
NOMINAIÓN ORIG N “JAMÓN TRUL”
348
UNIVERSIDAD DE EXTREMADURA DEPARTAMENTO DE PRODUCCIÓN ANIMAL Y CIENCIA DE LOS ALIMENTOS ESTUDIO DESCRIPTIVO-COMPARATIVO DE PRODUCTOS CÁRNICOS ASOCIADOS A LA DENOMINACIÓN DE ORIGEN “JAMÓN DE TERUEL” Memoria presentada por la Licenciada Raquel Reina Toribio Para optar al Grado de Doctor
Transcript of NOMINAIÓN ORIG N “JAMÓN TRUL”

UNIVERSIDAD DE EXTREMADURA
DEPARTAMENTO DE PRODUCCIÓN ANIMAL Y CIENCIA DE LOS
ALIMENTOS
ESTUDIO DESCRIPTIVO-COMPARATIVO
DE PRODUCTOS CÁRNICOS ASOCIADOS A LA
DENOMINACIÓN DE ORIGEN “JAMÓN DE TERUEL”
Memoria presentada por la Licenciada
Raquel Reina Toribio
Para optar al Grado de Doctor


Carmen García González, Profesora Titular del Área de Tecnología de Alimentos de la
Facultad de Veterinaria de la Universidad de Extremadura,
INFORMA:
Que la Tesis Doctoral titulada “Estudio descriptivo-comparativo de productos
cárnicos asociados a la Denominación de Origen “Jamón de Teruel” de la que es
autora la Licenciada en Química Dña. Raquel Reina Toribio, ha sido realizada en el
Departamento de Producción Animal y Ciencia de los Alimentos (área de Tecnología de
los Alimentos) bajo mi dirección y cumple las condiciones exigidas para optar al grado
de Doctor.
Para que así conste, lo firmo autorizando su presentación
Cáceres, 15 de junio de 2011
Carmen García González
DEPARTAMENTO
PRODUCCIÓN ANIMAL Y CIENCIA DE LOS ALIMENTOS
Tecnología de los Alimentos


Pascual López Buesa, Profesor Titular del Área de Tecnología de Alimentos de la
Facultad de Veterinaria de la Universidad de Zaragoza,
INFORMA:
Que la Tesis Doctoral titulada “Estudio descriptivo-comparativo de productos
cárnicos asociados a la Denominación de Origen “Jamón de Teruel” de la que es
autora la Licenciada en Química Dña. Raquel Reina Toribio, ha sido realizada en el
Departamento de Producción Animal y Ciencia de los Alimentos, (área de Tecnología de
los Alimentos) de la Universidad de Extremadura, bajo mi dirección y cumple las
condiciones exigidas para optar al grado de Doctor.
Para que así conste, lo firmo autorizando su presentación
Cáceres, 15 de junio de 2011
Pascual López Buesa
DEPARTAMENTO
PRODUCCIÓN ANIMAL Y CIENCIA DE LOS ALIMENTOS
Tecnología de los Alimentos


A mis padres, mi hermano, Manolo
y muy especialmente a mi tio Julio


AGRADECIMIENTOS
Quiero expresar mi agradecimiento a todas las personas que han hecho posible la
realización de esta tesis doctoral:
A la Dra. Carmen García Gonzalez y al Dr. Pascual López-Buesa, mis directores de tesis,
por haberme dado la oportunidad de realizar este trabajo; especialmente a Carmen, por todo el
tiempo y paciencia dedicados, por su interés mostrado, y por todo lo que he aprendido de ella.
A Jesus Ventanas, por la confianzada depositada en mi y darme la oportunidad, junto a
Carmen, en iniciarme en la investigación y poner a mi disposición todos los medios necesarios
para la elaboración de este trabajo.
A mis compañeros de las unidades de Tecnología e Higiene de los Alimentos de la
Facultad de Veterinaria, profesores, técnicos y becarios con los que he convivido y trabajado
estos años, porque de alguna manera han formado parte de mi trabajo, colaborando en las
numerosas catas realizadas o proporcionanadome ayuda cuando la he necesitado, especialmente
a Ana Antúnez, Verónica Fuentes y Fernando Lobo.
A mis “amigas”, por vuestro cariño y buenos momentos compartidos a lo largo de todos
estos años.
A mis padres, porque siempre han creido en mí y han valorado mi trabajo, por su cariño,
por su ayuda, su ánimo en los momentos de bajón, y por muchas cosas más… A mi hermano, por
el interés especial que siempre ha mostrado por mi trabajo y a Manolo, por valorarme y
entenderme, por su cariño y compañía, y en sobre todo por su infinita paciencia.
Y muy especialmente a mi tio Julio, por sus consejos, su preocupación y confianza, por
estar cuando lo necesito y ser una referencia para mi. Por haberme guiado siempre por el buen
camino y haberme ayudado tanto.
A todos vosotros……. GRACIAS!


INDICE DE CONTENIDOS


ABREVIATURAS...................................................................................................................................................................... III
RESUMEN ................................................................................................................................................................................ VII
1
1. INTRODUCCIÓN ................................................................................................................................................................. 3
1.1. SITUACIÓN ECONÓMICA ............................................................................................................................................. 5
1.1.1. IMPORTANCIA DEL JAMÓN Y LA PALETA EN EL MERCADO ........................................................... 5
1.1.2. IMPORTANCIA DE LA DOP EN EL MERCADO........................................................................................... 7
1.2. DENOMINACIÓN DE ORIGEN JAMÓN DE TERUEL ......................................................................................... 9
1.2.1. CARACTERÍSTICAS DE LA DOP TERUEL ................................................................................................. 10
1.2.2. LA DENOMINACIÓN PROTEGIDA JAMÓN DE TERUEL Y SU PROTAGONISMO EN LA
PROVINCIA.......................................................................................................................................................................... 15
1.3. LA GRASA DEL CERDO .............................................................................................................................................. 18
1.4. ANTECEDENTES BIBLIOGRÁFICOS SOBRE PRODUCTOS CURADOS.................................................. 24
1.5. ESTRUCTURA ANATÓMICA DE LA PALETA Y DEL JAMÓN CURADO ................................................. 27
1.5.1. COMPOSICIÓN QUÍMICA.................................................................................................................................. 27
1.5.2. PARÁMETROS RELACIONADOS CON LA CALIDAD DE LA CARNE .............................................. 30
2
2. PLANTEAMIENTOS Y OBJETIVOS ........................................................................................................................ 37
3
3. MATERIAL Y MÉTODOS ............................................................................................................................................. 41
3.1. MATERIAL BIOLÓGICO ............................................................................................................................................. 41
3.1.1. NATURALEZA DE LAS MUESTRAS ............................................................................................................. 41
3.1.2. NATURALEZA DEL ALIMENTO SUMINISTRADO ................................................................................. 41
3.1.3. PROCESADO DE LAS MUESTRAS ................................................................................................................. 42
3.1.3.1. PROCESADO DEL JAMÓN CURADO .......................................................................................... 42
3.1.3.2. PROCESADO DE LA PALETA CURADA .................................................................................... 43
3.1.3.3. PROCESADO DEL LOMO CURADO ............................................................................................ 43
3.2. TRATAMIENTO DE LAS MUESTRAS ................................................................................................................... 44
3.2.1. JAMÓN CURADO................................................................................................................................................... 44
3.2.2. PALETA CURADA ................................................................................................................................................ 46

3.2.3. LOMO CURADO ..................................................................................................................................................... 47
3.3. MÉTODOS DE ANÁLISIS............................................................................................................................................ 48
3.3.1. CONTENIDO DE HUMEDAD ........................................................................................................................... 48
3.3.2. CONTENIDO DE PROTEÍNAS ......................................................................................................................... 48
3.3.3. CONTENIDO EN GRASA INTRAMUSCULAR............................................................................................ 48
3.3.4. COLOR INSTRUMENTAL .................................................................................................................................. 49
3.3.5. COLOR QUÍMICO. DETERMINACIÓN DE PIGMENTOS RELACIONADOS CON EL COLOR:
MIOGLOBINA ..................................................................................................................................................................... 49
3.3.6. DETERMINACIÓN DEL CONTENIDO DE CLORUROS ......................................................................... 50
3.3.7. CUANTIFICACIÓN DE PRODUCTOS DERIVADOS DE LA OXIDACIÓN LIPÍDICA ................... 50
3.3.8. DETERMINACIÓN DE pH ................................................................................................................................. 51
3.3.9. ANÁLISIS DE PERFIL DE TEXTURA (TPA) .............................................................................................. 51
3.3.10. FRACCIONAMIENTO DE LA GRASA INTRAMUSCULAR Y DETERMINACIÓN DEL PERFIL
DE ÁCIDOS GRASOS DE CADA FRACCIÓN ........................................................................................................... 54
3.3.11. DETERMINACIÓN DE AMINOÁCIDOS LIBRES ................................................................................... 55
3.3.12. DETERMINACIÓN DE COMPUESTOS NITROGENADOS DE BAJO PESO MOLECULAR
DERIVADOS DEL ATP: NUCLEÓTIDOS .................................................................................................................. 57
3.3.13. ANÁLISIS DE COMPUESTOS VOLÁTILES .............................................................................................. 59
3.3.14. EVALUACIÓN SENSORIAL ............................................................................................................................ 60
3.3.15. ANÁLISIS ESTADÍTICO .................................................................................................................................. 62
4
4. RESULTADOS ................................................................................................................................................................... 65
4.1. JAMÓN CURADO ........................................................................................................................................................... 65
4.1.1. CARACTERIZACIÓN DEL JAMÓN CURADO ............................................................................................. 65
4.1.2. INFLUENCIA DE LA EXPRESIÓN DEL IGF2 EN LAS CARACTERÍSTICAS FÍSICO-QUÍMICAS
Y SENSORIALES DEL JAMÓN CURADO .................................................................................................................. 79
4.1.3. INFUENCIA DE LA EXPRESIÓN DEL IGF2 EN FUNCIÓN DEL SEXO SOBRE LAS
CARACTERÍSTICAS FÍSICO-QUÍMICAS Y SENSORIALES DEL JAMÓN CURADO ................................ 90
4.1.4. INFLUENCIA DE LA EXPRESIÓN DEL IGF2 EN FUNCIÓN DE LA CAMPAÑA SOBRE LAS
CARACTERÍSTICAS FÍSICO-QUÍMICAS Y SENSORIALES DEL JAMÓN CURADO ............................. 111
4.2. PALETA CURADA ...................................................................................................................................................... 136
4.2.1. CARACTERIZACIÓN DE LA PALETA CURADA .................................................................................... 136
4.2.2. INFLUENCIA DE LA EXPRESIÓN DEL IGF2 SOBRE LAS CARACTERÍSTICAS FÍSICO-
QUÍMICAS Y SENSORIALES DE LA PALETA CURADA ................................................................................. 148

4.2.3. INFLUENCIA DE LA EXPRESIÓN DEL IGF2 EN FUNCIÓN DEL SEXO SOBRE LAS
CARACTERÍSTICAS FÍSICO-QUÍMICAS Y SENSORIALES DE LA PALETA CURADA ....................... 158
4.2.4. INFLUENCIA DE LA EXPRESIÓN DEL IGF2 EN FUNCIÓN DE LA CAMPAÑA SOBRE LAS
CARACTERÍSTICAS FÍSICO-QUÍMICAS Y SENSORIALES DE LA PALETA CURADA ....................... 180
4.3. LOMO CURADO .......................................................................................................................................................... 198
4.3.1. CARACTERIZACIÓN DEL LOMO CURADO ............................................................................................ 198
4.3.2. INFLUENCIA DE LA EXPRESIÓN DEL IGF2 SOBRE LAS CARACTERÍSTICAS FÍSICO-
QUÍMICAS Y SENSORIALES DEL LOMO CURADO ......................................................................................... 211
4.3.3. INFLUENCIA DE LA EXPRESIÓN DEL IGF2 EN FUNCIÓN DEL SEXO SOBRE LAS
CARACTERÍSTICAS FÍSICO-QUÍMICAS Y SENSORIALES DEL LOMO CURADO ............................... 222
4.3.4. INFLUENCIA DE LA EXPRESIÓN DEL IGF2 EN FUNCIÓN DE LA CAMPAÑA SOBRE LAS
CARACTERÍSTICAS FÍSICO-QUÍMICAS Y SENSORIALES DEL LOMO CURADO ............................... 241
5
5. DISCUSIÓN GENERAL ............................................................................................................................................... 263
5.1. EFECTO DEL ALELO IGF2 ..................................................................................................................................... 264
5.1.1. CALIDAD DE LA GRASA ................................................................................................................................. 265
5.1.2. CALIDAD DE LOS PRODUCTOS CURADOS ........................................................................................... 268
5.1.3. CALIDAD SENSORIAL DE LOS PRODUCTOS CURADOS ................................................................. 271
5.2. EFECTO DE LA CAMPAÑA .................................................................................................................................... 287
5.2.1. CALIDAD DE LA GRASA ................................................................................................................................. 287
5.2.2. CALIDAD DE LOS PRODUCTOS CURADOS ........................................................................................... 288
5.2.3. CALIDAD SENSORIAL DE LOS PRODUCTOS CURADOS ................................................................. 290
6
6. CONCLUSIONES ............................................................................................................................................................ 299
7
7. BIBLIOGRAFÍA ............................................................................................................................................................. 303


ABREVIATURAS


Abreviaturas
III
ABREVIATURAS
Abreviaturas generales AOAC Association of Official Analytical Chemists (Asociación Oficial de
Química Analítica) BOA Boletín Oficial de Aragón CRA Capacidad de retención de agua CRDO Consejo Regulador de la Denominación de Origen Jamón de Teruel DOP Denominación de Origen ETG Especialidad tradicional garantizada FABP3 Fatty Acid Binding Proteins ICEX Instituto Español de Comercio Exterior IGF2 Insuline-like growth factor 2 IGP Indicación Geográfica Protegida ISO International Organization for Standarization Organización
Internacional para la Estandarización) HR Humedad relativa LEPR Receptor de leptina MARM Ministerio de Agricultura y Medio Rural y Marino MC4R Receptor de melanocortina-4
PSE Pale Soft Exudative (carnes pálidas, blandas y exudativas) QTL Quantitative trait loci RPM Revoluciones por minuto RN Rendement Napole RYR1 Gen del halotano WOK Web of Knowledge
Abreviaturas de parámetros físico-químicos a* Eje rojo-verde AQD Análisis cuantitativo-descriptivo b* Eje amarillo-azul BHT Butilato hidroxitolueno CG-EM Cromatografía de gases/Espectrometría de masas L* Coordenada de luminosidad f extracto fresco FID Detector de ionización de llama HPLC High Pressure Liquid Cromatography (Cromatógrafo de líquidos de alta resolución) N Newton
pH -log [H+] s Extracto seco sg Extracto seco y desengrasado SPME Solid Phase Microextraction (Microextracción en fase solida) SPSS Statistical Package for the Social Sciences
TBA Thio-barbituric acid reactive substances (Ácido tiobarbitúrico) TEP 1,1,3,3-tetraetoxipropano TPA Texture Profile Analysis (Análisis del Perfil de Textura)
UAA Unidades Arbitrarias de Area

Abreviaturas
IV
Abreviaturas de ácidos grasos LN Lípidos neutros AGL Ácidos grasos libres LP Lípidos polares LT Lípidos totales C10:0 Ácido cáprico C15:1 Ácido pentadecenoico C16:0 Ácido palmítico C16:1 Ácido palmitoleico C17:1 Ácido heptadecenoico C18:0 Ácido esteárico C18:1 Ácido oleico C18:2 (ω-6) Ácido linoleico C18:3 (ω-3) o (ω-6) Ácido linolénico C20:0 Ácido araquídico C20:2 Ácido eicosadienoico C20:3 (ω-3) o (ω-6) Ácido eicosatrienoico C20:4 (ω-6) Ácido araquidónico AGMI Ácidos grasos monoinsaturados AGS Ácidos grasos saturados AGPI Ácidos grasos poliinsaturados
Abreviaturas de aminoácidos Ala Alanina Arg Arginina Asn Asparagina Asp Ácido aspártico Cys Cisteína Gln Glutamina Glu Ácido Glutámico Gly Glicina Hys Histidina Iso Isoleucina Leu Leucina Met Metionina Phe Fenilalanina Pro Prolina Ser Serina Trh Treonina Trp Triptófano Tyr Tirosina Val Valina
Abreviaturas de nucleótidos ATP Adenosina 5´trifosfato ADP Adenosina 5´difosfato AMP Adenosina 5´monofosfato GMP Guanosina 5´monofosfato Ino Inosina IMP Inosina 5´monofosfato Hx Hipoxantina

RESUMEN


RESUMEN
VII
RESUMEN
La grasa intramuscular tiene una gran importancia, no solo desde un punto de vista
nutricional o tecnológico, sino también por su posible repercusión sobre algunas características
sensoriales de los productos curados, como puede ser la textura, el olor y el flavor. Desde el
punto de vista tecnológico, la grasa intramuscular es de enorme importancia puesto que
determina el grado de penetración al interior de los productos curados de la sal, aditivos y/o
especias empleadas durante el procesado así como el nivel de desecación del producto. La
infiltración grasa de la canal y de la carne son caracteres en cuya génesis hay un notable
componente genético, permitiendo por lo tanto su modulación mediante el cruce de los
individuos más adecuados para el propósito particular que se persiga. Desde hace algunos años,
se ha llevado a cabo la búsqueda de QTLs (Quantitative trait loci) o dicho de otro modo, la
localización de genes responsables de caracteres productivos o tecnológicos, dando buenos
resultados como herramienta de selección genética. El gen para el que se ha descrito mayor
influencia sobre la infiltración grasa es el IGF2 (insuline like growth factor 2) cuyos efectos
sobre la composición de la canal y de la carne, entre otros, son un mayor crecimiento de masa
muscular a expensas del tejido adiposo.
En este trabajo se estudiaron los efectos del IGF2 sobre distintas características físico-
químicas y sensoriales en productos curados asociados a la DOP “Jamón de Teruel”, (jamones,
paletas y lomos curados). Para ello se realizaron determinaciones físico químicas básicas
(porcentaje de humedad, grasa intramuscular, proteínas, sal, color instrumental, contenido de
mioglobina, TBA, pH, textura instrumental, perfil de ácidos grasos, aminoácidos y nucleótidos).
También se llevó a cabo un análisis sensorial utilizando un panel entrenad y una caracterización
del perfil de compuestos volátiles mediante una microextracción den fase sólida (SPME).
Además, el seguimiento del comportamiento del IGF2 en los productos curados obtenidos a lo
largo de tres generaciones, junto con la posibilidad de disponer de jamones, paletas y lomos de
los mismos animales, nos permitió obtener una amplia serie de resultados para realizar una
completa caracterización de los tres productos.
Las diferencias genotípicas para el gen del IGF2 tuvieron un gran efecto sobre espesor de
la grasa subcutánea en jamones y paletas, siendo en éstas mucho más llamativo, sin embargo, se
observó un mayor contenido de grasa intramuscular en los jamones procedentes de los lotes AG,
aunque este efecto no se manifestó en los lomos y en las paletas. No obstante, en los tres tipos de
piezas las puntuaciones alcanzadas en el descriptor de veteado en el análisis sensorial
mostraron valores superiores en los lotes AG. El resto de parámetros composicionales no se

RESUMEN
VIII
vieron afectados por las diferencias en el contenido de grasa subcutánea e intramuscular
mostradas, no obstante, si afectaron a otros parámetros como el perfil de ácidos grasos,
aminoácidos y nucleótidos, incluso en productos donde la grasa intramuscular no se vio afectada
por el IGF2, la paleta y el lomo.
Aunque se han observado diferencias relacionadas con la expresión del IGF2, son mucho
más importantes las diferencias relacionadas con la campaña, por lo que el factor procesado,
influye mucho más en las características de las piezas que otros factores como la genética o el
sexo.

CAPÍTULO I: INTRODUCCIÓN


CAPÍTULO I: INTRODUCCIÓN
3
1. INTRODUCCIÓN
España es tradicionalmente un país elaborador de productos cárnicos curados,
siendo el jamón el producto estrella y por lo tanto, un pilar fundamental de la economía
española. Actualmente, se ha convertido en uno de los alimentos más consumidos en
España (cada español consume al año cerca de 3,2 Kg. de jamón curado destacando
especialmente la demanda en el hogar (76% del total) (Martín, 2010)). Además de tener
un apreciado sabor y un aporte de nutrientes adecuado para la salud, es muy fácil de
combinar con todo tipo de comidas.
Dada la gran variedad de tipos y categorías de jamón y la complicada clasificación
del producto, conviene destacar aquellos jamones que han logrado el aval y
reconocimiento de la Unión Europea como: Denominación de Origen Protegida (DOP),
Indicación Geográfica Protegida (IGP) o Especialidad Tradicional Garantizada (ETG). Estas
son figuras de calidad que, respaldadas por un riguroso reglamento de elaboración y
control, se aplican a ciertos productos agrícolas y alimenticios.
A nivel nacional, son los Consejos Reguladores los organismos de vigilancia y
control del cumplimiento de las normas estipuladas por el reglamento de cada figura de
calidad. Las DOP garantizan que el producto cumple los requisitos para ser de una elevada
calidad. Para elaborar jamones es preciso conocer y controlar desde la cría del animal
hasta la comercialización del jamón, y se realizan diferentes controles, como salinidad,
tiempo de elaboración o etiquetado, que certifican que el jamón es de DOP. Por lo tanto,
todos los jamones deben de llevar una etiqueta y un precinto debajo de la pezuña, donde
se indicará el tipo de DOP a la que pertenece ese jamón.
Figura 1.1. Sellos indicadores de la Denominación de Origen Protegida

CAPÍTULO I: INTRODUCCIÓN
4
La DOP Jamón de Teruel fue la primera que se creó en España para amparar,
controlar y garantizar la producción de jamones. En Octubre de 1984 fue reconocida por la
Consejería de Agricultura del Gobierno de Aragón y en 1985 ratificada por el Ministerio de
Agricultura.
Seguidamente, Guijuelo fue la primera denominación de jamón y paleta procedente
de cerdo ibérico que obtuvo el marchamo de calidad en 1986. Más adelante, alcanzaron la
distinción de DOP Dehesa de Extremadura, en 1987, y Jamón de Huelva, en 1995. En 2004
se estableció la Denominación específica o IGP “Jamón de Trévelez” y finalmente en otoño
de 2010 el Jamón Ibérico de los Pedroches fue reconocido por la Unión Europea como
DOP.
Además de los sistemas de protección anteriormente citados, existe otro tipo de
sistema como es la ETG Jamón Serrano, registrada por la Unión Europea en 1999 con el
objetivo de proteger, tipificar y mejorar la imagen del Jamón Serrano. Dentro de esta
figura y con el impulso de Confecarne, en el año 2000 nació la Fundación del Jamón
Serrano como una entidad sin ánimo de lucro para gestionar y dirigir el proyecto de la
implantación y gestión de la ETG del Jamón Serrano, convirtiéndose de esta manera en el
centro y motor de la misma. La Fundación, como Órgano de Representación de la ETG del
Jamón Serrano, trabaja en la protección, mejora, proyección y promoción del Jamón
Serrano de calidad, diferenciándolo del resto de jamones curados que no se elaboran
conforme marca el Pliego de Condiciones y que no garantizan la calidad mínima exigible
que garantiza el cumplimiento de la ETG. Dicha fundación integra a 118 empresas,
ubicadas en las comunidades autónomas de Castilla y León, Castilla-La Mancha y Cataluña
principalmente, donde se encuentran mayoritariamente las industrias cárnicas. Además,
mucho antes de que surgiera la ETG, se creó dentro del sector otra asociación voluntaria
de empresas, el Consorcio del Jamón Serrano Español, cuyo objetivo primordial era y es la
producción de un Jamón Serrano exclusivo y de alta calidad para su comercialización en
los mercados exteriores.

CAPÍTULO I: INTRODUCCIÓN
5
1.1. SITUACIÓN ECONÓMICA
1.1.1. IMPORTANCIA DEL JAMÓN Y LA PALETA EN EL MERCADO
Tal como muestra la figura 1.2, desde comienzos de esta década, la producción de
jamones y paletas en España ha seguido un crecimiento continuo, aunque podemos decir
que en los últimos años se ha estancado. Las informaciones de 2007 nos muestran un
crecimiento del 1,8% en el número de piezas provenientes de cerdo blanco y del 3% en el
caso de las provenientes de cerdo Ibérico. Estos crecimientos son menores que los
registrados entre 2005 y 2006 en que se incrementaron las producciones en un 5,5% para
las piezas de cerdo blanco y en un 5,4% para las de Ibérico, lo que da idea del frenazo que
poco a poco se ha registrado en la producción (Cruz, 2009).
Figura 1.2. Evolución y composición de la producción de las piezas curadas
Las menores cifras de crecimiento en los últimos años muestran una tendencia
hacia la madurez del mercado, con el mantenimiento de la demanda. Del total de la
producción registrada, aproximadamente el 88% corresponden a la categoría de jamones
y paletas procedentes de cerdo blanco (figura 1.3), mientras que el resto (12%), en torno a
los 5 millones de piezas, son perniles curados de cerdo Ibérico. Los incrementos más
fuertes en los últimos años se han registrado en la producción de jamones procedentes de
cerdos ibéricos. Esto coincide con el fuerte incremento registrado en el número de cabezas
de ganado porcino Ibérico, obligado por una mayor demanda de este tipo de productos
curados y, sobre todo, de las carnes frescas de estos animales que han entrado con fuerza
en la restauración. (Cruz, 2007).

CAPÍTULO I: INTRODUCCIÓN
6
Figura 1.3. Composición de la producción de jamones y paletas curadas
De los 945.000 animales censados en 1997 hemos pasado a más de 2.043.000 de
cabezas de porcino extensivo en España en 2007, distribuidos fundamentalmente en la
zona de las dehesas que comprende Castilla y León, Extremadura, Andalucía y parte de
Castilla-La Mancha, aunque la cría de estos animales se da, en muchos casos, en
condiciones de estabulación. El incremento en el número de cabezas no ha sido uniforme
en cada una de estas comunidades autónomas y así en Extremadura ha sido donde menos
ha crecido, mientras que en las provincias de Castilla y León el incremento llega a
prácticamente el 31% (Cruz, 2007).
La producción de jamones curados de cerdo blanco sufrió una reducción de la
producción del 12% entre 2008 y 2009, cifras algo inferiores a las ya registradas en 2003.
Esta caída rompió la tendencia al alza continua desde comienzos de la década pasada. Para
encontrar una explicación habría que hablar de la caída de precio que se registró en los
jamones Ibéricos, especialmente en la categoría de cebo, que hizo que el precio de venta
del jamón Serrano estuviera, a veces, por encima del Ibérico, que también fue protagonista
de todo tipo de promociones comerciales (Cruz, 2010).
En lo referente a las paletas, la tendencia parece ser de crecimiento constante, con
una subida en la producción del 2,3% (4,30 millones de piezas). Incluso en 2009 se pudo
ver beneficiada de la crisis económica que, en muchos casos hizo sustituir la compra y
regalo de jamones curados (caso de cestas de Navidad, por ejemplo) por el de paletas, con
un menor precio de venta (Cruz, 2010).
El estancamiento de la producción en los últimos años, ha motivado que muchas
empresas se decidieran a conquistar nuevos mercados. Analizando la evolución de las
exportaciones, cabe destacar el fuerte crecimiento que se produjo entre 2007 y 2008, ya

CAPÍTULO I: INTRODUCCIÓN
7
que se incrementaron las ventas un 43% en lo referente a la cantidad. Sin embargo, dicho
crecimiento se redujo a tan sólo un 11,8% en valor, lo que puede dar muestra de la fuerte
competencia que tienen los exportadores españoles obligados a mantener e incluso
reducir los precios de sus productos (figura 1.4) (Cruz, 2009).
Figura 1.4. Principales destinos y evolución de las exportaciones españolas de Jamón Curado (2008)
1.1.2. IMPORTANCIA DE LA DOP EN EL MERCADO
De de entre todas las DOPs, la más comercializada en los últimos años ha sido la
DOP Jamón de Teruel, alcanzando en 2009 una comercialización de 519.365 piezas,
cantidad que correspondió al 44,83% (figura 1.5) de la producción anual con respecto al
resto de DOPs, doblando la producción de su inmediato seguidor, Guijuelo (24,14%).
Trevélez, Dehesa de Extremadura y Huelva les siguen en el número de piezas. Asimismo,
únicamente dos DOPs, Dehesa de Extremadura y Trevélez comercializan con países de la
Unión Europea y terceros (MARM, 2009).

CAPÍTULO I: INTRODUCCIÓN
8
Figura 1.5. DOPs e IGPs de Jamón. Distribución de la comercialización (MARM, 2009)
Por otro lado, la ordenación según el valor económico de las distintas
certificaciones varía considerablemente, debido a las diferencias en los costes unitarios de
las piezas, relegando a la DOP de Teruel al segundo puesto, por detrás de la de Guijuelo, y
seguida muy de cerca por la Dehesa de Extremadura que alcanza un 23,34% del valor
total. El último lugar, tanto en número de piezas como en valor económico, lo ocupa la
Denominación Específica Los Pedroches, con el 2,62% de la comercialización (Fandos et
al., 2007; MARM, 2009).
Respecto a la evolución de la comercialización, tal y como puede observarse en la
figura 1.6, lo más destacable sería el mayor incremento que se alcanzó en el periodo
comprendido entre el 2004 y 2006, con un aumento de más del 120% que suponía la
comercialización de 1.475.575 piezas. Sin embargo, en los últimos años se ha producido un
estancamiento, como se indicaba anteriormente, que también ha afectado a los productos
curados amparados por alguna denominación (MARM, 2009).
Figura 1.6. Evolución de la comercialización de piezas (MARM, 2009)

CAPÍTULO I: INTRODUCCIÓN
9
1.2. DENOMINACIÓN DE ORIGEN JAMÓN DE TERUEL
La DOP Jamón de Teruel fue reconocida en Octubre de 1984 por la Consejería de
Agricultura del Gobierno de Aragón y en 1985 ratificada por el Ministerio de Agricultura.
Posteriormente, el primer Reglamento fue modificado por el Departamento de Agricultura,
Ganadería y Montes del Gobierno de Aragón (Boletín Oficial de Aragón (BOA), 1993 (a)).
En 1997 se incluyó en la lista de productos de especial calidad aprobada por la Comisión
Europea, lo que implica su protección en el ámbito europeo.
Con el nacimiento de la DOP Jamón de Teruel concurrieron todos los sectores
vinculados a esta actividad productiva, desde ganaderos a secaderos, así como la
Administración pública. Esta condición pionera no sorprende si se tiene en cuenta la
tradición existente en la provincia en el consumo de productos derivados del cerdo y, más
concretamente, en la elaboración de jamón Serrano. Los objetivos perseguidos a la hora de
impulsar la Denominación fueron: contribuir al desarrollo de la provincia y en concreto al
del sector porcino; lograr que los cerdos sacrificados para la Denominación fueran criados
en Teruel; explotar al máximo las ventajas climáticas de la provincia para curar jamones y
productos de charcutería y utilizar la coyuntura ya existente para desarrollar una
industria próspera que aprovechara estos conocimientos y los aplicara en la elaboración
de jamón a gran escala, siempre sin perder de vista la tradición artesana conjugada con
métodos modernos de control y elaboración. La unificación de criterios entre los
productores, y la garantía del origen del jamón y de su alta calidad con el distintivo de
Jamón de Teruel fueron otros de los objetivos que se persiguieron a la hora de impulsar la
Denominación, junto con la voluntad de mejorar la comercialización e introducir el
producto en canales de distribución de toda España, especialmente en Levante, Aragón y
Cataluña.
La selección de los animales y la experiencia ancestral en la preparación de los
perniles tiene en Teruel dos inmejorables aliados: el relieve y el clima, óptimos ambos
para la curación natural de la carne, el paso fundamental para que el jamón desarrolle
plenamente todo su sabor y aroma. Tradicionalmente ha sido una zona ideal para la
curación natural de jamón por su altitud, ya que más del 62% de su superficie se
encuentra por encima de 1.000 m, y por su clima continental, con inviernos fríos, veranos
frescos y viento dominante de componente noroeste.

CAPÍTULO I: INTRODUCCIÓN
10
La zona de producción se extiende por toda la provincia de Teruel, pero la zona de
curación la forman solo las zonas de la provincia que tienen una altura mínima de 800
metros. Los cerdos aptos para la producción de jamones de DOP proceden de las razas
Landrace (tipo estándar) y Large White o cruce de ambas para la línea materna, y
Landrace (tipo estándar) o Duroc para la línea paterna. Deben ser sacrificados sobre los 8
o los 9 meses, y han de tener un peso entre 115 y 130 kilos y un espesor de tocino de entre
4 y 7 cm a la altura de la cuarta costilla. El tiempo de curación de estos jamones debe ser
mínimo de 14 meses en secaderos naturales.
Las características que deben cumplir los jamones una vez curados son: forma
alargada, perfilado y redondeado en sus bordes, conservando la pata. Puede presentarse
con toda la corteza o perfilado en corte tipo ‘V’ con un peso de entre 8 y 9 Kg, y nunca
inferior a 7 Kg. Sus características sensoriales deben ser color rojo y aspecto brillante al
corte, con grasa parcialmente infiltrada en la masa muscular. La carne debe tener un sabor
delicado y poco salado y la grasa debe tener consistencia untuosa, brillante, coloración
blanco-amarillenta, aromática y de sabor agradable.
1.2.1. CARACTERÍSTICAS DE LA DOP TERUEL
La DOP Jamón de Teruel es sinónimo de calidad, que se verifica con la implantación
de un buen sistema de trazabilidad y adecuadas prácticas de manejo y producción. La
“trazabilidad” se define como la posibilidad de encontrar y seguir el rastro a través de las
etapas de producción, transformación y distribución de un animal para la producción de
alimentos. Por tanto, es imprescindible tener un buen sistema de identificación mediante
el cual se puedan conocer todos los pasos, procesos y movimientos que ha sufrido el
alimento desde su producción hasta su puesta en circulación. Las fases que componen la
cadena de producción para el Jamón DOP Teruel son las siguientes:
Explotaciones ganaderas
Granjas
La zona de producción de cerdos aptos para la DOP Teruel está constituida por el
área geográfica de la provincia de Teruel, por lo que las granjas de producción de lechones
y cebo están enclavadas en dicha provincia. De las 441 explotaciones, el 62% se ubica en la
comarca del Bajo Aragón, el 23% en el Jiloca (Calamocha), el 10% en el Maestrazgo y el 5%
restante se reparten en la Hoya (Teruel), Albarracín y la cuenca minera.

CAPÍTULO I: INTRODUCCIÓN
11
Figura 1.7. Mapa de la Provincia de Teruel por comarcas
Razas
El tipo de ganado es el procedente de cruces entre: razas Landrace (tipo estándar),
Large White o cruce de ambas como línea materna y raza Landrace (tipo estándar) o
Duroc como línea paterna.
El cruce más empleado es Duroc x (Landrace x Large White). La raza Landrace se
caracteriza por ser una raza muy prolífica (con un promedio de 12 lechones) y versátil (se
usa como línea pura, materna o paterna). Esta raza está reconocida como de tipo magro, ya
que presenta bajos valores de engrasamiento. La raza Large White mejora la calidad de la
carne cuando es empleada en cruces. A nivel reproductivo destaca su elevada fertilidad,
prolificidad y la buena aptitud y actitud maternales (carácter tranquilo, cuidado de las
crías, capacidad lechera, etc.) que la hacen muy interesante tanto en cría en pureza como
en cruzamientos como línea materna. También es una de las razas que posee un alto índice
de crecimiento. La raza Duroc se caracteriza por su rusticidad y buen crecimiento e índice
de conversión, lo cual la hace adecuada para sacrificios superiores a 100 kg de peso vivo.
Además, su carne tiene una buena calidad tecnológica y presenta elevada capacidad de
retención de agua y contenido en grasa intramuscular (Latorre, 2003).
Cría de los animales
Los machos deben estar castrados antes de entrar en cebadero y las hembras no
deben estar en celo en el momento del sacrificio. La castración de los machos incrementa
la infiltración grasa, y evita el olor sexual de la carne (Diestre, 1990). El animal debe pesar

CAPÍTULO I: INTRODUCCIÓN
12
al sacrificio entre 115-130 kg y tener un mínimo de 8 meses de vida, de los cuales mes y
medio son de leche y seis meses y medio de cebo.
Alimentación
El 60-70% del pienso consumido procede de fábricas ubicadas en la provincia de
Teruel que se proveen de cereales de la zona: trigo, cebada, maíz, girasol, etc. El Consejo
Regulador puede dictar normas sobre la utilización de piensos de acuerdo con el avance
de las técnicas ganaderas y con el fin de mantener o mejorar la calidad del cerdo o de los
jamones producidos. En los 15 días anteriores al sacrificio los cerdos no reciben ningún
tipo de tratamiento ni alimentación medicamentosa.
Mataderos
Pre-sacrificio
Al final del cebo, los cerdos son transportados en camiones adecuados, de forma
que el animal no sufra alteraciones o molestias que puedan provocar estrés o dañar su
integridad física. Los mataderos y salas de despiece deben estar enclavados en el área
geográfica de la provincia de Teruel, reunir las condiciones técnico-sanitarias exigidas por
la legislación vigente y estar inscritos en el Registro del Consejo Regulador. En los corrales
de espera previa al sacrificio habrá clara separación física entre los cerdos amparados por
la DOP y el resto de partidas de cerdos. Asimismo, se separarán las partidas destinadas a la
obtención de perniles para la DOP según sus granjas de procedencia. Los cerdos
permanecerán en los corrales de recepción del matadero un tiempo no inferior a 12 h con
el fin de eliminar la fatiga del transporte y asegurar un nivel mínimo de reservas de
glucógeno muscular. Durante este tiempo se les proporcionará agua ad libitum con un
mínimo del 1% de azúcar.
Sacrificio
El sacrificio se llevará a cabo siguiendo las directrices de la legislación vigente. Se
realizará un aturdimiento previo exigiendo posteriormente un completo desangrado y no
pudiendo taladrar las extremidades posteriores del cerdo.
Post-sacrificio
Sólo podrán destinarse a la elaboración de jamón DOP Teruel aquellos perniles que
procedan de canales con un peso entre 115 y 130 kg y un espesor de tocino de entre 4 y 7
cm a la altura de la cuarta costilla. El oreo de las canales suele realizarse a una

CAPÍTULO I: INTRODUCCIÓN
13
temperatura inferior a 10ºC, con una duración mínima de 4 h y una humedad relativa (HR)
del 95% en la primera hora y 85% en las restantes. Una vez despiezada la canal, se
obtienen los perniles cortando la extremidad posterior del cerdo en su parte superior por
la sínfisis isquio-pubiana, son perfilados y puede dejarse íntegra la corteza o practicar el
“corte en V”, permitido desde el 1 de marzo de 2005 (BOA, 2005). A continuación suelen
mantenerse de 24 a 48 h entre -2 y 2 ºC (para conseguir una temperatura de 2 ºC en el
interior de la pieza), deben tener un espesor de tocino de 1,5-4 cm y un peso superior a
11,5 kg. En el momento del sacrificio debe estar presente un veedor, que trabaja para el
Consejo Regulador, encargado de comprobar la procedencia de los animales, espesor de
tocino de las canales y los parámetros mínimos que deben cumplir los perniles.
Los perniles aptos son identificados de forma indeleble con un sello de 10 dígitos
que indicarán: el nº de matadero, la semana del año de sacrificio, el nº de la granja y el nº
del cerdo.
Secaderos
La zona de elaboración está constituida por aquellos términos municipales de la
provincia de Teruel cuya altitud media no sea inferior a 800 m. Actualmente, de los 52
secaderos, 19 se ubican en la comarca de Gúdar-Javalambre, 10 en la de Teruel, 11 en la
del Jiloca (Calamocha), 7 en la del Maestrazgo y 5 entre el Matarraña, la Cuenca Minera y el
Bajo Aragón (figura 1.7).
El proceso se realizará exclusivamente en secaderos inscritos en el Registro del
Consejo Regulador y tendrá una duración mínima de 14 meses, obligatorio desde el 1 de
abril de 2005, por acuerdo interno del mismo Consejo. En la inscripción deben indicarse
las características de sus cámaras frigoríficas y de su secadero: número, superficie,
volumen y capacidad.
El transporte de los perniles desde el matadero al secadero se realizará en
vehículos frigoríficos, de forma que entren en la nave de salado con una temperatura
interior de entre 0 y 2 ºC.
Curación
Esta operación es llevada a cabo en secaderos naturales, controlando la
ventilación, que permitan las condiciones óptimas de HR y temperatura. Si se realiza
mediante métodos artificiales, las instalaciones deben contar con instrumentos que
garanticen un adecuado grado termo-higrométrico.

CAPÍTULO I: INTRODUCCIÓN
14
Salazón
Previo al salado, mediante presión manual o mecánica, se elimina la sangre
residual del jamón para evitar posibles problemas microbiológicos y de aspecto al corte.
La salazón se realiza por el denominado proceso en seco, enterrando los perniles en sal
gruesa, en contacto directo con las piezas, durante un periodo de tiempo proporcional a
los kilos que pesa el pernil, sin exceder nunca los 14 días, por orden expresa de la DOP.
Asimismo, se usan nitrificantes para favorecer la acción de la sal, inhibir la actividad de
determinados microorganismos y contribuir a la coloración roja del magro. Las
condiciones de esta fase suelen ser: temperatura 4ºC y HR entre 75 y 90%. Las mermas
varían entre 2 y 8%.
Terminada la salazón se lavan las piezas con agua templada, para eliminar la sal
adherida. A continuación se prensan, por métodos físicos o mecánicos, para darles una
forma más alargada y estrecha que es la que demanda el mercado.
Post-salado o asentamiento
Las mermas durante esta fase varían entre 10 y 15%. Se realiza en cámaras con
temperaturas entre 3 y 6 ºC y HR entre 80 y 90%. El tiempo de permanencia de las piezas
depende del peso de las mismas oscilando entre 45 y 90 días. En cualquier caso, el periodo
será suficiente para que la cantidad de NaCl al final no sea superior al 1% del peso fresco.
Secado
En esta fase, la temperatura irá aumentando aproximadamente desde los 10 hasta
los 30 ºC y la HR irá disminuyendo desde 85 hasta 70%. El tiempo necesario se estima en 6
meses, pudiendo prolongarse hasta los 10 meses en función del peso de la pieza. Las
mermas durante esta fase rondan el 35%.
Se lleva a cabo en secaderos naturales provistos de ventanas con apertura
regulable que permita el control de la ventilación para conseguir condiciones óptimas de
humedad relativa y temperatura. A tal efecto, los locales de secado pueden estar provistos
de aparatos idóneos para mantener el adecuado grado termohigrométrico
Maduración
La maduración se realiza en ambiente natural en naves y/o bodegas donde se
almacenan colgadas las piezas bajo condiciones especiales de humedad y temperatura del

CAPÍTULO I: INTRODUCCIÓN
15
medio natural. Deben permanecer el tiempo necesario para completar los 14 meses, como
mínimo, que dura todo el proceso de elaboración. Finalizada la maduración, un veedor
controlará los jamones que cumplan los requisitos, es decir, buen aspecto, ningún defecto
tras la cala y peso superior a 7 kg. A continuación, se marcarán a fuego con una estrella de
8 puntas y la palabra TERUEL en la corteza, y se identificarán con un brazalete numerado
que garantiza que el producto está protegido por la DOP.
Figura 1.9. Marca identificativa sobre DOP Jamón de Teruel
1.2.2. LA DENOMINACIÓN PROTEGIDA JAMÓN DE TERUEL Y SU PROTAGONISMO EN
LA PROVINCIA
Llegados a este punto, parece necesario destacar la relevancia que supone el sector
porcino, y más concretamente la DOP Jamón de Teruel, en dicha provincia. Para ello, nos
centraremos en las cifras que hacen referencia al producto en cuestión, así como en otras
cuestiones relativas a la DOP y su Consejo Regulador.
Al analizar la importancia del sector jamonero en la provincia de Teruel, cabría
destacar que éste supone la mayor actividad económica de dicha provincia, con más de
2.000 puestos de trabajo directos y unos 1.000 indirectos. En el año 2005 las ventas del
jamón con DOP llegaron a los 38 millones de euros (según el CRDO (Consejo Regulador de
la Denominación de Origen Jamón de Teruel), a las que se sumaron otros 50 millones de
euros procedentes de las ventas del resto del canal del cerdo (paletas, lomos y embutidos).
No obstante, como se ha indicado anteriormente, en los últimos años se ha
producido un estancamiento en la comercialización que ha afectado de la misma manera a
los jamones amparados por dicha denominación. Este hecho ha motivado que las
industrias busquen alternativas a esto, recurriendo a la exportación de sus productos y a
la promoción de otros, como son la paleta y el lomo curado, procedentes de cerdos
amparados por dicha denominación.

CAPÍTULO I: INTRODUCCIÓN
16
En cuanto al mercado al que se dirige, la mayoría de las ventas se realizan en
España y se centran en las comunidades autónomas de Valencia, Cataluña y Aragón. En lo
referente al mercado exterior, la evolución de las exportaciones de todo el jamón y la
paleta curada que se elabora en la provincia de Teruel (incluyendo no DOP) durante los
últimos años, puede considerarse algo inestable. Entre los años 2003 y 2007 se produjo un
crecimiento continuo, especialmente en los años 2006 y 2007, exportando 831.085 Kilos y
generando por lo tanto ingresos de 4.497.440 euros. Sin embargo, esta tendencia al alza se
vio afectada en 2007 descendiendo hasta 2008 en un 15% en cuanto a los Kilos
exportados, con un descenso del 10% del valor económico (Fuente: ICEX (Instituto
Español de Comercio Exterior)). Estos datos se pueden considerar algo mejores con
respecto al descenso que se produjo tanto en los kilos exportados como en el valor
económico del jamón curado en general producido en España. Respecto a los destinos a los
que se dirigen las exportaciones se encuentran fundamentalmente países de la Unión
Europea como Alemania, Reino Unido, Francia, Italia o Portugal, pero también llega a
países más lejanos como Argentina o Japón. (ICEX).
Con respecto a los productos curados relacionados con el Jamón de Teruel, se
encuentra la paleta curada, que posee características muy similares al jamón curado pero
que siempre se ha encontrado en un segundo plano. “Jamón de Teruel” es la única
Denominación que todavía hoy no ampara las paletas. Aunque estas piezas proceden de
cerdos controlados por el consejo regulador de la DOP y por lo tanto, tienen una elevada
calidad, son poco apreciadas por el consumidor que casi siempre se decanta por el Jamón
curado. A pesar de ello, datos mostrados anteriormente, demuestran el aumento de las
ventas de estos productos en los últimos años, en contra del descenso de los jamones
curados.
Las características tecnológicas de la paleta y la calidad sensorial del producto
final, al igual que las del jamón curado, dependen de muchos factores vinculados al
proceso productivo del animal del que proceden y al proceso de elaboración de las propias
piezas. El estudio de las paletas curadas resulta muy interesante bajo dos puntos de vista,
en primer lugar su caracterización permitirá transmitir al consumidor las características
de su calidad y promover su consumo, y en segundo lugar, debido a un proceso de
elaboración más corto, puede servir de modelo para estudiar los cambios que acontecen
en otras piezas similares de procesado más largo.
A pesar de que la paleta y el lomo curado no se encuentran amparados bajo la DOP
de Teruel, ambas poseen una mención de Calidad, por tratarse de productos que proceden
de cerdos controlados por el consejo regulador, garantizando por lo tanto su alta calidad.

CAPÍTULO I: INTRODUCCIÓN
17
La paleta curada fue amparada por la marca Calidad Alimentaria en Mayo de 2008,
pudiendo proceder de cerdos amparados por la DOP o no, aún así, aquellas procedentes de
la DOP llevarán además la palabra Teruel junto al logotipo “C”, como una garantía más de
la excelencia del producto. Las paletas deben tener un tiempo de curación mínimo de 7
meses, de los cuales, al menos los tres últimos deben ser de maduración en ambientes
naturales, alcanzando un peso final superior a los 4 kilos. Sus características serán
similares a las del Jamón de Teruel.
El lomo curado fue amparado por la marca Calidad Alimentaria en Octubre de
1993, y puede proceder o no de cerdos amparados por la denominación, al igual que las
paletas, añadiendo la palabra Teruel en función de su procedencia. Las piezas deben
curarse al aire durante un mínimo de 60 días, de las que al menos los 21 últimos deben ser
en ambiente natural. Este producto se caracteriza por su aspecto homogéneo, liso y de
color sonrosado. Destacando al gusto por un sabor delicado y poco salado, que el
Reglamento de utilización del nombre Calidad Alimentaria para el lomo embuchado define
como sabor característico.

CAPÍTULO I: INTRODUCCIÓN
18
1.3. LA GRASA DEL CERDO
Las características de los productos cárnicos curados están influenciadas por el
contenido y la composición de la grasa. La infiltración grasa de la canal y de la carne son
caracteres en cuya génesis hay un notable componente genético (Sellier, 1998). Este hecho
permite precisamente su modulación mediante el cruce de los individuos más adecuados
para el propósito particular que se persiga. En las últimas décadas este propósito ha sido,
por cuestiones económicas y nutricionales, la obtención de individuos con canales y carne
lo mas magras posible. Sin embargo, la elaboración de productos curados, y especialmente
de jamón curado, precisa de materias primas con mayor contenido graso. Para conseguir
mayores contenidos grasos en la canal y en la carne de los cerdos cuyo destino industrial
es la elaboración de productos curados caben varias estrategias, que suelen emplearse en
combinación:
1) Estrategias alimentarias.
2) El sacrificio a pesos altos y/o edades avanzadas.
3) La utilización de razas con niveles altos de engrasamiento.
Como ejemplo de la primera estrategia cabe citar el uso de dietas pobres en
proteína y ricas en carbohidratos que incrementan la síntesis endógena de grasas (Wood
et al., 1979; Wood et al., 2004).
La segunda estrategia se basa en el hecho de que el tejido adiposo es de deposición
tardía respecto al tejido magro (Hammond, 1952; Zembayashi, 1994). Las mayores
diferencias en torno al contenido de tejido adiposo con respecto al muscular pueden
explicarse a partir de trabajos clásicos de Hammond (1952), ya que establecieron con
bastante precisión que los tejidos del animal muestran una intensidad máxima de
crecimiento que varía a lo largo de distintas etapas de su vida, de manera que el tejido
adiposo es el que crece con mayor intensidad en las últimas etapas de vida, por lo que el
sacrificio de los animales con pesos elevados da lugar a mayor contenido de grasa en la
canal y en la carne. Dentro del tejido adiposo pueden diferenciarse tres partes, la grasa
subcutánea, la intramuscular y la intermuscular, donde la velocidad de crecimiento de
éstas también variará con la edad y peso del animal, de forma que la primera en
depositarse será la subcutánea, seguida de la intramuscular y finalmente la intermuscular.
Estos mismos depósitos grasos no tienen la misma importancia en la canal y en la calidad
de la carne. El contenido de grasa intermuscular no puede eliminarse, ya que mantiene
unidos los músculos y el de grasa subcutánea, puede ser fácilmente recortado del magro
en la carne fresca, aunque esto puede tener consecuencias sobre productos curados como

CAPÍTULO I: INTRODUCCIÓN
19
son el jamón y la paleta, ya que requieren de un espesor determinado para que el proceso
de secado sea adecuado (Monzoils et al., 2005; Kouba y Bonneau, 2009). Sin embargo, el
contenido de grasa intramuscular ha tenido un gran impacto sobre el consumidor, positivo
por la mejora de las características sensoriales (Gandemer, 2009), pero negativo por su
implicación nutricional (Jiménez-Colmenero et al., 2010). Es por ello, que no solo es
importante el contenido de grasa total, si no la deposición de ésta y su distribución (Fisher
et al., 2003; Kempster y Evans, 1979; Whitemore et al., 1982; Wood et al., 1983). El
contenido de grasa subcutánea e intramuscular también está relacionado con la edad y
peso del animal (Davies y Pryor, 1977). Estos autores encontraron que el crecimiento de la
grasa intramuscular era diferente del de la grasa subcutánea. Sin embargo, en un estudio
más reciente (Kouba y Bonneau, 2009), corroboraron datos de Fisher et al., (2003), que
encontraron valores similares en el coeficiente alométrico del tejido adiposo subcutáneo e
intramuscular en las paletas, mientras que el tejido adiposo subcutáneo crecía más rápido
que el intramuscular en el caso del jamón. De manera que, en nuestro caso, al aumentar el
peso del cerdo en vivo, deberíamos esperar menores diferencias entra la grasa
intramuscular y la subcutánea. Al aumentar el contenido de grasa intramuscular,
aumentaría en exceso el contenido de grasa subcutánea, por lo que la paleta tendría mayor
desperdicio.
La tercera estrategia, que contribuye a la variación sobre la composición del tejido
adiposo del cerdo, ha sido la variación genética determinada por las razas de cerdos
empleadas y los cruces de las mismas (Sellier, 1998). Uno de los pilares más importantes
en la producción cárnica es la selección genética, basada en el cruzamiento de aquellos
animales más aptos para los caracteres que se deseaba seleccionar. En el caso de las razas
de cerdos blancos se ha buscado aumentar el porcentaje de magro de la canal motivado
por las preferencias del consumidor por carnes magras y por la mejor eficacia productiva
de los animales más magros. De hecho, se produjo un descenso en la grasa dorsal de 0,4-
0,6 mm/año y un aumento en el porcentaje de magro de 0,6%/año en las canales porcinas
en España de 1990 a 1997 (Gispert et al., 1997). Este descenso de la grasa dorsal lleva
implícito un descenso de la grasa intramuscular que tiene un papel importante en la
calidad de los productos cárnicos curados.
El ejemplo paradigmático de esta estrategia lo constituye la utilización de razas no
sometidas a procesos de mejora genética como la raza Ibérica. Pero no se detiene ahí: la
utilización de la raza Duroc, generalmente como línea paterna, está muy extendida incluso
en la cría de animales para la producción de carne fresca, por su mayor capacidad de
infiltrar grasa que razas industriales como la Landrace, la Large White y, sobre todo, la

CAPÍTULO I: INTRODUCCIÓN
20
Pietrain. Precisamente la DOP “Jamón de Teruel” hace uso de esta raza como línea
finalizadora paterna (BOE, 2011). Hay que destacar en este punto, que existen en el
mercado diferentes lineas Duroc que aportan diferente grado de infiltración grasa
(Lonergan et al., 2001; Schwab et al., 2005); su influencia sobre la conformación y sobre la
calidad de productos curados ha sido estudiada recientemente por Cilla et al., (2006) (a).
Es obvio por lo tanto, que los productores de jamón curado pueden aumentar la
infiltración grasa utilizando simplemente razas o líneas con mayor capacidad de
infiltración, algo que no exige ninguna sofisticación tecnológica. Sin embargo, esta
respuesta es claramente simplista porque conseguir infiltrar mas grasa es muy costoso
desde el punto de vista económico, dado que la deposición de tejido adiposo es mucho
menos eficaz que la deposición de tejido magro. El aumento de la deposición de tejido
adiposo se traduce en el aumento consiguiente del índice de transformación, lo que pone
en peligro la rentabilidad económica del proceso. Teniendo en cuenta que alrededor del
70% del costo de producir un cerdo se debe a los costos de su alimentación. Si estos
aumentan, el delicado equilibrio entre costos de producción y precio de venta se desplaza
hacía el extremo negativo (como ha ocurrido tantas veces en el sector porcino en los
últimos años).
Ante esta situación, cabe pensar en la posibilidad de seleccionar, al menos dentro
de una raza o de una linea genética de una determinada raza, a aquellos individuos
portadores de alelos de genes con capacidad demostrada de influir de manera selectiva en
el contenido graso de la canal o de la carne, con la esperanza de modificar (aumentar)
directamente el carácter “infiltración grasa” al menor costo posible en términos de índice
de transformación.
El problema que se plantea a la hora de implementar esta práctica es doble: por un
lado, el contenido graso de la canal y/o de la carne es un carácter poligénico, es decir, está
influido por muchos genes, cada uno de los cuales (salvo excepciones que se citan mas
adelante) ejerce efectos muy pequeños sobre el carácter; por otro lado, el conocimiento de
genes que cumplan la premisa anterior es muy limitado. Los genes más relevantes, por la
magnitud de sus efectos y/o por la amplitud de los trabajos publicados, de los estudiados
hasta ahora se comentan a continuación.
Quizá el mas conocido y estudiado y con mayores efectos fenotípicos sea el gen
RYR1 o “gen del halotano” (Fujii et al., 1991), uno de cuyos alelos está asociado a la
aparición de carnes defectuosas (carnes con pH45 muy bajos que dan lugar a carnes PSE).
A pesar de este problema, el hecho de que este mismo alelo esté asociado a carnes y
canales mas magras (por mecanismos bioquímicos desconocidos pero asociados al

CAPÍTULO I: INTRODUCCIÓN
21
aumento de los niveles de calcio sarcoplásmico) ha hecho que haya sido seleccionado en
las últimas décadas y esté ampliamente introducido en razas como la Large White, la
Landrace y, sobre todo, la Pietrain. La aparición de un test de PCR para analizar la
mutación causal ha permitido eliminar esta mutación en las líneas en las que se ha
considerado conveniente. De hecho, en la DOP, se ha eliminado el alelo mutante, y todos
los animales reproductores son homocigotos para el alelo asociado a canales menos
magras y menos susceptibles de rendir carnes PSE.
En el gen RN (“Rendement Napole”) también hay descritas dos mutaciones (Milan
et al., 2000; Ciobanu et al., 2001) que influyen en el engrasamiento de la canal y de la
carne. Este gen, también llamado PRKAG3, codifica una subunidad de la cAMP kinasa que
está implicada en la deposición de glucógeno en el músculo y a través de ello en los niveles
de pH24. Por mecanismos bioquímicos también desconocidos, como en el caso del gen
RYR1, una mutación que hace aumentar los niveles de glucógeno aumenta también el
contenido magro de la canal (y disminuye los valores del pH 24 dando lugar a las carnes
“ácidas”), mientras que una mutación que hace disminuir el contenido en glucógeno,
disminuye a su vez los niveles de engrasamiento (y aumenta levemente los valores del
pH24). Mientras que la primera mutación parece restringida a la raza Hampshire (la gran
disminución del pH24 se conoce como “efecto Hampshire”), la segunda está ampliamente
distribuida en distintas razas.
Hay dos genes que actúan sobre el contenido graso y sobre la conformación de la
canal porque influyen en el comportamiento alimentario de los cerdos: son MC4R (el
receptor de melanocortina-4 y LEPR (el receptor de leptina). Tanto en MC4R (Kim et al.,
2000) como en LEPR (Ovilo et al., 2005) se han descubierto mutaciones cuya expresión
fenotípica es el aumento en el consumo de alimento. El mayor consumo de alimento
provoca mayores niveles de engrasamiento a diferentes niveles y cambios en la
conformación de la canal consistentes sobre todo en un aumento del espesor del tocino
dorsal y de las partes de la canal, como la panceta, con mayor contenido graso a expensas
de las piezas más magras, como el lomo.
En el gen FABP3 (también conocido como H-FABP), de la familia de las “Fatty Acid
Binding Proteins” también se han descrito polimorfismos asociados a aumentos en los
niveles de engrasamiento, tanto del tocino dorsal, como de la grasa intramuscular
(Gerbens et al., 1999; Gerbens et al., 2000). La proteína codificada por este gen es un
transportador de ácidos grasos en sangre por lo que se supone que los alelos asociados a
altos niveles de engrasamiento codifican proteínas más eficaces en esta tarea.

CAPÍTULO I: INTRODUCCIÓN
22
El gen para el que se ha descrito mayor influencia sobre la infiltración grasa es
IGF2 (insuline like growth factor 2). En 1999 descubrieron una mutación en el gen IGF2
cuyos efectos sobre la composición de la canal y de la carne (Nezer et al, 1999; Jeon et al,
1999) eran los siguientes:
1) Mayor masa muscular y mas carne a expensas de tejido adiposo
2) Mayor contenido magro en el jamón
3) Mayor superficie del L. dorsi
4) Menor espesor del tocino dorsal
5) Mayor tamaño del corazón
El gen IGF2 parece ser el responsable de un QTL mayor (quantitative trait loci) con
efectos sobre el crecimiento muscular y la composición corporal en el cerdo, responsable
de entre el 15 y el 30% de la variación fenotípica en el contenido en masa muscular y de
entre el 10 y el 20% del espesor del tocino dorsal, lo que se traduce entre un 3 y un 4%
más de masa muscular a expensas de tejido adiposo (Nezer et al., 1999).
El IGF2 codifica un factor de crecimiento que determina, en periodos tempranos
del desarrollo, la diferenciación celular. Modificando el número de células musculares y
adipocitos, modifica posteriormente el contenido graso de la canal (Van Laere et al., 2003.
El alelo salvaje está relacionado con fenotipos grasos, mientras que el alelo mutado lo está
con fenotipos magros. Van Laere et al (2003) suponían que el genotipo AA estaba
ampliamente extendido en cruces comerciales de cerdos que habían sido seleccionados
durante mucho tiempo por su capacidad de producir carne, mientras que detectaban que
el genotipo GG era mayoritario en jabalíes y en razas no seleccionadas. Detectaban sin
embargo una incidencia muy distinta en poblaciones de cerdos Large White.
La transmisión del genotipo de IGF2 se produce únicamente por vía paterna y tiene
la ventaja desde el punto de vista productivo de que no exige la selección, y llegado el caso,
el recambio de las madres, sino que los esfuerzos de mejora se pueden concentrar
únicamente en los machos.
No se puede hablar de manera general de una composición corporal óptima ya que
ésta dependerá del uso que se pretenda hacer de la canal y/o de la carne. Para la
producción de carne fresca, lo óptimo serían las canales bien conformadas con bajos
contenidos en grasa y abundancia de tejido muscular. Si los resultados de Nezer et al., Jeon
et al., y Van Laere et a.l, se confirman para poblaciones basadas en cruces de razas sería
muy conveniente seleccionar machos AA para que toda la F1 tenga el alelo A por vía

CAPÍTULO I: INTRODUCCIÓN
23
paterna. Sin embargo, para la elaboración de productos curados, especialmente de jamón y
de paleta, es muy recomendable partir de animales que presenten mayor grado de
infiltración grasa, ya que la calidad sensorial de estos productos está muy relacionada con
la cantidad de grasa que contienen y con la composición cualitativa de ésta.
El alelo mutante de IGF2 está ampliamente distribuido en diversas razas
(Carrodeguas et al., 2006), entre otras las que se utilizan para la elaboración de jamones
de Teruel. Por ello consideramos que la selección del alelo relacionado con fenotipos
grasos podría ser una buena alternativa para modificar (aumentar) el contenido graso de
aquellos animales criados por la DOP, con objeto de mejorar la aptitud de los perniles para
ser transformados en jamón curado.
La utilización de machos Duroc GG sería de gran interés para la DOP con un
objetivo distinto al que tiene en el cruce con razas Ibéricas. Este objetivo es mejorar la
infiltración grasa del jamón dado que ésta es muy baja en la genética Landrace/Large
White. Análisis realizados utilizando machos Duroc para la elaboración de Jamones
amparados por la DOP Teruel, encontraron que más del 94,8% de ellos eran
homocigóticos AA para el gen IGF2, un 5,0% eran heterocigóticos y solo un 0,1% eran
homocigóticos GG (Burgos et al., 2006; Burgos et al., 2008). La información del genotipo,
junto con el conocimiento ya existente de la deposición grasa favorecida en cerdos con el
alelo G, aumenta la posibilidad de incrementar el contenido de grasa en los cerdos de la
DOP por introgresión del alelo G del gen IGF2 en las líneas Duroc usadas por los criadores
para la DOP Jamón de Teruel. Para evaluar la viabilidad económica de la introgresión del
alelo G es necesario analizar los efectos de ambos alelos en el gen IGF2 sobre el
rendimiento de crecimiento del animal, en la conformación de la canal, o en la calidad y
composición de la carne.

CAPÍTULO I: INTRODUCCIÓN
24
1.4. ANTECEDENTES BIBLIOGRÁFICOS SOBRE PRODUCTOS CURADOS
Los primeros artículos científicos indexados (WOK) sobre el jamón curado datan
del año 1936, y tratan sobre la optimización de la técnica del procesado del jamón curado
(Miller y Ziegler, 1936). En los años 50 y 60, los trabajos publicados continúan
centrándose en la mejora del proceso de curación, ya sea mediante la optimización de la
adición de sales (Wistreich et al., 1958; Wistreich et al., 1959) y fosfatos (Madsen et al.,
1957) o la evaluación de las características de la materia prima (Heck y Brown, 1961);
Además empiezan a aparecer los primeros estudios de caracterización, teniendo en cuenta
dos músculos representativos de la profundidad y superficie del jamón: el bíceps femoris y
el semimembranosus (Briskey et al., 1959). En los artículos de los años 70 la materia prima
empieza a tener un mayor interés sobre la calidad del producto final (Kuntapan et al.,
1972), a pesar de ello, la optimización del proceso de curación con el uso de estufajes con
el fin de acortar el proceso, pero sin que la calidad del producto final se viera afectada,
sigue siendo importante para los investigadores (Kingsley et al., 1978). Al llegar a los años
90, empiezan a aparecer los primeros trabajos sobre jamón Ibérico (García et al., 1991;
Ventanas et al., 1992), aunque el número de publicaciones encontradas sobre jamón
curado sigue siendo superior a las de jamón Ibérico durante toda la década. A principios
del año 2000 el estudio del jamón Ibérico toma un mayor protagonismo, debido a que la
importancia económica de este producto va en aumento, incidiendo por lo tanto sobre un
incremento de la investigación.
En los últimos 50 años, podemos comprobar la gran variedad de estudios
realizados sobre el jamón curado. De los 395 trabajos revisados en la web, encontramos
73 que se centran en el efecto de la materia prima en la calidad del producto final, como
pueden ser la alimentación (Isabel et al., 1999), raza y sexo (Gou et al., 1995; Dumont y
Roy, 1975) o genética (Stalder et al., 2004) del animal; 75 se centran en la búsqueda de la
optimización del proceso de curación (Kemp et al., 1975; Arnau et al., 1997), sin tener en
cuenta la utilización de aditivos, como son los efectos de la sal, nitratos o fosfatos (Alino et
al., 2010; Palmia et al., 1992; Kamm y Coffin, 1968), aumentando así en un número de 125
trabajos. Otro de los temas muy investigados ha sido el análisis sensorial (Barbieri et al.,
1992; Piotrows et al., 1970; Rivas-Canedo et al., 2009), encontrando hasta 71 trabajos.
Existe un menor interés en el análisis de la de composición de la grasa subcutánea e
intramuscular (Kouba y Bonneau, 2009; Buscailhon et al., 1994) con 38 publicaciones,
contenido de proteínas (Flores et al., 1984; Toldrá et al., 1992) con 23 o textura (Ruiz-
Carrascal et al., 2000; Morales et al., 2010) con 16. Ya durante la última década empiezan a
ser más estudiados los efectos del envasado, tras la aparición de nuevos formatos en la
presentación del producto (Cilla et al., 2006 (a)).

CAPÍTULO I: INTRODUCCIÓN
25
En el caso del jamón Ibérico, a pesar de que hasta principios de los años 90 no
existen referencias bibliográficas, vemos como en los últimos 20 años su interés ha ido
creciendo, ya que encontramos hasta 109 trabajos, de los cuales 39 se centra en el análisis
de compuestos volátiles (García et al., 1991; Martin et al., 2000; Carrapiso et al., 2010 ) y
30 al análisis de la grasa subcutánea e intramuscular (Martin et al., 1999; Timón et al.,
2001; Narvaez-Rivas et al., 2007); dejando 40 relacionados con los efectos de la materia
prima sobre la calidad del producto final (de la Hoz et al., 1996; Carrapiso et al., 2007) o la
conservación (Andrés et al., 2006, Fuentes et al., 2010). En el caso del jamón curado, el
análisis de compuestos volátiles también se ha considerado importante sobre todo en los
últimos años, pero no tanto el de la grasa. Sin embargo, la caracterización del jamón
curado ha sido mucho más completa que la del Ibérico, donde se centran principalmente
en el estudio de la grasa y de los compuestos volátiles.
Al centramos en la DOP Jamón de Teruel, vemos que a pesar de tratarse de un
producto de elevada calidad, existe un gran vacío al revisar la bibliografía internacional,
encontrando tan solo 9 artículos indexados. La primera publicación encontrada data de
finales de los 90, donde se evalúa la calidad de este producto (Sierra et al., 1998), sin
embargo hasta finales de la década del 2000 no vuelve a encontrarse nada, a partir de
entonces los investigadores se centran especialmente en características del animal vivo
que influyen por lo tanto en la calidad del producto final (Daza et al., 2010; Latorre et al.,
2008) y en estudiar la aceptación de estos productos por parte del consumidor (Cilla et al.,
2006 (b); Resano et al., 2007). Una revisión en la bibliografía española muestra un poco
más de información, localizando hasta 13 artículos publicados en diferentes revistas
durante los últimos años. A pesar de esto, al revisarlos nos encontramos de nuevo con
poca información sobre la caracterización de estos productos, ya que se centran
fundamentalmente en datos de comercialización (Fandos et al., 2007), consumidores (San
Juan et al., 2004) o indicaciones sobre la trazabilidad de estos productos (Cárnica, 2000;
Latorre et al., 2005). Este hecho ha generado un desconocimiento del producto por parte
del consumidor debido a la falta de una completa caracterización que pudiera resaltar el
por qué de su alta calidad, en comparación con otros jamones curados.
Al revisar la información sobre otros productos, como el lomo o la paleta curada,
vemos como el número de publicaciones es bastante inferior que en el caso del jamón
curado, encontrando tan solo 24 trabajos sobre lomo curado durante los últimos años, 6
de los cuales se centran en lomos procedentes de cerdos controlados por el consejo
regulador de la DOP de Teruel y relacionados con características del animal vivo (Latorre
et al., 2008; Latorre et al., 2009). Con respecto al lomo Ibérico, encontramos una cifra muy

CAPÍTULO I: INTRODUCCIÓN
26
similar con 26 trabajos, la mayor parte de ellos a partir del año 2007 y relacionados
también con características del animal vivo (Muriel et al., 2004; Soto et al., 2010).
Con respecto a las paletas curadas, existen escasas referencias bibliográficas tanto
las procedentes de cerdo blanco como de cerdo Ibérico, encontrando tan solo alguna
publicación sobre la paleta en fresco (Barea et al., 2003; Barat et al., 2006) y una sobre la
paleta curada (Sarraga et al., 2007). La paleta curada es un producto muy similar al jamón
curado, por lo que desde el punto de vista científico resulta muy interesante, ya que una
caracterización podría utilizarse para predecir el comportamiento frente a posibles
cambios en productos de procesado más largo, ya que el proceso de curación de este
producto es más corto que el del jamón. Por lo tanto, estudios realizados en la paleta sobre
la materia prima, como pueden ser la influencia de la alimentación de los cerdos, la
genética, el cruce de razas, el peso de sacrificio o las características de procesado, pueden
ofrecernos información extrapolable al jamón curado en menos tiempo.

CAPÍTULO I: INTRODUCCIÓN
27
1.5. ESTRUCTURA ANATÓMICA DE LA PALETA Y DEL JAMÓN CURADO
Al revisar la bibliografía vemos que durante los 60 años en los que se ha estado
investigando el jamón curado, se ha conseguido que tengamos un gran conocimiento del
producto, desde su composición química hasta su composición anatómica (Ruiz, 2005).
Por otra parte, la paleta curada es la gran desconocida para el investigador, tanto su
composición química como anatómica, ya que se trata de un producto que siempre se ha
visto en un segundo plano, por lo que se considera necesario conocerlo, empezando por su
composición inicial.
La porción ósea de la extremidad anterior del cerdo (paleta) está compuesta por
los siguientes huesos: escápula, humero, radio, cúbito, carpianos y metacarpianos. Las
masas musculares correspondientes a los músculos son: latissimus dorsi, serratus ventralis,
serratus dorsalis anterior, interspinalis, rhomboideus, supraspinatus, infraspinatus, cutaneus
trunci, splenius, pectoralis profundi, semispinalis capitis, cutaneous faicei, ticeps brachii long,
medial y lateral head, teres major y minor, subscapularis, tensor fasciae antibrachii,
deltoideus, pectoralis superficialis, omo-transverarius, scalenus, coracobrachialis,
brachiocephalicus, cutaneus colli, brachialis, anconeus, extensor carpi radialis, obliquus y
ulnaris, bíceps brachii, retractor costae, superficial y profundi digital flexor thoracic,
common y lateral digital extensor thoracic, pronator teres y flexor carpi radialis y ulnaris y
la grasa subcutánea de cobertura. Debido a la gran variedad de músculos encontrados en
la paleta, hemos subrayado aquellos que se encuentran en mayor cantidad.
1.5.1. COMPOSICIÓN QUÍMICA
En 1991 Lawrie presentó los valores medios para un músculo típico de mamífero
adulto, siendo un 75% de agua, un 19% de proteína, un 2,5% de grasa, un 1,65% de
nitrógeno no proteico, un 1,20% de carbohidratos y un 0,60% de minerales. Unos años
después Gou (1993) se centró más en los valores medios de la carne de cerdo,
proponiendo un 13,9% de grasa subcutánea, un 68,2% de músculo, un 13,8% de grasa
intermuscular y un 8,7% de hueso.
En 1997 encontramos datos publicados por Belitz y Grosch, donde se presentaba la
composición química de la carne para el jamón y la paleta de cerdo (Belitz y Grosch, 1997).
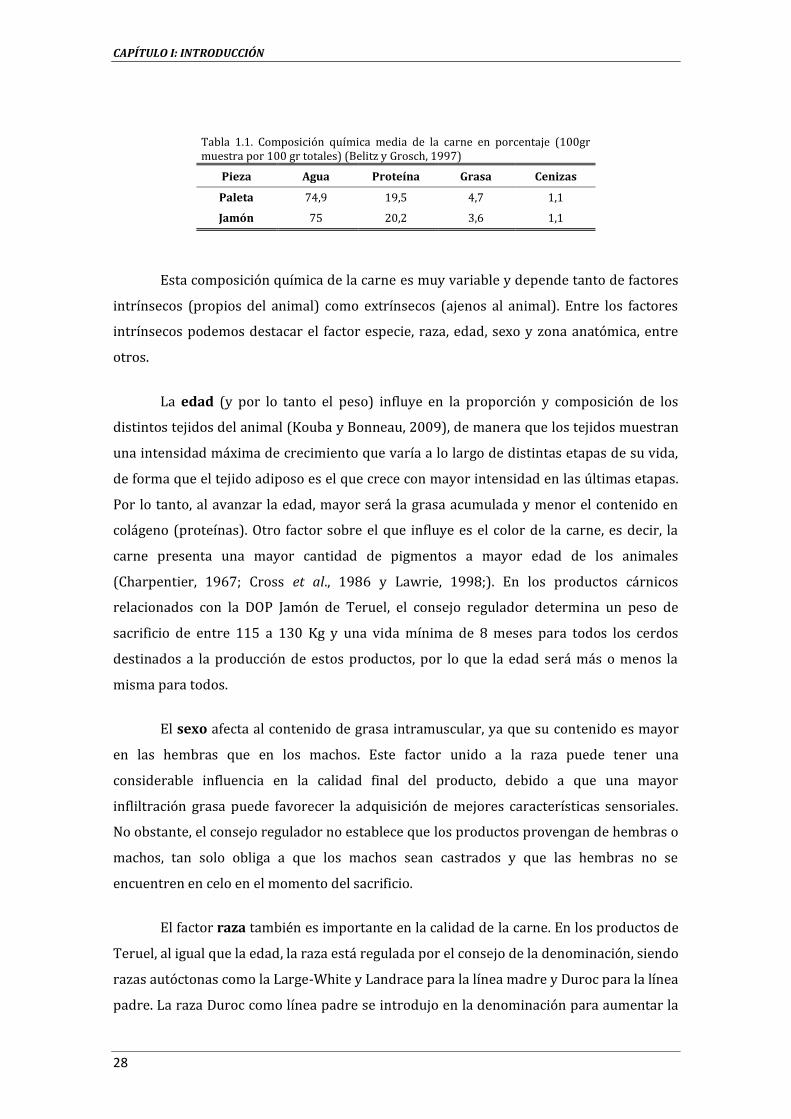
CAPÍTULO I: INTRODUCCIÓN
28
Tabla 1.1. Composición química media de la carne en porcentaje (100gr muestra por 100 gr totales) (Belitz y Grosch, 1997)
Pieza Agua Proteína Grasa Cenizas
Paleta 74,9 19,5 4,7 1,1
Jamón 75 20,2 3,6 1,1
Esta composición química de la carne es muy variable y depende tanto de factores
intrínsecos (propios del animal) como extrínsecos (ajenos al animal). Entre los factores
intrínsecos podemos destacar el factor especie, raza, edad, sexo y zona anatómica, entre
otros.
La edad (y por lo tanto el peso) influye en la proporción y composición de los
distintos tejidos del animal (Kouba y Bonneau, 2009), de manera que los tejidos muestran
una intensidad máxima de crecimiento que varía a lo largo de distintas etapas de su vida,
de forma que el tejido adiposo es el que crece con mayor intensidad en las últimas etapas.
Por lo tanto, al avanzar la edad, mayor será la grasa acumulada y menor el contenido en
colágeno (proteínas). Otro factor sobre el que influye es el color de la carne, es decir, la
carne presenta una mayor cantidad de pigmentos a mayor edad de los animales
(Charpentier, 1967; Cross et al., 1986 y Lawrie, 1998;). En los productos cárnicos
relacionados con la DOP Jamón de Teruel, el consejo regulador determina un peso de
sacrificio de entre 115 a 130 Kg y una vida mínima de 8 meses para todos los cerdos
destinados a la producción de estos productos, por lo que la edad será más o menos la
misma para todos.
El sexo afecta al contenido de grasa intramuscular, ya que su contenido es mayor
en las hembras que en los machos. Este factor unido a la raza puede tener una
considerable influencia en la calidad final del producto, debido a que una mayor
infliltración grasa puede favorecer la adquisición de mejores características sensoriales.
No obstante, el consejo regulador no establece que los productos provengan de hembras o
machos, tan solo obliga a que los machos sean castrados y que las hembras no se
encuentren en celo en el momento del sacrificio.
El factor raza también es importante en la calidad de la carne. En los productos de
Teruel, al igual que la edad, la raza está regulada por el consejo de la denominación, siendo
razas autóctonas como la Large-White y Landrace para la línea madre y Duroc para la línea
padre. La raza Duroc como línea padre se introdujo en la denominación para aumentar la

CAPÍTULO I: INTRODUCCIÓN
29
infiltración grasa (Peloso et al., 2010) dado que ésta era muy baja en la genética
Landrace/Large White.
La selección genética es importante porque como ya se ha mencionado en un
apartado anterior, puede mejorar la infiltración de la grasa. Esta herramienta nos permite
seleccionar, al menos dentro de una raza o de una genética de una determinada raza, a
aquellos individuos portadores de alelos de genes con capacidad demostrada para influir
de manera selectiva en el contenido graso de la canal o de la carne, con la esperanza de
aumentar directamente el carácter infiltración grasa con el menor costo posible en
términos de índice de transformación.
La zona anatómica también tiene influencia sobre la composición de la carne, la
grasa se deposita en distintas localizaciones anatómicas como es la subcutánea,
intermuscular e intramuscular. Estos mismos depósitos no tienen la misma importancia
en la canal y en la calidad de la carne, ya que un exceso de grasa subcutánea no es bueno
para el consumo de carne fresca, pero si necesario para la curación de la paleta y del
jamón curado. La velocidad de deposición también se verá influenciada por la zona
anatómica, ya que algunos autores han encontrado que la velocidad de deposición de la
grasa subcutánea será mayor que la de la grasa intramuscular en el caso del jamón curado,
pero igual en el caso de la paleta curada (Kouba y Bonneau, 2009).
Entre los factores extrínsecos el más importante es la alimentación, que incide
directamente sobre la composición de la carne, ya que en función de la dieta utilizada se
puede modificar el contenido de grasa, el color de la carne u otras propiedades sensoriales
de la misma, afectando por lo tanto a la calidad del producto final. Por ejemplo, si se
aumenta en la dieta el contenido de hidratos de carbono o de grasa, aumenta el
engrasamiento de las canales (Ordóñez et al., 1998). Mediante aportes grasos controlados
podemos mejorar la jugosidad y la terneza como consecuencia del incremento de grasa
intramuscular, pero por otra parte esto implicaría un descenso en el contenido de
proteínas. En el caso de los productos de la DOP de Teruel, la alimentación de los animales
también se encuentra regulada por el Consejo Regulador, de forma que las únicas
variaciones que se pueden producir vendrán determinadas por avances tecnológicos, que
pueden influir en animales criados en distintos años.

CAPÍTULO I: INTRODUCCIÓN
30
1.5.2. PARÁMETROS RELACIONADOS CON LA CALIDAD DE LA CARNE
La composición y estructura de la carne está íntimamente relacionada con su
calidad tanto nutritiva como sensorial, de ahí que cada uno de sus componentes tenga un
papel fundamental como responsables de la calidad final del producto. Desde el punto de
vista nutritivo y en función de la cantidad y calidad de sus componentes químicos, así
como en función de su utilización por el organismo, este producto pertenece al grupo
básico de “alimentos proteicos”. La carne además es una buena fuente de aminoácidos
esenciales, vitaminas (especialmente del grupo B) y minerales, entre los que destaca su
alto contenido en hierro. Este mineral tiene una excelente absorción y potencia la
asimilación procedente de otras fuentes como las leguminosas o los cereales (Godber,
1994). Otra de las propiedades de la carne es su contenido medio-alto en lípidos, que hace
que sea un alimento muy energético.
El agua en la carne es el componente más abundante, alcanzando entre un 65-75%
dependiendo del tipo de carne. A pesar de esto, se encuentra bien integrada formando
parte de la estructura cárnica. El agua de la canal se encuentra principalmente en el tejido
muscular magro; el tejido adiposo contiene poca agua. Muchas de las propiedades físicas
de la carne como el color, la textura y la firmeza dependen en parte de la capacidad de
retención de agua (CRA) de la carne, que está muy relacionada con el pH final de la misma.
El contenido en agua de la carne de cerdo es de aproximadamente 75% siendo muy similar
al de otras especies como pollo y vacuno (Fennema y Grosch, 1992; Primo et al., 1997).
En los productos cárnicos curados, el agua juega un papel muy importante en la
calidad, ya que el proceso de curación se basa principalmente en la pérdida gradual de
agua hasta alcanzar la merma adecuada en función del producto, ya que no será la misma
para jamón, paleta o lomo curado. Este parámetro se encuentra regulado en algunas
marcas registradas, por ejemplo, en la ETG el contenido máximo acuoso sobre el producto
desengrasado debe ser del 57% y en el caso de la DOP Jamón de Trévelez, los jamones
deben mermar a lo largo de todo el proceso un mínimo del 35% (MARM).
La fracción lipídica es la parte más variable en la carne y se encuentra en el tejido
adiposo subcutáneo, en el interior de la cavidad corporal o incluida en el tejido
intermuscular e intramuscular. Los lípidos están implicados en muchos aspectos de
calidad de la carne y productos cárnicos, tales como el valor nutricional, características
sensoriales y propiedades tecnológicas, y este estado va a depender de la composición en
ácidos grasos de la grasa, que a su vez están relacionados con factores como la raza
(Petrón et al., 2004), el sexo, la edad y el peso (Latorre et al., 2008; Latorre et al., 2009), la

CAPÍTULO I: INTRODUCCIÓN
31
alimentación (Daza et al., 2010), y otras como la localización anatómica (Kouba y Bonneau,
2009; Fisher et al., 2003).
En la etapa de crecimiento del animal la grasa se deposita en distintas
localizaciones anatómicas como es la subcutánea, intermuscular (entre músculos) e
intramuscular (dentro del músculo) a distinta velocidad (Fisher et al., 2003; Kouba y
Bonneau, 2009). Estos mismos depósitos, como se ha indicado anteriormente, no tendrán
la misma importancia en todos los productos.
La grasa subcutánea o de cobertura del cerdo tiene como principal función la de
reserva energética principalmente, y como aislante térmico. Este tejido se organiza en dos
capas, cuya composición en ácidos grasos ha sido motivo de numerosos estudios desde
finales de los 60, donde numerosos autores han demostrado diferencias entre las dos
capas, siendo la más externa más insaturada debido al mayor porcentaje de ácido linoleico
(C18:2(ω-6)) principalmente (Monziols et al., 2007). Esta cobertura tan solo puede
considerarse importante desde el punto de vista tecnológico en el caso de algunos
productos curados como la paleta o el jamón, dado que interviene en los procesos de
penetración de sal y de pérdida de agua.
La grasa intramuscular parece tener una gran importancia, no solo desde un
punto de vista nutricional o tecnológico, sino también por su posible repercusión sobre
algunas características sensoriales de los productos curados, como puede ser la textura, el
olor y el flavor. Desde el punto de vista tecnológico, la grasa intramuscular es de enorme
importancia puesto que determina el grado de penetración al interior de los productos
curados de la sal, aditivos y/o especias empleadas durante el procesado así como el nivel
de desecación del producto. Es por ello, que aquellos productos con un mayor contenido
de grasa intramuscular necesitarán un mayor periodo de maduración para conseguir la
pérdida de humedad adecuada relacionada con una óptima penetración de la sal. Esto se
debe a que el coeficiente de difusividad del agua y de la sal es menor en la grasa que en el
magro (Ventanas, 2006). En el jamón Ibérico, la grasa intramuscular visible (veteado) está
asociada a la imagen típica de calidad del producto (Ventanas et al. 2001) y parece que
tiene un efecto positivo sobre las características sensoriales (hay una relación positiva con
la jugosidad y el brillo y negativa con la fibrosidad, dureza y sequedad), aunque en otros
tipos de jamones curados su efecto ha sido contradictorio (Ruiz-Carrascal et al., 2000). El
contenido de grasa intramuscular de los cerdos influyen distintos factores, entre ellos el
desarrollo de la canal (Mayoral et al., 1999), la raza del cerdo y su cruzamiento (Berdagué
et al., 1993) y el sistema de alimentación (Pérez-Palacios et al., 2009).

CAPÍTULO I: INTRODUCCIÓN
32
Los compuestos nitrogenados de la carne están constituidos por proteínas,
péptidos, aminoácidos libres y otros compuestos. Las proteínas representan el
componente más abundante de la materia seca del músculo, influyendo por lo tanto en el
valor nutritivo de la carne, ya que su alto contenido y composición hace que sea una buena
fuente de aminoácidos esenciales y portadora de minerales, destacando el hierro, que
forma complejo con la miolobina.
Las proteínas se pueden clasificar atendiendo a su solubilidad en tres grupos
(Primo, 1997; Ordoñez et al., 1998); las proteínas del aparato contráctil (proteínas
miofibrilares), las proteínas solubles (proteínas sarcoplasmáticas), y las proteínas
insolubles (proteínas del tejido conjuntivo y proteínas de los orgánulos). Dentro de las
proteínas solubles podemos destacar la mioglobina. La cantidad de mioglobina muscular
se ve afectada por factores genéticos, la dieta del animal, el tipo de fibra muscular, la
especie, el ejercicio y por la edad. En cerdos se observa un descenso en el contenido de
mioglobina cuando los animales tienen una deficiencia en hierro, también se ha descrito
un aumento cuando los animales poseen una deficiencia en vitamina E y por el ejercicio
(Belitz y Grosch, 1997). El contenido de mioglobina también está relacionado con otro
atributo como es el color, no solo porque es el principal responsable en su formación, sino
porque el deterioro de la carne por su envejecimiento o mal almacenamiento cambia su
estado químico, por lo que se podría considerar como un marcador de la calidad. El color
es una característica de calidad muy importante que contribuye a la aceptación sensorial
de los productos cárnicos curados, por su estrecha relación con la mioglobina. Este
parámetro se encuentra descrito en las especificaciones de la ETG (MARM) y de las
denominaciones, por ejemplo, en el caso de la DOP de Teruel, se establece un color rojo y
de aspecto brillante al corte, pero no indica la cantidad de éste (BOE, 1993).
Dentro de la fracción nitrogenada también se incluyen los aminoácidos y los
nucleótidos, compuestos que muchos autores han descrito su relación con el sabor y flavor
final de la carne (Aristoy y Toldrá, 1998). Los aminoácidos son generados a partir de la
degradación de las proteínas como consecuencia del proceso de curación del jamón, paleta
o lomo curado, contribuyendo por lo tanto al desarrollo de su flavor (Larrea et al., 2006).
Las características del sabor de cada aminoácido han sido ampliamente estudiadas
individualmente y se ha caracterizado la naturaleza del sabor de los aminoácidos en cinco
sabores: dulce, amargo, ácido, salado y umami (Jurado, 2005), cuyo sabor estará
determinado por la concentración del aminoácido y por su umbral de reconocimiento
(concentración más baja a la que puede ser detectado) (Belitz y Grosch, 1997). Otro
aspecto conocido es la capacidad que tienen los aminoácidos de actuar como

CAPÍTULO I: INTRODUCCIÓN
33
potenciadores del sabor. Un ejemplo de esto es el efecto sinérgico del ácido glutámico
(Glu) con los 5 nucleótidos que provoca que el sabor umami sufra una clara potenciación
(Kawai et al., 2002). Además de su implicación con el sabor, los aminoácidos libres
también están relacionados con la formación de compuestos volátiles mediante reacciones
de Maillard, generando principalmente compuestos azufrados, nitrogenados y los
aldehídos de Strecker (Mottram, 1998; Jurado et al., 2009).
Los nucleótidos son compuestos nitrogenados de bajo peso molecular que se
originan por la degradación del ATP (Adenosina 5´trifosfato) que se encuentra en los
músculos tras el sacrificio (Shahidi et al., 1994; Saito et al., 2007). Estos compuestos
también se han descrito relacionados con la campaña del flavor, pero en este caso actúan
solo como potenciadores globales del flavor (Kawai et al., 2002). Es por ello, que tanto los
aminoácidos libres como los nucleótidos podrían potenciar el flavor característico de la
carne y de los productos cárnicos (Tovar, 2009).
Los compuestos volátiles, generados durante el proceso de secado-maduración,
derivan principalmente de la degradación de precursores como los compuestos
nitrogenados de bajo peso molecular (aminoácidos y nucleótidos) (Zhao et al., 2005; Mora
et al., 2009) y de los ácidos grasos que conforman la grasa (Elmore et al., 2002; Jurado et
al., 2009). La mayor parte de estos compuestos son liposolubles (Olivares et al., 2009), por
lo que se ha encontrado una mayor liberación de volátiles en productos con mayor
contenido en lípidos (Guinard et al., 2002; Seuvre et al., 2007). La presencia de algunos
compuestos volátiles o su concentración están íntimamente relacionados con la calidad
sensorial, mediante el aroma y el flavor (Pérez-Palacios et al., 2010; Ventanas et al., 2010).
Además, podrían utilizarse como indicadores del tiempo del proceso de curación, y como
indicadores del desarrollo del flavor (Jurado et al., 2009).
La textura es un atributo sensorial muy complejo que abarca las impresiones que
ocurren cuando el alimento contacta con la superficie de los dedos, la lengua y los dientes.
La textura de los productos curados (jamón, paleta y lomo) se debe por una parte al
proceso de elaboración que estos sufren, y por otra a las características de la materia
prima utilizada para su elaboración. La textura está relacionada con la grasa intramuscular
y con el contenido en agua. La grasa intramuscular afecta directamente a la jugosidad y
sabor, e indirectamente a la dureza (Jeremiah, 1978; Ruiz, 2005).
Uno de los principales defectos de textura en los productos curados es la textura
blanda y/o pastosa (Morales et al., 2008 (a); García-Rey et al., 2004). Este defecto está
relacionado con otras propiedades como son un pH elevado de la materia prima, por un

CAPÍTULO I: INTRODUCCIÓN
34
pH muy bajo (Sánchez, 2005), por la presencia de grasa intramuscular e intermuscular que
dificulte el salado y secado, por un contenido de sal bajo, o bien por una temperatura de
procesado elevada (Morales et al., 2008 (b)). En sentido contrario, la jugosidad es el
atributo que mayor influye positivamente sobre la aceptación de estos productos, ya que
está relacionado directamente con la secreción de saliva durante la masticación, o como
consecuencia de la liberación de sustancias aromáticas presentes en la grasa.
Por otra parte, actualmente, la calidad sensorial de los productos cárnicos es la
cualidad más importante, ya que de ella depende la decisión de su consumo. El hombre no
sólo consume un producto cárnico por su valor nutritivo sino por las sensaciones
sensoriales que se producen durante su ingestión, de manera que el consumidor aprecia
su calidad sensorial y decide si les gusta o no. Desde hace algunas décadas se ha
intentando cuantificar las sensaciones percibidas por los consumidores al probar un
alimento. Para ello se ha desarrollado una ciencia que se denomina evaluación sensorial.
La evaluación sensorial debe abarcar los aspectos más relevantes del alimento
que condicionan la aceptación o el rechazo del consumidor. Estas características son las
relacionadas con el aspecto, la textura, el olor, el sabor y el flavor, ya que todas ellas
muestran una relación más o menos marcada con la calidad que el consumidor percibe.
Actualmente, y ante la falta de métodos analíticos y rápidos utilizables en la propia
industria, la evaluación sensorial se ha convertido en una herramienta fundamental para
conocer las características de un producto y cómo serán percibidas y valoradas por el
consumidor. La evaluación sensorial permite establecer qué aspectos son positivos o
negativos, de forma que el fabricante puede modificar el procesado en función de aquellas
características que condicionan la elección del consumidor. Además, el estudio de la
calidad sensorial es importante para poder establecer relaciones con análisis físico-
químicos menos invasivos y laboriosos, que permitan la evaluación rápida y objetiva de un
determinado atributo sensorial.

CAPÍTULO II: PLANTEAMIENTOS Y OBJETIVOS


CAPÍTULO II: PLANTEAMIENTOS Y OBJETIVOS
37
2. PLANTEAMIENTOS Y OBJETIVOS
El grupo de investigación de Tecnología y calidad de los Alimentos de la UEX
(TECAL), centra sus esfuerzos, desde hace más de dos décadas, en el estudio de la calidad
de los productos cárnicos en sus más diversos aspectos. Los factores que condicionan
dicha calidad y los parámetros que permiten cuantificarla han sido objeto de numerosos
trabajos científicos entre los que destacan los dedicados al jamón Ibérico. Esta experiencia
ha permitido establecer relaciones con la Universidad de Zaragoza para desarrollar un
proyecto de investigación centrado en productos de gran importancia económica para
Aragón como son el Jamón de Teruel y otros productos asociados.
Esta investigación ha aprovechado las fortalezas de la Universidad de Zaragoza y
de la Universidad de Extremadura en el ámbito de la Genética y de la Tecnología de los
Alimentos y ha planteado el estudio de la influencia que tiene la expresión de distintos
“Quantitative Trait Loci” QTL en los cerdos, sobre los productos obtenidos a partir de
ellos.
El amplio seguimiento realizado, para el estudio genético, en varias generaciones
de animales, ha permitido contar con un importante número de muestras de las tres
piezas mas apreciadas y plantear una caracterización exhaustiva y un estudio comparativo
de las mismas que constituyen el objetivo principal de esta Tesis Doctoral.
Dentro de este objetivo principal se encuadran los siguientes objetivos
secundarios:
1. Caracterización físico-química y sensorial de productos cárnicos asociados a la D.O.
“Jamón de Teruel”:
Jamón curado
Paleta curada
Lomo curado
2. Evaluación del efecto del QTL IGF2 sobre productos cárnicos asociados a la D.O. “Jamón
de Teruel”.
3. Evaluación de la influencia de los alelos A y G de IGF2 sobre la calidad de los productos
cárnicos derivados de su progenie.


CAPÍTULO III. MATERIAL Y MÉTODOS


CAPÍTULO III: MATERIAL Y MÉTODOS
41
3. MATERIAL Y MÉTODOS
3.1. MATERIAL BIOLÓGICO
3.1.1. NATURALEZA DE LAS MUESTRAS
Para este estudio se utilizaron jamones, paletas y lomos curados procedentes de
hembras porcinas Large White x Landrace homocigóticas AA y cerdos heterocigóticos AG
procedentes de líneas Duroc. Las cerdas AA se cruzaron con las líneas Duroc AG para
asegurarse que cualquier alelo G detectado en su descendencia fuera de origen paterno. Se
seleccionaron 20 cerdos de su progenie, 10 AA y 10 AG y se criaron según las condiciones
establecidas por la DOP Jamón de Teruel (Latorre et al., 2005). Los cerdos se sacrificaron
con un peso entre 115-130 Kg y las piezas (jamones, paletas y lomos) se procesaron según
lo establecido por el consejo regulador de la DOP Jamón de Teruel (BOA, 1993 (a)).
3.1.2. NATURALEZA DEL ALIMENTO SUMINISTRADO
Los ingredientes empleados en la formulación del pienso suministrado a los
animales aparecen expuestos en la tabla 3.1.1.
Tabla 3.1.1. Materias primas para la alimentación del animal
Ingredientes (%)
Cebada 62,03%
Trigo 15,00%
Harina de soja 4,20%
Harina de colza 10,00% Salvado 4,17%
Melaza 1,50%
Grasa 0,50%
Carbonato cálcico 0,96% Lisina 50% liquida 0,61%
Cloruro sódico 0,45%
Fosfato dicálcico 0,27%
L-Treonina 0,04% Metionina hidroxianáloga 0,07%
La composición en ácidos grasos del alimento se indica en la tabla 3.1.2.

CAPÍTULO III: MATERIAL Y MÉTODOS
42
Tabla 3.1.2 Composición en ácidos grasos expresados en mg/100g muestra y en % de área de la grasa de los alimentos suministrados a los cerdos
PIENSO (mg ac graso/100 gr muestra) PIENSO (% Área)
% GRASA (f) 2,10 2,10
C14:0 27,16 0,49
C15:0 4,91 0,12
C16:0 878,00 23,10 C17:0 10,75 0,20
C18:0 201,60 4,99
C20:0 13,61 0,23
C22:0 5,28 0,13 C23:0 0,77 0,02
C16:1 7,54 0,17
C17:1 5,39 0,11
C18:1 391,54 28,54 C20:1 11,69 0,63
C22:1 5,47 0,14
C24:1 4,01 0,03
C18:2(ω-6) 1659,19 38,05 C18:3(ω-3) 141,90 3,13
C20:2 (ω-6) 1,96 0,10
C20:4 (ω-6) 2,28 0,05
C20:5(ω-3) 2,09 0,05 C22:6(ω-3) 1,31 0,06
AGS 1142,08 29,28 AGMI 425,63 29,63
AGPI 1808,72 41,45
ω-6 1663,42 38,20
ω-3 145,30 3,24 ω-6/ω-3 11,45 11,78
(f) extracto fresco
3.1.3. PROCESADO DE LAS MUESTRAS
3.1.3.1 PROCESADO DEL JAMÓN CURADO
El proceso de elaboración se inició con el salado en seco, donde los perniles fueron
frotados superficialmente con las sales de curado y a continuación se apilaron y fueron
cubiertos de sal, donde permanecieron durante 10 días. Las condiciones de esta fase
fueron 2,4ºC de temperatura y humedad relativa (HR) de 91%. Finalizada la fase de
salado, los perniles se lavaron con agua para eliminar los posibles restos de sal adheridos
y se trasladaron a cámaras para la fase de post-salado. En esta etapa los perniles
permanecieron a temperaturas de 3,5ºC y humedad relativa de 75% durante 66 días. En la
siguiente fase, la temperatura fue aumentando desde los 12 hasta los 30ºC y la HR fue
disminuyendo desde el 85 hasta 74% durante los 60 días que duró la etapa de secado. A
continuación, las piezas se maduraron colgadas en naves en ambiente natural durante 241
días, con una temperatura de 15ºC y una HR del 74% aproximadamente. Transcurrido ese
tiempo, se llevaron a unas cámaras durante 60 días en condiciones de estufaje, 28ºC de
temperatura y 65% de HR. Finalmente se almacenaron en condiciones de post-estufaje

CAPÍTULO III: MATERIAL Y MÉTODOS
43
hasta la finalización del procesado, con temperaturas entre 14 y 18ºC y HR entre 60 y
70%.
3.1.3.2 PROCESADO DE LA PALETA CURADA
El proceso de elaboración se inició con el salado en seco, donde las paletas fueron
frotadas superficialmente con las sales de curado y a continuación se apilaron y fueron
cubiertas de sal, donde permanecieron durante 5 días. Las condiciones de esta fase fueron
2,4ºC de temperatura y humedad relativa (HR) de 91%. Finalizada la fase de salado, las
paletas se lavaron con agua para eliminar los posibles restos de sal adheridos y se
trasladaron a cámaras para la fase de post-salado. En esta etapa las paletas permanecieron
a temperaturas de 3,5ºC y humedad relativa de 75% durante 45 días. En la siguiente fase,
la temperatura fue aumentando desde los 12 hasta los 30ºC y la HR fue disminuyendo
desde el 85 hasta 74% durante los 45 días que duró la etapa de secado. A continuación, las
piezas se maduraron colgadas en naves en ambiente natural durante 120 días, con una
temperatura de 15ºC y una HR del 74% aproximadamente. Transcurrido ese tiempo, se
llevaron a unas cámaras durante 60 días en condiciones de estufaje, 28ºC de temperatura
y 65% de HR. Finalmente se almacenaron en condiciones de post-estufaje hasta la
finalización del procesado, con temperaturas entre 14 y 18ºC y HR entre 60 y 70%.
3.1.3.3 PROCESADO DEL LOMO CURADO
Las cintas de lomo se adobaron mediante masaje en bombo durante 15 minutos
con una mezcla de adobo que incluye sales nitrificantes, especias y aditivos (tabla 3.1.1), y
se dejaron macerar en refrigeración entre 2-4ºC durante 4-5 días. Una vez concluido el
adobo, las piezas se embutieron en tripas naturales y se ataron manualmente los
extremos. El curado y secado-maduración, se realizó en varias etapas en cámaras con
condiciones controladas durante 70-75 días hasta que alcanzaron una merma de 37-38%.
Durante la primera semana la curación se llevó a cabo en frio con temperaturas entre 5 y
10ºC. Posteriormente se aumentó la temperatura hasta 12-14ºC. La HR cambió entre 72-
80%. Cada día se tocó el producto, y se variaron levemente la HR y la temperatura para
llegar hasta el citado 37-38% de merma. En los lomos procedentes a la segunda y tercera
campaña no se añadieron las especias.

CAPÍTULO III: MATERIAL Y MÉTODOS
44
Tabla 3.1.1. Ingredientes utilizados para el adobo del lomo curado
Ingredientes Composición (%)
Pieza lomo 94,00
Sal 3,00
Pimentón 1,80
Azúcar 0,60
Lactosa 0,30
Especias naturales 0,29
E-301 0,0001
E-252 0,0001
E-250 0,0001
3.2. TRATAMIENTO DE LAS MUESTRAS
3.2.1. JAMÓN CURADO
Para la caracterización del jamón curado asociado a la DOP Jamón de Teruel se
emplearon 50 muestras pertenecientes a tres campañas distintas, 20 muestras de la
primera campaña, 20 de la segunda y 10 de la tercera.
Tras la recepción de los jamones se tomaron medidas morfológicas iniciales, como
peso, longitud, anchura y perímetro de cada pieza, según se muestra en la figura 3.2.1:
Figura 3.2.1. Medidas de conformación del jamón curado
En lugar de realizar el tradicional muestreo en superficie y profundidad con los
músculos principales de estas zonas del jamón, se realizaron dos cortes tranversales a
cada pieza con ayuda de una sierra (MANCA, BM-170). Para utilizar siempre la misma
porción de la muestra se tomó como referencia la punta del jamón, por lo que se realizó el
primer corte a los 9 cm de ésta y el segundo a los 15 cm del primero corte, como se
muestra en la figura 3.2.2.

CAPÍTULO III: MATERIAL Y MÉTODOS
45
15 cm
9 cm
Figura 3.2.3
Figura 3.2.4
Figura 3.2.2. Cortes iniciales para Jamón Curado.
De esta forma, la porción utilizada para la caracterización del producto abarca una
gran cantidad de músculos (figura 3.2.3 y figura 3.2.4).
Figura 3.2.3. 1.-Semitendinosus, 2.-Semimembranosus, 3.-Gracilis, 4.-Soleus, 5.-Gastrocnemius, 6.- Biceps Femoris. Figura 3.2.4. 7.- Recto Femoral, 8.- Sartorio, 9.- Psoas Mayor, 10.- Gluteo Profundi, 11.- Gluteo Medio, 12.- Gluteo Biceps, 13.- Tensor de la Fascia Lata
Estos cortes permitieron visualizar y medir el espesor de la grasa subcutánea en
tres zonas (figura 3.2.5), para así poder poner de manifiesto las posibles diferencias entre
lotes.
Figura 3.2.5. Puntos de medida de la grasa subcutánea del jamón curado
A su vez, el fragmento obtenido se subdividió en tres cortes más, según se muestra
en la figura 3.2.6, para poder tomar las medidas de color instrumental en distintas caras de

CAPÍTULO III: MATERIAL Y MÉTODOS
46
cada loncha, de esta manera se dispone de una medida del color más representativa. Cada
loncha se reservaba siempre para las mismas determinaciones, utilizando siempre los
mismos músculos para cada análisis.
Figura 3.2.6. Distribución de medidas físico-químicas, instrumentales y sensoriales del jamón curado
Las muestras se conservaron en refrigeración hasta su posterior análisis.
3.2.2. PALETA CURADA
Para la caracterización de la paleta curada asociada a la DOP de Teruel se
emplearon 30 muestras pertenecientes a dos campañas distintas, 20 muestras de la
primera campaña y 10 de la tercera. La metodología para el muestreo fue muy similar a la
del jamón curado, con alguna pequeña variación. Inicialmente, al igual que con el jamón
curado, tras la recepción de las paletas se tomaron medidas morfológicas iniciales, como
peso, longitud, anchura y perímetro de cada pieza, tal como mostraba en la figura 3.2.1.
A continuación se realizaron dos cortes tranversales a cada pieza con ayuda de una
sierra (MANCA, BM-170). Para utilizar siempre la misma porción de la muestra, en el caso
de la paleta curada, se utilizó una porción de 12 cm , tomándose también como referencia
la punta de la paleta, realizándose el primer corte a los 9 cm de ésta y el segundo a los 12
cm del primero corte, tal como se muestra en la figura 3.2.7.

CAPÍTULO III: MATERIAL Y MÉTODOS
47
12 cm9 cm
Figura 3.2.9Figura 3.2.8
Figura 3.2.7. Corte trasversal para la paleta curada
De forma, al igual que para el jamón curado, la porción utilizada para la
caracterización del producto nos permitió abarcar una mayor cantidad de músculos
(figura 3.2.8 y figura 3.2.9).
Figura 3.2.8. 1.- Teres Major, 2.- Subscapularis, 3.- Supraspinatus, 4.- Pectoralis Profundi, 5.- Latissimus Dorsi, 6.- Triceps Brachi, 7.- Infraspinatus. Figura 3.2.9. 8.- Pectoralis Superficialis, 9.- Biceps Brachi, 10.- Triceps Brachi, 11.- Tensor Facial Antibrachi, 12.- Flexor Digitorum Superficialis - Thoracic, 13.- Brachiocephalicus, 14.- Brachialis, 15.- Extensor Carpi Radialis, 16.- Anconeus, 17.- Flexor Digitorum Profundi - Thoracic
El resto del procedimiento se realizó fue igual al que se descrito para el jamón
curado.
3.2.3. LOMO CURADO
Para la caracterización del lomo curado asociado a la DOP de Teruel se emplearon
45 muestras pertenecientes a tres campañas distintas, 20 muestras de la primera campaña
y 15 de la segunda y 10 de la tercera. La metodología que se siguió para el muestreo fue
muy simple en comparación con los otros dos productos. Inicialmente, tras la recepción
de los lomos se tomaron medidas morfológicas iniciales, como peso, longitud y diámetro
de cada pieza.Posteriormente se almacenó en refrigeración hasta su posterior análisis.

CAPÍTULO III: MATERIAL Y MÉTODOS
48
3.3. MÉTODOS DE ANÁLISIS
Los métodos utilizados se describirán en orden en el que se realizaron sobre las
muestras.
3.3.1. CONTENIDO DE HUMEDAD
La humedad se determinó utilizando el método ISO-1442 (I.S.O. 1997). Para ello se
mezcló una cantidad de muestra conocida (5 g) con arena de mar previamente desecada y
etanol sobre una cápsula de porcelana. La mezcla pesada se introdujo en una estufa a
102Cº hasta alcanzar un peso constante (4-5 horas). Finalmente las cápsulas se
introdujeron en un desecador con gel de sílice hasta alcanzar la temperatura ambiente. El
porcentaje de humedad se determinó por diferencia de pesada y se expresó en porcentaje
de humedad sobre el peso total de la muestra. Los análisis de cada muestra se realizaron
por triplicado.
3.3.2. CONTENIDO DE PROTEÍNAS
El contenido de proteínas se llevó a cabo según el método de Kjeldahl ISO-937 que
se basa en la determinación de nitrógeno total. Para ello, utilizando un digestor Büchi
(MOD II-425), se sometió a digestión 1 g de muestra con 25 ml de ácido sulfúrico y 15 g de
catalizador Kjeldahl. El resultado de la digestión se destiló con un destilador Büchi (MOD
426) utilizando 100 ml de NaOH al 30% y 100 ml de agua destilada y recogiéndose sobre
100 ml de ácido bórico al 2%. Finalmente el contenido de nitrógeno total se valoró por
titulación con HCl 0,1N utilizando como indicador el rojo de metilo-azul de metileno. Para
conocer la cantidad de proteínas a partir del contenido de nitrógeno total se multiplicó el
resultado por un factor (6.25). El contenido final en proteínas fue calculado en porcentaje
sobre el peso total de la muestra. Los análisis de cada muestra se realizaron por triplicado.
3.3.3. CONTENIDO EN GRASA INTRAMUSCULAR
La extracción y cuantificación de la grasa intramuscular se realizó utilizando el
método descrito por Bligh & Dyer (1959). Se homogeneizaron 40 g de muestra con 120 ml
de una mezcla de cloroformo:metanol (1:2) en un Sorvall Omnimixer (MOD. 17106). La
mezcla se centrifugó durante 10 minutos a 3000 rpm (revoluciones por minuto) y el
sobrenadante se filtró sobre un embudo de decantación. El resto de muestra se volvió a
centrifugar con 40 ml de cloroformo durante 10 minutos a 3000 rpm y se volvió a filtrar
uniéndose los sobrenadantes en el embudo. Se añadieron 40 ml de agua sobre la mezcla y
se agitó el embudo para que todas las fases se mezclaran bien. La mezcla se dejó en reposo
durante toda la noche para que las fases se separasen claramente. Finalmente, la fase

CAPÍTULO III: MATERIAL Y MÉTODOS
49
liposoluble se filtró con sulfato de sodio anhidro y se recogió sobre un matraz erlenmeyer
de boca esmerilada previamente pesado. El disolvente fue eliminado en un rotavapor y las
cantidades residuales con una corriente de nitrógeno. La grasa final se calculó por
diferencia de pesos. El contenido de grasa intramuscular se expresó en porcentaje sobre el
peso total de la muestra. Los análisis de cada muestra se realizaron por triplicado.
3.3.4. COLOR INSTRUMENTAL
Para realizar las medidas del color instrumental se utilizó un colorímetro portátil
Chroma Meter CR-300 (Minolta Camera Co., Japón), con un área de medición de 8 mm de
diámetro, una lámpara de xenón para la iluminación difusa de la muestra y un iluminante
CIE tipo D65. Las medidas de reflectancia se recogieron en un ángulo de visión de 0º. El
equipo fue calibrado mediante un patrón de calibración. Las mediciones se hicieron en el
espacio de color CIE L*a*b*, obteniéndose las coordenadas de cromaticidad L*
(luminosidad), a* (eje rojo-verde) y b* (eje amarillo-azul). Se realizaron 6 medidas sobre
distintos puntos de la superficie de cada muestra.
3.3.5. COLOR QUÍMICO. DETERMINACIÓN DE PIGMENTOS RELACIONADOS CON EL
COLOR: MIOGLOBINA
Para la extracción y cuantificación del contenido de mioglobina y hierro hemínico
se empleó el método descrito por Hornsey (1956) y modificado por Gorospe et al., (1986).
Se partió de 10 g de muestra previamente picada que se introdujo en un matraz
erlenmeyer de boca esmerilada, al que se añadieron 40 ml de acetona, 4 ml de agua (2 ml
para un 80% de humedad) y 1 ml de ácido clorhídrico al 35%. La mezcla se homogeneizó
en un Sorvall Omnimixer (MOD. 17106) durante dos minutos aproximadamente, se tapó y
se mantuvo en oscuridad durante 12 horas a temperatura de refrigeración.
Transcurrido ese tiempo, la mezcla se filtró sobre papel Whatman nº 5 y la
disolución restante se introdujo en cubetas de cuarzo de 1 cm de lado y se medió el
contenido de pigmentos con un espectrofotómetro U-2000 (UV VIS SCANNING
SPECTROPHOTOMETER, HITACHI, JAPÓN) a una longitud de onda de 640 nm.
Para realizar la calibración del equipo y restar la absorbancia de los reactivos
utilizados se utilizó un blanco que se preparó con las mismas proporciones de reactivos
pero sin añadir la muestra. Los análisis de cada muestra se realizaron por triplicado.

CAPÍTULO III: MATERIAL Y MÉTODOS
50
3.3.6. DETERMINACIÓN DEL CONTENIDO DE CLORUROS
Para la preparación del extracto se pasaron 10 g de muestra sobre un erlenmeyer
de boca esmerilada de 250 ml, se añadieron 150 ml de etanol al 40% y se tuvieron en
agitación durante una hora a 100ºC aproximadamente. La mezcla se filtró sobre un matraz
aforado de 250 ml y se añadieron consecutivamente 5 ml de cada uno de los reactivos
Carrez (solución acuosa de ferrocianuro potásico al 15% y solución acuosa de acetato de
zinc al 30%). La mezcla se enrasó con etanol al 40% y se dejó en reposo durante 10 min.
Pasado el tiempo se centrifugó a 3000 rpm durante 5 min y se filtró sobre otro matraz
aforado de 200 ml, enrasando si era necesario con agua destilada. El contenido del matraz
se trasvasó a un vaso de precipitados de 500 ml, se colocó en una placa calefactora y se
evaporó hasta que quedaron unos 100 ml del contenido. Se dejó enfriar y se enrasó sobre
un matraz aforado de 200 ml con agua.
El contenido en cloruro sódico del extracto se determinó mediante el método
Volhard (AOAC, 1984 (Association of Official Analytical Chemists)). Para ello se
introdujeron 10 ml del extracto en un erlenmeyer de 250 ml y se le añadió 10 ml de
solución de nitrato de plata 0,1 N, 1 ml de solución de ácido nítrico concentrado, 1 ml de
solución de sulfato férrico al 4 % y 50 ml de agua destilada. La mezcla se dejo en reposo
durante 10 min en oscuridad. El exceso de plata se valoró con una solución de sulfocianuro
potásico 0,1 N.
Los extractos se prepararon por triplicado para cada muestra y se realizaron dos
valoraciones por extracto.
3.3.7. CUANTIFICACIÓN DE PRODUCTOS DERIVADOS DE LA OXIDACIÓN LIPÍDICA
La cuantificación de los productos derivados de la oxidación lipídica se llevó a cabo
mediante la técnica de extracción de sustancias reactivas del ácido tiobarbitúrico (TBA) y
posterior análisis mediante espectrofotometría de acuerdo con el método de Salih et al.,
(1987).
Para ello se homogeneizaron 2,5g de muestra previamente picada con 7,.5 ml de
ácido percórico y 0,25 ml de BHT (Butilato hidroxitolueno) con un politrón (A/3.440
FUNGILAB). La mezcla resultante se centrifugó durante 10 min a 3000 rpm, se filtró sobre
un matraz aforado de 10 ml y se enrasó con agua destilada. Se tomaron 2 ml del extracto y
se mezclaron con 2 ml de una disolución de TBA 0,02M sobre tubos de ensayo con tapón
de rosca. Los tubos se cerraron y se introdujeron en un baño previamente calentado a
90ºC durante 30 min. Finalmente los tubos se dejaron enfriar a temperatura ambiente y se

CAPÍTULO III: MATERIAL Y MÉTODOS
51
centrifugaron durante 5 min a 3000 rpm. Los extractos se introdujeron en cubetas de
plástico de 1 cm de lado y se midieron con un espectrofotómetro U-2000 (UV VIS
SCANNING SPECTROPHOTOMETER, HITACHI, JAPÓN) a una longitud de onda de 532 nm.
Para la cuantificación se preparó una curva de calibrado utilizando distintas
concentraciones de una solución de 1,1,3,3-tetraetoxipropano (TEP) (0.2268 g) en 1 litro
preparada con ácido perclórico al 3,86%, sometida a las mismas condiciones que la
muestra. Los valores de TBA se calcularon a partir de la curva de calibrado y se expresaron
como mg de malondialdehído por Kg de muestra. Los análisis de cada muestra se
realizaron por triplicado.
3.3.8. DETERMINACIÓN DE pH
El pH se determinó mediante electrometría de electrodo selectivo (pH metro). Para
ello se tomaron 10 g de la muestra previamente picada y se mezcló con una cantidad igual
de agua destilada, que en este caso fueron 10 ml. Se dejó 10 minutos la mezcla en reposo,
se centrifugó a 3000 rpm durante 10 minutos y finalmente se filtró el sobrenadante, que
posteriormente se utilizó para la medida del pH.
Para medir el pH se utilizó un pH-metro CRISON, el cual fue calibrado previamente
con dos disoluciones tampón, una de pH 4 y otra de pH7.
Se realizaron 3 medidas de pH por muestra.
3.3.9. ANÁLISIS DE PERFIL DE TEXTURA (TPA)
La evaluación instrumental de la textura se realizó mediante el método descrito
por Bourne (1978). El ensayo es el “análisis de perfil de textura intramuscular” o “Texture
Profile Analysis” (TPA) y fue desarrollado mediante un texturómetro (Texture Analizer
Micro System TA.XT2).
Este método se basa en la realización de dos ciclos de compresión-descompresión
sucesivos sobre el alimento, imitando la acción de las mandíbulas, como se muestra en la
figura 3.3.1:

CAPÍTULO III: MATERIAL Y MÉTODOS
52
Figura 3.3.1. Gráfica producto del análisis de perfil de textura
Para ello se prepararon cubos de 1 cm de lado (unos 14 por muestra),
considerándose equivalente a la cantidad de alimento desprendida en un mordisco. Para
los ciclos de compresión-descompresión se empleó una sonda cilíndrica de 5 cm de
diámetro. La velocidad del ensayo fue de 1 mm/seg y el grado de compresión de la
muestra fue del 70% del grosor total de la misma.
A partir de la curva obtenida se calcularon los distintos parámetros que se
muestran a continuación, mostrando la definición sensorial y la instrumental,
respectivamente:
Dureza: Se expresa en unidades de fuerza, Kg, g, Newton (Rosenthal, 1999)
Fuerza requerida para comprimir un alimento entre los molares (Civille y
Szczesniak, 1973)
Fuerza máxima que tiene lugar en cualquier momento durante el primer ciclo de
compresión (Bourne, 1978)
Elasticidad: Adimensional. Una longitud dividida por otra longitud (Rosenthal, 1999)
Es una medida de cuanta estructura original de la muestra se ha roto por la
compresión inicial (Rosenthal, 1999)
Es la altura que recupera el alimento durante el tiempo que recorre durante el
primer ciclo y el segundo = L1/L2 (Bourne, 1978)

CAPÍTULO III: MATERIAL Y MÉTODOS
53
Adhesividad: No existen medidas reales de este parámetro, que se expresa en unidades del
integrador interno del ordenador Kg x s o g x s o Newton x s (Rosenthal, 1999)
El trabajo requerido para retirar el alimento de la superficie (Civille y Szczesniak,
1973)
Siguiendo el primer ciclo de compresión, se elimina la fuerza cuando la cruceta se
mueve hacia su posición original. Si el material es pegajoso o adhesivo, la fuerza se
convierte en negativa. El área de esta fuerza negativa se toma como una medida de
la adhesividad de la muestra (Rosenthal, 1999)
Cohesividad: Adimensional (Rosenthal, 1999)
La fuerza que los enlaces internos hacen sobre el alimento (Szczesniak, 1963)
Se mide tomando el trabajo total dado en la muestra durante el segundo ciclo y
dividiéndole por el trabajo dado durante el primer ciclo. El trabajo se mide como el
área por debajo de las respectivas curvas = B/A (Bourne, 1978)
Fracturabilidad: Se expresa en unidades de fuerza, Kg, g, Newton (Bourne, 1978)
La fuerza a la que el material se fractura
Es la primera caída significante de la curva durante el primer ciclo de compresión
(Bourne, 1978). Producto de un alto grado de dureza y de cohesividad (Szczesniak,
2002)
Masticabilidad: Kg o Newton
La energía requerida para desintegrar un alimento semisólido de modo que esté
listo para ser tragado (Civille y Szczesniak, 1973)
Dureza x Cohesividad x Elasticidad
Gomosidad: Kg o Newton
La energía requerida para desintegrar un alimento semisólido de modo que esté
listo para ser tragado (Civille y Szczesniak, 1973)
Dureza x Cohesividad x Elasticidad

CAPÍTULO III: MATERIAL Y MÉTODOS
54
3.3.10. FRACCIONAMIENTO DE LA GRASA INTRAMUSCULAR Y DETERMINACIÓN DEL
PERFIL DE ÁCIDOS GRASOS DE CADA FRACCIÓN
Fraccionamiento de la grasa intramuscular
El fraccionamiento de la grasa intramuscular en triglicéridos (lípidos neutros
(LN)), fosfolípidos (lípidos totales (LP)) y ácidos grasos libres (AGL) se llevó a cabo
mediante el método descrito por Kaluzny et al., (1985). El fraccionamiento se llevó a cabo
utilizando columnas de aminopropil activadas previamente con tres lavados de 2,5 ml de
hexano.
Se partió de 15 mg de grasa de la muestra y se introdujo en las columnas redisuelta
con dos lavados de 150 µl de la mezcla hexano:cloroformo:metanol (95:3:2). Se añadieron
5 ml de cloroformo para extraer la fracción de triglicéridos, 5 ml de ácido acético al 2%
con dietiléter para la fracción de ácidos grasos libres y finalmente 2,5 ml de
metanol:cloroformo (6:1) y 2,5 ml de acetato sódico 0,05M en metanol:cloroformo 6:1
para la extracción de los fosfolípidos.
Metilación de los ácidos grasos
Para la preparación de los ésteres metílicos de los ácidos grasos de cada una de las
fracciones se utilizó el método descrito por Sandler y Karo, (1992). Previamente se añadió
1 ml de ácido tridecanoico 0,04M a cada una de las fracciones y se evaporaron los
disolventes que se utilizaron para extraer cada fracción con corrientes de nitrógeno. Tras
la evaporación se añadió 1 ml de metilato sódico, la mezcla se agitó y se introdujo en la
estufa a 80ºC durante 30 min. Después se dejó enfriar, se añadió 1 ml de sulfúrico en
metanol al 5% y se volvió a introducir en la estufa a 80ºC durante 30 minutos más. Se dejó
la mezcla enfriar y se añadieron 1 ml de hexano y 1 ml de cloruro sódico sobresaturado
agitando manualmente entre cada adición. La mezcla se centrifugó 2 min a 4000 rpm y se
extrajo la capa más externa (hexano) con ayuda de una pipeta pasteur sobre un vial de 1,5
ml, se evaporó el disolvente con corrientes de nitrógeno y se rediluyó con hexano hasta un
volumen conocido de 1 ml.
Análisis cromatográfico
La separación y cuantificación de los ácidos grasos de cada una de las fracciones se
realizó mediante un cromatógrafo de gases HP 6890N, equipado con un detector on-
column y una columna capilar de polietilenglicol (60m x 0,32mm x 0,25µl) Supelcowax 10

CAPÍTULO III: MATERIAL Y MÉTODOS
55
(Supelco, EEUU). La detección de los compuestos se realizó mediante un detector de
ionización de llama (FID). Las condiciones cromatográficas fueron:
Flujo de gas portador (N2) 0,7 ml/min
Temperatura del horno (isoterma) 180ºC
Temperatura del inyector 250ºC
Temperatura del detector 250ºC
Identificación y cuantificación de los ácidos grasos
La identificación de los ácidos grasos se llevó a cabo por comparación de los
tiempos de retención de las muestras con patrones sometidos a las mismas condiciones
cromatográficas.
Para su cuantificación se utilizaron rectas de calibrado elaboradas para cada ácido
graso, relacionando el cociente de concentración del ácido graso problema/concentración
patrón interno frente al área del ácido graso problema/área del patrón interno.
El contenido total de cada ácido graso de cada fracción se calculó a partir del peso
de la muestra y se expresó en mg del ácido graso en 100 mg de la muestra. Los análisis de
cada muestra se realizaron por triplicado.
3.3.11. DETERMINACIÓN DE AMINOÁCIDOS LIBRES
La extracción y cuantificación de los aminoácidos libres se llevó a cabo según el
método descrito por Ruiz et al., (1999) (a).
Preparación del extracto
Se homogeneizaron 5 g de muestra picada con 1ml de patrón interno (10 mg/ml
de norleucina) y 49 ml de ácido sulfosalicílico al 10% con un politrón (Omni 5000). Este
extracto se mantuvo durante 17 horas a temperatura de refrigeración (1-2ºC).
Posteriormente la mezcla se centrifugó durante 10 minutos a 4000 rpm y se filtró
sobre un matraz erlenmeyer de 150 ml con papel Whatman nº 5. El filtrado resultante se
ajustó a pH 6 con NaOH 4N y se enrasó a 100 ml con agua miliq.
Se preparó un único extracto por muestra y se almacenó a -80ºC hasta su
derivatización.

