
República Bolivariana de Venezuela Universidad...
145
República Bolivariana de Venezuela Universidad Nacional Experimental de Guayana Coordinación de Investigación y Postgrado Coordinación Estudios de Postgrado de Ciencias Ambientales Variación espacial de la comunidad de macroinvertebrados en tres fases hidrológicas en el Bajo Caura Tesis de Maestría como requisito para optar al grado de Magister en Ciencias Ambientales Programa de Maestría en Ciencias Ambientales Mención Ecología Aplicada Autor: Lic. Ligia Blanco Belmonte Tutor: Dr. Hernán Castellanos Ciudad Guayana, Abril de 2006.
Transcript of República Bolivariana de Venezuela Universidad...

República Bolivariana de Venezuela
Universidad Nacional Experimental de Guayana Coordinación de Investigación y Postgrado
Coordinación Estudios de Postgrado de Ciencias Ambientales
Variación espacial de la comunidad de macroinvertebrados en tres fases
hidrológicas en el Bajo Caura
Tesis de Maestría como requisito para optar al grado de Magister en Ciencias Ambientales
Programa de Maestría en Ciencias Ambientales Mención Ecología Aplicada
Autor: Lic. Ligia Blanco Belmonte Tutor: Dr. Hernán Castellanos
Ciudad Guayana, Abril de 2006.

Universidad Nacional Experimental de Guayana Coordinación de Investigación y Postgrado
Coordinación Estudios de Postgrado de Ciencias Ambientales
Variación espacial de la comunidad de macroinvertebrados en tres fases
hidrológicas en el Bajo Caura
Ciudad Guayana 2006

Agradecimientos
“Los que han asumido las responsabilidades de este arte y se entregan al numen que se oculta detrás de él son descendientes directos de una inmensa y antigua comunidad de santos, trovadores, bardos, griots, poetas ambulantes, vagabundos, brujos y chiflados”
Clarissa Pinkola Estés
A la Energía Universal, A aquellos seres hermosos en mi vida que ya no están,
A mi familia, A mis amigos,
y a aquellos maestros que se cruzaron en mi camino para enseñarme a vivir con ideales y sueños.

Índice General
ANEXOS
LISTA DE TABLAS TABLAS 1. Lista de macroinvertebrados registrados en varios sustratos de los transectos estudiados en el Bajo Caura…………………………………………………………I
1A. Distribución, abundancia y biomasa de los Macroinvertebrados en el sedimento de nueve transectos del Bajo Caura en fase de aguas bajas...........IX
1B. Distribución, abundancia y biomasa de los Macroinvertebrados en el sedimento de nueve transectos del Bajo Caura en fase de subida de aguas.... XVII
1C. Distribución, abundancia y biomasa de los Macroinvertebrados en el sedimento de nueve transectos del Bajo Caura en fase de aguas altas............XXV
2. Distribución y abundancia de los Macroinvertebrados en el sustrato de maderos sumergidos en siete transectos del Bajo Caura durante la fase de aguas bajas.............................................................................................................. XXXVIII
3. Distribución y abundancia de los Macroinvertebrados en hojas verdes del bosque inundable de cinco transectos del Bajo Caura durante la fase de aguas altas.................................................................................................................. XL 4. Valores de variables físico-químicas en cada una de las estaciones de muestreo establecidas en 9 transectos a lo largo del Bajo Caura durante aguas altas (8) y aguas bajas (3)...................................................................................................XLIII

LISTA DE FIGURAS
FIGURAS
5. Ubicación de los transectos de estudio en el Bajo Caura...............................XLIV
6. Esquema del Transecto de estaciones en cada sitio de muestreo. ..............XLV
7. Materiales utilizados en el estudio. A. Nuecleador de sedimentos. B. Cuadrata y C. Cedazos de diferente tamaño de apertura. ............................................XLV
8. Composición porcentual media de los principales grupos de invertebrados encontrados en los diferentes sustratos colectados en los transectos de estudio en el Bajo Caura…………………………………………………………………………XLVI 9. Densidad media (Org/m²) de macroinvertebrados en el sedimento de todas las estaciones ubicadas en los 9 transectos del Bajo Caura durante las fases de aguas bajas (A) y aguas altas (B). ………………………………………………..XLVII
10. Composición porcentual del sedimento en las estaciones de muestreo en los 9 transectos tomados durante la fase de aguas bajas. …………………………..XLVIII 11. Análisis de conglomerados de la comunidad de invertebrados en el sedimento del transecto Patiquín (Caño Mato) durante aguas bajas (3) y durante aguas altas (8). …………………………………………………………………………………… IXL
12. Análisis de conglomerados de la comunidad de invertebrados en el sedimento del transecto Cejal (Caño Mato) durante aguas bajas (3) y durante aguas altas (8)…………………………………………………………………………………………IXL
13. Análisis de conglomerados de la comunidad de invertebrados en el sedimento del transecto Pozo Rico (Caño Mato) durante aguas bajas (3) y durante aguas altas (8). ………………………………………………………………………………... ...L
14. Análisis de conglomerados de la comunidad de invertebrados en el sedimento del transecto Brava (Río Caura) durante aguas bajas (3) y durante aguas altas (8)…………………………………………………………………………………………...L
15. Análisis de conglomerados de la comunidad de invertebrados en el sedimento del transecto Naparaico (Río Caura) durante aguas bajas (3) y durante aguas altas (8). …………………………………………………………………………………….. …LI

16. Análisis de conglomerados de la comunidad de invertebrados en el sedimento del transecto Aricagua (Río Caura) durante aguas bajas (3) y durante aguas altas (8). ……………………………………………………………………………………….. LI
17. Análisis de conglomerados de la comunidad de invertebrados en el sedimento del transecto Garzones (Caño Sipao) durante aguas bajas (3) y durante aguas altas (8). ……………………………………………………………………………….....LII
18. Análisis de conglomerados de la comunidad de invertebrados en el sedimento del transecto Chiribital (Caño Sipao) durante aguas bajas (3) y durante aguas altas (8)..……………………………………………………………………………………......LII 19. Análisis de conglomerados de la comunidad de invertebrados en el sedimento del transecto Caramacatico (Caño Sipao) durante aguas bajas (3) y durante aguas altas (8). ..……………………………………………………………………………......LIII
20. Análisis de Componentes Principales de las variables abióticas (Materia orgánica, Conductividad, Oxígeno Disuelto, Temperatura (ºC),Transparencia, y Profundidad) tomadas en cada sitio de muestreo de las 9 transectos en el Bajo río Caura durante la Transición (subida de aguas), aguas altas y aguas bajas………………………………………………………………………………..........LIV
21. Análisis de Componentes Principales de las variables abióticas (Materia orgánica, Conductividad, Oxígeno Disuelto, Temperatura (ºC),Transparencia, y Profundidad) tomadas cerca del sustrato de maderos sumergidos en el Bajo río Caura durante la fase de aguas bajas……………………………………………......LV
22. Gráfico de ordenación del Análisis de Correspondencia Canónica (ACC) mostrando la distribución de los taxones y estaciones en los nueve transectos en relación con las variables ambientales (189 taxones)………………………..........LVI
23. Gráfico de ordenación del Análisis de Correspondencia Canónica (ACC) mostrando la distribución de los taxones y estaciones en relación con las variables ambientales solo en las estaciones de maderos sumergidos (93 taxones).........LVII 24. Gráfico de ordenación del Análisis de Correspondencia Canónica (ACC) mostrando la distribución de los taxones y estaciones en relación con las variables ambientales solo en las estaciones de hojas verdes del bosque inundable (93 taxones)……………………………………………………………………………… LVIII
25. Cambios en la biomasa (g.Ind.m-²), riqueza e Índice de diversidad Shannon–Wienner (H`) media de los macroinvertebrados que viven en el sedimento a lo

largo de un gradiente de cuatro transectos de inundación del Caño Mato – río Caura– durante las fases de aguas bajas y aguas altas…………………………..LIX
26. Cambios en la biomasa (g.Ind.m-²), riqueza e Índice de diversidad Shannon–Wienner (H`) media de los macroinvertebrados que viven asociados a las hojas verdes del bosque de inundación del Caño Mato (tributario) y del río Caura durante la fase de aguas altas…………………………………………………………………..LX Resumen……………………………………………………………………………..........1 1. Introducción General
1.1. Fundamentos teóricos………………………………………………………… 2
1.2. Biodiversidad ………………………………………………………………… 5
1.3. Grupos Funcionales ………………………………………………………… 6
1.4. Especies claves ………………………………………………………………. 9
1.5. Variables……………………………………………………………………… 11
1.6. Sustratos……………………………………………………………………… 12
1.7. Morfología de las lagunas ………………………………………………….. 15
1.8. Geomorfología de los ríos ………………………………………………… 17
1.9. Antecedentes y objetivos ………………………………………………… 18
2. Estructura de la comunidad de macroinvertebrados 2.1. Introducción ………………………………………………………………….. 20
2.2. Material y métodos…………………………………………………………… 21
2.3. Resultados ……………………………………………………………………. 27
3. Comparación de las comunidades de macroinvertebrados encontradas en los
diferentes sustratos estudiados en las 3 fases hidrológicas 3.1. Introducción ………………………………………………………………….. 32 3.2. Material y métodos……………………………………………………………. 33 3.3. Resultados …………………………………………………………………… 34 4. Relación de las comunidades de macroinvertebrados con las variables físico-
químicas
4.1. Introducción………………………………………………………………….. 35 4.2. Material y métodos………………………………………………………….. 36 4.3. Resultados …………………………………………………………………… 37

5. El rol del sustrato en la variabilidad espacial de la comunidad de
macroinvertebrados 5.1. Introducción………………………………………………………………….. 39 5.2. Material y métodos…………………………………………………………. 41 5.3. Resultados …………………………………………………………………… 42 6. Discusión ……………………………………………………………………… 43 7. Conclusiones…………………………………………………………………… 49
8. Recomendaciones……………………………………………………………… 51 9. Referencias ……………………………………………………………………. 52

1
RESUMEN
La estructura de las comunidades de macroinvertebrados fue estudiada en
relación con el tipo de hábitat ribereño presente entre lagunas y caños tropicales y
algunas variables físico-químicas. Se trabajó en 9 transectos localizados en el
Bajo río Caura y en dos de sus principales tributarios (Caño Mato y Caño Sipao).
Los macroinvertebrados fueron recolectados en tres tipos de hábitat: sedimento,
maderos sumergidos y hojas verdes del bosque inundable. Las comunidades de
macroinvertebrados fueron dominadas por insectos de los ordenes Diptera
(Chironomus sp, Stenochironomus sp.) y Ephemeroptera (Brasilocaenis irmleri,
Asthenopus curtus, Callibaetis sp). Ademas de los insectos, otros grupos
dominantes fueron los Ostracoda, Cladocera y Annelida. La composición de los
grupos funcionales fue dominada por los Recolectores. En todos los transectos,
los sustratos temporales presentaron mayor abundancia y biomasa de
macroinvertebrados en comparación al sustrato sedimento. Los resultados
sugieren que los sustratos temporales fueron los tipos de hábitat más importantes
para las comunidades de invertebrados. La química del agua, la composición de
los sedimentos, la diversidad en los hábitat mostró una considerable variación
espacial a lo largo de la zona de confluencia del río Caura con los tributarios.

2
Capitulo 1
Introducción Fundamentos teóricos
Los fundamentos teóricos sobre la ecología de los ríos a menudo no reflejan
ampliamente el papel crucial de la complejidad espacial y de la dinámica fluvial en
los ecosistemas de ríos tropicales. La inclusión de las aguas superficiales (lóticas
y lénticas), de las subsuperficiales (hiporréicas y endorreicas = freáticas), de los
sistemas riparinos y de los ecotonos (manantiales) como componentes interactivos
que contribuyen a la biodiversidad total, es crucial para el desarrollo de un marco
holístico de los ríos como ecosistemas (Fig. 1).
Los patrones y procesos en el paisaje lótico están orientados al movimiento de
aguas abajo del río (Ward, 1989, Ward et al., 2002). La direccionalidad es
importante en los ecosistemas lóticos, tanto, que la estructura y la conexión
ecológica dentro del sistema se dividen en vectores verticales, longitudinales y
laterales (Ward, 1989; Ward et al., 2002; Ward y Stanford, 1995a; Petts y Amoros,
1996) (Fig. 2).
Wiens (2002) describe como la aplicabilidad de estos diferentes conceptos en
ecología de los ríos cambian con la magnitud relativa de los vectores de conexión:
el concepto de río contínuo (RCC) (Vannote et al., 1980) es aplicable a los ríos
restringidos, encajonados, donde las uniones longitudinales son fuertes. El

3
concepto de inundación de pulso (Junk et al., 1989; Malard et. al., 2000), el
concepto de corredor hiporréico y el de hidrosistemas fluviales (Petts y Amoros,
1996) son apropiados para aquellos
ríos abiertos donde la estructura
geomorfológica facilite los enlaces
laterales, verticales y longitudinales
(Fig. 2). Por último, el concepto de
discontinuidad serial (Ward y
Standford, 1995a, 1995b; Poole,
2002) puede ser integrado en la
jerarquía de cuenca porque la
discontinuidad está representada por
fronteras entre segmentos
adyacentes del río que difieren
marcadamente en su estructura
geomorfológica y así en su conexión
(Ward y Standford, 1983).
La conexión de un río y el intercambio de la materia, de la biota y de la energía
juegan un papel importante y poco entendido para explicar la diversidad del
paisaje ribereño. Las especies de macroinvertebrados registradas en él son un
producto de las comunidades terrestres y acuáticas que viven a lo largo de un
gradiente espacio-temporal. Las perturbaciones naturales promueven su
incremento a lo largo de él, como también lo limitan y excluyen. En consecuencia,
el paisaje del río da la base para conocer cuan complejas son las interacciones
entre estas perturbaciones naturales y los patrones de la estructura en la
productividad de la diversidad de las especies. (Ward et al., 2002).
Es así como el sistema ribereño, formado por un conjunto de mosaicos de parches
unidos por una diversidad de procesos, mantiene unas comunidades altamente
complejas. Ellas muestran a los invertebrados como componentes importantes del
sistema con una alta diversidad funcional y taxonómica y con patrones de

4
distribución, movimientos y efectos del flujo en varios procesos ecológicos del
paisaje (Malmqvist, 2002).
Uno de estos componentes, la distribución de los macroinvertebrados en los ríos,
está gobernado por un sin número de factores que típicamente actúan a diferentes
escalas (regionales, locales, hábitat). Entre ellos, está la estructura de la
comunidad local, la cual puede ser vista como el resultado de un proceso contínuo
de filtros ambientales. Tales filtros están enmarcados desde procesos regionales a
aquellos inmersos en una amplia cuenca, donde involucra la especiación, la
historia geológica, el clima y las características de los parches individuales a
pequeña escala como la depredación, el sustrato y la velocidad de la corriente
(Malmqvist, 2002).
Otro factor importante que maneja esta distribución de invertebrados es la
dispersión que está ligada a diferentes sistemas ecológicos a lo largo de las
diferentes fronteras, la cual ocurre en los sistemas acuáticos como también en los
alrededores terrestres y en las zonas terrestres de interconexión con otros cuerpos
de agua (Malmqvist, 2002).
En los ríos, hay un factor básico, el detrito, el cual juega un papel dominante como
base del flujo de energía (Fisher y Likens, 1973) y estructura de las cadenas
tróficas (Cummins, 1973; Wallace y Webster, 1996). Puede relacionarse
directamente con las densidades y las biomasas de los consumidores secundarios
como los invertebrados (Minshall 1967; Minshall y Minshall, 1977) y limitar la
biomasa y producción de los invertebrados detritívoros por el carbono en el
contenido. Además, al incrementarse la materia orgánica particulada gruesa
(MOPG), aumenta la densidad de algunas especies de invertebrados (Richardson,
1991) y al reducirse la hojarasca de las cabeceras de los ríos, disminuye la
producción secundaria de invertebrados (Wallace et al., 1997).

5
Este uso constante de la materia orgánica hace que los invertebrados afecten el
ciclo de nutrientes y el carbono convirtiéndose en intermediarios cruciales de los
productores primarios, del detrito o de los consumidores secundarios y de los
depredadores con una mayor jerarquía trófica (Malmqvist et al., 2001). Uno de los
tipos de carbono, el carbono bacterial, puede ser más importante como recurso
energético para algunos invertebrados que lo sugerido hasta ahora (Fuller et al.,
2004) y no sólo las condiciones ambientales sino también el sustrato y la oferta
alimentaria tienen que ver con el cambio de la composición de la comunidad de
invertebrados.
BIODIVERSIDAD
Un concepto amplio sobre la biodiversidad, acompañada de una heterogeneidad
espacio temporal, podría ser un tema unificador en la ecología de los ríos cuando
la diversidad es vista desde un punto funcional, donde los organismos sean
actores de una movilización energética. Particularmente en el trópico americano,
los esfuerzos para conocer la diversidad de la fauna de macroinvertebrados han
sido grandes pero no suficientes para la gran cantidad de información que aún
falta por identificarse (Báldi y McCollin, 2003; Bonetto y Ezcurra, 1964; Baptista et
al., 2001a, 2001b; Blanco-Belmonte, 1990; 1998; Callisto et al., 2001; César et al.,
2000; Fittkau, 1971, 2001; Galdean et al., 2001; García y Pereira, 2003; Goulding
et al., 1988; Junk, 1973, 1980; Poi de Neiff y Carignan, 1997; Trivinho et al, 2000,
entre otros.)
La diversidad de las especies es importante para el funcionamiento y resiliencia
del sistema (Aarts y Nienhuis, 1999) y, cuando se considera la restauración de
ambientes heterogéneos, el conocimiento de los mecanismos detrás de la relación
entre la heterogeneidad ambiental y la diversidad de las especies es necesaria.
Por lo tanto, estudios básicos como los que actualmente se manejan en los
trópicos son imprescindibles, tomando en cuenta la ayuda de nuevas tecnologías

6
como el PCR (Reacción en cadena de la Polimerasa), porque ayudarían más
rápidamente a estudios donde involucre la restauración de áreas que
aceleradamente están siendo fragmentadas.
Las medidas de diversidad de especies, incluyendo diversidad alpha, beta y
gamma, son un resultado de una historia de perturbaciones, fragmentación y
fenómenos sucesionales a lo largo del paisaje ribereño (Rosales, 2000). Por
ejemplo, al propender un enfoque jerárquico de la diversidad funcional en los
ecosistemas, del plano de inundación en ríos perturbados y naturales, aumentaría
el entendimiento del fenómeno ecológico que opera a diferentes escalas a lo largo
de los gradientes ambientales multidimensionales (Rosales, 2000).
Reestablecer la diversidad funcional (i.e. procesos sucesionales e hidrológicos) a
lo largo de un corredor ribereño, puede servir como foco para iniciativas de
conservación del río. Una vez que las iniciativas hayan sido reestablecidas, la
heterogeneidad de los hábitat incrementará seguidamente por un correspondiente
incremento de diversidad de especies de la biota acuática y riparina. (Ward y
Tockner, 2001). Aparte de la diversidad funcional del sistema, es importante la
visión de los grupos funcionales de organismos. Este concepto innovador sobre
diversidad funcional de organismos fue introducido a nivel más dinámico para los
invertebrados por Cummins (1974) en el concepto de río continuo publicado por
Vannote et al. (1980).
GRUPOS FUNCIONALES
Según la definición de Cummins y Klug (1979) y Merritt y Cummins (1996), los
invertebrados acuáticos pueden ser identificados funcionalmente de acuerdo al
tipo y tamaño de la materia orgánica procesada y son separados en Partidores,
Recolectores, Filtradores, Raspadores y Depredadores.

Los Partidores son los invertebrados que aprovechan la materia orgánica
particulada gruesa, principalmente vegetal (MOPG> 1mm) y procesan del 0,01 al
0,4 % del peso de su cuerpo diariamente (Henriques-Oliveira et al., 2003) (Fig. 3).
Fig. 3. Modelo esquemático de los partidores y las
interacciones microbianas en un sistema acuático.
(Extraído de la web del concepto de río contínuo).
Los consumidores del fondo también
llamados Recolectores consumen
materia orgánica particulada fina
(0,45 µm< MOPF <1 mm) que es una
mezcla originada por la
fragmentación de MOPG y por la
floculación de materia orgánica
disuelta (MOD<0,45 µm) junto con las
algas y organismos microscópicos.
Los Recolectores son diferenciados
en subcategorías basadas en la
ubicación del alimento (suspendido o
sedimentado) y por la forma de
obtenerlo (Filtradores FI, Atrapadores
AT y Minadores MI) (Fig. 4).
Los Filtradores (FI) capturan las
partículas finas suspendidas en el
agua (seston) construyendo filtros o
redes como los construidos por
algunas familias de Trichoptera
(Hydropsyche) o como otros ordenes
de insecto que modifican apéndices
de su cuerpo (i.e. Simulium,
Brachycentrus) o interacción de las
branquias de los invertebrados (i.e.
mejillón) para capturarlas desde el
sedimento suspendido (Cummins y
Klug, 1979).

Dentro de los Atrapadores (AT), hay organismos con cepillos y peines bucales con
los que recogen y capturan el sedimento particulado fino. Los Minadores (MI), como
los gusanos tubifícidos y un buen número de larvas dípteras, cavan dentro de
depósitos de sedimento fino y procesan por lo menos el peso de su cuerpo
diariamente pero descartan un alto % como seudo-heces. La tercera categoría
alimentaria son los Raspadores, los cuales se alimentan de una mezcla de material
microscópico adherido a la superficie de las rocas u otros sustrato, llamado perifiton,
formado predominantemente por algas o el llamado “biofilm” formado por bacterias
heterótrofas. Ellos crecen en el sustrato durante el proceso de descomposición,
observándose en el trópico relaciones iniciales de proto-cooperación con los
invertebrados (Bastardo, 1999; Blanco-Belmonte et al., 2003). Muchos organismos
incluidos en este grupo muerden el alimento de manera muy semejante al ramoneo
del ganado siendo más que un raspado o rasguño del sustrato.
Fig. 4. Modelo esquemático de los recolectores y la actividad microbiana. (Extraído de la web del
concepto de río contínuo).
Los Depredadores (DE), cuarta categoría de esos grupos funcionales, ingieren tejido
animal vivo o muerto recientemente (en particular de otros invertebrados acuáticos)
capturando toda la presa o parte de ella (Engullidores) o chupando los fluidos
(Chupadores).

9
A menudo, la taxa no puede ser asignada a un solo grupo funcional. Un ejemplo es el
observado en algunas especies de poliquetos que tienen la habilidad de cambiar de
Filtradores en suspensión a Recolectores de fondo, dependiendo de la disponibilidad
de las partículas en suspensión; otras especies de bivalvos despliegan
simultáneamente el mecanismo para alimentarse con las partículas en suspensión y
con las depositadas en el fondo. Los partidores a menudo ingieren MOPF mientras
consumen hojarasca (Cummins, 1973; Cummins y Klug, 1979; López y Levinton,
1987; Halse et al., 2002).
Hay hipótesis, como la de redundancia, que consideran que mientras haya una
especie de cada grupo funcional presente, los procesos de los ecosistemas
continuarán. Estos organismos son llamados especies claves.
ESPECIES CLAVES
Recientemente, una pregunta ha sido formulada cuestionando si, bajo que
condiciones, los roles funcionales de cada especie son necesarios para que los
procesos del ecosistema persistan. Esta pregunta basada en la “hipótesis de la
redundancia” predice que no todas las especies son igualmente necesarias en
cualquier momento para que los procesos del ecosistema continúen (Lawton y
Brown, 1994). Si algunas especies fueran “redundantes”, en términos de sus
relaciones funcionales, entonces su pérdida no resultaría en cambios observables en
el flujo de energía o en el ciclo de nutrientes. Este concepto de “redundancia
paralela”, usado en el análisis de ingeniería para la confiabilidad de los sistemas, es
aplicado en la comparación del rol de las especies en los ecosistemas. La “hipótesis
de la redundancia” puede ser desglosada en tres sub-hipótesis:

10
1. La hipótesis de los “grupos funcionales” predice que mientras haya una
especie de cada grupo funcional presente, los procesos de los ecosistemas
continuarán.
2. La hipótesis del “nivel trófico” predice que mientras la biomasa o el cambio de
organismos de cada nivel trófico permanece relativamente uniforme y sea
independiente de la composición de las especies, el flujo de energía y los
procesos en el ecosistema persistirán. Finalmente,
3. La hipótesis de la “especie clave (EC)” predice que no todas las especies
tienen igual importancia funcional; solamente son pocas especies las que
verdaderamente son necesarias para los procesos en el ecosistemas aunque
ellas no sean abundantes (Bond, 1993; 2001)
Esta última sub-hipótesis está basada en el término “especie clave,” que fue acuñado
por Paine (1969a) como una especie de un alto estatus trófico, cuyas actividades
ejercen una desproporcionada influencia en los patrones de la diversidad de las
especies en la comunidad. El mecanismo por el cual la especie clave regula la
diversidad de otras especies fue descrito por el mismo Paine (1969a) en la siguiente
hipótesis: “… la diversidad de las especies locales está directamente relacionada con
la eficiencia por la cual el depredador previene la monopolización de los mayores
requisitos ambientales de una especie”. La confirmación es dada en sus estudios
experimentales, donde la remoción del depredador reduce la diversidad de la presa
debido a una intensa competencia de la presa misma.
Paine (1969a) identificó así un proceso muy específico del ecosistema: la regulación
de la diversidad de especies por el efecto “top-down”: el cual consiste en
interacciones competitivas y la persistencia de la comunidad. Las especies que
juegan este papel de EC en los ecosistemas pueden ser generalistas tróficos o
especialistas, raros o comunes y no necesitarían ser importantes removedores de
energía (Paine, 1969b; Bond, 1993). Los experimentos de corto plazo (Paine, 1992;
Raffaelli y Hall, 1992) y los de largo plazo indican que el efecto de EC en el
ecosistema no es redundante, lo cual contribuye a su gran significancia (Bond, 1993,
2001; Schulze y Money, 1993).

11
Este concepto, ha sido redefinido por Davic (2003) para permitir predicciones de
estas especies dentro de los ecosistemas de la siguiente manera: Una “especie
clave” es considerada como aquella especie que mantiene una fuerte interacción con
otras especies cuyo efecto “top-down” en la diversidad de ellas y la competencia es
alta en relación con su biomasa dominante dentro de un grupo funcional. Esta
definición operativa enlaza la importancia de la comunidad de especies claves a un
proceso específico del ecosistema (i.e. la regulación de la diversidad de las especies)
dentro de los grupos funcionales a niveles tróficos bajos, porque están estructurados
para la competencia por un recurso alimentario limitado. Esta predicción a priori de
las especies claves tiene un valor aplicado para la conservación de las áreas
naturales.
Todas estas especies claves en la comunidad de invertebrados dependen de una
serie de variables bióticas y abióticas para su distribución y abundancia.
VARIABLES
Uno de los principales intereses de los ecólogos ha sido entender cuales factores
explican mejor los patrones observados en la distribución y la abundancia de
macroinvertebrados (Hynes, 1970). Ellos han sido considerados decisivos para
conocer la estructura del conjunto de macroinvertebrados en los ríos: el sustrato
(Reice, 1980; Rae, 1985, Baptista et al., 2001a,b), la temperatura (Vannote y
Sweeney, 1980; Ward y Stanford, 1983), la perturbación humana (Siegfried y Knight,
1977; Death y Winterbourn, 1995), las interacciones bióticas (Kohler, 1992).
El régimen hidrológico es considerado también uno de los componentes físicos más
importantes que influencian la comunidad de invertebrados (Wissinger, 1999; Whiles
y Goldowitz, 2001). Algunos autores han encontrado una relación entre la
permanencia del agua y el incremento de la riqueza (Driver, 1977; Williams y
Williams, 1999). Mientras que otros señalan a la prolongada permanencia del agua

12
como una limitante (Huener y Kadlec, 1992; Mallory et al., 1994), la cual puede
afectar la reproducción y levante de muchos invertebrados (Neckles et al., 1990)
aunque puede maximizar la productividad y riqueza en sitios con una intermitencia
hidrológica (Whiles y Goldowitz, 2001).
Por otro lado, los sustratos, que son una de las variables claves en la heterogeneidad
del hábitat para los invertebrados, pueden ser un importante recurso energético por
sí mismos o albergar una producción combinada de carbono suplementario para los
consumidores acuáticos que envuelve el carbón fijado por el fitoplancton (Hamilton y
Lewis, 1990) o por las bacterias (Fuller, et al., 2004). La producción basada en el
material béntico y material terrestre, combinados con la producción del fitoplancton
para las larvas de peces, parece ser un importante recurso energético en los ríos con
áreas de inundación (Fuller et al., 2004; Dangles et al., 2002).
SUSTRATOS
La heterogeneidad de los hábitat es un factor importante que influencia la distribución
de los macroinvertebrados en los ríos (Vinson y Hawkins, 1998; Junk, 1983, 1980).
Ella tiene que ver con la diversidad de sustratos (Baptista et al., 2001a), con la
variabilidad física en un gradiente (Rosi-Marchall y Wallace, 2002) y con la materia
orgánica gruesa y fina entre otros (Junk, 1973; Irmler y Junk, 1982). Las cadenas
tróficas de muchos sistemas, como las áreas de inundación, los suelos con alto
contenido de detrito y las cabeceras de los ríos (Polis y Hurd, 1996), basan sus
entradas en hojas muertas, maderos sumergidos, y microbios asociados con los
sustratos del bosque (Vannote et al. 1980; Rosemond et al. 2001).
El sustrato es un aspecto complejo del ambiente físico y cuando uno enfoca la
distribución de macroinvertebrados en diferentes hábitat a una amplia escala, los
cambios de los patrones ecológicos pueden ser registrados a lo largo del río o de la
cuenca, pero la mayoría de los patrones locales no son reproducibles en otros ríos

13
(Vinson y Hawkins, 1998). Es por esto que los procesos de estructurar los conjuntos
de insectos en diferentes tipos de sustratos, ya descritos y mejor evidenciados en
ríos de las zonas templadas (Rabeni y Minshall 1977; Minshall y Minshall, 1977),
deberían ser probados y comparados con los ríos tropicales y subtropicales
(Arunachalam et al., 1991; Dudgeon, 1994).
Un ejemplo es el encontrado por Baptista et al. (2001a), quienes observaron que los
conjuntos de insectos acuáticos a lo largo de la cuenca del río Macaé, en Brazil,
presentan una interrupción a lo largo del gradiente longitudinal, separando la cuenca
alta (4to orden, 700 m.s.n.m.) de la cuenca baja (5to y 6t0 orden). Por otro lado la
riqueza de sustratos durante aguas bajas son menos afectados por el flujo o entrada
de aguas, permitiendo un período más amplio para la colonización y procesamiento
de la materia orgánica béntica por los macroinvertebrados. De acuerdo a Ladle y
Ladle (1992), la preferencia de ellos por un sustrato es determinada primero por la
conducta de oviposición del organismo, pero esta distribución puede ser modificada
luego por un arrastre pasivo o una migración activa.
Las comunidades establecidas en los sustratos de roca, por ejemplo, son más
estables sirviendo como refugio para los organismos jóvenes (Merrit y Cummins,
1996). Sin embargo, la hojarasca como sustrato es preferido por muchos taxa ya que
ofrece un mejor refugio y condiciones alimentarias debido a la alta heterogeneidad y
al crecimiento de una flora perifítica rica. La falta de consumidores de hojarasca
(partidores) en trabajos realizados en el trópico, presentados en la 48 Reunión Anual
de la Sociedad Bentológica norteamericana en Keystone, Colorado, 29 Mayo– 1
Junio 2000, llevó a preguntarse de qué clase de alimento dependían los detritívoros
en el trópico y cuales organismos estaban procesando la hojarasca si los
invertebrados no lo hacían (Boyero, 2000). Recientemente, Blanco-Belmonte et al.
(2003) han evidenciado una proto-cooperación entre microcrustáceos y bacterias en
la descomposición de las hojas de matorrales en un experimento en un embalse de
aguas negras en Venezuela. Según Covich et al. (1999), una forma de adentrarse
conceptualmente en el rol de las especies en los ecosistemas de aguas dulces

14
puede ser el considerar un grupo de especies bénticas en hábitat adicionales como
un recurso de información del rol de una especie particular en los procesos
ecológicos, dada la carencia de trabajos empíricos y teóricos sobre este tema.
Rosemond et al. (2001) han demostrado experimentalmente que la limitación de un
recurso hace que la biomasa de detritívoros cambie como una respuesta de las
manipulaciones de la cantidad de materia orgánica particulada (Culp y Davies, 1985;
Richardson, 1991; Wallace et al., 1997; Batzer, 1998), o de carbón microbiano
disponible (Mikola y Setälä, 1998). En sistemas en los cuales el Carbón particulado
es utilizado, como en el trópico, la colonización y crecimiento de los detritívoros es
afectado por la cantidad y calidad de detrito (Cummins, 1974; Lawson et al., 1984).
En resumen, el detrito, como energía básica dominante en los ríos (Fisher y Likens,
1973), puede estar relacionado positivamente con las densidades y biomasas de los
consumidores (Egglishaw, 1964; Minshall, 1967; Culp y Davies, 1985).
Por otro lado, la disponibilidad de la materia orgánica es baja en los sustratos donde
el sedimento es arenoso y lleva a una riqueza y diversidad baja (Hawkins, 1984). En
los ecosistemas acuáticos, los detritívoros que viven en las arenas de agua o
sedimentos blandos son una de las principales fuentes de alimentos para los
depredadores, como los peces y aves, o para otros invertebrados carnívoros como
las larvas de Odonata (Healey, 1984; Walde y Davies, 1984; Hershey, 1987; Hershey
y Dodson, 1985; Wallace y Webster, 1996).
Muchos estudios han tratado de explicar el patrón de distribución de la fauna y la
flora a lo largo de una sección longitudinal del río (Vannote et al., 1980; Winterbourn
et al., 1981; Culp y Davies, 1982; Cushing et al., 1983; Statzner y Higler, 1985;
Naiman et al., 1987; Brussock y Brown, 1991; Palmer et al., 1994). Para explicar el
gradiente de la fauna observada a lo largo de un río en estos estudios, dos puntos de
vista han sido tomados: 1. Vannote et al. (1980) argumenta que las comunidades de
un río están ajustadas a un gradiente contínuo de cambios en las condiciones físicas
para asumir estrategias que involucran el mínimo costo energético de tal forma que
las comunidades río abajo capitalizan las ineficiencias de los procesos del río arriba.

