
Sara Maria Cunha RESPOSTA AO STRESS DE Listeria ... · o júri presidente Prof ... gastrointestinal...
62
Universidade de Aveiro Ano 2012/2013 Departamento de Biologia Sara Maria Cunha Oliveira Silva RESPOSTA AO STRESS DE Listeria monocytogenes
Transcript of Sara Maria Cunha RESPOSTA AO STRESS DE Listeria ... · o júri presidente Prof ... gastrointestinal...

Universidade de Aveiro Ano 2012/2013
Departamento de Biologia
Sara Maria Cunha Oliveira Silva
RESPOSTA AO STRESS DE Listeria monocytogenes

Universidade de Aveiro Ano 2012/2013
Departamento de Biologia
Sara Maria Cunha Oliveira Silva
STRESS RESPONSE OF Listeria monocytogenes
Dissertação apresentada à Universidade de Aveiro para cumprimento dos requisitos necessários à obtenção do grau de Mestre em Biologia Molecular e Celular, realizada sob a orientação científica da Professora Doutora Paula Cristina Maia Teixeira, Professora Auxiliar da Escola Superior de Biotecnologia da Universidade Católica Portuguesa e co-orientação do Professor Doutor António Carlos Matias Correia, Professor Catedrático do Departamento de Biologia da Universidade de Aveiro e Doutora Joana Gabriela Laranjeira Silva, investigadora da Escola Superior de Biotecnologia da Universidade Católica Portuguesa.

DECLARAÇÃO Declaro que este relatório é integralmente da minha autoria, estando devidamente referenciadas as fontes e obras consultadas, bem como identificadas de modo claro as citações dessas obras. Não contém, por isso, qualquer tipo de plágio quer de textos publicados, qualquer que seja o meio dessa publicação, incluindo meios eletrónicos, quer de trabalhos académicos.

Dedico este trabalho à minha família e amigos.

o júri
presidente Prof. Doutora Maria Adelaide de Pinho Almeida professora auxiliar do Departamento de Biologia da Universidade de Aveiro
Doutor Artur Jorge da Costa Peixoto Alves investigador auxiliar, Centro de Estudos do Ambiente e do Mar, Universidade de Aveiro
Prof. Doutora Paula Cristina Maia Teixeira professora auxiliar da Universidade Católica Portuguesa, Escola Superior de Biotecnologia

agradecimentos
Gostaria de deixar aqui o meu agradecimento à Escola Superior de Biotecnologia – Universidade Católica Portuguesa, pela oportunidade e condições proporcionadas para a realização deste Projeto de Dissertação.
À minha orientadora, Prof. Doutora Paula Teixeira, por me ter aceite como sua aluna de mestrado, pela sua paciência e constante disponibilidade, pelo conhecimento que me transmitiu e acima de tudo pela sábia contribuição para a minha formação profissional. Ao Prof. Doutor António Correia, meu co-orientador por me ter aceite igualmente como sua aluna de mestrado, pela sua disponibilidade, pelos conselhos prestados e valiosas contribuições. À Prof. Doutora Joana Silva, minha co-orientadora, pelo disponibilidade, acompanhamento e amizade ao longo deste período. A todos os colegas e amigos que fiz durante este projeto no Laboratório das Lácticas e do Pescado do Centro de Biotecnologia e Química Fina (CBQF), bem como nos Serviços de Microbiologia do Centro de Inovação e Apoio Empresarial (CINATE) pelo fantástico ambiente de trabalho, ajuda e paciência. À Prof. Doutora Teresa Brandão pela sua incansável disponibilidade para me ajudar e por tornar a análise estatística deste trabalho mais fácil. Ao Prof. Paul Gibbs por simpaticamente ter corrigido o inglês desta dissertação e pela partilha do seu conhecimento e sabedoria. À minha família e aos meus amigos, pelo incentivo, motivação e apoio dados durante este período.
Por fim, a todos aqueles que direta ou indiretamente contribuíram para que os objectivos deste trabalho pudessem ser alcançados.

palavras-chave
Listeria monocytogenes, stresse ácido, stresse alcalino, tracto gastro-intestinal, resistência a antibióticos.
resumo
Trinta e cinco isolados de Listeria monocytogenes provenientes de alimentos (n=20) e pacientes humanos com listeriose (n=15) e com diferentes perfis de resistência a antibióticos foram caracterizados e comparados com base na: (i) sua capacidade de sobrevivência à passagem pelo trato gastrointestinal simulado, (ii) sua capacidade de sobrivência a condições extremas de pH, (iii) potencial relação entre a resistência a antibióticos e a resistência às condições de stresse investigadas. A resposta às várias condições de stresse demonstrou ser estirpe- e stresse-dependente e não foi observada nenhuma relação entre isolados alimentares e clínicos (p > 0.05). Os resultados mostraram que L. monocytogenes sobrevive em condições ácidas e alcalinas extremas e não sobrevive quando submetida à passagem pelo trato gastrointestinal simulado, ou seja, à atividade dos sais biliares após ação conjunta do ácido clorídrico e pepsina. Não foi observada qualquer correlação entre a resistência a antibióticos e a resposta às condições de stresse aplicadas para os isolados estudados.

keywords
Listeria monocytogenes, acid stress, alkali stress, gastro-intestinal tract, antibiotic resistance.
abstract
Thirty-five Listeria monocytogenes isolates previously collected from food (n=20) and human patients suffering from listeriosis (n=15), with different antibiotic resistance profiles were characterized and compared based on: (i) their ability to survive through sequential conditions that parallel the digestive tract; (ii) their capacity to survive extreme pH values; (iii) the potential relationship, between antibiotic resistance and the resistance of L. monocytogenes isolates to the stress conditions investigated. The response was shown to be strain- and stress-dependent and no relation between food and clinical isolates was observed (p > 0.05). The results showed that L. monocytogenes is able to survive under extreme acid and alkaline conditions and did not survive when submitted to simulated sequential gastro-intestinal transit, i.e. the activity of bile salts after combined action of hydrochloric acid and pepsin. No correlation was observed between antibiotic resistance and response to the stress conditions applied to the isolates investigated.

i
Table of contents
List of abbreviations iii
List of figures v
List of tables vi
1. Listeria monocytogenes, a foodborne pathogen 1 1.1. History 1 1.2. Characteristics of the organism 1 1.3. Listeriosis 3 1.4. Background and occurrence 4 1.5. Stress resistance 6 1.6. Acid Stress 7
1.6.1. Induction of proteins 8 1.6.2. pH homeostasis 9 1.6.3. Glutamate decarboxylase system 9 1.6.4. Sigma factor (σB) 10 1.6.5. Two-component regulatory system 10
1.7. Alkali Stress 11 1.8. Antimicrobial resistance and stress resistance 12 1.9. Aim of the thesis 13
2. Material and Methods 15 2.1. Origin of isolates 15 2.2. Growth and storage conditions 16 2.3. Inoculum 17 2.4. Simulation of gastrointestinal conditions 17 2.5. Acid and alkaline treatments 18 2.6. Enumeration 18 2.7. Statistical analysis 19
3. Results and discussion 20 3.1. Effect of simulated digestion conditions 20 3.2. Effect of the pH in the survival of Listeria monocytogenes 26

ii
3.2.1. Acidic stress 26 3.2.2. Alkaline stress 30
3.3. Antimicrobial resistance and stress resistance 34
4. Conclusions 35
5. Proposals for future work 36
6. References 37

iii
List of abbreviations
AITR Alkali-Tolerance Response
ANOVA One-way analysis of variance
AR Antibiotic Resistant
AS Antibiotic Sensitive
ASP Alkali-Shock Proteins
ATP Adenosine triphosphate
ATR Acid Tolerance Response
BPW Buffered Peptone Water
CFU Colony Forming Units
DNA Deoxyribonucleic Acid
ECDC European Centre for Disease Prevention and Control
EFSA European Food Safety Authority
EU European Union
GABA Glutamate-γ-aminobutyrate
GAD Glutamate decarboxylase
GI Gastrointestinal
GSI Gastric and small intestinal
GSP General Stress Proteins

iv
HCl Hydrochloric acid
HPP High Pressure Processing
MA Massachusetts
MIC Minimum inhibitory concentration
NaHCO3 Sodium Bicarbonate
NaOH Sodium Hydroxide
PCR Polymerase Chain Reaction
PEF Pulsed Electric Field
RTE Ready-to-eat
TSAYE Tryptone Soy Agar supplemented with 0.6% Yeast Extract
TSBYE Tryptone Soy Broth supplemented with 0.6% Yeast Extract
UK United Kingdom
USA United States of America
UV Ultraviolet
VTEC Verocytotoxigenic Escherichia coli
WT Wild type

v
List of figures
Page Figure Title
4 1 Successive steps of human listeriosis (in Lecuit et al., 2007)
25 2 Survival (%) of food (A) and clinical (B) isolates of Listeria
monocytogenes resistant (C) or susceptible (D) to antibiotics during
stomach conditions simulation. Data are the mean of three independent
experiments; error bars represent the standard error of the mean.
29 3 Survival (%) of food (A) and clinical (B) isolates of Listeria
monocytogenes resistant (C) or susceptible (D) to antibiotics after
treatment with 10 mM HCl. Data are the mean of three independent
experiments; error bars represent the standard error of the mean.
33 4 Survival (%) of food (A) and clinical (B) isolates of Listeria
monocytogenes resistant (C) or susceptible (D) to antibiotics after
treatment with 100 mM NaOH. Data are the mean of three independent
experiments; error bars represent the standard error of the mean.

vi
List of tables
Page Table Title
15 1 Listeria monocytogenes isolates selected for the study and characterization
according to the serogroup and resistance to erythromycin, ciprofloxacin
and nitrofurantoin
18 2 Processing conditions used in each step of simulated digestion
23 3 Survival of thirty-five Listeria monocytogenes isolates when exposed to
simulated gastrointestinal conditions
28 4 Survival of thirty-five Listeria monocytogenes isolates when exposed to
acidic treatment with 10 mM HCl
32 5 Survival of thirty-five Listeria monocytogenes isolates when exposed to
alkaline treatment with 100 mM NaOH

1
1. Listeria monocytogenes, a foodborne pathogen
1.1. History Murray et al. first described Listeria monocytogenes in 1926. These researchers
isolated L. monocytogenes as the etiological agent of sudden death of young rabbits in the
animal-breeding establishment of the Department of Pathology of Cambridge, England
(Murray et al., 1926). In 1927, Pirie (Pirie, 1927), during investigations of unusual deaths
verified in gerbils in South Africa, isolated an identical bacterium.
The first documented case of human listeriosis was reported in 1929 in Denmark
and involved a soldier at the end of World War I (Nyfeldt, 1929; Allerberger, 2002). After
an increase in the number of reports on Listeria isolations in the late 70’s and early 80’s,
the first human listeriosis outbreak directly linked to the consumption of Listeria-
contaminated food was reported in 1983 (Schlech et al., 1983). Since then, several
epidemic outbreaks of listeriosis in humans occurred in North America and Europe
implicating L. monocytogenes as a causative agent (Farber and Peterkin, 1991) and
recognizing it as an important foodborne infection (Vazquez-Boland et al. 2001).
1.2. Characteristics of the organism Listeria spp. are Gram-positive; non spore-forming; non-capsulated, facultative
anaerobic; catalase positive; oxidase negative bacteria, with low G+C content (36 to 42%)
closely related to members of the genus Bacillus (Sallen et al., 1996), Clostridium,
Enterococcus, Streptococcus and Staphylococcus (Vazquez-Boland et al. 2001;
Swaminathan and Gerner-Smidt, 2007). At the time of writing, this genus is composed of
ten species: L. monocytogenes, L. ivanovii, L. welshimeri, L. innocua, L. seeligeri, L. grayi
(Vazquez-Boland et al. 2001), L. marthii (Graves et al., 2010), L. rocourtii (Leclercq et al.,
2010), L. fleischmannii (Bertsch et al., 2012) and L. weihenstephanensis (Halter et al.,
2012). Although species of the genus Listeria are environmental bacteria (Leclercq et al.,
2010), it also includes mammalian pathogenic species (i.e., L. monocytogenes, a human
and animal pathogen and L. ivanovii, an animal pathogen) (Orsi et al., 2008).
Listeria monocytogenes exhibits a wide range of temperature for growth, -1.5 to 45

2
ºC, with optimal growth between 30 ºC and 37 ºC (Gray and Killinger, 1966; Bell and
Kyriakides, 2005). It also grows in a wide range of pH values, i.e. 4.1 to 9.0 (Giotis et al.,
2007) and at high salt concentrations, e.g. 10% NaCl solutions (Duché et al., 2002).
Strains of Listeria monocytogenes species are divided into serotypes on the basis of
group-specific surface proteins, such as somatic (O) and flagellar (H) antigens
(Jeyaletchumi et al., 2010). Through examination of these particular proteins in slide
agglutination, at least 13 serotypes have been recognized (i.e. 1/2a, 1/2b, 1/2c, 3a, 3b, 3c,
4a, 4ab, 4b, 4c, 4d, 4e and 7) (Jeyaletchumi et al., 2010).
Molecular subtyping studies have established that L. monocytogenes can be divided
into four major evolutionary lineages, including lineages I and II, which are common, and
lineages III A/C and IIIB, which are rare (Roberts et al., 2006; Orsi et al., 2008). Isolates
from all four lineages have been associated with human listeriosis (Orsi et al., 2008).
Lineage I encompasses serotypes 1/2b, 3b, 3c, and 4b strains, whereas lineage II includes
serotypes 1/2a, 1/2c, and 3a (Nightingale et al., 2005; Liu et al., 2006). Most human
listeriosis cases and outbreaks have been associated with lineage I isolates, in particular
those of serotype 4b. On the other hand, lineage II isolates seem to be overrepresented
among isolates from ready-to-eat (RTE) and other food products, environmental sources
and animal listeriosis cases; and under-represented among human clinical cases
(Nightingale et al., 2005; Orsi et al., 2008; Ward et al., 2008).
Within lineage III, three different lineages have been identified (lineage IIIA, IIIB
and IIIC) (Liu et al., 2006) and represent serotypes 4a, 4b and 4c (Roberts et al., 2009).
These strains are mainly associated with isolates from animals with clinical listeriosis
(Roberts et al., 2009).
Serological and genetic typing schemes are clinically relevant since they
demonstrate that serotypes 1/2a, 1/2c, 1/2b, and 4b (lineages I and II) are involved with
around 98% of the documented human listeriosis cases and that serotypes 4a and 4c
(lineage III) are rarely associated with outbreaks of the infection (Liu et al., 2006).

