
(RRUBBI IAACCE EAE E )) BAAO O RLLOONNGGOO DDE UUMM … · 2018-09-06 · universidade federal da...
62
UNIVERSIDADE FEDERAL DA PARAÍBA CENTRO DE CIÊNCIAS EXATAS E DA NATUREZA PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS BIOLÓGICAS (ZOOLOGIA) RENATA MARINHO CRUZ ABELHAS VISITANTES FLORAIS DE RICHARDIA GRANDIFLORA (RUBIACEAE) AO LONGO DE UM GRADIENTE URBANO-RURAL JOÃO PESSOA 2013
Transcript of (RRUBBI IAACCE EAE E )) BAAO O RLLOONNGGOO DDE UUMM … · 2018-09-06 · universidade federal da...

UNIVERSIDADE FEDERAL DA PARAÍBA
CENTRO DE CIÊNCIAS EXATAS E DA NATUREZA
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS BIOLÓGICAS (ZOOLOGIA)
RENATA MARINHO CRUZ
AABBEELLHHAASS VVIISSIITTAANNTTEESS FFLLOORRAAIISS DDEE RRIICCHHAARRDDIIAA GGRRAANNDDIIFFLLOORRAA
((RRUUBBIIAACCEEAAEE)) AAOO LLOONNGGOO DDEE UUMM GGRRAADDIIEENNTTEE UURRBBAANNOO--RRUURRAALL
JOÃO PESSOA
2013

RENATA MARINHO CRUZ
AABBEELLHHAASS VVIISSIITTAANNTTEESS FFLLOORRAAIISS DDEE RRIICCHHAARRDDIIAA GGRRAANNDDIIFFLLOORRAA
((RRUUBBIIAACCEEAAEE)) AAOO LLOONNGGOO DDEE UUMM GGRRAADDIIEENNTTEE UURRBBAANNOO--RRUURRAALL
Dissertação apresentada ao Programa de
Pós-Graduação em Ciências Biológicas
(Zoologia), da Universidade Federal da
Paraíba, em cumprimento às exigências
para obtenção do título de Mestre em
Ciências Biológicas (Zoologia).
Orientador: Dr. Celso Feitosa Martins
JOÃO PESSOA
2013

C957a Cruz, Renata Marinho.
Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural / Renata Marinho Cruz.-- João Pessoa, 2013. 60f. : il. Orientador: Celso Feitosa Martins Dissertação (Mestrado) – UFPB/CCEN 1. Zoologia. 2. Polinização. 3. Polinizadores. 4. Urbanização. 5. Paisagem. 6. Sucesso reprodutivo.
UFPB/BC CDU: 59(043)

Dedico esta dissertação aos meus pais, pelo amor,
confiança e total apoio.

“Deus fez o mundo pela evolução e nós procuramos,
na medida do possível, reescrever a história desse
mundo.”
Padre Jesus Santiago Moure

AGRADECIMENTOS
Ao PPGCB/UFPB, pelo apoio estrutural para a realização deste projeto.
Ao CNPq, pela bolsa de estudos concedida.
Ao Prof. Celso Feitosa Martins, pelo apoio ao desenvolvimento deste estudo e pela
orientação sempre presente.
A Antonio Meira Leal, diretor da Vila Olímpica Ronaldo Marinho (Antigo DEDE/CIEF), pela
autorização para a realização das observações na área. Bem como, a todos os curiosos que
me fizeram descobrir o quanto uma bióloga trabalhando em área urbana deve estar pronta
para responder às mais estranhas perguntas.
A Creiton Vieira Magalhães, gerente de Administração da Secretaria de Estado da Segurança
e da Defesa Social (João Pessoa – PB), por permitir a realização do estudo nos jardins da
secretaria.
A Antonio Alberto Diniz de Medeiros (Chefe) e Odívio Francelino de Pontes (Chefe Adjunto)
por permitirem o livre acesso à Estação Experimental de Mangabeira (EMEPA-PB). Além
disso, aos funcionários da estação que colaboraram para a manutenção dos experimentos na
área e me presentearam com o revigorante suco das 11h00.
A Wálter Joaquim de Souza, pela permissão das observações no Sítio Ponta de Gramame e
pelos funcionários que colaboraram para a manutenção dos experimentos de polinização.
A Ricardo Lucena, pela autorização e apoio para a realização das observações na Granja
Pitumirim.
A José Adolfo Carniato, por permitir a realização do estudo no Sítio Olho D’água. Gostaria de
agradecer também a Ivanildo Pereira de Lima (Nildo), funcionário do sítio, pela atenção ao
nos receber e por sempre se preocupar em manter os indivíduos de Richardia grandiflora na
área.

À minha família, especialmente meus pais, Edson Jorge da Silva Cruz e Maria do Socorro
Marinho Cruz, e meu irmão, Edson Jorge da Silva Cruz Júnior, pelo amor, dedicação,
compreensão e motivação durante toda a minha trajetória. Gostaria de destacar, ainda, a
disposição e cuidado do meu pai, que me levou e acompanhou na maioria dos dias de
observação.
Ao meu namorado e músico predileto Antonio Luiz Drummond, pelo estímulo, carinho,
compreensão e bons momentos compartilhados.
A Carolina Nunes Liberal, minha amiga e “mãe bióloga”, pelo apoio, sugestões, conversas,
caronas e ajuda indispensável.
A Ana Carolina Toscano de Sousa, pelo auxílio nas atividades de campo e laboratório.
Aos amigos do laboratório de entomologia, Adriano Medeiros, Aline Lopes, Ana Cláudia
Alves, Anthony Bessa, Daniel Dal Bó, Everton Lorenzo, Jean Miguel, Liedson Tavares, Luiz
Paulo Araujo, Marcella Peixoto, Matilde Ernesto, Renato Barbosa, Virgínia Araujo, Valdemar
Neto e Wellington Santos pelo companheirismo nas atividades de laboratório e experiências
compartilhadas na hora do café.
Aos amigos Hannah Nunes, Carolina Costa e Danilo Cavalcanti, que embora não tenhamos
nos reunido com a frequência que gostaríamos, compartilhamos bons momentos
carregados de muitas recordações e risadas.
Ao amigo Raphael Seixas dos Santos, pela companhia nas aulas de tênis, que revigoraram
meu ânimo e melhoraram o meu desempenho com a rede entomológica (recomendo aulas
de tênis a todos os entomólogos).
A todos os professores e funcionários do Programa de Pós-Graduação em Ciências Biológicas
(Zoologia) da UFPB, pela contribuição e atenção ao longo do curso.

RESUMO
Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente
urbano-rural. A crescente urbanização tem sido um dos principais fatores causadores da
perda e fragmentação de habitats, que afetam as comunidades de polinizadores e
modificam as relações planta-polinizador. Embora tenha crescido o número de estudos
focando na influência da paisagem sobre os polinizadores, no Brasil esse conhecimento
ainda é incipiente. Neste estudo, as abelhas visitantes florais e o sucesso reprodutivo de
Richardia grandiflora (Cham. & Schltdl.) Steud foram analisados ao longo de um gradiente
urbano-rural, associando a possíveis alterações nas relações planta-polinizador. O estudo foi
desenvolvido em seis áreas localizadas no estado da Paraíba, divididas em três diferentes
categorias: urbanas, periurbanas e rurais. Para cada área foram estimados os percentuais de
cobertura vegetal (arbórea e herbáceo-arbustiva), superfícies impermeáveis, solo livre e
água, em duas escalas espaciais. As abelhas visitantes florais foram observadas
mensalmente, de fevereiro de 2012 a janeiro de 2013, sendo registrada a composição de
espécies, riqueza, frequência de ocorrência e número de visitas. Por fim, o sucesso
reprodutivo foi estimado a partir de experimentos de polinização (livre e cruzada manual
complementar) para obtenção da Eficácia Reprodutiva (ER) de R. grandiflora em cada área.
Nos agrupamentos a partir da caracterização da paisagem e composição de espécies, as
áreas urbanas se diferenciaram das demais, mas as áreas periurbanas e rurais não foram
diferenciadas entre si. A pouca alteração na riqueza das abelhas eussociais e no número de
visitas por meliponíneos, ao longo do gradiente urbano-rural, indicaram uma boa tolerância
à urbanização. As abelhas não eussociais apresentaram uma maior riqueza e frequência nas
áreas periurbanas e rurais. Além disso, a correlação positiva entre o número de visitas de
abelhas não eussociais e a vegetação herbáceo-arbustiva, na escala espacial menor, sugere
uma maior influência da paisagem local nessas espécies. A espécie introduzida Apis mellifera
apresentou uma maior frequência nas áreas urbanas, que pode estar relacionada à menor
disponibilidade de recursos, bem como à sua flexibilidade ecológica e poder competitivo.
Finalmente, as áreas rurais apresentaram as maiores taxas de Eficácia Reprodutiva,
enquanto as áreas urbanas, valores mais baixos. Nas áreas mais urbanizadas, possivelmente,
A. mellifera não foi um polinizador tão eficiente quanto as espécies nativas, o que pode ser
consequência da fragmentação das populações de Richardia grandiflora.
Palavras-chave: paisagem, urbanização, polinizadores, polinização, sucesso reprodutivo.

ABSTRACT
Bee visitors of the flowers of Richardia grandiflora (Rubiaceae) along an urban-rural
gradient. Increasing urbanization is among the main drivers of the loss and fragmentation of
habitats, that affect pollinator communities and change the plant-pollinator interactions.
Although studies focusing on the influence of landscape on pollinators have increased , in
Brazil this knowledge is incipient. Here, we analysed the bee visitors and reproductive
success of Richardia grandiflora (Cham. & Schltdl.) Steud along an urban-rural gradient,
relating to possible changes in the plant-pollinator interactions. This study was carried out in
six areas located in the state of Paraíba, belonging to three different categories: urban, peri-
urban and rural areas. We estimated the percentage of vegetation cover (woody and
herbaceous-shrub), impervious surfaces, bare soil and water, in two spatial scales.
Moreover, bee visitors of the flowers of R. grandiflora were observed monthly since
February 2012 until January 2013, and the species composition, richness, frequency of
occurrence and number of visits was recorded. Finally, we estimated the reproductive
success from pollination treatments (open-pollination and supplementary cross-pollination)
to obtain the Reproductive Efficacy of R. grandiflora in each area. In the clusters from the
landscape characterization and species composition the urban areas were differentiated
from the remaining, but peri-urban and country areas were not differentiated from each
other. The little change in the richness of eusocial bees and the number of visits by stingless
bees along the urban-rural gradient indicated good tolerance to urbanization. Non-eusocial
bees had higher richness and frequency in rural and peri-urban areas. Moreover, the positive
correlation between the number of non-eusocial bee visits and herbaceous-shrub
vegetation, in the smallest spatial scale, suggests a greater influence of the local landscape
on these species. The introduced species Apis mellifera had a higher frequency in urban
areas, which may be related to lower availability of resources, as well as its ecological
flexibility and competitive power. Finally, rural areas had the highest rates of Reproductive
Efficacy while urban areas showed lower values. Possibly, A. mellifera was not as efficient as
a native pollinator species, which may be a consequence of the fragmentation of the
populations of Richardia grandiflora in the most urbanized areas.
Keywords: landscape, urbanization, pollinators, pollination, reproductive success.

LISTA DE FIGURAS
Figura 1 Localização das áreas de estudo. (a) Localização dos municípios de João Pessoa, Conde e Alhandra no estado da Paraíba. (b) Localização das seis áreas de estudo: U1 (Vila Olímpica Ronaldo Marinho/DEDE); U2 (Secretaria de Estado da Segurança e Defesa Social), P1 (Estação Experimental de Mangabeira), P2 (Sítio Ponta de Gramame); R1 (Granja Pitumirim); R2 (Sítio Olho D’água). Fonte: Google Earth 2013.
20
Figura 2 Áreas de estudo. (a) U1 - Vila Olímpica Ronaldo Marinho/Dede. (b) U2 - Secretaria de Estado da Segurança e Defesa Social. (c) P1 - Estação Experimental de Mangabeira. (d) P2 - Sítio Ponta de Gramame. (e) R1 - Granja Pitumirim. (f) R2 - Sítio Olho D’Água.
21
Figura 3 Metodologia utilizada para estimativa das variáveis da paisagem. (a) Escalas espaciais utilizadas, quadrados com 1000m e 500m de lado, e divisão em 16 quadrantes. (b) Quadrante dividido em uma grade com 841 células. Fonte: Google Earth 2013.
22
Figura 4 Richardia grandiflora (Cham. & Schltdl.) Steud. (a) Inflorescência. (b) Flor (Foto: Carolina N. Liberal).
23
Figura 5 Quadrantes utilizados para a observação dos visitantes florais. (a) Esquema dos quadrantes (1m2) e distância mínima entre eles. (b) Marcação do quadrante em uma área de estudo.
24
Figura 6 Experimentos para análise do sucesso reprodutivo. (a) Inflorescências marcadas para acompanhamento até a maturação dos frutos. (b) Detalhe da marcação da inflorescência.
25
Figura 7 Abelhas visitantes florais de Richardia grandiflora. (a) Apis mellifera; (b) Augochlora sp.; (c) Callonychium brasiliense; (d) Ceratina sp.; (e) Dicranthidium arenarium; (f) Partamona littoralis.
30
Figura 8 Riqueza de espécies eussociais e não eussociais obtidas para as áreas urbanas, periurbanas e rurais.
31
Figura 9 Dendrograma de similaridade qualitativa (índice de Jaccard) entre as áreas urbanas, periurbanas e rurais.
31
Figura 10 Frequência relativa de ocorrência de visitas por Apis mellifera, abelhas eussociais nativas e abelhas não eussociais, nas áreas urbanas, periurbanas e rurais. * Não foram verificadas diferenças significativas entre as frequências de P2 e R1 (χ2 = 1,2054; gl = 2; p = 0,5473).
33
Figura 11 Frequência relativa de ocorrência de visitas, por espécies de abelhas, nas áreas urbanas, periurbanas e rurais.
33

Figura 12 Boxplots do número de visitas observados por dia em cada área. (a) Visitas realizadas por abelhas eussociais, incluindo Apis mellifera; (b) Visitas realizadas por A. mellifera; (c) Visitas realizadas apenas por abelhas eussociais nativas; (d) Visitas realizadas por abelhas não eussociais. As letras diferentes indicam diferenças significativas (p<0,05) entre as áreas, de acordo com os testes de Kruskall-Wallis e a posteriori de comparações múltiplas.
34
Figura 13 Análise de Correspondência entre as espécies de abelhas (número de visitas) e as áreas de estudo. As espécies eussociais estão representadas em azul e as não eussociais em preto. As áreas correspondem aos triângulos vermelhos.
35
Figura 14 Estimativa das variáveis da paisagem, utilizando o quadrado com 500m de lado (0,25km2), para as áreas urbanas, periurbanas e rurais.
36
Figura 15 Estimativa das variáveis da paisagem, utilizando o quadrado com de 1000m de lado (1km2), para as áreas urbanas, periurbanas e rurais.
37
Figura 16 Análise de Agrupamento (Cluster) entre as áreas urbanas, periurbanas e rurais, a partir das variáveis da paisagem. (a) 500m de lado; (b) 1000m de lado.
38

LISTA DE TABELAS
Tabela 1 Espécies de abelhas (Apidae) registradas nas áreas urbanas, periurbanas e rurais.
29
Tabela 2 Matriz de similaridade (índice de Jaccard) obtida para as áreas urbanas, periurbanas e rurais.
32
Tabela 3 Correlação entre as variáveis da paisagem para cada escala espacial (500m e 1000m). Os valores do r (Spearman) estão apresentados nas células cinza e os do p nas células brancas. Os valores de p significativos estão destacados em negrito.
37
Tabela 4 Correlações (r de Spearman) entre as variáveis da paisagem e o número de visitas e riqueza de abelhas. Os valores de p significativos estão destacados em negrito.
39
Tabela 5 Correlações significativas (r de Spearman) entre as variáveis da paisagem e o número de visitas por espécies. *p < 0,05; ** p < 0,01; *** p < 0,001.
40
Tabela 6 Sucessos reprodutivos para os experimentos de polinização e Eficácias Reprodutivas (ER) obtidas para cada área de estudo. As letras minúsculas, em cada linha, indicam diferenças significativas (p < 0,05) de acordo com o teste de χ2.
41
Tabela 7 Correlações (r de Spearman), com respectivos valores de p, entre a Eficácia Reprodutiva (ER) e as variáveis da paisagem e das abelhas (número de visitas e riqueza).
42

