Tema X. CATABOLISMO · 2018-09-08 · respiración celular aerobia, mientras que si esa sustancia...
19
Tema X. CATABOLISMO • Concepto de catabolismo • Reacciones de oxidación-reducción • Tipos de catabolismo • Respiración celular • Respiración celular aerobia - Oxidación de glúcidos: la glucosa - Oxidación de lípidos - Oxidación de proteínas • La fermentación • Fermentación láctica • Fermentación alcohólica CONCEPTO DE CATABOLISMO La reacción esquemática de todo fenómeno catabólico es la siguiente: AB A + B + Energía en la cual AB es una sustancia rica en energía. Dicho compuesto se descompone en los compuestos A y B, pobres en energía y la energía desprendida se almacena normalemente en forma de ATP. REACCIONES DE OXIDACIÓN - REDUCCIÓN Los electrones poseen diferentes cantidades de energía potencial dependiendo de su distancia al núcleo del átomo y de la atracción ejercida por el núcleo sobre ellos. Un ingreso de energía lanzará a un electrón a un nivel energético más alto, pero si no se añade energía, el electrón permanecerá en el nivel energético más bajo que encuentre disponible. Las reacciones químicas son, esencialmente, transformaciones de energía en virtud de las cuales la energía almacenada en los enlaces químicos se transfiere a otros enlaces químicos recién formados. En estas transferencias, los electrones se desplazan de un nivel de energía a otro. En muchas reacciones, los electrones pasan de un átomo o molécula a otro. Estas reacciones, que son de gran importancia en los sistemas vivos, se conocen como de oxidación-reducción -o redox-. La pérdida de un electrón se denomina oxidación y el átomo o molécula que pierde el electrón se dice que se ha oxidado. La razón de que la pérdida de electrones se conozca como oxidación es que el oxígeno, que atrae muy fuertemente a los electrones, es el que por lo general actúa como aceptor de electrones. La reducción es, por el contrario, la ganancia de un electrón. La oxidación y la reducción siempre ocurren simultáneamente, porque el electrón que pierde el átomo oxidado es aceptado por otro átomo que se reduce en el proceso. En las reacciones de oxidación-reducción se produce un movimiento de electrones de un átomo a otro. Un átomo o molécula que pierde electrones se oxida; el que los gana se reduce. Una reacción de oxidación-reducción. 1
Transcript of Tema X. CATABOLISMO · 2018-09-08 · respiración celular aerobia, mientras que si esa sustancia...

Tema X. CATABOLISMO
• Concepto de catabolismo • Reacciones de oxidación-reducción
• Tipos de catabolismo • Respiración celular
• Respiración celular aerobia - Oxidación de glúcidos: la glucosa - Oxidación de lípidos - Oxidación de proteínas
• La fermentación • Fermentación láctica • Fermentación alcohólica
CONCEPTO DE CATABOLISMO
La reacción esquemática de todo fenómeno catabólico es la siguiente: AB A + B + Energía
en la cual AB es una sustancia rica en energía. Dicho compuesto se descompone en los
compuestos A y B, pobres en energía y la energía desprendida se almacena normalemente en forma de ATP.
REACCIONES DE OXIDACIÓN - REDUCCIÓN
Los electrones poseen diferentes cantidades de energía potencial dependiendo de su distancia al núcleo del átomo y de la atracción ejercida por el núcleo sobre ellos. Un ingreso de energía lanzará a un electrón a un nivel energético más alto, pero si no se añade energía, el electrón permanecerá en el nivel energético más bajo que encuentre disponible.
Las reacciones químicas son, esencialmente, transformaciones de energía en virtud de las cuales la energía almacenada en los enlaces químicos se transfiere a otros enlaces químicos recién formados. En estas transferencias, los electrones se desplazan de un nivel de energía a otro. En muchas reacciones, los electrones pasan de un átomo o molécula a otro. Estas reacciones, que son de gran importancia en los sistemas vivos, se conocen como de oxidación-reducción -o redox-. La pérdida de un electrón se denomina oxidación y el átomo o molécula que pierde el electrón se dice que se ha oxidado. La razón de que la pérdida de electrones se conozca como oxidación es que el oxígeno, que atrae muy fuertemente a los electrones, es el que por lo general actúa como aceptor de electrones. La reducción es, por el contrario, la ganancia de un electrón. La oxidación y la reducción siempre ocurren simultáneamente, porque el electrón que pierde el átomo oxidado es aceptado por otro átomo que se reduce en el proceso. En las reacciones de oxidación-reducción se produce un movimiento de electrones de un átomo a otro. Un átomo o molécula que pierde electrones se oxida; el que los gana se reduce.
Una reacción de oxidación-reducción.
1