15
2. Statzner y Higler (1985) y Statzner et al. (1988) manejan una segunda idea, el
estrés hidráulico, asociado con la geomorfología de la cuenca del río, el cual
consideran que es el principal factor que estructura las comunidades lóticas, sin
embargo, este factor no siempre varía en forma predecible longitudinalmente.
Las características de un sistema lótico en una determinada área no pueden ser
vistas en una sola dimensión sino como la integración de varias dimensiones
(Cressa, 2003; Ward, 1989) (Fig. 3). Muchos de los estudios sobre invertebrados en
ambientes lóticos, realizados en la región, escasamente han tomado en cuenta la
dimensión lateral, la cual está formada por una zona de inundación diversa con
ambientes lénticos heterogéneos y que la secuencia hidrológica producto de los
pulsos de inundación es el principal proceso responsable de las adaptaciones que se
producen en la biota (Junk et al., 1989; Cressa, 2003).
MORFOLOGÍA DE LAS LAGUNAS
Las lagunas del área de inundación, que están en dimensión lateral, presentan una
variación química y ecológica considerable y además un factor común: el agua de río
que les inunda (Welcomme, 1985; Hamilton y Lewis, 1990). Mucha de esta variación
entre las lagunas es probablemente causada, como lo apunta Melack (1984), por
diferencias en la morfología de su cubeta. Pocos estudios existen sobre el grado en
que las características químicas y ecológicas de las lagunas de inundación puedan
ayudar a predecir su morfología. Hutchinson (1957) y Drago (1977; 1980; 1981)
presentan un sistema de clasificación para las lagunas basado en su origen, grados y
tipos de conexión. Por otro lado, Hamilton y Lewis (1990) basan la distribución y
dinámica de la biota en relación con la morfología de la cubeta y las características
químicas y ecológicas.
Las inundaciones anuales afectan en mayor o en menor grado los ambientes lénticos
dependiendo de su situación, origen, grado y tipo de conexión (Drago,1981). Por

16
ejemplo, las inundaciones anuales de la varzea e igapo del río Orinoco y del Bajo
Caura, respectivamente, siguen el hidrograma del canal principal que tiene una forma
unimodal con una duración de 4-6 meses (Hamilton y Lewis, 1990). La conexión
directa indica una mayor aproximación entre el cuerpo de agua (lagunas) y la
hidrometría del río; la indirecta, por el contrario, señala que debe existir una mayor
magnitud en los cambios hidrométricos de aquél para que los mismos sean acusados
por la laguna (Drago, 1981).
Las lagunas redondas como las encontradas para este estudio en el río Caura,
tienen una pendiente suave de la cubeta porque están formadas en áreas de
sedimentación (Pozo Rico, Patiquin, Chiribital, Garzones Naparaico) y una
comunicación directa. Las madres viejas (Cejal) o lagunas alargadas y dendríticas
como las lagunas Aricagua, Brava y Caramacatico, lato sensu, son a menudo mas
profundas que las lagunas anteriores y su comunicación varía entre una conexión
directa e indirecta.
Uno de los más comunes y mal entendidos en la ecología de los ríos tropicales ha
sido pensar ellos son ambientes físicos constantes, porque carecen de fluctuaciones
en la temperatura mayores de 4,5 ºC. Las fluctuaciones anuales e interanuales en la
lluvia y en el flujo o descarga de un río son importantes en los trópicos empero,
especialmente con el efecto de El Niño; estas son una de las mayores fuerzas que
manejan el ciclo de nutrientes y la dinámica de la cadena alimentaria. Esta
variabilidad anual en la lluvia con relación a la influencia de la entrada de hojarasca
en los macroinvertebrados ha sido tratado por Covich et al. (1999), un tópico muy
estudiado en la comunidad de peces tropicales pero desconocido en la comunidad
de macroinvertebrados (Boyero, 2000).
La perturbación física es una figura muy importante en los ríos porque interrumpe y
fractura las estructuras de las poblaciones, de las comunidades y del ecosistema
(Resh et al., 1988). Las inundaciones son ejemplos de perturbaciones naturales que
afectan las comunidades bénticas con una frecuencia e intensidad variable. Durante
estas perturbaciones, los sustratos pueden ser cambiados o movidos y su biota

17
agotada (Lake et al., 1994), creando parches desnudos que son recolonizados por un
grupo de colonos de los parches adyacentes, de la fauna de arrastre (Boyero et al.,
2001; 2002), o insectos adultos de otros ríos, tramos o cuencas. Estos cambios
rápidos podrían indicar una alta resiliencia en las comunidades de
macroinvertebrados tropicales (Boyero, 2000) igualmente encontrados por otros
autores para los ríos templados (Doeg et al., 1989). Como todos los procesos en
ecología, la perturbación y la recolonización de los hábitat bénticos ocurren a unas
escalas temporales (Peckarsky, 1987; Lake et al., 1994) y espaciales (Mackay, 1992)
diferentes.
Por otra parte, la dinámica geomorfológica en los ríos depende de su dinámica fluvial
y estos procesos físicos tienen fuertes implicaciones en la diversidad y la
composición de la biota (Rosales et al, 1999).
GEOMORFOLOGÍA EN LOS RÍOS
Los ríos están sujetos a umbrales que definen cambios significativos en los procesos
y morfología y delimitan el paisaje ribereño y los hábitat. Estos umbrales están
acondicionados por eventos que gobiernan los procesos y la forma en el canal del
río, entre los cuales los más importantes son el régimen fluvial, la cantidad y el
calibre del sedimento enviado por el canal y la topografía (la cual esta determinada
por el gradiente del canal) (Church, 2002). Estos factores determinan el régimen del
transporte del sedimento y el carácter de los depósitos aluviales a lo largo él. Los
cambios ocurren sistemáticamente a lo largo del sistema del drenaje como el flujo, el
gradiente y el carácter de cambio del sedimento, así una secuencia característica de
morfología y hábitat – por consiguiente el paisaje ribereño- puede ser descrito desde
las tierras altas a los canales distales. La secuencia es asociada con la eficiencia de
los ríos de mover el sedimento y con la estabilidad de los albardones (Church, 2002).

18
Un concepto inicial en esta sección, para considerar, es el rol de la dinámica del
canal de un río en relación con la biodiversidad, la cual depende de varios factores
incluyendo la riqueza y la abundancia relativa. La riqueza refleja el número de
especies por hábitat (diversidad α ) y el número de hábitat (N) en una región
(Whittaker, 1960). Si la “región” es un tramo de área de inundación, el número de
hábitat, la dinámica geomorfológica y la diversidad sedimentológica reflejará la
riqueza de las especies (Richards et al., 2002).
Richards et al. (2002) identificaron la dinámica de un canal como una clave control
para la biodiversidad y el patrón espacial porque: 1. La biodiversidad puede, a nivel
de tramo del río, estar muy relacionada con la diversidad del hábitat. 2. La diversidad
del hábitat refleja los procesos de migración y 3. La dinámica del canal integra las
influencias de los pulsos de inundación y la perturbación en la renovación y
regeneración de especies; hay perturbaciones creadas por los pulsos de inundación.
Comúnmente este término está asociado con la hidrometría, la humedad, la provisión
de nutrientes y sedimentos, los cuales juntos traen oportunidades de regeneración.
Sin embargo, ellos son también responsables de los procesos de sedimentación y
erosión, los cuales proveen oportunidades para la formación de parches a la escala
de hábitat. Entonces, la extensión del concepto de “pulso de inundación” pasaría al
concepto de “pulso de flujo” como lo explica Tockner et al. (2000), porque también
involucra a los geomorfólogos fluviales en la relación de estas perturbaciones con el
medio biótico.
ANTECEDENTES Y OBJETIVOS
En Sudamérica, son pocos los estudios que han enfocado gradientes longitudinales
(Illies, 1964; Domínguez y Ballesteros, 1992; Jacobsen et al.,1997). Entre ellos, Illies
(1964) efectuó muestreos de la fauna de macroinvertebrados en el río Huallaga en
Perú en forma cualitativa y sólo en un tipo de sustrato: rocas. Mientras que
Domínguez y Ballesteros (1992) trabajaron en Argentina en el río Cañas-Horcones
restringiendo el estudio sólo al orden Ephemeroptera. Más recientemente, Marchese

19
et al. (1992, 2002, 2005) analizaron las dimensiones laterales, longitudinales y
temporales de las comunidades bentónicas como también sus relaciones con las
características físicas, químicas y biológicas del río Paraná en Argentina sólo en el
sustrato sedimento; Jacobsen et al. (1997) compararon la estructura de la comunidad
de macroinvertebrados en un gradiente altitudinal en varios ríos del Ecuador pero
solo a nivel de familia. En Venezuela, específicamente en el río Caura, son escasos
los estudios realizados. Sin embargo, Rosales et al. (2002) analizaron la composición
relativa de los invertebrados en diferentes sustratos y la importancia de estos en la
dieta íctica de varias especies comerciales conseguidas en 9 transectos a lo largo del
Bajo Caura. García y Pereira (2003) realizaron un sondeo longitudinal rápido de la
diversidad de invertebrados en la cuenca del río Caura y en varios afluentes,
relacionándolos con algunas variables abióticas; Magalhães y Pereira (2003)
analizaron la riqueza de los crustáceos decápodos, el uso del hábitat y su
distribución en un sondeo logitudinal en el río Caura.
Estos estudios longitudinales en el río Caura son una base fundamental de
información. Sin embargo, la importancia del aporte lateral de la fauna de
macroinvertebrados en las zonas riparinas y de la dinámica estacional, tomando en
cuenta los nuevos paradigmas (Wiens, 2002; Tuckner et al., 2000; Petts y Amoros,
1996; Newbold et al., 1981, 1982; Elwood et al., 1983; Ward y Standford, 1983;
Statzner et al., 1988; Resh et al., 1988; Ward, 1989; Pringle et al, 1988; Junk et al.,
1989; Neiff, 1990, 1999) y enfoques que ya han sido utilizados en el río Caura por
Rosales (2000), son imprescindibles. Ellos sugieren que la diversidad alfa de la
vegetación incrementa hacia las márgenes de las zonas de confluencia de los
tributarios con el canal principal, indicando la presencia de ecotonos longitudinales,
los cuales dependen de la dinámica del flujo del río Caura y de la variabilidad de la
inundación. La relevancia de este estudio hace que sea importante conocer si esta
variabilidad igualmente ocurre en la fauna de macroinvertebrados evaluando la
estructura, densidad, diversidad, funcionalidad de la comunidad de
macroinvertebrados en dimensiones laterales, longitudinales y temporales como una

20
línea base de estudio sobre una visión que sea útil para el desarrollo de estrategias
que mantengan los valores de este ecosistema.
El objetivo general de este estudio es evaluar la variación espacial de la fauna de
macroinvertebrados acuáticos mediante el análisis de la composición, la densidad y
la biomasa de la comunidad en relación con algunas variables físicas y químicas
durante tres fases hidrológicas en el Bajo Caura. Los objetivos específicos son:
1. Caracterizar la comunidad de macroinvertebrados.
2. Comparar las comunidades de macroinvertebrados encontradas en los
diferentes sustratos estudiados en las 3 fases hidrológicas.
3. Relacionar las comunidades de macroinvertebrados con las variables físico-
químicas.
4. Examinar el rol del sustrato en la variabilidad espacial de la comunidad de
macroinvertebrados.
Esta tesis está organizada de la siguiente manera: cuatro capìtulos (2 – 5) donde se
introduce el tema, la metodología utilizada y los resultados de cada uno de los
objetivos planteados y dos capítulos donde se encuentran una discusión y una
conclusión general (6 y 7 respectivamente). El capítulo 8 es una serie de
recomendaciones generales derivadas de este estudio.
Capitulo 2: Composición de la comunidad de macroinvertebrados
2.1 Introducción
La composición taxonómica de la comunidad de macroinvertebrados en un río
cambia a lo largo de su cauce. Los cambios graduales (Ej. temperatura) y los
cambios abruptos (Ej. hidrometría) son factores importantes que afectan la
composición de una comunidad (Ramírez y Pringle, 2001).

21
Recientemente, la importancia del cambio de flujo en las descargas de agua han sido
reconocidas en la ecología de los ríos (Stanley et al., 1997; Tockner et al., 2000). La
extensión del área inundada puede aumentar por ordenes de magnitud durante una
inundación anual (Tockner et al, 2000) con efectos concomitantes en la distribución
de los organismos terrestres y acuáticos (Kohler et al., 1999). Kohler et al. (1999)
encontraron que los peces y los macroinvertebrados se redistribuían en cuerpos de
agua (temporales y permanentes) durante la fase de aguas altas.
Consecuentemente, la comunidad de invertebrados post-inundación era afectada
fuertemente por la depredación de peces y otros invertebrados depredadores. Ellos
sugirieron que el mosaico de estados sucesionales en la zona de inundación puede
reflejar interacciones bióticas como también fuerzas físicas estocásticas (Robinson et
al., 2002). Arscott et al. (2003) encontraron que, aunque la estructura de toda el área
de inundación era muy dinámica, la fracción de diferentes elementos del paisaje era
más bien constante. El fraccionamiento es importante porque la abundancia de
muchas especies depende de una formación contínua de nuevos hábitat.
El objetivo de este capítulo es examinar la composición de la comunidad de
macroinvertebrados bénticos en transectos transversales que toman en cuenta los
cuerpos lóticos y lénticos en el Bajo río Caura.
2.2. Material y métodos
2.2.1. Area estudio Este estudio fue conducido en el sector bajo del río Caura (aguas abajo del Salto
Pará), el segundo afluente más importante de la margen derecha del Río Orinoco. El
río Caura drena un área de 47.500 km2 (3º 37´-7º 47´ Lat. N, 63º23´-65º35´ Long. O),
que corresponde a un 5% del área total de Venezuela (Vargas y Rangel, 1996) (Fig.
5). La cuenca del río Caura es la cuarta cuenca más larga de Venezuela precedida
por los ríos Apure, Caroní, y Orinoco (Peña y Huber, 1996).

22
Peña (1996) divide la cuenca del Río Caura en dos subsectores hidrográficos con
características morfométricas diferentes, la margen derecha y la margen izquierda.
La margen derecha ocupa un 32% (14.650 Km2) y la margen izquierda un 68%, la
cual esta constituida por siete subcuencas y un número importante de intercuencas
entre las cuales están los caños Sipao y Mato. El área de estudio está localizada
entre una zona de Bosque seco en las planicies que bordean al caño Sipao y una
zona de Bosque Húmedo tropical que bordean las planicies del caño Mato (Holdridge
et al., 1971; Martínez, 1966). La precipitación media anual, tomando como fuente la
Estación Hidrometeorológica de CVG-EDELCA en Maripa (7º 25´ N, 65º10´ O, 35
m.s.n.m.) es de 1.980 mm/año sobre un período de 23 años (Vargas y Rangel,
1996). El período de lluvias va desde Abril hasta Octubre y el período de sequía
desde Enero hasta Marzo. Durante el período de lluvias, el Bajo Caura, hasta el nivel
de la población de las Trincheras, comienza a ser afectado por el fenómeno de
remanso, o represamiento de las aguas producido por el Orinoco en la época de
aguas altas. Según Rosales y Huber (1996), la planicie inundable del Bajo Caura se
extiende por debajo del Raudal La Mura y representa la mayor superficie de bosques
inundados por aguas mayormente oligotróficas de los ríos que drenan el Escudo
Guayanés.
Los sedimentos, por los alrededores de Maripa y Aripao, son de origen fluvio-deltáico
de la Formación Mesa, por sucesivas deposiciones del Río Orinoco, de finales del
terciario y del cuaternario reciente del Orinoco y el Caura (Rincón y Estanga, 1996;
Peña, 1996). Los suelos asociados con estos bosques inundables son considerados
por Rosales y Huber (1996) generalmente arcillosos con muy altos contenidos de
materia orgánica. Ellos comentan que estos bosques presentan un sotobosque ralo
con flora muy similar a aquella encontrada en el Río Mapire pero sin ningún elemento
distintivo claro de la flora asociada con aguas oligotróficas (“Igapó”) o eutróficas
(“várzea”), encontrando los bosques entremezclados en el Bajo Caura, como clasifica
Prance (1979) para el Amazonas. Knab-Vispo (2002) registró que las especies
alrededor de las lagunas del Bajo Caura y sus afluentes están caracterizados por una
mezcla de especies típicas del Igapó como Acosimum nitens, Macrolobium

23
multijugum, M. Acaciifolium, etc., y de especies típicas de la várzea como Maquira
coriacea, Gustavia augusta, Ruprechtia tenuifolia, Virola surinamensis.
2.2.3. Métodos
2.2.3.1. Trabajo de campo Se eligieron los siguientes sustratos para el estudio de macroinvertebrados: A) El
sedimento como el sustrato de mayor cobertura de los ambientes lótico y léntico, B)
Los troncos sumergidos como uno de los sustratos más frecuentemente encontrados
en todas los transectos durante la fase de aguas bajas y C) Las hojas verdes del
bosque inundado como el sustrato más frecuentemente encontrado durante la fase
de aguas altas. Se coleccionaron muestras en la sección baja de los caños Sipao,
Mato y río Caura. Cada transecto seleccionado tiene de referencia el nombre de la
laguna (Fig. 6). Las nueve lagunas seleccionadas pueden clasificarse según su
morfología, grado y tipo de conexión en: a) Lagunas pequeñas en formación con
conexión directa (Caño Sipao: Garzones, Chiribital y Caramacatico), b) lagunas
grandes con conexión indirecta, dendríticas y profundas (Rio Caura: Brava, Aricagua
y Naparaico) y c) lagunas pequeñas con conexión indirecta en forma de disco (Mato:
Pozo Rico y Patiquín) y en forma de herradura (Mato: Cejal) (Hutchinson, 1957;
Drago, 1981 y Hamilton y Lewis, 1990).
Sobre los transectos (Fig. 6), se establecieron 4 estaciones del sustrato sedimento,
CNT: en el centro de la laguna, ORI: en la orilla de la laguna próxima al Caño, BOS:
en el bosque inundado de la laguna y SIP o MAT: en la orilla del caño Mato o Sipao
próxima a la laguna estudiada y de los sustratos maderos sumergidos y hojas
verdes. Para las lagunas del río Caura, se estableció este mismo transecto sólo que
la estación ubicada a la orilla del río no fue colectada porque la fuerza del río no
permitió en ninguna fase la perfecta colección de la muestra. Se tomaron 4 replicas
de sedimento en cada estación de muestreo con un nucleador marca Wildco de 5 cm
de diámetro de apertura y 50 cm de altura (Fig. 7A). Una limpieza parcial de las
réplicas fue realizada en el campo colando parte de la muestra a través de una
batería de cedazos de 40, 250 y 500 µm de apertura (Fig. 7C). Cada replica fue

24
etiquetada y guardada en envases plásticos de 500 ml y preservada en formol al 4
%. Se realizaron tres campañas para la recolección de estas muestras: durante la
época de aislamiento de las lagunas del río: aguas bajas (AB), durante la época de
transición: subida de aguas (AT) y durante la época de conexión: aguas altas (AA)
(Downing y Rigler, 1984; Rosenberg, et al. 1997). La importancia de tomar estas
fases radica en que los organismos condicionan, adecuan o sincronizan sus ciclos de
vida (fertilidad, reproducción, dispersión de huevos, crecimiento, por ej.) a la época
en que ocurren los eventos hidrológicos.
La colección de muestras de maderos sumergidos se realizó por triplicado, en todas
las lagunas excepto en laguna Patiquín donde no se consiguieron en el transecto
estudiado. Se colectaron trozos o pedazos de madera de árboles sumergidos en las
orillas de las lagunas o cerca del bosque que estuvieran dentro de un marco de
madera de 625 cm² (Fig. 7B). Los trozos de madera fueron introducidos en una bolsa
o envase plástico, debidamente etiquetados y preservados en 4% de formol para su
transporte al laboratorio. Este sustrato sólo pudo muestrearse durante la fase de
aguas bajas porque es imposible utilizar la misma técnica de colección para las
demás fases estudiadas.
Las muestras de las hojas verdes del bosque inundado fueron coleccionadas
demarcando primero con una cuadrata de 625 cm2. Cuidadosamente se introducía
una bolsa plástica en el agua y se rodeaban rápidamente las ramas con las hojas
seleccionadas de un árbol sumergido, cortándose la sección vegetal elegida. Se
tomaron tres réplicas en cada estación del bosque. Cada replica fue etiquetada,
preservada en formol al 4 % y guardada en doble bolsa plástica y cerradas con un
fleje.
Para el sustrato sedimento, se tomó un nucleador adicional en cada estación del
transecto y el agua intersticial fue tomada para medir la conductividad con un
conductímetro marca Lamotte chemical, el pH con un pH-metro digital marca Orion,
la temperatura y el oxígeno disuelto con un oxigenómetro marca YSI. La profundidad
fue medida con el mismo nucleador y la transparencia con el disco de secchi. Para

25
los demás sustratos se tomaron igualmente muestras de agua del área muestreada.
Otra muestra adicional de sedimento fue tomada en la fase de aguas bajas para
conocer el aporte porcentual de arena, arcilla, limo y materia orgánica particulada
gruesa.
2.2.3.1. Trabajo de laboratorio La taxonomía de los macroinvertebrados acuáticos estuvo basada principalmente en
las formas inmaduras que son las más frecuentemente encontradas. Si el adulto en
las trampas de luz como las exuvias de las pupas y las exuvias del último estadio
larval se conseguían para los insectos holometabólicos (i.e. aquellos con
metamorfosis incompleta como la encontrada en los Ephemeroptera), entonces
lograba corroborarse desde el adulto las posibilidades de las formas inmaduras. No
fueron obtenidas crías en condiciones de laboratorio ni en el campo por la
disposición de tiempo para este estudio. Algunos grupos fueron identificados hasta el
nivel de familia o género separándolos en morfotipos, los cuales fueron codificados
(Roldán, 1988; Merritt y Cummins, 1996). Fue necesario ordenar las claves de
clasificación para cada grupo y cotejar la información taxonómica existente para
Venezuela (Hulbert et al. 1981 a,b). La identificación fue determinada usando un
microscopio estereoscópico marca Wild y algunos detalles con un microscopio óptico
marca Olympus a diferente niveles taxonómicos para cada grupo. Se utilizaron las
claves generales de Merritt y Cummins (1996), Pennak (1978), Edmondson (1959),
Usinger (1963), Lopretto y Tell (1995), Wiggins (1977), Epler (1995, 1996), Limongi
(1983), Peters (1971), Domínguez et al. (1992; 1997), Traver (1943), Demoulin
(1966), Berner (1978) y Puthz (1975). Las diferentes técnicas generales utilizadas
para la identificación de los invertebrados fueron consideradas según Pennak (1978).
La colección de referencia reposa en el Laboratorio de Macroinvertebrados acuáticos
de la Estación de Investigaciones Hidrobiológicas de Guayana (EDIHG) de
Fundación La Salle de Ciencias Naturales.

26
Cada taxon de los macroinvertebrados codificados fue asignado a un grupo funcional
según su alimentación basada en Jackson y Sweeney (1995), Merrit y Cummins
(1996) y Cummins y Klug (1979).
La abundancia de macroinvertebrados fue obtenida contando todos los individuos de
los sustratos de maderos sumergidos y hojas verdes del bosque inundable. En el
sedimento, el producto de la limpieza de la réplica a analizar fue teñido con una
solución acuosa de Rosa de bengala al 1% y dejado por 24 horas para su fijación.
Las muestras de sedimento del mes de junio (época de transición hidrológica) fueron
identificadas y contadas en su totalidad. Sin embargo, el tiempo, el conteo, la
desviación estándar entre las replicas y la energía consumida fue la base para
plantearse la realización de un submuestreo para el resto de las réplicas
provenientes de las otras fechas. El producto del primer cedazo fue analizado en su
totalidad y los siguientes cedazos fueron sometidos a un submuestreo.
La técnica de submuestreo en el sedimento consistió en adaptar un mezclador de
muestras microbiológicas que permitía regular la velocidad de mezcla (r.p.m.) similar
al utilizado en la técnica de submuestreo descrita por Blanco-Belmonte (1988) para
macroinvertebrados asociados a la vegetación acuática. El producto del segundo y
tercer cedazo de la limpieza de la réplica fue ubicado en un Elmneyer de 250 ml. El
frasco fue colocado en un centro fijo adaptado al mezclador; la boca del frasco
acoplada a un anillo metálico movible con acolchonamiento interno. Se efectuaron
varias pruebas consecutivas midiendo el tiempo, el número de cm3 a analizar y el
conteo. Los resultados fueron analizados mediante un ANOVA de una vía y el test t
del programa Statigraphic v.4. Se seleccionó un tiempo de 17 segundos para la
mezcla y se tomaron de 4 a 8 réplicas de 1 ml cada una, coleccionadas con una
micropipeta automática de boca ancha. Cuando las subréplicas presentaban
ausencia de organismos se analizaba toda la réplica.
El material de los otros sustratos fue lavado directamente sobre un cedazo de 250
µm, fue analizado y contado a 6x bajo un microscopio estereoscópico Marca Wild; los
organismos preservados en alcohol al 70%. Cada trozo de madera y hoja verde

27
fueron medidos para el cálculo del área y el peso seco calculado en g después de
secar el material en la estufa a 120ºC por 24 horas. La materia orgánica gruesa
proveniente de cada nucleador o sustrato fue secada a 120º C en la estufa, pesada y
estimada su media para cada estación.
El cálculo de la biomasa fue estimada de dos formas: volumétricamente y como peso
húmedo (g) sobre una balanza electrónica Mettler PC4400 (Error ± 0.01 mg). En los
organismos coleccionados en las maderas y en las hojas verdes del bosque
inundable se realizó una estimación total. Para la biomasa de los organismos que
habitan en el sedimento se realizó un estimación general de pesos medios de un
número indicado de organismos para cada grupo. Se utilizó la estimación de Junk
(1973) para aquellos grupos de tamaño pequeño con dificultad para el manejo y
colección del número deseado.
La densidad de la comunidad de macroinvertebrados de todos los sustratos es
expresada en el número de organismos en un metro cuadrado (Org.m-2).
2.3. Resultados La comunidad de macroinvertebrados en los sustratos analizados en el Bajo río
Caura (Tabla 1) fue dominado por insectos en un 70-75% en los sustratos maderos,
hojas verdes y rocas (sustrato no incluido), mientras que en un 5-22% en el detrito
flotante (sustrato no incluido), vegetación acuática (sustrato no incluido) y sedimento
(Fig. 8). La clase Insecta estuvo representada por 12 órdenes. Los órdenes más
frecuentes y abundantes en la mayoría de los sustratos fueron los Díptera y
Ephemeroptera. Otros macroinvertebrados, no insectos, que predominaron en todos
los sustratos fueron: los anélidos, los nematodos, los hidracáridos y los
microcrustaceos (Copepoda, Cladocera y Ostracoda).

28
El orden Díptera mostró una marcada predominancia dentro de la clase Insecta en la
mayoría de los sustratos. Los Chironomini, detritívoros, son más abundantes que los
Tanypodini, predadores. Se conoce en la literatura (Irmler, 1975) que este último
depreda los Chironomini. En el sedimento, se registraron frecuentemente masas
gelatinosas de Chironomini del primer estadio durante la fase de aguas altas; larvas
de estadios superiores a menudo se encontraron viviendo en estuches de arena muy
fina. En la madera, las larvas de Stenochironomus sp. (Diptera: Chironomini) minaron
finos surcos donde vivían en estuches sedosos. En la vegetación acuática y las hojas
verdes, otros géneros de dípteros se encontraron escondidos en el interior de las
raíces y en capas de limo. Es muy posible que existan especies endémicas de
Chironomidae como asegura Reiss (1974) para esta región y que su aporte a la
biodiversidad sea uno de los más altos de la clase Insecta como Vásquez et al
(1988) observaron para el Orinoco. Al igual que el orden Diptera, se encontraron los
Ephemeroptera como un componente frecuente en todos los sustratos de este
estudio (Tablas 1A-1C).
El orden Coleoptera no es tan frecuente como los ordenes anteriores, sin embargo,
es el que presenta el mayor número de familias en un orden de la clase Insecta. Los
ditiscidos e hydrofílidos son las familias con el mayor % de aparición en los maderos
sumergidos y hojarasca.
Dentro de la Clase Hydracarina, se registraron 42 codificaciones en su mayoría
localizadas en las hojas del bosque inundable. La frecuencia de los organismos en
algunas codificaciones es muy baja, registrándose la mayoría de las veces de 1 a 3
especímenes para una codificación durante las fechas estudiadas. La mayoría de los
hidracáridos encontrados durante aguas altas eran formas inmaduras (Dr. Rossi
com. per.)
Dentro de la Clase Crustácea, se registraron 5 órdenes. Los órdenes más frecuentes
y abundantes fueron los ostrácodos, los cladóceros y los copépodos. Dentro del
orden Cladocera, la familia Macrothricidae fue la más frecuentemente registrada para

29
el sedimento. La especie Illiocryptus sp2 (MAC 5) fue registrada sólo para las
estaciones de sedimento de los tributarios y para las orillas de la laguna Aricagua,
mientras que Illiocryptus sp1 (MAC3) es más frecuente en las demás estaciones del
sedimento. Del orden Ostracoda, hubo 14 codificaciones, las cuales en su mayoría
estuvieron en el sedimento. Pelocypris sp1 y Cypridae sp1 fueron las más
predominantes. El orden Decapoda sólo se registró entre la hojarasca con dos
especies: Macrobrachium amazonicum (JEL1) y Euryrhyncus amazoniensis (JEL2).
Este ùltimo figura como nuevo registro geográfico para Venezuela (identificación y
conocimiento de registro dado por el Dr. G. Pereira com. Pers.).
Los anélidos estuvieron representados por oligoquetos naiididos y por hirudíneos
principalmente del género Helobdella. Los primeros predominaron en el sedimento
principalmente en las estaciones del Bosque inundado (BOS) y en las orillas de las
lagunas, siendo Pristina sp1 (NAI 2) y Dero sp (NAI 6) los más abundantes y
frecuentes de este grupo. Entre los hirudíneos, Helobdella stagnalis (Hi 2) fue la
especie con más amplia aparición, no sólo en el sedimento sino también en los
maderos.
Los valores medios de la densidad (Org.m-2), riqueza, diversidad y biomasa
registrados en las muestras de sedimento para las cuatro estaciones de las nueve
lagunas en las diferentes épocas de estudio son mostrados en las Tablas 1A-1C. En
general, los transectos en el río Caura presentaron tendencias de mas baja densidad
durante la fase de aguas altas, mientras que los transectos de los tributarios
presentaron las mayores densidades en esta misma fase (Fig. 9). Los valores más
altos de densidad para los invertebrados en el sedimento durante el período de
estudio fueron registrados en Patiquín (PATBOS 64.031 ± 43.822,99 Org.m-2),
PATCNT (56.906 ± 36.029,68 Org.m-2) y PATORI (46.119 ± 7.562,54 Org.m-2)
durante la fase de aguas altas. Se observaron los valores más bajos en Chiribital
(CHIORI 53 ± 75,02 Org.m-2) durante la fase de subida de aguas y en Aricagua (CNT
372 ± 57,34 Org.m-2) durante la fase de aguas altas.

30
Se comprobó mediante el análisis de la varianza de clasificación doble (ANOVA) la
significación de las diferencias en las densidades observadas entre las muestras de
sedimento de las estaciones CNT, ORI y BOS para las diferentes épocas en los
transectos estudiados. La diferencia de las densidades entre los transectos durante
las fases de aguas altas y aguas bajas resultó ser significativa (F=9,82; g.l. 2,6;
P<0.005) para las estaciones centrales (CNT), (F=8,14; g.l. 2,6; P<0.005) para las
estaciones de las orillas de la laguna (ORI) y (F=8,17; g.l. 1,4; P<0.005) para las
estaciones del bosque inundado (BOS). Para los siguientes análisis de este estudio,
fueron omitidos los datos de la fase de subida de aguas o de transición por no
presentar diferencias significativas con las otras fases.
Brasilocaenis irmleri (B1), del grupo Ephemeroptera, apareció durante la fase de
aguas bajas solamente en dos estaciones del sedimento del Caño Sipao con unas
densidades entre 35 y 255 Org.m-2. Durante la fase de aguas altas, estuvo presente
en las estaciones del Bosque de Patiquín, Naparaico, Garzones y Caramacatico,
alcanzando en esta última la máxima densidad de 3.979 Org.m-2. Según los
resultados de Imler (1975) de experimentos realizados en laboratorio, Brasilocaenis
irmleri es un organismo que depende de la hojarasca más que del sedimento, tiene
un rápido desarrollo y es adaptable muy bien a los cambios. Los resultados que
consigue Irmler (1975) para Brasilocaenis en tres bosques de inundación del
Amazonas Central ubican a esta especie con una específica periodicidad y como
organismo característico de aguas altas al igual que este estudio.
Se comprobó mediante el análisis de la varianza de clasificación simple, la
significación de las diferencias en las densidades observadas en las muestras de
maderos sumergidos para las lagunas estudiadas. No hubo diferencia significativa de
las densidades registradas entre los sitios de estudio. Hubo densidades entre 22.164
y 28.955 Org.m-2 en los maderos sumergidos (Tabla 2). Un factor común en todas las
muestras de madera fue la presencia de Asthenopus curtus (Ephemeroptera),
representando más del 40 al 70% de la densidad y un 90% de la biomasa.

