
Libro Curso de Biología Molecular y...
71
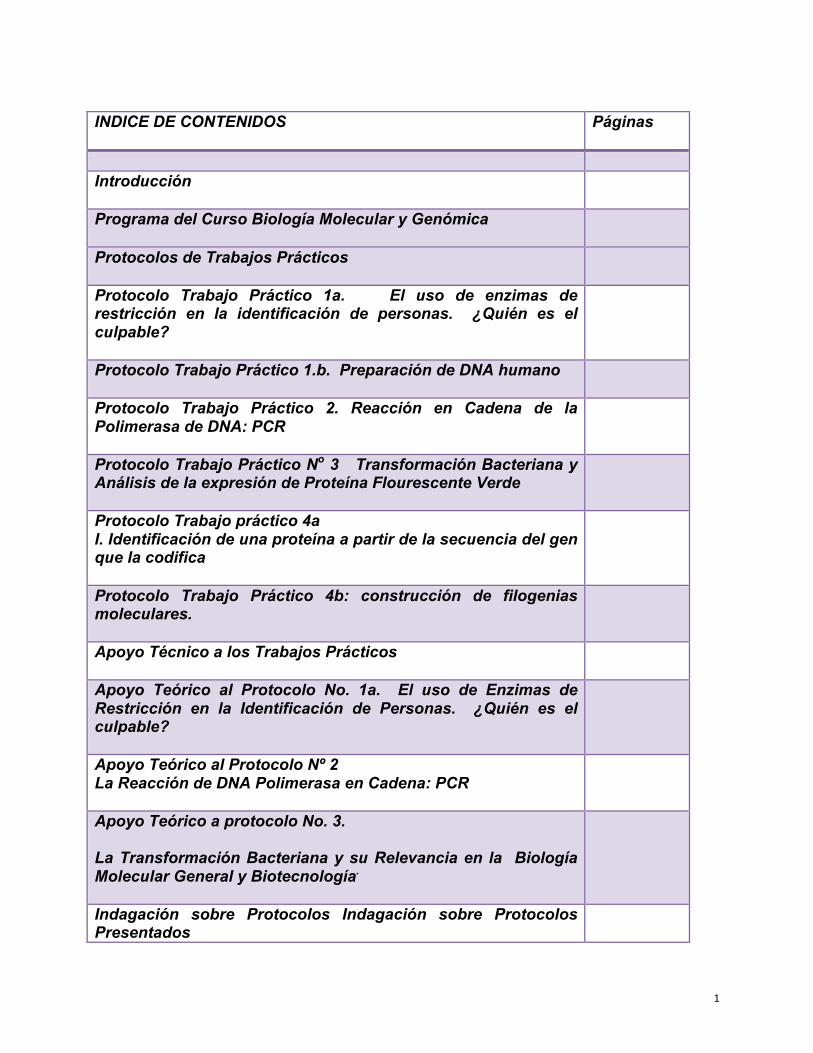
1 INDICE DE CONTENIDOS Páginas Introducción Programa del Curso Biología Molecular y Genómica Protocolos de Trabajos Prácticos Protocolo Trabajo Práctico 1a. El uso de enzimas de restricción en la identificación de personas. ¿Quién es el culpable? Protocolo Trabajo Práctico 1.b. Preparación de DNA humano Protocolo Trabajo Práctico 2. Reacción en Cadena de la Polimerasa de DNA: PCR Protocolo Trabajo Práctico N o 3 Transformación Bacteriana y Análisis de la expresión de Proteína Flourescente Verde Protocolo Trabajo práctico 4a I. Identificación de una proteína a partir de la secuencia del gen que la codifica Protocolo Trabajo Práctico 4b: construcción de filogenias moleculares. Apoyo Técnico a los Trabajos Prácticos Apoyo Teórico al Protocolo No. 1a. El uso de Enzimas de Restricción en la Identificación de Personas. ¿Quién es el culpable? Apoyo Teórico al Protocolo Nº 2 La Reacción de DNA Polimerasa en Cadena: PCR Apoyo Teórico a protocolo No. 3. La Transformación Bacteriana y su Relevancia en la Biología Molecular General y Biotecnología . Indagación sobre Protocolos Indagación sobre Protocolos Presentados
-
Upload
trinhxuyen -
Category
Documents
-
view
222 -
download
0
Transcript of Libro Curso de Biología Molecular y...
1
INDICE DE CONTENIDOS
Páginas
Introducción
Programa del Curso Biología Molecular y Genómica
Protocolos de Trabajos Prácticos
Protocolo Trabajo Práctico 1a. El uso de enzimas de restricción en la identificación de personas. ¿Quién es el culpable?
Protocolo Trabajo Práctico 1.b. Preparación de DNA humano
Protocolo Trabajo Práctico 2. Reacción en Cadena de la Polimerasa de DNA: PCR
Protocolo Trabajo Práctico No 3 Transformación Bacteriana y Análisis de la expresión de Proteína Flourescente Verde
Protocolo Trabajo práctico 4a I. Identificación de una proteína a partir de la secuencia del gen que la codifica
Protocolo Trabajo Práctico 4b: construcción de filogenias moleculares.
Apoyo Técnico a los Trabajos Prácticos
Apoyo Teórico al Protocolo No. 1a. El uso de Enzimas de Restricción en la Identificación de Personas. ¿Quién es el culpable?
Apoyo Teórico al Protocolo Nº 2 La Reacción de DNA Polimerasa en Cadena: PCR
Apoyo Teórico a protocolo No. 3. La Transformación Bacteriana y su Relevancia en la Biología Molecular General y Biotecnología.
Indagación sobre Protocolos Indagación sobre Protocolos Presentados

2
Indagación Protocolo 1. Trabajo Práctico I. El uso de las Enzimas de Restricción en la Identificación de personas. ¿Quien es el Culpable?
Indagación sobre Trabajo Práctico No. 2. La Reacción en Cadena de la DNA Polimerasa, PCR.
Trabajos Prácticos sin Laboratorios
I. Unidad de indagación sobre enzimas de restricción II. Parte Experimental. II.a. Duplicación de su DNA. II b. Sintetizar otra copia de su DNA de doble hebra
Lista Académicos Profesores Curso de Biología Molecular y Genómica
Lista de Profesores Curso de Biología Molecular y Genómica

3
INTRODUCCION
Este Curso Modelo Teórico- Práctico de Biología Molecular y Genómica fue diseñado por la Red Latinoamericana de Ciencias Biológicas (RELAB) y esta siendo patrocinado también por la Fundación “Wellcome Trust” del Reino Unido, por la Facultad de Medicina de la Universidad de Chile, por el Programa de Educación en Ciencias de la Red Interamericana de Academias de Ciencias (IANAS) y por la Fundación Allende Connelly. I.- Objetivos del Curso Objetivo General Contribuir al mejoramiento de la enseñanza – aprendizaje de la biología molecular y la genómica, mediante la capacitación teórica y práctica de profesores de biología de educación secundaria en cuatro conceptos fundamentales de esta áreas de la biología moderna mediante el acceso a materiales y a equipamiento sofisticado para que los profesores puedan compartir sus experiencias con sus alumnos usando el enfoque indagatorio. Objetivos Específicos
1) Profundizar el concepto fundamental de la biología molecular que nos plantea que la identidad de todos los seres vivos está determinada por la secuencia de deoxinucleotidos de su material genético. Por lo tanto, las diferencias y similitudes entre los seres vivos se pueden estudiar y definir mediante la lectura de esas secuencias.
2) Conocer y entender que las diferentes células de un organismo pluricelular contienen la
totalidad de la información genética de ese organismo. Por lo tanto, las diferencias en morfología, función y proteomas que existen entre las células de ese organismo se deben al hecho que la expresión de la información genética está regulada, solo expresándose una parte del total de la información. Esta regulación se hace a nivel de la transcripción (síntesis de RNA a partir del molde de DNA por las RNA polimerasas) y también a nivel post-transcripcional. Programas y mecanismos que regulan la expresión de la información genética juegan papeles esenciales en fenómenos biológicos tan importantes como la embriogénesis y la diferenciación celular. Los experimentos que demostraron la validez de este concepto, merecieron el Premio Nobel de Fisiología y Medicina del año 2012 al Dr. John Gurdon por sus hallazgos de transplantes nucleares en anfibios en 1962.
3) Entender los mecanismos y los procesos por medio de los cuales la información
genética genera otro tipo de macromoléculas, las proteínas. En la década de los 1960’s se dio un fundamental paso al aclararse la manera como la información genética escrita en el idioma de los nucleótidos puede dar origen a proteínas cuya estructura está constituida por 20 diferentes aminoácidos. Los laboratorios de Niremberg, Ochoa y Khorana, descifraron el código genético y donde hay combinaciones de tres nucleótidos (codón) que codifican para un aminoácido. En esa misma década en el laboratorio de Lipmann y otros laboratorios se aclararon los factores que intervienen en la síntesis de proteínas a partir del RNA mensajeros.

4
Esos resultados nos dijeron que: a) el código genético es universal (o casi universal) para todas las especies vivientes y b) que la maquinaria para traducir los RNA mensajeros a proteínas son también muy parecidas. Esta información tiene una enorme implicancia y nos dice que todos los seres vivos somos compatibles en cuanto podemos usar la información genética de cualquier otro organismo. En otras palabras que la ingeniería genética es posible. En la década de los 70’s se obtuvieron los avances técnicos que convirtieron a la ingeniería genética en una realidad e hicieron que la biotecnología se desarrollara enormemente.
4) Conocer y manejar el concepto que la evolución de las especies de los seres vivos está
impulsada por las mutaciones que sufre el DNA que se transmite a la descendencia de los organismos vivos. Estas mutaciones son alteraciones (cambios, deleciones, inserciones) en las secuencias de nucleótidos del material genético y que con cierta frecuencia altera la estructura y la función de sus productos génicos. De acuerdo a los postulado por Darwin, si los cambios causados por las mutaciones resultan en una ventaja competitiva en la propagación de las especies, esas mutaciones se perpetúan en la descendencia. Los genes y sus mutaciones, por lo tanto, reflejan el proceso evolutivo y pueden servir de relojes para medir el tiempo que ha intervenido en la separación de especies relacionadas. La genómica evolutiva se centra en el estudio de la evolución a nivel molecular y se nutre en la comparación entre genes similares y el número de mutaciones que los diferencia. Los bancos de datos con cientos de miles de genes y la bioinformática nos permiten ahora hacer esos estudios totalmente “in silico”.
5) Entregar a los Profesores que se capacitan en el curso la posibilidad de transmitirles a
sus alumnos el aprendizaje de los conceptos 1-4 mediante materiales didácticos con el enfoque ECBI y mediante la visita de laboratorios portátiles a sus establecimientos educacionales junto con estudiantes de postgrado en biología molecular que colaborarán en el montaje de los experimentos similares a los realizados en el curso.
Historia y Perspectivas Nuestro grupo de la Facultad de Medicina de la Universidad de Chile tiene una larga experiencia en organizar cursos teórico-prácticos en biología molecular. Inicialmente estos curso se centraron en alumnos de doctorado y post doctorado (1968-1996). En los últimos 15 años nos centramos en cursos de biología molecular para profesores de biología de la secundaria. La RELAB que nació en 1975, siguió esa misma ruta entrando en el siglo XXI a la organización de cursos para profesores secundarios que se hicieron en Chile, en Argentina y en Costa Rica. En el año 2011, el Consejo Directivo Regional RELAB reunido en Santiago en Agosto, decidió dar un giro a estos cursos teórico-prácticos para resolver un serio problema que limitaba su utilidad. Ese serio problema era que los profesores que tomaban esos cursos no podían compartir la importante experiencia de los experimentos con sus alumnos. Para resolver este problema se nos ocurrió la idea de los laboratorios portátiles que pudieran ser enviados a los establecimientos educacionales para que los estudiantes pudieran hacer también parte importante de los experimentos. Los profesores que podrían solicitar la visita de esos equipos científicos serían solo aquellos que ya habían aprobado el curso teórico-práctico en la Universidad. Para poder financiar una experiencia piloto, postulamos a un proyecto de la

5
Wellcome Trust del Reino Unido que nos aprobó un proyecto por USD$ 48.000 dólares. Con esos fondos RELAB decidió organizar un curso piloto en Enero del 2012 en Santiago y cursos similares en la Universidad Nacional Autonóma de México (UNAM), la Universidad de Costa Rica en San José y en la Universidad de Chile aquí en Santiago. Lo primero que decidimos era diseñar un curso que se focalizará en aspectos fundamentales de la biología molecular y la genómica. De esa discusión salieron los 4 conceptos que presentamos anteriormente. Luego diseñamos clases y experimentos que sirvieron para profundizar en estos conceptos. Ese fue el curso piloto que realizamos en Enero de este año en presencia de los observadores de México y Costa Rica. El curso fue muy exitoso según los profesores que lo tomaron y nos dio la idea de recortar algunos contenidos y algunos experimentos. Después de esto el mismo curso se repitió en Costa Rica en la primera semana de Julio y nuevamente en México en la última semana de ese mismo mes. Las organizadoras de esos cursos, la Dra. Ana Victoria Lizano en Costa Rica, y la Dra. Claudia Segal en México, nos han informado que esos cursos a lo que fueron becados profesores de otros países latinoamericanos fueron muy exitosos y que hay mucho entusiasmo por continuar con los cursos. Nuestra idea de estos cursos fue presentada en la reunión anual de la American Association for the Advancement of Science (AAAS). En conversaciones posteriores a esa presentación los Profesores Tim y Karen Spuck nos informaron que en Estados Unidos hay programas muy parecidos. En el Estado de Pennsylvania existe un Programa llamado Ciencia-en-Movimiento (www.science-in-motion.org) que tiene como participantes un grupo de 12 universidades que han recibido del Estado equipo científico sofisticado que las universidades envían a los establecimientos de educación secundaria acompañados de un docente que colabora en el montaje de los experimentos. Este programa tiene equipo para los laboratorios portátiles en todas las aéreas de las ciencias experimentales (física, química, biología, astronomía). La National Science Foundation que es como el CONICYT de Chile tiene un programa de becas de postgrado en ciencias (www.gk-12-org) que ha otorgado becas a más de 11000 estudiantes de Magister y Doctorado con la condición que estos becarios le dediquen un tiempo significativo de su tiempo para colaborar en la docencia pre-universitaria de las ciencias y la ingeniería. Por medio de la Embajada de Estados Unidos conseguimos el viaje de los esposos Spuck a Chile en Julio de este año lo que les permitió informar a autoridades del MINEDUC y de CONICYT y a los directivos de postgrado de las Universidades en Santiago, La Serena, Valparaíso y Concepción sobre estos proyectos. Las autoridades chilenas quedaron muy bien impresionadas con estos dos programas y se comprometieron a estudiar seriamente la posibilidad de apoyar un proyecto de laboratorios portátiles que ayuden a mejorar la educación en ciencias. Actualmente estamos trabajando para preparar un proyecto FONDEF que nos permita armar un piloto de proyecto “ciencia-en-movimiento” de Chile. Este curso es un eslabón importante para ese proyecto.
JORGE E. ALLENDE

6
Programa del Curso Biología Molecular y Genómica Instituto de Ciencias Biomédicas
Programa de Biología Celular y Molecular Facultad de Medicina - Universidad de Chile
Código CPEIP 12-0287 5 al 9 Noviembre 2013
Lunes Concepto Fundamental 1 La Identidad genética de especies e individuos se define por la información contenida en el DNA de cada célula 08:30 Bienvenida e Introducción al Curso
08:45 Evaluación Inicial
09:00-11:30 Clase 1 Dr. Jorge E. Allende
Una visión personal de la historia de la Biología Molecular
• DNA como material genético • El Dogma Central: de DNA a mRNA a Proteína
Café • El Proyecto del Genoma Humano
11:40-13:00 Clase 2 Dr. Juan Venegas
El Proceso de Duplicación de DNA
• Las DNA polimerasas • Introducción a la técnica de PCR
13:00-14:00 Almuerzo
14:00-17:00 Trabajo Práctico
• Dra. Germaine Jacob • Dra. Gittith Sánchez
1) Enzimas de Restricción 2) Preparación de DNA humano para usar en PCR
Martes Concepto Fundamental 2 Todas las células de un organismo multicelular tienen la información genética total del organismo, y, aún así, sus células son diferentes en morfología y función y expresan diferentes tipos de proteínas.

