Tesis Poliquetos Epibiontes de Mano de Leon FINAL 12 Marzo
137
Instituto Tecnológico de Los Mochis OPCION 1: TESIS PROFESIONAL Poliquetos (Annelida: Polychaeta) epibiontes de la almeja mano de león Nodipecten subnodosus Sowerby, 1835 en Laguna Ojo de Liebre, Guerrero Negro, Baja California Sur, México. Que presenta: Araujo Leyva Osmar Roberto
-
Upload
ana-cris-quezada -
Category
Documents
-
view
223 -
download
5
Transcript of Tesis Poliquetos Epibiontes de Mano de Leon FINAL 12 Marzo

Instituto Tecnológico de Los Mochis
OPCION 1: TESIS PROFESIONAL
Poliquetos (Annelida: Polychaeta) epibiontes de la almeja
mano de león Nodipecten subnodosus Sowerby, 1835 en
Laguna Ojo de Liebre, Guerrero Negro, Baja California
Sur, México.
Que presenta:
Araujo Leyva Osmar Roberto
Enero del 2013

i
DEDICATORIAS
A la vida y a Dios por crear una naturaleza tan hermosa y
llena de misterios, por brindarme salud y fortaleza para
seguir adelante en mis metas y seguir adquiriendo sabiduría.
A mi madre y hermana por cuidarme y volverme un hombre
fuerte lleno de virtudes y por todo el apoyo incondicional que
me dan día con día, las amo.
A mi novia Janeth por darme paciencia y serenidad en
momentos difíciles y por regalarme un amor tan puro y
sincero, Te amo.
A mis amigos que nunca me exigen y siempre me dan a pesar
de dedicarles muy poco de mí tiempo.
Y a todos mis profesores, que este trabajo es fruto de la suma
de sus enseñanzas y de su paciencia durante mi formación
como profesional.

ii
AGRADECIMIENTOS
Al Dr. Jesús Ángel de León por brindarme la oportunidad de trabajar y aprender este grupo
de animales un poco más a fondo, así como todas las enseñanzas como ser humano y el
apoyo recibido durante mi estancia en la UANL.
Al Dr. Jesús Manuel Díaz Gaxiola por sus valiosas críticas, sugerencias y todo el apoyo
durante la realización de este trabajo.
Al Dr. Gerardo Góngora Garza y a la Dr. Mariana Tovar Hernández por su apoyo durante
el análisis de las muestras.
A todo cuerpo académico del ITLM que crearon cimientos durante mi formación como
profesionista: Dr. Jesús Manuel Díaz Gaxiola, Biol. Dora Patricia Sánchez Vargas y Biol.
Gerardo Rangel Cota.
Y a todos mis compañeros de grupo que de algún modo aportaron estímulo y
competitividad, así como risas y lazos de amistad durante los cuatro años de grupo.
A Ángel de León Espinosa por su valiosa amistad y su mamá Elsy Espinosa Treviño por su
apoyo y calidez incondicional durante mi estancia en Monterrey.
A Jesús Horacio Flores Acosta y sus padres Horacio Flores Varela y Blanca Elva Acosta
por darme cobijo cuando más lo necesité así como su buena vibra y amistad incondicional.
A la música por hacer de mi vida un mundo de fantasía
A todos los biólogos por aportar datos a la ciencia y hacer mi labor investigativa un poco
menos compleja.

iii
“Si no deseas mucho, hasta las cosas más pequeñas te parecerán grandes”
Platón
RESUMEN GENERAL
Se presenta la información de la fauna de poliquetos epibiontes de la almeja mano de león
Nodipecten subnodosus de la Laguna Ojo de Liebre, Baja California Norte, México. Se
muestrearon cuatro bancos almejeros durante el mes de enero 2012. Se colecto un numero
de entre 20 y 30 almejas por zona, obteniendo biometrías de peso y longitud para la especie
N. subnodosus. Y se obtuvo un total de 376 poliquetos epibiontes agrupados en: ocho
órdenes, 16 familias, 36 géneros y 46 especies. Las especies con mayor abundancia fueron
Eunice chicasi, Platynereis bicanaliculata y Polycirrus californicus. Los análisis
estadísticos destacan a El Banco Chocolatero como la zona más afectada con una
correlación baja de; 0.6519 y a La Concha como la zona de mayor correlación (0.8677). La
relación entre los índices gonadosomáticos y hepatosomáticos se puede considerar normal
con variedad de picos tanto gonadosomáticos como hepatosomáticos en los diferentes
bancos, puesto que las almejas colectadas tuvieron tallas medias de 100 mm. No se
encontró una relación directa entre la fauna epibionte y estado de decremento poblacional
observado en los bancos almejeros en los últimos años en la comunidad pesquera de la
Laguna Ojo de Liebre. Se puede destacar a La Concha como el Banco con un coeficiente de
multi correlación de 0.8974 y la zona con mayor diversidad H1 =2.805. Se encontró una
estrecha relación la diversidad de poliquetos obtenidos en las cuatro áreas muestreadas, esto
se observa en el análisis clúster, en donde La Concha obtuvo los valores de diversidad
mayores y una similitud de 0.73 aislando a El Dátil, El Zacatoso, y Banco El Chocolatero
en un estrato distinto debido a sus valores bajos de diversidad y similitud.

iv
CONTENIDO
Tema PaginaAGRADECIMIENTOS iiRESUMEN GENERAL iiiINTRODUCCIÓN 1PLANTEAMIENTO DEL PROBLEMA 2JUSTIFICACIÓN 3OBJETIVOS 4ANTECEDENTES 5Características generales de Nodipecten subnodosus 5Anatomía y reproducción. 6Aspectos pesqueros 9Epibiontes sobre bivalvos 10Características generales clase Polychaeta 11Alimentación 13Reproducción 14Poliquetos asociados a otros organismos 15AREA DE ESTUDIO 16METODOLOGIA. 17Área de colecta. 17Colecta en campo. 18Separado eh identificación de organismos 19Toma de datos morfológicos de la almeja y Análisis estadísticos 21RESULTADOS 23Diagnosis de las especies 30Análisis de relación longitud peso 58Índice gonadosomático y hepatosomático. 61Factor de condición múltiple (KM). 63Índices ecológicos 68Análisis de componentes principales 69Diversidad Alfa 70Análisis Clúster 71Análisis SHE. 72

v
DISCUSIÓN 73CONCLUSIÓN 77LITERATURA CITADA 78
LISTA DE FIGURAS
Figura Tema PagináFigura 1. Conchas de N. subnodosus. 5Figura 2. Anatomía interna de un pectínido con sus órganos diferenciados 7Figura 3. Estructura corporal de un poliqueto 11
Figura 4. Parte Anterior de un poliqueto evidencian sus respectivos órganos de los sentidos 12
Figura 5. Estructura de dos parapodio birramio. 12Figura 6. Mapa: Laguna Ojo de Liebre. 16Figura 7. Buzo 18Figura 8. Muestra llenada con agua marina 18Figura 9. Almeja colectada 18Figura 10. Cubetas con muestras. 18Figura 11. CIBNOR Unidad G. Negro 19Figura 12. Retirando epibiontes de la concha. 19Figura 13. Lab. De biosistemática 20Figura 14. Identificación de poliquetos. 20Figura 15. Relación longitud peso banco Chocolatero. 58Figura 16. Relación longitud peso banco El Zacatoso. 59Figura 17. Relación longitud peso banco El Dátil. 59Figura 18. Relación longitud peso banco La Concha. 60Figura 19. Índice gonadosomático y hepatosomático Banco Chocolatero 61Figura 20. Índice gonadosomático y hepatosomático banco El Zacatoso 62Figura 21. Índice gonadosomático y hepatosomático banco El Dátil 62Figura 22. Índice gonadosomático y hepatosomático banco La Concha 63Figura 23. Longitud vs KM Banco Chocolatero 64Figura 24. Peso vs KM Banco Chocolatero 64Figura 25. Longitud vs KM El Zacatoso 65Figura 26. Peso vs KM El Zacatoso 65Figura 27. Longitud vs KM El Dátil 66Figura 28. Peso vs KM El Dátil 67Figura 29. Longitud vs KM La Concha 67Figura 30. Peso vs KM La Concha 68

vi
Fig 31. Análisis de componentes principales para las cuatro estaciones Banco Chocolatero, El Zacatoso, El Dátil y La Concha 69
Figura 32. Grafica de diversidad alfa para las cuatro zonas muestreadas. 70Figura 33 Análisis Cluster Laguna Ojo de Liebre. 71Figura 34 .
Análisis SHE de los cuatro bancos almejeros (Banco Chocolatero, El Zacatoso, El Dátil y la Concha 74
LISTA DE CUADROS
Cuadro Tema PagináCuadro 1. Abundancia de organismos por especies y estaciones de muestreo. 24-25Cuadro 2. Factor de condición múltiple y error típico bancos almejeros 63
Cuadro 3. Valores de diversidad de la poliqueto fauna epibionte para las cuatro estaciones. 68
LISTA DE LÁMINAS
Lámina Tema PagináLámina 1. Epibiontes de N. subnodosus parte uno 31Lámina 2 Epibiontes de N. subnodosus parte dos 32

1
INTRODUCCIÓN
Aunque ha habido numerosos estudios e informes aislados de relaciones simbióticas de
poliquetos y otros animales marinos, el único intento anterior para proporcionar una visión
general de estos fenómenos entre los poliquetos y otros organismos proviene de la década
de 1950, con no más de 70 especies simbiontes brevemente descritas (Martin y Britayev,
1998). En los últimos años se han realizado revisiones con un registro que incluye 292
especies de poliquetos comensales provenientes de 28 familias involucradas en 713
relaciones y 81 especies de poliquetos parásitos provenientes de 13 familias involucradas
con 253 relaciones simbióticas (Martin y Britayev, 1998).
Los estudios taxonómicos de poliquetos se enfocan a la fauna asociada a fondos blandos o
rocosos, (Díaz y Liñero 2003) muy pocos se han enfocado al análisis de la epifauna de
especies de interés comercial, tal es el caso de N. subnodosus en la Laguna Ojo de Liebre,
Guerrero Negro, Baja California Sur. México.
Dentro de las especies comerciales de moluscos, los pectínidos son bivalvos de gran
tamaño, a causa de esta cualidad son explotados en las costas del todo el mundo.
Nodipecten subnodosus o almeja mano de león es una especie grande con un promedio de
200 mm de ancho con hábitat epibentónico que normalmente se adhiere al sustrato duro
FAO (2005). Su caparazón es rígido y escamoso lo que facilita la colonización de gusanos
poliquetos u otros organismos en su superficie. Existen registros de cultivos acuícolas de
moluscos en algunas localidades de América donde la infestación por parte de algunas
especies de poliquetos han sido señaladas como responsables del retardo en crecimiento e
incremento en la mortalidad de los cultivos de moluscos. (Diaz y Liñero 2003)
.Este estudio pretende dar a conocer las especies epibentónicas de poliquetos que habitan
usualmente la valva dorsal (haz) de la almeja mano de león, así como análisis sobre
aspectos biológicos pesqueros de la almeja mano de león en cuatro de los principales
bancos almejeros del área: El Banco Chocolatero, El Zacatoso, El Dátil y La Concha,
además de la interacción de los organismos epibiontes y posibles alteraciones en el
hospedero.

2
PLANTEAMIENTO DEL PROBLEMA
La pesca de almeja mano león constituye una de las actividades de mayor importancia en el
poblado de Guerrero Negro, sin embargo los últimos años ha venido en decadencia, a causa
de altas mortandades en la población de N. subnodosus. Aunque se han encontrado algunas
de las causas de decremento poblacional como infecciones por vibriosis, actualmente hay
poco o nulo registro de estudios que aporten información sobre especies de poliquetos
responsables de la mortandad directa e indirectamente en el medio natural (Diaz y Liñero
2003). La cantidad de estudios ecológicos con índole de epibiosis en organismos anélidos
es muy escasa, el factor determinante es el enfoque en estudios de análisis biológico-
pesqueros con respecto a especies comerciales. La taxonomía es el segundo factor, a
menudo suelen realizarse trabajos en los cuales el objetivo es conocer la fauna de alguna
zona en específico. La almeja N. subnodosus es uno de los recursos pesqueros de mayor
importancia en el municipio de Mulegé, en los últimos años la sobreexplotación en bancos
almejeros ha obligado a que las poblaciones disminuyan drásticamente.

3
JUSTIFICACIÓN
El potencial económico de la Laguna Ojo de Libre depende significativamente de los
recursos biológicos de la zona costera. Por lo tanto, el entendimiento de las especies que se
explotan en los campos pesqueros debe de ser pleno para la extracción sustentable del
recurso y que reitere ganancias sin ningún factor de riesgo sobre el área de explotación.
Si bien es cierto N. subnodosus constituye poblaciones elevadas en todo Baja California
Sur, existen poco estudios sobre la comprensión de las poblaciones que la conforman y la
interacción entre especies. En este trabajo se pretende describir la estructura comunitaria de
la poliqueto fauna asociada a la almeja mano de león, una lista detallada de especies con las
que convive, la relación de estas especies con el tamaño, tallas e índices gonadosomático y
hepatosomático de la almeja y su papel como hábitat en el ambiente marino

4
OBJETIVOS
General:
Caracterizar la composición de poliquetos epibiontes asociados a la almeja mano de león,
así como determinar aspectos ecológicos entre su relación simbiótica con los poliquetos
epibiontes en la Laguna Ojo de Liebre, BCS.
Específicos:
Realizar un listado taxonómico de la anélidofauna presente en la almeja mano de león N.
subnodosus con sus respectivas descripciones, así como encontrar relaciones entre los
organismos en las cuatro zonas muestreadas dentro de la bahía Ojo de Liebre.
Evaluar el crecimiento de las poblaciones de N. subnodosus aplicando las regresiones de
relación longitud-peso, índices gonadosomáticos y hepatosomáticos así como el factor de
condición múltiple, enfocándose en el estado actual de los bancos almejeros de la Laguna
Ojo de Liebre.
Evaluar el estado actual de N. subnodosus mediante análisis estadístico haciendo énfasis en
la comparación de datos entre distintas áreas muestreadas y su fauna de poliquetos
asociada.
Analizar la población epibionte de N. subnodosus utilizando índices ecológicos que puedan
relacionarse con el estado actual en los bancos almejeros comparando los datos obtenidos
sobre la almeja mano de león.

5
ANTECEDENTES
Características generales de Nodipecten subnodosus
La almeja mano de león, N. subnodosus (Sowerby, 1935) pertenece al Phylum Mollusca,
Clase Bivalvia y a la familia Pectinidae (Morse y Zardus, 1997). Esta especie, al igual que
todos los pectínidos, tiene una concha con dos valvas comprimidas y articuladas
dorsalmente, pero a diferencia de las otras especies de la familia, N. subnodosus tiene una
concha muy grande, fuerte, gruesa, convexa y sólida. Es equivalva y la ornamentación
externa del disco está formada por costillas radiales anchas en forma redondeada
rectangular y uniformemente espaciadas, con nueve costillas en la valva izquierda, donde
tienen alternativamente una o más series de nódulos, mientras que la valva derecha posee
de nueve a 10 costillas que carecen de nódulos (Figura.1) (Peña, 2001).
Figura 1. Conchas de N. subnodosus.

6
Actualmente existen 400 especies de pectínidos distribuidas en el mundo, 67 de ellas están
distribuidas dentro de los mares iberoamericanos con 28 representantes en las costas del
Pacifico mexicano, con tres especies consideradas de mayor importancia comercial: Almeja
voladora (Pecten vogdesi Arnold, 1906), almeja catarina (Argopecten vertricosus, Sowerby
II, 1842) y la almeja mano de león (Nodipecten subnodosus, Sowerby, 1835) (Keen y Coan
1974).N. subnodosus generalmente habita lagunas, bahías y canales profundos de 6 m, con
corriente de agua elevada (González, 2003).En la laguna Ojo de Liebre, B.C.S., se
encuentran en densidades de 4 a 12 individuos por cada 10 m2 (Massó-Rojas et al., 2000) y
generalmente asociadas a “camas” de pastos marinos de la especie Zoostera marina.
Anatomía y reproducción.
Estos organismos presentan la cavidad del manto muy espaciosa y branquias grandes;
Benninger y Le Pennec (1991) mencionan que estas características le dan a los pectínidos la
capacidad de acumular alimento eficientemente, además de su capacidad respiratoria.
Tienen un pie reducido y comprimido en forma de hacha. Al igual que todos los pectínidos,
N. subnodosus carece del músculo aductor anterior, pero tiene un gran músculo aductor
posterior situado cerca del centro que, según Benninger y Le Pennec (1991), Chantler
(1991) y Dore (1991), este musculo le sirve a los pectínidos para cerrar las valvas con
fuerza y llevar a cabo su nado. En los pectínidos, el músculo aductor representa,
dependiendo de la especie, entre 5% y 20% del peso vivo del animal. Está compuesto por
dos tipos de tejido muscular, el estriado y el liso; juntos tienen forma cilíndrica,
normalmente de color blanco o crema y en ocasiones un poco café o rosa (Dore 1991). Por
otro lado, estas almejas presentan un ventrículo plegado en torno al intestino de manera
quela cavidad pericárdica no solo incluye al corazón sino también un breve segmento del
tubo digestivo. Debajo de esta cavidad se encuentran dos nefridios dispuestos en forma de
U. El sistema digestivo (Figura 2) se compone de boca, esófago, estómago, estilo cristalino,
glándula digestiva, intestino, recto y ano. El esófago conduce directamente al estómago,
situado dentro de la glándula digestiva. El intestino, después de atravesar la glándula
digestiva desciende ventralmente y entra en la gónada, donde da una vuelta y sale de ésta al
lado del músculo aductor y atraviesa el pericardio y el ventrículo.

