
SEMINARIOS GUÍA DE ESTUDIO DE CONTENIDOS … 1-Parte A... · Átomos de un mismo elemento pueden...
71
SEMINARIOS GUÍA DE ESTUDIO DE CONTENIDOS TEÓRICOS TEMAS: UNIONES QUÍMICAS. GRUPOS FUNCIONALES ESTRUCTURA DE GLÚCIDOS, LÍPIDOS, AMINOÁCIDOS Y NUCLEÓTIDOS DEPARTAMENTO DE BIOQUÍMICA, FACULTAD DE MEDICINA, UBA
Transcript of SEMINARIOS GUÍA DE ESTUDIO DE CONTENIDOS … 1-Parte A... · Átomos de un mismo elemento pueden...

SEMINARIOS
GUÍA DE ESTUDIO DE CONTENIDOS TEÓRICOS
TEMAS: UNIONES QUÍMICAS. GRUPOS FUNCIONALES ESTRUCTURA DE GLÚCIDOS, LÍPIDOS, AMINOÁCIDOS Y NUCLEÓTIDOS DEPARTAMENTO DE BIOQUÍMICA, FACULTAD DE MEDICINA, UBA

TEMA Pág. UNIONES QUÍMICAS .GRUPOS FUNCIONALES Introducción. Estructura atómica. Conceptos básicos------------------------- ----- 1 Concepto de electronegatividad------------------------------------------------------ - 6 Uniones químicas------------------------------------------------------------------------ 7 Uniones iónicas---------------------------------------------------------------- ---------- 7 Uniones covalentes----------------------------------------------------------------------- 8 Polaridad de las moléculas------------------------------------------------------------- 11 Uniones químicas y propiedades fisico-químicas de los compuestos-------- --- 12 Uniones intermoleculares ------------------------------------------------------ ------- 12 Grupos funcionales---------------------------------------------------------------------- 14 ESTRUCTURA DE LAS PRINCIPALES MOLÉCULAS DEL ORGANISMO ESTRUCTURA DE GLUCIDOS Monosacáridos -------------------------------------------------------------------------- 18 Asimetría--------------------------------------------------------------------------- ---- - 19 Estereoisómeros, enantiómeros, epímeros ------------------------------------- ----- 21 Estructura cíclica de monosacáridos ------------------------------------------------- 22 Derivados de azúcares ------------------------------------------------------------------ 26 Disacáridos, oligosacáridos y polisacáridos ----------------------------------------- 27 ESTRUCTURA DE LÍPIDOS Clasificación y estructura -------------------------------------------------------------- 30 Ácidos grasos ---------------------------------------------------------------------------- 31 Acilglicéridos------------------------------------------------------------------------ ---- 33 Lípidos complejos----------------------------------------------------------------------- 33 Lípidos isoprenoides -------------------------------------------------------------------- 38 Esteroides --------------------------------------------------------------------------------- 39 Prostaglandinas ---------------------------------------------------------------------- ---- 40 Comportamiento de los lípidos anfipáticos en agua --------------------------------- 41 ESTRUCTURA DE AMINOÁCIDOS Introducción ------------------------------------------------------------------------------ 43 Estereoquímica de los aminoácidos --------------------------------------------------- 46 Clasificación de aminoácidos ---------------------------------------------------------- 48 Propiedades ácido-base de los aminoácidos ------------------------------------- ---- 55 Reacciones químicas de los aminoácidos --------------------------------------------- 57 ESTRUCTURA DE NUCLEÓTIDOS Estructura general de nucleótidos. Bases púricas y pirimidínicas ----------------- 60 Propiedades fisicoquímicas de las bases púricas y pirimidínicas ------------------ 62 Nucleósidos --------------------------------------------------------------------------- --- 64 Nucleótidos ---------------------------------------------------------------------------- --- 65 ADN:Doble hélice ------------------------------------------------------------------------ 68

1
UNIONES QUIMICAS. GRUPOS FUNCIONALES
De un centenar de elementos químicos conocidos, sólo aproximadamente 20 de
ellos forman parte de los seres vivos. Más aún, sólo 6 elementos no metálicos (C, O, H, N,
P y S) constituyen más del 95 % de la masa total de las células. Los datos de la tabla
siguiente indican la composición elemental del cuerpo humano:
Composición elemental del cuerpo humano (En porcentaje respecto a peso seco):
ELEMENTO PORCENTAJE
Carbono 50 Oxígeno 20 Hidrógeno 10 Nitrógeno 8,5 Fósforo 2,5 Azufre 0,8
Estos pocos elementos forman la mayor parte de los diferentes compuestos químicos
presentes en los organismos vivos, desde los estructuralmente más simples (como agua,
dióxido de carbono, glucosa) hasta los más complejos, como proteínas, glucógeno,
celulosa, ácidos nucleicos, entre otros.
La molécula es la mínima porción de sustancia que puede existir conservando las
propiedades de ésta. La fórmula química de una sustancia establece qué elementos la
forman y en qué proporción atómica (átomo: es la mínima porción de materia que puede
existir en estado de libertad). Por ejemplo la sustancia química agua tiene una fórmula:
H2O, esto indica que está formada por oxígeno e hidrógeno. En este caso se unen dos
átomos de H y un átomo de O. Esta sustancia puede dividirse en átomos, pero los átomos
de H y de O en los que se disocia no tienen las mismas propiedades que la sustancia agua.
Incluso dos sustancias diferentes pueden estar formadas por los mismos elementos y
unidos en la misma proporción. Veamos el siguiente ejemplo:

2
CH3
CH3
C
C
C
H2
H3
C OH
O
Propanal Propanona
En ambos casos, la fórmula molecular es C3H6O, sin embargo estos átomos están unidos de
diferente forma lo cual determina que los dos compuestos señalados tengan propiedades
físicas y químicas diferentes y definan, por lo tanto compuestos químicos diferentes.
Por lo tanto, sustancias diferentes pueden estar formadas por elementos idénticos unidos en
la misma proporción. La diferencia se debe a la presencia de enlaces atómicos distintos en
cada uno de los compuestos señalados. Esto explica porqué, a partir de unos pocos
elementos, se pueden generar la diversidad de sustancias químicas presentes en los
organismos vivos.
Es importante conocer entonces cual es la fuerza que mantiene unidos a los átomos entre sí
para formar los distintos compuestos y como se relacionan las propiedades físicas y
químicas de las sustancias con el tipo de unión química que se establece entre los átomos
que la forman.
Para estudiar los distintos tipos de uniones químicas es necesario tener conocimientos
básicos acerca de estructura atómica y por lo tanto es altamente recomendable que leer este
tema antes de comenzar con esta unidad temática.
ESTRUCTURA ATOMICA: CONCEPTOS BÁSICOS
Los átomos están formados por un sistema de partículas que incluyen:
-electrones: Partículas con carga eléctrica negativa.
-protones: Partículas con carga eléctrica positiva.
-neutrones: Partículas sin carga eléctrica.
Los protones y neutrones constituyen el núcleo atómico, en tanto que los electrones giran
alrededor del núcleo.
El número atómico de un elemento, simbolizado por la letra Z, es igual al número de
protones del mismo. En el átomo neutro, el número de electrones es igual al número de

3
protones. El número másico, A, es igual al número de protones más neutrones que hay en
el átomo. Así por ejemplo, el elemento cloro tiene la siguiente composición de partículas:
Z= 17 →→→→ 17 protones y 17 electrones
17Cl A= 35 →→→→ Número de neutrones= 35-17= 18 neutrones
Átomos de un mismo elemento pueden diferir en el número de neutrones. Isótopos son
átomos de un mismo elemento con distinto número de neutrones. Por ejemplo 35Cl y 37Cl
son isótopos del cloro.
El siguiente texto en itálica es sólo para recordar algunos conceptos que pueden requerir
para una mejor comprensión de tema Uniones Químicas
Los electrones giran alrededor del núcleo. Sin embargo, existe una zona del espacio alrededor del
núcleo donde es mayor la probabilidad de encontrar a un electrón. A esta zona se la denomina
orbital.
Las funciones de onda son ecuaciones matemáticas que describen el movimiento y el estado
energético del electrón de un átomo. De la resolución de estas ecuaciones surgen valores
numéricos que se conocen como números cuánticos. Estos son cuatro: principal, azimutal,
magnético y de spin. El número cuántico principal, n, determina el nivel principal de energía del
electrón. Esta energía está vinculada con la distancia del electrón al núcleo, por lo tanto el
número cuántico principal permite estimar la distancia radial del electrón al núcleo. El número
cuántico principal toma valores desde n=1 hasta n=7. El nivel n=1 es el más cercano al núcleo y
tiene la energía más baja. A veces se usan las letras K, L, M, N, O, P, Q para designar los niveles
principales 1,2, 3,etc , respectivamente.
El número cuántico azimutal, l, puede asumir todos los valores enteros desde 0 hasta n-1, es decir:
0, 1, 2,..., (n-1). Este número designa el subnivel o subcapa dentro del nivel principal de energía, e
indica la forma del orbital o nube electrónica en el cual se mueve el electrón alrededor del núcleo.
Los números cuánticos azimutales se designan por las letras s, p, d ó f. Cuando un electrón tiene
un valor de l=0, el orbital se designa s, cuando l=1, el orbital se designa p, cuando l=2, d y
cuando l=3, f. La forma de los orbitales s es una esfera concéntrica con el núcleo, en tanto que los
orbitales p, por ejemplo tienen forma de pera. Los orbitales s pueden contener como máximo 2
electrones, los p, 6 ; los d, 10 y los f, 14 electrones .
Cada nivel energético está formado por uno ó más subniveles. Así por ejemplo el nivel
caracterizado por n=1 tiene un sólo subnivel, caracterizado por l=0 (subnivel s) (Si n=1, l sólo

4
puede tomar el valor n-1=0). En cambio el nivel n=2 tiene dos subniveles (l toma valores desde 0
hasta n-1, es decir 0 y 1), los cuales se caracterizan por las letras s y p respectivamente. El nivel 3
tiene tres subniveles, dado que los valores de l serán ( desde 0 hasta n-1) : 0, 1 y 2 ( s, p, d).
Con respecto a número cuántico magnético, m , y al número cuántico de espín, s , diremos que el
primero designa la orientación del orbital en el espacio (toma valores enteros, desde -l hasta +l).
En número de spin describe el sentido de giro del electrón sobre su eje y puede tener dos valores,
designados +1/2 y -1/2 (generalemte se simbolizan con flechas apuntando en sentido contrario; ↑↑↑↑
y ↓↓↓↓).
Aplicamos estos conceptos al átomo más sencillo, es decir el átomo de H (hidrógeno). Este
elemento tiene Z=1, por lo tanto tiene un solo electrón y el mismo se halla en el nivel energético
más bajo, es decir tiene n=1. El valor de l será 0 (toma el valor n-1=0) de modo que se halla en un
orbital s y ocupa una nube esférica alrededor del núcleo. Dado que l=0, el único valor posible de
m será 0 y por lo tanto el orbital tendrá una única orientación posible en el espacio como
corresponde para una esfera. El único electrón del átomo de hidrógeno gira alrededor de su eje
(puede hacerlo en los dos posibles sentidos diferentes, +1/2 o - 1/2.)
El elemento helio (He ) tiene número atómico Z=2, por lo tanto su configuración
electrónica será:
2He = 2 electrones
[Estos dos electrones se ubican en el nivel de menor energía, n=1.
Si n=1 ===> l= 0 ===> Los electrones están en un orbital s
ml = 0 ===> una única orientación espacial del orbital
ms = +1/2 y - -1/2 (Los dos electrones tienen distinto sentido de giro) ]
La estructura electrónica para este átomo será: 1 S2
Los dos electrones se disponen en el nivel energético más bajo, n=1, ocupando ambos un
orbital esférico, s, que tiene una única orientación espacial. Cada uno de ellos gira
alrededor de su eje en sentido opuesto ↑↑↑↑ y ↓↓↓↓ .
Para el elemento litio (Li), que tiene Z=3, el tercer electrón no puede ir al orbtal 1s por
estar lleno. Tiene que pasar al nivel de energía n=2. Ahora tenemos que decidir si el
electrón irá al orbital 2s o a uno de los tres orbitales p: Se ocupan primero los orbitales de
+ + + +

5
menor energía y el orbital 2s tiene menor energía que el 2p. En general el orden de llenado
de los orbitales es el siguiente: 1s, 2s, 2p, 3s, 3p, 4s, 3d, 4p....
El elemento carbono (C) tiene 6 electrones y su estructura electrónica será: 1s2 2s2 2p2 .
En el último nivel energético, n=2, tiene 4 electrones los que de acuerdo a lo expuesto
debieran estar 2 en un orbital s y 2 en los orbitales p (uno en el px y otro en el py). Sin
embargo, en el átomo de C ocurre un fenómeno que se conoce como hibridización de
orbitales: Estos cuatro electrones del último nivel se ubican en cuatro orbitales híbridos
denominados sp3 (formados por la hibridización de tres orbitales p y un orbital s), todos de
igual energía. Estos cuatro orbitales tienen forma de pera que se extienden a los cuatro
vértices de un tetraedro.
Hibridización de orbitales en el átomo de C:
**
*
**
*
sp3
sp3
sp3sp3
1s1s
sp3 sp3 sp3 sp3
Es importante destacar que elementos diferentes cuyos átomos tienen estructuras
electrónicas semejantes en sus capas externas o niveles de valencia tienen muchas
propiedades químicas en común. Esta idea que relaciona la semejanza en las estructuras
electrónicas con la semejanza en las propiedades físicas y químicas de los elementos es la
base de la ley periódica que establece:"Las propiedades de los elementos son función
periódica de sus números atómicos"
Comparando la distribución electrónica de los elementos 3Li y 11Na observamos:
3Li : 1 s2 2s1
11Na : 1 s2 2s2 2p6 3s1

