
Planeta vivo: El origen y la evolución temprana de la … · El estudio del origen y la evolución...
8
Enseñanza de las Ciencias de la Tierra, 2010 (18.1) – 25 ISSN: 1132-9157 – Pags. 25-32 FUNDAMENTOS CONCEPTUALES Y DIDáCTICOS Planeta vivo: El origen y la evolución temprana de la vida en la Tierra Living planet: the origin and early evolution of life on Earth Carlos Briones Llorente Laboratorio de Evolución Molecular. Centro de Astrobiología (CSIC-INTA, asociado al NASA Astrobiology Institute). Torrejón de Ardoz, Madrid Resumen Nuestro planeta es, por lo que sabemos hasta la fecha, el único lugar del cosmos donde existen seres vivos. El estudio del origen y la evolución temprana de la vida en la Tierra es un campo de investigación interdisciplinar muy activo, que está logrando responder a algunas de las muchas incógnitas planteadas desde hace un siglo y medio, cuando Charles R. Darwin sugirió que toda la biodiversidad observada podría ser el resultado de la evolución a través del tiempo a partir de una única forma de vida. Los primeros resultados de la química prebiótica iniciaron en 1953 el estudio sobre el origen de la vida como una disciplina experimental. Actualmente estudiamos en los laboratorios distintas etapas evolutivas desde las primeras biomoléculas hasta esa especie ancestral, que denominamos LUCA, de la que derivamos todas las demás. Diversas evidencias experimentales apoyan la hipótesis del “Mundo RNA”, según la cual esa molécula fue anterior al DNA y a las proteínas y podría postularse como el punto de partida de la evolución darwiniana. En cualquier caso, muchas son las preguntas que aún debemos responder en la investigación sobre la emergencia de la vida en la Tierra... y quizá fuera de ella. Palabras clave: Química prebiótica, superficies minerales, Mundo RNA, metabolismo, evolución, virus, células, astrobiología Abstract Our planet is, as far as we know, the only place in the universe where living beings exist. The study of the origin and early evolution of life on Earth is a very active, interdisciplinary research field. We are currently addressing some of the questions raised since, a century and a half ago, Charles R. Darwin suggested that the biodiversity observed could be the outcome of evolution from a single form of life. The first achievements of prebiotic chemistry initiated in 1953 the study of the origin of life as an experimental discipline. Nowadays, many laboratories investigate different evolutionary stages from the first biomolecules to LUCA, the common ancestor of all current species. Several experimental evidences support the “RNA World” hypothesis, which proposes that this molecule originated before DNA and proteins and that RNA could therefore represent the starting point of Darwinian evolution. In any case, there are still many open questions to be addressed in the study of the emergence of life on Earth and perhaps beyond. Keywords: Prebiotic chemistry, mineral surfaces, RNA World, metabolism, evolution, virus, cell, astrobiology INTRODUCCIóN La Astrobiología es una ciencia joven en la que convergen distintos campos de la física, la química, la geología, la biología y la ingeniería. A través de esta aproximación interdisciplinar se trata de responder a distintas cuestiones relacionadas con el origen, evo- lución y presencia de la vida en el Universo. El origen de la vida es en sí mismo un tema de investigación apasionante, que encierra preguntas complejas como cuándo ocurrió la transición de la materia inanimada a la viva, cómo se produjo ese salto cualitativo, si tuvo lugar en la Tierra o fuera de ella, y si el paso de la química a la biología puede haber sido un hecho frecuente en diferentes lugares del universo. El mundo molecular anterior a las células no deja fósiles que puedan ser analizados para re-
Transcript of Planeta vivo: El origen y la evolución temprana de la … · El estudio del origen y la evolución...

Enseñanza de las Ciencias de la Tierra, 2010 (18.1) – 25ISSN: 1132-9157 – Pags. 25-32
Fu
nd
am
ento
s
Co
nCe
ptu
ale
s
yd
idá
CtiC
osPlaneta vivo: El origen y la evolución temprana de la
vida en la Tierra
Living planet: the origin and early evolution of life on Earth
CarlosBrionesllorenteLaboratorio de Evolución Molecular. Centro de Astrobiología (CSIC-INTA, asociado al NASA Astrobiology Institute). Torrejón de Ardoz, Madrid
Resumen Nuestroplanetaes,porloquesabemoshastalafecha,elúnicolugardelcosmosdondeexistenseresvivos.ElestudiodelorigenylaevolucióntempranadelavidaenlaTierraesuncampodeinvestigacióninterdisciplinarmuyactivo,queestálograndoresponderaalgunasdelasmuchasincógnitasplanteadasdesdehaceunsigloymedio,cuandoCharlesR.Darwinsugirióquetodalabiodiversidadobservadapodríaserelresultadodelaevoluciónatravésdeltiempoapartirdeunaúnicaformadevida.Losprimerosresultadosdelaquímicaprebióticainiciaronen1953elestudiosobreelorigendelavidacomounadisciplinaexperimental.Actualmenteestudiamosenloslaboratoriosdistintasetapasevolutivasdesdelasprimerasbiomoléculashastaesaespecieancestral,quedenominamosLUCA,delaquederivamostodaslasdemás.Diversasevidenciasexperimentalesapoyanlahipótesisdel“MundoRNA”,segúnlacualesamoléculafueanterioralDNAyalasproteínasypodríapostularsecomoelpuntodepartidadelaevolucióndarwiniana.Encualquiercaso,muchassonlaspreguntasqueaúndebemosresponderenlainvestigaciónsobrelaemergenciadelavidaenlaTierra...yquizáfueradeella.
Palabrasclave:Químicaprebiótica,superficiesminerales,MundoRNA,metabolismo,evolución,virus,células,astrobiología
Abstract Our planet is, as far as we know, the only place in the universe where living beings exist. The study of the origin and early evolution of life on Earth is a very active, interdisciplinary research field. We are currently addressing some of the questions raised since, a century and a half ago, Charles R. Darwin suggested that the biodiversity observed could be the outcome of evolution from a single form of life. The first achievements of prebiotic chemistry initiated in 1953 the study of the origin of life as an experimental discipline. Nowadays, many laboratories investigate different evolutionary stages from the first biomolecules to LUCA, the common ancestor of all current species. Several experimental evidences support the “RNA World” hypothesis, which proposes that this molecule originated before DNA and proteins and that RNA could therefore represent the starting point of Darwinian evolution. In any case, there are still many open questions to be addressed in the study of the emergence of life on Earth and perhaps beyond.
