Osteología de Las Aves
14
OSTEOLOGÍA DE LAS AVES Muchas aves domésticas, particularmente las que pertenecen a los órdenes Galliformes (gallinas, pavos, etc.) y Anseriformes (patos, gansos, etc.), se han hecho terrestres dada la domesticación, pero poseen un esqueleto básicamente adaptado para el vuelo, como el de sus inmediatos antecesores. La evolución que han sufrido las aves se pone de manifiesto al observar las que son totalmente terrestres y apreciar en sus esqueletos (avestruces, emú o dromeos, etc.), que derivan secundariamente de los ancestrales voladores. La ligereza y la fuerza son los signos más importantes en los esqueletos de las aves. La fuerza y la rigidez se mantienen mediante la fusión (y a menudo la deleción concomitante) de los huesos. La fusión es menos importante en regiones como el cráneo y el tinturen pelviano, pero la fusión y la deleción ocurren en amplia extensión en los huesos de las alas y del miembro pelviano, donde los carpianos, metacarpianos y falanges de las alas y tarsianos y metatarsianos de las patas, la presentan muy manifiestamente. El ^squeleto es ligero por prolongación de gran cantidad de sacos aéreos en los huesos. Estos sacos aéreos sustituyen a la medula en muchos de los huesos de los miembros, y las extensiones adicionales puede que invadan partes del cráneo, de la columna vertebral y del cinturón pelviano. De forma arbitraria, el esqueleto se divide en parte axial, compuesto por el cráneo (incluido el aparato hioideo), columna vertebral, costillas y esternón; y un esqueleto apendicular constituido por los huesos de los miembros y los cinturones pectoral y pelviano. CRÁNEO Las aves poseen uno de los cráneos más altamente especializados entre los vertebrados vivientes. La fusión de los elementos más importantes a menudo hace que no existan las estructuras de las suturas originales, excepto en el embrión; la neumatización del cráneo puede originarse por las extensiones epiteliales de los sacos aéreos, en muchas zonas y, por tanto, produce unas estructuras extremadamente ligeras. Además de los hechos antes citados resulta que el cráneo es cinético. Todas las aves poseen un cráneo cinético (al menos en algún grado) de forma que la mandíbula superior es móvil, hacia arriba y abajo y se articula con la caja encefálica por medio de la articulación móvil, conocida como charnela frontonasal (cráneo-frontal). Como consecuencia de este mecanismo cinético el cuadrado es móvil en su articulación con la caja encefálica y está conectado indirectamente con la mandíbula superior por los huesos palatales, que son el pterigoideo y el palatino, los cuales se deslizan a lo largo del rostrum parasfenoidal. Un vómer simple se localiza en el plano medio de la región palatal rostral. Cuando el cuadrado rota rostralmente, los huesos palatales empujan rostralmente sobre la porción caudoventral de los maxilares y, por tanto, producen la rotación de la mandíbula superior hacia arriba, en su punta. Contrariamente, cuando el cuadrado rota caudalmente, se retrae el paladar y en consecuencia se deprime la mandíbula inferior. Una serie compleja de músculos operan este mecanismo cinético, que permite a las aves una extensión más ancha que la que normalmente sería posible y un mecanismo de cierre del pico más rápido; además, permite que el cráneo mantenga su eje primario aunque se mueva la mandíbula.
Transcript of Osteología de Las Aves