CAPÍTULO III: MATERIAL Y MÉTODOS
56
Derivatización de los aminoácidos
Para la derivatización se utilizó el método descrito por Yang y Sepúlveda, (1985).
Una vez el extracto alcanzó una temperatura ambiente (estaba almacenado a -80ºC), se
tomaron 50 µl y se mezclaron con 200 µl de una solución marcadora constituida por
etanol, trietilamina y fenilisotiocianato en una proporción 7:2:1.
La mezcla se mantuvo durante 10 minutos a temperatura ambiente para que se
produjera la reacción de derivatización y se evaporó hasta sequedad utilizando un Speed
Vac (Savant SVC 200) acoplado a una trampa refrigerada (Savant RT 4104) y una trampa
de vacío (Genevac CPV 100/2).
Finalmente el residuo se redisolvió en 200 µl de una disolución de tampón fosfato
(0,5 M y pH 7,4) y acetonitrilo al 5%.
Análisis cromatográfico
Se tomaron 20 µl de la disolución derivatizada del apartado anterior y se analizó
en un cromatógrafo de líquidos de alta resolución (HPLC) con un equipo HP series 1100
con detector ultravioleta.
Para la separación de los aminoácidos se utilizó una columna de fase reversa
Supelcosil LC-18 (Supelco) de 25 cm de longitud, 4,6 cm de diámetro interno y un tamaño
de partículas de 5 µm.
La fase móvil estaba compuesta por dos disoluciones tampón, una compuesta por
tampón acetato sódico 0,03M con 0,05% de trietilamina ajustado a un pH de 6,85 (A) y
otra compuesta por acetonitrilo (grado HPLC) al 90% en agua miliq (B).
El desarrollo cromatográfico que se llevó a cabo para la separación de los
aminoácidos fue por gradiente de elución, presentado en la tabla 3.3.2:
Tabla 3.3.2. Gradiente empleado en la determinación de aminoácidos
tiempo (min) flujo (ml/min) %A %B
0 1 96,8 3,2
0,5 1 95,5 4,5
5,5 1 91,5 8,5 15 1 88,5 11,5
24 1 73,5 26,5
46 1,5 1,0 99,0
58 1 96,8 3,2

CAPÍTULO III: MATERIAL Y MÉTODOS
57
La longitud de onda que se empleó para la detección de los aminoácidos fue
254nm.
Identificación y cuantificación de los aminoácidos
La identificación de los aminoácidos se llevó a cabo por comparación de los
tiempos de retención de patrones comerciales (Sigma-Aldrich), analizados en las mismas
condiciones que la muestra.
Para la cuantificación se prepararon rectas de calibrado de cada uno de los
aminoácidos, relacionando el cociente de concentración del aminoácido
problema/concentración patrón interno frente al área del aminoácido problema/área del
patrón interno, utilizando como patrón interno la norleucina, que se adicionaba a cada
muestra inicialmente durante la preparación del extracto.
El contenido total de cada aminoácido se calculó a partir del peso de la muestra y
se expresó en mg de aminoácido en 100 mg de muestra. Se realizó un único análisis por
muestra.
3.3.12. DETERMINACIÓN DE COMPUESTOS NITROGENADOS DE BAJO PESO
MOLECULAR DERIVADOS DEL ATP: NUCLEÓTIDOS
La extracción y cuantificación de los nucleótidos se llevó a cabo según el método
descrito por Watanabe et al., (1989).
Preparación del extracto
Se homogeneizaron 5 g de muestra picada con 20ml de HClO4 0,6N con un politrón
(Omni 5000). La mezcla se centrifugó durante 5 minutos a 4000 rpm y se filtró sobre un
matraz erlenmeyer de 150 ml con papel Whatman nº 5. El sólido restante se volvió a
mezclar con 20 ml de HClO4 y se repitió el proceso anterior mezclando finalmente ambos
sobrenadantes.
El filtrado conjunto se neutralizó a pH 6.5 con KOH 6N, introduciendo el matraz
sobre hielo picado para que las sales de KClO4 precipitasen, eliminándose posteriormente
por filtrado. El extracto final se enrasó a 100 ml con agua miliq.
Se preparó un único extracto por muestra y se almacenó a -80ºC hasta su análisis
cromatográfico.

CAPÍTULO III: MATERIAL Y MÉTODOS
58
Análisis cromatográfico
Se tomaron 20 µl del extracto anterior y se analizaron en un cromatógrafo de
líquidos de alta resolución (HPLC) con un equipo HP series 1100 con detector ultravioleta.
La longitud de onda que se utilizó en el análisis fue de 254nm.
Para la separación de los nucleótidos se utilizó una columna de fase reversa
Supelcosil LC-18 (Supelco) de 25 cm de longitud, 4,6 cm de diámetro interno y un tamaño
de partículas de 5 µm.
La fase móvil estaba compuesta por dos disoluciones tampón, una compuesta por
tampón fosfato potásico 0,1N a pH 6 (A) y otra compuesta por metanol de grado HPLC (B).
El desarrollo cromatográfico que se llevó a cabo para la separación de los
nucleótidos fue por gradiente de elución, presentado en la tabla 3.3.3:
Tabla 3.3.3. Gradiente empleado en la determinación de nucleótidos
Tiempo (min) Flujo (ml/min) %A %B 0 1 100 0 5 1 100 0
12 1 85 15 30 1 50 50 35 1 100 0
Identificación y cuantificación
La identificación de los nucleótidos se llevó a cabo por comparación de los tiempos
de retención de patrones comerciales (Sigma-Aldrich), analizados en las mismas
condiciones que la muestra.
La cuantificación se llevó a cabo elaborando rectas de calibrado de cada uno de los
nucleótidos a partir de los patrones comerciales, relacionando la concentración del
nucleótido frente al área de ese nucleótido.
El contenido total de cada nucleótido se calculó a partir del peso de la muestra y se
expresó en mg de nucleótido en 100 mg de muestra. Se realizó un único análisis por
muestra.

CAPÍTULO III: MATERIAL Y MÉTODOS
59
3.3.13. ANÁLISIS DE COMPUESTOS VOLÁTILES
Extracción de los compuestos volátiles
Para la extracción de los compuestos volátiles se pesó 1 g de muestra, previamente
picada, sobre un vial de vidrio y se selló con un tapón de plástico con septum de
teflón/silicona. Para este ensayo se utilizó una fibra de SPME (Solid Phase
Microextraction) de carboxenpolidimetilsiloxano (CARB/PDMS) (75 µm de grosor). Para
la activación de la fibra se empleó el mismo método que se utilizaría posteriormente para
la muestra durante 50 minutos, con el fin de reabsorber los posibles contaminantes.
Durante este tiempo, incubamos la muestra picada en el vial en un baño de metacrilato con
un sistema calefactor de la casa Selecta (Barcelona) a 37ºC durante 30 minutos.
Posteriormente se introdujo la fibra a través del septum del tapón y se mantuvo el vial con
la fibra en el baño durante 30 minutos más. Transcurrido el tiempo de extracción, la fibra
de SPME se transfirió de forma inmediata al inyector de un cromatógrafo de gases (modo
splitness, 250ºC) para la desorción de los compuestos capturados por la fibra de SPME y
su posterior separación, detección e identificación mediante CG-EM (Cromatografía de
gases/Espectrometría de masas).
Análisis cromatográfico (CG/EM)
El análisis se llevó a cabo en un cromatógrafo HP-6890 serie II acoplado a un
espectrómetro de masas Agilent 5973. La separación de los compuestos volátiles se realizó
en una columna capilar de sílice fundido con una fase estacionaria apolar de tipo HP-5 (5%
fenil-metil silicona, Hewlet-Packard, 50m x 0,32µm x 1,05 µm), operando a 6,5 psi de
presión de cabeza de columna, resultando un flujo de 1,3 mL min-1 de Helio.
El programa del horno utilizado para la separación de los compuestos volátiles fue
el siguiente: temperatura inicial de 40 ºC durante 10 minutos, posteriormente la
temperatura subió hasta 200ºC a razón de 5ºC/min, después tuvo otra subida de
temperatura hasta 250ºC a razón de 20ºC/min, permaneciendo en isoterma a 250ºC
durante 5 minutos más (tiempo total de la carrera cromatográgica 50 minutos
aproximadamente). La línea de transferencia del espectrómetro de masas se mantuvo a
280ºC. El espectro de masas se obtuvo mediante impacto electrónico a 70eV, con un
electrón multiplicador voltio de 1756V y la recogida de datos se realizó a una velocidad de
1scan/s en un rango de masas de 30 a 500m/z.

CAPÍTULO III: MATERIAL Y MÉTODOS
60
Identificación de los compuestos volátiles
Los compuestos volátiles detectados fueron identificados comparando sus
espectros de masas con los espectros incluidos en las bibliotecas Willey y NIST/EPA/NIH y
con los índices de Kovats descritos (Kondjoyan y Berdagué, 1996). Para calcular los
índices de Kovats se utilizó un patrón comercial de hidrocarburos (Sigma, St. Louis, USA)
que se sometió a las mismas condiciones que las muestras.
El contenido de compuestos volátiles se expresó en UAA (Unidades Arbitrarias de
Área), realizándose un análisis por cada muestra.
3.3.14. EVALUACIÓN SENSORIAL
Para la evaluación sensorial se realizó un análisis cuantitativo-descriptivo (AQD)
utilizando un panel entrenado compuesto por 12 jueces que analizaron 3 muestras por
sesión, en orden aleatorio y siempre comparando el mismo producto. Al tratarse de
distintos productos, los atributos que se emplearon para el análisis sensorial fueron
distintos, de forma que se evaluaron 19 características sensoriales para la paleta y el
jamón curado y 15 para el lomo curado, 4 menos ya que no se tuvieron en cuenta los
atributos relacionados con el aspecto y textura de la grasa.
La evaluación sensorial de los productos curados se llevó a cabo utilizando los
siguientes descriptores:
Color: Sensación resultante de estimular la retina por las ondas luminosas
comprendidas en la región visible del espectro.
Veteado: Medida de la cantidad de grasa infiltrada (intramuscular) en el tejido
muscular.
Brillo: Describe el aspecto de un producto que lo asemeja a una superficie pulida
que muestra reflejos brillantes.
Olor: Propiedad sensorial perceptible por el órgano olfativo cuando inspira
determinadas sustancias volátiles.
Dureza: Fuerza requerida para deformar el alimento durante la masticación.
Fluidez: Tendencia de la grasa a licuarse y deslizarse sobre la superficie

CAPÍTULO III: MATERIAL Y MÉTODOS
61
Masticabilidad: Tiempo necesario o número de masticaciones requeridas para
dejar un producto sólido en las condiciones adecuadas para su deglución.
Jugosidad: Sensación asociada a la liberación de agua y grasa por la masticación
así como por la secreción de jugos bucales provocados por la carne durante la masticación.
Pastosidad: Sensación percibida en la masticación asociada a la combinación de
bajos valores de dureza y valores relativamente altos de adhesividad y de gomosidad.
Salado: Describe el sabor elemental producido por soluciones acuosas tales como
el cloruro sódico.
Dulce: Describe el sabor elemental producido pos soluciones acuosas tales como la
sacarosa.
Intensidad de Flavor: Conjunto complejo de las propiedades olfativas y gustativas
que se perciben durante la degustación y que puede estar influido por las propiedades
táctiles, térmicas, dolorosas, e incluso por efectos cinestésicos.
Persistencia: Sensación olfato-gustativa raramente similar o muy próxima a la
sensación percibida cuando el producto estaba en la boca, que permanece localizada
durante un cierto tiempo y cuya duración se puede medir.
Curado: Conjunto complejo de características organolépticas olfato-gustativas que
se desarrollan como consecuencia de la adición de sales curantes y del posterior
secado/maduración del producto.
Rancidez: Olor y flavor pungente a grasa vieja
Para el desarrollo de las sesiones de catas se utilizó el programa informático FIZZ
versión 2.20 (Biosistemes, Francia). Las escalas utilizadas para las fichas fueron no
estructuradas de 10 cm de longitud siendo los extremos para los valores “poco” y “mucho”
para cada descriptor. A cada catador se le proporcionó dos trozos de jamón, paleta o lomo
de un espesor de 1 mm cortados con una máquina loncheadora (O.M.S. TGI-300),
presentados en platos transparentes y en orden aleatorio. Las sesiones de cata se
realizaron en una sala de catas estandarizada con seis cabinas con dotación informática
(donde se presentaban las fichas con los atributos), tornos para la presentación de las
muestras, luz fluorescente y totalmente independientes unas de otras.

CAPÍTULO III: MATERIAL Y MÉTODOS
62
3.3.15. ANÁLISIS ESTADÍSTICO
Los datos fueron analizados estadisticamente con ayuda del programa informatico
SPSS version 15.0. Para conocer el efecto de los distintos factores estudiados (genética,
sexo y campaña) en los diferentes parametros físico-quimicos, instrumentales y
sensoriales se empleó un análisis multivariante de la varianza mediante un modelo lineal
general y el test de Tukey para comparar las medias de los grupos con un nivel de
significación p ≤ 0,05.
Previo al tratamiento estadístico se llevó a cabo un tratamiento de datos anómalos
utilizando el test de Grubb, recomendado por las normas ISO.

CAPÍTULO IV: RESULTADOS


CAPÍTULO IV: RESULTADOS
65
4. RESULTADOS
4.1. JAMÓN CURADO
4.1.1. CARACTERIZACIÓN DEL JAMÓN CURADO
Características generales del jamón curado
La tabla 4.1.1 muestra los valores medios de las medidas morfológicas de todos los
jamones curados utilizados en el presente estudio (n=50).
Tabla 4.1.1. Medidas morfológicas generales del jamón curado (± desviación estándar de la media)
Peso (g) Longitud (cm) Anchura (cm) Perímetro (cm)
7,9 ± 0,7 75,4 ± 2,0 28,1 ± 1,4 65,2 ± 3,1
El Consejo Regulador de la DOP Jamón de Teruel tan solo establece que el peso de
los jamones debe encontrarse entre 8 y 9 Kg, nunca inferior de 7 Kg, por lo que todas las
piezas analizadas se encuentran dentro de los valores establecidos para el peso. El resto de
parámetros no están especificados en dicha normativa.
Las tablas 4.1.2 y 4.1.3 muestran el espesor de la grasa subcutánea de las zonas
proximal y distal de la porción utilizada para el análisis físico-químico, tal como se explicó
en la figura 3.2.2 del apartado material y métodos.
A
B
C
Figura 4.1.1. Zona proximal del jamón curado
Tabla 4.1.2. Medidas de la grasa subcutánea de la zona proximal del jamón curado (± desviación estándar de la media)
A (cm) B (cm) C (cm)
2,6 ± 1,1 2,4 ± 0,9 4,9 ± 1,5

CAPÍTULO IV: RESULTADOS
66
A
B
C
Figura 4.1.2. Zona distal del jamón curado
Perfil de textura instrumental
En el jamón curado, la textura es una característica que está directamente
relacionada con la estructura del músculo, especialmente con la degradación de las
proteínas miofibrilares y el colágeno, con el contenido de grasa intramuscular y con la
rampa de temperaturas utilizada durante el proceso de secado-maduración de las
muestras.
Los resultados obtenidos del TPA para el jamón curado se exponen en la tabla
4.1.4. Ruiz-Ramírez et al., (2006) observaron que los jamones curados con elevada
proteólisis y un bajo contenido en cloruros mostraban valores bajos de dureza y altos de
cohesividad y elasticidad, especialmente con bajos contenidos de agua. Al comparar los
resultados de la tabla 4.1.4 con otros productos similares como jamón Serrano, Ibérico o
de Bayona (Ventanas, 2006; Ruiz, 2005; Costa et al., 2008) podemos destacar el bajo valor
de la dureza y de la cohesividad, y el alto de la elasticidad.
Tabla 4.1.4. Análisis de perfil de textura (TPA) para el jamón curado (± desviación estándar de la media)
Fuerza (N) 10,13 ± 4,63
Dureza (N) 11,48 ± 5,14
Fracturabilidad (Adimensional) 9,29 ± 1,15 Adhesividad (N*seg) -0,24 ± 0,07
Elasticidad (cm) 0,59 ± 0,07
Cohesividad (Adimensional) 0,48 ± 0,03
Gomosidad (N) 5,15 ± 2,25 Masticabilidad (N) 3,08 ± 1,54
(N): Newton
Tabla 4.1.3. Medidas de la grasa subcutánea de la zona distal del jamón curado (± desviación estándar de la media)
A (cm) B (cm) C (cm)
7,1 ± 2,0 2,1 ± 0,6 3,3 ± 1,0

CAPÍTULO IV: RESULTADOS
67
Parámetros generales físico-químicos
Con el fin de obtener valores de referencia para la discusión de los resultados de
las distintas determinaciones, se midieron una serie de parámetros generales entre los
que se encuentran la humedad, proteínas y grasa intramuscular. Junto con estos, están
incluidos en la tabla 4.1.5 el resto de parámetros relacionados con la composición química
del producto.
Tabla 4.1.5. Parámetros físico-químicos generales del jamón curado (± desviación estándar de la media).
Humedad (g/100 g) 50,96 ± 4,45
Proteínas (f) (g/100 g) 33,16 ± 4,58
Proteínas (s,dg) (g/100 g) 76,54 ± 11,92
Grasa (f) (g/100 g) 4,36 ± 1,17 Grasa (s) (g/100 g) 8,92 ± 2,39
L (Adimensional) 37,96 ± 2,94
a (Adimensional) 15,05 ± 1,58
b (Adimensional) 8,68 ± 1,51 mg Mb/g m 2,39 ± 0,66
ppm Fe heminico 8,10 ± 2,24
Cloruros (f) (g/100 g) 3,49 ± 0,86
Cloruros (s) (g/100 g) 7,28 ± 1,77 pH (Adimensional) 6,08 ± 0,17
mg MDA/Kg 0,24 ± 0,08
(f): extracto fresco; (s): extracto seco; (s, dg): extracto seco y desengrasado
Como se observa en la tabla 4.1.5, la humedad media de los jamones fue de
50,96%, un valor muy similar a los descritos por otros autores para jamón Serrano
(48,5%) o jamón Ibérico (49%), e inferior para otros como el de Parma (61,8%), San
Daniel (60,4%) o Bayona (57%) (Toldrá, 2002). Las diferencias pueden deberse al
músculo analizado, al proceso de elaboración y al tamaño de las piezas, que en el caso de
los jamones de Parma o San Daniel son mayores.
El porcentaje medio de proteínas fue de 33,16%, igual al descrito para jamón
Serrano (33,1%) y superior al Ibérico (24,6%) (Toldrá, 2002).
El porcentaje medio de grasa intramuscular obtenido fue de 4,36%, algo inferior al
descrito para jamón Serrano (5,9%) y muy inferior al Ibérico (12,89-13,21%), sin embargo
este parámetro será más parecido al descrito en otros como el de Bayona (5%) y superior
al de San Daniel (3,6%) o al de Parma (3,5), debido a la utilización de animales de la misma
raza para la producción de los jamones (Toldrá, 2002; Ventanas et al., 2005).
El porcentaje de cloruros presente en los jamones fue de 3,49%, muy por debajo al
descrito para el resto de jamones, con un 8,7% para el Serrano o un 6,5% para el Ibérico
(Toldrá, 2002; Ventanas et al., 2005).

CAPÍTULO IV: RESULTADOS
68
El color es una de las características de apariencia más destacadas en el jamón
curado (Ramirez y Cava, 2008) y que dependerá de la humedad, del contenido de grasa
intramuscular y del contenido en mioglobina. El valor medio del contenido en mioglobina
de los jamones fue de 2,39 mg/g, un valor inferior al descrito por la bibliografía para
jamón curado o para jamón Ibérico. La concentración de mioglobina en el músculo de
cerdo se encuentra entre 4 y 9 mg/g. Sin embargo, la concentración de mioglobina
dependerá del tipo de músculo (Aristoy y Toldrá, 1998), de la edad del animal y del
ejercicio que realiza éste, incrementando el contenido de mioglobina con ambos. Es por
ello que los jamones Ibéricos presentan contenidos muy superiores comparados con los
jamones procedentes de cerdos blancos.
En cuanto al color instrumental, los jamones presentaron un valor de la
coordenada L de 37,96, inferior al descrito para jamones Ibéricos. El parámetro que
describe la coordenada a* siguió una tendencia contraria a la luminosidad, obteniendo un
valor de 15,05, superior al descrito para jamón Serrano o de Parma, con valores en torno a
12 (Costa et al., 2008) pero bastante inferior al descrito para jamón Ibérico (24,15-22,73)
(Muriel et al., 2004). El parámetro que describe la coordenada b* obtuvo un valor de 8,68,
muy superior al descrito para jamón Serrano y de Parma (2-3) (Costa et al., 2008) y muy
similar al descrito para jamón Ibérico (8,55-9,45) (Muriel et al., 2004).
Composición de la grasa intramuscular
La grasa intramuscular está constituida mayoritariamente por lípidos neutros (LN)
(triglicéridos, diglicéridos y monoglicéridos), ácidos grasos libres (AGL) y lípidos polares
(LP) (fosfolípidos).
En los jamones procedentes de cerdos blancos, al igual que en los Ibéricos, los LN
representan la mayoría de los lípidos de la grasa intramuscular, constituyendo
aproximadamente el 90% (Tejeda, 1999). Esta alta proporción hace que la composición de
ácidos grasos totales sea fiel reflejo, tal como se muestra en los resultados de la tabla 4.1.6,
donde nos encontramos con un 86,6%. Sin embargo, el contenido de AGL es diferente en
los jamones procedentes de cerdo blanco representando algo más del 20% en
comparación con los Ibéricos, donde esta fracción se reduce hasta niveles del 7-8%. Los
jamones de nuestro estudio presentaron una menor proporción de AGL en comparación
con otros jamones curados (12,8%). La fracción de LP representa la fracción minoritaria, y
puede verse alterada en función de la alimentación del animal. Los jamones de Teruel
mostraron una proporción de AGPI en torno al 0,6% (tabla 4.1.6).
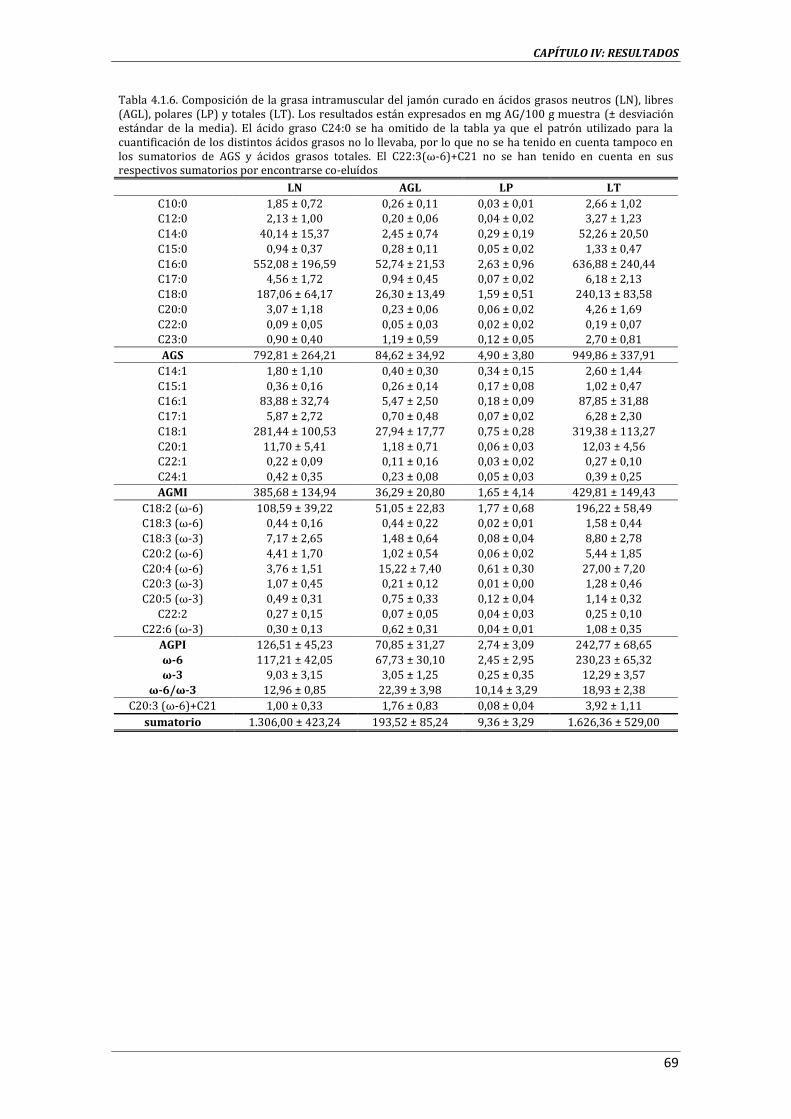
CAPÍTULO IV: RESULTADOS
69
Tabla 4.1.6. Composición de la grasa intramuscular del jamón curado en ácidos grasos neutros (LN), libres (AGL), polares (LP) y totales (LT). Los resultados están expresados en mg AG/100 g muestra (± desviación estándar de la media). El ácido graso C24:0 se ha omitido de la tabla ya que el patrón utilizado para la cuantificación de los distintos ácidos grasos no lo llevaba, por lo que no se ha tenido en cuenta tampoco en los sumatorios de AGS y ácidos grasos totales. El C22:3(ω-6)+C21 no se han tenido en cuenta en sus respectivos sumatorios por encontrarse co-eluídos
LN AGL LP LT
C10:0 1,85 ± 0,72 0,26 ± 0,11 0,03 ± 0,01 2,66 ± 1,02 C12:0 2,13 ± 1,00 0,20 ± 0,06 0,04 ± 0,02 3,27 ± 1,23
C14:0 40,14 ± 15,37 2,45 ± 0,74 0,29 ± 0,19 52,26 ± 20,50
C15:0 0,94 ± 0,37 0,28 ± 0,11 0,05 ± 0,02 1,33 ± 0,47
C16:0 552,08 ± 196,59 52,74 ± 21,53 2,63 ± 0,96 636,88 ± 240,44 C17:0 4,56 ± 1,72 0,94 ± 0,45 0,07 ± 0,02 6,18 ± 2,13
C18:0 187,06 ± 64,17 26,30 ± 13,49 1,59 ± 0,51 240,13 ± 83,58
C20:0 3,07 ± 1,18 0,23 ± 0,06 0,06 ± 0,02 4,26 ± 1,69
C22:0 0,09 ± 0,05 0,05 ± 0,03 0,02 ± 0,02 0,19 ± 0,07 C23:0 0,90 ± 0,40 1,19 ± 0,59 0,12 ± 0,05 2,70 ± 0,81
AGS 792,81 ± 264,21 84,62 ± 34,92 4,90 ± 3,80 949,86 ± 337,91
C14:1 1,80 ± 1,10 0,40 ± 0,30 0,34 ± 0,15 2,60 ± 1,44
C15:1 0,36 ± 0,16 0,26 ± 0,14 0,17 ± 0,08 1,02 ± 0,47 C16:1 83,88 ± 32,74 5,47 ± 2,50 0,18 ± 0,09 87,85 ± 31,88
C17:1 5,87 ± 2,72 0,70 ± 0,48 0,07 ± 0,02 6,28 ± 2,30
C18:1 281,44 ± 100,53 27,94 ± 17,77 0,75 ± 0,28 319,38 ± 113,27
C20:1 11,70 ± 5,41 1,18 ± 0,71 0,06 ± 0,03 12,03 ± 4,56 C22:1 0,22 ± 0,09 0,11 ± 0,16 0,03 ± 0,02 0,27 ± 0,10
C24:1 0,42 ± 0,35 0,23 ± 0,08 0,05 ± 0,03 0,39 ± 0,25
AGMI 385,68 ± 134,94 36,29 ± 20,80 1,65 ± 4,14 429,81 ± 149,43
C18:2 (ω-6) 108,59 ± 39,22 51,05 ± 22,83 1,77 ± 0,68 196,22 ± 58,49 C18:3 (ω-6) 0,44 ± 0,16 0,44 ± 0,22 0,02 ± 0,01 1,58 ± 0,44
C18:3 (ω-3) 7,17 ± 2,65 1,48 ± 0,64 0,08 ± 0,04 8,80 ± 2,78
C20:2 (ω-6) 4,41 ± 1,70 1,02 ± 0,54 0,06 ± 0,02 5,44 ± 1,85
C20:4 (ω-6) 3,76 ± 1,51 15,22 ± 7,40 0,61 ± 0,30 27,00 ± 7,20 C20:3 (ω-3) 1,07 ± 0,45 0,21 ± 0,12 0,01 ± 0,00 1,28 ± 0,46
C20:5 (ω-3) 0,49 ± 0,31 0,75 ± 0,33 0,12 ± 0,04 1,14 ± 0,32
C22:2 0,27 ± 0,15 0,07 ± 0,05 0,04 ± 0,03 0,25 ± 0,10
C22:6 (ω-3) 0,30 ± 0,13 0,62 ± 0,31 0,04 ± 0,01 1,08 ± 0,35
AGPI 126,51 ± 45,23 70,85 ± 31,27 2,74 ± 3,09 242,77 ± 68,65
ω-6 117,21 ± 42,05 67,73 ± 30,10 2,45 ± 2,95 230,23 ± 65,32 ω-3 9,03 ± 3,15 3,05 ± 1,25 0,25 ± 0,35 12,29 ± 3,57
ω-6/ω-3 12,96 ± 0,85 22,39 ± 3,98 10,14 ± 3,29 18,93 ± 2,38
C20:3 (ω-6)+C21 1,00 ± 0,33 1,76 ± 0,83 0,08 ± 0,04 3,92 ± 1,11
sumatorio 1.306,00 ± 423,24 193,52 ± 85,24 9,36 ± 3,29 1.626,36 ± 529,00

CAPÍTULO IV: RESULTADOS
70
En la tabla 4.1.7, en la fracción de triglicéridos (LN), podemos comprobar los altos
valores del ácido oleico (C18:1) con un 49,50%, seguido del palmítico (C16:0) con un
24,52% y del esteárico (C18:0) con un 11,71%. Estos valores se encuentran entre los
intervalos descritos para el jamón Ibérico, siendo entre 50-58% para el C18:1, entre el 22-
25% para el C16:0 y entre 8,5-13% para el C18:0. Estos resultados pueden explicarse por
la utilización de piensos enriquecidos con ácidos grasos esenciales.
Los AGMI (ácidos grasos monoinsaturados) fueron los mayoritarios en la fracción
de AGL, estos resultados difieren de los descritos por Jurado (2005) para jamón Ibérico,
que obtuvo una fracción mayoritaria de AGS (ácidos grasos saturados) o de otros trabajos
en los que se observó que los AGPI (ácidos grasos poliinsaturados) eran los mayoritarios
en dicha fracción (revisado por Toldrá et al., 1997).
Respecto al porcentaje de los sumatorios de ácidos grasos de los LT, destacamos el
39,41% de los AGS. Estos valores se encuentran entre los establecidos para jamones
procedentes de cerdos de razas blancas (38-40%), siendo superiores a los esperados para
jamones Ibéricos (36%), sin embargo los niveles de AGPI fueron superiores a los
esperados, ya que superaron a los descritos por otros autores en jamón Ibérico (6-7%).
Estos altos valores pueden deberse al tipo de alimento y a la cantidad ingerida del mismo,
por el uso de piensos enriquecidos, que influyen directamente en la cantidad de AGPI y
más concretamente en los valores de ácido linoleico (C18:2(ω-6)), ya que este ácido graso
es esencial. En jamones Ibéricos, este porcentaje se encuentra entre el 5-7%, sin embargo
en razas blancas presentan valores más elevados (10-12%).
Otro valor a destacar es la elevada relación existente entre ω-6/ ω-3 debido a la
pequeña cantidad del ácido linolénico (C18:3 (ω-3)) presente en un 0,40% y a la elevada
del linoleico (C18:2 (ω-6)) en un 9,31%. Desde el punto de vista nutritivo se recomiendan
valores por debajo de 6 (Cava et al., 1997).

CAPÍTULO IV: RESULTADOS
71
Tabla 4.1.7. Composición de la grasa intramuscular del jamón curado en ácidos grasos neutros (LN), libres (AGL), polares (LP) y totales (LT). Los resultados están expresados % (± desviación estándar de la media). El C22:3(ω-6)+C21 no se han tenido en cuenta en sus respectivos sumatorios por encontrarse co-eluídos.
LN AGL LP LT
C10:0 0,06 ± 0,01 0,06 ± 0,01 0,15 ± 0,07 0,05 ± 0,01
C12:0 0,06 ± 0,02 0,04 ± 0,01 0,22 ± 0,10 0,06 ± 0,01
C14:0 1,34 ± 0,46 0,68 ± 0,21 0,85 ± 0,26 1,51 ± 0,20
C15:0 0,04 ± 0,01 0,09 ± 0,03 0,27 ± 0,09 0,05 ± 0,01 C16:0 24,52 ± 3,64 18,39 ± 4,03 17,63 ± 1,68 26,72 ± 1,40
C17:0 0,23 ± 0,05 0,33 ± 0,07 0,59 ± 0,11 0,25 ± 0,05
C18:0 11,71 ± 1,84 11,34 ± 0,91 15,06 ± 2,07 10,54 ± 0,99
C20:0 0,17 ± 0,07 0,10 ± 0,03 0,24 ± 0,08 0,09 ± 0,01 C22:0 0,01 ± 0,01 0,03 ± 0,02 0,58 ± 0,49 0,01 ± 0,00
C23:0 0,09 ± 0,05 0,79 ± 0,32 1,81 ± 0,99 0,13 ± 0,03
C24:0 0,01 ± 0,01 0,08 ± 0,03 1,03 ± 0,89 0,01 ± 0,00
AGS 38,24 ± 3,36 31,93 ± 3,80 38,42 ± 4,08 39,41 ± 2,11
C14:1 0,05 ± 0,03 0,08 ± 0,04 1,53 ± 0,77 0,07 ± 0,03
C15:1 0,02 ± 0,01 0,10 ± 0,05 1,49 ± 1,23 0,05 ± 0,02
C16:1 3,39 ± 0,81 1,77 ± 0,49 1,19 ± 0,41 3,47 ± 0,62
C17:1 0,28 ± 0,06 0,21 ± 0,05 0,82 ± 0,44 0,27 ± 0,05 C18:1 49,50 ± 3,82 32,01 ± 3,89 19,39 ± 2,33 44,27 ± 2,29
C20:1 0,88 ± 0,38 0,64 ± 0,26 0,69 ± 0,26 0,55 ± 0,07
C22:1 0,02 ± 0,01 0,04 ± 0,01 0,69 ± 0,57 0,01 ± 0,00
C24:1 0,04 ± 0,03 0,17 ± 0,05 0,88 ± 0,68 0,03 ± 0,02
AGMI 54,18 ± 3,60 35,01 ± 4,14 26,70 ± 2,71 48,72 ± 2,76
C18:2 (ω-6) 6,11 ± 0,98 22,12 ± 2,20 16,04 ± 3,16 9,31 ± 1,62
C18:3 (ω-6) 0,02 ± 0,01 0,17 ± 0,03 0,12 ± 0,06 0,07 ± 0,01
C18:3 (ω-3) 0,39 ± 0,06 0,64 ± 0,07 0,71 ± 0,29 0,40 ± 0,07 C20:2 (ω-6) 0,33 ± 0,11 0,55 ± 0,14 0,65 ± 0,17 0,26 ± 0,04
C20:4 (ω-6) 0,24 ± 0,07 6,94 ± 1,49 5,74 ± 2,08 1,27 ± 0,29
C20:3 (ω-3) 0,07 ± 0,03 0,11 ± 0,04 0,09 ± 0,03 0,06 ± 0,01
C20:5 (ω-3) 0,04 ± 0,02 0,33 ± 0,05 1,20 ± 0,47 0,05 ± 0,01 C22:2 0,02 ± 0,01 0,05 ± 0,03 0,88 ± 0,78 0,01 ± 0,00
C22:6 (ω-3) 0,05 ± 0,03 0,74 ± 0,30 1,14 ± 0,51 0,10 ± 0,03
AGPI 7,26 ± 1,12 31,65 ± 3,09 26,56 ± 4,67 11,53 ± 1,95
ω-6 6,70 ± 1,04 29,78 ± 2,95 22,55 ± 4,95 10,91 ± 1,87
ω-3 0,55 ± 0,09 1,82 ± 0,35 3,13 ± 0,78 0,61 ± 0,10
ω-6/ω-3 12,25 ± 0,93 16,90 ± 3,29 7,91 ± 2,83 18,02 ± 1,61
C20:3 (ω-6) + C21 0,07 ± 0,02 0,99 ± 0,25 0,93 ± 0,23 0,19 ± 0,04

CAPÍTULO IV: RESULTADOS
72
Determinación del contenido en aminoácidos libres
El método de cromatografía utilizado permitió la identificación de 20 aminoácidos,
tal como se muestra en la tabla 4.1.8, que se mostraron en un total de 15 picos, tal como se
muestra en la figura 4.1.3.
Asp
Glu
Gly-Gln
Ser-Asn
Tyr
Val
Trp
Nor
Phe
His
Trh
Cys-Ala
Pro
Arg
Met
Iso
Leu
Lys
Figura 4.1.3. Cromatograma de aminoácidos libres obtenido mediante HPLC de magro de jamón curado.
A pesar de que los aminoácidos Pro (Prolina) y Arg (Arginina) se separaron en dos
picos, se cuantificaron conjuntamente ya que el cromatograma obtenido del patrón no
consiguió separarlos, apareciendo ambos en un único pico. Al igual que los aminoácidos
Tyr (Tirosina) y Val (Valina), que se cuantificaron conjuntos porque la mayor parte de las
muestras obtuvo un único pico.

CAPÍTULO IV: RESULTADOS
73
Tabla 4.1.8. Contenido en aminoácidos libres en jamón curado. Los resultados se expresan en mg aa/100 g muestra (± desviación estándar de la media).
E.H E.S
Ac. Aspártico (Asp) 264,56 ± 71,67 532,95 ± 136,01 Ac. Glutámico (Glu) 532,39 ± 125,04 1.075,36 ± 239,76
Serina + Asparagina (Ser-Asn) 368,12 ± 122,60 748,70 ± 254,14
Glicina + Glutamina (Gly-Gln) 353,70 ± 81,79 746,08 ± 210,06
Histidina (Hys) 217,29 ± 44,28 440,16 ± 87,53 Treonina (Trh) 105,41 ± 38,96 201,75 ± 58,38
Cisteína + Alanina (Cys-Ala) 1.119,58 ± 274,23 2.290,71 ± 611,04
Prolina + Arginina (Pro-Arg) 1.978,81 ± 310,25 4.158,62 ± 892,71
Tirosina + Valina (Tyr-Val) 476,21 ± 60,89 989,81 ± 165,93 Metionina (Met) 101,95 ± 20,84 207,83 ± 41,78
Isoleucina (Iso) 237,60 ± 47,78 484,98 ± 99,27
Leucina (Leu) 447,35 ± 91,98 911,51 ± 181,77
Fenilalanina (Phe) 230,74 ± 48,48 470,47 ± 97,66 Triptófano (Trp) 68,04 ± 9,29 141,83 ± 23,48
Lisina (Lys) 643,79 ± 133,92 1.318,98 ± 264,49
sumatorio 7.145,52 ± 1.184,86 14.719,77 ± 2.751,77
(EH): extracto húmedo; (ES): extracto seco
La Lys (lisina) y el Glu (ácido glutámico) fueron los aminoácidos que alcanzaron los
mayores niveles en el jamón curado sin tener en cuenta los que se encontraban co-eluidos,
seguidos de la Leu (leucina) y del Asp (ácido aspártico). Estos resultados coinciden con los
descritos por otros autores en jamón Ibérico (Martín et al., 2001; Ruiz et al., 1999), en
jamón curado (Toldrá et al., 2000), en jamón de Parma (Careri et al., 1993) y en jamón
curado francés (Buscailhon et al., 1994; Jurado, 2005).
El perfil de aminoácidos libres obtenido fue muy similar a los observados en otros
estudios sobre jamón curado (Toldrá et al., 2000) o jamón Ibérico (Jurado, 2005). Sin
embargo el contenido total de aminoácidos libres fue algo superior al obtenido en estudios
anteriores, con 12.575 mg/100g para jamón curado descrito por Toldrá et al., (2000) o
13.000 mg/100g descritos por Jurado (2005), a pesar de que los perfiles fueron similares.
(Los datos se expresan en extracto seco).

CAPÍTULO IV: RESULTADOS
74
Determinación de los compuestos nitrogenados derivados del ATP
El método de cromatografía utilizado permitió la identificación de 6 nucleótidos,
tal como se muestra en la tabla 4.1.9, que se presentaron en un total de 5 picos, tal como se
muestra en la figura 4.1.4.
IMP
GMP
ADP
AMP-Hx
Inosina
Figura 4.1.4. Cromatograma de nucleótidos libres obtenido mediante HPLC de magro de jamón curado. Tabla 4.1.9. Contenido en 5´nucleótidos del jamón curado. Los resultados se expresan en mg de nuc./100 g muestra (± desviación estándar de la media).
E.H. E.S.
Inosina 5´monofosfato (IMP) 8,35 ± 3,38 17,15 ± 7,18
Guanosina 5´monofosfato (GMP) 18,82 ± 4,38 38,66 ± 9,31 Adenosina 5´difosfato (ADP) 9,93 ± 3,40 20,47 ± 7,50
Adenosina 5´monofosfato(AMP)+ Hipoxantina(Hx) 120,00 ± 23,46 246,98 ± 54,17
Inosina (Ino) 24,94 ± 7,86 50,74 ± 14,92
sumatorio 182,04 ± 28,49 374,00 ± 65,73
(EH): extracto húmedo; (ES): extracto seco
Tal como muestra la bibliografía, tras el sacrificio del animal el ATP se degrada a
ADP (adenosina 5´difosfato) y AMP (adenosina 5´monofosfato) rápidamente, con la
subsiguiente acumulación de IMP (inosina 5´monofosfato). Por otra parte la IMP es
hidrolizada a Ino (Inosina), la cual es hidrolizada a Hx (Hipoxantina). Es por ello, que nos
encontramos con altos valores de AMP e Hx y bajos de IMP y ADP (tabla 4.1.9).

CAPÍTULO IV: RESULTADOS
75
Determinación de compuestos volátiles
Se detectaron un total de 123 compuestos volátiles en el total de jamones
analizados, de los cuales se presentan en la tabla 4.1.10 aquellos que aparecieron al menos
en dos de las tres campañas de jamones analizadas.
En la tabla 4.1.10 se muestran 113 compuestos pertenecientes a 12 familias
químicas diferentes: aldehídos (18), cetonas (15), alcoholes (12), hidrocarburos alifáticos
(18), ácidos (9), ésteres (8), compuestos azufrados (6), compuestos nitrogenados (8),
furanos (8), hidrocarburos aromáticos (6), terpenos (2) y compuestos clorados (2).
Entre los compuestos volátiles seleccionados, 63 han sido asociados a la oxidación
lipídica, 33 tienen su origen en aminoácidos a través de reacciones de Strecker (ej:
aldehídos ramificados) y de Maillard (ej: 2,6-dimetilpirazina), 11 de ellos han sido
relacionados tanto con la oxidación lipídica como con reacciones de Maillard y los 2
restantes están asociados a agentes externos del producto. El predominio de compuestos
derivados de las reacciones de oxidación coincidió con resultados obtenidos por Jurado
(2005) para los compuestos volátiles detectados en jamón Ibérico durante la fase de
secadero.
Tabla 4.1.10. Compuestos volátiles identificados en jamón curado agrupados por familia química. Los contenidos de cada compuesto se expresan en % de área y en UAA (106) (± desviación estándar de la media)
IK Posición Compuesto UAA (106) UAA (%) Posible origen
ALDEHIDOS 178,39 ± 76,63 13,76 ± 4,59 <500 2 acetaldehído 11,14 ± 4,23 0,96 ± 0,37 aa
550 12 2-metilpropanal 5,59 ± 3,30 0,42 ± 0,22 aa
649 22 3-metilbutanal 70,80 ± 50,57 5,52 ± 3,57 aa
660 24 2-metilbutanal 25,14 ± 16,44 1,75 ± 0,87 aa 698 27 pentanal 4,82 ± 2,55 0,39 ± 0,20 lipidos
744 37 2-metil-2-butenal 1,39 ± 0,63 0,11 ± 0,04 aa
800 51 hexanal 32,94 ± 18,02 2,53 ± 1,24 lipidos
902 65 heptanal 4,43 ± 1,88 0,37 ± 0,18 lipidos 908 66 3-(metiltio)propanal 2,75 ± 1,57 0,21 ± 0,11 aa
969 74 benzaldehído 3,11 ± 1,32 0,25 ± 0,10 aa
1006 83 octanal 2,96 ± 1,06 0,25 ± 0,12 lipidos
1053 87 benzenoacetaldehido 3,23 ± 1,52 0,27 ± 0,13 aa 1104 93 nonanal 8,35 ± 5,72 0,57 ± 0,29 lipidos
1157 94 2-nonenal 0,41 ± 0,14 0,04 ± 0,01 lipidos
1209 99 decanal 0,59 ± 0,37 0,05 ± 0,03 lipidos
1495 108 dodecanal 0,72 ± 0,52 0,06 ± 0,05 lipidos
CETONAS 218,10 ± 73,30 16,97 ± 5,12 <500 5 2-propanona + propanal 103,40 ± 44,02 8,39 ± 3,75 lipidos
586 16 2,3-butanodiona 6,36 ± 4,99 0,54 ± 0,41 lipidos
598 17 2-butanona + butanal 37,35 ± 33,44 2,31 ± 1,27 lipidos 656 23 1-hidroxi-2-propanona 4,98 ± 2,65 0,40 ± 0,21 lipidos
687 26 2-pentanona 31,34 ± 16,90 2,58 ± 1,52 lipidos
710 31 3-hidroxi-2-butanona 7,21 ± 4,47 0,61 ± 0,41 lipidos
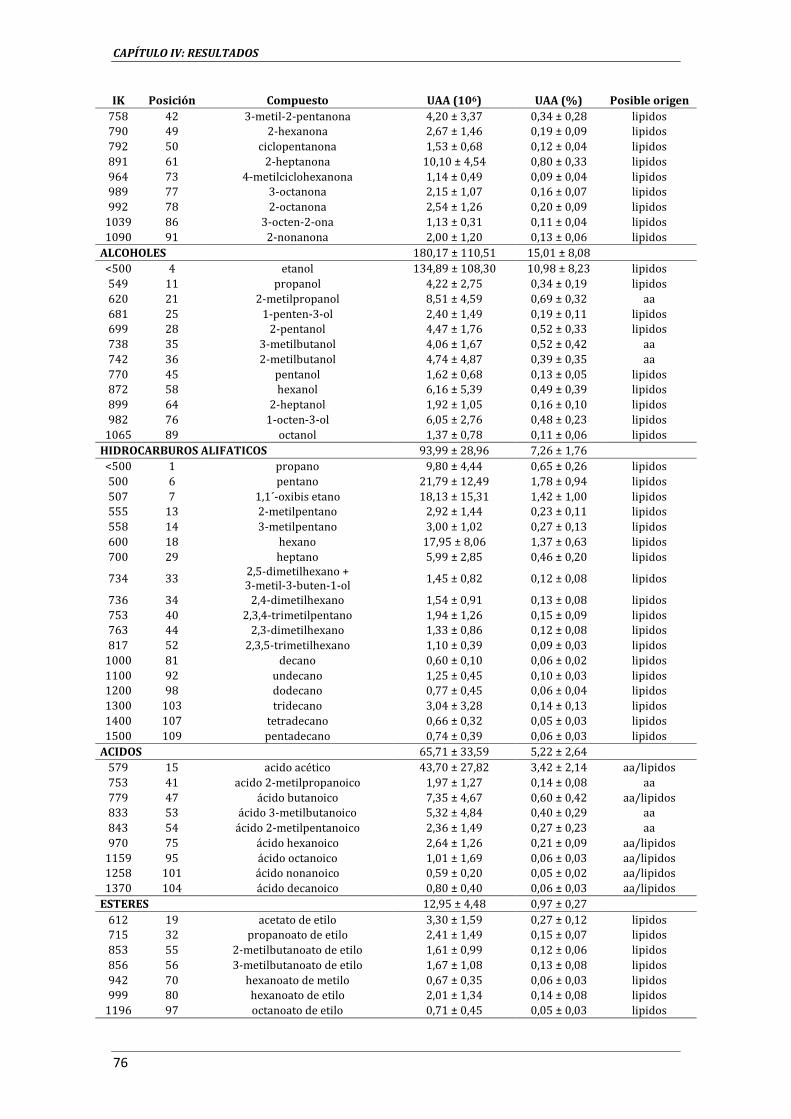
CAPÍTULO IV: RESULTADOS
76
IK Posición Compuesto UAA (106) UAA (%) Posible origen
758 42 3-metil-2-pentanona 4,20 ± 3,37 0,34 ± 0,28 lipidos 790 49 2-hexanona 2,67 ± 1,46 0,19 ± 0,09 lipidos
792 50 ciclopentanona 1,53 ± 0,68 0,12 ± 0,04 lipidos
891 61 2-heptanona 10,10 ± 4,54 0,80 ± 0,33 lipidos
964 73 4-metilciclohexanona 1,14 ± 0,49 0,09 ± 0,04 lipidos 989 77 3-octanona 2,15 ± 1,07 0,16 ± 0,07 lipidos
992 78 2-octanona 2,54 ± 1,26 0,20 ± 0,09 lipidos
1039 86 3-octen-2-ona 1,13 ± 0,31 0,11 ± 0,04 lipidos
1090 91 2-nonanona 2,00 ± 1,20 0,13 ± 0,06 lipidos
ALCOHOLES 180,17 ± 110,51 15,01 ± 8,08 <500 4 etanol 134,89 ± 108,30 10,98 ± 8,23 lipidos
549 11 propanol 4,22 ± 2,75 0,34 ± 0,19 lipidos
620 21 2-metilpropanol 8,51 ± 4,59 0,69 ± 0,32 aa
681 25 1-penten-3-ol 2,40 ± 1,49 0,19 ± 0,11 lipidos 699 28 2-pentanol 4,47 ± 1,76 0,52 ± 0,33 lipidos
738 35 3-metilbutanol 4,06 ± 1,67 0,52 ± 0,42 aa
742 36 2-metilbutanol 4,74 ± 4,87 0,39 ± 0,35 aa
770 45 pentanol 1,62 ± 0,68 0,13 ± 0,05 lipidos 872 58 hexanol 6,16 ± 5,39 0,49 ± 0,39 lipidos
899 64 2-heptanol 1,92 ± 1,05 0,16 ± 0,10 lipidos
982 76 1-octen-3-ol 6,05 ± 2,76 0,48 ± 0,23 lipidos
1065 89 octanol 1,37 ± 0,78 0,11 ± 0,06 lipidos
HIDROCARBUROS ALIFATICOS 93,99 ± 28,96 7,26 ± 1,76 <500 1 propano 9,80 ± 4,44 0,65 ± 0,26 lipidos
500 6 pentano 21,79 ± 12,49 1,78 ± 0,94 lipidos
507 7 1,1´-oxibis etano 18,13 ± 15,31 1,42 ± 1,00 lipidos 555 13 2-metilpentano 2,92 ± 1,44 0,23 ± 0,11 lipidos
558 14 3-metilpentano 3,00 ± 1,02 0,27 ± 0,13 lipidos
600 18 hexano 17,95 ± 8,06 1,37 ± 0,63 lipidos
700 29 heptano 5,99 ± 2,85 0,46 ± 0,20 lipidos
734 33 2,5-dimetilhexano + 3-metil-3-buten-1-ol
1,45 ± 0,82 0,12 ± 0,08 lipidos
736 34 2,4-dimetilhexano 1,54 ± 0,91 0,13 ± 0,08 lipidos
753 40 2,3,4-trimetilpentano 1,94 ± 1,26 0,15 ± 0,09 lipidos 763 44 2,3-dimetilhexano 1,33 ± 0,86 0,12 ± 0,08 lipidos
817 52 2,3,5-trimetilhexano 1,10 ± 0,39 0,09 ± 0,03 lipidos
1000 81 decano 0,60 ± 0,10 0,06 ± 0,02 lipidos
1100 92 undecano 1,25 ± 0,45 0,10 ± 0,03 lipidos 1200 98 dodecano 0,77 ± 0,45 0,06 ± 0,04 lipidos
1300 103 tridecano 3,04 ± 3,28 0,14 ± 0,13 lipidos
1400 107 tetradecano 0,66 ± 0,32 0,05 ± 0,03 lipidos
1500 109 pentadecano 0,74 ± 0,39 0,06 ± 0,03 lipidos
ACIDOS 65,71 ± 33,59 5,22 ± 2,64 579 15 acido acético 43,70 ± 27,82 3,42 ± 2,14 aa/lipidos
753 41 acido 2-metilpropanoico 1,97 ± 1,27 0,14 ± 0,08 aa
779 47 ácido butanoico 7,35 ± 4,67 0,60 ± 0,42 aa/lipidos 833 53 ácido 3-metilbutanoico 5,32 ± 4,84 0,40 ± 0,29 aa
843 54 ácido 2-metilpentanoico 2,36 ± 1,49 0,27 ± 0,23 aa
970 75 ácido hexanoico 2,64 ± 1,26 0,21 ± 0,09 aa/lipidos
1159 95 ácido octanoico 1,01 ± 1,69 0,06 ± 0,03 aa/lipidos 1258 101 ácido nonanoico 0,59 ± 0,20 0,05 ± 0,02 aa/lipidos
1370 104 ácido decanoico 0,80 ± 0,40 0,06 ± 0,03 aa/lipidos
ESTERES 12,95 ± 4,48 0,97 ± 0,27 612 19 acetato de etilo 3,30 ± 1,59 0,27 ± 0,12 lipidos
715 32 propanoato de etilo 2,41 ± 1,49 0,15 ± 0,07 lipidos
853 55 2-metilbutanoato de etilo 1,61 ± 0,99 0,12 ± 0,06 lipidos
856 56 3-metilbutanoato de etilo 1,67 ± 1,08 0,13 ± 0,08 lipidos
942 70 hexanoato de metilo 0,67 ± 0,35 0,06 ± 0,03 lipidos 999 80 hexanoato de etilo 2,01 ± 1,34 0,14 ± 0,08 lipidos
1196 97 octanoato de etilo 0,71 ± 0,45 0,05 ± 0,03 lipidos
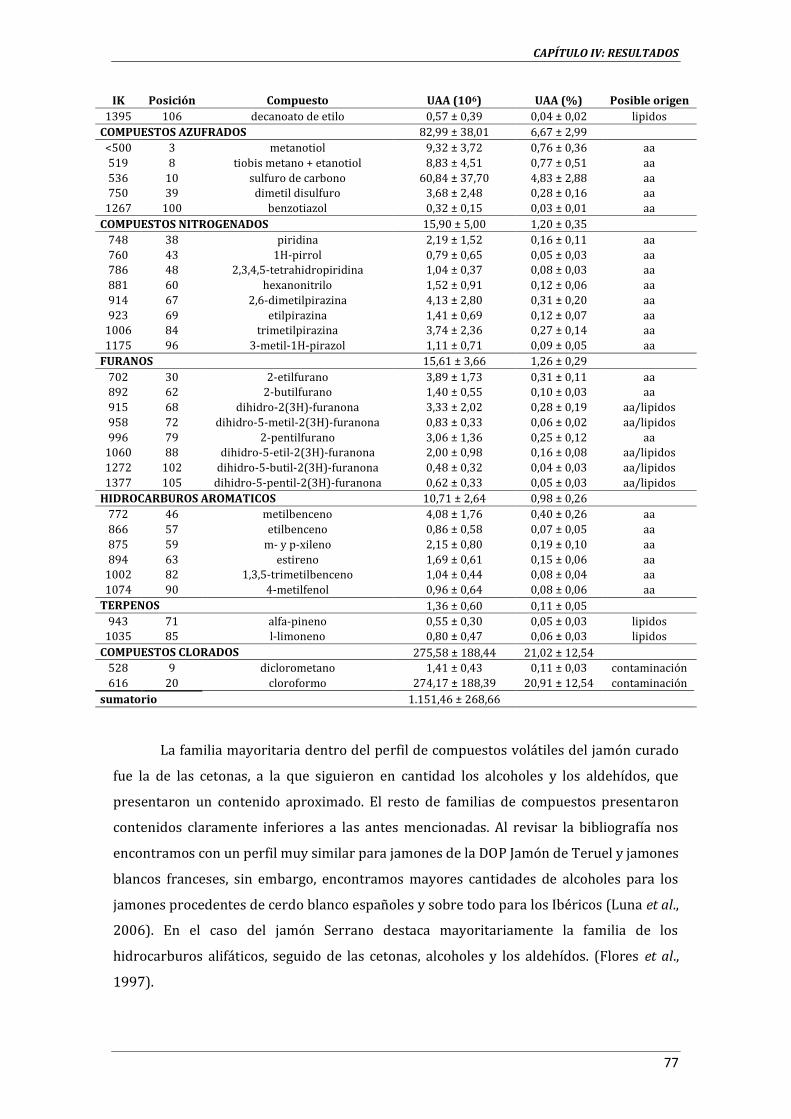
CAPÍTULO IV: RESULTADOS
77
IK Posición Compuesto UAA (106) UAA (%) Posible origen
1395 106 decanoato de etilo 0,57 ± 0,39 0,04 ± 0,02 lipidos
COMPUESTOS AZUFRADOS 82,99 ± 38,01 6,67 ± 2,99 <500 3 metanotiol 9,32 ± 3,72 0,76 ± 0,36 aa
519 8 tiobis metano + etanotiol 8,83 ± 4,51 0,77 ± 0,51 aa
536 10 sulfuro de carbono 60,84 ± 37,70 4,83 ± 2,88 aa 750 39 dimetil disulfuro 3,68 ± 2,48 0,28 ± 0,16 aa
1267 100 benzotiazol 0,32 ± 0,15 0,03 ± 0,01 aa
COMPUESTOS NITROGENADOS 15,90 ± 5,00 1,20 ± 0,35 748 38 piridina 2,19 ± 1,52 0,16 ± 0,11 aa
760 43 1H-pirrol 0,79 ± 0,65 0,05 ± 0,03 aa 786 48 2,3,4,5-tetrahidropiridina 1,04 ± 0,37 0,08 ± 0,03 aa
881 60 hexanonitrilo 1,52 ± 0,91 0,12 ± 0,06 aa
914 67 2,6-dimetilpirazina 4,13 ± 2,80 0,31 ± 0,20 aa
923 69 etilpirazina 1,41 ± 0,69 0,12 ± 0,07 aa 1006 84 trimetilpirazina 3,74 ± 2,36 0,27 ± 0,14 aa
1175 96 3-metil-1H-pirazol 1,11 ± 0,71 0,09 ± 0,05 aa
FURANOS 15,61 ± 3,66 1,26 ± 0,29 702 30 2-etilfurano 3,89 ± 1,73 0,31 ± 0,11 aa
892 62 2-butilfurano 1,40 ± 0,55 0,10 ± 0,03 aa
915 68 dihidro-2(3H)-furanona 3,33 ± 2,02 0,28 ± 0,19 aa/lipidos
958 72 dihidro-5-metil-2(3H)-furanona 0,83 ± 0,33 0,06 ± 0,02 aa/lipidos
996 79 2-pentilfurano 3,06 ± 1,36 0,25 ± 0,12 aa 1060 88 dihidro-5-etil-2(3H)-furanona 2,00 ± 0,98 0,16 ± 0,08 aa/lipidos
1272 102 dihidro-5-butil-2(3H)-furanona 0,48 ± 0,32 0,04 ± 0,03 aa/lipidos
1377 105 dihidro-5-pentil-2(3H)-furanona 0,62 ± 0,33 0,05 ± 0,03 aa/lipidos
HIDROCARBUROS AROMATICOS 10,71 ± 2,64 0,98 ± 0,26 772 46 metilbenceno 4,08 ± 1,76 0,40 ± 0,26 aa
866 57 etilbenceno 0,86 ± 0,58 0,07 ± 0,05 aa
875 59 m- y p-xileno 2,15 ± 0,80 0,19 ± 0,10 aa
894 63 estireno 1,69 ± 0,61 0,15 ± 0,06 aa 1002 82 1,3,5-trimetilbenceno 1,04 ± 0,44 0,08 ± 0,04 aa
1074 90 4-metilfenol 0,96 ± 0,64 0,08 ± 0,06 aa
TERPENOS 1,36 ± 0,60 0,11 ± 0,05 943 71 alfa-pineno 0,55 ± 0,30 0,05 ± 0,03 lipidos
1035 85 l-limoneno 0,80 ± 0,47 0,06 ± 0,03 lipidos
COMPUESTOS CLORADOS 275,58 ± 188,44 21,02 ± 12,54 528 9 diclorometano 1,41 ± 0,43 0,11 ± 0,03 contaminación
616 20 cloroformo 274,17 ± 188,39 20,91 ± 12,54 contaminación
sumatorio 1.151,46 ± 268,66
La familia mayoritaria dentro del perfil de compuestos volátiles del jamón curado
fue la de las cetonas, a la que siguieron en cantidad los alcoholes y los aldehídos, que
presentaron un contenido aproximado. El resto de familias de compuestos presentaron
contenidos claramente inferiores a las antes mencionadas. Al revisar la bibliografía nos
encontramos con un perfil muy similar para jamones de la DOP Jamón de Teruel y jamones
blancos franceses, sin embargo, encontramos mayores cantidades de alcoholes para los
jamones procedentes de cerdo blanco españoles y sobre todo para los Ibéricos (Luna et al.,
2006). En el caso del jamón Serrano destaca mayoritariamente la familia de los
hidrocarburos alifáticos, seguido de las cetonas, alcoholes y los aldehídos. (Flores et al.,
1997).

CAPÍTULO IV: RESULTADOS
78
Evaluación sensorial
Los valores de los diferentes atributos sensoriales utilizados para caracterizar el
jamón curado se incluyen en la tabla 4.1.11 y en el gráfico de araña representado en la
figura 4.1.5.
Tabla 4.1.11. Análisis cuantitativo-descriptivo de jamón curado (± desviación estándar de la media)
Color grasa Amarillo 1,4 ± 0,7
Rosa 1,1 ± 0,4
Color magro
Rojo 4,1 ± 0,8
Brillo 3,7 ± 0,7
Veteado 3,8 ± 1,2
Olor Intensidad 4,3 ± 0,7
Textura grasa Dureza 4,1 ± 1,0
Fluidez 4,4 ± 0,8
Textura magro
Dureza 2,6 ± 0,7
Sequedad 3,0 ± 0,8
Jugosidad 4,6 ± 0,6
Pastosidad 4,5 ± 1,3
Sabor Salado 4,9 ± 0,5 Dulce 1,0 ± 0,3
Amargo 1,0 ± 0,2
Flavor
Intensidad 4,8 ± 0,5
Persistencia 3,4 ± 0,7
Curado 1,3 ± 0,3
Rancidez 4,7 ± 0,5
Los valores emitidos por el panel sensorial para el jamón curado lo definen como
un jamón tierno, jugoso y con una baja sequedad, además de un flavor intenso y algo
salado, a pesar de que el contenido en cloruros de la tabla 4.1.4 indicaba que la
concentración de sal era baja.
0
1
2
3
4
5AMARILLO GRASA
ROSA GRASA
ROJO MAGRO
BRILLO MAGRO
VETEADO
INTENSIDAD OLOR
DUREZA GRASA
FLUIDEZ GRASA
DUREZA MAGROSEQUEDAD MAGROJUGOSIDAD MAGRO
PASTOSIDAD MAGRO
SALADO
DULCE
AMARGO
INTENSIDAD FLAVOR
PERSISTENCIA
CURADO
RANCIDEZ
Figura 4.1.5. Perfil sensorial del jamón curado

CAPÍTULO IV: RESULTADOS
79
4.1.2. INFLUENCIA DE LA EXPRESIÓN DEL IGF2 EN LAS CARACTERÍSTICAS FÍSICO-
QUÍMICAS Y SENSORIALES DEL JAMÓN CURADO
En este apartado se va a evaluar la influencia de los alelos A y G del QTL
(Quantitative Trait Loci) IGF2 (insuline-like growth factor 2) sobre todos los parámetros
analizados en los jamones curados. Para ello, tan solo van a incluirse aquellos parámetros
que presentaron diferencias significativas entre los dos lotes estudiados.
Características generales del jamón curado
Al evaluar las características generales del jamón curado: peso, longitud, anchura y
perímetro, no se encontraron diferencias significativas entre ambos lotes. No obstante, en
estudios realizados sobre otro lote de jamones procedentes de cerdos AA y AG analizados
en fresco, si se observaron diferencias significativas entre los lotes, presentando mayor
peso los jamones procedentes de los cerdos AG (Burgos et al., 2011). Aunque estos autores
señalaron diferencias significativas en materia fresca, a lo largo del procesado dichas
diferencias desaparecen.
El corte transversal de los jamones mostró diferencias significativas en cuanto al
espesor de la grasa subcutánea, que se percibieron claramente tanto en la zona proximal
(figura 4.1.6) como en la distal (figura 4.1.7), presentando diferencias significativas en dos
de las tres medidas tomadas en cada zona, tal y como se muestra en la tabla 4.1.12.
A
B
C
AG
AB
C
AA
Figura 4.1.6. Zona proximal del jamón curado

CAPÍTULO IV: RESULTADOS
80
A
B
C
AG
A
B
C
AA
Figura 4.1.7. Zona distal del jamón curado
Tabla 4.1.12. Espesor de la grasa subcutánea de las zonas proximal y distal del jamón curado (± desviación estándar de la media)
Zona proximal Zona distal
AA AG p
AA AG p
A (cm) 2,0 ± 0,7 3,2 ± 1,1 *** B (cm) 1,8 ± 0,4 2,3 ± 0,6 ***
B (cm) 2,0 ± 0,7 2,8 ± 1,0 ** C (cm) 3,2 ± 0,9 3,9 ± 1,6 *
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; Nivel de significación p<0,05
Los jamones procedentes de cerdos heterocigóticos (AG) presentaron un mayor
espesor de la grasa subcutánea (4,0 cm valor medio) en comparación con los jamones de
los cerdos homocigóticos (AA) (3,5 cm valor medio). Nuestros resultados confirman los de
Burgos et al,. (2010), que afirmaban que el alelo A era el responsable de aumentar el
contenido magro de la canal, aunque solo si era transmitido por vía paterna. Sin embargo,
el alelo G era el responsable de aumentar el contenido de grasa, independientemente de su
localización anatómica.
Perfil de textura instrumental
La tabla 4.1.13 muestra las diferencias significativas existentes entre los
parámetros de textura. Los cuatro parámetros mostrados en la tabla fueron
significativamente superiores en el lote AA, posiblemente debido a que son parámetros
relacionados inversamente con el contenido en grasa intramuscular, que fue inferior en
ese lote.
Tabla 4.1.13. Análisis de perfil de textura (TPA) del jamón curado (± desviación estándar de la media)
AA AG p
Fuerza (N) 11,46 ± 5,26 8,27 ± 2,78 *
Dureza (N) 12,97 ± 5,87 9,43 ± 3,10 *
Gomosidad (N) 6,42 ± 3,19 4,14 ± 1,32 **
Masticabilidad (N) 4,44 ± 2,85 2,41 ± 0,95 **
(N): Newton ** Nivel de significación p<0,01; * Nivel de significación p<0,05

CAPÍTULO IV: RESULTADOS
81
Parámetros generales físico-químicos
En cuanto a los parámetros generales, se encontraron diferencias significativas en
el contenido de grasa intramuscular y en el de proteínas (en extracto fresco), tal como se
muestra en la tabla 4.1.14.
Tabla 4.1.14. Parámetros físico-químicos generales del jamón curado (± desviación estándar de la media)
AA AG p
Proteínas (f) (g/100 g) 34,16 ± 5,42 31,61 ± 2,54 *
Grasa (f) (g/100 g) 3,85 ± 1,00 4,86 ± 1,11 **
Grasa (s) (g/100 g) 7,84 ± 2,16 9,99 ± 2,14 **
(f): extracto fresco; (s): extracto seco ** Nivel de significación p<0,01; Nivel de significación p<0,05
En el caso de los jamones, fue muy clara la diferencia existente entre el contenido
de grasa subcutánea y el de grasa intramuscular, siendo significativamente superiores los
valores del lote AG. En este caso, nuestros datos no coincidieron con los de Burgos et al.,
(2011), que no encontró diferencias significativas al estudiar el contenido de grasa
intramuscular de distintas piezas como el psoas major o el bíceps femoris. No obstante,
estos autores si observaron diferencias significativas cuando realizaron la determinación
en toda la masa del jamón.
En cuanto al contenido en proteínas en extracto fresco, el lote AA fue
significativamente superior al AG.
Composición de la grasa intramuscular
Como era de esperar, la fracción de triglicéridos (LN) fue la fracción mayoritaria de
la grasa intramuscular, con un porcentaje de 85,33% para el lote AA y de 88,01% para el
lote AG de la grasa total. La fracción de fosfolípidos (LP) fue la fracción minoritaria con un
0,65% para el lote AA y un 0,60% para el lote AG. La fracción de AGL fue de 14,02% para el
lote AA y de 11,39% para el lote AG.
Fracción de lípidos neutros
La tabla 4.1.15 muestra el contenido de los ácidos grasos identificados en la
fracción de LN de la grasa intramuscular extraída de los jamones curados que mostraron
diferencias significativas.
Existieron diferencias significativas en la mayoría de los ácidos grasos, y por lo
tanto en los sumatorios y contenido total, siendo superiores los valores del lote AG. Esta

CAPÍTULO IV: RESULTADOS
82
diferencia entre lotes coincidió con el mayor contenido de grasa intramuscular que
presentó el lote AG.
Tabla 4.1.15. Composición de la grasa intramuscular en la fracción de triglicéridos (LN) del jamón curado. Los resultados están expresados en mg de AG/100g muestra (± desviación estándar de la media)
LN AA LN AG P
C10:0 1,52 ± 0,67 2,06 ± 0,55 **
C12:0 1,79 ± 0,96 2,64 ± 1,13 **
C14:0 32,07 ± 12,61 48,20 ± 14,66 *** C15:0 0,78 ± 0,28 1,10 ± 0,40 **
C16:0 446,30 ± 155,66 635,21 ± 172,18 ***
C17:0 4,17 ± 1,62 5,68 ± 2,59 *
C18:0 167,65 ± 67,40 208,63 ± 56,57 * C20:0 2,20 ± 0,58 3,90 ± 1,39 ***
C22:0 0,09 ± 0,06 0,16 ± 0,12 *
C15:1 0,30 ± 0,13 0,44 ± 0,21 **
C16:1 66,39 ± 23,62 96,43 ± 31,33 *** C17:1 5,08 ± 2,06 6,65 ± 3,13 *
C18:1 247,77 ± 96,49 316,87 ± 96,88 *
C20:1 9,52 ± 4,31 13,00 ± 4,94 *
C22:1 0,19 ± 0,09 0,26 ± 0,08 ** C18:2 (ω-6) 92,49 ± 32,35 124,69 ± 40,76 **
C18:3 (ω-6) 0,40 ± 0,13 0,49 ± 0,17 *
C18:3 (ω-3) 6,08 ± 2,23 8,25 ± 2,73 **
C22:2 0,22 ± 0,12 0,32 ± 0,17 *
AGS 657,39 ± 228,50 908,62 ± 227,67 ***
AGMI 331,24 ± 118,98 436,00 ± 128,36 ** AGPI 108,25 ± 38,12 144,55 ± 46,66 **
ω-6 100,31 ± 35,37 134,13 ± 43,45 **
ω-3 7,72 ± 2,68 10,10 ± 3,23 **
C20:3 (ω-6) +C21 0,91 ± 0,34 1,09 ± 0,30 *
sumatorio 1.097,79 ± 407,56 1.490,27 ± 435,83 **
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05.
En cuanto a los valores expresados en %, las diferencias fueron menos marcadas
entre ambos lotes (tabla 4.1.16), aunque en este caso el lote AA presentó mayor
proporción de algunos ácidos grasos, destacando el valor de C18:3 (ω-6), que proviene
fundamentalmente de la alimentación del animal, y que por lo tanto parece ser ajeno al
efecto del IGF2.
Tabla 4.1.16. Composición de la grasa intramuscular en la fracción de triglicéridos (LN) del jamón curado. Los resultados están expresados en % (± desviación estándar de la media).
LN AA % LN AG % p
C18:3 (ω-6) 0,03 ± 0,01 0,02 ± 0,00 ** C20:5 (ω-3) 0,04 ± 0,02 0,17 ± 0,24 *
ω-3 0,56 ± 0,10 0,68 ± 0,23 * ω-6/ ω-3 12,38 ± 0,92 10,54 ± 2,54 **
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05

CAPÍTULO IV: RESULTADOS
83
Fracción de lípidos polares
Los ácidos grasos que componen la fracción de fosfolípidos (LP) que presentaron
diferencias significativas, se muestra en la tabla 4.1.17. A pesar de ser la fracción
minoritaria, se mantienen las diferencias significativas observadas en los ácidos grasos de
la fracción LN, existiendo diferencias significativas en más ácidos grasos que en la fracción
de AGL (tabla 4.1.19), siguiendo la misma tendencia que el resto de fracciones, es decir,
siendo superiores las cantidades en el lote AG, a excepción del ácido eicosatrienoico
(C20:3 (ω-3)), que fue significativamente superior en el lote AA.
Tabla 4.1.17. Composición de la grasa intramuscular en la fracción de fosfolípidos (LP) del jamón curado. Los resultados están expresados en mg de AG/100 g muestra (± desviación estándar de la media).
LP AA LP AG p
C10:0 0,03 ± 0,01 0,03 ± 0,01 **
C12:0 0,04 ± 0,02 0,05 ± 0,01 *
C15:0 0,04 ± 0,01 0,05 ± 0,02 ** C16:0 2,14 ± 0,56 2,94 ± 0,99 **
C17:0 0,06 ± 0,02 0,08 ± 0,02 **
C18:0 1,41 ± 0,51 1,77 ± 0,46 *
C20:0 0,05 ± 0,02 0,07 ± 0,02 ** C23:0 0,10 ± 0,04 0,14 ± 0,06 *
C20:1 0,05 ± 0,02 0,06 ± 0,03 *
C24:1 0,04 ± 0,02 0,05 ± 0,03 *
C18:3 (ω-3) 0,05 ± 0,02 0,10 ± 0,05 *** C20:2 (ω-6) 0,05 ± 0,02 0,06 ± 0,02 *
C20:3 (ω-3) 0,02 ± 0,01 0,01 ± 0,00 ***
C22:6 (ω-3) 0,03 ± 0,01 0,04 ± 0,01 *
AGS 4,16 ± 1,06 5,42 ± 1,45 **
AGMI 1,49 ± 0,45 1,83 ± 0,40 **
ω-3 0,22 ± 0,06 0,27 ± 0,06 *
sumatorio 8,37 ± 2,34 10,13 ± 2,35 *
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05
Los resultados expresados en porcentajes se muestran en la tabla 4.1.18. A
diferencia del resto de fracciones, excepto el ácido palmitoleico (C16:1), el resto de ácidos
grasos presentaron proporciones significativamente superiores en el lote AA, incluso la
fracción de AGPI, que no mostró diferencias significativas en ninguna de las fracciones.

CAPÍTULO IV: RESULTADOS
84
Tabla 4.1.18. Composición de la grasa intramuscular en la fracción de fosfolípidos (LP) del jamón curado. Los resultados están expresados en % (± desviación estándar de la media).
LP AA % LP AG % p
C16:0 18,12 ± 1,82 17,16 ± 1,43 * C17:0 0,63 ± 0,12 0,54 ± 0,08 **
C22:0 0,74 ± 0,62 0,40 ± 0,36 *
C17:1 1,05 ± 0,35 1,27 ± 0,37 *
C18:2 (ω-6) 17,06 ± 3,32 15,08 ± 2,80 * C20:4 (ω-6) 6,96 ± 1,60 4,52 ± 1,44 ***
C20:3 (ω-3) 0,10 ± 0,03 0,07 ± 0,01 ***
AGPI 28,79 ± 4,43 24,43 ± 3,66 ***
ω-6 24,82 ± 4,68 20,41 ± 4,08 **
C20:3 (ω-6) +C21 1,04 ± 0,23 0,82 ± 0,17 **
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05
Fracción de ácidos grasos libres
En cuanto a la composición de la fracción de AGL, al igual que el resto de
fracciones, las diferencias significativas siguieron la misma tendencia, siendo los valores
superiores en el lote AG, a excepción de la relación ω-6/ω-3 que fue superior en el lote AA,
tal como se muestra en la tabla 4.1.19. Sin embargo, al tratarse de una fracción minoritaria,
existieron menos diferencias significativas entre los lotes.
Tabla 4.1.19. Composición de la grasa intramuscular en la fracción de AGL del jamón curado. Los resultados están expresados en mg de AG/100 g muestra (± desviación estándar de la media).
AGL AA AGL AG p
C12 0,18 ± 0,05 0,23 ± 0,08 * C15 0,24 ± 0,06 0,31 ± 0,15 *
C22 0,04 ± 0,02 0,07 ± 0,04 *
C15:1 0,20 ± 0,08 0,27 ± 0,12 *
C20:5 (ω-3) 0,66 ± 0,22 1,16 ± 0,76 **
ω-3 2,86 ± 1,01 3,96 ± 1,83 *
ω-6/ ω-3 21,98 ± 2,82 18,67 ± 5,46 *
** Nivel de significación p<0,01; * Nivel de significación p<0,05
Al igual que en la fracción de LN, existieron diferencias significativas en el C18:3
(ω-6), siendo también superior en el lote AA, tal como se muestra en la tabla 4.1.20. Sin
embargo, en esta fracción si existieron diferencias significativas en el C18:0, que en este
caso fue superior en el lote AA.
Tabla 4.1.20. Composición de la grasa intramuscular en la fracción de AGL del jamón curado. Los resultados están expresados en % (± desviación estándar de la media)
AGL AA % AGL AG % p
C18:0 11,67 ± 0,84 11,00 ± 0,89 **
C18:3 (ω-6) 0,18 ± 0,03 0,16 ± 0,03 *
** Nivel de significación p<0,01; * Nivel de significación p<0,05

CAPÍTULO IV: RESULTADOS
85
Lípidos totales
En los resultados obtenidos en la tabla 4.1.21 se observa que el contenido total de
ácidos grasos fue superior significativamente en el lote AG, al igual que los sumatorios de
AGS y AGMI, no existiendo diferencias significativas en cuanto al contenido de AGPI de la
grasa intramuscular de los jamones. En cuanto a los ácidos grasos mayoritarios, el C16:0,
C18:1, C18:0 y C16:1, fueron significativamente superiores los valores del lote AG.
Tabla 4.1.21. Composición de la grasa intramuscular en la fracción de LT del jamón curado. Los resultados están expresados en mg de AG/100 g muestra (± desviación estándar de la media).
LT AA LT AG p
C10:0 2,11 ± 0,83 3,13 ± 0,94 ***
C12:0 2,66 ± 0,94 4,10 ± 1,56 ***
C14:0 44,75 ± 18,60 65,32 ± 26,03 ** C16:0 536,75 ± 195,87 737,01 ± 242,44 **
C18:0 211,96 ± 70,48 268,30 ± 87,41 *
C20:0 3,41 ± 1,22 4,93 ± 1,64 **
C22:0 0,17 ± 0,06 0,21 ± 0,07 * C23:0 2,47 ± 0,90 2,94 ± 0,65 *
C15:1 0,89 ± 0,41 1,22 ± 0,56 *
C16:1 75,46 ± 31,72 100,23 ± 27,38 **
C18:1 281,36 ± 117,21 357,39 ± 97,20 * C20:1 10,22 ± 4,51 13,83 ± 3,91 **
C18:3 (ω-3) 8,00 ± 2,95 9,60 ± 2,38 *
C20:3 (ω-3) 1,14 ± 0,51 1,41 ± 0,35 *
AGS 811,14 ± 272,57 1.094,13 ± 355,74 **
AGMI 376,57 ± 152,19 483,23 ± 128,63 *
ω-3 11,26 ± 3,94 13,25 ± 2,78 *
sumatorio 1.415,82 ± 472,06 1.836,42 ± 508,05 **
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05
La tabla 4.1.22 muestra los porcentajes de los ácidos grasos presentes en los
lípidos totales de la grasa intramuscular, donde las diferencias significativas fueron
mínimas, destacando los proporciones significativamente mayores del C18:3 (ω-6) y del
C20:4 ( ω-6) (ácido araquidónico) del lote AA.
Tabla 4.1.22. Composición de la grasa intramuscular en la fracción de LT del jamón curado. Los resultados están expresados en % (± desviación estándar de la media).
LT AA % LT AG % p
C16:0 26,22 ± 1,18 27,22 ± 1,44 *
C20:0 0,09 ± 0,01 0,10 ± 0,02 *
C20:1 0,53 ± 0,07 0,57 ± 0,05 *
C18:3 (ω-6) 0,08 ± 0,01 0,07 ± 0,01 ** C20:4 (ω-6) 1,41 ± 0,36 1,20 ± 0,30 *
AGS 38,84 ± 1,79 40,14 ± 2,50 * AGPI 12,14 ± 2,13 11,01 ± 1,79 *
ω-6 11,50 ± 2,02 10,41 ± 1,72 *
** Nivel de significación p<0,01; * Nivel de significación p<0,05
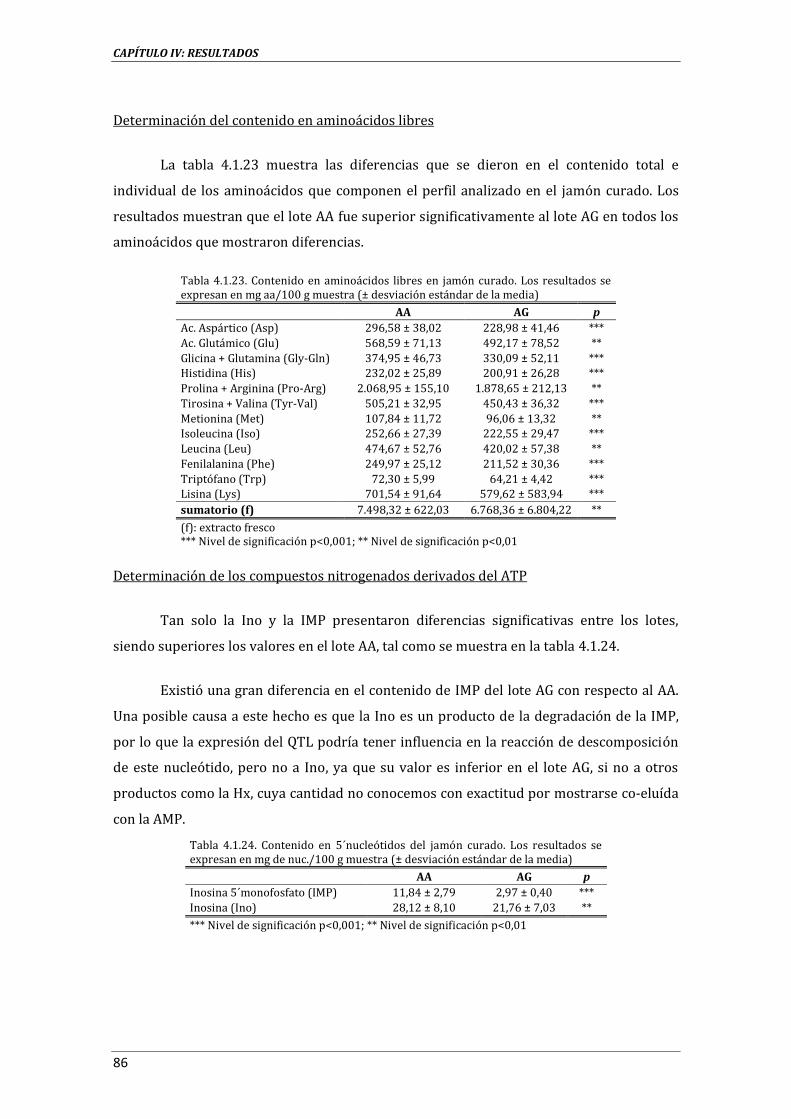
CAPÍTULO IV: RESULTADOS
86
Determinación del contenido en aminoácidos libres
La tabla 4.1.23 muestra las diferencias que se dieron en el contenido total e
individual de los aminoácidos que componen el perfil analizado en el jamón curado. Los
resultados muestran que el lote AA fue superior significativamente al lote AG en todos los
aminoácidos que mostraron diferencias.
Tabla 4.1.23. Contenido en aminoácidos libres en jamón curado. Los resultados se expresan en mg aa/100 g muestra (± desviación estándar de la media)
AA AG p
Ac. Aspártico (Asp) 296,58 ± 38,02 228,98 ± 41,46 ***
Ac. Glutámico (Glu) 568,59 ± 71,13 492,17 ± 78,52 **
Glicina + Glutamina (Gly-Gln) 374,95 ± 46,73 330,09 ± 52,11 *** Histidina (His) 232,02 ± 25,89 200,91 ± 26,28 ***
Prolina + Arginina (Pro-Arg) 2.068,95 ± 155,10 1.878,65 ± 212,13 **
Tirosina + Valina (Tyr-Val) 505,21 ± 32,95 450,43 ± 36,32 ***
Metionina (Met) 107,84 ± 11,72 96,06 ± 13,32 ** Isoleucina (Iso) 252,66 ± 27,39 222,55 ± 29,47 ***
Leucina (Leu) 474,67 ± 52,76 420,02 ± 57,38 **
Fenilalanina (Phe) 249,97 ± 25,12 211,52 ± 30,36 ***
Triptófano (Trp) 72,30 ± 5,99 64,21 ± 4,42 *** Lisina (Lys) 701,54 ± 91,64 579,62 ± 583,94 ***
sumatorio (f) 7.498,32 ± 622,03 6.768,36 ± 6.804,22 **
(f): extracto fresco *** Nivel de significación p<0,001; ** Nivel de significación p<0,01
Determinación de los compuestos nitrogenados derivados del ATP
Tan solo la Ino y la IMP presentaron diferencias significativas entre los lotes,
siendo superiores los valores en el lote AA, tal como se muestra en la tabla 4.1.24.
Existió una gran diferencia en el contenido de IMP del lote AG con respecto al AA.
Una posible causa a este hecho es que la Ino es un producto de la degradación de la IMP,
por lo que la expresión del QTL podría tener influencia en la reacción de descomposición
de este nucleótido, pero no a Ino, ya que su valor es inferior en el lote AG, si no a otros
productos como la Hx, cuya cantidad no conocemos con exactitud por mostrarse co-eluída
con la AMP.
Tabla 4.1.24. Contenido en 5´nucleótidos del jamón curado. Los resultados se expresan en mg de nuc./100 g muestra (± desviación estándar de la media)
AA AG p
Inosina 5´monofosfato (IMP) 11,84 ± 2,79 2,97 ± 0,40 ***
Inosina (Ino) 28,12 ± 8,10 21,76 ± 7,03 **
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01

CAPÍTULO IV: RESULTADOS
87
Determinación de compuestos volátiles
De los 113 compuestos volátiles detectados en los jamones (tabla 4.1.10), tan solo
30 presentaron diferencias significativas entre los lotes del estudio, 12 de ellos superiores
significativamente en el lote AG y 18 superiores significativamente en el lote AA. En cuanto
al contenido total, no existieron diferencias significativas entre los lotes. Las familias que
presentaron diferencias significativas entre lotes fue la de los hidrocarburos alifáticos, los
furanos y los terpenos, siendo solo el valor de los terpenos superior en el lote AG, debido a
la mayor proporción detectada del l-limoneno (tabla 4.1.25).
A pesar de observarse diferencias significativas en 30 compuestos volátiles, no
existieron diferencias en el contenido total de los compuestos volátiles entre los jamones
de ambos lotes (tabla 4.1.25). Este hecho puede verse influenciado por dos causas, una
asociada a las propias muestras y otra al procedimiento de extracción de los compuestos.
La primera de ellas no sería importante dada la similitud de las muestras, ya que los
animales se alimentaron con el mismo pienso y se criaron bajo las mismas condiciones
hasta la obtención del producto final, de forma que las diferencias encontradas pueden
deberse a la influencia del IGF2, que generó diferencias en el contenido de grasa
intramuscular, en la composición de ácidos grasos y en el contenido de aminoácidos libres.
Aún así, las pequeñas diferencias (no significativas) existentes en el contenido total de
compuestos volátiles entre los lotes pueden asociarse a la presencia del mayor contenido
de grasa intramuscular del lote AG, ya que existe una estrecha relación entre el contenido
y la composición de la grasa y la generación de compuestos volátiles (Estévez et al., 2003;
Ventanas et al., 2008).
El 2-butilfurano y el benzaldehído fueron significativamente superiores en el lote
AA. Dado que los furanos y los aldehídos son compuestos sintetizados a partir de
aminoácidos libres y péptidos mediante reacciones de Maillard, podemos asociar la mayor
cantidad de estos compuestos (tabla 4.1.25) con el mayor contenido en aminoácidos del
lote AA (tabla 4.1.23). Sin embargo, los hidrocarburos alifáticos y los alcoholes fueron
significativamente superiores en el lote AA, algo que esperaríamos del lote AG, ya que
estos compuestos proceden principalmente de los lípidos. Tan solo el grupo de los ácidos
destacó por ser superior significativamente en el lote AG.

CAPÍTULO IV: RESULTADOS
88
Tabla 4.1.25. Compuestos volátiles identificados en jamón curado agrupados por familia química. Los contenidos de cada compuesto se expresan en UAA (106) (± desviación estándar de la media).
IK Posición Compuesto AA AG p Posible origen
ALDEHIDOS
744 37 2-metil-2-butenal 1,12 ± 0,47 1,77 ± 0,91 ** aa
969 74 benzaldehído 3,42 ± 1,20 2,57 ± 1,02 * aa
CETONAS
586 16 2,3-butanodiona 8,03 ± 5,68 4,31 ± 2,90 ** lípidos 989 77 3-octanona 2,64 ± 1,65 1,89 ± 0,67 * lípidos
1039 86 3-octen-2-ona 1,20 ± 0,27 0,99 ± 0,28 * lípidos
ALCOHOLES
620 21 2-metilpropanol 9,90 ± 5,74 6,47 ± 2,69 * aa 699 28 2-pentanol 6,29 ± 3,25 3,32 ± 1,12 *** lípidos
738 35 3-metilbutanol 13,32 ± 16,90 4,25 ± 2,01 * aa
899 64 2-heptanol 2,15 ± 1,15 1,39 ± 0,57 ** lípidos
HIDROCARBUROS ALIFATICOS 93,97 ± 23,77 80,95 ± 19,14 *
558 14 3-metilpentano 4,78 ± 3,71 3,02 ± 1,09 * lípidos
600 18 hexano 20,81 ± 9,82 14,45 ± 5,24 ** lípidos
700 29 heptano 6,68 ± 2,79 4,02 ± 1,42 *** lípidos
1300 103 tridecano 2,57 ± 2,43 0,74 ± 0,16 ** lípidos
ACIDOS
753 41 acido 2-metilpropanoico 1,49 ± 0,50 2,21 ± 1,48 * aa
1258 101 ácido nonanoico 0,46 ± 0,12 0,66 ± 0,25 ** aa/lípidos
1370 104 ácido decanoico 0,55 ± 0,25 1,30 ± 0,72 *** aa/lípidos
ESTERES
715 32 propanoato de etilo 2,67 ± 2,09 1,13 ± 0,28 ** lípidos
853 55 2-metilbutanoato de etilo 1,26 ± 0,56 1,71 ± 0,83 * lípidos
999 80 hexanoato de etilo 2,23 ± 1,45 1,40 ± 0,73 * lípidos 1196 97 octanoato de etilo 0,55 ± 0,31 0,85 ± 0,49 * lípidos
1395 106 decanoato de etilo 0,43 ± 0,21 0,69 ± 0,51 * lípidos
COMPUESTOS NITROGENADOS
760 43 1H-pirrol 0,54 ± 0,23 0,86 ± 0,55 * aa 786 48 2,3,4,5-tetrahidropiridina 1,18 ± 0,36 0,90 ± 0,36 * aa
923 69 etilpirazina 1,17 ± 0,58 1,58 ± 0,74 * aa
FURANOS 16,12 ± 3,14 14,23 ± 3,33 *
892 62 2-butilfurano 1,58 ± 0,65 1,21 ± 0,40 * lípidos 996 79 2-pentilfurano 3,46 ± 1,64 2,56 ± 0,87 * lípidos
HIDROCARBUROS AROMATICOS
875 59 m- y p-xileno 1,81 ± 0,29 2,22 ± 0,91 * aa
894 63 estireno 1,60 ± 0,28 1,33 ± 0,36 ** aa
TERPENOS 1,12 ± 0,39 1,54 ± 0,69 *
1035 85 l-limoneno 0,56 ± 0,19 0,99 ± 0,54 ** lípidos
COMPUESTOS CLORADOS
528 9 diclorometano 1,18 ± 0,37 1,52 ± 0,46 ** contaminación
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05

CAPÍTULO IV: RESULTADOS
89
Evaluación sensorial
No se observaron diferencias significativas en las características sensoriales
evaluadas. La figura 4.1.8 muestra el perfil sensorial mediante un gráfico que pone de
manifiesto la gran similitud entre ambos lotes.
0
1
2
3
4
5AMARILLO GRASA
ROSA GRASA
ROJO MAGRO
BRILLO MAGRO
VETEADO
INTENSIDAD OLOR
DUREZA GRASA
FLUIDEZ GRASA
DUREZA MAGROSEQUEDAD JUGOSIDAD
PASTOSIDAD
SALADO
DULCE
AMARGO
INTENSIDAD …
PERSISTENCIA
CURADO
RANCIDEZ
AA
AG
Figura 4.1.8. Perfil sensorial del jamón curado

CAPÍTULO IV: RESULTADOS
90
4.1.3. INFUENCIA DE LA EXPRESIÓN DEL IGF2 EN FUNCIÓN DEL SEXO SOBRE LAS
CARACTERÍSTICAS FÍSICO-QUÍMICAS Y SENSORIALES DEL JAMÓN CURADO
En este apartado se va a evaluar la influencia de los alelos A y G del QTL
(Quantitative Trait Loci) IGF2 y su interacción con el sexo de los animales sobre todos los
parámetros analizados en los jamones curados. Para ello, tan solo van a incluirse aquellos
parámetros que presentaron diferencias significativas entre los cuatro lotes estudiados
(AA*H, AA*M, AG*H y AG*M, siendo H el factor hembra y M el factor macho).
Características generales del jamón curado
Al evaluar las características generales del jamón curado: peso, longitud, anchura y
perímetro, no se encontraron diferencias significativas entre los cuatro lotes, coincidiendo
con los resultados mostrados por la única influencia del IGF2. Estos resultados no
coincidieron con los de Burgos et al., (2011), ya que al evaluar el efecto sexo sobre la
conformación de otro lote de jamones analizados en fresco, si encontraron diferencias
significativas. Aunque estos autores señalaron diferencias significativas en materia fresca,
a lo largo del procesado dichas diferencias desaparecen.
El corte transversal de los jamones mostró diferencias en cuanto al espesor de la
grasa subcutánea, que se percibieron claramente tanto en la zona proximal (figura 4.1.9)
como en la distal (figura 4.1.10), presentando diferencias significativas en dos de las tres
medidas tomadas en cada zona, tal y como se muestra en las tablas 4.1.27 y 4.1.28.
Tabla 4.1.27. Medidas de la grasa subcutánea de la zona próximal del jamón curado (± desviación estándar de la media)
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
A (cm) 2,0a ± 0,6 1,9a ± 0,5 2,6ab ± 0,6 3,2b ± 0,7 ns *** ns ***
B (cm) 2,0ab ± 0,6 1,8a ± 0,5 2,5ab ± 1,1 2,8b ± 0,6 ns ** ns *
*** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo
Tabla 4.1.28. Medidas de la grasa subcutánea de la zona distal del jamón curado (± desviación estándar de la media)
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
B (cm) 1,8a ± 0,4 1,7a ± 0,4 2,1ab ± 0,3 2,4b ± 0,6 ns ** ns **
C (cm) 3,1a ± 0,9 3,0a ± 0,7 2,9a ± 0,4 4,3b ± 1,6 * ns * **
** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo
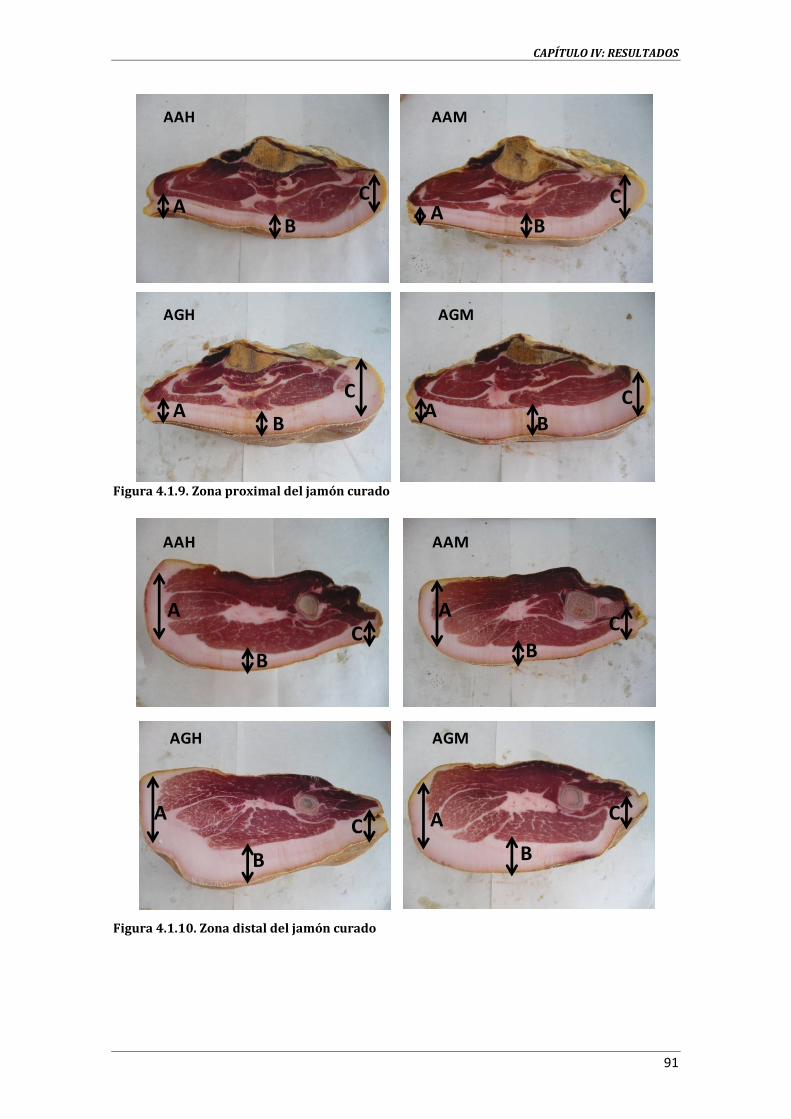
CAPÍTULO IV: RESULTADOS
91
AAH AAM
AGMAGH
AB
AC
B
C
AB
CA
B
C
Figura 4.1.9. Zona proximal del jamón curado
AAH AAM
AGH AGM
A
BC
A
A
B
C
B
CACB
Figura 4.1.10. Zona distal del jamón curado

CAPÍTULO IV: RESULTADOS
92
Los jamones procedentes de cerdos heterocigóticos (AGH y AGM) presentaron un
mayor espesor de la grasa subcutánea, siendo superior el efecto del macho (4,1 cm valor
medio) en comparación con la hembra (3,7 cm valor medio). Los jamones procedentes de
cerdos homocigóticos (AAH y AAM) presentaron menor espesor de grasa subcutánea en
comparación con los otros dos lotes, no existiendo diferencias debidas al efecto sexo, ya
que ambos lotes AA mostraron el mismo espesor de grasa (3,4 cm valor medio).
Perfil de textura instrumental
La tabla 4.1.29 muestra las diferencias significativas existentes entre los
parámetros de textura. De los ocho parámetros evaluados, seis presentaron diferencias
significativas por el efecto genético, siendo las muestras pertenecientes a los lotes AA las
que presentaron mayores valores. El efecto sexo no ejerció ninguna influencia sobre los
parámetros de textura evaluados, a pesar de que en la mayor parte de los casos, los lotes
pertenecientes a los machos mostraron mayores valores.
Parámetros generales físico-químicos
En cuanto a los parámetros generales, el contenido de grasa intramuscular y el pH
estuvieron claramente influenciados por el efecto genético, siendo superiores los valores
de los lotes AG. La coordenada a* y el porcentaje de cloruros estuvieron influenciados por
el efecto sexo, siendo superiores los valores de los lotes pertenecientes a los machos. El
resto de parámetros que mostraron diferencias se debieron a la interacción de ambos
factores, siendo superior el porcentaje de proteínas del lote AAH y de mg MDA/Kg del lote
AGH, tal como se muestra en la tabla 4.1.30.
Las diferencias significativas producidas en el contenido de grasa subcutánea e
intramuscular estuvieron claramente influenciadas por el efecto genético y no por el
efecto sexo, a pesar de que el sexo se ha descrito como un factor influyente en el contenido
de grasa intramuscular, siendo en el caso de las hembras superior (Latorre et al., 2008;
Latorre et al., 2009).

Tabla 4.2.29. Análisis de perfil de textura (TPA) del jamón curado (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
Fuerza (N) 10,70ab ± 5,73 12,31b ± 4,33 7,57a ± 1,12 8,38ab ± 3,25 ns ** ns * Dureza (N) 12,20ab ± 6,51 13,83b ± 4,70 8,62a ± 1,26 9,60ab ± 3,63 ns ** ns *
Adhesividad (N*seg) -0,26ab ± 0,04 -0,20b ± 0,05 -0,22ab ± 0,08 -0,27a ± 0,02 ns ns ** *
Elasticidad (cm) 0,62 ± 0,08 0,58 ± 0,06 0,58 ± 0,05 0,62 ± 0,03 ns ns * ns
Gomosidad (N) 6,12ab ± 3,63 6,75b ± 2,43 3,80a ± 0,67 4,70ab ± 2,04 ns ** ns * Masticabilidad (N) 4,08 ± 2,91 4,07 ± 1,70 2,03 ± 0,52 2,93 ± 1,42 ns ** ns *
(N): Newton ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo
Tabla 4.1.30. Parámetros físico-químicos generales del jamón curado (± desviación estándar de la media)
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
Proteínas (f) (g/100 g) 35,40 ± 6,23 31,58 ± 1,88 31,38 ± 2,23 32,96 ± 3,84 ns ns * ns
Proteínas (s,dg) (g/100 g) 80,64 ± 14,88 70,38 ± 5,48 72,94 ± 7,17 78,49 ± 10,45 ns ns ** ns Grasa (f) (g/100 g) 3,56a ± 0,95 4,16ab ± 0,91 4,85b ± 1,27 4,87b ± 0,92 ns ** ns **
Grasa (s) (g/100 g) 6,85a ± 1,50 8,43ab ± 1,88 9,94b ± 2,40 10,04b ± 1,82 ns *** ns ***
a (Adimensional) 14,40 ± 1,45 15,88 ± 1,62 14,68 ± 1,09 15,22 ± 1,57 * ns ns ns
Cloruros (g/100 g) (s) 3,23ab ± 0,69 3,24ab ± 0,33 3,14a ± 0,69 4,04b ± 1,01 * ns ns * Cloruros (f) (g/100 g) 6,81ab ± 1,50 7,22ab ± 1,30 6,55a ± 1,55 8,53b ± 1,96 * ns ns *
pH (Adimensional) 6,10ab ± 0,19 6,13b ± 0,18 5,94a ± 0,02 6,06ab ± 0,17 ns * ns *
mg MDA/Kg 0,23ab ± 0,05 0,24ab ± 0,07 0,31b ± 0,15 0,21a ± 0,07 ns ns * ns
(f): extracto fresco; (s): extracto seco; (s,dg): extracto seco y desengrasado *** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
94
Composición de la grasa intramuscular
La fracción de triglicéridos (LN) fue la fracción mayoritaria de la grasa
intramuscular en los cuatro lotes estudiados, con un porcentaje del 83,6% para el lote
AAH, 85,4% para el AAM, 89,8% para el AGH y de 88,3% para el lote AGM de la grasa total.
La fracción de fosfolípidos (LP) fue la fracción minoritaria con un 0,8, 0,6, 0,7 y 0,5%
respectivamente. La fracción de AGL representó el 15,6, 13,9, 9,5 y 11,1%
respectivamente.
Fracción de lípidos neutros
La tabla 4.1.31 muestra el contenido en mg/g de ácido graso y la tabla 4.1.32 el
porcentaje de ácidos grasos identificados en la fracción de triglicéridos (LN) de la grasa
intramuscular extraída de los jamones curados.
Existieron diferencias significativas en la mayoría de los ácidos grasos, y por lo
tanto, en los sumatorios y contenido total, cuatro de ellos como consecuencia del efecto
sexo, nueve por el efecto genético y el resto por el efecto de ambos factores. El contenido
total presentó diferencias debidas el efecto de los dos factores y su interacción, siendo
superior el valor del lote AGM.
En cuanto a los distintos sumatorios, tan solo el contenido de AGS mostró una
influencia por los dos factores, siendo el lote AGM el que mayor contenido de esta fracción
presentó, coincidiendo con el mayor contenido de este lote de los ácidos grasos
mayoritarios C16:0 y C18:0 y el contenido total. El resto de sumatorios, AGMI, AGPI, ω-3 y
ω-6 mostraron una clara influencia por el efecto genético, siendo superiores los valores de
los lotes AG, coincidiendo también con el mayor contenido de los ácidos monoinsaturados
mayoritarios, C16:1 y C18:1, que mostraron ambos lotes (AG).
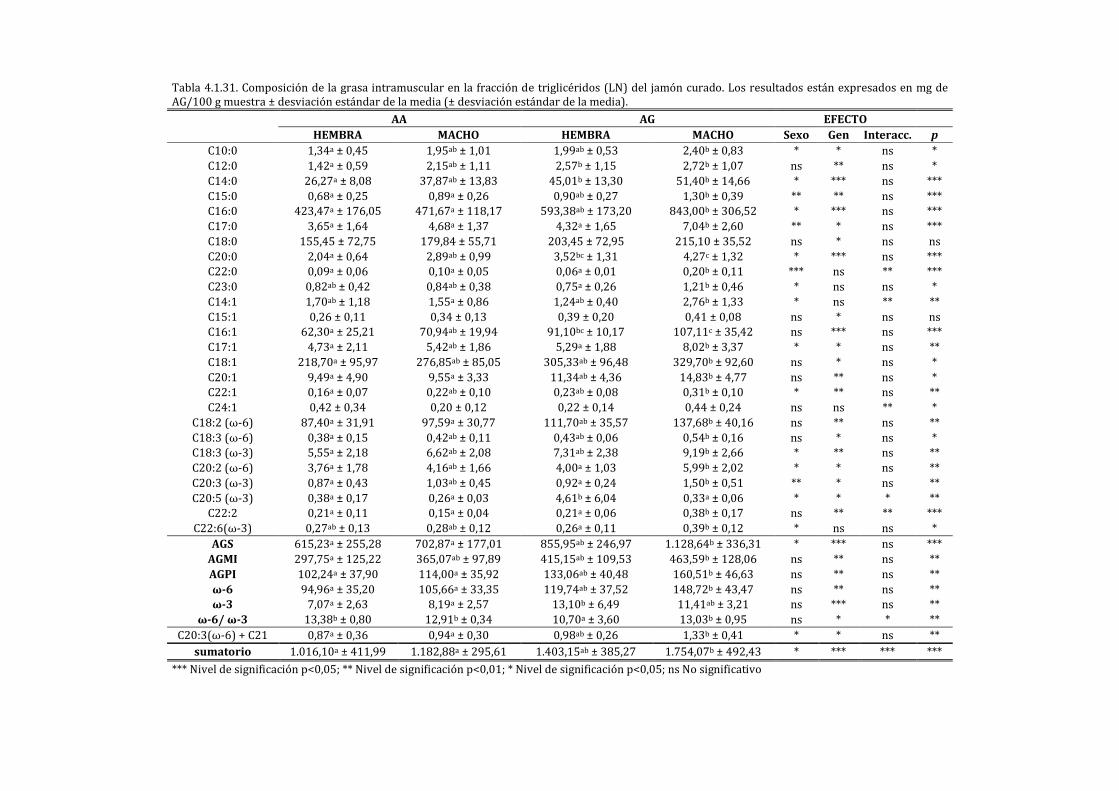
Tabla 4.1.31. Composición de la grasa intramuscular en la fracción de triglicéridos (LN) del jamón curado. Los resultados están expresados en mg de AG/100 g muestra ± desviación estándar de la media (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
C10:0 1,34a ± 0,45 1,95ab ± 1,01 1,99ab ± 0,53 2,40b ± 0,83 * * ns *
C12:0 1,42a ± 0,59 2,15ab ± 1,11 2,57b ± 1,15 2,72b ± 1,07 ns ** ns *
C14:0 26,27a ± 8,08 37,87ab ± 13,83 45,01b ± 13,30 51,40b ± 14,66 * *** ns ***
C15:0 0,68a ± 0,25 0,89a ± 0,26 0,90ab ± 0,27 1,30b ± 0,39 ** ** ns *** C16:0 423,47a ± 176,05 471,67a ± 118,17 593,38ab ± 173,20 843,00b ± 306,52 * *** ns ***
C17:0 3,65a ± 1,64 4,68a ± 1,37 4,32a ± 1,65 7,04b ± 2,60 ** * ns ***
C18:0 155,45 ± 72,75 179,84 ± 55,71 203,45 ± 72,95 215,10 ± 35,52 ns * ns ns
C20:0 2,04a ± 0,64 2,89ab ± 0,99 3,52bc ± 1,31 4,27c ± 1,32 * *** ns *** C22:0 0,09a ± 0,06 0,10a ± 0,05 0,06a ± 0,01 0,20b ± 0,11 *** ns ** ***
C23:0 0,82ab ± 0,42 0,84ab ± 0,38 0,75a ± 0,26 1,21b ± 0,46 * ns ns *
C14:1 1,70ab ± 1,18 1,55a ± 0,86 1,24ab ± 0,40 2,76b ± 1,33 * ns ** **
C15:1 0,26 ± 0,11 0,34 ± 0,13 0,39 ± 0,20 0,41 ± 0,08 ns * ns ns C16:1 62,30a ± 25,21 70,94ab ± 19,94 91,10bc ± 10,17 107,11c ± 35,42 ns *** ns ***
C17:1 4,73a ± 2,11 5,42ab ± 1,86 5,29a ± 1,88 8,02b ± 3,37 * * ns **
C18:1 218,70a ± 95,97 276,85ab ± 85,05 305,33ab ± 96,48 329,70b ± 92,60 ns * ns *
C20:1 9,49a ± 4,90 9,55a ± 3,33 11,34ab ± 4,36 14,83b ± 4,77 ns ** ns * C22:1 0,16a ± 0,07 0,22ab ± 0,10 0,23ab ± 0,08 0,31b ± 0,10 * ** ns **
C24:1 0,42 ± 0,34 0,20 ± 0,12 0,22 ± 0,14 0,44 ± 0,24 ns ns ** *
C18:2 (ω-6) 87,40a ± 31,91 97,59a ± 30,77 111,70ab ± 35,57 137,68b ± 40,16 ns ** ns **
C18:3 (ω-6) 0,38a ± 0,15 0,42ab ± 0,11 0,43ab ± 0,06 0,54b ± 0,16 ns * ns * C18:3 (ω-3) 5,55a ± 2,18 6,62ab ± 2,08 7,31ab ± 2,38 9,19b ± 2,66 * ** ns **
C20:2 (ω-6) 3,76a ± 1,78 4,16ab ± 1,66 4,00a ± 1,03 5,99b ± 2,02 * * ns **
C20:3 (ω-3) 0,87a ± 0,43 1,03ab ± 0,45 0,92a ± 0,24 1,50b ± 0,51 ** * ns **
C20:5 (ω-3) 0,38a ± 0,17 0,26a ± 0,03 4,61b ± 6,04 0,33a ± 0,06 * * * ** C22:2 0,21a ± 0,11 0,15a ± 0,04 0,21a ± 0,06 0,38b ± 0,17 ns ** ** ***
C22:6(ω-3) 0,27ab ± 0,13 0,28ab ± 0,12 0,26a ± 0,11 0,39b ± 0,12 * ns ns *
AGS 615,23a ± 255,28 702,87a ± 177,01 855,95ab ± 246,97 1.128,64b ± 336,31 * *** ns ***
AGMI 297,75a ± 125,22 365,07ab ± 97,89 415,15ab ± 109,53 463,59b ± 128,06 ns ** ns **
AGPI 102,24a ± 37,90 114,00a ± 35,92 133,06ab ± 40,48 160,51b ± 46,63 ns ** ns **
ω-6 94,96a ± 35,20 105,66a ± 33,35 119,74ab ± 37,52 148,72b ± 43,47 ns ** ns ** ω-3 7,07a ± 2,63 8,19a ± 2,57 13,10b ± 6,49 11,41ab ± 3,21 ns *** ns **
ω-6/ ω-3 13,38b ± 0,80 12,91b ± 0,34 10,70a ± 3,60 13,03b ± 0,95 ns * * **
C20:3(ω-6) + C21 0,87a ± 0,36 0,94a ± 0,30 0,98ab ± 0,26 1,33b ± 0,41 * * ns **
sumatorio 1.016,10a ± 411,99 1.182,88a ± 295,61 1.403,15ab ± 385,27 1.754,07b ± 492,43 * *** *** ***
*** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo
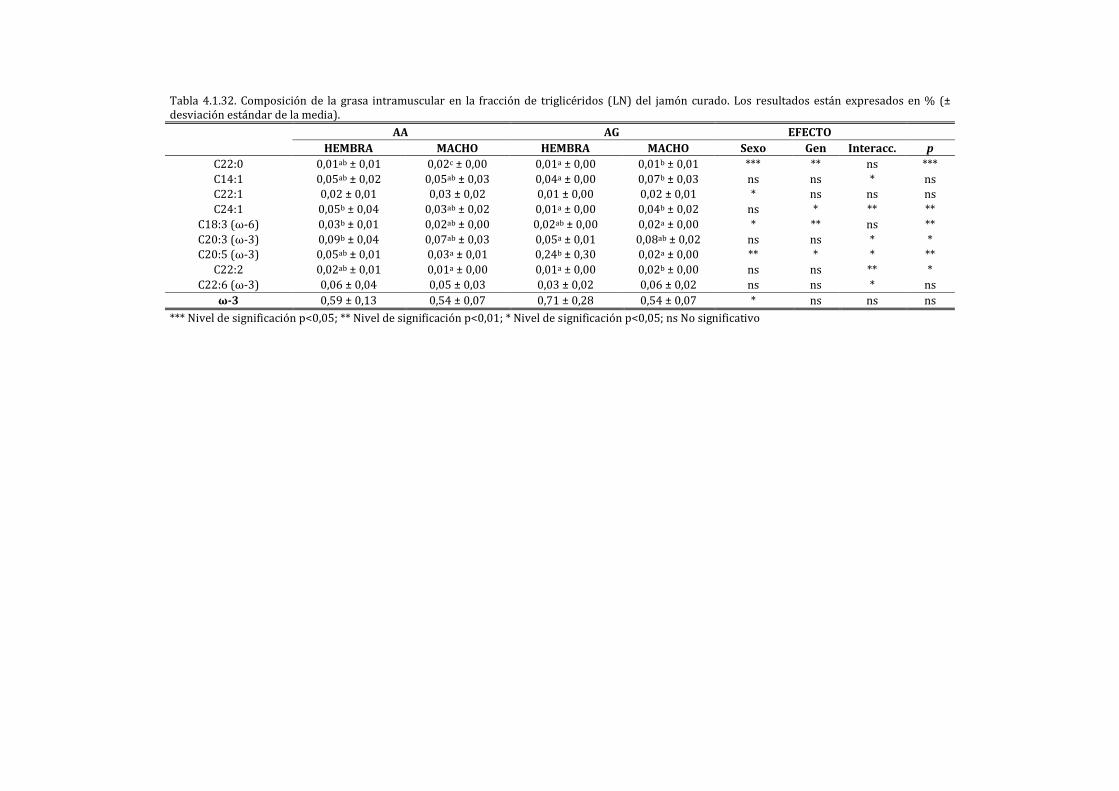
Tabla 4.1.32. Composición de la grasa intramuscular en la fracción de triglicéridos (LN) del jamón curado. Los resultados están expresados en % (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
C22:0 0,01ab ± 0,01 0,02c ± 0,00 0,01a ± 0,00 0,01b ± 0,01 *** ** ns ***
C14:1 0,05ab ± 0,02 0,05ab ± 0,03 0,04a ± 0,00 0,07b ± 0,03 ns ns * ns C22:1 0,02 ± 0,01 0,03 ± 0,02 0,01 ± 0,00 0,02 ± 0,01 * ns ns ns
C24:1 0,05b ± 0,04 0,03ab ± 0,02 0,01a ± 0,00 0,04b ± 0,02 ns * ** **
C18:3 (ω-6) 0,03b ± 0,01 0,02ab ± 0,00 0,02ab ± 0,00 0,02a ± 0,00 * ** ns **
C20:3 (ω-3) 0,09b ± 0,04 0,07ab ± 0,03 0,05a ± 0,01 0,08ab ± 0,02 ns ns * * C20:5 (ω-3) 0,05ab ± 0,01 0,03a ± 0,01 0,24b ± 0,30 0,02a ± 0,00 ** * * **
C22:2 0,02ab ± 0,01 0,01a ± 0,00 0,01a ± 0,00 0,02b ± 0,00 ns ns ** *
C22:6 (ω-3) 0,06 ± 0,04 0,05 ± 0,03 0,03 ± 0,02 0,06 ± 0,02 ns ns * ns
ω-3 0,59 ± 0,13 0,54 ± 0,07 0,71 ± 0,28 0,54 ± 0,07 * ns ns ns
*** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
97
En cuanto a los valores expresados en % (tabla 4.1.32), las diferencias fueron
menos marcadas entre los lotes en comparación con la tabla anterior, donde los resultados
estaban influenciados por el contenido de grasa intramuscular de cada lote. Tan solo
existieron diferencias en la proporción de ω-3 debidas a la influencia del sexo, siendo
superiores las proporciones de los lotes pertenecientes a las hembras, al comparar dentro
de una misma genética.
El resto de ácidos que mostraron diferencias fueron minoritarios, por lo que el
efecto de ambos factores no fue muy marcado.
Fracción de lípidos polares
El análisis del perfil de fosfolípidos (LP) de la grasa intramuscular se muestra en la
tabla 4.1.33. De los ácidos grasos que mostraron diferencias significativas, cuatro de ellos
fueron como consecuencia del efecto sexo, dieciséis por el efecto genético y dos por el
efecto de ambos factores. Además de existir diferencias significativas en la mayor parte de
los ácidos grasos que componen la fracción, también existieron diferencias en los distintos
sumatorios, excepto en el contenido de AGPI. El contenido de AGS, AGMI y ω-3 mostraron
diferencias debidas al efecto genético, siendo superior el contenido de los lotes AG. Tan
solo el contenido de ω-6 mostró diferencias por el efecto sexo, siendo superiores los
valores de los lotes pertenecientes a las hembras.

Tabla 4.1.33. Composición de la grasa intramuscular en la fracción de fosfolípidos (LP) del jamón curado. Los resultados están expresados en mg de AG/100 g muestra (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
C10:0 0,03a ± 0,01 0,025a ± 0,009 0,03ab ± 0,01 0,04b ± 0,01 ns ** ns *
C12:0 0,03a ± 0,01 0,04ab ± 0,02 0,05b ± 0,02 0,05b ± 0,01 ns ** ns ** C14:0 0,24 ± 0,17 0,32 ± 0,19 0,33 ± 0,19 0,17 ± 0,05 ns ns * ns
C15:0 0,03a ± 0,01 0,04ab ± 0,01 0,05b ± 0,01 0,06b ± 0,02 ns ** ns **
C16:0 2,07a ± 0,68 2,22ab ± 0,36 2,90ab ± 1,06 2,97b ± 0,90 ns ** ns **
C17:0 0,06 ± 0,02 0,06 ± 0,02 0,08 ± 0,02 0,08 ± 0,03 ns ** ns * C18:0 1,25a ± 0,42 1,57ab ± 0,54 1,90b ± 0,46 1,51ab ± 0,27 ns * * **
C20:0 0,04a ± 0,02 0,05a ± 0,02 0,08b ± 0,02 0,06ab ± 0,02 ns *** ns **
C22:0 0,03ab ± 0,02 0,02a ± 0,01 0,02ab ± 0,02 0,05b ± 0,03 ns ns * *
C23:0 0,11ab ± 0,04 0,08a ± 0,01 0,15b ± 0,07 0,13b ± 0,04 ns ** ns ** C15:1 0,16a ± 0,05 0,15a ± 0,09 0,26b ± 0,12 0,16a ± 0,05 * * ns **
C16:1 0,11a ± 0,05 0,15a ± 0,06 0,28b ± 0,14 0,21ab ± 0,08 ns *** ** ***
C17:1 0,07a ± 0,01 0,08a ± 0,04 0,26b ± 0,20 0,07a ± 0,02 ** ** ** ***
C18:1 0,66 ± 0,32 0,65 ± 0,07 0,82 ± 0,20 0,81 ± 0,28 ns * ns ns C20:1 0,04a ± 0,02 0,04a ± 0,01 0,08b ± 0,03 0,05a ± 0,02 ** *** ns ***
C22:1 0,03ab ± 0,02 0,03ab ± 0,01 0,02a ± 0,01 0,05b ± 0,03 ns ns ** *
C24:1 0,03a ± 0,02 0,04ab ± 0,02 0,05ab ± 0,04 0,07b ± 0,04 ns ** ns *
C18:3 (ω-6) 0,02ab ± 0,01 0,02ab ± 0,01 0,01ab ± 0,00 0,02b ± 0,01 * ns ** ** C18:3 (ω-3) 0,04a ± 0,02 0,06ab ± 0,02 0,09bc ± 0,05 0,10c ± 0,02 ns *** ns ***
C20:2 (ω-6) 0,05ab ± 0,02 0,05a ± 0,02 0,06ab ± 0,02 0,07b ± 0,02 ns ** ns *
C20:4 (ω-6) 0,72 ± 0,35 0,49 ± 0,17 0,67 ± 0,28 0,50 ± 0,27 * ns ns ns
C20:3 (ω-3) 0,01a ± 0,01 0,01a ± 0,00 0,01a ± 0,00 0,02b ± 0,01 * ns ** ** C22:2 0,03ab ± 0,02 0,04ab ± 0,02 0,01a ± 0,00 0,07b ± 0,05 ** ns * **
C22:6 (ω-3) 0,04ab ± 0,02 0,03a ± 0,01 0,04ab ± 0,01 0,05b ± 0,01 ns * ns *
AGS 3,88a ± 1,05 4,42ab ± 0,93 5,60b ± 1,56 5,12b ± 1,16 ns ** ns **
AGMI 1,38a ± 0,34 1,44a ± 0,29 2,15b ± 0,54 1,79ab ± 0,33 ns *** ns ***
ω-6 2,60 ± 1,15 1,98 ± 0,50 2,75 ± 0,95 2,20 ± 0,54 * ns ns ns
ω-3 0,22 ± 0,08 0,22 ± 0,04 0,25 ± 0,07 0,28 ± 0,04 ns * ns ns ω-6/ ω-3 11,93 ± 4,19 8,98 ± 1,29 11,50 ± 4,67 8,03 ± 2,13 ** ns ns *
C20:3 (ω-6)+C21 0,07 ± 0,02 0,07 ± 0,02 0,10 ± 0,04 0,08 ± 0,04 ns * ns ns
sumatorio 8,18a ± 2,44 8,17a ± 1,52 10,86b ± 2,71 9,53ab ± 1,69 ns ** ns **
*** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

Tabla 4.1.34. Composición de la grasa intramuscular en la fracción de fosfolípidos (LP) del jamón curado. Los resultados están expresados en % (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
C10:0 0,13 ± 0,03 0,14 ± 0,06 0,11 ± 0,04 0,16 ± 0,07 * ns ns ns
C15:0 0,25 ± 0,04 0,27 ± 0,05 0,29 ± 0,08 0,24 ± 0,06 ns ns * ns C17:0 0,60ab ± 0,10 0,65b ± 0,13 0,53a ± 0,10 0,58ab ± 0,09 ns * ns ns
C22:0 0,64ab ± 0,61 0,84b ± 0,59 0,08a ± 0,02 1,08b ± 0,97 ** ns ns **
C24:0 0,82ab ± 0,67 1,24b ± 1,01 0,09a ± 0,01 1,19b ± 0,79 ** ns ns **
C15:1 2,74b ± 2,25 0,63a ± 0,14 1,31a ± 0,63 0,84a ± 0,18 ** ns * *** C16:1 0,86a ± 0,19 1,27b ± 0,49 1,36b ± 0,43 1,17ab ± 0,28 ns ns ** **
C17:1 1,47b ± 1,08 0,67a ± 0,36 0,99ab ± 0,52 0,47a ± 0,12 ** ns ns **
C22:1 0,52ab ± 0,29 0,91b ± 0,70 0,13a ± 0,01 1,29b ± 0,99 *** ns ns **
C18:2 (ω-6) 18,41b ± 2,82 15,55ab ± 3,19 15,05a ± 2,80 15,10ab ± 2,70 ns * ns * C18:3 (ω-6) 0,16b ± 0,05 0,09a ± 0,06 0,11ab ± 0,03 0,16b ± 0,05 ns ns *** **
C20:2 (ω-6) 0,68 ± 0,13 0,64 ± 0,16 0,55 ± 0,17 0,70 ± 0,19 ns ns * ns
C20:4 (ω-6) 7,65b ± 1,41 5,67a ± 2,03 4,75a ± 1,60 4,32a ± 1,21 * *** ns ***
C20:3 (ω-3) 0,10ab ± 0,04 0,12b ± 0,02 0,06a ± 0,01 0,11b ± 0,05 ** ns ns ** C22:2 0,77ab ± 0,66 0,74ab ± 0,59 0,10a ± 0,03 1,20b ± 0,88 * ns ** **
AGS 37,72ab ± 3,49 40,82b ± 2,82 36,40a ± 4,18 38,66ab ± 3,16 * ns ns * AGPI 30,98b ± 3,48 25,79a ± 4,71 23,46a ± 4,18 24,84a ± 2,86 ns ** ** ***
ω-6 26,90b ± 3,65 21,95a ± 4,90 20,47a ± 4,21 20,28a ± 3,79 ns ** * **
C20:3 (ω-6)+C21 1,10b ± 0,21 0,98ab ± 0,23 0,82a ± 0,24 0,83a ± 0,09 ns ** ns **
*** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
100
Los resultados expresados en porcentaje se muestran en la tabla 4.1.34. Al igual
que ocurría con los triglicéridos (LN), los fosfolípidos mostraron diferencias en menos
ácidos grasos al expresarlos en porcentaje. Tan solo las proporciones de AGS, AGPI y ω-6
mostraron diferencias. Los AGS mostraron una clara influencia por el efecto sexo, siendo
superiores los lotes procedentes de machos y los ω-6 por el efecto genético, siendo
superiores los lotes AA, a diferencia de la tabla 4.1.33, cuyos sumatorios tuvieron una
influencia diferente en ambos casos. La fracción de AGPI también mostró diferencias
atribuidas al efecto genético, siendo superiores los porcentajes de los lotes AA,
especialmente el lote AAH.
Fracción de ácidos grasos libres
A diferencia de las otras dos fracciones, en la composición de la fracción de AGL, las
diferencias existentes entre los ácidos grasos estuvieron más influenciadas por el efecto
sexo que por el efecto genético, tal como se muestra en la tabla 4.1.35. En cuanto a los
sumatorios, tan solo la fracción de AGMI mostró diferencias significativas influidas por el
sexo, siendo superiores los valores de los lotes pertenecientes a los machos. Los ácidos
grasos saturados y monoinsaturados mayoritarios (C16:0, C18:0, C6:1 y C18:1) también se
mostraron claramente influidos por el efecto del sexo, siendo superiores los valores de los
lotes procedentes de machos.
La tabla 4.1.36 muestra las diferencias significativas existentes entre las
proporciones de ácidos grasos dentro de cada lote. El número de ácidos grasos que
mostraron diferencias significativas fue inferior al mostrado en la tabla anterior (tabla
4.1.35), coincidiendo así con los resultados mostrados para las otras dos fracciones
estudiadas, triglicéridos (LN) y fosfolípidos (LP). Tan solo existieron diferencias en dos
ácidos mayoritarios, el C18:0 y C16:1, pero no existieron diferencias en ningún sumatorio.
El C18:0 estuvo influenciado por el efecto genético y por la interacción de ambos factores,
siendo superior el valor de los lotes AA. Por otra parte, el C16:1 mostró una influencia por
la interacción de ambos factores, siendo superior el valor del lote AGH.

