Musculo Cardiaco y Liso
85
4. Músculo cardiaco. Este tipo de tejido muscular forma la mayor parte del corazón de los vertebrados. Las células presentan estriaciones longitudinales y transversales imperfectas y difieren del músculo esquelético sobre todo en la posición central de su núcleo y en la ramificación e interconexión de las fibras. El músculo cardiaco carece de control voluntario. Está inervado por el sistema nervioso vegetativo, aunque los impulsos procedentes de él sólo aumentan o disminuyen su actividad sin ser responsables de la contracción rítmica característica del miocardio vivo. El mecanismo de la contracción cardiaca se basa en la generación y transmisión automática de impulsos. El músculo cardíaco (miocardio) es un tipo de músculo estriado encontrado en el corazón. Su función es bombear la sangre a través del sistema circulatorio por contracción. El músculo cardíaco generalmente funciona involuntaria y rítmicamente, sin tener inervación (estimulación nerviosa. Es un músculo miogénico, es decir autoexcitable. Las fibras estriadas y con ramificaciones del músculo cardíaco forman una red interconectada en la pared del corazón. El músculo cardíaco se contrae automáticamente a su propio ritmo, unas 100.000 veces al día. No se puede controlar conscientemente, sin embargo, su ritmo de contracción está regulado por el sistema nervioso autónomo dependiendo de que el cuerpo esté activo o en reposo. 4.1. Estructura. El músculo cardíaco (Fig 1) está formado por células musculares ramificadas, que poseen 1 o 2 núcleos y que se unen entre sí a través Figura 1 Figura 2
-
Upload
horacio-sanabria-monjardin -
Category
Documents
-
view
316 -
download
0
Transcript of Musculo Cardiaco y Liso

4. Músculo cardiaco.
Este tipo de tejido muscular forma la mayor parte del corazón de los vertebrados. Las células presentan estriaciones longitudinales y transversales imperfectas y difieren del músculo esquelético sobre todo en la posición central de su núcleo y en la ramificación e interconexión de las fibras. El músculo cardiaco carece de control voluntario. Está inervado por el sistema nervioso vegetativo, aunque los impulsos procedentes de él sólo aumentan o disminuyen su actividad sin ser responsables de la contracción rítmica característica del miocardio vivo. El mecanismo de la contracción cardiaca se basa en la generación y transmisión automática de impulsos.
El músculo cardíaco (miocardio) es un tipo de músculo estriado encontrado en el corazón. Su función es bombear la sangre a través del sistema circulatorio por contracción.
El músculo cardíaco generalmente funciona involuntaria y rítmicamente, sin tener inervación (estimulación nerviosa. Es un músculo miogénico, es decir autoexcitable.
Las fibras estriadas y con ramificaciones del músculo cardíaco forman una red interconectada en la pared del corazón. El músculo cardíaco se contrae automáticamente a su propio ritmo, unas 100.000 veces al día. No se puede controlar conscientemente, sin embargo, su ritmo de contracción está regulado por el sistema nervioso autónomo dependiendo de que el cuerpo esté activo o en reposo.
4.1. Estructura.
El músculo cardíaco (Fig 1) está formado por células musculares ramificadas, que poseen 1 o 2 núcleos y que se unen entre sí a través de un tipo de unión propia del músculo cardíaco
llamada disco intercalar (Fig 2). A diferencia del músculo esquelético, las fibras musculares cardíacas corresponden a un conjunto de células cardíacas unidas entre sí en disposición lineal.
Figura 1 Figura 2
Figura 3 Figura 4
Figura 5 Figura 6

Las células musculares cardíacas, de unos 15 m m de diámetro y unos 100 mm de largo, tienen el núcleo ubicado al centro del citoplasma (Fig 3) y presentan estriaciones transversales similares a las del músculo esquelético (Fig 4 y 5)). El retículo sarcoplásmico no es muy desarrollado y se distribuye irregularmente entre las miofibrillas, que no aparecen claramente separadas. Sin embargo, las mitocondrias, que son extremadamente numerosas, están distribuidas regularmente dividiendo a las células cardíacas en miofibrillas aparentes. En el sarcoplasma hay numerosas gotas de lípido y partículas de glicógeno. Con frecuencia las células musculares cardíacas presentan pigmentos de lipofuscina cerca de los polos nucleares. Las células están rodeadas por una lámina externa, comparable a la lámina basal de los epitelios.
Existen ciertas diferencias estructurales entre el músculo de los ventrículos y de las aurículas. Las células musculares de las aurículas son mas pequeñas y vecinos al núcleo, en asociación con complejos de Golgi presentes en esa zona, se observan gránulos de unos 0.4 m m de diámetro que contienen el factor natriurético auricular, auriculina o atriopeptina.
Estructuralmente, las miofibrillas del músculo cardíaco, son esencialmente iguales a la del miofibrillas del músculo esquelético. Por otra parte, los túbulos T del músculo cardíaco son de mayor diámetro que los del músculo esquelético y se ubican a nivel del disco Z (Fig 6). Los túbulos se asocian generalmente con una sola expansión de las cisternas del retículo sarcoplasmico. De manera que lo característico del músculo cardíaco son las díadas, compuestas de un túbulo T y de una cisterna de retículo endoplásmico.
Discos intercalares
Los discos intercalares son los sistemas de unión que asocian a las células musculares cardíacas para formar las fibras del miocardio (Fig 1). Estas estructuras se encuentran en regiones de la membrana donde los extremos de dos células se enfrentan y se ubican en lugar de un disco Z. Su nombre deriva del hecho que en cortes longitudinales aparecen como estructuras escaleriformes.
Los discos intercalares presentan: (Fig 2 y 3).

una porción transversa, en la cuál se ubican dos tipos de unión intercelular : fascia adherens y mácula adherens una la porción lateral, que corre paralela a los miofilamentos, en la cuál se ubican uniones de comunicación (nexos o gap junctions).
La fascia adherens es un tipo de unión propia del corazón, pero su estructura es semejante a la de las zonas de adhesión de los epitelios. Estas estructuras anclan filamentos de actina a la membrana plasmática y también unen las membranas de células adyacentes. De esta manera, asocian el aparato contráctil de cada célula con el de la célula vecina. Su organización molecular es similar a la descrita en el capítulo de epitelios.
La mácula adherens corresponde a desmosomas típicos que se ubican en las porciones transversas y paralelas del disco. Estas estructuras anclan los filamentos intermedios de desmina de la fibra cardíaca y participan, junto con la fascia adherens, en la adhesión de las membranas plasmáticas de células vecinas.
Las uniones de comunicación (nexos), corresponden a sitios que permiten el paso de iones y moléculas pequeñas desde el citoplasma de una célula a la célula vecina. Su ultraestructura y composición molecular es similar a la de las uniones de comunicación descritas en el capítulo de epitelios.
Figura 1
Figura 2 Figura 3

4.2. Características eléctricas.
El sistema cardiovascular o aparato circulatorio está formado por el corazón y los vasos sanguíneos. Su función es llevar a cabo la circulación de la sangre por todo el organismo, para distribuir el oxígeno y demás nutrientes a las células del organismo, y recoger sus productos metabólicos de desecho para su eliminación.
La importancia de este sistema en un organismo pluricelular radica en que constituye el nexo de comunicación entre las células y el medio externo, distribuyendo sustancias que son esenciales para el metabolismo y la supervivencia celular.
La bomba cardiaca está formada por cuatro cavidades (dos aurículas y dos ventrículos) que funcionan como dos bombas en serie, trabajando al unísono y manteniendo cada una de ellas un circuito. El corazón derecho, junto con las arterias, capilares y venas pulmonares, forman el circuito menor o circulación pulmonar; mientras que el corazón izquierdo, junto con las arterias, capilares y venas sistémicas forman el circuito mayor o circulación sistémica.
El movimiento de la sangre en este sistema cerrado sería:
a) Tomando como punto de inicio el ventrículo izquierdo, recorrería todos los vasos sistémicos hasta volver a la aurícula derecha.
b) Pasa al ventrículo derecho que la bombea a los vasos pulmonares, retornando a la aurícula izquierda para pasar al ventrículo y cerrar de esta forma todo el circuito.
Histología cardiaca
La pared cardiaca, al igual que la pared vascular, está formada por tres capas de tejidos. La más interna recibe el nombre de endocardio y es una capa de células epiteliales muy planas con uniones muy fuertes entre ellas, y en contacto continuo con la sangre.
La capa media es la más importante y se denomina miocardio, está formada por fibras musculares estriadas cardíacas que superficialmente se disponen de forma oblicua, en la porción central delmiocardio adoptan una disposición circular y las más profundas se sitúan longitudinalmente. Su contracción garantiza un acortamiento en todos los ejesdel espacio y justifica la función de bomba que tiene esta estructura. La tercera capa de la pared cardiaca y la más externa es el epicardio, formado por tejido conectivo laxo con redes de fibras elásticas, vasos y nervios.
El pericardio es una bolsa que recubre todo el corazón. El pericardio permite los movimientos del corazón con un rozamiento mínimo, funciona como lubricante, disminuyendo la fricción en los continuos movimientos cardíacos, e impide los llenados excesivos.
Miocardiocito o fibra muscular cardiaca
Las fibras cardíacas son fibras musculares estriadas, mucho más cortas que las esqueléticas. Tienen unas 150 μ de longitud y unas 15-20 μ de ancho. Son células uninucleadas con un

contenido en mitocondrias mucho mayor que en las esqueléticas. Las miofibrillas presentan estriaciones con el mismo bandeado que en el músculo esquelético.
A diferencia de las fibras esqueléticas el retículo sarcoplásmico está menos desarrollado y los túbulos T, que presentan una anchura mayor, se sitúan sobre las líneas Z. Por otro lado, la agrupación de una cisterna del retículo y el túbulo T da lugar a una diada más que a una triada.
Las fibras cardíacas no presentan un contorno uniforme sino que tienen prolongaciones o ramificaciones por donde se unen unas con otras formando una especie de red o malla. La zona de contacto entre las fibras cardíacas da lugar a unas regiones especializadas a nivel de la membrana plasmática denominadas discos intercalares. Los discos intercalares son un tipo especial de unión intercelular, gap que garantiza la comunicación eléctrica ente estas células; y por otro lado, proporciona lugares de adhesión y anclaje de una célula con otra. Los discos intercalares proporcionan la base estructural que permite que el corazón se comporte como unsincitio funcional, ya que no morfológico porque cada célula mantiene su individualidad.
Funcionamiento del corazón
El corazón podría considerarse como una bomba electromecánica; es decir, un sistema que genera de forma automática el impulso cardíaco y lo transmite a todas las células de trabajo.
El músculo cardíaco es un músculo excitable, los miocardiocitos presentan las siguientes características exclusivas:
a) Son células automáticas capaces de contraerse sin ningún estímulo externo.

b) Son células rítmicas, lo cual permite que mantengan una frecuencia de contracción suficiente para mantener la actividad de bombeo sin detenciones que pudieran poner en riesgo la supervivencia del organismo. Estas dos características no son observables en la mayor parte de las fibras cardíacas pero subyacen en todas ellas.
Existen dos sincitios musculares, uno el auricular y otro el ventricular, aislados uno del otro por una barrera de tejido fibroso, que constituye el plano donde asientan las válvulas cardíacas.
Propiedades eléctricas del corazón
Los miocardiocitos son autoexcitables o automáticos, lo que significa que no requieren la presencia de un estímulo externo para generar una respuesta contráctil. Esta capacidad de despolarizarse y contraerse rítmicamente sin inervación, se denomina ritmicidad miogénica y es responsable del automatismo cardíaco.
Desde el punto de vista de sus propiedades eléctricas se pueden distinguir dos tipos de fibras cardíacas:
a) Fibras automáticas o de respuesta lenta. b) Fibras de trabajo o de respuesta rápida. Las primeras se caracterizan por ser
capaces de generar y conducir el potencial de acción; las segundas, en condiciones normales, no son automáticas y requieren un estímulo para su excitación; sin embargo, en algunas ocasiones, pueden funcionar de forma automática aunque con un ritmo más lento que las primeras.