OSTEOLOGÍA DE LAS AVES
Muchas aves domésticas, particularmente las que pertenecen a los órdenes Galliformes (gallinas, pavos, etc.) y Anseriformes (patos, gansos, etc.), se han hecho terrestres dada la domesticación, pero poseen un esqueleto básicamente adaptado para el vuelo, como el de sus inmediatos antecesores. La evolución que han sufrido las aves se pone de manifiesto al observar las que son totalmente terrestres y apreciar en sus esqueletos (avestruces, emú o dromeos, etc.), que derivan secundariamente de los ancestrales voladores.
La ligereza y la fuerza son los signos más importantes en los esqueletos de las aves. La fuerza y la rigidez se mantienen mediante la fusión (y a menudo la deleción concomitante) de los huesos. La fusión es menos importante en regiones como el cráneo y el tinturen pelviano, pero la fusión y la deleción ocurren en amplia extensión en los huesos de las alas y del miembro pelviano, donde los carpianos, metacarpianos y falanges de las alas y tarsianos y metatarsianos de las patas, la presentan muy manifiestamente. El ^squeleto es ligero por prolongación de gran cantidad de sacos aéreos en los huesos. Estos sacos aéreos sustituyen a la medula en muchos de los huesos de los miembros, y las extensiones adicionales puede que invadan partes del cráneo, de la columna vertebral y del cinturón pelviano.
De forma arbitraria, el esqueleto se divide en parte axial, compuesto por el cráneo (incluido el aparato hioideo), columna vertebral, costillas y esternón; y un esqueleto apendicular constituido por los huesos de los miembros y los cinturones pectoral y pelviano.
CRÁNEO
Las aves poseen uno de los cráneos más altamente especializados entre los vertebrados vivientes. La fusión de los elementos más importantes a menudo hace que no existan las estructuras de las suturas originales, excepto en el embrión; la neumatización del cráneo
puede originarse por las extensiones epiteliales de los sacos aéreos, en muchas zonas y, por tanto, produce unas estructuras extremadamente ligeras. Además de los hechos antes citados resulta que el cráneo es cinético.
Todas las aves poseen un cráneo cinético (al menos en algún grado) de forma que la mandíbula superior es móvil, hacia arriba y abajo y se articula con la caja encefálica por medio de la articulación móvil, conocida como charnela frontonasal (cráneo-frontal). Como
consecuencia de este mecanismo cinético el cuadrado es móvil en su articulación con la caja encefálica y está conectado indirectamente con la mandíbula superior por los huesos palatales, que son el pterigoideo y el palatino, los cuales se deslizan a lo largo del rostrum parasfenoidal. Un vómer simple se localiza en el plano medio de la región palatal rostral. Cuando el cuadrado rota rostralmente, los huesos palatales empujan rostralmente sobre la porción caudoventral de los maxilares y, por tanto, producen la rotación de la mandíbula superior hacia arriba, en su punta. Contrariamente, cuando el cuadrado rota caudalmente, se retrae el paladar y en consecuencia se deprime la mandíbula inferior. Una serie compleja de músculos operan este mecanismo cinético, que permite a las aves una extensión más ancha que la que normalmente sería posible y un mecanismo de cierre del pico más rápido; además, permite que el cráneo mantenga su eje primario aunque se mueva la mandíbula.

Todas las aves gallifor caudoventral de los maxilares y, por tanto, producen la rotación de la mandíbula superior hacia arriba, en su punta. Contrariamente, cuando el cuadrado rota caudalmente, se retrae el paladar y en consecuencia se deprime cinético, que permite a las aves una extensión más ancha que la que normalmente sería posible y un mecanismo de cierre del pico más rápido; además, permite que el cráneo mantenga su eje primario aunque se mueva la mandíbula. Todas las aves galliformes y anseriformes tienen la cinética craneal bien desarrollada.
Las aves se caracterizan también por sus ojos extraordinariamente desarrollados, que se acomodan a las grandes órbitas del cráneo. Los dos huesos orbitales están separados por un tabique óseo, llamado septum interorbital, que deriva de los elementos me-setmoideos. El foramen óptico, que es bastante grande, se localiza en el borde caudal del septum. La pared

caudal de la órbita deriva de varios elementos esfenoidales que se unen y hacen inseparables, en el adulto. El globo ocular, propiamente dicho, también posee un esqueleto óseo, el anillo esclerótico, esto es, una serie de placas óseas solapadas que forman un anillo en la esclerótica del ojo.
Los lagrimales (prefrontales), que se localizan en los bordes rostrales de las órbitas, se articulan fundamentalmente con las ramas laterales de los nasales, y en menor grado con los frontales. En varios grupos de aves pueden separarse los huesos lagrimal y etmoidal. Puede que exista en algunas aves un solo hueso lagrimal, pero haya uno o dos, éstos se unen con otros elementos en la pared orbital rostral. En la mayor parte de las anátidas, los etmoides no están presentes, pero los lagrimales están bien desarrollados. En las aves galliformes existen ambos huesos, pero el etmoides es muy pequeño, frágil y, normalmente, se destruye al preparar el esqueleto; el lagrimal, en la gallina, es un hueso muy importante del borde rostral de la órbita.