Tabla 4.1.35. Composición de la grasa intramuscular en la fracción de AGL del jamón curado. Los resultados están expresados en mg de AG/100 g muestra (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
C12:0 0,17 ± 0,04 0,20 ± 0,06 0,24 ± 0,08 0,22 ± 0,08 ns * ns ns
C16:0 48,53ab ± 16,48 57,54ab ± 19,90 39,49ab ± 15,42 62,75b ± 25,36 ** ns ns *
C18:0 22,58 ± 8,86 24,00 ± 7,04 18,17 ± 8,89 28,49 ± 12,65 * ns ns ns
C20:0 0,19a ± 0,04 0,24ab ± 0,05 0,28b ± 0,10 0,24ab ± 0,05 ns * * * C22:0 0,04 ± 0,02 0,05 ± 0,03 0,07 ± 0,05 0,07 ± 0,03 ns * ns ns
C14:1 0,40 ± 0,27 0,30 ± 0,21 0,24 ± 0,12 0,50 ± 0,32 ns ns * ns
C15:1 0,25ab ± 0,11 0,18a ± 0,08 0,26ab ± 0,13 0,38b ± 0,22 ns * * *
C16:1 4,96 ± 1,47 6,16 ± 2,48 4,53 ± 1,89 6,96 ± 3,97 * ns ns ns C17:1 0,54ab ± 0,23 0,66ab ± 0,36 0,35a ± 0,12 0,79b ± 0,49 ** ns ns *
C18:1 22,27 ± 8,85 27,54 ± 13,23 17,92 ± 9,28 30,63 ± 15,26 * ns ns ns
C22:1 0,05 ± 0,02 0,08 ± 0,04 0,06 ± 0,02 0,09 ± 0,05 ** ns ns *
C20:3 (ω-3) 0,20ab ± 0,09 0,18ab ± 0,06 0,11a ± 0,03 0,26b ± 0,14 * ns ** ** C20:5 (ω-3) 0,66a ± 0,26 0,61a ± 0,13 1,42b ± 0,90 0,73a ± 0,25 * ** * **
C22:2 0,06ab ± 0,04 0,05a ± 0,03 0,02a ± 0,01 0,10b ± 0,06 * ns *** ***
AGMI 29,64 ± 11,00 36,41 ± 16,82 24,70 ± 11,99 37,61 ± 21,43 * ns ns ns
ω-6/ ω-3 22,74b ± 2,84 21,26b ± 2,34 15,45a ± 6,84 23,19b ± 2,71 * * *** ***
*** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo
Tabla 4.1.36. Composición de la grasa intramuscular en la fracción de AGL del jamón curado. Los resultados están expresados en % (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
C12:0 0,04a ± 0,00 0,04ab ± 0,01 0,05b ± 0,01 0,05ab ± 0,00 ns * ns * C18:0 11,79b ± 0,70 11,56b ± 0,93 10,60a ± 0,92 11,40ab ± 0,69 ns ** * **
C22:0 0,03a ± 0,01 0,03ab ± 0,02 0,03a ± 0,01 0,05b ± 0,03 * ns ns *
C24:0 0,07 ± 0,03 0,09 ± 0,04 0,06 ± 0,00 0,09 ± 0,04 ** ns ns *
C15:1 0,12 ± 0,05 0,08 ± 0,05 0,12 ± 0,07 0,09 ± 0,02 * ns ns ns C16:1 1,64 ± 0,48 1,86 ± 0,50 2,05 ± 0,45 1,65 ± 0,22 ns ns * ns
C20:5 (ω-3) 0,31 ± 0,03 0,34 ± 0,06 0,35 ± 0,02 0,32 ± 0,04 ns ns * ns
*** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
102
Lípidos totales
En la tabla 4.1.37 se presenta la composición de los ácidos grasos presentes en los
lípidos totales de la grasa intramuscular de los cuatro lotes en estudio. Todos los
sumatorios mostraron diferencias significativas, influenciadas sobre todo por el efecto
genético y en alguno de ellos, como los sumatorios de AGS, AGMI y ω-3 también por el
efecto sexo. A pesar de ello, todos los sumatorios fueron superiores en los lotes de genética
AG, independientemente del sexo de los animales.
Los ácidos grasos mayoritarios C16:0 y C18:1 fueron los responsables de la
tendencia de los sumatorios, sin embargo el C18:0 fue superior significativamente en el
lote AGM (donde aquí el efecto sexo si tuvo influencia) y el C16:1 no mantuvo diferencias
entre tres de los cuatro lotes, siendo el lote AAH el único que presentó diferencias, siendo
inferior su valor en comparación con el resto de lotes.
La tabla 4.1.38 muestra los porcentajes de los ácidos grasos presentes en los
lípidos totales de la grasa intramuscular, donde tan solo tres mostraron diferencias
significativas entre los lotes, aún así se trata de ácidos grasos minoritarios, por lo que su
repercusión no fue importante.
El número de ácidos grasos que mostraron diferencias significativas fue inferior al
mostrado en la tabla anterior (tabla 4.1.37), coincidiendo así con los resultados mostrados
para las tres fracciones estudiadas, triglicéridos (LN), fosfolípidos (LP) y AGL.

Tabla 4.1.37. Composición de la grasa intramuscular en la fracción de lípidos totales del jamón curado. Los resultados están expresados en mg de AG/100 g muestra (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
C10:0 1,70a ± 0,74 2,738ab ± 0,671 3,05b ± 0,96 3,20b ± 0,88 ns * ns * C12:0 2,26a ± 0,82 3,40ab ± 1,22 4,04ab ± 1,12 4,15b ± 1,78 ns ** ns *
C14:0 36,50a ± 12,40 53,69ab ± 19,28 64,66ab ± 22,52 65,83b ± 27,61 * ** ns **
C15:0 1,02a ± 0,33 1,53b ± 0,32 1,41ab ± 0,40 1,47b ± 0,48 ** ns ns **
C16:0 457,75a ± 151,13 622,32ab ± 194,69 736,18b ± 217,07 737,66b ± 252,46 * ** ns ** C17:0 4,78a ± 1,74 6,56b ± 2,06 5,98ab ± 1,78 7,02b ± 1,58 ** * ns **
C18:0 182,65a ± 62,22 243,71ab ± 61,67 255,25ab ± 76,05 278,56b ± 91,19 * ** ns *
C20:0 3,07a ± 0,98 3,82ab ± 1,28 5,03b ± 1,59 4,86b ± 1,62 ns *** ns ***
C22:0 0,14a ± 0,04 0,21b ± 0,09 0,22ab ± 0,07 0,19ab ± 0,06 * ns * * C14:1 1,75a ± 0,62 2,64ab ± 1,23 2,96ab ± 1,00 2,80b ± 1,84 * * ns *
C16:1 60,61a ± 21,87 91,55b ± 31,57 101,59b ± 23,63 99,16b ± 29,05 * ** ns **
C17:1 4,58a ± 1,47 6,86b ± 2,17 6,31ab ± 1,92 7,34b ± 2,34 ** * ns **
C18:1 235,99a ± 78,72 330,52ab ± 126,95 358,26b ± 93,29 356,70b ± 96,74 * ** ns ** C20:1 8,30a ± 2,60 12,30b ± 5,01 12,94b ± 3,76 14,53b ± 3,74 ** ** ns ***
C22:1 0,22 ± 0,08 0,29 ± 0,13 0,28 ± 0,04 0,28 ± 0,05 ns ns * ns
C18:2(ω-6) 166,28a ± 61,37 195,50ab ± 55,96 209,83b ± 28,79 220,71b ± 55,78 ns ** ns **
C18:3(ω-3) 6,82a ± 2,31 9,26b ± 2,93 9,48b ± 2,27 9,23b ± 1,72 * * * ** C20:2 (ω-6) 4,22a ± 1,18 5,63ab ± 1,89 5,37ab ± 1,68 6,14b ± 1,68 * * ns *
C20:3(ω-3) 0,90a ± 0,24 1,09ab ± 0,28 1,37b ± 0,33 1,44b ± 0,35 * *** ns ***
C20:5(ω-3) 0,94 ± 0,28 1,17 ± 0,36 1,14 ± 0,21 1,24 ± 0,23 * ns ns *
C22:2 0,18a ± 0,04 0,31b ± 0,13 0,27b ± 0,09 0,24ab ± 0,06 ns ns ** **
AGS 692,14a ± 219,73 940,69ab ± 254,05 1.078,67b ± 317,29 1.105,98b ± 370,34 * ** ns **
AGMI 312,46a ± 100,45 445,61b ± 161,58 483,81b ± 121,28 482,60b ± 129,50 * ** ns ** AGPI 206,78a ± 72,49 242,70ab ± 67,66 257,92b ± 33,81 269,57b ± 60,50 ns ** ns **
ω-6 197,06a ± 70,01 229,82ab ± 64,08 244,68b ± 31,91 256,44b ± 59,41 ns ** ns *
ω-3 9,54a ± 2,72 12,57b ± 3,59 12,96b ± 2,69 12,89b ± 2,01 * * * **
sumatorio 1.214,88a ± 373,74 1.632,84ab ± 450,69 1.824,52b ± 448,86 1.862,41b ± 534,92 * ** ns **
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

Tabla 4.1.38. Composición de la grasa intramuscular en la fracción de lípidos totales del jamón curado. Los resultados están expresados en % (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
C20:0 0,09 ± 0,01 0,09 ± 0,01 0,10 ± 0,02 0,10 ± 0,01 ns * ns ns C15:1 0,06b ± 0,04 0,05ab ± 0,01 0,06ab ± 0,03 0,03a ± 0,01 ** ns ns *
C18:3(ω-6) 0,08 ± 0,01 0,07 ± 0,01 0,07 ± 0,01 0,07 ± 0,01 ns * ns ns
** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
105
Determinación del contenido en aminoácidos libres
La tabla 4.1.39 muestra las diferencias significativas en el contenido total e
individual de la composición de aminoácidos libres analizada en el jamón curado.
Al estudiar el efecto de los dos factores, sexo y genética, en la composición de
aminoácidos libres vemos claramente como la genética tuvo un efecto más marcado, ya
que el sexo solo tuvo influencia en cuatro aminoácidos en conjunto con la genética. Se
observó como los lotes pertenecientes a la genética AA presentaron mayores cantidades
de aminoácidos libres y contenido total, siendo en algunos de los casos el lote
perteneciente a la hembra el que mostró mayores cantidades.
Determinación de los compuestos nitrogenados derivados del ATP
A diferencia de los resultados obtenidos en la tabla 4.1.24, al estudiar el efecto de
ambos factores, existieron diferencias significativas en todos los nucleótidos analizados
(tabla 4.1.40). El efecto sexo fue el causante de las difererencias generadas entre lotes,
observándose mayores valores en los lotes pretenecientes a los machos, aunque el factor
genético tuvo influencia sobre algún nucleótido, como la IMP y la Ino, destacando los
valores de los lotes pertenecientes a la genética AA.

Tabla 4.1.39. Contenido en aminoácidos libres en jamón curado en extracto fresco. Los resultados se expresan en mg aa/100 g muestra (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Inter p
Asp 294,09b ± 48,33 299,07b ± 18,88 212,61a ± 20,31 237,16a ± 50,36 ns *** ns *** Glu 561,52b ± 83,21 575,66b ± 50,76 467,33a ± 8,16 504,58ab ± 101,59 ns *** ns **
Ser-Asp 398,26b ± 69,22 375,11b ± 36,57 285,74a ± 34,79 333,85ab ± 80,35 ns *** ns ***
Gly-Gln 379,76b ± 50,09 370,14b ± 40,39 308,28a ± 10,30 340,99ab ± 66,47 ns ** ns **
Hys 234,97c ± 30,76 229,08bc ± 17,55 193,74a ± 6,26 204,50ab ± 33,72 ns *** ns ** Trh 115,27b ± 25,91 97,01ab ± 14,04 90,62a ± 14,20 93,96a ± 12,15 ns * * **
Pro-Arg 2.024,81b ± 191,44 2.113,08b ± 82,37 1.798,35a ± 42,82 1.918,80ab ± 271,35 ns *** ns **
Met 110,70b ± 11,81 104,97b ± 10,81 104,35b ± 11,58 90,52a ± 12,89 ** ** ns **
Iso 265,43c ± 18,33 239,88bc ± 31,63 217,78ab ± 16,77 210,31a ± 25,47 * *** ns *** Leu 492,87c ± 38,41 456,48bc ± 60,78 418,02ab ± 36,42 399,94a ± 57,31 ns *** ns ***
Phe 260,69c ± 21,03 239,25bc ± 26,22 213,84ab ± 22,24 197,74a ± 26,46 * *** ns ***
Trp 79,16c ± 8,54 71,95bc ± 7,83 66,69ab ± 4,47 62,97a ± 3,96 ** *** ns ***
Lys 745,34b ± 87,11 657,74ab ± 83,53 591,18a ± 47,69 590,31a ± 74,81 ns *** ns ***
sumatorio (f) 7.497,84c ± 833,34 7.449,59bc ± 427,05 6.491,80a ± 165,09 6.715,52ab ± 934,64 ns *** ns ***
sumatorio (s) 15.052,31bc ± 1.745,42 15.209,75c ± 934,59 13.079,37a ± 489,45 13.733,52ab ± 1.320,79 ns *** ns **
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo
Tabla 4.1.40. Contenido en 5´nucleótidos del jamón curado en extracto fresco. Los resultados se expresan en mg de nuc/100 g muestra (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
IMP 10,20c ± 3,44 13,88d ± 0,91 2,63a ± 0,24 5,70b ± 2,85 *** *** ns ***
GMP 14,60a ± 6,41 21,50b ± 4,42 19,42b ± 1,32 19,71b ± 1,26 ** ns ** **
ADP 8,99 ± 1,49 11,56 ± 5,22 11,62 ± 2,53 8,21 ± 2,59 ns ns ** *
AMP-Hx 96,27a ± 25,71 131,97b ± 25,85 111,03ab ± 11,38 123,75b ± 8,78 *** ns ns *** Ino 29,96b ± 5,52 26,29ab ± 10,03 26,33ab ± 2,57 20,77a ± 8,38 * * ns *
sumatorio (f) 160,02a ± 24,64 205,19b ± 41,28 171,04a ± 16,00 178,15ab ± 8,22 ** ns ** ** sumatorio (s) 320,03a ± 46,18 417,01b ± 80,60 353,95a ± 38,12 366,46ab ± 27,56 ** ns * **
(f): extracto fresco; (s): extracto seco *** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
107
Determinación de compuestos volátiles
De los 113 compuestos volátiles detectados en los jamones (tabla 4.1.10), 59
presentaron diferencias significativas entre los lotes del estudio. Sin embargo, es difícil
explicar dichas diferencias en base a los efectos achacables a los distintos factores
considerados. Individualmente, las diferencias observadas no muestran un
comportamiento que se pueda normalizar, por esta razón hemos preferido representar
una visión conjunta que recoja los sumatorios de cada familia (figura 4.1.11 y 4.1.12). Esta
representación permite apreciar que solo existieron diferencias significativas en algunas
familias, como los compuestos azufrados, los ésteres y los furanos, donde en todos los
casos el efecto genético fue el más marcado. El contenido total de compuestos volátiles, a
pesar de que no aparece representado en las figuras, no presentó diferencias significativas
entre los lotes.
Algunos de los compuestos a destacar son la 2,3-butanodiona que fue superior en
los dos lotes pertenecientes a los machos y el 2-propanol en los pertenecientes a las
hembras. Sin embargo, el efecto genético fue más marcado destacando el 2-pentanol y el
propanoato de etilo en los lotes AA y el ácido decanoico, el 1H-pirrol, el etilbenceno y el l-
limoneno en los lotes AG.
0
50
100
150
200
250
AAH
AAM
AGH
AGM
**
Figura 4.1.11. Diagrama de barras correspondientes a la abundancia total de las
principales familias químicas, expresadas en UAA (106) en los lotes de jamones estudiados (** Nivel de significación p<0,01)

CAPÍTULO IV: RESULTADOS
108
02468
101214161820
AAH
AAM
AGH
AGM
*
Figura 4.1.12. Diagrama de barras correspondientes a la abundancia total de las principales familias químicas, expresadas en UAA (106) en los lotes de jamones estudiados (** Nivel de significación p<0,01; * Nivel de significación p<0,05)
Evaluación sensorial
De las 19 características sensoriales evaluadas, 12 presentaron diferencias
significativas entre los lotes estudiados (tabla 4.1.42). Destacaron los elevados parámetros
de los lotes pertenecientes a los machos, especialmente el AGM, como son el veteado, la
fluidez de la grasa, la dureza del magro y el sabor dulce; por otra parte, la persistencia del
flavor y el color amarillo de la grasa fueron superiores significativamente en el lote AAH.
Por último, destacó la elevada pastosidad del lote AGH y las menores puntuaciones en
general del lote AAH. No existió una clara influencia por ninguno de los dos factores en
particular, contribuyendo ambos de forma más general.

CAPÍTULO IV: RESULTADOS
109
0
1
2
3
4
5
6AMARILLO GRASA
ROSA GRASA
ROJO MAGRO
BRILLO MAGRO
VETEADO
INTENSIDAD OLOR
DUREZA GRASA
FLUIDEZ GRASA
DUREZA MAGRO
SEQUEDAD JUGOSIDAD
PASTOSIDAD
SALADO
DULCE
AMARGO
INTENSIDAD FLAVOR
PERSISTENCIA
CURADO
RANCIDEZ
AAH
AAM
AGH
AGM
Figura 4.1.11. Perfil sensorial del jamón curado
El gráfico de araña muestra claramente como el lote AGM presentó un mayor
veteado y fluidez de la grasa, sin embargo el lote AAM mostró una mayor intensidad de
olor y persistencia, aunque las diferencias con el resto de lotes fueron visiblemente
pequeñas. El resto de lotes no mostró ninguna diferencia pronunciada.

Tabla 4.1.42. Análisis cuantitativo-descriptivo del jamón curado (± desviación estándar de la media)
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
Color grasa Amarillo 1,11a ± 0,33 1,80b ± 0,88 1,14a ± 0,31 1,46ab ± 0,50 ** ns ns *
Color magro Veteado 3,66ab ± 0,66 3,13a ± 0,50 2,86a ± 0,85 4,51b ± 0,96 * ns *** ***
Textura grasa Fluidez 3,75a ± 0,67 4,52bc ± 0,36 4,01ab ± 0,48 5,12c ± 0,70 *** * ns ***
Textura magro
Dureza 2,43ab ± 0,79 2,16a ± 0,46 2,58ab ± 0,86 2,98b ± 0,37 ns * ns *
Sequedad 2,98 ± 0,81 3,56 ± 1,03 2,65 ± 0,78 3,17 ± 0,75 * ns ns ns
Pastosidad 4,69ab ± 1,48 4,06a ± 0,88 5,44b ± 1,20 3,79a ± 1,11 ** ns ns *
Sabor
Salado 4,88ab ± 0,37 4,94b ± 0,49 5,12b ± 0,50 4,50a ± 0,41 ns ns ** **
Dulce 1,15b ± 0,23 0,94ab ± 0,21 0,68a ± 0,11 0,95b ± 0,33 ns ** ** **
Amargo 1,10 ± 0,27 1,04 ± 0,28 0,90 ± 0,19 0,95 ± 0,19 ns * ns ns
Flavor Rancidez 4,56 ± 0,33 4,96 ± 0,43 4,69 ± 0,76 4,48 ± 0,41 ns ns * ns
Persistencia 3,42ab ± 0,71 3,87b ± 0,35 3,18a ± 0,78 3,33ab ± 0,55 ns * ns *
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
111
4.1.4. INFLUENCIA DE LA EXPRESIÓN DEL IGF2 EN FUNCIÓN DE LA CAMPAÑA SOBRE LAS CARACTERÍSTICAS FÍSICO-QUÍMICAS Y SENSORIALES DEL JAMÓN CURADO
En este apartado se va a evaluar la influencia del QTL (Quantitative Trait Loci)
IGF2 (insuline-like growth factor 2), del sexo y la campaña sobre algunos parámetros
físico-químicos en jamones procedentes de cerdos de distintas generaciones. Para ello se
evaluarán las diferencias existentes entre tres lotes, el primero perteneciente a los cerdos
de la primera generación y dos lotes más, que corresponden con las dos generaciones
posteriores. A cada lote se le denominará, primera, segunda y tercera campaña.
Características generales del jamón curado
Las características generales del jamón curado mostraron diferencias significativas
en los cuatro parámetros estudiados (tablas 4.1.43 y 4.1.44). Estas diferencias estuvieron
influenciadas claramente por el efecto campaña, ya que como se vio en el apartado
anterior, al evaluar el efecto de la genética y del sexo, no existieron diferencias
significativas en ningún parámetro morfológico, llegando a la conclusión de que la
selección de los cerdos, aunque presentaba diferencias en la materia prima achacables a la
genética o al sexo, estas diferencias se perdían durante el proceso de curación o eran
enmascaradas por las diferencias entre campañas. Por lo general, los jamones de la tercera
campaña resultaron los más grandes, en especial los pertenecientes al lote AGH, ya que los
cerdos pertenecientes a esta campaña presentaron mayores pesos de sacrificio, generando
por lo tanto jamones de mayor tamaño. Sin embargo, los jamones pertenecientes a la
segunda campaña resultaron ser los más pequeños, especialmente los pertenecientes al
lote AAH, a pesar de que presentaron altas medidas de longitud.

Tabla 4.1.43. Medidas morfológicas generales del jamón curado (± desviación estándar de la media). Parte I.
J1 J2
AA AG AA AG
HEMBRA MACHO HEMBRA MACHO HEMBRA MACHO HEMBRA MACHO
Peso (G) 8,5bc ± 0,4 7,7abc ± 0,3 7,5ab ± 0,1 8,3bc ± 0,6 7,1a ± 0,3 8,0abc ± 0,1 7,9abc ± 0,2 7,7abc ± 0,4
Longitud (cm) 74,2abc ± 0,2 76,2cdef ± 0,2 75,3abcde ± 1,8 74,9abcd ± 0,4 78,0f ± 0,6 75,7bcdef ± 0,4 75,3abcde ± 1,1 73,3ab ± 1,5
Anchura (cm) 29,2cd ± 0,2 29,2cd ± 0,2 28,0bcd ± 1,6 28,0bcd ± 0,0 25,9a ± 0,8 28,2bcd ± 0,2 27,2ab ± 0,7 29,0bcd ± 0,0 Perímetro (cm) 66,9b ± 0,9 65,3ab ± 2,8 63,5ab ± 0,4 66,9b ± 0,9 62,1a ± 0,4 64,1ab ± 0,1 63,5ab ± 1,5 64,9ab ± 0,6
Tabla 4.1.44. Medidas morfológicas generales del jamón curado (± desviación estándar de la media). Parte II.
J3
AA AG
EFECTO
HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
Peso (G) 8,0abc ± 0,0 7,4ab ± 0,9 8,7c ± 0,0 8,3bc ± 0,4 * ns * *** ***
Longitud (cm) 77,0def ± 0,8 73,0a ± 0,0 78,0f ± 0,0 77,3ef ± 0,4 ns *** ** ** *** Anchura (cm) 29,0bcd ± 0,0 27,5abc ± 0,5 29,5d ± 0,0 28,5bcd ± 1,1 ns ns *** ns ***
Perímetro (cm) 66,0b ± 0,0 66,1b ± 2,4 67,0b ± 0,0 64,7ab ± 0,6 ns ns *** ** ***
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
113
El corte transversal de los jamones mostró diferencias significativas, en cuanto al
espesor de la grasa subcutánea, en la zona proximal (figura 4.1.12), presentando
diferencias significativas en las tres medidas tomadas en esa zona, tal y como se muestra
en las tablas 4.1.45 y 4.1.46. Por lo general, los jamones de la segunda campaña tuvieron
un mayor espesor de la grasa subcutánea en el punto C, influenciado únicamente por el
efecto genético, pero los de la tercera fueron superiores significativamente en los otros
dos puntos, influenciados por el efecto genético y además por el efecto campaña. No
obstante, al hacer un promedio de todas las medidas de la zona proximal se vió como los
jamones de la tercera campaña mostraron un mayor espesor de grasa subcutánea (3,5
cm), seguido de los jamones de la segunda campaña (3,3 cm) y finalmente los de la
primera (3,0 cm).
Figura 4.1.12. Zona proximal del jamón curado
La zona distal también mostró diferencias significativas entre las tres campañas
(tablas 4.1.47 y 4.1.48). Por lo general, los jamones de la primera campaña mostraron un
mayor espesor en el punto A, y los de la tercera en los otros dos puntos. Sin embargo, al
hacer las medias de nuevo se observaron resultados distintos a los de la zona proximal.
Los jamones de la segunda campaña mostraron un mayor espesor (4,5 cm), seguido de los
jamones de la segunda (4,2 cm) y después los de la tercera (4,1 cm).
Figura 4.1.13. Zona distal del jamón curado

4.1.45. Espesor de la grasa subcutánea de la zona proximal del jamón curado (± desviación estándar de la media). Parte I.
J1 J2
AA AG AA AG
HEMBRA MACHO HEMBRA MACHO HEMBRA MACHO HEMBRA MACHO
A (cm) 2,5bcde ± 0,5 1,9ab ± 0,1 2,2abc ± 0,2 2,4abcd ± 0,3 1,3a ± 0,3 1,5ab ± 0,3 2,6bcde ± 0,7 3,1cde ± 0,1
B (cm) 2,3abcd ± 0,5 2,3abcd ± 0,3 2,7bcd ± 0,1 2,9cd ± 0,6 1,4a ± 0,3 1,4a ± 0,3 2,0abc ± 0,3 2,0abc ± 0,0
C (cm) 4,7ab ± 0,2 5,0ab ± 0,5 4,5ab ± 0,4 5,5b ± 0,7 4,4ab ± 0,2 5,2b ± 0,2 5,5b ± 0,0 5,7b ± 0,5
4.1.46. Espesor de la grasa subcutánea de la zona proximal del jamón curado (± desviación estándar de la media). Parte II.
J3
AA AG
EFECTO
HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
A (cm) 1,9ab ± 0,1 2,4abcde ± 0,1 3,5e ± 0,0 3,4de ± 0,6 *** ns ** ns ***
B (cm) 2,7bcd ± 0,2 1,8ab ± 0,3 4,2e ± 0,0 3,2d ± 0,2 *** * *** ns ***
C (cm) 5,5b ± 0,4 3,6a ± 1,6 4,5ab ± 0,0 5,1b ± 0,3 * ns ns ** **
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo
4.1.47. Espesorde la grasa subcutánea de la zona distal del jamón curado (± desviación estándar de la media). Parte I.
J1 J2
AA AG AA AG
HEMBRA MACHO HEMBRA MACHO HEMBRA MACHO HEMBRA MACHO
A (cm) 6,4abc ± 0,4 6,6abc ± 0,4 8,3bc ± 0,6 6,4abc ± 0,6 8,5bc ± 0,9 7,5bc ± 1,6 7,0abc ± 2,4 7,7bc ± 0,8 B (cm) 1,7ab ± 0,1 2,1bc ± 0,2 2,5c ± 0,0 2,6c ± 0,4 2,0bc ± 0,0 2,0bc ± 0,0 2,0bc ± 0,3 2,3bc ± 0,6
C (cm) 3,9abc ± 0,1 2,9ab ± 0,2 3,0ab ± 0,1 3,2abc ± 0,5 3,8abc ± 0,2 3,5abc ± 0,7 2,9a ± 0,4 5,0bc ± 1,9
4.1.48. Espesor de la grasa subcutánea de la zona distal del jamón curado (± desviación estándar de la media). Parte II.
J3
AA AG
EFECTO
HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
A (cm) 3,8a ± 1,4 9,5c ± 2,0 8,0bc ± 0,0 5,8ab ± 1,0 ns ns ns *** ** B (cm) 2,1bc ± 0,2 1,2a ± 0,2 2,5c ± 0,0 2,5c ± 0,1 ns ns *** * *** C (cm) 3,0ab ± 0,2 3,1ab ± 0,5 2,5a ± 0,0 5,3c ± 1,3 ns * ns ns **
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
115
Parámetros generales físico-químicos
En cuanto a los parámetros generales, se encontraron diferencias significativas en
todos los parámetros evaluados, tal como se muestra en las tablas 4.1.49 y 4.1.50.
Las tablas muestran como los factores campaña y sexo ejercieron una interacción
significante sobre todos los parámetros excepto sobre el contenido de grasa
intramuscular, cuyas diferencias se vieron afectadas únicamente por el factor genético. Los
jamones de la primera campaña, especialmente los pertenecientes al lote AGH, mostraron
un mayor contenido de grasa intramuscular, a pesar de que los jamones de esta campaña
presentaron menor espesor de la grasa subcutánea tanto en la zona distal como en la
proximal.
La humedad y la coordenada L también fueron superiores significativamente en los
jamones de la primera campaña, especialmente en los pertenecientes al lote AGM. Los
jamones de la segunda campaña presentaron un mayor valor de las coordenadas a* y b*.
Sin embargo, a pesar del elevado valor de la coordenada a* de los jamones de la segunda
campaña, el contenido en mioglobina fue superior en los jamones pertenecientes a la
tercera campaña.
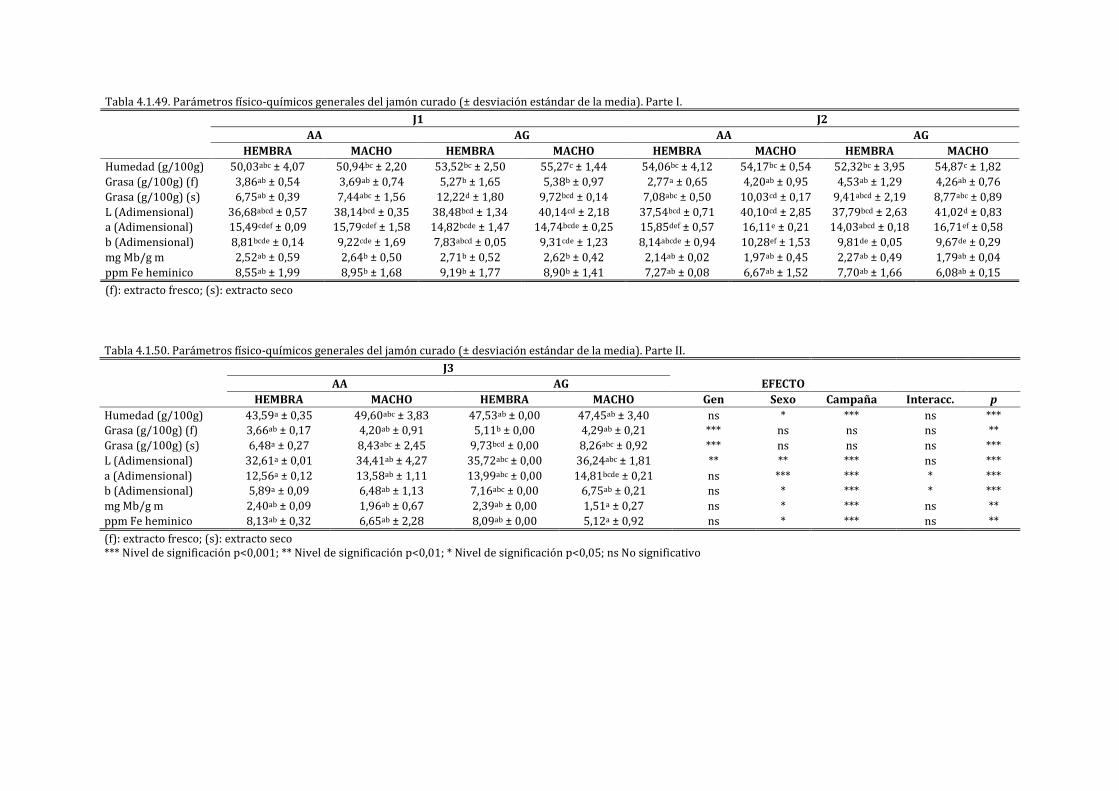
Tabla 4.1.49. Parámetros físico-químicos generales del jamón curado (± desviación estándar de la media). Parte I.
J1 J2
AA AG AA AG
HEMBRA MACHO HEMBRA MACHO HEMBRA MACHO HEMBRA MACHO
Humedad (g/100g) 50,03abc ± 4,07 50,94bc ± 2,20 53,52bc ± 2,50 55,27c ± 1,44 54,06bc ± 4,12 54,17bc ± 0,54 52,32bc ± 3,95 54,87c ± 1,82
Grasa (g/100g) (f) 3,86ab ± 0,54 3,69ab ± 0,74 5,27b ± 1,65 5,38b ± 0,97 2,77a ± 0,65 4,20ab ± 0,95 4,53ab ± 1,29 4,26ab ± 0,76
Grasa (g/100g) (s) 6,75ab ± 0,39 7,44abc ± 1,56 12,22d ± 1,80 9,72bcd ± 0,14 7,08abc ± 0,50 10,03cd ± 0,17 9,41abcd ± 2,19 8,77abc ± 0,89
L (Adimensional) 36,68abcd ± 0,57 38,14bcd ± 0,35 38,48bcd ± 1,34 40,14cd ± 2,18 37,54bcd ± 0,71 40,10cd ± 2,85 37,79bcd ± 2,63 41,02d ± 0,83 a (Adimensional) 15,49cdef ± 0,09 15,79cdef ± 1,58 14,82bcde ± 1,47 14,74bcde ± 0,25 15,85def ± 0,57 16,11e ± 0,21 14,03abcd ± 0,18 16,71ef ± 0,58
b (Adimensional) 8,81bcde ± 0,14 9,22cde ± 1,69 7,83abcd ± 0,05 9,31cde ± 1,23 8,14abcde ± 0,94 10,28ef ± 1,53 9,81de ± 0,05 9,67de ± 0,29
mg Mb/g m 2,52ab ± 0,59 2,64b ± 0,50 2,71b ± 0,52 2,62b ± 0,42 2,14ab ± 0,02 1,97ab ± 0,45 2,27ab ± 0,49 1,79ab ± 0,04
ppm Fe heminico 8,55ab ± 1,99 8,95b ± 1,68 9,19b ± 1,77 8,90b ± 1,41 7,27ab ± 0,08 6,67ab ± 1,52 7,70ab ± 1,66 6,08ab ± 0,15
(f): extracto fresco; (s): extracto seco
Tabla 4.1.50. Parámetros físico-químicos generales del jamón curado (± desviación estándar de la media). Parte II.
J3
AA AG
EFECTO
HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
Humedad (g/100g) 43,59a ± 0,35 49,60abc ± 3,83 47,53ab ± 0,00 47,45ab ± 3,40 ns * *** ns *** Grasa (g/100g) (f) 3,66ab ± 0,17 4,20ab ± 0,91 5,11b ± 0,00 4,29ab ± 0,21 *** ns ns ns **
Grasa (g/100g) (s) 6,48a ± 0,27 8,43abc ± 2,45 9,73bcd ± 0,00 8,26abc ± 0,92 *** ns ns ns ***
L (Adimensional) 32,61a ± 0,01 34,41ab ± 4,27 35,72abc ± 0,00 36,24abc ± 1,81 ** ** *** ns ***
a (Adimensional) 12,56a ± 0,12 13,58ab ± 1,11 13,99abc ± 0,00 14,81bcde ± 0,21 ns *** *** * *** b (Adimensional) 5,89a ± 0,09 6,48ab ± 1,13 7,16abc ± 0,00 6,75ab ± 0,21 ns * *** * ***
mg Mb/g m 2,40ab ± 0,09 1,96ab ± 0,67 2,39ab ± 0,00 1,51a ± 0,27 ns * *** ns **
ppm Fe heminico 8,13ab ± 0,32 6,65ab ± 2,28 8,09ab ± 0,00 5,12a ± 0,92 ns * *** ns **
(f): extracto fresco; (s): extracto seco *** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
117
Composición de la grasa intramuscular
El contenido total de grasa intramuscular mostró diferencias significativas, siendo
superior el contenido en los jamones pertenecientes a la primera campaña, seguido de los
jamones de la tercera y finalmente de los de la segunda (tablas 4.1.49 y 4.1.50),
coincidiendo así con el contenido total de ácidos grasos mostrado en las tablas 4.1.51 y
4.1.52.
En cuanto a los ácidos grasos mayoritarios, el C16:0, C16:1 y C18:1 fueron
significativamente superiores en los jamones de la primera campaña, especialmente los
jamones pertenecientes al lote AGH, coincidiendo con el lote que presentó mayor
contenido de AGS, AGMI y el sumatorio total de ácidos grasos. Otros ácidos grasos
mayoritarios que mostraron diferencias fueron el C18:0, superior en los jamones de la
tercera campaña, o el C18:2 (ω-6) superior en la segunda campaña, especialmente los
pertenecientes al lote AGM, coincidiendo con el lote que presentó mayor contenido de
AGPI y de ω-6.
Por lo general, las diferencias significativas generadas entre lotes se debieron
sobre todo al factor campaña, seguido del factor genético. El factor sexo fue el que menos
repercusión tuvo sobre los resultados, al igual que la interacción entre los tres factores,
que tan solo mostraron diferencias en seis ácidos grasos.

Tabla 4.1.51. Composición de la grasa intramuscular en la fracción de lípidos totales del jamón curado. Los resultados están expresados en mg de AG/100 g muestra (± desviación estándar de la media). Parte I.
J1 J2
AA AG AA AG
HEMBRA MACHO HEMBRA MACHO HEMBRA MACHO HEMBRA MACHO
C10:0 1,58abc ± 0,05 2,66bcde ± 0,60 2,84cde ± 1,17 2,45bcd ± 0,28 1,38ab ± 0,41 3,09de ± 0,55 2,88cde ± 0,67 2,69bcde ± 0,25 C12:0 2,81abc ± 0,07 3,59bc ± 0,02 5,81d ± 0,06 3,64bc ± 0,44 2,08ab ± 0,80 3,47bc ± 0,62 3,68bc ± 0,86 3,94c ± 1,64 C14:0 47,65bcd ± 8,28 58,18de ± 7,19 100,91f ± 0,30 63,38de ± 6,58 33,40abc ± 1,48 54,09cde ± 11,36 57,99de ± 16,19 75,37e ± 9,33 C15:0 1,33bcd ± 0,26 1,71cde ± 0,28 2,11e ± 0,11 1,77de ± 0,42 0,67a ± 0,05 1,69cde ± 0,10 1,25bcd ± 0,09 1,22bc ± 0,06 C16:0 579,97abcd ± 142,80 646,45bcd ± 21,73 1.072,17e ± 47,17 724,00cd ± 56,73 360,25ab ± 113,87 545,56abcd ± 49,35 693,62cd ± 160,36 763,89d ± 262,07 C17:0 5,83ab ± 0,68 7,29b ± 0,24 6,33b ± 3,02 8,03b ± 2,28 2,43a ± 0,16 7,37b ± 0,47 5,49ab ± 0,80 6,12b ± 0,39 C18:0 207,72abc ± 53,66 251,94abcd ± 53,89 367,45cd ± 1,93 313,86bcd ± 97,93 114,97a ± 23,42 167,05ab ± 14,26 236,85abc ± 58,39 287,59bcd ± 104,59 C20:0 3,62ab ± 0,04 3,78ab ± 0,24 5,67c ± 0,45 5,86c ± 1,24 2,49a ± 0,24 3,33ab ± 0,03 4,85bc ± 1,14 5,87c ± 0,87 C22:0 0,16abc ± 0,01 0,16abc ± 0,00 0,23cd ± 0,01 0,19bc ± 0,01 0,14ab ± 0,01 0,28d ± 0,08 0,23cd ± 0,01 0,20bcd ± 0,07 C23:0 2,34ab ± 0,34 2,53ab ± 0,31 3,10bc ± 0,08 3,00bc ± 0,13 1,18a ± 0,13 2,69bc ± 1,00 3,06bc ± 0,84 4,04c ± 1,06 AGS 853,01ab ± 204,40 978,30abc ± 67,12 1.469,43d ± 201,63 1.302,90cd ± 381,93 519,00a ± 128,90 788,61ab ± 56,55 1.009,90abc ± 233,19 1.150,93cd ± 377,89
C14:1 2,14abc ± 0,51 3,70c ± 0,62 2,34abc ± 0,55 3,61c ± 2,18 1,86abc ± 0,63 2,95abc ± 0,19 3,41bc ± 0,24 2,42abc ± 0,62 C15:1 1,01ab ± 0,51 0,86a ± 0,03 0,89a ± 0,03 1,05ab ± 0,25 0,68a ± 0,19 1,14ab ± 0,58 1,25ab ± 0,40 1,77b ± 0,43 C16:1 71,09ab ± 0,74 88,26abc ± 2,71 133,93e ± 0,36 120,63cd ± 25,53 47,70a ± 2,62 87,78abc ± 22,17 97,12bcd ± 18,18 82,25abc ± 8,37 C17:1 5,39abc ± 1,08 7,93bcd ± 0,30 9,41d ± 0,30 8,76cd ± 2,41 2,65a ± 0,65 6,13bcd ± 1,45 5,71abc ± 0,99 5,59abc ± 0,79 C18:1 235,96ab ± 25,16 322,66ab ± 60,51 394,43b ± 24,04 406,69b ± 101,59 145,73a ± 34,65 256,20ab ± 13,47 376,11b ± 3,07 347,23b ± 113,01 C20:1 9,47ab ± 1,06 12,28ab ± 0,09 14,37ab ± 1,21 16,76c ± 4,39 5,53a ± 0,19 8,79ab ± 0,61 11,82abc ± 2,48 14,19ab ± 2,47 C22:1 0,20ab ± 0,07 0,23ab ± 0,07 0,29abc ± 0,03 0,25abc ± 0,03 0,14a ± 0,02 0,31abc ± 0,11 0,27abc ± 0,05 0,42c ± 0,17 C24:1 0,24a ± 0,16 0,27a ± 0,13 0,11a ± 0,00 0,13a ± 0,00 0,21a ± 0,08 0,47a ± 0,03 0,56a ± 0,21 0,38a ± 0,32 AGMI 325,50ab ± 26,28 436,19bc ± 60,35 587,27c ± 88,18 558,16c ± 132,66 204,51a ± 36,87 363,77abc ± 22,93 496,25bc ± 21,40 454,25bc ± 119,69
C18:2(ω-6) 155,98ab ± 16,34 186,14abc ± 21,60 193,36abc ± 67,14 202,81bc ± 45,05 98,72a ± 10,31 227,37bc ± 14,13 206,00bc ± 8,32 256,58c ± 86,08 C18:3(ω-6) 1,61b ± 0,30 1,57b ± 0,01 1,64b ± 0,24 1,68b ± 0,30 0,88a ± 0,05 2,00b ± 0,07 1,48ab ± 0,18 1,60b ± 0,26 C18:3(ω-3) 6,68ab ± 0,53 8,48bc ± 1,42 12,34d ± 0,84 8,94bcd ± 1,24 3,78a ± 0,27 10,36cd ± 1,81 9,29bcd ± 1,63 9,48bcd ± 0,58
C20:2 4,31ab ± 0,12 5,40bcd ± 0,28 4,44abc ± 1,22 5,63bcd ± 0,09 2,53a ± 0,07 4,96abcd ± 1,08 5,51bcd ± 0,40 7,13d ± 2,34 C20:4 23,81abc ± 3,13 26,54abc ± 3,17 21,09ab ± 0,46 25,96abc ± 5,60 19,28a ± 8,11 27,28abc ± 10,77 30,66abc ± 6,96 34,41bc ± 2,72
C20:3(ω-3) 1,06abc ± 0,03 1,07abcd ± 0,05 1,65cde ± 0,28 1,49bcde ± 0,03 0,54a ± 0,00 0,91ab ± 0,06 1,27bcde ± 0,21 1,58cde ± 0,52 C20:5(ω-3) 0,88ab ± 0,05 1,09ab ± 0,00 1,16ab ± 0,24 1,00ab ± 0,08 0,77a ± 0,33 0,90ab ± 0,10 1,15ab ± 0,24 1,38c ± 0,20
C22:2 0,18ab ± 0,05 0,25ab ± 0,10 0,20ab ± 0,10 0,26ab ± 0,09 0,17ab ± 0,01 0,36ab ± 0,09 0,27ab ± 0,01 0,27ab ± 0,13 C22:6(ω-3) 0,92bc ± 0,08 0,97bc ± 0,22 0,83ab ± 0,03 1,04bcd ± 0,15 0,48a ± 0,09 0,75ab ± 0,09 0,97bc ± 0,05 1,34d ± 0,25
AGPI 195,44ab ± 19,22 231,52abc ± 23,23 240,28bc ± 75,66 249,95bc ± 54,06 127,16a ± 14,83 274,87bc ± 20,56 256,60bc ± 12,25 313,77c ± 89,73 ω-6 185,71ab ± 18,83 219,65abc ± 22,02 220,54abc ± 68,19 236,08bc ± 50,12 121,41a ± 14,67 261,61bc ± 19,48 243,66bc ± 12,28 299,72bc ± 89,21 ω-3 9,55b ± 0,50 11,62bc ± 1,41 15,99d ± 1,10 12,46bcd ± 1,29 5,58a ± 0,42 12,91bcd ± 1,93 12,67bcd ± 1,73 13,78cd ± 0,95
ω-6/ω-3 19,43ab ± 1,38 18,98a ± 1,33 13,71a ± 3,69 18,85a ± 3,07 21,79ab ± 2,61 20,56ab ± 2,83 19,56ab ± 3,03 21,65ab ± 5,71 C20:3 (ω-6)+C21 3,23ab ± 0,29 3,80bc ± 0,27 4,43bcd ± 0,03 4,56bcd ± 0,16 1,81a ± 0,28 3,10ab ± 0,52 4,45bcd ± 0,85 5,86d ± 1,44
sumatorio 1.377,17abc ± 228,85 1.649,81bc ± 140,34 2.363,52d ± 103,74 1.937,42cd ± 302,10 852,48a ± 159,79 1.430,36abc ± 85,18 1.767,20bcd ± 256,78 1.924,81cd ± 587,42
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo
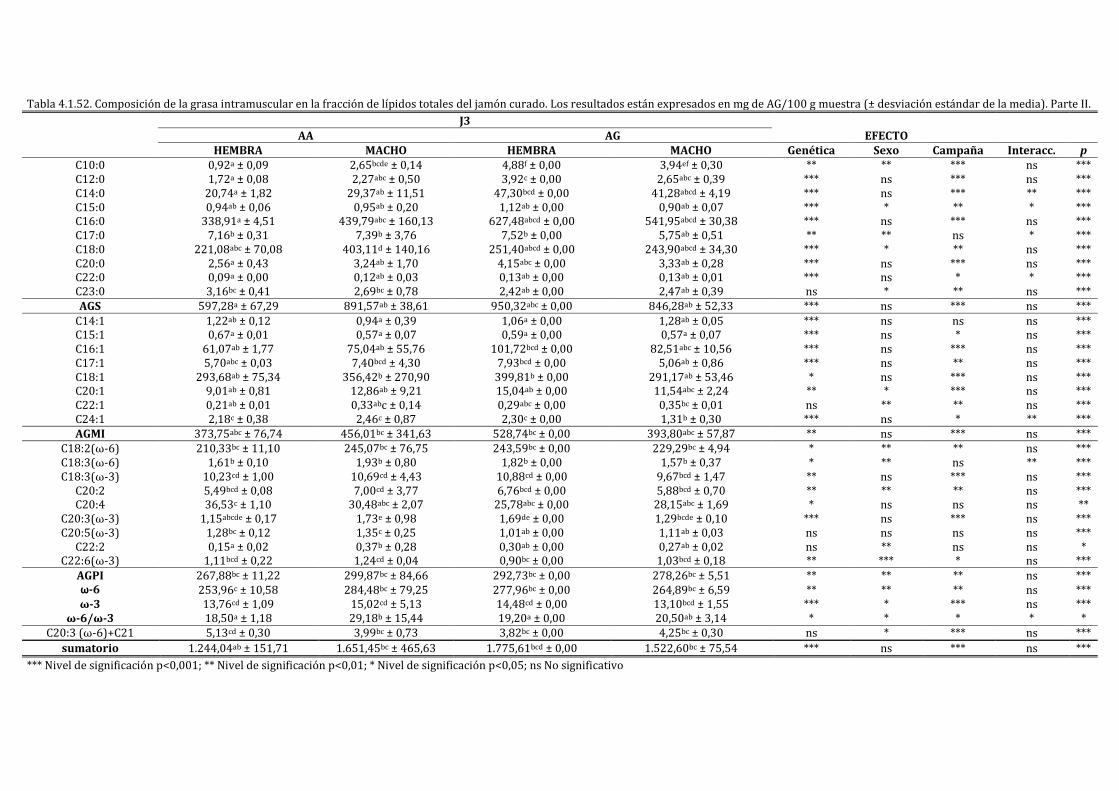
Tabla 4.1.52. Composición de la grasa intramuscular en la fracción de lípidos totales del jamón curado. Los resultados están expresados en mg de AG/100 g muestra (± desviación estándar de la media). Parte II.
J3
AA AG
EFECTO
HEMBRA MACHO HEMBRA MACHO Genética Sexo Campaña Interacc. p
C10:0 0,92a ± 0,09 2,65bcde ± 0,14 4,88f ± 0,00 3,94ef ± 0,30 ** ** *** ns *** C12:0 1,72a ± 0,08 2,27abc ± 0,50 3,92c ± 0,00 2,65abc ± 0,39 *** ns *** ns *** C14:0 20,74a ± 1,82 29,37ab ± 11,51 47,30bcd ± 0,00 41,28abcd ± 4,19 *** ns *** ** *** C15:0 0,94ab ± 0,06 0,95ab ± 0,20 1,12ab ± 0,00 0,90ab ± 0,07 *** * ** * *** C16:0 338,91a ± 4,51 439,79abc ± 160,13 627,48abcd ± 0,00 541,95abcd ± 30,38 *** ns *** ns *** C17:0 7,16b ± 0,31 7,39b ± 3,76 7,52b ± 0,00 5,75ab ± 0,51 ** ** ns * *** C18:0 221,08abc ± 70,08 403,11d ± 140,16 251,40abcd ± 0,00 243,90abcd ± 34,30 *** * ** ns *** C20:0 2,56a ± 0,43 3,24ab ± 1,70 4,15abc ± 0,00 3,33ab ± 0,28 *** ns *** ns *** C22:0 0,09a ± 0,00 0,12ab ± 0,03 0,13ab ± 0,00 0,13ab ± 0,01 *** ns * * *** C23:0 3,16bc ± 0,41 2,69bc ± 0,78 2,42ab ± 0,00 2,47ab ± 0,39 ns * ** ns *** AGS 597,28a ± 67,29 891,57ab ± 38,61 950,32abc ± 0,00 846,28ab ± 52,33 *** ns *** ns ***
C14:1 1,22ab ± 0,12 0,94a ± 0,39 1,06a ± 0,00 1,28ab ± 0,05 *** ns ns ns *** C15:1 0,67a ± 0,01 0,57a ± 0,07 0,59a ± 0,00 0,57a ± 0,07 *** ns * ns *** C16:1 61,07ab ± 1,77 75,04ab ± 55,76 101,72bcd ± 0,00 82,51abc ± 10,56 *** ns *** ns *** C17:1 5,70abc ± 0,03 7,40bcd ± 4,30 7,93bcd ± 0,00 5,06ab ± 0,86 *** ns ** ns *** C18:1 293,68ab ± 75,34 356,42b ± 270,90 399,81b ± 0,00 291,17ab ± 53,46 * ns *** ns *** C20:1 9,01ab ± 0,81 12,86ab ± 9,21 15,04ab ± 0,00 11,54abc ± 2,24 ** * *** ns *** C22:1 0,21ab ± 0,01 0,33abc ± 0,14 0,29abc ± 0,00 0,35bc ± 0,01 ns ** ** ns *** C24:1 2,18c ± 0,38 2,46c ± 0,87 2,30c ± 0,00 1,31b ± 0,30 *** ns * ** *** AGMI 373,75abc ± 76,74 456,01bc ± 341,63 528,74bc ± 0,00 393,80abc ± 57,87 ** ns *** ns ***
C18:2(ω-6) 210,33bc ± 11,10 245,07bc ± 76,75 243,59bc ± 0,00 229,29bc ± 4,94 * ** ** ns *** C18:3(ω-6) 1,61b ± 0,10 1,93b ± 0,80 1,82b ± 0,00 1,57b ± 0,37 * ** ns ** *** C18:3(ω-3) 10,23cd ± 1,00 10,69cd ± 4,43 10,88cd ± 0,00 9,67bcd ± 1,47 ** ns *** ns ***
C20:2 5,49bcd ± 0,08 7,00cd ± 3,77 6,76bcd ± 0,00 5,88bcd ± 0,70 ** ** ** ns *** C20:4 36,53c ± 1,10 30,48abc ± 2,07 25,78abc ± 0,00 28,15abc ± 1,69 * ns ns ns **
C20:3(ω-3) 1,15abcde ± 0,17 1,73e ± 0,98 1,69de ± 0,00 1,29bcde ± 0,10 *** ns *** ns *** C20:5(ω-3) 1,28bc ± 0,12 1,35c ± 0,25 1,01ab ± 0,00 1,11ab ± 0,03 ns ns ns ns ***
C22:2 0,15a ± 0,02 0,37b ± 0,28 0,30ab ± 0,00 0,27ab ± 0,02 ns ** ns ns * C22:6(ω-3) 1,11bcd ± 0,22 1,24cd ± 0,04 0,90bc ± 0,00 1,03bcd ± 0,18 ** *** * ns ***
AGPI 267,88bc ± 11,22 299,87bc ± 84,66 292,73bc ± 0,00 278,26bc ± 5,51 ** ** ** ns *** ω-6 253,96c ± 10,58 284,48bc ± 79,25 277,96bc ± 0,00 264,89bc ± 6,59 ** ** ** ns *** ω-3 13,76cd ± 1,09 15,02cd ± 5,13 14,48cd ± 0,00 13,10bcd ± 1,55 *** * *** ns ***
ω-6/ω-3 18,50a ± 1,18 29,18b ± 15,44 19,20a ± 0,00 20,50ab ± 3,14 * * * * * C20:3 (ω-6)+C21 5,13cd ± 0,30 3,99bc ± 0,73 3,82bc ± 0,00 4,25bc ± 0,30 ns * *** ns ***
sumatorio 1.244,04ab ± 151,71 1.651,45bc ± 465,63 1.775,61bcd ± 0,00 1.522,60bc ± 75,54 *** ns *** ns ***
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
120
Las tablas 4.1.53 y 4.1.54 muestran los porcentajes de los ácidos grasos presentes
en los lípidos totales de la grasa intramuscular. A diferencia de los porcentajes de ácidos
grasos que se evaluaron en capítulos anteriores, al estudiar el efecto de los tres factores
por separado, vemos como existieron diferencias significativas en gran parte de los ácidos
grasos. Sin embargo, a diferencia de las tablas 4.1.51 y 4.1.52, el factor que más efecto tuvo
sobre los porcentajes fue la campaña, ya que el factor sexo y genético no mostraron un
efecto muy marcado.
Existieron diferencias significativas en la proporción de AGS, siendo en este caso
superior en los jamones de la segunda campaña, especialmente los pertenecientes al lote
AGM. Los AGMI no mostraron diferencias significativas y el resto de sumatorios fueron
superiores en los jamones de la tercera campaña, influenciados todos ellos especialmente
por el efecto campaña. Estos resultados siguieron una tendencia contraria a los mostrados
en las tablas 4.1.51 y 4.1.52, que presentaron por lo general mayores cantidades de ácidos
grasos los jamones de la primera campaña, influenciados posiblemente por el mayor
contenido de grasa intramuscular que presentaron los jamones pertenecientes a esa
campaña.

Tabla 4.1.53. Composición de la grasa intramuscular en la fracción de lípidos totales del jamón curado. Los resultados están expresados en % (± desviación estándar de la media). Parte I.
J1 J2
AA AG AA AG
HEMBRA MACHO HEMBRA MACHO HEMBRA MACHO HEMBRA MACHO
C10:0 0,05ab ± 0,00 0,04ab ± 0,01 0,05ab ± 0,00 0,05ab ± 0,01 0,05ab ± 0,00 0,05ab ± 0,00 0,04ab ± 0,00 0,05ab ± 0,00 C12:0 0,06bcd ± 0,00 0,06cd ± 0,00 0,06cd ± 0,01 0,07d ± 0,01 0,06d ± 0,01 0,06cd ± 0,01 0,06cd ± 0,01 0,06bcd ± 0,00 C14:0 1,50cd ± 0,14 1,50bcd ± 0,02 1,58cd ± 0,01 1,70d ± 0,09 1,54cd ± 0,05 1,51cd ± 0,13 1,65cd ± 0,05 1,41bc ± 0,07 C15:0 0,05 ± 0,00 0,05 ± 0,01 0,05 ± 0,00 0,05 ± 0,01 0,05 ± 0,00 0,05 ± 0,00 0,05 ± 0,01 0,05 ± 0,00 C16:0 25,14ab ± 0,26 26,39abc ± 0,83 27,38bc ± 0,25 27,24bc ± 1,56 26,78abc ± 0,17 26,48abc ± 0,78 26,80abc ± 1,44 27,52bc ± 0,42 C17:0 0,25 ± 0,00 0,23 ± 0,04 0,22 ± 0,05 0,25 ± 0,07 0,21 ± 0,01 0,27 ± 0,06 0,21 ± 0,01 0,27 ± 0,01 C18:0 10,96 ± 0,11 9,68 ± 0,09 9,98 ± 0,15 11,20 ± 0,23 11,22 ± 0,11 10,45 ± 1,63 10,34 ± 0,99 11,27 ± 0,75 C20:0 0,08a ± 0,01 0,09abc ± 0,01 0,09a ± 0,00 0,10abc ± 0,01 0,08a ± 0,00 0,09ab ± 0,01 0,09abc ± 0,01 0,09abc ± 0,00 C22:0 0,01ab ± 0,00 0,01a ± 0,00 0,01a ± 0,00 0,01ab ± 0,00 0,01abc ± 0,00 0,01c ± 0,00 0,01bc ± 0,00 0,01abc ± 0,00 C23:0 0,12abc ± 0,00 0,11ab ± 0,01 0,11ab ± 0,03 0,10ab ± 0,02 0,11ab ± 0,02 0,10a ± 0,01 0,14bc ± 0,01 0,16c ± 0,01 C24:0 0,01abcd ± 0,00 0,01a ± 0,00 0,01ab ± 0,00 0,01a ± 0,00 0,01abcd ± 0,00 0,01abc ± 0,00 0,01de ± 0,00 0,02e ± 0,00 AGS 38,23abc ± 0,30 38,17abc ± 0,93 39,53abc ± 0,36 40,78bc ± 1,60 40,12abc ± 0,09 39,08abc ± 2,21 39,42abc ± 2,11 40,89c ± 1,08
C14:1 0,09d ± 0,03 0,08bcd ± 0,00 0,06abcd ± 0,02 0,07abcd ± 0,02 0,07bcd ± 0,01 0,08cd ± 0,00 0,08bcd ± 0,00 0,07bcd ± 0,02 C15:1 0,07ab ± 0,00 0,05ab ± 0,00 0,04ab ± 0,00 0,03ab ± 0,01 0,09b ± 0,05 0,06ab ± 0,01 0,08ab ± 0,03 0,08ab ± 0,04 C16:1 3,21ab ± 0,09 3,52b ± 0,06 3,80b ± 0,01 3,50b ± 0,17 3,59b ± 0,18 2,97ab ± 0,18 3,34b ± 0,01 3,14ab ± 0,06 C17:1 0,28 ± 0,00 0,26 ± 0,02 0,26 ± 0,07 0,30 ± 0,08 0,25 ± 0,01 0,29 ± 0,01 0,27 ± 0,03 0,27 ± 0,01 C18:1 44,27 ± 1,44 44,92 ± 0,66 43,92 ± 0,04 45,55 ± 1,46 44,72 ± 0,19 45,64 ± 1,93 44,14 ± 1,58 41,89 ± 0,69 C20:1 0,51abc ± 0,03 0,60cd ± 0,05 0,57bcd ± 0,02 0,59bcd ± 0,05 0,45a ± 0,01 0,47a ± 0,03 0,52abc ± 0,04 0,51ab ± 0,01 C22:1 0,01ab ± 0,00 0,01abc ± 0,00 0,01abc ± 0,00 0,01a ± 0,00 0,01bcde ± 0,00 0,01bcdef ± 0,00 0,01abcd ± 0,00 0,01cdef ± 0,00 C24:1 0,02abc ± 0,01 0,01a ± 0,00 0,01a ± 0,00 0,01ab ± 0,00 0,03cdef ± 0,00 0,04def ± 0,01 0,04def ± 0,01 0,03bcd ± 0,02 AGMI 48,46 ± 1,44 49,46 ± 0,57 48,67 ± 0,05 50,07 ± 1,38 49,22 ± 0,35 49,56 ± 1,86 48,48 ± 1,56 46,01 ± 0,67
C18:2(ω-6) 9,96bcd ± 0,08 8,73abc ± 0,19 8,50ab ± 0,03 7,69a ± 0,09 8,75abc ± 0,26 8,08ab ± 0,11 9,72bcd ± 1,49 10,45cd ± 0,20 C18:3(ω-6) 0,09c ± 0,01 0,07ab ± 0,01 0,06a ± 0,01 0,06a ± 0,01 0,07ab ± 0,00 0,06ab ± 0,00 0,07abc ± 0,00 0,08bc ± 0,00 C18:3(ω-3) 0,40ab ± 0,00 0,42ab ± 0,00 0,40ab ± 0,01 0,34a ± 0,03 0,40ab ± 0,06 0,41ab ± 0,10 0,40ab ± 0,02 0,43ab ± 0,02
C20:2 0,24ab ± 0,00 0,25abc ± 0,03 0,26bcde ± 0,00 0,23a ± 0,01 0,24ab ± 0,02 0,23a ± 0,00 0,30de ± 0,01 0,29d ± 0,01 C20:4 1,57cd ± 0,11 1,07ab ± 0,02 0,91a ± 0,12 0,92a ± 0,09 1,27abc ± 0,20 1,00a ± 0,06 1,36abcd ± 0,33 1,36abcd ± 0,13
C20:3(ω-3) 0,05abc ± 0,00 0,05ab ± 0,00 0,06cde ± 0,00 0,06bcd ± 0,00 0,05ab ± 0,01 0,05a ± 0,00 0,05abc ± 0,01 0,06cde ± 0,00 C20:5(ω-3) 0,07de ± 0,00 0,05abcd ± 0,00 0,04ab ± 0,00 0,04a ± 0,00 0,05abcd ± 0,01 0,04abc ± 0,00 0,05abcd ± 0,01 0,06cd ± 0,01
C22:2 0,01ab ± 0,00 0,01abc ± 0,00 0,01a ± 0,00 0,01ab ± 0,00 0,01abcd ± 0,00 0,02cde ± 0,00 0,02de ± 0,00 0,01bcde ± 0,00 C22:6(ω-3) 0,11ab ± 0,02 0,09a ± 0,03 0,09a ± 0,03 0,07a ± 0,02 0,08a ± 0,01 0,07a ± 0,01 0,10ab ± 0,01 0,11ab ± 0,01
AGPI 12,50cdef ± 0,11 10,75abcd ± 0,22 10,32abc ± 0,18 9,41a ± 0,18 10,92abcd ± 0,19 9,95ab ± 0,14 12,07bcdef ± 1,76 12,85def ± 0,27 ω-6 11,86cde ± 0,10 10,13abcd ± 0,20 9,73abc ± 0,15 8,89a ± 0,14 10,33abcd ± 0,18 9,37ab ± 0,09 11,45bcde ± 1,73 12,18de ± 0,29 ω-3 0,64abc ± 0,02 0,61ab ± 0,03 0,58ab ± 0,03 0,51a ± 0,06 0,57a ± 0,05 0,57a ± 0,11 0,61ab ± 0,04 0,65abc ± 0,02
ω-6/ω-3 18,67 ± 0,64 16,62 ± 0,54 16,75 ± 0,77 17,71 ± 1,96 18,18 ± 1,62 16,90 ± 3,16 18,83 ± 2,02 18,65 ± 0,95 C20:3 (ω-6)+C21 0,23cd ± 0,01 0,17abc ± 0,01 0,16ab ± 0,04 0,14a ± 0,02 0,18abc ± 0,01 0,17abc ± 0,04 0,21bcd ± 0,00 0,24d ± 0,02

Tabla 4.1.54. Composición de la grasa intramuscular en la fracción de lípidos totales del jamón curado. Los resultados están expresados en % (± desviación estándar de la media). Parte II.
J3
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Genética Sexo Campaña Interacc. p C10:0 0,02a ± 0,00 0,04b ± 0,01 0,05c ± 0,00 0,05ab ± 0,00 *** * ns * *** C12:0 0,03a ± 0,00 0,05abc ± 0,01 0,06bcd ± 0,00 0,04ab ± 0,00 ns ns *** * *** C14:0 1,16a ± 0,02 1,26ab ± 0,04 1,52cd ± 0,00 1,50cd ± 0,19 *** ns *** * *** C15:0 0,06 ± 0,00 0,05 ± 0,01 0,05 ± 0,00 0,05 ± 0,00 ns ns ns ns ns C16:0 24,71a ± 0,05 24,66a ± 1,22 26,00abc ± 0,00 28,06c ± 1,31 *** * * * *** C17:0 0,28 ± 0,01 0,25 ± 0,02 0,22 ± 0,00 0,25 ± 0,04 ns ns ns ns ns C18:0 10,59 ± 0,43 9,85 ± 0,59 9,46 ± 0,00 9,39 ± 1,22 ns ns ** ns ** C20:0 0,10abc ± 0,00 0,10abc ± 0,01 0,11bc ± 0,00 0,11cd ± 0,01 * ns *** ns *** C22:0 0,01ab ± 0,00 0,01a ± 0,00 0,01a ± 0,00 0,01a ± 0,00 ns ns *** * *** C23:0 0,12abc ± 0,02 0,16c ± 0,01 0,14bc ± 0,00 0,13abc ± 0,02 * ns *** * *** C24:0 0,01bcd ± 0,00 0,01cd ± 0,00 0,01abcd ± 0,00 0,01a ± 0,00 ns ns *** ** *** AGS 37,09ab ± 0,38 36,44a ± 1,92 37,64abc ± 0,00 39,60abc ± 2,34 ** ns ** ns **
C14:1 0,03a ± 0,00 0,04ab ± 0,00 0,04abc ± 0,00 0,05abcd ± 0,00 ns ns *** ns *** C15:1 0,03a ± 0,00 0,03a ± 0,01 0,03a ± 0,00 0,03a ± 0,00 ns ns *** ns *** C16:1 2,27a ± 0,05 3,54b ± 1,31 3,92b ± 0,00 3,76b ± 1,20 ** ns ns * ** C17:1 0,22 ± 0,03 0,28 ± 0,01 0,26 ± 0,00 0,24 ± 0,04 ns ns ns * ns C18:1 43,59 ± 0,69 45,12 ± 6,50 45,50 ± 0,00 43,69 ± 3,57 ns ns ns ns ns C20:1 0,62d ± 0,02 0,59bcd ± 0,08 0,64d ± 0,00 0,59bcd ± 0,02 * ns *** ns *** C22:1 0,02def ± 0,00 0,02ef ± 0,00 0,01cdef ± 0,00 0,02f ± 0,00 ns * *** ns *** C24:1 0,05f ± 0,00 0,05f ± 0,00 0,05ef ± 0,00 0,03bcd ± 0,00 ** ns *** ** *** AGMI 46,83 ± 0,66 49,67 ± 7,87 50,46 ± 0,00 48,39 ± 4,82 ns ns ns ns ns
C18:2(ω-6) 10,74d ± 0,10 11,09d ± 3,22 9,38abcd ± 0,00 9,64bcd ± 0,21 ns ns *** ns *** C18:3(ω-6) 0,09c ± 0,01 0,08abc ± 0,01 0,06ab ± 0,00 0,06a ± 0,00 *** ** ns ns *** C18:3(ω-3) 0,41ab ± 0,01 0,47b ± 0,11 0,41ab ± 0,00 0,38ab ± 0,08 ns ns ns ns ns
C20:2 0,39f ± 0,02 0,33e ± 0,02 0,28cd ± 0,00 0,27bcd ± 0,01 * *** *** ** *** C20:4 1,53bcd ± 0,04 1,82d ± 0,60 1,35abcd ± 0,00 1,21abc ± 0,18 ** ns *** * ***
C20:3(ω-3) 0,09g ± 0,00 0,08f ± 0,00 0,07e ± 0,00 0,06de ± 0,00 ns ** *** ns *** C20:5(ω-3) 0,06cde ± 0,00 0,08e ± 0,04 0,06bcd ± 0,00 0,05abcd ± 0,00 ** ns *** ** ***
C22:2 0,01ab ± 0,00 0,02e ± 0,00 0,02e ± 0,00 0,01bcde ± 0,00 ns ns *** ** *** C22:6(ω-3) 0,17c ± 0,00 0,15bcd ± 0,06 0,10ab ± 0,00 0,11ab ± 0,01 ** ns *** ns ***
AGPI 13,49ef ± 0,08 14,12f ± 4,07 11,73abcde ± 0,00 11,80bcdef ± 0,30 * ns *** ns *** ω-6 12,75e ± 0,08 13,32e ± 3,86 11,08abcde ± 0,00 11,18bcde ± 0,33 ns ns *** ns *** ω-3 0,73bc ± 0,00 0,78c ± 0,21 0,63abc ± 0,00 0,61ab ± 0,08 ** ns *** ns ***
ω-6/ω-3 17,39 ± 0,10 17,07 ± 0,27 17,51 ± 0,00 18,74 ± 2,86 ns ns ns ns ns C20:3 (ω-6)+C21 0,22bcd ± 0,00 0,23cd ± 0,05 0,17abc ± 0,00 0,20abcd ± 0,00 ns ns ** ns ***
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
123
Determinación de compuestos volátiles
Como hemos señalado en apartados anteriores, dadas las particularidades del
análisis de compuestos volátiles, que suponen una gran variabilidad de los resultados,
hemos utilizado las tablas 4.1.55, 4.1.56 y 4.1.57 para poner de manifiesto (dado que se
cuenta con un número relativamente alto de muestras perfectamente identificadas) que a
pesar de la variación, los aldehídos, las cetonas y los alcoholes fueron los principales
representantes de estos compuestos y que el procesado es uno de los factores más
directamente implicados en su generación.
De los 113 compuestos volátiles detectados en los jamones (tabla 4.1.10), 105
presentaron diferencias significativas entre lotes, 35 de ellos superiores
significativamente en los jamones de la primera campaña, 24 superiores en la segunda, 29
en la tercera y el resto no siguieron una tendencia clara según el efecto en estudio.
Existieron diferencias significativas en el contenido total de compuestos, debidas
únicamente al efecto genético, destacando principalmente el lote AAH de la primera
campaña por su mayor contenido y el lote AGM de la segunda campaña por su menor
contenido.
En los jamones de la primera campaña destacaron los altos valores de compuestos
como el 2-metil-2-butenal, 3-hidroxi-2-butanona, etanol, octanol, dodecano, la mayor
parte de los ácidos, la dihidro-5-etil-2(3H)-furanona, el metil- y etilbenceno y el alfa-
pineno, cuyos altos valores contribuyeron a que estos jamones presentaran un mayor
sumatorio de todos los compuestos; en los de la segunda campaña destacaron algunos
compuestos como el 2-nonenal, la ciclopentanona, el 2-heptanol, el 3-metilpentano, el
tridecano, la mayor parte de los ésteres y el 2-etilfurano. En los jamones de la tercera
campaña destacaron el 3-(metiltio)propanal, la 2-heptanona, el 1-penten-3-ol, el 2-
pentanol, el pentano, el decanoato de etilo, el sulfuro de carbono, el hexanonitrilo y el
etilfurano.
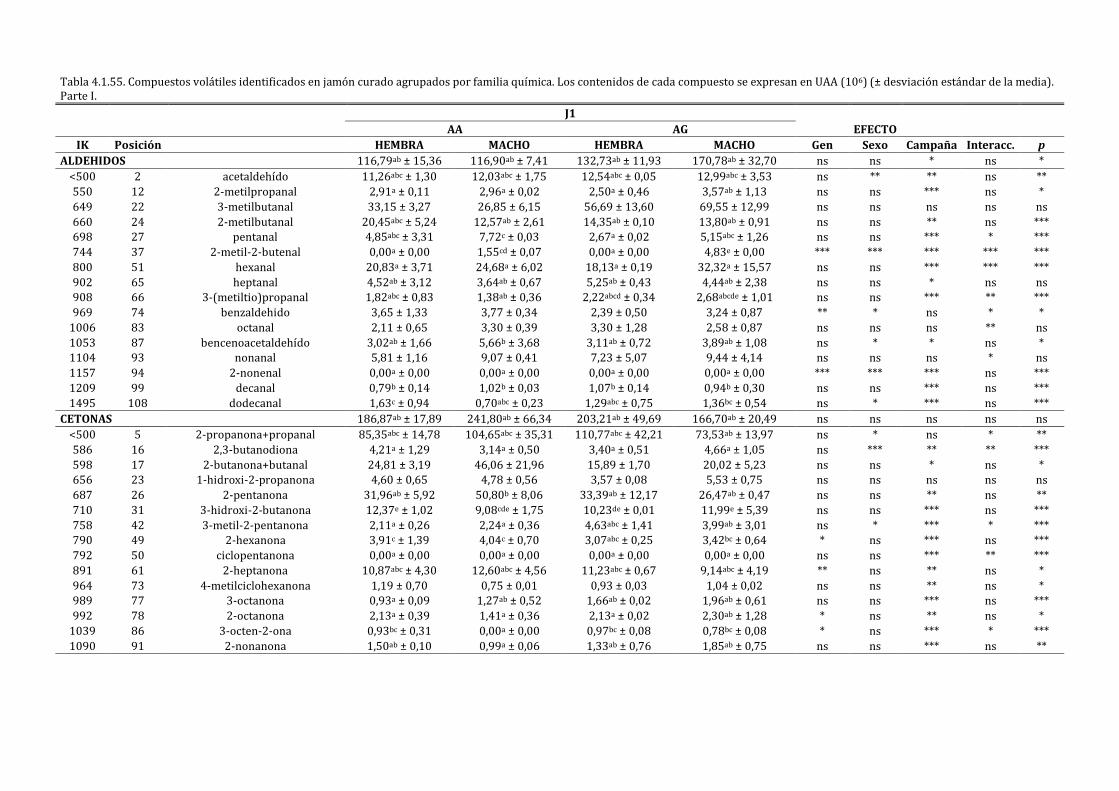
Tabla 4.1.55. Compuestos volátiles identificados en jamón curado agrupados por familia química. Los contenidos de cada compuesto se expresan en UAA (106) (± desviación estándar de la media). Parte I.
J1
AA AG
EFECTO
IK Posición
HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
ALDEHIDOS 116,79ab ± 15,36 116,90ab ± 7,41 132,73ab ± 11,93 170,78ab ± 32,70 ns ns * ns *
<500 2 acetaldehído 11,26abc ± 1,30 12,03abc ± 1,75 12,54abc ± 0,05 12,99abc ± 3,53 ns ** ** ns **
550 12 2-metilpropanal 2,91a ± 0,11 2,96a ± 0,02 2,50a ± 0,46 3,57ab ± 1,13 ns ns *** ns *
649 22 3-metilbutanal 33,15 ± 3,27 26,85 ± 6,15 56,69 ± 13,60 69,55 ± 12,99 ns ns ns ns ns
660 24 2-metilbutanal 20,45abc ± 5,24 12,57ab ± 2,61 14,35ab ± 0,10 13,80ab ± 0,91 ns ns ** ns *** 698 27 pentanal 4,85abc ± 3,31 7,72c ± 0,03 2,67a ± 0,02 5,15abc ± 1,26 ns ns *** * ***
744 37 2-metil-2-butenal 0,00a ± 0,00 1,55cd ± 0,07 0,00a ± 0,00 4,83e ± 0,00 *** *** *** *** ***
800 51 hexanal 20,83a ± 3,71 24,68a ± 6,02 18,13a ± 0,19 32,32a ± 15,57 ns ns *** *** ***
902 65 heptanal 4,52ab ± 3,12 3,64ab ± 0,67 5,25ab ± 0,43 4,44ab ± 2,38 ns ns * ns ns 908 66 3-(metiltio)propanal 1,82abc ± 0,83 1,38ab ± 0,36 2,22abcd ± 0,34 2,68abcde ± 1,01 ns ns *** ** ***
969 74 benzaldehido 3,65 ± 1,33 3,77 ± 0,34 2,39 ± 0,50 3,24 ± 0,87 ** * ns * *
1006 83 octanal 2,11 ± 0,65 3,30 ± 0,39 3,30 ± 1,28 2,58 ± 0,87 ns ns ns ** ns
1053 87 bencenoacetaldehído 3,02ab ± 1,66 5,66b ± 3,68 3,11ab ± 0,72 3,89ab ± 1,08 ns * * ns * 1104 93 nonanal 5,81 ± 1,16 9,07 ± 0,41 7,23 ± 5,07 9,44 ± 4,14 ns ns ns * ns
1157 94 2-nonenal 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 *** *** *** ns ***
1209 99 decanal 0,79b ± 0,14 1,02b ± 0,03 1,07b ± 0,14 0,94b ± 0,30 ns ns *** ns ***
1495 108 dodecanal 1,63c ± 0,94 0,70abc ± 0,23 1,29abc ± 0,75 1,36bc ± 0,54 ns * *** ns ***
CETONAS 186,87ab ± 17,89 241,80ab ± 66,34 203,21ab ± 49,69 166,70ab ± 20,49 ns ns ns ns ns
<500 5 2-propanona+propanal 85,35abc ± 14,78 104,65abc ± 35,31 110,77abc ± 42,21 73,53ab ± 13,97 ns * ns * **
586 16 2,3-butanodiona 4,21a ± 1,29 3,14a ± 0,50 3,40a ± 0,51 4,66a ± 1,05 ns *** ** ** ***
598 17 2-butanona+butanal 24,81 ± 3,19 46,06 ± 21,96 15,89 ± 1,70 20,02 ± 5,23 ns ns * ns * 656 23 1-hidroxi-2-propanona 4,60 ± 0,65 4,78 ± 0,56 3,57 ± 0,08 5,53 ± 0,75 ns ns ns ns ns
687 26 2-pentanona 31,96ab ± 5,92 50,80b ± 8,06 33,39ab ± 12,17 26,47ab ± 0,47 ns ns ** ns **
710 31 3-hidroxi-2-butanona 12,37e ± 1,02 9,08cde ± 1,75 10,23de ± 0,01 11,99e ± 5,39 ns ns *** ns ***
758 42 3-metil-2-pentanona 2,11a ± 0,26 2,24a ± 0,36 4,63abc ± 1,41 3,99ab ± 3,01 ns * *** * *** 790 49 2-hexanona 3,91c ± 1,39 4,04c ± 0,70 3,07abc ± 0,25 3,42bc ± 0,64 * ns *** ns ***
792 50 ciclopentanona 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 ns ns *** ** ***
891 61 2-heptanona 10,87abc ± 4,30 12,60abc ± 4,56 11,23abc ± 0,67 9,14abc ± 4,19 ** ns ** ns *
964 73 4-metilciclohexanona 1,19 ± 0,70 0,75 ± 0,01 0,93 ± 0,03 1,04 ± 0,02 ns ns ** ns * 989 77 3-octanona 0,93a ± 0,09 1,27ab ± 0,52 1,66ab ± 0,02 1,96ab ± 0,61 ns ns *** ns ***
992 78 2-octanona 2,13a ± 0,39 1,41a ± 0,36 2,13a ± 0,02 2,30ab ± 1,28 * ns ** ns *
1039 86 3-octen-2-ona 0,93bc ± 0,31 0,00a ± 0,00 0,97bc ± 0,08 0,78bc ± 0,08 * ns *** * ***
1090 91 2-nonanona 1,50ab ± 0,10 0,99a ± 0,06 1,33ab ± 0,76 1,85ab ± 0,75 ns ns *** ns **
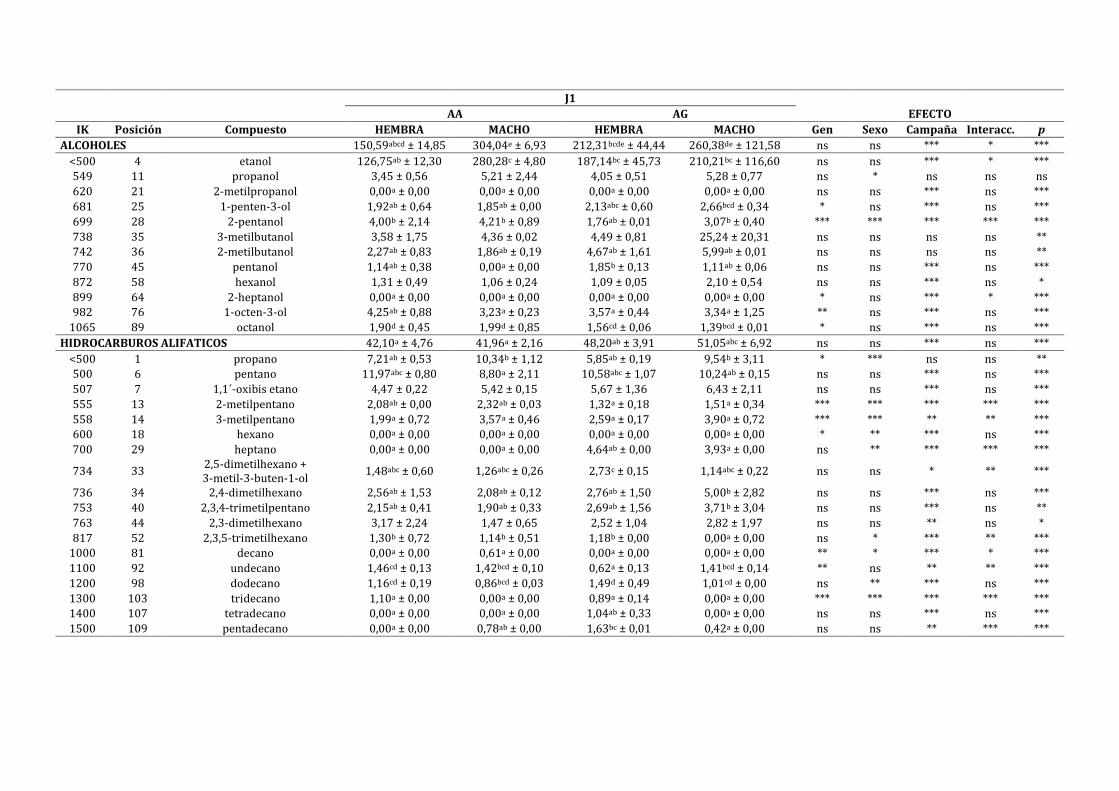
J1
AA AG EFECTO
IK Posición Compuesto HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
ALCOHOLES 150,59abcd ± 14,85 304,04e ± 6,93 212,31bcde ± 44,44 260,38de ± 121,58 ns ns *** * ***
<500 4 etanol 126,75ab ± 12,30 280,28c ± 4,80 187,14bc ± 45,73 210,21bc ± 116,60 ns ns *** * *** 549 11 propanol 3,45 ± 0,56 5,21 ± 2,44 4,05 ± 0,51 5,28 ± 0,77 ns * ns ns ns
620 21 2-metilpropanol 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 ns ns *** ns ***
681 25 1-penten-3-ol 1,92ab ± 0,64 1,85ab ± 0,00 2,13abc ± 0,60 2,66bcd ± 0,34 * ns *** ns ***
699 28 2-pentanol 4,00b ± 2,14 4,21b ± 0,89 1,76ab ± 0,01 3,07b ± 0,40 *** *** *** *** ***
738 35 3-metilbutanol 3,58 ± 1,75 4,36 ± 0,02 4,49 ± 0,81 25,24 ± 20,31 ns ns ns ns ** 742 36 2-metilbutanol 2,27ab ± 0,83 1,86ab ± 0,19 4,67ab ± 1,61 5,99ab ± 0,01 ns ns ns ns **
770 45 pentanol 1,14ab ± 0,38 0,00a ± 0,00 1,85b ± 0,13 1,11ab ± 0,06 ns ns *** ns ***
872 58 hexanol 1,31 ± 0,49 1,06 ± 0,24 1,09 ± 0,05 2,10 ± 0,54 ns ns *** ns *
899 64 2-heptanol 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 * ns *** * *** 982 76 1-octen-3-ol 4,25ab ± 0,88 3,23a ± 0,23 3,57a ± 0,44 3,34a ± 1,25 ** ns *** ns ***
1065 89 octanol 1,90d ± 0,45 1,99d ± 0,85 1,56cd ± 0,06 1,39bcd ± 0,01 * ns *** ns ***
HIDROCARBUROS ALIFATICOS 42,10a ± 4,76 41,96a ± 2,16 48,20ab ± 3,91 51,05abc ± 6,92 ns ns *** ns ***
<500 1 propano 7,21ab ± 0,53 10,34b ± 1,12 5,85ab ± 0,19 9,54b ± 3,11 * *** ns ns ** 500 6 pentano 11,97abc ± 0,80 8,80a ± 2,11 10,58abc ± 1,07 10,24ab ± 0,15 ns ns *** ns ***
507 7 1,1´-oxibis etano 4,47 ± 0,22 5,42 ± 0,15 5,67 ± 1,36 6,43 ± 2,11 ns ns *** ns ***
555 13 2-metilpentano 2,08ab ± 0,00 2,32ab ± 0,03 1,32a ± 0,18 1,51a ± 0,34 *** *** *** *** ***
558 14 3-metilpentano 1,99a ± 0,72 3,57a ± 0,46 2,59a ± 0,17 3,90a ± 0,72 *** *** ** ** *** 600 18 hexano 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 * ** *** ns ***
700 29 heptano 0,00a ± 0,00 0,00a ± 0,00 4,64ab ± 0,00 3,93a ± 0,00 ns ** *** *** ***
734 33 2,5-dimetilhexano + 3-metil-3-buten-1-ol
1,48abc ± 0,60 1,26abc ± 0,26 2,73c ± 0,15 1,14abc ± 0,22 ns ns * ** ***
736 34 2,4-dimetilhexano 2,56ab ± 1,53 2,08ab ± 0,12 2,76ab ± 1,50 5,00b ± 2,82 ns ns *** ns ***
753 40 2,3,4-trimetilpentano 2,15ab ± 0,41 1,90ab ± 0,33 2,69ab ± 1,56 3,71b ± 3,04 ns ns *** ns **
763 44 2,3-dimetilhexano 3,17 ± 2,24 1,47 ± 0,65 2,52 ± 1,04 2,82 ± 1,97 ns ns ** ns *
817 52 2,3,5-trimetilhexano 1,30b ± 0,72 1,14b ± 0,51 1,18b ± 0,00 0,00a ± 0,00 ns * *** ** *** 1000 81 decano 0,00a ± 0,00 0,61a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 ** * *** * ***
1100 92 undecano 1,46cd ± 0,13 1,42bcd ± 0,10 0,62a ± 0,13 1,41bcd ± 0,14 ** ns ** ** ***
1200 98 dodecano 1,16cd ± 0,19 0,86bcd ± 0,03 1,49d ± 0,49 1,01cd ± 0,00 ns ** *** ns ***
1300 103 tridecano 1,10a ± 0,00 0,00a ± 0,00 0,89a ± 0,14 0,00a ± 0,00 *** *** *** *** *** 1400 107 tetradecano 0,00a ± 0,00 0,00a ± 0,00 1,04ab ± 0,33 0,00a ± 0,00 ns ns *** ns ***
1500 109 pentadecano 0,00a ± 0,00 0,78ab ± 0,00 1,63bc ± 0,01 0,42a ± 0,00 ns ns ** *** ***

J1
AA AG EFECTO
IK Posición Compuesto HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
ACIDOS 78,69cd ± 6,29 64,79bcd ± 7,56 97,07d ± 58,46 66,39bcd ± 17,46 ns ns *** * ***
579 15 ácido acético 54,30cd ± 2,05 43,37bcd ± 3,88 55,17cd ± 42,30 41,48bcd ± 13,25 ** ns *** *** *** 753 41 ácido 2-metilpropanoico 1,76ab ± 0,35 1,97ab ± 0,20 3,45b ± 2,17 2,23ab ± 1,08 ns ns *** ns ***
779 47 ácido butanoico 9,80ab ± 1,00 11,16ab ± 4,53 16,72b ± 13,23 8,52ab ± 3,20 ns ns *** ns ***
833 53 ácido 3-metilbutanoico 5,43abc ± 2,06 1,64ab ± 0,21 9,07c ± 0,54 5,65abc ± 3,65 ns ns *** ns ***
843 54 ácido 2-metilpentanoico 2,56ab ± 1,48 0,96a ± 0,29 6,19c ± 2,17 2,69ab ± 0,92 ns ns ** ns ***
970 75 ácido hexanoico 2,75ab ± 0,94 3,20ab ± 0,05 2,71ab ± 0,30 2,62ab ± 0,45 * *** *** ns *** 1159 95 ácido octanoico 0,84abc ± 0,32 1,05bc ± 0,21 0,91abc ± 0,39 1,13c ± 0,41 ns ns *** ns ***
1258 101 ácido nonanoico 0,45bc ± 0,14 0,30ab ± 0,03 0,82d ± 0,40 0,74cd ± 0,19 *** ** *** ** ***
1370 104 ácido decanoico 0,79b ± 0,09 1,14bc ± 0,16 2,03d ± 0,18 1,32c ± 0,44 *** *** *** *** ***
ESTERES 11,36ab ± 3,23 8,84ab ± 0,50 10,81ab ± 2,38 9,69ab ± 2,87 ns ns ** ns **
612 19 acetato de etilo 2,88a ± 1,40 2,99a ± 0,05 3,05a ± 2,04 2,14a ± 1,07 ns ns ns * *
715 32 propanoato de etilo 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 * ns *** ns ***
853 55 2-metilbutanoato de etilo 1,96ab ± 0,10 0,80ab ± 0,06 1,87ab ± 0,02 1,75ab ± 0,91 ns ns * ns *
856 56 3-metilbutanoato de etilo 3,18ab ± 0,26 1,28ab ± 0,28 2,64ab ± 0,10 1,33ab ± 0,13 ns * ns ns ** 942 70 hexanoato de metilo 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 ns ** *** * ***
999 80 hexanoato de etilo 2,11abc ± 1,19 2,46abc ± 0,26 1,14abc ± 0,05 2,17abc ± 0,68 ns ns ns * ***
1196 97 octanoato de etilo 0,72a ± 0,37 0,84a ± 0,07 1,08ab ± 0,97 1,13ab ± 0,44 * ns ns ** **
1395 106 decanoato de etilo 0,52abc ± 0,30 0,47abc ± 0,03 1,04bcd ± 0,53 1,17cd ± 0,42 *** ** ** ** ***
COMPUESTOS AZUFRADOS 111,55ab ± 55,32 81,18ab ± 26,92 67,75ab ± 10,60 104,75ab ± 30,78 ** ** *** ** ***
<500 3 metanotiol 7,34ab ± 0,19 7,10a ± 0,77 8,34ab ± 0,15 7,70ab ± 1,11 ns ns *** * ***
519 8 metano tiobis + etanotiol 5,40abc ± 1,64 2,91a ± 0,17 3,60ab ± 1,35 9,43bcde ± 0,54 ns *** *** ns ***
536 10 sulfuro de carbono 94,77bc ± 55,66 67,86abc ± 26,67 52,30abc ± 9,50 84,29abc ± 28,66 ** ** *** ** *** 750 39 dimetil disulfuro 3,54ab ± 0,30 2,84ab ± 0,00 2,99ab ± 1,50 2,93ab ± 0,85 * ns * ns **
1267 100 benzotiazol 0,49cd ± 0,16 0,46bcd ± 0,23 0,52d ± 0,19 0,40abcd ± 0,02 ns * *** ns ***
COMPUESTOS NITROGENADOS 13,87ab ± 1,90 13,33ab ± 2,05 16,36ab ± 1,56 16,08ab ± 3,47 ns ns *** ns **
748 38 piridina 4,02cd ± 1,80 3,13bcd ± 1,08 2,94bcd ± 0,07 4,46d ± 0,52 ns ns *** ns *** 760 43 1H-pirrol 0,66a ± 0,32 0,37a ± 0,01 2,57b ± 1,00 1,15a ± 0,65 ** ns *** ns ***
786 48 2,3,4,5-tetrahidropiridina 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 ** ns *** ns ***
881 60 hexanonitrilo 1,18ab ± 0,34 1,48abc ± 0,38 1,99bc ± 0,42 1,16ab ± 0,33 ** ** *** ns ***
914 67 2,6-dimetilpirazina 1,92a ± 0,03 2,51a ± 0,31 2,40a ± 0,05 2,00a ± 0,52 ns ns *** ns *** 923 69 etilpirazina 2,51ab ± 1,08 1,65a ± 0,39 2,39ab ± 0,35 2,85ab ± 1,07 * *** *** *** ***
1006 84 trimetilpirazina 2,71ab ± 0,43 2,71ab ± 1,23 2,27ab ± 0,45 2,54ab ± 1,31 ns ns *** ns *
1175 96 3-metil-1H-pirazol 0,87abc ± 0,09 1,49bc ± 0,31 1,80c ± 0,87 1,90c ± 0,51 * ns *** ns ***

J1
AA AG EFECTO
IK Posición Compuesto HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
FURANOS 12,31abcd ± 1,50 11,52abc ± 0,63 12,04abcd ± 1,43 10,26ab ± 1,49 *** *** *** ns ***
702 30 2-etilfurano 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 ns ns *** ns *** 892 62 2-butilfurano 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 ** ** *** * ***
915 68 dihidro-2(3H)-furanona 4,09c ± 0,45 4,75c ± 0,31 4,25c ± 1,13 3,67bc ± 0,80 ns ns *** ns ***
958 72 dihidro-5-metil-2(3H)-furanona 1,03b ± 0,10 0,85ab ± 0,02 0,83ab ± 0,03 0,93ab ± 0,03 ns ns ** ns ns
996 79 2-pentilfurano 2,51ab ± 1,08 1,65a ± 0,39 2,39ab ± 0,35 2,85ab ± 1,07 * *** *** *** ***
1060 88 dihidro-5-etil-2(3H)-furanona 3,41e ± 0,16 3,20de ± 0,04 3,03cde ± 0,64 1,95abcd ± 0,26 *** ns *** * *** 1272 102 dihidro-5-butil-2(3H)-furanona 0,30 ± 0,02 0,31 ± 0,07 0,77 ± 0,52 0,29 ± 0,06 ns * ns * **
1377 105 dihidro-5-pentil-2(3H)-furanona 0,97b ± 0,04 0,75ab ± 0,36 0,77ab ± 0,41 0,57ab ± 0,19 * ns ns * ns
HIDROCARBUROS AROMATICOS 15,78bc ± 2,13 8,78abc ± 0,58 10,69abc ± 2,94 9,47abc ± 1,98 ** * ns ns **
772 46 metilbenceno 9,80b ± 1,00 3,06ab ± 0,53 3,81ab ± 1,74 4,05ab ± 2,03 * ** ns ns ** 866 57 etilbenceno 1,46bcd ± 0,82 1,56cd ± 0,60 2,06d ± 0,49 0,75abc ± 0,24 ns ns *** * ***
875 59 m- y p- xileno 1,64a ± 0,14 1,57a ± 0,25 1,64a ± 0,02 1,61a ± 0,37 * * *** * ***
894 63 estireno 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 *** ** *** ns ***
1002 82 1,3,5-trimetilbenceno 1,22bc ± 0,25 1,35c ± 0,17 0,99abc ± 0,22 1,21bc ± 0,17 ** ns *** ns *** 1074 90 4-metilfenol 1,65abc ± 0,59 1,24abc ± 0,01 2,20c ± 1,51 1,86bc ± 0,55 ns ns *** ns ***
TERPENOS 1,21bcd ± 0,25 1,50cde ± 0,50 1,76de ± 0,50 2,27e ± 0,33 * ** *** ns ***
943 71 alfa-pineno 0,44abc ± 0,21 0,87bcd ± 0,43 1,19d ± 0,47 0,80bcd ± 0,33 ns ns *** ns ***
1035 85 l-limoneno 0,76b ± 0,16 0,63ab ± 0,13 0,57ab ± 0,03 1,47d ± 0,25 ** ** *** *** ***
COMPUESTOS CLORADOS 190,53ab ± 172,37 433,67b ± 335,62 70,21a ± 4,12 135,17ab ± 115,00 ** ns ns ns **
528 9 diclorometano 1,09bc ± 0,66 1,08bc ± 0,12 1,08bc ± 0,09 1,21bc ± 0,02 *** ** *** ns ***
616 20 cloroformo 189,45ab ± 171,96 432,58b ± 335,62 69,14a ± 4,11 133,95ab ± 114,98 ** ns ns ns **
sumatorio 923,98ab ± 220,04 1.329,61b ± 318,46 882,59ab ± 133,20 1.001,18ab ± 132,56 * ns ns ns *
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo
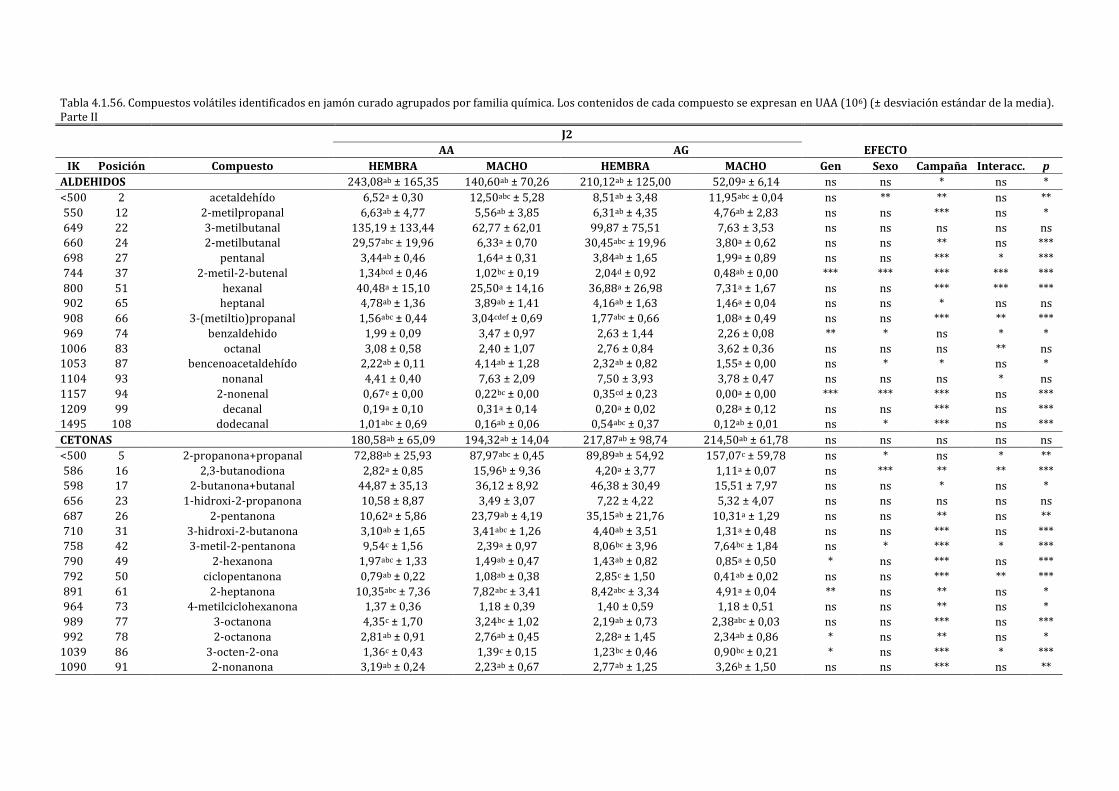
Tabla 4.1.56. Compuestos volátiles identificados en jamón curado agrupados por familia química. Los contenidos de cada compuesto se expresan en UAA (106) (± desviación estándar de la media). Parte II
J2
AA AG
EFECTO
IK Posición Compuesto HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
ALDEHIDOS 243,08ab ± 165,35 140,60ab ± 70,26 210,12ab ± 125,00 52,09a ± 6,14 ns ns * ns *
<500 2 acetaldehído 6,52a ± 0,30 12,50abc ± 5,28 8,51ab ± 3,48 11,95abc ± 0,04 ns ** ** ns **
550 12 2-metilpropanal 6,63ab ± 4,77 5,56ab ± 3,85 6,31ab ± 4,35 4,76ab ± 2,83 ns ns *** ns *
649 22 3-metilbutanal 135,19 ± 133,44 62,77 ± 62,01 99,87 ± 75,51 7,63 ± 3,53 ns ns ns ns ns 660 24 2-metilbutanal 29,57abc ± 19,96 6,33a ± 0,70 30,45abc ± 19,96 3,80a ± 0,62 ns ns ** ns ***
698 27 pentanal 3,44ab ± 0,46 1,64a ± 0,31 3,84ab ± 1,65 1,99a ± 0,89 ns ns *** * ***
744 37 2-metil-2-butenal 1,34bcd ± 0,46 1,02bc ± 0,19 2,04d ± 0,92 0,48ab ± 0,00 *** *** *** *** ***
800 51 hexanal 40,48a ± 15,10 25,50a ± 14,16 36,88a ± 26,98 7,31a ± 1,67 ns ns *** *** *** 902 65 heptanal 4,78ab ± 1,36 3,89ab ± 1,41 4,16ab ± 1,63 1,46a ± 0,04 ns ns * ns ns
908 66 3-(metiltio)propanal 1,56abc ± 0,44 3,04cdef ± 0,69 1,77abc ± 0,66 1,08a ± 0,49 ns ns *** ** ***
969 74 benzaldehido 1,99 ± 0,09 3,47 ± 0,97 2,63 ± 1,44 2,26 ± 0,08 ** * ns * *
1006 83 octanal 3,08 ± 0,58 2,40 ± 1,07 2,76 ± 0,84 3,62 ± 0,36 ns ns ns ** ns 1053 87 bencenoacetaldehído 2,22ab ± 0,11 4,14ab ± 1,28 2,32ab ± 0,82 1,55a ± 0,00 ns * * ns *
1104 93 nonanal 4,41 ± 0,40 7,63 ± 2,09 7,50 ± 3,93 3,78 ± 0,47 ns ns ns * ns
1157 94 2-nonenal 0,67e ± 0,00 0,22bc ± 0,00 0,35cd ± 0,23 0,00a ± 0,00 *** *** *** ns ***
1209 99 decanal 0,19a ± 0,10 0,31a ± 0,14 0,20a ± 0,02 0,28a ± 0,12 ns ns *** ns *** 1495 108 dodecanal 1,01abc ± 0,69 0,16ab ± 0,06 0,54abc ± 0,37 0,12ab ± 0,01 ns * *** ns ***
CETONAS 180,58ab ± 65,09 194,32ab ± 14,04 217,87ab ± 98,74 214,50ab ± 61,78 ns ns ns ns ns
<500 5 2-propanona+propanal 72,88ab ± 25,93 87,97abc ± 0,45 89,89ab ± 54,92 157,07c ± 59,78 ns * ns * **
586 16 2,3-butanodiona 2,82a ± 0,85 15,96b ± 9,36 4,20a ± 3,77 1,11a ± 0,07 ns *** ** ** *** 598 17 2-butanona+butanal 44,87 ± 35,13 36,12 ± 8,92 46,38 ± 30,49 15,51 ± 7,97 ns ns * ns *
656 23 1-hidroxi-2-propanona 10,58 ± 8,87 3,49 ± 3,07 7,22 ± 4,22 5,32 ± 4,07 ns ns ns ns ns
687 26 2-pentanona 10,62a ± 5,86 23,79ab ± 4,19 35,15ab ± 21,76 10,31a ± 1,29 ns ns ** ns **
710 31 3-hidroxi-2-butanona 3,10ab ± 1,65 3,41abc ± 1,26 4,40ab ± 3,51 1,31a ± 0,48 ns ns *** ns *** 758 42 3-metil-2-pentanona 9,54c ± 1,56 2,39a ± 0,97 8,06bc ± 3,96 7,64bc ± 1,84 ns * *** * ***
790 49 2-hexanona 1,97abc ± 1,33 1,49ab ± 0,47 1,43ab ± 0,82 0,85a ± 0,50 * ns *** ns ***
792 50 ciclopentanona 0,79ab ± 0,22 1,08ab ± 0,38 2,85c ± 1,50 0,41ab ± 0,02 ns ns *** ** ***
891 61 2-heptanona 10,35abc ± 7,36 7,82abc ± 3,41 8,42abc ± 3,34 4,91a ± 0,04 ** ns ** ns * 964 73 4-metilciclohexanona 1,37 ± 0,36 1,18 ± 0,39 1,40 ± 0,59 1,18 ± 0,51 ns ns ** ns *
989 77 3-octanona 4,35c ± 1,70 3,24bc ± 1,02 2,19ab ± 0,73 2,38abc ± 0,03 ns ns *** ns ***
992 78 2-octanona 2,81ab ± 0,91 2,76ab ± 0,45 2,28a ± 1,45 2,34ab ± 0,86 * ns ** ns *
1039 86 3-octen-2-ona 1,36c ± 0,43 1,39c ± 0,15 1,23bc ± 0,46 0,90bc ± 0,21 * ns *** * *** 1090 91 2-nonanona 3,19ab ± 0,24 2,23ab ± 0,67 2,77ab ± 1,25 3,26b ± 1,50 ns ns *** ns **
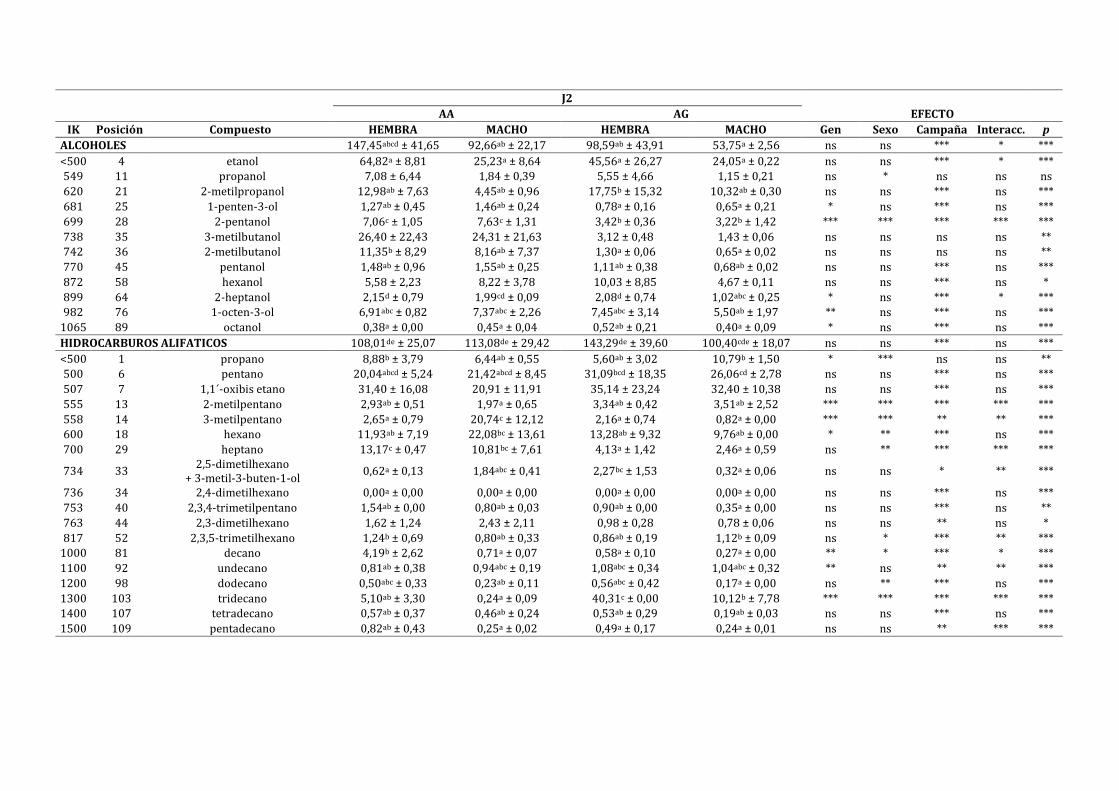
J2
AA AG EFECTO
IK Posición Compuesto HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
ALCOHOLES 147,45abcd ± 41,65 92,66ab ± 22,17 98,59ab ± 43,91 53,75a ± 2,56 ns ns *** * ***
<500 4 etanol 64,82a ± 8,81 25,23a ± 8,64 45,56a ± 26,27 24,05a ± 0,22 ns ns *** * *** 549 11 propanol 7,08 ± 6,44 1,84 ± 0,39 5,55 ± 4,66 1,15 ± 0,21 ns * ns ns ns
620 21 2-metilpropanol 12,98ab ± 7,63 4,45ab ± 0,96 17,75b ± 15,32 10,32ab ± 0,30 ns ns *** ns ***
681 25 1-penten-3-ol 1,27ab ± 0,45 1,46ab ± 0,24 0,78a ± 0,16 0,65a ± 0,21 * ns *** ns ***
699 28 2-pentanol 7,06c ± 1,05 7,63c ± 1,31 3,42b ± 0,36 3,22b ± 1,42 *** *** *** *** ***
738 35 3-metilbutanol 26,40 ± 22,43 24,31 ± 21,63 3,12 ± 0,48 1,43 ± 0,06 ns ns ns ns ** 742 36 2-metilbutanol 11,35b ± 8,29 8,16ab ± 7,37 1,30a ± 0,06 0,65a ± 0,02 ns ns ns ns **
770 45 pentanol 1,48ab ± 0,96 1,55ab ± 0,25 1,11ab ± 0,38 0,68ab ± 0,02 ns ns *** ns ***
872 58 hexanol 5,58 ± 2,23 8,22 ± 3,78 10,03 ± 8,85 4,67 ± 0,11 ns ns *** ns *
899 64 2-heptanol 2,15d ± 0,79 1,99cd ± 0,09 2,08d ± 0,74 1,02abc ± 0,25 * ns *** * *** 982 76 1-octen-3-ol 6,91abc ± 0,82 7,37abc ± 2,26 7,45abc ± 3,14 5,50ab ± 1,97 ** ns *** ns ***
1065 89 octanol 0,38a ± 0,00 0,45a ± 0,04 0,52ab ± 0,21 0,40a ± 0,09 * ns *** ns ***
HIDROCARBUROS ALIFATICOS 108,01de ± 25,07 113,08de ± 29,42 143,29de ± 39,60 100,40cde ± 18,07 ns ns *** ns ***
<500 1 propano 8,88b ± 3,79 6,44ab ± 0,55 5,60ab ± 3,02 10,79b ± 1,50 * *** ns ns ** 500 6 pentano 20,04abcd ± 5,24 21,42abcd ± 8,45 31,09bcd ± 18,35 26,06cd ± 2,78 ns ns *** ns ***
507 7 1,1´-oxibis etano 31,40 ± 16,08 20,91 ± 11,91 35,14 ± 23,24 32,40 ± 10,38 ns ns *** ns ***
555 13 2-metilpentano 2,93ab ± 0,51 1,97a ± 0,65 3,34ab ± 0,42 3,51ab ± 2,52 *** *** *** *** ***
558 14 3-metilpentano 2,65a ± 0,79 20,74c ± 12,12 2,16a ± 0,74 0,82a ± 0,00 *** *** ** ** *** 600 18 hexano 11,93ab ± 7,19 22,08bc ± 13,61 13,28ab ± 9,32 9,76ab ± 0,00 * ** *** ns ***
700 29 heptano 13,17c ± 0,47 10,81bc ± 7,61 4,13a ± 1,42 2,46a ± 0,59 ns ** *** *** ***
734 33 2,5-dimetilhexano
+ 3-metil-3-buten-1-ol 0,62a ± 0,13 1,84abc ± 0,41 2,27bc ± 1,53 0,32a ± 0,06 ns ns * ** ***
736 34 2,4-dimetilhexano 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 ns ns *** ns ***
753 40 2,3,4-trimetilpentano 1,54ab ± 0,00 0,80ab ± 0,03 0,90ab ± 0,00 0,35a ± 0,00 ns ns *** ns **
763 44 2,3-dimetilhexano 1,62 ± 1,24 2,43 ± 2,11 0,98 ± 0,28 0,78 ± 0,06 ns ns ** ns *
817 52 2,3,5-trimetilhexano 1,24b ± 0,69 0,80ab ± 0,33 0,86ab ± 0,19 1,12b ± 0,09 ns * *** ** *** 1000 81 decano 4,19b ± 2,62 0,71a ± 0,07 0,58a ± 0,10 0,27a ± 0,00 ** * *** * ***
1100 92 undecano 0,81ab ± 0,38 0,94abc ± 0,19 1,08abc ± 0,34 1,04abc ± 0,32 ** ns ** ** ***
1200 98 dodecano 0,50abc ± 0,33 0,23ab ± 0,11 0,56abc ± 0,42 0,17a ± 0,00 ns ** *** ns ***
1300 103 tridecano 5,10ab ± 3,30 0,24a ± 0,09 40,31c ± 0,00 10,12b ± 7,78 *** *** *** *** *** 1400 107 tetradecano 0,57ab ± 0,37 0,46ab ± 0,24 0,53ab ± 0,29 0,19ab ± 0,03 ns ns *** ns ***
1500 109 pentadecano 0,82ab ± 0,43 0,25a ± 0,02 0,49a ± 0,17 0,24a ± 0,01 ns ns ** *** ***
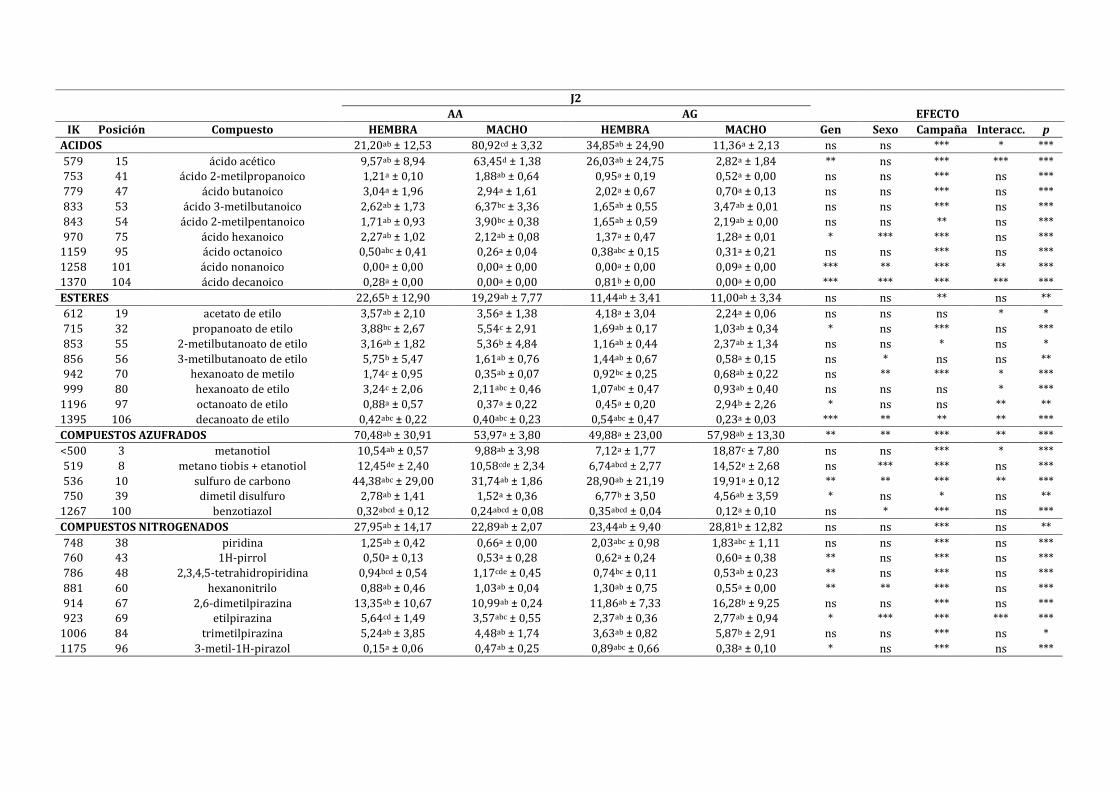
J2
AA AG EFECTO
IK Posición Compuesto HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
ACIDOS 21,20ab ± 12,53 80,92cd ± 3,32 34,85ab ± 24,90 11,36a ± 2,13 ns ns *** * ***
579 15 ácido acético 9,57ab ± 8,94 63,45d ± 1,38 26,03ab ± 24,75 2,82a ± 1,84 ** ns *** *** *** 753 41 ácido 2-metilpropanoico 1,21a ± 0,10 1,88ab ± 0,64 0,95a ± 0,19 0,52a ± 0,00 ns ns *** ns ***
779 47 ácido butanoico 3,04a ± 1,96 2,94a ± 1,61 2,02a ± 0,67 0,70a ± 0,13 ns ns *** ns ***
833 53 ácido 3-metilbutanoico 2,62ab ± 1,73 6,37bc ± 3,36 1,65ab ± 0,55 3,47ab ± 0,01 ns ns *** ns ***
843 54 ácido 2-metilpentanoico 1,71ab ± 0,93 3,90bc ± 0,38 1,65ab ± 0,59 2,19ab ± 0,00 ns ns ** ns ***
970 75 ácido hexanoico 2,27ab ± 1,02 2,12ab ± 0,08 1,37a ± 0,47 1,28a ± 0,01 * *** *** ns *** 1159 95 ácido octanoico 0,50abc ± 0,41 0,26a ± 0,04 0,38abc ± 0,15 0,31a ± 0,21 ns ns *** ns ***
1258 101 ácido nonanoico 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,09a ± 0,00 *** ** *** ** ***
1370 104 ácido decanoico 0,28a ± 0,00 0,00a ± 0,00 0,81b ± 0,00 0,00a ± 0,00 *** *** *** *** ***
ESTERES 22,65b ± 12,90 19,29ab ± 7,77 11,44ab ± 3,41 11,00ab ± 3,34 ns ns ** ns **
612 19 acetato de etilo 3,57ab ± 2,10 3,56a ± 1,38 4,18a ± 3,04 2,24a ± 0,06 ns ns ns * *
715 32 propanoato de etilo 3,88bc ± 2,67 5,54c ± 2,91 1,69ab ± 0,17 1,03ab ± 0,34 * ns *** ns ***
853 55 2-metilbutanoato de etilo 3,16ab ± 1,82 5,36b ± 4,84 1,16ab ± 0,44 2,37ab ± 1,34 ns ns * ns *
856 56 3-metilbutanoato de etilo 5,75b ± 5,47 1,61ab ± 0,76 1,44ab ± 0,67 0,58a ± 0,15 ns * ns ns ** 942 70 hexanoato de metilo 1,74c ± 0,95 0,35ab ± 0,07 0,92bc ± 0,25 0,68ab ± 0,22 ns ** *** * ***
999 80 hexanoato de etilo 3,24c ± 2,06 2,11abc ± 0,46 1,07abc ± 0,47 0,93ab ± 0,40 ns ns ns * ***
1196 97 octanoato de etilo 0,88a ± 0,57 0,37a ± 0,22 0,45a ± 0,20 2,94b ± 2,26 * ns ns ** **
1395 106 decanoato de etilo 0,42abc ± 0,22 0,40abc ± 0,23 0,54abc ± 0,47 0,23a ± 0,03 *** ** ** ** ***
COMPUESTOS AZUFRADOS 70,48ab ± 30,91 53,97a ± 3,80 49,88a ± 23,00 57,98ab ± 13,30 ** ** *** ** ***
<500 3 metanotiol 10,54ab ± 0,57 9,88ab ± 3,98 7,12a ± 1,77 18,87c ± 7,80 ns ns *** * ***
519 8 metano tiobis + etanotiol 12,45de ± 2,40 10,58cde ± 2,34 6,74abcd ± 2,77 14,52e ± 2,68 ns *** *** ns ***
536 10 sulfuro de carbono 44,38abc ± 29,00 31,74ab ± 1,86 28,90ab ± 21,19 19,91a ± 0,12 ** ** *** ** *** 750 39 dimetil disulfuro 2,78ab ± 1,41 1,52a ± 0,36 6,77b ± 3,50 4,56ab ± 3,59 * ns * ns **
1267 100 benzotiazol 0,32abcd ± 0,12 0,24abcd ± 0,08 0,35abcd ± 0,04 0,12a ± 0,10 ns * *** ns ***
COMPUESTOS NITROGENADOS 27,95ab ± 14,17 22,89ab ± 2,07 23,44ab ± 9,40 28,81b ± 12,82 ns ns *** ns **
748 38 piridina 1,25ab ± 0,42 0,66a ± 0,00 2,03abc ± 0,98 1,83abc ± 1,11 ns ns *** ns *** 760 43 1H-pirrol 0,50a ± 0,13 0,53a ± 0,28 0,62a ± 0,24 0,60a ± 0,38 ** ns *** ns ***
786 48 2,3,4,5-tetrahidropiridina 0,94bcd ± 0,54 1,17cde ± 0,45 0,74bc ± 0,11 0,53ab ± 0,23 ** ns *** ns ***
881 60 hexanonitrilo 0,88ab ± 0,46 1,03ab ± 0,04 1,30ab ± 0,75 0,55a ± 0,00 ** ** *** ns ***
914 67 2,6-dimetilpirazina 13,35ab ± 10,67 10,99ab ± 0,24 11,86ab ± 7,33 16,28b ± 9,25 ns ns *** ns *** 923 69 etilpirazina 5,64cd ± 1,49 3,57abc ± 0,55 2,37ab ± 0,36 2,77ab ± 0,94 * *** *** *** ***
1006 84 trimetilpirazina 5,24ab ± 3,85 4,48ab ± 1,74 3,63ab ± 0,82 5,87b ± 2,91 ns ns *** ns *
1175 96 3-metil-1H-pirazol 0,15a ± 0,06 0,47ab ± 0,25 0,89abc ± 0,66 0,38a ± 0,10 * ns *** ns ***