31
Al igual que el sustrato anterior, la significación de las diferencias en las densidades
observadas entre las muestras de hojas del bosque inundable para los sitios
estudiados fue comprobada mediante el análisis de la varianza de clasificación
simple. No hubo diferencia significativa entre las densidades encontradas en las
hojas verdes del bosque inundable. Las densidades más altas fueron registradas en
Aricagua (376.110 Org.m-2, Tabla 3) y las más bajas en Naparaico (4.676 Org.m-2,
Tabla 3). Las densidades estimadas en las hojas verdes del bosque inundable fueron
las más altas registradas en un metro cuadrado de sustrato (Tabla 3). La densidad
de los quironómidos es muy alta, sin embargo, es en este sustrato donde la
diversidad de hydracáridos es más alta.
Dentro de los grupos funcionales, los recolectores predominan de un 60 a un 90 %
en el sedimento. No hubo partidores en los sedimentos; sin embargo, tanto en las
hojas como en los maderos, estos grupos estuvieron presentes (Fig. 9A).
Los valores más altos de biomasa de macroinvertebrados en un metro cuadrado en
el bajo Caura fueron registradas en los sustratos temporales: maderos y hojas verdes
del bosque inundable (Tabla 2 y 3) obtenidos en las fases de aguas bajas y aguas
altas respectivamente. En los maderos sumergidos, se obtuvieron los valores más
altos de biomasa de macroinvertebrados (26,78 y 94,6 g.m-2), muy por encima de
todos los sustratos estudiados, siendo el valor más alto encontrado en Brava (Río
Caura). En el sustrato de las hojas verdes del bosque inundable, se obtuvieron
valores de biomasa entre 13,10 – 41,24 g.m-2, siendo el más alto en Chiribital (Caño
Sipao) (Tabla 3). En el sedimento, los valores mas altos registrados durante la fase
de aguas altas fueron en el bosque de Caramacatico (Caño Sipao) y orilla de Pozo
Rico, los cuales alcanzaron 24,92 y 23,10 g.m-2 respectivamente. Esto valores fueron
muy altos en comparación a los valores comúnmente encontrado en todos los
transectos (<0,01 – 15,0 g.m-2) (Tabla 1A – 1C).
En general, la composición de los sedimentos en los centros y orillas de las lagunas
ubicadas en los transectos de Caño Mato fueron limo-arcillosos, mientras que en los

32
transectos del río Caura, las orillas de las lagunas Brava y Aricagua fueron más
arcillo-arenosas y los centros conservaron la misma composición de los transectos
del Caño Mato. La composición del sedimento en las lagunas en los transectos del
Caño Sipao tendió a ser más areno-arcillo-limosos (Fig. 10).
Capitulo 3: Comparación de las comunidades de macroinvertebrados
3.1 Introducción Los macroinvertebrados acuáticos juegan un papel clave en los ecosistemas de ríos
debido a su posición intermedia en la cadena alimentaria ligada a la producción
alóctona/auctóctona con los niveles tróficos altos (Munn y Brusven, 1991). Algunos
hábitat son físicamente complejos y proveen cuantitativamente diferentes espacios
de vida disponibles en hábitat físicamente simples. Tal diversidad de espacios
favorece el aumento del número de nichos y consecuentemente aumenta la riqueza.
Dean y Connell (1987) y Bell et al., (1991) han observado que hábitat complejos
contienen más especies que los hábitat simples. Downes et al., (1998) y Vinson y
Hawkins (1998) señalan que cuantificar la cantidad y variedad del espacio disponible
para los organismos es difícil y que la relación de la abundancia y riqueza de
macroinvertebrados con respecto a la heterogeneidad del sustrato no es consistente.
Es importante acotar que la mayoría de los estudios han medido estos espacios con
métodos que son algo subjetivos a un solo hábitat o que resulta en números que no
pueden ser comparados con cualquier medida de acuerdo a la medida de McCoy y
Bell (1991). Además, la complejidad per se depende de la escala espacial a la que es
examinada según refiere Kolasa y Pickett, (1991). Estas dificultades han sido
significativas para las predicciones específicas de cómo un espacio de nicho puede
incrementarse por la complejidad del hábitat. Para resolver esta controversia, McCoy
y Bell (1991) propusieron un esquema tridimensional para describir la estructura del
hábitat, siendo el eje de su complejidad referida a la abundancia (ej. de aberturas, de
huecos): más complejos son los hábitat mientras más espacios diferentes halla.

33
1998). Boyero (2003) considera una clara necesidad de utilizar medidas métricas
para la heterogeneidad del sustrato, la cual proveerá información más consistente de
la distribución espacial del habitat y su relación con los macroinvertebrados. El objetivo de este capitulo es comparar la densidad de morfoespecies o códigos de
macroinvertebrados registrada en los diferentes estaciones y sustratos a lo largo de 9
transectos en el bajo Caura, usando un espacio de medida métrica (1 m2) para cada
sustrato.
3.2. Material y métodos
Un Análisis de varianza de uno y dos factores para varias muestras por grupos fue
aplicado a los datos de densidad de las muestras del sustrato sedimento entre las
diferentes estaciones y diferentes fechas con el propósito de comprobar si el
conjunto de datos son significativamente diferentes. Los datos fueron transformados
a log+1 para normalizar la frecuencia de distribución de los conteos (binomial
negativos) y eliminar la dependencia de la varianza con la media aritmética (s2 > X) y
así asegurar que los componentes de la varianza sean aditivas (ver Elliot, 1979).
Para examinar las posibles diferencias de las comunidades de invertebrados entre
los diferentes transectos y entre las diferentes fases estacionales (Aguas altas y
aguas bajas, fue aplicado un análisis de conglomerados usando el coeficiente de
disimilitud Euclideano y el método UPGMA, el cual le da igual peso a las estaciones
provenientes de diferentes sitios y sustratos para una misma época (Kovach
Computing Service 1985-2001; programa MSVP versión 3.12d).
Las distancias euclideanas entre las estaciones de los transectos, ubicadas en un
espacio multidimensional en cada eje representando la densidad transformada de las
especies, son ilustradas en dendrogramas individuales para cada transecto.

34
3.3. Resultados
Los dendrogramas ilustran el resultado de la comparación de las diferentes
estaciones ubicadas en los 9 transectos estudiados (Figs. 11-19). El contraste de las
estructuras de las comunidades entre las estaciones durante las fases de aguas altas
y aguas bajas es distintivo en diferentes grados a lo largo de los transectos.
En la Fig. 11, puede observarse claramente el conglomerado consecutivo formado
por las estaciones de sedimento en la fase de aguas altas, uniéndoles una similitud
de sólo un 20%. Un segundo conglomerado es formado por las estaciones de
sedimento en la fase de aguas bajas y hojas verdes. La similitud de esta última
estación con la comunidad de macroinvertebrados en la orilla de la laguna Patiquín
es de un 40%, el más alto conseguido en esta laguna, pero ocurriendo en diferentes
fases estacionales.
La Fig. 12 ilustra el dendrograma para una laguna de herradura (Cejal) y se observa
claramente el 100% de similitud en la estación de la orilla de la laguna en las
diferentes fases estacionales. Sin embargo, la similitud entre la comunidad del centro
de la laguna para las diferentes fases es de un 40%, evidenciándose el 100% de
disimilitud entre los maderos sumergidos y la orilla de la laguna para la fase de aguas
bajas.
En la Fig. 13, se muestra el análisis de conglomerados en la comunidad de
invertebrados de la laguna Pozo Rico (Mato). En este diagrama, puede observarse
claramente la disimilitud mayor en un 70% de todas las estaciones tomadas en este
transecto durante la fase de aguas altas y la fase de aguas bajas.
Uno de los casos más particulares es el del transecto Brava, donde las estaciones
centro y orilla de la laguna presentaron una mayor similitud con respecto a las otras

35
estaciones (fase de aguas bajas: 38%, fase de aguas altas: 41%) (Fig. 14). La
particularidad no es sólo la similitud entre las estaciones, sino que esta laguna es
una de las más profundas con una pendiente pronunciada en la orilla. En
consecuencia, el sustrato de maderos sumergidos no guarda ninguna relación con la
estación de la orilla de la laguna.
Los análisis de conglomerados realizados en las comunidades de invertebrados del
resto de los transectos (Naparaico, Aricagua, Chiribital, Garzones y Caramacatico)
son ilustrados en los dendrogramas de las Figs. 15-19. Al igual que el transecto
anterior, las comunidades de invertebrados en las estaciones del centro y orilla de las
lagunas guardan mayor similitud que las otras estaciones en el transecto. La
diferencia estacional es marcada y la separación entre los sustratos, aún habiendo
sido tomados en los mismos espacios o áreas, muestran una comunidad de
invertebrados diferenciada.
Capitulo 4:
Relación de las comunidades de macroinvertebrados con las variables físico-químicas
4.1 Introducción Los factores abióticos y los cambios hidrológicos en particular influyen fuertemente
en la dinámica de los paisajes ribereños creando y manteniendo la heterogeneidad
(dinámica de parches, mosaicos en el paisaje, ecotonos), los patrones sucesionales
(ecología de las islas) y los moderadores ambientales (clima, temperatura) (Robinson
et al., 2002).
La estructura de las comunidades de invertebrados está determinada por factores
abióticos (perturbaciones naturales y antrópicas, química del agua y tipo de sustrato)
y bióticos (productividad, competencia, depredación) (Peckarsky, 1983). Tales
perturbaciones son consideradas como uno de las fuerzas controladoras más
importantes en las comunidades de macroinvertebrados que viven en un río (Resh et
al., 1988; Reice et al., 1990; Lake, 2000; Zimmermann y Death, 2002).

36
Las inundaciones, uno de los factores abióticos más importantes en el trópico, han
sido el motivo de las controversias generadas por el RCC (River Continuum Concept)
debido a que se ingnoró el efecto de estas fuerzas en el concepto. Por otro lado, la
escasa información específica para el río/área de inundación/ecosistema
considerado en las comunidades bióticas generó otra controversia para el concepto
de flujo. Por esto, el objetivo de este capítulo es comparar las densidades de las
comunidades de macroinvertebrados provenientes de las estaciones ubicadas en los
transectos (río/área de inundación) en diferentes regiones del Bajo Caura durante las
fases de aguas altas y aguas bajas con respecto a las variables tomadas para cada
estación.
4.2. Material y métodos
Para los análisis de este capítulo, las densidades de los macroinvertebrados y los
valores de la variables abióticas fueron transformados a log (x+1) y log (x)
respectivamente para estabilizar las varianzas de la densidad de las especies a lo
largo de las transectos y para conformar mejor los datos por las asunciones
estadísticas.
Un Análisis de Componentes Principales (ACP) fue realizado con el programa MSVP
versión 3.12d usando los datos coleccionados durante aguas altas y aguas bajas. El
ACP fue utilizado antes del Análisis de Correspondencia Canónica (ACC) para
reducir las variables abióticas redundantes que mejor respondieran a un modelo
lineal (i.e. se omitieron aquellas variables que mostraron mayor inflación de la
varianza). Las asociaciones estadísticas entre la estructura de las comunidades y las
variables ambientales fueron cuantificadas usando el Análisis de Correspondencia
Canónica (ACC), una técnica de ordenamiento no lineal de autovectores
especialmente diseñado para un análisis de gradientes directo de las relaciones
entre un conjunto de datos ecológicos multivariados. (Kovach Computing Service
1985-2001; programa MSVP versión 3.12d). Se realizó un ACC por separado

37
(maderos sumergidos y hojas verdes) para los sustratos temporales y para todas las
muestras combinadas. El ACC genera un gráfico de ordenación (biplot) que dibuja
más claramente las diferentes clasificaciones que pueden darse entre las relaciones
de la densidad de las comunidades y las variables abióticas.
4.3. Resultados
El Análisis de componentes principales (ACP) para las variables abióticas tomadas
en todas las estaciones de sedimentos en los 9 transectos del estudio muestra que
75,73% de la variación total es explicada por los tres primeros componentes (Fig.
20). El primer componente explica 35.53% de la variación total con tres variables
(Oxígeno disuelto OD, Transparencia TRANS y Profundidad PROF) con autovalores
>0,500. El segundo eje explica 20,64% con una sola variable (Materia orgánica
particulada gruesa MO) y el tercer eje (17,56%) con las variables: Conductividad
(COND) y pH. En la Fig. 20 se ve claramente que esta variación separa claramente
las fases de aguas altas y aguas bajas. La única variable que no explica la diferencia
estacional entre las variables ambientales es la Temperatura. (Fig. 20)
El Análisis de componentes principales (ACP) para las variables abióticas tomadas
en los maderos sumergidos (7 variables x 8 casos) muestra que 85,34% de la
variación total es explicada por los tres primeros componentes (Fig. 21). El primer
componente explica un 38,67% de la variación total con una variable, pH, con
autovalores >0,500. El segundo componente explica 66,7% con una sola variable,
PROF, y el tercer componente con las variables COND y TRANS. En la Fig. 21 se
observa la separación química de las subregiones estudiadas.
Para los análisis de Correspondencia Canónica (ACC), se omitieron aquellas
variables como T(ºC) y MO, que no explican claramente las diferentes subregiones
escogidas para este estudio, a partir de los análisis de ACP para las variables
abióticas (ACP) y aquellas estaciones de muestreo que presentan alta similitud en el
análisis de las variables bióticas (Análisis de conglomerados Fig. 12 CEJO).

38
El análisis de Correspondencia Canónica (ACC) para todas las muestras combinadas
(sedimento, madera y hojas verdes del bosque de inundación) mostró las diferentes
relaciones entre la distribución de las especies, a lo largo de los gradientes de los
factores ambientales extraidos en los tres primeros ejes y los cambios estacionales
en la estructura de la comunidad de invertebrados (Fig. 22). Los gradientes de
PROF, pH y TRANS incrementaron a lo largo del primer y cuarto eje de ordenación,
los cuales coinciden con la ordenación de las estaciones en la mayoría de los
transectos encontrados en aguas altas sobre la media (PROF:738,1 cm, pH: 5,88 y
TRANS: 59,9 cm; Fig. 22). Los gradientes OD y COND incrementaron a lo largo de
los ejes 2 y 3 de ordenación, los cuales coinciden con la ordenación de las
estaciones en la mayoría de los transectos encontrados en aguas bajas sobre la
media (OD 6,33 y COND: 17,65 mS/cm) o por debajo de la media (OD: 3,7 y COND
23,26 mS/cm; Fig. 22).
Las especies registradas en los maderos sumergidos están claramente distribuidas a
lo largo del tercer eje (Ej. Asthenopus curtus, más abundante y Climacia sp,
específica) mientras que las especies registradas en las hojas verdes del bosque de
inundación están dentro del agrupamiento de la fase de aguas altas. Las especies
como Hellobdella stagnalis y Chaoborus sp. registradas en los sedimentos de las
lagunas están ubicadas dentro del agrupamiento horizontal de la fase de aguas altas
(Fig. 22).
Un Análisis de Correspondencia Canónico en los sustratos maderos sumergidos y
hojas verdes del bosque de inundación son ilustrados en las Figs. 23 y 24
respectivamente. En este análisis, en el sustrato de maderos sumergidos, se
separan claramente las tres principales regiones: Caura, Mato y Sipao. La mayoría
de las especies caen dentro del agrupamiento del Caura que estuvo alineado con la
transparencia y conductividad (Fig 23). Las especies comunes para todos los
maderos sumergidos, como Asthenopus curtus, están en la intersección de los tres
círculos y las especies registradas sólo en las otras subregiones, se encontraron

39
fuera de esta intersección. La separación de las regiones estuvo fuertemente
alineada con el gradiente de la transparencia y el pH; sin embargo, en este caso, el
gradiente de la conductividad proporcionó en el tercer eje una separación más clara
de las regiones (Fig. 23).
El ACC en el sustrato hojas verdes del bosque inundado igualmente separa
claramente las dos regiones muestreadas: Caura y Mato. Al igual que en los análisis
anteriores, la separación estuvo fuertemente alineada con los gradientes de la
transparencia, la conductividad y el pH principalmente.
Capítulo 5: El rol del sustrato en la variabilidad espacial de la comunidad de
macroinvertebrados 5.1. Introducción
Una cantidad desproporcionada de investigaciones sobre la estructura de la
comunidad béntica en los ríos ha sido enfocado particularmente en hábitat como los
guijarros o saltos de agua, comparados con otros tipos de hábitat, aunque en un
número de estudios en las regiones templadas se ha investigado la estructura de la
comunidad béntica con respecto a otros tipos de sustratos (Egglishaw, 1969; Rabeni
y Minshall, 1977; Minshall y Minshall, 1977; Huryn y Wallace, 1987).
A pesar de su importancia, son aún escasos los estudios que han sido publicados
sobre la distribución de los macroinvertebrados encontrados en diferentes hábitat en
un río (Dudgeon, 1994; Arunachalam et al., 1991; Ramírez et al., 1998; Magalhães y
Pereira, 2003). Es ahora cuando se reconoce que los ríos contienen una variedad de
hábitat con diferentes condiciones físico-químicas (Ramírez et al., 1998; Pringle et
al., 1988). La distribución de macroinvertebrados en varios hábitat en los ríos refleja,
hasta cierto grado, una distribución de recursos (ej. Alimento, oxígeno, predadores,

40
Rabeni y Minshall, 1977; Ramírez et al., 1998), y provee información acerca de cómo
las comunidades podrían responder ante los cambios en los parámetros ambientales.
La alta diversidad biológica registrada en el trópico (Welcomme, 1985, 1990;
McConnell, 1987) está asociada con una alta dinámica natural de los hábitat
acuáticos, ecotonales y terrestres (Ward et al., 2002; Robinson et al., 2002; Rosales
et al, 1999). Esta heterogeneidad espacial en los sistemas lóticos está caracterizada
por una amplia variedad de sustratos, los cuales experimentan una variabilidad
estacional y temporal a diferentes niveles hidrométricos y con una entrada de
nutrientes que pueden influir en la supervivencia de los organismos. La rápida
adaptación de los macroinvertebrados a esta heterogeneidad espacial es la
condición que les aventaja con respecto a otros organismos.
Los macroinvertebrados bénticos están asociados a diferentes sustratos
permanentes (rocas, grava, sedimento) y sustratos temporales (maderos
sumergidos, hojas verdes del bosque de inundación, hojarasca, carpetas de material
alóctono flotante) en un río. Sustratos investigados en este estudio (maderos
sumergidos y hojas) son los que Benke et al. (1984) consideran particularmente
importantes para los ríos de aguas negras ya que al entrar en las cadenas
alimentarias como detrito, la mayor parte es refractaria y procesada vía “microbial
loop”.
La naturaleza del sustrato es muy importante para determinar la estructura de
comunidad ecológica presente. Algunas especies requieren ciertos tipos de
sustratos. Por ejemplo, los invertebrados como algunos tricópteros necesitan grava
para fijar sus estuches y crear redes de filtro para su alimentación. Otros organismos
necesitan limo y áreas someras donde el material orgánico es acumulado. Las
grandes rocas proveen protección a los organismos de la corriente y a su vez
soportan un crecimiento algal para su alimentación.

41
Una evidencia considerable (Harper et al. 1997) muestra que los macroinvertebrados
prefieren diferentes tallas de sustratos en diferentes parches: desde arcillas, maderos
sumergidos, vegetación hasta rocas y que esta variabilidad de sustratos tiene un
efecto acumulativo sobre la biodiversidad. Rosales et al. (1999) sugieren que un
gradiente longitudinal sumado a una alta variación lateral, bosques inundables en
una zona de confluencia de un tributario en un gran río tropical con un pulso de
inundación significativo, tiene una alta diversidad beta en términos de números de
hábitat, más que cualquier otro paisaje en el perfil longitudinal de un río.
Tuckner et al. (2000) encontraron grandes cambios en la curvas de la riqueza de
especies de diferentes grupos de organismos, incluidos los macroinvertebrados
bénticos, a lo largo de un transecto transversal del área de inundación, sugiriendo
que una pérdida de conectividad hidrológica afectaría a los grupos diferentemente,
aunque se redujera toda la biodiversidad. Por consiguiente, el hábitat del área de
inundación puede ser usado alternativamente por los animales terrestres y acuáticos,
dependiendo del ciclo hidrológico, aumentando así toda la productividad del sistema
(Robinson et al., 2002). Estas áreas sometidas a cambios estacionales proveen de
alimento abundante y de alta calidad a los organismos terrestres durante la fase de
aguas bajas y a los organismos acuáticos, una diversidad de sustratos colonizables,
el forraje y la reproducción que es clave durante la fase de aguas altas.
El objetivo de este capítulo es examinar el rol del sustrato en la variabilidad espacial
de la comunidad de macroinvertebrados mediante la diversidad, riqueza y biomasa
de los macroinvertebrados en tres diferentes sustratos debido al escaso
conocimiento que existe sobre los macroinvertebrados en los ambientes leníticos y
lóticos del Bajo Caura.
5.2. Material y métodos La diversidad fue medida mediante el Índice de Shannon - Wienner (H´)

42
Indice de Shannon - Wienner: H´ = - Σ pi log Pi donde pi = Probabilidad de aparición de la especie o código I para todas las
estaciones en los 9 transectos de estudio.
La riqueza fue medida contando el número de las especies o codificaciones de las
morfoespecies para todas las estaciones en los 9 transectos de estudio.
5.3. Resultados La diversidad (Shannon-Wiener, H'), biomasa (g m-2) y riqueza de los taxa de
macroinvertebrados registrados en el sedimento en tres transectos en el Caño Mato
(Patiquin, Cejal, y Pozo Rico) y Naparaico en el río Caura son ilustradas en la Figura
25 durante las fases de aguas altas y aguas bajas. Los resultados sugieren un
incremento lateral similar en diversidad, biomasa y riqueza de taxa para todos los
puntos de muestreo en la zona de confluencia. Las flechas de la Fig. 25 indican las
estaciones ubicadas en el bosque de inundación adyacente con la más alta
diversidad y riqueza de taxa, en todas los transectos. Estos datos sugieren que las
zonas de confluencia del caño Mato con el rìo Caura presenta un gradiente
comparable, en biodiversidad, biomasa y riqueza, como el modelo propuesto por
Rosales et al. (1999).
Se ilustra tambien en la Fig. 26 los resultados en riqueza de taxa, diversidad de
especies (H') y biomasa (g.m-²) de los macroinvertebrados asociados a las hojas
verdes del bosque inundado, el cual es considerado en este estudio como un
sustrato temporal encontrado en el sitio del bosque inundable durante la fase de
aguas altas. Estos resultados representan los primeros datos generados de este tipo
de sustrato en el bosque de inundación del río Caura. Los resultados indican que la
abundancia relativa de especies a menudo está caracterizada por pocas especies
dominantes y por un gran número de especies raras: 66% de la abundancia
representadas por tres familias: Chironomidae, Staphylinidae y Naididae; mientras
que 24 familias (que comprenden 44% de la riqueza total) aparecen sólo una vez en

43
la zona de confluencia. En los otros sitios (Patiquin, Naparaico y Brava) 70-82% de
la abundancia total estuvo representada por dos familias: Chironomidae y Naididae
de donde 13-20 familias (18-30% de la riqueza total) aparece sólo una vez. Estos
datos provienen de hábitat escasamente estudiados en el trópico, y coincidiendo con
estudios generados en otras áreas de investigación. Vinson y Hawkins (1998) han
encontrado que la riqueza de los invertebrados acuáticos puede variar
marcadamente en relación con la estructura del hábitat. Un sustrato físicamente
complejo, como las hojas verdes del bosque de inundación posiblemente soporta
más taxa que aquellos estructuralmente simples como la arena o las rocas.
De la misma forma cuando se examinan datos de sur America tropical, América
central y Africa del Oeste, Claro et al. (2004) y Tedesco et al. (2005) registran que las
más altas riquezas de especies de peces están asociadas con los canales que tienen
conexión directa con refugios en los bosques de inundación, por lo que también
parece encontrarse una fuerte asociación entre una más alta riqueza y los bosques
de inundación.
6. Discusión
La suma del número presente de taxa de invertebrados en el bajo Caura encontrado
entre los sustratos temporales y permanentes (sedimento, pozos de roca, vegetación
acuática, maderos sumergidos, hojarasca y hojas verdes del bosque de inundación)
en los 9 transectos, es más alto que los reportados por otros autores para el Orinoco
y el Caura (Blanco-Belmonte, 1990; 1998; García y Pereira, 2003). Los escasos
estudios en estas regiones no permiten realizar conclusiones aún definitivas al
respecto, más aún conociendo la debilidad en identificaciones taxonómicas que el
trópico tiene con varios grupos de invertebrados.
La aparición esporádica de órdenes como Neuroptera (Climacia sp1, NEU1) no sólo
tiene que ver con el desarrollo de esponjas en la superficie de algunos de los

44
maderos sumergidos encontrados en algunas lagunas (Pennak, 1978), sino que
muestra igualmente los numerosos factores que pueden contribuir a una alta
diversidad observada en el bajo Caura. Es decir, una amplia variedad de hábitat
permite que diferentes especies tomen ventaja de diferentes nichos (Rodríguez-
Olarte et al., 2003)
El número de hábitat en los que aparecen las especies de Ephemeroptera revela,
además de una diversidad alta de este grupo en el Bajo Caura, la condición
generalista de algunas ninfas como Callibaetis sp1 (CAL1) y Brasilocaenis irmleri
(B1); mientras que otras revelan una condición clave como Asthenopus spp (E4) en
los maderos sumergidos, Campsurus spp. (CAM1) en el sedimento, Simothraulopsis
demerara (E9), Leentvaria palpalis (E10) y el nuevo género de Leptophlebiidae g.n.
(E13) en las rocas. La conducta minadora de Asthenopus curtus ha sido observada
anteriormente en los pecíolos de Eichhornia crassipes en las lagunas de inundación
del Orinoco por el autor. Sin embargo, desempeña un papel equivalente al de Adusta
povilla en el Caura observada también por Pert (1970) en las maderos sumergidos
del lago Volta en África. Este género puede considerarse especializado en la
explotación y beneficio del sustrato invadido, aunque accidentalmente esté en otros
hábitat como en las astillas de madera del detrito flotante. Se prestó atención
taxonómica a este orden, ya que a menudo es utilizado en trabajos de biomonitoreo
por la alta diversidad de hábitat en donde habitan y especialmente a la sensibilidad
de respuesta de algunas especies a las alteraciones de algunas variables abióticas
en los ecosistemas acuáticos (Leal y Esteves, 1999; Rosenberg et al., 1997).
Dentro de los organismos que mostraron un patrón claro de abundancia durante las
fases estudiadas están los Chaoboridae (CHA 1, Diptera) (Máx aguas altas:1273 Org
/m-2) y Brasilocaenis irmleri (B1, Ephemeroptera). Ellos, en general, aparecen con
mayor frecuencia y densidad en casi todas las lagunas durante las fases de subida y
aguas altas y fueron un componente menor, pero importante, de los
macroinvertebrados durante la fase de aguas bajas en Aricagua. Una dinámica
similar presentada por los Chaoboridae fue evidenciada por Reiss (1977) para una

45
laguna de aguas negras en el Amazonas Central. Para las lagunas del Bajo Caura,
las máximas densidades obtenidas durante aguas altas son casi tres veces mayores
que las registradas por Reiss (op.cit.) en la laguna Tupé (445 individuals/m2) en esta
misma época. A diferencia del neotrópico, en los lagos de Norte América, la larva de
Chaoborus sp. es un componente importante de la zona profunda, migrando a las
aguas abiertas durante la noche y depredando selectivamente el zooplancton
(Wetzel, 1983).
Las densidades extremadamente altas, registradas en la laguna Caramacatico y
Naparaico durante las épocas de subida de aguas y aguas altas, posiblemente
puedan deberse al producto de un incendio durante 1999 en la zona del bosque
estudiado. El efecto a corto plazo por un incendio, tala y quema en estos bosques
equivaldría a una pérdida permanentemente de la capa orgánica por volatilización,
lixiviación o escorrentía de los nutrientes (Dezzeo, 1994; Hernández, 1999). Se
pregunta entoces si fue posiblemente ésta disposición de nutrientes la que permitió
una colonización rápida, artificial de quironómidos, Brasilocaenis irmleri y nematodos
durante la época siguiente y que bajó anormalmente la densidad en la fase de aguas
bajas del siguiente año. Los efectos de actividades antropogénicas en las
comunidades de macroinvertebrados bénticos han sido reportadas por varios autores
(Leal y Esteves, 1999; Nakamura, 2000). Ellos han demostrado que una
perturbación tanto física como química (fuerte influencia de las concentraciones de
nitrógeno, fósforo y materia orgánica en el sedimento) son responsables de alterar la
diversidad, la densidad y la biomasa de las comunidades bénticas en el Amazonas
(Junk y Furch, 1985; Reiss, 1977).
Los resultados de este estudio revelan una fuerte e inesperada relación entre la
densidad de la comunidad de invertebrados y cincos variables abióticas ambientales
en los 9 transectos estudiados del Bajo Caura: TRANS medida con el disco de
secchi, PROF, pH, OD y COND (Fig. 22). Relaciones similares con la composición
íctica y algunas variables abióticas en las lagunas del río Orinoco han sido
registradas por Rodríguez y Lewis (1997). La mayoría de las estaciones en los

46
transectos mostraron claramente trayectorias de cambios estacionales. Estos
cambios involucran, en general, un aumento en la abundancia de los
macroinvertebrados durante aguas altas para las estaciones de sedimento en los
transectos de los tributarios y Naparaico concomitantemente con el aumento de la
profundidad, la transparencia y el pH (Figs. 7 y 22). Resh et al., (1988) y Lake (2000)
comentan que la inundación en los ríos ejerce una fuerte influencia en la fauna de
macroinvertebrados, causando un aumento típico en densidad y diversidad durante
la subida y un declive en aguas altas. Sin embargo, en el trópico existen factores que
pueden limitar esta generalización.
Las estaciones ubicadas en los transectos de Aricagua y Brava sobre el río Caura
presentan un incremento opuesto a los encontrados en los otros transectos y esto
posiblemente pueda deberse a la morfología de las lagunas y a la formación del
bosque de inundación. Las lagunas en los transectos de los tributarios (Mato y Sipao)
y Naparaico (Caura) presentan forma de disco, más pequeñas y menos profundas
que las lagunas Aricagua y Brava, las cuales son alargadas, dendríticas y más
profundas. Las lagunas en los transectos del tributario Mato son abiertas con
conexión directa al río, mientras que los de Aricagua y Brava poseen albardones
más altos. Los hábitat como los maderos sumergidos parecen influenciar claramente
la estructura del conjunto de invertebrados durante la fase de aguas bajas aun y
cuando comparten un espacio reducido con estaciones de sedimento de orilla,
hojarasca, etc.
Dentro de cada tipo de sustrato conviven un reducido número de especies que
tienden a especializarse. Pérez, (1984) en su estudio sobre el uso del hábitat y la
estructura de la comunidad de peces de un río tropical asociado a un bosque en
Venezuela, encontró una relación similar entre las características del hábitat y la
estructura de la comunidad de peces midiendo la complejidad del hábitat y
correlacionándola con la diversidad, riqueza y equidad.

47
Los valores de biomasa y densidad en los diferentes sustratos permitieron considerar
igualmente la importancia de las orillas durante la fase de aguas bajas, donde los
efemerópteros y tricopteros son un componente importante de estos microhábitat y
de la dieta de los peces insectívoros del río Caura (Gonzalez y Vispo, 2003). De igual
manera, Goulding et al. (1988) observaron la importancia de las orillas,
indirectamente en sus estudios sobre la diversidad y la ecología alimentaria vista
desde las comunidades ícticas en el río Negro en Brasil. Naiman et al. (1988), Pringle
et al. (1988) y Rosales (2000) señalan que la potencial importancia de estas zonas
está relacionada con la calidad de las mismas y que el estudio de mosaico de
sustratos provee una importante información para el funcionamiento de un sistema.
En general, los valores de diversidad de la comunidad de macroinvertebrados en el
bajo Caura son bajos comparados con otros ambientes de aguas blancas (Irmler,
1975; Fittkau et al., 1975) pero relativamente altos comparados con otros ambientes
de aguas negras del Amazonas (Reiss, 1977; Nessimian et al., 1998). Los valores de
diversidad obtenidos para este estudio son bajos en valores de equitatividad que
indican una distribución desequilibrada en donde, dominan dos o tres especies o
codificaciones.
Muchos de los trabajos recientes sobre los patrones de diversidad biótica a gran
escala han enfocado como los factores locales y los procesos históricos regionales
operan como filtros del paisaje para influenciar la ocurrencia y persistencia de
especies en hábitat (Tonn, 1990; Ricklefs y Latham, 1993; Poff, 1997). La maxima
riqueza local por sitio parece estar relacionada con una riqueza regional (Caley y
Schluter, 1997), la cual pareciera estar restringida por las condiciones ambientales
regionales y la historia biogeográfica (Ricklefs, 1987). Una aproximación para poder
estructurar la diversidad de las especies en la zona de confluencia (ecotonal) podría
dividirse de una manera similar a lo descrito por Lamb y Mallik (2003) para la zona
riparina/ecotono de un bosque:

48
1. Las especies características de hábitat específicos (sustratos temporales
como los maderos sumergidos, las hojas verdes del bosque de inundación, la
hojarasca, los sustratos permanentes como los sedimentos (arena, grava,
arcilla, rocas, entre otros) encontradas en baja abundancia en la zona de
confluencia: Nematoda en el sedimento, Tanypodinae en las hojas verdes,
Asthenopus curtus (Ephemeroptera) y Pristina sp (Annelida) en los maderos
sumergidos,
2. Las especies que no cambian en abundancia a lo largo de la zona de
confluencia: Cyrnellus spp (Trichoptera) en los maderos sumergidos y
Cypridae sp6 (Ostracoda OS9) en el sedimento, Ceratopogonidae spp. en las
hojas verdes,
3. Las especies especialistas en las zonas de confluencia, las cuales son raras
en las comunidades adyacentes pero pueden estar presentes con abundancia
relativamente alta en la dinámica de la zona de confluencia: Stenochironomus
sp., en el sedimento, Tenagobia sp, Hydrochus spp y Helobdella stagnalis en
los maderos sumergidos.
El cambio en la prevalencia de la abundancia de las especies puede representar una
respuesta a un cambio en las condiciones ambientales encontradas en la zona de
confluencia (Lamb y Mallik, 2003). Sin embargo, los datos obtenidos de este estudio
sugieren que las zonas de confluencia de los tributarios del río Caura presentan
gradientes comparables, en términos de diversidad de macroinvertebrados, al
modelo propuesto por Rosales et al. (1999).