3
1.3. Listeriosis
On one hand, L. monocytogenes is well-adjusted as a saprophyte for peaceful
survival in soil, water and decaying vegetation (Schuchat et al., 1991), but on the other
hand, it is a facultative intracellular pathogen responsible for causing listeriosis (Gray et
al., 2006), a severe foodborne, opportunistic infection, both in humans and animals (Ripio
et al., 2007).
Although L. monocytogenes is considered as a significant human and animal
pathogen responsible for listeriosis, L. innocua, L. ivanovii, L. welshimeri and L. seeligeri
have also been associated with occasional human infections (Rocourt et al., 1986; Andre
and Genicot, 1987; Cummins et al., 1994; Perrin et al., 2003; Guillet et al., 2010). Though
some cases of cutaneous listeriosis have been reported in veterinarians and farmers, who
have had direct contact with infected animals (Cain et al., 1986; Schuchat et al.,
1991; McLauchlin et al., 1994; Swaminathan and Gerner-Smidt, 2007), the major source
of human infections is due to ingestion of contaminated food (Farber and Peterkin, 1991;
Vázquez-Boland et al., 2001). Human invasive listeriosis can occur in healthy individuals,
but manifests primarily in pregnant women and neonates, the elderly (55 to 60 years and
older) and immunocompromised or debilitated individuals with underlying diseases
(Vázquez-Boland et al., 2001). The virulence of the infecting organism, the susceptibility
of the host and the size of the inoculum can influence the severity of the disease (Schuchat
et al., 1991).
The different steps of human listeriosis are shown schematically in Figure 1. After
the ingestion of Listeria-contaminated food, the bacteria are able to survive the stomach’s
acid conditions and reach the gastrointestinal tract. There, they have the capacity to cross
the intestinal barrier and are thought to disseminate from the mesenteric lymph nodes to
the spleen and liver (Lecuit et al., 2007). If not controlled properly by the immune system,
L. monocytogenes infection may reach the brain or placenta, resulting in infections of the
central nervous system mostly in immunocompromised patients and intrauterine or
cervical infections in pregnant women, which may lead to spontaneous abortion (2nd/3rd
trimester), stillbirth and neonatal infection with high mortality (Vázquez-Boland et al.,
2001).

4
Figure 1. Successive steps of human listeriosis (in Lecuit et al., 2007)
In healthy individuals, the ingestion of highly contaminated food products may lead
to a self-limited gastroenteritis (Lecuit et al., 2007). In groups at risk for invasive
listeriosis, general signs of infection often occur after an incubation period that can vary
between 10 and 70 days, making it difficult to identify the contaminated source in sporadic
cases (Lecuit et al., 2007).
1.4. Background and occurrence Listeria monocytogenes has been isolated from a wide range of raw foods and RTE
meats, poultry, dairy products, seafood and vegetables and from processed food that
becomes contaminated after processing such as soft cheeses (Lou and Yousef, 2007). From 2010 until 2012 a survey was carried out on L. monocytogenes with the aim
to estimate the prevalence of the pathogen in certain RTE foods at retail in the European
Union (EU) (European Food Safety Authority, 2013). 13,088 food samples including fish,
meat products and soft or semi-soft cheeses were collected and examined for the presence
of L. monocytogenes (European Food Safety Authority, 2013). The prevalence across the
entire European Union in fish samples was of 10.4% at the time of sampling, and of 10.3%
at the end of shelf-life. For meat and cheese samples, at the end of shelf-life, the
prevalence was of 2.07% and 0.47% respectively (European Food Safety Authority, 2013).
The data provided by this survey, gathered in all EU, will be useful in assessing the
exposure of EU consumers to L. monocytogenes via the specific three RTE food categories

5
tested that are thought to pose a particular risk for L. monocytogenes (European Food
Safety Authority, 2013).
In 2006, listeriosis was reported in 23 EU Member States as well as in Bulgaria (in
EU since 2007) and Norway and was the fifth most common zoonotic infection in Europe
after Campylobacter, Salmonella, Yersinia and verocytotoxigenic Escherichia coli (VTEC)
infections (Denny and McLauchlin, 2008). Listeriosis occurs infrequently (0.3 cases per
year per 100,000 of the population in the whole EU) (Denny and McLauchlin, 2008). On
one hand, from the 1,583 cases reported from the Member States in 2006, 64% were
accounted for in Germany, France and United Kingdom (Denny and McLauchlin, 2008).
On the other hand, Denmark, Finland and Luxembourg reported the highest incidence rates
of t 0.9 cases per 100,000 population (Denny and McLauchlin, 2008).
Listeriosis is not a reportable disease in Portugal nor has an active surveillance
system been established, thus available data are scarce (Almeida et al., 2006).
Nevertheless, some studies have been helping to understand the disease in Portugal
(Almeida et al., 2006; Almeida et al., 2009; Almeida et al., 2010). Almeida et al. (2006)
performed a retrospective study on listeriosis in Portugal and gathered all the data available
from 1994 until 2003. During that period, at least 35 cases of listeriosis were registered,
with a death rate of 37.5% for the 16 cases for which full case histories were available
(Almeida et al., 2006). According to the data collected in the study, the year with most
cases was 2003 with an estimated incidence of at least 1.4 cases per million inhabitants
(Almeida et al., 2006). These results must be assessed carefully since they are only an
initial estimate: not all the country's healthcare units were contacted, clinical symptoms
were sometimes not evident, thus making diagnosis difficult, and due to the fact that in
Portugal the notification of listeriosis is not legally mandatory (Almeida et al., 2006).
Despite the fact that there are no official reports of listeriosis cases in Portugal, this study
showed that L. monocytogenes is a threat to public health in a similar dimension to what
happens in other countries (Almeida et al., 2006). The only official data are from 2004,
recorded in the report of the European Food Safety Authority (EFSA)/European Centre for
Disease Prevention and Control (ECDC), reporting 38 human cases that year (EFSA,
2007). This authority has not reported data before or after 2004 for Portugal (EFSA, 2007;
EFSA, 2011; EFSA 2013). Between 2004 and 2007, Almeida et al. (2009) gathered cases
of listeriosis diagnosed in Portugal: 67 cases were identified with a death rate above 40%

6
in 24 cases of which full cases histories were available (Almeida et al., 2009). The number
of cases has increased over the years with an annual rate of 2.3 cases per million
inhabitants, roughly double the value reported in 2003 (Almeida et al., 2009; Almeida et
al., 2010).
For the presence of L. monocytogenes in foods, Mena et al. (2004) investigated its
presence on products sold in Portugal (collected from Portuguese producers and retailers).
From the 1035 samples, 7.0% were positive for L. monocytogenes (Mena et al., 2004). The
occurrence of L. monocytogenes in raw meat, raw chicken, raw milk, raw fish, flour and
frozen vegetables reflects the ubiquity of this organism (Mena et al., 2004). These products
showed the highest incidence of the pathogen with 17.7%, 60%, 16.7%, 12%, 18.5% and
12.9% of positive samples respectively (Mena et al., 2004). Although values are high, the
presence of L. monocytogenes in these products is not as problematic as its presence in
RTE products as these raw foods are typically cooked or pasteurized before consumption
(Mena et al., 2004). In RTE products, an occurrence of 25% in ham, 8.3% in dried fruits,
4.1% in pastry and 4% in fresh cheese was recorded; foods that are part of the eating habits
of Portuguese people in general (Mena et al., 2004).
1.5. Stress resistance Bacteria are adapted for optimal functioning in their normal physiological
environments (Beales, 2004). Although changing environments inflict stress on an
organism (Beales, 2004), bacteria have developed adaptive networks to face the challenges
and survive (Abee and Wouters, 1999).
Yousef and Courtney (2003) defined microbial stress as the imposition of
detrimental nutritional conditions, toxic chemicals and suboptimal physical conditions that
not necessarily cause physical damage but alter organism behaviour (Wesche et al., 2009).
Biological stressors could include competitive microorganisms (e.g., lactic acid bacteria),
microbial metabolites and antagonism (Yousef and Courtney, 2003). Hence, traditional or
novel food preservation techniques such as use of brine, high pressure processing (HPP),
pulsed electric field (PEF), ionizing radiation and ultraviolet (UV) irradiation can be
considered as microbial stressors (Lado and Yousef, 2002). These “hurdles” can cause
different degrees of stress in different bacteria that will influence the physiology, function

7
and activity of the microorganisms (Donnelly, 2002).
Once microorganisms sense stress, the cells respond in various ways in order to
survive: promote changes in membrane fluidity (e.g. cold shock), alter cell protein
structure or disrupt ribosomes (e.g. heat) or harm nucleic acids (e.g. γ radiation) (Yousef
and Courtney, 2003). At a molecular level, stress response includes transcription leading to
the synthesis of regulatory proteins that cope with the imposed stress (Yousef and
Courtney, 2003). Depending on the magnitude of the stress involved, stress can be
distinguished as either “sub-lethal” or “lethal or severe” (Donnelly, 2002). Sub-lethal stress
implies damage to structures within cells, the expression of which involves some loss of
cell function that may be transient or permanent (Wesche et al., 2009). It can result in
microbial “injury” and can be manifested as either retarded growth or complete prevention
of growth (Donnelly, 2002). When microorganisms are exposed to sub-lethal stress, it is
generally thought that this exposure can induce a transient adaptation (adaptive response)
accompanied by a temporary physiological change that often results in increased tolerance
to subsequent applications of the same type of stress or at times to a different stress (Hill et
al., 2002; Wesche et al., 2009). However, lethal or severe stress causes irreversible damage
to the microbial cells (Lou and Yousef, 1997) killing some, but not necessarily all,
bacterial cells (Wesche et al., 2009). When only a fraction of the population experiences
lethality, additional gene responses and adaptive mutations may actually improve survival
of the overall population (Wesche et al., 2009).
1.6. Acid Stress A common method of food preservation is its acidification and it is used worldwide
(Gahan et al., 1996). It is achieved either via fermentation or by addition of specific acid
food preservatives such as acetic, lactic and propionic acids (Hazan et al., 2004). The weak
acids in non-dissociated form permeate the cell membrane and, once inside the cytoplasm,
the acid dissociates (Phan-Thanh et al., 2000; Hazan et al., 2004). In this way, the acid
cannot diffuse out, decreasing the intracellular pH to dramatic values, which disrupts the
metabolic activities of the cell (Phan-Thanh et al., 2000; Hazan et al., 2004). Strong acids
completely dissociate into H+ and anions (Phan-Thanh et al., 2000). Because cell
membranes have a very low permeability to H+, protons enter and exit cells by interacting

8
with systems that control H+ transport, such as F0F1-ATPase, Na+/H+ antiporters, electron
transport systems (respiratory chains) (Phan-Thanh et al., 2000).
The acid tolerance response (ATR) is an induced protective response in
microorganisms against acid stress (Foster and Hall, 1990). The microbial response to acid
stress includes changes in membrane composition; increase in proton efflux; increase in
amino acid catabolism and induction of DNA repair enzymes (Yousef and Courtney,
2003). ATR is dependent on growth phase (exponential and stationary phase cells) and
differs among different bacterial species (Jordan et al., 1999; Yousef and Courtney, 2003).
The variations in intracellular or extracellular pH can be a signal for induction of acid
shock or adaptation proteins (Yousef and Courtney, 2003). Membrane bound proteins may
sense external or periplasmic pH (Yousef and Courtney, 2003). Internal pH may affect
gene expression or modulate a regulatory element involved in gene expression (Yousef and
Courtney, 2003).
Listeria monocytogenes is a neutrophile that is not able to grow at a pH below 4.5
(Walecka and Bania, 2012). During its life cycle, the pathogen faces a low-pH
environment at a number of stages, including in acidic foods, during gastric passage,
following exposure to fatty acids in the intestine and in the phagosome of the macrophages
(Cotter and Hill, 2003). In order to survive, Listeria possesses some stress adaptation
mechanisms (Cotter and Hill, 2003) such as,
x Induction of proteins;
x pH homeostasis;
x Glutamate decarboxylase system;
x Sigma factor (σB);
x Two-component regulatory systems.
1.6.1. Induction of proteins
In order to study the expression of proteins, Phan-Thanh and Mahouin (1999)
examined cells exposed to lethal acid pH (acid stress) and non-lethal acid pH (acid
adaptation). Although they observed that more proteins were induced in cells exposed to
lethal pH, the majority of induced proteins were common to both pH conditions used
(Phan-Thanh and Mahouin, 1999). Along with protein GroEL, which is also induced

9
during low temperatures (cross-protection), various transcriptional regulators and ATP
synthase were induced (Phan-Thanh and Mahouin, 1999). These results suggest that acid
adaptation also provides cross-protection against other stress factors (Phan-Thanh and
Mahouin, 1999; Gandhi and Chikindas, 2006). This phenomenon has great implications for
the food industry (Gandhi and Chikindas, 2006).
1.6.2. pH homeostasis
pH homeostasis allows microorganisms to maintain their intracytoplasmic pH via
proton transport across the cell membrane (Gandhi and Chikindas, 2006).
Listeria monocytogenes is a facultative anaerobic bacterium that can use the active
transport of H+ coupled with electron transport in respiratory chains (aerobic organisms) or
carry out H+ transport via H+-ATPase molecules using energy from ATP hydrolysis
(Shabala et al., 2002). Cotter et al. (2000) decided to study a multi-subunit enzyme called
F0F1-ATPase, highly conserved that had been extensively studied in other bacteria, namely
Escherichia coli. This enzyme serves as a channel for proton translocation across the cell
membrane by using ATP (Cotter et al., 2000). F0 rotation is caused by proton gradient and
followed by rotation of portion F1 that leads to ATP synthesis. Cotter et al. (2000)
observed a 3 log reduction under acid stress in the presence of proton inhibitor (DCCD-
N,N’-dicyclohexylcarbodiimide) concluding that F0F1-ATPase has a role in acid adaptation
of Listeria (Cotter et al., 2000).
1.6.3. Glutamate decarboxylase system
Glutamate decarboxylase (GAD) system is another mechanism that L.
monocytogenes also uses to survive acid stress (Gandhi and Chikindas, 2006). This system
is composed of three genes gadA, gadB and gadC (Gandhi and Chikindas, 2006). The first
two genes encode for two glutamate decarboxylases and gadC codes for a glutamate-γ-
aminobutyrate (GABA) antiporter (Gandhi and Chikindas, 2006). It is thought that
glutamate is taken up by the cell via a specific transporter followed by its decarboxylation,
which happens in the cytoplasm (Gandhi and Chikindas, 2006). In the process, γ-

10
aminobutyrate is produced resulting in the use of an intracellular proton (Gandhi and
Chikindas, 2006). Afterwards, an antiporter located in the cell membrane exports γ-
aminobutyrate from the cell (Gandhi and Chikindas, 2006). Due to proton loss from the
cell, the pH in the cytoplasm increases (Gandhi and Chikindas, 2006). Cotter et al. (2001)
studied the role of the GAD system in the acid resistance of L. monocytogenes during
gastric passage and found that the addition of glutamate increased the survival of the wild
strain in gastric fluid (Cotter et al., 2001). Deletion of the genes gadA, gadB and gadC
resulted in an increased sensitivity to low pH (Cotter et al., 2001). These results
demonstrated the importance of a functional GAD system in acid resistance in Listeria
(Cotter et al., 2001).
1.6.4. Sigma factor (σB)
Wiedmann et al. (1998) studied the role of general stress transcription factor (σB)
on the acid resistance of L. monocytogenes. The acid resistance of a sigB mutant, in
stationary phase, was lower than in the wild type (WT) (Wiedmann et al., 1998). This
study suggested that the rate of survival of L. monocytogenes is dependent on the
expression of σB-dependent proteins upon exposure to an acidic environment (Wiedmann
et al., 1998). Kazmierczak et al. (2003) observed that the alternative σB factor regulates the
expression of gadB gene involved in acid stress survival and regulates virulence gene
expression in L. monocytogenes.
1.6.5. Two-component regulatory system
Cotter et al. (1999) identified the role of a two-component regulatory system
consisting of lisR and lisK, which encode the response regulator and histidine kinase,
respectively in L. monocytogenes. This study by Cotter et al. (1999) showed that these
two-component signal transduction systems can sense alterations in the environment such
as low pH by a membrane-associated histidine kinase and the response regulator enables
the cell to respond by altering gene expression. Thus, the signal LisRK transduction system
is involved in response to acidic stress and regulates the virulence gene expression in L.