Los orificios externos de la nariz de las aves anseriformes y galliformes se denominan «holorhinales»; caudalmente, se extienden a la parte correspondiente de la charnela nasofrontal*.
Las ramas laterales de los nasales (ventral-mente llamadas apófisis maxilares de los nasales) se unen ventralmente con los maxilares. Los maxilares, en las aves, están muy reducidos si se comparan con otros grupos de vertebrados y son huesos, generalmente, pequeños que se extienden medialmente como las apófisis mediales de los maxilares. Lateralmente, los maxilares dan origen a huesos delgados, semejantes a palillos, que son las apófisis cigomáticas, unidas a los huesos yugales para formar la porción rostral de los arcos cigomáticos. La parte caudal de cada arco cigomático está formada por otro hueso también semejante a un palillo, el cuadrato-yugal, que se articula caudalmente con el hueso cuadrado. El hueso cuadrado es complejo en todas las aves y sirve, no solamente de suspensor de la mandíbula, sino que se articula con la mandíbula inferior y, además, los cóndilos del cuadrado se articulan con la mandíbula inferior (mandíbula), medialmente con el pterigoideo y, dorsalmente, mediante una apófisis ótica, con el hueso escamoso, rostral al meato auditivo externo (cavidad timpánica). Además, una apófisis orbital del hueso cuadrado se extiende dentro de la órbita y sirve para el origen de un músculo importante en el cierre de la mandíbula inferior.
Se observan distintos centros «óticos» de osificación que rodean y protegen el oído interno aviar, y una columna auris fija en la ventana oval, que se localiza entre el proótico y el opistótico. Estos componentes óticos no se observan con facilidad desde el exterior del cráneo. La membrana timpánica está conectada por una serie de ligamentos y cartílagos extracolumnares a los estribos óseos. Las ondas sonoras se transmiten, por tanto, desde el tímpano del oído a través de la columna auris a su lámina de sostén, donde se transforman en ondas de presión que entran en la perilinfa del oído interno.
La parte caudal del cráneo está formada por una serie de huesos que se unen en el adulto. Inmediatamente caudal al parietal está el supraoccipital mediano; surge de los centros de osificación bilateral que se unen en un simple elemento en el adulto. El supraoccipital forma el borde dorsal del foramen magnum, que está bordeado lateralmente por un par de exoccipitales y ventralmente por un basoccipital. Contribuye, fundamentalmente, con un cóndilo occipital que caracteriza a las aves y a la mayoría de los reptiles. Cada uno de los exoccipitales contribuye ligeramente a formar el cóndilo occipital y los bordes caudoventral y caudales de la cavidad timpánica. Los exoccipitales se unen rostralmente con el basisfenoides mediano, que aquí está representado por una lámina basitemporal, hueso plano que forma la base caudal del cráneo.
Muchos de los huesos del neurocráneo se unen, generalmente, en el adulto y pueden tener incluso pocos vestigios de las estructuras de suturas originales; sin embargo, regiones muy generalizadas son fácilmente discernibles, como un par de huesos frontales, extensos dorsalmente, y unos parietales más pequeños, caudalmente que, como los frontales, constituyen el suelo de la cavidad. El borde total dorsal de la órbita está formado por la prolongación lateral de los huesos frontales. Los huesos nasales se articulan con el borde rostral de los huesos frontales en la charnela nasofrontal. Los nasales son huesos complejos en todas las aves. Constituyen el techo de la cavidad nasal y su porción rostral es redonda, extendida rostroventralmente como una rama lateral para formar el borde caudal de la nariz externa. Rostralmente, los nasales se unen a las apófisis nasales de los premaxilares,