Las fibras de cada región cardiaca se caracterizan por poseer un ritmo distinto, las ventriculares son las más lentas, mientras que un grupo de auriculares poseen el ritmo más elevado. Este grupo de fibras auriculares se denomina nodo sinusal, y al presentar la ritmicidad más alta, su actividad es la que marca la frecuencia básica del corazón, denominándoselas por ello células marcapaso.
La generación y propagación de un potencial de acción cardíaco es posible por la existencia de un sistema especializado de excitación y conducción.
La generación y propagación de un potencial de acción cardíaco es posible por la existencia de un sistema especializado de excitación y conducción.
Sistema eléctrico del corazón.
Las fibras que componen este sistema son el nodo sinusal, el nodo aurículo-ventricular, el haz de His y las fibras de Purkinje. La propagación del potencial de acción en las células cardíacas se produce a través de uniones de tipo gap, como si fuese una membrana continua. La despolarización iniciada en el nodo sinusal se expande por todas las fibras auriculares de arriba abajo. Al alcanzar el plano fibroso, sólo dispone de un punto para continuar la propagación, el nodo auriculo-ventricular, en este punto se produce un enlentecimiento (retraso de 0,1 seg) de la propagación debido a la geometría de las fibras que forman este nodo. Se caracteriza este nodo por ser un haz estrecho con pocas uniones gap donde la

velocidad de conducción del impulso eléctrico es más baja y, por lo tanto da lugar a ese retraso. A continuación, el potencial se desplaza rápidamente por el resto del sistema de conducción, alcanzando casi al unísono todas las fibras ventriculares.
Potencial de acción cardiaco
Para que se desarrolle una respuesta contráctil, lo primero que ha de generarse es una respuesta eléctrica en la membrana. Esta respuesta se denomina potencial de acción cardíaco.
La morfología del potencial de acción varía de una célula a otra dependiendo de su localización. Una característica general es su larga duración, a diferencia del músculo esquelético y de las neuronas, cuya duración es de 1-5 ms, el potencial de las fibras cardíacas oscila entre 150 y 300 ms.
Este tiempo tan prolongado tiene importantes consecuencias funcionales, ya que van a superponerse en el tiempo el potencial de acción (fenómeno eléctrico) con la contracción de la fibra (fenómeno mecánico). Consecuencia de este tiempo tan elongado es que los periodos refractarios también se extienden, garantizando que el músculo no pueda reexcitarse en ningún momento, excepto muy al final de la contracción.
Las dos variedades más diferenciadas de potencial de acción son, el potencial marcapasos o potencial de acción de las células de respuesta lenta, y el potencial de acción de las fibras de trabajo o células de respuesta rápida.
Potencial marcapasos o respuesta lenta
En las células del nodo sinusal y nodo aurículo-ventricular, el potencial de membrana en reposo no se mantiene en un valor estable, sino que presenta una serie de fluctuaciones rítmicas que van a dar lugar a la generación automática y rítmica de potenciales de acción. Las fases en que se desarrolla esta actuación son:
a) Fase de reposo inestable. El potencial de membrana no se mantiene constante, sino que va despolarizándose hasta generar el potencial de acción. La apertura de canales

para cationes permite que entren cargas positivas y que la célula se despolarice lentamente hasta alcanzar el umbral (-50 mV).Esta lenta despolarización que precede al potencial de acción se conoce como prepotencial, potencial marcapasos o despolarización diastólica , y su desarrollo temporal es un factor clave para la frecuencia cardíaca. A este tipo de potenciales se les describe con el término de respuestas lentas debido a esta fase de pendiente poco pronunciada.
b) Fase de despolarización. Debido a la entrada de iones de Ca++ del exterior. c) Fase de repolarización.
El potencial de acción se propaga por las fibras auriculares dando lugar a los potenciales de acción de dichas fibras y llega al nodo AV antes de que el potencial marcapasos de la células del nodo hayan alcanzado por sí solas el umbral. Posteriormente se propaga por el haz de His, y llega a las fibras ventriculares.
Potencial de acción ventricular o respuesta rápida
En el resto de fibras cardíacas, auriculares y ventriculares, el potencial de acción se desarrolla en las siguientes fases:
Fase 0 o fase de despolarización rápida. Fase 1 o de repolarización breve. Fase 2 o de meseta. Es la fase más característica de los potenciales de acción
cardíacos. Durante la misma se produce una apertura de canales lentos de Ca. Fase 3 o fase de repolarización. Fase 4 o potencial de membrana en reposo. En condiciones basales estas fibras
presentan una gran permeabilidad al potasio, lo que hace que su valor en reposo esté próximo a su punto de equilibrio (-90 mv).

Periodo refractario
El periodo refractario absoluto abarca el tiempo desde que se inicia la fase 0 hasta casi la mitad de la fase 3. Por término medio es de unos 200 ms. El periodo refractario relativo abarca el tiempo restante hasta que la membrana se ha repolarizado por completo. Dura unos 50 ms, y hay una cierta recuperación de la excitabilidad ya que se pueden generar potenciales de acción si el estímulo es muy fuerte.
Propagación delpotencial de acción cardiaco
El potencial de acción iniciado en el nodo sinusal se extiende por todas las fibras cardiacas según la secuencia coordinada descrita en el sistema de conducción. Puede apreciarse el retraso temporal producido en el nodo AV y la rapidez con que el potencial se inicia en todas las fibras ventriculares. Esta despolarización, prácticamente al unísono, permite que la contracción se desarrolle de igual manera y el corazón pueda funcionar como una bomba.

4.3. Características contráctiles.
La función cardiaca esta central izada en la capacidad del corazón de bombear sangre oxigenada hacia los tej idos peri féricos, pero si estudiamos al corazón de esta forma tan amplia es dif íci l determinar donde se encuentra la anomalía responsable de la fal la cardiaca. Por el lo es conveniente estudiar al corazón como músculo y no como una bomba y apl icar los principios de la f isiología cardiaca.
Las células musculares cardíacas consti tuyen el 75% del volumen total del corazón, siendo los componentes principales de los miocitos las miofibri l las y en un menor porcentaje las mitocondrias. El resto de los componentes son: el sistema T, el retículo sarcoplásmico, el núcleo, el sarcoplasma, el sarcolema y los l isosomas.
El sarcolema es la membrana celular de la f ibra muscular. El la se invagina en el interior celular para tomar más contacto con las miof ibri l las, formando una red de paredes gruesas que recibe el nombre de túbulos T. En sectores del túbulo T muy di latados que toman estrecha relación con el retículo endoplásmatico penetra el potencial de acción que provocará la l iberación de calcio necesaria para la contracción muscular. Estos túbulos también mediarán la recolección del mismo para provocar la relajación.
En el músculo cardíaco se puede dist inguir uniones entre las células ( los discos intercalares ) que unen los miocitos por sus extremos haciendo que el corazón funcione en forma sincronizada como un sincit io. La función del miocito es la contracción y detallaremos como se produce la misma a nivel molecular.

La maquinaria contráctil esta representada por la miofibri l las . Las mismas están compuestas por unidades contráct i les denominadas sarcómeras de 2,2 um de longitud y un ancho equivalente a la miof ibri l la. Con microscopio electrónico se puede ver una estructura electro densa denominada disco Z que separa una sarcómera de otra. Este disco se encuentra ubicado en una región poco densa l lamada la banda I (por isotrópica) en donde solamente hay f i lamentos f inos. Estas bandas alternan con otras denominadas bandas A (por anisotrópica) donde se hal lan f i lamentos gruesos y f inos. En la parte media de las bandas A se encuentra la banda H de menor densidad aun (donde solo hay f i lamentos gruesos). Fig. 1. Estas dist intas bandas sufren variaciones periódicas que se deben a la superposición de las proteínas cito esquelét icas.
En la sarcómera pueden dist inguirse los f i lamentos de act ina (f i lamento f ino) que nacen de los discos Z, donde existe la a act inina que es la proteína que une la act ina y la t i t ina , esta ult ima es una proteína elást ica ( la más grande del organismo). La t i t ina posee dos funciones:
mantiene a la miosina en su posición y, debido a que tiene una parte elástica,
actúa como resorte recuperando la longitud de la miof ibri l la después de la contracción muscular. Fig. 2
La miosina, proteína que forma el f i lamento grueso, esta formada por dos cadenas l igeras (muy parecidas a la calmodulina y troponina c pero que han perdido la af inidad por el calcio) , que forman la cola y dos cadenas pesadas que formas la cabeza. Cada cadena pesada esta compuesta por tres dominios: uno el terminal NH2, el segmento central y el extremo terminal COOH, estos dos últ imos son los que interaccionan con la act ina. La ATP asa que proporciona la energía necesaria a part ir de la hidról isis del ATP se encuentra en un sit io diferente al de la unión a la act ina. Fig. 1
El f i lamento f ino esta formado por act ina G proteína globular la cual se va pol imerizando para formar un f i lamento de act ina F que luego se combinara con otro para formar el f i lamento de act ina. Fig. 2
Cada siete pares de act ina G encontramos un complejo de proteínas reguladoras formado por la troponina T que se une a la tropomiosina , la troponina C que es la que t iene af inidad por el calcio y la Troponina I que t iene función inhibidora. Todas estas proteínas t ienen forma globular.

Fig. 1
A su vez este complejo esta unido al f i lamento f ino por la proteína tropomiosina, de forma alargada y acintada que se ubica entre los f i lamentos de act ina cada siete pares de act ina G.
En estado de reposo la unión de la tropomiosina con la troponinaT mantienen un estado inhibitorio que evita la interacción de la act ina y la miosina.
Cuando ingresa el Ca+ este se une a la troponina C formando un complejo Ca+ troponina C que provoca un cambio de posición en la tropomiosina. Este cambio deja l ibre el si t io del f i lamento de act ina para que se una con la cabeza de miosima. Así, la contracción muscular consiste en la unión y desunión cíclica de la cabeza de miosina con el fi lamento de actina con la concomitante hidrólisis de una molécula de APT por la ATPasa presente en la cabeza de la miosina.
Mecanismo de la contracción propiamente dicho:
Ante la l legada del est imulo apropiado la tropomiosina deja l ibre el si t io de la act ina para que interactúe con la miosina, formando fuertes puentes transversales que actúan a modo de remo, desplazando cícl icamente los f i lamentos de act ina.

La cabeza de la miosina se adhiere al f i lamento de act ina arrastrando el disco Z hacia el centro de la sarcómera. Luego la miosina se desconecta del f i lamento de act ina y recupera su posición de reposo momento en que encontramos a los puentes cruzados débilmente unidos. A continuación el f i lamento de miosina se une nuevamente al f i lamento de act ina pero en un punto más cercano al disco Z, con lo cual el f i lamento de act ina se corre un poco más hacia el centro de la sarcómera. Estos episodios se suceden varias veces, lo que provoca que
Fig.2

Potencial de acción calcio citoplasmático
Formación del complejo Ca+ troponina C
Liberación de la inhibición de la actina
Interacción de miosina y actina
Contracción muscular
Esquema 1
los f i lamentos de act ina se acerquen mutuamente y la sarcómera acorta su longitud durante la contracción como resultado del desl izamiento intersección de los f i lamentos de act ina sobre la miosina. Las bandas I y H se acortan mientras que la banda A permanece sin modif icaciones.

Se ha comprobado que en miocardios defectuosos está presente una regulación anómala del calcio que hace fracasar la función contráct i l . Recientes investigaciones han encontrado fármacos( Vetmedin) que incrementan la af inidad de los f i lamentos de troponina por el Ca+ mejorando la contract i l idad cardiaca ( no requir iendo más energía si no aumentando su ef iciencia)
SISTEMA NERVIOSO AUTÓNOMO ;
El corazón esta regulado por el sistema nervioso autónomo. El simpático lo modula posit ivamente y el parasimpático ejerce una modulación negativa.Los neurotransmisores del sistema adrenérgico adrenalina y a noradrenalina, van a actuar en dist intos receptores: los α y los β1 y β2. Aunque los mismos t ienen mecanismos intrínsecos diferentes pero van regular la función del músculo cardiaco en solo sentido.
Los receptores α t ienen mayor af inidad por la adrenalina y los receptores b1 t ienen igual af inidad para cualquiera de los dos neurotransmisores mientras que los B2 t iene mayor af inidad por la adrenalina.
Fig 3

Esquema 2
El receptor β tanto el subtipo 1 como 2,cuando se unen al agonista adrenérgico, est imula a la proteína G s que es un hetero trímero de membrana formado por tres subunidades la α, β y g . A su vez, esta proteína est imula a la adenil ciclasa produciendo el segundo mensajero AMP cícl ico a part ir de ATP. El aumento de la concentración de este segundo mensajero act iva a la proteína quinasa A que fosfori la varias proteínas del miocito .
Sus acciones son las siguientes:
fosfori lación de la fosfolamban , proteína que est imula la Ca+ ATPasa para que introduzca el Ca+ al retículo sarcoplásmico,
fosfori lación de los canales de Ca+ permit iendo que ingrese más Ca+ (lo que aumentara la act ividad enzimática de la cabeza de la miosina),
fosfori lación de los canales de Na+ para cerrarlos cuando la célula este despolarizada. ( ver esquema 2)

Las acciones β agonista aumentarán la fuerza y la velocidad de contracción, pero también faci l i tarán la relajación, acción mediada por la fosfolamban.
El receptor α cuando es est imulado por la noradrenalina, act iva a la proteína Gs de membrana que a su vez est imula posit ivamente la proteína fosfol ipasa c que va a actuar sobre el fosfat idi l inositol bifosfato originando inositol tr i fosfato (IP3) y diaci l gl icerol (DAG).
El (IP3) provoca l iberación de calcio del retículo sarcoplásmico y el DAG activa a la proteína quinasa c la cual produce:
un preacondicionamiento de los canales de K+,
interviene en la contract i l idad y
pone en marcha factores de crecimiento celular que podrían estar relacionados con la hipertrof ia cardíaca en aquellos pacientes con sobre est imulación adrenérgica.
En conclusión el sistema simpático provocará:
un aumento de la frecuencia,
aumento de la contract i l idad y
un aumento de la conducción en las aurículas y los ventrículos, aumentando el automatismo con posibi l idad de aparición de marcapasos ectópicos
El parasimpático t iene como neurotransmisor la aceti l col ina la que va a actuar sobre los receptores col inérgicos. Los que se encuentran en el corazón son los receptores M2 un subtipo de los receptores muscarínicos. La act ivación de los mismos provoca la act ivación de la proteína G i que inhibe a la adenil ciclasa con lo que disminuyen las concentraciones de AMP. Esta disminución provoca el cierre de los canales de Ca+ y la apertura de los canales de K+ produciendo una hiperpolarización en el nódulo sinoauricular y auriculo ventr icular (efecto inotrópico negativo).
TEORIA DE LA LIBERACIÓN DE CALCIO INDUCIDA POR CALCIO:
La concentración de Ca+ ionizado en el medio extracelular es de 10-3 nM. La del medio intracelular estando la célula relajada es de 10-7 nM. Estos valores se elevan cuando la célula toma el estado de contracción alcanzando un valor de 10-5 nM.