11
monocytogenes (Cotter et al., 1999).
Because acidic pH of foods helps to prevent the growth of foodborne pathogens,
the ATR observed in L. monocytogenes is noteworthy, since the exposure of the pathogen
to mild acidic conditions could confer resistance to more severe acidic conditions (Gandhi
and Chikindas, 2006).
1.7. Alkali Stress The resistance of L. monocytogenes to moderate and severe alkaline pH conditions
is of particular concern especially when alkali cleaners and sanitizers are used in cleaning
and decontamination within the food industry (Sharma et al., 2003; Giotis et al., 2007;
Giotis et al., 2008a). Such exposure can induce alkaline adaptation and it may account for
this bacterium’s persistence in food-processing environments (Giotis et al., 2008b). These
adaptations enable the pathogen to resist high pH conditions related to human defence
mechanisms, e.g. alkali conditions in phagolysosomes and pancreatic secretions in the
small intestine (Sharma et al., 2003; Giotis et al., 2007; Giotis et al., 2008a). Strongly
alkaline cleaners, like sodium hydroxide (NaOH), are used, for example, to remove heavy
soils (proteins and fats) in meat processing plants (Sharma et al., 2003).
The ability of Listeria to induce an alkali-tolerance response (AlTR) could be a
significant factor in the persistence and clinical impact of this pathogen in alkaline food-
processing systems and pathogen’s virulence within human gastrointestinal tract (Giotis et
al., 2008a).
Little is known about the factors or mechanisms by which AlTR initiates (Giotis et
al., 2008a). In fact, knowledge about the mechanisms used by Gram-positive bacteria to
adapt and grow at alkali pHs comes mainly from studies of alkaliphilic strains of Bacillus
species (Giotis et al., 2008b). The alkali stress promotes a specific response in these
organisms resulting in a transient enhanced expression of a subset of genes that increase
metabolic acid production, changes in cell surface properties and an increase of expression
and activity of monovalent- cation/proton antiporters (Padan et al., 2005). These response
mechanisms allow the maintenance of neutral cytoplasmic pH and therefore enable the
growth of Bacillus in alkali conditions (Padan et al., 2005). On the other hand, other
bacteria, such as Enterococcus faecalis, use other response mechanisms different from the

12
maintenance of neutral cytoplasmic pH (Mugikura et al., 1990; Flahaut et al., 1997). Their
adaptation to alkali environments is mainly accomplished due to de novo protein synthesis
by induction of proteins/enzymes capable of remaining active at high pH values (Giotis et
al., 2008b). These proteins provide either a specific protective function to alkali pH
(Alkali-Shock Proteins, ASP) (Giotis et al., 2008b) or provide a rather non-specific
protective function (General Stress Proteins, GSP), regardless of the type of stress (i.e. heat
shock proteins and proteins involved in the SOS response) (Giotis et al., 2008b).
Giotis et al. (2008b), in order to enlighten the response of L. monocytogenes
towards alkali stress, attempted to identify the contribution of protein synthesis in the
induction of AlTR. They concluded that alkali pH provides L. monocytogenes with non-
specific multiple-stress resistance and AlTR functions to minimize excess alkalization and
energy expenditures while mobilizing available carbon sources (Giotis et al., 2008b).
1.8. Antimicrobial resistance and stress resistance
Antibiotics were, originally, developed to treat human infectious diseases (Barbosa
and Levy, 2000). However, soon after, several were applied into animal husbandry where
they have proven to be highly effective in treating bacterial infections of significant animal
health consequence in possibly every food animal species, including poultry and
aquaculture (Doyle et al., 2013). Broad use and misuse of antibiotics created a strong
selective pressure, which consistently has resulted in the survival and spread of resistant
bacteria (Barbosa and Levy, 2000), which constitutes a major public health threat (Zhang
et al., 2013). Nevertheless, the occurrence of resistance among targeted pathogens in food
animals is not what raises most public-health and food safety concerns (Doyle et al., 2013).
Instead, it is the unintentional consequences of the effects of the antibiotics on the bacteria
that often reside in the gastrointestinal tracts of the food animals (Doyle et al., 2013).
Other implication of antibiotic resistant pathogens is that the acquisition of
antibiotic resistance may influence the behaviour of such organisms during typical food
processing conditions (Duffy et al., 2006). There are reports in the literature that suggest
that bacteria resistant to antibiotics may display different resistance to stresses, such as
acid or heat (McGee, 2003; Walsh et al., 2005). McGee (2003) suggested a possible link
between the acquisition of antibiotic resistance and survival of Escherichia coli O157:H7
at a low pH. This study is however not in accordance with other published study by Duffy

13
et al. (2006) that showed that there were no differences in acid resistance between
antibiotic-resistant and –susceptible E. coli strains. Other reports in the literature also
propose a possible link between the acquisition of antibiotic resistance and pathogen
thermal resistance (Walsh et al., 2001; Walsh et al., 2005). Walsh et al. (2001) reported
that the presence or absence of antibiotic resistance genes did not modulate the heat
resistance of the L. monocytogenes strains studied. Later, in 2005, Walsh et al. concluded
that no significant differences were observed in the heat resistance of sensitive strains of
Salmonella Enteritidis and Salmonella Typhimurium, and their laboratory-acquired
antibiotic-resistant mutant strains. However, a potential link between multiantibiotic
resistance and heat resistance in S. Typhimurium DT104 was suggested (Walsh et al.,
2005).
Considering the continuing increase, albeit slow, of the antimicrobial resistance of
L. monocytogenes particularly of strains isolated from food and production environments
(Barbosa et al., 2013; Conter et al., 2009), further investigation on the stress responses of
such strains, in particular for multiple antibiotic resistant strains, is justified (Walsh et al.,
2001).
1.9. Aim of the thesis
Since L. monocytogenes was recognized as a foodborne pathogen in the 1980s,
extensive research on the pathogen’s cell biology, virulence mechanisms and epidemiology
has been done (Walecka and Bania, 2012). Before causing infection, L. monocytogenes has
to overcome numerous stresses encountered both in the environment as well as in the host
(Walecka and Bania, 2012). Thus, understanding microbial stress resistance and response
to sub-lethal stresses may be of great value for food safety (Walecka and Bania, 2012). It
is possible that knowledge about the role of the factors affecting stress responses could
influence the design of modern food preservation methods (Walecka and Bania, 2012). In
this study, 35 L. monocytogenes isolates from different origins (clinical and food) were
characterized with the following objectives:
x To assess the survival of L. monocytogenes isolates based on their capacity to
survive through conditions that are typically encountered in the gastrointestinal
tract;

14
x To evaluate the survival of L. monocytogenes isolates when submitted to different
stress conditions (acidic and alkaline);
x To investigate the potential relationship, if any, between antibiotic resistance and
the resistance of L. monocytogenes isolates to the stress conditions investigated.
15
2. Material and Methods
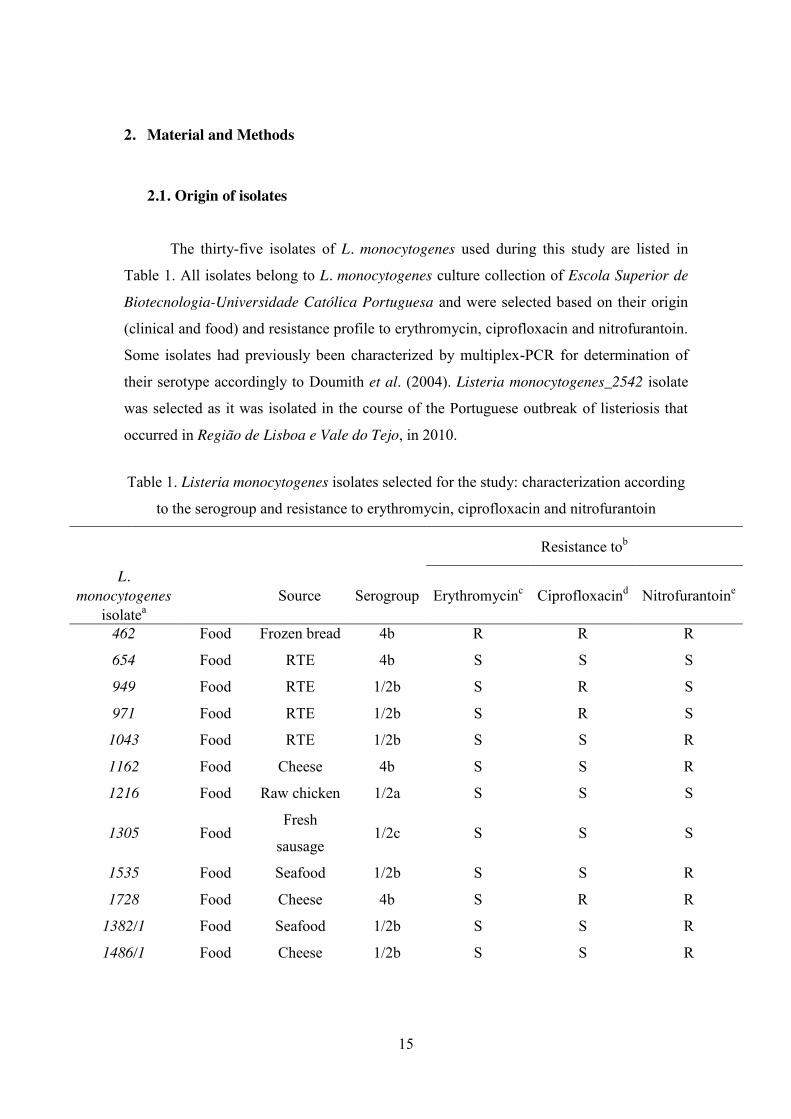
2.1. Origin of isolates The thirty-five isolates of L. monocytogenes used during this study are listed in
Table 1. All isolates belong to L. monocytogenes culture collection of Escola Superior de
Biotecnologia-Universidade Católica Portuguesa and were selected based on their origin
(clinical and food) and resistance profile to erythromycin, ciprofloxacin and nitrofurantoin.
Some isolates had previously been characterized by multiplex-PCR for determination of
their serotype accordingly to Doumith et al. (2004). Listeria monocytogenes_2542 isolate
was selected as it was isolated in the course of the Portuguese outbreak of listeriosis that
occurred in Região de Lisboa e Vale do Tejo, in 2010.
Table 1. Listeria monocytogenes isolates selected for the study: characterization according
to the serogroup and resistance to erythromycin, ciprofloxacin and nitrofurantoin
Resistance tob
L. monocytogenes
isolatea Source Serogroup Erythromycinc Ciprofloxacind Nitrofurantoine
462 Food Frozen bread 4b R R R
654 Food RTE 4b S S S
949 Food RTE 1/2b S R S
971 Food RTE 1/2b S R S
1043 Food RTE 1/2b S S R
1162 Food Cheese 4b S S R
1216 Food Raw chicken 1/2a S S S
1305 Food Fresh
sausage 1/2c S S S
1535 Food Seafood 1/2b S S R
1728 Food Cheese 4b S R R
1382/1 Food Seafood 1/2b S S R
1486/1 Food Cheese 1/2b S S R

16
1604/2 Food Cheese 4b S S S
1852/3 Food Alheira 1/2b R R R
1940/1 Food Fermented
sausage 4b S R S
800/2 Food Bread 1/2b S S R
841/2 Food Garlic bread 4b R S R
863/1 Food Garlic bread 1/2c S S S
925/1 Food Cheese 1/2b S S S
969/3 Food Garlic bread 1/2b S S R
1062 Clinical Blood 1/2b S S R
1543 Clinical Blood 4b S S S
1547 Clinical Blood 1/2b S S S
2065 Clinical Blood 1/2b S S S
2086 Clinical Blood 1/2a S S S
2103 Clinical Blood 1/2a S R S
2104 Clinical Blood 4b S S R
2264 Clinical Blood 4b S R S
2265 Clinical Blood 4b S R S
2388 Clinical Peritoneal
fluid 1/2a S R S
2542 Clinical Placenta 4b S S S
2571 Clinical CSF 4b S R S
2658 Clinical Blood 1/2b S R R
3390 Clinical CSF 4b S R R
3391 Clinical Blood 1/2a S R R aCodes of L. monocytogenes isolates of the Listeria culture collection of Escola Superior de
Biotecnologia-Universidade Católica Portuguesa. bGrowth (R, resistant) or no growth (S,
susceptible) on Mueller-Hinton agar with 3% (v/v) lysed horse blood. cMinimum inhibitory
concentration (MIC, μg/mL) ≥ 8; dMIC (μg/mL) ≥ 4; eMIC (μg/mL) ≥ 128
2.2. Growth and storage conditions Stock cultures were grown on Tryptic Soy Agar (Pronadisa, Madrid, Spain)