J2
AA AG EFECTO
IK Posición Compuesto HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
FURANOS 16,40d ± 1,86 13,74abcd ± 2,17 15,39bcd ± 3,23 9,49a ± 1,07 *** *** *** ns ***
702 30 2-etilfurano 6,01d ± 0,19 3,99cd ± 1,21 4,41cd ± 2,17 3,34bc ± 0,13 ns ns *** ns *** 892 62 2-butilfurano 1,06bc ± 0,25 1,01bc ± 0,35 1,51cd ± 0,54 0,60ab ± 0,06 ** ** *** * ***
915 68 dihidro-2(3H)-furanona 1,10a ± 0,16 1,03a ± 0,09 3,26abc ± 2,15 1,24ab ± 0,42 ns ns *** ns ***
958 72 dihidro-5-metil-2(3H)-furanona 0,78ab ± 0,41 0,79ab ± 0,39 0,84ab ± 0,27 0,55ab ± 0,24 ns ns ** ns ns
996 79 2-pentilfurano 5,64cd ± 1,49 3,57abc ± 0,55 2,37ab ± 0,36 2,77ab ± 0,94 * *** *** *** ***
1060 88 dihidro-5-etil-2(3H)-furanona 1,17ab ± 0,12 2,06abcde ± 0,92 1,61ab ± 1,16 0,70a ± 0,00 *** ns *** * *** 1272 102 dihidro-5-butil-2(3H)-furanona 0,13 ± 0,04 0,48 ± 0,31 0,67 ± 0,45 0,13 ± 0,07 ns * ns * **
1377 105 dihidro-5-pentil-2(3H)-furanona 0,50ab ± 0,28 0,81ab ± 0,32 0,73ab ± 0,46 0,17a ± 0,08 * ns ns * ns
HIDROCARBUROS AROMATICOS 13,37abc ± 5,85 7,83ab ± 0,45 12,01abc ± 5,01 6,98a ± 0,88 ** * ns ns **
772 46 metilbenceno 8,33ab ± 5,74 2,73ab ± 0,27 6,42ab ± 4,49 2,42a ± 0,41 * ** ns ns ** 866 57 etilbenceno 0,29a ± 0,01 0,50abc ± 0,03 0,62abc ± 0,39 0,39ab ± 0,00 ns ns *** * ***
875 59 m- y p- xileno 1,87a ± 0,14 1,75a ± 0,27 2,16a ± 0,98 1,79a ± 0,61 * * *** * ***
894 63 estireno 1,52cd ± 0,12 1,50c ± 0,11 1,63c ± 0,43 1,39c ± 0,38 *** ** *** ns ***
1002 82 1,3,5-trimetilbenceno 0,99abc ± 0,11 0,86abc ± 0,29 0,73ab ± 0,12 0,75ab ± 0,01 ** ns *** ns *** 1074 90 4-metilfenol 0,36a ± 0,02 0,49ab ± 0,26 0,45ab ± 0,20 0,24a ± 0,05 ns ns *** ns ***
TERPENOS 0,86abc ± 0,31 1,24bcd ± 0,13 1,23bcd ± 0,24 1,20bcd ± 0,09 * ** *** ns ***
943 71 alfa-pineno 0,41abc ± 0,17 0,89cd ± 0,13 0,53abc ± 0,22 0,64abcd ± 0,10 ns ns *** ns ***
1035 85 l-limoneno 0,45ab ± 0,19 0,35ab ± 0,03 0,70b ± 0,07 0,57ab ± 0,04 ** ** *** *** ***
COMPUESTOS CLORADOS 342,74ab ± 49,53 157,09ab ± 101,38 364,01ab ± 97,72 232,41ab ± 9,13 ** ns ns ns **
528 9 diclorometano 0,00a ± 0,00 0,69b ± 0,00 1,80cd ± 0,66 2,09d ± 0,03 *** ** *** ns ***
616 20 cloroformo 342,74ab ± 49,53 156,39ab ± 101,38 362,21ab ± 97,65 230,32ab ± 9,12 ** ns ns ns **
sumatorio 1.195,85ab ± 264,57 896,09ab ± 199,24 1.182,30ab ± 347,46 778,88a ± 89,97 * ns ns ns *
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

Tabla 4.1.57. Compuestos volátiles identificados en jamón curado agrupados por familia química. Los contenidos de cada compuesto se expresan en UAA (106) (± desviación estándar de la media). Parte III.
J3
AA AG EFECTO
IK Posición Compuesto HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
ALDEHIDOS 191,09ab ± 11,26 293,38b ± 10,01 213,70ab ± 0,00 223,54ab ± 61,12 ns ns * ns *
<500 2 acetaldehído 16,12bc ± 0,34 17,46c ± 4,14 9,83abc ± 0,00 13,67abc ± 5,13 ns ** ** ns **
550 12 2-metilpropanal 9,69ab ± 0,71 10,51b ± 0,20 6,22ab ± 0,00 7,19ab ± 1,50 ns ns *** ns *
649 22 3-metilbutanal 70,31 ± 1,73 107,27 ± 37,62 68,91 ± 0,00 101,35 ± 52,76 ns ns ns ns ns
660 24 2-metilbutanal 25,55abc ± 3,19 45,44c ± 3,78 20,78abc ± 0,00 34,80bc ± 9,36 ns ns ** ns *** 698 27 pentanal 4,17abc ± 1,02 6,58bc ± 2,19 6,55bc ± 0,00 3,10ab ± 0,51 ns ns *** * ***
744 37 2-metil-2-butenal 0,47ab ± 0,02 1,05bc ± 0,55 0,98abc ± 0,00 0,89abc ± 0,23 *** *** *** *** ***
800 51 hexanal 39,81a ± 8,46 79,35b ± 35,94 78,00b ± 0,00 35,86a ± 11,03 ns ns *** *** ***
902 65 heptanal 4,05ab ± 0,37 5,39ab ± 0,73 6,26b ± 0,00 5,62ab ± 2,52 ns ns * ns ns 908 66 3-(metiltio)propanal 3,93ef ± 0,83 4,38f ± 0,11 2,85bcdef ± 0,00 3,55def ± 0,00 ns ns *** ** ***
969 74 benzaldehido 3,72 ± 0,17 3,58 ± 1,00 1,83 ± 0,00 3,19 ± 0,59 ** * ns * *
1006 83 octanal 3,01 ± 0,16 2,97 ± 0,68 3,43 ± 0,00 3,33 ± 0,04 ns ns ns ** ns
1053 87 bencenoacetaldehído 2,17ab ± 0,02 3,79ab ± 1,48 2,84ab ± 0,00 3,34ab ± 1,51 ns * * ns * 1104 93 nonanal 7,15 ± 0,55 4,93 ± 0,40 4,70 ± 0,00 6,86 ± 2,62 ns ns ns * ns
1157 94 2-nonenal 0,29bcd ± 0,07 0,44d ± 0,05 0,17ab ± 0,00 0,34bcd ± 0,10 *** *** *** ns ***
1209 99 decanal 0,37a ± 0,17 0,24a ± 0,04 0,34a ± 0,00 0,22a ± 0,01 ns ns *** ns ***
1495 108 dodecanal 0,29ab ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,24ab ± 0,04 ns * *** ns ***
CETONAS 190,97ab ± 10,86 259,46b ± 36,04 113,93a ± 0,00 165,31ab ± 18,38 ns ns ns ns ns
<500 5 2-propanona+propanal 103,24abc ± 11,50 142,83bc ± 18,22 55,77a ± 0,00 70,05ab ± 0,85 ns * ns * **
586 16 2,3-butanodiona 0,00a ± 0,00 14,22b ± 0,00 2,62a ± 0,00 16,14b ± 2,10 ns *** ** ** ***
598 17 2-butanona+butanal 12,53 ± 6,08 14,33 ± 0,67 7,54 ± 0,00 28,54 ± 9,40 ns ns * ns * 656 23 1-hidroxi-2-propanona 4,13 ± 0,15 5,04 ± 0,46 1,54 ± 0,00 4,40 ± 0,16 ns ns ns ns ns
687 26 2-pentanona 36,39ab ± 1,64 42,35ab ± 46,24 19,93ab ± 0,00 21,85ab ± 11,30 ns ns ** ns **
710 31 3-hidroxi-2-butanona 4,12abc ± 0,03 5,55abcd ± 2,05 5,97abcd ± 0,00 7,71bcde ± 2,87 ns ns *** ns ***
758 42 3-metil-2-pentanona 0,70a ± 0,02 1,25a ± 1,07 0,56a ± 0,00 0,41a ± 0,10 ns * *** * *** 790 49 2-hexanona 3,07abc ± 0,99 3,20bc ± 1,22 2,35abc ± 0,00 2,07abc ± 0,87 * ns *** ns ***
792 50 ciclopentanona 1,41abc ± 0,02 1,86bc ± 0,76 1,09ab ± 0,00 1,28abc ± 0,39 ns ns *** ** ***
891 61 2-heptanona 16,72bc ± 0,41 16,86c ± 6,51 10,87abc ± 0,00 6,48ab ± 0,08 ** ns ** ns *
964 73 4-metilciclohexanona 1,10 ± 0,05 0,83 ± 0,28 0,41 ± 0,00 0,54 ± 0,02 ns ns ** ns * 989 77 3-octanona 1,26ab ± 0,06 1,90ab ± 0,28 0,97a ± 0,00 1,49ab ± 0,61 ns ns *** ns ***
992 78 2-octanona 3,65ab ± 1,90 4,59b ± 2,28 2,53ab ± 0,00 2,17ab ± 0,83 * ns ** ns *
1039 86 3-octen-2-ona 1,26c ± 0,02 1,36c ± 0,83 0,57ab ± 0,00 0,83bc ± 0,14 * ns *** * ***
1090 91 2-nonanona 1,37ab ± 0,07 3,27b ± 2,38 1,21ab ± 0,00 1,36ab ± 0,50 ns ns *** ns **

J3
AA AA EFECTO
IK Posición Compuesto HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
ALCOHOLES 84,28a ± 8,34 111,50abc ± 5,87 230,14cde ± 0,00 66,76a ± 4,28 ns ns *** * ***
<500 4 etanol 42,71a ± 3,16 50,48a ± 21,79 195,98bc ± 0,00 40,68a ± 0,23 ns ns *** * *** 549 11 propanol 3,06 ± 1,64 2,06 ± 0,00 5,23 ± 0,00 1,27 ± 0,05 ns * ns ns ns
620 21 2-metilpropanol 5,20ab ± 0,53 5,89ab ± 1,48 0,52a ± 0,00 4,29ab ± 0,67 ns ns *** ns ***
681 25 1-penten-3-ol 3,77cd ± 0,30 4,12d ± 3,54 1,89ab ± 0,00 2,95bcd ± 0,25 * ns *** ns ***
699 28 2-pentanol 3,39b ± 0,16 13,96d ± 0,00 0,00a ± 0,00 0,00a ± 0,00 *** *** *** *** ***
738 35 3-metilbutanol 2,57 ± 0,13 6,12 ± 0,89 9,10 ± 0,00 3,60 ± 1,65 ns ns ns ns ** 742 36 2-metilbutanol 1,00a ± 0,65 2,93ab ± 0,97 3,70ab ± 0,00 1,18a ± 0,56 ns ns ns ns **
770 45 pentanol 2,07bc ± 0,47 3,87c ± 3,09 1,75ab ± 0,00 2,10bc ± 0,98 ns ns *** ns ***
872 58 hexanol 7,48 ± 3,23 11,38 ± 12,26 6,27 ± 0,00 2,23 ± 0,35 ns ns *** ns *
899 64 2-heptanol 1,16abcd ± 0,01 0,86ab ± 0,46 0,49ab ± 0,00 0,91ab ± 0,32 * ns *** * *** 982 76 1-octen-3-ol 10,85c ± 0,17 8,64bc ± 5,03 4,59ab ± 0,00 6,71abc ± 1,54 ** ns *** ns ***
1065 89 octanol 1,01abc ± 0,31 1,19abcd ± 0,87 0,62ab ± 0,00 0,82abc ± 0,12 * ns *** ns ***
HIDROCARBUROS ALIFATICOS 83,92abcd ± 17,65 149,36e ± 29,14 83,66abcd ± 0,00 94,31bcd ± 7,00 ns ns *** ns ***
<500 1 propano 9,05b ± 0,76 9,94b ± 3,09 3,12a ± 0,00 8,62ab ± 0,53 * *** ns ns ** 500 6 pentano 27,75d ± 9,18 27,78d ± 0,44 23,60abcd ± 0,00 30,10d ± 2,55 ns ns *** ns ***
507 7 1,1´-oxibis etano 17,43 ± 2,55 27,27 ± 6,96 13,75 ± 0,00 19,32 ± 8,71 ns ns *** ns ***
555 13 2-metilpentano 4,73b ± 1,73 14,48c ± 2,75 2,90ab ± 0,00 2,49ab ± 1,07 *** *** *** *** ***
558 14 3-metilpentano 5,92a ± 2,43 17,44bc ± 2,27 4,28a ± 0,00 8,84ab ± 2,61 *** *** ** ** *** 600 18 hexano 9,66ab ± 1,18 36,28c ± 9,90 11,87ab ± 0,00 14,10ab ± 1,08 * ** *** ns ***
700 29 heptano 0,00a ± 0,00 4,96ab ± 0,00 16,66c ± 0,00 3,08a ± 0,59 ns ** *** *** ***
734 33 2,5-dimetilhexano
+ 3-metil-3-buten-1-ol 1,04ab ± 0,00 1,38abc ± 0,28 0,56a ± 0,00 0,84ab ± 0,59 ns ns * ** ***
736 34 2,4-dimetilhexano 0,41a ± 0,05 0,68a ± 0,03 0,50a ± 0,00 0,29a ± 0,00 ns ns *** ns ***
753 40 2,3,4-trimetilpentano 0,77a ± 0,08 0,93ab ± 0,11 1,05ab ± 0,00 0,63a ± 0,06 ns ns *** ns **
763 44 2,3-dimetilhexano 0,55 ± 0,01 0,42 ± 0,11 1,20 ± 0,00 0,32 ± 0,07 ns ns ** ns *
817 52 2,3,5-trimetilhexano 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 ns * *** ** *** 1000 81 decano 1,16a ± 0,67 0,43a ± 0,00 0,70a ± 0,00 0,60a ± 0,05 ** * *** * ***
1100 92 undecano 1,79d ± 0,13 1,14abc ± 0,30 1,02abc ± 0,00 1,21abcd ± 0,09 ** ns ** ** ***
1200 98 dodecano 0,88bcd ± 0,09 0,90cd ± 0,25 0,80abc ± 0,00 0,97cd ± 0,25 ns ** *** ns ***
1300 103 tridecano 0,94a ± 0,10 1,09a ± 0,56 0,46a ± 0,00 0,87a ± 0,15 *** *** *** *** *** 1400 107 tetradecano 1,13b ± 0,48 2,25c ± 2,06 0,62ab ± 0,00 1,10b ± 0,02 ns ns *** ns ***
1500 109 pentadecano 0,73ab ± 0,09 2,00c ± 1,97 0,57a ± 0,00 0,93ab ± 0,18 ns ns ** *** ***

J3
AA AA EFECTO
IK Posición Compuesto HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
ACIDOS 52,53abcd ± 7,06 69,98bcd ± 8,14 33,32abc ± 0,00 66,13bcd ± 7,24 ns ns *** * ***
579 15 ácido acético 39,63bcd ± 7,61 43,52bcd ± 0,53 21,88abc ± 0,00 44,35bcd ± 7,85 ** ns *** *** *** 753 41 ácido 2-metilpropanoico 0,79a ± 0,02 1,29a ± 0,30 0,75a ± 0,00 0,99a ± 0,04 ns ns *** ns ***
779 47 ácido butanoico 6,56ab ± 0,30 11,20ab ± 2,08 6,19ab ± 0,00 10,65ab ± 4,03 ns ns *** ns ***
833 53 ácido 3-metilbutanoico 1,41ab ± 0,03 3,93ab ± 0,15 1,27a ± 0,00 2,38ab ± 1,08 ns ns *** ns ***
843 54 ácido 2-metilpentanoico 1,14a ± 0,13 2,32ab ± 0,44 0,91a ± 0,00 1,52ab ± 0,54 ns ns ** ns ***
970 75 ácido hexanoico 2,16ab ± 0,33 6,95c ± 4,81 1,57a ± 0,00 4,60bc ± 1,92 * *** *** ns *** 1159 95 ácido octanoico 0,51abc ± 0,16 0,56abc ± 0,32 0,41ab ± 0,00 0,42ab ± 0,04 ns ns *** ns ***
1258 101 ácido nonanoico 0,00a ± 0,00 0,00a ± 0,00 0,22ab ± 0,00 1,06d ± 0,00 *** ** *** ** ***
1370 104 ácido decanoico 0,34a ± 0,00 0,21a ± 0,11 0,14a ± 0,00 0,16a ± 0,04 *** *** *** *** ***
ESTERES 5,82a ± 0,92 6,57a ± 0,03 16,22ab ± 0,00 7,70a ± 0,54 ns ns ** ns **
612 19 acetato de etilo 1,77a ± 0,61 3,90ab ± 0,54 6,88b ± 0,00 3,42a ± 0,37 ns ns ns * *
715 32 propanoato de etilo 0,79ab ± 0,00 0,00a ± 0,00 1,27ab ± 0,00 0,59ab ± 0,18 * ns *** ns ***
853 55 2-metilbutanoato de etilo 0,47a ± 0,00 0,00a ± 0,00 0,80ab ± 0,00 2,21ab ± 0,00 ns ns * ns *
856 56 3-metilbutanoato de etilo 0,64a ± 0,02 0,75a ± 0,11 1,37ab ± 0,00 0,43a ± 0,02 ns * ns ns ** 942 70 hexanoato de metilo 0,66ab ± 0,00 0,44ab ± 0,00 0,24ab ± 0,00 0,29ab ± 0,02 ns ** *** * ***
999 80 hexanoato de etilo 0,70ab ± 0,15 0,96ab ± 0,19 2,81bc ± 0,00 0,42a ± 0,03 ns ns ns * ***
1196 97 octanoato de etilo 0,46a ± 0,32 0,17a ± 0,05 1,29ab ± 0,00 0,18a ± 0,00 * ns ns ** **
1395 106 decanoato de etilo 0,34ab ± 0,13 0,34ab ± 0,22 1,56d ± 0,00 0,17a ± 0,03 *** ** ** ** ***
COMPUESTOS AZUFRADOS 114,39ab ± 12,39 77,36ab ± 15,53 262,69c ± 0,00 132,21b ± 43,42 ** ** *** ** ***
<500 3 metanotiol 15,12bc ± 0,98 11,74abc ± 0,37 10,07ab ± 0,00 11,69abc ± 4,55 ns ns *** * ***
519 8 metano tiobis + etanotiol 10,02cde ± 1,62 11,44de ± 6,60 4,67abc ± 0,00 12,17de ± 2,54 ns *** *** ns ***
536 10 sulfuro de carbono 87,01abc ± 12,43 51,80abc ± 9,66 245,06d ± 0,00 106,20c ± 42,30 ** ** *** ** *** 750 39 dimetil disulfuro 2,02a ± 0,02 2,21a ± 0,32 2,69ab ± 0,00 1,99a ± 0,05 * ns * ns **
1267 100 benzotiazol 0,23abcd ± 0,01 0,16ab ± 0,04 0,20abc ± 0,00 0,15a ± 0,00 ns * *** ns ***
COMPUESTOS NITROGENADOS 16,22ab ± 0,57 16,15ab ± 4,41 13,07ab ± 0,00 9,99a ± 0,69 ns ns *** ns **
748 38 piridina 0,38a ± 0,02 0,90ab ± 0,30 0,60a ± 0,00 0,58a ± 0,15 ns ns *** ns *** 760 43 1H-pirrol 0,42a ± 0,00 0,49a ± 0,11 0,23a ± 0,00 0,38a ± 0,01 ** ns *** ns ***
786 48 2,3,4,5-tetrahidropiridina 1,42de ± 0,02 1,65e ± 0,18 1,13bcde ± 0,00 1,17bcde ± 0,01 ** ns *** ns ***
881 60 hexanonitrilo 3,82e ± 0,51 2,64cd ± 1,52 2,00bc ± 0,00 1,44abc ± 0,16 ** ** *** ns ***
914 67 2,6-dimetilpirazina 2,33a ± 0,41 2,35a ± 0,36 1,48a ± 0,00 2,17a ± 0,40 ns ns *** ns *** 923 69 etilpirazina 4,41bcd ± 0,50 3,62abc ± 0,02 5,98d ± 0,00 1,58a ± 0,27 * *** *** *** ***
1006 84 trimetilpirazina 2,34ab ± 0,07 3,46ab ± 1,73 0,94a ± 0,00 1,74ab ± 0,19 ns ns *** ns *
1175 96 3-metil-1H-pirazol 1,10abc ± 0,22 1,04abc ± 0,23 0,72ab ± 0,00 0,94abc ± 0,13 * ns *** ns ***

J3
AA AA EFECTO
IK Posición Compuesto HEMBRA MACHO HEMBRA MACHO Gen Sexo Generación Interacc. p
FURANOS 16,07cd ± 1,18 16,58d ± 3,27 14,58bcd ± 0,00 11,76abc ± 1,41 *** *** *** ns ***
702 30 2-etilfurano 1,62ab ± 0,59 2,32bc ± 0,64 3,07bc ± 0,00 1,83abc ± 0,95 ns ns *** ns *** 892 62 2-butilfurano 2,49e ± 0,43 2,05de ± 0,38 1,59cd ± 0,00 1,43cd ± 0,09 ** ** *** * ***
915 68 dihidro-2(3H)-furanona 3,14abc ± 0,31 5,21c ± 0,62 1,28ab ± 0,00 3,61bc ± 0,03 ns ns *** ns ***
958 72 dihidro-5-metil-2(3H)-furanona 0,73ab ± 0,00 0,68ab ± 0,09 0,42a ± 0,00 0,68ab ± 0,01 ns ns ** ns ns
996 79 2-pentilfurano 4,41bcd ± 0,50 3,62abc ± 0,02 5,98d ± 0,00 1,58a ± 0,27 * *** *** *** ***
1060 88 dihidro-5-etil-2(3H)-furanona 2,28bcde ± 0,60 1,71abc ± 0,87 1,36ab ± 0,00 1,75abc ± 0,66 *** ns *** * *** 1272 102 dihidro-5-butil-2(3H)-furanona 0,65 ± 0,01 0,45 ± 0,36 0,41 ± 0,00 0,30 ± 0,01 ns * ns * **
1377 105 dihidro-5-pentil-2(3H)-furanona 0,75ab ± 0,07 0,53ab ± 0,34 0,47ab ± 0,00 0,58ab ± 0,31 * ns ns * ns
HIDROCARBUROS AROMATICOS 12,81c ± 1,58 16,39abc ± 4,72 8,48abc ± 0,00 10,09abc ± 1,45 ** * ns ns **
772 46 metilbenceno 6,46ab ± 0,52 5,18ab ± 1,65 3,92ab ± 0,00 3,08ab ± 0,50 * ** ns ns ** 866 57 etilbenceno 0,43ab ± 0,03 0,86abc ± 0,52 0,22a ± 0,00 0,86abc ± 0,46 ns ns *** * ***
875 59 m- y p- xileno 3,19a ± 0,84 6,55b ± 2,01 2,96a ± 0,00 3,41a ± 1,59 * * *** * ***
894 63 estireno 1,30c ± 0,18 2,22d ± 1,13 0,57ab ± 0,00 1,15bc ± 0,41 *** ** *** ns ***
1002 82 1,3,5-trimetilbenceno 0,86abc ± 0,27 0,82ab ± 0,42 0,50a ± 0,00 0,64a ± 0,01 ** ns *** ns *** 1074 90 4-metilfenol 0,56ab ± 0,06 0,76ab ± 0,02 0,31a ± 0,00 0,95abc ± 0,55 ns ns *** ns ***
TERPENOS 0,71ab ± 0,05 0,66ab ± 0,10 0,38a ± 0,00 0,86abc ± 0,36 * ** *** ns ***
943 71 alfa-pineno 0,26abc ± 0,01 0,24ab ± 0,04 0,14a ± 0,00 0,23ab ± 0,00 ns ns *** ns ***
1035 85 l-limoneno 0,45ab ± 0,04 0,43ab ± 0,06 0,24a ± 0,00 0,63ab ± 0,36 ** ** *** *** ***
COMPUESTOS CLORADOS 365,14ab ± 81,86 306,87ab ± 2,33 50,49a ± 0,00 243,37ab ± 142,25 ** ns ns ns **
528 9 diclorometano 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 *** ** *** ns ***
616 20 cloroformo 365,14ab ± 81,86 306,87ab ± 2,33 50,49a ± 0,00 243,37ab ± 142,25 ** ns ns ns **
sumatorio 1.130,44ab ± 85,25 1.321,32ab ± 88,49 1.035,78ab ± 0,00 1.031,13ab ± 135,44 * ns ns ns *
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
136
4.2. PALETA CURADA
4.2.1. CARACTERIZACIÓN DE LA PALETA CURADA
Debido a la escasez de trabajos publicados sobre paleta curada, los resultados de la
caracterización de las paletas de Teruel se compararán con los jamones de Teruel del
mismo estudio y con paletas Ibéricas de cebo analizadas en el mismo lugar y bajo las
mismas condiciones en un proyecto paralelo al nuestro (Ref,. RTA 2008-00026). El
Consejo Regulador de la DOP Jamón de Teruel no ampara dichos productos, aún así desde
2008 poseen una marca diferenciada de calidad denominada “C´alial” regulada por el
Departamento de Agricultura y Alimentación del Gobierno de Aragón (BOA, 2008).
Características generales de la paleta curada
La tabla 4.2.1 muestra los valores medios de las medidas morfológicas de todas las
paletas curadas analizadas en el presente estudio (n=30).
Tabla 4.2.1. Medidas morfológicas generales de la paleta Curada (± desviación estándar de la media)
Peso (g) Longitud (cm) Anchura (cm) Perímetro (cm)
4,5 ± 0,4 62,9 ± 1,2 24,3 ± 1,1 55,0 ± 1,6
El peso de las paletas fue prácticamente la mitad que el de los jamones, con una
longitud y perímetro inferiores, siendo la anchura el parámetro más parecido entre ambas
piezas.
La marca “C´alial” establece un peso superior de 4 Kg para las paletas curadas, por
lo que todas las piezas analizadas se encuentraron dentro de los valores establecidos para
el peso. El resto de parámetros no se encuentran especificados en dicha normativa (BOA,
2008).
Las tablas 4.2.2 y 4.2.3 muestran el espesor de la grasa subcutánea de las zonas
proximal y distal de la porción utilizada para el análisis físico-químico, tal como se explicó
en la Figura 3.2.9 del apartado material y métodos.

CAPÍTULO IV: RESULTADOS
137
Figura 4.2.1. Zona proximal de la paleta curada
Figura 4.2.2. Zona distal de la paleta curada
Perfil de textura instrumental
La tabla 4.2.4 muestra los resultados obtenidos del análisis de perfil de textura. Los
valores de dureza fueron superiores en la paleta curada (19,97 N) en comparación con el
jamón (11,48 N) (tabla 4.1.5). Si consideramos que la textura es una característica que está
directamente relacionada con la estructura del músculo, con el contenido de grasa
intramuscular y con la rampa de temperaturas utilizada durante el proceso de secado-
maduración de las muestras, estos valores serían esperados dada la menor musculatura y
el menor grado de humedad de las paletas. Además se encontraron valores superiores en
el porcentaje de cloruros, parámetros relacionados también con la dureza, tal como
observó Ruiz-Ramírez et al., (2006). Otros parámetros a destacar son la mayor gomosidad
y masticabilidad de las paletas (9,36 N; 5,60 N) frente a los jamones (5,15 N; 3,08 N) (tabla
4.1.5). El resto de parámetros mostraron valores similares en ambos productos. Al
comparar la paleta curada con la Ibérica se encontraron valores inferiores en la paleta
curada en los parámetros de dureza (31,27 N (paleta Ibérica)), gomosidad (14,90N (paleta
Ibérica)) y masticabilidad (9,22 N (paleta Ibérica)) (Reina et al., 2010).
Tabla 4.2.2. Medidas de la grasa subcutánea de la zona proximal de la paleta curada (± desviación estándar de la media)
A (cm) B (cm) C (cm)
2,7 ± 0,7 2,9 ± 0,7 4,1 ± 0,8
Tabla 4.2.3. Medidas de la grasa subcutánea de la zona distal de la paleta curada (± desviación estándar de la media)
A (cm) B (cm) C (cm)
3,5 ± 1,2 1,3 ± 0,4 1,7 ± 0,7

CAPÍTULO IV: RESULTADOS
138
Tabla 4.2.4. Análisis de perfil de textura (TPA) para la paleta curada (± desviación estándar de la media)
Fuerza (N) 18,09 ± 2,99 Dureza (N) 19,97 ± 3,93
Fracturabilidad (Adimensional) 10,01 ± 0,56
Adhesividad (N*seg) -0,20 ± 0,05
Elasticidad (cm) 0,59 ± 0,04 Cohesividad 0,49 ± 0,02
Gomosidad (N) 9,36 ± 1,82
Masticabilidad (N) 5,60 ± 1,15
(N): Newton
Parámetros generales físico-químicos
Con el fin de obtener valores de referencia para la discusión de los resultados de
las distintas determinaciones, se midieron una serie de parámetros generales entre los
que se encuentran la humedad, proteínas y grasa intramuscular. Junto con estos, están
incluidos en la tabla 4.2.5 el resto de parámetros relacionados con la composición química
del producto.
Tabla 4.2.5. Parámetros físico-químicos generales de la paleta curada (± desviación estándar de la media).
Humedad (g/100 g) 46,34 ± 6,57
Proteínas (f) (g/100 g) 38,70 ± 0,83 Proteínas (s, dg) (g/100 g) 87,30 ± 2,58
Grasa (f) (g/100 g) 7,04 ± 1,14
Grasa (s) (g/100 g) 12,76 ± 3,63
L (Adimensional) 33,98 ± 2,35 a (Adimensional) 12,74 ± 1,63
b (Adimensional) 5,71 ± 0,74
mg Mb/g m 4,42 ± 0,89
ppm Fe heminico 14,88 ± 2,97 Cloruros (f) (g/100 g) 4,34 ± 0,71
Cloruros (s) (g/100 g) 8,95 ± 1,05
pH (Adimensional) 5,88 ± 0,29
mg MDA/Kg 0,14 ± 0,07
(f): extracto fresco; (s): extracto seco; (s, dg): extracto seco y desengrasado
Como se observa en la tabla 4.2.5, la humedad media de las paletas fue de 46,34%,
un valor inferior al obtenido en el jamón curado y en las paletas Ibéricas (50,49%), con
medidas morfológicas muy similares. Los resultados se encuentran por debajo de la norma
de calidad, que establece un máximo de un 55%.
El porcentaje medio de proteínas fue de 38,70%, superior al mostrado para el
jamón de Teruel (33,16%) (tabla 4.1.5) y al de las paletas Ibéricas (34,29%) (Reina et al.,
2010). El porcentaje medio de grasa intramuscular obtenido fue de 7,04%, muy superior
al que encontramos en el jamón (4,36%) (tabla 4.1.5), pero algo inferior al de las paletas
Ibéricas (7,49%) (Reina et al., 2010). El porcentaje de cloruros presente en las paletas fue

CAPÍTULO IV: RESULTADOS
139
de 4,34%, superior al descrito por el jamón, con un 3,49% (tabla 4.1.5) pero muy inferior
al encontrado en las paletas Ibéricas, con un valor de 5,45% (Reina et al., 2010). El
contenido en materia seca fue de 8,95%, muy por debajo al límite establecido por la norma
de calidad, que indica un máximo de un 15%.
El color es también una de las características de apariencia más destacadas en la
paleta curada y que dependerá, al igual que en el jamón, de la humedad, del contenido de
grasa intramuscular y del contenido en mioglobina. El valor medio del contenido en
mioglobina de las paletas fue de 4,42 mg/g, un valor muy superior al que encontramos en
el jamón. La concentración de mioglobina depende del tipo de músculo (Aristoy y Toldrá,
1998), además de otros factores ya citados, como la edad del animal o el ejercicio que
realiza éste. Las paletas de Teruel mostraron un valor superior a las Ibéricas (Reina et al.,
2010).
En cuanto al color instrumental, las paletas presentaron un valor de la coordenada
L de 33,98, inferior al descrito para el jamón (tabla 4.1.5). Sin embargo, la coordenada a*
fue de 12,74, inferior al del jamón, que presentó un valor de 15,01 (tabla 4.1.5), a pesar de
presentar un contenido en mioglobina superior. Al comparar con las paletas Ibéricas, éstas
presentaron un valor de luminosidad superior (37,52), pero también de la coordenada a*
(15,87), valores más acordes con lo esperado por su elevado contenido en mioglobina
(Reina et al., 2010). La coordenada b* obtuvo un valor de 5,71, inferior al descrito para
jamón y para las paletas Ibéricas (6,55) (Reina et al., 2010). En cuanto al valor de mg de
malonaldehído, resultó inferior en las paletas (0,14 mg) en comparación con los jamones
(0,24 mg) (tabla 4.1.5), a pesar de que el contenido de grasa intramuscular fue superior.
Composición de la grasa intramuscular
En las paletas curadas, al igual que en los jamones, los triglicéridos (LN)
representaron la mayoría de los lípidos de la grasa intramuscular, constituyendo
aproximadamente el 78,7% (tabla 4.2.6) frente al 86,6% del jamón curado (tabla 4.1.6).
Sin embargo, el contenido de fosfolípidos (LP) y AGL fue superior en las paletas,
constituyendo para la fracción de AGL el 20% frente al 12,8% (tabla 4.1.6) descrito en el
jamón y el 1,3% de la fracción de fosfolípidos frente al 0,6% del jamón (tabla 4.1.6).
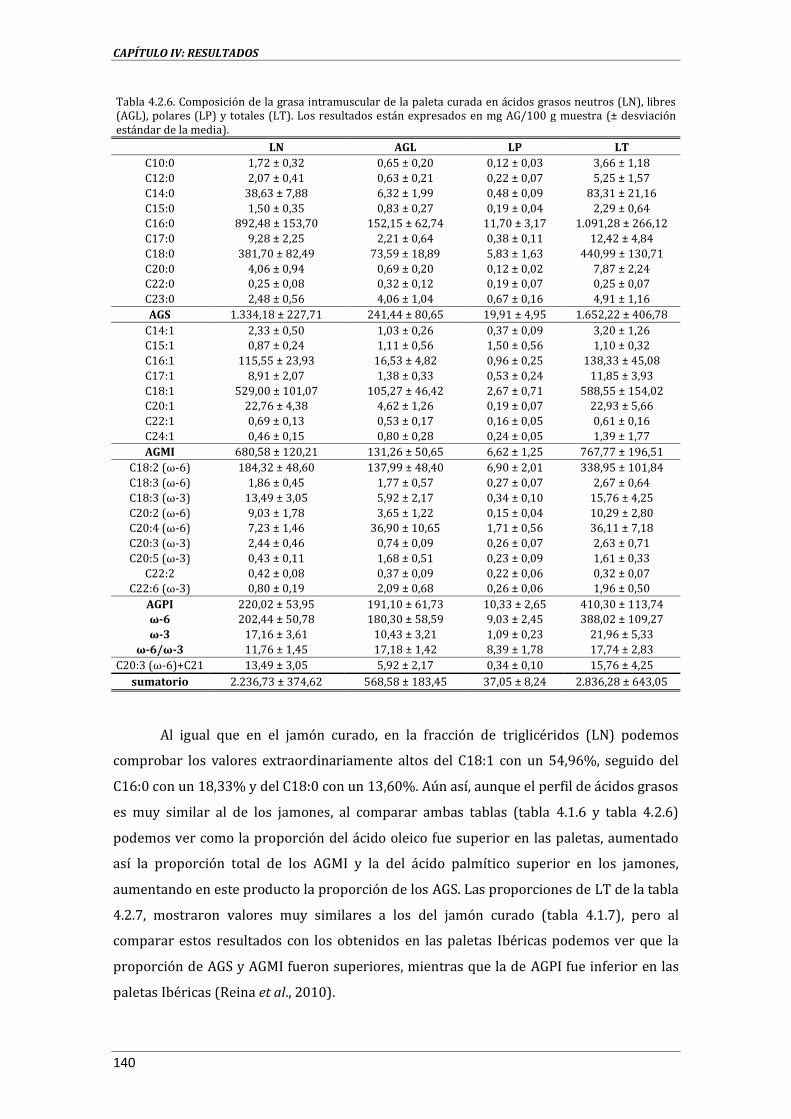
CAPÍTULO IV: RESULTADOS
140
Tabla 4.2.6. Composición de la grasa intramuscular de la paleta curada en ácidos grasos neutros (LN), libres (AGL), polares (LP) y totales (LT). Los resultados están expresados en mg AG/100 g muestra (± desviación estándar de la media).
LN AGL LP LT
C10:0 1,72 ± 0,32 0,65 ± 0,20 0,12 ± 0,03 3,66 ± 1,18
C12:0 2,07 ± 0,41 0,63 ± 0,21 0,22 ± 0,07 5,25 ± 1,57
C14:0 38,63 ± 7,88 6,32 ± 1,99 0,48 ± 0,09 83,31 ± 21,16
C15:0 1,50 ± 0,35 0,83 ± 0,27 0,19 ± 0,04 2,29 ± 0,64 C16:0 892,48 ± 153,70 152,15 ± 62,74 11,70 ± 3,17 1.091,28 ± 266,12
C17:0 9,28 ± 2,25 2,21 ± 0,64 0,38 ± 0,11 12,42 ± 4,84
C18:0 381,70 ± 82,49 73,59 ± 18,89 5,83 ± 1,63 440,99 ± 130,71
C20:0 4,06 ± 0,94 0,69 ± 0,20 0,12 ± 0,02 7,87 ± 2,24 C22:0 0,25 ± 0,08 0,32 ± 0,12 0,19 ± 0,07 0,25 ± 0,07
C23:0 2,48 ± 0,56 4,06 ± 1,04 0,67 ± 0,16 4,91 ± 1,16
AGS 1.334,18 ± 227,71 241,44 ± 80,65 19,91 ± 4,95 1.652,22 ± 406,78
C14:1 2,33 ± 0,50 1,03 ± 0,26 0,37 ± 0,09 3,20 ± 1,26 C15:1 0,87 ± 0,24 1,11 ± 0,56 1,50 ± 0,56 1,10 ± 0,32
C16:1 115,55 ± 23,93 16,53 ± 4,82 0,96 ± 0,25 138,33 ± 45,08
C17:1 8,91 ± 2,07 1,38 ± 0,33 0,53 ± 0,24 11,85 ± 3,93
C18:1 529,00 ± 101,07 105,27 ± 46,42 2,67 ± 0,71 588,55 ± 154,02 C20:1 22,76 ± 4,38 4,62 ± 1,26 0,19 ± 0,07 22,93 ± 5,66
C22:1 0,69 ± 0,13 0,53 ± 0,17 0,16 ± 0,05 0,61 ± 0,16
C24:1 0,46 ± 0,15 0,80 ± 0,28 0,24 ± 0,05 1,39 ± 1,77
AGMI 680,58 ± 120,21 131,26 ± 50,65 6,62 ± 1,25 767,77 ± 196,51
C18:2 (ω-6) 184,32 ± 48,60 137,99 ± 48,40 6,90 ± 2,01 338,95 ± 101,84
C18:3 (ω-6) 1,86 ± 0,45 1,77 ± 0,57 0,27 ± 0,07 2,67 ± 0,64
C18:3 (ω-3) 13,49 ± 3,05 5,92 ± 2,17 0,34 ± 0,10 15,76 ± 4,25
C20:2 (ω-6) 9,03 ± 1,78 3,65 ± 1,22 0,15 ± 0,04 10,29 ± 2,80 C20:4 (ω-6) 7,23 ± 1,46 36,90 ± 10,65 1,71 ± 0,56 36,11 ± 7,18
C20:3 (ω-3) 2,44 ± 0,46 0,74 ± 0,09 0,26 ± 0,07 2,63 ± 0,71
C20:5 (ω-3) 0,43 ± 0,11 1,68 ± 0,51 0,23 ± 0,09 1,61 ± 0,33
C22:2 0,42 ± 0,08 0,37 ± 0,09 0,22 ± 0,06 0,32 ± 0,07 C22:6 (ω-3) 0,80 ± 0,19 2,09 ± 0,68 0,26 ± 0,06 1,96 ± 0,50
AGPI 220,02 ± 53,95 191,10 ± 61,73 10,33 ± 2,65 410,30 ± 113,74 ω-6 202,44 ± 50,78 180,30 ± 58,59 9,03 ± 2,45 388,02 ± 109,27
ω-3 17,16 ± 3,61 10,43 ± 3,21 1,09 ± 0,23 21,96 ± 5,33
ω-6/ω-3 11,76 ± 1,45 17,18 ± 1,42 8,39 ± 1,78 17,74 ± 2,83
C20:3 (ω-6)+C21 13,49 ± 3,05 5,92 ± 2,17 0,34 ± 0,10 15,76 ± 4,25
sumatorio 2.236,73 ± 374,62 568,58 ± 183,45 37,05 ± 8,24 2.836,28 ± 643,05
Al igual que en el jamón curado, en la fracción de triglicéridos (LN) podemos
comprobar los valores extraordinariamente altos del C18:1 con un 54,96%, seguido del
C16:0 con un 18,33% y del C18:0 con un 13,60%. Aún así, aunque el perfil de ácidos grasos
es muy similar al de los jamones, al comparar ambas tablas (tabla 4.1.6 y tabla 4.2.6)
podemos ver como la proporción del ácido oleico fue superior en las paletas, aumentado
así la proporción total de los AGMI y la del ácido palmítico superior en los jamones,
aumentando en este producto la proporción de los AGS. Las proporciones de LT de la tabla
4.2.7, mostraron valores muy similares a los del jamón curado (tabla 4.1.7), pero al
comparar estos resultados con los obtenidos en las paletas Ibéricas podemos ver que la
proporción de AGS y AGMI fueron superiores, mientras que la de AGPI fue inferior en las
paletas Ibéricas (Reina et al., 2010).
CAPÍTULO IV: RESULTADOS
141
Tabla 4.2.7. Composición de la grasa intramuscular de la paleta curada en ácidos grasos neutros (LN), libres (AGL), polares (LP) y totales (LT). Los resultados están expresados % (± desviación estándar de la media).
LN AGL LP LT
C10:0 0,04 ± 0,01 0,07 ± 0,02 0,19 ± 0,04 0,04 ± 0,01 C12:0 0,04 ± 0,02 0,04 ± 0,01 0,23 ± 0,06 0,05 ± 0,02
C14:0 0,70 ± 0,46 0,48 ± 0,06 0,67 ± 0,10 1,37 ± 0,20
C15:0 0,03 ± 0,01 0,07 ± 0,01 0,29 ± 0,04 0,05 ± 0,01
C16:0 18,83 ± 3,64 11,93 ± 0,74 18,07 ± 1,14 26,33 ± 1,86 C17:0 0,24 ± 0,05 0,27 ± 0,03 0,74 ± 0,14 0,27 ± 0,05
C18:0 13,60 ± 1,84 11,28 ± 0,68 13,91 ± 1,06 10,85 ± 1,07
C20:0 0,22 ± 0,07 0,16 ± 0,03 0,34 ± 0,15 0,11 ± 0,02
C22:0 0,02 ± 0,01 0,09 ± 0,03 0,65 ± 0,10 0,01 ± 0,00 C23:0 0,14 ± 0,05 0,95 ± 0,18 2,79 ± 0,42 0,13 ± 0,02
C24:0 0,05 ± 0,01 0,24 ± 0,09 2,35 ± 0,26 0,01 ± 0,00
AGS 33,90 ± 1,26 25,58 ± 1,02 40,23 ± 1,83 39,20 ± 2,67
C14:1 0,05 ± 0,03 0,08 ± 0,01 0,51 ± 0,09 0,05 ± 0,02 C15:1 0,02 ± 0,01 0,09 ± 0,04 1,16 ± 0,28 0,03 ± 0,01
C16:1 2,43 ± 0,81 1,64 ± 0,23 1,45 ± 0,19 3,22 ± 0,56
C17:1 0,28 ± 0,06 0,23 ± 0,05 0,97 ± 0,34 0,30 ± 0,08
C18:1 54,96 ± 3,82 40,00 ± 1,67 20,13 ± 1,02 45,54 ± 2,15 C20:1 1,22 ± 0,38 0,96 ± 0,09 0,71 ± 0,12 0,64 ± 0,08
C22:1 0,04 ± 0,01 0,14 ± 0,04 0,59 ± 0,13 0,02 ± 0,00
C24:1 0,05 ± 0,03 0,38 ± 0,12 1,84 ± 0,46 0,01 ± 0,01
AGMI 59,05 ± 1,19 43,52 ± 1,66 27,36 ± 1,25 49,80 ± 2,55
C18:2 (ω-6) 5,94 ± 0,98 19,32 ± 0,98 16,47 ± 1,84 8,76 ± 1,47
C18:3 (ω-6) 0,06 ± 0,01 0,22 ± 0,05 0,38 ± 0,17 0,07 ± 0,01
C18:3 (ω-3) 0,38 ± 0,06 0,74 ± 0,04 0,78 ± 0,12 0,41 ± 0,08
C20:2 (ω-6) 0,42 ± 0,11 0,66 ± 0,08 0,55 ± 0,09 0,29 ± 0,05 C20:4 (ω-6) 0,30 ± 0,07 6,44 ± 1,02 5,33 ± 1,01 1,00 ± 0,25
C20:3 (ω-3) 0,11 ± 0,03 0,21 ± 0,05 0,62 ± 0,23 0,07 ± 0,01
C20:5 (ω-3) 0,02 ± 0,02 0,27 ± 0,06 0,76 ± 0,25 0,04 ± 0,01
C22:2 0,03 ± 0,01 0,10 ± 0,02 0,88 ± 0,12 0,01 ± 0,00 C22:6 (ω-3) 0,09 ± 0,03 0,90 ± 0,18 2,18 ± 0,36 0,10 ± 0,02
AGPI 7,34 ± 0,68 28,86 ± 1,84 27,96 ± 2,24 10,75 ± 1,83 ω-6 6,72 ± 0,65 26,64 ± 1,61 22,74 ± 2,43 10,12 ± 1,72
ω-3 0,60 ± 0,06 2,12 ± 0,27 4,35 ± 0,45 0,62 ± 0,11
ω-6/ω-3 11,36 ± 1,05 12,82 ± 1,41 5,36 ± 0,98 16,36 ± 1,25
C20:3 (ω-6)+C21 0,10 ± 0,02 0,93 ± 0,16 0,64 ± 0,07 0,16 ± 0,03
Determinación del contenido en aminoácidos libres
El método de cromatografía utilizado permitió la obtención de cromatogramas con
el mismo perfil al obtenido para los jamones curados, identificándose 20 aminoácidos, tal
como se muestra en la tabla 4.2.8, que se presentaron en 15 picos, mostrados en la figura
4.2.3.

CAPÍTULO IV: RESULTADOS
142
Asp
Glu
Gly-Gln
Ser-Asn Tyr-Val
Trp
Nor
Phe
His
Trh
Cys-Ala
Pro
Arg
Met
Iso
Leu
Lys
Figura 4.2.3. Cromatograma de aminoácidos libres obtenido mediante HPLC de magro de paleta curada.
A pesar de que los aminoácidos Pro y Arg se separaron en dos picos, se
cuantificaron conjuntamente, ya que el cromatograma obtenido del patrón no consiguió
separarlos, apareciendo ambos en un único pico.
Tabla 4.2.8. Contenido en aminoácidos libres en la paleta curada. Los resultados se expresan en mg aa/100 g muestra (± desviación estándar de la media).
E.H E.S
Ac. Aspártico (Asp) 165,69 ± 25,18 335,40 ± 51,04 Ac. Glutámico (Glu) 424,35 ± 75,02 858,71 ± 153,56
Serina + Asparagina (Ser-Asp) 244,17 ± 45,67 494,55 ± 94,46
Glicina + Glutamina (Gly-Gln) 229,94 ± 33,28 465,25 ± 67,25
Histidina (Hys) 149,05 ± 22,47 302,16 ± 47,90 Treonina (Trh) 149,88 ± 51,23 302,78 ± 102,45
Cisteína + Alanina (Cys-Ala) 776,54 ± 195,19 1.574,93 ± 415,49
Prolina + Arginina (Pro-Arg) 1.491,64 ± 201,40 3.021,43 ± 423,03
Tirosina + Valina (Tyr-Val) 327,99 ± 48,03 664,53 ± 100,89 Metionina (Met) 54,39 ± 9,79 110,31 ± 20,82
Isoleucina (Iso) 120,24 ± 17,82 243,79 ± 38,74
Leucina (Leu) 314,03 ± 41,97 635,28 ± 83,35
Fenilalanina (Phe) 147,02 ± 19,96 297,92 ± 42,25 Triptófano (Trp) 56,48 ± 7,93 114,59 ± 17,58
Lisina (Lys) 482,31 ± 55,55 976,45 ± 113,17
sumatorio 5.133,73 ± 649,16 10.398,09 ± 1.369,54
(EH): Extracto húmedo; (ES): Extracto seco
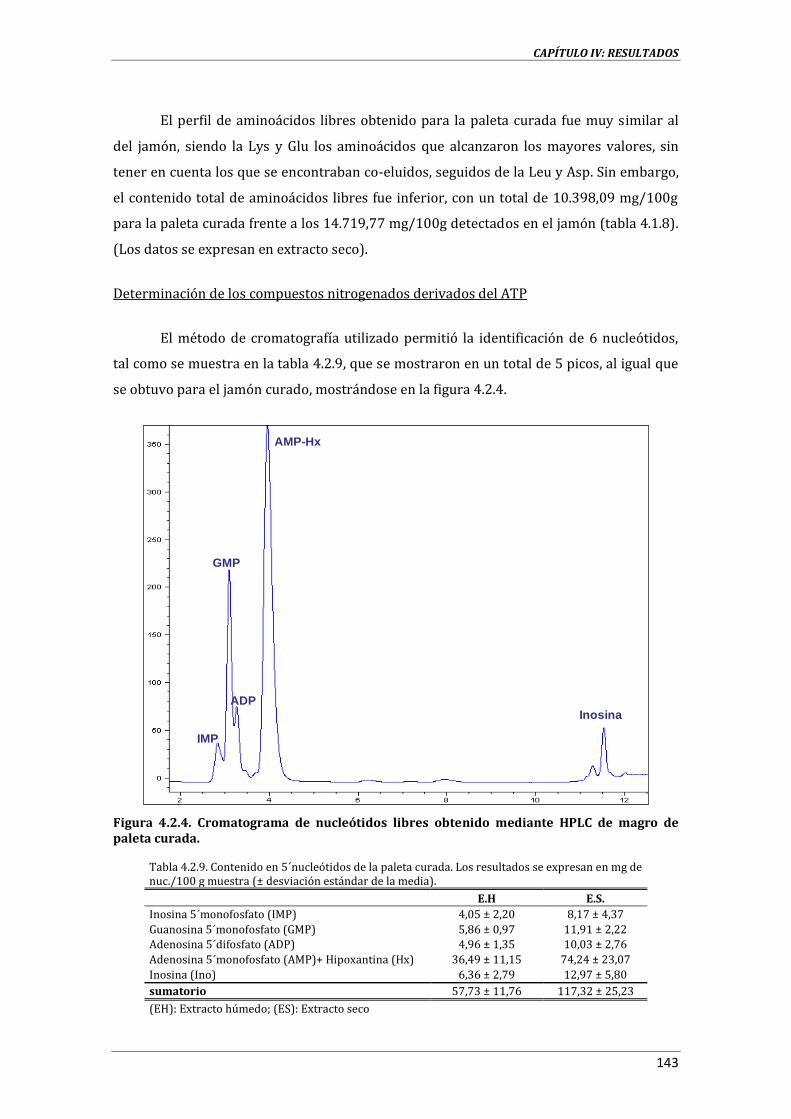
CAPÍTULO IV: RESULTADOS
143
El perfil de aminoácidos libres obtenido para la paleta curada fue muy similar al
del jamón, siendo la Lys y Glu los aminoácidos que alcanzaron los mayores valores, sin
tener en cuenta los que se encontraban co-eluidos, seguidos de la Leu y Asp. Sin embargo,
el contenido total de aminoácidos libres fue inferior, con un total de 10.398,09 mg/100g
para la paleta curada frente a los 14.719,77 mg/100g detectados en el jamón (tabla 4.1.8).
(Los datos se expresan en extracto seco).
Determinación de los compuestos nitrogenados derivados del ATP
El método de cromatografía utilizado permitió la identificación de 6 nucleótidos,
tal como se muestra en la tabla 4.2.9, que se mostraron en un total de 5 picos, al igual que
se obtuvo para el jamón curado, mostrándose en la figura 4.2.4.
IMP
Inosina
AMP-Hx
ADP
GMP
Figura 4.2.4. Cromatograma de nucleótidos libres obtenido mediante HPLC de magro de paleta curada.
Tabla 4.2.9. Contenido en 5´nucleótidos de la paleta curada. Los resultados se expresan en mg de nuc./100 g muestra (± desviación estándar de la media).
E.H E.S.
Inosina 5´monofosfato (IMP) 4,05 ± 2,20 8,17 ± 4,37
Guanosina 5´monofosfato (GMP) 5,86 ± 0,97 11,91 ± 2,22 Adenosina 5´difosfato (ADP) 4,96 ± 1,35 10,03 ± 2,76
Adenosina 5´monofosfato (AMP)+ Hipoxantina (Hx) 36,49 ± 11,15 74,24 ± 23,07
Inosina (Ino) 6,36 ± 2,79 12,97 ± 5,80
sumatorio 57,73 ± 11,76 117,32 ± 25,23
(EH): Extracto húmedo; (ES): Extracto seco

CAPÍTULO IV: RESULTADOS
144
El perfil de nucleótidos obtenido para la paleta curada fue muy similar al del
jamón, siendo la AMP co-eluída con la Hx los nucleótidos que alcanzaron los mayores
niveles. Sin embargo, el contenido total de nucleótidos fue muy inferior, con un total de
117,32 mg/100g para la paleta curada frente a los 374,00 mg/100g detectados en el
jamón (tabla 4.1.9). (Los datos se expresan en extracto seco).
Determinación de compuestos volátiles
Se detectaron un total de 128 compuestos volátiles en las 30 paletas curadas
analizadas, de los cuales se presentan en la tabla 4.2.10 los 97 compuestos que
aparecieron en las dos campañas analizadas.
En la tabla 4.2.10 se muestran 97 compuestos pertenecientes a 13 familias
químicas diferentes: aldehídos (15), cetonas (14), alcoholes (15), hidrocarburos alifáticos
(14), ácidos (7), ésteres (6), compuestos azufrados (5), compuestos nitrogenados (5),
furanos (7), hidrocarburos aromáticos (4), terpenos (2), éteres (1) y compuestos clorados
(2).
Entre los compuestos volátiles seleccionados, 55 han sido asociados a la oxidación
lipídica, 30 tienen su origen en aminoácidos a través de reacciones de Strecker (ej:
aldehídos ramificados) y de Maillard (ej: 2,6-dimetilpirazina), 8 de ellos han sido
relacionados tanto con la oxidación lipídica como con reacciones de Maillard y los 2
restantes están asociados a agentes externos del producto.
Tabla 4.2.10. Compuestos volátiles identificados en la paleta curada agrupados por familia química. Los contenidos de cada compuesto se expresan en % de área y en UAA (106) (± desviación estándar de la media).
IK Posicion Compuesto UAA (106) UAA (%) Posible origen
ALDEHIDOS 192,16 ± 90,47 15,11 ± 4,21
<500 2 acetaldehído 6,72 ± 3,36 0,52 ± 0,25 aa
550 12 2-metilpropanal 11,59 ± 8,00 0,81 ± 0,47 aa
649 23 3-metilbutanal 59,42 ± 36,78 3,96 ± 1,84 aa 660 24 2-metilbutanal 27,76 ± 15,24 2,01 ± 0,95 aa
698 28 pentanal 11,08 ± 6,07 1,00 ± 0,73 lipidos
800 46 hexanal 42,53 ± 29,93 4,07 ± 2,74 lipidos
902 57 heptanal 6,65 ± 4,06 0,48 ± 0,27 lipidos 908 59 3-(metiltio)propanal 2,51 ± 1,15 0,23 ± 0,15 aa
969 67 benzaldehído 3,34 ± 2,20 0,27 ± 0,18 aa
1006 75 octanal 6,19 ± 5,79 0,59 ± 0,53 lipidos
1053 79 bencenoacetaldehido 1,71 ± 1,00 0,20 ± 0,28 aa 1104 86 nonanal 10,26 ± 9,66 0,75 ± 0,58 lipidos
1157 87 2-nonenal 0,88 ± 0,43 0,07 ± 0,04 lipidos
1209 88 decanal 0,91 ± 0,80 0,08 ± 0,09 lipidos
1495 93 dodecanal 0,62 ± 0,31 0,06 ± 0,04 lipidos
CETONAS 217,61 ± 115,75 19,31 ± 8,98
<500 5 2-propanona + propanal 95,87 ± 62,28 8,26 ± 5,82 lipidos
586 14 2,3-butanodiona 11,18 ± 8,64 1,01 ± 0,77 lipidos

CAPÍTULO IV: RESULTADOS
145
IK Posicion Compuesto UAA (106) UAA (%) Posible origen
598 16 2-butanona + butanal 18,25 ± 9,13 1,55 ± 0,95 lipidos 687 27 2-pentanona 56,81 ± 43,94 4,34 ± 3,01 lipidos
710 30 3-hidroxi-2-butanona 4,55 ± 2,35 1,59 ± 1,60 lipidos
790 44 2-hexanona 3,80 ± 1,98 0,29 ± 0,13 lipidos
792 45 ciclopentanona 3,27 ± 1,83 0,40 ± 0,41 lipidos 891 55 2-heptanona 17,66 ± 12,56 1,31 ± 0,81 lipidos
964 66 4-metilciclohexanona 1,16 ± 0,56 0,11 ± 0,07 lipidos
989 70 3-octanona 1,81 ± 1,01 0,16 ± 0,08 lipidos
992 71 2-octanona 1,88 ± 0,90 0,17 ± 0,12 lipidos
1090 83 2-nonanona 1,36 ± 0,66 0,14 ± 0,10 lipidos
ALCOHOLES 144,24 ± 57,50 12,66 ± 5,09
<500 4 etanol 65,50 ± 40,67 5,86 ± 4,54 lipidos
549 11 propanol 4,99 ± 3,93 0,43 ± 0,28 lipidos
604 18 2-metil-3-buten-1-ol 1,88 ± 0,73 0,16 ± 0,06 aa 620 21 2-metilpropanol 10,13 ± 5,16 0,86 ± 0,44 aa
673 25 1-metoxi-2-propanol 6,67 ± 6,31 0,55 ± 0,43 aa
681 26 1-penten-3-ol 6,52 ± 4,33 0,49 ± 0,30 lipidos
699 29 2-pentanol 13,27 ± 7,20 1,36 ± 0,93 lipidos 738 33 3-metilbutanol 5,82 ± 3,19 0,65 ± 0,74 aa
742 34 2-metilbutanol 2,39 ± 1,43 0,23 ± 0,19 aa
770 40 pentanol 7,58 ± 5,47 0,54 ± 0,44 lipidos
872 52 hexanol 6,11 ± 3,77 0,47 ± 0,32 lipidos 899 56 2-heptanol 2,05 ± 1,18 0,20 ± 0,17 lipidos
906 58 2-butoxietanol 2,89 ± 1,67 0,23 ± 0,13 aa
982 69 1-octen-3-ol 6,80 ± 5,14 0,46 ± 0,29 lipidos
1065 82 octanol 1,65 ± 1,09 0,15 ± 0,14 lipidos
HIDROCARBUROS ALIFÁTICOS 89,89 ± 20,70 9,10 ± 4,89
<500 1 propano 5,66 ± 2,87 0,52 ± 0,33 lipidos
500 6 pentano 18,08 ± 9,51 1,63 ± 1,05 lipidos
507 7 1,1´-oxibis etano 9,05 ± 5,26 0,66 ± 0,45 lipidos 555 13 2-metilpentano 3,48 ± 1,56 0,34 ± 0,22 lipidos
600 17 hexano 13,64 ± 7,92 1,24 ± 0,76 lipidos
624 22 metilciclopentano 8,04 ± 2,97 1,06 ± 0,99 lipidos
734 32 2,5-dimetilhexano + 3-metil-3-buten-1-ol
2,21 ± 1,42 0,19 ± 0,12 lipidos
753 38 2,3,4-trimetilpentano 4,35 ± 2,96 0,53 ± 0,49 lipidos
763 39 2,3-dimetilhexano 1,74 ± 1,55 0,18 ± 0,24 lipidos
785 43 2,2,5-trimetilhexano 2,14 ± 1,39 0,28 ± 0,27 lipidos 1000 74 decano 18,53 ± 17,24 2,19 ± 2,40 lipidos
1100 84 undecano 2,52 ± 1,69 0,22 ± 0,17 lipidos
1400 92 tetradecano 0,44 ± 0,21 0,04 ± 0,03 lipidos
ACIDOS 66,59 ± 45,45 5,08 ± 2,73
579 15 acido acético 35,37 ± 31,59 2,87 ± 2,27 aa/lipidos
753 37 acido 2-metilpropanoico 2,21 ± 1,38 0,15 ± 0,09 aa
779 42 ácido butanoico 11,26 ± 9,92 0,88 ± 0,72 aa/lipidos
833 47 ácido 3-metilbutanoico 8,74 ± 8,65 0,53 ± 0,46 aa 843 48 ácido 2-metilpentanoico 2,61 ± 1,43 0,21 ± 0,11 aa
970 68 ácido hexanoico 5,39 ± 4,49 0,35 ± 0,24 aa/lipidos
1258 89 ácido nonanoico 1,02 ± 0,57 0,09 ± 0,07 aa/lipidos
ESTERES 11,71 ± 4,74 1,28 ± 0,74
612 19 acetato de etilo 2,43 ± 1,35 0,35 ± 0,48 lipidos
728 31 2-hidroxipropanoato de metilo 1,22 ± 0,83 0,10 ± 0,06 lipidos
853 49 2-metilbutanoato de etilo 3,87 ± 3,18 0,44 ± 0,42 lipidos
856 50 3-metilbutanoato de etilo 2,08 ± 1,71 0,20 ± 0,15 lipidos 942 63 hexanoato de metilo 0,69 ± 0,42 0,06 ± 0,04 lipidos
999 73 hexanoato de etilo 1,41 ± 0,78 0,12 ± 0,10 lipidos
COMPUESTOS AZUFRADOS 120,36 ± 82,08 11,61 ± 9,48
<500 3 metanotiol 6,60 ± 3,53 0,51 ± 0,30 aa 519 8 tiobis metano + etanotiol 5,45 ± 3,07 0,41 ± 0,22 aa

CAPÍTULO IV: RESULTADOS
146
IK Posicion Compuesto UAA (106) UAA (%) Posible origen
536 10 sulfuro de carbono 106,25 ± 79,78 10,53 ± 9,35 aa 750 36 dimetil disulfuro 2,05 ± 1,27 0,17 ± 0,11 aa
COMPUESTOS NITROGENADOS 12,85 ± 5,16 0,96 ± 0,37
748 35 piridina 2,92 ± 2,45 0,27 ± 0,20 aa
881 54 hexanonitrilo 3,16 ± 1,90 0,26 ± 0,16 aa 914 60 2,6-dimetilpirazina 3,28 ± 2,01 0,20 ± 0,10 aa
923 62 etilpirazina 0,92 ± 0,48 0,06 ± 0,05 aa
1006 76 trimetilpirazina 2,56 ± 1,04 0,17 ± 0,08 aa
FURANOS 17,34 ± 7,76 1,53 ± 0,76
915 61 dihidro-2(3H)-furanona 4,82 ± 2,12 0,37 ± 0,17 aa/lipidos 958 65 dihidro-5-metil-2(3H)-furanona 1,44 ± 0,85 0,12 ± 0,06 aa/lipidos
996 72 2-pentilfurano 4,96 ± 2,82 0,49 ± 0,40 aa
1060 80 dihidro-5-etil-2(3H)-furanona 2,64 ± 1,57 0,20 ± 0,10 aa/lipidos
1068 81 dihidro-5-metil-5-etil-2(3H)-furanona 2,03 ± 1,55 0,18 ± 0,12 aa/lipidos 1272 90 dihidro-5-butil-2(3H)-furanona 0,75 ± 0,63 0,07 ± 0,06 aa/lipidos
1377 91 dihidro-5-pentil-2(3H)-furanona 0,69 ± 0,46 0,09 ± 0,09 aa/lipidos
HIDROCARBUROS AROMÁTICOS 11,87 ± 5,90 1,05 ± 0,51
772 41 metilbenceno 5,84 ± 2,96 0,51 ± 0,29 aa 866 51 etilbenceno 1,06 ± 0,64 0,10 ± 0,07 aa
875 53 m- y p-xileno 3,10 ± 2,24 0,26 ± 0,19 aa
1103 85 nitro benceno 1,87 ± 1,29 0,17 ± 0,14 aa
TERPENOS 2,48 ± 1,33 0,22 ± 0,12
943 64 alfa-pineno 0,84 ± 0,49 0,07 ± 0,04 lipidos
1035 77 l-limoneno 1,65 ± 1,01 0,15 ± 0,10 lipidos
ETERES 1,96 ± 1,31 0,17 ± 0,12
1042 78 1,8-cineol 1,96 ± 1,31 0,17 ± 0,12 lipidos
COMPUESTOS CLORADOS 348,48 ± 295,85 21,94 ± 13,26
528 9 diclorometano 1,56 ± 0,77 0,20 ± 0,15 contaminación
616 20 cloroformo 346,91 ± 295,93 21,74 ± 13,31 contaminación
sumatorio 1.237,54 ± 479,97
La familia mayoritaria dentro del perfil de compuestos volátiles de la paleta curada
fue la de las cetonas, a la que siguieron en cantidad los aldehídos y los alcoholes, que
presentaron contenido aproximado. El perfil de compuestos volátiles fue muy similar al
mostrado para los jamones curados de Teruel, con algunas diferencias como la mayor
proporción de las cetonas y los aldehídos en las paletas y la mayor proporción de los
alcoholes en los jamones. Sin embargo el contenido total de compuestos volátiles fue
superior en las paletas que en los jamones, encontrando un valor de 1237,54 UAA (106)
frente a los 1151,46 UAA (106) que se detectaron en el jamón (tabla 4.1.10).

CAPÍTULO IV: RESULTADOS
147
Evaluación sensorial
Los valores de los diferentes atributos sensoriales utilizados para caracterizar la
paleta curada se incluyen en la tabla 4.2.11 y en el gráfico de araña de la figura 4.2.5.
Tabla 4.2.11. Análisis cuantitativo-descriptivo de la paleta curada (± desviación estándar de la media).
Color grasa Amarillo 0,8 ± 0,1
Rosa 1,6 ± 0,3
Color magro
Rojo 3,9 ± 0,7
Brillo 3,9 ± 0,9
Veteado 3,5 ± 0,8
Olor Intensidad 3,3 ± 0,6
Textura grasa Dureza 4,0 ± 0,7
Fluidez 3,9 ± 0,5
Textura magro
Dureza 2,4 ± 0,7
Sequedad 2,6 ± 0,5
Jugosidad 5,3 ± 0,6
Pastosidad 2,2 ± 0,8
Sabor
Salado 5,3 ± 0,5
Dulce 1,0 ± 0,2 Amargo 0,7 ± 0,2
Flavor
Intensidad 4,8 ± 0,6
Curado 2,8 ± 0,7
Rancidez 0,8 ± 0,2
Persistencia 4,8 ± 0,6
Al comparar los valores emitidos por el panel sensorial para la paleta curada con el
jamón curado, vemos como la paleta se describió como un producto menos rancio y
pastoso, con un mayor flavor “a curado” y más persistente que el jamón. Sin embargo, la
fluidez de la grasa y la intensidad del olor fueron mayores en el jamón que en la paleta.
0
1
2
3
4
5
6AMARILLO GRASA
ROSA GRASA
ROJO MAGRO
BRILLO MAGRO
VETEADO
INTENSIDAD OLOR
DUREZA GRASA
FLUIDEZ GRASA
DUREZA MAGROSEQUEDAD MAGROJUGOSIDAD MAGRO
PASTOSIDAD MAGRO
SALADO
DULCE
AMARGO
INTENSIDAD …
PERSISTENCIA
CURADO
RANCIDEZ
JAMON CURADO PALETA CURADA
Figura 4.2.5. Perfil sensorial de la paleta curada

CAPÍTULO IV: RESULTADOS
148
4.2.2. INFLUENCIA DE LA EXPRESIÓN DEL IGF2 SOBRE LAS CARACTERÍSTICAS
FÍSICO-QUÍMICAS Y SENSORIALES DE LA PALETA CURADA
En este apartado se va a evaluar la influencia de los alelos A y G del IGF2 (insuline-
like growth factor 2) del QTL (Quantitative Trait Loci) sobre todos los parámetros
analizados en las paletas curadas. Para ello, tan solo van a incluirse aquellos parámetros
que presentaron diferencias significativas entre los dos lotes estudiados.
Características generales de la paleta curada
Al evaluar las características generales de la paleta curada: peso, longitud, anchura
y perímetro, no se encontraron diferencias significativas entre ambos lotes. Sin embargo,
si se observaron diferencias significativas en el peso de paletas analizadas en fresco
procedentes de los mismos lotes de cerdos, presentando mayor peso las paletas
procedentes de los cerdos AA (Burgos et al., 2011).
El corte transversal de las paletas mostró diferencias significativas en cuanto al
espesor de la grasa subcutánea, tanto en la zona proximal (figura 4.2.6) como en la distal
(figura 4.2.7), presentando diferencias significativas en cinco de las seis medidas tomadas
en cada zona, tal y como se muestra en la tabla 4.2.12.
AGAA
A BC
AB
C
Figura 4.2.6. Zona proximal de la paleta curada
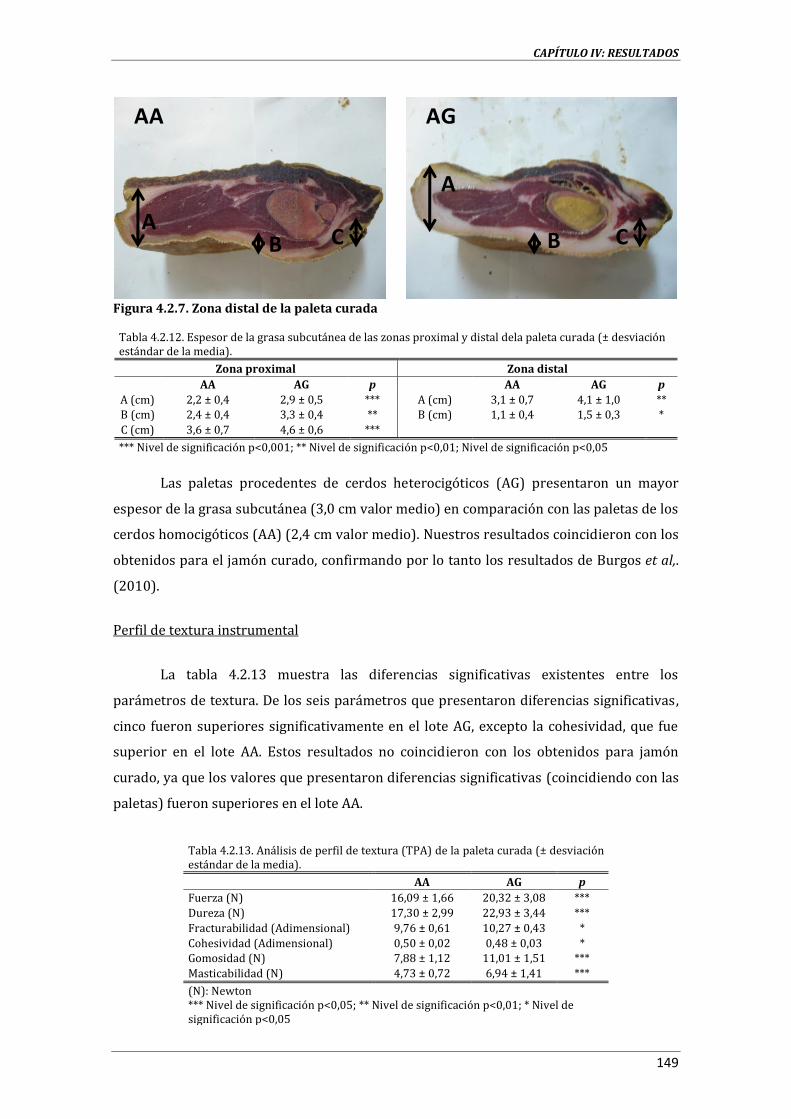
CAPÍTULO IV: RESULTADOS
149
AGAA
A
B CA
B C
Figura 4.2.7. Zona distal de la paleta curada
Tabla 4.2.12. Espesor de la grasa subcutánea de las zonas proximal y distal dela paleta curada (± desviación estándar de la media).
Zona proximal Zona distal
AA AG p
AA AG p
A (cm) 2,2 ± 0,4 2,9 ± 0,5 *** A (cm) 3,1 ± 0,7 4,1 ± 1,0 ** B (cm) 2,4 ± 0,4 3,3 ± 0,4 ** B (cm) 1,1 ± 0,4 1,5 ± 0,3 *
C (cm) 3,6 ± 0,7 4,6 ± 0,6 ***
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; Nivel de significación p<0,05
Las paletas procedentes de cerdos heterocigóticos (AG) presentaron un mayor
espesor de la grasa subcutánea (3,0 cm valor medio) en comparación con las paletas de los
cerdos homocigóticos (AA) (2,4 cm valor medio). Nuestros resultados coincidieron con los
obtenidos para el jamón curado, confirmando por lo tanto los resultados de Burgos et al,.
(2010).
Perfil de textura instrumental
La tabla 4.2.13 muestra las diferencias significativas existentes entre los
parámetros de textura. De los seis parámetros que presentaron diferencias significativas,
cinco fueron superiores significativamente en el lote AG, excepto la cohesividad, que fue
superior en el lote AA. Estos resultados no coincidieron con los obtenidos para jamón
curado, ya que los valores que presentaron diferencias significativas (coincidiendo con las
paletas) fueron superiores en el lote AA.
Tabla 4.2.13. Análisis de perfil de textura (TPA) de la paleta curada (± desviación estándar de la media).
AA AG p
Fuerza (N) 16,09 ± 1,66 20,32 ± 3,08 ***
Dureza (N) 17,30 ± 2,99 22,93 ± 3,44 ***
Fracturabilidad (Adimensional) 9,76 ± 0,61 10,27 ± 0,43 *
Cohesividad (Adimensional) 0,50 ± 0,02 0,48 ± 0,03 * Gomosidad (N) 7,88 ± 1,12 11,01 ± 1,51 ***
Masticabilidad (N) 4,73 ± 0,72 6,94 ± 1,41 ***
(N): Newton *** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05

CAPÍTULO IV: RESULTADOS
150
Parámetros generales físico-químicos
En cuanto a los parámetros generales, tan solo se encontraron diferencias
significativas en los mg MDA/Kg, tal como se muestra en la tabla 4.2.14.
Tabla 4.2.14. Parámetros físico-químicos generales de la paleta curada (± desviación estándar de la media).
AA AG p
mg MDA/Kg 0,08 ± 0,01 0,15 ± 0,09 **
(f): extracto fresco ** Nivel de significación p<0,01
En el caso de las paletas, al igual que en los jamones, existieron diferencias
significativas en el contenido de grasa subcutánea, siendo significativamente superiores
los resultados pertenecientes al lote AG, pero no existieron diferencias en el contenido de
grasa intramuscular. Nuestros datos coincidieron con los de Burgos et al., (2011), que no
encontraron diferencias significativas al estudiar el contenido de grasa intramuscular de
distintas piezas como el psoas major o el bíceps femoris.
El contenido en mg MDA/Kg fue significativamente superior en el lote AG,
coincidiendo con el lote que presentó mayor espesor de grasa subcutánea.
Composición de la grasa intramuscular
La fracción de triglicéridos (LN) fue la fracción mayoritaria de la grasa
intramuscular en ambos lotes, con un porcentaje de 77,6% para el lote AA y de 83,0% para
el lote AG de la grasa total. La fracción de fosfolípidos (LP) fue la fracción minoritaria con
un 1,4% para el lote AA y un 1,1% para el lote AG. La fracción de AGL fue de 21,0% para el
lote AA y de 16,0% para el lote AG.
Fracción de lípidos neutros
La tabla 4.2.15 muestra el contenido de los ácidos grasos identificados en la
fracción de triglicéridos (LN) de la grasa intramuscular extraída de las paletas curadas.
Existieron diferencias significativas en tan solo cuatro ácidos grasos y en el
contenido total de los AGS, siendo superiores los valores del lote AG. Esta diferencia entre
lotes coincidió con el mayor contenido de grasa intramuscular presente en el lote AG.
CAPÍTULO IV: RESULTADOS
151
Tabla 4.2.15. Composición de la grasa intramuscular en la fracción de triglicéridos (LN) de la paleta curada. Los resultados están expresados en mg de AG/100 g muestra (± desviación estándar de la media).
LN AA LN AG p
C16:0 843,16 ± 102,86 1.017,96 ± 251,44 *
C17:0 7,98 ± 1,09 9,95 ± 2,44 * C20:0 3,47 ± 0,71 4,37 ± 0,76 **
C14:1 2,62 ± 0,31 2,03 ± 0,54 **
AGS 1.254,39 ± 175,28 1.491,97 ± 310,73 *
** Nivel de significación p<0,01; * Nivel de significación p<0,05
En cuanto a los valores expresados en %, las diferencias fueron más marcadas
entre ambos lotes, aunque en este caso el lote AA presentó mayor proporción de algunos
ácidos grasos, destacando el valor del C18:1, de los AGMI y los AGPI.
Tabla 4.2.16. Composición de la grasa intramuscular en la fracción de triglicéridos (LN) de la paleta curada. Los resultados están expresados en % (± desviación estándar de la media).
LN AA % LN AG % p
C18:0 12,82 ± 1,12 14,06 ± 0,72 **
C23:0 0,15 ± 0,02 0,13 ± 0,01 ** C24:0 0,05 ± 0,02 0,04 ± 0,02 *
C14:1 0,05 ± 0,01 0,04 ± 0,00 **
C18:1 55,98 ± 1,34 54,65 ± 0,96 **
C20:1 1,30 ± 0,25 1,14 ± 0,10 * C22:1 0,05 ± 0,01 0,04 ± 0,01 **
C18:2 (ω-6) 6,26 ± 0,52 5,61 ± 0,60 **
C18:3 (ω-6) 0,06 ± 0,01 0,05 ± 0,00 **
C20:4 (ω-6) 0,33 ± 0,03 0,28 ± 0,03 *** C20:3 (ω-3) 0,11 ± 0,01 0,10 ± 0,01 **
C22:2 0,03 ± 0,00 0,02 ± 0,00 **
C22:6 (ω-3) 0,10 ± 0,01 0,09 ± 0,01 ***
AGMI 60,18 ± 1,50 58,56 ± 0,94 **
AGPI 7,73 ± 0,57 6,96 ± 0,65 **
ω-6 7,08 ± 0,54 6,36 ± 0,62 **
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05
Fracción de lípidos polares
El análisis del perfil de fosfolípidos (LP) de la grasa intramuscular se muestra en la
tabla 4.2.17. La mayoría de los ácidos grasos de la fracción de fosfolípidos mostró valores
más altos en el lote AA y un mayor número de diferencias significativas entre ellos. Solo el
C10:0 (ácido cáprico) fue significativamente superior en el lote AG.

CAPÍTULO IV: RESULTADOS
152
Tabla 4.2.17. Composición de la grasa intramuscular en la fracción de fosfolípidos (LP) de la paleta curada. Los resultados están expresados en mg de AG/100 g muestra (± desviación estándar de la media).
LP AA LP AG p
C14:0 0,51 ± 0,06 0,41 ± 0,06 ***
C16:0 12,39 ± 2,43 9,90 ± 2,46 *
C17:0 0,42 ± 0,09 0,31 ± 0,07 ** C18:0 6,57 ± 1,49 4,68 ± 1,14 **
C14:1 0,41 ± 0,08 0,31 ± 0,07 **
C16:1 1,05 ± 0,21 0,79 ± 0,18 **
C17:1 0,65 ± 0,25 0,40 ± 0,18 ** C18:1 3,04 ± 0,67 2,27 ± 0,62 **
C18:2 (ω-6) 5,32 ± 0,11 6,48 ± 1,81 *
C20:2 (ω-6) 0,17 ± 0,05 0,12 ± 0,03 **
C20:3 (ω -3) 0,24 ± 0,06 0,30 ± 0,08 * C20:5 (ω-3) 0,26 ± 0,10 0,17 ± 0,06 *
AGS 21,38 ± 3,80 16,70 ± 3,71 ** AGMI 7,24 ± 0,89 5,67 ± 1,07 ***
sumatorio 37,57 ± 4,90 32,09 ± 6,86 *
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05
Los resultados expresados en porcentajes se muestran en la tabla 4.2.18. A
diferencia de la tabla anterior, excepto el C15:1 (ácido penatdecenoico), C17:1 (ácido
heptadecenoico) y el C20:4 (ω-6) (ácido araquidónico), que fueron significativamente
superiores en el lote AA, el resto de ácidos grasos presentaron proporciones
significativamente superiores en el lote AG.
Tabla 4.2.18. Composición de la grasa intramuscular en la fracción de fosfolípidos (LP) de la paleta curada. Los resultados están expresados en % (± des viación estándar de la media).
LP AA % LP AG % p
C15:0 0,28 ± 0,04 0,32 ± 0,03 *
C23:0 2,49 ± 0,33 3,08 ± 0,35 ***
C24:0 2,30 ± 0,21 2,68 ± 0,52 * C14:1 0,48 ± 0,10 0,55 ± 0,07 *
C15:1 1,27 ± 0,29 0,95 ± 0,19 **
C17:1 1,14 ± 0,28 0,69 ± 0,22 ***
C24:1 1,75 ± 0,47 2,14 ± 0,17 ** C18:3 (ω-3) 0,72 ± 0,09 0,81 ± 0,09 *
C20:4 (ω-6) 5,85 ± 1,22 4,73 ± 1,19 *
C20:3 (ω-3) 0,65 ± 0,15 1,04 ± 0,37 **
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05
Fracción de ácidos grasos libres
En cuanto a la composición de la fracción de AGL, al igual que la fracción de
fosfolípidos, las diferencias significativas siguieron la misma tendencia, siendo los valores
superiores en el lote AA, a excepción del C10:0, que fue superior en el lote AG, tal como se
muestra en la tabla 4.2.19.

CAPÍTULO IV: RESULTADOS
153
Tabla 4.2.19. Composición de la grasa intramuscular en la fracción de AGL de la paleta curada. Los resultados están expresados en mg de AG/100 g muestra (± desviación estándar de la media).
AGL AA AGL AG p
C10:0 0,55 ± 0,14 0,75 ± 0,21 **
C17:0 2,49 ± 0,79 1,59 ± 0,06 ***
C18:0 82,42 ± 26,97 66,20 ± 11,60 * C23:0 4,57 ± 1,05 3,49 ± 0,82 **
C15:1 1,30 ± 0,61 0,69 ± 0,24 **
C17:1 1,40 ± 0,29 1,08 ± 0,06 ***
C18:1 103,10 ± 42,06 77,55 ± 11,57 * C20:2 (ω-6) 3,93 ± 1,19 2,95 ± 0,58 *
C20:4 (ω-6) 42,08 ± 12,28 31,13 ± 6,19 **
C20:5 (ω-3) 1,80 ± 0,48 1,38 ± 0,28 **
C22:6 (ω-3) 2,12 ± 0,44 1,65 ± 0,42 **
AGMI 132,37 ± 51,64 101,76 ± 14,79 *
ω-3 10,80 ± 2,88 8,77 ± 1,73 *
C20:3 (ω-6)+C21 5,04 ± 1,45 4,00 ± 0,85 *
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05
La tabla 4.2.20 muestra las diferencias significativas existentes entre las
proporciones de ácidos grasos dentro de cada lote. Existieron mayores proporciones en el
lote AA, excepto el C10:0, el C16:0 y el C16:1, que fueron significativamente superiores en
el lote AG, por lo que la proporción de los AGS fue significativamente superior en ese lote.
Tabla 4.2.20. Composición de la grasa intramuscular en la fracción de AGL de la paleta curada. Los resultados están expresados en % (± desviación estándar de la media).
AGL AA % AGL AG % p
C10:0 0,06 ± 0,01 0,07 ± 0,01 *
C16:0 11,30 ± 0,60 13,45 ± 1,80 ***
C24:0 0,26 ± 0,08 0,16 ± 0,03 *** C16:1 1,55 ± 0,23 1,74 ± 0,21 *
C20:1 1,00 ± 0,04 0,95 ± 0,06 **
C18:3 (ω-6) 0,24 ± 0,04 0,20 ± 0,05 *
C20:2 (ω-6) 0,69 ± 0,09 0,61 ± 0,03 ** C20:3 (ω-3) 0,24 ± 0,06 0,16 ± 0,01 ***
C20:5 (ω-3) 0,29 ± 0,02 0,25 ± 0,06 *
AGS 25,00 ± 0,98 26,96 ± 1,58 **
ω-3 2,23 ± 0,23 2,00 ± 0,25 *
C20:3 (ω-6)+C21 5,04 ± 1,45 4,00 ± 0,85 *
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05
Lípidos totales
En los resultados obtenidos en la tabla 4.2.21, al igual que en la fracción de LN, se
observó que el contenido total del C16:0, del C20:0 (ácido araquídico) y del sumatorio de
AGS fueron significativamente superiores en el lote AG, no existiendo diferencias
significativas entre el resto de ácidos grasos.

CAPÍTULO IV: RESULTADOS
154
Tabla 4.2.21. Composición de la grasa intramuscular en la fracción de LT de la paleta curada. Los resultados están expresados en mg de AG/100 g muestra (± desviación estándar de la media).
LT AA LT AG p
C16:0 969,90 ± 207,98 1.212,65 ± 262,53 *
C20:0 6,73 ± 1,32 8,60 ± 2,23 *
AGS 1.474,27 ± 328,55 1.807,46 ± 373,97 *
* Nivel de significación p<0,05
La tabla 4.2.22 muestra los porcentajes de los ácidos grasos presentes en los LT de
la grasa intramuscular, donde destacaron las proporciones significativamente superiores
del C18:0 del lote AG, ya que el resto de proporciones fueron significativamente superiores
en el lote AA.
Tabla 4.2.22. Composición de la grasa intramuscular en la fracción de LT de la paleta curada. Los resultados están expresados en % (± desviación estándar de la media).
LT AA % LT AG % p
C16:0 25,49 ± 1,51 27,17 ± 1,79 *
C20:0 0,10 ± 0,01 0,12 ± 0,02 *
C18:2 (ω-6) 9,44 ± 1,55 8,27 ± 0,73 *
C18:3 (ω-6) 0,07 ± 0,01 0,06 ± 0,01 ** C18:3 (ω-3) 0,45 ± 0,07 0,37 ± 0,06 **
C20:2 (ω-6) 0,31 ± 0,05 0,27 ± 0,04 *
C20:4 (ω-6) 1,13 ± 0,25 0,88 ± 0,16 **
C20:3 (ω-3) 0,07 ± 0,01 0,06 ± 0,01 * C20:5 (ω-3) 0,05 ± 0,01 0,04 ± 0,01 *
C22:6 (ω-3) 0,11 ± 0,02 0,09 ± 0,01 *
AGS 38,11 ± 1,75 40,59 ± 3,28 * AGPI 11,65 ± 1,89 10,05 ± 0,89 **
ω-6 10,95 ± 1,79 9,47 ± 0,83 **
ω-3 0,68 ± 0,10 0,56 ± 0,08 **
C20:3 (ω-6)+C21 0,18 ± 0,02 0,15 ± 0,02 **
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05
Determinación del contenido en aminoácidos libres
El único aminoácido que mostró diferencias significativas fue la treonina, que fue
superior (p<0,05) en el lote AA, coincidiendo con los resultados mostrados para jamón
curado, a pesar de que en este producto se dieron diferencias significativas en todos los
aminoácidos analizados.

CAPÍTULO IV: RESULTADOS
155
Determinación de los compuestos nitrogenados derivados del ATP
A diferencia de los resultados obtenidos para jamón curado, cuatro de los seis
nucleótidos analizados presentaron diferencias significativas entre los lotes (tabla 4.2.24).
La IMP fue significativamente superior en el lote AG, no coincidiendo con los resultados
del jamón curado, sin embargo, el resto de los nucleótidos fueron significativamente
superiores en el lote AA.
Tabla 4.2.24. Contenido en 5´nucleótidos de la paleta curada. Los resultados se expresan en mg de nuc./100 g muestra (± desviación estándar de la media).
AA AG p
Inosina 5´monofosfato (IMP) 2,99 ± 1,33 6,68 ± 4,33 ** Adenosina 5´monofosfato (AMP) + Hipoxantina (Hx) 41,07 ± 7,70 31,91 ± 12,71 *
Inosina (Ino) 8,10 ± 2,45 4,63 ± 2,35 **
sumatorio (f) 63,15 ± 8,56 53,73 ± 14,02 *
sumatorio (s) 130,18 ± 19,68 103,61 ± 29,44 **
(f): extracto fresco; (s): extracto seco ** Nivel de significación p<0,01; * Nivel de significación p<0,05
Determinación de compuestos volátiles
De los 97 compuestos volátiles detectados en las paletas (tabla 4.2.10), tan solo 17
presentaron diferencias significativas entre los lotes del estudio, 7 de ellos superiores
significativamente en el lote AA y 10 superiores significativamente en el lote AG. En cuanto
al contenido total, no existieron diferencias significativas entre los lotes, coincidiendo con
el jamón curado (tabla 4.1.25).
La no existencia de diferencias significativas en el contenido total de compuestos
volátiles coincidió con la ausencia de diferencias significativas en el contenido de grasa
intramuscular que mostraron las paletas, a diferencia del jamón, que a pesar de mostrar
diferencias en el contenido de grasa intramuscular, no mostró diferencias en el contenido
de compuestos volátiles.
Las únicas familias que presentaron diferencias entre los lotes fue la de los ácidos,
siendo superior el valor obtenido en el lote AA, y la de los éteres, siendo superior el
contenido en el lote AG.

CAPÍTULO IV: RESULTADOS
156
Tabla 4.2.25. Compuestos volátiles identificados en la paleta curada agrupados por familia química. Los contenidos de cada compuesto se expresan en UAA (106) (± desviación estándar de la media).
IK Posición Compuesto AA AG p Posible origen
ALDEHIDOS
660 24 2-metilbutanal 25,45 ± 16,16 42,38 ± 26,15 * aa
698 28 pentanal 12,29 ± 7,35 8,08 ± 2,08 * lípidos
969 67 benzaldehído 2,29 ± 1,27 4,38 ± 2,94 * aa
1006 75 octanal 6,68 ± 5,80 3,13 ± 1,30 * lípidos 1053 79 bencenoacetaldehido 1,36 ± 0,70 2,11 ± 1,15 * aa
CETONAS
710 30 3-hidroxi-2-butanona 3,77 ± 1,21 21,22 ± 22,98 ** lípidos
1090 83 2-nonanona 0,97 ± 0,42 1,56 ± 0,69 ** lípidos
ALCOHOLES
673 25 1-metoxi-2-propanol 3,73 ± 3,20 9,85 ± 7,45 ** aa
699 29 2-pentanol 9,94 ± 2,84 15,04 ± 8,47 * lípidos
738 33 3-metilbutanol 7,83 ± 4,47 4,58 ± 2,90 * aa 770 40 pentanol 4,40 ± 2,73 8,25 ± 4,38 ** lípidos
HIDROCARBUROS ALIFÁTICOS
624 22 metilciclopentano 4,09 ± 0,29 9,36 ± 3,90 *** lípidos
ACIDOS
1258 89 ácido nonanoico 2,92 ± 2,62 1,04 ± 0,72 * aa/lípidos
ESTERES
853 49 2-metilbutanoato de etilo 5,14 ± 4,36 0,70 ± 0,10 *** lípidos
856 50 3-metilbutanoato de etilo 1,50 ± 0,64 3,29 ± 2,96 * lípidos
COMPUESTOS NITROGENADOS
1006 76 trimetilpirazina 4,11 ± 2,65 2,43 ± 0,99 * aa
FURANOS
1272 90 dihidro-5-butil-2(3H)-furanona 0,45 ± 0,29 0,90 ± 0,63 * aa/lípidos
COMPUESTOS CLORADOS
528 9 diclorometano 2,49 ± 1,34 1,40 ± 0,82 * contaminación
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05
Algunos de los compuestos a destacar son el 1-metoxi-2-propanol, el 1-pentanol o
el metilciclopentano, que fueron significativamente superiores en el lote AG. Por otra
parte, el pentanal, el ácido nonanoico y el ácido 2-metilbutanoato de etilo fueron
significativamente superiores en el lote AA, compuestos que proceden principalmente de
los lípidos.
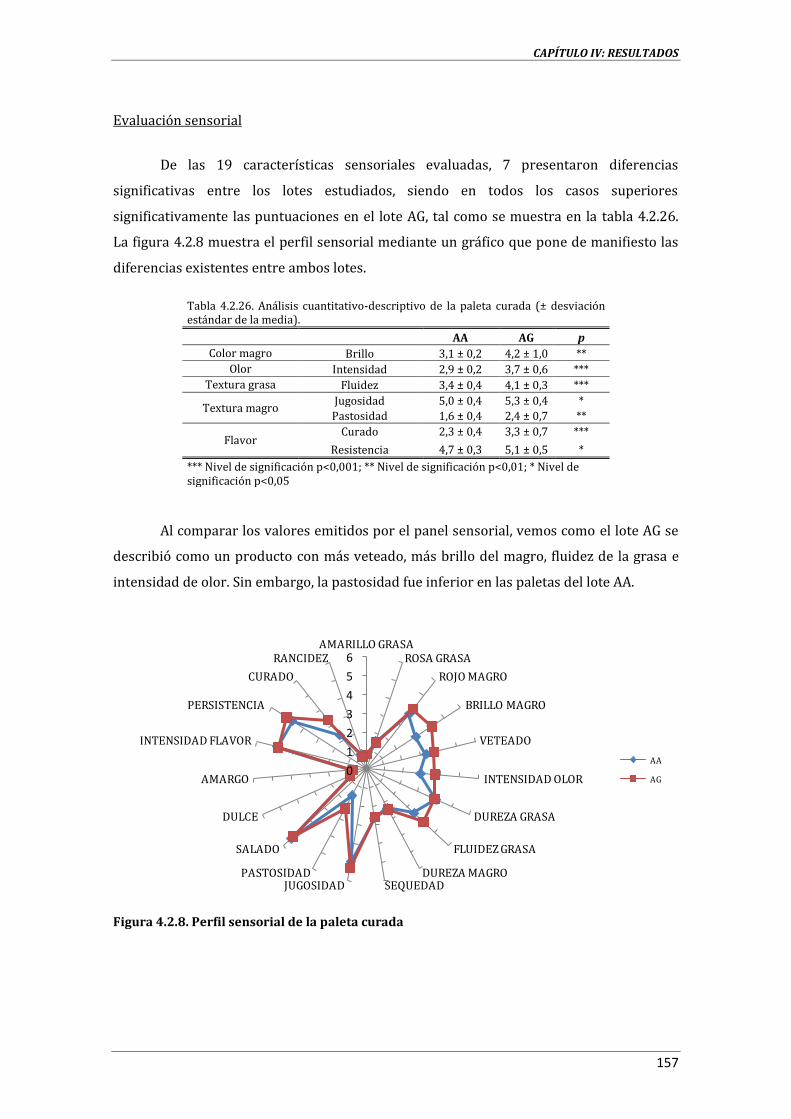
CAPÍTULO IV: RESULTADOS
157
Evaluación sensorial
De las 19 características sensoriales evaluadas, 7 presentaron diferencias
significativas entre los lotes estudiados, siendo en todos los casos superiores
significativamente las puntuaciones en el lote AG, tal como se muestra en la tabla 4.2.26.
La figura 4.2.8 muestra el perfil sensorial mediante un gráfico que pone de manifiesto las
diferencias existentes entre ambos lotes.
Tabla 4.2.26. Análisis cuantitativo-descriptivo de la paleta curada (± desviación estándar de la media).
AA AG p
Color magro Brillo 3,1 ± 0,2 4,2 ± 1,0 **
Olor Intensidad 2,9 ± 0,2 3,7 ± 0,6 ***
Textura grasa Fluidez 3,4 ± 0,4 4,1 ± 0,3 ***
Textura magro Jugosidad 5,0 ± 0,4 5,3 ± 0,4 *
Pastosidad 1,6 ± 0,4 2,4 ± 0,7 **
Flavor Curado 2,3 ± 0,4 3,3 ± 0,7 ***
Resistencia 4,7 ± 0,3 5,1 ± 0,5 *
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05
Al comparar los valores emitidos por el panel sensorial, vemos como el lote AG se
describió como un producto con más veteado, más brillo del magro, fluidez de la grasa e
intensidad de olor. Sin embargo, la pastosidad fue inferior en las paletas del lote AA.
0
1
2
3
4
5
6AMARILLO GRASA
ROSA GRASA
ROJO MAGRO
BRILLO MAGRO
VETEADO
INTENSIDAD OLOR
DUREZA GRASA
FLUIDEZ GRASA
DUREZA MAGROSEQUEDAD JUGOSIDAD
PASTOSIDAD
SALADO
DULCE
AMARGO
INTENSIDAD FLAVOR
PERSISTENCIA
CURADO
RANCIDEZ
AA
AG
Figura 4.2.8. Perfil sensorial de la paleta curada

CAPÍTULO IV: RESULTADOS
158
4.2.3. INFLUENCIA DE LA EXPRESIÓN DEL IGF2 EN FUNCIÓN DEL SEXO SOBRE LAS
CARACTERÍSTICAS FÍSICO-QUÍMICAS Y SENSORIALES DE LA PALETA CURADA
En este apartado se va a evaluar la influencia de los alelos A y G del IGF2 (insuline-
like growth factor 2) y su interacción con el sexo de los animales sobre todos los
parámetros analizados en las paletas curadas. Para ello, tan solo van a incluirse aquellos
parámetros que presentaron diferencias significativas entre los cuatro lotes estudiados
(AA*H, AA*M, AG*H y AG*M, siendo H el factor hembra y M el factor macho).
Características generales de la paleta curada
Al evaluar las características generales de la paleta curada: peso, longitud, anchura
y perímetro, no se encontraron diferencias significativas entre los cuatro lotes,
coincidiendo con los resultados mostrados por la influencia del IGF2. Estos resultados
coincidieron con los de Burgos et al., (2011), ya que al evaluar el efecto sexo, la
conformación de las paletas no presentó diferencias significativas.
El corte transversal de las paletas mostró diferencias significativas en cuanto al
espesor de la grasa subcutánea, tanto en la zona proximal (figura 4.2.9) como en la distal
(figura 4.2.10), presentando diferencias significativas en cinco de las seis medidas
tomadas en cada zona debidas al efecto genético y tan solo tres debidas al efecto sexo, tal y
como se muestra en las tablas 4.2.27 y 4.2.28.
Tabla 4.2.27. Medidas de la grasa subcutánea de la zona proximal en la paleta curada (± desviación estándar de la media)
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
A (cm) 2,0a ± 0,3 2,4ab ± 0,3 3,5c ± 0,0 2,7b ± 0,4 ns *** *** ***
B (cm) 2,5a ± 0,4 2,2a ± 0,2 3,6b ± 0,4 3,1b ± 0,2 ** *** ns ***
C (cm) 3,0a ± 0,2 3,8b ± 0,5 4,5c ± 0,0 4,5c ± 0,5 * *** * ***
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo
Tabla 4.2.28. Medidas de la grasa subcutánea de la zona distal en la paleta curada (± desviación estándar de la media)
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
A (cm) 2,5a ± 1,2 3,1ab ± 0,1 3,4ab ± 1,3 4,4b ± 0,7 * ** ns **
B (cm) 1,2ab ± 0,4 0,9a ± 0,2 1,6b ± 0,1 1,5b ± 0,4 ns *** ns **
C (cm) 1,6 ± 0,4 1,9 ± 0,8 2,0 ± 0,8 1,4 ± 0,6 ns ns ns ns
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo
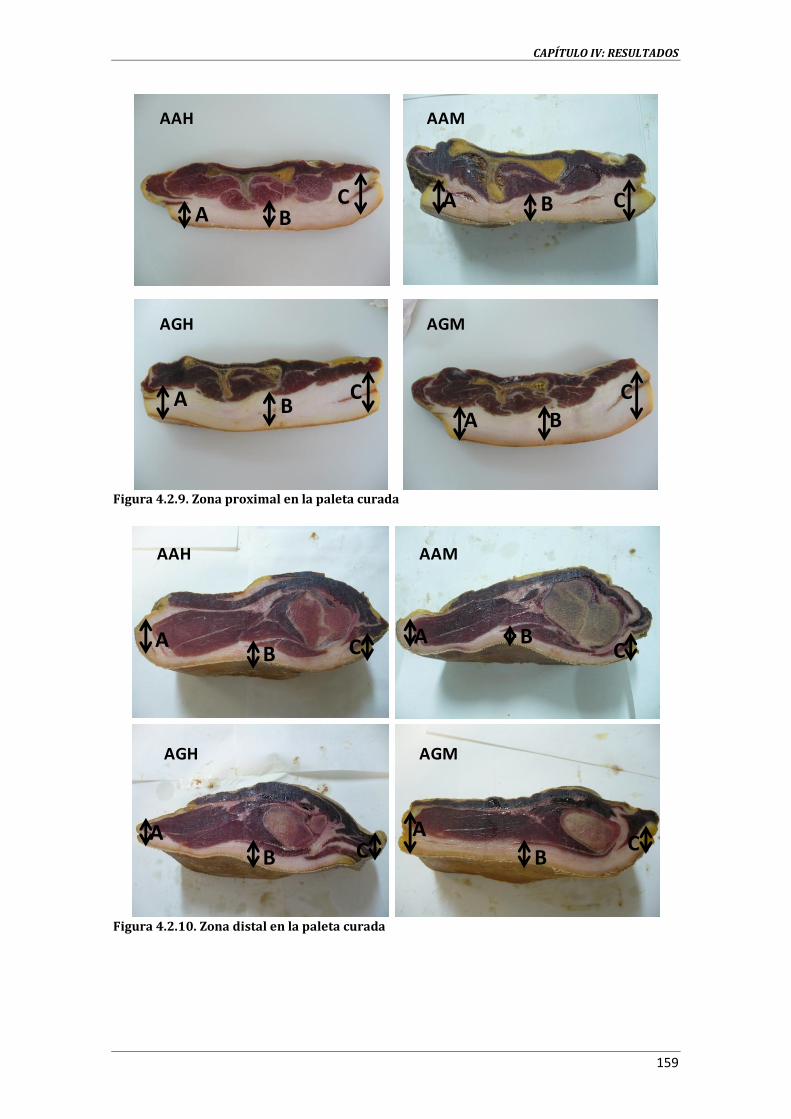
CAPÍTULO IV: RESULTADOS
159
AGH
AAMAAH
AGM
A BC
BA C
A B C
B
C
A
Figura 4.2.9. Zona proximal en la paleta curada
AAH AAM
AGH AGM
A A
AA
B
B
B
B
C C
C C
Figura 4.2.10. Zona distal en la paleta curada

CAPÍTULO IV: RESULTADOS
160
Las paletas procedentes de cerdos heterocigóticos (AGH y AGM) presentaron un
mayor espesor de la grasa subcutánea siendo superior el espesor de las muestras
procedentes de hembras (3,1 cm valor medio) en comparación con las procedentes de
machos (2,9 cm valor medio), siguiendo una tendencia contraria a los jamones, que
mostraron mayor espesor los lotes procedentes de machos que los de hembras. Las
paletas procedentes de los cerdos homocigóticos (AAH y AAM) presentaron menor
espesor de grasa subcutánea en comparación con los otros dos lotes, además la influencia
del sexo no fue la misma, presentando mayor contenido las paletas procedentes de
machos (2,4 cm valor medio) en comparación con las procedentes de hembras (2,1 cm
valor medio).
Perfil de textura instrumental
La tabla 4.2.29 muestra las diferencias significativas existentes entre los
parámetros de textura. De los ocho parámetros evaluados, tres presentaron diferencias
significativas por el efecto sexo y cinco por el efecto genético. Los parámetros de fuerza,
dureza, gomosidad y masticabilidad fueron superiores en las paletas AG, e inferiores en las
AA independientemente del sexo, resultando las AGM más duras, gomosas y masticables
pero menos elásticas y cohesivas, al contrario que las AA, a pesar de que las AAM
obtuvieron mayores parámetros de elasticidad y cohesividad.
Parámetros generales físico-químicos
En cuanto a los parámetros generales, mostraron diferencias significativas el
contenido de grasa intramuscular debidas al efecto genético y los mg Mb/g m, el pH y los
mg MDA/ Kg debidas al efecto sexo, tal como se muestra en la tabla 4.2.30. Se observaron
diferencias significativas que podrían ser achacables a la genética, en el contenido de grasa
subcutánea e intramuscular, siendo significativamente superiores los resultados
pertenecientes al lote AGH en ambos casos. Sin embargo, el efecto sexo mostró diferencias
en el contenido de grasa subcutánea en algún punto, pero no en el de grasa intramuscular,
a pesar de que entre los lotes pertenecientes a las paletas AG si existió un mayor contenido
en el lote H con respecto al lote M. El efecto sexo mostró diferencias significativas en el
contenido de mioglobina (mg Mb/g), siendo superiores los valores de los lotes
pertenecientes a las hembras en comparación con los machos, independientemente de la
genética. También existieron diferencias en el valor del pH y en el contenido de MDA, pero
en este caso la tendencia no fue determinante solo por el efecto sexo, sino que el efecto
genético también tubo parte.

Tabla 4.2.29. Análisis de perfil de textura (TPA) de la paleta curada (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
Fuerza (N) 16,84a ± 1,65 16,09a ± 0,00 17,77a ± 3,41 21,67b ± 2,35 ns ** * *** Dureza (N) 17,58a ± 1,17 17,66a ± 0,34 20,09a ± 4,01 24,27b ± 2,48 * *** * ***
Fracturabilidad (Adimensional) 10,17ab ± 0,49 9,64a ± 0,19 10,31ab ± 0,30 10,74b ± 1,06 ns * ns *
Adhesividad (N*seg) -0,18b ± 0,01 -0,20b ± 0,01 -0,27a ± 0,07 -0,17b ± 0,06 * ns ** **
Elasticidad (cm) 0,56a ± 0,04 0,64b ± 0,01 0,59ab ± 0,06 0,56a ± 0,03 ns ns ** ** Cohesividad (Adimensional) 0,49ab ± 0,02 0,51b ± 0,00 0,51b ± 0,02 0,47a ± 0,02 ns ns *** **
Gomosidad (N) 8,09a ± 0,32 6,61a ± 0,58 10,23b ± 1,71 11,38b ± 1,29 ns *** ** ***
Masticabilidad (N) 4,94a ± 0,59 4,09a ± 0,33 7,48b ± 1,91 6,57b ± 0,98 * *** ns ***
(N): Newton *** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo
Tabla 4.2.30. Parámetros físico-químicos generales de la paleta curada (± desviación estándar de la media)
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
Humedad (g/100 g) 45,60a ± 7,57 52,36b ± 0,62 50,26ab ± 1,44 49,55ab ± 0,81 ns ns * ns Proteínas (f) (g/100 g) 39,12 ± 0,38 38,80 ± 0,29 38,21 ± 1,29 38,61 ± 0,93 ns ns * ns Proteínas (s,dg) (g/100 g) 85,96 ± 2,43 88,54 ± 1,91 88,45 ± 0,03 86,88 ± 3,25 ns ns ** ns
Grasa (f) (g/100 g) 6,21a ± 0,67 6,71ab ± 0,64 7,80b ± 1,67 7,32ab ± 0,68 ns ** ns *
Grasa (s) (g/100 g) 10,76a ± 2,37 12,93ab ± 3,00 13,97b ± 0,23 12,91ab ± 4,04 ns * * * L (Adimensional) 33,51 ± 2,20 35,31 ± 0,44 35,83 ± 0,65 33,86 ± 2,61 ns ns * ns
mg Mb/g m 4,75b ± 0,80 3,35a ± 0,10 4,70b ± 0,60 4,42ab ± 0,99 * ns ns **
Cloruros (s) (g/100 g) 4,76 ± 0,74 3,98 ± 0,42 4,20 ± 0,01 4,44 ± 0,61 ns ns * ns
pH (Adimensional) 5,92bc ± 0,28 5,78b ± 0,25 5,46a ± 0,09 6,22c ± 0,06 ** ns *** *** mg MDA/Kg 0,09ab ± 0,01 0,13ab ± 0,05 0,07a ± 0,02 0,16b ± 0,08 ** ns ns ns
(f): extracto fresco; (s): extracto seco; (s,dg): extracto seco y desengrasado *** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
162
Composición de la grasa intramuscular
La fracción de triglicéridos (LN) fue la fracción mayoritaria de la grasa
intramuscular en los cuatro lotes estudiados, con un porcentaje de 76,6% para el lote AAH,
82,8% para el AAM, 86,6% para el AGH y de 81,1% para el lote AGM de la grasa total. La
fracción de fosfolípidos (LP) fue la fracción minoritaria con un 1,3, 1,6, 1,0 y 1,2%
respectivamente. La fracción de AGL representó el 22,2, 15,6, 12,4 y 17,7%
respectivamente.
Fracción de lípidos neutros
La tabla 4.2.31 muestra el contenido de los ácidos grasos identificados en la
fracción de triglicéridos (LN) de la grasa intramuscular extraída de las paletas curadas.
Existieron diferencias significativas en doce ácidos grasos, cinco de ellos como
consecuencia del efecto sexo, otros cinco por el efecto genético y dos por el efecto de
ambos factores. El sumatorio de AGS y AGMI se mostró influenciado por el efecto genético,
siendo las muestras del lote AG las que presentaron los mayores niveles de estos ácidos
grasos, coincidiendo con el contenido de C18:1. Otro ácido graso mayoritario, como es el
C16:0, mostró diferencias causadas por el efecto de ambos factores, sexo y genética, siendo
superior el valor del lote AGH e inferior el lote AAH.
En cuanto a los valores expresados en % (tabla 4.2.32), las diferencias fueron más
marcadas entre los lotes, donde cuatro ácidos mostraron diferencias por el efecto sexo,
ocho por el efecto genético y tres por el efecto de ambos. Los AGS y AGPI mostraron
diferencias debidas al efecto de ambos factores, siendo superior el valor del lote AGM y
AAH respectivamente. Para los AGMI el efecto sexo no dio lugar a diferencias, pero si el
efecto genético, siendo superior el valor del lote AAM.
De los ácidos grasos mayoritarios que mostraron diferencias significativas, el
C16:0 fue superior en el lote AG y el C18:1 superior en el lote AA, ambos influenciados por
el efecto genético. El C18:0 estuvo influenciado por ambos factores, siendo superior
significativamente el valor del lote AGM.

Tabla 4.2.31. Composición de la grasa intramuscular en la fracción de triglicéridos (LN) de la paleta curada. Los resultados están expresados en mg de AG/100 g muestra (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
C14:0 37,28a ± 0,99 36,56a ± 5,46 52,13b ± 0,97 40,50a ± 9,46 ** *** ns **
C16:0 912,09a ± 22,73 828,35a ± 115,75 1.202,67b ± 12,40 959,60ab ± 279,84 * ** ns *
C17:0 8,92ab ± 2,35 8,31a ± 1,12 11,13b ± 0,10 9,70ab ± 2,54 ns * ns ns C20:0 3,73ab ± 0,91 3,14a ± 0,19 4,55ab ± 0,95 5,00b ± 1,47 ns ** ns *
C14:1 2,47b ± 0,02 2,57b ± 0,39 1,82a ± 0,12 2,00a ± 0,59 ns *** ns **
C15:1 0,89b ± 0,19 0,54a ± 0,08 0,91b ± 0,27 1,00b ± 0,17 ns ** * **
C18:1 518,01 ± 87,68 566,52 ± 93,64 716,96 ± 7,69 591,99 ± 174,05 ns * ns ns C18:3 (ω-6) 2,12b ± 0,38 1,77ab ± 0,22 2,30c ± 0,00 1,65a ± 0,38 *** ns ns **
C20:4 (ω-6) 8,35b ± 1,69 5,91a ± 0,15 7,32ab ± 1,69 5,75a ± 0,30 *** ns ns **
C20:5 (ω-3) 0,48b ± 0,03 0,29a ± 0,00 0,46b ± 0,06 0,41b ± 0,10 *** ns * **
AGS 1.337,51 ± 107,20 1.304,47 ± 113,29 1.710,67 ± 67,18 1.485,93 ± 432,71 ns * ns ns
AGMI 662,60 ± 90,17 699,64 ± 92,21 875,77 ± 33,92 745,89 ± 205,13 ns * ns ns
C20:3 (ω-6)+C21 2,21b ± 0,50 1,90ab ± 0,31 1,89ab ± 0,35 1,65a ± 0,06 * ns ns *
sumatorio 2.212,55 ± 202,51 2.218,20 ± 229,79 2.821,27 ± 167,59 2.445,28 ± 696,91 ns * ns ns
*** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

Tabla 4.2.32. Composición de la grasa intramuscular en la fracción de triglicéridos (LN) de la paleta curada. Los resultados están expresados en % (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
C12:0 0,03ab ± 0,01 0,03ab ± 0,00 0,04b ± 0,01 0,03a ± 0,00 ns ns * *
C15:0 0,03 ± 0,00 0,03 ± 0,00 0,03 ± 0,00 0,03 ± 0,00 * ns ns ns
C16:0 18,29 ± 0,94 18,55 ± 0,63 19,09 ± 0,17 19,12 ± 0,30 ns ** ns * C18:0 13,13b ± 0,00 12,68a ± 0,54 13,17b ± 0,10 14,77c ± 0,22 *** *** *** ***
C20:0 0,20ab ± 0,04 0,24b ± 0,05 0,19a ± 0,02 0,22ab ± 0,01 * ns ns ns
C23:0 0,14a ± 0,01 0,16b ± 0,02 0,13a ± 0,02 0,13a ± 0,01 ns ** * **
C24:0 0,04b ± 0,01 0,05b ± 0,01 0,02a ± 0,01 0,04b ± 0,01 ** ** ns ** C14:1 0,05 ± 0,01 0,05 ± 0,01 0,04 ± 0,00 0,04 ± 0,00 ns ** ns *
C18:1 55,31 ± 1,42 55,63 ± 0,18 54,45 ± 0,14 54,46 ± 0,64 ns * ns ns
C20:1 1,05a ± 0,06 1,15ab ± 0,00 1,05a ± 0,09 1,20b ± 0,08 *** ns ns ***
C22:1 0,04a ± 0,01 0,06b ± 0,01 0,04a ± 0,01 0,04a ± 0,01 ns *** *** *** C18:2 (ω-6) 6,53b ± 0,36 6,00ab ± 0,58 5,92ab ± 0,84 5,41a ± 0,34 * ** ns **
C18:3 (ω-3) 0,46b ± 0,11 0,39ab ± 0,07 0,41ab ± 0,03 0,35a ± 0,02 * ns ns *
C20:2 (ω-6) 0,39a ± 0,00 0,42ab ± 0,01 0,44b ± 0,05 0,39a ± 0,02 ns ns *** **
C20:4 (ω-6) 0,32b ± 0,02 0,34b ± 0,04 0,32b ± 0,02 0,27a ± 0,02 * * ** *** C20:3 (ω-3) 0,10ab ± 0,01 0,11b ± 0,00 0,11ab ± 0,01 0,10a ± 0,00 ns ns * *
C20:5 (ω-3) 0,02a ± 0,00 0,02ab ± 0,00 0,02b ± 0,00 0,02ab ± 0,00 ns ns ** *
C22:2 0,02ab ± 0,00 0,03b ± 0,01 0,03ab ± 0,01 0,02a ± 0,00 ns ns ** *
C22:6 (ω-3) 0,09 ± 0,01 0,09 ± 0,03 0,09 ± 0,01 0,08 ± 0,01 ns * ns ns
AGS 32,85ab ± 0,98 32,67a ± 1,01 33,68b ± 0,15 35,32c ± 0,39 * *** ** ***
AGMI 59,15 ± 1,52 59,80 ± 0,42 58,50 ± 0,17 58,39 ± 0,64 ns * ns ns AGPI 7,99b ± 0,48 7,46ab ± 0,59 7,39ab ± 0,86 6,69a ± 0,37 ** ** ns **
ω-6 6,59b ± 0,36 6,06ab ± 0,58 5,97ab ± 0,84 5,47a ± 0,35 * ** ns **
ω-3 0,67b ± 0,12 0,61ab ± 0,08 0,63ab ± 0,04 0,55a ± 0,02 * ns ns *
C20:3 (ω-6)+C21 0,10b ± 0,01 0,11c ± 0,00 0,10b ± 0,00 0,09a ± 0,00 ns *** *** ***
*** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
165
Fracción de lípidos polares
El análisis del perfil de fosfolípidos (LP) de la grasa intramuscular se muestra en la
tabla 4.2.33. De los ácidos grasos que mostraron diferencias significativas cinco de ellos
fueron como consecuencia del efecto sexo, otros ocho por el efecto genético y cuatro por el
efecto de ambos factores.
No existieron diferencias significativas en ningún sumatorio como consecuencia
del efecto sexo, excepto la relación ω-6/ ω-3, cuya diferencia fue más marcada entre los
lotes AAH y AAM. Existieron diferencias en los sumatorios AGS y AGMI como consecuencia
del efecto genético, siendo superiores los valores del lote AA, coincidiendo con los
mayores valores de los ácidos grasos mayoritarios C16:0 y C18:1.
Los resultados expresados en porcentajes se muestran en la tabla 4.2.34. A
diferencia de la tabla anterior, solo el porcentaje de AGPI presentó diferencias
significativas por el efecto de ambos factores, siendo superior el valor del lote AAH.

Tabla 4.2.33. Composición de la grasa intramuscular en la fracción de fosfolípidos (LP) de la paleta curada. Los resultados están expresados en mg de AG/100 g muestra (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
C14:0 0,63b ± 0,16 0,48a ± 0,04 0,40a ± 0,07 0,41a ± 0,05 ns *** * ***
C15:0 0,17ab ± 0,04 0,22b ± 0,00 0,14a ± 0,03 0,22b ± 0,04 *** ns ns ** C16:0 10,96ab ± 1,88 13,81b ± 2,39 9,41a ± 1,37 10,15a ± 2,84 ns ** ns *
C17:0 0,44a ± 0,05 0,33b ± 0,05 0,22a ± 0,01 0,36b ± 0,07 ns *** *** ***
C18:0 6,81a ± 1,71 6,32a ± 1,15 4,26a ± 0,09 4,88ab ± 1,37 ns ** ns **
C20:0 0,13ab ± 0,03 0,13ab ± 0,01 0,15bc ± 0,00 0,12a ± 0,01 * ns * * C22:0 0,16 ± 0,02 0,21 ± 0,05 0,23 ± 0,13 0,15 ± 0,03 ns ns * ns
C23:0 0,59ab ± 0,12 0,75b ± 0,18 0,50a ± 0,06 0,68ab ± 0,10 ** ns ns *
C14:1 0,43b ± 0,07 0,39ab ± 0,10 0,31a ± 0,11 0,32ab ± 0,04 ns ** ns *
C15:1 2,27c ± 0,94 1,20ab ± 0,31 1,79bc ± 0,04 0,71a ± 0,14 *** * ns *** C16:1 0,77ab ± 0,02 1,14c ± 0,19 0,63a ± 0,13 0,87b ± 0,17 *** ** ns ***
C17:1 0,71b ± 0,24 0,59ab ± 0,24 0,63bc ± 0,16 0,29a ± 0,10 ** * ns **
C18:1 2,52ab ± 0,26 3,16b ± 0,64 1,90a ± 0,14 2,45ab ± 0,71 * ** ns **
C20:1 0,20ab ± 0,04 0,25b ± 0,07 0,13a ± 0,00 0,16a ± 0,07 ns ** ns ** C22:1 0,17 ± 0,04 0,13 ± 0,01 0,17 ± 0,03 0,16 ± 0,04 * ns ns ns
C24:1 0,23 ± 0,04 0,31 ± 0,12 0,30 ± 0,11 0,25 ± 0,03 ns ns * ns
C18:2 (ω-6) 5,21a ± 0,06 8,51b ± 2,53 6,80ab ± 1,76 6,26ab ± 1,81 ns ns * *
C18:3 (ω-3) 0,36b ± 0,05 0,29ab ± 0,06 0,24a ± 0,01 0,29ab ± 0,07 ns * * * C20:2 (ω-6) 0,18a ± 0,04 0,12b ± 0,01 0,13a ± 0,00 0,12a ± 0,03 ** ns ns **
C20:4 (ω-6) 1,62ab ± 0,22 2,42b ± 0,88 1,46a ± 0,10 1,36a ± 0,55 ns * ns **
C20:3 (ω-3) 0,23a ± 0,05 0,32ab ± 0,00 0,34bc ± 0,00 0,28ab ± 0,10 ns ns ** *
AGS 20,26ab ± 3,74 22,60b ± 3,60 15,62a ± 1,66 17,30ab ± 4,40 ns ** ns *
AGMI 7,29b ± 1,00 7,17b ± 1,24 5,86ab ± 0,18 5,21a ± 1,09 ns *** ns **
AGPI 8,61a ± 0,43 12,55b ± 3,36 9,78ab ± 1,99 9,24ab ± 2,46 ns ns * * ω-6 5,49a ± 0,06 8,77b ± 2,60 7,04ab ± 1,79 6,50ab ± 1,81 ns ns * *
ω-6/ ω-3 5,04a ± 0,76 8,49b ± 2,35 7,10ab ± 1,12 6,33ab ± 1,59 * ns ** **
C20:3 (ω-6)+C21 0,17 ± 0,01 0,22 ± 0,08 0,23 ± 0,06 0,15 ± 0,04 ns ns ** *
sumatorio 36,34 ± 3,36 42,54 ± 7,68 31,48 ± 3,73 31,90 ± 7,84 ns ** ns *
*** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

Tabla 4.2.34. Composición de la grasa intramuscular en la fracción de fosfolípidos (LP) de la paleta curada. Los resultados están expresados en % (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
C12:0 0,19 ± 0,02 0,21 ± 0,06 0,27 ± 0,04 0,28 ± 0,11 ns * ns ns C14:0 0,73ab ± 0,06 0,62a ± 0,08 0,84b ± 0,21 0,61a ± 0,10 ** ns ns **
C17:0 0,81b ± 0,12 0,61a ± 0,04 0,67ab ± 0,01 0,80b ± 0,14 ns ns *** **
C20:0 0,50b ± 0,04 0,38ab ± 0,09 0,42ab ± 0,15 0,25a ± 0,12 ** * ns **
C23:0 2,45a ± 0,40 2,54ab ± 0,22 3,05c ± 0,43 2,95bc ± 0,09 ns *** ns ** C24:0 2,40 ± 0,26 2,20 ± 0,07 2,74 ± 0,69 2,64 ± 0,41 ns * ns ns
C14:1 0,53bc ± 0,09 0,37a ± 0,02 0,61c ± 0,02 0,51b ± 0,06 *** *** ns ***
C17:1 1,10ab ± 0,23 1,18b ± 0,32 1,09ab ± 0,49 0,64a ± 0,17 ns * * **
C22:1 0,51a ± 0,07 0,59ab ± 0,12 0,48a ± 0,12 0,71b ± 0,12 ** ns ns ** C24:1 1,71 ± 0,50 2,24 ± 0,00 2,08 ± 0,25 1,83 ± 0,49 ns ns * ns
C18:2 (ω-6) 15,78ab ± 0,49 19,10c ± 0,00 14,65a ± 1,50 16,28b ± 0,59 *** ***
***
C18:3 (ω-6) 0,52b ± 0,07 0,35a ± 0,05 0,62b ± 0,01 0,33a ± 0,17 *** ns ns ***
C18:3 (ω-3) 0,73a ± 0,04 0,71a ± 0,13 0,92b ± 0,02 0,75a ± 0,08 * **
** C20:2 (ω-6) 0,52 ± 0,08 0,60 ± 0,02 0,61 ± 0,15 0,50 ± 0,04 ns ns ** *
C20:4 (ω-6) 5,43ab ± 0,75 6,02b ± 0,25 5,34ab ± 0,79 4,37a ± 0,91 ns ** * **
C20:3(ω-3) 0,52a ± 0,24 0,63ab ± 0,18 0,83b ± 0,00 0,73ab ± 0,17 ns ** ns *
C22:2 0,78ab ± 0,09 0,95c ± 0,03 0,73a ± 0,02 0,86bc ± 0,06 *** ** ns *** C22:6(ω-3) 2,13ab ± 0,34 1,85a ± 0,06 2,49b ± 0,48 2,19ab ± 0,30 * * ns *
AGPI 27,08a ± 0,68 30,96b ± 0,43 26,98a ± 1,85 26,83a ± 0,75 *** *** *** *** ω-6 16,30ab ± 0,43 19,45c ± 0,06 15,28a ± 1,49 16,61b ± 0,58 *** *** ** ***
ω-3 4,05a ± 0,54 3,95a ± 0,37 5,02b ± 0,62 4,49ab ± 0,23 ns *** ns **
ω-6/ ω-3 4,10b ± 0,52 4,96c ± 0,45 3,12a ± 0,68 3,71ab ± 0,26 ** *** ns ***
C20:3 (ω-6)+C21 0,64a ± 0,05 0,65a ± 0,11 0,92b ± 0,22 0,62a ± 0,07 ** * ** **
*** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
168
Fracción de ácidos grasos libres
En cuanto a la composición de la fracción de AGL, a diferencia de las otras dos
fracciones, las diferencias existentes entre los ácidos grasos se debieron más a la
interacción entre los dos factores del estudio, que su influencia por separado, tal como se
muestra en la tabla 4.2.35. En cuanto a los sumatorios, las tres fracciones, AGS, AGMI y
AGPI fueron superiores en el lote AAH seguidas del lote AGM.
La tabla 4.2.36 muestra las diferencias significativas existentes entre las
proporciones de ácidos grasos dentro de cada lote. Existieron mayores proporciones
(superiores en el lote AG) de los ácidos grasos mayoritarios C16:0 y el C16:1 debidas al
efecto genético y del C18:1 (superiores en el lote AGH) debidas al efecto sexo.
La proporción de AGS estuvo influenciada por el efecto genético, siendo superior
significativamente el valor del lote AGM y la proporción de AGMI y AGPI, influenciada por
el efecto sexo, superior en el lote AGH y AAM, respectivamente.