SUMÁRIO
1 INTRODUÇÃO..................................................................................................................... 12
1.1 Abelhas e polinização..................................................................................................... 12
1.2 Paisagem e o gradiente urbano-rural...................................................................... 13
1.3 Efeitos da urbanização e fragmentação de habitats nas comunidades de abelhas e nas relações planta-polinizador.............................................................
14
2 OBJETIVOS............................................................................................................................ 16
2.1 Objetivo geral..................................................................................................................... 16
2.2 Objetivos específicos....................................................................................................... 16
3 HIPÓTESES............................................................................................................................ 17
4 METODOLOGIA................................................................................................................. 18
4.1 Áreas de estudo................................................................................................................. 18
4.1.1 Áreas urbanas e periurbanas.................................................................................... 18
4.1.2 Áreas rurais...................................................................................................................... 19
4.1.3 Estimativas das variáveis da paisagem................................................................ 22
4.2 Richardia grandiflora (Cham. & Schltdl.) Steud.................................................. 23
4.3 Observação, coleta e identificação das abelhas................................................. 24
4.4 Sucesso reprodutivo........................................................................................................ 25
4.5 Análises estatísticas......................................................................................................... 27
5 RESULTADOS .................................................................................................................... 28
5.1 Abelhas visitantes florais nas áreas urbanas, periurbanas e rurais.......... 28
5.2 Variáveis da paisagem.................................................................................................... 35
5.3 Relações entre as variáveis da paisagem e as abelhas.................................. 38
5.4 Sucesso reprodutivo........................................................................................................ 40
6 DISCUSSÃO.......................................................................................................................... 43
6.1 Polinizadores e paisagem.............................................................................................. 43
6.2 Sucesso reprodutivo........................................................................................................ 46
7 CONCLUSÕES...................................................................................................................... 50
REFERÊNCIAS.......................................................................................................................... 51

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
12
1 INTRODUÇÃO
1.1 Abelhas e polinização
A polinização constitui um dos principais processos biológicos, sem o qual a maioria
das espécies de angiospermas não se reproduz (Buchmann & Nabhan 1996) e, por isso, é
considerada um serviço ecossistêmico – serviço proporcionado pelo ambiente, indispensável
para a espécie humana (Klein et al. 2006, Kremen et al. 2002). Do ponto de vista animal, a
polinização é um produto secundário da coleta de um recurso (pólen, néctar, resina, óleo)
que é fornecido nas flores, enquanto para as plantas é a maneira de aumentar o fluxo gênico
entre os indivíduos (Janzen 1980). Estima-se que dentre as 250 mil espécies de
angiospermas, cerca de 90% são polinizadas por animais, principalmente por insetos
(Buchmann & Nabhan 1996, Ollerton 1999, Raven et al. 2007). Segundo Baker & Hurd
(1968), a diversificação das angiospermas coincide com um aumento pronunciado na
diversidade dos grupos de insetos, cuja ecologia está intimamente ligada à dessas plantas.
Dentre os insetos, as abelhas (Hymenoptera: Apidae sensu Melo & Gonçalves 2005) são
consideradas os principais polinizadores de plantas cultivadas e silvestres (Freitas & Pereira
2004, Kevan & Imperatriz-Fonseca 2002, Kremen et al. 2012, Michener 2007). Dessa forma,
constituem um grupo chave para a manutenção de muitos ecossistemas terrestres (Neff &
Simpson 1993) e para a produção agrícola (FAO 2004).
Todas as espécies de abelhas existentes dependem, de alguma forma, dos produtos
provenientes das angiospermas (Danforth et al. 2006, Michener 2007, Wcislo & Cane 1996).
Além desses recursos, para a maioria dos polinizadores, os habitats devem apresentar
condições adequadas de temperatura, bem como vegetação e substratos apropriados para a
nidificação (Cane 2001, Liow 2001). O aumento na degradação da vegetação nativa para a
implantação e/ou expansão das cidades ou áreas agrícolas, e a consequente redução de
fontes de recurso e sítios de nidificação, tem sido apontada como uma das principais causas
do declínio na diversidade de polinizadores (Dias et al. 1999, Freitas et al. 2009). Esse
declínio tem sido evidenciado em diversos estudos realizados nas regiões temperadas
(Biesmeijer et al. 2006, Kearns & Inouye 1997, Kevan & Phillips 2001, Klein et al. 2007,

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
13
Ricketts et al. 2008) e a discussão acerca dele se tornou pertinente em várias regiões do
globo.
1.2 Paisagem e o gradiente urbano-rural
Segundo Metzger (2001), paisagem pode ser definida como um mosaico
heterogêneo, composto por unidades interativas, sendo a heterogeneidade existente em
pelo menos um fator, segundo um observador e numa certa escala de observação. No
contexto ecológico, esse mosaico pode ser considerado como um conjunto de habitats que
apresentam condições mais ou menos favoráveis para a espécie ou comunidade estudada
(Metzger 2001). A escala espacial utilizada deve ser escolhida considerando a capacidade de
deslocamento e dispersão das espécies analisadas.
As atividades antrópicas têm modificado os ecossistemas naturais, gerando paisagens
modificadas. Dentre os tipos dominantes de paisagens modificadas estão as áreas urbanas e
as terras agrícolas (Koh & Gardner 2010). Os ambientes urbanos constituem áreas
espacialmente heterogêneas, sob intensa atividade humana, que apresentam uma grande
quantidade de superfícies impermeáveis e estruturas artificiais (pavimentos, construções,
etc) (McIntyre et al. 2001). Em contraposição aos ambientes urbanos, as áreas rurais
constituem regiões com predominância de cobertura vegetal, que podem estar destinadas a
atividades agropecuárias, turismo natural, silvicultura, extrativismo ou conservação
ambiental. Da mesma forma que a urbanização, a intensificação da atividade agrícola,
primariamente, degrada o habitat através da redução da cobertura vegetal nativa (Freitas et
al. 2009, McIntyre et al. 2001).
A partir do início do século XX, a dicotomia “urbano” e “rural” vem sendo
amplamente discutida, destacando-se a importância de considerar uma transição menos
abrupta entre esses dois tipos de ambientes (Reis 2006). Nesse contexto, enquadram-se as
áreas periurbanas, que constituem uma zona de transição entre os ambientes urbanos e
rurais, nas quais é possível encontrar características de ambos os tipos de ambientes na
partilha pelo uso do solo (Vale & Gerardi 2006). O espaço periurbano, portanto, apresenta
um nível de urbanização intermediário entre os extremos urbano e rural. Dessa forma, ao
utilizar áreas urbanas, periurbanas e rurais, está sendo realizada uma abordagem em

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
14
gradiente, que pode ser definido através da redução da urbanização (redução de superfícies
impermeáveis e aumento da cobertura vegetal) das áreas urbanas para as rurais.
1.3 Efeitos da urbanização e fragmentação de habitats nas comunidades de abelhas e nas relações planta-polinizador
A urbanização é um processo crescente no mundo, constituindo uma das principais
atividades antrópicas causadoras de alterações drásticas e persistentes nos habitats
(McKinney 2006). Através desse processo, ocorre o aumento da quantidade de superfícies
impermeáveis (prédios, estradas, áreas industriais), com a correspondente redução das
áreas verdes (Ahrné et al. 2009). Nas áreas urbanas, portanto, os habitats adequados (áreas
ruderais, parques, jardins, fragmentos, etc) estão incorporados numa matriz altamente
inadequada (superfícies impermeáveis), gerando uma fragmentação extrema de habitats
(Andrieu et al. 2009). Além disso, nesses ambientes também podem ser verificados, mais
intensamente, fatores como poluição, maior número de espécies vegetais exóticas e
alterações climáticas locais (Kim 1992, McIntyre et al. 2001, Rebele 1994).
Diversos estudos têm demonstrado o efeito da ocupação humana, e a consequente
redução de habitats adequados e da disponibilidade de recursos, bem como aumento do
isolamento de ambientes favoráveis, nas comunidades de polinizadores (Melo et al. 2006,
Viana et al. 2012). A maioria desses estudos aponta uma diminuição na riqueza e abundância
de espécies de polinizadores, seja em resposta à fragmentação ambiental (Steffan-Dewenter
& Tscharntke 1999, à distância de habitats naturais (Chachoff & Aizen 2006) ou à
urbanização (Bates et al. 2011, Zanette et al. 2005). Entretanto, nem todos os ambientes
alterados são inviáveis para os polinizadores, podendo abrigar ambientes alternativos que
criam conectividade entre fragmentos remanescentes e fornecem habitats e recursos
valiosos para as abelhas (Chacoff & Aizen 2006, Fetridige et al. 2008, McFrederick & LeBuhn
2006, Saure 1996). Dessa forma, mesmo nas cidades é possível encontrar um número
considerável de espécies de abelhas (Banaszak-Cibicka & Zmihorski 2011, Taura & Laroca
1991).
É importante considerar que os grupos de abelhas podem responder de maneira
diferenciada aos impactos das ações antrópicas. Em alguns casos, pode ser verificada pouca
alteração na abundância total, porém modificação na abundância relativa de cada espécie

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
15
em resposta a distúrbios antrópicos (Samejima et al. 2004). Espécies generalistas, por
exemplo, podem estar presentes em abundância em áreas com diferentes níveis de
urbanização (Zanette et al. 2005). Por outro lado, espécies que apresentam requerimentos
de habitat mais específicos e pequena capacidade de deslocamento tendem a ser mais
sensíveis à fragmentação ambiental e aumento da urbanização (Bates et al. 2011, Biesmeijer
et al. 2006, Gathmann et al. 1994, Geslin et al. 2013, Greenleaf et al. 2007, Steffan-Dewenter
et al. 2002). Além disso, espécies exóticas, que geralmente apresentam maior flexibilidade
ecológica e poder de competição, podem apresentar suas populações aumentadas em
ambientes antropizados, em detrimento das espécies nativas (Aizen & Feinsinger 1994a,
Aizen & Feinsinger 1994b, Melo et al. 2006, Steffan-Dewenter et al. 2002).
A redução no tamanho e na conectividade entre os habitats, como influência da
urbanização, pode provocar, ainda, alterações nas relações planta-polinizador (Aguilar et al.
2006, Biesmeijer et al. 2006, Harris & Johnson 2004, Tscharntke & Brandl 2004). A escassez
de habitats e recursos adequados, bem como a pouca atratividade das pequenas ou pouco
densas populações de plantas, podem promover a queda na densidade populacional e
riqueza de espécies de polinizadores (Andrieu et al. 2009, Goverde et al. 2002, McIntyre &
Hostetler 2001, Steffan-Dewenter & Tscharntke 1999). Ao mesmo tempo, a quantidade de
recursos disponíveis e o seu arranjo espacial podem modificar o comportamento de
forrageio dos polinizadores, como uma forma de manter o ganho energético nesses
ambientes fragmentados (Andrieu et al. 2009, Charnov 1976, Goverde et al. 2002). Essas
alterações na abundância, riqueza e comportamento dos polinizadores geram efeitos
negativos na produção de sementes e no fluxo gênico entre as populações de plantas (Kunin
1997, Steffan-Dewenter & Tscharntke 1999).
Embora se observe um crescente de estudos analisando os efeitos da paisagem na
disponibilidade e comportamento dos polinizadores, no Brasil esse conhecimento ainda é
incipiente de modo que pouco se conhece sobre as relações entre a estrutura da paisagem e
os polinizadores (Viana et al. 2012). Essa realidade reflete a necessidade de investigar a
composição, frequência, riqueza e número de visitas de abelhas ao longo de um gradiente
urbano-rural, associando a variáveis da paisagem e possíveis interferências nas relações
planta-polinizador.

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
16
2 OBJETIVOS
2.1 Objetivo geral
Estudar os efeitos das paisagens modificadas ao longo de um gradiente urbano-rural
sobre as abelhas visitantes florais e sucesso reprodutivo de Richardia grandiflora (Cham. &
Schltdl.) Steud.
2.2 Objetivos específicos
Avaliar a composição de espécies, riqueza, frequência e número de visitas das
abelhas visitantes florais de Richardia grandiflora em cada área;
Estimar os percentuais de cobertura vegetal (arbórea e herbáceo-arbustiva),
superfícies impermeáveis, solo livre e água para cada área;
Relacionar as variáveis da paisagem (cobertura vegetal, superfícies impermeáveis,
solo livre e água) com o número de visitas e a riqueza das abelhas visitantes florais de
Richardia grandiflora;
Analisar o sucesso reprodutivo de Richardia grandiflora, nas diferentes áreas,
associando às variáveis da paisagem, número de visitas e riqueza de abelhas.

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
17
3 HIPÓTESES
(1) A redução da urbanização ao longo do gradiente urbano-rural (diminuição no
número de edificações, pavimentações e aumento de superfícies livres e vegetação)
altera a composição e frequência dos polinizadores, bem como aumenta a riqueza e
número de visitas das abelhas nativas;
(2) Áreas mais urbanizadas apresentam uma maior frequência da espécie exótica Apis
mellifera;
(3) A Eficácia Reprodutiva é afetada positivamente ao longo do gradiente urbano-rural.

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
18
4 METODOLOGIA
4.1 Áreas de estudo
O estudo foi desenvolvido nos municípios de João Pessoa, Conde e Alhandra
localizados no estado da Paraíba, Brasil (Figura 1a). O clima da região, de acordo com as
divisões climáticas de Köppen, está inserido dentro do domínio tropical quente-úmido (As’).
Caracteriza-se por apresentar um período chuvoso que se inicia no mês de fevereiro/março
e prolonga-se até julho/agosto. O período seco começa no mês de setembro e estende-se
até fevereiro. As médias térmicas anuais são de 22 - 26° C e as precipitações pluviométricas
situam-se em torno de 1500 mm/ano. Os solos da região são arenosos ou argilosos de baixa
fertilidade, lixiviados (podzólicos ou latossolos) (Feliciano & Melo 2003).
Foram utilizadas seis áreas de estudo (Figura 1b), distribuídas em três diferentes
categorias: urbanas, periurbanas e rurais.
4.1.1 Áreas urbanas e periurbanas
As áreas urbanas (U) utilizadas neste estudo estão localizadas na cidade de João
Pessoa (3421,30 hab/Km2) (IBGE 2010a) e foram os jardins da (1) Vila Olímpica Ronaldo
Marinho (DEDE/CIEF) (7°6'45"S, 34°51'3"W) (Figura 2a), localizada no bairro dos Estados, e
da (2) Secretaria de Estado da Segurança e Defesa Social (7°9'42"S, 34°50'12"W) (Figura 2b),
localizada no bairro de Mangabeira. Ambos os bairros estão inseridos na matriz urbana da
cidade de João Pessoa.
As áreas periurbanas (P) estão localizadas na margem da cidade João Pessoa e foram
a (1) Estação Experimental de Mangabeira (07°11'52"S, 34°48'42"W) (Figura 2c), pertencente
à Empresa Estadual de Pesquisa Agropecuária da Paraíba (EMEPA-PB), localizada no bairro
Costa do Sol, e o (2) Sítio Ponta de Gramame, localizado no bairro de Gramame (7°12'26"S,
34°51'55"W) (Figura 2d). Embora a ocupação do bairro Costa do Sol tenho aumentado nos
últimos anos, com intensificação da especulação imobiliária, construção civil e atividade
turística, a área apresenta uma baixa densidade populacional, com grande parte destinada

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
19
ao uso agrícola (Barbosa 2005). Da mesma forma, no Bairro de Gramame apenas parte do
espaço foi assimilado pela expansão urbana, permanecendo a outra com elementos rurais
(Leite 2011).
4.1.2 Áreas rurais
As áreas rurais (R) utilizadas neste estudo estão localizadas nos municípios de Conde
e Alhandra, que são integrantes da região metropolitana de João Pessoa. As cidades de
Conde (123,74 hab/Km2) e Alhandra (98,58 hab/Km2) possuem aproximadamente 1/3 da
população vivendo na zona rural (IBGE 2010a, IBGE 2010b). Nessas cidades, os
estabelecimentos agropecuários desempenham um papel de destaque, ocupando uma área
aproximada de 7278 hectares no Conde e 10811 hectares em Alhandra (IBGE 2006). As áreas
que fizeram parte deste estudo foram a (1) Granja Pitumirim, localizada no município do
Conde (7°16'24"S, 34°53'21"W) (Figura 2e), e o (2) Sítio Olho D’água (07°25'31"S,
34°53'21"W) (Figura 2f), localizado no assentamento Tapuiu no município de Alhandra.