Keywords: Prebiotic chemistry, mineral surfaces, RNA World, metabolism, evolution, virus, cell, astrobiology
IntroduccIón
La Astrobiología es una ciencia joven en la queconvergendistintoscamposdelafísica,laquímica,lageología,labiologíaylaingeniería.Atravésdeestaaproximacióninterdisciplinarsetrataderesponderadistintascuestionesrelacionadasconelorigen,evo-luciónypresenciadelavidaenelUniverso.Elorigende lavidaesensímismountemade investigación
apasionante,queencierrapreguntascomplejascomocuándoocurriólatransicióndelamateriainanimadaa la viva, cómo se produjo ese salto cualitativo, situvolugarenlaTierraofueradeella,ysielpasodelaquímicaa labiologíapuedehabersidounhechofrecuenteendiferenteslugaresdeluniverso.
El mundo molecular anterior a las células nodeja fósiles que puedan ser analizados para re-

26 – Enseñanza de las Ciencias de la Tierra, 2010 (18.1)
construir el origen de la vida. Así, lo que hoy sa-bemossobreaquellaépocadehacemásde3.500millonesdeañoseselresultadodelaspruebasin-directasderivadasdedosaproximacionescomple-mentarias.Laprimera líneade investigaciónes ladenominadabottom-upo“deabajohaciaarriba”,y consiste en intentar llegar a la biología a partirde una química progresivamente más compleja.Paraellosesigueelcaminoiniciadoporelfamosoexperimento de química prebiótica de Stanley L.Miller, con el que demostró que los aminoácidosque constituyen las proteínas podían obtenerseabióticamenteapartirdelosgasespotencialmentepresentes en la atmósfera terrestre primitiva (Mi-ller,1953).
Lasegundavíaeslaconocidacomotop-downo“dearribahaciaabajo”,ysebasaenlacompara-ciónde losgenomasymetabolismosde losorga-nismosactualesenbuscadecaracterísticascomu-nesquepudieronexistirenunaespeciecelulardelaquehemosderivadotodaslasdemás,denomina-da“progenote”oLUCA–del inglésLast Universal Common Ancestor–.Podemosconsiderarqueestasegunda aproximación fue iniciada por Charles R.Darwin hace un siglo y medio. En efecto, a pesarde que en esa época se desconocía la naturalezadelmaterialgenéticoy losmecanismosmolecula-resqueoperanenlosseresvivos,tantoenelpá-rrafofinalde“Elorigende lasespecies” (Darwin,1859) como en la correspondencia que mantuvoconotroscientíficosdesuépoca,Darwinyasugi-rióquelabiodiversidadqueobservabapodríaserel resultadode laevoluciónatravésdeltiempoapartirdeunaúnicaformadevida.
Para investigar sobre el origen resulta nece-sario,enprimer lugar,definirdeuna formaope-rativa lo que entendemos por vida. A pesar dequetodaslasdefinicionesdevidaresultanincom-pletasycuestionables,unadelasmásutilizadasen investigaciónes laquehapropuestoelNASAAstrobiologyInstitute:“Unservivoesunsistemaquímicoautomantenido,queevolucionacomore-sultadodesuinteracciónconelmedio”.Estadefi-niciónincluyelascaracterísticasprincipalesdelavidayenglobaa todos losseresvivoscelulares.Noobstanteestadefinición–yorasmuchas–dejaa entidades replicativas acelulares muy relevan-tesenbiología,comolosvirusylosviroides,enellímiteentrelovivoylonovivo.Unafrontera,pre-cisamente,muyinteresanteparainvestigarsobreelorigende lavida.Porotraparte,unacuestiónadicional que debemos introducir es que en losseresvivossedistingueentreelfenotipo–lasca-racterísticasvisiblesocuantificablesdelosorga-nismos,comosuformaoeltipodemetabolismoqueposeen–yelgenotipo–la informacióngené-ticaqueesresponsabledelfenotipoobservado–.
Los materIaLes de partIda: La químIca prebIótIca y Los mundos mIneraLes
Desde la formación de la Tierra, hace aproxi-madamente 4.550 millones de años (Ma), hastaque hace entre 4.200 y 3.900 Ma aparecieron lasprimeras rocas sedimentarias, la actividad geoló-gicadenuestroplaneta, juntoconel intensobom-bardeometeoríticoalqueestabasometido,produ-cía temperaturas y presiones incompatibles con laexistenciadeagua líquida.Por tanto,seconsideraquelavidadifícilmentepudooriginarseenlaTierraduranteeleónHadeano,hacemásde3.900Ma.Undato relevante es que el desbalance isotópico delcarbonopresenteenrocassedimentariasde3.800MaencontradasenIsua,Groenlandia,podríaindicarunsistema“biológico”defijacióndeCO2 (Mojzsisy col., 1996). Si esto fuera así, algún tipo de vidafotosintéticasehabríadesarrolladoentansolo100Ma,unintervaloquevariosautoresconsiderande-masiadocorto.Encualquiercaso,sesabequehace3.500 Ma la vida ya existía, y además había dadolugaraunagranvariedaddemorfologíascelularesy relaciones ecológicas. Esto lo podemos afirmarporquesehandetectadoycaracterizadomicrofósi-lesconesaantigüedadenlossílexdeApex–grupoWarrawoona,AustraliaOccidental–cuyamorfologíaescompatibleconladelascianobacteriasfilamen-tosasactuales(SchopfyPacker,1987).TambiénenWarrawoona,yenrocasdelamismaépoca,sehanencontrado estromatolitos fósiles similares a lascomunidadesdeespeciesunicelularesque formanactualmentelostapetesmicrobianos.