7
La parte final del intestino está unida al lado posterior del músculo aductor, quedando libre
en la región del recto y del ano. Se ha comprobado que, en algunas especies de pectínidos,
el estómago y el estilo cristalino son responsables de la digestión extracelular, mientras que
la glándula digestiva es el principal sitio de digestión intracelular (Benninger y
Le Pennec 1991).El sistema nervioso se ha descrito sólo para Pecten
maximus y se ha observado que es bilateral, y que se compone de tres
pares de ganglios y dos pares de cordones nerviosos (Benninger y
LePennec 1991).
Figura 2. Anatomía interna de un pectínido con sus órganos diferenciados.
No se ha descrito la anatomía del sistema reproductor de N. subnodosus; sin embargo, se
han hecho descripciones detalladas en numerosas especies de pectínidos y se ha visto que
no difiere entre ellas. Así, se ha observado que en los pectínidos la glándula reproductiva
deriva del complejo celómico, y las células germinales primarias provienen de células del
mesodermo (Raven 1966, Wada 1968). La gónada es una glándula acinosa enracimo,
limitada externamente por un tegumento más o menos plegado, que se infiltra a veces hacia
el interior en forma de trabéculas intersticiales. Contiene gonoductos que sirven de
conductos evacuadores, vasos sanguíneos, fibras musculares y la atraviesa parte del
intestino (Román et al., 2000a). La gónada está anatómicamente bien diferenciada y se

8
localiza en la parte anterior de la masa visceral, asociada dorsalmente a la glándula
digestiva y unida en su borde medio posterior al músculo.
De acuerdo con Barber y Blake (1991) el ciclo reproductivo de los pectínidos tiene seis
etapas perfectamente diferenciables: vegetativa, diferenciación, crecimiento citoplasmático,
maduración, desove y reabsorción de gametos no desovados. Dentro de los pectínidos,
dependiendo de la especie y el lugar, se puede presentar un ciclo reproductivo anual
(generalmente con mucha sincronización de las etapas mencionadas anteriormente) como
en Zygochlamys patagonica en el Canal de Beagle, Chile (Collado et al., 2000), semianual
como en Z. patagonica en Chile, (Jaramillo et al., 1993) o continuo en especies que tienen
desoves sucesivos prolongados durante todo el año (Román et al. 2000a) como es el caso de
Aequipecten tehuelchus en el Golfo de San José, Argentina (Christiansen et al., 1974) y A.
ventricosus en la Bahía de La Paz, B.C.S., México (Luna- González et al., 2000). En las
últimas modalidades hay poca sincronización en la época reproductiva, ya que suelen
registrarse desoves parciales y es frecuente la presencia de atresias ovocitarias y lisis como
parte de la estrategia reproductiva (Román et al., 2000a).
Se ha comprobado que en aguas frías, es común registrar una marcada estacionalidad, la
cual disminuye en aguas templadas y es casi imperceptible en aguas cálidas (Sastry 1979,
Newell et al., 1982, Barber y Blake 1991, Román et al. 2000a). Por otro lado, también se ha
comprobado que en una misma especie e incluso en una misma población pueden existir
diferencias en el ciclo reproductivo y se ha visto que esto depende de la latitud y de las
diferencias ambientales particulares (Sastry 1979, Barber y Blake1991, Román et al.
2000a). Estas variaciones han sido evidenciadas en algunas especies de pectínidos. Por
ejemplo, en diferentes trabajos realizados en las Islas Británicas sobre Aequipecten
opercularis se han observado variaciones en cuanto al inicio y duración del desove (Broom
y Mason 1978, Taylor y Venn 1979, Brand et al. 1980, Allison 1994). En el Atlántico en
Galicia, España, A. opercularis desova cuatro o cinco veces al año (Román et al. 2000a),
mientras que en el Mediterráneo Occidental el ciclo gametogénico muestra una marcada
estacionalidad con dos periodos de maduración (Canales 1998).

9
Aspectos pesqueros
La almeja mano de león, N. subnodosus, es un recurso pesquero de importancia en Baja
California Sur. Debido al sabor y al tamaño de su músculo aductor y a las tallas que alcanza
(longitud máxima de 218 mm (Félix-Pico et al., 1999), esta especie tiene una gran
demanda a nivel internacional; de ahí su importancia económica. Lo anterior ha provocado
una sobreexplotación de este recurso en los diferentes cuerpos de agua de la Península de
Baja California. Debido a ello, su abundancia ha disminuido, al grado que su pesquería en
la actualidad se restringe (a nivel nacional) a su límite norte de distribución, en la laguna
Ojo de Liebre, B.C.S.
Es una especie hermafrodita funcional que presenta un rápido crecimiento, pudiendo
alcanzar una longitud máxima de 220 mm. Esta especie es altamente apreciada en el
mercado nacional e internacional gracias al buen sabor que presenta el músculo abductor
comúnmente conocido como “callo” que puede pesar hasta 260 gr (Gonzales 2003).
Las ventajas de la especie sobre otros pectínidos son notables. Por ejemplo, un kilogramo
de callo de almeja catarina se obtiene a partir de 170 organismos de 5 cm de altura de la
concha, en aproximadamente 12 meses de cultivo, y tiene un valor comercial de2.9 dólares
en el mercado norteamericano al mayoreo. En cambio, un kilogramo de callos de almeja
mano de león se produce a partir de 40 ejemplares de 12 cm de altura de la concha en 18
meses de cultivo, vendiéndose a 6.25 dólares en el mercado norteamericano al mayoreo
(González 2003). Estas características hacen de esta especie un organismo de gran interés
en la acuicultura. Lo anterior ha estimulado la investigación básica para lograr su cultivo.

10
Epibiontes sobre Bivalvos
Epibiosis es la asociación entre epibiontes (organismos que crecen unido a una superficie
viva) y basibiosis (organismos que proporcionan sustrato para los epibiontes). Esta
asociación crea una compleja red de beneficios ventajas y desventajas de ambos
organismos (Wahl, 1989). Los bivalvos se asocian a menudo con epibiontes incrustantes
(ver Feifarek, 1987), (Díaz y Liñero 2003 y Liñero y Díaz 2006), ya que la estructura rígida
de sus valvas proporciona un área óptima para el ensamblaje de muchos organismos
bentónicos. En el ensamblaje de epibiones sobre pectínidos es frecuente encontrar
foraminiferos, esponjas, hidroides, crustáceos, briozoarios, ascidias y poliquetos (Schejter
y Bremec, 2007).
Este último grupo constituye uno de los más importantes debido a su abundancia,
diversidad, capacidad de colonización y distribución (Liñero y Díaz 2006), por lo anterior
se han realizado estudios sobre la epibiosis de poliquetos sobre bivalvos principalmente de
importancia económica que de alguna manera tiene repercusiones en tallas, pesos y ciclos
reproductivos, eh incluso la muerte en algunas especies de organismos cultivados (Díaz y
Liñero 2003, Liñero y Díaz 2006).
Los poliquetos perforadores infestan frecuentemente las conchas de los bivalvos
cultivados, suelen causar graves daños a las conchas de moluscos, que afectan a la aptitud
de sus anfitriones y causando a menudo una pérdida financiera para acuicultores (Avilés et
al., 2007).

11
Características generales clase Polychaeta
Los poliquetos, aunque son animales marinos muy comunes pasan generalmente
inadvertidos a los observadores casuales debidos a sus hábitos ocultos. En las 8000 especies
descritas de lo poliquetos existe una gran diversidad de formas corporales y estilos de vida
que se refleja en el establecimiento de 86 familias. (Ruppert y Barnes 2000). La mayor
parte tienen menos de 10 cm de largo y un diámetro que oscila de 2 a 10 mm, pero existen
algunas formas intersticiales menores de 1 mm; de la misma manera, no son raras las
formas de 70 o más cm de longitud. (Ruppert y Barnes 2000).
Figura 3. Estructura corporal de un poliqueto
El cuerpo de los poliquetos se divide básicamente en tres regiones (Figura. 3). La región
anterior “Acron” (Figura. 4) está formada por el prostomio y peristomio. El peristomio es el
segmento pre oral donde se hallan los glangios cerebrales y órganos sensoriales como ojos,
palpos y cirros, mientras el peristomio o segmento oral es donde se abre la boca y
generalmente cuenta con órganos de captura y órganos sensoriales relacionados con la
alimentación como pueden ser tentáculos(de León Gonzales et al., 2009).

12
Figura 4. Parte Anterior de un poliqueto evidencian sus respectivos órganos de los sentidos
El soma o metastomio, cuenta con una serie de segmentos que comúnmente son muy
parecidos entre sí, las estructuras más características con las que cuenta el soma son los
pies laterales llamados parapodios o parápodos. Típicamente los parápodos se dividen en
dos ramas: notopodios como rama dorsal y neuropodios como rama ventral (Figura. 5), (de
León Gonzales et al., 2009).En el extremo posterior se encuentra el último segmento que es
denominado pigidio y en algunos grupos porta uritos.
Figura 5. Estructura de dos parapodios birramio.

13
Los gusanos poliquetos se distribuyen básicamente en todas las aguas saladas del planeta y
solamente un pequeño porcentaje en aguas dulceacuícolas. Actualmente se conocen unas 12
mil especies en todo el mundo, mientras que nuestro país tiene registradas más de mil, aún
que se estima que falta una gran cantidad de especies no descritas (de León Gonzales et al.,
2009).
Alimentación
El tipo de alimentación de los poliquetos depende de sus costumbres. Básicamente se puede
catalogar dentro de los siguientes tipos:
Depredadores: Se incluyen aquí miembros de varias familias bentónicas, pelágicas y
habitantes de tubos de comportamiento activo. No todos los poliquetos con mandíbulas son
depredadores; esta variedad de hábitos se refleja con claridad en el género Nereis, que
tiene una faringe también eversible provistas de mandíbulas. En este género hay especies
herbívoras, que emplean sus mandíbulas para rascar la cobertura de algas de la superficie
de las rocas, omnívoras, que se alimentan de algas, de detritos y otros invertebrados;
existen también especies comensales y carnívoras por lo que no todas las especies
mandibuladas son depredadoras obligadas (Hickman et al, 2002).
Filtradores: Se trata siempre de poliquetos sedentarios en los que en la cabeza o segmento
posterior se dispone de estructuras filtradoras. En los gusanos abanicó como los serpúlidos,
las radiolas forman una corona que retiene las partículas que lleva en suspensión una
corriente que producen los sirios de las mismas. Las partículas recorren el eje de la radiola
por un canal ciliado hasta su base a la vez que se produce en un proceso de selección de las
mismas en función del tamaño, de forma que las más pequeñas circulan por la parte
inferior del canal y las más grandes por encima algunas de ellas se dedican a la formación
del tubo y el resto que no son desechadas se dirigen a la boca (Hickman et al, 2002).

14
Reproducción
Los poliquetos muestran una alta capacidad de regeneración; son capaces de regenerar el
cuerpo entero desde un único segmento, especialmente en aquellas especies que tienen el
cuerpo indiferenciado. En este proceso tiene una gran importancia el sistema nervioso. La
reproducción asexual por gemación o fragmentación del cuerpo no es muy abundante pero
existe en algunos grupos. Lo más común en los poliquetos es la reproducción sexual
(Ruppert y Barnes 2000). Son dioicos (es muy raro el hermafroditismo), y tienen gónadas
diferenciadas asociadas a septos o vasos sanguíneos; en los más primitivos cada segmento
tiene gónadas, pero en los grupos más recientes las gónadas aparecen sólo en los segmentos
genitales, casi siempre en la región abdominal del animal cuando se encuentran
diferenciados regionalmente.
Estructuralmente, las gónadas no son órganos bien definidos, sino que son unas masas de
gametos en desarrollo formadas por gametogonias o gametocitos que se expulsan al
celoma. De esta forma, al alcanzar el gusano la madurez el celoma está completamente
lleno de gametos que pueden dar un color característico al gusano. La expulsión de todos
estos gametos puede realizarse por medio de celomoductos asociados a los nefridios, por
gonoductos o incluso directamente por la rotura del cuerpo, lo que conlleva la muerte del
animal. En la fotografía superior se muestra un corte transversal de Nereis, donde se
observa, además del vaso ventral, el cordón nervioso y los músculos ventrales, una serie de
huevos dispersos por el celoma. (Ruppert y Barnes, 2000 y Hickman et al., 2002)

15
Poliquetos asociados a otros organismos
Los poliquetos se caracterizan por habitar la zona del bentos aunque de la misma manera
pueden adherirse a cualquier superficie solida por lo que también son considerados como
organismos epibiontes de otros seres vivos. Tal es el caso de Liñero y Díaz (2006)
encontrando 43 especies y 15 familias de poliquetos sobre el bivalvo Spondylus
americanus. Los estudios asociados a la epibiosis de poliquetos se han enfocado a los
bivalvos de interés comercial, sin embargo el estudio está centrado en tres principales
familias de poliquetos Spionidae, Sabellidae y Cirratulidae, responsables de estragos y
pérdidas millonarias en grandes cultivos de moluscos (Avilés et al, 2007).
A su vez existen trabajos de índole ecológica en diferentes grupos de moluscos
comerciales; Pinctata imbricata, Liñero y Díaz (2006) Haliotis discus, Avilés et al., (2007)
Crassostrea rhizhophorae, Díaz, et al (2009) Isognomon alatus, Díaz y Liñero (2003)
Perna viridis Liñero (1999) Fissurella latimarginata, Olivares et al., (1998) Familia
Mytilidae, Zaixso (2004) entre otros estudios vinculados a lo anterior, que cuyo objetivo
respecta a la composición de poliquetos
La epibiosis de poliquetos no solo es común es moluscos, si no en cualquier organismo que
pueda sustentar un sustrato con las cualidades óptimas de adhesión, tal es el caso de la
tortuga verde, Chelonia mydas registrando un porcentaje de 6.9% de ocurrencia sobre su
caparazón (Alonso 2007).
Algunas especies no solo actúan como patógenos, si no como simbiontes benéficos, la
especie Spirobranchus giganteus ayuda a la fijación de carbonato de calcio en corales
placoideos y se encuentra en 80% de la especies de corales placoideos como epibionte
(Rowley 2008).

16
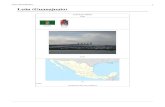
AREA DE ESTUDIO
La Laguna Ojo de Liebre (Figura 6) se localiza en la costa occidental del estado de Baja
California Sur 27° 45′ 0″ N, 114° 15′ 0″ W, y se extiende 40 km tierra adentro con una
superficie de 366,6 km². Forma parte del Complejo Lagunar de Guerrero Negro B.C.S. en
la costa del pacifico a la Bahía de Sebastián Vizcaíno. Dentro de la laguna se encuentran
tres tipos de sustratos: rocoso, arenoso-fino y lodos, en los que el gradiente de energía del
oleaje se presentan en forma descendiente, (Sarur-Zanatta et al., 1984).
Figura 6. Mapa: Laguna Ojo de Liebre proporcionado por Salinera Guerrero Negro.
La Laguna Ojo de Liebre se caracteriza por ser un cuerpo de agua somero con varios
canales bifurcados. Gran parte de los declives de los canales y las zonas someras están
cubiertos por densos lechos de pastos marinos que albergan poblaciones de invertebrados.
Estos lechos se extienden hasta formar marismas.

17
El cuerpo de agua se comunica al mar a través de una boca estrecha. Aunque de umbral
profundo, lo cual permite un flujo permanente de masas de agua oxigenada, ricas en
nutrientes, así como el reflujo de masas de agua con mayor concentración salina por efecto
de la evaporaci6n (Contreras, 1985).
El clima en el área de la laguna es árido, con una temperatura media anual entre los 18 y
22°C y una baja precipitación promedio de 3 cm/año, por lo que los escurrimientos hacia la
laguna son considerados de muy baja escala (Marinone y Lizárraga, 1982).
La laguna presenta una elevada salinidad, debido al poco aporte de aguadulce,
consecuencia de la precipitación irregular que tiene un máximo de 5.0cm/año, habiendo
períodos prolongados de sequía. De la misma manera la laguna no recibe escurrimientos de
áreas agrícolas y prácticamente ningún agua de desecho (Aradit y Llinas, 1991).Cabe
mencionar que en el extremo interno de la laguna se encuentran grandes zonas de
evaporación y por consiguiente una gran producción de sal.
METODOLOGIA.
Área de colecta.
El estudio se realizó en las zonas con gran acumulación de almejas (Bancos): Banco
Chocolatero con coordenadas: 27°52´06´´N, 114°14´53´´W El Zacatoso , 27° 52′ 21″ N,
114° 07′ 07″ O, Punta El Dátil, 27° 46′ 34″ N, 114° 12′ 17″ O, e Isla La Concha,
27° 49′ 17″ N, 114° 12′ 56″ O

18
Colecta en campo.
Se realizaron cuatro colectas durante los días 10-13 de enero del 2012. Los ejemplares de
N. subnodosus fueron colectados realizando un muestreo diario: en sustrato blando
arenosos con la ayuda de buzos (Figura. 7) a una profundidad de entre 5 y 15 m, una vez
recolectado el material se etiquetó y depositó en bolsas ziploc de 25 x 35 cm con agua
marina (Figura. 8 y 9) depositadas en cubetas (Figura. 10) para su traslado al laboratorio.
Figura 7. Buzo Figura 8. Muestra llenada con agua marina
Figura 9. Almeja colectada Figura 10. Cubetas con muestras.