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
20
Figura 1 – Localização das áreas de estudo. (a) Localização dos municípios de João Pessoa, Conde e Alhandra no estado da Paraíba. (b) Localização das seis áreas de estudo: U1 (Vila Olímpica Ronaldo Marinho/DEDE); U2 (Secretaria de Estado da Segurança e Defesa Social), P1 (Estação Experimental de Mangabeira), P2 (Sítio Ponta de Gramame); R1 (Granja Pitumirim); R2 (Sítio Olho D’água). Fonte: Google Earth 2013.
a
b

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
21
Figura 2 – Áreas de estudo. (a) U1 - Vila Olímpica Ronaldo Marinho/Dede. (b) U2 - Secretaria de Estado da Segurança e Defesa Social. (c) P1 - Estação Experimental de Mangabeira. (d) P2 - Sítio Ponta de Gramame. (e) R1 - Granja Pitumirim. (f) R2 - Sítio Olho D’Água.
a b
c d
e f

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
22
4.1.3 Estimativas das variáveis da paisagem
As seis áreas de estudo foram caracterizadas a partir de imagens de satélite (Google
Earth 2013), utilizando duas escalas espaciais diferentes: um quadrado com 1000m de lado e
outro com 500m de lado, inscrito no quadrado maior (Figura 3a). O quadrado maior foi
divido em um total de 16 quadrantes, cada um com 250m de lado, que foram subdivididos
em uma grade contendo 841 subquadrantes (células com ≅74,32 m2) (Figura 3b).
Figura 3 – Metodologia utilizada para estimativa das variáveis da paisagem. (a) Escalas espaciais utilizadas, quadrados com 1000m e 500m de lado, e divisão em 16 quadrantes. (b) Quadrante dividido em uma grade com 841 células. Fonte: Google Earth 2013.
Cada célula foi analisada e classificada nas seguintes categorias: (1) vegetação arbórea,
(2) vegetação herbáceo-arbustiva, (3) solo livre, (4) superfícies impermeáveis (construções e
pavimentos) e (5) água. A categoria solo livre foi incluída devido à importância para abelhas
que nidificam no solo (Michener 2007, Zanette et al. 2005). A partir da contagem do número
de células por categoria foi possível estimar a porcentagem de cada variável da paisagem.
1000m
250m
250m
1000m
500m
500m
a b

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
23
4.2 Richardia grandiflora (Cham. & Schltdl.) Steud.
Richardia grandiflora (Cham. & Schltdl.) Steud. (Figura 4) é uma erva ruderal,
pertencente à família Rubiaceae (subfamília Rubioidae, tribo Spermacoceae) conhecida
popularmente como “ervanço”, “poaia” ou “ipeca-mirim” (Agra et al. 2007). A palavra
ruderal provém de ruderes, que significa ruínas, é utilizada para qualificar aquelas plantas
que durante o processo evolutivo se adaptaram a ambientes antropizados, ocupando beiras
de calçadas, terrenos baldios e outros tipos de ambientes modificados pelo homem (Lorenzi
1991). R. grandiflora apresenta distribuição neotropical (Andersson 1992), sendo nativa da
Argentina, Bolívia, Brasil, Uruguai e Paraguai (Lewis & Oliver 1974). No Brasil, pode ser
encontrada em todas as regiões (Pereira & Barbosa 2006).
Figura 4 –Richardia grandiflora (Cham. & Schltdl.) Steud. (a) Inflorescência. (b) Flor (Foto: Carolina N. Liberal).
Embora estudos sobre os visitantes florais de Richardia grandiflora sejam escassos, a
importância dessa espécie como fonte de recursos para abelhas pode ser evidenciada em
levantamentos da flora utilizada por insetos e em análises polínicas de mel (Boff 2008,
Carvalho 2013, Kiill et al. 2000, Moura 2008, Nascimento 2011, Pires et al. 2009, Pott & Pott
1986, Santos et al. 2006, Silva 2006, Silva et al. 2008, Vidal et al. 2008). No único estudo
direcionado à biologia floral, sistema reprodutivo e polinizadores de Richardia grandiflora,
realizado por Cruz (2011), a espécie foi considerada como autoincompatível e as abelhas
a b

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
24
foram o grupo de visitantes mais rico e frequente, sendo apontadas como os principais
polinizadores.
R. grandiflora foi escolhida como ferramenta para a realização deste estudo devido à
importante relação entre suas flores e as abelhas, bem como à sua característica ruderal, o
que possibilitou que indivíduos pudessem ser encontrados nas seis áreas de estudo.
4.3 Observação, coleta e identificação das abelhas
Em cada área de estudo foram observados três grupos de 50 flores. Cada grupo
estava contido dentro de um quadrante de 1m2 e distando, no mínimo, 3m do outro (Figura
5).
Figura 5 – Quadrantes utilizados para a observação dos visitantes florais. (a) Esquema dos quadrantes (1m2) e distância mínima entre eles. (b) Marcação do quadrante em uma área de estudo.
As observações foram realizadas mensalmente de fevereiro/2012 a janeiro/2013,
com um total de 72 dias de observação, 12 em cada área. Em cada dia de observação, as
flores foram monitoradas das 7h00min às 12h00min, durante 15min a cada meia hora,
sendo 5min por quadrante. Assim, foi obtido um total de 30min de observação por hora,
totalizando 2h30min por dia de observação.
Exemplares foram coletados para identificação com rede entomológica e frascos
coletores. Em seguida, foram colocadas em câmara mortífera contendo acetato de etila,
montadas e depositadas na Coleção Entomológica do Departamento de Sistemática e
Ecologia da Universidade Federal da Paraíba.
a b

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
25
No processo de identificação, primeiramente foi utilizada a chave taxonômica de
gêneros de abelhas ocorrentes no Brasil apresentada em Silveira et al. (2002). O nível
específico foi atingido através do auxílio de chaves taxonômicas de grupos específicos e
comparação com a coleção de referência da Universidade Federal da Paraíba. Foi utilizada a
classificação proposta por Melo & Gonçalves (2005) e seguida por Moure et al. (2007), na
qual todas as abelhas então agrupadas na família Apidae. Nessa classificação, as famílias
consideradas por Michener (2007) e Silveira et al. (2002) são tratadas como subfamílias.
4.4 Sucesso reprodutivo
Para a análise do sucesso reprodutivo, foram realizados os experimentos de
polinização livre e polinização cruzada manual complementar. Em ambos, 50 inflorescências
foram escolhidas em cada área, totalizando 300 inflorescências por experimento. As
inflorescências foram marcadas com palitos de madeira (Figura 6) e acompanhadas até a
maturação dos frutos. As flores utilizadas nos experimentos foram marcadas no cálice com
caneta permanente, para posterior reconhecimento.
Figura 6 – Experimentos para análise do sucesso reprodutivo. (a) Inflorescências marcadas para acompanhamento até a maturação dos frutos. (b) Detalhe da marcação da inflorescência.
a b

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
26
Em seguida, as inflorescências foram levadas para o laboratório para contagem do
número de sementes produzidas. A partir desses dados, o sucesso reprodutivo (SR) foi
calculado utilizando a fórmula:
Na polinização livre, as inflorescências foram mantidas sob condições naturais. Na
polinização cruzada manual, os grãos de pólen de flores de uma planta foram transportados
para o estigma de flores de outra planta, com o auxílio de uma pinça. Para assegurar o
cruzamento entre indivíduos diferentes, os mesmos foram feitos utilizando a distância
mínima de 3m entre as flores doadoras e receptoras. O experimento foi realizado pela
manhã (7h00min – 9h00min), que corresponde ao horário com maior frequência dos
polinizadores naturais (Cruz 2011). Após a realização dos cruzamentos, as inflorescências
não foram ensacadas, de modo que permaneceram disponíveis para os visitantes florais.
Portanto, a polinização cruzada manual foi realizada de forma complementar (Guedes et al.
2011).
Para fins de análise, foi considerada a razão entre os sucessos da polinização livre e
polinização cruzada manual complementar, denominada de Eficácia Reprodutiva (ER)
(Zapata & Arroyo 1978), que, para espécies xenogâmicas, mede o percentual de flores que
foram polinizadas com pólen proveniente de outro indivíduo:
Os valores de ER mais próximos de 0 indicam um maior déficit de polinização,
enquanto os mais próximos de 1 sugerem uma maior taxa de polinização natural.

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
27
4.5 Análises estatísticas
Todos os cálculos foram realizados utilizando o pacote estatístico R versão 2.12.1 (R
Development Core Team 2010). Neste estudo, as análises foram feitas considerando os dois
grupos básicos: abelhas eussociais e abelhas não eussociais. Além disso, a espécie eussocial
introduzida Apis mellifera foi analisada separadamente.
Para analisar a similaridade entre as áreas quanto à composição de espécies, foi
utilizado o índice qualitativo de Jaccard (presença e ausência). Para facilitar a visualização
dos resultados, as relações estabelecidas entre as áreas foram agrupadas empregando o
método de ligação média UPGMA, produzindo ao final um dendrograma.
O teste de Qui-Quadrado (χ2), com correção de Yates, foi utilizado para comparar a
frequência relativa de ocorrência dos grupos de visitantes nas áreas urbanas, periurbanas e
rurais. Esse teste foi aplicado, ainda, para comparar os sucessos reprodutivos obtidos para
cada experimento de polinização nas diferentes áreas.
Para comparar o número de visitas obtidas em cada área, foi utilizado o teste de
Kruskal-Wallis. Em sequência foi aplicado o teste a posteriori de comparação múltipla
usando a função “kruskalmc” do pacote pgirmess (Giraudoux 2011). A aceitação ou rejeição
da hipótese nula foi realizada através da obtenção de um p-valor, adotando o nível de
significância = 0,05 (Siegel & Castellan Jr. 2006).
A Análise de Correspondência (AC) foi utilizada para analisar o nível de associação
entre as espécies de abelhas e as áreas, através do número de visitas realizadas pelos
indivíduos.
A Análise de Agrupamento (Cluster), utilizando a distância Euclidiana e o método de
ligação média UPGMA, foi aplicada para verificar se as seis áreas, previamente categorizadas
como urbanas, periurbanas e rurais, poderiam ser também diferenciadas a partir das
estimativas das variáveis da paisagem. A análise de agrupamento foi realizada para as duas
escalas espaciais: 500m e 1000m.
Por fim, a correlação de Spearman foi aplicada para analisar a relação entre as
variáveis da paisagem (vegetação arbórea, vegetação herbáceo-arbustiva, solo livre,
superfícies impermeáveis e água) e as abelhas (número de visitas e riqueza). A correlação foi
empregada, ainda, para relacionar as variáveis da paisagem, número de visitas e riqueza de
abelhas com a Eficácia Reprodutiva (ER) obtida para cada área.

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
28
5 RESULTADOS
5.1 Abelhas visitantes florais nas áreas urbanas, periurbanas e rurais
Foram registradas um total de 5304 visitas realizadas por 29 espécies de abelhas,
pertencentes a 17 gêneros e quatro subfamílias (Tabela 1). As subfamílias Apinae e
Halictinae foram as melhores representadas, com 13 espécies cada uma. Para as subfamílias
Megachilinae e Andreninae foram encontradas, respectivamente, 2 e 1 espécie. Dentre as
espécies observadas, apenas duas foram registradas nas seis áreas, Apis mellifera (Figura 7a)
e Trigona spinipes. Onze espécies foram registradas apenas em uma única área: Ancylocelis
apiformis, Augochlora sp. 6, Augochlora sp. 7, Augochloropsis sp., Dialictus sp.1, Exomalopsis
cf. analis, Exomalopsis (E.) sp., Frieseomelitta doederleini, Megachile (A.) sp., Thectochlora
alaris e Trigona fuscipennis.
As áreas urbanas apresentaram uma riqueza de espécies inferior às demais (S=3 em
U1 e S=7 em U2). Os maiores valores foram obtidos paras as áreas periurbanas (S= 20 em P1
e S=18 em P2), seguida pelas áreas rurais (S=14 em R1 e S=17 em R2). Considerando
separadamente os grupos de abelhas, percebeu-se que a riqueza de espécies eussociais
variou pouco nos três tipos de área (Figura 8), com o menor valor obtido para a área U2 (2
espécies) e o maior para as áreas P1, P2 e R1 (4 espécies). Por outro lado, a riqueza de
espécies de abelhas não eussociais foi superior nas áreas periurbanas e rurais, com valores
que variaram de 10 (R1) a 16 (P1).
A análise do dendrograma baseado no índice qualitativo de Jaccard permitiu observar
que a composição de espécies de abelhas foi mais similar entre as áreas periurbanas e rurais
(Figura 9). A maior similaridade foi obtida entre a área periurbana P2 e a rural R1 (0,60)
(Figura 9 e Tabela 2). As áreas urbanas mostraram uma menor similaridade na composição
de espécies de abelhas em relação aos outros tipos de área, sendo o menor valor obtido
entre as áreas U1 e R2 (0,11).

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
29
Tabela 1 – Espécies de abelhas (Apidae) registradas nas áreas urbanas, periurbanas e rurais.
Espécies de abelhas (Apidae) U1 U2 P1 P2 R1 R2
ANDRENINAE
Calliopsini Callonychium brasiliense (Ducke, 1907)
APINAE Apini
Apina Apis mellifera Linnaeus, 1758
Meliponina Frieseomelitta doederleini (Friese, 1900)
Partamona littoralis Pedro & Camargo, 2003
Plebeia flavocincta (Cockerell, 1912)
Trigona spinipes (Fabricius, 1793)
Trigona fuscipennis Friese 1900
Emphorini Ancyloscelis apiformis (Fabricius, 1793)
Eucerini Melissoptila uncicornis (Ducke, 1910)
Exomalopsini Exomalopsis (Exomalopsis) cf. analis Spinola, 1853
Exomalopsis (Exomalopsis) sp.
Xylocopini
Ceratinina Ceratina (Ceratinula) manni Cockerell , 1912
Ceratina (Crewella) sp. 1
Ceratina (Crewella) sp. 2
HALICTINAE Halictini Dialictus opacus (Moure, 1940)
Dialictus sp. 1
Dialictus sp. 2
Augochlorini Augochlora sp. 1
Augochlora sp. 2
Augochlora sp. 3
Augochlora sp. 4
Augochlora sp. 5
Augochlora sp. 6
Augochlora sp. 7
Augochlorella sp.
Augochloropsis sp.
Thectochlora alaris (Vachal, 1904)
MEGACHILINAE
Anthidiini Dicranthidium arenarium (Ducke, 1907)
Megachilini Megachile (Acentron) sp.

