
Nature artículo
6
LETTERS The infl ammasome reco gn iz es cytoso li c mi cr ob ia l and host DNA and triggers an innate immune response Daniel A. Muruve 1 *, Virginie Pe ´trilli 3 *, Anne K. Zaiss 2 , Lindsay R. White 1 , Sharon A. Clark 1 , P. Joel Ross 4 , Robin J. Parks 4 & Jurg Tschopp 3 The innate immune system recognizes nucleic acids during infec- tion and tissue damage. Whereas viral RNA is detected by endo- somal toll-like receptors (TLR3, TLR7, TLR8) and cytoplasmic RIG-I and MDA5, endosomal TLR9 and cytoplasmic DAI bind DNA 1 , resulting in the activation of nuclear factor-kB and inter- fero n regul atory factor transcription factor s. Howev er, viruse s also trigger pro-inflammatory responses 2 , which remain poorly defined. Here we show that internalized adenoviral DNA induces matur ation of pro-in terle ukin- 1b in mac rophag es, whi ch is dependenton NALP3 and ASC, compon ent s of theinnatecytos oli c molecular complex termed the inflammasome. Correspondingly, NALP3- and ASC-deficient mice display reduced innate inflam- matory responses to adenovirus particles. Inflammasome activa- tion also occurs as a result of transfected cytosolic bacterial, viral and mammalian (host) DNA, but in this case sensing is dependent on ASC but not NALP3. The DNA-se nsi ng pro-infla mma tor y pathway functions independently of TLRs and interferon regula- tory factors. Thus, in addition to viral and bacterial components or danger signals in general, inflammasomes sense potentially dangerous cytoplasmic DNA, strengthening their central role in innate immunity. CpG-ri ch DNA fro m bac ter ia and vir uses sti mul ate s dendri tic cel ls andmacro phag es to releas e considerableamounts of type I int erf eron (IFN) by act iva ting the DNA-se nsi ng protein TLR9 (re f. 1).Howev er, there is ample evidence that DNA, including host (self) DNA, can be recognized independently of TLR9 (refs 2, 3). A double-stranded (ds)DNA-binding protein designated DAI was recently identified that detects both microbial and host DNA 4 . In the absence of DAI, the type I IFN response triggered by viral DNA is severely hampered, suggesting an important role of the DAI signalling pathway in the antiviral response. In addition to type I IFN, a successful anti-viral res ponse is als o depe nde nt on a str ong pro -in fla mmator y component controlled by two alarm cytokines, TNF and interleukin (IL)-1 b (ref. 5). Although the mechanism of the type I IFN response has been intensively investigated, little is known about the mechanism of the virus-mediated inflammatory response. Adenov iruses are non-e nvelo ped DNA virus es that cause respir - atory and gastrointestinal disease in humans. In addition, recom- binant adenovirus vectors are studied and developed for gene and oncolytic therapy 6 . Adenovirus vectors trigger the innate immune system, which results in the rapid induction of inflammator y cyto- kines such as IL-1b and TNF 6 and acute inflammation in vivo 7,8 . In contrast to most RNA viruses, the signalling pathways that mediate this effect have not been fully defined. The NALP proteins are cytoplasmic NOD-like receptors (NLRs) 9 . Thebest unders too d is NALP3 (al so cal led cry opy rin , NLRP3) , whi ch senses exogeno us and host ligan ds such as bacter ial peptidoglyc an, ATP or uric acid 10 . NALP3 recruits, via the adaptor protein ASC, the inf lammat ory caspase-1 int o a molecular comple x ter med the inflammasome 10 . Onc e act iva ted, cas pas e-1 proces ses pro -IL-1b and pro-IL-18 to their active and secreted forms. Other NLRs that ar e known to formIL-1b-processing inflammaso mes includ e NALP1 and IPAF, the latter of which directly activates caspase-1 in response to bacterial flagellin 11 . Given the limited information on the inflam- matory response to DNA viruses and DNA in general, we have explored here the role of the inflammasome in this pathway. To test whethe r adenovi rus can activ ate the inflammasome, differ- entiated human THP-1 cells were infected with wild-type human adenovirus (serotype 5) and the processing and secretion of caspase- 1 and IL-1b protein was determined (Fig. 1a). Because antiviral anti- bodies enhance the interaction and internalization of adenovirus with leukocytes 12 , aden ovi rusinfectio ns werealso perf orme d in thepresence of human serum. Adenovirus infection induced robust processing of the 35-kDa pro-IL-1b protein to the mature, secreted 17-kDa IL-1b cytokine within 2 h, which increased over time. Similar results were obtained after infection with serotype 3 human adenovirus (Fig. 1b). To deter mine whet her vira l repl icat ion was nece ssar y to acti vate IL-1 b, THP-1 cells were incubated with the non-repl icating adenovirus vec- tors AdLacZ or AdGFP. Both adenovirus vectors efficiently activated pro-IL-1b maturation (Fig. 1c). To examine further the role of the aden ovi rus viri on in infl ammasome acti vat ion, THP -1 cell s were chal - lenged with ultraviolet/psoralen-inactivated adenovirus, adenovirus vect ors lac king CAR and inte grin -bin din g caps id doma ins, or a helper-de pendent adenovirus vector (Fig. 1d). All of the recombinant adenoviru s vectors effectively induced IL-1 b processing and secretio n (Fig. 1d and Supplementary Fig. 1), confirming that the adenovirus virion activates the inflammasome, independent of the transgene or viral gene expression. IL-1b secretion on adenovirus transduction was not restricted to human cells, as cytokine releas e f rom murine macro- phages was almost as efficient as that triggered by lipopolysaccharide (LPS) and ATP (Fig. 1e). Moreover, infection with HSV-1, a DNA virus of the Herpesviridae family, also caused potent caspase-1 and pro-IL-1b maturation, suggesting that DNA viruses in general may activate an IL-1b-based inflammato ry response (Fig. 1f). To dete rmi ne mor e pre cis ely the vir ion compon ent res pon sible for IL-1b activation, THP-1 cells were incubated with infectious empty adenovirus capsids or adenovirus DNA. An increasing titre of empty viral capsids was incapable of inducing IL-1b processing (Fig. 2a). Because empty viral capsids retain the binding and internalization properties of mature adenovirus virions 13 , these results suggested that the capsid protein s or the mere intern aliza tion event are prob- ably not responsible for inflammaso me activa tion. To test whether *These authors contributed equally to this work. 1 Department of Medicine, 2 Biochemistry and Molecular Biology, University of Calgary, Alberta T2N 4N1, Canada. 3 Department of Biochemistry, University of Lausanne, Epalinges, Switzerland. 4 Molecular Medicine Program, Ottawa Health Research Institute, Ottawa, Ontario K1Y 4E9, Canada. Vol 452 | 6 March 2008 | doi:10.1038/nature06664 103 Nature PublishingGroup ©2008
-
Upload
anonymous-hxd6pjl -
Category
Documents
-
view
214 -
download
0
Transcript of Nature artículo

8/10/2019 Nature artículo
http://slidepdf.com/reader/full/nature-articulo 1/6
LETTERS
The inflammasome recognizes cytosolic microbial andhost DNA and triggers an innate immune responseDaniel A. Muruve 1*, Virginie Pe trilli3 *, Anne K. Zaiss 2 , Lindsay R. White 1 , Sharon A. Clark 1 , P. Joel Ross 4 ,Robin J. Parks 4 & Jurg Tschopp 3
The innate immune system recognizes nucleic acids during infec-tion and tissue damage. Whereas viral RNA is detected by endo-somal toll-like receptors (TLR3, TLR7, TLR8) and cytoplasmicRIG-I and MDA5, endosomal TLR9 and cytoplasmic DAI bindDNA1 , resulting in the activation of nuclear factor- kB and inter-feron regulatory factor transcription factors. However, virusesalso trigger pro-inflammatory responses 2 , which remain poorly defined. Here we show that internalized adenoviral DNA inducesmaturation of pro-interleukin-1 b in macrophages, which isdependenton NALP3 andASC, components of theinnatecytosolicmolecular complex termed the inflammasome. Correspondingly,NALP3- and ASC-deficient mice display reduced innate inflam-matory responses to adenovirus particles. Inflammasome activa-tion also occurs as a result of transfected cytosolic bacterial, viraland mammalian (host) DNA, but in this case sensing is dependenton ASC but not NALP3. The DNA-sensing pro-inflammatory pathway functions independently of TLRs and interferon regula-tory factors. Thus, in addition to viral and bacterial componentsor danger signals in general, inflammasomes sense potentially dangerous cytoplasmic DNA, strengthening their central role ininnate immunity.
