
TÁCTICAS REPRODUCTIVAS DE Haliotis fulgens … · Acrosoma: Estructura en forma de casquete que...
108
INSTITUTO POLITECNICO NACIONAL CENTRO INTERDISCIPLINARIO DE CIENCIAS MARINAS TÁCTICAS REPRODUCTIVAS DE Haliotis fulgens PHILIPPI, 1845 Y Haliotis corrugata W. WOOD, 1828 (GASTROPODA: ARCHAEOGASTROPODA) EN DOS ZONAS DE PESCA EN LA COSTA OCCIDENTAL DE BAJA CALIFORNIA SUR. TESIS QUE PARA OBTENER EL GRADO DE DOCTORADO EN CIENCIAS MARINAS PRESENTA MÓNICA NURENSKAYA VÉLEZ ARELLANO LA PAZ, B.C.S., JULIO DE 2016
Transcript of TÁCTICAS REPRODUCTIVAS DE Haliotis fulgens … · Acrosoma: Estructura en forma de casquete que...

INSTITUTO POLITECNICO NACIONAL CENTRO INTERDISCIPLINARIO DE CIENCIAS MARINAS
TÁCTICAS REPRODUCTIVAS DE Haliotis fulgens PHILIPPI, 1845 Y Haliotis corrugata W.
WOOD, 1828 (GASTROPODA: ARCHAEOGASTROPODA) EN DOS ZONAS DE PESCA EN LA COSTA OCCIDENTAL DE BAJA
CALIFORNIA SUR.
TESIS
QUE PARA OBTENER EL GRADO DE DOCTORADO EN CIENCIAS MARINAS
PRESENTA
MÓNICA NURENSKAYA VÉLEZ ARELLANO
LA PAZ, B.C.S., JULIO DE 2016



DEDICATORIA
Resulta difícil agradecer a todos los que merecen parte del crédito por este trabajo
sin excluir a alguno, pues se complica delinear la medida en que cada uno de mis
logros es consecuencia de lo vivido con cada persona, de cada consejo, de cada
regaño, de cada palabra de aliento. Pero no omito a nadie cuando pienso en ello.
Por eso, brindo el mérito correspondiente a todos los que me acompañaron a lo
largo de este camino, el cual no hubiera podido recorrer sin marchar al lado de mi
hermosa familia, pues me apoyó siempre y, en aras de enorgullecerlos, he llegado
hasta aquí.
A ellos dedico con especial énfasis este trabajo: Mamá, Papá, Karen y Prisca, sin
ustedes y sus llamadas continuas mi estancia en La Paz sería más nostálgica.
A Tony, que nunca perdiste cuenta de este trabajo y, aunque no lo viste
culminado, estoy segura de que en el cielo celebras con nosotros.
A mis abuelitos, Santiago y María de Jesús, tías, primas, primos… valga siempre
su recuerdo en mi mente, agradezco saber que creen en mí y que puedo contar
con ustedes en todo momento.
A los amigos entrañables, compañeros de escuela, cómplices de aventuras y
colegas que hacen más ameno el andar y el trabajo, porque sin ustedes los
momentos divertidos, las horas compartidas en el laboratorio y en el campo,
terminar este trabajo hubiera sido tortuoso.
Finalmente quiero dedicar este trabajo a mi compañero de vida con quien hoy
cumplo uno de tantos sueños, uno de los muchos que nos faltan consumar:
gracias por tu apoyo y esmero, sin duda eres mi brújula, le das dirección a mi vida.
Te amo Milton A. Spanopoulos Zarco.

AGRADECIMIENTOS
Al Centro Interdisciplinario de Ciencias Marinas del Instituto Politécnico Nacional, por todas las facilidades otorgadas para la realización de este trabajo. A la Secretaria de Investigación y Posgrado del Instituto Politécnico Nacional por el apoyo suministrado a través de los proyectos SIP-IPN, claves 20161106, 20151089, 20141323, 20130751 y 20121218 y al Programa Institucional de Formación de Investigadores a través de la Comisión de Operación y Fomento de Actividades Académicas del IPN por el apoyo económico otorgado durante mis estudios. Al Consejo Nacional de Ciencia y Tecnología (CONACyT) por el proyecto Extinción local, adaptación o compensación en organismos de vida larga ante el calentamiento global; Panopea sp. en las costas del noroeste de México. (clave 241603) y por el apoyo económico otorgado durante mis estudios. Al Centro Regional de Investigaciones Pesqueras, unidad La Paz por el apoyo otorgado a través del proyecto abulón del CRIP La Paz/INAPESCA. Un distinguido agradecimiento al Dr. Federico Andrés García Domínguez y al Dr. Daniel Lluch Cota por su apoyo y confianza en mi trabajo, y su capacidad para guiar mis ideas, lo que ha sido un aporte invaluable, no solamente en el desarrollo de esta tesis, sino también en mi formación como investigadora, además por brindarme la oportunidad de ser parte de sus proyectos y por su participación activa en la dirección de este trabajo. Al Dr. Daniel Lluch Belda, por su apoyo incondicional y por sus atinados comentarios en los primeros meses del desarrollo de este trabajo, como miembro del comité. Al comité revisor, Dr. Federico Andrés García Domínguez, Dr. Daniel Lluch Cota, Dr. Oscar Holguin Quiñones, Dr. Mauricio Ramírez Rodríguez y Dr. José Luis Gutiérrez González, por sus observaciones y aportaciones a esta tesis. A la Dra. Ma. Teresa Sicard González por recibirme y facilitarme lo necesario durante mi estancia en el Laboratorio de Ecofisiología del CIBNOR y a la M. en C. Rosa Linda Salgado García por la disposición mostrada durante mi estancia en el laboratorio, y el apoyo recibido en el procesamiento de muestras, al instruirme en la técnica pero también por su compañerismo y solidaridad. Al Tec. Ariel Arturo Cruz Villacorta, por la toma de imágenes en el microscopio electrónico de barrido, también por su asesoría en la técnica de microscopía electrónica de barrido, uso del microscopio, así como su agradable compañía, durante este proceso, gracias.

A la Dra. Fiorenza Micheli por recibirme y otorgarme todas las facilidades durante mi estancia en Hopkins Marine Station, también a su asistente Cheryl Butner, por todas las atenciones dentro y fuera del laboratorio. A los miembros del Laboratorio 2 del CINVESTAV-México principalmente a la Dra. Mineko Shibayama Salas y al Dr. Víctor Tsusumi por recibirme y permitirme participar en las actividades como otro miembro del laboratorio. A la M. en C. Karla Montserrat Gil Becerril y a la QFB Silvia Galindo Gómez por el apoyo técnico recibido durante mi estancia y muchas gracias a la M. en C. Dora Angélica Silva Olivares por el apoyo técnico en el procesamiento de las muestras para microscopía electrónica de transmisión, así como en la observación de estas. A la Dra. Rebeca Sánchez Cárdenas, quien me recibió en el Laboratorio de Ecología de Pesquerías de la FACIMAR, también quiero agradecer su cariño y amistad brindada. Al personal del INAPESCA que me apoyaron durante los muestreos Biol. Pesq. Carlos Monroy, Dr. Eduardo Quiroz, Biol. Mar. Mauro Guadarrama y Dra. Verónica Fernández, que hicieron más corto los viajes con su amable compañía y a Nancy Catillo, que fue parte primordial en el procesamiento histológico, además de su incondicional amistad. A los directores y miembros de las cooperativas “Bahía Tortugas”, “Emancipación”, “California de San Ignacio” y “Progreso”, quienes apoyaron en la toma de muestras y sin su ayuda este trabajo no se hubiera podido realizar. A todos los que colaboraron en la disección de muestras en el laboratorio, Biol. Alma Rosa Rivera Camacho, al M. en C. Edgardo Camacho, la Dra. Liliana Carvalho Saucedo, a la Dra. Uri Argumedo Hernández, a la M. en C. Cintya Nevarez, a Nancy Castillo, que sin su ayuda el estrés hubiera sido mayor. A los estudiantes de verano científico Mareli Cahua Fernández, Ma. Concepción López Ojeda y Aritzel Aramara Morfin Jiménez, quienes me apoyaron en el procesamiento de algunas muestras. A los miembros del Depto. de Servicios Escolares del CICIMAR, Lic. Humberto Ceseña Amador y C. Cesar Casas Núñez, por la atención, amabilidad, disposición, dedicación e interés mostrada día a día, a cada uno de los estudiantes e investigadores de este centro de investigación. Un muy especial agradecimiento a los miembros de la unidad de informática Lic. Manuel Pacheco Hoyo, M. en C. Roberto Aguilera Angulo y C. Susana Cárdenas Chávez, por su amable disponibilidad y eficiencia para resolver mis problemas computacionales. A mis maestros: Dr. Arturo Tripp Quezada, M. en C. Marcial Villalejo Fuerte, Dr. Enrique Nava Sánchez y Dr. Héctor Villalobos Ortiz, que dentro y fuera de las aulas me han transmitido su conocimiento y que formarán parte de mi quehacer profesional.

PREFACIO A partir de la presente tesis se derivaron los siguientes trabajos: Artículos como primer autor (enfocados a la reproducción de Haliotis spp.) Vélez-Arellano, N. F. A. García-Domínguez, D. B. Lluch-Cota, J. L. Gutiérrez-
González & R. Sánchez-Cárdenas. 2015. Histological validation of Morphochromatically-defined gonadal maturation stages of green Abalone (Haliotis fulgens) Philippi, 1845 and pink abalone (Haliotis corrugata) Wood, 1828. Int. J. Morphol. 33(3):1054-1059.
Vélez-Arellano, N, F. A. García-Domínguez, D. Lluch-Cota, J. L. Gutiérrez-
González, O. Holguin-Quiñones y M. Ramírez-Rodríguez. 2016. Biochemical changes during the reproductive cycle of Haliotis fulgens Philippi 1845 (Gastropoda: Archaeogastropoda) in the Baja California Sur West Coast. J. Shellfish Res. 35(1): 199-207.
Vélez-Arellano, N, F. A. García-Domínguez, D. Lluch-Cota, J. L. Gutiérrez-
González, L. A. Salcido-Guevara & R. Sánchez-Cárdenas. 2016. Morphophysiological indices of the Green abalone Haliotis fulgens Philippi, 1845 at mexican Ocean Pacific coast. Turk. J. Fish. Aquat. S. C. En revisión.
Vélez-Arellano N., R. L. Salgado García, F. A. García-Domínguez, D. B. Lluch-
Cota & J. L. Gutiérrez-González. Fecundidad de Haliotis corrugata W. Wood 1828 y de Haliotis fulgens Philippi 1845 en la costa occidental central de la península de Baja California, México. En preparación.
Artículos como coautor (enfocados a la alimentación de Haliotis spp.)
Vega-García, D., A. Piñón-Gimate, N. Vélez-Arellano & S. E. Lluch-Cota. 2015. Differences in diet of Green (Haliotis fulgens) and Pink (Haliotis corrugata) wild abalone along the pacific coast of the Baja California Peninsula, using stable isotope analyses. J. Shellfish Res. 34(3):879-884.
Siqueiros-Beltrones, D., U. Argumedo-Hernández, N. Vélez-Arellano & F. A. García-Domínguez. 2015. Diatom species diversity in the diet of healthy and sick specimens of adult Haliotis fulgens and Haliotis corrugata. Rev. Biol. Mar. Oceanog. 50(2): 271-281.

Ponencias: Vélez-Arellano, N., F. A. García-Domínguez, J. L. Gutiérrez-González y D. B.
Lluch-Cota. 2013. Relación del ciclo reproductivo del abulón azul y la temperatura en B.C.S., México. IV Congreso Mexicano de Ecología. Villahermosa, Tab. 18 a 22 de marzo de 2013.
Vélez-Arellano, N., F. A. García-Domínguez, D. B. Lluch-Cota y J. L. Gutiérrez-
González. 2013. Comparación del estudio del ciclo reproductivo de Haliotis fulgens (Gastropoda) utilizando una escala morfocromática y un análisis histológico. XXXVI Congreso Nacional de Histología, Cuernavaca, Mor. 16 a 18 de octubre.
Vélez-Arellano, M. N., F. A. García-Domínguez, M, Shibayama, A. Silva-Olivares,
D. B. Lluch-Cota y J. L. Gutiérrez-González. 2014. Descripción de la ovogénesis del abulón azul Haliotis fulgens. Mollusca 2014. Encuentro de las Américas. México, D. F. 22 al 27 de junio.
Siqueiros Beltrones, D., U. Argumedo-Hernández, N. Vélez-Arellano y F. A. García-Domínguez. 2014. Diversidad de especies de diatomeas en la dieta de adultos de Haliotis fulgens Philippi, 1945 y Haliotis corrugata Swainson, 1822, incluyendo especímenes con síndrome de deshidratación. VXIII Congreso Nacional de Oceanografía. La Paz, B. C. S. 4 a 6 de junio.

ÍNDICE
GLOSARIO ............................................................................................................................ i
LISTA DE FIGURAS............................................................................................................ v
LISTA DE TABLAS ........................................................................................................... viii
RESUMEN ........................................................................................................................... ix
ABSTRACT ........................................................................................................................... x
INTRODUCCIÓN ................................................................................................................. 1
ANTECEDENTES ................................................................................................................ 6
OBJETIVO GENERAL ........................................................................................................ 9
OBJETIVOS ESPECÍFICOS .......................................................................................... 9
MATERIALES Y MÉTODOS ............................................................................................ 10
ÁREA DE ESTUDIO ...................................................................................................... 10
METODOLOGÍA DE CAMPO ...................................................................................... 11
METODOLOGÍA DE LABORATORIO ........................................................................ 11
Técnica de microscopía electrónica de transmisión ............................................ 11
Técnica de microscopía electrónica de barrido .................................................... 12
Técnica histológica .................................................................................................... 12
Proporción de sexos .................................................................................................. 13
Ciclo reproductivo ...................................................................................................... 13
Talla de primera madurez ......................................................................................... 13
Fecundidad ................................................................................................................. 14
Análisis bioquímico de los tejidos............................................................................ 15
Índices morfofisiológicos de peso ........................................................................... 17
Índices morfofisiológicos de área ............................................................................ 18
Análisis de la variación del diámetro de los ovocitos (AVDO) ............................ 18
Factores ambientales ................................................................................................ 19
Análisis estadísticos .................................................................................................. 19
RESULTADOS ................................................................................................................... 19
Proporción de sexos ...................................................................................................... 19
Haliotis fulgens ........................................................................................................... 19
Haliotis corrugata ....................................................................................................... 20
FASES DEL CICLO GONÁDICO ................................................................................ 21
Desarrollo .................................................................................................................... 21
Madurez ....................................................................................................................... 23

Desove ......................................................................................................................... 24
Posdesove .................................................................................................................. 25
Indiferenciación .......................................................................................................... 26
CICLO REPRODUCTIVO ............................................................................................. 26
Haliotis fulgens ........................................................................................................... 26
Haliotis corrugata ....................................................................................................... 28
TALLA DE PRIMERA MADUREZ ............................................................................... 30
Haliotis fulgens ........................................................................................................... 30
Haliotis corrugata ....................................................................................................... 31
FECUNDIDAD ................................................................................................................ 32
Haliotis fulgens ........................................................................................................... 32
Haliotis corrugata ....................................................................................................... 33
CICLO DE ALMACENAMIENTO DE NUTRIENTES ............................................... 33
Índices morfofisiológicos de peso ........................................................................... 45
Índices morfofisiológicos de área ............................................................................ 50
ANÁLISIS DEL DIÁMETRO PROMEDIO MENSUAL DE LOS OVOCITOS ........ 54
FACTORES AMBIENTALES ....................................................................................... 55
DESCRIPCIÓN DE LA GAMETOGÉNESIS.............................................................. 56
Espermatogénesis ..................................................................................................... 56
Ovogénesis ................................................................................................................. 60
DISCUSIÓN .................................................................................................................... 63
CONCLUSIONES .............................................................................................................. 74
REFERENCIAS .............................................................................................................. 76

i
GLOSARIO
Acrosoma: Estructura en forma de casquete que cubre la cabeza del
espermatozoide y que contiene enzimas hidrolíticas, que le permiten fecundar el
óvulo (Villee, 1996).
Ciclo gametogénico: Secuencia de desarrollo y maduración por los que pasa el
gameto dentro de la gónada en una línea celular (ovogénesis y espermatogénesis)
(Kraeuter y Castagna, 2001).
Ciclo gonádico: Conjunto de acontecimientos histofisiológicos que se producen
en la gónada (cada estadio de desarrollo gonádico), a intervalos de tiempo cíclicos
(Romo-Piñera, 2010).
Ciclo reproductivo: Intervalo de tiempo entre periodos reproductivos en una
población (Hahn, 1989)
Corion: Cubierta mucosa que cubre, da protección y flotabilidad al ovocito (Ortíz-Ordoñez et al., 2009). Cromatina: Sustancia que se encuentra en el núcleo de la célula formando el
material cromosómico durante la interface; está compuesto de ADN unido a
proteínas (Villee, 1996).
Desove: Expulsión de los gametos por el organismo (Kraeuter y Castagna, 2001).
Dioico: Se dice del organismo que tiene los sexos masculino y femenino en
distintos individuos o colonias (Villee, 1996).
Electrodensos: Se refiere al material oscuro observado en un microscopio
electrónico, zonas gruesas que no permite atravesar los electrones (Najmudeen,
2008).

ii
Época reproductiva: Temporada en la que los organismos de una población se
encuentran en etapa de madurez o desove (Gurney y Mundy, 2004).
Espermatocito: Los espermatocitos primarios son células diploides derivadas del
proceso de espermatogénesis formadas por el aumento en tamaño de las
espermatogonias. Los espermatocitos secundarios son células haploides
originadas tras la meiosis I; cada espermatocito secundario se diferencia en una
espermátida (De Robertis et al., 1977).
Espermatogonias: Células reproductoras masculinas primarias y diploides (De
Robertis et al., 1977).
Espermatozoides: Células reproductoras masculinas haploides y maduras, que
han pasado por una serie de modificaciones que la hacen viable para alcanzar y
perforar la membrana del óvulo, y se caracteriza por su movilidad por medio de un
flagelo (De Robertis et al., 1977).
Estrategia reproductiva: Conjunto de características de una especie para la
reproducción (Wootton, 1989).
Eucromatina: Es una forma de la cromatina ligeramente compactada con una
gran concentración de genes, y a menudo se encuentra en transcripción activa
(De Robertis et al., 1977).
Euritermo: Aquellos organismos que son capaces de soportar las oscilaciones de
temperatura (Bayard, 1974).
Fases de desarrollo gonádico: Se refiere a la división artificial del proceso de
desarrollo gonádico (Romo-Piñera, 2010).

iii
Gametogénesis: Proceso de formación de gametos o células reproductores
(Kraeuter y Castagna, 2001).
Gametos: Células reproductoras ovulo o espermatozoides, cuya unión, en la
reproducción sexual, inicial el desarrollo de un nuevo individuo (Villee, 1996).
Glándula digestiva: Glándula encargada de producir los jugos digestivos
necesarios para la digestión química de los alimentos (Kraeuter y Castagna 2001).
Gonocórico: Individuo que posee estructuras reproductoras únicamente
femeninas o masculinas (Romo-Piñera, 2010).
Hepatopáncreas: Glándula secretora de jugos digestivos presente en crustáceos,
arácnidos y moluscos; su función es similar a la que realizan el hígado y el
páncreas en los vertebrados (Kraeuter y Castagna, 2001).
Heterocromatina: Son regiones de la cromatina compactas, condensadas,
empaquetadas que se tiñen fuertemente con coloraciones para ADN (De Robertis
et al., 1977).
Intensidad de desove: Término referido a la frecuencia en la que se presenta la
etapa de desove en un periodo determinado (Gurney y Mundy, 2004).
Ovocito: Gameto o célula sexual femenina (Kraeuter y Castagna, 2001).
Ovogonias: Células reproductoras femeninas primarias y diploides (Kraeuter y
Castagna, 2001).
Periodo reproductivo: Momento dentro del ciclo reproductivo en el que se
presentan organismos en desove y en madurez (Gurney y Mundy, 2004).

iv
Táctica reproductiva: Variaciones en las características reproductivas debidas a
cambios ambientales (Wootton, 1989)
Tejido conjuntivo o conectivo: Tejido que provee soporte estructural y
transporte de nutrientes (Leeson et al., 1987).
Temperatura superficial del mar (TSM): Temperatura medida en el océano
dentro de los primeros centímetros de profundidad (Levinton, 2001).
Vitelo: Material nutritivo de reserva de los ovocitos que se utiliza durante el
desarrollo del embrión (Romo-Piñera, 2010).
Vitelogénesis: Formación y acumulación del vitelo en el citoplasma de las células
(Kraeuter y Castagna, 2001).
Zona tropical: Zona latitudinal incluida entre los dos trópicos (23º 27´ norte y sur)
(Levinton, 2001).
Zona templada: Zona latitudinal incluida entre 23º 27´y 66º 33´ (norte y sur)
(Levinton, 2001).

v
LISTA DE FIGURAS
Figura 1. Límites de distribución reportados para las especies de abulón de la
costa noroeste del pacífico.. .............................................................................................. 2
Figura 2. Zonas geográficas reglamentadas para la captura de abulón en la
península de Baja California y ubicación de las zonas de estudio .............................. 4
Figura 3. Etapa de desarrollo en Haliotis fulgens. . ..................................................... 22
Figura 4. Etapa de madurez en Haliotis fulgens. . ....................................................... 23
Figura 5. Etapa de desove en Haliotis fulgens. ........................................................... 24
Figura 6. Etapa de posdesove en Haliotis fulgens.. ..................................................... 25
Figura 7. Gónada de en etapa de indiferenciación de Haliotis fulgens. ................... 26
Figura 8. Ciclo reproductivo de Haliotis fulgens en Baja California Sur, México..... 28
Figura 9. Ciclo reproductivo de Haliotis corrugata en Baja California Sur, México. 29
Figura 10. Histograma de frecuencia de tallas, señala cuando el 50% de la
población se encuentra madura. ..................................................................................... 30
Figura 11. Histograma de frecuencia de tallas, señala cuando el 50% de la
población se encuentra madura. ..................................................................................... 31
Figura 12. Comportamiento de la concentración de carbohidratos durante un ciclo
anual en el pie, manto, gónada y hepatopáncreas de Haliotis fulgens en Baja
California Sur, México. ...................................................................................................... 34
Figura 13. Comportamiento de la concentración de carbohidratos en las distintas
etapas del ciclo reproductivo en el pie, manto, gónada y hepatopáncreas de
Haliotis fulgens en Baja California Sur, México. ........................................................... 35

vi
Figura 14. Comportamiento de la concentración de carbohidratos durante un ciclo
anual en el pie, manto, gónada y hepatopáncreas de Haliotis corrugata I en Baja
California Sur, México. ...................................................................................................... 36
Figura 15. Comportamiento de la concentración de carbohidratos en las distintas
etapas del ciclo reproductivo en pie, manto, gónada y hepatopáncreas de Haliotis
corrugata en Baja California Sur, México. ..................................................................... 37
Figura 16. Comportamiento de la concentración de lípidos durante un ciclo anual
en pie, manto, gónada y hepatopáncreas de Haliotis fulgens en Baja California
Sur, México. ........................................................................................................................ 38
Figura 17. Concentración de lípidos en las distintas etapas del ciclo reproductivo
en pie, manto, gónada y hepatopáncreas de Haliotis fulgens en Baja California
Sur, México. ........................................................................................................................ 39
Figura 18. Comportamiento de la concentración de lípidos durante un ciclo anual
en pie, manto, gónada y hepatopáncreas de Haliotis corrugata en Baja California
Sur, México. ........................................................................................................................ 40
Figura 19. Comportamiento de la concentración de lípidos en las distintas etapas
del ciclo reproductivo en pie, manto, gónada y hepatopáncreas de Haliotis
corrugata en la zona de pesca III en Baja California Sur, México. ............................ 41
Figura 20. Comportamiento de la concentración de proteínas durante un ciclo
anual en pie, manto, gónada y hepatopáncreas de Haliotis fulgens en Baja
California Sur, México. ...................................................................................................... 42
Figura 21. Comportamiento de la concentración de proteínas en las distintas
etapas del ciclo reproductivo en pie, manto, gónada y hepatopáncreas de Haliotis
fulgens en Baja California Sur, México. ......................................................................... 43

vii
Figura 22. Comportamiento de la concentración de proteínas durante un ciclo
anual en pie, manto, gónada y hepatopáncreas de Haliotis corrugata en Baja
California Sur, México. ...................................................................................................... 44
Figura 23. Comportamiento de la concentración de proteínas en las distintas
etapas del ciclo reproductivo en pie, manto, gónada y hepatopáncreas de Haliotis
corrugata en Baja California Sur, México. ..................................................................... 45
Figura 24. Comportamiento mensual de los índices morfofisiológicos de Haliotis
fulgens en la zona de pesca II en Baja California Sur, México. ................................. 46
Figura 25. Comportamiento mensual de los índices morfofisiológicos de Haliotis
fulgens en la zona de pesca III en Baja California Sur, México. ................................ 47
Figura 26. Comportamiento mensual de los índices morfofisiológicos de Haliotis
corrugata en la zona de pesca II en Baja California Sur, México. ............................. 48
Figura 27. Comportamiento mensual de los índices morfofisiológicos de Haliotis
corrugata en la zona de pesca III en Baja California Sur, México. ............................ 49
Figura 28. Índices de área de Haliotis fulgens en Baja California Sur, México. ...... 51
Figura 29. Índices de área de Haliotis corrugata en Baja California Sur, México. 53
Figura 30. Diámetro promedio mensual de los ovocitos de Haliotis fulgens en Baja
California Sur, México. ...................................................................................................... 54
Figura 31. Diámetro promedio mensual de los ovocitos de Haliotis corrugata en
Baja California Sur, México. ............................................................................................. 55
Figura 32. Variación en la Temperatura Superficial del Mar y fotoperiodo en la
zona de pesca II y en la zona de pesca III en Baja California Sur, México. ............ 56
Figura 33. Vista panorámica de la espermatogénesis en Haliotis corrugata. .......... 57
Figura 34. Microfotografía de espermatozoide de Haliotis fulgens............................ 59

viii
Figura 35. Corte transversal del arreglo mitocondrial de un espermatozoide de
Haliotis corrugata. .............................................................................................................. 59
Figura 36. Células de la ovogénesis.. ............................................................................ 61
Figura 37. Ovocitos atrésicos ......................................................................................... 62
LISTA DE TABLAS
Tabla 1. Número de hembras, machos y ejemplares indiferenciados de Haliotis
fulgens.. ............................................................................................................................... 20
Tabla 2. Número de hembras, machos y ejemplares indiferenciados de Haliotis
corrugata. ............................................................................................................................ 21
Tabla 3. Fecundidad de Haliotis fulgens en las zonas de pesca II y III en enero de
2013. .................................................................................................................................... 32
Tabla 4. Fecundidad de Haliotis fulgens y Haliotis corrugata por organismo en la
zona de pesca III en agosto de 2012. ............................................................................ 33
Tabla 5. Valores máximos y mínimos de los índices morfofisiológicos de Haliotis
fulgens y Haliotis corrugata en la costa occidental de Baja California Sur, México.
.............................................................................................................................................. 50
Tabla 6. Frecuencias en porcentaje, iguales o mayores al 50% de desove y
madurez, reportados en Haliotis fulgens en trabajos antecedentes. ........................ 65
Tabla 7. Talla de primera madurez L50 de H. corrugata y H. fulgens, estimada en
las cuatro zonas de pesca, en trabajos antecedentes ................................................. 70
Tabla 8. Fecundidad estimada en trabajos antecedentes para Haliotis fulgens. ... 72

ix
RESUMEN Para conocer las tácticas reproductivas del abulón azul, Haliotis fulgens y del abulón
amarillo, Haliotis corrugata en dos zonas de pesca de la costa occidental de Baja
California Sur, se determinaron sus ciclos reproductivo, gonádico y de
almacenamiento de nutrientes, así como su relación con factores ambientales locales.
Se realizaron muestreos mensuales en la zona de pesca II (Bahía Tortugas-Bahía y
Asunción) y en la zona de pesca III (La Bocana), de septiembre de 2011 a agosto de
2013. Se obtuvieron muestras de gónada y masa visceral. Para conocer el ciclo
reproductivo y de almacenamiento de nutrientes, se aplicaron técnicas histológicas y
bioquímicas a porciones de gónada, hepatopáncreas, manto y pie. Para describir la
gametogénesis se utilizó microscopia electrónica de barrido y de transmisión. Cómo
indicadores del ambiente se utilizaron datos mensuales del fotoperiodo y de la
temperatura superficial del mar, obtenidos a partir de imágenes de satélite de la región
de estudio. Los resultados indican que en H. fulgens y H. corrugata, el desove, el
desarrollo y la reabsorción de gametos se presentaron durante todo el ciclo
reproductivo. Se encontró que para el crecimiento de la gónada también presentan
una táctica oportunista, debido a que utilizan los nutrientes disponibles en el medio,
los almacenados en el hepatopáncreas, los obtenidos por la reabsorción de gametos
residuales o una combinación de todo lo anterior. Las variaciones en las tácticas se
relacionan con cambios interanuales de las condiciones ambientales. Por ejemplo,
que ambas especies desovan durante todo el año, pero H. fulgens presenta un pico
reproductivo de junio a diciembre en las dos zonas, mientras que H. corrugata, no
presenta ningún pico reproductivo. La fecundidad de H. fulgens varía en cada zona de
pesca de dos a cuatro millones de huevos. Se concluye que en ambas especies la
estrategia reproductiva consiste en un conjunto de tácticas oportunistas que incluyen
desoves euritérmicos (con afinidad a desovar con mayor intensidad alrededor de los
20ºC), desarrollo continuo de la gónada, cambios en la proporción de sexos y en la
fecundidad que les permiten maximizar el aprovechamiento de los recursos
ambientales disponibles para su reproducción, y no existen diferencias en la
estrategia reproductiva de ambas especies entre las zonas estudiadas, pero si en
cuanto a las respuestas oportunistas de las tácticas reproductivas a la variaciones
ambientales temporales, interanuales y climatológicos.

x
ABSTRACT To better understand the reproductive tactics of green abalone, Haliotis fulgens,
and pink abalone, Haliotis corrugata in two fishing zones off the west coast of Baja
California Sur their reproductive, gonadal and storage of nutrient cycles were
determined, as well as their related local environmental factors. Monthly samples
were taken in fishing zone II (Bahía Tortugas- Bahía Asunción) and fishing zone III
(La Bocana), from September 2011 to August 2013. Gonad and visceral mass
samples were obtained. To understand the reproductive and storage nutrient
cycles, histological and biochemical techniques were applied to portions of the
gonads, hepatopancreas, mantle and foot. To describe gametogenesis, scanning
electron microscopy and transmission was used. Monthly photoperiod data and
sea surface temperature were used as environmental indicators, which were
obtained from satellite images of the study area. The results indicate that H.
fulgens and H. corrugata spawning, development, and resorption of gametes were
presented throughout the reproductive cycle. It was found that the growth of the
gonad also has an opportunistic tactic, because the nutrients available in the
environment are used, stored in the hepatopancreas, obtained by reabsorption of
residual gametes, or a combination of the above. Variations in strategies relate to
interannual changes in environmental conditions. For example, both species
spawn throughout the year, but H. fulgens has a reproductive peak from June to
December in the two zones, while H. corrugata has no reproductive peak. H.
fulgens fecundity varies in each fishing zone between two to four million eggs. It is
concluded that in both species, the reproductive strategy is a set of opportunistic
tactics that include eurythermal spawns (with the affinity to spawn more intensely
around 20 ° C), continued development of the gonad, changes in sex ratio and
fecundity that allow them to maximize the use of environmental resources available
for reproduction, and there are no differences in reproductive strategy of both
species among the studied zones, other than the opportunistic responses to
reproductive tactics to temporary and annual environmental variations.