7
09:00-11:30 Clase 3 Dr. Miguel Allende Regulación de la Expresión Génica
• Regulación a nivel de la transcripción en procariontes y eucariontes.
• Células madres Café
• Embriogénesis
11:30-13:00 Clase 4 Dr. Héctor Toledo Regulación post-transcripcional y post-traduccional
• Regulación génica a nivel post-transcripcional • Procesamiento de mRNA • Regulación génica a nivel post-traduccional • Proteómica
13:00-14:00 Almuerzo
14:00-17:30 Trabajo Práctico
• Dra. Nora Riveros
1) Reacción en Cadena de la DNA Polimerasa
Miércoles Concepto Fundamental 3 El código genético que traduce la información genética para generar proteínas con secuencias aminoacídicas especiales es casi universal para todos los organismos vivos. El mecanismo de transducción basado en los ribosomas también es muy similar. Estos dos hechos son la base para la ingeniería genética por medio de la cual el material genético de una especie diferente puede ser expresado correctamente en una célula que recibe este material. 09:00-11:30 Clase 5 Dr. Marcelo Antonelli
La maquinaria de la transducción
• El código genético • La síntesis de proteínas
Café • Ingeniería genética y los vectores de expresión • Genotecas
11:40-13:00 Clase 6 Dr. Marcelo Antonelli
Biotecnología y sus Aplicaciones

8
13:00-14:00 Almuerzo
14:00-17:00 Trabajo Práctico
• Dr. Marcelo Antonelli
Herramientas para Ingeniería Genética
• Transformación de una bacteria con el gen de la proteína GFP (proteína verde fluorescente).
Jueves Concepto Fundamental 4 Se puede observar la evolución darwiniana a nivel molecular mediante el estudio de las secuencias de genes y de sus productos proteicos en diferentes especies. La evolución molecular y su ocurrencia en el tiempo que permiten estimar cuando se separaron las diferentes ramas de los seres vivos, se pueden analizar observando las diferencias en las secuencias de genes y proteínas. 09:00-10:30 Clase 7 Dr. Andrés Iriarte
Introducción a la Bioinformática • Aplicaciones de la informática en el genoma humano
Café • Aplicaciones de la informática en medicina
11:00-13:00 Clase 8 Dr. Mauricio Moraga
Filogenia y Evolución
13:00-14:00 Almuerzo
14:00-17:30 Trabajo Práctico • Dr. Sergio Flores
Introducción a la Bioinformática
1) Identificación de una proteína a partir de la secuencia del gen que la codifica
2) Estudios de Filogenia en el computador
Viernes 09:00-13:00 Discusión y Análisis
Evaluación del curso : sugerencias y críticas
• Discusión General en que intervienen todos los participantes del curso.

9
• Biología Molecular y Genómica: Impactos en la Salud, Agricultura, la Sociedad y la Ética
• Mesa Redonda: Dres. Jorge E. Allende, Hector Toledo, Juan Venegas, Nora Riveros, Andrés Iriarte, Marcelo Antonelli, Catherine Connelly, Eugenio Spencer
14:00-15:45 Discusión y Análisis
• Propuestas de cómo introducir la metódica de la
indagación en cursos de Biología de enseñanza media. Sugerencias de esquemas en Biología Molecular y Genética. Dra. Pilar Reyes, Profesora Filomena Vargas
16:00-16:30 Evaluación Final
16:30-17:00 Despedida

10
PROTOCOLOS DE TRABAJOS PRACTICOS

11
Protocolo Trabajo Práctico 1a. El uso de enzimas de restricción en la identificación de personas. ¿Quién es el culpable? En este trabajo práctico se utiliza la técnica de "fingerprinting" o "huellas" de DNA como herramienta forense para identificar al culpable de un crimen. Cada grupo recibirá 6 muestras de DNA para utilizar en la identificación de un "culpable." El protocolo 1A. tiene dos partes: parte A, la digestión del DNA con tres enzimas de restricción y una parte B, el análisis de los fragmentos de DNA producidos por esta digestión mediante el proceso de electroforesis en un gel de agarosa. Una presentación sobre los mecanismos de las reacciones de las enzimas de restricción y su importancia para la identificación del DNA de las personas, se encuentra en la sección Apoyo Teórico al Protocolo 1. Los protocolos presentados han sido adaptados para este trabajo práctico un protocolo comercial presentado por Bio Rad en su "Kit" Biotechnology Explorer DNA Fingerprinting, Bio Rad Catalog Number 166-0007-EDU.
***** Materiales disponibles para cada estación de trabajo. Se dispone de muestras de DNA obtenidas en el lugar del crimen y de 5 sospechosos. EcoR1 /Pst 1, enzimas de restricción 1 tubo Puntas de pipetas automáticas 15 puntas Micropipeta automática 1 de 10 microlitros Microtubos Eppendorf de colores 6 tubos Soporte para microtubos 1 Un marcador de tubos y hielo El laboratorio proporcionará: Incubadora 37ºC o baño de agua a 37ºC 1 Geles de Agarosa al 8% preparados 1 Cámaras de electroforesis 3 Fuente de Poder 1 Transiluminador 1 Centrífuga de alta velocidad (6000 rpm) 1 Centrífuga de baja velocidad (minifuga) 2

12
Protocolo Experimental: PARTE A. Digestión de las muestras de DNA con una mezcla de enzimas de restricción. l. La mezcla de 3 enzimas de restricción marcada como ENZ se maneja en hielo. 2. Marque los tubos Eppendorf de colores de la siguiente manera: EC = escena del crimen (verde) Sl = sospechoso I (azul) S2 = sospechoso 2 (naranja) S4 = sospechoso 3 (violeta) S3 = sospechoso 4 (rojo) S5 = sospechoso 5 (amarillo) Marque los tubos con el nombre de su grupo y coloque los tubos en el soporte de espuma. 3. Pipetee l0 µl de cada muestra de DNA y transfiéralo al tubo Eppendorf que corresponda. Utilice una punta nueva para cada muestra de DNA. Asegúrese que cada muestra quede en el fondo del tubo. 4. Pipetee l0 µl de la mezcla de enzimas, ENZ y agréguela cada tubo, depositándola en el fondo de cada tubo. Utilice una punta nueva para cada tubo. 5. Tape los tubos y mezcle los componentes agitándolos con golpes de dedo. Centrifugue los tubos durante 5 segundos a máxima velocidad para recolectar toda la solución al fondo de los tubos. 6. Coloque los tubos en el soporte/flotador de espuma e incúbelos durante 45 minutos a 37º C en un baño termoregulado o hasta el día siguiente a temperatura ambiente. 7. A los 45 minutos de incubación a 37ºC, retire los tubos del baño termo regulado y colóquelos en el refrigerador hasta la etapa siguiente.

13
PROTOCOLO 1a. RESUMIDO RECUERDE DE USAR UNA PUNTA DE PIPETA NUEVA CADA VEZ QUE PIPETEE Y ANOTE CADA ACCION CON UN CHEQUEO: ! " Marcar los microtubos Eppendorf con lápiz negro permanente: EC, S1, S2, S3, S4, S5 " Poner los microtubos en un soporte sobre hielo. " Adicionar muestras de DNA, uno por uno al tubo indicado: 10 µl. " Adicionar la muestra de enzimas de restricción (ENZ) a cada tubo: 10 µl. " Tapar y agitar cada tubo usando un "Vortex" o con golpes de dedo. " Centrifugar a máxima velocidad por 5 segundos en una minifuga. " Poner cada tubo en el soporte para la incubación. " Incubar los tubos durante 45 minutos a 37ºC en baño termo regulado. " Retirar los tubos a los 45 minutos y colocarlos en hielo o en el refrigerador.
Parte B. Separación de los fragmentos por electroforesis en geles de agarosa. l. Saque del refrigerador o hielo las muestras de DNA digeridas. En cualquier caso, centrifugue por 5 segundos a máxima velocidad para bajar toda la solución de los tubos hasta el fondo. 2. Utilizando una punta nueva para cada muestra, coloque 5 µl del amortiguador de carga (loading dye "LD') en cada tubo. Tape los tubos y mezcle golpeando ligeramente con el dedo. 3. Ud. dispondrá de un gel de agarosa al 0,8 %, colocado en la cámara de electroforesis con el amortiguador TAE, el cual debe cubrir el gel. ¡ATENCION! El amortiguador TAE utilizado para preparar los geles de agarosa contiene bromuro de etidio, un potente agente mutagénico, por lo tanto, debe usar guantes en todas las operaciones siguientes. 4. Asegúrese que los pocillos del gel estén próximos al electrodo negativo (negro). Los fragmentos de DNA (cargados negativamente) migran hacia el polo positivo (electrodo rojo).

14
5. Utilizando una punta nueva en cada caso, cargue primero la muestra de estándares de peso molecular de DNA y a continuación, cargue las muestras en el gel en el siguiente orden: Pocillo Muestra ❏ carril l: estandares de pesos moleculares, l0 µl ❏ carril 2: EC 20 µl ❏ carril 3: S1 20 µl ❏ carril 4: S2 20 µl ❏ carril 5: S3 20 µl ❏ carril 6: S4 20 µl ❏ carril 7: S5 20 µl 6. Coloque la tapa de la cámara de electroforesis y conecte los terminales a la fuente de poder. 7- Dé el paso de corriente y corra la electroforesis a 100 volts. 8. Cuando termine la electroforesis, apague la fuente de poder y remueva la tapa de la cámara. Con cuidado, saque la bandeja que contiene el gel. iCUIDADO! EI gel es MUY resbaloso. Empuje suavemente el gel con el pulgar y deje que resbale hacia una bandeja plástica para su posterior manipulación. 9. Exponga el gel a la luz ultravioleta en un transiluminador y obtenga una imagen de él. 10. Discuta los resultados obtenidos y decida quien es el culpable

15
Protocolo Trabajo Práctico 1.b. Preparación de DNA humano Para la preparación de DNA humano se eligió usar las células de la mucosa bucal de los mismos participantes del curso. El proceso de extracción requiere el uso de un detergente suave para romper la membrana celular y una solución salina (nace al 0,9% P/V) para limitar las interacciones iónicas entre las moléculas. El protocolo también incluye la adición de un producto llamado Instagene que es una resina de intercambio iónico (producto de BioRad ) que remueve metales pesados de la preparación que podrían inhibir el proceso de PCR. Este protocolo simple, indicado a continuación, permite obtener un DNA relativamente intacto para su uso posterior en el Trabajo Práctico de PCR. 1. Cada miembro del equipo debe tener:
• 1 tubo con tapa que contiene 200 µl de Instagene, (marcarlo con sus iniciales)
• 1 microtubo de 1,5 ml (marcarlo con sus iniciales) • 1 taza (copa) que contiene 10 ml de solución salina 0,9% (marcarlo con sus
iniciales) 2.. Vierta la solución salina de la copa a la boca
• No botar la solución salina después de completar este etapa. • Enjuague vigorosamente durante 30 segundos. • Expulsar el enjuague de la solución salina (que ya tiene células de la
mucosa bucal) de vuelta a la taza. 3. Fije el volumen de una micropipeta P-1000 a un volumen de 1000 microlitros (1 ml) y transfiera 1 ml de la solución de enjuague al microtubo con sus iniciales. Si no tiene una P-1000, vierta 1 ml de la solución salina con cuidado en el microtubo (use de las marcas de volumen del mismo microtubo para medir 1 ml. 4. Centrifuge su tubo en una centrífuga equilibrada durante 2 minutos a velocidad máxima. Cuando la centrífuga se haya detenido completamente, retire el tubo. Usted debe ser capaz de ver un sedimento de células blancas (una bolita) en el fondo del tubo. Lo ideal sería que el sedimentado sea el tamaño de la "cabeza" de un fósforo. Si usted no puede ver la bolita, o es demasiada pequeña, vierta el sobrenadante salino y adicione más de su enjuague salino, y centrifugue de nuevo. 5. Bote el sobrenadante y descártelo. Teniendo cuidado de no perder su bolita o pelet de células, cuidadosamente limpie el exceso de líquido en el microtubo invertido con un pañuelo o toalla de papel. Es aceptable que una pequeña cantidad de solución salina (50 µl) quede sobre el pelet en el fondo del tubo. 6. Resuspender el pelet totalmente por agitación con la mano o con un vortex hasta que no haya grumos de células.

16
7. Usando una micropipeta de volumen variable fijada en 20 µl, transfiera sus células resuspendidas a un tubo con tapón de rosca que contiene el InstaGene con sus iniciales. Puede ser necesario utilizar la pipeta varias veces para la transferencia de todas sus células. 8. Atornille las tapas de los tubos fuertemente. Agite o utilice un vortex para mezclar el contenido. 9. Coloque los tubos en el soporte de espuma. Cuando todos los miembros de su equipo hayan recogido sus muestras, deje flotar el soporte flotar en el baño de agua a 56ºC durante 10 minutos. En el punto medio (5 minutos), agite los tubos varias veces. Coloque los tubos de nuevo en el baño de agua para los otros 5 minutos. El total de tiempo de incubación en el baño de agua a 56° C es de 10 min. 10. Retire los tubos del baño de agua y agite varias veces. Ahora coloque los tubos en un baño de agua a 100º C durante 5 minutos. 11. Retire los tubos del baño de agua y agite varias veces para volver a suspender la muestra. Ubique ocho de los tubos en una centrifuga Deje sedimentar el pelet por centrifugación durante 5 minutos a 6.000 x g (o 10 minutos a 2.000 x g). 12. Guarde su tubo tapado en el refrigerador hasta el período siguiente de laboratorio, o vaya al paso 2 de la lección 2 si su profesor le indica que debe hacerlo.

17
PROTOCOLO 1b. RESUMIDO
ANOTAR EN EL RESUMEN CADA ACCIÓN AL TERMINARLA: !
" Marcar un tubo de 1,5 ml, un tubo con tapa y el vaso con sus iniciales. o " Colectar las células de la mucosa bucal mediante un lavado de la boca con solución salina (se llamará 'enjuague'). o " Transferir 1 ml del enjuague al tubo de 1,5 ml o " Centrifugar a máxima velocidad por 2 minutos para obtener una bolita de células en el fondo del tubo. Botar el sobrenadante, cuidadosamente. o " Agitar el tubo (prefiera un Vortex) hasta dispersar las células y que no haya grumos. o " Transferir 20 µl de las células al tubo con tapa que contiene “Instagen”. Tapar el tubo " Agitar el tubo usando preferencialmente un Vortex. o " Cuando todos los grupo tengan listos sus tubos con tapas se inicia la incubación a 56°C por 10 minutos. (Completados los primeros 5 minutos, agitar el tubo con un Vortex. A continuación, calentar el tubo en un baño de 100°C por 5 minutos. " Retirar el tubo, agitarlo, y centrifugar a 6000 x g por 5 minutos. Guardar el tubo en el refrigerador hasta el siguiente trabajo práctico.

18
Protocolo Trabajo Práctico 2. Reacción en Cadena de la Polimerasa de DNA: PCR En este trabajo práctico se analizarán el DNA humano preparado anteriormente por cada participante del curso (Protocolo 1.b). La misma preparación de su DNA se analizará en este práctico por la presencia o ausencia de un gen específico y posteriormente se comentará la distribución del gen dentro de este mismo grupo de personas. El gen en estudio se llama ALU PV92 y está ubicado en el cromosoma 16. La presencia o ausencia de este gen no refleja una ventaja o desventaja genética para las personas pero, como es un gen muy estudiado en humanos, su análisis le da un interés adicional al trabajo práctico que pretende demostrar la capacidad analítica del método de PCR. La cantidad del DNA preparado ayer para este experimento representa una cantidad muy, muy pequeña de su DNA y es insuficiente para una análisis químico típico. Para analizar su DNA se requiere amplificarlo y para está amplificación es necesario utilizar un instrumento llamado "Termociclador." Este instrumento termociclador ha revolucionado los estudios científicos, médicos y forenses debido a la amplificación de trazas de DNA para su posterior análisis en el laboratorio. Una descripción detallada de la técnica de PCR y de los genes ALU esta presentada en sección Apoyo Teórico de este guía. Durante todo el proceso de la amplificación de un DNA en el termociclador, las reacciones realizadas en forma cíclica por este instrumento son: 1. - denaturación del DNA 1 minuto a 94ºC 2. - hibridación de los partidor, 1 minuto a 60ºC 3. - polimerización 2 minutos a 72ºC En la etapa 1. se separan las hebras de DNA para formar monohebras y esto ocurre en forma óptima a 94ºC. Para copiar una hebra simple de DNA, se requiere un "partidor" que permite iniciar la síntesis de la hebra nueva en la posición 3´ del DNA molde (etapa 2.). En la etapa de polimerización, la temperatura es de 72ºC, la temperatura óptima para la enzima Taq polimerasa que sintetiza una cadena nueva del DNA (etapa 3.). Se repite 40 veces los pasos de denaturación, hibridación y polimerización, cada paso a la temperatura apropiada y finalmente, se realiza una polimerización continua por 10 minutos a 72ºC. Este protocolo está basado en un Kit de PCR Informática diseñado por Biotechnology Explorer™ de Bio Rad, "Chromosome 16: PV92," Catalog #166-2100EDU.

