Señalización celular
40
Señalización Celular Julio 17, 2007 4.2.1. Neurotransmisor. El grupo de sustancias químicas cuya descarga, a partir de vesículas existentes en la neurona pre-sináptica, hacia la brecha sináptica, produce un cambio en el potencial de acción de la neurona post-sináptica se conocen como neurotransmisores. Cualquiera de los compuestos químicos que se liberan en la superficie presináptica y se ligan a los correspondientes receptores de la superficie postsináptica. Son numerosos los ya descubiertos y de naturaleza química muy distinta de unos a otros. Los primeros conocidos fueron la adrenalina y la acetilcolina. El neurotransmisor es el que permite el paso del impulso nervioso a través de la sinapsis. Algunos neurotransmisores se sintetizan directamente en las terminaciones nerviosas por acción de los enzimas que se han sintetizado en el soma y se han transportado a las terminaciones nerviosas. En función de la naturaleza del neurotransmisor se puede producir en el soma o en las terminaciones nerviosas. La sinapsis es una estructura densa, red o rejilla presináptica, y las vesículas sinápticas se disponen en los huecos que les ofrece dicha rejilla. Para el pasaje de los neurotransmisores participan las proteínas de transmembrana, tanto a nivel del extremo pre sináptico como post sináptico. Los neurotransmisores son sustancias sintetizadas a nivel neuronal, donde se encuentran sus precursores y se liberan por despolarización pre sináptica a nivel del intersticio sináptico, actuando sobre los receptores post sinápticos específicos. Clasificación de los aminoácidos neurotransmisores. Los aminoácidos neurotransmisores han sido clasificados en inhibitorios y excitatorios. Entre los inhibitorios tenemos: el gama amino butírico o GABA, la taurina, la glicina y la alanina. Actúan sobre receptores asociados a canales iónicos, abren canales de cloro, producen una hiperpolarización de la membrana post sináptica y disminuyen la actividad neuronal. Entre los excitatorios tenemos: el homocisteico, el aspártico y el glutámico. También actúan sobre
-
Upload
claudia-montes-de-guerrero -
Category
Documents
-
view
65 -
download
0
Transcript of Señalización celular

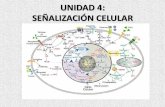
Señalización Celular
Julio 17, 2007
4.2.1. Neurotransmisor. El grupo de sustancias químicas cuya descarga, a partir de vesículas existentes en la neurona pre-sináptica, hacia la brecha sináptica, produce un cambio en el potencial de acción de la neurona post-sináptica se conocen como neurotransmisores.
Cualquiera de los compuestos químicos que se liberan en la superficie presináptica y se ligan a los correspondientes receptores de la superficie postsináptica. Son numerosos los ya descubiertos y de naturaleza química muy distinta de unos a otros. Los primeros conocidos fueron la adrenalina y la acetilcolina. El neurotransmisor es el que permite el paso del impulso nervioso a través de la sinapsis. Algunos neurotransmisores se sintetizan directamente en las terminaciones nerviosas por acción de los enzimas que se han sintetizado en el soma y se han transportado a las terminaciones nerviosas. En función de la naturaleza del neurotransmisor se puede producir en el soma o en las terminaciones nerviosas.
La sinapsis es una estructura densa, red o rejilla presináptica, y las vesículas sinápticas se disponen en los huecos que les ofrece dicha rejilla. Para el pasaje de los neurotransmisores participan las proteínas de transmembrana, tanto a nivel del extremo pre sináptico como post sináptico. Los neurotransmisores son sustancias sintetizadas a nivel neuronal, donde se encuentran sus precursores y se liberan por despolarización pre sináptica a nivel del intersticio sináptico, actuando sobre los receptores post sinápticos específicos.
Clasificación de los aminoácidos neurotransmisores. Los aminoácidos neurotransmisores han sido clasificados en inhibitorios y excitatorios. Entre los inhibitorios tenemos: el gama amino butírico o GABA, la taurina, la glicina y la alanina. Actúan sobre receptores asociados a canales iónicos, abren canales de cloro, producen una hiperpolarización de la membrana post sináptica y disminuyen la actividad neuronal. Entre los excitatorios tenemos: el homocisteico, el aspártico y el glutámico. También actúan sobre receptores asociados a canales iónicos, abren los canales de sodio, producen una despolarización de la membrana post sináptica y aumentan la actividad neuronal.
Orígen de los aminoácidos. Las fuentes principales de glutamato y aspartato son el ciclo de Krebs y sus intermediarios alfa oxoglutarato y oxalacetato. A través de la acción de enzimas transaminasas el alfa oxoglutarato se convierte en

glutamato y el oxalacetato en aspartato. Estas reacciones ocurren en todas las células.
Características de los aminoácidos. Tanto el aspártico como el glutámico son aminoácidos no esenciales, no atraviesan la barrera hematoencefálica, son sintetizados a partir de la glucosa y de precursores como la glutamina.El mensajero al actuar sobre un receptor post sináptico específico desencadena cambios de la permeabilidad de la membrana y cambios de la actividad enzimática, participando los neuromediadores como el AMPc y el GMPc.
Los eventos químicos asociados con la neurotransmisión son:
Síntesis del neurotransmisor, donde a veces participan las neuroglias. Almacenamiento de moléculas de neurotransmisor en vesículas sinápticas. Liberación de transmisores por exocitosis, lo cual es Ca+2-dependiente. A la neurona presináptica llega un impulso nervioso y abre los canales de Ca+2. El Ca+2 entra y el neurotransmisor es vertido en el espacio sináptico. El neurotransmisor se une al neuroreceptor. Activación del receptor de la membrana plasmática de la neurona postsináptica. Iniciación de las acciones del segundo mensajero. El receptor de la neurona postsináptica envía unas respuestas intracelulares que pueden desencadenar diferentes respuestas. Inactivación del transmisor, ya sea por degradación química o por readsorción en las membranas. El neurotransmisor tiene que desaparecer por: la presencia de un enzima específico que inactive el neurotransmisor o a través de la recaptación (proceso mediante el que la célula presináptica vuelve a coger el neurotransmisor y lo guarda en su interior (implica un ahorro).
Algunas moléculas son la acetilcolina, la norepinefrina y la dopamina. El gas óxido nítrico es también un neurotransmisor, con un especial mecanismo que no cumple en todos los términos con la definición dada. Las drogas de acción cerebral actúan en alguna o algunas de estas etapas.

En una misma neurona pueden existir diferentes tipos de neurotransmisores:
Aminoácidos y aminas. Aminoácidos y péptidos. Aminas y péptidos. Péptidos. Purinas y aminas. Aminas.
Normalmente hay coexistencia entre neurotransmisores. Es extraño que haya neuronas que sólo eliminen 1 único neurotransmisor. Lo más normal y frecuente es la coliberación de neurotransmisores. Los neuromoduladores son sustancias peptídicas que se originan fuera de la sinapsis y que modifican la excitabilidad neuronal, siendo algunos de ellos los neuropeptidos, la sustancia P, y las prostaglandinas. Los neuromoduladores son sustancias secretadas de manera natural que actúa de manera similar a un neurotransmisor, con la diferencia de que no queda restringido al espacio sináptico sino que se difunde por el fluido extracelular. Un neuromodulador es un producto liberado en una sinapsis que influye directamente en las consecuencias postsinápticas de la neurotransmisión, por ejemplo, el ATP, la adenosina, el GTP, la feniletilamina, etc. Los neuromediadores son transmisores químicos que aumentan las respuestas post sinápticas y se los ha denominado segundos mensajeros. Un neuromediador proteíco de membrana que específicamente es un ligando, media la acción de una sustancia sobre un determinado receptor. Por ejemplo, el ligando endógeno para las benzodiacepinas o los barbitúricos en el receptor GABA. Y la familia de proteínas G. No es exactamente lo mismo que un neuromodulador ya que el segundo mensajero es habitualmente el AMPc. Se define la neurosecreción como la específica capacidad de segregar un producto de síntesis neuronal al medio extracelular, con la propiedad de llevar a cabo un efecto fisiológico en un efector u órgano diana. Los neurotransmisores y neuromoduladores actúan como transmisores o propagadores de mensajes específicos o primeros mensajeros, que al activar al sistema trasductor de la adenilato ciclasa membranal, activan al sistema amplificador que transforma al ATP o GTP en AMPc o GMPc o segundo mensajero, que al activar diversos componentes enzimáticos, dan orígen a la respuesta celular. Los neurotransmisores se pueden clasificar en colinérgicos, adrenérgicos depresores y estimulantes, derivados de la fenil alanina o derivados del triptofano. Los adrenérgicos depresores son la noradrenalina, la dopamina y la serotonina. Los adrenérgicos estimulantes son la fenil etil amina. Los derivados de la fenilalanina son la fenil etil amina, la noradrenalina, la dopamina y la tiramina. Los receptores adrenérgicos conocidos son el alfa 1 para NA y AD, el alfa 2 para AD y NA, el beta 1 para AD, NA y DA, y el beta 2 para AD.E l sistema noradrenérgico está centrado en el locus coeruleus (LC) y subcerúleo (SC) siendo éstos sus principales núcleos de almacenaje. Del LC parten dos haces de proyección ascendente, el haz noradrenérgico ventral (HNAV) y el haz noradrenérgico dorsal (HNAD). La noradrenalina facilita la atención y la capacidad de aprendizaje.
El sistema dopaminérgico posee sus reservorios principales a nivel del locus niger (LN). Los receptores dopaminérgicos se agrupan en dos subtipos, los D1 con alta afinidad agonista, que poseen una proteína (NS) de acople entre el receptor y la adenilato ciclasa, incrementando la síntesis de AMPc, y los D2 de baja afinidad agonista, inhiben la adenil

