
Metabolismo ii catabolismo
56
CATABOLISMO Profesor: Adán Gonçalves
-
Upload
adan-goncalves-conselleria-de-educacion-xunta-de-galicia -
Category
Education
-
view
57 -
download
1
Transcript of Metabolismo ii catabolismo

CATABOLISMO
Profesor: Adán Gonçalves

1. O CATABOLISMO LIBERA ENERXÍA GRAZAS A REACCIÓNS DE TRANSFERENCIA DE ELECTRÓNS
O catabolismo é a fase degradativa ou oxidativa do metabolismo na que se libera enerxía.Nas vías catabólicas os compostos orgánicos complexos iniciais son transformados (oxidados) sucesivamente en moléculas máis sinxelas ata converterse en produtos finais; moitos deles denomínanse produtos de excreción.A enerxía liberada nestas reaccións acumúlase en forma de ATP que logo se emprega en distintas actividades celulares.
NOTA: Hai rutas que participan tanto no catabolismo como no
anabolismo e denomínanase rutas anfibólicas. Por exemplo a ruta das pentosas fosfato.

UTILIZACIÓN CELULAR DA ENERXÍA PARA:1. Síntetizar biomoléculas e macromoléculas a
partir de precursores simples (anabolismo)2. Transportar activamente ións e moléculas a
través da súa membrana.3. Realizar traballo mecánico na contracción
muscular e en outros movementos celulares4. Producir calor para mellorar as reaccións5. Impulsar os procesos de transcrición, tradución e
duplicación do ADN.

A LIBERACIÓN DE ENERXÍA NO CATABOLISMO
Nas vías catabólicas, pásase dos reactivos cara os produtos que teñen menor enerxía libre (G), polo que se libera enerxía ao exterior (G.Son polo tanto reaccións exergónicas.
AS REACCIÓNS CATABÓLICAS SON REACCIÓNS REDOX
As reaccións de oxidación-redución (redox) son reaccións de transferencia de electróns, nas que a substancia que se oxida perde electróns e a que se reduce os capta.O axente oxidante é quen acepta os electróns e o axente redutor e quen cede os electróns.As reaccións catabólicas son reaccións de oxidación de moléculas ricas en enerxía. Un exemplo é a respiración celular da glicosa (C6H12O6) nos eucariotas, que se oxida en presenza de O2 (axente oxidante) e libera enerxía. Os C da glicosa pasan ao seu máximo grao de oxidación, o CO2.
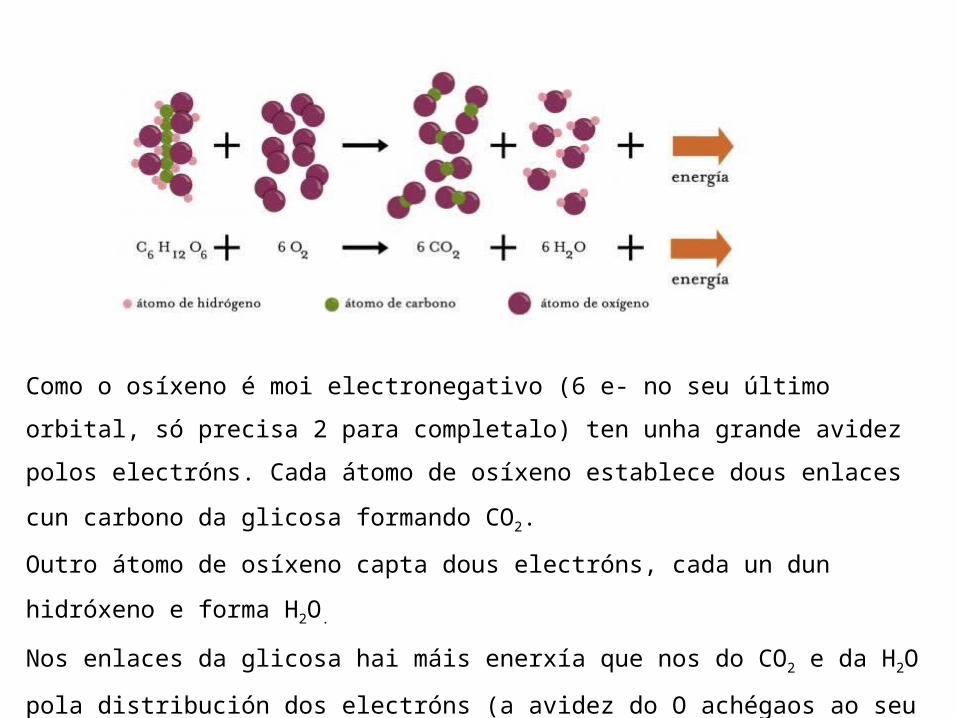
Como o osíxeno é moi electronegativo (6 e- no seu último orbital, só precisa 2 para completalo) ten unha grande avidez polos electróns. Cada átomo de osíxeno establece dous enlaces cun carbono da glicosa formando CO2.Outro átomo de osíxeno capta dous electróns, cada un dun hidróxeno e forma H2O. Nos enlaces da glicosa hai máis enerxía que nos do CO2 e da H2O pola distribución dos electróns (a avidez do O achégaos ao seu núcleo pasando a un estado enerxético menor) por iso liberase enerxía.
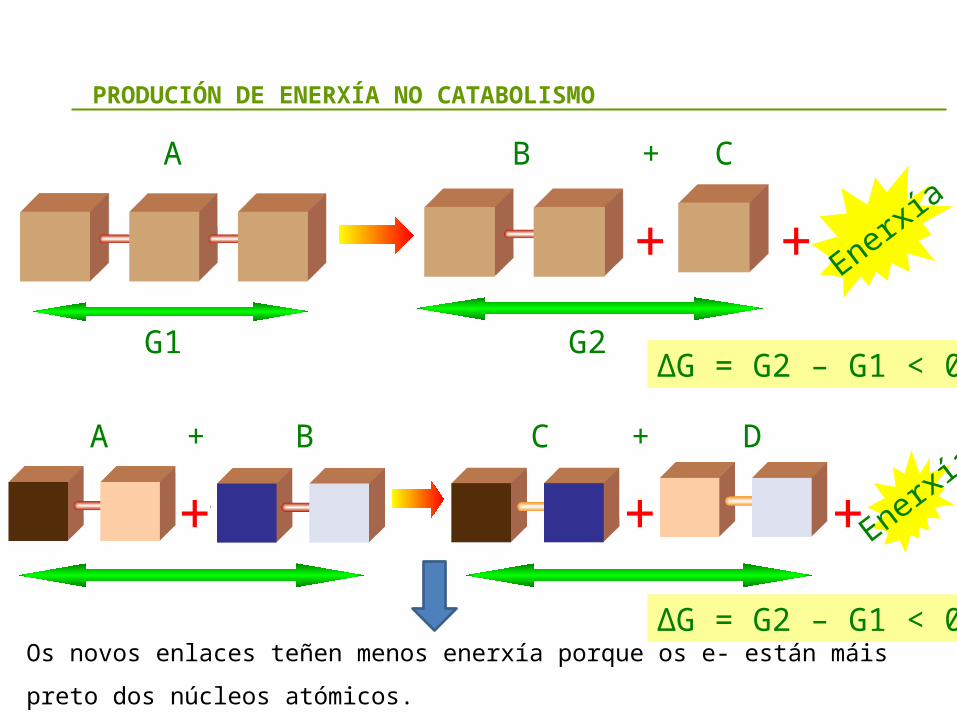
PRODUCIÓN DE ENERXÍA NO CATABOLISMO
+
A B C
Enerxía
A
+
+ + +B C D
G1 G2ΔG = G2 – G1 < 0
ΔG = G2 – G1 < 0
+
++
Enerxía
Os novos enlaces teñen menos enerxía porque os e- están máis preto dos núcleos atómicos.

LIBERACIÓN GRADUAL DE ENERXÍA NO CATABOLISMO POR RESPIRACIÓN
A liberación de enerxía na respiración sucede de xeito gradual. Isto débese a varios factores: Hai reaccións sucesivas Ocorre un transporte de H. Os e- da glicosa non pasan directamente
ao O, senón que nas primeiras etapas viaxan xunto a protóns formando átomos de H que pasan a un coenzima (xeralmente NAD+) que actúa como transportador de H. Unha deshidroxenasa elimina da glicosa 2 e- e 2 H+ , é dicir, 2 átomos de H, e cede 2 e- e un H+ ao NAD+ que se reduce a NADH, deixando un protón libre no medio (exprésase NADH + H+).
A cadea de transporte electrónico. O NADH non pasa directamente os e- ao O, senón que llos transfire a unha cadea transportadora de e- formada por citocromos (unhas proteínas) nas membranas internas da mitocondria. Finalmente chegan ao O e tras a unión aos H+ libres se reduce a H2O. Asociado a este transporte hai un fluxo de H+ que permite sintetizar ATP a expensas de ADP + Pi mediante ATP sintasas.
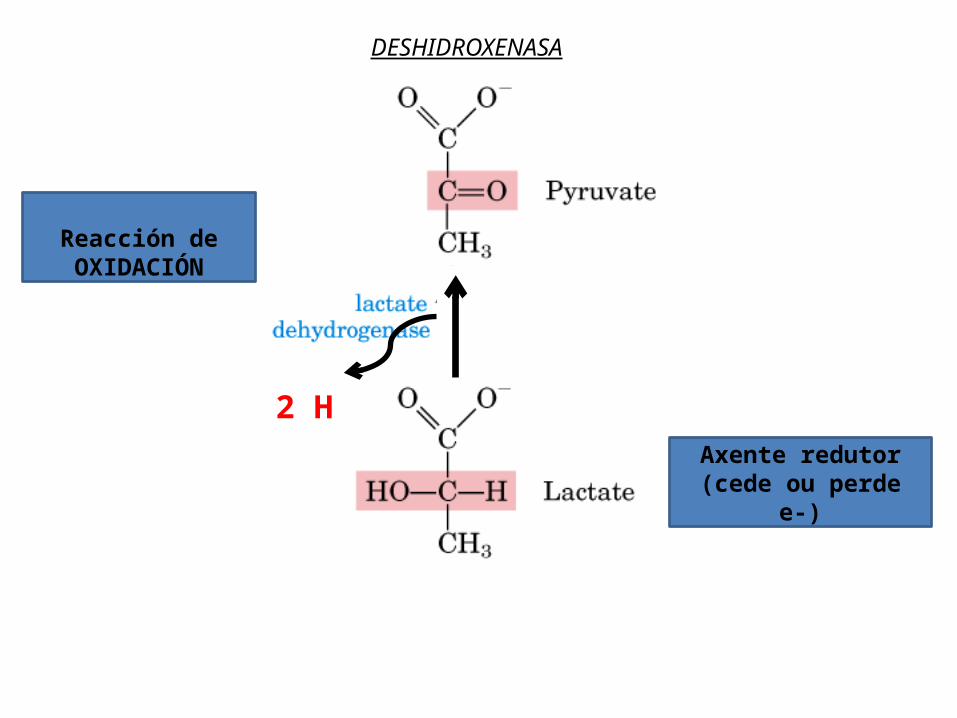
Reacción de OXIDACIÓN
Axente redutor(cede ou perde
e-)
2 H
DESHIDROXENASA

REACCIÓNS REDOX COMPOSTO OXIDADO
COMPOSTO REDUCIDO
Reaccións redox
A BO+ AO B+AH B+ A BH+
A B+ A+ B- +e-
BBH
B-
AOA
A+
OXIDACIÓN
REDUCIÓN
HIDRÓXENO ELECTRÓNS ENERXÍA
ELIMINACIÓN
ADICIÓN
ELIMINACIÓN
ADICIÓN
LIBERACIÓN
ALMACENAMENTO
CARACTERÍSTICAS DAS REACCIÓNS REDOX