19
Registro de epibiontes
La separación de organismos epibiontes se realizó en el Centro de Investigaciones
Biológicas del Noroeste unidad Guerrero Negro (CIBNOR) (Figura 11). Los organismos
fueron extraídos realizando un cuidadoso raspado de la concha con el uso pinzas de
disección (Fig. 12), después depositadas en bolsas ziploc y viales con formaldehido al 10%
con su respectiva etiqueta, tomando en cuenta datos como, lugar, fecha, estación de
muestreo y el número de la almeja.
Figura 11. CIBNOR Unidad G. Negro Figura 12. Retirando epibiontes de la concha.

20
Separado e identificación de organismos
Los organismos epibiontes fueron trasladados al laboratorio de Biosistemática en la
Facultad de Ciencias Biológicas, Universidad Autónoma de Nuevo León (Figura 13). Una
vez ahí los poliquetos se separaron de los diferentes grupos como, crustáceos,
equinodermos y moluscos. Utilizando para esto un estereoscopio Olimpus SZG1 y un
microscopio Olimpus BX51 con óptica DIC, pinzas de disección y cajas de petri (Figura
14), los organismos fueron identificados hasta el taxa menor posible con la ayuda de claves
taxonómicas actuales (De León Gonzales et al., 2009).
Figura 13. Lab. de Biosistematica Figura 14. Identificacion de poliquetos.

21
Toma de datos morfológicos de la almeja y Análisis estadísticos
Se tomaron datos morfológicos de la almeja correspondientes a la altura, anchura, longitud,
con la ayuda de un vernier electrónico y el peso blando total con la ayuda de una balanza
electrónica.
Se tomaron datos morfométricos como: longitud total (LT); altura total (AT), anchura
(ANC) en milímetros. También datos gravimétricos en (g) como: Peso (W) de glándula
digestiva y gónadas. Con los datos obtenidos se realizaron los siguientes cálculos:
Relación longitud- peso
Se estimó la relación longitud peso con el fin de obtener datos de correlación entre ellos,
utilizando la fórmula:
W= aL b(2-3)
Donde:
W: Peso en gramos (Peso blando)
a: intercepto al origen
L: longitud elevada a un exponente (2 a 3 y multiplicado por una constante).
Índice gonadosomático:
Wg/Wt (100)
Donde:
Wg: Peso de la gónada
Wt: Peso total

22
Índice hepatosomático:
Wh/Wt (100)
Donde:
Wh: Peso del glándula digestiva
Wt: Peso total
Factor de condición múltiple
KM= W/LbAc
Donde:
W: peso en gramos
L: Longitud en centímetros
A: Anchura
b y c: pendientes de la relación múltiple peso longitud anchura en donde b es la pendiente de longitud y c la
de altura.
Índice de diversidad Shannon-Wiener
H' = Σ pi ln p
Donde:
H’ :es el índice de diversidad,
ln: es el logaritmo natural
pi: es la proporción de la muestra, que representa el número total de individuos de una especie (ni)
dividido para el número de individuos de todas las especies (n).

23
Análisis de datos en PAST
Para realizar, los cálculos de diversidad alfa, Análisis SHE, Análisis Clúster, Análisis de
componentes principales, se utilizó el programa PAST en el cual se procesaron los datos y
se generaron gráficas.
RESULTADOS
Durante el presente estudio se revisaron las muestras de cuatro estaciones en la bahía Ojo
de Liebre, Guerrero Negro, BCS. Se identificaron 376 organismos que fueron agrupados en
8 órdenes, 16 familias, 36 géneros y 46 especies las familias mejor representadas fueron,
Syllidae, Nereididae y Eunicidae y Terebellidae. Las familias con el mayor número de
especies fueron: Syllidae, (10 especies), Nereididae (11 especies), Polynoidae (5 especies)
y Sabellidae (4 especies) (cuadro 1). Las cuatro especies más ampliamente distribuidas
fueron Eunice chicasi, Polycirrus californicus, y Platynereis bicanaliculata y quienes
constituyeron el 36% de la abundancia total.
El Banco Chocolatero fue la estación con el mayor número de familias 14, mientras que en
El Dátil solo se obtuvieron siete familias colocándose como el área más pobre. En lo que
respecta a géneros, el banco La Concha presentó el número más elevado con 26, seguido
por el Banco Chocolatero con 21 y el Zacatoso aportando la menor cantidad con 10. La
Zona con mayor número de especies fue La Concha, aportando 33 especies del total
muestreado, así mismo El Zacatoso arrojó la cifra menor con un total de 12 especies.
A continuación se muestra una listado de especies con sus respectivas descripciones
biológicas para cada especie ordenadas en arreglo filogenético siguiendo el acomodo
propuesto por Fauchald, 1977.

24
Cuadro 1.- Abundancia de organismos por especies y estaciones de muestreo.
Especies FamiliaBanco
Chocolatero El Zacatal El DátilLa
Concha
Flabelligella macrochaeta Fauchald, 1972 Acrocirridae Banse, 1969 1 0 0 0
Axiothella mucosa Andrews, 1891 Maldanidae Malmgren, 1867 6 0 0 10
Nereiphylla mimica Eibye-Jacobsen, 1992 Phyllodocidae Örsted, 1843 2 1 0 9
Lepidametria commensalis Webster, 1879 Polynoidae Malmgren, 1867 2 4 2 9
Harmothoe crassicirrata Johnson, 1897 Polynoidae Malmgren, 1867 1 0 0 1
Harmothoe sp 1 Polynoidae Malmgren, 1867 0 0 0 1
Lepidonotus spiculus Treadwell, 1906 Polynoidae Malmgren, 1867 0 0 0 5
Mamgreniella sp 1 Polynoidae Malmgren, 1867 0 0 0 1
Sthenelais fusca Johnson, 1897 Sigalionidae Malmgren, 1867 2 0 0 0
Paleanotus purpurea Rioja, 1947 Chrysopetalidae Ehlers, 1864 1 0 0 0
Exogone lourei Berkeley & Berkeley, 1938 Syllidae Grube, 1850 1 0 0 14
Paraehlersia ferrugina Langerhans, 1881. Syllidae Grube, 1850 0 0 0 2
Salvatoria brevicirra Rioja, 1941 Syllidae Grube, 1850 0 0 0 4
Salvatoria sp 1 Syllidae Grube, 1850 1 0 0 0
Syllis aciculate Treadwell, 1945 Syllidae Grube, 1850 2 0 0 4
Syllis gracilis Grube, 1840 Syllidae Grube, 1850 0 0 1 1
Syllis cf prolifera Krohn1852 Syllidae Grube, 1850 0 1 0 4
Opisthosyllis sp 1 Syllidae Grube, 1850 0 0 1 0
Sphaerosyllis sp 1 Syllidae Grube, 1850 0 0 0 1
Branchiosyllis sp 1 Syllidae Grube, 1850 0 1 1 14
Platynereis bicanaliculata Baird, 1863 Nereididae Blainville, 1818 5 21 3 3
Ceratonereis singularis Treadwell, 1929 Nereididae Blainville, 1818 0 0 1 1
Neanthes acuminata Ehlers 1868 Nereididae Blainville, 1818 0 0 0 2
Eunereis eugeniae Leon-Gonzalez y Solis-Weiss, 2000 Nereididae Blainville, 1818 0 0 0 1

25
Especies FamiliaBanco
Chocolatero El Zacatal El DátilLa
Concha
Nereis grubei Kinberg, 1866 Nereididae Blainville, 1818 0 0 0 4
Nereis sp 1 Nereididae Blainville, 1818 0 0 1 0
Nereis sp 2 Nereididae Blainville, 1818 0 0 1 0
Nereis sp 3 Nereididae Blainville, 1818 0 0 0 1
Pareurythoe spirocirrata Essenberg, 1917 Amphinomidae Savigny in Lamarck, 1818 4 0 23 1Eunice chicasi de León-González, Rivera y Romero, 2004
Eunicidae Berthold, 1827 6 10 7 38
Eunice sp 1 Eunicidae Berthold, 1827 0 0 2 1
Marphysa sanguinea Montagu, 1815 Eunicidae Berthold, 1827 0 1 0 1
Lumbrineris latreilli Audouin y Milne Edwards, 1834 Lumbrineridae Schmarda, 1861 1 0 1 8
Lumbrineris japonica Marenzeller, 1879 Lumbrineridae Schmarda, 1861 0 2 0 4
Lumbrineris inflata Moore, 1911 Lumbrineridae Schmarda, 1861 0 0 0 1
Arabella iricolor Montagu, 1804 Oenonidae Kinberg, 1865 0 0 0 6
Trophoniella salazarae Salazar Vallejo 2012 Flabelligeridaede Saint-Joseph, 1894 2 0 5 1
Piromis sp Flabelligeridaede Saint-Joseph, 1894 1 0 1 0
Polycirrus californicus Moore, 1909 Terebellidae Grube, 1851 1 8 7 27
Eupolymnia nebulosa Montagu, 1818 Terebellidae Grube, 1851 0 3 0 0
Amphitrite cirrata O. F. Müller, 1771 in 1776 Terebellidae Grube, 1851 0 4 7 0
Parasabella pallida Moore, 1923 Sabellidae 0 0 0 5
Branchiomma bairdi McIntosh, 1885 Sabellidae 0 0 0 4
Chone mollis Bush in Moore, 1904 Sabellidae 3 0 13 1
Paradialychone ecaudata Moore, 1903 Sabellidae 6 0 4 1
Crucigera websteri Benedict, 1887 Serpulidae Johnston, 1865 1 0 0 0
Total de individuos 48 56 81 191

26
Lista sistemática de especies:
Phylum Annelida
Clase Polychaeta
Orden Spionida
Super Orden Cirratuliformia
Familia: Acrocirridae Banse, 1969
Flabelligella macrochaeta Fauchald, 1972
Orden Capitellida
Familia Maldanidae Malmgren, 1867
Axiothella mucosa Andrews, 1891
Orden Phyllodocida
Super Orden Phyllodociformia
Familia Phyllodocidae Örsted, 1843
Nereiphylla mimica Eibye-Jacobsen, 1992
Super Orden Aphroditiformia
Super Familia Aphroditoidea
Familia Polynoidae Malmgren, 1867
Lepidametria commensalis Webster, 1879
Harmothoe crassicirrata Johnson, 1897
Harmothoe sp 1
Malmgreniella sp 1

27
Lepidonotus spiculus Treadwell, 1906
Familia Sigalionidae Malmgren, 1867
Sthenelais fusca Johnson, 1897
Super Familia Chrysopetalacea
Familia Chrysopetalidae Ehlers, 1864
Paleanotus purpurea Rioja, 1947
Super Orden Nereidiformia
Familia Syllidae Malmgren, 1867
Branchiosyllis sp 1
Exogone lourei Berkeley y Berkeley, 1938
Paraehlersia ferrugina Langerhans, 1881
Opisthosyllis sp 1
Salvatoria sp 1
Sphaerosyllis sp 1
Salvatoria brevicirra Rioja, 1941
Syllis aciculate Treadwell, 1945
Syllis gracilis Grube, 1840
Syllis cf prolifera
Familia Nereididae Blainville, 1818

28
Platynereis bicanaliculata Baird, 1863
Ceratonereis singularis Treadwell, 1929
Neanthes acuminata Ehlers (1868)
Nereis grubei Kinberg, 1866
Nereis sp 1
Nereis sp 2
Nereis sp 3
Orden Amphinomida
Familia Amphinomidae Lamarck, 1818
Pareurythoe spirocirrata Essenberg, 1917
Orden Eunicida
Super Orden Eunicea
Familia Eunicidae Berthold, 1827
Eunice chicasi de León-González, Rivera y Romero, 2004
Eunice sp 1
Marphysa sanguinea Montagu, 1815
Familia Lumbrineridae Schmarda, 1861
Lumbrineris latreilli Audouin and Milne Edwards, 1834
Lumbrineris japonica Marenzeller, 1879
Lumbrineris inflata Moore, 1911

29
Familia Oenonidae Kinberg, 1865
Arabella iricolor Montagu, 1804
Orden Flabelligerida
Familia Flabelligeridae de Saint-Joseph, 1894
Trophoniella salazarae Salazar Vallejo, 2012
Piromis sp 1
Orden Terebellida
Familia Terebellidae Malmgren, 1867
Polycirrus californicus, Moore, 1909
Eupolymnia nebulosa, Montagu, 1818
Amphitrite cirrata, O. F. Müller, 1771 in 1776
Orden Sabellida
Familia Sabellidae Latreille, 1825
Parasabella pallida Moore, 1923
Branchiomma bairdi McIntosh, 1885
Chone mollis Bush in Moore, 1904
Paradialychone ecaudata Moore, 1903
Familia Serpulidae Rafinesque, 1815
Crucigera websteri Benedict, 1887

30
Diagnosis de las especies.
Lámina 1. Epibiontes de N. Subnodosus parte uno. A-B. Lepidametria commensalis, C.
Lepidonotus spiculus, D-E. Sthenelais fusca, F-G. Platynereis bicanaliculata, H.
Ceratonereis singularis, I. Neanthes acuminata, J. Eunereis eugeniae, K. Nereis grubei, L.
Pareurythoe spirocirrata.

31
Lámina 2. Epibiontes de N. Subnodosus parte dos. A. Eunice chicasi, B. Marphysa
sanguinea, C. Arabella iricolor, D. Trophoniella salazarae, E. Amphitrite cirrata, F.
Eupolymnia nebulosa, G. Branchiomma bairdi, H. Chone mollis, I. Paradialychone
ecaudata, J. Crucigera websteri
Acrocirridae Banse, 1969

32
Flabelligella macrochaeta Fauchald, 1972
Flabelliderma macrochaeta Fauchald, 1972: 222-223.
Diagnosis: Prostomio reducido, en forma de palpo: con un par de manchas oculares fácil de
distinguir, faringe parcialmente eversible. Notosetas con vainas anulares que llevan
numerosas espinas, una seta capilar por rama; Neurosetas con falcígeros compuestos largos
con puntas redondeadas terminalmente formando una V.
Material examinado: un ejemplar: Guerrero Negro, Laguna Ojo de Liebre, Banco
Chocolatero (1)
Distribución.- Atlántico Centro Oriental y el Pacífico. Se conoce su localidad tipo para el
Pacifico Mexicano.
Maldanidae Malmgren, 1867
Clymenella mucosa (Andrews, 1891)
Axiothea mucosa (Andrews, 1891). 14 (852): 277-302
Material examinado: 16 ejemplares: Banco Chocolatero (6); La Concha (10).
Diagnosis: Prostomio ovalado, un poco más largo que ancho. Órganos nucales casi
paralelos, rectos y anchos, casi de la misma longitud que la cabeza. Setígero cuarto con
collar. Parte anterior cuatro a ocho setígeros provistos de bandas completas glandulares, los
segmentos posteriores con zonas glandulares en todo el parapodio. Sin espinas. Pigidio
formado por un embudo con 22-26 cirros.
Distribución.- Pacífico Oriental y el Atlántico Centro-Occidental.
Phyllodocidae Örsted, 1843

33
Nereiphylla mimica Eibye-Jacobsen, 1992
Nereiphylla mimica Eibye-Jacobsen, 1992: 589-613.
Material examinado: 12 ejemplares: Banco Chocolatero (2); El Zacatal (1); La Concha (9).
Diagnosis: Prostomio oval margen dorsoposterior recto más ancho que largo: cuatro
antenas fusiformes tan largas como el prostomio, par dorsal insertas justo debajo del borde
frontal: Ojos grandes en lente de color rojo, oscuro, papila nucal ausente. Segundo
segmento con una o dos setas, primer segmento dorsal reducido, segmento ligeramente
separado por el lado, Cirro tentacular ventral del segmento dos cirriforme. Todos los cirros
dorsales delgados con forma ovalada. Lóbulos neuropodiales no divididos en lóbulos sub y
supra aciculares, con un cirro anal que disminuye su grosor distalmente.
Distribución.- Centro América: desde Belice al norte de Brasil. Posiblemente se trate de
una sinonimia entre Nereiphylla mimica y Nereiphylla paretti registrada en el Pacifico.
Polynoidae Malmgren, 1867
Lepidametria commensalis Webster, 1879
(Lámina 1 A-B)
Antinoe parasitica Webster, 1879. 9: 202-272. Augener, H. 1928: 647-834.
Lepidasthenia commensalis Fiege, 2004: 863-876.
Material examinado: 17 ejemplares: Banco Chocolatero (2); El Zacatal (4); El Dátil (2); La
Concha (9).
Diagnosis: Longitud hasta 100 mm. Con un ancho de 9 mm. Incluyendo setas. Con hasta
80 segmentos, cuerpo alargado plano dorsoventralmente. Entre 30-50 pares de élitros
continuando hasta la parte posterior, tubérculos ovales, lisos y carentes de franjas papilares.
La disposición de los élitros en la región posterior es irregular. Los élitros de los ejemplares
jóvenes tienen una peculiar zona cónica levantó justo medial a los elitroforos.
Pocas notosetas delgadas de tipo capilar, neurosetas gruesas bidentadas de color ámbar.
Cuerpo color púrpura rojizo color oscuro a gris oscuro o negro; cirros dorsales con una