Al elevarse las concentraciones de Ca+ , aumenta la interacción entre el Ca+ y la troponina C lo que gati l la el proceso contráct i l .
En cada onda de despolarización ingresan pequeñas concentraciones de Ca+ del medio extracelular a través de canales de calcio voltaje dependiente , que act ivan la l iberación del Ca+ del retículo sarcoplasmático.
Esta teoría t iene una correlación molecular demostrada por la existencia de un receptor en retículo sarcoplasmático que l ibera calcio hacia los túbulos T. La l iberación del calcio esta en relación con la duración del potencial de acción.
Cada canal de calcio voltaje dependiente del sarcolema controla a un grupo de canales de l iberación de calcio del retículo sarcoplasmático. Fig. 4. Esto se debe a la proximidad anatómica de los canales de calcio del sarcolema ubicados en los túbulos T y los canales de l iberación de calcio del retículo sarcoplasmático(RS).
Los canales de l iberación de calcio del retículo sarcoplasmático son parte de una compleja estructura proteica l lamado receptor Ryanodine ( RR) cuya densidad es de 800 receptores por micrómetro cuadrado de RS. Este receptor se ext iende de la membrana del RS hasta los túbulos T (TT) contr ibuyendo una región l lamada pie o canal de unión.
Este receptor t iene dos porciones: la más grande es el pie que l iga los túbulos T con el RS y una más pequeña en la región C terminal que consti tuye el canal poro
que puede ser act ivado por fosfori lación ante estímulos ß adrenérgicos o cambios de voltajes.
En la contracción, la onda de despolarización act iva los canales L de calcio de los TT lo que permite la entrada de pequeñas cantidades de calcio al ci tosol. Este Ca+ interactúa con el pie del RR, y provoca un cambio conformacional en dicho receptor que abre los canales de calcio del RS sal iendo el Ca+ hacia el ci tosol.En el miocito debe exist ir un balance de Ca*. es decir que la misma cantidad de este ion que ingresa debe sal ir . Esto se logra por dos mecanismos: 1) por intercambio Na+/Ca+; 2) por la regulación de la bomba deCa+.

Fig. 4
INCREMENTO DEL CALCIO EN EL RETÍCULO SARCOPLASMATICO POR LA BOMBA DE Ca++/ATPasa
Los iones de Ca+ ingresan al RS por la acción de la bomba Ca++/ATPasa (también l lamada SERCA), Esta bomba es una proteína de membrana que representa mas del 90% de las proteínas del RS.
Existen tres genes que codif ican cinco isoformas de la misma ,predominando en el miocardio la isoforma 2ª.
Por cada mol de ATP hidrol izado por la enzima dos moles de calcio son traídos al RS.
La fosfolamban ( FL) l lamada así por ser receptor de fosfatos es el principal regulador de la bomba Ca++/ATPasa, y se encuentra en la misma relación molar con dicha bomba.
La fosfolamban es una proteína pentamérica que se encuentra en la membrana del RS y cuya act ivación depende de su estado de fosfori lación. Normalmente inhibe la bomba Ca++/ATPasa cuando no esta fosfori lado. Cada una de las cinco subunidades de la fosfolamban puede ser fosfori lada en dos sit ios diferentes por dos o tres proteínas quinasas (PK). Una de las más importantes es la act ivada por el AMPc en respuesta a la est imulación ß adrenérgica del miocito, incrementando el calcio en el RS.

El calcio incorporado al RS por la bomba de Ca++/ ATP asa es almacenado l igado a la proteína calsecuestr ina que se encuentra cerca de los túbulos T. El calcio almacenado por la calcecuestr ina esta disponible para el proceso de l iberación.
Existe otra proteína de almacenamiento que es la calrectul ina .
Durante la relajación la bomba de calcio y el intercambiador Na+/Ca+ del RS compiten para ret irar el calcio citosól ico.
El equi l ibr io del balance de calcio se l leva a cabo por una serie de intercambiadores iónicos principalmente por el Na+/Ca+.
La act ividad de este intercambiador depende del potencial de membrana y de la concentración de Na* y Ca+ a ambos lados de la membrana.
ESTRUCTURA DE LOS CANALES DE CALCIO
Los canales de calcio dependiente de voltaje o canales L son proteínas macromoleculares que atraviesan la bicapa l ipidica .
Todos los modelos de acople electro- mecánico atr ibuyen un papel crucial a la apertura de estos canales, para el inicio de la contracción muscular cardíaca. Los canales iónicos t ienen dos propiedades: bloqueo y permeabil idad y protegiendo a cada canal existen dos o más puertas hipotét icas.
Los iones pueden pasar solamente cuando ambas puertas están abiertas. En el potencial de reposo la puerta de act ivación esta cerrada y la de inactivación esta abierta y en la despolarización la puerta de act ivación se abre permit iendo el pasaje ionico.
Existe una semejanza estructural entre los canales de Na+ y Ca+ operados por voltajes. Ambos canales t ienen una subunidad a1 importante con cuatro dominios transmembrana, también t ienen otras subunidades a2 ,ß y d .

Cada uno de los cuatro dominios transmenbrana de la subunidad a1 se compone de seis hél ices exist iendo en cada dominio un segmento hel icoidal especif ico cargado posit ivamente que consti tuye el sensor de voltaje.
La act ivación de la compuerta se debe a un cambio de carga en la subunidad a1, adquir iendo esta subunidad carga posit iva.
El canal real del poro se encuentra en la subunidad a1, entre las hél ices 5 y 6 donde los iones de calcio transcurren.
La subunidad a1 puede ser fosfori lada en varios sit ios, especialmente en el extremo carboxi lo terminal. Los grupos fosfatos del ATP son transferidos a la subunidad a1 y en esta misma subunidad se produce una alteración de las cargas lo que provoca una mayor probabil idad de apertura del canal.
La función de las subunidad b es la de aumentar el f lujo de la subunidad a1.
INTERCAMBIO SODIO/ PROTON Y EQUILIBRIO ACIDO BASE:
El pH intracelular es el resultado del equi l ibr io entre los cambiadores alcal inizantes , acidif icantes y de la producción metabólica de ácidos .
Si el interior la célular t iene un pH ácido debido al aumento de la concentración de protones se pone en marcha el intercambiador Na/ H sacando el protón de la célula y provocando el ingreso de Na+, (1-1). Como resultado aumenta el pH intracelular y la concentración de sodio, este ult imo se equil ibra por la acción de el intercambiador Na/ Ca++ o por la bomba Na+/K+.
Esta bomba también puede funcionar para extraer Na+ a expensas de un aumento de la concentración de protón disminuyendo la act ividad del intercambiador COOH-/Cl-.
El cotransportador Na+/COOH- sirve también para corregir la acidosis debido al transporte interno del bicarbonato.
El intercambiador COOH-/Cl- acidif ica el medio cuando el mismo se encuentra con un pH elevado.
REGULACIÓN MEDIADA POR TRIFOSFATO DE INOSITOL(IP3 ):

Es un sistema totalmente diferente implicado en la regulación del calcio, est imulando el intercambio sodio calcio directamente. Existe un receptor para el IP3, que t iene un alto grado de homología con el RR.
El ( IP3) es un mensajero intracelular que se forma, como di j imos anteriormente, luego de la act ivación de la fosfol ipasa C, mecanismo que se desencadena luego de la act ivación del receptor α adrenérgico. Una vez formado se une a el receptor de (IP3) y media la l iberación de Ca+ del retículo sarcoplasmático
El IP3 también part icipa en la transducción de señales inducida por angiotensina II , endotel ina y a1 agonista.
Tiene un papel importante en el paro cardiaco, ya que aumenta su regulación para ayudar a mantener la l iberación de Calcio del RS.
BOMBA DE NA++/K ATPasa
Todo el Na+ que ingresó, ya sea por la despolarización temprana o por el intercambiador Na/ Ca+ debe sal ir del interior celular por la bomba Na+ / K ATPasa.
Por cada molécula de ATP que se consume se el iminan 3 moléculas de Na+ y 2 de K+ ingresan al interior celular. Por lo tanto se pierde una carga posit iva y esta es una bomba electrogénica contr ibuyendo en – 10 mV al potencial de reposo.
CAMBIOS IÓNICOS NECESARIOS PARA LA CONTRACCIÓN MUSCULAR:
La actividad eléctr ica de los miocitos se relaciona con la difusión de iones a través de la membrana.
Durante el estado de reposo se detecta en la célula un potencial de negativo de 80 a 90 mV que se l lama potencial de reposo, atr ibuido a la difusión de K+ y a la bomba de Na+./ K+ ATPasa que saca tres moléculas de Na+ al exterior y envía dos moléculas de K+ al interior.
En una célula ventr icular t ípica podemos describir dist intas corr ientes iónicas que van a determinar las fases que van a l levar a la despolarización celular.
La fase 0 del potencial de acción de respuesta rápida, característ ico de las f ibras musculares y del sistema de conducción, es bien empinada, se debe a la apertura de los canales de sodio operados por voltaje.
Estos canales están consti tuidos por subunidades α y β, a su vez cada subunidad α esta consti tuido por cuatro dominios, cada dominio consta de seis hél ices y en este ult imo se local iza el sensor de voltaje.

Durante esta fase se abren la compuerta de act ivación y de inactivación del canal permit iendo que entre masivamente el sodio.
Este cambio l leva a que el interior se vuelva más posit ivo y al cierre de la compuerta de inactivación con lo que el sodio ingresa pero en pequeñas concentraciones.
La fase 1 se caracteriza por que se act iva una corr iente de K+, l lamada corr iente transitoria de K+, que vuelve menos posit ivo al interior celular.
La fase que continua se caracteriza por tener una alta resistencia a los iones. Está representada fundamentalmente por una corriente de entrada de calcio y en esta fase también aparecen: una corriente de entrada de Na+ y una corriente sostenida de K+ , la cual necesita de la entrada de calcio para act ivarse y f inalmente la corriente rect i f icadora tardía de K+. La fase 2 se representa en la curva como la meseta.
La fase 3 esta dada por las corr ientes de K+, que como todas las corr ientes son de sal ida de K+, l lamadas corr iente rect i f icadora y corr iente rect i f icadora anómala.
En la fase 4 se evidencia la actividad de la bomba de Na+/K+, la de Na+/ Ca+ y la aparición de una corr iente de K+.
En el Nódulo sinusal y el Nódulo aurículo Ventr icular que son células marcapasos estas fases no son tan del imitadas y se caracterizan por tener en la fase 4 la l lamada corriente marcapaso que es una corriente de entrada de Na+ acompañada de una menor permeabil idad para el K+ lo que faci l i ta alcanzar rápidamente en valor umbral y disparar el potencial de acción. Fig. 5
¿ CÓMO SE PRODUCE EL ACOPLE ELECTRO MECANICO?
Con el potencial de acción ingresa calcio a la célula fundamentalmente en la fase 2, esta cantidad de calcio no es suficiente para que se produzca el proceso contráct i l , si no que esta pequeña concentración induce a su vez a la l iberación de calcio inducida por calcio.
El calcio que ingresa se une a la troponina C l iberando la inhibición de la act ina y se produce la contracción muscular.
Todo este mecanismo desde que l lega el potencial de acción hasta que se produce la contracción muscular se l lama acople electromecánico .
MECANISMO DE FRANK STARLING :

No se puede hablar de contracción del músculo cardíaco y no mencionar a Frank Starl ing, quien se dedicó a estudiar los factores que r igen el volumen de sangre impulsado por el corazón .
La ley de Frank Starling afirma que cuanto más se llene el corazón durante la diástole más será el volumen expulsado durante la sístole, y dentro de los limites fisiológicos expulsara toda la sangre que le l legue .
Cuando el retorno venoso aumenta ,el músculo cardiaco se est ira y aumenta su longitud, lo que hace que el corazón se contraiga con más fuerza y expulse automáticamente toda la sangre.
Esta capacidad de est iramiento muscular hacia una longitud optima para contraer con mayor fuerza es característ ica de los músculos cardíacos y esquelét icos.
Básicamente lo que propone esta ley es que a mayor precarga( retorno venoso) la f ibra muscular aumentara su longitud lo que provocara una mayor fuerza de contracción muscular
MECANISMO DE LA INSUFICIENCIA CARDIACA:
Se ha comprobado que en los miocardiocitos anormales no se cumple la Ley de Frank Starl ing en períodos avanzados de la enfermedad.
En preparaciones de t iras musculares de pacientes con insuficiencia cardíaca las curvas de longitud tensión se encontraban alteradas.
La falta de respuesta en la insuficiencia cardíaca se debe a que la sensibi l idad al Ca+ ya es máxima y no aumenta con la elongación, auque otros estudios atr ibuyen a alteraciones de las proteínas reguladoras de la act ina.
En la insuficiencia cardíaca se han encontrado diversas alteraciones del calcio relacionadas con alteraciones de la función sistól ica y diastól ica.
A pesar de la aparente incapacidad de movil ización del calcio, la concentración pico durante la sístole es la misma que la del miocito normal lo que signif icaría que en la mayoría de los casos la alteración no se debe a una disponibi l idad disminuida de calcio sino una baja sensibi l idad a este ion por los f i lamentos de act ina, lo que l levaría a un retraso en la act ivación y disminución de la fuerza contráct i l .
Sin embargo está demostrado un muy importante aumento del calcio citosól ico en reposo, aumento debido a una regulación en baja de los genes de la ATPasa del calcio del retículo sarcoplásmico (SERCA) y del fosfolambán que act iva al SERCA. Ambos genes están disminuidos en la insuficiencia cardíaca.