ciclasa y disminuyen la síntesis de AMPc, presentando una proteína reguladora (NI). El sistema dopaminérgico controla la motilidad voluntaria y las funciones cognitivas. Su aumento produce hiperactividad psicomotora e hiperactividad psíquica propia de la manía. Favorece la secreción de prolactina y paratohormona, facilita el esfuerzo y la motivación para el aprendizaje.Mientras el déficit de dopamina es responsable del Parkinson su exceso produce esquizofrenia. En deprimidos disminuye en forma secundaria a la disminución de NA. Los derivados del triptofano son la serotonina (5-HT), la triptamina, la bufotenina, la dimetil triptamina y la N acetil serotonina. La secreción de monoaminas (NA y DA) e indolaminas (5-HT) depende del sistema hipotálamo hipofisario. La serotonina o 5 hidroxi triptamina posee sus reservorios y fuentes principales a nivel del núcleo dorsal del rafe (NRD) y del núcleo del rafe mediano (NRM). Posee dos agrupaciones de fibras ascendentes y una descendente. El haz procencefálico medioventral, el haz procencefálico mediodorsal, y el haz rafespinal. La biosíntesis de serotonina disminuye con la adrenalectomía y aumenta con la administración de glucocorticoides. La serotonina favorece la consolidación y recuperación de la memoria. Al parecer la serotonina aumenta con la ansiedad, las fobias y el pánico. La bufotenina o N-dimetil serotonina es una amina secundaria con actividad psicotrópica. Inyectada por vía intravenosa produce despersonalización, alucinaciones visuales y alteraciones similares a la esquizofrenia.
Los colinérgicos son la acetil colina y el GABA. El sistema colinérgico (ACo), se distribuye el 5% en el pirenóforo, el 25% en los axones y el 70% en los terminales pre sinápticos. La acetilcolina (ACo) almacenada en las vesículas es estable o ligada mientras que la recién sintetizada es lábil o disponible. Al despolarizarse la membrana, entra Ca++ y Na+, pasando la ACo a la hendidura sináptica. La acetilcolina se sintetiza merced a la colina acetiltransferasa (CAT) y se inactiva a nivel de sus receptores post sinápticos por la acetil colinesterasa (ACoE) en colina y acetato. Los receptores de la ACo son de dos tipos, nicotínicos o de respuesta rápida y muscarínicos, pre y post sinápticos, que actúan a largo plazo trabajando como segundos mensajeros y poniendo en marcha al sistema GTP – GMPc. La ACo predomina en la neocorteza, el sistema límbico, la retina y el hipotálamo. Proporciona la plasticidad para el aprendizaje y la memoria. La colina acetil transferasa predomina en el estriado, en neocorteza e hipocampo. Su disminución con la edad es responsable de las pérdidas de atención y memoria. La ACo regula el ritmo circadiano vigilar-dormir y participa del aprendizaje y la memoria.
El sistema de la monoamino oxidasa (MAO) es una enzima que se localiza en la membrana mitocondrial. Cataboliza a la NA, DA y 5-HT, y se clasifica en MAO A con sustrato en NA y 5-HT y MAO B con sustrato en FEA.
El sistema aminoacidérgico está constituído por el principal sistema inhibitorio representado por el GABA y la glicina y por los sistemas estimulantes representados por los glutamatos y aspartatos. El ácido gamma amino butírico (GABA) procede de la neocorteza inhibidora 4S y 8S y del sistema estrio palidal (SEP). Los receptores post sinápticos para el GABA son el GABA-A que abriendo los ionóforos o canales de Cl-, hiperpolarizan la membrana post sináptica con el bloqueo de la transmisión sináptica, y el GABA-B que actúan a través de los canales de Ca++. El GABA posee manifestaciones anticonvulsivas y antiagresivas, aumenta la liberación de PRL y de GH.

La histamina (HA) se forma por decarboxilación del aminoácido histidina catalizada por la histidina decarboxilasa. La histamina cerebral se inactiva por metilación catalizada por la histamina N metil transferasa. La histamina se encuentra en vesículas sinápticas y se libera por concentraciones despolarizantes de K+. Se conocen dos tipos de receptores histaminérgicos, los H1 y los H2 que son antagonistas de muchos antidepresivos. Las neuronas histaminérgicas controlan el estado de vigilancia. Colibera con la sustancia P y la somatostatina favoreciendo a través de la primera la trasmisión del dolor y de la segunda la estimulación de los linfocitos T que participan del fenómeno inmunitario.
La fenil etil amina (FEA) neocortical predomina durante la vigilia, actuando como una anfetamina biológica, dependiendo de ella el índice de vigilancia.
Los neuropéptidos son péptidos complejos que actúan como comensajeros, coliberando con el neurotransmisor y modulando su acción. Los neuropéptidos se clasifican en opioides y no opioides. Por un lado actúan como neurohormonas (TRH, CRF, LHRH, PRL), y por el otro como moduladores de neurotransmisores. Se sintetizan en el retículo endoplásmico rugoso como una molécula precursora, la propiomelanocortina, y luego en el complejo de Golgi en forma molecular de encefalinas, endorfinas, ACTH y MSH.
Los opioides se denominan así por su acción similar al opio o a sus derivados como la morfina, y son ellos las encefalinas, las endorfinas y las dinorfinas. Con respecto a las encefalinas, tanto la metencefalina como la leuencefalina actúan de preferencia sobre receptores delta, estimulando la secreción neurohipofisaria, siendo indispensables para la memoria y siendo moduladoras de la actividad afectiva instintual.La escotofobina provoca miedo a la oscuridad. Las endorfinas y en particular las beta endorfinas actúan de preferencia sobre los receptores Mu, inhibiendo la secreción adenohipofisaria de LH, siendo consideradas una sustancia antishock, provocando analgesia y somnolencia. Las dinorfinas actúan de preferencia sobre los receptores kappa naurohipofisarios. Las encefalinas y las endorfinas actuarían inhibiendo las sinapsis dopamínicas o de placer, la dinorfina endógena regularía la libido, las beta endorfinas favorecen las reacciones rápidas de defensa. La actividad analgésica de las beta endorfinas contrarrestan la agresión, favoreciendo la lucha o la retirada. Las endorfinas producen sed, hambre y analgesia y las dinorfinas promueven la alimentación, mientras que la calcitonina y la bombecina inducen a la saciedad. Las beta endorfinas aumentan el apetito favoreciendo la obesidad y la naloxona lo contrarresta. Los opioides se comportan como neurotransmisores post sinápticos a través de los receptores Mu y como neuromoduladores a nivel pre sináptico, favoreciendo la liberación de DA. Producen una brusca disminución de la actividad neuronal con caída del AMPc, desencadenando el bloqueo de la transmisión. Ejercen actividades multifuncionales tendientes a lograr la homeostasis y la supervivencia, regulan los estados emocionales, regulan la reacciones caracterológicas y regulan los procesos cognitivos.
Los neuropéptidos no opioides son la sustancia P, la colecistoquinina, el polipéptido intestinal vasoactivo y el neuropéptido YY. La sustancia P es un transmisor del dolor. La colecistoquinina (CCK) se libera conjuntamente con la DA. Su elevación post prandial es la responsable de la somnolencia post alimentaria. La progesterona incrementa la secreción de CCK. El polipéptido intestinal vasoactivo (VIP) y el péptido histidina isoleucina (PHI) incrementan unas cien veces la acción de la adenilato ciclasa, activando el AMPc. El péptido YY inhibe ésta acción, participa de la vida

emocional y de la algesia, aumenta el apetito sexual. El neuropéptido YY, coliberador adrenérgico, es un potente vasoconstrictor cerebral y mediando la NA mantiene contraídas las arterias cerebrales e inhibe el AMPc. El potencial de acción moviliza al Ca++, que desencadena la reacción entre la neurina de la membrana sináptica y la stenina de las vesículas sinápticas, originando la proteína contráctil neurostenina que contrae a las vesículas sinápticas. El Mg++ bloquea su acción.
El monóxido de nitrógeno (NO) es un neuromediador que se difunde libremente a través de las neuromembranas sinápticas. El óxido nítrico es considerado un nuevo neurotransmisor, descubierto en la neuroglía y destruído en segundos por la óxido nítrico sintetasa. El receptor sería el hierro. El glutamato provoca la entrada de Ca++ membranal, interactuando con la calmodulina, iniciando la síntesis de óxido nítrico. La acción de la ACo está mediada por el NO que estimula a los receptores del glutamato, con aumento del GMPc, merced a la NO-sintetasa. El NO se forma a partir del oxígeno y la arginina y la NO-sintetasa, que activa a la calmodulina (CM) sensible al Ca++. El NO participa de la plasticidad sináptica como segundo mensajero en el aprendizaje.
Criterios para establecer si una sustancia es un
neurotransmisor.
1. Presencia del neurotransmisor (Bioquímica, Anatomía, Histología).
1.1. El agente químico debe estar localizado en los elementos presinápticos y probablemente distribuido en todo el cerebro.1.2. La neurona debe contener los precursores, las enzimas selectivas o un mecanismo de transporte específico para el
neurotransmisor.1.3. En la sinapsis debe existir receptores para el neurotransmisor.La animación muestra el torrente axconal que lleva el precusor Triptofano a la neurona serotoninérgica. La enzima triptofano-hidroxilasa transforma al triptofano en 5-hidroxitriptofano. Una descarboxilasa la transforma en Serotonina.La Serotonina se almacena en vesículas para ser liberada cuando sea necesario.Al unirse al receptor la Serotonina cumple su efecto transmisor. Para que termine su acción la Serotonina es recapturada. La Monoaminoxidasa transforma a la Serotonina en ácido 5-hidroxiindolacético para su eliminación.
2. Liberación (Fisiología) La estimulación de las aferencias debe producir liberación del neurotransmisor en cantidades fisiológicas.
3. Identidad de acción (Fisiología, Farmacología)3.1. La aplicación directa del neurotransmisor a la sinapsis debe producir efectos idénticos a los producidos por estimulación eléctrica.3.2. La interacción del neurotransmisor con su receptor debe inducir