En los sistemas vivos las reacciones que capturan energía (fotosíntesis) y las reacciones que liberan energía (glucólisis y respiración), son reacciones de oxidación-reducción. La oxidación completa de un mol de glucosa libera 686 kilocalorías de energía libre; de modo inverso, la reducción del dióxido de carbono para formar un mol de glucosa almacena 686 kilocalorías de energía libre en los enlaces químicos de la glucosa. Si esta energía fuera liberada de una sola vez, la mayor parte se disiparía como calor. Esto no solamente no sería útil para la célula, sino que la alta temperatura resultante sería letal. Sin embargo, la vida ha evolucionado adquiriendo mecanismos que regulan la marcha de estas reacciones químicas y una multitud de otras, de modo tal que la energía se almacena en enlaces químicos particulares de los que puede ser liberada en pequeñas cantidades cuando la célula lo necesite.
La transferencia de electrones se realiza en las reacciones orgánicas en átomos de H (1H es 1e-+1H+). Una sustancia se oxida cuando pierde electrones y se deshidrogena. Una sustancia se reduce al ganar electrones y al aceptar protones (H+), hidrogenándose. Los hidrógenos (en los que van los electrones) desprendidos por un compuesto, antes de llegar a su aceptor final van a ser transportados por compuestos como el NAD+, NADP+ (nicotina adenina dinucleótido fosfato), FAD+ (flavín adenin dinucleótido). Se trata de nucleótidos no nucleicos que van a ser además, coenzimas de las enzimas deshidrogenasas.
Los electrones, antes de llegar a la molécula aceptora final de electrones, van a ser captados por unas moléculas transportadoras de electrones, que son los citocromos. Se va perdiendo energía cuando los electrones pasan de un transportador a otro. Esa energía se utilizará para sintetizar ATP a partir de ADP + Pi. TIPOS DE CATABOLISMO
Existen dos tipos de catabolismo:
• La RESPIRACIÓN CELULAR (aerobia y anaerobia). El dador de electrones suele ser un compuesto orgánico, pero el aceptor final de electrones es una molécula inorgánica. Si esa molécula inorgánica es el O2, estaremos hablando de la respiración celular aerobia, mientras que si esa sustancia inorgánica aceptora final de electrones es otro compuesto como los iones nitrato (NO3) o sulfato (SO4
2- ), estaremos hablando de una respiración celular anaerobia.
• La FERMENTACIÓN. Tanto el dador de electrones como el aceptor final de electrones son compuestos orgánicos. Otra diferencia entre la respiración y la fermentación, es que en esta última, la fosforilación de ATP a partir de ADP solo se hace a nivel de sustrato, mientras que en la respiración también se lleva a cabo en la cadena respiratoria.
RESPIRACIÓN CELULAR RESPIRACIÓN CELULAR AEROBIA
Al alimentarnos, las proteínas, lípidos y polisacáridos han de ser degradados a moléculas
menores que utilizará la célula. La digestión mediante enzimas de los alimentos pasa las proteínas a aminoácidos, los lípidos a ac. grasos y glicerol y los polisacáridos a monosacáridos.
Para obtener energía estas moléculas son degradadas a compuestos que pueden ingresar en las vías centrales de la respiración celular aerobia -glucólisis y ciclo de Krebs- en diferentes pasos.
2

Glucólisis
Oxidación de glúcidos: la glucosa Estudiaremos con detalle la oxidación de la glucosa porque es una fuente principal de energía en
la mayoría de las células. Cuando la glucosa se degrada en una serie de pequeños pasos por medio de enzimas, una proporción significativa de la energía contenida en la molécula vuelve a empaquetarse en los enlaces fosfato de las moléculas de ATP.
Es un proceso en cuatro fases:
• Degradación de la glucosa es la glucólisis1 que se efectúa en el citoplasma de la célula. En él la glucosa se transformará en ac pirúvico. Es común a la respiración celular y a las fermentaciones.
• El ácido pirúvico penetra en las mitocondrias transformándose en Acetil-Coenzima A
(Acetil-CoA). • El Acetil-CoA se degrada completamente hasta CO2 y H2O en la matriz mitocondrial
mediante el Ciclo de Krebs en el que se va a generar una gran cantidad de poder reductor (NADH , NADPH) y algo de ATP.
• Estos compuestos cederán sus electrones a la cadena respiratoria o cadena de transporte electrónico, transfiriendo sus electrones a sus transportadores hasta el aceptor final de electrones que es el O2. Estos moléculas transportadoras se encargarán de crear un gradiente osmótico de H+ que servirá para la síntesis de ATP (fosforilación oxidativa).
1 La glucólisis en realidad es un proceso ANAEROBIO y lo llevan a cabo tanto los organismos que usan el oxígeno como receptor final de electrones como los que no lo hacen. Lo incluimos aquí para dar una continuidad al proceso de oxidación de la glucosa.
3
Todos estos procesos están íntimamente relacionados. En condiciones anaeróbicas que veremos más adelante, el proceso de fermentación transforma
al ácido pirúvico producido por la glucólisis o en etanol, o en ácido láctico.
La oxidación consiste en la pérdida de un electrón y la reducción es la ganancia de un electrón. Dado que en las reacciones de oxido-reducción espontáneas, los electrones van de niveles de energía mayores a niveles de energía menores, cuando una molécula se oxida, habitualmente libera energía cuando. En la oxidación de la glucosa, los enlaces carbono-carbono (C-C), carbono-hidrógeno (C-H) y oxígeno-oxígeno (O-O) se cambian por enlaces carbono-oxígeno (C-O) e hidrógeno-oxígeno (H-O), a medida que los átomos de oxígeno atraen y acaparan electrones. La ecuación resumida de este proceso es:
Glucosa + Oxígeno => Dióxido de Carbono + Agua + Energía
o bien,
C6H12O6 + 6O2=> 6CO2 + 6H2O *G = -686 kcal/mol
Los sistemas vivos son expertos en conversiones energéticas. Su organización les permite atrapar
esta energía libre, de modo que no se disipe al azar, sino que pueda usarse para hacer el trabajo de la célula. Aproximadamente el 40% de la energía libre desprendida por la oxidación de la glucosa se conserva en la conversión de ADP a ATP.
4