En resumen se puede af irmar que en el miocardio insuficiente el potencial para generar fuerza es comparable al miocardio normal y no hay disminución del calcio citosól ico. En cambio, las propiedades diastól icas alteradas en la insuficiencia cardíaca podrían deberse a un aumento del calcio citosól ico
Fig. 5

SISTEMA DE CONDUCCIÓN DEL CORAZÓN
El sistema especializado de excitación y conducción está formado por: el Nódulo Sinusal (SA, de Keith-Flack), las vías internodulares, el Nódulo Auriculo-Ventricular (AV, de Aschoff-Tawara), el Haz Aurículo-Ventricual (AV, de His, que se bifurca en ramas derecha e izquierda) y las fibras de Purkinje.
El sistema de conducción del corazón coordina el ciclo cardíaco ya que genera impulsos rítmicos que permiten la contracción ordenada de las aurículas y los ventrículos. Dicho impulso se genera en el nódulo sinusal.
NÓDULO SINUSAL
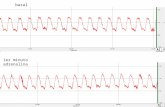
Las fibras cardíacas del nódulo sinusal tienen la capacidad de autoexcitarse y producir descargas que provocan las contracciones rítmicas automáticas del corazón. Es por ello que habitualmente el nódulo SA controla la frecuencia del latido de todo el corazón (60-100 latidos/min).¿Cómo es el mecanismo de la ritmicidad del nódulo sinusal? El potencial de membrana en reposo del nódulo sinusal es de aproximadamente -60 mV4. Dicho nódulo presenta una actividad eléctrica espontánea que se denomina Despolarización Diastólica Espontánea (DDE) o Prepotencial (fase 4) y es un proceso que se repite indefinidamente durante toda la vida de una persona.
Tres son las corrientes iónicas que median la génesis de la DDE: una corriente entrante,

Iƒ, inducida por hiperpolarización, una corriente entrante de Ca2+, ICa, y una corriente saliente de K+, IK (corriente rectificadora tardía).La corriente entrante Iƒ (denominada tipo “ƒunny”) se activa cerca del final de la repolarización y está mediada principalmente por iones Na+ debido a la presencia de canales lentos de sodio en las fibras del nódulo sinusal. Cuanto más negativo sea el potencial de membrana mayor será la activación de la corriente Iƒ. La segunda corriente entrante, ICa, se activa al final de la fase 4 y acelera la despolarización diastólica que lleva al disparo del potencial de acción. Esta corriente está mediada por iones Ca2+ debido a la presencia de canales lentos de calcio.
Cuando el potencial de membrana de las fibras del nódulo sinusal alcanza un valor liminal (umbral) de aproximadamente -40 mV, otros canales de calcio6 dependientes de voltaje se “activan” (abren), despolarizando la célula (fase 0) y desencadenando, de esta manera, el potencial de acción. Al mismo tiempo, se abren una gran cantidad de canales de potasio. Aproximadamente 100-150 ms después de su apertura, los canales lentos calcio se “inactivan” (cierran) por lo que se interrumpe el flujo de entrada de los iones positivos de Ca2+ mientras que comienzan a difundir iones K+ hacia el exterior de la fibra. Esta corriente saliente de K+, IK, tiende a repolarizar la célula (fase 3) luego de la despolarización y produce un exceso de negatividad dentro de la fibra (hiperpolarización). Esta acción “repolarizante” de la corriente IK se opone a los efectos “despolarizantes” de las corrientes Iƒ e ICa que se activan durante la fase 4. Sin embargo, los canales de potasio comienzan a cerrarse durante la fase 4 por lo que el eflujo de K+ disminuye durante esta fase. A medida que la corriente IK disminuye, su oposición a los efectos “despolarizantes” de las dos corrientes entrantes (Iƒ e ICa) disminuye gradualmente desencadenando, de esta manera, la Despolarización Diastólica Espontánea.El impulso generado en el nódulo sinusal viaja a través del músculo auricular, despolarizando las fibras y provocando su contracción, hasta el nódulo aurículo-ventricular.
NÓDULO Y HAZ AURÍCULO-VENTRICULAR
Aquí se produce un retraso en la conducción del impulso (de aproximadamente 0,13 segundos) para que las aurículas tengan tiempo de vaciar su contenido hacia los ventrículos antes de que comience la contracción ventricular. La causa del retraso se debe a la disminución de las uniones gap entre las células del nódulo y del haz AV. La conducción a través del haz AV es unidireccional, evitando, de esta manera, que los potenciales de acción viajen retrógradamente desde los ventrículos hacia las aurículas.
TRANSMISIÓN RÁPIDA EN PURKINJE Y MÚSCULO VENTRICULAR
Las fibras de Purkinje son grandes y presentan muchas uniones gap muy permeables, lo que permite una transmisión casi instantánea (0,03-0,06 seg.) del impulso a todo el músculo ventricular. Esto permite que todas las porciones de los ventrículos comiencen a contraerse casi al mismo tiempo (tipo sincrónico de contracción), hecho fundamental para que el corazón pueda cumplir su rol de bomba eficaz.
MARCAPASOS
El nódulo sinusal es prácticamente siempre el marcapasos (pacemaker) del corazón. Otras partes del corazón pueden presentar una excitación rítmica intrínseca como, por ejemplo, las fibras del nódulo AV y las fibras de Purkinje. Sin embargo, como la frecuencia

de descarga del nódulo SA es mayor que la frecuencia de descarga tanto del nódulo AV como de Purkinje, el nódulo sinusal produce siempre una nueva descarga antes de que el nódulo AV o Purkinje puedan alcanzar sus propios umbrales de autoexcitación. En el caso de que el nódulo SA no sea el marcapasos, se estará en presencia de un marcapasos ectópico, que puede producir una debilidad significativa en el bombeo cardíaco.
MÚSCULO CARDÍACO AURICULAR Y VENTRICULAR
El músculo cardíaco, al igual que el músculo esquelético, es estriado y posee miofibrillas con filamentos de actina y miosina. Además, el músculo cardíaco funciona como un sincicio debido a las uniones gap presentes en los discos intercalados de las membranas celulares. Gracias a este sincicio, los potenciales de acción viajan fácilmente de una célula muscular a la otra.
La despolarización (fase 0) ocurre antes de la aparición de la fuerza de contracción. Se continúa de un período breve de repolarización temprana (fase 1) y es seguida por una meseta (plateau, fase 2). Luego, la membrana se repolariza (fase 3) hasta alcanzar el potencial de membrana en reposo (fase 4). La finalización de la repolarización (fase 4) coincide aproximadamente con la fuerza pico. La relajación del músculo sucede principalmente durante la fase 4 del potencial de acción. La duración de la contracción suelen ser similar a la duración del potencial de acción.
POTENCIAL DE ACCIÓN
¿Cómo se generan los potenciales de acción en el músculo cardíaco? Cualquier acontecimiento que produzca una elevación del potencial de membrana en reposo (-90 mV) hasta un valor umbral puede ser responsable de la génesis de un potencial de acción (ej.: excitación mecánica, química o eléctrica). En este caso, el impulso eléctrico generado y propagado por el sistema de conducción es el responsable de generar un potencial de acción en el músculo cardíaco.
FASES DEL POTENCIAL DE ACCIÓN
La despolarización se produce gracias a la presencia de canales rápidos de sodio en la membrana del miocito. El impulso eléctrico mencionado eleva el potencial de membrana lo suficiente para provocar la apertura de dichos canales rápidos de sodio voltaje dependiente9. En el valor umbral (aproximadamente -65 mV), el número de iones Na+ que entran en la fibra supera al número de iones K+ que salen de la misma. Así, se produce un círculo vicioso de retroalimentación positiva en el que la entrada de Na+ produce una elevación adicional del potencial de membrana y abre aún más canales rápidos de sodio activados por el voltaje y permite que se produzca una mayor entrada de Na+ al interior de la fibra. Los canales rápidos de sodio voltaje dependientes tienen dos compuertas: de activación (o M3) y de inactivación (o H). Además, presentan tres estados distintos: reposo, activado e inactivado.
El mismo voltaje (umbral) que dispara la apertura de M3 (estado activado) provoca el cierre de H pero, como las compuertas presentan diferente cinética de activación, H se cierra unas milésimas de segundo después de la apertura de M3 (estado inactivado). Durante el estado activado del canal, fluyen grandes cantidades de Na+ hacia el interior de la fibra produciendo la despolarización de la célula (fase 0). Por otra parte, el mismo

voltaje que había provocado la apertura de estos canales también genera la activación de canales de potasio voltaje dependientes. Sin embargo, estos canales son de “activación lenta”10, por lo que se abren milésimas de segundo después de la apertura de la compuerta de activación (M3) de los canales de sodio. La apertura de los canales de potasio genera la salida de K+ hacia el exterior de la fibra al mismo tiempo que comienzan a cerrarse los canales de sodio. Las corrientes responsables del eflujo de K+ se denominan IK (rectificadora tardía), IK1 (rectificadora hacia dentro) e Ito (transitoria hacia afuera). El aumento de la salida de K+ en simultáneo con el descenso de la entrada de Na+ recuperan el potencial de reposo normal de la célula (repolarización).
¿Por qué se produce la meseta? Nuevamente, el valor umbral alcanzado por la célula miocárdica ocasiona la apertura de canales de calcio (predominantemente, de canales tipo L) voltaje dependientes, que permanecen abiertos varias décimas de segundo. Durante su apertura, fluyen hacia el interior de la célula grandes cantidades de iones calcio11 manteniendo prolongado el proceso de despolarización. Este influjo de cargas positivas mediadas por iones calcio queda equilibrado por el eflujo de cargas positivas en forma de iones potasio a través de las corrientes IK, IK1 e Ito (esta última responsable de generar la fase de repolarización temprana y que no se inactiva completamente hasta el final de la fase 2).
¿Por qué la repolarización es lenta? Las corrientes de K+ (IK, IK1 e Ito) aumentan con el tiempo en tanto las corrientes de Ca2+ se inactivan. Cuando el eflujo de K+ excede el influjo de Ca2+ comienza la repolarización. Las corrientes de K+ transitoria hacia afuera (Ito) y rectificadora tardía (IK) ayudan al inicio de la repolarización pero no así la corriente rectificadora hacia adentro (IK1) debido a que la conductancia de los canales responsables de esta corriente es muy baja para los valores del potencial de membrana durante la meseta. Sin embargo, una vez que la fase 3 ha comenzado, la contribución de la corriente rectificadora hacia adentro (IK1) aumenta considerablemente debido a que el eflujo de iones K+ disminuye el valor del potencial de membrana provocando un aumento de la conductancia de los canales que median la corriente IK1.
¿Qué ocurre durante la fase 4? Durante esta fase predominan los mecanismos para conservar la homeostasis intracelular. El exceso de Na+ que ingresa a la célula durante la fase 0 del potencial de acción se elimina por acción de la Na+,K+-ATPasa. Esta enzima elimina 3 iones Na+ y los intercambia por 2 iones K+ (que salieron de la célula durante las fases 2 y 3 del potencial de acción). La eliminación del Ca2+ intracelular (que ingresó durante la fase 2) ocurre por medio de una bomba de Ca2+ dependiente de ATP (que mueve iones Ca2+ de regreso hacia los sitios de almacenamiento en el retículo sarcoplásmico – ver apéndice) y un mecanismo de intercambio Na+/Ca2+, que intercambia 3 iones Na+ desde el exterior por cada ion Ca2+ expulsado.
PERÍODOS REFRACTARIOS
El músculo cardíaco requiere un período de tiempo luego del potencial de acción para poder ser reexcitado por un estímulo. El período refractario absoluto está definido por la porción del potencial de acción en la que ningún estímulo (lo fuerte que sea) puede generar una nueva respuesta celular. Esto se debe a que la compuerta de inactivación de los canales de sodio no se abre de nuevo hasta que el potencial vuelva hasta (casi) los valores de reposo. Luego, durante el período refractario efectivo se puede generar una respuesta local, no propagada, sólo con una señal excitadora muy intensa (estímulo supra-umbral). El período refractario relativo se extiende desde el final del período

refractario efectivo hasta el momento en el que la célula recupera su potencial de membrana de reposo.
Cuando una fibra rápida muscular cardíaca es estimulada durante el período refractario relativo del potencial de acción evocado previamente, las características del nuevo potencial de acción variarán según el valor de potencial de membrana que exista en el momento de la nueva estimulación. Cuanto más tardía sea la estimulación durante el período refractario relativo, mayor será la amplitud de la respuesta y la pendiente de la despolarización debido a que el número de canales de sodio que se han recuperado del estado de inactivación es mayor a medida que la repolarización progresa durante la fase 3. A consecuencia de la mayor amplitud y pendiente de la despolarización de la respuesta evocada, la velocidad de propagación también aumenta cuanto más tardíamente se estimule la fibra durante el período refractario relativo. Una vez que la fibra se repolarizado completamente, la respuesta será siempre la misma sin importar en qué momento de la fase 4 se aplique un estímulo. A medida que se estimula más tardíamente a las fibras lentas durante el período refractario relativo, la amplitud y la pendiente de la despolarización aumentan progresivamente, al igual que lo observado con las fibras rápidas. Además, el mismo efecto sobre la velocidad de conducción se manifiesta en ambos tipos de fibras. De todas maneras, la recuperación completa de la excitabilidad ocurre mucho más lentamente en las fibras lentas que en las fibras rápidas.
EFECTOS DEL SISTEMA NERVIOSO AUTÓNOMO SOBRE EL CORAZÓN
La eficacia de la función de bomba del corazón está controlada por los nervios simpáticos y parasimpáticos (vagos). Los nervios vagos se distribuyen principalmente a los nódulos SA y AV y tienen poca representación en el músculo auricular y casi nula en el músculo ventricular. Por otra parte, los nervios simpáticos se distribuyen en todas las regiones del corazón, con una intensa representación en el músculo ventricular. SIMPÁTICO El sistema nervioso simpático aumenta todas las propiedades cardíacas. Los nervios simpáticos estimulan al corazón mediante la liberación de noradrenalina desde las terminales nerviosas. Dicho neurotransmisor actúa, principalmente, sobre receptores β1 presentes en el músculo cardíaco. Los receptores β1 están ligados a la proteína Gs. La