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
30
Figura 7 – Abelhas visitantes florais de Richardia grandiflora. (a) Apis mellifera; (b) Augochlora sp.; (c) Callonychium brasiliense; (d) Ceratina sp.; (e) Dicranthidium arenarium; (f) Partamona littoralis.
a
c
e
d
b
f

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
31
Figura 8 – Riqueza de espécies eussociais e não eussociais obtidas para as áreas urbanas, periurbanas e rurais.
Figura 9 – Dendrograma de similaridade qualitativa (índice de Jaccard) entre as áreas urbanas, periurbanas e rurais.
0
2
4
6
8
10
12
14
16
18
U1 U2 P1 P2 R1 R2
Nú
me
ro d
e e
spé
cie
s
Eussociais Não Eussociais

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
32
Tabela 2 – Matriz de similaridade (índice de Jaccard) obtida para as áreas urbanas, periurbanas e rurais.
U1 U2 P1 P2 R1
U2 0,25
P1 0,15 0,35
P2 0,17 0,32 0,58
R1 0,21 0,31 0,48 0,60
R2 0,11 0,33 0,42 0,52 0,48
Ao comparar a frequência relativa de ocorrência de visitas por grupo de abelhas
(Figura 10), o teste χ2 apontou uma igualdade estatística apenas entre as áreas P2 e R1 (χ2 =
1,2054; gl = 2; p = 0,5473). As demais áreas diferiram estatisticamente entre si (p < 0,05).
Nas áreas urbanas, observou-se uma predominância dos indivíduos da espécie introduzida
Apis mellifera, apresentando uma frequência de mais de 70% nas duas áreas (U1 e U2).
Tanto nas áreas periurbanas quanto nas rurais, a maior frequência foi registrada para as
abelhas não eussociais, sendo de 45,5%, 56,5%, 50,5% e 79,4%, respectivamente, em P1, P2,
R1 e R2 (Figura 10). Dentre as abelhas não eussociais é importante destacar as espécies de
Augochlorini (Figura 7b), que estiveram presentes em todas essas áreas e foram as que
ocorreram com mais frequência em P2 e R2 (32% e 35,4%) (Figura 11). Na área periurbana
P1 a frequência registrada para A. mellifera ainda foi uma das maiores (32,17%) e as
espécies de abelhas eussociais nativas apresentaram uma frequência de 22,4%. Na área
periurbana P2 e nas áreas rurais (R1 e R2) a segunda maior frequência foi registrada para as
espécies eussociais nativas (39,7%, 46,7% e 16,7%, respectivamente) e A. mellifera
apresentou uma frequência inferior a 4%. Dentre as espécies de abelhas eussociais nativas,
pode-se destacar Partamona littoralis (Figura 7f) na área P2 (25,1%) e Plebeia flavocincta na
área R1 (29,6%).
Comparando o número de visitas obtido por grupo de abelhas em cada área (Figura
12), observou-se um maior número de visitas por abelhas eussociais (incluindo A. mellifera)
nas áreas urbanas que nas áreas periurbanas e rurais (Figura12a). Entretanto, as visitas das
áreas P1, P2 e R2 não diferiram significativamente das obtidas na área U2 (p > 0,05).
Analisando separadamente as visitas pelos indivíduos da espécie Apis mellifera (Figura12b),
foi obtida uma diferença significativa (p < 0,05) entre as áreas urbanas e as demais, com
exceção da área periurbana P1. Para as abelhas eussociais nativas, as únicas diferenças

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
33
0
10
20
30
40
50
60
70
80
90
100
U1 U2 P1 P2 R1 R2
Frq
uên
cia
rela
tiva
de
oco
rrên
cia
(%)
Outras
Dicrathidium arenarium
Ceratina spp.
Callonychium brasiliense
Augochlorini spp.
Trigona spinipes
Plebeia flavocincta
Partamona littoralis
Apis mellifera
significativas obtidas foram entre a área U2 e as áreas P1, P2 e R1 (Figura12c). Por fim, para
as espécies não eussociais, o número de visitas nas áreas P2 e R2 diferiram
significativamente das áreas urbanas. Embora os resultados mostrem um maior número de
visitas nas áreas P1 e R1 em relação às áreas urbanas, a diferença entre a área U2 não foi
estatisticamente significativa. Da mesma forma, não foi constatada diferença entre as áreas
periurbanas e rurais (Figura12d).
Figura 10 – Frequência relativa de ocorrência de visitas por Apis mellifera, abelhas eussociais nativas e abelhas não eussociais, nas áreas urbanas, periurbanas e rurais. * Não foram verificadas diferenças significativas entre as frequências de P2 e R1 (χ2 = 1,2054; gl = 2; p = 0,5473).
Figura 11 – Frequência relativa de ocorrência de visitas, por espécies de abelhas, nas áreas urbanas, periurbanas e rurais.
0
10
20
30
40
50
60
70
80
90
100
U1 U2 P1 P2 R1 R2
Freq
uên
cia
rela
tiva
de
oco
rrê
nci
a (
%)
Não eussociais
Eussociais nativas
Apis mellifera
*
*

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
34
Figura 12 – Boxplots do número de visitas observados por dia em cada área. (a) Visitas realizadas por abelhas eussociais, incluindo Apis mellifera; (b) Visitas realizadas por A. mellifera; (c) Visitas realizadas apenas por abelhas eussociais nativas; (d) Visitas realizadas por abelhas não eussociais. As letras diferentes indicam diferenças significativas (p<0,05) entre as áreas, de acordo com os testes de Kruskall-Wallis e a posteriori de comparações múltiplas.
Através da Análise de Correspondência foi demonstrado que as áreas urbanas estão
fortemente associadas à espécie Apis mellifera (Figura 13). A área P1, desconsiderando as
espécies registradas unicamente nessa área (Frieseomelitta doederleini, Exomalopsis spp. e
Augochloropsis sp.), apresentou uma grande associação com a espécie eussocial Trigona
b
d
a
c
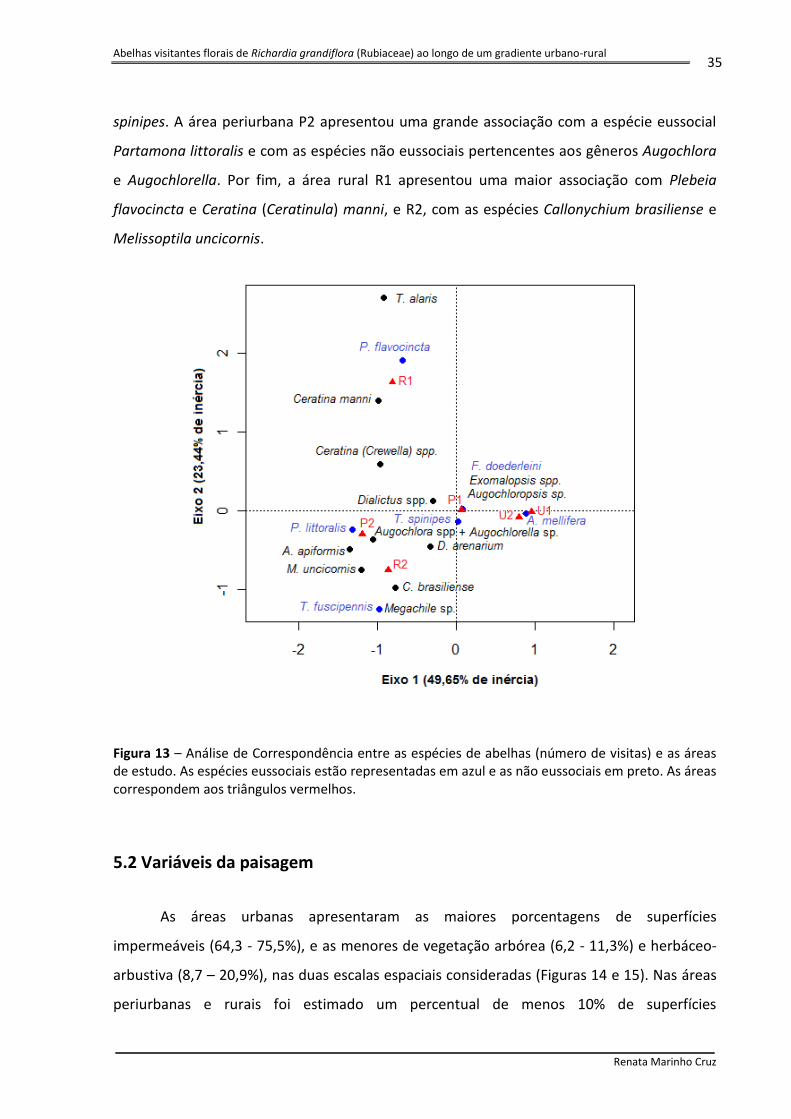
Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
35
spinipes. A área periurbana P2 apresentou uma grande associação com a espécie eussocial
Partamona littoralis e com as espécies não eussociais pertencentes aos gêneros Augochlora
e Augochlorella. Por fim, a área rural R1 apresentou uma maior associação com Plebeia
flavocincta e Ceratina (Ceratinula) manni, e R2, com as espécies Callonychium brasiliense e
Melissoptila uncicornis.
Figura 13 – Análise de Correspondência entre as espécies de abelhas (número de visitas) e as áreas de estudo. As espécies eussociais estão representadas em azul e as não eussociais em preto. As áreas correspondem aos triângulos vermelhos.
5.2 Variáveis da paisagem
As áreas urbanas apresentaram as maiores porcentagens de superfícies
impermeáveis (64,3 - 75,5%), e as menores de vegetação arbórea (6,2 - 11,3%) e herbáceo-
arbustiva (8,7 – 20,9%), nas duas escalas espaciais consideradas (Figuras 14 e 15). Nas áreas
periurbanas e rurais foi estimado um percentual de menos 10% de superfícies

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
36
0
20
40
60
80
100
120
U1 U2 P1 P2 R1 R2
Po
rce
nta
gem
da
área
(%
)
500m
Água
Impermeável
Solo
Herbáceo-arbustiva
Arbórea
impermeáveis. Os menores valores para essa variável foram obtidos nas áreas rurais (0,4% -
2,7%) e nas periurbanas variaram de 3,5 a 9,3%. De uma maneira geral, o percentual de
vegetação arbórea e herbáceo-arbustiva foi similar entre as áreas periurbanas e rurais. A
exceção foi a área P1 que apresentou um maior percentual de vegetação arbórea (40,8% e
56,2%). Em geral, as porcentagens de solo livre e água foram relativamente baixas em todas
as áreas, nas duas escalas. A única exceção foi o percentual de solo livre em R2, que foi de
16,5 e 23,5%.
Relacionando as variáveis da paisagem (Tabela 3) foi possível observar que, em
ambas as escalas espaciais, houve uma forte correlação negativa entre a vegetação
herbáceo-arbustiva e as superfícies impermeáveis (r = – 1; p < 0,0001 e r = – 0,89; p =
0,0188). Por esse motivo, essas duas variáveis serão consideradas em conjunto na discussão.
Com exceção da relação entre água e solo livre na escala de 1000m (r = – 0,83; p = 0,0416),
as demais não foram significativas.
Além disso, ao correlacionar cada variável na escala de 500m, com a sua
correspondente na escala de 1000m, observou-se uma forte correlação entre os percentuais
de vegetação arbórea (r = 0,89; p = 0,0188). Da mesma forma, foi observada uma forte
correlação entre os valores vegetação herbáceo-arbustiva (r = 0,94; p = 0,0048) e superfícies
impermeáveis (r = 0,94; p = 0,0048). Apenas para as variáveis água e solo livre não foram
verificadas correlação significativas entre os valores obtidos para cada escala (p > 0,05).
Figura 14 – Estimativa das variáveis da paisagem, utilizando o quadrado com 500m de lado (0,25km2), para as áreas urbanas, periurbanas e rurais.

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
37
Figura 15 – Estimativa das variáveis da paisagem, utilizando o quadrado com de 1000m de lado (1km2), para as áreas urbanas, periurbanas e rurais.
Tabela 3 – Correlação entre as variáveis da paisagem para cada escala espacial (500m e 1000m). Os valores do r (Spearman) estão apresentados nas células cinza e os do p nas células brancas. Os valores de p significativos estão destacados em negrito.
Variáveis da paisagem (%)
Água Arbórea Herbáceo-arbustiva
Solo Impermeável
500m
Água - 0,5441 0,8717 0,8717 0,8717
Arbórea 0,31 - 0,8717 0,3965 0,8717
Herbáceo-arbustiva 0,09 0,09 - 0,3287 p < 0,0001
Solo –0,09 –0,43 0,49 - 0,3287
Impermeável –0,09 –0,09 –1 –0,49 -
1000m
Água - 0,5441 0,4685 0,0416 0,704
Arbórea 0,31 - 0,2657 0,2657 0,2657
Herbáceo-arbustiva –0,37 0,54 - 0,5451 0,0188
Solo –0,83 –0,54 0,31 - 0,5441
Impermeável 0,2 –0,54 –0,89 –0,31 -
A partir da Análise de Agrupamento (Cluster) foi evidenciada uma separação entre as
áreas urbanas e as demais. Entretanto, não foi verificada uma separação clara das áreas
periurbanas e rurais. Além disso, foi apresentada uma maior proximidade entre as áreas R1
e P2 (Figura 16).
0
20
40
60
80
100
120
U1 U2 P1 P2 R1 R2
Po
rce
nta
gem
da
área
(%
)
1000m
Água
Impermeável
Solo
Herbáceo-arbustiva
Arbórea