medialmente. Estas apófisis se unen a lo largo de la línea media dorsal de la región nasal. Las apófisis rostromediales de los nasales (apófisis premaxilares) forman, por tanto, los bordes dorsocaudales de la nariz externa. Sus ramas laterales se extienden rostroventralmente hasta encontrarse con las apófisis maxilares de los premaxilares y formar el borde rostro-ventral de la nariz externa.
MANDÍBULA INFERIOR
Cinco huesos, generalmente, son los que contribuyen a la formación de la mandíbula inferior; en el adulto sus suturas están enmascaradas por la osificación. El hueso mayor y más rostral de la mandíbula inferior es el dentario, que forma una sínfisis mandibular, distalmente. Otros componentes pueden incluir los huesos esplenio, angular, sub-angular, prearticular y el articular que, como su nombre indica, se articula con los cóndilos del cuadrado. En las aves como en sus predecesores los reptiles, existe un foramen mandibular caudal al hueso dentario.
APARATO HIOIDES
El aparato hioides se describe con detalle en el capítulo 63 y, por tanto, no se repetirá aquí.
COLUMNA VERTEBRAL
El único cóndilo occipital del cráneo se articula con el atlas, la I de las vértebras cervicales. El hueso atlas tiene forma de anillo, presenta la superficie craneal profundamente cóncava para articularse con el con-
dilo occipital; caudalmente, existen tres caritas articulares que se conectan con el axis o II vértebra cervical. El axis, como en todos los vertebrados, envía su apófisis odontoides
cranealmente para ponerse en contacto con el cóndilo occipital y el atlas. Con la excepción del atlas el resto de la columna vertebral está formada de vértebras heterocelas, con las llamadas superficies articulares en forma de silla de montar. Hay excepciones, como, por ejemplo, los loros que tienen vértebras opistocelas (con superficies caudales cóncavas) en la región torácica. Del mismo modo que la mayor parte de los vertebrados, las vértebras se articulan una con otra por medio de articulaciones sinoviales, entre las cigapófisis, y por una articulación con los centros sucesivos. No es usual que las aves tengan numerosas vértebras cervicales fusionadas; comúnmente las tienen en la región torácica y siempre en las regiones sacra y caudal.
Las vértebras son algo más arbitrariamente asignadas a las regiones que en los mamíferos. En general, las vértebras cervicales se pueden considerar como las que empiezan con el atlas y terminan con una costilla esternal para comenzar la I torácica. En la gallina las dos últimas vértebras cervicales poseen los segmentos vertebrales de las costillas.

Las vértebras torácicas, dorsales o de la espalda, son normalmente cuatro o seis. Pueden
ser definidas como vértebras que poseen costillas formadas de partes vertebral y esternal. Estas últimas se articulan con las apófisis esternocostales del esternón, excluyendo las que se unen con el sinsacro. Ocasionalmente, existen dos o más vértebras torácicas unidas para formar un simple notarium u os do fsale. Muchas aves galliformes poseen una estructura como la descrita; sin embargo, las aves anseriformes no.
Las restantes vértebras presentan aún mayores problemas en su definición, como las vértebras de las regiones lumbar y sacra, que están típicamente unidas en un simple sinsacrum en todas las aves. Este sinsacro está a menudo inseparablemente unido lateralmente con los ilios.
Caudal al sinsacro se localizan de cuatro a nueve vértebras caudales libres pero hay una notable variación en cuanto a este número, aun dentro de las mismas especies.
La parte final de la columna vertebral está formada por la fusión de probablemente cuatro a ocho huesos (el número preciso es muy discutible y puede variar), o vértebras caudales embrionarias, para formar una estructura única llamada pigostilo («corcusilla» o
«rabadilla»), a la que se unen varios músculos caudales y también fascias que van a