Con ello, la pregunta que surge es: ¿cómo sepudo pasar, en relativamente poco tiempo, de unplanetaconvulsoeinhóspitoaunavidaplenamenteestablecida? Siguiendo la escuela de pensamientoiniciadaporDarwin,AlexandrI.OparinyJohnB.S.Haldanepublicaronenlosaños20delsiglopasadosendas obras que trataron científicamente al pro-blemadelorigendelavida(Oparin,1924;Haldane,1929).Tresdécadasdespuésseprodujoelsiguientehito fundamental: la primera aproximación experi-mentalalorigendelavida.Elexperimentoqueinau-guraríaelcampodelaquímicaprebióticafuereali-zadoporStanleyL.MillerenellaboratoriodeHaroldUrey, y permitió demostrar que varios monómerosbiológicos–incluidoslosaminoácidosqueconstitu-yenlasproteínas–podíanobtenerseapartirdelosgasesprovenientesdelvulcanismoqueformabanlaatmósferaterrestreprimitiva(Miller,1953).Así,gra-ciasalaportedeenergíasuministradopordescar-gaseléctricas,CH4,NH3,H2OyH2reaccionabanparaformar,enausenciadecualquieractividadbiológi-ca,unamezcladelosmonómerosesencialesparalavidaquecomenzóadenominarse“sopaprebiótica”.Eléxitoypopularidaddelasdiferentesvariantesdeesteexperimentofueenorme,sobretodocuandosecomprobóqueelmeteoritoMurchison–unacondri-

Enseñanza de las Ciencias de la Tierra, 2010 (18.1) – 27
tacarbonáceacaídaenVictoria,Australia,en1969–conteníaentresumateriaorgánicalosaminoácidosque Miller había sintetizado en el laboratorio. Noobstante,sehademostradoquesilaatmósferapri-mitivanoeratanreductora,yenconcretoconteníaCOoCO2,laeficienciadelasíntesisdeaminoácidosen “experimentos tipo Miller” disminuye notable-mente.Por tanto,hoyendíaelpapeldeestepro-cesoenelorigendelavidasecuestionaenfunciónde las discrepancias existentes en los modelos deatmósferaprimitiva.
En la década siguiente, otro experimento re-levante de química prebiótica permitió al españolJoan Oró sintetizar abióticamente la adenina, unadelascuatrobasesnitrogenadasqueformanlosnu-cleótidos–monómerosdelosdosácidosnucleicosnaturales,DNAyRNA–.Desdeentonces,diferenteslíneasde investigaciónhan llevadoaobtenergrancantidaddemoléculaspequeñasybio-monómerosa partir de precursores químicos sencillos. En mu-chosdeestosexperimentoshasidoimprescindiblecontarconcatalizadoresinorgánicosenelproceso,desde iones metálicos en disolución hasta las su-perficies de distintos minerales o rocas. Con ellosehapuestodemanifiestoanecesidadderealizarabordajes experimentales en la intersección entrela geología, la química inorgánica y la bioquímica.Enunadeestaslíneasdetrabajosehademostradoque filosilicatos como la montmorillonita permitencatalizarlaformacióndecadenasdeRNAdehasta50nucleótidosde longitudapartirdeprecursoresmonoméricosactivados(HuangyFerris,2006).
Larelevanciadelamontmorillonita–yenmenormedidade lamicay lapirita–para laaparicióndelas macromoléculas biológicas enlaza los trabajosactualesconlahipótesisdeGrahamCairns-Smithenlosaños60sobreunMundo de arcilla.Enél,uncier-to tipo de “herencia” primitiva anterior al materialgenéticopodríahaberestadobasadaenlatransmi-sión,durantesucrecimiento,delasirregularidadespresentes en cristales de silicatos (Cairns-Smith,1965). En este contexto, también resultan relevan-teslostrabajosdeGunterWächtershäusersobreelMundo de hierro y azufre,enelcuallassuperficiesde pirita y otros sulfuros metálicos pudieron cata-lizar lapolimerizacióndepéptidosapartirdeami-noácidos,yademáshaberalbergadoalgunaformademetabolismoprimitivo(Wächtershäuser,1988).
eL orIgen de La vIda: mundo rna y mun-dos pre-rna
Apesardeloséxitosexperimentalesobtenidosenelámbitodelaquímicaprebiótica,desdelasínte-sisdelosprimerosmonómerosobio-polímeroscor-toshastaelorigendelprimersistemaquepudieraserdefinidocomoun“servivo”huboderecorrerseun largo camino. Durante las últimas décadas se
ha trabajado intensamente para intentar entendercómopudoserlaevoluciónquellevódelasprime-ras moléculas biológicas a LUCA, es decir, desdelasraícesdelárboldelavidahastaesepuntodesutroncoenquesesepararonlasramasquehanori-ginado toda la biodiversidad actual. A lo largo deltroncocomúndelavidadebieronproducirsevariosprocesos,cadaunodeloscualesconstituyeunim-portanteretocientífico:elfinaldelasíntesisdemo-nómerosbiológicos,laeleccióndelaquiralidad,elorigendelasmembranasbiológicas,laaparicióndelaprimerapoblacióndemoléculascapacesdeauto-replicarse,elestablecimientodelcódigogenético,laconstrucciónderedesmetabólicas,olaprogresivaformacióndeungenomasuficientementecomplejocomoparapermitir laevolucióndeuna“célulamí-nima”viable.