Tabla 4.2.35. Composición de la grasa intramuscular en la fracción de AGL de la paleta curada. Los resultados están expresados en mg de AG/100 g muestra (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
C10:0 0,59ab ± 0,17 0,50a ± 0,08 0,74b ± 0,10 0,67ab ± 0,18 ns * ns ns
C12:0 0,56 ± 0,13 0,48 ± 0,04 0,75 ± 0,31 0,68 ± 0,21 ns * ns ns C14:0 6,50 ± 1,83 4,68 ± 0,48 5,42 ± 0,99 5,99 ± 1,39 ns ns * ns
C15:0 0,82b ± 0,24 0,48a ± 0,00 0,51a ± 0,01 0,97b ± 0,22 ns ns *** ***
C16:0 162,04c ± 40,70 92,00ab ± 18,51 88,72a ± 1,81 129,59bc ± 20,68 ns ns *** ***
C17:0 2,80ab ± 0,72 1,54a ± 0,06 1,57a ± 0,04 3,12b ± 1,30 ns ns *** ** C18:0 93,85 ± 25,95 70,98 ± 24,55 61,97 ± 0,78 78,79 ± 18,68 ns ns * ns
C23:0 4,67b ± 1,14 4,48ab ± 0,93 3,06a ± 0,63 3,71ab ± 0,85 ns ** ns *
C15:1 1,61b ± 0,65 0,79a ± 0,25 0,45a ± 0,10 1,12ab ± 0,45 ns * *** **
C16:1 21,23b ± 6,54 11,51a ± 1,73 12,86a ± 0,50 16,98ab ± 3,01 ns ns *** ** C17:1 1,90b ± 0,71 1,28a ± 0,13 1,06a ± 0,09 1,30ab ± 0,21 ns * * **
C18:1 118,07b ± 41,35 66,09a ± 11,38 71,11a ± 9,98 81,41ab ± 11,45 * ns ** **
C20:1 5,36b ± 1,34 3,81a ± 0,50 4,72ab ± 0,11 4,28ab ± 0,89 * ns ns *
C22:1 0,48ab ± 0,03 0,59b ± 0,18 0,35a ± 0,02 0,45ab ± 0,14 * * ns * C18:2 (ω-6) 161,58b ± 56,82 103,12a ± 15,02 97,12a ± 7,89 127,03ab ± 23,16 ns ns ** **
C18:3 (ω-3) 6,62b ± 2,34 4,40a ± 0,60 4,12a ± 0,29 5,46ab ± 1,14 ns ns ** *
C20:2 (ω-6) 4,32b ± 1,08 2,86a ± 0,40 2,22a ± 0,02 3,06a ± 0,57 ns ** *** ***
C20:4 (ω-6) 44,88b ± 13,13 33,42ab ± 4,08 33,80ab ± 1,29 32,38a ± 6,16 ns ns ns * C20:3 (ω-3) 1,47b ± 0,51 0,67a ± 0,01 0,66a ± 0,03 0,76a ± 0,12 ** ** *** ***
C20:5 (ω-3) 1,99b ± 0,48 1,61ab ± 0,42 1,56ab ± 0,02 1,39a ± 0,30 ns * ns *
C22:6 (ω-3) 2,52b ± 0,65 2,04ab ± 0,40 1,48a ± 0,35 1,74a ± 0,44 ns ** ns **
AGS 272,76b ± 63,48 176,18a ± 38,50 163,62a ± 1,97 224,49ab ± 36,55 ns ns *** **
AGMI 150,52b ± 48,91 85,97a ± 13,57 92,35a ± 9,65 107,18ab ± 15,20 * ns ** **
AGPI 225,74b ± 73,34 150,21a ± 20,26 143,05a ± 9,48 173,77ab ± 31,69 ns ns ** * ω-6 163,56b ± 57,15 104,73a ± 15,04 98,85a ± 8,38 128,62ab ± 23,57 ns ns ** *
ω-3 12,60b ± 3,49 8,71a ± 1,12 7,81a ± 0,64 9,35ab ± 1,89 ns * ** **
sumatorio 654,12b ± 183,08 416,65a ± 69,36 403,22a ± 9,20 509,61ab ± 80,89 ns ns ** **
*** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

Tabla 4.2.36. Composición de la grasa intramuscular en la fracción de AGL de la paleta curada. Los resultados están expresados en % (± desviación estándar de la media)
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
C10:0 0,04a ± 0,01 0,07b ± 0,00 0,07b ± 0,00 0,07b ± 0,01 *** * *** ***
C12:0 0,03a ± 0,00 0,05b ± 0,01 0,05b ± 0,01 0,04ab ± 0,01 ns ns ** * C15:0 0,06a ± 0,01 0,07ab ± 0,01 0,06a ± 0,01 0,08b ± 0,01 ** ns ns **
C16:0 11,16a ± 0,59 11,15a ± 0,27 12,98b ± 0,24 12,53b ± 0,43 ns *** ns ***
C17:0 0,25a ± 0,02 0,27ab ± 0,01 0,26ab ± 0,04 0,29b ± 0,02 ** ns ns **
C18:0 11,05 ± 0,72 11,59 ± 0,73 11,37 ± 0,08 11,71 ± 0,15 * ns ns ns C22:0 0,06a ± 0,02 0,08ab ± 0,01 0,12b ± 0,04 0,06a ± 0,01 * ns *** ***
C23:0 0,79a ± 0,12 1,14c ± 0,15 1,01bc ± 0,08 0,91ab ± 0,17 * ns *** **
C24:0 0,22ab ± 0,07 0,31b ± 0,08 0,15a ± 0,03 0,17a ± 0,03 * *** ns ***
C14:1 0,07a ± 0,01 0,14b ± 0,05 0,08a ± 0,01 0,07a ± 0,02 ** * ** *** C16:1 1,49 ± 0,27 1,61 ± 0,17 1,70 ± 0,21 1,76 ± 0,21 ns * ns ns
C18:1 39,94a ± 0,34 39,86a ± 0,64 41,99b ± 1,97 38,97a ± 1,11 ** ns ** **
C22:1 0,13ab ± 0,00 0,17b ± 0,04 0,13ab ± 0,02 0,12a ± 0,04 ns * ns *
C18:3 (ω-6) 0,19a ± 0,01 0,27b ± 0,03 0,21a ± 0,02 0,21a ± 0,05 * ns ** ** C18:3 (ω-3) 0,71a ± 0,04 0,79b ± 0,01 0,72a ± 0,08 0,74ab ± 0,02 ** ns ns *
C20:2 (ω-6) 0,70 ± 0,12 0,69 ± 0,03 0,61 ± 0,04 0,61 ± 0,02 ns ** ns *
C20:4 (ω-6) 6,07a ± 0,86 7,78b ± 0,90 5,89a ± 0,06 6,28a ± 0,63 ** ** * ***
C20:3 (ω-3) 0,33c ± 0,00 0,20ab ± 0,02 0,21b ± 0,06 0,16a ± 0,00 *** *** ** *** C20:5 (ω-3) 0,25ab ± 0,03 0,31b ± 0,01 0,27ab ± 0,06 0,24a ± 0,05 ns ns * *
C22:2 0,07a ± 0,01 0,12c ± 0,02 0,10bc ± 0,00 0,09b ± 0,01 ** ns *** ***
AGS 24,26a ± 0,82 25,39b ± 0,99 26,73c ± 0,25 26,49c ± 0,44 ns *** * *** AGMI 43,38a ± 0,47 43,51a ± 0,80 45,54b ± 2,13 42,56a ± 1,15 ** ns ** **
AGPI 28,33 ± 1,40 30,35 ± 1,89 28,42 ± 1,90 29,13 ± 1,07 * ns ns ns
ω-3 2,38 ± 0,42 2,34 ± 0,18 2,09 ± 0,36 1,95 ± 0,21 ns * ns *
ω-6/ ω-3 8,29a ± 1,49 8,33a ± 0,52 9,68ab ± 1,76 10,46b ± 1,07 ns ** ns **
C20:3 (ω-6)+C21 0,82a ± 0,09 1,09b ± 0,11 0,91ab ± 0,18 0,88ab ± 0,15 * ns * **
*** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
171
Lipidos totales
En la tabla 4.1.37 se presenta la composición en ácidos grasos de los lípidos totales
presentes en la grasa intramuscular de los cuatro lotes en estudio. Tan solo los sumatorios
de AGS y AGMI mostraron diferencias significativas. El contenido de AGS mostró
diferencias debidas al efecto genético, siendo los lotes AG superiores significativamente
seguido de los lotes AA, coincidiendo con el contenido del ácido graso mayoritario C16:0.
El contenido de AGMI mostró diferencias debido a la interacción de ambos factores, siendo
superior el valor del lote AGH, coincidiendo en este caso con el contenido de los dos ácidos
monoinsaturados mayoritarios, el C16:1 y el C18:1.
Ni el sexo ni la genética parecen afectar al contenido total de ácidos grasos, no
presentando diferencias significativas entre los cuatro lotes estudiados.
La tabla 4.2.38 muestra los porcentajes de los ácidos grasos presentes en los
lípidos totales de la grasa intramuscular, donde destacaron las proporciones
significativamente superiores de los ácidos C16:0 y C18:0, influenciados por la genética y
del C6:1, influenciado por el sexo.
En cuanto a los sumatorios, el efecto genético tan solo dio lugar a diferencias en la
proporción de los AGS, siendo superior el valor de los lotes AG, sin embargo, el resto de
sumatorios que presentaron diferencias se vieron influenciados por ambos factores,
destacando la mayor proporción del lote AAH.

Tabla 4.2.37. Composición de la grasa intramuscular en la fracción de lípidos totales de la paleta curada. Los resultados están expresados en mg de AG/100 g muestra (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
C10:0 2,63a ± 0,77 4,568b ± 0,831 4,10b ± 1,23 3,37ab ± 0,85 * ns ** ** C12:0 3,97a ± 1,01 5,42ab ± 1,03 6,25b ± 2,10 5,29ab ± 1,19 ns * ns *
C14:0 71,12a ± 22,00 78,14ab ± 7,61 84,94ab ± 13,93 88,08b ± 18,36 ns * ns *
C15:0 1,86 ± 0,34 2,44 ± 0,59 1,88 ± 0,13 2,02 ± 0,15 * ns ns *
C16:0 897,73a ± 194,76 1.052,37ab ± 191,28 1.289,48b ± 257,40 1.174,24ab ± 256,57 ns ** ns * C20:0 6,25a ± 1,21 7,38ab ± 1,18 7,51ab ± 0,78 8,16b ± 1,84 ns * ns *
C23:0 4,50 ± 0,70 5,85 ± 1,17 5,11 ± 1,51 4,47 ± 0,74 ns ns * ns
C15:1 1,01 ± 0,20 0,98 ± 0,21 1,36 ± 0,35 1,12 ± 0,31 ns * ns ns
C16:1 96,76a ± 40,58 138,08ab ± 19,84 142,01b ± 6,76 130,12ab ± 6,81 ns ns * * C17:1 10,80ab ± 2,96 13,22ab ± 3,79 16,27b ± 0,53 10,82a ± 4,27 ns ns ** *
C18:1 469,64 ± 84,56 622,44 ± 180,05 624,28 ± 100,07 574,06 ± 92,57 ns ns * ns
C22:1 0,50a ± 0,11 0,53ab ± 0,10 0,70b ± 0,11 0,65ab ± 0,13 ns ** ns *
C24:1 0,21b ± 0,06 0,16ab ± 0,01 0,18ab ± 0,05 0,11a ± 0,00 ** ns ns ** C22:2 0,28 ± 0,04 0,37 ± 0,08 0,27 ± 0,04 0,34 ± 0,08 * ns ns ns
AGS 1.371,65a ± 302,92 1.550,30ab ± 270,08 1.849,86b ± 268,41 1.712,84ab ± 302,22 ns ** ns * AGMI 600,11 ± 114,17 804,99 ± 199,63 811,43 ± 103,99 744,34 ± 97,28 ns ns * *
C20:3(ω-6)+C21 5,86 ± 1,02 6,38 ± 0,78 6,16 ± 0,10 5,79 ± 1,25 ns ns * ns
*** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo
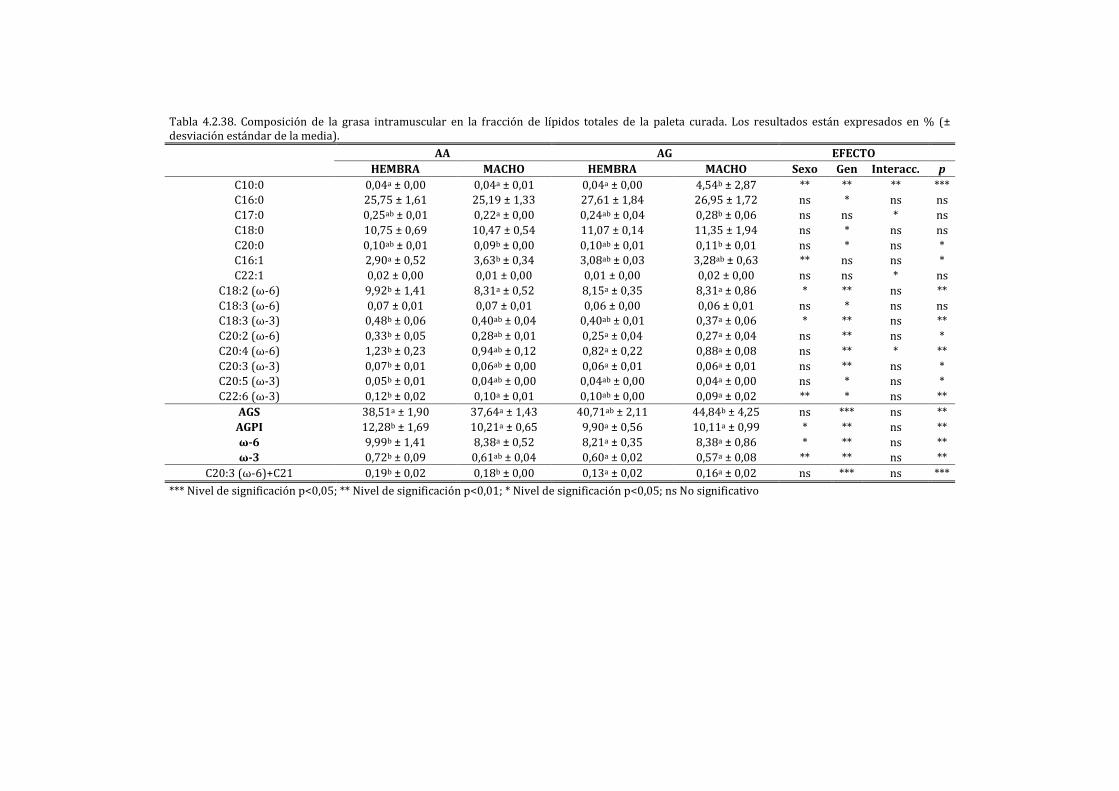
Tabla 4.2.38. Composición de la grasa intramuscular en la fracción de lípidos totales de la paleta curada. Los resultados están expresados en % (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
C10:0 0,04a ± 0,00 0,04a ± 0,01 0,04a ± 0,00 4,54b ± 2,87 ** ** ** *** C16:0 25,75 ± 1,61 25,19 ± 1,33 27,61 ± 1,84 26,95 ± 1,72 ns * ns ns
C17:0 0,25ab ± 0,01 0,22a ± 0,00 0,24ab ± 0,04 0,28b ± 0,06 ns ns * ns
C18:0 10,75 ± 0,69 10,47 ± 0,54 11,07 ± 0,14 11,35 ± 1,94 ns * ns ns
C20:0 0,10ab ± 0,01 0,09b ± 0,00 0,10ab ± 0,01 0,11b ± 0,01 ns * ns * C16:1 2,90a ± 0,52 3,63b ± 0,34 3,08ab ± 0,03 3,28ab ± 0,63 ** ns ns *
C22:1 0,02 ± 0,00 0,01 ± 0,00 0,01 ± 0,00 0,02 ± 0,00 ns ns * ns
C18:2 (ω-6) 9,92b ± 1,41 8,31a ± 0,52 8,15a ± 0,35 8,31a ± 0,86 * ** ns **
C18:3 (ω-6) 0,07 ± 0,01 0,07 ± 0,01 0,06 ± 0,00 0,06 ± 0,01 ns * ns ns C18:3 (ω-3) 0,48b ± 0,06 0,40ab ± 0,04 0,40ab ± 0,01 0,37a ± 0,06 * ** ns **
C20:2 (ω-6) 0,33b ± 0,05 0,28ab ± 0,01 0,25a ± 0,04 0,27a ± 0,04 ns ** ns *
C20:4 (ω-6) 1,23b ± 0,23 0,94ab ± 0,12 0,82a ± 0,22 0,88a ± 0,08 ns ** * **
C20:3 (ω-3) 0,07b ± 0,01 0,06ab ± 0,00 0,06a ± 0,01 0,06a ± 0,01 ns ** ns * C20:5 (ω-3) 0,05b ± 0,01 0,04ab ± 0,00 0,04ab ± 0,00 0,04a ± 0,00 ns * ns *
C22:6 (ω-3) 0,12b ± 0,02 0,10a ± 0,01 0,10ab ± 0,00 0,09a ± 0,02 ** * ns **
AGS 38,51a ± 1,90 37,64a ± 1,43 40,71ab ± 2,11 44,84b ± 4,25 ns *** ns **
AGPI 12,28b ± 1,69 10,21a ± 0,65 9,90a ± 0,56 10,11a ± 0,99 * ** ns **
ω-6 9,99b ± 1,41 8,38a ± 0,52 8,21a ± 0,35 8,38a ± 0,86 * ** ns **
ω-3 0,72b ± 0,09 0,61ab ± 0,04 0,60a ± 0,02 0,57a ± 0,08 ** ** ns **
C20:3 (ω-6)+C21 0,19b ± 0,02 0,18b ± 0,00 0,13a ± 0,02 0,16a ± 0,02 ns *** ns ***
*** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
174
Determinación del contenido en aminoácidos libres
La tabla 4.2.39 muestra las diferencias significativas en el contenido total e
individual de la composición de aminoácidos libres analizada en la paleta curada.
A pesar de que al evaluar el efecto genético por si solo tan solo la treonina mostró
diferencias significativas (tabla 4.2.23), al estudiar el efecto de los dos factores, sexo y
genética, existieron diferencias significativas en un mayor número de aminoácidos libres.
En este caso la Trh también mostró diferencias debidas al efecto de la interacción de
ambos factores, siendo el lote AAH el superior, coincidiendo con otros aminoácidos como
la co-elución Ser-Asp y Phe, donde el lote AGH mostró mayor cantidad. El resto de
aminoácidos que mostraron diferencias significativas fueron superiores en el lote AAM,
influidos por el efecto genético o por ambos factores.
Determinación de los compuestos nitrogenados derivados del ATP
A diferencia de los resultados obtenidos en la tabla 4.2.24, al estudiar ambos
factores, existieron diferencias significativas en todos los nucleótidos analizados (tabla
4.2.40). Existieron diferencias significativas en el contenido total de nucleótidos
influenciados por la interacción de ambos factores, destacando la mayor cantidad de estos,
tanto en extracto fresco como seco, del lote AAH. El resto de nucleótidos fueron superiores
en el lote AAH excepto la IMP, que fue significativamente superior en el lote AGM.

Tabla 4.2.39. Contenido en aminoácidos libres en la paleta curada en extracto fresco. Los resultados se expresan en mg aa/100 g muestra (± desviación estándar de la media)
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
Asp 158,52ab ± 12,00 180,20b ± 29,81 124,94a ± 5,03 167,65b ± 24,99 ** * ns **
Ser-Asp 198,74a ± 24,84 270,07a ± 43,28 259,59b ± 55,78 230,29ab ± 31,31 ns ns ** *
Gly-Gln 215,63 ± 23,36 250,97 ± 37,46 246,98 ± 2,05 229,11 ± 33,56 ns ns * ns
Trh 210,40b ± 76,73 102,83a ± 11,20 114,27a ± 3,37 157,20ab ± 50,08 ns ns ** ** Leu 302,77a ± 35,08 373,02b ± 8,75 307,00a ± 62,94 307,16a ± 29,32 * * * **
Phe 143,23 ± 16,07 154,57 ± 17,13 159,14 ± 27,88 135,80 ± 14,99 ns ns * ns
Lys 482,91ab ± 39,48 538,55b ± 20,10 448,02a ± 27,54 451,38a ± 54,92 ns ** ns **
** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo
Tabla 4.2.40. Contenido en 5´nucleótidos de la paleta curada en extracto fresco. Los resultados se expresan en mg de nuc./100 g muestra (± desviación estándar de la media)
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
IMP 2,88b ± 0,13 3,33b ± 1,86 1,24a ± 0,15 7,95c ± 0,79 *** ** *** ***
GMP 7,19b ± 1,27 5,56a ± 0,24 6,14ab ± 0,70 4,98a ± 0,82 *** * ns *** ADP 5,40b ± 0,62 4,27ab ± 1,61 2,72a ± 0,02 5,30b ± 1,00 ns ns *** **
AMP + Hx 43,05 ± 7,77 34,83 ± 0,65 30,95 ± 15,77 38,46 ± 3,56 ns ns * ns
Ino 8,38b ± 2,52 7,82b ± 2,34 7,14ab ± 0,80 3,89a ± 2,19 * ** ns **
sumatorio (f) 66,69 ± 10,15 55,81 ± 5,36 54,04 ± 18,09 60,57 ± 6,54 ns ns * ns
sumatorio (s) 138,27 ± 19,38 114,01 ± 12,04 107,16 ± 35,88 119,50 ± 13,22 ns ns * ns
(f): extracto fresco; (s): extracto seco *** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
176
Determinación de compuestos volátiles
Nuevamente se repite el comportamiento observado en el caso de los jamones, de
los 97 compuestos volátiles detectados en las paletas, tan solo 58 presentaron diferencias
significativas entre los lotes del estudio. Aún así las diferencias no mostraron una
tendencia en función del factor estudiado por lo que en lugar de presentar una tabla con
todos los valores, tan solo se incluyen dos figuras con los sumatorios de cada familia
(figuras 4.2.11 y 4.1.12).
En cuanto al contenido total, a pesar de no verse representado en las figuras,
presentó diferencias significativas entre los cuatro lotes estudiados, destacando el elevado
valor del lote AGH en comparación con el resto, a pesar de que el contenido de cloroformo
fue también superior en ese lote. Las diferencias existentes en el contenido total de
compuestos volátiles podrían estar asociadas a la presencia del mayor contenido de grasa
intramuscular del lote AGH, ya que el segundo lote que presentó mayor contenido (sin
tener en cuenta el cloroformo) fue el lote AGM, coincidiendo también con el lote que
mostró mayor contenido de grasa intramuscular en segundo lugar.
A pesar de que al observar las figuras puede parecer que existieron diferencias
significativas en la mayor parte de las familias, la gran variabilidad de las muestras dentro
de un mismo lote proporcionó desviaciones de la media tan elevadas, que tan solo tres
familias mostraron diferencias significativas, los compuestos azufrados que fueron
superiores en el lote AGM, los ésteres en el AGH y los éteres en el AGH, donde en todos los
casos el efecto genético fue el que generó las diferencias.

CAPÍTULO IV: RESULTADOS
177
0
50
100
150
200
250
300
AAH
AAM
AGH
AGM
*
Figura 4.2.11. Diagrama de barras correspondiente a la abundancia total de las principales familias química, expresadas en UAA (106) en los lotes de paletas estudiados (* Nivel de significación p<0,05)
0
5
10
15
20
25
AAH
AAM
AGH
AGM
***
*
Figura 4.2.12. Diagrama de barras correspondiente a la abundancia total de las principales familias química, expresadas en UAA (106) en los lotes de paletas estudiados (*** Nivel de significación p<0,001; * Nivel de significación p<0,05)

CAPÍTULO IV: RESULTADOS
178
Evaluación sensorial
De las 19 características sensoriales evaluadas, 12 presentaron diferencias
significativas entre los lotes estudiados (tabla 4.2.42). Destacaron las elevadas
puntuaciones de los atributos en los lotes pertenecientes a la genética AG, a pesar de que
el efecto genético tuvo una influencia similar al efecto sexo. La dureza de la grasa, el sabor
dulce y el flavor “a curado” fueron superiores en el lote AGH; por otra parte, el brillo del
magro, la intensidad del olor y la pastosidad fueron superiores en el lote AGM. La figura
4.2.11 muestra el perfil sensorial mediante un gráfico que pone de manifiesto las
diferencias existentes entre los cuatro lotes.
El gráfico de araña muestra claramente las diferencias de los lotes AGM y AGH
anteriormente citadas. El resto de lotes no mostraron ninguna diferencia pronunciada.
0
1
2
3
4
5
6AMARILLO GRASA
ROSA GRASA
ROJO MAGRO
BRILLO MAGRO
VETEADO
INTENSIDAD OLOR
DUREZA GRASA
FLUIDEZ GRASA
DUREZA MAGROSEQUEDAD JUGOSIDAD
PASTOSIDAD
SALADO
DULCE
AMARGO
INTENSIDAD FLAVOR
PERSISTENCIA
CURADO
RANCIDEZ
AAH
AAM
AGH
AGM
Figura 4.2.11. Perfil sensorial de la paleta curada

Tabla 4.2.42. Análisis cuantitativo-descriptivo de la paleta curada (± desviación estándar de la media)
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
Color grasa Rosa 1,33 ± 0,40 1,81 ± 0,37 1,69 ± 0,24 1,43 ± 0,18 ns ns ** *
Color magro Rojo 3,85b ± 0,39 2,67a ± 0,01 4,18b ± 0,92 3,99b ± 0,49 ** ** * ***
Brillo 3,19a ± 0,05 3,10a ± 0,33 2,89a ± 0,01 4,70b ± 0,88 *** ** *** ***
Olor Intensidad 2,91a ± 0,09 2,82a ± 0,29 3,10a ± 0,65 4,03b ± 0,43 * *** ** ***
Textura grasa Dureza 3,53a ± 0,13 4,35ab ± 0,75 4,51b ± 0,32 3,95ab ± 0,63 ns ns ** * Fluidez 3,77 ± 0,76 3,47 ± 0,33 4,24 ± 0,33 4,05 ± 0,27 ns * ns ns
Textura magro
Dureza 2,49 ± 0,60 1,71 ± 0,32 1,98 ± 0,30 2,48 ± 0,49 ns ns ** *
Sequedad 3,14b ± 0,26 2,12a ± 0,24 2,93b ± 0,03 2,66ab ± 0,49 *** ns * ***
Pastosidad 1,87ab ± 0,66 1,39a ± 0,06 2,09ab ± 0,67 2,59b ± 0,64 ns ** * **
Sabor Dulce 1,09ab ± 0,15 0,84a ± 0,15 1,17b ± 0,01 0,89a ± 0,19 *** ns ns **
Flavor Curado 2,31a ± 0,38 2,08a ± 0,15 4,54c ± 0,09 3,01b ± 0,54 *** *** *** ***
Rancidez 0,97 ± 0,51 0,96 ± 0,25 0,72 ± 0,01 0,65 ± 0,09 ns * ns ns
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
180
4.2.4. INFLUENCIA DE LA EXPRESIÓN DEL IGF2 EN FUNCIÓN DE LA CAMPAÑA SOBRE
LAS CARACTERÍSTICAS FÍSICO-QUÍMICAS Y SENSORIALES DE LA PALETA CURADA
En este apartado se va a evaluar la influencia del QTL (Quantitative Trait Loci)
IGF2 (insuline-like growth factor 2), del sexo y la campaña sobre algunos parámetros
físico-químicos en paletas procedentes de cerdos de distintas generaciones. Para ello se
evaluarán las diferencias existentes entre dos lotes, el primero perteneciente a los cerdos
de la primera generación y otro lote que corresponde a cerdos de la tercera generación. A
cada lote se le denominará, primera y tercera campaña.
Características generales de la paleta curada
Las características generales de la paleta tan solo mostraron diferencias
significativas en la medida de la anchura (tablas 4.2.43 y 4.2.44). Esta diferencia estuvo
influenciada únicamente por el efecto campaña, que también generó diferencias en la
longitud de las paletas, pero no suficientes como para originar una diferencia significativa
global. Estos datos confirman lo que sucedía con los jamones curados, ya que al evaluar el
efecto de la genética y del sexo, no existieron diferencias significativas en ningún
parámetro morfológico, llegando a la conclusión de que la selección de los cerdos no
influía sobre estos parámetros en curado, aunque sí en fresco. Por lo general, las paletas de
la tercera campaña resultaron las más grandes, en especial las pertenecientes a los lotes
AG, ya que los cerdos pertenecientes a esta campaña presentaron mayores pesos de
sacrificio, generando por lo tanto paletas de mayor tamaño.
El corte transversal de las paletas mostró diferencias significativas en cuanto al
espesor de la grasa subcutánea en la zona proximal (figura 4.2.12) en las tres medidas
tomadas en ese corte, tal y como se muestra en la tabla 4.2.44. Por lo general, las paletas
de la primera campaña tuvieron un mayor espesor de la grasa subcutánea en los tres
puntos medidos, cuya influencia fue únicamente el efecto genético, coincidiendo así con el
jamón curado. Al hacer un promedio de todas las medidas de la zona proximal se vio como
las paletas de la primera campaña mostraron un mayor espesor de grasa subcutánea (3,3
cm) en comparación con las paletas de la tercera campaña (3,1 cm).
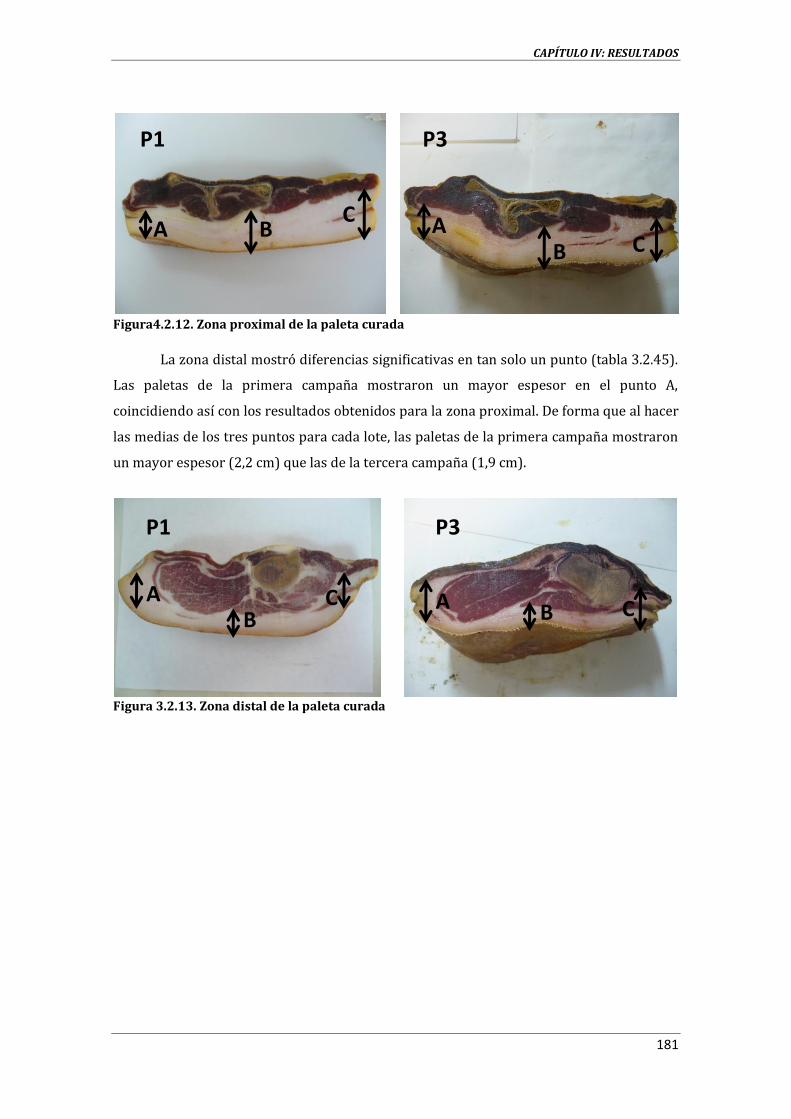
CAPÍTULO IV: RESULTADOS
181
P1 P3
A BC A
B C
Figura4.2.12. Zona proximal de la paleta curada
La zona distal mostró diferencias significativas en tan solo un punto (tabla 3.2.45).
Las paletas de la primera campaña mostraron un mayor espesor en el punto A,
coincidiendo así con los resultados obtenidos para la zona proximal. De forma que al hacer
las medias de los tres puntos para cada lote, las paletas de la primera campaña mostraron
un mayor espesor (2,2 cm) que las de la tercera campaña (1,9 cm).
ACB
CB
A
P1 P3
Figura 3.2.13. Zona distal de la paleta curada

Tabla 4.2.43. Medidas morfológicas generales de la paleta curada (± desviación estándar de la media).
P1 P3
AA AG AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
Peso (G) 4,6 ± 0,3 4,8 ± 0,4 4,6 ± 0,8 4,5 ± 0,4 4,2 ± 0,4 4,0 ± 0,4 4,6 ± 0,0 4,3 ± 0,5 ns ns ns ns ns
Longitud (cm) 62,8 ± 0,8 62,9 ± 1,3 61,9 ± 0,9 62,6 ± 0,9 64,7 ± 1,2 63,0 ± 0,0 64,0 ± 0,0 63,0 ± 1,6 ns ns ** ns ns
Anchura (cm) 23,9a ± 0,7 24,0a ± 0,4 23,6a ± 0,9 24,0a ± 1,1 25,3a ± 1,2 25,0a ± 0,0 28,0b ± 0,0 25,0b ± 1,8 ns ns ** ns **
Perímetro (cm) 54,9 ± 1,1 55,5 ± 0,8 55,8 ± 1,9 55,5 ± 1,4 54,3 ± 2,1 53,3 ± 3,0 54,2 ± 0,0 54,8 ± 2,4 ns ns ns ns ns
4.2.44. Espesor de la grasa subcutánea de la zona proximal de la paleta curada (± desviación estándar de la media).
P1 P3
AA AG AA AG
EFECTO
HEMBRA MACHO HEMBRA MACHO HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
A (cm) 2,2ab ± 0,3 2,3ab ± 0,2 3,6c ± 0,2 2,5ab ± 0,3 2,0a ± 0,4 2,8abc ± 0,3 2,5ab ± 0,0 3,0bc ± 0,4 *** ns ns ns ***
B (cm) 2,7abcd ± 0,4 2,4abc ± 0,4 3,5d ± 0,5 3,2cd ± 0,2 2,1ab ± 0,3 2,0a ± 0,0 3,2cd ± 0,0 3,2bcd ± 0,5 *** ns ns ns *** C (cm) 3,5ab ± 0,9 3,8ab ± 0,5 5,0b ± 0,6 4,2ab ± 0,5 3,2a ± 0,2 3,6ab ± 0,6 4,5ab ± 0,0 4,9b ± 0,3 *** ns ns ns **
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo
4.2.45. Espesor de la grasa subcutánea de la zona distal de la paleta curada (± desviación estándar de la media).
P1 P3
AA AG AA AG
EFECTO
HEMBRA MACHO HEMBRA MACHO HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
A(cm) 2,4 ± 1,4 3,4 ± 0,4 3,7 ± 1,3 4,5 ± 0,7 2,7 ± 0,9 3,0 ± 0,0 2,4 ± 0,0 4,3 ± 0,6 * * ns ns *
B (cm) 1,1 ± 0,5 1,1 ± 0,4 1,7 ± 0,7 1,6 ± 0,4 1,2 ± 0,2 0,9 ± 0,2 1,5 ± 0,0 1,4 ± 0,3 * ns ns ns ns
C (cm) 1,7 ± 0,7 1,9 ± 1,0 2,3 ± 0,6 1,3 ± 0,5 1,8 ± 0,2 1,8 ± 0,3 0,8 ± 0,0 1,6 ± 0,6 ns ns ns ns ns
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
183
Parámetros generales físico-químicos
En cuanto a los parámetros generales, se encontraron diferencias significativas en
algunos de los parámetros evaluados, tal como se muestra en las tablas 4.2.46 y 4.2.47.
La tabla muestra como el único factor que influyó sobre las características físico-
químicas fue la campaña, generando diferencias en parámetros como la humedad, la
coordenada L y la coordenada b*, siendo superiores los valores de las paletas
pertenecientes a la primera campaña. La grasa intramuscular también mostró diferencias
entre lotes, siendo también superiores los valores de las paletas de la primera campaña,
coincidiendo con el mayor contenido de grasa subcutánea que presentaron esas paletas.

Tabla 4.2.46. Parámetros físico-químicos generales de la paleta curada (± desviación estándar de la media). Parte I.
P1
AA AG
EFECTO
HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
Humedad (g/100 g) 51,21b ± 2,52 51,30b ± 2,23 50,26b ± 1,61 49,55b ± 1,05 ns ns *** ns ***
Grasa (f) (g/100 g) 6,32 ± 0,75 7,30 ± 1,12 8,28 ± 1,54 7,37 ± 0,47 ns ns * ns ns Grasa (s) (g/100 g) 13,00ab ± 2,85 15,05ab ± 2,00 16,66b ± 2,06 14,60ab ± 2,68 ns ns *** ns ** L 35,06bc ± 1,01 35,27c ± 0,51 35,66c ± 0,79 34,73bc ± 0,85 ns ns *** ns *** a 13,37 ± 1,58 12,78 ± 0,91 12,36 ± 0,28 12,64 ± 0,61 ns ns ns ns ns
b 6,30d ± 0,52 6,24d ± 0,41 5,92bc ± 0,63 5,61abc ± 0,56 * * *** ns ***
mg Mb/g m 4,27 ± 0,55 3,67 ± 0,45 4,51 ± 0,54 4,09 ± 0,36 ns ns ** ns *
ppm Fe heminico 13,39 ± 1,42 13,48 ± 1,66 14,43 ± 1,22 14,19 ± 1,04 ns ns ** ns ns
(f): extracto fresco; (s): extracto seco Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo
Tabla 4.2.47. Parámetros físico-químicos generales de la paleta curada (± desviación estándar de la media). Parte II.
P3
AA AG
EFECTO
HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
Humedad (g/100 g) 36,25a ± 1,47 40,02a ± 3,31 39,99a ± 0,00 37,68a ± 5,09 ns ns *** ns ***
Grasa (f) (g/100 g) 6,02 ± 0,45 6,44 ± 0,44 5,89 ± 0,00 6,75 ± 0,65 ns ns * ns ns Grasa (s) (g/100 g) 9,46a ± 1,41 10,80a ± 0,63 9,82a ± 0,00 10,95ab ± 0,92 ns ns *** ns ** L 30,94a ± 0,76 34,32abc ± 1,18 31,03a ± 0,00 31,76ab ± 2,40 ns ns *** ns *** a 11,04 ± 1,39 13,96 ± 1,54 12,16 ± 0,00 12,44 ± 2,46 ns ns ns ns ns
b 4,64ab ± 0,29 5,84bc ± 0,29 4,24a ± 0,00 5,17abc ± 0,37 * * *** ns ***
mg Mb/g m 5,56 ± 0,42 4,34 ± 0,97 5,43 ± 0,00 5,67 ± 1,70 ns ns ** ns *
ppm Fe heminico 18,85 ± 1,43 14,73 ± 3,30 18,43 ± 0,00 18,85 ± 6,40 ns ns ** ns ns
(f): extracto fresco; (s): extracto seco *** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
185
Composición de la grasa intramuscular
A pesar de que exitieron diferencias significativas en el contenido de grasa
intramuscular, el contenido total de ácidos grasos no mostró diferencias significativas, tal
como se muestra en las tablas 4.2.48 y 4.2.49.
En cuanto a los ácidos grasos mayoritarios, el C18:1 y el C18:2 (ω-6) presentaron
diferencias significativas, debidas al efecto campaña, siendo superior en las paletas de la
tercera campaña, especialmente en las pertenecientes al lote AAM. El resto de ácidos
grasos mayoritarios no presentaron diferencias significativas entre las dos campañas.
Como consecuencia de esto, el sumatorio de AGS no mostró diferencias significativas, pero
si el resto de sumatorios, AGMI, AGPI, ω-6 y ω-3, siendo superiores los valores de las
paletas de la tercera campaña, especialmente las pertenecientes al lote AAM, todas ellas
influídas por el efecto campaña.
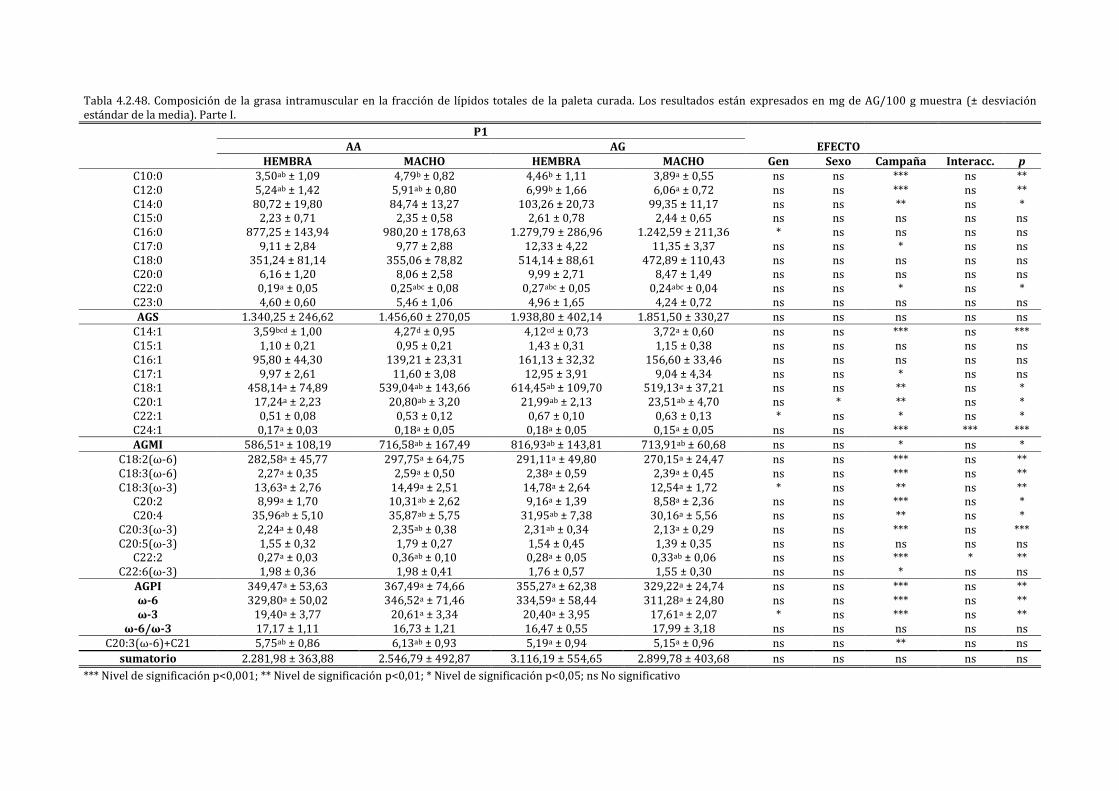
Tabla 4.2.48. Composición de la grasa intramuscular en la fracción de lípidos totales de la paleta curada. Los resultados están expresados en mg de AG/100 g muestra (± desviación estándar de la media). Parte I.
P1
AA AG
EFECTO
HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
C10:0 3,50ab ± 1,09 4,79b ± 0,82 4,46b ± 1,11 3,89a ± 0,55 ns ns *** ns ** C12:0 5,24ab ± 1,42 5,91ab ± 0,80 6,99b ± 1,66 6,06a ± 0,72 ns ns *** ns ** C14:0 80,72 ± 19,80 84,74 ± 13,27 103,26 ± 20,73 99,35 ± 11,17 ns ns ** ns * C15:0 2,23 ± 0,71 2,35 ± 0,58 2,61 ± 0,78 2,44 ± 0,65 ns ns ns ns ns C16:0 877,25 ± 143,94 980,20 ± 178,63 1.279,79 ± 286,96 1.242,59 ± 211,36 * ns ns ns ns C17:0 9,11 ± 2,84 9,77 ± 2,88 12,33 ± 4,22 11,35 ± 3,37 ns ns * ns ns C18:0 351,24 ± 81,14 355,06 ± 78,82 514,14 ± 88,61 472,89 ± 110,43 ns ns ns ns ns C20:0 6,16 ± 1,20 8,06 ± 2,58 9,99 ± 2,71 8,47 ± 1,49 ns ns ns ns ns C22:0 0,19a ± 0,05 0,25abc ± 0,08 0,27abc ± 0,05 0,24abc ± 0,04 ns ns * ns * C23:0 4,60 ± 0,60 5,46 ± 1,06 4,96 ± 1,65 4,24 ± 0,72 ns ns ns ns ns AGS 1.340,25 ± 246,62 1.456,60 ± 270,05 1.938,80 ± 402,14 1.851,50 ± 330,27 ns ns ns ns ns
C14:1 3,59bcd ± 1,00 4,27d ± 0,95 4,12cd ± 0,73 3,72a ± 0,60 ns ns *** ns *** C15:1 1,10 ± 0,21 0,95 ± 0,21 1,43 ± 0,31 1,15 ± 0,38 ns ns ns ns ns C16:1 95,80 ± 44,30 139,21 ± 23,31 161,13 ± 32,32 156,60 ± 33,46 ns ns ns ns ns C17:1 9,97 ± 2,61 11,60 ± 3,08 12,95 ± 3,91 9,04 ± 4,34 ns ns * ns ns C18:1 458,14a ± 74,89 539,04ab ± 143,66 614,45ab ± 109,70 519,13a ± 37,21 ns ns ** ns * C20:1 17,24a ± 2,23 20,80ab ± 3,20 21,99ab ± 2,13 23,51ab ± 4,70 ns * ** ns * C22:1 0,51 ± 0,08 0,53 ± 0,12 0,67 ± 0,10 0,63 ± 0,13 * ns * ns * C24:1 0,17a ± 0,03 0,18a ± 0,05 0,18a ± 0,05 0,15a ± 0,05 ns ns *** *** *** AGMI 586,51a ± 108,19 716,58ab ± 167,49 816,93ab ± 143,81 713,91ab ± 60,68 ns ns * ns *
C18:2(ω-6) 282,58a ± 45,77 297,75a ± 64,75 291,11a ± 49,80 270,15a ± 24,47 ns ns *** ns ** C18:3(ω-6) 2,27a ± 0,35 2,59a ± 0,50 2,38a ± 0,59 2,39a ± 0,45 ns ns *** ns ** C18:3(ω-3) 13,63a ± 2,76 14,49a ± 2,51 14,78a ± 2,64 12,54a ± 1,72 * ns ** ns **
C20:2 8,99a ± 1,70 10,31ab ± 2,62 9,16a ± 1,39 8,58a ± 2,36 ns ns *** ns * C20:4 35,96ab ± 5,10 35,87ab ± 5,75 31,95ab ± 7,38 30,16a ± 5,56 ns ns ** ns *
C20:3(ω-3) 2,24a ± 0,48 2,35ab ± 0,38 2,31ab ± 0,34 2,13a ± 0,29 ns ns *** ns *** C20:5(ω-3) 1,55 ± 0,32 1,79 ± 0,27 1,54 ± 0,45 1,39 ± 0,35 ns ns ns ns ns
C22:2 0,27a ± 0,03 0,36ab ± 0,10 0,28a ± 0,05 0,33ab ± 0,06 ns ns *** * ** C22:6(ω-3) 1,98 ± 0,36 1,98 ± 0,41 1,76 ± 0,57 1,55 ± 0,30 ns ns * ns ns
AGPI 349,47a ± 53,63 367,49a ± 74,66 355,27a ± 62,38 329,22a ± 24,74 ns ns *** ns ** ω-6 329,80a ± 50,02 346,52a ± 71,46 334,59a ± 58,44 311,28a ± 24,80 ns ns *** ns ** ω-3 19,40a ± 3,77 20,61a ± 3,34 20,40a ± 3,95 17,61a ± 2,07 * ns *** ns **
ω-6/ω-3 17,17 ± 1,11 16,73 ± 1,21 16,47 ± 0,55 17,99 ± 3,18 ns ns ns ns ns C20:3(ω-6)+C21 5,75ab ± 0,86 6,13ab ± 0,93 5,19a ± 0,94 5,15a ± 0,96 ns ns ** ns ns
sumatorio 2.281,98 ± 363,88 2.546,79 ± 492,87 3.116,19 ± 554,65 2.899,78 ± 403,68 ns ns ns ns ns
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

Tabla 4.2.49. Composición de la grasa intramuscular en la fracción de lípidos totales de la paleta curada. Los resultados están expresados en mg de AG/100 g muestra (± desviación estándar de la media). Parte II.
P3
AA AG
EFECTO
HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
C10:0 2,05a ± 0,54 2,99ab ± 0,47 2,65ab ± 0,00 2,60ab ± 0,61 ns ns *** ns ** C12:0 3,09a ± 0,72 4,20ab ± 0,13 3,28a ± 0,00 4,14ab ± 0,72 ns ns *** ns ** C14:0 67,06 ± 3,00 75,11 ± 2,64 62,56 ± 0,00 71,17 ± 13,39 ns ns ** ns * C15:0 2,05 ± 0,13 2,10 ± 0,04 1,88 ± 0,00 2,09 ± 0,21 ns ns ns ns ns C16:0 931,88 ± 254,47 1.232,79 ± 52,05 1.328,23 ± 0,00 1.071,72 ± 282,92 * ns ns ns ns C17:0 12,40 ± 3,62 18,51 ± 2,96 13,53 ± 0,00 14,64 ± 4,36 ns ns * ns ns C18:0 412,05 ± 106,56 576,53 ± 62,41 456,85 ± 0,00 489,53 ± 192,71 ns ns ns ns ns C20:0 6,39 ± 1,21 8,35 ± 0,71 7,39 ± 0,00 7,70 ± 2,19 ns ns ns ns ns C22:0 0,21ab ± 0,03 0,35bc ± 0,06 0,38c ± 0,00 0,26abc ± 0,04 ns ns * ns * C23:0 4,33 ± 0,82 6,83 ± 0,80 5,72 ± 0,00 4,83 ± 0,62 ns ns ns ns ns AGS 1.441,52 ± 361,38 1.927,78 ± 121,82 1.882,48 ± 0,00 1.668,68 ± 477,05 ns ns ns ns ns
C14:1 1,22a ± 0,13 2,20abc ± 0,27 1,81ab ± 0,00 1,99ab ± 0,31 ns ns *** ns *** C15:1 0,89 ± 0,13 1,22 ± 0,10 0,86 ± 0,00 1,09 ± 0,14 ns ns ns ns ns C16:1 98,36 ± 33,43 177,00 ± 44,54 139,14 ± 0,00 150,65 ± 46,63 ns ns ns ns ns C17:1 12,19 ± 2,98 17,27 ± 1,87 15,35 ± 0,00 13,46 ± 2,37 ns ns * ns ns C18:1 580,81ab ± 150,09 830,94b ± 31,66 663,59ab ± 0,00 644,93ab ± 101,64 ns ns ** ns * C20:1 20,59ab ± 3,75 30,06b ± 1,24 26,51ab ± 0,00 25,79ab ± 5,17 ns * ** ns * C22:1 0,48 ± 0,14 0,82 ± 0,24 0,84 ± 0,00 0,84 ± 0,26 * ns * ns * C24:1 3,62b ± 0,18 4,91c ± 0,31 4,69c ± 0,00 3,11b ± 0,85 ns ns *** *** *** AGMI 718,16ab ± 190,50 1.064,43b ± 75,87 852,79ab ± 0,00 841,85ab ± 139,66 ns ns * ns *
C18:2(ω-6) 460,98ab ± 114,34 572,85b ± 152,41 377,01ab ± 0,00 418,12ab ± 93,38 ns ns *** ns ** C18:3(ω-6) 2,96ab ± 0,48 4,00b ± 0,36 3,54ab ± 0,00 2,89ab ± 0,37 ns ns *** ns ** C18:3(ω-3) 21,70ab ± 5,62 25,97b ± 7,58 14,84a ± 0,00 18,10ab ± 3,71 * ns ** ns **
C20:2 13,33ab ± 2,84 16,93b ± 2,41 11,53ab ± 0,00 11,92ab ± 2,83 ns ns *** ns * C20:4 41,74ab ± 2,63 41,30ab ± 0,84 47,99ab ± 0,00 41,40b ± 6,63 ns ns ** ns *
C20:3(ω-3) 3,58b ± 0,03 3,98b ± 0,16 3,16bc ± 0,00 3,59b ± 0,12 ns ns *** ns *** C20:5(ω-3) 1,67 ± 0,05 1,73 ± 0,02 1,80 ± 0,00 1,72 ± 0,25 ns ns ns ns ns
C22:2 0,30a ± 0,05 0,59b ± 0,18 0,59b ± 0,00 0,46ab ± 0,18 ns ns *** * ** C22:6(ω-3) 2,23 ± 0,17 2,17 ± 0,05 2,31 ± 0,00 2,20 ± 0,15 ns ns * ns ns
AGPI 548,48ab ± 125,11 669,51b ± 163,62 462,79ab ± 0,00 500,39ab ± 104,33 ns ns *** ns ** ω-6 519,00ab ± 119,26 635,08b ± 156,02 440,07ab ± 0,00 474,33ab ± 100,58 ns ns *** ns ** ω-3 29,18ab ± 5,80 33,85b ± 7,77 22,12ab ± 0,00 25,60ab ± 3,84 * ns *** ns **
ω-6/ω-3 17,65 ± 0,67 18,69 ± 0,32 19,90 ± 0,00 18,35 ± 1,64 ns ns ns ns ns C20:3(ω-6)+C21 6,04ab ± 1,23 8,35b ± 1,52 6,34ab ± 0,00 6,75ab ± 0,99 ns ns ** ns ns
sumatorio 2.714,19 ± 665,21 3.670,07 ± 211,09 3.204,40 ± 0,00 3.017,67 ± 638,94 ns ns ns ns ns
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
188
Las tablas 4.2.50 y 4.2.51 muestran los porcentajes de los ácidos grasos presentes
en los lípidos totales de la grasa intramuscular. A diferencia de las tablas anteriores, vemos
como al expresar los resultados en porcentajes aparecen diferencias significativas debidas
al efecto campaña y además al efecto genético.
Las proporciones de AGPI, ω-6 y ω-3, fueron significativamente superiores en las
paletas de la tercera campaña, especialmente las pertenecientes al lote AAH. Los AGMI y
AGS no mostraron diferencias significativas, junto con los ácidos grasos mayoritarios de
ambas fracciones. Estos resultados siguieron una tendencia diferente a los resultados
mostrados en las tablas anteriores, donde las paletas pertenecientes al lote AAM
presentaron por lo general mayores cantidades de ácidos grasos. Esta diferencia puede
deberse a que al expresar los ácidos grasos en porcentaje existe un efecto genético y de
campaña, frente al único efecto genético de los ácidos grasos de las tablas 4.2.48 y 4.2.49.

Tabla 4.2.50. Composición de la grasa intramuscular en la fracción de lípidos totales de la paleta curada. Los resultados están expresados en % (± desviación estándar de la media). Parte I.
P1
AA AG
EFECTO
HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
C10:0 0,05b ± 0,00 0,05b ± 0,01 0,04b ± 0,00 0,04b ± 0,01 * ns *** ns *** C12:0 0,06b ± 0,01 0,06b ± 0,01 0,06b ± 0,00 0,06b ± 0,01 ns ns *** ns *** C14:0 1,51b ± 0,19 1,45ab ± 0,13 1,46ab ± 0,13 1,46ab ± 0,13 ns ns ** ns ** C15:0 0,05 ± 0,01 0,05 ± 0,01 0,05 ± 0,01 0,05 ± 0,01 ns ns ns ns ns C16:0 26,11 ± 1,89 25,38 ± 1,52 28,05 ± 1,81 27,16 ± 1,79 * ns ns ns ns C17:0 0,27 ± 0,06 0,26 ± 0,05 0,27 ± 0,06 0,29 ± 0,07 ns ns ns ns ns C18:0 10,32 ± 0,52 10,46 ± 0,43 11,56 ± 1,26 11,52 ± 1,06 ns ns ns ns ns C20:0 0,10a ± 0,01 0,10ab ± 0,02 0,10ab ± 0,01 0,11ab ± 0,01 * ns * ns * C22:0 0,01 ± 0,00 0,01 ± 0,00 0,01 ± 0,00 0,01 ± 0,00 ns ns ns ns ns C23:0 0,14 ± 0,02 0,14 ± 0,01 0,12 ± 0,03 0,12 ± 0,02 ns ns ns ns ns C24:0 0,01 ± 0,00 0,01 ± 0,00 0,01 ± 0,00 0,01 ± 0,00 ns ns ns ns ns AGS 38,62 ± 2,36 37,98 ± 1,42 41,72 ± 2,89 40,82 ± 2,72 * ns ns ns ns
C14:1 0,06bc ± 0,01 0,07c ± 0,01 0,05abc ± 0,01 0,06abc ± 0,01 ns ns *** ns *** C15:1 0,03 ± 0,01 0,04 ± 0,01 0,05 ± 0,03 0,04 ± 0,02 ns ns ns ns ns C16:1 3,29 ± 0,36 3,51 ± 0,28 3,25 ± 0,20 3,26 ± 0,20 ns ns ns ns ns C17:1 0,32 ± 0,05 0,33 ± 0,07 0,32 ± 0,09 0,29 ± 0,11 ns ns ns ns ns C18:1 45,79 ± 1,69 46,48 ± 1,40 44,86 ± 1,84 45,30 ± 1,82 ns ns ns ns ns C20:1 0,61 ± 0,05 0,62 ± 0,02 0,58 ± 0,06 0,64 ± 0,08 ns ns ns ns ns C22:1 0,02 ± 0,00 0,01 ± 0,00 0,01 ± 0,00 0,02 ± 0,00 ns ns * ns * C24:1 0,01a ± 0,00 0,01a ± 0,00 0,01a ± 0,00 0,01a ± 0,00 ns * *** ns *** AGMI 50,13 ± 1,63 51,07 ± 1,11 49,13 ± 1,83 49,61 ± 1,97 ns ns ns ns ns
C18:2(ω-6) 9,16ab ± 0,86 8,41ab ± 0,55 7,46a ± 1,19 7,81a ± 0,63 ** ns ** ns ** C18:3(ω-6) 0,08 ± 0,01 0,07 ± 0,01 0,06 ± 0,01 0,06 ± 0,00 * ns * ns * C18:3(ω-3) 0,46ab ± 0,05 0,42ab ± 0,03 0,37ab ± 0,06 0,36ab ± 0,06 ** ns * ns *
C20:2 0,31ab ± 0,04 0,29ab ± 0,02 0,24a ± 0,04 0,25a ± 0,04 ** ns * ns * C20:4 1,18ab ± 0,19 0,97ab ± 0,12 0,75ab ± 0,18 0,86a ± 0,07 ** ns * ns *
C20:3(ω-3) 0,07 ± 0,01 0,06 ± 0,00 0,06 ± 0,01 0,06 ± 0,01 * ns * ns * C20:5(ω-3) 0,05 ± 0,01 0,04 ± 0,00 0,03 ± 0,01 0,04 ± 0,00 * ns ns ns ns
C22:2 0,01a ± 0,00 0,01a ± 0,00 0,01a ± 0,00 0,01a ± 0,00 * ns *** * *** C22:6(ω-3) 0,12 ± 0,02 0,10 ± 0,01 0,08 ± 0,02 0,09 ± 0,01 * ns ns ns ns
AGPI 11,41ab ± 1,10 10,37ab ± 0,67 9,05a ± 1,50 9,53a ± 0,75 ** ns ** ns ** ω-6 10,72ab ± 1,05 9,74ab ± 0,65 8,51a ± 1,41 8,98a ± 0,68 ** ns ** ns ** ω-3 0,69 ± 0,08 0,62 ± 0,03 0,54 ± 0,09 0,54 ± 0,08 ** ns * ns *
ω-6/ω-3 15,66 ± 1,36 15,69 ± 0,82 15,75 ± 0,38 16,83 ± 1,56 ns ns ns ns ns C20:3(ω-6)+C21 0,19b ± 0,03 0,17ab ± 0,02 0,13a ± 0,02 0,15ab ± 0,02 ** ns * ns *
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

Tabla 4.1.51. Composición de la grasa intramuscular en la fracción de lípidos totales de la paleta curada. Los resultados están expresados en % (± desviación estándar de la media). Parte II.
P3
AA AG
EFECTO
HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
C10:0 0,01a ± 0,00 0,02a ± 0,00 0,02a ± 0,00 0,02a ± 0,00 * ns *** ns *** C12:0 0,03a ± 0,00 0,03a ± 0,00 0,03a ± 0,00 0,03a ± 0,00 ns ns *** ns *** C14:0 1,12ab ± 0,11 1,13ab ± 0,03 1,06a ± 0,00 1,25ab ± 0,14 ns ns ** ns ** C15:0 0,05 ± 0,00 0,04 ± 0,00 0,04 ± 0,00 0,05 ± 0,00 ns ns ns ns ns C16:0 25,14 ± 0,57 24,71 ± 0,36 25,85 ± 0,00 25,81 ± 1,03 * ns ns ns ns C17:0 0,26 ± 0,02 0,26 ± 0,03 0,24 ± 0,00 0,26 ± 0,02 ns ns ns ns ns C18:0 11,46 ± 0,04 11,32 ± 0,07 11,39 ± 0,00 11,38 ± 0,03 ns ns ns ns ns C20:0 0,11ab ± 0,01 0,10ab ± 0,01 0,13ab ± 0,00 0,12b ± 0,01 * ns * ns * C22:0 0,01 ± 0,00 0,01 ± 0,00 0,01 ± 0,00 0,01 ± 0,00 ns ns ns ns ns C23:0 0,13 ± 0,00 0,13 ± 0,01 0,15 ± 0,00 0,13 ± 0,01 ns ns ns ns ns C24:0 0,01 ± 0,00 0,01 ± 0,00 0,01 ± 0,00 0,01 ± 0,00 ns ns ns ns ns AGS 38,32 ± 0,64 37,76 ± 0,38 38,92 ± 0,00 39,06 ± 1,02 * ns ns ns ns
C14:1 0,03a ± 0,00 0,03a ± 0,00 0,03a ± 0,00 0,03ab ± 0,00 ns ns *** ns *** C15:1 0,02 ± 0,00 0,02 ± 0,00 0,02 ± 0,00 0,02 ± 0,00 ns ns ns ns ns C16:1 2,40 ± 0,27 3,31 ± 0,90 3,10 ± 0,00 3,30 ± 0,96 ns ns ns ns ns C17:1 0,26 ± 0,02 0,27 ± 0,02 0,25 ± 0,00 0,27 ± 0,05 ns ns ns ns ns C18:1 44,38 ± 1,71 46,28 ± 3,15 47,77 ± 0,00 45,05 ± 2,96 ns ns ns ns ns C20:1 0,61 ± 0,05 0,61 ± 0,05 0,72 ± 0,00 0,70 ± 0,08 ns ns ns ns ns C22:1 0,02 ± 0,00 0,02 ± 0,00 0,02 ± 0,00 0,02 ± 0,00 ns ns * ns * C24:1 0,04c ± 0,01 0,03bc ± 0,00 0,04bc ± 0,00 0,03b ± 0,01 ns * *** ns *** AGMI 47,74 ± 1,81 50,58 ± 4,08 51,94 ± 0,00 49,44 ± 4,03 ns ns ns ns ns
C18:2(n-6) 11,19b ± 1,23 10,06ab ± 2,28 8,28ab ± 0,00 9,08a ± 0,52 ** ns ** ns ** C18:3(n-6) 0,07 ± 0,01 0,06 ± 0,01 0,06 ± 0,00 0,06 ± 0,01 * ns * ns * C18:3(n-3) 0,51b ± 0,06 0,46ab ± 0,12 0,32a ± 0,00 0,40ab ± 0,03 ** ns * ns *
C20:2 0,37b ± 0,04 0,33ab ± 0,03 0,29ab ± 0,00 0,31ab ± 0,01 ** ns * ns * C20:4 1,32ab ± 0,26 1,15b ± 0,36 1,13ab ± 0,00 0,97ab ± 0,11 ** ns * ns *
C20:3(n-3) 0,08 ± 0,01 0,07 ± 0,01 0,07 ± 0,00 0,07 ± 0,00 * ns * ns * C20:5(n-3) 0,05 ± 0,01 0,05 ± 0,02 0,04 ± 0,00 0,04 ± 0,01 * ns ns ns ns
C22:2 0,01a ± 0,00 0,01a ± 0,00 0,02b ± 0,00 0,01ab ± 0,00 * ns *** * *** C22:6(n-3) 0,13 ± 0,02 0,11 ± 0,03 0,10 ± 0,00 0,10 ± 0,01 * ns ns ns ns
AGPI 13,72b ± 1,48 12,31ab ± 2,85 10,32ab ± 0,00 11,04ab ± 0,57 ** ns ** ns ** ω-6 12,94b ± 1,41 11,60ab ± 2,68 9,77ab ± 0,00 10,42ab ± 0,54 ** ns ** ns ** ω-3 0,77 ± 0,08 0,69 ± 0,17 0,53 ± 0,00 0,61 ± 0,05 ** ns * ns *
ω-6/ω-3 16,90 ± 0,32 16,83 ± 0,34 18,44 ± 0,00 17,00 ± 0,94 ns ns ns ns ns C20:3 (ω-6)+C21 0,19ab ± 0,02 0,16b ± 0,02 0,16ab ± 0,00 0,17ab ± 0,02 ** ns * ns *
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
191
Determinación de compuestos volátiles
Como en ocasiones anteriores, al analizar los resultados de los compuestos
volátiles, hemos utilizado las tablas 4.2.52 y 4.2.53 para poner de manifiesto (dado que se
cuenta con un número relativamente alto de muestras perfectamente identificadas) que a
pesar de la variación, los aldehídos, las cetonas y los alcoholes fueron los principales
representantes de estos compuestos y que el procesado es uno de los factores más
directamente implicados en su generación.
No existieron diferencias significativas en el contenido total de compuestos
volátiles entre las paletas de las dos campañas (Tablas 4.2.52 y 4.2.53), aunque si
existieron en la mayor parte de los compuestos volátiles detectados. De los 97 compuestos
volátiles detectados en las paletas (tabla 4.2.10), 67 presentaron diferencias significativas
entre los lotes, 39 de ellos superiores significativamente en las paletas de la primera
campaña y 28 superiores en las de la tercera, existiendo influencia de los tres factores por
lo general.
En las paletas de la primera campaña destacaron los elevados valores de algunos
compuestos como el pentanal, el hexano, el 2,3,4-trimetilpentano y la mayor parte de las
cetonas, alcoholes, ácidos y ésteres que mostraron diferencias. Sin embargo en las paletas
de la tercera campaña destacaron el 2-metilpropanal, el metanotiol, el hexanonitrilo y los
compuestos clorados.

Tabla 4.2.52. Compuestos volátiles identificados en la paleta curada agrupados por familia química. Los contenidos de cada compuesto se expresan en UAA (106) (± desviación estándar de la media). Parte I.
P1
AA AG
EFECTO
IK Posición Compuesto HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p ALDEHIDOS 175,46ab ± 29,96 104,81a ± 49,89 249,36ab ± 108,85 335,69ab ± 207,86 * ns ns * *
<500 2 acetaldehído 5,44 ± 1,99 6,35 ± 3,14 6,87 ± 3,39 8,16 ± 4,83 ns ns ns ns ns 550 12 2-metilpropanal 2,20a ± 1,45 2,71a ± 0,91 6,76ab ± 0,09 12,30abc ± 9,09 ns ns *** ns *** 649 23 3-metilbutanal 54,94 ± 3,65 37,20 ± 31,65 60,18 ± 32,99 171,87 ± 152,15 ns ns ns ns ns 660 24 2-metilbutanal 34,33 ± 3,56 6,52 ± 1,91 28,78 ± 17,94 48,08 ± 42,89 ns ns ns ns ns 698 28 pentanal 17,69ab ± 4,70 10,15ab ± 5,38 24,28b ± 11,90 12,05ab ± 6,66 ns ns ** ns ** 800 46 hexanal 20,27a ± 13,54 19,85a ± 12,48 62,66a ± 44,03 51,33a ± 3,90 *** ** *** *** *** 902 57 heptanal 6,40 ± 5,32 3,93 ± 1,42 9,27 ± 5,76 7,22 ± 4,33 * ns ns ns ns 908 59 3-(metiltio)propanal 2,77 ± 0,31 1,76 ± 0,62 1,70 ± 0,38 2,79 ± 1,18 ns ns ns * ns 969 67 benzaldehído 1,75 ± 1,11 4,49 ± 2,99 3,57 ± 2,00 6,58 ± 3,60 ns * * ns *
1006 75 octanal 14,06b ± 0,71 3,86ab ± 2,07 13,26ab ± 11,65 2,80ab ± 1,70 ns ** ** ns ** 1053 79 bencenoacetaldehido 1,17 ± 0,82 0,80 ± 0,31 2,79 ± 0,02 1,86 ± 1,51 * * ns ns * 1104 86 nonanal 11,68a ± 9,11 2,34a ± 0,01 25,12b ± 4,51 5,99a ± 4,31 ** *** ** ns *** 1157 87 2-nonenal 1,12 ± 0,68 0,76 ± 0,31 1,67 ± 0,94 2,85 ± 2,25 ns ns * ns ns 1209 88 decanal 0,90ab ± 0,55 0,71ab ± 0,38 1,79b ± 1,25 1,06ab ± 0,63 ns ns ** ns * 1495 93 dodecanal 0,74a ± 0,09 3,37b ± 2,05 0,66a ± 0,03 0,76a ± 0,39 ns * ** * ***
CETONAS 181,89a ± 61,27 235,81ab ± 113,81 327,57ab ± 148,07 409,05b ± 67,05 ns ns ** ns ** <500 5 2-propanona+propanal 39,31a ± 22,42 114,41ab ± 84,57 100,40ab ± 66,65 169,26b ± 36,81 ns ns ns ns ** 586 14 2,3-butanodiona 14,16ab ± 8,95 9,06ab ± 4,95 26,26b ± 13,28 9,19ab ± 5,68 ns ** ** ns ** 598 16 2-butanona+butanal 19,50 ± 4,62 28,25 ± 2,18 21,58 ± 0,84 18,78 ± 16,89 ns ns ** ns ns 687 27 2-pentanona 39,64 ± 33,57 29,02 ± 15,82 110,10 ± 66,49 86,44 ± 27,97 ns ns ns ns ** 710 30 3-hidroxi-2-butanona 53,70b ± 6,26 24,68ab ± 23,06 27,83ab ± 22,69 45,71b ± 16,22 ns ns *** ns *** 790 44 2-hexanona 2,83 ± 1,74 3,15 ± 1,76 5,32 ± 3,05 3,78 ± 2,23 ns ns ns ns ns 792 45 ciclopentanona 3,44a ± 0,69 7,03a ± 4,36 6,53a ± 0,72 51,33b ± 3,90 *** *** *** *** *** 891 55 2-heptanona 3,75 ± 1,24 11,05 ± 4,97 21,39 ± 8,30 18,28 ± 16,33 ns ns ns ns * 964 66 4-metil-ciclohexanona 1,40 ± 0,77 1,34 ± 0,68 1,36 ± 0,12 1,69 ± 0,22 ns ns ** ns * 989 70 3-octanona 1,82a ± 0,70 2,06a ± 1,16 2,45a ± 1,35 1,76a ± 1,49 *** *** *** *** *** 992 71 2-octanona 1,31 ± 0,67 1,98 ± 0,61 2,30 ± 0,23 1,42 ± 1,04 ns ns ns ns ns
1090 83 2-nonanona 1,03a ± 0,55 3,75b ± 1,73 2,05ab ± 0,06 1,43a ± 0,19 ns * *** * *** ALCOHOLES 167,08 ± 41,65 144,35 ± 62,71 185,66 ± 39,92 165,72 ± 67,39 ns ns ** ns ns
<500 4 etanol 105,00 ± 27,99 86,43 ± 51,64 60,09 ± 26,80 78,35 ± 30,37 ns ns ** ns ns 549 11 propanol 3,60 ± 2,78 7,82 ± 6,29 3,71 ± 0,25 1,28 ± 0,32 ns ns ns ns ns 604 18 2-metil-3-buten-1-ol 3,06ab ± 1,35 1,97a ± 0,74 4,35b ± 1,17 2,22ab ± 0,48 ns * ** ns *** 620 21 2-metilpropanol 0,00a ± 0,00 0,00a ± 0,00 17,78d ± 0,00 8,92c ± 4,23 *** ** ** *** *** 673 25 1-metoxi-2-propanol 4,15 ± 3,78 4,25 ± 4,13 13,09 ± 8,80 9,99 ± 9,01 ns ns ** ns ns 681 26 1-penten-3-ol 1,73 ± 0,94 4,54 ± 3,40 8,05 ± 5,18 6,82 ± 5,94 ns ns ns ns ns 699 29 2-pentanol 17,35bc ± 7,29 10,68abc ± 3,63 30,99d ± 1,94 10,77abc ± 6,36 * *** *** ns ***

P1
AA AG EFECTO
IK Posición Compuesto HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p 738 33 3-metilbutanol 9,06b ± 2,41 9,49b ± 0,91 4,29a ± 0,46 8,63b ± 0,99 ** ns *** ns *** 742 34 2-metilbutanol 3,60b ± 1,18 3,73b ± 1,35 2,81ab ± 0,10 2,66ab ± 0,99 ns ns *** ns *** 770 40 pentanol 2,93 ± 2,21 1,44 ± 0,47 12,84 ± 0,01 14,97 ± 11,93 ns ns ns ns * 872 52 hexanol 4,56 ± 2,03 6,84 ± 2,85 10,89 ± 4,80 6,03 ± 4,55 ns ns ns ns ns 899 56 2-heptanol 2,87b ± 1,32 2,62ab ± 0,87 2,42ab ± 0,19 2,15ab ± 1,04 ns ns *** ns ** 906 58 2-butoxietanol 2,57a ± 1,05 1,60a ± 0,21 4,03ab ± 1,79 2,48a ± 1,45 ns *** ns ** *** 982 69 1-octen-3-ol 5,02 ± 3,58 2,14 ± 0,34 7,62 ± 0,45 8,34 ± 7,14 ns ns ns ns ns
1065 82 octanol 1,57 ± 1,10 0,80 ± 0,33 2,72 ± 0,55 2,11 ± 1,88 ns ns ns ns ns HIDROCARBUROS ALIFÁTICOS 92,17bc ± 11,80 86,47abc ± 21,12 130,09c ± 37,80 91,51bc ± 18,96 ns ns *** * **
<500 1 propano 3,72a ± 1,19 5,36ab ± 2,39 6,68ab ± 2,51 5,89ab ± 3,46 ns ns ns ns ns 500 6 pentano 17,27abc ± 0,97 13,58ab ± 5,92 33,10bc ± 24,16 9,46a ± 1,53 ns ns * ns ** 507 7 1,1´-oxibis etano 9,20ab ± 1,36 6,47ab ± 3,52 10,19ab ± 7,02 3,07a ± 2,32 ns ns * * ** 555 13 2-metilpentano 3,73 ± 1,79 3,36 ± 1,91 3,95 ± 0,05 10,10 ± 8,15 ns ns ns ns ns 600 17 hexano 6,67ab ± 0,24 0,00a ± 0,00 16,95c ± 0,41 8,42abc ± 7,00 ns * ns ns *** 624 22 metilciclopentano 14,82bc ± 9,87 6,91abc ± 2,72 0,00a ± 0,00 16,99c ± 0,00 ns ns ** * ***
734 32 2,5-dimetilhexano
+ 3-metil-3-buten-1-ol 2,93a ± 0,52 1,28a ± 0,45 2,46a ± 0,46 2,79a ± 1,95 *** *** ns *** ***
753 38 2,3,4-trimetilpentano 3,03abc ± 1,46 5,32bc ± 3,23 5,47c ± 1,27 2,49abc ± 0,23 ns ns *** * *** 763 39 2,3-dimetilhexano 1,62ab ± 0,15 4,05b ± 2,53 1,88ab ± 0,54 1,26ab ± 0,37 ns ns *** ns ** 785 43 2,2,5-trimetilhexano 4,12 ± 2,11 4,31 ± 3,28 4,32 ± 0,01 2,59 ± 1,78 ns ns *** ns *
1000 74 decano 21,36ab ± 5,81 29,83ab ± 19,35 41,83b ± 21,98 24,53ab ± 15,02 ns ns *** ns ** 1100 84 undecano 3,01 ± 2,63 3,40 ± 1,72 1,05 ± 0,00 3,15 ± 2,32 ns ns ns ns ns 1400 92 tetradecano 0,69ab ± 0,15 2,60b ± 1,71 2,21ab ± 1,25 0,80ab ± 0,12 ns ns *** * **
ACIDOS 77,45ab ± 19,31 63,12ab ± 24,23 48,05ab ± 18,53 108,92b ± 38,93 ns ns ** * ** 579 15 acido acético 37,12ab ± 2,47 30,48a ± 21,18 10,09a ± 4,22 71,01b ± 24,33 ns ns * ** *** 753 37 acido 2-metilpropanoico 7,03b ± 3,43 1,80a ± 1,03 2,92ab ± 0,90 1,60a ± 1,30 * ** ** ns *** 779 42 ácido butanoico 26,63 ± 17,41 2,09 ± 0,42 14,92 ± 8,08 16,21 ± 12,61 ns ns * ns * 833 47 ácido 3-metilbutanoico 1,92 ± 0,99 14,87 ± 11,26 7,97 ± 4,15 8,92 ± 5,12 ns ns * ns * 843 48 ácido 2-metilpentanoico 2,76 ± 1,02 3,08 ± 1,43 1,87 ± 0,25 2,30 ± 1,44 ns ns ns ns ns 970 68 ácido hexanoico 0,68 ± 0,26 4,73 ± 4,26 9,12 ± 6,28 7,20 ± 6,16 ns ns ns ns ns
1258 89 ácido nonanoico 1,30a ± 0,16 6,08b ± 3,39 1,16a ± 0,63 1,68a ± 0,63 * ns ** * *** ESTERES 26,13bc ± 15,67 27,43c ± 12,51 13,13abc ± 3,43 11,54abc ± 2,84 * ns *** ns **
612 19 acetato de etilo 19,67b ± 15,44 1,79a ± 0,50 2,54a ± 0,18 3,51ab ± 1,73 ns ns * ns ** 728 31 2-hidroxipropanoato de metilo 1,19ab ± 0,40 1,19ab ± 0,89 1,17ab ± 0,02 1,53ab ± 1,20 ns ** ns ns * 853 49 2-metilbutanoato de etilo 1,09a ± 0,00 11,66b ± 5,56 0,96a ± 0,00 0,45a ± 0,00 ** * ** ** *** 856 50 3-metilbutanoato de etilo 1,89 ± 0,85 9,10 ± 8,79 6,26 ± 3,35 3,73 ± 2,46 ns ns ** ns * 942 63 hexanoato de metilo 1,32b ± 0,00 0,97ab ± 0,50 0,39ab ± 0,25 0,92ab ± 0,68 ns ns ** ns **

P1
AA AG EFECTO
IK Posición Compuesto HEMBRA MACHO HEMBRA MACHO Genética Sexo Campaña Interacc. p 999 73 hexanoato de etilo 0,97abc ± 0,36 2,71d ± 0,68 1,81cd ± 0,48 1,40bc ± 0,36 ns ** *** ** ***
COMPUESTOS AZUFRADOS 67,20ab ± 21,38 48,04a ± 16,92 68,30ab ± 21,13 61,62ab ± 9,90 ns ** *** ns *** <500 3 metanotiol 3,45a ± 0,23 3,12a ± 1,30 5,90ab ± 3,67 6,04ab ± 3,01 ns ** *** * *** 519 8 tiobis metano + etanotiol 5,14ab ± 1,69 2,54a ± 1,27 7,14ab ± 4,36 2,75a ± 1,74 ns ns *** ns ** 536 10 sulfuro de carbono 56,84ab ± 20,81 38,18a ± 17,65 52,65ab ± 19,01 50,82ab ± 7,33 ns *** *** ns *** 750 36 dimetil disulfuro 1,77ab ± 0,74 4,20b ± 1,94 2,61ab ± 1,53 2,01ab ± 1,28 ns ns ** ns *
COMPUESTOS NITROGENADOS 12,84a ± 5,03 15,24a ± 3,60 153,84b ± 0,77 13,28a ± 7,31 *** *** *** *** *** 748 35 piridina 4,14 ± 3,64 2,86 ± 1,79 3,34 ± 0,30 4,11 ± 2,85 ns ns ** ns ns 881 54 hexanonitrilo 1,46a ± 0,22 2,47ab ± 0,39 4,81b ± 0,03 2,88ab ± 1,82 *** *** ns ns *** 914 60 2,6-dimetilpirazina 1,61 ± 0,29 2,43 ± 0,43 2,14 ± 0,62 3,21 ± 2,50 ns ns ns ns ns 923 62 etilpirazina 0,90a ± 0,63 0,99a ± 0,34 143,55b ± 0,00 1,13a ± 1,03 *** *** *** *** ***
1006 76 trimetilpirazina 4,73bc ± 2,54 6,49c ± 3,12 0,00a ± 0,00 1,95ab ± 0,96 ** ns ns ns ** FURANOS 19,28 ± 7,32 20,11 ± 8,39 24,01 ± 7,25 19,20 ± 6,83 ns ns ** ns *
915 61 dihidro-2(3H)-furanona 4,66 ± 1,74 6,14 ± 3,22 3,95 ± 0,15 5,16 ± 3,24 ns ns ns ns ns 958 65 dihidro-5-metil-2(3H)-furanona 1,02 ± 0,62 1,65 ± 1,09 2,59 ± 1,62 1,65 ± 1,10 ns ns * ns ns 996 72 2-pentilfurano 5,13ab ± 2,50 5,28ab ± 2,06 8,50b ± 2,42 5,64ab ± 1,92 ns ns *** ns **
1060 80 dihidro-5-etil-2(3H)-furanona 3,24 ± 2,36 2,35 ± 1,54 3,06 ± 1,67 2,20 ± 0,89 ns ns ns ns ns 1068 81 dihidro-5-metil-5-etil-2(3H)-furanona 3,84 ± 2,67 2,21 ± 1,15 3,96 ± 1,87 2,66 ± 2,02 ns ns ** ns * 1272 90 dihidro-5-butil-2(3H)-furanona 0,65ab ± 0,39 0,50a ± 0,20 1,11b ± 0,17 0,66ab ± 0,23 ns * *** ns *** 1377 91 dihidro-5-pentil-2(3H)-furanona 0,73ab ± 0,40 1,99b ± 1,28 0,82ab ± 0,09 1,22ab ± 0,55 ns ns *** ns **
HIDROCARBUROS AROMÁTICOS 11,48 ± 5,56 15,51 ± 2,46 14,21 ± 2,37 12,96 ± 6,99 ns ns ** ns * 772 41 metilbenceno 6,04 ± 3,07 6,72 ± 0,20 8,07 ± 1,69 7,13 ± 4,01 ns ns ** ns ns 866 51 etilbenceno 0,98abc ± 0,73 1,78c ± 0,71 1,63bc ± 0,45 1,17abc ± 0,69 ns ns *** ns ** 875 53 m- y p- xileno 2,07 ± 1,08 5,02 ± 2,54 1,63 ± 0,45 3,03 ± 1,98 ns * ns ns *
1103 85 nitro benceno 2,39 ± 1,55 1,99 ± 0,96 2,87 ± 2,28 1,63 ± 0,96 ns ns * ns ns TERPENOS 3,17ab ± 1,09 2,11ab ± 1,01 4,38b ± 1,27 2,86ab ± 1,36 ns ns *** ns **
943 64 alfa-pineno 1,06ab ± 0,48 0,85a ± 0,37 1,86b ± 0,14 0,85a ± 0,55 ns * *** ns *** 1035 77 l-limoneno 2,11 ± 0,77 1,26 ± 0,87 2,52 ± 1,26 2,01 ± 0,84 ns ns ** ns ns
ETERES 2,05ab ± 1,82 0,96a ± 0,58 3,84b ± 0,80 1,39ab ± 0,71 ns ns ns ns * 1042 78 1,8-cineol 2,05ab ± 1,82 0,96a ± 0,58 3,84b ± 0,80 1,39ab ± 0,71 ns ns ns ns *
COMPUESTOS CLORADOS 409,30ab ± 227,35 99,18a ± 40,40 410,41ab ± 243,05 220,21a ± 124,38 ns ** *** ** *** 528 9 diclorometano 5,10c ± 0,93 1,82b ± 0,62 1,93b ± 0,23 1,69ab ± 1,03 ** ** *** ns *** 616 20 cloroformo 404,19ab ± 226,96 97,37a ± 40,68 408,48ab ± 243,20 218,53a ± 123,89 ns ** *** ** ***
sumatorio 1.330,67 ± 513,42 827,23 ± 215,71 1.804,10 ± 860,60 1.143,65 ± 325,84 ns ns ns ns *
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

Tabla 4.2.53. Compuestos volátiles identificados en la paleta curada agrupados por familia química. Los contenidos de cada compuesto se expresan en UAA (106) (± desviación estándar de la media). Parte II.
P3
AA AG EFECTO IK Posición Compuesto HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
ALDEHIDOS 206,57ab ± 18,79 229,56ab ± 94,33 405,99b ± 0,00 210,28ab ± 45,18 * ns ns * * <500 2 acetaldehído 4,87 ± 1,56 3,31 ± 1,41 6,54 ± 0,00 4,59 ± 0,92 ns ns ns ns ns 550 12 2-metilpropanal 18,10bc ± 0,12 20,32c ± 3,10 18,94bc ± 0,00 20,13c ± 5,74 ns ns *** ns *** 649 23 3-metilbutanal 83,44 ± 21,04 75,63 ± 51,35 116,80 ± 0,00 64,51 ± 22,18 ns ns ns ns ns 660 24 2-metilbutanal 45,27 ± 0,25 39,09 ± 7,09 41,52 ± 0,00 41,99 ± 0,49 ns ns ns ns ns 698 28 pentanal 4,37a ± 0,68 8,14a ± 3,21 8,14a ± 0,00 8,40a ± 0,84 ns ns ** ns ** 800 46 hexanal 32,75a ± 8,43 62,46a ± 22,70 182,54b ± 0,00 47,85a ± 21,08 *** ** *** *** *** 902 57 heptanal 3,58 ± 0,45 5,46 ± 2,44 11,11 ± 0,00 5,68 ± 2,99 * ns ns ns ns 908 59 3-(metiltio)propanal 2,49 ± 0,39 2,73 ± 1,28 2,35 ± 0,00 2,31 ± 0,01 ns ns ns * ns 969 67 benzaldehído 1,54 ± 0,04 2,50 ± 0,86 1,63 ± 0,00 2,94 ± 0,83 ns * * ns *
1006 75 octanal 2,25a ± 0,06 2,82ab ± 1,63 4,39ab ± 0,00 3,05ab ± 1,51 ns ** ** ns ** 1053 79 bencenoacetaldehido 1,80 ± 0,05 1,74 ± 0,52 2,70 ± 0,00 1,36 ± 0,48 * * ns ns * 1104 86 nonanal 4,63a ± 0,27 4,58a ± 1,53 8,35a ± 0,00 6,03a ± 3,02 ** *** ** ns *** 1157 87 2-nonenal 0,79 ± 0,24 0,40 ± 0,10 0,63 ± 0,00 0,73 ± 0,30 ns ns * ns ns 1209 88 decanal 0,42ab ± 0,25 0,20a ± 0,13 0,23ab ± 0,00 0,26ab ± 0,07 ns ns ** ns * 1495 93 dodecanal 0,27a ± 0,16 0,19a ± 0,00 0,13a ± 0,00 0,47a ± 0,21 ns * ** * ***
CETONAS 255,67ab ± 17,63 189,23a ± 24,85 160,73a ± 0,00 162,54a ± 19,93 ns ns ** ns ** <500 5 2-propanona+propanal 110,06ab ± 18,07 55,90ab ± 16,72 42,99a ± 0,00 75,64ab ± 19,14 ns ns ns ns ** 586 14 2,3-butanodiona 4,88a ± 0,00 4,33a ± 0,00 10,80ab ± 0,00 2,76a ± 0,65 ns ** ** ns ** 598 16 2-butanona+butanal 10,71 ± 1,07 11,87 ± 4,88 12,23 ± 0,00 15,43 ± 0,07 ns ns ** ns ns 687 27 2-pentanona 94,00 ± 0,60 79,15 ± 0,00 52,18 ± 0,00 31,16 ± 7,77 ns ns ns ns ** 710 30 3-hidroxi-2-butanona 4,20a ± 0,41 3,78a ± 1,41 3,27a ± 0,00 4,62a ± 0,83 ns ns *** ns *** 790 44 2-hexanona 4,69 ± 0,09 4,34 ± 3,06 4,12 ± 0,00 3,31 ± 1,13 ns ns ns ns ns 792 45 ciclopentanona 2,46a ± 0,13 1,64a ± 0,72 1,75a ± 0,00 1,74a ± 0,03 *** *** *** *** *** 891 55 2-heptanona 21,00 ± 2,05 21,81 ± 9,56 15,10 ± 0,00 22,54 ± 1,51 ns ns ns ns * 964 66 4-metil-ciclohexanona 0,69 ± 0,00 0,87 ± 0,30 0,74 ± 0,00 0,69 ± 0,19 ns ns ** ns * 989 70 3-octanona 1,13a ± 0,11 1,90a ± 0,53 14,97a ± 0,00 1,31b ± 0,75 *** *** *** *** *** 992 71 2-octanona 1,14 ± 0,28 2,68 ± 0,90 1,87 ± 0,00 2,51 ± 1,51 ns ns ns ns ns
1090 83 2-nonanona 0,69a ± 0,03 0,98a ± 0,40 0,70a ± 0,00 0,84a ± 0,12 ns * *** * *** ALCOHOLES 103,34 ± 22,91 98,79 ± 24,61 84,77 ± 0,00 86,71 ± 36,19 ns ns ** ns ns
<500 4 etanol 45,41 ± 19,72 38,09 ± 0,73 30,99 ± 0,00 46,18 ± 29,93 ns ns ** ns ns 549 11 propanol 1,91 ± 0,13 2,37 ± 1,36 3,47 ± 0,00 3,34 ± 0,36 ns ns ns ns ns 604 18 2-metil-3-buten-1-ol 1,68a ± 0,06 2,36ab ± 1,40 1,25a ± 0,00 0,98a ± 0,10 ns * ** ns *** 620 21 2-metilpropanol 8,99c ± 0,00 5,24bc ± 0,00 0,00a ± 0,00 1,68ab ± 0,00 *** ** ** *** *** 673 25 1-metoxi-2-propanol 1,35 ± 0,00 0,70 ± 0,00 0,00 ± 0,00 2,94 ± 0,48 ns ns ** ns ns 681 26 1-penten-3-ol 7,09 ± 0,65 6,85 ± 4,58 4,98 ± 0,00 6,58 ± 0,22 ns ns ns ns ns 699 29 2-pentanol 5,83ab ± 1,26 11,18abc ± 0,00 20,25cd ± 0,00 2,73a ± 0,00 * *** *** ns *** 738 33 3-metilbutanol 3,19a ± 0,32 2,69a ± 0,17 2,61a ± 0,00 2,22a ± 0,94 ** ns *** ns ***
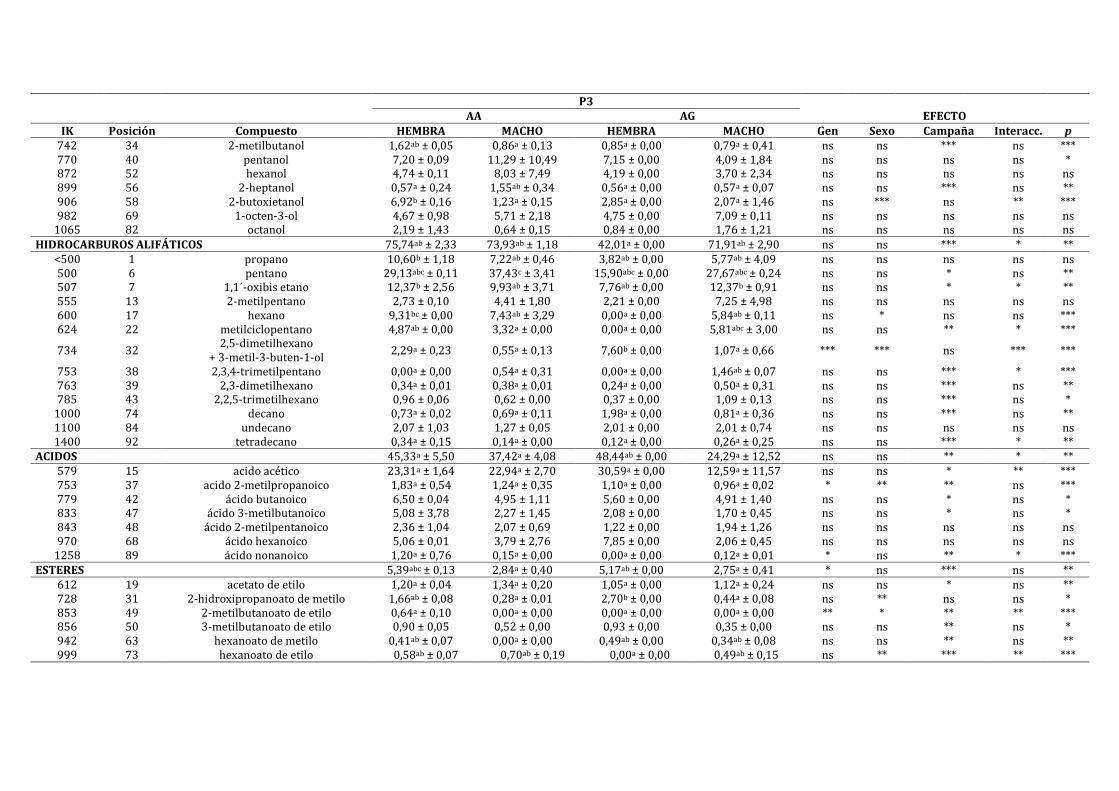
P3
AA AG EFECTO IK Posición Compuesto HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
742 34 2-metilbutanol 1,62ab ± 0,05 0,86a ± 0,13 0,85a ± 0,00 0,79a ± 0,41 ns ns *** ns *** 770 40 pentanol 7,20 ± 0,09 11,29 ± 10,49 7,15 ± 0,00 4,09 ± 1,84 ns ns ns ns * 872 52 hexanol 4,74 ± 0,11 8,03 ± 7,49 4,19 ± 0,00 3,70 ± 2,34 ns ns ns ns ns 899 56 2-heptanol 0,57a ± 0,24 1,55ab ± 0,34 0,56a ± 0,00 0,57a ± 0,07 ns ns *** ns ** 906 58 2-butoxietanol 6,92b ± 0,16 1,23a ± 0,15 2,85a ± 0,00 2,07a ± 1,46 ns *** ns ** *** 982 69 1-octen-3-ol 4,67 ± 0,98 5,71 ± 2,18 4,75 ± 0,00 7,09 ± 0,11 ns ns ns ns ns
1065 82 octanol 2,19 ± 1,43 0,64 ± 0,15 0,84 ± 0,00 1,76 ± 1,21 ns ns ns ns ns HIDROCARBUROS ALIFÁTICOS 75,74ab ± 2,33 73,93ab ± 1,18 42,01a ± 0,00 71,91ab ± 2,90 ns ns *** * **
<500 1 propano 10,60b ± 1,18 7,22ab ± 0,46 3,82ab ± 0,00 5,77ab ± 4,09 ns ns ns ns ns 500 6 pentano 29,13abc ± 0,11 37,43c ± 3,41 15,90abc ± 0,00 27,67abc ± 0,24 ns ns * ns ** 507 7 1,1´-oxibis etano 12,37b ± 2,56 9,93ab ± 3,71 7,76ab ± 0,00 12,37b ± 0,91 ns ns * * ** 555 13 2-metilpentano 2,73 ± 0,10 4,41 ± 1,80 2,21 ± 0,00 7,25 ± 4,98 ns ns ns ns ns 600 17 hexano 9,31bc ± 0,00 7,43ab ± 3,29 0,00a ± 0,00 5,84ab ± 0,11 ns * ns ns *** 624 22 metilciclopentano 4,87ab ± 0,00 3,32a ± 0,00 0,00a ± 0,00 5,81abc ± 3,00 ns ns ** * ***
734 32 2,5-dimetilhexano
+ 3-metil-3-buten-1-ol 2,29a ± 0,23 0,55a ± 0,13 7,60b ± 0,00 1,07a ± 0,66 *** *** ns *** ***
753 38 2,3,4-trimetilpentano 0,00a ± 0,00 0,54a ± 0,31 0,00a ± 0,00 1,46ab ± 0,07 ns ns *** * *** 763 39 2,3-dimetilhexano 0,34a ± 0,01 0,38a ± 0,01 0,24a ± 0,00 0,50a ± 0,31 ns ns *** ns ** 785 43 2,2,5-trimetilhexano 0,96 ± 0,06 0,62 ± 0,00 0,37 ± 0,00 1,09 ± 0,13 ns ns *** ns *
1000 74 decano 0,73a ± 0,02 0,69a ± 0,11 1,98a ± 0,00 0,81a ± 0,36 ns ns *** ns ** 1100 84 undecano 2,07 ± 1,03 1,27 ± 0,05 2,01 ± 0,00 2,01 ± 0,74 ns ns ns ns ns 1400 92 tetradecano 0,34a ± 0,15 0,14a ± 0,00 0,12a ± 0,00 0,26a ± 0,25 ns ns *** * **
ACIDOS 45,33a ± 5,50 37,42a ± 4,08 48,44ab ± 0,00 24,29a ± 12,52 ns ns ** * ** 579 15 acido acético 23,31a ± 1,64 22,94a ± 2,70 30,59a ± 0,00 12,59a ± 11,57 ns ns * ** *** 753 37 acido 2-metilpropanoico 1,83a ± 0,54 1,24a ± 0,35 1,10a ± 0,00 0,96a ± 0,02 * ** ** ns *** 779 42 ácido butanoico 6,50 ± 0,04 4,95 ± 1,11 5,60 ± 0,00 4,91 ± 1,40 ns ns * ns * 833 47 ácido 3-metilbutanoico 5,08 ± 3,78 2,27 ± 1,45 2,08 ± 0,00 1,70 ± 0,45 ns ns * ns * 843 48 ácido 2-metilpentanoico 2,36 ± 1,04 2,07 ± 0,69 1,22 ± 0,00 1,94 ± 1,26 ns ns ns ns ns 970 68 ácido hexanoico 5,06 ± 0,01 3,79 ± 2,76 7,85 ± 0,00 2,06 ± 0,45 ns ns ns ns ns
1258 89 ácido nonanoico 1,20a ± 0,76 0,15a ± 0,00 0,00a ± 0,00 0,12a ± 0,01 * ns ** * *** ESTERES 5,39abc ± 0,13 2,84a ± 0,40 5,17ab ± 0,00 2,75a ± 0,41 * ns *** ns **
612 19 acetato de etilo 1,20a ± 0,04 1,34a ± 0,20 1,05a ± 0,00 1,12a ± 0,24 ns ns * ns ** 728 31 2-hidroxipropanoato de metilo 1,66ab ± 0,08 0,28a ± 0,01 2,70b ± 0,00 0,44a ± 0,08 ns ** ns ns * 853 49 2-metilbutanoato de etilo 0,64a ± 0,10 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 ** * ** ** *** 856 50 3-metilbutanoato de etilo 0,90 ± 0,05 0,52 ± 0,00 0,93 ± 0,00 0,35 ± 0,00 ns ns ** ns * 942 63 hexanoato de metilo 0,41ab ± 0,07 0,00a ± 0,00 0,49ab ± 0,00 0,34ab ± 0,08 ns ns ** ns ** 999 73 hexanoato de etilo 0,58ab ± 0,07 0,70ab ± 0,19 0,00a ± 0,00 0,49ab ± 0,15 ns ** *** ** ***

P3
AA AG EFECTO IK Posición Compuesto HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
999 73 hexanoato de etilo 0,58ab ± 0,07 0,70ab ± 0,19 0,00a ± 0,00 0,49ab ± 0,15 ns ** *** ** *** COMPUESTOS AZUFRADOS 193,01bc ± 28,09 291,18cd ± 33,06 137,68ab ± 0,00 387,84d ± 134,12 ns ** *** ns ***
<500 3 metanotiol 10,01bc ± 0,47 9,45bc ± 0,73 14,17c ± 0,00 6,08ab ± 0,10 ns ** *** * *** 519 8 tiobis metano + etanotiol 7,36ab ± 0,08 8,49b ± 1,82 6,55ab ± 0,00 8,88b ± 1,62 ns ns *** ns ** 536 10 sulfuro de carbono 174,24bc ± 27,69 272,02cd ± 31,53 115,95ab ± 0,00 371,45d ± 131,95 ns *** *** ns *** 750 36 dimetil disulfuro 1,41ab ± 0,16 1,22ab ± 1,02 1,00a ± 0,00 1,43ab ± 0,73 ns ns ** ns *
COMPUESTOS NITROGENADOS 10,66a ± 0,70 8,92a ± 0,08 13,81a ± 0,00 7,29a ± 1,20 *** *** *** *** *** 748 35 piridina 0,60 ± 0,06 0,33 ± 0,00 0,60 ± 0,00 0,43 ± 0,00 ns ns ** ns ns 881 54 hexanonitrilo 3,27ab ± 0,60 1,65a ± 0,02 7,36c ± 0,00 1,83a ± 0,94 *** *** ns ns *** 914 60 2,6-dimetilpirazina 3,35 ± 0,03 3,35 ± 0,41 3,18 ± 0,00 1,92 ± 0,02 ns ns ns ns ns 923 62 etilpirazina 0,77a ± 0,00 0,98a ± 0,07 0,72a ± 0,00 0,80a ± 0,16 *** *** *** *** ***
1006 76 trimetilpirazina 2,66abc ± 0,10 2,62abc ± 0,54 1,96ab ± 0,00 2,31abc ± 0,22 ** ns ns ns ** FURANOS 10,41 ± 0,21 11,53 ± 3,03 12,11 ± 0,00 10,70 ± 2,32 ns ns ** ns *
915 61 dihidro-2(3H)-furanona 4,78 ± 0,25 4,78 ± 0,41 3,80 ± 0,00 4,44 ± 0,62 ns ns ns ns ns 958 65 dihidro-5-metil-2(3H)-furanona 0,80 ± 0,16 1,03 ± 0,44 1,14 ± 0,00 0,88 ± 0,10 ns ns * ns ns 996 72 2-pentilfurano 1,45a ± 0,06 2,38a ± 0,19 3,13a ± 0,00 2,93a ± 1,31 ns ns *** ns **
1060 80 dihidro-5-etil-2(3H)-furanona 2,31 ± 0,02 2,23 ± 1,38 2,76 ± 0,00 1,68 ± 0,51 ns ns ns ns ns 1068 81 dihidro-5-metil-5-etil-2(3H)-furanona 0,72 ± 0,02 0,63 ± 0,24 0,84 ± 0,00 0,50 ± 0,00 ns ns ** ns * 1272 90 dihidro-5-butil-2(3H)-furanona 0,25a ± 0,01 0,24a ± 0,24 0,26a ± 0,00 0,15a ± 0,05 ns * *** ns *** 1377 91 dihidro-5-pentil-2(3H)-furanona 0,11a ± 0,02 0,23a ± 0,14 0,18a ± 0,00 0,11a ± 0,02 ns ns *** ns **
HIDROCARBUROS AROMÁTICOS 6,80 ± 2,89 9,23 ± 5,49 6,45 ± 0,00 5,67 ± 0,91 ns ns ** ns * 772 41 metilbenceno 3,63 ± 2,18 4,93 ± 4,19 3,84 ± 0,00 2,77 ± 0,91 ns ns ** ns ns 866 51 etilbenceno 0,00a ± 0,00 0,57abc ± 0,04 0,31ab ± 0,00 0,25ab ± 0,02 ns ns *** ns ** 875 53 m- y p- xileno 1,46 ± 0,01 3,00 ± 1,26 1,71 ± 0,00 1,28 ± 0,00 ns * ns ns *
1103 85 nitro benceno 1,71 ± 1,13 0,73 ± 0,00 0,59 ± 0,00 1,36 ± 0,01 ns ns * ns ns TERPENOS 2,15ab ± 1,69 0,78a ± 0,04 0,82a ± 0,00 1,39a ± 0,44 ns ns *** ns **
943 64 alfa-pineno 0,43a ± 0,03 0,41a ± 0,04 0,46a ± 0,00 0,45a ± 0,06 ns * *** ns *** 1035 77 l-limoneno 1,72 ± 1,69 0,38 ± 0,00 0,36 ± 0,00 0,95 ± 0,42 ns ns ** ns ns
ETERES 1,41ab ± 0,64 1,24ab ± 0,36 1,25ab ± 0,00 1,82ab ± 1,15 ns ns ns ns * 1042 78 1,8-cineol 1,41ab ± 0,64 1,24ab ± 0,36 1,25ab ± 0,00 1,82ab ± 1,15 ns ns ns ns *
COMPUESTOS CLORADOS 625,42b ± 62,62 682,00b ± 0,00 682,00b ± 0,00 182,98a ± 131,87 ns ** *** ** *** 528 9 diclorometano 0,90ab ± 0,30 0,00a ± 0,00 0,00a ± 0,00 0,32ab ± 0,02 ** ** *** ns *** 616 20 cloroformo 624,53b ± 62,77 682,00b ± 0,00 682,00b ± 0,00 182,66a ± 131,86 ns ** *** ** ***
sumatorio 1.541,90 ± 75,30 1.636,63 ± 65,79 1.601,22 ± 0,00 1.156,16 ± 60,42 ns ns ns ns *
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
198
4.3. LOMO CURADO
4.3.1. CARACTERIZACIÓN DEL LOMO CURADO
Al igual que en el caso de la paleta curada, el Consejo Regulador de la DOP Jamón
de Teruel no ampara dichos productos, aún así desde 1993 poseen una marca diferenciada
de calidad denominada “Aragón Calidad Alimentaria” regulada por el Departamento de
Agricultura y Alimentación del Gobierno de Aragón (BOA, 1993 (b)).
Características generales del lomo curado
La tabla 4.3.1 muestra los valores medios de las medidas morfológicas de todos los
lomos curados utilizados en el presente estudio (n=45).
Tabla 4.3.1. Medidas morfológicas generales del lomo curado (± desviación estándar de la media).
Peso (g) Longitud (cm) Perímetro (cm) Diámetro (cm)
1,7 ± 0,1 51,2 ± 3,4 21,0 ± 0,7 6,7 ± 0,2
La marca “Aragón Calidad Alimentaria” establece un calibre mínimo de 60 mm, por
lo que todas las piezas analizadas se encuentran por encima de los valores establecidos
para el calibre. El resto de datos morfológicos no están especificados en dicha normativa
(BOA, 1993).
Perfil de textura instrumental
Los resultados obtenidos del análisis de perfil de textura (TPA) para el lomo
curado se exponen en la tabla 4.3.2. Estos resultados son muy diferentes a los descritos
para lomos Ibéricos por Ramírez (2008), que encontró mayores valores en todos los
parámetros de textura descritos a continuación.
Tabla 4.3.2. Análisis de perfil de textura (TPA) para el lomo curado (± desviación estándar de la media).
Fuerza (N) 28,00 ± 2,84
Dureza (N) 30,66 ± 3,07 Fracturabilidad (Adimensional) 10,53 ± 0,55
Adhesividad (N*seg) -0,16 ± 0,01
Elasticidad (cm) 0,78 ± 0,02
Cohesividad (Adimensional) 0,60 ± 0,01 Gomosidad (N) 18,29 ± 1,88
Masticabilidad (N) 14,21 ± 1,48
(N): Newton

CAPÍTULO IV: RESULTADOS
199
Parámetros generales físico-químicos
Los resultados de las determinaciones de humedad, proteínas y grasa
intramuscular se muestran en la tabla 4.3.3, junto con otros parámetros relacionados con
la composición química del producto.
Tabla 4.3.3. Parámetros físico-químicos del lomo curado (± desviación estándar de la media).
Humedad (g/100 g) 50,97 ± 2,06
Proteínas (f) (g/100 g) 39,49 ± 2,84
Proteínas (s, dg) (g/100 g) 83,69 ± 8,48 Grasa (f) (g/100 g) 4,80 ± 2,02
Grasa (s) (g/100 g) 8,96 ± 3,24
L (Adimensional) 38,04 ± 2,29
a (Adimensional) 14,74 ± 1,28 b (Adimensional) 9,94 ± 1,82
mg Mb/g m 1,85 ± 0,51
ppm Fe heminico 6,26 ± 1,73
Cloruros (f) (g/100 g) 2,20 ± 0,74 Cloruros (s) (g/100 g) 4,47 ± 1,50
pH (Adimensional) 5,96 ± 0,03
mg MDA/Kg 0,25 ± 0,06
(f): extracto fresco; (s): extracto seco; (s, dg): extracto seco y desengrasado
Como se observa en la tabla 4.3.3, la humedad media de los lomos fue de 50,97%,
un valor algo superior al indicado en la norma de calidad, que establece que la humedad
sea inferior al 50% (BOA, 1993 (b)). Estos resultados son similares a los descritos por
otros autores para lomo curado procedentes de cerdos alimentados con piensos
enriquecidos (Martín, 2008) que encontraron valores entre un 46,8% y un 49,6%. Sin
embargo, nuestros resultados fueron superiores a los descritos por Ramírez (2008) para
lomos procedentes de cerdos cruzados con Duroc al 50%, que encontraron porcentajes de
humedad de entre 33,97 y 38,66%, respectivamente. Las diferencias pueden deberse a
efectos de procesado de las piezas.
En relación al contenido en grasa intramuscular de los lomos curados, los valores
estuvieron comprendidos en torno a un 4,80%. Estos valores son ligeramente inferiores a
los descritos por Ramírez (2008) en lomos de cerdos cruzados (en torno al 7% en materia
fresca), por Muriel (2003) en lomos de cerdos Ibéricos de montanera y pienso (superior al
7% en extracto fresco), o por los descritos por Ventanas (2006) en lomos procedentes de
cerdos Ibéricos alimentados con piensos enriquecidos con alto oleico (en torno a un 8,29%
en extracto fresco). Sin embargo nuestros valores fueron similares a los descritos por
Martin (2008) para lomo curado procedente de cerdos alimentados con piensos
enriquecidos (entre un 4,7% y un 6% en extracto fresco). Las diferencias encontradas

CAPÍTULO IV: RESULTADOS
200
pueden deberse a diversos factores como la genética del animal, la alimentación recibida
por este y por la zona del lomo considerada para el estudio por los distintos autores, ya
que en nuestro caso las muestras empleadas para la cuantificación de la grasa
intramuscular fueron tomadas en todos los casos de la zona central del lomo, donde el
contenido graso es menor que el existente en otras zonas como la cabezada.
El porcentaje medio de proteínas fue de 39,49%, inferior al descrito para lomos
Ibéricos cruzados con Duroc con un 48% aproximadamente (Ramírez, 2008) o a lomos
Ibéricos puros de montanera que presentaron un 42,85% o alimentados con piensos
enriquecidos con un 47,79% (Ventanas, 2006). Las diferencias pueden deberse al distinto
contenido en grasa intramuscular y al distinto grado de hidratación de los lomos.
El porcentaje de cloruros presente en los lomos fue de 2,20%, inferior al descrito
por Ventanas (2006) que encontró valores entre 2,68% y un 3,29% para lomos Ibéricos
llegando a la conclusión de que ni la genética ni la alimentación eran agentes influyentes
en el contenido de cloruros. Sin embargo al expresar los resultados en extracto seco los
resultados son más similares, ya que los lomos Ibéricos de ese estudio presentaron mucho
menor contenido en humedad (en torno al 28%).
El valor obtenido para la concentración de mioglobina (mg/g muestra) fue de 1,85,
inferior al descrito por Ventanas (2006), que mostró valores de concentración de
mioglobina (mg/ g muestra) entre 2,73 y 4,69. La actividad física realizada por los cerdos
Ibéricos explicaría el mayor contenido en mioglobina de sus músculos y en definitiva en
los productos curados derivados.
En cuanto al color instrumental, los lomos presentaron un valor de luminosidad de
38,04, superior al descrito por Ventanas (2006) que mostró valores entre 30,3 y 34,5,
aunque similar al descrito por Ramírez (2008), que mostró valores comprendidos entre
36,0 y 40,5. Estas diferencias con respecto a los datos del primer autor pueden deberse a
factores como el % de humedad, la capacidad de retención de agua (CRA) que estaría
estrechamente relacionada con el pH y con la integridad de las proteínas musculares, el
nivel de engrasamiento, etc. El parámetro obtenido para la coordenada a* obtuvo un valor
de 14,74, superior al descrito por Ventanas (2006) que obtuvo valores entre 8,8 y 10,9,
pero inferior al descrito por Ramírez (2008) que propuso valores entre 16,5 y 18,2.
Algunos autores han relacionado valores altos de a* en jamón con una intensa desecación
del producto a lo largo del procesado (Arnau et al., 2003) y otros lo han relacionado con
procesados a temperaturas más elevadas en la fase de secadero (Andrés et al., 2005). El
valor obtenido para la coordenada b* fue de 8,45, superior a los descritos por Ventanas

CAPÍTULO IV: RESULTADOS
201
(2006) y Ramírez (2008). El valor de la coordenada b* viene determinada por las
características de la grasa, que es la que contribuye con “coloraciones amarillentas” a la
matización del color. Del mismo modo la cantidad de nitrosomioglobina oxidada, en este
caso del magro, podría contribuir a estas coloraciones amarillentas (Skibsted, 1992). En
cuanto al valor de mg de MDA, resultó inferior en los lomos curados (0,25 mg) en
comparación con los lomos Ibéricos estudiados por Ramírez (2008) que encontró valores
de entre 1,76 y 1,85 mg o por Ventanas (2006) que encontró valores de entre 0,67 y 1,24
mg, teniendo en cuneta que el contenido de grasa intramuscular fue superior en las piezas
Ibéricas.
Composición de la grasa intramuscular
Los triglicéridos (LN) representaron la fracción mayoritaria de la grasa
intramuscular, constituyendo aproximadamente el 86,8% (tabla 4.3.4), seguido de la
fracción de AGL con un 11,6% y por último de la fracción de fosfolípidos (LP) con un 1,6%.
En la tabla 4.3.5, podemos comprobar las altas proporciones del C18:1 con un
54,09%, seguido muy por debajo del C16:0 con un 19,34% y del C18:0 con un 14,26% en
la fracción de triglicéridos (LN). Estos valores difieren de los obtenidos por Martín (2008),
que obtuvo proporciones inferiores del C18:1 (43,83%) y del C18:0 (13,50%) y superiores
del C16:0 (26,10%). Al comparar con lomos Ibéricos descritos por Ventanas (2006), la
proporción del C16:0 descrita fue muy similar a la mostrada (51,43%), sin embargo el
contenido de C18:0 y C18:1 fueron inferiores (11,03% y 51,43%).
Respecto a las proporciones de la fracción de fosfolípidos (LP) podemos destacar la
mayor proporción de los AGS con un 29,64%, en comparación con los 31,81% descritos
por Martín (2008) y los 33,65% descritos por Ventanas (2006).
En cuanto a las proporciones de los sumatorios de la fracción de AGL, destacamos
el 26,01% de los AGS frente a los 32,51% en lomos Ibéricos, los 35,54% de los AGMI frente
a los 41,21% en lomo Ibérico y los 36,11% de los AGPI frente a los 31,91% de los lomos
Ibéricos (Ventanas, 2006).

CAPÍTULO IV: RESULTADOS
202
Tabla 4.3.4. Composición de la grasa intramuscular del lomo curado en ácidos grasos neutros (LN), libres (AGL), polares (LP) y totales (LT). Los resultados están expresados en mg AG/100 g muestra (± desviación estándar de la media).
LN AGL LP LT
C10:0 1,55 ± 0,28 0,27 ± 0,07 0,06 ± 0,01 2,97 ± 1,44
C12:0 1,63 ± 0,27 0,30 ± 0,08 0,11 ± 0,03 3,33 ± 1,88
C14:0 25,49 ± 4,11 1,97 ± 0,53 0,27 ± 0,05 51,64 ± 26,18 C15:0 0,75 ± 0,19 0,24 ± 0,04 0,09 ± 0,02 1,00 ± 0,35
C16:0 500,09 ± 85,99 43,09 ± 8,74 6,69 ± 1,12 663,05 ± 326,08
C17:0 3,43 ± 0,68 0,75 ± 0,18 0,17 ± 0,03 6,18 ± 3,51
C18:0 241,04 ± 53,65 22,15 ± 3,90 3,17 ± 0,45 278,53 ± 140,89 C20:0 3,20 ± 0,62 0,24 ± 0,06 0,07 ± 0,02 4,71 ± 2,94
C22:0 0,22 ± 0,05 0,13 ± 0,03 0,13 ± 0,04 0,14 ± 0,06
C23:0 0,89 ± 0,16 1,85 ± 0,31 0,33 ± 0,06 2,26 ± 0,71
AGS 778,28 ± 142,00 70,99 ± 12,74 11,10 ± 1,59 1.013,82 ± 482,16
C14:1 1,95 ± 0,43 0,45 ± 0,12 0,22 ± 0,05 2,41 ± 1,12
C15:1 0,43 ± 0,13 0,47 ± 0,14 0,73 ± 0,09 1,22 ± 0,63
C16:1 70,27 ± 11,01 5,69 ± 1,57 0,48 ± 0,09 103,84 ± 58,21
C17:1 3,27 ± 0,68 0,40 ± 0,10 0,26 ± 0,05 6,26 ± 3,41 C18:1 309,78 ± 70,77 20,23 ± 3,20 1,93 ± 0,47 368,05 ± 189,79
C20:1 13,03 ± 3,24 1,19 ± 0,32 0,11 ± 0,03 12,24 ± 6,18
C22:1 0,49 ± 0,12 0,13 ± 0,02 0,08 ± 0,02 0,30 ± 0,17
C24:1 0,26 ± 0,06 0,44 ± 0,06 0,22 ± 0,07 1,26 ± 1,04
AGMI 399,49 ± 81,71 28,99 ± 4,94 4,04 ± 0,62 495,56 ± 246,77
C18:2 (ω-6) 82,47 ± 17,05 43,51 ± 6,41 5,16 ± 0,97 168,66 ± 66,70
C18:3 (ω-6) 0,44 ± 0,10 0,57 ± 0,12 0,11 ± 0,03 1,53 ± 0,59
C18:3 (ω-3) 5,65 ± 1,19 1,46 ± 0,26 0,19 ± 0,04 6,79 ± 2,92 C20:2 (ω-6) 4,33 ± 0,97 0,99 ± 0,18 0,18 ± 0,03 4,33 ± 2,05
C20:4 (ω-6) 3,50 ± 0,67 18,74 ± 3,80 1,71 ± 0,27 22,33 ± 6,28
C20:3 (ω-3) 1,19 ± 0,26 0,32 ± 0,06 0,22 ± 0,12 1,11 ± 0,51
C20:5 (ω-3) 0,36 ± 0,07 0,89 ± 0,17 0,21 ± 0,06 0,99 ± 0,28 C22:2 0,27 ± 0,06 0,18 ± 0,03 0,17 ± 0,06 0,25 ± 0,15
C22:6 (ω-3) 0,29 ± 0,06 1,12 ± 0,19 0,17 ± 0,02 1,05 ± 0,37
AGPI 98,52 ± 20,21 67,77 ± 10,40 8,12 ± 1,32 207,04 ± 77,63
ω-6 90,75 ± 18,68 63,81 ± 9,81 7,16 ± 1,21 196,85 ± 73,83
ω-3 7,50 ± 1,50 3,79 ± 0,60 0,79 ± 0,17 9,94 ± 3,85
ω-6/ω-3 12,10 ± 0,37 16,88 ± 0,84 9,33 ± 2,15 19,99 ± 2,18
C20:3 (ω-6)+C21 0,96 ± 0,20 2,28 ± 0,44 0,23 ± 0,05 3,45 ± 0,98
sumatorio 1.277,25 ± 238,97 170,02 ± 27,06 23,49 ± 3,36 1.719,88 ± 770,57

CAPÍTULO IV: RESULTADOS
203
Tabla 4.3.5. Composición de la grasa intramuscular del lomo curado en ácidos grasos neutros (LN), libres (AGL), polares (LP) y totales (LT). Los resultados están expresados en % (± desviación estándar de la media).
LN AGL LP LT
C10:0 0,06 ± 0,01 0,09 ± 0,01 0,15 ± 0,04 0,04 ± 0,02
C12:0 0,05 ± 0,01 0,06 ± 0,01 0,17 ± 0,03 0,05 ± 0,02
C14:0 0,85 ± 0,11 0,49 ± 0,08 0,47 ± 0,07 1,49 ± 0,20 C15:0 0,02 ± 0,00 0,07 ± 0,01 0,14 ± 0,03 0,04 ± 0,01
C16:0 19,34 ± 1,26 12,05 ± 0,98 14,86 ± 1,66 27,59 ± 1,81
C17:0 0,14 ± 0,01 0,26 ± 0,03 0,45 ± 0,05 0,19 ± 0,05
C18:0 14,26 ± 1,12 11,18 ± 0,29 11,37 ± 0,95 11,17 ± 1,31 C20:0 0,29 ± 0,03 0,16 ± 0,02 0,17 ± 0,07 0,11 ± 0,01
C22:0 0,02 ± 0,00 0,12 ± 0,03 0,74 ± 0,18 0,01 ± 0,00
C23:0 0,09 ± 0,01 1,29 ± 0,18 1,89 ± 0,17 0,11 ± 0,03
C24:0 0,04 ± 0,01 0,29 ± 0,05 1,32 ± 0,29 0,01 ± 0,00
AGS 35,17 ± 2,07 26,06 ± 1,16 31,73 ± 2,53 40,80 ± 2,84
C14:1 0,06 ± 0,01 0,11 ± 0,02 0,40 ± 0,06 0,07 ± 0,03
C15:1 0,01 ± 0,00 0,12 ± 0,03 1,47 ± 0,32 0,04 ± 0,02
C16:1 2,60 ± 0,21 1,50 ± 0,19 1,02 ± 0,15 3,80 ± 0,71 C17:1 0,17 ± 0,01 0,18 ± 0,02 1,18 ± 0,37 0,22 ± 0,05
C18:1 54,09 ± 2,03 32,19 ± 2,62 18,79 ± 1,09 45,43 ± 2,64
C20:1 1,12 ± 0,13 0,78 ± 0,07 0,55 ± 0,08 0,55 ± 0,07
C22:1 0,05 ± 0,01 0,13 ± 0,04 0,36 ± 0,07 0,01 ± 0,00 C24:1 0,05 ± 0,01 0,53 ± 0,10 1,41 ± 0,43 0,02 ± 0,02
AGMI 58,14 ± 2,04 35,54 ± 2,68 25,18 ± 1,63 50,15 ± 3,14
C18:2 (ω-6) 4,66 ± 0,45 21,51 ± 1,13 17,26 ± 2,38 7,05 ± 2,33
C18:3 (ω-6) 0,02 ± 0,00 0,25 ± 0,03 0,20 ± 0,06 0,06 ± 0,01 C18:3 (ω-3) 0,28 ± 0,03 0,71 ± 0,11 0,51 ± 0,06 0,28 ± 0,10
C20:2 (ω-6) 0,32 ± 0,03 0,66 ± 0,06 0,94 ± 0,12 0,19 ± 0,07
C20:4 (ω-6) 0,26 ± 0,03 10,52 ± 1,44 6,56 ± 1,00 1,08 ± 0,41
C20:3 (ω-3) 0,09 ± 0,01 0,24 ± 0,05 0,51 ± 0,18 0,05 ± 0,02 C20:5 (ω-3) 0,03 ± 0,00 0,50 ± 0,06 0,83 ± 0,19 0,05 ± 0,01
C22:2 0,03 ± 0,00 0,13 ± 0,02 0,97 ± 0,21 0,01 ± 0,00
C22:6 (ω-3) 0,06 ± 0,01 1,58 ± 0,21 1,86 ± 0,30 0,10 ± 0,03
AGPI 5,75 ± 0,51 36,11 ± 2,49 29,64 ± 3,42 8,88 ± 2,92 ω-6/ω-3 4,69 ± 0,45 21,76 ± 1,14 17,46 ± 2,38 8,39 ± 2,77
ω-3 0,46 ± 0,04 3,04 ± 0,31 3,71 ± 0,54 0,48 ± 0,15 ω-6/ω-3 10,20 ± 0,79 7,23 ± 0,67 4,81 ± 1,17 17,39 ± 1,47
C20:3 (ω-6)+C21 0,08 ± 0,01 1,44 ± 0,18 1,19 ± 0,10 0,17 ± 0,05

CAPÍTULO IV: RESULTADOS
204
Determinación del contenido en aminoácidos libres
El método de cromatografía utilizado permitió la identificación de 20 aminoácidos,
tal como se muestra en la tabla 4.3.6, que se presentaron en un total de 15 picos, tal como
se muestra en la figura 4.3.1.
Asp
GluGly-Gln
Ser-Asn
TyrTrp
Nor
Phe
His
Trh
Cys-Ala
Pro
Arg
Met
Iso
Leu
Lys
Val
Figura 4.3.1. Cromatograma de aminoácidos libres obtenido mediante HPLC de magro de
lomo curado.
A pesar de que los aminoácidos Pro y Arg se separaron en dos picos, se
cuantificaron conjuntamente ya que el cromatograma obtenido del patrón no consiguió
separarlos, apareciendo ambos en un único pico. Al igual que los aminoácidos Tyr y Val,
que se cuantificaron conjuntos porque la mayor parte de las muestras obtuvo un único
pico.