19
Protocolo Experimental: Cada estudiante de cada grupo debe tener: un tubo de PCR un tubo sin tapa que funciona como adaptador un tubo con su DNA (guardado del Trabajo Práctico 1b.) 1. Marque cada tubo con sus iniciales y ponga el tubo de PCR en el tubo sin tapa, el que funciona como un adaptador que sostiene el tubo de PCR y enseguida, ponga el tubo de PCR con su adaptador en la gradilla. 2. Localice ahora la solución PCR Master Mix (etiquetada "Master") que debe estar en el recipiente con hielo. Transfiera 10 µl del Master Mix a su tubo de PCR. 3. Coloque 5 µl de su DNA, guardado del Trabajo Práctico 1b., en el tubo de PCR ya con la solución Master Mix. En seguida, adicione 5 µl de agua. Agite golpeando suavemente el fondo del tubo. 4. Centrifugue en la Minifuga para asegurar que toda la solución esté en el fondo del tubo. Coloque el tubo de PCR en el termociclador. 5. Cuando todos los tubos PCR estén en el Termociclador, el Instructor iniciará el termociclador para que comiencen los ciclos térmicos de la reacción de PCR. Se programarán 40 ciclos de amplificación, lo cual tomará aproximadamente 2 horas. El programa completo de amplificación que se usará será el siguiente: Programa de Amplificación
Ciclo Paso Función Temperatura Tiempo 1 1 Pre-denaturalización 94º C 2 minutos 2* 1 Denaturalización 94º C 1 minuto 2 Hibridización 60º C 1 minuto 3 Extensión 72º C 2 minutos 3 1 Extensión final 72º C 10 minutos *Repetir 30 a 40 veces los pasos 1, 2, y 3 del ciclo 2 (Este valor depende del largo del DNA en estudio).

20
Al término de la amplificación remueva las muestras de PCR del termociclador y póngalas en una gradilla. A continuación se realizará un análisis de los productos por electroforesis en geles. Procedimiento para la electroforesis en geles de Agarosa 1. Remueva las muestras PCR-del Termociclador y pongalas en una gradilla 2. En un gel de agarosa previamente preparado, colocado dentro de una cámara de electroforesis con alrededor de 275 ml de buffer de electroforesis, coloque las muestras en el orden indicado (el Master Mix ya tiene buffer de muestra): Marcar sus adiciones !
Carril Muestra Volumen " 1 Master Mix con DNA Standard 5 µl " 2 Estudiante 1 20 µl " 3 Estudiante 2 20 µl " 4 Estudiante 3 20 µl " 5 Estudiante 4 20 µl 3. Realice la electroforesis de las muestras. Use 100 V con la fuente de poder y deje correr el gel por 30 minutos. 4. Cuando termine la electroforesis, coloque el gel en una bandeja y ponga 60 ml del colorante de DNA Bio-Safe u otro colorante para teñir el DNA. Cubra con plástico adhesivo y deje teñir el gel toda la noche o el tiempo indicado por su instructor. 5. La presencia o ausencia del inserto Alu puede ser detectado por la reacción de PCR seguida por la electroforesis en geles de agarosa. Los casos de DNA que llevan un gen ALU migrarán más atrás que los genomas sin este inserto. Analice los resultados de la electroforesis dentro su grupo y posteriormente con los grupos vecinos.

21
Protocolo Trabajo Práctico No 3 Transformación Bacteriana y Análisis de la expresión de Proteína Flourescente Verde Introducción. En este laboratorio, los alumnos realizarán un procedimiento conocido como transformación genética de bacterias. La transformación ocurre cuando a una célula bacteriana se le introduce un fragmento de material genético contenido en una molécula de DNA. Desde un punto de visto molecular, la transformación consiste en un cambio genético estable producido por la incorporación de un DNA de un organismo procarionte o eucarionte en una célula bacteriana. Este procedimiento puede modificar las características genéticas de la bacteria recipiente y, a menudo, el DNA introducido proporciona a la bacteria un nuevo rasgo o carácter que puede ser identificado de manera posterior a la transformación. El fragmento de DNA utilizado en la transformación generalmente contiene en su secuencia al menos un gen o un grupo de genes. En este trabajo práctico se estudiará la transformación bacteriana mediante la utilización de un fragmento de DNA conocido como plásmido. En esta molécula de DNA de origen bacteriano se pueden introducir genes derivados de otros organismos. Esta construcción híbrida llamada también DNA recombinante puede ser introducida dentro de la bacteria o célula hospedante a ser modificada. Los plásmidos son pequeños segmentos de DNA que tienen una forma circular y que han sido obtenidos a partir de las bacterias. Los plásmidos poseen genes que codifican para una o más características heredables que pueden ser beneficiosas para la supervivencia de las bacterias. Una característica importante de los plásmidos es que éstos replican su DNA en forma independiente del DNA cromosomal de la célula hospedante. En la naturaleza, las bacterias pueden transferir los plásmidos entre sí, lo que les permite compartir estos genes beneficiosos. Este mecanismo permite que las bacterias de manera natural puedan adaptarse a nuevos ambientes. La reciente aparición de resistencia bacteriana a los antibióticos se debe a la transferencia de DNA plásmidial de una bacteria a otra. En el Trabajo Práctico No 3. se utilizarán un "Kit" de la Companía Bio Rad catálogo No BRLSI-1660003EDU para la transformación de bacterias con el plásmido pGLO. Transformación con el plasmido pGLO En este trabajo práctico los estudiantes utilizan un procedimiento sencillo para transformar bacterias con el plásmido pGLO que contiene un gen que codifica para una proteína fluorescente verde, la Green Fluorescent Protein o GFP (pGLO-GFP). El plásmido contiene además otro gen que le da resistencia al antibiótico ampicilina. La fuente natural del gen GFP es la medusa bioluminiscente Aequorea victoria. El producto del gen GFP es una proteína fluorescente, que hace que la medusa fluoresca y brille en la oscuridad. Las bacterias transformadas con el plásmido que contiene este gen, adquieren la propiedad de fluorescer con un color verdoso intenso que se hace visible al ser expuestas éstas a la luz ultravioleta. En el trabajo práctico, se procederá a la transformación de la bacteria no patógena Eschericia coli con el plásmido pGLO. Posteriormente las bacterias son crecidas sobre

22
un medio de cultivo sólido conteniendo el antibiótico ampicilina, por 16 hrs. a 37ºC. Aquellas bacterias que han incorporado el plásmido pGLO pueden expresar el gen de la proteína fluorescente verde GFP, lo que les permite producir un color verde brillante fluorescente. A continuación de la tranformación, se realizarán un experimento de separación de las proteínas de la bacteria mediante la técnica de electroforesis en geles desnaturantes de proteínas, con la finalidad de identificar la proteína GFP expresada por las bacterias transformadas con el plásmido pGLO.. A continuación se presenta una breve explicación del mecanismo de expresión de proteínas utilizado en el plásmido pGLO. El pGLO, (ver imagen mas abajo) es un plásmido producido por la empresa de biotecnología Bio-Rad (www.bio-rad.com). Este plásmido incorpora un sistema especial de genes que permiten realizar la expresión regulada de la proteína fluorescente verde en las bacterias transformadas. El gen de la proteína fluorescente verde puede ser activado en las células transformadas con sólo agregar el azúcar arabinosa al medio de cultivo de las células bacterianas. La selección de las células que han sido transformadas con el DNA de pGLO se lleva a cabo mediante el crecimiento de las bacterias en placas de agar que contienen el medio de cultivo y el antibiótico de selección ampicilina. Las células transformadas aparecen de color blanco (fenotipo silvestre) en las placas que no contienen arabinosa, y de color verde fluorescente cuando la arabinosa ha sido incluida en el agar nutritivo. El diseño único del plásmido pGLO permite a educadores y estudiantes, explorar fácilmente los mecanismos de regulación de la expresión génica y los procedimientos experimentales de la selección genética. Todo el proceso se observa con una lámpara UV de onda larga relativamente barato. Para una discusión más detallada de estos conceptos, refiérase al Sector Apoyo Teórico Trabajo Practico 3. El plásmido pGLO Protocolo del Trabajo Práctico No 3.
1. Rotule un microtubo de ensayo (con tapa) "+ DNA pGLO" y otro "-DNA pGLO." Marque ambos tubos con el nombre de sus iniciales. Coloque ambos tubos en una gradilla en su mesón de trabajo.

23
2. Abra ambos tubos y mediante una pipeta de transferencia estéril, transfiera 250 µl de solución de Transformación (CaCl2)., Traspase inmediatamente ambos tubos al hielo (figura a continuación).
3. Use una asa estéril para tomar (picar) una colonia de bacterias de la placa de partida del experimento. Tome el tubo rotulado "+ DNA pGLO" y sumerja el asa en la solución de transformación hasta la base del tubo. Gire el asa en círculo con el dedo índice y el pulgar hasta que toda la colonia se disperse en la solución de transformación (no debe observar fragmentos flotantes). Vuelva a colocar el tubo en la gradilla en el hielo. Utilizando una nueva asa estéril repita el procedimiento con el tubo rotulado -DNA pGLO (figura a continuación).
4. Examine la solución de DNA del pGLO con la lámpara UV. Anote sus observaciones. A continuación, sumerja una asa estéril en el tubo que contiene el DNA del plásmido pGLO. Retire el asa de la solución. Debe ver una película de la solución de plásmido sobre el asa. Introduzca el asa en el tubo que contiene la suspensión de bacterias en el tubo rotulado + DNA. Cierre el tubo y colóquelo en la gradilla para tubos en el hielo. Cierre el tubo rotulado -DNA. No agregue el DNA del plásmido en el tubo –DNA. ¿Por qué no debe agregar el DNA del pGLO en este tubo? 5. Incube los tubos en hielo durante 10 minutos. Asegúrese de empujar la gradilla con los tubos hasta el fondo de la cubeta que contiene el hielo, de manera que los tubos hagan contacto con el hielo.

24
6. Mientras que los microtubos están incubándose en el hielo, rotule las cuatro placas de agar en la base (no en la cubierta) de la siguiente manera: +pGLO/ LB / amp +pGLO/ LB / amp / ara -pGLO LB /amp -pGLO/ LB / amp / ara
7. “Shock Térmico”: Coloque los tubos en una gradilla de poliestireno y transfiera los tubos a un baño de agua a 42°C. Incube los tubos durante 50 segundos exactos. Asegúrese de que los tubos hacen el contacto con el agua calentada a 42oC. Al cabo de los 50 segundos transfiera los dos tubos de vuelta en el hielo. Para mejorar el resultado de la transformación, se recomienda que los cambios de temperatura se hagan lo más rápido posible. Incube entonces los tubos en hielo durante 2 minutos.
8. Retire la gradilla con los tubos del hielo y colóquela en su mesón de trabajo. Abra un tubo y, con una pipeta estéril nueva, añada 250 µl de medio LB en el tubo y ciérrelo nuevamente. Repita el procedimiento con una nueva pipeta estéril para el otro tubo. Incube los tubos durante 10 minutos a temperatura ambiente. 9. Golpee los tubos cerrados con su dedo índice para mezclar el contenido. Con una pipeta estéril nueva para cada tubo, tome 100 µl de cada una de las suspensiones de transformación y agréguelas sobre las placas de agar correspondientes (incluyendo las placas control).

25
10. Disemine a través de toda la superficie del agar la suspensión de bacterias. (Figura arriba) Utilice para este propósito un asa estéril diferente para cada placa de agar. Con el asa se debe rastrillar rápidamente la superficie del agar en cada placa de modo de dispersar lo más ampliamente posible sobre el agar la suspensión de bacterias (Figura inferior). 11. Apile las placas con una cinta adhesiva. Coloque su nombre y el del grupo en la parte inferior de la pila de placas. Coloque el apilamiento boca abajo en la incubadora a 37 ° C hasta el día siguiente. Observación de las bacterias Al día siguiente los alumnos procederán a mirar las características fenotípicas de las bacterias transformadas que crecieron en las placas. Anote sus resultados. Observe las bacterias transformadas bajo la lámpara UV. Debe observar una coloración intensa verdosa, producto de la fluorescencia de la proteína GFP. Analice la implicancia de este resultado. Análisis de la expresión de la proteína GFP mediante electroforesis desnaturante de acrilamida. La electroforesis de proteínas permite separar y analizar las proteínas presentes en un extracto celular. Combinada con otras técnicas permite inclusive identificarlas. -Para realizar este procedimiento se inocula una colonia de cada una de las placas de la transformación bacteriana (realizada el día anterior), en un tubo estéril cónico de 50

26
ml, que contiene 3 ml de medio líquido LB (Luria Bertani), ampicilina a una concentración final de 100 µg/ml y 5 mg/ml de arabinosa. -Las bacterias se crecen durante al menos ocho horas en tubos cónicos de 50 ml conteniendo cada uno las bacterias obtenidas a partir de las condiciones de transformación siguiente:
+pGLO/ LB / amp +pGLO/ LB / amp / ara
-pGLO LB /amp
-pGLO/ LB / amp / ara Discuta porqué en el medio LB se debe adicionar ampicilina. -Luego de las ocho horas de crecimiento, extraiga 50 µl de bacteria de cada uno de los tubos de crecimiento y colóquelos en microtubos de 1 ml previamente rotulados. -Agregue a cada tubo 50 µl de solución de Laemmli 2x (solución de preparación de muestras proteicas) y caliente 5 minutos a ebullición para romper las células. -Cargue 5 µl de esta solución en un gel de poliacrilamida. Junto con las muestras bacterianas cargue en el gel 5 µl de una muestra que contiene una mezcla de proteínas de peso molecular conocido que servirán para estimar el peso molecular de la GFP. -Someta las muestras a electroforesis durante 1 hora a 100-120 Volt. -Terminada la electroforesis, tiña el gel con una solución de tinción que contiene Azul de Coomassie en ácido acético y metanol. -Destiña el gel con ácido acético y metanol. Observe el patrón de bandeo obtenido en el gel. ¿Logra identificar la proteína GFP de entre todas las proteínas observadas? -Sacar foto de su gel para documentar sus resultados.