interacción entre la noradrenalina y el receptor adrenérgico β1 desencadena la estimulación de la adenilato ciclasa por la Gs con la consiguiente formación de AMPc y activación de la proteín quinasa dependiente de AMPc (PKA). ¿Cómo se estimulan las propiedades cardíacas? Los efectos simpáticos sobre el corazón se explican por un aumento de la permeabilidad de la membrana a los iones calcio y sodio. Más específicamente, la señalización intracelular mediada por el receptor β1 provoca la activación de la PKA lo que determina una serie de efectos que influyen sobre cada una de las propiedades cardíacas.
- Cronotropismo, Dromotropismo y Batmotropismo: la PKA fosforila los canales de calcio y sodio involucrados en la génesis de la Despolarización Diastólica Espontánea facilitando, de esta manera, su apertura. Esto provoca un aumento de la permeabilidad de la membrana de la fibra (en este caso, la fibra del marcapaso) al Ca2+ y al Na+ produciendo un aumento de la velocidad del ascenso (pendiente) del potencial de membrana hacia el valor umbral. Por todo esto, se acelera la autoexcitación espontánea de las células del nódulo sinusal. Este efecto se traduce como un aumento en la frecuencia cardíaca (efecto cronotrópico positivo). A su vez, la mayor permeabilidad al Ca2+ y al Na+ provoca una mayor amplitud de la respuesta de despolarización. Cuanto mayor es la amplitud del potencial de acción, más efectiva se torna la despolarización de las partes adyacentes de la membrana (efecto batmotrópico positivo) y mas rápida es la onda de despolarización que se propaga por la fibra (efecto dromotrópico positivo).
- Inotropismo: los iones calcio tienen una función importante en la excitación del proceso contráctil de las miofibrillas (acoplamiento excitación-contracción). Sin embargo, la cantidad de calcio que ingresa a la célula en cada despolarización es muy inferior a la necesaria para producir la contracción por lo que el calcio, que se unirá a las proteínas del sarcómero12, proviene principalmente de depósitos intracelulares ubicados en el retículo sarcoplásmico. El calcio almacenado en el retículo sarcoplásmico es liberado por el calcio que ingresa a la célula, constituyendo un mecanismo de liberación inducido por el mismo ion (liberación de calcio inducida por el calcio). Los canales por donde egresa el calcio del retículo sarcoplásmico se denominan receptores de rianodina y se encuentran agrupados en las regiones del retículo sarcoplásmico cercanas a los túbulos T del sarcolema. La fosforilación de los canales de calcio (tipo L) mediada por PKA es responsable de una mayor entrada de Ca2+ a la fibra muscular miocárdica. Además, la PKA fosforila proteínas contráctiles del músculo (troponina I). Ambos efectos determinan un aumento de la contractilidad miocárdica (efecto inotrópico positivo).
- Lusitropismo: el calcio citoplasmático es recapturado activamente por el retículo sarcoplásmico mediante una bomba calcio-ATPasa13 y es retirado del sarcolema por un intercambiador de sodio-calcio (saca 1 ion Ca2+ y mete 3 Na+). Esta bomba está regulada por una proteína denominada fosfolamban. La activación de la PKA provoca fosforilación de dicha proteína. La fosfolamban fosforilada aumenta la velocidad con la que la bomba calcio-ATPasa del retículo sarcoplásmico secuestra calcio14 acelerando, de esta manera, la relajación del músculo miocárdico (efecto lusitrópico positivo).
PARASIMPÁTICO
El sistema nervioso parasimpático produce los efectos contrarios a los del simpático sobre el corazón: disminuye todas las propiedades cardíacas. La estimulación de los nervios vagos que inervan al corazón hace que se libere acetilcolina en las terminales nerviosas. La acetilcolina actúa sobre receptores M2 presentes en el miocardio. Estos receptores

están ligados a la proteína Gi. La proteína Gi disminuye la producción de AMPc (a través de la inhibición de la adenilato ciclasa por la subunidad α), aumenta la permeabilidad al potasio (la subunidad βγ abre canales de K+) y disminuye la disponibilidad de calcio en el sarcolema (por supresión de la actividad de canales lentos calcio – tipo L– sensibles a voltaje).
¿Cómo se inhiben las propiedades cardíacas? Los efectos parasimpáticos sobre el corazón se explican por un aumento de la permeabilidad de la membrana al potasio. Además, la disminución de la producción de AMPc hace que no se active la PKA. De esta manera, se inhiben todos los efectos simpáticos que se producían por activación de la PKA (ver arriba).
- Cronotropismo, Dromotropismo y Batmotropismo: la apertura de canales de potasio (regulados por la Gi) y el consecuente aumento de permeabilidad a ese ion conducen a hiperpolarización del músculo miocárdico15. De esta manera, se retrasa la llegada al valor umbral (menor pendiente de despolarización), se disminuye la amplitud del potencial de acción y se incrementa la corriente repolarizante de potasio en las fibras musculares. Estos efectos se traducen como una disminución en la frecuencia cardíaca (efecto cronotrópico negativo) y del nivel de excitabilidad (efecto batmotrópico negativo) y reducción de la velocidad de conducción a través del tejido especializado (efecto dromotrópico negativo). -Inotropismo y Lusitropismo: la proteína Gi disminuye la actividad de los canales lentos calcio (tipo L). Así, disminuye el acoplamiento excito-contráctil del músculo cardíaco (efecto inotrópico negativo). Además, como la activación de Gi disminuye la producción de AMPc y, por ende, la actividad de la PKA, no se produce fosforilación de fosfolamban por lo que se reduce la actividad de la bomba calcio-ATPasa presente en el retículo sarcoplásmico (efecto lusitrópico negativo).

Por lo tanto los efectos del simpático sobre el corazón se deben tanto a modificaciones de la frecuencia como a modificaciones de la fuerza contráctil (aumento). En cambio, los efectos del parasimpático se basan principalmente en modificaciones de la frecuencia disminución). Esta diferencia se debe fundamentalmente a la distinta distribución anatómica de las terminales nerviosas de ambas divisiones del sistema nervioso autónomo.
4.3.1. Importancia del Ca2+ en la fibra cardiaca.
Fibra muscular CARDÍACA: CARDIOMIOCITOS
El TJ muscular cardíaco es una variedad del TJ muscular especializada en atender la función cardiovascular mediante la diferenciación de una clase de células, los CARDIOMIOCITOS, que tienen similitudes tanto con la fibra muscular lisa como con la estriada esquelética.
Similitudes con la fibra muscular lisa:

- Regulados por el SN Vegetativo, es decir, son de contracción involuntaria.
- Son independientes, es decir, son células uni-binucleadas que poseen una “frontera” con respecto a las células vecinas; NO forman un sincitio, a diferencia de las C. musculares esqueléticas (estriadas), que forman un magma citoplasmático con muchos núcleos.
- Sus células se comunican por Gap Junctions o uniones de comunicación directa (es lo mismo), lo que permite la propagación del estímulo eléctrico a todo el entramado celular para la posterior activación de la contracción (Ca2+).
Similitudes con la fibra muscular esquelética/estriada:
- Citoesqueleto contráctil estriado de miofibrillas constituidas por SARCÓMERAS (que son las que dan la estriación al citoplasma).
¡¡¡OJO!!! El citoesqueleto de las fibras musculares LISAS NO está constituido por sarcómeras, sino que forma una malla contráctil trenzada o un compactado celular que da lugar a una contracción mucho menos potente.
ESTRUCTURA DE LA C. MUSCULAR CARDÍACA o CARDIOMIOCITO
- Adopta una morfología fusiforme, algo estrellada y con prolongaciones celulares que le sirven para unirse a las células vecinas (a través de discos intercalares), creándose una malla tridimensional de células en el
corazón que forma el miocardio.
Razón por la que forman esta estructura en malla: esta disposición en malla les proporciona una mayor fuerza/potencia contráctil.
- En los huecos entre las C. musculares cardíacas o entre cardiomiocitos se alojan CAPILARES, por lo que la fibra muscular cardíaca está tremendamente VASCULARIZADA (oxigenada).

Contenido de los cardiomiocitos:
1. Un núcleo central.
2. Una Mb plasmática o SARCOLEMA.
3. Una estructura intracelular de miofibrillas contráctiles: cintas de moléculas contráctiles que forman las MIOFIBRILLAS de la C. muscular, que a su vez están constituidas por SARCÓMERAS.
Las miofibrillas se extienden desde los extremos de las células hasta las del lado opuesto, formando cuerdas cuya contracción determina el cambio de tamaño del músculo. En estas cuerdas podemos observar la estriación y las sarcómeras. Es decir, las miofibrillas están enganchadas de lado a lado y se contraen (NO se rompen porque el extremo de las células forma complejos de unión que evitan que se desgarren al contraerse).
Entre las miofibrillas encontramos el resto de orgánulos:
I. Mitocondrias (llamadas sarcosomas en su descubrimiento debido a que se observaron en cuerpos de estas C. musculares). El corazón exagera al máximo el potencial contráctil de sus células, por eso tiene muchas más mitocondrias que la célula muscular esquelética estriada.

II. RETÍCULO SARCOPLÁSMICO (RS) = retículo endoplasmático. Es muy diferente al RE de cualquier célula ya que, en vez de formar cisternas alineadas cuya acumulación da lugar al AG, forma envoltorios sobre las miofibrillas que están especializados en acumular el calcio cerca de las sarcómeras.
Adaptación Sarcómera/Retículo Sarcoplásmico/Túbulos-T: cada sarcómera tiene su retículo sarcoplásmico pegado/acoplado, que será el que le proporcione el Ca2+ iniciador de la contracción gracias a la activación de la liberación de este Ca2+ por los túbulos-T
- El retículo sarcoplásmico tiene almacenado el Ca2+ que necesita la sarcómera para contraerse, pero NO lo libera directamente, sino que requiere de una activación.
- La activación del RS para la liberación del Ca2+ viene dada por estructuras de la Mb plasmática, que son entrantes tubulares (de la Mb) – a modo de tuberías - que se dirigen hacia la sarcómera y se acoplan al RS, los TÚBULOS-T (equivalentes a la caveolina-3 de las c. musculares lisas).
- Los túbulos-T entran hasta la intimidad de las cisternas del RS pegadas a las sarcómeras y llevan hasta cada sarcómera el POTENCIAL DE ACCIÓN de la estimulación eléctrica de la célula muscular.
Activación nerviosa despolarización de la Mb Túbulos-T (intimidad) estímulo eléctrico hasta c/ una de las sarcómeras activación de la contracción.
SISTEMA DE TÚBULOS-T: conductos ramificados conectados con el exterior de la célula por invaginaciones de la Mb plasmática o sarcolema que alcanzan a c/ una de las sarcómeras pegándose al retículo sarcoplásmico que tiene c/ una de estas sarcómeras. Llevan la excitación eléctrica, activando la liberación del Ca2+, el cual provoca cambios de conformación en la sarcómera (actina-miosina) e inicia la contracción.
En resumen: el RS se pega a c/sarcómera para acercarle el Ca2+, pero primero al RS le entra el túbulo-T, que es el que acerca la excitación eléctrica a toda la zona del RS y activa la liberación del Ca2+. Retículo Sarcoplásmico/Túbulo-T = DIADAS. Como cada RS tiene un túbulo-T forman parejas.
*En la C. muscular esquelética estriada se forman TRIADAS (1 túbulo-T por c/2 RS). III. Gran acúmulo de moléculas: glucógeno, alta concentración de lípidos y lipofuscina (puede llegar a representar el 20% del cardiomiocito en el anciano).
La presencia de lipofuscina en el cardiomiocito es un signo de envejecimiento.
UNIONES INTERCELULARES con las células “vecinas”: DISCOS INTERCALARES o ESCALERIFORMES
Representan complejos de unión entre los cardiomiocitos, en los cuales distinguimos 2 zonas: 1) Verticales (transversales con respecto al eje del cardiomiocito) = DESMOSOMAS + FASCIA ADHERENS (uniones intermedias)

Función: unión firme de las células entre sí y fijación de las miofibrillas. - Función de los desmosomas: fijan, unen firmemente a las células musculares cardíacas evitando su deslizamiento.
Cuando la célula se contrae estira de la propia Mb y de su unión a la célula vecina. - Función de la fascia adherens: fijan miofibrillas y también se unen entre ellas. Acompaña a la fijación de los cardiomiocitos por los desmosomas y engancha las miofibrillas.
Las miofibrillas se pueden unir porque la fascia adherens tiene puntos de unión para la alfa-actinina (de las miofibrillas). Cuando se contraen todos los cardiomiocitos a la vez el corazón se hace más “pequeño”.
2) Horizontales (longitudinales respecto al eje de la célula; donde pisamos en la “escalera”) = GAP Junctions o uniones de comunicación directa + desmosomas
- Función de las Gap Junctions: comunicar a los cardiomiocitos entre sí para que sea posible el intercambio de iones. No tienen fuerza para la fijación, por lo que su única función es la de comunicar.
DIFERENCIAS ENTRE LAS C. MUSCULARES DEL CORAZÓN SEGÚN SU LOCALIZACIÓN

ESPECÍFICA (aurícula o ventrículo) No todas las células musculares del corazón son iguales, sino que presentan diferencias según se sitúen en la aurícula o en el ventrículo: 1. CARDIOMIOCITOS AURICULARES = más pequeños.
2. CARDIOMIOCITOS VENTRICULARES = más grandes.
Poseen características metabólicas distintas (que no vamos a tratar este año).
Además, también se distinguen cardiomiocitos modificados o grupos heterogéneos de cardiomiocitos que han perdido la capacidad de contracción, y en su lugar han diferenciado la capacidad de autoexcitación, como por ejemplo: 3. CÉLULAS MARCAPASOS: son ritmogénicas, es decir, marcan el ritmo contráctil del corazón.
4. CÉLULAS DE PURKINJE: forman el sistema de conducción del corazón. Permiten la propagación rápida del estímulo eléctrico a través del miocardio.
Otros cardiomiocitos que también están en la aurícula (derecha) pero asociados a los nódulos sinusales, donde están las células endocrinas del corazón: 5. CARDIOMIOCITOS CON FUNCIÓN ENDOCRINA: están en la aurícula y producen polipéptidos hormonales que contribuyen a la regulación hidroelectrolítica de nuestro cuerpo.
REGENERACIÓN DEL CORAZÓN (Lunes, 16 de Mayo a las 11:30)

