Marilys D. Milián Jiménez (1) T. González Arencibia ... morfologica.pdf · y Colocasia), ha sido...
44
Caracterización de la variabilidad…………. CARACTERIZACIÓN MORFOAGRONÓMICA DEL GERMOPLASMA DE Xanthosoma EN CUBA Marilys D. Milián Jiménez (1) , Xonia Xiqués Martín (2) , María I. Román Gutiérrez (1) , Clara T. González Arencibia (2) , Irelio Sánchez Ramos (1) , Margarita Nadal (2) , Yoel Beovides García (1) , Dablys Guerra Hernández (1) , Daviel Guerra Hernández (1) . (1) Instituto de Investigaciones en Viandas Tropicales (INIVIT), Apdo 6, Santo Domingo, Villa Clara Cuba. (2) Facultad de Biología, Universidad de La Habana, Cuba. Palabras clave: caracterización, variabilidad, germoplasma, Xanthosoma, descriptores. RESUMEN Se realizó la caracterización de la variabilidad del germoplasma del género Xanthosoma perteneciente al Banco de Germoplasma de Aráceas del Instituto de Investigaciones en Viandas Tropicales (INIVIT), ubicada en Santo Domingo, Villa Clara, Cuba, compuesto por 71 clones. De acuerdo a la evaluación morfoagronómica y con el empleo de análisis multivariados, se pudieron diferenciar los genotipos y formar grupos afines, así como determinar las variables más importantes para la caracterización, con las que quedó definido un listado de 13 descriptores mínimos.. Los estudios realizados permiten sugerir la presencia en la colección cubana de las especies Xanthosoma sagittifolium (L.) Schott, donde aparecen los clones de los grupos Blanco y Amarillo , Xanthosoma violaceum Schott, donde están incluidos los clones del grupo Morado , Xanthosoma brasilense Engl., cuyo único representante es el clon ‘Belembe’, Xanthosoma atrovirens Koch y Bouché, representada por el clon ‘Amarilla Trinidad’ y Alocasia macrorrhiza Schott, donde se incluye el clon ‘Picante’. I. INTRODUCCION La malanga, como se conoce en Cuba a las diferentes especies comestibles de Aráceas ( Xanthosoma y Colocasia), ha sido tradicionalmente un cultivo de subsistencia, esto explica su marginalización. Aunque es un producto principal para millones de personas en los trópicos, se tiene poca información disponible sobre su cultivo y requerimientos.
Transcript of Marilys D. Milián Jiménez (1) T. González Arencibia ... morfologica.pdf · y Colocasia), ha sido...

Caracterización de la variabilidad………….
CARACTERIZACIÓN MORFOAGRONÓMICA DEL GERMOPLASMA DE Xanthosoma EN CUBA
Marilys D. Milián Jiménez(1), Xonia Xiqués Martín(2), María I. Román Gutiérrez(1), Clara T. González Arencibia(2), Irelio Sánchez Ramos(1), Margarita Nadal(2), Yoel Beovides García(1), Dablys Guerra Hernández(1), Daviel Guerra Hernández (1). (1) Instituto de Investigaciones en Viandas Tropicales (INIVIT), Apdo 6, Santo Domingo,
Villa Clara Cuba. (2) Facultad de Biología, Universidad de La Habana, Cuba. Palabras clave: caracterización, variabilidad, germoplasma, Xanthosoma, descriptores. RESUMEN Se realizó la caracterización de la variabilidad del germoplasma del género Xanthosoma perteneciente al Banco de Germoplasma de Aráceas del Instituto de Investigaciones en Viandas Tropicales (INIVIT), ubicada en Santo Domingo, Villa Clara, Cuba, compuesto por 71 clones. De acuerdo a la evaluación morfoagronómica y con el empleo de análisis multivariados, se pudieron diferenciar los genotipos y formar grupos afines, así como determinar las variables más importantes para la caracterización, con las que quedó definido un listado de 13 descriptores mínimos.. Los estudios realizados permiten sugerir la presencia en la colección cubana de las especies Xanthosoma sagittifolium (L.) Schott, donde aparecen los clones de los grupos Blanco y Amarillo, Xanthosoma violaceum Schott, donde están incluidos los clones del grupo Morado, Xanthosoma brasilense Engl., cuyo único representante es el clon ‘Belembe’, Xanthosoma atrovirens Koch y Bouché, representada por el clon ‘Amarilla Trinidad’ y Alocasia macrorrhiza Schott, donde se incluye el clon ‘Picante’. I. INTRODUCCION
La malanga, como se conoce en Cuba a las diferentes especies comestibles de Aráceas (Xanthosoma y Colocasia), ha sido tradicionalmente un cultivo de subsistencia, esto explica su marginalización. Aunque es un producto principal para millones de personas en los trópicos, se tiene poca información disponible sobre su cultivo y requerimientos.

Caracterización de la variabilidad………….
2
Esta situación está cambiando con las áreas de consumo, especialmente en la costa Atlántica de las Estados Unidos, donde millones de Latinoamericanos consumen malanga y otros cultivos tropicales, un hecho que ha estimulado la producción comercial en las Antillas y en la América Central (Giacometti y León, 1994).
El género Xanthosoma es de origen americano desde México hasta Brasil, pero su cultivo se concentra en la zona del Caribe; en Puerto Rico constituye uno de los cultivos más antiguos heredados de los aborígenes Arawak (Barret, 1930; Montaldo, 1977).
La producción mundial de malanga Xanthosoma se estima en 4 000 000 t, las zonas de producción más importantes son la central y occidental de África Tropical, Las Antillas, Venezuela y Oceanía (Cuba, MINAGRI, 1998).
En Cuba, el género Xanthosoma es el de mayor importancia en la preferencia de la población con relación a otros de las aráceas comestibles. Su producción ha aumentado en los últimos años en detrimento de las áreas de Colocasia sobre todo porque esta última requiere un mayor gasto de agua. Los valores nutricionales y su fácil cocción unidas a sus cualidades digestivas, hacen de las especies del género Xanthosoma, un producto de alta demanda en el mercado nacional, así como, en la dieta de hospitales, hogares de ancianos y círculos infantiles, es por ello que el Ministerio de la Agricultura, plantea obtener un aumento significativo en la producción de este cultivo en los próximos años, con la finalidad de satisfacer las demandas crecientes del mismo (Cuba, MINAGRI, 1998).
Para lograr una recuperación de la producción de este renglón alimenticio es necesario fortalecer un aspecto importante como es el mejoramiento genético del cultivo, para lo cual es necesario contar con un banco de germoplasma que garantice la clasificación y conservación de los recursos fitogenéticos los cuales representan la fuente de variabilidad que facilitará los programas de mejoramiento. Los bancos de germoplasma constituyen una acumulación de material genético mediante la introducción de plantas de una zona, región o país, con el objetivo de enriquecer la fuente genética para trabajos de mejoramiento, ellos facilitan los procesos de cruzamiento, selección, evaluación y multiplicación de materiales, de manera integral y de gran beneficio para el desarrollo agrícola de un país (Gómez 1983).
El Plan de Acción Mundial para la Conservación y la Utilización Sostenible de los Recursos Fitogenéticos tiene entre las actividades prioritarias la conservación y mejoramiento in situ, la conservación ex situ y dentro de las actividades destinadas a mejorar la utilización de los recursos fitogenéticos, está el incremento de los estudios sobre caracterización, evaluación y la formación de colecciones núcleos, ya que son importantes para el ordenamiento eficaz y global de las colecciones y permiten a los usuarios tener una respuesta rápida a sus necesidades informativas (FAO, 1996b).
A nivel mundial se han realizado varios intentos de clasificación e identificación del germoplasma de malanga, uno de los primeros realizado en Cuba fue el desarrollado por Roig (1913) el que logró identificar diferentes clones de Xanthosoma y Colocasia. Varios autores hacen énfasis en que no está clara la posición taxonómica de las especies cultivadas de Xanthosoma, aunque se han reconocido cuatro: X. atrovirens Koch y Bouché , X. caracu Koch y Bouché, X. nigrum (X. violaceum Schott ) y X. sagittifolium Schott; Hay algunos cultivares que no son asignables a ninguna de éstas; sin embargo, casi todos los autores consultados (León, 1979; Gómez, 1983;

Caracterización de la variabilidad………….
3
León, 1987; Purseglove, 1988; Giacometti y León, 1994) coinciden en que para Xanthosoma es preferible hablar de clones del género y agrupar la mayoría en una especie polimórfica, Xanthosoma sagittifolium, en razón de las deficiencias de la clasificación existente, hasta que una revisión moderna del género, aclare la situación taxonómica de la especie mencionada.
Los recursos genéticos en Cuba reciben una especial atención y entre ellos se encuentran las raíces y tubérculos tropicales, por el significativo papel que desempeñan en la alimentación de la población (Milián et al., 1993b).
En el Instituto de Investigaciones en Viandas Tropicales (INIVIT) se mantiene desde 1967, una colección de clones de malanga del género Xanthosoma procedentes de prospecciones, de introducciones y del programa de mejoramiento genético del cultivo. En esta colección se han desarrollado estudios morfológicos, que han permitido la caracterización y evaluación preliminar del germoplasma (Milián et al., 1993a; Milián et al., 1993b). Estos resultados han demostrado que existen grandes diferencias entre clones de una misma especie y similitud entre clones de especies diferentes (Milián et al., 1998), por lo que deben ser ampliados y complementados con el empleo de análisis citogenéticos y genético-bioquímicos que ayuden a dilucidar la situación taxonómica del género. Por otra parte resulta necesario mejorar la estructura de clones en este cultivo y seleccionar cultivares más rústicos y resistentes al ataque de plagas y enfermedades. En este contexto, es de gran importancia el estudio detallado de los recursos fitogenéticos del género Xanthosoma disponibles en el país.
Teniendo en cuenta los antecedentes y las recomendaciones de otros autores, el presente trabajo se ha trazado el siguiente objetivo:
♦ Seleccionar los caracteres morfoagronómicos más importantes para la evaluación de los clones.
II. MATERIALES Y MÉTODOS
II.1. MATERIAL VEGETAL
El trabajo se desarrolló en el Banco de Germoplasma de Aráceas del Instituto de Investigaciones en Viandas Tropicales (INIVIT), ubicado en el municipio de Santo Domingo, provincia de Villa Clara, Cuba.
La colección se plantó, de acuerdo a un Diseño Completamente Aleatorizado, en el mes de abril, durante los años 1994 a 1999 sobre un suelo Pardo sialítico con carbonatos (Cuba, MINAG, 1995; Hernández et al., 1995) y se aplicaron las normas de cultivo recomendadas en el Instructivo Técnico para la Malanga (Ministerio de la Agricultura, 1984).
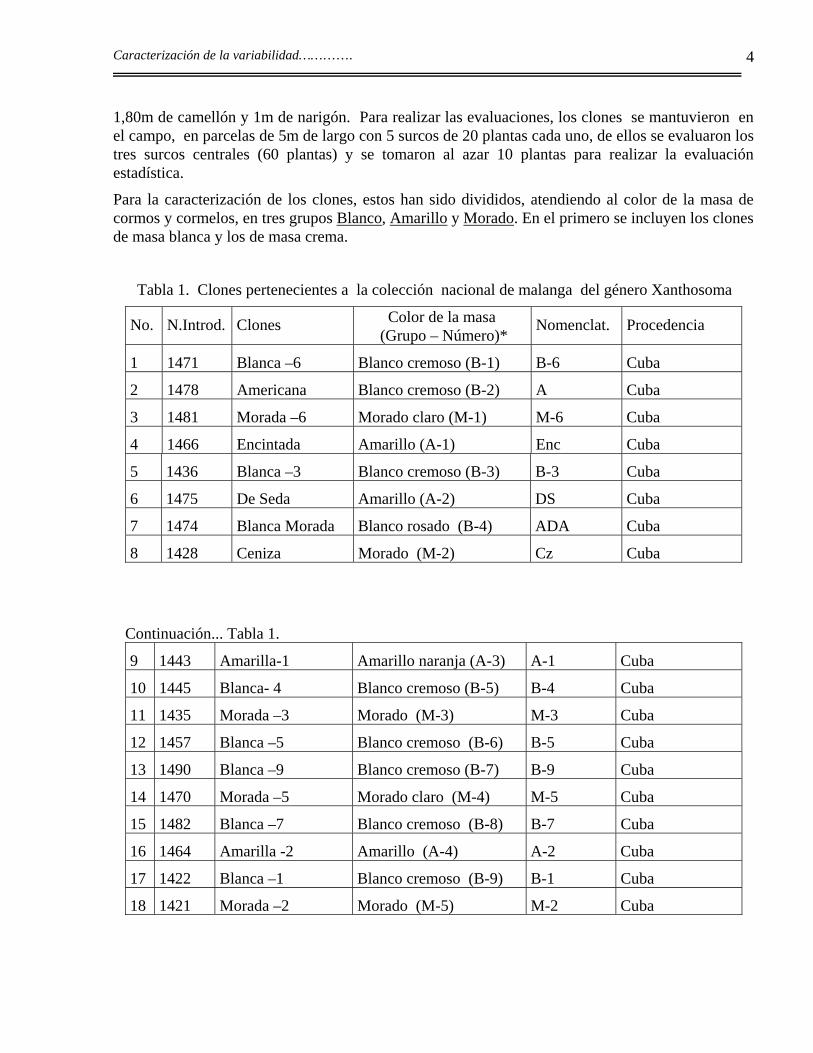
Se analizaron 71 clones pertenecientes a la colección nacional de malanga del género Xanthosoma (Tabla 1). Los clones se conservan ex situ: en condiciones de campo e in vitro, 56 de ellos son cubanos y 15 corresponden a introducciones foráneas (Milián et al., 1993b).
Como material de reproducción se utilizaron fracciones de cormos en el área objeto de evaluación, los cuales se plantaron a una distancia de 0,90m x 0.35m. Las parcelas estuvieron separadas a
Caracterización de la variabilidad………….
4
1,80m de camellón y 1m de narigón. Para realizar las evaluaciones, los clones se mantuvieron en el campo, en parcelas de 5m de largo con 5 surcos de 20 plantas cada uno, de ellos se evaluaron los tres surcos centrales (60 plantas) y se tomaron al azar 10 plantas para realizar la evaluación estadística.
Para la caracterización de los clones, estos han sido divididos, atendiendo al color de la masa de cormos y cormelos, en tres grupos Blanco, Amarillo y Morado. En el primero se incluyen los clones de masa blanca y los de masa crema.
Tabla 1. Clones pertenecientes a la colección nacional de malanga del género Xanthosoma
No. N.Introd. Clones Color de la masa (Grupo – Número)*
Nomenclat. Procedencia
1 1471 Blanca –6 Blanco cremoso (B-1) B-6 Cuba
2 1478 Americana Blanco cremoso (B-2) A Cuba
3 1481 Morada –6 Morado claro (M-1) M-6 Cuba
4 1466 Encintada Amarillo (A-1) Enc Cuba
5 1436 Blanca –3 Blanco cremoso (B-3) B-3 Cuba
6 1475 De Seda Amarillo (A-2) DS Cuba
7 1474 Blanca Morada Blanco rosado (B-4) ADA Cuba
8 1428 Ceniza Morado (M-2) Cz Cuba
Continuación... Tabla 1.
9 1443 Amarilla-1 Amarillo naranja (A-3) A-1 Cuba
10 1445 Blanca- 4 Blanco cremoso (B-5) B-4 Cuba
11 1435 Morada –3 Morado (M-3) M-3 Cuba
12 1457 Blanca –5 Blanco cremoso (B-6) B-5 Cuba
13 1490 Blanca –9 Blanco cremoso (B-7) B-9 Cuba
14 1470 Morada –5 Morado claro (M-4) M-5 Cuba
15 1482 Blanca –7 Blanco cremoso (B-8) B-7 Cuba
16 1464 Amarilla -2 Amarillo (A-4) A-2 Cuba
17 1422 Blanca –1 Blanco cremoso (B-9) B-1 Cuba
18 1421 Morada –2 Morado (M-5) M-2 Cuba

