INTERACCION ENTRE HORMONAS TIROIDEAS Y EL SISTEMA ...
262
UNIVERSIDAD COMPLuTENSE DE MADRID FACuLTAD DE FARMACIA DEPARTAMENTO DE BIOQUIMICA Y BIOLOGIA MOLECULAR II INTERACCION ENTRE HORMONAS TIROIDEAS Y EL SISTEMA IGFs/IGFBPs: MECANISMOS MOLECULARES TESIS DOCTORAL SONIA RAMOS RIVERO Madrid, 1999
Transcript of INTERACCION ENTRE HORMONAS TIROIDEAS Y EL SISTEMA ...

UNIVERSIDAD COMPLuTENSEDE MADRID
FACuLTAD DE FARMACIA
DEPARTAMENTO DE BIOQUIMICA Y BIOLOGIA MOLECULAR II
INTERACCION ENTRE HORMONAS TIROIDEAS
Y EL SISTEMA IGFs/IGFBPs:
MECANISMOS MOLECULARES
TESIS DOCTORAL
SONIA RAMOS RIVERO
Madrid, 1999

DEPARTAMENTO DE BIOQUíMICA Y BIOLOGíA MOLECULAR II
UNIVERSIDAD COMPLUTENSEDE MADRID
FACULTAD DE FARMACIA
INTERACCION ENTRE HORMONAS TIROIDEAS
Y El SISTEMA IGFs/IGFBPs:
MECANISMOS MOLECULARES
Memoria que presentaSONIA RAMOS RIVERO para
optar al gradode Doctor en Farmaciapor la
UniversidadComplutensede Madrid
Madrid, 1999

Estetrabajoseha llevadoa caboen el Grupode Endocrinología
y MetabolismoPerinatal,en el Institutode Bioquímica(Centro
Mixto: F. Farmacia - C.S.I.C.), ubicadoen la Facultadde
Farmaciade la UniversidadComplutensede Madrid.
Ha sido realizadopor Sonia RamosRivero y dirigido por la
Dra. Ana M~’ Pascual-LeonePascual.

Deseoexpresarmi agradecimientoal ProfesorD. ManuelRuiz Amil, Director
del Departamentode Bioquímica y Biología Molecular de la Facultad de
Farmacia,asícomo a la Dra. EvangelinaPalaciosAlaiz, actualDirectoradel
Instituto de Bioquímica,por suacogidaen el mismo.
A la Dra. Ana M8 Pascual-LeonePascual,Directorade estaTesis,mí mayor
y másprofundagratitudpor su orientacióny apoyocientífico constantes,así
comoporel esfuerzoy entusiasmopuestosen la realizaciónde estetrabajode
investigación.
Expresotambiénmi agradecimientoa la Dra. GabrielaMorrealede Escobar
porhaberfacilitadoensu laboratoriola dosificaciónde lashormonastiroideas.
A mis compañerasdel Grupode Endocrinologíay MetabolismoPerinatal:M~
Angeles Martín, María Agote y Ana de la Puentepor su continuo apoyo,
ayuday sumás sinceraamistad.
Agradezco igualmente a mis compañeros Fernando Escrivá, Carmen
Alvarez y, en especiala Luis Goya, sucolaboraciónen el aprendizajede las
técnicasde ‘Western inmunoblot’, dosificaciónde GH y la metodologíade
biología molecular,así como su constanteayuda y amistad.
De la misma forma, agradezcola inestimablelabor de Angel García en el
animalario de] Centro, y la de AdoraciónUrrea y CarmenCarrizosaen la
Biblioteca del Centro.
Porúltimo, quiero hacerconstarmi mássinceroagradecimientoa mi familia
y a mis amigosporsu apoyoincondicionaly muestrasconstantesde ánimo.

INDICE
ABREVIATURAS 17
1. INTRODUCCION 20
1.1. HORMONAS TIROIDEAS 21
1.1.1. Síntesisy secreción 21
1.1.2. Regulacióndel axis tiroideo 21
1.1.2.1. Tirotropina 22
1.1.2.2. Control neuroendocrino 22
1.1.2.2.1.Hormonade liberaciónde tirotropina 22
1.1.2.2.2.Otros factoreshipotalámicos 23
1.1.2.2.3.Hormonastiroideas 24
1 . 1.3. Metabolismoy accionesde las hormonastiroideas 24
1.1.3.1. Metabolismo 24
1.1.3.2. Accionesbiológicasde las hormonastiroideas 25
1.1.4. Interacciónentrehormonastiroidease insulina 25
1.2. FACTORES DE CRECIMIENTO SIMILARES A LA INSULINA
(IGEs) 27
1.2.1. Estructura,síntesisy transporte 27
1.2.1.1. Estructura 27
1.2.1.2. Síntesisy transporte 28
1.2.2. Accionesbiológicasde los IGFs 29
1.2.2.1.Iii vitro 29
1.2.2.2. In vivo 30
1.2.2.3. Acciones autocrinas, paracrinasy endocrinas 31
1.3. PROTEíNASTRANSPORTADORASDE IGFs (IGFBPs) 32
1.3.1. Estructura,síntesisy regulación 32
1.3.1.1.Estructura 32
1.3.1.2. Síntesis 32
1.3.1.3. Regulación 33
1.3.1.3.1. Variaciones diurnas 33
1.3.1.3.2. Nutrición 33
5