17
supplemented with 0.6% (w/v) of Yeast Extract (Lab M, Lancashire, United Kingdom)
(TSAYE) at 37ºC for 24h and preserved at -20ºC in Tryptic Soy Broth supplemented with
0.6% (w/v) of Yeast Extract (TSBYE) containing 30% (v/v) of glycerol (Sigma,
Steinheim, Germany).
2.3. Inoculum The preparation of the inoculum was done according to Ramalheira et al. (2010).
One pure colony of each isolate was transferred from TSAYE incubated at 37 ºC for 24 h
to 10 mL of TSBYE and incubated in the same conditions. This culture was then
subsequently diluted 1:100 in TSBYE and incubated at 37 ºC for 24h to yield cells in the
stationary phase of growth. Cells were harvested by centrifugation (7000 g, 10 min, 4 ºC;
Rotina 35R, Hettich, Germany), re-suspended in 10 mL of sterile Ringer’s solution (Lab
M, Lancashire, United Kingdom) and mixed to obtain an inoculum of approximately 107
CFU/mL.
2.4. Simulation of gastrointestinal conditions The simulated gastrointestinal pathway was performed according to Madureira et
al. (2011) in buffered peptone water (BPW; Lab M) and is described in Table 2 – including
the compounds and concentrations used, the period of time and the intensities of stirring in
all steps (stirring was used to somehow simulate the peristaltic movements).
0.5 mL aliquots of inoculum (prepared as described in section 2.3) were added to
glass flasks with 49.5 mL of BPW adjusted to pH 2.5 with Hydrochloric Acid (1 M HCl,
Pronalab, Lisbon, Portugal) and 25 mg/mL of pepsin (Sigma), prepared in 0.1 N HCl. The
glass flasks were kept at 37 ºC and samples were taken just before inoculation, and after 15
min, 45 min, 70 min and 90 min (esophagus-stomach step). Following exposure to
stomach conditions, the duodenum step simulation used 12 g/L bile salts (Pronadisa)
diluted in 0.1 M Sodium Bicarbonate (NaHCO3). This solution was added at the beginning
of this step at a rate of 0.25 mL/mL and a sample was taken at time 110 min (intestinal
phase). Finally, the ileum step was brought about by an increase of pH to 6.5 using 0.1 M
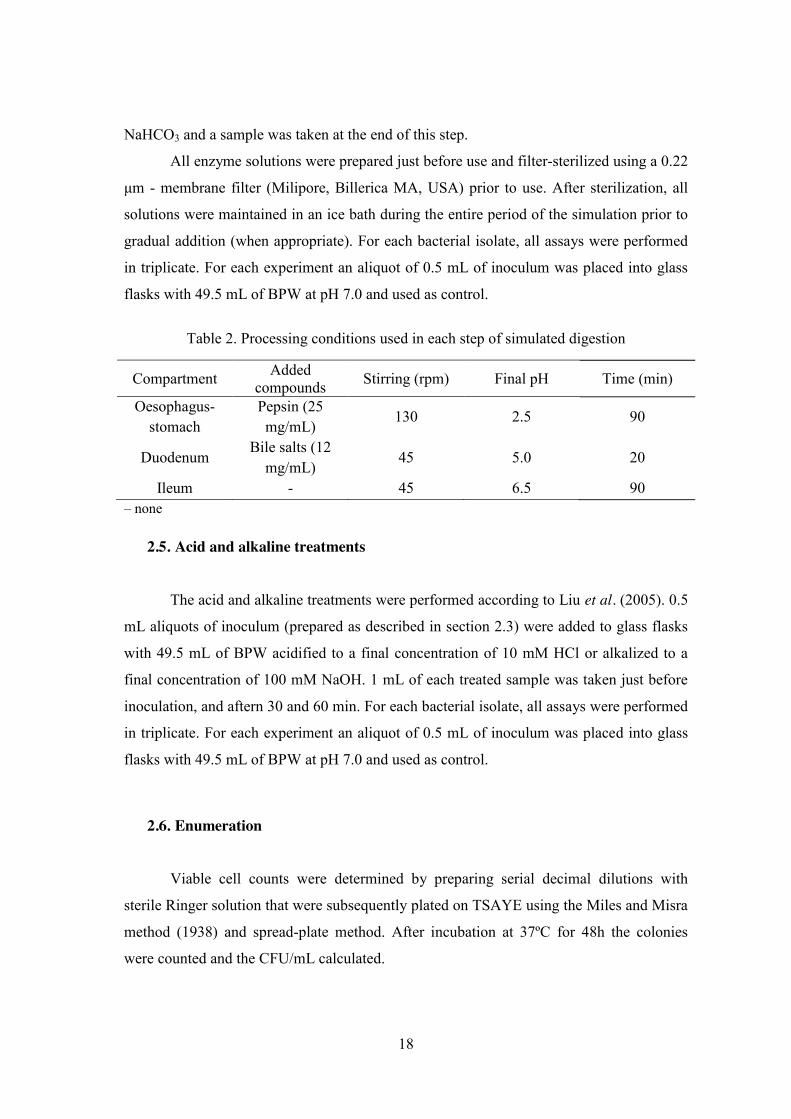
18
NaHCO3 and a sample was taken at the end of this step.
All enzyme solutions were prepared just before use and filter-sterilized using a 0.22
μm - membrane filter (Milipore, Billerica MA, USA) prior to use. After sterilization, all
solutions were maintained in an ice bath during the entire period of the simulation prior to
gradual addition (when appropriate). For each bacterial isolate, all assays were performed
in triplicate. For each experiment an aliquot of 0.5 mL of inoculum was placed into glass
flasks with 49.5 mL of BPW at pH 7.0 and used as control.
Table 2. Processing conditions used in each step of simulated digestion
Compartment Added compounds Stirring (rpm) Final pH Time (min)
Oesophagus-stomach
Pepsin (25 mg/mL)
130 2.5 90
Duodenum Bile salts (12
mg/mL) 45 5.0 20
Ileum - 45 6.5 90 – none
2.5. Acid and alkaline treatments The acid and alkaline treatments were performed according to Liu et al. (2005). 0.5
mL aliquots of inoculum (prepared as described in section 2.3) were added to glass flasks
with 49.5 mL of BPW acidified to a final concentration of 10 mM HCl or alkalized to a
final concentration of 100 mM NaOH. 1 mL of each treated sample was taken just before
inoculation, and aftern 30 and 60 min. For each bacterial isolate, all assays were performed
in triplicate. For each experiment an aliquot of 0.5 mL of inoculum was placed into glass
flasks with 49.5 mL of BPW at pH 7.0 and used as control.
2.6. Enumeration
Viable cell counts were determined by preparing serial decimal dilutions with
sterile Ringer solution that were subsequently plated on TSAYE using the Miles and Misra
method (1938) and spread-plate method. After incubation at 37ºC for 48h the colonies
were counted and the CFU/mL calculated.

19
2.7. Statistical analysis Microbial counts were transformed to logarithmic reduction using the equation: log
(N/N0), where N is the microbial cell density at a particular sampling time and N0 is the
initial cell density. One-way analysis of variance (ANOVA) was carried out to assess
statistically any significant differences among all experimental data on the survival of L.
monocytogenes in simulated gastrointestinal (GI) tract and acid and alkaline treatments.
All tests were performed to a 5% significance level, using Kaleidagraph software (version
4.4, Synergy Software, Reading, USA).

20
3. Results and discussion
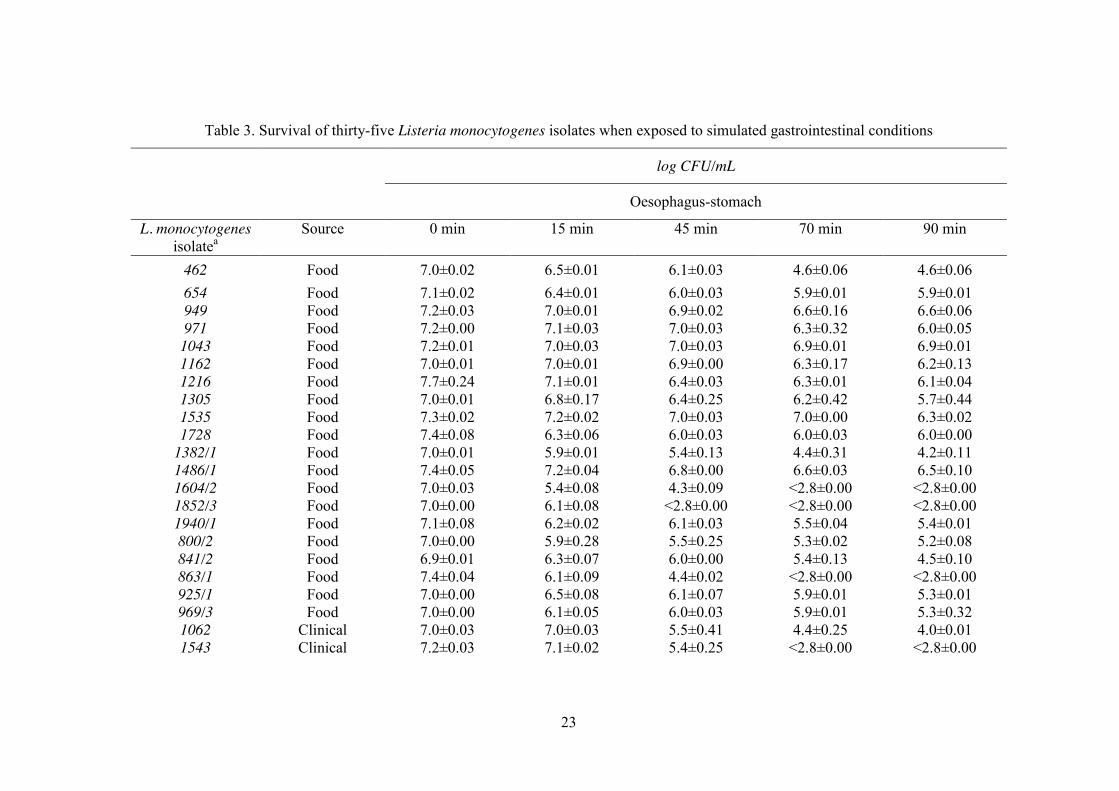
3.1. Effect of simulated digestion conditions
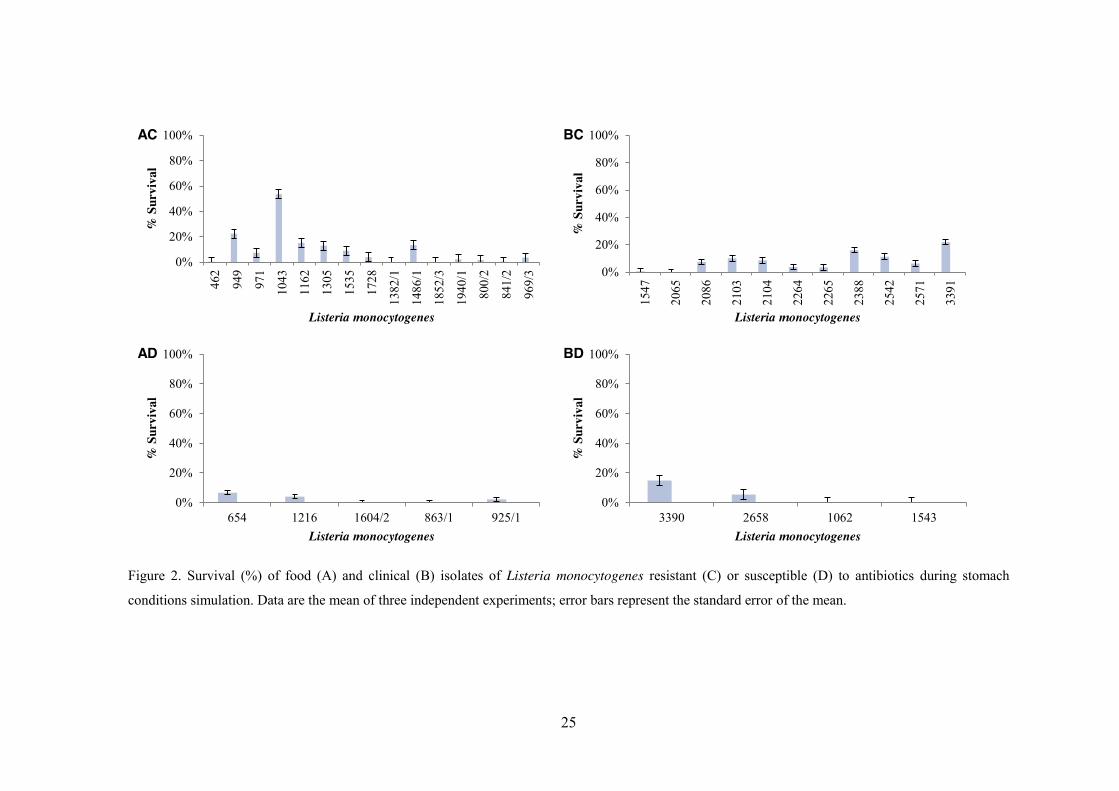
The decreases in viability of the thirty-five isolates of L. monocytogenes tested in
the simulated gastrointestinal model is plotted in figure 2; a minimum threshold of 3 log
CFU/mL was take into account, owing to the intrinsic sensitivity limitation of Miles and
Misra’s plating method of enumeration. The survival in each step of digestion is tabulated
in Table 3.
Examination of Figure 2, for stomach conditions, indicates that the majority of the
isolates were susceptible to the treatment showing a survival of 15% or lower. The isolate,
L. monocytogenes_1043, from a RTE food, showed the highest resistance to the decrease
in pH in the presence of pepsin, with approximately 53% survivors (Figure 2). Upon
addition of bile salts (110 min, Table 1), all the isolates that resisted until that moment
(stomach conditions), were reduced to values below the detection limit of the enumeration
technique. This step was dramatically detrimental: the viable cell counts dropped more
than 4 log cycles from their initial value, even for isolate L. monocytogenes_2542, clinical
4b, associated with the deadly Portuguese outbreak of listeriosis in 2010. No correlation
was found between origin (clinical or food), antibiotic resistance profile and survival of L.
monocytogenes (p>0.05) in simulated gastric conditions. During exposure to oesophagus-stomach simulated conditions, five of the thirty-
five L. monocytogenes isolates (L. monocytogenes_1604/2, L. monocytogenes_1852/3, L.
monocytogenes_863/1, L. monocytogenes_1543, L. monocytogenes_2065) were incapable
to survive to the simulated gastric juice, which corresponds to pH 2.5. Upon exposure to
duodenum conditions (i.e. bile salts), viable cell numbers of all isolates decreased abruptly.
Apparently, L. monocytogenes cells were in a fragile state when they reached this stage of
digestion with several injuries caused by acid before exposure to bile salts. It is important
to recall that bile salts are natural detergents that facilitate the digestion and absorption of
hydrophobic components of the diet (Sánchez et al., 2007). However, their antimicrobial
nature makes them strongly inhibitory for bacteria (their detergent property dissolves
bacterial membranes), thus, restricting their survival throughout the GI tract (Sánchez et
al., 2007). In contrast to this deleterious action of acid and pepsin conjugated with bile

21
salts, Melo et al. (2013) and Olier et al. (2004) reported that all L. monocytogenes strains
were able to survive the exposure to the gastric fluid as well to the intestinal fluid.
However, it is important to point out that Melo et al. (2013) exposed their L.
monocytogenes strains to a cheese-simulated medium previously to the GI stress. In fact, a
simulated or a real food matrix confer resistance to GI passage due to the protection
conferred by food components as other studies have demonstrated (Barmpalia-Davis et al.,
2009; Peterson et al., 2007). Nevertheless, our results are in accordance with those
reported from Barbosa et al. (2012), Jiang et al. (2010), King et al. (2003) and Ramalheira
et al. (2010) (studies also carried out in broth) who proved that an increase in bile salts
susceptibility was observed when cells had first been exposed to the low pH environment
of the gastric system.
It is important to note that in the current study two of the three L. monocytogenes
isolates with the highest percentages of resistance to GI passage were recovered from RTE
foods. Note also that RTE foods do not undergo any treatment to ensure their safety before
consumption and therefore the risk of foodborne disease must be considered if L.
monocytogenes cells are present in the food.
Comparison of survival patterns to different stress conditions of clinical versus
food isolates is well documented in the literature (Avery and Buncic, 1997; Barmpalia-
Davis et al., 2008; Dykes and Moorhead, 2000; Lianou et al., 2006; Olier et al., 2004;
Ramalheira et al., 2010; Viallete et al., 2003; Werbrouck et al., 2008). Avery and Buncic
(1997) investigated the effect on pathogenicity and growth characteristics of thirty isolates
of L. monocytogenes (fifteen clinical and fifteen from meat) after long-term storage at 4 ºC.
Clinical isolates demonstrated higher resistance to the effects of unfavourable storage
conditions, concerning pathogenicity and lag-phase duration at 37 ºC (Avery and Buncic,
1997). Barmpalia-Davis et al. (2008a) compared the ability to survive in a dynamic GI
model of thirteen strains of L. monocytogenes (outbreak origin/food associated and
human). Survival of GI tract conditions was demonstrated to be growth phase and strain-
dependent (Barmpalia-Davis et al. (2008a). The highest acid susceptibility was observed
for two strains of clinical origin (Barmpalia-Davis et al. (2008a). In the current study, the
survival of the acidic phase also proved to be strain-dependent as had been demonstrated in
other studies (Faleiro et al., 2003;; Francis and O’Beirne, 2005;; Lianou et al., 2006). Dykes
and Moorhead (2000) tested thirty L. monocytogenes strains of different origins (clinical