6
Ambos elementos tienen un electrón en su último nivel energético (niveles 2 s y 3 s para el
Li y el Na respectivamente), y tienen propiedades químicas similares, lo cual está
relacionado con la similitud en la estructura electrónica. Además conviene recordar que
ambos elementos están ubicados en el mismo grupo en la tabla periódica, Grupo I, dado
que ambos tienen 1 electrón en el último nivel.
Las estructuras electrónicas en los elementos 7N y 15P indican que ambos elementos
tienen 5 electrones de valencia:
7N 1 s2 2s2 2p3 = 5 electrones en el último nivel (nivel n=2).
15P 1 s2 2s2 2p6 3s2 3p3 = 5 electrones en el último nivel,(nivel n=3)
Ambos elementos tienen comportamiento químico similar y pertenecen al grupo V de la
tabla periódica (ver tabla periódica). El elemento N (nitrógeno) pertenece al segundo
período (n=2) en tanto que el P (fósforo ) pertenece al tercero ( n= 3 ).
Si se analizan las estructuras de los llamados gases inertes o nobles, los cuales se
caracterizan por su escasa reactividad química, se comprueba que todos ellos tienen 8
electrones en su último nivel energético, excepto el helio (He), que tiene 2. Ellos se ubican
en el Grupo VIII de la tabla periódica.
Concepto de electronegatividad
La electronegatividad de un elemento expresa la tendencia del mismo a atraer electrones. A
cada elemento se le asigna un número positivo que indica la electronegatividad del mismo.
Linus Pauling desarrolló una escala de electronegatividades (relativa) en la cual asignó al
fluor (F ), que es el elemento con mayor capacidad para atraer electrones (es decir , es el
más electronegativo ), el valor de 4. Al cesio (Cs ), que junto con el francio (Fr ) son los
elementos que sostienen más debilmente a los electrones, se les asigna un valor más bajo,
0,7. De acuerdo a la ubicación de los elementos en la tabla periódica, se puede decir que la
electronegatividad aumenta al recorrer un período de izquierda a derecha y en un grupo de
abajo hacia arriba.
Sentido de aumento de la electronegatividad en la T.P.

7
UNIONES QUIMICAS
Cuando se acercan dos átomos se ejercen varias fuerzas electrostáticas entre ellos.
Algunas de éstas tratan de mantener los átomos unidos, otras tienden a separarlos (recordar
que cargas eléctricas del mismo signo se repelen, cargas de signo contrario se atraen: los
electrones se repelen entre sí, los protones atraen a los electrones, entre otros ejemplos).
Cuando las fuerzas de atracción superan a las de repulsión, se forma un enlace químico.
La regla del octeto establece que, cuando se forma un enlace químico, los átomos ceden o
comparten electrones, de manera tal que la capa más externa o de valencia de cada átomo
contenga 8 electrones. La inercia química de los gases nobles se atribuye a esta
configuración electrónica. Puede decirse por lo tanto que cuando dos átomos se unen entre
sí lo hacen tratando de adquirir cada uno de ellos la estructura de un gas noble, logrando
así una estructura estable.
El átomo de sodio es un buen ejemplo para ilustrar la regla del octeto:
11Na : 1s2 2s2 2p6 3s1
Cuando este elemento se une a otro elemento, tiende a adquirir configuración de gas noble.
Este elemento tenderá a perder el electrón de su último nivel y adquirir la estructura de un
gas noble, en este caso el 10Ne (neón), más que a ganar 7 electrones y adquirir la
estructura del 18Ar (argón). El sodio tiene una electronegatividad baja (no tiene tendencia
a ganar electrones, tiene tendencia a perder electrones, es un elemento muy
electropositivo).
Las uniones químicas son principalmente de dos clases:
a) Uniones iónicas
b) Uniones covalentes
Uniones iónicas
Se forman por transferencia de electrones de un átomo (electropositivo) a otro
(electronegativo). El que pierde los electrones se convierte en un ión positivo (catión) y el
que los gana se convierte en un ión negativo (anión). Entre ambas especies se genera una
atracción electrostática, que constituye la unión iónica.
Este tipo de unión está presente por ejemplo en el compuesto NaCl:
17Cl : 1s2 2s2 2p6 3s2 3p5 ⇒ 7 electrones de valencia

8
11Na : 1s2 2s2 2p6 3s1 ⇒ 1 electrón de valencia
El cloro, al adquirir 1 electrón toma la configuración electrónica de un gas noble,
por lo tanto se forma la especie Cl- . El sodio tenderá a perder su electrón de valencia para
adquirir la estructura de gas noble (8 electrones en su último nivel ) y se convertirá en Na+:
Los iones cloruro se rodean de iones sodio y éstos de iones cloruro para formar la
red cristalina. Otros ejemplos son el cloruro de magnesio y el oxido de calcio.
En este tipo de unión química existe transferencia de electrones desde un átomo a otro, por
lo tanto se da entre elementos de muy diferente electronegatividad, frecuentemente entre
elementos del Grupo I o II con los elementos del grupo VI u VII.
Los compuestos con este tipo de unión se caracterizan por: poseer elevado punto de fusión
y de ebullición, presentar estructura cristalina iónica, ser solubles en agua y poco solubles
en solventes orgánicos y por ser buenos conductores de la corriente eléctrica en solución
acuosa o en estado fundido. Este tipo de unión se presenta en sales y bases
Uniones covalentes
Enlace covalente es aquel que se verifica cuando los átomos, para adquirir
estructura electrónica de gas noble, comparten pares de electrones. Ambos elementos
unidos por este tipo de unión tienen electronegatividades similares.
En el compuesto metano, CH4 , las uniones son covalentes. El carbono (C), que tiene un
valor de electronegatividad de 2,5, se une con el H (hidrógeno) cuyo valor de
electronegatividad es 2,5:
1H : 1s1 6C : 1s2 2s2 2p2
Cl Na Cl NaCl NaClCl Na Cl NaCl NaClCl Na
ClCl Mg ClCl MgClCl Mg ClClCl Mg ClCl Mg ClClCl Mg

9
Compartiendo pares de electrones, ambos elementos adquieren estructura
electrónica de gas noble: 8 electrones el C y 2 electrones el H (éste adquiere la
configuración del He, el gas noble más cercano en la T.P).
En la molécula de nitrógeno (N2 ), se comparten entre ambos átomos 3 pares de electrones.
15 N 1s2 2s2 2p6 3s 2 3p3
Unión covalente triple
En el dióxido de carbono, CO2, el átomo de carbono comparte con cada átomo de oxígeno
un par electrónico:
Uniones covalentes dobles En la molécula de agua, H2O, el átomo de oxígeno comparte con cada átomo de H un par
electrónico:
El elemento S (azufre) tiene la siguiente configuración electrónica:
16S : 1s2 2s2 2p6 3s2 3p4
Sólo necesita 2 electrones para adquirir la configuración de gas noble (ocho electrones en
el último nivel). En el compuesto SO3 cada átomo de S se une a tres átomos de O, de
manera que los electrones se distribuyen como se ilustra:
C H CH
H
HC H CH
H
H
N N N N NNN N NN NN N NN N
CO O C OOCO OCOO OO C OO C OO
H HO OH H H O HH HOHH HOO OH HOOH H H O HH O H

10
uniones covalentes dativas
Esta molécula presenta una unión covalente doble y dos uniones covalentes dativa. En la
unión covalente dativa el par electrónico que establece la unión entre los dos átomos
pertenece a uno solo de ellos, en este caso el átomo de S . Se simboliza con una flecha →,
dirigida hacia el átomo que acepta el par electrónico.
Los compuestos con uniones covalentes se caracterizan por: poseer bajo punto de fusión y
ebullición, ser solubles en solventes orgánicos (los que poseen uniones covalentes), ser
insolubles en agua, no conducir la corriente eléctrica. La unión covalente es la más
generalizada entre los compuestos de la llamada química orgánica.
Es importante destacar que el par electrónico que se comparte por dos átomos no se halla
equidistante de ambos núcleos, sino que la mayor parte del tiempo se halla desplazado
hacia el elemento más electronegativo. Evidentemente esto no ocurre en las moléculas
diatómicas homonucleares:
Molécula de hidrógeno molécula de agua
Como se ve en los ejemplos anteriores, en la molécula de H2O los electrones de unión se
hallan más desplazados hacia el O, elemento más electronegativo que el H. Si bien la
diferencia de electronegatividades entre ambos no es suficiente para generar iones, se
genera sobre los núcleos de ambos átomos una densidad de carga positiva ( + ) sobre el H y
negativa ( - ) sobre el O.
En el caso de la molécula de hidrógeno (H2) (molécula homonuclear), el par electrónico
está igualmente compartido por ambos átomos. Cuando se genera cierta carga sobre los
átomos que se unen mediante uniones covalentes, se dice que el enlace covalente es polar,
para diferenciarlo de los enlaces HH, ClCl, O========O , etc., que son covalentes no
polares .
SO O
O
S
O
OOSO O
O
SOO OO
OO
S
O
OO S
O
OO
H H OH H
δ-δ+ δ+
H HH H OH H
δ-δ+ δ+
OH HOH H
δ-δ+ δ+

11
Polaridad de las moléculas
Debido a la polaridad de los enlaces individuales, la totalidad de la molécula puede tener
centros de carga positiva y negativa separados. Dicha molécula constituye un dipolo
(sistema formado por dos centros con cargas opuestas separados una cierta distancia), lo
cual puede simbolizarse con una flecha apuntando al elemento más electronegativo.
Las moléculas diatómicas formadas por un mismo elemento son no polares (por ejemplo
O2) , en tanto que las moléculas diatómicas formadas por elementos diferentes pueden
tener cierto grado de polaridad (ejemplo C=O) y la polaridad de las mismas será mayor
cuanto mayor sea la diferencia de electronegatividad entre los dos átomos que se unen. En
las moléculas poliatómicas, la presencia de enlaces polares no es suficiente para que la
misma resulte polar. Los ejemplos siguientes ilustran el concepto: H2O y Cl4C.
Ambos dipolos O-H presentes en la molécula de agua no se anulan, ambos contribuyen a
la polaridad de la molécula. En cambio, los cuatro dipolos C-Cl se anulan mutuamente (En
este sistema formado por distintos dipolos se crean fuerzas que se anulan mutuamente). La
molécula, si bien tiene enlaces polares, como un todo, resulta no polar. Por lo tanto,
además de la presencia de enlaces polares, es necesario tener presente la geometría de la
molécula para determinar si la misma es o no polar: la presencia de grupos polares en
una molécula es condición necesaria pero no suficiente para que la misma resulte
polar.
H HH Hδ-
CCl Cl
Cl
Clδ- δ-
δ-
δ-
CCl Cl
Cl
Clδ- δ-
δ-
CClCl ClCl
ClCl
ClClδ- δ-
δ-H
H
O105
H
H
O105105 C
Cl
ClCl
Cl
C
Cl
ClCl
Cl

12
Uniones químicas y propiedades fisicoquímicas de los compuestos
Los compuestos químicos con uniones iónicas son sólidos cristalinos con alto punto de
fusión debido a la fuerza de unión entre iones. Son compuestos muy solubles en agua y
solventes polares. En cambio, los compuestos covalentes tienen bajo punto de fusión y
ebullición y la solubilidad en agua y solventes polares aumenta al aumentar la polaridad de
la molécula. Por ejemplo:
A : CH4 B: CH3 OH
Considerando las moléculas A y B, esta última será más soluble en agua dado que B es
una molécula polar en tanto A, no lo es.
UNIONES INTERMOLECULARES
Enlace electrostático: Un grupo cargado en una molécula puede atraer un grupo con carga
opuesta presente en otra molécula y consecuentemente se crea entre ambas moléculas una
fuerza de atracción. Esta fuerza es directamente proporcional a la cargas de los grupos
involucrados e inversamente proporcional a la distancia que las separa.
Uniones puente de hidrógeno: Muchas sustancias hidrogenadas presentan propiedades
anormales como un punto de ebullición o de fusión anormalmente elevados, tal es el caso
del agua. Esto se debe a que las moléculas de agua, en el estado líquido o sólido tienden a
asociarse por medio de uniones intermoleculares en las que el H actúa como puente, y por
eso se llama enlaces puente de hidrógeno. El oxígeno, muy electronegativo, tiene gran
tendencia a atraer el par electrónico compartido dejando al H con una cierta densidad de
carga positiva. Este hidrógeno es atraído por otro oxígeno de otra molécula, formándose
una unión puente de hidrógeno:
H
CH H
H
H
CH H
H
H
CH H
OH
H
CH H
OH
H H H H
O
O O
H H
Uniones puente de hidrogeno
Uniones puente de hidrogeno entre moleculas de agua
H H H H
O
O O
H H
Uniones puente de hidrogeno
Uniones puente de hidrogeno entre moleculas de agua