34
banda subterminal oscura, los élitros con manchas pequeñas de pigmento de color gris a
negro con una mancha clara cerca de la elitroforo.
Distribución.- Océano Atlántico Norte, América Central, con su localidad tipo en el Golfo
de México y
Discusión: debido a que esta especie no está registrada para el pacifico, puede que se trate
de un sinónimo u otra especie similar
Harmothoe crassicirrata Johnson, 1897
Harmothoe crassicirrata Johnson, 1897: 153-199
Material examinado: dos ejemplares: Banco Chocolatero (1); La Concha (1).
Diagnosis: Prostomio con lóbulos laterales bien definidos. Picos anteriores prominentes y
agudos, fisura media amplia más o menos un medio del tamaño del prostomio. Tentáculos
con un segmento basal cónico y ancho, alcanzando la altura de los lóbulos prostomiales.
Tentáculos más cortos que los cirros prostomiales, pero casi el doble que las antenas. Esta
última inserta debajo al nivel del tentáculo, el segmento basal grueso, todos los demás
similares a este. Cuatro ojos, par anterior mayor, colocado aproximadamente en la mitad
del prostomio, par posterior más o menos la mitad del tamaño del par anterior colocado más
atrás cerca de la base del prostomio. Cirro peristomial similar al tentáculo pero mucho más
grueso y largo, las articulaciones basales de los cirros dorsales y ventrales están fusionadas
en su totalidad. Cirro dorsal claviforme y mucho más grueso, todos los cirros dorsales
papilados a diferencia de los ventrales. Parapodios divididos en dos ramas desiguales,
ambas largas y cilíndricas. Entre ocho y 16 setas dorsales aserradas y muy sólidas. Setas
ventrales más numerosas y finas (55 o más). Con cinco pares de élitros con coloración café
oscura a rojiza.
Distribución.- Océano Pacifico; Bahía de Monterrey California
Lepidonotus spiculus Treadwell, 1906

35
(Lámina 1 C)
Polynoe specula: Treadwell, 1906:1151-1152,
Lepidonotus caelorus: Moore, 1903, 55: 401-490
Material examinado: cinco ejemplares: La Concha (5).
Diagnosis: Prostomio con picos anteriores largos y un cetaforo medio robusto; Antena
lateral inserta en la parte terminal. Las escamas están adornadas con espinas y una franja
marginal. Setas neuropodiales un diente grueso bien definido en el extremo distal de la
región dentada. Parapodios romos, truncados. Con 12 pares de élitros, élitros con espinas
conspicuas, veteadas y de color gris aunque Existe una gran variación en la pigmentación
de esta especie así como en el tamaño de las antenas
Distribución.- Se encuentra desde Japón hacia el norte alrededor de la Cuenca del Pacífico
y la costa oeste de América del Norte hasta el sur de California.
Sigalionidae Malmgren, 1867
Sthenelais fusca Johnson, 1897
(Lámina 1 D-E)
Sthenelais fusca: Johnson 1897:185-186, pl. 9, figs. 60-61, pl.
Material examinado: dos ejemplares: Banco Chocolatero (2)
Diagnosis: Prostomio redondeado, dos pares de ojos, el par anterior situado hacia delante,
casi oculto bajo la antena si se desde vista dorsal. Tentáculo cortó aplanado y folioso. Un
par de segmentos peristomiales dirigidos hacia adelante. Setígeros en parapodios de forma
y tamaño similar a los tentáculos, mientras el setígero ventral de tamaño mucho menor.
Palpos largos, cilíndricos y lisos, generalmente enrollados y retorcidos en especímenes ya
preservados. Primer par de élitros blanquecinos romboidales, el resto reniforme con
segmentos anchos (2-27).
Parapodios alargados, parte anterior redondeada con un apéndice cilíndrico ligeramente
redondeado. Setas dorsales de tipo capilar serradas. Setas ventrales de cuatro tipos: simples,

36
articuladas moderadamente largas, con apéndice dentado y de tamaño corto y setas cortas
articuladas basalmente. Cirros ventrales no alcanza la porción final del parapodio pero
presentan una expansión dorsal en forma de ala moniliforme cerca de la parte distal.
Distribución.- Océano Pacifico; California
Chrysopetalidae Ehlers, 1864
Paleanotus purpurea Rioja, 1947
Paleonotus purpurea Rioja, 1947b: 517-519
Material examinado: un ejemplar: Banco Chocolatero (1).
Diagnosis: Prostomio oval visible entre las paleas de los primeros segmentos. Paleas cubren
todo el dorso incluso la línea media dorsal. Antena media inserta detrás del par de ojos
anteriores. Paleas principales simétricas, con puntas redondeadas; neurosetas espinígeros y
falcígeras.
Distribución.- Océano Pacifico, Oeste de México
Syllidae Malmgren, 1867

37
Exogone lourei Berkeley & Berkeley, 1938
Exogone lourei Berkeley y Berkeley, 1938: 44,; Rioja, 1941: 703, pl. 3,; Hartman, 1968:
425,; Banse, 1972: 200,; Perkins, 1981: 1092; Uebelacker,1984 (in part): 30–39.
Exogone (Exogone) lourei.–San Martín, 1991a: 728-735; Núñez et al., 1992: 45; Kudenov
y Harris, 1995: 15,.
Material examinado: 15 ejemplares: Banco Chocolatero (1); La Concha (14).
Diagnosis: Cuerpo largo y delgado, filiforme 5,2 mm de largo, 0.25 mm de ancho, con 45
setígeros. Prostomio pentagonal; Con dos pares de ojos en el arreglo trapezoidal, antenas
inserta cerca una de otra, en fila delante de los ojos anteriores, antena media cilíndrica con
punta estrecha, mucho más larga que las antenas laterales, pero ligeramente más corta que
el prostomio, palpos juntos, antenas laterales ovaladas, cerca de un quinto o sexto de la
longitud de las antena media. Palpos largo, fusionado a lo largo de su de longitud, de forma
triangular con una muesca distal. Peristomio tan largo como los siguientes segmentos;
cirros tentaculares similares a las antenas laterales, pero más cortos. Cirros dorsales
ovales a ligeramente piriformes, ligeramente más largo que las antenas laterales pero más
corto que los lóbulos parapodiales, presentes en todos los segmentos. Parapodios anteriores
cada uno con un espinígero compuestos por seta con hoja unidentada, larga y delgada,
además de cuatro a cinco falcígeros con un diente subdistal largo y un diente distal corto
más largo que las espinas marginales. Espinígero provisto de un proceso triangular, con
espinas diminutas en la superficie con la hoja distal bífida. Setas dorsales sencillas desde
una vista anterior, parapodios con las puntas redondeadas y finamente espinosos. Setas
ventrales simples, parapodios posteriores sigmoides bidentados, con un diente subdistal
más largo y ancho y diente distal pequeño.
Acícula solitaria, delgada, distalmente redondeada. Faringe larga y delgada, a través de seis
a siete segmentos, con dientes faríngeos en el margen anterior, proventrículo más corto que

38
la faringe, a través de tres segmentos, con alrededor de 23 hileras de células musculares.
Pigidio con dos cirros anales de largos.
Distribución. Pacífico, las costas de la Columbia Británica a Panamá. Zona del Caribe: La
Florida, el Golfo de México, Belice, Cuba. Atlántico oriental (Islas Canarias). Australia
(Australia Occidental).
Paraehlersia ferrugina Langerhans, 1881.
Paraehlersia ferrugina Langerhans, San Martin, G. 2003: 1-554
Ehlersia ferruginea Langerhans, 1881, 42(3): p 95-124.
Material examinado: dos ejemplares: La Concha (2).
Diagnosis: Cuerpo largo y delgado, con hasta 10 mm de longitud, 0.3 de anchura, y 72
setígeros, sin patrón de coloración. Una banda ciliada provista de largos cilios y de poros en
cada segmento. Prostomio pentagonal, con dos pares de ojos gruesos en disposición
trapezoidal y dos manchas oculares anteriores por lo común bien patentes. Tres antenas
claramente moniliformes, al menos al extremo aunque las bases pueden ser lisas o
irregularmente articuladas; antena central dos veces más grande que las laterales; antenas
laterales también moniliformes, ligeramente más largas que el prostomio incluyendo los
palpos, insertas por delate de los ojos anteriores y por detrás de las manchas oculares.
Palpos gruesos, robustos más largos que el prostomio, unidos a la base. Segmento bucal
dorsalmente cubierto por el primer setígero: con dos pares de cirros tentaculares
moniliformes, los dorsales tan largos como la antena central, los ventrales más cortos.
Cirros dorsales largos y delgados, los de la parte anterior claramente moniliformes y
progresivamente se van haciendo lisos y delgados, alternando largos y cortos, pasando por
cirros ligeramente articulados. Parápodios sub rectangulares: cirros ventrales largos,
digitiformes.

39
Los parápodos posteriores son más pequeños y alargados, con cirros ventrales más largos y
delgados. Parápodios anteriores con 3-6 setas compuestas pseudoespínigeras, con extremos
bífidos y espinulación corta. Parápodios medios con tres setas similares a los de los
parápodos anteriores pero más largas. Los parápodios posteriores llevan una seta
pseudoespínigera aparentemente unidentada y cinco falcígeros compuestos. Faringe ancha,
de longitud similar al proventrículo, armada de un grueso diente agudo en parte anterior y
un círculo distal de papilas. Proventrículo rectangular, a través de unos a seis a siete
segmentos con al menos 25 hileras de células musculares.
Distribución: Es una especie cosmopolita generalmente de mares templados, aunque podría
tratarse de un complejo de especies.
Salvatoria brevicirra Rioja, 1941
Brania heterocirra: Rioja, 1941: 700, pl. 3,
Syllis sp 7, Góngora 2011 : 116
Material examinado: cuatro ejemplares: La Concha; (4).
Diagnosis: De 20 a 23 segmentos. Prostomio transverso con sus bordes laterales
redondeados. Palpos bien desarrollados, redondeados anteriormente, fusionados en su
mayor parte, existiendo entre ellos un surco perfectamente acusado. Con dos pares de ojos:
Los anteriores mayores y más separados entre sí. En el borde anterior del prostomio existe
un par de manchas oculares muy pequeñas. Las antenas laterales se insertan anteriormente
y a los lados. Las tres antenas son similares en forma y longitud: La antena media casi
alcanza el borde anterior que forman los palpos. El peristomio es tan largo como el
prostomio. Con dos pares de cirros tentaculares: El par superior mayo el inferior. Los cirros
dorsales de igual forma que las antenas y los cirros tentaculares: los del primer setígero
claramente mayores al resto. Cirros ventrales digitiformes con su extremo redondeado.
Falcígeros con un artejo terminal bidentado.

40
En los setígeros posteriores existen en cada haz dos setas simples aciculares, sinuosas y
unidentadas: dorsal y otra ventral. Pigidio redondeado, provisto de dos cirros anales mucho
más largos que los cirros dorsales. Faringe desde el segundo hasta el quinto setígero, el
proventrículo ocupa de dos a dos y medio segmentos, a partir del quinto setígero.
Distribución.- Océano pacifico, Oeste de México
Syllis aciculata Treadwell, 1945
Typosyllis aciculata orientalis, Imajima, 1966 26: 1-452.Imajima, 2003; 23: 1-221
Material examinado: seis ejemplares: Banco Chocolatero (2); La Concha (4).
Diagnosis: Color café, con surcos ínter segméntales obscuros. Prostomio pentagonal
ligeramente más ancho que largo, un par de palpos, tres antenas y dos pares de ojos. Los
palpos unidos en sus bases por una tercera parte de su longitud. Antena media inserta detrás
del punto medio de los ojos posteriores. Antenas laterales justo enfrente del par de ojos
anteriores. Articuladas con 13 artejos. Dos pares de ojos en arreglo trapezoidal: el par
anterior en lentes, el doble de tamaño que el par posterior. Peristomio con dos pares de
cirros tentaculares moniliformes; el par dorsal compuesto por 17 artejos y el par ventral
compuesto por 10 artejos. Faringe con el margen anterior liso, de seis segmentos de largo y
se extiende desde el cuarto al décimo setígero. Es simple, sin color y con un diente anterior.
El proventrículo se origina en el décimo segmento y se extiende hasta el segmento 17. Los
lóbulos parapodiales anteriores son de forma rectangular, cada uno con un cirro tentacular
articulado, lóbulo cónico con 10 setas delgadas, en lóbulos ventrales. Los lóbulos
posteriores son más pequeños, con una acícula delgada de color amarillo y de cinco a seis
setas compuestas.
Distribución: Norte de California hasta Panamá.

41
Syllis gracilis Grube, 1840
Syllis gracilis Grube, 1840:77; Góngora-Garza, 1984:39
Material examinado: dos ejemplares: Banco El Dátil (1); La Concha (1).
Diagnosis: Prostomio redondeado, con tres antenas articuladas, la media con 8 a 13
artículos, las laterales con 9 a 18 artículos. Cuatro ojos pequeños. Palpos largos,
triangulares separados en casi toda su longitud. La faringe se extiende hasta el setígero 10;
presenta el margen liso y rodeado por diez papilas suaves. El proventrículo abarca los
setígeros 8 a16, con 36 a 38 hileras de células musculares. Cuatro cirros tentaculares
formados por 11 a 14 artejos; cirros dorsales con 6 a 13 artículos; cirros ventrales con 6 a
14 artículos. Setas incluyen falcígeros compuestos bidentados y ligeramente aserrados,
reemplazados por setas simples robustas en forma de "Y". Setas simples superiores e
inferiores delgadas y bidentadas. Pigidio con dos cirros anales, con 6 a 12 artículos.
Distribución: Japón; mar Mediterráneo; islas Canarias; Sudáfrica; costa atlántica y pacífica
de Panamá; Nueva Inglaterra y Carolina del Norte E.U.; Norte del golfo de México y golfo
de California; Cuba.
Syllis cf prolifera Krohn 1852
Typosyllis (Typosyllis) prolifera San Martín G (2003), p 1-554
Material examinado: cinco ejemplares: Guerrero Negro, Laguna Ojo de Liebre, El Zacatal
(1); La Concha (4).
Diagnosis: Prostomio redondeado, rectangular, con cuatro ojos y con o sin manchas
oculares cerca de la base de los palpos. Faringe con diez papilas distales, un diente
mediodorsal en posición sub-terminal, proventrículo entre los setígeros 8-13 y 10-20.
Palpos triangulares. Antena media con 12-31 artejos, la antena lateral con 9-23. Cirros
tentaculares dorsales con 13-29 artejos, los ventrales con 8-16. Parapodios anteriores con
cirros dorsales largos 8-42 artejos, 6-45 en la región media, cirros ventrales largos y
digitiformes.

42
Las setas son falcígeros bidentados con los dientes más o menos redondeados y
asenaciones gruesas; en la parte posterior del cuerpo existen setas simples bífidas y setas
simples delgadas, también bífidas.
Distribución: Circumtropical, Golfo de California, Nayarit.
Discusión: La descripción corresponde a la especie Syllis prolifera ya que presenta la
mayoría de las características de esta especie sin embargo es necesario realizar un estudio
adicional para saber si realmente se trata de Syllis prolifera.
Nereididae Blainville, 1818
Platynereis bicanaliculata Baird, 1863
(Lámina 1 F-G)
Nereis bicanaliculata: Baird, 1863 106-110
Platynerieis bicanaliculata: Imajima y Hartman 1964: 152:
Material examinado: 32 ejemplares: Banco Chocolatero (5); El Zacatal (21); El Dátil (3);
La Concha (3).
Diagnosis: El cuerpo es pardo oscuro, con parches cerca de los parapodios, con entre 95-
100 segmentos. Prostomio con dos pares de ojos en arreglo trapezoidal, con pigmentación
oscura excepto en la parte media. Antenas frontales delgadas, palpos gruesos. Faringe
carece de paragnatos en los anillos I, II y V, el anillo III tiene parches ovales varias con
filas de conos, área IV con una media luna de 7 a 10 parches de conos, en el anillo oral VI
tiene de dos a tres líneas de pectinas, áreas VII y VII con dos filas de pectinas. Peristomio
ligeramente más largo que el siguiente segmento, con 4 pares de cirros tentaculares de
largos, el par dorsal posterior es aproximadamente 10 veces más largo. Parapodios
dorsalmente triangulares, lóbulos medio y ventral. Notopodios con espinígeros simples.
Neuropodios con falcígeros y espinígeros cortos y gruesos.
Distribución.- Anfipacífico en aguas subtropicales a templadas. Costa oeste de Canadá
hasta el oeste de México

43
Ceratonereis singularis Treadwell, 1929
(Lámina 1 H)
Ceratonereis singularis Treadwell, 1929: 1-3, figs. 1-8.
Material examinado: dos ejemplares: Banco El Dátil (1); La Concha (1).
Diagnosis: Cuerpo pálido, con dos bandas pardo-amarillentas transversas en cada segmento
anterior, los medios y posteriores sin pigmento. Prostomio hendido en la parte medio
frontal, con dos pares de ojos negros en arreglo trapezoidal, los anteriores ligeramente
mayores y más separados entre sí. Palpos delgados y largos; antenas piriformes rebasan el
borde distal de los palpostílos .Peristomio tan largo como el siguiente segmento, cirros
tentaculares delgados, el mayor alcanza el setígero 18. Faringe con paragnatos sólo en el
anillo maxilar, área II: 11-13 en dos líneas; área III: 11 en grupo oval; área IV: 16 en grupo
oval. Parapodios anteriores con cuatro lóbulos, el notopodial superior y neuropodial
inferior digitiformes, el notopodial inferior y neuropodial superior subtriangulares. Cirro
dorsal largo, inserto medialmente, cirro ventral corto, basal. Parapodios posteriores
similares a los anteriores, pero con el lóbulo notopodial superior y neuropodial inferior
reducidos; cirro dorsal delgado y largo, dehiscente, el ventral largo pero menor que el
dorsal. Notosetas espinígeros homogonfos y falcígeros sesquigonfos unidentados, con el
borde interior finamente serrado; las neuropodiales espinígeros y falcígeros heterogonfos
unidentados Pigidio con ano terminal, sin cirros anales.
Distribución.- Pacifico Norte, Baja California, México

44
Neanthes acuminata Ehlers 1868
(Lámina 1 I)
Nereis (Neanthes) acuminata:.- Day 1973: 41; Gardiner 1976
Nereis (Neanthes) arenaceodentata: Pettibone 1963: 162
Neanthes acuminata:.-Taylor 1984: 31-15
Material examinado: dos ejemplares: Banco La Concha (2).
Diagnosis: Prostomio más ancho que largo con dos pares de ojos negros bien desarrollados;
Cirros tentaculares mayores alcanzan el setígeros tres a cuatro: Faringe con paragnatos
cónicos; área I con 17 conos ovales, área II con 25 conos curvos, área III con 35
paragnatos, área IV con 30 conos, Áreas V, VI, VII-VIII fusionados en una banda continua
de paragnatos cónicos, de tres a cuatro filas irregulares. Lígula notopodial dorsal y media
presentes, lígula notopodial con región basal presente y región posterior ligeramente más
amplia, setas notopodiales espinígeros homogonfos y setas neuropodiales con setas
espinígeros homogonfos y falcígeros heterogonfos en los paquetes sub aciculares.
Distribución.- Océano Pacifico y Golfo de México.
Eunereis eugeniae de León-González y Solis-Weiss, 2000
(Lámina 1 J)
Eunereis eugeniae de León-González y Solís-Weiss, 2000: 550-552, figs. 1a-g.
Material examinado: un ejemplar: Banco La Concha (1).
Diagnosis: Cuerpo completo sin pigmentación evidente, con más de 90 setígeros.
Prostomio pentagonal con un par de antenas frontales digitiformes que rebasan el borde
anterior de los palpos. Dos pares de ojos en arreglo cuadrangular, los anteriores en lente.
Palpos biarticulados con el palpostílo cónico. Peristomio tan ancho como los dos primeros
setígeros, con cuatro pares de cirros tentaculares cortos, el par mayor llega hasta el setígero
cuatro. Faringe con paragnatos sólo en el anillo oral: Área VII-VIII: 7 conos transparentes
en una línea.