1
INTRODUCCIÓN
Los abulones son gasterópodos marinos pertenecientes a la subclase
Vetigastropoda, a la familia Haliotidae y al género monotípico Haliotis, que incluye
aproximadamente 200 especies (Gofas, 2013). Las especies del género Haliotis
son generalmente organismos dioicos, sin embargo existe como caso incidental, la
presencia de hermafroditismo en H. gigantea (Murayama, 1935 citado en Cox,
1962), en H. tuberculata (Girard, 1972) y en H. fulgens (M. Ortiz, comunicación
personal, citado en Guzmán del Próo, 1992).
Los abulones maduros presentan una tasa de crecimiento promedio anual
de 26 mm, aunque H. laevigata llega a 45 mm por año. El crecimiento es
altamente dependiente del consumo de alimento y por lo tanto de su disponibilidad
(Mazón-Suástegui et al., 1996).
Los abulones son organismos herbívoros que durante su fase larvaria se
alimentan de plancton que ingresa en la boca por acción de los cilios orales.
Durante la etapa juvenil se alimentan de diatomeas bentónicas y al alcanzar la
talla adulta, lo que coincide con la aparición del primer poro respiratorio,
comienzan a incluir diferentes especies de macroalgas en su dieta (Cox, 1962;
Haaker et al., 1998). El hábitat de las especies de abulón, lo constituyen las zonas
intermareal y submareal de las costas rocosas, sobre las que se desarrolla la
comunidad bentónica asociada a macroalgas (Dayton y Tegner, 1984).
El género Haliotis, se distribuye en aguas tropicales y templadas (Geiger y
Owen, 2012, citado en Gordon y Petter, 2013), principalmente en las costas del
Océano Pacífico de Australia, Japón, Canadá, Sudáfrica, California. En México se
encuentra en las partes norteña y central de costa occidental de la Península de
Baja California (Dybas, 1994) y se reportan siete especies: H. fulgens Philippi
1845 (abulón azul), H. corrugata Gray 1828 (abulón amarillo), H. cracherodii Leach
1817 (abulón negro), H. rufescens Swainson 1822 (abulón rojo), H. sorenseni
Bartsch 1940 (abulón rosa o chino), H. assimilis (Linnaeus 1758) (abulón rayado) y
H. walallensis Stearns 1899 (abulón aplanado) (Fig. 1).
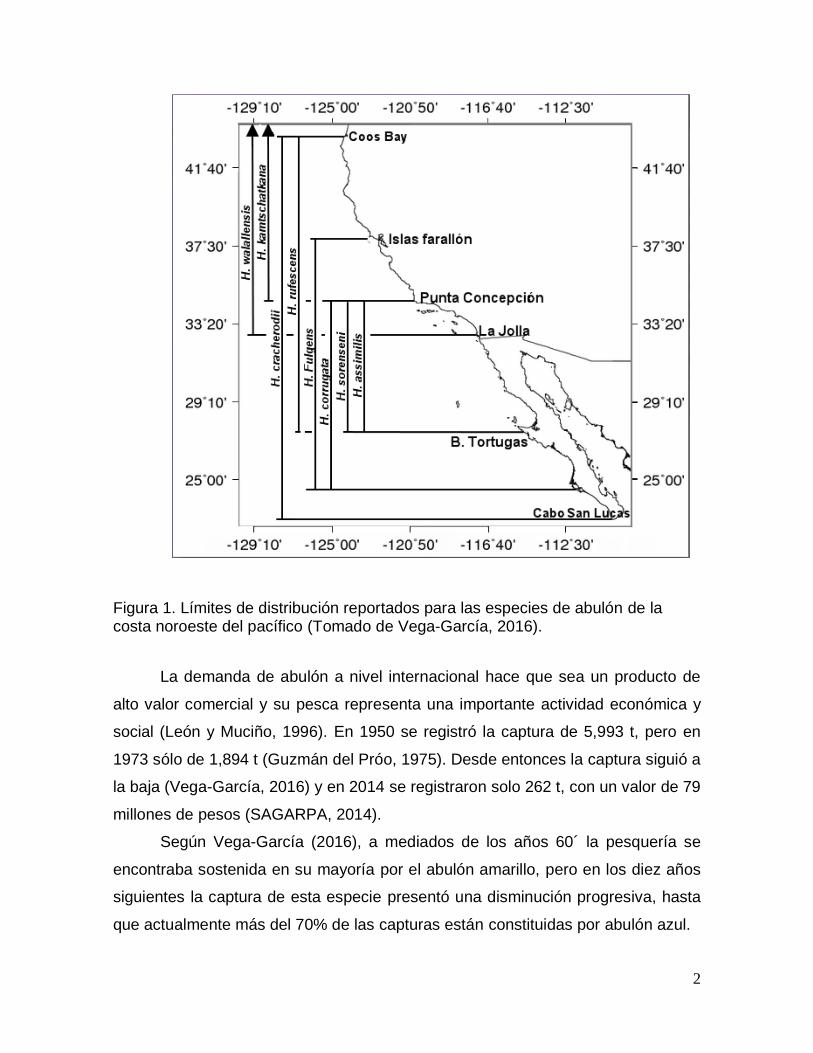
2
Figura 1. Límites de distribución reportados para las especies de abulón de la costa noroeste del pacífico (Tomado de Vega-García, 2016).
La demanda de abulón a nivel internacional hace que sea un producto de
alto valor comercial y su pesca representa una importante actividad económica y
social (León y Muciño, 1996). En 1950 se registró la captura de 5,993 t, pero en
1973 sólo de 1,894 t (Guzmán del Próo, 1975). Desde entonces la captura siguió a
la baja (Vega-García, 2016) y en 2014 se registraron solo 262 t, con un valor de 79
millones de pesos (SAGARPA, 2014).
Según Vega-García (2016), a mediados de los años 60´ la pesquería se
encontraba sostenida en su mayoría por el abulón amarillo, pero en los diez años
siguientes la captura de esta especie presentó una disminución progresiva, hasta
que actualmente más del 70% de las capturas están constituidas por abulón azul.

3
En México, la administración o regulación de las pesquerías de
invertebrados bentónicos se basa principalmente en tres medidas regulatorias,
todas fundamentadas en el conocimiento del ciclo reproductivo: 1) el
establecimiento de dos tipos de vedas: la veda reproductiva, que impide la captura
durante la época de máxima actividad reproductiva, y la veda prohibitiva, que no
permite la captura en ningún momento; 2) la determinación de la talla mínima de
captura, que se fija con base en la talla de primera reproducción y 3) el
establecimiento de zonas de reserva, que se aplica a recursos con hábitos de
agregación reproductora (Baqueiro, 1992). Todas estas medidas ya han sido
implementadas desde 1956 a favor de la mejora en el manejo del abulón. Así se
implementó en la costa del Pacífico de Baja California una veda, del 16 de enero
al 15 de marzo, para la captura de abulón amarillo, rojo, azul y negro, y se
establecieron tallas mínimas de captura por especie. En 1972 la veda se cambió a
julio y agosto (León y Muciño, 1995). Actualmente el aprovechamiento de las
distintas especies de abulón se ajusta a las Normas Oficiales Mexicanas NOM-
005-PESC-1993 (SEPESCA, 1993) y NOM-009-SAG/PESC-2015 (SAGARPA,
2016), en las que se define el procedimiento para la determinar las épocas y
zonas de veda para la captura de las diferentes especies de la flora y fauna
acuáticas.

4
Figura 2. Zonas geográficas reglamentadas para la captura de abulón en la península de Baja California y ubicación de las zonas de estudio.
La figura 2, muestra la división por zonas de captura y cooperativas en
B.C.S. Para cada una de las zonas de pesca de abulón se implantan periodos de
veda diferenciales con duración de cinco meses. Así, en la zonas II y III la veda es
del 1º de agosto al 31 de diciembre; estas zonas, junto con la cooperativa de la
Isla Cedros, que pertenece a la zona de pesca I, se consideran como la zona
centro, que aporta más del 95% de la producción total (Keyl, 2004). En 1998, en
esta zona habitaban aproximadamente 10,000 personas, cuyas actividades

5
económicas estaban sustentadas básicamente en la pesquería de abulón y
langosta, pues de esta población 9,644 personas eran miembros de las
cooperativas pesqueras (Ponce-Díaz et al., 1998).
Para que estas pesquerías sean sustentables a largo plazo, es necesario
efectuar algunos estudios biológicos y pesqueros cuyos resultados permiten
administrarlas adecuadamente. Uno de los más importantes es el del ciclo
reproductivo, que en los invertebrados marinos puede estar afectado por
interacciones entre factores exógenos, como el fotoperiodo, el nivel de mareas y la
temperatura (Rose et al., 1990; López et al., 2005), y los factores endógenos de
tipo endocrino y neuronal (López et al., 2005), lo que puede provocar distinta
estacionalidad de la época reproductiva, cambios en la intensidad y frecuencia de
desove, fecundidad y talla de primera madurez, así como modificaciones en el
almacenamiento y transferencia de nutrientes, etc.
Estas características en conjunto, están determinadas por su genoma en
respuesta a los cambios ambientales y la combinación de los rasgos reproductivos
de los individuos pertenecientes al mismo conjunto genético, puede ser
considerada como la estrategia reproductiva de esos individuos (Saborido-Rey,
2004). Es decir que la estrategia reproductiva de una población comprende el
modelo reproductivo total, que consiste en un conjunto de tácticas que varían en
respuesta a las fluctuaciones ambientales y le permiten reproducirse exitosamente
(Calvo et al., 1998). Por esto es que las historias de vida de los organismos no
pueden basarse en predicciones generales, por lo que es necesario realizar
estudios particulares para cada población, y realizar ajustes de acuerdo a la
información específica de estas, de lo contrario, la información insuficiente sobre la
historia de vida de un organismo, dará como resultado correlaciones negativas
que no se ajusten al modelo teórico (Calvo et al., 1998). Con base a lo anterior, en
el presente trabajo se pretende describir las tácticas reproductivas de Haliotis
fulgens y de Haliotis corrugata en Baja California Sur, México. Ya que no se
cuenta con algún trabajo que describa simultáneamente, tanto el ciclo
reproductivo, como el ciclo de almacenamiento de nutrientes, ni que describa o

6
discuta las tácticas reproductivas que utilizan H. fulgens y H. corrugata como
estrategia para lograr reproducirse exitosamente
ANTECEDENTES
Existen numerosos trabajos en relación con la biología reproductiva del
género Haliotis, que describen épocas de desove para varias especies y muestran
la variabilidad en la periodicidad del desove entre especies, o en una sola especie
entre los años y sitios (Boolootian et al., 1962; Webber y Giese, 1969; Young y De
Martini, 1970; Tutschulte y Connell, 1981; Litaay y De Silva, 2003; Najmudeen y
Victor, 2004). Por ejemplo, para H. cracheroidii, Boolootian et al. (1962) reportaron
la época de desove entre primavera y otoño en Monterey, California, pero años
después en el mismo sitio, Webber y Giese (1969) reportaron como época de
desove la primavera y el verano. Otro ejemplo es la variación en la época de
desove de H. varia en diferentes localidades (Najmudeen y Victor, 2004).
En México los primeros trabajos sobre reproducción de abulones estudiaron
H. fulgens en Isla Cedros, Baja California (Sevilla, 1971) y en Bahía Tortugas Baja
California Sur (Belmar-Pérez y Guzmán del Próo, 1991). En el primero, efectuado
de enero a diciembre de 1965, se reportó que la mayor parte de los organismos se
reprodujeron de junio a septiembre y sólo unos cuantos en octubre, y que el
desove (junio-septiembre) coincidió con ascensos en la temperatura del agua
(valores próximos a 20ºC). En el segundo trabajo, realizado con muestreos
estacionales entre noviembre de 1985 y noviembre de 1988, se reportó que los
periodos de máxima actividad reproductiva coincidieron con el otoño y la
primavera, aunque en invierno y verano pueden presentarse expulsiones totales,
pero menos frecuentes.
Sobre la reproducción de H. corrugata, en Punta Eugenia y Punta Abreojos,
Andrade (1971) determinó tres grados de madurez de la gónada (inmaduro,
madurando y maduro) y reporta que el máximo grado de madurez ocurrió a
mediados de verano, en coincidencia con el aumento de la temperatura; también
observó que los organismos de Punta Eugenia, alcanzaron su máxima madurez,
un mes antes que los estudiados en Punta Abreojos, 150 Km al sur.

7
En otras especies de Haliotis se ha reportado que los desoves pueden
ocurrir cuando las temperaturas son las más altas, durante los descensos o en las
temperaturas mínimas. Por ejemplo, H. cracheroidii desova cerca de las
temperaturas máximas (mayores a 14°C) (Webber y Giese, 1969), mientras que H.
iris y H. australis lo hacen después de una disminución de la temperatura (menor a
8°C) (Wilson y Schiel, 1995), pero H. asinina desova todo el año, excepto en
verano, cuando la temperatura del agua es más cálida (Singhagraiwan y Doi,
1992; Capinpin et al., 1998). Capinpin et al. (1998) y Counihan et al. (2001),
aseguran que el desove de H. asinina puede ser inducido por cambios bruscos de
la temperatura causados por las mareas.
También se ha reportado que la temperatura no determina por sí sola la
reproducción, sino que es la combinación de dos o más factores externos (como
fotoperiodo, ciclo de mareas, alimentación y viento), lo que interviene en el
proceso reproductivo. Esto se ha reportado en poblaciones silvestres de H. discus
hannai, H. rufescens (Young y De Martini, 1970), H. iris, H. australis (Poore, 1973),
H. diversicolor diversicolor (Takashima et al., 1978), H. asinina (Jebreen et al.,
2000), H. rubra y H. laevigata (Grubert et al., 2005).
Por otro lado en especies cultivadas como H. tuberculata, H. discus hannai,
H. rufescens y H. varia se ha observado que el control de la temperatura y la dieta,
estimulan la maduración de las gónadas y el desove (Najmudeen y Victor, 2004;
Fukazawa et al., 2007; Manganaro et al., 2008; Muñoz et al., 2012). En cuanto a la
nutrición, Capinpin et al. (1998), observaron en el laboratorio que con
disponibilidad constante de alimento, los individuos de H. asinina presentaron
intervalos de desoves cortos; esto les sugirió que el alimento interviene en la
maduración más rápida de los óvulos. Otros autores coinciden con esta hipótesis
(Webber y Giese, 1969; Shepherd y Laws, 1974; Capinpin et al., 1998; Counihan
et al., 2001).
El fotoperiodo, es otro factor externo que puede influir en el proceso
reproductivo de Haliotis. Webber y Giese (1969) sugieren que los días con más de
12 horas luz, pueden estimular la gametogénesis de H. cracheroidii, pero los datos
no fueron totalmente concluyentes, ya que el aumento de la duración del día no se

8
correlacionó con el aumento del tamaño de las gónadas. Para H. asinina, se
encontró una correlación de la maduración de la gónada con el aumento de horas
luz, y otra correlación negativa entre las mayores temperatura del aire y los
desoves (Setyono, 2006).
Con referencia a la relación entre el ciclo de almacenamiento de nutrientes
y el ciclo reproductivo, se ha observado que en H. cracherodii, H. rufescens
(Boolootian et al., 1962), H. rubra (Litaay y De Silva, 2003) y H. varia (Najmudeen,
2007), el índice gonádico y el índice hepático están inversamente correlacionados,
lo que sugiere que el hepatopáncreas almacena nutrientes esenciales para el
desarrollo de los gametos durante el ciclo reproductivo y se refleja en la variación
de proteínas y lípidos en la gónada y en la glándula digestiva (Najmudeen, 2007).
Los análisis proximales de sustancias de reserva en relación con la reproducción
en H. cracherodii (Webber, 1970), H. rubra (Litaay y De Silva, 2003; Litaay, 2005)
y H. varia (Najmudeen, 2007), señalan que durante la reproducción existe una
demanda metabólica para la producción de gametos. El único estudio de este tipo
realizado en H. fulgens es el de Nelson et al. (2002) quienes demostraron que los
lípidos actúan como moléculas esenciales en el crecimiento y en la maduración
gonadal.
En cuanto a estudios sobre fecundidad, existen trabajos que reportan la
fecundidad de algunas especies de Haliotis en condiciones de cultivo. Por
ejemplo, en H. rubra y H. laevigata se encontró que la cantidad de huevos varía de
acuerdo a la temperatura del agua (Grubert et al., 2005), y en H. rufescens se
reportan valores de 6.19 x106 a 12.6 X106 huevos en organismos de 134 a 198.5
mm de longitud (Giorgi y De Martini, 1977).
En poblaciones silvestres de H. iris se estimaron de 11.25 X 106 a 12.86
X106 huevos en individuos de 68 a 155 mm (Poore, 1973); para H. asinina de 1.5 x
105 a 6 x 105 huevos en longitudes de 58.1 a 69 mm; para H. rubra de 1.09 X 106
a 7.5 X 106 huevos en individuos de 120 a 145 mm (Litaay y De Silva, 2003) y para
H. tuberculata coccinea de 85,749 a 691,546 huevos en longitudes de 39 a 62 mm
(Bilbao et al., 2010). En todos los trabajos se ha observado que existe una
variación de acuerdo a la especie y talla.

9
En el abulón amarillo H. corrugata, Ortiz-Quintanilla et al. (1987) estimaron
de 1.3 x 106 a 10.4 x 106 huevos en longitudes de 134 a 186 mm, y atribuyen la
diferencia al estadio de madurez de los organismos. Para H. fulgens, Guzmán del
Próo et al. (2001) estimaron de 0.99 x 106 a 22.3 x 106 de huevos en individuos de
115 a 198 mm. Para esta especie, Ortiz-Quintanilla et al. (1987) reportan valores
de 3.34 x 106, 5.16 x 106 y 7.24 x 106 huevos en organismos de 111 a 200 mm.
Resumiendo, existen varios trabajos, realizados entre 1970 y 1992, que
describen los aspectos reproductivos de H. fulgens y de H. corrugata, efectuados
de forma puntual, con el objetivo de determinar la época de desove o la
fecundidad. Sin embargo, no se dispone de información sobre posibles relaciones
entre el ciclo reproductivo y el ciclo de transferencia de nutrientes, ni sobre los
efectos que sobre ellos tienen factores como la temperatura y el fotoperiodo, la
fecundidad, la proporción de sexos y la talla de primera madurez. La existencia de
estas relaciones supone la existencia de procesos de adaptación reproductiva
(tácticas), asociados al éxito de la sobrevivencia de las especies. En el caso de los
abulones amarillo y azul, conocer esas tácticas contribuirá a la comprensión de
sus comportamientos ante fenómenos asociados a cambios ambientales.
OBJETIVO GENERAL
Determinar las tácticas reproductivas del abulón azul Haliotis fulgens y del abulón
amarillo Haliotis corrugata en la costa noroccidental de Baja California Sur.
OBJETIVOS ESPECÍFICOS
Describir, analizar y comparar el ciclo reproductivo de H. fulgens y H.
corrugata.
Caracterizar el ciclo de almacenamiento y utilización de nutrientes y su relación
con el ciclo reproductivo de H. fulgens y H. corrugata.
Determinar la relación de la temperatura y el fotoperiodo con el ciclo
reproductivo de H. fulgens y H. corrugata.
Estimar la fecundidad en H. fulgens y H. corrugata.

10
MATERIALES Y MÉTODOS
ÁREA DE ESTUDIO
El área de estudio se encuentra ubicada en la costa noroccidental de Baja
California Sur, México; esta región pertenece a la zona de transición templado-
tropical, que se caracteriza por tener un gradiente estacional importante debido a
la influencia de la Corriente de California y de la Corriente Noroecuatorial, la
primera impone características templadas durante el invierno-primavera y la
segunda características tropicales durante el verano-otoño. En la zona se presenta
una amplia variabilidad climática a diferentes escalas, desde semanas hasta largo
plazo, evidenciadas en variables físicas como la temperatura del mar, nivel medio
de mar, actividad de surgencias y los patrones de circulación (Chelton, 1981;
Lluch-Belda, 1999). También forma parte del Centro de Actividad Biológica del
Pacífico mexicano (BAC), caracterizado por niveles de concentración pigmentaria,
volúmenes de desplazamiento macrozooplanctónico y abundancia relativa de
huevos y larvas de peces neríticos claramente superiores al promedio de la zona
costera (Lluch-Belda et al., 2000). Destaca la presencia de macroalgas como
Macrocystis pyrifera, especie dominante en la zona de Bahía Tortugas y Eisenia
arborea dominante en La Bocana (Vega-García, 2016) y en ella se localizan dos
de las áreas de pesca de abulón definidas en la NOM-009-SAG/PESC-2015.
La zona de pesca II, abarca desde el primer tercio noroeste de Bahía
Asunción, a partir del punto localizado a 27º 09' N; 114º 13' W, hacia el sur hasta
Punta Holcomb (26º 42' 20" N; 113º 15' 50" W) y hasta el límite del Mar Territorial
(Fig. 2). En esta zona los muestreos se realizaron en Bahía Asunción y en Bahía
Tortugas, en donde destacan por su interés en el manejo pesquero del abulón las
cooperativas “Bahía Tortugas” y “Emancipación” en Bahía Tortugas y “California
de San Ignacio” en Bahía Asunción.
La zona de pesca III se encuentra limitada en la parte norte por Punta
Malarrimo (27º 47' 30" N; 114º 32' 20" W y 27º 57" N, 114º 32' 20" W), y por la
parte sur a partir del punto localizado a 27º 09' N; 114º 13' W, hasta el límite del

11
Mar Territorial (Fig. 2). La Bocana es una de las zonas de mayor productividad
pesquera, en donde la cooperativa “Progreso” destaca por su interés en la
investigación sobre abulón.
METODOLOGÍA DE CAMPO
Con la colaboración de las Cooperativas “Bahía Tortugas”, “Emancipación”
de Bahía Tortugas, “California de San Ignacio” de Bahía Asunción y “Progreso” de
La Bocana, se realizaron muestreos mensuales de septiembre de 2011 a agosto
de 2013, de aproximadamente 30 ejemplares por especie de cada zona de pesca.
Los abulones fueron capturados por miembros de dichas cooperativas, mediante
buceos autónomos a una profundidad de entre cuatro y 12 m.
En condiciones de laboratorio cada ejemplar fue medido, pesado y
disectado para pesar la gónada, el callo y las vísceras y, obtener un trozo de 3 cm
de la porción central de gónada que se fijó en formol al 10%, para posteriormente
describir y determinar histológicamente las fases del ciclo reproductivo.
Adicionalmente, entre febrero de 2012 y febrero de 2013, cada mes se
transportaron vivos diez organismos de cada especie, desde La Bocana, en la
zona de pesca III, al Laboratorio de Invertebrados Marinos del CICIMAR, en la
Ciudad de La Paz. Los individuos se transportaron en hieleras cerradas rellenas
con geles congelados y varias capas de frondas de Macrocystis.
METODOLOGÍA DE LABORATORIO
Técnica de microscopía electrónica de transmisión
Para describir la ultraestructura y morfología de la gametogénesis, se obtuvo un
fragmento de gónada de la parte apical del complejo hepatogonadal de
aproximadamente 5 mm3 de los organismos transportados vivos al laboratorio y se
fijó en glutaraldehído al 2.5%. Posteriormente se lavaron con solución fisiológica
para después realizar una posfijación con tetraróxido de Osmio al 2%. A
continuación se deshidrataron con etanol y se colocaron en óxido de propileno,
con la finalidad de proporcionar un medio entre el alcohol y la resina. Finalmente
se incluyeron en resina spur utilizando moldes cilíndricos para colocarlos en una

12
incubadora a 65 ºC, con el fin de que la resina polimerizara. Una vez polimerizada
la resina, se procedió a tallar el bloque hasta obtener una forma de pirámide. Se
obtuvieron cortes semifinos de 0.3 μm de espesor utilizando un ultramicrotomo con
una cuchilla de vidrio, estos se recogieron con ayuda de un asa para su tinción
con azul de toluidina.
Posteriormente se seleccionaron áreas representativas de la
gametogénesis para tomar fotografías y realizar cortes finos (50-80 nm de
espesor). Estos se colocaron sobre rejillas de cobre de 200 mallas, para ser
contrastados y observados con un microscopio electrónico de transmisión. Se
obtuvieron microfotografías y se realizaron las descripciones ultraestructurales,
correspondientes a cada fase de la gametogénesis de cada sexo.
Técnica de microscopía electrónica de barrido
Se fijaron fragmentos de 5 mm3 de la gónada en glutaraldehído al 2.5% y se
lavaron utilizando cambios decrecientes de solución salina de 37 ups a agua
destilada. Posteriormente se posfijaron en tetraóxido de Osmio al 2% y se realizó
una pre-deshidratación con etanol; se secaron a punto crítico en CO2. Las
muestras fueron depositadas en un evaporador de metal para recibir un baño de
paladio y finalmente se observaron y capturaron imágenes utilizando un
microscopio electrónico de barrido. A partir de éstas, se efectuaron descripciones
morfológicas correspondientes a cada una de las fases de la gametogénesis de
cada sexo.
Técnica histológica
Las porciones de las gónadas fijadas en formol al 10% se deshidrataron con
etanol. Después se aclararon con CitriSolv y se incluyeron en parafina. Los cortes
histológicos se hicieron de 5 μm de grosor, se tiñeron con la técnica Hematoxilina-
Eosina (Humason, 1979) y se montaron con resina sintética, para su observación
en el microscopio. Posteriormente se describieron las fases del ciclo reproductivo
de acuerdo al criterio de Vélez-Arellano et al. (2015).

13
Proporción de sexos
El sexo de cada ejemplar se determinó macroscópicamente mediante la
coloración de la gónada (verde en hembras y crema para machos), y se confirmó
con el análisis histológico (Sevilla, 1971; Vélez-Arellano et al., 2015). La
proporción sexual se estimó dividiendo el número total de hembras entre el
número total de machos. Para conocer si esta proporción difiere de una proporción
1:1, se aplicó una prueba de 2 (Zar, 2010).
Ciclo reproductivo
Una vez obtenidas las preparaciones histológicas, el método consistió en
dividir el ciclo completo en categorías arbitrarias con base en la presencia y
número de las células gametogénicas, así como el grosor y coloración de la
gónada (indiferenciada, desarrollo, madurez, desove o posdesove) (Sevilla 1971;
Belmar-Pérez y Guzmán-del Próo, 1992; Vélez-Arellano et al., 2015). Con los
datos se estimaron las frecuencias relativas de cada categoría por mes. Los
meses con mayor frecuencia de madurez y desove indicaron la época
reproductiva.
Talla de primera madurez
La talla de primera madurez o de reclutamiento a la reproducción se calculó
simultáneamente para ambos sexos y se definió como la longitud en la cual el
50% de los organismos se encuentran sexualmente maduros, es decir los que
presenten estadios gonádicos de madurez y desove. Para estimarla se utilizó el
modelo logístico:
Donde: Y = Proporción de organismos sexualmente maduros x = Longitud de los organismos a, b = Constantes

14
Fecundidad
La fecundidad se estimó utilizando únicamente las hembras que se
encontraron en estadio de madurez. En la zona de pesca III, se analizaron cuatro
hembras maduras de abulón azul y dos de abulón amarillo, recolectadas en
agosto de 2012, y dos de abulón azul en enero de 2013. En la zona de pesca II, se
utilizaron cuatro hembras maduras de abulón azul recolectadas en enero de 2013.
En cada caso se realizó un corte longitudinal del saco gonadal para eliminar
la porción de hepatopáncreas y se obtuvo el peso húmedo de la gónada con una
balanza analítica (Precisa TX200). Se recolectaron muestras de los ovocitos
desprendidos de las trabéculas (Bilbao et al., 2010) y se colocaron en cajas de
Petri de vidrio para realizar lavados consecutivos con agua marina filtrada (1µm) y
eliminar los residuos de tejido conectivo. Los ovocitos se recolectaron con pipetas
de vidrio y se colocaron en tubos eppendorf de 1.5 mL. El exceso de agua de las
muestras se eliminó por decantación y absorción con papel absorbente.
Las muestras de ovocitos (de 4 g) se diluyeron en 50 mL de agua marina
filtrada (con una malla de 1µm) para obtener una solución de ovocitos concentrada
(0.08 g-1 mL-1) que se mantuvo homogenizada en un agitador automático (Corning)
para preparar alícuotas diluidas (0.005 g-1 mL-1, 200 mL) y realizar el conteo de
ovocitos, en un contador de partículas Multisizer 3 (Beckmann Coulter) con
apertura de 560 µm, usando como diluyente agua marina filtrada (1µm). La
calibración del equipo se realizó de acuerdo a las instrucciones del fabricante. La
lectura de las muestras se hizo en agitación constante para evitar la
sedimentación de los ovocitos.
La concentración de ovocitos por muestra se estimó considerando un
intervalo de talla de ovocitos maduros de 80 a 200 µm, determinada con las
observaciones histológicas de las mismas gónadas. La tasa de dilución óptima de
las alícuotas se estableció realizando diluciones seriales con muestras de ovocitos
del mismo organismo. Se registró coeficiente de determinación, R2 de 0.9376 para
una dilución 0.005 g mL. El flujo de la muestra en el contador de partículas se
estableció midiendo el volumen de diluyente desplazado a través de la apertura

15
(560 µm) por minuto. El flujo resultante se utilizó para estimar la concentración de
ovocitos (σT) de acuerdo a la siguiente fórmula:
σT (cel−1 mL−1) =σ(VT)
f
Donde: σ = número de ovocitos en la muestra (80 a 200 µm)
VT = volumen total de la muestra (mL)
f = flujo (mL-1 min-1)
El número de huevos por unidad de peso (N) se calculó con base en la
concentración de ovocitos (σT) y el peso húmedo de los ovocitos utilizados para
preparar la muestra (0.005 g).
N (cel−1 g−1) = σT(1 g)
g
Donde g = peso húmedo de ovocitos por muestra (gramos).
La fecundidad (F) de cada organismo se estimó con base en el peso
húmedo total de la gónada (G) y al número de ovocitos por unidad de peso (N) de
acuerdo a la siguiente fórmula:
F = N G
Análisis bioquímico de los tejidos
Se obtuvieron fragmentos de aproximadamente 2 cm3 de gónada, músculo
del pie y manto, para determinar la composición bioquímica de estos tejidos
(proteínas, lípidos y carbohidratos). Los tejidos se almacenaron a -54°C en un
ultracongelador. Posteriormente se tomaron submuestras de 0.3 g, que fueron
homogenizadas en 1.5 mL de solución salina fría (NaCl a 35 ups), para obtener el
extracto crudo para realizar los análisis bioquímicos.
Las técnicas de bioquímica utilizadas fueron previamente estandarizadas
para cada tejido y las concentraciones de cada componente bioquímico se

16
calcularon a partir de curvas de calibración de los estándares respectivos. Para las
proteínas se utilizó suero de albúmina, para los carbohidratos una solución de
dextrosa y para los lípidos una mezcla de colesterol y triglicéridos.
Proteínas
Para la determinación de proteínas se utilizó el método de Bradford (1976),
usando un agente cromógeno comercial y suero de albúmina bovina como
solución estándar. A 10 μL de la muestra homogenizada se le agregaron 200 μL
de NaOH 0.1 N para digerir durante dos horas. De la muestra homogenizada y
digerida se tomaron 10 μL y se adicionó 1 mL de solución reactiva de Bradford
para leer la absorbancia a 595 nm de longitud de onda.
Lípidos
La determinación de los lípidos totales se realizó por el método de la
sulfofosfovainillina (Barnes y Blackstock, 1973). Se utilizó un reactivo comercial y
solución estándar. A 25 μL de extracto crudo, se le agregaron 250 μL de ácido
sulfúrico concentrado, y se incubó la solución a 90°C durante 10 minutos e
inmediatamente se detuvo la reacción en un baño frío a 4°C. Posteriormente se
mezclaron 20 μL de extracto crudo-ácido con 200 μL de solución de
sulfofosfovainillina en una microplaca. Esta mezcla se incubó durante 40 minutos y
con un lector de microplaca se leyó la absorbancia a 540 nm.
Carbohidratos
Los carbohidratos totales se determinaron por el método de la antrona
(Roe, 1955). Se tomaron 100 μL de extracto crudo y se adicionaron 100 μL de
ácido tricloroacético al 20% (para la remoción de proteínas en la muestra). Esta
mezcla se centrifugó a 4000 rpm a 5°C durante 10 minutos. Del sobrenadante
obtenido se tomaron 100 μL, y se agregó 1 mL de solución de antrona (0.1%
disuelta en ácido sulfúrico concentrado), después se incubaron a 90°C durante
cuatro minutos y se detuvo la reacción en un baño frío a 4°C; posteriormente se
leyó la absorbancia a 620 nm.