Caracterización de la variabilidad………….
5
19 1427 Tricolor Blanco cremoso (B-10) T Cuba
20 1460 Morada Ceniza Blanco cremoso (B-11) MCz Cuba
21 1437 Picante Blanco cremoso (B-12) P Cuba
22 1480 Chopo Amarillo Amarillo (A-5) ChA Cuba
23 1450 Morada –4 Morado (M-6) M-4 Cuba
24 1446 Sergio Cuarentena Blanco rosado (M-7) SC Cuba
25 1486 Blanca –8 Blanco (B-13) B-8 Cuba
26 1485 Morada –7 Morado claro (B-8) M-7 Cuba
27 1244 Stoupan Blanco (B-14) St Guadaloupe
28 1242 Amarilla Criolla-1 Blanco rosado (A-1) AC-1 Cuba
29 1243 Amarilla Amarillo (A-7) A Cuba
30 1255 Amarilla Ceniza Amarillo naranja (A-8) ACz Cuba
31 1238 Amarilla Especial Amarillo naranja (A-9) AE Cuba
32 1232 Amarilla Criolla Amarillo naranja(A-10) AC Cuba
33 1261 Amarilla Especial-4
Amarillo (A-11) AE-4 Cuba
34 1245 Belembe Amarillo claro (A-12) B Guadaloupe
35 1239 Javánica Blanco cremoso (B-15) J Cuba
36 1229 Amarilla Trinidad Amarillo (A-13) AT Trin. Tobago
Continuación... Tabla 1.
37 1249 Morada1727 Rosado (M-9) M1727 Cuba
38 1250 México-1 Blanco rosado (M-10) Mj-1 México
39 1251 México-27 Morado claro (M-11) Mj -27 México
40 1252 México-16 Morado claro (M-12) Mj -16 México
41 1236 Jardín Morado claro (M-13) Jd Cuba
42 1253 México-2 Morado claro (M-14) Mj-2 México
43 1230 Riza Blanco (B-16) R Cuba
44 1262 Blanca Baracoa Blanco (B-17) BB Cuba
45 1265 Cuarentena Morado claro (M-15) Cuarent Saot. y Príncipe
46 1231 Blanca Blanco (B-18) Bca Cuba
Caracterización de la variabilidad………….
6
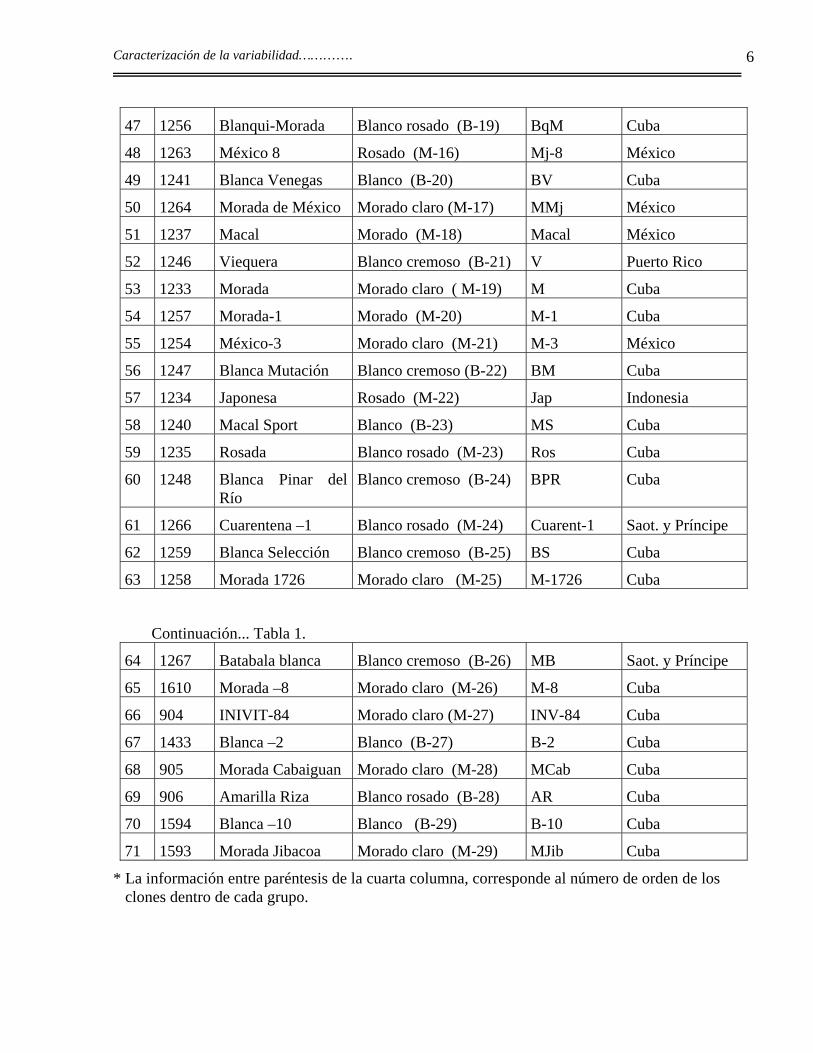
47 1256 Blanqui-Morada Blanco rosado (B-19) BqM Cuba
48 1263 México 8 Rosado (M-16) Mj-8 México
49 1241 Blanca Venegas Blanco (B-20) BV Cuba
50 1264 Morada de México Morado claro (M-17) MMj México
51 1237 Macal Morado (M-18) Macal México
52 1246 Viequera Blanco cremoso (B-21) V Puerto Rico
53 1233 Morada Morado claro ( M-19) M Cuba
54 1257 Morada-1 Morado (M-20) M-1 Cuba
55 1254 México-3 Morado claro (M-21) M-3 México
56 1247 Blanca Mutación Blanco cremoso (B-22) BM Cuba
57 1234 Japonesa Rosado (M-22) Jap Indonesia
58 1240 Macal Sport Blanco (B-23) MS Cuba
59 1235 Rosada Blanco rosado (M-23) Ros Cuba
60 1248 Blanca Pinar del Río
Blanco cremoso (B-24) BPR Cuba
61 1266 Cuarentena –1 Blanco rosado (M-24) Cuarent-1 Saot. y Príncipe
62 1259 Blanca Selección Blanco cremoso (B-25) BS Cuba
63 1258 Morada 1726 Morado claro (M-25) M-1726 Cuba
Continuación... Tabla 1.
64 1267 Batabala blanca Blanco cremoso (B-26) MB Saot. y Príncipe
65 1610 Morada –8 Morado claro (M-26) M-8 Cuba
66 904 INIVIT-84 Morado claro (M-27) INV-84 Cuba
67 1433 Blanca –2 Blanco (B-27) B-2 Cuba
68 905 Morada Cabaiguan Morado claro (M-28) MCab Cuba
69 906 Amarilla Riza Blanco rosado (B-28) AR Cuba
70 1594 Blanca –10 Blanco (B-29) B-10 Cuba
71 1593 Morada Jibacoa Morado claro (M-29) MJib Cuba
* La información entre paréntesis de la cuarta columna, corresponde al número de orden de los clones dentro de cada grupo.
Caracterización de la variabilidad………….
7
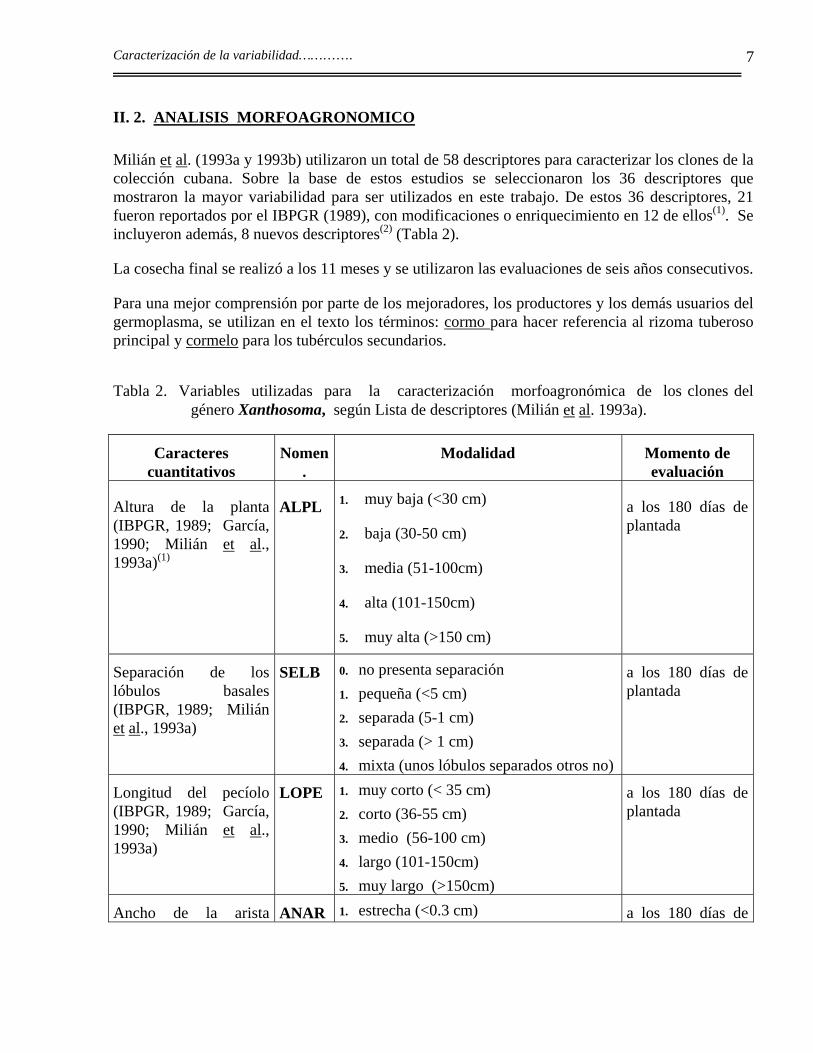
II. 2. ANALISIS MORFOAGRONOMICO
Milián et al. (1993a y 1993b) utilizaron un total de 58 descriptores para caracterizar los clones de la colección cubana. Sobre la base de estos estudios se seleccionaron los 36 descriptores que mostraron la mayor variabilidad para ser utilizados en este trabajo. De estos 36 descriptores, 21 fueron reportados por el IBPGR (1989), con modificaciones o enriquecimiento en 12 de ellos(1). Se incluyeron además, 8 nuevos descriptores(2) (Tabla 2).
La cosecha final se realizó a los 11 meses y se utilizaron las evaluaciones de seis años consecutivos.
Para una mejor comprensión por parte de los mejoradores, los productores y los demás usuarios del germoplasma, se utilizan en el texto los términos: cormo para hacer referencia al rizoma tuberoso principal y cormelo para los tubérculos secundarios.
Tabla 2. Variables utilizadas para la caracterización morfoagronómica de los clones del género Xanthosoma, según Lista de descriptores (Milián et al. 1993a).
Caracteres cuantitativos
Nomen.
Modalidad Momento de evaluación
Altura de la planta (IBPGR, 1989; García, 1990; Milián et al., 1993a)(1)
ALPL 1. muy baja (<30 cm)
2. baja (30-50 cm)
3. media (51-100cm)
4. alta (101-150cm)
5. muy alta (>150 cm)
a los 180 días de plantada
Separación de los lóbulos basales (IBPGR, 1989; Milián et al., 1993a)
SELB 0. no presenta separación 1. pequeña (<5 cm) 2. separada (5-1 cm) 3. separada (> 1 cm) 4. mixta (unos lóbulos separados otros no)
a los 180 días de plantada
Longitud del pecíolo (IBPGR, 1989; García, 1990; Milián et al., 1993a)
LOPE 1. muy corto (< 35 cm) 2. corto (36-55 cm) 3. medio (56-100 cm) 4. largo (101-150cm) 5. muy largo (>150cm)
a los 180 días de plantada
Ancho de la arista ANAR 1. estrecha (<0.3 cm) a los 180 días de
Caracterización de la variabilidad………….
8
(García, 1990; Milián et al., 1993a)(2) 2. estrecha media (0.3-1cm)
3. ancha (>1cm)
plantada
Continuación... Tabla 2.
Ancho de la hoja (IBPGR, 1989; García, 1990; Milián et al., 1993a)
ANHO 1. muy estrecha (<15cm) 2. estrecha (15-20cm) 3. media (20-30cm) 4. ancha (30-40cm) 5. muy ancha (>40cm)
a los 180 días de plantada
Longitud de la hoja (IBPGR, 1989; García, 1990; Milián et al., 1993a)
LOHO 1. muy corta (<50cm) 2. corta (15-30cm) 3. media (31-40cm) 4. larga (41-50cm) 5. muy larga (>50cm)
a los 180 días de plantada
Número de cormos/ planta (IBPGR, 1989; García, 1990; Milián et al., 1993a)
NRCO 1. bajo (<2) 2. medio (2-4) 3. alto (>4)
a los 360 días de plantada
Número de cormelos/ planta (IBPGR, 1989; García, 1990; Milián et al., 1993a)
NCOR 1. bajo (<4) 2. medio (4-8) 3. alto (>8)
a los 360 días de plantada
Distancia internerval de la hoja (García, 1990; Milián et al., 1993a)(2)
DIIN 1. corta (<2 cm) 2. media (2-4 cm) 3. larga (>4 cm)
a los 180 días de plantada
Relación Largo/Ancho de la hoja (García, 1990; Milián et al., 1993a) (2)
RLAH 1. baja (<0.6)
2. media (0.6-1)
3. alta (>1)
a los 180 días de plantada
Relación longitud de la hoja/ longitud del pecíolo (García, 1990; Milián et al., 1993a) (2)
RLHP 1. baja (<0.3)
2. media (0.3-0.48)
3. alta (> 0.48)
a los 180 días de plantada
Relación longitud del pecíolo/ Ancho de la
LPHA 1. baja (<1.8) a los 180 días de plantada

Caracterización de la variabilidad………….
9
hoja (García, 1990; Milián et al., 1993a) (2)
2. media (1.8-2.60)
3. alta (>2.6)
Continuación... Tabla 2.
Número de hijos (IBPGR, 1989; García, 1990; Milián et al., 1993a) (1)
NROH 1. ninguno (0)
2. poco (1-2)
3. medio (3-5)
4. alto (6-10)
5. muy alto (>10)
a los 360 días de plantada
Porcentaje de Materia Seca en el follaje (Milián et al., 1993a) (2)
PMSF 1. bajo (<6%)
2. medio (6-10%)
3. alto (>10%)
a los 360 días de plantada.
Porcentaje de Materia Seca en los cormos (García, 1990; Milián et al., 1993a) (1)
PMSC 1. bajo (<12%)
2. medio (12-18%)
3. alto (>18%)
a los 360 días de plantada.
Porcentaje de Materia Seca en los cormelos (García, 1990; Milián et al., 1993a) (1)
PMSL 1. bajo (<15%)
2. medio (15-25%)
3. alto (>25%)
a los 360 días de plantada.
Contenido de Almidón IBPGR, 1989; Milián et al., 1993a) (1)
COAL 1. bajo (<15%)
2. medio (15-25%)
3. alto (>25%)
a los 360 días de plantada.
Continuación... Tabla 2.
Caracteres cualitativos Nomen. Modalidad Momento de evaluación

Caracterización de la variabilidad………….
10
Número de cromosomas NRCR 1. 26 cromosomas
2. 24 cromosomas
Raíces en metafase mitótica
Continuación... Tabla 2.
Hábito de crecimiento (IBPGR, 1989; García, 1990; Milián et al., 1993a) (1)
HACR 1. acaulescente cerrado
2. acaulescente semicerrado
3. acaulescente abierto
4. erecto superficial cerrado
5. erecto superficial semicerrado
6. erecto superficial abierto
7. Reclinado superficial cerrado
8. Reclinado superficial semicerrado
9. Reclinado superficial abierto
a los 180 días de plantada
Inserción del pecíolo en la lámina de la hoja (IBPGR, 1989; Milián et al., 1993a) (1)
IPLH 1. peltada
2. subpeltada
3. no peltada o sagitada
a los 180 días de plantada
Orientación de la lámina (IBPGR, 1989; Milián et al., 1993a) (1)
ORLM 1. erecta
2. inclinada
3. horizontal
a los 180 días de plantada
Margen de la hoja (IBPGR, 1989; Milián et al., 1993a)
MAHO 1. entero liso 3. lobulado
2. entero ondulado 4. dividido
a los 180 días de plantada
Forma de la hoja (IBPGR, 1989; Milián et al., 1993a)
FOHO sin lóbulo basal
hastada
sagitada
a los 180 días de plantada
Color de las nervaduras por el haz (IBPGR, 1989; Milián et al., 1993a) (1)
CNHA 1. verde 5.morado
2. verde claro 6.morado verdoso
3. verde oscuro 7.morado oscuro
a los 180 días de plantada

Caracterización de la variabilidad………….
11
4. verde morado 8.policromado
9.otro
Continuación... Tabla 2.
Color de las nervaduras por el envés (IBPGR, 1989; Milián et al., 1993a) (1)
CNEN 1. verde 5.morado
2. verde claro 6.morado verdoso
3. verde oscuro 7.morado oscuro
4. verde morado 8.policromado
9.otro
a los 180 días de plantada
Color del pecíolo (IBPGR, 1989; García, 1990; Milián et al., 1993a)
COPE 1. verde 5. morado
2. verde claro 6. morado verdoso
3. verde oscuro 7. morado oscuro
4. verde morado 8. policromado
a los 180 días de plantada
Color de la parte basal pecíolo (IBPGR, 1989; García, 1990; Milián et al., 1993a) (1)
CPBP 1. blanco 5. rosado
2. blanco verdoso 6. amarillo rosáceo
3. verde 7. amarillo verdoso
4. verde claro 8. rojo
9. morado
a los 180 días de plantada
Color de la arista del pecíolo (García, 1990; Milián et al., 1993a) (1)
COAP 1. verde 3. morado
2. rosado 4. morado oscuro
a los 180 días de plantada
Presencia de cera en el pecíolo (García, 1990; Milián et al., 1993a) (2)
PCPE 1. ninguna
2. escasa
3. media
4. abundante
a los 180 días de plantada
Color de la hoja por el haz (IBPGR, 1989;
COHO 1. verde a los 180 días de

Caracterización de la variabilidad………….
12
García, 1990; et al., 1993a)
2. verde claro
3. verde oscuro
4. verde oscuro c/borde morado
plantada
Continuación... Tabla 2.
Color de la epidermis de cormos y cormelos (García, 1990; Milián et al., 1993a)
CECC 1. castaño claro
2. castaño oscuro
a los 360 días de plantada
Color de las yemas de cormos y cormelos (García, 1990; Milián et al., 1993a) (2)
CYCC 1. blanco 5. amarillo rojizo
2. blanco oscuro 6. rosado
3. blanco amarillento7. rosado oscuro
4. amarillo 8. rojo
9. morado
a los 360 días de plantada.
Color de la masa de cormos y cormelos (IBPGR, 1989; García, 1990; Milián et al., 1993a)
CMCC 0. blanco 5. amarillo naranja
1. blanco cremoso 6. rosado
2. blanco rosado 7. rojo claro
3. amarillo 8 morado
4. amarillo claro 9 morado claro
a los 360 días de plantada.
Forma de los cormos y cormelos (IBPGR, 1989; García, 1990; Milián et al., 1993a)
FRCC 1. cónico
2. cilíndrico
3. clíptico
a los 360 días de plantada.
Floración (García, 1990; Milián et al., 1993a)
FLOR 1. ninguna
2. baja (< 5% de plantas florecidas)
3. media (entre 5- 15% plantas florecidas)
4. alta (entre 15-20% plantas florecidas)
Después de los seis meses