341.3.1.3.3. Ejercicio
1.3.1.3.4. Ontogenia
1.3.1.3.5. Hormonas
1.3.2. Tipos de IGFBPs y acciones biológicas
1.3.2.1. Tipos de IGFBPs
1.3.2.2. Acciones biológicas
1.4. FACTORESENDOCRINOSQUEREGULANEL DESARROLLO:
HORMONAS TIROIDEAS E IGFs, SU INTERACCION
1.4.1. Interacciónentre las hormonastiroideasy el axi39 IGFs/IGFBPs.
1.5. TRANSPORTADORESDE GLUCOSA (GLUTs)
1.5.1. Estructura,funcióny expresióntisular
1.5.2. Efecto de] estadotiroideo sobrelos transportadoresde glucosa
1.6. PLANTEAMIENTO DEL TEMA.
2. MATERIAL Y METODOS
2.1. ANIMALES
2.2. COMPOSICIONDE LA DIETA GENERAL
2.3. HIPOTIROIDISMO
2.4. MODELOS DE HIPOTIROIDISMO
2.4.1. Animaleshipotiroideosneonatales
2.4.1.1. Ratasneonatalestiroidectomizadas
2.4.1.2. Ratasneonatalestratadascon MMI
2.4.1.3. Ratasneonatalestiroidectomizadastratadascon STZ
2.4.2. Animaleshipotiroideosdestetados
2.4.2.1. Ratasdestetadastiroidectomizadas
2.4.2.2. Ratasdestetadastiroidectomizadastratadascon insulina
2.4.3. Animaleshipotiroideosadultos
2.4.3.1. Ratasadultastiroidectomizadas
2.4.3.2.Ratasadultastiroidectonizadastratadascon insulina
2.5. MODELOS DE HIPOTIROIDISMO CON REI-LABILITACION
2.5.1. Animaleshipotiroideostratadoscon tiroxina
2.5.1.1.Rehabilitaciónmedianteinyecciónde ‘[4
2.5.1.2.Rehabilitaciónmedianteimplantaciónde pellets
34
34
34
35
3,7
38
39
41
41
42
44
48
49
49
51
51
51
51
53
53
53
53
54
54
54
54
55
55
55
55
6

552.6. MODELO DE DIABETES POR ESTREPTOZOTOCINA
2.6.1. Tratamientode animalesdiabéticoscon ‘[4
(GLUTs)2.7. ESTUDIO DE LOS TRANSPORTADORESDE GLUCOSA
EN RATAS NEONATALES HIPOTIROIDEAS
2.8. RECOGIDADE MUESTRAS
2.9. DETERMINACION RADIOINMUNOLOGICA DE IGF-I.
2.9.1. Fundamentodel radioininunoanálisis
2.9.2. Filtraciónen gel
2.9.2.1. Fundamento
2.9.2.2.Material y reactivos
2.9.2.3.Calibraciónde la columna
2.9.2.4. Filtraciónde los sueros
2.9.3. Marcajeradiactivode IGF-I y II
2.9.3.1. Fundamento
2.9.3.2. Material y reactivos
2.9.3.3. Procedimiento
2.9.3.4. Cálculo del rendimientoy la actividadespecífica
2.9.3.5. Perfil de elución de radiactividad
2.9.3.6. Repurificacióndel IGF marcado
2.9.4. Desarrollodel RIA de IGF-I
2.9.4.1. Material y reactivos
2.9.4.2. Preparaciónde las muestras
2.9.4.3.Procedimiento
2.9.4.4. Cálculos
2.10. DETERMINACION RADIOINMUNOLOGICA DE T, Y ~.
2.10.1.Marcajeradiactivo de T3 y T4
2.10.1.1.Material y reactivos
2.10.1.2.Procedimiento
2.10.1.3.Perfil de eluciónde radiactividad
2.10.2.Desarrollodel RIA de ‘[3
2.10.2.1.Material y reactivos
2.10.2.2.Procedimiento
2.10.2.3.Cálculos
58
58
59
59
59
60
60
61
61
62
62
62
63
63
63
64
64
64
64
66
66
66
67
67
67
67
68
68
68
69
69
7