TIPOS DE CATABOLISMO
Hai dous tipos: Respiración e fermentación. Respiración: intervén a cadea de transporte de electróns que
permite transferir e- dende un composto orgánico inicial a un aceptor final que é un composto inorgánico. En función do aceptor final falamos de:
Respiración aeróbica: o aceptor final (axente oxidante) é o O2 que se reduce a auga ao aceptar e- e protóns.
Respiración anaeróbica: o aceptor final non é o O2, senón outra substancia inorgánica por exemplo NO-
2 (nitrito) ou NO-3
(nitrato).As reaccións catabólicas da respiración varían dependendo dos substrato de partida. Os glícidos e lípidos son oxidados ata o final como fonte principal para obter enerxía. Porén as proteínas e ácidos nucleicos habitualmente son degradados para obter aa e nt respectivamente e soamente en condicións excepcionais de extrema necesidade empréganse para obter enerxía.
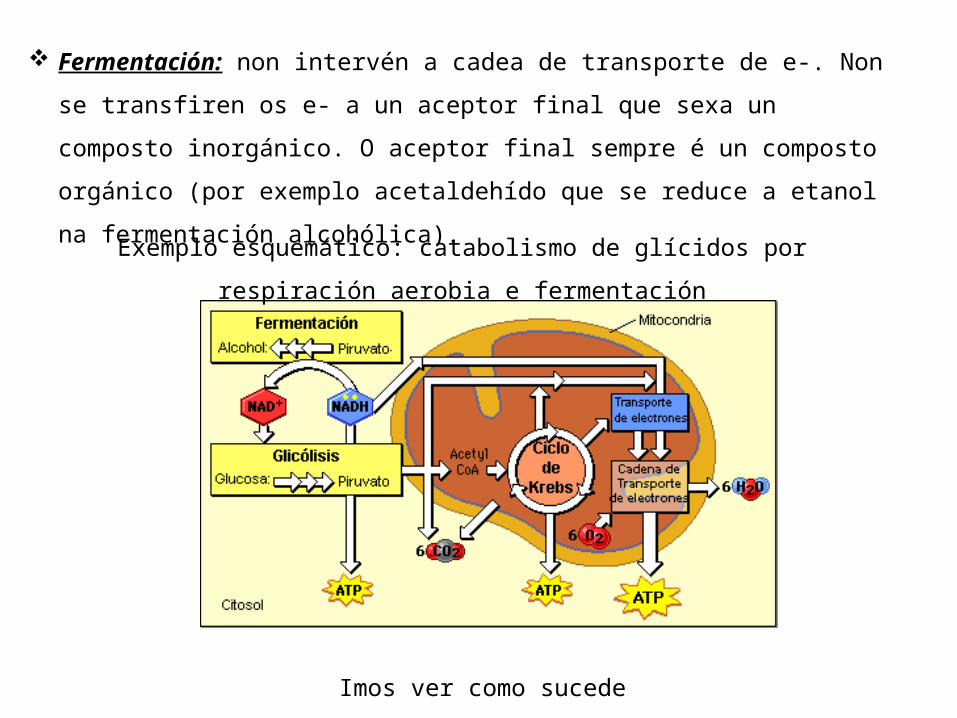
Fermentación: non intervén a cadea de transporte de e-. Non se transfiren os e- a un aceptor final que sexa un composto inorgánico. O aceptor final sempre é un composto orgánico (por exemplo acetaldehído que se reduce a etanol na fermentación alcohólica).
Exemplo esquemático: catabolismo de glícidos por respiración aerobia e fermentación
Imos ver como sucede

2. O CATABOLISMO PODE SUCEDER POR RESPIRACIÓN OU FERMENTACIÓN
Os polisacáridos contidos nos alimentos son degradados na dixestión a disacáridos e despois monosacáridos.O glicóxeno, almacenado no músculo (reserva propia) e no fígado tamén pode ser degradado a glicosa cando sexa preciso.Do mesmo xeito do amidón dos vexetais pode obterse glicosa.Un proceso clave do metabolismo dos glícidos é a glicolise mediante a cal a partir da oxidación da glicosa obtemos 2 moléculas de ácido pirúvico ou piruvato (Pyr). A finalidade do proceso é liberar a enerxía dos enlaces –C-C- da glicosa e almacenala en forma de ATP. Ademais obtense poder redutor en forma de NADH+H+. A partir do Pyr pódese iniciar a fermentación ou a respiración segundo o O2 dispoñible.
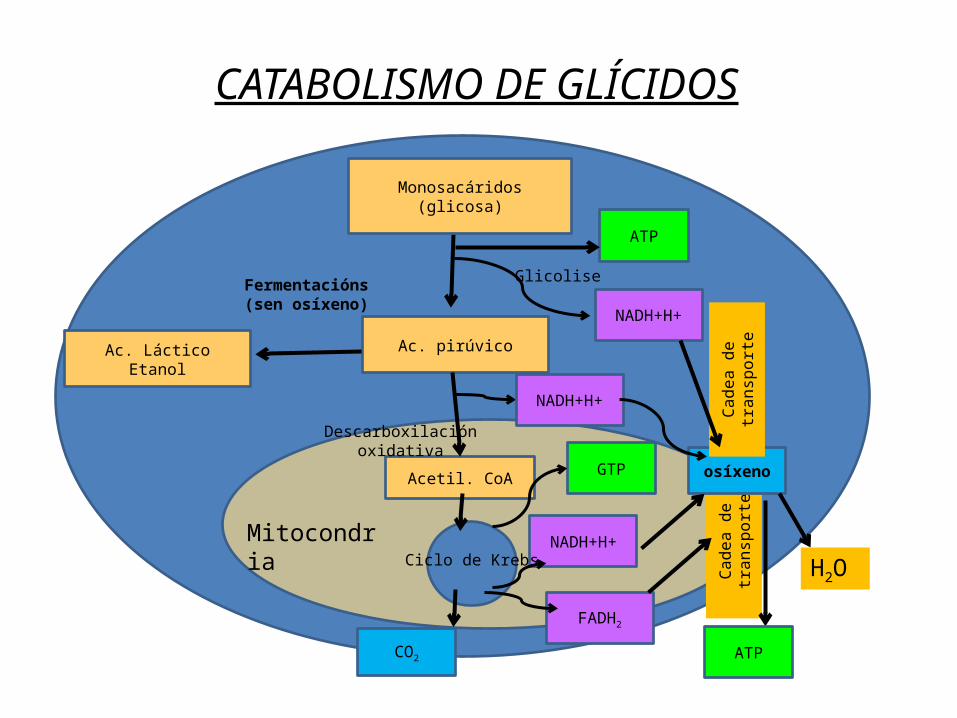
CATABOLISMO DE GLÍCIDOS
Moléculas sencillas digeridas(glucosa)
Monosacáridos(glicosa)
Ac. pirúvico
Acetil. CoA
Ac. LácticoEtanol
Fermentacións(sen osíxeno)
Glicolise
Descarboxilación oxidativa
Cade
a de
tra
nspo
rte
ATP
NADH+H+
NADH+H+
GTP osíxeno
ATP
Ciclo de Krebs
CO2
H2OMitocondria
NADH+H+
FADH2
Cade
a de
tra
nspo
rte

3. A GLICOLISE: OBTENCIÓN DE ENERXÍA A PARTIR DE GLICOSA
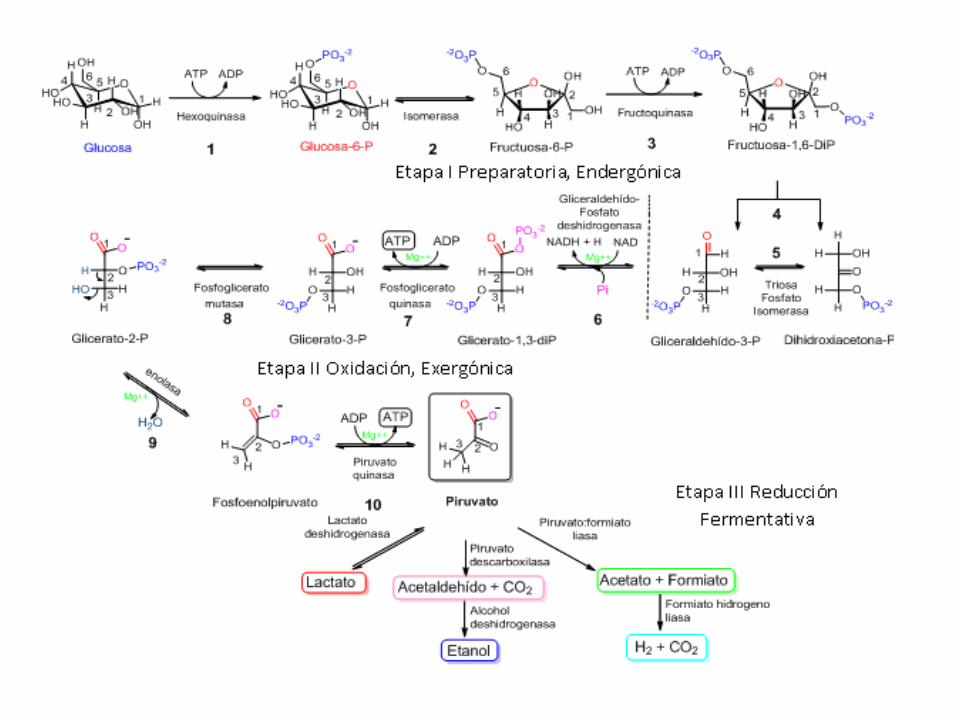
A glicolise ou ruta Embden-Meyerhof é considerada a ruta máis antiga empregada polos seres vivos para obter enerxía. É a ruta central do catabolismo de glícidos en plantas, animais e microorganismos.Prodúcese no citosol, escinde a glicosa (C6H12O6) en 2 moléculas de
Pyr (CH3-CO-COOH) e a enerxía liberada no proceso permite obter de xeito neto 2 ATP.Adoita falarse de dúas fases: Fase preparativa: gástanse 2 ATP. Dende a glicosa ata o
gliceraldehído- 3P Fase oxidativa: prodúcense 4 ATP (netos 2 ATP) + 2 NADH+H+ +
2 Piruvato
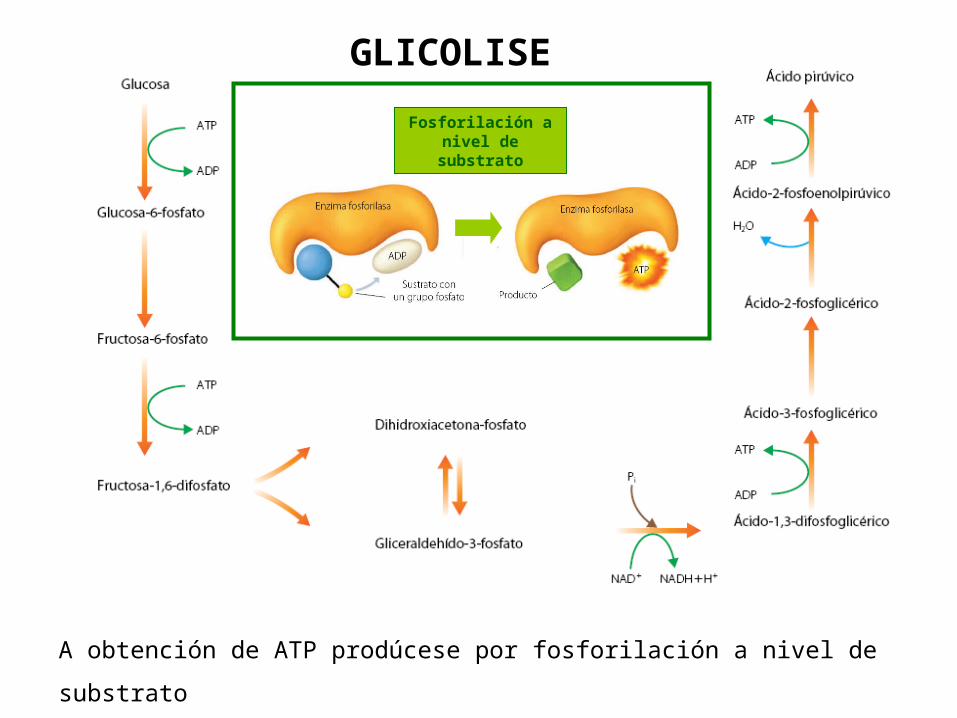
Fosforilación a nivel de substrato
GLICOLISE
A obtención de ATP prodúcese por fosforilación a nivel de substrato
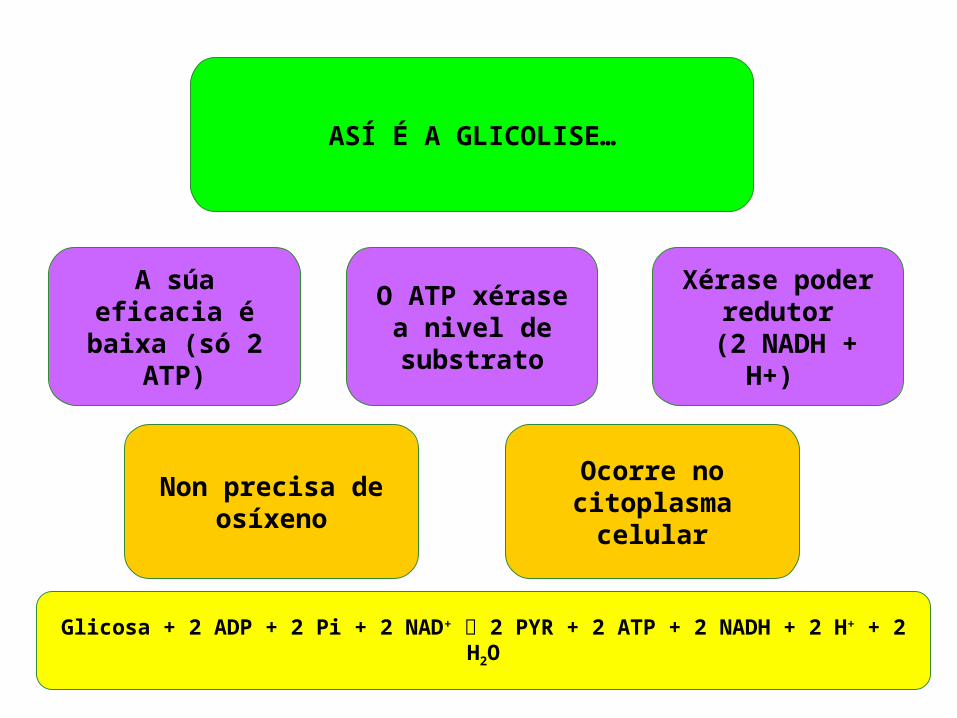
Non precisa de osíxeno
A súa eficacia é baixa (só 2 ATP)
ASÍ É A GLICOLISE…
Ocorre no citoplasma celular
Xérase poder redutor
(2 NADH + H+)
O ATP xérase a nivel de
substrato
Glicosa + 2 ADP + 2 Pi + 2 NAD+ 2 PYR + 2 ATP + 2 NADH + 2 H+ + 2 H2O