En presencia de oxígeno, el ácido pirvico entra en el ciclo de Krebs donde se sintetiza más ATP y se transfieren más electrones y protones a las coenzimas. Estas coenzimas aceptoras de electrones transfieren su carga a la cadena transportadora de electrones a lo largo de la cual, paso a paso, los electrones caen a niveles inferiores de energía. A medida que esto ocurre, se fabrica mucho más ATP. Al final de la cadena transportadora, los electrones se reúnen con los protones y se combinan con el oxígeno, formándose agua. En ausencia de oxígeno, el ácido pirúvico puede convertirse en ácido láctico o etanol. Este proceso, llamado fermentación, no produce ATP, pero regenera las moléculas de coenzima aceptoras de electrones, necesarias para que la glucólisis continúe.
Glucólisis La glucólisis es un proceso que sucede en el citoplasma celular, en el cual una molécula de
glucosa de 6 carbonos se escinde en dos moléculas de 3 carbonos de ácido pirúvico. Este proceso da como resultado un rendimiento neto de dos moléculas de ATP (a partir de ADP y fosfato inorgánico) y dos moléculas de NADH (a partir de NAD+).
La glucólisis comienza con una molécula de glucosa. En este proceso tiene dos fases, en la primera se invierte energía (2ATPs) por transferencia de un grupo fosfato desde una molécula de ATP, una por cada paso, a la molécula de azúcar. La molécula de 6 carbonos en la segunda fase se escinde y, de allí en adelante, la secuencia produce energía (4 ATPs). En cierto momento se reduce una molécula de NAD+ a NADH y H+ almacenandose parte de la energía producida por la oxidación de la glucosa. En los pasos finales las moléculas de ADP toman energía del sistema, fosforilándose, como yahemos dicho 4 ATPs.
GLUCÓLISIS
1ª ETAPA 2ª ETAPA5

Resumiendo: para iniciar la secuencia glucolítica es necesaria la energía de los enlaces fosfato de
dos moléculas de ATP. Posteriormente se producen dos moléculas de NADH a partir de dos de NAD+ y cuatro de ATP a partir de cuatro de ADP:
Glucosa + 2ATP + 4ADP + 2Pi + 2NAD+ => 2 Ácido pirúvico + 2ADP + 4ATP + 2NADH + 2H+ + 2H2O
De esta forma, una molécula de glucosa se convierte en dos moléculas de ácido pirúvico. La
ganancia neta, la energía recuperada, es dos moléculas de ATP y dos moléculas de NADH por molécula de glucosa. Las dos moléculas de ácido pirúvico contienen todavía una gran parte de la energía que se encontraba almacenada en la molécula de glucosa original. La serie de reacciones que constituyen la glucólisis se lleva a cabo virtualmente en todas las células vivas, desde las células procarióticas hasta las células eucarióticas de nuestros propios cuerpos.
Respiración aerobia: continuación de la oxidación de la glucosa La respiración aerobia se después de la glucólisis contiúa con etapas más:
1. Obtención de acetil Co-A a partir de Ac. Pirúvico. 2. Oxidación de acetil Co-A en la matriz mitocondrial en el Ciclo de Krebs o ciclo del
ácido cítrico. 3. Transporte de electrones: la cadena respiratoria.Fosforilación oxidativa.
1. Obtención de acetil Co-A a partir de Ac. Pirúvico.
En el curso de la respiración, las moléculas de tres carbonos de ácido pirúvico producido por la glucólisis son degradadas a grupos acetilo de dos carbonos, que luego entran ayudados por el CoA al ciclo de Krebs.
Antes de entrar al ciclo de Krebs, el ácido pirúvico sufre varias transformaciones hasta convertirse en acetil CoA:
• Pérdida de C y O en forma de CO2 (descarboxilación). • Unión del grupo acetilo a la CoA.
6