Caracterización de la variabilidad………….
13
5. Muy alta (> 30% plantas florecidas)
Porción masculina de la inflorescencia (García, 1990; Milián et al., 1993a)
POMI 1. no hay flores
2. encerrada
3. expuesta
Después de los seis meses
Se calculó la frecuencia absoluta de valores de cada carácter los cuales fueron expresados en porcentaje (%).
Con los 36 descriptores señalados se realizó un análisis de Componentes Principales a partir de una matríz de correlaciones de Spearman entre las variables cualitativas y cuantitativas (Linares, 1988), empleando el sistema Statistica, versión 5. Para seleccionar las correlaciones significativas se tuvo en cuenta la probabilidad de p<0.01 (Sigarroa, 1985).
El listado de descriptores mínimos se obtuvo seleccionando en el análisis general de todos los clones, aquellas variables originales que tuvieron mayor peso en los componentes C1, C2 y C3, teniendo en cuenta el siguiente criterio: se seleccionaron los autovectores mayor y menor, y se utilizó el promedio de esos valores como umbral a partir del cual efectuar la selección de las variables de mayor contribución. También se tomaron en cuenta las correlaciones de las variables con los respectivos ejes y sus coeficientes de determinación en el mismo (Fundora et al., 1992). Los descriptores mínimos seleccionados para continuar el estudio de la colección son los siguientes:
• Separación de los lóbulos basales de las hojas • Longitud del pecíolo • Número de hijos • Contenido de materia seca en los cormos • Contenido de materia seca en los cormelos • Color del pecíolo • Color de la base del pecíolo • Color de la arista del pecíolo • Presencia de cera en el pecíolo • Color de las hojas • Color de las yemas de los cormos y los cormelos • Color de la masa o pulpa de los cormos y los cormelos • Número de cromosomas
Los 71 clones estudiados fueron agrupados de acuerdo a un análisis de Conglomerados (cluster analysis) a partir de una matriz de distancia Euclidiana (Daviers, 1973).
Para cada uno de los grupos de clones formados de acuerdo al color de la masa de los cormos y los cormelos y después en conjunto, se realizaron análisis de Componentes Principales (Linares, 1988), con el empleo de los 13 descriptores mínimos. De acuerdo a la contribución de los componentes se hicieron los agrupamientos teniendo en cuenta los caracteres que más aportaron a la variabilidad en
Caracterización de la variabilidad………….
14
la matriz de valores y vectores propios para los componentes C1, C2 y las correlaciones de Spearman significativas entre las variables. Para facilitar el análisis de Componentes Principales los clones fueron identificados por números según el grupo (Tabla 1).
Para verificar el agrupamiento de los 71 clones, realizado a partir del análisis de Componentes Principales, se utilizó un análisis Factorial Discriminante (Linares, 1988).
III. RESULTADOS Y DISCUSIÓN
III.1. ANÁLISIS MORFOAGRONÓMICO
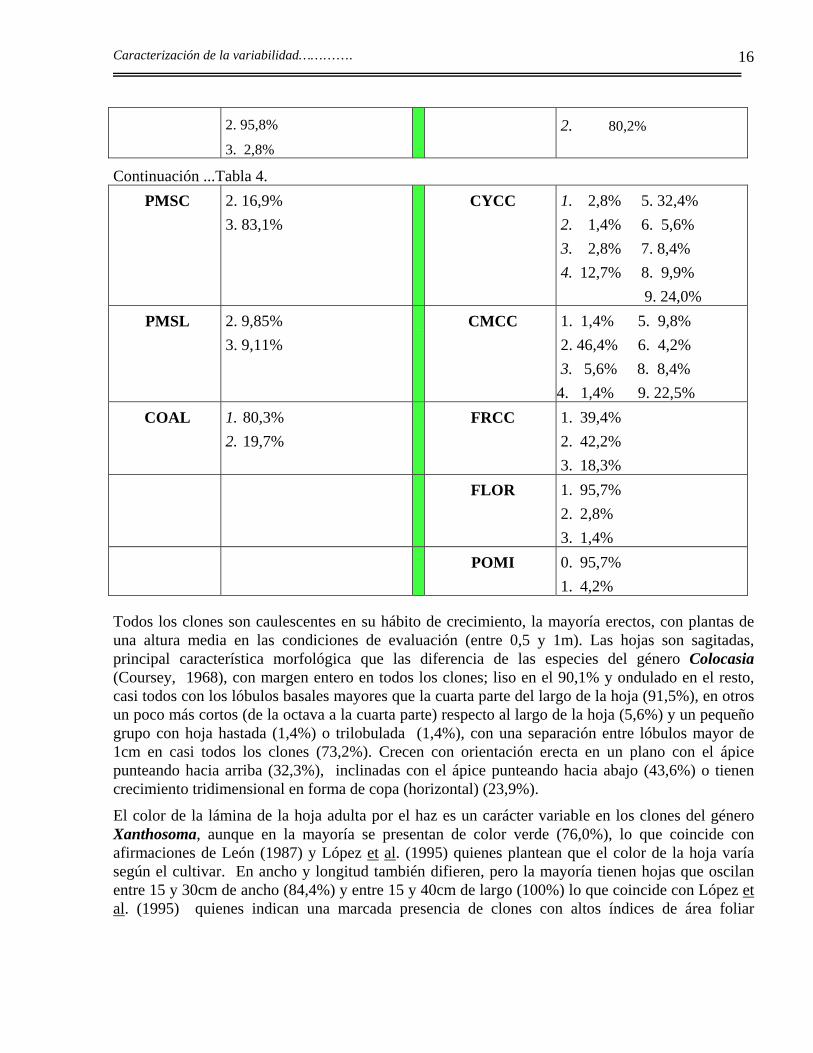
El análisis del porcentaje, realizado con la utilización de las variables cualitativas y cuantitativas de la Lista de Descriptores de Xanthosoma (Milián et al., 1993a) muestra una amplia variabilidad del germoplasma conservado (Tabla 4).
Tabla 4. Tabla de frecuencias absolutas de los valores de cada carácter expresado en porcentaje.
Caracteres cuantitativos
Porcentaje de clones por modalidad
Caracteres cualitativos
Porcentaje de clones por modalidad
ALPL 1. 1,4% 2. 29,5% 3. 69,0%
NRCR 1. 55,0% 2. 45.0%
SELB 0. 1,4% 1. 2,8% 2. 19,7% 3. 73,2% 4. 2,8%
HACR 1. 35,2% 2. 32,3% 3. 32,3%
LOPE 1. 1,4% 2. 38,0% 3. 60,5%
IPLH 3. 100%
ANAR 1. 56,3% 2. 42,2% 3. 1,4%
ORLM 0. 32,3% 1. 43,6% 2. 23,9%
ANHO 1. 1,4% 3. 59,1% 2. 25,3% 4. 14,0%
MAHO 1. 90,1% 2. 9,8%
LOHO 2. 87,3% 3. 12,6%
FOHO 2. 1,4% 4. 5,6%

Caracterización de la variabilidad………….
15
5. 91,5% 6. 1,4%
Continuación ...Tabla 4.
NRCO 1. 15,4%
2. 77,5%
3. 7,0%
CNHA 1. 88,8%
2. 8,4%
3. 2,8%
NCOR 1. 56,3%
2. 36,6%
3. 7,0%
CNEN 1. 11,2% 6. 1,4%
2. 84,5% 8. 1,4%
4. 1,4%
DIIN 1. 71,8%
2. 28,2%
COPE 1. 59,1% 6. 1,4%
2. 5,6% 7. 2,8%
3. 2,8% 8. 1,4%
4. 26,7%
RLAH 1. 5,63%
2. 50,7%
3. 43,66%
CPBP 1. 5,6% 5. 29,5%
2. 18,3% 6. 15,4%
3. 5,6% 7. 12,6%
4. 5,6% 8. 2,8%
9. 4,2%
RLHP 1. 36,61%
2. 32,39%
3. 30,98%
COAP 1. 7,0%
2. 15,4%
3. 73,2%
4. 4,2% LPHA 1. 8.54%
2. 44,8%
3. 46,66%
PCPE 1. 2,8%
2. 18,3%
3. 23,9%
4. 54,9%
NROH 2. 39,4%
3. 45,0%
4. 15,4%
COHO 1. 76,0%
2. 5,6%
3. 14,0%
4. 4,2%
PMSF 1. 1,4% CECC 1. 19,7%
Caracterización de la variabilidad………….
16
2. 95,8%
3. 2,8% 2. 80,2%
Continuación ...Tabla 4. PMSC 2. 16,9%
3. 83,1%
CYCC 1. 2,8% 5. 32,4% 2. 1,4% 6. 5,6% 3. 2,8% 7. 8,4% 4. 12,7% 8. 9,9% 9. 24,0%
PMSL 2. 9,85% 3. 9,11%
CMCC 1. 1,4% 5. 9,8% 2. 46,4% 6. 4,2% 3. 5,6% 8. 8,4% 4. 1,4% 9. 22,5%
COAL 1. 80,3% 2. 19,7%
FRCC 1. 39,4% 2. 42,2% 3. 18,3%
FLOR 1. 95,7% 2. 2,8% 3. 1,4%
POMI 0. 95,7% 1. 4,2%
Todos los clones son caulescentes en su hábito de crecimiento, la mayoría erectos, con plantas de una altura media en las condiciones de evaluación (entre 0,5 y 1m). Las hojas son sagitadas, principal característica morfológica que las diferencia de las especies del género Colocasia (Coursey, 1968), con margen entero en todos los clones; liso en el 90,1% y ondulado en el resto, casi todos con los lóbulos basales mayores que la cuarta parte del largo de la hoja (91,5%), en otros un poco más cortos (de la octava a la cuarta parte) respecto al largo de la hoja (5,6%) y un pequeño grupo con hoja hastada (1,4%) o trilobulada (1,4%), con una separación entre lóbulos mayor de 1cm en casi todos los clones (73,2%). Crecen con orientación erecta en un plano con el ápice punteando hacia arriba (32,3%), inclinadas con el ápice punteando hacia abajo (43,6%) o tienen crecimiento tridimensional en forma de copa (horizontal) (23,9%).
El color de la lámina de la hoja adulta por el haz es un carácter variable en los clones del género Xanthosoma, aunque en la mayoría se presentan de color verde (76,0%), lo que coincide con afirmaciones de León (1987) y López et al. (1995) quienes plantean que el color de la hoja varía según el cultivar. En ancho y longitud también difieren, pero la mayoría tienen hojas que oscilan entre 15 y 30cm de ancho (84,4%) y entre 15 y 40cm de largo (100%) lo que coincide con López et al. (1995) quienes indican una marcada presencia de clones con altos índices de área foliar

Caracterización de la variabilidad………….
17
obtenidos en la etapa de máximo crecimiento del follaje de los que dependen los mayores rendimientos. Las nervaduras en casi todos los clones son de color verde por el haz (88,8%) y verde claro por el envés (84,5%), mientras que unos pocos clones presentan otras coloraciones.
Se observan diferencias en cuanto a la longitud del pecíolo entre los diferentes clones, la que varía desde muy cortos (35cm) en unos clones (1,4%) hasta pecíolos de longitud media (entre 56 y 100cm) donde se agrupan la mayoría de los clones (60,5%). Su coloración también muestra una marcada diversidad; más de la mitad de los clones tienen pecíolos verdes (59,1%), pero el resto presenta coloraciones entre el verde morado (26,7%), verde claro (5,6%), verde oscuro (2,8%), morado oscuro (2,8%), morado verdoso (1,4%) y policromado (1,4%). Según López et al.(1995) el pecíolo surge a partir de una contracción gradual del extremo superior de la vaina y junto con ésta sirve de sostén al limbo; su longitud y coloración cambian con la variedad.
La parte basal del pecíolo tiene una coloración característica en los diferentes clones y también cambia con la variedad, según López et al. (1995). La mayor parte de los clones la tienen rosada (29,5%), blanco verdoso (18,3%), amarillo rosáceo (15,4%) y amarillo verdoso (12,6 %), el resto es blanco (5,6%), verde (5,6 %), verde claro (5,6 %), morado (4,2%) o rojo (2,8%).
La arista del pecíolo varía igualmente en su ancho y coloración entre los diferentes clones; más de la mitad la tienen estrecha (< 0,3 cm) (56,3 %), mientras que el 24,2% tiene un ancho medio (entre 0,3-1,0cm) y el 1,4% la tienen ancha (> 1,0 cm). Su coloración es morada en la mayoría de los clones (73,2%), aunque también aparecen algunos con la arista verde (7,0%), rosada (15,4%) o morado oscuro (4,2%).
El pecíolo también se caracteriza por presentar cera en su superficie. En más de la mitad de los clones, ésta es abundante, o sea, cubre todo el pecíolo (54,9%), en otros cubre sólo sus dos terceras partes (23,9%), o cubre solo 1/3 (18,3%) y un pequeño grupo(2,8%) no presenta cera.
En el presente trabajo se ha encontrado un 42.2% de clones con cormos y cormelos cilíndricos, 39.4% cónicos y 18.3% con cormos y cormelos elipsoidales, lo que coincide con López et al. (1995) cuando plantea que el cormo, el cual para la planta es una reserva de nutrientes y agua, y los tubérculos o cormelos que se forman del cormo y constituyen la parte comestible de mayor calidad, son la parte útil de la planta y que su forma varía según el clon, entre cilíndrico, elipsoidal o cónico y se ramifica en cormelos laterales.
La mayoría de los clones evaluados tienen la pulpa o masa de cormos y cormelos con tonalidades entre el blanco y el blanco rosado (47.8%), el 35.1% son interiormente entre rosado claro y morado; mientras que sólo el 16.8% presentan tonalidades amarillas en la pulpa de sus cormos y cormelos lo que coincide con López et al. (1995) cuando plantea que el color de la pulpa en clones Xanthosoma es por lo general blanca pero que puede ser amarilla o morada.
La coloración de las yemas también es un carácter que distingue a los clones dentro del género. Según López et al. (1995) las cicatrices foliares en la malanga forman un anillo completo en torno al cormo; en cada vaina sucesiva se distingue una yema cada vez más pequeña pero siempre presente. En su mayoría, los clones presentan las yemas de sus cormos y cormelos, con una gran variabilidad en su coloración, aunque predominan las tonalidades rosadas, rojas o moradas y amarillas (47.9%).

Caracterización de la variabilidad………….
18
Está demostrado que el cormo desempeña una función muy importante en el desarrollo de la planta, por las reservas energéticas que contienen. Además, en las especies de Xanthosoma la propagación se realiza fundamentalmente, por medio de secciones o pedazos del cormo primario (López et al., 1995) y los cormelos son la parte comestible de mayor calidad aunque también pueden ser usados como material de propagación (León, 1987) especialmente en los clones de masa amarilla. En esta colección casi todos los clones producen entre dos y cuatro cormos (77.5%) y pocos cormelos (>4) (56.3%); sin embargo, hay un 43,6% de clones que producen más de cuatro cormelos por planta y dentro de éstos, un 7% produce más de ocho, lo que supone la presencia de genotipos de gran valor para los programas de mejoramiento o la introducción directa en condiciones de producción.
En cuanto a su coloración la mayoría de los clones tienen cormos y cormelos con epidermis de color castaño oscuro (80.2%) lo que coincide con observaciones realizadas por López et al. (1995).
Los clones mostraron un contenido de materia seca en el follaje, relativamente bajo debido a que la evaluación se realizó a los 360 días de plantada. La mayoría presentó entre el 6 y el 10% destacándose los clones ‘Morada 2’ y ‘Morada 3’ con los valores más elevados (10, 11 y 10, 28% respectivamente) lo que indica que estos clones tienen un ciclo más largo y ‘Belembe’ con el porcentaje más bajo (5.57%). La materia seca en el follaje comienza a declinar a partir del máximo desarrollo foliar probablemente a causa del traslado de las sustancias de reserva de estos órganos hacia los cormos que crecen con rapidez a medida que decrece la materia seca en el follaje (López et al. (1995); de ahí la importancia del momento en que se han realizado estas evaluaciones.
El momento de la evaluación de la materia seca (%) en los cormos y los cormelos, también se hizo coincidir con la cosecha donde precisamente se presentan los niveles más elevados; se detectó la presencia de un alto contenido en la totalidad de los clones el que oscila entre 15 y 35% en los cormos y entre 21y 40% en los cormelos. Se observó, además, una tendencia en los clones de masa amarilla a presentar los valores más altos en correspondencia con el nivel de materia seca del follaje de la planta madre y el de los cormos totales por plantón, lo que coincide con López et al. (1995).
Las especies de Xanthosoma comestibles tienen un contenido apreciable de almidón en sus cormos y cormelos, el que está en dependencia del porcentaje de peso seco de cada sección y varía según la especie y las condiciones del cultivo. Sus granos esféricos, de tamaño pequeño y uniforme, unido al contenido de vitaminas y minerales hacen que esta planta sea una fuente de alimentos nutritivos y de alta digestibilidad (López et al, 1995). El alto contenido de almidón en algunos clones facilita su uso industrial (Piedrahita, 1979).
Los resultados de la Tabla 4 muestran un 19,7% de los clones con un porcentaje de almidón en sus cormos y cormelos por encima del 15%, independientemente de la especie.
No todos los clones florecen en las condiciones climáticas de Cuba; en el período de evaluación donde las temperaturas estuvieron por encima de los 25oC solo lo hizo el 5,6%. Según López et al. (1995) para que se produzca la floración se requieren de temperaturas medias inferiores a este valor cuando las plantas tienen entre 8 y 10 meses de plantadas. Este autor plantea además, que la floración en Xanthosoma es muy característica; en la inflorescencia, las flores masculinas se presentan en la parte superior, las estériles en el centro y en la parte inferior las femeninas.
Como se pudo apreciar, el análisis de frecuencia de aparición de los caracteres, los clones muestran una gran diversidad (Tabla 4), lo que puede estar dado por sus diferencias en la procedencia