A glicolise produce NADH+H+, é dicir poder redutor, pero precisa NAD+ (o coenzima oxidado) para ter lugar. Isto implica que é un proceso que non pode suceder só indefinidamente e necesita dunha vía metabólica que rexenere os coenzimas oxidados que son reducidos cando ten lugar.Esta vía pode ser a respiración ou a fermentación.Existe polo tanto unha interdependencia entre a glicolise e estas vías que ven dada por esta coenzima.
DESTINO DO PIRUVATO, PRODUCTO FINAL DE LA GLICOLISE
1) EN CÉLULAS ANAEROBIAS ESTRICTASRealizan fermentacións ou respiracións incompletas: só lles aportan o 5% da enerxía (2 ATP por molécula de glicosa), pero rexeneran o NAD+.Exemplo: fermentación láctica (por exemplo bacterias Lactobacillus) pasa a ácido láctico (ácido 2-hidroxi-propanoico) (CH3-CHOH-COOH)

2) EN CÉLULAS ANAEROBIAS FACULTATIVASRealizan excepcionalmente fermentacións. Ex. lévedos do xénero Saccharomyces (eucariotas) realizan a fermentación
alcohólica pola que a glicosa pasa a etanol e CO2. Só produce 2 ATP, pero rexenera o poder redutor.
Ex. As células musculares esqueléticas en condiciones anaeróbicas (sen O2) realizan unha fermentación láctica. Produce tamén 2 ATP e rexenera o poder redutor. Cando sometemos aos nosos músculos a un sobreesforzo físico, as células musculares poden non recibir o suficiente O2 para levar a cabo a respiración aerobia do Pyr e o degradan por fermentación a ácido láctico. Posteriormente o ácido láctico será transportado ao fígado, onde en condición aerobias (con O2) será reconvertido a Pyr.
3) EN CÉLULAS AEROBIASLevan a cabo unha respiración aeróbica onde o Pyr pasa dentro da mitocondria onde será degradado a CO2 e H2O.
DESTINO DO PIRUVATO, PRODUCTO FINAL DE LA GLICOLISE

RESPIRACIÓN CELULAR AEROBIA
1. Descarboxilación oxidativa do piruvato. Produce ácido acético en forma de acetil coenzima A. Matriz mitocondrial
2. Ciclo de Krebs. O ácido acético é oxidado totalmente para formar CO2. Matriz mitocondrial.3. Transporte de electróns. Ata o O2. Participa a cadea respiratoria. Na membrana mitocondrial interna.
4. A enerxía almacenada nos coenzimas liberase e é empregada para sintetizar ATP.(fosforilación oxidativa)
DESPOIS DA GLICOLISE:
CO2
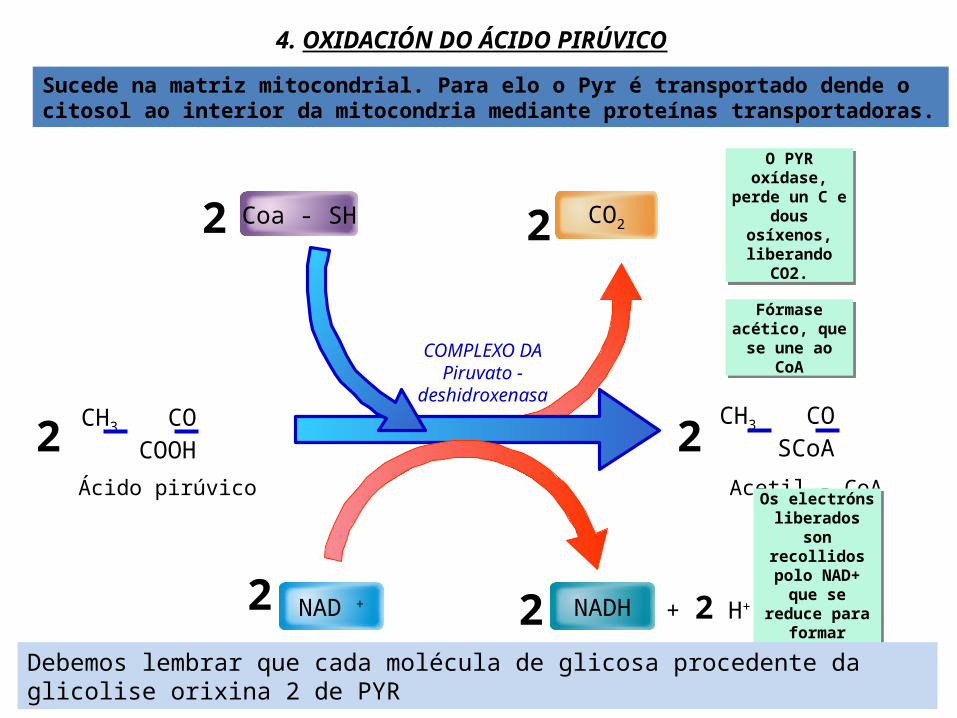
4. OXIDACIÓN DO ÁCIDO PIRÚVICO
Coa - SH
NAD + NADH + 2 H+
CH3 CO COOH CH3 CO SCoA
Ácido pirúvico Acetil - CoA
COMPLEXO DA Piruvato -
deshidroxenasa
2 2
2 2
2 2O PYR oxídase,
perde un C e dous osíxenos, liberando CO2.
Fórmase acético, que se
une ao CoA
Os electróns liberados son
recollidos polo NAD+ que se reduce para
formar NADH+H+
Debemos lembrar que cada molécula de glicosa procedente da glicolise orixina 2 de PYR
Sucede na matriz mitocondrial. Para elo o Pyr é transportado dende o citosol ao interior da mitocondria mediante proteínas transportadoras.
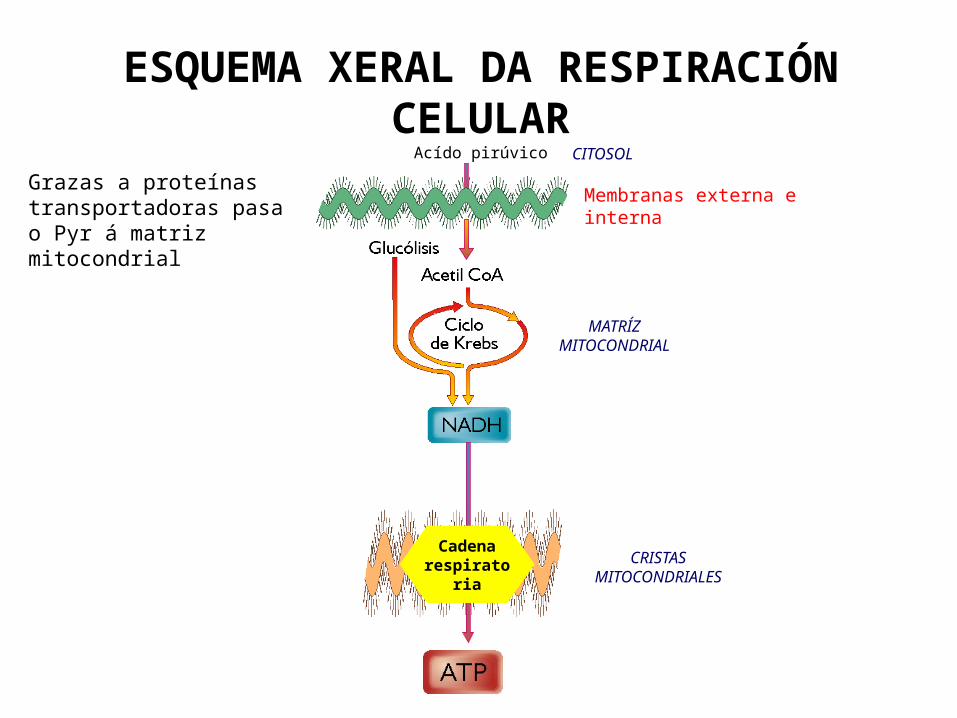
ESQUEMA XERAL DA RESPIRACIÓN CELULAR
Cadena respiratoria
Acído pirúvico CITOSOL
MATRÍZ MITOCONDRIAL
CRISTAS MITOCONDRIALES
Membranas externa e internaGrazas a proteínas transportadoras pasa o Pyr á matriz mitocondrial
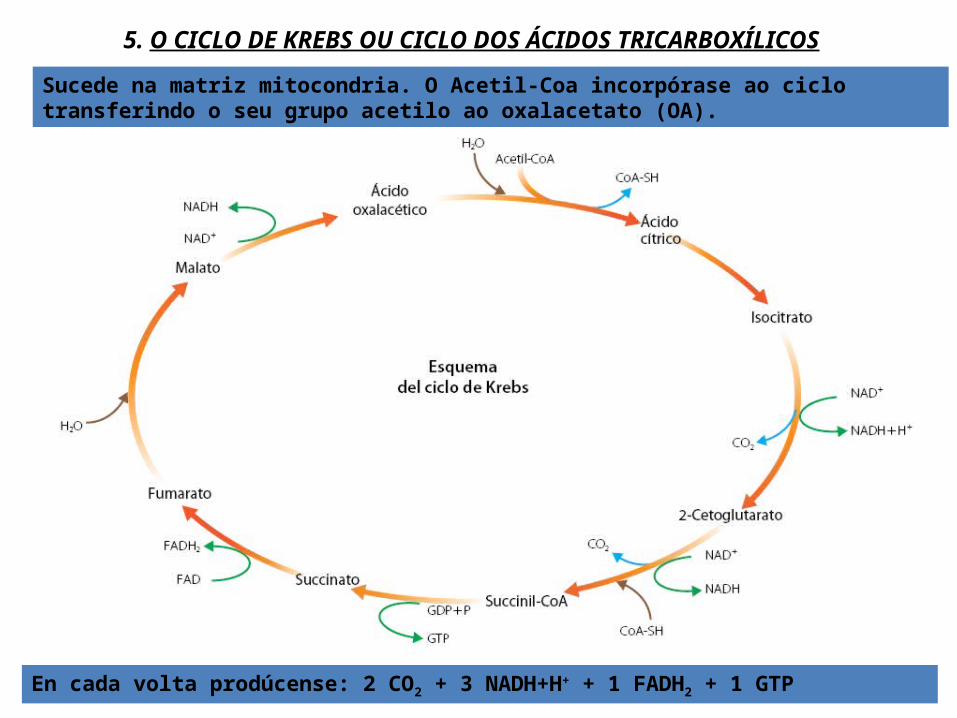
5. O CICLO DE KREBS OU CICLO DOS ÁCIDOS TRICARBOXÍLICOS
Sucede na matriz mitocondria. O Acetil-Coa incorpórase ao ciclo transferindo o seu grupo acetilo ao oxalacetato (OA).
En cada volta prodúcense: 2 CO2 + 3 NADH+H+ + 1 FADH2 + 1 GTP
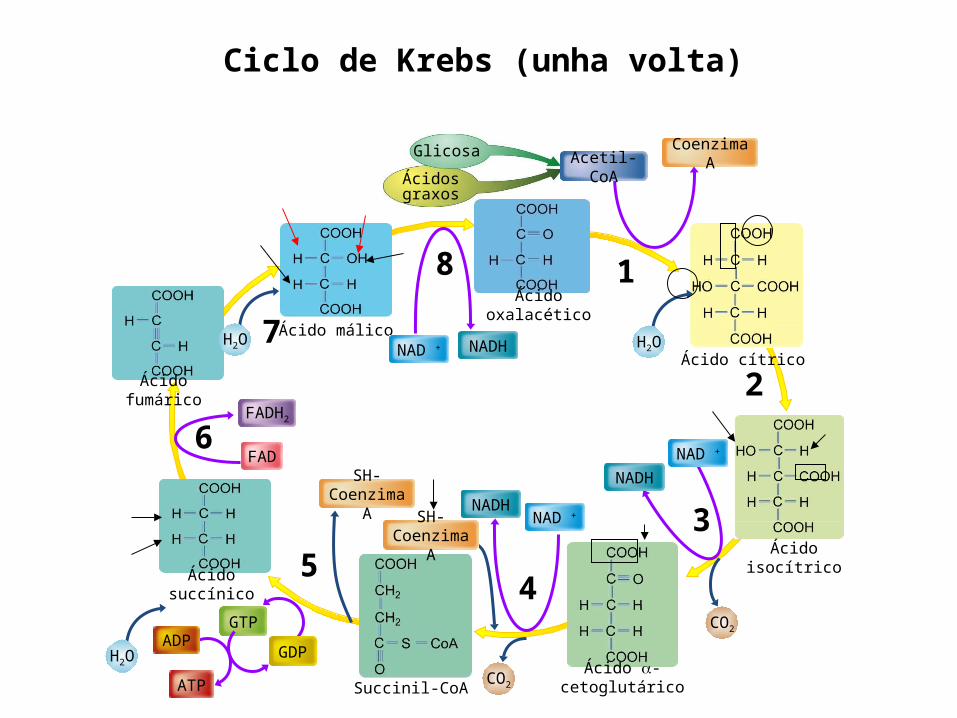
Ciclo de Krebs (unha volta)
Ácido cítrico
Ácido isocítrico
Ácido -cetoglutáricoSuccinil-CoA
Ácido succínico
Ácido fumárico
Ácido málico
Glicosa
Ácidos graxos
Ácido oxalacético
H2O
SH-Coenzima A
SH-Coenzima A
FAD
FADH2
NADHNAD +
NADHNAD +
Coenzima AAcetil-CoA
NAD + NADH
GDP
GTP
ATP
ADP
CO2
CO2
H2O H2O
1
4
2
35
6
7
8
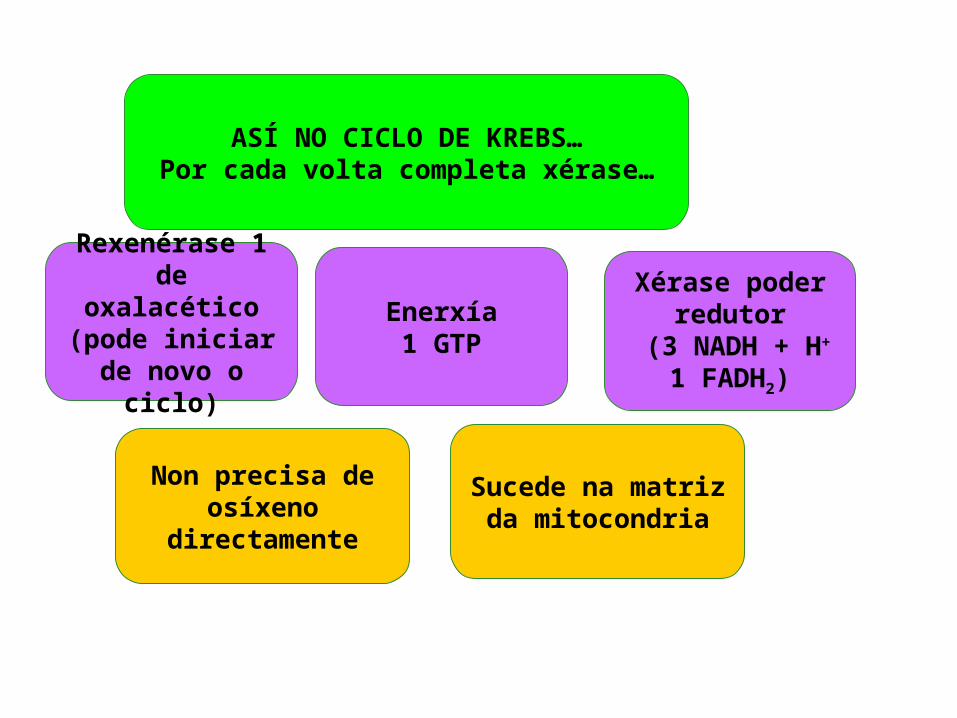
Non precisa de osíxeno
directamente
Rexenérase 1 de oxalacético(pode iniciar
de novo o ciclo)
ASÍ NO CICLO DE KREBS…Por cada volta completa xérase…
Sucede na matriz da
mitocondria
Xérase poder redutor
(3 NADH + H+
1 FADH2)
Enerxía1 GTP