CAPÍTULO IV: RESULTADOS
205
Tabla 4.3.6. Contenido en aminoácidos libres en lomo curado. Los resultados se expresan en mg aa/100 g muestra (± desviación estándar de la media).
E.H E.S
Ac. Aspártico (Asp) 161,60 ± 35,84 330,52 ± 74,76 Ac. Glutámico (Gln) 538,08 ± 39,37 1.099,41 ± 91,69
Serina + Asparagina (Ser-Asn) 319,33 ± 26,17 652,74 ± 63,52
Glicina + Glutamina (Gly-Gln) 272,32 ± 16,58 556,46 ± 41,84
Histidina (His) 145,78 ± 9,98 298,04 ± 26,23 Treonina (Trh) 129,85 ± 21,84 265,40 ± 46,04
Cisteína + Alanina (Cys-Ala) 1.121,19 ± 132,20 2.291,68 ± 296,05
Prolina + Arginina (Pro-Arg) 2.774,34 ± 197,44 5.667,87 ± 461,87
Tirosina + Valina (Tyr-Val) 526,78 ± 21,12 1.076,44 ± 65,23 Metionina (Met) 136,28 ± 5,76 278,52 ± 17,89
Isoleucina (Iso) 234,12 ± 9,96 478,44 ± 30,31
Leucina (Leu) 322,23 ± 15,39 658,71 ± 47,08
Fenilalanina (Phe) 225,79 ± 8,73 461,46 ± 28,86 Triptófano (Trp) 86,52 ± 3,15 176,82 ± 10,67
Lisina (Lys) 626,71 ± 28,74 1.280,78 ± 84,45
sumatorio 7.620,94 ± 420,80 15.573,29 ± 1.120,44
(EH): Extracto húmedo; (ES): Extracto seco
El perfil de aminoácidos libres mostrado en la tabla 4.3.6 fue muy similar a los
obtenidos para jamón y para paleta curada, sin embargo existió alguna variación en algún
aminoácido como fue el caso de la Phe, que se encontró en mayor cantidad que el Asp,
contrario a lo que sucedía con la paleta y el jamón curado.
En cuanto al contenido total de aminoácidos libres fue algo superior al obtenido
para jamón curado (14.719 mg/100g en extracto seco) y considerablemente más alto al
descrito en la paleta curada (10.398 mg/100 g en extracto seco).
Determinación de los compuestos nitrogenados derivados del ATP
El método de cromatografía utilizado permitió la identificación de 6 nucleótidos,
tal como se muestra en la tabla 4.3.7, que se presentaron en un total de 5 picos, tal como se
muestra en la figura 4.3.2.

CAPÍTULO IV: RESULTADOS
206
IMP
GMP
ADP
AMP-Hx
Inosina
Figura 4.3.2. Cromatograma de nucleótidos libres obtenido mediante HPLC de magro de lomo curado.
Tabla 4.3.7. Contenido en 5´nucleótidos del lomo curado. Los resultados se expresan en mg de nuc./100 g muestra (± desviación estándar de la media).
E.H. E.S.
Inosina 5´monofosfato (IMP) 1,76 ± 0,42 3,57 ± 0,86 Guanosina 5´monofosfato (GMP) 3,45 ± 1,08 6,98 ± 2,12
Adenosina 5´difosfato (ADP) 1,63 ± 0,28 3,30 ± 0,57
Adenosina 5´monofosfato (AMP)+Hipoxantina (Hx) 41,58 ± 2,80 84,49 ± 6,84
Inosina (Ino) 6,34 ± 1,86 12,87 ± 3,76
sumatorio 54,75 ± 4,69 111,21 ± 10,36
(EH): Extracto húmedo; (ES): Extracto seco
El perfil de nucleótidos mostrado en la tabla 4.3.7 fue similar al descrito para el
jamón y la paleta curada. El contenido total fue similar al descrito para la paleta curada
(117,32 mg/g muestra en extracto seco) pero muy inferior al descrito para el jamón
curado (374,0 mg/g muestra en extracto seco). Las diferencias descritas pudieron deberse
a las diferencias existentes en los tiempos de curación de cada producto.
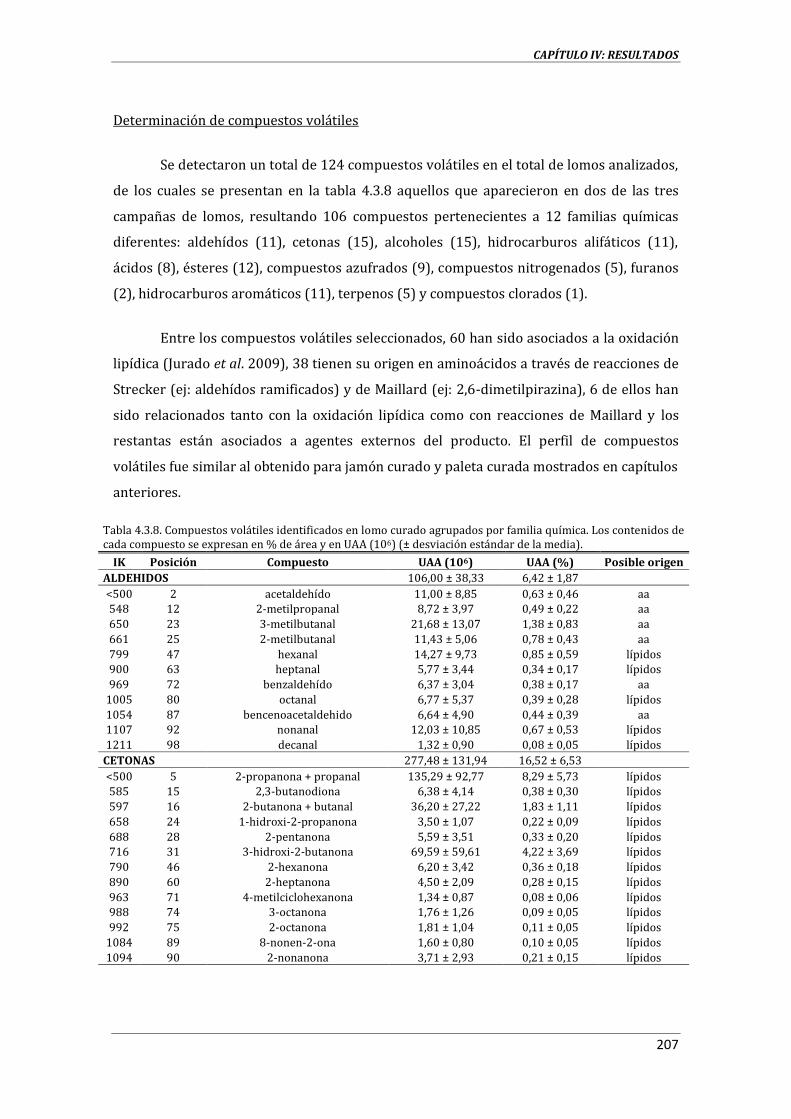
CAPÍTULO IV: RESULTADOS
207
Determinación de compuestos volátiles
Se detectaron un total de 124 compuestos volátiles en el total de lomos analizados,
de los cuales se presentan en la tabla 4.3.8 aquellos que aparecieron en dos de las tres
campañas de lomos, resultando 106 compuestos pertenecientes a 12 familias químicas
diferentes: aldehídos (11), cetonas (15), alcoholes (15), hidrocarburos alifáticos (11),
ácidos (8), ésteres (12), compuestos azufrados (9), compuestos nitrogenados (5), furanos
(2), hidrocarburos aromáticos (11), terpenos (5) y compuestos clorados (1).
Entre los compuestos volátiles seleccionados, 60 han sido asociados a la oxidación
lipídica (Jurado et al. 2009), 38 tienen su origen en aminoácidos a través de reacciones de
Strecker (ej: aldehídos ramificados) y de Maillard (ej: 2,6-dimetilpirazina), 6 de ellos han
sido relacionados tanto con la oxidación lipídica como con reacciones de Maillard y los
restantas están asociados a agentes externos del producto. El perfil de compuestos
volátiles fue similar al obtenido para jamón curado y paleta curada mostrados en capítulos
anteriores.
Tabla 4.3.8. Compuestos volátiles identificados en lomo curado agrupados por familia química. Los contenidos de cada compuesto se expresan en % de área y en UAA (106) (± desviación estándar de la media).
IK Posición Compuesto UAA (106) UAA (%) Posible origen
ALDEHIDOS 106,00 ± 38,33 6,42 ± 1,87 <500 2 acetaldehído 11,00 ± 8,85 0,63 ± 0,46 aa
548 12 2-metilpropanal 8,72 ± 3,97 0,49 ± 0,22 aa
650 23 3-metilbutanal 21,68 ± 13,07 1,38 ± 0,83 aa
661 25 2-metilbutanal 11,43 ± 5,06 0,78 ± 0,43 aa
799 47 hexanal 14,27 ± 9,73 0,85 ± 0,59 lípidos 900 63 heptanal 5,77 ± 3,44 0,34 ± 0,17 lípidos
969 72 benzaldehído 6,37 ± 3,04 0,38 ± 0,17 aa
1005 80 octanal 6,77 ± 5,37 0,39 ± 0,28 lípidos
1054 87 bencenoacetaldehido 6,64 ± 4,90 0,44 ± 0,39 aa 1107 92 nonanal 12,03 ± 10,85 0,67 ± 0,53 lípidos
1211 98 decanal 1,32 ± 0,90 0,08 ± 0,05 lípidos
CETONAS
277,48 ± 131,94 16,52 ± 6,53 <500 5 2-propanona + propanal 135,29 ± 92,77 8,29 ± 5,73 lípidos
585 15 2,3-butanodiona 6,38 ± 4,14 0,38 ± 0,30 lípidos
597 16 2-butanona + butanal 36,20 ± 27,22 1,83 ± 1,11 lípidos
658 24 1-hidroxi-2-propanona 3,50 ± 1,07 0,22 ± 0,09 lípidos
688 28 2-pentanona 5,59 ± 3,51 0,33 ± 0,20 lípidos 716 31 3-hidroxi-2-butanona 69,59 ± 59,61 4,22 ± 3,69 lípidos
790 46 2-hexanona 6,20 ± 3,42 0,36 ± 0,18 lípidos
890 60 2-heptanona 4,50 ± 2,09 0,28 ± 0,15 lípidos
963 71 4-metilciclohexanona 1,34 ± 0,87 0,08 ± 0,06 lípidos 988 74 3-octanona 1,76 ± 1,26 0,09 ± 0,05 lípidos
992 75 2-octanona 1,81 ± 1,04 0,11 ± 0,05 lípidos
1084 89 8-nonen-2-ona 1,60 ± 0,80 0,10 ± 0,05 lípidos
1094 90 2-nonanona 3,71 ± 2,93 0,21 ± 0,15 lípidos

CAPÍTULO IV: RESULTADOS
208
IK Posición Compuesto UAA (106) UAA (%) Posible origen
ALCOHOLES
584,07 ± 285,79 32,72 ± 9,67 <500 4 etanol 378,78 ± 252,58 21,88 ± 10,60 lípidos
548 11 propanol 21,58 ± 18,60 0,62 ± 0,39 lípidos 602 18 2-butanol 24,47 ± 21,89 1,34 ± 1,16 lípidos
621 22 2-metilpropanol 22,02 ± 11,89 1,31 ± 0,68 aa
669 26 1-metoxi-2-propanol 2,60 ± 1,05 0,18 ± 0,08 aa
683 27 1-penten-3-ol 2,08 ± 1,05 0,12 ± 0,04 lípidos 736 33 2-metil-3-buten-2-ol 4,56 ± 3,67 0,25 ± 0,19 lípidos
739 34 3-metilbutanol 31,86 ± 21,58 1,66 ± 0,81 aa
743 35 2-metilbutanol 15,30 ± 12,74 0,73 ± 0,45 aa
771 41 pentanol 1,78 ± 1,12 0,11 ± 0,06 lípidos 790 44 2,3-butanodiol 46,84 ± 30,60 2,67 ± 2,14 lípidos
869 57 hexanol 4,61 ± 3,17 0,25 ± 0,18 lípidos
897 62 2-heptanol 8,63 ± 5,00 0,52 ± 0,33 lípidos
905 64 2-butoxietanol 14,17 ± 11,41 0,81 ± 0,62 lípidos
980 73 1-octen-3-ol 4,78 ± 3,02 0,27 ± 0,14 lípidos
HIDROCARBUROS ALIFÁTICOS 91,62 ± 30,64 4,93 ± 1,16 <500 1 propano 8,31 ± 5,62 0,27 ± 0,13 lípidos
500 6 pentano 19,51 ± 9,88 1,21 ± 0,67 lípidos
505 7 1,1´-oxibis etano 12,60 ± 8,72 0,66 ± 0,34 lípidos 550 13 2-metilpentano 4,86 ± 1,87 0,29 ± 0,15 lípidos
559 14 3-metilpentano 3,89 ± 2,05 0,25 ± 0,14 lípidos
600 17 hexano 20,79 ± 8,98 1,21 ± 0,65 lípidos
759 39 2,3,4-trimetilpentano 5,12 ± 3,31 0,32 ± 0,18 lípidos 778 43 4-metilheptano 1,29 ± 0,23 0,35 ± 0,28 lípidos
1000 78 decano 12,70 ± 19,53 0,23 ± 0,14 lípidos
1200 97 dodecano 1,52 ± 0,78 0,09 ± 0,05 lípidos
1300 101 tridecano 1,03 ± 0,69 0,05 ± 0,04 lípidos
ACIDOS
236,86 ± 257,81 9,07 ± 4,37 583 19 acido acético 198,71 ± 247,67 6,55 ± 3,83 aa/lípidos
685 29 ácido propanoico 2,47 ± 1,17 0,16 ± 0,08 aa/lípidos
759 38 acido 2-metilpropanoico 5,20 ± 4,16 0,25 ± 0,17 aa 784 45 ácido butanoico 13,09 ± 4,96 0,65 ± 0,42 aa/lípidos
837 50 ácido 3-metilbutanoico 5,17 ± 3,29 0,71 ± 0,73 aa
846 51 ácido 2-metilpentanoico 8,33 ± 7,76 0,50 ± 0,48 aa
1165 94 ácido octanoico 2,72 ± 2,95 0,16 ± 0,19 aa/lípidos 1256 100 ácido nonanoico 1,17 ± 0,60 0,07 ± 0,04 aa/lípidos
ESTERES
92,49 ± 56,75 6,20 ± 4,08 524 9 acetato de metilo 4,16 ± 3,14 0,26 ± 0,21 lípidos
612 20 acetato de etilo 48,43 ± 40,85 3,48 ± 3,07 lípidos 722 32 propanoato de etilo 4,11 ± 2,91 0,24 ± 0,18 lípidos
763 40 2-metilpropanoato de etilo 6,59 ± 4,97 0,40 ± 0,30 lípidos
800 48 butanoato de etilo 7,75 ± 1,45 0,47 ± 0,16 lípidos
815 49 2-hidroxipropanoato de etilo 4,30 ± 2,91 0,26 ± 0,18 lípidos 852 52 2-metilbutanoato de etilo 2,42 ± 1,17 0,15 ± 0,06 lípidos
854 53 3-metilbutanoato de etilo 7,60 ± 9,40 0,47 ± 0,56 lípidos
926 67 hexanoato de metilo 1,67 ± 1,33 0,10 ± 0,08 lípidos
998 77 hexanoato de etilo 2,32 ± 1,08 0,16 ± 0,09 lípidos 1196 96 octanoato de etilo 1,91 ± 1,07 0,12 ± 0,07 lípidos
1395 102 decanoato de etilo 1,24 ± 0,73 0,07 ± 0,05 lípidos
COMPUESTOS AZUFRADOS 152,59 ± 119,40 5,30 ± 3,80 <500 3 metanotiol 6,65 ± 3,94 0,41 ± 0,26 aa
517 8 tiobis metano + etanotiol 6,12 ± 3,39 0,38 ± 0,22 aa
534 10 sulfuro de carbono 52,58 ± 83,16 2,98 ± 3,58 aa
702 30 3-metiltio-1-propene + 1-metiltio-1-propene 81,32 ± 106,02 1,13 ± 0,65 aa
763 37 dimetil disulfuro 3,22 ± 1,07 0,22 ± 0,10 aa
875 55 3,3´-tiobis-1-propene 1,81 ± 1,06 0,12 ± 0,08 aa
1251 99 benzotiazol 0,89 ± 0,58 0,06 ± 0,04 aa
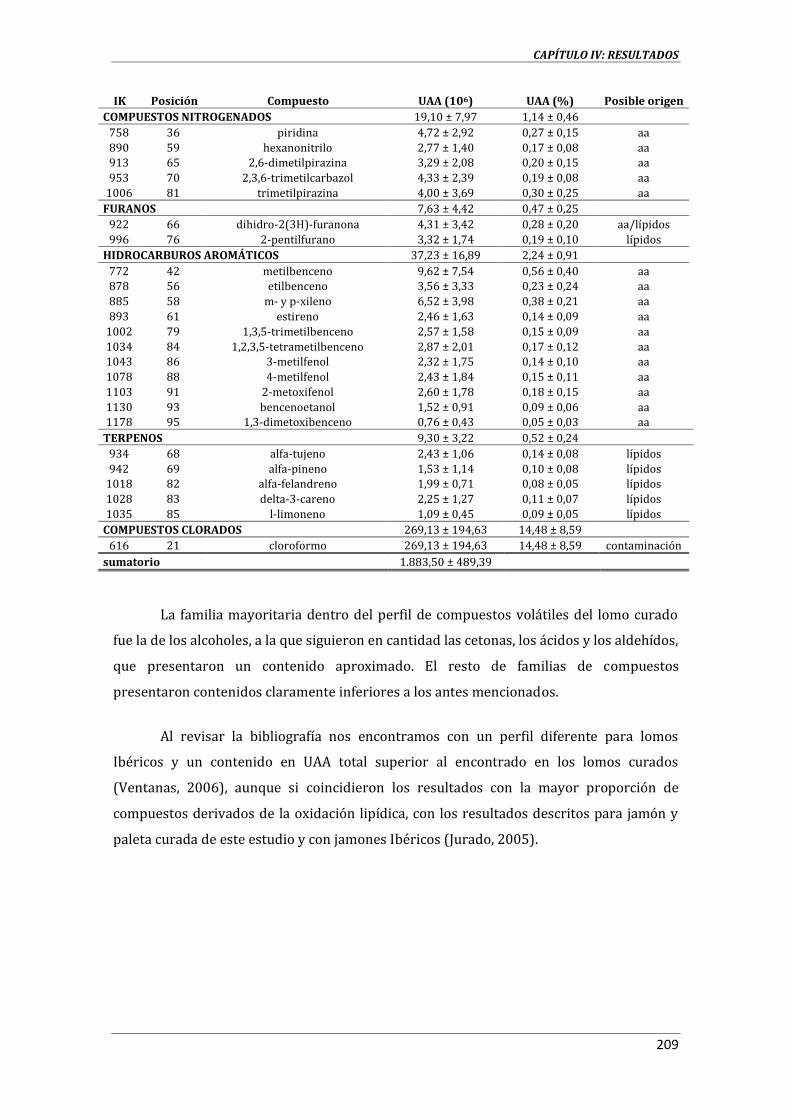
CAPÍTULO IV: RESULTADOS
209
IK Posición Compuesto UAA (106) UAA (%) Posible origen
COMPUESTOS NITROGENADOS 19,10 ± 7,97 1,14 ± 0,46 758 36 piridina 4,72 ± 2,92 0,27 ± 0,15 aa
890 59 hexanonitrilo 2,77 ± 1,40 0,17 ± 0,08 aa 913 65 2,6-dimetilpirazina 3,29 ± 2,08 0,20 ± 0,15 aa
953 70 2,3,6-trimetilcarbazol 4,33 ± 2,39 0,19 ± 0,08 aa
1006 81 trimetilpirazina 4,00 ± 3,69 0,30 ± 0,25 aa
FURANOS
7,63 ± 4,42 0,47 ± 0,25 922 66 dihidro-2(3H)-furanona 4,31 ± 3,42 0,28 ± 0,20 aa/lípidos
996 76 2-pentilfurano 3,32 ± 1,74 0,19 ± 0,10 lípidos
HIDROCARBUROS AROMÁTICOS 37,23 ± 16,89 2,24 ± 0,91 772 42 metilbenceno 9,62 ± 7,54 0,56 ± 0,40 aa
878 56 etilbenceno 3,56 ± 3,33 0,23 ± 0,24 aa
885 58 m- y p-xileno 6,52 ± 3,98 0,38 ± 0,21 aa
893 61 estireno 2,46 ± 1,63 0,14 ± 0,09 aa
1002 79 1,3,5-trimetilbenceno 2,57 ± 1,58 0,15 ± 0,09 aa
1034 84 1,2,3,5-tetrametilbenceno 2,87 ± 2,01 0,17 ± 0,12 aa 1043 86 3-metilfenol 2,32 ± 1,75 0,14 ± 0,10 aa
1078 88 4-metilfenol 2,43 ± 1,84 0,15 ± 0,11 aa
1103 91 2-metoxifenol 2,60 ± 1,78 0,18 ± 0,15 aa
1130 93 bencenoetanol 1,52 ± 0,91 0,09 ± 0,06 aa 1178 95 1,3-dimetoxibenceno 0,76 ± 0,43 0,05 ± 0,03 aa
TERPENOS
9,30 ± 3,22 0,52 ± 0,24 934 68 alfa-tujeno 2,43 ± 1,06 0,14 ± 0,08 lípidos
942 69 alfa-pineno 1,53 ± 1,14 0,10 ± 0,08 lípidos 1018 82 alfa-felandreno 1,99 ± 0,71 0,08 ± 0,05 lípidos
1028 83 delta-3-careno 2,25 ± 1,27 0,11 ± 0,07 lípidos
1035 85 l-limoneno 1,09 ± 0,45 0,09 ± 0,05 lípidos
COMPUESTOS CLORADOS 269,13 ± 194,63 14,48 ± 8,59 616 21 cloroformo 269,13 ± 194,63 14,48 ± 8,59 contaminación
sumatorio 1.883,50 ± 489,39
La familia mayoritaria dentro del perfil de compuestos volátiles del lomo curado
fue la de los alcoholes, a la que siguieron en cantidad las cetonas, los ácidos y los aldehídos,
que presentaron un contenido aproximado. El resto de familias de compuestos
presentaron contenidos claramente inferiores a los antes mencionados.
Al revisar la bibliografía nos encontramos con un perfil diferente para lomos
Ibéricos y un contenido en UAA total superior al encontrado en los lomos curados
(Ventanas, 2006), aunque si coincidieron los resultados con la mayor proporción de
compuestos derivados de la oxidación lipídica, con los resultados descritos para jamón y
paleta curada de este estudio y con jamones Ibéricos (Jurado, 2005).

CAPÍTULO IV: RESULTADOS
210
Evaluación sensorial
Los valores de los diferentes atributos sensoriales utilizados para caracterizar el
lomo curado se incluyen en la tabla 4.3.9 y en el gráfico de araña de la figura 4.3.1.
Tabla 4.3.9. Análisis cuantitativo-descriptivo del lomo curado (± desviación estándar de la media).
Color magro
Rojo 2,4 ± 0,4
Brillo 1,8 ± 0,2
Veteado 2,0 ± 0,2
Olor Intensidad 2,7 ± 0,3
Textura magro
Dureza 3,8 ± 0,3
Sequedad 4,1 ± 0,3
Jugosidad 3,4 ± 0,3
Fibrosidad 2,7 ± 0,6
Sabor
Salado 4,1 ± 0,3
Dulce 1,2 ± 0,1
Amargo 0,9 ± 0,1
Flavor
Intensidad 4,5 ± 0,4
Curado 2,8 ± 0,2
Rancidez 1,0 ± 0,1
Persistencia 3,8 ± 0,4
Los valores emitidos por el panel sensorial para el lomo curado lo definen como un
lomo de gran intensidad de flavor y persistencia, algo seco y salado. Al comparar con el
análisis sensorial realizado sobre lomos Ibéricos (Ventanas, 2006) vemos como las
puntuaciones de todos los atributos fueron superiores a las mostradas en la tabla 4.3.9.
0
1
2
3
4
5ROJO
BRILLO
VETEADO
INTENSIDAD OLOR
DUREZA
SEQUEDAD
JUGOSIDAD
FIBROSIDADSALADO
DULCE
AMARGO
INTENSIDAD FLAVOR
CURADO
RANCIDEZ
PERSISTENCIA
Figura 4.3.1. Perfil sensorial del lomo curado

CAPÍTULO IV: RESULTADOS
211
4.3.2. INFLUENCIA DE LA EXPRESIÓN DEL IGF2 SOBRE LAS CARACTERÍSTICAS
FÍSICO-QUÍMICAS Y SENSORIALES DEL LOMO CURADO
En este apartado se va a evaluar la influencia de los alelos A y G del IGF2 (insuline-
like growth factor 2) del QTL (Quantitative Trait Loci) sobre todos los parámetros
analizados en los lomos curados. Para ello, tan solo van a incluirse aquellos parámetros
que presentaron diferencias significativas entre los dos lotes estudiados.
Características generales del lomo curado
Al evaluar las características generales del lomo curado: peso, longitud, perímetro
y diámetro, tan solo la longitud presentó diferencias significativas entre ambos lotes,
siendo mayor el valor de los lomos del lote AA (tabla 4.3.10). A pesar de las escasas
diferencias significativas en el resto de parámetros morfológicos, los lomos pertenecientes
al lote AA presentaron mayores valores. Sin embargo, si se observaron diferencias
significativas en el peso de lomos analizados en fresco procedentes de los mismos lotes de
cerdos, presentando mayor peso los lomos procedentes de los cerdos AA (Burgos et al.,
2011).
Tabla 4.3.10. Medidas morfológicas generales del lomo curado (± desviación estándar de la media).
AA AG p
Longitud (cm) 52,5 ± 3,5 48,9 ± 1,7 ***
*** Nivel de significación p<0,001
Perfil de textura instrumental
La tabla 4.3.11 muestra las diferencias significativas existentes entre los
parámetros de textura. De los seis parámetros que presentaron diferencias significativas,
cuatro fueron superiores significativamente en el lote AA, excepto la adhesividad y la
elasticidad, que fueron superiores significativamente en el lote AG. Estos resultados
coincidieron con los obtenidos para jamón curado (tabla 4.2.13), ya que aquellos valores
que presentaron diferencias significativas fueron superiores en el lote AA.

CAPÍTULO IV: RESULTADOS
212
Tabla 4.3.11. Análisis de perfil de textura (TPA) del lomo curado (± desviación estándar de la media).
AA AG p
Fuerza (N) 29,27 ± 2,70 26,89 ± 2,82 **
Dureza (N) 32,05 ± 3,04 29,45 ± 2,91 **
Adhesividad (N*seg) -0,17 ± 0,02 -0,16 ± 0,01 **
Elasticidad (cm) 0,77 ± 0,01 0,81 ± 0,01 *** Gomosidad (N) 19,02 ± 0,01 17,66 ± 1,96 **
Masticabilidad (N) 15,04 ± 0,25 13,91 ± 1,62 **
(N): Newton *** Nivel de significación p<0,001; ** Nivel de significación p<0,01
Parámetros generales físico-químicos
En cuanto a los parámetros generales, se encontraron diferencias significativas en
5 de los catorce parámetros mostrados en la tabla 4.3.2, tal como se muestra en la tabla
4.3.12.
Tabla 4.3.12. Parámetros físico-químicos generales del lomo curado (± desviación estándar de la media).
AA AG p
a (Adimensional) 14,36 ± 1,34 15,15 ± 1,10 *
mg Mb/g m 1,62 ± 0,30 1,87 ± 0,33 *
ppm Fe heminico 5,49 ± 1,02 6,35 ± 1,11 * pH (Adimensional) 5,94 ± 0,00 5,96 ± 0,04 *
mg MDA/Kg 0,21 ± 0,06 0,28 ± 0,07 ***
*** Nivel de significación p<0,001; * Nivel de significación p<0,05
A diferencia de los jamones y las paletas curadas, el contenido de grasa
intramuscular no presentó diferencias significativas entre ambos lotes, sin embargo la
coordenada a* y el contenido en mioglobina si mostraron diferencias significativas, siendo
superiores los valores del lote AG.
El contenido en mg MDA/Kg fue significativamente superior en el lote AG, siendo a
su vez el lote que mostró mayor contenido en grasa intramuscular, a pesar de que no
presentó diferencias significativas entre ambos lotes (4,66 vs 4,94%).
Composición de la grasa intramuscular
La fracción de triglicéridos (LN) fue superior significativamente en el lote AG con
un valor de 87,8% frente al 84,5% del lote AA. La fracción de fosfolípidos (LP) también fue
superior en el lote AG con un 1,7% frente al 1,6% del lote AA. Sin embargo, la fracción de
AGL fue superior en el lote AA con un 13,8% frente al 10,6% del lote AG.

CAPÍTULO IV: RESULTADOS
213
Fracción de lípidos neutros
La tabla 4.3.13 muestra el contenido de los ácidos grasos identificados en la
fracción de triglicéridos (LN) de la grasa intramuscular extraída de los lomos curados.
Existieron diferencias significativas en el contenido total de ácidos grasos y en los
sumatorios de AGS y AGMI, siendo significativamente superiores los valores
pertenecientes al lote AG, a pesar de que el contenido de grasa intramuscular no mostró
diferencias significativas, coincidiendo por ello con los resultados mostrados para la paleta
curada.
En cuanto al resto de ácidos grasos, existieron diferencias significativas en algunos
de ellos, destacando los mayoritarios como son el C16:0, el C16:1 y el C18:1, siendo su
contenido significativamente superior en el lote AG.
Tabla 4.3.13. Composición de la grasa intramuscular en la fracción de triglicéridos (LN) del lomo curado. Los resultados están expresados en mg de AG/100 g muestra.
LN AA LN AG p
C10:0 1,27 ± 0,11 1,63 ± 0,32 ***
C12:0 1,43 ± 0,18 1,94 ± 0,18 *** C14:0 22,82 ± 2,44 27,49 ± 4,95 ***
C16:0 453,31 ± 64,55 535,19 ± 98,09 **
C14:1 1,59 ± 0,25 1,84 ± 0,30 **
C15:1 0,32 ± 0,07 0,46 ± 0,15 *** C16:1 66,93 ± 9,10 81,94 ± 18,52 **
C18:1 272,63 ± 52,60 337,64 ± 81,37 **
C20:1 7,60 ± 0,52 13,67 ± 2,97 ***
C24:1 0,24 ± 0,05 0,30 ± 0,06 *** C20:5(ω-3) 0,36 ± 0,06 0,43 ± 0,12 *
C22:2 0,24 ± 0,04 0,35 ± 0,10 ***
AGS 727,14 ± 116,20 816,73 ± 158,08 *
AGMI 352,86 ± 57,40 439,78 ± 103,82 **
sumatorio 1.180,55 ± 188,40 1.354,41 ± 276,64 *
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05
En cuanto a los valores expresados en %, las diferencias fueron más marcadas
entre ambos lotes, aunque en este caso el lote AA presentó mayor proporción de algunos
ácidos grasos, destacando el valor de C18:3 (ω-3) que fue superior significativamente en el
lote AA.

CAPÍTULO IV: RESULTADOS
214
Tabla 4.3.14. Composición de la grasa intramuscular en la fracción de triglicéridos (LN) del lomo curado. Los resultados están expresados en % (± desviación estándar de la media).
LN AA % LN AG % p
C10:0 0,07 ± 0,01 0,06 ± 0,00 ***
C12:0 0,05 ± 0,01 0,05 ± 0,00 *
C15:0 0,03 ± 0,00 0,02 ± 0,00 *** C17:0 0,14 ± 0,00 0,15 ± 0,01 *
C20:0 0,27 ± 0,03 0,30 ± 0,03 **
C22:0 0,03 ± 0,00 0,02 ± 0,00 **
C23:0 0,09 ± 0,01 0,08 ± 0,01 ** C14:1 0,05 ± 0,00 0,06 ± 0,00 ***
C16:1 2,22 ± 0,10 2,64 ± 0,14 ***
C17:1 0,15 ± 0,00 0,17 ± 0,01 ***
C20:1 1,00 ± 0,11 1,15 ± 0,08 *** C24:1 0,05 ± 0,01 0,05 ± 0,01 *
C18:2 (ω-6) 3,99 ± 0,03 4,60 ± 0,32 ***
C18:3 (ω-6) 0,03 ± 0,00 0,02 ± 0,00 ***
C18:3 (ω-3) 0,29 ± 0,03 0,27 ± 0,02 * C20:2 (ω-6) 0,29 ± 0,02 0,33 ± 0,03 ***
C20:4 (ω-6) 0,27 ± 0,03 0,25 ± 0,03 **
C20:3 (ω-3) 0,07 ± 0,00 0,08 ± 0,00 ***
C20:5 (ω-3) 0,03 ± 0,01 0,03 ± 0,00 *** C22:2 0,02 ± 0,00 0,02 ± 0,00 **
C22:6 (ω-3) 0,06 ± 0,01 0,06 ± 0,01 ***
AGMI 57,03 ± 2,25 58,73 ± 1,66 **
AGPI 5,07 ± 0,08 5,66 ± 0,38 ***
ω-6 4,58 ± 0,05 5,19 ± 0,36 ***
ω-3 0,46 ± 0,04 0,44 ± 0,03 * ω-6/ ω-3 10,00 ± 0,85 11,84 ± 0,55 ***
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05
Fracción de lípidos polares
El análisis del perfil de fosfolípidos (LP) de la grasa intramuscular se muestra en la
tabla 4.3.15. A pesar de ser la fracción minoritaria, al igual que la fracción de triglicéridos
(LN), existió un contenido total significativamente superior en el lote AG, sin embargo,
existió una mayor cantidad de ácidos grasos libres minoritarios que presentaron mayores
valores significativamente en el lote AA.

CAPÍTULO IV: RESULTADOS
215
Tabla 4.3.15. Composición de la grasa intramuscular en la fracción de fosfolípidos (LP) del lomo curado. Los resultados están expresados en mg de AG/100 g muestra (± desviación estándar de la media).
LP AA LP AG p
C10:0 0,06 ± 0,01 0,07 ± 0,01 ***
C12:0 0,11 ± 0,01 0,08 ± 0,02 ***
C14:0 0,28 ± 0,04 0,25 ± 0,05 * C20:0 0,12 ± 0,01 0,05 ± 0,01 ***
C22:0 0,13 ± 0,02 0,09 ± 0,03 ***
C15:1 0,70 ± 0,06 0,77 ± 0,11 *
C16:1 0,46 ± 0,07 0,58 ± 0,17 ** C17:1 0,28 ± 0,04 0,25 ± 0,06 *
C24:1 0,14 ± 0,02 0,16 ± 0,01 **
C18:2 (ω-6) 4,56 ± 0,48 5,69 ± 1,20 ***
C18:3 (ω-6) 0,13 ± 0,02 0,07 ± 0,01 *** C20:4 (ω-6) 1,89 ± 0,07 1,65 ± 0,34 **
C20:3 (ω-3) 0,332 ± 0,125 0,031 ± 0,003 ***
C20:5 (ω-3) 0,15 ± 0,02 0,18 ± 0,04 *
ω-6 6,76 ± 0,48 7,59 ± 1,57 *
ω-3 0,82 ± 0,15 0,58 ± 0,11 ***
ω-6/ ω-3 8,48 ± 1,57 13,00 ± 1,82 ***
C20:3 (ω-6)+C21 0,19 ± 0,01 0,24 ± 0,05 ***
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05
Los resultados expresados en porcentajes se muestran en la tabla 4.3.16. Destacó
la proporción significativamente superior del C18:0 y C18:1 en el lote AA, haciendo que la
proporción de AGMI fuera superior significativamente en el lote AA.
Tabla 4.3.16. Composición de la grasa intramuscular en la fracción de fosfolípidos (LP) del lomo curado. Los resultados están expresados en %.
LP AA % LP AG % p
C10:0 0,10 ± 0,01 0,16 ± 0,05 ***
C16:0 14,07 ± 1,96 16,15 ± 0,68 ***
C17:0 0,47 ± 0,03 0,39 ± 0,00 ***
C18:0 11,88 ± 0,06 10,53 ± 0,63 *** C20:0 0,26 ± 0,11 0,12 ± 0,04 ***
C14:1 0,40 ± 0,02 0,31 ± 0,03 ***
C15:1 1,26 ± 0,17 1,67 ± 0,40 ***
C16:1 0,96 ± 0,12 1,07 ± 0,17 * C17:1 0,88 ± 0,15 1,24 ± 0,38 ***
C18:1 20,57 ± 0,31 17,98 ± 0,97 ***
C20:1 0,61 ± 0,02 0,44 ± 0,05 ***
C22:1 0,35 ± 0,06 0,42 ± 0,05 *** C24:1 1,65 ± 0,43 1,21 ± 0,40 **
C18:3 (ω-6) 0,28 ± 0,04 0,16 ± 0,06 ***
C20:3 (ω-3) 0,67 ± 0,13 0,15 ± 0,03 ***
C22:6 (ω-3) 1,84 ± 0,01 2,56 ± 0,87 ***
AGMI 26,674 ± 0,657 24,323 ± 1,738 ***
C20:3 (ω-6)+C21 1,133 ± 0,080 1,194 ± 0,082 *
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05
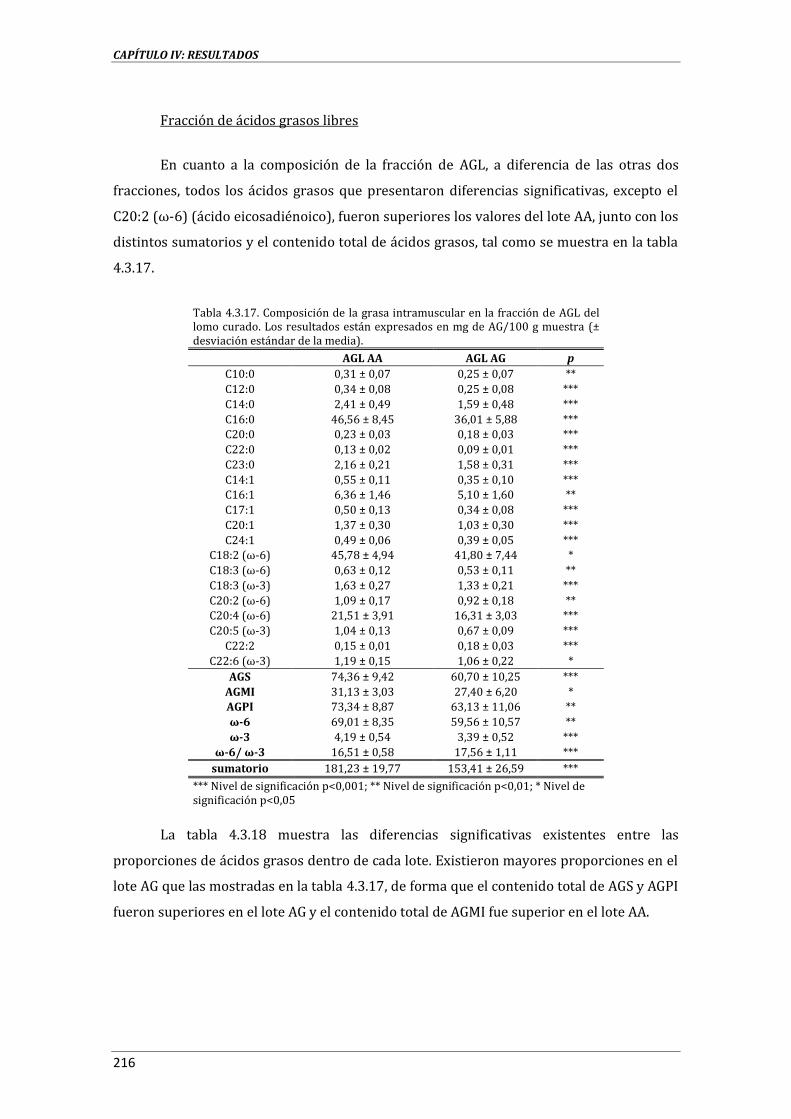
CAPÍTULO IV: RESULTADOS
216
Fracción de ácidos grasos libres
En cuanto a la composición de la fracción de AGL, a diferencia de las otras dos
fracciones, todos los ácidos grasos que presentaron diferencias significativas, excepto el
C20:2 (ω-6) (ácido eicosadiénoico), fueron superiores los valores del lote AA, junto con los
distintos sumatorios y el contenido total de ácidos grasos, tal como se muestra en la tabla
4.3.17.
Tabla 4.3.17. Composición de la grasa intramuscular en la fracción de AGL del lomo curado. Los resultados están expresados en mg de AG/100 g muestra (± desviación estándar de la media).
AGL AA AGL AG p
C10:0 0,31 ± 0,07 0,25 ± 0,07 **
C12:0 0,34 ± 0,08 0,25 ± 0,08 ***
C14:0 2,41 ± 0,49 1,59 ± 0,48 ***
C16:0 46,56 ± 8,45 36,01 ± 5,88 *** C20:0 0,23 ± 0,03 0,18 ± 0,03 ***
C22:0 0,13 ± 0,02 0,09 ± 0,01 ***
C23:0 2,16 ± 0,21 1,58 ± 0,31 ***
C14:1 0,55 ± 0,11 0,35 ± 0,10 *** C16:1 6,36 ± 1,46 5,10 ± 1,60 **
C17:1 0,50 ± 0,13 0,34 ± 0,08 ***
C20:1 1,37 ± 0,30 1,03 ± 0,30 ***
C24:1 0,49 ± 0,06 0,39 ± 0,05 *** C18:2 (ω-6) 45,78 ± 4,94 41,80 ± 7,44 *
C18:3 (ω-6) 0,63 ± 0,12 0,53 ± 0,11 **
C18:3 (ω-3) 1,63 ± 0,27 1,33 ± 0,21 ***
C20:2 (ω-6) 1,09 ± 0,17 0,92 ± 0,18 ** C20:4 (ω-6) 21,51 ± 3,91 16,31 ± 3,03 ***
C20:5 (ω-3) 1,04 ± 0,13 0,67 ± 0,09 ***
C22:2 0,15 ± 0,01 0,18 ± 0,03 ***
C22:6 (ω-3) 1,19 ± 0,15 1,06 ± 0,22 *
AGS 74,36 ± 9,42 60,70 ± 10,25 ***
AGMI 31,13 ± 3,03 27,40 ± 6,20 * AGPI 73,34 ± 8,87 63,13 ± 11,06 **
ω-6 69,01 ± 8,35 59,56 ± 10,57 **
ω-3 4,19 ± 0,54 3,39 ± 0,52 ***
ω-6/ ω-3 16,51 ± 0,58 17,56 ± 1,11 ***
sumatorio 181,23 ± 19,77 153,41 ± 26,59 ***
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05
La tabla 4.3.18 muestra las diferencias significativas existentes entre las
proporciones de ácidos grasos dentro de cada lote. Existieron mayores proporciones en el
lote AG que las mostradas en la tabla 4.3.17, de forma que el contenido total de AGS y AGPI
fueron superiores en el lote AG y el contenido total de AGMI fue superior en el lote AA.

CAPÍTULO IV: RESULTADOS
217
Tabla 4.3.18. Composición de la grasa intramuscular en la fracción de AGL del lomo curado. Los resultados están expresados en % (± desviación estándar de la media).
AGL AA % AGL AG % p
C10:0 0,06 ± 0,00 0,09 ± 0,01 ***
C16:0 11,40 ± 0,37 13,57 ± 2,02 ***
C18:0 11,36 ± 0,26 11,01 ± 0,29 *** C22:0 0,11 ± 0,03 0,09 ± 0,00 ***
C16:1 1,26 ± 0,02 1,46 ± 0,18 ***
C17:1 0,17 ± 0,01 0,15 ± 0,01 ***
C18:1 35,06 ± 1,50 30,95 ± 2,54 *** C24:1 0,44 ± 0,01 0,55 ± 0,11 ***
C18:2 (ω-6) 20,70 ± 0,77 22,22 ± 1,25 ***
C20:2 (ω-6) 0,61 ± 0,02 0,69 ± 0,08 ***
C20:3 (ω-3) 0,22 ± 0,04 0,26 ± 0,06 ** C22:2 0,14 ± 0,03 0,12 ± 0,01 *
C22:6 (ω-3) 1,41 ± 0,14 1,61 ± 0,19 ***
AGS 25,49 ± 0,53 27,01 ± 2,21 **
AGMI 38,03 ± 1,44 34,24 ± 2,59 ***
AGPI 35,05 ± 1,58 36,90 ± 3,05 *
ω-6 32,11 ± 1,40 33,66 ± 2,89 * ω-3 2,81 ± 0,18 3,11 ± 0,30 ***
ω-6/ ω-3 11,47 ± 0,47 10,86 ± 0,88 **
C20:3 (ω-6)+C21 1,37 ± 0,17 1,50 ± 0,18 *
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05
Lípidos totales
No existieron diferencias significativas en ningún ácido graso de los que componen
los lípidos totales, a pesar de que el contenido total de los ácidos grasos mayoritarios
C16:0, C18:0, C16:1 y C18:1 fueron superiores en el lote AG, al igual que los sumatorios de
AGS, AGMI y el contenido total.
La tabla 4.3.19 muestra los porcentajes de los ácidos grasos presentes en los
lípidos totales de la grasa intramuscular, donde destacaron las proporciones
significativamente superiores del C16:0 del lote AG.
Tabla 4.3.19. Composición de la grasa intramuscular en la fracción de lípidos totales del lomo curado. Los resultados están expresados en % (± desviación estándar de la media).
LT AA % LT AG % p
C14:0 1,42 ± 0,21 1,59 ± 0,14 **
C16:0 26,76 ± 1,67 28,42 ± 1,55 **
AGS 39,80 ± 2,53 41,83 ± 2,77 *
** Nivel de significación p<0,01; * Nivel de significación p<0,05
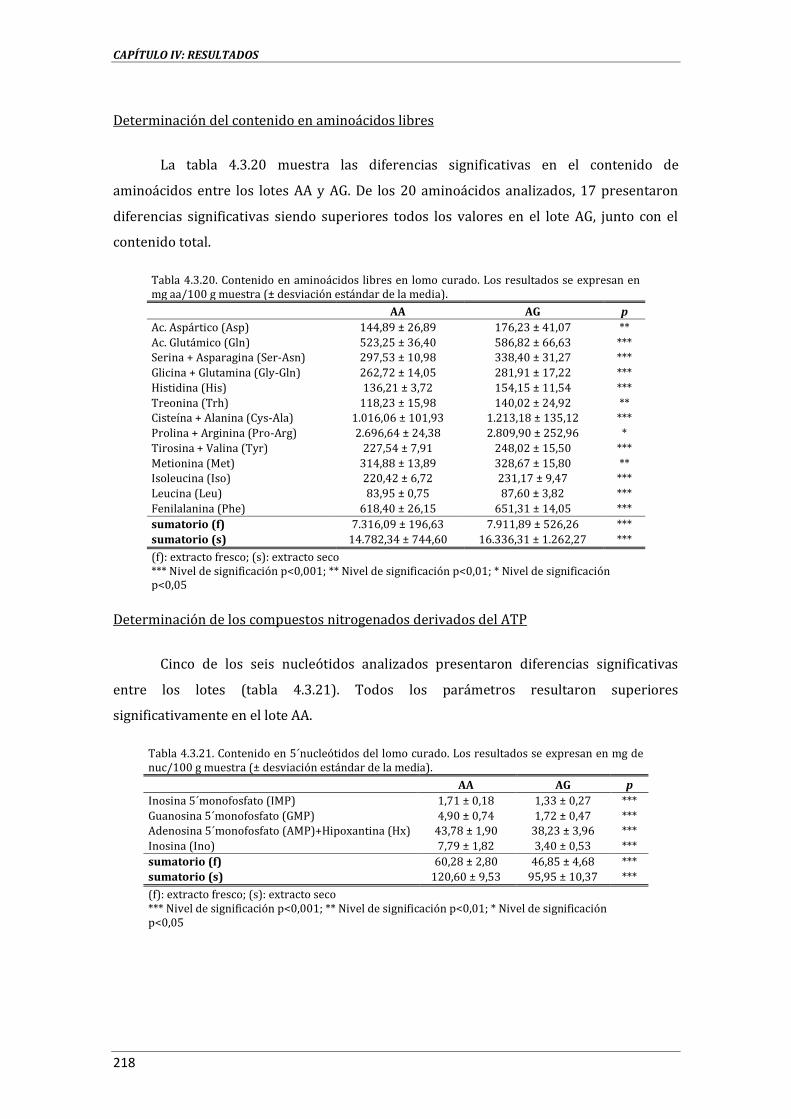
CAPÍTULO IV: RESULTADOS
218
Determinación del contenido en aminoácidos libres
La tabla 4.3.20 muestra las diferencias significativas en el contenido de
aminoácidos entre los lotes AA y AG. De los 20 aminoácidos analizados, 17 presentaron
diferencias significativas siendo superiores todos los valores en el lote AG, junto con el
contenido total.
Tabla 4.3.20. Contenido en aminoácidos libres en lomo curado. Los resultados se expresan en mg aa/100 g muestra (± desviación estándar de la media).
AA AG p
Ac. Aspártico (Asp) 144,89 ± 26,89 176,23 ± 41,07 **
Ac. Glutámico (Gln) 523,25 ± 36,40 586,82 ± 66,63 *** Serina + Asparagina (Ser-Asn) 297,53 ± 10,98 338,40 ± 31,27 ***
Glicina + Glutamina (Gly-Gln) 262,72 ± 14,05 281,91 ± 17,22 ***
Histidina (His) 136,21 ± 3,72 154,15 ± 11,54 ***
Treonina (Trh) 118,23 ± 15,98 140,02 ± 24,92 ** Cisteína + Alanina (Cys-Ala) 1.016,06 ± 101,93 1.213,18 ± 135,12 ***
Prolina + Arginina (Pro-Arg) 2.696,64 ± 24,38 2.809,90 ± 252,96 *
Tirosina + Valina (Tyr) 227,54 ± 7,91 248,02 ± 15,50 ***
Metionina (Met) 314,88 ± 13,89 328,67 ± 15,80 ** Isoleucina (Iso) 220,42 ± 6,72 231,17 ± 9,47 ***
Leucina (Leu) 83,95 ± 0,75 87,60 ± 3,82 ***
Fenilalanina (Phe) 618,40 ± 26,15 651,31 ± 14,05 ***
sumatorio (f) 7.316,09 ± 196,63 7.911,89 ± 526,26 ***
sumatorio (s) 14.782,34 ± 744,60 16.336,31 ± 1.262,27 ***
(f): extracto fresco; (s): extracto seco *** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05
Determinación de los compuestos nitrogenados derivados del ATP
Cinco de los seis nucleótidos analizados presentaron diferencias significativas
entre los lotes (tabla 4.3.21). Todos los parámetros resultaron superiores
significativamente en el lote AA.
Tabla 4.3.21. Contenido en 5´nucleótidos del lomo curado. Los resultados se expresan en mg de nuc/100 g muestra (± desviación estándar de la media).
AA AG p
Inosina 5´monofosfato (IMP) 1,71 ± 0,18 1,33 ± 0,27 ***
Guanosina 5´monofosfato (GMP) 4,90 ± 0,74 1,72 ± 0,47 *** Adenosina 5´monofosfato (AMP)+Hipoxantina (Hx) 43,78 ± 1,90 38,23 ± 3,96 ***
Inosina (Ino) 7,79 ± 1,82 3,40 ± 0,53 ***
sumatorio (f) 60,28 ± 2,80 46,85 ± 4,68 ***
sumatorio (s) 120,60 ± 9,53 95,95 ± 10,37 ***
(f): extracto fresco; (s): extracto seco *** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05

CAPÍTULO IV: RESULTADOS
219
Determinación de compuestos volátiles
De los 105 compuestos volátiles detectados (tabla 4.3.8), tan solo 33 presentaron
diferencias significativas entre los lotes del estudio, 16 de ellos superiores
significativamente en el lote AA y 17 superiores significativamente en el lote AG. En cuanto
al contenido total, no existieron diferencias significativas entre los lotes (tabla 4.3.22),
coincidiendo con los resultados obtenidos para el jamón curado y la paleta curada. Las
únicas familias que presentaron diferencias significativas entre los lotes fueron la de los
ésteres y los hidrocarburos alifáticos, resultando las UAA del lote AA superiores en ambos
casos.
Algunos de los compuestos a destacar son el 3-hidroxi-2-butanona, el pentano, el
acetato de etilo y el dimetil disulfuro, que fueron significativamente superiores en el lote
AA. Por otra parte, el 2-butanol y el ácido 2-metilpropanoico fueron significativamente
superiores en el lote AG.

CAPÍTULO IV: RESULTADOS
220
Tabla 4.3.22. Compuestos volátiles identificados en el lomo curado agrupados por familia química. Los contenidos de cada compuesto se expresan en UAA (106) (± desviación estándar de la media).
IK Posición Compuesto AA AG p Posible origen
ALDEHIDOS
1211 98 decanal 0,98 ± 0,61 1,68 ± 1,02 * lípidos
CETONAS
585 15 2,3-butanodiona 9,35 ± 5,78 4,87 ± 3,39 * lípidos
658 24 1-hidroxi-2-propanona 3,03 ± 0,98 4,06 ± 1,07 ** lípidos 716 31 3-hidroxi-2-butanona 85,81 ± 65,78 34,27 ± 21,91 ** lípidos
992 75 2-octanona 1,26 ± 0,47 2,05 ± 1,14 * lípidos
1084 89 8-nonen-2-ona 1,15 ± 0,39 1,85 ± 1,00 * lípidos
1094 90 2-nonanona 2,12 ± 1,15 3,97 ± 2,70 * lípidos
ALCOHOLES
602 18 2-butanol 9,37 ± 6,74 32,20 ± 22,70 *** lípidos
621 22 2-metilpropanol 22,55 ± 10,47 15,75 ± 6,12 * aa
771 41 pentanol 1,92 ± 1,09 1,17 ± 0,71 * lípidos
HIDROCARBUROS ALIFÁTICOS 86,16 ± 14,44 72,61 ± 17,61 *
500 6 pentano 23,60 ± 8,37 15,33 ± 9,92 * lípidos
600 17 hexano 24,80 ± 8,57 18,40 ± 8,69 * lípidos
759 39 2,3,4-trimetilpentano 1,96 ± 1,05 5,57 ± 3,14 *** lípidos 1000 78 decano 5,06 ± 3,46 2,07 ± 1,12 ** lípidos
1300 101 tridecano 0,67 ± 0,45 1,24 ± 0,82 ** lípidos
ACIDOS
685 29 ácido propanoico 1,93 ± 0,83 2,93 ± 1,30 * aa/lípidos 759 38 acido 2-metilpropanoico 4,93 ± 3,60 13,21 ± 12,10 * aa
784 45 ácido butanoico 12,52 ± 3,22 18,79 ± 11,16 * aa/lípidos
1256 100 ácido nonanoico 0,91 ± 0,41 1,42 ± 0,69 * aa/lípidos
ESTERES 106,63 ± 61,27 52,94 ± 17,33 **
612 20 acetato de etilo 64,64 ± 51,01 16,12 ± 7,66 *** lípidos
722 32 propanoato de etilo 4,84 ± 3,04 2,47 ± 1,71 ** lípidos
815 49 2-hidroxipropanoato de etilo 4,50 ± 3,17 3,84 ± 2,79 ** lípidos
852 52 2-metilbutanoato de etilo 3,37 ± 2,12 2,14 ± 0,88 * lípidos 854 53 3-metilbutanoato de etilo 4,43 ± 1,84 2,79 ± 0,98 ** lípidos
926 67 hexanoato de metilo 0,60 ± 0,24 1,40 ± 0,88 ** lípidos
1395 102 decanoato de etilo 0,82 ± 0,31 1,42 ± 0,78 ** lípidos
COMPUESTOS AZUFRADOS
763 37 dimetil disulfuro 8,34 ± 7,75 3,11 ± 1,01 ** aa
1251 99 benzotiazol 0,65 ± 0,35 1,35 ± 1,06 * aa
HIDROCARBUROS AROMÁTICOS
893 61 estireno 3,08 ± 1,90 0,82 ± 0,05 *** aa 1043 86 3-metilfenol 2,82 ± 2,09 1,28 ± 0,48 ** aa
1178 95 1,3-dimetoxibenceno 0,38 ± 0,10 1,01 ± 0,55 *** aa
TERPENOS
934 68 alfa-tujeno 2,87 ± 1,05 2,10 ± 1,02 * lípidos 1035 85 l-limoneno 0,77 ± 0,25 1,38 ± 0,64 ** lípidos
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo
Evaluación sensorial
De las 15 características sensoriales evaluadas, 13 presentaron diferencias
significativas entre los lotes estudiados, 6 superiores significativamente en el lote AG y el
resto en el lote AA, tal como se muestra en la tabla 4.3.23. La figura 4.3.3 muestra el perfil
sensorial mediante un gráfico que pone de manifiesto las diferencias existentes entre
ambos lotes.

CAPÍTULO IV: RESULTADOS
221
Tabla 4.3.23. Análisis cuantitativo-descriptivo del lomo curado (± desviación estándar de la media).
AA AG p
Magro Veteado 1,8 ± 0,1 2,1 ± 0,1 ***
Olor Intensidad 2,3 ± 0,1 2,9 ± 0,4 ***
Magro
Dureza 4,1 ± 0,0 3,7 ± 0,4 ***
Sequedad 4,5 ± 0,1 3,9 ± 0,2 ***
Jugosidad 3,2 ± 0,1 3,4 ± 0,4 **
Fibrosidad 3,1 ± 0,6 2,1 ± 0,3 ***
Sabor
Salado 4,0 ± 0,3 4,3 ± 0,2 ***
Dulce 1,3 ± 0,0 1,2 ± 0,0 ***
Amargo 0,8 ± 0,0 1,0 ± 0,2 ***
Flavor
Intensidad 4,3 ± 0,3 4,9 ± 0,2 ***
Curado 3,1 ± 0,5 2,7 ± 0,1 ***
Rancidez 1,0 ± 0,1 1,0 ± 0,0 **
Persistencia 3,5 ± 0,3 4,0 ± 0,5 **
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo
Al comparar los valores emitidos por el panel sensorial, vemos como el lote AA se
describió como un producto más duro, seco y fibroso. Sin embargo, la intensidad del flavor
y la persistencia fueron superiores en las paletas del lote AA.
0
1
2
3
4
5ROJO
BRILLO
VETEADO
INTENSIDAD OLOR
DUREZA
SEQUEDAD
JUGOSIDAD
FIBROSIDADSALADO
DULCE
AMARGO
INTENSIDAD FLAVOR
CURADO
RANCIDEZ
PERSISTENCIA
AA
AG
Figura 4.3.3. Perfil sensorial del lomo curado

CAPÍTULO IV: RESULTADOS
222
4.3.3. INFLUENCIA DE LA EXPRESIÓN DEL IGF2 EN FUNCIÓN DEL SEXO SOBRE LAS
CARACTERÍSTICAS FÍSICO-QUÍMICAS Y SENSORIALES DEL LOMO CURADO
En este apartado se va a evaluar la influencia de los alelos A y G del QTL
(Quantitative Trait Loci) IGF2 en función del sexo de los animales sobre todos los
parámetros analizados en los lomos curados. Para ello, tan solo van a incluirse aquellos
parámetros que presentaron diferencias significativas entre los cuatro lotes estudiados
(AA*H, AA*M, AG*H y AG*M, siendo H el factor hembra y M el factor macho).
Características generales del lomo curado
Al evaluar las características generales del lomo curado: peso, longitud, perímetro
y diámetro, todos los parámetros presentaron diferencias significativas entre los lotes,
mostrando una clara influencia por el efecto sexo, ya que tan solo el parámetro de longitud
estuvo influido por la genética, coincidiendo con los resultados mostrados en la tabla
4.3.10. Los lomos procedentes de los machos fueron los más gruesos y pesados y los
pertenecientes a los lotes AA los más largos.
Perfil de textura instrumental
La tabla 4.3.25 muestra las diferencias significativas existentes entre los
parámetros de textura. Todos los parámetros evaluados mostraron diferencias
significativas, dos de ellos influenciados por el efecto genético, dos por el efecto sexo y el
resto por el efecto de ambos factores. El lote AAM fue el lote que presentó mayores valores
por lo general, coincidiendo en la mayor parte de los casos con el lote AGM, por lo que en
estos parámetros fue más marcado el efecto sexo. Otros parámetros superiores en el lote
AAH fueron la dureza y la fracturabilidad, éste último superior también en el lote AGH.

Tabla 4.3.24. Medidas morfológicas generales del lomo curado (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
Peso (G) 1,7ab ± 0,1 1,8b ± 0,1 1,6a ± 0,0 1,7b ± 0,1 ** ns ns *
Longitud (cm) 53,1c ± 3,8 51,9bc ± 3,0 48,7a ± 1,7 49,0ab ± 1,6 ns *** ns ** Perímetro (cm) 20,7ab ± 0,3 21,2bc ± 0,7 20,5a ± 0,5 21,5c ± 0,6 *** ns ns ***
Diámetro (cm) 6,6ab ± 0,1 6,8bc ± 0,2 6,5a ± 0,1 6,9c ± 0,2 *** ns ns ***
*** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo
Tabla 4.3.25. Análisis de perfil de textura (TPA) del lomo curado (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
Fuerza (N) 31,11b ± 2,30 26,83a ± 2,58 26,06a ± 3,41 28,28ab ± 1,95 ns * *** ***
Dureza (N) 31,35ab ± 4,15 32,99b ± 0,36 28,62a ± 3,45 30,83ab ± 2,15 * * ns *
Fracturabilidad (Adimensional) 11,10b ± 0,33 9,88a ± 0,29 10,97b ± 0,64 9,69a ± 0,34 *** ns ns *** Adhesividad (N*seg) -0,19ab ± 0,01 -0,14c ± 0,01 -0,16b ± 0,01 -0,15c ± 0,01 *** ** *** ***
Elasticidad (cm) 0,76ab ± 0,02 0,78b ± 0,01 0,80b ± 0,03 0,78b ± 0,01 ns ** *** ***
Cohesividad (Adimensional) 0,59ab ± 0,01 0,60b ± 0,01 0,60ab ± 0,01 0,61b ± 0,01 ** * ns **
Gomosidad (N) 18,58ab ± 2,33 19,56b ± 0,25 17,03a ± 2,23 18,70ab ± 1,53 * * ns * Masticabilidad (N) 14,10ab ± 1,72 15,13b ± 0,32 13,40a ± 1,73 14,77ab ± 1,41 * ns ns *
*** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo
Tabla 4.3.26. Parámetros físico-químicos generales del lomo curado (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
a (Adimensional) 14,39 ± 1,55 14,32 ± 1,06 15,30 ± 0,57 15,00 ± 1,41 ns * ns ns
pH (Adimensional) 5,90a ± 0,10 5,93ab ± 0,01 5,97b ± 0,03 5,94ab ± 0,04 ns * ns *
mg MDA/Kg 0,20a ± 0,06 0,23a ± 0,05 0,22a ± 0,06 0,38b ± 0,02 *** *** *** ***
(s,dg): extracto seco y desengrasado *** Nivel de significación p<0,05; Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
224
Parámetros generales físico-químicos
En cuanto a los parámetros generales, la coordenada a*, el pH y los mg MDA/Kg
fueron los únicos parámetros que mostraron diferencias significativas, influenciados en la
mayor parte de los casos por el efecto genético, siendo superiores los valores de los lotes
AG. La coordenada a* y el pH estuvieron influenciados también por el efecto genético,
siendo superiores los valores del lote AGH y los mg MDA/Kg, influenciados por ambos
factores, fueron superiores en el lote AGM, tal como se muestra en la tabla 4.3.26.
A diferencia de los otros dos productos estudiados, el contenido de grasa
intramuscular no se mostró influenciado por ninguno de los dos factores evaluados, no
presentando diferencias significativas, a pesar de que el IGF2 y el sexo se han descrito
como factores influyentes en el contenido de grasa intramuscular, por lo que el factor
posición anatómica podría haber influido en este caso.
Composición de la grasa intramuscular
La fracción de triglicéridos (LN) fue la fracción mayoritaria de la grasa
intramuscular en los cuatro lotes estudiados, con un porcentaje de 80,2% para el lote AAH,
85,7% para el AAM, 86,8% para el AGH y de 89,4% para el lote AGM de la grasa total. La
fracción de fosfolípidos (LP) fue la fracción minoritaria con un 1,9, 1,4, 1,6 y 2,2%
respectivamente. La fracción de AGL representó respectivamente.el 17,9, 12,4, 11,6 y 8,4%
Fracción de lípidos neutros
La tabla 4.3.27 muestra el contenido de los ácidos grasos identificados en la
fracción de triglicéridos (LN) de la grasa intramuscular extraída de los lomos curados. A
pesar de no exisitir diferencias en el contenido de grasa intramuscular, si existieron
diferencias significativas en la mayoría de los ácidos grasos, y por lo tanto en los
sumatorios y contenido total, doce de ellos como consecuencia del efecto sexo y el resto
por el efecto de ambos factores, pero ninguno por el efecto único de la genética. El
contenido total presentó diferencias debidas el efecto de la genética y a la interacción de
ambos factores, siendo superior el valor del lote AGH.
En cuanto a los distintos sumatorios, el contenido de AGS mostró influencia por el
efecto genético, siendo el lote AGH el que mayor contenido de esta fracción presentó,
coincidiendo con el mayor contenido del ácido graso mayoritario C16:0 y el contenido
total de ese lote. Los sumatorios AGPI, ω-3 y ω-6 mostraron una clara influencia por el

CAPÍTULO IV: RESULTADOS
225
efecto sexo, siendo superiores los valores del lote AAM, coincidiendo también con el mayor
contenido del ácido poliinsaturado mayoritario C18:2 (ω-6) que mostró ese lote. Por
último, el contenido de AGMI fue superior en el lote AGH, influenciado por ambos factores
y coincidiendo con el mayor contenido del C18:1.
En cuanto a los valores expresados en % (tabla 4.3.28), las diferencias también
fueron marcadas entre los lotes, donde tanto el efecto genético como el sexo mostraron
influencia sobre los distintos lotes. La proporción de AGS estuvo influenciada por el efecto
genético, coincidiendo con la mayor proporción del C18:0, que fue superior en todos los
lotes excepto el AAM. La proporción de AGMI fue superior en los lotes AAM y AGH,
influenciada por la interacción de ambos factores y coincidiendo también con las mayores
proporciones del C16:1 y C18:1 de ambos lotes. Por último, la proporción de ω-3, estuvo
influenciada por ambos factores, siendo superior el lote AAM. El resto de ácidos que
mostraron diferencias fueron minoritarios, por lo que su repercusión no fue importante.

Tabla 4.3.27. Composición de la grasa intramuscular en la fracción de triglicéridos (LN) del lomo curado. Los resultados están expresados en mg de AG/100 g muestra (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
C10:0 1,21a ± 0,13 2,38c ± 0,40 1,53ab ± 0,43 1,79b ± 0,13 *** ns *** ***
C12:0 1,26a ± 0,14 2,31c ± 0,41 1,73b ± 0,43 1,87b ± 0,12 *** ns *** *** C14:0 21,00a ± 2,22 26,45b ± 1,87 27,48b ± 6,69 27,53b ± 2,44 * ** * ***
C15:0 0,54a ± 0,10 0,91c ± 0,15 0,84bc ± 0,27 0,71ab ± 0,10 * ns *** ***
C16:0 410,02a ± 57,76 410,82a ± 0,00 563,20b ± 135,67 488,50ab ± 26,37 ns *** ns ***
C17:0 2,99a ± 0,65 5,28b ± 0,96 3,61a ± 0,96 3,23a ± 0,40 *** ** *** *** C18:0 199,29a ± 34,64 294,53b ± 57,93 252,38ab ± 74,92 224,29a ± 13,91 * ns *** **
C20:0 2,46a ± 0,35 3,96b ± 0,43 3,58b ± 0,90 2,81a ± 0,21 * ns *** ***
C22:0 0,17a ± 0,04 0,28b ± 0,01 0,26b ± 0,06 0,18a ± 0,02 ns ns *** ***
C23:0 0,77a ± 0,13 1,11b ± 0,15 0,88a ± 0,19 0,82a ± 0,09 ** * *** *** C14:1 1,43a ± 0,26 2,37b ± 0,38 2,22b ± 0,62 1,77a ± 0,05 * ns *** ***
C15:1 0,33a ± 0,09 0,48bc ± 0,13 0,53c ± 0,19 0,34ab ± 0,04 ns ns *** **
C16:1 55,03a ± 3,99 90,74b ± 1,13 90,51b ± 24,71 67,66a ± 4,02 ns ns *** ***
C17:1 2,60a ± 0,45 5,36c ± 1,03 3,76b ± 1,05 2,86a ± 0,30 *** ** *** *** C18:1 249,59b ± 35,90 174,12a ± 0,00 373,77c ± 108,85 277,43b ± 19,47 *** *** ns ***
C20:1 9,31a ± 1,81 16,28c ± 3,70 14,75bc ± 4,08 11,87ab ± 0,63 * ns *** ***
C22:1 0,36a ± 0,09 0,64b ± 0,06 0,60b ± 0,17 0,35a ± 0,01 ns ns *** ***
C24:1 0,20a ± 0,05 0,29b ± 0,04 0,29b ± 0,06 0,33b ± 0,06 *** *** ns *** C18:2 (ω-6) 68,23a ± 14,03 104,80c ± 15,99 86,09b ± 21,82 73,10ab ± 5,24 * ns *** ***
C18:3 (ω-6) 0,36a ± 0,03 0,67b ± 0,09 0,42a ± 0,12 0,37a ± 0,03 *** *** *** ***
C18:3 (ω-3) 4,80a ± 0,98 7,37b ± 1,15 5,67a ± 1,49 5,04a ± 0,41 ** * *** ***
C20:2 (ω-6) 3,33a ± 0,62 5,87c ± 0,88 4,60b ± 1,22 3,67ab ± 0,34 ** ns *** *** C20:4 (ω-6) 2,86a ± 0,49 4,57b ± 0,68 3,50a ± 0,70 3,30a ± 0,39 *** ns *** ***
C20:3 (ω-3) 0,89a ± 0,14 1,45b ± 0,16 1,39b ± 0,39 0,98a ± 0,08 ns ns *** ***
C20:5 (ω-3) 0,29a ± 0,06 0,45b ± 0,04 0,40b ± 0,10 0,48b ± 0,13 *** * ns ***
C22:2 0,20a ± 0,04 0,52b ± 0,16 0,30a ± 0,08 0,44b ± 0,11 *** ns * *** C22:6 (ω-3) 0,25a ± 0,05 0,37b ± 0,05 0,30a ± 0,07 0,29a ± 0,06 ** ns ** ***
AGS 639,72a ± 94,38 748,02ab ± 61,56 855,48b ± 220,31 751,72ab ± 42,53 ns ** ** ** AGMI 318,85a ± 41,61 290,27a ± 4,82 486,42b ± 139,30 362,61a ± 23,82 ** *** * ***
AGPI 81,21a ± 16,07 126,07c ± 19,08 102,67b ± 25,80 87,68ab ± 6,68 * ns *** ***
ω-6 74,79a ± 14,92 115,91c ± 17,58 94,61b ± 23,82 80,45ab ± 5,99 * ns *** ***
ω-3 6,23a ± 1,14 9,64b ± 1,37 7,76a ± 1,94 6,79a ± 0,60 ** ns *** ***
C20:3 (ω-6)+C21 0,77a ± 0,17 1,16b ± 0,07 1,06b ± 0,30 0,83a ± 0,10 ns ns *** ***
sumatorio 1.040,55a ± 147,40 1.166,18a ± 83,66 1.445,63b ± 380,53 1.202,84ab ± 72,22 ns ** ** **
*** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo
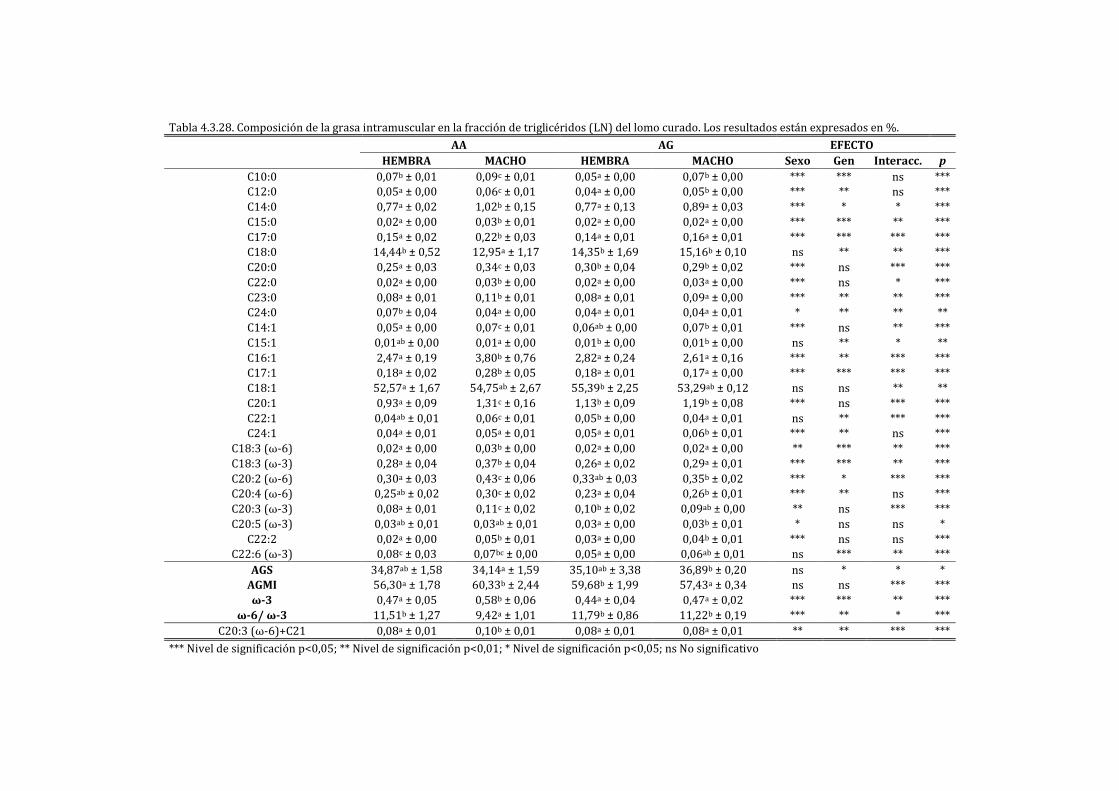
Tabla 4.3.28. Composición de la grasa intramuscular en la fracción de triglicéridos (LN) del lomo curado. Los resultados están expresados en %.
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
C10:0 0,07b ± 0,01 0,09c ± 0,01 0,05a ± 0,00 0,07b ± 0,00 *** *** ns *** C12:0 0,05a ± 0,00 0,06c ± 0,01 0,04a ± 0,00 0,05b ± 0,00 *** ** ns ***
C14:0 0,77a ± 0,02 1,02b ± 0,15 0,77a ± 0,13 0,89a ± 0,03 *** * * ***
C15:0 0,02a ± 0,00 0,03b ± 0,01 0,02a ± 0,00 0,02a ± 0,00 *** *** ** ***
C17:0 0,15a ± 0,02 0,22b ± 0,03 0,14a ± 0,01 0,16a ± 0,01 *** *** *** *** C18:0 14,44b ± 0,52 12,95a ± 1,17 14,35b ± 1,69 15,16b ± 0,10 ns ** ** ***
C20:0 0,25a ± 0,03 0,34c ± 0,03 0,30b ± 0,04 0,29b ± 0,02 *** ns *** ***
C22:0 0,02a ± 0,00 0,03b ± 0,00 0,02a ± 0,00 0,03a ± 0,00 *** ns * ***
C23:0 0,08a ± 0,01 0,11b ± 0,01 0,08a ± 0,01 0,09a ± 0,00 *** ** ** *** C24:0 0,07b ± 0,04 0,04a ± 0,00 0,04a ± 0,01 0,04a ± 0,01 * ** ** **
C14:1 0,05a ± 0,00 0,07c ± 0,01 0,06ab ± 0,00 0,07b ± 0,01 *** ns ** ***
C15:1 0,01ab ± 0,00 0,01a ± 0,00 0,01b ± 0,00 0,01b ± 0,00 ns ** * **
C16:1 2,47a ± 0,19 3,80b ± 0,76 2,82a ± 0,24 2,61a ± 0,16 *** ** *** *** C17:1 0,18a ± 0,02 0,28b ± 0,05 0,18a ± 0,01 0,17a ± 0,00 *** *** *** ***
C18:1 52,57a ± 1,67 54,75ab ± 2,67 55,39b ± 2,25 53,29ab ± 0,12 ns ns ** **
C20:1 0,93a ± 0,09 1,31c ± 0,16 1,13b ± 0,09 1,19b ± 0,08 *** ns *** ***
C22:1 0,04ab ± 0,01 0,06c ± 0,01 0,05b ± 0,00 0,04a ± 0,01 ns ** *** *** C24:1 0,04a ± 0,01 0,05a ± 0,01 0,05a ± 0,01 0,06b ± 0,01 *** ** ns ***
C18:3 (ω-6) 0,02a ± 0,00 0,03b ± 0,00 0,02a ± 0,00 0,02a ± 0,00 ** *** ** ***
C18:3 (ω-3) 0,28a ± 0,04 0,37b ± 0,04 0,26a ± 0,02 0,29a ± 0,01 *** *** ** ***
C20:2 (ω-6) 0,30a ± 0,03 0,43c ± 0,06 0,33ab ± 0,03 0,35b ± 0,02 *** * *** *** C20:4 (ω-6) 0,25ab ± 0,02 0,30c ± 0,02 0,23a ± 0,04 0,26b ± 0,01 *** ** ns ***
C20:3 (ω-3) 0,08a ± 0,01 0,11c ± 0,02 0,10b ± 0,02 0,09ab ± 0,00 ** ns *** ***
C20:5 (ω-3) 0,03ab ± 0,01 0,03ab ± 0,01 0,03a ± 0,00 0,03b ± 0,01 * ns ns *
C22:2 0,02a ± 0,00 0,05b ± 0,01 0,03a ± 0,00 0,04b ± 0,01 *** ns ns *** C22:6 (ω-3) 0,08c ± 0,03 0,07bc ± 0,00 0,05a ± 0,00 0,06ab ± 0,01 ns *** ** ***
AGS 34,87ab ± 1,58 34,14a ± 1,59 35,10ab ± 3,38 36,89b ± 0,20 ns * * * AGMI 56,30a ± 1,78 60,33b ± 2,44 59,68b ± 1,99 57,43a ± 0,34 ns ns *** ***
ω-3 0,47a ± 0,05 0,58b ± 0,06 0,44a ± 0,04 0,47a ± 0,02 *** *** ** ***
ω-6/ ω-3 11,51b ± 1,27 9,42a ± 1,01 11,79b ± 0,86 11,22b ± 0,19 *** ** * ***
C20:3 (ω-6)+C21 0,08a ± 0,01 0,10b ± 0,01 0,08a ± 0,01 0,08a ± 0,01 ** ** *** ***
*** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
228
Fracción de lípidos polares
El análisis del perfil de fosfolípidos (LP) de la grasa intramuscular se muestra en la
tabla 4.3.29. De los ácidos grasos que mostraron diferencias significativas seis de ellos
fueron como consecuencia del efecto sexo, siete por el efecto genético y nueve por el efecto
de ambos factores. Además de existir diferencias significativas en la mayor parte de los
ácidos grasos que componen la fracción, también existieron diferencias en los distintos
sumatorios, excepto en el contenido de AGS. El contenido de AGMI estuvo influenciado por
el efecto sexo, siendo superiores los valores de los lotes pertenecientes a los machos. El
resto de sumatorios que mostraron diferencias fueron debidas a ambos factores, siendo
superiores los valores del lote AGM, excepto el contenido de ω-3, que fue superior en el
lote AAM.
Los resultados expresados en porcentajes se muestran en la tabla 4.3.30. A
diferencia de lo que ocurría con los triglicéridos, los fosfolípidos mostraron diferencias en
menos ácidos grasos al expresarlos en porcentajes. Tan solo las proporciones de AGS, AGPI
y ω-6 mostraron diferencias. Los AGS mostraron una clara influencia del efecto sexo
siendo superior el lote AAM y los AGPI y ω-6 por el efecto genético, siendo superior el lote
AAH, a diferencia de la tabla 4.3.29, donde las fracciones de AGPI y ω-6 mostraron
diferencias atribuidas a ambos factores, siendo superiores los porcentajes del lote AGM.

Tabla 4.3.29. Composición de la grasa intramuscular en la fracción de fosfolípidos (LP) del lomo curado. Los resultados están expresados en mg de AG/100 g muestra.
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
C10:0 0,12b ± 0,03 0,048a ± 0,002 0,07a ± 0,02 0,06a ± 0,01 *** ** *** *** C12:0 0,12b ± 0,02 0,11b ± 0,01 0,14b ± 0,05 0,08a ± 0,00 *** ns ** ***
C15:0 0,11b ± 0,00 0,08a ± 0,00 0,09a ± 0,03 0,09a ± 0,01 ** ns ** ***
C16:0 5,94 ± 0,53 7,04 ± 0,50 7,18 ± 2,05 6,53 ± 0,38 ns ns * ns
C20:0 0,14c ± 0,00 0,10b ± 0,00 0,05a ± 0,02 0,05a ± 0,01 *** *** *** *** C22:0 0,13b ± 0,02 0,18c ± 0,04 0,14bc ± 0,06 0,07b ± 0,02 ns *** *** ***
C23:0 0,43ab ± 0,12 0,83c ± 0,37 0,32a ± 0,12 0,68bc ± 0,24 *** ns ns ***
C14:1 0,19a ± 0,03 0,29b ± 0,01 0,21a ± 0,07 0,21a ± 0,03 *** * ** ***
C15:1 1,40ab ± 0,67 0,66a ± 0,08 1,54b ± 1,05 1,99b ± 0,87 ns ** * ** C16:1 0,46a ± 0,09 0,46a ± 0,05 0,62b ± 0,23 0,51ab ± 0,06 ns ** ns *
C17:1 0,52b ± 0,18 0,22a ± 0,02 0,47b ± 0,31 0,53b ± 0,21 * * * **
C18:1 1,47a ± 0,16 2,26b ± 0,10 1,93ab ± 0,68 2,21b ± 0,50 *** ns ns ***
C20:1 0,12 ± 0,03 0,12 ± 0,01 0,11 ± 0,03 0,10 ± 0,02 ns * ns ns C24:1 2,61b ± 1,45 0,28a ± 0,07 1,06a ± 0,81 0,16a ± 0,01 *** ** * ***
C18:2 (ω-6) 3,84a ± 0,19 5,52b ± 0,11 5,14b ± 1,36 6,61c ± 0,86 *** *** ns ***
C18:3 (ω-6) 0,14c ± 0,03 0,09ab ± 0,00 0,09a ± 0,04 0,10b ± 0,00 ns *** *** ***
C20:2 (ω-6) 0,14a ± 0,02 0,22c ± 0,02 0,17ab ± 0,05 0,19bc ± 0,03 *** ns ** *** C20:4 (ω-6) 1,76ab ± 0,21 1,82b ± 0,04 1,53a ± 0,38 1,85b ± 0,28 * ns ns *
C20:3 (ω-3) 0,23a ± 0,13 0,47b ± 0,08 0,12a ± 0,07 0,14a ± 0,08 *** *** *** ***
C20:5 (ω-3) 0,20 ± 0,05 0,23 ± 0,06 0,24 ± 0,09 0,17 ± 0,02 ns ns * *
C22:2 0,15a ± 0,04 0,22a ± 0,06 0,13a ± 0,04 2,43b ± 1,56 *** *** *** *** C22:6 (ω-3) 0,15a ± 0,01 0,18a ± 0,01 0,24b ± 0,08 0,16a ± 0,03 * ** *** ***
AGMI 6,86b ± 2,16 4,36a ± 0,10 6,02b ± 1,91 5,80ab ± 0,70 ** ns * ** AGPI 6,79a ± 0,43 8,93b ± 0,21 7,86ab ± 2,05 11,84c ± 2,46 *** ** * ***
ω-6 5,89a ± 0,33 7,65bc ± 0,13 6,92ab ± 1,78 8,75c ± 1,16 *** ** ns ***
ω-3 0,75a ± 0,17 1,06b ± 0,12 0,80a ± 0,25 0,67a ± 0,06 * *** *** ***
ω-6/ ω-3 8,18a ± 1,72 7,31a ± 0,82 8,93a ± 1,30 13,34b ± 2,63 ** *** *** ***
C20:3 (ω-6)+C21 0,23ab ± 0,05 0,19a ± 0,01 0,22a ± 0,05 0,28b ± 0,05 ns ** *** ***
sumatorio 24,48a ± 2,97 26,36ab ± 0,98 25,92ab ± 6,68 29,42b ± 2,76 * ns ns *
*** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

Tabla 4.3.30. Composición de la grasa intramuscular en la fracción de fosfolípidos (LP) del lomo curado. Los resultados están expresados en %.
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
C10:0 0,13 ± 0,03 0,14 ± 0,06 0,11 ± 0,04 0,16 ± 0,07 * ns ns ns
C15:0 0,25 ± 0,04 0,27 ± 0,05 0,29 ± 0,08 0,24 ± 0,06 ns ns * ns
C17:0 0,60ab ± 0,10 0,65b ± 0,13 0,53a ± 0,10 0,58ab ± 0,09 ns * ns ns
C22:0 0,64ab ± 0,61 0,84b ± 0,59 0,08a ± 0,02 1,08b ± 0,97 ** ns ns ** C24:0 0,82ab ± 0,67 1,24b ± 1,01 0,09a ± 0,01 1,19b ± 0,79 ** ns ns **
C15:1 2,74b ± 2,25 0,63a ± 0,14 1,31a ± 0,63 0,84a ± 0,18 ** ns * ***
C16:1 0,86a ± 0,19 1,27b ± 0,49 1,36b ± 0,43 1,17ab ± 0,28 ns ns ** **
C17:1 1,47b ± 1,08 0,67a ± 0,36 0,99ab ± 0,52 0,47a ± 0,12 ** ns ns ** C22:1 0,52ab ± 0,29 0,91b ± 0,70 0,13a ± 0,01 1,29b ± 0,99 *** ns ns **
C18:2 (ω-6) 18,41b ± 2,82 15,55ab ± 3,19 15,05a ± 2,80 15,10ab ± 2,70 ns * ns *
C18:3 (ω-6) 0,16b ± 0,05 0,09a ± 0,06 0,11ab ± 0,03 0,16b ± 0,05 ns ns *** **
C20:2 (ω-6) 0,68 ± 0,13 0,64 ± 0,16 0,55 ± 0,17 0,70 ± 0,19 ns ns * ns C20:4 (ω-6) 7,65b ± 1,41 5,67a ± 2,03 4,75a ± 1,60 4,32a ± 1,21 * *** ns ***
C20:3 (ω-3) 0,10ab ± 0,04 0,12b ± 0,02 0,06a ± 0,01 0,11b ± 0,05 ** ns ns **
C22:2 0,77ab ± 0,66 0,74ab ± 0,59 0,10a ± 0,03 1,20b ± 0,88 * ns ** **
AGS 37,72ab ± 3,49 40,82b ± 2,82 36,40a ± 4,18 38,66ab ± 3,16 * ns ns *
AGPI 30,98b ± 3,48 25,79a ± 4,71 23,46a ± 4,18 24,84a ± 2,86 ns ** ** ***
ω-6 26,90b ± 3,65 21,95a ± 4,90 20,47a ± 4,21 20,28a ± 3,79 ns ** * **
C20:3 (ω-6)+C21 1,10b ± 0,21 0,98ab ± 0,23 0,82a ± 0,24 0,83a ± 0,09 ns ** ns **
*** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
231
Fracción de ácidos grasos libres
En cuanto a la composición de la fracción de AGL, a diferencia de las otras dos
fracciones, las diferencias existentes entre los ácidos grasos estuvieron influenciadas en la
mayor parte de los casos por ambos factores, tal como se muestra en la tabla 4.3.31. En
cuanto a los sumatorios, todos ellos mostraron diferencias significativas influidas por
ambos factores, siendo superiores los valores del lote AAH. Los ácidos grasos saturados y
monoinsaturados mayoritarios (C16:0, C18:0, C6:1 y C18:1) también se mostraron
claramente influidos por ambos factores, siendo superiores también los valores del lote
AAH.
La tabla 4.3.32 muestra las diferencias significativas existentes entre las
proporciones de ácidos grasos dentro de cada lote. Al igual que la tabla anterior, todos los
ácidos grasos mostraron diferencias significativas. Existieron diferencias en todos los
sumatorios, siendo superiores los valores del lote AGM en todos los casos, excepto en la
proporción de AGMI, cuya porporción coincidió en tres de los cuatro, siendo el lote AGM
inferior, coincidiendo con la menor proporción del ácido monoinsaturado C18:1 de ese
lote.
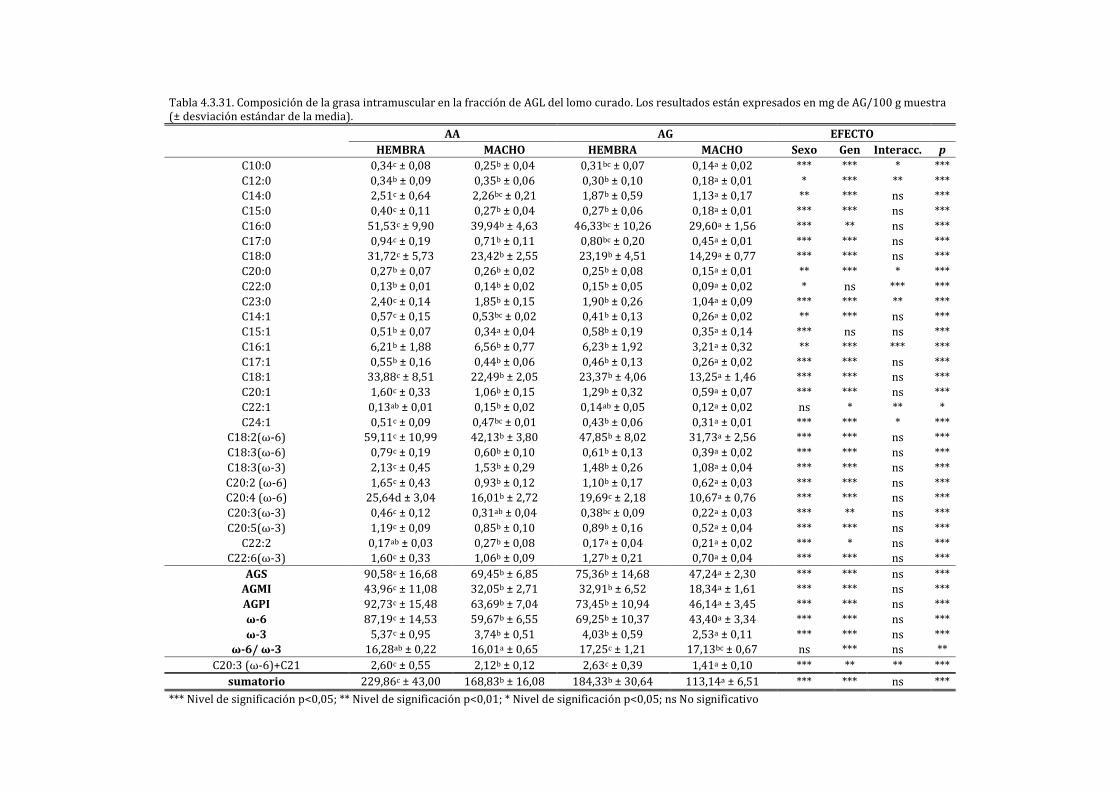
Tabla 4.3.31. Composición de la grasa intramuscular en la fracción de AGL del lomo curado. Los resultados están expresados en mg de AG/100 g muestra (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
C10:0 0,34c ± 0,08 0,25b ± 0,04 0,31bc ± 0,07 0,14a ± 0,02 *** *** * ***
C12:0 0,34b ± 0,09 0,35b ± 0,06 0,30b ± 0,10 0,18a ± 0,01 * *** ** *** C14:0 2,51c ± 0,64 2,26bc ± 0,21 1,87b ± 0,59 1,13a ± 0,17 ** *** ns ***
C15:0 0,40c ± 0,11 0,27b ± 0,04 0,27b ± 0,06 0,18a ± 0,01 *** *** ns ***
C16:0 51,53c ± 9,90 39,94b ± 4,63 46,33bc ± 10,26 29,60a ± 1,56 *** ** ns ***
C17:0 0,94c ± 0,19 0,71b ± 0,11 0,80bc ± 0,20 0,45a ± 0,01 *** *** ns *** C18:0 31,72c ± 5,73 23,42b ± 2,55 23,19b ± 4,51 14,29a ± 0,77 *** *** ns ***
C20:0 0,27b ± 0,07 0,26b ± 0,02 0,25b ± 0,08 0,15a ± 0,01 ** *** * ***
C22:0 0,13b ± 0,01 0,14b ± 0,02 0,15b ± 0,05 0,09a ± 0,02 * ns *** ***
C23:0 2,40c ± 0,14 1,85b ± 0,15 1,90b ± 0,26 1,04a ± 0,09 *** *** ** *** C14:1 0,57c ± 0,15 0,53bc ± 0,02 0,41b ± 0,13 0,26a ± 0,02 ** *** ns ***
C15:1 0,51b ± 0,07 0,34a ± 0,04 0,58b ± 0,19 0,35a ± 0,14 *** ns ns ***
C16:1 6,21b ± 1,88 6,56b ± 0,77 6,23b ± 1,92 3,21a ± 0,32 ** *** *** ***
C17:1 0,55b ± 0,16 0,44b ± 0,06 0,46b ± 0,13 0,26a ± 0,02 *** *** ns *** C18:1 33,88c ± 8,51 22,49b ± 2,05 23,37b ± 4,06 13,25a ± 1,46 *** *** ns ***
C20:1 1,60c ± 0,33 1,06b ± 0,15 1,29b ± 0,32 0,59a ± 0,07 *** *** ns ***
C22:1 0,13ab ± 0,01 0,15b ± 0,02 0,14ab ± 0,05 0,12a ± 0,02 ns * ** *
C24:1 0,51c ± 0,09 0,47bc ± 0,01 0,43b ± 0,06 0,31a ± 0,01 *** *** * *** C18:2(ω-6) 59,11c ± 10,99 42,13b ± 3,80 47,85b ± 8,02 31,73a ± 2,56 *** *** ns ***
C18:3(ω-6) 0,79c ± 0,19 0,60b ± 0,10 0,61b ± 0,13 0,39a ± 0,02 *** *** ns ***
C18:3(ω-3) 2,13c ± 0,45 1,53b ± 0,29 1,48b ± 0,26 1,08a ± 0,04 *** *** ns ***
C20:2 (ω-6) 1,65c ± 0,43 0,93b ± 0,12 1,10b ± 0,17 0,62a ± 0,03 *** *** ns *** C20:4 (ω-6) 25,64d ± 3,04 16,01b ± 2,72 19,69c ± 2,18 10,67a ± 0,76 *** *** ns ***
C20:3(ω-3) 0,46c ± 0,12 0,31ab ± 0,04 0,38bc ± 0,09 0,22a ± 0,03 *** ** ns ***
C20:5(ω-3) 1,19c ± 0,09 0,85b ± 0,10 0,89b ± 0,16 0,52a ± 0,04 *** *** ns ***
C22:2 0,17ab ± 0,03 0,27b ± 0,08 0,17a ± 0,04 0,21a ± 0,02 *** * ns *** C22:6(ω-3) 1,60c ± 0,33 1,06b ± 0,09 1,27b ± 0,21 0,70a ± 0,04 *** *** ns ***
AGS 90,58c ± 16,68 69,45b ± 6,85 75,36b ± 14,68 47,24a ± 2,30 *** *** ns *** AGMI 43,96c ± 11,08 32,05b ± 2,71 32,91b ± 6,52 18,34a ± 1,61 *** *** ns ***
AGPI 92,73c ± 15,48 63,69b ± 7,04 73,45b ± 10,94 46,14a ± 3,45 *** *** ns ***
ω-6 87,19c ± 14,53 59,67b ± 6,55 69,25b ± 10,37 43,40a ± 3,34 *** *** ns ***
ω-3 5,37c ± 0,95 3,74b ± 0,51 4,03b ± 0,59 2,53a ± 0,11 *** *** ns *** ω-6/ ω-3 16,28ab ± 0,22 16,01a ± 0,65 17,25c ± 1,21 17,13bc ± 0,67 ns *** ns **
C20:3 (ω-6)+C21 2,60c ± 0,55 2,12b ± 0,12 2,63c ± 0,39 1,41a ± 0,10 *** ** ** ***
sumatorio 229,86c ± 43,00 168,83b ± 16,08 184,33b ± 30,64 113,14a ± 6,51 *** *** ns ***
*** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

Tabla 4.3.32. Composición de la grasa intramuscular en la fracción de AGL del lomo curado. Los resultados están expresados en % (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Genética Interacc. p
C10:0 0,10 ± 0,03 0,09 ± 0,01 0,08 ± 0,02 0,10 ± 0,01 ns ns *** ns C12:0 0,06a ± 0,01 0,08b ± 0,02 0,06a ± 0,01 0,07ab ± 0,01 *** ns *** ns
C14:0 0,47a ± 0,05 0,56b ± 0,10 0,46a ± 0,10 0,49ab ± 0,03 * ns *** ns
C15:0 0,07a ± 0,01 0,07a ± 0,02 0,06a ± 0,01 0,09b ± 0,00 ** ns *** **
C16:0 11,43a ± 0,37 11,70a ± 0,61 12,78a ± 2,77 14,89b ± 0,29 ** *** *** * C17:0 0,25a ± 0,02 0,28ab ± 0,04 0,27ab ± 0,06 0,31b ± 0,03 * * *** ns
C18:0 11,35 ± 0,26 11,37 ± 0,27 11,33 ± 0,67 11,13 ± 0,16 ns ns *** ns
C20:0 0,15a ± 0,03 0,17ab ± 0,01 0,15a ± 0,02 0,18b ± 0,02 ** ns *** ns
C22:0 0,11 ± 0,04 0,11 ± 0,01 0,12 ± 0,03 0,13 ± 0,03 ns ns *** ns C23:0 1,19a ± 0,19 1,49b ± 0,15 1,30ab ± 0,22 1,18a ± 0,06 ns ns *** ***
C24:0 0,24a ± 0,03 0,37bc ± 0,05 0,31ab ± 0,06 0,39c ± 0,12 *** * *** ns
C14:1 0,10a ± 0,01 0,15b ± 0,03 0,10a ± 0,02 0,12a ± 0,02 *** ns *** *
C15:1 0,10a ± 0,02 0,13ab ± 0,03 0,15ab ± 0,04 0,16b ± 0,07 ns ** *** ns C16:1 1,40a ± 0,16 1,72b ± 0,19 1,62ab ± 0,43 1,63ab ± 0,14 * ns *** *
C17:1 0,18a ± 0,01 0,20ab ± 0,03 0,18a ± 0,05 0,23b ± 0,02 ** ns *** ns
C18:1 33,36b ± 3,36 33,90b ± 0,74 32,41b ± 2,58 28,52a ± 1,97 * *** *** **
C20:1 0,72a ± 0,04 0,82b ± 0,02 0,83b ± 0,11 0,73a ± 0,06 ns ns *** *** C22:1 0,10a ± 0,04 0,16bc ± 0,03 0,12ab ± 0,05 0,18c ± 0,03 *** ns *** ns
C24:1 0,42a ± 0,03 0,66b ± 0,10 0,51a ± 0,11 0,61b ± 0,09 *** ns *** *
C18:2(ω-6) 20,66a ± 1,03 20,75a ± 0,28 21,35a ± 1,07 23,67b ± 1,05 *** *** *** ***
C18:3(ω-6) 0,23a ± 0,02 0,26ab ± 0,02 0,24a ± 0,04 0,28b ± 0,03 ** ns *** ns C18:3(ω-3) 0,69a ± 0,05 0,67a ± 0,09 0,68a ± 0,15 0,84b ± 0,10 * * *** **
C20:2 (ω-6) 0,60a ± 0,02 0,67ab ± 0,03 0,67b ± 0,09 0,71b ± 0,05 ** ** *** ns
C20:4 (ω-6) 11,02 ± 1,25 9,94 ± 0,65 9,94 ± 2,03 11,40 ± 1,33 ns ns *** **
C20:3(ω-3) 0,21a ± 0,05 0,23ab ± 0,03 0,26ab ± 0,06 0,27b ± 0,05 ns ** *** ns C20:5(ω-3) 0,49 ± 0,08 0,51 ± 0,03 0,49 ± 0,07 0,51 ± 0,06 ns ns *** ns
C22:2 0,14a ± 0,04 0,33b ± 0,14 0,12a ± 0,02 0,34b ± 0,07 *** ns *** ns
C22:6(ω-3) 1,63 ± 0,28 1,44 ± 0,16 1,68 ± 0,25 1,50 ± 0,09 ** ns *** ns
AGS 25,43a ± 0,52 26,30a ± 0,93 26,93a ± 3,10 28,98b ± 0,50 ** *** *** ns
AGMI 36,37b ± 3,42 37,73b ± 0,75 35,92b ± 2,90 32,17a ± 1,93 ns *** *** **
AGPI 35,66a ± 2,18 34,80a ± 0,24 35,44a ± 3,15 39,53b ± 2,61 * ** *** ** ω-6 32,51a ± 1,87 31,62a ± 0,34 32,22a ± 2,94 36,07b ± 2,39 * ** *** **
ω-3 3,01 ± 0,43 2,86 ± 0,05 3,11 ± 0,39 3,12 ± 0,19 ns ns *** ns
ω-6/ ω-3 10,95ab ± 1,16 11,07b ± 0,07 10,45a ± 1,10 11,56ab ± 0,46 * ns *** ns
C20:3 (ω-6)+C21 1,27a ± 0,19 1,50b ± 0,10 1,54b ± 0,25 1,44ab ± 0,07 ns ns *** **
*** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
234
Lípidos totales
A diferencia del resto de fracciones estudiadas, no existieron diferencias
significativas en ningún ácido graso de los lípidos totales, por lo que tampoco existieron
diferencias en ningún sumatorio. A pesar de la gran cantidad de diferencias encontradas al
estudiar cada una de las fracciones por separado, estas se compensaron cuando se
analizaron en conjunto.
Al expresar los ácidos grasos en porcentajes, existieron diferencias en tres ácidos y
en la proporción de AGS (tabla 4.3.33). Tan solo la proporción de AGS mostró diferencias
debidas al efecto genético, coincidiendo así con la proporción del C16:0 y siendo superior
en ambos casos las proporciones de los lotes pertenecientes a la genética AG. Los otros dos
ácidos que mostraron diferencias fueron minoritarios, por lo que su repercusión no fue
importante.
Tabla 4.3.33. Composición de la grasa intramuscular en la fracción de lípidos totales del lomo curado. Los resultados están expresados en % (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
C14:0 1,35a ± 0,11 1,45ab ± 0,23 1,63c ± 0,10 1,55bc ± 0,16 ns *** ns **
C16:0 27,19 ± 1,62 26,29 ± 1,60 28,44 ± 1,60 28,72 ± 0,11 ns *** ns ** C15:1 0,04ab ± 0,01 0,04a ± 0,01 0,04b ± 0,02 0,03b ± 0,00 * ns ns *
AGS 40,69ab ± 2,14 38,97a ± 2,29 41,50b ± 2,01 42,16b ± 1,60 ns ** ns **
*** Nivel de significación p<0,05; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
235
Determinación del contenido en aminoácidos libres
La tabla 4.3.34 muestra las diferencias significativas en el contenido total e
individual de la composición de aminoácidos libres analizada en el lomo curado.
Al estudiar el efecto de los dos factores, sexo y genética, en la composición de
aminoácidos libres vemos claramente como la genética tuvo un efecto más marcado, ya
que el sexo por si solo no tuvo influencia en ningún aminoácido. A diferencia de los otros
dos productos estudiados (jamones y paletas) los lotes pertenecientes a la genética AG
presentaron mayores cantidades de aminoácidos libres y contenido total.
Determinación de los compuestos nitrogenados derivados del ATP
Al igual que los resultados obtenidos en la tabla 4.3.21, existieron diferencias
significativas en todos los nucleótidos analizados (tabla 4.3.35), influenciados claramente
por el efecto genético. Destacaron las mayores cantidades por lo general de los lotes
pertenecientes a la genética AA en todos los casos.

Tabla 4.3.34. Contenido en aminoácidos libres en lomo curado en extracto fresco. Los resultados se expresan en mg aa/100 g muestra (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
Asp 1,48a ± 0,35 1,39a ± 0,08 1,88b ± 0,51 1,50a ± 0,15 * * ns ** Glu 5,63b ± 0,35 4,88a ± 0,25 6,01b ± 0,94 5,92b ± 0,30 * *** * ***
Ser-Asp 4,32a ± 0,14 4,20a ± 0,16 4,73b ± 0,59 5,02b ± 0,16 ns *** * ***
Gly-Gln 4,24a ± 0,26 4,32a ± 0,10 4,72b ± 0,58 4,71b ± 0,15 ns *** ns **
His 2,72a ± 0,07 2,91b ± 0,03 3,08c ± 0,27 3,16c ± 0,09 ** *** ns *** Trh 1,40a ± 0,18 1,55a ± 0,16 1,59a ± 0,24 1,86b ± 0,27 ** ** ns ***
Cys-Ala 14,50b ± 0,42 11,73a ± 0,60 15,53bc ± 0,28 15,79c ± 1,55 *** *** *** ***
Pro-Arg 28,75b ± 0,23 27,65a ± 0,55 27,09a ± 1,62 27,89ab ± 0,34 ns * ** **
Tyr-Val 5,73 ± 0,22 5,55 ± 0,04 5,95 ± 0,70 5,92 ± 0,05 ns * ns ns Met 1,30ab ± 0,07 1,27a ± 0,00 1,34b ± 0,09 1,33ab ± 0,04 ns ** ns *
Iso 2,74a ± 0,11 2,71a ± 0,01 2,90b ± 0,23 2,93b ± 0,03 ns *** ns ***
Leu 4,71a ± 0,27 4,71a ± 0,01 4,80ab ± 0,20 4,99b ± 0,23 ns ** ns *
Phe 2,89ab ± 0,10 2,84a ± 0,02 3,12b ± 0,42 3,04ab ± 0,09 ns ** ns * Trp 1,04a ± 0,01 1,09ab ± 0,03 1,14b ± 0,13 1,08ab ± 0,01 ns * * *
Lys 8,52ab ± 0,47 8,45a ± 0,12 8,51ab ± 0,50 8,90b ± 0,17 ns ns * *
sumatorio (s) 89,96b ± 1,75 85,25a ± 1,10 92,37bc ± 5,56 94,02c ± 1,44 ns *** ** ***
sumatorio (f) 174,65a ± 4,78 172,89a ± 5,88 187,98b ± 11,11 197,30c ± 4,88 ns *** * **
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo
Tabla 4.3.35. Contenido en 5´nucleótidos del lomo curado en extracto fresco. Los resultados se expresan en mg de nuc./100 g muestra (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
IMP 2,22c ± 0,46 1,63ab ± 0,15 1,36a ± 0,39 1,94bc ± 0,40 ns * *** ***
GMP 4,65c ± 0,64 5,19c ± 0,81 1,85a ± 0,56 2,76bc ± 0,88 ** *** ns ***
ADP 1,49a ± 0,18 2,34b ± 0,44 2,17b ± 0,75 1,62a ± 0,31 ns ns *** *** AMP + HX 44,37b ± 2,43 46,32b ± 1,19 39,11a ± 0,39 37,23a ± 2,69 ns *** ** ***
Ino 8,18 ± 2,41 7,27 ± 0,68 6,85 ± 3,52 5,60 ± 1,85 ns * ns ns
sumatorio (s) 60,91b ± 3,57 62,74b ± 1,24 51,35a ± 3,87 49,16a ± 4,43 ns *** ns ***
sumatorio (f) 118,28b ± 7,54 127,14c ± 4,04 104,43a ± 7,24 102,68a ± 7,99 ns *** * ***
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
237
Determinación de compuestos volátiles
De los 105 compuestos volátiles detectados en los lomos (tabla 4.3.8), tan solo 63
presentaron diferencias significativas entre los lotes del estudio. Aún así las diferencias no
mostraron una tendencia en función del factor estudiado por lo que en lugar de presentar
una tabla con todos los valores, tan solo se incluyen dos figuras con los sumatorios de cada
familia (figuras 4.3.4 y 4.3.5).
En cuanto al contenido total, existieron diferencias significativas entre los cuatro
lotes, destacando el menor valor del lote AGM en comparación con el resto de lotes, no
obstante no se incluirá en las figuras 4.3.4 y 4.3.5. Las diferencias existentes en el
contenido total de compuestos volátiles no pueden estar asociadas al contenido de grasa
intramuscular, ya que los lomos no mostraron diferencias significativas en dicho
contenido. Al agrupar los compuestos volátiles por familias nos encontramos con
diferencias significativas en la mayor parte de éstas, influenciadas únicamente por el
efecto genético, pero sin seguir un patrón determinado (figuras 4.3.4 y 4.3.5).
Algunos de los compuestos a destacar son el 2-butanol, el hexano, el ácido 3-
metilbutanoico, el acetato de etilo, el 2-metilpropanoato de etilo, y especialmente la co-
elución formada por el 3-metiltio-1-propeno + 1-metiltio-1-propeno que fueron
significativamente superiores en el lote AHH. Por otra parte, el 2,3,4-trimetilpentano, el
ácido acético y especialmente el sulfuro de carbono fueron superiores significativamente
en el lote AAM. El acetaldehído, la 2-pentanona, el 2-metilbutanol, el hexanol, el decano, el
butanoato de etilo y la dihidro-2(3H)-furanona en el lote AGH, y tan solo el decanoato de
etilo en el lote AGM.

CAPÍTULO IV: RESULTADOS
238
0
100
200
300
400
500
600
700
800
AAH
AAM
AGH
AGM
*
*
* * *
Figura 4.3.4. Diagrama de barras correspondiente a la abundancia total de las principales familias química, expresadas en UAA (106) en los lotes de lomos estudiados (* Nivel de significación p<0,05)
0
5
10
15
20
25
30
35
40
45
50
COMP. NITROGENADOS
FURANOS HID. AROMATICOS
TERPENOS
AAH
AAM
AGH
AGM
*
Figura 4.3.5. Diagrama de barras correspondiente a la abundancia total de las principales familias química, expresadas en UAA (106) en los lotes de lomos estudiados (* Nivel de significación p<0,05)

CAPÍTULO IV: RESULTADOS
239
Evaluación sensorial
De las 15 características sensoriales evaluadas, 13 presentaron diferencias
significativas entre los lotes estudiados (tabla 4.3.37), sin embargo las diferencias no
siguieron una tendencia hacia un lote determinado, ya sea por efecto del sexo como por el
efecto de la genética.
0
1
2
3
4
5ROJO
BRILLO
VETEADO
INTENSIDAD OLOR
DUREZA
SEQUEDAD
JUGOSIDAD
FIBROSIDADSALADO
DULCE
AMARGO
INTENSIDAD FLAVOR
CURADO
RANCIDEZ
PERSISTENCIA
AAH
AAM
AGH
AGM
Figura 4.3.4. Perfil sensorial del lomo curado
El gráfico de araña muestra claramente como el lote AAH presentó una mayor
dureza y sequedad del magro junto una menor fibrosidad, sin embargo el lote AAM mostró
una mayor fibrosidad y flavor “a curado” y el lote AGH una mayor persistencia, aunque las
diferencias de estos lotes con respecto al resto fueron visiblemente menores.

Tabla 4.3.37. Análisis cuantitativo-descriptivo del lomo curado (± desviación estándar de la media).
AA AG EFECTO
HEMBRA MACHO HEMBRA MACHO Sexo Gen Interacc. p
Color magro
Rojo 2,69c ± 0,38 2,21ab ± 0,12 2,53bc ± 0,57 1,86a ± 0,17 * *** ns ***
Brillo 1,56a ± 0,34 1,89b ± 0,09 1,88b ± 0,21 1,79ab ± 0,23 ns ns ** **
Veteado 1,71a ± 0,13 1,97b ± 0,07 2,13c ± 0,18 2,18c ± 0,05 *** *** ** ***
Olor Intensidad 2,70ab ± 0,24 2,43a ± 0,25 2,96b ± 0,43 2,76ab ± 0,30 ** * ns **
Textura magro
Dureza 4,03b ± 0,08 3,49a ± 0,40 3,75ab ± 0,49 3,64a ± 0,13 ns ** * **
Sequedad 4,63b ± 0,26 3,99a ± 0,40 3,92a ± 0,22 3,98a ± 0,09 *** ** *** ***
Fibrosidad 2,90ab ± 0,70 3,33b ± 0,29 2,36a ± 0,68 2,38a ± 0,26 *** ns ns ***
Sabor Dulce 1,22ab ± 0,20 1,61c ± 0,26 1,31b ± 0,12 1,08a ± 0,02 *** ns *** ***
Amargo 0,74a ± 0,05 0,79a ± 0,03 0,84a ± 0,16 1,21b ± 0,06 *** *** *** ***
Flavor
Intensidad 4,01a ± 0,42 4,57bc ± 0,08 4,97c ± 0,26 4,25ab ± 0,46 ** ns *** ***
Curado 3,10ab ± 0,59 3,20b ± 0,21 2,80a ± 0,18 2,76a ± 0,08 ** ns ns **
Rancidez 1,06c ± 0,08 0,89a ± 0,10 0,96ab ± 0,02 1,01bc ± 0,04 ns ** *** ***
Persistencia 3,56a ± 0,34 3,45a ± 0,17 4,41b ± 0,43 3,20a ± 0,36 ** *** *** ***
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
241
4.3.4. INFLUENCIA DE LA EXPRESIÓN DEL IGF2 EN FUNCIÓN DE LA CAMPAÑA SOBRE
LAS CARACTERÍSTICAS FÍSICO-QUÍMICAS Y SENSORIALES DEL LOMO CURADO
En este apartado se va a evaluar la influencia del QTL (Quantitative Trait Loci)
IGF2 (insuline-like growth factor 2), del sexo y la campaña sobre algunos parámetros
físico-químicos en lomos procedentes de cerdos de distintas generaciones. Para ello se
evaluarán las diferencias existentes entre tres lotes, el primero perteneciente a los cerdos
de la primera generación y dos lotes más, que corresponden con las dos generaciones
posteriores. A cada lote se le denominará, primera, segunda y tercera campaña.
Características generales del lomo curado
En este apartado tan solo se compararán los resultados de los lomos procedentes
de la segunda y la tercera campaña debido a que los lomos de la primera habían sido
utilizados con anterioridad al inicio del presente estudio, tratándose además de lomos
adobados. A estos lomos se les realizaron las mismas determinaciones excepto las
medidas morfológicas iniciales, por lo que no se dispone de ellas. La tabla 4.3.37 muestra
las diferencias significativas entre dos lotes, pertenecientes a la segunda y la tercera
campaña, donde se dieron diferencias significativas en todos los parámetros morfológicos.
Los lomos de la tercera campaña resultaron más alargados que los de la segunda. Estas
diferencias estuvieron claramente influenciadas en primer lugar por el efecto campaña y
en segundo lugar por el efecto sexo, ya que como se vio anteriormente, al evaluar el efecto
de la genética, tan solo la longitud mostró diferencias significativas, llegando a la
conclusión de que la selección de los cerdos no influía sobre estos parámetros en el
producto curado, aunque sí en fresco. Por lo general, los lomos de la tercera campaña
resultaron los más grandes, siendo los pertenecientes al lote AA más pesados y largos y los
AG más anchos, coincidiendo así con los resultados mostrados para el jamón curado, ya
que los cerdos pertenecientes a esta campaña presentaron mayores pesos de sacrificio,
generando por lo tanto lomos de mayor tamaño.

CAPÍTULO IV: RESULTADOS
242
Parámetros generales físico-químicos
En cuanto a los parámetros generales, se encontraron diferencias significativas en
todos los parámetros evaluados, tal como se muestra en las tablas 4.3.38 y 4.3.39.
Las tablas muestran como el factor campaña actúo sobre todos los parámetros
físico-químicos, excepto aquellos relacionados con el contenido de mioglobina,
influenciados por el factor genético. A diferencia de los resultados mostrados para el
jamón curado, el contenido de grasa intramuscular no se vio influenciado por el factor
genético, sino por el factor campaña. Los lomos de la primera campaña presentaron
mayores valores en todos los parámetros evaluados excepto en el contenido de humedad,
que fue superior en los lomos de la tercera campaña, especialmente los pertenecientes al
lote AAH. El contenido de grasa intramuscular fue superior en los lomos de la primera
campaña, a pesar de que no existieron diferencias significativas entre los lotes de esa
misma campaña, ya que como se ha mencionado anteriormente el efecto fue claramente
de la campaña y no del sexo o de la genética. El resto de parámetros relacionados con el
color fueron superiores en los lomos pertenecientes a los lotes AG, donde la tabla muestra
como su influencia fue claramente genética.

Tabla 4.3.37. Medidas morfológicas generales del lomo curado (± desviación estándar de la media).
L2 L3
AA AG AA AG
EFECTO
HEMBRA MACHO HEMBRA MACHO HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p Peso (G) 1,8ab ± 0,0 1,7ab ± 0,1 1,6a ± 0,1 1,9ab ± 0,1 1,9ab ± 0,0 2,0b ± 0,0 1,8ab ± 0,4 1,8ab ± 0,1 ns ns * ns * Longitud (cm) 51,9ab ± 0,4 47,1a ± 1,5 48,3a ± 0,8 49,0ab ± 0,0 60,0c ± 0,0 56,5bc ± 1,5 55,5bc ± 7,8 46,5a ± 1,5 *** *** *** ** *** Anchura (cm) 20,6abc ± 0,0 21,2cd ± 0,1 20,2a ± 0,1 21,6d ± 0,0 20,3ab ± 0,2 22,4e ± 0,2 21,0bcd ± 0,9 22,5e ± 0,2 ns *** *** ** *** Perímetro (cm) 6,6abc ± 0,0 6,7cd ± 0,0 6,4a ± 0,0 6,9d ± 0,0 6,5ab ± 0,1 7,1e ± 0,1 6,7bcd ± 0,3 7,2e ± 0,1 ns *** *** ** ***
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo
Tabla 4.3.38. Parámetros físico-químicos generales del lomo curado (± desviación estándar de la media). Parte I.
L1 L2
AA AG
AA AG
HEMBRA MACHO HEMBRA MACHO HEMBRA MACHO HEMBRA MACHO
Humedad (g/100g) 50,94bc ± 0,04 47,72a ± 0,45 50,77bc ± 0,31 52,45cd ± 1,15 48,54ab ± 0,02 51,89cd ± 0,95 50,64bc ± 0,50 51,26c ± 0,46 Grasa (f) (g/100g) 6,45b ± 0,29 7,66b ± 0,68 7,68b ± 1,43 6,77b ± 1,33 3,49a ± 0,08 3,40a ± 0,00 3,23a ± 0,42 3,27a ± 0,00 Grasa (s) (g/100g) 13,14b ± 0,25 14,66b ± 0,88 15,59b ± 1,97 14,28b ± 1,78 6,79a ± 0,03 7,07a ± 0,21 6,55a ± 0,34 6,71a ± 0,59 L (Adimensional) 39,36bc ± 0,41 39,08bc ± 0,84 41,08c ± 0,26 39,60bc ± 1,91 36,16ab ± 0,12 37,60abc ± 0,18 36,97ab ± 3,02 38,03abc ± 0,78 a (Adimendional) 13,64abc ± 0,25 14,68bcd ± 0,95 15,50cd ± 0,02 15,60d ± 1,41 14,67bcd ± 0,40 14,24abcd ± 0,26 15,79d ± 0,26 14,44abcd ± 0,67 b (Adimensional) 8,88bc ± 0,28 9,62cd ± 0,10 10,48d ± 0,01 10,47d ± 0,86 8,03ab ± 0,01 7,76ab ± 0,20 7,73ab ± 0,62 8,30ab ± 0,52 mg Mb/g m 1,50ab ± 0,22 1,33a ± 0,13 2,40c ± 0,74 1,85abc ± 0,12 1,96abc ± 0,16 1,98abc ± 0,01 1,86abc ± 0,04 1,80abc ± 0,01 ppm Fe heminico 5,10ab ± 0,75 4,50a ± 0,44 8,15c ± 2,50 6,27abc ± 0,42 6,65abc ± 0,53 6,71abc ± 0,02 6,31abc ± 0,15 6,10abc ± 0,04
(f): extracto fresco; (s): extracto seco
Tabla 4.3.39. Parámetros físico-químicos generales del lomo curado (± desviación estándar de la media). Parte II.
L3
AA AG
EFECTO
HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
Humedad (g/100g) 54,10d ± 1,23 51,00bc ± 0,27 50,93bc ± 3,85 50,19abc ± 0,60 ns ns * *** *** Grasa (f) (g/100g) 2,54a ± 0,16 3,26a ± 0,68 3,59a ± 0,37 3,97a ± 0,23 ns ns *** ns *** Grasa (s) (g/100g) 5,53a ± 0,20 6,65a ± 1,23 7,38a ± 1,33 7,97a ± 0,24 ns ns *** ns *** L (Adimensional) 37,00ab ± 0,12 39,10bc ± 0,43 36,49ab ± 3,49 34,88a ± 1,26 ns ns *** ns *** a (Adimendional) 13,06ab ± 0,17 12,71a ± 0,67 14,75bcd ± 0,83 13,15ab ± 0,47 *** ns *** ns *** b (Adimensional) 7,63a ± 0,24 8,57abc ± 0,39 7,61a ± 0,22 7,71ab ± 0,39 * * *** * *** mg Mb/g m 1,65abc ± 0,03 1,23a ± 0,18 2,10bc ± 0,85 2,21bc ± 0,07 *** ns ns ns *** ppm Fe heminico 5,60abc ± 0,09 4,18a ± 0,60 7,11bc ± 2,90 7,48bc ± 0,24 *** ns ns ns ***
(f): extracto fresco; (s): extracto seco *** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
244
Composición de la grasa intramuscular
El contenido total de grasa intramuscular mostró diferencias significativas, siendo
superior en los lomos pertenecientes a la primera campaña, seguido de los lomos de la
tercera y finalmente los de la segunda (tablas 4.3.40 y 4.3.41), coincidiendo así con los
resultados mostrados para el jamón curado (tablas 4.1.49 y 4.1.50), a pesar de que en el
caso del jamón, la genética también influyó sobre el contenido de ácidos grasos y en el
lomo no, donde el efecto campaña tuvo una mayor influencia.
Como era de esperar, los ácidos grasos mayoritarios como el C16:0, C18:0, 16:1,
C18:1 y C18:2 (ω-6) fueron significativamente superiores en los lomos de la primera
campaña, especialmente los lomos pertenecientes al lote AGH, coincidiendo con el lote que
presentó mayor contenido de todos los sumatorios y ácidos grasos totales.
Por lo general, las diferencias significativas generadas entre lotes se debieron
sobre todo al factor campaña, seguido del factor genético y sexo, aunque ambos tuvieron
menos repercusión, al igual que la interacción entre los tres factores, que mostraron
diferencias en dieciocho ácidos grasos.