17
Índices morfofisiológicos de peso
Índices del pie y de la masa visceral
Como una medida para evaluar la condición de los organismos se calculó el
índice del pie propuesto por Arellano-Martínez (2004) (IP = peso del pie / peso
total del organismo sin concha).También se calculó el índice de la masa visceral
propuesto por Romo-Piñera (2010) (IMV = peso de la masa visceral / peso del
organismo sin concha), ambos suponen que los valores altos indican que los
organismos presentan una buena condición, mientras que los valores bajos
indican que los organismos han tenido un desgaste en su condición nutricional.
Índice hepatogonadal
Como una medida de la madurez gonádica se calculó el índice
hepatogonadal (IHG = peso de la glándula hepatogonadal / peso total del
organismo sin concha X 100). De acuerdo al criterio de Villalejo-Fuerte y Ceballos-
Vázquez (1996), los valores bajos de IHG, se interpretaron como evidencia de
inactividad gametogénica o posdesove, mientras que un aumento sostenido del
IHG indica actividad gametogénica. Los puntos máximos en la curva fueron
considerados como evidencia de madurez y una disminución sostenida, como
evidencia de población en desove.
Índice de condición
Se calculó el índice de condición utilizado por Villalejo-Fuerte y Ceballos-Vázquez
(1996): IC = (peso del organismo sin concha / peso total del organismo) X100. Los
valores más bajos indican la etapa de desarrollo gonadal y los valores más altos
coinciden con las de madurez y desove.

18
Índices morfofisiológicos de área
Índice gonádico
Como un indicador de la época reproductiva se calculó el índice propuesto
por Poore (1973) (IG = área de la gónada / área del complejo hepatogonadal).
Las áreas se midieron a partir de imágenes digitales obtenidas de
preparaciones histológicas de cortes transversales del complejo hepatogonadal,
utilizando el programa Sigma Scan Pro versión 5.
Índice hepático
Como un indicador de la transferencia de nutrientes durante el ciclo
reproductivo, se calculó un índice hepático (IH= área del hepatopáncreas/ área del
complejo hepatogonadal). Las áreas se midieron a partir de imágenes digitales,
obtenidas de preparaciones histológicas de cortes transversales del complejo
hepatogonadal, utilizando el programa Sigma Scan Pro versión 5.
Análisis de la variación del diámetro de los ovocitos (AVDO)
Con base en el estudio histológico, se efectuó el seguimiento del diámetro
promedio mensual de los ovocitos (Grant y Tyler, 1983). De las preparaciones
histológicas, se seleccionaron cinco por mes, que correspondieran a hembras en
etapas de desarrollo o madurez y se tomaron fotografías con un fotomicroscopio.
Utilizando el programa de cómputo Sigma Scan Pro Versión 5, se midió y
promedio el diámetro mayor y el diámetro menor de alrededor 50 ovocitos de cada
preparación, seleccionando sólo aquellos que presentaban el nucléolo. Con los
datos obtenidos se determinó el diámetro promedio. Cuando el tamaño promedio
de los ovocitos fue mayor, se consideró que la gónada se encontraba en su fase
de máxima madurez; cuando se observó una caída brusca del tamaño promedio
de los ovocitos, se consideró que se ha efectuado un desove.

19
Factores ambientales
Para saber si existe una relación entre el ciclo reproductivo y el ciclo de
almacenamiento de nutrientes con algunos factores ambiéntales. Se obtuvieron
registros de temperatura de septiembre de 2011 a agosto de 2013 en las zonas de
pesca II y III, a partir de imágenes de satélite de la NOAA, e información del
fotoperiodo de la página del Observatorio Naval de los Estados Unidos (USNO,
2015).
Análisis estadísticos
Para determinar la relación entre la frecuencia relativa de organismos en
cada fase del ciclo reproductivo con la temperatura, el fotoperiodo, los diferentes
índices y el contenido de nutrientes en cada uno de los tejidos, se realizó un
análisis de correlación de rangos de Spearman.
Para comparar la diferencia en el diámetro de los ovocitos en cada uno de
los meses muestreados, así como la diferencia en los índices calculados para
cada uno de los meses se realizaron análisis de variancia de una vía. Para
determinar la diferencia del contenido de nutrientes en cada uno de los tejidos
durante las distintas fases del ciclo reproductivo, se aplicó un análisis de variancia
de una vía, seguido por una prueba de Tukey para la comparación de medias y así
detectar en qué meses o fases del ciclo se observaron esas diferencias (Sokal y
Rohlf, 1979).
RESULTADOS
Proporción de sexos
Haliotis fulgens
Se analizaron 1011 ejemplares de Haliotis fulgens, 420 de la zona de pesca
II y 591 de la III (Tabla 1). Una prueba de X2 demostró que existe diferencia
significativa en la proporción de sexos de 1:1, de una hembra por cada 0.6
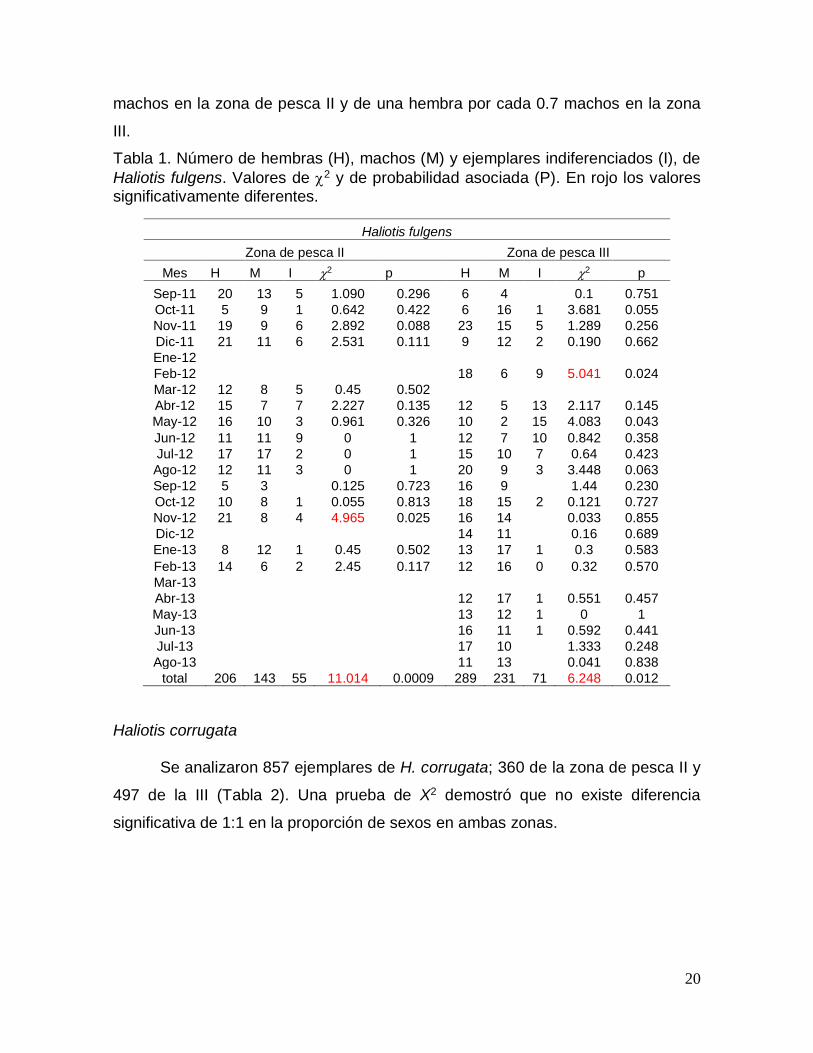
20
machos en la zona de pesca II y de una hembra por cada 0.7 machos en la zona
III.
Tabla 1. Número de hembras (H), machos (M) y ejemplares indiferenciados (I), de
Haliotis fulgens. Valores de 2 y de probabilidad asociada (P). En rojo los valores significativamente diferentes.
Haliotis fulgens
Zona de pesca II Zona de pesca III
Mes H M I 2 p H M I 2 p
Sep-11 20 13 5 1.090 0.296 6 4
0.1 0.751
Oct-11 5 9 1 0.642 0.422 6 16 1 3.681 0.055
Nov-11 19 9 6 2.892 0.088 23 15 5 1.289 0.256
Dic-11 21 11 6 2.531 0.111 9 12 2 0.190 0.662
Ene-12
Feb-12
18 6 9 5.041 0.024
Mar-12 12 8 5 0.45 0.502
Abr-12 15 7 7 2.227 0.135 12 5 13 2.117 0.145
May-12 16 10 3 0.961 0.326 10 2 15 4.083 0.043
Jun-12 11 11 9 0 1 12 7 10 0.842 0.358
Jul-12 17 17 2 0 1 15 10 7 0.64 0.423
Ago-12 12 11 3 0 1 20 9 3 3.448 0.063
Sep-12 5 3
0.125 0.723 16 9
1.44 0.230
Oct-12 10 8 1 0.055 0.813 18 15 2 0.121 0.727
Nov-12 21 8 4 4.965 0.025 16 14
0.033 0.855
Dic-12
14 11
0.16 0.689
Ene-13 8 12 1 0.45 0.502 13 17 1 0.3 0.583
Feb-13 14 6 2 2.45 0.117 12 16 0 0.32 0.570
Mar-13
Abr-13
12 17 1 0.551 0.457
May-13
13 12 1 0 1
Jun-13
16 11 1 0.592 0.441
Jul-13
17 10
1.333 0.248
Ago-13
11 13
0.041 0.838
total 206 143 55 11.014 0.0009 289 231 71 6.248 0.012
Haliotis corrugata
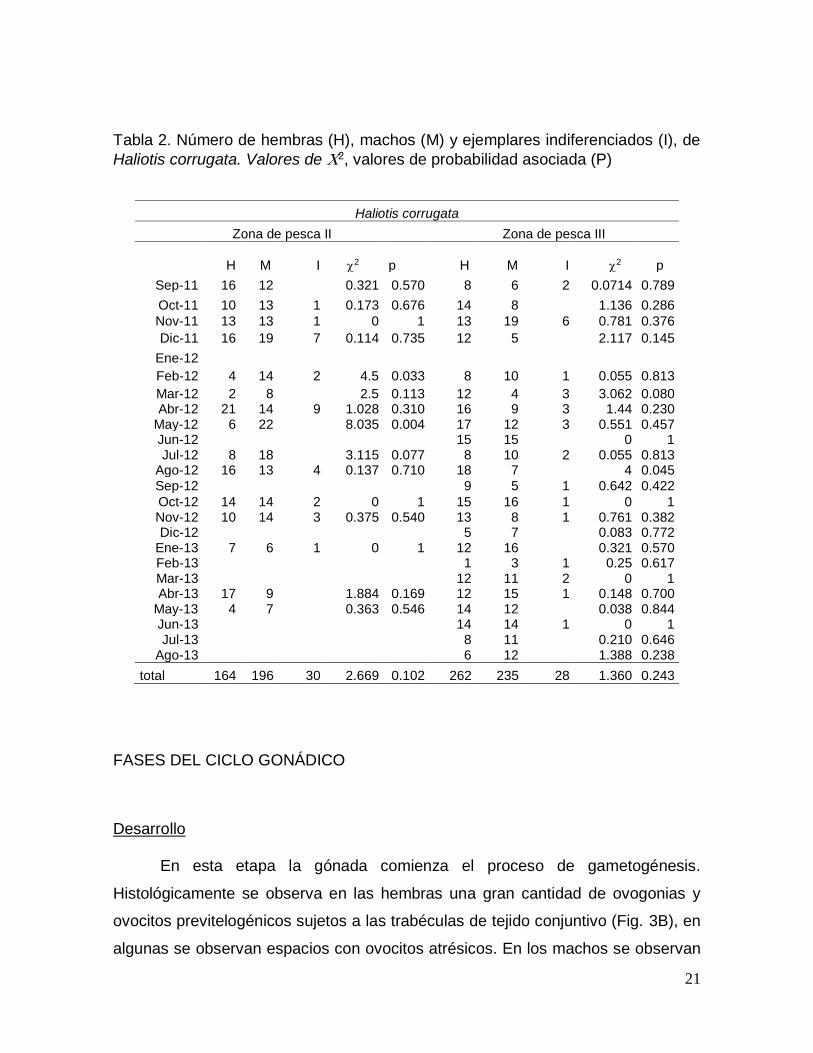
Se analizaron 857 ejemplares de H. corrugata; 360 de la zona de pesca II y
497 de la III (Tabla 2). Una prueba de X2 demostró que no existe diferencia
significativa de 1:1 en la proporción de sexos en ambas zonas.
21
Tabla 2. Número de hembras (H), machos (M) y ejemplares indiferenciados (I), de
Haliotis corrugata. Valores de 2, valores de probabilidad asociada (P)
Haliotis corrugata
Zona de pesca II Zona de pesca III
H M I 2
p H M I 2 p
Sep-11 16 12
0.321 0.570 8 6 2 0.0714 0.789
Oct-11 10 13 1 0.173 0.676 14 8
1.136 0.286
Nov-11 13 13 1 0 1 13 19 6 0.781 0.376
Dic-11 16 19 7 0.114 0.735 12 5
2.117 0.145
Ene-12 Feb-12 4 14 2 4.5 0.033 8 10 1 0.055 0.813
Mar-12 2 8
2.5 0.113 12 4 3 3.062 0.080 Abr-12 21 14 9 1.028 0.310 16 9 3 1.44 0.230
May-12 6 22
8.035 0.004 17 12 3 0.551 0.457 Jun-12
15 15
0 1
Jul-12 8 18
3.115 0.077 8 10 2 0.055 0.813 Ago-12 16 13 4 0.137 0.710 18 7
4 0.045
Sep-12
9 5 1 0.642 0.422
Oct-12 14 14 2 0 1 15 16 1 0 1 Nov-12 10 14 3 0.375 0.540 13 8 1 0.761 0.382 Dic-12
5 7
0.083 0.772
Ene-13 7 6 1 0 1 12 16
0.321 0.570 Feb-13
1 3 1 0.25 0.617
Mar-13
12 11 2 0 1 Abr-13 17 9
1.884 0.169 12 15 1 0.148 0.700
May-13 4 7
0.363 0.546 14 12
0.038 0.844 Jun-13
14 14 1 0 1
Jul-13
8 11
0.210 0.646 Ago-13
6 12
1.388 0.238
total 164 196 30 2.669 0.102 262 235 28 1.360 0.243
FASES DEL CICLO GONÁDICO
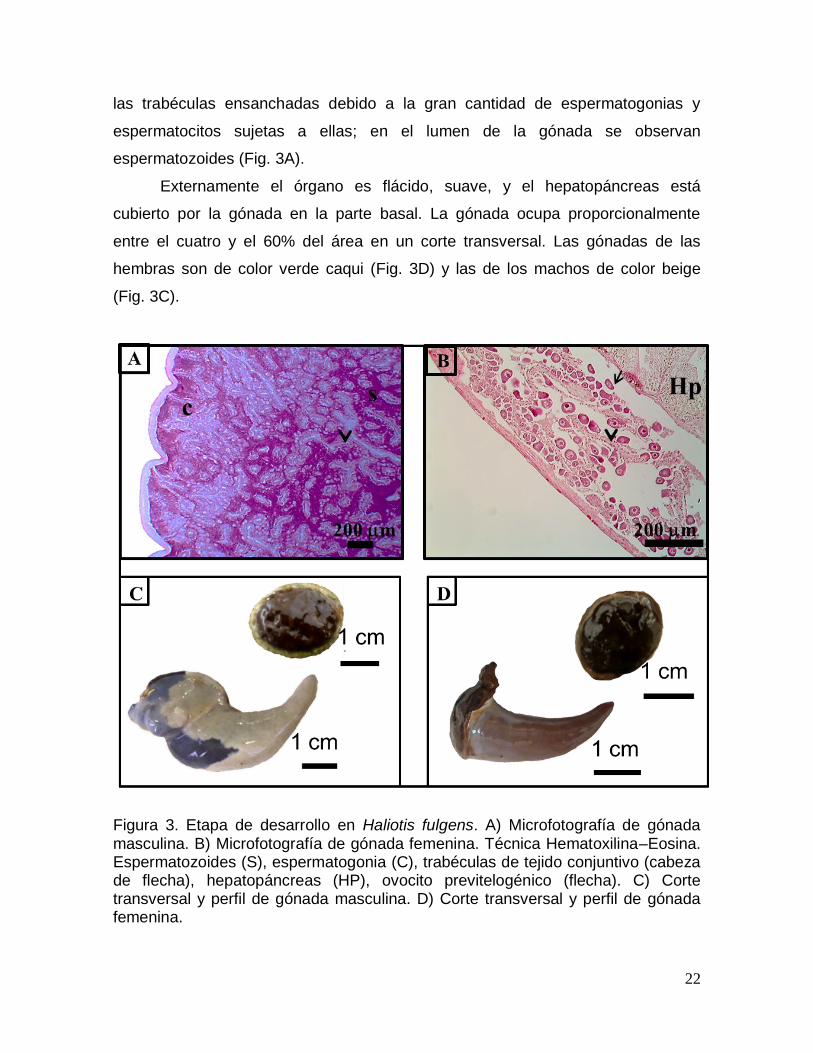
Desarrollo
En esta etapa la gónada comienza el proceso de gametogénesis.
Histológicamente se observa en las hembras una gran cantidad de ovogonias y
ovocitos previtelogénicos sujetos a las trabéculas de tejido conjuntivo (Fig. 3B), en
algunas se observan espacios con ovocitos atrésicos. En los machos se observan
22
las trabéculas ensanchadas debido a la gran cantidad de espermatogonias y
espermatocitos sujetas a ellas; en el lumen de la gónada se observan
espermatozoides (Fig. 3A).
Externamente el órgano es flácido, suave, y el hepatopáncreas está
cubierto por la gónada en la parte basal. La gónada ocupa proporcionalmente
entre el cuatro y el 60% del área en un corte transversal. Las gónadas de las
hembras son de color verde caqui (Fig. 3D) y las de los machos de color beige
(Fig. 3C).
Figura 3. Etapa de desarrollo en Haliotis fulgens. A) Microfotografía de gónada masculina. B) Microfotografía de gónada femenina. Técnica Hematoxilina–Eosina. Espermatozoides (S), espermatogonia (C), trabéculas de tejido conjuntivo (cabeza de flecha), hepatopáncreas (HP), ovocito previtelogénico (flecha). C) Corte transversal y perfil de gónada masculina. D) Corte transversal y perfil de gónada femenina.
23
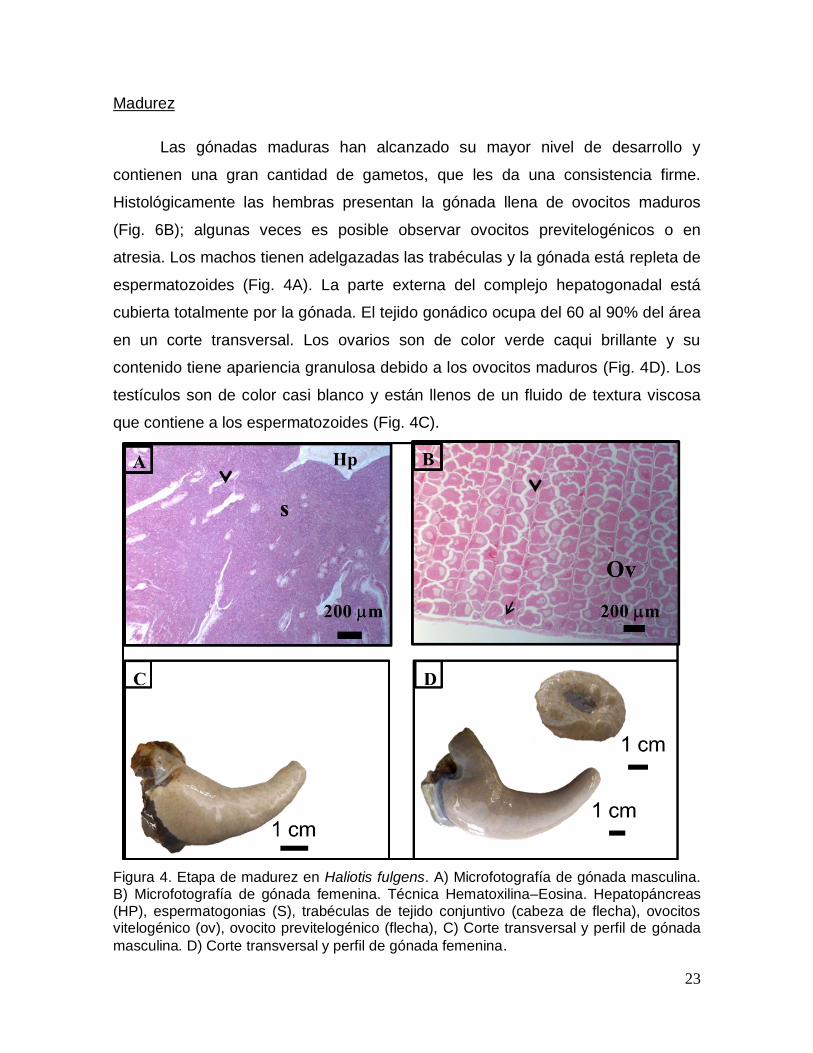
Madurez
Las gónadas maduras han alcanzado su mayor nivel de desarrollo y
contienen una gran cantidad de gametos, que les da una consistencia firme.
Histológicamente las hembras presentan la gónada llena de ovocitos maduros
(Fig. 6B); algunas veces es posible observar ovocitos previtelogénicos o en
atresia. Los machos tienen adelgazadas las trabéculas y la gónada está repleta de
espermatozoides (Fig. 4A). La parte externa del complejo hepatogonadal está
cubierta totalmente por la gónada. El tejido gonádico ocupa del 60 al 90% del área
en un corte transversal. Los ovarios son de color verde caqui brillante y su
contenido tiene apariencia granulosa debido a los ovocitos maduros (Fig. 4D). Los
testículos son de color casi blanco y están llenos de un fluido de textura viscosa
que contiene a los espermatozoides (Fig. 4C).
Figura 4. Etapa de madurez en Haliotis fulgens. A) Microfotografía de gónada masculina. B) Microfotografía de gónada femenina. Técnica Hematoxilina–Eosina. Hepatopáncreas (HP), espermatogonias (S), trabéculas de tejido conjuntivo (cabeza de flecha), ovocitos vitelogénico (ov), ovocito previtelogénico (flecha), C) Corte transversal y perfil de gónada
masculina. D) Corte transversal y perfil de gónada femenina.
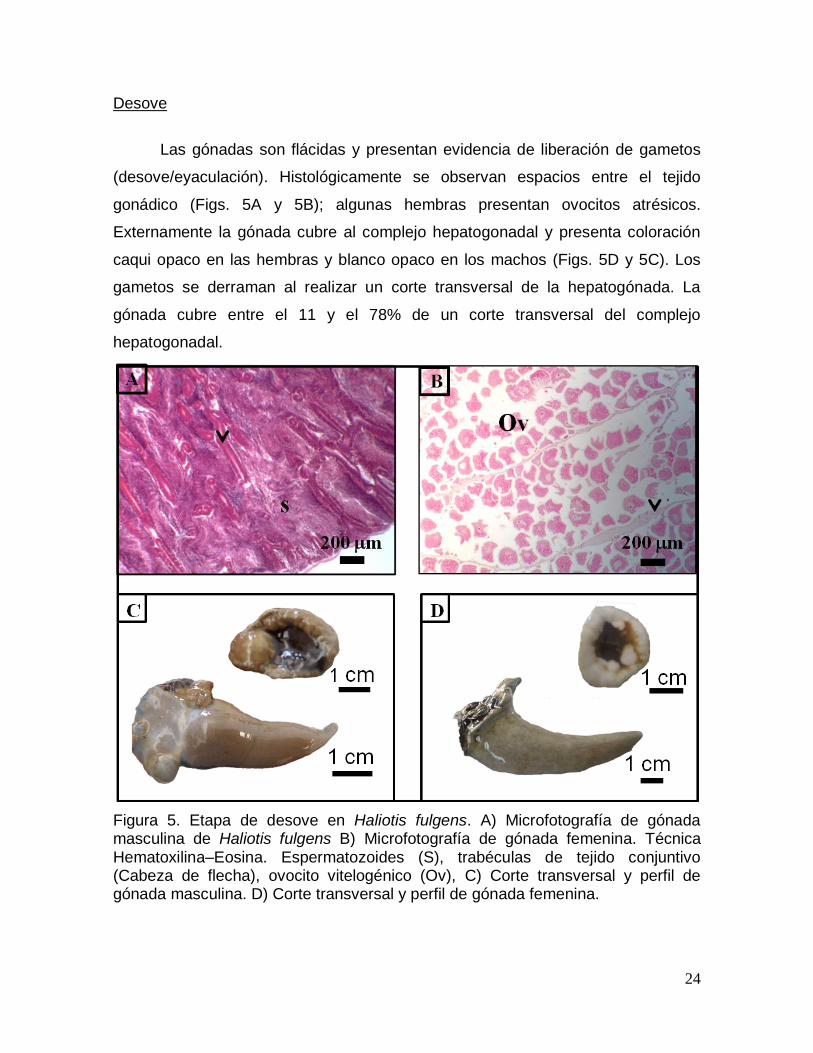
24
Desove
Las gónadas son flácidas y presentan evidencia de liberación de gametos
(desove/eyaculación). Histológicamente se observan espacios entre el tejido
gonádico (Figs. 5A y 5B); algunas hembras presentan ovocitos atrésicos.
Externamente la gónada cubre al complejo hepatogonadal y presenta coloración
caqui opaco en las hembras y blanco opaco en los machos (Figs. 5D y 5C). Los
gametos se derraman al realizar un corte transversal de la hepatogónada. La
gónada cubre entre el 11 y el 78% de un corte transversal del complejo
hepatogonadal.
Figura 5. Etapa de desove en Haliotis fulgens. A) Microfotografía de gónada masculina de Haliotis fulgens B) Microfotografía de gónada femenina. Técnica Hematoxilina–Eosina. Espermatozoides (S), trabéculas de tejido conjuntivo (Cabeza de flecha), ovocito vitelogénico (Ov), C) Corte transversal y perfil de gónada masculina. D) Corte transversal y perfil de gónada femenina.
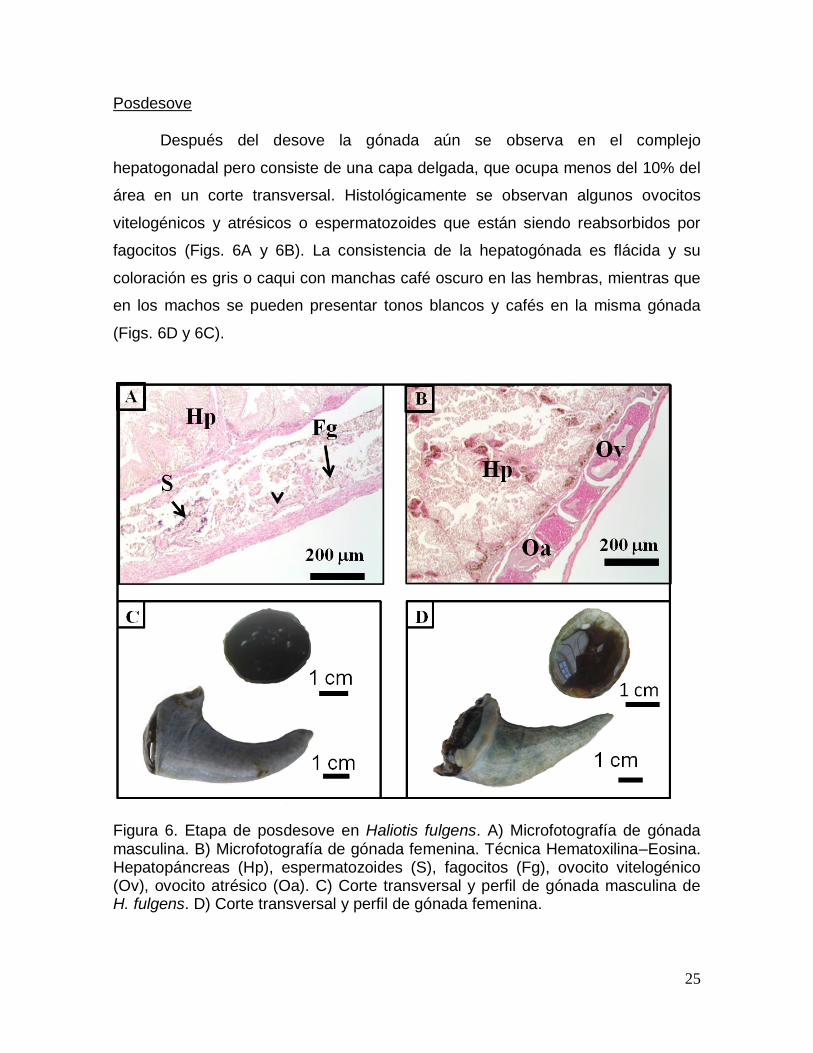
25
Posdesove
Después del desove la gónada aún se observa en el complejo
hepatogonadal pero consiste de una capa delgada, que ocupa menos del 10% del
área en un corte transversal. Histológicamente se observan algunos ovocitos
vitelogénicos y atrésicos o espermatozoides que están siendo reabsorbidos por
fagocitos (Figs. 6A y 6B). La consistencia de la hepatogónada es flácida y su
coloración es gris o caqui con manchas café oscuro en las hembras, mientras que
en los machos se pueden presentar tonos blancos y cafés en la misma gónada
(Figs. 6D y 6C).
Figura 6. Etapa de posdesove en Haliotis fulgens. A) Microfotografía de gónada masculina. B) Microfotografía de gónada femenina. Técnica Hematoxilina–Eosina. Hepatopáncreas (Hp), espermatozoides (S), fagocitos (Fg), ovocito vitelogénico (Ov), ovocito atrésico (Oa). C) Corte transversal y perfil de gónada masculina de H. fulgens. D) Corte transversal y perfil de gónada femenina.

26
Indiferenciación
La gónada no presenta evidencia de gametogénesis, es decir, las células
germinales en el epitelio que envuelve al complejo hepatogonadal, no se han
diferenciado a gametos. El complejo hepatogonadal está compuesto casi
completamente por hepatopáncreas y es posible observar un espacio vacío entre
el epitelio que cubre al complejo y el hepatopáncreas (Fig. 7A). Externamente el
órgano es de consistencia suave, flácido al tacto; el epitelio que lo cubre es
delgado y tiene una apariencia blanquecina casi transparente, que permite ver la
coloración externa casi negra del hepatopáncreas. Presenta tamaños entre 2 y 6
cm y en un corte transversal se observa el hepatopáncreas de color café (Fig. 7B).
Figura 7. Gónada de en etapa de indiferenciación de Haliotis fulgens. A) Corte transversal con la técnica histológica. Técnica Hematoxilina–Eosina. Hepatopáncreas (hp), espacio gonadal vacío (cabeza de flecha). B) Morfología externa del complejo hepatogonadal y corte transversal en fresco.
CICLO REPRODUCTIVO
Haliotis fulgens
El proceso de diferenciación de etapas de desarrollo gonadal y su
frecuencia relativa de aparición por mes permite apreciar el desarrollo del ciclo
reproductivo.