• Captura del NAD+ de un electrón del pirúvico y un hidrógeno de la CoA transformándose en NADH + H+
7
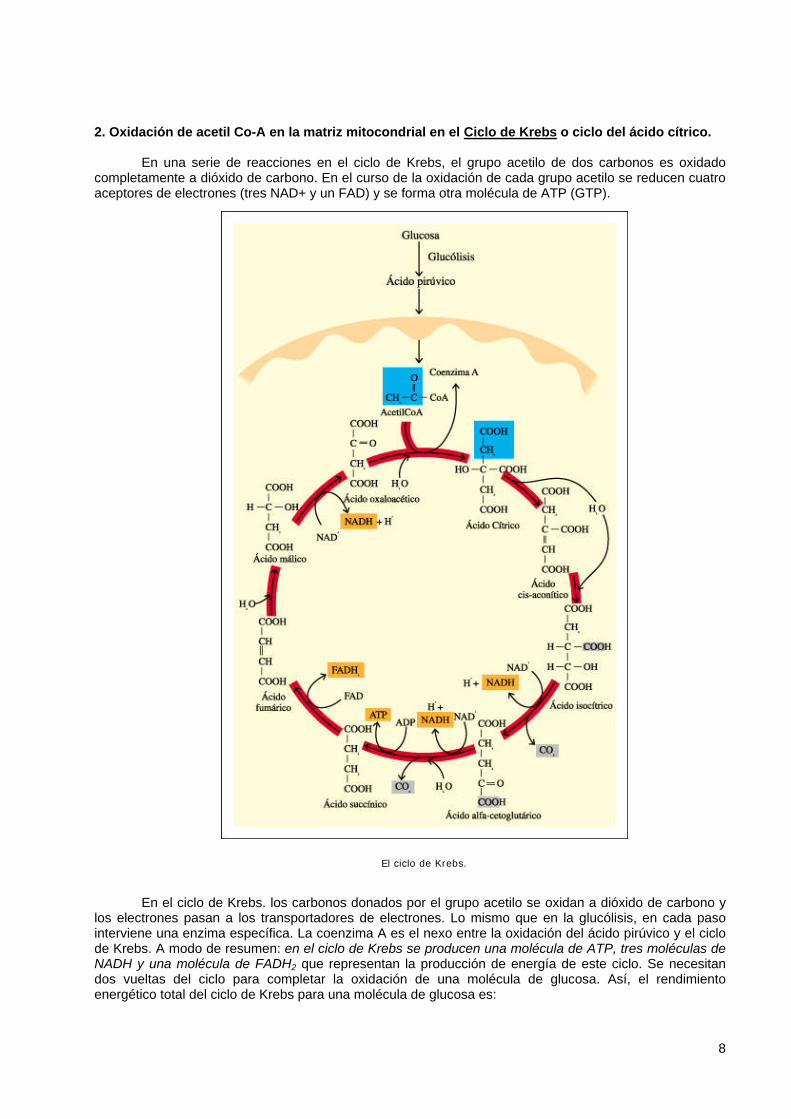
2. Oxidación de acetil Co-A en la matriz mitocondrial en el Ciclo de Krebs o ciclo del ácido cítrico.
En una serie de reacciones en el ciclo de Krebs, el grupo acetilo de dos carbonos es oxidado completamente a dióxido de carbono. En el curso de la oxidación de cada grupo acetilo se reducen cuatro aceptores de electrones (tres NAD+ y un FAD) y se forma otra molécula de ATP (GTP).
El ciclo de Krebs. En el ciclo de Krebs. los carbonos donados por el grupo acetilo se oxidan a dióxido de carbono y
los electrones pasan a los transportadores de electrones. Lo mismo que en la glucólisis, en cada paso interviene una enzima específica. La coenzima A es el nexo entre la oxidación del ácido pirúvico y el ciclo de Krebs. A modo de resumen: en el ciclo de Krebs se producen una molécula de ATP, tres moléculas de NADH y una molécula de FADH2 que representan la producción de energía de este ciclo. Se necesitan dos vueltas del ciclo para completar la oxidación de una molécula de glucosa. Así, el rendimiento energético total del ciclo de Krebs para una molécula de glucosa es:
8
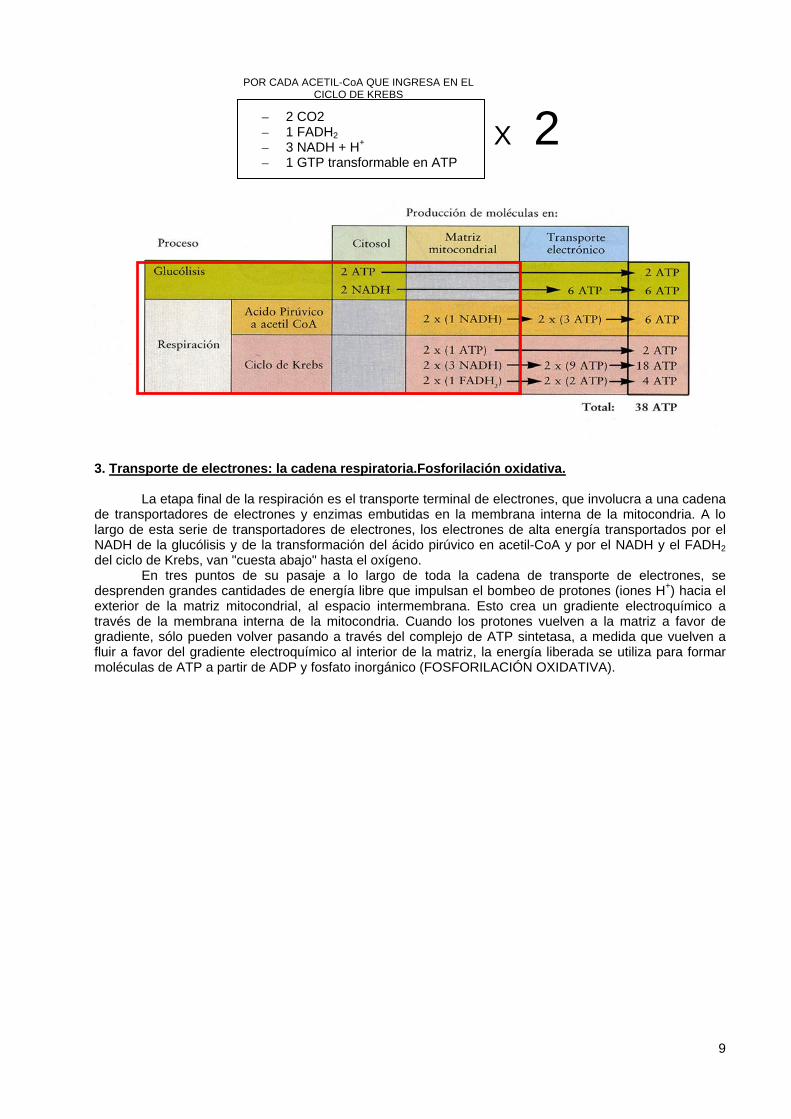
– 2 CO2 – 1 FADH2 – 3 NADH + H+ – 1 GTP transformable en ATP
POR CADA ACETIL-CoA QUE INGRESA EN EL CICLO DE KREBS
X 2
3. Transporte de electrones: la cadena respiratoria.Fosforilación oxidativa.
La etapa final de la respiración es el transporte terminal de electrones, que involucra a una cadena de transportadores de electrones y enzimas embutidas en la membrana interna de la mitocondria. A lo largo de esta serie de transportadores de electrones, los electrones de alta energía transportados por el NADH de la glucólisis y de la transformación del ácido pirúvico en acetil-CoA y por el NADH y el FADH2 del ciclo de Krebs, van "cuesta abajo" hasta el oxígeno.
En tres puntos de su pasaje a lo largo de toda la cadena de transporte de electrones, se desprenden grandes cantidades de energía libre que impulsan el bombeo de protones (iones H+) hacia el exterior de la matriz mitocondrial, al espacio intermembrana. Esto crea un gradiente electroquímico a través de la membrana interna de la mitocondria. Cuando los protones vuelven a la matriz a favor de gradiente, sólo pueden volver pasando a través del complejo de ATP sintetasa, a medida que vuelven a fluir a favor del gradiente electroquímico al interior de la matriz, la energía liberada se utiliza para formar moléculas de ATP a partir de ADP y fosfato inorgánico (FOSFORILACIÓN OXIDATIVA).
9