2.10.3. Desarrollo del MAde 14. . . . 69
2.10.3.1. Material y reactivos. . 69
2. 10.3.2. Procedimiento 70
2.10.3.3. Cálculos 70
2.11. DETERMINACION RADIOINMUNOLOGICA DE INSULINA. ... 70
2.11.1.Material y reactivos 70
2.11.2.Procedimento 71
2.11.3.Cálculos 71
2.12. DETERMINACION RADIOINMUNOLOGICA DE HORMONA DE
CRECIMIENTO (GH) 71
2.12.1. Preparacionde las hipófisis parael MA 71
2.12.2. Marcajeradiactivo de Gil 71
2.12.2.1.Material y reactivos 72
2.12.2.2.Procedimiento 72
2.12.2.3.Cálculo del rendimientoy la actividadespecífica 72
2.12.2.4. Perfil de elución de radiactividad tras el marcaje radiactivo de
Gil 72
2.12.2.5.Repurificaciónde la GEl marcada 73
2.12.3.Desarrollodel RíA de GH 73
2.12.3.1.Material y reactivos 73
2.12.3.2.Procedimiento 73
2.12.3.3.Cálculos. 74
2.13. DETERMINACION DE LA CONCENTRACION SERICADE IGF-II
POR ANALISIS DE RADIORECEPTOR(RRA) 74
2.13.1. Fundamento 74
2.13.2.Preparaciónde membranaplasmáticaparticuladade hígadode rata. 74
2.13.3.Desarrollodel RRA de IGF-II 75
2.13.3.1,Material y reactivos 75
2.13.3.2.Procedimiento 75
2.13.3.3.Cálculos 76
TECNICAS ELECTROFORETICAS 76
2.14. ANALISIS DELAS PROTEíNASLIGADORAS DE IGF: WESTERN
BLOT 76
8

2.14.1. Fundamento
2.14.2. Electroforesis
2.14.2.1.Preparacióndel gel
2.14.2.2.Preparaciónde las muestras.
2.14.2.3.Condicionesde electroforesis.
2.14.3. Electrotransferencia
2.14.4. Westernligand blot
2.14.4.1.Reactivos
2.14.4.2.Preparaciónde la membrana. .
2.14.4.3.Incubacióncon el radioligando.
2.14.4.4.Lavado y detecciónde las IGFBPs.
2.14.5.Westerninmunoblot
2. 14.5.1. Reactivos
2.14.5.2. Preparaciónde la membrana
2.14.5.3.Incubaciónconel primer anticuerpo.
2.14.5.4.Incubacióncon el anticuerposecundario.
2.14.5.5.Detecciónde las IGFBPs
2.14.6. Westerninmunoblot
2.14.6.1.Reactivos
2.14.6.2.Preparaciónde la membrana
2.14.6.3.Incubaciánconel anticuerpoprimario
2.14.6.4.Incubacióncon el anticuerposecundario
2.14.6.5. Detecciónde las IGFBPs
2.15. ANALISIS DE LOS TRANSPORTADORESDEGLUCOSA(GLUTs):
WESTERNINMUNOBLOT
2.15.1. Preparaciónde las muestras
2. 15.1.1. Reactivos
2.15.1.2.Extracciónde las membranasde tejidos muscularesy corazon.
2. 15.1.3. Extracciónde las membranashepáticas
2.15.2.Westernimnunoblot
2.15.2.1.Reactivos
2.15.2.2.Preparaciónde la membrana
2.15.2.3.Incubacióncon el anticuerpoprimario
76
76
77
77
77
77
79
79
79
79
79
80
80
80
81
81
81
81
81
82
82
82
82
83
83
83
84
84
84
84
86
86
9