27
Protocolo Trabajo práctico 4a I. Identificación de una proteína a partir de la secuencia del gen que la codifica
La bacteria patógena Vibrio cholera produce fuertes diarreas provocando deshidratación severa. En un laboratorio de microbiología clínica se aisló una cepa bacteriana que producía una patología suave (S), la cual se volvía virulenta al ser transformada con DNA cromosomal de la cepa agresiva. A continuación se indica la secuencia del DNA de Vibrio que provoca el cambio
>DNA de vibrio ATGATTAAATTAAAATTTGGTGTTTTTTTTACAGTTTTACTATCTTCAGCATATGCACAT GGAACACCTCAAAATATTACTGATTTGTGTGCAGAATACCACAACACACAAATATATACG CTAAATGATAAGATATTTTCGTATACAGAATCTCTAGCTGGAAAAAGAGAGATGGCTATC ATTACTTTTAAGAATGGTGCAATTTTTCAAGTAGAAGTACCAGGTAGTCAACATATAGAT TCACAAAAAAAAGCGATTGAAAGGATGAAGGATACCCTGAGGATTGCATATCTTACTGAA GCTAAAGTCGAAAAGTTATGTGTATGGAATAATAAAACGCCTCATGCGATTGCCGCAATT AGTATGGCAAAT
OBJETIVOS: Identificar la proteína causante de la recuperación del fenotipo virulento y posiblemente de la patología en humanos. Análisis de la secuencia de DNA de este gen mediante bioinformática.
1. Identifique los marcos de lectura (ORF) codificados. 2. Una vez definidos los marcos de lectura, identifique los que corresponden a
una proteína de función conocida. 3. Deduzca cual de ellas puede estar relacionada con la patología de acuerdo a
su función. 4. Analice las relaciones estructurales y filogenéticas entre las proteínas
encontradas. Instrucciones para efectuar este análisis: Ingrese al sitio web: http://www.ncbi.nlm.nih.gov/ . Una vez en la página, vaya a Sequence Analysis y de ahí a Tool. Ingresar a la página ORF FINDER. Estando aquí, ingresar la secuencia del DNA en el cuadro (incluyendo >DNA de vibrio). Pinchar la casilla orf finder. Una vez que aparecen los posibles marcos de lectura, hacer click en cada uno y visualizar la proteína codificada. En cada caso ingresar a BLAST (en la misma página en que aparecen los marcos de lectura identificados). Hacer click sobre el botón format para encontrar, en orden de aparición, las proteínas que estén archivadas hasta el momento en las bases de datos que tengan mayor similitud con la cadena de aminoácidos encontrada. La proteína con mayor score es la que más probablemente se correlaciona con la patología de su caso. Se puede visualizar un alineamiento múltiple entre las secuencias de proteínas detectadas (Other reports: multiple alingment) o un árbol de distancia filogenética entre las secuencias de proteínas encontradas (Other reports: distance tree report). De las secuencias incluidas

28
en el alineamiento múltiple elija cinco diferentes (de diferentes organismos o propiedades) haciendo click en la descripción y vuelva a realizar el alineamiento. En la misma página Reports haga click en la imagen rosada debajo del esquema de la proteína (título: putative conserved domains…..) Aparecerá una página titulada Conserved Domains on …. Aparecerá un esquema similar al anterior. Posicionando la “mano” (cursor) sobre uno de las imágenes rosadas aparecerá una figura que muestra la estructura de un dominio correspondiente al indicado. Se puede abrir ese ícono y se podrá visualizar la estructura de la proteína (se debe contar con el programa Cn3D 4.3 que es de dominio público). Alternativamente se puede elegir unas 5-6 secuencias del alineamiento original en formato FASTA y archivarlas en su documento junto a la secuencia usada en el problema. Luego en Google ingresar CLUSTAL W. Aparecerá en primer lugar ClustalW EBI. Ingresar las secuencias guardadas y realizar el alineamiento múltiple. Analizar las similitudes entre las secuencias. Revise si en las bases de datos de PDB o PDBsum (buscar en Google) existe algún archivo con esta(s) proteína(a). Si es así, analice las propiedades estructurales de la proteína (estructura secundaria, terciaria, dominios, similitudes con proteínas de otros organismos). II. Genoma humano y Medicina
La fibrosis quística es una enfermedad genética recesiva que causa severas alteraciones en la absorción de nutrientes y/o en el aparato respiratorio, provocando en muchos casos la muerte de los pacientes a edades tempranas. En el año 1989 se identificó el gen alterado en esta patología , el que codifica una proteína de membrana denominada CFTR (cystic fibrosis transmembrane regulator). En relación con este gen, investigue en las bases de datos computacionales de la secuencia del genoma humano:
a. la localización cromosómica del gen CFTR b. la organización y tamaño del gen. c. la identidad de los genes aledaños d. el tamaño del mRNA y de la proteína CFTR e. la función del CFTR f. la función alterada de la mutante ΔF508
Una fuente de información, pero no la única, se puede encontrar en la página web: http://www.ncbi.nlm.nih.gov. Una vez ubicada esta página, ir a Genomes and Maps y dentro de ésta seleccionar Genome, buscar Human Genome Chromosome 7, buscar CFTR. Hacer clic en cromosoma 7, aparecen el mapa de la región de gen y se puede manipular el tamaño del segmento y localizar SNPs.

29
III. Análisis de genomas bacterianos completos. En esta actividad se revisarán algunas de las herramientas que proporciona este servidor del Craig Venter Institute http://www.jcvi.org. Particularmente en la Microbial and environmental Genomics IV. Análisis de vias metabólicas. En esta actividad de analizarán algunas de las herramientas que proporciona este servidor. En particular la orientada a analizar vías metabólicas en los genomas secuenciados http://www.genome.jp/kegg/ V. Visualización de estructuras de proteínas. Para realizar esta actividad se utilizarán los programas computacionales RASWIN y MAGE. El primero es un programa que permite visualizar archivos tipo pdb (protein data bank), es decir archivos que tienen datos de estructuras de proteínas reales o predichas en formato pdb. El segundo programa está diseñado para educación sobre estructura de proteínas y es muy didáctico. Utiliza archivos en formato kin (kinimage). Se empleará la colección de archivos que pondré a su disposición. Algunas páginas que pueden ser útiles para educación. http://www.neb.com/nebecomm/enzymefinder.asp http://poltaco.blogspot.com/2008/07/laboratorio-virtual-digestin-con.html http://learn.genetics.utah.edu/content/labs/extraction/ http://biomodel.uah.es/lab/inicio.htm#cibertorio http://learn.genetics.utah.edu/es/ http://biomodel.uah.es/lab/inicio.htm

30
Protocolo Trabajo Práctico 4b: construcción de filogenias moleculares. La historia evolutiva de un grupo de organismos se denomina filogenia y la representación gráfica de esta historia, árbol filogenético. Este árbol rescata la secuencia de eventos de especiación, la relación de ancestría de los diversos taxones y en algunos casos los tiempos transcurridos en el proceso. La reconstrucción de esta filogenia es siempre indirecta y en muchos casos más de un árbol filogenético puede representar la historia evolutiva de un grupo. Esta actividad pretende que usted aplique sus conocimientos relativos al análisis filogenético a un caso particular de un gen eucarionte, y basado en el carácter homólogo de éste, reconstruya la filogenia para un grupo de taxones. Esta actividad pretende además que se familiarice con el uso de MEGA, un software de uso libre que puede ser bajado de la red y utilizado para realizar alineamientos y análisis filogenético mediante diversas aproximaciones (distancia, máxima parsimonia, máxima verosimilitud (versión 5)). 1. ALINEAMIENTO En la construcción de una filogenia molecular debe contarse con un alineamientos de secuencias que permita garantizar que la comparación está siendo realizada de manera adecuada, es decir, carácter a carácter, entre caracteres homólogos. Para este procedimiento se puede recurrir a diversos software disponibles y plataformas online gratuitas. En esta ocasión utilizaremos el motor de alineamiento incluido en el software MEGA. En este primer paso abra Mega
Seleccione Alignment Seleccione Alignment Explorer Seleccione Retrieve sequences
Para mayor comodidad se dispone de un archivo que contiene las secuencias a analizar en formato fasta (Carpeta ejercicios). Este formato es reconocido por la mayoría de los software de alineamiento.
Ir a carpeta EJERCICIOS Elegir alineamiento.fas
Ahora usted tiene en pantalla las secuencias y puede proceder a alinearlas. Antes de eso si lo desea explore las secuencias, moviéndose en sentido vertical y horizontal. Puede además traducir la secuencia a proteína y utilizar otras funciones disponibles.

31
Para alinear las secuencias
Selecione Edit Select all
Seleccione Alignment Align by Clustal Seleccione Ok en el cuadro sin modificar los parámetros.
Explore el alineamiento, moviéndose en sentido horizontal y vertical. Localice las regiones donde hay mayor o menor similitud (guíese por los asteriscos sobre las secuencias que indican identidad entre todas las secuencias). Localice ahora las regiones donde hay gaps. Mire los bordes de las regiones con gaps y determine la calidad del alineamiento en estas zonas. 2. ANALISIS FILOGENETICO Para el análisis filogenético el programa MEGA cuenta con diversas herramientas que permiten determinar distancias, construir filogenias bajo diferentes supuestos y modelos, y determinar la solidez de éstas. En este práctico nos restringiremos a la construcción de filogenias mediante métodos de distancia (Neighbor-Joining). Para exportar a mega ir a
Data Export Alignment Mega Format Seleccione guardar. En el siguiente cuadro apriete Ok En el siguiente cuadro elija Yes. Cerrar y aceptar abrir en Mega. Yes.
Explorar la secuencia utilizando los botones bajo la barra de menú. Ver codones, sitios variables, sitios parsimoniosos, etc. Con sitios variables seleccionado, evalúe de manera cualitativa si estos se presentan con mayor frecuencia en primera, segunda o tercera posición. Ir a Phylogeny en ventana principal (la ventana principal podría estar oculta bajo la ventana de exploración de secuencia). Para construir una filogenia con las secuencias alineadas proceda de la siguiente manera: En phylogeny seleccione Construct Phylogeny
Neighbor-Joining En la primera oportunidad acepte los parámetros (clic en compute).

32
Aparece una ventana nueva con el dendrograma construido. Revise la posición de las distintos taxones. Discuta con sus compañeros la consistencia de esta filogenia con su conocimiento previo respecto a la relación de dichos taxones. Explore los distintos botones bajo la barra de menú. Repita el procedimiento anterior para evaluar otros parámetros:
En phylogeny seleccione Construct Phylogeny
Neighbor-Joining
En esta oportunidad explore las pestañas y opciones disponibles y según las instrucciones del profesor:
Pruebe diversos modelos de sustitución. Evalúe considerar, 1ra, y 2da posición. Evalúe considerar, solo 3ra posición. Realice test de filogenia (Bootstrap).
Compare los árboles obtenidos. ¿Donde se ubican los genes en el árbol? ¿Puede decir algo sobre la evolución del gen en estas especies? ¿Puede utilizar este gen para evaluar la historia evolutiva de estos taxones? Si lo desea puede explorar en el menú principal las otras aproximaciones teórico-metodológicas para obtener árboles filogenéticos (máxima parsimonia y máxima verosimilitud).

33
APOYO TECNICO A LOS TRABAJOS PRACTICOS

34
Apoyo Teórico al Protocolo No. 1a. El uso de Enzimas de Restricción en la Identificación de Personas. ¿Quién es el culpable? Las Huellas Genéticas en el DNA Los seres humanos tienen una gran parte de su secuencia de DNA en común pero también existen grandes variaciones que permiten su fragmentación con enzimas de restricción. Cada enzima de restricción se dirige a una parte del DNA donde hay una secuencia que reconoce. Estas secuencias se encuentran en diferentes posiciones en el DNA de diferentes personas, resultando que el DNA de cada individuo se cortará en una manera específica y no da un conjunto de productos comparables con otros, es decir, resulta en fragmentos de longitud variable del DNA. (Una excepción a esta regla son los gemelos idénticos que tienen DNA idéntico.) Estas diferencias son conocidas como polimorfismo de longitud de fragmentos de restricción (RFLP). Esta técnica también se utiliza a menudo para determinar el genotipo de un individuo para un determinado gen, por ejemplo, para detectar la presencia o ausencia de una mutación que confiere un trastorno genético determinado. Las Enzimas de Restricción. Las enzimas de restricción conforman un grupo grande de enzimas que cortan el DNA de doble hebra en sitios muy específicos. Más de 3000 enzimas de restricción han sido descubiertas hasta la fecha y muchas de ellas están disponibles comercialmente. Su descubrimiento en bacterias y su caracterización en los años 70 contribuyeron al desarrollo de muchísimas tecnologías de la biología molecular moderna. Los investigadores Daniel Nathans, Werner Arber, y Hamilton O. Smith recibieron el premio Nobel en 1978 por este trabajo. La primera aplicación de la técnica “Huella genética” (DNA Fingerprinting) la realizó Alec Jeffrys en 1984. La determinación de huella genética de DNA es una técnica utilizada para distinguir a un individuo de otros individuos de la misma especie, usando solamente muestras de su DNA. El proceso comienza con la extracción de DNA de las células, usando una pequeña muestra de sangre, saliva, semen u otro tejido. Las enzimas de restricción cortan por reconocimiento de secuencias específicas, por ejemplo, la enzima de restricción Sma1 reconoce la secuencia 5´CCCGGG3´ apareada con 3´GGGCCC5´de la otra hebra y corta el DNA de doble hebra entre la C central y la base G de ambas hebras (5´CCC #GGG3´), dejando dos fragmentos del DNA con sus segmentos terminales “romos”. Otras enzimas de restricción cortan el DNA de doble hebra dejando sus segmentos terminales “escalonados”, es decir, con bases nucleotídicas no apareadas en cada extremo. Por ejemplo, la enzima de restricción EcoR1 reconoce la secuencia 5´GAATTC, se une a este fragmento (y a todos los fragmentos con esta secuencia en la muestra de DNA) y hace un corte entre las bases G y A generando extremos cohesivos de la siguiente manera: (X representa cualquier base):

35
A continuación se muestra un DNA de doble hebra sin cortar, con el sitio del corte en rojo:
5´XXXXXXGAATTCXXXXXX-3´ 3´XXXXXXCTTAAGXXXXXX-5´
Después de incubar con la enzima de restricción EcoR1, los productos son:
5´XXXXXXG- AATTCXXXXXX-3´ 3´XXXXXXCTTAA -GXXXXXX-5´
Estos fragmentos de doble hebra, cada uno con un extremo no apareado, se conocen como “extremos cohesivos” (“pegagosos”) porque se pueden volver a reasociar por apareamiento de las bases libres entre sí o con un DNA de otra especie previamente cortado con la misma enzima. Para hacer un estudio de huellas, normalmente se utiliza más de una enzima de restricción para generar más fragmentos cortados asegurando perfiles de cortes diferentes. Identificación de DNA humano usando los Segmentos Terminales Repetidos. Otro método común de realizar huellas genéticas de DNA es la que explora secuencias repetidas altamente variables llamadas “repeticiones cortas en tándem,” denominadas con la abreviatura STR. (La sigla STRs en inglés significa Short Terminal Repeats y se traduce al español como, Segmentos (cortos) Terminales Repetidos). Diferentes personas tienen distinto número de unidades de repetición. Por ejemplo, un gen llamado CSF contiene una secuencia AGAT repetida entre 6 a 16 veces. Dos personas no relacionadas serán propensas a tener diferente número de repeticiones de esta secuencia AGAT. Alrededor del 5 al 20% de las personas tienen un número variable de repeticiones de un STR específico compartido. Analizar varios STR al mismo tiempo así como estudiar otros tipos de regiones STR se transforma en una técnica con poder increíble para identificar personas por análisis de su DNA. Entre más regiones de STR se analicen, más discriminatoria es la prueba. En los Estados Unidos de América se utilizan actualmente 13 loci diferentes (repeticiones frecuentes en el DNA) para la discriminación de personas. Este número ha demostrado la generación de la probabilidad de identidad de uno en un trillón ( 1 X 1018) o más. Por lo tanto, es posible establecer que es muy poco probable que una identidad haya surgido por casualidad, excepto en el caso de gemelos idénticos que tienen idénticos perfiles genéticos.

36
Electroforesis en Geles Los fragmentos de DNA cortados por las enzimas de restricción se pueden separar mediante electroforesis en geles. Este método coloca la muestra de DNA en un pocillo en un extremo de un gel de agarosa. Un gel de agarosa es muy parecido una fina capa de gelatina puesta en una cajita plástica. Un campo eléctrico se aplica al gel y, dado que el DNA está cargado negativamente, es atraído por el extremo positivo del campo y comienza a moverse a través del gel. El gel de agarosa tiene poros que permiten que las moléculas de DNA puedan moverse hacia el polo negativo. Cuanto más grande sea el DNA, los fragmentos ya no pueden viajar a través de la matriz del gel tan rápido como los fragmentos cortos y se quedan atrapados cerca del inicio del gel donde la muestra se ha cargado. Los fragmentos más cortos se mueven con más facilidad y así viajarán más lejos. Esto da lugar a un patrón único de bandas para el DNA de cada individuo. La imagen de un gel de agarosa típica se visualiza con los productos de un análisis de DNA tratado con tres enzimas de restricción, EcoR1, Pst1 y BamIII. El lado izquierdo muestra el gel después del tratamiento con un colorante específico, (compuesto que se intercala entremedio de las hebras del DNA) y permite observar los fragmentos en luz visible o (dependiente del colorante) en luz UV. El lado derecho del gel muestra un autoradiografía del gel donde las bandas representan el DNA que ha sido marcado previamente en el nucleótido 5’ terminal de las cadenas con fosforo-32P- fósforo (radioactividad).