45
Mandíbulas quitinosas con ocho dientes. Parapodios anteriores con lóbulos triangulares,
sólo el lóbulo neuropodial superior redondeado distalmente. Cirro dorsal y ventral
subiguales. Parapodios medios con el lóbulo notopodial superior triangular, el notopodial
inferior subulado; lóbulo neuropodial superior mamiliforme, el inferior subulado. Cirro
dorsal inserto medialmente, mayor que el ventral, el cual está inserto basalmente.
Parapodios posteriores con el notopodio formado por el lóbulo notopodial superior delgado
y redondeado distalmente; el inferior triangular. Neuropodio con el lóbulo superior
mamiliforme, el inferior subulado. Cirro dorsal y ventral subiguales . Parapodios anteriores
con espinígeros homogonfos notopodiales supraciculares; neurosetas supraciculares
espinígeros homogonfos y falcígeros heterogonfos, las infraciculares un espinígero y
numerosos falcígeros heterogonfos, estos últimos similares a los supraciculares, con el
apéndice delgado, un pequeño diente apical delgado , dirigido hacia abajo, finamente
dentado en su margen interno . Notosetas supraciculares en parapodios medios un
espinígero y dos falcígeros homogonfos, estos últimos con el apéndice corto, con cuatro
pequeños dientes en su margen interno, el manubrio de la seta con una membrana
crenulada unida a las aristas. Neurosetas sólo en posición supracicular, con un falcígero
heterogonfo con el apéndice pequeño redondeado distalmente, fuertemente dentado en su
margen interno. Parapodios posteriores con un falcígero homogonfo notopodial
supracicular, similar al de parapodios medios. Neurosetas sólo en posición infracicular,
estas son un espinígero y un falcígero heterogonfo. Pigidio con ano terminal y dos cirros
anales largos.
Distribución.- Océano Pacifico Norte, Baja California Sur, Punta Eugenia

46
Nereis grubei Kinberg, 1866
(Lámina 1 K)
Heteronereis grubei Kinberg, 1866: 337-357.
Material examinado: cuatro ejemplares: Banco La Concha (4).
Diagnosis: Prostomio más largo que ancho, con un par de antenas frontales subuladas
gruesas. Dos pares de ojos, los anteriores en lente, ovales, los posteriores de menor tamaño,
redondos. Palpos globosos, palpostílos cónicos. Peristomio tan ancho como los primeros
dos setígeros, con cuatro pares de cirros tentaculares cortos, el par mayor alcanza hasta el
primer setígero. Faringe con paragnatos en el siguiente arreglo: área I: dos conos en línea ;
área II: 12-14 en dos líneas; área III: 20 conos en tres líneas; área IV: 25 conos en arreglo
creciente; área V: 15 conos en un parche; área VI: cuatro conos grandes en cruz; áreas VII-
VIII: 65 en cuatro líneas. Mandíbulas con cinco dientes. Parapodios anteriores con dos
lóbulos redondeados, el neuropodial superior mamiliforme, el inferior subulado. cirro
dorsal y ventral subiguales. Parapodios medios similares a los anteriores, con el cirro dorsal
inserto medialmente. Parapodios posteriores con el lóbulo notopodial superior expandido,
cirro dorsal distal. Setación en parapodios anteriores con el siguiente arreglo: Notosetas
espinígeros homogonfos; neurosetas supraciculares espinígeros homogonfos y falcígeros
heterogonfos, estas últimas con el apéndice delgado. Parapodios medios y posteriores con
setación similar, notosetas falcígeros homogonfos con lámina corta, presentan tres dientes
pequeños en el lado interno; las neuropodiales en arreglo similar al de parapodios
anteriores, pero los falcígeros infraciculares con el apéndice ancho
Distribución.- Océano Pacifico. Se conoce para su localidad tipo, Chile.

47
Amphinomidae Lamarck, 1818
Pareurythoe spirocirrata Essenberg, 1917
(Lámina 1 L)
Eurythoe spirocirrata Essenberg, 1917: 71-74.
Material examinado: 26 ejemplares: Banco Chocolatero (4); El Dátil (23); La Concha (1).
Diagnosis: Cuerpo largo y delgado, gris en el alcohol. Superficie dorsal ligeramente
convexa. Superficie ventral fuertemente convexa. Branquias marginales. Dos cirros en cada
parapodio. Carúnculo unido al prostomio, amplio y liso, que se extiende hasta el cuarto
segmento. El cuerpo es largo, algo uniforme en anchura, atenuado gradualmente hacia el
extremo posterior con ambos extremos obtusamente redondeado. La superficie dorsal es
casi recta ligeramente arqueada. La superficie ventral es convexa. Con un tentáculo corto.
El extremo posterior de la carúncula es estrecho y hendido. Prostomio tiene dos pares de
cirros. Puede o no presentar ojos. Las setas dorsales son de dos tipos, una aserrada en la
zona terminal y la otra aserrada débilmente en la zona distal.
Distribución.- Pacifico Este: México, Perú e Islas Galápagos
Eunicidae Berthold, 1827
Eunice chicasi de León-González, Rivera y Romero, 2004
(Lámina 2 A)
Eunice chicasi de León-González et al., 2004: 93, Fig. 1 A-G.
Material examinado: 61 ejemplares: Banco Chocolatero (6); El Zacatal (10); El Dátil (7);
La Concha (38).
Diagnosis Prostomio de menor tamaño que el peristomio, palpos redondeados, con un
surco medio dorsal. Antenas prostomiales en semicírculo, equidistantes, palpos alcanzan el
borde posterior del quinto setígero. Ceratóforos cortos, ceratostilos con articulación mixta,
palpos con 15 artejos, Ojos redondos.

48
Primer anillo peristomial tres a cuatro veces más grande que el segundo, separación visible
dorsal y ventral. Cirros peristomiales lisos, digitiformes alcanzan la mitad del peristomio.
Aparato maxilar con cuatro pares de maxilas y una impar: Formula maxilar= 1+1, 5+6,
6+0, 7+8, 1+1; Mx-III formando parte del arco distal. Branquias pectinadas desde el sexto
setígero al 98 ; el mayor desarrollo se observa en el setígero 14 con 10 filamentos.
Branquias más grandes que el cirro dorsal, filamentos branquiales de similar longitud que el
cirro dorsal. Cirro dorsal liso, digitiforme. Cirro ventral de los setígeros 1-4 digitiformes,
del quinto al 36 con la base inflada de forma alargada, posteriores digitiformes. Lóbulo
presetal truncado, lóbulo postsetal redondeado decreciendo en tamaño en los setígeros
posteriores. Setas pectinadas heterodontas, con hasta 10 dentículos; falcígeros compuestos
bidentados en los setígeros anteriores con los dientes de similar tamaño, en setígeros
posteriores diente proximal triangular más grande que el distal; falcígeros tridentados en los
últimos seis setígeros, con el diente distal pequeño. Acículas amarillas, en setígeros
anteriores mucronadas o romas, en setígeros medios y posteriores en forma de “T”. Pigidio
con dos pares de cirros anales lisos, los dorsales mucho más grandes, ventrales muy cortos
digitados.
Distribución.- Pacifico Oriental Tropical. Se conoce para su localidad tipo, Isla Manguera,
El Salvador, América Central. En México, para Puerto Ángel y Huatulco, Oaxaca. Esta es
la cita más norteña de E. chicasi.
Marphysa sanguinea Montagu, 1815
(Lámina 2 B)
Nereis sanguinea Montagu, 1815: 11: 18-21
Material examinado: dos ejemplares: Banco El Zacatal (1); La Concha (1).
Diagnosis: Setígeros anteriores cilíndricos, por lo general aplanados dorsoventralmente; la
epidermis es iridiscente anterodorsalmente. Prostomio con una hendidura anteroventral
profunda. Posee cinco apéndices; Tres antenas y dos palpos laterales más cortos dispuestos
en línea curva cerca del borde posterior.

49
Ojos presentes. Peristomio dividido en dos anillos, el anterior más grande que el posterior:
cirros peristomiales ausentes. Mandíbulas fuertemente esclerotizadas con formula maxilar:
Mx I = 1 +1; MX II = 3-4 +4-5, los dientes anteriores más grandes; III Mx = 4.6 (todos los
dientes pequeños y uniformes en tamaño), Mx IV = 03.05 + 05.08 (todos los dientes de
tamaño uniforme; Mx V = 1.1.Branquias presentes desde setígero 13-27, setígeros
posteriores, inicialmente formados por un solo filamento aumentando hasta 6 dispuestos a
lo largo de eje único. Notosetas de setígeros uno y dos de dos tipos: de tipo delgada y
dentada y gruesa y dentada. Setas ventral de tipo compuesta espinígeras con espina sub
acicular bífida. Pigidio con dos cirros anales.
Distribución.- Pacifico Oriental Tropical, Océano Pacifico Sur. Gran Caribe, Océano
Atlántico, Mar Rojo Mar del Norte y Mar Europeo. Se conoce para su localidad tipo,
Inglaterra, Cornwall, Polpero. Debido a que ha sido ampliamente registrada en el mundo se
cree que pertenece a un complejo de especies. Para la región del Pacifico existen tres
especies con el complejo sanguínea registrando como única diferencia el número de
filamentos branquiales y el tamaño de los organismos por lo que su registro en Baja
California sería cuestionable.
Lumbrineridae Schmarda, 1861
Lumbrineris latreilli Audouin y Milne Edwards, 1834
Lumbrineris latreilli: Audouin y Milne Edwards, 1834: 168, pl. 38, Fauvel, 1911.
1-41.Hartman, 1938. (1): 3-31. Bellan, 2001: pp. 214-231.
Lumbriconereis tingens Keferstein, 1862.:1-147
Lumbriconereis nardonis Grube, 1840. 1-92.
Material examinado: 10 ejemplares: Banco Chocolatero (1); El Dátil (1); La Concha (8).
Diagnosis: El color es rojo pálido a marrón. Con 90 segmentos o más. Prostomio cónico y
ligeramente más largo que ancho.

50
Parapodios anteriores tienen setas limbadas y falcígeros compuestos, este último a través
del setígeros 18 a 23, cada uno es multidentado; posteriormente se sustituyen por falcígeros
simples. Parapodios posteriores tienen lóbulos postsetal no más largos que los de delante.
Acícula son de color amarillo, número de 2 a 3 en un fascículo. Las Maxilas están bien
desarrolladas, los portadores de una incisión lateral, ti; La maxila II tiene cinco o cuatro
dientes, III tiene dos dientes y IV un solo diente. Mandíbulas son compactas, y se funden en
sentido medial.
Discusión: Se asemeja a Lumbrineris japonica ya que la única diferencia es el color de la
acícula que en este caso es de color amarillo en vez de poseer una acícula negra como en L.
japonica.
Distribución.- Océano Pacifico, Golfo de México y Mar Mediterráneo. Su localidad tipo es
en Francia.
Lumbrineris japonica Marenzeller, 1879
Lumbriconereis japonica Marenzeller; 1879; 137-138, pl. 5,; Isuka, 1912:139-140, pl. 14,
Material examinado: seis ejemplares: Banco El Zacatal (2); La Concha (4).
Diagnosis: Cuerpo cortó pálido Prostomio globular. Lóbulopostsetal alargado en setígeros
anteriores Falcígeros compuestos cubiertos en los primeros siete setígeros. Setas limbadas
en todos los setígeros Falcígeros cubiertos simples en setígeros medios. Ganchos
aciculares de color negro.
Distribución.- Océano Pacifico Occidental; Del oeste de Canadá al oeste de México. Se
conoce su localidad tipo Eno-Sima, Japón

51
Lumbrineris inflata Moore, 1911
Lumbrineris cervicalis: Treadwell, 1922:176; Uschakov, 1955:239
Material examinado: un ejemplar: Guerrero Negro, Laguna Ojo de Liebre, La Concha (1).
Diagnosis: Cuerpo alargado pálido, prostomio globular. Maxila III con tres dientes la IV
con dos dientes Lóbulo postsetal alargado en todos los parapodios. Acículas amarillas.
Falcígeros compuestos cubiertos en los primeros 20 a 25 setígeros. Setas limbadas en todos
los setígeros. Falcígeros cubiertos simples en setígeros medios y posteriores
Distribución.- De Columbia británica hasta el Oeste de México; Golfo de México, Mar de
Bering, Mar Amarillo, Japón.
Oenonidae Kinberg, 1865
Arabella iricolor Montagu, 1804
(Lámina 2 C)
Nereis iriscolor: Montagu, 1804:82.
Material examinado: seis ejemplares: Banco La Concha (6).
Diagnosis: Cuerpo alargado y cilíndrico, hasta 20 mm de longitud. La coloración es pardo
rosada muy iridiscente. El prostomio es cónico, desprovisto de apéndices sensitivos y con 4
ojos transversales situados en la parte posterior. La fórmula mandibular es la siguiente: M I:
I+9, M II: 6-7 + 12-14, M III: 6+ 4-6, M IV: 5-6 + 6, M V: I+ I. Los parápodos llevan un
pequeño cirro dorsal, sedas simples geniculadas y limbadas.
Distribución.- De los Estados Unidos, Reino Unido, Australia, México, Grecia, Sudáfrica,
Italia, Canadá, Francia, Turquía, Tanzania, Albania, Isla de Man, Madagascar, Croacia,
Sri Lanka, Guernsey, Jersey.

52
Flabelligeridae de Saint-Joseph, 1894
Trophoniella salazarae Salazar Vallejo 2012: 512, fig. 26
(Lámina 2 D)
Material examinado: ocho ejemplares: Banco Chocolatero (2); El Dátil (5); La Concha (1)
Diagnosis: Cuerpo cilíndrico anteriormente, atenuándose posteriormente. Túnica rugosa,
maculada sin partículas de sedimento. Papilas del cuerpo erosionadas sin partículas de
sedimento y de tamaño diminuto. Prostomio un poco pálido, ojos negros, carúnculo
negruzco bien desarrollado, extendido a lo largo del prostomio. Palpos de color marrón
rojizo claro, ligeramente más largos que la placa branquial y un poco deprimido con un
tallo estrecho y grande, con alas laterales muy notables. Caja cefálica con setas conspicuas
con proyección triangulada. Parapodios poco desarrollados, las setas salen de la pared del
cuerpo, notopodio con dos grandes papilas una basal y la otra lateral, neuropodio con
papilas similares pero de menor longitud. Notosetas medias en series cortas y por paquetes,
multiarticuladas, neurosetas falcadas, articuladas. Pigidio sin cirros anales.
Distribución.- Pacifico Mexicano; Baja California- Nayarit.
Terebellidae Malmgren, 1867
Amphitrite cirrata Müller, 1771 in 1776
(Lámina 2 E)
Amphitrite radiata Moore, 1923: 75:193.
Material examinado: 11 ejemplares: Banco El Zacatal (3); El Dátil (7).
Diagnosis: Color pálido tórax con 17 setígeros de los cuales 16 son uncinígeros. Tres pares
de branquias filamentosas y sésiles Cojinetes laterales reducidos Con escudos ventrales.
Notosetas torácicas ligeramente limbadas con las puntas marginalmente serradas. Uncinos
torácicos en hileras dobles a partir del séptimo setígero.