2.15.2.4. Incubación con el anticuerpo secundario. 86
2.15.2.5. Detección de los GLUTs
E IGFBPs: ENSA2.16. ANALISIS DE EXPRESION DE IGFs
PROTECCIONDE RIBONUCLEASAS. ...
2.16.1. Extracción del ARNtotal
2.16.1.1. Preparación de las muestras
2.16.2. Ensayo de protección de ribonucleasas.
2.16.2.1.Fundamento
2.16.2.2.Preparaciónde las muestras
2.16.2.3. Síntesisde la ribosonda
2.16.2.3.1.Reactivos
2.16.2.3.2.Obtencióndel plásmido
2.16.2.3.3.Linearizacióndel plásmido
2. 16.2.3.4.Marcajede la ribosonda
2.16.2.3.5.Hibridación en solucion
2.16.2.3.5.1.Reactivos
2.16.2.3.5.2.Hibridaciónde la ribosonda
2. 16.2.3.5.3.Digestiónde la ARNasa
2. 16.2.3.5.4.Electroforesis
2.16.2.3.5.5.Cuantificacióndensitométrica
2.17. OTRAS DETERMINACIONESANALíTICAS.
2.17.1.Análisis de la glucemia
2.17.1.1. Glucosa en sangre
2.17.1.2. Glucosa en sangre desproteinizada
2.17.2. Determinación de las proteínas
2.18. CALCULOSESTADíSTICOS
2.18.1. Análisis de la varianza
2.18.2. Regresión lineal y correlación
3. RESULTADOS
3.1. TIROIDECTOMIA (T> Y TRATAMIENTO
EN PERIODONEONATAL
3.1.1. Evolución del pesocorporal
YODE
87
87
87
88
88
88
88
88
89
90
91
92
92
92
92
93
93
93
93
93
94
94
95
95
96
CON METIMAZOL (MMI>
97
98
98
86
10

3.1.2. Niveles séricosde ‘[3 y ‘[4 y plasmáticose hipofisariosdeGil 98
3. 1.3. Glucemiae insulinemia 101
3.1.4. Factoresde crecimientosimilaresa la insulina (IGFs): nivelesséricos
y expresióndel ARNm hepático 101
3.1.4.1. Niveles séricos 101
31.4.2. Expresiónde] ARNm hepático 103
3.1.5. Estudio de regresiónlineal y correlación 105
3. 1.6. Proteínasligadorasde IGFs (IGFBPs): nivelesséricosy expresióndel
ARNm hepático 105
3.1.6.1. Niveles séricos 105
3.1.6.1.1.Westernligand blot 105
3.1.6.1.2.Westerninmunoblot 108
3.1.6.2.Expresióndel ARNm hepático 108
3.2. TRATAMIENTO CON ESTREPTOZOTOCINA(STZ) DE RATAS
NEONATALES TIROIDECTOMIZADAS 111
3.2.1. Evolución del pesocorporal 111
3.2.2. Niveles séricosde T3 y ‘[4 y plasmáticose hipofisariosde Gil 111
3.2.3. Glucemiae insulinemia 113
3,2.4. IGFs: niveles séricosy expresióndel ARNm hepático 113
3.2.4.1.Niveles séricos 113
3.2.4.2. Expresióndel ARNm hepático 113
3.2.5. Estudiode regresiónlineal y correlación 115
3.2.6. IGFBPs:nivelesséricosy expresióndel ARNm hepático 115
3.2.6.1. Niveles séricos 115
3.2.6.1.1.Westernligand blot 115
3.2.6.1.2.Westerninmunoblot 115
3.2.6.2.Expresióndel ARNm hepático 115
3.2.7. Resumende los resultadosdel estudiode la tiroidectomiay la
administración de MMI en ratas neonatalesy de las ratas
neonatalestiroidectomizadastratadascon STZ 119
3.3. TIROIDECTOMIA EN PERIODODE DESTETEY ADULTO 121
3.3.1. Evolucióndel pesocorporal 121
3.3.2. Niveles séricosde T3 y T4 y plasmáticose hipofisariosde Gil 121
11

3.3.3. Glucemiae insulinemia 125
3.3.4. IGFs: nivelesséricosy expresióndel ARNm hepático 125
3.3.4.1.Niveles séricos 125
3.3.4.2.Expresióndel ARNm hepático 125
3.3.5. Estudiode regresiónlineal y correlación 125
3.3.6. IGFBPs: nivelesséricosy expresióndel ARNm hepático 128
3.3.6.1. Niveles séricos 128
3.3.6.1.1.Westernligand blot 128
3.3.6.1.2.Westerninmunoblot 128
3.3.6.2. Expresióndel ARiNm hepático 131
3.4. TRATAMIENTO CON INSULINA DE RATAS
TIROIDECTOMIZADAS DESTETADAS Y ADULTAS 131
3.4.1. Evolución del pesocorporal 131
3.4.2. Niveles séricosde ‘[3 y T4 y plasmáticose hipofisariosde Gil 134
3.4.3. Glucemiae insulinemia 134
3.4.4. IGFs: nivelesséricosy expresióndel ARNm hepático 134
3.4.4.1.Niveles séricos 134
3.4.4.2.Expresióndel ARNm hepático 137
3.4.5. Estudiode regresiónlineal y correlación 137
3.4.6. IGFBPs: nivelesséricosy expresióndel ARNm hepático 137
3.4.6.1. Niveles séricos 137
3.4.6.1.1. Westernligand blot 137
3.4.6.2. Expresióndel ARNm hepático 140
3.4.7. Resumende los resultadosdel estudiode la tiroidectomíaenratas
destetadasy adultasy de animalesdestetadosy adultosoperados
tratadoscon insulina 140
3.5. TRATAMIENTO CON ‘[4 DE RATAS TIROIDECTOMIZADAS
NEONATALES, DESTETADAS Y ADULTAS 142
3.5.1. Tratamiento con ‘[4 de las ratas neonatales tiroidectomizadas.
Comparaciónentredosis inyectadasy pellets 143
3.5.1.1. Evolucióndel pesocorporal 143
3.5.1.2. Niveles séricosde ‘[3 y ‘[4 y plasmáticose hipofisariosde Gil. . . . 143
3.5.1.3.Glucemiae insulinemia 145
12