37
Apoyo Teórico al Protocolo Nº 2 La Reacción de DNA Polimerasa en Cadena: PCR Introducción. La Reacción en Cadena de la Polimerasa (PCR) es una técnica enzimática que permite amplificar millones de veces una molécula de DNA. Esta técnica, descubierta en el año 1983 por Kary Mullis, transformó en pocos años los estudios de la biología molecular hasta entonces arduos y exclusivos, proporcionando una técnica accesible y manejable por investigadores de todos las áreas de la biología. El objetivo de técnica PCR es producir “in vitro” (o sea, sin el uso de microorganismos) un gran número de copias de una secuencia específica de DNA, a partir de una cantidad inicial de DNA pequeñísima. En un tiempo corto, mediante esta técnica pueden sintetizarse millones de copias de un DNA o de un segmento específico de DNA, los que pueden ser utilizados en investigación básica o aplicada. El impacto y aplicación de la PCR tanto en biología básica como en biología aplicada son gigantescos, particularmente en el proceso de clonamiento de genes, y revolucionarios en muchos otros campos como en el diagnóstico clínico , farmacología y medicina forense. ¿Qué es la PCR? PCR es una sigla del nombre en inglés de una técnica muy usada en la biología molecular: Polymerase Chain Reaction , traducida como "Reacción en cadena de la Polimerasa." El sistema de PCR se basa en repetidos ciclos térmicos que permiten proceder independientemente para realizar reacciones que ocurren a tres temperaturas diferentes: un calentamiento a 95ºC, que provoca la denaturación del DNA "molde," seguido de una disminución de la temperatura a cerca de 60ºC , que permite la hibridación de oligonucleótidos (partidores) al DNA molde y luego un calentamiento a 72ºC que permite la polimerización de muchas copias de este DNA. La síntesis del DNA son catalizada por una DNA polimerasa especial llamada TAQ polimerasa (aislada de la bacteria Thermus aquaticus), que es termoestable y resiste hasta temperaturas sobre 90 ºC y con una actividad óptima cerca de los 70 ºC. La mayoría de las DNA polimerasas se inactivan a esas altas temperaturas. Para entender este proceso de reacciones en cadena necesitamos recordar brevemente detalles de la estructura de una molécula de DNA. Este repaso sobre su estructura ayudará a apreciar por qué la reacción de PCR tiene que proceder en una forma cíclica, alternando ordenadamente tres reacciones que ocurren a tres diferentes temperaturas. La extraordinaria estabilidad del DNA helicoidal surge como consecuencia de las interacciones hidrofóbicas entre las bases de cada cadena y los puentes de hidrógeno entre las cadenas complementarias. En sus estudios previos posiblemente han

38
analizado o construido un modelo de DNA, y observado que la superficie exterior de esta molécula tiene cargas negativas debido a la presencia de grupos fosfatos que dan el carácter ácido al polímero. Además, cada fosfato se une a un azúcar desoxiribosa, formando largas cadenas alternando azúcar-fosfato-azúcar, conectadas por la llamada unión fosfodiéster. Las bases nitrogenadas A,T,G y C están unidas a los azucares, quedando en el centro de la molécula cilíndrica, orientadas perpendicularmente a la cadena fosfodiéster, parecida a los peldaños de una escalera tipo caracol. Las bases A,T,G y C de una cadena están posicionadas opuestas a sus bases complementarias T, A, C, G, como consecuencia de la especificidad de la DNA polimerasa durante la síntesis de las cadenas. Las fuerzas sumadas de todas las interacciones 'base-base' (A-T y G-C) dan gran estabilidad a la molécula debido al número enorme de puentes de hidrógeno ( -H- -+- -H- ) presentes entre las dos cadenas de miles de pares de bases. Merece mención el hecho de que un DNA con alto contenido de pares de bases G-C es significativamente más estable que un DNA rico en pares A-T, debido a la mayor cantidad de puentes de hidrógeno (3 en G-C y 2 en A-T). El conjunto de todas estas interacciones que dan estabilidad se resumen en la formación de una estructura cilíndrica compacta que ha sido descrita frecuentemente como una "pila de monedas.
Modelo de la doble hélice de DNA
El DNA, a pesar de su estabilidad molecular, para su funcionamiento en el ambiente fisiológico requiere una flexibilidad grande para exponer y separar segmentos específicos, por ejemplo, durante la expresión de genes específicos. En los animales y plantas superiores, el DNA nuclear se encuentra envuelto en proteínas básicas llamadas histonas que se sueltan de los segmentos específicos de estar “activadas” durante la expresión de genes particulares. La activación se inicia y coordina por la asociación al DNA de otras proteínas reguladores que despejan las histonas y apoyan la separación de las hebras de la doble hélice en segmentos de 20, 40 o más pares de bases. El segmento desenrollado o 'abierto' parecería una burbuja

39
han analizado o construido un modelo de DNA, y observado que la superficie exterior de esta molécula tiene cargas negativas debido a la presencia de grupos fosfatos que dan el carácter ácido al polímero. Además, cada fosfato se une a un azúcar desoxiribosa, formando largas cadenas alternando azúcar-fosfato-azúcar, conectadas por la llamada unión fosfodiéster. Las bases nitrogenadas A,T,G y C están unidas a los azucares, quedando en el centro de la molécula cilíndrica, orientadas perpendicularmente a la cadena fosfodiéster, parecida a los peldaños de una escalera tipo caracol. Las bases A,T,G y C de una cadena están posicionadas opuestas a sus bases complementarias T, A, C, G, como consecuencia de la especificidad de la DNA polimerasa durante la síntesis de las cadenas. Las fuerzas sumadas de todas las interacciones 'base-base' (A-T y G-C) dan gran estabilidad a la molécula debido al número enorme de puentes de hidrógeno ( -H- -+- -H- ) presentes entre las dos cadenas de miles de pares de bases. Merece mención el hecho de que un DNA con alto contenido de pares de bases G-C es significativamente más estable que un DNA rico en pares A-T, debido a la mayor cantidad de puentes de hidrógeno (3 en G-C y 2 en A-T). El conjunto de todas estas interacciones que dan estabilidad se resumen en la formación de una estructura cilíndrica compacta que ha sido descrita frecuentemente como una "pila de monedas.
Modelo de la doble hélice de DNA
El DNA, a pesar de su estabilidad molecular, para su funcionamiento en el ambiente fisiológico requiere una flexibilidad grande para exponer y separar segmentos específicos, por ejemplo, durante la expresión de genes específicos. En los animales y plantas superiores, el DNA nuclear se encuentra envuelto en proteínas básicas llamadas histonas que se sueltan de los segmentos específicos de estar “activadas” durante la expresión de genes particulares. La activación se inicia y coordina por la asociación al DNA de otras proteínas reguladores que despejan las histonas y apoyan la separación de las hebras de la doble hélice en segmentos de 20, 40 o más pares de bases. El segmento desenrollado o 'abierto' parecería una burbuja

40
que distorsiona la forma regular de la hélice. Estos cambios en la hélice ocurren mediante este tipo de interacciones durante los procesos de replicación, transcripción, reparación, y metilación de las bases en el DNA. Al liberarse de éstas proteínas reguladoras, el DNA logra reformar la doble hélice. Este dinamismo y flexibilidad para asociarse y disociarse específicamente es la base de la expresión génica. La alteración de la estructura secundaria del DNA purificado en solución es también dinámica en condiciones en el laboratorio, por ejemplo, a temperaturas sobre 45 – 50 grados, en la presencia de sales concentradas, o la urea. se observan alteraciones radicales en la estructura secundaria del DNA, favoreciendo la separación total de las hebras. Al sacar la urea por diálisis, se vuelve a tener la forma apareada. La gran estabilidad de la molécula de DNA en forma de doble hélice depende totalmente de la temperatura ambiente. Al subir sobre las temperaturas fisiológicas, a 50º, 60º C, la molécula empieza a abrirse poco a poco, en diferentes segmentos, hasta que a temperaturas mas altas, 75º a 90ºC, se logra la separación total con la formación de las monohebras. Esta reacción se llama denaturación de DNA y es un proceso totalmente reversible con la formación progresiva de las hebras reasociadas (renaturadas) al bajar la temperatura a 30ºC. La reasociación de DNA de gran tamaño (miles de bases de largo) requiere un tiempo relativamente largo y depende de la concentración y complejidad de la muestra. Una solución de DNA genómica, con todo su multiplicidad de secuencias primarias, recuperará totalmente su carácter de doble hélice durante un proceso de re-naturación. Esta propiedad es justamente la que se aprovecha durante el proceso de la reacción en cadena de las polimerasas PCR. Como lo indica el nombre PCR, este proceso involucra una secuencia específica de reacciones distintas (en “cadena” o secuencial), que requieren de diferentes temperaturas. Aumentando gradualmente la temperatura, desde 60º, el DNA va perdiendo su forma de hélice y hay separación parcial de las hebras. Entre 80 y 95ºC, se produce la separación rápida y total de las hebras. Este último cambio de estructura del DNA, su denaturación, constituye la primera etapa o ciclo de una reacción de PCR. Primer ciclo del PCR: Denaturación del DNA a 95º, con la separación de las dos hebras. La segunda etapa de PCR, al cambiar la temperatura a 55º - 65º, (las hebras larguísimas de DNA no alcanzan re-asociarse eficientemente a estas temperaturas), fragmentos monohebras pequeños que se llaman “partidores” logran asociarse a sitios

41
complementarios. Los partidores normalmente son de aproximadamente 24 desoxinucleótidos de largo y están preparados por síntesis química en el laboratorio, según el diseño del investigador que busca la amplificación de un gen específico. Uno de estos partidores reconoce una secuencia específica y corta (20-24 bases de largo) ubicada en una hebra de DNA, y el otro partidor reconoce otro segmento específico ubicado en la otra hebra. La temperatura que permite la unión de los partidores es determinada por su secuencia primaria (número de bases G-C y de A-T) y generalmente está cerca de 55º y 65º. Segundo ciclo del PCR: Fragmentos "partidores" se unen al DNA a temperaturas entre 55 y 65º, por apareamiento específico de bases La tercera reacción del PCR es el proceso de síntesis de hebras complementarias a cada una de las monohebras generadas en la etapa 1, que ya tienen asociados los partidores como se indica en la figura del ciclo 2. La enzima TAQ polimerasa, usada universalmente, tiene su actividad máxima a 72ºC y la temperatura de reacción se cambia para efectuar el tercer ciclo y permitir una síntesis óptima. Tercer ciclo de PCR: Elongación de las cadenas por la TAQ polimerasa, a 72º C, incorporando paso por paso los desoxinucleotidos (ver flechitas). La unión de DNA polimerasa a ambas hebras del DNA resultará en la síntesis de dos moléculas nuevas desde los partidores. Las moléculas sintetizadas toman la forma de doble hélice después del paso de la Taq polimerasa. En resumen, para este ciclo, se requiere la síntesis del DNA nuevo en la presencia del DNA "molde" y una

42
enzima DNA polimerasa termoestable, que tiene actividad máxima enzimática alrededor de 72ºC, pero también requiere que la enzima no se inactiva a la temperatura del ciclo de denaturación del DNA, a 90-95ºC. La reacción de síntesis requiere también la presencia de los sustratos de la enzima desoxinucleótidos trifosfatos, dNTP, los dos partidores de oligonucleotidos de 20 a 24 nucleótidos de largo y condiciones adecuados de pH y sales (Mgh o Mnh). Es evidente que existe un gran desafío técnico para lograr ejecutar estos tres ciclos de reacciones en forma repetida, miles de veces. Las tres reacciones independientes requieren una temperatura específica y dos de los ciclos duran solamente algunos pocos segundos mientras la reacción de elongación requiere varios minutos (el tiempo del ciclo tres depende del largo del DNA para amplificar). El instrumento que permite realizar estos ciclos de temperatura con precisión se "Termociclador" y contiene un sistema de programación donde se puede especificar las condiciones para cada caso experimental (temperatura y tiempo para cada ciclo). La variedad de instrumentos que hoy están disponibles en el comercio tienen amplias capacidades para ejecutar varias reacciones simultáneas, según la necesidad del laboratorio. Las incubaciones normalmente se hacen en un volumen total de 10 a 200 microlitros y la programación del tiempo la diseña el investigador. Por ejemplo, el primer ciclo podría ser a 92º por 30 segundos, el segundo ciclo de la incubación para la asociación de los partidores podría ser de 20 segundos y la incubación para el tercer ciclo de elongación puede ser de varios minutos (10 a 15 minutos). A continuación hay una representación del número y tipos de fragmentos que aparecen en cada ciclo de PCR. Se aprecia que la cantidad de moléculas amplificadas para formar el producto deseado (el "target" de los partidores específicos) sobrepasa muchísimo la cantidad de todos los otros productos no deseados. Al analizar los productos en un sistema de electroforesis en geles de agarosa, se aprecia una banda fuerte y preponderante debido a que la cantidad de los otros productos son relativamente muy menores.

43
Distribución del tamaño de productos de PCR después de varios ciclos de amplificación
. Tabla Inserto: La población de moléculas de DNA al final de los ciclos .
DNA molde!
DNA intermediario! Cantidad DNA a 20 ciclos !

44
Los Genes "ALU' ¿Qué genes estamos buscando? Introducción a las inserciones de DNA no codificantes en el genoma. En este trabajo práctico se analizará el DNA humano preparado anteriormente por cada uno de los mismos participantes del curso. Las 20 preparaciones de DNA obtenidas, se analizarán para la presencia o ausencia de este segmento de DNA específico y posteriormente se comentará la distribución de Alu dentro de este mismo grupo de personas. La secuencia en estudio se llama ALU PV92, ubicado en cromosoma 16. Se estima que 500-2,000 secuencias tipo Alu se encuentran en el genoma humano, pero en este estudio se analizará secuencias Alu del cromosoma 16, ubicado en el locus PV92. La presencia o ausencia de Alu en particular no refleja de ningún modo una ventaja o desventaja genética para las personas pero, como es un segmento muy estudiado en humanos, su análisis da un interés adicional al trabajo práctico, que pretende demostrar la capacidad analítica de la técnica de PCR. Una descripción breve de ALU se presenta a continuación. Se estima que los 46 cromosomas que forman el DNA genómico humano contienen aproximadamente 35,000 genes. Cada cromosoma contiene una serie de genes específicos. Los cromosomas más grandes contienen más DNA, por lo tanto tienen más genes en comparación a los cromosomas más pequeños. Cada uno de los cromosomas homólogos (pares) tienen genes similares. La mayoría de los genes tienen el código de una proteína particular y algunos codifican una molécula de RNA. Sorprendentemente, los 35,000 genes codificantes constituyen sólo el 5% del DNA cromosómico. Hay muchas regiones en el DNA que no se sabe su función, pero estudios recientes (2012) han mostrado que un 85% o más de DNA tiene un papel regulatorio en la expresión de nuestros genes. La vasta mayoría de las inserciones Alu ocurren en regiones no codificantes para genes determinados. Las secuencias ALU están esparcidas en bloques entre segmentos funcionales de genes, algunas veces están insertos dentro de los genes codificantes, dividiéndolos en segmentos. Hay dos tipos de DNA no codificante: a las secuencias de DNA no codificante que se encuentra dentro de los genes se les llaman intrones y al DNA no codificante que se encuentra fuera de los genes, separándolos entre sí, se le denomina secuencias intergénicas. La función del DNA no codificante intergénico no es conocida totalmente, aunque se sabe que una parte de estas secuencias intervienen en la regulación de la expresión génica de las regiones codificantes. El grupo más grande de secuencias intergénicas se conocen como transposones y entre ellos, se encuentran los retrotransposones que han sido introducidos en los cromosomas por retrovirus. Entre los tipos de DNA retrotransposones se encuentran los miles de elementos ALU que tienen secuencias de DNA de cerca de 300 pares de bases nucleotídicas y por esto son clasificados como segmentos cortos (SINE). El nombre “Alu” viene de un sitio de restricción para la enzima Alu I que se encuentra dentro de estas secuencias.

45
ALGUNOS TIPOS DE SECUENCIAS INTERGENICAS TRANSPOSONES (DNA introducido por diferentes mecanismos) $ % RETROTRANSPOSONES + TRANSPOSONES DE DNA (DNA introducido por retrovirus) (Transposición no involucra RNA) $ % LINES + SINES segmentos largos segmentos cortos % ALU Los elementos ALU se encuentran sólo en primates. Por lo tanto, los cientos de miles de copias de ALU se han acumulado en los primates desde su separación del resto de los vertebrados, hace aproximadamente 65 millones de años. Se piensa que por cada 200 nacimientos ocurre una nueva transposición de un elemento ALU. En el cromosoma 16 humano hay un locus designado PV92 donde, en algunas personas, hay un inserto de DNA llamado ALU. No todas las personas tienen este inserto.
La mayoría de las inserciones Alu se han “fijado” en un lugar, lo que significa que ambos cromosomas homólogos en toda la población humana tienen una inserción en el mismo locus o posición cromosómica. Sin embargo, un número de inserciones ALU son polimórficas, lo que significa que una inserción puede estar presente o ausente en cada uno de los pares de cromosomas en cada individuo. Existen personas que tienen el inserto Alu en uno de sus cromosomas 16 (heterocigotas pv92+/pv92-), otros tienen el inserto Alu en los dos cromosomas 16 (homocigotas pv92+/pv92+ ).
Otras personas no tienen ninguna copia Alu en ninguno de sus cromosomas, es decir, son homocigotos.(pv92-/pv92-). La presencia o ausencia de este inserto puede ser detectada usando la reacción de PCR seguida por una electroforesis en gel de agarosa.
Dado que la inserción del elemento Alu está en un segmento no codificante, no produce ningún efecto en el fenotipo y su presencia o ausencia no tiene ninguna relevancia para la salud de la persona.