CpG-rich DNA from bacteria and viruses stimulates dendritic cellsandmacrophages to release considerableamounts of type I interferon(IFN) by activating theDNA-sensing protein TLR9 (ref. 1).However,there is ample evidence that DNA, including host (self) DNA, can berecognized independently of TLR9 (refs 2, 3). A double-stranded(ds)DNA-binding protein designated DAI was recently identifiedthat detects both microbial and host DNA 4 . In the absence of DAI,the type I IFN response triggered by viral DNA is severely hampered,suggesting an important role of the DAI signalling pathway in theantiviral response. In addition to type I IFN, a successful anti-viralresponse is also dependent on a strongpro-inflammatory componentcontrolled by two alarm cytokines, TNF and interleukin (IL)-1 b(ref. 5). Although the mechanism of the type I IFN response has beenintensively investigated, little is known about the mechanism of thevirus-mediated inflammatory response.
Adenoviruses are non-enveloped DNA viruses that cause respir-atory and gastrointestinal disease in humans. In addition, recom-binant adenovirus vectors are studied and developed for gene andoncolytic therapy 6. Adenovirus vectors trigger the innate immunesystem, which results in the rapid induction of inflammatory cyto-kines such as IL-1b and TNF 6 and acute inflammation in vivo 7,8 . Incontrast to most RNA viruses, the signalling pathways that mediatethis effect have not been fully defined.
The NALP proteins are cytoplasmic NOD-like receptors (NLRs) 9 .Thebest understood is NALP3 (also calledcryopyrin, NLRP3), which
senses exogenous and host ligands such as bacterial peptidoglycan,ATP or uric acid 10 . NALP3 recruits, via the adaptor protein ASC, theinflammatory caspase-1 into a molecular complex termed theinflammasome 10 . Once activated, caspase-1 processes pro-IL-1 band pro-IL-18 to their active and secreted forms. Other NLRs thatare known to form IL-1 b-processing inflammasomes include NALP1and IPAF, the latter of which directly activates caspase-1 in responseto bacterial flagellin 11 . Given the limited information on the inflam-matory response to DNA viruses and DNA in general, we haveexplored here the role of the inflammasome in this pathway.
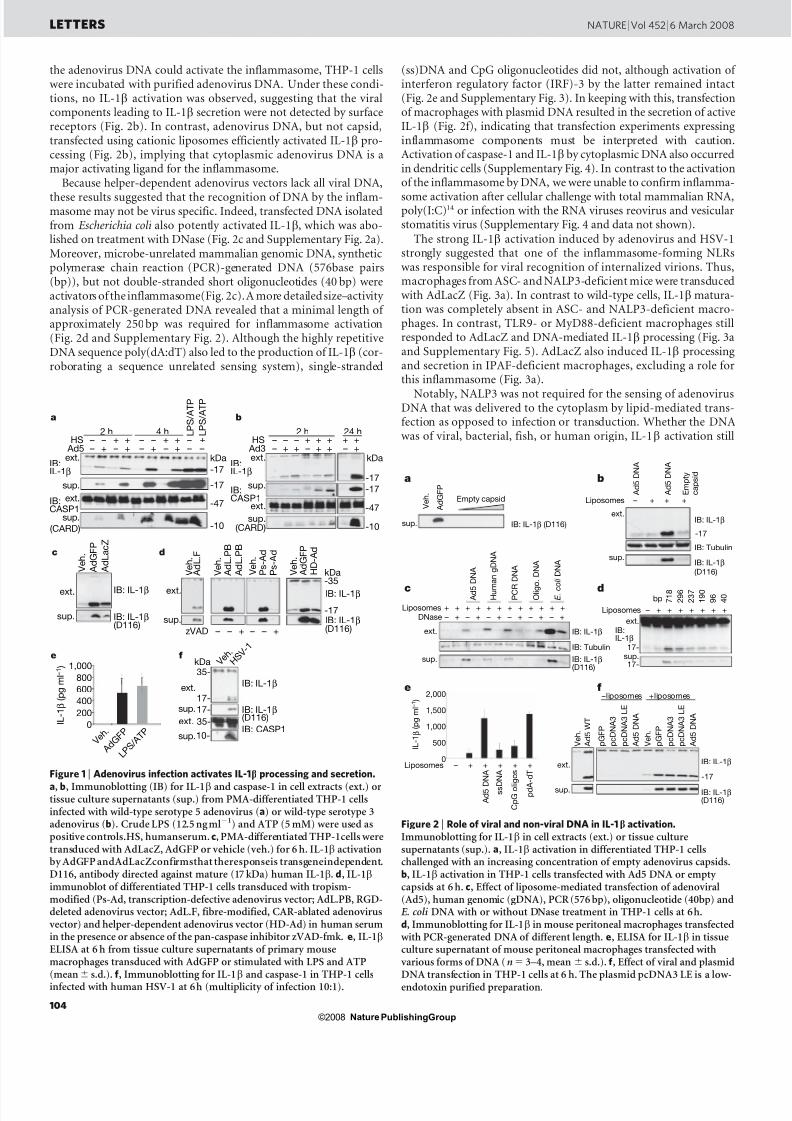
To test whether adenovirus can activate the inflammasome, differ-entiated human THP-1 cells were infected with wild-type humanadenovirus (serotype 5) and the processing and secretion of caspase-1 and IL-1b protein was determined (Fig. 1a). Because antiviral anti-bodies enhance the interaction and internalization of adenovirus withleukocytes 12 , adenovirusinfectionswerealsoperformed in thepresenceof human serum. Adenovirus infection induced robust processing of the 35-kDa pro-IL-1 b protein to the mature, secreted 17-kDa IL-1 bcytokine within 2 h, which increased over time. Similar results wereobtained after infection with serotype 3 human adenovirus (Fig. 1b).To determinewhetherviral replicationwasnecessary to activateIL-1 b,THP-1 cells were incubated with the non-replicating adenovirus vec-tors AdLacZ or AdGFP. Both adenovirus vectors efficiently activatedpro-IL-1 b maturation (Fig. 1c). To examine further the role of theadenovirus virion in inflammasomeactivation,THP-1 cells werechal-lenged with ultraviolet/psoralen-inactivated adenovirus, adenovirusvectors lacking CAR and integrin-binding capsid domains, or ahelper-dependent adenovirus vector (Fig. 1d). All of the recombinantadenovirus vectors effectively induced IL-1 b processing and secretion(Fig. 1d and Supplementary Fig. 1), confirming that the adenovirusvirion activates the inflammasome, independent of the transgene orviral gene expression. IL-1b secretion on adenovirus transduction wasnot restricted to human cells, as cytokine release from murine macro-phages was almost as efficient as that triggered by lipopolysaccharide(LPS) and ATP (Fig. 1e). Moreover, infection with HSV-1, a DNAvirus of the Herpesviridae family, also caused potent caspase-1 andpro-IL-1 b maturation, suggesting that DNA viruses in general may activate an IL-1b-based inflammatory response (Fig. 1f).