Caracterización de la variabilidad………….
19
geográfica y por considerarse Cuba, sino dentro del centro de origen del cultivo, como un centro de dispersión secundaria.
En la Tabla 5 se muestran las correlaciones existentes entre los 13 caracteres analizados para el grupo Blanco, de las cuales seis fueron significativas. Caracteres como el color de la hoja, color del pecíolo, presencia de cera en el pecíolo, porcentaje de materia en los cormos y los cormelos, color de la parte basal del pecíolo y la separación de los lóbulos basales estuvieron involucrados en la correlaciones significativas obtenidas.
El carácter porcentaje de materia seca en cormos se correlaciona positivamente con el color de las yemas de los cormos y cormelos y con la separación de los lóbulos basales de las hojas; el porcentaje de materia seca en cormelos, con el color de la parte basal de pecíolo y con la presencia de cera en el pecíolo, lo que significa que los clones con alto contenido de materia seca en los cormos (%) tendrán una coloración rosado oscuro, rojo o morado en las yemas de sus cormos o cormelos y mayor separación de los lóbulos basales. Como el porcentaje de materia seca en los cormos se correlaciona positivamente con el de los cormelos, se puede afirmar que los clones con alto contenido en los primeros, lo tendrán también en los segundos y presentarán coloraciones amarillo verdoso, rojo o morado en la parte basal del pecíolo y abundante cera a lo largo de éste. Lo anterior confirma la importancia de este descriptor ya que permite utilizarlo como criterio de selección en los programas de mejoramiento genético en este cultivo, permitiendo pronosticar el comportamiento de atributos de calidad, como el contenido de materia seca, antes del momento de la cosecha lo cual en Xanthosoma es doblemente importante dado su prolongado ciclo.
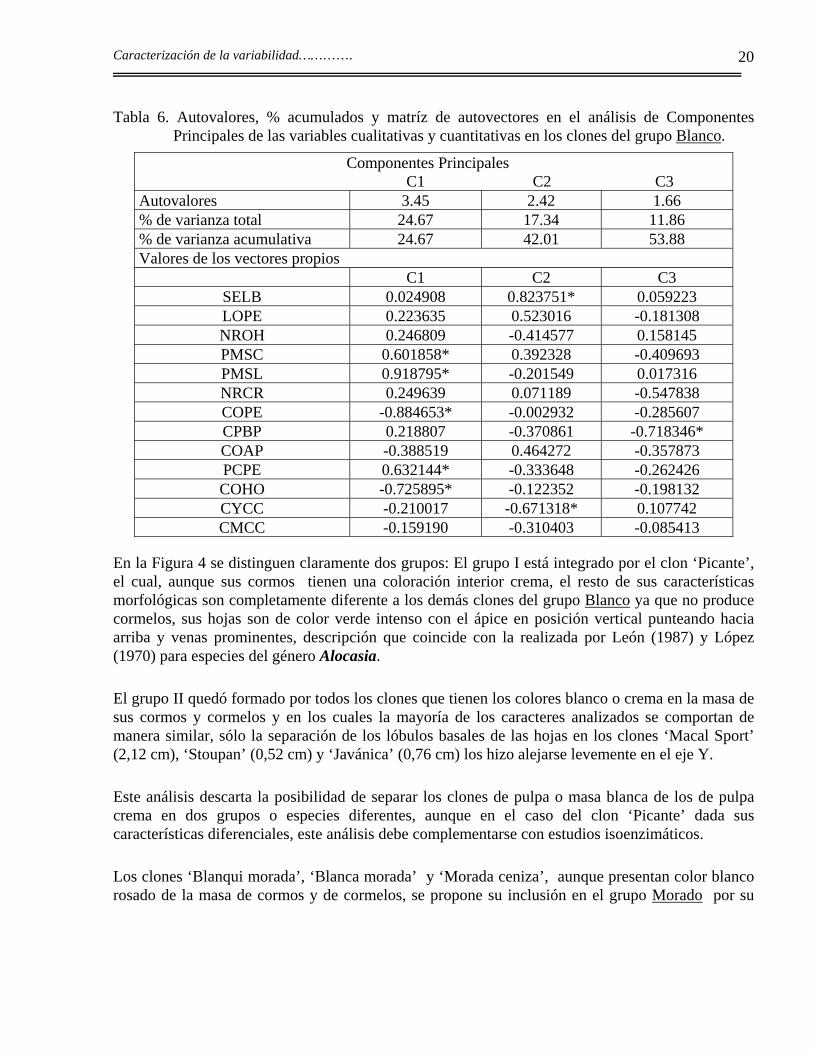
En el grupo Blanco, los resultados de los análisis de Componentes Principales se presentan en la Tabla 6. Con tres componentes se explica el 53,88% de la variación total. Las variables más importantes para la caracterización de los clones para los tres componentes fueron:
C1: porcentaje de materia seca en cormos (PMSC ) y en cormelos (MSCL), color del pecíolo (COPE ) y de la hoja (COHO );
C2: separación de los lóbulos (SELB) y el color de las yemas (CYCC);
C3: el color de la base del pecíolo (CPBP).
Los caracteres cualitativos relacionados con el pecíolo son, en este grupo, los que más aportan a la variabilidad de los clones; el color de las yemas también es importante. Estos caracteres están poco influenciados por el ambiente y están determinados por pocos genes lo que los hace más estables y por tanto más útiles en la caracterización de los clones.
El porcentaje de materia seca en los cormelos tuvo un alto valor en la diferenciación de los clones, esto puede estar determinado por los diferentes calibres que presentan los cormelos ya que, según Rodríguez Manzano (1998), el contenido de materia seca varía con el calibre y con los diferentes orígenes de los cormelos, los últimos en madurar son los cormelos terciarios. En los cormos este carácter es más estable y es también importante en la diferenciación de los clones, lo mismo que la separación de los lóbulos basales en las hojas.
Caracterización de la variabilidad………….
20
Tabla 6. Autovalores, % acumulados y matríz de autovectores en el análisis de Componentes Principales de las variables cualitativas y cuantitativas en los clones del grupo Blanco.
Componentes Principales C1 C2 C3 Autovalores 3.45 2.42 1.66 % de varianza total 24.67 17.34 11.86 % de varianza acumulativa 24.67 42.01 53.88 Valores de los vectores propios C1 C2 C3
SELB 0.024908 0.823751* 0.059223 LOPE 0.223635 0.523016 -0.181308 NROH 0.246809 -0.414577 0.158145 PMSC 0.601858* 0.392328 -0.409693 PMSL 0.918795* -0.201549 0.017316 NRCR 0.249639 0.071189 -0.547838 COPE -0.884653* -0.002932 -0.285607 CPBP 0.218807 -0.370861 -0.718346* COAP -0.388519 0.464272 -0.357873 PCPE 0.632144* -0.333648 -0.262426 COHO -0.725895* -0.122352 -0.198132 CYCC -0.210017 -0.671318* 0.107742 CMCC -0.159190 -0.310403 -0.085413
En la Figura 4 se distinguen claramente dos grupos: El grupo I está integrado por el clon ‘Picante’, el cual, aunque sus cormos tienen una coloración interior crema, el resto de sus características morfológicas son completamente diferente a los demás clones del grupo Blanco ya que no produce cormelos, sus hojas son de color verde intenso con el ápice en posición vertical punteando hacia arriba y venas prominentes, descripción que coincide con la realizada por León (1987) y López (1970) para especies del género Alocasia.
El grupo II quedó formado por todos los clones que tienen los colores blanco o crema en la masa de sus cormos y cormelos y en los cuales la mayoría de los caracteres analizados se comportan de manera similar, sólo la separación de los lóbulos basales de las hojas en los clones ‘Macal Sport’ (2,12 cm), ‘Stoupan’ (0,52 cm) y ‘Javánica’ (0,76 cm) los hizo alejarse levemente en el eje Y.
Este análisis descarta la posibilidad de separar los clones de pulpa o masa blanca de los de pulpa crema en dos grupos o especies diferentes, aunque en el caso del clon ‘Picante’ dada sus características diferenciales, este análisis debe complementarse con estudios isoenzimáticos.
Los clones ‘Blanqui morada’, ‘Blanca morada’ y ‘Morada ceniza’, aunque presentan color blanco rosado de la masa de cormos y de cormelos, se propone su inclusión en el grupo Morado por su

Caracterización de la variabilidad………….
21
número de cromosomas (2n=24), por presentar el pecíolo de color verde, la arista de color morado y las yemas de color rosado.
Figura 4. Agrupamientos formados a partir del análisis de Componentes Principales en el grupo
Blanco.
De las correlaciones existentes entre los 12 caracteres analizados para el grupo Amarillo, cinco fueron significativas (Tabla 7). El carácter porcentaje de materia seca en cormelos, como componente del rendimiento, está correlacionado con la presencia de cera en el pecíolo y éste a su vez, con el color de la yemas de los cormos y los cormelos lo que significa que en la malangas amarillas la presencia de abundante cera a lo largo del pecíolo indica que los cormelos presentan un alto porcentaje de materia seca Los clones de este grupo generalmente presentan abundante cera en le pecíolo y también los más altos valores de materia seca en cormos y cormelos, en comparación con el resto de los clones estudiados.
En el grupo Amarillo, con tres componentes se explica el 68,59 % de la variabilidad total (Tabla 8).
Las variables que más aportaron para la diferenciación de los clones fueron:
C1: Presencia de cera (PCPE), color de la hoja (COHO), color de la arista del pecíolo (COAP).
C2: Número de hijos (NROH), color del pecíolo (COPE).
C3: % de materia seca en cormos (PMSC) y color de las yemas (CYCC).
1
2
3
4
5
6
7
8
910
11
12
13
1415
16
17
18
19
20
21
22
23
24
25
2627
28
29
2.5
1.5
0.5
0.5
1.5
2.5
3.5
-5.5 -4.5 -3.5 -2.5 -1.5 -0.5 0.5 1.5
Componente 1
Com
pone
nte
2
I
II

Caracterización de la variabilidad………….
22
Tabla 8. Autovalores, % acumulados y matríz de autovectores en el análisis de Componentes Principales de las variables analizadas en los clones del grupo Amarillo.
Componentes Principales C1 C2 C3 Autovalores 3.28 2.97 1.97 % de varianza total 27.33 24.81 16.43 % de varianza acumulativa 27.33 52.15 68.59 Valores de los vectores propios C1 C2 C3
SELB -0.335713 -0.109760 0.174695 LOPE 0.565461 -0.573989 -0.236150 NROH -0.018989 0.791256* -0.300806 PMSC -0.273212 0.392285 -0.818692* PMSL 0.566390 0.580309 0.184754 COPE 0.431320 -0.711273* -0.464815 CPBP 0.513204 0.535935 -0.522619 COAP 0.653479* -0.559069 -0.187021 PCPE 0.753228* 0.534036 0.127299 COHO 0.680160* 0.129932 -0.131717 CYCC 0.552181 0.209520 0.725592* CMCC -0.490663 0.260861 -0.085811
El carácter presencia de cera en el pecíolo, es uno de los que más contribuyó a la variabilidad en el Componente C1. Según Milián et al. (1993b) en el grupo Amarillo, éste es un carácter importante a la hora de diferenciar los clones ya que, a pesar de que la mayoría de los mismos presentan abundante cera a lo largo del pecíolo, existen diferencias entre ellos. El número de hijos también es un carácter altamente valorado en la distinción de los clones ya que, dentro de las especies de Xanthosoma comestibles, este grupo es el que mayor ahijamiento presenta, carácter en el que, según Rodríguez Nodals (1979), se asemejan al género Colocasia. También, la coloración del pecíolo mostró importancia con su contribución en el Componente C2, lo que sugiere valorarlo para integrar la Lista de los Descriptores más importantes en Xanthosoma.
Este grupo sobresale por los valores más altos de materia seca en cormos y cormelos entre todos los evaluados, sin embargo dentro del propio grupo Amarillo este carácter también evidencia una gran contribución a la variabilidad en el Componente C3.
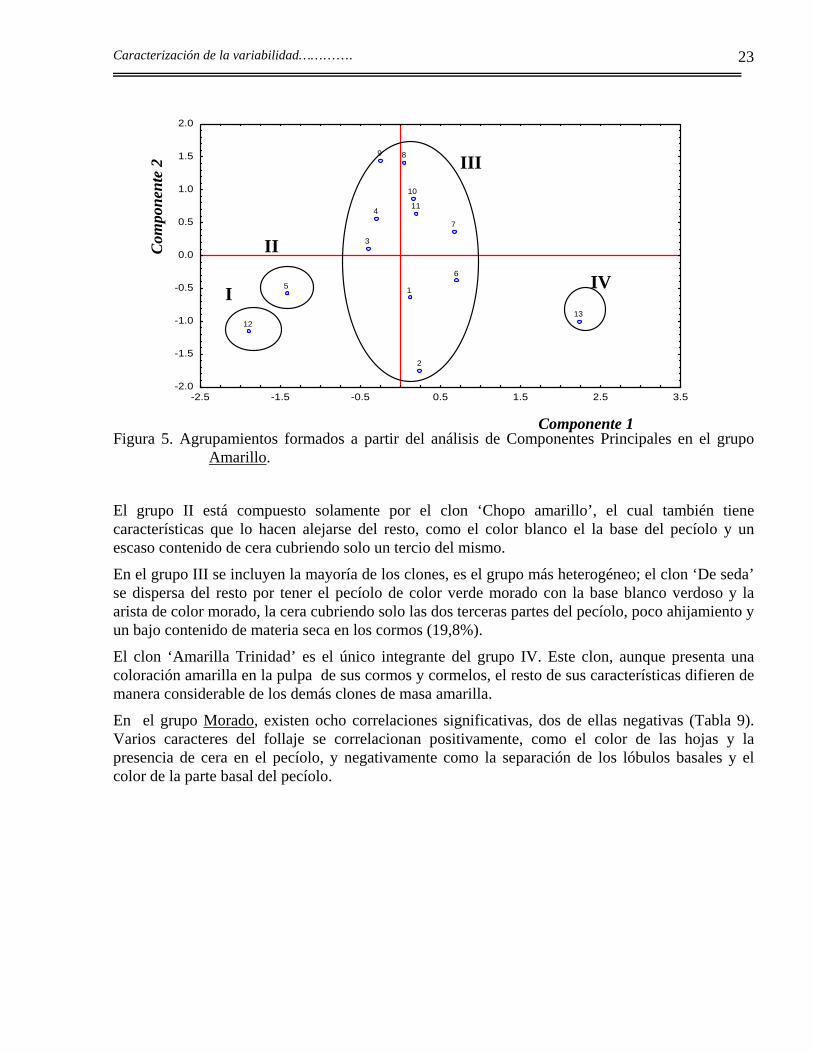
La Figura 5 muestra la presencia de cuatro agrupamientos. Como se puede apreciar los clones están bastante dispersos. En el grupo I se incluye el clon ‘Belembe’ que, aunque comparte la similitud de algunos caracteres con el clon ‘Chopo amarillo’, es diferente de éste y del resto de los clones en cualidades como el color del pecíolo, color de la arista y de la base del pecíolo, color de las yemas, presencia de cera, número de hijos, color de la masa, color de las nervaduras por el envés de las hojas, altura de la planta y tiene la característica muy distintiva de presentar hojas trilobuladas con el ápice de los lóbulos punteando hacia arriba.
Caracterización de la variabilidad………….
23
Figura 5. Agrupamientos formados a partir del análisis de Componentes Principales en el grupo Amarillo.
El grupo II está compuesto solamente por el clon ‘Chopo amarillo’, el cual también tiene características que lo hacen alejarse del resto, como el color blanco el la base del pecíolo y un escaso contenido de cera cubriendo solo un tercio del mismo.
En el grupo III se incluyen la mayoría de los clones, es el grupo más heterogéneo; el clon ‘De seda’ se dispersa del resto por tener el pecíolo de color verde morado con la base blanco verdoso y la arista de color morado, la cera cubriendo solo las dos terceras partes del pecíolo, poco ahijamiento y un bajo contenido de materia seca en los cormos (19,8%).
El clon ‘Amarilla Trinidad’ es el único integrante del grupo IV. Este clon, aunque presenta una coloración amarilla en la pulpa de sus cormos y cormelos, el resto de sus características difieren de manera considerable de los demás clones de masa amarilla.
En el grupo Morado, existen ocho correlaciones significativas, dos de ellas negativas (Tabla 9). Varios caracteres del follaje se correlacionan positivamente, como el color de las hojas y la presencia de cera en el pecíolo, y negativamente como la separación de los lóbulos basales y el color de la parte basal del pecíolo.
1
2
3
4
56
7
89
10
11
1213
-2.0
-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
2.0
-2.5 -1.5 -0.5 0.5 1.5 2.5 3.5
Com
pone
nte
2
Componente 1
I
II
III
IV

Caracterización de la variabilidad………….
24
El color de la masa de cormos y cormelos se correlaciona negativamente con la separación de los lóbulos basales y positivamente con el color de la arista del pecíolo, lo que significa que los clones con una pequeña o nula separación de los lóbulos y una arista de color morado o morado oscuro van a presentar cormos y cormelos con masa de color también morado o morado claro.
El porcentaje de materia seca en los cormos se correlaciona positivamente con el color de las yemas directamente, e indirectamente con el color del pecíolo y de su arista, por lo que también pueden utilizarse como criterios de selección. Es decir, que cuando las yemas tienen una coloración roja o morada y el pecíolo presenta tonalidades morado verdoso o morado oscuro, con la arista también de este color, el porcentaje de materia seca en los cormos será mayor.
Landa et al. (1996) encontraron correspondencia entre el color de la masa rosada o morada de los cormos y cormelos y el número cromosómico 2n=24 en un grupo de clones de malanga morada, lo que lleva a plantear que la observación del carácter mencionado, puede indicar la existencia de un número cromosómico dado.
La matriz de los valores y vectores propios (Tabla 10) muestra los caracteres que más incidieron en el 54.4% de la variabilidad total acumulada hasta el tercer componente en el grupo Morado; los caracteres más importantes en la diferenciación de los clones son:
C1: color de la arista del pecíolo (COAP), color de las yemas de cormos y cormelos (CYCC), contenido de materia seca de cormos (PMSC), color del pecíolo (COPE) y color de la pulpa o masa (CMCE)
C2: color de la base del pecíolo (CPBP), separación de los lóbulos basales (SELB), presencia de cera en el pecíolo (PCPE) y color de la hoja (COHO)
C3: longitud del pecíolo (LOPE).
El color de la pulpa o masa de los cormos y cormelos (CMCC) tiene un alto valor en la diferenciación de los clones aún dentro del mismo grupo (Morado), donde se supone que la variación existente sea mínima.
El color de la yemas de cormos y cormelos, a pesar de no estar incluido en los descriptores para Xanthosoma del IBPGR (1989), es el segundo en aportar la mayor variabilidad al Componente C1 por lo que se propone incorporarlo al listado de descriptores mínimos. La mayoría de los caracteres seleccionados en los dos primeros componentes son cualitativos, y esto garantiza una mayor estabilidad genética de los clones. Para Poey (1978) y Milián et al (1998), los caracteres cualitativos están determinados por uno o pocos genes, poco afectados por el ambiente, aunque en algunos caracteres su expresión puede verse alterada por la acción de genes modificadores
El porcentaje de materia seca en los cormos (PMSC) también tuvo un valor alto en el Componente C1. Este carácter puede ser mucho más estable que el contenido de materia seca en los cormelos; ya que éste último varía según el calibre de los mismos (Rodríguez Manzano, 1998).