46
A continuación se representa un esquema para el caso de una persona heterocigota para el segmento ALU en el cromosoma 16 y los sitios de unión de los partidores para iniciar el PCR.
Gen materno, locus PV92+, con un inserto del segmento ALU
____//Sitio del Partidor1.____Inserto del gen ALU _____._____________//___ Sitio del Partidor 2
Gen paterno, locus PV92-, sin un inserto:
___//Sitio del Partidor 1_________________//____ Sitio del Partidor 2
Como se muestra en la Figura anterior, los partidores para realizar la reacción de PCR en este trabajo práctico están diseñados para colocarse a ambos lados del producto de PCR del gen paterno. Sin la inserción ALU daría origen a una banda que migra como un DNA de 641 bp de largo. En el caso de DNA con el inserto, como el caso materno, la banda producto de PCR sería de un tamaño de 941 pares de bases. Este incremento en el tamaño es debido a la presencia de los 300 pares de bases del inserto ALU. Cuando todos los productos de PCR son analizados mediante electroforesis en gel de agarosa, hay 3 tipos de patrones que pueden ser visualizados en el gel. Si para el DNA de una persona ambos cromosomas tienen insertos ALU, se producirá un solo tipo de banda, de 941 pares de bases (homocigoto pv92+/pv92+). Si en otra muestra de DNA ninguno de sus dos cromosomas contiene el inserto ALU entonces cada producto de PCR tendrá 641 pb y migrarán como una banda de 641 pb (homocigoto pv92-/pv92-). Si la muestra tiene un inserto ALU en un cromosoma pero no en el otro, entonces habrá un producto de la PCR de 641 pb y otro de 941 pb.y el gel se visualizarán dos bandas de 941 y 641 pb (heterocigoto pv92+/pv92-).

47
Apoyo Teórico a protocolo No. 3. La Transformación Bacteriana y su Relevancia en la Biología Molecular General y Biotecnología. La transformación biológica tiene una historia interesante. En 1928, Frederick Griffith, un médico de Londres que trabajaba en un laboratorio de patología, llevó a cabo un experimento que él nunca fue capaz de interpretar por completo durante su vida. Griffith modificó permanentemente (transformó) una cepa no patógena de Diplococcus pneumonía en una cepa bacteriana de neumococo mortalmente patógena. Griffith realizó esta sorprendente modificación de la patogenicidad de las bacterias mediante el tratamiento de las bacterias no patógenas con bacterias que habían sido muertas por acción del calor. En este experimento, las cepas de bacterias (no patógenas) que habían sido puestas en contacto con los restos no vivos de la cepa virulenta, fueron capaces de matar a ratones inyectados con ellas. Griffith repitió el experimento muchas veces, siempre con los mismos resultados. Él y muchos de sus colegas estaban muy perplejos. ¿Qué estaba sucediendo que transformaba bacterias no patógenas en asesinos mortales? Muchos años después, éste vendría a ser conocido como el primer caso registrado de transformación biológica llevada a cabo en un laboratorio. Griffith no sabía de nada del DNA y su función, pero se dio cuenta que la transformación se podía heredar. Los experimentos de Griffith pueden hoy día ser visto como el nacimiento de la manipulación genética que condujeron al desarrollo de la tecnología del DNA recombinante y biotecnología, con las perspectivas de poder manipular genes humanos. Dieciséis años después del experimento de Griffith, en 1944, un grupo de investigación del Rockefeller Institute, dirigido por Oswald T. Avery, publicó un artículo que se derivaba directamente de la obra de Griffith. El trabajo de Avery pretendía dilucidar cuál era la sustancia responsable de la modificación fenotípica de las bacterias de neumonía. Avery y sus colaboradores demostraron que la sustancia es el DNA, y que la transformación biológica es producida cuando las células incorporan y expresan el DNA extraño. A pesar de que pasaron muchos años antes de que el crédito de este descubrimiento se le atribuyese a Avery, hoy él es universalmente reconocido como el autor indiscutido del desarrollo de este conocimiento biológico. A partir del trabajo de Avery y otros, Douglas Hanahan desarrolló la técnica de transformación utilizada hoy en día en todos los laboratorios del mundo. ¿Cómo se produce la regulación de la expresión de la arabinosa? Los organismos regulan la expresión de sus genes y, finalmente, las cantidades y tipos de proteínas presentes en sus células debido a varias razones, entre ellas la adaptación al medio. La regulación de los genes no sólo permite la adaptación de los seres vivos a las diferentes condiciones ambientales, sino que también evita la sobre expresión de proteínas en condiciones no deseadas y un derroche energético innecesario que pondría al organismo en una desventaja competitiva. Los genes implicados en el transporte y degradación (catabolismo) de los alimentos son buenos ejemplos de genes altamente regulados. Por ejemplo, el azúcar arabinosa es

48
una fuente de energía y una fuente de carbono para la bacteria E. coli. Esta bacteria producen tres enzimas (proteínas) necesarias para degradar la arabinosa como fuente de alimento. Los genes que codifican estas enzimas no se expresan cuando la arabinosa está ausente en el medio, pero se expresan cuando la arabinosa está presente en su entorno. La regulación de la expresión de las proteínas ocurre a menudo a nivel de la transcripción del DNA en ARN. Esta regulación se lleva a cabo en una secuencia muy específica en el DNA, llamada promotor, donde la ARN polimerasa se une al DNA y comienza la transcripción del gen. El segmento promotor se encuentra generalmente justo antes de los segmentos de DNA que codifican los genes. En el esquema a continuación, el promotor se indica con la letra P (PBAD en el caso en la figura). En bacterias, es frecuente observar que grupos de genes, relacionados metabólicamente, se agrupan y se transcribe en un sólo ARN mensajero a partir de un promotor. Estos grupos de genes controlados por un único promotor son llamados operones. Los tres genes (araB, araA y araD) que codifican para las tres enzimas que participan en la degradación de la arabinosa se encuentran agrupados en lo que se conoce como el operón arabinosa y donde la RNA polimerasa transcribe los genes de este operón (ver Figura Regulación del Operon Arabinosa).
Regulación del Operón Arabinosa La expresión de estos tres genes depende de la iniciación de la transcripción en un único promotor, llamado PBAD. La transcripción de estos tres genes requieren de la presencia simultánea de:

49
El DNA templado que contiene el promotor PBAD ,del operón arabinosa (que lleva las secuencias que codifican a los ARN mensajeros para las proteínas araB, AraA y araD) la ARN polimerasa, la proteína araC que une al promotor PBAD, la azúcar arabinosa. Cuando la arabinosa está presente en el medio ambiente, las bacterias la ocupan como nutriente. Una vez dentro de la bacteria, la arabinosa interactúa directamente con araC, la proteína que se une al DNA en la secuencia promotora. Esta interacción hace que araC cambie su conformación. Este cambio conformacional de araC promueve la unión de la ARN polimerasa en la secuencia promotora del operón arabinosa y permite la transcripción del único ARN que codifica para los tres genes, araB, AraA y araD. Cuando las tres enzimas se sintetizan, se degrada la arabinosa. En ausencia de arabinosa, araC vuelve a su conformación original y la transcripción del operón arabinosa se inhibe. El plásmido pGLO es construido en el laboratorio e incorpora parte de la estructura del operón arabinosa. Tanto el promotor PBAD como el gen araC están presentes. Sin embargo, los genes que codifican para el catabolismo de arabinosa, araB, araA y araD, han sido sustituidos por el gen que codifica para la proteína fluorescente verde (GFP) (ver Figura Expresión de la proteína fluorescente verde en el plásmido). Por lo tanto, en la presencia de arabinosa, la proteína araC promueve la unión de la ARN polimerasa al promotor y la transcripción del ARN que codifica para la proteína fluorescente verde GFP. Las células que expresan esta proteína en su citoplasma fluorescente de un color verde brillante, en la medida que produzcan más y más GFP fluorescente. En ausencia de arabinosa, araC no facilita la unión de la ARN polimerasa y el gen GFP no se transcribe. Cuando la proteína GFP no se sintetiza, las colonias de bacterias presentan el fenotipo silvestre, es decir, colonias de color blanco, sin fluorescencia.

50
Este es un excelente ejemplo del dogma central de la biología molecular en acción: DNA→ ARN→PROTEINA→ FENOTIPO, CARÁCTER O RASGO GENÉTICO.
Electroforesis Denaturante de proteínas. Esta técnica permite separar las moléculas de una mezcla mediante la aplicación de un campo eléctrico. Las proteínas presentan propiedades ácido-base que están determinadas por el número y tipo de sus radicales ionizables, junto con los grupos amino y carboxilo terminal. Las proteínas que han sido modificadas post-traduccionalmente, ya sea mediante glicosilación, fosforilación, sulfonación, acetilación etc., poseen cargas adicionales. A un pH diferente al de su punto isoeléctrico, las proteínas presentan carga y al ser sometidas a un campo eléctrico migran a diferente velocidad según esta carga. Cuando una molécula cualquiera es sometida a un campo eléctrico, la velocidad con que migra depende de dos fuerzas opuestas, una impulsora y otra de resistencia. La primera depende fundamentalmente de la magnitud del campo eléctrico aplicado y de la carga neta de la molécula. En la segunda influyen, entre otros factores, el tamaño y la forma de la molécula y la viscosidad del medio a través del cual se mueve. Al respecto, la velocidad de migración resulta ser directamente proporcional a la carga neta de la molécula y al potencial aplicado e inversamente proporcional al tamaño de la molécula y a la viscosidad del medio. En relación a este último punto, un factor
EXPRESIÓN DE LA PROTEÍNA FLUORESCENTE VERDE EN EL PLÁSMIDO pGLO

51
importante en una electroforesis es el soporte o matriz en el cual está migrando la mezcla de proteínas en solución. La matriz óptima para electroforesis es aquella que es estable, que disminuye o elimina la convección y que no reacciona químicamente con la muestra. Se ha utilizado una variedad de matrices o soportes para la separación de moléculas mediante electroforesis, entre las cuales se encuentran la agarosa y la poliacrilamida. Poliacrilamida: es un polímero que proviene de la polimerización de la acrilamida en conjunto con la N,N, metilen-bis-acrilamida, que actúa como reactivo entrecruzador. Mediante variaciones en las concentraciones de ambos reactivos se puede lograr distintos tamaños de poros en los geles. Para iniciar la polimerización se adiciona un catalizador que generalmente es una amina cuaternaria, N,N,N',N'-tetrametil etilen-diamina (TEMED), y el iniciador, persulfato de amonio, que genera radicales libres de oxígeno. Los poros del gel son de tamaño pequeño, lo cual retarda la migración de las proteínas de gran tamaño, permitiendo separar con gran resolución aquellas que tengan densidad de carga (razón entre la carga y la masa de la proteína) similares. Como resultado de la electroforesis se obtienen bandas de las proteínas separadas, las cuales se pueden visualizar por tinción u otros medios. Existen varios sistemas para efectuar electroforesis en geles de poliacrilamida, entre ellos se pueden mencionar: i. Electroforesis en condiciones nativas: en estas condiciones las proteínas se
separan principalmente de acuerdo a la carga neta que presentan a un pH determinado, a la forma y al tamaño de ellas.
ii. Electroforesis en condiciones denaturantes en geles de poliacrilamida en
presencia de dodecil sulfato de sodio (SDS): en este sistema las proteínas se separan según su tamaño. Para realizar una electroforesis en estas condiciones, las proteínas se incuban previamente en presencia de SDS, que es un detergente aniónico que rodea a la proteína en solución, evitando la formación de agrupaciones de la proteína. El tratamiento de las proteínas con este detergente y con agentes reductores de grupos sulfhidrilos (generalmente β-mercaptoetanol), cambia su forma tridimensional compacta a una de tipo filamentoso. El SDS se une a las regiones hidrofóbicas de las proteínas confiriéndoles una importante carga negativa, haciendo que la carga intrínseca de la proteína no sea significativa. Así, se eliminan las diferencias en cuanto a forma y a carga eléctrica y las velocidades de migración de las proteínas de la mezcla se correlacionan en forma muy precisa con el peso molecular.
Por esto, este método se usa frecuentemente para determinar el peso molecular de las proteínas, por comparación con la movilidad electroforética de proteínas patrones de peso molecular conocido. El tratamiento con SDS permite además la disociación de las subunidades de las proteínas oligoméricas.

52
Uso de antibióticos como marcador de selección de bacterias que contienen el gen de interés El plásmido pGLO utilizado en el trabajo práctico contiene el gen de la β-lactamasa que le proporciona a la bacteria que lo posee resistencia al antibiótico ampicilina. La proteína β-lactamasa es producida y secretada por bacteria que han sido transformadas con el plásmido que contiene el gen para la β-lactamasa. La β-lactamasa secretada inactiva a la ampicilina presente en el LB/ agar, lo que permite el crecimiento bacteriano. Sólo las bacterias que contienen el plásmido pGLO pueden crecer y formar colonias en las placas que contienen ampicilina. Las células no transformadas (aquellas bacterias que no contienen el plásmido) no puede crecer en las placas de selección con ampicilina. Glosario de algunos términos usados en este Trabajo Práctico. Agar Proporciona una matriz sólida para permitir el crecimiento de bacterias. Contiene además mezclas de nutrientes con carbohidratos, aminoácidos, nucleótidos, sales y vitaminas. Arabinosa Un hidrato de carbono aislado de plantas, que es normalmente utilizado como nutriente para bacterias β-lactamasa La β-lactamasa es una proteína con actividad enzimática que proporciona resistencia al antibiótico ampicilina. La proteína β-lactamasa es producida y secretada por bacterias que han sido transformadas con un plásmido que contiene el gen de la β-lactamasa (por ejemplo el pGLO). La proteína β-lactamasa secretada por la bacteria inactiva a la ampicilina presente en el LB/agar, lo que permite a la bacteria dividirse, crecer y expresar los genes contenidos en el plásmido recientemente adquirido en la transformación (en nuestro caso la proteína GFP ). Biotecnología Es la manipulación de los organismos vivos a nivel genético, para producir productos potencialmente beneficiosos. Clonación o clonamiento Es el proceso de generar copias idénticas de un individuo. Por ejemplo, cuando a partir de la división de una célula única se origina una población de células, todas las células de la población serán genéticamente iguales. Esta población se llama clon o población clonal. El proceso de generación de una población clonal se llama "clonación". Luego de la transformación de una bacteria con un DNA plasmidial (pGLO), cada vez que la bacteria se divida, va a producir copias idénticas del plásmido que son heredadas por las células hijas. Se generan por tanto copias idénticas de esta secuencia específica de DNA que constituyen una población clonal o clon de moléculas de DNA.

53
Ingeniería Genética Consiste en la manipulación del material genético de un organismo (DNA) mediante la introducción o eliminación de genes específicos. Regulación de la expresión Génica La expresión génica es cuidadosamente regulada en todos los organismos que se conocen para permitir su adaptación a las diferentes condiciones ambientales y evitar la sobreproducción de proteínas no utilizadas en determinadas condiciones fisiológicas. Los genes implicados en la degradación de diferentes fuentes de alimentos son buenos ejemplos de genes altamente regulados. Por ejemplo, la arabinosa es una azúcar simple, que es a la vez una fuente de energía y una fuente de carbono para las bacterias. Los genes de bacterias que codifican para las enzimas necesarias para la degradación de la arabinosa no se expresan cuando la arabinosa no está presente en el medio ambiente. Sin embargo, cuando la arabinosa se encuentra presente en el medio, estos genes se activan. Cuando la arabinosa se agota en el medio, los genes se desactivan nuevamente. Proteína fluorescente verde El gen que codifica para la proteína fluorescente verde (GFP) fue aislado originalmente a partir de la medusa bioluminiscente Aequorea victoria. El gen de la GFP ha sido recientemente clonado. La conformación tridimensional única de la proteína GFP permite que emita energía en forma de luz verde cuando se expone a la luz ultravioleta. Plásmido o plasmidio Es una molécula de DNA circular, capaz de replicarse de manera autónoma, que porta además uno o varios genes de resistencia a antibióticos y permite expresar secuencias de DNA codificantes para genes foráneos (como el GFP). pGLO Plásmido que contiene la secuencia codificante para la proteína GFP. Además contiene el gen de resistencia a ampicilina, β-lactamasa. Tecnología del DNA recombinante Consiste en el proceso de cortar y recombinar fragmentos de DNA con la finalidad de aislar genes o alterar su estructura y función. Detección o búsqueda de genes (“screening”) Conjunto de técnicas diseñadas para identificación y aislamiento de genes a partir de una genoteca. Vector Molécula de DNA con capacidad de replicarse de manera autónoma, en la que fragmentos de DNA foráneos se insertan y son propagados en una célula hospedante (por ejemplo un plásmido).