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
38
Figura 16 – Análise de Agrupamento (Cluster) entre as áreas urbanas, periurbanas e rurais, a partir das variáveis da paisagem. (a) 500m de lado; (b) 1000m de lado.
5.3 Relações entre as variáveis da paisagem e as abelhas
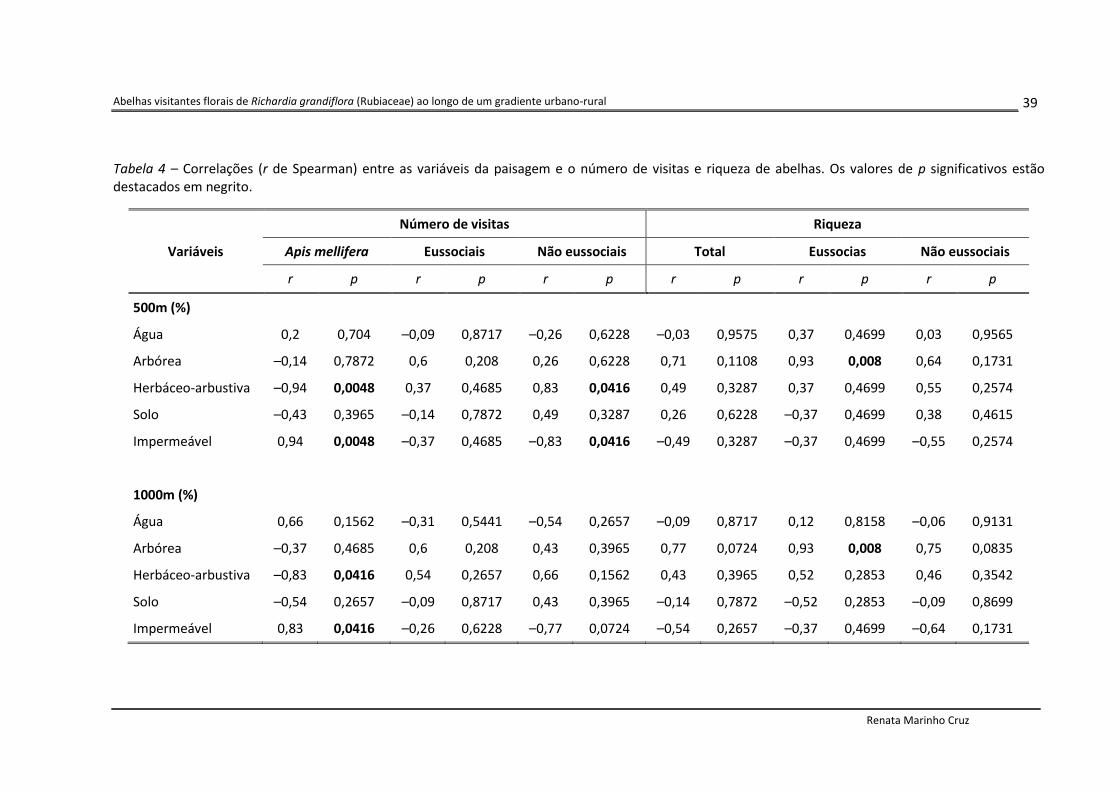
Ao relacionar as variáveis da paisagem com o número de visitas obtido para cada
grupo de abelha (Tabela 4), foi verificada uma forte correlação entre as visitas por Apis
mellifera e os percentuais de vegetação herbáceo-arbustiva e superfícies impermeáveis, nas
duas escalas espaciais. Para a vegetação herbáceo-arbustiva a correlação foi negativa (r = –
0,94; p < 0,0048; r = – 0,83; p = 0,0416) enquanto que para as superfícies impermeáveis foi
positiva (r = 0,94; p = 0,0048; r = 0,83; p = 0,0416). Da mesma forma, essas duas variáveis se
correlacionaram fortemente, na escala de 500m, com o número de visitas por abelhas não
eussociais. Entretanto, diferentemente do observado para A. mellifera, a correlação foi
positiva para a vegetação herbáceo-arbustiva (r = 0,83; p = 0,0416) e negativa para as
superfícies impermeáveis (r = – 0,83; p = 0,0416).
A única correlação significativa entre as variáveis da paisagem e a riqueza de espécies
foi obtida ao relacionar a porcentagem de vegetação arbórea com o número de espécies
eussociais encontradas em cada área (Tabela 4). A correlação foi verificada para as duas
escalas espaciais, sendo forte e positiva (r = 0,93; p = 0,008).
a b
Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
39
Tabela 4 – Correlações (r de Spearman) entre as variáveis da paisagem e o número de visitas e riqueza de abelhas. Os valores de p significativos estão destacados em negrito.
Número de visitas Riqueza
Variáveis Apis mellifera Eussociais Não eussociais Total Eussocias Não eussociais
r p r p r p r p r p r p
500m (%)
Água 0,2 0,704 –0,09 0,8717 –0,26 0,6228 –0,03 0,9575 0,37 0,4699 0,03 0,9565
Arbórea –0,14 0,7872 0,6 0,208 0,26 0,6228 0,71 0,1108 0,93 0,008 0,64 0,1731
Herbáceo-arbustiva –0,94 0,0048 0,37 0,4685 0,83 0,0416 0,49 0,3287 0,37 0,4699 0,55 0,2574
Solo –0,43 0,3965 –0,14 0,7872 0,49 0,3287 0,26 0,6228 –0,37 0,4699 0,38 0,4615
Impermeável 0,94 0,0048 –0,37 0,4685 –0,83 0,0416 –0,49 0,3287 –0,37 0,4699 –0,55 0,2574
1000m (%)
Água 0,66 0,1562 –0,31 0,5441 –0,54 0,2657 –0,09 0,8717 0,12 0,8158 –0,06 0,9131
Arbórea –0,37 0,4685 0,6 0,208 0,43 0,3965 0,77 0,0724 0,93 0,008 0,75 0,0835
Herbáceo-arbustiva –0,83 0,0416 0,54 0,2657 0,66 0,1562 0,43 0,3965 0,52 0,2853 0,46 0,3542
Solo –0,54 0,2657 –0,09 0,8717 0,43 0,3965 –0,14 0,7872 –0,52 0,2853 –0,09 0,8699
Impermeável 0,83 0,0416 –0,26 0,6228 –0,77 0,0724 –0,54 0,2657 –0,37 0,4699 –0,64 0,1731

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
40
Relacionando as espécies com as variáveis da paisagem, para algumas foram obtidas
correlações significativas (Tabela 5). Dentre elas, Augochlorini spp., Ceratina (Crewella) spp.
e Ceratina (Ceratinula) manni apresentaram uma correlação forte e positiva com a
vegetação herbáceo-arbustiva (r de 0,83 a 0,94). Plebeia flavocinta e Dialictus sp. 2 se
correlacionaram fortemente com a vegetação arbórea (r de 0,81 a 0,93), nas duas escalas.
Por fim, Callonychium brasiliense apresentou uma forte correlação positiva com solo livre (r
= 0,81), na escala de 500m, e negativa com superfícies impermeáveis (r = – 0,84), na escala
de 1000m.
Tabela 5 – Correlações significativas (r de Spearman) entre as variáveis da paisagem e o número de visitas por espécies. *p < 0,05; ** p < 0,01; *** p < 0,001.
Eussociais Não eussociais
P.
flavocincta Augochlorini
spp. C.
brasiliense Ceratina (C.) spp.
Ceratina (C.) manni
Dialictus sp.2
500m (%) Água
Arbórea 0,81*
0,88*
Herbáceo-arbustiva
0,83*
0,83* 0,88* Solo
0,81*
Impermeável
–0,83*
–0,83* –0,88*
1000m (%) Água
-
Arbórea 0,84*
0,93**
Herbáceo-arbustiva
0,94** 0,94** Solo
Impermeável
–0,84*
5.4 Sucesso reprodutivo
Não foi verificada diferença significativa (p > 0,05) entre os sucessos reprodutivos
obtidos nos experimentos de polinização livre das áreas urbanas (U1 e U2) (Tabela 5). Por
sua vez, os sucessos reprodutivos da polinização livre das áreas periurbanas e rurais
diferiram das urbanas (p < 0,05). A única exceção foi a igualdade estatística entre os
resultados da área urbana U1 e a rural R1 (χ2 = 3,3722; gl = 1; p = 0,0663). Essa igualdade nas

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
41
áreas U1 e R1 foi registrada, ainda, para o sucesso reprodutivo oriundo da polinização
cruzada manual complementar (χ2 = 0,5685; gl = 1; p = 0,4508). As áreas rurais foram as
únicas em que não foi verificada diferença significativa entre o sucessos reprodutivos da
polinização livre e cruzada manual complementar (R1: χ2 = 2,3717; gl = 1; p = 0,2415 e R2: χ2
= 2,1298; gl = 1; p = 0,1445). Além disso, os valores da Eficácia Reprodutiva (ER) dessas áreas
foram os maiores obtidos (0,9 e 0,93).
Tabela 6 – Sucessos reprodutivos para os experimentos de polinização e Eficácias Reprodutivas (ER) obtidas para cada área de estudo. As letras minúsculas, em cada linha, indicam diferenças significativas (p < 0,05) de acordo com o teste de χ2.
Experimento Sucesso Reprodutivo (%)
U1 U2 P1 P2 R1 R2
Polinização livre 42,3 ab
41,3 b
13,5 c 74,6
d 52,1
a* 75,9
d*
Polinização cruzada manual complementar
61,4 a 77,8
b 27,6
c 92
d 57,6
a* 81,3
e*
ER 0,69 0,53 0,49 0,81 0,9 0,93
* Não foi verificada diferença significativa entre a polinização livre e cruzada. R1: χ2 = 2,3717; gl = 1; p = 0,2415
e R2: χ2 = 2,1298; gl = 1; p = 0,1445.
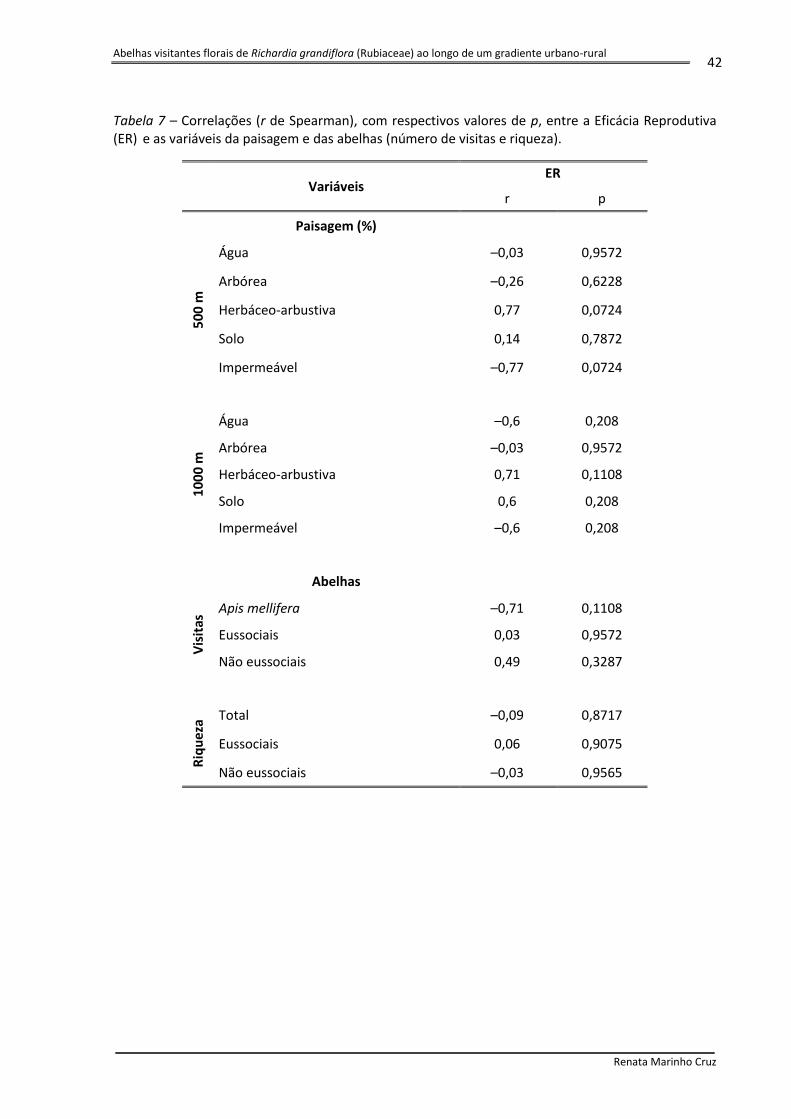
Não foram obtidas correlações significativas entre as variáveis da paisagem e a Eficácia
Reprodutiva (ER) (Tabela 6). Contudo, os índices de correlação obtidos para a vegetação
herbáceo-arbustiva e superfícies impermeáveis foram altos e quase atingiram o nível de
significância. Foi obtido um índice positivo para a vegetação herbáceo-arbustiva (500m: r =
0,77; 1000m: r = 0,71) e negativo para as superfícies impermeáveis (500m: r = – 0,77;
1000m: r = – 0,6). Da mesma forma, não foram verificadas correlações significativas entre as
abelhas (número de visitas e riqueza) e a ER. Dentre os valores de correlação obtidos, o
único que se destacou foi o número de visitas por Apis mellifera (r = – 0,71).
Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
42
Tabela 7 – Correlações (r de Spearman), com respectivos valores de p, entre a Eficácia Reprodutiva (ER) e as variáveis da paisagem e das abelhas (número de visitas e riqueza).
Variáveis
ER
r p
Paisagem (%)
50
0 m
Água –0,03 0,9572
Arbórea –0,26 0,6228
Herbáceo-arbustiva 0,77 0,0724
Solo 0,14 0,7872
Impermeável –0,77 0,0724
10
00
m
Água –0,6 0,208
Arbórea –0,03 0,9572
Herbáceo-arbustiva 0,71 0,1108
Solo 0,6 0,208
Impermeável –0,6 0,208
Abelhas
Vis
itas
Apis mellifera –0,71 0,1108
Eussociais 0,03 0,9572
Não eussociais 0,49 0,3287
Riq
uez
a Total –0,09 0,8717
Eussociais 0,06 0,9075
Não eussociais –0,03 0,9565

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
43
6 DISCUSSÃO
6.1 Polinizadores e paisagem
As áreas urbanas puderam ser diferenciadas das demais através das variáveis da
paisagem e composição de espécies, mas o mesmo não foi obtido para as áreas periurbanas
e rurais. As abelhas eussociais, excluindo Apis mellifera, e não eussociais apresentaram
respostas diferentes à urbanização, sendo as últimas aparentemente mais sensíveis a essas
alterações. Além disso, a hipótese de que as áreas mais urbanizadas apresentam uma maior
frequência da espécie introduzida A. mellifera foi confirmada.
A partir das estimativas das variáveis da paisagem e da Análise de Agrupamento, as
áreas urbanas foram nitidamente diferenciadas das demais, devido a sua paisagem
composta em grande parte por superfícies impermeáveis e com pouca cobertura vegetal
(arbórea e herbáceo-arbustiva). Contudo, as áreas periurbanas e rurais não puderam ser
diferenciadas entre si pelas características da paisagem. De fato, em termos de percentual
de superfícies impermeáveis, áreas periurbanas são mais próximas a rurais que urbanas
(Radford & James 2013). Possivelmente, uma melhor diferenciação seria obtida apenas com
a utilização de escalas espaciais maiores, que, entretanto, não seriam úteis para
compreender os efeitos sobre as abelhas, por não serem compatíveis com os raios de voo da
maioria das espécies. Da mesma forma, a partir da composição de espécies foi reafirmada
uma maior semelhança entre as áreas urbanas e a não diferenciação das áreas periurbanas e
rurais.
A riqueza de abelhas eussociais pouco se alterou ao longo do gradiente urbano-rural.
As abelhas eussociais possuem a grande capacidade de encontrar e estocar alimento, o que
pode fazer com que elas tenham um melhor sucesso em áreas onde os recursos são
escassos, como as áreas urbanas (Taura & Laroca 1991, Velthuis 1997, Zanette et al. 2005).
Considerando apenas as abelhas eussociais nativas (Meliponina), observou-se que o número
de visitas foi similar entre as áreas urbanas, periurbanas e rurais. Os meliponíneos
apresentam uma variedade de locais de nidificação, incluindo ninhos aéreos e em cavidades
preexistentes (ocos de árvore, solo, ninhos de cupins), o que pode refletir o seu potencial de

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
44
adaptação (Nogueira-Neto 1997, Velthuis 1997, Sousa et al. 2002). Além disso, algumas
espécies são bastante tolerantes às condições dos ambientes urbanos, nidificando, inclusive,
em frestas de construções e muros (Pedro & Camargo 1999, Taura & Laroca 1991).
Analisando o efeito do distúrbio humano em florestas, Samejima et al. (2004) obtiveram que
o distúrbio humano reduziu a densidade de ninhos, mas teve pouco efeito sobre o número
total de abelhas sem ferrão. Zanette et al. (2005), por sua vez, reportaram que a abundância
de meliponíneos foi diretamente afetada pela perda da cobertura vegetal e aumento das
superfícies impermeáveis associadas com a urbanização. Em ambos os estudos, entretanto,
foi verificada uma alteração na abundância relativa por espécie, que pode variar de acordo
com os requerimentos de nidificação de cada espécie (Samejima et al. 2004, Zanette et al.
2005). A espécie Trigona spinipes merece destaque, pois esteve presente em todas as áreas
e com uma frequência semelhante. T. spinipes é uma espécie generalista, que constrói
ninhos aéreos, frequentemente encontrada em ambientes urbanos e que parece não ser
sensível às mudanças relacionadas à urbanização (Zanette et al. 2005). Outra espécie que
deve ser destacada é Plebeia flavocincta que, embora possa ser encontrada nidificando em
substratos antrópicos (muros, construções), foi mais frequente nas áreas periurbanas e
rurais e apresentou correlação positiva com a vegetação arbórea. Os indivíduos
pertencentes a esse gênero nidificam, originalmente, em cavidades preexistentes, como
ocos de árvore e troncos (Roubik 2006). Por fim, foi verificada uma correlação positiva entre
a vegetação arbórea e a riqueza de abelhas eussociais, corroborando o fato de que embora
possa ser observada uma grande densidade de colônias em áreas urbanas e fragmentos,
geralmente se encontra uma baixa diversidade de espécies, relacionada, sobretudo, à baixa
densidade de árvores (Cane 2001, Sousa & Kleinert 2010).
Assim como Trigona spinipes, Apis mellifera também foi encontrada em todas as seis
áreas de estudo. A. mellifera é uma espécie eussocial introduzida, superabundante em
diversos levantamentos realizados no Brasil, que pode ser encontrada em todos os
ambientes (urbanos, agrícolas e naturais) (Minussi & Alves-dos-Santos 2007, Silveira et al.
2002). Entretanto, diferentemente dos meliponíneos, a frequência e o número de visitas
pelos indivíduos de A. mellifera foram superiores nas áreas urbanas. A predominância dessa
espécie nas áreas mais urbanizadas foi evidenciada, ainda, através da correlação negativa
entre o número de visitas e a vegetação herbáceo-arbustiva (e consequentemente positiva
para superfícies impermeáveis). Situação semelhante foi observada por Steffan-Dewenter et