13
Cualquier molécula que tenga un átomo de H unido a un átomo pequeño y electronegativo
como el N, O o F puede formar uniones puente de hidrógeno. En los compuestos de
importancia biológica grupos N-H y O-H son los cuantitativamente importantes.
Una molécula para ser soluble en agua tiene que tener la capacidad de romper la fuerte
cohesión entre moléculas de agua dada por las uniones puente de hidrógeno. Aquellos
compuestos con múltiples grupos -OH, -NH2, podrán solubilizarse en agua porque los
mismos tendrán capacidad de desestabilizar las uniones puente de hidrógeno entre las
moléculas de agua y establecer nuevas uniones puente de hidrógeno entre el O del agua y
ellas mismas. La energía del enlace por puente de hidrógeno oscila entre 3 y 7 kcal/mol.
Este tipo de fuerza intermolecular es la más intensa. La relación entre las energías de
enlace es, aproximadamente: 100:10:1 (enlaces covalentes: enlace puente de hidrógeno:
otras fuerzas intermoleculares)
Fuerzas de Van Der Waals: También son fuerzas de atracción molecular. Dado que la
distribución de la carga electrónica alrededor de un átomo cambia con el tiempo, se
establece en determinado tiempo una distribución asimétrica de cargas la cual induce una
similar asimetría en la distribución de cargas en los átomos vecinos, lo cual lleva a la
formación de dipolos que se atraen mutuamente.
Puede darse por la atracción entre una molécula polar y otra molécula que aislada no es
polar. Al aproximarse el dipolo a la molécula no polar, su carga parcial atrae o repele los
electrones de la otra molécula induciendo en ésta un dipolo (dipolo inducido).
También es posible la atracción entre dos moléculas no polares, por ejemplo la molécula
de H2. Si los electrones de ambos H se encuentran en un determinado momento en uno de
los extremos de su orbital molecular puede quedar el núcleo expuesto por corto tiempo y
adquirir una carga parcial positiva. Esta carga positiva en un átomo atraerá los electrones
de otra molécula de H2 y determinará la atracción transitoria entre ambas moléculas.
La magnitud de las fuerzas de Van der Waals varía con la distancia entre dipolos: Sólo son
efectivas a distancias muy cortas. La energía de una unión de este tipo es de alrededor de 1
kcal/mol, considerablemente más débil que las uniones electrostáticas y las puente de
hidrógeno.
Cuando dos moléculas tienen superficies complementarias, como por ejemplo una enzima
y su sustrato o un antígeno y su anticuerpo, ser posible que ambas estructuras se acerquen
lo suficiente como para que se establezcan múltiples uniones de Van der Waals, que a
cortas distancias serán efectivas y de este modo se producirá una unión estable entre ambas
estructuras. Por ejemplo:

14
GRUPOS FUNCIONALES
Las propiedades químicas de los compuestos dependen de la estructura de los
mismos. Sin embargo, ciertas propiedades son atribuibles fundamentalmente a un grupo
atómico ordenado de manera particular, más que al conjunto de la molécula. Así por
ejemplo las propiedades químicas de los ácidos orgánicos se atribuyen al grupo ,
llamado grupo carboxilo, el cual caracteriza a los mismos. Tales grupos de átomos que
determinan las propiedades químicas de los distintos compuestos se denominan grupos
funcionales.
Un determinado grupo funcional, por ejemplo el grupo –OH puede estar en un
carbono primario, secundario o terciario. Es conveniente entonces recordar que un carbono
primario es aquel que tiene sólo una de sus valencias implicadas en una unión con otro
átomo de C. Un carbono secundario tiene dos de sus valencias involucradas en uniones con
átomos de carbono. Un carbono terciario tiene tres uniones C-C.
A continuación están indicados los grupos funcionales que frecuentemente se
encuentran en las moléculas de importancia biológica. Las valencias libres en cada grupo
funcional (indicada por una línea, --) en las diferentes moléculas representan una unión a
una cadena hidrocarbonada.
Enzima Sustrato Complejo Enzima-SustratoEnzima Sustrato Complejo Enzima-Sustrato
C OH
O
C OH
O

15
GRUPOS FUNCIONALES
R-OH
Grupo Funcional Fórmula Sufijo Ejemplo
Hidroxilo R-Ol CH3-CH2-OH
etanol (Alcohol primario)
CH3-CH-CH3
OHCH3-C-CH3
OH
CH3
2-propanol 2 metil 2 propanol
(Alcohol secundario) (Alcohol terciario)
Alcoxi R-O-R´ R-R´eter CH3-O- CH3
dimetilcetona
R-OH
Grupo Funcional Fórmula Sufijo Ejemplo
Hidroxilo R-Ol CH3-CH2-OH
etanol (Alcohol primario)
CH3-CH-CH3
OH
CH3-CH-CH3
OHCH3-C-CH3
OH
CH3
CH3-C-CH3
OH
CH3
OH
CH3
2-propanol 2 metil 2 propanol
(Alcohol secundario) (Alcohol terciario)
Alcoxi R-O-R´ R-R´eter CH3-O- CH3
dimetilcetona
Cetona C
O
R R´ R-R´cetona C
O
CH3 CH3
Dimetilcetona opropanona
Carboxilo CO
OHR R-oico CO
OHCH3
Acido etanoico
Aldehido CO
HR R- al CH3-COH
etanal
Cetona C
O
R R´C
O
R R´ R-R´cetona C
O
CH3 CH3C
O
CH3 CH3
Dimetilcetona opropanona
Carboxilo CO
OHR CO
OHR R-oico CO
OHCH3C
O
OHCH3
Acido etanoico
Aldehido CO
HR CO
HR R- al CH3-COH
etanal
CH3-COHCH3-C
OH
etanal
Grupo Funcional Fórmula Sufijo EjemploGrupo Funcional Fórmula Sufijo EjemploGrupo Funcional Fórmula Sufijo EjemploGrupo Funcional Fórmula Sufijo Ejemplo

16
Los ésteres se consideran como el producto de reacción entre un ácido carboxílico y un
alcohol. En tanto que los anhídridos son el producto de reacción entre dos ácidos. Ambos
compuestos se hidrolizan (reaccionan en medio acuosos) y generan los compuestos que
los constituyen:
ester + agua <========> ácido + alcohol
anhídrido + agua < ========> ácido + ácido
Las amidas son el producto de deshidratación entre un ácido carboxílico y el amoníaco. De
este modo la reacción de hidrólisis de las amidas rinde un ácido carboxílico y amoníaco:
amida + agua < ======= > ácido carboxílico + amoníaco
Las amidas N sustituídas son aquellas en los que los átomos de H unidos al N de las
amidas son reemplazados por cadenas hidrocarbonadas: uno o los dos H (amidas
monosustituídas o disustituídas). Por ejemplo:
Ester C
O
R O R´ R-ato de R´de ilo C
O
CH3 CH3O
Etanoato de metilo
Anhidrido C
O
R O C
O
R CH3 C
O
CH3O C
O
Anhidrido acético
Amida C
O
R O NH2 C
O
CH3 NH2
Propanamida
Grupo Funcional Fórmula Sufijo Ejemplo
Amina R- NH2 R-amina CH3-CH2-NH2
R-amida
etanamina
Ester C
O
R O R´C
O
R O R´ R-ato de R´de ilo C
O
CH3 CH3OC
O
CH3 CH3O
Etanoato de metilo
Anhidrido C
O
R O C
O
RC
O
R O C
O
R CH3 C
O
CH3O C
O
CH3 C
O
C
O
CH3O C
O
C
O
Anhidrido acético
Amida C
O
R O NH2C
O
R O NH2 C
O
CH3 NH2C
O
CH3 NH2
Propanamida
Grupo Funcional Fórmula Sufijo Ejemplo
Amina R- NH2 R-amina CH3-CH2-NH2
R-amida
etanamina
C
O
CH3 NH CH3
C
O
CH3 N CH3
CH2-CH3
Amida monosustituída Amida disustituída
C
O
CH3 NH CH3
C
O
CH3 N CH3
CH2-CH3
C
O
CH3 N CH3
CH2-CH3
Amida monosustituída Amida disustituída

17
Las aminas pueden ser clasificadas en aminas primarias, secundarias y terciarias. Por
ejemplo:
CH3-CH2- CH2- NH2 amina primaria grupo funcional : NH2
H3C−−−− NH −−−−CH3 amina secundaria grupo funcional : NH
H3C N CH2−−−−CH3 amina terciaria grupo funcional : N
Un grupo con S es el grupo tiol, -SH, similar al grupo hidroxilo. Por ejemplo el etanotiol
tiene la siguiente estructura:
CH3 CH2 SHCH3 CH2 SH

18
ESTRUCTURA DE LOS PRINCIPALES COMPUESTOS DEL ORGANI SMO
Nuestro organismo está formado por compuestos de estructuras muy diversas,
desde moléculas relativamente simples como los azúcares y aminoácidos hasta polímeros
enormemente complejos como las proteínas y los ácidos nucleicos. Las estructuras de estos
compuestos se relacionan íntimamente con sus funciones fisiológicas.
A continuación analizaremos brevemente las estructuras de glúcidos, lípidos,
aminoácidos y nucleótidos.
GLÚCIDOS
Los glúcidos previamente denominados hidratos de carbono o carbohidratos o
azúcares pueden clasificarse según el número de subunidades en MONOSACÁRIDOS,
DISACÁRIDOS, OLIGOSACÁRIDOS Y POLISACÁRIDOS. Los monosacáridos, como
glucosa, fructosa y galactosa, son los más sencillos. Son nutrientes esenciales para el
organismo y a partir de ellos se sintetizan otros compuestos. Se unen por enlaces
glicosídicos para formar los otros tipos de carbohidratos. Los disacáridos, maltosa,
sacarosa, lactosa, contienen 2 unidades de monosacáridos unidos por un enlace glicosídico.
Los oligosacáridos, como por ejemplo la porción de carbohidratos de las glicoproteínas y
los glicolípidos, contienen de 3 a 10 unidades de monosacáridos. Los polisacáridos, como
el almidón y el glucógeno contienen hasta miles de unidades de monosacárido y además de
constituir la reserva de monosacáridos celulares, junto con los oligosacáridos están
involucrados en procesos de reconocimiento entre células y en el transporte de proteínas
entre distintos compartimientos subcelulares.
MONOSACÁRIDOS
Los glúcidos más sencillos son los monosacáridos y pueden definirse como
polihidroxialdehídos o polihidroxicetonas. Su fórmula molecular es (C-H2O)n con n ≥3
y es por eso que se denominaron hidratos de carbono. Son cadenas carbonadas donde un
carbono es un grupo carbonilo y los otros generalmente presentan grupos hidroxilo. Si el
grupo carbonilo es un aldehído, el monoscárido es una aldosa (glucosa, etc.) y si es una
cetona es una cetosa (fructosa, etc.). En la mayoría de los monosacáridos la cadena de
carbonos presenta de 3 (triosa) a 7 (heptosa) carbonos. Los monosacáridos más simples
son de 3 carbonos: GLICERALDEHÍDO (aldotriosa) y DIHIDROXIACETONA
(cetotriosa). Es decir, dependiendo del número de carbonos del monosacárido es:

19
MONOSACARID
O
Nº DE
CARBON
OS
ALDEHIDO
CETONA EJEMPLO
TRIOSA 3 ALDOTRIOS
A
CETOTRIOS
A
GLICERALDEHÏ
DO
TETROSA 4 ALDOTETRO
SA
CETOTETRO
SA
ERITRULOSA
PENTOSA 5 ALDOPENTO
SA
CETOPENTO
SA
RIBOSA
HEXOSA 6 ALDOHEXOS
A
CETOHEXOS
A
GALACTOSA
HEPTOSA 7 ALDOHEPTO
SA
CETOHEPTO
SA
SEDOHEPTULOS
A
En la siguiente figura se muestran las estructuras de dos hexosas muy comunes: la glucosa,
que es una ALDOSA por tener una función aldehído, y la fructosa, que es una CETOSA
por poseer una función cetona. O CH2OH H-C C=O H-C-OH HO-C-H HO-C-H H-C-OH H-C-OH H-C-OH H-C-OH H-C-OH CH2OH CH2OH
Glucosa Fructosa
Otro azúcar importante, no ya desde el punto de vista energético es la ribosa. Esta
pentosa forma parte de los ácidos nucleidos (del ARN como ribosa y del ADN como
desoxirribosa). La ribosa es una aldosa, por poseer la función aldehído.
Asimetría
La mayoría de los monosacáridos presenta una estructura asimétrica. Es importante
tener esto en cuenta porque las enzimas generalmente reconocen una sola de las formas
isoméricas.