Caracterización de la variabilidad………….
25
Tabla 10. Autovalores, porcentajes acumulados y matríz de autovectores en el análisis de Componentes Principales de las variables analizadas en los clones del grupo Morado.
Componentes Principales C1 C2 C3 Autovalores 3.17 2.08 1.27 % de varianza total 26.45 17.33 10.60 % de varianza acumulativa 26.45 43.79 54.40 Valores de los vectores propios C1 C2 C3
SELB -0.505400 0.630050* -0.048932 LOPE -0.304918 0.086496 0.609381* NROH 0.163779 -0.049452 0.485630 PMSC 0.604502* -0.009087 0.458116 PMSL -0.306304 -0.219627 -0.326080 COPE 0.605431* 0.410585 0.055267 CPBP 0.359591 -0.705625* -0.108920 COAP 0.836424* 0.236522 0.033164 PCPE 0.148543 0.614833* -0.492705 COHO 0.247127 0.695185* 0.128649 CYCC 0.749077* -0.203127 -0.183419 CMCC 0.707323* -0.017710 -0.195749
.
La Figura 6 muestra la distribución de los clones de acuerdo al análisis de Componentes Principales; permite apreciar la formación de tres grupos con la combinación de los caracteres de los Componentes C1 y C2.
El grupo I está representado por los clones ‘México 1’, ‘México 8’, ‘Japonesa’, ‘Cuarentena 1’, los cuales presentan las aristas del pecíolo (COAP) de color rosado, las yemas de los cormos y los cormelos (CYCC) de color rosado oscuro, el pecíolo de color verde (COPE) y tiene valores medios en el contenido de materia seca en los cormos (PMSC). Dichos clones fueron introducidos desde México y Saotomé y Príncipe y tienen la pulpa de color blanco rosado o rosado.
Los clones ubicados en el grupo II tienen una coloración morada en la arista del pecíolo y conforman el grupo más heterogéneo. Los clones presentaron mayor variabilidad fenotípica, supuestamente atribuible a la acumulación de mutaciones espontáneas en la evolución del género (León, 1987), ya que en él es muy difícil la obtención de semilla botánica (León, 1987; López et al., 1995; Milián et al., 1993a), dado por la rareza y la especial característica que presenta la flor de la malanga (sus óvulos están cubiertos por una sustancia gelatinosa y una bráctea que impiden la autofecundación). Además, en Xanthosoma existe una diferencia de 48 h entre la actividad del polen y la receptividad del óvulo (López et al., 1995).

Caracterización de la variabilidad………….
26
Existen en este grupo seis clones que tiene una coloración en la base del pecíolo diferente del rosado: el clon ‘Morada de México’ y el ‘México-3’ tienen la base del pecíolo de color amarillo verdoso, los clones ‘Morada-8’, ‘INIVIT-84’ y ‘Morada Cabaiguán’ la tienen de color morado y el clon ‘Morada Jibacoa’ tiene el pecíolo con la base de color rojo.
La fuerte incidencia del color morado oscuro del pecíolo y de su arista hicieron al clon ‘Jardín’ alejarse en el grupo III; la base del pecíolo es rosada y presenta abundante cera a lo largo del mismo, as hojas son de un color verde intenso con las nervaduras de color morado verdoso por el envés, también sus yemas son de color morado y la masa de los cormos y los cormelos es de color morado claro, aunque en estos caracteres es similar a otros clones.
Figura 6. Agrupamientos formados a partir del análisis de Componentes Principales en el grupo Morado.
La Figura 7 muestra el dendrograma obtenido de los clones de la colección cubana de malanga del género Xanthosoma donde se puede apreciar la formación de tres grupos: el I incluye los clones del grupo Morado, en el II se ubica el clon ‘Picante’, mientras que en el III se agrupan los clones de los grupos Blanco y Amarillo mostrando coincidencia con la distribución por grupos propuesta.
Debemos señalar que los clones ‘Sergio cuarentena’, ‘Morada 1727’, ‘Cuarentena-1’, ‘Rosada’, ‘México-1’, ‘Blanqui morada’, ‘Blanca morada’ y ‘Morada ceniza’, aunque pertenecen al grupo Morado aparecen en el agrupamiento de los clones de masa amarilla y blanca, debido fundamentalmente, a que la coloración de la masa de los cormos y los cormelos es de color blanco rosado y presentan, en comparación con los clones del grupo Morado, valores altos para el número
1
2
3
4
5
67
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
-3
-2
-1
0
1
2
3
-3 -2 -1 0 1 2 3
Componente 1
Com
pone
nte
2
I
II
III

Caracterización de la variabilidad………….
27
de hijos y para el porcentaje de materia seca en los cormos y en los cormelos, hojas largas y abundante cera a lo largo del pecíolo.
Figura 7. Dendrograma obtenido del análisis de Conglomerados (Cluster analysis) de los clones de la colección cubana del género Xanthosoma con los caracteres cuantitativos y cualitativos. Los resultados de las Correlaciones por Rangos de Spearman entre las variables analizadas en los 71 clones de la colección se muestran en la Tabla 11. Se destaca el número de cromosomas, que tuvo una correlación positiva con el color del pecíolo, de su parte basal y de su arista, con el color de las yemas y de la masa de los cormos y cormelos. Es decir, los clones con número cromosómico de 2n=24 generalmente tendrán coloraciones morada o morado oscuro en el pecíolo, rojo o morado en su parte basal y la arista también de color morado; mientras que las yemas se presentarán de color rosado oscuro o morado, lo mismo que la masa de cormos y cormelos
Por su parte los clones con 2n=26 cromosomas presentarán pecíolos de color verde, con su parte basal y sus aristas también verde o con tonalidades rosadas estas últimas, las yemas de color blanco, blanco amarillento, amarillo o amarillo rojizo. Los cormos y cormelos tendrán coloración interior desde blanco hasta amarillo naranja lo cual se corresponde fielmente con el análisis morfoagronómico realizado e indica que es posible conocer la ploidía en los clones de esta colección de Xanthosoma a través del registro de los caracteres mencionados. Katsiotis y Forsberg (1995) y García et al. (1996) plantearon la posibilidad de distinguir el nivel de ploidía mediante mediciones del grano de polen en diversos cultivos.
De manera general, se puede afirmar que en Xanthosoma es posible predecir atributos de calidad como el contenido de materia seca en los cormos y los cormelos teniendo en cuenta el número de hijos, el color de la parte basal del pecíolo, la presencia de cera a lo largo de éste, el color de las hojas y el color de las yemas de los cormos y los cormelos ya que dichos caracteres están
I II III
Distancia Euclidiana
0
2
4
6
8
10
71
68
65
57
48
51
39
40
63
26
41
66
55
50
23
18
11
14
54
8
53
42
45
3
21
28
24
37
61
60
13
36
31
30
29
33
16
9
32
35
4
58
44
43
64
19
67
25
49
10
52
70
62
15
5
47
59
27
56
38
46
20
17
12
7
2
34
22
6
69
1
Caracterización de la variabilidad………….
28
correlacionados con el mismo. Así, generalmente los clones con mayor número de hijos, con abundante cera en el pecíolo, con hojas de color verde oscuro completa o con el borde morado, con la parte basal del pecíolo de colores claros entre el blanco y el amarillo rosáceo o verdoso y con las yemas de los cormos y de los cormelos, también entre el blanco y el amarillo rojizo, presentarán los mayores porcentajes de materia seca en sus órganos de reserva.
Para Xanthosoma no existen reportes relacionados con el estudio de las asociaciones entre los diferentes caracteres; pero en Colocasia, Pandey et al. (1996) estudiaron las correlaciones entre ocho caracteres subterráneos con incidencia en el rendimiento y señalaron que el peso de los cormos madres y cormelos puede ser usado como criterio de selección. También en este cultivo Rodríguez Manzano (1998) obtuvo significación para 20 correlaciones entre los caracteres de los órganos foliares y subterráneos importantes para realizar las selecciones indirectas en los programas de mejoramiento genético.
En la Tabla 12 se presentan los resultados del análisis de Componentes Principales en los 71 clones; es posible apreciar los caracteres que más incidieron en el 56,45% de la variabilidad total acumulada hasta el tercer componente. C1: Color de la masa de los cormos y los cormelos (CMCC), color de las yemas (CYCC), número
de cromosomas (NRCR), color de la arista del pecíolo (COAP), % de materia seca en los cormos (PMSC)
C2: Color de la base del pecíolo(CPBP), número de hijos (NROH) C3: Color del pecíolo (COPE), % de materia seca en los cormelos (PMSL)
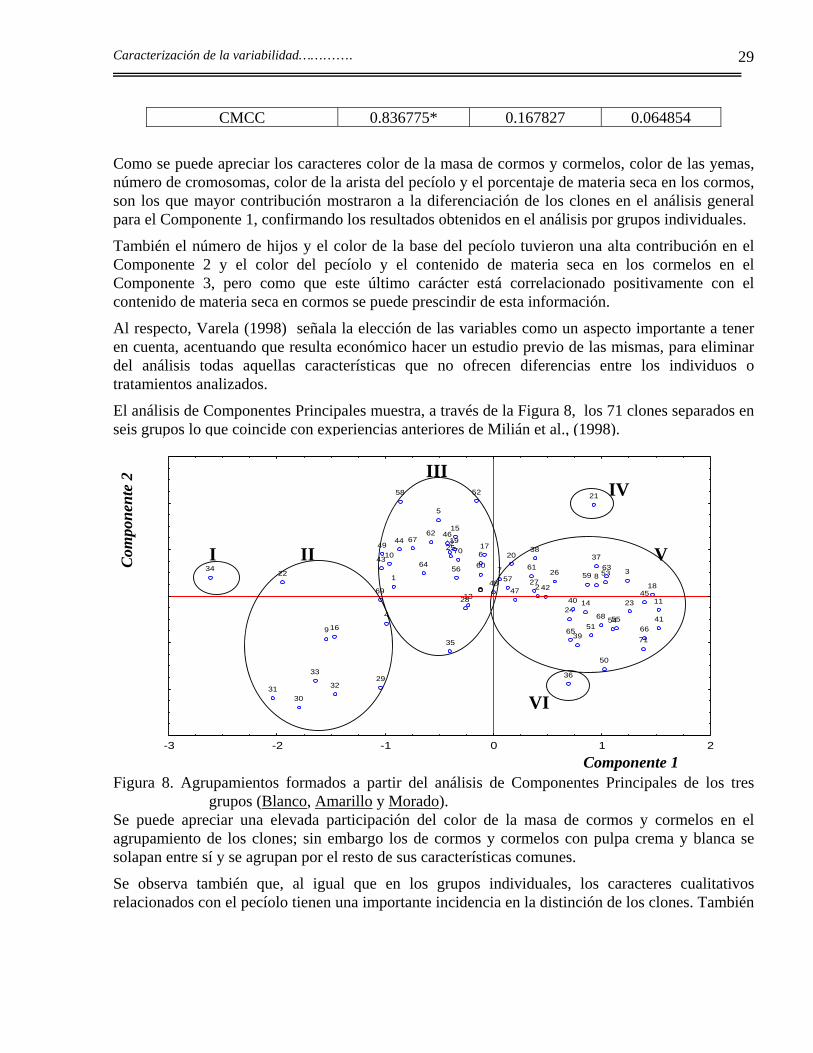
Tabla 12. Autovalores, % acumulado y matriz de autovectores en el análisis de Componentes Principales de las variables cualitativas y cuantitativas en la colección de Xanthosoma.
Componentes Principales C1 C2 C3 Autovalores 2.74 2.44 1.58 % de varianza total 22.90 20.35 13.19 % de varianza acumulativa 22.90 43.26 56.45 Valores de los vectores propios
C1 C2 C3 SELB 0.019393 -0.575480 0.031929 LOPE 0.304196 -0.428127 0.101985 NROH -0.542286 -0.590586* -0.131011 PMSC -0.601407* -0.460895 -0.254881 PMSL 0.042681 -0.344792 0.728480* NRCR 0.779292* -0.226296 0.089211 COPE 0.396631 -0.286067 -0.749545* CPBP 0.304400 -0.709425* 0.182304 COAP 0.637506* 0.310060 -0.221947 PCPE 0.106865 -0.479174 0.314858 COHO 0.270666 -0.439239 -0.434829 CYCC 0.806106* -0.305096 0.137388
Caracterización de la variabilidad………….
29
CMCC 0.836775* 0.167827 0.064854
Como se puede apreciar los caracteres color de la masa de cormos y cormelos, color de las yemas, número de cromosomas, color de la arista del pecíolo y el porcentaje de materia seca en los cormos, son los que mayor contribución mostraron a la diferenciación de los clones en el análisis general para el Componente 1, confirmando los resultados obtenidos en el análisis por grupos individuales.
También el número de hijos y el color de la base del pecíolo tuvieron una alta contribución en el Componente 2 y el color del pecíolo y el contenido de materia seca en los cormelos en el Componente 3, pero como que este último carácter está correlacionado positivamente con el contenido de materia seca en cormos se puede prescindir de esta información.
Al respecto, Varela (1998) señala la elección de las variables como un aspecto importante a tener en cuenta, acentuando que resulta económico hacer un estudio previo de las mismas, para eliminar del análisis todas aquellas características que no ofrecen diferencias entre los individuos o tratamientos analizados.
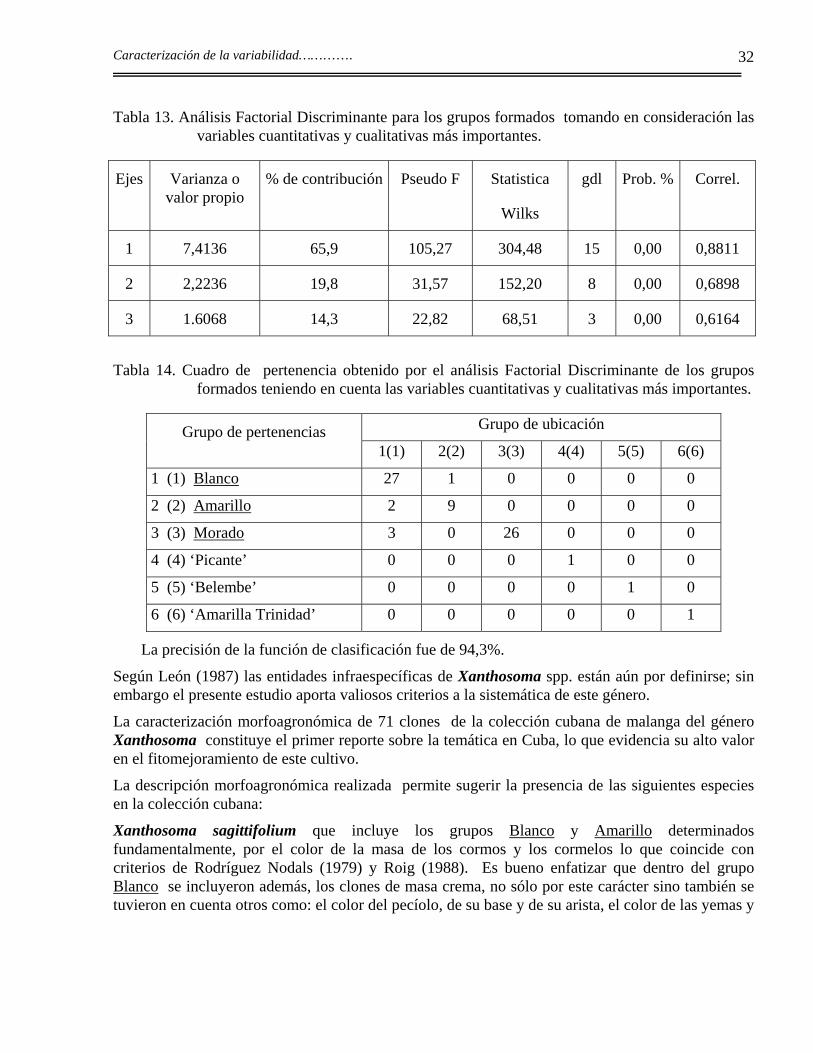
El análisis de Componentes Principales muestra, a través de la Figura 8, los 71 clones separados en seis grupos lo que coincide con experiencias anteriores de Milián et al., (1998).
Figura 8. Agrupamientos formados a partir del análisis de Componentes Principales de los tres grupos (Blanco, Amarillo y Morado).
Se puede apreciar una elevada participación del color de la masa de cormos y cormelos en el agrupamiento de los clones; sin embargo los de cormos y cormelos con pulpa crema y blanca se solapan entre sí y se agrupan por el resto de sus características comunes.
Se observa también que, al igual que en los grupos individuales, los caracteres cualitativos relacionados con el pecíolo tienen una importante incidencia en la distinción de los clones. También
12
3
4
5
6
78
9
10
11
12
1314
15
16
17
18
19
20
21
22
2324
25
2627
28
29
3031 32
33
34
35
36
3738
39
40
41
42
43
44
45
46
4748
49
50
51
52
53
5455
5657
58
5960 61
62
6364
65 66
67
68
69
70
71
-3 -2 -1 0 1 2
Com
pone
nte
2
Componente 1
III
VII
IV
I
VI