Representación esquemática de la cadena transportadora de electrones
En esta representación de la cadena respiratoria, las moléculas que se indican: flavina mononucleótido (FMN), coenzima Q (CoQ) y los citocromos b, c, a y a3, son los principales transportadores de electrones de la cadena. Al menos otras nueve moléculas transportadoras funcionan como intermediarias además de las que se muestran aquí.
Los electrones transportados por la NADH entran en la cadena cuando son transferidos a la FMN, que entonces se reduce (azul). Casi instantáneamente, el FMN cede los electrones al CoQ. El FMN vuelve así a su forma oxidada (naranja), listo para recibir otro par de electrones, y la CoQ se reduce. CoQ entonces pasa los electrones al siguiente aceptor, y vuelve a su forma oxidada. El proceso se repite en sentido descendente. Los electrones, al pasar por la cadena respiratoria, van saltando a niveles energéticos sucesivamente inferiores.
Los electrones que son transportados por el FADH2 se encuentran en un nivel energético ligeramente inferior que los del NADH. En consecuencia, entran en la cadena de transporte más abajo, a la altura de la CoQ. Los electrones finalmente son aceptados por el oxígeno, que se combina con protones (iones hidrógeno) en solución, y forman agua.
10

La fosforilación oxidativa.
De acuerdo con la teoría quimiosmótica, los protones son bombeados hacia afuera de la matriz
mitocondrial, a medida que los electrones descienden a lo largo de la cadena de transporte electrónico, que se encuentra en la membrana mitocondrial interna. El movimiento de protones a favor del gradiente electroquímico, a medida que pasan a través del complejo de la ATP sintetasa, suministra la energía por medio de la cual se regenera el ATP a partir del ADP y el fosfato inorgánico.
Parece que los protones se bombean a través de la membrana interna mitocondrial hacia el espacio intermembrana con la energía que pierden los electrones, pero sólo en determinados puntos de la cadena respiratoria (sistemas I, III y IV)
Se bombean 2 H+ en cada uno de los tres puntos. La matriz entonces queda cargada negativamente con respecto al espacio intermembrana.
El NADH libera los electrones con un nivel energético muy alto y tienen energía suficiente para bombear H+ en los tres puntos de la cadena, un par de protones por cada uno de ellos, por tanto bombea 6 H+ en total.
El FAD en cambio cede sus electrones algo más abajo en la cadena respiratoria, a la CoQ, y sólo ceden energía suficiente para bombear protones en dos puntos de la cadena y por tanto 4 H+.
Al salir protones de la matriz, esta queda cargada negativamente con respecto al espacio intermembrana. Esto provoca un flujo de protones que intentan volver a la matriz para igualar la diferencia de potencial, pero como la membrana interna es impermeable a los H+, sólo pueden volver por una proteína de membrana, la ATP-sintetasa. Esta enzima utiliza la energía acumulada en el gradiente de H+ para sintetizar ATP (1 ATP por cada 2 protones que pasan). A este proceso se le llama, como se ha dicho: FOSFORILACIÓN OXIDATIVA.
Por cada NADH por tanto se forman 3 ATPs porque sus electrones tienen la energía suficiente para bombear 6 electrones en los tres puntos posibles de la cadena (2 H+ en cada uno).
11

Por cada FADH2 sin embargo solo se forman 2 ATPs porque sus electrones sólo tienen energía suficiente para bombear electrones en dos puntos de la cadena (2 H+ en cada uno), y por tanto bombean 4 H+ que vuelven luego por la ATP sintetasa.
La fosforilación oxidativa.
Rendimiento energético global de una molécula de glucosa
La glucólisis produce dos moléculas de ATP directamente y dos moléculas de NADH. La conversión de ácido pirúvico en acetil CoA, que ocurre dentro de la mitocondria, produce dos
moléculas de NADH por cada molécula de glucosa y rinde, de esta forma, seis moléculas de ATP. El ciclo de Krebs, que también se desarrolla dentro de la mitocondria, produce dos moléculas de
ATP, seis de NADH y dos de FADH2, o un total de 24 moléculas de ATP por cada molécula de glucosa. La producción total a partir de una molécula de glucosa es un máximo de 38 moléculas de ATP. Las moléculas de ATP, una vez formadas, son exportadas a través de la membrana de la
mitocondria por un sistema de cotransporte que al mismo tiempo ingresa una molécula de ADP por cada ATP exportado.
12