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
45
al. 2002, que registraram um aumento na densidade de visitas de A. mellifera em resposta à
redução do percentual de áreas seminaturais. Similarmente, Aizen & Feinsinger (1994a e
1994b) observaram um aumento da população da espécie A. mellifera, em detrimento das
abelhas nativas, com o aumento da fragmentação florestal. O aumento do grau de
impermeabilização, e a consequente redução da cobertura vegetal, implica em um menor
número de recursos disponíveis, o que pode ter provocado o aumento da densidade de
visitas por A. mellifera nas áreas urbanas (Steffan-Dewenter et al. 2002). Ao mesmo tempo,
espécies exóticas, como A. mellifera, geralmente apresentam grande flexibilidade ecológica
e poder de competição, de modo que podem ter sido favorecidas pela influência antrópica
(Buchmann 1996, Melo et al. 2006, Roubik 1996). Esse favorecimento pode ter gerado,
ainda, uma pressão competitiva sobre as abelhas nativas, provavelmente minimizada nas
áreas menos urbanizadas devido a uma maior disponibilidade de fontes alimentares (Aizen &
Feinsinger 1994b, Pedro & Camargo 1991, Schaffer et al. 1983, Steffan-Dewenter et al. 2002,
Wilms et al. 1996).
Por outro lado, as abelhas não eussociais apresentaram uma riqueza e frequência
maior nas áreas periurbanas e rurais. Além disso, foi verificada uma correlação positiva entre
o número de visitas e a vegetação herbáceo-arbustiva. De forma semelhante, Hennig &
Ghazoul (2011) observaram, em áreas urbanas de Zürich, um aumento da diversidade e
visitas de abelhas relacionada à extensão de áreas verdes. Espécies não eussociais possuem
requerimentos de habitat mais específicos e restritos que as abelhas eussociais, o que pode
fazer com que suas populações tendam mais a declinar (Biesmeijer et al. 2006). Diversos
estudos apontam que espécies não eussociais são mais sensíveis à fragmentação de habitat
e ao aumento da urbanização (Bates et al. 2011, Gathmann et al. 1994, Geslin et al. 2013,
Greenleaf et al. 2007, Steffan-Dewenter et al. 2002). Geslin et al. 2013, por exemplo,
observou uma redução na frequência de abelhas solitárias em ambientes urbanos e
suburbanos, quando comparada à obtida nas áreas seminaturais e rurais.
A correlação entre o número de visitas das abelhas não eussociais e a vegetação
herbáceo-arbustiva foi obtida apenas na escala menor (500m). Analisando a relação entre
abelhas e a porcentagem de habitas seminaturais, Steffan-Dewenter et al. 2002 demonstram
que a distribuição de abelhas solitárias é melhor explicada utilizando escalas espaciais
menores. Provavelmente, isso se deve ao fato de que as abelhas não eussociais apresentam
raios de forrageio menores que as eussociais, de modo que são mais afetadas por

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
46
modificações locais na paisagem. Nas abelhas eussociais o raio de forrageio pode chegar a 2
– 3km ou mais (Buchmann & Shipman 1991, Osborne et al. 2008, Steffan-Dewenter &
Tscharntke 2000, Zurbuchen et al. 2010), enquanto que em espécies solitárias pequenas,
varia de poucas centenas de metros (100-200m) até aproximadamente 1,3km (Beil et al.
2008, Gathmann & Tscharntke 2002, Zurbuchen et al. 2010). Geralmente, espécies com
pequena capacidade de deslocamento ou dispersão percebem a paisagem num contexto
mais local, ao contrário de espécies com maior capacidade de deslocamento, que tendem a
perceber a paisagem num contexto mais amplo (Metzger 2001). Entretanto, ao relacionar as
variáveis vegetação arbórea e herbáceo-arbustiva com o número de visitas por espécie de
abelha, foram obtidas correlações significativas nas duas escalas. Isso pode ter ocorrido
devido à correlação significativa entre as porcentagens dessas variáveis das duas escalas
espaciais, ou porque o raio de voo de algumas espécies abrange as duas escalas espaciais
estudadas. Considerando que não foi observada correlação significativa entre a variável solo
livre nas duas escalas, a correlação obtida entre a espécie Callonychium brasiliense e a
variável solo livre, na menor escala, deve ser destacada. C. brasiliense é uma espécie que
nidifica no solo e possui entre 4 e 7mm de comprimento (Silveira et al. 2002). Espécies com
menor tamanho corporal geralmente apresentam raios de voo também menores (Gathmann
et al. 1994, Greenleaf et al. 2007, Steffan-Dewenter & Tscharntke 1999) o que, juntamente
com a correlação obtida na escala de 500m, pode evidenciar a importância da
disponibilidade local de solo livre para a manutenção dessa espécie.
6.2 Sucesso reprodutivo
A hipótese de que o sucesso reprodutivo é influenciado positivamente ao longo do
gradiente urbano-rural foi confirmada. As áreas menos urbanizadas apresentaram uma
maior frequência de abelhas eussociais nativas e não eussociais. Além disso, o menor
sucesso reprodutivo obtido nas áreas mais urbanizadas pode ter resultado do maior número
de visitas de A. mellifera e/ou da fragmentação das populações de Richardia grandiflora
nessas áreas.
A produção de frutos pode estar relacionada não apenas à eficiência dos
polinizadores, mas também à limitação de recursos (Horvitz & Schemske 1988) ou outros

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
47
fatores que independam das abelhas. Dessa forma, outros fatores das áreas, como
disponibilidade de nutrientes e água no solo, microclima etc., podem estar influenciando os
resultados do sucesso reprodutivo oriundo da polinização livre. A partir do experimento de
polinização cruzada manual complementar se obtém a taxa de frutificação máxima para as
condições locais (Guedes et al. 2011). Assim, pode-se admitir que a Eficácia Reprodutiva
(ER), para espécies xenogâmicas, como Richardia grandiflora (Cruz 2011), fornece uma
indicação indireta da eficiência dos polinizadores (Zapata & Arroyo 1978). Essa situação foi
evidenciada, neste estudo, através da igualdade estatística entre os resultados da
polinização livre para as áreas U1 e R1, sendo, entretanto, verificado um maior valor de ER
para a área rural R1.
A igualdade estatística entre os resultados da polinização livre e cruzada das áreas
rurais, bem como os altos valores de Eficácia Reprodutiva (ER), demonstraram a ocorrência
de uma alta taxa de polinização natural. Em uma das áreas periurbanas (P2), embora os
resultados da polinização livre e cruzada tenham diferido estatisticamente, também foi
verificado um alto valor de ER. Essas áreas possuem em comum uma baixa frequência de
visitas por Apis mellifera e maiores frequências de abelhas não eussociais e eussociais
nativas (Meliponina). Diversos estudos apontam os meliponíneos como importantes
polinizadores da flora silvestre e cultivada (Imperatriz-Fonseca et al. 2006), dentre eles,
espécies de Frieseomelitta, Partamona, Plebeia e Trigona, gêneros que foram registrados
neste estudo (Ex. Alves & Freitas 2007, Kiill & Simão-Bianchini 2011, Maués et al. 2000,
Schlindwein & Medeiros 2006). Embora meliponíneos sejam generalistas, estudos
demonstram uma grande fidelidade floral - operárias visitam uma única espécie de planta
em uma mesma viagem – o que pode conferir uma maior eficiência na polinização (Heard
1999, Ramalho et al. 1994). Da mesma forma, as abelhas não eussociais também são
consideradas como importantes polinizadoras de muitas plantas nativas (Freitas 2004,
Nunes-Silva et al. 2010). Para a subfamília Halictinae, uma das mais ricas e frequentes neste
estudo, também existem referências de espécies atuando como polinizadoras (Dikmen 2007,
Nunes-Silva et al. 2010, Venturieri et al 2005). Entretanto, neste estudo, não foi obtida
correlação significativa entre as abelhas não eussociais ou eussociais nativas e a ER. Para as
abelhas não eussociais foi sugerida, apenas, uma correlação positiva moderada.
Não foram obtidas correlações significativas entre as abelhas (riqueza e número de
visitas) e a ER. Entretanto, a alta frequência e número de visitas por A. mellifera nas áreas

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
48
mais urbanizadas, sugerem que os indivíduos dessa espécie não polinizaram as flores de R.
grandiflora tão eficientemente quanto os nativos. Esse fato poderia explicar os menores
valores de ER obtidos para as áreas urbanas e a área periurbana P1. Garibaldi et al. 2013
verificaram uma associação entre as visitas de insetos nativos e o aumento no sucesso
reprodutivo de 41 sistemas de cultivo em todo mundo, enquanto que para Apis mellifera foi
obtido um aumento significativo em apenas 14% dos sistemas analisados. Alguns estudos
apontam que A. mellifera geralmente visita muitas flores pertencentes a um mesmo
indivíduo, o que favorece a geitonogamia (Aizen & Feinsinger 1994a, Celebrezze & Paton
2004, Dupont et al. 2004, Goulson 2003). Além disso, os indivíduos dessa espécie forrageiam
mais cedo do que a maioria das abelhas nativas, deixando as anteras praticamente vazias
(Carmo et al. 2004, Carneiro & Martins 2012, Goulson 2003), e depositam poucos grãos de
pólen nos estigmas (Jacobi & Del Sarto 2007). Os indivíduos de A. mellifera podem
apresentar, ainda, baixa fidelidade floral (Westerkamp 1991), o que pode gerar o depósito
de grãos de pólen heteroespecíficos e dificultar a germinação daqueles provenientes de
indivíduos da mesma espécie (Neves & Viana 2011). Contudo, no estudo de polinização de
Richardia grandiflora, realizado por Cruz (2011), A. mellifera foi a espécie de abelha mais
frequente, tocou as estruturas reprodutivas florais em todas as visitas e, ainda, foi verificada
uma alta taxa de polinização natural. Da mesma forma, diversos estudos demonstraram a
eficiência dos indivíduos da espécie introduzida A. mellifera na polinização de espécies
vegetais cultivadas e silvestres (ex. Alves & Freitas 2007, Carneiro & Martins 2012, Freitas &
Paxton 1998, Nadia et al. 2007, Neves & Viana 2011, Santos et al. 2005).
Além disso, o menor sucesso reprodutivo obtido nas áreas mais urbanizadas pode ter
sido influenciado pela fragmentação de habitat. Da mesma forma que para as abelhas, não
foram verificadas correlações significativas entre as variáveis da paisagem e a Eficácia
Reprodutiva, mas os resultados das correlações sugerem uma influência positiva da
vegetação herbáceo-arbustiva (e negativa das superfícies impermeáveis) no sucesso
reprodutivo de Richardia grandiflora. Nas áreas mais urbanizadas, as populações de R.
grandiflora estão cercadas por uma matriz de superfícies impermeáveis, constituindo,
portanto, em um caso extremo de fragmentação de habitat (Andrieu et al. 2009). Diversos
estudos têm demonstrado que populações de plantas mais isoladas geralmente sofrem mais
com a erosão genética (Fischer & Matthies 1997, Oostermeijer et al. 1994) e apresentam
uma redução no sucesso reprodutivo (Aizen & Feinsinger 1994a, Kéry et al. 2000, Steffan-

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
49
Dewenter & Tscharntke 1999). Em ambientes fragmentados são, geralmente, observadas
alterações nas relações planta-polinizador (Goverde et al. 2002, Tscharntke & Brandl 2004).
Em um estudo analisando os efeitos da fragmentação de habitat em populações da
Asteraceae ruderal Crepis sancta (L.) Bornm (Andrieu et al. 2009), foram observadas
modificações no comportamento dos indivíduos da espécie Apis mellifera entre áreas
urbanas e rurais. Nas populações urbanas, os indivíduos de A. mellifera apresentaram uma
maior duração nas visitas e, ainda, visitaram inflorescências mais próximas. A partir da teoria
do forrageamento ótimo (Charnov 1976), é possível compreender essa modificação como
sendo uma maneira de compensar a energia gasta para chegar até as inflorescências
(Andrieu et al. 2009, Goulson 1999, Zimmerman 1982). Essas alterações no comportamento
dos polinizadores podem conduzir a uma alta geitonogamia e, consequentemente, reduzir o
sucesso reprodutivo (Andrieu et al. 2009). Além disso, Geslin et al. 2013 verificaram que
para alguns grupos de abelhas pode haver um aumento do comportamento generalista ao
longo do gradiente urbano-rural, o que também reduz o sucesso reprodutivo das espécies
vegetais.
Dessa forma, a fragmentação de habitats encontrada nas áreas urbanas pode ter
modificado o comportamento dos polinizadores, o que poderia reduzir a eficiência total dos
polinizadores nas áreas mais urbanizadas. Os resultados deste estudo mostraram que
Richardia grandiflora apresenta maior sucesso reprodutivo em áreas menos urbanizadas,
sugerindo, ainda, uma relação com a maior disponibilidade de abelhas nativas. Entretanto,
para essas relações serem melhor compreendidas são necessários testes específicos de
eficiência de polinizadores ao longo de gradientes urbano-rurais.

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
50
7 CONCLUSÕES
As áreas urbanas se diferenciaram das demais através da caracterização da paisagem
e composição de espécies. Para as áreas periurbanas e rurais não houve uma clara
diferenciação, devido à similaridade que elas apresentam nas escalas espaciais consideradas.
De uma maneira geral, a pouca alteração na riqueza das abelhas eussociais e no
número de visitas por meliponíneos indica uma boa tolerância desse grupo às modificações
ocasionadas pela urbanização. Para a espécie introduzida Apis mellifera, corroborando o que
foi encontrado em estudos sobre fragmentação de habitat, foi verificada uma maior
frequência nas áreas urbanas, o que pode estar relacionada com menor disponibilidade de
recursos, assim como sua flexibilidade ecológica e poder competitivo.
As abelhas não eussociais apresentaram uma maior riqueza e frequência nas áreas
periurbanas e rurais. Além disso, a correlação positiva entre o número de visitas e a
vegetação herbáceo-arbustiva, na escala espacial menor, sugere uma maior influência da
paisagem local nessas espécies. Essa condição destaca a importância de se preservarem
diferentes habitats dentro das áreas urbanas, como, por exemplo, áreas para abelhas que
nidificam no solo, como Callonychium brasiliense.
Por fim, o sucesso reprodutivo foi afetado positivamente ao longo do gradiente
urbano-rural, pois as áreas rurais apresentaram as maiores taxas de polinização natural,
enquanto as urbanas, valores mais baixos. Em relação às áreas periurbanas foi observada
uma situação mista, reafirmando a condição intermediária, entre o urbano-rural, que essas
áreas representam. Possivelmente, A. mellifera não foi um polinizador tão eficiente quanto
as abelhas nativas, o que pode ser consequência da fragmentação das populações de
Richardia grandiflora nas áreas mais urbanizadas. Entretanto, são necessárias novas
observações e testes para obter conclusões mais precisas.