Caracterización de la variabilidad………….
30
la coloración de las yemas y el contenido de materia seca en cormos y cormelos juega un importante papel.
En el grupo I se separa el clon ‘Belembe’, ya que presenta caracteres muy particulares.
El grupo II presenta cierta dispersión entre los clones que lo conforman, pero caracteres como el propio color amarillo de la pulpa, la coloración también amarilla de las yemas, el alto número de hijos, el elevado contenido de materia seca en los cormos en comparación con el resto de los clones, los hace conformar el mismo grupo. El clon ‘De Seda’ tiene la masa de sus cormos y cormelos de color amarillo, sin embargo se integra al grupo Blanco, ya que caracteres como el bajo contenido de materia seca en sus cormos y cormelos, así como, su escaso ahijamiento, entre otros, lo hacen separarse del grupo Amarillo. Lo mismo sucede con el clon ‘Amarilla criolla-1’ para dichos caracteres, pero este clon tiene además, la masa de sus cormos y cormelos de color blanco rosado.
Los clones de pulpa blanca y crema integran el grupo III; se distinguen por esta característica y además, porque tienen una coloración amarillo rojiza de las yemas, verde del pecíolo y morado de su arista y un contenido medio de materia seca en los cormos en comparación con el resto de los clones. El clon ‘Javánica’ es el de mayor dispersión entre los de masa blanca, lo que puede estar motivado por su coloración amarillo verdoso en la base del pecíolo, su abundante contenido de cera a lo largo del mismo y sus yemas de color amarillo rojizo, todo lo cual coincide con características del grupo Amarillo.
En el grupo IV se separa el clon ‘Picante’ que, aunque la pulpa de sus cormos es de color crema, sólo en este carácter es similar a los demás clones de este grupo. Se caracteriza por tener sus hojas con el ápice punteando hacia arriba en posición casi vertical, las nervaduras son prominentes y no produce cormelos. Además, el pecíolo es policromado donde se puede distinguir una coloración blanca verdosa con pigmentación morada en todo el pecíolo que se extiende a las nervaduras por el envés y en la base presenta una coloración blanca; la arista es de color morado oscuro, las hojas presentan un color verde intenso y las nervaduras por el haz son de color verde oscuro. Tiene, casi siempre, una alta presencia de flores.
El grupo V lo integran los clones con la pulpa de cormos y cormelos de color rosado o morado y que tienen otras características comunes y diferentes del resto de los clones como la coloración verde o verde morada del pecíolo con su parte basal rosada, cubierto de abundante cera en la mayoría de los clones, las yemas son de color morado y sus cormos presentan los porcentajes más bajos de materia seca.
En este grupo se ubicaron los clones ‘Blanca morada’, ‘Blanqui morada’ y ‘Morada ceniza’ los que, a pesar de tener una coloración blanca rosada de la pulpa de sus cormos y cormelos, presentan un número cromosómico 2n=24 que los diferencia del resto de los clones ubicados en el grupo Blanco. Este resultado unido a la separación de los lóbulos basales de las hojas, la longitud y color del pecíolo, número de hijos, contenido de materia seca en los cormos y también en los cormelos, color de la base y de la arista del pecíolo, presencia de cera, color de las hojas y de las yemas llevaron a reubicar este material en el grupo Morado.
El grupo VI es para el clon ‘Amarilla Trinidad’ que, a pesar de tener pulpa amarilla, no forma parte de este grupo ya que tiene características propias muy diferentes del resto de los clones. Tiene la

Caracterización de la variabilidad………….
31
hoja hastada de color verde intenso con borde morado, el pecíolo tiene coloración morada con la base de color rojo y la arista morado oscuro.
Según Roig (1913), quien caracterizó e identificó clones de malanga (Xanthosoma y Colocasia), es de destacar la importancia que tienen los caracteres de los cormos, cormelos, hojas y pecíolos en las observaciones y evaluaciones para la diferenciación de los clones. La inflorescencia es un carácter también importante, pero no todos los clones florecen.
El análisis Factorial Discriminante confirmó un 94,3% de precisión de la función de clasificación de los clones en estos seis grupos. Se pudo apreciar que las dos primeras funciones discriminantes fueron altamente significativas y por tanto, válidas para el análisis (Tabla 13), indicando que en general, el agrupamiento de los clones fue el correcto de acuerdo a los resultados del análisis de Componentes Principales.
En la Tabla 14 puede observarse que el 94,3% del total de clones estaban bien ubicados. Del grupo Blanco es separado el clon ‘Javánica’ y reubicado en en el grupo Amarillo, lo cual debe estar motivado por su coloración amarillo verdoso en la base del pecíolo, su abundante contenido de cera a lo largo del mismo y sus yemas de color amarillo rojizo, todo lo cual coincide con características del grupo Amarillo. Este clon fue el de mayor dispersión entre los de masa blanca agrupados por el análisis de Componentes Principales (Figura 8).
Los clones ‘De Seda’ y ‘Amarilla criolla-1’, que estaban dentro del grupo Amarillo, se pasan para el grupo Blanco, lo que puede explicarse porque ‘De Seda’ tiene poca cera en el pecíolo, el color del pecíolo es verde morado, con la base blanco verdoso; mientras que ‘Amarilla criolla-1’ tiene la masa de los cormos y los cormelos de color blanco rosado, con la base del pecíolo blanco verdoso. Ambos clones presentan los valores más bajos del número de hijos y porcentaje de materia seca en los cormos y los cormelos, todo lo cual los hacen alejarse de los clones del grupo Amarillo y situarse en el Blanco, lo que también ocurrió en el análisis de Componentes Principales (Figura 8).
Existen un grupo de variables como: número de cromosomas, color del pecíolo, color de las hojas y color de las yemas que acercan los clones de los grupos Blanco y Amarillo, por lo que algunos autores como León (1976), García (1979), Rodríguez Nodals (1979), León (1987), Roig (1988) y Giacometti y León (1994) los reunen en una sola especie: Xanthosoma sagittifolium, lo que apoya la propuesta de reubicación de los clones de un grupo en otro.
El análisis Factorial Discriminante comprobó que 26 de los clones del grupo Morado están bien clasificados en éste; sin embargo ‘México-1’, ‘México-8’ y ‘Cuarentena-1’, los saca de su grupo de pertenencia para ubicarlos en el grupo Blanco, debido a que la masa de sus cormos y cormelos es de color blanco rosado y rosado, el color de las hojas es verde, la arista del pecíolo es de color rosado y presenta abundante cera, cualidades que los acercan al nuevo grupo de ubicación propuesto.
Debe señalarse que en el análisis de Componentes Principales los clones mencionados, aunque están dentro del grupo Morado, se sitúan en los límites del grupo de pertenencia con el grupo de ubicación. A pesar de estos resultados, se propone mantener estos tres clones dentro del grupo Morado, debido a que su número cromosómico es de 2n=24 y la mayoría de los caracteres cualitativos y cuantitativos analizados se corresponden con los clones de este grupo.
‘Picante’, ‘Belembe’ y ‘Amarilla Trinidad’ son colocados en grupos diferentes pues, como ya fue analizado, presentan fenotipos muy característicos.
Caracterización de la variabilidad………….
32
Tabla 13. Análisis Factorial Discriminante para los grupos formados tomando en consideración las variables cuantitativas y cualitativas más importantes.
Ejes Varianza o valor propio
% de contribución Pseudo F Statistica
Wilks
gdl Prob. % Correl.
1 7,4136 65,9 105,27 304,48 15 0,00 0,8811
2 2,2236 19,8 31,57 152,20 8 0,00 0,6898
3 1.6068 14,3 22,82 68,51 3 0,00 0,6164
Tabla 14. Cuadro de pertenencia obtenido por el análisis Factorial Discriminante de los grupos formados teniendo en cuenta las variables cuantitativas y cualitativas más importantes.
Grupo de ubicación Grupo de pertenencias 1(1) 2(2) 3(3) 4(4) 5(5) 6(6)
1 (1) Blanco 27 1 0 0 0 0
2 (2) Amarillo 2 9 0 0 0 0
3 (3) Morado 3 0 26 0 0 0
4 (4) ‘Picante’ 0 0 0 1 0 0
5 (5) ‘Belembe’ 0 0 0 0 1 0
6 (6) ‘Amarilla Trinidad’ 0 0 0 0 0 1
La precisión de la función de clasificación fue de 94,3%.
Según León (1987) las entidades infraespecíficas de Xanthosoma spp. están aún por definirse; sin embargo el presente estudio aporta valiosos criterios a la sistemática de este género.
La caracterización morfoagronómica de 71 clones de la colección cubana de malanga del género Xanthosoma constituye el primer reporte sobre la temática en Cuba, lo que evidencia su alto valor en el fitomejoramiento de este cultivo.
La descripción morfoagronómica realizada permite sugerir la presencia de las siguientes especies en la colección cubana:
Xanthosoma sagittifolium que incluye los grupos Blanco y Amarillo determinados fundamentalmente, por el color de la masa de los cormos y los cormelos lo que coincide con criterios de Rodríguez Nodals (1979) y Roig (1988). Es bueno enfatizar que dentro del grupo Blanco se incluyeron además, los clones de masa crema, no sólo por este carácter sino también se tuvieron en cuenta otros como: el color del pecíolo, de su base y de su arista, el color de las yemas y

Caracterización de la variabilidad………….
33
de la masa de sus cormos y sus cormelos, la presencia de cera, el contenido de materia seca en los cormos y en los cormelos, entre otros.
Autores como Cordero (1986), León (1987), García (1990), Milián (1993b), Giacometti y León (1994) y Rodríguez et al. (1999) habían clasificado en la especie Xanthosoma sagittifolium solamente los clones de masa blanca, mientras que los de pulpa crema los ubicaron en Xanthosoma caracu y en Xanthosoma atrovirens los de masa amarilla. El presente estudio ha demostrado que este carácter por sí solo no puede distinguir especies y que es necesario incluir el número cromosómico y otros descriptores cualitativos y cuantitativos de alta contribución a la diferenciación de los clones, unido a la caracterización isoenzimática que ofrece una mayor información acerca de los genotipos y que, según Simpson y Withers (1986) juega un papel importante en la caracterización y evaluación de los recursos genéticos vegetales. Además, es importante que la clasificación de especies se realice en ejemplares vivos, ya que según León (1987) dicha clasificación y la descripción se hizo en ejemplares de herbario, basada principalmente en caracteres foliares muy variables.
Xanthosoma violaceum donde se incluyen los clones cuya coloración de la masa o pulpa de sus cormos y cormelos es rosada o morada. Fueron incluidos además, en esta especie los clones ‘Blanca morada’, ‘Blanqui morada’ y ‘Morada ceniza’ quienes, a pesar de tener la masa de color blanco rosado, presentaron un número cromosómico igual a los clones de masa rosada o morada (2n=24). Los clones de esta especie presentan coloraciones morado y morado oscuro en el pecíolo, rojo o morado en la base y morado en la arista. Las yemas y la masa de los cormos y los cormelo son de color rosado oscuro o morado.
Esta ubicación propuesta resultó ser más adecuada ya que dichos clones, en el análisis de Componentes Principales siempre se ubicaron en el límite entre ambos grupos y el Análisis Discriminante confirmó la precisión de la clasificación. De ahí que estos resultados corroboren lo planteado anteriormente sobre la importancia de incluir varios caracteres en la clasificación de las especies en lugar de uno solo.
Existen en la colección clones muy diferentes del resto como ‘Belembe’ introducido desde la Isla de Guadalupe cuya descripción coincide con la reportada por León (1987) y Purseglove (1988) para Xanthosoma brasilense Engl. crece en macollas hasta de un metro, generalmente de unos 50cm de alto, con varios tallos que provienen de un cormo pequeño de pulpa amarillenta que no se utiliza como alimento. Las hojas tienen pecíolos largos y delgados, de 20 a 40 cm de longitud; la lámina es hastada, con dos lóbulos basales triangulares, a menudo asimétricos, y un lóbulo basal más grande de ápice agudo. Se come las hojas más tiernas, a menudo con parte del pecíolo; como en otras Aráceas, la cocción elimina los cristales de oxalato de calcio y el látex que se encuentra en las hojas.
El clon ‘Picante’ también difiere en muchas de sus características de los clones de las demás especies de Xanthosoma. Este clon fue prospectado en la región oriental cubana y sus características morfológicas lo hacen muy diferente del resto del material estudiado; sin embargo, por la coincidencia de su descripción con la reportada por López (1970) para las especies del género Alocasia y por León (1987) para la Alocasia macrorrhiza se considera conveniente su ubicación como un clon de esta especie o de otra perteneciente a este género. Este último autor plantea que la A. macrorrhiza es una especie de cultivo primitivo en la que la parte utilizable es el

Caracterización de la variabilidad………….
34
tallo aéreo, cilíndrico hasta de cuatro metros de longitud. Por el alto contenido de oxalato, los troncos se limpian, cortan en tajadas y se someten a cocción prolongada. El tallo aéreo lleva hojas lanceoladas con láminas colocadas en el mismo plano que el pecíolo.
Por su parte, López (1970) añade que en Cuba las especies del género Alocasia no se cultivan como clones comestibles, siendo, en cambio, muy comunes en los jardines como plantas de adorno. En otros países se distinguen por la gran cantidad de almidón que contienen sus rizomas que además, presentan gran número de cristales de oxalato. La especie más común es A. macrorrhiza Schott de grandes hojas verdes y lustrosas, gran rizoma acre y cáustico y numerosísimos tuberculitos no comestibles, a la que también se le llama malanga de jardín (Roig, 1988).
El clon ‘Amarilla Trinidad’ presenta la hoja hastada y de color verde oscuro con borde morado, el pecíolo es de color morado oscuro con la base de color rojo y la arista morado oscuro, además, los cormos y cormelos son pequeños y tienen forma irregular, descripción que coincide con la expuesta por Cordero (1986) para la especie Xanthosoma atrovirens.
El presente estudio, al incluir por primera vez en esta colección, análisis morfoagronómicos, facilitaría la selección de las de mejores características para su explotación comercial, además de ayudar al esclarecimiento de la situación taxonómica de los clones dentro del género Xanthosoma, ya que según FAO (1996 a), se considera la falta de caracterización y evaluación como un obstáculo importante para utilizar los recursos fitogenéticos en programas de mejoramiento.
V. CONCLUSIONES
A partir de los 13 descriptores mínimos seleccionados, se caracterizaron los clones de la colección de germoplasma del género Xanthosoma y se pudieron conocer las particularidades de los grupos que lo conforman.
Los estudios realizados han permitido sugerir que en la colección cubana de germoplasma del género Xanthosoma, están representadas las especies Xanthosoma sagittifolium (L.) Schott, donde aparecen los clones de los grupos Blanco y Amarillo, Xanthosoma violaceum Schott, donde están incluidos los clones del grupo Morado, Xanthosoma brasilense Engl., cuyo único representante es el clon ‘Belembe’, Xanthosoma atrovirens Koch y Bouché, representada por el clon ‘Amarilla Trinidad’ y Alocasia macrorrhiza Schott, donde está incluido el clon ‘Picante’.
VI. RECOMENDACIONES
Emplear esta metodología en la caracterización de otras colecciones de germoplasma de raíces
y tubérculos.