Resumen del máximo rendimiento energético a partir de la oxidación de una molécula de glucosa
Oxidación de lípidos
La célula, a parte de la glucosa, va a utilizar otras moléculas como combustibles respiratorios como lípidos y proteínas. Los lípidos tienen un papel muy importante como combustible rico en energía ya que pueden acumularse en grandes cantidades en las células en forma de triglicéridos. Éstos poseen una gran cantidad de energía almacenada (alrededor de 9 kcal/gr.) y se pueden acumular en forma casi anhidra, ocupando muy poco volumen. En contraste con ello, el glucógeno y el almidón están muy hidratados y no pueden acumularse en forma tan concentrada y además encierran menor cantidad de energía (aproximadamente 4 kcal/gr.).
Los triglicéridos se disocian en glicerina y ácidos grasos y cada componente va a tener un catabolismo propio.
Aunque la glicerina puede ser transportada al hígado donde se utiliza para sintetizar glucosa, también puede transformarse fácilmente en 3 fosfoglideraldehido incorporándose así al metabolismo general de glúcidos.
Los ácidos grasos cubren hasta el 40 % de las necesidades totales de combustible en el hombre. Forman parte de los lípidos saponificables, algunos de los cuales tienen como función la reserva de energía. La hidrólisis de estos lípidos llevada a cabo por las lipasas suministra grandes cantidades de ácidos grasos, que son distribuidos por la sangre desde el tejido adiposo al resto de los tejidos, excepto al cerebro que sólo utiliza glucosa como carburante metabólico.
Los ácidos grasos son moléculas que almacenan en sus enlaces una gran cantidad de energía
química. La oxidación de los ácidos grasos se produce en la mitocondria y su entrada en este orgánulo está precedida de la unión entre el ácido graso y una molécula de coenzima A en el citosol. Algunos tejidos, como es el caso del músculo cardiaco, obtienen la mayor parte de la energía que necesitan mediante la oxidación de ácidos grasos.
Activación de los ácidos grasos.
Previamente a su oxidación, los ácidos grasos de cadena larga son activados en el citosol mediante su unión con el coenzima A para formar moléculas de acil graso-CoA. Esta reacción es catalizada por el enzima acil-CoA sintetasa de la membrana externa de la mitocondria, y en ella se consume 2 moléculas de ATP. El paso a través de la membrana interna se hace por medio de una
13

proteína específica de transporte, pero para ello, el ácido graso ha de ser transferido desde el coenzima A a una molécula mediadora del transporte, la carnitina. Una vez que el complejo acil-carnitina ha atravesado la membrana interna, el ácido graso es transferido al coenzima A de la matriz. Los ácidos grasos de cadena corta entran libres hasta la matriz y allí son activados por acción de acil-CoA sintetasas específicas.
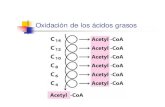
La β-oxidación o hélice de Lynen: una oxidación por etapas
La β-oxidación consiste en la degradación de los ácidos grasos para formar moléculas de acetil-CoA que pueden ser oxidadas posteriormente en el ciclo de Krebs. En las células animales este proceso se desarrolla en la matriz de la mitocondria y en los peroxisomas. Sin embargo, en las células vegetales y en las levaduras solo tiene lugar en los peroxisomas. La β-oxidación es un proceso que se realiza por etapas.
En cada una de las etapas de la β-oxidación se libera una molécula de acetil-CoA. Previamente, el átomo de carbono β del ácido graso debe ser oxidado para formar un grupo cetónico (C = O).
El resultado de cada una de las etapas es la formación de un ácido graso con dos átomos de carbono menos que el anterior y de dos coenzimas reducidos (FADH2 y NADH).
La repetición de estas reacciones rompe las moléculas de los ácidos grasos en moléculas de acetil-CoA y produce electrones de alta energía que pueden ser transferidos a la cadena respiratoria. En el ejemplo que puedes ver a continuación, una molécula de ácido palmítico (16 C) libera 8 moléculas de acetil-CoA tras 7 etapas de oxidación que producirán 7 moléculas de NADH y 7 de FADH2.
14