54
Referencias 1. Hanahan, Douglas, Studies on transformation of Escherichia coli with plasmids. J. Mol. Biol., 166, 557 (1983). 2. Hanahan, Douglas, Techniques for transformation of E. coli . In DNA cloning: A practical approach (Ed. D. M. Glover), vol. 1. IRL Press, Oxford (1987). 3. Two positively regulated systems, ara and mal, Robert Schleif, In Escherichia coli and Salmonella, Cellular and Molecular Biology, Frederick Neidhardt, Editor in Chief, ASM Press, Washington, D.C. 1996.

55
PAGINA CON FOTOGRAFIA DE INDAGACION SOBRE PROTOCOLOS PRESENTADOS

56
Indagación Protocolo 1. La metodología de trabajo será con modalidad grupal. Idealmente cada grupo debe tener 3 a 4 integrantes máximo. Es recomendable que cada grupo tenga su identidad. Trabajo Práctico I. El uso de las Enzimas de Restricción en la Identificación de personas. ¿Quien es el Culpable? Orientación general previa al trabajo práctico. En la escena de un crimen se obtiene una muestra de DNA del posible culpable. También se dispone de muestras de DNA de 5 o mas sospechosos. El desafío es resolver las siguientes preguntas: ¿Cómo identificar al culpable de un crimen usando las muestras de DNA? (Consultar a los participantes que discutan los diferentes métodos disponibles para analizar el DNA.) ¿Qué precauciones debe tomar en la escena del crimen para obtener resultados confiables? ¿Cuáles métodos son los mas usados en la forense para identificar un culpable)? Luego, invítelos a compartir sus creencias con el resto de los grupos. Simultáneamente se va desarrollando un protocolo que permitiría identificar una persona entre varias. ¿Cuál protocolo es lo más cercano a la técnica experimental propuesta para este trabajo práctico? Indagación sobre la parte experimental: identificación de personas. ¿Qué precauciones debe tomar en la escena del crimen? ¿Qué controles prácticos son importantes mientras se toman muestras en el sitio del crimen para obtener resultados confiables? ¿Qué son las enzimas de restricción? Discuta las características de las enzimas de restricción, considerando su origen (fuente biológica) y su modo de alterar el DNA. ¿Por qué colocar las muestras en un baño termo regulado durante la digestión? ¿Por qué dejarlas en el refrigerador o en un ambiente de baja temperatura antes y después de la digestión? ¿Podría diseñar un estudio (una serie de análisis) para determinar la estabilidad enzimática de una enzima de restricción, por ejemplo, de la enzima EcoR1? ¿Cómo se benefician los microorganismos al poseer enzimas de restricción?

57
¿Como es posible que los microorganismos no corten su propio DNA endógeno con las enzimas de restricción que invaden su medio interior? Nota: La bacteria se protege de invasores como fagos destruyendo el DNA foráneo con enzimas de restricción, pero no destruye su DNA propio. La bacteria modifica su DNA en el momento de su síntesis. La modificación consiste en la metilación de bases a través de la acción de una metiltransferasa (una enzima que agrega un grupo metilo a purinas o pirimidinas). Así, las bases metiladas de la bacteria no son reconocidas como sitios de restricción y como consecuencia, el DNA no sufre cortes en su propio DNA en estos sitios. Esencialmente, la bacteria disfrazan su DNA en una manera que lo protege de la digestión y solamente el DNA de un fago "invasor" sufre cortes. ¿Por qué conviene usar mas que de enzima de restricción en la digestión para realizar una identificación de DNA? ¿Qué tan confiable es este método de la "huellas genéticas" o cortes en el DNA para la identificación de un "culpable", si no hay otra evidencia disponible (por ejemplo, como testigos o huellas dactilares)? ¿Es posible que en el caso donde los patrones de dos DNA no son distinguibles entre sí con tres enzimas de restricción, se pudieran lograr diferenciarlos con otros métodos? Comentar. ¿Qué pasaría si el "culpable" tiene un gemelo idéntico? ¿Podría distinguir entre los dos gemelos con este método? En el análisis de fragmentos de DNA en geles de agarosa . Posterior a la incubación del DNA con enzimas de restricción, se analiza los fragmentos mediante el uso de electroforesis en geles de agarosa. Si todos los fragmentos tienen la misma carga eléctrica negativa, ¿porque no llegan todos juntos al polo positivo? ¿Hasta qué punto los diferentes fragmentos de DNA se mueven hacia el final del gel? Entre todos los fragmentos que podría tener en un caso de DNA fragmentado, cuál tipo migrará mas rápido hacia el final de gel: el más grande o el más chico? En los experimentos de electroforesis se ha usado agarosa al 3% (3 gramos en 100 de volumen final de agua o tampón). ¿La separación de las bandas de DNA habría sido igual utilizando una preparación al 5% de agarosa?, al 1% de agarosa? Discuta su respuesta.

58
El uso de las enzimas de restricción en la ingeniería genética. En el trabajo práctico 1a. y 1b. se utilizaron preparaciones de su propio DNA total, aislado de su saliva y se analizó posteriormente mediante el uso de enzimas de restricción y la separación de los fragmentos formados por electroforesis. El uso de las enzimas de restricción se utiliza muchísimo también en la ingeniería genética. ¿Tiene un concepto claro sobre lo que es la ingeniería genética? En el trabajo práctico 3 se realiza un experimento típico de este proceso, insertando (transfectando) en una bacteria un plásmido construido para expresar dos genes foráneos. Describa el objetivo de Trabajo Práctico 3, indicando como representativa de un protocolo de ingeniería genética. Discuta sus ideas sobre lo que es la ingeniería genética. ¿Ha leído comentarios en la prensa o textos científicos sobre este tema? Nota: En la sección Apoyo Teórico para el Trabajo Practico No 1. se describe las enzimas de restricción que cortan DNA en forma escalonada, dejando fragmentos con terminales llamados "cohesivos." Los investigadores de Biología Molecular aprovechan este tipo de enzimas para cortar un DNA que les interesa modificar mediante la combinación de fragmentos de DNA de diferentes orígenes. Realizando cortes en dos DNA con la misma enzima de restricción, dejaría a ambos fragmentos con lo mismos terminales cohesivos y con la probabilidad de lograr "pegar" un fragmento nuevo un este sitio, con el resultado de la formación de una "inserción" de un tipo de DNA diferente en el DNA en estudio. Este tipo de procesamiento permite "construir" un DNA nuevo por tener una secuencia diferente insertado donde no existían en el gen natural. Este resumen indica muy superficialmente lo que es la base de la formación de genes transgénicas por las técnicas de ingeniería genética. ¿Ha escuchado a personas expresar temores sobre la ingeniería genética? ¿Ha escuchado hablar sobre el peligro de plantas transgénicas? ¿Ha leído información en la prensa o visto películas sobre estos temas? ¿Ha investigado en Internet qué es la ingeniería genética y algunos de sus beneficios? ¿Sabía que toda la insulina usada para diabéticos es manufacturada en laboratorios usando técnicas de ingeniería genética y esta insulina es idéntica a la insulina producida en su cuerpo?

59
¿Sabía que las mejoras realizadas en muchas plantas para resistir las heladas se deben a estudios de ingeniería genética? ¿Sabía que las mejoras realizadas en muchas plantas para resistir la salinidad permiten producir cultivos a la orilla del mar y esto se debe a estudios de ingeniería genética? Comparte sus creencias sobre este tema con el resto de los grupos.

60
Indagación sobre Trabajo Práctico No. 2. La Reacción en Cadena de la DNA Polimerasa, PCR. 1. Conociendo La TAQ Polimerasa. ¿Qué son las enzimas? ¿En el caso de hombre, cuales temperaturas podrían contribuir favorablemente a la actividad de enzimas típicas encontradas en los diferentes tejidos del cuerpo? ¿Cuáles temperaturas podrían denaturar (destruir su actividad) una enzima típica en solución? ¿A cual temperatura actúa la enzima TAQ polimerasa? ¿A qué temperatura tiene máxima actividad la TAQ Polimerasa? (Indica un rango probable). Dibuje en un gráfico (temperatura vs actividad enzimática) su concepto de los resultados esperados de un estudio de la actividad enzimática de TAQ Polimerasa versus temperatura (entre 25º y 80ºC). ¿Es la TAQ polimerasa una enzima típica? ¿De donde aislaron esta enzima? ¿Usted tenía conocimientos de la presencia de "vida" o sistemas biológicos en ambientes tan poco amigable como dentro de un géiser? ¿Cree que se podría encontrar una bacteria con una actividad enzimática similar a la TAQ Polimerasa en Los Salares de Atacama? ¿Qué rango de temperaturas se usan en este trabajo práctico de PCR? Indica que reacción esta en progreso en cada fase del incubación en el termociclador. 2. La Polimeración de DNA específico a partir de Partidores. En 1953, después de muchos estudios de científicos cristalógrafos, se definió la estructura bicoidial ordenada de DNA: un doble hélice con el apareamiento de bases que conocemos hoy. En general, se considera que el DNA es una molécula muy estable en condiciones fisiológicos. ¿Cómo esta estructurado el DNA recién aislado de la saliva (como preparada para esta experimento) cuando lo tiene a temperaturas entre 20ºC y 37ºC? ¿Qué pasa con el DNA de doble hebra a temperaturas altas (sobre 45ºC hasta 60 o 70ºC)?

61
¿Qué pasa con el DNA en solución cuando se reajusta la temperatura de 70-80ºC y vuelve a la temperatura ambiente? ¿Qué rangos de temperaturas se usan en el trabajo práctico de PCR? Indica la forma de DNA en cada una de estas temperaturas (¿doble hebra o hebra simple?) Basando en sus conocimientos de las bases A,G, C y T, y los pares que se forman en el DNA de doble hebra, ¿podría la estructura primaria de un DNA afectar la temperatura de la denaturación un DNA de doble hebra? Justifique su respuesta. Nota: La separación de las hebras de una molécula DNA de doble hebra por altas temperaturas es consecuencia de las agitaciones moleculares de los puentes de hidrógeno entre los pares de bases, eventualmente logrando su separación. De bajar la temperatura de nuevo, las hebras logran reasociarse para formar doble hebras. La secuencia primaria entonces puede afectar la temperatura de reformar el doble hélice. Se ha encontrado moléculas de DNA en muchos lugares "extremos" en el mundo, como en tumbas milenarias, en el interior de pedazos de cuarzo (donde hay insectos encapsulados) después de miles de años de su formación. Además, se ha aislado fragmentos de DNA mitocondrial de huesos de momias encontradas en el norte de Chile en estudios antropológicos. Las secuencias primarias de estas muestras de DNA han sido determinadas y indican que durante muchísimos años, muchas moléculas de DNA no se han degradado. Secuencias descifradas de estos DNA indican que su origen era mitocondrial y no nuclear. ¿Qué nos dicen estos acontecimientos en relación a la estabilidad de DNA? Nota: Denaturación de DNA o "Melting." El este trabajo práctico hay dos componentes que son sensibles a la temperatura tanto para mantener una estructura específica como tener función biológica. La estructura helicoidial de DNA se mantiene como doble hebra porque las bases están complementarias (donde hay un A en una cadena, hay un T en la cadena complementaria, y donde un G, un C.) y se forma las cadenas bicoidial muy estables debido a la formación de miles de 'puentes de hidrogeno. La separación de las hebras de una molécula DNA de doble hebra por altas temperaturas es consecuencia de las agitaciones moleculares entre estos puentes de hidrogeno entre las bases, eventualmente logrando su separación. De bajar la temperatura de nuevo, las hebras logran reasociarse para formar doble hebras nuevamente, y recuperan su estructura de doble hélice original. De bajar la temperatura muy rápidamente, no todas las hebras logran re-asociarse perfectamente. En el caso de las proteínas, estas están también estructuradas tridimensionalmente y poseen una conformación espacial que determina su actividad catalítica (en el caso de proteínas enzimas).

62
Consideraciones de la ética del análisis de DNA de personas. Discute los problemas que podría surgir de este tipo de información. Nota: En los años recientes, las técnicas disponibles para secuenciar el DNA total de una persona han mejoradas muchísimas y se puede determinar la secuencia de todo el DNA de una persona en un día. Las muestras de DNA humano pueden quedar guardadas en una "genoteca" para diferentes usos, según el caso y la secuencia del DNA particular se pueden guardar en un "banco de datos." Algunos países mantienen genotecas de DNA de personas, almacenando información relacionadas con crímenes o personas sospechosas, pero generalmente no de toda la población. ¿Sería muy útil tener archivos que identifican a todas las personas? ¿Quienes tendrán acceso a tales archivos? ¿Tendría utilidad esta información para el médico de un paciente con una enfermedad de difícil diagnóstico? Las mutaciones en la secuencia de DNA de un gen pueden o no afectar a la expresión del gen o la secuencia de aminoácidos en una proteína codificada. ¿Esta información seria ventajosa para su médico? ¿Es posible que una genoteca de DNA sea base de abusos realizados por la policía? ¿o por un gobierno? ¿o por un empleador? ¿Cree Ud. que la existencia de una genoteca de toda la población sería una violación de los derechos de privacidad?

63
LA INDAGACIÓN EN LOS CURSOS DE BIOLOGÍA DE ENSEÑANZA MEDIA:
Sugerencias de esquemas en Biología Molecular y
Genómica

64
Trabajos Prácticos sin laboratorios 1. Unidad de indagación sobre enzimas de restricción***1
INTRODUCCIÓN En esta actividad los estudiantes utilizarán facsímiles de los protocolos usados para determinar una "huella" de DNA. La actividad permite entender y visualizar el proceso de análisis de DNA usando enzimas de restricción y posteriormente, realizando un modelo del proceso de "electroforesis en geles," los participantes pueden comparar su propio DNA con los de sus compañeros de curso. El ejercicio ilustra el proceso usado en estudios forenses para la identificación de personas y en otros estudios genéticos. El protocolo se distingue por su simplicidad y claridad y además, utiliza materiales convencionales, siendo papel y tijeras las protagonistas principales. Para identificar personas se utilizan enzimas de restricción (tijeras) para analizar una muestra del DNA (cinta de papel) de ellos mismos. Todos los estudiantes inventarán una secuencia de 100 bases de DNA para usarla como una hebra de "su" DNA. Esta "molécula" será duplicada para formar una doble hebra escrito en cintas de papel que se cortarán en sitios específicos con sus "enzimas de restricción." Finalmente, se analizarán su perfil o 'huella genética" usando un proceso que representa la técnica de electroforesis en "geles." Cada alumno puede comparar la migración de su DNA con los obtenidos de sus compañeros de curso. De tener un DNA de uno o más "sospechosos" en el estudio, podrían incluir en el estudio la determinación de "quien es el culpable" del supuesto crimen. Objetivo General Aplicar una técnica de análisis de DNA para identificar personas usando materiales aplicables en diferentes etapas de la educación básica. Objetivos Específicos. 1. Reconocer que la existencia de las ideas iniciales de los participantes, orientan las posibles explicaciones frente a solución del problema y que son modificables a la luz de la evidencia. 2. Formular predicciones fundamentadas en las ideas iniciales proveniente de las experiencias personales o del mundo científico. 3. Aclarar sus conocimientos sobre la puesta en marcha del protocolo de la técnica “Huella genética” (DNA Fingerprinting) de DNA, específicamente, explicar lo que hacen las enzimas de restricción y entender el sistema de electroforesis en geles.