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
51
REFERÊNCIAS
Agra, M. F.; Freitas, P. F. & Barbosa-Filho, J. M. 2007. Synopsis of the plants known as medicinal and poisonous in Northeast of Brazil. Brazilian Journal of Pharmacognosy, 17(1) : 114-140.
Aguilar, R.; Ashworth, L.; Galetto, L. & Aizen, M. A. 2006. Plant reproductive susceptibility to habitat fragmentation: review and synthesis through a meta-analysis. Ecology Letters, 9: 968–980.
Ahrné, K.; Bengtsson, J. & Elmqvist, T. 2009. Bumble bee (Bombus spp) along a gradient of increasing urbanization. PLoS ONE, 4(5): e5574
Aizen, M. A. & Feinsinger, P. 1994a. Forest fragmentation, pollination, and plant reproduction in a chaco dry forest, Argentina. Ecology, 75: 330–351.
Aizen, M. A. & Feinsinger, P. 1994b. Habitat fragmentation, native insect pollinators, and feral Honey bees in argentine "chaco serrano". Ecological Applications, 4(2): 378-392.
Alves, J. E. & Freitas, B. M. 2007. Requerimentos de polinização da goiabeira. Ciência Rural, 37(5): 1281-1286.
Andersson, L. 1992. A provisional checklist of neotropical Rubiaceae. Scripta Botanica Belgica, 1: 1-199.
Andrieu, E.; Dornier, A.; Rouifed, S.; Schatz, B. & Cheptou, P. O. 2009. The town Crepis and the country Crepis: How does fragmentation affect a plant–pollinator interaction? Acta Oecologica, 35: 1-7.
Baker, H. G. & Hurd, P. D, JR. 1968. Intrafloral ecology. Annual Review of Entomology, 13: 385-414.
Banaszak-Cibicka W. & Z mihorski, M. 2011.Wild bees along an urban gradient: winners and losers. Journal of Insect Conservation, 16: 331-343.
Barbosa, A. G. 2005. Produção do Espaço e Transformações Urbanas no Litoral Sul de João Pessoa – PB. Dissertação (Mestrado em Geografia). Universidade Federal do Rio Grande do Norte. 192p.
Bates, A. J.; Sadler, J. P.; Fairbrass, A. J.; Falk, S. J.; Hale, J. D. & Matthews, T. J. 2011. Changing bee and hoverfly pollinator assemblages along an urban-rural gradient. . PloS ONE, 6 (8): e23459.
Beil, M.; Horn, H.; Schwabe A. 2008. Analysis of pollen loads in a wild bee community (Hymenoptera: Apidae), a method for elucidating habitat use and foraging distances. Apidologie,39: 456-467.

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
52
Biesmeijer, J. C.; Roberts, S. P. M.; Reemer, M.; Ohlemuller, R.; Edwards, M.; Peeters, T. ; Schaffers, A. P.; Potts, S. G.; Kleukers, R.; Thomas, C. D.; Settele, J. & Kuni, W. E. 2006. Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science, 313: 351-354.
Boff, S. V. Flora de capões e Hymenoptera (abelhas e vespas) visitantes florais de flores no pantanal do Miranda-abobral. 2008. Dissertação (Mestrado em Biologia Vegetal).Universidade Federal de Mato Grosso do Sul. 97p.
Buchmann, S. L. & Nabhan, G. P. 1996. The forgotten pollinators. Island Press, 292p.
Buchmann, S. L. & Shipman, C .W. 1991. Foraging distances flown by honey bee colonies: analyses using Mathematica software. American. Bee Journal, 131: 771.
Buchmann, S. L. 1996. Competition between honey bees and native bees in the Sonoran Desert and global bee conservation issues, p. 125-142. In: Matheson, A.; Buchmann, S. L.; O’toole, C.; Westrich, P. & Willians, I. (Eds.). The Conservation of Bees. Academic Press, 254p.
Cane, J. H. 2001. Habitat fragmentation and native bees: a premature verdict? Conservation Ecology, 5(1): 3. Disponível em < http://www.consecol.org/vol5/iss1/ art3/ >. (Acesso em: 20 /02/2012).
Carmo, R. M.; Franceschinelli, E. V. & Silveira, F. A. 2004. Introduced honeybees (A. mellifera) reduce pollination success without affecting the floral resource taken by native pollinators. Biotropica, 36 (3): 371-376.
Carneiro, L. T. & Martins, C. F. 2012. Africanized honey bees pollinate and preempt the pollen of Spondias mombin (Anacardiaceae) flowers. Apidologie, 43 (4): 474-486.
Carvalho, D. M. 2013. Interação abelha-planta em sistemas agrícolas: forças de interação e nichos tróficos. Dissertação (Mestrado em Zoologia). Universidade Estadual de Feira de Santana. 48p.
Celebrezze, T. & Paton, D. C. 2004. Do introduced honeybees (Apis mellifera, Hymenoptera) provide full pollination service to bird-adapted Australian plants with small flowers? An experimental study of Brachyloma ericoides (Epacridaceae). Austral Ecology, 29: 129-136.
Chacoff, N. P. & Aizen, M. A. 2006. Edge effects on flower-visiting insects in grapefruit plantations bordering premontane subtropical forest. Journal of Applied Ecology, 43: 18-27.
Charnov, E.L.1976. Optimal foraging, marginal value theorem.Theoretical. Population Biology, 9: 129–136.
Cruz, R. M. 2011. Biologia floral e visitantes florais de Richardia grandiflora (Rubiaceae). Monografia (Graduação em Ciências Biológicas). Universidade Federal da Paraíba. 53p.
Danforth, B. N.; Sipes, S.; Fang, J. & Brady, S. G. 2006. The history of early bee diversification based on five genes plus morphology. Proceedings of the National Academy of Sciences, 103(41): 15118–15123.

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
53
Dias, B. S. F.; Raw, A. & Imperatriz-Fonseca, V. L. 1999. International pollinators initiative: the São Paulo Declaration on Pollinators - Report on the Recommendations of the Workshop on the Conservation and Sustainable Use of Pollinators in Agriculture with Emphasis on Bees. Brazilian Ministry of the Environment (MMA), University of Sao Paulo, Brazilian Corporation for Agricultural Research (EMBRAPA).
Dikmen, F. 2007. The role and the importance of the family halictidae (apiformes: apoidea) in pollination of natural and agricultural vegetation. Mellifera, 7(13-14): 16-19.
Dupont, Y. L.; Hansen, D. M.; Valido, A. & Olesen, J. M. 2004. Impact of introduced honey bees on native pollination interactions of the endemic Echium wildpretii (Boraginaceae) on Tenerife, Canary Islands. Biological Conservation, 118: 301-311.
FAO (FOOD AND AGRICULTURE ORGANIZATION). 2004. Conservation and management of pollinators for sustainable agriculture - the international response, p. 19-25. In: Freitas, B. M. & Portela, J. O. B. (Eds.). Solitary bees: conservation, rearing and management for pollination. Imprensa Universitária/UFC. 285p.
Feliciano, M. L. M. & Melo, R. B. (Org.) 2003. Atlas do Estado da Paraíba: Informação para gestão do patrimônio natural (mapas). 1 ed. SEPLAN/IDEME. 58p.
Fetridge, E. D.; Ascher, J. S. & Langellotto, G. A. 2008. The bee fauna of residential gardens in a suburb of New York City (Hymenoptera: Apoidea). Annales of the Entomological Society of America, 101(6): 1067-1077.
Fischer, M. & Matthies, D. 1997. Mating structure and inbreeding and outbreeding depression in the rare plant Gentianella germanica (Gentianaceae). American Journal of Botany, 84(12): 1685-1692.
Freitas, B. M. & Paxton, R. J. 1998. A comparison of two pollinators: the introduced honey bee Apis mellifera and an indigenous bee Centris tarsata on cashew Anacardium occidentale in its native range of NE Brazil. Journal Applied Ecology, 35: 109-121.
Freitas, B. M. & Pereira, J. O. P. (Eds) 2004. Solitary bees: conservation, rearing and management for pollination. Imprensa Universitária/UFC. 285p.
Freitas, B. M. 2004. Solitary Bees: conservation, rearing and management for pollination. Imprensa Universitária, 285p.
Freitas, B. M.; Imperatriz-Fonseca, V. L.; Medina, L. M.; Kleinert, A .M. P.; Galleto, L.; Nates-Parra, G. & Quezadaeuán, J. J.G. 2009. Diversity, threats and conservation of native bees in the Neotropics. Apidologie, 40: 332-346.
Garibaldi, L. A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M. A.; Bommarco, R.; Cunningham, S. A.; Kremen, C.; Cavalheiro, L. G.; Harder, L. D.; Afik, O.; Bartomeus, I.; Benjamin, F.; Boreux, V.; Cariveau, D.; Chacoff, N.; Dudenhöffer; Freitas, B. M.; Ghazoul, J.; Greenleaf, S.; Hipólito, J.; Holzschuh, A.; Howlett, B.; Issacs, R.; Javorek, S. K.; Kennedy, C. M.; Krewenka, K. M., Krishnan, S.; Mandelik, Y.; Mayfield, M. M.; Motzke, I.; Munyuli, T.; Nault, B. A.; Otieno, M.; Petersen, J.; Pisanty, G.; Potts, S. G.; Rader, R.; Ricketts, T. H.; Rundlöf, M.; Seymour, C. L.; Shüepp, C.; Szentgyörgýi, H.; Taki, H.; Tscharntke, T.; Vergara, C. H.; Viana, B. F.; Wanger, T. C; Westphal, C.; Williams, N. & Klein, A. M. 2013. Wild pollinators enhance fruit set crops regardless of honey bee abundance. Science, 339: 1608-1611.

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
54
Gathmann, A. & Tscharntke, T. 2002. Foraging ranges of solitary bees. Journal of Animal Ecology, 71: 757-764.
Gathmann, A.; Greiler, H. J. & Tscharntke, T. 1994. Trapnesting bees and wasps colonizing set-aside fields: succession and body size, management by cutting and sowing. Oecologia, 98: 8–14.
Geslin, B.; Gauzens, B.; Thébault, E. & Dajoz, I. 2013. Plant Pollinator Networks along a Gradient of Urbanisation. PloS ONE, 8(5): e63421.
Giraudoux, P. 2011. Pgirmess: Data analysis in ecology. R package version 1.5.1. Disponível em < http://CRAN.R-project.org/package=pgirmess>.
Google Earth (Versão 7.1.1.1580). 2013. Software. Google Inc., Mountain View, USA.
Goulson, D. 1999. Foraging strategies of insects for gathering nectar and pollen, and implications for plant ecology and evolution. Perspectives in Plant Ecology, 2: 185–209.
Goulson, D. 2003. Effects of introduced bees on native ecosystems. Annual Review of Ecology and Systematics, 34: 1-26.
Goverde, M.; Schweizer, K.; Baur, B. & Erhardt, A. 2002. Small-scale fragmentation effects on pollinator behaviour: experimental evidence from the bumblebee Bombus veteranus on calcareous grasslands. Biological Conservation, 104: 293–299.
Greenleaf S. S.; Williams N. M.; Winfree R. & Kremen C. 2007. Bee foraging ranges and their relationship to body size. Oecologia, 153: 589–596.
Guedes, R. S.; Zanella, F. C. V.; Martins, C. F & Schlindwein, C. 2011. Déficit de polinização da aceroleira no período seco no semiárido paraibano. Revista Brasileira de Fruticultura, 33(2): 465-471.
Harris, L. F.; Johnson, S. D. 2004. The consequences of habitat fragmentation for plant pollinator mutualisms. International Journal of Tropical Insect Science, 24: 29–43.
Heard, T. A. 1999. The role of stingless bees in crop polinnation. Annual Review of Entomology, 44: 183-206.
Hennig, E. I.; Ghazoul, J. 2011. Plant–pollinator interactions within the urban environment. Perspectives in Plant Ecology, Evolution and Systematics, 13: 137–150.
Horvitz, C. C. & Schemske, D. W. 1988. A test of the pollinator limitation hypothesis for a Neotropical herb. Ecology, 69(1): 200-206.
IBGE. 2006. Censo Agropecuário 2006: Condição do produtor – Total – Masculino e Feminino – Área dos estabelecimentos agropecuários. IBGE Cidades@. Disponível em < http://www.ibge.gov.br/cidadesat/ topwindow.htm?1>. (Acesso em 08/04/2013).
IBGE. 2010a. Densidade demográfica. IBGE Cidades@. Disponível em < http://www. ibge.gov.br/ cidadesat/ topwindow.htm?1>. (Acesso em 08/04/2013).
IBGE. 2010b. Resultados do Censo 2010. Disponível em < http://www.ibge.gov.br/ home/estatistica/populacao/censo2010/tabelas_pdf/total_populacao_paraiba.pdf>. (Acesso em 08/04/2013).

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
55
Imperatriz-Fonseca, V. L.; Saraiva, A. M & De Jong, D. (Eds.) 2006. Bees as pollinators in Brazil: assessing the status and suggesting best practices. 112p.
Jacobi, C. M. & Del Sarto, M. C. L. 2007. Pollination of two species of Vellozia (Velloziaceae) from high-altitude quartzitic grassland, Brazil. Acta Botanica Brasilica, 21(2): 325-333.
Janzen, D. H. 1980. Ecologia vegetal nos trópicos. EPU/USP. 79p.
Kearns, C. A. & Inouye, D. W. 1997. Pollinators, flowering plants, and conservation biology. BioScience, 47(5): 297-307.
Kéry, M.: Matthies, D. & Spillmann, H. H. 2000. Reduced fecundity and offspring performance in small populations of the declining grassland plants Primula veris and Gentiana lutea. Journal of Ecology, 88: 17–30.
Kevan, P. & Imperatriz-Fonseca, V. L. (Eds) 2002. Pollinating Bees: the conservation link between agriculture and nature. Ministério do Meio Ambiente. 313p.
Kevan, P. G. & Phillips, T. P. 2001. The economic impacts of pollinator declines: an approach to assessing the consequences. Conservation Ecology, 5(1):8. Disponível em < http://www.consecol.org/vol5/iss1/art8/ >. (Acesso em: 10 /04/2013).
Kiill, L. H. P. & Simão-Bianchini, R. 2011. Biologia reprodutiva e polinização de Jacquemontia nodiflora (Desr.) G. Don (Convolvulaceae) em Caatinga na região de Petrolina, PE, Brasil. Hoehnea, 38(4): 511-520.
Kill, L. H. P.; Haji, F. N. P. & Lima, P. C. F. 2000. Visitantes florais de plantas invasoras de áreas com fruteiras irrigadas. Scientia Agricola, 57(3): 575- 580.
Kim, H. H. Urban heat island. 1992. International Journal of Remote Sensing 13(12): 2319-2336.
Klein, A. M.; Steffan-Dewenter, I. & Tscharntke, T. 2006. Rainforest promotes trophic interactions and diversity of trap-nesting Hymenoptera in adjacent agroforestry. Journal of Animal Ecology, 75: 315-323.
Klein, A. M.; Vaissière, B. E.; Cane, J. H.; Steffan-Dewenter, I.; Cunningham, S. A.; Kremen, C. & Tscharntke, T. 2007. Importance of pollinators in changing landscapes for world crops. Proceedings of the Royal Society B-Biological Sciences, 274: 303-313.
Koh, L. P. & Gardner, T. A. 2010. Conservation in human-modified landscapes, p. 236-261. In: Sodhi, N. S & Ehrlich, P. E. (Eds.) Conservation biology for all. Oxford University Press. 344p.
Kremen, C., Williams, N. M. & Thorp, R. W. 2002. Crop pollination from native bees at risk from agricultural intensification. Proceedings of the National Academy of Sciences of the United States of America. 99: 16812-16816.
Kunin, W. E. 1997. Population size and density effects in pollination: pollinator foraging and plant reproductive success in experimental arrays of Brassica kaber. Journal of Ecology, 85:225–234.
Leite, A. P. A. 2011. A produção de habitações populares no espaço periurbano da cidade de João Pessoa – PB (2006 – 2011): O conjunto habitacional Gervásio Maia. Dissertação (Mestrado em Arquitetura e Urbanismo). Universidade Federal da Paraíba. 128p.