To determine more precisely thevirioncomponent responsible forIL-1b activation, THP-1 cells were incubated with infectious empty adenovirus capsids or adenovirus DNA. An increasing titre of empty viral capsids was incapable of inducing IL-1 b processing (Fig. 2a).Because empty viral capsids retain the binding and internalizationproperties of mature adenovirus virions 13 , these results suggestedthat the capsid proteins or the mere internalization event are prob-ably not responsible for inflammasome activation. To test whether
* These authors contributed equally to this work.
1Department of Medicine, 2 Biochemistry and Molecular Biology, University of Calgary, Alberta T2N 4N1, Canada. 3 Department of Biochemistry, University of Lausanne, Epalinges,Switzerland. 4 Molecular Medicine Program, Ottawa Health Research Institute, Ottawa, Ontario K1Y 4E9, Canada.
Vol 452 |6 March 2008 |doi:10.1038/nature06664
103
Nature PublishingGroup©2008
8/10/2019 Nature artículo
http://slidepdf.com/reader/full/nature-articulo 2/6
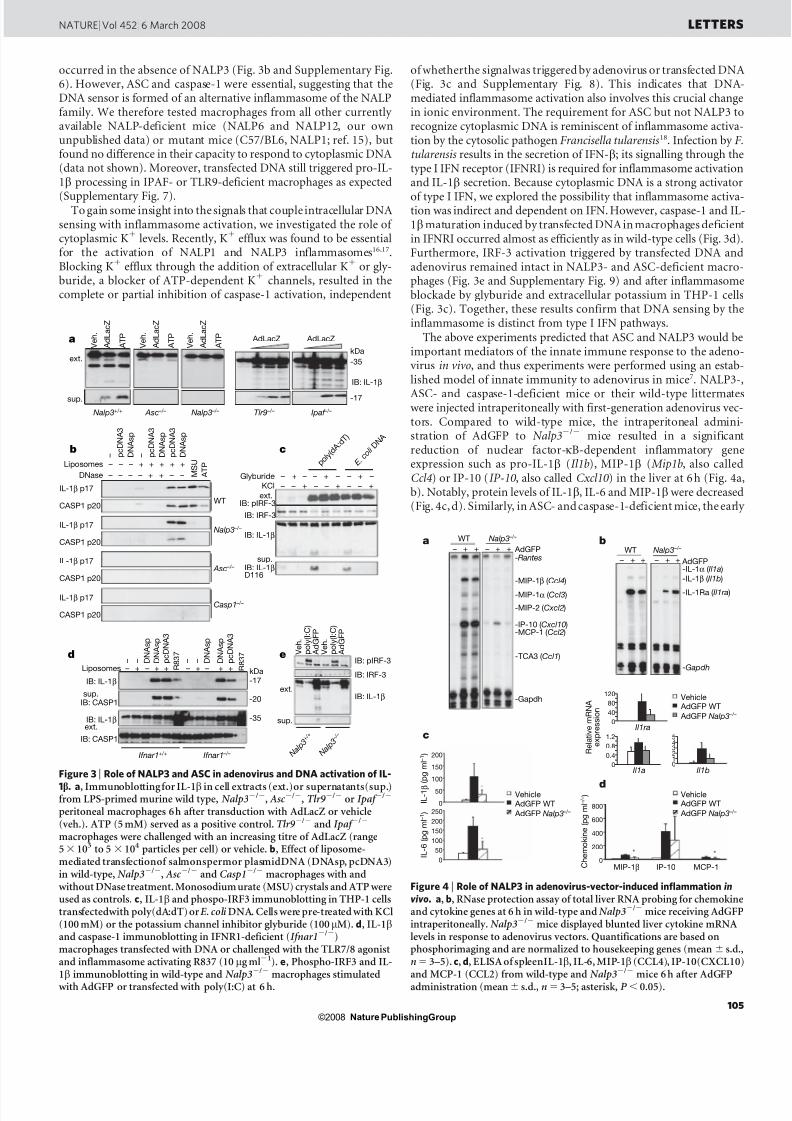
the adenovirus DNA could activate the inflammasome, THP-1 cellswere incubated with purified adenovirus DNA. Under these condi-tions, no IL-1 b activation was observed, suggesting that the viralcomponents leading to IL-1 b secretion were not detected by surfacereceptors (Fig. 2b). In contrast, adenovirus DNA, but not capsid,transfected using cationic liposomes efficiently activated IL-1 b pro-cessing (Fig. 2b), implying that cytoplasmic adenovirus DNA is amajor activating ligand for the inflammasome.
Because helper-dependent adenovirus vectors lack all viral DNA,these results suggested that the recognition of DNA by the inflam-masome may not be virus specific. Indeed, transfected DNA isolatedfrom Escherichia coli also potently activated IL-1 b , which was abo-lished on treatment with DNase (Fig. 2c and Supplementary Fig. 2a).Moreover, microbe-unrelated mammalian genomic DNA, syntheticpolymerase chain reaction (PCR)-generated DNA (576base pairs(bp)), but not double-stranded short oligonucleotides (40 bp) wereactivators of the inflammasome(Fig. 2c).A more detailedsize–activity analysis of PCR-generated DNA revealed that a minimal length of approximately 250bp was required for inflammasome activation(Fig. 2d and Supplementary Fig. 2). Although the highly repetitiveDNA sequence poly(dA:dT) also led to the production of IL-1 b (cor-roborating a sequence unrelated sensing system), single-stranded
(ss)DNA and CpG oligonucleotides did not, although activation of interferon regulatory factor (IRF)-3 by the latter remained intact(Fig. 2e and Supplementary Fig. 3). In keeping with this, transfectionof macrophages with plasmid DNA resulted in the secretion of activeIL-1b (Fig. 2f), indicating that transfection experiments expressinginflammasome components must be interpreted with caution.Activation of caspase-1 and IL-1 b by cytoplasmic DNA also occurredin dendritic cells (Supplementary Fig. 4). In contrast to the activationof the inflammasome by DNA, we were unable to confirm inflamma-
some activation after cellular challenge with total mammalian RNA,poly(I:C) 14 or infection with the RNA viruses reovirus and vesicularstomatitis virus (Supplementary Fig. 4 and data not shown).
The strong IL-1 b activation induced by adenovirus and HSV-1strongly suggested that one of the inflammasome-forming NLRswas responsible for viral recognition of internalized virions. Thus,macrophages from ASC- and NALP3-deficient mice were transducedwith AdLacZ (Fig. 3a). In contrast to wild-type cells, IL-1 b matura-tion was completely absent in ASC- and NALP3-deficient macro-phages. In contrast, TLR9- or MyD88-deficient macrophages stillresponded to AdLacZ and DNA-mediated IL-1 b processing (Fig. 3aand Supplementary Fig. 5). AdLacZ also induced IL-1 b processingand secretion in IPAF-deficient macrophages, excluding a role forthis inflammasome (Fig. 3a).