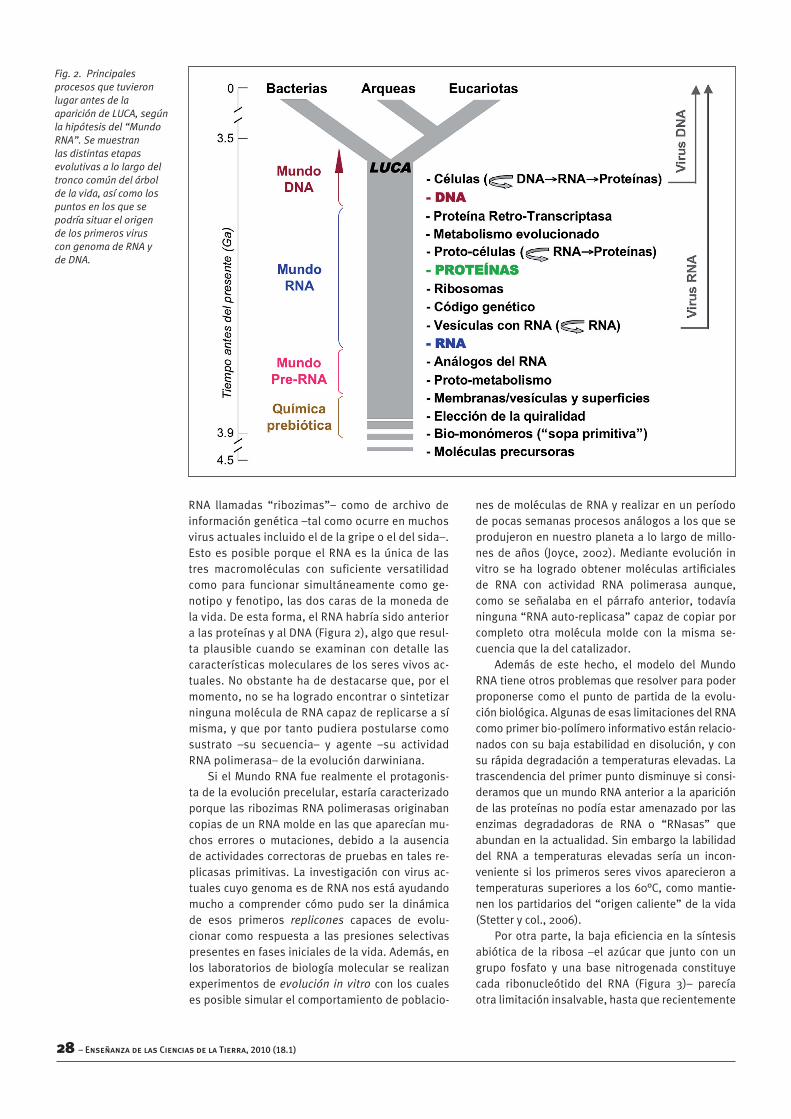
Enestecampoexisteunaparadojacentralqueresultadifícilderesolver.Entodaslascélulasac-tuales, la información genética está almacenadaenelDNA,elflujodeinformacióngenéticasepro-duceensentidoDNA→RNA→Proteína,ylamayorparte del metabolismo y funciones celulares larealizanlasproteínas.Lasproteínasestáncodifi-cadasporelDNA,yasuvezlareplicacióndelDNAnopuedellevarseacabosinproteínasconactivi-dad“DNApolimerasa”(Figura1).Entonces,¿quéaparecióprimero,elDNAolasproteínas?Estaeraunapreguntasinrespuestahastaelcomienzodela década de 1980. Desde entonces, diferentesevidencias experimentales apuntan a que la pri-meramacromoléculabiológicanofueningunadeesasdos,sino laqueen labiologíaactualactúacomo intermediaria en la expresión del mensajegenético: el RNA. La hipótesis del “Mundo RNA”(Gilbert,1986)mantienequeantesdelaapariciónde las proteínas y el DNA, el RNA podría haberdesempeñandotantofuncionescatalíticas–comohacen en las células determinadas enzimas de
Fig. 1. Representación esquemática del flujo de información genética en los seres vivos. Los procesos que tienen lugar en las células se muestran con flechas continuas, mientras que los realizados sólo por ciertos virus se indican con trazo discontinuo. La replicación del DNA y del RNA, la transcripción y la retrotranscripción son catalizadas por proteínas con la correspondiente actividad enzimática, mientras que la traducción se realiza por complejos agregados de RNA y proteínas denominados ribosomas.
28 – Enseñanza de las Ciencias de la Tierra, 2010 (18.1)
RNA llamadas “ribozimas”– como de archivo deinformacióngenética–talcomoocurreenmuchosvirusactualesincluidoeldelagripeoeldelsida–.EstoesposibleporqueelRNAes laúnicade lastres macromoléculas con suficiente versatilidadcomo para funcionar simultáneamente como ge-notipoyfenotipo,lasdoscarasdelamonedadelavida.Deestaforma,elRNAhabríasidoanterioralasproteínasyalDNA(Figura2),algoqueresul-ta plausible cuando se examinan con detalle lascaracterísticasmolecularesdelosseresvivosac-tuales.Noobstantehadedestacarseque,porelmomento,nosehalogradoencontrarosintetizarningunamoléculadeRNAcapazdereplicarseasímisma,yqueportantopudierapostularsecomosustrato –su secuencia– y agente –su actividadRNApolimerasa–delaevolucióndarwiniana.
SielMundoRNA fue realmenteelprotagonis-tadelaevoluciónprecelular,estaríacaracterizadoporque lasribozimasRNApolimerasasoriginabancopiasdeunRNAmoldeenlasqueaparecíanmu-chos errores o mutaciones, debido a la ausenciadeactividadescorrectorasdepruebasentalesre-plicasas primitivas. La investigación con virus ac-tualescuyogenomaesdeRNAnosestáayudandomucho a comprender cómo pudo ser la dinámicade esos primeros replicones capaces de evolu-cionar como respuesta a las presiones selectivaspresentesenfasesinicialesdelavida.Además,enlos laboratorios de biología molecular se realizanexperimentosdeevolución in vitro con loscualesesposiblesimularelcomportamientodepoblacio-
nesdemoléculasdeRNAyrealizarenunperíododepocassemanasprocesosanálogosalosqueseprodujeronennuestroplanetaalolargodemillo-nes de años (Joyce, 2002). Mediante evolución invitro se ha logrado obtener moléculas artificialesde RNA con actividad RNA polimerasa aunque,como se señalaba en el párrafo anterior, todavíaninguna“RNAauto-replicasa”capazdecopiarporcompleto otra molécula molde con la misma se-cuenciaqueladelcatalizador.