Caracterización de la variabilidad………….
35
VI. REFERENCIAS BIBLIOGRAFICAS
ACOSTA, A. 1991. Identificación y descripción de las variedades de Yautía (Xanthosoma spp.), en
la colección de subestación en Gurabo. Universidad de Puerto Rico, Recinto de Mayagüez. ACOSTA, R. 1999. Caracterización citogenética, morfoagronómica y genético-bioquímica de diez
clones de plátano Burro (Musa spp.) grupo ABB. Tesis de Diploma, Lic. E n Biología, UH. AFZA, R; D., KAEMMER; J., DOLEZEL; J., KOENIG; M., VAN DUREN; G., KAHL; F.J.,
NOVAK. 1992. The potential of nuclear DNA flow-cytometry and DNA fingerprinting for Musa improvement programmes.International Symposium on Genetic Improvement of Bananas for Resistance to Deseases and Pest, Montpellier, 7-9 sept.
ALCÁZAR, M.D. 1995. Peroxidase isozymes in the defence response of Capsicum annoum to Phytophtora capsici. Physiol. Plantarum. 94: 736-742.
ANGEL, F.; D. ARIAS; J. THOME; C. IGLESIAS and W.M. ROCA. 1993. Towards the construction a molecular map of cassava (Manihot esculenta Crantz): Comparison of restriction enzymes and probe sources in detecting RFLPs. J. Biotech., 31:103-113.
AMANO, M; TSUNODA, S. NASUTION, R. NAKAMURA, R. NAKAMURA, T. HIRAI, Y.. 1989. Chromosome studies on some species. In genus Musa In: The 6 th International Congress of Sabrao. Tsukuba, Japan. 4 p.
ANDERSON,W.R;FAIRBANKS, D.J. 1990. Molecular markers important tols for plants genetic resources characterization diversity .V 6(3,4).
ARIAS, G., I. 1998a. Araceae. Fascículo 1/1. En: Flora de la República de Cuba. Serie A. Plantas vasculares. Koeltz Scientific Books. Koenigstern/ Federal Republic of Germany.—46p.
ARIAS, E. 1998b. Utilización de las isoenzimas en la caracterización de plantas de piña (Ananas sp.) de interés para el mejoramiento. Tesis para optar por el grado de Maestro en Ciencias Biológicas. Facultad de Biología. UH. 51 p.
BAI, K. V. 1982. Cytogenetics. Root and tubers 23. IITA, Nigeria. Pág. 114-116. BARRET, O. W. 1930. Los cultivos tropicales. La Habana cultural p.469-471. BLACKBURN, E. H. 1991. Structure and function of telomeres. Nature, 350: 569-573. BLACKBURN, E. H. 1992. Telomerases. Ann. Rev. Biochen., 61: 113-129. BLACKBURN, E. H.; SZOSTAK, J. W. 1984 The molecular structure of centromeres and
telomeres. Ann. Rev. Biochen., 53: 163-194. BONNER, J, W. WARNER, R. M. Y BREWBAKER, J.L. 1994. A chemosystematic study of Musa
cultivars. Hortsience, 9: 325-328. BREWER, G. J. y C. F. SINGH. 1971. Introduction to isozymes techniques. Acad. Pres. New York.
186 pp. BROWN, A. Y BERTKE, A. 1969. Citology. Edición revolucionaria. Instituto Cubano del Libro,
607 p.. BROWN, A.H.D. Y M.T. CLEGG. 1983. Isozymes of plant genetic resources. In: Isozymes:
Current topics in biological and medical research. Proc. 4th. Intern. Cong., Austin. 1982. Vol. 2: 285-295.

Caracterización de la variabilidad………….
36
CASPERSSON, T,; FARBER, S.; FOLEY, G. E.; KUDYNOWSKI, J.; MODEST, E. J.;SIMONSSON, E.; WAGH, U.; ZECH, L. 1968. Chemical differentiation along metaphase chromosomes. Exp. Cell Res., 49: 219-222.
CHAKRAVORTI, A. 1951. Origen of cultivated bananas of South East Asia. Indian J.G.. Plant Breeding II: 34-46.
CHAPEL, J., L. IGLESIAS, A. BARRETO, F. BAISNE y J. P. SIMÓN. 1974. A simplified apparatus for vertical slab electrophoresis in polyacrilamide gel. Laboratory practice. No. 23: 311 – 312.
CHETELAT, R. T. TAKAHASHI, M.DE VERNA, J.W. 1993. Maplocation Diaphorrase-3 (Día-3) a newisozyme marker on 9s. TGC Report. 43:10-11.
CLARKE, L. Y J., CARBON. 1980. Isolation of a yeast centromere and construction of functional small circular chromosome. Nature, 287: 504-509
CLARKE, L.; J., CARBON. 1985. The structure and function of yeast centromeres. Ann. Rev. Genet., 19: 29-56.
CONWUERME, I.C. 1978. The tropical Tuber Crops Jonh Wiley E Sons, Chicheter, New York CORDERO GARCÍA, M. 1975. Origen, distribución y clasificación botánica de la Yautía. En:
Curso de adiestramiento en el cultivo de la yautía. Sec. De Estado de Agricultura, Dpto de Producción. Dpto de Investigaciones Agropecuarias. Centro Agronómico Tropical de investigación y Enseñanza (CATIE)-Centro Norte de Desarrollo Agropecuario (CENDA).
CORDERO, M. 1986. Origen, distribución y clasificación botánica de la Yautía. En: Curso Nacional de Yautía. 20-24 de mayo de 1985. FAO. La Herradura. Santiago de los Caballeros, R. D. Pág 1-5. – 90p.
CORNIDE, M. T. y O, LEÓN. 1998. Principales aplicaciones de los marcadores moleculares. Uso de los marcadores moleculares en la genética y la selección de las plantas. CNIC.18-25 .
CUBA, MINAGRIG. 1984. Instructivo Técnico del cultivo de la malanga .-La Habana: CIDA, -p.23.
CUBA, MINAGRIG. Instituto de Suelos. 1995. Nueva versión de la clasificación genética de los suelos de Cuba. Ciudad de La Habana. 26 p.
CUBA, CONSEJO NACIONAL DE CIENCIA Y TÉCNICA. 1976. Situación y perspectivas de la genética vegetal en Cuba. I Parte. Primera reunión Nacional de Genética Vegetal. La Habana, Cuba. 39p.
CUBA, MINISTERIO DE LA AGRICULTURA. 1998. Instructivo Técnico sobre el cultivo de la malanga. Ciudad de la Habana.
CUENCO, A.; C. DE ARMAS Y H. GÁLVEZ. 1987. Comparación de siete clones de malanga (Xanthosoma spp.) Tesis para optar por el título de Técnico Agrónomo, curso 1986-1987. IPA “Martín Torres”-INIVIT.
DANTAS, J. ; SHEPHERD, K. ; SOARES FILHO, W.; CORDEIRO, Z.; SILVA, S.; SOUZA, A. 1993. Citogénética e melhoramento genético da bananeira (Musa spp.) EMBRAPA-CNPMF. Documentos 48. 61p.
DARLINGTON, C. D.1951. Mendel and the determinants. In. “Genetics in the 20 th Century”. New York.
DARLINGTON, C. D. Y A. P. WYLLIE. 1956. Chromosome atlas of flowering plants. London. Allen. Unwir. Pág. 374-393.
DAVIERS, J. C. 1973. Stadistics and date analysis in geology. I.Winley. Sans Inc. 536 pp

Caracterización de la variabilidad………….
37
DAVIS, B. J. 1964. Disc electrophoresis II. Methods and applications to human serum proteins. Ann. Ny. Acad. Sci. No. 121: 404 – 422. .
DEBOUK G. DANIAL. 1981. Proyectos de Recolección de Germoplasma de Phaseolus en México. Informe CIAT-INIA, 1978-1979.
ELLIS,.R.H; A. R. WALKINSON, R. J. SUMMERFIELD. 1995. Evaluation descriptors of times to floqering to the genetic characteristic.Plants Genetic Resources Newsletter. N103.
ENGLER, S. 1879. Araceae. In. Candolle, A. de, Monographie phanerogamarum.—Paris V.I. ENRIQUEZ, G. A. 1991. Relación de los recursos fitogenéticos con otras ciencias. Pág. 3-14. En:
Castillo, R.; J. Estrella; C. Tapia (Eds.) 1991. Técnicas para el manejo y uso de los recursos fitogenéticos. Dpto de Recursos Fitogenéticos. Instituto Nacional de Investigaciones Agropecuarias, Quito, Ecuador.—248p.
ESQUINAS ALCAZAR, J. T.. 1981.Los recursos fitogenéticos, una inversión segura para el futuro. Consejo Internacional de Recursos fitogenéticos – División de Producción y Protección de Plantas. Organización para la Agricultura y la Alimentación de las Naciones Unidas – FAO/Instituto Nacional de Investigaciones Agrarias. Ministerio de Agricultura y Pesca, España. Madrid. España, pág 11-16.
FAO. 1991. Production year book. Volume 45p. 67-184.Rome. Italy. FAO. 1996 a. Conservación y utilización sostenible de los recursos fitogenéticos para la
Alimentación y la Agricultura. Plan de Acción Mundial. Informe sobre el estado de los Recursos Fitogenéticos en el Mundo.
FAO, 1996b. Informe sobre el Estado de los Recursos Fitogenéticos en el Mundo. Preparado para la Conf. Técn.. Internac. sobre los Recursos Fitogenéticos. Leipzig, Alemania, junio-1996. 75p,
FAO-INIVIT. 1990. Manual de Intercambio y manejo de germoplasma in vitro de Yautía. Xanthosoma spp.. Oficina Regional para América Latina y el Caribe – Instituto de Investigaciones de Viandas Tropicales. Red de Cooperación Técnica en Producción de Cultivos Alimenticios de la oficina Regional de la FAO para América Latina y el Caribe (FAO/RLAC) en Cooperación con el Instituto de Investigaciones en Viandas Tropicales (INIVIT), Santo Domingo, Villa Clara, Cuba- Santiago de Chile.—25p.
FLORIDO, M. 1999. Caracterización de variedades y especies silvestres de tomate atendiendo a características morfobioquímicas y tolerancia al calor. Tesis de Maestría. 87 pp.
FRAGENE, M.; F. ANGEL; R. GOMEZ; F. RODRIGUEZ; M. BONIERBALE; J. THOME; C. IGLESIAS y W. M. ROCA. 1998. A linkage map of cassava (Manihot esculenta Crantz) based on RFLP and RAPD markers. In: Proceedings of Second International Scientific Meeting of Cassava Biotechnology Network, Bogor, Indonesia, 22-26 August, 1994. Working document No. 150.-- Centro Internacional de Agricultura Tropical (CIAT), Cali, Colombia, p. 49-61.
FUNDORA, Z.; R., VERA; E., YABER; O., BARRIOS. 1992. La estadística multivariada en la sanidad vegetal. INIFAT-MINAGRI, La Habana, 47 pp.
GALL, J.G Y M.L., PARDUE. 1969. Formation and detection of RNA-DNA hybrid molecules in cytological preparations. Proc. Nac. Acad. Sci., 63: 378-383.
GARCÍA, G. M. 1980. Prospección sobre Recursos Genéticos de Raíces y Tubérculos Tropicales. Centro de Mejoramiento de Semillas Agámicas (CEMSA) Santo Domingo, Villa Clara, Cuba.
GARCÍA, M. 1979. La conservación de los recursos genéticos: algunos apuntes sobre la importancia del germoplasma cubano en Viandas Tropicales, pág. 204-247. En: CEMSA, Ministerio de la

Caracterización de la variabilidad………….
38
Agricultura. Centro de Mejoramiento de Semillas Agámicas “Fructuoso Rodríguez” Memorias 1969-1975, 282 p. Santo Domingo, Villa Clara. Cuba.
GARCÍA, M.1990. Generalidades sobre el cultivo de la malanga Xanthosoma. Villa Clara, INIVIT.—6p. (Conferencia mimeografiada).
GARCÍA, D.; M.E., GONZÁLEZ; O., SAM. 1996. Algunas características asociadas con la ploidía en papa. Cultivos Tropicales. 17(1): 65-66.
GIACOMETTI, D.C. Y J. LEÓN. 1994. Tannia, Yautía (Xanthosoma sagittifolium) En: J. E. Hernando Bermejo and J. León. Neglected crops: 1492 from a Different Perspective. Plant Production and Protection Series N0 26 FAO, Rome, Italy p.253-258.
GOODING, H. J. y CAMPBELL, J. S. 1961. Preliminary trials of West Indiam Xanthosoma cultivars. Tropical Agriculture (Trinidad) 38: 145-152).
GOGORCENA, Y. Y L. M. ORTIZ. 1988. Identificación y caracterización de mandarinas y mandarinos híbridos mediante caracteres bioquímicos. Serie Prod. Vegetal: 1-18.
GOLA N., C. 1969. Tratado de Botánica. Edit. Rev. Instituto del Libro. GÓMEZ ARIAS, N. 1983. Germoplasma de Aráceas Alimenticias en Colombia. Universidad del
Valle. Facultad de Ingeniería. Departamento de Procesos Químicos y Biológicos. Sección de alimentos. Cali. Colombia. González, C. 1989. Comportamiento genético - bioquímico de la lima persa SRA - 58 (Citrus latifolia Tan.) sobre diferentes patrones en Cuba. 100p. Tesis Candidato a Doctor en Ciencias Biológicas. Esc. De Biología de La Habana.
GONZÁLEZ, C.; M. LAVISTA Y J. GONZÁLEZ. 1984. Estudio de patrones de isoenzimas para la lima persa. Prsc. Soc. Citricultura.—Brasil,
GONZÁLEZ, C.1989. Comportamiento genético - bioquímico de la Lima persa _ SRA-58 (Citrus latifolia Tan.) sobre diferentes patrones en Cuba. Tesis presentada en opción del grado de Candidato a Doctor en Ciencias Biológicas.
GREIDER, C.W. Y, E.H., BLACKBURN. 1985. Identification of a specific telomere terminal transference activity in Tetrahymena extracts. Cells, 43:405-413.
GUEDES, M. Y C.I., RODRÍGUEZ. 1974. Disc electrophoretic pattern of phenoloxidase from leaves of coffee cultivars. Sep. De Portugail. Acta Biológica. Serie A., Vol XIII:169-177.
HANSON, J. 1986. Methods of storing tropical root crop germoplasm with special reference to yam. Plant Genetic Resources Newsletter, 64:-24-32p., IBPGR-FAO.
HAUDRICOURT, A. 1941. Les colocaseis alimentaires. Rewee internatinale de Botanique Appliquie et agriculture tropical. 21(233-234): 40-65.
HAYWARD, M.D.; N.J., MCADAM; J.G., JONES; C., EVANS; G.M., EVANS; J.W., FORSTER; A., USTIN; K.G., HOSSAIN; B., QUADER; M., STAMMERS; J.K., WILL. 1994. Genetics markers and the selection of quantitative traits in forage grasses. Euphytica. 77: 269-275.
HERNÁNDEZ, M.; O. MILIÁN Y A. RODRÍGUEZ. 1991. Instructivo Técnico del cultivo de la malanga Xanthosoma (grupo amarillo). Instituto de Investigaciones en Viandas Tropicales (INIVIT) 6p.
HERNÁNDEZ, A.; J. M. PÉREZ; M. MORALES y R. LÓPEZ. 1995. Correlación de la nueva clasificación genética de los suelos de Cuba, con clasificaciones internacionales (Soil taxonomy y FAO-UNESCO) y clasificaciones nacionales (Segunda clasificación genética y Clasificación de series de suelos).
HOPKINS, A. A.; CH. M. TALIAFERRO. 1996. Chromosome numbers and nuclean DNA.V:36(5).

Caracterización de la variabilidad………….
39
HUSSAIN ,.A, H. RAMÍREZ, W .BUHUK. 1987. Identificación de cultivos de leguminosas forrajeras por métodos electroforéticos Agrotecnia V:67,713-717.
HYAMS, J.S.; B.R., BRINKLEY. 1989. Mitosis. Molecules and mechanisms. Academic Press, London, XII,350 p.
IBPGR. 1989. Descriptors for Xanthosoma. International Board for Plant Genetic Resources, Rome. IGLESIAS, L. 1988. Aplicación de la técnica electroforética en el manejo de los recursos
fitogenéticos. Conferencia: 6 pp. IGLESIAS, L. 1994. Utilización de marcadores bioquímicos y moleculares en el mejoramiento
genético de la papa. Cultivos Tropicales. 15(2): 108-121. IITA. 1982. Land preparation and planting in cocoyams. In: Tuber and Root Crops Production
Manual. Manual Series No. 0, May p. 199-201. IPGRI. 1995. Molecular genetic techniques for plant genetic resources. Report of IPGRI Workshop,
Roma, Italy, 9-11 octubre. Ayad, W. G., T. Hodgkin, A. Janadat y V. R. Rao, ed. 137 pp. IPGRI-INIBAP/CIRAD. 1996. Descriptores para el banano (Musa spp.). Instituto Internacional de
Recursos Fitogenéticos, Roma, Italia; Red Internacional para el Mejoramiento de Banano y el Plátano, Montpellier, Francia, y el Centre de cooperation internationale en recherche agronomique pour le développement, Montpellier, Francia. 55 pp. ISBN 92-9043-307-8
JACOBS, M.. 1974. Controle genetique des isozyme de leuciminopectidase chez Arabidopsis thaliana. Bull. Sec Reg. Bot. Belg. Inf. Lab. voor Planten-genetica. Brusel: 76-94.
JARRET, R. 1990. Molecular methods for detecting genetic diversity in Musa. In identification of genetic diversity the genus Musa. (R. L. Jarret. Ed.). INIBAP, France. Pp 8-16.
JO GARCIA, M.; R. HERNÁNDEZ; E. HERNÁNDEZ; H. DIAZ. 1992. Aplicación del análisis multivariadoen el estudio de la diversidad genética del frijol (Phaseolus vulgaris L.). Centro Agrícola, Año 19, No. 2-3, pág. 83-87.
KARP, A., K. V., SKRESOVICH, W. G. A., BHAT Y T., HODKIN. 1997. Molecular tools in plant genetic resources conservation: a guide to the tehnologies. IPGRI Technical Bulltin, No. 2, 47 pp.
KASEMSAK, P. 1992. Genome classification of bananas by chemotaxonomy. Bangkok. 137 p. KAT, T. A. 1978. La investigación básica en el plasma germinal. En: Recursos genéticos disponibles
en México, SOME FI, Boletín, Chapingo, México, s/p. KATSIOTIS, A.; R.A., FORSBERG. 1995. Pollen grain size in four ploidy levels of genus Avena.
Euphytica. 83: 103-108. LACADENA, J.R. 1970. Genética Vegetal. Fundamentos de su aplicación. (2da Edición). LACADENA, J.R.1995.Cytogenetic: yesterday, today and forever. A conceptual and historical view
Chromosomal Today. Vol 12. 842 pp. LACADENA, J.R. 1996. Citogenética. Ed. Complutense, Madrid, 931pp. LEBOT, V. Y ARADHYA, K.M. 1992 .Collecting and evaluating taro (Colocasia esculenta ) for
isozyme variation. Plant Genetic Resources Newsletter. No. 90. pp 47 - 49. LEON H. 1946. Flora de Cuba. Vol. 1 – Contr. Ocas. Mus. Hist. Nat. Colegio “De la Salle”. 8. LEÓN, J. 1968. Fundamentos Botánicos de los Cultivos Tropicales. San José, C.R. Inst.
Panamericano de Ciencias Agrícolas. OEA. Edit. IICA. Serie textos y materiales de enseñanza 18: 136-138.
LEÓN, J. 1976. Procceding of the fourth. Symposium of the International Society for Tropical Root Crops. Edited by James Cock, MacIrtyre and Graham. CIAT, Cali, Colombia, 1-7 August.