Notably, NALP3 was not required for the sensing of adenovirusDNA that was delivered to the cytoplasm by lipid-mediated trans-fection as opposed to infection or transduction. Whether the DNAwas of viral, bacterial, fish, or human origin, IL-1 b activation still
a
c
V e h .
A d G F P
A d L a c Z
d
zVAD – – + – – +
V e h .
A d L . P B
A d L . P B
V e h .
P s - A
d
P s - A
d
V e h .
A d L . F
V e h .
A d G F P
H D - A
d
-17
-35
IB: IL-1 β
IB: IL-1 β IB: IL-1 β
(D116) IB: IL-1 β(D116)
L P S / A T P
HS L P S / A T P
Ad5HS
Ad3––
–+
–+
+–
++
++
+–
++
ext.
ext.
ext.sup.
sup.
ext.
ext.sup.
sup.
ext.
sup.
sup.(CARD)
– – + + – – –+ + +– + – + – + –– + –
2 h 4 h
-17
-17kDa
kDa
kDaIB:
IB:
IL-1β
CASP1
ext.
ext.
sup.
sup.(CARD)
IB:
IB:
IL-1β
CASP1-47
-10
b
2 h 24 h
-17-17
-47
-10
0200400600800
1,000
I L - 1 β ( p g
m l – 1 )
e
V e h .
A d G F P
L P S / A T P
17-
35-kDa
17-
V e h .
H S V - 1f
IB: IL-1 β
IB: IL-1 β(D116) 35-
10-IB: CASP1
Figure 1 | Adenovirus infection activates IL-1 b processing and secretion.
a , b , Immunoblotting (IB) for IL-1 b and caspase-1 in cell extracts (ext.) ortissue culture supernatants (sup.) from PMA-differentiated THP-1 cellsinfected with wild-type serotype 5 adenovirus ( a ) or wild-type serotype 3adenovirus ( b). Crude LPS (12.5 ng ml 2 1 ) and ATP (5 mM) were used aspositive controls.HS, humanserum. c, PMA-differentiated THP-1cells weretransduced with AdLacZ, AdGFP or vehicle (veh.) for 6 h. IL-1 b activationby AdGFP andAdLacZconfirmsthat theresponseis transgeneindependent.D116, antibody directed against mature (17 kDa) human IL-1 b . d , IL-1bimmunoblot of differentiated THP-1 cells transduced with tropism-modified (Ps-Ad, transcription-defective adenovirus vector; AdL.PB, RGD-deleted adenovirus vector; AdL.F, fibre-modified, CAR-ablated adenovirus vector) and helper-dependent adenovirus vector (HD-Ad) in human serumin the presence or absence of the pan-caspase inhibitor zVAD-fmk. e, IL-1bELISA at 6 h from tissue culture supernatants of primary mousemacrophages transduced with AdGFP or stimulated with LPS and ATP(mean 6 s.d.). f, Immunoblotting for IL-1 b and caspase-1 in THP-1 cellsinfected with human HSV-1 at 6h (multiplicity of infection 10:1).
a
V e h .
A d G F P
Empty capsid
sup.
sup.
sup.
2,000
1,500
1,000
500
0
ext.
ext.
ext.ext.
sup.
sup.
IB: IL-1 β (D116)
V e h .
A d 5 W T
p G F P
A d 5 D N A
p c D N A 3
p c D N A 3 L E
p G F P
A d 5 D N A
p c D N A 3
p c D N A 3 L E
V e h .
f
-17
–liposomes +liposomes
E m p t y
c a p s i db
IB: IL-1 β
IB: Tubulin
Liposomes – + + +
-17
A d 5 D N A
A d 5 D N A
c
LiposomesDNase
+ + + + + + + + + + + +– + – + – + – + – + – +
A d 5
D N A
H u m
a n g D
N A
P C R
D N A
O l i g o .
D N A
E . c
o l i D N A
d 7 1 8
2 9 6 2 3 7
1 9 0 9 6 4 0bp
Liposomes – + + + + + + +
17-
17-
I L - 1 β ( p g
m l – 1 )
– + + + + +
A d 5 D N A
s s D N A
C p G o l
i g o s
p d A - d T
e
Liposomes
IB: IL-1 β (D116)
IB: IL-1 β
IB: IL-1 β (D116)
IB: IL-1 β IB:IL-1β
IB: TubulinIB: IL-1 β (D116)
Figure 2 | Role of viral and non-viral DNA in IL-1 b activation.Immunoblotting for IL-1 b in cell extracts (ext.) or tissue culturesupernatants (sup.). a , IL-1b activation in differentiated THP-1 cellschallenged with an increasing concentration of empty adenovirus capsids.b , IL-1b activation in THP-1 cells transfected with Ad5 DNA or empty capsids at 6 h. c, Effect of liposome-mediated transfection of adenoviral(Ad5), human genomic (gDNA), PCR (576 bp), oligonucleotide (40bp) andE. coli DNA with or without DNase treatment in THP-1 cells at 6h.d , Immunoblotting for IL-1 b in mouse peritoneal macrophages transfected with PCR-generated DNA of different length. e , ELISA for IL-1b in tissueculture supernatant of mouse peritoneal macrophages transfected with various forms of DNA ( n 5 3–4, mean 6 s.d.). f, Effect of viral and plasmidDNA transfection in THP-1 cells at 6 h. The plasmid pcDNA3 LE is a low-endotoxin purified preparation.
LETTERS NATURE |Vol 452 |6 March 2008
104
Nature PublishingGroup©2008
8/10/2019 Nature artículo
http://slidepdf.com/reader/full/nature-articulo 3/6
occurred in the absence of NALP3 (Fig. 3b and Supplementary Fig.6). However, ASC and caspase-1 were essential, suggesting that theDNA sensor is formed of an alternative inflammasome of the NALPfamily. We therefore tested macrophages from all other currently available NALP-deficient mice (NALP6 and NALP12, our ownunpublished data) or mutant mice (C57/BL6, NALP1; ref. 15), butfound no difference in their capacity to respond to cytoplasmic DNA(data not shown). Moreover, transfected DNA still triggered pro-IL-1b processing in IPAF- or TLR9-deficient macrophages as expected
(Supplementary Fig. 7).To gain some insight into thesignals that couple intracellular DNA
sensing with inflammasome activation, we investigated the role of cytoplasmic K
1levels. Recently, K
1efflux was found to be essential
for the activation of NALP1 and NALP3 inflammasomes 16,17 .Blocking K
1efflux through the addition of extracellular K
1or gly-
buride, a blocker of ATP-dependent K1
channels, resulted in thecomplete or partial inhibition of caspase-1 activation, independent
of whetherthe signalwas triggered by adenovirus or transfected DNA(Fig. 3c and Supplementary Fig. 8). This indicates that DNA-mediated inflammasome activation also involves this crucial changein ionic environment. The requirement for ASC but not NALP3 torecognize cytoplasmic DNA is reminiscent of inflammasome activa-tion by the cytosolic pathogen Francisella tularensis 18 . Infection by F.tularensis results in the secretion of IFN- b ; its signalling through thetype I IFN receptor (IFNRI) is required for inflammasome activationand IL-1b secretion. Because cytoplasmic DNA is a strong activator
of type I IFN, we explored the possibility that inflammasome activa-tion was indirect and dependent on IFN. However, caspase-1 and IL-1b maturation induced by transfected DNAin macrophages deficientin IFNRI occurred almost as efficiently as in wild-type cells (Fig. 3d).Furthermore, IRF-3 activation triggered by transfected DNA andadenovirus remained intact in NALP3- and ASC-deficient macro-phages (Fig. 3e and Supplementary Fig. 9) and after inflammasomeblockade by glyburide and extracellular potassium in THP-1 cells(Fig. 3c). Together, these results confirm that DNA sensing by theinflammasome is distinct from type I IFN pathways.