insertarse en la cola. El movimiento del pigostilo en las especies que vuelan tiene un papel muy importante. La primera ave conocida, el Archaeopteryx, carecía de pigostilo y tenía una serie numerosa de vértebras caudales libres, a cada una de las cuales se unía un par de plumas de la cola.
Además de servir de inserción de las plumas de la cola, el pigostilo también posee un par de glándulas oleosas (glándula uropigia). Aunque muchas aves carecen de esta glándula (p. ej., algunos loros y las palomas), es muy útil para las aves acuáticas, puesto que impermeabilizan las plumas y éstas no se mojan, lo cual permite el buceo de estas especies.
TÓRAX COSTILLAS
Típicamente, existen siete pares de verdaderas costillas, que están formadas por un segmento dorsal (costilla vertebral) y un segmento ventral (costilla esternal). Las costillas dorsales poseen dos articulaciones con sus correspondientes vértebras; la cabeza superior (capitulum) se articula con la apófisis transversa de la vértebra y la cabeza inferior o ventral (tuberculum) lo hace con el cuerpo vertebral. Las dos primeras costillas, de los siete pares, carecen por lo general de segmento ventral y no se articulan con el esternón. El resto tiene costillas esternales que se articulan dorsalmente con las costillas vertebrales y, ventralmente, con las apófisis esternocostales del esternón. El último par de costillas posee segmentos vertebrales y esternales. Excepto para la primera y última, las costillas vertebrales poseen apófisis uncinadas que solapan las costillas sucesivas y proporcionan rigidez a la caja
torácica. Las apófisis uncinadas son características solamente de unos cuantos grupos aviares.
ESTERNÓN
El esternón (hueso del pecho) es un hueso extenso que presenta un quilla dirigida ventralmente, superficie ósea para el origen de la mayor parte de los músculos pectorales y supracoracoideos*. El término metaesternón se utiliza generalmente para aplicarlo a la región caudal del esternón; el término esternón costal para la zona de inserción de las costillas esternales. Caudalmente, el esternón se caracteriza por poseer unas aberturas o ventanas que están limitadas por membranas ligamentosas, con zonas para el origen de porciones de los músculos pectorales. El esternón de la gallina se denomina esternón de cuatro escotaduras; existen apófisis caudolaterales (pars lateralis) que forman los límites laterales de las escotaduras laterales, y apófisis metasternales laterales (pars medicáis), que forman los contornos laterales de las escotaduras medias. Hay una gran variación en cuanto al borde caudal del esternón en las aves; algunas poseen solamente dos escotaduras (p. ej., los patos) y algunas tienen unos bordes caudales sólidos con pocas aberturas (p. ej., algunos loros) o son totalmente sólidos (p. ej., avestruces). En la superficie dorsal del esternón existen varios forámenes, a través de los cuales los sacos aéreos se comunican con el interior. Cranealmente, el esternón se proyecta lateralmente con un par de apófisis esternocoracoidales y, medialmente, existe una proyección ósea simple que es la espina manu-brial. El punto más craneal de la quilla del esternón se conoce con el nombre de vértice carinal.

CINTURÓN PECTORAL Y MIEMBROS
El cinturón pectoral está compuesto por tres pares de huesos que soportan las alas. Se
forman por las clavículas unidas (llamadas fúrculas), los huesos coracoides y las escápulas.
El hueso más robusto del cinturón pectoral es el coracoides, dirigido ventral y caudalmente
para articularse con el esternón por el sulcus coracoidal. El hueso coracoides está hueco e invadido por los sacos aéreos claviculares. Las clavículas son pequeños huesos semejantes
a bastones que ventral-mente se unen en una lámina aplanada, llamada hypocleidium (cara furcular); se conecta al vértice carinal del esternón por un ligamento, llamado hipocleidal, que representa el rafe del músculo mayor del vuelo.

La escápula es plana, larga y extendida caudalmente, paralela a la columna vertebral,
engrosada ligeramente en su extremo próxima!, con un foramen neumático cerca de la apófisis acromium. La escápula se articula cranealmente con el coracoides y el furculum, participando junto con el primero en la formación de la fosa glenoidea, fosa articular para la cabeza del húmero. Estos tres huesos —escápula, coracoides y furculum — se juntan dorsalmente dejando un canal trióseo (foramen triosseum) a través del cual pasa el tendón del músculo supracoracoides, para insertarse en el húmero como una parte importante del mecanismo de vuelo; actúa elevando el húmero y el ala.
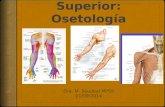
El húmero es el mayor de los huesos del ala, se articula proximalmente con el cinturón
torácico en la fosa glenoidea por su convexa y grande cabeza. El foramen neumático en el extremo proximal del húmero permite la penetración del saco aéreo clavicular, con lo cual se

neumatiza el interior de este hueso largo. Sobre la superficie ventral de la cresta deltoides del húmero, los músculos pectorales que son los más importantes en el vuelo, se insertan y actúan deprimiendo el húmero. Distalmente, el húmero se articula con el radio y el cubito mediante dos cóndilos, el mayor de los cuales se articula con el cubito.
El radio y cubito constituyen los huesos del antebrazo. El cubito es considerablemente
mayor que el radio, pero ambos huesos parecen aproximadamente de la misma longitud. Proximalmente, el cubito y el radio se articulan con los dos cóndilos del húmero; distalmente, con el carpo. El extremo proximal del radio tiene una carita articular que se articula con el más pequeño de los cóndilos humerales. Proximalmente, el cubito presenta una superficie cóncava para articularse con el mayor de los cóndilos del húmero y una elevación llamada olécranon. Igualmente existe en el cubito un foramen nutricio localizado en la mitad de su longitud y sobre la superficie externa una serie de proyecciones óseas pequeñas que representan los puntos de inserción para las plumas secundarias del ala. En las aves galliformes estas proyecciones están un tanto reducidas; sin embargo, son muy manifiestas en las aves muy voladoras. La extremidad distal del radio posee una carita para la articula-ción con el cubito; también se articula con el carpo. Distalmente, el cubito tiene dos caritas articulares, una para articularse con el hueso carporradial (radial o escafolunar) y otra para el hueso carpocubital (ulnar o cuneiforme).
Los huesos carpometacarpianos son tres elementos unidos: metacarpianos II, III y IV
(alguna vez son considerados representantes de los dedos I, II y III). El metacarpiano II está representado por una pequeña proyección sobre el lado radial del carpometacarpo; el III y IV son elementos largos que se unen en su extremidad distal y que dejan un amplio espacio interóseo entre ellos. El III dedo es el mayor y normalmente contiene dos falanges, mientras
que el II y el IV contienen una, en la mayoría de las aves. Sin embargo, en el embrión se