65
4. Visualizar como se obtienen e interpretan evidencias a través de la detección de diferencias de secuencias de DNA analizado en un posible estudio legal (aspectos éticos, forenses, casos de paternidad, herencias). 5. Comunicar en forma oral y escrita sus resultados, análisis de estos y conclusión que permite dar solución al problema planteado inicialmente. Profundización Temática. Duplicación de DNA. Las reglas del apareamiento de las bases nucleótidicos y una sólida compresión de los mecanismos que gobiernan la síntesis de DNA son necesarias para la apreciación y ejecución de este módulo. Se presenta en forma muy resumida a continuación. Para iniciar el proceso de duplicación, una enzima llamada DNA helicasa desenvuelve el hélice de DNA, separando las dos hebras previamente unidas por puentes de hidrógeno entre los pares de bases, dejando camino para la maquinaria de duplicación. A continuación, la enzima DNA polimerasa se une a una de los dos cadenas de DNA y usando un pequeño "partidor," una secuencia de 20 nucleótidos de largo requerida para iniciar) y copia la secuencia complementaria a la cadena matriz, para formar finalmente una cadena complementaria nueva. El partidor se sintetiza otra enzima. El proceso de duplicación en la mayoría de los organismos ocurre con la molécula del DNA en la forma de doble hélice, donde las hebras de DNA están orientadas en direcciones opuestas entre sí, una de orientación 5' hacia 3', indicado la hebra matriz o líder y la hebra llamada complementario, orientada con su término 3' hacia 5'. El proceso de duplicación se complica por el hecho de que la DNA polimerasa sólo puede trabajar en una sola dirección a lo largo de la cadena azúcar-fosfato. La DNA polimerasa además requiere un partidor nucleotídico para iniciar todo el proceso. Así, mientras que la DNA polimerasa puede ejecutarse de forma continua a lo largo de una hebra, elongando en la orientación 5' hacia 3', dando un producto conocido como la cadena "líder" o "adelantada." La otra cadena , llamada la cadena "complementaria" o "atrasada" se forma por segmentos (Fragmentos de Okasaki) de una manera gradual en etapas. Las enzimas de restricción y las huellas genéticas. Las Enzimas de Restricción conforman un grupo grande de enzimas encontrado en bacteria, virus, y otros organismos y que cortan el DNA de doble hebra en sitios muy específicos. Su presencia en la bacteria, por ejemplo, permite que este bacteria podría defenderse contra infección por un DNA foránea, realizando cortes en el DNA de invasor. Su descubrimiento el bacteria y su caracterización en los años setenta contribuyeron al desarrollo de muchísimas tecnologías de la biología molecular moderna y los investigadores Daniel Nathans, Werner Arber, and Hamilton O. Smith. recibieron el premio Nobel en 1978 por este trabajo. La primera aplicación de la técnica "Huellas genéticas" (DNA Fingerprinting) se realizó Alec Jeffrys en 1984.

66
Mas de 3000 enzimas de restricción has sido descubiertas hasta la fecha y muchas de éstas están disponibles comercialmente para comprar. Determinación de huellas genéticas del DNA de una persona u otras especies es una técnica utilizada para distinguir entre los individuos de la misma especie usando solamente muestras de su DNA. El proceso comienza con la extracción de DNA de las células de un individuo usando una muestra de sangre, saliva, semen, u otro tejido apropiado. Los seres humanos tienen una gran parte de su secuencia de DNA en común pero existen también grandes variaciones que permitan su fragmentación con enzimas de restricción en una forma específica para cada individuo. Cada enzima de restricción pueden dirigirse a una parte del DNA donde hay una secuencia que reconoce. Estas secuencias se encuentran en diferentes posiciones en el DNA de diferentes personas, resultando que el DNA cada individuo se cortará en una manera especial, y no da un conjunto de productos comparables con otros, es decir, resulta en fragmentos de longitud variable del DNA. Una excepción a esta regla son los gemelos idénticos que tienen DNA idéntico. Estas diferencias son conocidas como polimorfismos de longitud de fragmentos de restricción. Esta técnica también se utiliza a menudo para determinar el genotipo de un individuo para un determinado gen, por ejemplo, para detectar la presencia o ausencia de una mutación que confiere un trastorno genético determinado. Las enzimas de restricción cortan, como indicado, por reconocimiento de secuencias específicas. Por ejemplo, la enzima de restricción conocida como Sma1 busca la 5'CCCGGG3' apareada con 3'CCCGGG5' de otra hebra y corta el DNA de doble hebra entre la C central (5'CCCIGGG3' )y la base G de ambas hebras. Otras corten escalonado, y quedan bases no apareadas en cada extremo. Por ejemplo, la enzima de restricción EcoR1 reconoce la secuencia 5'-GAATTC- (donde X es cualquier base desoxinucleotídico) y hace un corte escalonado dejando los productos indicados, con extremos no-apareados: 5'-(X)nG- y -AATTC(X)n- 3' 3'-(X)nCTTAA- -G(X)n-5'. Estos fragmentos tienen lo que se conocen como extremos pegajosos porque pueden volver a asociar una secuencia que tiene la secuencia complementaria de DNA de una hebra.. Normalmente se utilizan más que una enzima de restricción (2 o 3) para hacer un estudio de huellas. Otro método común de realizar huellas genéticas de DNA explota secuencias cortas repetidas, altamente variable llamada repeticiones cortas en tándem (STRs, sigla en ingles para Short Terminal Repeats). Diferente personas tienen distintos números de unidades de repetición. Por ejemplo, un gen llamado CSF contiene una secuencia -AGAT- repetida entre 6 a 16 veces. Dos personas no relacionadas serán propensos a tener diferentes números de repeticiones de esta secuencia AGAT.

67
Mientras que el número variable de repeticiones de un STR especifico será compartida por alrededor de 5 - 20% de las personas, si nos fijamos en otros tipos de regiones STR o examinen varias STR al mismo tiempo, este método de analizar varios SRT se convierte en un poder increíble para identificar personas por análisis de su ADN. Lo más regiones STR que se analizan de una persona, lo más discriminatorio la prueba. En los Estados Unidos de América, utilicen actualmente 13 loci diferentes (repeticiones frecuentes en el ADN) para la discriminación. Esto ha resultado en la capacidad de generar probabilidades de identidad de 1 en un trillón (1 con 18 ceros detrás) o más. Por lo tanto, es posible establecer una identidad que muy poco probable haya surgido por casualidad, excepto en el caso de gemelos idénticos, que tienen idénticos perfiles genéticos. Electroforesis en geles. Los fragmentos de DNA cortadas por las enzimas de restricción se pueden separar mediante electroforesis en geles. Este método coloca la muestra de DNA en un pozo en un extremo de un gel de agarosa. Un gel de agarosa es muy parecida a una fina capa de gelatina puesto en una cajita plástica. Un campo eléctrico se aplica al gel y, dado que el DNA está cargado negativamente, es atraído por el extremo positivo del campo y comienza a moverse a través del gel. Cuanto más grande sea, los fragmentos ya no puede viajar a través de la matriz del gel tan rápidos como los fragmentos cortos y se quedan atrapados cerca el inicio del gel donde la muestra se ha cargado. Los fragmentos más cortos mueven adelante con más facilidad y así viajarán más lejos. Esto da lugar a un patrón único de bandas para el DNA de cada individuo. El imagen en la Figura 1 de un gel de agarosa típica se visualiza un análisis de DNA previamente tratada con tres enzimas de restricción, EcoR1, Pst1 y BamH1. Por el lado derecho se ve una autoradiografía del mismo gel donde usaron DNA radioactivo.
Figura 1. Electroforesis en un gel de agarosa de DNA cortada por 3 enzimas de restricción.

68
Después de escribir su propia secuencia de DNA, los "experimentos" que realizan los alumnos son los siguientes: Usando la misma tira de papel con su propio DNA de una hebra, confeccionan la hebra complementaria en la misma cinta y entonces, usan "tijeras" (para representar la enzima Helicasa de DNA), separan las dos hebras, formando un sitio de iniciación de la síntesis de ADN. Posteriormente, se asocia la DNA Polimerasa al término 3' de una de las hebras y se inicia la duplicación de esta hebra del DNA, para tener una nueva molécula de DNA de doble hebra. La otra molécula de DNA de una hebra que quedó se duplica también por la polimerasa y este DNA se guarda aparte en una Genoteca que representa el DNA de este grupo de alumnos. La otra molécula de DNA recién duplicada se "incuba" con una "Enzima de Restricción" que corta en sitios específicos para dejar varios fragmentos más chicos. En el experimento de este unidad cada alumno debería tener 6 a 7 fragmentos si usa una enzima de restricción simple (corta entre cada A y T). Los fragmentos son de diferentes tamaños y el largo total de cada fragmento equivale el número de bases del segmento. Finalmente, cada alumno (o grupo) se confeccionan un "gel de electroforesis" (el gel es una hoja grande de papel de envolver) y marcan este papel usando un plumón con los resultados de la electroforesis, donde las distancias de migración de cada fragmento representa el perfil de su huella genética. Inspección visual de las variadas bandas presentados en los diferentes geles permite que los alumnos podrán apreciar la gran utilidad de la técnica para demostrar diferencias entre los DNA de las personas. La investigación de una escena del crimen podría ser agregado como parte de las actividades de esta unidad pero aumentaría el tiempo dedicado al tema. El profesor de la unidad entregará un DNA tomado de la "escena del crimen" para análisis junto con los DNA de los alumnos y comparar su perfil de migración con otros DNA. Otro profesor o voluntario podría representar el culpable y discretamente poner el perfil de este DNA cortada con la misma enzima de restricción en un gel para análisis. Alternativamente, estos aspectos relacionados con la identificación de un "culpable" podrían quedar solamente para la discusión final de la práctica (Objetivo 4). Dado que tanto las huellas dactilares de DNA y la cromatografía de proteínas en papel se basan en los mismos principios de electroforesis (separación de las moléculas por tamaño) se presta para discutir algunos paralelos conceptuales interesantes entre estas dos técnicas (Objetivo 2) Vocabulario Duplicación del DNA, DNA polimerasa, enzimas de restricción, la huella de ADN, polimorfismos mostrados por longitud de fragmentos, repeticiones cortas, en tándem, reacción en cadena (PCR), electroforesis en geles. Grupos de trabajo. Cada alumno se "crea" y "replica" una cadena de ADN suya. Para los análisis de restricción el ideal sería que cada alumno realiza la corrida de su gel. Si el espacio y materiales no permiten, se recomiendan forman grupos de 3 a 4 alumnos. En el análisis de los geles todos los grupos deberían participar juntos.

69
Materiales. • Tijeras de niño; • Cinta de papel de máquina de sumar (1 metro por persona o por muestra de
ADN si trabajan en grupos); • Reglas de madera de un metro; • lápices de mina y dos de color; • plumones para marcar los posos y la migración de fragmentos, • papel de envolver o espació sólido (suelo) de un metro por 90 cm por cada tres
personas (cada corrida de ADN requiere 30 cm). • Opción de trabajo: en lugar de papel sobre el piso podría usar el mismo piso de
la sala, un pasillo, ventana o el gimnasio, enmarcando el "gel" con cinta adhesiva (de los pintores de casas) o cinta de color) para definir sus limites y para marcar los posos para cargar las muestras.
Secuencia de la Lección. I: Para comenzar. El propósito de esta actividad es la creación de una secuencia de ADN, escrita sobre una cinta de papel como los de las máquinas de sumar, replicar este ADN, y luego utilizar estas secuencias de ADN para realizar un análisis de restricción. Este análisis permite determinar las "huellas de un ADN." Una sólida comprensión de la estructura del ADN y las reglas de apareamiento de las bases nucleotídicos para formar la doble hebra . I.a. ¿Que necesito? Una cinta de maquina de sumar de 1 metro de largo, una tira para cada participante o grupo. Estas tiras van a usar para anotar la secuencia de una hebra de su "DNA" (cada participante o un alumno representando el grupo). Cada persona o grupo requiere 3 lápices (negro, rojo y otro color); un plumón,1 tijeras y 1 regla en cm de un metro. Cada cinta con un DNA usará una espacio de 30 cm de ancho por 1 metro de largo en un papel de envolver con medidas de 1 cm indicadas por una orilla (para guiar en la ubicación en el papel la distancia de migración de los fragmentos). I.b. ¿Como es el "gel" de agarosa? Los geles usados en los laboratorios de biología molecular son formados usando agarosa gelificada en la presencia de un tampón para ajustar de pH y fuerza iónica. Estos geles, por su textura y suavidad, parecen una jalea sumergida en un liquido. Usando electrodos y un voltaje controlado, se pasa corriente por el gel para efectuar la migración del DNA hacia el polo positivo, según su tamaño. En esta Unidad, el 'gel' será un papel grande donde los alumnos pondrán una marca o banda representando cada fragmento de DNA que resultó como producto de un corte por la enzima de restricción. El profesor anunciará la secuencia que corta la enzima de restricción que usarán todo los participantes. Se cortará las tiras en estos sitios usando tijeras que representan las enzimas de restricción.

70
I.c. ¿Que voy a usar para representar mi DNA? Cada persona debe poner una regla de un metro en el centro de la cinta de un metro de largo. Dibujar una línea recta por el largo de la cinta a aproximadamente 3 cm desde el borde del papel, usando un lápiz de mina y la regla para guía. Sobre esta línea y con la regla fija sobre el papel, escribe al azar, una tras otra, las bases nucleotídicas (A, T, C o G), espaciándolas un centímetro cada una. Escribe 100 bases (letras) a la larga del papel para crear una cadena de DNA. Nota: su ADN puede tener bases repetidas 2,3 4 veces, como TTGGGCAAGGGATAATTTTCGA y totalmente al azar. Recuerde que todos los alumnos (o grupos) están creando su propia secuencia de DNA individual de doble hebra y el DNA de cada persona es diferente, por lo que deben trabajar en forma independiente para componer su secuencia de DNA II. Parte Experimental. II.a. Duplicación de su DNA. A continuación, escribe por debajo de la regla, en la misma manera de mantener las bases un cm de otra, las bases complementarias a cada base dibujada arriba de la regla, hasta completar las 100 bases de esta cadena. Ahora, ha preparado su DNA de doble hebra de 100 pares de bases. La Figura 2. representa la distribución de una base y su base complementario, en un papel de 6 a 7 cm. de ancho. (M) (la regla aquí) (C) I________base_______I_______ base________I Papel visto 0______________________________________7 cm = al ancho de la cinta, desde el canto. El primer sitio de corte sería a las 3,5 cm. Figura 2. La anotación del DNA en la cinta de papel, con la regla de madera centrada sobre el papel. La línea horizontal inferior representa el ancho de la cinta de papel (7 cm de ancho, aprox.), visto desde el canto y donde se ve la posición de la primera base de la hebra matriz (M) que es complementaria a otra base (C), y con espacio libre 1 a 2 cm en el centro donde se ubica la regla de un metro. La cinta tiene un metro de largo con las bases separadas por un cm, todo el largo de la cinta. Conviene marcar la cinta con pequeñas marcas, indicando los centímetros a la larga del papel, para guiar cuando necesita cortar entre dos bases. Para el primer paso de la duplicación del DNA recién dibujado en sus cintas se utiliza unas tijeras para representar a la enzima DNA Helicasa. La DNA Helicasa desenrrolla las hebras del DNA apareados, formando una burbuja (espacio) dentro las hebras entrelazadas para permitir la asociación de la maquinaria de la síntesis de DNA .

71
Por lo tanto, cada estudiante, con sus tijeras que representan la Helicasa, debe cortar entre las dos hebras de su DNA a la larga del papel. La Figura 3 (página aparte) permite visualizar todo el proceso de este experimento de la preparación de las cadenas de DNA de doble hebra. II b. Sintetizar otra copia de su DNA de doble hebra. La asociación de la DNA polimerasa al termino 3' de una de las hebras permite la duplicación de las hebras originales, que permite tener dos molécula de DNA de doble hebra (ver Figura 3 donde las hebras rojo y azul son los últimos sintetizadas). Para hacer otra copia de su DNA, con su lápiz rojo escribe el complemento de uno de las hebras (su lápiz rojo es la DNA polimerasa y la síntesis es el acto de escribir ahora el complemento de una hebras). En seguida, escribe con lápiz azul el complemento del DNA en la otra tira de papel. Ahora, su lápiz azul es otra molécula de la DNA polimerasa realizando la síntesis de otra copia. Cada persona (grupo) debe terminar con dos copias exactas de su DNA. La molécula de ADN con colores rojo y azul se guarda aparte en una Genoteca que representa el ADN de este grupo de alumnos. La otra molécula de ADN (negro, hebras) se "incuba" con una "Enzima de Restricción" que corta en sitios específicas para dejar varios fragmentos más chicos. Ver sector II.c. y Figura 3.
















![Biología Molecular e Ingeniería Genética [libro].pdf](https://static.fdocuments.ec/doc/165x107/55cf9dcd550346d033af4466/biologia-molecular-e-ingenieria-genetica-libropdf.jpg)
![Biología Molecular e Ingeniería Genética [libro]LUQUE - copia.pdf](https://static.fdocuments.ec/doc/165x107/577c7e7c1a28abe054a15e3f/biologia-molecular-e-ingenieria-genetica-libroluque-copiapdf.jpg)