Caracterización de la variabilidad………….
40
LEÓN, J. 1987. Botánica de los cultivos tropicales. IICA. San José, Costa Rica, 445p. C 1968. XXI. Colección Libros y materiales educativos. IICA; No 84.
LEWONTIN,.R.C. 1979. La base genética de la evolución. Ediciones Omega .S.A. Barcelona, España.350p.
LINARES, G. 1988. Análisis de datos. La Habana. Pueblo y Educación, 590 pp. LIMA, G. H. Establecimiento de métodos y técnicas auxiliares al mejoramiento Genético de los
cítricos Tesis de doctorado. Univ. Habana, 1983 LÓPEZ ZADA, M. 1970. El cultivo de la malanga. Conferencia mimeografiada. Facultad de
Ciencias Agropecuarias, Universidad Central de Las Villas, Cuba.—39p. LÓPEZ ZADA, M.; E. VÁZQUEZ Y R. LÓPEZ FLEITES. 1995. Raíces y tubérculos. Ciudad de La
Habana. Ed. Pueblo y Educación. 2. Malanga, pág. 98-162.Cuba.—312p. LUGO, Y. 1996. Caracterización citogenética e isoenzimática de clones de la colección cubana de
malanga Xanthosoma. Tesis de Diploma. Universidad de Oriente, Santiago de Cuba.- INIVIT, Santo Domingo, Cuba.—41p.
MARCHANT, C. J. 1973. Chromosome variation in Araceae: V. Acoreae to Lasieae.—Kew Bull. 28: 199-210.
MARKERT, C. L. y F. MOLLER. 1959. Multiple forms of enzymes tissue, ontogenetics and species especif patterns. Proc. Nat. Acad. Sci. USA. 45: 753-763.
MARTÍN, F. W..1973. El almacenamiento de plasma germinativo en forma negativa de raíces y tubérculos tropicales. In. Conf. Téc. de la FAO sobre recursos genéticos de plantas de cultivo. Resúmenes. Roma. Italia.
MARTÍNEZ W., O.; L. M. REYES C.; M. BELTRÁN. 1998. Quimiovariabilidad en el género Musa: relaciones de similitud y diferenciación. Rev. INFOMUSA, La Revista Internacional sobre Bananos y Plátanos, Vol. 7, No.2, dic.
MILIÁN J. M.; I. SÁNCHEZ; M. GARCÍA. 1993a. Observaciones sobre la Lista de descriptores para la caracterización de los recursos genéticos en Xanthosoma spp.. Instituto de Investigaciones en Viandas Tropicales (INIVIT), Santo Domingo, Villa Clara, Cuba. 14p.
MILIÁN J. M.; I. SÁNCHEZ; M. GARCÍA. 1993b. Caracterización y evaluación de la colección cubana de malanga Xanthosoma spp. – Instituto de Investigaciones en Viandas Tropicales. INIVIT., Santo Domingo, Villa Clara, Cuba. 35p.
MILIÁN JIMÉNEZ, M.; I. SÁNCHEZ RAMOS; M. GARCÍA GARCÍA; D. GUERRA HERNÁNDEZ Y D. GUERRA HERNÁNDEZ. 1998. Variabilidad genética del género Xanthosoma en Cuba. Edición Especial VIII Jornada Científica. – Instituto de Investigaciones en Viandas Tropicales. INIVIT., Santo Domingo, Villa Clara, Cuba. 19p.
MILLER, O.L; BEATTY, B.R. 1969. Visualization of nucleolar genes. Science, 164:955-957. MONTALDO, A. 1977. Cultivo de raíces y tubérculos tropicales. Instituto Interamericano de
Ciencias Agrícola de la OEA, San José Costa Rica. MOSS, D. W. 1982. Isoenzimas. Capman and Hall, London and New York. NEAL, M. C. 1948. In Gardens of Hawai. Bernice P. Bishop Museum Spec. Publ. No. 40.—805 p. NGUYEN, V. X; H. YOSTINO; M. TAHANA. 1999. Genetic analysis of 12 polymorph isozime in
Colocasia esculenta V:40(3). NURSE, P. 1994. Ordering S phase and M phase in the cell cycle. Cell, 79: 547-550. O’HAIR, S. K. 1990. Tropical Root and Tuber Crops pág 424-428 In: J. Janick and J. E. Simon
(eds.) Advances in new crops. Timber Press, Portland, OR. Ohnishi, O. Matsuoka, Y. (1996)

Caracterización de la variabilidad………….
41
Search for buckwheat.2. taxonomy of Fagopyrum (Polygonaceae) pecies based on morphology, isozymes and ep ANA variability. Genes & Genetic Systems. 71 (6) 383-390.
ORNSTEIN, l. 1964. Disc electrophoreses 1. Bakground and theory. Ann. N. Y. Acad. Sci. No. 121-349.
ORTIZ, J.M. 1998a. Las isoenzimas y la resolución de problemas filogenéticos y evolutivos. Curso de Especialización en Recursos Fitogenéticos. España.
PANDEY, G.; V. K. DOBHAL y R. L. SAPRA. 1996. Genetic variability, correlation and path análisis in taro Colocasia esculenta. Journal of Hill research (India) V: 9(2), 299-302.
PEREA, D.M. 1995. Estrategias para el mejoramiento de bananos y plátanos. Augura. Vol. 18 (1): 56-65.
PÉREZ DE LA VEGA, M. 1997. El uso de marcadores moleculares en genética vegetal y mejora. Invest. Agr: Prod. Veg. 12 (1,2y3): 30-60.
PERRET, P.M. 1990. Background paper on field genebank, Musa Conservation and Documentation Proceedings of a Worshop. INIBAP. International Network for the Improvement of Banana and Plantains. Montpellier: 19-20.
PHILLIPS, P. L. 1993. Plant cytogenetics. En: Botanical Sciences (S.I.) : (S.N.) –p. 341-359. PIEDRAHITA G. C. 1979. Utilización de Aráceas comestibles en la alimentación e industria. Rev.
Esso Agrícola #34, Agosto – Septiembre. POEY, F. R. 1978. El mejoramiento integral del maíz: Valor nutritivo y rendimiento, hipótesis y
métodos. Colegio de post graduados, México.-- 206p. PURSEGLOVE, J. W. 1988. Tropical Crops Monocotyledons. Longman Cientific y Technical.
Copublished in the United States with John Willey y Sons, Inc., New York. QUIRÓS, C.F. 1991. Exine pattern of a hybrid between Lycopersicon esculentum and Solanum
pennelli. J. Hered. 66: 45-47. RAMIREZ, H.; A. HUSSAIN; W. M. ROCA and W. BUSHUK. 1987. Isozyme electrophoregrams
of sixteen enzymes in five tissues of cassava (Manihot esculenta Crantz) varieties. Euphytica, 36: 39-48.
RAMIREZ, H.; A. CALDERON Y W. M. ROCA. 1993. Técnicas moleculares para evaluar y mejorar el germoplasma vegetal.En: Cultivo de tejidos en la Agricultura: Fundamentos y aplicaciones. W. M. Roca y L. A. Mroginski (eds.). Centro Internacional de Agricultura Tropical (CIAT), Cali, Colombia, Parte C, Cap. 36, p. 825-856.
RAYMOND, S. 1964. Acrilamide gel electrophoresis. Ann. Ny. Acad. Sci. No. 121: 350–361. RITOSSA, F.M.; S., SPIELGERMAN. 1965. Localization of DNA complementary to RNA in the
nucleous organizer region of D. Melanogaster. Proc. Nat. Acad. Sci. 53: 737-745. ROIG, J. T.1913. Las especies y variedades de malanga cultivadas en Cuba.—Santiago de la Vegas.
E. A. 21. ROIG, J. T.1988. Diccionario Botánico de nombres vulgares cubanos. LL-Z. – Tomo II. 3 ra Edición.
Editorial Científico Técnica. La Habana. ROMÁN, M. I.; A., RODRÍGUEZ; X., XIQUÉS; C., GONZÁLEZ;. 1997. Caracterización
isoenzimática de 17 clones diploides de plátano fruta Musa spp. Biología, Vol. 11(1): 61-70. RODRÍGUEZ NODALS, A.A. 1973. Las colecciones de germoplasma en la Dirección General de
Semillas de la República de Cuba. Trabajo presentado en la Primera Reunión sobre Recursos Genéticos de la Región del Caribe. FAO-CATIE.- Turrialba, Costa Rica, p-5.

Caracterización de la variabilidad………….
42
RODRÍGUEZ NODALS, A.A. 1979. Botánica aplicada. Pág. 198. En: CEMSA, Ministerio de la Agricultura. Centro de Mejoramiento de Semillas Agámicas “Fructuoso Rodríguez” Memoria 1969-1975. 282 p. Marzo.
RODRÍGUEZ, A. 1991. Caracterización y evaluación de la colección nacional de Malanga (Colocasia esculenta) .Tesis para optar el título de ingeniera agrónoma, VCLV, Villa Clara,100p.
RODRÍGUEZ,.A. 1998. Caracterización del germoplasma Colocasia esculenta L. Tesis para optar por el título de maestro en Ciencias Biológicas. Universidad de La Habana.
RODRÍGUEZ MORALES, S,; M. GARCÍA GARCÍA; M. MILIÁN JIMÉNEZ AND I. SÁNCHEZ RAMOS. 1999. Establishment, maintenance and use under field conditions of Cuban germplasm of tropical root and tuber crops and banana and plantain. p. 25-29. En: Engelman, Florent (editor). Management of field and in vitro germplasm collections. Proceedings of a Consultation Meeting, 15-20 January 1996, CIAT, Cali, Colombia. International Plant Genetic Resources Institute, Rome, Italy. 162p. ISBN 92-9043-432-5
ROSERO, 1975. El cultivo de la yautía. (X. sagittifolium Schott) en la zona de Uraba, Departamento de Antioquia. Colombia. – 26p.
SÁNCHEZ RAMOS, I; MILIÁN JIMÉNEZ, M.; A. RAYAS; S. RODRÍGUEZ MORALES. 1998. Caracterización de la variabilidad en el germoplasma de ñame (Dioscorea spp.) en Cuba. Rev. Agrotecnia de Cuba. Edición Especial VIII Jornada Científica del INIFAT.
SANDOVAL, J.A. Y J., ESCOUTE. 1996. Aspectos citológicos y descripción de una metodología para el conteo de los cromosomas en el género Musa. CORBANA. Corporación Bananera Nacional 21(45): 51-56. San José Costa Rica.
SCANDALIOS, J. G. 1974. Isozimes in development and differentiation. Ann. Rev. Plant. Physical l. 25: 225-258.
SCHNELL, R. J.; R. GOENAGA; C. T. OLANO. 1999. Genetic similarities among cocoyam cultivars based on randomly amplified polymorphic DNA (RAPD) analysis National Germplasm Repository USDA-ARS. Scientia-Horticulturae, 80:3-4.—267-276.
SHEPHERD, K.. 1990. Observations on the conservation of Musa germplam. In: Musa Conservation and Documentation: Proceedings of a Workshop. INIBAP. International Network for the Improvement of Bananas and Plantains: 35-36.
SIGARROA, A. 1985.Biometría y Diseño Experimetal. La Habana. Ed. Pueblo y Educación 734pp. SIGARROA, A. Y M.T., CORNIDE. 1995. Marcadores moleculares para la selección de variedades
vegetales. Avances en Biotecnología Moderna. Vol. 3:17-47. SILAYOI, B. ; P. SOMPEN. 1991.Chromosome numbers and Karyotypes of some thai bananas.
Journal of Kasetsart. Science 25: 400-407. SIMPSON, M. J. A. y L. A. WITHERS. 1986. Characterization of plant genetic resources using
isozyme electrophoresis: a guide to the literature. A Technical Report Commissioned by the IBPGR Advisory Committee on in vitro storage. IBPGR. Rome.—102p.
STALKER, H. T.; T.D. PHILLIPS; J.P. MURPHY Y T.M. JONES. 1994. Variation of isozyme patterns among Arachis spp.. Theor Appl. Genet. 87: 746-755.
STEBBINS,.L.G. 1971. Chromosomal evolution in higher plants.LTD. London 216p. SWANSON, C.P. 1960. Cytology and Cytogenetics. Macmillan & Co., Ltd., London, X, 596 pp. TANDEHNJIE, J. 1990. Comparison of the crude protein and moisture contents of leaves and
petioles among cocoyam (Xanthosoma sagittifolium) accessions. Research Report submitted to ITA, University Center, Dschang, June.

Caracterización de la variabilidad………….
43
TANKSLEY, S.D. Y T. J., ORTOS. 1983. Isozymes in Plant genetic and Breeding. Part A (Eds.) Amrsterdan Elsevier. 109-138.
TAMBONG, J. T.; X. NDZANA; J. G. WUTCH; R. DADSON. 1997. Variability and germplasm loss in the Cameroon national collection of cocoyam (Xanthosoma sagittifolium Schott (L.)). Roots and Tuber Research Project (ROTREP), Institute of Agricultural Research. Cameroon. Plant Genetic Resources-Newsletter, No. 112.—49-54.
TEZENAS DU’ MONTCEL, H. Y P. PERRET. 1990. Proposals for an efficient network on International . Musa conservation. In: Musa Conservation and Documentation: Proceeding of a Workshop INIBAP. International Network for the Improvement of Banana and Plantains:31-34.
TOYAO, T. N. HEWSTONE Y C. MUÑOZ. 1991.Técnicas de recuento de cromosomas en plantas agrícolas. INIA. Boletíon Técnico No 191.
TRIEST, L. 1992. The role of isozymes in studies of plant population: several considerations of data obtained in water plants. Belg Joum. Bot. 125 (2) 262-269.
USTIMENKO, G. V. BAKUMOVSKI. 1982. El cultivo de plantas tropicales y subtropicales. Capítulo 3. Tubérculos feculentos. Editorial MIR, Moscú. Pág. 162- 224.—425p.
VALLS, J.F.; MASS, B..L.; LÓPEZ, C.R.. 1995.Recursos genéticos de Arachis silvestre y diversidad genética. En: Biología, Agronomía de especies forrajeras de Arachis. Edd. Kerridge, P.C. .CIAT.
WARNE, R. Y M. S. STRAUSS. 1985.Use of isoenzymes for the determination of genetic variability from tissue culture and between cultivars of taro (Colocasia esculenta) VII Symposium Guadaloupe.
WITHERS, L. A. AND WILLIAMS, J.T. 1998. “In vitro” conservation research highlights, International Board for plant Genetic Resourse, Rome. Italy. 91p.
WITHERS, L. A. 1988. Tissue culture for genetic conservation IBPGR Report, Rome. Italy. 91p. ZHENG, P.; CHENG, Y.D. 1993. Peroxidase isoenzymes analysis of Guang dong sweet potato
germplasm. In: Proceeding of the Asian Sweet Potato Germplasm. Network Meeting; Gianzhru.China. 25 February- 2 March.1991. Manila Filipines, International Potato Center, 114-119.
MORPHOAGRONOMIC CHARACTERIZATION OF GERMPLASM Xanthosoma GENUS IN CUBA Key words: characterization, variability, germplasm, Xanthosoma, descriptors, ABSTRACT The variability of the Xanthosoma germplasm made up of 71 clones which belong to the Araceas
germplasm bank in the Research Institute of Tropical Root and Tuber Crops (INIVIT) situated in
Santo Domingo, Villa Clara, Cuba was characterized. Based on morphoagronomic evaluations and
using multivariate analysis. It was possible to make a difference between genotypes to form affinity

Caracterización de la variabilidad………….
44
groups, as well as, to determine the most important variables for characterizations. Where a list of
13 minimum descriptors was defined. Surveys permitted to suggest the presence of Xanthosoma
sagittifolium (L.) Schott in the Cuban collection where clones from white and yellow groups are
available, Xanthosoma violaceum Schott involving clones from the purple group, Xanthosoma
brasilense Engl. Represented by ´Belembe´clone, Xanthosoma atrovirens Koch and Bouche based
on ´Amarilla Trinidad´ clone and Alocasia macrorrhiza Schott where ´Picante´ clone is included.