27
En las preparaciones histológicas de H. fulgens en la zona de pesca II fue posible
observar organismos indiferenciados, en desarrollo y desove durante todo el
periodo de muestreo, lo que indica reproducción continua durante todo el año (Fig.
8A). La presencia de organismos en desove y madurez fue más frecuente de
septiembre de 2011 a diciembre de 2011 y de octubre de 2012 a febrero de 2013
(mayor al 40%). La etapa de posdesove no se presentó en septiembre de 2011,
octubre de 2012 y febrero de 2013 (Fig. 8A).
En la zona de pesca III, los resultados muestran la creciente presencia de
individuos indiferenciados a partir de septiembre de 2011 y su predominancia
hasta agosto de 2012. La etapa de desarrollo se presentó en todo el periodo,
excepto en octubre de 2011, con mayor frecuencia de febrero a agosto de 2013,
que en los mismos meses en 2012 (mayor al 30%). La madurez y el desove se
observaron de febrero a mayo, se incrementaron de junio a septiembre y
predominaron hasta febrero de 2013, lo que sugiere la presencia de cambios en
factores ambientales que influyen en el proceso de maduración. Finalmente el
posdesove se presentó durante todo el periodo de estudio, con excepción de
febrero, julio y agosto de 2013; la mayor frecuencia se observó en septiembre de
2011 (30%) (Fig. 8B).
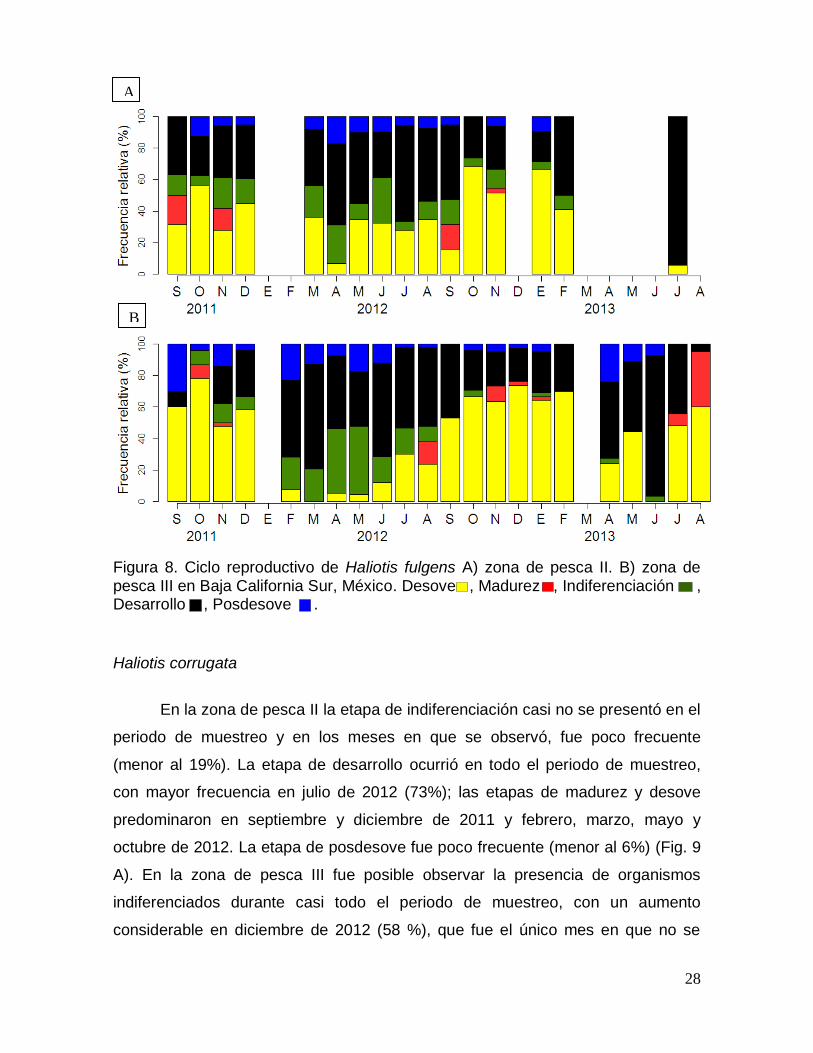
28
Figura 8. Ciclo reproductivo de Haliotis fulgens A) zona de pesca II. B) zona de pesca III en Baja California Sur, México. Desove , Madurez , Indiferenciación , Desarrollo , Posdesove .
Haliotis corrugata
En la zona de pesca II la etapa de indiferenciación casi no se presentó en el
periodo de muestreo y en los meses en que se observó, fue poco frecuente
(menor al 19%). La etapa de desarrollo ocurrió en todo el periodo de muestreo,
con mayor frecuencia en julio de 2012 (73%); las etapas de madurez y desove
predominaron en septiembre y diciembre de 2011 y febrero, marzo, mayo y
octubre de 2012. La etapa de posdesove fue poco frecuente (menor al 6%) (Fig. 9
A). En la zona de pesca III fue posible observar la presencia de organismos
indiferenciados durante casi todo el periodo de muestreo, con un aumento
considerable en diciembre de 2012 (58 %), que fue el único mes en que no se
A
B
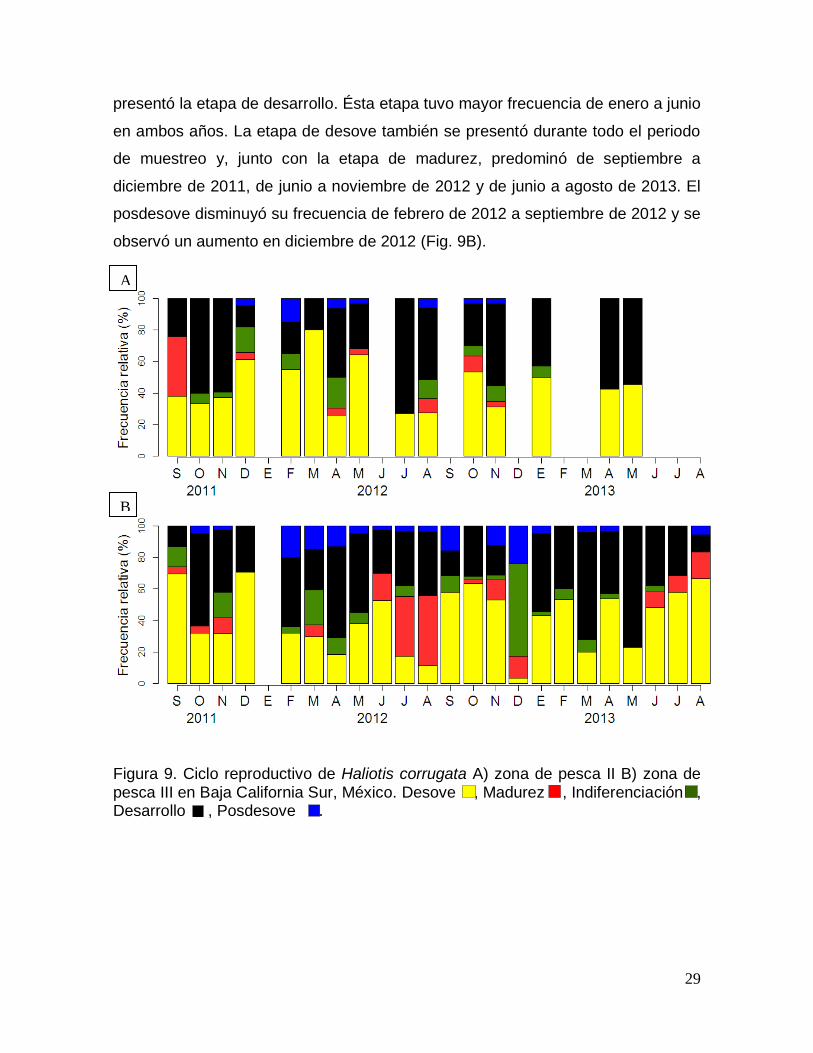
29
presentó la etapa de desarrollo. Ésta etapa tuvo mayor frecuencia de enero a junio
en ambos años. La etapa de desove también se presentó durante todo el periodo
de muestreo y, junto con la etapa de madurez, predominó de septiembre a
diciembre de 2011, de junio a noviembre de 2012 y de junio a agosto de 2013. El
posdesove disminuyó su frecuencia de febrero de 2012 a septiembre de 2012 y se
observó un aumento en diciembre de 2012 (Fig. 9B).
Figura 9. Ciclo reproductivo de Haliotis corrugata A) zona de pesca II B) zona de pesca III en Baja California Sur, México. Desove , Madurez , Indiferenciación , Desarrollo , Posdesove . .
A
B
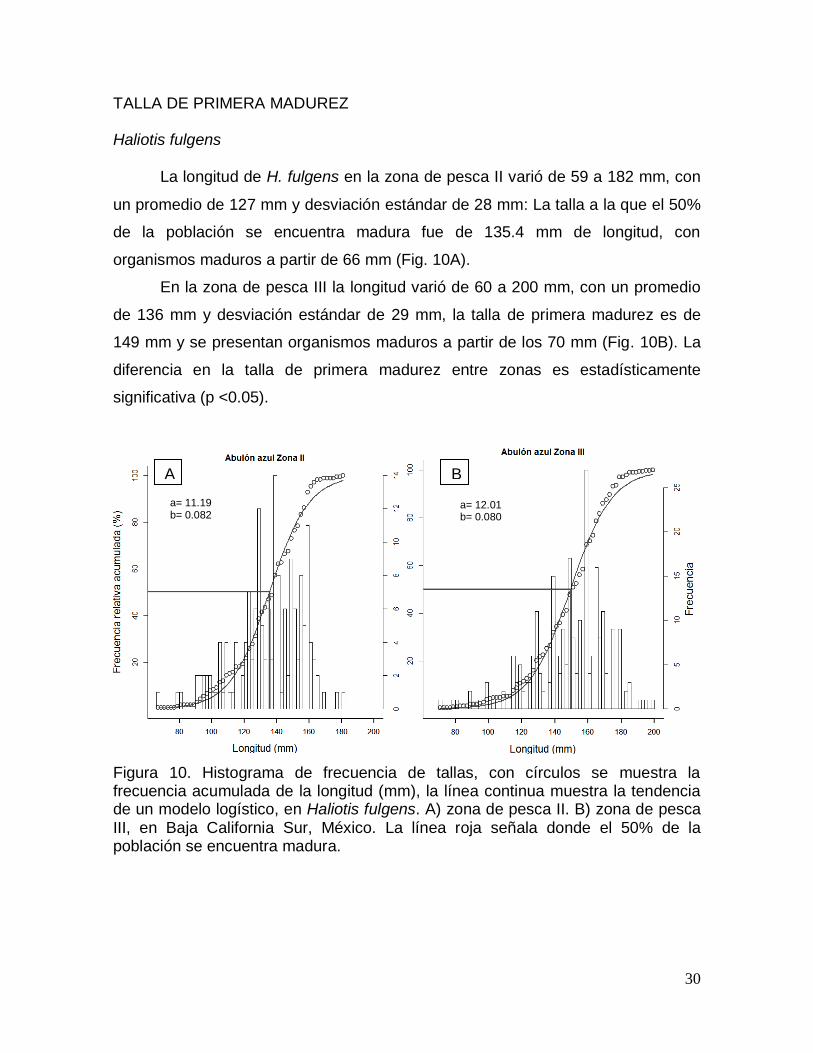
30
TALLA DE PRIMERA MADUREZ
Haliotis fulgens
La longitud de H. fulgens en la zona de pesca II varió de 59 a 182 mm, con
un promedio de 127 mm y desviación estándar de 28 mm: La talla a la que el 50%
de la población se encuentra madura fue de 135.4 mm de longitud, con
organismos maduros a partir de 66 mm (Fig. 10A).
En la zona de pesca III la longitud varió de 60 a 200 mm, con un promedio
de 136 mm y desviación estándar de 29 mm, la talla de primera madurez es de
149 mm y se presentan organismos maduros a partir de los 70 mm (Fig. 10B). La
diferencia en la talla de primera madurez entre zonas es estadísticamente
significativa (p <0.05).
Figura 10. Histograma de frecuencia de tallas, con círculos se muestra la frecuencia acumulada de la longitud (mm), la línea continua muestra la tendencia de un modelo logístico, en Haliotis fulgens. A) zona de pesca II. B) zona de pesca III, en Baja California Sur, México. La línea roja señala donde el 50% de la población se encuentra madura.
A B
a= 11.19 b= 0.082
a= 12.01 b= 0.080
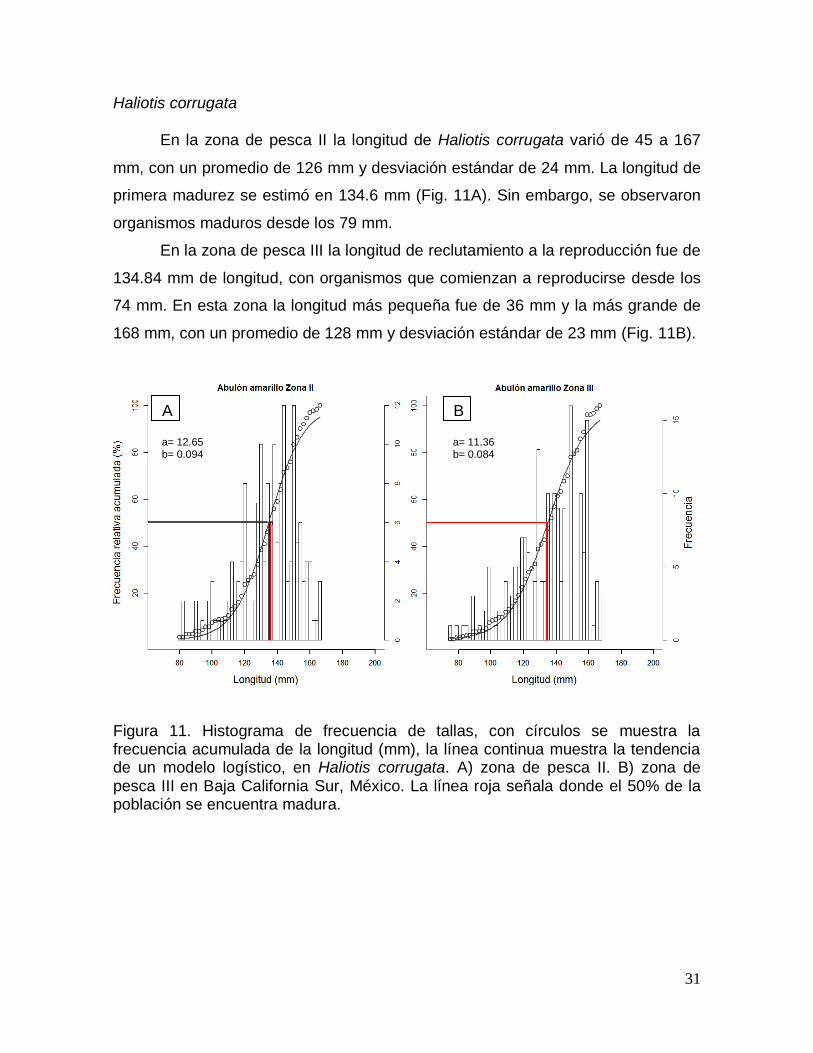
31
Haliotis corrugata
En la zona de pesca II la longitud de Haliotis corrugata varió de 45 a 167
mm, con un promedio de 126 mm y desviación estándar de 24 mm. La longitud de
primera madurez se estimó en 134.6 mm (Fig. 11A). Sin embargo, se observaron
organismos maduros desde los 79 mm.
En la zona de pesca III la longitud de reclutamiento a la reproducción fue de
134.84 mm de longitud, con organismos que comienzan a reproducirse desde los
74 mm. En esta zona la longitud más pequeña fue de 36 mm y la más grande de
168 mm, con un promedio de 128 mm y desviación estándar de 23 mm (Fig. 11B).
Figura 11. Histograma de frecuencia de tallas, con círculos se muestra la frecuencia acumulada de la longitud (mm), la línea continua muestra la tendencia de un modelo logístico, en Haliotis corrugata. A) zona de pesca II. B) zona de pesca III en Baja California Sur, México. La línea roja señala donde el 50% de la población se encuentra madura.
A B
a= 12.65 b= 0.094
a= 11.36 b= 0.084
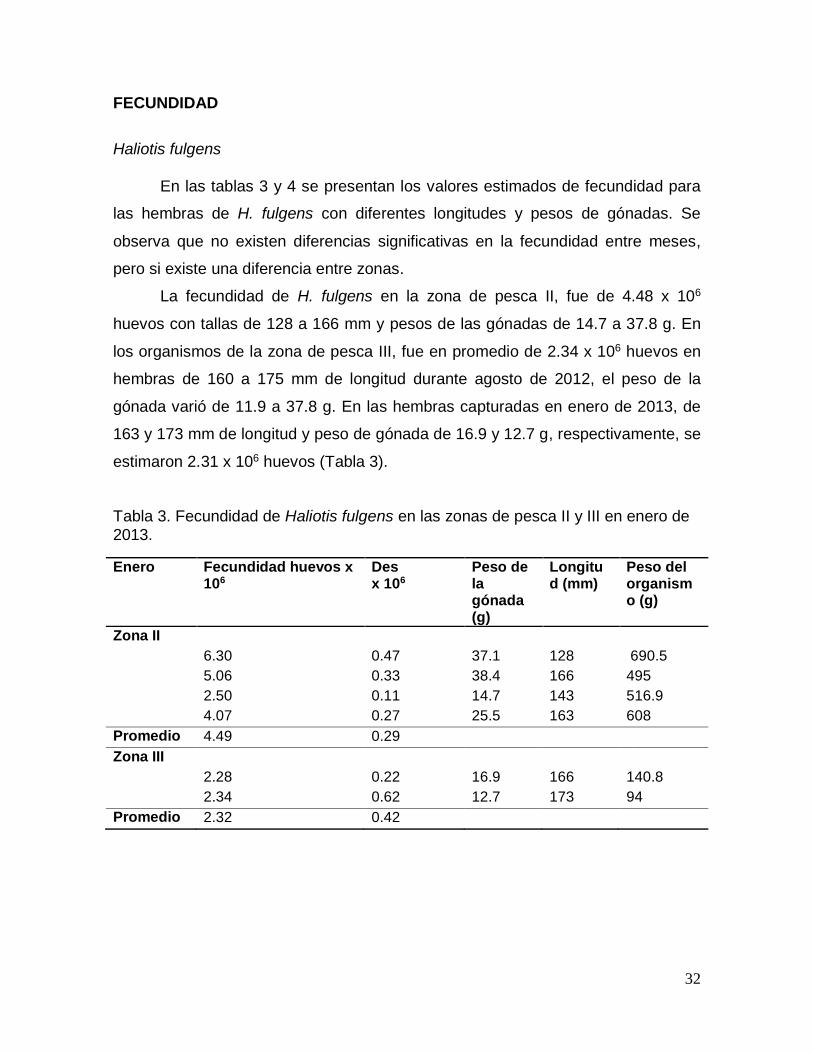
32
FECUNDIDAD
Haliotis fulgens
En las tablas 3 y 4 se presentan los valores estimados de fecundidad para
las hembras de H. fulgens con diferentes longitudes y pesos de gónadas. Se
observa que no existen diferencias significativas en la fecundidad entre meses,
pero si existe una diferencia entre zonas.
La fecundidad de H. fulgens en la zona de pesca II, fue de 4.48 x 106
huevos con tallas de 128 a 166 mm y pesos de las gónadas de 14.7 a 37.8 g. En
los organismos de la zona de pesca III, fue en promedio de 2.34 x 106 huevos en
hembras de 160 a 175 mm de longitud durante agosto de 2012, el peso de la
gónada varió de 11.9 a 37.8 g. En las hembras capturadas en enero de 2013, de
163 y 173 mm de longitud y peso de gónada de 16.9 y 12.7 g, respectivamente, se
estimaron 2.31 x 106 huevos (Tabla 3).
Tabla 3. Fecundidad de Haliotis fulgens en las zonas de pesca II y III en enero de 2013.
Enero Fecundidad huevos x 106
Des x 106
Peso de la gónada (g)
Longitud (mm)
Peso del organismo (g)
Zona II
6.30 0.47 37.1 128 690.5
5.06 0.33 38.4 166 495
2.50 0.11 14.7 143 516.9
4.07 0.27 25.5 163 608
Promedio 4.49 0.29
Zona III
2.28 0.22 16.9 166 140.8
2.34 0.62 12.7 173 94
Promedio 2.32 0.42
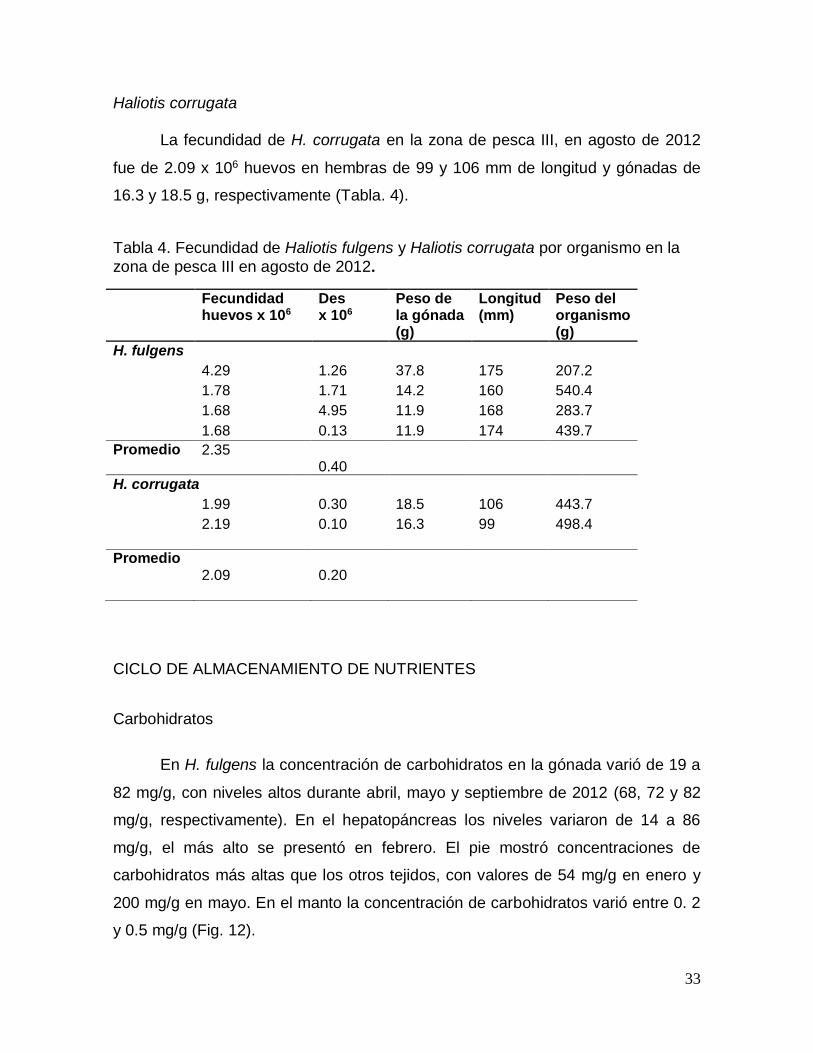
33
Haliotis corrugata
La fecundidad de H. corrugata en la zona de pesca III, en agosto de 2012
fue de 2.09 x 106 huevos en hembras de 99 y 106 mm de longitud y gónadas de
16.3 y 18.5 g, respectivamente (Tabla. 4).
Tabla 4. Fecundidad de Haliotis fulgens y Haliotis corrugata por organismo en la
zona de pesca III en agosto de 2012.
Fecundidad huevos x 106
Des x 106
Peso de la gónada (g)
Longitud (mm)
Peso del organismo (g)
H. fulgens
4.29 1.26 37.8 175 207.2
1.78 1.71 14.2 160 540.4
1.68 4.95 11.9 168 283.7
1.68 0.13 11.9 174 439.7
Promedio 2.35 0.40
H. corrugata
1.99 0.30 18.5 106 443.7
2.19 0.10 16.3 99
498.4
Promedio 2.09
0.20
CICLO DE ALMACENAMIENTO DE NUTRIENTES
Carbohidratos
En H. fulgens la concentración de carbohidratos en la gónada varió de 19 a
82 mg/g, con niveles altos durante abril, mayo y septiembre de 2012 (68, 72 y 82
mg/g, respectivamente). En el hepatopáncreas los niveles variaron de 14 a 86
mg/g, el más alto se presentó en febrero. El pie mostró concentraciones de
carbohidratos más altas que los otros tejidos, con valores de 54 mg/g en enero y
200 mg/g en mayo. En el manto la concentración de carbohidratos varió entre 0. 2
y 0.5 mg/g (Fig. 12).

34
Figura 12. Comportamiento de la concentración de carbohidratos en (mg/g) de
febrero de 2012 a febrero de 2013 en el pie, manto, gónada y hepatopáncreas de
Haliotis fulgens en la zona de pesca III en Baja California Sur, México.
La concentración de carbohidratos en H. fulgens en las diferentes etapas
del ciclo reproductivo no presentó una diferencia significativa (p <0.05) (Fig. 13).
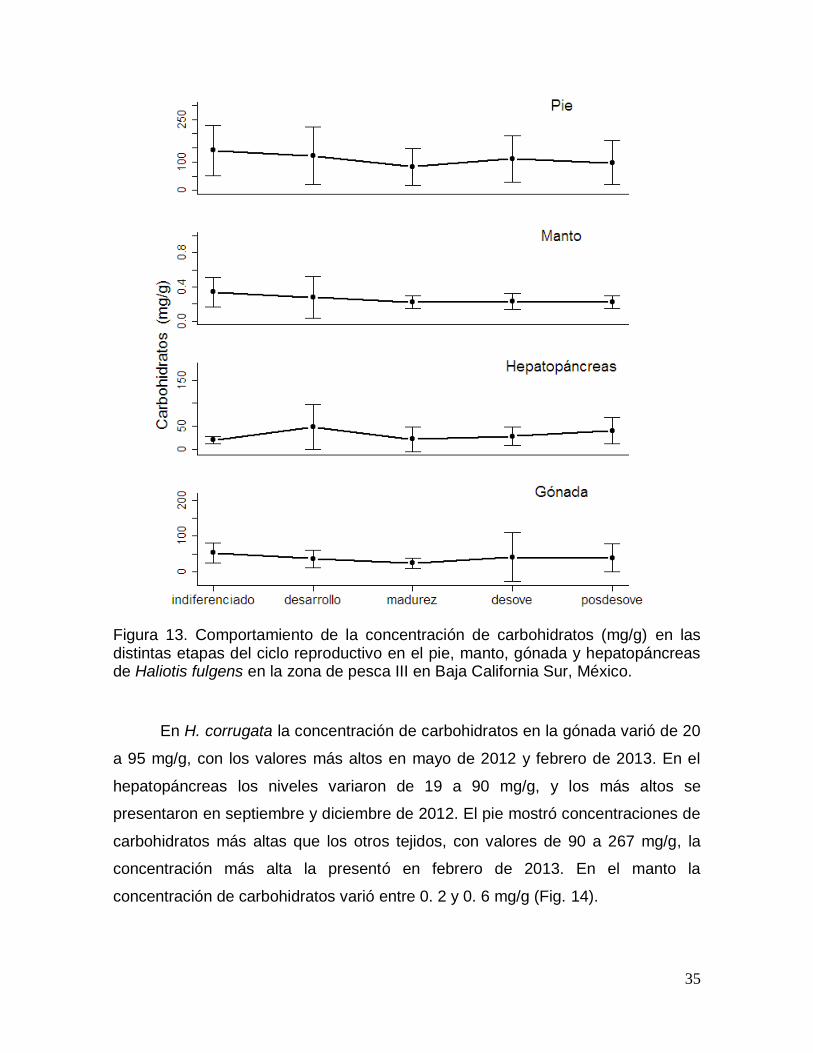
35
Figura 13. Comportamiento de la concentración de carbohidratos (mg/g) en las distintas etapas del ciclo reproductivo en el pie, manto, gónada y hepatopáncreas de Haliotis fulgens en la zona de pesca III en Baja California Sur, México.
En H. corrugata la concentración de carbohidratos en la gónada varió de 20
a 95 mg/g, con los valores más altos en mayo de 2012 y febrero de 2013. En el
hepatopáncreas los niveles variaron de 19 a 90 mg/g, y los más altos se
presentaron en septiembre y diciembre de 2012. El pie mostró concentraciones de
carbohidratos más altas que los otros tejidos, con valores de 90 a 267 mg/g, la
concentración más alta la presentó en febrero de 2013. En el manto la
concentración de carbohidratos varió entre 0. 2 y 0. 6 mg/g (Fig. 14).