22
and meat) for their capacity to resist acid stress. Although they reported an increased acid
resistance in all clinical L. monocytogenes strains, no significant differences were apparent
based on source (Dykes and Moorhead, 2000). Lianou et al. (2006) characterized and
compared twenty-five L. monocytogenes strains (clinical and food) representing several
serotypes, based on growth rates and heat and acid resistance. These authors reported that
though no clear origin-related trends were observed under the test conditions (heating at 55
ºC or pH 3), outbreak-related isolates of serotype 4b demonstrated the highest acid
resistance (Lianou et al., 2006). In the current study and probably due to the unequal
distribution of each serotype investigated, no relationship between the serotype and
simulated GI passage was observed (p> 0.05). Olier et al. (2004) reported that there was no
relationship between the origin (human, food or food processing environment) of fifty
strains of L. monocytogenes and capacity to degrade bile salts. Ramalheira et al. (2010)
showed that clinical isolates were more resistant than food isolates to GI passage
performed in BPW. Vialette et al. (2003) investigated the growth rates of eight L.
monocytogenes strains (four clinical associated to fish products and four from other
seafoods) with respect to stress conditions that can occur during food processing. Clinical
strains displayed better adaptation to pH and osmotic stress than seafood strains in
stationary phase Vialette et al. (2003). Werbrouck et al. (2008) did not find any clear
correlation between the origin (clinical and meat) and survival of L. monocytogenes
serotype 4b in synthetic gastric fluid.
Although static gastric and small intestinal (GSI) models do not reproduce the
dynamic processes occurring during human digestion such as gastric emptying or
continuous changes in pH and secretion flow rates, the generated information would be a
valuable contribution for further studies in the context of microbiological risk assessments.
23
Table 3. Survival of thirty-five Listeria monocytogenes isolates when exposed to simulated gastrointestinal conditions
log CFU/mL
Oesophagus-stomach
L. monocytogenes isolatea
Source
0 min
15 min
45 min
70 min
90 min
462 Food 7.0±0.02 6.5±0.01 6.1±0.03 4.6±0.06 4.6±0.06 654 Food 7.1±0.02 6.4±0.01 6.0±0.03 5.9±0.01 5.9±0.01 949 Food 7.2±0.03 7.0±0.01 6.9±0.02 6.6±0.16 6.6±0.06 971 Food 7.2±0.00 7.1±0.03 7.0±0.03 6.3±0.32 6.0±0.05
1043 Food 7.2±0.01 7.0±0.03 7.0±0.03 6.9±0.01 6.9±0.01 1162 Food 7.0±0.01 7.0±0.01 6.9±0.00 6.3±0.17 6.2±0.13 1216 Food 7.7±0.24 7.1±0.01 6.4±0.03 6.3±0.01 6.1±0.04 1305 Food 7.0±0.01 6.8±0.17 6.4±0.25 6.2±0.42 5.7±0.44 1535 Food 7.3±0.02 7.2±0.02 7.0±0.03 7.0±0.00 6.3±0.02 1728 Food 7.4±0.08 6.3±0.06 6.0±0.03 6.0±0.03 6.0±0.00
1382/1 Food 7.0±0.01 5.9±0.01 5.4±0.13 4.4±0.31 4.2±0.11 1486/1 Food 7.4±0.05 7.2±0.04 6.8±0.00 6.6±0.03 6.5±0.10 1604/2 Food 7.0±0.03 5.4±0.08 4.3±0.09 <2.8±0.00 <2.8±0.00 1852/3 Food 7.0±0.00 6.1±0.08 <2.8±0.00 <2.8±0.00 <2.8±0.00 1940/1 Food 7.1±0.08 6.2±0.02 6.1±0.03 5.5±0.04 5.4±0.01 800/2 Food 7.0±0.00 5.9±0.28 5.5±0.25 5.3±0.02 5.2±0.08 841/2 Food 6.9±0.01 6.3±0.07 6.0±0.00 5.4±0.13 4.5±0.10 863/1 Food 7.4±0.04 6.1±0.09 4.4±0.02 <2.8±0.00 <2.8±0.00 925/1 Food 7.0±0.00 6.5±0.08 6.1±0.07 5.9±0.01 5.3±0.01 969/3 Food 7.0±0.00 6.1±0.05 6.0±0.03 5.9±0.01 5.3±0.32 1062 Clinical 7.0±0.03 7.0±0.03 5.5±0.41 4.4±0.25 4.0±0.01 1543 Clinical 7.2±0.03 7.1±0.02 5.4±0.25 <2.8±0.00 <2.8±0.00

24
1547 Clinical 7.0±0.01 6.5±0.07 6.1±0.01 5.3±0.31 4.4±0.36 2065 Clinical 6.9±0.01 6.2±0.00 5.3±0.14 4.0±0.00 <2.8±0.00 2086 Clinical 7.1±0.01 7.0±0.01 6.9±0.01 6.3±0.33 5.9±0.01 2103 Clinical 7.0±0.00 6.9±0.16 6.4±0.00 6.1±0.11 6.0±0.00 2104 Clinical 7.1±0.01 7.0±0.00 6.6±0.04 6.3±0.00 6.0±0.02 2264 Clinical 7.5±0.01 7.3±0.04 6.5±0.02 6.4±0.01 6.0±0.01 2265 Clinical 7.4±0.01 7.3±0.02 6.5±0.04 6.3±0.01 6.0±0.01 2388 Clinical 7.1±0.05 7.0±0.01 6.5±0.19 6.3±0.00 6.3±0.00 2542 Clinical 7.2±0.03 7.1±0.01 7.0±0.01 6.4±0.10 6.2±0.06 2571 Clinical 7.4±0.01 7.3±0.03 6.7±0.20 6.5±0.05 6.2±0.10 2658 Clinical 7.2±0.03 7.2±0.03 7.0±0.00 6.1±0.02 6.0±0.03 3390 Clinical 6.9±0.01 6.6±0.03 6.4±0.02 6.3±0.03 6.1±0.08 3391 Clinical 7.0±0.00 6.6±0.02 6.4±0.03 6.4±0.02 6.3±0.03
aCodes in L. monocytogenes collection of Escola Superior de Biotecnologia-Universidade Católica Portuguesa All the isolates were reduced to values
below the detection limit of the enumeration technique in duodenum and ileum steps.
25
Figure 2. Survival (%) of food (A) and clinical (B) isolates of Listeria monocytogenes resistant (C) or susceptible (D) to antibiotics during stomach
conditions simulation. Data are the mean of three independent experiments; error bars represent the standard error of the mean.
0%
20%
40%
60%
80%
100%
462
949
971
1043
1162
1305
1535
1728
1382
/1
1486
/1
1852
/3
1940
/1
800/
2
841/
2
969/
3
% S
urvi
val
Listeria monocytogenes
AC
0%
20%
40%
60%
80%
100%
1547
2065
2086
2103
2104
2264
2265
2388
2542
2571
3391
% S
urvi
val
Listeria monocytogenes
BC
0%
20%
40%
60%
80%
100%
654 1216 1604/2 863/1 925/1
% S
urvi
val
Listeria monocytogenes
AD
0%
20%
40%
60%
80%
100%
3390 2658 1062 1543
% S
urvi
val
Listeria monocytogenes
BD

26
3.2. Effect of the pH in the survival of Listeria monocytogenes Concerning exposure to HCl and NaOH, Figures 3 and 4 demonstrate that survival
in the conditions studied, was strain dependent.
The ability of L. monocytogenes strains to tolerate and survive under harsh
environmental conditions such as extreme pH values (<4.0 or >9.0) is well documented in
literature (Cotter and Hill, 2003; Dykes and Moorhead, 2000; Giotis et al., 2008b; Giotis et
al., 2008a, Lianou et al., 2006; Liu et al., 2005; Metselaar et al., 2013). External stresses
often result in inhibition of growth and once the stress factors are removed, L.
monocytogenes is capable of multiplying (Giotis et al., 2007; Liu et al., 2005). The
mechanisms underlying L. monocytogenes response to environmental stress conditions,
such as acid, alkali, oxidative and carbon-starvation stresses have become clearer,
following the identification of stress response genes (Gandhi and Chikindas, 2007). In
particular, an alternative sigma factor gene σB that plays a vital role in the general stress
response of L. monocytogenes to acid and alkali hurdles (Giotis et al., 2008a; Sue et al.,
2004).
3.2.1. Acidic stress
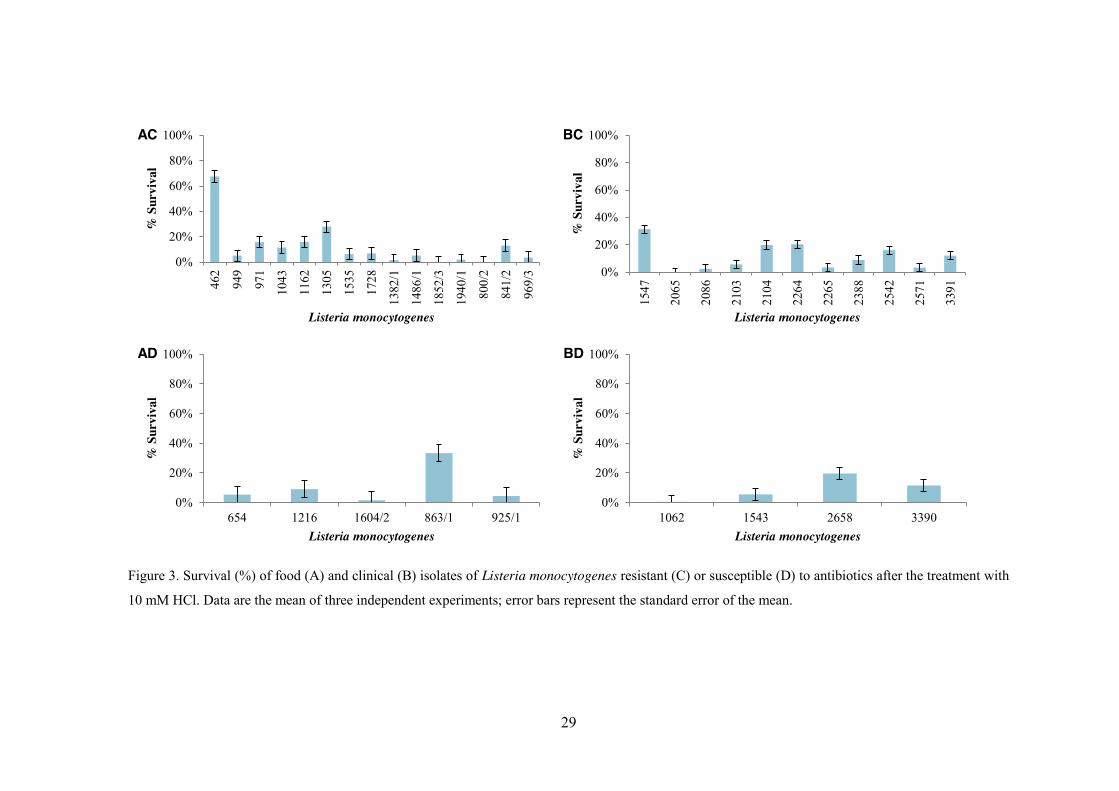
Regarding the 10 mM HCl challenge, after 1 h treatment, pH 1.5, all L.
monocytogenes isolates displayed reductions in their mean viable cell numbers (Table 4).
Of thirty-five L. monocytogenes isolates, only three, two food (L. monocytogenes_1852/3
and L. monocytogenes_800/2) and one clinical (L. monocytogenes_2065), all belonging to
serogrup 1/2b, were not detected after 1 h treatment (Table 4). In fact, viable cell numbers
decreased to values below the detection limit of the enumeration technique following
inoculation for the isolates L. monocytogenes_1852/3 and L. monocytogenes_2065, while
for the isolate L. monocytogenes_800/2 this was observed after 30 min of treatment. In
contrast, L. monocytogenes_462, isolated from frozen bread, 4b, showed, alongside garlic
bread isolate L. monocytogenes_863/1, 1/2c, and L. monocytogenes_1547, isolated from
blood, 1/2b, the highest resistance to acid treatment with 68% and approximately 30%
survivors respectively (Figure 4). Regarding this challenge, isolate L.

27
monocytogenes_2542, clinical 4b, associated with the Portuguese outbreak, showed a
survival of 16% (Figure 4). As in the previous challenge, no significant differences in the
resistance to both treatments were detected between food and clinical isolates or between
antibiotic sensitive and antibiotic resistant isolates (p > 0.05).
Although, a number of researchers have reported the inhibitory effects of low pH
on L. monocytogenes (Conner et al., 1990; Ita and Hutkins, 1991; Phan-Thanh et al., 2000;
Vasseur et al., 1999), the results of the current study demonstrated that L. monocytogenes
isolates possibly evolved adaptive networks that allow them to tolerate low pH as
previously reported in other studies (Barmpalia-Davis et al., 2008; Dykes and Moorhead,
2000; Francis and O’Beirne, 2005;; Liu et al., 2005). Indeed, of thirty-five L.
monocytogenes isolates, only two (L. monocytogenes_1852/3; L. monocytogenes_2065)
were not detected after 30 minutes exposure and one (L. monocytogenes_800/2) after one
hour. Nonetheless the pH tested in this treatment was very low, pH 1.5, Ita and Hutkins
(1991) observed that low intracellular pH was not the major factor in the inhibition of L.
monocytogenes at acid pH. In fact, cells treated with organic acids or HCl at pH values as
low as 3.5 were able to maintain their cytoplasmic pH at a value near 5 (Ita and Hutkins,
1991). Budde and Jakobsen (2000), Shabala et al. (2002) and Siegumfeldt et al. (1999)
also demonstrated that L. monocytogenes maintains its intracellular pH within a narrow
range of 7.6 to 8.0 at extracellular pH values of 5.0 to 8.0 and at pH 4.0 in the presence of
glucose. Confronted with acidic conditions, bacterial cells attempted to resist by
maintaining its intracellular homeostasis whose failure leads to loss of cell viability
(Chitarra et al., 2000). Listeria, a facultative bacterium, may use aerobic or anaerobic
processes to maintain its intracellular pH homeostasis (Phan-Thanh et al., 2000); indeed, in
aerobic bacteria, the active transport of H+ is coupled to the process of electron transport in
respiratory chains; in anaerobic bacteria, H+ transport is carried out through a specific H+
channel in the F0F1-ATPase molecule (proton pump) using energy from ATP hydrolysis
(Cotter et al., 2000).