The above experiments predicted that ASC and NALP3 would beimportant mediators of the innate immune response to the adeno-virus in vivo , and thus experiments were performed using an estab-lished model of innate immunity to adenovirus in mice 7 . NALP3-,ASC- and caspase-1-deficient mice or their wild-type littermateswere injected intraperitoneally with first-generation adenovirus vec-tors. Compared to wild-type mice, the intraperitoneal admini-stration of AdGFP to Nalp32 / 2 mice resulted in a significantreduction of nuclear factor- kB-dependent inflammatory geneexpression such as pro-IL-1 b (Il1b ), MIP-1 b (Mip1b , also calledCcl4 ) or IP-10 ( IP-10 , also called Cxcl10 ) in the liver at 6h (Fig. 4a,b). Notably, protein levels of IL-1 b, IL-6 and MIP-1 b were decreased(Fig. 4c,d). Similarly, in ASC- andcaspase-1-deficient mice, the early
a
Asc –/–
V e h .
A d L a c Z
A T P
ext.
ext.
ext.
ext.
sup.
sup.
sup.
sup.
V e h .
A d L a c Z
A T P
V e h .
A d L a c Z
A T P
-17
-35kDa
AdLacZ AdLacZ
Nalp3 +/+ Nalp3 –/–
N a l p 3
+ / +
N a l p 3
– / –
Tlr9 –/– Ipaf –/–
Liposomes – – – + + + + +– – – – + + –DNase
IL-1β p17
WT
Nalp3 –/–
Asc –/–
Casp1 –/–
CASP1 p20
IL-1β p17
CASP1 p20
IL-1β p17
CASP1 p20
IL-1β p17
CASP1 p20
Liposomes –– + – + + + +
D N A s p
p c D N A 3
D N A s p
p c D N A 3
D N A s p
– –– + –
D N A s p
– R 8 3 7
R 8 3 7
-17kDa
-20
-35
Ifnar1 +/+ Ifnar1 –/–
d
Glyburide – + – – + – – + –– – + – – + – – +KCl
IB: pIRF-3IB: IRF-3
IB: IL-1 β
IB: IL-1 β
IB: CASP1
IB: IL-1 β
IB: CASP1
IB: IL-1 βD116
p o l y ( d A : d T
)
E . c o l i D
N A
IB: pIRF-3
IB: IRF-3
IB: IL-1 β
V e h .
p o l y ( I : C )
A d G F P
V e h .
p o l y ( I : C )
A d G F P
e
IB: IL-1 β
– A T P
M S U
p c D N A 3
D N A s p
D N A s p
– – D N A s p
p c D N A 3
p c D N A 3
b c
Figure 3 | Role of NALP3 and ASC in adenovirus and DNA activation of IL-
1b. a , Immunoblottingfor IL-1 b in cell extracts (ext.)or supernatants(sup.)from LPS-primed murine wild type, Nalp3 2 / 2 , Asc 2 / 2 , Tlr9 2 / 2 or Ipaf 2 / 2
peritoneal macrophages 6h after transduction with AdLacZ or vehicle(veh.). ATP (5 mM) served as a positive control. Tlr9 2 / 2 and Ipaf 2 / 2
macrophages were challenged with an increasing titre of AdLacZ (range5 3 103 to 5 3 104 particles per cell) or vehicle. b , Effect of liposome-mediated transfectionof salmonspermor plasmidDNA (DNAsp, pcDNA3)in wild-type, Nalp3 2 / 2 , Asc 2 / 2 and Casp12 / 2 macrophages with and without DNase treatment. Monosodiumurate (MSU) crystals and ATP wereused as controls. c, IL-1b and phospo-IRF3 immunoblotting in THP-1 cellstransfectedwith poly(dA:dT)or E. coliDNA. Cells were pre-treated with KCl(100 mM) or the potassium channel inhibitor glyburide (100 mM). d , IL-1band caspase-1 immunoblotting in IFNR1-deficient ( Ifnar1 2 / 2 )macrophages transfected with DNA or challenged with the TLR7/8 agonistand inflammasome activating R837 (10 mg ml2 1 ). e , Phospho-IRF3 and IL-1b immunoblotting in wild-type and Nalp3
2 / 2macrophages stimulated
with AdGFP or transfected with poly(I:C) at 6 h.
b
AdGFP
-IL-1 β ( Il1b )-IL-1 α ( Il1a )
-IL-1Ra ( Il1ra )
-Gapdh
a WT
-Rantes
-MIP-1 β ( Ccl4 )
-MIP-2 ( Cxcl2 )
-IP-10 ( Cxcl10 )
-TCA3 ( Ccl1 )
-Gapdh
200
150
100
50
0
200250
150100
500
AdGFP– + + – + +– + + – + +
-MIP-1 α ( Ccl3 )
-MCP-1 ( Ccl2 )
c
Vehicle AdGFP WT AdGFP Nalp3 –/–
Vehicle AdGFP WT AdGFP Nalp3 –/–
Vehicle AdGFP WT AdGFP Nalp3 –/–
I L - 1 β ( p g
m l – 1 )
I L - 6 ( p g m
l – 1 )
d
MCP-1IP-10MIP-1 β
012345
0
1.20.80.4
0
800
600
400
200
0
4080
120
Il1a
Il1ra
Il1b
R e l a t i v e m
R N A
e x p r e s s i o n
C h e m o k i n e
( p g m
l – / – )
Nalp3 –/–
WT Nalp3 –/–
Figure 4 | Role of NALP3 in adenovirus-vector-induced inflammation in
vivo . a , b, RNase protection assay of total liver RNA probing for chemokineand cytokine genes at 6 h in wild-type and Nalp3 2 / 2 mice receiving AdGFPintraperitoneally. Nalp3 2 / 2 mice displayed blunted liver cytokine mRNAlevels in response to adenovirus vectors. Quantifications are based onphosphorimaging and are normalized to housekeeping genes (mean 6 s.d.,n 5 3–5). c, d , ELISA of spleenIL-1b , IL-6, MIP-1 b (CCL4), IP-10(CXCL10)and MCP-1 (CCL2) from wild-type and Nalp3 2 / 2 mice 6 h after AdGFPadministration (mean 6 s.d., n 5 3–5; asterisk, P , 0.05).
NATURE |Vol 452 |6 March 2008 LETTERS
105
Nature PublishingGroup©2008

8/10/2019 Nature artículo
http://slidepdf.com/reader/full/nature-articulo 4/6
inflammatory response in the liver was blunted after intraperitonealadministration of AdLacZ compared to wild-type controls (Supple-mentary Fig. 9). These results show that the NALP3 inflammasomeand IL-1b are mediators of the innate immune response to the ade-novirus virion in vivo . However, the partial reduction in the inflam-matory response observed in mice deficient in inflammasomecomponents is consistent with redundant mechanisms that exist inthe innate response to adenovirus 19 .