Además de este hecho, el modelo del MundoRNAtieneotrosproblemasqueresolverparapoderproponerse como el punto de partida de la evolu-ciónbiológica.AlgunasdeesaslimitacionesdelRNAcomoprimerbio-polímeroinformativoestánrelacio-nadosconsubajaestabilidadendisolución,yconsurápidadegradaciónatemperaturaselevadas.Latrascendenciadelprimerpuntodisminuyesiconsi-deramosqueunmundoRNAanterioralaaparicióndelasproteínasnopodíaestaramenazadoporlasenzimas degradadoras de RNA o “RNasas” queabundanenlaactualidad.Sinembargolalabilidaddel RNA a temperaturas elevadas sería un incon-veniente si los primeros seres vivos aparecieron atemperaturas superiores a los 60°C, como mantie-nen lospartidariosdel“origencaliente”de lavida(Stetterycol.,2006).
Porotraparte, labajaeficienciaen lasíntesisabiótica de la ribosa –el azúcar que junto con ungrupo fosfato y una base nitrogenada constituyecada ribonucleótido del RNA (Figura 3)– parecíaotralimitacióninsalvable,hastaquerecientemente
Fig. 2. Principales procesos que tuvieron lugar antes de la aparición de LUCA, según la hipótesis del “Mundo RNA”. Se muestran las distintas etapas evolutivas a lo largo del tronco común del árbol de la vida, así como los puntos en los que se podría situar el origen de los primeros virus con genoma de RNA y de DNA.

Enseñanza de las Ciencias de la Tierra, 2010 (18.1) – 29
sehadescritounsistemaalternativoparaobtenerlosmonómerosdelRNAapartirdeunúnicoprecur-sor sencillo (Powner y col., 2009). Otro problematienequeverconelhechodequemediantepoli-merización abiótica sobre arcillas, o en sistemasquecontienenlípidos,sehanobtenidooligómerosdeRNAdehasta40-50nucleótidos,mientrasquelasribozimasmássencillasconcapacidadRNApo-limerasaseleccionadasmedianteevolucióninvitroson relativamente complejas y tienen 180 nt delongitud.Comounposiblemecanismoquemedieen ese “salto evolutivo” necesario para el origendel Mundo RNA se ha desarrollado un modelo deevolución modular basado en la unión o ligacióninicialdemoléculascortasydesecuenciaaleatoriadeRNA.MedianteestemecanismosepermitiríalaprogresivaadicióndemódulosfuncionalesdeRNAaunamoléculaprogresivamentemáslargaycom-plejaquellegaríaaadquiriractividadRNApolime-rasa(Brionesycol.,2009).
Enparalelo,antelasaparenteslimitacionesdelRNAcomoprimeramacromoléculacapazdeportarinformaciónheredable,ydadoqueelDNAproba-blementesurgióconposterioridadaél,durantelasúltimasdécadasunalíneadeinvestigaciónimpor-tanteenquímicaorgánicaybioquímicahasidolasíntesisdeanálogosdelosácidosnucleicosnatu-rales.Entreestospolímeros,resultaránrelevantesparaelorigendelavidalosqueseanmásestablesqueelDNAyelRNA,yqueademásesténformadospor monómeros más fácilmente sintetizables quelos nucleótidos en condiciones prebióticas. En al-gunosdeelloselesqueletoazúcar-fosfatodelRNAydelDNAsehasustituidoporpolímerosdeglicerolodeciertosazúcaresmáspequeñosque laspen-tosas. Otro análogo muy investigado desde quese sintetizó a principios de la década de 1990 esel denominado ácido nucleico peptídico o “PNA”,queposeeunesqueletolongitudinalparecidoaldelas proteínas del cual sobresalen las bases nitro-genadas en la conformación adecuada para inte-raccionarespecíficamenteconlosácidosnucleicosnaturales(Figura3).
Por tanto, tal vez algún polímero informativosimilar al PNA u otros análogos protagonizó un“Mundo pre-RNA” capaz de resistir las rigurosaspresiones selectivas de hace unos 3.800 Ma, yposteriormente transferir al RNA la informaciónseleccionadahastaentonces(Joyce,2002;DelayeyLazcano,2005).Aligualqueenotrasetapasdelcaminohacialavida,enelámbitodelosanálogosdeácidosnucleicosestentadorpensarendistin-tos orígenes más o menos simultáneos de otrastantas moléculas con información heredable. Noobstante,haderecordarsequeningunodeestosposiblespolímerospre-RNAprevaleceenlabiolo-gíaactual,porloqueseasumequesudecliveyase había producido mucho antes de la aparicióndeLUCA.
Los compartImentos y eL metaboLIsmo
ComoyademostróC.R.Darwin,lavidaconllevaevolución.ElgenetistaTheodosiusDobzhanskysin-tetizóestehechoenelenunciadoquedatítuloaunodesustrabajos:“Nadaenbiologíatienesentidosinoesalaluzdelaevolución”(Dobzhansky,1973).Talevidenciasólolacuestionanhoyendíalosparti-dariosdelcreacionismoo“diseñointeligente”,conprejuicios basados en sus propias creencias y noenargumentocientíficoalguno.Así,elorigendelavidadebiósuponertambiénelpuntodepartidadela evolución. Y, tal como hemos indicado, quizá elRNAfuelaprimeramoléculacapazdeevolucionar.Pero, para intentar entender el origen de la vida yde laevolución,necesitamosdos ingredientesadi-cionales,tannecesarioscomolareplicacióndeunamoléculainformativa:laexistenciadeuncomparti-mentocapazdediferenciarelindividuodesuentor-no,yladisponibilidaddeunmetabolismograciasalcual el “ser vivo” intercambiaría materia y energíaconelmedio.
Lavidaactualestásiemprecompartimentada,yessabidoquelacélulaconstituyelaunidadestruc-turalyfuncionaldelosorganismos.Así,asumimosqueyaenlasprimerasetapasdelavidaalgúntipode compartimento debió permitir la individualiza-ción de la entidad capaz de evolucionar frente almedio en el que lo hacía. Esto permitiría un com-portamiento termodinámico muy diferente dentrodelservivo–enelcuallamateriaseordenabayla
Fig. 3: Representación esquemática del RNA y el PNA, mostrando su capacidad para interaccionar específicamente a través de puentes de hidrógeno entre sus bases nitrogenadas. Se señala un monómero de cada uno de los polímeros: un ribonucleótido del RNA –que se une a sus vecinos mediante enlaces éster-fosfato– y una unidad de N-(2-aminoetil)glicina del PNA –que se enlaza mediante enlaces peptídicos a los monómeros contiguos–.