3.5.1.4. IGFs: niveles séricos y expresión del ARNmhepático 145
3.5.1.4.1. Niveles séricos 145
3.5.1.4.2. Expresión del ARNmhepático 147
3.5.1.5. Estudio de regresión lineal y correlación 147
3.5.1.6. IGFBPs: niveles séricos y expresión del ARNmhepático 147
3.5.1.6.1. Niveles sericos 147
3.5.1.6.1.1. Western ligand blot 147
3.5.1.6.1.2. Western inmunoblot 150
3.5.1.6.2. Expresión del ARNmhepático 150
3.5.1.7. Resumen de los resultados del estudio del tratamiento con ‘[4
en ratas neonatales tiroidectomizadas 152
3.5.2.Tratamiento con ‘[4 de las ratas destetadasy adultastiroidectomizadas.
Comparaciónde dosisinyectadasy pellets 153
3.5.2.1. Evolucióndel pesocorporal 153
3.5.2.2. Niveles séricosde T3 y ‘[4 y plasmáticose hipofisariosde Gil. . . . 155
3.5.2.3. Glucemiae insulinemia 157
3.5.2.4.IGFs: nivelesséricosy expresióndel ARNm hepático 157
3.5.2.5. Estudiode regresiónlineal y correlación 159
3.5.2.6. IGFBPs: nivelesséricosy expresióndel ARNm hepático 159
3.5.2.6.1.Niveles serícos 159
3.5.2.6.1.1.Western ligand blot 159
3.5.2.6.1.2. Western inmunoblot 163
3.5.2.6.2.Expresióndel ARNm hepático 163
3.5.2.7. Resumende los resultadosdel estudiodel tratamientocon T4
en ratasdestetadasy adultastiroidectomizadas 165
3.6. DIABETES EN PERIODO NEONATAL Y ADULTO Y
REHABILITACION CONT4 166
3.6.1. Evolución del peso corporal 166
3.6.2. Niveles séricos de ‘[3 y T4 y plasmáticos e hipofisarios de Gil 168
3.6.3. Glucemia e insulinemia 169
3.6.4. IGEs: niveles séricos y expresióndel ARNm hepático 169
3.6.4.1. Niveles séricos 169
3.6.4.2. Expresión del ARNmhepático 169
13

3.6.5. IGFBPs: niveles séricos y expresión del ARNmhepático 171
3.6.5.1. Niveles séricos 171
3.6.5.1.1. Western ligand blot 171
3.6.5.1.2. Western imnunoblot 171
3.6.5.2. Expresión del ARNmhepático 171
3.6.6. Resumen de los resultados del estudio de la diabetes y el
tratamiento con 14 en ratas neonatales y adultas 174
3.7. HIPOTIROIDISMO, IGFs, IGFBPs Y TRANSPORTADORESDE
GLUCOSA(GLUTs) EN PERIODONEONATAL 175
3.7.1. Evolución del peso corporal 176
3.7.2. Niveles séricos de T3 y ‘[4 y plasmáticos e hipofisariosde GH 176
3.7.3. Glucemia e insulinemia 176
3.7.4. IGFs: niveles séricos y expresión del ARNmhepático 178
3.7.4.1. Niveles séricos 178
3.7.4.2. Expresión del ARNmhepático 178
3.7.5. Estudio de regresiónlineal y correlación 180
3.7.6. IGFBPs:nivelesséricosy expresióndel ARNm hepático 180
3.7.6.1. Niveles séricos 180
3.7.6.1.1.Westernligand blot 180
3.7.6.1.2.Westerninmunoblot 183
3.7.6.2. Expresiónhepática 183
3.7.7. Niveles de GLUTs en membranacruda 185
3.7.7.1. Corazón 185
3.7.7.2. Hígado 185
3.7.7.3.Músculosesqueléticos:cuadricepsy gastrocnemius 187
3.7.8. Resumende los resultadosdel estudio del hipotiroidismo en ratas
neonatales 187
3.7.9. Resumende los resultadosde las variacionesde los transportadoresde
glucosa(GLUTs) en ratashipotiroideasneonatales 189
4. DISCUSION 190
4.1. ESTUDIO DE LAS ALTERACIONESDEL AXIS IGFs/IGFBPsEN
SITUACIONES DE HIPOTIROIDISMO DESDE LA ETAPA
14