20
Un átomo de carbono al cual se unen cuatro grupos químicos diferentes
constituye un centro asimétrico (o quiral). La triosa gliceraldehído tiene 1 (UN) carbono
asimétrico y por lo tanto tiene 2 (DOS) isómeros ópticos activos (ENANTIÓMEROS ): el
D-gliceraldehído y el L-gliceraldehído. Los monosacáridos de más carbonos se asignan
como D o L según la configuración del carbono asimétrico más alejado del grupo carbonilo
(el aldehido o la cetona).
Aquellos que tienen la misma configuración que el D-gliceraldehído (el OH hacia
la derecha) pertenecen a la serie D y los que tienen la configuración del L-gliceraldehído
(el OH hacia la izquierda) pertenecen a la serie L. Dado que la mayor parte de los azúcares
de los organismos vivos pertenecen a la serie D, se asume que los azúcares son D a menos
que se indique específicamente. Cuando decimos glucosa, entonces, nos referimos a la D-
glucosa.
D-gliceraldehido L-gliceraldehido
Imagen especular

21
ESTEREOISOMEROS, ENANTIOMEROS Y EPIMEROS
Los ESTEREOISÓMEROS presentan la misma fórmula química pero difieren en la
posición del grupo hidroxilo en uno o más de los carbonos asimétricos. Un azúcar con
n centros asimétricos presentará 2n estereoisómeros, a menos que tenga un plano de
simetría. Dentro de los estereoisómeros, los enantiómeros son imágenes
especulares entre sí. El D-gliceraldehído y el L-gliceraldehído son enantiómeros, lo
mismo que la D- y la L-glucosa, es decir, los monosacáridos D y L con el mismo nombre
son enantiómeros y los sustituyentes de todos los carbonos asimétricos son los opuestos.
No importa de qué forma se roten éstos azúcares, sus átomos no pueden superponerse.
D-Gliceraldehído D-GlucosaD-Gliceraldehído D-Glucosa

22
Diasteroisómeros son estereoisómeros que no son imágenes especulares y pueden
no presentar carbonos asimétricos.
Epímeros son diastereoisómeros que contienen más de un C asimétrico y que
difieren en la posición del grupo hidroxilo de sólo uno de los carbonos asimétricos. La
D-glucosa y la D-galactosa son epímeros, que difieren solo en la posición 4.
En resumen:
ESTEREOISOMEROS
ENANTIOMEROS DIASTERÓMEROS
EPIMEROS
ESTRUCTURAS CÍCLICAS DE LOS MONOSACÁRIDOS
Aunque generalmente los monosacáridos se representan como cadenas lineales (fórmula de
Fischer), en solución existen principalmente como anillos en los cuales el grupo carbonilo
(aldehído o cetona) ha reaccionado químicamente con un grupo hidroxilo de la misma
molécula. Las hexosas, como la aldohexosa glucosa y la cetohexosa fructosa, forman
anillos de 6 miembros (piranosas) o de 5 miembros (furanosas). Estas representaciones
cíclicas llevan el nombre de fórmulas de Haworth.
D-glucosa D-galactosa
EPIMEROS

23
OH aldehído | R-CHO + R'-OH → R - C - OR' hemiacetal | H OH cetona | R1-CO-R2 + R'-OH → R1 - C - OR' hemicetal | R2
H
Proyección de Fischer
Fórmula de Haworth
El carbono carbonílico forma un hemiacetal (si es aldehído) o hemicetal (si es una
cetona), generando un nuevo centro asimétrico. El carbono del aldehído o cetona originales
que ahora se une a un oxígeno en el anillo se denomina carbono anomérico, y entonces se
los llama anómeros a los isómeros que se generan: ALFA y BETA.
D-glucosa D-fructosa
α-D-glucopiranosa α-D-fructofuranosa

24
Se designa αααα (alfa), al anómero que presenta los grupos -OH y -CH2OH (que sustituyen a
los carbonos unidos por el oxígeno) en trans (apuntan en sentidos opuestos), y se designa
ββββ (beta) si los grupos son cis (apuntan hacia el mismo sentido). También se pueden
escribir ésas estructuras como las proyecciones de Fischer: α (alfa), si el OH del carbono
anomérico está del mismo lado que el anillo y β (beta) si está del lado opuesto.
Las fórmulas de Haworth se usan más frecuentemente:
α-D-Glucosa β-D-Glucosaα-D-Glucosa β-D-Glucosa
α-D-glucosa
β-D-glucosa
α-D-glucosa
β-D-glucosa

25
El azúcar se dibuja con el anillo perpendicular al plano del papel, el oxígeno se escribe
atrás a la derecha, el C-1 a la derecha y los sustituyentes por encima o debajo del plano del
papel. El grupo hidroxilo unido al carbono anomérico es αααα si está por debajo del plano del
anillo o si el -OH y el -CH2OH que se encuentran en los dos carbonos que están unidos por
el oxígeno son trans, y en cambio es ββββ si está por encima o si el -OH y el CH2OH son cis.
Lo que se escribe a la derecha en la proyección de Fischer se escribe hacia abajo en la
fórmula de Haworth. Las formas alfa y beta del mismo monosacárido son formas
especiales de epímeros denominadas anómeros que difieren sólo en la configuración del
carbono acetálico (o cetálico).
Los azúcares pueden dibujarse más exactamente con las formas silla o bote
predominando en solución la forma silla.
En solución acuosa, el anillo puede abrirse y cerrarse. Cuando se abre, se forma la cadena
lineal, y cuando se cierra, el hidroxilo puede adoptar ambas configuraciones alfa o beta. El
proceso mediante el cual se equilibran las formas α y β en solución se denomina
mutarrotación. Dentro de las células la mutarrotación es catalizada enzimáticamente, por
Forma silla
Forma bote
Forma silla
Forma bote

26
lo que ocurre muy rápido. Sin embargo, cuando el carbono anomérico une el azúcar a otro
compuesto, el enlace se fija en una configuración. Las enzimas pueden diferenciar esas
configuraciones y son específicas para solamente una de ellas.
DERIVADOS DE AZÚCARES
Aminoazúcares
Son aquellos que contienen grupos amino, como la glucosamina y la
galactosamina. El grupo amino en general está acetilado.
Azúcares sulfatados
Algunos azúcares contienen grupos sulfato, en particular los componentes de los
glicosaminoglicanos como el condroitín sulfato y la heparina, componentes principales del
tejido conectivo.
Azúcares fosforilados
Dentro de las células los azúcares a menudo incorporan grupos fosfato, lo que
evita que atraviesen las membranas debido a sus cargas negativas. Cuando la glucosa entra
a las células se fosforila en la posición 6. Los grupos fosfato también pueden unir azúcares
a nucleósidos. Por ejemplo, la glucosa 1-fosfato puede reaccionar con UTP para formar
UDP-glucosa.
Glucosamina
N-Acetil-β-D-glucosamina
N-Acetilglucosamina 6-sulfato

27
OXIDACIÓN Y REDUCCIÓN DE CARBOHIDRATOS
El grupo aldehído o el carbono 6 de las hexosas a veces se oxida a ácido
carboxílico. Por ejemplo, la glucosa puede oxidarse en el carbono 1 para formar ácido
glucónico, o en el carbono 6 para formar ácido glucurónico.
Los azúcares que se pueden oxidar en el carbono 1 se denominan azúcares
reductores.
Si el grupo aldehído de un azúcar se reduce, el azúcar se convierte en un
polialcohol (poliol). Por ejemplo, la glucosa puede reducirse a sorbitol y la galactosa a
galactitol.
DISACÁRIDOS-OLIGOSACÁRIDOS Y POLISACÁRIDOS
Los monosacáridos pueden unirse mediante reacciones donde se remueve H2O
(deshidratación) formándose enlaces glicosídicos, entre un átomo de carbono de un

28
monosacárido y el átomo de carbono de otro monosacárido. Cuando el grupo hidroxilo del
carbono anomérico de un monosacárido reacciona con un -OH o un -NH de otro
compuesto se forman glicósidos. El enlace puede ser alfa o beta dependiendo de la
posición del átomo unido al carbono anomérico. Los monosacáridos se unen por enlaces
O-glicosídicos para formar disacáridos, oligosacáridos y polisacáridos.
Si sólo se unen unos pocos monosacáridos, el polímero resultante es un
oligosacárido. Si se une un gran número (cientos o miles) el polímero resultante es una
macromolécula denominada polisacárido. Dos polisacáridos comunes son el glucógeno y
el almidón, formas de almacenamiento de los glúcidos en las células animales y vegetales
respectivamente. Ambos están compuestos enteramente de moléculas de glucosa unidas
entre el C 1 de una glucosa y el C 4 de otra. El glucógeno y la amilopectina (una forma del
almidón) también tienen uniones donde el C 1 de una glucosa está unido al C 6 de una
segunda glucosa. Estos enlaces producen la formación de ramas que están presentes tanto
en el glucógeno como en la amilopectina pero no en la amilosa.
Sacarosa (Glucosa-α(1→2)-fructosa)
Maltosa (Glucosa-α(1→4)-glucosa)
Lactosa (Galactosa-β(1→4)-glucosa)

29
GLUCOGENO
enlace α1→4
La celulosa, por otra parte, tiene una función bien diferente como el principal componente
estructural de la pared de las células vegetales. Se compone enteramente de moléculas de
glucosa unidas por enlaces de tipo ββββ y es un polisacárido no ramificado. Este tipo de
uniones hace que forme cadenas muy extendidas que constituyen fibras de gran resistencia
mecánica.
Otro tipo de enlaces con azúcares
Se encuentran enlaces N-glicosídicos en los nucleótidos. Por ej. en la adenosina del
ATP, la adenina está unida por el N al C1 de la ribosa, a través de un enlace N-glicosídico.
Adenina
Enlace N-glicosídico
Ribosa
enlace α 1→6

30
ESTRUCTURA DE LÍPIDOS
Clasificación y estructura
Se incluye dentro de este grupo a compuestos químicos de naturaleza diversa que
tienen una característica común: su insolubilidad en agua y solventes polares y su
solubilidad en solventes no polares como cloroformo, benceno, etc. Están formados por
carbono e hidrógeno y en menor medida por oxígeno. También pueden contener fósforo,
nitrógeno y azufre. Los lípidos son mayormente una forma de almacenamiento de
combustible metabólico, también forman parte de la estructura de las membranas, otros
lípidos particulares son hormonas, vitaminas, etc.
Por tratarse de un grupo de compuestos muy heterogéneo es difícil su clasificación.
Un criterio posible considera a los lípidos según contengan ácidos grasos en su estructura y
que mediante una reacción química (saponificación) pueden liberarlos. Según este criterio
se clasifica a los lípidos en saponificables y no saponificables.
Dentro del grupo de los lípidos saponificables se encuentran:
• Lípidos simples: Son aquellos lípidos que sólo contienen carbono, hidrógeno y
oxígeno. Estos lípidos simples se subdividen a su vez en:
1. Acilglicéridos o grasas: Cuando los acilglicéridos son sólidos se les llama grasas y
cuando son líquidos a temperatura ambiente se llaman aceites.
2. Céridos o ceras.
• Lípidos complejos: Son los lípidos que además de contener en su molécula
carbono, hidrógeno y oxígeno, también contienen otros elementos como nitrógeno,
fósforo, azufre u otra biomolécula como un glúcido. Los lípidos que forman parte de
las biomembranas son los lípidos complejos, razón por la cual también se les llama
lípidos de membrana. Los lípidos complejos comprende diferentes subgrupos de
compuestos
Los lípidos insaponificables son aquellos que NO poseen ácidos grasos en su estructura y no
producen reacciones de saponificación. Entre los lípidos insaponificables encontramos a: • Terpenos. • Esteroides. • Prostaglandinas.
31
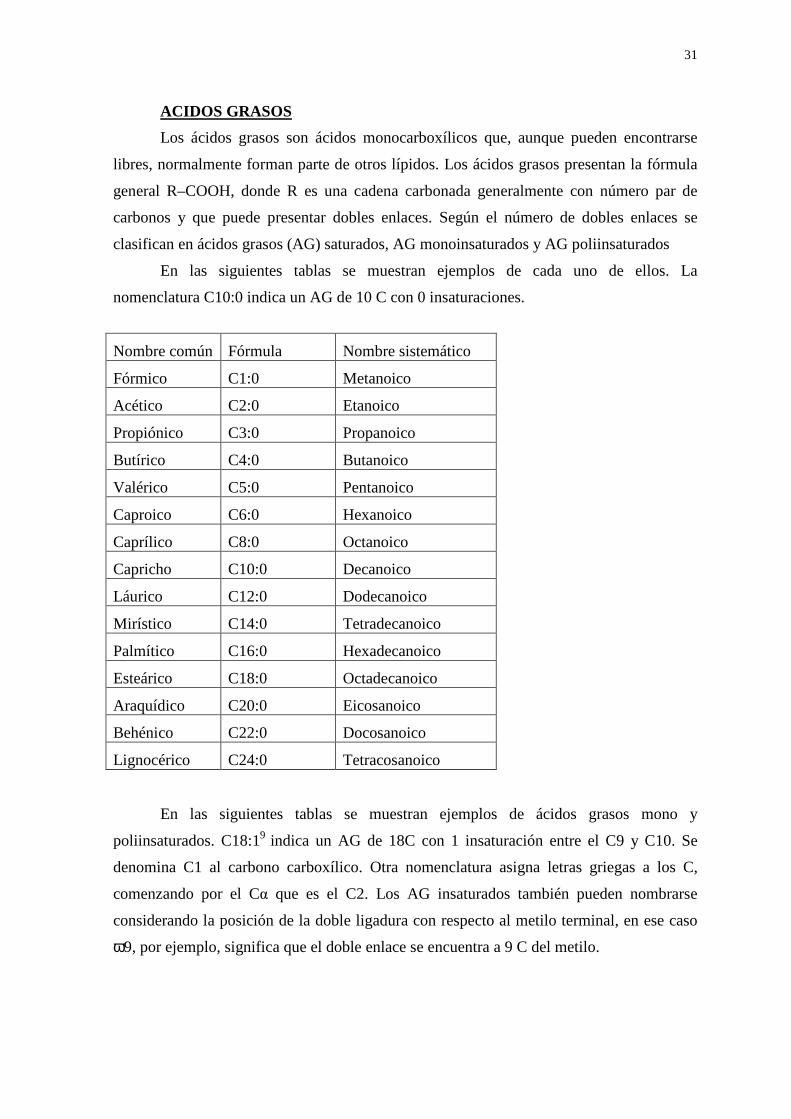
ACIDOS GRASOS
Los ácidos grasos son ácidos monocarboxílicos que, aunque pueden encontrarse
libres, normalmente forman parte de otros lípidos. Los ácidos grasos presentan la fórmula
general R–COOH, donde R es una cadena carbonada generalmente con número par de
carbonos y que puede presentar dobles enlaces. Según el número de dobles enlaces se
clasifican en ácidos grasos (AG) saturados, AG monoinsaturados y AG poliinsaturados
En las siguientes tablas se muestran ejemplos de cada uno de ellos. La
nomenclatura C10:0 indica un AG de 10 C con 0 insaturaciones.
Nombre común Fórmula Nombre sistemático
Fórmico C1:0 Metanoico
Acético C2:0 Etanoico
Propiónico C3:0 Propanoico
Butírico C4:0 Butanoico
Valérico C5:0 Pentanoico
Caproico C6:0 Hexanoico
Caprílico C8:0 Octanoico
Capricho C10:0 Decanoico
Láurico C12:0 Dodecanoico
Mirístico C14:0 Tetradecanoico
Palmítico C16:0 Hexadecanoico
Esteárico C18:0 Octadecanoico
Araquídico C20:0 Eicosanoico
Behénico C22:0 Docosanoico
Lignocérico C24:0 Tetracosanoico
En las siguientes tablas se muestran ejemplos de ácidos grasos mono y
poliinsaturados. C18:19 indica un AG de 18C con 1 insaturación entre el C9 y C10. Se
denomina C1 al carbono carboxílico. Otra nomenclatura asigna letras griegas a los C,
comenzando por el Cα que es el C2. Los AG insaturados también pueden nombrarse
considerando la posición de la doble ligadura con respecto al metilo terminal, en ese caso
ω9, por ejemplo, significa que el doble enlace se encuentra a 9 C del metilo.