49
7. Conclusiónes
1. 242 codificaciones/especies para la fauna de macroinvertebrados han sido
registradas hasta ahora, siendo este trabajo un aporte significativo al acervo
científico de la región.
2. La comunidad de macroinvertebrados estuvo dominada por los insectos de
los ordenes Diptera y Ephemeroptera. Ademas de los insectos, otros grupos
dominantes fueron los Ostracoda, Cladocera y Annelida.
3. La composición de los grupos funcionales fue dominada por los recolectores.
4. En todos los transectos, los sustratos temporales, tanto en aguas bajas como
altas, presentaron mayor abundancia, diversidad, riqueza y biomasa de
macroinvertebrados en comparación al sustrato sedimento.
5. Los valores de biomasa, riqueza, diversidad y densidad en los diferentes
estaciones del sedimento permitieron considerar la importancia de las orillas
durante la fase de aguas bajas y del bosque durante la fase de aguas altas.
6. Durante el momento de transición (subida de aguas), sólo en las estaciones
de bosque hubo aumento de la densidad, la diversidad y la biomasa. Los
valores de biomasa obtenidos para este estudio son bajos en valores de
equitatividad que indican una distribución desequilibrada en donde dominan
dos o tres especies o codificaciones.
7. El análisis de similitud con base en las codificaciones/especies registradas
revelaron una diferencia de una estructura definida y diferenciada de los
macroinvertebrados entre los sustratos temporales y permanentes durante
aguas altas y bajas.
8. Los análisis de Correlación Canónica revelan una fuerte relación entre la
estructura de los macroinvertebrados durante las fases de aguas altas y bajas
y cinco variables ambientales abióticas (Oxígeno disuelto, Transparencia,
Profundidad, Conductividad y pH).
9. Un reducido número de especies conviven dentro de cada tipo de sustrato,
pero tienden a especializarse en determinados sustratos o microhábitat (ej.

50
Asthenopus spp. en los maderos sumergidos, Campsurus spp. en el
sedimento, Brasilocaenis irmleri en la hojarasca) y comparten con otras
especies que muestran capacidad para utilizar varios de estos (Callibaetis
sp.). 10. La comparación de varios transectos en un perfil longitudinal del río, sugiere
un gradiente donde las zonas de confluencia presentan altos valores de
biomasa, diversidad y riqueza. Esta misma comparación sugiere una
constante y alta variación lateral de los bosques de inundación.

51
8. Recomendaciones
De los resultados obtenidos, se estima que un control del flujo del agua en el río
Caura durante aguas bajas afectaría fuertemente las orillas de las lagunas y con esto
la comunidad de macroinvertebrados, especialmente efemerópteros y tricópteros,
que desaparecerían casi inmediatamente como se ha visto en casi todas las
represas hechas en los sistemas de grandes ríos. Esto afectaría directamente a las
comunidades que viven de la pesca por ser los invertebrados un ítem importante en
la dieta de los peces.
Dada la alta riqueza de macroinvertebrados y los escasos estudios taxónomicos de
muchos de los grupos de invertebrados en una de las cuencas más pristinas de
latinoamérica, es importante reactivar el interes en la sistemática en las nuevas
generaciones de biólogos. Las acciones de protección ambiental no tendrían éxito si
se desconoce la importancia de la biodiversidad en regiones tan especiales en el
trópico como lo es la cuenca del Caura.
Es importante desarrollar estudios más detallados donde variables como el nitrógeno
total, disponibilidad del fósforo y la materia orgánica lábil en el sedimento permitan
relacionar la densidad, riqueza, biomasa y diversidad de la fauna de
macroinvertebrados bénticos.

52
9. REFERENCIAS
Aarts, B. G. W y P. H. Nienhuis 1999. Ecological sustainability and biodiversity. The International Journal of Sustainable Development and World. Ecology: 1-29.
Arscott, D. B., Tockner, K. y J. V. Ward 2003. Spatio-temporal patterns of benthic
invertebrates along the river continuum of a braided Alpine river (Tagliamento River, NE Italy). Archiv für Hydrobiologie 158:431-460.
Arunachalam, M., Madhusoodanan Nair, K. C., Vijverberg, J. y K. Kortmulder 1991.
Substrate selection and seasonal variation in densities of invertebrates in stream pools of a tropical river. Hydrobiologia 213: 141-148.
Báldi, A., y D. McCollin 2003. Island ecology and contingent theory: the role of spatial
scale and taxonomic bias. Global Ecology & Biogeography 12:1-3. Baptista, D. F., Buss, D. F., Dorville, L. F. M. y J. L. Nessimian 2001a. Diversity and
habitat preference of aquatic insects along the longitudinal gradient of the Macaé River basin, Rio de Janeiro. Brazil. Rev. Bras. Biol. 61:249-258.
Baptista, D. F., Dorville, L. F. M. y D. F. Buss 2001b. Spatial and temporal
organization of aquatic insects assemblages in the longitudinal gradient of a tropical river. Rev. Bras. Biol. 61:295-304.
Bastardo, H. 1999. El proceso de descomposición y su importancia ambiental.
Trabajo de ascenso a titular en el escalafón universitario de la ilustre Universidad Central de Venezuela.
Batzer, D. P. 1998. Trophic interactions among detritus, benthic midges, and
predatory fish in a freshwater marsh. Ecology 79: 1688–1698.
Bell, S. B., McCoy, E. D. y H. R. Mushinsky 1991. (Eds.) Habitat Structure: The physical arrangements of objects in space. Chapman and Hall, London. 438 p.
Benke, A. C., VanArsdall, T. C. Jr., Gillespie, D. M. y F. K. Parrish 1984. Invertebrate
productivity in a subtropical blackwater river: The importance of habitat and life history. Ecological Monographs 54: 25-63.
Berner, L. 1978. The status of Asthenopus curtus (Hagen) (Ephemeroptera:
Polymitarcidae). Acta Amazonica 8(1):103-105.
Blanco-Belmonte, L. 1988. Técnica para la obtención de submuestras de organismos micropleustónicos. Mem. Congreso Iberoamericano y del Caribe. Mem. Soc. Cienc. Nat. La Salle, Suppl. 2. 48:89-96.

53
Blanco-Belmonte, L. 1990. Estudio de las comunidades de invertebrados asociados a
las macrofitas acuáticas de tres lagunas de inundación de la sección baja del río Orinoco, Venezuela. Mem. Soc. Cienc. Nat. La Salle. 49/50:71-107.
Blanco-Belmonte, L., Neiff, J. J. y A. Poi de Neiff 1998. Invertebrate fauna associated
with floating macrophytes in the floodplain lakes of the Orinoco (Venezuela) and Paraná (Argentina). Verh. Internat. Verein. Limnol. 26:2030-2034.
Blanco-Belmonte, L., Bastardo, Asmine y J. Rosales 2003. Contribución funcional de
los invertebrados, bacterias y hongos en la descomposición de hojas en un río de aguas negras tropical. V Congreso Venezolano de Ecología. Isla de Margarita, 3 al 7 de Noviembre de 2003.
Bond, W. J. 1993. Keystone species. 237-253. En: E. D. Schulze y H. A. Mooney
(Eds.). Biodiversity and ecosystem function. Springer-Verlag, Berlin, Germany.
Bond, W. J. 2001. Keystone species: hunting the shark?. Science 292:63-64.
Bonetto, A. A. e I. de Ezcurra 1964. La fauna bentónica de algunas aguas rápidas del Paraná Medio. Physis 24:311-316.
Boyero, L. 2000. Towards a global stream ecology. Tree 15:390-391.
Boyero, Luz y Robert C. Bailey 2001. Organization of macroinvertebrate communities
at a hierarchy of spatial scales in a tropical stream. Hydrobiologia 464: 219–225.
Boyero, L., y J. L. De Lope 2002. Short-term recolonization of stones in a tropical
island stream. Marine and Freshwater Research 53:993-996.
Brussock, P. P. y A. V. Brown 1991. Riffle-pool geomorphology disrupts longitudinal patterns of stream benthos. Hydrobiologia 220: 109-117.
Caley M. J. y D. Schluter 1997. The relationship between local and regional diversity.
Ecology 78: 70-80.
Callisto, M., Moreno, P. y F. A. R. Barbosa 2001. Habitat diversity and benthic functional trophic groups at Serra do Cipó, Southeast Brazil. Rev. Bras. Biol. 61:59-266.
César, I. I., Ocox, C., Paggi, A. C., Spaccesi, R. C. A., F. Tangorra, M. y M. P.
Tassara 2000. Diversidad de invertebrados bentónicos del río de la Plata. Biología Acuática 19:27-63.
Church, M. 2002. Geomorphic thresholds in riverine landscapes. Freshwater Biology
47:541-557.

54
Claro-Jr., Ferreira, Luiz L., E., Zuanon, J. y C. Araujo-Lima 2004. O efeito da
floresta alagada na alimentação de três espécies de peixes onívoros em lagos de várzea da Amazônia Central, Brasil. Acta Amazonica 34(1): 133 – 137.
Covich, Alan P., Palmer, Margaret A. y Todd A. Crowl 1999. The Role of Benthic
Invertebrate Species in Freshwater Ecosystems Zoobenthic species influence energy flows and nutrient cycling. BioScience 49 (2):119-127.
Cressa, Claudia 2003. Comparación entre estudios realizados en sub-cuencas y
cuencas hidrográficas de Venezuela. Ecotropicos 16(2):64-74.
Culp, J. M. y R. W. Davies 1982. Analysis of longitudinal zonation and river continuum concept in the Oldman-South Saskatchewan river system. Can. J. Fish. Aquat. Sci. 39: 1258-1266.
Culp, J. M. y R. W. Davies 1985. Responses of benthic macroinvertebrate species to
manipulation of interstitial detritus in Carnation Creek, B.C., Can. J. Fish. Aquat. Sci. 42:139-146.
Cummins, K. W. 1973. Trophic relations of aquatic insects. Ann. Rev. Entomol. 18:
183-206.
Cummins, K. W. 1974. Structure and function of stream ecosystems. BioScience 24:631-641.
Cummins, K. W., y M. J. Klug 1979. Feeding ecology of stream invertebrates. Ann.
Rev. Ecol. System. 10: 147-172.
Cushing, C. E., McIntire, C. D., Cummins, K. W., Minshall, G. W., Petersen, R. C., Sedell, J. R. y R. L. Vannote 1983. Relationships among chemical, physical, and biological indices along river continua based on multivariate analyses. Arch. Hydrobiol. 98: 317-326.
Dangles, O., Jonsson, M. y B. Malmqvist 2002. The importance of detritivore species
diversity for maintaining stream ecosystem functioning following the invasion of a riparian plant. Biological Invasions. 4:441-446.
Davic, R. D. 2003. Linking keystone species and functional groups: a new operational
definition of the keystone species concept. Conservation Ecology 7(1): r11.
Dean, R. L. y J. H. Connell 1987. Marine invertebrates in an algal succession. I. Variations in abundance and diversity with succession. J. Exp. Mar. Biol. Ecol. 109:195-215.

55
Death, R. G. y M. J. Winterbourn 1995. Diversity patterns in stream benthic invertebrate communities: the influence of habitat stability. Ecology 76: 1446-1460.
Demoulin, G. 1966. Contribution á l´etude des Ephéméroptéres du Surinam. Bull.
Inst. R. Sci. Nat. Belg. 42(37):1-22.
Dezzeo, N. 1994. (Ed.). Ecología de la altiplanicie de la Gran Sabana Guayana Venezolana). I. Investigaciones sobre la dinámica bosque-sabana en el sector SE, subcuencas de los ríos Yuruaní, Arabopó y Alto Kukenán. Scientia Guayanae 4: 1-205.
Doeg, T. J., Lake, P. S. y R. Marchant 1989. Colonization of experimentally disturbed
patches by stream macroinvertebrates in the Acheron River Victoria. Australian Journal of Ecology 14:207-220.
Domínguez, E. y J. M. Ballesteros Valdez 1992. Altitudinal replacement of
Ephemeroptera in a subtropical river. Hydrobiologia 246: 83-88.
Domínguez, E., Hubbard, M. D. y W. L. Peters 1992. Clave para ninfas y adultos de las familias y géneros de Ephemeroptera (Insecta) sudamericanos. Biología Acuática 16:1-38.
Domínguez, E., Peters, W. L., Peters, J. G. y H. M. Savage 1997. The imago of
Simothraulopsis Demoulin with redescription of the nymph (Ephemeroptera: Leptophlebiidae: Atalophlebiinae). Aquatic Insects 19(3):141-150.
Downes, B. J., Lake, P. S. Schreiber, E. S. G. y A. Glaister 1998. Habitat structure
and regulation of local species diversity in a stony, upland stream. Ecological Monographs 68: 237-257.
Downing, J. A. y F. H. Rigler 1984. (Eds.). A manual on methods for the assessment
of secondary productivity in fresh waters. Blackwell Scientific Publications. Oxford, England.
Drago, E. C. 1977. Erosión y sedimentación en un tramo de cauce del río Paraná
medio (República Argentina). Asociación Geológica Argentina Revista. 32:277-290.
Drago, E. C. 1980. Aspectos limnológicos de ambientes próximos a la ciudad de
Santa Fe (Paraná Medio): Comportamiento hidrológico y sólidos suspendidos. Ecología (Argentina) 5:31-48.
Drago, E. C. 1981. Grados de conexión y fases hidrológicas en ambientes leníticos
de la llanura aluvial del río Paraná (Argentina). Ecología (Argentina). 6:27-33.

56
Driver, E. H. 1977. Chironomid communities in small prairie ponds: some characteristics and controls. Freshwater Biology 7:121-133.
Dudgeon, D. 1994. The influence of riparian vegetation on macroinvertebrate
community structure and functional organization in six new Guinea streams. Hydrobiologia 294: 65-85.
Edmondson, W. T. 1959. Fresh-Water Biology. John Wiley & Sons, Inc. New York.
1248 p.
Egglishaw, H. J. 1964. The distributional relationship between the bottom fauna and plant detritus in streams. Journal of Anim. Ecology 38: 19-33.
Egglishaw, H. J. 1969. The distribution of benthic invertebrates on substrata in fast
flowing streams. J. Anim. Ecol. 38: 19—33.
Elliot, J. M. 1979. Some methods for the Statistical analysis of samples of benthic invertebrates. Freshwater Biological Association. Scientific Publication Nº25. UK. 160 p.
Elwood, J. W., J. D. Newbold, R. V. O'Neill, and W. Van Winkle. 1983. Resource
spiraling: an operational paradigm for analyzing lotic ecosystems. Pages 3-27 in T. D. Fontaine and S. M. Bartell, editors. Dynamics of lotic ecosystems. Ann Arbor Science, Ann Arbor, Michigan, USA.
Epler, J. H. 1995. Identification manual for the larval Chironomidae (Diptera) of
Florida. Final Report DEP Contract Number WM579. Department of Environmental Protection. Florida State, Tallahassee. 317 p.
Epler, J. H. 1996. Identification manual for the water beetles of Florida (Coleoptera:
Dryopidae, Dytiscidae, Elmidae, Gyrinidae, Haliplidae, Hydraenidae, Hidrophilidae, Noteridae, Psephenidae, Ptilodactylidae, Scirtidae). Final Report DEP Contract Number WM621. Department of Environmental Protection. Florida State, Tallahassee.
Fisher, S. G. y G. E. Likens 1973. Energy Flow in Bear Brook, New Hampshire: An
integrative approach to stream ecosystem metabolism. Ecol. Monogr. 43:421-439.
Fittkau, E. J. 1971. Distribution and ecology of amazonian chironomids (Diptera). The
Canadian Entomologist 103:407-413.
Fittkau, E. J., Irmler, U., Junk, W. J., Reiss, F. y G. W. Schmidt 1975. Productivity, biomass and population dynamics in Amazonian water bodies. 289-311. En: (Eds.) F. B. Golley y E. Medina: Tropical ecological systems. Trends in terrestrial and aquatic research. New York Springer.

57
Fittkau, E. J. 2001. Amazonian Chironomidae (Diptera, Chironomidae): A contribution
to chironomid research in the Neotropics. Amazoniana 16:313-323.
Fuller, Randall L., Kennedy, Brian y Carl Nielsen 2004. Macroinvertebrate responses to algal and bacterial manipulations in streams. Hydrobiologia 52: 113-126.
Galdean, N., Callisto, M. y F. A. R. Barbosa 2001. Biodiversity assessment of benthic
macroinvertebrates in altitudinal lotic ecosystems of Serra do Cipó (MG, Brazil). Rev. Bras. Biol. 61:239-248.
García, José V. y Guido Pereira 2003. Diversity of Benthic macroinvertebrates from
the Caura River Basin, Bolívar State, Venezuela. Rap Bulletin of Biological Assesment 28:144-150.
Goulding, M., Leal Carvalho, M. y E. G. Ferreira 1988. Rio Negro: rich life in poor
water: Amazonian diversity and food chain ecology as seen through fish communities. SPB Academic Publishing, The Hague, The Netherlands. 189 p.
Halse, S. A., Cale, D. J., Jasinska, E. J. y R. J. Shiel 2002. Monitoring change in
aquatic invertebrate biodiversity: sample size, faunal elements and analytical methods. Aquatic Ecology 36:395–410.
Hamilton, S. K., y Lewis, Jr. W. M. 1990. Basin morphology in relation to chemical
and ecological characteristics of lakes on the Orinoco River floodplain, Venezuela. Arch. Hydrobiol. 119:393-425.
Harper, D., Mekotova, J., Hulme, S., White, J. y J. Hall 1997. Habitat heterogeneity
and aquatic invertebrate diversity in floodplain forests. Global Ecology and Biogeography Letters 6:275-285.
Hawkins, C. P. 1984. Substrate associations and longitudinal distributions in species
of Ephemerellidae (Ephemeroptera: Insecta) from western Oregon. Freshw. Invertebr. Biol. 3(4): 181-188.
Healey, M. 1984. Fish predation on aquatic insects. 255-288. En: The ecology of
aquatic insects. (Eds.). V. H. Resh y D. M. Rosenberg. Praeger Press, New York. 625 p.
Henriques-Oliveira, A. L., Nessimian, J. L. y L. F. M. Dorville 2003. Feeding habitats
of chironomid larvae (Insecta: Diptera) from a stream the Floresta da Tijuca, Rio de Janeiro, Brazil. Braz. J. Biol. 63:269-281.
Hernández, L. 1999. Ecología de la altiplanicie de la Gran Sabana, II. Estructura,
diversidad, crecimiento y adaptación en los bosques de las subcuencas de los ríos Yuruaní y Alto Kukenán. Scientia Guayanae 9: 1-160.

58
Hershey, A. E. 1987. Tubes and foraging behavior in larval Chironomidae:
implications for predator avoidance. Oecologia 73: 236-241.
Hershey, A. E. y S. I. Dodson 1985. Selective predation by a sculpin and a stonefly on two chironomids in laboratory feeding trails. Hydrobiol. 124: 269-273.
Holdridge, L. R., Grenke, W. C., Hatheway, W. H., Liang, T. y J. A. Tosi Jr. 1971.
Forest environments in tropical life zones: A pilot study. Pergamon, Oxford. England.
Huener, J. D. y J. A. Kadlec 1992. Macroinvertebrate response to marsh
management strategies in Utah: Wetlands. 12 72–78.
Hulbert, S. H., Rodríguez, G. y N. W. Dos Santos 1981a. (Eds.). Aquatic Biota of Tropical South America. Parte1. Arthropoda. San Diego University Press, San Diego. 323 p.
Hulbert, S. H., Rodríguez, G. y N. W. Dos Santos 1981b. (Eds.). Aquatic Biota of
Tropical South America. Parte 2. Anarthropoda. San Diego University Press, San Diego. 298 p.
Huryn, A. D y J. B. Wallace 1987. Local geomorphology as a determinant of
macrofaunal production in a mountain stream. Ecology 68:1932-1942. Hutchinson, G. E. 1957. A treatise on limnology, v. 1. Geography, Physics and
Chemistry. Wiley. 1015 p.
Hynes, H. B. N. 1970. The ecology of running waters. University of Toronto Press, Toronto.
I l l ies, J. 1964. The invertebrate fauna of the Huallaga, a Peruvian tributary
of the Amazon River, from the sources down to Tingo Maria. Verh. Int. Verein. Limnol. 15: 1077-1083.
Irmler, U. 1975. Ecological studies of the aquatic soil invertebrates in three inundation
forests of central Amazonia. Amazoniana. 5:337-409.
Irmler, U. y W. J. Junk 1982. The inhabitation of artificially exposed leaf samples by aquatic macro-invertebrates at the margin of amazonian inundation forests. Tropical Ecology 23:64-75.
Jackson, J. K. y B. W. Sweeney 1995. Egg and larval development times for 35
species diversity of tropical stream insects from Costa Rica. J. North Amer. Benthol. Soc. 14:115-130.

59
Jacobsen, D., Schultz, R. y A. Encalada 1997. Structure and diversity of stream invertebrate assemblages: the influence of temperature with altitude and latitude. Freshwater Biology 38:247-261.
Junk, W. J. 1973. Investigations on the ecology and production-biology of the "floating
meadows" (Paspalo-Echinochloetum) on the Middle Amazon. II. The aquatic fauna in the root system of the floating vegetation. Amazoniana 4(1): 9-102.
Junk, W. J. 1980. Areas inundáveis-Um desafio para limnologia. Acta Amazonica
10(4):775-795.
Junk, W. J. 1983. Aquatic Habitats in Amazonia. The Environmentalist. 3 Suppl. 5:24-34.
Junk, W. J. y K. Furch 1985. The Physical and Chemical Properties of Amazonian
Waters and their Relationships with the Biota. 3-17. En: Key Environments: Amazonia. (Eds.) G. T. Prance y T. E. Lovejoy. Pergamon Press, Oxford.
Junk, W. J., Bayley, P. B. y R. E. Sparks 1989. The flood pulse concept in river-
floodplain systems. Canadian Special Publication of Fisheries and Aquatic Sciences 106:110-127.
Knab-Vispo, C. 2002. Vegetación. Informe Técnico entregado a Fundacite-Guayana
como un capítulo del Proyecto Ecología Integral del Bajo Caura. 45 p.
Kohler, S. L. 1992. Competition and the structure of a benthic stream community. Ecol. Monog. 62: 165-188.
Kohler, S. L., Corti, D., Slamecka, M. C. y D. W. Schneider 1999. Prairie floodplain
ponds: mechanisms affecting invertebrate community structure. 711–730. En: Invertebrates in Freshwater Wetlands of North America: Ecology and Management. (Eds.) D. P. Batzer, R. B. Rader y S. A. Wissinger. John Wiley and Sons, New York.
Kolasa, J., y S. T. A. Pickett 1991. (Eds.). Ecological Heterogeneity. Springer-Verlag
New York, Inc. 332 p.
Ladle, M. y R. J. Ladle 1992. Life history patterns of river invertebrates. Hydrobiologia 248: 31-37.
Lake, P. S., Schreiber, E. S. G., Milne, B. J. y R. G. Pearson 1994. Species richness
in streams: patterns over time, with stream size and with latitude. Verh. int. Ver. Limnol. 25: 1822–1826.
Lake, P. S. 2000. Disturbance, patchiness, and diversity in streams. J. North Amer.
Benthol. Soc. 19 :573-592.

60
Lamb, E. G. y A. U. Mallik 2003. Plant species traits across a riparian - zone/forest ecotone. Journal of Vegetation Science 14:853-858.
Lawson, D. L., Klug, M. J. y R. W. Merritt 1984. The influence of the physical,
chemical,and microbiological characteristics of decomposing leaves on the growth of the detritivore Tipula abdominalis (Diptera:Tipulidae). Can. J. Zool. 62:2339-2343.
Lawton, J. H. y V. K. Brown 1994. Redundancy in ecosystems. En: Biodiversity and
ecosystem function. 255-270. (Eds.) E. D. Schulze y H. A. Mooney. Springer-Verlag, London.
Leal, J. J. F. y F. A. Esteves 1999. Density and biomass of Campsurus sp.
(Ephemeroptera) and other macroinvertebrates in an Amazonian lake impacted by bauxite tailings (Lago Batata, Pará, Brazil). Amazoniana 15(3/4):193-209.
Limongi, J. 1983. Estudio morfo-taxonomico de náyades en algunas especies de
Odonata (Insecta) en Venezuela. Trabajo de Grado para la obtención del título de Licenciatura, Universidad Central de Venezuela. 100 p.
López, G. R., y J. S. Levinton 1987. Ecology of deposit-feeding animals in marine
sediments. Quart. Rev. Biol. 62: 235-260.
Lopretto, E. y G. Tell 1995. Ecosistemas de aguas continentales. Metodologías para su estudio. Ediciones Sur. La Plata. Tomo III. 895 p.
Mackay, R. J. 1992. Colonization by lotic invertebrates. A review of processes and
patterns. Can. J. Fish. Aqua. Sci. 49:617-628.
Marchese, M. e I. Ezcurra de Drago 1992. Benthos of the lotic environmets in the Middle Paraná River system. Transverse zonation. Hydrobiologia 237: 1-14.
Marchese, M., Ezcurra de Drago, I. y E. Drago 2002. Benthic Macroinvertebrates and
Physical habitat relationships in the Paraná river-floodplain system. 111-132. En: The ecohydrology of Southamerican Rivers and Wetlands. (Ed.) M. Mc.Clain. Publicación Especial #6 de International Association of Hydrological Sciences.
Marchese, M., Wantzen, Karl M., e I. Ezcurra de Drago 2005. Benthic invertebrate
assemblages and species diversity patterns of the Upper Paraguay River. Tropical ecology 21(5):485-499.
McConnell, R. H. L. 1987. Ecological Studies in Tropical Fish Communities.
Cambridge University Press, 382 p.

61
McCoy, E. D. y Bell, S. S. 1991. Habitat structure: the evolution and diversification of a complex topic. 3-27. En: Habitat structure: the physical arrangements of objects in space. (Eds.) E. D. McCoy, S. S. Bell y H. R. Mushinsky. Chapman and Hall.
Magalhães, C. y G. Pereira 2003. Survey of decapod crustaceans in the Caura River
Basin, Bolívar State, Venezuela: Species Richness, Habitat, Zoogeographical Aspects and Conservation Implications. Rap Bulletin of Biological Assesment 28:151-159.
Malard, F., Tockner, K., Dole-Olivier, M. J. y J. V. Ward 2002. A landscape
perspective of surface-subsurface hydrological exchanges in river corridors. Freshwater Biology 47:621-640.
Mallory, M. L., Blancher, P. J., Weatherhead, P. J. y D. K. McNicol 1994.
Presence or absence of fish as a cue to macroinvertebrate abundance in boreal wetlands. Hydrobiol. 279/280: 345-351.
Malmqvist, B. 2002. Aquatic invertebrates in riverine landscapes. Freshwater Biology
47:679-694.
Malmqvist, B., Wottom, R. S. y Y. Zhang. 2001. Suspension feeders transform massive amounts of seston in large northern rivers. Oikos 92:35-43.
Martínez, María 1966. Clima: Gradientes bioclimáticos. En: Ecología de la Cuenca
del Río Caura, Venezuela. I. Caracterización general. (Eds.) J. Rosales y O. Huber. Scientia Guaianae 6:
Merritt R. W. y Cummins, K. W. 1996. An introduction to the aquatic insects of North
America. Kendall/Hunt Publishing Company. Iowa. 862 p.
Mikola, J. y H. Setälä 1998. Productivity and trophic-level biomasses in a microbial-based soil food web. Oikos 82: 158-168.
Minshall, G. W. y J. N. Minshall 1977. Microdistribution of benthic invertebrates in a
rocky mountain (USA) stream. Hydrobiologia 55: 231-249.
Minshall, J. N. 1967. Role of allochthonous detritus in the trophic structure of a woodland spring brook community. Ecology 48:139-144.
Munn, M. D. y M. A. Brusven 1991. Benthic macroinvertebrate communities in
nonregulated waters of the Clearwater River, Idaho, U.S.A. Regulated rivers: Research & Management 6:1-11.

62
Naiman, R. J., Melillo, J. M., Lock, M. A., Ford, T. E. y S. R. Reice 1987. Longitudinal patterns of ecosystem processes and community structure in a subarctic river continuum. Ecology 68:1139-1156.
Naiman, R. J., Décamps, H., Pastor, J. y C. A. Johnston 1988. The potential
importance of boundaries to fluvial ecosystems. J. North Am. Benthological Soc. 7:289-306.
Nakamura, K. 2000. Comparación preliminar de la diversidad íctica en tres
morichales con distintos niveles de intervención humana, cuenca baja del río Caura, Estado Bolívar, Venezuela. Tesis para la obtención del grado de Licenciatura de la Escuela de Biología de la Universidad Central de Venezuela. 157 p.
Neckles, H. A., Murkin, H. R. y J. A. Cooper 1990. Influences of seasonal flooding on
macroinvertebrate abundance in wetland habitats. Freshwater Biology 23:311-322.
Neiff, J. J. 1990. Ideas para la interpretación ecológica del Paraná. Interciencia
15:424-441.
Neiff, J. J. 1999. El régimen de pulsos en ríos y grandes humedales de Sudamérica. 97-146. En: Trópicos sobre humedales subtropicales y templados de Sudamérica (Eds.). A. I. Malvárez, UNESCO, Montevideo, Uruguay.
Nessimian, J. L., Dorville, L. F. M., Sanseverino, A. M. y D. F. Baptista 1998.
Relation between flood pulse and functional composition of the macroinvertebrate benthic fauna in the lower Rio Negro, Amazonas, Brazil. Amazoniana 15: 35–50.
Newbold, J. D., O´Neill, R. V y W. Van Winkle 1981. Measuring nutrient spiraling in
streams. Can. J. Fish. Aqua. Sci. 38:860-863.
Newbold, J. D., O´Neill, R. V., Elwood, J. W. y W. Van Winkle 1982. Nutrient spiraling in streams: implications for nutrient limitation and invertebrate activity. American Naturalist 120: 628-652.
Paine, R. T. 1969a. A note on trophic complexity and community stability. American
Naturalist 103:91-93.
Paine, R. T. 1969b. The Pisaster-Tegula interaction: prey patches, predator food preference, and intertidal community structure. Ecology 50: 950-961.
Paine, R. T. 1992. Food-web analysis through field measurement of per capita
interaction strength. Nature 355:73-75.

63
Peckarsky, B. L. 1987. Mayfly cerci as defense against stonefly predation: deflection
and detection. Oikos 48: 161–170.
Peckarsky, B. L. 1983. Biotic interactions or abiotic limitations?. A model of lotic community structure. 303-323. En: Dynamics of Lotic Ecosystems. (Eds.) T. D. Fontaine III, y S. M. Bartell. Ann Arbor Science, Ann Arbor, MI.
Pennak, Robert W. 1978. Fresh-Water Invertebrates of the United States. A wiley-
Interscience Publication. 2 da. Edición. 803 p.
Peña, O. 1996. Hidrografía. En: Ecología de la Cuenca del Río Caura, Venezuela. I. Caracterización general. (Eds.) J. Rosales y O. Huber. Scientia Guaianae 6:29-33.
Peña, O. y O. Huber. 1996. Características geográficas generales. En: Ecología de la
Cuenca del Río Caura. (Eds.) Rosales, J. y O. Huber. Scientia Guaianae 6: 4–10.
Pérez, L. E. 1984. Uso del habitat por la comunidad de peces de un río tropical
asociado a un bosque. Mem. Soc. Cienc. Nat. La Salle 121(44):143-162.
Petr, T., 1970. Macroinvertebrates of flooded trees in the man-made Volta Lake (Ghana) with special reference to the burrowing mayfly Povilla adusta Navas. Hydrobiologia 36: 373–98.
Peters, W. L. 1971. A revision of the Leptophlebiidae of the West Indies
(Ephemeroptera). Smithsonian Contributions to Zoology 62:1-48.
Petts, G. E. y Amoros, C. (Eds.) 1996. Fluvial Hydrosystems. Chapman & Hall. Londres, 306 p.
Poff, N. L. 1997. Landscape filters and species traits: towards mechanistic
understanding and prediction in stream ecology. J. North Amer. Benthol. Soc. 16: 391–409.
Poi de Neiff, A., y R. Carignan 1997. Macroinvertebrates on Eichhornia crassipes
roots in two lakes of the Paraná River floodplain. Hidrobiologia 345:185-196.
Polis, G. A. y S. D. Hurd 1996. Linking marine and terrestrial food webs: allochthonous input from the Ocean supports high secondary productivity on small islands and coastal land communities. American Naturalist 147: 396-423.
Poole, G. C. 2002. Fluvial landscape ecology: addressing uniqueness within the river
discontinuum. Freshwater Biology 47:641-660.

64
Pringle, C. M., Naiman, R. J., Bretschko, G., Karr, J. R., Oswood, M. A., Webster, J. R., Welcomme, R. L. y M. J. Winterbourn 1988. Patcht dynamics of lotic systems: the stream as a mosaic. Journal North American Benthological Society 7:503-524.
Puthz, V. 1975. Eine neue Caenidengattung aus dem Amazonasgebiet (Insecta:
Ephemeroptera: Caenidae). Amazoniana 5(3):411-415.
Rabeni, C. F. y G. W. Minshall 1977. Factors affecting micro-distribution of stream benthic insects. Oikos 29: 33-43.
Rae, J. G. 1985. A multivariate study of resource partitioning in soft bottom lotic
Chironomidae. Hydrobiologia 126: 275-285.
Raffaelli, D. G. y S. J. Hall 1992. Compartments and predation in estuarine food web. J. Anim. Ecol. 61: 551-560.
Ramírez, A., Paaby, P., Pringle, C. M. y G. Aguero 1998. Effect of habitat type on
benthic macroinvertebrates in a tropical lowland stream, Costa Rica. Revista de Biología Tropical 46: 201-213.
Ramírez A. y C. M. Pringle 2001. Spatial and temporal patterns of invertebrate drift in
streams draining a Neotropical landscape. Freshwater Biology 46:47-62.
Reice, S. R. 1980. The role of substratum in benthic macroinvertebrate microdistribution and litter decomposition in a woodland stream. Ecology 61:580–590.
Reice, S. R., Wissmar, R. C. y R. J. Naiman 1990. Disturbance regimes, resilience,
and recovery of animal communities and habitats in lotic ecosystems. Environmental Biology 14(5):647-59.
Resh, V. H., Brown, A. V., Covich, A. P., Gurtz, M. E., Li, H. W., Minshall, G. W.,
Reice, S. R., Sheldom, A. L., Wallace, J. B. y R. C. Wissmar 1988. The role of disturbance in stream ecology. J. North Am. Benthol. Soc. 7:435-455.
Reiss, F. 1974. Vier neue Chironomus-Arten (Chironomidae, Diptera) und ihre
ökologische Bedeutung für die Benthosfauna zentralamazonischer Seen und Überschwemmungswälder. Amazoniana 5: 3–23.
Reiss, F. 1977. Qualitative and quantitative investigations on the macrobenthic fauna
of Central Amazon lakes. I. Lago Tupé, a black water lake on the lower Rio Negro. Amazoniana 6(2):203-236.
Richards, K., Brasington, J. y F. Hughes 2002. Geomorphic dynamics of floodplains:
ecological implications and a potential modeling strategy. Freshwater Biology 47:559-579.