28
Table 4. Survival of thirty-five Listeria monocytogenes isolates when exposed to acidic
treatment with 10 mM HCl
log cfu/mL
L. monocytogenes isolatea Source 0 min 30 min 60 min 462 Food 7.2±0.06 7.1±0.06 7.1±0.02 654 Food 6.7±0.43 5.8±0.36 5.0±0.01 949 Food 6.9±0.19 6.3±0.37 5.6±0.01 971 Food 6.4±0.39 5.6±0.24 5.2±0.02
1043 Food 6.1±0.40 5.2±0.15 4.9±0.02 1162 Food 6.1±0.26 5.4±0.05 5.2±0.02 1216 Food 7.0±0.05 6.6±0.01 5.9±0.02 1305 Food 6.9±0.16 6.4±0.03 6.2±0.01 1535 Food 6.7±0.20 5.6±0.02 5.4±0.01 1728 Food 6.9±0.36 6.3±0.34 5.4±0.01
1382/1 Food 6.7±0.52 4.7±0.21 4.3±0.01 1486/1 Food 6.9±0.02 6.4±0.11 5.6±0.02 1604/2 Food 7.1±0.03 5.9±0.02 5.3±0.02 1852/3 Food 6.3±0.13 <2.8±0.00 <2.8±0.00 1940/1 Food 7.0±0.25 5.3±0.01 5.2±0.03 800/2 Food 5.9±0.41 4.0±0.01 <2.8±0.00 841/2 Food 7.1±0.23 6.5±0.32 6.1±0.04 863/1 Food 5.9±0.27 5.4±0.09 5.2±0.07 925/1 Food 5.9±0.32 4.5±0.05 4.4±0.01 969/3 Food 5.7±0.23 4.3±0.04 4.1±0.03 1062 Clinical 6.8±0.14 4.7±0.10 4.4±0.01 1543 Clinical 6.8±0.15 6.1±0.09 5.5±0.02 1547 Clinical 6.1±0.28 5.6±0.33 5.5±0.33 2065 Clinical 7.2±0.09 <2.8±0.00 <2.8±0.00 2086 Clinical 6.9±0.12 5.6±0.15 5.3±0.01 2103 Clinical 7.1±0.02 6.5±0.14 5.9±0.00 2104 Clinical 6.9±0.13 6.3±0.01 6.1±0.01 2264 Clinical 6.9±0.13 6.4±0.07 6.2±0.05 2265 Clinical 6.8±0.18 5.8±0.38 5.2±0.33 2388 Clinical 6.7±0.24 5.8±0.33 5.2±0.33 2542 Clinical 7.0±0.03 6.2±0.15 6.2±0.02 2571 Clinical 6.8±0.13 5.8±0.26 5.3±0.02 2658 Clinical 6.9±0.04 6.3±0.16 6.2±0.02 3390 Clinical 7.0±0.02 6.4±0.12 6.1±0.09 3391 Clinical 7.3±0.19 6.6±0.04 6.2±0.18
aCodes in L. monocytogenes collection of Escola Superior de Biotecnologia-Universidade Católica
Portuguesa.
29
Figure 3. Survival (%) of food (A) and clinical (B) isolates of Listeria monocytogenes resistant (C) or susceptible (D) to antibiotics after the treatment with
10 mM HCl. Data are the mean of three independent experiments; error bars represent the standard error of the mean.
0%
20%
40%
60%
80%
100%
462
949
971
1043
1162
1305
1535
1728
1382
/1
1486
/1
1852
/3
1940
/1
800/
2
841/
2
969/
3
% S
urvi
val
Listeria monocytogenes
AC
0%
20%
40%
60%
80%
100%
1547
2065
2086
2103
2104
2264
2265
2388
2542
2571
3391
% S
urvi
val
Listeria monocytogenes
BC
0%
20%
40%
60%
80%
100%
654 1216 1604/2 863/1 925/1
% S
urvi
val
Listeria monocytogenes
AD
0%
20%
40%
60%
80%
100%
1062 1543 2658 3390
% S
urvi
val
Listeria monocytogenes
BD

30
3.2.2. Alkaline stress
With reference to 100 mM NaOH treatment, after 1 h at pH 12.0, L. monocytogenes
isolates displayed reductions in their mean viable cell numbers (Table 5). Although this
noteworthy capacity to tolerate this challenging condition, two isolates, L.
monocytogenes_863/1, garlic bread 1/2c, and L. monocytogenes_1547, blood 1/2b, failed
to recover after 30 min of such treatment (Table 5). On the other hand, the alheira isolate
L. monocytogenes_1852/3, 1/2b, and L. monocytogenes_2542, clinical 4b, associated with
the Portuguese outbreak, demonstrated the highest resistance to alkaline treatment with
approximately 38% and 31% survivors respectively (Figure 5). One hour of exposure to
pH 12.0 resulted in 0.8 log cycle reduction for isolate L. monocytogenes_2542 (Table 5).
No significant differences or clear trends in overall reduction were apparent based on
source (clinical or food) or antibiotic resistance profile (p > 0.05).
The use of alkaline cleaners and sanitizers is widespread in food processing
environments, thus L. monocytogenes may be exposed to alkaline conditions in a variety of
pre- and post- processing environments (Sharma et al., 2002). Alkaline solutions, such as
NaOH are generally used in detergents to remove heavy soils (proteins and fats), eliminate
carbonized sediment, oil or grease since they facilitate protein denaturation, fats
saponification and have a bactericidal activity (Sharma et al., 2002; Vasseur et al., 1999).
Our results have shown a great tolerance to high pH values since only two (L.
monocytogenes_863/1, L. monocytogenes_1547) of thirty-five L. monocytogenes isolates
were not detected after one hour of exposure to such conditions. Growth of L.
monocytogenes at pH > 9.5 is unusual (Gray and Killinger, 1966), but a number of recent
studies demonstrated that the expression of factor σB-dependent genes plays a fundamental
role in cellular survival of L. monocytogenes under adverse conditions, such as alkaline
environments (Becker et al., 2000; Ferreira et al., 2001; Giotis et al., 2008b; Giotis et al.,
2008a; Sharma et al., 2002; Wemekamp-Kamphuis et al., 2004; Wiedmann et al., 1998).
Cheroutre-Vialette et al. (1998) investigated the effects of water activity and pH shifts on
the growth of three L. monocytogenes strains using different osmotic solutes among them
NaOH. They concluded that cells were not affected by the presence of NaOH (Cheroutre-
Vialette et al., 1998). Vasseur et al. (1999) performed a similar study and evaluated the
responses of five L. monocytogenes strains to an alkaline treatment by the addition of

31
NaOH in order to achieve different pH values. They saw that increased alkalinity induced
an increase in lag phases for all strains (Vasseur et al., 1999). However, at pH 11 (highest
pH value tested) three of five strains were more tolerant to alkaline conditions (Vasseur et
al., 1999).
From a food safety point of view, an obvious implication from this study is that use
of acid or alkali in a food processing environment may not be sufficient to eliminate all
cells of L. monocytogenes.
The absence of a clear association of acid or alkaline resistance with origin of the
isolate (clinical or food) and antibiotic resistance profile could be due to the limited
statistical power of the analysis resulting from the small number of clinical and AS
isolates.
32
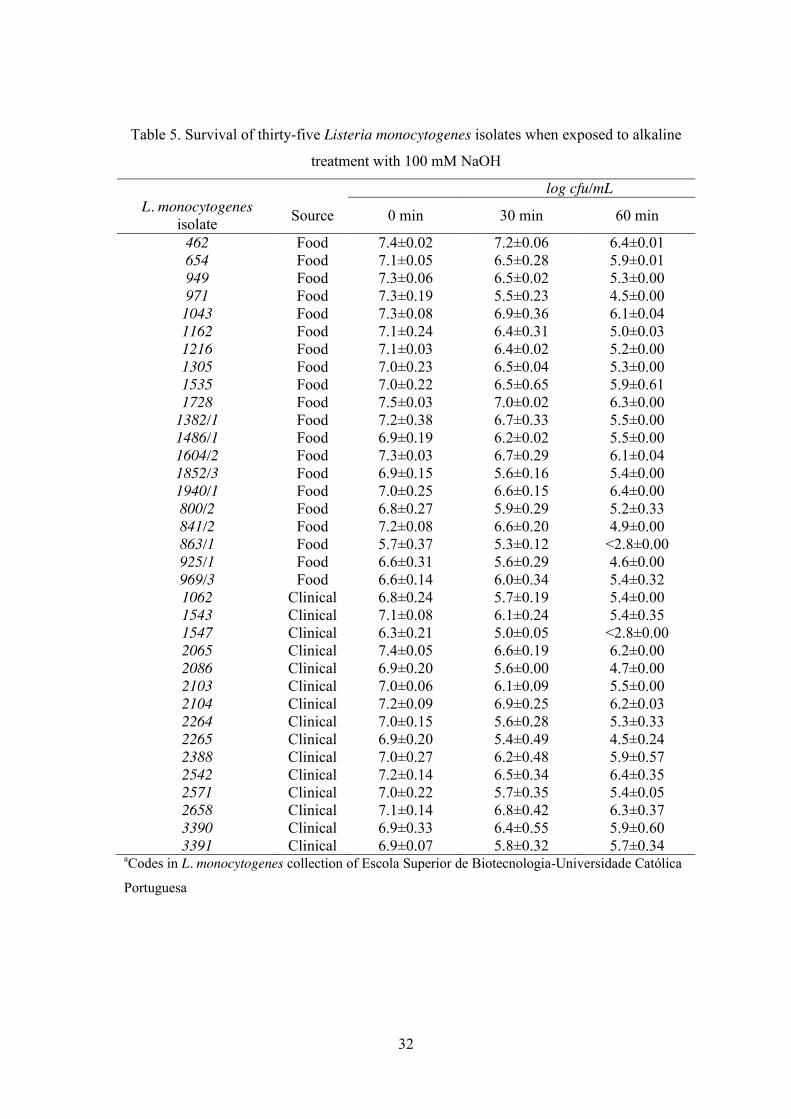
Table 5. Survival of thirty-five Listeria monocytogenes isolates when exposed to alkaline
treatment with 100 mM NaOH
log cfu/mL L. monocytogenes
isolate Source 0 min 30 min 60 min
462 Food 7.4±0.02 7.2±0.06 6.4±0.01 654 Food 7.1±0.05 6.5±0.28 5.9±0.01 949 Food 7.3±0.06 6.5±0.02 5.3±0.00 971 Food 7.3±0.19 5.5±0.23 4.5±0.00
1043 Food 7.3±0.08 6.9±0.36 6.1±0.04 1162 Food 7.1±0.24 6.4±0.31 5.0±0.03 1216 Food 7.1±0.03 6.4±0.02 5.2±0.00 1305 Food 7.0±0.23 6.5±0.04 5.3±0.00 1535 Food 7.0±0.22 6.5±0.65 5.9±0.61 1728 Food 7.5±0.03 7.0±0.02 6.3±0.00
1382/1 Food 7.2±0.38 6.7±0.33 5.5±0.00 1486/1 Food 6.9±0.19 6.2±0.02 5.5±0.00 1604/2 Food 7.3±0.03 6.7±0.29 6.1±0.04 1852/3 Food 6.9±0.15 5.6±0.16 5.4±0.00 1940/1 Food 7.0±0.25 6.6±0.15 6.4±0.00 800/2 Food 6.8±0.27 5.9±0.29 5.2±0.33 841/2 Food 7.2±0.08 6.6±0.20 4.9±0.00 863/1 Food 5.7±0.37 5.3±0.12 <2.8±0.00 925/1 Food 6.6±0.31 5.6±0.29 4.6±0.00 969/3 Food 6.6±0.14 6.0±0.34 5.4±0.32 1062 Clinical 6.8±0.24 5.7±0.19 5.4±0.00 1543 Clinical 7.1±0.08 6.1±0.24 5.4±0.35 1547 Clinical 6.3±0.21 5.0±0.05 <2.8±0.00 2065 Clinical 7.4±0.05 6.6±0.19 6.2±0.00 2086 Clinical 6.9±0.20 5.6±0.00 4.7±0.00 2103 Clinical 7.0±0.06 6.1±0.09 5.5±0.00 2104 Clinical 7.2±0.09 6.9±0.25 6.2±0.03 2264 Clinical 7.0±0.15 5.6±0.28 5.3±0.33 2265 Clinical 6.9±0.20 5.4±0.49 4.5±0.24 2388 Clinical 7.0±0.27 6.2±0.48 5.9±0.57 2542 Clinical 7.2±0.14 6.5±0.34 6.4±0.35 2571 Clinical 7.0±0.22 5.7±0.35 5.4±0.05 2658 Clinical 7.1±0.14 6.8±0.42 6.3±0.37 3390 Clinical 6.9±0.33 6.4±0.55 5.9±0.60 3391 Clinical 6.9±0.07 5.8±0.32 5.7±0.34
aCodes in L. monocytogenes collection of Escola Superior de Biotecnologia-Universidade Católica
Portuguesa

33
Figure 4. Survival (%) of food (A) and clinical (B) isolates of Listeria monocytogenes resistant (C) or susceptible (D) to antibiotics after the treatment with
100 mM NaOH. Data are the mean of three independent experiments; error bars represent the standard error of the mean.
0%
20%
40%
60%
80%
100%
462
949
971
1043
1162
1305
1535
1728
1382
/1
1486
/1
1852
/3
1940
/1
800/
2
841/
2
969/
3
% S
urvi
val
Listeria monocytogenes
AC
0%
20%
40%
60%
80%
100%
1547
2065
2086
2103
2104
2264
2265
2388
2542
2571
3391
% S
urvi
val
Listeria monocytogenes
BC
0%
20%
40%
60%
80%
100%
654 1216 1604/2 863/1 925/1
% S
urvi
val
Listeria monocytogenes
AD
0%
20%
40%
60%
80%
100%
1062 1543 2658 3390
% S
urvi
val
Listeria monocytogenes
BD

34
3.3. Antimicrobial resistance and stress resistance
The antibiotic-resistant profiles of the thirty-five L. monocytogenes isolates used
are shown in Table 1. In this study, the results obtained for antibiotic resistant (AR)
isolates did not differ from those obtained for antibiotic sensitive (AS) isolates under any
of the conditions examined (Figures 2-4), suggesting that antibiotic resistance mechanisms
did not confer any cross-resistance to gastrointestinal passage, acid or alkaline stresses.

35
4. Conclusions
Once L. monocytogenes is ingested by a mammalian host through contaminated
foods, its survival within that host depends upon the bacterium’s ability to survive the acid
environment of the stomach, the secretion of proteolytic enzymes and the inhibitory action
of bile salts in the small intestine. In this study the survival of different isolates of L.
monocytogenes through simulated GI conditions was evaluated. It was demonstrated that
L. monocytogenes was able to survive in extreme acid conditions. However, it did not
survive when submitted to sequential GI transit, i.e., presence of bile salts. Although GI
conditions do not simulate the physiological environment of digestion in the human GI
tract, they allow us to understand the microorganism’s fate in the digestive environment.
From a food safety point of view the results of this study renders many of the
conventional food manufacturing processes ineffective against L. monocytogenes cells, as a
certain number of this species will survive at very extreme pH conditions.
It was also concluded in the present study that antibiotic-resistant isolates of L.
monocytogenes are not more GI-, acid- or alkaline-tolerant than antibiotic-susceptible
isolates, but, at the same time, are not less GI-, acid- or alkaline-tolerant either. It is
important to point out that an unequal number of AR and AS isolates was used in this
study and this may have contributed to bias in the results. The continuing increase in the
prevalence of AR L. monocytogenes in foods justifies further investigation of the stress
responses of such strains, in particular for multiple antibiotic-resistant strains.
The response of thirty-five L. monocytogenes isolates was shown to be strain- and
stress-dependent and no relation between food and clinical isolates was observed.