Nun principio o balance enerxético do ciclo de Krebs parece moi pobre, só un GTP que equivale a un ATP, pero como veremos os coenzimas reducidos producidos neste ciclo de reaccións e nas anteriores (descarboxilación do PYR e glicolise) liberan enerxía cando se incorporan á cadea respiratoria e provocan a fosforilación oxidativa.5. CADEA DE TRANSPORTE ELECTRÓNICA OU CADEA RESPIRATORIA
A cadea respiratoria está formada por seis compoñentes: Os complexos proteicos I, II, III e IV englobados na membrana
mitocondrial interna. A ubiquinona ou CoQ capaz de trasladarse pola bicapa lipídica. O citocromo c, unha pequena heteroproteína localizada na
membrana que traslada e- (Fe+3/Fe+2) dende o complexo III ao IV (estes tamén conteñen citocromos).
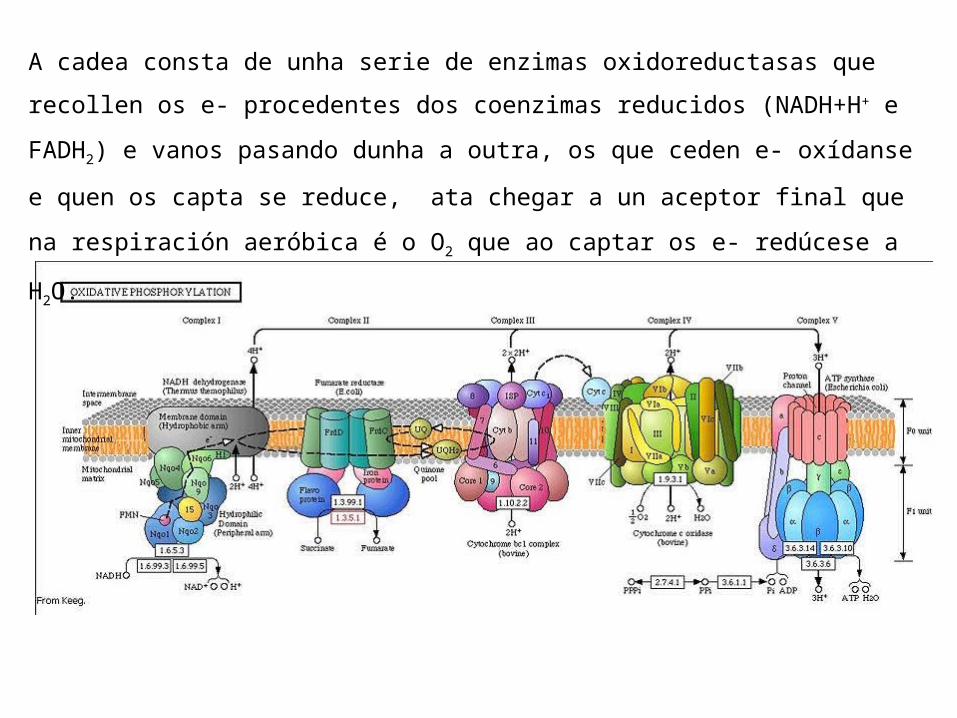
A cadea consta de unha serie de enzimas oxidoreductasas que recollen os e- procedentes dos coenzimas reducidos (NADH+H+ e FADH2) e vanos pasando dunha a outra, os que ceden e- oxídanse e quen os capta se reduce, ata chegar a un aceptor final que na respiración aeróbica é o O2 que ao captar os e- redúcese a H2O.

O fluxo de e- é a favor de gradiente dende o NADH de potencial redox máis baixo ata o O2 de potencial redox máis alto. A medida que os e- se desprazan pola cadea pasan a ocupar unha posición máis próxima ao núcleo da molécula que os transporta e polo tanto teñen menor enerxía (lembrar que a maior electronegatividade máis avidez polos e- me que o potencial redox expresa a tendencia a captar e-).
NOTA: Algúns microorganismos, como xa dixemos, levan a cabo unha respiración anaeróbica na que o aceptor último dos e- non é o O2 senón outra molécula. Este é o caso das bacterias reductoras do nitrato (NO-
3) que empregan este nitrato como aceptor último.

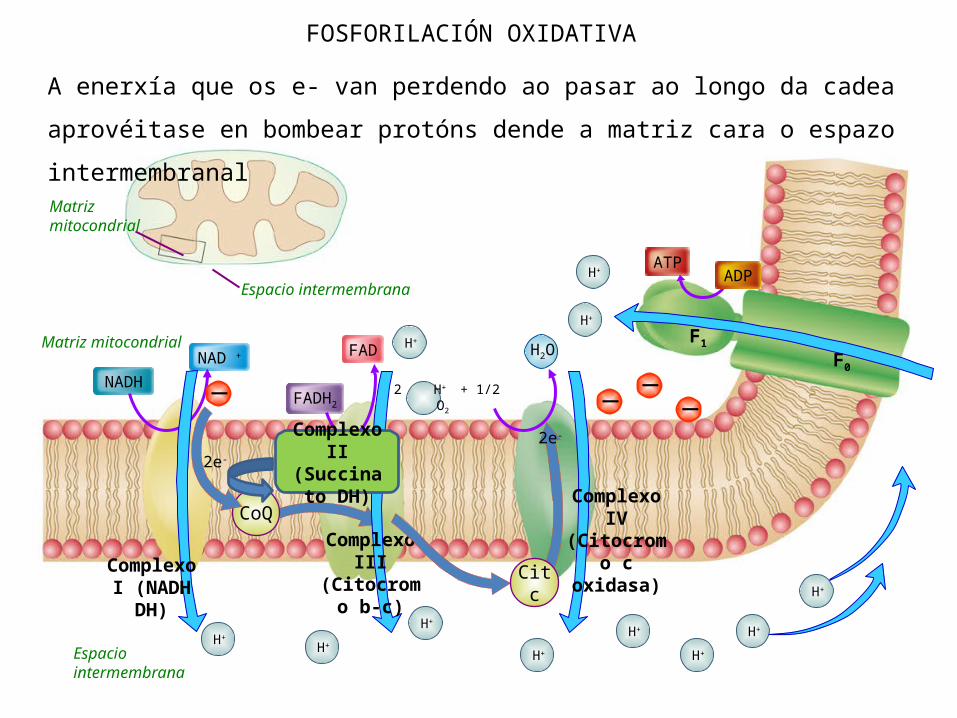
6. FOSFORILACIÓN OXIDATIVA
É a síntese de ATP mediante ATP-asas acoplada ao transporte de e- na cadea respiratoria. Esta síntese é posible porque asociado ao transporte de e- prodúcese un fluxo de H+ en tres puntos da cadea dende a matriz mitocondrial ata o espazo intermembranal.Hai fluxo de H+ a nivel do complexo I, complexo III e do complexo IV. O NADH, debido ao seu potencial redox, cede os seus e- a nivel do complexo I polo que participa dende o principio; o FADH2 os cede dende o complexo II. Esta é a razón pola que o NADH (3/2,5 ATP) xera máis enerxía que o FADH2
(2/1,5 ATP).