cambios en la membrana que produzcan potenciales postsinápticos excitatorios o inhibitorios.3.3. La estimulación aferente o la aplicación directa de la sustancia debe producir efectos semejantes a los producidos por la aplicación de agentes farmacológicos.
4. Inactivación (Bioquímica, Histología) Deben existir mecanismos de inactivación (difusión, enzimas metabólicas, sistemas de recaptación) que hagan concluir la interacción del neurotransmisor con el receptor.
4.2.2. Mecanismos receptoriales El receptor recibe el impacto del neurotransmisor y lleva a cabo la transducción (recibe una señal y transmite otra). El ATP necesario para la síntesis de neurotransmisor es proporcionado por las mitocondrias de la terminal presináptica. Esta síntesis debe ser muy rápida, debido a que la cantidad del mismo almacenada en las vesículas se halla limitada para unos segundos o minutos de actividad plena. Los receptores son proteínas bajo control genético.
Los receptores tienen dos componentes importantes:
1. Componente de fijación, que protruye al exterior de la membrana en el surco sináptico, y fija el neurotransmisor liberado de la terminal presináptica.2. Componente ionóforo, que penetra de la membrana al interior de la neurona y puede ser de 2 tipos: Un conducto de iones activado químicamente -conductos activados por ligando-, cuyo paradigma es el NMDA -N-metil-D-aspartato-, verdadero conglomerado o complejo de canales iónicos, que pueden ser de 3 tipos principales: Canales de sodio, Canales de potasio y Canales de cloruro. También pertenecen a esta categoría los receptores denominados AMPA/Kainate; y una enzima que activa un sistema metabólico interno de las células y son receptores metabotrópicos que se unen a proteínas G en el interior de las células.
Las neuronas regulan su actividad por mecanismos de retroalimentación que involucran a receptores de la terminación nerviosa, conocidos como autoreceptores. Ejemplo de autoreceptor es el receptor b2-adrenérgico en las terminaciones noradrenérgicas, que media muchas de las acciones fisiológicas de las catecolaminas endógenas Adrenalina y Noradrenalina, y es el blanco de varios agentes terapéuticos. Pese a que el mecanismo de recaptura de neurotransmisores se conoce desde hace unos 30 años. La disponibilidad de fármacos selectivos que actúan sobre tales receptores es bastante reciente. La clonación de receptores y la posibilidad de expresarlos en células no neuronales está permitiendo un mayor conocimiento del mecanismo de acción de los diferentes transportadores, asi como la posibilidad de encontrar nuevos fármacos selectivos.Los receptores pueden ser:
Receptores sensoriales Hay cinco tipos mayores de receptores sensoriales en el cuerpo humano: mecanoreceptores, que detectan la deformación mecánica; termoreceptores, que detectan los cambios de temperatura; fotoreceptores en la retina; nociceptores, que detectan el daño tisular y quimioreceptores. Algunos tipos de receptores, como los mecanoreceptores periféricos, son esencialmente transductores que convierten el estímulo detectado en un impulso eléctrico. Cuando el cambio detectado es tan grande que supera el umbral, se genera un

potencial de acción que es conducido del sistema nervioso periférico hacia el Sistema nervioso central. Como ya se mencionó, un incremento en el estímulo es convertido en un incremento en la frecuencia de disparo de la neurona eferente. La mayoría de los receptores sensoriales experimentan el fenómeno de adaptación cuando reciben continuamente en forma prolongada un estímulo apropiado, resultando en una disminución progresiva de la frecuencia de disparo en la parte del receptor estimulada. En algunos tipos de receptores (receptores fásicos) la frecuencia de disparo se detiene completamente; y en otros (receptores tónicos) la frecuencia cae a un nivel de mantenimiento. Ejemplos de mecanoreceptores fásicos incluyen los receptores de los folículos pilosos y los Corpúsculos de Paccini. Mientras los receptores de las cápsulas de unión y los musculares, son ejemplos de mecanoreceptores tónicos.
Quimiorreceptores Los quimiorreceptores están contenidos en neuronas especializadas y son capaces de responder a pequeños cambios químicos en el espacio extracelular. Tipos de quimioreceptores incluyen, los receptores del gusto, los receptores del olfato, varios receptores hipotalámicos que pueden detectar la osmolalidad sanguínea y concentraciones sanguínea de aminoácidos, ácidos grasos y glucosa; y los receptores de la aorta y las carótidas capaces de detectar cambios en las concentraciones de oxígeno y bióxido de carbono de la sangre.
4.2.2.1. Receptores ionotróficos Determinan la apertura o cierre de canales y producen despolarizaciones (génesis de potenciales de respuesta excitatorios) o hiperpolarizaciones (génesis de potenciales de respuesta inhibitorios). Es una respuesta rápida. El mecanismo de acción de estos receptores puede ser de dos formas, via señalización extracelular a través de la acción de un neurotrasmisor que induce, al unirse al receptor la apertura del canal, algunos canales pueden necesitar la unión de dos neurotransmisores como es el caso del receptor de Acetilcolina o el receptor NMDA que necesita glutamato y glicina. La señalización tambien puede ser intracelular, generalmente fosforilando en la cara citoplasmica del canal el receptor, induciendo la apertura del canal.
4.2.2.2. Receptores metabotróficos Liberan mensajeros intracelulares (AMPcíclico, CA y fosfolípidos). Cuando el receptor recibe el neurotransmisor, pone en funcionamiento la adenilatociclasa y el ATP se transforma en AMPcíclico. Una vez ha actuado, es destruido por la fosfodiesterasa. EL AMPcíclico activa una proteinquinasa, que fosforila 1 proteína.
Los receptores de Ca+2, cuando reciben el neurotransmisor, abren 1 canal de Ca+2, entra Ca+2 extracelular y se junta a la proteína calmodulina, formando la calmodulina-Ca, que activa una proteinquinasa que fosforila una proteína. El receptor de membrana, cuando recibe el neurotransmisor, activa la PLC (fosfolipasa C). La PLC actúa sobre los fosfolípidos de membrana

(concretamente sobre el fosfotidilinositol) y se derivan 2 productos (inositol trifosfato [IP3] y diacilglicerol). El inositol trifosfato actúa sobre el retículo endoplasmático liberando el Ca+2intracelular. El diacilglicerol, en presencia de Ca+2, activa la proteinquinasa que fosforila la proteína y da lugar a la respuesta postsináptica.
1. Si el receptor es inotrópico sólo abre o cierra canales. Hay receptores que ponen en marcha proteínas G. El neurotransmisor actúa sobre el
receptor, que activa una proteína G, que activa la adenil ciclasa y que transforma el ATP en AMPcíclico. Este AMPcíclico puede actuar sobre el canal de membrana, abriéndolo. Son canales iónicos operados por vías metabólicas activadas por proteína G. Cuando se fosforila el canal, se abre.
2. El neurotransmisor y el receptor provocan que la proteína G active la PLC y active el fosfatidil inositol dando (IP3 y diacilglicerol), que libera el Ca+2 y la proteinquinasa fosforila la proteína del canal y se
abre.
3. El neurotransmisor estimula el receptor, que provoca que la proteína G abra el canal.

Los recientes avances sobre el conocimiento de la farmacología de los receptores de los aminoácidos excitatorios permite la aplicación del conocimiento fino de su papel en la etiología de las enfermedades neurodegenerativas y
su tratamiento. Los receptores ionotrópicos de los aminoácidos excitatorios pueden ser divididos en dos largas familias: la familia del receptor NMDA y la familia de los receptores AMPA y KAINATO. Los estudios de clonaje de receptores han mostrado que hay un largo número de potenciales subtipos de receptores en ambas familias. Han sido desarrollados antagonistas para los receptores NMDA los cuales pueden interactuar como mínimo con cuatro sitios del receptor, reconocidos como drogas independientes. Para los receptores AMPA y KAINATO, dos clases de antagonistas han sido bien identificados. Razonable potencia, selectividad y penetración cerebral son las propiedades fundamentales que presentan los antagonistas que se conocen actualmente para éstos sitios y comprenden también la inhibición
de la liberación del ácido glutámico presináptico, como puede ser el Riluzole. La capacidad del ácido glutámico para matar neuronas por su excitotoxicidad ha sido ampliamente demostrada. La acetilcolina como neurotransmisor, actúa sobre los receptores nicotínicos (actúa igual que la nicotina del tabaco) y sobre los receptores muscarínicos (actúa por setas). Los receptores colinérgicos son Muscarínico (M1), Muscarínico (M2) y el nicotínico. Los receptores muscarínicos se pueden bloquear con atropina (se extrae de la Atropa belladona). El curare (dextrotubocuranina) bloquea la sinapsi colinérgica entre el músculo esquelético. El mismo neurotransmisor, a
veces polariza y, a veces, despolariza dependiendo del receptor y los canales que operen el receptor. En la sinapsis colinérgica, se coge ácido acético y se esterifica. La colina + Co-A + ácido acético dan acetilcolina, que se libera en el espacio sináptico y actúa sobre el receptor nicotínico y muscarínico (M1-M2). El enzima acetilcolinesterasa hidroliza el éster de acetilcolina y libera colina y acetato. Después es recaptado y se vuelve a formar acetilcolina. De todas las sinapsis, se tiene que conocer el neurotransmisor, la biosíntesis y la degradación del neurotransmisor. La dopamina, noradrenalina y adrenalina provienen de la fenilalanina que, mediante la fenilalaninahidroxilasa, le introduce un OH y forma la tiroxina, que mediante la tiroxinahidroxilasa le introduce otro OH y forma la dihidroxifenilalanina. Se descarboxila (se saca COO mediante la carboxilasa) y se forma la dopamina (neurotransmisor de las neuronas dopaminérgicas). Si se le introduce otro OH, se forma la noradrenalina o

norepinefrina (neurotransmisor de las neuronas noradrenilaninérgicas). Por acción de una N-metil transferasa se forma la epinefrina o adrenalina (neurotransmisor de las neuronas adrenérgicas).
4.2.3. Señalización sináptica
La sinapsis es el proceso esencial en la comunicación neuronal y constituye el lenguaje básico del sistema nervioso. Afortunadamente, las semejanzas de los mecanismos sinápticos son mucho más amplias que las diferencias, asociadas éstas a la existencia de distintos neurotransmisores con características particulares. Elliot en 1904 fue el primero que sugirió la posibilidad de que la información era transferida de una neurona a otra por la liberación de una sustancia química desde las fibras nerviosas; Loewi es, sin embargo, el
primero que mostró la existencia de una sustancia química en el líquido perfundido con la estimulación del nervio vago y fue su colaborador Navratil quien más tarde demostró que esta sustancia era la acetilcolina. La sinapsis es un punto de machimbre o de enlace entre dos neuronas, la presináptica y la postsináptica. Las fibras nerviosas actúan como terminales de bujías eléctricas de los motores de explosión. Hay una luz o una brecha sináptica entre los terminales, brecha sináptica donde descargan vesículas sinápticas que difunden, ayudan a que ocurran reacciones físicas y químicas, recapturan los neurotransmisores ya usados y propagan potencial eléctrico desde una pared o membrana de la brecha o hendidura, la de la neurona presináptica, a la pared o membrana de la otra, la postsináptica.En su extremo, el axon de los nervios se ramifica en muchos terminales pequeños que llegan a estar en contacto estrecho con las dendritas de otras neuronas. Al contacto entre dos neuronas se le llama sinapsis. El axon y la dendrita nunca se tocan. Siempre hay un pequeño vacio llamado hendidura sinaptica. Cuando la señal electrica llega a un terminal nervioso, hace que el nervio libere neurotransmisores. Los neurotransmisores son agentes quimicos que viajan una corta distancia hasta las dendritas mas próximas. A la neurona que libera el neurotransmisor se le llama neurona presinaptica. A la neurona receptora de la señal se le llama neurona postsinaptica. Dependiendo del tipo de neurotransmisor liberado, las neuronas postsinapticas son estimuladas (excitadas) o desestimuladas (inhibidas). Cada neurona se comunica con muchas otras al mismo tiempo. Puesto que una neurona puede enviar o no un estimulo, su comportamiento siempre se basa en el equilibrio de influencias que la excitan o la inhiben en un momento dado. Las neuronas son capaces de enviar estimulos varias veces por segundo.La sinapsis es un hecho comunicativo entre dos neuronas, una presináptica y otra postsináptica. Es imprescindible la conducción previa del impulso nervioso en la neurona