36
Figura 14. Comportamiento de la concentración de carbohidratos (mg/g) de febrero de 2012 a febrero de 2013 en el pie, manto, gónada y hepatopáncreas de Haliotis corrugata en la zona de pesca III en Baja California Sur, México.
En H. corrugata en el pie y manto no se registró diferencia significativa (p<
0.05) durante las distintas etapas del ciclo reproductivo. Únicamente se notó una
diferencia en la concentración de carbohidratos en la gónada durante la etapa de
indiferenciación donde fue mayor que en las otras etapas. En el hepatopáncreas
se presentó una fuerte disminución de la concentración de carbohidratos durante
el desarrollo y un fuerte aumento en la madurez (Fig. 15).
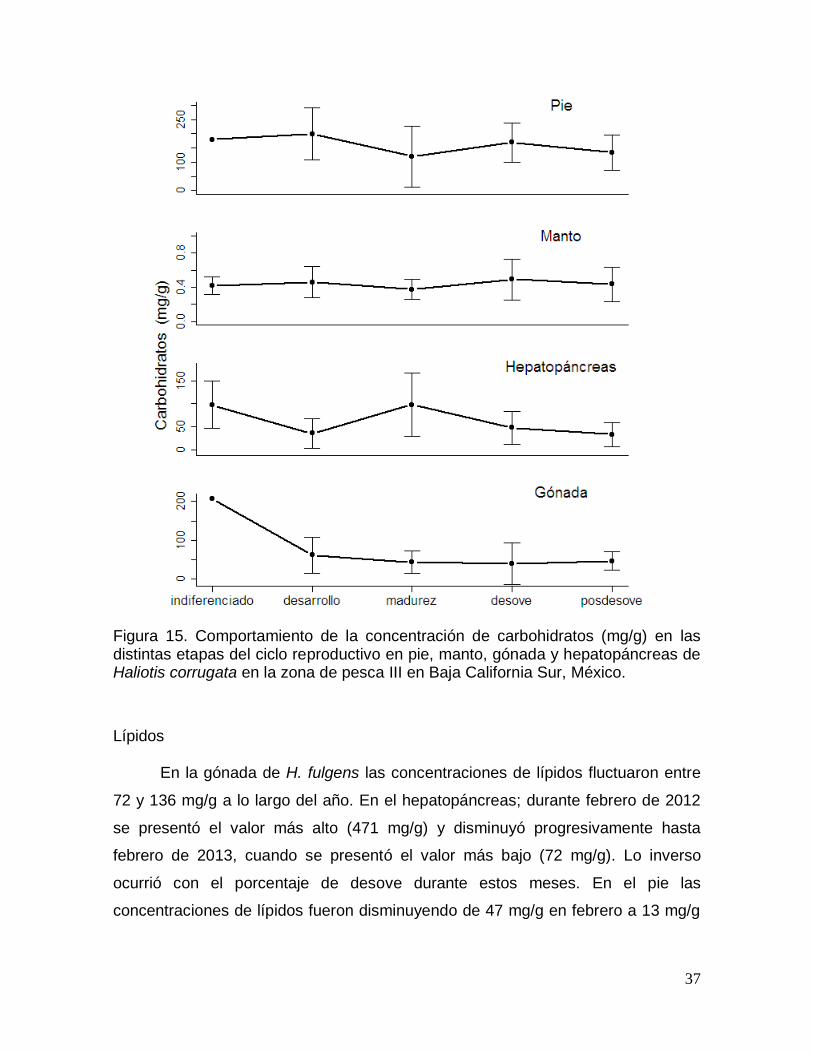
37
Figura 15. Comportamiento de la concentración de carbohidratos (mg/g) en las distintas etapas del ciclo reproductivo en pie, manto, gónada y hepatopáncreas de Haliotis corrugata en la zona de pesca III en Baja California Sur, México.
Lípidos
En la gónada de H. fulgens las concentraciones de lípidos fluctuaron entre
72 y 136 mg/g a lo largo del año. En el hepatopáncreas; durante febrero de 2012
se presentó el valor más alto (471 mg/g) y disminuyó progresivamente hasta
febrero de 2013, cuando se presentó el valor más bajo (72 mg/g). Lo inverso
ocurrió con el porcentaje de desove durante estos meses. En el pie las
concentraciones de lípidos fueron disminuyendo de 47 mg/g en febrero a 13 mg/g
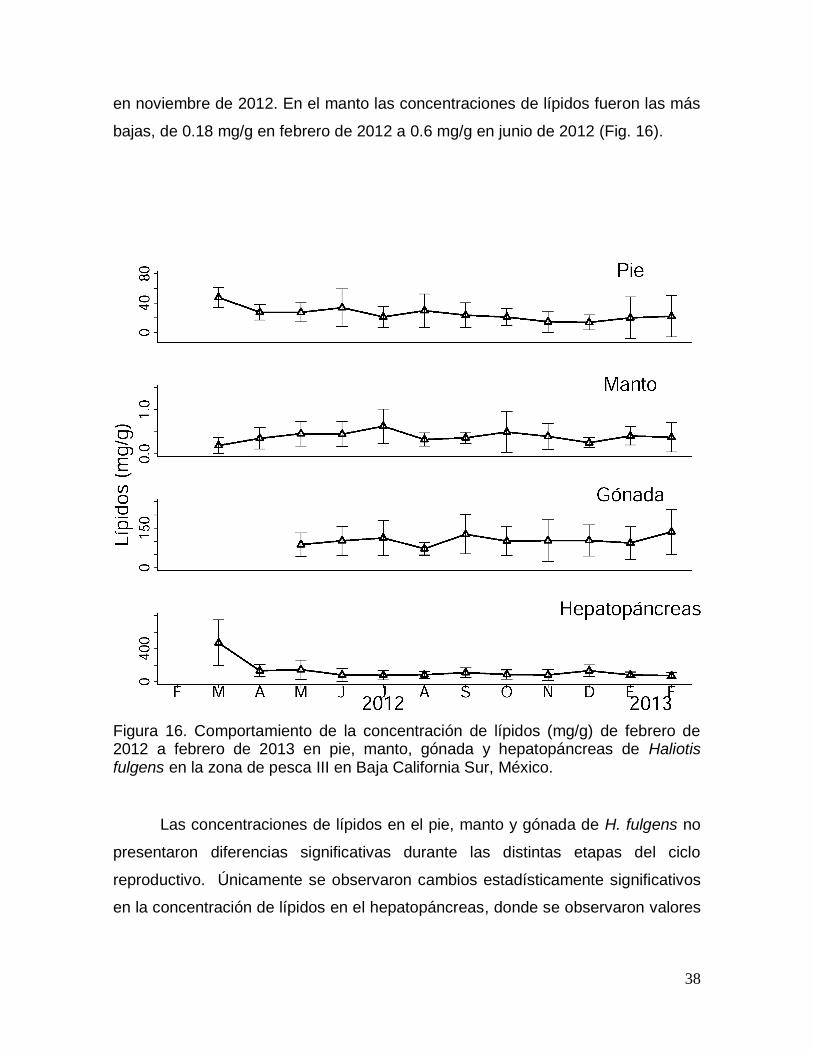
38
en noviembre de 2012. En el manto las concentraciones de lípidos fueron las más
bajas, de 0.18 mg/g en febrero de 2012 a 0.6 mg/g en junio de 2012 (Fig. 16).
Figura 16. Comportamiento de la concentración de lípidos (mg/g) de febrero de 2012 a febrero de 2013 en pie, manto, gónada y hepatopáncreas de Haliotis fulgens en la zona de pesca III en Baja California Sur, México.
Las concentraciones de lípidos en el pie, manto y gónada de H. fulgens no
presentaron diferencias significativas durante las distintas etapas del ciclo
reproductivo. Únicamente se observaron cambios estadísticamente significativos
en la concentración de lípidos en el hepatopáncreas, donde se observaron valores
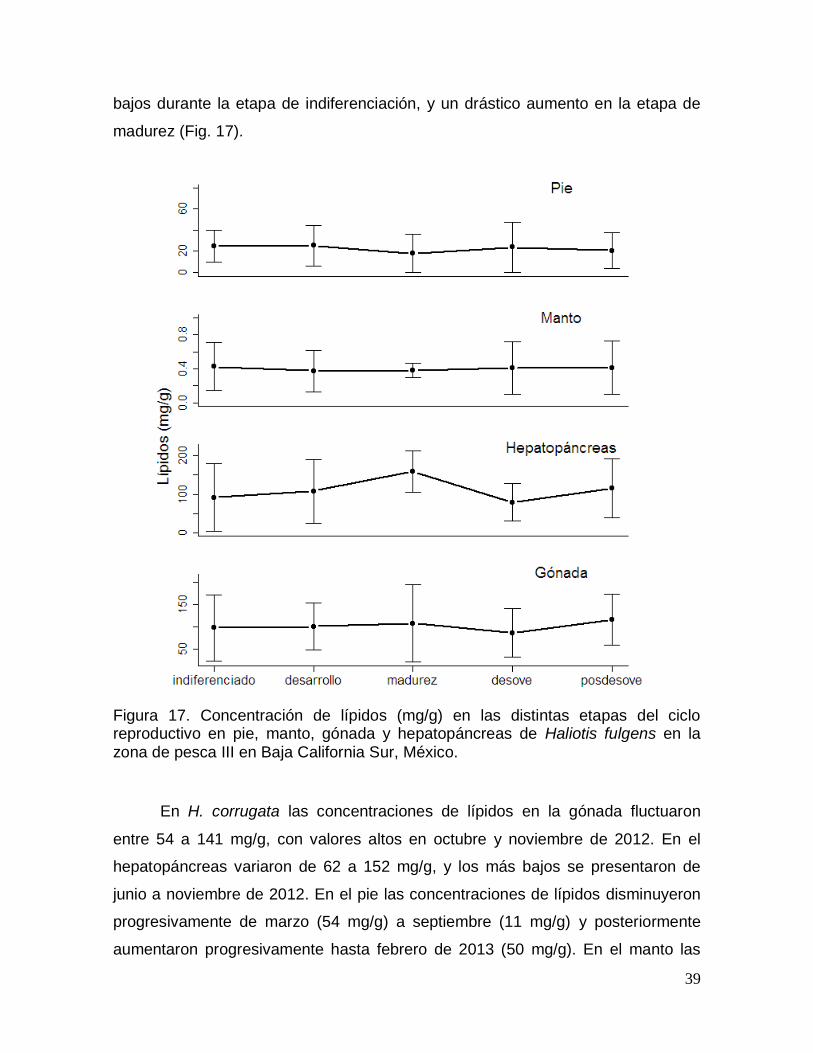
39
bajos durante la etapa de indiferenciación, y un drástico aumento en la etapa de
madurez (Fig. 17).
Figura 17. Concentración de lípidos (mg/g) en las distintas etapas del ciclo reproductivo en pie, manto, gónada y hepatopáncreas de Haliotis fulgens en la zona de pesca III en Baja California Sur, México.
En H. corrugata las concentraciones de lípidos en la gónada fluctuaron
entre 54 a 141 mg/g, con valores altos en octubre y noviembre de 2012. En el
hepatopáncreas variaron de 62 a 152 mg/g, y los más bajos se presentaron de
junio a noviembre de 2012. En el pie las concentraciones de lípidos disminuyeron
progresivamente de marzo (54 mg/g) a septiembre (11 mg/g) y posteriormente
aumentaron progresivamente hasta febrero de 2013 (50 mg/g). En el manto las
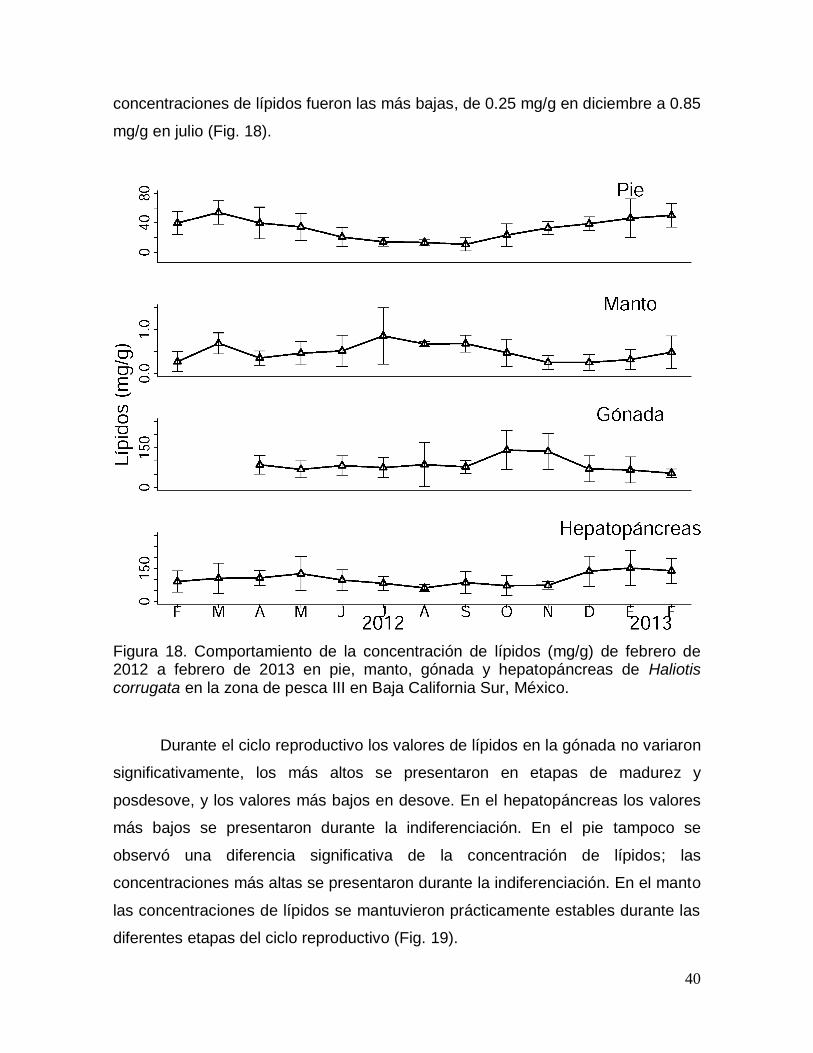
40
concentraciones de lípidos fueron las más bajas, de 0.25 mg/g en diciembre a 0.85
mg/g en julio (Fig. 18).
Figura 18. Comportamiento de la concentración de lípidos (mg/g) de febrero de 2012 a febrero de 2013 en pie, manto, gónada y hepatopáncreas de Haliotis corrugata en la zona de pesca III en Baja California Sur, México.
Durante el ciclo reproductivo los valores de lípidos en la gónada no variaron
significativamente, los más altos se presentaron en etapas de madurez y
posdesove, y los valores más bajos en desove. En el hepatopáncreas los valores
más bajos se presentaron durante la indiferenciación. En el pie tampoco se
observó una diferencia significativa de la concentración de lípidos; las
concentraciones más altas se presentaron durante la indiferenciación. En el manto
las concentraciones de lípidos se mantuvieron prácticamente estables durante las
diferentes etapas del ciclo reproductivo (Fig. 19).
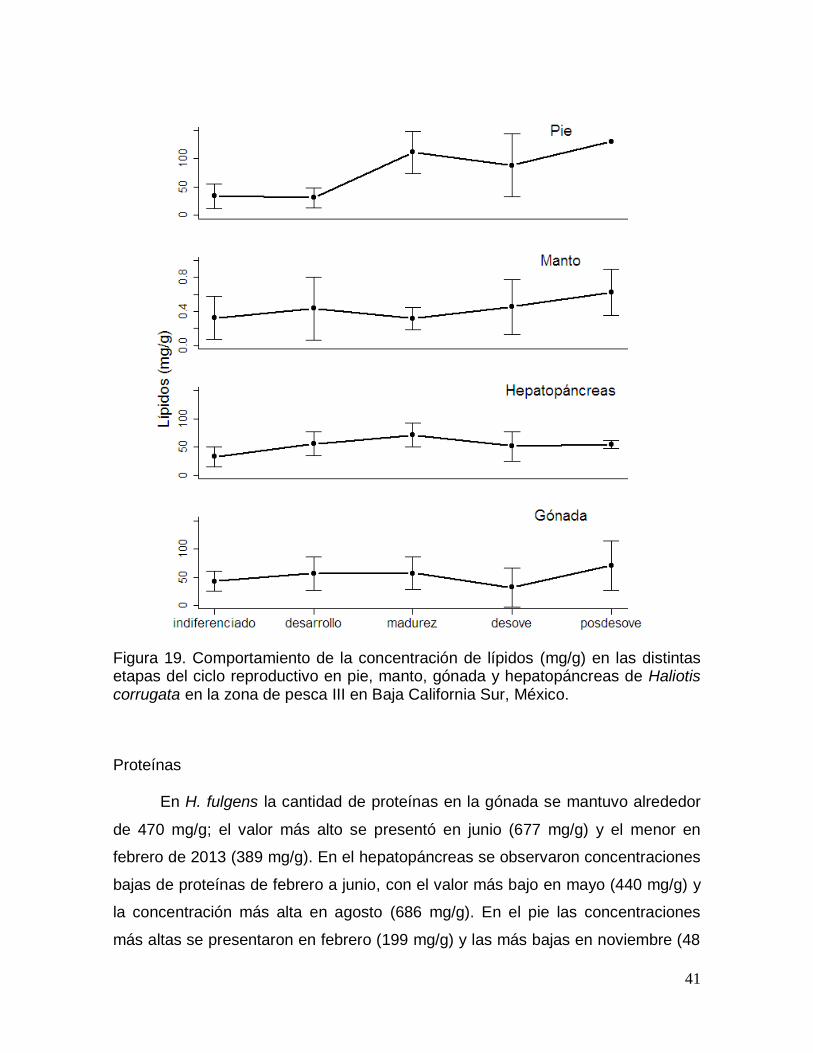
41
Figura 19. Comportamiento de la concentración de lípidos (mg/g) en las distintas etapas del ciclo reproductivo en pie, manto, gónada y hepatopáncreas de Haliotis corrugata en la zona de pesca III en Baja California Sur, México.
Proteínas
En H. fulgens la cantidad de proteínas en la gónada se mantuvo alrededor
de 470 mg/g; el valor más alto se presentó en junio (677 mg/g) y el menor en
febrero de 2013 (389 mg/g). En el hepatopáncreas se observaron concentraciones
bajas de proteínas de febrero a junio, con el valor más bajo en mayo (440 mg/g) y
la concentración más alta en agosto (686 mg/g). En el pie las concentraciones
más altas se presentaron en febrero (199 mg/g) y las más bajas en noviembre (48
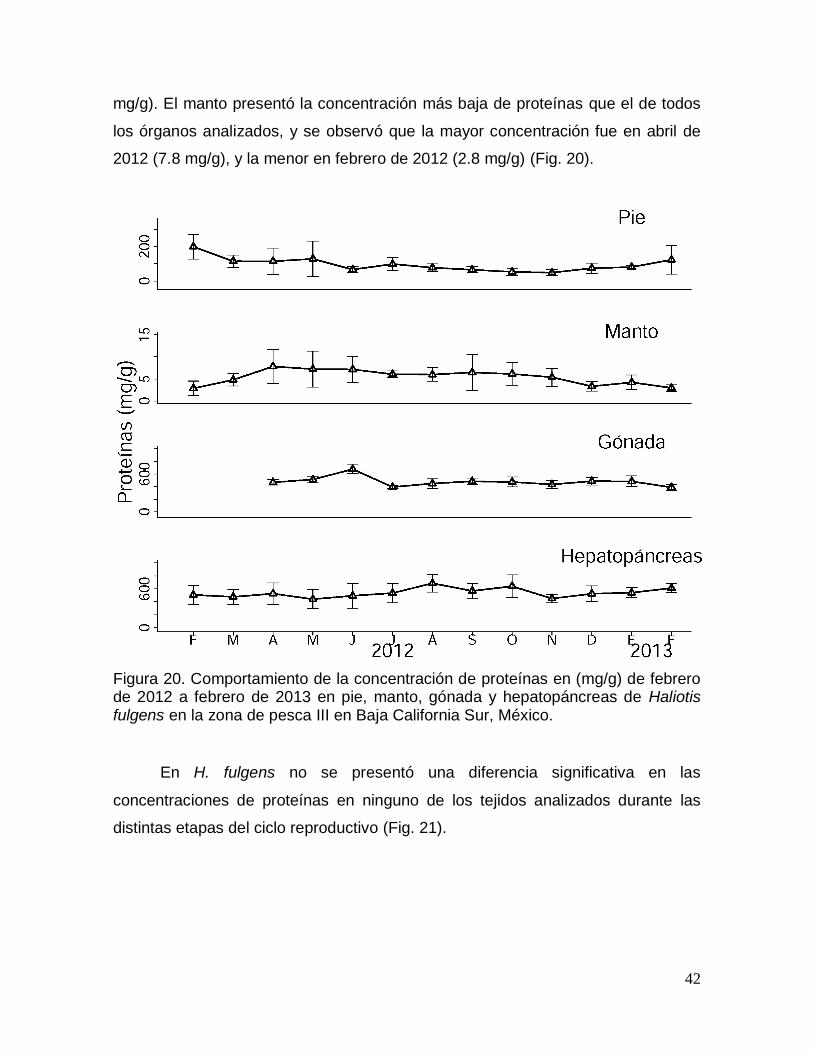
42
mg/g). El manto presentó la concentración más baja de proteínas que el de todos
los órganos analizados, y se observó que la mayor concentración fue en abril de
2012 (7.8 mg/g), y la menor en febrero de 2012 (2.8 mg/g) (Fig. 20).
Figura 20. Comportamiento de la concentración de proteínas en (mg/g) de febrero de 2012 a febrero de 2013 en pie, manto, gónada y hepatopáncreas de Haliotis fulgens en la zona de pesca III en Baja California Sur, México.
En H. fulgens no se presentó una diferencia significativa en las
concentraciones de proteínas en ninguno de los tejidos analizados durante las
distintas etapas del ciclo reproductivo (Fig. 21).
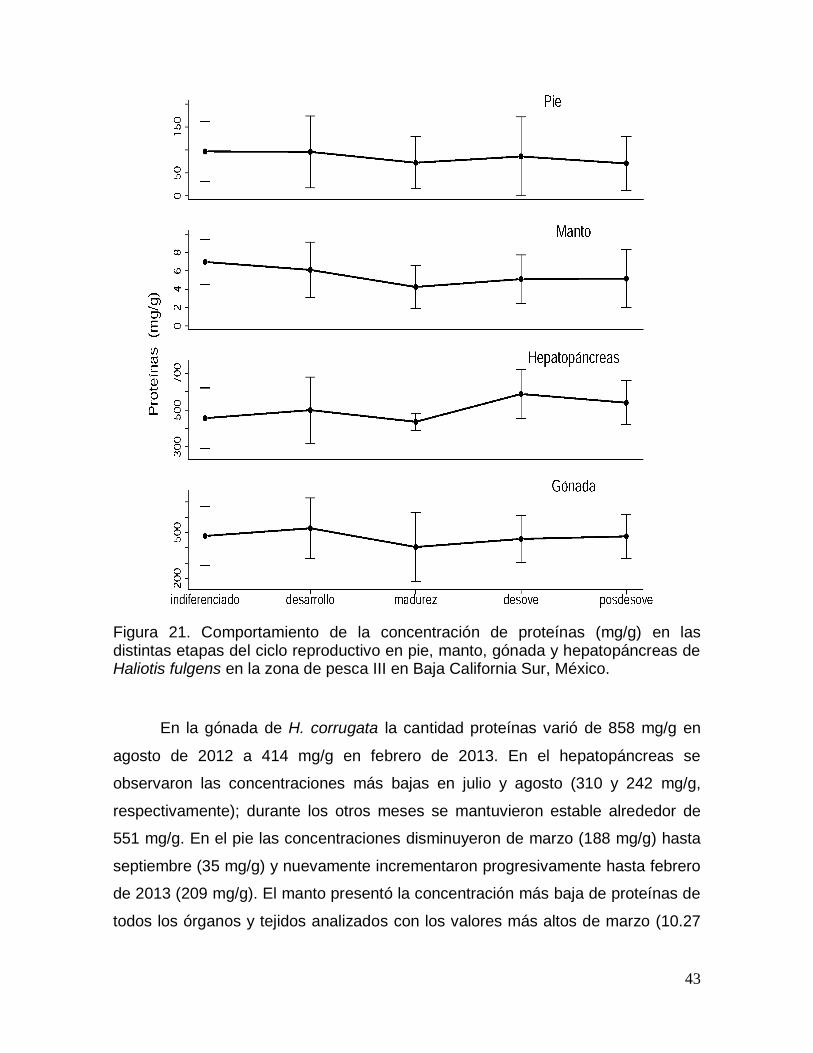
43
Figura 21. Comportamiento de la concentración de proteínas (mg/g) en las distintas etapas del ciclo reproductivo en pie, manto, gónada y hepatopáncreas de Haliotis fulgens en la zona de pesca III en Baja California Sur, México.
En la gónada de H. corrugata la cantidad proteínas varió de 858 mg/g en
agosto de 2012 a 414 mg/g en febrero de 2013. En el hepatopáncreas se
observaron las concentraciones más bajas en julio y agosto (310 y 242 mg/g,
respectivamente); durante los otros meses se mantuvieron estable alrededor de
551 mg/g. En el pie las concentraciones disminuyeron de marzo (188 mg/g) hasta
septiembre (35 mg/g) y nuevamente incrementaron progresivamente hasta febrero
de 2013 (209 mg/g). El manto presentó la concentración más baja de proteínas de
todos los órganos y tejidos analizados con los valores más altos de marzo (10.27
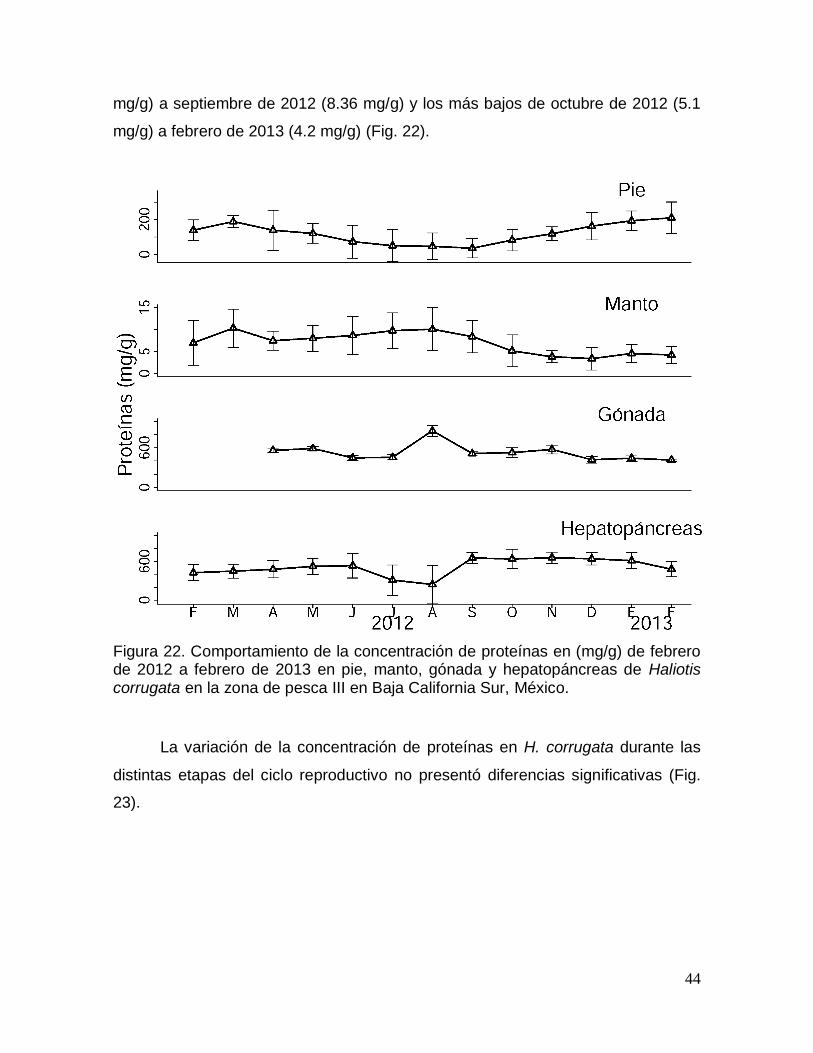
44
mg/g) a septiembre de 2012 (8.36 mg/g) y los más bajos de octubre de 2012 (5.1
mg/g) a febrero de 2013 (4.2 mg/g) (Fig. 22).
Figura 22. Comportamiento de la concentración de proteínas en (mg/g) de febrero de 2012 a febrero de 2013 en pie, manto, gónada y hepatopáncreas de Haliotis corrugata en la zona de pesca III en Baja California Sur, México.
La variación de la concentración de proteínas en H. corrugata durante las
distintas etapas del ciclo reproductivo no presentó diferencias significativas (Fig.
23).
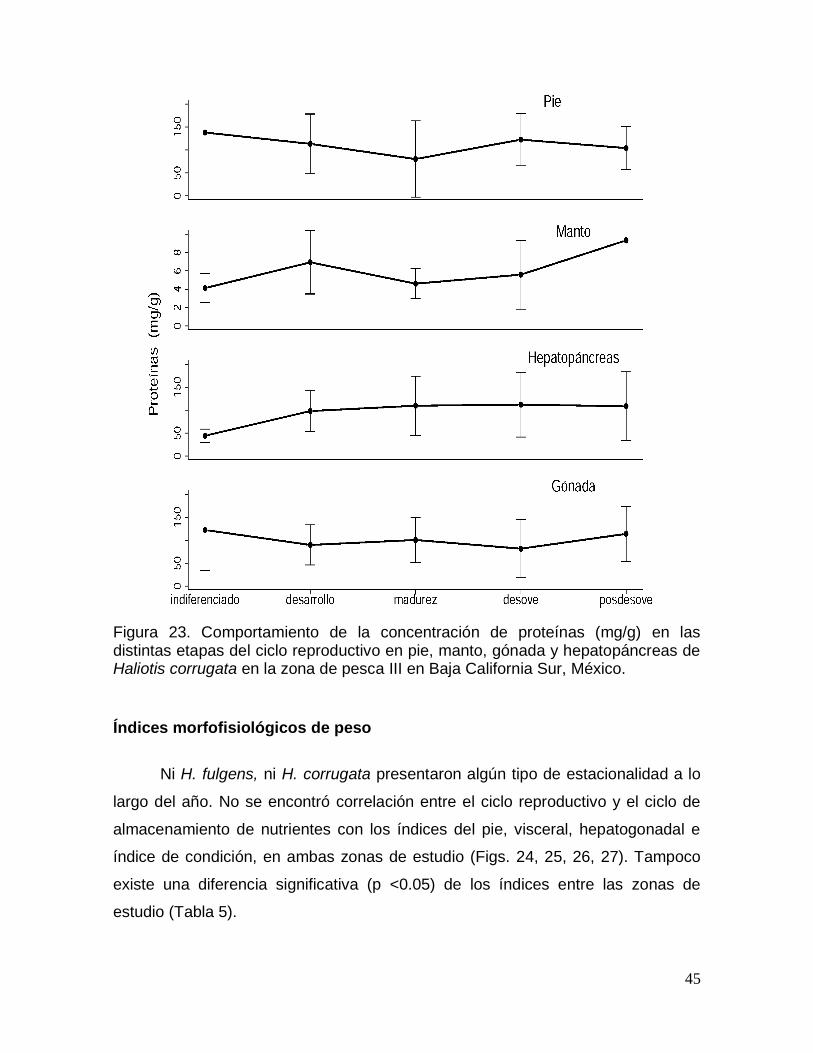
45
Figura 23. Comportamiento de la concentración de proteínas (mg/g) en las distintas etapas del ciclo reproductivo en pie, manto, gónada y hepatopáncreas de Haliotis corrugata en la zona de pesca III en Baja California Sur, México.
Índices morfofisiológicos de peso
Ni H. fulgens, ni H. corrugata presentaron algún tipo de estacionalidad a lo
largo del año. No se encontró correlación entre el ciclo reproductivo y el ciclo de
almacenamiento de nutrientes con los índices del pie, visceral, hepatogonadal e
índice de condición, en ambas zonas de estudio (Figs. 24, 25, 26, 27). Tampoco
existe una diferencia significativa (p <0.05) de los índices entre las zonas de
estudio (Tabla 5).
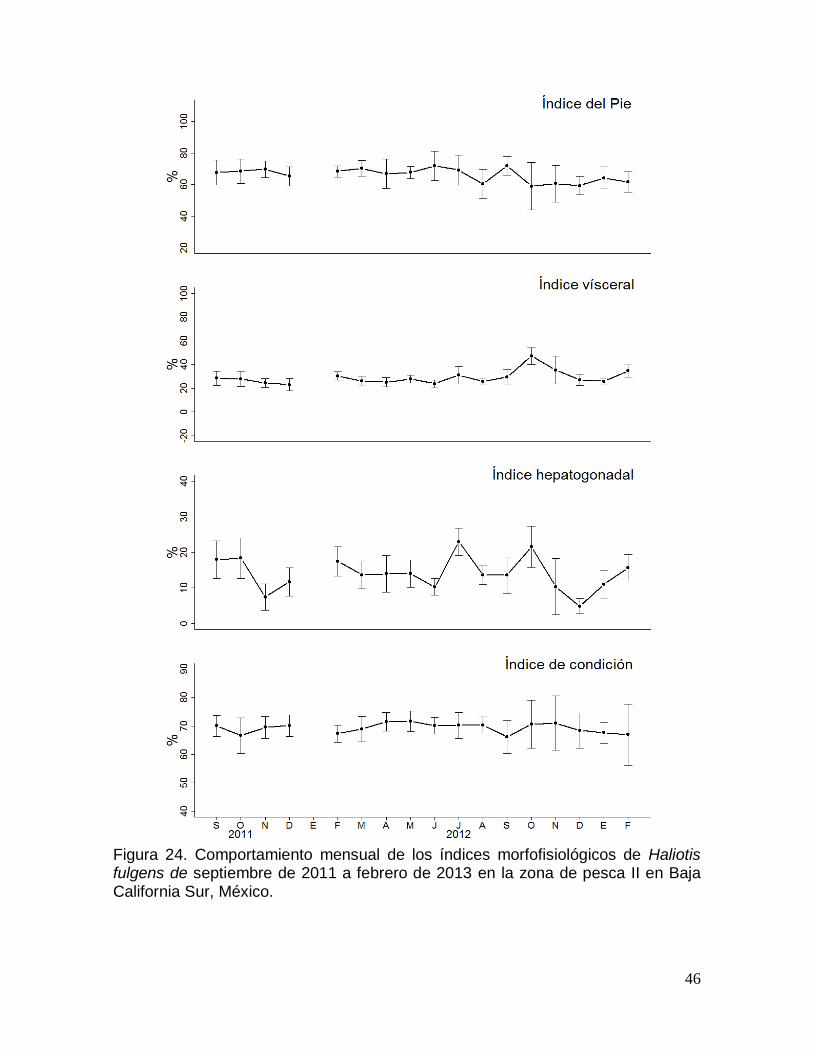
46
Figura 24. Comportamiento mensual de los índices morfofisiológicos de Haliotis fulgens de septiembre de 2011 a febrero de 2013 en la zona de pesca II en Baja California Sur, México.
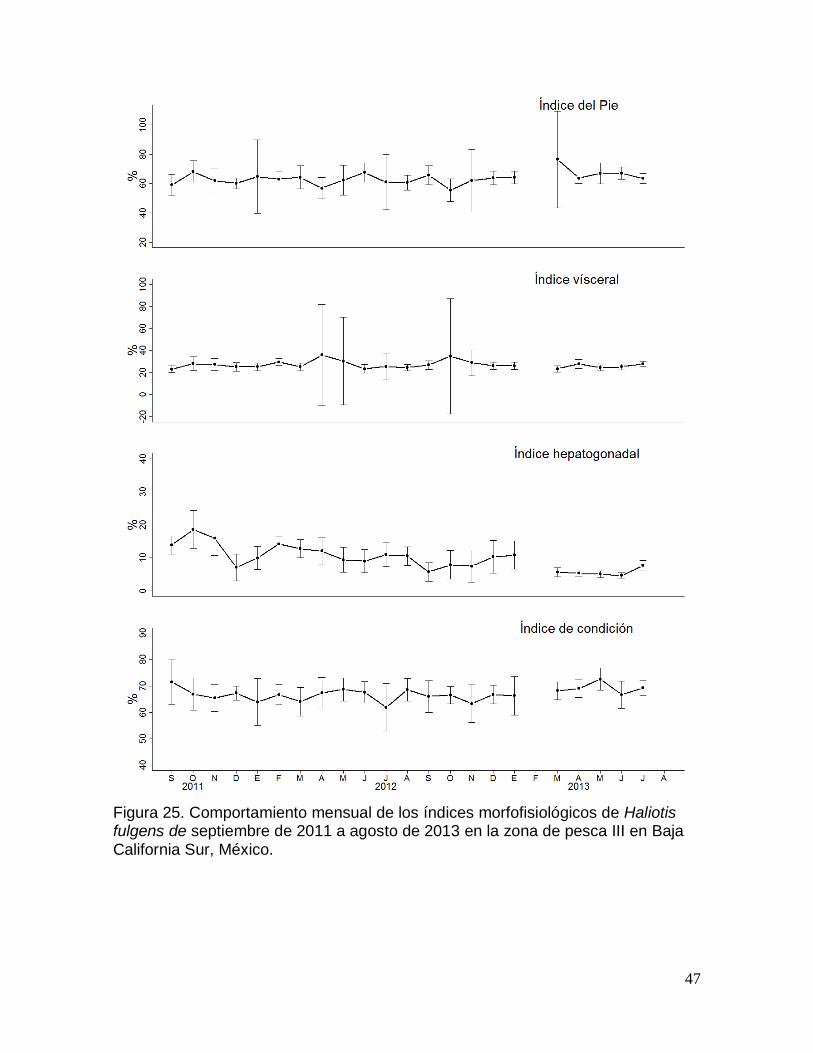
47
Figura 25. Comportamiento mensual de los índices morfofisiológicos de Haliotis fulgens de septiembre de 2011 a agosto de 2013 en la zona de pesca III en Baja California Sur, México.
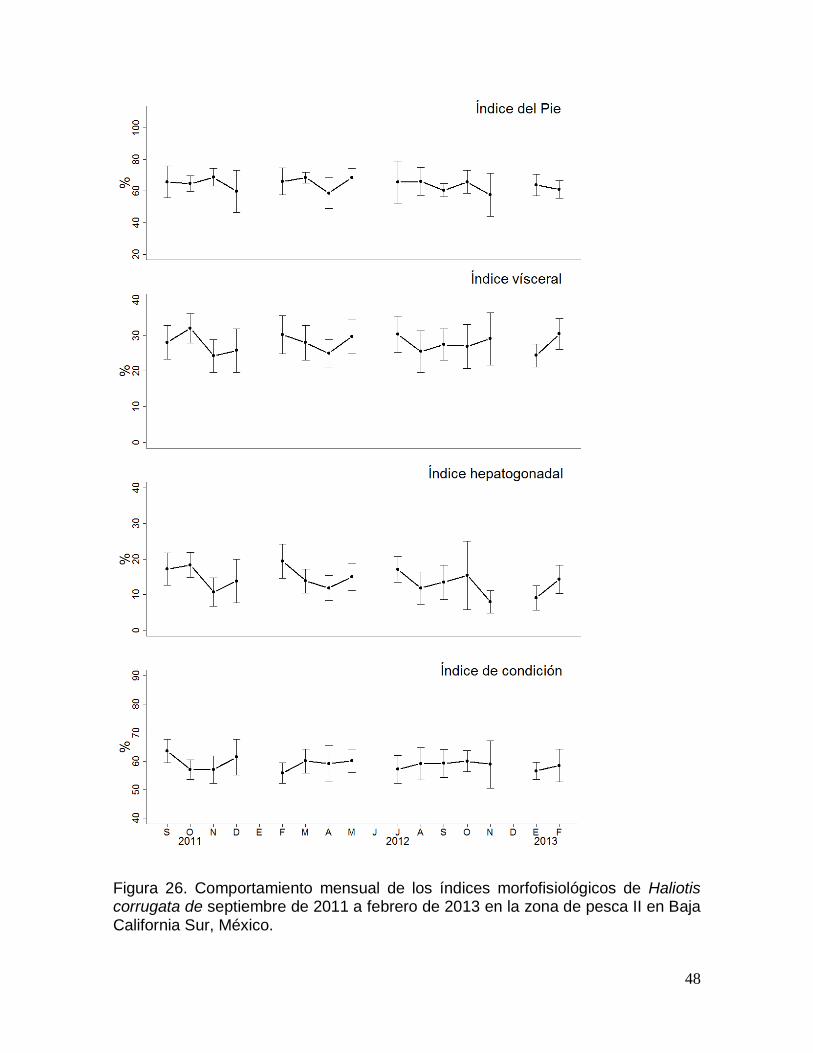
48
Figura 26. Comportamiento mensual de los índices morfofisiológicos de Haliotis corrugata de septiembre de 2011 a febrero de 2013 en la zona de pesca II en Baja California Sur, México.
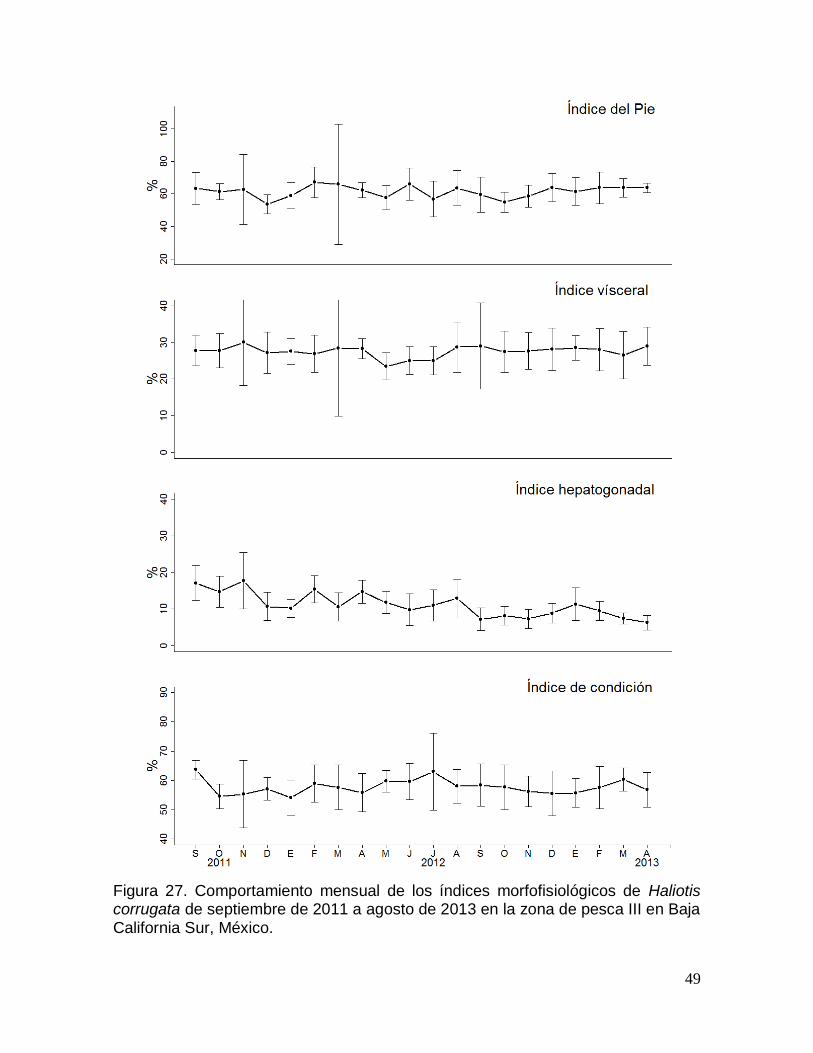
49
Figura 27. Comportamiento mensual de los índices morfofisiológicos de Haliotis corrugata de septiembre de 2011 a agosto de 2013 en la zona de pesca III en Baja California Sur, México.
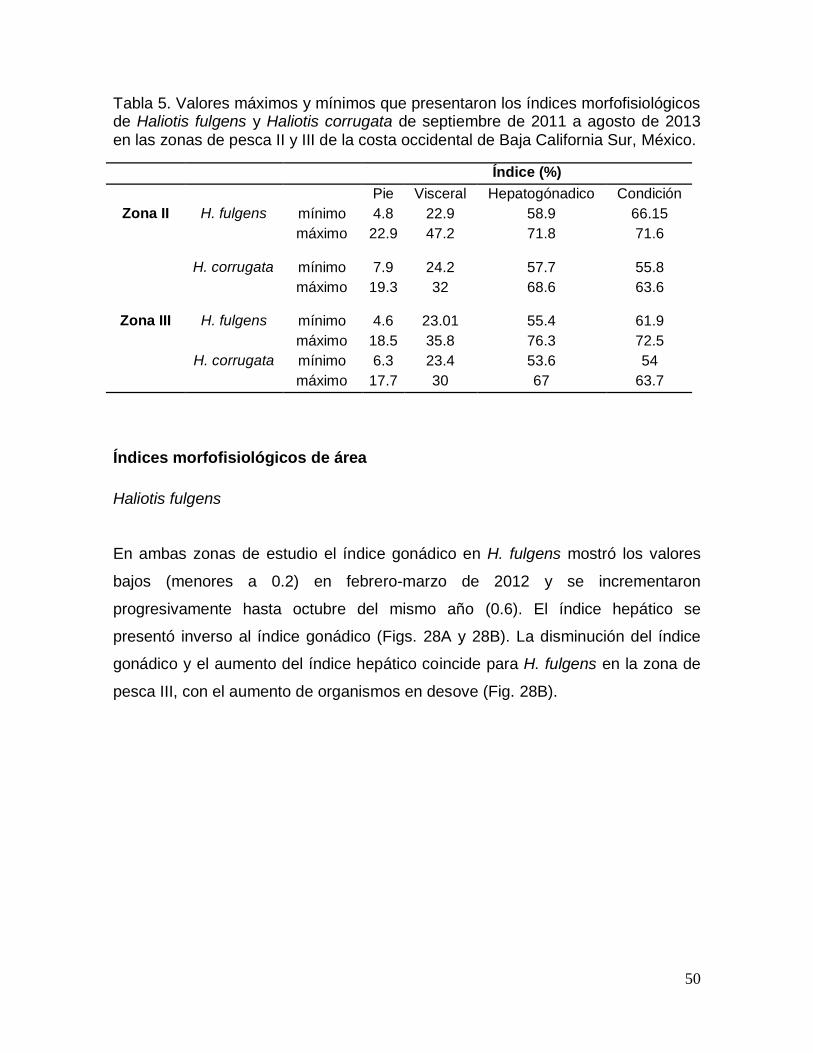
50
Tabla 5. Valores máximos y mínimos que presentaron los índices morfofisiológicos de Haliotis fulgens y Haliotis corrugata de septiembre de 2011 a agosto de 2013 en las zonas de pesca II y III de la costa occidental de Baja California Sur, México.
Índice (%)
Pie Visceral Hepatogónadico Condición
Zona II H. fulgens mínimo 4.8 22.9 58.9 66.15
máximo 22.9 47.2 71.8 71.6
H. corrugata mínimo 7.9 24.2 57.7 55.8
máximo 19.3 32 68.6 63.6 Zona III H. fulgens mínimo 4.6 23.01 55.4 61.9
máximo 18.5 35.8 76.3 72.5
H. corrugata mínimo 6.3 23.4 53.6 54
máximo 17.7 30 67 63.7
Índices morfofisiológicos de área
Haliotis fulgens
En ambas zonas de estudio el índice gonádico en H. fulgens mostró los valores
bajos (menores a 0.2) en febrero-marzo de 2012 y se incrementaron
progresivamente hasta octubre del mismo año (0.6). El índice hepático se
presentó inverso al índice gonádico (Figs. 28A y 28B). La disminución del índice
gonádico y el aumento del índice hepático coincide para H. fulgens en la zona de
pesca III, con el aumento de organismos en desove (Fig. 28B).
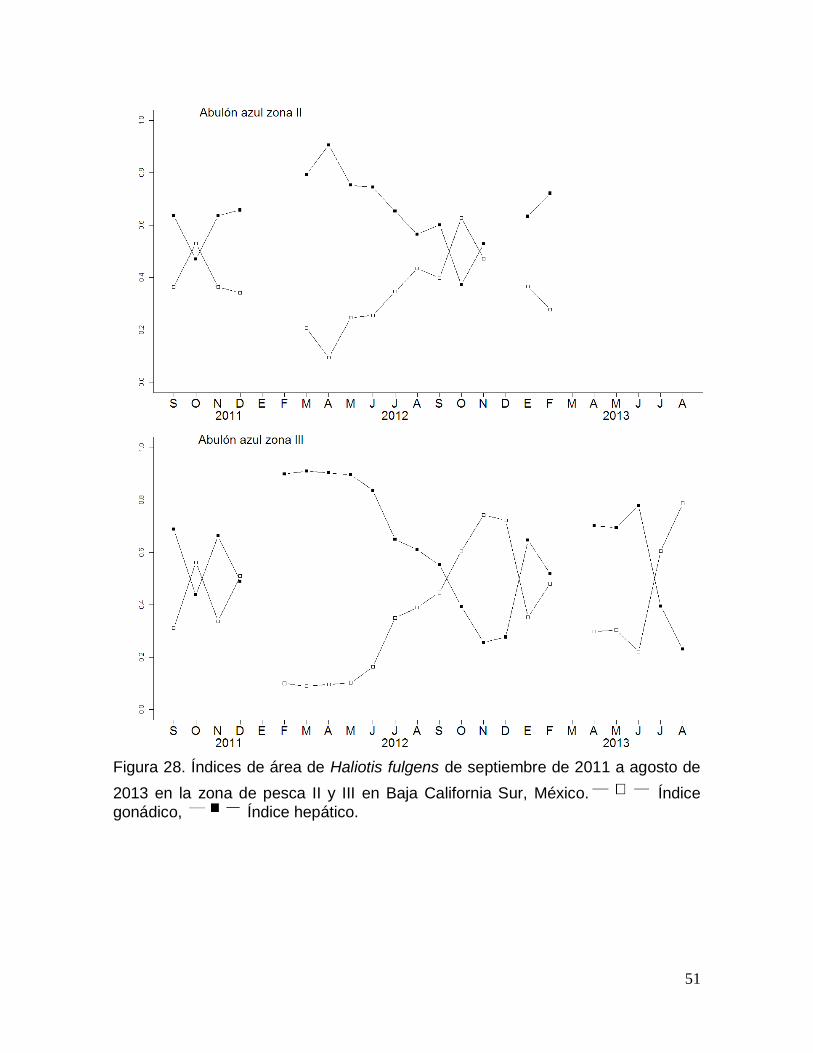
51
Figura 28. Índices de área de Haliotis fulgens de septiembre de 2011 a agosto de
2013 en la zona de pesca II y III en Baja California Sur, México. Índice
gonádico, Índice hepático.