QUIMIÓSMOSE
Forma parte da fosforilación oxidativa xa que é o proceso polo cal se produce o retorno dos H+ dende o espazo intermembranal cara a matriz mitocondrial. Este retorno é o verdadeiro responsable da síntese de ATP nas ATP-asas.Cando hai un exceso de H+ no espazo intermembranoso. Éstes regresan a través de canais que hai nas ATP-asas situadas na membrana. Ese fluxo de retorno move un rotor existente na ATP-asa que permite a síntese de ATP a expensas de ADP + Pi.
FOSFORILACIÓN OXIDATIVA
CoQ
FADH2
NADHNAD + FAD
H+
H+
H+
2 H+ + 1/2 O2
H+
H+H+
H+
H+
H+
H+
H+
ATPADP
H2O
2e-
2e-
_ _ __
F1
F0
Complexo I (NADH DH)
Complexo III
(Citocromo b-c)
Complexo IV
(Citocromo c oxidasa)
Matriz mitocondrial
Espacio intermembrana
Matriz mitocondrial
Espacio intermembrana
Cit c
A enerxía que os e- van perdendo ao pasar ao longo da cadea aprovéitase en bombear protóns dende a matriz cara o espazo intermembranal
Complexo II (Succinato
DH)
ATP32
Ciclode
Krebs
NADH2
NADH2
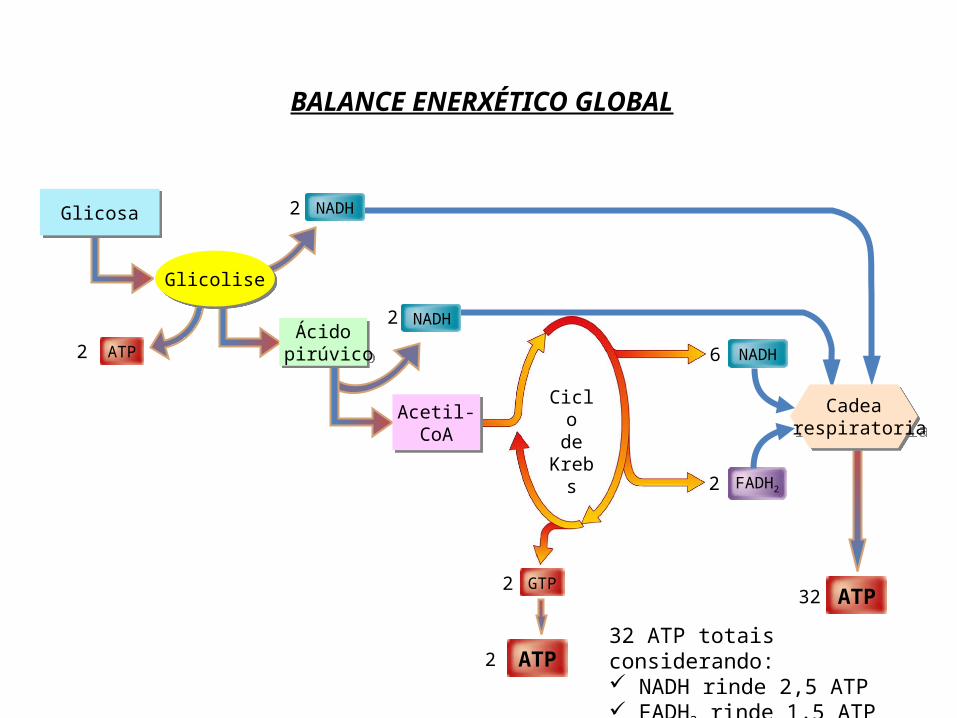
BALANCE ENERXÉTICO GLOBAL
Acetil-CoA
Glicosa
Ácido pirúvico
FADH22
NADH6
Cadea respiratoria
GTP2
ATP2
Glicolise
ATP232 ATP totais considerando: NADH rinde 2,5 ATP FADH2 rinde 1,5 ATP
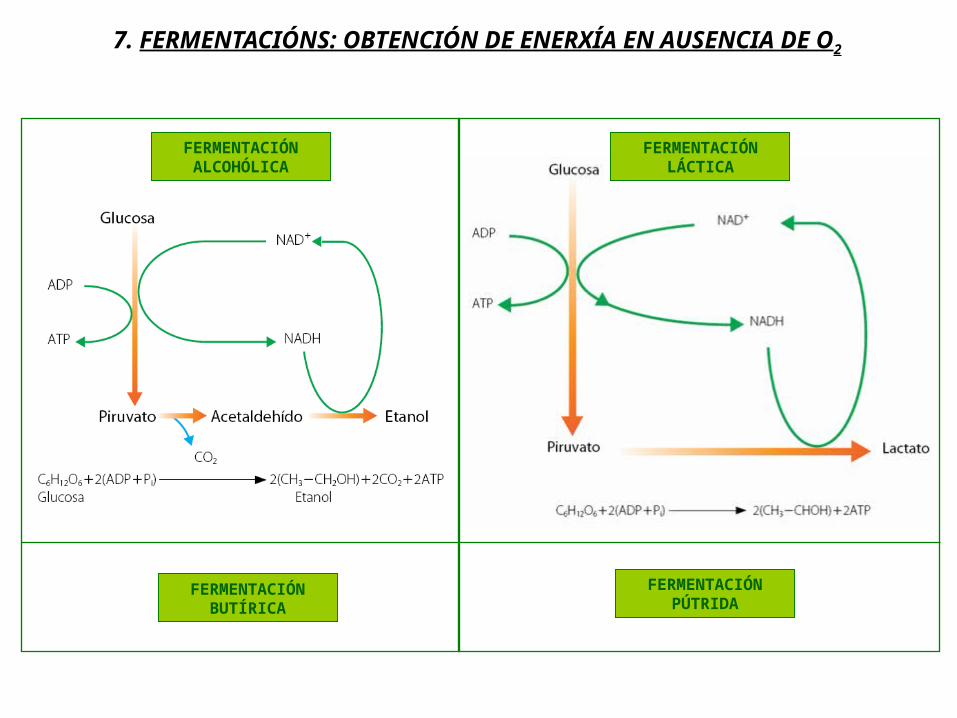
FERMENTACIÓN ALCOHÓLICA
FERMENTACIÓN LÁCTICA
FERMENTACIÓN BUTÍRICA
FERMENTACIÓN PÚTRIDA
7. FERMENTACIÓNS: OBTENCIÓN DE ENERXÍA EN AUSENCIA DE O2
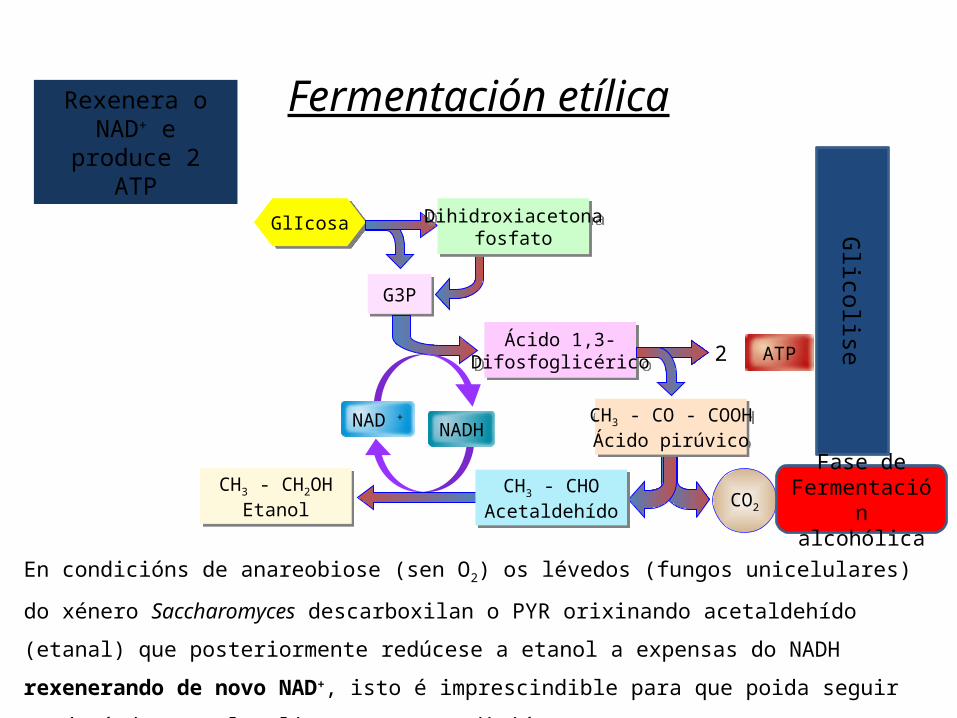
CH3 - CH2OHEtanol
Fermentación etílica
G3P
GlIcosa Dihidroxiacetonafosfato
Ácido 1,3-Difosfoglicérico ATP2
NADH
CO2
CH3 - CHOAcetaldehído
NAD + CH3 - CO - COOHÁcido pirúvico
Glicolise
Fase de Fermentación alcohólica
En condicións de anareobiose (sen O2) os lévedos (fungos unicelulares) do xénero Saccharomyces descarboxilan o PYR orixinando acetaldehído (etanal) que posteriormente redúcese a etanol a expensas do NADH rexenerando de novo NAD+, isto é imprescindible para que poida seguir producíndose a glucolise nestas condicións.
Rexenera o NAD+ e
produce 2 ATP

APLICACIÓNS DA FERMENTACIÓN ALCOHÓLICA
A fabricación de bebidas alcohólicas (viño, cervexa, whisky, ron...) basease na fermentación alcohólica a partir de diversas fontes: o etanol do viño procede da glicosa da uva; o etanol da cervexa procede da glicosa da cebada...As especies máis importantes son S. Cerevisae e S. Elipsoidens.S. Cerevisae tamén é responsable da fermentación alcohólica que nos permite elaborar pan. O CO2 forma as burbullas que son retidas polo gluten permitindo que levede o pan.
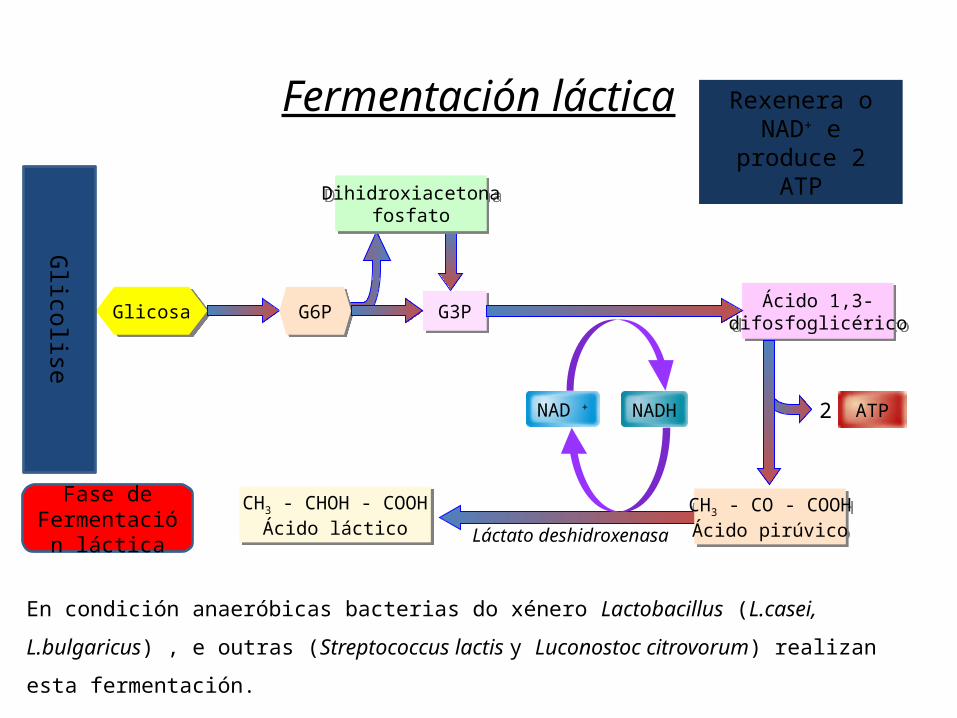
CH3 - CHOH - COOHÁcido láctico
Fermentación láctica
Glicosa
Dihidroxiacetonafosfato
ATP2NADHNAD +
G6P G3P Ácido 1,3-difosfoglicérico
CH3 - CO - COOHÁcido pirúvicoLáctato deshidroxenasa
Glicolise
Fase de Fermentació
n láctica
En condición anaeróbicas bacterias do xénero Lactobacillus (L.casei, L.bulgaricus) , e outras (Streptococcus lactis y Luconostoc citrovorum) realizan esta fermentación.
Rexenera o NAD+ e
produce 2 ATP