NEONATALA LA ADULTA 191
4.1.1. Niveles de lOEs en suero y expresión de su ARNmen hígado en
animales neonatales hipotiroideos 191
4.1.2. IGFs en animales detetadosy adultos tiroidectomizados 195
4. 1.3. IGFs en animales tiroidectomizados neonatales tratados con
estreptozotocina (STZ) y en detetados y adultos tratados con
insulina 195
4.1.4. Consideracionesfinalesdel estudiode las variacionesde los IGFs. . . 196
4.1.5. IGFBPs: nivelesséricosy expresióndel ARNm hepáticoen animales
hipotiroideosneonatales 197
4. 1.5.1. IGFBPs en los animales neonataleshipotiroideos y tratados con
STZ 198
4.1.5.2. Consideracionesfinales del estudio de las variaciones de las
IGFBPs 200
4.1.6. IGFBPsen animalesdestetadosy adultostiroidectomizadosy tratados
con insulina 201
4.1.6.1.Consideracionesfinalesdel estudiode las variacionesde las IGFBPs
en animaleshipotiroideos 202
4.2. ESTUDIO DE LAS ALTERACIONES DEL AXIS IGFs/IGFBPsEN
SITUACIONES DE HIPOTIROIDISMO (TIROIDECTOMIA) DESDE
LA ETAPA NEONATAL A LA ADULTA DESPUES DE LA
REHABILITACION CON ‘[4 202
4.2.1. lOEs e IGFBPs en neonatos 204
4.2.2. lOEs e IGFBPs en destetadosy adultos 206
4.3. ESTUDIO DE LA REilABILITACION CON T4 EN SITUACIONES
DE DIABETES EN PERIODONEONATAL Y ADULTO 208
4.3.1. lOEs e IGFBPs: nivelesséricosy expresióndel ARNm hepático. . . . 209
4.4. ESTUDIODE LAS VARIACIONES DE LOS TRANSPORTADORES
DE GLUCOSA (GLUTs) EN ANIMALES NEONATALES
HIPOTIROIDEOS. POSIBLE CORRELACION ENTRE LAS
ACCIONES DE LAS HORMONAS TIROIDEAS, GLUTs Y EL
AXIS IGFs/IGFBPs 213
4.4.1. IGFs e IGFBPs: nivelesséricosy expresióndel ARNm hepático. . . . 214
15

4.4.2. GLUTs: nivelesen membranacrudade corazón,hígado,cuadricepsy
gastroenemius 216
CONCLUSIONES 219
BIBLIOGRAFíA 224
16

ABREVIATURAS
ACTI-I
ADN
ADNasa
ALS
ANOVA
APS
ARN
ARNasa
B
BCIP
BSA
DA
DEPC(il20)
DIT
DS
DTT
ECL
EDTA
FG
FGF
FSil
Gil
GLUT
HEPES
Hormona corticotrópica
Acido desoxirribonucleico. Sufijos: c, complementario
Desoxirribonucleasa
Subunidad ácido lábil (Acid labile-subunit)
Análisis de la varianza
Persulfatoamónico
Acido ribonucleico.Sufijos: m, mesanjero;t, transportador
Ribonucleasa
Concentraciónde ligandounido a su receptor
Ligamiento inespecífico
Fosfato de 5-bromo-4-cloro-3-indolilo
Albúmina séricabovina
Dopamina
Dietilpirocarbamato
Diyodotirosina
Desviaciónestándar
Ditiotreitol
Técnica de quimioluminiscencia (TMEnhanced
chemiluminiscence)
Acido etilenaminotetracético
Filtración engel
Factor de crecimiento de los fibroblastos
growth factor)
Hormona estimulante del folículo (Folicle
hormone)
Hormonade crecimiento(‘Growth hormone’).Prefijo: r,
rata
Tansportadorde glucosa(‘Glucosetransporte?)
ácido N-2-hidroxietilpiperazina-N’-2-etanosulfónico
( Fibroblast
stimulating
17