32
Palmitoleico C16:19;ω7 cis-9-hexadecanoico
Oleico C18:19; ω9 cis-9-octadecanoico
Elaídico C18:19; ω9 trans-9-octadecanoico
Erúcico C22:113; ω9 cis-13-dococenoico
Nervónico C24:113; ω9 cis-15-tetracosenoico
Linoleico C18:29,12; ω6 todos cis-9,12-octadecadienoico
γ-Linolénico C18:36,9,12; ω6 todos cis-6,9,12-octadecatrienoico
α-Linolénico C18:39,12,15; ω3 Todos cis-9,12,15-octadecatrienoico
Araquidónico C20:45,8,11,14; ω6 todos cis-5,8,11,14-eicosatetraenoico
Timnodónico C20:55,8,11,14,17; ω3 todos cis-5,8,1,14,17-eicosapentenoico
Clupanodónico C22:57,10,13,16,19; ω3 todos cis-7,10,13,16,19-docosapentenoico
Cervónico C22:64,7,10,13,16,19; ω3 todos cis-4,7,10,13,16,19-docosahexenoico
Propiedades de los ácidos grasos
Los ácidos grasos poseen una zona hidrófilica, el grupo carboxilo (-COOH) y una
zona lipófilica, la cadena hidrocarbonada que presenta grupos metileno (-CH2-) y grupos
metilo (-CH3) terminales. Por eso las moléculas de los ácidos grasos son anfipáticas, pues
por una parte, la cadena alifática es no polar y por tanto, soluble en disolventes orgánicos
(lipófilica), y por otra, el grupo carboxilo es polar y soluble en agua (hidrófilico).
Desde el punto de vista químico, los ácidos grasos son capaces de formar enlaces
éster con los grupos alcohol de otras moléculas. Cuando estos enlaces se hidrolizan con un
álcali, se rompen y se obtienen las sales de los ácidos grasos correspondientes,
denominados jabones, mediante el proceso denominado saponificación.

33
ACILGLICEROLES O ACILGLICÉRIDOS O GRASAS NEUTRAS
Los acilgliceroles son ésteres de glicerol con ácidos grasos. Como el glicerol puede
fomar tres enlaces éster, los acilglicéridos pueden presentar uno, dos o tres ácidos grasos
en su estructura, por lo que hablamos de mono, di o triacilglicéridos (o triglicéridos).
CERAS
Las ceras son ésteres de los ácidos grasos con alcoholes primarios de cadena larga y
responden a la formula general: O || R-O-C-R’
En general son sólidas y totalmente insolubles en agua. Su función está relacionada
con su impermeabilidad al agua y con su firme consistencia. Así las plumas, el pelo, la
piel, las hojas, frutos, están cubiertas de una capa cérea protectora. Una de las ceras más
conocidas es la que segregan las abejas para confeccionar su panal.
LÍPIDOS COMPLEJOS
Los lípidos compuestos son aquellos cuya molécula presenta dos o más
componentes claramente diferenciados de los cuales uno de ellos una vez separado
presenta propiedades de lípido. Dentro de este grupo están los fosfolípidos y los
glucolípidos.
Fosfolípidos
Este grupo de lípidos se caracteriza por presentar en su estructura un grupo fosfato
esterificado con un alcohol. El alcohol puede ser glicerol, que es precursor de los
fosfoglicéridos o esfingol (esfingomielinas). Los fosfolípidos de glicerol o
fosfoacilgliceroles son lípidos de membrana que presentan en su estructura glicerol que se
encuentra esterificado con ácido fosfórico y ácidos grasos.

34
sn-glicerol-3-fosfato Ácido fosfatídico (1,2-diacil-sn-glicerol-3-fosfato).
El grupo fosfato a su vez puede estar esterificado con un alcohol sencillo, de
manera que el grupo fosfato forma un puente con dos uniones ester entre dos alcoholes:
puente fosfodiester. Dependiendo de la naturaleza del alcohol sencillo que aporta el
residuo A que se une al grupo fosfato (ver esquema siguiente) tenemos diferentes familias
de fosfolípidos.

35
Los esfingolípidos derivan del esfingol. El esfingol es un alcohol de 18 C con un
grupo -OH en el C1, una doble ligadura entre C4 y C5 y un grupo amino en el C2 (adenás
tiene otro OH- en el C3). Los ácidos grasos se unen al esfingol y forman las ceramidas. En
las ceramidas el ácido graso se une al grupo amino del esfingol mediane una unión
AMIDA.
Esfingol (o Esfingosina) (Ceramida)
En las ceramidas, el grupo hidroxilo del C1 proveniente del esfingol se puede
unir a otros compuestos. Puede unirse a un grupo fosfato y a través de este se puede unir a
FAMILIA GRUPO A
FOSFATIDILCOLINAS
(LECITINAS) COLINA
FOSFATIDILETANOLAMINAS
(CEFALINAS) ETANOLAMINA
FOSFATIDILSERINAS
SERINA
FOPSFATIDILINOSITOL
MIOINOSITOL
FOSFATIDILGLICEROLES
GLICEROL
CARDIOLIPINAS FOSFATIDILGLICEROL

36
la colina, formando la esfingomielina. Este compuesto es un importante componente de las
membranas celulares
Estructura general de las esfingomielinas
En la estructura de los esfingolípidos se distinguen dos colas hidrofóbicas: una la aporta el
esfingol (desde C3 a C18) y la otra cola la aporta el ácido graso (el ácido graso y el
esfingol se unen para formar la ceramida mediante una union amida). En las
esfingomielinas el grupo fosfato y la colina forman la cabeza polar de la molécula. Por lo
tanto la esfingomielina es una molécula anfipática con 2 colas hidrofóbicas. Cuando la
molécula anfipática tiene dos colas hidrofóbicas, esa molécula en un medio acuoso se
ordena formando bicapas. Es por eso que los esfingolípidos son constituyentes
mayoritarios de las membranas, por ejemplo de las membranas de neuronas.
Glucolípidos
Son lípidos complejos que contienen monosacáridos o sus derivados (glucosa,
galactosa, Nacetilglucosamina, etc). El monosacárido se une al C1 de la ceramida. Por lo
tanto son moléculas anfipáticas, con un cabeza polar que la aporta el o los glúcidos que se
unen al C1 de la ceramida, y 2 colas hidrofóbicas. Esto explica porqué son parte de las
bicapas lipídicas de las membranas de todas las células.
En los glucolípidos, un glúcido se une al C1 de la ceramida. Los más sencillos
contienen una única molécula de azúcar, aunque también pueden presentar cadenas de 2 a
10 residuos de azúcares. Cuando los glucolípidos contienen en su estructura azúcares
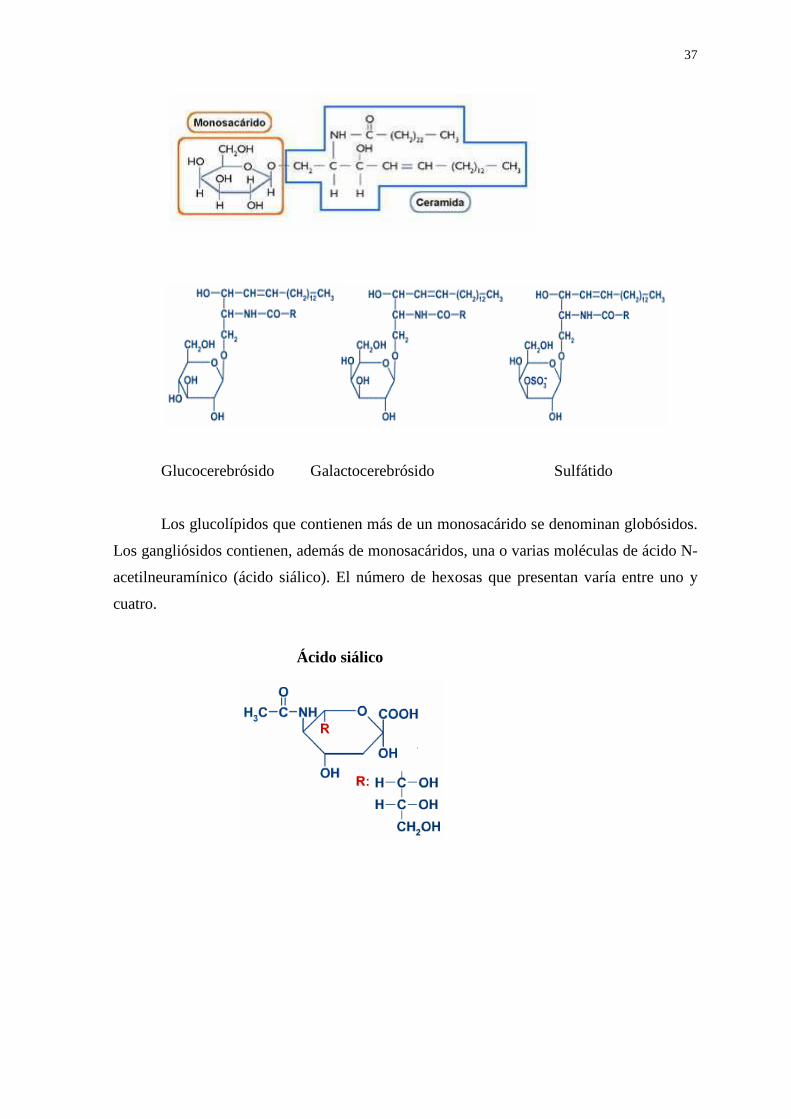
neutros como glucosa o galactosa (sólo una molécula), se los denomina CEREBRÓSIDOS
(glucocerebrósido o galactocerebrósido). Los sulfátidos son los ésteres sulfúricos de los
cerebrósidos (el sulfato esterifica el C3 de la hexosa).
37
Glucocerebrósido Galactocerebrósido Sulfátido
Los glucolípidos que contienen más de un monosacárido se denominan globósidos.
Los gangliósidos contienen, además de monosacáridos, una o varias moléculas de ácido N-
acetilneuramínico (ácido siálico). El número de hexosas que presentan varía entre uno y
cuatro.
Ácido siálico

38
Estructura de un gangliósido
LÍPIDOS ISOPRENOIDES
Este grupo de lípidos se caracteriza por ser derivados del isopreno.
A partir de la condensación de varias unidades de isopreno activo (isopreno
fosforilado) se sintetizan los diferentes lípidos isoprenoides. Cada dos unidades de
isopreno dan lugar a un terpeno, de manera que hay monoterpenos, sesquiterpenos,
diterpenos, etc. según contengan, respectivamente, dos, tres, cuatro, etc. isoprenos. Hay
terpenos acíclicos, ramificados y cíclicos que pueden contener otros grupos funcionales
(cetona, alcohol). Entre los monoterpenos figuran diversos compuestos volátiles con
aromas característicos, como el limoneno del limón y el alcanfor.
Limoneno Vitamina A o retinol
El retinol o vitamina A1 y el deshidro-3-retinol o vitamina A2 son diterpenos
parcialmente ciclados, mientras el fitol es un diterpeno lineal. La vitamina A tiene un
grupo –OH en el extremo de la cadena hidrocarbonada. Ese hidroxilo se puede esterificar
con un ácido graso.
Son también terpenoides la vitamina E o α-tocoferol y los derivados quinónicos,
como las ubiquinonas, la plastoquinona y las vitaminas K.
Ceramida
Ácido siálico
Galactosa
Glucosa
N-Acetil Galactosamina
Ceramida
Ácido siálico
Galactosa
Glucosa
N-Acetil Galactosamina

39
Vitamina E o α-tocoferol
Los carotenoides son derivados poliisoprénicos de 40 átomos de carbono
(tetraterpenos). Por ejemplo, el β-caroteno:
El β-caroteno es un pigmento presente en las frutas y hortalizas como las zanahorias y el
tomate. Cuando el β-caroteno se cliva genera la Vitamina A. (compare las estructuras de
la vitamina A y del β-caroteno
ESTEROIDES
Los esteroides pueden considerarse también lípidos isoprenoides, puesto que en
último término provienen del isopentenilpirofosfato. Su estructura se relaciona con la del
anillo esterano o ciclopentano-perhidrofenantreno.
Los esteroles presentan un grupo hidroxilo en el C-3 y diferentes sustituyentes en el
C-17. En la figura pueden verse diversos esteroles.

