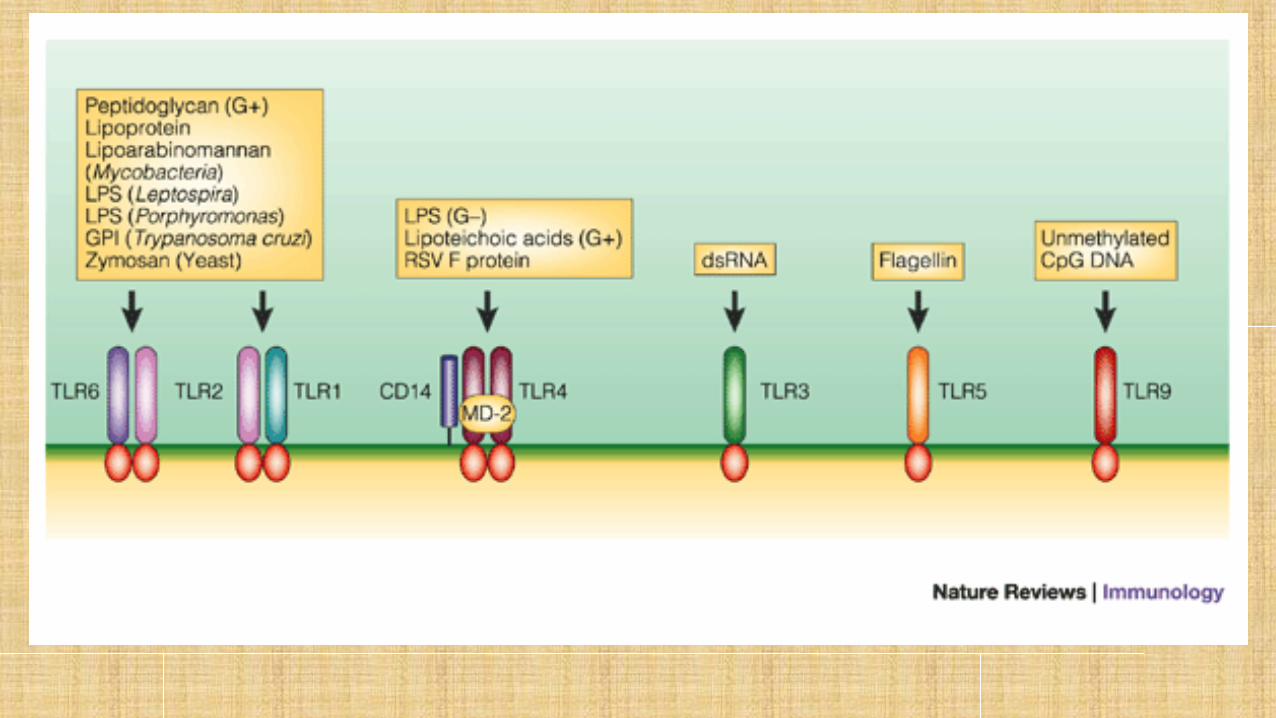
LIGANDOS

Ligandos
Acorde a la localización los TLR’s han sido divididos en dos categorías:
◦ a) los que se localizan en la superficies de membrana (TLR-1, -2, -4,-5, -6, -10, -11, -12 y -13) ◦ b) los que encuentran principalmente en las membranas de los endosomas (TLR-3, -7, -8 y -9).
El más reciente descubrimiento de un miembro de la familia TLR, es el TLR-15, el cual ha sido asociado con el reconocimiento de los componentes de Salmonella spp

TLR1 Reconoce lipopéptidos de la membrana celular de Borrelia burgdorferi y también se asocia con TLR4 e inhibe su señalización en células endoteliales.
el TLR1 puede tener una función más importante de regulación a través de la interacción con diferentes TLR

TLR2 El TLR2 reconoce diferentes productos bacterianos como lipoproteínas de bacterias gramnegativas, peptidoglicano de bacterias grampositivas, ácido lipoteicoico de bacterias grampositivas,lipoarabinomanano de micobacterias, una modulina soluble en fenol de Staphylococcus epidermidis,glicoinositolfosfolípidos de Trypanosoma cruzi,glicolípidos de Treponema maltophilum,porinas de Neisseria, zymosan de hongos

TLR3 Este receptor reconoce ácido ribonucleico (ARN) de doble cadena, un patrón molecular asociado con posibles infecciones por virus.
Cabe destacar que el ARN de doble cadena es un inductor muy potente de los interferones tipo I, que tiene propiedades antivirales, además de que promueve la maduración de células dendríticas.

TLR 4 TLR4 distingue
LPS de bacterias gramnegativas,
taxol de plantas,
proteína de fusión del virus respiratorio sincitial,
proteínas de la cubierta del virus del tumor mamario de ratón y virus de la leucemia murina de Molones
proteína 60 de choque térmico (HSP60) derivada de Chlamydia pneumoniae,
HSP60 y HSP70 del huésped,
extradominio A de fibronectina,
oligosacáridos del ácido hialurónico del hospedero,
fragmentos de polisacárido de sulfato de heparan del hospedero,
fibrinógeno del hospedero.
Mientras que TLR2 reconoce LPS cilíndrico, el LPS cónico estimula a las células sólo mediante TLR

TLR5 Reconoce la flagelina de bacterias gramnegativas.
Se identificó al observar que la flagelina—proteína epiteliales basolaterales del intestino, estimula la producción de respuestas inmunitarias inflamatorias; cuando aquélla hace contacto con la superficie apical no hay tal respuesta.
Las investigaciones con inmunolocalización demostraron que TLR5 se expresa sólo en la superficie basolateral del epitelio intestinal y al contacto con flagelina de Salmonella desencadena una cascada de señalización que media el factor de trascripción proinflamatorio NF-κβ.