APLICACIÓNS DA FERMENTACIÓN LÁCTICA
Esta fermentación emprégase na industria para a fabricación do iogur, queixo, callada (=cuajada)...En tódolos casos as bacterias responsables desta fermentación atópanse no leite e o ácido láctico que producen acidifica o leite provocando a precipitación das súas proteínas, como a caseina e fórmase o callo (=cuajo) separandose do soro. Ambolos dous son a materia prima da que se obteñen os derivados lácteos. NOTA: prodúcese tamén fermentación láctica nos músculos cando non
hai aporte suficiente de O2. Isto débese a que ao facer un exercicio moi intenso, sobre todo con falta de entrenamento, ao cabo dun tempo a cantidade de glicosa oxidada é superior ao aporte de O2 que recibe o sangue e polo tanto o músculo, por iso o Pyr non é quen de completar a súa oxidación; nestas condicións anaeróbicas o NADH formado na glicolise oxídase a expensas da redución de Pyr a ácido láctico.
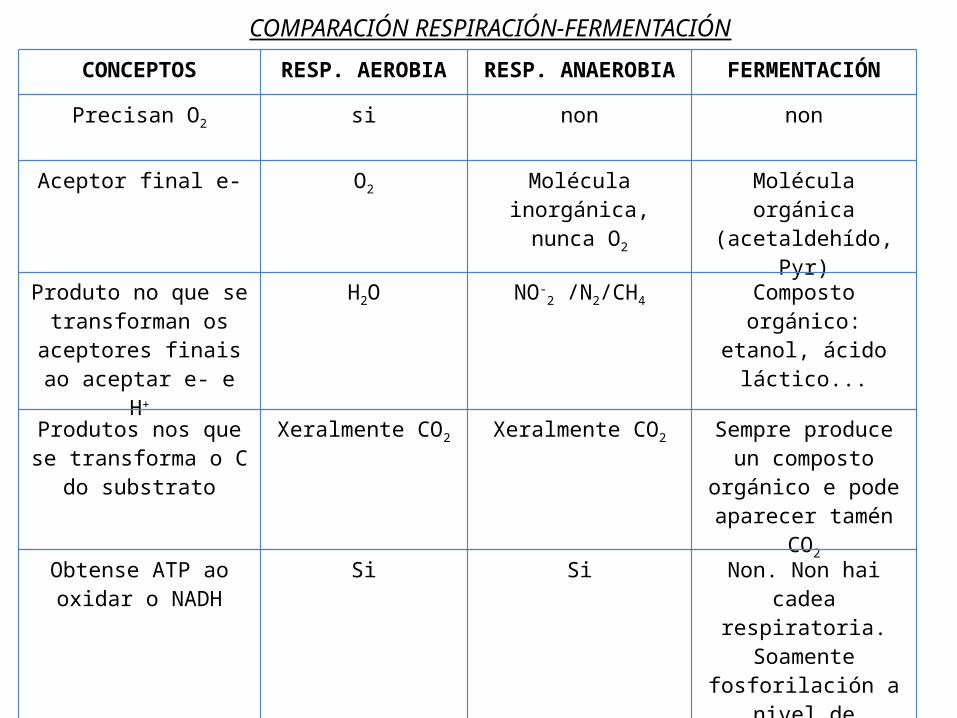
COMPARACIÓN RESPIRACIÓN-FERMENTACIÓNCONCEPTOS RESP. AEROBIA RESP.
ANAEROBIAFERMENTACIÓN
Precisan O2 si non non
Aceptor final e- O2 Molécula inorgánica, nunca
O2
Molécula orgánica (acetaldehído, Pyr)
Produto no que se transforman os
aceptores finais ao aceptar e- e H+
H2O NO-2 /N2/CH4 Composto
orgánico: etanol, ácido láctico...
Produtos nos que se transforma o C do
substrato
Xeralmente CO2 Xeralmente CO2 Sempre produce un composto
orgánico e pode aparecer tamén
CO2
Obtense ATP ao oxidar o NADH
Si Si Non. Non hai cadea respiratoria.
Soamente fosforilación a
nivel de substrato.Enerxía obtida 32 ATP 32 ATP Adoita ser 2 ATP

8. CATABOLISMO DOS LÍPIDOS
Os triglicéridos constitúen unha reserva enerxética de grande importancia debido ao seu alto valor enerxético. Como xa comentamos no tema adicado aos lípidos, as graxas proporcionan en media unhas 9,4 Kcal/g , mentres que os glícidos rinden só 4,3 Kcal/g. Aínda que tamén almacenamos glícidos (o glicóxeno no figado e músculos), o xeito máis habitual que ten o noso corpo de almacenar enerxía é en forma de graxas.A principal vía metabólica para a obtención de enerxía é a oxidación dos ácidos graxos que se obteñen basicamente da hidrólise dos triglicéridos (TAG).O catabolismo dos TAG consta de varias fases: Hidrólise dos TAG a glicerina e ácidos graxos por unha lipasa no citosol. A glicerina transfórmase en dihidroxiacetona fosfato (no citosol) que é
un intermediario da glicolise onde será degradada ou pode seguir unha vía anabólica e dar glicosa.
Os ácidos graxos activados sofren a β-oxidación nas mitocondrias.
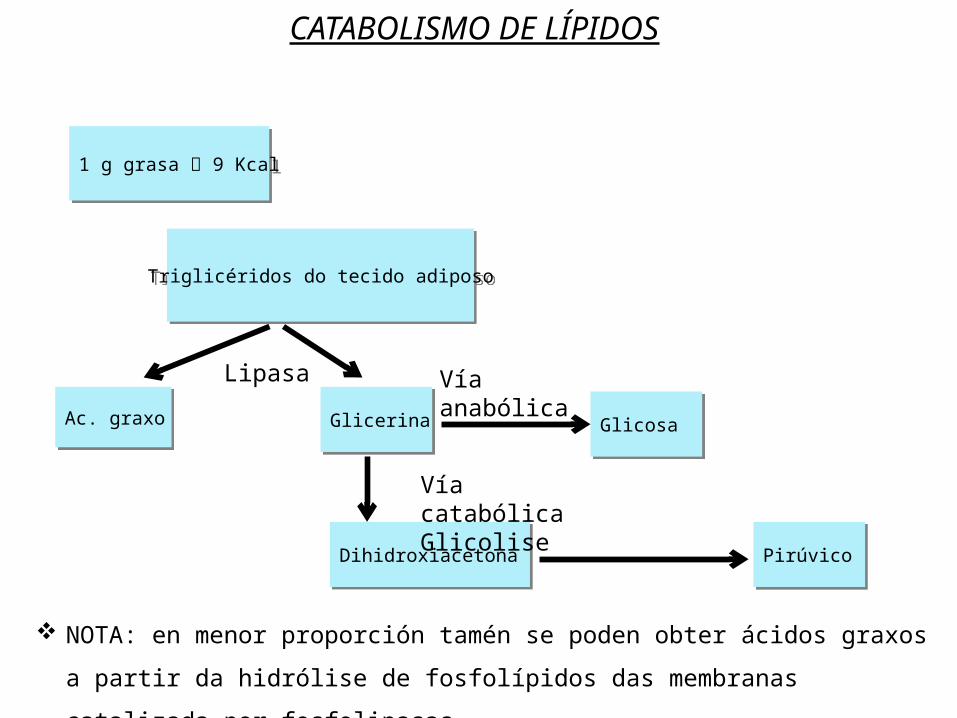
CATABOLISMO DE LÍPIDOS
Triglicéridos do tecido adiposo
1 g grasa 9 Kcal
Ac. graxo Glicerina Glicosa
Dihidroxiacetona Pirúvico
Lipasa Vía anabólica
Vía catabólicaGlicolise
NOTA: en menor proporción tamén se poden obter ácidos graxos a partir da hidrólise de fosfolípidos das membranas catalizada por fosfolipasas.
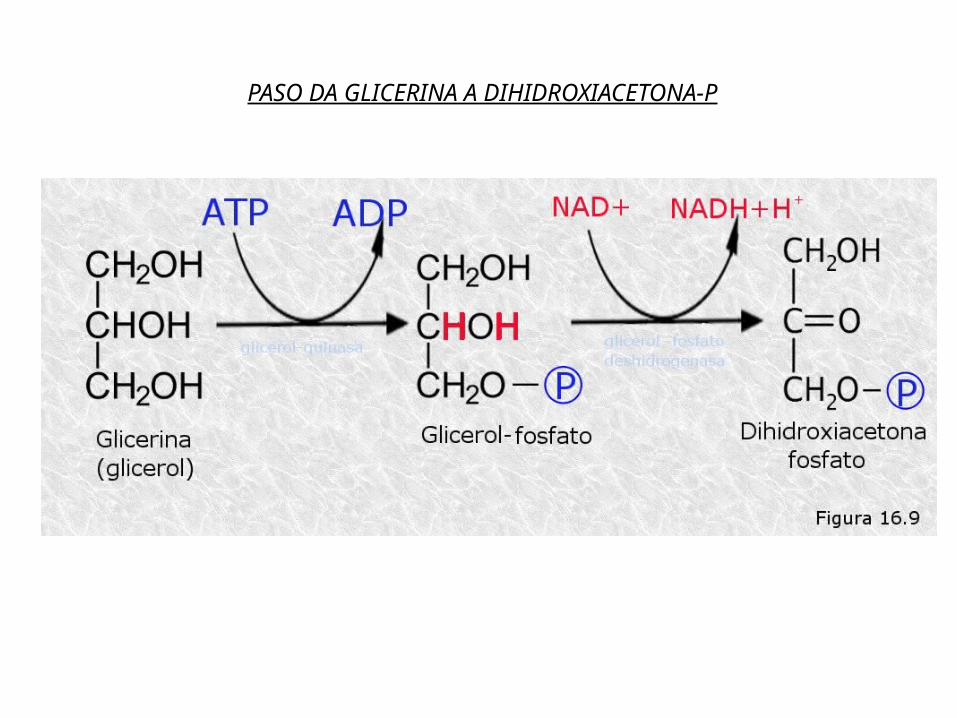
PASO DA GLICERINA A DIHIDROXIACETONA-P