40
Colesterol Estradiol Progesterona
Testosterona Aldosterona Cortisol
El colesterol (o 3-hidroxi-5,6-colesteno) es un esterol de 27 átomos de carbono,
cuyo grupo hidroxilo adopta la configuración β. El colesterol puede ser esterificado por
ácidos grasos, donde el OH- del C3 se une con el grupo carboxilo del ácido graso.
El colecalciferol o vitamina D3, los ácidos biliares y sus sales, y las diversas
hormonas esteroideas (corticoides, andrógenos y estrógenos) son derivados del
colesterol.
Vitamina D3 o colecalciferol
PROSTAGLANDINAS
Las prostaglandinas son lípidos cuya molécula básica está constituida por 20
átomos de carbono que forman un anillo ciclopentano y dos cadenas alifáticas.

41
La figura muestra la estructura de la prostaglandina A1 y A2. El número 1 indica
que en las cadenas laterales hay sólo 1 doble ligadura. En la PGA2, hay 2 dobles ligaduras
en las cadenas laterales.
Al igual que los leucotrienos, las prostaglandinas derivan del ácido araquidónico.
Ambos tipo de compuestos tienen diversas funciones.
(Este tema será desarrollado con más detalle en clases proximas.)
COMPORTAMIENTO DE LOS LÍPIDOS ANFIPÁTICOS EN AGUA
En un entorno acuoso, las cabezas polares de los lípidos anfipáticos tienen a
orientarse hacia su entorno polar, mientras que las colas hidrofóbicas o no polares tienden
a minimizar el contacto con el agua. Las colas no polares de los lípidos, al no tener
afinidad por el agua, son excluidas del entorno acuoso. En estas condiciones las colas
hidrofóbicas quedan muy juntas favoreciendo las interacciones por fuerzas de Vand der
Waals entre moléculas. Por lo tanto las moléculas anfipáticas en medio acuoso se ordenan
formando una bicapa lipídica (si es una molécula con dos colas hidrofóbicas), o una
micela (si es una estructura con sólo una cola hidrofóbica). Las micelas son esferas de una
sola capa y solamente pueden llegar hasta cierto tamaño, mientras que las bicapas pueden
ser considerablemente más largas. También pueden formar túbulos (pequeños tubos).
Las micelas y las bicapas se separan del ambiente polar mediante un proceso
conocido como “efecto hidrofóbico”. Cuando se disuelve una sustancia no polar en un
micela
bicapa

42
entorno polar, las moléculas polares (i.e. agua en una solución acuosa) se acomodan de
manera más ordenada alrededor de la sustancia no polar disuelta debido a que las
moléculas polares no pueden formar puentes de hidrógeno con las moléculas no polares. Es
por esto que, en un entorno acuoso, las moléculas polares del agua forman una caja
ordenada de “clatrato” alrededor de la molécula no polar disuelta. De cualquier manera,
cuando la molécula no polar se separa del líquido polar, la entropía (el estado de desorden)
de la molécula polar en el líquido se incrementa. Esto es esencialmente una forma de fase
de separación, similar a la separación espontánea que ocurre cuando se ponen juntos agua
y aceite.

43
ESTRUCTURA DE AMINOÁCIDOS
Introducción
Las proteínas son las macromoléculas más abundantes en las células y constituyen
alrededor del 50% de su peso seco. Dentro de las células exhiben una enorme diversidad de
funciones biológicas: como constituyentes de las membranas biológicas (glicoproteínas de
identificación, transportadores, canales, etc.), como catalizadores de reacciones químicas
(enzimas), interactuando con los ácidos nucleicos (histonas, ribosomas), interactuando con
hormonas y neurotransmisores (receptores), etc.
Para entender los aspectos estructurales y las características químicas de las
proteínas, es fundamental primero analizar los componentes bioquímicos que las
constituyen, los aminoácidos.
Los aminoácidos son moléculas relativamente pequeñas compuestas por carbono,
oxígeno, hidrógeno y nitrógeno. La hidrólisis ácida o básica (es decir la ruptura en
unidades constitutivas por acción de un ácido o una base) de las proteínas y/o péptidos mas
comunes origina unos veinte aminoácidos que presentan una característica común dada por
la presencia de un grupo carboxilo (-COOH) y un grupo amino (-NH2) ligados a un
mismo carbono, que es llamado carbono alfa (Cαααα). Este carbono presenta generalmente
sus otros dos enlaces compartidos con un hidrógeno y un grupo (serie de átomos unidos)
denominado grupo radical (-R). La diferencia entre aminoácidos, está dada entonces por
las características del grupo radical unido a dicho Cα. Esto a su vez provoca que de
acuerdo a las características del grupo radical, varíe la estructura general, el tamaño, la
carga eléctrica y su solubilidad. Todos los aminoácidos pueden ser cristalizados y su
estructura estudiada mediante la difracción de rayos X.
Carbono alfa
Grupo radical
Los aminoácidos que constituyen normalmente las proteínas comunes son
denominados aminoácidos primarios o comunes. Estos se diferencian de todos los otros
aminoácidos que están en los organismos vivos y que forman parte de proteínas especiales,

44
como componentes de éstas, o bien se encuentran en forma libre participando de vías
metabólicas particulares. De entre los veinte aminoácidos comunes existe uno, la prolina
en el que el carbono alfa y el nitrógeno forman parte de un anillo del grupo radical, es decir
no se observa un grupo amino sino que en este caso en particular se habla de un grupo
imino, por lo que se suele denominar a la prolina un iminoácido.
Cada organismo vivo sintetiza sus propias proteínas a partir de los aminoácidos.
Las plantas superiores sintetizan a su vez todos los aminoácidos necesarios. Hay que
destacar que los animales carecemos de esa capacidad. Cada especie animal puede
sintetizar sólo algunos aminoácidos que necesita y, por lo tanto, depende de la dieta para
incorporar aquellos aminoácidos que debe sintetizar para formar proteínas. Esos
aminoácidos se los considera esenciales y no porque sean los únicos necesarios para la
vida de la especie, sino porque deben estar incluidos en la dieta. Cada especie, tiene su
grupo de aminoácidos esenciales propios.
La mayoría de los aminoácidos que ingerimos se encuentran en forma de proteínas,
sin embargo sólo los aminoácidos pueden incorporarse a las diferentes rutas metabólicas.
Para ello, las proteínas y péptidos ingeridos sufren un proceso de hidrólisis por medio de
enzimas proteolíticas (secretadas por el estómago, páncreas e intestino delgado) en el
tracto gastrointestinal. Después de la acción de las enzimas los aminoácidos quedan libres,
son absorbidos por el intestino y transportados a la corriente sanguínea, por medio de la
cual llegan al hígado donde ocurre su metabolismo y distribución. Las proteínas endógenas
también se degradan después de un tiempo, adquieren señales que indican a las enzimas de
degradación cuando deben comenzar su proceso.
Como se mencionó anteriormente, a los aminoácidos comunes se los suele
clasificar en una primera aproximación en aminoácidos esenciales y no esenciales, de
acuerdo a la capacidad de ser o no sintetizados, por la célula, tejido u organismo en
consideración. En general se le asigna una abreviatura de tres letras a cada aminoácido
primario o bien una sola letra (en mayúscula), la cual se emplea cuando se desean expresar
a los aminoácidos constituyentes en una cadena peptídica o proteica.
Carbono alfa
Prolina
Carbono alfa
Prolina

45
Los aminoácidos comunes se pueden clasificar en:
Esenciales (deben ser suministrados por la dieta)
Arginina Arg R Histidina His H Leucina Leu L Isoleucina Ile I Lisina Lys K Metionina Met M Fenilalanina Phe F Treonina Thr T Triptofano Trp W Valina Val V
No esenciales (podemos sintetizarlos)
Alanina Ala A Asparagina Asn N Ac. Aspártico Asp D Cisteína Cys C Glutamina Gln Q Ac. Glutámico Glu E Glicina Gly G Prolina Pro P Serina Ser S Tirosina Tyr Y
Dentro de esta clasificación de aminoácidos esenciales, cabe destacar que en el
caso de la arginina y la histidina son esenciales durante el crecimiento en el niño, pero no
para el adulto.
En los períodos en que el organismo atraviesa crisis funcionales (desnutrición
aguda o crónica, traumatismos o trastornos articulares y/o musculares, alteraciones en el
tracto gastrointestinal, hepatitis, afecciones renales, deficiencias cerebrales o nerviosas,
etc.) o demandas extra por razones mecánicas (atletas, etc.) o cerebrales (estrés, exámenes,
etc.) se produce aumento en el consumo de aminoácidos, por lo que muchas veces
conviene completar la dieta habitual por medio de la administración exógena de
aminoácidos. Por las mismas razones, la ingesta de alimentos que contienen aminoácidos
significa además un extraordinario recurso preventivo, por cuanto incorpora al organismo
mecanismos de fortalecimiento ante previsibles compromisos extra, sean éstos derivados
de circunstancias especiales o de inevitables decadencias de las funciones orgánicas
derivadas de la edad.

46
Como ya hemos comentado de los veinte aminoácidos proteicos conocidos, ocho
son esenciales para la vida humana y dos semiesenciales. Estos diez aminoácidos son los
que requieren ser incorporados al organismo con la alimentación, especialmente, cuando se
da una situación de disfunción o enfermedad.
El organismo no almacena el exceso de aminoácidos que provienen de la dieta, los
transforma en intermediarios metabólicos comunes como el piruvato, oxalacetato y α-
cetoglutarato, es decir, que los aminoácidos van a ser precursores de la glucosa, ácidos
grasos y cuerpos cetónicos, por lo tanto, actúan como combustible y precursores
metabólicos.
Estereoquímica de los Aminoácidos
Por sus características estructurales los aminoácidos son compuestos ópticamente
activos. Si se consideran los veinte aminoácidos comunes, a excepción de la glicina, todos
los demás presentan al menos un átomo de carbono asimétrico, el Carbono alfa, al cual se
encuentran unidos el grupo carboxilo, el grupo amino, un hidrógeno y un grupo radical. Al
ser estos cuatro sustituyentes diferentes, genera como ya hemos visto para los glúcidos,
que exista para cada aminoácido por lo menos dos formas isómeras, las cuales son
idénticas en todas sus propiedades químicas y físicas a excepción de la dirección a la que
pueden rotar el plano de un haz de luz polarizada. Entre ambos isómeros hay que distinguir
entre los que rotan el plano hacia la izquierda, levorrotatorios, levógiros [-], y los que lo
hacen hacia la derecha, dextrorrotatorios, dextrógiros [+]. Estos isómeros son llamados
isómeros ópticos, enantiómeros o estereoisómeros.
Como en el caso de los hidratos de carbono, existe una segunda denominación en
estos isómeros ópticos, que está basada en la configuración de los cuatro sustituyentes del
carbono asimétrico, esta convención es la más empleada en la literatura y utiliza como
compuesto de referencia al gliceraldehído. Al igual que en los hidratos de carbono existen
entonces L-aminoácidos y D-aminoácidos.
Los isómeros ópticos de los aminoácidos que presenten en su carbono asimétrico
una configuración similar a la del L-gliceraldehido son designados como L-aminoácidos
y los que presenten una configuración similar a la del D-gliceraldehido, son designados D-
aminoácidos, independientemente de la dirección a la que roten el plano de un haz de luz
polarizada. En la naturaleza encontramos una mezcla de ambos que se denomina racémica,
pero los aminoácidos que forman las proteínas son L-estereoisómeros, a excepción de
la glicina, que no posee átomos de carbono asimétricos.

47
D-Gliceraldehido
C O
H
OHC
H C2 OH
H
H
HC
H C2 OH
HO
L-Gliceraldehido
C O
COO-
C
R
H NH3+
D- -Aminoácidoαααα
COO -
H N3
+H
R
C
L- -Aminoácidoαααα
Asimismo al igual que en los azúcares, cuando un aminoácido posea más de un átomo de
carbono asimétrico habrá 2n posibles estereoisómeros. La mayoría de los aminoácidos
comunes, posee un solo carbono asimétrico, las excepciones son treonina e isoleucina que
poseen 2 átomos de carbono asimétricos; esto origina 4 posibles estereoisómeros los cuales
son llamados diasteroisómeros; sin embargo solo uno de cada uno de ellos se encuentra
formando parte de las proteínas.