65
Richardson, J. S. 1991. Seasonal food limitation of detritivores in a montane stream:
an experimental test. Ecology 72:873-887.
Ricklefs, R. E. 1987. Community diversity: relative roles of local and regional processes. Science 235:167-171.
Ricklefs, R. E. y R. E. Latham, 1993. Global diversity patterns in mangrove floras.
215-219. En: Species diversity in ecological communities. (Eds.) R. E. Ricklefs y D. Schluter. Univ. of Chicago Press.
Rincón, H. y Y. Estanga 1996. Geología. En: Ecología de la Cuenca del Río Caura,
Venezuela. I. Caracterización general. (Eds.) J. Rosales y O. Huber. Scientia Guaianae 6: 20-24.
Robinson, C. T., Tockner, K. y J. V. Ward 2002. The fauna of dynamic riverine
landscapes. Freshwater Biology 47:661-677. Rodríguez , M. A. y W. M. Lewis, Jr. 1997. Structure of fish assemblages along
environmental gradients in floodplain lakes of the Orinoco River. Ecological Monographs, 67(1):109–128.
Rodríguez-Olarte, D., Taphorn B. D., Lasso-Alcalá, O. y C. Vispo 2003. Fishes of the
Lower Caura River, Orinoco Basin, Venezuela. En: Plants and vertebrates of the Caura´s riparian corridor: their biology, use and conservation. (Eds.) C. Vispo y C. Knab-Vispo. Scientia Guaianae 12:181-222.
Roldán, G. 1988. Guía para el estudio de los macroinvertebrados acuáticos del
Departamento de Antioquia. Universidad de Antioquia, Fondo FEN, Colombia. Bogotá. 217 p.
Rosales, J. 1996. Vegetación: Bosques ribereños. En: Ecología de la Cuenca del Río
Caura, Venezuela I. Caracterización General. (Eds.) J. Rosales, y O. Huber. Scientia Guaianae 6:66-69.
Rosales, J. 2000. An ecohydrological approach for riparian forest biodiversity
conservation in large tropical rivers. Unpublished PhD Thesis, the University of Birmingham.
Rosales, J., Bradley, C., Petts, G. y D. Gilvear. 1999. A conceptual model of
hydrogeomorphological-vegetation interactions within confluence zones of large tropical rivers, Orinoco, Venezuela. Hydrological and Geochemical Processes in Large Scale River: 1-7.
Rosales, J., Conrad, V., Dezzeo, N., Blanco-Belmonte, L., Knab-Vispo, C., González,
N., Daza, F., Bradley, C., Gilvear, D., Escalante, G., Chacón N. y Geoffrey Petts 2002. Ecohydrology of Riparian Forest in the Orinoco River Basin. 93-

66
110. En: The Ecohydrology of South America Rivers and Wetlands. (Ed.) M. E. McClain. and Wetlands. (Ed.) M. Mc.Clain. Publicación Especial #6 de International Association of Hydrological Sciences.
Rosemond, Amy D., Pringle, C. M., Ramírez, Alonso y Michael J. Paul 2001. A test of
top-down and bottom-up control in a detritus-based food web. Ecology 82(8): 2279–2293.
Rosenberg, D. M., Davies, I. J., Cobb, D. G. y A. P. Wiens 1997. (unpublished).
Protocols for Monitoring Biodiversity: Benthic Macronivertebrates in Fresh Waters. Fisheries and Oceans Canada. http://www.eman-rese.ca/eman/ecotools/protocols/freshwater/benthics/benthic_fresh_e.pdf
Rosi-Marshall, E. J. y J. B. Wallace 2002. Invertebrate food webs along a stream
resource gradient. Freshwater Biology 47:129-141.
Schulze, E. D. y H. A. Mooney 1993. Ecosystem function of biodiversity: a summary. 497-510. En: (Eds.) E.-D. Schulze y H. A. Mooney. Biodiversity and ecosystem function. Springer-Verlag, Berlin, Germany.
Siegfried, C. A. y A. W. Knight 1977. The effects of a washout in a sierra foothill
stream. Am. Midl. Nat. 98: 200-207.
Stanley, E. H., Fisher, S. G. y N. B. Grimm 1997. Ecosystem expansion and contraction in streams. BioScience 47:427-435.
Statzner, B. y B. Higler 1985. Questions and comments on the River Continuum
Concept. Can J. Fish. Aquat. Sci. 42:1038-1044.
Statzner, B., Gore, J. A. y V. Resh 1988. Hydraulic streams ecology: observed patterns and potential applications. J. North Amer. Benthol. Soc. 7:307-360.
Tedesco, P. A., Oberdorff, T., Lasso, Carlos A., Zapata, Milton y B. Hugueny 2005.
Evidence of history in explaining diversity patterns in tropical riverine fish. J. Biogeogr. 32: 1899–1907.
Tockner, K., F. Malard y J. V. Ward 2000. An extension of the flood pulse concept.
Hydrological Process 14:2861-2883. Tonn, W. M., Magnuson, J. J., Rask, M. y J. Toivonen 1990. Intercontinental
comparison of small-lake fish assemblages: the balance between local and regional processes. Am. Nat. 136: 345–375.
Traver, J. R. 1943. New Venezuelan mayflies. Boletín de Entomología Venezolana
2:79-98.

67
Trivinho-Strixino, S., Correia, L. C. S. y K. Sonoda 2000. Phytophilous chironomidae (diptera) and other macroinvertebrates in the ox-bow Infernão Lake (Jataí Ecological Station, Luiz Antônio, SP, Brazil). Rev. Bras. Biol. 60: 527-535.
Tuckner, K., Mallard, F. y J. V. Ward 2000. An extension of the flood pulse concept.
Hydrological Process 14:2861-2883.
Vannote, R. L., Minshall, G. W., Cummins, K. W., Sedell, J. R. y C. E. Cushing 1980. The river continuum concept. Can. J. Fish. Aqua. Sci. 37:130-137.
Vannote, R. L. y B. W. Sweeney 1980. Geographic Analysis of Thermal Equilibria: A
conceptual model for evaluating the effect of natural and modified thermal regimes on aquatic insect communities. American Naturalist 115(5):667-695.
Vargas, H. y J. Rangel 1996. Hidrología y sedimentos. En: Ecología de la Cuenca del Río Caura, Venezuela. I. Caracterización general. (Eds.) J. Rosales y O. Huber. Scientia Guaianae 6: 34-39.
Vinson M. R. y C. P. Hawkins 1998. Biodiversity of stream insects: variation at local,
basin and regional scales. Annu. Rev. Entomol. 43: 271-93.
Vásquez, E., Sánchez, L. Pérez, L. E. y Blanco, L. 1990. Estudios hidrobiológicos y piscicultura en algunos cuerpos de agua (ríos, lagunas, embalses) de la cuenca baja del Río Orinoco. En: El Río Orinoco como Ecosistema. (Eds.) F. Weibezahn., H. Alvarez y Lewis Jr., W. M. Galac, Impresos Rubel. Caracas, 450 p.
González, N. y C. Vispo 2003. Aspects of the diets and feeding ecologies of fish
from nine floodplain lakes of the Lower Caura, Venezuelan Guayana. En: Plants and vertebrates of the Caura´s riparian corridor: their biology, use and conservation. (Eds.) C. Vispo y C. Knab-Vispo. Scientia Guaianae 12: 329-366.
Usinger, Robert L. 1963. Aquatic Insects of California. University of California Press.
California. 508 p.
Walde, S. J. y R. W. Davies 1984. Invertebrate predation and lotic prey communities: Evaluation of in situ enclosure/exclosure experiments. Ecology 65: 1206-1213.
Wallace J. B., Eggerton, S. L, Meyer, J. L. y J. R. Webster 1997. Multiple trophic
levels of a forest stream linked to terrestrial litter inputs. Science 277: 102–104.
Wallace, J. B. y J. R. Webster 1996. The role of macroinvertebrates in stream ecosystem function. Ann. Rev. Entomol. 41: 115-139.

68
Ward, J. V. 1989. The four dimensional nature of lotic ecosystems. J. North Amer. Benthol. Soc. 8:2-8.
Ward, J. V. y K. Tockner 2001. Biodiversity: towards a unifying theme for river
ecology. Freshwater Biology 46:807-819.
Ward, J. V., Tockner, K., Arscott, D. B. y C. Claret 2002. Riverine landscape diversity. Freshwater Biology 47:517-539.
Ward, J. V. y J. A. Standford 1983. The serial discontinuity concept of lotic
ecosystems. 347-356. En: Dynamic of Lotic Ecosystems. (Eds.) T.D. Fontaine y S.M. Bartell. Ann Arbor Sciences Publishers, Ann Arbor, MI.
Ward, J. V. y J. A. Standford 1995a. Ecological connectivity in alluvial river
ecosystems and its disruption by flow regulation. Regulated Rivers: Research and Management 10: 159-168.
Ward, J. V. y J. A. Standford 1995b. Ecological connectivity in alluvial river
ecosystems and its disruption by flow regulation. Regulated Rivers: Research and Management 11: 105-119.
Welcomme, R. L. 1985. Pesca fluvial. FAO Fish. Tech. Pap. 265. Roma, 330 p.
Welcomme, R. L. 1990. Status of fisheries in South American rivers. Interciencia 15:
337–345
Wetzel, R. G. 1983. Limnology. 2nd ed. Saunders College Publishing. 767 p.
Whiles, M. R. y B. S. Goldowitz 2001. Hydrologic influences on insect emergence production from central Platte River wetlands. Ecological Applications 11: 1829-1842.
Whittaker, R. H. 1960. Vegetation of the Siskyou Mountains, Oregon and California.
Ecological Monographs 30:279-338.
Wiens, J. A. 2002. Riverine landscapes: taking landscape ecology into the water. Freshwater Biology 47:501-515.
Wiggins, G. B. 1977. Larvae of the North American Caddisfly Genera. Toronto
University Press, Toronto. 401 p.
Winterbourn, M. J. 1981: The use of aquatic invertebrates in studies of stream water quality. Water and Soil Publication 22: 5–16.
Williams, D. D. y B. W. Feltmate 1992. Aquatic Insects. CAB International. 358 p.

69
Williams, D. D. y N. E. Williams 1999. Canadian Springs: postglacial development of the invertebrate fauna. 447-467. En: Invertebrates in Freshwater Wetlands of North America: Ecology and Management (Eds.) D. P. Batzer, R. Rader, y S. A. Wissinger. John Wiley & Sons, N.Y. 1100 p.
Wissinger S. A. 1999. Ecology of wetland invertebrates: synthesis and applications
for conservation and management. 1043–1086. En: Invertebrates in Freshwater Wetlands of North America: Ecology and Management (Eds.) D.P. Batzer, R.B. Rader y S.A. Wissinger. John Wiley and Sons, New York.
Zimmermann, E. M. y R. G. Death 2002. Effect of substrate stability and canopy
cover on stream invertebrate communities. New Zealand Journal of Marine and Freshwater Research 36: 537-545.

ANEXOS

TABLAS

Tabla 1. Lista de macroinvertebrados registrados en varios sustratos de los transectos estudiados en el Bajo Caura. (Abreviaciones: PAT= Patiquin, CEJ= Cejal, POZ= Pozo Rico, GAR= Garzones, CHI= Chiribital, CAR= Caramacatico, NAP= Naparaico, BRA= Brava, ARI= Aricagua; A=Sedimento, B=Madera, C=Hojas verdes del bosque de inundaciòn, roc=Pozo de rocas, vga=Vegetación acuática, hoj=Hojarasca, X=Presencia, - =muestra no tomada. *=Nuevos registros para Venezuela, **=Nuevos registros geográficos para Venezuela y ***=Nuevo registro para el mundo).
MATO SIPAO CAURA Otros substratos
PAT CEJ POZ GAR CHI CAR NAP BRA ARI roc vga hoj TAXA COD A B C A B C A B C A B C A B C A B C A B C A B C A B C Arthropoda (Insecta) Ephemeroptera Baetidae Callibaetis sp1 CAL 1 - - - X - - X Caenidae Brasilocaenis irmleri Puthz B 1 X - - X X X - X - X - X X X X X X Polymitarcyidae Campsurus notatus CAM 1 - - - X - X - X X X Asthenopus curtus E 4 - X X X - X - X - X X X X X Leptophebiidae Simothraulopsis demerara Peters, Peters &Savage E 9 - - - - - X Leentvaria palpalis Demoulin* E 10 - - - - - X Microphlebia surinamensis Peters E 12 - - - - - X Hydrosmilodon sp.n* E 11 - - - - - X Leptophlebiidae g.n.*** E 13 - - - - - X Trichoptera Odontoceridae sp1 BRA 1 - - X - X - X - X Odontoceridae sp1 BRA 3 X Hydroptilidae sp1 HYT 1 - - - X - - X X X X X X Hydroptilidae sp2 HYT 2 - - - X - - X X X X Hydroptilidae sp3 HYT 3 - - - - - X Hydroptilidae sp4 HYT 4 - - - - - X Hydroptilidae sp5 HYT 5 X Hydropsychidae Macronema sp1 TRO 1 - - - X - - X X X Macronema sp2 TRO 2 - - - - - X Synoestropsis sp1 TRO 3 - - - - - X Synoestropsis sp2 TRO 4 - - - - - X Polycentropidae Cyrnellus sp1 CYR 1 - X - X X X - X - X - X X X X Cyrnellus sp2 CYR 2 - X X - X X X - X - X - X X X X X Plecoptera
(Continuación Tabla 1)
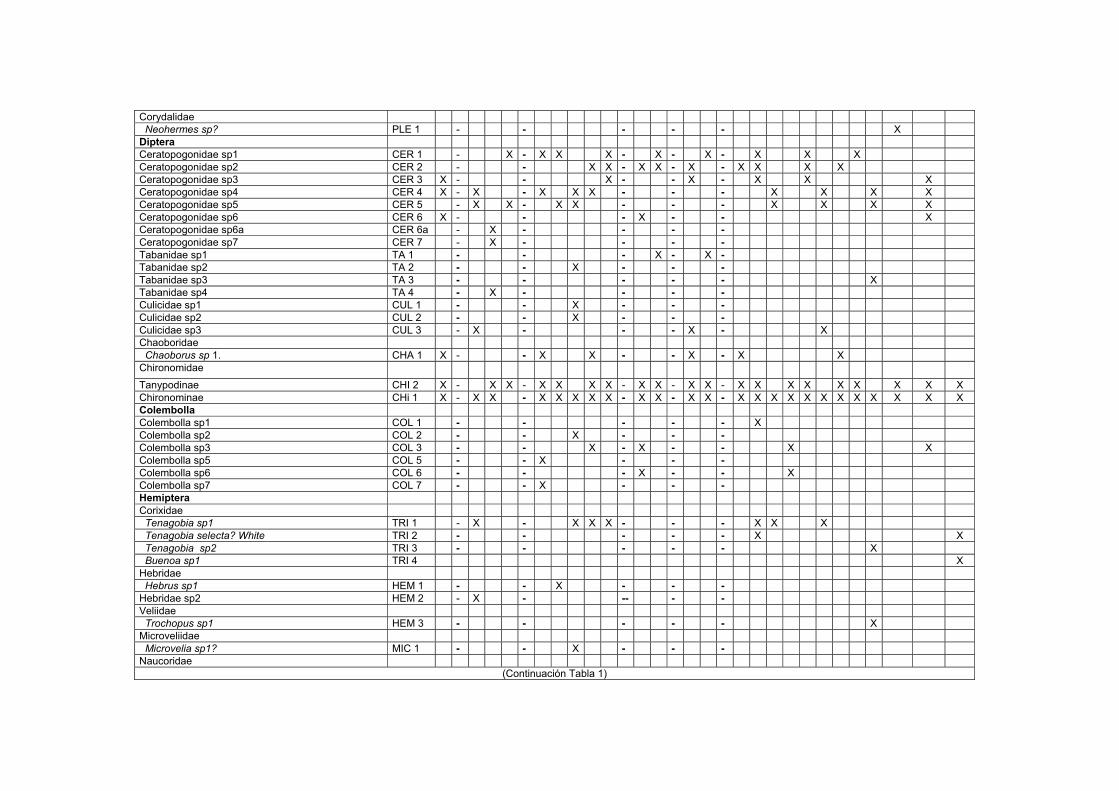
Corydalidae Neohermes sp? PLE 1 - - - - - X Diptera Ceratopogonidae sp1 CER 1 - X - X X X - X - X - X X X Ceratopogonidae sp2 CER 2 - - X X - X X - X - X X X X Ceratopogonidae sp3 CER 3 X - - X - - X - X X X Ceratopogonidae sp4 CER 4 X - X - X X X - - - X X X X Ceratopogonidae sp5 CER 5 - X X - X X - - - X X X X Ceratopogonidae sp6 CER 6 X - - - X - - X Ceratopogonidae sp6a CER 6a - X - - - - Ceratopogonidae sp7 CER 7 - X - - - - Tabanidae sp1 TA 1 - - - X - X - Tabanidae sp2 TA 2 - - X - - - Tabanidae sp3 TA 3 - - - - - X Tabanidae sp4 TA 4 - X - - - - Culicidae sp1 CUL 1 - - X - - - Culicidae sp2 CUL 2 - - X - - - Culicidae sp3 CUL 3 - X - - - X - X Chaoboridae Chaoborus sp 1. CHA 1 X - - X X - - X - X X Chironomidae
Tanypodinae CHI 2 X - X X - X X X X - X X - X X - X X X X X X X X X Chironominae CHi 1 X - X X - X X X X X - X X - X X - X X X X X X X X X X X X Colembolla Colembolla sp1 COL 1 - - - - - X Colembolla sp2 COL 2 - - X - - - Colembolla sp3 COL 3 - - X - X - - X X Colembolla sp5 COL 5 - - X - - - Colembolla sp6 COL 6 - - - X - - X Colembolla sp7 COL 7 - - X - - - Hemiptera Corixidae Tenagobia sp1 TRI 1 - X - X X X - - - X X X Tenagobia selecta? White TRI 2 - - - - - X X Tenagobia sp2 TRI 3 - - - - - X Buenoa sp1 TRI 4 X Hebridae Hebrus sp1 HEM 1 - - X - - - Hebridae sp2 HEM 2 - X - -- - - Veliidae Trochopus sp1 HEM 3 - - - - - X Microveliidae Microvelia sp1? MIC 1 - - X - - - Naucoridae
(Continuación Tabla 1)

Pelocoris sp1 PEC 1 - - X - - - X Coleoptera Dytiscidae Pachydrus sp1 PAC 1 X Desmopachria sp1 DES 1 - - - - - X Laccophilus sp1 (A) LAC 1 - - - - - X X Laccophilus sp2(Ll) LAC 2 X Laccophilus sp3 (L) LAC 3 X Dytiscidae sp1 (A) DYS 1 - - X - - - Dytiscidae sp2 (L) DYC 1 - - X - - - Dytiscidae sp3 (A) DYC 2 - X - - - - Noteridae Hydroanthus sp1 HYR 1 - - - - - X Mesonoterus sp1 NOT 1 - X - - - - X Noteridae sp1 NOT 2 - - - - - X Hydrophilidae Gyriinus sp1 (L) GYR 1 - - - - - X Gyretes sp1 (L) GYR 2 X Helochares sp 1 (A) HEL 1 - - - - - X Hydrochus sp1 HUC 1 - X - X X - - - X Hydrochus sp2 HUC 2 - X - X X - - - X Hydrochus sp3 HUC 3 - - X X - - - Paracymus sp1? PAR 1 - - - - - X Paracymus sp2? PAR 2 - - - - - X Paracymus sp3? PAR 3 - - - - - X Hydrophilidae sp1 n.i. (A) HYP 1 - X - X - X - - X Hydrophilidae sp2 n.i. (A) HYP 2 - - X - - - Hydrophilidae sp3 n.i. (A) HYP 3 - - X - - - Hydrophilidae sp4 n.i. (A) HYP 4 - - X - - - Hydrophilidae sp5 n.i. (A) HYP 5 - X - - - - X Dryopidae sp1 (L) DRYO 1 X - - X - - - X X Elmidae sp1 (A) ELM 1 - - X - X - X - X X X X Elmidae sp2 (L) ELM 2 - - - - - X X Elmidae sp3 (A) ELM 3 - - - - - X Elmidae sp4 (L) ELM 4 Elmidae sp5 (L) ELM 5 X X Elmidae sp6 (L) ELM 6 - X - - - - Helodidae sp1 (L) HED 1 - X - X X X - X - - X Staphilinidae sp1 (A) STA 1 - X - X X - - - X Staphilinidae sp2 (A) STA 2 - X - - - - Staphilinidae sp3 (A) STA 3 - - X - - - Staphilinidae sp4 (A) STA 4 - X - - - - Thysanura sp1 THY 1 - - - - X - Neuroptera -
(Continuación Tabla 1)
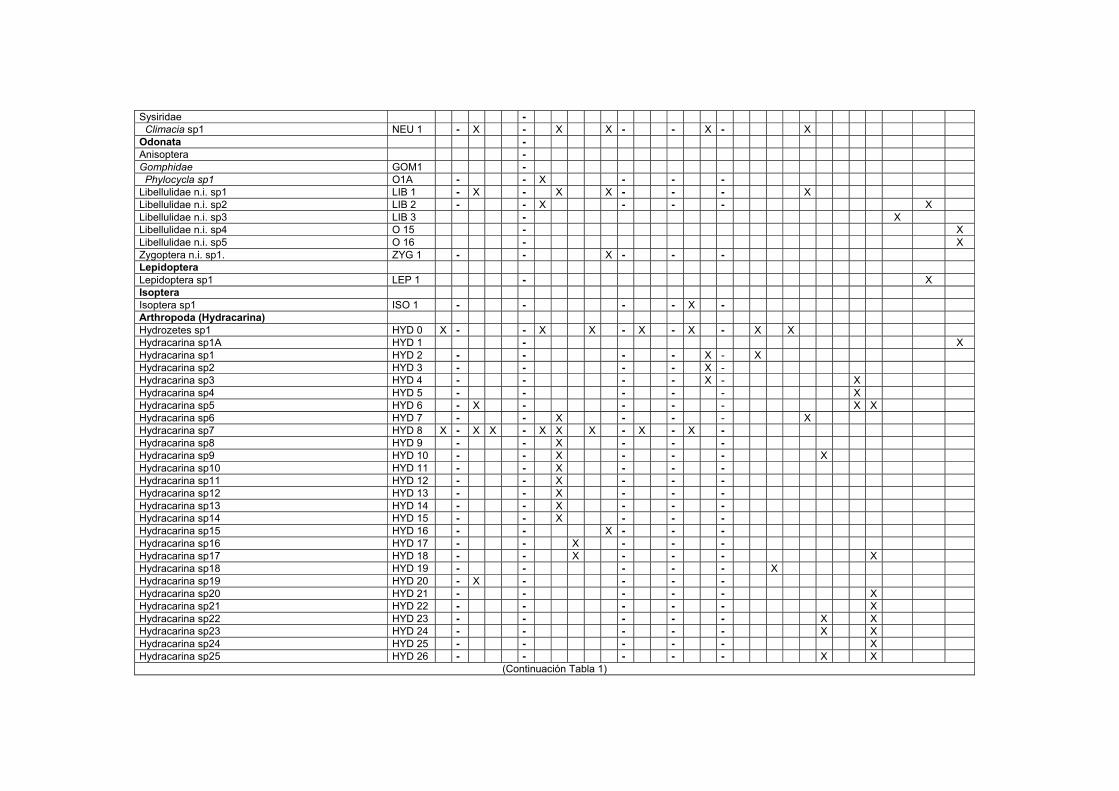
Sysiridae - Climacia sp1 NEU 1 - X - X X - - X - X Odonata - Anisoptera - Gomphidae GOM1 - Phylocycla sp1 O1A - - X - - - Libellulidae n.i. sp1 LIB 1 - X - X X - - - X Libellulidae n.i. sp2 LIB 2 - - X - - - X Libellulidae n.i. sp3 LIB 3 - X Libellulidae n.i. sp4 O 15 - X Libellulidae n.i. sp5 O 16 - X Zygoptera n.i. sp1. ZYG 1 - - X - - - Lepidoptera Lepidoptera sp1 LEP 1 - X Isoptera Isoptera sp1 ISO 1 - - - - X - Arthropoda (Hydracarina) Hydrozetes sp1 HYD 0 X - - X X - X - X - X X Hydracarina sp1A HYD 1 - X Hydracarina sp1 HYD 2 - - - - X - X Hydracarina sp2 HYD 3 - - - - X - Hydracarina sp3 HYD 4 - - - - X - X Hydracarina sp4 HYD 5 - - - - - X Hydracarina sp5 HYD 6 - X - - - - X X Hydracarina sp6 HYD 7 - - X - - - X Hydracarina sp7 HYD 8 X - X X - X X X - X - X - Hydracarina sp8 HYD 9 - - X - - - Hydracarina sp9 HYD 10 - - X - - - X Hydracarina sp10 HYD 11 - - X - - - Hydracarina sp11 HYD 12 - - X - - - Hydracarina sp12 HYD 13 - - X - - - Hydracarina sp13 HYD 14 - - X - - - Hydracarina sp14 HYD 15 - - X - - - Hydracarina sp15 HYD 16 - - X - - - Hydracarina sp16 HYD 17 - - X - - - Hydracarina sp17 HYD 18 - - X - - - X Hydracarina sp18 HYD 19 - - - - - X Hydracarina sp19 HYD 20 - X - - - - Hydracarina sp20 HYD 21 - - - - - X Hydracarina sp21 HYD 22 - - - - - X Hydracarina sp22 HYD 23 - - - - - X X Hydracarina sp23 HYD 24 - - - - - X X Hydracarina sp24 HYD 25 - - - - - X Hydracarina sp25 HYD 26 - - - - - X X
(Continuación Tabla 1)

Hydracarina sp26 HYD 27 - - - - - X X Hydracarina sp27 HYD 28 - - - - - X X Hydracarina sp28 HYD 29 - - - - - X X Hydracarina sp29 HYD 30 - X - - - - Hydracarina sp30 HYD 31 - - - - - X X Hydracarina sp31 HYD 32 - - - - - X Hydracarina sp32 HYD 33 - - - - - X Hydracarina sp33 HYD 34 - - - - - X X Hydracarina sp34 HYD 35 - - - - - X Hydracarina sp35 HYD 36 - - - - - X X Hydracarina sp36 HYD 37 - - - - - X Hydracarina sp37 HYD 38 X - - - - - X Hydracarina sp38 HYD 39 - - - - - X Hydracarina sp39 HYD 40 - - X - - X - X Hydracarina sp30 HYD 41 - X X - - X - - Arthropoda (Arachnida) - Arachnidae sp1 ARA 1 - X - - - - Arachnidae sp2 ARA 2 - - X - - - Arachnidae sp3 ARA 3 - - - - - X Arachnidae sp4 ARA 4 - X X X - - X - Arthropoda (Crustacea) Ostracoda Darwinula sp1 OS 0 X - X Ostracoda sp1 OS 1 X - - X - X - - X Ostracoda sp2 OS 2 - - X - - - X Pelocypris sp1? OS 3 X - X - X X - X - X - X X Clamydotheca sp1? OS 3a - - X X - X - - X X Cypridae sp1 OS 4 X - X - X X - X - X - X X X Cypridae sp2 OS 5 - - X X - X - X - X X Ostracoda sp3 OS 6 - - - - X Ostracoda sp4 OS 7 - - - - - X Ostracoda sp5 OS 8 X - - - X - - X X Ostracoda sp6 OS 9 X - - - X - X - X X Ostracoda sp7 OS 11 X - - X - - - Ostracoda sp8 OS 12 - X - - - - Ostracoda sp9 OS 13 - - - - - X Copepoda Cyclopoidea COP 1 X - X X - X X X X X - X X - X X - X X X X X X X X Calanoidea COP 2 - X - X X - X - X X - X X X Branchiopoda Cyclestheria hislopi Baird CYC 1 - X - X X - - - X X X Branchiopoda sp1 EUL 1 - - X - - - Cladocera Macrothricidae
(Continuación Tabla 1)

Macrothricidae sp1 MAC 2 - X - - - - Illyocryptus sp1 MAC 3 X - X X - X - X - X - X X X X X Macrothricidae sp2 MAC 4 - X - - - - X Illyocryptus sp2 MAC 5 X - - X X - - - X Bosminidae Bosmina sp1 BOS 1 X - - X X X - - - X X X Bosminopsis sp1 BOS 2 - X Bosminopsis sp1 BOS 3 - - - X - - Moinidae Moina sp1 MOI 1 - - X - - - X Chydoridae Alona sp1 ALO 1 - - X - - - Chydoridae sp1 CHY 1 X - - X - - - X Chydoridae sp2 CHY 2 - - - - - X Chydoridae sp3 CHY 3 - X - X - - - X X X X Chydoridae sp4 CHY 3a - - - - - X Chydoridae sp5 CHY 4 - - - - - X X Chydoridae sp6 CHY 5 - - X - - - X X Chydoridae sp7 CHY 6 X - - X - - - X Chydoridae sp8 CHY 7 - X X - - - - Chydoridae sp9 CHY 8 - - - - - X Chydoridae sp10 CHY 9 - - - - - X X Chydoridae sp10A CHY 10 - X Camptocercus sp1 CHY 11 - X - - - - Chydoridae sp11 CHY 12 - - X - - - X Chydoridae sp12 CHY 13 X - - - - - Chydoridae sp13 CHY 15 - X - - - - Chydoridae sp14 CHY 16 - X - X - - - Sididae Diaphanosoma sp1 DIA 1 X - X - X X - X - - X X X X Daphnidae Daphnidae sp1 DAP 1 - X - - - - Dapnidae sp2 DAP 2 - - X - - - Daphnidae sp3 DAP 3 X - X - - - - Daphnidae sp4 DAP 4 - - - - - X Daphnidae sp5 DAP 5 - - - - - X Decapoda Palaemonidae Macrobrachium amazonicum Heller JEL 1 - - - - - X Euryrrhyncus amazoniensis?** JEL 2 - - - - - X Nematoda Nematoda sp1 ME 1 X - X X X - X X X X X - X X - X X - X X X X X X X X X Nematoda sp2 ME 2 X - X X - X X X - X X - X X - X X X X X X X X X X Nematoda sp3 ME 3 X
(Continuación Tabla 1)

Annelida (Oligochaeta) Naididae sp1A NAI 1 X X Pristina sp1 NAI 2 X - X X X - X X X X X - X - X X - X X X X X X X X X Naididae sp1 NAI 3 - - - - - X X X X X Naididae sp2 NAI 4 - - - X - X - X X X Naididae sp3 NAI 5 X - X - X - - - X X X Naididae sp4 NAI 6 X - X X X - X X X X X - X X - X X - X X X X X X X X X X Naididae sp5 NAI 6a X - X - X X - X - X - X X X Dero sp1 NAI 7 - X - X X - - - X Naididae sp6 NAI 8 - - X - - - X Dero sp2 NAI 9 - - X X - - - X X Naididae sp7 NAI 10 - - X - - - Naididae sp8 NAI 11 - X Dero sp3 NAI 12 - X Dero vagus? NAI 13 - X Naididae sp9 NAI 14 - X Lumbriculidae sp1 LUM 1 - X - - - - Annelida (Hirudinea) Helobdella sp1 Hi 1 - X - X - X X - X - X X Helobdella stagnalis (Limnaeus) Hi 2 - X - X X - X - X - X X X X Helodeblla sp2A Hi 3 X X Helodeblla sp2 Hi 4 - - X - X - X - X X Helobdella sp3 Hi 5 - - - - X - X X Helobdella sp4 Hi 6 - - - X - - X Helobdella sp5 Hi 7 X X Helobdella triserialis? (Blanchard) Hi 8 X Platyhelminthes (Turbellaria) Turbellaria sp 1 TU 1 - X - X X - X X - X - X X X X Turbelaria sp2 TU 2 - - - X - - Turbelaria sp3 TU 3 X Coelenterata Hydra sp1 COE 1 - X - X - - - X Mollusca Gasteropoda n.i. GAS 1 X Ancylidae sp1 ANC 1 - X - X X X - X - - Pelecypoda sp1 PEL1 1

Tabla 1A. Distribución, abundancia y biomasa de los Macroinvertebrados en el sedimento de nueve transectos del Bajo Caura en aguas bajas. (Abreviaciones: COD= Códigos, PAT= Laguna Patiquin, CEJ= L. Cejal, POZ= L. Pozo Rico, NAP= L. Naparaico, BRA= L. Brava, ARI= L. Aricagua, GAR= L. Garzones, CHI= L. Chiribital, CAR= L. Caramacatico. CNT= Centro de la laguna, ORI= Orilla de la laguna, MAT= Orilla Caño Mato, SIP= Orilla Caño Sipao).
MATO CAURA EPOCA DE SEQUÍA
PAT CEJ POZ NAP BRA ARI TAXA COD CNT ORI MAT CNT ORI CNT ORI MAT CNT ORI CNT ORI CNT ORI
Insecta Ephemeroptera Polymitarcyidae Campsurus sp CAM 1 Caenidae Brasilocaenis irmleri B 1 Diptera Chironominae CHI 1 x x x x x x x Stenochironomus sp CER 1 Tanypodinae CHI 2 x x x Chaoborus sp1 CHA 1 x x x x x Ceratopogonidae sp2 CER 2 x Ceratopogonidae sp3 CER 3 x Ceratopogonidae sp4 CER 4 x x Ceratopogonidae sp6 CER 6 x Ceratopogonidae sp6a CER 6a Ceratopogonidae sp7 CER 7 Tabanidae sp4 TAB 4 Collembola Colembolla sp3 COL 3 Colembolla sp5 COL 5 x Colembolla sp6 COL 6 x Colembolla sp7 COL 7 Hemiptera Tenagobia sp1 TRI 1 Coleoptera Elmidae sp4 (L) ELM 4
(continuación Tabla 1A...)