Os ácidos graxos son transportados aos tecidos onde poden ser metabolizados (fígado, riles e tecido muscular aerobio). O proceso de degradación dos ácidos graxos pode dividirse nas seguintes fases:1. Activación do ác graxo: únese a CoA-SH formando un Acil-CoA ou Acil
graxo CoA (forma activada) no citosol. Precisa 1 ATP a AMP+PPi2. O Acil-CoA precisa unirse á carnitina e de proteínas transportadoras
para introducirse na mitocondria atravesando as membranas mitocondriais,
3. O Acil-Coa sofre na matriz mitocondrial un ciclo de catro reaccións denominado β-oxidación dos ácidos graxos ou hélice de Lynen que permite a súa degradación completa a Acetil-CoA (excepto nº impar de C) que ingresa no ciclo de Krebs.
NOTA: En cada volta do ciclo o ac graxo perde 2 C en forma de Acetil-CoA excepto en ac graxos cun nº impar de C que na última volta dará lugar a un resto de 3C (propionil-CoA) que é convertido en succinil-CoA e ingresa no Ciclo de Krebs.
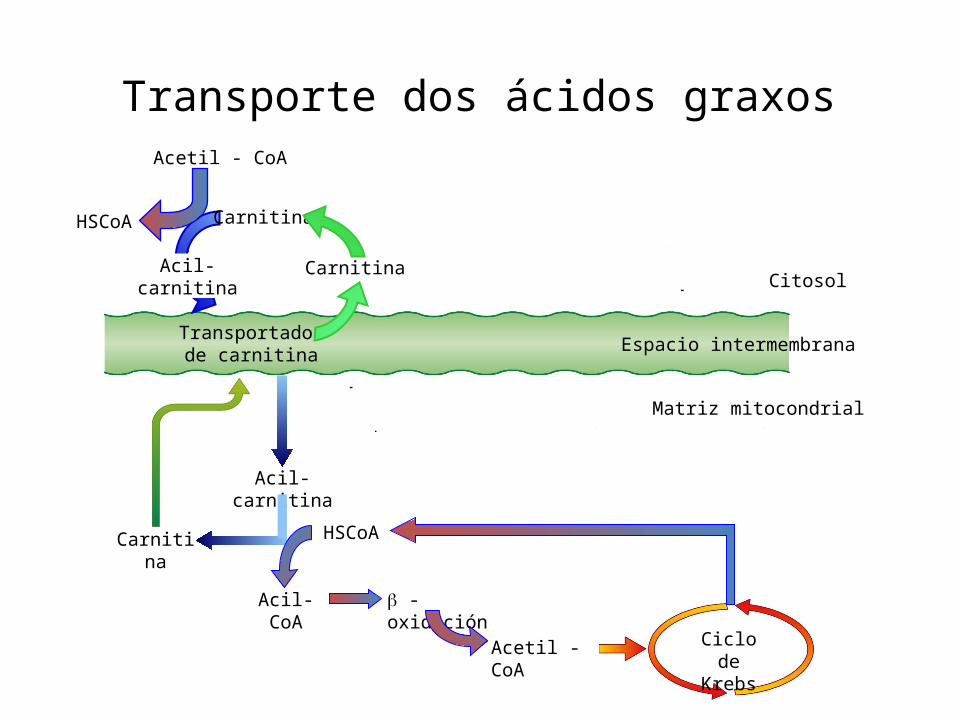
Transporte dos ácidos graxos
Ciclo de Krebs
Transportadorde carnitina
Acil-carnitina
Carnitina HSCoA
Acil-CoA - oxidación
Acetil - CoA
Acil-carnitina
Carnitina
Carnitina
Espacio intermembrana
Citosol
Matriz mitocondrial
Acetil - CoA
HSCoA
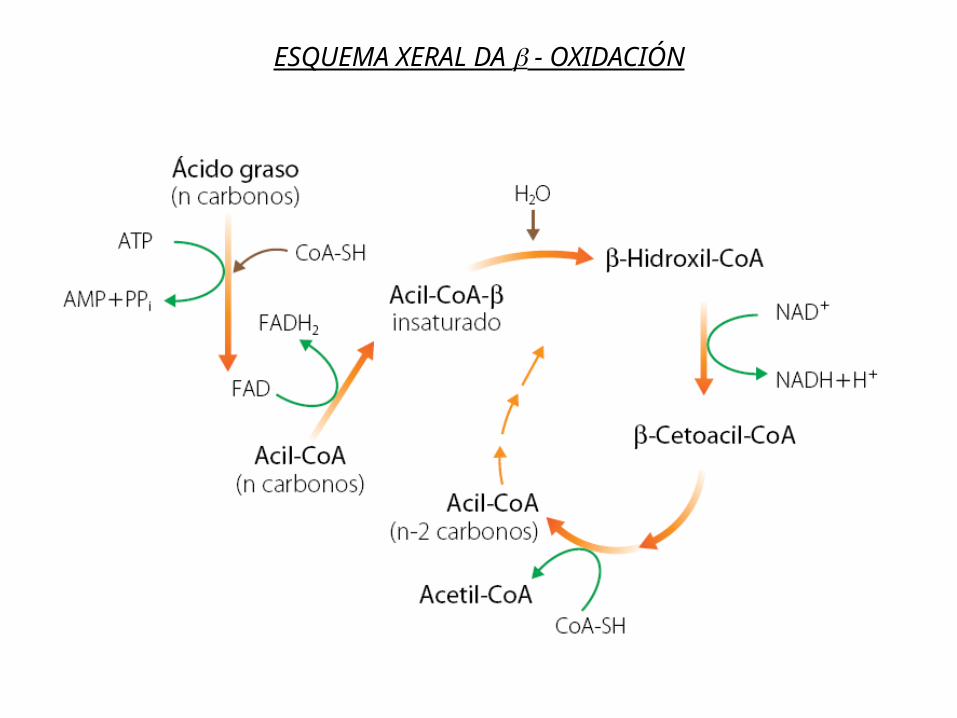
ESQUEMA XERAL DA - OXIDACIÓN
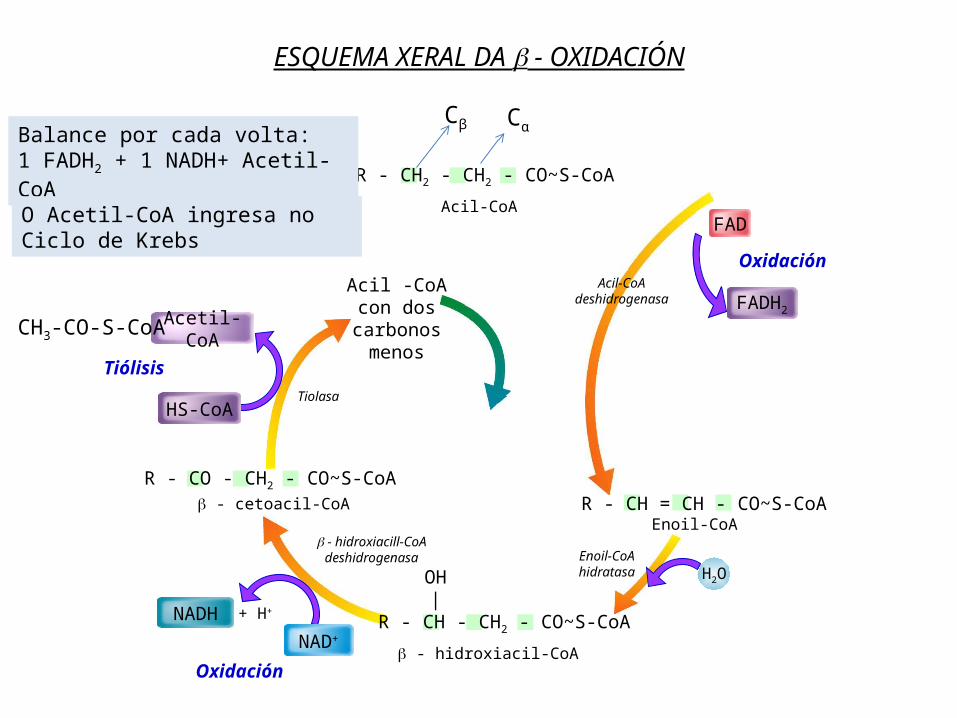
ESQUEMA XERAL DA - OXIDACIÓN
Acil -CoAcon dos
carbonosmenos
NADH + H+
NAD+
Oxidación
FADH2
FAD
Oxidación
R - CH2 - CH2 - CO~S-CoA
Acil-CoA
Acetil-CoA
HS-CoA
Tiólisis
R - CO - CH2 - CO~S-CoA - cetoacil-CoA
R - CH - CH2 - CO~S-CoA
OH|
- hidroxiacil-CoA
R - CH = CH - CO~S-CoAEnoil-CoA
- hidroxiacill-CoAdeshidrogenasa
Acil-CoAdeshidrogenasa
Tiolasa
Enoil-CoAhidratasa H2O
CαCβ
CH3-CO-S-CoA
Balance por cada volta: 1 FADH2 + 1 NADH+ Acetil-CoAO Acetil-CoA ingresa no Ciclo de Krebs

9. CATABOLISMO DE PROTEÍNAS
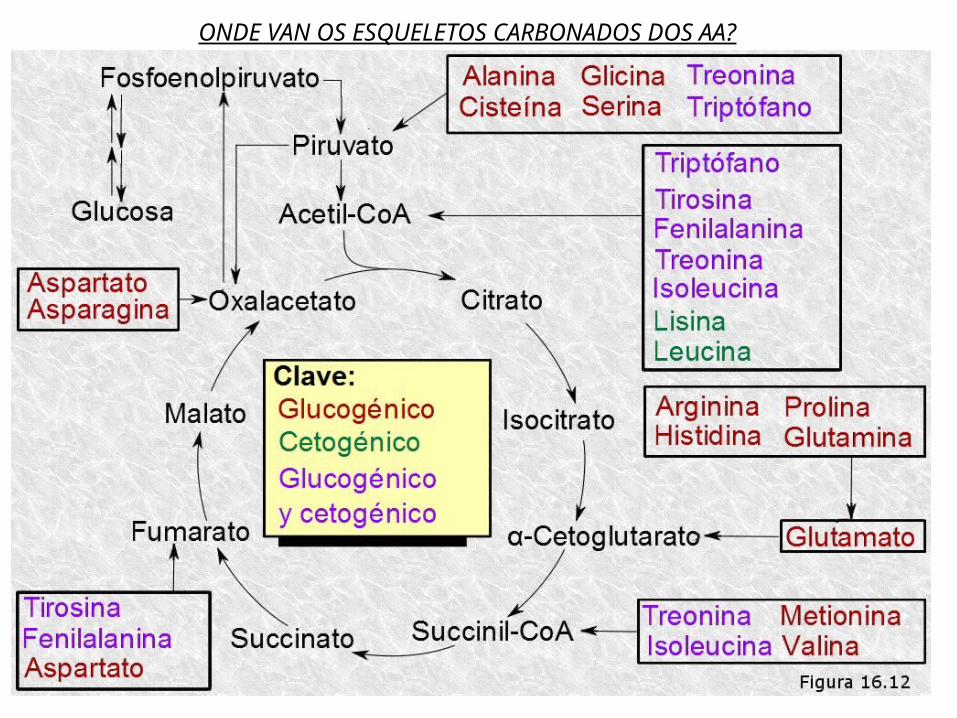
Os aa son empregado xeralmente para construir proteínas, pero cando se atopan en exceso poden ser utilizados excepcionalmente para obter enerxía separando os grupos aminos que se poden excretar.As proteínas son hidrolizadas a aa e éstos poden ser degradados en rutas oxidativas acopladas a síntese de ATP, aínda que non son tan bos combustibles como os lípidos e glícidos.Para os 20 aa proteicos existen 20 rutas multienzimáticas de oxidación, todas elas converxen nunhas poucas rutas terminais que conducen ao Pyr, ó Acetil-CoA ou a intermediarios do Ciclo de Krebs.En liñas xerais se pode falar de dous procesos fundamentais na degradación de aa: Transaminación Desaminación oxidativa