30 – Enseñanza de las Ciencias de la Tierra, 2010 (18.1)
entropía descendía– y en su entorno. Además, laexistenciadeuncompartimentoevitabaladilucióndelasbiomoléculasdeesaproto-célulaenelmedio,conloquesemejorabanlosparámetroscinéticosysegarantizabaelmantenimientode laredde inte-raccionesquímicasnecesariasparalasupervivenciadelsistemareplicativo.
Lacompartimentacióndelossistemasquímicosprogresivamentemáscomplejospudolograrsegra-ciasalaexistenciademicroambientesenlasuperfi-cieoelinteriordedeterminadosmineralesorocas.Enestesentido,resultasugerente lapropuestadeunsistemadecompartimentosmineralesformadosporsulfurosdehierroyasociadosalassurgenciashidrotermalessubmarinas,quepodríanhaberalber-gadolasreaccionesmetabólicasnecesariasparalasprimerasetapasdelaevoluciónprecelular(MartinyRusell,2003).
Alternativamente,losprimeroscompartimentospudieronestar formadosporsustanciasanfifílicas,que en un medio acuoso originarían espontánea-mente vesículas capaces de encerrar al sistemaevolutivotalcomoocurreenlabiosferaactualconlos lípidos que forman las membranas biológicas.Moléculasmássimplesqueloslípidosdenuestrasmembranas, como por ejemplo ciertos ácidos gra-sosdecadenacorta,sehanpropuestocomocandi-datosplausiblesparaformarlasprimerasbicapasyvesículas(Segréycol.,2001).Laexistenciadeunamembrana –en la cual podrían insertarse algunasmoléculascapacesdeaumentarsupermeabilidad,comopéptidoscortosdesecuenciaaleatoria–per-mitiríaelintercambiodesustanciasentreelservivoysuentorno,yconellolaaparicióndelosprimerosgradientes moleculares utilizables para la produc-cióndeenergíaquímica.
Un sistema semipermeable de separación entreel ser vivo y su entorno lleva consigo, por tanto, laposibilidaddeacoplar losflujosdemateriaconsis-temas de obtención de energía (Mitchell, 1961). Sepermitiríaasíelestablecimientodeunproto-metabo-lismocapazdegarantizar laviabilidadyautonomíadelsistemaevolutivo.Dehecho,lapropiareplicaciónde una molécula informativa como el RNA requieredealgúntipodemetabolismobasalquegaranticeelaporteconstantehaciaelinteriordelcompartimentode los ribonucleótidos necesarios para que actuaralahipotéticaribozimaRNA-polimerasadelaqueha-blábamosenelapartadoanterior.Laaparicióndelasprimeras rutasmetabólicasen lasqueseasociaranfuncionalmentedistintasreaccionesquímicasiríaha-ciendo progresivamente más compleja y robusta lareddeinteraccionesnecesariasparaextraerenergíade los distintos tipos de moléculas presentes en elmedio,ytambiénparagarantizarladisponibilidaddemonómerosrequeridosenelmantenimientodelflujodelainformacióngenética(Shapiro,2007).
Durantedécadas,unaprofundacontroversiahamantenidoseparadasdosescuelascientíficasenel
ámbitodelorigende lavida: lospartidariosde la“replicacióntemprana”,conelmodelodelMundoRNAcomobandera,ylosquepostulabanun“me-tabolismo primordial” anterior a la evolución demoléculasconinformaciónheredable.Afortunada-mente,enlosúltimosañosseestáproduciendolaesperada síntesis entre ambas aproximaciones, yactualmenteseinvestigaconvesículasquecontie-nenmoléculasdeRNAyposeenpotencialidadparaser metabólicamente viables (Ricardo y Szostak,2009).
Luca y La evoLucIón de Los prImeros or-ganIsmos
Sipudiéramostrazar lagenealogíade todas lasespecies que constituyen la biodiversidad actualcomprobaríamosquelatotalidaddelosseresvivoscompartimoselprimerodenuestrosancestros:LUCA.AunquenuncapodremosconoceraLUCAendetalle,sísabemosqueesaespecieeraunmicroorganismoconmembranalipídicaysinnúcleodefinido.Enellayasehabíaestablecidoelsentidodelflujodeinfor-mación genética como DNA→RNA→Proteína, y sumetabolismocentraleraelmismoquecompartimostodoslosorganismosactuales.