48
Clasificación de los Aminoácidos
Además de las características generales de los aminoácidos, existen diferentes
grupos radicales unidos al carbono alfa, que hacen que un aminoácido presente por
ejemplo otro grupo ácido más, lo que da lugar a los denominados aminoácidos ácidos, que
presente algún grupo básico, produciendo los denominados aminoácidos básicos, o que
incluso incorpore en su estructura otros elementos, como el azufre (S), que se denominan
aminoácidos azufrados. Por otro lado puede suceder que en lugar de tener una estructura
molecular lineal tenga una configuración cíclica, aromática, y los denominados
aminoácidos de cadena simple o ramificada. Finalmente pueden encontrarse aminoácidos
en que la posición del grupo amino no se encuentre en el carbono alfa sino en otro más
alejado del grupo funcional carboxilo, generando los llamados beta y gama aminoácidos.
Una forma de realizar la clasificación es la de agrupar a los aminoácidos según las
características físico-químicas que les confieren los grupos radicales (R) que se
encuentran ligados al Cα. De acuerdo a las propiedades de éstos grupos R y en particular
de la polaridad que éstos presentan a un pH fisiológico, los aminoácidos se pueden
clasificar en:
1.- Aminoácidos no polares o hidrofóbicos
2.- Aminoácidos polares pero sin carga neta efectiva
3.- Aminoácidos negativamente cargados, o aminoácidos ácidos, y
4.- Aminoácidos positivamente cargados o aminoácidos básicos
Aminoácidos no polares
Dentro de los ocho aminoácidos con grupos no polares y con características
hidrofóbicas, se incluyen cinco aminoácidos cuyos grupos radicales son cadenas alifáticas
hidrocarbonadas: L-alanina (Ala) ; L-valina (Val) ; L-leucina (Leu) ; L-isoleucina (Ile); L-
prolina (Pro) ;

49
COO -
C H
H N+
2 CH 2
CH 2H C2CH 2
COO -
H N3
+C H
CH CH 3
CH 3
COO -
H N3
+C H
CH 2
CH
H C3 CH 3
COO-
H N3
+C H
CH
H C3 CH 3
COO -
CH 3
H N3+
C H
L-Alanina L-Valina L-Leucina L-Isoleucina L-Prolina
dos aminoácidos cuyos grupos radicales poseen anillos aromáticos: L-fenilalanina (Phe) y
L-triptofano (Trp)
COO -
H N3+
C H
CH 2
COO -
H N3
+C H
CH 2
C CH
NH
L-Fenilalanina L-Triptofano
y uno el cual contiene azufre en su grupo radical en forma de éter de azufre: L-metionina
(Met)
COO -
H N3+
C H
CH 2
CH 2
CH 3
S
L-Metionina
Aminoácidos polares
Los aminoácidos polares pero sin carga neta efectiva, son aminoácidos mas
hidrofílicos que los anteriores y poseen grupos funcionales capaces de formar uniones
puente de hidrógeno con el agua (por lo tanto más solubles). Esta categoría incluye siete
aminoácidos: L-serina (Ser) ; L-treonina (Thr) ; L-tirosina (Tyr) ; L-asparagina (Apn) ; L-
glutamina (Gln) L-cisteína (Cys) y glicina (Gly).

50
COO -
H N3+
C H
CH OH2
COO -
H N3+
C H
CH OH
CH 3
COO -
H N3+
C H
CH 2
OH
L-Serina L-Treonina L-Tirosina
La polaridad de los tres primeros (Ser, Thr, Tyr) está dada por sus grupos hidroxilo
presentes en el R, la de los dos siguientes (Asn, Gln) por sus grupos amida; en el caso de
la Cys la polaridad de este aminoácido está dada por el grupo tiol o sulfhidrilo.
NH2
COO-
H N3+
C H
CH 2
C O
COO -
H N3+
C H
CH 2
NH2
C O
CH 2
COO-
H N3+
C H
CH 2
SH
COO -
H N3
+C H
H
L-Asparagina L-Glutamina L-Cisteina Glicina
En el caso de la cisteína en particular es importante mencionar que este aminoácido
puede presentarse en las proteínas en forma dimérica unido covalentemente a través de un
puente disulfuro. Al aminoácido formado se lo suele denominar cistina, y se conoce
que es importante para las características estructurales de ciertas proteínas como la
insulina y anticuerpos. En éstas proteínas la cistina sirve de nexo entre dos cadenas
polipeptídicas formando lo que se denomina un enlace cruzado (cross-linking) a través de
la unión disulfuro.

51
C O O H
H N2 C H
H C2 S H H S
C O O H
H N2 C H
C H 2
C O O H
H N2 C H
SH C2
C O O H
H N2 C H
C H 2S
C y s C y s
C is t in a
F or m a ci ón d e C i st i n a
L os g ru p o s tio l d e d o s m o lé c u la s deL -C is te in a so n r ap ida m e n te o x id a d o sp a ra d a r un en lac e d isu lfu ro
En cuanto a la glicina el grupo R (que es un hidrógeno) es demasiado pequeño
para alterar la polaridad de los grupos carboxilo y amino del aminoácido.
Aminoácidos con carga negativa
Los dos aminoácidos comunes cuyos grupos R poseen una carga neta negativa
a pH 7 (aminoácidos ácidos) son el ácido L-aspártico (Asp) y el ácido L-glutámico (Glu).
COO-
H N3
+C H
CH 2
COO -
COO -
H N3
+C H
CH 2
CH 2
COO -
Acido L- Aspártico Acido L-Glutámico
Ambos son derivados de L-asparagina (Asn) y L-glutamina (Gln) por hidrólisis de
sus grupos amino ubicados en el carbono gamma y delta respectivamente.
Aminoácidos con carga positiva
Los aminoácidos en los cuales la carga neta aportada por sus grupo R a pH 7 es
positiva (aminoácidos básicos), son: L-lisina (Lys) ; L-arginina (Arg) y L-histidina (His)

52
COO-
H N3
+C H
CH 2
NHC
CH
NH+H
C
COO-
H N3+
C H
CH 2
CH 2
CH 2
NH2
C NH 2+
NH
COO-
H N3
+C H
CH 2
CH 2
NH3+
CH 2
CH 2
L-Lisina L-Arginina L-Histidina
La lisina, posee un segundo grupo amino en el carbono epsilon (ε) de su cadena alifática;
la arginina presenta un grupo guanidino positivamente cargado mientras que la histidina
tiene en su estructura un grupo imidazol ionizado.
Otros Aminoácidos de importancia
Además de los 20 aminoácidos comunes hallados en las proteínas existen otros
aminoácidos menos frecuentes que son componentes de ciertas proteínas
COOH
C H
HN CH 2
CHH C2
OH
4-Hidroxiprolina
2
1
3
45
Derivado de la L-ProlinaForma parte de las proteínasfibrosas del Colágeno
COOH
H N2 C H
CH 2
CH 2
NH 2
C
CH 2
5-Hidroxilisina
H OH
2
1
3
4
5
6
Derivado de la L-LisinaForma parte de las proteínasfibrosas del Colágeno
COOH
H N2 C H
CH 2
CH 2
NH
CH 2
2
1
3
4
5
6
CH 2
CH 3
6- N-Metilisina
Derivado de la L-LisinaForma parte de la Miosinaque interviene en lacontracción muscular

53
y que en general provienen de aminoácidos comunes modificados. Entre ellos podemos
nombrar: 4-hidroxi-L-prolina y 5-hidroxi-L-lisina que se encuentran en el colágeno
formando parte de las proteínas fibrosas del tejido conectivo. Por otro lado, la N-metil-L-
lisina que se encuentra en la miosina (proteína que actúa en los procesos de contracción
muscular), el ácido γ-carboxi-L-glutámico que forma parte de la proteína de coagulación
protrombina y de otras proteínas con capacidad de unir el ión calcio, y la desmosina un
derivado de lisina que se encuentra en la elastina, otra proteína de característica
fibrosa.
N
CHOOC
COOH
NH2
CH 2
CH 2
CH 2
C
CH 2
C
CH 2
CH 2
H N2 H
H C2H CH 2
H C2 C
NH2
H
COOH
H C2
H C2
NH2
COOHH
Desmosina
Derivado de la L-LisinaSe encuentra solamente comounidad constitutiva de laproteína Elastina
C H
CH 2
HC
COOH
COOH
COOH
-Carboxi-Glutámicoγγγγ
Derivado del Acidos L-GlutámicoForma parte de la Protrombina asicomo de otras proteínas que regulanla concentración de ion Ca++
H N2
Ciertos
aminoácidos no proteicos actúan como precursores o intermediarios en el metabolismo,
como: β-alanina que es el precursor del ácido pantoténico que a su vez es parte constitutiva
de la Coenzima A (CoA); y además ornitina y citrulina que intervienen en el ciclo de la
urea.
CH 2
CH 2
NH2
COOH
-Alaninaββββ
Forma parte de laPanteteína y de laCoenzima A
C H
COOH
H N2
CH 2
CH 2
CH 2
NH2
Ornitina
Intermediario delCiclo de la Urea
C H
COOH
H N2
CH 2
CH 2
CH 2
NH2
C O
NH
Citrulina
Intermediario delCiclo de la Urea

54
La homocisteína, es un aminoácido precursor de la cisteína Cys; y la taurina es un caso
particular de aminoácido no carboxílico el cual se combina con los ácidos y sales biliares;
CH 2
CH 2
NH2
SO H3
Taurina
Aminoácido libre dentro de lascélulas o combinado con losácidos biliares
C H
COOH
H N2
CH 2
CH 2
SH
Homoscisteína
Metabolito de la síntesisde L-Metionina
Existe otra serie de aminoácidos que intervienen en la síntesis de agentes químicos para la
transmisión de impulsos nerviosos. Este es el caso de la L-dopa y el 5-hidroxi-triptofano
conocido con el nombre de serotonina. Otros aminoácidos que actúan como
neurotransmisores son el ácido γ amino butírico o GABA, la glicina, el ácido glutámico y
el ácido aspártico.
CH 2
CH 2
NH2
CH 2
COOH
-AminobutíricoγγγγGABA
Neurotransmisor InhibitorioSe lo encuentra en tejido nervioso5-Hidrixi-Triptofano
C H
CH 2
C CH
NH
HO
H N2
COOH
5
Aminoácido precursor dela Serotonina (5HT)
C H
CH 2
COOH
H N2
HOOH
L-DOPA
Aminoácido precursor de laMelatonina y la Dopamina
Finalmente,
debemos también considerar aquellos aminoácidos relacionados con una acción hormonal
como es el caso de los aminoácidos asociados a la glándula tiroides:

55
C H
CH 2
COOH
H N2
HO
I 3
3-Yodotirosina
Se lo encuentra entejido tiroideo y ensuero
C H
CH 2
COOH
H N2
HO
I 3 5 I
3, 5 -Diyodotirosina
Se lo encuentra enasociación con laTiroglobulina
O
C H
CH 2
COOH
H N2
I I
I
OH
3, 5, 3'-Triyodotironina
Se lo encuentra entejido tiroideo
O
C H
CH 2
COOH
H N2
I I
I
OH
I
Tiroxina
Se lo encuentra enasociación con laTiroglobulina
Propiedades Ácido/Base de los Aminoácidos
Los aminoácidos se encuentran en las disoluciones acuosas formando iones
dipolares o iones híbridos. Esta característica hace que un aminoácido sencillo, mono-
amino y mono-carboxílico, como por ejemplo alanina, se comporte como un ácido dibásico
cuando se encuentra totalmente protonado a pH bajo. Si se realiza una valoración con
hidróxido de sodio de este aminoácido, se observará que la curva de valoración será
bifásica: en un comienzo tendremos la molécula con los dos protones (-COOH y -NH3+)
y a medida que aumenta la alcalinidad del medio, el aminoácido cederá primero el H+ del
grupo carboxilo, y más tarde a mayor alcalinidad cederá el H+ del grupo amino. Existe un
valor de pH en el cual la molécula del aminoácido no posee carga eléctrica neta (es decir
cuando la forma preponderante sea la del ión bipolar), a este pH se lo denomina punto
isoeléctrico. El punto isoeléctrico es una propiedad característica de la molécula
aminoacídica y depende de sus constantes de disociación ácido/ base.
En general el grupo carboxilo de los ácidos monoamino-monocarboxílicos es un
ácido mas fuerte que el grupo carboxilo de los ácidos alifáticos comparables (el grupo
carboxilo de Ala es mas ácido que el del ácido acético o el del ácido láctico), mientras que
el grupo amino es un ácido mas fuerte que el grupo amino de las aminas alifáticas
comparables. Ninguno de los aminoácidos monoamino-monocarboxílicos que poseen
grupos radicales sin carga neta efectiva, presentan capacidad amortiguadora significativa
en el rango de pH fisiológico, solo lo hacen en las cercanías de los valores de sus

56
constantes de disociación que son similares para todos ellos (zonas de pH 1,3-3 y pH 8,6-
10,6).
Valoración de la Glicina.
Ión dipolar
De todos los aminoácidos comunes solamente la histidina presenta una capacidad
amortiguadora significativa entre pH 6-8 (La hemoglobina, proteína transportadora de O2
y CO2 es rica en este aminoácido).
El grupo γ-carboxilo del ácido glutámico y δ-carboxilo del ácido aspártico poseen
valores de constantes de disociación más cercanos a los ácidos carboxílicos sencillos como
el ácido acético. El grupo sulfhidrilo de la cisteína y el grupo hidroxilo de la tirosina son
ácidos muy débiles. Los grupos ε-amino de la lisina y el grupo guanidino de la arginina,
son fuertemente básicos y pierden sus protones a valores de pH muy elevados.