MATO CAURA EPOCA DE SEQUÍA
PAT CEJ POZ NAP BRA ARI TAXA COD CNT ORI MAT CNT ORI CNT ORI MAT CNT ORI CNT ORI CNT ORI
Elmidae sp5 (L) ELM 5 x x Elmidae sp6 (L) ELM 6 Dytiscidae sp3 (A) DYC 2 Odonata Anisoptera Gomphidae Phylocycla sp1 O1A x Libellulidae sp1 n.i. LIB 1 Isoptera ISO 1 Hydracarina Hydrozetes sp1 HYD 0 Hydracarina sp8 HYD 8 x Hydracarina sp16 HYD 16 x Hydracarina sp34 HYD 34 x Hydracarina sp35 HYD 35 Hydracarina sp36 HYD 36 x Hydracarina sp37 HYD 37 Hydracarina sp38 HYD 38 x x x Hydracarina sp39 HYD 39 x Hydracarina sp40 HYD 40 13 Hydracarina sp41 HYD 41 Arachnida Arachnidae sp4 ARA 4 Crustacea Ostracoda Ostracoda sp1 OS 1 Ostracoda sp2 OS 2 Pelocypris sp1 OS 3 x x x x x Clamydotheca sp1 OS 3a x x Cypridae sp1 OS 4 Cypridae sp2 OS 5 x x x Ostracoda sp3 OS 6
(continuación Tabla 1A...)

MATO CAURA EPOCA DE SEQUÍA
PAT CEJ POZ NAP BRA ARI TAXA COD CNT ORI MAT CNT ORI CNT ORI MAT CNT ORI CNT ORI CNT ORI
Ostracoda sp4 OS 7 Ostracoda sp5 OS 8 Ostracoda sp6 OS 9 x x x Ostracoda sp7 OS 11 Ostracoda sp8 OS 12 Ostracoda sp9 OS 13 Copepoda Cyclopoidea COP 1 x x x x x x x Calanoidea COP 2 x x x x x Cladocera Macrothricidae Illyocryptus sp1 MAC 3 Illyocryptus sp2 MAC 5 x x Bosminidae Bosmina sp.1 BOS1 x x x x x x Bosminopsis sp.1 BOS3 Moinidae Moina sp1 MOI 1 x x Chydoridae Alona sp1 ALO 1 Chydoridae sp1 CHY 1 x Chydoridae sp3 CHY 3 x x x Chydoridae sp4 CHY3a x Chydoridae sp6 CHY 5 x Chydoridae sp7 CHY 6 Chydoridae sp8 CHY 7 Camptocercus sp1 CHY 11 Chydoridae sp11 CHY12 Chydoridae sp12 CHY13 Chydoridae sp13 CHY15 Chydoridae sp14 CHY16 Sididae Diaphanosoma sp1 DIA 1 x x x x x x x
(continuación Tabla 1A...)
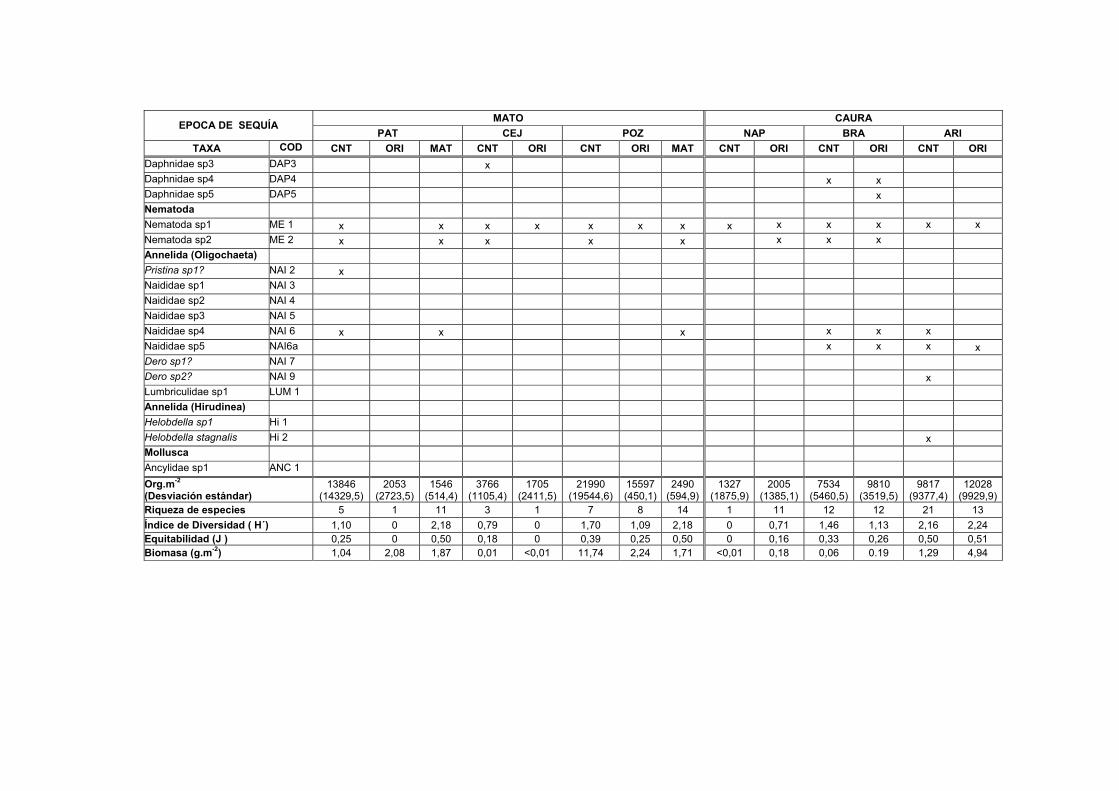
MATO CAURA EPOCA DE SEQUÍA
PAT CEJ POZ NAP BRA ARI TAXA COD CNT ORI MAT CNT ORI CNT ORI MAT CNT ORI CNT ORI CNT ORI
Daphnidae sp3 DAP3 x Daphnidae sp4 DAP4 x x Daphnidae sp5 DAP5 x Nematoda Nematoda sp1 ME 1 x x x x x x x x x x x x x Nematoda sp2 ME 2 x x x x x x x x Annelida (Oligochaeta) Pristina sp1? NAI 2 x Naididae sp1 NAI 3 Naididae sp2 NAI 4 Naididae sp3 NAI 5 Naididae sp4 NAI 6 x x x x x x Naididae sp5 NAI6a x x x x Dero sp1? NAI 7 Dero sp2? NAI 9 x Lumbriculidae sp1 LUM 1 Annelida (Hirudinea) Helobdella sp1 Hi 1 Helobdella stagnalis Hi 2 x Mollusca Ancylidae sp1 ANC 1 Org.m-2 (Desviación estándar)
13846 (14329,5)
2053 (2723,5)
1546 (514,4)
3766 (1105,4)
1705 (2411,5)
21990 (19544,6)
15597 (450,1)
2490 (594,9)
1327 (1875,9)
2005 (1385,1)
7534 (5460,5)
9810 (3519,5)
9817 (9377,4)
12028 (9929,9)
Riqueza de especies 5 1 11 3 1 7 8 14 1 11 12 12 21 13 Índice de Diversidad ( H´) 1,10 0 2,18 0,79 0 1,70 1,09 2,18 0 0,71 1,46 1,13 2,16 2,24 Equitabilidad (J ) 0,25 0 0,50 0,18 0 0,39 0,25 0,50 0 0,16 0,33 0,26 0,50 0,51 Biomasa (g.m-2) 1,04 2,08 1,87 0,01 <0,01 11,74 2,24 1,71 <0,01 0,18 0,06 0.19 1,29 4,94

(Continuación .....Tabla 1A. Distribución, abundancia y biomasa de los Macroinvertebrados en el sedimento de nueve transectos del Bajo Caura en aguas bajas. (Abreviaciones: COD= Códigos, PAT= Laguna Patiquin, CEJ= L. Cejal, POZ= L. Pozo Rico, NAP= L. Naparaico, BRA= L. Brava, ARI= L. Aricagua, GAR= L. Garzones, CHI= L. Chiribital, CAR= L. Caramacatico. CNT= Centro de la laguna, ORI= Orilla de la laguna, MAT= Orilla Caño Mato, SIP= Orilla Caño Sipao).
SIPAO EPOCA DE SEQUÍA
GAR CHI CAR TAXA COD CNT ORI CNT ORI SIP CNT ORI SIP
Insecta Ephemeroptera Polymitarcyidae Campsurus spp CAM 1 x x Caenidae Brasilocaenis irmleri? B 1 Diptera Chironominae CHI 1 x x x x x x Stenochironomus sp CER 1 Tanypodinae CHI 2 x x x Chaoborus sp1 CHA 1 Ceratopogonidae sp2 CER 2 x Ceratopogonidae sp3 CER 3 Ceratopogonidae sp4 CER 4 Ceratopogonidae sp6 CER 6 x Ceratopogonidae sp6a CER 6a Ceratopogonidae sp7 CER 7 Tabanidae sp4 TAB 4 Collembola Colembolla sp3 COL 3 Colembolla sp5 COL 5 Colembolla sp6 COL 6 x Colembolla sp7 COL 7 Hemiptera Tenagobia sp1 TRI 1 Coleoptera Elmidae sp4 (L) ELM 4 Elmidae sp5 (L) ELM 5
(continuación Tabla 1A....)

SIPAO EPOCA DE SEQUÍA
GAR CHI CAR TAXA COD CNT ORI CNT ORI SIP CNT ORI SIP
Elmidae sp6 (L) ELM 6 Dytiscidae sp3 (A) DYC 2 Odonata Anisoptera Gomphidae Phylocycla sp1 O1A Libellulidae sp1 n.i. LIB 1 Isoptera ISO 1 Hydracarina Hydrozetes sp1 HYD 0 Hydracarina sp8 HYD 8 x x x x Hydracarina sp16 HYD 16 Hydracarina sp34 HYD 34 Hydracarina sp35 HYD 35 Hydracarina sp36 HYD 36 Hydracarina sp37 HYD 37 Hydracarina sp38 HYD 38 Hydracarina sp39 HYD 39 Hydracarina sp40 HYD 40 Hydracarina sp41 HYD 41 (Arachnida Arachnidae sp4 ARA 4 (Crustacea Ostracoda Ostracoda sp1 OS 1 x Ostracoda sp2 OS 2 Pelocypris sp1 OS 3 x x x Clamydotheca sp1 OS 3a Cypridae sp1 OS 4 Cypridae sp2 OS 5 x x x Ostracoda sp3 OS 6 Ostracoda sp4 OS 7
(continuación Tabla 1A...)

SIPAO EPOCA DE SEQUÍA
GAR CHI CAR TAXA COD CNT ORI CNT ORI SIP CNT ORI SIP
Ostracoda sp5 OS 8 x Ostracoda sp6 OS 9 x x x Ostracoda sp7 OS 11 Ostracoda sp8 OS 12 Ostracoda sp9 OS 13 Copepoda Cyclopoidea COP 1 x Calanoidea COP 2 x x Cladocera Macrothricidae Illyocryptus sp1 MAC 3 Illyocryptus sp2 MAC 5 Bosminidae Bosmina sp.1 BOS1 x Bosminopsis sp.1 BOS3 Moinidae Moina sp1 MOI 1 Chydoridae Alona sp1 ALO 1 Chydoridae sp1 CHY 1 Chydoridae sp3 CHY 3 Chydoridae sp4 CHY3a Chydoridae sp6 CHY 5 Chydoridae sp7 CHY 6 Chydoridae sp8 CHY 7 Camptocercus sp1 CHY 11 Chydoridae sp11 CHY12 Chydoridae sp12 CHY13 Chydoridae sp13 CHY15 Chydoridae sp14 CHY16 Sididae Diaphanosoma sp1 DIA 1
(continuación Tabla 1A...)
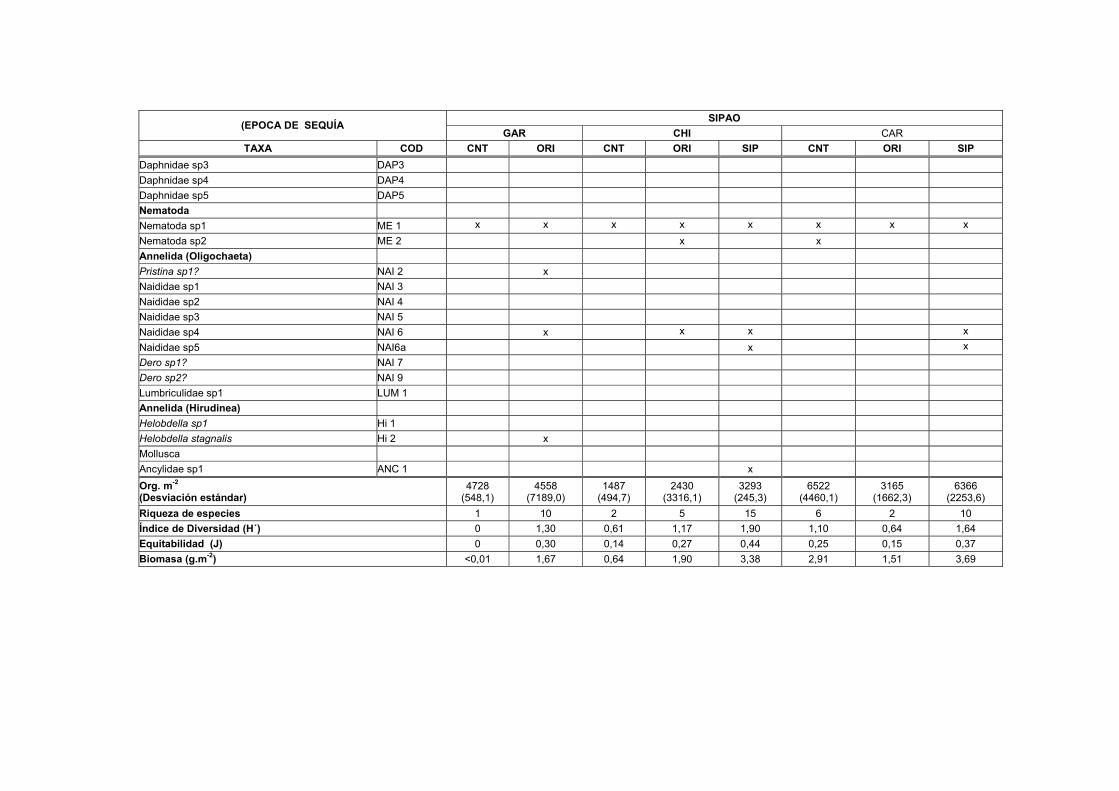
SIPAO (EPOCA DE SEQUÍA
GAR CHI CAR TAXA COD CNT ORI CNT ORI SIP CNT ORI SIP
Daphnidae sp3 DAP3 Daphnidae sp4 DAP4 Daphnidae sp5 DAP5 Nematoda Nematoda sp1 ME 1 x x x x x x x x Nematoda sp2 ME 2 x x Annelida (Oligochaeta) Pristina sp1? NAI 2 x Naididae sp1 NAI 3 Naididae sp2 NAI 4 Naididae sp3 NAI 5 Naididae sp4 NAI 6 x x x x Naididae sp5 NAI6a x x Dero sp1? NAI 7 Dero sp2? NAI 9 Lumbriculidae sp1 LUM 1 Annelida (Hirudinea) Helobdella sp1 Hi 1 Helobdella stagnalis Hi 2 x Mollusca Ancylidae sp1 ANC 1 x Org. m-2 (Desviación estándar)
4728 (548,1)
4558 (7189,0)
1487 (494,7)
2430 (3316,1)
3293 (245,3)
6522 (4460,1)
3165 (1662,3)
6366 (2253,6)
Riqueza de especies 1 10 2 5 15 6 2 10 Índice de Diversidad (H´) 0 1,30 0,61 1,17 1,90 1,10 0,64 1,64 Equitabilidad (J) 0 0,30 0,14 0,27 0,44 0,25 0,15 0,37 Biomasa (g.m-2) <0,01 1,67 0,64 1,90 3,38 2,91 1,51 3,69

Tabla 1B. Distribución, abundancia y biomasa de los Macroinvertebrados en el sedimento de nueve transectos del Bajo Caura en la fase de subida de aguas. (Abreviaciones: COD= Códigos, PAT= Laguna Patiquin, CEJ= L. Cejal, POZ= L. Pozo Rico, NAP= L. Naparaico, BRA= L. Brava, ARI= L. Aricagua, GAR= L. Garzones, CHI= L. Chiribital, CAR= L. Caramacatico. CNT= Centro de la laguna, ORI= Orilla de la laguna, MAT= Orilla Caño Mato, SIP= Orilla Caño Sipao).
MATO CAURA EPOCA DE TRANSICION
PAT CEJ POZ NAP BRA TAXA COD CNT ORI BOS MAT CNT ORI BOS CNT ORI BOS MAT CNT ORI BOS CNT (Insecta Ephemeroptera Polymitarcyidae Campsurus spp CAM 1 x Caenidae Brasilocaenis irmleri? B 1 Diptera Chironominae CHI 1 x x x x x x x x Stenochironomus sp CER 1 Tanypodinae CHI 2 x x x x Chaoborus sp1 CHA 1 x x x Ceratopogonidae sp2 CER 2 Ceratopogonidae sp3 CER 3 Ceratopogonidae sp4 CER 4 Ceratopogonidae sp6 CER 6 Ceratopogonidae sp6a CER 6a Ceratopogonidae sp7 CER 7 x Tabanidae sp4 TAB 4 Collembola Colembolla sp3 COL 3 Colembolla sp5 COL 5 Colembolla sp6 COL 6 Colembolla sp7 COL 7 Hemiptera Tenagobia sp1 TRI 1 Coleoptera Elmidae sp4 (L) ELM 4 Elmidae sp5 (L) ELM 5
(Continuación Tabla 1B...)

MATO CAURA EPOCA DE TRANSICION
PAT CEJ POZ NAP BRA TAXA COD CNT ORI BOS MAT CNT ORI BOS CNT ORI BOS MAT CNT ORI BOS CNT Elmidae sp6 (L) ELM 6 x Dytiscidae sp3 (A) DYC 2 x Odonata Anisoptera Gomphidae Phylocycla sp1 O1A Libellulidae sp1 n.i. LIB 1 x Isoptera ISO 1 Hydracarina Hydrozetes sp1 HYD 0 Hydracarina sp8 HYD 8 x x x Hydracarina sp16 HYD 16 Hydracarina sp34 HYD 34 Hydracarina sp35 HYD 35 Hydracarina sp36 HYD 36 Hydracarina sp37 HYD 37 Hydracarina sp38 HYD 38 x Hydracarina sp39 HYD 39 Hydracarina sp40 HYD 40 x Hydracarina sp41 HYD 41 x Arachnida Arachnidae sp4 ARA 4 Crustacea Ostracoda Ostracoda sp1 OS 1 x x x x Ostracoda sp2 OS 2 x Pelocypris sp1 OS 3 x x x Clamydotheca sp1 OS 3a Cypridae sp1 OS 4 Cypridae sp2 OS 5 Ostracoda sp3 OS 6 x Ostracoda sp4 OS 7 Ostracoda sp5 OS 8
(Continuación Tabla 1B...)
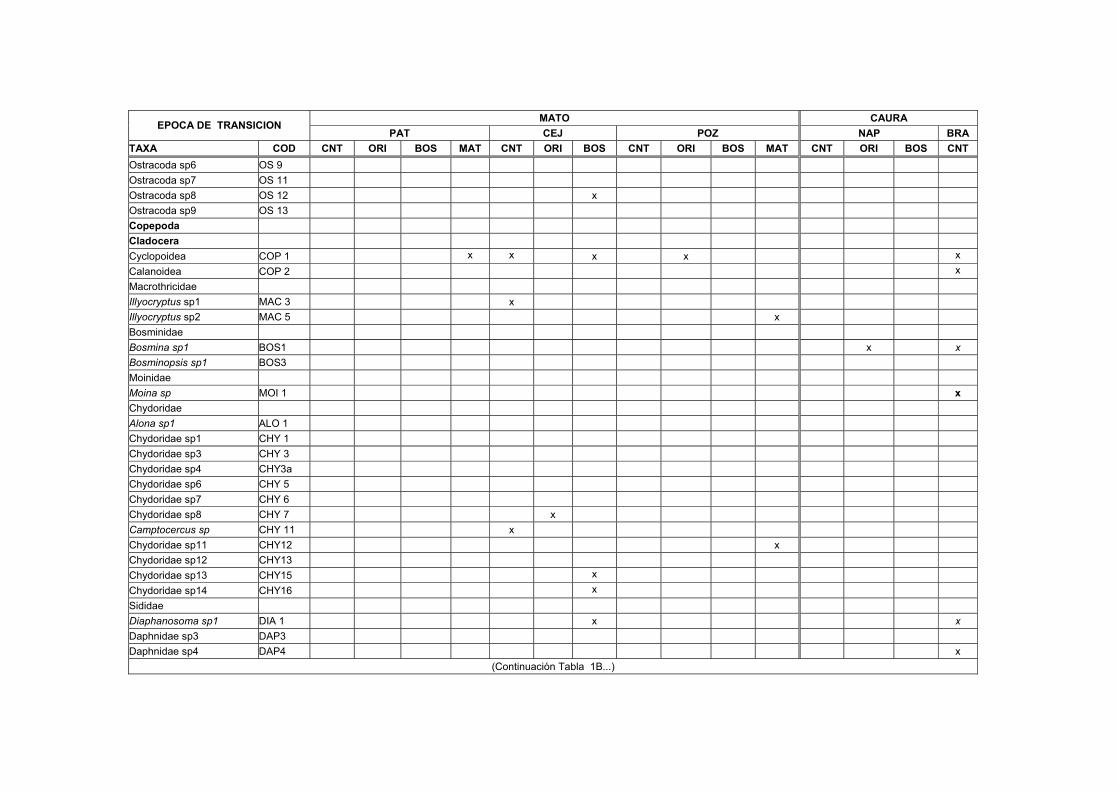
MATO CAURA EPOCA DE TRANSICION
PAT CEJ POZ NAP BRA TAXA COD CNT ORI BOS MAT CNT ORI BOS CNT ORI BOS MAT CNT ORI BOS CNT Ostracoda sp6 OS 9 Ostracoda sp7 OS 11 Ostracoda sp8 OS 12 x Ostracoda sp9 OS 13 Copepoda Cladocera Cyclopoidea COP 1 x x x x x Calanoidea COP 2 x Macrothricidae Illyocryptus sp1 MAC 3 x Illyocryptus sp2 MAC 5 x Bosminidae Bosmina sp1 BOS1 x x Bosminopsis sp1 BOS3 Moinidae Moina sp MOI 1 x Chydoridae Alona sp1 ALO 1 Chydoridae sp1 CHY 1 Chydoridae sp3 CHY 3 Chydoridae sp4 CHY3a Chydoridae sp6 CHY 5 Chydoridae sp7 CHY 6 Chydoridae sp8 CHY 7 x Camptocercus sp CHY 11 x Chydoridae sp11 CHY12 x Chydoridae sp12 CHY13 Chydoridae sp13 CHY15 x Chydoridae sp14 CHY16 x Sididae Diaphanosoma sp1 DIA 1 x x Daphnidae sp3 DAP3 Daphnidae sp4 DAP4 x
(Continuación Tabla 1B...)
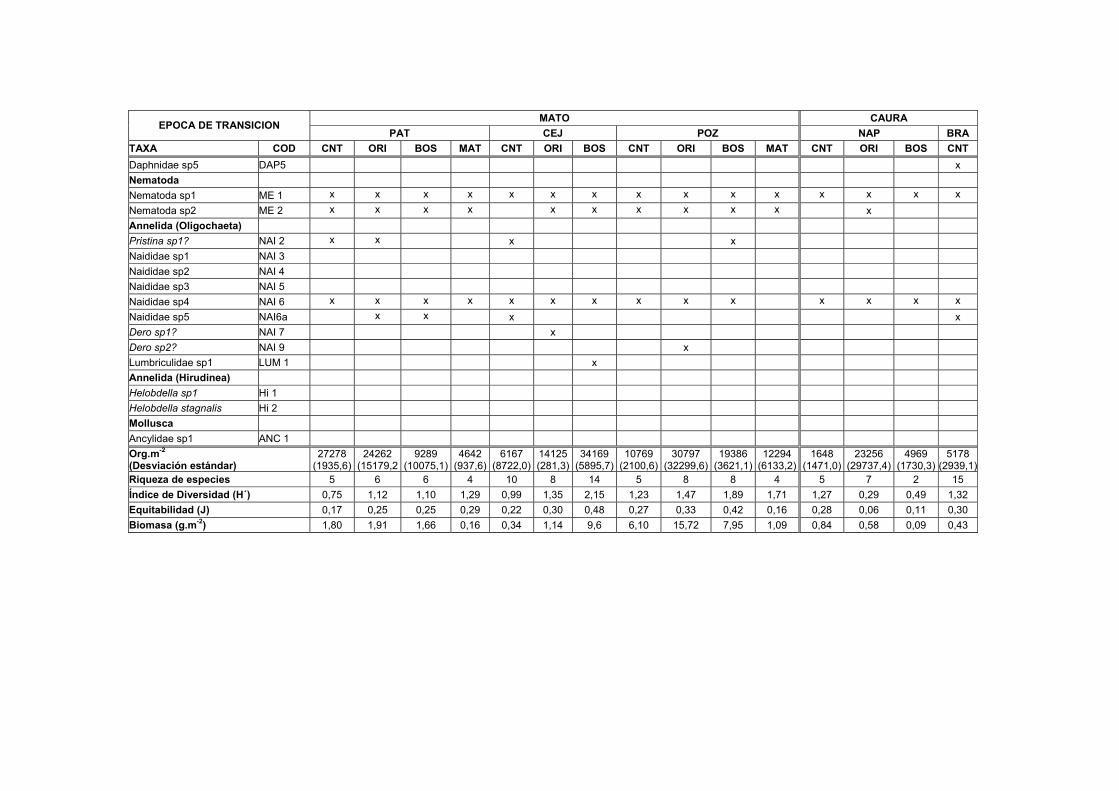
MATO CAURA EPOCA DE TRANSICION
PAT CEJ POZ NAP BRA TAXA COD CNT ORI BOS MAT CNT ORI BOS CNT ORI BOS MAT CNT ORI BOS CNT Daphnidae sp5 DAP5 x Nematoda Nematoda sp1 ME 1 x x x x x x x x x x x x x x x Nematoda sp2 ME 2 x x x x x x x x x x x Annelida (Oligochaeta) Pristina sp1? NAI 2 x x x x Naididae sp1 NAI 3 Naididae sp2 NAI 4 Naididae sp3 NAI 5 Naididae sp4 NAI 6 x x x x x x x x x x x x x x Naididae sp5 NAI6a x x x x Dero sp1? NAI 7 x Dero sp2? NAI 9 x Lumbriculidae sp1 LUM 1 x Annelida (Hirudinea) Helobdella sp1 Hi 1 Helobdella stagnalis Hi 2 Mollusca Ancylidae sp1 ANC 1 Org.m-2 (Desviación estándar)
27278 (1935,6)
24262 (15179,2
9289 (10075,1)
4642 (937,6)
6167 (8722,0)
14125(281,3)
34169 (5895,7)
10769 (2100,6)
30797 (32299,6)
19386 (3621,1)
12294 (6133,2)
1648 (1471,0)
23256 (29737,4)
4969 (1730,3)
5178 (2939,1)
Riqueza de especies 5 6 6 4 10 8 14 5 8 8 4 5 7 2 15 Índice de Diversidad (H´) 0,75 1,12 1,10 1,29 0,99 1,35 2,15 1,23 1,47 1,89 1,71 1,27 0,29 0,49 1,32 Equitabilidad (J) 0,17 0,25 0,25 0,29 0,22 0,30 0,48 0,27 0,33 0,42 0,16 0,28 0,06 0,11 0,30 Biomasa (g.m-2) 1,80 1,91 1,66 0,16 0,34 1,14 9,6 6,10 15,72 7,95 1,09 0,84 0,58 0,09 0,43
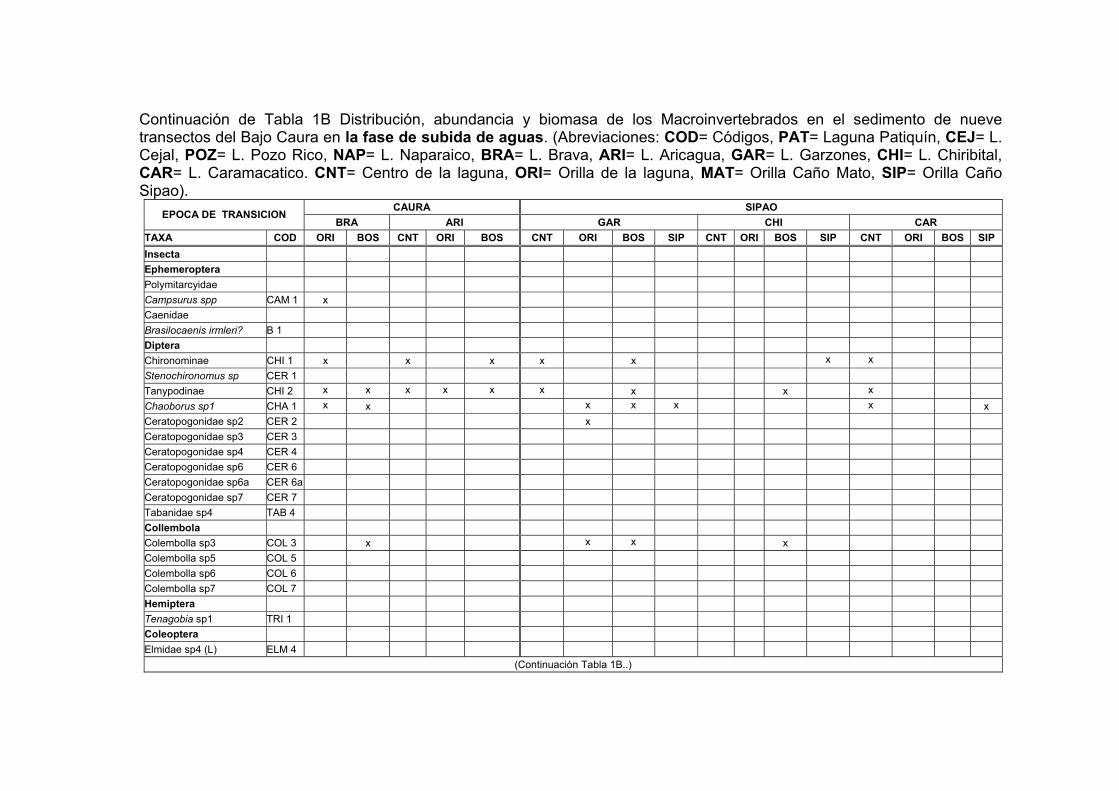
Continuación de Tabla 1B Distribución, abundancia y biomasa de los Macroinvertebrados en el sedimento de nueve transectos del Bajo Caura en la fase de subida de aguas. (Abreviaciones: COD= Códigos, PAT= Laguna Patiquín, CEJ= L. Cejal, POZ= L. Pozo Rico, NAP= L. Naparaico, BRA= L. Brava, ARI= L. Aricagua, GAR= L. Garzones, CHI= L. Chiribital, CAR= L. Caramacatico. CNT= Centro de la laguna, ORI= Orilla de la laguna, MAT= Orilla Caño Mato, SIP= Orilla Caño Sipao).
CAURA SIPAO EPOCA DE TRANSICION
BRA ARI GAR CHI CAR TAXA COD ORI BOS CNT ORI BOS CNT ORI BOS SIP CNT ORI BOS SIP CNT ORI BOS SIP Insecta Ephemeroptera Polymitarcyidae Campsurus spp CAM 1 x Caenidae Brasilocaenis irmleri? B 1 Diptera Chironominae CHI 1 x x x x x x x Stenochironomus sp CER 1 Tanypodinae CHI 2 x x x x x x x x x Chaoborus sp1 CHA 1 x x x x x x x Ceratopogonidae sp2 CER 2 x Ceratopogonidae sp3 CER 3 Ceratopogonidae sp4 CER 4 Ceratopogonidae sp6 CER 6 Ceratopogonidae sp6a CER 6a Ceratopogonidae sp7 CER 7 Tabanidae sp4 TAB 4 Collembola Colembolla sp3 COL 3 x x x x Colembolla sp5 COL 5 Colembolla sp6 COL 6 Colembolla sp7 COL 7 Hemiptera Tenagobia sp1 TRI 1 Coleoptera Elmidae sp4 (L) ELM 4
(Continuación Tabla 1B..)