La β oxidación se puede representar mediante la hélice de Lynen. En ella cada vuelta representa una etapa de oxidación en la que se libera una molécula de acetil-CoA y se genera una de FADH2 y otra de NADH.
Oxidación de proteínas En los seres vivos, las proteínas tienen otras funciones diferentes a la energética. La función
primordial de los aminoácidos es la de ser los componentes de las proteínas. Aun así, también pueden ser degradados y oxidados para producir energía. Hay varias situaciones que justifican su degradación: por una parte la célula debe efectuar el recambio de sus proteínas; por otra parte, las células animales no pueden almacenar aminoácidos libres por lo que un aporte elevado de proteínas en la dieta hace que una parte importante de los aminoácidos sea degradada. Esto mismo sucede en condiciones de ayuno, en las que las células no disponen de otras moléculas para satisfacer sus necesidades energéticas. La hidrólisis de las proteínas y la consiguiente liberación de aminoácidos tiene lugar en el citoplasma y puede hacerse en los lisosomas.
Previamente a la oxidación de sus cadenas carbonadas, los aminoácidos se desprenden de sus grupos amino, por un proceso de transaminación o de desaminación oxidativa. También pueden perder el grupo carboxilo mediante un proceso de descarboxilación.
• Transaminación. Es una reacción de interconversión en la que un aminoácido cede su grupo α-amino a un cetoácido (generalmente el α-cetoglutárico, que se transforma en ácido glutámico). Los enzimas que catalizan estas reacciones son las aminotransferasas (transaminasas), que están presentes en el citosol y en la matriz mitocondrial. Los grupos amino se almacenan así en forma de ácido glutámico, que puede cederlos para la síntesis de nuevos aminoácidos o bien combinarse con un nuevo grupo NH4
+ para formar glutamina y así transportar dicho grupo por la sangre.
15
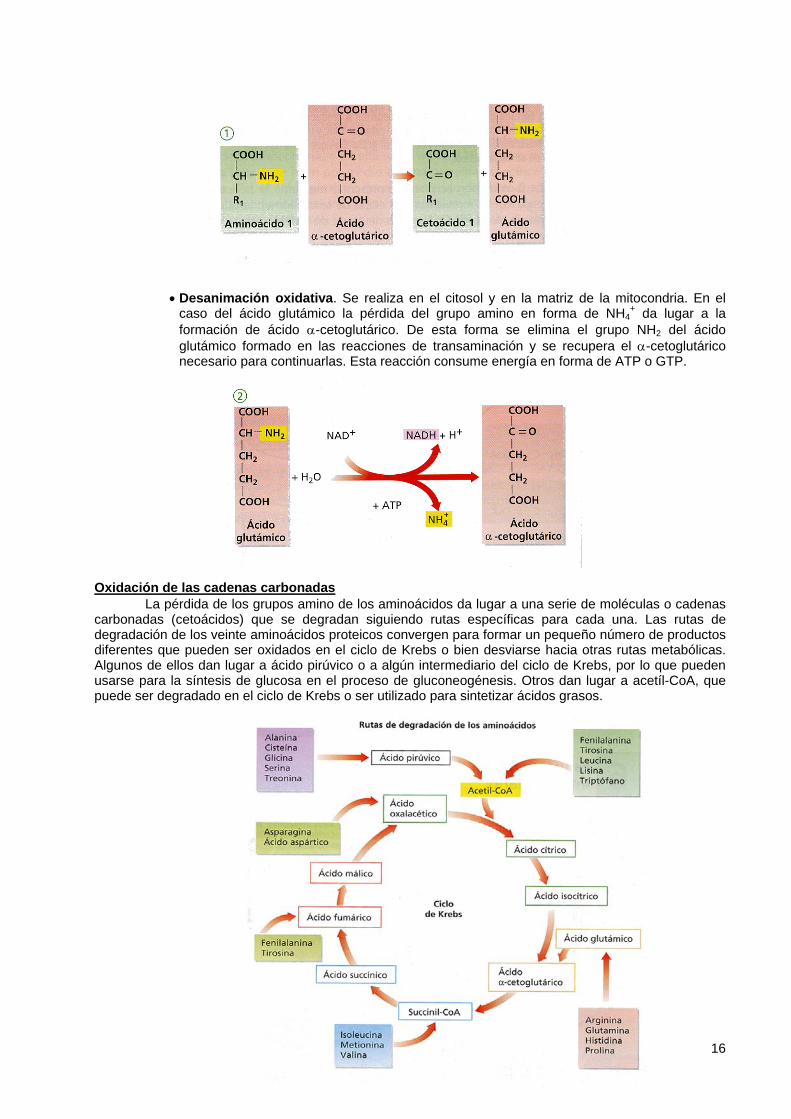
• Desanimación oxidativa. Se realiza en el citosol y en la matriz de la mitocondria. En el caso del ácido glutámico la pérdida del grupo amino en forma de NH4
+ da lugar a la formación de ácido α-cetoglutárico. De esta forma se elimina el grupo NH2 del ácido glutámico formado en las reacciones de transaminación y se recupera el α-cetoglutárico necesario para continuarlas. Esta reacción consume energía en forma de ATP o GTP.
Oxidación de las cadenas carbonadas
La pérdida de los grupos amino de los aminoácidos da lugar a una serie de moléculas o cadenas carbonadas (cetoácidos) que se degradan siguiendo rutas específicas para cada una. Las rutas de degradación de los veinte aminoácidos proteicos convergen para formar un pequeño número de productos diferentes que pueden ser oxidados en el ciclo de Krebs o bien desviarse hacia otras rutas metabólicas. Algunos de ellos dan lugar a ácido pirúvico o a algún intermediario del ciclo de Krebs, por lo que pueden usarse para la síntesis de glucosa en el proceso de gluconeogénesis. Otros dan lugar a acetíl-CoA, que puede ser degradado en el ciclo de Krebs o ser utilizado para sintetizar ácidos grasos.
16

La solución a un problema de residuos La eliminación del grupo amino de los aminoácidos da lugar a la formación de amoniaco (NH3) en
forma de ion amonio (NH4+), parte del cual puede ser recuperado en una reacción inversa a la
desaminación oxidativa para formar ácido glutámico. Sin embargo una parte importante del amoniaco formado en el catabolismo de los aminoácidos debe ser eliminado. Esta sustancia tiene una gran toxicidad y la respuesta de los organismos para su eliminación es un claro ejemplo de adaptación al medio. Hay diferentes estrategias:
• Animales amoniotélicos. Son animales acuáticos que pueden eliminar directamente los iones amonio, ya que la abundancia de agua les permite disminuir toxicidad de esta sustancia por dilución. Entre los más importantes están la mayor parte de los invertebrados, muchos peces y anfibios.
• Animales ureotélicos. La mayor parte de los vertebrados terrestres y muchos de los acuáticos excretan principalmente urea, una sustancia formada por la combinación de una molécula de CO2 y dos de amoniaco, cuya toxicidad es menor que la de éste. La urea se sintetiza en las células del hígado, en una serie de reacciones que transcurren en el citosol y en la mitocondria y que constituyen el ciclo de la urea.
• Animales uricotélicos. Algunos animales eliminan el nitrógeno procedente del catabolismo en forma de ácido úrico, una molécula con menor toxicidad, por lo que puede ser almacenada durante cierto tiempo. Es una sustancia difícilmente soluble, que tiende a cristalizar y formar suspensiones sólidas, lo que constituye una ventaja para los reptiles y las aves, cuyo consumo de agua es escaso. Estos animales son ovíparos y el embrión no puede eliminar los productos de excreción, por lo que se acumulan en el huevo. La acumulación de excrementos de ciertas aves en algunas zonas del litoral marino da lugar a un excelente abono nitrogenado que se denomina guano.
RESPIRACIÓN CELULAR ANAEROBIA La respiración anaerobia se diferencia de la aerobia en que la sustancia aceptora final de
electrones que se reduce es diferente al oxígeno. Se puede tratar de los iones nitrato (NO3) o sulfato (SO4
2- ). El dador de electrones suele ser un compuesto orgánico. En la respiración celular anaerobia también participan transportando protones y electrones los coenzimas NAD y FAD y también el sustrato se transforma finalmente en CO2, aunque la oxidación también puede ser incompleta. Este tipo de respiración es igual de rentable que la aerobia ( a partir de una glucosa, 38 ATPs) pero se da en ambientes anaerobios y la realizan procariotas como Escherichia coli, Agrobacterium... (suelen ser microorganismos aerobios facultativos). LA FERMENTACIÓN
La fermentación es un proceso catabólico en el cual tanto el dador de electrones como el aceptor final son compuestos orgánicos. Son rutas metabólicas mediante las cuales los organismos consiguen energía a partir de combustibles orgánicos y en ausencia de oxígeno molecular. En las fermentaciones, por tanto, el aceptor final de electrones no es el oxígeno, sino una molécula orgánica que varía según el tipo de fermentación.
Se trata de un proceso anaerobio en el que no interviene ni el ciclo de Krebs ni la cadena respiratoria.
El ácido pirúvico permanece en el citoplasma (no entra en la mitocondria como sucede para la respiración aerobia).
En ellas el NADH de la glucólisis cede sus electrones a una molécula orgánica que quedará más reducida, el NAD+ y que entrará otra vez en glucolísis.
Muchos microorganismo usan la fermentación como única fuente de energía (anaerobios estrictos) o sólo cuando les falta oxígeno (anaerobios facultativos). Los seres vivos superiores pueden fermentar la glucosa en ausencia de O2 pasándola a ácido láctico.
Hay dos tipos: fermentación láctica y alcohólica.
17