4.3.2. Tejido marcapaso y de conducción.
(ARCHIVO PDF)

5. Músculo liso.
El músculo liso está formado por fibras musculares lisas que corresponden a células uninucleadas, delgadas y aguzadas en los extremos, cuya longitud varía entre 20 y 500 mm (Figs 1 y 2). Este tipo de músculo forma la porción contráctil de la pared de diversos órganos tales como tubo digestivo (Fig 3) y vasos sanguíneos (Fig 4), que requieren de una contracción lenta y sostenida. Las células se organizan en grupos, formando haces, rodeados de tejido conjuntivo fibroso que contiene vasos sanguíneos.
El núcleo de las fibras musculares lisas se ubica en el centro de la fibra y los organelos citoplasmáticos tales como mitocondrias, aparato de Golgi, retículo endoplásmico rugoso y ribosomas libres se localizan, mayoritariamente, en la vecindad de los polos nucleares (Fig 5). El resto del citoplasma está ocupado por abundantes miofilamentos finos de actina, una proporción menor de miofilamentos gruesos de miosina, y un citoesqueleto de filamentos intermedios formados por desmina. Existen, también, numerosos cuerpos densos, estructuras que anclan filamentos finos (Fig 6).
Las fibras musculares lisas se disponen desplazadas una respecto de la otra, de manera que el extremo delgado de una fibra se ubica vecino a la parte ancha de la fibra vecina. Esta disposición de las fibras y la localización del núcleo en el centro, explica el aspecto del músculo liso en corte transversal (Figs 7 y 8).
Las fibras musculares lisas están rodeadas por una lámina basal (lámina externa) comparable a la lámina basal de los epitelios (Fig 9). Por fuera de la lámina externa, se dispone una trama de fibras reticulares.
En sitios discretos, las células adyacentes están asociadas por uniones de comunicación ("nexos"), de estructura y función similares a la explicada en tejidos epiteliales.
Figura 1 Figura 2

Figura 3 Figura 4
Figura 5 Figura 6
Figura 7 Figura 8
Figura 9

Bases estructurales de la contracción del músculo liso. El aparato contráctil del músculo liso se contrae mas lentamente que el del músculo estriado, pero permite un acortamiento mayor de las fibras musculares lisas (Fig 1). El mecanismo de contracción, en esta variedad de músculo, también se basa en el deslizamiento de los filamentos finos sobre los filamentos gruesos. Los filamentos de actina de las fibras musculares lisas son fáciles de detectar a nivel ultraestructural (Fig 2); en cambio la visualización de los filamentos gruesos requiere de condiciones de fijación especiales, que demuestran que en el músculo liso por cada filamento grueso hay una proporción mucho mayor de filamentos finos (1:14) que la que se observa en el músculo esquelético (1:6).En estas células, la contracción es regulada también por alza en las concentraciones citosólicas de Ca++. Sin embargo, la regulación de la contracción está asociada a miosina y no a actina. Un alza en las concentraciones citosólicas de Ca++ induce la fosforilación de las cadenas livianas de la miosina lo que:
produce una modificación en la cola de la molécula que permite la formación de filamentos gruesos (Fig 3) y
genera un cambio conformacional en la cabeza que permite su interacción con actina. Los filamentos gruesos preparados in vitro, a partir de miosina de músculo liso, aparecen polarizados en una sola dirección en un lado del filamento y en la dirección opuesta a lo largo del otro lado (Fig 3). En esta configuración no existe una zona libre de puentes, como la que se ve en el filamento grueso del músculo esquelético. Esta disposición tiene la ventaja que actina y miosina pueden interactuar sin interrupción a lo largo de todo el filamento grueso. Cuando la cabeza de la miosina se defosforila, los filamentos se desensamblan y la miosina se disocia de la actina. La fosforilación es catalizada por una enzima (quinasa de la cadena liviana de la miosina) cuya acción requiere de la presencia del complejo Ca-calmodulina.
El modelo aceptado de contracción de las fibras musculares lisas (Fig 4) establece que manojos de filamentos finos de actina, asociados a filamentos gruesos de miosina, se anclan por un extremo a cuerpos densos adheridos a la membrana plasmática y por el otro a filamentos intermedios no contráctiles a través de cuerpos densos citoplasmáticos. La a-actinina es uno de los componentes de los cuerpos densos. El rol de los cuerpos densos es similar al de los discos Z de las miofibrillas del músculo estriado. Los manojos contráctiles se orientarían oblicuos respecto del eje mayor de la célula (Fig 5), lo que explicaría el acortamiento que experimentan las fibras musculares lisas durante su contracción. En la superficie de las células musculares lisas existen numerosas vesículas membranosas o cavéolas (Figs 7 y 8), vecinas a cisternas o túbulos de retículo endoplásmico liso. Se cree que este sistema membranoso juega un papel en la captura y liberación de calcio, similar al que desempeña el retículo sarcoplásmico en el músculo estriado.
Además de su actividad contráctil, las células musculares lisas tienen la capacidad de sintetizar colágena tipo III, elastina y proteoglucanos.

Figura 1 Figura 2
Figura 3 Figura 4
Figura 5 Figura 6
Figura 7 Figura 8

Figura 9
Bases estructurales de la contracción del músculo liso. El aparato contráctil del músculo liso se contrae mas lentamente que el del músculo estriado, pero permite un acortamiento mayor de las fibras musculares lisas (Fig 1). El mecanismo de contracción, en esta variedad de músculo, también se basa en el deslizamiento de los filamentos finos sobre los filamentos gruesos. Los filamentos de actina de las fibras musculares lisas son fáciles de detectar a nivel ultraestructural (Fig 2); en cambio la visualización de los filamentos gruesos requiere de condiciones de fijación especiales, que demuestran que en el músculo liso por cada filamento grueso hay una proporción mucho mayor de filamentos finos (1:14) que la que se observa en el músculo esquelético (1:6).En estas células, la contracción es regulada también por alza en las concentraciones citosólicas de Ca++. Sin embargo, la regulación de la contracción está asociada a miosina y no a actina. Un alza en las concentraciones citosólicas de Ca++ induce la fosforilación de las cadenas livianas de la miosina lo que:
produce una modificación en la cola de la molécula que permite la formación de filamentos gruesos (Fig 3) y
genera un cambio conformacional en la cabeza que permite su interacción con actina. Los filamentos gruesos preparados in vitro, a partir de miosina de músculo liso, aparecen polarizados en una sola dirección en un lado del filamento y en la dirección opuesta a lo largo del otro lado (Fig 3). En esta configuración no existe una zona libre de puentes, como la que se ve en el filamento grueso del músculo esquelético. Esta disposición tiene la ventaja que actina y miosina pueden interactuar sin interrupción a lo largo de todo el filamento grueso. Cuando la cabeza de la miosina se defosforila, los filamentos se desensamblan y la miosina se disocia de la actina. La fosforilación es catalizada por una enzima (quinasa de la cadena liviana de la miosina) cuya acción requiere de la presencia del complejo Ca-calmodulina.
El modelo aceptado de contracción de las fibras musculares lisas (Fig 4) establece que manojos de filamentos finos de actina, asociados a filamentos gruesos de miosina, se anclan por un extremo a cuerpos densos adheridos a la membrana plasmática y por el otro a filamentos intermedios no contráctiles a través de cuerpos densos citoplasmáticos. La a-actinina es uno de los componentes de los cuerpos densos. El rol de los cuerpos densos es similar al de los discos Z de las miofibrillas del músculo estriado. Los manojos contráctiles se orientarían oblicuos respecto del eje mayor de la célula (Fig 5), lo que explicaría el acortamiento que experimentan las fibras musculares lisas durante su contracción. En la superficie de las células musculares lisas existen numerosas vesículas membranosas o cavéolas (Figs 7 y 8), vecinas a cisternas o túbulos de retículo endoplásmico liso. Se cree que este sistema membranoso juega un papel en la captura y

liberación de calcio, similar al que desempeña el retículo sarcoplásmico en el músculo estriado.
Además de su actividad contráctil, las células musculares lisas tienen la capacidad de sintetizar colágena tipo III, elastina y proteoglucanos.
Figura 1 Figura 2
Figura 3
Figura 6
Características y funciones:
Los músculos lisos forman las paredes de las vísceras y no están bajo el control de la
voluntad. Sus fibras no contienen estrías.

Este músculo tiene similitud con el músculo estriado o esquelético. La diferencia es que
no posee línea Z como lo posee el músculo estriado, sino que posee bolas densas que
reemplazan a estas líneas Z.
Este puede ser unitario o multiunitario. Se le llama unitario cuando existe entre cada fibra
de este músculo una unión (los llamados gap junctions); se les llama multiunitario si no
están enlazados por uniones, sino que funcionan de manera independiente.
Este músculo y su función es muy importante, por ejemplo, los seres humanos presentan
musculatura lisa en todo el tracto gastrointestinal, el cual, es importante porque interviene
en lo que son las contracciones de peristaltismo.
El funcionamiento de la contracción es mucho más duradero que la del músculo
esquelético debido a que no consume tanta energía como lo hace el mismo. La fase de
contracción de este tipo de músculo es duradera, puesto que cuando la acción de unión
de miosina y actina -mismos pasos de contracción que el músculo esquelético-, gasta
menor cantidad de energía (la misma cantidad de ATP, pero menor consumo de energía),
es decir, el metabolismo de gasto de energía de ATP es más lento que el del músculo
esquelético.
No solo el tiempo de la contracción es una diferencia del músculo esquelético con el
músculo liso (la distancia que se contrae es mucho mayor que la del músculo
esquelético).
TIPO DE MUSCULOS LISOS:
§ Músculo liso multiunitario: Compuesto de fibras musculares lisas separadas. Cada
fibra puede contraerse independientemente de las otras, su control es ejercida
principalmente por señales nerviosas. Rara vez muestran contracciones espontáneas.
Ejemplos: músculo ciliar del ojo, el iris del ojo, la membrana nictitante de algunos
animales inferiores.
§ Músculo liso visceral: Se disponen de forma tubular en las paredes de las arterias y
venas pequeñas, así como en los órganos huecos como el estómago, intestino, útero y
vejiga.

CELULAS DEL TEJIDO MUSCULAR LISO:
El músculo liso está formado por células con las siguientes características:
·Son células fusiformes, delgadas.
·Núcleo: central, alargado, cromatina laxa, con uno o mas nucléolos, en forma de “puro”,
uno por cada célula.
·Citoplasma: uniforme, levemente eosinófilo, sin estriaciones (contiene microfilamentos de
actina y miosina en desorden).
El músculo liso es involuntario, lento y forzado, no sujeto a la “ley del todo o nada”.
Se localiza en órganos huecos, excepto corazón, como:
Aparato respiratorio, aparato digestivo, aparato urinario, vasos sanguíneos, etc.
CONTRACCION DEL MUSCULO LISO
Aunque los iones de calcio en las células del músculo liso causan la contracción como en
el músculo estriado, el control del movimiento de los iones de calcio es diferente. En el
músculo liso relajado los iones de calcio están en el retículo sarcoplasmático por toda la
célula. Cuando se produce la excitación de la membrana, los iones de calcio se liberan al
citoplasma y se unen a una proteína llamada CALMODULINA. El complejo calcio-
calmodulina activa a una enzima que fosforila la miosina y permite su unión a la actina.
Luego se produce el deslizamiento de la misma forma que en las fibras musculares
estriadas. Las fibras musculares lisas pueden excitarse por terminales nerviosas o por
hormonas sanguíneas.
GENERALIDADES:
MUSCULO LOCALIZACION
APARIENCIA DE LAS CELULAS
TIPO DE ACTIVIDAD
ESTIMULACION
MUSCULO LISO (NO ESTRIADO)
Se encuentra en las paredes de las vísceras huecas y vasos
Fibras fusiforme pequeñas, independiente
Contracción débil, lenta rítmica o de tono sostenido; actúa
INVOLUNTARIA A TRAVES EL SISTEMA NERVIOSO

sanguíneos, iris y cuerpo ciliar del ojo; se une a los folículos pilosos de la piel (musculo erector del pelo)
s o aglomeradas, sin estrías; núcleo único, central.
fundamentalmente para impulsar sustancias (peristalsis) y restringir la corriente (vasoconstricción y actividad esfinteriana)
AUTONOMO.
CLINICAMENTE:
HIPERTROFIA E HIPERPLASIA DEL MUSCULO LISO:
Las células musculares lisas experimentan una hipertrofia compensadora en respuesta al
aumento de demandas. Las células musculares lisas de la pared uterina aumentan
durante el embarazo, no solo en tamaño sino también en número (hiperplasia), dado que
conservan su capacidad de dividirse. Ademas pueden desarrollarse nuevas células
musculares lisas a partir de células incompletamente diferenciadas (pericitos) localizadas
a lo largo de los vasos sanguíneos de pequeño calibre.
MUSCULO LISO EN LAS VIAS RESPIRARTORIAS:
Los músculos lisos pueden contraerse isométricamente, isotónicamente, o de manera
intermedia entre 2 extremos, en vivo. El musculo de la vía respiratoria no se contrae
isométricamente, porque la vía respiratoria no es rígida. Es probable que el musculo liso
de la vía respiratoria se contraiga semi- isotónicamente, haciéndose más corto hasta que
se limita por fuerzas opuestas. Ejemplo: Elastancia de la pared de la vía respiratoria y
fuerza de retracción elástica del pulmón. La fuerza desarrollada por los músculos lisos de
la vía respiratoria cuando son estimulados dependen de la longitud inicial.