presináptica y particularmente, en los denominados botones terminales, que son las últimas estructuras de la ramificación y diversificación axónica de la neurona presináptica. Esta circunstancia es el primer punto de acción para los fármacos y drogas que afectan a la sinapsis, pues en concreto, la modificación de la conductibilidad, aun no siendo un fenómeno tan asequible como otras etapas de la sinapsis, es uno de los caminos para la intervención de anestésicos que infiltrados a distintas concentraciones bloquean o modifican la conductibilildad. Algunos neurotransmisores como acetilcolina (ACh), glicina, glutamato, aspartato y ácido gamma-amino butírico (GABA), tienen una actividad biológica directa aumentando la conductancia a ciertos iones por adherencia a canales iónicos activados en la membrana postsináptica. Otros neurotransmisores, como la noradrenalina (NA), dopamina (DA) y serotonina (5-HT), no tienen actividad directa pero actúan indirectamente vía sistemas de segundo mensajero para causar la respuesta postsináptica. Estos sistemas implican adenosín-monofosfato-cíclico (AMPc), guanidín-monofosfato-cíclico (GMPc), inositol trifosfato (ITP), diacil glicerol (DAG), prostaglandinas (Pgs), leucotrienos, epóxidos y Ca++.
4.2.3.1. Clases de sinapsis. La función de la neurona es la comunicación y la función del SN es generar un comportamiento, ambos en virtud de las conexiones interneuronales. Una neurona ejerce su influencia para excitar a otras neuronas mediante los puntos de unión o sinapsis. Cada unión sináptica está formada por una parte de una neurona (terminal sináptico) que conduce un impulso a la sinapsis y por otra, de otra neurona (estructura postsináptica) que recibe el impulso en la sinapsis (Barr, 1994).
El impulso nervioso debe atravesar un espacio muy pequeño (20nm), denominado hendidura sináptica que separa las estructuras pre y postsinápticas y puede propagarse en cualquier dirección por la superficie de la neurona; sin embargo la dirección que toma en condiciones fisiológicas se determina por una polarización constante que se hace en la sinapsis, donde la transmisión se efectúa del axón de una neurona a la superficie de
otra neurona (Darnell, 1993). La dinámica estructural y funcional para que se lleve a cabo una sinapsis entre dos neuronas esta dada por el movimiento, descarga, recaptación y reformación (resíntesis) de un neurotransmisor. Algunos neurotransmisores, como los péptidos, por ejemplo, son producidos en el soma, empaquetados en las vesículas que migran a través del axón mediante flujo axoplásmico por medio de los microtúbulos hacia el terminal presináptico donde se conocen con el nombre de vesículas sinápticas. La naturaleza del contenido de la vesícula sináptica varía dependiendo de la región cerebral de donde esta proceda. Cuando un impulso llega al terminal sináptico esta acompañado por la entrada de iones calcio en el citoplasma neuronal. El calcio proviene de los fluidos tisulares que están fuera de la neurona (espacio extracelular), estos iones, que han atravesado la membrana celular se unen a la molécula transportadora. Los iones calcio impulsan la migración de alguna de las vesículas sinápticas hacia la membrana presináptica, la membrana de cada vesícula sufre un proceso de fusión como la membrana presináptica, lo cual esta seguido por una expulsión rápida exocitosis del neurotransmisor libre en la hendidura sináptica (Meyer, 1985).

El neurotransmisor liberado en la hendidura sináptica interaccciona directamente con las moléculas del receptor en la membrana postsináptica. Mediante este tipo de interacción se abren un gran número de canales iónicos específicos que permiten el flujo de una corriente eléctrica, transportada por iones cargados a través de la membrana postsináptica lo que afecta al estado electroquímico de la membrana en el área inmediata al canal. De esta forma la excitabilidad eléctrica de esta pequeña porción de membrana puede aumentar o disminuir mediante despolarización o hiperpolarización de la misma. Las alteraciones eléctricas individuales de la membrana postsináptica ejercen un efecto en el potencial de membrana de la neurona, que puede llevar a la generación del impulso nervioso (Barr, 1994). En algunos casos cuando se libera el neurotransmisor, este es inactivado generalmente por hidrólisis, los fragmentos que resultan del neurotransmisor se eliminan del sistema o se reciclan por endocitosis, en cuyo caso, los fragmentos se reincorporan a una nueva vesícula formada en la membrana presináptica. Este tipo de vesícula se conoce como vesícula encapsulada y tiene apariencia diferente a la vesícula sináptica. Los fragmentos que entran en la neurona por endocitosis son utilizados para la resíntesis del neurotransmisor (Cooper, 1994). Existen algunos principios generales para la identificación de dos tipo de sinapsis: excitadoras e inhibidoras. Datos electrofisiológicos, muestran la distribución de las sinapsis excitadoras a nivel de la porción superior del árbol dendrítico de las neuronas centrales y de las sinapsis inhibidoras que habitualmente están unidas a los segmentos iniciales de las dendritas o a los cuerpos celulares (Bradford, 1988). A partir de la excitabilidad de las neuronas, que es su propiedad específica, se desencadenan distintos tipos de mecanismos que trascienden a la propia neurona y que establecen una clara comunicación entre las mismas. Esto es a lo que denominamos sinapsis, una región celular clara, concreta y muy estructurada definida por el mantenimiento de un espacio interneural, y cuyo significado final es el de la comunicación interneural a la que nos referimos en términos generales como sinapsis eléctrica y sinapsis química. En el primer caso, se habla siempre de una comunicación excitatoria con la continuidad de la conducción de la excitabilidad o el impulso nervioso; en el segundo caso se habla de una comunicación excitatoria o inhibitoria mediada por una sustancia química que no es otra cosa que el neurotransmisor. La sinapsis es, sin lugar a dudas, la estructura más lábil y accesible a las distintas sustancias químicas como fármacos y drogas y, por tanto, el lugar y término de referencia más importante en los mecanismos de acción de los distintos psicofármacos. La sumatoria de los impulsos exitatorios e inhibitorios que llegan por todas las sinapsis que se relacionan con a cada neurona( 1000 a 200.000) determina si se produce o no la descarga del potencial de acción por el axón de esa neurona Las sinapsis suelen clasificarse en dos tipos según la transmisión del impulso: sinapsis eléctrica y sinapsis química.4.2.3.2. Sinapsis
eléctrica.En este tipo de sinapsis los procesos pre y postsináptico son continuos (2 nm entre ellos) debido a la unión citoplasmática por moléculas de proteínas tubulares a través de las cuales transita libremente el agua, pequeños iones y moléculas por esto el estímulo es capaz de pasar

directamente de una célula a la siguiente sin necesidad de mediación química (Barr, 1994). Corresponden a uniones de comunicación entre las membranas plasmáticas de los terminales presináptico y postsinápticos, las que al adoptar la configuración abierta permiten el libre flujo de iones desde el citoplasma del terminal presinático hacia el citoplasma del terminal postsináptico. La sinapsis eléctrica ofrece una vía de baja resistencia entre neuronas, y hay un retraso mínimo en la transmisión sináptica porque no existe un mediador químico. En este tipo de sinapsis no hay despolarización y la dirección de la transmisión está determinada por la fluctuación de los potenciales de membrana de las células interconectadas (Bradford, 1988).
4.2.3.3. Sinapsis química.
La mayoría de las sinapsis son de tipo químico, en las cuales una sustancia, el neurotransmisor hace de puente entre las dos neuronas, se
difunde a través del estrecho espacio y se adhiere a los receptores, que son moléculas especiales de proteínas que se encuentran en la membrana postsináptica (Bradford, 1988). La energía requerida para la liberación de un neurotransmisor se genera en la mitocondria del terminal presináptico. La unión de neurotransmisores a receptores de la membrana postsinápticas produce cambios en la permeabilidad de la membrana. La naturaleza del neurotransmisor y la molécula del
receptor determina si el efecto producido será de excitación o inhibición de la neurona postsináptica (Barr, 1994). Se han descrito varias formas de sinapsis según las estructuras implicadas. (Bradford, 1988). Axosomática: Sinapsis entre un axón y un soma. Axodendrítica: Sinapsis ocurrida entre un axón y una dendrita. Axoespinodendrítica: Sinapsis entre un axón y una espina dendrítica. Axoaxónica: Sinapsis entre dos axones. Dendrodendrítica: Sinapsis ocurrida entre dos dendritas. Somatosomática: Sinapsis entre dos somas. Dendrosomática: Sinapsis entre un soma y una dendrita.
Existen dos clases de sinapsis química: la sinapsis asimétrica o tipo I se caracteriza por la diferencia en densidad de las membranas presináptica y postsináptica, siendo más gruesa la última. Esta densidad consiste de un material proteico que puede estar asociado al receptor postsináptico; la sinapsis simétrica o tipo II se caracteriza porque las membranas presináptica y postsináptica poseen un grosor semejante (Bradford, 1988).
4.2.3.4. Mecanismos de exitación e inhibición sinaptica.