52
Haliotis corrugata
En H. corrugata en la zona de pesca II, tanto el índice gonádico, como el
índice hepático mostraron grandes fluctuaciones a lo largo del periodo de estudio.
El índice gonádico presentó valores altos en septiembre de 2011 (0.7) y mayo de
2012 (0.6), lo que concuerda con una frecuencia mayor de madurez y desove;
durante los otros meses los índices se mantuvieron en niveles medios de 0.5 (Fig.
29).
En la zona de pesca III se presentaron variaciones en los valores de los
índices gonádico y hepático de septiembre a diciembre de 2011. De febrero a abril
de 2012 el índice gonádico presentó valores bajos (menores a 0.2) que
aumentaron progresivamente hasta agosto de 2012 (0.7), y disminuyeron
nuevamente hasta mayo de 2013 (0.2), posteriormente se recupera en julio y
agosto de 2013, cuando se presentaron valores mayores de 0.7, estos aumentos
del índice gonádico concordaron con el aumento de organismos en etapa de
desove (Fig. 29).

53
Figura 29. Índices de área de Haliotis corrugata de septiembre de 2011 a agosto
de 2013 en la zona de pesca II y III en Baja California Sur, México. Índice
gonádico, Índice hepático.
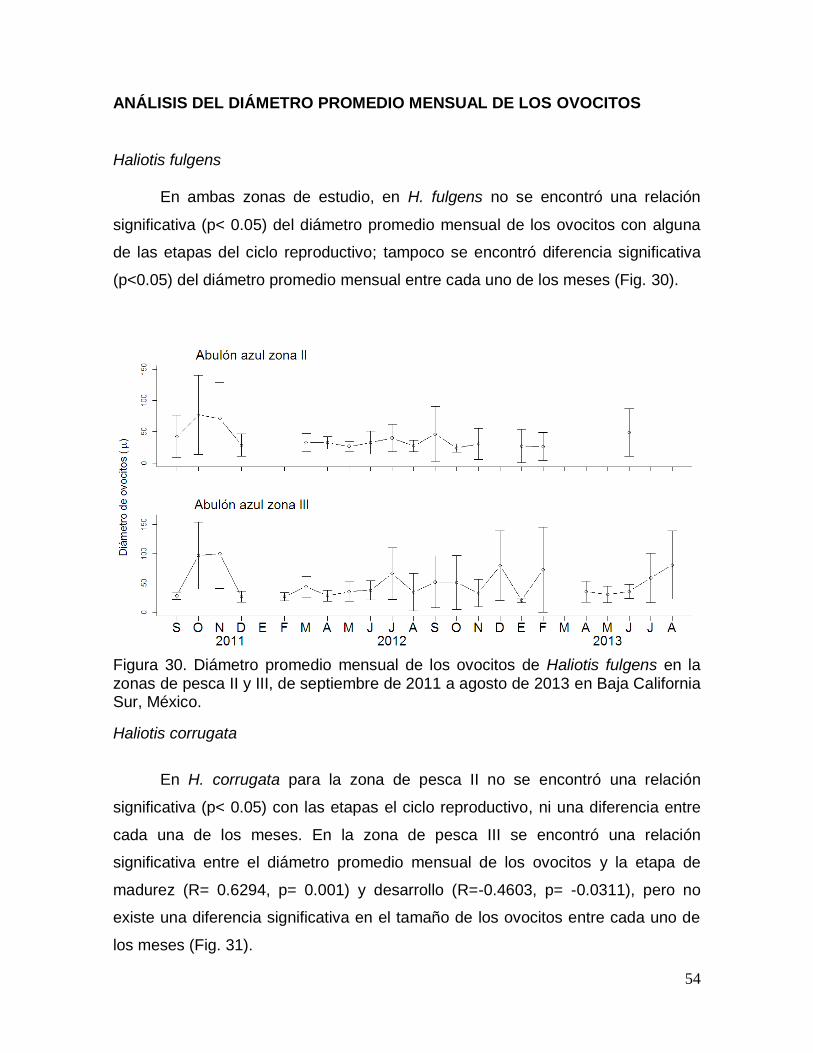
54
ANÁLISIS DEL DIÁMETRO PROMEDIO MENSUAL DE LOS OVOCITOS
Haliotis fulgens
En ambas zonas de estudio, en H. fulgens no se encontró una relación
significativa (p< 0.05) del diámetro promedio mensual de los ovocitos con alguna
de las etapas del ciclo reproductivo; tampoco se encontró diferencia significativa
(p<0.05) del diámetro promedio mensual entre cada uno de los meses (Fig. 30).
Figura 30. Diámetro promedio mensual de los ovocitos de Haliotis fulgens en la zonas de pesca II y III, de septiembre de 2011 a agosto de 2013 en Baja California Sur, México.
Haliotis corrugata
En H. corrugata para la zona de pesca II no se encontró una relación
significativa (p< 0.05) con las etapas el ciclo reproductivo, ni una diferencia entre
cada una de los meses. En la zona de pesca III se encontró una relación
significativa entre el diámetro promedio mensual de los ovocitos y la etapa de
madurez (R= 0.6294, p= 0.001) y desarrollo (R=-0.4603, p= -0.0311), pero no
existe una diferencia significativa en el tamaño de los ovocitos entre cada uno de
los meses (Fig. 31).
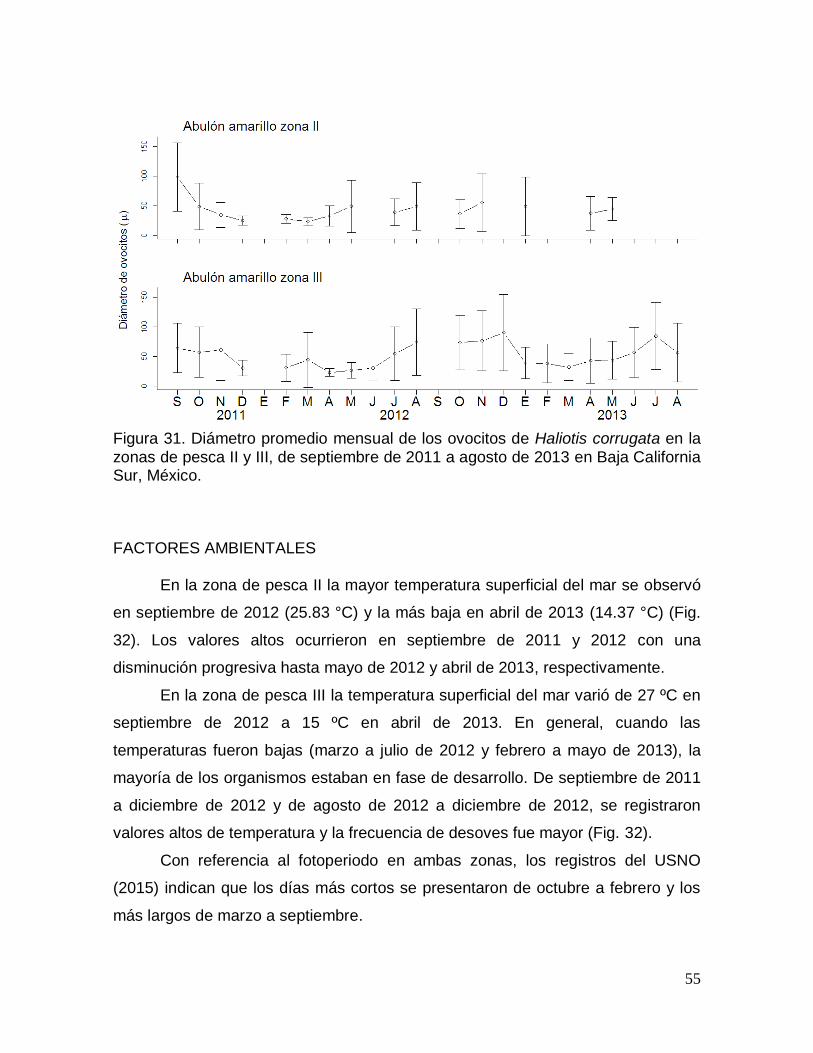
55
Figura 31. Diámetro promedio mensual de los ovocitos de Haliotis corrugata en la zonas de pesca II y III, de septiembre de 2011 a agosto de 2013 en Baja California Sur, México.
FACTORES AMBIENTALES
En la zona de pesca II la mayor temperatura superficial del mar se observó
en septiembre de 2012 (25.83 °C) y la más baja en abril de 2013 (14.37 °C) (Fig.
32). Los valores altos ocurrieron en septiembre de 2011 y 2012 con una
disminución progresiva hasta mayo de 2012 y abril de 2013, respectivamente.
En la zona de pesca III la temperatura superficial del mar varió de 27 ºC en
septiembre de 2012 a 15 ºC en abril de 2013. En general, cuando las
temperaturas fueron bajas (marzo a julio de 2012 y febrero a mayo de 2013), la
mayoría de los organismos estaban en fase de desarrollo. De septiembre de 2011
a diciembre de 2012 y de agosto de 2012 a diciembre de 2012, se registraron
valores altos de temperatura y la frecuencia de desoves fue mayor (Fig. 32).
Con referencia al fotoperiodo en ambas zonas, los registros del USNO
(2015) indican que los días más cortos se presentaron de octubre a febrero y los
más largos de marzo a septiembre.
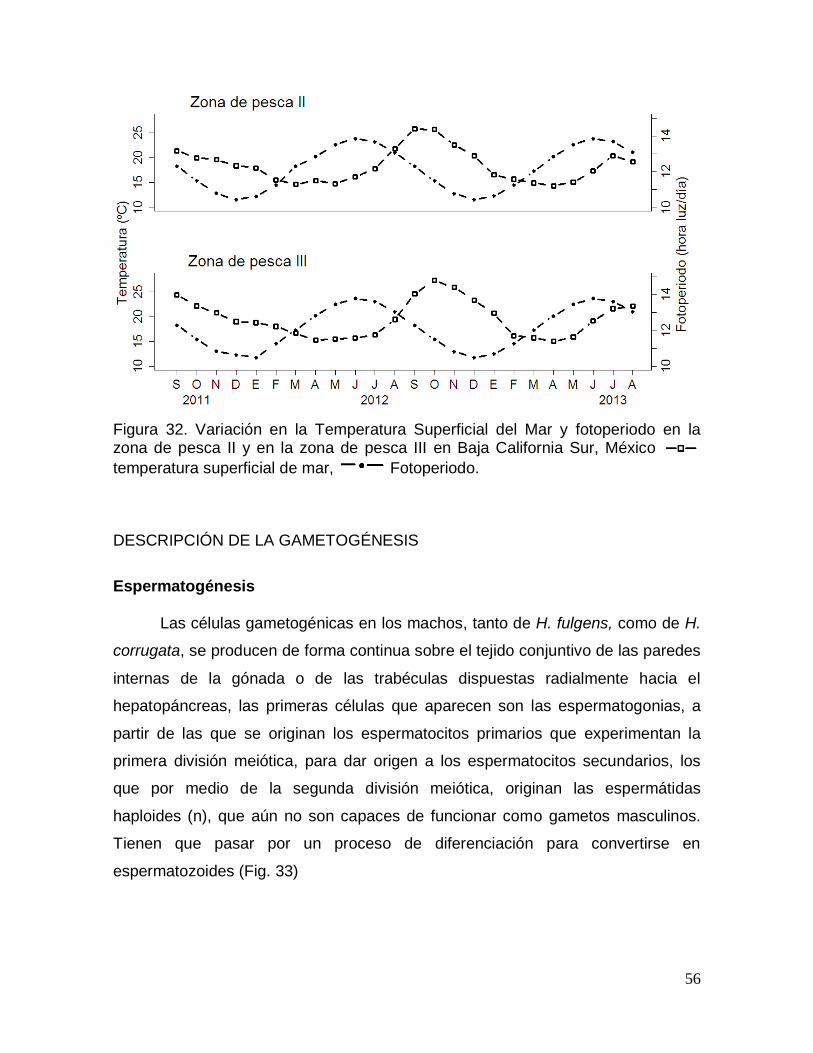
56
Figura 32. Variación en la Temperatura Superficial del Mar y fotoperiodo en la zona de pesca II y en la zona de pesca III en Baja California Sur, México
temperatura superficial de mar, Fotoperiodo.
DESCRIPCIÓN DE LA GAMETOGÉNESIS
Espermatogénesis
Las células gametogénicas en los machos, tanto de H. fulgens, como de H.
corrugata, se producen de forma continua sobre el tejido conjuntivo de las paredes
internas de la gónada o de las trabéculas dispuestas radialmente hacia el
hepatopáncreas, las primeras células que aparecen son las espermatogonias, a
partir de las que se originan los espermatocitos primarios que experimentan la
primera división meiótica, para dar origen a los espermatocitos secundarios, los
que por medio de la segunda división meiótica, originan las espermátidas
haploides (n), que aún no son capaces de funcionar como gametos masculinos.
Tienen que pasar por un proceso de diferenciación para convertirse en
espermatozoides (Fig. 33)
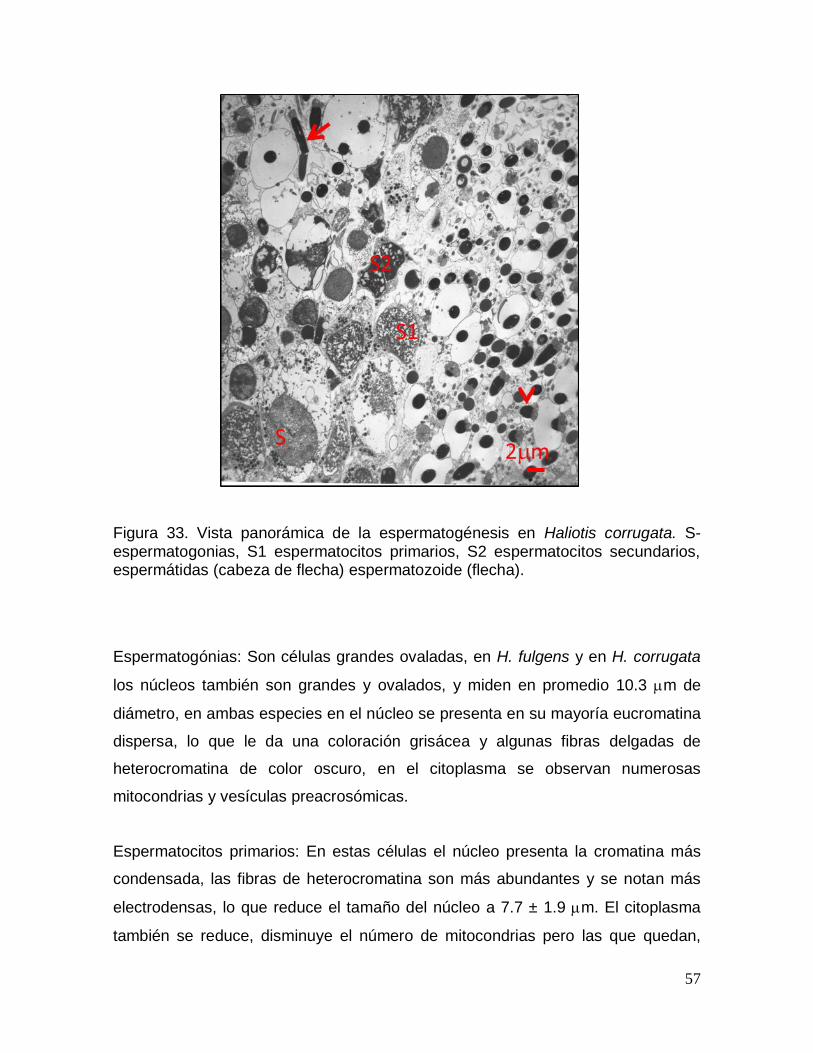
57
Figura 33. Vista panorámica de la espermatogénesis en Haliotis corrugata. S- espermatogonias, S1 espermatocitos primarios, S2 espermatocitos secundarios, espermátidas (cabeza de flecha) espermatozoide (flecha).
Espermatogónias: Son células grandes ovaladas, en H. fulgens y en H. corrugata
los núcleos también son grandes y ovalados, y miden en promedio 10.3 m de
diámetro, en ambas especies en el núcleo se presenta en su mayoría eucromatina
dispersa, lo que le da una coloración grisácea y algunas fibras delgadas de
heterocromatina de color oscuro, en el citoplasma se observan numerosas
mitocondrias y vesículas preacrosómicas.
Espermatocitos primarios: En estas células el núcleo presenta la cromatina más
condensada, las fibras de heterocromatina son más abundantes y se notan más
electrodensas, lo que reduce el tamaño del núcleo a 7.7 ± 1.9 m. El citoplasma
también se reduce, disminuye el número de mitocondrias pero las que quedan,

58
aumentan su volumen, las vesículas preacrosómicas se acumulan hacia un lado
de la célula.
Espermatocitos secundarios: El núcleo en estas células es de menor tamaño que
en las anteriores, 5.9 ± 1.5 m, aquí se presentan gruesas fibras de
heterocromatina que desplazan a la eucromatina, el número de mitocondrias ha
disminuido y aumentaron en tamaño, estas comienzan a desplazarse hacia uno de
los polos, las vesículas preacrosómicas comienzan a fusionarse.
Espermátidas: Estas células presentan la heterocromatina completamente
condensada, el núcleo empieza a alargarse y las mitocondrias se localizan en la
parte basal de la célula, en la parte apical se observa el acrosoma que comienza a
alargarse.
Espermatozoides: Son células alargadas con una longitud de 10.8 ± 1 m, ha
perdido prácticamente el citoplasma, solo se observa el núcleo con la
heterocromatina completamente condensada, se observan algunas lagunas
nucleares, se observa un acrosoma alargado que mide 3.5 ± 0.8 m de longitud
(Fig. 34), en la parte basar presenta cinco mitocondrias que rodean un centriolo,
las mitocondrias tienen un diámetro promedio de 1.3 ± 0.4 m (Figs. 34 y 35)
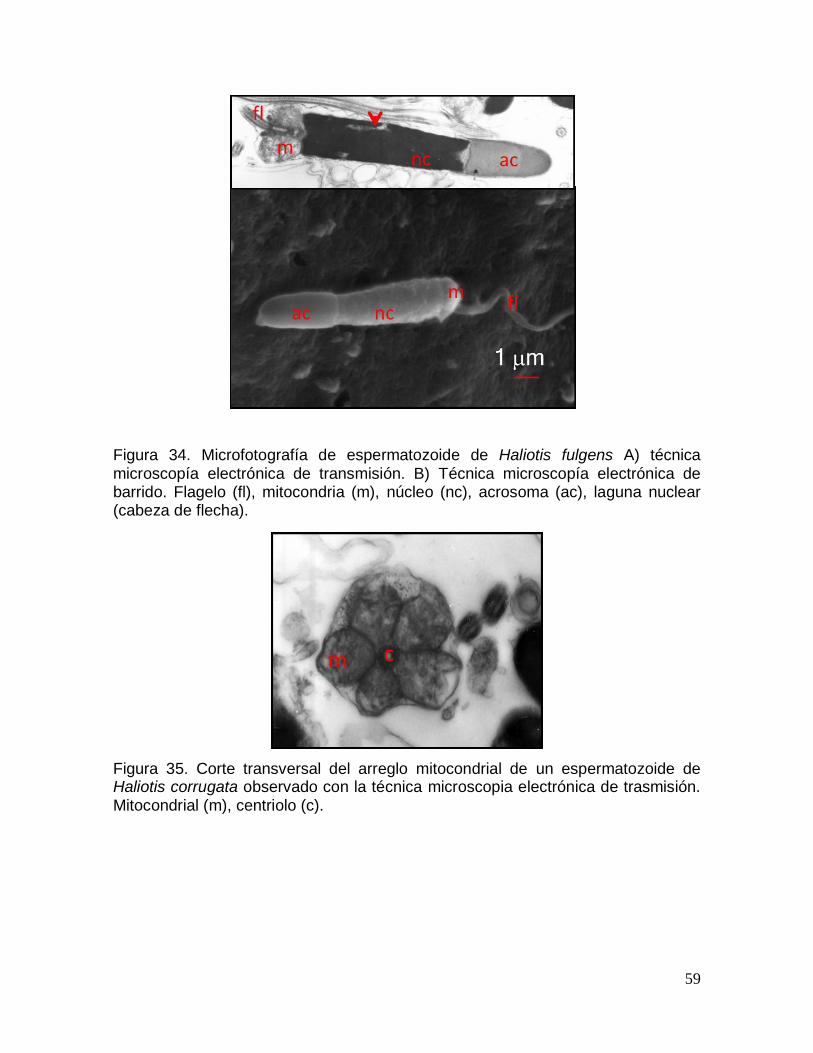
59
Figura 34. Microfotografía de espermatozoide de Haliotis fulgens A) técnica microscopía electrónica de transmisión. B) Técnica microscopía electrónica de barrido. Flagelo (fl), mitocondria (m), núcleo (nc), acrosoma (ac), laguna nuclear (cabeza de flecha).
Figura 35. Corte transversal del arreglo mitocondrial de un espermatozoide de Haliotis corrugata observado con la técnica microscopia electrónica de trasmisión. Mitocondrial (m), centriolo (c).