Tabla 4.3.40. Composición de la grasa intramuscular en la fracción de lípidos totales del lomo curado. Los resultados están expresados en mg de AG/100 g muestra (± desviación estándar de la media). Parte I.
L1 L2
AA AG AA AG
HEMBRA MACHO HEMBRA MACHO HEMBRA MACHO HEMBRA MACHO
C10:0 3,25ab ± 0,73 7,09b ± 4,94 3,99ab ± 0,25 3,52ab ± 0,35 2,43ab ± 0,83 3,23ab ± 0,66 1,26a ± 0,17 1,30a ± 0,12 C12:0 3,35abc ± 0,54 5,27c ± 2,84 4,06abc ± 0,31 4,43bc ± 0,80 2,92abc ± 1,06 4,52bc ± 0,04 2,36abc ± 1,28 1,90ab ± 0,14 C14:0 59,75bc ± 2,51 65,11bc ± 34,14 73,81c ± 11,14 71,35c ± 5,85 44,61abc ± 1,12 60,98bc ± 0,60 37,53abc ± 17,56 29,79ab ± 1,02 C15:0 1,08cde ± 0,02 0,93cd ± 0,05 1,43e ± 0,27 1,31de ± 0,26 0,76abc ± 0,02 0,89bcd ± 0,03 0,67abc ± 0,24 0,48ab ± 0,04 C16:0 957,65cde ± 29,48 872,94bcde ± 462,34 1.118,95e ± 16,06 1.010,70de ± 278,24 543,19abcd ± 2,45 598,60abcde ± 5,70 429,71abc ± 201,16 289,38a ± 18,70 C17:0 6,97cdef ± 0,24 4,41abcd ± 0,45 10,13f ± 1,64 7,97ef ± 2,97 3,27abc ± 0,81 3,84abcd ± 0,04 1,81a ± 0,30 2,18ab ± 0,02 C18:0 414,51bc ± 56,50 227,36a ± 65,38 488,36c ± 13,51 436,69bc ± 141,18 215,50a ± 4,68 223,62a ± 5,75 145,70a ± 76,64 112,87a ± 6,16 C20:0 4,62cde ± 0,26 3,00abc ± 1,01 5,09de ± 0,53 5,30e ± 1,35 4,61cde ± 0,12 5,12de ± 0,15 4,43bcde ± 0,44 2,28a ± 0,03 C22:0 0,29 ± 0,02 0,13 ± 0,01 0,37 ± 0,20 0,18 ± 0,02 0,10 ± 0,00 0,16 ± 0,00 0,12 ± 0,01 0,08 ± 0,00 C23:0 2,99ef ± 0,37 2,13bcd ± 0,35 2,62def ± 0,37 2,28cde ± 0,13 1,95bcd ± 0,33 1,79abc ± 0,23 1,44ab ± 0,40 1,39ab ± 0,03 AGS 1.454,46cde ± 79,62 1.188,37bcde ± 524,70 1.708,80e ± 28,87 1.543,73de± 414,40 819,33abcd ± 3,57 902,75abcd ± 10,41 686,43ab ± 326,17 441,64a ± 12,14
C14:1 2,72d ± 0,42 1,96c ± 0,21 3,13de ± 0,08 2,75d ± 0,24 3,35e ± 0,07 2,74d ± 0,22 1,67bc ± 0,22 1,50abc ± 0,12 C15:1 1,47ef ± 0,43 0,94bcd ± 0,14 1,35def ± 0,02 1,31cdef ± 0,07 1,71fg ± 0,07 0,87bc ± 0,02 1,02bcde ± 0,09 1,96g ± 0,27 C16:1 125,99defg ± 12,47 92,21cdef ± 17,40 161,30g ± 4,59 141,05fg ± 30,88 80,97bcd ± 2,38 137,84efg ± 12,23 62,54abc ± 32,62 39,17ab ± 1,66 C17:1 7,54gh ± 0,28 5,19de ± 0,54 8,63h ± 0,29 6,86fg ± 0,62 3,53bc ± 0,08 4,23cd ± 0,23 1,78a ± 0,19 2,10a ± 0,08 C18:1 530,28cd ± 54,95 335,08b ± 78,93 638,93d ± 10,45 557,62d ± 138,95 250,73ab ± 13,92 293,78ab ± 18,56 116,43a ± 22,62 130,91a ± 5,03 C20:1 15,76de ± 0,79 9,91abcd ± 2,78 15,27de ± 1,55 18,51e ± 5,05 8,11abc ± 0,01 11,84bcde ± 0,47 6,72abc ± 3,39 4,90ab ± 0,30 C22:1 0,45d ± 0,11 0,20abc ± 0,05 0,69e ± 0,04 0,41d ± 0,14 0,16ab ± 0,00 0,30abcd ± 0,02 0,14ab ± 0,02 0,11a ± 0,00 C24:1 2,09bcd ± 0,65 1,14b ± 0,23 3,09e ± 0,03 1,64bcd ± 0,63 0,07a ± 0,00 0,07a ± 0,00 0,06a ± 0,02 0,05a ± 0,00 AGMI 686,29de ± 65,27 446,62c ± 99,44 832,40e ± 10,62 730,15e ± 169,02 348,62abc ± 15,17 451,66c ± 16,43 190,35ab ± 47,72 180,69a ± 2,75
C18:2(ω-6) 197,99cd ± 5,12 151,52bc ± 38,06 243,85d ± 14,17 184,19cd ± 33,53 102,70ab ± 10,50 113,73ab ± 6,49 90,21ab ± 32,82 87,39a ± 1,48 C18:3(ω-6) 2,23f ± 0,19 1,56de ± 0,17 2,43f ± 0,00 1,75e ± 0,02 1,13bc ± 0,01 1,30cd ± 0,07 0,89ab ± 0,05 0,67a ± 0,02 C18:3(ω-3) 7,91ef ± 0,34 5,99cde ± 2,18 9,34fg ± 0,11 6,73de ± 0,42 4,97bcd ± 1,21 4,97abcd ± 0,20 2,63a ± 0,39 3,09ab ± 0,04
C20:2 4,66bcd ± 0,04 3,46abc ± 1,74 5,75cd ± 1,65 4,22abcd ± 0,21 2,91ab ± 0,12 3,93abcd ± 0,33 2,43ab ± 1,00 2,14a ± 0,06 C20:4 26,38cd ± 3,51 19,31ab ± 0,62 23,06bc ± 1,62 22,07bc ± 0,75 19,57ab ± 0,17 17,44ab ± 0,83 15,56a ± 4,70 15,30a ± 0,32
C20:3(ω-3) 1,32cd ± 0,03 0,96abcd ± 0,39 1,56d ± 0,45 1,26bcd ± 0,37 0,75abc ± 0,07 1,04abcd ± 0,07 0,50a ± 0,14 0,53a ± 0,01 C20:5(ω-3) 1,27d ± 0,13 0,86abc ± 0,10 1,29d ± 0,01 1,23d ± 0,05 0,87abc ± 0,02 0,81ab ± 0,09 0,65a ± 0,23 0,63a ± 0,04
C22:2 0,36d ± 0,05 0,28cd ± 0,11 0,64e ± 0,01 0,31d ± 0,06 0,12a ± 0,01 0,13ab ± 0,00 0,08a ± 0,03 0,11a ± 0,00 C22:6(ω-3) 1,51e ± 0,02 0,90abc ± 0,09 1,06c ± 0,01 1,14c ± 0,27 0,86abc ± 0,00 1,04c ± 0,08 0,67a ± 0,06 0,66a ± 0,04
AGPI 243,64cd ± 8,45 184,85bc ± 42,88 288,98d ± 17,31 222,91cd ± 35,07 133,59ab ± 11,98 144,39ab ± 7,01 109,57a ± 39,72 110,52a ± 1,80 ω-6 231,27cd ± 8,29 175,85bc ± 40,11 275,09d ± 16,93 212,22cd ± 34,23 126,02ab ± 11,11 136,40ab ± 7,19 109,09ab ± 37,91 105,50a ± 1,78 ω-3 12,01efg ± 0,43 8,71cd ± 2,69 13,25fg ± 0,53 10,38def ± 0,96 7,44bcd ± 1,23 7,85bcd ± 0,23 4,45a ± 0,69 4,91ab ± 0,10
ω-6/ω-3 19,27ab ± 0,72 20,63abc ± 1,91 20,76abc ± 0,92 20,39ab ± 2,09 17,15a ± 1,98 17,39ab ± 1,37 24,11bc ± 6,18 21,51abc ± 0,53 C20:3 (ω-6)+C21 4,41de ± 0,09 3,48bcd ± 0,70 3,48bcd ± 0,01 3,76cd ± 0,55 2,71abc ± 0,02 3,43bcd ± 0,11 2,44ab ± 0,85 2,16a ± 0,03
sumatorio 2.388,80cde ± 136,65 1.823,32bcd ± 607,83 2.833,65e ± 49,05 2.500,55de ± 595,85 1.304,24ab ± 19,81 1.502,23abc ± 20,46 931,44ab ± 377,21 735,00a ± 8,49
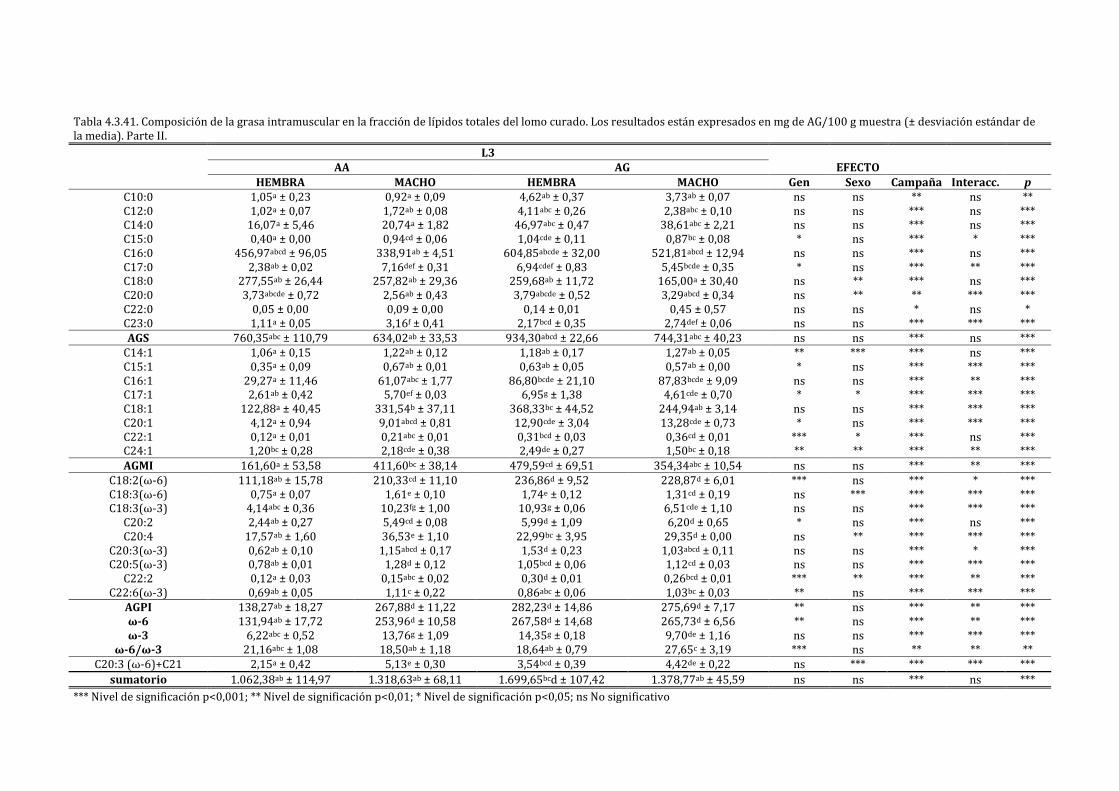
Tabla 4.3.41. Composición de la grasa intramuscular en la fracción de lípidos totales del lomo curado. Los resultados están expresados en mg de AG/100 g muestra (± desviación estándar de la media). Parte II.
L3
AA AG
EFECTO
HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
C10:0 1,05a ± 0,23 0,92a ± 0,09 4,62ab ± 0,37 3,73ab ± 0,07 ns ns ** ns ** C12:0 1,02a ± 0,07 1,72ab ± 0,08 4,11abc ± 0,26 2,38abc ± 0,10 ns ns *** ns *** C14:0 16,07a ± 5,46 20,74a ± 1,82 46,97abc ± 0,47 38,61abc ± 2,21 ns ns *** ns *** C15:0 0,40a ± 0,00 0,94cd ± 0,06 1,04cde ± 0,11 0,87bc ± 0,08 * ns *** * *** C16:0 456,97abcd ± 96,05 338,91ab ± 4,51 604,85abcde ± 32,00 521,81abcd ± 12,94 ns ns *** ns *** C17:0 2,38ab ± 0,02 7,16def ± 0,31 6,94cdef ± 0,83 5,45bcde ± 0,35 * ns *** ** *** C18:0 277,55ab ± 26,44 257,82ab ± 29,36 259,68ab ± 11,72 165,00a ± 30,40 ns ** *** ns *** C20:0 3,73abcde ± 0,72 2,56ab ± 0,43 3,79abcde ± 0,52 3,29abcd ± 0,34 ns ** ** *** *** C22:0 0,05 ± 0,00 0,09 ± 0,00 0,14 ± 0,01 0,45 ± 0,57 ns ns * ns * C23:0 1,11a ± 0,05 3,16f ± 0,41 2,17bcd ± 0,35 2,74def ± 0,06 ns ns *** *** *** AGS 760,35abc ± 110,79 634,02ab ± 33,53 934,30abcd ± 22,66 744,31abc ± 40,23 ns ns *** ns ***
C14:1 1,06a ± 0,15 1,22ab ± 0,12 1,18ab ± 0,17 1,27ab ± 0,05 ** *** *** ns *** C15:1 0,35a ± 0,09 0,67ab ± 0,01 0,63ab ± 0,05 0,57ab ± 0,00 * ns *** *** *** C16:1 29,27a ± 11,46 61,07abc ± 1,77 86,80bcde ± 21,10 87,83bcde ± 9,09 ns ns *** ** *** C17:1 2,61ab ± 0,42 5,70ef ± 0,03 6,95g ± 1,38 4,61cde ± 0,70 * * *** *** *** C18:1 122,88a ± 40,45 331,54b ± 37,11 368,33bc ± 44,52 244,94ab ± 3,14 ns ns *** *** *** C20:1 4,12a ± 0,94 9,01abcd ± 0,81 12,90cde ± 3,04 13,28cde ± 0,73 * ns *** *** *** C22:1 0,12a ± 0,01 0,21abc ± 0,01 0,31bcd ± 0,03 0,36cd ± 0,01 *** * *** ns *** C24:1 1,20bc ± 0,28 2,18cde ± 0,38 2,49de ± 0,27 1,50bc ± 0,18 ** ** *** ** *** AGMI 161,60a ± 53,58 411,60bc ± 38,14 479,59cd ± 69,51 354,34abc ± 10,54 ns ns *** ** ***
C18:2(ω-6) 111,18ab ± 15,78 210,33cd ± 11,10 236,86d ± 9,52 228,87d ± 6,01 *** ns *** * *** C18:3(ω-6) 0,75a ± 0,07 1,61e ± 0,10 1,74e ± 0,12 1,31cd ± 0,19 ns *** *** *** *** C18:3(ω-3) 4,14abc ± 0,36 10,23fg ± 1,00 10,93g ± 0,06 6,51cde ± 1,10 ns ns *** *** ***
C20:2 2,44ab ± 0,27 5,49cd ± 0,08 5,99d ± 1,09 6,20d ± 0,65 * ns *** ns *** C20:4 17,57ab ± 1,60 36,53e ± 1,10 22,99bc ± 3,95 29,35d ± 0,00 ns ** *** *** ***
C20:3(ω-3) 0,62ab ± 0,10 1,15abcd ± 0,17 1,53d ± 0,23 1,03abcd ± 0,11 ns ns *** * *** C20:5(ω-3) 0,78ab ± 0,01 1,28d ± 0,12 1,05bcd ± 0,06 1,12cd ± 0,03 ns ns *** *** ***
C22:2 0,12a ± 0,03 0,15abc ± 0,02 0,30d ± 0,01 0,26bcd ± 0,01 *** ** *** ** *** C22:6(ω-3) 0,69ab ± 0,05 1,11c ± 0,22 0,86abc ± 0,06 1,03bc ± 0,03 ** ns *** *** ***
AGPI 138,27ab ± 18,27 267,88d ± 11,22 282,23d ± 14,86 275,69d ± 7,17 ** ns *** ** *** ω-6 131,94ab ± 17,72 253,96d ± 10,58 267,58d ± 14,68 265,73d ± 6,56 ** ns *** ** *** ω-3 6,22abc ± 0,52 13,76g ± 1,09 14,35g ± 0,18 9,70de ± 1,16 ns ns *** *** ***
ω-6/ω-3 21,16abc ± 1,08 18,50ab ± 1,18 18,64ab ± 0,79 27,65c ± 3,19 *** ns ** ** ** C20:3 (ω-6)+C21 2,15a ± 0,42 5,13e ± 0,30 3,54bcd ± 0,39 4,42de ± 0,22 ns *** *** *** ***
sumatorio 1.062,38ab ± 114,97 1.318,63ab ± 68,11 1.699,65bcd ± 107,42 1.378,77ab ± 45,59 ns ns *** ns ***
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
247
Las tablas 4.3.42 y 4.3.43 muestran los porcentajes de los ácidos grasos presentes
en los lípidos totales de la grasa intramuscular. A diferencia de los porcentajes de ácidos
grasos que se evaluaron en capítulos anteriores, al estudiar el efecto de los tres factores
por separado vemos como existieron diferencias significativas en una gran parte de los
ácidos grasos. Coincidiendo con las tablas 4.3.40 y 4.3.41, el factor más influyente sobre las
diferencias generadas entre los porcentajes fue la campaña, ya que el factor genético y el
sexo no mostraron un efecto muy marcado, no obstante, las diferencias no siguieron la
misma tendencia.
Existieron diferencias significativas en la proporción de AGS, siendo por lo general
superiores en los lomos de la primera campaña y en el lote AAH de la segunda, por su
elevada proporción del C18:0. La proporción de AGMI mostraron diferencias significativas,
siendo superiores en los lomos de la segunda campaña pertenecientes al lote AGH, por su
mayor proporción de los ácidos monoinsaturados mayoritarios C16:1 y C18:1. Además, las
proporciones de AGPI, ω-3 y ω-6 fueron superiores también en los lomos de la segunda
campaña, especialmente los lomos pertenecientes al lote AAM.
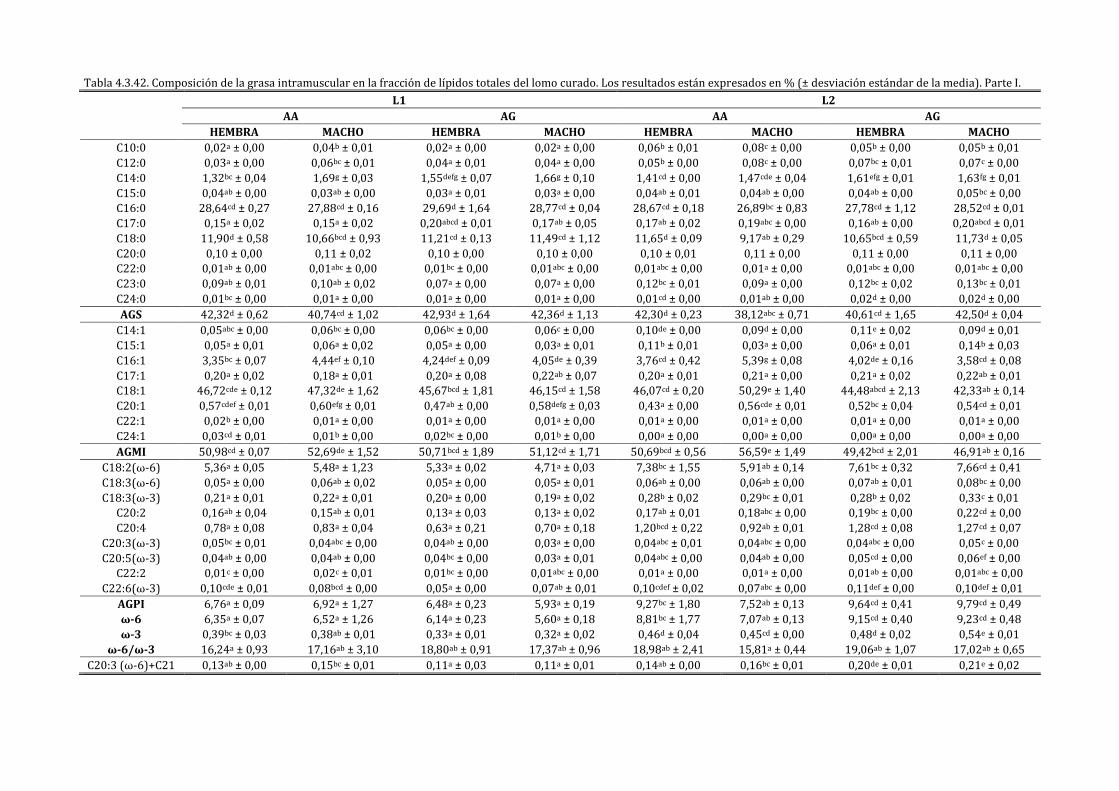
Tabla 4.3.42. Composición de la grasa intramuscular en la fracción de lípidos totales del lomo curado. Los resultados están expresados en % (± desviación estándar de la media). Parte I.
L1 L2
AA AG AA AG
HEMBRA MACHO HEMBRA MACHO HEMBRA MACHO HEMBRA MACHO
C10:0 0,02a ± 0,00 0,04b ± 0,01 0,02a ± 0,00 0,02a ± 0,00 0,06b ± 0,01 0,08c ± 0,00 0,05b ± 0,00 0,05b ± 0,01
C12:0 0,03a ± 0,00 0,06bc ± 0,01 0,04a ± 0,01 0,04a ± 0,00 0,05b ± 0,00 0,08c ± 0,00 0,07bc ± 0,01 0,07c ± 0,00
C14:0 1,32bc ± 0,04 1,69g ± 0,03 1,55defg ± 0,07 1,66g ± 0,10 1,41cd ± 0,00 1,47cde ± 0,04 1,61efg ± 0,01 1,63fg ± 0,01
C15:0 0,04ab ± 0,00 0,03ab ± 0,00 0,03a ± 0,01 0,03a ± 0,00 0,04ab ± 0,01 0,04ab ± 0,00 0,04ab ± 0,00 0,05bc ± 0,00 C16:0 28,64cd ± 0,27 27,88cd ± 0,16 29,69d ± 1,64 28,77cd ± 0,04 28,67cd ± 0,18 26,89bc ± 0,83 27,78cd ± 1,12 28,52cd ± 0,01
C17:0 0,15a ± 0,02 0,15a ± 0,02 0,20abcd ± 0,01 0,17ab ± 0,05 0,17ab ± 0,02 0,19abc ± 0,00 0,16ab ± 0,00 0,20abcd ± 0,01
C18:0 11,90d ± 0,58 10,66bcd ± 0,93 11,21cd ± 0,13 11,49cd ± 1,12 11,65d ± 0,09 9,17ab ± 0,29 10,65bcd ± 0,59 11,73d ± 0,05
C20:0 0,10 ± 0,00 0,11 ± 0,02 0,10 ± 0,00 0,10 ± 0,00 0,10 ± 0,01 0,11 ± 0,00 0,11 ± 0,00 0,11 ± 0,00 C22:0 0,01ab ± 0,00 0,01abc ± 0,00 0,01bc ± 0,00 0,01abc ± 0,00 0,01abc ± 0,00 0,01a ± 0,00 0,01abc ± 0,00 0,01abc ± 0,00
C23:0 0,09ab ± 0,01 0,10ab ± 0,02 0,07a ± 0,00 0,07a ± 0,00 0,12bc ± 0,01 0,09a ± 0,00 0,12bc ± 0,02 0,13bc ± 0,01
C24:0 0,01bc ± 0,00 0,01a ± 0,00 0,01a ± 0,00 0,01a ± 0,00 0,01cd ± 0,00 0,01ab ± 0,00 0,02d ± 0,00 0,02d ± 0,00
AGS 42,32d ± 0,62 40,74cd ± 1,02 42,93d ± 1,64 42,36d ± 1,13 42,30d ± 0,23 38,12abc ± 0,71 40,61cd ± 1,65 42,50d ± 0,04
C14:1 0,05abc ± 0,00 0,06bc ± 0,00 0,06bc ± 0,00 0,06c ± 0,00 0,10de ± 0,00 0,09d ± 0,00 0,11e ± 0,02 0,09d ± 0,01
C15:1 0,05a ± 0,01 0,06a ± 0,02 0,05a ± 0,00 0,03a ± 0,01 0,11b ± 0,01 0,03a ± 0,00 0,06a ± 0,01 0,14b ± 0,03
C16:1 3,35bc ± 0,07 4,44ef ± 0,10 4,24def ± 0,09 4,05de ± 0,39 3,76cd ± 0,42 5,39g ± 0,08 4,02de ± 0,16 3,58cd ± 0,08
C17:1 0,20a ± 0,02 0,18a ± 0,01 0,20a ± 0,08 0,22ab ± 0,07 0,20a ± 0,01 0,21a ± 0,00 0,21a ± 0,02 0,22ab ± 0,01 C18:1 46,72cde ± 0,12 47,32de ± 1,62 45,67bcd ± 1,81 46,15cd ± 1,58 46,07cd ± 0,20 50,29e ± 1,40 44,48abcd ± 2,13 42,33ab ± 0,14
C20:1 0,57cdef ± 0,01 0,60efg ± 0,01 0,47ab ± 0,00 0,58defg ± 0,03 0,43a ± 0,00 0,56cde ± 0,01 0,52bc ± 0,04 0,54cd ± 0,01
C22:1 0,02b ± 0,00 0,01a ± 0,00 0,01a ± 0,00 0,01a ± 0,00 0,01a ± 0,00 0,01a ± 0,00 0,01a ± 0,00 0,01a ± 0,00
C24:1 0,03cd ± 0,01 0,01b ± 0,00 0,02bc ± 0,00 0,01b ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00
AGMI 50,98cd ± 0,07 52,69de ± 1,52 50,71bcd ± 1,89 51,12cd ± 1,71 50,69bcd ± 0,56 56,59e ± 1,49 49,42bcd ± 2,01 46,91ab ± 0,16
C18:2(ω-6) 5,36a ± 0,05 5,48a ± 1,23 5,33a ± 0,02 4,71a ± 0,03 7,38bc ± 1,55 5,91ab ± 0,14 7,61bc ± 0,32 7,66cd ± 0,41
C18:3(ω-6) 0,05a ± 0,00 0,06ab ± 0,02 0,05a ± 0,00 0,05a ± 0,01 0,06ab ± 0,00 0,06ab ± 0,00 0,07ab ± 0,01 0,08bc ± 0,00
C18:3(ω-3) 0,21a ± 0,01 0,22a ± 0,01 0,20a ± 0,00 0,19a ± 0,02 0,28b ± 0,02 0,29bc ± 0,01 0,28b ± 0,02 0,33c ± 0,01 C20:2 0,16ab ± 0,04 0,15ab ± 0,01 0,13a ± 0,03 0,13a ± 0,02 0,17ab ± 0,01 0,18abc ± 0,00 0,19bc ± 0,00 0,22cd ± 0,00
C20:4 0,78a ± 0,08 0,83a ± 0,04 0,63a ± 0,21 0,70a ± 0,18 1,20bcd ± 0,22 0,92ab ± 0,01 1,28cd ± 0,08 1,27cd ± 0,07
C20:3(ω-3) 0,05bc ± 0,01 0,04abc ± 0,00 0,04ab ± 0,00 0,03a ± 0,00 0,04abc ± 0,01 0,04abc ± 0,00 0,04abc ± 0,00 0,05c ± 0,00
C20:5(ω-3) 0,04ab ± 0,00 0,04ab ± 0,00 0,04bc ± 0,00 0,03a ± 0,01 0,04abc ± 0,00 0,04ab ± 0,00 0,05cd ± 0,00 0,06ef ± 0,00 C22:2 0,01c ± 0,00 0,02c ± 0,01 0,01bc ± 0,00 0,01abc ± 0,00 0,01a ± 0,00 0,01a ± 0,00 0,01ab ± 0,00 0,01abc ± 0,00
C22:6(ω-3) 0,10cde ± 0,01 0,08bcd ± 0,00 0,05a ± 0,00 0,07ab ± 0,01 0,10cdef ± 0,02 0,07abc ± 0,00 0,11def ± 0,00 0,10def ± 0,01
AGPI 6,76a ± 0,09 6,92a ± 1,27 6,48a ± 0,23 5,93a ± 0,19 9,27bc ± 1,80 7,52ab ± 0,13 9,64cd ± 0,41 9,79cd ± 0,49
ω-6 6,35a ± 0,07 6,52a ± 1,26 6,14a ± 0,23 5,60a ± 0,18 8,81bc ± 1,77 7,07ab ± 0,13 9,15cd ± 0,40 9,23cd ± 0,48
ω-3 0,39bc ± 0,03 0,38ab ± 0,01 0,33a ± 0,01 0,32a ± 0,02 0,46d ± 0,04 0,45cd ± 0,00 0,48d ± 0,02 0,54e ± 0,01
ω-6/ω-3 16,24a ± 0,93 17,16ab ± 3,10 18,80ab ± 0,91 17,37ab ± 0,96 18,98ab ± 2,41 15,81a ± 0,44 19,06ab ± 1,07 17,02ab ± 0,65
C20:3 (ω-6)+C21 0,13ab ± 0,00 0,15bc ± 0,01 0,11a ± 0,03 0,11a ± 0,01 0,14ab ± 0,00 0,16bc ± 0,01 0,20de ± 0,01 0,21e ± 0,02

Tabla 4.3.43. Composición de la grasa intramuscular en la fracción de lípidos totales del lomo curado. Los resultados están expresados en % (± desviación estándar de la media). Parte II.
L3
AA AG
EFECTO
HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
C10:0 0,02a ± 0,01 0,02a ± 0,00 0,06b ± 0,00 0,05b ± 0,00 ns ** *** ns ***
C12:0 0,03a ± 0,00 0,03a ± 0,00 0,05b ± 0,01 0,04a ± 0,00 ns *** *** ns ***
C14:0 1,26ab ± 0,03 1,16a ± 0,02 1,62fg ± 0,15 1,50def ± 0,08 *** ** *** ** *** C15:0 0,04ab ± 0,01 0,06c ± 0,00 0,05bc ± 0,00 0,05bc ± 0,00 ns * *** ** ***
C16:0 24,66a ± 0,86 24,71a ± 0,05 27,60bcd ± 2,27 25,52ab ± 0,71 *** ** *** ** ***
C17:0 0,25de ± 0,01 0,28e ± 0,01 0,21bcd ± 0,01 0,24cde ± 0,00 ns ns *** ns ***
C18:0 9,85abc ± 0,42 10,59bcd ± 0,43 10,28bcd ± 1,16 8,53a ± 0,15 ns ** *** *** *** C20:0 0,10 ± 0,00 0,10 ± 0,00 0,11 ± 0,01 0,11 ± 0,01 ns ns ns ns ns
C22:0 0,01ab ± 0,00 0,01c ± 0,00 0,01abc ± 0,00 0,01abc ± 0,00 ** ns ns ns **
C23:0 0,16c ± 0,01 0,12bc ± 0,02 0,12bc ± 0,02 0,14c ± 0,01 ns ns *** ** ***
C24:0 0,01ab ± 0,00 0,01cd ± 0,00 0,01abc ± 0,00 0,01ab ± 0,00 ns * *** *** ***
AGS 36,39a ± 1,33 37,09ab ± 0,39 40,14bcd ± 3,54 36,19a ± 0,95 ** ** *** *** ***
C14:1 0,04ab ± 0,00 0,03a ± 0,00 0,04abc ± 0,00 0,05abc ± 0,00 * ns *** ns ***
C15:1 0,04a ± 0,00 0,03a ± 0,00 0,03a ± 0,00 0,03a ± 0,00 ns ns *** *** ***
C16:1 2,91ab ± 0,30 2,27a ± 0,05 3,59cd ± 0,47 4,72f ± 0,32 *** *** *** *** *** C17:1 0,28ab ± 0,01 0,22ab ± 0,03 0,24ab ± 0,03 0,32b ± 0,03 ns ns *** ns **
C18:1 41,52a ± 0,99 43,59abc ± 0,69 43,93abcd ± 2,23 46,44cd ± 1,37 * ** *** ** ***
C20:1 0,51bc ± 0,02 0,62fg ± 0,02 0,61efg ± 0,05 0,64g ± 0,01 ns *** *** *** ***
C22:1 0,02b ± 0,00 0,02b ± 0,00 0,02b ± 0,00 0,02b ± 0,00 * ns *** ** *** C24:1 0,05e ± 0,00 0,05e ± 0,00 0,04e ± 0,00 0,03d ± 0,00 *** *** *** *** ***
AGMI 45,37a ± 1,26 46,83ab ± 0,66 48,50abc ± 2,78 52,24cd ± 1,67 ns *** *** *** ***
C18:2(ω-6) 9,01cde ± 0,19 10,74f ± 0,10 9,37def ± 0,02 9,79ef ± 0,05 ns ns *** * ***
C18:3(ω-6) 0,08bc ± 0,01 0,09c ± 0,01 0,06ab ± 0,00 0,06ab ± 0,00 * * *** ns *** C18:3(ω-3) 0,37d ± 0,02 0,41de ± 0,01 0,43e ± 0,02 0,31bc ± 0,03 ns ns *** *** ***
C20:2 0,33f ± 0,02 0,39g ± 0,02 0,28e ± 0,01 0,27de ± 0,01 *** * *** * ***
C20:4 1,38cd ± 0,01 1,53de ± 0,04 1,18bc ± 0,24 1,19bcd ± 0,02 ns ns *** ns ***
C20:3(ω-3) 0,08e ± 0,00 0,09f ± 0,00 0,07de ± 0,00 0,06d ± 0,00 *** ns *** ** *** C20:5(ω-3) 0,06ef ± 0,01 0,06f ± 0,00 0,05de ± 0,01 0,06ef ± 0,00 ** ns *** *** ***
C22:2 0,01c ± 0,00 0,01abc ± 0,00 0,02c ± 0,00 0,01abc ± 0,00 ns ns *** ns ***
C22:6(ω-3) 0,13f ± 0,02 0,17g ± 0,00 0,09cde ± 0,01 0,11ef ± 0,01 *** * *** ** ***
AGPI 11,45de ± 0,20 13,49f ± 0,08 11,55de ± 0,26 11,87ef ± 0,03 ns ns *** ** ***
ω-6 10,80de ± 0,20 12,75f ± 0,08 10,89def ± 0,27 11,31ef ± 0,04 ns ns *** * ***
ω-3 0,63f ± 0,01 0,73f ± 0,00 0,64g ± 0,01 0,54e ± 0,04 *** ns *** *** *** ω-6/ω-3 17,02ab ± 0,16 17,39ab ± 0,10 16,96ab ± 0,78 20,94b ± 1,66 * ns ns * **
C20:3 (ω-6)+C21 0,20de ± 0,00 0,22e ± 0,00 0,17cd ± 0,00 0,20de ± 0,00 ns *** *** ns ***
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

CAPÍTULO IV: RESULTADOS
250
Determinación de compuestos volátiles
En las Tablas 4.3.44, 4.3.45 y 4.3.46 se muestra que a pesar de la variación
observada, los alcoholes y las cetonas fueron los principales representantes de los
compuestos volátiles y que el procesado es uno de los factores más directamente
implicados en su generación.
Existieron diferencias significativas en el contenido total de compuestos volátiles
entre los lomos de las tres campañas (tablas 4.3.44, 4.3.45 y 4.3.46) y en la mayor parte de
los compuestos volátiles detectados. Este hecho puede verse influenciado por las causas
explicadas anteriormente en jamones y paletas, como pueden ser las diferencias en el
procesado de las piezas y los tiempos de muestreo. No obstante, en los lomos curados
debemos tener en cuenta la gran diferencia que existió en el contenido de grasa
intramuscular entre las tres campañas y que los lomos de la primera campaña tuvieron un
procesado algo diferente al resto de campañas, ya que esos lomos fueron adobados. Esto
podría explicar el hecho de que el factor campaña fue determinante sobre las diferencias
generadas en la mayor parte de los compuestos, ya que el resto de factores influyeron
levemente.
De los 105 compuestos volátiles analizados en los lomos, 97 presentaron
diferencias significativas entre los lotes, 52 de ellos superiores significativamente en los
lomos de la primera campaña, 20 superiores en la segunda, 9 en la tercera y el resto no
siguieron una tendencia clara según el efecto en estudio. En cuanto al contenido total de
compuestos volátiles no destacó ninguna campaña en concreto, a pesar del adobo de los
lomos de la primera campaña, sino más bien destacaron dos lotes en particular, los lomos
pertenecientes al lote AGH de la primera campaña y los AAM de la segunda campaña.
En los lomos de la primera campaña destacaron los altos valores de algunos
compuestos como el hexanal, 3-hidroxi-2-butanona, 2-butanol, 2-heptanol y la mayor
parte de los ácidos, ésteres, compuestos nitrogenados y terpenos cuyos altos valores
pueden deberse al adobo realizado sobre el producto. En los lomos de la segunda campaña
destacaron el acetaldehído, la 2-pentanona, etanol, dodecano, sulfuro de carbono y el
metilbenceno. En los lomos de la tercera campaña el 2-metilpropanal, el 4-metilpentano,
etilbenceno y m- y p- xileno.

Tabla 4.3.44. Compuestos volátiles identificados en lomo curado agrupados por familia química. Los contenidos de cada compuesto se expresan en UAA (106) (± desviación estándar de la media). Parte I.
L1
AA
AG
EFECTO
IK Posición Compuesto HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p ALDEHIDOS 89,00a ± 13,85 109,47ab ± 70,67 132,22ab ± 57,71 93,84ab ± 21,73 ** * *** ns *
<500 2 acetaldehído 2,48a ± 0,45 3,39a ± 1,41 9,25ab ± 6,19 3,38a ± 0,36 *** *** *** *** *** 548 12 2-metilpropanal 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 3,28b ± 0,00 * ns ns ns *** 650 23 3-metilbutanal 26,53ab ± 8,12 35,61ab ± 29,68 61,33ab ± 69,75 39,33ab ± 15,87 ns ns ns ns ns 661 25 2-metilbutanal 6,82a ± 0,40 8,39a ± 5,53 8,11a ± 1,08 9,10a ± 0,29 ns ns * * * 799 47 hexanal 18,06 ± 14,73 8,86 ± 0,38 8,16 ± 0,33 3,40 ± 1,79 ns ns *** ns * 900 63 heptanal 8,20bcd ± 0,83 9,12d ± 5,09 8,40cd ± 1,88 7,14abcd ± 3,80 ** ns *** ns *** 969 72 benzaldehído 3,70ab ± 1,92 3,90ab ± 3,47 9,24b ± 3,72 9,10b ± 1,34 * ns *** ns ***
1005 80 octanal 3,59abc ± 0,04 7,92abcd ± 0,00 3,78abc ± 0,12 3,18abc ± 0,00 ns * *** ns *** 1054 87 bencenoacetaldehido 9,42ab ± 2,38 8,63ab ± 5,82 12,72b ± 4,64 9,27ab ± 1,03 ns ns ** ns *** 1107 92 nonanal 8,80ab ± 2,13 22,18b ± 19,31 8,81ab ± 2,73 5,01a ± 2,97 * ns *** ns *** 1211 98 decanal 1,39ab ± 0,57 1,47ab ± 0,73 2,43b ± 1,61 1,67ab ± 1,49 ns ns *** * **
CETONAS 166,96ab ± 36,48 547,97bc ± 290,91 510,55abc ± 460,70 226,85ab ± 23,25 * * *** ** ** <500 5 2-propanona+propanal 39,66a ± 8,19 58,02a ± 20,68 44,11a ± 15,84 127,77a ± 5,96 ns ns *** ns *** 585 15 2,3-butanodiona 7,47 ± 0,28 6,91 ± 4,67 2,04 ± 2,88 4,89 ± 0,68 ns ns * * ** 597 16 2-butanona+butanal 23,55 ± 14,38 262,73 ± 316,84 365,04 ± 445,97 23,92 ± 4,64 ns * *** ns ns 658 24 1-hidroxi-2-propanona 2,71ab ± 0,00 2,21ab ± 0,00 0,00a ± 0,00 5,08b ± 0,00 ns ns *** ns *** 688 28 2-pentanona 7,03abc ± 1,05 4,90abc ± 1,19 5,47abc ± 2,32 5,15abc ± 0,66 *** * *** *** *** 716 31 3-hidroxi-2-butanona 73,83bc ± 16,17 197,26d ± 40,40 70,19bc ± 15,26 46,20ab ± 20,29 ns ns ** ns *** 790 46 2-hexanona 0,00 ± 0,00 0,00 ± 0,00 0,00 ± 0,00 0,00 ± 0,00 * * ** ns * 890 60 2-heptanona 4,46a ± 1,44 6,03a ± 3,41 15,04b ± 8,33 5,10a ± 0,06 ns ns *** * ** 963 71 4-metilciclohexanona 1,77bcd ± 0,53 3,23d ± 1,53 3,34d ± 0,00 2,10cd ± 1,16 ns * *** * *** 988 74 3-octanona 3,34d ± 0,17 3,04bcd ± 1,03 0,00a ± 0,00 3,21cd ± 1,91 ns ns *** ** *** 992 75 2-octanona 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 * ns *** ns ***
1084 89 8-nonen-2-ona 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 ** ns ** ns *** 1094 90 2-nonanona 3,15 ± 1,27 3,64 ± 0,50 5,33 ± 1,78 3,44 ± 0,43 ns *** *** *** **
ALCOHOLES 355,48a ± 57,09 339,78a ± 14,07 570,90b ± 69,13 377,86a ± 20,54 ns *** *** *** *** <500 4 etanol 139,50ab ± 33,15 120,73a ± 19,75 200,61ab ± 140,67 166,84ab ± 5,94 * *** ** ns *** 548 11 propanol 14,54a ± 6,02 14,08a ± 3,72 27,64ab ± 35,21 5,25a ± 2,06 *** ns *** *** *** 602 18 2-butanol 22,27e ± 0,00 48,11f ± 0,00 81,72h ± 0,00 54,63g ± 0,00 ns ns ** ns *** 621 22 2-metil-1-propanol 23,83ab ± 0,00 13,72a ± 5,06 50,41b ± 0,00 28,70ab ± 0,00 ns ns ns ns ** 669 26 1-metoxi-2-propanol 12,06b ± 10,00 1,95ab ± 0,00 4,55ab ± 0,00 2,93ab ± 0,89 ns ns ns ns * 683 27 1-penten-3-ol 1,93 ± 0,25 2,58 ± 0,00 2,86 ± 1,17 1,87 ± 0,16 *** ns *** *** ns 736 33 2-metil-3-buten-2-ol 5,01de ± 2,39 3,58bcd ± 1,06 4,20cd ± 2,51 3,71bcd ± 0,13 ns ns ns ns *** 739 34 3-metilbutanol 21,80 ± 1,34 26,24 ± 2,40 27,59 ± 4,40 19,58 ± 2,44 ns * * ns ns 743 35 2-metilbutanol 7,67 ± 3,31 8,55 ± 0,84 6,10 ± 2,48 6,31 ± 0,33 ns * *** * * 771 41 pentanol 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 ns ** *** *** *** 790 44 2,3-butanodiol 72,69b ± 5,71 73,72b ± 16,37 120,96c ± 23,68 56,71b ± 14,42 ns ** ** ns ***

L1
AA AG EFECTO
IK Posición Compuesto HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p 790 44 2,3-butanodiol 72,69b ± 5,71 73,72b ± 16,37 120,96c ± 23,68 56,71b ± 14,42 ns ** ** ns *** 869 57 hexanol 6,01abc ± 0,33 3,78abc ± 0,08 9,63c ± 5,52 4,89abc ± 0,21 ns * *** ** ** 897 62 2-heptanol 12,30d ± 1,75 8,31cd ± 3,17 19,02e ± 3,53 5,41bc ± 3,64 ns ns *** ns *** 905 64 2-butoxi etanol 10,72abc ± 4,60 9,91ab ± 4,07 11,29abc ± 2,93 16,93abc ± 2,53 ns ns *** ns *** 980 73 1-octen-3-ol 5,15abc ± 0,98 4,52abc ± 0,00 4,32abc ± 0,50 4,10abc ± 1,97 ns ns ns *** ***
HIDROCARBUROS ALIFÁTICOS 86,89ab ± 22,53 37,57a ± 1,77 48,67ab ± 30,97 76,00ab ± 1,26 ns ns *** ns * <500 1 propano 2,13a ± 0,46 1,92a ± 0,41 0,00a ± 0,00 3,37a ± 0,00 ** ns *** * *** 500 6 pentano 18,12abc ± 0,97 17,12abc ± 0,80 12,32ab ± 3,42 5,61a ± 0,61 ns ns ns ns ** 505 7 1,1´-oxibis etano 10,71 ± 0,48 7,59 ± 0,62 11,24 ± 5,55 7,49 ± 0,72 ns ns ns * ns 550 13 2-metilpentano 9,90 ± 5,48 4,37 ± 1,04 3,11 ± 2,16 2,73 ± 0,71 ns ns ** ns * 559 14 3-metilpentano 7,10ab ± 0,19 2,80a ± 0,28 18,25b ± 22,01 1,82a ± 0,01 ns * *** *** * 600 17 hexano 34,20b ± 18,94 0,00a ± 0,00 0,00a ± 0,00 51,22b ± 0,00 *** *** *** *** *** 759 39 2,3,4-trimetilpentano 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 * *** *** ** *** 778 43 4-metilheptano 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 ns ns *** * ***
1000 78 decano 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 *** ns *** * *** 1200 97 dodecano 1,51abcd ± 0,10 1,60abcd ± 0,13 1,60abcd ± 0,90 3,57de ± 0,49 ns ns *** ns *** 1300 101 tridecano 3,21c ± 2,07 2,17abc ± 0,00 2,15abc ± 1,25 0,19ab ± 0,00 ns ns *** ns ***
ACIDOS
404,31bc ± 35,51 441,50cd ± 285,60 415,31bc ± 62,50 239,00abc ± 151,70 ns ns *** ns *** 583 19 acido acético 149,64abc ± 28,73 188,28bc ± 86,29 220,99c ± 0,00 130,64abc ± 85,17 *** * ns * ** 685 29 ácido propanoico 0,00a ± 0,00 0,00a ± 0,00 5,12d ± 0,00 2,70abcd ± 2,14 * ns *** ns *** 759 38 acido 2-metilpropanoico 49,62b ± 0,10 51,63b ± 36,57 30,27ab ± 17,29 22,50ab ± 19,30 ns ns *** ns *** 784 45 ácido butanoico 12,05ab ± 0,17 7,38a ± 0,49 29,86b ± 25,75 5,10a ± 3,27 * ns *** ns ** 837 50 ácido 3-metilbutanoico 150,53b ± 8,02 144,82b ± 144,93 56,62ab ± 25,29 61,26ab ± 58,63 ns ns *** * *** 846 51 ácido 2-metilpentanoico 30,64abc ± 6,01 38,90bc ± 29,19 61,03c ± 45,10 7,42ab ± 5,85 ns ns *** ns ***
1165 94 ácido octanoico 11,29d ± 0,57 8,59bcd ± 10,90 9,29cd ± 1,17 9,38d ± 0,00 ns ns *** *** *** 1256 100 ácido nonanoico 0,54abcd ± 0,00 1,90de ± 0,00 2,12e ± 0,82 0,00a ± 0,00 ns ns *** ns ***
ESTERES
227,06bc ± 30,03 147,27ab ± 24,77 373,33c ± 207,99 160,08ab ± 53,81 ns ns *** *** *** 524 9 acetato de metilo 11,74d ± 0,65 6,95c ± 2,37 6,52c ± 3,60 12,66d ± 2,17 ns ns *** ** *** 612 20 acetato de etilo 95,65b ± 21,02 86,27ab ± 17,65 187,80c ± 95,40 61,77ab ± 28,39 *** * *** ns *** 722 32 propanoato de etilo 9,29e ± 0,84 7,51de ± 0,80 5,49cd ± 3,58 1,88ab ± 1,14 ns ns *** ns *** 763 40 2-metilpropanoato de etilo 14,23ab ± 2,78 18,05ab ± 0,00 37,87b ± 38,30 17,43ab ± 0,00 ns ** *** ** ** 800 48 butanoato de etilo 12,59cd ± 0,15 7,18bc ± 0,44 14,50d ± 9,53 6,68abc ± 1,80 ns *** *** * *** 815 49 2-hidroxipropanoato de etilo 10,55c ± 0,75 3,26a ± 1,06 9,03bc ± 0,46 4,95ab ± 0,08 ns ** *** ns *** 852 52 2-metilbutanoato de etilo 23,80c ± 0,00 0,00a ± 0,00 30,89c ± 11,57 18,01bc ± 15,02 ns * *** ns *** 854 53 3-metilbutanoato de etilo 36,82ab ± 8,95 8,72a ± 0,00 68,32b ± 50,28 23,30a ± 15,82 ns ns *** ns *** 926 67 hexanoato de metilo 3,34cd ± 2,36 2,00abc ± 1,19 2,77bcd ± 0,37 4,91d ± 1,28 ns * *** * *** 998 77 hexanoato de etilo 6,20c ± 1,27 2,56ab ± 0,92 5,45bc ± 2,94 3,59abc ± 0,26 ns ns *** ns ***
1196 96 octanoato de etilo 1,64abcd ± 0,48 2,96cd ± 1,63 2,68abcd ± 1,24 2,83bcd ± 0,56 ns ns *** ns *** 1395 102 decanoato de etilo 1,22abc ± 0,03 1,81abc ± 1,97 2,01bc ± 0,43 2,07c ± 0,11 ns ns *** ns **
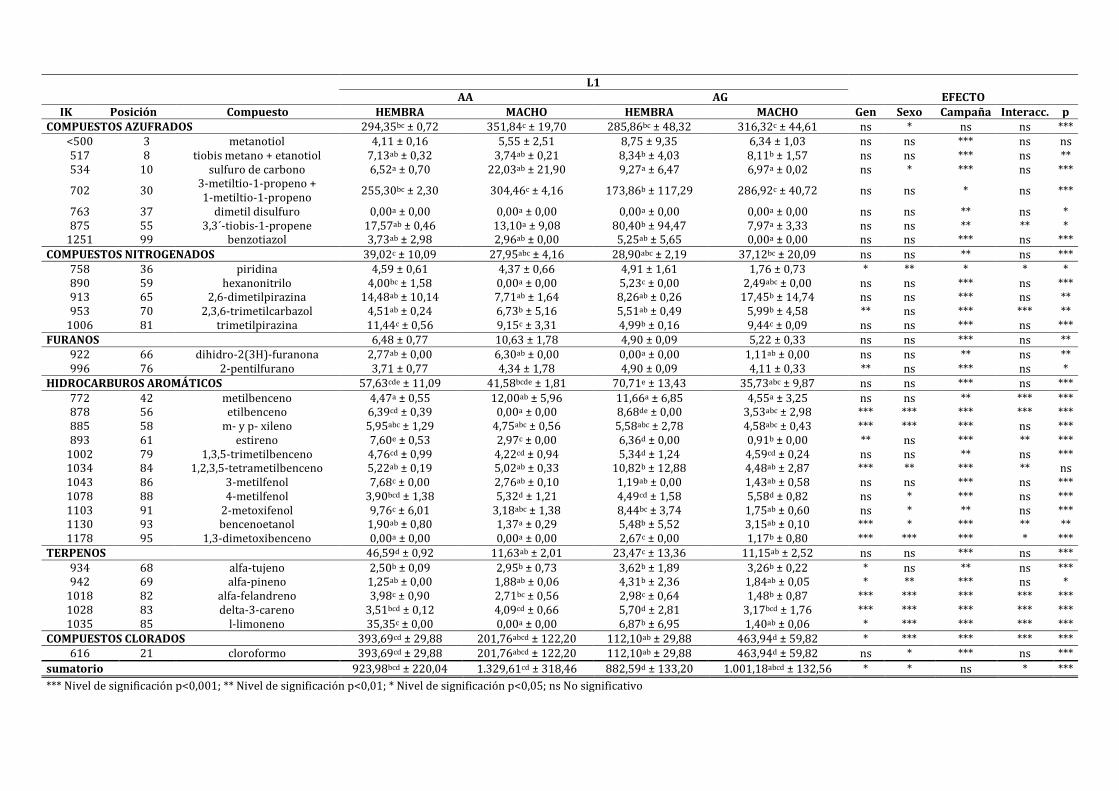
L1
AA AG EFECTO IK Posición Compuesto HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p
COMPUESTOS AZUFRADOS 294,35bc ± 0,72 351,84c ± 19,70 285,86bc ± 48,32 316,32c ± 44,61 ns * ns ns *** <500 3 metanotiol 4,11 ± 0,16 5,55 ± 2,51 8,75 ± 9,35 6,34 ± 1,03 ns ns *** ns ns 517 8 tiobis metano + etanotiol 7,13ab ± 0,32 3,74ab ± 0,21 8,34b ± 4,03 8,11b ± 1,57 ns ns *** ns ** 534 10 sulfuro de carbono 6,52a ± 0,70 22,03ab ± 21,90 9,27a ± 6,47 6,97a ± 0,02 ns * *** ns ***
702 30 3-metiltio-1-propeno + 1-metiltio-1-propeno
255,30bc ± 2,30 304,46c ± 4,16 173,86b ± 117,29 286,92c ± 40,72 ns ns * ns ***
763 37 dimetil disulfuro 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 ns ns ** ns * 875 55 3,3´-tiobis-1-propene 17,57ab ± 0,46 13,10a ± 9,08 80,40b ± 94,47 7,97a ± 3,33 ns ns ** ** *
1251 99 benzotiazol 3,73ab ± 2,98 2,96ab ± 0,00 5,25ab ± 5,65 0,00a ± 0,00 ns ns *** ns *** COMPUESTOS NITROGENADOS 39,02c ± 10,09 27,95abc ± 4,16 28,90abc ± 2,19 37,12bc ± 20,09 ns ns ** ns ***
758 36 piridina 4,59 ± 0,61 4,37 ± 0,66 4,91 ± 1,61 1,76 ± 0,73 * ** * * * 890 59 hexanonitrilo 4,00bc ± 1,58 0,00a ± 0,00 5,23c ± 0,00 2,49abc ± 0,00 ns ns *** ns *** 913 65 2,6-dimetilpirazina 14,48ab ± 10,14 7,71ab ± 1,64 8,26ab ± 0,26 17,45b ± 14,74 ns ns *** ns ** 953 70 2,3,6-trimetilcarbazol 4,51ab ± 0,24 6,73b ± 5,16 5,51ab ± 0,49 5,99b ± 4,58 ** ns *** *** **
1006 81 trimetilpirazina 11,44c ± 0,56 9,15c ± 3,31 4,99b ± 0,16 9,44c ± 0,09 ns ns *** ns *** FURANOS
6,48 ± 0,77 10,63 ± 1,78 4,90 ± 0,09 5,22 ± 0,33 ns ns *** ns **
922 66 dihidro-2(3H)-furanona 2,77ab ± 0,00 6,30ab ± 0,00 0,00a ± 0,00 1,11ab ± 0,00 ns ns ** ns ** 996 76 2-pentilfurano 3,71 ± 0,77 4,34 ± 1,78 4,90 ± 0,09 4,11 ± 0,33 ** ns *** ns *
HIDROCARBUROS AROMÁTICOS 57,63cde ± 11,09 41,58bcde ± 1,81 70,71e ± 13,43 35,73abc ± 9,87 ns ns *** ns *** 772 42 metilbenceno 4,47a ± 0,55 12,00ab ± 5,96 11,66a ± 6,85 4,55a ± 3,25 ns ns ** *** *** 878 56 etilbenceno 6,39cd ± 0,39 0,00a ± 0,00 8,68de ± 0,00 3,53abc ± 2,98 *** *** *** *** *** 885 58 m- y p- xileno 5,95abc ± 1,29 4,75abc ± 0,56 5,58abc ± 2,78 4,58abc ± 0,43 *** *** *** ns *** 893 61 estireno 7,60e ± 0,53 2,97c ± 0,00 6,36d ± 0,00 0,91b ± 0,00 ** ns *** ** ***
1002 79 1,3,5-trimetilbenceno 4,76cd ± 0,99 4,22cd ± 0,94 5,34d ± 1,24 4,59cd ± 0,24 ns ns ** ns *** 1034 84 1,2,3,5-tetrametilbenceno 5,22ab ± 0,19 5,02ab ± 0,33 10,82b ± 12,88 4,48ab ± 2,87 *** ** *** ** ns 1043 86 3-metilfenol 7,68c ± 0,00 2,76ab ± 0,10 1,19ab ± 0,00 1,43ab ± 0,58 ns ns *** ns *** 1078 88 4-metilfenol 3,90bcd ± 1,38 5,32d ± 1,21 4,49cd ± 1,58 5,58d ± 0,82 ns * *** ns *** 1103 91 2-metoxifenol 9,76c ± 6,01 3,18abc ± 1,38 8,44bc ± 3,74 1,75ab ± 0,60 ns * ** ns *** 1130 93 bencenoetanol 1,90ab ± 0,80 1,37a ± 0,29 5,48b ± 5,52 3,15ab ± 0,10 *** * *** ** ** 1178 95 1,3-dimetoxibenceno 0,00a ± 0,00 0,00a ± 0,00 2,67c ± 0,00 1,17b ± 0,80 *** *** *** * ***
TERPENOS
46,59d ± 0,92 11,63ab ± 2,01 23,47c ± 13,36 11,15ab ± 2,52 ns ns *** ns *** 934 68 alfa-tujeno 2,50b ± 0,09 2,95b ± 0,73 3,62b ± 1,89 3,26b ± 0,22 * ns ** ns *** 942 69 alfa-pineno 1,25ab ± 0,00 1,88ab ± 0,06 4,31b ± 2,36 1,84ab ± 0,05 * ** *** ns *
1018 82 alfa-felandreno 3,98c ± 0,90 2,71bc ± 0,56 2,98c ± 0,64 1,48b ± 0,87 *** *** *** *** *** 1028 83 delta-3-careno 3,51bcd ± 0,12 4,09cd ± 0,66 5,70d ± 2,81 3,17bcd ± 1,76 *** *** *** *** *** 1035 85 l-limoneno 35,35c ± 0,00 0,00a ± 0,00 6,87b ± 6,95 1,40ab ± 0,06 * *** *** *** ***
COMPUESTOS CLORADOS 393,69cd ± 29,88 201,76abcd ± 122,20 112,10ab ± 29,88 463,94d ± 59,82 * *** *** *** *** 616 21 cloroformo 393,69cd ± 29,88 201,76abcd ± 122,20 112,10ab ± 29,88 463,94d ± 59,82 ns * *** ns ***
sumatorio 923,98bcd ± 220,04 1.329,61cd ± 318,46 882,59d ± 133,20 1.001,18abcd ± 132,56 * * ns * ***
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

Tabla 4.3.45. Compuestos volátiles identificados en lomo curado agrupados por familia química. Los contenidos de cada compuesto se expresan en UAA (106) (± desviación estándar de la media). Parte II.
L2
AA
AG
EFECTO
IK Posición Compuesto HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p ALDEHIDOS 121,57ab ± 25,77 88,01a ± 4,60 132,55ab ± 31,84 122,96ab ± 11,42 ** * *** ns *
<500 2 acetaldehído 17,80bc ± 2,42 13,39abc ± 2,06 24,18c ± 5,77 21,38c ± 6,16 *** *** *** *** *** 548 12 2-metilpropanal 7,35f ± 0,01 4,08cd ± 1,16 3,82bc ± 0,69 6,02e ± 0,00 * ns ns ns *** 650 23 3-metilbutanal 25,34ab ± 18,26 7,07a ± 2,59 21,72ab ± 14,41 15,41ab ± 0,44 ns ns ns ns ns 661 25 2-metilbutanal 17,86ab ± 10,58 10,18a ± 0,98 16,58ab ± 9,80 11,37a ± 7,45 ns ns * * * 799 47 hexanal 22,34 ± 11,44 15,29 ± 5,62 16,76 ± 6,28 22,40 ± 7,12 ns ns *** ns * 900 63 heptanal 2,99abc ± 0,68 5,43abcd ± 0,71 7,14abcd ± 1,50 6,76abcd ± 0,75 ** ns *** ns *** 969 72 benzaldehído 9,43b ± 0,05 7,20ab ± 0,20 9,35b ± 0,84 9,49b ± 2,80 * ns *** ns ***
1005 80 octanal 9,05bcd ± 5,01 12,14d ± 0,31 9,99cd ± 0,67 4,18abc ± 3,88 ns * *** ns *** 1054 87 bencenoacetaldehido 1,70a ± 1,07 2,67a ± 0,01 3,31a ± 1,15 1,49a ± 1,18 ns ns ** ns *** 1107 92 nonanal 6,79ab ± 0,24 9,84ab ± 1,71 17,72ab ± 5,03 22,04b ± 4,78 * ns *** ns *** 1211 98 decanal 0,91ab ± 0,04 0,72ab ± 0,01 1,99ab ± 0,92 2,42b ± 0,05 ns ns *** * **
CETONAS 373,52abc ± 52,30 788,87c ± 38,31 381,12abc ± 168,77 324,71abc ± 84,02 * * *** ** ** <500 5 2-propanona+propanal 209,27a ± 39,41 577,40b ± 25,93 217,49a ± 133,00 188,02a ± 57,74 ns ns *** ns *** 585 15 2,3-butanodiona 20,94 ± 2,39 14,51 ± 3,35 11,05 ± 2,94 24,03 ± 22,88 ns ns * * ** 597 16 2-butanona+butanal 44,92 ± 16,34 41,53 ± 1,45 46,88 ± 17,63 46,81 ± 8,89 ns * *** ns ns 658 24 1-hidroxi-2-propanona 3,27ab ± 1,90 3,24ab ± 1,48 3,60ab ± 2,12 4,87b ± 0,00 ns ns *** ns *** 688 28 2-pentanona 15,26cd ± 8,97 8,37abcd ± 1,13 12,43bcd ± 1,83 19,36d ± 0,00 *** * *** *** *** 716 31 3-hidroxi-2-butanona 64,71abc ± 1,15 123,38c ± 15,39 61,77abc ± 32,46 22,01ab ± 21,45 ns ns ** ns *** 790 46 2-hexanona 6,32 ± 1,75 5,24 ± 0,17 8,63 ± 7,99 4,13 ± 3,86 * * ** ns * 890 60 2-heptanona 3,90a ± 0,87 6,04a ± 2,43 4,89a ± 2,09 3,68a ± 3,09 ns ns *** * ** 963 71 4-metilciclohexanona 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 ns * *** * *** 988 74 3-octanona 0,69abcd ± 0,06 1,61abcd ± 0,05 1,83abcd ± 0,79 1,71abcd ± 1,43 ns ns *** ** *** 992 75 2-octanona 1,81ab ± 0,59 3,04b ± 1,40 3,34b ± 0,99 1,68ab ± 1,43 * ns *** ns ***
1084 89 8-nonen-2-ona 0,99ab ± 0,38 2,37bc ± 0,00 3,34c ± 1,00 2,69c ± 0,00 ** ns ** ns *** 1094 90 2-nonanona 1,43 ± 0,15 2,15 ± 0,77 5,88 ± 3,82 5,72 ± 0,55 ns *** *** *** **
ALCOHOLES 357,24a ± 74,18 1.024,96e ± 138,61 766,40cd ± 68,01 920,69de ± 90,84 ns *** *** *** *** <500 4 etanol 222,11ab ± 4,35 895,05e ± 130,94 605,69d ± 102,08 824,45e ± 96,93 * *** ** ns *** 548 11 propanol 6,84a ± 0,37 5,93a ± 0,33 8,83a ± 4,03 4,25a ± 0,53 *** ns *** *** *** 602 18 2-butanol 2,36ab ± 0,10 2,61ab ± 0,55 3,01abc ± 0,86 0,00a ± 0,00 ns ns ** ns *** 621 22 2-metil-1-propanol 9,32a ± 3,88 26,21ab ± 0,67 17,15ab ± 9,54 8,56a ± 3,22 ns ns ns ns ** 669 26 1-metoxi-2-propanol 3,18ab ± 0,34 2,39ab ± 0,99 3,82ab ± 1,34 5,68ab ± 2,77 ns ns ns ns * 683 27 1-penten-3-ol 2,72 ± 1,25 2,01 ± 0,21 3,27 ± 1,92 2,62 ± 2,14 *** ns *** *** ns 736 33 2-metil-3-buten-2-ol 4,96de ± 0,00 7,47e ± 0,00 3,51bcd ± 0,22 0,00a ± 0,00 ns ns ns ns *** 739 34 3-metilbutanol 43,84 ± 41,70 30,15 ± 3,55 51,58 ± 29,38 34,05 ± 5,08 ns * * ns ns 743 35 2-metilbutanol 24,05 ± 18,46 15,75 ± 0,30 25,32 ± 12,17 13,83 ± 9,21 ns * *** * * 771 41 pentanol 2,82bc ± 1,64 2,23abc ± 0,02 3,71c ± 0,95 0,22a ± 0,00 ns ** *** *** *** 790 44 2,3-butanodiol 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 ns ** ** ns ***

L2
AA AG EFECTO
IK Posición Compuesto HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p 869 57 hexanol 7,16abc ± 3,84 4,65abc ± 1,85 8,91bc ± 3,03 0,35a ± 0,00 ns * *** ** ** 897 62 2-heptanol 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 ns ns *** ns *** 905 64 2-butoxi etanol 21,68bc ± 4,14 23,27c ± 14,56 24,04bc ± 11,96 18,42abc ± 0,79 ns ns *** ns *** 980 73 1-octen-3-ol 6,21abc ± 3,04 7,24bc ± 0,03 7,55bc ± 2,51 8,27c ± 1,51 ns ns ns *** ***
HIDROCARBUROS ALIFÁTICOS 60,53ab ± 2,19 92,70ab ± 7,72 103,40b ± 40,61 50,10ab ± 5,27 ns ns *** ns * <500 1 propano 17,35b ± 2,20 15,33b ± 2,03 16,45b ± 5,40 19,82b ± 0,00 ** ns *** * *** 500 6 pentano 13,61abc ± 1,83 22,45abc ± 0,39 19,72abc ± 13,07 5,12a ± 0,00 ns ns ns ns ** 505 7 1,1´-oxibis etano 11,99 ± 0,36 20,27 ± 10,44 22,20 ± 17,84 10,98 ± 1,13 ns ns ns * ns 550 13 2-metilpentano 2,92 ± 0,03 3,97 ± 0,24 7,74 ± 4,31 1,53 ± 1,09 ns ns ** ns * 559 14 3-metilpentano 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 ns * *** *** * 600 17 hexano 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 *** *** *** *** *** 759 39 2,3,4-trimetilpentano 3,52ab ± 1,87 21,03c ± 4,22 6,62b ± 2,66 3,46ab ± 3,01 * *** *** ** *** 778 43 4-metilheptano 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 ns ns *** * ***
1000 78 decano 8,49ab ± 3,58 9,65ab ± 1,82 26,06b ± 14,32 2,16a ± 1,86 *** ns *** * *** 1200 97 dodecano 2,65bcde ± 0,00 0,00a ± 0,00 3,10cde ± 0,00 4,59e ± 2,36 ns ns *** ns *** 1300 101 tridecano 0,00a ± 0,00 0,00a ± 0,00 1,52abc ± 0,00 2,45bc ± 0,93 ns ns *** ns ***
ACIDOS 165,51ab ± 8,14 174,42abc ± 38,22 179,30abc ± 65,08 88,92a ± 40,59 ns ns *** ns *** 583 19 acido acético 128,65abc ± 0,46 136,42abc ± 37,63 130,36abc ± 60,77 47,28ab ± 46,62 *** * ns * ** 685 29 ácido propanoico 2,93abcd ± 0,06 1,83abc ± 0,07 2,68abcd ± 1,34 3,28bcd ± 0,95 * ns *** ns *** 759 38 acido 2-metilpropanoico 8,04a ± 4,16 2,48a ± 0,07 11,16a ± 4,19 0,00a ± 0,00 ns ns *** ns *** 784 45 ácido butanoico 11,26ab ± 3,92 16,07ab ± 0,65 13,37ab ± 5,36 18,42ab ± 10,28 * ns *** ns ** 837 50 ácido 3-metilbutanoico 7,12a ± 4,12 9,38a ± 0,88 10,15a ± 8,32 7,90a ± 4,26 ns ns *** * *** 846 51 ácido 2-metilpentanoico 4,64ab ± 2,09 6,00ab ± 0,16 8,16ab ± 0,16 6,70ab ± 2,40 ns ns *** ns ***
1165 94 ácido octanoico 1,82abc ± 0,49 1,15ab ± 0,00 1,56ab ± 0,09 3,80abcd ± 0,00 ns ns *** *** *** 1256 100 ácido nonanoico 1,04abcde ± 0,33 1,10abcde ± 0,50 1,86cde ± 0,20 1,54bcde ± 1,24 ns ns *** ns ***
ESTERES 33,18a ± 8,57 121,02ab ± 3,36 52,33a ± 12,43 40,18a ± 5,06 ns ns *** *** *** 524 9 acetato de metilo 2,47ab ± 0,10 4,41abc ± 0,39 5,43bc ± 0,72 4,66abc ± 1,06 ns ns *** ** *** 612 20 acetato de etilo 10,91a ± 6,92 87,81b ± 1,51 19,88ab ± 10,51 11,16a ± 1,51 *** * *** ns *** 722 32 propanoato de etilo 3,41abc ± 0,77 4,21bc ± 1,05 3,26abc ± 0,47 3,62bc ± 0,00 ns ns *** ns *** 763 40 2-metilpropanoato de etilo 4,78a ± 1,73 3,24a ± 0,34 4,65a ± 1,14 8,65a ± 2,55 ns ** *** ** ** 800 48 butanoato de etilo 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 ns *** *** * *** 815 49 2-hidroxipropanoato de etilo 2,99a ± 0,50 4,51ab ± 1,37 4,89ab ± 3,28 2,70a ± 0,58 ns ** *** ns *** 852 52 2-metilbutanoato de etilo 1,91ab ± 0,38 5,14ab ± 0,64 3,14ab ± 0,96 1,61ab ± 0,79 ns * *** ns *** 854 53 3-metilbutanoato de etilo 2,85a ± 0,10 5,78a ± 1,74 3,32a ± 0,37 2,18a ± 0,77 ns ns *** ns *** 926 67 hexanoato de metilo 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 ns * *** * *** 998 77 hexanoato de etilo 1,44a ± 0,44 3,34abc ± 0,03 2,98abc ± 0,70 2,51ab ± 1,96 ns ns *** ns ***
1196 96 octanoato de etilo 1,40abcd ± 0,57 1,69abcd ± 0,25 3,40d ± 0,45 1,56abcd ± 1,07 ns ns *** ns *** 1395 102 decanoato de etilo 1,03abc ± 0,01 0,88abc ± 0,00 1,39abc ± 0,10 1,53abc ± 1,05 ns ns *** ns **
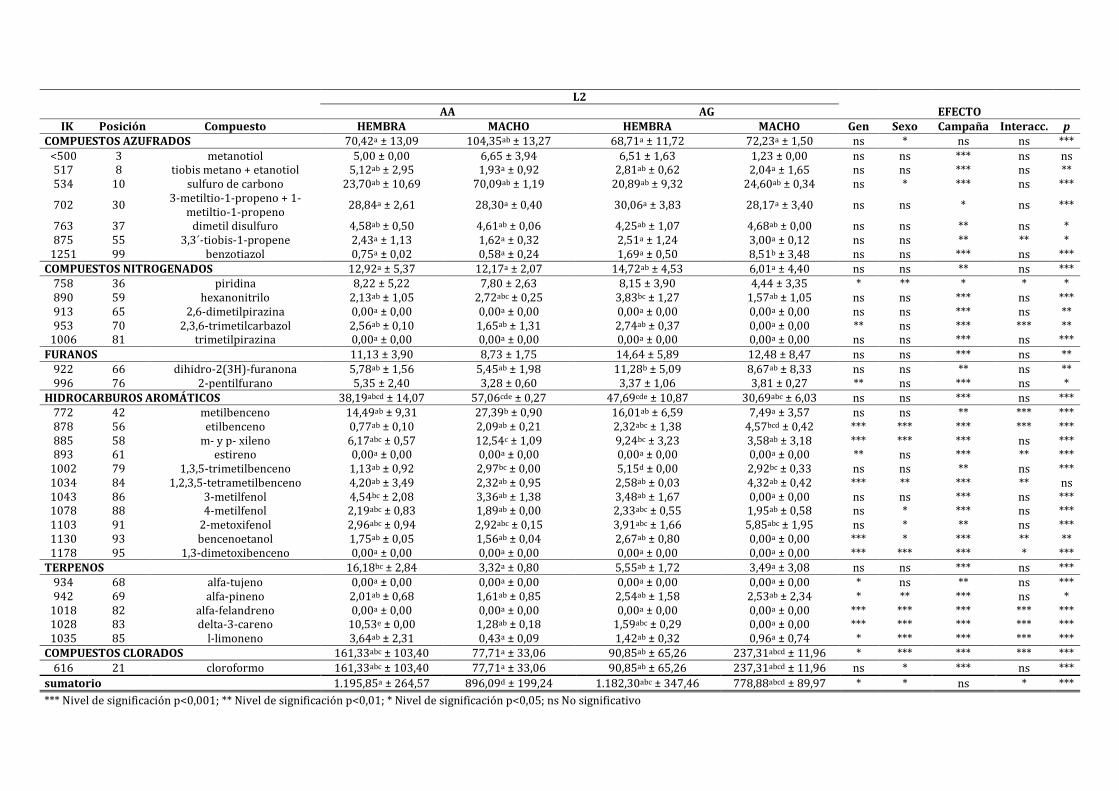
L2
AA AG EFECTO
IK Posición Compuesto HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p COMPUESTOS AZUFRADOS 70,42a ± 13,09 104,35ab ± 13,27 68,71a ± 11,72 72,23a ± 1,50 ns * ns ns ***
<500 3 metanotiol 5,00 ± 0,00 6,65 ± 3,94 6,51 ± 1,63 1,23 ± 0,00 ns ns *** ns ns 517 8 tiobis metano + etanotiol 5,12ab ± 2,95 1,93a ± 0,92 2,81ab ± 0,62 2,04a ± 1,65 ns ns *** ns ** 534 10 sulfuro de carbono 23,70ab ± 10,69 70,09ab ± 1,19 20,89ab ± 9,32 24,60ab ± 0,34 ns * *** ns ***
702 30 3-metiltio-1-propeno + 1-
metiltio-1-propeno 28,84a ± 2,61 28,30a ± 0,40 30,06a ± 3,83 28,17a ± 3,40 ns ns * ns ***
763 37 dimetil disulfuro 4,58ab ± 0,50 4,61ab ± 0,06 4,25ab ± 1,07 4,68ab ± 0,00 ns ns ** ns * 875 55 3,3´-tiobis-1-propene 2,43a ± 1,13 1,62a ± 0,32 2,51a ± 1,24 3,00a ± 0,12 ns ns ** ** *
1251 99 benzotiazol 0,75a ± 0,02 0,58a ± 0,24 1,69a ± 0,50 8,51b ± 3,48 ns ns *** ns *** COMPUESTOS NITROGENADOS 12,92a ± 5,37 12,17a ± 2,07 14,72ab ± 4,53 6,01a ± 4,40 ns ns ** ns ***
758 36 piridina 8,22 ± 5,22 7,80 ± 2,63 8,15 ± 3,90 4,44 ± 3,35 * ** * * * 890 59 hexanonitrilo 2,13ab ± 1,05 2,72abc ± 0,25 3,83bc ± 1,27 1,57ab ± 1,05 ns ns *** ns *** 913 65 2,6-dimetilpirazina 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 ns ns *** ns ** 953 70 2,3,6-trimetilcarbazol 2,56ab ± 0,10 1,65ab ± 1,31 2,74ab ± 0,37 0,00a ± 0,00 ** ns *** *** **
1006 81 trimetilpirazina 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 ns ns *** ns *** FURANOS 11,13 ± 3,90 8,73 ± 1,75 14,64 ± 5,89 12,48 ± 8,47 ns ns *** ns **
922 66 dihidro-2(3H)-furanona 5,78ab ± 1,56 5,45ab ± 1,98 11,28b ± 5,09 8,67ab ± 8,33 ns ns ** ns ** 996 76 2-pentilfurano 5,35 ± 2,40 3,28 ± 0,60 3,37 ± 1,06 3,81 ± 0,27 ** ns *** ns *
HIDROCARBUROS AROMÁTICOS 38,19abcd ± 14,07 57,06cde ± 0,27 47,69cde ± 10,87 30,69abc ± 6,03 ns ns *** ns *** 772 42 metilbenceno 14,49ab ± 9,31 27,39b ± 0,90 16,01ab ± 6,59 7,49a ± 3,57 ns ns ** *** *** 878 56 etilbenceno 0,77ab ± 0,10 2,09ab ± 0,21 2,32abc ± 1,38 4,57bcd ± 0,42 *** *** *** *** *** 885 58 m- y p- xileno 6,17abc ± 0,57 12,54c ± 1,09 9,24bc ± 3,23 3,58ab ± 3,18 *** *** *** ns *** 893 61 estireno 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 ** ns *** ** ***
1002 79 1,3,5-trimetilbenceno 1,13ab ± 0,92 2,97bc ± 0,00 5,15d ± 0,00 2,92bc ± 0,33 ns ns ** ns *** 1034 84 1,2,3,5-tetrametilbenceno 4,20ab ± 3,49 2,32ab ± 0,95 2,58ab ± 0,03 4,32ab ± 0,42 *** ** *** ** ns 1043 86 3-metilfenol 4,54bc ± 2,08 3,36ab ± 1,38 3,48ab ± 1,67 0,00a ± 0,00 ns ns *** ns *** 1078 88 4-metilfenol 2,19abc ± 0,83 1,89ab ± 0,00 2,33abc ± 0,55 1,95ab ± 0,58 ns * *** ns *** 1103 91 2-metoxifenol 2,96abc ± 0,94 2,92abc ± 0,15 3,91abc ± 1,66 5,85abc ± 1,95 ns * ** ns *** 1130 93 bencenoetanol 1,75ab ± 0,05 1,56ab ± 0,04 2,67ab ± 0,80 0,00a ± 0,00 *** * *** ** ** 1178 95 1,3-dimetoxibenceno 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 *** *** *** * ***
TERPENOS 16,18bc ± 2,84 3,32a ± 0,80 5,55ab ± 1,72 3,49a ± 3,08 ns ns *** ns *** 934 68 alfa-tujeno 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 * ns ** ns *** 942 69 alfa-pineno 2,01ab ± 0,68 1,61ab ± 0,85 2,54ab ± 1,58 2,53ab ± 2,34 * ** *** ns *
1018 82 alfa-felandreno 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 *** *** *** *** *** 1028 83 delta-3-careno 10,53e ± 0,00 1,28ab ± 0,18 1,59abc ± 0,29 0,00a ± 0,00 *** *** *** *** *** 1035 85 l-limoneno 3,64ab ± 2,31 0,43a ± 0,09 1,42ab ± 0,32 0,96a ± 0,74 * *** *** *** ***
COMPUESTOS CLORADOS 161,33abc ± 103,40 77,71a ± 33,06 90,85ab ± 65,26 237,31abcd ± 11,96 * *** *** *** *** 616 21 cloroformo 161,33abc ± 103,40 77,71a ± 33,06 90,85ab ± 65,26 237,31abcd ± 11,96 ns * *** ns ***
sumatorio 1.195,85a ± 264,57 896,09d ± 199,24 1.182,30abc ± 347,46 778,88abcd ± 89,97 * * ns * ***
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo

Tabla 4.3.46. Compuestos volátiles identificados en lomo curado agrupados por familia química. Los contenidos de cada compuesto se expresan UAA (106) (± desviación estándar de la media). Parte III.
L3
AA
AG
EFECTO
IK Posición Compuesto HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p ALDEHIDOS 63,07a ± 3,32 71,96a ± 2,02 216,14b ± 132,75 70,72a ± 6,84 ** * *** ns *
<500 2 acetaldehído 6,56ab ± 0,07 3,40a ± 0,04 7,04ab ± 1,51 4,41a ± 0,21 *** *** *** *** *** 548 12 2-metilpropanal 10,80h ± 0,00 4,95de ± 0,11 22,45i ± 0,00 9,04g ± 0,00 * ns ns ns *** 650 23 3-metilbutanal 17,15ab ± 4,12 13,18ab ± 0,44 100,72b ± 92,41 17,66ab ± 0,67 ns ns ns ns ns 661 25 2-metilbutanal 10,33a ± 1,73 9,30a ± 0,01 52,80b ± 47,49 11,91a ± 0,36 ns ns * * * 799 47 hexanal 6,14 ± 1,13 13,32 ± 0,79 17,11 ± 10,29 4,88 ± 0,95 ns ns *** ns * 900 63 heptanal 1,93a ± 0,37 3,01abc ± 0,05 2,54ab ± 0,07 2,06a ± 0,33 ** ns *** ns *** 969 72 benzaldehído 3,13a ± 1,34 3,77ab ± 0,03 5,07ab ± 4,36 3,57ab ± 0,51 * ns *** ns ***
1005 80 octanal 1,25a ± 0,18 2,18ab ± 0,03 1,41a ± 0,07 1,68a ± 0,27 ns * *** ns *** 1054 87 bencenoacetaldehido 2,12a ± 0,00 12,71b ± 1,35 3,33a ± 0,00 12,24b ± 5,40 ns ns ** ns *** 1107 92 nonanal 3,51a ± 0,42 5,88a ± 0,56 3,69a ± 0,44 2,97a ± 0,29 * ns *** ns *** 1211 98 decanal 0,16ab ± 0,00 0,27ab ± 0,13 0,00a ± 0,00 0,29ab ± 0,10 ns ns *** * **
CETONAS 182,76ab ± 91,77 88,65a ± 4,77 156,51ab ± 94,44 110,85ab ± 12,71 * * *** ** ** <500 5 2-propanona+propanal 131,95a ± 101,39 71,12a ± 2,69 119,44a ± 88,81 68,98a ± 13,68 ns ns *** ns *** 585 15 2,3-butanodiona 2,86 ± 0,00 1,55 ± 0,00 1,75 ± 0,00 2,29 ± 0,25 ns ns * * ** 597 16 2-butanona+butanal 12,65 ± 0,00 5,46 ± 2,18 6,97 ± 0,00 8,79 ± 0,70 ns * *** ns ns 658 24 1-hidroxi-2-propanona 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 ns ns *** ns *** 688 28 2-pentanona 2,65ab ± 2,54 0,96a ± 0,14 4,16abc ± 1,99 2,23ab ± 0,52 *** * *** *** *** 716 31 3-hidroxi-2-butanona 26,51ab ± 13,68 5,22a ± 1,64 12,42ab ± 5,77 19,33ab ± 0,71 ns ns ** ns *** 790 46 2-hexanona 0,75 ± 0,00 0,00 ± 0,00 1,84 ± 1,63 3,34 ± 0,41 * * ** ns * 890 60 2-heptanona 2,52a ± 1,43 1,14a ± 0,41 5,80a ± 0,79 2,15a ± 0,17 ns ns *** * ** 963 71 4-metilciclohexanona 0,23ab ± 0,07 0,60abc ± 0,40 0,44abc ± 0,39 0,42abc ± 0,02 ns * *** * *** 988 74 3-octanona 0,48ab ± 0,28 0,39a ± 0,05 0,59abc ± 0,21 0,79abcd ± 0,04 ns ns *** ** *** 992 75 2-octanona 0,76a ± 0,11 0,61a ± 0,03 0,82a ± 0,16 0,76a ± 0,08 * ns *** ns ***
1084 89 8-nonen-2-ona 0,36a ± 0,00 1,05ab ± 0,00 0,71a ± 0,63 0,51a ± 0,03 ** ns ** ns *** 1094 90 2-nonanona 1,04 ± 0,24 0,57 ± 0,27 1,57 ± 0,11 1,26 ± 0,29 ns *** *** *** **
ALCOHOLES 585,25bc ± 18,57 441,08ab ± 64,90 365,79a ± 46,71 372,03a ± 12,14 ns *** *** *** *** <500 4 etanol 446,76cd ± 24,58 344,50bc ± 41,09 231,44ab ± 46,73 302,34abc ± 9,14 * *** ** ns *** 548 11 propanol 11,40a ± 0,00 6,72a ± 0,73 51,52b ± 0,00 10,79a ± 4,70 *** ns *** *** *** 602 18 2-butanol 7,70cd ± 0,00 5,67bc ± 4,37 4,89bc ± 0,00 11,80d ± 1,96 ns ns ** ns *** 621 22 2-metil-1-propanol 43,47ab ± 0,69 50,04b ± 33,62 14,92ab ± 2,00 14,52a ± 0,89 ns ns ns ns ** 669 26 1-metoxi-2-propanol 2,45ab ± 1,28 2,89ab ± 0,00 0,96a ± 0,00 1,07a ± 0,05 ns ns ns ns * 683 27 1-penten-3-ol 1,15 ± 0,20 0,85 ± 0,13 1,65 ± 0,83 1,92 ± 0,03 *** ns *** *** ns 736 33 2-metil-3-buten-2-ol 1,56abc ± 1,01 0,80ab ± 0,02 1,03ab ± 0,42 1,38abc ± 0,02 ns ns ns ns *** 739 34 3-metilbutanol 43,93 ± 8,91 11,34 ± 1,78 34,73 ± 3,02 15,61 ± 0,01 ns * * ns ns 743 35 2-metilbutanol 20,65 ± 3,84 2,71 ± 0,11 15,97 ± 2,16 3,47 ± 0,10 ns * *** * * 771 41 pentanol 0,73ab ± 0,00 0,42a ± 0,09 0,64ab ± 0,41 0,55ab ± 0,01 ns ** *** *** *** 790 44 2,3-butanodiol 0,56a ± 0,00 2,82a ± 1,09 2,82a ± 0,00 2,18a ± 0,34 ns ** ** ns ***

L3
AA AG EFECTO
IK Posición Compuesto HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p 790 44 2,3-butanodiol 0,56a ± 0,00 2,82a ± 1,09 2,82a ± 0,00 2,18a ± 0,34 ns ** ** ns *** 869 57 hexanol 1,26a ± 0,45 2,72abc ± 0,25 1,78ab ± 1,08 1,52ab ± 0,06 ns * *** ** ** 897 62 2-heptanol 1,16ab ± 0,55 7,91cd ± 0,85 1,55ab ± 0,34 3,39abc ± 0,30 ns ns *** ns *** 905 64 2-butoxi etanol 0,39a ± 0,00 0,69a ± 0,11 0,57a ± 0,00 0,52a ± 0,04 ns ns *** ns *** 980 73 1-octen-3-ol 2,09ab ± 0,71 1,00a ± 0,37 1,31a ± 0,25 0,98a ± 0,16 ns ns ns *** ***
HIDROCARBUROS ALIFÁTICOS 65,06ab ± 15,94 59,46ab ± 2,65 47,72ab ± 11,54 69,60ab ± 9,21 ns ns *** ns * <500 1 propano 3,12a ± 0,00 3,63a ± 0,33 3,21a ± 0,83 2,64a ± 0,21 ** ns *** * *** 500 6 pentano 33,10c ± 11,69 28,71bc ± 0,88 22,51abc ± 5,12 22,88abc ± 0,74 ns ns ns ns ** 505 7 1,1´-oxibis etano 13,53 ± 6,89 11,54 ± 1,49 12,16 ± 4,61 17,17 ± 9,09 ns ns ns * ns 550 13 2-metilpentano 6,25 ± 3,82 5,43 ± 0,68 2,91 ± 1,18 8,04 ± 0,24 ns ns ** ns * 559 14 3-metilpentano 2,99a ± 1,11 1,12a ± 0,16 1,63a ± 0,97 6,95ab ± 0,58 ns * *** *** * 600 17 hexano 0,00a ± 0,00 6,00a ± 0,00 0,00a ± 0,00 6,64a ± 0,19 *** *** *** *** *** 759 39 2,3,4-trimetilpentano 0,94a ± 0,00 0,54a ± 0,00 1,14ab ± 0,00 1,18ab ± 0,00 * *** *** ** *** 778 43 4-metilheptano 2,29d ± 0,00 1,02b ± 0,33 1,63c ± 0,00 1,03b ± 0,24 ns ns *** * ***
1000 78 decano 1,92a ± 1,88 0,70a ± 0,02 1,48a ± 0,00 1,53a ± 0,15 *** ns *** * *** 1200 97 dodecano 0,71abc ± 0,30 0,61ab ± 0,05 0,61ab ± 0,50 1,12abc ± 0,08 ns ns *** ns *** 1300 101 tridecano 0,21ab ± 0,11 0,16ab ± 0,02 0,44ab ± 0,00 0,41ab ± 0,05 ns ns *** ns ***
ACIDOS 33,15a ± 0,54 37,02a ± 13,33 71,85a ± 59,87 89,25a ± 8,81 ns ns *** ns *** 583 19 acido acético 28,37a ± 0,00 31,67a ± 13,99 63,50ab ± 57,87 80,33abc ± 9,69 *** * ns * ** 685 29 ácido propanoico 0,92ab ± 0,00 0,58ab ± 0,21 4,23cd ± 0,00 1,70abc ± 0,96 * ns *** ns *** 759 38 acido 2-metilpropanoico 1,71a ± 0,21 1,12a ± 0,25 1,42a ± 0,30 1,42a ± 0,12 ns ns *** ns *** 784 45 ácido butanoico 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 * ns *** ns ** 837 50 ácido 3-metilbutanoico 0,85a ± 0,08 1,80a ± 0,36 1,22a ± 0,63 3,62a ± 0,09 ns ns *** * *** 846 51 ácido 2-metilpentanoico 0,94a ± 0,31 1,03a ± 0,29 1,15a ± 0,90 1,65ab ± 0,62 ns ns *** ns ***
1165 94 ácido octanoico 0,23a ± 0,06 0,37a ± 0,26 0,33a ± 0,18 0,37a ± 0,12 ns ns *** *** *** 1256 100 ácido nonanoico 0,13ab ± 0,00 0,44abc ± 0,00 0,00a ± 0,00 0,16ab ± 0,04 ns ns *** ns ***
ESTERES 53,86a ± 5,27 49,57a ± 4,46 20,99a ± 1,21 69,67a ± 4,86 ns ns *** *** *** 524 9 acetato de metilo 2,20ab ± 1,01 1,18a ± 0,07 1,06a ± 0,15 1,10a ± 0,02 ns ns *** ** *** 612 20 acetato de etilo 24,26ab ± 0,00 40,52ab ± 4,90 11,77a ± 1,53 53,50ab ± 4,36 *** * *** ns *** 722 32 propanoato de etilo 0,67ab ± 0,00 0,36a ± 0,08 0,67ab ± 0,42 0,49a ± 0,16 ns ns *** ns *** 763 40 2-metilpropanoato de etilo 4,62a ± 2,75 1,13a ± 0,03 1,15a ± 0,19 1,66a ± 0,20 ns ** *** ** ** 800 48 butanoato de etilo 7,39bc ± 0,14 0,00a ± 0,00 0,00a ± 0,00 4,90ab ± 0,50 ns *** *** * *** 815 49 2-hidroxipropanoato de etilo 4,88ab ± 0,44 1,23a ± 0,05 1,69a ± 0,16 1,67a ± 0,11 ns ** *** ns *** 852 52 2-metilbutanoato de etilo 2,21ab ± 0,69 1,04a ± 0,31 0,90a ± 0,14 1,64ab ± 0,02 ns * *** ns *** 854 53 3-metilbutanoato de etilo 3,04a ± 1,09 2,07a ± 0,43 1,34a ± 0,14 2,16a ± 0,08 ns ns *** ns *** 926 67 hexanoato de metilo 0,36ab ± 0,00 0,20ab ± 0,04 0,37ab ± 0,22 0,34ab ± 0,01 ns * *** * *** 998 77 hexanoato de etilo 2,45ab ± 0,60 0,99a ± 0,10 1,08a ± 0,47 1,03a ± 0,15 ns ns *** ns ***
1196 96 octanoato de etilo 1,18abc ± 0,85 0,62a ± 0,07 0,65a ± 0,12 0,75ab ± 0,03 ns ns *** ns *** 1395 102 decanoato de etilo 0,62abc ± 0,00 0,24a ± 0,01 0,31ab ± 0,16 0,42abc ± 0,01 ns ns *** ns **

L3
AA AG EFECTO
IK Posición Compuesto HEMBRA MACHO HEMBRA MACHO Gen Sexo Campaña Interacc. p COMPUESTOS AZUFRADOS 220,37abc ± 69,73 116,10ab ± 52,60 299,63bc ± 234,94 328,15c ± 58,39 ns * ns ns ***
<500 3 metanotiol 6,84 ± 2,69 4,69 ± 2,70 6,50 ± 4,31 2,61 ± 0,83 ns ns *** ns ns 517 8 tiobis metano + etanotiol 6,46ab ± 1,30 8,11b ± 2,09 7,71ab ± 3,03 7,05ab ± 0,01 ns ns *** ns ** 534 10 sulfuro de carbono 198,05bc ± 71,50 83,93ab ± 37,42 278,43c ± 239,61 312,94c ± 58,30 ns * *** ns ***
702 30 3-metiltio-1-propeno + 1-metiltio-1-propeno
6,55a ± 0,00 3,38a ± 1,13 4,18a ± 3,08 1,71a ± 0,30 ns ns * ns ***
763 37 dimetil disulfuro 1,54a ± 0,00 15,22b ± 14,02 1,52a ± 0,18 2,93ab ± 0,13 ns ns ** ns * 875 55 3,3´-tiobis-1-propene 0,73a ± 0,38 0,51a ± 0,14 1,06a ± 0,22 0,68a ± 0,26 ns ns ** ** *
1251 99 benzotiazol 0,20a ± 0,00 0,23a ± 0,15 0,24a ± 0,00 0,22a ± 0,01 ns ns *** ns *** COMPUESTOS NITROGENADOS 5,16a ± 0,26 5,55a ± 1,27 5,66a ± 1,32 6,09a ± 1,49 ns ns ** ns ***
758 36 piridina 2,95 ± 0,31 1,27 ± 0,41 3,19 ± 0,54 1,56 ± 0,16 * ** * * * 890 59 hexanonitrilo 1,00ab ± 0,30 2,08ab ± 0,12 1,01ab ± 0,74 2,24abc ± 1,13 ns ns *** ns *** 913 65 2,6-dimetilpirazina 0,50ab ± 0,21 1,12ab ± 0,39 0,60ab ± 0,02 0,88ab ± 0,29 ns ns *** ns ** 953 70 2,3,6-trimetilcarbazol 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 ** ns *** *** **
1006 81 trimetilpirazina 0,72a ± 0,05 1,08a ± 0,40 0,86a ± 0,01 1,41a ± 0,65 ns ns *** ns *** FURANOS 2,30 ± 0,44 2,38 ± 0,54 2,03 ± 1,46 5,25 ± 2,89 ns ns *** ns **
922 66 dihidro-2(3H)-furanona 1,08ab ± 0,01 1,26ab ± 0,59 1,03ab ± 0,53 0,93ab ± 0,09 ns ns ** ns ** 996 76 2-pentilfurano 1,22 ± 0,45 1,12 ± 0,45 1,00 ± 0,92 4,32 ± 2,94 ** ns *** ns *
HIDROCARBUROS AROMÁTICOS 8,78a ± 1,93 66,07de ± 5,71 9,53a ± 2,99 16,70ab ± 4,59 ns ns *** ns *** 772 42 metilbenceno 2,66a ± 0,49 6,57a ± 0,25 2,59a ± 0,97 3,15a ± 0,10 ns ns ** *** *** 878 56 etilbenceno 0,47ab ± 0,10 12,39e ± 2,08 0,47ab ± 0,29 1,83ab ± 1,00 *** *** *** *** *** 885 58 m- y p- xileno 1,14a ± 0,07 42,47d ± 3,79 1,44ab ± 0,65 5,97abc ± 3,98 *** *** *** ns *** 893 61 estireno 0,73b ± 0,19 0,96b ± 0,35 0,88b ± 0,06 0,71b ± 0,06 ** ns *** ** ***
1002 79 1,3,5-trimetilbenceno 0,73a ± 0,31 0,81a ± 0,01 0,76a ± 0,25 0,74a ± 0,00 ns ns ** ns *** 1034 84 1,2,3,5-tetrametilbenceno 0,92ab ± 0,03 0,67a ± 0,09 0,88ab ± 0,29 1,29ab ± 0,72 *** ** *** ** ns 1043 86 3-metilfenol 0,48a ± 0,14 0,72a ± 0,30 0,55a ± 0,31 1,11ab ± 0,22 ns ns *** ns *** 1078 88 4-metilfenol 0,40a ± 0,16 0,31a ± 0,12 0,55a ± 0,10 0,52a ± 0,04 ns * *** ns *** 1103 91 2-metoxifenol 0,41a ± 0,06 0,30a ± 0,06 0,50a ± 0,18 0,56a ± 0,12 ns * ** ns *** 1130 93 bencenoetanol 0,59a ± 0,32 0,34a ± 0,17 0,57a ± 0,00 0,45a ± 0,02 *** * *** ** ** 1178 95 1,3-dimetoxibenceno 0,25a ± 0,12 0,52ab ± 0,19 0,34a ± 0,00 0,36a ± 0,00 *** *** *** * ***
TERPENOS 1,42a ± 0,18 1,31a ± 0,10 1,28a ± 0,69 3,69a ± 0,16 ns ns *** ns *** 934 68 alfa-tujeno 0,17a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,45a ± 0,02 * ns ** ns *** 942 69 alfa-pineno 0,30a ± 0,01 0,25a ± 0,07 0,42a ± 0,15 0,47a ± 0,01 * ** *** ns *
1018 82 alfa-felandreno 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 0,00a ± 0,00 *** *** *** *** *** 1028 83 delta-3-careno 0,32a ± 0,08 0,30a ± 0,01 0,26a ± 0,00 0,91ab ± 0,06 *** *** *** *** *** 1035 85 l-limoneno 0,64a ± 0,12 0,76a ± 0,11 0,59a ± 0,54 1,86ab ± 0,23 * *** *** *** ***
COMPUESTOS CLORADOS 321,44abcd ± 106,24 873,01e ± 51,96 335,56bcd ± 260,33 336,96bcd ± 21,39 * *** *** *** *** 616 21 cloroformo 321,44abcd ± 106,24 873,01e ± 51,96 335,56bcd ± 260,33 336,96abc ± 21,39 ns * *** ns ***
sumatorio 1.130,44ab ± 85,25 1.321,32abc ± 88,49 1.035,78ab ± 0,00 1.031,13ab ± 135,44 * * ns * ***
*** Nivel de significación p<0,001; ** Nivel de significación p<0,01; * Nivel de significación p<0,05; ns No significativo


CAPÍTULO V: DISCUSIÓN GENERAL


CAPÍTULO V: DISCUSIÓN GENERAL
263
5. DISCUSIÓN GENERAL
Como se señala en la introducción de esta memoria, son numerosos los trabajos
dedicados a la investigación de los productos curados del cerdo, tanto de razas blancas
como Ibéricas. Todos estos estudios han permitido avanzar en el conocimiento de estos
alimentos de gran importancia económica en nuestro país. Se ha mejorado
sustancialmente la tecnología de su elaboración y se conocen mucho mejor los cambios
que acontecen durante ella. Sin embargo, la particularidad de estos productos, con una
gran variabilidad entre piezas, tiempos de curado o maduración relativamente largos o,
incluso muy largos y precios elevados, hace que generalmente, al realizar los distintos
estudios, el número de muestras por lote sea pequeño. Este reducido número unido a la
variabilidad antes mencionada y al hecho de que se trata de productos “vivos” en los que
los cambios bioquímicos y microbiológicos son constantes conduce en muchas ocasiones a
resultados de discusión complicada, cuya razón de ser es difícil de explicar.
En este estudio se nos brindó la oportunidad de realizar una investigación sobre
productos elaborados procedentes de cerdos que se habían genotipado para diferentes
QTLs (RYR1, RN, MC4R, LEPR, FABP3, IGF2) y cuyo seguimiento durante tres generaciones
podría poner de manifiesto el efecto de la expresión de los diferentes alelos en los
productos curados. La posibilidad de disponer de jamones, paletas y lomos procedentes de
los mismos animales y procesados en los mismos secaderos nos permitió obtener una
amplia serie de resultados para realizar una caracterización de los productos.
Sin embargo, contar con esta importante cantidad de datos necesita una
interpretación y colocarlos en el contexto adecuado para convertirlos en información útil,
es decir, el objetivo de la quimiometría. Aunque inicialmente no entraba dentro de los
objetivos de esta tesis su tratamiento quimiométrico y nuestra experiencia se centra más
en la discusión de los resultados en función de la experiencia previa propia y de otros
autores, nos dimos cuenta de la utilidad que podrían tener unos resultados
cuidadosamente obtenidos y verificados de un número relativamente alto de piezas
cárnicas, de ahí que en el apartado de resultados de esta memoria se recojan de manera
exhaustiva todos los datos analíticos presentados en función de todos los aspectos
estudiados. Como hemos indicado, un tratamiento quimiométrico de estos datos podría
aportar información extremadamente útil que esperamos poder desarrollar a corto plazo,
pero que actualmente no podemos abordar. Por esta razón, abordaremos la discusión de
los resultados de forma tradicional, conscientes de la dificultad de explicar algunos de
ellos en función de los parámetros estudiados, de forma que intentaremos discutir los

CAPÍTULO V: DISCUSIÓN GENERAL
264
efectos del IGF2 en los distintos productos curados y su comportamiento a lo largo de las
generaciones.
La discusión se ha dividido en dos apartados. El primer apartado discute los
resultados obtenidos en referencia al efecto del QTL IGF2, evaluando la calidad del jamón,
paleta y lomo curado según el contenido lipídico. El segundo apartado pretende
profundizar si los cambios producidos por el efecto del QTL IGF2 se mantienen a lo largo
de las distintas campañas de los tres productos estudiados.
5.1. EFECTO DEL ALELO IGF2
Tanto la grasa subcutánea, como la grasa intermuscular y la intramuscular, son
muy importantes para determinar la calidad de los productos curados, ya que contribuyen
a la calidad tecnológica y sensorial (Girard et al., 1986; Antequera et al., 1992). Debido a
las nuevas tendencias nutricionales actuales, los elaboradores de productos cárnicos se
han visto obligados a utilizar razas cada vez más magras, seleccionadas con vistas al
consumo de su carne en fresco, reduciendo así la calidad de los productos curados
necesitados de una mayor infiltración y cobertura grasa. La utilización de la raza Duroc
para la producción de Jamón de Teruel fue una de las primeras iniciativas que se tomaron
para resolver este problema, consiguiendo así un aumento de la grasa intramuscular, ya
que los cruces que se utilizaban hasta el momento, Landrace x Large White tenían una
infiltración muy baja. Un mayor nivel de grasa intramuscular tiene una influencia positiva
sobre las cualidades sensoriales de la carne de cerdo y de los productos derivados, aunque
la respuesta del consumidor es variable, no exenta de discrepancias, como revela la
literatura científica (Wood et al., 1988; Edwards, 1992).
Otra herramienta de selección genética que ha mostrado buenos resultados es la
búsqueda y utilización de QTLs, entre los que se encuentra una mutación en el gen IGF2
cuyos efectos sobre la composición de la canal y de la carne eran entre otros: una mayor
masa muscular y más carne a expensas de tejido adiposo (Nezer et al., 1999; Jeon et al.,
1999); Además, la transmisión del genotipo de IGF2 se producía únicamente por vía
paterna. Se ha encontrado que mientras que el alelo A era más frecuente en razas
ampliamente seleccionadas, el alelo G aparecía con mayor frecuencia en razas menos
sometidas a selección (Van Laere et al., 2003). Por este motivo resulta interesante la
localización de sementales Duroc que contengan el alelo G de IGF2 y evaluar así la
influencia de este alelo sobre la calidad de los productos curados derivados de su
progenie.

CAPÍTULO V: DISCUSIÓN GENERAL
265
5.1.1. CALIDAD DE LA GRASA
Tanto desde el punto de vista de su reducción para su consumo en fresco, como de
su incremento para favorecer el procesado de los productos curados, la grasa siempre ha
estado en el objetivo de la selección genética. En este sentido, se observaron diferencias
significativas en el contenido graso, intramuscular y subcutáneo, siendo superior en
aquellos productos (jamón y paleta curada) pertenecientes al lote AG, donde el alelo G fue
definido por Burgos et al., (2010) como el alelo responsable de aumentar el contenido de
grasa, independientemente de su localización anatómica, en comparación con el lote AA,
cuyo alelo se ha descrito como el responsable de aumentar el contenido magro de la canal.
En el caso de los jamones, fue muy clara la diferencia existente en el contenido de
grasa subcutánea, existiendo diferencias significativas en cuatro de los seis puntos de
medida y en el contenido total de grasa intramuscular (p<0,01), presentando diferencias
significativas tanto en el contenido expresado en extracto fresco como en seco. En el caso
de las paletas curadas, existió una mayor diferencia en el contenido de grasa subcutánea,
donde cinco de los seis puntos de medida mostraron diferencias significativas, pero no
existieron diferencias significativas en el contenido de grasa intramuscular, a pesar de que
las paletas procedentes del lote AG mostraron un mayor contenido que las del AA (6,42 vs
7,48% en extracto fresco). Parece claro que los efectos son más evidentes sobre la grasa
subcutánea que sobre la intramuscular, y sobre el jamón más que sobre la paleta. Esto
podría deberse a que la grasa subcutánea se deposita antes que la grasa intramuscular y
que a la edad de sacrificio de los cerdos utilizados para este estudio, que por otra parte es
la edad habitual de los cerdos destinados a chacinería, aún no se ha completado el
depósito de grasa intramuscular. Nuestros datos no coinciden con los resultados
obtenidos por Kouba y Bonneau (2009), que encontraron valores similares en la velocidad
de deposición del tejido adiposo subcutáneo e intramuscular en las paletas, mientras que
el tejido adiposo subcutáneo crecía más rápido que el intramuscular en el caso del jamón.
Por otra parte, estos resultados no coinciden con los de Burgos et al., (2011), que no
encontraron diferencias significativas al estudiar el contenido de grasa intramuscular de
distintas piezas como el psoas major o el bíceps femoris. No obstante, estos autores si
observaron diferencias significativas cuando realizaron la determinación en toda la masa
del jamón.
Las diferencias en el espesor de la grasa subcutánea de los jamones fueron
similares a las que se dieron en las paletas. Sin embargo, el incremento de grosor de la
grasa subcutánea no se percibe tanto en el jamón como en la paleta, debido a que en el
primero la proporción magro/grasa es mayor. Las diferencias en el contenido de grasa

CAPÍTULO V: DISCUSIÓN GENERAL
266
intramuscular solo se dieron en el jamón, pudiéndose apreciar un mayor veteado en toda
la loncha. Sensorialmente, el aspecto al corte de los jamones AG, con un mayor veteado y
un espesor de grasa subcutánea razonable será mejor valorado que el de las paletas del
mismo grupo, que presentaron un espesor de la grasa subcutánea excesivo con el mismo
veteado.
Con respecto al lomo curado, tan solo se tomaron medidas del contenido de grasa
intramuscular, no encontrando diferencias significativas entre ambos lotes, a pesar de que
los lomos procedentes del lote AG mostraron un mayor contenido que los del AA (4,9 vs
4,7% en extracto fresco).
Las diferencias en el contenido y composición de ácidos grasos entre lotes, pueden
estar ya presentes en el perfil de ácidos grasos de la materia prima (Ramírez y Cava,
2007), o deberse a cambios producidos en el perfil de ácidos grasos durante el procesado.
Sin embargo, dado que el procesado es el mismo según el producto estudiado, las
diferencias en la composición de ácidos grasos se deberán principalmente al contenido
graso de la canal y del músculo (Gandemer, 1998).
La fracción de ácidos grasos más importante es la de los triglicéridos (LN), ya que
representa la forma química en la que se encuentran los ácidos en el músculo,
representando el 87% en el jamón y el lomo curado y el 79% en la paleta curada respecto
al contenido total de ácidos grasos, por lo que existe una gran correlación con la
composición de ácidos grasos totales, a pesar de que las diferencias son más marcadas en
la fracción de triglicéridos (LN). Es por ello que esta fracción es la más eficiente en la
búsqueda de diferencias entre los productos curados (Viera-Alcaide et al., 2007 y 2008).
Se han descrito numerosos factores que afectan al contenido intramuscular de los
triglicéridos (LN), de entre los cuales los más importantes son la localización anatómica, la
raza, el sexo, la edad y la alimentación (Tejeda, 1999; Gandemer, 2002).
La composición en ácidos grasos de la fracción de triglicéridos (LN) se caracteriza
por tener una elevada proporción de AGS y AGMI y baja de AGPI (Tejeda et al., 2002; Raes
et al., 2004). Por otra parte, la fracción de fosfolípidos (LP) se caracteriza por tener
elevadas proporciones de AGPI, por lo que es la fracción más susceptible a la oxidación
lipídica (Gandemer, 1999; Raes et al., 2004). Teniendo en cuenta estos resultados, la
influencia de las posibles diferencias en la composición de los ácidos grasos de esta
fracción, será muy pequeña independientemente de la materia prima (genotipo o sistema
de explotación de los cerdos), lo que sugiere que la sensibilidad a la oxidación lipídica será

CAPÍTULO V: DISCUSIÓN GENERAL
267
muy similar independientemente de la materia prima utilizada (Gandemer, 1997;
Gandemer, 2002).
En los tres productos de este estudio, al existir mayores cantidades en el contenido
de grasa intramuscular en el lote AG, el contenido de lípidos totales fue superior
significativamente en ese lote y por lo tanto, el contenido de triglicéridos (LN), por lo que
las cantidades de las otras dos fracciones, AGL y fosfolípidos (LP), se vieron reducidas en
general y sus cantidades fueron en algunos casos superiores significativamente en el lote
AA. Es por ello que el efecto que puede haber ejercido el IGF2 en la composición de la
grasa intramuscular quedará más reflejado al evaluar las diferencias que existieron entre
las proporciones de los ácidos grasos (%) dentro de cada lote.
La composición en ácidos grasos de los lípidos totales y de cada una de las
fracciones (LN, AGL y LP) presentó escasas diferencias entre los dos lotes estudiados en
cada uno de los productos. En el caso del jamón curado, se observaron diferencias
significativas en el contenido total de AGS (debido a las diferencias del ácido graso
mayoritario C16:0) y AGPI de la fracción de lípidos totales, siendo el contenido de AGS
superior y el de AGPI inferior en el lote AG. La paleta curada siguió una tendencia muy
similar al jamón curado, con la diferencia de que en este caso la fracción de triglicéridos
(LN) también presentó diferencias significativas en el contenido de AGMI, coincidiendo
con la mayor proporción del ácido graso mayoritario C18:1 y en el de AGPI, debido a las
diferencias del C18:2(ω-6), (mayoritario en esa fracción), siendo superiores
significativamente los valores de las paletas del lote AA, coincidiendo así con la fracción de
lípidos totales, donde la proporción de los AGPI también fue significativamente superior
en el lote AA. En el lomo curado destacaron las diferencias mostradas en el contenido de
AGMI y AGPI de la fracción de triglicéridos (LN), que a diferencia de los otros dos
productos analizados, fueron superiores significativamente las proporciones del lote AG,
coincidiendo con las mayores proporciones de ese lote de los ácidos grasos C16:1 y
C18:2(ω-6).
La composición de la grasa intramuscular es importante porque está relacionada
con la calidad sensorial (Ventanas et al., 2007 (a) y (b)) y con la calidad nutricional de los
productos curados (Ventanas et al., 2007 (b); Jiménez-Colmenero et al., 2010). Desde el
punto de vista nutricional, es importante tener en cuenta la proporción de AGS, ya que se
asocian al incremento del riesgo de padecer obesidad, hipercolesterolemia, algunos tipos
de cáncer (Wood et al., 2004) y enfermedades inflamatorias y autoinmunes (Simopoulos,
2002; Jiménez-Colmenero et al., 2010). También la relación de ω-6/ω-3 es importante en
la dieta, ya que estos ácidos grasos son esenciales; aún así, estudios científicos han

CAPÍTULO V: DISCUSIÓN GENERAL
268
demostrado que no solo es necesario ingerir una cantidad adecuada de estos ácidos sino
que es necesario hacerlo en un proporción correcta (Simopoulos, 2002; Jiménez-
Colmenero et al., 2010), estableciendo por ello que la relación entre ellos sea de entre 3 y
10, o que se encuentren por encima de un tercio del total de ácidos grasos (Burgos et al.,
2010). Los tres productos mostraron diferencias significativas en la proporción de AGS,
siendo en los tres casos superiores en el lote AG. Sin embargo, la relación ω-6/ω-3 mostró
diferencias significativas tan solo en la fracción de triglicéridos (LN) del jamón y del lomo,
siendo superiores significativamente en el lote AA en el jamón y en el lote AG en el lomo.
Estos resultados están relacionados con la proporción de AGPI, cuyas diferencias en
ambos productos seguían la misma tendencia.
Desde el punto de vista nutricional las paletas procedentes del lote AA presentaron
ventajas por su menor proporción en AGS y mayor de AGMI. En el caso de los lomos, serían
también más apropiados los procedentes del lote AA, por presentar menores niveles de
AGS y ω-6/ω-3. El jamón curado, no puede diferenciarse entre lotes, ya que los jamones
del lote AA presentaron menor contenido de AGS, pero mayor de la relación ω-6/ω-3,
teniendo valores por encima de 10.
5.1.2. CALIDAD DE LOS PRODUCTOS CURADOS
El mayor o menor contenido de grasa subcutánea e intramuscular puede influir en
el resto de parámetros composicionales y sensoriales estrechamente relacionados con la
calidad (Antequera et al., 1992; Carrapiso et al., 2003). Además el contenido de grasa
intramuscular es uno de los principales factores que afectan a la aptitud tecnológica de los
productos curados durante la etapa de secado-maduración, ya que un adecuado nivel de
grasa es esencial para regular la difusión de la sal/agua y el proceso de secado (Arnau,
1998; Gandemer, 2002).
Las medidas morfológicas, que también pueden estar condicionadas por los
depósitos grasos, no mostraron diferencias significativas en ninguno de los tres productos,
excepto la longitud del lomo curado, que fue superior significativamente en el lote AA, no
coincidiendo con el resto de medidas, por lo que las posibles diferencias no se atribuyen al
efecto del QTL, sino más bien a la toma de muestra. Estos resultados difieren de los
obtenidos por Burgos et al., (2011) sobre otros lotes procedentes de cerdos AA y AG
analizados en fresco, donde si observaron diferencias significativas entre ambos lotes,
presentando mayor peso los productos procedentes de los cerdos AG. Aunque estos
autores señalaron diferencias significativas en materia fresca, a lo largo del procesado
dichas diferencias desaparecen.

CAPÍTULO V: DISCUSIÓN GENERAL
269
En general no existieron diferencias significativas en los parámetros físico-
químicos causadas por las diferencias en el contenido de grasa subcutánea e
intramuscular. A pesar de la existencia de diferencias significativas en el espesor de la
grasa subcutánea en la paleta y en el jamón, no existieron diferencias en el contenido de
humedad o de cloruros, aunque sí existieron diferencias en el contenido de proteínas en el
jamón curado, siendo superior el contenido del lote menos graso, es decir, el AA. Esta
única diferencia en la composición puede deberse principalmente a la ya comentada
mayor relación músculo/grasa del jamón curado en comparación con la paleta curada. Al
comparar con el lomo curado, la ausencia de diferencias significativas en los parámetros
composicionales fueron más esperadas, ya que el contenido de grasa intramuscular no
presentó diferencias significativas.
Por otra parte, los procesos oxidativos guardan una estrecha relación con el
contenido y la composición de la grasa intramuscular, por ejemplo, altos valores de AGPI
aumentan la susceptibilidad de los procesos oxidativos de la carne, y por lo tanto la
rancidez (Cava et al., 1999; Ramírez y Cava, 2007). Según estos autores, los productos más
insaturados mostraron una mayor intensidad en la oxidación lipídica que se reflejaba con
un alto contenido de TBA (mg MDA/Kg).
Al consultar las tablas de los tres productos, vemos como la proporción de AGPI
fue superior significativamente en el lote AA en el jamón y la paleta. En el caso del lomo las
diferencias no fueron significativas en la fracción de lípidos totales, pero si en la fracción
de triglicéridos (LN), siendo superior el contenido de AGPI en el lote AG. En este sentido, el
lote AA perteneciente a los jamones y paletas mostró mayor proporción de AGPI, pero
menor contenido de TBA (significativamente superior en el lote AG en las paletas, ya que
en los jamones no existieron diferencias significativas). Nuestros resultados no
coincidieron con lo expuesto anteriormente, ya que los productos más oxidados fueron los
que tuvieron una menor proporción de AGPI (lote AG). El comportamiento de los lomos
fue contrario al de las paletas y jamones, coincidiendo así con autores como Cava et al.,
(1999) y Ramírez y Cava (2007), ya que la fracción de triglicéridos (donde existieron
diferencias significativas) tuvo mayor proporción de AGPI en el lote AG, coincidiendo con
el mayor contenido de TBA que presentó ese lote.
La textura instrumental es otro parámetro relacionado con el contenido y
composición de la grasa intramuscular, aun asi el conocimiento que se tiene sobre el efecto
del contenido y composición de la grasa sobre la textura de jamones curados es escaso
(Ruiz-Carrascal et al., 2000; Muriel et al., 2004). Las medidas del perfil de textura
realizadas sobre nuestras piezas mostraron diferencias significativas en parámetros como

CAPÍTULO V: DISCUSIÓN GENERAL
270
dureza, fuerza, gomosidad y masticabilidad en los tres productos analizados, sin embargo
no siguieron la misma tendencia, ya que en el caso del jamón y del lomo estos parámetros
fueron superiores significativamente en el lote AA, pero en el caso de la paleta fueron
superiores en el AG.
Ruiz-Carrascal et al., (2000) encontraron una correlación negativa entre la dureza
del jamón y el contenido de grasa intramuscular, en concordancia con Parolari et al.,
(1988) y coincidiendo con nuestros resultados para jamón curado, ya que el lote AG
presentó mayor cantidad de grasa intramuscular y menor dureza. La paleta curada, al
igual que el lomo, no presentaron diferencias significativas en el contenido de grasa
intramuscular, por lo que las diferencias mostradas en el perfil de textura no pueden
relacionarse con este parámetro.
Por otra parte, Ruiz-Ramírez et al., (2006) observaron que los jamones curados
con elevada proteolisis y un bajo contenido en cloruros mostraban valores bajos de dureza
y altos de cohesividad y elasticidad, especialmente con bajos contenidos de agua. La
tendencia que siguieron los parámetros de la textura propuesta por estos autores si se
corresponde con los obtenidos en nuestros productos, ya que las muestras más duras,
fueron las menos elásticas. Además esta sería una posible explicación a la menor dureza
instrumental y sensorial de los lomos del lote AG, ya que presentaron mayor contenido de
aminoácidos libres, pero no explicaría la mayor dureza de las paletas AG y de los jamones
AA. En primer lugar porque las paletas no mostraron diferencias significativas en el
contenido general de aminoácidos libres (tan solo mostraron diferencias significativas en
el contenido de treonina) y en segundo lugar porque los jamones AA (más duros)
presentaron un mayor contenido de aminoácidos libres, aunque un menor contenido de
grasa intramuscular, como ya se ha mencionado anteriormente.
Recientemente, Morales et al., (2007) encontraron una correlación negativa entre
la dureza y el contenido en humedad en jamón curado, aunque esta relación dependía de
las temperaturas aplicadas al final de la etapa de secado-maduración. Además,
encontraron que jamones curados con un contenido de grasa intramuscular por encima
del 4% o con valores de NaCl por debajo del 2% era más fácil que presentaran texturas
blandas. Estos resultados confirman en cierta manera la contradicción de las paletas
curadas con respecto al jamón curado, ya que a pesar de que tuvieron un mayor contenido
de grasa intramuscular que el jamón, el contenido de humedad fue inferior, pudiendo por
lo tanto aumentar la dureza. Sin embargo, esto no explicaría el hecho de que las paletas
procedentes del lote AA fueran más blandas que las del lote AG, ya que tanto el contenido

CAPÍTULO V: DISCUSIÓN GENERAL
271
de grasa intramuscular como el porcentaje de humedad no mostraron diferencias
significativas entre lotes.
En cuanto a la cohesividad y adhesividad, Ramírez y Cava, (2008) encontraron
correlaciones positivas entre las dos medidas y el contenido de grasa intramuscular,
coincidiendo con los lomos de nuestro estudio, siendo superiores significativamente
ambos parámetros en el lote AG. Los jamones no mostraron diferencias en esos
parámetros de textura y las paletas en el contenido de grasa intramuscular.
5.1.3. CALIDAD SENSORIAL DE LOS PRODUCTOS CURADOS
El contenido de grasa, en especial el de grasa intramuscular, es considerado como
una de las características de calidad más importantes en los productos curados, afectando
a parámetros sensoriales como apariencia, jugosidad, intensidad o persistencia de flavor
(Gandemer, 2009).
Ramírez y Cava, (2008) afirmaban que una baja proporción de AGPI y una alta de
AGMI influía positivamente sobre determinados atributos sensoriales en jamón Ibérico,
aumentando el veteado y la jugosidad y disminuyendo la rancidez, incrementando por lo
tanto la preferencia de los consumidores (Ruiz et al., 2002(a)). Los jamones no
presentaron diferencias significativas en ninguno de los atributos nombrados entre ambos
lotes, a pesar de que el lote AG presentó menor proporción de AGPI. Las paletas curadas
tan solo presentaron diferencias significativas en el atributo de jugosidad, que fue superior
en el lote AG, independientemente de que el contenido de AGMI fuera significativamente
inferior en ese lote (en la fracción de triglicéridos (LN)). El lomo curado del lote AG mostró
puntuaciones significativamente superiores en los atributos de veteado y jugosidad e
inferiores en el atributo rancidez, coincidiendo con el mayor contenido de AGMI, a pesar
de que el contenido de grasa intramuscular no mostró diferencias significativas.
Además, el atributo rancidez también está relacionado con los procesos oxidativos
que tienen lugar durante el procesado de los productos curados (Cava et al., 1999;
Ramírez y Cava, 2007). Según estos autores, como ya se ha mencionado, los productos más
insaturados mostraron una mayor intensidad en la oxidación lipídica que se reflejaba con
un alto contenido de TBA. Nuestros resultados no coinciden con estos autores, ya que el
atributo rancidez tan solo mostró diferencias significativas en el lomo curado, siendo
superior en el lote AA, a pesar de que ese lote mostró valores significativamente inferiores
en el contenido de TBA (mg MDA/Kg) y en la proporción de los AGMI y AGPI.