El líquido extracelular tiene una gran cantidad de concentración de iones de sodio en cambio de la concentración de iones potasio es baja, por otra parte en el citoplasma de la neurona hay una alta concentración de iones potasio y una baja concentración de iones sodio. En estado de reposo los iones sodio pueden salir de la célula por difusión mediante los canales de la membrana. Unicamente pequeñas cantidades de sodio se difunden a través de la membrana, la cual, en
reposo, es poco permeable a estos iones, grandes cantidades de iones sodio penetran en el momento de la conducción de los impulsos la entrada de sodio y la pérdida de potasio intracelular encuentran la oposición de distintas proteínas de la membrana constituyendo la bomba de sodio-potasio. La bomba la produce la molécula rica en energía, el ATP, para transportar iones a través de la membrana contra un gradiente de concentración, así las concentraciones de iones en el citoplasma se mantienen mediante gasto de energía que se producen de manera importante por consecuencia de la actividad de la bomba. La diferencia que resulta de las concentraciones de iones producen en la membrana un potencial de reposo, con el interior de la neurona que alcanza aproximadamente menos 70 mV con respecto al exterior (Zimmermann, 1993). Durante la excitación que se atribuye a una variedad de estímulos ya sean químicos o físicos, se presenta una reducción del potencial de membrana y se dice que dicha membrana se despolariza. La reducción del potencial se extiende en dirección lateral en la membrana disminuyendo su magnitud con la mayor distancia a partir del punto de iniciación. Este cambio en el grado de potencial, es el tipo único de señalización en las dendritas y en el cuerpo de la neurona (Darnell, 1993). La producción de estímulos en cantidad e intensidad suficiente puede reducir el potencial de membrana en el segmento inicial del axón hasta 10 a 15 mV. Este es un valor que desencadena la abertura de los canales de sodio regulado por el voltaje de la membrana del axón, los iones aparecen localmente en la superficie exterior procedentes de la superficie interna

disminuyendo el gradiente y la concentración y atraídos por el exceso de carga negativa en el axoplasma. El interior del axón es temporalmente de +40 mV respecto al exterior. Este cambio se llama el potencial de acción. Una vez que se generó este potencial se propaga por la membrana, en circuitos locales de corriente eléctrica, que abren los canales cercanos de sodio. El potencial de acción al desplazarse produce un impulso nervioso (Meyer, 1985). Se dice que los estímulos que despolarizan la membrana de la neurona son excitatorios, debido a que cantidad suficiente de ellos iniciarán el potencial de acción. Algunos estímulos originan el efecto opuesto de hiperpolarización, en este caso el potencial de membrana excede la cifra del potencial de reposo -70mV. Los impulsos que causan hiperpolarización inhiben la generación de potenciales de acción, debido a que se oponen a los efectos de los estímulos despolarizantes (Zimmermann, 1993).
Primera etapa: Regulación de la síntesis del neurotransmisor. Un botón terminal en equilibrio mantiene una polarización de su membrana y una regulación de su capacidad de síntesis del neurotransmisor o de las proteínas asociadas a la consolidación de las vesículas sinápticas, de acuerdo con las exigencias funcionales de la neurona y de su propio control metabólico sobre la disponibilidad de neurotransmisor. El neurotransmisor puede almacenarse en vesículas sinápticas, pequeños reservorios globulares que contienen receptores en su pared exterior y permiten que el neurotransmisor se libere en sitios específicos de la terminal presináptica. Se ha hecho la analogía de la terminal presináptica como un espacio donde las vesículas sinápticas, así como las mitocondrias y otras estructuras subcelulares, están flotando. Sólo en un lugar determinado de este espacio se localizan los sitios por donde el neurotransmisor puede liberarse hacia el exterior. Como si las vesículas fueran huevos que sólo pudieran acomodarse en los huecos de sus cajas, que se encuentran en el piso de este espacio. Y sólo a través de los huecos de estas cajas se puede descargar el contenido hacia el exterior. En este caso, los huecos tienen receptores que reconocen los componentes de la cáscara del huevo. La presencia de estas vesículas y de receptores en sus membranas, y de moléculas de el neurotransmisor significa que existe todo lo necesario en el interior de la terminal para sintetizar todos estos componentes, y de mecanismos para la regulación de esta síntesis y de la liberación sináptica. La terminal presináptica tiene autorreceptores que le informan sobre los niveles del neurotransmisor en el exterior de la terminal. Si sus niveles son elevados, la terminal puede fabricar o liberar menos. Si éstos son bajos, puede hacer lo contrario. La activación, pues, de los autorreceptores puede tener efectos de estimulación o, generalmente, de inhibición de la liberación sináptica. La membrana de la terminal presináptica también posee moléculas transportadoras del mismo neurotransmisor que ellas liberan. Es un mecanismo de recaptación que sirve no sólo para ahorrar neurotransmisor al reutilizarlo, sino también contribuye a limitar sus efectos postsinápticos. Asumiendo esta condición, la conducción de un impulso nervioso, que rompe el esfuerzo fisiológico para el mantenimiento de la polarización, produce una significativa movilización de las vesículas sinápticas que almacenan neurotransmisores, los cuales llegan a unirse a la propia membrana plasmática; y la entrada de Ca++, como consecuencia de la despolarización, que abre los canales de Ca++, rompe la anastomosis vesícula-membrana y libera al espacio sináptico el neurotransmisor. El

aumento de la superficie de membrana que supone la ruptura de las vesículas sinápticas, es posteriormente restaurado por la pinocitosis interna, es decir, el desprendimiento interno de estas membranas que van a ser, desde luego, precursoras para la formación de nuevas vesículas sinápticas. Asimismo, el Ca++ que ha intervenido en el proceso es transportado extracelularmente por las proteínas transportadoras de Ca++ similares a la bomba de sodio-potasio, con lo que se vuelve al equilibrio en la neurona presináptica.
Segunda etapa: Liberación del neurotransmisor. Una vez que el neurotransmisor ha sido liberado al espacio sináptico, se difunde por el mismo y puede seguir las siguientes rutas:
Fijación en los lugares específicos de membrana tanto presináptica como postsináptica, que son los receptores.
Dispersión en el espacio sináptico y actuación fuera de la sinapsis como un neuromodulador.
Recaptación presináptica del neurotransmisor. Catabolización enzimática del neurotransmisor y por tanto
degradación de la estructura.
Tercera etapa: Activación de receptores.Receptores postsinápticos Las estructuras de unión del neurotransmisor en la membrana postsináptica son proteínas específicas de membrana que constituyen los receptores postsinápticos y que son la clave del reconocimiento de la liberación de neurotransmisores presinápticos. Es decir, los receptores postsinápticos son la estructura básica para interpretar la comunicación interneuronal, constituyen, por tanto, auténticas estructuras de diferenciación neuronal a la vez que son un claro exponente de la diversidad específica. La afinidad del neurotransmisor como ligando por el receptor postsináptico es una característica inherente a la diferenciación y especialización de las propias neuronas, hasta tal punto que un neurotransmisor determinado no produce efectos, por muy constante e intensa que sea su liberación, si la membrana postsináptica no contiene en su diferenciación los receptores específicos para ese neurotransmisor concreto. Una vez que el receptor potsináptico y neurotransmisor se unen, aparece inmediatamente una consecuencia en la membrana postsináptica. Indudablemente, hay una alteración constitucional que puede ser suficiente para modificar los canales iónicos de polarización de esa membrana, generando rápidamente una despolarización o una hiperpolarización, lo que conllevaría un aumento de la activación o inhibición, respectivamente, de la membrana postsináptica. Normalmente, la activación de los receptores postsinápticos por un neurotransmisor ejerce su efecto, es decir, es interpretada postsinápticamente, mediante la utilización de un neuromediador o de un segundo mensajero. En el primer caso, lo que ocurre es que la acción neurotransmisor-receptor es indirecta y necesita de una proteína de membrana mediadora ante la modificación de su permeabilidad iónica, o lo que es lo mismo, hacia la alteración de su polarización. En el segundo caso, bien por la neuromediación, bien por el complejo específico neurotransmisor-receptor, se activa un sistema enzimático de membrana, normalmente la adenilato-ciclasa, cuya actuación inmediata se realiza sobre el adenosín trifosfato (ATP) que se transforma en adenosín monofosfato cíclico (AMPc). El AMPc no sólo activa la apertura de los canales iónicos sino que también conduce la información de la actividad

de membrana de la que procede, hacia las estructuras somáticas y nucleares de la neurona, por lo que las consecuencias de una comunicación interneuronal pueden perfectamente trascender a mecanismos bioquímicos que impliquen cambios estructurales y/o funcionales a partir de la utilización del segundo mensajero. Al igual que ocurría en la segunda etapa, es decir, la dependencia del Ca++ en la liberación del neurotransmisor, todavía aquí nos encontramos con una incuestionable dependencia del Ca++ que es tanto más acusada cuanto mayor es la repercusión metabólica del AMPc. Como resultado final de esta etapa, lo que ocurre localmente es una movilización de los canales iónicos que afectan al Na+, K+, Cl- y Ca++. Como sabemos, la polarización de la membrana establece particularmente unas concentraciones de Na+ extracelular y de K+ intracelular. Si se abre el canal de Na+, aparece una única entrada que genera automáticamente una despolarización, es decir crea un potencial postsináptico excitador. Si lo que se activa es el canal de K+, se establece una acumulación excesiva de K+ intracelular cuya consecuencia es la contraria, es decir, una hiperpolarización de la membrana y por tanto un potencial postsináptico inhibidor. La acción sobre el canal de Cl- tiene efecto cuando la membrana está ligeramente despolarizada, en cuyo caso la apertura de este canal se opone a la aparición de un potencial postsináptico excitador. Cuando la membrana está polarizada, cuyo resultado final es una clara electronegatividad intracelular, la apertura de un canal de Cl- no tiene mayor repercusión al reafirmar la electronegatividad, y además es rápidamente reequilibrada por difusión. Por su parte los iones Ca++ situados extracelularmente actúan igual que los iones Na+; es decir, despolarizan la membrana y producen potenciales postsinápticos excitatorios, además de ser un potente activador enzimático.Receptores presinápticos o autorreceptores Es también enormemente transcendente la activación de los receptores específicos de membrana, situados en la neurona presináptica, que responden a la liberación del neurotransmisor con la misma eficacia y afinidad que los receptores postsinápticos. Ahora bien, el resultado de este mecanismo es un servomecanismo, es decir, la autorregulación de la liberación del neurotransmisor con el fin de que la comunicación neuronal sea eficaz, ajustada y esté adecuada a los niveles de excitabilidad que han provocado los potenciales de acción en los botones terminales. Estos receptores, a diferencia de los postsinápticos, no controlan canales iónicos. Sin embargo, sí que regulan la movilización de las vesículas, intervienen esencialmente en la disponibilidad del neurotransmisor y, por tanto, en su síntesis y en su liberación. Normalmente, su control es por retroalimentación y su activación favorece los mecanismos de inhibición presináptica que, lógicamente, se asientan en el funcionamiento de los segundos mensajeros.
Cuarta etapa: Inactivación del neurotransmisor Una vez cumplido el objetivo sináptico, es decir la comunicación a la neurona postsináptica, se produce la inactivación del neurotransmisor. El mecanismo de inactivación más importante es la recaptación presináptica, que se produce no por la utilización de receptores específicos para su fijación, sino por auténticos transportadores de membrana que vuelven a incorporarlos al espacio presináptico. Estos transportadores de membrana actúan bien sobre el neurotransmisor íntegramente liberado y reconocido por sus receptores o bien sobre metabolitos intermediarios fruto de la acción catabólica de sistemas enzimáticos postsinápticos. Indudablemente, la recaptación o la degradación catabólica del neurotransmisor termina con