Our data demonstrate that the internalized adenovirus DNA acti-
vates intracellular NALP3 and downstream signalling that involvesASC and caspase-dependent activation of IL-1 b. This is in line withfindings that IL-1 b and theIL-1RI have a significant role in the innateimmune response to adenovirus 8 , and is also consistent with therequirement for adenoviral DNA to mediate the innate response 20 .Similarly, the inflammasome-dependent IL-18 was shown to have amajor role in the protection against HSV-1 (ref. 21). The identifica-tion of the inflammasome as an antiviral innate mechanism is alsoconsistent with data demonstrating a PYD-containing protein(M13L) encoded by myxoma virus that disrupts the NALP–ASCinteraction and caspase-1 activation 22 , an important immunomodu-latory strategy to circumvent host antiviral responses.
In addition to detecting viral DNA, the inflammasome alsoresponded to cytoplasmic mammalian genomic DNA, bacterial
DNA and synthetic DNA. Unexpectedly, the response to transfectedcytoplasmic DNA occurred in a NALP3-independent manner, butrelied on ASC and caspase-1. Because ASC is part of the NALPinflammasome complex, it is likely that NALP inflammasomes con-stitute the sensing platform of cytoplasmic DNA. Alternatively, aNALP-independent mechanism, requiring only ASC for the activa-tion of the inflammasome as recently proposed, cannot beexcluded 23 . How the DNA is sensed is currently not clear. We haveno evidence that theonly characterized cytosolic DNAsensor, DAI,isinvolved (data not shown).
Our data clearly identify the NALP/ASC inflammasomes and IL-1b as an alternative innate mechanism to type I IFN capable of sens-ingcytoplasmic DNAandof triggeringa pro-inflammatory response.This has a number of implications. First, it is likely that the NALP3
inflammasome has an important role in the host response to DNAvirus infection in general. Second, our data raise the possibility thatautoimmune diseases such as systemic lupus erythematosus (SLE)might involve the dysregulation of the inflammasome. Because many patients of such diseases form immune complexes to dsDNA by means of autoreactive antibodies, the elevated IL-1 b levels observedduring active disease might be explained via this mechanism 24 .Similarly, chronic arthritis caused by mammalian DNA that escapesfrom degradation is associated with increased levels of IL-1 b and IL-18. Indeed, one form of SLE and rheumatoid arthritis was recently found to be associated with mutations in the NALP1 gene25 . Theidentification of the inflammasome as a general sensorof cytoplasmicDNA may therefore lead to new antiviral therapies and to new insights and treatments for nucleic-acid-dependent autoimmune
diseases.METHODS SUMMARYViruses and viralvectors. Wild-type serotype 5, serotype 3 adenovirus, serotype5 adenovirus vectors and helper-dependent adenovirus vectors were propagatedand purified as previously described 26–28 . Empty serotype 5 adenovirus capsidswere generated using the helper-dependent adenovirus vector system and 293-Cre cells, as previously described 13 .Animal studies. Mice genetically deficient for caspase-1 ( Casp1), Asc , Nalp3,Ipaf , interferon receptor 1 ( Ifnar1) or Tlr9 were on a C57BL/6 background 29,30 .For in vivo studies, mice were injected intraperitoneally with 2 3 1011 particles of adenovirus vectors.Cell culture. Human THP-1 cells were differentiated for 24 h with PMA(100 nM). Mouseperitoneal macrophages were isolated using 4–10% thioglycol-late solution andstimulated with 1–10 ngml 2 1 ultra-pure LPS (Invivogen).Viralinfection/transduction was performed using 10 4 particles per cell for adenovirusvectors or multiplicity of infection 100:1 for wild-type adenovirus 3 and 5.
DNA isolation, plasmids and transfections. Human and viral genomic DNAwas isolated from THP-1 cells and adenovirus by standard protocol. DNA of varying length was amplified in standard PCR reactions using oligonucleotidesspecific to the mouse Gapdh gene and to regions of the human RANTES (Ccl5 )and mouse IP-10 ( Cxcl10 ) promoters. PCR products were pooled, ethanol pre-cipitated and re-suspended in water. Complimentary sense and antisense oligo-nucleotides (40 bp) were synthesized and annealed to obtain dsDNA fragments.Bacterial DNA was isolated from E. coli . To remove contaminating LPS from theDNA preparation, E. coli DNA was incubated with polymyxin B (50 mg ml2 1 ) atroom temperature for 30 min. DNA was ethanol precipitated twice and re-
suspended in endotoxin-free water. Single-stranded DNA, poly(deoxyadenosine:deoxythymidine) (poly(dA:dT)) DNA, CpG oligonucleotides, poly(inosine:cytosine) (poly(I:C)) and R837 were obtained from commercial sources.Transfections were performed using Lipofectamine 2000 (4 ml ml2 1 ) as per themanufacturer’s protocol (Invitrogen). All transfections were performed using1–2 mg of DNA.
Full Methods and any associated references are available in the online version ofthe paper at www.nature.com/nature.
Received 9 June 2007; accepted 4 January 2008.Published online 20 February 2008.
1. Kawai, T. & Akira, S. TLR signaling. Cell Death Differ. 13, 816–825 (2006).2. Ishii, K. J.& Akira, S.Innate immunerecognition of, andregulation by,DNA. Trends
Immunol. 27, 525 –532 (2006).3. Stetson, D. B. & Medzhitov, R. Recognition of cytosolic DNA activates an IRF3-
dependent innate immune response. Immunity 24, 93 –103 (2006).4. Takaoka,A. etal. DAI(DLM-1/ZBP1)is a cytosolicDNA sensor andan activatorof
innate immune response. Nature 448, 501–505 (2007).5. Sergerie, Y., Rivest, S. & Boivin, G. Tumor necrosis factor- a and interleukin-1- b
play a critical role in the resistance against lethal herpes simplex virusencephalitis. J. Infect. Dis. 196, 853 –860 (2007).
6. Kay, M. A., Glorioso, J. C. & Naldini, L. Viral vectors for gene therapy: the art ofturning infectious agents into vehicles of therapeutics. Nature Med. 7, 33–40(2001).
7. Muruve, D. A., Barnes, M. J., Stillman, I. E. & Libermann, T. A. Adenoviral genetherapy leads to rapid induction of multiple chemokines and acute neutrophil-dependent hepatic injury in vivo. Hum. Gene Ther. 10, 965 –976 (1999).
8. Shayakhmetov, D. M., Li, Z. Y., Ni, S. & Lieber, A. Interference with the IL-1-signaling pathway improves thetoxicityprofileof systemically appliedadenovirusvectors. J. Immunol. 174, 7310 –7319 (2005).
9. Fritz, J. H., Ferrero, R. L., Philpott, D. J. & Girardin, S. E. Nod-like proteins inimmunity, inflammation and disease. Nature Immunol. 7, 1250 –1257 (2006).
10. Petrilli, V., Dostert, C., Muruve, D. A. & Tschopp, J. The inflammasome: a dangersensing complex triggering innate immunity. Curr. Opin. Immunol. 19, 615–622(2007).
11. Franchi, L.etal. IntracellularNOD-like receptorsin innate immunity, infection anddisease. Cell. Microbiol. 10, 1–8 (2008).