5.1. Estructura.
Las células musculares lisas poseen un aparato contráctil de filamentos finos y gruesos y
un citoesqueleto de filamentos intermedios de desmina y vimentina.
El resto del sarcoplasma está repleto de filamentos finos que forman parte del aparato de
contráctil. Los filamentos gruesos de miosina están dispersos por todo el sarcoplasma de
la célula muscular lisa. Los filamentos finos de una célula muscular lisa están adheridos a
densidades citoplasmáticas o cuerpos densos que se ven entre los filamentos. Estas
estructuras se hallan distribuidas por todo el sarcoplasma en una red de filamentos
intermedios de la proteína desmina.
Los componentes del aparato contráctil de las células musculares lisas son:
Filamentos finos que contienen actina, la isoforma muscular lisa de la tropomiosina, y dos
proteínas específicas del musculo liso, la caldesmona y la calponina. Con la
tropomiosisna muscular lisa no hay troponina asociada. la investigación indica que la
posición de la tropomiosina sobre el filamento de actina está regulada por la fosforilación
de las cabezas de la miosina. La caldesmona y la calponina son proteínas fijadoras de
actina que bloquean el sitio de unión para la miosina. La acción de estas proteínas es
dependiente del Ca2+ y también está controlada por la fosforilación de las cabezas de
miosina.
Filamentos gruesos que contienen miosina II y que son un poco diferentes de los que hay
en el musculo esquelético. Esta miosina también está compuesta por dos cadenas
polipeptídicas pesadas y cuatro ligera. Sin embargo, la estructura de los filamentos
gruesos de la célula muscular lisa es diferentes de la de los filamentos gruesos de la
célula muscular esquelética. las moléculas de miosina están escalonadas en paralelo
entre dos vecinas inmediatas y también están unidas a una compañera antiparalela
mediante una superposición breve en el extremo distal de sus colas.
Los cuerpos densos proveen un sitio de fijación para los filamentos finos y filamentos
intermedios.
Los cuerpos densos contienen una variedad de proteínas de placa - actinina, que fijan
filamentos tanto finos comoade adhesión, incluida la intermedios de sarcolema en forma
directa e indirecta desempeñan un papel importante en la trasmisión de las fuerzas
1364 1ecfc2457dfcb5f

contráctiles generadas en el interior de la célula hacia la superficie celular, lo que altera la
forma de la célula. Estos cuerpos densos son análogos de intracelulares de las líneas z
del musculo estriado.
La contracción del músculo liso es iniciada por una gran variedad de impulsos que
incluyen estímulos mecánicos, eléctricos y químicos.
Los mecanismos que causan la contracción de las células musculares lisas son muy
diferentes de los de las células musculares estriadas. El músculo liso tiene diversos
mecanismos de transducción de señales que inician y modulan la contracción de sus
células. Todos conducen a la elevación de la concentración intracelular del Ca2+ , que es
la responsable directa de la contracción muscular. Así, la contracción del músculo liso
puede ser desencadenada por:
Impulsos mecánicos, como el estiramiento pasivo del músculo liso vascular. Los impulsos
mecánicos activan canales iónicos mecanosensibles que conducen a la iniciación de la
contracción muscular espontanea.
Despolarizaciones eléctricas, como las que ocurren durante la estimulación nerviosa del
musculo liso. La liberación de los neurotransmisores acetilcolina y noradrelanina desde
sus terminaciones nerviosas sinápticas estimula receptores ubicados en la membrana
plasmática de la célula muscular y cambia el potencial de la membrana. Esto causa
apertura de canales de Ca2+ sensibles al voltaje.
ASPECTOS FUNCIONALES DEL MÚSCULO LISO
El músculo liso se especializa en la contracción lenta y prolongada. Las células
musculares lisas pueden permanecer contraídas por periodos prolongados sin fatigarse.
Se pueden contraer a la manera de una onda y producir movimientos peristálticos como
los del tubo digestivo y la vía espermática del varón o la contracción puede ocurrir en todo
el músculo al mismo tiempo para producir movimientos expulsivos como los de la vejiga,
la vesícula biliar y el útero. El músculo liso tiene una actividad contráctil espontánea en
ausencia de estímulos nerviosos.
La contracción del musculo liso suele ser regulada por neuronas posganglionares del
sistema nervioso automático (SNA); la mayor parte del músculo liso esta inervada en
forma directa tanto por nervios simpáticos como parasimpáticos. En el tubo digestivo el

tercer componente del SNA, la división entérica, es la fuente primaria de nervios para las
capas musculares.
Aunque la mayor parte del Ca2+ entra en el citoplasma durante la despolarización a
través de canales de Ca2+ activados por voltaje, algunos canales de Ca2+ , llamados
canales de Ca2+ activados por ligando, son activados por hormonas a través de sus
mecanismos de segundo mensajero. En consecuencia la contracción del musculo liso
también puede ser iniciada por ciertas hormonas secretadas desde el lóbulo posterior de
la glándula pituitaria, por ejemplo la oxitocina y en menor medida, la hormona
antidiurética. Además las células musculares lisas pueden ser estimuladas o inhibidas por
hormonas secretadas por la hormona suprarrenal (p. ej., adrenalina y noradrenalina). La
oxitocina es un poderoso estimulante de la contracción del musculo liso y su liberación
desde la neurohipofisis desempeña un papel esencial en la contracción uterina durante el
parto.
Las células musculares lisas de diferencian del mesodermo ubicado en las paredes de órganos huecos del sistema cardiovascular, del digestivo, del urinario y del reproductor. Las fibras musculares lisas del iris se diferencian a partir del ectodermo.
Las células musculares lisas maduras presentan una forma fusiforme con un núcleo oval central. Pueden medir desde 20 µm en las paredes de los vasos sanguíneos hasta 400 - 500 µm en el útero.
Observe en este esquema 3D los dos tipos de cortes de una fibra.
Cada célula se halla rodeada de una membrana basal rica en proteoglicanos, glicoproteínas y colágeno tipo III. El colágeno funciona como una red que sostiene y organiza el paquete muscular.
Dependiendo del sitio, las células musculares lisas producen colágeno, elastina y otros componentes de la matriz extracelular.

Músculo liso al MO
Paquetes de músculo liso y su relacioón con el epimisio y elperimisio. Masson 100x.

Corte longitudinal de fibras musculares lisas de la pared del estómago. Masson 400x.
Sección transversal de las mismas fibras del preparado anterior. Masson 400x.
CONTRACCION DEL MUSCULO LISO
Aunque los iones de calcio en las células del músculo liso causan la contracción como en el músculo estriado, el control del movimiento de los iones de calcio es diferente. En el músculo liso relajado los iones de calcio están en el retículo sarcoplasmático por toda la célula. Cuando se produce la excitación de la membrana, los iones de calcio se liberan al citoplasma y se unen a una proteína llamada CALMODULINA. El complejo calcio-calmodulina activa a una enzima que fosforila la miosina y permite su unión a la actina. Luego se produce el deslizamiento de la misma forma que en las fibras musculares estriadas. Las fibras musculares lisas pueden excitarse por terminales nerviosas o por hormonas sanguíneas.
REGENERACION
Las fibras musculares lisas presentan un alto poder de regeneración.
5.2. Músculo liso visceral.

Se llama músculo liso por presentar sus miofilamentos de actina y miosina con una ordenación distinta a la del esquelético, es decir sin estriaciones.
CARACTERÍSTICAS PRINCIPALES:
1. MÚSCULO UNITARIO:
El músculo liso se divide en dos tipos morfológicos: músculo liso unitario o visceral y músculo liso multiunitario.
El primero está formado por células musculares agrupadas de forma compacta, formando láminas o haces rodeadas de tejido conectivo. Es típico formando las paredes musculares de de las estructuras viscerales huecas.
El segundo, está formado por células musculares aisladas, aunque pueden estar próximas entre sí. Forman parte de las trabéculas del bazo, etc.(1)
2. TAMAÑO:
Son más pequeñas que las estriadas. Entre 2 y 10 µm de diámetro por 20 a 60 µm de longitud (2).
3. CONEXIONES INTERCELULARES:
En las unitarias se encuentran puentes de baja resistencia, que les dá características funcionales de músculos sincitial. En estos puntos de baja resistencia (GAP JUNCTIONS)(UNIONES ESTRECHAS)(DESMOSOMAS), se encuentran pequeños poros por donde pasan iones y pequeñas moléculas. Estas uniones permiten el acople eléctrico de estas células.
4. FILAMENTOS CONTRÁCTILES:
Están formadas por filamentos de actina y miosina, los cuales se disponen más o menos paralelamente al eje longitudinal, aunque no muestran estriaciones como en el esquelético. Relación filamentos delgados (fd) y gruesos (fg) es de 16/1 (m.e.: 6/1). Los filamentos delgados tienden a rodear a los filamentos gruesos.
La principal proteína del fd es la actina y la del fg es la miosina (3). El fd tiene la misma proporción que en m.e. de tropomiosina y actina, pero no posee troponina (esto indica que la regulación de la actividad ATPasa de la actomiosina es diferente.
En el m.l. el fg no contiene "zona desnuda" (4), por estar las moléculas de miosina orientadas en direcciones opuestas a cada lado del filamento. La relación actina/miosina es de 31/1 (en el m.e. es 3/1). Esta disminución de miosina en el m.l. plantea la interrogante de cómo desarrolla tanta fuerza.

Químicamente la actina y la miosina son distintas a las del m.e., siendo además la actividad enzimática de la actomiosina mucho más lenta que la encontrada en el m.e.
La actividad ATPasa de la miosina es 10 veces menor que en el m.e. y su actividad está sujeta a una regulación más directa del Ca2+. Esta miosina es capaz de interactuar con filamentos de actina y causar contracción sólo cuando su cadena ligera está fosforilada, cuando no lo está la miosina no puede interactuar con la actina y produce relajación muscular. Estas cadenas ligeras son un conjunto de proteínas asociadas al extremo globular de la molécula de miosina. En el m.l. hay dos clases de proteínas de cadena ligera (en el m.e. hay tres). Una de estas proteínas tiene un p.m de 20kD (LC1) y la otra de 17 kD (LC2). La preoteína de 20 kD participa en la regulación de la actividad ATPasa de la actomiosina.
5. FIJACIÓN DE LOS MIOFILAMENTOS:
En este tipo de músculo liso no se distinguen líneas Z, y en consecuencia no se pueden definir sarcomeras. Los filamentos delgados se fijan a áreas electrodensas de la membrana celular, constituídas por -actina. También se observan cuerpos densos de -actina homólogos a las líneas Z del músculo esquelético.
6. OTRAS CARACTERÍSTICAS MORFOLÓGICAS:
Presentan una capa de glucoproteínas, excepto en los puntos de uniones estrechas. Presentan un número elevado de vesículas pinocíticas de unos 600 de . El retículo endoplásmico está poco desarrollado. Pero si presentan retículo sarcoplásmico en contacto con numerosas invaginaciones denominadas cavéolas, las cuales posiblemente sirven para transportar sustancias desde la membrana celular hacia la proximidad del retículo sarcoplásmico.
7. CARACTERÍSTICAS FUNCIONALES: (comunes a los dos tipos de m.l.)
a) Su inervación motora es exclusivamente autónoma. Control neurogénico.
b) Pequeño potencial de membrana en reposo.
c) En el unitario es fácil que presenten actividad eléctrica espontánea, originándose zonas marcapasos cuya despolarización propagan al resto de las células adyacentes.
d) Presentan potenciales de acción de gran amplitud.
e) Exhiben cierto grado de tono intrínseco, es decir, una tensión basal de reposo a la que superponen las contracciones. ACTIVIDAD MIÓGENA.
f) Desarrollan contracciones lentas y prolongadas, con un gasto mínimo de energía (requieren de cinco a diez veces menos hidrólisis de ATP que el m.e. para realizar la misma tarea).
g) En el unitario es normal encontrar una respuesta de relajamiento frente a estiramiento lento de sus fibras. Propiedad debida a las características viscoelásticas de este tipo de

músculo, que le permite una adaptación de la longitud de sus fibras, sin generación de tensión.
h) En el unitario, y en algunos casos, también se observa la capacidad de éste para desarrollar tensión activa como consecuencia de una distensión fuerte. CONTRACCIÓN REACTIVA AL ESTIRAMIENTO.
INERVACION: CONTROL NEUROGÉNICO
La inervación del músculo liso es siempre autonómica, a partir de fibras postganglionares simpáticas y parasimpáticas.
Las terminaciones colinérgicas (todas las preganglionares, postganglionares anatómicamente parasimpáticas, postganglionares anatómicamente simpáticas que inervan a las glándulas sudoríparas y las postganglionares anatómicamente simpáticas que inervan a los vasos sanguíneos del músculo esquelético (vasodilatadoras)) tienen vesículas claras, mientras que las adrenérgicas tienen vesículas de núcleo denso, y las purinérgicas tienen vesículas opacas.
Estas terminaciones presentan como característica la formación de varicosidades (figura) dispuestas a distintos intervalos, las cuales presentan vesículas con neurotransmisores. Éstas actúan como uniones neuromusculares poco especializadas, con una hendidura sináptica que varía dependiendo del tipo de músculo liso. Así en el multiunitario la unión se aproxima más a la del músculo esquelético, con una aproximación media de 6 a 20 µm ("contacto directo"), mientras que en el unitario va de 80 a 120 µm o más ("contacto indirecto o por difusión").
Los receptores para la ACh en el m.l. gastrointestinal son de tipo muscarínico y dentro de éstos del tipo M2, la activación de los mismo determina:
- apertura de un conjunto de canales catiónicos voltaje dependientes- modulación de las propiedades de canales voltaje dependientes que normalmente operan en respuesta a las alteraciones del Vm. (canales M o canales de Ca2+ sensitivos al potencial)- liberan el Ca2+ almacenado.- incrementan el IP3 u otro análogo por acelerar la actividad de la diesterasa fosfolipasa C (PLC).- incremento de la permeabilidad al K+ causado por la apertura de canales de K activados por el calcio.- una disminución del AMPc- y un incremento del GMPc
La excitación es principalmente por vía colinérgica, aunque no es la única.
Los receptores para las terminaciones adrenérgicas son de dos tipos los alfa-adrenérgicos y los beta-adrenérgicos. Los alfa están asociados con la vasoconstricción e inhibición de la motilidad gástrica. Los beta están asociados con la vasodilatación y relajación en el digestivo. Los receptores alfa presentes en el músculo liso gástrico son del tipo 1. Hay excepciones en el sentido de que la NA causa contracción del EEI (esfínter esofágico