la activación de los receptores de membrana y desde luego termina con la actividad de los neurotransmisores.
Quinta etapa: Difusión del neurotransmisor. La ultima etapa que podemos definir en la sinapsis es la difusión del neurotransmisor. Esto nos permite incluir el concepto de neuromodulador, que produce sus efectos a grandes distancias del lugar de secreción, y que afecta a grupos neuronales y otras áreas de intervención. Este concepto es fundamental en la secreción de neuropéptidos más que de neurotransmisores clásicos, y aunque los propios neuropéptidos de secreción sináptica pueden actuar como auténticos neurotransmisores, la propia estructura de la molécula favorece su difusión y alejamiento de su actuación local, por lo que pueden ser considerados como neuromoduladores. La naturaleza de la materia transmisora no es en realidad quien define el efecto postsináptico de la neurotransmisión, sino que es la naturaleza de los canales iónicos controlados por los receptores de membrana postsinápticos la que define el efecto excitatorio o inhibitorio; de ahí que podamos encontrarnos con efectos claramente inhibitorios de un determinado neurotransmisor que en su generalidad es excitatorio.

5.2.5.3. Impulso nervioso La membrana plasmática forma el límite externo continuo del cuerpo celular y sus prolongaciones y en la neurona es el sitio de iniciación y conducción del impulso nervioso. Su espesor es de aproximadamente 8 nm lo cuál la hace demasiado delgada para poder ser observada por un microscopio óptico. Con microscopio electrónico se observa una campa interna y otra externa de moléculas dispuestas muy laxamente (cada capa aproximadamente de 2,5 nm) y separadas por una capa intermedia de lípidos. Moléculas de hidrato de carbono se encuentran adheridas al exterior de la capa plasmática y se unen con proteínas o lípidos formando lo que se conoce como cubierta celular o glucocálix. La membrana plasmática y la cubierta celular juntas forman una membrana semipermeable que permite la difusión de ciertos iones a través de ella pero limita otras. En estado de reposo los iones de K+ difunden a través de la membrana plasmática desde el citoplasma celular hacia el líquido tisular. La permeabilidad de la membrana a los iones de K+ es mucho mayor que el influjo de Na+. Esto da como resultado una diferencia de potencial estable de alrededor de -80 mv que pueden medirse a través de la membrana ya que el interior es negativo en relación al exterior. Este potencial se conoce como potencial de reposo. Cuando una célula nerviosa es excitada (estimulada) por un medio eléctrico, mecánico o químico, ocurre un rápido cambio de permeabilidad de la membrana a los iones de Na+, estos iones difunden desde el liquido tisular a través de la membrana plasmática hacia el citoplasma celular. Esto induce a que la membrana se despolarise progresivamente. La súbita entrada de iones Na+ seguida por la polaridad alterada produce determinado potencial de acción que es de aproximadamente +40 mv. Este potencial es muy breve (5 nseg) ya que muy pronto la mayor permeabilidad de la membrana a los iones de Na+ cesa y aumenta la permeabilidad de los iones K+, de modo
que estos comienzan a fluir desde el citoplasma celular y así el área localizada de la célula retorna al estado de reposo. Una vez generado el potencial de acción se propaga por la membrana plasmática, alejándose del sitio de iniciación y es conducido a lo largo de las neuritas como el impulso nervioso. Una vez que el impulso nervioso se ha difundido por una región de la membrana plasmática, no puede provocarse
otro potencial en forma inmediata. La duración de este estado no excitable se denomina período refractario. La transmisión, que no es más que un desplazamiento de cargas eléctricas por la membrana neuronal, constituye el impulso nervioso. Este impulso es la base de todas las funciones nerviosas, incluidas las superiores. Debido a esto, y empleando instrumentos especiales de

medición, se puede detectar la actividad nerviosa en forma de pequeñas corrientes eléctricas, tal es el caso de la electroencefalografía.
Un impulso nervioso es una onda de electronegatividad que recorre toda la neurona y que se origina como consecuencia de un cambio transitorio de la permeabilidad en la membrana celular, secundario a un estímulo. La excitabilidad de las neuronas depende de la existencia de distintas concentraciones de iones a ambos lados de la membrana celular y de la capacidad de transporte activo a través de estas membranas. La excitación neuronal se acompaña de un flujo de partículas cargadas a través de la membrana, lo cual
genera una corriente eléctrica. La membrana de las células está polarizada, debido a que hay un reparto desigual de cargas eléctricas entre el interior y el exterior de la célula. Esto crea una diferencia de potencial, siendo el exterior positivo respecto al interior. En el exterior, en el líquido intersticial, el anión más abundante es el de cloro. En el citoplasma, los aniones más abundantes son las proteínas, que en el pH celular se ionizan negativamente. El catión más abundante en el líquido intersticial es el de sodio, y en el citoplasma el de potasio. El desequilibrio iónico que produce la polarización de la membrana es debido a la distinta permeabilidad que presenta frente a cada uno de los iones. El ión de potasio atraviesa la membrana libremente. La permeabilidad para el sodio es menor, y además es expulsado por medio de un transporte activo llamado bomba de sodio. Las proteínas, debido a su tamaño, no pueden atravesar libremente la membrana. Toda esta dinámica establece una diferencia de potencial en condiciones de reposo, de unos -70 mV.Es lo que se denomina potencial de membrana. Cuando se aplica un estímulo adecuado a la membrana de la neurona, se altera su permeabilidad, permitiendo la entrada de iones de sodio a favor de su gradiente de concentración. Este tránsito es tan intenso que la bomba de sodio resulta ineficaz. El flujo de sodio invierte la diferencia de voltaje pasando el exterior a ser negativo y el interior positivo (+30 mV). Conforme se iguala el gradiente de concentración, el flujo de sodio decrece, mientras que el potasio sale de la célula para neutralizar la electronegatividad del exterior. El tránsito de potasio se produce un milisegundo después que el de sodio. La salida de potasio es mayor que la necesaria para restablecer el potencial de reposo, por lo que la membrana queda hiperpolarizada, con mayor electronegatividad en el interior.
La representación gráfica de la variación de potencial respecto al tiempo es el potencial de acción. La cantidad de estímulo necesario para provocar la actividad de una neurona, se denomina umbral de excitabilidad. Alcanzado este umbral, la respuesta es efectiva, independientemente de la interrupción o aumento del estímulo. Es decir, sigue la ley del todo o nada. Durante la despolarización, la neurona no es excitable, es decir, está en periodo refractario. Durante la hiperpolarización subsiguiente, la neurona es parcialmente excitable, parcialmente refractaria, es decir, que

necesitamos un estímulo más intenso para provocar un nuevo potencial de acción, ya que ha aumentado el umbral de excitabilidad.contacto entre células contiguas esa cubierta queda interrumpida, recibiendo esos lugares el nombre de NODOS DE RANVIER. En las fibras que poseen cubierta de mielina, dispuesta en torno a las células de Schwan, separadas por los denominados nódulos de Ranvier, la onda de electronegatividad se propaga saltando de nódulo en nódulo. Esta propagación saltatoria es más rápida, al no tener que despolarizar todos los puntos de la fibra nerviosa. Además permite un importante ahorro energético, ya que la bomba de sodio tiene que movilizar menor cantidad de iones. En el SNC los axones están rodeados por la mielina de los oligodendrocitos (fibras nerviosas mielínicas del SNC), mientras que en el SNP pueden estar rodeados, ya sea, por prolongaciones citoplasmáticas de las células de Schwann (fibras amielínicas) o por la mielina las células se Schwann (fibras nerviosas mielínicas del SNP)
AMIELÍNICAS o desnudas, son las fibras que no están recubiertas por vaina de mielina. En las fibras nerviosas amielínicas el impulso se conduce, como una onda continua de inversión de voltaje hasta los botones terminales de los axones. La velocidad que es proporcional al diámetro del

axón y varía entre 1 a 100 m/s.
¿A que llamamos Osmosis?
Si dos soluciones de distintas concentración se hallan separadas por una membrana fácilmente permeable para las moléculas del solvente (agua por ejemplo) y difícilmente permeable osmosis. El agua tiende a pasar desde la solución que tiene menor concentración hacia la que tiene mayor concentración.
La osmosis se define como la difusión de agua o cualquier solvente a través de una membrana semipermeable que separa a dos soluciones de distinta concentración propiciando que el solvente de la solución que tiene menor concentración se desplace hacia la solución que tiene mayor concentración con el fin de que la concentración de las dos soluciones se equilibren.
INSULINA
La Insulina (latín insula”isla”) es una hormona polipeptídica formada por 51 aminoácidos. Es segregada por las células beta de los islotes de Langerhans del páncreas, en forma de precursor inactivo (proinsulina), el cual pasa al aparato de Golgi, donde se modifica, eliminando una parte y uniendo los dos fragmentos restantes mediante puentes disulfuro.