53
Distribución.- Oriental del Atlántico Norte a las Islas Azores, Mediterráneo, América del
Norte del Atlántico, Groenlandia Occidental, el Ártico canadiense y Alaska, Siberia Ártico,
el Mar de Bering, Mar de Okhotsk, el Mar de Japón, América del Norte del Pacífico.
Eupolymnia nebulosa Montagu, 1818
(Lámina 2 F)
Amphiro nebulosa Montagu, 1808: 108-111
Terebella nebulosa Montagu, 1818: 12: 340-344
Terebella tuberculata Dalyell, 1853:.1-359.
Material examinado: tres ejemplares: Banco El Zacatal (3)
Diagnosis: Lóbulo tentacular con una doble hilera de manchas oculares sobre la solapa
prostomial .Pequeñas solapas laterales presentes sobre los segmentos II-IV. Branquias
irregularmente ramificadas con pedúnculo bien desarrollado. Notosetas simples con velo
asimétrico estriado diagonalmente. Uncinos desde el segundo setígero, dispuestos en una
sola hilera hasta el cuarto setígero, a partir del quinto los uncinos se disponen en doble
hilera, cara a cara. Fórmula dentaria de los uncinos 1:2:3-5. Apéndice de inserción del
ligamento bien desarrollado tanto en los uncinos torácicos como en los abdominales. Los
ejemplares preservados son de color crema claro.
Distribución.- Océano Atlántico Norte, Golfo de México y Golfo de California.

54
Polycirrus californicus Moore, 1909
Polycirrus perpleux: Moore, 1923: 198-199. Hartman, 1969:631-632.Fide Banse, 1980
Material examinado: 44 ejemplares: Banco Chocolatero (1); El Zacatal (8); El Dátil (7); La
Concha (27).
Diagnosis: Color en vida es rojo. El cuerpo consiste en 55 a 65 segmentos con alrededor de
28 setígeros. El vientre tiene un profundo surco neural en todo el cuerpo. Prostomio grande,
con proyecciones arqueadas más o menos el doble de largos que el ancho, el largo es
similar al tamaño de los primeros cuatro segmentos. Ojos ausentes. Tentáculos orales muy
números formando una masa. Tentáculos de dos tipos, delgados y filiformes distalmente
expandidos e insertados lateralmente, mientras otros mucho más gruesos insertados en el
centro. Labio inferior liso, triangular, formando el primer escudo ventral; los siguientes 8
de menor tamaño, medialmente divididos por un surco neural, escudos abruptamente
reducidos desde allí. Notoseta actual de los segmentos dos a 29, en pequeños fáculos
laterales, cada seta ligeramente limbada, suave, con punta puntiaguda. Uncinos presentes
alrededor del segmento 14, en una sola fila corta. Cada uncino una placa plana, más largo
que ancho, con una base prolongada, estrecha, un gran colmillo y dientes dos, visto de
perfil.
Distribución.- Océano Pacifico; San Diego California.

55
Sabellidae Latreille, 1825
Branchiomma bairdi McIntosh, 1885
(Lámina 2 G)
Dasychone bairdi McIntosh, 1885:, 12: 1-554.
Material examinado: cuatro ejemplares: Banco La Concha (4).
Diagnosis: Tubos cimentados con arena fina. El cuerpo alcanza hasta 40 mm de longitud;
verde olivo con marcas pardas, manchas oculares al costado de cada segmento disminuyen
hacia la parte posterior. Corona branquial con 16 pares de radiolos; bandas alternadas verde
olivo y blancas: bandas anaranjadas en el eje del radiolo. Macroestilodo en forma de correa
principalmente en la parte media del radiolo, el resto digitiformes cortos. Ojos rojos o
anaranjados en los radiolos. Collar separado dorsalmente, lóbulos ventrales subtriangulares,
con ápices redondeados. Tórax usualmente con ocho segmentos. Uncinos en forma de
gancho, con 2 o 3 hileras de dientes, ocupan un tercio de la longitud.
Distribución.- Golfo de México, Océano Pacifico; Baja California.
Chone mollis Bush in Moore, 1904
(Lámina 2 H)
Metachonemollis Bush, 1905:56:175: 216.
Material examinado: 18 ejemplares: Banco Chocolatero (3); El Dátil (13); La Concha (1).
Diagnosis: Sin escudos ventrales. Base branquial oculta por nivel collar. Aproximadamente
15 pares de radiolos conectados más o menos dos tercios de la longitud de la membrana
palmada, con conexión filiforme en el extremo intermedio de la longitud. Primer paquete
de setas pequeña, insertada en un surco al mismo nivel que todas las notosetas siguientes.
Setas espatuladas sin puntas. Anterior abdominal uncinos con rostro sobre cómo cerca de
las bases, pocos dientes gruesos en dos columnas; coronas algo agrandadas en segmentos
posteriores.
Distribución.- Alaska, Estados Unidos y Pacifico Mexicano.

56
Paradialychone ecaudata Moore, 1903
(Lámina 2 I)
Chone ecaudata Moore, 1903: Hartman, 1942: 101-140.
Jasmineira ecaudata Moore, 1923 75: 179-259.
Material examinado: 11 ejemplares, Banco Chocolatero (6); El Dátil (4); La Concha (1).
Diagnosis: Escudos ventrales presentes- De seis a ocho radiolos conectados más o menos a
la mitad de la longitud de la membrana palmada. Collar bajo, casi al mismo nivel que cubre
la base branquial, con la superposición de los extremos dorsales. Primer paquete de
notosetas grandes. Labios notopodiales conspicuos en tórax. Setas espatuladas con grandes
y puntiagudas puntas. Uncinos anteriores abdominales con rostro.
Distribución.- Canadá, Estados Unidos, México, Perú y las Islas Galápagos.
Parasabella pallida Moore, 1923
Demonax pallidus Moore, 1923: Tovar-Hernández,:60: 13–19.
Material examinado: cinco ejemplares: Banco La Concha (5).
Diagnosis: Los tubos son gruesos, exterior cubierto con granos de arena fina. El cuerpo es
de color blanco o amarillo pálido, la corona tentacular marrón rojizo, muy moteada para
parecer aterciopelada. El collar se compone de un par de lóbulos bajos, dorsales oblicuos, y
los lóbulos laterales de terminación ventral en un par de solapas triangulares. Notosetas
torácicas son más largas y más estrechas en la parte superior del haz; neurosetas consisten
en una línea anterior de peines y una línea posterior de los ganchos aviculares. Parapodios
abdominales son provistos de ganchos avicular en los notopodios y setas limbadas en
neuropodios.
Distribución.- Océano Pacifico; Bahía de Monterrey California

57
Serpulidae Rafinesque, 1815
Crucigera websteri Benedict, 1887
(Lámina 2 J)
Crucigera websteri Benedict, 1887: 85:101.
Material examinado: un ejemplar: Banco Chocolatero (1)
Diagnosis: Branquias compuesta por 18 filamentos. El opérculo tiene forma de campana,
con un pedúnculo fuerte, debajo del pedúnculo surge cuatro procesos digitales en forma de
cruz, situados justo debajo de la copa. Los radios, en sus superficies interiores, están
armados con dientes cónicos, en forma de corona. Setas del collar son largas y robustas.
Uncinos torácicos similares a los abdominales, pero mucho más grandes. Los tubos, en sus
superficies superiores, tienen dos o tres crestas delgadas muy irregulares.
Distribución.- Atlántico Centro Oriental y el Pacífico. Se conoce su localidad tipo para el
Golfo de México.

58
Análisis de relación longitud peso
Se colectaron un total 99 almejas en cuatro bancos distintos:29especímenes en Banco
Chocolatero; 30 en banco El Zacatoso; 20 en banco El Dátil y 20 banco La Concha. Se
realizaron las ecuaciones para la relación longitud peso de manera independiente para los
organismos de cada zona de colecta. Los resultados son los siguientes: En el banco
Chocolatero (Figura 15) se obtiene una ecuación de 0.597 L1.1218con un coeficiente de
alometría de 1.1218 y un coeficiente de determinación r2= 0.4251 (r= 0.6519).
40 60 80 100 120 140 1600
50
100
150
200
250
f(x) = 0.596993207179672 x^1.12177923084866R² = 0.425074408750254
Longitud (mm)
Peso
(gr)
Figura 15. Relación longitud peso Banco Chocolatero.
Mientras en banco El Zacatoso (Figura 16) se obtiene una ecuación de la relación longitud-
peso igual a 0.0002 L2.8147 con un coeficiente de determinación (r2) = 0.8247 y un
coeficiente de correlación de 0.9077.

59
70 80 90 100 110 120 130 1400
20406080
100120140160180
f(x) = 0.00017920349279605 x^2.81473784187463R² = 0.824668474566857
Longitud (mm)
Peso
(gr)
Figura 16. Relación longitud peso banco El Zacatoso.
Para el banco El Dátil (Figura 17) se obtuvo una relación longitud-peso de 0.0003 L2.6769,
con un coeficiente de determinación de (R2) = 0.699 y un coeficiente de correlación de
0.8360.
90 100 110 120 130 140 150 1600
50
100
150
200
250
f(x) = 0.000321019219889142 x^2.67688550494191R² = 0.699028626848662
Longitud (mm)
Peso
(gr)
Figura 17. Relación longitud peso banco El Dátil.

60
En el Banco La Concha (Figura 18), la ecuación de longitud-peso fue de 0.0075 L2.0294 con
un coeficiente de determinación de 0.753 y uno de correlación de 0.8677.
70 80 90 100 110 120 130 140 1500
20
40
60
80
100
120
140
160
180
f(x) = 0.00750876907599204 x^2.02943427823079R² = 0.753043108280109
Longitud (mm)
Peso
(gr)
Figura 18 . Relación longitud peso banco La Concha.

61
Índice gonadosomático y hepatosomático.
El índice gonadosomático se basa en el peso de la gónada con respecto al peso total del
organismo guarda una relación directa que explica el estadio de desarrollo gonádico, ya
que, alcanza un valor máximo inmediatamente antes del desove, de tal manera que por
ejemplo, al evaluar una población durante un ciclo anual, el valor de máximo se alcanza
antes de la reproducción. Mientras que se conoce como índice hepatosomático a la relación
que existe entre el peso del hígado y el peso del ejemplar. Esta relación, es específica para
hembras ya que el hígado- glándula digestiva en este caso segrega vitelogenina exógena
que van a ser captadas por el óvulo en desarrollo. Por lo tanto, es directamente proporcional
al ciclo reproductivo y decae justo antes del desove, siendo así un indicador de la puesta.
En el caso de la especie N. subnodosus se observa un solo pico gonadosomático en el banco
Chocolatero (Figura 19).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 150
2
4
6
8
10
12
14
Ind. Gona-dosomático
Ind. Hep-atosomático
Figura 19. Índice gonadosomático y hepatosomático Chocolatero
El banco El Zacatoso mostro Tres picos gonadosomáticos en banco (Figura 20).

62
1 2 3 4 5 6 7 8 9 1011 12 1314 1516 17 180
2
4
6
8
10
12
14
16
18
Ind. GonadosomáticoInd. Hepatosomático
Figura 20. Índice gonadosomático y hepatosomático El Zacatoso
Mientras que en el banco El Dátil se observaron dos picos gonadosomático marcados
(Figura 21).
1 3 5 7 9 11 13 15 17 190
5
10
15
20
25
30
35
Ind GonadosomáticoInd Hepatosomático
Figura 21. Índice gonadosomático y hepatosomático El Dátil
Se observa actividad hepatosomática marcada en la Concha con dos picos hepatosomáticos
y uno gran gonadosomáticos (Figura 22).

63
1 2 3 4 5 6 7 8 9 10 11 12 13 14 150
2
4
6
8
10
12
Indice GonadosomaticoIndice Hepatosomatico
Figura 22. Índice gonadosomático y hepatosomático La Concha
Factor de condición múltiple (KM).
Los siguientes gráficos muestran el comportamiento de la especie N. subnodosus y sus
cambios respecto al tiempo, explicando sus relaciones morfométricas. Se realizaron las
ecuaciones para el factor de condición múltiple de manera independiente para los
organismos de cada zona de colecta. Los resultados son los siguientes: El Banco
Chocolatero arrojó el factor de condición de condición múltiple más bajo KM= 0.6673 y
un error típico = 0.1195, El Zacatoso con un valor de Km= 0.6473 y un error típico de
0.863, El Dátil presentó un KM= 0.7865 y un error típico de 0.0711 y La Concha un KM=
0.8974 y error típico de 0.0656 (Cuadro 2).
Cuadro 2. Factor de condición múltiple y error típico bancos almejeros
Bancos Almejeros KM Error típicoBanco Chocolatero 0.6673 0.1195El Zacatoso 0.6473 0.0863El Dátil 0.7865 0.0711La Concha 0.8974 0.0656

64
En el Banco Chocolatero (Longitud vs KM) (Figura 23) se obtiene una ecuación de
4.6463X + 138.2 con un coeficiente de determinación der2= 0.153 y un coeficiente de
correlación de = 0.3911. Mientras que la ecuación de relación Peso vs KM (Figura 24) de
4.377X + 124.4 con un coeficiente de determinación de r2= 0.696 y un coeficiente de
correlación de = 0.8342
40 60 80 100 120 140 1600.00
200.00
400.00
600.00
800.00
1000.00
1200.00
1400.00
f(x) = 4.64628560520026 x + 138.203885087192R² = 0.153808767684656
Longitud (mm)
KM
Figura 23. Longitud vs KM Banco Chocolatero
0 50 100 150 200 2500.0000
200.0000
400.0000
600.0000
800.0000
1000.0000
1200.0000
1400.0000
f(x) = 4.37723413105992 x + 124.411551572577R² = 0.696052395420122
Peso (gr)
KM
Figura 24. Peso vs KM Banco Chocolatero

65
En el banco El Zacatoso la ecuación de Longitud vs KM (Figura 25) arrojó el siguiente
resultado de 0.203X - 12.23 con un coeficiente de determinación der2= 0.745 y un
coeficiente de correlación de = 0.8631. Por su parte la ecuación de Peso vs KM (Figura
26) fue de 1.1138X – 2.004 con un coeficiente de determinación de r2= 0.240 y un
coeficiente de correlación de = 0.4898.
70 80 90 100 110 120 130 1400.0000
2.0000
4.0000
6.0000
8.0000
10.0000
12.0000
14.0000
16.0000
f(x) = 0.203309806659414 x − 12.2314117225734R² = 0.745932186113929
Longitud (mm)
KM
Figura 25. Longitud vs KM El Zacatoso
3.45 3.5 3.55 3.6 3.65 3.7 3.75 3.80.0000
0.5000
1.0000
1.5000
2.0000
2.5000
f(x) = 1.11375285563088 x − 2.00416171791037R² = 0.240881122872515
Peso (gr)
KM
Figura 26. Peso vs KM El Zacatoso

66
Para el banco el Dátil la ecuación de Longitud vs KM (Figura 27) mostró un resultado de
0.3585X - 22.94 con un coeficiente de determinación der2= 0.545 y un coeficiente de
correlación de = 0.7382. Mientras que la ecuación de Peso vs KM (Figura 28) de
0.1494X+ 1.39 con un coeficiente de determinación de r2= 0.994 y un coeficiente de
correlación de = 0.9969.
90 100 110 120 130 140 150 1600
5
10
15
20
25
30
35
40
f(x) = 0.358525610671469 x − 22.9459457556801R² = 0.545701436425949
Longitud (mm)
KM
Figura 27. Longitud vs KM El Dátil

67
0 50 100 150 200 2500
5
10
15
20
25
30
35
40
f(x) = 0.149445259366074 x + 1.38997510154134R² = 0.994132509782332
Peso (gr)
KM
Figura 28. Peso vs KM El Dátil
En el lugar de colecta de La Concha la ecuación de Longitud vs KM (Figura 29) arrojó el
siguiente resultado de 0.0726X - 1.921 con un coeficiente de determinación der2= 0.537 y
un coeficiente de correlación de = 0.7328. Por otro lado la ecuación de Peso vs KM
(Figura 30) fue de 0.0441X+ 1.237 con un coeficiente de determinación de r 2= 0.970 y un
coeficiente de correlación de = 0.9848.
70 80 90 100 110 120 130 140 1500
1
2
3
4
5
6
7
8
9
10
f(x) = 0.0725531368114529 x − 1.92167612905794R² = 0.537191661677963
Longitud (mm)
KM
Figura 29. Longitud vs KM La Concha

68
20 40 60 80 100 120 140 160 1800
1
2
3
4
5
6
7
8
9
10
f(x) = 0.0440854816816833 x + 1.23701610262171R² = 0.97022459048934
Peso (gr)
KM
Figura 30. Peso vs KM La Concha
Índices ecológicos
Las cuatro zonas de recolecta tuvieron valores bajos de diversidad, siendo La Concha la
estación con el número más alto de especies con un H= a 2.805 y El Zacatal con un H=
1.894 representando a la estación rica en especies. El Banco Chocolatero se observó como
la estación con mayor equidad entre especies con un valor de 0.9171, mientras en las demás
estaciones no se apreció un rango de diferencia alto entre la diversidad y dominancia
(Cuadro 3).
Cuadro 3. Valores de diversidad de la poliqueto fauna epibionte para las cuatro estaciones.
Estación Shannon-Weiner Índice de Equidad Margalef SimpsonBanco Chocolatero 2.748 0.9171 4.882 0.9221El Zacatal 1.894 0.7898 2.484 0.7915El Dátil 2.308 0.7984 3.925 0.8501La Concha 2.805 0.8022 6.162 0.9027

69
Análisis de componentes principales
El análisis de componentes principales permite reducir la dimensionalidad de datos cuando
las masas de datos son muy similares entre sí, destacando los más importantes y los menos
relevantes. En el caso de la Laguna de Ojo de Liebre el mayor número de especies, familias
y géneros se recolectaron en el banco de La Concha (Figura 31), mientras las demás
estaciones se presentaron valores bajos de familias, géneros y especies. El ACP 1 tuvo un
valor de 61.524 donde destacan géneros, especies y el número total de organismos (0.5021,
0.5053 y 0.455) y el ACP 2 registró un valor de 25.027 donde destacan talla promedio y
número de almejas (0.5359 y 0.2491).
Figura 31. Análisis de componentes principales para las cuatro estaciones Banco
Chocolatero, El Zacatoso, El Dátil y La Concha

70
Diversidad Alfa
La diversidad alfa es considerada como la riqueza de especies dentro de una comunidad
homogénea. La zona con mayor diversidad alfa en la laguna Ojo de Liebre corresponde a
La Concha, seguida de El Banco Chocolatero y el Dátil con una diversidad similar y El
Zacatal representando a la zona de menor diversidad (Figura 32).
Figura32. Grafica de diversidad alfa para las cuatro zonas muestreadas.