60
Ovogénesis
La ovogénesis en H. fulgens y en H. corrugata comienza a partir de la formación
de ovogonias que crecen sujetas al tejido conjuntivo, sobre las paredes internas
de la gónada o en las trabéculas que se forman de manera radial hacia el
hepatopáncreas, conforme avanza la gametogénesis estas células crecen,
acumulando nutrientes en el citoplasma hasta formar el ovocito vitelogénico (Fig.
36).
Ovogonias: Las ovogonias son células piriformes que crecen sujetas al tejido
conjuntivo, presentan tallas en promedio de 35.38 ± 10.22 m, presentan un
núcleo grande redondo central. Con histología es posible observar que son de tipo
acidófilo.
Ovocitos previtelogénicos: Son células redondas que aún se encuentran
ligeramente unidas al tejido conjuntivo, tienen un diámetro en promedio de 71.9 ±
32.27 m, presentan un núcleo grande redondo central, con histología se puede
observar que es de tipo acidófilo. Con microscopía electrónica de transmisión es
posible observar algunas mitocondrias (Figs. 36A y 36C).
Ovocitos vitelogénicos: llegan a medir hasta 200 m, presentan dentro del
citoplasma una gran cantidad de plaquetas vitelinas y vacuolas lipídicas, alrededor
del citoplasma presentan un espacio previtelogénicos donde contiene una gran
cantidad de microvellocidades rodeadas por una envoltura vitelina. Con
microscopía electrónica de barrido la superficie se ve aparentemente lisa, debido a
la envoltura mucosa llamada corion, pero al desprenderse este, se observa que
contiene una gran cantidad de poros en la superficie (Figs. 36B, 36D y 36F).
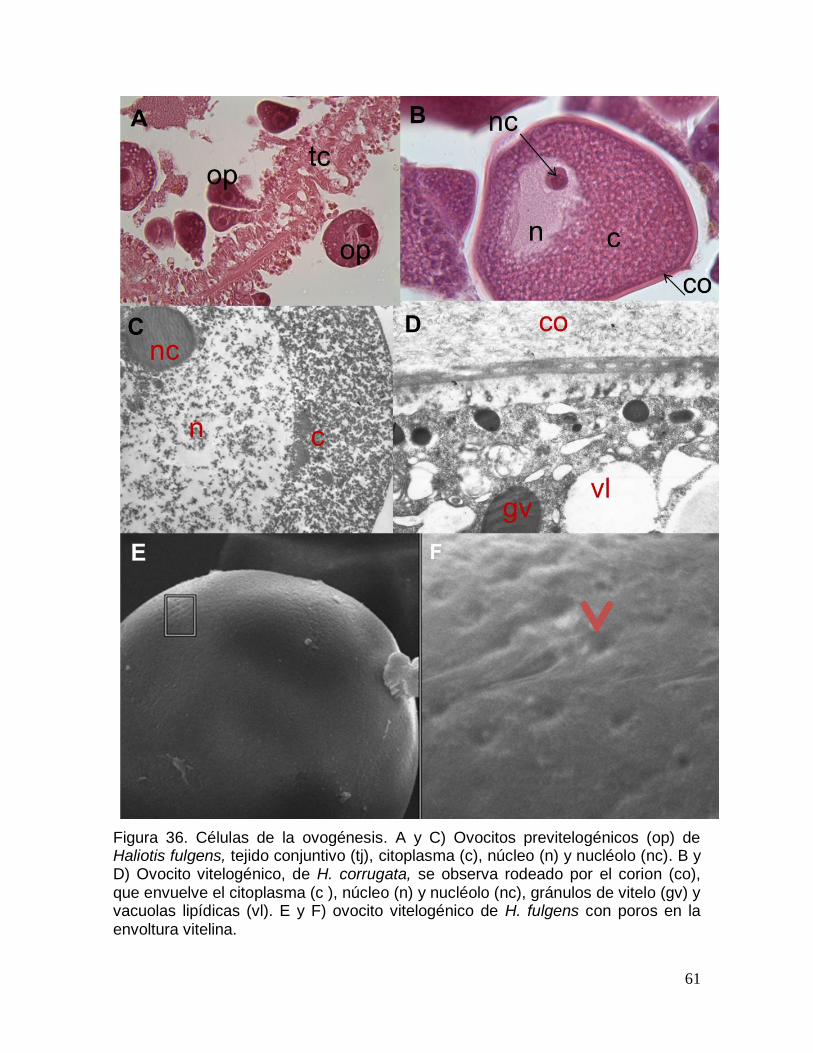
61
Figura 36. Células de la ovogénesis. A y C) Ovocitos previtelogénicos (op) de Haliotis fulgens, tejido conjuntivo (tj), citoplasma (c), núcleo (n) y nucléolo (nc). B y D) Ovocito vitelogénico, de H. corrugata, se observa rodeado por el corion (co), que envuelve el citoplasma (c ), núcleo (n) y nucléolo (nc), gránulos de vitelo (gv) y vacuolas lipídicas (vl). E y F) ovocito vitelogénico de H. fulgens con poros en la envoltura vitelina.
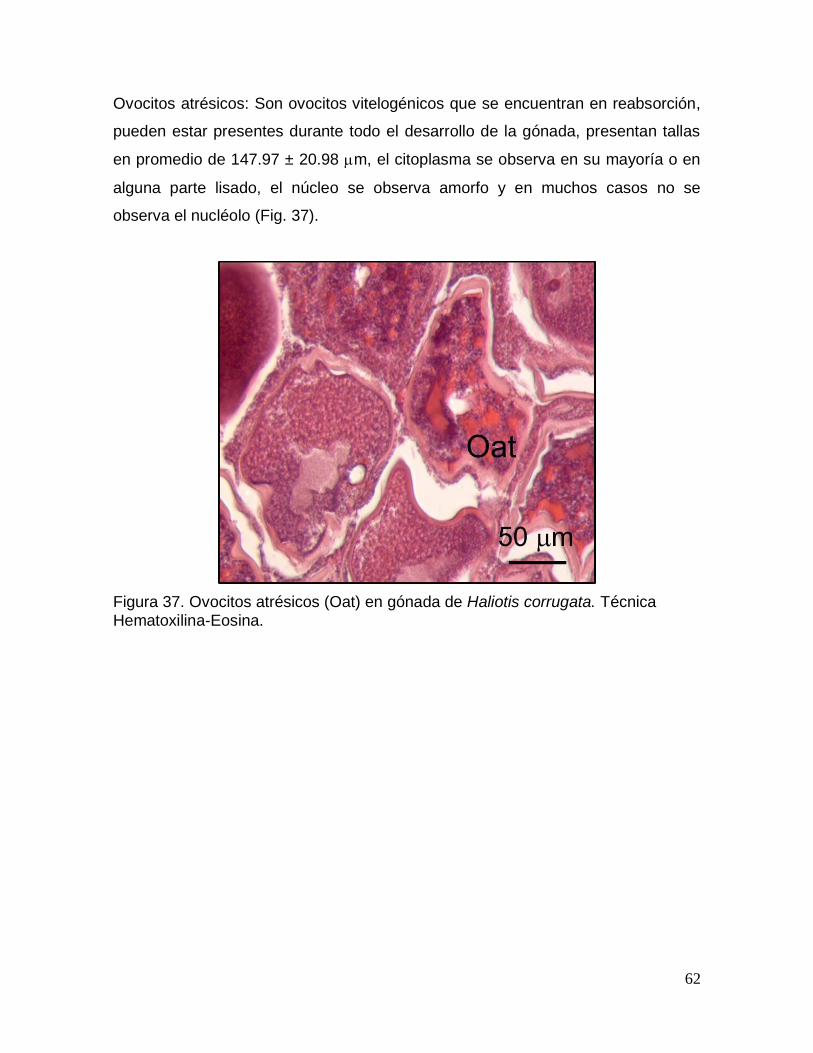
62
Ovocitos atrésicos: Son ovocitos vitelogénicos que se encuentran en reabsorción,
pueden estar presentes durante todo el desarrollo de la gónada, presentan tallas
en promedio de 147.97 ± 20.98 m, el citoplasma se observa en su mayoría o en
alguna parte lisado, el núcleo se observa amorfo y en muchos casos no se
observa el nucléolo (Fig. 37).
Figura 37. Ovocitos atrésicos (Oat) en gónada de Haliotis corrugata. Técnica Hematoxilina-Eosina.

63
DISCUSIÓN
El análisis histológico y ultramicroscópico de las gónadas de H. fulgens y H.
corrugata, demostró que las dos especies presentan desoves parciales durante
todo el ciclo y que el tamaño de la gónada no es una característica que distinga a
los organismos en desove, ya que este se presentó tanto en organismos con
gónadas gruesas, como en organismos con gónadas delgadas. Esto explica la
presencia de desoves continuos, lo que se confirma con la carente relación entre
el índice gonádico (basado en peso) y la época reproductiva. Webber y Giese
(1969), Zetina et al. (2000) y Litaay y De Silva (2003) en otros haliótidos también
encontraron que el índice gonádico no revela con certeza las fases de madurez.
No obstante, también se ha probado que el índice gonádico permite destacar las
fases clave de maduración gonádica (liberación de gametos) y es buen indicador
de la tendencia reproductiva (Moraisa et al., 2003; Najmudeen et al., 2004,
Najmudeen, 2007; Elhasni et al., 2013). La presencia de desoves parciales
durante todo el ciclo, dificulta la posibilidad de utilizar indicadores para describir el
ciclo reproductivo, por lo que el método histológico o las observaciones directas de
la gónada son las herramientas más precisas para este tipo de organismos.
Por otro lado los índices calculados con base en el área de la gónada, han
tenido mejores resultados, como es el caso del presente trabajo, en el que se
encontraron valores altos en el índice gonádico calculado con el área durante la
época reproductiva, que señalan que durante esta época, en promedio el área de
la gónada de los organismos, era mayor que durante los meses en los que los
desoves parciales fueron menos intensos. En estudios con Bolinus brandaris
(Abidli et al., 2012) y H. rubra (Litaay y De Silva, 2003) se afirma que estos índices
son precisos para mostrar cambios asociados a los desoves y desoves parciales,
pero Ault (1985) menciona que podría ocurrir una sobreestimación del área de la
gónada en el estadio de desove, cuando el área de la gónada no cambia debido a
que las trabéculas no se contraen después de la expulsión de los gametos, lo que
no se observó en las gónadas de las dos especies estudiadas en este trabajo.

64
Los resultados histológicos del presente trabajo, indican que H. fulgens y H.
corrugata presentan desarrollo de gametos y desoves parciales prácticamente
todo el año. Esto fue confirmado porque no hubo diferencias en la variación del
diámetro promedio mensual de los ovocitos, por la presencia de todos los estadíos
celulares del ciclo gametogénico en todas las gónadas analizadas y por la
presencia de ovocitos atrésicos en todas las fases del ciclo gonádico.
Además de desoves continuos, ambas especies presentaron un periodo
con mayor frecuencia de organismos en fase de madurez y desove, que señala
una época reproductiva de más intensidad que podría considerarse como una de
sus tácticas reproductivas.
En el presente trabajo en el caso de H. fulgens el desove y la madurez se
presentaron durante todo el año y el periodo de mayor intensidad fue de
septiembre a diciembre de 2011 y de octubre de 2012 a febrero de 2013, en
ambas zonas de estudio. En trabajos anteriores efectuados en H. fulgens en Bahía
Tortugas se reportaron ejemplares con signos de madurez sexual en cualquiera de
las cuatro estaciones del año, con una mayor intensidad de desove durante el
otoño-invierno de 1985 y durante la primavera de 1987 (Belmar-Pérez y Guzmán
del Próo, 1991), por lo que los autores caracterizan a la especie como asincrónica
y poco especializada en su ciclo reproductivo. También Sevilla (1971) reporta que
en Isla Cedros, los ejemplares examinados desovaron de junio a octubre de 1965,
y la producción de gametos continúa durante la etapa reproductiva, por lo que
supone que algunos ejemplares se reproducen más de una vez en una sola
temporada. García-Juárez y Ortiz-Quintanilla (1991), en la misma zona de estudio,
coinciden en señalar la misma época reproductiva y un desarrollo continuo de la
gónada, además de un segundo pico del 58% en diciembre de 1989 y del 73% en
diciembre de 1990, Andrade (1971) reporta que en Punta Eugenia, Punta Abreojos
y San Pablo, B.C.S., la época reproductiva es de junio a octubre, tanto para
hembras, como para machos y que el estadío de desarrollo se presenta durante
todo el ciclo, Astudillo (1993) reporta la época reproductiva de septiembre a
febrero en La Bocana, San Juanico y Las Pocitas, B.C.S. y Luna (1993) señala
que en Bahía Magdalena, la época reproductiva se presentó de octubre a enero.

65
Esto indica que H. fulgens tiene la capacidad de reproducirse durante todo el año
con variaciones interanuales en la intensidad del desove.
En resumen, la presencia de desove y madurez durante todo el año, con
una intensidad mayor o igual al 50% de septiembre a febrero, coincide con lo
encontrado por otros autores que realizaron análisis histológicos utilizando
muestreos mensuales (Tabla 6), lo que corrobora que la época reproductiva más
intensa es en este periodo, independientemente de las variaciones ambientales
interanuales y las debidas a los fenómenos de El Niño y La Niña.
Tabla 6. Frecuencias en porcentaje, iguales o mayores al 50% de desove y
madurez, en Haliotis fulgens reportadas por otros autores. * Año El Niño fuerte. +
Año El Niño débil. 1) Isla Cedros (Sevilla, 1971), 2) Punta Eugenia y Punta
Abreojos (Andrade, 1971), 3) San Juanico (Astudillo, 1973), 4) La Bocana
(Astudillo, 1973), 5) Isla Cedros (García-Juárez y Ortiz-Quintanilla, 1991), 6) Bahía
Magdalena (Luna, 1989), 7, 8, 9) Bahía Tortugas, Bahía Asunción (Presente
estudio), 10, 11, 12) La Bocana (Presente estudio).
No Año de
muestreo
E F MAMJJ A S O N D
1* 1965 100 80 60
2+ 1969-1970 57 82 100
3 1990 82
4 1990 62 63 91 84 100 72
5 1989-1990 73
6 1990-1991 71 76 71
7 2011 50 56
8 2012 68 51
9 2013 66
10 2011 60 78 50 58
11 2012 67 66 63 82
12 2013 64 60

66
En el presente trabajo en H. corrugata los desoves también ocurrieron
durante todos los meses de muestreo, pero la época reproductiva, varió de
acuerdo a la zona y año de estudio. En la zona II de septiembre a diciembre de
2011, de febrero a mayo y en octubre de 2012. En la zona III en septiembre y
diciembre de 2011, de junio a noviembre de 2012 y de junio a agosto de 2013. El
único trabajo previo que reporta la época reproductiva de H. corrugata es el de
Andrade (1971) en Punta Eugenia y Punta Abreojos, que fue de junio a octubre, y
reporta que los organismos en Punta Eugenia alcanzaron su madurez un mes
antes que los de Punta Abreojos.
Esto indica que H. corrugata y H. fulgens presentan un desarrollo gonádico
continuo que les permite presentar desoves durante todo el año y cuando las
condiciones ambientales son más favorables aumentar la intensidad, como una
táctica oportunista. Las variaciones del ciclo gonádico de acuerdo a la zona
geográfica y condiciones ambientales, explicarían los cambios en la estacionalidad
de la época reproductiva. Este tipo de cambios también se han registrado en
poblaciones de H. asinina (Counihan et al., 2001), H. madaka (Horiguchi et al.,
2005) y H. varia (Najmudeen y Victor, 2004), y sugieren que la táctica reproductiva
oportunista es propia del género Haliotis.
La presencia de desoves durante todo el año, con un pico de actividad, es
un comportamiento normal reportado para especies tropicales como H. tuberculata
coccinea (Bilbao et al., 2010), pero Webber y Giese (1969), Capinpin et al. (1998)
y Wilson y Schiel (1995), reportan que en especies de Haliotis de zonas templadas
y subtropicales, la maduración gonádica y los desoves presentan una marcada
estacionalidad, relacionada con los cambios de la temperatura.
Este comportamiento lo presentan también otras tres especies de
gasterópodos del orden Vetigastropoda, que coexisten con H. fulgens y H.
corrugata. Una de ellas es Tegula aureotincta, que presenta desoves durante todo
el año, con dos eventos principales de evacuación de gametos, uno en otoño-
invierno y el otro en primavera (Vélez-Arellano et al., 2011). La segunda es Tegula
eiseni, en la que los machos presentaron desoves durante todo el año y en las

67
hembras, aunque no se reportan desoves, los autores sugieren desoves parciales
por la presencia de gametogénesis y maduración durante todo el año, además
para ambos sexos se reportan dos picos con grandes eventos de desove uno en
primavera y otro en otoño (Vélez-Arellano et al., 2009). La última es Astraea
undosa, que también se considera una especie que se reproduce durante todo el
año y en la que las expulsiones de primavera y otoño son los principales picos
reproductivos (Belmar-Pérez y Guzmán del Próo, 1991).
Esta combinación de comportamientos en el desove, como especies
tropicales y templadas, encontrada en algunas especies de gastrópodos,
incluyendo H. fulgens y H. corrugata, en la zona de estudio, posiblemente se debe
a la complejidad del comportamiento climático, ya que esta zona se considera
como de transición templado-tropical, donde existe un gradiente estacional
importante debido a la influencia de la Corriente de California, que le da
características templadas durante el invierno-primavera y la de la Corriente
Noroecuatorial, que aporta características tropicales durante el verano-otoño.
En el presente trabajo no fue posible relacionar la temperatura superficial
del mar con la época reproductiva de H. fulgens y H. corrugata, debido a los
desoves continuos y a la variabilidad de la época reproductiva entre años y zonas.
Sin embargo, se observó que los desoves fueron más intensos a temperaturas
cercanas a 20°C, en coincidencia con lo reportado por Sevilla (1971) y Luna
(1993). Las dos especies tuvieron un amplio intervalo de temperaturas en el que
fueron capaces de desovar, lo que posiblemente es parte de la táctica
reproductiva que se complementa con el hecho de que ambas especies desovan
en pausas, cuando hay condiciones ambientales adecuadas. Al respecto Vélez-
Arellano et al. (2009), mencionan que en la zona de estudio existen cambios
drásticos de disminución o aumento de temperatura de 1 a 3°C, que ocurren en
cuestión de días o de una semana a la siguiente, por lo que estos cambios pueden
influir en las expulsiones parciales.
En cuanto al posible efecto del fotoperiodo en el ciclo reproductivo de H.
fulgens y H. corrugata, en el presente trabajo no se encontró una correlación
estadísticamente significativa, pero es posible observar el aumento del desove

68
durante los meses en los que los días comienzan a ser más cortos. Al respecto el
efecto del fotoperiodo sobre la capacidad reproductiva en los haliótidos ha sido
poco estudiado y los resultados son poco concluyentes. Es el caso de H.
cracherodii, especie en la que el tamaño de las gónadas de los reproductores, no
presenta una correlación estadística con la duración del día, aunque los periodos
de luz mayores a 12 horas estimulan la gametogénesis (Webber y Giese, 1969).
En contraste Setyono (2006) encontró que en H. asinina existe una relación entre
la maduración gonádica y el incremento de la duración del día, pero esta
maduración parece estar más asociada al incremento de la temperatura del aire y
a las mareas. Por otra parte Peña (1986), observó en condiciones de laboratorio,
que en H. discus un fotoperiodo extremadamente largo (24 horas de luz), tiene un
efecto negativo para la maduración gonadal y concluye que para que la
maduración gonadal suceda, se requiere una variación cíclica del fotoperiodo y de
la temperatura.
Por otra parte, los ciclos de reproducción de los haliótidos pueden ser
afectados por la disponibilidad de alimento. En sitios donde hay abundancia de
alimento durante todo el año, los organismos pueden completar dos ciclos
gonádicos por año, en tanto que una disponibilidad de alimento variable resulta en
bajo desarrollo gonadal (Ault, 1985). En el caso de H. fulgens, Vega-García et al.
(2015) reportan un claro patrón estacional de contenidos altos de nitrógeno en
isótopos estables durante el invierno, sin embargo para H. corrugata no se
encontró el mismo patrón y sugieren que los cambios se relacionan con la
variabilidad estacional de las fuentes de alimento. Lo anterior apoya que los
abulones tienen una táctica reproductiva oportunista, que favorece un aumento de
la intensidad del desove cuando existe abundancia de alimento.
En el mismo sentido la relación inversa entre el índice gonádico y el índice
hepático de H. fulgens y de H. corrugata en el presente estudio, apunta hacia una
transferencia de nutrientes desde el hepatopáncreas hacia la gónada. Esto
también ocurre en H. crachedorii, H. rufescens (Boolootian et al., 1962), H. rubra
(Litaay y De Silva, 2003) y H. varia (Najmudeen, 2007), y sugiere que la glándula
hepática reserva nutrientes esenciales para el desarrollo de los gametos durante

69
el ciclo reproductivo. Sin embargo, en el presente estudio en H. fulgens y H.
corrugata no fue posible observar cambios en los componentes bioquímicos,
posiblemente debido al desarrollo gonádico continuo.
Según Darriba et al. (2005) la relación entre el ciclo reproductivo y el ciclo
de almacenamiento de nutrientes define la estrategia reproductiva de la especie.
La táctica puede variar y ser de tipo conservativa u oportunista. En la conservativa,
la gametogénesis se realiza utilizando las reservas almacenadas en el
hepatopáncreas (Bayne, 1976) como se observó que lo hicieron H. fulgens y H.
corrugata en el presente trabajo. Esto también ha sido reportado para otras
especies de Haliotis (Boolootian et al., 1962; Litaay y De Silva, 2003; Najmudeen,
2007). Incluso, Litaay y De Silva (2003) y Litaay (2005) reportan que el
hepatopáncreas es el principal proveedor de reservas de nutrientes,
especialmente lípidos, durante la maduración.
En la táctica oportunista la gametogénesis se presenta cuando hay
disponibilidad suficiente de alimento para soportar los requerimientos energéticos
del proceso (Bayne, 1976). En ese sentido en el presente trabajo se observó que
H. fulgens y H. corrugata también se comportaron como oportunistas, puesto que
presentaron desarrollo gonádico, utilizando los nutrientes obtenidos del medio.
Otro órgano considerado como almacén de nutrientes en los moluscos es el pie
(Webber, 1970; Brousseau, 1987; Rodríguez-Astudillo et al., 2005; Ramesh y
Ravichandran, 2008). En H. cracherodii, el pie disminuye de tamaño conforme
aumenta el volumen de la gónada (Webber, 1970). En el presente trabajo no fue
posible observar la acumulación o transferencia de nutrientes del pie a algún otro
órgano, posiblemente debido a la condición oportunista de los organismos. Por lo
anterior es posible concluir que la táctica reproductiva utilizada por H. fulgens y H.
corrugata es oportunista, puesto que aprovechan los nutrientes del medio cuando
existe disponibilidad de alimento, mientras que por otro lado reabsorben sus
gametos y utilizan las reservas acumuladas en el hepatopáncreas cuando las
condiciones del medio no son favorables.
Varios autores reportan que la proporción de sexos en Haliotis no difiere
significativamente de la proporción esperada de 1:1 (Boolootian et al., 1962;

70
Webber y Giese, 1969; Young y De Martini, 1977; Poore, 1973; Tutschulte y
Connell, 1981; Capinpin et al., 1998; Litaay y De Silva, 2003; Najmudeen, 2004).
Sin embargo, en el presente trabajo la proporción de sexos de H. fulgens, varió
significativamente sesgada hacia las hembras en la zona de pesca II, de una
hembra por cada 0.6 machos y en la zona de pesca III, de una hembra por cada
0.7 machos. Para H. australis, se ha reportado que los machos son más
frecuentes y para H. iris que la proporción por sexos varía con la localidad, en una
con mayor proporción de hembras y en otra, con mayor proporción de machos
(Poore, 1973).
Sin embargo, existe la posibilidad de que los individuos jóvenes se hayan
clasificados como hembras, pues podrían formar primero ovocitos antes de
convertirse en organismos maduros. Este fenómeno se presenta en H. lamellosa
(Bolognari, 1953 en Mgaya, 1995) y en H. tuberculata (Girard, 1972 en Mgaya,
1995; Hayashi, 1980), esto podría dar una explicación para el aumento de
hembras en H. fulgens, ya que esta especie incluso tiene reportes de
hermafroditismo (Ortiz, comunicación personal, citado en Guzmán del Próo, 1992).
Morton (1991) propuso que la plasticidad fenotípica en bivalvos puede ser
una táctica reproductiva para maximizar el potencial reproductivo cuando la
proporción de sexos se encuentra sesgada. En Megapitaria squalida se encontró
una proporción de sexos sesgada hacia las hembras y además la presencia de
organismos hermafroditas (Romo-Piñera, 2010). Con lo referido anteriormente es
posible que la mayor proporción de hembras en la población de H. fulgens, es una
táctica que hace más eficiente la reproducción.
La talla de primera madurez refleja un compromiso evolutivo entre los
beneficios y los costos para la capacidad de reproducirse, comparativamente
temprano o tarde en la vida (Hart y Reynolds, 1992). En la Tabla 7 se presentan
los valores estimados en distintos años por diferentes autores, de la talla de
primera madurez de H. corrugata y de H. fulgens.