IGF
IGFBP
ip
iv
Kav
LBB
MIT
MMI
MOPS
N
NBT
Nonidet-P40
NT
PAGE
PBS
PDGF
PEG
PMSF
PTU
PVDF
lilA
rpm
RRA
sc
SCN
Factor de crecimiento similar a la insulina (“Insulin-like
growth factor”). Prefijos: r, rata; h, humano; br, humano-
recombinante; tr, des(1-3) IGF-I (IGF-I truncado)
Proteína ligadora de IGF (“Insulin-like growth factor
binding protein”). Prefijos: r, rata; h, humano; b, bovino;
rh, humano-recombinante
Administración intraperitoneal
Administración intravenosa
Coeficientede disponibilidad
Tampónde “ligand blotting”
Monoyodotirosina
2-mercapto-1 -metilimidazol (metimazol)
Acido 3-(N-morfonilol)propinosulfónico
Ligamiento inespecífico
Nitroazul de tetrazoilo
Oxido de octilfenol-etileno
Neurotensina
Electroforesisen geldepoliacrilamida(“polyacrylamidegel
electrophoresis”)
Tampónfosfato (‘Phosphatebuffer salme”)
Factorde crecimientode derivadode plaquetas(“Platelet-
derivedgrowth factor”)
Polietilenglicol
Fluoruro de fenil-metil-sulfonio
Propiltiouracilo
Difluoruro de polivinilideno
Radioinmunoanálisis(“Radioimmunoassay9Revolucionespor minuto
Radioanálisisde receptor(‘Radiorreceptorassay‘9
Administraciónsubcutánea
Isotiocianato
18

SDS Dodecilsulfatode sodio
SRJH Somatostatina
STZ Estreptozotocina
T Concentración de ligando total
Diyodotironina
Triyodotironina. Prefijos: r, rata; r: rT3 (‘[3 inversa,
“reverse-T3”)
‘[4 Tetrayodotironinao tiroxina. Prefijo: r, rata
‘[BA Albúmina fijadora de tiroxina (“Thyroxine-binding
albumin”)
TBG Globulina fijadora de tiroxina (“Thyroxine-binding
globulin)
TBPA Proalbúminafijadora de tiroxina (II Thyroxine-bindingpre-
albumin)
TBS SoluciónsalinatamponadaconTris (‘Tris-bufferedsalme’)
TE Tris basey EDTA
TEMED N, N, N’, N’-tetrametiletilendiamina
Tg Tiroglobulina
TPO Tiroperoxidasa
TRH ilormomadeliberacióndetirotropina(“Thyrotropin-release
hormone)
Tris Tris(hidroximetil)an-iinometano
TSH Tirotropina (‘ Thyroid-stimulatinghormone‘)
Tween-20 Monolaurato de polioxoetilenosorbitano
UI Unidades internacionales
V Volumen de elución de una sustanciadeterminadopor
filtración en gel
19

iNTRODUCCION

Introducción
1. INTRODUCCION.1.1. HORMONAS TIROIDEAS.
1.1.1. Síntesisy secrecion.
Las células productoras de las hormonas tiroideas 3, 5, 3’-L-triyodotironina
(T3) y 3, 5, 3’, 5’ -L-tetrayodotironina (‘[4) se organizan en folículos en el interior de
la glándula tiroidea; dichos folículos rodeanuna cavidadque contieneuna sustancia
viscosa denominadacoloide y que constituye el almacénpara la glicoproteina
tiroglobulina(Tg). Así pues,la síntesisde las hormonastiroideasseinicia mediante
la captaciónde yoduro por partedel tiroides medianteun mecanismode transporte
activo, a continuación,seproducela yodaciónen los gruposde tirosina de la Tg y,
mediante un acoplamiento intramolecular en una reacción catalizada por la enzima
tiroperoxidasa (TPO), se forman unos yodoaminoácidos precursores de las hormonas
tiroideas:monoyodo-y diyodotirosina(MIT y DIT, respectivamente).La Tg yodada
se almacenaen el coloide,y sirve de reservade ‘[3, ‘[4 y de yodo, perono pasacomo
tal a la sangrecirculante, sino que la secreciónde las hormonastiroideasconlíeva
una degradaciónde la Tg por partede la yodotirosinadesyodasa(hidrolasaácida),
que libera ‘[3, ‘[4, MIT y DIT y yodo, que se reciclaenel folículo (Guyton, 1986).
El productomayoritariode la secrecióntiroideaes la tiroxina, aunquela formaactiva
es la triyodada (‘[3), y estase generatanto en la glándula (en baja proporciónen
condicionesnormales)comoen tejidos extracelularespor pérdidade un átomo de
yodo de la T4 (desyodación).La ‘[3 tiene unaafinidad 10 vecesmayorporel receptor
que la T4 (St Germain, 1994).
Las hormonastiroideas,debido a su bajasolubilidaden solucionesacuosas,
circulanpor la sangreen una pequeñafracción (menosdel 1 %) en forma libre, y la
mayor parte de ellas lo hacenunidas a unas proteínastransportadorasque se
sintetizanen el hígado: globulina fijadora de tiroxina (TBG), albúmina (‘[BA) y
proalbúmina(TBPA>.
1.1.2. Regulacióndel axis tiroideo.
La síntesis y secreciónde las hormonastiroideas, expuestasde un modo
simplista, estánreguladasporun sistemacomplejoen el cual sepuedenconsiderar
tres nivelesdiferentes.La hipófisis sintetizay segregala hormonatirotropa (‘[Sil),
principal estimulo de la función tiroidea. Dicha secreción está sujeta a dos
influencias,dadoqueel hipotálamoproducedoshormonas,la hormonaliberadorade
21