12. Cotter, M. J., Zaiss, A. K. & Muruve, D. A. Neutrophils interact with adenovirusvectors via Fc receptors and complement receptor 1. J. Virol. 79, 14622 –14631(2005).
13. Stilwell, J. L., McCarty, D. M., Negishi, A., Superfine, R. & Samulski, R. J.Development and characterization of novel empty adenovirus capsids and theirimpact on cellular gene expression. J. Virol. 77, 12881–12885 (2003).
14. Kanneganti, T. D. et al. Critical role for cryopyrin/Nalp3 in activation of caspase-1in response to viral infection and double-stranded RNA. J. Biol. Chem. 281,36560 –36568 (2006).
15. Boyden, E. D. & Dietrich, W. F. Nalp1b controls mouse macrophage susceptibilityto anthrax lethal toxin. Nature Genet. 38, 240 –244 (2006).
16. Franchi, L., Kanneganti, T. D., Dubyak, G. R. & Nunez, G. Differential requirement
of P2X7 receptor and intracellular K1
for caspase-1 activation induced byintracellular and extracellular bacteria. J. Biol. Chem. 282, 18810–18818 (2007).17. Petrilli, V. et al. Activation of the NALP3 inflammasome is triggered by low
intracellular potassium concentration. Cell Death Differ. 14, 1583–1589 (2007).18. Henry, T., Brotcke, A., Weiss, D. S., Thompson, L. J. & Monack, D. M. Type I
interferon signaling is required for activation of the inflammasome duringFrancisella infection. J. Exp. Med. 204, 987 –994 (2007).
19. Zhu, J., Huang, X. & Yang, Y. Innate immune response to adenoviral vectors ismediated by both TLR-dependent and -independent pathways. J. Virol. 81,3170 –3180 (2007).
20. Nociari, M., Ocheretina, O., Schoggins, J. W. & Falck-Pedersen, E. Sensinginfection by adenovirus: toll-like receptor-independent viral DNA recognitionsignals activation of theinterferon regulatory factor 3 master regulator. J. Virol. 81,4145 –4157 (2007).
21. Fujioka, N. et al. Interleukin-18 protects mice against acute herpes simplex virustype 1 infection. J. Virol. 73, 2401 –2409 (1999).
22. Johnston,J. B. etal. A poxvirus-encodedpyrin domain protein interactswith ASC-1 to inhibit host inflammatory and apoptotic responses to infection. Immunity 23,587 –598 (2005).
LETTERS NATURE |Vol 452 |6 March 2008
106
Nature PublishingGroup©2008

8/10/2019 Nature artículo
http://slidepdf.com/reader/full/nature-articulo 5/6
23. Fernandes-Alnemri,T. etal. Thepyroptosome:a supramolecular assembly ofASCdimers mediating inflammatory cell death via caspase-1 activation. Cell DeathDiffer. 14, 1590 –1604 (2007).
24. Sun, K.H., Yu,C. L.,Tang, S.J. & Sun, G. H.Monoclonal anti-double-strandedDNAautoantibody stimulates the expression and release of IL-1 b , IL-6, IL-8, IL-10 andTNF-a fromnormalhumanmononuclearcellsinvolving in thelupus pathogenesis.Immunology 99, 352 –360 (2000).
25. Jin, Y. et al. NALP1 in vitiligo-associated multiple autoimmune disease. N. Engl. J.Med. 356, 1216–1225 (2007).
26. Tibbles, L. A. et al. Activation of p38 and ERK signaling during adenovirus vectorcellentry leadto expression of theC-X-C chemokine IP-10. J. Virol. 76, 1559 –1568
(2002).27. Cotten, M. et al. Psoralen treatment of adenovirus particles eliminates virus
replication and transcription while maintaining the endosomolytic activity of thevirus capsid. Virology 205, 254 –261 (1994).
28. Muruve, D. A. et al. Helper-dependent adenovirus vectors elicit intact innate butattenuated adaptive host immune responses in vivo. J. Virol. 78, 5966 –5972(2004).
29. Mariathasan, S. et al. Differential activation of the inflammasome by caspase-1adaptors ASC and Ipaf. Nature 430, 213–218 (2004).
30. Martinon, F., Petrilli,V., Mayor, A., Tardivel,A. & Tschopp, J. Gout-associated uricacid crystals activate the NALP3 inflammasome. Nature 440, 237 –241 (2006).
Supplementary Information is linked to the online version of the paper atwww.nature.com/nature .
Acknowledgements This work was supported by the Alberta Heritage Foundationfor Medical Research, operating and group grants from the Canadian Institutes forHealth Research, the Swiss Committee for Technology and Innovation andMUGEN. D.A.M. is the recipient of an AHFMR Scholar Award; A.K.Z. is therecipient of a Heart and Stroke Foundation of Canada Studentship; V.P. is therecipient of a Marie Curie training award. We thank R. Castillo for technical
assistance, V.Dixit, A.Trumpp andS. Akirafor knockout mice andP. Vandenabeelefor the caspase-1 antibody.
Author Contributions D.A.M., V.P., A.K.Z., L.R.W., S.A.C., P.J.R. and R.J.P.conceived the research and conducted the experiments. J.T. oversaw the wholeproject.
Author Information Reprints and permissions information is available atwww.nature.com/reprints. Correspondence and requests for materials should beaddressed to J.T. ([email protected]) .
NATURE |Vol 452 |6 March 2008 LETTERS
107
Nature PublishingGroup©2008

8/10/2019 Nature artículo
http://slidepdf.com/reader/full/nature-articulo 6/6
METHODSViruses and viral vectors. Wild-type serotype 5 and serotype 3 adenovirus(ATCC)was propagated on HeLa cells andpurifiedby caesium chloride gradientcentrifugation. Viral infectious titres were determined on HeLa cells. The type 5,E1/E3-deletedadenovirusencodingthe GFP or LacZtransgene underthe controlof a cytomegalovirus promoter was generated using the Ad-EASY system(Stratagene), plaque purified twice and then propagated on human embryonickidney 293 cells and purified as previously described 26 . The particle titre wasdetermined by measuring the optical density at 260 nm described as particlesper cell. Plaque assay on HeLa cells and PCR was used to test for replication-competent adenovirus that remained consistently less than 1 per 10 10 particles.Ps-Ad is AdGFP, which was rendered transcription defective by UV/psoralentreatment as previously described 27 . AdL.PB and AdL.F are tropism-modifiedE1-E3-deleted Ad5-based vectors carrying the luciferase transgene in the E1region under the control of the CMV promoter 26 . The helper-dependent ade-novirus vector expressing the LacZ transgene under control of the CMV pro-moter was produced using the Cre/ loxP system as described 28 . Empty serotype 5adenovirus capsids were generated using the helper-dependent adenovirus vec-tor system and 293-Cre cells as previously described 13 and titred according toprotein content compared to an adenovirus vector virion standard. Reovirus(provided by P. Forsyth) was propagated on Vero cells. Purified herpes simplex virus-1 (HSV-1) was obtained from Advanced Biotechnologies. Low endotoxintissue culture reagents and buffers were used for vector production and experi-ments. Adenovirus vectors were routinely tested for the presence of endotoxinusing the Limulus Amebocyte Lysate kit. All vectors contained less then 0.1endotoxin units ml 2 1 .Animal studies. All animal studies were performed in accordance with theAnimal Care Committee guidelines at the University of Calgary and theUniversity of Lausanne. Mice, genetically deficient for caspase-1, Asc , Nalp3,Ipaf , interferon receptor 1 ( Ifnar1) or Tlr9 on a C57BL/6 background or wild-type littermates, were generated as previously described 29,30 . Mice were age- andsex-matched for the experiments. All mice were between 25 and 30 g and8–10weeks of age. Adenovirus vectors were injected intraperitoneally with2 3 1011 particles in a total volume of 100 ml (vector plus vehicle). Controlanimals received vehicle alone (3% sucrose, 150 mM NaCl, 10 mM Tris pH 7.4and 1mM MgCl 2 ).Cell culture. HumanTHP-1promonocytic leukaemiacellswere grown in RPMI1640 media, supplemented with 10% FBS, 1% penicillin-streptomycin, 1%MEM sodium pyruvate (100 mM) and 0.01% b -mercaptoethanol. THP-1 celldifferentiation was achieved by overnight stimulation with 100 nM phorbol-12-myristate-13-acetate (PMA). Cells were periodically tested for mycoplasma
contamination.Mouse peritoneal macrophages were isolated by peritoneal lavage 72h afterintraperitoneal injection of 10% thioglycollate solution (BD Biosciences). Cellswere platedat thedensity of8 3 105 cellsin 12-well dishesand non-adherent cellswere removed after 3 h. Cells were cultured in RPMI medium complementedwith 10% FBS, sodium pyruvate, penicillin/streptomycin and L-glutamine.Twenty-four hours after isolation, the cells were transduced with adenovirusvectors. Vector and control transductions were performed in the presence of 10ngml 2 1 ultra-pure LPS (Invivogen).