71
Análisis Clúster
Se realizo un análisis clúster con el fin de agrupar las zonas muestreadas según su similitud
en este caso, se puede apreciar dos conglomerados con una similitud de 0.73, destacando de
manera aislada el banco La Concha, representando la zona con el mayor número de
especies (Figura 33), por otro lado, el grupo dos contiene a las tres estaciones restantes, de
las cuales El Dátil tiene una similitud de 0.88 acercándose a las El Zacatoso y El Banco
Chocolatero mientras estas últimas comparten similitud de 0.91.
Figura 33 Análisis Clúster Laguna Ojo de Liebre.

72
Análisis SHE.
Con respecto al análisis SHE S: Riqueza, H: Diversidad y E: uniformidad. El Presente
estudio arroja valores de riqueza y diversidad similares en las estaciones de El Banco
Chocolatero, El Zacatal y El Dátil, siendo La Concha la cual mostro los valores de riqueza
y diversidad más altos, así mismo se muestran valores de uniformidad bajos en todas las
estaciones, debido a un número de especies dominantes (Figura 34).
Figura 34 .Análisis SHE de los cuatro bancos almejeros (Banco Chocolatero, El Zacatoso,
El Dátil y la Concha).

73
DISCUSIÓN
Se identificaron un total de 376 organismos correspondientes a 46 especies dentro de las
cuales se puede destacar a cuatro especies más ampliamente distribuidas: Eunice chicasi,
Polycirrus californicus, y Platynereis bicanaliculata quienes constituyeron el 36% de la
abundancia total.
En las cuatro áreas de estudio, la estación más representativa fue La Concha con el mayor
número de especies (33), mientras que la estacion de el Zacatal es considerada la estación
menos diversa en especies con un número de 12.
En este estudio la especie más abundante fue Eunice chicasi (Eunicidae) conformando el
16% del total de los organismo, seguida de Polycirrus californicus (Terebellidae) con 11%
y Platynereis bicanaliculata (Nereididae) con 9%, cada una de las especies con presencia
todas las estaciones. Los demás grupos solo representaron cifras de 8% o menos de
abundancia.
Eunice chicasi, se conoce para el Pacífico sur como epibionte del molusco Spondylus
calcifer, reiterando en este estudio que su hábitat predominante es sobre la concha de
moluscos. E. chicasi se distribuye principalmente en el Pacifico sur con su localidad tipo
en El Salvador según reporto Ruiz-Cancino (2010), siendo este el registro más reciente de
E. chicasi para el norte del Pacífico.
Platynereis bicanaliculata es una especie común en el Pacifico que suele habitar fondos
rocosos y sustratos duros (De León González, 1999). Sin embargo no existen estudios
específicos sobre el papel que desempeña como epibionte de otros organismos, pues es
común que representantes de la familia Nereididae aparezcan con altas prevalencias y
abundancias en la un gran número de los trabajos, lo anterior es concordante con el hecho
de que han sido descritos como habitantes por excelencia de sustratos duros y calcáreos
(Ruppert y Barnes, 2000). Polycirrus californicus habita sobre sustrato arenoso grueso
(Rodríguez Valencia 2004), no obstante no existe ningún reporte que indique su hábitat
como epibionte potencial de algún molusco bivalvo, convirtiéndose este en el primer
registro como epibionte.

74
En la infracomunidad de poliquetos asociados N. subnodosus se pueden destacar las
especies de polynoides, registradas como epibiontes de equinodermos y otros grupos de
moluscos identificándose como organismos con gran capacidad de epibiosis, actividad
registrada con anterioridad por Pettibone (1993) y Rozbaczylo y Cañete (1993).
El número de especies registrado en estudios similares es inferior al obtenido en éste.
Liñero-Arana (1999) registró 11 especies asociadas a Perna viridis; Díaz y Liñero-Arana
(2003, b) para P. imbricata e I. alatus encontraron a 26 y 38 especies respectivamente;
Keough (1984) identificó tres especies de espirórbidos en Pinna bicolor Gmelin, 1791. De
León-González et al. (1993), registraron 31 especies de poliquetos epibiontes de Spondylus
princeps unicolor Sowerby, 1847, en el Pacífico americano, Liñero y Díaz (2006) encontró
43 especies epibiontes de Spondylus americanus en Venenzuela.En el presente estudio se
registran tres especies más, de las cuales las más comunes son Syllis gracilis, Lumbrineris
inflata y Piromis sp. Los poliquetos son un grupo de gran abundancia y capacidad de
colonización, familias como Terebellidae, Sabellidae y Serpulidae frecuentemente
colonizan las superficies de diferentes grupos bentónicos como moluscos y crustáceos
siendo sus caparazones estructuras de fácil colonización Hernández, Muñoz, y Rozbaczylo.,
(2001), es el caso de cultivos de Crassostrea gigas en Sinaloa donde se registraron
nereididos, sabelidos y polinoides como posibles comensales (Estrada 2011). Cabe destacar
algunas familias errantes como Nereididae, Eunicidae, Flabelligeridae y Syllidae a menudo
se encuentra en sustratos fangosos que es el hábitat natural de N. subnodosus (Liñero y
Díaz 2006).
En lo que respecta a los análisis estadísticos de relación longitud peso de N. subnodosus en
todas las áreas de muestreo se observo una correlación de tipo positiva con una factor de
determinación (r); (Banco Chocolatero; 0.6519 El Zacatoso; 0.9077. El Dátil; 0.8360, La
Concha; 0.8677, donde la los coeficientes de correlación son medio- altos, fenómeno ya
observado por Gonzales (2010) para la especie. Se obtuvo un índice de alometría negativo
con valores de L inferiores a tres. El Banco Chocolatero se encontró una correlación regular
(r= 0.6519), área donde se registraron las almejas con más daño anatómico: concha
quebradiza, algunas muertas y otras vacías.

75
La relación entre los índices gonadosomáticos y hepatosomáticos presentó variación,
evidenciando actividades fisiológicas sin ningún patrón fijo de desove. De acuerdo a los
datos recaudados se puede inferir que los organismos de las tres primeras estaciones; Banco
Chocolatero, banco El Zacatoso y banco El Dátil, estaban en por madurar y/o por desovar,
mientras que los organismos de banco La Concha expresaron niveles hepatosomáticos
altos, lo que indicaría una ligera proximidad al proceso de desove, como consecuencia los
individuos guardaban reservas vitelinas para la siguiente temporada reproductiva,
comportamiento observado por Sastry (1979), Barber y Blake (1991), Román et al. (2000a)
quienes comprobaron que en una misma especie de pectínido e incluso en una misma
población existen diferencias en el ciclo reproductivo y se ha visto depende de la latitud y
de diferencias ambientales particulares de cada zona.
El análisis de factor de condición múltiple exhibe correlación de tipo positiva en todas las
zonas. El Banco Chocolatero obtuvo un KM de 0.66730817 y al Zacatoso con un KM de
0.64730158 (Tabla 3). Estaciones con un factor de crecimiento medio-bajo, con poca
relación entre el peso y la longitud; El Banco Chocolatero presentó un coeficiente de
correlación de 0.399 entre la longitud y el KM y el Zacatoso mostró un coeficiente de
correlación de 0.4898 entre el peso y KM . La Concha fue la estación con mayor KM
0.8974, siendo los organismos del área los de mejor tamaño y peso expresado en KM
elevado, cabe destacar que La Concha fue la estación en el mayor número de individuos y
de especies registradas y El Zacatoso el área con menor diversidad y abundancia por lo que
puede existir una relación entre la cantidad de individuos epibiontes y el crecimiento, ya
que un factor de estrés en común puede alterar el hábitat completo de una comunidad; el
bajo aprovechamiento de nutrientes ocurre por la influencia y cambios en factores abióticos
así como la baja disponibilidad de alimento (Barber y Blake 1991) y al igual que el
hospedero, comensales, simbiontes y parásitos pueden verse afectados de igual manera o
bien beneficiar al parásito si se trata de un patógeno.
Todas las zonas obtuvieron valores de diversidad bajos, se puede resaltar a La Concha
como la zona con el índice H´ más elevado de 2.805, no obstante las cuatro zonas arrojaron
un DSi alto con poca diversidad y elevada dominancia, resultado del dominio de Eunice
chicasi, Polycirrus californicus y Platynereis bicanaliculata con el número más alto de

76
individuos, comportamiento observado en los análisis de componentes principales donde
agrupó al número mayor de familias, géneros y especies en La Concha, misma zona con
los picos más altos en los gráficos de diversidad alfa. El análisis clúster separa las áreas en
dos grupos, uno es conformado por La Concha dominando en abundancia y diversidad y el
segundo grupo obedece a las estaciones restantes quienes por su poca diversidad formaron
otro conglomerado. SHE resalta lo antes mencionado siendo La Concha donde los valores
de riqueza y diversidad fueron elevados.
Así mismo se muestran valores de uniformidad bajos en todas las estaciones, debido a la
dominancia de un grupo reducido de especies. De forma general se aprecia que cuando los
valores de S (riqueza) aumenta la uniformidad (E) se ve disminuida y viceversa, este
comportamiento ya fue observado antes por Hayek y Buzas (1997).

77
CONCLUSIÓN
Se identificaron 376 organismos con 46 especies como epibiontes de N. subnodosus. El
área con el mayor número de especies fue La Concha con 33 especies y el Zacatoso como
el número menor con 12 especies. Dentro de la comunidad de poliquetos que habitan en la
almeja mano de león se destacaron tres especies por ser las de mayor abundancia: Eunice
chicasi, Polycirrus y Platynereis bicanaliculata además dentro del estudio se obtuvieron
especies de Nereidos, Eunicidos y Silidos como potenciales a ser nuevos registros.
La relación longitud peso de los organismos no fue óptima para los individuos de la Laguna
Ojo de Liebre, únicamente La Concha obtuvo valores óptimos de correlación, mientras las
demás zonas presentaron valores medio-bajos
No se observó un patrón marcado de maduración en los individuos de N. Subnodosus con
una variación hepatosomatica-gonadosomatica normal, de acuerdo a la biología de la
especie
No se obtuvieron pruebas de una relación entre los poliquetos que habitan sobre la valva
dorsal de N. subnodosus, siendo el comportamiento de las variaciones entre las talla y peso
ya observado en otros estudios con valores de correlación mayores de 60%.
Se encontró poca relación entre la diversidad y similitud de especies en las diferencias áreas
de colecta, siendo La Concha el área con mayor número de especies e individuos, además
hubo poca similitud entre la diversidad de las cuatro zonas.

78
LITERATURA CITADA
Andrews, E. A. 1891. Report upon the Annelida Polychaeta of Beaufort, North Carolina.
Proceedings of the United States National Museum, 14(852): 277-302
Alonso L. 2007. Epibiontes Asociados a la Tortuga Verde Juvenil (Chelonia mydas) en el Área
de Alimentación y Desarrollo de Cerro Verde, Uruguay. Tesis de Licenciatura en
Ciencias Biológicas. Facultad de Ciencias Exactas y Naturales. Universidad de Buenos
Aires, Argentina.
Allison, E. H. 1993. The dynamics of exploited populations of scallops (Pecten maximus L.)
and queens (Chlamys opercularisL.) in the north Irish Sea. PhD thesis, University of
Liverpool, UK. 254 pp.
Aradist-Castellanos, A y J. Llinas. 1991. Aves migratorias: Patos y Gansos. En La reserva de la
biosfera el Vizcaíno en la Península de Baja California. (Eds. ORTEGA, A. Y
ARRIAGA L). Centro de Investigaciones Biológicas de Baja California Sur A.C. México.
231-246.
Augener, H. 1928. Die Polychaeten von Spitzbergen. Fauna Arctica, Jena, 5: 647-834. Webster,
Henry E. 1879. The Annelida Chaetopoda of the Virginian coast. Transactions Albany
Institute New York, 9: 202-272.
Audouin, J.V. and Milne Edwards, H. 1833. Classification des Annélides et description de celles
qui habitent les côtes de la France. Annales des sciences naturelles, Paris, sér. 1, 29:
195-269.
Aviles F., N. Rozbaczylo, M. Herve and M. Godoy. 2007. First report of Polychaetes from
genus Oriopsis (Polychaeta: Sabellidae) associated with the Japanese abalone, Haliotis
discus hannai and other native mollusk in Chile. Journal of Shellfish Research, Vol. 26,
No. 3, 863–867, 2007.
Barber, B. J. y N. J. Blake. 1991. Reproductive physiology. Pp. 377-409.En: Shumway, S. E.
(ed.), Scallops: Biology, Ecology and Aquaculture. Amsterdam: Elsevier.

79
Baird, William 1863. Descriptions of several new species of worms belonging to the Annelida
Errantia and Sedentaria or Tubicola of Milne Edwards. Proceedings of the Zoological
Society of London, 1863: 106-110.
Barnich, R.; Fiege, D. (2004). Revision of the genus Lepidastheniella Monro, 1924
(Polychaeta: Polynoidae: Lepidastheniinae) with notes on the subfamily
Lepidastheniinae and the description of a new species. Journal of Natural History. 38:
863-876.
Benninger, P. G. y M. Le Pennec. 1991. Fundamental anatomy of scallops. Pp. 133-224.En:
Shumway, S. E. (ed.). Scallops: Biology Ecology an Aquaculture. Elsevier Science, New
york.
Bellan, G. (2001). Polychaeta, in: Costello, M.J. et al. (Ed.) (2001). European register of
marine species: a check-list of the marine species in Europe and a bibliography of
guides to their identification. Collection Patrimoines Naturels, 50: pp. 214-231.
Brand, A. R., Paul, J. D., Hoogesteger, J. N. (1980). Spat of the scallops Chlamys opercularis
(L.) and Pecten maximus (L.) on artificial collectors. J. mar. biol. Ass. U. K. 60: 379-
390
Broom, M. J. y J. Mason. 1978. Growth and spawning in the pectinid Chlamys opercularis in
relation to temperature and phytoplankton concentration. Mar. Biol. 47:277-285.
Bush, Katharine J. 1904. On Spirorbis from Japan. In: Moore, J.P. Report on Sabellidae and
Serpulidae. Proceedings of the Academy of Natural Sciences, Philadelphia 56:
175.page(s): 216.
Canales, J. 1998. Estudio comparativo del ciclo reproductivo de dos poblaciones de
Aequipecten opercularis (L.) (Bivalvia: Pectinidae)Tesis de Doctorado, Universidad de
Valencia. 222 pp.
Chantler, P. D. 1991. The structure and function of scallop adductor muscles. Pp 225-304.En:
Shumway, S. E. (ed.), Scallops: Biology, Ecology and Aquaculture. Elsevier,Amsterdam.

80
Christieansen, H. E., E. Cabrera y S. R. Brodsky. 1974. Ecología de las poblaciones de vieiras
Chlamys tehuelcha (d’Orb 1846) en el golfo de San Matías (Río Negro, Argentina).
Estudio histológico del ciclo reproductivo. Inst. Biol. Marina Mar del Plata. Contrib. No
225, 17 pp.
Collado, G., C. Betancourt, K. Valderrama, L. Cocas, R. Ramírez, M. González, L.Guzmán y D.
Brown. 2000. Ciclo reproductivo de los ostiones Chlamys vítrea (King y Broderip,
1831) y Chlamys patagonica (King y Broderip, 1831) (Mollusca: Bivalvia) en el
extremo sur de Chile. Pp. 99. En: Memorias del XX Congreso de Ciencias del Mar.
Concepción, Chile.
Contreras, F. ,1985. Las lagunas costeras mexicanas. Centro de Ecodesarrollo-Secretaria de
pesca, Mexico 263 pp.
Day, J.H. 1973. New Polychaeta from Beaufort, with a key to All Species Recorded from North
Carolina. NOAA Technical Rep NMFS Circ 375: 53 pp.
Dauvin, J.-C.; Dewarumez, J.-M.; Gentil, F. (2003).Liste actualisée des espèces d’Annélides
Polychètes présentes en Manche [An up to date list of polychaetous annelids from the
English Channel]. Cah. Biol. Mar. 44(1): 67-95.
de León González, Jesús Ángel and Solis-Weiss, Vivianne 2000. A review of the polychaete
family Nereididae from Western Mexico. Bulletin of Marine Science, 67(1): 549-570.
de León González, J.A., A. Leija-Tristán & S. Salazar-Vallejo. 1993. Epifauna del ostión
espinoso Spondylus princeps unicolor (Mollusca: Bivalvia) de Puerto Escondido, Golfo
de California, México. Rev. Biol. Trop. 41: 877-881.
de León-González, J.A.; Giovanni R, Carlos; and Yaneth R, Mayra 2004. Sublittoral Eunicidae
and Onuphidae (Polychaeta) from soft bottom off El Salvador, eastern Pacific. Journal
of the Marine Biological Association of the United Kingdom, 84(1): 93-101.