inferior) vía receptor . Sin embargo la representación adrenérgica intestinal es pobre, por lo que se baraja la posibilidad de que las fibras inhibidoras sean tipo PIV(péptido intestinal vasoactivo) y de nucleótidos de adenosina. En definitiva nervios ni colinérgicos ni adrenérgicos.
POTENCIAL DE MEMBRANA DE LAS CÉLULAS MUSCULARES LISAS:
POTENCIAL DE REPOSO: principal característica es su pequeño valor (5).
La resistencia por unidad de área de la célula muscular lisa (13-21 k cm2) es más alta que la del m.e. (rápido de la rana)(1,5-4,0 k cm2) y que la fibra de Purkinje cardiaca)(1,2-2,0 k cm2). Esta alta resistencia refleja una baja permeabilidad pasiva al potasio y al cloro. Los coeficientes de permeablidad son: 8,3x10-8 cm seg-1 y 5,4x10-8 cm seg-1, respectivamente. Para el PNa es de 1,6x10-8 cm seg-1. De acuerdo con las concentraciones de estos iones dentro y fuera, los potenciales de equilibrio según la ecuación de Nernst son +62mV para el sodio, -89mV para el potasio y -22 mV para el cloro. Ninguno de estos potenciales están cercanos a los valores medidos de Vm que es de -40 a -75 mV. Es decir ninguno de los iones están en equilibrio al valor del Vm y ninguno se distribuye pasivamente a través de la membrana, por lo tanto, la célula muscular lisa debe gastar energia para mantener su estado de reposo. De acuerdo con la ecuación de Goldman, el Vm debería ser de -35 mV. La diferencia entre el valor teórico y el real se puede explicar por la existencia de bombas electrogénicas. Hay evidencias de una bomba de Na/K 3:1, y también de una bomba de cloro electrogénica, dado que la concentración de cloro en el interior es más alta que la esperada por su difusión pasiva, utilizando un cotransporte con el sodio.
Además de la difusión pasiva, y las bombas indicadas, juegan un importante papel la difusión por intercambio, en el mantenimiento de las concentraciones iónicas. Se conocen las de Na-Na, Na-Ca, y Na-Mg, y co-transporte de Na-Cl. Aquí el sodio se mueve siguiendo su gradiente electroquímico, mientras que el ión intercambiado o cotransportado lo hace en contra de su gradiente electroquímico. La energía necesaria proviene del gradiente electroquímico del sodio.
El intercambio Na-Na se hace al parecer con el retículo sarcoplásmico. Su importancia está bajo debate.
El intercmabio Na-Cl. El Cl entraría siguiendo al Na de acuerdo con su gradiente. Este sodio sería sacado por la bomba Na-K, dejando Cl en el interior celular. También es posible otro mecanismo de intercambio Cl-CO3H-.
Intercambio Na-Ca. Parece participar en el control del Ca iónico libre, aunque no la única, ya que sin gradiente de Na se puede conseguir relajar al músculo. Posiblemente exista una bomba de Ca.
ACTIVIDAD MARCAPASOS: En el músculo unitario es muy característico encontrar células que presentan un Vm inestable en reposo de forma que presentan una despolarización creciente (prepotenciales fundamentalmente de calcio) en el tiempo, que lleva a la célula a su umbral de excitabilidad, seguida de una repolarización y vuelta a empezar. Esta despolarización, gracias al diseño de sincitio funcional de este músculo, se

propaga de forma electrotónica en todas direcciones a través de las uniones de baja resistencia, generando un ritmo de despolarización que se conoce como RITMO ELÉCTRICO BÁSICO (ONDAS ELÉCTRICAS LENTAS), con una frecuencia variable en cuanto al número de ondas de despolarización por minuto. Esta propiedad además parece depender del conjunto celular, ya que no ha podido ser evidenciada en células lisas aisladas.
La duración de estos ritmos viene determinada por la velocidad de despolarización de los prepotenciales y por la distancia existente entre el nivel umbral y el potencial de membrana. Este último parámetro puede ser regulado por la presencia de fármacos como la ACh la cual disminuye la distancia incrementado el número de potenciales de acción y en consecuencia el incremento del tono basal. La NA tiene un efecto contrario. También hay que tener encuenta la posible participación de hormonas entéricas y no entéricas, así como de factores miógenos no nerviosos (histamina, adenosina, prostaglandinas y leucotrienos).
Estas ondas lentas son la respuesta a señales originadas en plexos de axones que contienen células intersticiales de Cajal, las cuales forman uniones estrechas con el músculo liso, y no son como se pensaba, producto de las características funcionales propias de las células musculares.
POTENCIALES DE ACCIÓN:
Los potenciales de acción suelen aparecer sobre estas ondas de despolarización propagadas electrotónicamente, ondas que sufren decremento en su propagación. Así mismo, las posibles espigas no alteran ni la frecuencia, ni la amplitud de estas ondas lentas de despolarización.
El mecanismo iónico implicado no está aún bien establecido, aunque si sabe que existe un incremento inicial de la conductancia al sodio y otra conductancia catiónica activada a -50 mV (posiblemente el calcio).
El que la célula alcance su umbral de excitabilidad con estas ondas de despolarización y genere potenciales de acción, depende de su estado de excitabilidad, el cual viene gobernado por factores importantes como: el influjo nervioso extrínseco o intrínseco, el influjo hormonal y el influjo de factores miogénicos. Además de estos factores en algunos casos también interviene el grado de estiramiento. (La ACh despolariza la membrana del músculo liso, mientras que la NA la hiperpolariza).
Estos potenciales de acción son muy variables en el músculo liso (ver figura), yendo de una espiga rápida a una muy lenta, a veces estos potenciales pueden unirse dando origen a una fase de meseta larga. Hay acuerdo de que la espiga del potencial de acción se debe a una corriente de entrada de calcio. No obstante existe una corriente de salida de potasio que se solapa con la corriente de entrada de calcio, la cual es causa de la primera, haciendo que la espiga sea menor de lo esperado, y que rara vez llegue a potencial cero.
Si hay poco calcio, la corriente de entrada viene soportada por el sodio . La repolarización se debe a la salida de potasio, y ésta puede ocurrir de forma temprana, durante la fase de despolarización, evitando así la inversión del potencial (el ECa es de +100 mV).

Estos potenciales de acción se encuentran siempre superpuestos a la región ascendente de las ondas lentas de despolarización, aunque estas ondas no necesariamente van acompañadas de potenciales de acción (Ver figura).
CONDUCCIÓN DE LA EXCITACIÓN:
La propagación tanto de las ondas lentas de despolarización, como de los potenciales de acción, es lenta pero uniforme gracias a las características sincitiales funcional del músculo liso unitario.
ACOPLAMIENTO EXCITACIÓN-CONTRACCIÓN
El factor acoplador es el calcio iónico libre en el citoplasma. No obstante la célula muscular lisa tiene escaso retículo endoplásmico, principal lugar de almacenamiento de calcio en el m.e.. Por esta razón se han sugerido otros lugares de almacenamiento, como por ejemplo las vesículas encontradas y próximas a la membrana celular, las mitocondrias (también escasas) y el movimiento de calcio a través de la membrana citoplasmática, mediante canales de calcio voltaje dependientes o a través de canales operados por receptores.
La despolarización de la membrana provocada por el influjo neurógeno determina la apertura de canales de calcio voltaje dependientes que favorecen las corrientes de entrada de calcio iónico libre. Así mismo esta despolarización propagada al retículo sarcoplásmico permite liberación de calcio almacenado en el mismo. Este tipo de respuesta se conoce como ACOPLAMIENTO EXCITACIÓN-CONTRACCIÓN, dado que los incrementos intracelulares de calcio iónico libre permiten su unión a la proteína citoplasmátrica calmodulina (6) la cual forma parte de una proteína más grande denominada miosina cinasa de cadena ligera (MCL-k). Esta fijación al calcio permite que la calmodulina catalice la fosforilación de la cadena ligera de miosina. Cuanto más calcio se ligue a la calmodulina, más fosforilación se produce de la ATPasa de actomiosina, y en consecuenia mayor interacción se produce entre la miosina y la actina (esta fosforilación es causa de la mayor lentitud en la contracción, ya que en el m.e. se produce sólo reorientación que es más rápido). Por el contrario la disminución en la concentración de calcio iónico libre citoplasmático, determina mayor defosforilación y disminuye la interacción miosina-actina, por activación de una fosfatasa de cadena ligera. La unión actina-miosina dura más en el músculo liso, por lo que la hidrólisis del ATP es más lenta y en consecuencia supone menor consumo de energía.
Junto a este acople determinado por la despolarización de la membrana, existe otro acople determinado por los agentes neurohormonales, ACOPLE FARMACOMECÁNICO, gracias al cual se produce incremento de calcio iónico intracelular sin despolarización de la membrana, sino mediado por receptores a distintos neuromoduladores.
La ACh através de su receptor M1 activa a la fosfolipasa C la cual a su vez produce IP3 y diacilglicerol. El IP3 libera el calcio iónico secuestrado, mientrás que el diaciglicerol activa a una fosfokinasa C (PKC) encargada de fosforilar distintas proteínas intracelulares.
La modulación inhibidora de la contracción que parece realizar el AMPc consiste en la facilitación que éste determina, en la fosforilación mediante una proteína kinasa de la MCL-k, haciéndose insensible la MCL-k-P al complejo Calmodulina-Ca2+.

CARACTERÍSTICAS CONTRÁCTILES DEL MÚSCULO LISO (UNITARIO)
La primera característica es el retardo que aparece entre el pico del potencial de acción y el pico de contracción, provocado por los eventos bioquímicos de fosforilación comentados (latencia de unos 200 mseg). La máxima contracción puede aparecer a los 500 mseg del pico de la espiga del potencial de acción.
Cada potencial de acción genera una onda de contracción, y la contracción final es el resultado de la suma de contracciones individuales. No obstante se puede observar contracción sin la presencia de potenciales de acción, como ocurre en el colon. La contracción no requiere la presencia de potenciales de acción, aunque siempre que se produce un potencial de acción hay una contracción. Esta contracción es debida a las ondas de despolarización. Las ondas lentas establecen los aspectos del peristaltismo.
Dada la lentitud de los potenciales de acción y la lentitud de las ondas de contracción éstas pueden superponerse de forma que se forme una contracción tetánica de baja frecuencia, con características muy distintas a las del músculo esquelético. Este fenómeno es debido a la lentitud conque se varía la concentración intracelular del calcio iónico libre, el cual mientras esté presente en concentraciones mayores a 10-7 mol/l produce efecto contráctil.
No obstante es normal encontrar al músculo liso unitario con un tono basal de contracción generado normalmente más que por el influjo nervioso, que también participa, por la modulación humoral y hormonal de la excitabilidad. Este tono es debido a un incremento mantenido de la concentración de calcio iónico libre intracelular, probablemente causado por un bajo recambio.
La relajación de tensión, es una característica también del músculo liso, por la cual se pueden producir cambios de longitud sin cambios intensos de tensión. Esta es una propiedad debida a las características viscoelásticas de éstos.
Por otro lado el estiramiento intenso de la fibra muscular lisa determina una respuesta de tensión independiente al ambiente nervioso y humoral de la misma CONTRACCIÓN REACTIVA AL ESTIRAMIENTO. No obstante la distensión mantenida determina a su vez una respuesta de relajación muscular, que favorece la disminución de la tensión. Esta respuesta puramente miógena, afecta seguramente a los cambios en la permeabilidad al calcio que se producen como consecuencia del estiramiento rápido.

Músculo liso
Es el músculo involuntario. Está compuesto de células con forma de huso con un núcleo central, que carecen de estrías transversales aunque muestran débiles estrías longitudinales. Su movimiento o contracción lo determina el sistema nervioso vegetativo. Se localiza en la piel, órganos internos, aparato reproductor, grandes vasos sanguíneos y aparato excretor.
El músculo liso humano, también llamado músculo visceral o involuntario, está constituido por células delgadas con forma de huso. Controladas por el sistema nervioso autónomo, las células del músculo liso ayudan a formar la estructura de la piel, los vasos sanguíneos y los órganos internos.
5.3. Músculo liso multiunitario.
Las células del tercer tipo de músculo no poseen estriaciones; por esta razón conforman
el músculo liso. Además estas células carecen de un sistema de túbulos T. El músculo
liso está en las paredes de vísceras huecas, paredes de vasos sanguíneos, conductos
más grandes de glándulas compuestas, vías respiratorias y haces pequeñas de la dermis
de la piel. El músculo liso no está controlado por la voluntad; lo regulan el sistema
nervioso central autónomo, hormonas y condiciones fisiológicas locales. Por este motivo,
el músculo liso también se conoce como músculo involuntario.

Existen dos tipos de músculo liso:
-Células de músculo liso multiunitarias, que pueden contraerse de manera independiente una de la otra porque cada célula muscular tiene su inervación propia.
-Músculo liso unitario (unidad única, vascular), cuyas membranas celulares forman uniones de intersticio con las células musculares lisas contiguas y las fibras nerviosas sólo hacen sinapsis con unas cuantas de las fibras musculares. En consecuencia, las células del músculo liso unitario no pueden contraerse de manera independiente unas de otras.
Además de sus funciones contráctiles, parte del músculo liso es capaz de sintetizar proteínas exógenas. Entre las sustancias que elaboran las células de músculo liso para su uso extracelular se encuentran colágena, elastina, glucosaminoglucanos, proteoglucanos y factores de crecimiento.
Microscopia óptica de fibras de músculo liso
La microscopia óptica revela que las fibras de músculo liso tienen forma de huso, son cortas y poseen un núcleo colocado en el centro.

Durante el acortamiento muscular, el núcleo adopta un aspecto en "sacacorchos" característico, como resultado del método de contracción del músculo liso.
Control de la contracción del músculo liso
La contracción de las fibras del músculo liso se lleva a cabo de la siguiente manera:
1. Los iones calcio, liberados del retículo sarcoplásmico y provenientes de las membranas, se unen con la calmodulina y esto altera su conformación. El complejo calcio-calmodulina se une con la caldesmona, lo que induce su liberación del sitio activo de la actina F, y luego activa a la cinasa de la cadena ligera de miosina.
2. La cinasa de la cadena ligera de miosina fosforila una de las cadenas más ligeras de miosina, conocida como la cadena reguladora, y permite el desdoblamiento de la molécula de meromiosina ligera para formar la molécula de de miosina II parecida a un "palo de golf" típica.
3. La cadena ligera fosforilada posibilita la interacción entre el sitio de unión de actina de la miosina II y hace posible la interacción entre la actina y el subfragmento S1 de la miosina II, lo que lleva a la contracción.
5.4. Actividades eléctricas y mecánicas del músculo liso.