FERMENTACIÓN LÁCTICA
Consiste en la transformación de la glucosa en dos moléculas de ácido láctico. El aceptor final de electrones es el ácido pirúvico.
Reacción enzimática que produce ácido láctico anaeróbicamente a partir de ácido pirúvico en las células musculares.
En el curso de esta reacción, el NADH se oxida y el ácido pirúvico se reduce. Las moléculas de
NAD+ producidas en esta reacción vuelven a la glucólisis. Sin estas moléculas, la glucólisis no puede seguir adelante.
Se da en Lactobacillus y en Streptococcus usadas para hacer queso, yogur, kefir, etc. Transforma la lactosa en glucosa y luego ésta en ácido láctico. El cambio de pH hace que la caseína de la leche precipite y forme el cuajo que se separa del suero.
El músculo esquelético trabaja normálmente en aerobiosis y el ácido pirúvico se oxida en la mitocondria hasta CO2 y H2O. Si el ejercicio es fuerte no hay bastante CO2 para oxidar el pirúvico y obtener así ATP. En estas condiciones, las células convieten pirúvico en láctico y así consiguen NAD+ que usan en la glucólisis consiguiendo algo de ATP (2 ATPs por glucosa en lugar de 38 ATPs que se consiguen en la respiración aerobia).
La acumulación de ácido láctico da como resultado dolor y fatiga muscular. Por ejemplo, se produce en las células musculares de los vertebrados durante ejercicios intensos, como en el caso de una carrera. Cuando corremos rápido, aumentamos la frecuencia respiratoria, incrementando de este modo el suministro de oxígeno, pero incluso este incremento puede no ser suficiente para satisfacer los requerimientos inmediatos de las células musculares. Sin embargo, las células pueden continuar trabajando y acumular lo que se conoce como deuda de oxígeno. La glucólisis continúa, utilizando la glucosa liberada por el glucógeno almacenado en el músculo, pero el ácido pirúvico resultante no entra en la vía aeróbica de la respiración sino que se convierte en ácido láctico que, a medida que se acumula, disminuye el pH del músculo y reduce la capacidad de las fibras musculares para contraerse, produciendo la sensación de fatiga muscular. El ácido láctico se difunde en la sangre y es llevado al hígado. Posteriormente, cuando el oxígeno es más abundante (como resultado de la inspiración y espiración profunda que siguen al ejercicio intenso) y se reduce la demanda de ATP, el ácido láctico se resintetiza en ácido pirúvico y nuevamente en glucosa o glucógeno.
¿Por qué el ácido pirúvico se convierte en ácido láctico sólo para volver a convertirse en ácido pirúvico? La función de la conversión inicial es simple: usa el NADH y regenera el NAD+, sin el cual la glucólisis no podría continuar. FERMENTACIÓN ALCOHÓLICA
El ácido pirúvico puede convertirse en etanol (alcohol etílico). El producto de reacción depende del tipo de célula. Por ejemplo, las levaduras, presentes en el hollejo de las uvas, pueden crecer con o sin oxígeno. Cuando los jugos azucarados de las uvas y de otras frutas se extraen y se almacenan en condiciones anaeróbicas, las levaduras transforman el mosto en vino, convirtiendo la glucosa en etanol. Cuando el azúcar se agota, las levaduras dejan de funcionar; en este momento, la concentración de alcohol es entre 12% y 17% dependiendo de la variedad de uvas y de la estación en la cual fueron cosechadas.
En el primer paso de la glucólisis se desprende dióxido de carbono. En el segundo, se oxida el NADH y se reduce el acetaldehído. La mayor parte de la energía química de la glucosa permanece en el alcohol, que es el producto final de la secuencia. Sin embargo, regenerando NAD+, estos pasos permiten
18

que la glucólisis continúe, con su pequeño (2ATPs), pero en algunos casos vitalmente necesario, rendimiento de ATP.
Pasos por los cuales el ácido pirúvico, formado en la glucólisis, se convierte anaeróbicamente en etanol.
Cada molécula de glucosa se transforma en dos moléculas de etanol. La realizan levaduras del género Saccharomyces y según la especie producen cerveza, whisky,
sidra, vino, etc. La fabricación del pan es un proceso de fermentación de los polisacáridos de la harina. El CO2
ahueca la masa de pan y tanto él como el alcohol desaparecen en la cocción.
19


