TLR6 El TLR6 de humano tiene 69% de identidad con TLR1,
TLR6 reconoce sólo lipoproteínas diacetiladas como las producidas por mitoplasmas, que son potentes activadores de macrófagos.
TLR6 actúa de manera sinérgica con TLR2 para reconocer peptidoglicanos, que son componentes de bacterias grampositivas.

TLR7
Este receptor reconoce compuestos sintéticos considerados inmunomoduladores como los derivados de la imidazoquinolina, además de loxoribina y bropirimina, que tienen capacidad antiviral debido a que inducen la producción de citoquinas proinflamatorias, en especial INF-

TLR8 El TLR8 pertenece a la subfamilia del TLR9 y TLR7 también reconoce compuestos antivirales.

TLR9 Es esencial para el reconocimiento de motivos CpG no metilados de ADN
En humanos, TLR9 es expresado por células dendríticas plasmacitoideas y células B, y esto correlaciona con la capacidad de estas células de responder a la presencia de oligonucleótidos CpG; mientras que en ratones, el TLR9 se encuentra en células dendríticas mieloideas, macrófagos y células B.

TLR10 El TLR10 está filogenéticamente relacionado con el TLR1 y TLR6.
Se desconoce cuál es el ligando de este receptor que se expresa preferentemente en linfocitos B de tejidos inmunitarios, como bazo, nódulos linfáticos y timo

TLR11 Este receptor se ha identificado recientemente en ratones, se expresa en células epiteliales urinarias y regula la resistencia a infecciones por bacterias uropatógenas. Sin embargo el ligando específico no se ha identificado

los TLR1; 2; 4; 5; 6, y 10 se expresan en la superficie celular y migran a los fagosomas (vesícula rodeada de membrana en un fagocito que se forma por la invaginación de la membrana celular y del material fagocitado) tras activarse al reconocer el ligando.
Los TLR3; 7; 8, y 9 se expresan en compartimientos intracelulares, principalmente en el endosoma S y el retículo endoplasmático.

El reconocimiento de los ligandos por parte de los TLR, presentes en las células dendríticas y macrófagos, conduce a la rápida producción de citocinas y quimiocinas que indican la presencia del patógeno.
Esta respuesta inicia un reclutamiento rápido de células del sistema inmunitario al lugar de la infección y la activa, con lo que se inicia una respuesta inmediata frente al patógeno.
Las señales originadas por los TLR promueven la expresión de moléculas de adherencia, tanto en las células epiteliales como en las células hematopoyéticas circulantes.

Como ya se ha mencionado, se han identificado 15 tipos de TLR en mamíferos, 13 de ellos, del TLR1 al TLR13, en humanos y ratones.
Otras formas equivalentes se han aislado en otras especies de mamíferos; sin embargo, algunos TLR encontrados en humanos no están presentes en otros mamíferos y, por el contrario, otros mamíferos pueden expresar TLR que no se han aislado en humanos.
Por ejemplo, los TLR11; 12, y 13 sólo se expresan en ratones. Por el momento sólo se han descrito 10 TLR funcionales en humanos.
Esta circunstancia condiciona o limita el empleo de animales de experimentación como modelos de inmunidad innata extrapolables a la especie humana.

Receptores Ligandos exógenos Ligandos endógenos Localización
TLR 2/1 Pam3CSK4 Biglicano β defensina-3 Membrana celularTLR 2/6 Péptidoglicano
PorinasLipoarabinomananaFosfolipomananasGlucosil fosfatidil inositolHemaglutininaTriacil lipopéptidosDiacillipopéptidosÁcido lipoteicoicoZymosan
TLR4
Lipopolisacárido (LPS) Biglicano tenacina-C Membrana celularManana HSP60 y 70 Glicosilinositol fosfolípidos Fibrinógeno Proteínas de cápside Fibronectina Proteínas de fusión Àcido hialurónico del virus sinsicial respiratorio Heparan sulfato elastasa del neutrófilo
Pneumolisinas estreptococcicasPaclixatelTaxol
LDL
TLR10 Desconocido Desconocido Membrana celular
TLR5 Flagelina Desconocido Membrana celular
TLR3Poly I:CRNA cadena doble
RNA mensajero Endosomas
TLR7/8
ResiquimodR848LoxoribinassRNA siRNA
RNA mensajero Endosomas
TLR9 DNA con motivos CpG Complejos inmunes Endosomas Hemozoina del Plasmodium de IgG-cromatina


Bibliografía Akira S, Hemmi H. 2003.
Recognition of pathogen-associated molecular pattern by TLR family. Immunol Lett 85(2): 85-95.
Hallman M, Ramet M, Ezekowitz RA. 2001.
Toll-like receptors as sensors of pathogens. Pediatr Res 50: 315-321.
Los receptores tipo Toll en el desarrollo y función del sistema hematopoyético,
Eduardo Vadillo,*,** Rosana Pelayo*Unidad de Investigación Médica en Enfermedades Oncológicas, Hospital de Oncología, Centro Médico Nacional Siglo XXI, IMSS.Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional.
http://www.archbronconeumol.org/es/papel-los-receptores-toll-like-las/articulo/13147573/
Abbas A., Lichtman A. (2004). Inmunología celular y molecular. Elsevier España S.A. - Madrid, España
http://www.elsevier.es/es-revista-seminarios-fundacion-espanola-reumatologia-274-articulo-abc-los-toll-like-receptors-relacion-13184459


