71
Tabla 7. Talla de primera madurez L50 de Haliotis corrugata y Haliotis
fulgens, estimada por otros autores en las cuatro zonas de pesca (SAGARPA,
2016). 1) Guzmán del Próo et al., 1980, 2) Lelevier et al., 1989, 3) Vega et al., 1994, 4)
Presente estudio, 5) Vega y Michel 1992, 6) Astudillo, 1993.
No Zona I
Zona II Zona III Zona IV
1 H. corrugata 140 135 125 110 2 130 135 132 110 3 121 4 79-167
L50=135 74-169 L50=135
1 H. fulgens 150 145 130 120 5 145 6 102-192
L50=129 77-180 L50=101
2 148 148 147 122 4 59-182
L50=135 60-200 L50=149
Con los datos publicados en los diferentes trabajos previamente realizados
(Tabla 7), debido a que únicamente se reportan las tallas como dato puntual, sin
mencionar la metodología, ni el intervalo de tallas utilizado, no es posible efectuar
una comparación con los datos obtenidos en este trabajo para saber si existen
diferencias. Sin embargo la talla de primera madurez de H. fulgens reportada en el
presente trabajo para las zonas II y III si es significativamente diferente, al
respecto en otros haliótidos como en H. rufescens, se sugiere que la talla de
primera madurez se reduce tras el suministro continuo de una dieta óptima y en H.
tuberculata coccinea la disminución se atribuye a la degradación de las costas y la
sobreexplotación (Bilbao, 2010). Button (2008) propone que la talla de primera
madurez no se ve afectada por la reducción de la población, sino que al realizar
una evaluación de poblaciones deterioradas, el porcentaje de organismos
maduros de tallas pequeñas aumenta al disminuir la talla promedio de la
población. Por lo anterior, con la información existente y la generada en el
presente trabajo, no es posible concluir si la disminución de la talla de primera
madurez de H. fulgens en la zona de pesca II, es una táctica que le permite

72
apresurar la reproducción, o es debida a la influencia de la pesquería, por la que
han disminuido los organismos de tallas grandes.
En el caso de la fecundidad, en el presente trabajo se estimaron valores
que difieren por zona, posiblemente esto se debe a las diferencias en las
condiciones ambientales y a la presión pesquera, ya que se sabe que la gran
variabilidad en el tamaño de los desoves entre poblaciones de la misma especie, o
bien, en un mismo organismo a lo largo de uno o varios ciclos anuales, pueden
atribuirse a factores biológicos, ambientales y antropogénicos (Ault, 1985; Giorgi y
De Martini, 1977; Molina, 1983). En este sentido, Kasyanov (2001), menciona que
el nivel de máxima fecundidad de una especie, se presenta en las condiciones de
hábitat más favorables, cuando los organismos gastan la energía máxima
disponible para la reproducción y Ault (1985), demostró que la fecundidad de
organismos silvestres recién capturados de H. rufescens, fue tres veces menor
que en organismos mantenidos en el laboratorio en condiciones ideales durante
70 a 90 días con longitudes semejantes.
Al comparar la fecundidad calculada en el presente trabajo con los reportes
anteriores (Tabla 8), se observa que existen diferencias con la calculada en el
presente estudio, posiblemente debido a la diferencia en las metodologías
empleadas. Por lo que no es posible comparar los resultados.
Tabla 8. Fecundidad de Haliotis fulgens estimada por otros autores 1) Ortiz-Quintanilla et al., 1987, 2) Guzmán del Próo et al., 2001, 3) Presente estudio.
No Especie Mes año Talla Huevos x 106
1 H. fulgens Jun 82 182-166 5.16 1 Jun 85 172-153 3.34 1 Sep 84 200-111 7.24 2 Oct 97 115-199 5.95 3 Ene 2013/ Zona II 128-166 4.48 3 Ago 2012 / Zona III 160-175 2.34 3 Ene 2013/ Zona III 163-173 2.3 1 H. corrugata Jun 82 186-134 10.4 1 Jun 85 142 1.34 1 Sep 84 167-120 3.75 3 Ago 2012 99 -106 2.09

73
Así mismo, Poore (1973) reportó que en enero de 1973, H. australis
presentó una fecundidad entre 46,000 y 2,910,000 huevos, en ejemplares cuya
longitud de la concha varió de 62 a 91 mm, respectivamente y menciona que fue
menor a la observada en julio de 1968, cuando fue de 195,000 a 1,380,000, a
longitudes de 67 mm a 87 mm, respectivamente, el autor lo atribuye a la etapa del
ciclo en la que se encontraban los organismos. De manera similar Ortiz-Quintanilla
et al. (1987), atribuyeron al grado de madurez los valores de fecundidad diferentes
que encontraron entre meses. Por el contrario, en este trabajo la fecundidad no
varió entre meses, pero esto puede ser debido a que únicamente se analizaron
hembras en estadio de madurez (determinado mediante el análisis histológico).
De acuerdo a Wootton (1989), la suma de las tácticas reproductivas
conforma la estrategia reproductiva de una especie. En resumen, de acuerdo a
esta definición, la estrategia reproductiva de H. fulgens y H. corrugata consiste en
un conjunto de tácticas compartidas como son la presencia de desoves parciales
euritérmicos durante todo al año, con afinidad a desovar con más intensidad
alrededor de los 20ºC, un comportamiento oportunista en el aprovechamiento de
nutrientes durante la gametogénesis, ya sea utilizando los nutrientes en el medio,
los nutrientes almacenados en el hepatopáncreas, los nutrientes reabsorbidos de
los gametos que no son expulsados, o una combinación de todo lo anterior.
También se presentaron tácticas distintas entre especies, como la presencia de un
pico reproductivo en H. fulgens de junio a diciembre en las dos zonas, mientras
que H. corrugata, no presenta ningún pico reproductivo, H. fulgens presentó una
mayor proporción de hembras como una táctica para asegurar la reproducción,
mientras que en H. corrugata la proporción de sexos fue de 1:1. Adicionalmente,
en H. fulgens la fecundidad varió de acuerdo a las condiciones del medio.
Desafortunadamente, no fue posible conocer como se comportó la fecundidad en
H. corrugata, debido a que en una de las localidades de estudio no fue posible
obtener muestras, por lo que no se pudo efectuar la comparación correspondiente.
A pesar de estas variaciones oportunistas de las tácticas reproductivas, se
concluye que la estrategia reproductiva de H. fulgens y de H. corrugata, es la

74
misma en ambas zonas de estudio, ya que las tácticas son oportunistas y variarán
de acuerdo a las fluctuaciones ambientales temporales propias de cada localidad,
ya sean las debidas a la variación interanual, o a las causadas por fenómenos
climatológicos.
CONCLUSIONES
Las tácticas reproductivas de H. fulgens y H. corrugata son semejantes y
oportunistas, se modifican temporalmente, de acuerdo al año y a la localidad a
causa de las condiciones ambientales que se presenten, todos estos cambios
a favor de la reproducción, por lo que se concluye que no hay diferencias entre
las zona de pesca II y III
El desarrollo gonádico continuo de H. fulgens y de H. corrugata, permite
que estas especies puedan desovar en cualquier momento en que las
condiciones sean adecuadas, por lo que emplean una táctica oportunista,
debido a que utilizan el alimento disponible para soportar los requerimientos
energéticos de estos procesos, o en los momentos en que las condiciones no
son adecuadas reabsorben sus gametos para aprovechar los nutrientes y
utilizan las reservas almacenadas en el hepatopáncreas, lo que les permite
realizar estos procesos cuando el alimento no está disponible.
La variación de la proporción de sexos (sesgada a las hembras) y la
diferencia por zonas en la fecundidad de H. fulgens, son modificaciones en
estas tácticas que les permite maximizar el aprovechamiento de los recursos
ambientales disponibles o limitantes para su reproducción, en este caso debido
al alimento.
Ambas especies son desovadoras euritérmicas. Sin embargo, Haliotis
fulgens presentó un periodo de intensa actividad reproductiva de principios de
verano a finales de invierno tanto en la zona II, como en la zona III y en Haliotis
corrugata, el periodo donde se observó una intensa actividad reproductiva varió
de acuerdo a la zona de estudio y al año, en la zona II fue de septiembre a
diciembre de 2011, y de febrero a mayo y en octubre de 2012, para la zona III

75
fue en septiembre y diciembre de 2011, de junio a noviembre de 2012 y de
junio a agosto de 2013.
No se encontró una clara relación entre el ciclo reproductivo y el ciclo de
almacenamiento de nutrientes, debido a la gametogénesis continua de la
gónada.
No se encontró una clara relación del ciclo reproductivo con el fotoperiodo
en las zonas de estudio.

76
REFERENCIAS
Abidli, S., Y. Lahbib & N. T. El Menif. 2012. Relative growth and reproductive cycle
in two populations of Bolinus brandaris (Gastropoda: Muricidae) from
northern Tunisia (Bizerta Lagoon and small Gulf of Tunis). Biologia.
67(4):751-761.
Andrade, P. M. 1971. Algunos aspectos sobre madurez gonadal en Haliotis
fulgens y Haliotis corrugata (phylum Mollusca, clase Gastropoda) por
observación directa. Tesis de licenciatura. Universidad Autónoma de Baja
California Sur. La Paz, México. 52 p.
Arellano-Martínez, M. 2004. Características ecofisiológicas de la reproducción de
Nodipecten subnodosus (Sowerby, 1835) (Pectinidae) en la laguna Ojo de
Liebre, B.C.S., México, Tesis de doctorado. Centro Interdisciplinario de
Ciencias Marinas, Instituto Politécnico Nacional. La Paz, México. 167 p.
Astudillo, J. M. 1993. Algunos aspectos reproductivos del abulón azul Haliotis
fulgens Philippi, 1845 (Mollusca: Haliotidae) en tres áreas: La Bocana, San
Juanico y La Poza, B.C.S., México en un ciclo anual. Tesis de licenciatura.
Universidad Autónoma de Baja California Sur. La Paz, México. 71 p.
Ault, J. S. 1985. Some quantitative aspects of reproduction and growth of the red
abalone, Haliotis rufescens Swainson. J. World Aquacult. Soc. 16:398-425.
Baqueiro, C. E., A. Avilés-Quevedo, J. A. Masso-Rojas, M. Muciño-Díaz, P.
Rogers-Nieto & A. Vélez-Barajas. 1992. Manual de métodos de muestreo y
evaluación de poblaciones de moluscos y otros recursos bentónicos.
Secretaría de Pesca, México, D.F. 73 p.
Barnes, H. & J. Blackstock. 1973. Estimation of lipids in marine animals and
tissues: detailed investigation of the sulphophosphovainillin method for
“total” lipids. J. Exp. Mar. Biol. Ecol. 12:103-118.
Bayard, H. 1974. Introducción a la Biología Marina. Acribia Zaragoza. 455 p.

77
Bayne, R. D. 1976. Aspects of reproduction in bivalve molluscs, 432–448. En: M.
Wiley, editor. Estuarine Processes. Academic Press. New York.
Belmar-Pérez, J. & S. A. Guzmán-del Próo. 1991. Madurez sexual y ciclo gonádico
de Haliotis fulgens y Astraea undosa en Bahía Tortugas B.C.S. (1986-
1988).121-129. En: S. Guzmán del Próo, editor. Memorias del Taller
México-Australia sobre reclutamiento de recursos bentónicos de Baja
California. SEPESCA-IPN, México. 212 p.
Bilbao, A., V. Tuset, M. Viera, G. Courtois de Vicose, H. Fernández-Palacios, R.
Haroun & M. Izquierdo. 2010. Reproduction, fecundity and growth of
abalone (Haliotis tuberculata coccinea, Reeve 1846) in the Canary Islands.
J. Shellfish Res. 29:959-967.
Bolognari, A., 1953. Ricerche sulla sessualita di Haliotis lamellose Lam. Arch. Zool.
Ital. 38:361-402.
Boolootian, R. A., A. Farmanfarmaian & A. C. Giese. 1962. On the reproductive
cycle and breeding habits of two western species of Haliotis. Biol. Bull.
122:183-193.
Bradford, M. M. 1976. A rapid and sensitive method for the quantification of
microgram quantities of protein utilizing the principle of protein-dye binding.
Anal. Biochem. 72:248-253.
Brousseau, D. J. 1987. A comparative study of the reproductive cycle of the soft-
shell clam, Mya arenaria in Long Island Sound. J. Shellfish Res. 6:7-15.
Button, C. A. 2008. The influence of density-dependent aggregation characteristics
on the population biology of benthic broadcast-spawning gastropods: pink
abalone (Haliotis corrugata), red abalone (Haliotis rufescens), and wavy
turban snails (Megastraea undosa). Tesis de doctorado. University of
California at San Diego. 209 p.
Calvo, J., E. Morriconi & P. M. Orler. 1998. Estrategias reproductivas de moluscos
bivalvos y equinoideos. 195-231. En: E. E. Boschi, editor. El Mar Argentino
y sus recursos pesqueros. Tomo 2. Los Moluscos de interés pesquero.

78
Cultivo y estrategias reproductivas de bivalvos y equinoideos, Instituto
Nacional de Investigación y Desarrollo Pesquero. Secretaría de Agricultura,
Ganadería, Pesca y Alimentación, Mar del Plata. 231 p.
Capinpin, E. C., V. C. Encena & N. C. Bayon. 1998. Studies on the reproductive
biology of the Donkey’s ear abalone, Haliotis asinina Linne. Aquaculture.
166:141–150.
Chelton, D. B. 1981. Interannual variability of the California Current - Physical
Factors. CALCOFI Rep. 22:34-48.
Counihan, R. T., D. C. McNamara, D. C. Souter, E. J. Jebreen, N. P. Preston, C.
R. Johnson & B. M. Degnan. 2001. Pattern, synchrony and predictability of
spawning of the tropical abalone Haliotis asinina from Heron Reef, Australia.
Mar. Ecol. Prog. Ser. 213:193–202.
Cox, K. W. 1962. California Abalones, Family Haliotidae. Calif. Dept. Fish Game.
Fish. Bull. 118:1-133.
Darriba, S., F. San Juan & A. Guerra. 2005. Energy storage and utilization in
relation to the reproductive cycle in the razor clam Ensis arcuatus (Jeffreys,
1865). ICES J. Mar Sci. 62:886-896.
Dayton, P. K & M. J. Tegner. 1984. Catastrophic Storms, El Niño, and Patch
Stability in a Southern California Kelp Community. Science. 224:283-285.
De Robertis, E. D. P., F. A. Saez & E. M. F. De Robertis. 1977. Biología Celular.
9.° ed. Librería El Ateneo Editorial. Buenos Aires. 528 p.
DOF. 1993. Norma Oficial Mexicana 004-PESC-1993, para regular el
aprovechamiento de la almeja catarina, en aguas de jurisdicción federal de
los estados de Baja California y Baja California Sur, 21 de diciembre 1993.
Dybas, C. L. 1994. Tough but tender abalone stage a comeback. Sea - Front.
40(3):12-13.
Elhasni, K., P. Vasconcelos, M. Ghorbel & O. Jarboui. 2013. Reproductive cycle of
Bolinus brandaris (Gastropoda: Muricidae) in the Gulf of Gabes (southern
Tunisia). Medit. Mar. Sci. 14(1):24-35.

79
Fukazawa, H., T. Kawamura, H. Takami & Y. Watanabe. 2007. Oogenesis and
relevant changes in egg quality of abalone Haliotis discus hannai during a
single spawning season. Aquaculture. 270:265-275.
García-Juárez, A. R. & M. Ortiz-Quintanilla.1991. Madurez sexual en abulón azul,
Haliotis fulgens (Mollusca: Gastropoda) de Isla de Cedros, Baja California,
65-75. En: S. Guzmán del Próo, editor. Memorias del Taller México-
Australia sobre reclutamiento de recursos bentónicos de Baja California.
SEPESCA-IPN, México. 212 p.
Geiger, D. L. & B. Owen. 2012. Abalone: World-wide Haliotidae. Onch
Books. Hackenheim. Germany. 361 p.
Giorgi, A. E. & J. D. De Martini. 1977. A study of the reproductive biology of the red
abalone, Haliotis rufescens Swainson, near Mendocino, California. Calif.
Fish. and Game. 63(2):80-94.
Girard, A. 1972. La reproduction de l´ormeau, Haliotis tuberculata L. Rev. Trav.
Inst. Peches Marit. 36(2):163-184.
Gofas, S. 2013. Prosobranchia. Accessed through: World Register of Marine
Species at http://www.marinespecies.org/aphia.php?p=taxdetails&id=102 on
2014-03-18.
Gordon, H. R. & A. C. Petter. 2013. World abalone supply, markets, and pricing:
2011 update. J. Shellfish Res. 32(1):5–7.
Grant, A. & P. A. Tyler.1983. The analysis of data in studies of invertebrate
reproduction. II. The analysis of oocytes size / frequency data, and
comparison of different types of data. Int. J. Invertebr. Reprod. 6:259-269.
Grubert, M. A., C. N. Mundy & A. J. Ritar. 2005. The effects of sperm density and
gamete contact time on the fertilization success of blacklip (Haliotis rubra;
Leach, 1814) and greenlip (H. laevigata; Donovan, 1808) Abalone. J.
Shellfish Res. 24(2):407-413.
Gurney, L. J. & C. N. Mundy. 2004. Reproductive biology: Identifying spawning
cycles in the genus Haliotis. TAFI. 23:1-17

80
Guzmán del Próo, S. A., J. Pineda, B. J. Molina, F. Uribe, R. Aguilar, M. Andrade,
G. León, V. Marin & C.E. Castro. 1980. Análisis de la pesquería del abulón
de Baja California: fundamentos biológicos para un nuevo régimen de
explotación del recurso. Informé Técnico del Instituto Nacional de la Pesca
(documento interno no publicado). México. 294 p.
Guzmán-del Próo, S. A. 1992. A review of the biology of abalone and its fishery in
México, 341-360. En: S. A. Shepherd, M. J. Tegner & S. A. Guzmán-del
Próo, editores. Abalone of the World: Their Biology, Fisheries and Culture.
Blackwell, Oxford. 608 p.
Guzmán-del Próo, S. A., J. Carrillo-Laguna, J. Belmar-Pérez & E. Martínez-Villeda.
2001. Fecundidad de abulón azul (Haliotis fulgens) en Bahía Tortugas, BCS
En El Niño de 1997. Informe Técnico. ENCB-IPN. México.
Haaker, P. L., D. O. Parker, K. C. Barsky & C. S. Y. Chun. 1998. Growth of the red
abalone, Haliotis rufescens (Swainson), at Johnsons Let, Santa Rosa
Island, California. J. Shellfish Res. 17(3):747-753.
Hahn, K. O. 1989. Gonad Reproductive Cycles, 13-39. En: K.O. Hahn, editor.
CRC Handbook of Culture of Abalone and other Marine Gastropods. CRC
Press, Boca Raton, FL. 348 p.
Hart, P. J. & J. D. Reynolds. 2002. Handbook of Fish Biology and Fisheries.
Volume 1. Blackwell Publishing, Oxford Fish Biology. 413 p.
Hayashi, I. 1980. The reproductive biology of the ormer, Haliotis tuberculate. J.
Mar. Biol. Ass. U. K. 60:415-430.
Horiguchi, T., M. Kojima, N. Takiguchi, M. Kaya, H. Shiraishi & M. Morita. 2005.
Continuing observation of disturbed reproductive cycle and ovarian
spermatogenesis in the giant abalone, Haliotis madaka from an organotin-
contaminated site of Japan. Mar. Pollut. Bull. 51:817-822.
Humason, L. G. 1979. Animal Tissue Techniques. 4.º ed. W. H. Freeman, San
Francisco. 661 p.

81
Jebreen, E. J., R. T. Counihan, D. R. Fielder & B. M. Degnan. 2000. Synchronous
oogenesis during the semilunar spawning cycle of the tropical abalone
Haliotis asinina. J. Shellfish Res. 19:845-851.
Kasyanov, V. L. 2001. Reproductive Strategy of Marine Bivalves and Echinoderns.
Science Publishers, Enfield. 229 p.
Keyl, F. 2004. Evaluación y manejo del recurso marino Haliotis spp. (abulón) en la
península de Baja California, México mediante un modelo pesquero-
climático. Tesis de maestría. Universidad de Costa Rica, San Pedro, Costa
Rica. 121 p.
Kraeuter, J. N. & M. Castagna. 2001. Biology of the hard clam. 1.º ed. Elsevier.
Oxford. 751 p.
Leeson, C. R., T. S. Leeson & A. A. Paparo. 1987. Histología. 5.º ed. Nueva
Editorial Interamericana, S. A. de C. V. México, D.F. 618 p.
Lelevier, G. A., G. León, M. Ortíz, J. R. Turrubiates, J. González. & M. A.
Reinecke. 1989. Análisis biológico pesquero del stock abulón en la
península de Baja California, durante las temporadas de pesca 1981-1988.
Centro Regional de Investigaciones Pesqueras en Ensenada. Informe
Técnico del Instituto Nacional de la Pesca (inédito).
León, G. & M. Muciño. 1995. Nota informativa sobre la pesquería de abulón en
Baja California. Centro Regional de Investigaciones Pesqueras en La Paz.
Informe Técnico del Instituto Nacional de la Pesca (inédito).
León, G. & M. Muciño. 1996. Pesquería de abulón, 15-41. En: M. Casas-Valdez &
G. Ponce-Díaz, editores. Estudio de Potencial Pesquero y Acuícola de Baja
California Sur. Vol. I. SEMARNAP, Gobierno del estado de BCS, FAO,
INAPESCA, UABCS, CIB, CICIMAR, CET DEL MAR, México. 350 p.
Levinton, J. S. 2001. Marine Biology Function, Biodiversity, Ecology. 2.º ed.
Oxford University press. New York. 515 p.
Litaay, M. & S. S. De Silva. 2003. Spawning season, fecundity and proximate
composition of the gonads of wild-caught blacklip abalone (Haliotis rubra)

82
from Port Fairy waters, southeastern Australia. Aquat. Living Resour.
16:353-361.
Litaay, M. 2005. The Blacklip abalone (Haliotis rubra L), proximate profiles of the
gonad & the digestive gland in relation to maturation of the female. Mar.
Chimica Act. 6(2):2-7.
Lluch-Belda, D. 1999. The interdecadal climatic change signal in the temperate
Large Marine Ecosystems of the Pacific, 42-47. En: K. Sherman & Q. Tang,
editores. Large Marine Ecosystems of the Pacific Rim. Assesment,
Sustainability and Management. Blackwell Science, Qingdao. 468 p.
Lluch-Belda, D., J. Elorduy-Garay, S. E. Lluch-Cota & G. Ponce-Díaz. 2000. B A C
Centros de Actividad Biológica del Pacífico mexicano. Centro de
Investigaciones Biológicas del Noroeste, México. 365 p.
López, J., C. Rodríguez & J. F. Carrasco. 2005. Comparación del ciclo reproductor
de Solen marginatus (Pulteney, 1799) (Mollusca: Bivalvia) en la rías del Eo
y Villaviciosa (Asturias, noroeste de España): relación con las variables
ambientales. Bol. Inst. Esp. Oceanogr. 21(1-4):317-327.
Luna, G., A. 1993. Ciclo reproductivo del abulón azul Haliotis fulgens, Philippi
1845. (Mollusca: Haliotidae) en la Bahía Magdalena, Baja California Sur,
México. Tesis de licenciatura. Universidad Autónoma de Baja California Sur,
La Paz, México. 63 p.
Manganaro, M., F. Marino, G. Lanteri, A. Manganaro & B. Macri. 2008.
Reproduction trials under experimental conditions of Mediterranean abalone
(Haliotis tuberculata, L. 1758). Chem. Ecol. 24(1):159-164.
Mazón-Suástegui, J., M. Muciño-Diaz & A. L. Bazúa-Sicre. 1996. Cultivo de abulón
Haliotis spp., 475-512. En: M. Casas-Valdez & G. Ponce-Díaz, editores.
Estudio de Potencial Pesquero y Acuícola de Baja California Sur. Vol. II.
SEMARNAP, Gobierno del estado de BCS, FAO, INAPESCA, UABCS, CIB,
CICIMAR, CET DEL MAR, México. 693 p.

83
Mgaya, Y. D. 1995. Synopsis of biological data on the European abalone (ormer),
Haliotis tuberculata Linnaeus 1758 (Gastropoda: Haliotidae). No. 156 FAO
Fisheries Synopsis, FAO, Roma. 28 p.
Molina, M.J. 1983. Estimaciones de fecundidad en Haliotis fulgens de El Bajo,
Bahía Rosario, Baja California. Cienc. Pesq. 4:77-95.
Moraisa, S., D. Boaventura, L. Narcisoa, P. Re & S. J. Hawkinsc. 2003. Gonad
development and fatty acid composition of Patella depressa Pennant
(Gastropoda: Prosobranchia) populations with different patterns of spatial
distribution, in exposed and sheltered sites. J. Exp. Mar. Biol. Ecol. 294:6-
80.
Morton, B. 1991. Do the Bivalvia demonstrate environment-specific sexual
strategies? A Hong Kong model. J. Zool. 223:131–142.
Muñoz, P., R. Ambler & C. Bulboa. 2012. Settlement, survival, and post-Larval
growth of Red abalone, Haliotis rufescens, on polycarbonate plates treated
with germlings of Ulva sp. J. World. Aquacult. Soc. 43(6):890-895.
Murayama, S. 1935. On the development of the Japanese abalone Haliotis
gigantea. Tokyo Imper. Univ., Coll. Agric., J. 13(3):227–232.
Najmudeen, T. M. 2008. Ultrastructural studies of oogenesis in the variable
abalone Haliotis varia (Vetigastropoda: Haliotidae). Aquat. Biol. 2:143-151.
Najmudeen, T. M. & A. C. C. Victor. 2004. Reproductive biology of the tropical
abalone Haliotis varia from Gulf of Mannar. J. Mar. Biol. Ass. India.
46(2):156-161.
Najmudeen, T. M. 2007. Variation in biochemical composition during gonad
maturation of the tropical abalone Haliotis varia Linnaeus 1785
(Vetigastropoda: Haliotidae). Mar. Biol. Res. 3:454-461.
Nelson, M. M., D. L. Leighton, C. F. Phleger & D. Nichols. 2002. Comparison of
growth and lipid composition in the green abalone Haliotis fulgens provided
specific macroalgal diets. C.B.P. Part B. 131:695-712.

84
Ortíz-Ordoñez, E., E. L. Mendoza-Santana, J. Belmar-Pérez & T. D. Padilla-
Benavides. 2009. Histological description of the male and female gonads in
Tegula eiseni, T. funebralis, T. aureotincta, T. gallina and T. regina from
Bahia Tortugas, B.C.S., Mexico. Int. J. Morphol. 27(3): 691-697
Ortiz-Quintanilla, M., A. R. García-Juárez & F. Salgado-Hernández. 1987.
Madurez, índice gonadal y fecundidad en dos especies de abulón Haliotis
spp. (Mollusca: Gastropoda) de la Isla de Cedros, Baja California. VII
Congreso Nacional de Oceanografía. Ensenada, B.C. 86-92.
Peña, J. B. 1986. La gónada de Haliotis discus (Reeve, 1946) (Gastropoda;
Prosobranchia) y los factores que influyen en su maduración. IBERUS.
6(2):229-235.
Ponce-Díaz, G., A. Vega-Velázquez, M. Ramade-Villanueva, G. León-Carballo &
R. Franco-Santiago. 1998. Socioeconomic characteristic of the abalone
fishery along the west coast of the Baja California Peninsula, México, J.
Shellfish Res. 17(3):853-857.
Poore, G. C. B. 1973. Ecology of New Zealand abalones, Haliotis species
(Mollusca: Gastropoda). New Zeal. J. Mar. Fresh. 7(1-2):67-84.
Ramesh, R. & S. Ravichandran. 2008. Seasonal variation on the proximate
composition of Turbo brunneus. Int. J. Zool. Res. 4(1):28-34.
Rodríguez-Astudillo, S., M. Villalejo-Fuerte, F. García-Domínguez & R. Guerrero-
Caballero. 2005. Biochemical composition and its relationship with the
gonadal index of the black oyster Hyotissa hyotis (Linnaeus, 1758) at
Espiritu Santo Gulf of California. J. Shellfish Res. 24(4):975-978.
Roe, J. H. 1955. The determination of sugar in blood and spinal fluid with antrone
reagent. J. Biol. Chem. 212:335-343.
Romo-Piñera, A. K. 2010. Estrategia reproductiva de Megapitaria squalida
(Sowerby, 1835) en dos zonas de Baja California Sur, México. Tesis de
doctorado. Centro Interdisciplinario de Ciencias Marinas, Instituto
Politécnico Nacional. La Paz, México. 120 p.

85
Rose, R., R. Dybdalh & S. Harders. 1990. Reproductive cycle of the estern
Australian silverlip pearl oyster Pinctada maxima (Jameson) (Mollusca:
Pteriidae). J. Shellfish Res. 9(2):261-272.
Saborido-Rey, F. 2004. Ecología de la reproducción y potencial reproductivo en las
poblaciones de peces marinos. Curso de doctorado, 2002-2004.
Universidad de Vigo. Inst. de Invest. Mar. Vigo, España. 71 p.
SAGARPA (Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y
Alimentación). 2016. Norma Oficial Mexicana NOM-009-SAG/PESC-2015,
que establece el procedimiento para determinar las épocas y zonas de veda
para la captura de las diferentes especies de la flora y fauna acuáticas, en
aguas de jurisdicción federal de los Estados Unidos Mexicanos. Diario
Oficial (México). 12 de febrero de 2016.
SAGARPA. 2014. Anuario Estadístico de Pesca. Secretaría de Agricultura,
Ganadería, Desarrollo Rural, Pesca y Alimentación. CONAPESCA, México.
220 p.
SEPESCA (Secretaría de Pesca). 1993. Norma Oficial Mexicana NOM 005-PESC-
1993, para regular el aprovechamiento de las poblaciones de las distintas
especies de abulón, en aguas de jurisdicción federal de la península de
Baja California. Diario Oficial (México). 21 de diciembre de 1993.
Setyono, D. D. E. 2006. Reproductive aspects of the tropical abalone, Haliotis
asinina, from southern Lombok waters, Indonesia. MRI. 30:1-14.
Sevilla, M. L. 1971. Desarrollo gonádico del abulón azul Haliotis fulgens Phillippi.
Rev. Soc. Mex. Hist. Nat. XXXII:129–139.
Shepherd, S. A. & H. M. Laws. 1974. Studies on southern Australian abalone
(Genus Haliotis). II. Reproduction of five species. Aust. J. Mar. Fresh. Res.
25(1):49-62.
Singhagraiwan, T. & M. Doi. 1992. Spawning pattern and fecundity of the donkey´s
ear abalone, Haliotis asinina Linne, observed in captivity. Thai. Mar. Fish.
Res. Bull. 3:61-69.

86
Sokal, R. & F. Rohlf. 1979. Biometría. Principios y métodos estadísticos en la
investigación biológica. Blume Ediciones, Madrid. 832 p.
Takashima, F., M. Okuno, K. Nishimura & M. Nomura. 1978. Gametogenesis and
reproductive cycle in Haliotis diversicolor diversicolor. Reeve. J. Tokyo Univ.
Fish. 65:1-8.
Tutschulte, T. & J. H. Connell. 1981. Reproductive biology of three species of
abalone (Haliotis) in Southern California. The Veliger. 23(3):195-206.
USNO. 2015. The United States Naval Observatory. Consultado el 29 de enero de
2015, http://aa.usno.navy.mil/data/
Vega, V. A. & E. Michel. 1992. Contribución al conocimiento de la biología
reproductiva del abulón azul (Haliotis fulgens Philippi 1845), en el litoral de
Punta Eugenia, B.C.S. México. Res. IX Simp. Biol. Mar. UABCS. La Paz,
México.
Vega, V. A., G. León & M. Muciño. 1994. Sinopsis de información biológica,
pesquera y acuacultural de los abulones (Haliotis spp.) de la península de
Baja California, México. Centro Regional de Investigación Pesquera en La
Paz. Informe Técnico del Instituto Nacional de la Pesca (inédito).
Vega-García, D. 2016. Cambios históricos en las poblaciones de abulón azul y
amarillo en la Península de Baja. Tesis de doctorado. Centro de
Investigaciones Biológicas del Noroeste, S.C. La Paz, México. 70 p.
Vega-García, D., A. Piñón-Gimate, N. Vélez-Arellano & S. E. Lluch-Cota. 2015.
Differences in diet of Green (Haliotis fulgens) and Pink (Haliotis corrugata)
wild abalone along the pacific coast of the Baja California Peninsula, using
stable isotope analyses. J. Shellfish Res. 34(3):879-884.
Vélez-Arellano, N., F. García-Domínguez, D. B. Lluch-Cota, J. L. Gutiérrez-
González & R. Sánchez-Cárdenas. 2015. Histological validation of
morphochromatically-defined gonadal maturation stages of green abalone
(Haliotis fulgens) Philippi, 1845 and pink abalone (Haliotis corrugata) Wood,
1828. Int. J. Morphology. 33(3):1054-1059.

87
Vélez-Arellano, N., l. E. Mendoza-Santana, E. Ortiz -Ordoñez & S. A. Guzmán del
Próo. 2011. Ciclo gonádico de Tegula aureotincta (Mollusca: Gastropoda)
en Bahía Asunción, Baja California Sur, México. Hidrobiologica. 21(2):178-
184.
Vélez-Arellano, N., S. Guzmán del Próo & E. Ortiz-Ordoñez. 2009. Gonadal cycle
of Tegula eiseni (Jordan, 1936) (Mollusca: Gastropoda) in Bahía Asunción,
Baja California Sur, México. J. Shellfish Res. 28(3):577–580.
Villalejo-Fuerte, M. & B. P. Ceballos-Vázquez. 1996. Variación de los índices de
condición general, gonádico y de rendimiento muscular en Argopecten
circularis (Bivalvia: Pectinidae). Rev. Biol. Trop. 44:591-594.
Villee, C. A., 1996. Biología. 8.° ed. Mc Graw Hill. México D.F. 944 p.
Webber, H. H. & A. C. Giese. 1969. Reproductive cycle and gametogenesis in the
black abalone Haliotis cracheroidii (Gastropoda: Prosobranchiata). Marine
Biology. 4:152-159.
Webber, H. H., 1970. Changes in metabolite composition during the reproductive
cycle of the abalone Haliotis cracheroidii (Gastropoda: Prosobranchiata).
Physiol. Zool. 43(3):213-231.
Wilson, N. H. F. & D. R. Schiel. 1995. Reproduction in two species of abalone
(Haliotis iris and H. australis) in southern New Zealand. Mar. Freshwater
Res. 46:629-637.
Wootton, R. J., 1989, Introduction: strategies and tactics in fish reproduction.1-12.
En: G. W. Potts & M. N. Wootton, editores. Fish Reproduction: Strategies
and Tactics. Academic Press, London, 410p.
Young, J. S. & J. D. De Martini. 1970. The Reproductive cycle, gonadal histology,
and gametogenesis of the red abalone Haliotis rufescens (Swainson). Calif.
Fish and Game. 56(4):298-309.
Zar, H. J. 2010. Biostatistical analysis. Prentice-Hall. Englewood Cliffs. 620 p.

88
Zetina, Z. A., D. Aldana, T. Brule & E. Baqueiro. 2000. Ciclo reproductor e índices
de condición usados en Melongena corona (Mollusca: Gastropoda). Rev.
Biol. Trop. 48(1):77-86.
