Myeloid dendritic cells were derived from bone marrow cells. Isolated bonemarrow cells were differentiated for 8 days in IMDM medium supplementedwith 2 mM L-glutamine, 100U ml 2 1 penicillin, 100 mg ml2 1 streptomycin,50 mM 2-mercaptoethanol (all from Invitrogen), 10% heat-inactivated FBS(IMDM complete medium), and 10% supernatant of J558 cells (GM-CSF-producing cells).
Viral infections and transductions in THP-1 cells were performed in six-wellplates or 24-well plates with 2 3 106 cells per well and 2 3 105 cells per well,respectively. Cells were incubated for the specified time periods in 1 ml of medium (6-well) or 0.3ml of medium (24-well) containing viruses or viralvectors (10 4 particles per cell for adenovirus vectors; multiplicity of infection100:1for wild-type adenovirus 3 and 5). For selected experiments, the FBS in thecell culture medium was replaced by 5–20% pooled human serum. Humanserum was collected from ten healthy donors, pooled, and filtered through a
sterile 0.45-mm mesh. In experiments involving the analysis of IL-1 b processing,crude LPS (12.5 mg ml2 1 ) and ATP (5 mM) were used as positive controls. Toinhibit caspase-dependent IL-1 b processing, cells were treated with 0.05mMzVAD-fmk (Sigma), 100 mM glyburide (glibencamide, Sigma) and 100 mMKCl for 15–30min before viral infection/transduction. Sphingosine-1-phosphate (S1P, 10 nM, Sigma), an unrelated modulator of cation channels,was used as a control.Immunoblotting. For immunoblotting cells were lysed with buffer (10 mM TrispH 7.5, 1% NP-40, 150 mM NaCl, and protease inhibitor cocktail), proteinsseparated on sodium dodecyl sulphate-polyacrylamide gels and then transferredonto polyvinylidene difluoride or nitrocellulose membranes. The membraneswere blocked with 5% milk proteins in 1 3 PBS and 0.5% Tween-20 and thenprobedwith primary antibodiesas follows:polyclonal rabbit anti-humanmature(17kDa)IL-1 b (D116,Cell Signaling), polyclonalrabbit anti-humanIL-1 b (CellSignaling), polyclonalsheep anti-human IL-1 b , polyclonalsheep anti-mouse IL-1b , polyclonalrabbit anti-humancaspase-1 (Santa Cruz), polyclonalrabbit anti-mouse caspase-1 (gift from P. Vandenabeele) and polyclonal rabbit anti-IRF-3(phospho-specific and total, Cell Signaling). Appropriate HRP-conjugatedsecondary antibodies were used and proteins detected using ECL reagent(Amersham).DNA isolation, plasmids and transfections. To obtain genomic DNA, THP-1cells were lysed in a buffer containing 100 mM NaCl, 10mM Tris pH 8.0, 0.5%SDS, 10 mM EDTA and 100 mg ml2 1 proteinase K at 56 u C. The lysate was cen-trifuged and the supernatant extracted with phenol:chloroform:isoamyl alcohol(25:24:1). The aqueous layer was ethanol precipitated and the DNA re-
suspended in ddH 2O. Viral DNA was obtained from wild-type serotype 5 ade-novirus as described above. Plasmids used in this study were pcDNA3.1 andpeGFP. pcDNA3.1 LE was purified using a low endotoxin purification protocol(Qiagen). PCR DNA and DNA of varying lengths were amplified in standardPCRreactions using oligonucleotides specific to a 576-bpfragmentof themouseGapdh gene and to regions of the human RANTES and mouse IP-10 promoters.PCR products were pooled, ethanol precipitated and re-suspended in water.Complimentary sense and antisense oligonucleotides (40 bp) were synthesizedand annealed to obtain dsDNA fragments. Bacterial DNA was isolated from E.coli usingthe DNeasy kit(Qiagen).To removecontaminating LPSfromthe DNApreparation, E. coli DNA was incubated with polymyxin B (50 mg ml2 1 ) at roomtemperature for 30 min. DNA was ethanol precipitated twice and re-suspendedin endotoxin-free water. Single-stranded DNA and poly(dA:dT) were obtainedfrom Sigma. CpG oligonucleotides, poly(I:C) and R837 were obtained fromInvivogen. DNase treatments were performed using RQ1 RNase-free DNase asper the manufacturer’s protocol (Promega). Equivalent volumes of DNase-treated and untreatedDNA were used in transfection studies.Transfections wereperformed using Lipofectamine 2000 (4 ml ml
2 1 ) as per the manufacturer’s pro-tocol (Invitrogen). All transfections were performed using 1–2 mg of DNA andRNA.RNase protection assay and ELISA. Total RNA from liver tissues was isolatedusing the RNeasy kit (Qiagen) according to the manufacturer’s protocol. RNaseprotection assays were carried out using the RiboQuant MultiProbe RNaseprotection assay system and the mCK2b and mCK5c probe sets (BDPharmingen) as per the manufacturer’s protocol. Gels were visualized by auto-radiography and quantified using a Personal F/X Molecular Imager and theQuantity One software (BioRad). The signal intensity of individual genes wasnormalized to the intensity of the housekeeping gene Gapdh or L32 within thesame sample.
ELISAs for mouse IL-1b and IL-6 were performed on tissue culture super-natants or spleen extracts using OptiEIA ELISA kits as per the manufacturer’s
protocol (BD Bioscience). Analysis of mouse IP-10 (CXCL10), MIP-1 b (CCL4)and MCP-1 (CCL2) proteins in spleen extracts was performed using theMultiplex Antibody BeadAssay and Luminex as per the manufacturer’sprotocol(Biosource). Results were normalized to protein content within individual sam-ples as determined by the Bradford assay.Statistics. All values are expressed as the mean 6 s.d. of individual samples.Samples were analysed using the student’s t -test.
doi:10.1038/nature06664
Nature PublishingGroup©2008


