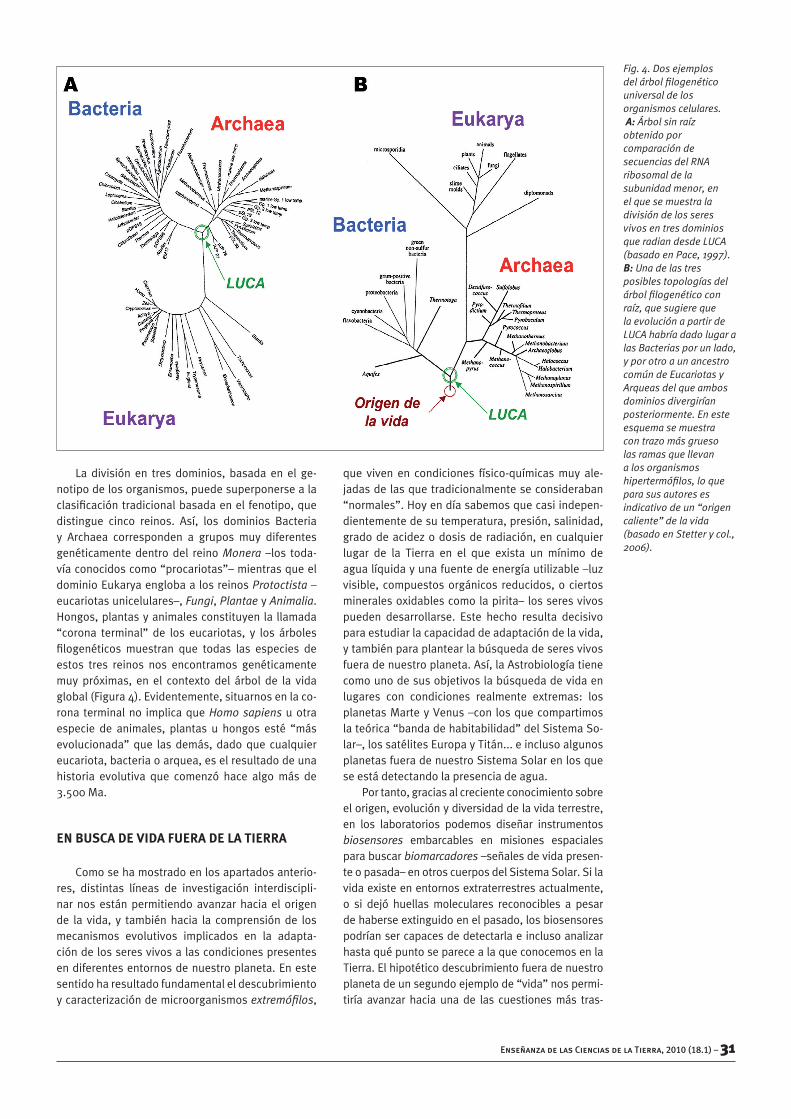
Laexistenciadenuestroantepasadocomúnsedemostrógraciasalacomparacióndelasecuenciadenucleótidosdedeterminadosgenes–fundamen-talmente, el que codifica el RNA de la subunidadmenordelosribosomas–entodaslasespeciesco-nocidas(Woeseycol.,1990;Pace,1997).Estameto-dología,quefueposiblehacesólotresdécadasgra-cias a los avances producidos en la secuenciaciónautomática de ácidos nucleicos, permitió determi-narsinningúngénerodedudasquetodoslosorga-nismoscompartimoseseancestrocomúnsugeridoporDarwin,apartirdelcualhemosevolucionadoentres grandes ramas diferentes correspondientes alosgrandesdominiosfilogenéticosactuales:Bacte-ria,ArchaeayEukarya(Figura4).
Bacteriasyarqueassonseresvivosunicelularesquecarecendenúcleodefinidoyposeenorganiza-cióndetipo“procariótico”.EldominioEukaryaeng-lobaa todos loseucariotas:organismosunicelula-resopluricelularescuyainformacióngenéticaestáalmacenadaenunnúcleocelulardiferenciado,yqueengeneralposeenuncomplejosistemademembra-nasyorgánulosespecializados.Hoyendíaseasu-mequeelnúcleoeucarióticoseoriginóporinterna-lizacióndeunaarqueadentrodeunabacteria.Porsu parte, el origen de las mitocondrias –orgánulosfundamentales para el metabolismo presentes enprácticamentetodosloseucariotas–yloscloroplas-tos–orgánulosfotosintéticosdeplantasyalgas–sedebióaeventosposterioresde“endosimbiosis”dedistintos tiposdebacteriasenel citoplasmaeuca-riótico(MargulisyChapman,1998).
Enseñanza de las Ciencias de la Tierra, 2010 (18.1) – 31
La división en tres dominios, basada en el ge-notipodelosorganismos,puedesuperponersealaclasificacióntradicionalbasadaenelfenotipo,quedistingue cinco reinos. Así, los dominios Bacteriay Archaea corresponden a grupos muy diferentesgenéticamente dentro del reino Monera –los toda-víaconocidoscomo“procariotas”–mientrasqueeldominioEukaryaenglobaalosreinosProtoctista–eucariotasunicelulares–,Fungi,PlantaeyAnimalia.Hongos,plantasyanimalesconstituyenla llamada“corona terminal” de los eucariotas, y los árbolesfilogenéticos muestran que todas las especies deestos tres reinos nos encontramos genéticamentemuy próximas, en el contexto del árbol de la vidaglobal(Figura4).Evidentemente,situarnosenlaco-rona terminalno implicaqueHomo sapiensuotraespecie de animales, plantas u hongos esté “másevolucionada” que las demás, dado que cualquiereucariota,bacteriaoarquea,eselresultadodeunahistoria evolutiva que comenzó hace algo más de3.500Ma.
en busca de vIda fuera de La tIerra
Comosehamostradoenlosapartadosanterio-res, distintas líneas de investigación interdiscipli-nar nos están permitiendo avanzar hacia el origende la vida, y también hacia la comprensión de losmecanismos evolutivos implicados en la adapta-cióndelosseresvivosalascondicionespresentesendiferentesentornosdenuestroplaneta.Enestesentidoharesultadofundamentaleldescubrimientoycaracterizacióndemicroorganismosextremófilos,
que viven en condiciones físico-químicas muy ale-jadasdelasquetradicionalmenteseconsideraban“normales”.Hoyendíasabemosquecasiindepen-dientementedesutemperatura,presión,salinidad,gradodeacidezodosisderadiación,encualquierlugar de la Tierra en el que exista un mínimo deagualíquidayunafuentedeenergíautilizable–luzvisible, compuestos orgánicos reducidos, o ciertosmineralesoxidablescomolapirita–losseresvivospueden desarrollarse. Este hecho resulta decisivoparaestudiarlacapacidaddeadaptacióndelavida,ytambiénparaplantearlabúsquedadeseresvivosfueradenuestroplaneta.Así,laAstrobiologíatienecomounodesusobjetivoslabúsquedadevidaenlugares con condiciones realmente extremas: losplanetasMarteyVenus–con losquecompartimoslateórica“bandadehabitabilidad”delSistemaSo-lar–,lossatélitesEuropayTitán...einclusoalgunosplanetasfueradenuestroSistemaSolarenlosqueseestádetectandolapresenciadeagua.