pueden presentar falanges rudimentarias distales que, subsiguientemente, se unen con el elemento más proximal.
CINTURÓN Y MIEMBROS PELVIANOS
El cinturón pelviano está formado por la fusión de tres huesos separados en el embrión: ilion, isquion y pubis. Los iliones también se unen con el sinsacro. Los huesos púbicos
están dirigidos caudalmente y unidos cranealmente con el isquion; se quedan libres caudalmente pero se unen al isquion mediante una membrana fuerte. Cranealmente, hay un espacio de separación entre los dos huesos, llamado foramen obturador. El foramen ilioisquiático está presente entre el isquion y el ilion. El acetábulo, que asienta entre el ilion y el isquion, está perforado y se acomoda a la cabeza del fémur (hueso del muslo). El extremo proximal del fémur tiene un trocánter muy prominente, lateral a su cabeza. En el extremo distal del fémur posee un surco grande (surco rotuliano) que acomoda al hueso sesamoideo patelar (hueso de la rodilla). Distalmente, el fémur tiene dos cóndilos que se articulan con el tibiotarso y el peroné.
El tibiotarso es el mayor de los huesos del miembro pelviano. Está provisto, proximalmente, de dos cóndilos para la articulación con el fémur y de crestas, para los orígenes de ciertos músculos extensores importantes.
Igual que en el fémur, el tibiotarso tiene un foramen nutricio en su eje caudal. La parte lateral del eje superior presenta una cresta para la inserción del peroné. El gallo presenta la característica aviar de tener un peroné muy reducido; en algunas razas el peroné es de mayor tamaño y extensión.

La extremidad distal del tibiotarso presenta cóndilos prominentes para articularse con el tarsometatarso. También existen depresiones para la inserción de los ligamentos colaterales de la articulación tibiotarso-tarso-metatarsiana, presente a los lados de la extremidad distal. Como quiera que alguno de los huesos tarsianos proximales se unen con el extremo distal de la tibia en el embrión para formarse el tibiotarso, los elementos distales se unen con el metatarso alargado (tres en número) para formar así el tarsometatarso de las aves. Los metatarsianos presentes son probablemente del II al IV.
El tarsometatarsiano del adulto se caracteriza, probablemente, por la presencia de caritas internas y externas para articularse con el tibiotarso. En la extremidad proximal, plantarmente, existe una serie de crestas calcáneas que forman canales tendinosos para la inserción de los tendones de los dedos; esta zona del tarsometatarso se considera a menudo como hipotarso. En el extremo proximal del tarsometatarso existen varios forámenes, dorsalmente. En el gallo hay un núcleo óseo que surge del lado más interno del tarsometatarso, aproximadamente dos tercios distalmente a lo largo del eje. Distal-mente, el tarsometatarso presenta tres trócleas grandes para articularse con los dedos II III y IV.

Entre las trócleas para los dedos III Y IV existe un foramen grande, llamado foramen distal, que indica la zona de fusión entre los metatarsianos III y IV. Plantarmente, en el extremo distal del tarsometatarso, se localiza (aunque no está unido al tarsometatarsiano propiamente dicho) el I metatarsiano, que sirve para articularse con el dedo I, o dedo gordo y se dirige posteriormente provisto de dos falanges.

Los dedos II, III y IV, se corresponden con las respectivas trócleas; tienen tres, cuatro y cinco falanges, respectivamente. La falange distal de cada dedo es una garra, que en vida está provista de un recubrimiento córneo exteriormente.