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
56
Lewis, W. H.; & Oliver, R. L. 1974. Revision of Richardia (Rubiaceae). Brittonia, 26: 271-301.
Liow, L. H. 2001. Bee diversity along a gradient of disturbance in tropical lowland forests of Southeast Asia. CBM:s skriftserie, 3: 101-130.
Lorenzi, H. 1991. Plantas daninhas do Brasil: terrestres, aquáticas, parasitas, tóxicas e medicinais. 2.ed. Plantarum, 440p.
Maués, M. M.;Souza, L. A. & Miyanaga, R. 2000. Insetos polinizadores do cupuaçuzeiro (Theobroma grandiflorum Willd. Ex Sprengel) Schum. Sterculiaceae) no estado do Pará, Brasil. Embrapa CPATU, 19p.
McFrederick, Q. S. & LeBuhn, G. 2006. Are urban parks refuges for bumble bees Bombus spp. (Hymenoptera: Apidae)? Biological Conservation, 129: 372–382.
McIntyre, N. E. & Hostetler, M. E. 2001. Effects of urban land use on pollinator (Hymenoptera: Apoidea) communities in a desert metropolis. Basic Appl. Ecol. 2, 209–218.
McIntyre, N. E; Rango, J.; Fagan, W. F. & Faeth, S. H. 2001. Ground arthropod community structure in a heterogeneous urban environment. Landscape and Urban Planning, 52: 257-274.
McKinney, M. L. 2006. Urbanization as a major cause of biotic homogenization. Biological Conservation, 127: 247-260.
Melo, G. A. R. & Gonçalves, R. B. 2005. Higher-level bee classifications (Hymenoptera, Apoidea, Apidae sensu lato). Revista Brasileira de Zoologia, 22(1): 153-159.
Melo, G. A. R.; Martins, A. C. & Gonçalves, R. B. 2006. Alterações de longo prazo na estrutura de assembléias de abelhas: conhecimento atual e perspectivas, p. 150-155. In: Santana, W. C.; Lobo, C. H. & Hartfelder, K. H. (Eds.). Anais do VII Encontro sobre Abelhas. Ribeirão Preto, FFCLRP-USP, FMRP-USP. CD-ROM.
Metzger, J. P. 2001. O que é ecologia de paisagens? Biota Neotropica, 1(1-2): 1-9.
Michener, C. D. 2007. The bees of the world. 2 ed. Johns Hopkins University Press. 953p.
Minussi, L. C. & Alves-dos-Santos, I. 2007. Abelhas nativas versus Apis mellifera Linnaeus, espécie exótica (Hymenoptera: Apidae). Bioscience Journal, 23 (1): 58-62.
Moura, D. C. 2008. Interações entre plantas e abelhas nas matas ciliares do rio São Francisco. Tese (Doutorado em Biologia Vegetal). Universidade Federal de Pernambuco. 156p.
Moure, J. S.; Urban, D. & Melo, G. A. R. 2007. Catalogue of Bees (Hymenoptera, Apoidea) in the Neotropical Region. Sociedade Brasileira de Entomologia. 1058p.
Nadia, T. L.; Machado, I. C. & Lopes, A. V. 2007. Polinização de Spondias tuberosa Arruda (Anacardiaceae) e análise da partilha de polinizadores com Ziziphus joazeiro Mart. (Rhamnaceae), espécies frutíferas e endêmicas da caatinga. Revista Brasileira de Botânica, 30(1): 89-100.

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
57
Nascimento, A. S. 2011. Caracterização botânica e geográfica do mel de Apis mellifera L. produzido no território do Recôncavo da Bahia. 2011. Dissertação (Mestrado em Ciências Agrárias). Universidade Federal do Recôncavo da Bahia. 68p.
Neff, J. L. & Simpson, B. B. 1993. Bees, pollination systems and plant diversity. In: Lasalle, J. & Gauld, L.D. (Eds.). Hymenoptera and Biodiversity. CAB Internacional. 348p.
Neves, E. L. & Viana, B. F. 2011. Pollination efficiency of Apis mellifera Linnaeus, 1758 (Hymenoptera, Apidae) on the monoecious plants Jatropha mollissima (Pohl) Baill. And Jatropha mutabilis (Pohl) Baill. (Euphorbiaceae) in a semi-arid Caatinga area, northeastern Brazil. Brazilian Journal of Biology, 71(1): 107-113.
Nogueira-Neto, P. 1997. Vida e criação de abelhas indígenas sem ferrão. Nogueirapis, 445p.
Nunes-Silva, P.; Hrncir, M. & Imperatriz-Fonseca, V. L. 2010. A polinização por vibração. Oecologia Australis, 14(1): 140-151.
Ollerton, J. 1999. La evolución de las relaciones polinizador-planta en los Artrópodes. Boletín de la Sociedad Entomologica Aragonesa, 26: 741-758.
Oostermeijer, J. G. B.; van Eijck, M. W. & den Nijs, J. C. M.1994. Offspring fitness in relation to population size and genetic variation in the rare perennial plant species Gentiana pneumonanthe (Gentianaceae). Oecologia 97: 289–296.
Osborne, J. L.; Martin, A. P.; Carreck, N. L.; Swain, J. L.; Knight, M. E.; Goulson, D.; Hale, R. J. & Sanderson, R. A. 2008. Bumblebee flight distances in relation to the forage landscape. Journal of Animal Ecology, 77: 406-415.
Pedro, S. R. & Camargo, J. M. F. 1991. Interactions on floral resources between the Africanized honey bee Apis mellifera L. and the native bee community (Hymenoptera: Apoidea) in a natural "cerrado" ecosystem in southeast Brazil. Apidologie, 22: 397-415.
Pedro, S. R. M. & Camargo, J. M. F. 1999. Apoidea apiformes, p. 193 – 211. In: Brandão, C. R. F. & Canello, E. M. (Eds.) Biodiversidade do Estado de São Paulo: síntese do conhecimento ao final do século XX, Invertebrados terrestres (Vol. 5). FAPESP, 279p.
Pereira, M. S. & Barbosa, M. R. V. 2006. A família Rubiaceae na Reserva Biológica Guaribas, Paraíba, Brasil. Subfamília Rubioideae. Acta Botanica Brasilica, 20(2): 455-470.
Pires, J. M.; Carrer, C. C.; Carvalho, M. C.; Orlandelli, C. R.; Carrer; P. L. M & Pires, L. C. 2009. Diagnóstico do pasto apícola numa região de caatinga no município de Caiçara do Rio dos Ventos/RN. In: XIX Congresso Brasileiro de Zootecnia. Anais... ABZ e FZEA/USP.
Pott, A. & Pott, V. J. 1986. Inventário da flora apícola do Pantanal do Mato Grosso do Sul. Boletim Informativo da Empresa Brasileira de Pesquisa Agropecuária (EMBRAPA), 3: p. 1-18.
R Development Core Team. 2010. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Disponível em < http://www.R-project.org/.>.

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
58
Radford, K. G. & James, P. 2013. Changes in the value of ecosystem services along a rural–urban gradient: A case study of Greater Manchester, UK. Landscape and Urban Planning, 109: 117-127.
Ramalho, M.; Giannini, T. C.; Malagodibraga, K. S. & Imperatriz-Fonseca, V. L. 1994. Pollen harvest by stingless bee foragers (Hymenoptera, Apidae, Meliponinae). Grana, 33: 239–44.
Raven, P. H.; Evert, R. F. & Eichhorn, S. E. 2007. Biologia Vegetal. 7 ed. Guanabara Koogan.
Rebele, F. 1994. Urban ecology and special features of urban ecosystems. Global Ecology and Biogeography Letters, 4(6): 173-187.
Reis, D. S. 2006. O rural e urbano no Brasil. In: XV Encontro nacional de estudos populacionais – ABEP. Anais... ABEP. Disponível em <http://www.abep.nepo.unicamp. br/encontro2006/docspdf/abep2006_777.pdf>. (Acesso em: 16/06/2013).
Ricketts, T. H.; Regetz, J.; Steffan-Dewenter, I.; Cunningham, S. A.; Kremen, C.; Bogdanski, A.; Gemmill-Herren, B.; Greenleaf, S. S.; Klein, A. M.; Mayfield, M. M.; Morandin, L. A.; Ochieng, A. & Viana, B. F. 2008. Landscape effects on crop pollination services: are there general patterns? Ecology Letters, 11: 499-515.
Roubik, D. W. 1996. Measuring the meaning of honey bees, p. 163-172. In: Matheson, A.; Buchmann, S. L.; O’toole, C.; Westrich, P. & Willians, I. (Eds.). The Conservation of Bees. Academic Press, 254p.
Roubik, D. W. Stingless bee nesting biology. 2006. Apidologie, 37: 124-143.
Samejima, H.; Marzuki, M.; Nagamitsu, T. & Nakasizuka, T. 2004.The effects of human disturbance on a stingless bee community in a tropical rainforest. Biological Conservation , 120(4): 577-587.
Santos, M. J. S.; Machado, I. C. & Lopes, A. V. 2005. Biologia reprodutiva de duas espécies de Jatropha L. (Euphorbiaceae) em Caatinga, Nordeste do Brasil. Revista Brasileira de Botânica, 28(2): 361-373.
Santos, R. F.; Kiill, L. H. P. & Araújo, J. L. P. 2006. Levantamento da flora melífera de interesse apícola no município de Petrolina-PE. Caatinga, 19(3): 221-227.
Saure, C. 1996. Urban habitats for bees: the example of the city of Berlin, p. 47-53. In: Matheson, A.; Buchmann, S. L.; O’toole, C.; Westrich, P. & Willians, I. (Eds.). The Conservation of Bees. Academic Press, 254p.
Schaffer, W. M.; Zeh, D. W.; Buchmann, S. L.; Kleinhaus, S.; Schaffer, M. V.; Antrim, M. 1983. Competition for nectar between introduced honeybees and native North American bees and ants. Ecology, 64: 564-577.
Schlindwein, C. & Medeiros, P. C. R. 2006. Pollination in Turnera subulata (Turneraceae): Unilateral reproductive dependence of the narrowly oligolectic bee Protomeliturga turnerae (Hymenoptera, Andrenidae). Flora, 201: 178-188.
Siegel, S. & Castellan Jr., J. 2006. Estatística Não-paramétrica para Ciências do Comportamento. 2. ed. Artmed. 448p.

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
59
Silva, M. A.; Barbosa, J. S. & Albuquerque, H. N. 2010. Levantamento das plantas espontâneas e suas propriedades fitoterapêuticas: um estudo no complexo Aluízio Campos – Campina Grande – PB. Revista Brasileira de Informações Científicas, 1(1): 52-66.
Silva, R. A. 2006. Caracterização da flora apícola e do mel produzido por Apis mellifera L., 1758 (Hymenoptera: Apidae) no estado da Paraíba.Tese (Doutorado em Zootecnia). Universidade Federal da Paraíba. 86p.
Silva, R. A.; Evangelista-Rodrigues, A.; Aquino, I. S.; Felix, L. P.; Mata, M. F. & Peronico, A. A. 2008. Caracterização da flora apícola do semi-árido da Paraíba. Archivos de Zootecnia, 57: 427-438.
Silveira; F. A.; Melo, G. A. R. & Almeida, E. A. B. 2002. Abelhas brasileiras: sistemática e identificação. 1 ed. Fernando A. Silveira. 253p.
Sousa, L. A.; Pereira, T. O.; Prezoto, F. & Faria-Mucci, G. M. 2002. Nest foundation and diversity of Meliponini (Hymenoptera, Apidae) in a urban area of the municipality of Juiz de Fora, MG, Brazil. Bioscience Journal, 18(2): 59-65.
Sousa, V. C.; Kleinert, A. M. P. 2010. Identificação de ninhos de meliponíneos (Apidae, Meliponini) em área aberta do Parque de Ciência e Tecnologia da USP. In: IX Encontro sobre Abelhas. Resumos...USP/RP.
Steffan-Dewenter, I. & Tscharntke, T. 1999. Effects of habitat isolation on pollinator communities and seed set. Oecologia 121: 432–440.
Steffan-Dewenter, I. & Tscharntke, T. 2000. Resource overlap and possible competition between honey bees and wild bees in central Europe. Oecologia, 122:288-296.
Steffan-Dewenter, I.; Münzenberg, U.; Bürger, C.; Thies, C. & Tschartke, T. 2002. Scale-dependent effects of landscape context on three pollinator guilds. Ecology, 83(5): 1421-1432.
Taura, H. M. & Laroca, S. 1991. Abelhas altamente sociais de uma área restrita em Curitiba (Brasil): Distribuição dos ninhos e abundância relativa. Acta Biológica Paranaense, 20(1-4): 85-101.
Tscharntke, T. & Brandl, R. 2004. Plant-insect interactions in fragmented landscapes. Annual Review Entomology,49: 405-430.
Vale, A. R & Gerardi, L. H. O. 2006. Crescimento urbano e as teorias sobre o espaço periurbano: analisando o caso do município de Araraquara (SP), p. 231-246. In: Gerardi, L. H. O. & Carvalho, P. F. (Org.) Geografia: ações e reflexões. UNESP-Rio Claro/AGETEO. 434p.
Velthuis, H. H. W. 1997. Biologia das abelhas sem ferrão. Edusp, 33p.
Venturieri G. C.; Rodrigues S. T. & Pereira C. A. B. 2005. As abelhas e as flores do açaizeiro (Euterpe olecea Mart. – Arecaceae). Mensagem Doce, 80: 32- 33.
Viana, B. F.; Boscolo, D.; Mariano-Neto, E.; Lopes, L. E.; Lopes, A. V.; Ferreira, P. A; Pigozzo, C. M. & Primo, L. 2012. A polinização no contexto da paisagem: o que de fato sabemos e o que precisamos saber?, p. 67-102. In: Imperatriz-Fonseca, V. L.; Canhos, D. A. L.; Alves, D. A. & Saraiva, A. M. Polinizadores no Brasil. EDUSP. 488p.

Abelhas visitantes florais de Richardia grandiflora (Rubiaceae) ao longo de um gradiente urbano-rural
Renata Marinho Cruz
60
Vidal, M. G.; Santana, N. S. & Vidal, D. 2008. Flora apícola e manejo de apiários na região do Recôncavo Sul da Bahia. Revista Acadêmica: Ciências Agrárias Ambientais, 6(4): 503-509.
Wcislo, W. T. & Cane, J. H. 1996. Floral resource utilization by solitary bees (Hymenoptera: Apoidea) and exploitation of their stored foods by natural enemies. Annual Review of Entomology, 41: 257-286.
Westerkamp, C. H. 1991. Honey bees are poor pollinators - Why? Plant Systematic and Evolution, 177(1): 71-75.
Wilms, W.; Imperatriz-Fonseca, V. L. & Engels, W. 1996. Resource partitioning between highly eusocialbees and possible impact of the introduced Africanized honey bee on native stingless bees in the Brazilian Atlantic Rainforest. Studies on the Neotropical Fauna Environmental, 31: 137-151.
Zanette, L. R. S.; Martins, R. P. & Ribeiro, S. P. 2005. Effects of urbanization on Neotropical wasp and bee assemblages in a Brazilian metropolis. Landscape and Urban Planning, 71: 105-121.
Zapata, T. R. & Arroyo, M. T. K. 1978. Plant Reproductive Ecology of a Secondary Deciduous Tropical Forest in Venezuela. Biotropica, 10(3): 221-230.
Zimmerman, M., 1982. Optimal foraging – random movement by pollen collecting bumblebees. Oecologia, 53: 394–398.
Zurbuchen, A.; Landert, L.; Klaiber, J.; Müller, A.; Hein, S. & Dorn, S. 2010. Maximum foraging ranges in solitary bees: only few individuals have the capcapability to cover long foraging distances. Biological Conservation, 143: 669-676.


