Portanto,graciasalcrecienteconocimientosobreelorigen,evoluciónydiversidaddelavidaterrestre,en los laboratorios podemos diseñar instrumentosbiosensores embarcables en misiones espacialesparabuscarbiomarcadores–señalesdevidapresen-teopasada–enotroscuerposdelSistemaSolar.Silavidaexisteenentornosextraterrestresactualmente,o si dejó huellas moleculares reconocibles a pesardehaberseextinguidoenelpasado,losbiosensorespodríansercapacesdedetectarlaeinclusoanalizarhastaquépuntoseparecealaqueconocemosenlaTierra.Elhipotéticodescubrimientofueradenuestroplanetadeunsegundoejemplode“vida”nospermi-tiría avanzar hacia una de las cuestiones más tras-
Fig. 4. Dos ejemplos del árbol filogenético universal de los organismos celulares. A: Árbol sin raíz obtenido por comparación de secuencias del RNA ribosomal de la subunidad menor, en el que se muestra la división de los seres vivos en tres dominios que radian desde LUCA (basado en Pace, 1997). B: Una de las tres posibles topologías del árbol filogenético con raíz, que sugiere que la evolución a partir de LUCA habría dado lugar a las Bacterias por un lado, y por otro a un ancestro común de Eucariotas y Arqueas del que ambos dominios divergirían posteriormente. En este esquema se muestra con trazo más grueso las ramas que llevan a los organismos hipertermófilos, lo que para sus autores es indicativo de un “origen caliente” de la vida (basado en Stetter y col., 2006).

32 – Enseñanza de las Ciencias de la Tierra, 2010 (18.1)
cendentales que están implícitas en el estudio delorigen:¿laemergenciadelavidapuedeserunhechofrecuente en los lugares cuyas condiciones físico-químicas lo permiten? O, expresado de otra forma:¿habitamosenunode losmuchos“planetasvivos”queexistenenelUniverso?
bIbLIografía
Briones,C.,Stich,M.,Manrubia,S.C.(2009).ThedawnoftheRNAworld:Towardfunctionalcomplexitythroughli-gationofrandomRNAoligomers.RNA,15,743-749.
Cairns-Smith,A.G.(1965).Theoriginoflifeandthena-tureoftheprimitivegene.J. Theoret. Biol.,10,53-88.
Darwin,C.R.The origin of species by means of natural selection, Londres, John Murray, 1859. [Versión recienteenespañol:C.R.Darwin,El origen de las especies,EspasaCalpe,Madrid,2009]
Delaye, L., Lazcano, A. (2005). Prebiological evolutionandthephysicsoftheoriginoflife.Phys. Life Rev.,2,47-64.
Dobzhansky, T. (1973). Nothing in biology makessenseexceptinthelightofevolution.The American Biol-ogy Teacher,35,125-129.
Gilbert, W. (1986). Origin of life: The RNA world. Na-ture,319,618.
Haldane,J.B.S.(1929).Theoriginoflife.Rationalist An-nual,148,3-10.
Huang,W.,Ferris,J.P.(2006).One-step,regioselectivesynthesisofupto50-mersofRNAoligomersbymontmo-rillonitecatalysis.J. Am. Chem. Soc.,128,8914-8919.
Joyce,G.F.(2002).TheantiquityofRNA-basedevolu-tion.Nature,418,214-221.
Martin,W.,Rusell,M.J.(2003).Ontheoriginsofcells:ahypothesis for theevolutionarytransitions fromabioticgeochemistrytochemoautotrophicprokaryotes,andfromprokaryotes to nucleated cells. Phil. Trans. R. Soc. Lond. B,358,59-85.
Margulis,L.,Chapman,M.J.(1998).Endosymbioses:cycli-calandpermanentinevolution.TrendsMicrobiol.6:342-346.
Miller,S.L.(1953).AproductionofaminoacidsunderpossibleprimitiveEarthconditions.Science,117,528-529.
Mitchell, P. (1961). Coupling of phosphorylation toelectronandhydrogentransferbyachemi-osmotictypeofmechanism.Nature,191,144-148.
Mojzsis, S.J., Arrhenius, G., McKeegan, K.D., Harrison,T.M., Nutman, A.P., Friend, C.R.L. (1996). Evidence for lifeonEarthbefore3,800millionyearsago.Nature,384,55-59.
Oparin,A.I.Proskhodenie zhisni,MoscovksiiRabotch-ii,Moscú,1924.[Primeratraducciónalinglés:A.I.Oparin,The origin of life,MacMillan,NewYork,1938;Versiónenespañol: A.I. Oparin, El origen de la vida, Akal, Madrid,1989]
Pace,N.(1997).Amolecularviewofmicrobialdiversityandthebiosphere.Science,276,734-740.
Powner, M.W., Gerland, B., Sutherland, J.D. (2009).Synthesisofactivatedpyrimidineribonucleotidesinprebi-oticallyplausibleconditions.Nature,459,239-242.
Ricardo,A.,Szostak,J.W.(2009).Elorigendelavida.Investigación y Ciencia398:38-46.
Segré,D.,Ben-Eli,D.,Deamer,D.W.,Lancet,D.(2001).Thelipidworld.Orig. Life Evol. Biosph.,31,119-145.
Schopf,W.,Packer,B.M.(1987).EarlyArchean(3.3-bil-lionto3.5-billion-year-old)microfossilsfromWarrawoonagroup,Australia.Science,237,70-73.
Shapiro,R.(2007).Elorigendelavida.Investigación y Ciencia,371,18-25.
Stetter,K.O.(2006).Hyperthermophilesinthehistoryoflife.Phil. Trans. R. Soc. Lond. B,361,1837-1842.
Wächtershäuser,G.(1988).Beforeenzymesandtem-plates:theoryofsurfacemetabolism.Microbiol. Rev.,52,452-484.
Woese, C.R., Kandler, O., Wheelis, M.L. (1990). To-wardsanaturalsystemoforganisms:Proposalforthedo-mainsArchaea,BacteriaandEucarya.Proc. Natl. Acad. Sci. USA,87,4576-4579.n
Este artículo fue solicitado desde E.C.T. el día 12 de enero de 2010 y aceptado definitivamente para su publicación el 1 de junio de 2010.


