36
5. Proposals for future work
The present work demonstrated that L. monocytogenes is able to survive under
extreme acid and alkaline conditions, but not through sequential GI tract, i.e. the activity of
bile salts after the combined action of acid and pepsin. Also, the presence or absence of
antibiotic resistance genes did not modulate the resistance response of the isolates
examined towards the stress conditions investigated. In order to investigate and understand
the mechanisms that underly these answers is important to comprehend the bacterial
strategies that effectively allow resistance development as well as overcoming some
limitations inherent to the technique used to simulate GI tract digestion. Therefore,
suggestions for future work include:
1. Improvement in the artificial digestive systems, extending their potential and
approach the in vivo human situation more closely. Always bearing in mind that it
is still impossible to mimic fully the overall digestive parameters in vivo in a single
in vitro model;
2. Evaluation of genotypic and phenotypic characteristics of L. monocytogenes
isolates, concerning:
x Resistance, adaptation and cross-adaptation to conventional food preservatives;
x Resistance to different stress conditions (e.g., salt, pH, etc.);
x Acquisition of antibiotic resistance genes by L. monocytogenes and possible
linkage to resistance to food processing stresses.

37
6. References
Abee, T., and Wouters, J. A. (1999). Microbial stress response in minimal processing.
International Journal of Food Microbiology, 50, 65–91.
Allerberger, F. (2003). Listeria: growth, phenotypic differentiation and molecular
microbiology. FEMS Immunology and Medical Microbiology, 35, 183–189.
Almeida, G. N., Gibbs, P. A, Hogg, T. A, and Teixeira, P. C. (2006). Listeriosis in
Portugal: an existing but under reported infection. BMC Infectious Diseases, 6, 153.
Almeida, G. N., Magalhães, R. B., Barbosa, J. B., Hogg, T. A., and Teixeira, P. C. (2009).
Listeriose em Portugal: 2004 - 2007. Revista Portuguesa de Doenças Infecciosas, 6,
90–94.
Almeida, G., Schlech, A., Magalhães, R., Santos, I., Hogg, T., Leclercq, A., and Teixeira,
P. (2010). Distribution and characterization of Listeria monocytogenes clinical
isolates in Portugal, 1994-2007. European Journal of Clinical Microbiology and
Infectious Diseases, 29, 1219–1227.
Andre, P., and Genicot, A. (1987). First isolation of Listeria welshimeri in a human.
Zentralblatt für Bakteriologie, Mikrobiologie und Hygiene, 263, 605–606.
Avery, S. M., and Buncic, S. (1997). Differences in pathogenicity for chick embryos and
growth kinetics at 37 ºC between clinical and meat isolates of Listeria
monocytogenes previously stored at 4 ºC. International Journal of Food
Microbiology, 34, 319–27.
Barbosa, J., Borges, S., Magalhães, R., Ferreira, V., Santos, I., Silva, J., Almeida, G.,
Gibbs, P., and Teixeira, P. (2012). Behaviour of Listeria monocytogenes isolates
through gastro-intestinal tract passage simulation, before and after two sub-lethal
stresses. Food Microbiology, 30, 24–28.
Barbosa, J., Magalhães, R., Santos, I., Ferreira, V., Brandão, T. R. S., Silva, J., Almeida,
G., and Teixeira, P. (2013). Evaluation of Antibiotic Resistance Patterns of Food
and Clinical Listeria monocytogenes Isolates in Portugal. Foodborne Pathogens
and Disease, 10, 861-866.
Barbosa, T. M., and Levy, S. B. (2000). The impact of antibiotic use on resistance
development and persistence. Drug resistance updates, 3, 303–311.

38
Barmpalia-Davis, I. M., Geornaras, I., Kendall, P. A., and Sofos, J. N. (2008a). Survival of
Listeria monocytogenes in a simulated dynamic gastrointestinal model during
storage of inoculated bologna and salami slices in vacuum packages. Journal of
Food Protection, 71, 2014–2023.
Barmpalia-Davis, I. M., Geornaras, I., Kendall, P. A., and Sofos, J. N. (2009). Effect of fat
content on survival of Listeria monocytogenes during simulated digestion of
inoculated beef frankfurters stored at 7 degrees C. Food Microbiology, 26, 483–490.
Barmpalia-Davis, I. M., Geornaras, I., Kendall, P. A., and Sofos, J. N. (2008). Differences
in survival among 13 Listeria monocytogenes strains in a dynamic model of the
stomach and small intestine. Applied and Environmental Microbiology, 74, 5563–
5567.
Beales, N. (2004). Adaptation of Microorganisms to Cold Temperatures, Weak Acid
Preservatives, Low pH, and Osmotic Stress: A Review. Comprehensive Reviews in
Food Science and Food Safety, 3, 1–20.
Becker, L. A., Cetin, M. S., Hutkins, R. W., and Benson, A. K. (1998). Identification of the
Gene Encoding the Alternative Sigma Factor sigma B from Listeria monocytogenes
and Its Role in Osmotolerance. Journal of Bacteriology, 180, 4547–4554.
Bell, C., and Kyriakides, A. (2005). Listeria: A Practical Approach to the Organism and
its Control in Foods (Practical Food Microbiology) (2nd ed., pp. 296–300). Oxford:
Blackwell Publishing.
Bertsch, D., Rau, J., Eugster, M. R., Haug, M. C., Lawson, P. A., Lacroix, C., and Meile,
L. (2013). Listeria fleischmannii sp. nov., isolated from cheese. International
Journal of Systematic and Evolutionary Microbiology, 63, 526–532.
Bowman, J. P., Lee Chang, K. J., Pinfold, T. and Ross, T. (2010). Transcriptomic and
phenotypic responses of Listeria monocytogenes strains possessing different growth
efficiencies under acidic conditions. Applied and Environmental Microbiology, 4836–
4850.
Budde, B.B., and Jakobsen, M. (2000). Real-time measurements of the interaction between
single cells of Listeria monocytogenes and nisin on a solid surface. Applied and
Environmental Microbiology, 66, 3586–3591.
Cain, D. B., and McCann, V. L. (1986). An unusual case of cutaneous listeriosis. Journal
of Clinical Microbiology, 23, 976–977.

39
Cheroutre-Vialette, M., Lebert, I., Hebraud, M., Labadie, JC. and Lebert, A. (1998).
Effects of pH or aw stress on growth of Listeria monocytogenes. International
Journal of Food Microbiology, 42, 71–77.
Chitarra, L. G., Breeuwer, P., van den Bulk, R. W., and Abee, T. (2000). Rapid
fluorescence assessment of intracellular pH as a viability indicator of Clavibacter
michiganensis subsp michiganensis. Journal of Applied Microbiology, 88, 809–
816.
Conner, D.E., Scott, V.N. and Bernard, D.T. (1990). Growth, inhibition and survival of
Listeria monocytogenes as affected by acidic conditions. Journal of Food
Protection, 55, 652–655.
Conter, M., Paludi, D., Zanardi, E., Ghidini, S., Vergara, A., and Ianieri, A. (2009).
Characterization of antimicrobial resistance of foodborne Listeria monocytogenes.
International Journal of Food Microbiology, 128, 497–500.
Cotter, P. D., and Hill, C. (2003). Surviving the Acid Test : Responses of Gram-Positive
Bacteria to Low pH. Microbiology and Molecular Biology Reviews, 67, 429–453.
Cotter, P. D., Emerson, N., Gahan, C. G. M., Hill, C., Cotter, P. D., Emerson, N., and
Gahan, C. G. M. (1999). Identification and Disruption of lisRK, a Genetic Locus
Encoding a Two-Component Signal Transduction System Involved in Stress
Tolerance and Virulence in Listeria monocytogenes. Journal of Bacteriology, 181,
6840–6843.
Cotter, P. D., Gahan, C. G., and Hill, C. (2000). Analysis of the role of the Listeria
monocytogenes F0F1-ATPase operon in the acid tolerance response. International
Journal of Food Microbiology, 60, 137–146.
Cotter, P. D., Gahan, C. G., and Hill, C. (2001). A glutamate decarboxylase system
protects Listeria monocytogenes in gastric fluid. Molecular Microbiology, 40, 465–
75.
Cummins, A. J., Fielding, A. K., and McLauchlin, J. (1994). Listeria ivanovii infection in a
patient with AIDS. The Journal of infection, 28, 89–91.
Denny, J., and McLauchlin, J. (2008). Human Listeria monocytogenes infections in Europe
- an opportunity for improved european surveillance. Surveillance and Outbreak
Reports, 13, 1–5.
Doherty, A.M., McMahon, C.M.M., Sheridan, J.J., Blair, I.S., McDowell, D.A. and

40
Hegarty, T. (1998) Thermal resistance of Yersinia enterocolitica and Listeria
monocytogenes in meat and potato substrates. Journal of Food Safety, 18, 69-83.
Dombroski, C.S., Jaykus, L.A., Green, D.P. and Farkas, B.E. (1999) Use of mutant strain
for evaluating processing strategies to inactivate Vibrio vulnificus in oysters.
Journal of Food Protection, 62, 592-600.
Donnelly, C. W. (2002). Detection and isolation of Listeria monocytogenes from food
samples: implications of sublethal injury. Journal of AOAC International, 85, 495–
500.
Doumith, M., Buchrieser, C., Glaser, P., Jacquet, C., and Martin, P. (2004). Differentiation
of the major Listeria monocytogenes serovars by multiplex PCR. Journal of
Clinical Microbiology, 42, 3819–3822. Doyle, M. P., Loneragan, G. H., Scott, H. M., and Singer, R. S. (2013). Antimicrobial
Resistance: Challenges and Perspectives. Comprehensive Reviews in Food Science
and Food Safety, 12, 234–248.
Duché, O., Trémoulet, F., Glaser, P., and Labadie, J. (2002). Salt Stress Proteins Induced
in Listeria monocytogenes. Applied and Environmental Microbiology, 68, 1491–1498.
Duffy, G., Walsh, C., Blair, I. S., and McDowell, D. A. (2006). Survival of antibiotic
resistant and antibiotic sensitive strains of E. coli O157 and E. coli O26 in food
matrices. International Journal of Food Microbiology, 109, 179–186.
Dykes, G. A., and Moorhead, S. M. (2000). Survival of osmotic and acid stress by Listeria
monocytogenes strains of clinical or meat origin. International Journal of Food
Microbiology, 56, 161–6.
European Food Safety Authority (2007). The Community Summary Report on Trends and
Sources of Zoonoses, Zoonotic Agents, Antimicrobial resistance and Foodborne
outbreaks in the European Union in 2006. EFSA Journal 2007, 130, 1-352.
European Food Safety Authority, European Centre for Disease Prevention and Control
(2011). The European Union Summary Report on Trends and Sources of Zoonoses,
Zoonotic Agents and Food-borne Outbreaks in 2009. EFSA Journal 2011, 9, 2090.
European Food Safety Authority, European Centre for Disease Prevention and Control
(2013). The European Union Summary Report on Trends and Sources of Zoonoses,
Zoonotic Agents and Food-borne Outbreaks in 2011. EFSA Journal 2013, 11,
3129.

41
Farber, J. M., and Peterkin, P. I. (1991). Listeria monocytogenes, a Food-Borne Pathogen.
Microbiological Reviews, 55, 476–511.
Ferreira, A., O’Byrne, C. P., and Boor, K. J. (2001). Role of B in Heat, Ethanol, Acid, and
Oxidative Stress Resistance and during Carbon Starvation in Listeria
monocytogenes. Applied and Environmental Microbiology, 67, 4454–4457.
Flahaut, S., Hartke, A., Giard, J. C., and Auffray, Y. (1997). Alkaline stress response in
Enterococcus faecalis: adaptation, cross-protection, and changes in protein synthesis.
Applied and Environmental Microbiology, 63, 812–814.
Francis, G. A., and O’Beirne, D. (2005). Variation among strains of Listeria
monocytogenes: differences in survival on packaged vegetables and in response to
heat and acid conditions. Food Control, 16, 687–694.
Francis, G. A., and O’Beirne, D. (2005). Variation among strains of Listeria
monocytogenes: differences in survival on packaged vegetables and in response to
heat and acid conditions. Food Control, 16, 687–694.
Gahan, C., O’Driscoll, B., and Hill, C. (1996). Acid adaptation of Listeria monocytogenes
can enhance survival in acidic foods and during milk fermentation. Applied and
Environmental Microbiology, 62, 3128–3132.
Gandhi, M., and Chikindas, M. L. (2007). Listeria: A foodborne pathogen that knows how
to survive. International Journal of Food Microbiology, 113, 1–15.
Giotis, E. S., Blair, I. S., and McDowell, D. A. (2007). Morphological changes in Listeria
monocytogenes subjected to sublethal alkaline stress. International Journal of Food
Microbiology, 120, 250–258.
Giotis, E. S., Julotok, M., Wilkinson, B. J., Blair, I. S., and McDowell, D. (2008a). Role of
sigma B factor in the alkaline tolerance response of Listeria monocytogenes 10403S
and cross-protection against subsequent ethanol and osmotic stress. Journal of Food
Protection, 71, 1481–1485.
Giotis, E. S., Muthaiyan, A., Blair, I. S., Wilkinson, B. J., and McDowell, D. A. (2008b).
Genomic and proteomic analysis of the Alkali-Tolerance Response (AlTR) in Listeria
monocytogenes 10403S. BMC Microbiology, 8, 102.
Graves, L. M., Helsel, L. O., Steigerwalt, A. G., Morey, R. E., Daneshvar, M. I., Roof, S.
E., Orsi, R. H., Fortes, E. D., Milillo, S. R., den Bakker, H.C., Wiedmann, M.,
Swaminathann, B., and Sauders, B. D. (2010). Listeria marthii sp. nov., isolated from

42
the natural environment, Finger Lakes National Forest. International Journal of
Systematic and Evolutionary Microbiology, 60, 1280–1288.
Gray, M. J., Freitag, N. E., and Boor, K. J. (2006). How the Bacterial Pathogen Listeria
monocytogenes Mediates the Switch from Environmental Dr. Jekyll to Pathogenic Mr.
Hyde. Infection and Immunity, 74, 2505–2512.
Gray, M. L. and Killinger, A. H. (1966). Listeria monocytogenes and listeric infections.
Bacteriology Reviews, 30, 309–382.
Gray, M. L., and Killinger, A. H. (1966). Listeria monocytogenes and Listeric Infections.
Bacteriological Reviews, 30, 309–382.
Guillet, C., Join-Lambert, O., Le Monnier, A., Leclercq, A., Mechaï, F., Mamzer-Bruneel,
M. F., Bielecka, M. K., Scortti, M., Disson, O., Berche, P., Vazquez-Boland, J.,
Lortholary, O., and Lecuit, M. (2010). Human Listeriosis caused by Listeria ivanovii.
Emerging Infectious Diseases, 16, 136–138.
Hazan, R., Levine, A., and Abeliovich, H. (2004). Benzoic Acid, a Weak Organic Acid
Food Preservative, Exerts Specific Effects on Intracellular Membrane Trafficking
Pathways in Saccharomyces cerevisiae. Applied and Environmental Microbiology,
70, 4449–4457.
Hill, C., Cotter, P. D., Sleator, R. D., and Gahan, C. G. M. (2002). Bacterial stress response
in Listeria monocytogenes : jumping the hurdles imposed by minimal processing.
International Dairy Journal, 12, 273–283.
Ita, P.S. and Hutkins, R.W. (1991). Intracellular pH and survival of Listeria
monocytogenes Scott A in tryptic soy broth containing acetic, lactic, citric and
hydrochloric acids. Journal of Food Protection, 54, 15–19.
Jeyaletchumi, P., Tunung, R., Margaret, S. P., Son, R., Farinazleen, M. G., and Cheah, Y.
K. (2010). Detection of Listeria monocytogenes in foods. International Food
Research Journal, 11, 1–11.
Jiang, L., Olesen, I., Andersen, T., Fang, W., and Jespersen, L. (2010). Survival of Listeria
monocytogenes in Simulated Gastrointestinal System and Transcriptional Profiling.
Foodborne Pathogens and Disease, 7, 1–8.
Jordan, K. N., Oxford, L., and Byrne, C. P. O. (1999). Survival of Low-pH Stress by
Escherichia coli O157:H7: Correlation between Alterations in the Cell Envelope and
Increased Acid Tolerance. Applied and Environmental Microbiology, 65, 3048–3055.