Interviene en el aprovechamiento metabólico de los nutrientes, sobre todo con el anabolismo de los hidratos de carbono. Su déficit provoca la diabetes mellitus y su exceso provoca hiperinsulinismo con hipoglucemia.
La insulina es la hormona “anabólica” por excelencia, permite disponer a las células del aporte necesario de glucosa para los procesos de síntesis con gasto de energía, que luego por glucólisis y respiración celular se obtendrá la energía necesaria en forma de ATP (mononucleótido de adenosina trifosforilado) que usa el metabolismo como unidad de energía transportable para dichos procesos.Mantiene la concentración de glucosa en nuestra sangre. Lo consigue porque cuando el nivel de glucosa es elevada el páncreas lo libera a la sangre. Su función es favorecer la absorción celular de la glucosa.Es una de las 2 hormonas que produce el pancreas junto con el glucagón (al contrario de la insulina, cuando el nivel de glucosa disminuye es liberado a la sangre). La insulina se produce en el Páncreas en los “Islotes de Langerhans”, mediante unas células llamadas Beta.Una manera de detectar si las Células beta producen insulina, es haciendo una prueba, para ver si existe péptido C en sangre. El péptido C se libera a la sangre cuando las células Beta procesan la proinsulina, convirtiéndola en insulina. Cuando sólo entre un 10% y un 20% de las células Beta están en buen estado, comienzan a aparecer los síntomas de la diabetes.
Diabetes tipo 1Característicamente se da en la época temprana de la vida y se debe a un deficit absoluto de insulina, dado por la destrucción de las células beta del páncreas por procesos autoinmunes o idiopáticos. Sólo cerca de 1 entre cada 20 personas diabéticas tiene diabetes tipo 1, la cual se presenta más frecuentemente en jóvenes y niños. Este tipo de diabetes se conocía como diabetes mellitus insulinodependiente o diabetes juvenil. En ella, las células beta del páncreas no producen insulina o apenas producen. En los primeros años de la enfermedad suelen quedar reservas pancreáticas que permiten una secreción mínima de insulina (este período se denomina luna de miel).

Diabetes tipo 2Se caracteriza por un complejo mecanismo fisiopatológico, cuyo rasgo principal es el déficit relativo de producción de insulina y una deficiente utilización periférica por los tejidos de glucosa (resistencia a la insulina). Se desarrolla a menudo en etapas adultas de la vida, y es muy frecuente la asociación con la obesidad; anteriormente llamada diabetes del adulto, diabetes relacionada con la obesidad, diabetes no insulino dependiente. Varios fármacos y otras causas pueden, sin embargo, causar este tipo de diabetes.
Hay cuatro tipos fundamentales de Insulina, basados en:* Cuán rápido la insulina empieza su trabajo (arranque)* Cuándo trabaja más fuertemente (hora de su pico)* Cuánto tiempo se mantiene en actividad en su cuerpo (duración).
La Insulina de acción rápida (Lispro) ingresa a la sangre en los primeros 15 minutos después de la inyección. Su pico se produce más tarde entre 30 y 90 minutos y se puede mantener a lo largo de aproximadamente 5 horas.
La Insulina de acción corta (regular) usualmente ingresa a la sangre en los primeros 30 minutos después de la inyección. Su pico es alcanzado entre 2 y 4 horas más tarde y permanece en la sangre por aproximadamente 4 a 8 horas.
La Insulina de acción intermedia (NPH y lenta) ingresa a la sangre 2 a 6 horas después de la inyección. Tienen las dos Insulina un pico de 4 a 14 horas más tarde y permanecen en la sangre por aproximadamente 14 a 20 horas.
La Insulina de acción prolongada (ultralenta) se toma de 6 a 14 horas para iniciar su trabajo. Ella no tiene pico de acción o es un pico muy corto entre 10 a 16 horas después de la inyección. Permanece en la sangre entre 20 y 24 horas.
fibrosis quística
La fibrosis quística (abreviatura FQ), originalmente denominada fibrosis quística del páncreas, y también conocida como mucoviscidosis (del lat. muccus, "moco", y viscōsus, "pegajoso"), es una enfermedad frecuente que afecta al organismo en forma generalizada, causando muerte prematura. La dificultad para respirar es el síntoma más común, emergente de infecciones pulmonares crónicas, las cuales pueden mostrarse resistentes al tratamiento con antibióticos y otros fármacos. La FQ es un trastorno multisistémico que causa la formación y acumulación de un moco espeso y pegajoso, afectando fundamentalmente a pulmones, intestinos, páncreas e hígado. Asimismo, se caracteriza por la presencia de una alta concentración de sal (NaCl) en el sudor, lo que sentó las bases de la prueba estándar para este diagnóstico: el examen de electrolitos del sudor. El mismo evalúa, entre otros iones, los niveles de cloruro excretados. Una variedad de síntomas, incluyendo infecciones sinusales, disminución del crecimiento, diarrea son el resultado de los efectos de la FQ sobre los distintos órganos. Se trata de una de las enfermedades fatales más comunes. Su prevalencia es mayor entre caucásicos; una de cada 25 personas de ascendencia europea es portadora asintomática de un gen para FQ, siendo la enfermedad genética no hereditable más frecuente entre esta población. Y aproximadamente una de cada 25 personas es portador sano heterocigoto. Los afectados pueden ser diagnosticados mediante pruebas genéticas

prenatales; también por screnning neonatal o, durante la infancia temprana, por la mencionada prueba del sudor. No existe cura para la FQ,sin embargo hoy en día existen tratamientos en los que se puede tener una vida completamente normal alargando la misma un 25 %.La supervivencia media para estos pacientes se estima en 35 años, alcanzando valores más altos en algunos países (57,8 en EE.UU.).[1] [2] En casos severos, el empeoramiento de la enfermedad puede imponer la necesidad de un trasplante de pulmón.
La FQ es causada por una mutación en un gen llamado regulador de la conductancia transmembrana de la fibrosis quística (CFTR, por sus siglas en inglés). Este gen interviene en la producción de sudor, jugos gástricos, y moco. Aunque la mayoría de las personas sanas tienen dos copias funcionales del gen, sólo una es necesaria para impedir el desarrollo de fibrosis quística. La FQ se desarrolla cuando ninguno de estos genes opera normalmente. En consecuencia, se la considera una enfermedad autosómica recesiva. El nombre fibrosis quística se refiere a los procesos característicos de cicatrización (fibrosis) y formación de quistes dentro del páncreas, reconocidos por primera vez en los años 1930.[3]
Síntomas y signos [editar]
La sintomatología de la fibrosis quística varía en función de la edad del individuo, el grado en que se ven afectados órganos específicos, la terapéutica instituida previamente, y los tipos de infecciones asociadas. Esta enfermedad compromete al organismo en su totalidad y muestra su impacto sobre el crecimiento, la función respiratoria, la digestión. El periodo neonatal se caracteriza por un pobre aumento de peso y por obstrucción intestinal producida por heces densas y voluminosas. Otros síntomas aparecen, más tarde, durante la niñez y al inicio de la adultez. Éstos incluyen retardo del crecimiento, advenimiento de la enfermedad pulmonar, y dificultades crecientes por la malabsorción de vitaminas y nutrientes en el tracto gastrointestinal.
A la mayoría de los niños se les diagnostica fibrosis quística antes del primer año de vida, cuando la mucosidad pegajosa que afecta pulmones y páncreas, comienza a mostrar su impacto. En el tracto respiratorio, esas secreciones sirven como caldo de cultivo para diversas bacterias responsables de infecciones crónicas, con deterioro progresivo y permanente del parénquima pulmonar. Conforme se agrava la condición respiratoria, los pacientes sufren hipertensión pulmonar. Por otra parte, en el páncreas, el moco obstruye el tránsito de las enzimas sintetizadas por la glándula e impide que lleguen hasta los intestinos para digerir y absorber el alimento.
Otros síntomas incluyen expectoración de sangre o esputo sanguinolento, dilatación crónica de los bronquios o bronquiolos (bronquiectasia), elevación de la presión sanguínea en el pulmón, insuficiencia cardiaca, sensación de no estar recibiendo suficiente oxígeno o disnea, insuficiencia respiratoria y atelectasia; podría requerirse soporte ventilatorio.[4] Además de las infecciones bacterianas más comunes, las personas con FQ desarrollan con mayor facilidad otros tipos de enfermedades respiratorias. Entre éstas se encuentra la aspergilosis broncopulmonar alérgica, caracterizada por una respuesta de hipersensibilidad ante un hongo (moho) ordinario del género Aspergillus (Aspergillus fumigatus), que agudiza los problemas respiratorios. Otro ejemplo es la infección con el complejo Mycobacterium avium (MAC), grupo de actinobacterias

emparentadas con Mycobacterium tuberculosis, que puede ocasionar daños mayores al pulmón, y que no responde a la terapéutica con antibióticos convencionales.
El moco en los senos paranasales es igualmente denso y pegajoso, y también puede causar oclusión de los orificios por donde los senos habitualmente drenan, lo cual hace que se acumulen secreciones que actúan como caldo de cultivo para los patógenos antes mencionados. En estos casos, se pueden presentar dolor facial, fiebre, secreción nasal profusa y cefaleas. En las personas con FQ, a menudo se observa crecimiento sobreabundante de tejido nasal (pólipos), a consecuencia de la inflamación por infección sinusal crónica. Estos pólipos pueden agravar la obstrucción de las vías respiratorias superiores e intensificar las dificultades respiratorias.[5] [6]
CANALES DE ESCURRIMIENTO DE POTASIO Y SODIO
CANAL DE POTASIO
Cuando se abre el canal de potasio el potencial de la membrana se hace más negativo (hiperpolarización).
El potasio está más concentrado en el interior de la célula, por ese motivo cuando se abren canales de potasio este ion tiende a salir por gradiente de concentración. Esto extrae cargas eléctricas positivas del interior de la célula, y deja el potencial de ésta más negativo. El potencial de equilibrio del potasio es de aproximadamente –100 mV, y cuando aumenta la permeabilidad a este ión el potencial de la membrana se acerca a este potencial de equilibrio.
CANAL DE SODIO
La apertura del canal de sodio lleva el potencial de membrana a un valor muy positivo (+66 mV). El sodio tiende a entrar en la célula por gradiente de concentración y por atracción electrostática, con lo que introduce en la célula cargas positivas y produce depolarización. Durante el potencial de acción, la apertura de los canales de sodio dependientes de voltaje hace que el potencial de la membrana se haga positivo (+30 mV), aunque en este caso queda algo más bajo que el potencial de equilibrio del sodio, porque los canales están abiertos durante un tiempo muy corto y no da tiempo a que se equilibren las cargas.
Mecanismo mediado q permite la entrada de soluto
En biología celular se denomina transporte de membrana al conjunto de mecanismos que regulan el paso de solutos, como iones y pequeñas moléculas, a través de membranas plasmáticas, esto es, bicapas lipídicas que poseen proteínas embebidas en ellas. Dicha propiedad se debe a la selectividad de membrana, una característica de las membranas celulares que las faculta como agentes de separación específica de