CAURA SIPAO EPOCA DE TRANSICION
BRA ARI GAR CHI CAR TAXA COD ORI BOS CNT ORI BOS CNT ORI BOS SIP CNT ORI BOS SIP CNT ORI BOS SIP Elmidae sp5 (L) ELM 5 Elmidae sp6 (L) ELM 6 Dytiscidae sp3 (A) DYC 2 Odonata Anisoptera Gomphidae Phylocycla sp1 O1A Libellulidae n.i. LIB 1 Isoptera ISO 1 x Hydracarina Hydrozetes sp1 HYD 0 x x x x Hydracarina sp8 HYD 8 x x Hydracarina sp16 HYD 16 Hydracarina sp34 HYD 34 Hydracarina sp35 HYD 35 Hydracarina sp36 HYD 36 Hydracarina sp37 HYD 37 Hydracarina sp38 HYD 38 Hydracarina sp39 HYD 39 Hydracarina sp40 HYD 40 Hydracarina sp41 HYD 41 Arachnida Arachnidae sp4 ARA 4 Crustacea Ostracoda x Ostracoda sp1 OS 1 x x x Ostracoda sp2 OS 2 x x x Pelocypris sp1 OS 3 x x x x x Clamydotheca sp1 OS 3a x x x Cypridae sp1 OS 4 x x x Cypridae sp2 OS 5 x Ostracoda sp3 OS 6
(Continuación Tabla 1B..)
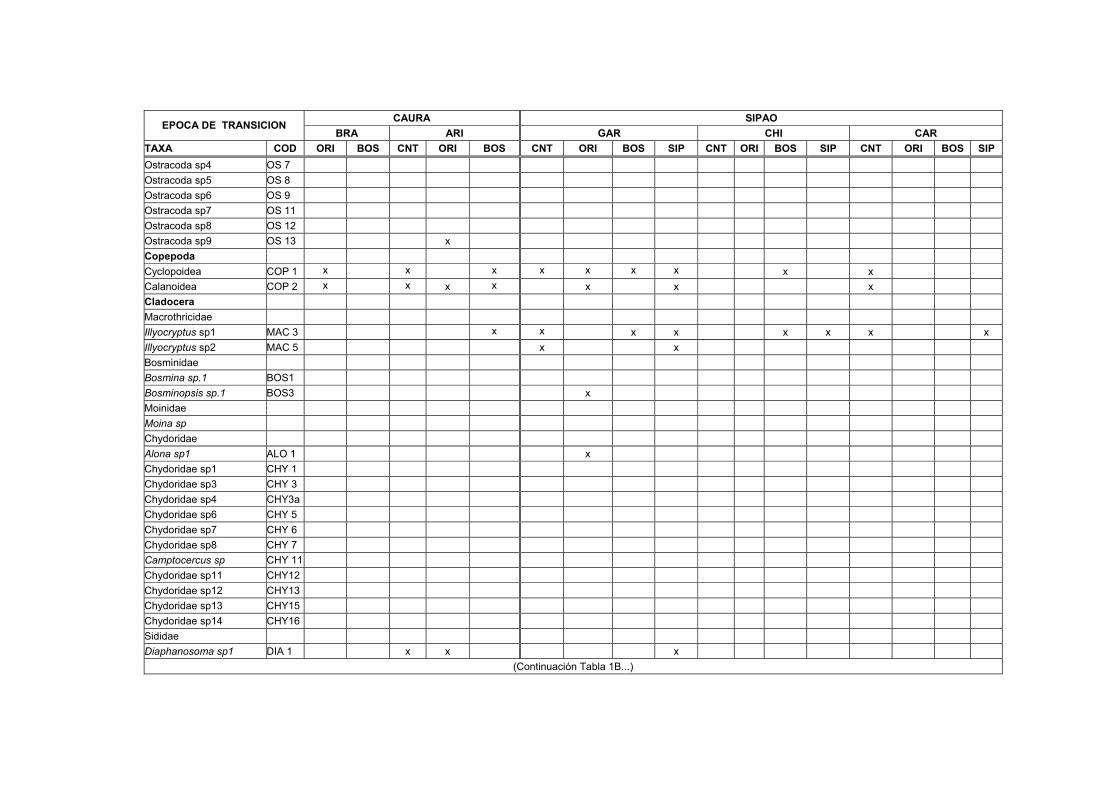
CAURA SIPAO EPOCA DE TRANSICION
BRA ARI GAR CHI CAR TAXA COD ORI BOS CNT ORI BOS CNT ORI BOS SIP CNT ORI BOS SIP CNT ORI BOS SIP Ostracoda sp4 OS 7 Ostracoda sp5 OS 8 Ostracoda sp6 OS 9 Ostracoda sp7 OS 11 Ostracoda sp8 OS 12 Ostracoda sp9 OS 13 x Copepoda Cyclopoidea COP 1 x x x x x x x x x Calanoidea COP 2 x x x x x x x Cladocera Macrothricidae Illyocryptus sp1 MAC 3 x x x x x x x x Illyocryptus sp2 MAC 5 x x Bosminidae Bosmina sp.1 BOS1 Bosminopsis sp.1 BOS3 x Moinidae Moina sp Chydoridae Alona sp1 ALO 1 x Chydoridae sp1 CHY 1 Chydoridae sp3 CHY 3 Chydoridae sp4 CHY3a Chydoridae sp6 CHY 5 Chydoridae sp7 CHY 6 Chydoridae sp8 CHY 7 Camptocercus sp CHY 11 Chydoridae sp11 CHY12 Chydoridae sp12 CHY13 Chydoridae sp13 CHY15 Chydoridae sp14 CHY16 Sididae Diaphanosoma sp1 DIA 1 x x x
(Continuación Tabla 1B...)
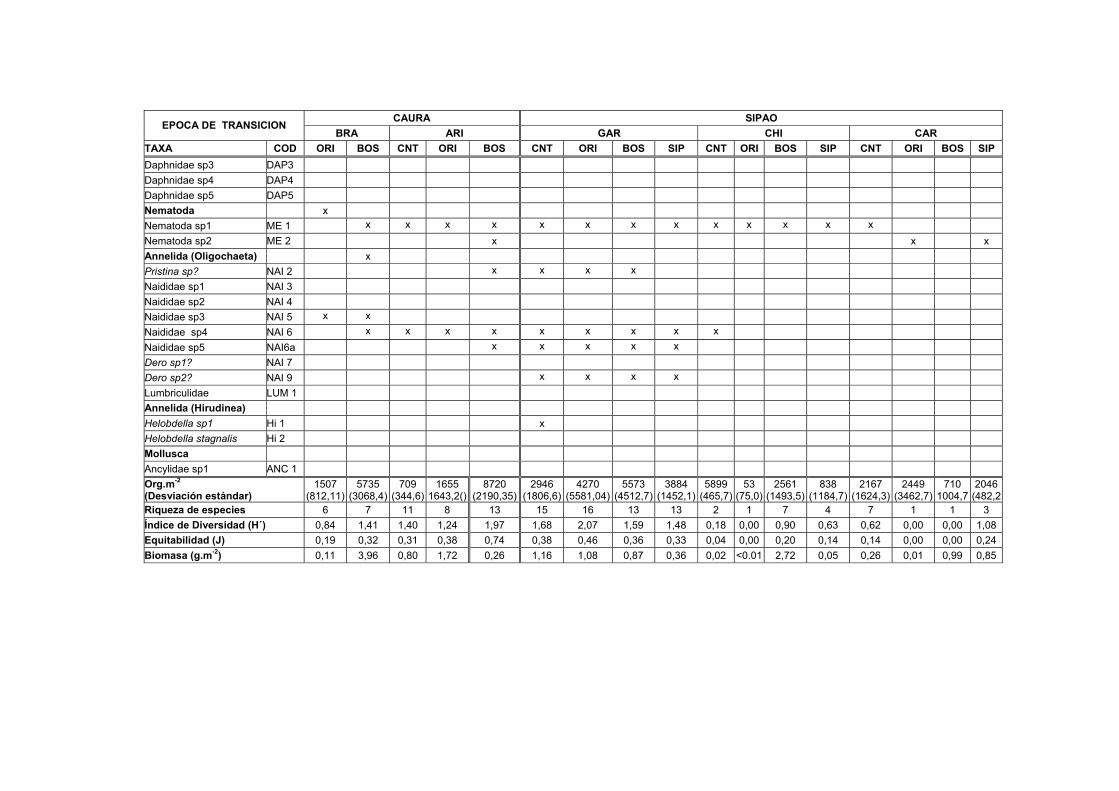
CAURA SIPAO EPOCA DE TRANSICION
BRA ARI GAR CHI CAR TAXA COD ORI BOS CNT ORI BOS CNT ORI BOS SIP CNT ORI BOS SIP CNT ORI BOS SIP Daphnidae sp3 DAP3 Daphnidae sp4 DAP4 Daphnidae sp5 DAP5 Nematoda x Nematoda sp1 ME 1 x x x x x x x x x x x x x Nematoda sp2 ME 2 x x x Annelida (Oligochaeta) x Pristina sp? NAI 2 x x x x Naididae sp1 NAI 3 Naididae sp2 NAI 4 Naididae sp3 NAI 5 x x Naididae sp4 NAI 6 x x x x x x x x x Naididae sp5 NAI6a x x x x x Dero sp1? NAI 7 Dero sp2? NAI 9 x x x x Lumbriculidae LUM 1 Annelida (Hirudinea) Helobdella sp1 Hi 1 x Helobdella stagnalis Hi 2 Mollusca Ancylidae sp1 ANC 1 Org.m-2 (Desviación estándar)
1507 (812,11)
5735 (3068,4)
709 (344,6)
1655 1643,2()
8720 (2190,35)
2946 (1806,6)
4270 (5581,04)
5573 (4512,7)
3884 (1452,1)
5899 (465,7)
53 (75,0)
2561 (1493,5)
838 (1184,7)
2167 (1624,3)
2449 (3462,7)
710 1004,7
2046 (482,2
Riqueza de especies 6 7 11 8 13 15 16 13 13 2 1 7 4 7 1 1 3 Índice de Diversidad (H´) 0,84 1,41 1,40 1,24 1,97 1,68 2,07 1,59 1,48 0,18 0,00 0,90 0,63 0,62 0,00 0,00 1,08 Equitabilidad (J) 0,19 0,32 0,31 0,38 0,74 0,38 0,46 0,36 0,33 0,04 0,00 0,20 0,14 0,14 0,00 0,00 0,24 Biomasa (g.m-2) 0,11 3,96 0,80 1,72 0,26 1,16 1,08 0,87 0,36 0,02 <0.01 2,72 0,05 0,26 0,01 0,99 0,85

Tabla 1C Distribución, abundancia y biomasa de los Macroinvertebrados en el sedimento de nueve transectos del Bajo Caura en fase de aguas altas. (Abreviaciones: COD= Códigos, PAT= Laguna Patiquín, CEJ= L. Cejal, POZ= L. Pozo Rico, NAP= L. Naparaico, BRA= L. Brava, ARI= L. Aricagua, GAR= L. Garzones, CHI= L. Chiribital, CAR= L. Caramacatico. CNT= Centro de la laguna, ORI= Orilla de la laguna, MAT= Orilla Caño Mato, SIP= Orilla Caño Sipao).
MATO EPOCA DE CONEXION PAT CEJ POZ
TAXA COD CNT ORI BOS MAT CNT ORI BOS CNT ORI BOS MAT Insecta Ephemeroptera Polymitarcyidae Campsurus sp CAM 1 Caenidae Brasilocaenis irmleri? B 1 x Diptera Chironominae CHI 1 x x x x x x Stenochironomus sp CER 1 x x Tanypodinae CHI 2 x x x x x x Chaoborus sp1 CHA 1 Ceratopogonidae sp2 CER 2 Ceratopogonidae sp3 CER 3 Ceratopogonidae sp4 CER 4 Ceratopogonidae sp6 CER 6 Ceratopogonidae sp6a CER 6a x Ceratopogonidae sp7 CER 7 Tabanidae sp4 TAB 4 x Collembola Colembolla sp3 COL 3 Colembolla sp5 COL 5 Colembolla sp6 COL 6 Colembolla sp7 COL 7 x Hemiptera Tenagobia sp1 TRI 1 Coleoptera Elmidae sp4 (L) ELM 4 Elmidae sp5 (L) ELM 5
(Continuación Tabla 1C...)

MATO EPOCA DE CONEXION PAT CEJ POZ
TAXA COD CNT ORI BOS MAT CNT ORI BOS CNT ORI BOS MAT Elmidae sp6 (L) ELM 6 Dytiscidae sp3 (A) DYC 2 Odonata Anisoptera Gomphidae Phylocycla sp1 O1A Libellulidae sp1 n.i. LIB 1 Isoptera ISO 1 Hydracarina Hydrozetes sp1 HYD 0 x x x Hydracarina sp8 HYD 8 x x Hydracarina sp34 HYD 34 Hydracarina sp35 HYD 35 Hydracarina sp36 HYD 36 Hydracarina sp37 HYD 37 Hydracarina sp38 HYD 38 Hydracarina sp39 HYD 39 Hydracarina sp40 HYD 40 Hydracarina sp41 HYD 41 Arachnida Arachnidae sp4 ARA 4 x Crustacea Ostracoda Ostracoda sp1 OS 1 x Ostracoda sp2 OS 2 Pelocypris sp1 OS 3 x x x Clamydotheca sp1 OS 3a Cypridae sp1 OS 4 x x x x x Cypridae sp2 OS 5 x Ostracoda sp3 OS 6 Ostracoda sp4 OS 7 Ostracoda sp5 OS 8 x x Ostracoda sp6 OS 9 x x x x x
(Continuación Tabla 1C...)

MATO EPOCA DE CONEXION PAT CEJ POZ
TAXA COD CNT ORI BOS MAT CNT ORI BOS CNT ORI BOS MAT Ostracoda sp7 OS 11 x x Ostracoda sp8 OS 12 Ostracoda sp9 OS 13 Copepoda Cyclopoidea COP 1 x x Calanoidea COP 2 Cladocera Macrothricidae Illyocryptus sp1 MAC 3 x x x Illyocryptus sp2 MAC 5 x Bosminidae Bosmina sp.1 BOS1 Bosminopsis sp.1 BOS3 Moinidae Moina sp MOI 1 Chydoridae Alona sp1 ALO 1 Chydoridae sp1 CHY 1 x x Chydoridae sp3 CHY 3 Chydoridae sp4 CHY3a Chydoridae sp6 CHY 5 Chydoridae sp7 CHY 6 x Chydoridae sp8 CHY 7 Camptocercus sp CHY 11 Chydoridae sp11 CHY12 Chydoridae sp12 CHY13 x Chydoridae sp13 CHY15 Chydoridae sp14 CHY16 Sididae Diaphanosoma sp1 DIA 1 x x Daphnidae sp3 DAP3 x Daphnidae sp4 DAP4 Daphnidae sp5 DAP5
(Continuación Tabla 1C...)

MATO EPOCA DE CONEXION PAT CEJ POZ
TAXA COD CNT ORI BOS MAT CNT ORI BOS CNT ORI BOS MAT Nematoda Nematoda sp1 ME 1 x x x x x x x x x x Nematoda sp2 ME 2 x x x x x x x Annelida (Oligochaeta) Pristina sp? NAI 2 x x x Naididae sp1 NAI 3 Naididae sp2 NAI 4 Naididae sp3 NAI 5 x x Naididae sp4 NAI 6 x x x x x x x x x Naididae sp5 NAI6a x Dero sp1? NAI 7 x x Dero sp2? NAI 9 Lumbriculidae LUM 1 Annelida (Hirudinea) Helobdella sp1 Hi 1 Helobdella stagnalis Hi 2 Platyhelminthes (Turbellaria) Turbellaria sp1 TU 1 Mollusca Ancylidae sp1 ANC 1 x Org. m-2
(Desviación estándar) 56907
(36029,6) 47119
(7562,5) 64031
(43822,9) 21512
(3412,5) 5453
(284,9) 1722
(1027,8) 13487
(2609,1) 33210
(24456,7) 27097 (337,6)
12080 (2612,4)
4621 (1905,1)
Riqueza de especies 12 8 18 8 2 1 7 8 11 8 8 Índice de Diversidad (H´) 1,43 1,17 2,03 1,49 0,29 0,00 1,26 1,75 1,85 1,91 0,62 Equitabilidad (J) 0,32 0,26 0,45 0,33 0,07 0,00 0,28 0,39 0,41 0,43 0,14 Biomasa (g.m-2) 13,61 3,75 15,94 5,09 0,04 <0.01 8,58 21,01 23,36 10,31 0,19
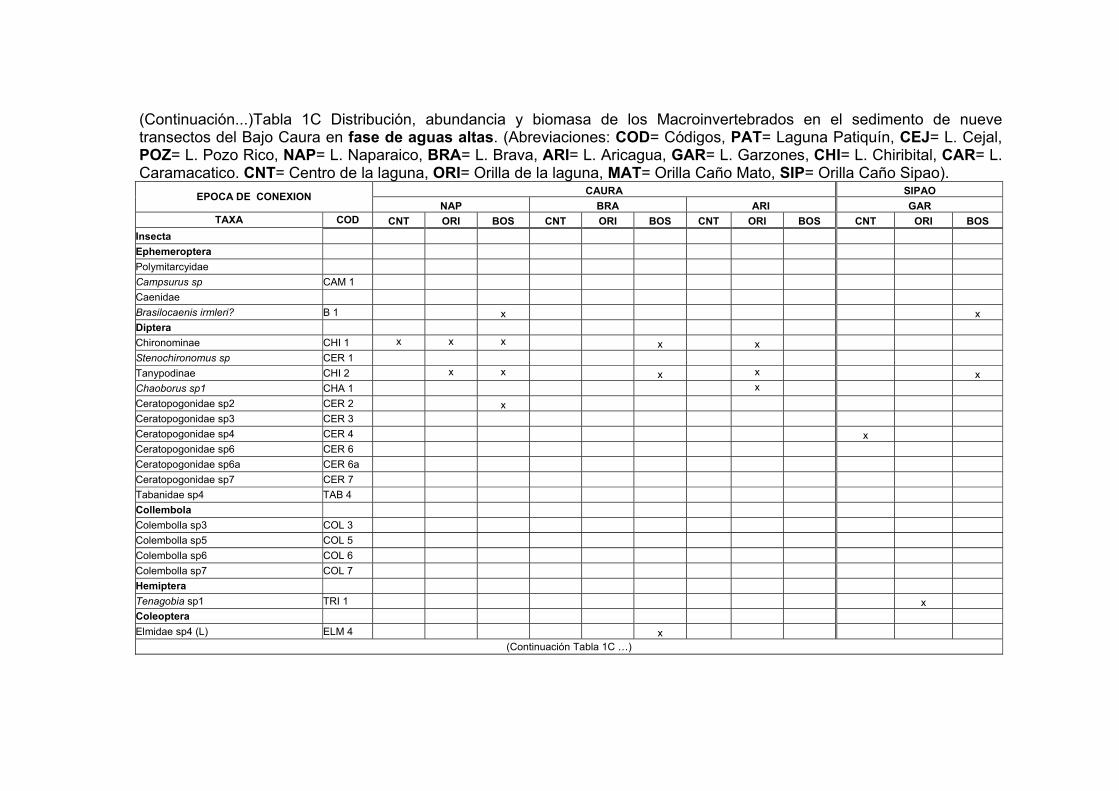
(Continuación...)Tabla 1C Distribución, abundancia y biomasa de los Macroinvertebrados en el sedimento de nueve transectos del Bajo Caura en fase de aguas altas. (Abreviaciones: COD= Códigos, PAT= Laguna Patiquín, CEJ= L. Cejal, POZ= L. Pozo Rico, NAP= L. Naparaico, BRA= L. Brava, ARI= L. Aricagua, GAR= L. Garzones, CHI= L. Chiribital, CAR= L. Caramacatico. CNT= Centro de la laguna, ORI= Orilla de la laguna, MAT= Orilla Caño Mato, SIP= Orilla Caño Sipao).
CAURA SIPAO EPOCA DE CONEXION NAP BRA ARI GAR
TAXA COD CNT ORI BOS CNT ORI BOS CNT ORI BOS CNT ORI BOS Insecta Ephemeroptera Polymitarcyidae Campsurus sp CAM 1 Caenidae Brasilocaenis irmleri? B 1 x x Diptera Chironominae CHI 1 x x x x x Stenochironomus sp CER 1 Tanypodinae CHI 2 x x x x x Chaoborus sp1 CHA 1 x Ceratopogonidae sp2 CER 2 x Ceratopogonidae sp3 CER 3 Ceratopogonidae sp4 CER 4 x Ceratopogonidae sp6 CER 6 Ceratopogonidae sp6a CER 6a Ceratopogonidae sp7 CER 7 Tabanidae sp4 TAB 4 Collembola Colembolla sp3 COL 3 Colembolla sp5 COL 5 Colembolla sp6 COL 6 Colembolla sp7 COL 7 Hemiptera Tenagobia sp1 TRI 1 x Coleoptera Elmidae sp4 (L) ELM 4 x
(Continuación Tabla 1C …)

CAURA SIPAO EPOCA DE CONEXION
NAP BRA ARI GAR TAXA COD CNT ORI BOS CNT ORI BOS CNT ORI BOS CNT ORI BOS
Elmidae sp5 (L) ELM 5 Elmidae sp6 (L) ELM 6 Dytiscidae sp3 (A) DYC 2 Odonata Anisoptera Gomphidae Phylocycla sp1 O1A Libellulidae sp1 n.i. LIB 1 Isoptera ISO 1 Hydracarina Hydrozetes sp1 HYD 0 x x x x Hydracarina sp8 HYD 8 x Hydracarina sp34 HYD 34 x Hydracarina sp35 HYD 35 x Hydracarina sp36 HYD 36 x Hydracarina sp37 HYD 37 x Hydracarina sp38 HYD 38 x Hydracarina sp39 HYD 39 Hydracarina sp40 HYD 40 Hydracarina sp41 HYD 41 (Arachnida Arachnidae sp4 ARA 4 Crustacea Ostracoda Ostracoda sp1 OS 1 Ostracoda sp2 OS 2 Pelocypris sp1 OS 3 x x x x x Clamydotheca sp1 OS 3a x x x Cypridae sp1 OS 4 x Cypridae sp2 OS 5 x x x Ostracoda sp3 OS 6
(Continuación Tabla 1C..)

CAURA SIPAO EPOCA DE CONEXION
NAP BRA ARI GAR TAXA COD CNT ORI BOS CNT ORI BOS CNT ORI BOS CNT ORI BOS
Ostracoda sp4 OS 7 x Ostracoda sp5 OS 8 x x Ostracoda sp6 OS 9 x Ostracoda sp7 OS 11 Ostracoda sp8 OS 12 Ostracoda sp9 OS 13 Copepoda Cyclopoidea COP 1 x x x Calanoidea COP 2 Cladocera Macrothricidae Illyocryptus sp1 MAC 3 x Illyocryptus sp2 MAC 5 Bosminidae Bosmina sp.1 BOS1 Bosminopsis sp.1 BOS3 x Moinidae Moina sp MOI 1 Chydoridae Alona sp1 ALO 1 Chydoridae sp1 CHY 1 Chydoridae sp3 CHY 3 Chydoridae sp4 CHY3a Chydoridae sp6 CHY 5 Chydoridae sp7 CHY 6 Chydoridae sp8 CHY 7 Camptocercus sp CHY 11 Chydoridae sp11 CHY12 x Chydoridae sp12 CHY13 Chydoridae sp13 CHY15 Chydoridae sp14 CHY16 Sididae Diaphanosoma sp1 DIA 1 x
(Continuación Tabla 1C...)

CAURA SIPAO EPOCA DE CONEXION
NAP BRA ARI GAR TAXA COD CNT ORI BOS CNT ORI BOS CNT ORI BOS CNT ORI BOS
Daphnidae sp3 DAP3 Daphnidae sp4 DAP4 Daphnidae sp5 DAP5 Nematoda Nematoda sp1 ME 1 x x x x x x x x x x x Nematoda sp2 ME 2 x x x x x x Annelida (Oligochaeta) Pristina sp? NAI 2 x x x Naididae sp1 NAI 3 x Naididae sp2 NAI 4 x Naididae sp3 NAI 5 x Naididae sp4 NAI 6 x x x x x x x x x Naididae sp5 NAI6a x Dero sp1? NAI 7 Dero sp2? NAI 9 Lumbriculidae LUM 1 Annelida (Hirudinea) Helobdella sp1 Hi 1 Helobdella stagnalis Hi 2 Platyhelminthes (Turbellaria) Turbellaria sp1 TU 1 x Mollusca Ancylidae sp1 ANC 1 x x Org- m-2 (Desviación estándar)
2419 (45,01)
9309 (3141,5)
7321 (630,2)
1990 (649,0)
4929 (3849,9)
2311 (952,5)
186 (217,4)
5038 (4223,99
1990 (2813,5)
37917 (36743,0)
25984 (17450,9)
6886 (4583,9)
Riqueza de especies 3 6 18 3 6 11 3 12 2 7 7 7 Índice de Diversidad (H´) 1,02 0,74 2,22 0,86 0,66 1,63 0,79 1,75 0,69 0,53 1,43 1,72 Equitabilidad (J) 0,23 0,17 0,50 0,19 0,15 0,36 0,18 0,39 0,15 0,12 0,32 0,38 Biomasa (g.m-2) 1,71 1,24 4,57 0,21 0,04 2,56 <0,01 3,13 0,11 3,91 10,88 7,80

(Continuación...)Tabla 1C. Distribución, abundancia y biomasa de los Macroinvertebrados en el sedimento de nueve transectos del Bajo Caura en fase de aguas altas. (Abreviaciones: COD= Códigos, PAT= Laguna Patiquín, CEJ= L. Cejal, POZ= L. Pozo Rico, NAP= L. Naparaico, BRA= L. Brava, ARI= L. Aricagua, GAR= L. Garzones, CHI= L. Chiribital, CAR= L. Caramacatico. CNT= Centro de la laguna, ORI= Orilla de la laguna, MAT= Orilla Caño Mato, SIP= Orilla Caño Sipao).
SIPAO EPOCA DE CONEXION CHI CAR TAXA COD CNT ORI BOS SIP CNT ORI BOS SIP
Insecta Ephemeroptera Polymitarcyidae Campsurus sp CAM 1 Caenidae Brasilocaenis irmleri? B 1 x Diptera Chironominae CHI 1 x x Stenochironomus sp CER 1 Tanypodinae CHI 2 x x x Chaoborus sp1 CHA 1 Ceratopogonidae sp2 CER 2 x x Ceratopogonidae sp3 CER 3 x Ceratopogonidae sp4 CER 4 Ceratopogonidae sp6 CER 6 Ceratopogonidae sp6a CER 6a Ceratopogonidae sp7 CER 7 Tabanidae sp4 TAB 4 Collembola Colembolla sp3 COL 3 x Colembolla sp5 COL 5 Colembolla sp6 COL 6 Colembolla sp7 COL 7 Hemiptera Tenagobia sp1 TRI 1 Coleoptera
(Continuación Tabla 1C...)

SIPAO EPOCA DE CONEXION CHI CAR TAXA COD CNT ORI BOS SIP CNT ORI BOS SIP
Elmidae sp4 (L) ELM 4 Elmidae sp5 (L) ELM 5 Elmidae sp6 (L) ELM 6 Dytiscidae sp3 (A) DYC 2 Odonata Phylocycla sp1 O1A Libellulidae sp1 n.i. LIB 1 Isoptera ISO 1 Hydracarina Hydrozetes sp1 HYD 0 x x x Hydracarina sp8 HYD 8 x x Hydracarina sp34 HYD 34 Hydracarina sp35 HYD 35 Hydracarina sp36 HYD 36 Hydracarina sp37 HYD 37 Hydracarina sp38 HYD 38 Hydracarina sp39 HYD 39 Hydracarina sp40 HYD 40 x Hydracarina sp41 HYD 41 Arachnida Arachnidae sp4 ARA 4 Crustacea Ostracoda Ostracoda sp1 OS 1 Ostracoda sp2 OS 2 Pelocypris sp1 OS 3 x x x x x x x Clamydotheca sp1 OS 3a x Cypridae sp1 OS 4 x x Cypridae sp2 OS 5 x x x x x x Ostracoda sp3 OS 6 Ostracoda sp4 OS 7 Ostracoda sp5 OS 8
(Continuación Tabla 1C...)

SIPAO EPOCA DE CONEXION CHI CAR TAXA COD CNT ORI BOS SIP CNT ORI BOS SIP
Ostracoda sp6 OS 9 Ostracoda sp7 OS 11 Ostracoda sp8 OS 12 Ostracoda sp9 OS 13 Copepoda Cyclopoidea COP 1 x x Calanoidea COP 2 x x Cladocera Macrothricidae Illyocryptus sp1 MAC 3 x x Illyocryptus sp2 MAC 5 Bosminidae Bosmina sp.1 BOS1 Bosminopsis sp.1 BOS3 Moinidae Moina sp MOI 1 Chydoridae Alona sp1 ALO 1 Chydoridae sp1 CHY 1 Chydoridae sp3 CHY 3 Chydoridae sp4 CHY3a Chydoridae sp6 CHY 5 Chydoridae sp7 CHY 6 Chydoridae sp8 CHY 7 Camptocercus sp CHY 11 Chydoridae sp11 CHY12 Chydoridae sp12 CHY13 Chydoridae sp13 CHY15 Chydoridae sp14 CHY16 Sididae Diaphanosoma sp1 DIA 1 x Daphnidae sp3 DAP3
(Continuación Tabla 1C...)

SIPAO EPOCA DE CONEXION CHI CAR TAXA COD CNT ORI BOS SIP CNT ORI BOS SIP
Daphnidae sp4 DAP4 Daphnidae sp5 DAP5 Nematoda Nematoda sp1 ME 1 x x x x x x x x Nematoda sp2 ME 2 x x x Annelida (Oligochaeta) Pristina sp? NAI 2 x Naididae sp1 NAI 3 Naididae sp2 NAI 4 Naididae sp3 NAI 5 Naididae sp4 NAI 6 x x x x x Naididae sp5 NAI6a Dero sp1? NAI 7 Dero sp2? NAI 9 Lumbriculidae LUM 1 Annelida (Hirudinea) Helobdella sp1 Hi 1 x Helobdella stagnalis Hi 2 Platyhelminthes (Turbellaria) Turbellaria sp1 TU 1 x Mollusca Ancylidae sp1 ANC 1 Org.m-2 (Desviación stándard)
14589 (15004,8)
23999 (5506,1)
18717 (11345,1)
3051 (1776,5)
16579 (4689,9)
14851 (12224,4)
16870 (3421,1)
967 (432,0)
Riqueza de especies 7 4 8 11 2 7 9 10 Indice de Diversidad (H´) 1,58 1,03 1,78 1,36 0,35 0,73 1,94 0,83 Equitabilidad (J) 0,35 0,23 0,40 0,30 0,08 0,16 0,43 0,19 Biomasa (g.m-2) 3,77 11,15 8,38 0,74 2,55 3,58 24,92 0,15

Tabla. 2. Distribución y abundancia de los Macroinvertebrados en maderos sumergidos en siete transectos del Bajo Caura durante la fase de aguas bajas. (Abreviaciones: COD= Códigos, CEJ= L. Cejal, POZ= L. Pozo Rico, NAP= L. Naparaico, BRA= L. Brava, ARI= L. Aricagua, GAR= L. Garzones, CHI= L. Chiribital, CAR= L. Caramacatico, y *=Densidad Media < 0,5)
MATO CAURA SIPAO TAXA COD CEJ POZ NAP ARI BRA GAR CHI CAR
Insecta Ephemeroptera Caenidae Callibaetis sp CAL 1 x Brasilocaenis irmleri B 1 x x x x x Polymitarcyidae Asthenopus spp E4 x x x x x x x x Trichoptera Odontoceridae sp1 BRA 1 x x x x Hydroptilidae sp1 HYT 1 x x x Hydroptilidae sp2 HYT 2 x x x Hydropsychidae sp1 TRO 1 x x Polycentropididae Cyrnellus sp 1 CYR 1 x x x x x x x Cyrnellus sp 2 CYR 2 x x x x x x x x Diptera Tanypodinae CHI 1 x x x x x x x x Chironomidae CHI 2 x x x x x x x Stenochironomus sp1 CER 1 x x x x x x x Ceratopogonidae sp2 CER 2 x x x x Ceratopogonidae sp3 CER 3 x x x Ceratopogonidae sp5 CER 5 x x Tabanidae sp1 TA 1 x x Collembola Collembola sp1 COL 1 x Hemiptera Corixidae Tenagobia sp1 TRI 1 x x Tenagobia selecta? TRI 2 x Hebridae Hebrus sp1? HEM 1 x Naucoridae Pelocoris sp1 PEC 1 x Coleoptera Dytiscidae Pachydrus sp1 PAC 1 x Desmopachria sp1 DES 1 x Laccophilus sp1 LAC 1 x Dytiscidae sp1 (A) DYS 1 x Dytiscidae sp2 (L) DYC 1 x Noteridae Hydrochantus sp1 HYR 1 x

(Continuación Tabla 2...) MATO CAURA SIPAO
TAXA COD CEJ POZ NAP ARI BRA GAR CHI CAR Mesonoterus sp1 NOT 1 x Gyrinidae Gyrinus sp1 (L) GYR 1 x Hydrophilidae Helochares sp1 (A) HEL 1 x Hydrochus sp1 (A) HUC 1 x x x x Hydrochus sp2 (A) HUC 2 x x x Hydrochus sp3 (A) HUC 3 x Paracymus sp1 (A) PAR 1 x Paracymus sp2 (A) PAR 2 x Paracymus sp3 (A) PAR 3 x Hydrophilidae sp1 n.i (A) HYP 1 x x x x Hydrophilidae sp2 n.i (A) HYP 2 x Hydrophilidae sp3 n.i (A) HYP 3 x Hydrophilidae sp4 n.i (A) HYP 4 x Hydrophilidae sp5 n.i (A) HYP 5 x Dryopidae sp1 (L) DRYO 1 x Elmidae sp1 (A) ELM 1 x x x x x x Elmidae sp2 (L) ELM 2 x x Elmidae sp3 (A) ELM 3 x Helodidae sp1 (L) HED 1 x x x x x Staphilinidae sp1(A) STA 1 x x x Staphilinidae sp2 (A) STA 2 x Thysanura Thysanura sp1 (L) THY 1 x Neuroptera Sysyridae Climacia sp? (L) NEU 1 x x x x Odonata Libellulidae sp1 n.i. LIB 1 x x x Zygoptera sp1 n.i ZYG 1 x Hydracarina Hydracarina sp2 HYD 2 x x Hydracarina sp3 HYD 3 x Hydracarina sp4 HYD 4 x x Hydracarina sp5 HYD 5 x Hydracarina sp6 HYD 6 x Hydracarina sp7 HYD 7 x x Hydracarina sp8 HYD 8 x Hydracarina sp9 HYD 9 x Hydracarina sp10 HYD 10 x Hydracarina sp11 HYD 11 x Hydracarina sp12 HYD 12 x Hydracarina sp13 HYD 13 x Hydracarina sp14 HYD 14 x Hydracarina sp15 HYD 15 x Hydracarina sp17 HYD 16 x Arachnida
(Continuación Tabla 2...)