43
Kazmierczak, M. J., Mithoe, S. C., Boor, K. J., and Wiedmann, M. (2003). Listeria
monocytogenes σB Regulates Stress Response and Virulence Functions. Journal of
Bacteriology, 185, 5722–5734.
King, T., Ferenci, T., and Szabo, E. A. (2003). The effect of growth atmosphere on the
ability of Listeria monocytogenes to survive exposure to acid, proteolytic enzymes
and bile salts. International Journal of Food Microbiology, 84, 133–143.
Lado, B. H., and Yousef, A. E. (2002). Alternative food-preservation technologies:
efficacy and mechanisms. Microbes and Infection, 4, 433–440.
Lang Halter, E., Neuhaus, K., and Scherer, S. (2013). Listeria weihenstephanensis sp. nov.,
isolated from the water plant Lemna trisulca taken from a freshwater pond.
International Journal of Systematic and Evolutionary Microbiology, 63, 641–647.
Leclercq, A., Clermont, D., Bizet, C., Grimont, P. A. D., Le Flèche-Matéos, A., Roche, S.
M., Buchrieser, C., Cadet-Daniel, V., Le Monnier, A., Lecuit, M., and Allerberger, F.
(2010). Listeria rocourtiae sp. nov.. International Journal of Systematic and
Evolutionary Microbiology, 60, 2210–2214.
Lecuit, M. (2007). Human listeriosis and animal models. Microbes and Infection, 9, 1216–
1225.
Lianou, A., Stopforth, J. D., Yoon, Y., Wiedmann, M., and Sofos, J. N. (2006). Growth
and stress resistance variation in culture broth among Listeria monocytogenes strains
of various serotypes and origins. Journal of Food Protection, 69, 2640–2647.
Liu, D., Lawrence, M. L., Ainsworth, a J., and Austin, F. W. (2005). Comparative
assessment of acid, alkali and salt tolerance in Listeria monocytogenes virulent and
avirulent strains. FEMS Microbiology Letters, 243, 373–378.
Liu, D., Lawrence, M. L., Wiedmann, M., Gorski, L., Mandrell, R. E., Ainsworth, A. J.,
and Austin, F. W. (2006). Listeria monocytogenes subgroups IIIA, IIIB, and IIIC
delineate genetically distinct populations with varied pathogenic potential. Journal of
Clinical Microbiology, 44, 4229–4233.
Lou, Y., and Yousef, A. E. (1999). Characteristics of Listeria monocytogenes Important
to Food Processors in Listeria, Listeriosis and Food Safety, (2nd ed., pp. 157-214).
New York: Marcel Dekker, Inc.

44
Lou, Y., and Yousef, A. E. (1997). Adaptation to sublethal environmental stresses protects
Listeria monocytogenes against lethal preservation factors. Applied and
Environmental Microbiology, 63, 1252–1255.
Madureira, A. R., Amorim, M., Gomes, A. M., Pintado, M. E., and Malcata, F. X. (2011).
Protective effect of whey cheese matrix on probiotic strains exposed to simulated
gastrointestinal conditions. Food Research International, 44, 465–470.
McGee, P. J. (2003). The relationship between antibiotic resistance and acid tolerance of
E. coli O157:H7. Escherichia Coli O157: H7 in Cattle Production Systems - a
Food Safety Perspective. PhD Thesis, University College Dublin.
McLauchlin, J., and Low, J. C. (1994). Primary cutaneous listeriosis in adults: an
occupational disease of veterinarians and farmers. The Veterinary Record, 135, 615–
617.
Melo, J., Schrama, D., Hussey, S., Andrew, P. W., and Faleiro, M. L. (2013). Listeria
monocytogenes dairy isolates show a different proteome response to sequential
exposure to gastric and intestinal fluids. International Journal of Food Microbiology,
163, 51–63.
Mena, C. (2004). Incidence of Listeria monocytogenes in different food products
commercialized in Portugal. Food Microbiology, 21, 213–216.
Metselaar, K. I., den Besten, H. M. W., Abee, T., Moezelaar, R., and Zwietering, M. H.
(2013). Isolation and quantification of highly acid resistant variants of Listeria
monocytogenes. International Journal of Food Microbiology, 166, 508–514.
Miles, O., and Misra, S. S., (1938). The estimation of the bactericidal power of the blood.
Journal of Hygiene, 45, 41−45.
Murray, E., Webb, R., and Swann, M. (1926). A disease of rabbits characterised by a large
mononuclear leukocytosis, caused by a hitherto undescribed bacillus, Bacterium
monocytogenes. Journal of Pathology and Bacteriology, 29, 407-439.
Nightingale, K. K., Windham, K., and Wiedmann, M. (2005). Evolution and Molecular
Phylogeny of Listeria monocytogenes Isolated from Human and Animal Listeriosis
Cases and Foods. Journal of Bacteriology, 187, 5537–5551.
Nyfeldt, A. (1929). Etiologie de la mononucleose infectieuse. Comptes rendus des Seances
de la Societe de Biologie, 101, 590-591. 1929.

45
Olier, M., Rousseaux, S., Piveteau, P., Lemaître, J.P., Rousset, A., and Guzzo, J. (2004).
Screening of glutamate decarboxylase activity and bile salt resistance of human
asymptomatic carriage, clinical, food, and environmental isolates of Listeria
monocytogenes. International Journal of Food Microbiology, 93, 87–99.
Orsi, R. H., Sun, Q., and Wiedmann, M. (2008). Genome-wide analyses reveal lineage
specific contributions of positive selection and recombination to the evolution of
Listeria monocytogenes. BMC Evolutionary Biology, 8, 233.
Padan, E., Bibi, E., Ito, M., and Krulwich, T. A. (2005). Alkaline pH homeostasis in
bacteria: new insights. Biochimica et Biophysica Acta, 1717, 67–88.
Perrin, M., Bemer, M., and Delamare, C. (2003). Fatal Case of Listeria innocua
Bacteremia. Journal of Clinical Microbiology, 41, 5308–5310.
Peterson, L.D., Faith, N.G., and Czuprynski, C.J (2007). Resistance of Listeria
monocytogenes F2365 cells to synthetic gastric fluid is greater following growth on
ready-to-eat deli turkey meat than in brain heart infusion broth. Journal of Food
Protection, 70, 2589–2595.
Phan-Thanh, L., and Mahouin, F. (1999). A proteomic approach to study the acid response
in Listeria monocytogenes Proteomics and 2-DE. Electrophoresis, 20, 2214–2224.
Phan-Thanh, L., Mahouin, F., and Aligé, S. (2000). Acid responses of Listeria
monocytogenes. International Journal of Food Microbiology, 55, 121–126.
Pirie, J. H. H. (1927). A new disease of veld rodents “Tiger River Disease”. Publication of
the South African Institute of Medical Research, 3, 163-186.
Ramalheira, R., Almeida, M., Azeredo, J., Branda, T. R. S., Almeida, G., Silva, J., and
Teixeira, P. (2010). Survival of Clinical and Food Isolates of Listeria monocytogenes
Through Simulated Gastrointestinal Tract Conditions. Foodborne Pathogens and
Disease, 7, 121–128.
Ripio, A., Domi, G., Lara, A., and Sua, N. (1997). A Gly145Ser substitution in the
transcriptional activator PrfA causes constitutive overexpression of virulence factors
in Listeria monocytogenes. Journal of Bacteriology, 179, 1533–1540.
Roberts, A. J., Williams, S. K., Wiedmann, M., and Nightingale, K. K. (2009). Some
Listeria monocytogenes outbreak strains demonstrate significantly reduced invasion,
inlA transcript levels, and swarming motility in vitro. Applied and Environmental
Microbiology, 75, 5647–5658.

46
Roberts, A., Nightingale, K., Jeffers, G., Fortes, E., Kongo, J. M., and Wiedmann, M.
(2006). Genetic and phenotypic characterization of Listeria monocytogenes lineage
III. Microbiology, 152, 685–693.
Rocourt, J., H. Hof, A. Schrettenbrunner, R. Malinverni and J. Bille. (1986). Acute
purulent Listeria seelingeri meningitis in an immunocompetent adult. Schweizerische
medizinische. Wochenschrift, 116, 248-251.
Ross, R. P., Morgan, S., and Hill, C. (2002). Preservation and fermentation: past, present
and future. International Journal of Food Microbiology, 79, 3–16.
Sallen, B., Rajoharison, A., Desvarenne, S., Quinn, F., and Mabilat, C. (1996).
Comparative Analysis of 16S and 23S rRNA Sequences of Listeria Species.
International Journal of Systematic Bacteriology, 46, 669–674.
Sánchez, B., Champomier-Vergès, M.-C., Stuer-Lauridsen, B., Ruas-Madiedo, P.,
Anglade, P., Baraige, F., de los Reyes-Gavilán, C., Johansen, E., Zagorec, M., and
Margolles, A. (2007). Adaptation and response of Bifidobacterium animalis subsp.
lactis to bile: a proteomic and physiological approach. Applied and environmental
microbiology, 73, 6757–6767.
Schlech 3rd, W. F., P. Lavigne, R. Bortolussi, A. Allen, E. Haldane, A. Wort, A.
Hightower, S. Johnson, S. King and E. Nicholls. (1983). Epidemic listeriosis -
evidence for transmission by food. New England Journal of Medicine, 308, 203- 205.
Schuchat, A., Swaminathan, B., and Broome, C. V. (1991). Epidemiology of Human
Listeriosis. Clinical Microbiology Reviews, 4, 169–183.
Shabala, L., Budde, B., Ross, T., Siegumfeldt, H., and McMeekin, T. (2002). Responses of
Listeria monocytogenes to acid stress and glucose availability monitored by
measurements of intracellular pH and viable counts. International Journal of Food
Microbiology, 75, 89–97.
Sharma, M., Taormina, P., and Beuchat, L. (2003). Habituation of Foodborne Pathogens
Exposed to Extreme pH Conditions : Genetic Basis and Implications in Foods and
Food Processing Environments. Food Science Technology Research, 9, 115–127.
Siegumfeldt, H., Rechinger, K.B., and Jakobsen, M. (1999). Use of fluorescence ratio
imaging for intracellular pH determination of individual bacterial cells in mixed
cultures. Microbiology, 145,1703–1709.

47
Sue, D., Fink, D., Wiedmann, M., and Boor, K. J. (2004). sigmaB-dependent gene
induction and expression in Listeria monocytogenes during osmotic and acid stress
conditions simulating the intestinal environment. Microbiology, 150, 3843–55.
Swaminathan, B., and Gerner-Smidt, P. (2007). The epidemiology of human listeriosis.
Microbes and Infection, 9, 1236–1243.
Vasseur, C., Baverel, L., Hébraud, M., and Labadie, J. (1999). Effect of osmotic, alkaline,
acid or thermal stresses on the growth and inhibition of Listeria monocytogenes.
Journal of Applied Microbiology, 86, 469–76.
Vázquez-Boland, J. A., Kuhn, M., Berche, P., Chakraborty, T., Domínguez-Bernal, G.,
Goebel, W., Lez-Zorn, B. G., Wehland, J., Goebel, W., and Kreft, J. (2001). Listeria
Pathogenesis and Molecular Virulence Determinants. Clinical Microbiology REviews,
14, 584–640.
Vialette, M., Pinon, A., Chasseignaux, E., and Lange, M. (2003). Growths kinetics
comparison of clinical and seafood Listeria monocytogenes isolates in acid and
osmotic environment. International Journal of Food Microbiology, 82, 121–131.
Walecka, E. and Bania, J. (2012). Stress Response in Microbiology. (1st ed., pp. 91–114).
Norflok: Caister Academic Press.
Walsh, C., Duffy, D., Sheridan, J. J., Fanning, S., Blair, I. S., and McDowell, D. A. (2005).
Thermal resistance of antibiotic resistant and antibiotic sensitive Salmonella spp. on
chicken meat. Journal of Food Safety, 25, 288–302.
Walsh, D., Sheridan, J. J., Duffy, G., Blair, I. S., McDowell, D. A., and Harrington, D.
(2001). Thermal resistance of wild-type and antibiotic-resistant Listeria
monocytogenes in meat and potato substrates. Journal of Applied Microbiology, 90,
555–560.
Ward, T. J., Ducey, T. F., Usgaard, T., Dunn, K. a, and Bielawski, J. P. (2008). Multilocus
genotyping assays for single nucleotide polymorphism-based subtyping of Listeria
monocytogenes isolates. Applied and Environmental Microbiology, 74, 7629–7642.
Wemekamp-Kamphuis, H. H., Wouters, J. A., de Leeuw, P. P. L. A., Hain, T.,
Chakraborty, T., and Abee, T. (2004). Identification of sigma factor sigma B-
controlled genes and their impact on acid stress, high hydrostatic pressure, and
freeze survival in Listeria monocytogenes EGD-e. Applied and Environmental
Microbiology, 70, 3457–66.

48
Werbrouck, H., Botteldoorn, N., Ceelen, L., Decostere, A., Uyttendaele, M., Herman, L.,
and Van Coillie, E. (2008). Characterization of virulence properties of Listeria
monocytogenes serotype 4b strains of different origins. Zoonoses and Public Health,
55, 242–248.
Wesche, A. M., Gurtler, J. B., Marks, B. P., and Ryser, E. T. (2009). Stress, sublethal
injury, resuscitation, and virulence of bacterial foodborne pathogens. Journal of Food
Protection, 72, 1121–1138.
Wiedmann, M., Arvik, T. J., Hurley, R. J., and Boor, J. (1998). General Stress
Transcription Factor B and Its Role in Acid Tolerance and Virulence of Listeria
monocytogenes. Journal of Bacteriology, 180, 3650–3656.
Wiedmann, M., Arvik, T. J., Hurley, R. J., and Boor, K. J. (1998). General Stress
Transcription Factor sigma B and Its Role in Acid Tolerance and Virulence of
Listeria monocytogenes. Journal of Bacteriology, 180, 3650–3656.
Yousef, A. E. and Courtney, P. D. (2003). Basics of stress adaptation and implications in
new-generation foods in Microbial stress adaptation and food safety, (1st ed., pp. 256-
310). Florida: CRC Press.
Zhang, L., Huang, Y., Zhou, Y., Buckley, T., and Wang, H. H. (2013). Antibiotic
administration routes significantly influence the levels of antibiotic resistance in gut
microbiota. Antimicrobial Agents and Chemotherapy, 57, 3659–3666.

![Proteomes of Lactobacillus delbrueckii subsp. bulgaricus LBB.B5 … › content › msys › 2 › 5 › e00027-17.full.pdf · “translation (J)” (21 proteins [15.91%]) were significantly](https://static.fdocuments.ec/doc/165x107/5f27274997fd4b72907bdf00/proteomes-of-lactobacillus-delbrueckii-subsp-bulgaricus-lbbb5-a-content-a.jpg)
