sustancias de distinta índole química; es decir, la posibilidad de permitir la permeabilidad de ciertas sustancias pero no de otras.[1]
Los movimientos de casi todos los solutos a través de la membrana están mediados por proteínas transportadoras de membrana, más o menos especializadas en el transporte de moléculas concretas. Puesto que la diversidad y fisiología de las distintas células de un organismo está relacionada en buena medida con su capacidad de captar unos u otros elementos externos, se postula que debe existir un acervo de proteínas transportadoras específico para cada tipo celular y para cada momento fisiológico determinado;[1] dicha expresión diferencial se encuentra regulada mediante: la transcripción diferencial de los genes codificantes para esas proteínas y su traducción, es decir, mediante los mecanismos genético-moleculares, pero también a nivel de la biología celular: dichas proteínas pueden requerir de activación mediada por rutas de señalización celular, activación a nivel bioquímico o, incluso, de localización en vesículas del citoplasma.[2
Termodinámicamente, el flujo de sustancias de un compartimento a otro puede realizarse a favor o en contra de un gradiente, ya sea de concentración, o electroquímico. Si el intercambio de sustancias se realiza a favor del gradiente, esto es, en el sentido de los potenciales decrecientes, el requerimiento de energía externo al sistema es nulo; si, en cambio, el transporte se hace en contra del gradiente, se requiere el aporte de energía, energía metabólica en nuestro caso.[3] Por ejemplo, un mecanismo químico de separación clásico que no requiere un aporte de energía externo es la diálisis: en ella, una membrana semipermeable separa dos soluciones que difieren en la concentración de un mismo soluto. Si la membrana permite el paso de agua pero no el del soluto, sucede que el agua fluye hacia el compartimento más concentrado en soluto, a fin de establecer un equilibrio en el cual la energía del sistema sea mínima. Para que suceda este flujo, puesto que el agua se desplaza de un lugar muy concentrado a uno muy diluido en disolvente (en cuanto a soluto, se da la situación opuesta), y, por ello, lo hace a favor de gradiente, no se requiere un aporte de energía externo.
Diagrama de una membrana plasmática 1. fosfolípido 2. colesterol 3. glicolípido 4. glúcido 5. proteína transmembrana 6. glicoproteína 7. proteína anclada a un fosfolípido 8. glicoproteína periférica.
La naturaleza de las membranas biológicas, especialmente la de sus lípidos, es anfipática, lo que se traduce en que forman una bicapa que alberga una parte interna hidrofóbica y una externa hidrofílica, permite que surja una posibilidad de transporte, la difusión simple o difusión pasiva, que consiste en la difusión de sustancias a su través sin gasto de energía metabólica y sin ayuda de proteínas transportadoras. En el caso de

que la sustancia a transportar posea una carga neta, difundirá no sólo en respuesta a un gradiente de concentración, sino también al potencial de membrana, esto es, al gradiente electroquímico.
Permeabilidad relativa de una bicapa de fosfolípidos a distintas sustancias[1]
Tipo de sustancia Ejemplos Comportamiento
Gases CO2, N2, O2 Permean
Moléculas polares pequeñas sin carga
Urea, agua, etanolPermean, total o parcialmente
Grandes moléculas polares sin carga
glucosa, fructosa No permean
Iones K+, Na+, Cl-, HCO3- No permean
Moléculas polares cargadasATP, aminoácidos, glucosa-6-fosfato
No permean
Puesto que son pocas las moléculas que son capaces de difundir a través de una membrana lipídica, la mayoría de los procesos de transporte involucran a proteínas de transporte. Se trata de proteínas transmembrana que poseen multitud de hélices alfa inmersas en la matriz lipídica o bien, al menos en bacterias, beta láminas.[4] Dicha estructura probablemente implique una vía de entrada a través de entornos hidrofílicos proteicos que causarían una disrupción en el medio altamente hidrofóbico constituido por los lípidos.[1] Las proteínas intervienen de diversas formas en el transporte: actúan tanto como bombas impulsadas por ATP, esto es, por energía metabólica, o como canales de difusión facilitada.
Difusión simple [editar]
Una membrana semipermeable separa dos compartimentos con concentraciones distintas de un soluto: con el paso del tiempo, el soluto difundirá hasta alcanzar el equilibrio a ambos lados.

Como se mencionó anteriormente, la difusión pasiva es un fenómeno espontáneo puesto que suceden incrementando la entropía del sistema, y disminuyendo la energía libre.[5] No requiere de la intervención de proteínas de membrana, pero sí de las características de la sustancia a transportar y de la naturaleza de la bicapa. Para el caso de una membrana fosfolipídica pura, la velocidad de difusión de una sustancia depende de su:
gradiente de concentración, hidrofobicidad, tamaño, carga, si la molécula posee carga neta.
Estos factores afectan de diversa manera a la velocidad de difusión pasiva:
a mayor gradiente de concentración, mayor velocidad de difusión, a mayor hidrofobicidad, esto es, mayor coeficiente de partición, mayor
solubilidad en lípido y por tanto mayor velocidad de difusión, a mayor tamaño, menor velocidad de difusión, dado un potencial de membrana, es decir, la diferencia de potencial entre la cara
exoplasmática y la endoplasmática de la membrana, y un gradiente de concentración se define un gradiente electroquímico que determina las direcciones de transporte energéticamente favorables de una molécula cargada, dependiendo de la naturaleza de ésta y del signo del potencial, si bien la mayor parte de las células animales poseen carga negativa en su exterior.[6]
La difusión simple a través de la membrana lipídica muestra una cinética de no saturación, esto es, que, puesto que la tasa neta de entrada está determinada sólo por la diferencia en el número de moléculas a cada lado de la membrana, la entrada aumenta en proporción a la concentración de soluto en el fluido extracelular. Esta característica distingue la difusión simple de los mecanismos de penetración por canales de transporte mediado.[6]
Difusión facilitada [editar]
La difusión facilitada involucra el uso de un proteína para facilitar el movimiento de moléculas a través de la membrana. En algunos casos, las moléculas pasan a través de canales con la proteína. En otros casos, la proteína cambia su forma, permitiendo que las moléculas pasen a través de ella.
Bajo el mismo principio termodinámico que en el caso de la difusión simple, es decir, que el soluto a transportar lo hace a favor de gradiente, la difusión facilitada opera de modo similar, pero está facilitada por la existencia de proteínas canal, que son las que facilitan el transporte de, en este caso, agua o algunos iones y moléculas hidrófilas.

Estas proteínas integrales de membrana conforman estructuras en forma de poro inmersas en la bicapa, que dejan un canal interno hidrofílico que permite el paso de moléculas altamente lipófobas como las mencionadas anteriormente. La apertura de este canal interno puede ser constitutiva, es decir, continua y desregulada, en los canales no regulados, o bien puede requerir una señal que medie su apertura o cierre: es el caso de los canales regulados.[1]
Transporte activo y cotransporte [editar]
En él se efectúa un transporte en contra del gradiente de concentración o electroquímico y, para ello, las proteínas transportadoras implicadas consumen energía metabólica (comúnmente adenosín trifosfato). La hidrólisis del compuesto que actúa como moneda energética puede ser muy evidente, como en el caso de los transportadores que son ATPasas, o puede tener un origen indirecto: por ejemplo, los cotransportadores emplean gradientes de determinados solutos para impulsar el transporte de un determinado compuesto en contra de su gradiente, a costa de la disipación del primer gradiente mencionado. Pudiera parecer que en este caso no interviene un gasto energético, pero no es así puesto que el establecimiento del gradiente de la sustancia transportada colateralmente al compuesto objetivo ha requerido de la hidrólisis de ATP en su generación mediante unos determinados tipos de proteínas denominados bombas.[2] Por ello, se define transporte activo primario como aquél que hidroliza ATP de forma directa para transportar el compuesto en cuestión, y transporte activo secundario como aquél que utiliza la energía almacenada en un gradiente electroquímico.
El descubrimiento de la existencia de este tipo de transportadores se produjo al estudiar cinéticamente la transferencia de moléculas a través de las membranas: para algunos solutos, se observó que la velocidad de entrada alcanza una meseta a partir de cierta concentración externa a partir de la cual no se produce un incremento significativo de velocidad de captación, esto es, surge una respuesta tipo curva logística. Se interpretó que el transporte aquí se produce por la formación de un complejo sustrato-transportador, conceptualmente idéntico al complejo enzima-sustrato de la cinética enzimática. Por ello, cada proteína transportadora posee una constante de afinidad por el soluto que es igual a la concentración del soluto cuando la velocidad de transporte es la mitad de su valor máximo (equivaldría, para el caso de un enzima, a la constante de Michaelis-Menten).[6]
Algunos rasgos importantes del transporte activo, además de su capacidad de intervenir aun en contra de gradiente, su cinética y el empleo de ATP, son: su elevado grado de selectividad y su facilidad de inhibición farmacológica selectiva.[6]
Transportadores [editar]
Uniporte, simporte y antiporte de moléculas a través de transportadores de membrana.

Un transportador puede movilizar diversos iones y moléculas; según la direccionalidad, se distinguen:
antiportadores: aquéllos que transportan un tipo de molécula en contra de su gradiente al mismo tiempo que desplazan uno o más iones diferentes a favor del suyo, siendo ambos gradientes contrapuestos,
simportadores: los que desplazan el compuesto a transportar en contra de su gradiente acoplando este trasiego al desplazamiento de uno o más iones diferentes a favor del suyo, que, en este caso, es equivalente al de la molécula a transportar.
Ambos reciben el nombre de contransportadores.