81
de León González, J. A. 1999. Nereididae (Annelida: Polychaeta) de México. Universidad
Autónoma de Nuevo León. Facultad de Ciencias Biológicas. Informe final SNIB-
CONABIO proyecto No. H011. México D. F.
de León-González J.A., Bastida-Zavala J.R., Carrera-Parra L.F., García-Garza M.E., Peña-Rivera
A., Salazar-Vallejo S.I., Solís-Weiss V. (Eds). 2009. Poliquetos (Annelida: Polychaeta)
de México y América Tropical. Tomo I. Universidad Autónoma de Nuevo León.
Monterrey, Nuevo León, México. 737 p.
Díaz, O. 1999. Poliquetos asociados a sustratos artificiales sumergidos en la costa nororiental
de Venezuela. Tesis MSc., Universidad de Oriente, Cumaná, Venezuela. 166 p.
Díaz-Díaz, O.; Liñero-Arana, I. Poliquetos asociados a Isognomon alatus (Gmelin, 1791)
(Bivalvia: Isognomonidae) en la costa nororiental de Venezuela. Iberus 21(2): 31-35.
2003b.
Díaz- Díaz y Liñero Arana, Poliquetos epibiontes de Pinctata imbricata Roding. 1798 (Bivalvia:
Pteriidae) en el Golfo de Cariago Venezuela. Universidad Autónoma de Mexico. Sucre,
Venezuela. 2003
Díaz, O.D., I. Liñero-Arana, S. Villafranca and T. Allen 2009 Epizoic polychaetes (Annelida:
Polychaeta) on Crassostrea rhizhophorae (Guilding, 1828) from La Restinga Lagoon,
Margarita Island, Venezuela [Poliquetos (Annelida: Polychaeta) epizoicos
de Crassostrea rhizhophorae(Guilding, 1828) de La Laguna Ecotrópicos 22(1):13-22.
Dore, I. 1991. Shellfish: a guide to oysters, mussels, scallops, clams and similar products for
the commercials used. Van Nostrand Reinhold New York, 239 pp.
Eibye-Jacobsen, D. 1992.Phyllodocids annelida polychaeta of Belize with the description of
three new species. Proceedings of the Biological Society of Washington, 105(3): 589-613.
Essenberg, Christine 1917. New species of Amphinomidae from the Pacific coast.University of
California Publications in Zoology, 18(4): 71-74.

82
Estrada C. Aidé. 2011 Relación entre parámetros físico-químicos y biológicos en el cultivo de
Crassostrea gigas, informe final de residencia profesional. Instituto tecnológico de Los
Mochis, Sinaloa. Mexico.
FAO. 2005. The state of World fisheries and aquaculture. Editorial Group, FAO Inf. Div.
ISSN1020-5489. FAO, Rome, Italy
Fauvel, P. 1911. Troisième note préliminaire sur les polychètes provenant des campagnes de
l'Hirondelle et de la Princesse-Alice, ou déposées dans la Musée Océanographique de
Monaco. Bulletin de l'Institute océanographique, 194: 1-41.
Feifarek, B. P. 1987. Spines and epibionts as antipredator defenses in the thorny oyster
Spondylus americanus Hermann. Journal of Experimental Marine Biology and Ecology
105: 39-56.
Félix-Pico, E., M. Villalejo-Fuerte, A. Tripp-Quezada, & O. Holguin-Quiñones. 1999. Growth
and survival of Lyropecten subnodosus (Sowerby, 1835) in suspended culture at the
national marinepark of Bahia de Loreto, B.C.S., Mexico. In: Strand, Ø (Ed.) Book of
abstractas 12th Internacional Pectinid Workshop, Bergen, Norway.pp. 39-40.
Góngora G. 2011. Diversidad en la fauna de Sílinos (Polychaeta: Syllidae: Syllinae) de las
costas del Noroeste de México. Universidad Autónoma de Nuevo León. Mexico.
González J. A. 2010 Efecto combinado de temperatura y dieta en el crecimiento, composición
bioquímica y respiración de juveniles de almeja mano de león Nodipecten subnodosus
(Sowerby1835). Universidad del mar campus Oaxaca
González E. F. 2003. Temperaturas letales y temperatura óptima para crecimiento en una
población de almeja mano de león (Nodipecten subnodosus Sowerby, 1835), CIBNOR,
La Paz, B. C. S.,
Grube, A.E. 1840. Actinien, Echinodermen und Würmer des Adriatischen- und Mittelmeers,
nach eigenen Sammlungen beschrieben.1-92. J.H. Bon. Königsberg.
Hartman, O. 1938.Annotated list of the types of polychaetous annelids in the Museum of
Comparative Zoology. Bulletin of the Museum of Comparative Zoology, 85(1): 3-31.

83
Hartman, O. 1942: The identity of some marine annelid worms in the United States National
Museum. Proceedings of the United States National Museum, 92(3142): 101-140.
Hayek C L, M A Buzas (1997) Surveying Natural Populations. Columbia University Press. New
York, N.Y., USA. 563 p.
Hernández, C.E., G. Muñoz & N. Rozbaczylo (2001). Poliquetos asociados con
Austromegabalanus psittacus (Molina, 1782) (Crustacea: Cirripedia) en Península
Gualpén, Chile central: Biodiversidad y efecto del tamaño del sustrato biológico,
Scientific Electronic Library Online - Chile. Retrieved at November 18, 2012, from the
website temoa : Open Educational Resources (OER) Portal
at http://www.temoa.info/node/585741
Hickman, C. P., Roberts, L. S. y Larson, A.2002. Principios integrales de Zoología. Editorial
Interamericana.(Undécima Edición Española).
Imajima, Minoru and Hartman, O. 1964.The polychaetous annelids of Japan. Occasional Papers
of the Allan Hancock Foundation, 26: 1-452.
Imajima, Minoru 2003 Polychaetous Annelids from Sagami Bay and Sagami Sea collected by
the Emperor Showa of Japan and deposited at the Showa Memorial Institute,
National Science Museum, Tokyo (II), Orders included within the Phyllodocida,
Amphinomida, Spintherida and Enicida. National Science Museum Monographs, 23: 1-
221
Rodríguez Valencia, J.A. 2004 Respuesta de los poliquetos bentónicos a la variabilidad
ambiental y condiciones el niño en bahía Petacalco (Guerrero, México) Ciencias
Marinas, diciembre, año/vol. 30, número 004Universidad Autónoma de Baja California
Ensenada, México pp. 515–526
Jaramillo, R., J. Winter, J. Valencia y A. Rivera. 1993. Gametogenic cycle of the Chiloé
Scallop(Chlamys amandi).J. Shellfish Res. 12:59-64.

84
Johnson, Herbert P. 1897. A preliminary account of the marine annelids of the Pacific coast,
with descriptions of new species. Proceedings of the California Academy of Sciences, Ser.
3, 1(5): 153-199.
Keen, A. y E. Coan. 1974. Marine mollusca genera of western north America, an illustrated
key. Stanford university press. 208 pp.
Keough, M. 1984. Dynamics of the epifauna of the bivalve Pinna bicolor: interaction among
recruitment, predation and competition. Ecology 63: 677-688.
Laura Schejter & Claudia S. Bremec (2007)."Epibionts on Flexopecten felipponei (Dall, 1922),
an uncommon scallop from Argentina". American Malacological Bulletin 22 (1): 75–82.
Liñero-Arana, I. 1999. Poliquetos (Annelida: Polychaeta) asociados al mejillón
verde Perna viridis en la Península de Araya, Venezuela. Bol. Inst. Oceanogr.
Venezuela. Univ. Oriente, Cumaná.
Liñero y Díaz, Poliquetos (Annelida: Polychaeta) epibiontes de Spondylus americanus
(Bivalvia: Spondylidae) en el Parque Nacional Mochima, Venezuela, Instituto
Oceanográfico de Venezuela, Venezuela 2006.
Luna-González, A., C. Cáceres-Martínez, C. Zúñiga-Pacheco, S. López-López y B. P. Ceballos-
Vázquez. 2000. Reproductive cycle of Argopecten ventricosus (Sowerby II, 1842)
(Bivalvia: Pectinidae) in the Rada of Puerto de Pichilingue, B.C.S., Mexico and its
relation to temperature, salinity, and quality of food. J. Shellfish Res. 19:107-112.
Marinone, L.S.G. y L. Lizarraga A. 1982.Transporte litoral sobre las barras de Guerrero
Negro y Ojo de Liebre. Ciencias Marinas Val. 8 (1).
Martin, D. & Britayev, T. A., 1998 Symbiotic Polychaetes: review of known species. Oceanogr.
Mar. Biol. Ann. Rev. 36, 217-340.
Massó-Rojas, J. A., E. Morales-Bojorques, J. Talavera-Mayer, M. Fajardo-León y R. Hernández-
Valenzuela. 2000. La Pesquería de Almeja Mano de León, Baja California, pp. 349-
366. En: Instituto Nacional de Pesca, SAGARPA, México (ed.), Sustentabilidad y Pesca
Responsable en México, Evaluación y manejo.

85
Marenzeller, Emil von 1879.Südjapanische Anneliden.I. Amphinomea, Aphroditea,
Lycoridea, Phyllodocea, Hesionea, Syllidea, Eunicea, Glycerea, Sternaspidea,
Chaetopterea, Cirratulea, Amphictenea. Denkschriften der Mathematisch-naturwissen
schaftlichen classe der Kaiserliche Akademie der Wissenschaften, Wien, 41(2): 109-154.
McIntosh, W.C. 1885. Report on the Annelida Polychaeta collected by H.M.S. Challenger
during the years 1873-1876. Report on the Scientific Results of the Voyage of H.M.S.
Challenger during the years 1872-76, Ser. Zoology, 12: 1-554.
Montagu G. (1804). Description of several marine animals found on the south coast of
Devonshire. Transactions of the Linnean Society, London, 7, pp. 61-85;pls. 6-7.
Montagu, George 1808. II. Description of several Marine Animals [Polychaeta p.108-111, pl.6-
8] found on the South Coast of Devonshire. Transactions of the Linnean Society of
London, 9: 108-111.
Montagu, George 1815. Descriptions of several new or rare animals principally marine, found
on the south coast of Devonshire. Transactions of the Linnean Society of London, 11: 1-
26.
Montagu, G. 1818. Descriptions of five British species of the genus Terebella. Transactions of
the Linnean Society of London, 12: 340-344.
Moore, 1923 The polychaetous annelids dredged by the U.S.S. 'Albatross' off the coast of
southern California in 1904. IV. Spionidae to Sabellariidae. Proceedings of the
Academy of Natural Sciences, Philadelphia, 75: 179-259.
Moore, J.P. 1903. Polychaeta from the coastal slope of Japan and from Kamchatka and Bering
Sea. Proceedings of the Academy of Natural Sciences, Philadelphia, 55: 401-490.

86
Newell, R.I.E., T.J. Hilbish, R.K. Koehn and C.J. Newell. – 1982.Temporal variation in the
reproductive cycles of Mytilus edulisL. (Bivalvia, Mytilidae) from localities on the east
coast of the United States. Biol. Bull., 162: 299-310
Olivares, A., M. Bretos, R. Chihuailaf & A. Zenis.1998. Biometría, hábitat y epibiontes en
Fissurella latimarginata (Mollusca: Prosobranchia) en el norte de Chile. Estud.
Oceanol.
Peña, J. 2001. Taxonomía, morfología, distribución y hábitat de los pectínidos
iberoamericanos. Pp. 1-25. En: Maeda-Martínez, A. N. (ed.), Los moluscos pectínidos de
iberómerica: Ciencia y Acuicultura. Editorial Limusa S.A. de C.V. México.
Pettibone, M.H. 1963. Marine polychaete worms of the New England region, 1.Aphroditidae to
Trochochaetidae. Bull. US Natl. Mus., 227: 1-356.
Pettibone M.H.1993 Scaled polychaetes (Polynoidae) associated with ophiuroids and other
invertebrates and review of species referred to Malmgrenia McIntosh and replaced by
Malmgreniella Hartman, with descriptions of new taxa /
Raven, C. P. 1966. Morphogenesis: the analysis of molluscan development. 2nd edition,
Pergamon Press, Oxford.
Rioja, Enrique 1941. Estudios Anelidologicos. III. Datos para el conocimiento de la fauna de
poliquetos de las costas del pacifico de Mexico. Anales del Instituto de Biología, Mexico,
12(2): 669-746.
Rodríguez-Valencia, J.A. 2004.Response of benthic polychaetes to environmental variability
and El Niño conditions at Petacalco Bay (Mexico). Ciencias Marinas. 30(4): 515-526.
Román, G., M. J. Campos y J. Cano. 2000a. Viabilidad del cultivo de la viera (Pecten maximus)
en mar abierto en la costa de Málaga. Informe final.
Rowley, S. 2008. A critical evaluation of the symbiotic association between tropical tube-
dwelling Polychaetes and their Hermatypic coral hosts, with a focus on
Spirobranchus giganteus(Pallas, 1766). The Plymouth Student Scientist.

87
Rozbaczylo N & JI Cañete. 1993. A new species of scaleworm, Harmothoe commensalis
(Polychaeta: Polynoidae),from mantle cavities of two Chilean clams. Proceedings of the
Biological Society of Washington 106 (4): 666-672.
Ruiz-Cancino, G., L.F. Carrera-Parra & R. Bastida-Zavala. 2011(2010). Eunícidos (Polychaeta:
Eunicidae) del Pacífico sur de México. Ciencia y Mar 14(40:( 2760.)
Ruppert–Barnes, 2000 "Zoología de los Invertebrados", Sexta Ed. Interamericana, México, 1994.
Salazar-Vallejo S.I. 2012. Revisión of trophoniella hartman, 1959 (Polychaeta,
Flabelligeridae). Zoosystema. 34(3): 453-519.
San Martin, G. 2003. Fauna Ibérica. Vol. 21. Annelida Polychaeta II. Syllidae. Museo Nacional
den Ciencias Naturales, Madrid.
Sastry, A. N. 1979. Pelecypoda (excluding Ostreidae).En: Giese, A.C.y J. S. Pearse, (eds.),
Reproduction of Marine Invertebrates. New York: Academic Press, pp.113-292.
Sarur-Zanata, J. C., R. Millan Nuñez., C.A. Sigala Gutiérrez y C.A. Small Mattox-Sheahen. 1984.
Variation and similarity in three zones with different type of substrate in Laguna Ojo
de Liebre, B. C. S., México. Cienc. Mar.
Schejter L y C Bremec (2007) Benthic richness in the Argentine continental shelf: The role of
Zygochlamys patagonica (Mollusca, Bivalvia,Pectinidae) as settlement substrate.
Journal of the Marine Biological Association UK 87: 917-925.
Taller Técnico Regional de la FAO, Estado actual del cultivo y manejo de moluscos bivalvos y
su proyección futura Factores que afectan su sustentabilidad en América Latina, 20-
24 de agosto de 2007, Puerto Montt, Chile.
Taylor, A.e. & T,J. Venn. 1979. Seasonal variation in weight and biochemical composition of
the tissues of . the queen scallop, Chlamys opercularis from the Clyde Sea area. J.
Mar. Biol. Ass. U. K. 59:605-621.

88
Tovar-Hernández, María Ana; Harris, Leslie, H. 2010: Parasabella Bush, 1905, replacement
name for the polychaete genus Demonax Kinberg, 1867 (Annelida, Polychaeta,
Sabellidae).ZooKeys 60: 13–19.
Treadwell, Aaron L. 1906. Polychaetous annelids of the Hawaiian Islands collected by the
steamer Albatross in 1902.Bulletin of the United States Fish Commission, 23(3): 1145-
1181.
Treadwell, Aaron L. 1922. Polychaetous annelids collected at Friday Harbor, State of
Washington, in February and March, 1920.Carnegie Institute of Washington
Publication, 312: 171-181.
Treadwell, Aaron L. 1929. New species of polychaetous annelids in the collections of the
American Museum of Natural History from Porto Rico, Florida, Lower California, and
British Somaliland. American Museum Novitates, 392: 1-13.
Verrill, A.E. 1873. Report upon the invertebrate animals of Vineyard Sound and the adjacent
waters, with an account of the physical characters of the region. Report of the United
States Commission for Fisheries, 1871-72: 295-778
Wada, S. K. 1968. Mollusca. I. Amphineura, Gastropoda, Scaphopoda, Pelecypoda. Pp.485-
525. En: Kume, M. y K. Dan (eds.),.Invertebrate Embriology, Nolit Publishing House,
Belgrade.
Wahl, M. 1989. Marine epibiosis. I. Fouling and antifouling: Some basic aspects. Marine
Ecology Progress Series 58: 175-189.
Zaixso, H. E. 2004. . Bancos de cholga Aulacomya atra atra (Molina) (Bivalvia: Mytilidae) del
golfo San José (Chubut, Argentina): Diversidad y relaciones con facies afines Revista de
Biología Marina y Oceanografía

89
OPCION DE TITULACIÓN 1 TESIS
PROFESIONAL
OSMAR ROBERTO ARAUJO LEYVA
LIC. BIOLOGÍA
No. De control: 07440449
REALIZADO EN:

90
GUERRERO NEGRO, BAJA CALIFORNIA SUR.
MEXICO.
TEMA:
Poliquetos (Annelida: Polychaeta) epibiontes de la almeja
mano de león Nodipecten subnodosus Sowerby, 1835 en
Laguna Ojo de Liebre, Guerrero Negro, Baja California
Sur, México.