CAPÍTULO V: DISCUSIÓN GENERAL
272
Numerosos autores han encontrado una contribución destacada en el sabor de la
carne y de los productos cárnicos resultantes de procesos proteolíticos, en concreto,
péptidos y aminoácidos. No hay estudios anteriores sobre el efecto del contenido de grasa
intramuscular en la concentración de aminoácidos libres de los productos curados (Martín
et al., 2001). Aunque siempre se ha concedido más importancia a las condiciones de
procesado en la generación de aminoácidos libres, también parece influir la calidad de la
materia prima (Toldrá y Flores, 1998; Jurado et al., 2007) y el tipo de músculo (Aristoy y
Toldrá, 1998; Cornet et al., 1999; Okrouhla et al., 2006).
Martín et al., 2001 observaron que las condiciones de procesado (temperatura y
contenido de sal) determinaban no solo el nivel, sino también el tipo de compuestos
procedentes de la degradación de proteínas durante el procesado de los jamones Ibéricos.
Estos mismos autores observaron que la formación de aminoácidos libres aumentaba
significativamente en los primeros 240 días y posteriormente la concentración se
mantenía prácticamente constante, coincidiendo con los resultados de Jurado (2005). Este
incremento de aminoácidos en la etapa de secadero fue considerado por el aumento de
temperatura, característico de dicha etapa, estimulando la actividad enzimática (Toldrá et
al., 1997; Toldrá et al., 2000; Jurado et al., 2007).
Por lo tanto, durante la etapa de secadero confluyen las mejores condiciones para
la formación masiva de aminoácidos libres, por una parte porque los perniles no
presentan una deshidratación excesiva y la sal todavía no se ha distribuido uniformemente
por toda la pieza. Esto explicaría el por qué numerosos autores han encontrado que el
contenido de aminoácidos libres estaba correlacionado negativamente con el contenido de
sal (Virgili et al., 1999; Toscani et al., 2000; Virgili et al., 2006); por otra parte, la
temperatura alcanzada durante esa etapa parece favorecer la formación de aminoácidos
libres (Armero et al., 1999; Jurado, 2005; Virgili et al., 2007).
Esto puede explicar en cierta medida las diferencias que se produjeron en el
contenido total de aminoácidos libres al comparar los tres productos estudiados. El jamón
curado obtuvo cantidades muy similares al lomo, ya que a pesar de que el proceso de
secadero del lomo fue más corto que el del jamón, el contenido final de cloruros del lomo
fue también muy inferior, por lo que la actividad enzimática durante la etapa de secadero
en el jamón fue inferior a la del lomo, disminuyendo así las diferencias entre ambos.
Por otra parte, la paleta curada presentó menores cantidades de aminoácidos
libres al comparar con los otros productos estudiados. Menores al comparar con el jamón
por su etapa de secadero más corta y por su similar contenido en cloruros; e inferior al

CAPÍTULO V: DISCUSIÓN GENERAL
273
comparar con el lomo curado, por su mayor contenido en cloruros, independientemente
del mayor tiempo en secadero que se mantuvieron las paletas curadas.
En cuanto a las diferencias en el contenido de aminoácidos libres producidas por
las diferencias en el contenido de grasa intramuscular son difíciles de explicar, teniendo en
cuenta además que estos aminoácidos son aminoácidos no esenciales (es decir,
aminoácidos que pueden ser sintetizados por el organismo) (Belitz y Gross, 1997; Jurado,
2005). Por una parte, existieron diferencias significativas en la mayor parte de los
aminoácidos en el jamón curado, siendo significativamente superiores los valores del lote
AA; en el lomo curado también existieron diferencias significativas en la mayor parte de
los compuestos, pero en este caso fueron superiores los valores del lote AG, a pesar de que
el contenido de grasa intramuscular no presentó diferencias significativas. Por último, la
paleta curada tan solo presentó diferencias significativas en el contenido de treonina, que
fue superior en el lote AA, sin embargo no mostró diferencias significativas en el contenido
de grasa intramuscular.
Estas diferencias no pueden deberse ni al tiempo de secadero, ya que fue el mismo
en los distintos lotes de cada pieza, ni al contenido en cloruros, ya que este parámetro no
presentó diferencias significativas entre lotes en ninguno de los tres productos.
Algunos autores encontraron una estrecha relación entre la concentración de
determinados aminoácidos libres con el tipo metabólico de las fibras musculares (Aristoy
y Toldrá, 1998; Cournet y Bousset, 1999). Ellos encontraron que la concentración de
determinados aminoácidos libres no esenciales y el total de aminoácidos libres
presentaron una concentración significativamente más elevada en los músculos oxidativos
que en los glicolíticos, clasificando como músculo glicolítico el longissimus dorsi y el
semimembranosus y el masseter y trapezious como oxidativo.
Estos resultados no explican las diferencias encontradas en el contenido de
aminoácidos entre lotes, ya que según estos autores el comportamiento de los lomos y de
los jamones debería haber sido el mismo, por tratarse de músculos con una misma
actividad enzimática, no obstante, los lomos AG y los jamones AA presentaron mayor
contenido.
Rosell y Toldrá, (1998) observaron una mayor actividad proteolítica en general en
jamones procedentes de cerdos blancos al comparar con jamones procedentes de cerdos
Ibéricos, ya que las enzimas que mostraron una mayor actividad proteolítica en los
jamones Ibéricos estaban relacionadas con la edad de matanza de los cerdos y con la

CAPÍTULO V: DISCUSIÓN GENERAL
274
mayor duración del procesado de los jamones, por lo que ambas causas no tienen efecto al
comparar ambos lotes en nuestros productos. Coincidiendo con estos autores, en el caso
del jamón curado, podemos pensar que el menor contenido de grasa intramuscular que
presentó el lote AA pudo haber influido sobre la actividad proteolítica, facilitando la
generación de aminoácidos libres en ese lote, sin embargo, la ausencia de diferencias
significativas en el contenido de grasa intramuscular en la paleta curada, puede explicar
también la ausencia de diferencias significativas en el contenido de aminoácidos libres.
No obstante, los lomos curados presentaron una tendencia contraria a los jamones,
ya que los lomos del lote AG presentaron mayores cantidades. En este sentido, nuestros
resultados no coincidieron con los de Cava et al., (2004), al comparar lomos procedentes
de cerdos Ibéricos de distintas razas con lomos procedentes de razas blancas (Large
White-Landrace x Large White), que sí coincidían con Rosell y Toldrá, (1998) en cuanto a
la mayor actividad proteolítica de las líneas más magras. En este sentido no es de extrañar
la tendencia contraria de nuestros resultados, ya que los lomos, a diferencia de los
jamones, no presentaron diferencias significativas en el contenido de grasa intramuscular.
Sin embargo, Zhao et al., (2005) observaron que la temperatura y el contenido de
cloruros inhibían la actividad proteolítica, pero que el pH la aumentaba, de forma que al
aumentar el pH disminuía el efecto de la temperatura y la sal. Estos resultados pueden
explicar las mayores cantidades de aminoácidos obtenidas en el lote AG de los lomos, ya
que el valor del pH fue superior significativamente en los lomos del lote AG, a pesar de que
el contenido en mioglobina fue también superior en los lomos de ese lote, no coincidiendo
con los resultados de Aristoy y Toldrá, (1998), que indicaba que el contenido de
mioglobina ejercía un efecto inhibidor en la actividad enzimática.
La mayor cantidad de aminoácidos en general en el lote AA en el jamón curado y en
el lote AG en el lomo curado podría tener una implicación importante en las características
sensoriales, más concretamente en el sabor y flavor, ya que a pesar de que el umbral de
percepción de algunos aminoácidos es muy alto, puede tener una implicación muy
importante en el sabor debido a un efecto sinérgico sobre la percepción de otros
compuestos (Kato y Nishimura, 1987). En este sentido, que una mayor cantidad de
aminoácidos libres influiría en las características sensoriales del producto final y por lo
tanto en el grado de aceptabilidad de estos productos. Nuestros resultados mostraron una
mayor intensidad de flavor y persistencia en el lote de lomo que presentó una mayor
cantidad de aminoácidos libres, sin embargo, el lote AA del jamón presentó una mayor
cantidad de aminoácidos, pero la intensidad de flavor no mostró diferencias significativas,
posiblemente por el menor contenido de grasa intramuscular (relacionado con

CAPÍTULO V: DISCUSIÓN GENERAL
275
parámetros sensoriales como intensidad o persistencia de favor (Gandemer, 2009) que
mostraron los jamones de ese lote.
Los aminoácidos libres han sido descritos como sabor-activos, pudiendo ejercer
una fuerte influencia en el flavor final de los productos curados (Kato et al., 1989; Toldrá y
Flores, 1998; Hidalgo y Zamora, 2004; Jurado et al., 2007). En la actualidad los jamones
Ibéricos de alta calidad más apreciados son aquellos que presentan un ligero sabor a
dulce, relacionado estrechamente con un menor contenido en sal y con la mayoría de
aminoácidos libres con sabor dulce (Ruiz et al., 1999 (a); Jurado, 2005).
Jurado, (2005) observó una relación entre el contenido de glicina-glutamina y
prolina con el sabor dulce, estos resultados coinciden con los obtenidos en los jamones,
que presentaron mayores puntuaciones en el gusto dulce los jamones delo lote AA,
coincidiendo con el mayor contenido de los aminoácidos implicados, sin embargo, los
resultados del lomo curado no coinciden, ya que el lote AA obtuvo mayor puntuación del
sabor dulce, teniendo menor contenido de aminoácidos.
Por otro lado, Jurado (2005) no encontró una influencia de los aminoácidos libres
con el sabor salado, pero Careri et al. 1993 en cambio, si encontraron una estrecha
relación entre la cantidad de ácido glutámico y aspártico libre y el sabor salado, no
coincidiendo con nuestros resultados para jamón, ya que a pesar de que el contenido de
aminoácidos fue superior en el lote AA, el contenido de cloruros y el sabor salado no
mostraron diferencias significativas entre lotes; Sin embargo, el lomo curado del lote AG
presentó mayor contenido de estos aminoácidos, mostrando también mayor puntuación
en el sabor salado, aunque el contenido de cloruros tampoco mostró diferencias
significativas.
Otro aminoácido que se ha relacionado con el flavor ha sido la lisina, que mediante
reacciones de Maillard dan lugar a la formación de piracinas (Hwang et al., 1994; Hwang et
al., 1995), compuestos implicados en el flavor “a curado”. Jurado (2005) encontró una
correlación negativa entre el contenido de lisina y el flavor “a curado”, mientras que los
resultados de Careri et al., (1993) mostraron correlaciones positivas ente el contenido de
tirosina y valina y el sabor a producto cárnico curado. Nuestros resultados coinciden con
los de Jurado (2005), ya que los lotes que mostraron mayor contenido de lisina, mostraron
menor puntuación en el flavor “a curado”.
A pesar de que el mayor contenido de aminoácidos libres está relacionado con una
mayor intensidad de flavor (Kato et al., 1989; Toldrá y Flores, 1998; Hidalgo y Zamora,

CAPÍTULO V: DISCUSIÓN GENERAL
276
2004; Jurado et al., 2007), una actividad proteolítica en exceso puede afectar a la textura
final de los productos curados, que a su vez guarda una estrecha relación con el contenido
de grasa intramuscular (Virgili et al., 1995; Ruiz-Carrascal et al., 2000; Ruiz-Ramírez et al.,
2006) y debido a los procesos lipolíticos y oxidativos que tienen lugar durante la curación,
influye también en el desarrollo del aroma (López et al., 1992; Ruiz_Carrascal et al., 2000).
Existen otros compuestos, derivados de compuestos nitrogenados, distintos a los
aminoácidos pero que también afectan al flavor de la carne, como son los nucleótidos. Al
observar nuestros resultados vemos como en general, aquellos nucleótidos que
presentaron diferencias significativas en los tres productos, mostraron mayores
contenidos en el lote AA, coincidiendo con el menor contenido de grasa intramuscular de
ese lote, excepto el contenido de IMP en la paleta curada, que fue superior
significativamente en el lote AG.
No hay estudios anteriores que expliquen el efecto del contenido de grasa
intramuscular en la concentración de nucleótidos en los productos curados, ya que
siempre se ha concedido más importancia a las condiciones de procesado en la generación
y evolución de éstos (Tikk et al., 2006; Aristoy y Toldrá, 2009; Hernández-Cázares et al.,
2011), junto con su efecto como potenciadores en el flavor de los productos curados
(Jurado, 2005; Tikk et al., 2006). Por ejemplo, se ha demostrado que la IMP actúa como
potenciador del sabor, contribuyendo al sabor umami (Aristoy y Toldrá, 2009), mientras
que la Hx junto con algún aminoácido y péptido puede contribuir al sabor amargo de la
carne (Tikk et al., 2006).
Hernández-Cázares et al., (2011) mostraron que el contenido de cloruros actuaba
como catalizador de la reacción de descomposición de ATP a IMP, pero que el paso de IMP
a Ino no se veía afectado. También observaron que la reducción del contenido de IMP tras
el post-salado disminuía la intensidad de flavor del producto final, sin embargo, este efecto
potenciador podía verse reemplazado por otras sustancias tales como las adición de sales
o la generación de determinados aminoácidos acumulados durante el procesado.
Nuestros resultados siguieron una tendencia contraria a los resultados observados
por estos autores, ya que los jamones no mostraron diferencias significativas en la
intensidad de flavor, a pesar de que el lote AA mostró mayor contenido de IMP y de
aminoácidos, esperando por lo tanto un efecto potenciador y por ello una intensidad de
flavor mayor. Por otro lado, los lomos del lote AG presentaron mayor puntuación de este
atributo, coincidiendo con el mayor contenido de aminoácidos de ese lote, pero el menor
de IMP. Sin embargo, en la paleta curada el lote AG presentó mayor contenido de IMP, a

CAPÍTULO V: DISCUSIÓN GENERAL
277
pesar de la no existencia de diferencias significativas en el contenido de aminoácidos, de
grasa intramuscular y de intensidad de flavor.
Estos resultados nos hacen pensar que el efecto potenciador de la IMP no tiene
influencia en la intensidad del flavor en general, que se encuentra influenciada por el
contenido de aminoácidos libres y por el de grasa intramuscular, ya que en el caso del
jamón curado, el contenido de aminoácidos fue superior en el lote AA y el contenido de
grasa intramuscular inferior, no mostrando diferencias significativas en la intensidad de
flavor, independientemente de que el contenido de IMP fuera superior o inferior. En el
caso del lomo, al no presentar diferencias significativas en el contenido de grasa
intramuscular y si en el de aminoácidos, siendo superior el lote AG, la intensidad de flavor
resultó superior en ese lote, independientemente de que el contenido de IMP fuera
inferior. Por último, la paleta curada mostró diferencias en el contenido de IMP, pero no
las mostró en la intensidad de flavor, debido a la no existencia de diferencias en el
contenido de grasa intramuscular y aminoácidos libres.
En cuanto a la relación existente entre el contenido de Hipoxantina y el sabor
amargo observada por Tikk, M. et al., (2006), nuestros resultados no son muy
concluyentes, ya que la hipoxantina apareció coeluída con la AMP, producto intermedio
entre la ATP y la IMP, por lo que desconocemos el contenido real de la hipoxantina.
Nuestros resultados mostraron que el sabor amargo estuvo influenciado por el contenido
de aminoácidos independientemente del mayor o menor contenido de hipoxantina co-
eluída. En el caso del lomo, que fue el único que mostró diferencias significativas en el
contenido de hipoxantina y el sabor amargo, ambos parámetros mostraron una
correlación negativa, ya que los lomos del lote AA presentaron más contenido de
hipoxantina pero menos sabor amargo, coincidiendo con el menor contendido de
aminoácidos en ese lote. En el caso del jamón curado, presentaron mayor sabor amargo los
jamones del lote AA, coincidiendo con el mayor contenido de aminoácidos, a pesar de que
el contenido de hipoxantina no presentó diferencias significativas. Sin embargo, la paleta
curada, aunque presentó diferencias significativas en el contenido de hipoxantina, siendo
superior en el lote AA, el sabor amargo no mostró diferencias entre lotes, coincidiendo con
la no existencia de diferencias en el contenido de aminoácidos.
El olor y flavor de los productos curados también guardan una estrecha relación
con los compuestos volátiles, además su estudio podría proporcionar una buena
herramienta para la clasificación del producto final. Existen numerosos estudios que han
señalado la variabilidad en el contenido de volátiles, la cual puede deberse a que los
productos curados no son homogéneos (jamón y paleta especialmente) y que los músculos

CAPÍTULO V: DISCUSIÓN GENERAL
278
y grasa subcutánea de las muestras pueden ser diferentes (García-González et al., 2005;
Luna et al., 2006; Narváez-Rivas et al., 2010), pudiendo por ello generar diferencias
significativas sin guardar relación con el mayor o menor contenido de grasa
intramuscular.
En los últimos años se han llevado a cabo varios trabajos en distintos músculos
asociados a productos curados Ibéricos, como jamón y lomo, confirmando la influencia del
contenido de grasa, particularmente el contenido de grasa intramuscular, en la mayoría de
las características sensoriales (Muriel et al., 2004; Ruiz et al., 2002 (b); Ventanas et al.,
2005; Ventanas et al., 2008). Sin embargo, aunque el efecto de la composición de la grasa
intramuscular en la generación de compuestos volátiles en jamones curados ha sido
ampliamente estudiada (Ruiz. et al., 2002(b)), poco se sabe sobre el efecto del contenido
de grasa intramuscular en la generación y liberación de compuestos aromáticos durante el
proceso de maduración (Ventanas et al. 2008). No obstante, la presencia de un elevado
contenido de grasa intramuscular se asocia con una mayor formación de compuestos
volátiles y por tanto con un óptimo desarrollo del aroma y el flavor durante el procesado
(Ventanas et al., 2008). Por otra parte, de los componentes del alimento que pueden influir
en la liberación de los compuestos volátiles al espacio de cabeza, los lípidos son los que
presentan un mayor efecto en el reparto de los compuestos volátiles entre el producto y la
fase gaseosa (de Roos, 1997).
Dentro de los compuestos responsables del aroma se han identificado decenas de
ellos, siendo los grupos más abundantes los hidrocarburos, aldehídos y alcoholes, aunque
también existen otros compuestos como cetonas, ácidos grasos libres, γ-lactonas
(furanonas) y ésteres (Möhler, 1982; Pérez, 2006). En particular, el aroma del curado ha
sido relacionado con la presencia de aldehídos ramificados con grupos metilo, alcoholes
secundarios, metil-cetonas, etil ésteres y el dimetil trisulfuro (Hansen-Moller et al, 1997;
Carrapiso et al., 2002; Carrapiso et al., 2010).
El jamón curado presentó diferencias significativas en 30 compuestos volátiles. Las
familias que mostraron diferencias fueron las de los hidrocarburos alifáticos y los furanos,
superiores significativamente en el lote AA (p<0,05) y la de los terpenos en el lote AG
(p<0,05). Las paletas curadas presentaron diferencias significativas en 18 compuestos, no
existiendo diferencias en ninguna familia. Por último, los lomos presentaron diferencias en
33 compuestos, donde tan solo la familia de los ésteres presentó diferencias, siendo
superior el valor del lote AA (p<0,01).

CAPÍTULO V: DISCUSIÓN GENERAL
279
Los hidrocarburos alifáticos proceden de la oxidación lipídica (Möhler, 1982; Ruiz
et al., 2002(b)), mientras que aquellos cuya cadena principal tiene hasta 10 átomos de
carbono, así como los hidrocarburos ramificados, parecen tener su origen en su
acumulación en los tejidos grasos del animal, posiblemente procedentes de la
alimentación que reciben durante el periodo de cebo (Meynier et al., 1999; Ruiz et al.,
2002(b)). En el jamón curado, los cuatro hidrocarburos alifáticos que presentaron
diferencias significativas fueron superiores en el lote AA, de los cuales dos de ellos (el
hexano (p<0,001) y el heptano (p<0,01)) fueron alcanos cuya cadena principal tiene
menos de 10 átomos de carbono, por lo que su origen se debe a la oxidación lipídica
(Möhler, 1982; Ruiz et al., 2002(b)). Sin embargo, también mostraron diferencias un
alcano lineal de más de 10 átomos de carbono (tridecanoico (p<0,01) y un hidrocarburo
ramificado (3-metilpentano (p<0,05)), cuya mayor cantidad en el lote AA podría deberse a
la baja polaridad de estos compuestos, estando más retenidos por lo tanto en la fase grasa.
En la paleta curada tan solo el metilciclopentano presentó diferencias (p<0,001), pero en
este caso fue superior el valor en el lote AG, como era de esperar. El lomo curado mostró
diferencias en cinco hidrocarburos, tres de ellos superiores en el lote AA (el pentano
(p<0,05), el hexano (p<0,05) y el decano (p<0,01)), cuyo origen es lipídico (la polaridad no
influiría es este caso porque el contenido de grasa intramuscular no presentó diferencias)
y dos en el lote AG (el 2,3,4-trimetilpentano (p<0,001) y el tridecano (p<0,01)) cuyo origen
procede de la alimentación. Aún así, la presencia de estos compuestos no parece tener una
influencia sobre las características sensoriales de los productos curados debido a sus
elevados umbrales de olfacción (García y Timón, 2001).
Sin embargo, en el jamón Serrano los hidrocarburos aromáticos como el m- y p-
xileno poseen un aroma descrito como “ahumado fenólico”, mientras que el o-xileno se
describe como aroma a “caramelo-frutal” (Flores et al., 1997; Flores et al., 1998; Ventanas,
2006; Pérez, 2006). Los hidrocarburos aromáticos son productos procedentes de la
pirolisis de la lignina y presentan bajo umbral de olfacción con característicos aromas a
humo (Stahnke, 1998; Muriel, 2003). En los jamones, tan solo el m- y p- xileno presentó
diferencias significativas, siendo superior el área del lote AG. El origen de este compuesto
en productos curados no se ha relacionado con la pirolisis de la lignina, sino más bien con
la alimentación recibida por los animales, y su presencia en los jamones se debería a su
deposición directa en los tejidos grasos del animal (Berdagué et al., 1993; Ruiz et al., 1999
(b)), de ahí su mayor contenido en el lote AG. En la paleta curada no existieron diferencias
significativas en ningún hidrocarburo aromático, sin embargo en el lomo tres compuestos
mostraron diferencias, dos superiores en el lote AA (estireno (p<0,001) y 3-metilfenol
(p<0,01)) y uno en el lote AG (1,3-dimetoxibenceno (<0,001)). En el lomo curado, cuyo

CAPÍTULO V: DISCUSIÓN GENERAL
280
procesado no incluye fases de ahumado, los fenoles pueden proceder de ingredientes
ahumados como el pimentón, en el que se han descrito numerosos compuestos fenólicos
(Mateo y Zumalacarregui, 1996). La acumulación de este tipo de compuestos en los
jamones y lomos que presentaron mayor contenido graso explicarían las mayores
cantidades del xileno y del 1,3-dimetoxibenceno. No obstante, habría que tener en cuenta
las características de estos compuestos que presentan un marcado carácter apolar (Jo y
Ahn, 1999) y por tanto liposolubles, por lo que tendrían una mayor afinidad por la fase
grasa, lo que podría haber dificultado la liberación al espacio de cabeza de muchos de
estos compuestos en los productos de mayor contenido de grasa, como se observa en
aquellos hidrocarburos aromáticos y alifáticos que fueron superiores en el lote AA.
Otra familia que fue más abundante en el lote AA en el jamón fue la de los furanos.
La presencia de los furanos en los alimentos se relaciona con procesos de calentamiento,
sin embargo son compuestos habituales en el jamón curado y en embutidos crudos
curados (Muriel, 2003) y se ha demostrado su formación a partir de la oxidación del ácido
linoleico (Grosh, 1987; Shahidi et al., 1986; Pérez, 2006). Nuestros resultados muestran
mayor proporción de estos compuestos en aquellos jamones que presentaron menor
contenido de grasa intramuscular (jamones procedentes del lote AA), y por lo tanto, mayor
proporción de AGPI, más susceptibles a la oxidación. En el jamón curado, tan solo dos
furanos de los ocho identificados mostraron diferencias significativas, el 2-pentilfurano
(p<0,05) y el 2-butilfurano (p<0,05), ambos superiores en el lote AA. El 2-pentilfurano es
un compuesto asociado a la oxidación más concretamente del ácido linoleico (Grosch,
1987; Ruiz et al., 2002 (b); Elmore et al., 2005; Ventanas, 2006), sin embargo los jamones
no presentaron diferencias significativas en dicho ácido graso. Este compuesto no se ha
identificado como olor-activo del jamón Ibérico (Carrapiso et al., 2002, 2007 y 2010),
aunque sí en jamón Serrano, describiéndose como un aroma a “jamón” (Flores et al, 1997;
Pérez, 2006). En la paleta curada, tan solo la dihidro-5-butil-2(3H)-furanona presentó
diferencias significativas (p<0,05), siendo superior el valor del lote AG. Las lactonas
resultan de la degradación de lípidos por la formación de compuestos cíclicos a partir de
los correspondientes hidroxiácidos, que son subproductos del metabolismo de los ácidos
grasos (Forrs, 1972), aunque no se descarta la intervención de reacciones de Maillard
(Ruiz et al., (2002) (b)). Estos compuestos se caracterizan por aportar olores agradables,
definidos como a coco, a mantequilla o a frutas (Belitz y Gross, 1997; Pérez, 2006) y por
tener bajos umbrales de olfacción, influyendo de forma importante en el aroma del jamón
curado. No obstante, en los trabajos llevados a cabo por Carrapiso et al. (2002, 2007 y
2010) no se incluye ninguna lactona entre los compuestos olor-activos del jamón Ibérico.

CAPÍTULO V: DISCUSIÓN GENERAL
281
La única familia que mostró un valor significativamente superior en el lote AG en el
jamón curado fue la de los terpenos, por el mayor contenido de limoneno (p<0,01). En las
paletas no existieron diferencias significativas entre estos compuestos, mientras que los
lomos también mostraron un mayor contenido de limoneno (p<0,01) en el lote AG, pero
además presentaron mayor contenido de alfa-tujeno (p<0,05) en el lote AA. La presencia
de compuestos terpénicos en productos cárnicos elaborados sin especias, como el jamón o
la paleta curada, pueden provenir de la alimentación del animal y acumularse en el tejido
graso (Buscailhon et al., 1993; Ruiz et al., 1999 (b); Andrés et al., 2002) explicando así el
mayor contenido del lote AG, sin embargo, es en los productos especiados en los que este
tipo de compuestos se encuentran en mayor número y cantidad. Meynier et al., (1999)
detectaron en salami de Milán que el 50% de los compuestos volátiles eran terpenos
aportados por la pimienta utilizada en su elaboración. El aroma que caracteriza al
limoneno es el limón (Belitz y Gross, 1997; Acree y Arn, 1997).
Los ésteres se forman a partir de la interacción de los ácidos grasos libres y los
alcoholes generados durante la oxidación lipídica en el tejido intramuscular (Shahidi et al,
1986), aunque algunos como el acetato de etilo, también se han descrito como productos
del metabolismo microbiano (Stahnke, 1994; Edwards et al., 1999; Bruna et al., 2001). En
el jamón curado, de los cinco esteres que mostraron diferencias significativas, dos fueron
superiores en el lote AA (el etil propanoato (p<0,01) y el etil hexanoato (p<0,05)) y tres en
el lote AG (el etil 2-metilbutanoato (p<0,05), el etil octanoato (p<0,05) y el etil decanoato
(p<0,05)). En la paleta curada tan solo dos mostraron diferencias, el etil 2-metilbutanoato
(p<0,001) superior en el AA y el etil 3-metilbutanoato (p<0,05) en el lote AG. Sin embargo,
siete de los doce ésteres identificados en los lomos mostraron diferencias, 5 de ellos
superiores en el lote AA (el acetato de etilo (0<0,001), el etil propanoato (p<0,01), el etil 2-
hidroxipropanoato (p<0,01), el etil 2-metilbutanoato (p<0,05) y el etil 3-metilbutanoato
(p<0,01)) y dos en el AG (el metil hexanoato (p<0,01) y el etil decanoato (p<0,01)). Los
ésteres procedentes de ácidos de cadena corta son importantes para el aroma y sabor, ya
que son los responsables de las notas dulces afrutadas que se detectan en la carne de
porcino (Baines y Mlotkiewicz, 1984; Pérez, 2006), como el etil 2-metilbutanoato
(superior en el lote AG del jamón y en el AA de la paleta y el lomo) que se ha descrito como
un aroma a “caramelo-dulce” (Flores et al. 1997). Por otra parte, el metil hexanoato
(superior en el lote AG del lomo) es el único éster encontrado en jamón Serrano con aroma
a “carne cocida” (Flores et al., 1997). También los ésteres de cadena larga, como el etil
decanoato (superior en el lote AG del jamón y del lomo), se han descrito por poseer notas
etéreas y a uva (Miranda-López et al., 1992; Ruiz et al., 1998).

CAPÍTULO V: DISCUSIÓN GENERAL
282
Los aldehídos constituyen una de las familias más importantes en el análisis
cuantitativo de compuestos volátiles presentes en el jamón curado (Sabio y Vidal-Aragón,
1998; García et al., 1991; López et al., 1992) debido a su bajo umbral de olfacción (Shahidi
et al., 1986). En cuanto a su origen, los aldehídos lineales provienen principalmente de la
degradación oxidativa de los ácidos grasos (López et al., 1992) y los aldehídos ramificados
y aromáticos podrían provenir de la degradación de Strecker de los aminoácidos (García et
al., 1991; López et al., 1992). El benzaldehído, aldehído de Strecker formado por la
degradación de la fenilalanina (Pripis-Nicolau et al., 2000), fue superior significativamente
en el lote AA (p<0,05) en el jamón curado, coincidiendo con el mayor contenido de
aminoácidos libres detectados en ese lote, entre ellos la fenilalanina (p<0,001). Sin
embargo, en la paleta curada fue significativamente superior (p<0,05) en el lote AG,
aunque el contenido de fenilalanina no mostró diferencias significativas. No obstante, su
implicación en el flavor del producto final no se ha descrito. La paleta curada presentó
diferencias significativas en otros aldehídos, entre ellos el pentanal (p<0,05) y el octanal
(p<0,05), superiores en el lote AA, cuyo origen lipídico puede explicarse por su mayor
proporción de AGPI en ese lote; además, su contribución con el aroma se ha descrito como
“verde”, “aceite” o “grasa” (Forss, 1972; Carrapiso et al., 2002). También dos compuestos
fueron superiores en el lote AG, el 2-metilbutanal (p<0,05) y el bencenoacetaldehido
(p<0,05). Existen distintos argumentos en cuanto al origen del 2-metilbutanal, algunos
autores lo relacionan con el metabolismo de los microorganismos presentes en el jamón
(Hinrichsen y Andersen, 1994 y 1995; Jacobsen y Hinrichsen, 1997); Otros sostienen que
la principal vía de formación de estos compuestos son las reacciones de Strecker y las
reacciones de Maillard (Berdagué et al., 1991; García et al., 1991; Ventanas et al., 1992;
Flores et al., 1997; Toldrá et al., 2000, revisado por Ruiz et al, 2002 (b)). El 2-metilbutanal
es importante por su contribución en el aroma de jamón curado (Hinrichsen y Pedersen,
1995) y, concretamente en el jamón Serrano; además este compuesto se ha encontrado
entre los compuestos más olor-activos del jamón Ibérico y su olor se ha descrito como
tostado y frutal (Carrapiso et al., 2002 y 2010) o como herbal, con notas de sudor y rancio
(Sánchez del Pulgar, 2011). Sin embargo, el bencenoacetaldehido no se ha encontrado
entre los compuestos volátiles más olor-activos del jamón Ibérico en estudios anteriores
(Carrapiso et al., 2002 y 2010; García-González et al., 2008), a excepción de los jamones
alterados o “calados” (Carrapiso et al., 2010), aunque se ha descrito como un olor a miel
(Sanchez del Pulgar, 2011). El lomo tan solo mostró diferencias en el contenido de decanal,
superior en el lote AG (p<0,05), coincidiendo con la mayor proporción de AGPI que se
detectaron en ese lote.

CAPÍTULO V: DISCUSIÓN GENERAL
283
Las cetonas deben su origen a la oxidación lipídica de ácidos grasos insaturados
(Sahidi et al., 1986; revisado por Ruiz et al., 2002 (b)). El jamón curado mostró diferencias
significativas en la 2,3-butanodiona (p<0,001), la 3-octanona (p<0,05) y la 3-octen-2-ona
(p<0,05), superiores en el lote AA, coincidiendo con la mayor proporción de AGPI más
susceptibles a la oxidación en ese lote. Sin embargo, la paleta curada mostró diferencias
significativas en dos compuestos, la 3-hidroxi-2-butanona (p<0,01) y la 2-nonanona
(p<0,01), siendo en este caso superiores en el lote AG, a pesar de la menor proporción de
AGPI que presentó este lote. El lomo curado presentó diferencias en seis compuestos de
los quince identificados, dos superiores en el lote AA (2,3-butanodiona (p<0,05) y la 3-
hidroxi-2-butanona (p<0,01)), coincidiendo con la mayor proporción de AGPI y cuatro
superiores en el AG (1-hidroxi-2-propanona (p<0,01), 2-octanona (p<0,05), 8-nonen-2-
ona (p<0,05) y la 2-nonanona (p<0,05)). La 2,3-butanodiona tiene su origen en la
oxidación de los ácidos grasos insaturados, aunque también se ha relacionado con
reacciones de Maillard a partir de un aminoácido y un azúcar reductor (Mottram, 1994;
Song and Cadwallader, 2008). Se ha identificado como uno de los compuestos volátiles
más olor-activos del jamón Ibérico en los trabajos realizados hasta la fecha (Flores et al.,
1997; Carrapiso et al., 2002; García-González et al., 2008; Carrapiso et al., 2010) y se ha
encontrado en el perfil de compuestos volátiles de la mayoría de los jamones curados
estudiados (Sánchez del Pulgar, 2011). En todos estos trabajos su olor se ha descrito como
olor a mantequilla, si bien en los estudios de Carrapiso et al., (2002), García-González et al.,
(2008) y Carrapiso et al., (2010), el olor de este compuesto también se ha relacionado con
el de la vainilla y caramelo. La 3-hidroxi-2-butanona ha sido identificada como
responsable del olor a “fresa” (Flores et al., 1997), mientras que las cetonas con
insaturaciones, como la 3-octen-2-ona o la 8-nonen-2-ona, son responsables de impartir
notas aromáticas particulares a grasas animales y vegetales (Forss, 1972; Ventanas, 2006).
El hecho de que los procesos oxidativos se hayan visto frenados por la presencia de
especias en ajos en los lomos, podría haber favorecido la formación de compuestos
volátiles derivados de la actividad de mohos y levaduras presentes principalmente en la
superficie de las piezas, lo que podría explicar el mayor contenido de ésteres y cetonas en
los lomos del lote AG. Por el contrario, en los jamones y paletas, productos procesados sin
especias, ajo y pimentón, predominarían los compuestos derivados de la oxidación
lipídica, por lo que el mayor contenido de algunos ésteres y cetonas o incluso de otros
compuestos derivados de la oxidación lipídica en el lote AG del jamón y la paleta, podría
deberse a la polaridad de estos productos, aportándoles por lo tanto un carácter lipofílico
(Ventanas, 2006).

CAPÍTULO V: DISCUSIÓN GENERAL
284
Los alcoholes representaron la segunda familia más abundante del total de los
compuestos volátiles identificados en el jamón y lomo curado y la tercera en las paletas.
Los alcoholes son en su mayor parte productos derivados de la oxidación de los ácidos
grasos insaturados (Forss, 1972; Shahidi et al., 1986). No obstante, los alcoholes de bajo
peso molecular y los ramificados pueden proceder de la fermentación microbiana de los
glúcidos (Kandler, 1983; Sunensen et al., 2001). Dentro de este grupo se identificaron 12
compuestos en los jamones y 15 en las paletas y en los lomos. En jamón curado cuatro
compuestos presentaron diferencias significativas, siendo superior el valor del lote AA (2-
metilpropanol (p<0,05), 3-metilbutanol (p<0,05), 2-pentanol (p<0,001) y el 2-heptanol
(0,01)). La paleta curada mostró diferencias en cuatro compuestos, tres superiores en el
lote AG (1-metoxi-2-propanol (p<0,01), 2-pentanol (p<0,05) y 1-pentanol (p<0,01)) y uno
en el AA (3-metilbutanol (p<0,05)), este último coincidiendo con el jamón. Sin embargo,
los lomos mostraron diferencias en tres compuestos, uno superior en el lote AA (2-butanol
(p<0,001)) y dos en el AG (2-metilpropanol (p<0,05) y 1-pentanol (0,05)). Los alcoholes
no ramificados proceden en su mayor parte de la oxidación lipídica de los ácidos grasos
insaturados (Forss, 1972; Shahidi et al., 1986), como el 2-heptanol y el 2-pentanol, este
último relacionado con el ácido linoleico (Forrs, 1972), coincidiendo el mayor contenido
de estos volátiles con el lote que mostró mayor proporción de AGMI o AGPI, excepto la
paleta curada, que siguió una tendencia contraria. En cambio, los alcoholes ramificados
que presentaron diferencias, como el 2-metilpropanol (p<0,05) y el 3-metilbutanol
(p<0,05) deben su origen a la degradación de los aminoácidos (Ruiz et al., 1999 (b)).
Concretamente, el 2-metilpropanol procede de la reducción de su correspondiente
aldehído, el 2-metilpropanal (Stahnke et al., 1994) que a su vez deriva del aminoácido
valina, via degradación de Strecker. Por otro lado el 3 y 2-metilbutanol son productos de
las reacciones de reducción que sufren el 3 y 2-metilbutanal (Hertz y Chang, 1970), que
derivan de la degradación de aminoácidos leucina e isoleucina vía degradación de Strecker
o bien como producto del metabolismo microbiano (Bailey et al., 1992; Muriel, 2003). El
lote AA mostró un mayor contenido de volátiles ramificados en los tres productos
analizados, coincidiendo con el mayor contenido de aminoácidos en el jamón, sin embargo
las paletas no presentaron diferencias significativas en el contenido de aminoácidos y los
lomos tuvieron un mayor contenido en el lote AG. En cuanto a su influencia en el aroma,
ésta es menor que la de otros compuestos carbonilo como los aldehídos, ya que el umbral
de olfacción de los alcoholes es más elevado (Drum and Spanier, 1991). Sin embargo, en
carne de cerdo se considera que los alcoholes de cadena lineal producen ciertas notas
“verdes”, “a madera” y a “grasa-floral” y tienen una contribución importante en el aroma
de jamón (Barbieri et al., 1992).

CAPÍTULO V: DISCUSIÓN GENERAL
285
Los ácidos carboxílicos provienen de la hidrólisis de triglicéridos y fosfolípidos y las
enzimas responsables de estos procesos permanecen activas durante el proceso de curado
(Girard y Bucharles, 1991; Motilva et al., 1993; revisado por Ruiz et al., 2002 (b)). Timón et
al., (1998) encontraron altos porcentajes de estos compuestos en jamones Ibéricos,
esperados por el elevado contenido de grasa intramuscular que presentaron los jamones.
De los 9 ácidos identificados en el jamón, tan solo 3 (ácido 2-metilpropanoico (p<0,05),
ácido nonanoico (p<0,001) y el ácido decanoico (p<0,001)) mostraron diferencias
significativas, todos ellos superiores en el lote AG. Los lomos mostraron diferencias en
cuatro ácidos, todos ellos superiores en el lote AG (ácido propanoico (p<0,05), ácido 2-
metilpropanoico (p<0,05), ácido butanoico (p<0,05) y ácido nonanoico (p<0,05))
coincidiendo con el jamón, ya que ambos productos mostraron una mayor proporción de
triglicéridos. Sin embargo, las paletas tan solo mostraron diferencias en el ácido nonanoico
(p<0,05), que fue superior en el lote AA, al contrario que los jamones y los lomos, pero
coincidiendo con Ventanas, (2006), que encontró valores del ácido nonanoico superiores
en los lomos con menor contenido graso, atribuyendo este hecho al marcado carácter
apolar de este compuesto, probablemente debido a la longitud de la cadena
hidrocarbonada, lo que podría atribuirse a la mayor afinidad y por tanto “retención” por la
fase grasa, teniendo más efecto en la paleta que en el jamón por su mayor contenido de
grasa intramuscular. De los ácidos grasos de cadena lineal, aquellos que presentan menos
de seis átomos de carbono, como el ácido butanoico, superior en los lomos del lote AG, son
los que pueden contribuir en mayor medida al aroma del producto curado, debido a que
presentan bajos niveles de olfacción y poseen notas aromáticas relacionadas con un aroma
fuerte a “queso” (Ansorena et al., 2001; Carrapiso et al., 2002; García-González et al., 2008;
Sánchez del Pulgar, 2011). El resto de ácidos identificados se caracterizan por presentar
bajos umbrales de olfacción y notas aromáticas dulces (Mateo y Zumalacarregui, 1996).
La mayor parte de los compuestos nitrogenados detectados en los productos
cárnicos proceden de reacciones de tipo Maillard que tienen lugar entre productos de la
oxidación lipídica y determinados aminoácidos (Stahnke, 1994; Hwang et al., 1995; Ruiz,
et al., 2002(b)). Aunque estas reacciones se ven favorecidas por un medio ligeramente
básico, temperaturas elevadas y actividad de agua entre 0,4 y 0,8, los compuestos
derivados de estas reacciones, como las piracinas y los pirroles, se encuentran descritos
entre los compuestos volátiles de los productos curados (Mateo y Zumalacárregui, 1996;
Ruiz et al., 1998; Schmidt y Berger, 1998; Andrés et al., 2002). De los ocho compuestos
nitrogenados identificados en jamón curado, tan solo tres mostraron diferencias
significativas, dos de ellos superiores en el lote AG (1H-pirrol (p<0,05) y la etilpirazina
(p<0,05)) y uno en el lote AA (2,3,4,5-tetrahidropiridina (p<0,05)). El elevado contenido

CAPÍTULO V: DISCUSIÓN GENERAL
286
de compuestos nitrogenados en el lote AG del jamón, a pesar de tratarse del lote con
menor contenido de aminoácidos, puede explicarse por el carácter polar de dichos
compuestos, aumentando por lo tanto su liberación en el lote más graso. En la paleta
curada tan solo la trimetilpirazina mostró diferencias, siendo superior el valor en el lote
AA, a pesar de que las paletas no mostraron diferencias significativas en el contenido de
aminoácidos. El lomo, sin embargo, no mostró diferencias significativas en ningún
compuesto de esta familia. La potencia aromática de los compuestos cíclicos nitrogenados
es muy importante en la percepción total, ya que la mayoría de ellos tienen un bajo umbral
de percepción (Ruiz et al., 2002(b)). En carne se han hallado más de 50 compuestos
(Möhler, 1982) siendo los grupos fundamentales el pirrol, pirazol y pirazina. Las pirazinas
son consideradas como componentes clave en los alimentos por impartir ciertas notas de
aroma a “frutos secos” y “tostado” (Barbieri et al., 1992).
Los compuestos volátiles con azufre se forman vía Strecker a partir de la reacción
entre aminoácidos azufrados como la cisteína, la cistina y la metionina y compuestos
carbonilos derivados de la oxidación lipídica (Shahidi et al., 1986; Pripis-Nicolau et al.,
2000; revisado por Ruiz et al., 2002 (b)). Estos compuestos son los más sobresalientes del
aroma de la carne (Möhler, 1982) porque poseen un bajo umbral de olfacción (Chang y
Petersen, 1977; Drumm y Spanier, 1991). El lomo curado fue el único producto que
presentó diferencias en la familia de los compuestos azufrados, existiendo diferencias en
dos, uno superior en el lote AA (dimetil disulfuro (p<0,01)) y otro superior en el lote AG
(benzotiazol (p<0,05)). El dimetil disulfuro no ha sido incluido entre los compuestos más
olor-activos del jamón Ibérico (Carrapiso et al., 2002), aunque sí parece estar implicado en
el olor de jamón Serrano (Flores et al., 1997 y 1998). Las posibles diferencias encontradas
en los lomos en comparación con los otros productos pueden atribuirse a la utilización del
ajo como parte del adobo de los lomos curados, ya que muchos de los compuestos
azufrados detectados han sido identificados previamente en distintos tipos de embutidos
crudos curados que incluían el ajo entre las especias del adobo (Mateo y Zumalacárregui,
1996; Edwards et al., 1999; Ansorena et al., 2001).
Finalmente, sólo se detectaron dos compuestos clorados, el 2-diclorometano y el
cloroformo, aunque tan solo el diclorometano mostró diferencias significativas en el jamón
y la paleta. A pesar de ello, el origen de estos compuestos podría ser atribuido a los
residuos de pesticidas ingeridos por los cerdos (Buscailhon et al., 1993) o a contaminación
de tipo laboratorial (Rui et al., 1999 (b); Ventanas, 2006)

CAPÍTULO V: DISCUSIÓN GENERAL
287
5.2. EFECTO DE LA CAMPAÑA
Tras evaluar los efectos producidos por la incidencia del alelo IGF2 en los tres
productos estudiados, parece interesante comprobar si dichos efectos se mantienen a los
largo de tres campañas sucesivas, donde se ha llevado a cabo la misma selección de los
lotes (AA y AG) en los tres productos, para así poder descartar un efecto casual de los
resultados del apartado anterior. Es por ello que en este apartado tan solo se han evaluado
las características que más diferencias se dieron entre lotes.
5.2.1. CALIDAD DE LA GRASA
En el jamón curado se observaron diferencias significativas en el espesor de la
grasa subcutánea en los tres puntos de medida tanto de la zona proximal como de la distal.
Las diferencias generadas en la zona distal fueron debidas por el efecto genético, el sexo y
por la interacción de ambos factores, pero en ningún caso por el efecto de la campaña. La
zona proximal presentó diferencias debidas al efecto genético pero también al efecto
campaña, de foma que los jamones de la tercera campaña presentaron un mayor espesor
en dos de los tres puntos medidos. Por lo tanto, al hacer la media de las seis medidas
tomadas en ambas zonas nos encontramos con una ausenia de diferencias significativas al
comparar las tres campañas, por lo que las diferencias generadas se debieron
principalmente al factor genético, ya que al evaluar los factores sexo y genética en un
apartado anterior, el sexo no generó influencia sobre este parámetro. Por otra parte, el
contenido de grasa intramuscular no presentó diferencias significativas entre las
campañas ni entre sexos, ya que las diferencias se debieron únicamente al efecto genético.
Las paletas mostraron diferencias significativas en el contenido de grasa
subcutánea en ambas zonas (proximal y distal) únicamente debidas al efecto genético,
pero no por el efecto sexo ni campaña. Por el contrario, el contenido de grasa
intramuscular mostró diferencias significativas únicamente debidas al efecto campaña y
no al efecto sexo y genético, siendo superior el valor de las paletas de la primera campaña,
coincidiendo así con los lomos. Una posible explicación a las diferencias generadas en el
lomo curado sería el adobo utilizado en los lomos de la primera campaña y la zona de
toma de muestra para la cuantificación de la grasa intramuscular, ya que se realizó en los
extremos, en comparación con los lomos de la segunda y tercera campaña, donde se
realizó en el centro de las piezas, zona que se ha descrito como menos grasa (Ventanas,
2006).

CAPÍTULO V: DISCUSIÓN GENERAL
288
El contenido de grasa intramuscular del lomo tuvo un comportamiento similar al
de la paleta, ya que las diferencias significativas se debieron únicamente al factor genético,
siendo superiores los valores de los lomos de la primera campaña. Las diferencias
encontradas entre campañas pueden deberse a qu los lomos de la primera campaña
fueron adobados y a la toma de muestra para la cuantificación del contenido de grasa
intramuscular, explicado anteriormente.
Las diferencias en el contenido graso, también pueden verse reflejadas en la
composición de ácidos grasos, ya que las posibles diferencias en la composición se deben
principalmente al contenido graso de la canal y del músculo (Gandemer, 1998). Como ya
se ha indicado anteriormente, el contenido de ácidos grasos está influenciado por el
contenido de grasa intramuscular, por lo que las posibles diferencias entre lotes debidas a
los factores estudiados serán más evidentes al estudiar la composición de ácidos grasos
expresada en porcentajes de area.
El jamón y la paleta mostraron un comportamiento muy similar, ya que existieron
diferencias significativas en la proporción de AGS y AGPI. En el jamón curado la
proporción de AGS fue superior en los jamones de la primera campaña (que presentó
mayor contenido de grasa intramuscular) y la de AGPI en los jamones de la tercera, en
ambos casos influenciados por el efecto de la campaña y la genética. La paleta curada
presentó mayores proporciones de AGS en las de la primera campaña, debidas únicamente
al efecto genético y mayores proporciones de AGPI en las de la tercera, coincidiendo con el
jamón. En el lomo curado, al existir mayores cantidades en el contenido de grasa
intramuscular en los lomos de la primera campaña, el contenido de lípidos totales fue
significativamente superior en ese lote, pero además las proporciones de AGS y AGMI
también lo fueron, donde aquí también el factor sexo tuvo influencia. Sin embargo, la
proporción de AGPI fue superior en los lomos de la tercera campaña, coincidiendo así con
los otros dos productos analizados, aunque en este caso solo el efecto campaña tuvo
repercusión sobre las diferencias. En este sentido, el factor campaña fue el determinante
en la mayor parte de las diferencias generadas en la composición de la grasa
intramuscular, influenciados por la materia prima (las tres campañas presentaron
distintos pesos de sacrificio) y por el procesado de cada uno de los productos.
5.2.2. CALIDAD DE LOS PRODUCTOS CURADOS
Las medidas morfológicas mostraron diferencias significativas en los tres
productos, aunque no en todas las medidas. De forma general, los productos
pertenecientes a la tercera campaña resultaron más grandes. No obstante, al extudiar el

CAPÍTULO V: DISCUSIÓN GENERAL
289
efecto del IGF2, observamos que no existieron diferencias significativas en los parámetros
morfológicos, por lo que las diferencias obtenidas al comparar campañas se debían
únicamente al efecto campaña, y por lo tanto al efecto canal, ya que los cerdos
pertenecientes a la tercera campaña fueron los más pesados, influyendo por lo tanto en las
medidas morfológicas y sobre el resto de parámetros físico químicos.
Existieron en general diferencias significativas en los parámetros físico-químicos,
pudiendo estar causadas por diversos facotres como las diferencias morfológicas o las
diferencias en el contenido de grasa subcutánea e intramuscular.
Al evaluar las características físico-químicas, nos encontamos con que las
diferencias mostradas no estuvieron influenciadas por el contenido graso de las distintas
piezas. Los jamones no mostraron diferencias en el contenido de humedad, siendo inferior
en los jamones de la tercera campaña, a pesar de tratarse de los más grandes (los animales
se sacrificaron con mayor peso), aunque el contenido de grasa subcutánea e intramuscular
no mostró diferencias entre campañas. Las diferencias se debieron por lo tanto al
procesado de las piezas, ya que el único factor que influyó sobre este parámetro fue el de
la campaña. Estas diferencias pudieron influir sobre el resto de parámetros físico-
químicos analizados, influenciados también por el efecto campaña, como es la
luminosidad, de forma que los jamones de la tercera campaña mostraron menores valores
de luminosidad.
La paleta curada mostró diferencias en todos los parámetros físico-químicos
evaluados, achacables únicamente al efecto campaña, a pesar de que en este caso la grasa
intramuscular sí mostró diferencias, que pudieron influir también sobre el resto de
parámetros. Por ejemplo, la humedad fue mayor en las piezas más grasas (primera
campaña), que mostraron también un valor de luminosidad mayor.
El lomo curado tuvo un comportamiento similar a la paleta, ya que las diferencias
se debieron principalmente al efecto campaña y en algunos casos al efecto genético. Sin
embargo, a pesar de las grandes diferencias mostradas en el contenido de grasa
intramuscular entre la primera campaña y el resto, la humedad no mostró diferencias
entre campañas, aunque si el valor de luminosidad, que resultó superior en las paletas más
grasas (primera campaña).
Es por ello, que al evaluar los resultados vemos como el alelo IGF2 se comporta de
la misma manera a los largo de las distintas generaciones de animales, ya que las

CAPÍTULO V: DISCUSIÓN GENERAL
290
diferencias producidas en los tres productos se debieron principalmente al factor
campaña.
Nuestros resultados no coinciden con la bibliografía, ya que en primer lugar Kouba
y Bonneau, (2009) demostraron que la edad del animal (y por lo tanto, el peso) influía
sobre la composición y proporción de los distintos tejidos del animal, de forma que el
tejido adiposo es el que crecía con mayor intensidad en las últimas etapas. Nuestros
resultados muestran que los productos de la tercera campaña, procedentes de cerdos
sacrificados con un mayor peso, proporcionaban muestras de mayor tamaño pero no de
mayor contenido graso en ninguno de los productos estudiados. En segundo lugar, estos
mismos autores (Kouba y Bonneau, 2009) afirmaban que la deposición de la grasa
subcutánea era mayor que la intramuscular en el jamón, e igual en la paleta, no
coincidiendo con nuestros resultados, ya que los jamones no mostraron diferencias en el
espesor de la grasa subcutánea entre campañas, pero si en el contenido de grasa
intramuscular siendo superior en los jamones de la primera campaña, que resultaron por
lo general más pequeños que el resto, sin embargo, las paletas más pequeñas (tan solo en
el parámetro de anchura) contenían mayor cantidad de grasa intramuscular y de
subcutánea.
5.2.3. CALIDAD SENSORIAL DE LOS PRODUCTOS CURADOS
Como ya se ha indicado anteriormente, la generación de compuestos volátiles es
muy variable, ya que los productos curados son muy homogéneos y además los músculos y
la grasa de las muestras pueden ser diferentes (Luna et al., 2006), pudiendo por ello
generar diferencias significativas. Además en este apartado existe una variable más que se
debe tener en cuenta; en primer lugar cada campaña se analizó en el momento de la
recepción de las muestras, transcurriendo unos meses entre campañas en cada producto,
por lo que las diferencias en el procesado de las piezas y los tiempos de muestreo pueden
generar diferencias entre campañas, ajenas a las características sensoriales del producto.
En segundo lugar, los lomos de la primera campaña fueron adobados, por lo que parte de
las diferencias significativas entre los lomos pueden deberse al adobo.
Dada la gran cantidad de compuestos volátiles identificados y la gran variedad de
lotes comparados, debido a que la mayor parte de las diferencias generadas entre lotes se
debieron al efecto campaña, la discusión se llevará a cabo evaluando tan solo las
diferencias más marcadas entre campañas, que pueden haber influido en el contenido
total de compuestos volátiles.

CAPÍTULO V: DISCUSIÓN GENERAL
291
El jamón curado presentó diferencias significativas en 105 compuestos volátiles,
siendo superior el contenido total de los jamones de la segunda campaña. Las paletas
curadas presentaron diferencias significativas en 67 compuestos, siendo superior el
contenido total de las paletas de la tercera campaña. Por último, los lomos presentaron
diferencias en 97 compuestos, donde los lomos de la segunda campaña presentaron mayor
contenido de compuestos volátiles (sin tener en cuenta, en todos los lotes, el contenido en
UAA del cloroformo, a pesar de encontrarse recogido en las tablas).
Los hidrocarburos alifáticos fueron superiores en los jamones de la segunda y
tercera campaña en comparación con los de la primera, debido generalmente por la gran
proporción de alcanos saturados con menos de 10 átomos de carbono (procedentes de la
oxidación lipídica (Ruiz et al., 2002(b)) relacionados además con la mayor proporción de
AGPI que presentaron estos lotes. Además también mostraron mayores cantidades de
alcanos ramificados y de tridecano (los jamones de la segunda campaña) cuyo origen es su
acumulación en los tejidos grasos del animal, posiblemente procedentes de la
alimentación que recibieron durante el periodo de cebo (Meynier et al., 1999; Ruiz et al.,
2002(b)). Las paletas de la primera campaña también mostraron un mayor contenido de
estos compuestos, sobre todo por las mayores proporciones de alcanos ramificados y
saturados con más de diez átomos de carbono, coincidiendo con el mayor contenido de
grasa intramuscular de ese lote. Además, las paletas de la tercera mostraron un mayor
contenido de alcanos saturados de menos de 10 átomos de carbono, coincidiendo también
con la mayor proporción de AGPI de ese lote. Los lomos de la segunda campaña mostraron
mayor contenido de estos compuestos, a pesar de que las diferencias fueron menores que
en el resto de productos. Destacó el elevado contenido de hexano en los lomos de la
primera campaña, cuyo valor no es relevante por tratarse de un compuesto relacionado
con la contaminación del ambiente (Ventanas, 2006). En los lomos de la segunda y tercera
campaña destacaron principalmente los altos valores de hidrocarburos saturados con
menos de 10 átomos de carbono, relacionados también con la mayor proporción de AGPI.
Aún así, como ya se ha indicado, la presencia de estos compuestos no influye sobre las
características sensoriales de los productos curados debido a sus elevados umbrales de
olfacción (García y Timón, 2001). Las diferencias generadas en estos compuestos se
debieron únicamente al efecto que desempeñó el factor campaña sobre los tres productos.
Los hidrocarburos aromáticos presentaron diferencias significativas entre los
jamones de las distintas campañas, debidas principalmente al mayor contenido de m- y p-
xileno en los jamones de la tercera campaña. En la paleta curada existieron diferencias
significativas, siendo superiores los valores de las paletas de la primera campaña, debido

CAPÍTULO V: DISCUSIÓN GENERAL
292
al elevado contenido de todos los hidrocarburos aromáticos que mostraron diferencias,
relacionados con la alimentación recibida por los animales y su mayor deposición en el
tejido graso (Berdagué et al., 1993; Ruiz et al., 1999(b)) y con el mayor contenido de grasa
intramuscular de ese lote. Los lomos mostraron un mayor contenido en la primera y
segunda campaña, en comparación con la tercera. El mayor contenido de estos
compuestos en la primera campaña está relacionado con el adobo (Mateo y
Zumalacarregui, 1996), sin embargo, el mayor contenido en los lomos de la segunda se
debe principalmente al mayor contenido de metilbenceno, que podría atribuirse a una
contaminación laboratorial (Muriel et al., 2004).
Los furanos, que provienen de la oxidación lipídica (Sahidi et al., 1986),
presentaron diferencias significativas entre los jamones, de forma que los de la segunda y
tercera campaña mostraron mayor contenido que los de la primera, principalmente
debido a la ausencia de compuestos como el 2-penilfurano o el 2-butilfurano en los
jamones de la primera campaña; sin embargo, las paletas de la primera campaña
mostraron mayor contenido que las de la tercera, por su mayor proporción de 2-
pentifurano y 5-metildihidro-5-etil-2(3H)-furanona, coincidiendo así con los lomos, que
presentaron mayor contenido los lomos de la primera y segunda campaña comparados
con los de la tercera. Estos compuestos están relacionados con la oxidación lipídica, más
concretamente el 2-pentilfurano, que está relacionado con la oxidación del ácido linoleico,
explicando por ello el mayor contenido de los lomos de la segunda campaña en
comparación con la primera, ya que presentaron mayor proporción de éste ácido graso,
pero no explica el mayor contenido de las paletas de la primera, que mostraron mayor
contenido de grasa pero menor proporción del ácido linoleico.
La familia de los terpenos fue superior significativamente en los jamones y las
paletas de la primera campaña, por su mayor contenido de alfa-pineno y l-limoneno. No
obstante, los lomos presentaron un mayor contenido de terpenos, siendo superior
significativamente el contenido en los lomos adobados de la primera campaña, ya que se
trata de productos especiados, en los que este tipo de compuestos se encuentran en mayor
número y cantidad (Meynier et al., 1999).
Los ésteres fueron superiores en los jamones de la primera y segunda campaña,
coincidiendo con las paletas y los lomos de la primera. Los jamones no mostraron
diferencias en el contenido de grasa intramuscular, sin embargo los lomos y las paletas de
la primera campaña presentaron mayor contenido, pero menor proporción de AGPI. A
pesar de que el origen de estos compuestos está relacionado con la oxidación lipídica en el
tejido intramuscular (Shahidi et al., 1986), su mayor liberación en los productos más

CAPÍTULO V: DISCUSIÓN GENERAL
293
grasos podría estar determinada por el carácter lipofílico de estos compuestos, explicando
por ello su mayor proporción en los lotes más grasos de las paletas y los lomos, pero no en
los jamones.
Los aldehídos constituyen una de las familias más abundantes en el análisis
cuantitativo de compuestos volátiles. A pesar de que exitieron diferencias significativas en
los jamones y las paletas, si tenemos en cuenta únicamente el efecto campaña, estas
diferencias desaparecen. Los lomos si mostraron diferencias, siendo superior los valores
de la segunda campaña, debido a los elevados valores del acetaldehído. Este compuesto
puede provenir del aminoácido cisteína por una reacción de descarboxilación que conduce
a la liberación de sulfuro de hidrógeno y la formación de una base de Schiff, que por
hidrólisis da lugar a una base de amoníaco y acetaldehído (Mottram, 1998; Pripis-Nicolau,
2000). También fueron superiores los valores de hexanal, octanal y nonanal, que
provienen principalmente de la degradación oxidativa de los ácidos grasos (Lopez et al.,
1992), a pesar de que la mayor proporción de AGPI se mostró en los lomos de la tercera
campaña.
Las cetonas deben su origen a la oxidación lipídica (Sahidi et al., 1986).
Coincidiendo con los aldehídos, al evaluar únicamente el efecto campaña, solo mostraron
diferencias significativas los lomos, siendo también el contenido de los lomos de la
segunda campaña superior, debido al elevado valor de la 2-propanona, que se mostró co-
eluída con el propanal. A pesar de no conocer el contenido exacto de cada compuesto, el
mayor contenido de aldehídos lineales de ese lote nos hace pensar que el valor de la 2-
propanona fue inferior. Aún así, la mayor parte de las cetonas fueron superiores en los
lomos de la segunda campaña, coincidiendo así con la familia de los aldehídos
El hecho de que los procesos oxidativos se hayan visto frenados por la presencia de
especias, como el ajo en los lomos, podría haber favorecido la formación de compuestos
volátiles derivados de la actividad de mohos y levaduras presentes principalmente en la
superficie de las piezas, lo que podría explicar el mayor contenido de ésteres y cetonas en
los lomos de la segunda generación. Por el contrario, los jamones y paletas, productos
procesados sin especias, ajo y pimentón, no mostraron diferencias significativas entre
campañas.
Los alcoholes representaron la segunda familia más abundante del total de los
compuestos volátiles identificados en el jamón y lomo curado y la tercera en las paletas.
Estos compuestos mostraron diferencias significativas en los tres productos, siendo
superiores los jamones y paletas de la primera campaña y los lomos de la segunda. En los

CAPÍTULO V: DISCUSIÓN GENERAL
294
jamones de la primera campaña destacó el mayor contenido de etanol, coincidiendo con
las paletas de la primera y los lomos de la segunda, producto vinculado a la fermentación
de hidratos de carbono por acción bacteriana (Montel et al., 1998), mientras que Stahnke
et al., (2002) establecieron que la formación de este compuesto podría tener lugar por
diferentes rutas. No obstante, junto con el resto de alcoholes saturados, presentaron un
umbral de olfacción alto (Drumm y Spanier, 1991), por lo que carecen de interés sensorial.
Por otro lado, tres alcoholes ramificados fueron superiores en los jamones de la segunda
campaña, coincidiendo de nuevo con las paletas de la primera y con dos de los lomos de la
segunda. Estos compuestos son productos de la reducción de sus correspondientes
aldehídos, que derivan de la degradación de los aminoácidos (Muriel, 2003). Además
destacaron los elevados valores del 2-butanol y 2,3-butanodiol en los lomos de la primera
campaña. Se ha descrito el origen del 2,3-butanodiol a partir de la fermentación por acción
de los microorganismos (Meynier et al., 1999), pudiendo ser superior por el adobo que
llevaban estos lomos.
Timón et al., (1998) encontraron altos porcentajes de ácidos carboxílicos en
jamones Ibéricos, esperados por el elevado contenido de grasa intramuscular que
presentaron los jamones, coincidiendo con nuestros resultados, ya que los productos más
grasos (paletas y lomos de la primera campaña) mostraron mayor contenido de ácidos
carboxílicos, incluso los jamones de la primera campaña, a pesar de no presentar
diferencias significativas en el contenido de grasa intramuscular.
La mayor parte de los compuestos nitrogenados detectados en los productos
cárnicos proceden de reacciones de tipo Maillard, que tienen lugar entre productos de la
oxidación lipídica y determinados aminoácidos (Ruiz et al., 2002(b)). Se mostraron
mayores cantidades de estos compuestos en los jamones de la segunda campaña y en las
paletas y lomos de la primera. El carácter polar de dichos compuestos, disminuye su
carácter lipofílico, aumentando así la liberación de estos compuestos en los lotes más
grasos, de ahí su mayor contenido en las paletas y los lomos de la primera campaña. Los
jamones no mostraron diferencias significativas en el contenido de grasa intramuscular y
la proporción de AGMI fue superior en los jamones de la primera y la de AGPI en los de la
tercera, por lo que el mayor contenido de compuestos nitrogenados en los jamones de la
segunda campaña se debieron a otras causas, como pueden ser efectos de procesado de las
muestras, ya que las diferencias se debieron únicamente al efecto campaña.
Los compuestos volátiles con azufre se forman a partir de aminoácidos azufrados
como la cisteína, la cistina y la metionina por la reacción de degradación de Strecker
(Shahidi et al., 1986). Estos compuestos fueron superiores en los jamones de la primera y

CAPÍTULO V: DISCUSIÓN GENERAL
295
tercera campaña, en las paletas de la segunda y los lomos de la tercera. Estas diferencias se
debieron especialmente al elevado valor del sulfuro de carbono, que fue el compuesto
mayoritario de esta familia en los tres productos. El dimetil disulfuro se forma por
reacciones de Strecker a partir de la metionina (Pripis-Nicalau et al., 2000), implicado en
el olor del jamón Serrano (Fores et al., 1997) por su bajo umbral de olfacción.
Finalmente, los compuestos clorados mostraron diferencias significativas en los
tres productos. A pesar de ello, el origen de estos compuestos podría ser atribuido a los
residuos de pesticidas ingeridos por los cerdos (Buscailhon et al., 1993) o a contaminación
de tipo laboratorial (Rui et al., 1999 (b); Ventanas, 2006)
Con esto podemos concluir que el factor campaña fue el mayor responsable de las
diferencias significativas que se dieron entre lotes en los tres productos estudiados; sin
embargo, no por ello el factor genético o el factor sexo no tuvieron influencia sobre la
generación de compuestos volátiles, ya que como se vio en apartados anteriores, al
estudiar la influencia de estos factores por separado, el número de diferencias
significativas seguía siendo elevado. Con esto concluimos que el efecto genético puede ser
importante en la generación de estos compuestos, por su gran relación con el contenido de
grasa intramuscular, pero estas diferencias pueden verse enmascaradas por las
diferencias generadas por otros efectos imposibles de evitar y producidos sobre las
muestras a lo largo de las distintas campañas, como son pequeños cambios en la materia
prima o especialmente en los tiempos de procesado, que son los más influyentes en la
generación de compuestos volátiles.


CAPÍTULO VI. CONCLUSIONES


CAPÍTULO VI: CONCLUSIONES
299
6. CONCLUSIONES
1. Como era de esperar las características sensoriales de la paleta y el jamón
mostraron valores muy próximos, solamente la pastosidad mostró valores
significativamente más bajos en la paleta. Así mismo, el lomo se diferenció
significativamente de los dos productos anteriores, alcanzando peores puntuaciones en
variables relacionadas directamente con la calidad, como la jugosidad o la intensidad de
olor.
2. La expresión del IGF2 se traduce en un marcado incremento del espesor de la
grasa subcutánea en jamones y paletas, siendo en éstas mucho más llamativo.
3. Se observó un mayor contenido de grasa intramuscular en los jamones
procedentes de los lotes AG, aunque este efecto no se manifestó en los lomos y en las
paletas. No obstante, en los tres tipos de piezas las puntuaciones alcanzadas en el
descriptor de veteado en el análisis sensorial mostraron valores superiores en los lotes
AG.
4. El análisis sensorial realizado sobre las paletas utilizando un panel entrenado,
mostró que las paletas procedentes de cerdos AG obtuvieron mejores puntuaciones en
distintos descriptores relacionados directamente con la calidad, como una mayor
jugosidad, veteado y brillo.
5. Aunque se han observado diferencias relacionadas con la expresión del IGF2,
son mucho más importantes las diferencias relacionadas con la campaña, por lo que el
factor procesado, influye mucho más en las características de las piezas que otros factores
como la genética o el sexo.
6. Las diferencias significativas observadas en el contenido de aminoácidos libres
en los distintos productos no siguieron una pauta uniforme. Esto hace pensar que la
proteólisis producida durante la maduración de estos productos se ve afectada por
numerosos factores que pueden interferir entre sí: como el pH, la sal, la genética, el
contenido de grasa intramuscular y la duración del procesado. Así, la mayor duración de
procesado y el menor contenido en grasa intramuscular puede favorecer la generación de
la mayor cantidad de aminoácidos que se observa en los jamones del lote AA.
7. Se han caracterizado por primera vez los volátiles presentes en lomo y paleta
curada asociados a la DOP T y se ha ampliado la identificación de los volátiles presentes en

CAPÍTULO VI: CONCLUSIONES
300
el jamón de Teruel. Las diferencias observadas entre los compuestos volátiles de los
distintos lotes son difíciles de explicar en función de los factores estudiados. De la misma
forma que ocurre en el caso de los aminoácidos, pensamos que los numerosos factores que
pueden afectar a la proteólisis y lipolisis interfieren entre si favoreciendo o no la
generación de distintos compuestos.

CAPÍTULO VII. BIBLIOGRAFÍA GENERAL


CAPÍTULO VII: BIBLIOGRAFÍA GENERAL
303
7. BIBLIOGRAFÍA
Acree, T. and Arn, H. (1997). “Flavornet Gas chromatography-olfatometry (GCO) of natural
products”. Cornell University. http://www.nysaes.cornell.edu/flavornet.
Alino, M., Grau, R., Toldrá, F. and Barat, J.M. (2010). Physicochemical changes in dry-cured
hams salted with potassium, calcium and magnesium chloride as a partial
replacement for sodium chloride. Meat Science, 86, 331-336
Andrés, A.I., Cava, R. and Ruiz, J. (2002). Monitoring volatile compounds during dry-cured
ham ripening by solid-phase microextraction coupled to a new direct extraction
decive. Journal of Chromatography A, 963, 83-88.
Andrés, A.I., Ventanas, Ventanas, S., Ventanas, J., Cava, R. and Ruiz, J. (2005).
Physicochemical changes throughout the ripening of dry cured hams with different
salt content and processing conditions. European Food Research and Technology,
221, 30-35.
Andrés, A.I., Adamsen, C.E., Moller, J.K.S., Ruiz, J. and Skibsted, L.H. (2006). High-pressure
treatment of dry-cured Iberian ham. Effect on colour and oxidative stability during
chill storage packed in modified atmosphere. European Food Research and
Technology, 222, 486-491.
Ansorena, D., Gimeno, O., Astiasarán, I. and Bello, J. (2001). Analysis of volatile compounds
of GC-MS of a dry-fermented sausage: chorizo de Pamplona. Meat Science, 34, 67-
75.
Antequera, T., López-Bote, C.J., Córdoba, J.J., García, C., Asensio, M.A., Ventanas, J., García-
Regueiro, J.A. and Díaz, I. (1992). Lipid oxidative changes in the processing of
Iberian pig hams. Food Chemistry, 45, 105-110.
AOAC (1984). Method 24010. Official Methods of Analysis. S. Williams (ed.), Arlintgton,
V.A. Washinton, DC, Association of Official Analytical Chemist.
Aristoy, M.C. and Toldrá, F. (1998). Concentration on free amino acids and dipeptides in
porcine skeletal muscles with different oxidative patterns. Meat Science, 50, 327-
332.
Aristoy, M.C. and Toldrá, F. (2009). Nucleotides and its derived compounds. In M.L. Nollet
& F. Toldrá (Eds.), Handbook of muscle foods analysis, 279−288. Boca Raton, FL,
USA: CRC Press.
Armero, E., Basega, M., Aristoy, M. C. and Toldrá, F. (1999). Effects of sire type and sex on
pork muscle exopeptidase activity, natural dipeptides and free amino acids. Journal
of the Science of Food and Agriculture, 79, 1280–1284.

CAPÍTULO VII: BIBLIOGRAFÍA GENERAL
304
Arnau, J., Guerrero, L. and Gou, P. (1997). Effects of temperature during the last month of
ageing and of salting time on dry-cured ham aged for six months. Journal of the
Science of Food and Agriculture, 74, 193-198.
Arnau, J. (1998). Principales problemas tecnológicos en la elaboración de jamón curado.
Special Symposium – 44th ICoMST, Barcelona, pp. 72–86
Arnau, J., Gou, P. and Comaposada, J. (2003). Effect of the relative humidity of drying air
during the resting period on the composition and appearance of dry-cured ham
surface. Meat Science, 65, 1275-1280.
Baines, D.A. and Mlotkiewicz, J.A. (1984). The chemistry of meat flavor. En Recent
advances in the chemistry of meat; Bailey, A.J. Ed. The Royal Society of Chemistry,
Burlington House, London. Pp 119-164.
Bailey, M.E., Rourke, T.J., Gutheil, R.A. and Wang, C.Y.J. (1992).En: Off-flavors in Foods and
Beverages. Ed: G. Charalambous. Elsevier Science Publishers B.V. Amsterdam, 7.
Barat, J.M., Vidal-Brotons, D., Lopez-Pascual, E. and Grass, M.L. (2006). Quantifications and
kinetics of the residual brine generation during ham and shoulder pile salting.
Meat Science, 73, 576-580.
Barbieri, G., Bolzoni, L., Parolari, G., Virgili, R., Buttini, R., Careri, M. and Mangia, A. (1992).
Flavor compounds of dry-cured ham. Journal of Agricultural and Food Chemistry,
40, 2389-2394.
Barea, R., Nieto, R., Lara, L., García, M.A., Vilchez, M.A. and Aguilera, J.F. (2003). Effects of
dietary protein content and feeding level on carcass characteristics and organ
weights of Iberian pigs growing between 50 and 100 kg live weight. Animal
Science, 82, 405-413.
Belitz, H.D. & Grosch, W. (1997). “Química de los alimentos”. Ed. Acribia, Zaragoza
Berdague, J.L., Denoyer, J.L., Le Quere, J.L. and Semon, E. (1991). Volatile components of
dry-cured ham. Journal of Agricultural and Food Chemistry, 39, 1257-1261.
Berdagué, J.L., Bonnaud, N., Rousset, S. and Touraille, C. (1993). Influence of pig crossbreed
on the composition, volatile compound content and flavor of dry cured ham. Meat
Science, 34, 119-129.
Bligh, E.G. & Dyer, W.J. (1959). A rapid method of total lipid extraction and purification.
Canadian Journal of Biochemistry and Phisiology, 37, 911-917.
BOA (1993) (a). Orden de 29 de julio de 1993, del Departamento de Agricultura, Ganadería
y Montes, por la que se aprueba el Reglamento de la Denominación de Origen
“Jamón de Teruel” y su Consejo Regulador. Boletín Oficial de Aragón 93, 3168-3177
BOA (1993) (b). ORDEN de 19 de octubre de 1993, del Departamento de Agricultura,
Ganadería y Montes, por la que se aprueba el Reglamento de utilización de la

CAPÍTULO VII: BIBLIOGRAFÍA GENERAL
305
marca. "Aragón Calidad Alimentaria" para el lomo embuchado. Boletín Oficial de
Aragón, 125, 3855-3856.
BOA (2005). Orden de 18 de febrero de 2005, del Departamento de Agricultura y
Alimentación, por la que se modifica el Reglamento de la Denominación de Origen
Protegida “Jamón de Teruel” y su Consejo Regulador. Boletín Oficial de Aragón, 28,
2985
BOA (2008). ORDEN de 19 de mayo 2008, del Consejero de Agricultura y Alimentación,
por la que se aprueba el Reglamento Técnico para la utilización de la marca
«C’alial» en la «paleta curada o paletilla». Boletín Oficial de Aragón,87
BOE (2011). Boletín Oficial del Estado, 20, pag. 6676
http://www.boe.es/boe/dias/2011/01/24/pdfs/BOE-B-2011-2392.pdf
Bourne, M.C. (1978). Texture profile Analysis. Food Technology, 62-66
Briskey, E. J., Bray, R.W., Hoekstra, W.G., Philips, P.H. and Grummer, R.H. (1959). The
chemical and physical characteristics of various pork ham muscle classes. Journal
of Animal Science, 18, 146-152
Bruna, J.M., Hierro, E.M., de la Hoz, L., Mottram, D.S., Fernández, M. and Ordoñez, J.A.
(2001). The contribution of Penicillium aurantiogriseum to the volatile
composition and sensory quality of dry fermented sausages. Meat Science, 59, 97-
107.
Burgos, C., Carrodeguas, J.A., Moreno, C., Altarriba, J.,Tarrafeta, L., Barcelona, J. A. and
López-Buesa, P.(2006). Allelic incidence in several pig breeds of a missense variant
of pig melanocortin-4 receptor (MC4R) gene associated with carcass and
productive traits; its relation to IGF2 genotype. Meat Science, 73, 144-150.
Burgos, C., Carrodeguas, J.A., Tarrafeta, L y López-Buesa, P. (2008). Genética molecular y
selección porcina; el caso de IGF2 y MC4R. Eurocarne, 170
Burgos, C., Moreno, C., Carrodeguas, J.A., Barcelona, J.A.,Tarrafeta, L. and López-Buesa, P.
(2010). The influence of diet, sex, IGF2 and RYR1 genotypes and anatomical
location on pork loin composition. Journal of Food Composition and Analysis, 23,
307-313
Burgos, C., Galve, A, Moreno, C., Altarriba, J., Reina, R., García, C. and López-Buesa, P.
(2011). The effectiveness of IGF2 to enhance fat content in pig carcasses and pork
to improve cured ham quality (En revisión).
Buscailhon, S., Berdagué, J.L. and Monin, G. (1993). Time related changes in volatile
compounds of lean tissue during processing of French dry-cured ham. Journal of
Agricultural and Food Chemistry 63, 69-75.

CAPÍTULO VII: BIBLIOGRAFÍA GENERAL
306
Buscailhon, S., Monin, G., Cornet, M. and Bousset, J. (1994). Time-related changes in
nitrogen fractions and free amino acids of lean tissue of French dry-cured. Meat
Science, 37, 449-456.
Buscailhon, S., Gandemer, G. and Monin, G. (1994). Time-related changes in intramuscular
lipids of French dry-cured ham. Meat Science, 37, 245-255.
Careri, M., Alessandro, M., Barbieri, G., Bolzoni, L., Virgili, R. and Parolari, G. (1993).
Sensory property relationships to chemical-data of Italian-type dry-cured ham.
Journal of Food Science, 58, 968-972.
Cárnica 2000 (2000). Implantación de un sistema de control de la trazabilidad del Jamón
de Teruel. Cárnica 2000, 27, 87
Careri, M., Alessandro, M., Barbieri, G., Bolzoni Virgili, R. and Parolari, G. (1993). Sensory
property relationships to chemical data of Italian type dry-cured ham. Journal of
Food Science, 58, 968–972.
Carrapiso, A.I., Jurado, A., Timon, M.L. and García, C. (2002). Odor-active compounds of
Iberian hams with different aroma characteristics. Journal of Agricultural and Food
Chemistry, 50, 6453-6458.
Carrapiso, A.I., Bonilla, F. and García, C. (2003). Effect of crossbreeding and rearing system
on sensory characteristics of Iberian ham. Meat Science, 65, 623–629.
Carrapiso, A.I., Jurado, A., Martin, L. and García, C. (2007). The duration of the outdoor
rearing period of pigs influences Iberian ham characteristics. Irish Journal of
Agricultural and Food Research, 46, 105-115.
Carrapiso, A.I., Martin, L., Jurado, A. and García, C. (2010). Characterisation of the most
odour-active compounds of bone tainted of dry-cured Iberian ham. Meat Science,
85, 54-58.
Carrodeguas, J.A., Burgos, C., Moreno, C., Sánchez, A.C., Ventanas, S., Tarrafeta, L.,
Barcelona, J.A., López, M.O., Oria, R. and López-Buesa, P. (2005). Incidence in
diverse pig populations of an IGF2 mutation with potential influence on meat
quality and quantity. An assay based on real time PCR (RT-PCR). Meat Science, 71,
577-582.
Cava, R., Ruiz, J., López-Bote, C., Martín, L., García, C., Ventanas, J. and Antequera, T. (1997).
Influence of finishing diet on fatty acid profiles of intramuscular lipids,
tryglicerides, and phospholipids in muscles of the Iberian pig. Meat Science, 45, 2,
263-270.
Cava, R., Ruiz, J., Ventanas, J. and Antequera, T. (1999). Oxidative and lipolytic changes
during ripening of Iberian hams as affected by feeding system: extensive feeding
and alpha-tocopheryl acetate supplementation. Meat Science, 52, 165-172.

CAPÍTULO VII: BIBLIOGRAFÍA GENERAL
307
Cava, R., Ferrer, J.M., Estévez, M., Morcuende, D. and Todrá, F. (2004). Composition and
proteolytic and lypolitic enzyme activities in muscle Longissimus Dorsi from
Iberian pigs and industrial genotype pigs. Food Chemistry, 88, 25-33.
Chang S.S. and Petersen, R.J. (1977). Recent developments in the flavour meat. Journal of
Food Science 42, 298-305.
Charpentier, J. (1967). Calidad de la canal y de la carne. En: CNRZ. Jouy-en-Josas, Francia.
Cilla, I., Martinez, L, Beltran, J.A. and Roncales, P. (2006) (a). Dry-cured ham quality and
acceptability as affected by the preservation system used for retail sale. Meat
Science, 73, 581-589.
Cilla, I., Martinez, L, Guerrero, L., Guardia, M.D., Arnau, J., Altarriba, J. and Roncales, P.
(2006) (b). Consumer beliefs and attitudes towards dry-cured ham and protected
designation of origin teruel ham in two Spanish regions differing in product
knowledge. Food Science and Technology Internathional, 12, 229-240.
Cilla, I., Altarriba, J., Guerrero, L., Gispert, M., Martínez, L., Moreno, C., Beltrán, J.A., Guardia,
M.D., Diestre, A., Arnau, J. and Roncalés, P. (2006). Effect of different Duroc line
sires on carcass composition, meat quality and dry-cured ham acceptability. Meat
Science, 72, 252-260.
Ciobanu, D., Bastiaansen, J., Malek, M., Helm, J., Woollard, J., Plastow, G. and Rothschild, M.
(2001). Evidence for New Alleles in the Protein Kinase Adenosine Monophosphate-
Activated 3-Subunit Gene Associated With Low Glycogen Content in Pig Skeletal
Muscle and Improved. Meat Quality Genetics, 159, 1151–1162.
Civille, G.V. and Szczesniak, A.S. (1973). Guidelines to training a texture profile panel.
Journal of Texture Studies, 4, 204-233.
Costa, M.D., Bergamin, W., Silveira, E.T.F. and de Felicio, P.E. (2008). Colour and texture
profiles of boneless reestructured dry-cured hams compared to traditional hams.
Scientia Agricola, 65, 169-173.
Cournet, M. and Bousset, J. (1999). Free amino acids and dipeptides in porcine muscles:
differences between 'red' and 'white' muscles. Meat Science, 51, 215-219.
Cross, H.R., Durland, P.R. and Seideman, S.C. (1986). Sensory Qualities of Meat. En: Muscle
as Food. Food Science and Technology. Ed. P.J. Bechtel. Seires ed. B.S. Schweigert.
Academic Press, Inc. New York.
Cruz, J. (2007). La evolución del sector productor de jamón curado en España. Eurocarne,
154
Cruz, J. (2009). El sector jamonero busca en las exportaciones una salida ante el
estancamiento y la competencia en el mercado nacional. Eurocarne, 174

CAPÍTULO VII: BIBLIOGRAFÍA GENERAL
308
Cruz, J. (2010). El crecimiento del consumo y las exportaciones hacen augurar cierta
recuperación del sector jamonero. Eurocarne, 189
Davies, A.S. and Pryor, W.J. (1977). Growth changes in the distribution of dissectable and
intramuscular fat in pigs. Journal of Agricultural Science, 89, 257-266.
Daza, A., Latorre, M.A. and Lopez-Bote, C.J. (2010). The use of barley as single ingredient in
the diet provided during the finishing period may improve the meat quality of
heavy pigs from PO Teruel ham (Spain). Spanish Journal of Agricultural Research, 8,
607-616.
De la Hoz, L., López, M.O., Hierro, E., Cambero, M.I. and Ordoñez, J.A. (1996). Effect of diet
on the fatty acid composition of intramuscular and intermuscular fat in Iberian pig
cured ham. Food Science and Technology International, 2, 391-397.
De Roos, K.B. (1997). How lipids influence food flavor. The Chemistry of Flavor
Interactions. Food Technology, 51, 60-62.
Diestre, A., Oliver, M.A., Gispert, M., Arpa, I. and Arnau, J. (1990). Consumer responses to
fresh meat and meat products from barrows and boars with different levels of boar
Saint. Animal Production, 50,519-530.
Drumm, T.D. and Spanier, A.M. (1991). Changes in the content of lipid autoxidation and
sulphur-containing compounds in cooked beef storage. Journal of Agricultural and
Food Chemistry, 39, 336-343.
Dumont, B.L. and Roy, G. (1975). Comparative studies of ham characteristics in Landrace-
Francais, Landrace-Belge and Pietrain pigs. Annales de Zootechnie, 24, 803-803.
Edwards, S.A., Wood, J.D., Moncrieff, C.B. and Porter, S.A. (1992). Composition of the Duroc
and Large White as a terminal sire breeds and their effect on pigment quality.
Animal Production, 54, 289–297
Edwars, R.A., Ordóñez, J.A., Dainty, R.H., Hierro, E.M. and de la Hoz, L. (1999).
Characterization of the headspace volatile compounds of selected Spanish dry
fermented sausages. Food Chemistry, 64, 461-465.
Elmore, J.S., Campo, M.M., Enser, M. and Mottram, D. (2002). Effect of lipid composition on
meat-like model system containing cysteine, Ribose,and polyunsaturated fatty
acids. Journal of Agricultural Food Chemistry, 50, 1126-1132.
Elmore, J.S., Cooper, S.L., Enser, M., Mottram, D.S., Sinclair, L.A., Wilkinson, R.G. and Wood,
J.D. (2005). Dietary manipulation of fatty acid composition in lamb and meats and
its effect on the volatile aroma compounds on grilled lamb. Meat Science, 69, 233-
242.
Estévez, M., Morcuende, D., Ventanas, S. and Cava, R. (2003). Analysis of volatiles in meat
from Iberian pigs and lean pigs after refrigeration and cooking by using SPME-GC-
MS. Journal of Agricultural and Food Chemistry, 51, 3429-3435.

CAPÍTULO VII: BIBLIOGRAFÍA GENERAL
309
Fandos, C., Espejel, J. y Flavián, C. (2007). La importancia de la Denominación de Origen
Protegida Jamón de Teruel para su consolidación en el mercado. Eurocarne, 159
Fennema, O.R. & Grosch, W. (1992). “Química de los alimentos”. Ed. Acribia, Zaragoza
Fisher, A.V., Greenb, D.M., Whittemoreb, C.T., Wood, J.D. and Schofield, C.P. (2003). Growth
of carcass components and its relation with conformation in pigs of three types. 65,
639-650
Flores, J., Bermell, S., Nieto, P. and Costell, E. (1984). Chemical-changes in protein during
the slow and rapid maturation processes of dry-cured ham and its relation with
quality. Revista de Agroquímica y Tecnología de los Alimentos, 24, 503-509.
Flores, M., Grimm, C., Toldrá, F. and Spanier, A. (1997). Correlations of sensory and volatile
compounds of Spanish “Serrano” dry-cured ham as a function of two processing
times. Journal of Agricultural of Food Chemistry, 45, 2178-2186.
Flores, M., Biron, C., Izquierdo, L. and Nieto, P. (1998). Characterization of green hams
from Iberian pigs by fast analysis of subcutaneous fat. Meat Science, 23, 253-262.
Forss, D.A. (1972). Odor and flavor compounds from lipids. Progress in the Chemistry of
Fats and Other Lipids, 13, 181-258
Fuentes, V., Ventanas, J., Morcuende, D., Estévez, M. and Ventanas, S. (2010). Lipid and
protein oxidation and sensory properties of vacuum-packaged dry-cured ham
subjected to high hydrostatic pressure. Meat Science, 85, 506-514.
Fujii, J., Otsu, K., Zorzat, F., de León, S., Khanna, V.K., Weiler, J.E., O´Brien, P.J. and Mac
Lennan, D.H. (1991). Identification of a mutation in porcine ryanodine receptor
associated with malignant hyperthermia. Science, 253, 448-451.
Gandemer, G. (1997). Phospholipides, lipolyse, oxydation et flaveur des produits carnés.
OCL, 4, 1-7.
Gandemer, G. (1998). Lipids and meat quality. Lipolysis, oxidation, and flavor. In the
proceedings of the 44th ICoMST, 30-August-4 September, Barcelona, Spain
Gandemer, G. (1999). Lipids and meat quality: lipolysis, oxidation, maillard reaction and
flavour. Science of Aliments, 19, 439-458.
Gandemer, G. (2002). Lipids in muscles and adipose tissues, changes during processing
and sensory properties of meat products. Meat Science, 62, 309–321
Gandemer, G. (2009). Dry cured ham quality as related to lipid quality of raw material and
lipid changes during processing: A review. Grasas y Aceites, 60, 297–307
García, C., Berdagué, J.J., Antequera, T., López Bote, C., Córdoba, J.J. and Ventanas, J. (1991).
Volatile components of dry-cured Iberian ham. Food Chemistry, 41, 23-32
García, C. and Timón, M.L. (2001). Los compuestos responsables del “flavor” del jamón
Ibérico. Variaciones en los distintos tipos de jamones. En: Tecnología del Jamón

CAPÍTULO VII: BIBLIOGRAFÍA GENERAL
310
Ibérico: de los sistemas tradicionales a la explotación racional del sabor y el aroma.
J. Ventanas, ed., Ediciones Mundi-Prensa, Madrid, 15-43
García-González, D.L., Luna, G., Morales, M.T. and Aparicio-Ruiz, R. (2005). Chemical
analysis of dry cured ham aroma by electronic nose. In TYPIC final public
conference (pp. 16–26), December 2005, Clermont-Ferrand: France.
García-González, D.L.; Tena, N.; Aparicio-Ruiz, R. and Morales, M.T. (2008). Relationship
between sensory attributes and volatile compounds qualifying dry-cured hams.
Meat Science, 80, 315-325.
García-Rey, R., Quiles-Zafra, R., Tapiador, J. and Luque de Castro, M. (2004). Relationship
between pH before salting and dry-cured ham quality. Meat Science, 67, 625-632.
Gerbens, F., van Erp, A.J.M., Harders, F.L., Verburg, F.J., Meuwissen, T.H.E., Veerkamp, J.H.
and te Pas, M.F. (1999). Effect of genetic variants of the Heart Fatty Acid-Binding
Protein gene on intramuscular fat and performance traits in pigs. Journal of Animal
Science, 77, 846-852.
Gerbens, F., de Koning, D.J., Harders, F.L., Meuwissen, T.H.E., Janss, L.L.G., Groenen, M.A.M.,
Veerkamp, J.H., Van Arendonk, J.A.M. and te Pas, M.F. (2000). The effect of
adypocite and heart fatty acid-binding protein genes on intramuscular fat and
backfat content in Meishan crossbreed pigs. Journal of Animal Science, 78, 552-559.
Girard, J.P., Goutefongea, R., Monin, G. and Touraille, C. (1986). Qualities des viands de
porc. In J. M. Perz, P. Mornet, & A. M. Rerat (Eds.), Le Porc et son elevage (pp. 461–
480).
Girard, J.P. and Bucharles, C. (1991). La acidificación. En: Tecnología de la carne y los
productos cárnicos. Girard J.P. Zaragoza, España. Acribia S.A.
Gispert, M., Valero, A., Oliver, M. A. y Diestre, A. (1997). Problemas asociados a la falta de
grasa en canales porcinas. Eurocarne, 61, 27-32.
Godber, J.S. (1994). Nutritional Value of Muscle Foods. En: Muscle Foods. Meat Poultry and
Seafood Technology. Eds. D.M. Kinsman, A.W. Kotula, B.C. Breidenstein. Chapman &
Hall, New York.
Gorospe, O., Sanchez-Monje, J.M. & Bello, J. (1986). Eliminación de interferencias en el
estudio químico del color de productos cárnicos comerciales, provocadas por
sustancias colorantes presentes. Alimentaria, 25-32.
Gou, P. (1993). Comparación de las características de crecimiento, canal, carne fresca y
jamón curado de cinco cruzamientos en porcino. Tesis Doctoral. Universidad
Politécnica de Valencia.
Gou, P., Guerrero, L. and Arnau, J. (1995). Sex and crossbreed effects on the characteristics
of dry-cured ham. Meat Science, 40, 21-31.

CAPÍTULO VII: BIBLIOGRAFÍA GENERAL
311
Grosch, W. (1987). Reactions of hydroperoxide products of low molecular weights. En:
Autooxidation of Unsaturated Lipids. Ed.: Chan, H.S.W. Academis Press. London.
Guinard, J.X., Wee, C., McSunas, A. and Fritter, D. (2002). Flavour release from salad
dressing varying in fat and garlic flavor. Food Quality and Preference 13, 129-137.
Hammond, J.A. (1952). Physiological limits to intense production in animals. British
Agricultural, Bulletin 4, 222-224.
Hansen-Moller, J. (1997). Evaluation of peptides generated in Italian-style dry cured ham
during processing. Journal of Agricultural and Food Chemistry, 45, 3123-3128.
Heck, M.C. and Brown, C.J. (1961). Relathionship between carcass measurements and
organoleptic quality in commercial and Country-Style cured ham. Journal of Animal
Science, 20, 589-592.
Hernández-Cázares, A.S., Aristoy, M.C. and Toldrá, F. (2011). Nucleotides and their
degradation products during processing of dry-cured ham, measured by HPLC and
an enzyme sensor. Meat Science, 87, 125-129.
Hertz, K.O. and Chang, S.S. (1970). Meat flavor. Advances in Food Research, 18, 1-83
Hidalgo, F.J. and Zamora, R. (2004). Strecker-type degradation produced by the lipid
oxidation products 4,5-epoxy-2-alkenals. Journal of Agricultural and Food
Chemistry, 52, 7126–7131.
Hinrichsen, L.L. and Andersen, H.J. (1994). Volatile compounds and chemical changes in
cured pork: role of three halotolerant bacteria. Journal of Agricultural and Food
Chemistry, 42, 1537-1542.
Hinrichsen, L.L. and Pedersen, S.B. (1995). Relationship Among Flavor, Volatile
Compounds, Chemical-Changes, and Microflora in Italian-Type Dry-Cured Ham
During Processing. Journal of Agricultural and Food Chemistry 43, 2932-2940.
Hornsey, H.C. (1956). The color of cooked cured pork. 1. Estimation of the nitric oxide ham
pigments. Journal of the Science of Food and Agriculture, 7,534-541.
Hwang, H.I., Hartman, T.G., Rosen, R.T., Lech, J. and Ho, C.T. (1994). Formation of Pyrazines
from the Maillard reaction of glucose and lysine-amine-15N. Journal of Agricultural
and Food Chemistry, 42, 1000-1004.
Hwang, H.I., Hartman, T.G. and Ho, C.T. (1995). Relative Reactivities of Amino Acids in
Pyrazine formation. Journal of Agricultural of Food Chemistry, 43, 179-184.
ICEX. Instituto Español de Comercio Exterior. www.icex.es
Isabel, B., Timón, M., Cava, R., García, C., Ruiz, J., Carmona, J.M., Soares, M. and Bote, C.J.L.
(1999). Dietary alpha-tocopheryl acetate supplementation modifies volatile
aldehyde and sensory properties of dry-cured hams. Irish Journal of Agricultural
and Food Research, 38, 137-142.

CAPÍTULO VII: BIBLIOGRAFÍA GENERAL
312
I.S.O. 937 (1978). Meat and meat product. Determination of nitrogen content.
Organización Internacional para la estandarización.
I.S.O. 1442 (1997). Meat and meat product. Determination of moisture content.
Organización Internacional para la estandarización.
Jacobsen, T. and Hinrichsen, L. (1997). Bioformation of flavour by Penicillium candidum,
Penicillium nalgiovense and Geotrichum candidum on glucose, peptone, maize oil
and meat extract. Food Chemistry, 60, 409-416.
Jeon, J.T., Carlborg, Ö., Törnsten, A., Giuffra, E., Amarger, V., Chardon, P., Andersson-Eklund,
L., Anderson, K., Hansson, I., Ludström, K. and Anderson, L. (1999). A paternally
expressed QTL affecting skeletal and cardiac muscle mass in pigs maps to the IGF2
locus. Nature Genetics, 21, 157-158.
Jeremiah, L.E. (1978). A review of factors affecting meat quality. Lacombe Research Station
Technical, Bulletin Number 1, Lacombe, Alberta, Canada.
Jiménez-Colmenero, F., Ventanas, J. and Toldrá, F. (2010). Nutritional composition of dry-
cured ham and its role in a healthy diet. Meat science, 84, 585-593.
Jo, C. and Ahn, D.U. (1999). Fat reduces volatiles production in oil emulsion system
analyzed by purge-and-trap dynamic headspace/gas chromatography. Journal of
Food Science, 64, 641-643.
Jurado, A. (2005). Evolución de compuestos relacionados con el sabor y el aroma del
jamón ibérico de distintas calidades durante su procesado. Tesis Doctoral.
Universidad de Extremadura.
Jurado, A., García, C., Timón, M. and Carrapiso, A.I. (2007). Effect of ripening time and
rearing system on amino acid-related flavour compounds of Iberian ham. Meat
Science, 75, 585-594
Jurado, A., Carrapiso, A.I., Ventanas, J. y García, G. (2009). Changes in SPME-extracted
volatile compounds from Iberian ham during ripening. Grasa y aceites, 60(3), 262-
270.
Kaluzny, M.A., Duncan, L.A., Merrit, M.V. and Epps, D.V. (1985). Rapid separation of lipid
classes in high yield and purity using bonded phase columns. Journal of lipid
research, 26, 135-140.
Kamm, L. and Coffin, E. (1968). An evaluation of method for determination of added
phosphate in ham. Journal of the Association of Official Analytical Chemist, 51, 968
Kandler, O. (1983). Carbohydrate metabolism in lactic acid bacteria. Antoine van
Leeuwenhoek, 49, 209-224.
Kato, H and Nishimura, T. (1987). Taste compounds and conditioning beef, pork and
chicken. En: Kawamura, Y.; Kare, M.R, ed. Umami: A basic taste. New York: Marcel
Dekker: 289-306.

CAPÍTULO VII: BIBLIOGRAFÍA GENERAL
313
Kato, H., Ra Rhue, M. and Nishimura, T. (1989). Role of free amino acids and peptides in
food taste. In R. Teranishi, R. G. Buttery & F. Shahidi (Eds.), Flavor chemistry, trends
and developments. ACS Symposium series 388. Washington, ACS, pp 158–17
Kawai, M., Okiyama, A. and Ueday, U. (2002). Taste enhancements between various amino
acids and IMP. Chemical senses, 27, 739-745.
Kemp, J.D., Langlois, B.E., Fox, J.D. and Varney, W.Y. (1975). Effects of curing ingredients
and holding times and temperatures on organoleptic and microbiological
properties of dry-cured sliced ham. Journal of Food Science, 40, 634-636.
Kempster, J.A. and Evans, D.G. (1979). The effects of genotype, sex and feeding regimen on
pig carcass development. 2. Tissue weight distribution and fat partition between
depots. Journal of Agricultural Science, 93, 349-358.
Kim, K.S., Larsen, N., Short, T., Plastow, G. and Rothschild, M.F. (2000). A missense variant
of the melanocortin-4-receptor (MC4R) gene is associated with fatness, growth,
and feed intake traits. Mammalian Genome, 11, 131-135.
Kingsley, G.R., Graham, P.P. and Young, R.W. (1978). Effects of frozen storage and dry-
curing on ham trygliceride fatty acids. Journal of Food Science, 43, 479-498.
Kondjoyan, N & Berdagué, J.L. (1996). A compilation of relative retention índices for the
analisys of aromatic compounds. INRA de theix. Saint Genes Champanelle. Francia.
Kouba, M. and Bonneau, M. (2009). Compared development of intramuscular and
subcutaneous fat in carcass and primal cuts of growing pigs from 30 to 140 Kg
body weight. Meat Science, 81, 270-274
Kuntapan, C., Fox, J.D., Moody, W.G. and Kemp, J.D. (1972). Cured ham properties as
affected by fresh ham quality. Journal of Animal Science, 35, 204
Larrea, V., Hernando, I., Quiles, A., Lluch, M.A. and Pérez-Munuera, I. (2006). Changes in
protein during Teruel dry-cured ham processing. Meat Science, 74, 586-593.
Latorre, M.A. (2003). Efecto del sexo, el tipo genético y el peso al sacrificio sobre la
productividad, la calidad de la canal y de la carne de cerdos pesados. Tesis Doctoral.
Universidad Politécnica de Madrid
Latorre, M.A., Mocé, M.L. y Fernández, C. (2005). El jamón de la D.O.P. Jamón de Teruel.
Eurocarne, 139, 139-146.
Latorre, M.A., García-Belenguer, E. and Arino, L. (2008). The effects of sex and slaughter
weight on growth performance and carcass traits of pigs intended for dry-cured
ham from Teruel (Spain). Journal of Animal Science, 86, 1993-1942.
Latorre, M.A., Ripoll, G., García-Belenguer, G. and Arino, L. (2009). The effect of gender and
slaughter weight on loin and fat characteristics of pigs intended for Teruel dry-
cured ham production. Spanish Journal of Agricultural Research, 7, 407-416.

CAPÍTULO VII: BIBLIOGRAFÍA GENERAL
314
Lawrie, R.A. (1991). Meat Science. Pergamon Press.
Lawrie, R.A. (1998). Ciencia de la carne. Ed. Acribia, Zaragoza
Lonergan, S.M., Huff-Lonergan, E., Rowe, L. J., Kuhlers, D. L. and Jungst, S.B. (2001).
Selection for lean growth efficiency in Duroc pigs influences pork quality. Journal of
Animal Science, 79, 2075-2085.
López, M.O., de la Hoz, L., Cambero, M.I., Gallardo, E., Reglero, G. and Ordóñez, J.A. (1992).
Volatile compounds of dry hams from Iberian pigs. Meat Science, 31, 267–277.
Luna, G., Aparicio, R. and García-Gonzalez, D. (2006). A tentative characterization of white
dry-cured hams from Teruel (Spain) by SPME-GC. Food Chemistry, 97, 621-630.
Madsen, O. Wilson, G.D. and Weir, C.E. (1957). Relationships between polyphosphates and
salt impurities in ham curing. Food Technology, 11, 15-15
MARM (2009). Datos de las Denominaciones de Origen Protegidas (D.O.P.) e Indicaciones
Geográficas Protegidas (I.G.P.) de Productos Agroalimentarios
(http://www.marm.es/es/alimentacion/temas/calidad-
agroalimentaria/INFORME_FINAL_VINCULADO_A_EXCEL_2009_tcm7-152858.pdf)
MARM. Ministerio de Medio Ambiente y Medio Rural y Marino.
http://www.marm.es/es/alimentacion/temas/calidad-agroalimentaria/calidad-
diferenciada/
Martin, L., Córdoba, J.J., Ventanas, J. and Antequera, T. (1999). Changes in intramuscular
lipids during ripening of Iberian dry-cured ham. Meat Science, 51, 129-134.
Martin, L., Timón, M.L., Petron, M.J., Ventanas, J. and Antequera, T. (2000). Evolution of
volatile aldehydes in Iberian ham matured under different processing conditions.
Meat Science, 54, 333-337.
Martín, L., Antequera, T., Ventanas, J., Benítez-Donoso, R. and Córdoba, J.J. (2001). Free
amino acids and other non-volatile compounds formed during processing of
Iberian ham. Meat Science, 59, 363-368.
Martín, D. (2008). Advances in the study of conjugated lin0leic acid (CLA) in swine feeding
and meat products: effect of combination with monounsaturated fatty acids. Tesis
Doctoral. Universidad de Extremadura.
Martín, V. (2010). Consumo de Jamón en España. Distribución y consumo, 110.
(http://www.marm.es)
Mateo, J. and Zumalacarregui, J.M. (1996). Volatile compounds in chorizo and their
changes during ripening. Meat Science, 44, 255-273
Mayoral, A.I., Dorado, M., Guillén, M.T., Robina, A., Vivo, J.M., Vázquez, C. and Ruiz, J.
(1999). Development of meat and carcass quality characteristics in Iberian pigs
reared outdoors. Meat Science, 52, 315-324.

CAPÍTULO VII: BIBLIOGRAFÍA GENERAL
315
Meynier, A., Genot, C. and Gandemer, G. (1999). Oxidation of muscle phospholipids in
relation to their fatty acid composition with emphasis on volatile compounds.
Journal Science of Food Agricultural, 79, 797-804.
Milan, D., Jeon, J.T., Looft, C., Amager, V., Thelander, M., Robic, A., Rogel-Gaillard C., Paul, S.,
Iannuccelli, N., Rask, L., Ronne, H., Lundström, K., Reinsch, N., Gellin, J., Kalm, E.,
Roy, P.L., Chardon, P. and Andersson, L. (2000). A mutation in an AMP-activated
protein kinase subunit is associated with excess glycogen in pig skeletal muscle.
Science, 288, 1248-1251.
Miller, R.C. and Ziegler, P.T. (1936). The progress of the distribution of the salt in ham
during the curing process. Journal of Agricultural Research, 52, 225-232
Miranda-Lopez, R.; Bibbey, L.M.; Watson, B.T. and McDaniel, M.R. (1992). Odor analysis of
pinot noir wines from grapes of different maturities by a gas chromatography-
olfactometry technique (Osme). Journal of Food Science, 57, 985-993.
Möhler, K. (1982). Ciencia y tecnología de la carne. Teoría y práctica. El curado. Editorial
Acribia. Zaragoza. pp 67-81.
Montel, M.C., Masson, F. and Talon, R. (1998). Bacterial role in flavour development. Meat
Science, 49, S111-S123.
Monziols, M., Bonneau, M., Davenel, A. and Kouba, M. (2005). Tissue distribution in pig
carcasses exhibiting large differences in their degree of leanness, with special
emphasis on intermuscular fat. Livestock Production Science 97, 267–274
Monziols, M., Bonneau, M., Davenel, B. and Kouba, M. (2007). Comparison of the lipid
content and fatty acid composition of intermuscular and subcutaneous adipose
tissues in pig carcasses. Science Direct, 76, 54-60.
Mora, L., Sentandreu, M.A., Koistinen, K.M., Fraser, P.D., Toldrá, F. and Bramley, P.M.
(2009). Naturally generated small peptides derived from myofibrillar proteins in
Serrano dry-cured ham. 57, 3228-3234.
Morales, R., Serra, X., Guerrero, L. and Gou, P. (2007). Softness in dry-cured porcine biceps
femoris muscles in relation to meat quality characteristics and processing
conditions. Meat Science, 77, 662-669.
Morales, R., Guerrero, A., Claret, A., Guardia, M.D. and Gou, P. (2008) (a). Beliefs and
attitudes of butchers and consumers towards dry-cured ham. Meat Science, 80,
1005-1012.
Morales, R., Arnau, J., Serra, X., Guerrero, L. and Gou, P. (2008) (b). Texture changes in dry-
cured ham pieces by mild thermal treatments at the end of the drying process.
Meat Science, 80, 231-238.

CAPÍTULO VII: BIBLIOGRAFÍA GENERAL
316
Morales, R., Arnau, J., Serra, X., Guerrero, L. and Gou, P. (2010). Texture changes in dry-
cured ham pieces by mild thermal treatments and the end of the drying process.
Meat Science, 80, 231-238.
Motilva, M.J., Toldrá, F., Nieto, P. and Flores, J. (1993). Muscle lipolysis phenomena in the
processing of dry-cured ham. Food Chemistry, 48, 121-125.
Mottram, D.S. (1994). Capítulo 2: The chemistry of meat flavor. En: Flavor of meat, meat
products and seafood. Editor: Shahidi, F. Blackie Academic & Professional, Londres,
61-83.
Mottram, D.S. (1998). Flavor formation in meat and meat products: a review. Food
Chemistry, 62, 415-424.
Muriel, E. (2003). Estudio comparativo de parámetros físico-químicos y sensoriales de
lomo Ibérico. Tesis Doctoral. Universidad de Extremadura.
Muriel, E., Ruiz, J., Martin, D., Petron, M.J. and Antequera, T. (2004). Physico-chemical and
sensory characteristics of dry-cured loin from different Iberian pigs lines. Food
Science and Technology International, 10, 117-123.
Narvaez-Rivas, M., Vicario, I.M., Constante, E.G. and León-Camacho, M. (2007). Changes in
the concentrations on free fatty acids, monoacylglycerol, and diacylglycerol in the
subcutaneous fat of Iberian ham during the dry-curing process. Journal of
Agricultural and Food Chemistry, 55, 10953-10961.
Narváez-Rivas, M., Gallardo, E., Rios, J.J. and León-Camacho, M. (2010). A tentative
characterization of volatile compounds from Iberian Dry-Cured Ham according to
different anatomical locations. A detailed study. Grasas y aceites, 61, 369-377.
Nezer, C., Moreau, I., Karim, L., Brouwers, B., Coppieters, W., Detilleux, J., Hanset, R., Kvasz,
A., Leroy, P. and Georges, M. (1999). An imprented QTL with major effects on
muscle mass and fat deposition maps to de IGF locus in pigs. Nature Genetics, 21,
155-156.
Okrouhla, M., Stupka, R., Čitek, J., Šprysl, M., Kluzakova, E., Trnka, M. and Štolc, L. (2006).
Amino acid composition of pig meat in relation to live weight and sex. Czech
Journal of Animal Science, 51, 529-534.
Olivares, A., Navarro, J.L. and Flores, M. (2009). Distribution of volatile compounds in lean
and subcutaneous fat tissues during processing of dry fermented sausages. Meat
Science, 42, 1303-1308.
Ordóñez, J., Cambero, I., Fernández, L., García, L., García de Fernando, G., De la Hoz, L and
Selgas, M.D. (1998). “Tecnología de los alimentos.” Vol.II. Alimentos de origen
animal. Ed. Síntesis, S.A.
Ovilo, C., Fernández, A., Noguera, J.L., Barragán, C., Letón, R., Rodríguez, C., Mercadé, A.,
Alves, E., Folch, J.M., Varona, L. and Toro, M. (2005). Fine mapping of porcine

CAPÍTULO VII: BIBLIOGRAFÍA GENERAL
317
chromosome 6 QTL and LEPR effects on body composition in multiple generations
of an Iberian by Landrace intercross. Genetics Research, 85, 57–67.
Palmia, F., Mazoyer, C., Diaferia, C., Baldini, P. and Porretta, A. (1992). Salt and water
distribution in typical Italian hams. Revista Española de Ciencia y Tecnología de los
Alimentos, 32, 71-83.
Parolari, G., Rivaldi, R., Leonelli, C., Bellatti, M. and Bovis, N. (1988). Colore e consistenza
del prosciutto crudo in rapporto alla material prima e alla tecnica di stagionatura.
Industria Conserve, 63, 45–49.
Peloso, J.V., Lopes, P.S., Gomide, L.A.M., Guimaraes, S.E.F. and Carneiro, P.L.S. (2010).
Carcass and ham quality characteristics of heavy pigs from different genetic groups
intended for the production of dry-cured hams. Meat Science, 86, 371-376.
Pérez, M. (2006). Interacción de los compuestos aromáticos del Jamón Curado con matriz
proteica. Tesis Doctoral. Universidad Politécnica de Valencia.
Pérez-Palacios, Ruiz, J., Tejeda, J.F. and T., Antequera (2009). Subcutaneous and
intramuscular lipid traits as tools for classifying Iberian pigs as a function of their
feeding background. Meat Science, 81, 632-640.
Pérez-Palacios, T., Antequera, T., Molano, R., Rodríguez, P.G. and Palacios, R. (2010).
Sensory traits prediction in dry-cured hams from fresh product via MRI and lipid
composition. Journal of Food Engineering, 101, 152-157.
Petrón, M.J., Muriel, E., Timón, M.L., Martín, L. and Antequera, T. (2004). Fatty acids and
triacylglycerols profiles from different types of Iberian dry-cured hams. Meat
Science, 68, 71-77.
Piotrows, E.G., Zaika, L.L. and Wasserma, A.E. (1970). Studies on aroma of cured ham.
Journal of Food Science, 35, 321-325.
Primo Yufera, E. (1997). “Química de los alimentos.” Ed. Síntesis S.A., Madrid.
Pripis-Nicolau, L., de Revel, G., Bertrand, A. and Maujean, A. (2000). Formation of Flavour
Components by the Reaction of Amino Acid and Carbonyl Compounds in Mild
Conditions. Journal of Agricultural and Food Chemistry, 48, 3761-3766.
Raes, K., De Smet, S. and Demeyer, D. (2004). Effect of dietary fatty acids on incorporation
of long chain polyunsaturated fatty acids and conjugated linoleic acid in lamb, beef
and pork meat: a review. Animal Feed Science and Technology, 113, 199–221
Ramírez, M.R. and Cava, R. (2007). Carcass composition and meat quality of 3 different
Iberian × Duroc genotype pigs. Meat Science, 75, 388–396
Ramírez, M.R. and Cava, R. (2008). Effect of Iberian × Duroc genotype on composition and
sensory properties of dry-cured ham. Journal of the Science of Food and Agriculture,
88, 667-675.

CAPÍTULO VII: BIBLIOGRAFÍA GENERAL
318
Ramírez, M.R. (2008). Assessment of the quality and technological characteristics of meat
and dry-cured products from Iberian pigs crossed with different Duroc genetic
lines. Tesis Doctoral. Universidad de Extremadura.
Reina, R., Ventanas, J., García-Casco, J.M. and García, C. (2010). Characterization of dry-
cured shoulders: Quality traits. En VII Congreso Internacional del cerdo
mediterraneo. Córdoba, España
Resano, H., Sanjuan, A.I. and Albisu, L.M. (2007). Consumer´s acceptability of cured ham in
Spain and the influence of information. Food Quality and Preference, 18, 1064-
1076.
Rivas-Canedo, A. Fernández-García, E. and Núñez, M. (2009). Volatile compounds in dry-
cured Serrano ham subjected to high pressure processing. Effect of the packaging
material. Meat Science, 82, 162-169.
Rosell, C.M. and Toldrá, F. (1998). Comparison of muscle proteolytic and lipolytic enzyme
levels in raw hams from Iberian and White pigs. Journal of the Science of Food and
Agriculture, 76, 117–122.
Rosenthal, A. (1999). Food texture, Measurements and Perception. Editorial Aspen
Publishers, INC. Maryland, USA.
Ruiz, J., Cava, R., Ventanas, J. and Jensen, T. (1998). Headspace solid phase microextraction
for the analysis of volatiles in a meat product: Dry-Cured Iberian Ham. Journal
Agricultural and Food Chemistry, 46, 4688-4694.
Ruiz, J., García, C., Díaz, M.C., Cava, R., Tejeda, J.F. and Ventanas, J. (1999) (a). Dry cured
Iberian ham non-volatile components as affected by the length of the curing
process. Food Research International, 32, 643-651.
Ruiz, J., Ventanas, J., Cava, R., Andrés, A.I. and García, C. (1999) (b). Volatile compounds of
dry-cured Iberian ham as affected by the lenght of the curing process. Meat Science,
52, 19-27.
Ruiz, J., García, C., Muriel, E., Andrés, A.I. and Ventanas, J. (2002) (a). Influence of sensory
characteristics on the acceptability of dry-cured ham. Meat Science, 61, 347–354
Ruiz, J., Muriel, E. and Ventanas, J. (2002) (b). The flavour of Iberian ham. En Research
advances in the quality of meat and meat products. Ed. F.Toldrá. Research
Signpost, Trivandrum, India. Pp 289-309.
Ruiz, J (2005).Textura de músculos de cerdo y de jamón curado con distintos niveles de
NaCl, pH y contenido de agua. Tesis Doctoral. Universidad Autónoma de Barcelona.
Ruiz-Carrascal, J., Ventanas, J., Cava, R., Andrés, A.I. and García, C. (2000). Texture and
appearance of dry-cured ham as affected by fat content and fatty acid composition.
Food Research International, 33, 91-95.

CAPÍTULO VII: BIBLIOGRAFÍA GENERAL
319
Ruiz-Ramírez, J., Arnau, J., Serra, X. and Gou, P. (2006). Effect of pH24, NaCl content and
proteolysis index on the relationship between water content and texture
parameters in biceps femoris and Semimembranosus muscles in dry-cured ham.
Meat Science, 72, 185-194
Sabio, E. and Vidal-Aragón, M.C. (1998). Volatile compounds present in six types of dry-
cured ham south European countries. Food Chemistry, 51, 660-672.
Saito, K., Ahhmed, A., Takeda, H., Kawahara, S., Irie, M. and Muguruma, M. (2007). Effects of
humidity-stabilizing sheet on the color and K value of beef stored at cold
temperatures. Meat Science, 75, 265-272
Salih, A. M., Smith, D.M., Price, J.F. and Dawson, L.E. (1987). Modified extraction 2-
thiobarbituric acid method for measuring lipid oxidation in poultry. Poultry
Science, 66, 1483-1489.
Sánchez, F. (2005). Modificaciones tecnológicas para mejorar la seguridad y calidad del
jamón curado. Tesis doctoral. IRTA.
Sánchez del Pulgar, J. (2011). Estudio en productos cárnicos mediante técnicas
olfatométricas. Tesis Doctoral. Universidad de Extremadura.
Sandler, S.R and Karo, W. (1992). Source book of advances organic laboratory
preparations. Academic Press, San Diego.
San Juan, A.I., Albisu, L.M., Aguelo, V. y Gracia, A. (2004). El Jamón de Teruel con
Denominación de Origen y los consumidores. Surcos de Aragón, 86, 26-30.
Sarraga, C., Guardia, M.D., Diaz, I., Guerrero, L., Regueiro, J.A.G. and Arnau, J. (2007).
Nutritional and sensory quality of porcine raw meat, cooked ham and dry-cured
shoulder as affected by dietary enrichment with docosahexaenoic acid (DHA) and
alpha-tocopheryl acetate. Meat Science, 76, 377-384.
Schwab, C.R., Baas, T.J., Berry, N.L., Mote, B.E. and Stalder, K.J. (2005). Selection for
Intramuscular Fat in Duroc Swine – An Update. In:
http://www.nsif.com/conferences/2005/pdf%5CSelectionIntramuscularFat.pdf
Shahidi, F. Rubin, L.J. and D’Souza, L.A. (1986). Meat flavor volatiles: a review of the
composition techniques of analysis, and sensory evaluation. Critical Reviews in
Food Science and Nutrition 24, 141-243.
Shahidi, F, Syowiecki, E., Dunajski, E. and Chong, X. (1994). Nonprotein nitrogen
compounds in harp seal (Phoca groenlandica) meat. Food Chemistry, 46, 407-413.
Sellier, P. (1998). Genetics of meat and carcass traits. In M. F. Rothschild & A. Ruvinsky
(Eds.). The genetics of the pig (pp. 463–510). New York, USA: CAB International.

CAPÍTULO VII: BIBLIOGRAFÍA GENERAL
320
Seuvre, A.M., Philippe, E., Rochard, S. and Voilley, A. (2007). Kinetic study of the release of
aroma compounds in different model food systems. Food Research International,
40, 480-492.
Schmidt, S. and Berger, R.G. (1998). Aroma compounds in fermented sausage of different
origins. Lebensmittel Wissenschaft und Technologie, 31, 559-567.
Shahidi, F. Rubin, L.J. and D’Souza, L.A. (1986). Meat flavor volatiles: a review of the
composition techniques of analysis, and sensory evaluation. Critical Reviews in
Food Science and Nutrition 24, 141-243.
Sierra, I., Campo, M.M., Olleta, J.L., Panea, B., Sanudo, C. and Pardos, J.J. (1998). Quality in
the denomination of origin “Jamon de Teruel”. Basis of the Quality of Typical
Mediterranean Animal Products, 365-369.
Simopoulos, A.P. (2002). The importance of the ratio of omega-6/omega-3 essential fatty
acids. Biomedicine and Pharmacotherapy, 56, 365–379.
Skibsted, L.H. (1992). Cured meat products and their Oxidative Stability. En: The Chemistry
on muscle-based Foods, Ed: Ledward D.A., Johnson D.E. y Knight M.K. The Royal
society of Chemistry 266, 287.
Song, H. and Cadwallader, K.R. (2008). Aroma components of American country ham.
Journal of Food Science, 73, 29-35.
Soto, E., Hoz, L., Ordoñez, J.A., Hierro, E., Herranz, B., López-Bote, P. and Cambero, M.I.
(2010). Volatile profile and sensory characteristics of dry-cured loins as affected
by feeding level in the period previous to the late fattening phase and by rearing
system of Iberian pigs. Journal of Muscle Foods, 21, 636-657.
Stahnke, L.H. (1994). Aroma components from dried sausages fermented with
Staphylococcus-Xylosus. Meat Science, 38, 39-53.
Stahnke, L.H. (1998). Character impact aroma compounds in fermented sausage.
Proceeding of the 44th International Congress of Food Science and Technology,
Barcelona, 786-787.
Stalder, K.J., Moeller, S.J. and Rothschild, M.F. (2004). Evidence of two QTL affecting fresh
and dry-cured ham processing quality characteristics. Journal of Animal Science, 82,
42-42
Sunesen, L.O., Dorigoni, V., Zanardi, E. and Stahnke, L. (2001). Volatile compounds released
during ripening in Italian dried sausage. Meat Science, 58, 93-97.
Szczesniak, A. (1963). Objective measurements of food texture. Journal of Food Science, 28,
410-428.
Szczesniak, A. (2002). Texture is a sensory property. Food Quality and Preferences, 13, 215-
225.

CAPÍTULO VII: BIBLIOGRAFÍA GENERAL
321
Tejeda, J.F. (1999). Estudio de la influencia de la raza y la alimentación sobre la fracción
lipídica intramuscular del cerdo Ibérico. Tesis Doctoral, Universidad de
Extremadura.
Tejeda, J.F., Gandemer, G., Antequera, T., Viau, M. and García, C. (2002). Lipid traits of
muscles as related to genotype and fattening diet in Iberian pigs: total
intramuscular lipids and triacylglycerols. Meat Science, 60, 357-363.
Timón, M.L., Ventanas, J., Martín, L., Tejeda, J.F. and García, C. (1998). Volatile compounds
in supercritical carbon dioxide extracts of Iberian ham. Journal of agricultural and
Food Chemistry, 46, 5143-5150.
Timón, M.L., Ventanas, J., Carrapiso, A.I., Jurado, A. and García, C. (2001). Subcutaneous and
intermuscular fat characterisation of dry-cured Iberian hams. Meat Science, 58, 85-
91.
Tikk, M., Tikk, K., Torngren, M.A., Meinert, L., Aaslyng, M.D. and Karlsson, A.H. (2006).
Development of inosine monophosphate and its degradation products during aging
of pork of different qualities in relation to basic taste and retronasal flavor
perception of the meat. Journal of Agricultural and Food Chemistry, 54, 7769−7777.
Toldrá, F., Miralles, M.C. and Flores, J. (1992). Protein extractability in dry-cured ham. Food
Chemistry, 44, 391-394.
Toldrá, F., Flores, M. and Sanz, Y. (1997). Dry-cured ham flavour: enzymatic generation
and process influence. Food Chemistry, 59, 523-530.
Toldrá, F. and Flores, M. (1998). The role of muscle proteases and lipases in flavor
development during the processing of dry-cured ham. Critical review s in food
science, 38, 331-352.
Toldrá, F., Aristoy, M.C. and Flores, M. (2000). Contribution of muscle aminopeptidases to
flavor development in dry-cured ham. Food ResearchInternational, 33, 181-185.
Toldrá, F. (2002). Manufacturing of dry-cured ham. En: Dry-cured meat products. Food &
Nutrition Press, INC. Trumbull, Connecticut, pp. 52.
Toscani, T., Virgili, R., Corbari, G. and Calzolari, L. (2000). Effects of different processing
plants on proximate composition, proteolysis and hardness of Parma ham.
Industria Conserve, 75, 259–270.
Tovar, J. (2009). Estudio comparativo de las rutas de formación de compuestos
relacionados con la actividad aromática en carne de distintas especies. Tesis
Doctoral. Universidad de Extramadura.
Van Laere, A.S., Nguyen, M., Braunschweig, M., Nezer, C., Collette, C., Moreau, L., Archibald,
A.L., Haley, C.S., Buys, N., Tally, M., Andersson, G., Georges, M. and Andersson, L.

CAPÍTULO VII: BIBLIOGRAFÍA GENERAL
322
(2003). A regulatory mutation in IGF2 causes a major QTL effect on muscle growth
in the pig. Nature, 425, 832-836.
Ventanas, J., Córdoba, J.J., Antequera, T., García, C., López-Bote, C. and Asensio, M.A. (1992).
Hydrolysis and Maillard reactions during ripening of Iberiam ham. Journal of Food
Science, 57, 813-815.
Ventanas, J., Tejeda, J.F. and Petrón, M.J. (2001). El jamón ibérico: de una imagen de calidad
a una calidad definida y contrastada. En: Tecnología del Jamón Ibérico: de los
sistemas tradicionales a la explotación racional del sabor y el aroma. J. Ventanas,
ed., Ediciones Mundi-Prensa, Madrid, 15-43
Ventanas, S., Ventanas, J., Ruiz, J. and Estévez, M. (2005). Iberian pigs for the development
of high-quality cured products. In: Recent Research Developments in Agricultural
& Food Chemistry. Signpost, Trivandrum, India. pp: 27-53.
Ventanas, S. (2006). Influencia de la raza y de la alimentación sobre el contenido y
características de la grasa intramuscular del lomo de cerdo ibérico: efecto sobre
parámetros determinantes de la calidad. Tesis Doctoral. Universidad de
Extremadura.
Ventanas, S., Ventanas, J. and Ruiz, J. (2007) (a). Sensory characteristics of Iberian dry-
cured loins: Influence of crossbreeding and rearing system. Meat Science, 75, 211-
219.
Ventanas, S., Ruiz, J., García, C. and Ventanas, J. (2007) (b). Preference and juiciness of
Iberian dry-cured loin as affected by intramuscular fat content, crossbreeding and
rearing system. Meat Science, 77, 324-330.
Ventanas, S., Ventanas, J., Estévez, M. and Ruiz, J. (2010). Analysis of volatile molecules in
Iberian dry-cured loins as affected by genetic, feeding systems and ingredients.
European Food Research and Technology, 231, 225-235.
Ventanas, S., Estevez, M., Andrés, A.I. and Ruiz, J. (2008). Analysis of volatile compounds of
Iberian dry-cured loins with different intramuscular fat contents using SPME–DED.
Meat Science, 79, 172-180
Viera-Alcaide, I., Vicario, I.M., Graciani Constente, E., Léon-Camacho, M. (2007).
Authentication of fattening diet of Iberian pig according to their triacylglycerol
profile from subcutaneous fat. Analytica Chimica Acta. 596, 319-324.
Viera-Alcaide, I., Vicario, I.M., Escudero-Gilette, M.L., Graciani Constente, E. and Léon-
Camacho, M. (2008). A multivariate study of the triacylglycerols composition of the
subcutaneous adipose tissue of Iberian pig in relation to the fattening diet and
genotype. Grasas y Aceites, 59, 327-336.

CAPÍTULO VII: BIBLIOGRAFÍA GENERAL
323
Virgili, R., Parolari, G., Schivazappa, C., Soresi Bordini, C. and Borri, M. (1995). Sensory and
texture quality of dry-cured ham as affected by endogenous cathepsin B activity
and muscle composition. Journal of Food Science, 60, 1183-1186.
Virgili, R., Parolari, G., Soresi-Bordini, C., Schivazappa, C., Cornet, M. and Monin, G. (1999).
Free amino acids and dipeptides in dry cured ham. Journal of Muscle Foods, 10,
119–130.
Virgili, R., Saccani, G., Gabba, L., Tanzi, E. and Soresi Bordini, C. (2007). Changes of free
amino acids and biogenic amines during extended ageing of Italian dry-cured ham.
LTW-Food Science and Technology, 40, 871-878.
Watanabe, A., Tsuneishi, E. and Takimoto, Y. (1989). Analysis of ATP and its breakdown
products in beef by reversed-phase HPLC. Journal of Food Science, 54, 1169-1172
Wistreich, H.E., Morse, R.E. and Kenyon, L.J. (1958). Curing of ham a study of sodium
chloride diffusion. Food Technology, 12, 45-55
Wistreich, H.E., Morse, R.E. and Kenyon, L.J. (1959). Curing of ham – A study of sodium
chloride accumulation. 1. Methods, effect of temperature, cations, muscles and
solution concentration. Food Technology, 13, 441-443
Whitemore, C.T., Henderson, R., Ellis, M., Smith, W.C., Laird, R., Wood, J.D. (1982). Influence
of test regime on the success of identification of different rates of lean and fat
deposition in pigs. Animal Production, 34, 380-381.
WOK. Web of Knowledge. http://www.accesowok.fecyt.es/
Wood, J.D., Lodge, G.A. and Lister, D. (1979). Response to different rates of energy intake
by Gloucester Old Spot and Large White boars and gilts given the same total feed
allowance. Animal Production, 28, 371–380.
Wood, J.D., Whelehan, O.P., Ellis, M., Smith, W.C. and Laird, R. (1983). Effects of selection
for low backfat thickness in pigs on the sites of tissue deposition in the body.
Animal Production. 36, 389-397.
Wood, J.D., Edwards, S.A. and Bichard, M. (1988). Influence of the Duroc breed on pig meat
quality. In Proceedings of the 34th ICoMST (pp. 571–572). Brisbane, Australia.
Wood, J.D., Nute, G.R., Richardson, R.I., Whittington, F.M., Southwood, O., Plastow, G.,
Mansbridge, da Costa, N. and Chang, K.C. (2004). Effects of breed, diet and muscle
on fat deposition and eating quality in pig. Meat Science, 67, 651–667.
Yang, C. and Sepúlveda, F. (1985). Separation of phenyltiocarbamyl amino acids by high-
performance liquid chromatography of spherisorb octadecysilane columns. Journal
of Cromatography , 346, 413-416.
Zembayashi, M. (1994). Effects of nutritional planes and breeds on intramuscular lipid in
m-longissimus-dorsi of steers. Meat Science, 38, 367-374.

CAPÍTULO VII: BIBLIOGRAFÍA GENERAL
324
Zhao, G.M., Zhou, G.H. Tian, W., Xu, X.L., Wang, Y.L. and Luo, X. (2005). Changes in alanyl
aminopeptidase activity and free amino acid contents in biceps femoris during
processing in Jinhua Ham. Meat Science, 71, 612-619.


