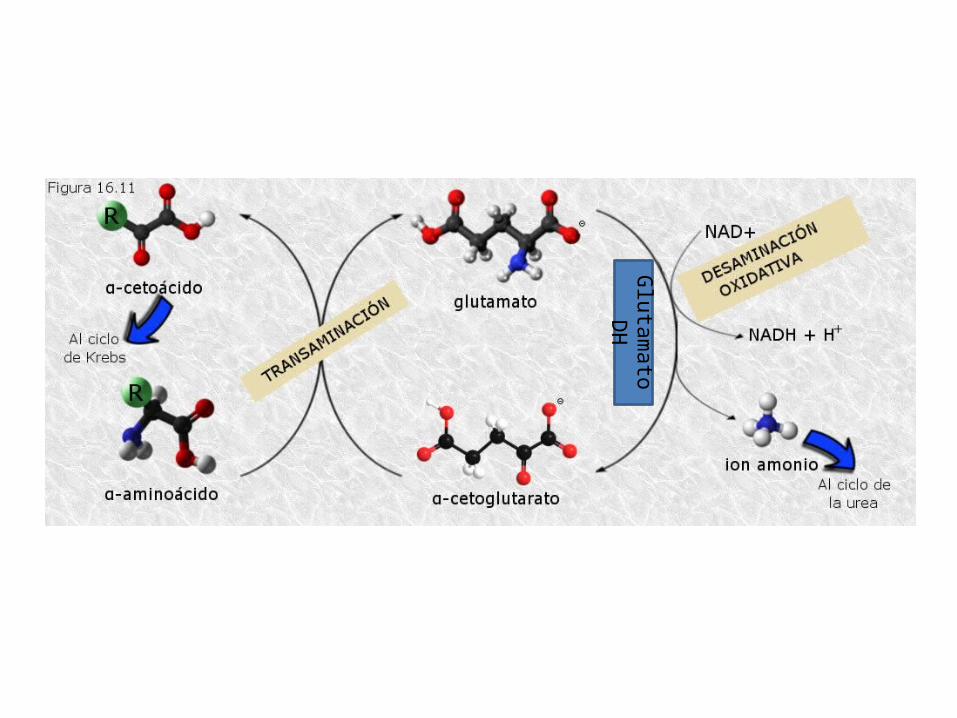
TRANSAMINACIÓN
Os aa, habitualmente, transfiren o seu grupo α-amino (-NH2) ao α-cetoglutarato ou ácido α-cetoglutárico que se transforma en ácido glutámico. Deste xeito todos os grupos amino dos distintos aa son recollidos en forma de ácido glutámico. Os aa ao ceder o grupo –NH2 quedan convertidos en α-cetoácidos (como o Pyr). O α-cetoácido ingresa no ciclo de Krebs. A reacción está catalizada por transaminasas. Sobre todo no fígado. DESAMINACIÓN OXIDATIVA
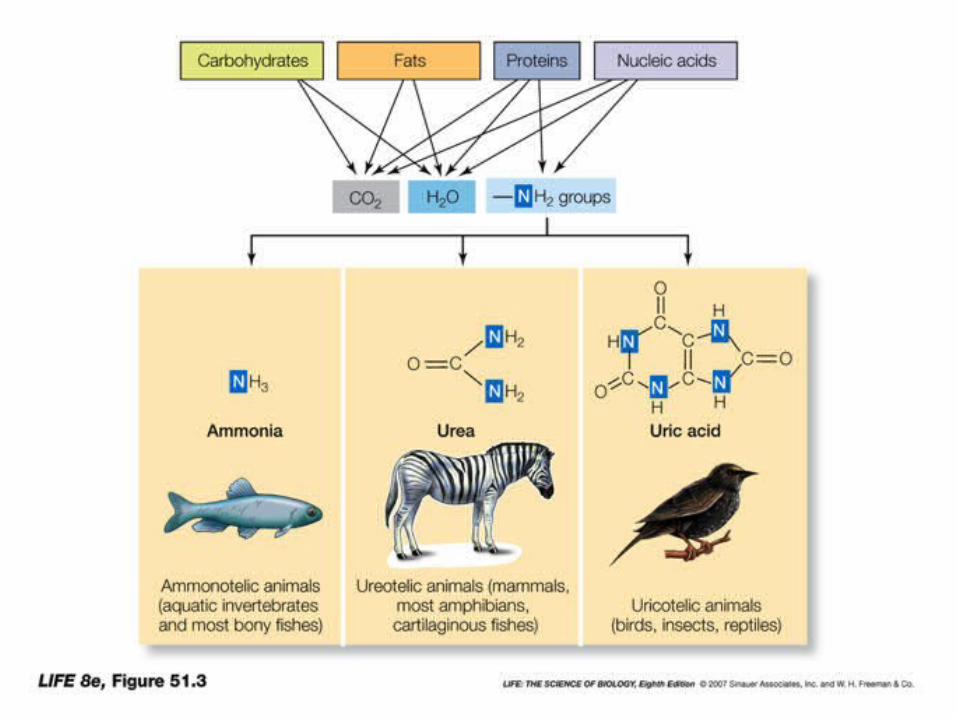
Ocorre no fígado e nos riles, o grupo amino sepárase do glutámico en forma de amonio. O amonio é moi tóxico e ten que ser excretado. Dependendo como se excrete os animais poden ser: Amoniotélicos: excretan amonio. Peixes óseos e invertebrados
mariños. Ureotélicos: expulsan amonio en forma de urea. Anfibios e
mamíferos. Uricotélicos: expulsan amonio en forma de ácido úrico. Réptiles,
moitos insectos e aves.
Glutamato DH
ONDE VAN OS ESQUELETOS CARBONADOS DOS AA?
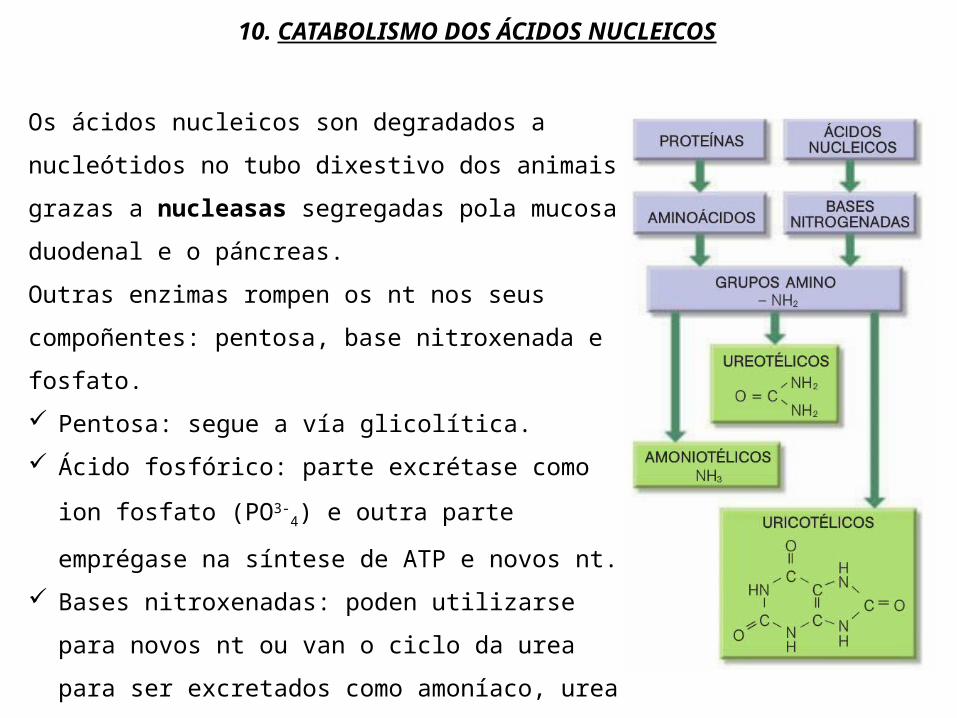
10. CATABOLISMO DOS ÁCIDOS NUCLEICOS
Os ácidos nucleicos son degradados a nucleótidos no tubo dixestivo dos animais grazas a nucleasas segregadas pola mucosa duodenal e o páncreas.Outras enzimas rompen os nt nos seus compoñentes: pentosa, base nitroxenada e fosfato. Pentosa: segue a vía glicolítica. Ácido fosfórico: parte excrétase como ion
fosfato (PO3-4) e outra parte emprégase na
síntese de ATP e novos nt. Bases nitroxenadas: poden utilizarse para
novos nt ou van o ciclo da urea para ser excretados como amoníaco, urea ou ácido úrico.
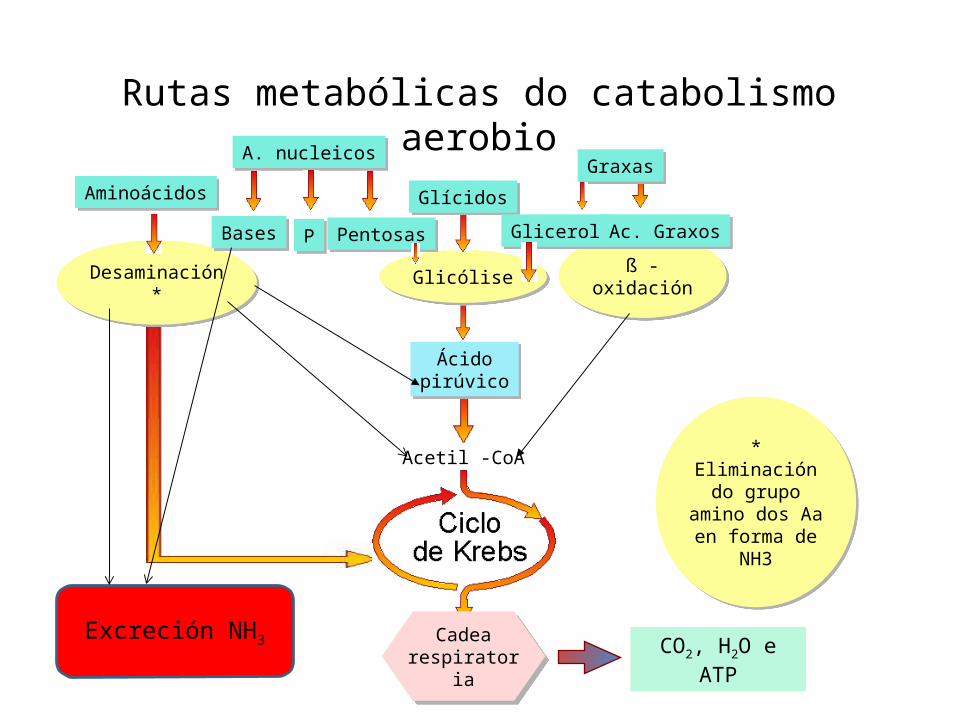
Rutas metabólicas do catabolismo aerobio
Cadearespiratoria
Desaminación * Glicólise ß -oxidación
Ácidopirúvico
Acetil -CoA
Aminoácidos Glícidos
Graxas
CO2, H2O e ATP
Glicerol Ac. Graxos
* Eliminación do grupo amino dos Aa en forma de
NH3
A. nucleicos
Bases P Pentosas
Excreción NH3
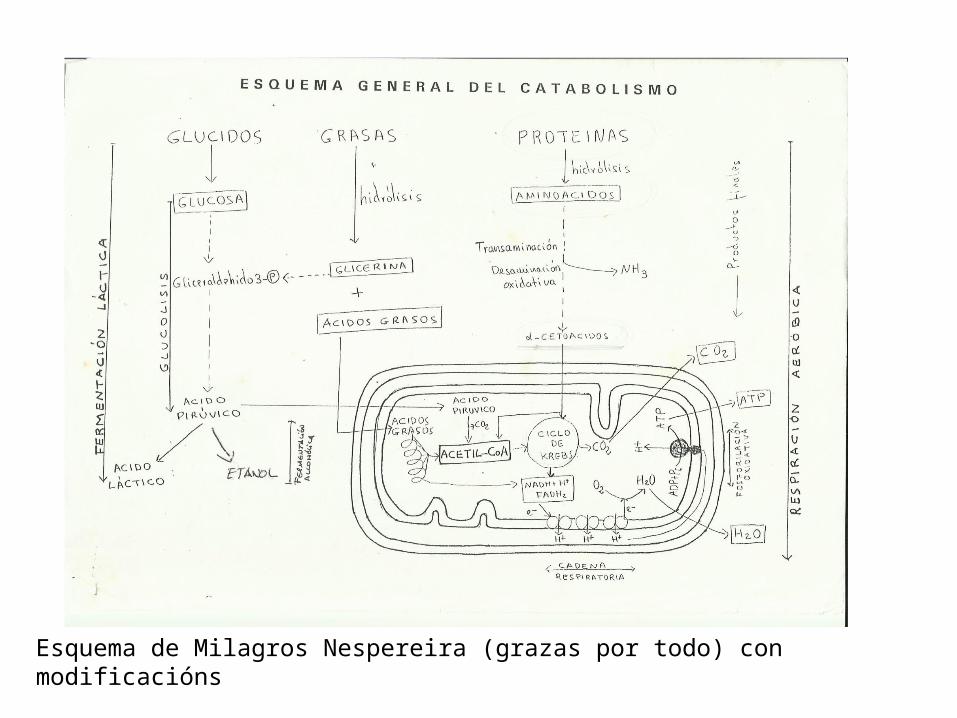
Esquema de Milagros Nespereira (grazas por todo) con modificacións

GRAZAS POR ATENDERME

WEBGRAFÍA https://biomedlabteziutlanletter.blogspot.com.es/2015/07/visualizacion-panoramica-
de-la.html
http://www.bionova.org.es/biocast/tema16.htm
http://
cienciasdejoseleg.blogspot.com.es/2013/08/diferentes-versiones-de-la-cadena-de.ht
ml
http://www.culturagalega.org/noticia.php?id=15702
https://mobile.twitter.com/hashtag/carrabouxo
https://es.slideshare.net/biologiahipatia/ud10-metabolismo-i


