57
Reacciones químicas de los aminoácidos
Los grupos carboxilo y amino de los aminoácidos presentan todas las reacciones
químicas que se pueden esperar de éstos, como ser la formación de sales, esterificación y
acetilación. Así también, presentan las reacciones químicas propias del metabolismo de
los aminoácidos: transaminación, deaminación y decarboxilación.
Existen reacciones colorimétricas específicas para los aminoácidos basadas
específicamente en los grupos funcionales R unidos al carbono alfa. Estas reacciones son
útiles tanto para la identificación cualitativa como para la valoración cuantitativa de los
mismos.
Además de lo mencionado en el párrafo anterior, existe una reacción específica
para visualizar tanto aminoácidos libres como proteínas, que se basa en la presencia de
NH3 en el medio de reacción. La ninhidrina es un poderoso agente oxidante que causa la
decarboxilación oxidativa de los α-aminoácidos produciendo CO2 , NH3 y un aldehído
con un átomo de carbono menos. La ninhidrina reducida reacciona luego con el NH3
liberado, produciendo un compuesto de color púrpura que absorbe luz visible permitiendo
de esa manera determinar la cantidad presente del aminoácido en solución. Otras aminas
diferentes a los aminoácidos reaccionan de forma similar con la ninhidrina, pero no
producen desprendimiento de CO2. En el caso de la L-prolina que es una imina, la
reacción que se produce con la ninhidrina genera una coloración amarilla.
Otra reacción de importancia en los aminoácidos es la que se produce entre el
grupo amino de los aminoácidos con un grupo carbonilo de los aldehídos formándose
lo que se denomina una base de Schiff. Esta es una reacción en la cual se produce una
condensación de la molécula del aldehído con el aminoácido formándose una unión
covalente entre el átomo de carbono del aldehído y el átomo de nitrógeno del aminoácido
(>C=N-) con pérdida de agua. El producto de esta reacción es de vida media corta y
vuelve a disociarse rápidamente al menos que exista una reacción química subsiguiente
que utilice a la base de Schiff como sustrato o bien que las concentraciones de los reactivos
sea elevada favoreciendo el sentido de la reacción hacia la formación de producto.

58
C O
H
R'
H2N-CH
COOH
R
R' C N C R
COOH
H H
H2O+ +
Aldehido(azúcar)
Amino(aminoácido o proteína) Base de Schiff
Cuando la glucemia se mantiene elevada por períodos prolongados, el aldehído de la
glucosa reacciona con grupos amino libres de las proteínas, formando bases de Schiff que
mediante una reacción química subsecuente producirá la glicosilación de la proteína. El
producto de esta última reacción química, a diferencia de la formación de la base de Schiff,
será permanente en la proteína hasta tanto se produzca el recambio de la misma. La
continua glicosilación de proteínas producida en la Diabetes Mellitus es el factor principal
de las alteraciones secundarias provocadas por esta patología.
La reacción química principal de los aminoácidos es aquella en la que se establecen
enlaces covalentes entre dos moléculas de aminoácidos. Esto se produce por la interacción
entre el grupo carboxilo unido al C alfa de un aminoácido con el grupo amino unido al
C alfa de otro aminoácido. La unión que se forma se denomina ENLACE PEPTÍDICO .
Este tipo de enlace covalente es de características rígidas, es decir las uniones entre los
átomos de C y N no pueden girar libremente. Esto hace que una serie de aminoácidos
unidos por estos tipos de enlaces presenten una estructura particular la cual va a depender
del tipo de aminoácidos que constituyan la cadena, como se verá en clases teóricas
posteriores.
Formación de enlaces peptídicos

59
En general todos los aminoácidos cuyos grupos radicales son de característica
aromática como Trp, Tyr, His y Phe absorben luz ultravioleta (UV). Dada la incidencia
de estos grupos aromáticos, la absorbancia de una solución de proteína está dentro del
espectro ultravioleta (289 nm). Todos los aminoácidos absorben luz en el UV lejano, lo
que se emplea como un método de determinación simple (<220 nm).

60
ESTRUCTURA DE NUCLEÓTIDOS
Una característica esencial de los seres vivos es su capacidad para reproducirse.
Para ello cada individuo debe contener una descripción completa de sí mismo, que además
ha de ser capaz de transmitir a sus descendientes para que ellos puedan construir otro
individuo con esas características. A nivel celular, una célula ha de disponer de esas
instrucciones para construir una réplica idéntica de sí misma. En una célula, esa
información se encuentra en el ácido desoxirribonucleico (ADN). El ADN tiene la
particularidad de que posee información también para hacer copias de sí mismo. Para que
la información contenida en el ADN se pueda expresar hace falta otra sustancia que es el
ácido ribonucleico (ARN).
Los ácidos nucleicos son polímeros de nucleótidos (polinucleótidos) y a su vez los
nucleótidos son moléculas constituidas por una base nitrogenada, un monosacárido de
cinco carbonos (pentosa) y de uno a tres fosfatos. Este tipo de estructuras participa en una
amplia variedad de procesos biológicos:
1. Constituyen las unidades estructurales de los ácidos nucleicos (ADN y ARN)
2. Forman parte de coenzimas de óxido reducción (FAD, NAD+, NADP+)
3. Forman parte de compuestos que actúan como segundos mensajeros en el mecanismo de
acción de determinadas hormonas y neurotransmisores (AMP cíclico, GMP cíclico).
4. Los nucleótidos de purina actúan en general como compuestos reservorios de energía
(ATP, GTP).
5. Los nucleótidos de pirimidina también son utilizados como intermediarios de alta
energía en procesos biosintéticos (UDP, CDP que forman UDP-glucosa y CDP-acilglicerol
intermediarios en la síntesis de glucógeno y lípidos respectivamente). ESTRUCTURA GENERAL DE LOS NUCLEÓTIDOS
Los nucleótidos son los ésteres fosfóricos de los nucleósidos. Un nucleósido resulta
de la unión β-glicosídica entre una pentosa (D-ribosa en el ARN o D-desoxirribosa en el
ADN) y una base nitrogenada que puede ser una purina o una pirimidina. Un nucleótido
está formado por la unión de un grupo fosfato al carbono 5’ de una pentosa. A su vez la
pentosa lleva unida al carbono 1’ una base nitrogenada

61
Bases púricas y pirimidínicas

62
Las bases nitrogenadas se asemejan estructuralmente a sustancias cíclicas de seis átomos
(dos de éstos átomos son nitrógeno) como en las pirimidinas o bien a un anillo de
pirimidina fusionado con un imidazol (estructura cíclica con dos anillos y cuatro átomos de
nitrógeno). Dentro del grupo de las bases púricas se encuentran la Adenina (A), y la
Guanina (G) mientras que Timina (T), Citosina (C), Uracilo (U) son bases pirimidínicas.
Estos derivados de la purina y la pirimidina son las bases que se encuentran con mayor
frecuencia en los ácidos nucleicos.
La hipoxantina y la xantina también son bases púricas e intermediarias en el
catabolismo de la adenina y la guanina. El producto final del catabolismo de las purinas en
los humanos es el ácido úrico, una base púrica completamente oxidada.
Las bases nombradas son las que se encuentran mayoritariamente en los ácidos
nucleicos. Sin embargo, también hay otras bases menos frecuentes, llamadas bases raras o
menores como la 5-metil-citosina, 5-hidroxi-metil-citosina, 2-metil-guanina y 6-metil-
adenina. Propiedades fisicoquímicas de las bases púricas y pirimidínicas
Solubilidad: En general son moléculas muy poco solubles en agua. A pH neutro, la
guanina es la menos soluble de las bases, seguida por la xantina. El ácido úrico,
constituyente normal de la orina, a pH alcalino forma uratos solubles, pero en medio ácido
es insoluble.
Absorción de luz ultravioleta: Las bases nitrogenadas, debido a su estructura
heterocíclica aromática, tienen la propiedad de absorber radiaciones en la zona UV del
espectro. Presentan un máximo de absorción a una longitud de onda de 260 nm. Esta

63
propiedad es importante pues puede utilizarse para la determinación cuali y cuantitativa de
las bases y sus derivados, incluyendo a los ácidos nucleicos.
Tautomerismo: Los compuestos con función cetona (>C=O) y con un hidrógeno unido en
el Cα a este grupo funcional, presentan un tipo de isomería especial que se denomina
tautomería ceto-enólica de modo que pueden existir dos isómeros posibles llamados
tautómeros. El compuesto en cuya estructura se presenta el grupo funcional cetona se
denomina lactama y aquel que presenta el grupo funcional alcohol se lo denomina
lactima.

64
Los compuestos que presentan una estructura HN-C=O, es decir la presencia de un
hidrógeno en el átomo de nitrógeno adyacente al grupo funcional cetona, también son
capaces de presentar este tipo de isomería. Las bases pirimidínicas que hemos nombrado,
presentan este tipo de estructura y por lo tanto dos tautómeros posibles. Para el caso de las
purinas, sólo la guanina presenta un grupo ceto (C6) en condiciones similares. La
importancia del predominio de una u otra forma en condiciones fisiológicas radica en el
apareamiento de las bases por uniones puente de hidrógeno entre las diferentes bases,
como en el ADN.
Formación de uniones puente de hidrógeno
Los átomos de hidrógeno de los grupos –NH2 y >NH de las bases nitrogenadas pueden
formar uniones puente de hidrógeno con los átomos de oxígeno >C=O de la posición 2 de
la timina, citosina y uracilo o de la posición 6 de la guanina. Este tipo de unión entre
moléculas, cuya característica se estudiará en clases posteriores, está íntimamente
relacionada con la estructura de los ácidos nucleicos.
NUCLEÓSIDOS
Un nucleósido está formado por una base púrica o pirimidínica unida a través del
N9 o el N1 a un azúcar, generalmente ribosa o 2-deoxirribosa por una unión beta-
glucosídica. La numeración de los carbonos de los azúcares es la habitual pero colocando
un tilde sobre el número, para distinguirlo del número correspondiente a la base. Los
nucleósidos que contienen ribosa se denominan ribonucleósidos y los que contienen 2-
deoxirribosa, desoxirribonucleósidos.
En la siguiente tabla se indica la nomenclatura corriente de los ribonucleósidos y
desoxirribonucleósidos de diferentes bases nitrogenadas:

65
BASE RIBONUCLEÓSIDO DESOXIRRIBONUCLEÓSIDO
Adenina Adenosina Desoxiadenosina
Guanina Guanosina Desoxiguanosina
Citosina Citidina Desoxicitidina
Uracilo Uridina
Timina Desoxitimidina
NUCLEÓTIDOS
Los nucleótidos, ribonucleótidos y/o desoxirribonucleótidos, son esteres
fosfofóricos de los nucleósidos. Cualquiera de los grupos hidroxilo del azúcar del
nucleósido puede ser fosforilado, pero en general se fosforila el hidroxilo del C 5´. El ácido
fosfórico se una a un grupo hidroxilo del azúcar formando un éster fosfórico (en general, el
grupo hidroxilo implicado en la unión es generalmente el de la posición 5’ sin embargo
también existen nucleótidos con grupos fosfatos unidos a los hidroxilos en 2’ o 3’).
A su vez, un segundo y tercer grupo fosfato se pueden unir al primero de ellos por uniones
anhídrido fosfórico. Los nucleótidos con un solo grupo fosfato se denominan nucleósido
monofosfato, con leósido. Se muestra a continuación la estructura del nucleótido ATP (ó
adenosín trifosfato) :

66
El ATP es el principal compuesto en el almacenamiento y transporte de energía para
distintos procesos metabólicos. También el GTP y el CTP cumplen esa función.
La presencia de restos de ácido fosfórico en los nucleótidos determina el carácter ácido de
éstas moléculas. A pH neutro, los grupos fosfato están ionizados y presentan cargas
negativas. La figura muestra la estructura de nucleósidos y nucleótidos: Otros nucleótidos de importancia biológica
El AMPc (3’,5’-monofosfato de adenosina) es mediador de diversas señales
extracelulares, al igual que el GMPc (3’,5’-monofosfato de guanina). A continuación se
indica la estructura del AMPc.
Nucleótidos

67
En distintos procesos biosintéticos también
intervienen nucleótidos como transportadores
activados de las unidades que formarán parte de
moléculas más complejas. Por ejemplo el UTP
participa de la síntesis de glucógeno, polímero de
glucosa que se utiliza como reserva de combustible
metabólico en el organismo, formando UDP-
glucosa. Siendo este último la especie que
transporta los restos de glucosa para la síntesis de
glucógeno por acción de la glucógeno sintasa.
Polinucleótidos
Existen dos clases de nucleótidos, los
ribonucleótidos en cuya composición encontramos
la pentosa ribosa y que forman parte del ARN y los
desoxirribonucleótidos, en donde participa la
desoxirribosa y que constituyen el ADN.
Las uniones entre nucleótidos son uniones
fosfodiéster. El grupo fosfato de un nucleótido se
une con el hidroxilo del carbono 5’ de otro
nucleótido, de este modo en la cadena quedan dos
extremos libres, de un lado el carbono 5’ de la
pentosa unido al fosfato y del otro el carbono 3’ de
la pentosa.
UDP-GlucosaUDP-Glucosa

68
ADN: Doble hélice
La molécula de ADN está constituída por dos largas cadenas de nucleótidos unidas entre sí
formando una doble hélice (antiparalelas: el extremo 5´ de una se enfrenta al extremo 3´de
la otra hebra). Las dos cadenas de nucleótidos que constituyen una molécula de ADN se
mantienen unidas entre sí porque se forman enlaces entre las bases nitrogenadas de ambas
cadenas que quedan enfrentadas.
La unión de las bases se realiza mediante puentes de hidrógeno, y este apareamiento está
condicionado químicamente de forma que la adenina (A) sólo se puede unir con la Timina
(T) y la Guanina (G) con la Citosina (C).

69
Las interacciones entre las cadenas polinuclotídicas se muestran en el esquema siguiente:


