MATO CAURA SIPAO TAXA COD CEJ POZ NAP ARI BRA GAR CHI CAR
Arachnidae sp1 ARA 1 x Crustacea Ostracoda Ostracoda sp2 OS 2 x Cladocera Illyocryptus sp 1 MAC 3 x Daphnidae sp 1 DAP 1 1 Copepoda Cyclopoidea COP 1 x x x x x Calanoidea COP 2 x Branchiopoda Cyclestheria hislopi CYC 1 x x Branchiopoda sp1 EUL 1 x Nematoda Nematoda sp1 ME 1 x x x x x x x x Nematoda sp 2 ME 2 x x x x x x x x Annelida (Oligochaeta) Prístina sp1 NAI 2 x x x x x x x x Naididae sp3 NAI 3 x x Naididae sp4 NAI 4 x x x Naididae sp5 NAI 5 x x Naididae sp6 NAI 6 x x x x x x x x Naididae sp6 NAI 8 x Dero sp2 NAI 9 x Naididae sp7 NAI 10 x Annelida (Hirudinea) Helobdella sp1 Hi 1 x x x x x Helobdella stagnalis? Hi 2 2 x x x x x Helobdella sp2 Hi 4 x x x x Helobdella sp3 Hi 5 x x x Helobdella sp4 Hi 6 x Platyhelminthes (Turbellaria)
Turbellaria sp1 TU 1 x x x x x x x Turbellaria sp2 TU 2 x Mollusca Pelecypoda sp1 PEL 1 x x x
Total ± Desviación estándar 128 ± 106,1
229 ± 209,3
593 ± 241,8
366 ± 270,8
274 ± 42,4
366 ± 155,1
397 ± 101,1
408 ± 117,3
Peso maderos (g) 33,0 23,12 51,67 63,95 80,84 111,7 111,8 217,59 Area maderos (cm2) 158,6 103,3 204,8 205,6 346,3 329,8 326,5 340,4 Org./100 g de madera 388 990 1148 583 339 327 355 188 Org.m-2 de madera 8068 22164 28955 18144 7912 11067 12159 11984 Biomasa (g) ± Desviación estándar
0,43 ± 0,25
0,42 ± 0,07
1,24 ± 1,72
1,57 ± 1,27
1,12± 1,40
1,1 ± 0,62
3,09 ± 1,72
1,16 ± 0,01
Biomasa (g) . 100g madera 1,29 1,81 2,41 2,46 1,39 0,98 2,76 0,53 Biomasa (g.m-2) 27,11 40,65 60,54 76,28 32,34 98,47 94,64 34,07 Riqueza de especies 23 39 41 20 23 34 28 26 Índice de diversidad (H´) 2,39 2,23 2,29 1,56 2,23 2,39 2,09 2,16 Equitabilidad (J) 0,53 0,50 0,51 0,35 0,50 0,53 0,46 0,48

Tabla 3. Distribución y abundancia de los Macroinvertebrados en hojas del bosque inundable de cinco transectos del Bajo Caura durante la fase de aguas altas. (Abreviaciones: PAT= Laguna Patiquín, POZ= L. Pozo Rico, NAP= L. Naparaico, ARI= L. Aricagua, BRA= L. Brava, y *=Densidad Media< 0,5).
MATO CAURA TAXA Código PAT POZ NAP ARI BRA
Insecta Ephemeroptera Polymitarcyidae Asthenopus curtus E4 x Trichoptera Hydroptilidae sp1 HYT 1 x x x Hydroptilidae sp2 HYT 2 x x Hydroptilidae sp3 HYT 3 x Hydroptilidae sp4 HYT 4 x Polycentropididae Cyrnellus sp1? CYR 1 x x x Cyrnellus sp 2 CYR 2 x x x x Diptera Chironomidae CHI 1 x x x x x Tanypodinae CHI 2 x x x x x Ceratopogonidae sp4 CER 4 x x x x Ceratopogonidae sp5 CER 5 x x x x x Tabanidae sp2 TA 2 x Tabanidae sp3 TA 3 x Culicidae sp1 CUL 1 x Culicidae sp2 CUL 2 x Culicidae sp3 CUL 3 x x Collembola Collembola sp2 COL 2 x Hemiptera Corixidae Tenagobia sp1 TRI 1 x x x x Corixidae sp1 TRI 3 x Hebridae sp2 HEM 2 x Veliidae Trochopus sp1 HEM 3 x Microveliidae Microvelia sp1 MIC 1 x Coleoptera Noteridae Mesonoterus sp1? NOT 1 x Noteridae sp1 NOT 2 x Hydrophilidae Hydrochus sp1? HUC 1 x Hydrochus sp2? HUC 2 x
(Continuación Tabla 3...)
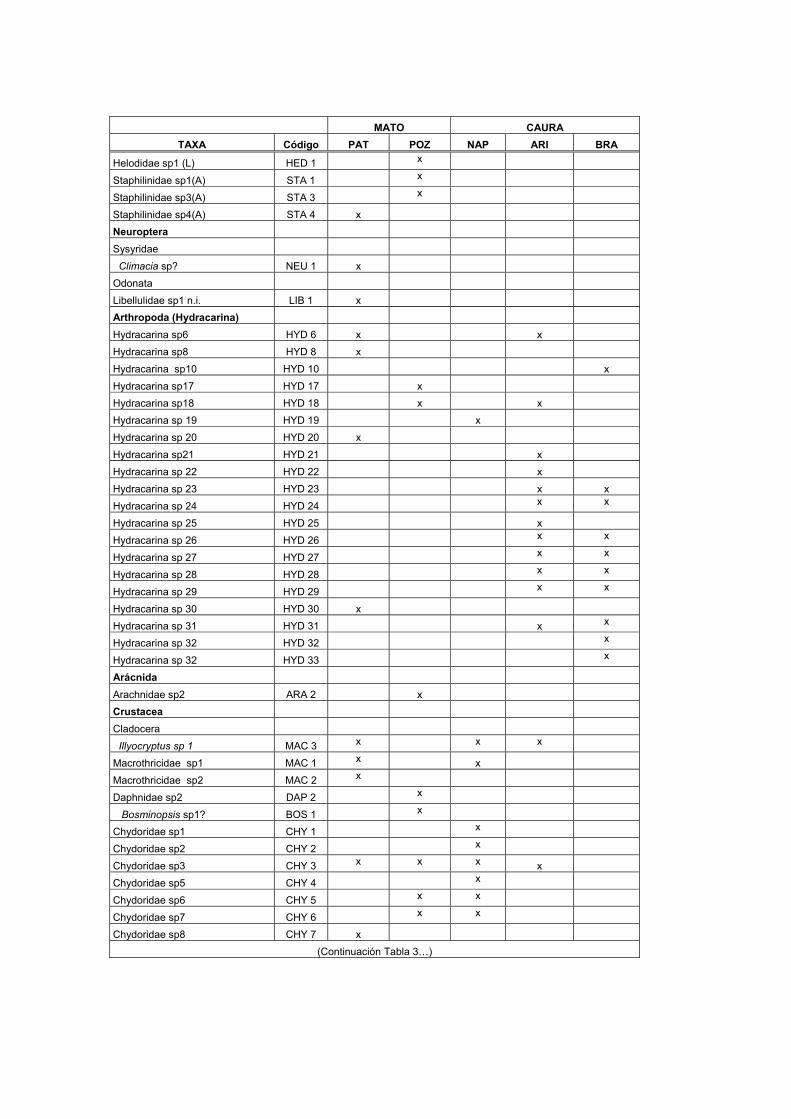
MATO CAURA TAXA Código PAT POZ NAP ARI BRA
Helodidae sp1 (L) HED 1 x
Staphilinidae sp1(A) STA 1 x Staphilinidae sp3(A) STA 3 x Staphilinidae sp4(A) STA 4 x Neuroptera Sysyridae Climacia sp? NEU 1 x Odonata Libellulidae sp1 n.i. LIB 1 x Arthropoda (Hydracarina) Hydracarina sp6 HYD 6 x x Hydracarina sp8 HYD 8 x Hydracarina sp10 HYD 10 x Hydracarina sp17 HYD 17 x Hydracarina sp18 HYD 18 x x Hydracarina sp 19 HYD 19 x Hydracarina sp 20 HYD 20 x Hydracarina sp21 HYD 21 x Hydracarina sp 22 HYD 22 x Hydracarina sp 23 HYD 23 x x Hydracarina sp 24 HYD 24 x x Hydracarina sp 25 HYD 25 x Hydracarina sp 26 HYD 26 x x Hydracarina sp 27 HYD 27 x x Hydracarina sp 28 HYD 28 x x Hydracarina sp 29 HYD 29 x x Hydracarina sp 30 HYD 30 x Hydracarina sp 31 HYD 31 x x Hydracarina sp 32 HYD 32 x Hydracarina sp 32 HYD 33 x Arácnida Arachnidae sp2 ARA 2 x Crustacea Cladocera Illyocryptus sp 1 MAC 3 x x x Macrothricidae sp1 MAC 1 x x Macrothricidae sp2 MAC 2 x Daphnidae sp2 DAP 2 x Bosminopsis sp1? BOS 1 x Chydoridae sp1 CHY 1 x Chydoridae sp2 CHY 2 x Chydoridae sp3 CHY 3 x x x x Chydoridae sp5 CHY 4 x Chydoridae sp6 CHY 5 x x Chydoridae sp7 CHY 6 x x Chydoridae sp8 CHY 7 x
(Continuación Tabla 3…)

MATO CAURA TAXA Código PAT POZ NAP ARI BRA
Chydoridae sp9 CHY 8 x
Copepoda Cyclopoidea COP 1 x x x x x Calanoidea COP 2 x x Branchiopoda Cyclestheria hislopi Baird CYC 1 x x x Nematoda Nematoda sp1 ME 1 x x x x Nematoda sp2 ME 2 x x Annelida (Oligochaeta) Prístina sp2 NAI 2 x x x x Naididae sp4 NAI 6 x x x x x Dero sp2 NAI 7 x Coelenterata Hydra sp1 COE 1 x x Mollusca Ancylidae sp1 ANC 1 x x Total ± Desviación estándar 372 ± 153,4 283 ± 142,6 118 ± 133,6 2354 ± 475,8 639 ± 50,91 Peso seco Hoja (g) 1,82 2,59 1,90 1,83 0,58 Área Hoja (cm2) 84,66 197,20 163,88 153,32 54,52 Org.m-2 hoja 45951 17495 5431 447698 117215 Org./100 g hojas secas 21374 13338 4676 376110 110172 Biomasa (g) ± Desviación estándar 0,25 ± 0,02 0,26 ± 0,04 0,22 ± 0,01 0,22 ± 0,03 0,22 ± 0,01 Biomasa (g) . 100g hojas 13,50 9,99 11,62 11,85 38,77 Biomasa (g.m-2) 29,52 13,18 13,42 14,34 40.35 Riqueza de Especies 29 33 21 35 22 Índice de diversidad (H´) 1,83 2,10 1,80 0,80 1,91 Equitabilidad (J) 0,44 0,50 0,43 0,19 0,46

Tabla 4. Valores de variables físico-químicas en cada una de las estaciones de muestreo establecidas en 9 transectos a lo largo del Bajo Caura durante la fase de aguas altas (8) y aguas bajas (3). Variables
Estaciones MO (g) OD
T (ºC)
COND (mS/cm) pH
TRANS(cm)
PROF (cm)
PATC8 0,172 3,0 25 22 5,73 100 460 PATO8 0,161 4,0 25 14 5,99 90 440 PATB8 0,726 3,2 25 25 5,70 110 210 PATM8 0,157 1,8 24,5 30 6,09 110 480 CEJC8 0,305 3,4 25 28 6,01 100 860 CEJO8 0,000 2,8 26 16 5,91 100 680 CEJB8 0,766 3,2 26 17 5,57 90 320 POZC8 0,000 3,4 25 10 5,62 120 610 POZO8 0,176 3,2 27 24 5,65 100 600 POZB8 0,260 3,4 27 16 5,76 90 330 POZM8 0,157 1,8 26,5 22 5,97 100 820 NAPC8 0,140 4,6 25 16 6,21 140 925 NAPO8 0,721 4,6 26 13 6,21 130 792,5 NAPB8 0,517 4,6 27 26 5,91 140 545 BRAC8 0,000 4,8 26,5 21 5,96 140 885 BRAO8 0,000 5,0 26,5 19 6,05 160 815 BRAB8 0,719 3,0 27 12 5,82 130 460 ARIC8 0,000 3,8 29 10 6,00 130 1145 ARIO8 0,745 4,3 29,5 15,5 6,09 135 1155 ARIB8 0,903 4,6 29 10 6,02 120 680 CARC8 0,000 2,8 28 12 5,79 150 960 CARO8 0,411 5,0 28,5 14 5,95 130 1020 CARB8 0,654 4,2 31 16 5,85 140 750 CARS8 0,638 4,0 31 15 5,79 130 910 CHIC8 0,000 3,4 27,5 12 5,82 150 930 CHIO8 0,000 4,8 28 10 6,05 130 900 CHIB8 0,902 3,8 29 10 5,93 140 710 CHIS8 0,369 4,0 29 14 5,76 160 980 GARC8 0,735 3,2 29 12 5,77 120 970 GARO8 0,737 3,8 29 24 5,77 110 930 GARB8 0,634 5,4 26 24 5,73 150 610 PATC3 0,21 6,6 34 12 6,97 110 150 PATO3 0,18 6,6 34 14 6,67 130 130 PATM3 0,11 7,2 28 14 6,80 60 330 CEJC3 0,29 5,6 27 20 6,73 70 200 CEJO3 0,3784 6,4 27 14 6,73 80 200 POZC3 0,6 7,0 26 14 6,36 50 180
POZO3 0,23 7,0 26 14 6,36 50 180 POZM3 0,22 7,4 26 14 6,80 70 100 NAPC3 0,07 7,8 29 20 6,39 80 185 NAPO3 7,9 7,8 29 20 6,39 60 145 BRAC3 0,155 7,4 30 7 6,18 40 250 BRAO3 0,62 7,4 30 7 6,18 40 350 ARIC3 4,065 8,0 31 10 5,63 60 215 ARIO3 6,93 8,0 31 10 5,60 40 122 CARC3 0,21 6,9 30 9 6,30 40 90 CARO3 4,9 6,9 30 9 6,30 30 82 CHIC3 1,779 6,9 30 9,5 6,20 20 90 CHIO3 15,42 6,9 30 9,5 6,20 20 80 CHIS3 5,04 6,9 30 9,5 6,20 70 90 GARC3 3,62 6,6 29 9,1 6,21 30 90 GARO3 0,9741 6,0 26 9 6,20 40 50 CEJMAD 33 4,0 22 26 6,03 78 340 POZ MAD 23,12 4,4 24,5 14 6,36 58 530 NAPMAD 51,67 3,4 26 18 5,78 110 263 BRA MAD 80,84 2,4 25 16 5,29 63 63 ARIMAD 63,95 3,2 24 16 5,81 96 200 CARMAD 217,59 3,0 28 18 5,62 90 465 CHIMAD 111,8 3,6 26 16 5,75 80 387 GARMAD 111,7 4,4 27 21 5,44 99 500 PATHOJ 1,82 3,2 25 25 5,70 110 210 POZHOJ 2,59 3,4 27 16 5,76 90 330 NAPHOJ 1,9 4,6 27 26 5,91 140 545 BRAHOJ 1,83 3,0 27 12 5,82 130 460 ARIHOJ 0,58 4,6 29 10 6,02 120 680

FIGURAS

Fig. 5. Ubicación de los transectos de estudio en el Bajo Caura. Caño Sipao: 1.Caramacatico, 2. Chiribital, 3. Garzones.; Río Caura: 4. Aricagua, 5. Brava, 6. Naparaico, Caño Mato: 7. Pozo Rico, 8. Cejal, 9. Patiquín

Fig. 6. Transecto de estaciones en cada sitio de muestreo. Los sustratos temporales fueron colectados entre las estaciones 2 y 3 (muestras de madera) y en los alrededores de la estación 3 fueron colectadas las hojas verdes del bosque inundable.
Fig. 7. Materiales utilizados en el estudio. A. Nucleador de sedimentos. B. Cuadrata y C. Cedazos de diferente tamaño de apertura.
A
B
C


Invertebrados en el Bajo Caura
0%10%20%30%40%50%60%70%80%90%
100%
maderossumergidos
Hojas verdes Hojarasca Vegetaciónacuática
sedimento
SustatosCollembola Coelenterata Turbellaria Nematoda Oligochaeta Hirudinea
Crustacea Hydracarina Insecta Mollusca
Fig. 8. Composición porcentual media de los principales grupos de invertebrados encontrados en los diferentes sustratos colectados en los transectos de estudio en el Bajo Caura.
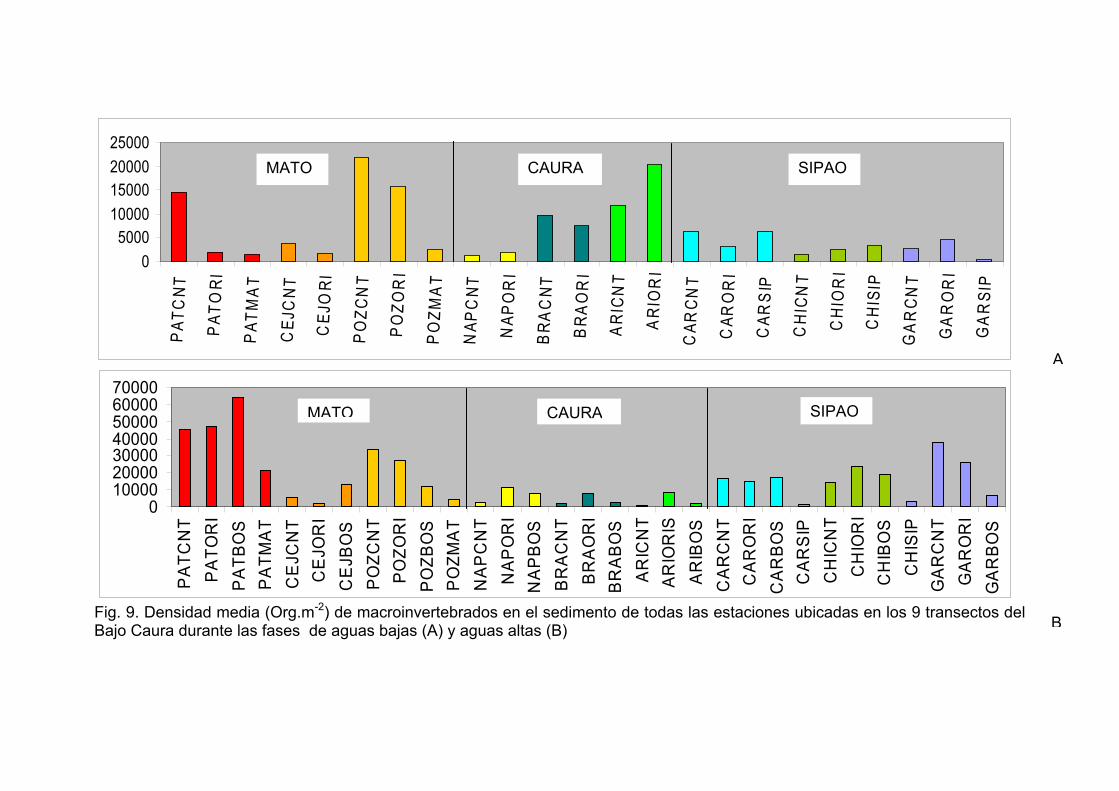
05000
10000150002000025000
PAT
CN
T
PAT
OR
I
PAT
MA
T
CEJ
CN
T
CEJ
OR
I
PO
ZCN
T
PO
ZOR
I
PO
ZMA
T
NAP
CN
T
NAP
OR
I
BR
AC
NT
BR
AO
RI
AR
ICN
T
AR
IOR
I
CAR
CN
T
CAR
OR
I
CAR
SIP
CH
ICN
T
CH
IOR
I
CH
ISIP
GA
RC
NT
GA
RO
RI
GA
RSI
P
010000200003000040000500006000070000
PA
TCN
TP
ATO
RI
PA
TBO
SP
ATM
AT
CE
JCN
TC
EJO
RI
CE
JBO
SP
OZC
NT
PO
ZOR
IP
OZB
OS
PO
ZMA
TN
AP
CN
TN
AP
OR
IN
AP
BO
SB
RA
CN
TB
RA
OR
IB
RA
BO
SA
RIC
NT
AR
IOR
ISA
RIB
OS
CA
RC
NT
CA
RO
RI
CA
RB
OS
CA
RS
IPC
HIC
NT
CH
IOR
IC
HIB
OS
CH
ISIP
GA
RC
NT
GA
RO
RI
GA
RB
OS
Fig. 9. Densidad media (Org.m-2) de macroinvertebrados en el sedimento de todas las estaciones ubicadas en los 9 transectos del Bajo Caura durante las fases de aguas bajas (A) y aguas altas (B)
MATO
MATO
CAURA SIPAO
CAURA SIPAO
A
B


Composición % del sedimento en Mato
0%20%40%60%80%
100%
patcnt patori patmat cejcnt cejori pozcnt pozori pozmat
Composición % del sedimento en Caura
0%20%40%60%80%
100%
napcnt napori bracnt braori aricnt ariori
Composición % del sedimento en Sipao
0%
50%
100%
chicnt chiori chisip carcnt carori garcnt garorimo arena arcilla limo
Fig. 10. Composición porcentual del sedimento en las estaciones de muestreo en los 9 transectos tomados durante la fase de aguas bajas.

Fig. 11. Análisis de conglomerados de la comunidad de invertebrados en el sedimento del transecto Patiquín (Caño Mato) durante aguas bajas (3) y durante aguas altas (8). PAT=Patiquín; B=Bosque inundable; C=Centro de la laguna; O=Orilla de la laguna; T=Orilla del Caño; H=Hojas. Basado en el coeficiente de disimilitud Euclidiana.
Fig. 12. Análisis de conglomerados de la comunidad de invertebrados en el sedimento del transecto Cejal (Caño Mato) durante aguas bajas (3) y durante aguas altas (8). CEJ=Cejal; B=Bosque inundable; C=Centro de la laguna; O=Orilla de la laguna; T=Orilla del Caño; M=Maderos. Basado en el coeficiente de disimilitud Euclidiana.

Fig. 13. Análisis de conglomerados de la comunidad de invertebrados en el sedimento del transecto Pozo Rico (Caño Mato) durante aguas bajas (3) y durante aguas altas (8). POZ=Pozo Rico; B=Bosque inundable; C=Centro de la laguna; O=Orilla de la laguna; T=Orilla del Caño; H=Hojas verdes del bosque de inundación; M=Maderos. Basado en el coeficiente de disimilitud Euclidiana.
Fig. 14. Análisis de conglomerados de la comunidad de invertebrados en el sedimento del transecto Brava (Río Caura) durante aguas bajas (3) y durante aguas altas (8). BRA=Brava; B=Bosque inundable; C=Centro de la laguna; O=Orilla de la laguna; T=Orilla del Caño; H=Hojas verdes del bosque de inundación; M=Maderos. Basado en el coeficiente de disimilitud Euclidiana.

Fig. 15. Análisis de conglomerados de la comunidad de invertebrados en el sedimento del transecto Naparaico (Río Caura) durante aguas bajas (3) y durante aguas altas (8). NAP=Naparaico; B=Bosque inundable; C=Centro de la laguna; O=Orilla de la laguna; T=Orilla del Caño; H=Hojas verdes del bosque de inundación; M=Maderos. Basado en el coeficiente de disimilitud Euclidiana.
Fig. 16. Análisis de conglomerados de la comunidad de invertebrados en el sedimento del transecto Aricagua (Río Caura) durante aguas bajas (3) y durante aguas altas (8). ARI=Aricagua; B=Bosque inundable; C=Centro de la laguna; O=Orilla de la laguna; T=Orilla del Caño; H=Hojas verdes del bosque de inundación; M=Maderos. Basado en el coeficiente de disimilitud Euclidiana.

Fig. 17. Análisis de conglomerados de la comunidad de invertebrados en el sedimento del transecto Garzones (Caño Sipao) durante aguas bajas (3) y durante aguas altas (8). GAR=Garzones; B=Bosque inundable; C=Centro de la laguna; O=Orilla de la laguna; T=Orilla del Caño; M=Maderos. Basado en el coeficiente de disimilitud Euclidiana.
Fig. 18. Análisis de conglomerados de la comunidad de invertebrados en el sedimento del transecto Chiribital (Caño Sipao) durante aguas bajas (3) y durante aguas altas (8). CHI=Chiribital; B=Bosque inundable; C=Centro de la laguna; O=Orilla de la laguna; T=Orilla del Caño; M=Maderos. Basado en el coeficiente de disimilitud Euclidiana.

Fig. 19. Análisis de conglomerados de la comunidad de invertebrados en el sedimento del transecto Caramacatico (Caño Sipao) durante aguas bajas (3) y durante aguas altas (8). CAR=Caramacatico; B=Bosque inundable; C=Centro de la laguna; O=Orilla de la laguna; T=Orilla del Caño; M=Maderos. Basado en el coeficiente de disimilitud Euclidiana.



PCA case scores
Axi
s 2
Axis 1
-0.2
-0.4
0.00.20.40.60.8
-0.07-0.15-0.22-0.29-0.37
0.000.070.150.220.290.37
-0.07-0.15-0.22-0.29-0.37
0.00 0.070.15
0.220.29
0.37
Fig. 20. Análisis de Componentes Principales de las variables abióticas (Materia orgánica, Conductividad, Oxígeno Disuelto, Temperatura (ºC),Transparencia, y Profundidad) tomadas en cada sitio de muestreo de las 9 transectos en el Bajo río Caura durante la Transición (subida de aguas), aguas altas y aguas bajas.
ANALISIS DE COMPONENTES PRINCIPALES de las variable abióticas Análisis de 7 variables x 84 casos Tolerancia de auto análisis set a 1E-7 Datos estandarizados Eje 1 Eje 2 Eje 3 Autovalores 2,627 1,445 1,229 Porcentaje 37,530 20,642 17,561 Porcentaje Acum. 37,530 58,172 75,733
Variables cargadas del ACP MO 0,080 -0,580 -0,438 OD 0,547 0,197 -0,123 T. (ºC) 0,325 0,413 -0,124 COND -0,220 -0,380 0,600 pH 0,307 0,196 0,627 TRANS -0,438 0,416 -0,155 PROF -0,505 0,312 -0,030
Aguas bajas Aguas
altas
Transición

Fig. 21. Análisis de Componentes Principales de las variables abióticas (Materia orgánica, Conductividad, Oxígeno Disuelto, Temperatura (ºC),Transparencia, y Profundidad) tomadas cerca del sustrato de maderos sumergidos en el Bajo río Caura durante la fase de aguas bajas. ANÁLISIS DE COMPONENTES PRINCIPALES Análisis de 7 variables x 8 casos Tolerancia de eigenanálisis set a 1E-7 Datos estandardizados Eje 1 Eje 2 Eje 3 Autovalores 2,708 1,964 1,303 Percentaje 38,679 28,059 18,610 Percentaje Acum. 38,679 66,738 85,348
ACP carga de variables MO -0,481 0,315 -0,080 OD 0,425 0,463 0,068 T. (ºC) -0,463 0,399 -0,216 COND 0,144 0,128 0,754 pH 0,516 0,121 -0,228 TRANS -0,234 0,246 0,537 PROF 0,176 0,660 -0,183

Fig. 22. Gráfico de ordenación del Análisis de Correspondencia Canónica (ACC) mostrando la distribución de los taxones y estaciones en los nueve transectos en relación con las variables ambientales (189 taxones). Cada símbolo representa un taxón individual. Cada símbolo de representa una estación individual. La proyección ortogonal del punto de las especies en las flechas ambientales representa el centro aproximado de la distribución de las especies a lo largo de un gradiente particular ambiental. Las flechas indican la dirección a la que más cambia una variable ambiental. Los valores medios de las variables ambientales son encontrados en el origen de las flechas y los valores numéricos de las variables ambientales incrementan del origen hacia la punta de la flecha. El origen corresponde a valores de 59,9 cm para la transparencia (TRANS), 17,65 mS/cm para la conductividad (COND), 5,88 (pH) y 738,1 cm (PROF) durante las fases de aguas altas y/o aguas bajas. Eje 1 Eje 2 Eje 3 Eje 4 Eje 5 Autovalores 0,359 0,203 0,175 0,143 0,134 Porcentaje 4,502 2,546 2,193 1,793 1,683 Porcentaje Acum 4,502 7,048 9,241 11,034 12,718 Porcentaje Acum. Constr. 35,398 55,419 72,662 86,763 100,000 Espec.-amb. correlaciones 0,879 0,798 0,823 0,796 0,799 Coeficientes canónicos para variables ambientales OD 0,676 0,348 -0,149 0,340 -0,053 COND -0,603 0,051 0,536 0,163 -0,194 pH 0,628 -0,140 0,322 0,319 0,305 TRANS -0,343 -0,613 0,052 0,153 -0,372 PROF 0,072 -0,474 0,199 -0,203 -0,575
Asthenopus
Climacia
Helobdella stagnalis
Chaoborus
POZO3
BRAB8
GARB8
ARIO3 BRAMAD

Eje 1 Eje 2 Eje 3 Eje 4 Eje 5 Autovalores 0,470 0,297 0,242 0,214 0,164 Porcentaje 26,461 16,717 13,608 12,028 9,222 Porcentaje Acum 26,461 43,178 56,786 68,813 78,036 Porcentaje Acum. Constr. 33,908 55,331 72,769 88,182 100,000 Espec.-amb. correlaciones 0,988 0,986 0,960 0,988 0,997 Coeficientes canónicos para variables ambientales OD 0,567 0,431 0,077 0,328 0,601 COND -0,281 0,599 0,678 0,182 0,130 pH 0,769 0,443 -0,302 -0,261 -0,156 TRANS -0,797 0,426 0,008 -0,355 0,185 PROF 0,419 0,269 0,239 -0,762 0,297 Fig. 23. Gráfico de ordenación del Análisis de Correspondencia Canónica (ACC) mostrando la distribución de los taxones y estaciones en relación con las variables ambientales (93 taxones) solo en las estaciones de maderos sumergidos. Cada símbolo representa un taxón individual. Cada símbolo representa una estación individual de los maderos. La proyección ortogonal del punto de las especies en las flechas ambientales representa el centro aproximado de la distribución de las especies a lo largo de un gradiente particular ambiental. Las flechas indican la dirección a la que más cambia una variable ambiental. Los valores medios de las variables ambientales son encontrados en el origen de las flechas y los valores numéricos de las variables ambientales incrementan del origen hacia la punta de la flecha. El origen corresponde a valores de 84,25 cm para la transparencia (TRANS), 18,1 mS/cm para la conductividad (COND), 3,5 para el pH, 63 cm para la profundidad (PROF), durante las fases de aguas bajas.

Eje 1 Eje 2 Eje 3 Eje 4 Autovalores 0,531 0,356 0,246 0,181 Porcentaje 40,405 27,082 18,745 13,768 Porcentaje Acum. 40,405 67,487 86,232 100,000 Porcentaje Acum. Constr. 40,405 67,487 86,232 100,000 Espec.-amb. correlaciones 1,000 1,000 1,000 1,000 Coeficientes canónicos para variables ambientales MO 0,657 -0,338 0,671 0,060 OD -0,193 0,176 -0,874 0,410 T. (ºC) -0,515 -0,340 -0,775 0,136 COND 0,659 0,552 0,430 0,275 pH -0,581 -0,034 -0,764 0,278 TRANS -0,733 0,306 0,168 0,583 PROF -0,649 -0,131 -0,678 0,319 Fig. 24. Gráfico de ordenación del Análisis de Correspondencia Canónica (ACC) mostrando la distribución de los taxones y estaciones en relación con las variables ambientales solo en las estaciones de hojas verdes del bosque inundable (93 taxones). Cada símbolo representa un taxón individual. Cada símbolo representa una estación individual de las hojas verdes del bosque inundado. La proyección ortogonal del punto de las especies en las flechas ambientales representa el centro aproximado de la distribución de las especies a lo largo de un gradiente particular ambiental. Las flechas indican la dirección a la que más cambia una variable ambiental. Los valores medios de las variables ambientales son encontrados en el origen de las flechas y los valores numéricos de las variables ambientales incrementan del origen hacia la punta de la flecha. El origen corresponde a valores de 118 cm para la transparencia (TRANS), 17,8 mS/cm para la conductividad (COND), 5,8 para el pH y 445 cm para la profundidad (PROF), durante las fases de aguas altas.


Fig. 25. Cambios en la biomasa (g.Org./m-²), riqueza e Índice de diversidad Shannon–Wienner (H`) media de los macroinvertebrados que viven en el sedimento a lo largo de un gradiente de cuatro transectos de inundación del Caño Mato – río Caura– durante las fases de aguas bajas y aguas altas- . LC: Centro de la laguna, LS: Orilla de la laguna, FOR: Bosque inundado.

Fig. 26. Cambios en la biomasa (g.Org.m-²), riqueza e Índice de diversidad Shannon–Wienner (H`) media de los macroinvertebrados que viven asociados a las hojas verdes del bosque de inundación del Caño Mato (tributario) y del río Caura durante la fase de aguas altas.















![PROPUESTA DE PROCEDIMIENTO FÍSICO …cidar.uneg.edu.ve/DB/bcuneg/EDOCS/TESIS/TESIS_POSTGRADO/...las soluciones de aluminato en función de la [ Na 2O] a diferentes temperaturas 22](https://static.fdocuments.ec/doc/165x107/5b5f171f7f8b9af90c8d1b0a/propuesta-de-procedimiento-fisico-cidarunegeduvedbbcunegedocstesistesispostgradolas.jpg)