Iniroducck5n
tirotropina (TRH), que estimula la síntesis y secreciónde TSil en la hipófisis, y la
somatostatina (SRIH), que la inhibe. Sin embargo el sistemade contrarregulación
positivo más importante se establecea un tercernivel, mediantela acciónsupresora
que las hormonastiroideasejercensobrela síntesisy secreciónde TSil.
Este esquemaelemental se ve complicado por las diferentes acciones,
inhibidoras o estimuladoras,que a los distintos niveles ejercenotras hormonasy
neurotransmisores.
1.1.2.1.Tirotropina.(TSH, ‘thyroid-stimulatinghormone).
La TSil esuna glucoproteinay su regulaciónpor las hormonastiroideases
un sistemade retroalimentaciénnegativaa nivel de la hipófisis; numerososestudios
han demostradoque un aumentoen los nivelesde las hormonastiroideasdisminuye
la secreciónde TSil y que el déficit de hormonastiroideasaumentasu secreción
(Belchetz, 1978). Así pues, se ha visto que en situacionesde hipotiroidismoa corto
plazo, la administraciónde ‘[3 puede inhibir la síntesis y liberación de ‘[Sil de
maneradosis-dependiente,vía retroalimentación. Porotro lado, en situacionesde
hipotiroidismo prolongado, cuando la síntesis proteica está considerablemente
disminuida, la administraciónde hormonatiroideaestimulala síntesisde proteínas,
incluida la ‘[Sil. Este efecto predominacuando la dosis de ‘[3 dadaes pequeña,
mientras que dosis altas de dicha hormonaharíanmás evidente la inhibición por
retroalimentación(Magner, 1990).
Además,estaregulacióntambiénocurrevía hipotalámica,porhormonasque
estimulano inhiben(TRH y DA, respectivamante)los nivelesde TSH. Así pues,las
hormonas tiroideas e hipotalámicas interactúanpara modular tanto los niveles
circulantesde ‘[Sil como la transcripciónde los genescodificadoresde la hormona
(Morley, 1981; Gershengorn,1986).
1.1.2.2.Controlneuroendocrino.
1.1.2.2.1. Hormona de liberación de tirotropina. (‘[RU, ‘thyrotropin-
releasehormone”).
El ‘[RU esun tripéptidoque poseeun efectopositivo enel hipotálamosobre
la función tiroidea, y aunquese encuentraampliamentedistribuido por el cuerpo,
pareceserque la regióndel núcleoparaventricularestáparticularmenteinvolucrada
en el axis TRH-TSH (de Greef, 1992). No obstante,estarespuestapositiva en el
hipotálamo puede ser abolida por lesiones hipotalámicasy estímulos externos
22

Introducción
(cambiosen la temperaturaambiente,stress,ciclo día-noche).
La liberacióndel ‘[RU estáreguladade forma negativapor las hormonas
tiroideas, y aunque sobre esta retroalimentación existen grandes lagunas, se sabe que:
en las ratas hipertiroideas,la liberaciónde ‘[RU está disminuida en un 30-40%,
mientras que el hipotiroidismo inducido por tiroidectomíao la administraciónde
drogas tirostáticas se asocia con un aumentoen el contenidohipotalámicodel ARNm
de pro-’[RH (de Greef, 1992). No obstante, el tratamiento con MMI, que induce
también un aumento del contenido de ARNmde pro-TRU en el hipotálamo, no altera
la liberación de ‘[Ril a nivel de este órgano (Rondeel, 1988; Rondeel, 1990) y sólo
cuando el hipotiroidismo se induce mediante tiroidectomía o la administración de
PTU, se aumenta moderadamente el nivel de ‘[RU en la sangre portal hipofisal
(Rondeel, 1992).
1.1.2.2.2. Otros factores hipotalámicos.
1. Somatostatina (SRIH).
La somatostatina inhibe la secreción de Gil, pero también la liberación de
‘[Sil. En la rata, la somatostatinano afectaa los nivelesbasalesplasmáticosde TSR,
pero sí inhibe la respuestade ‘[Sil a TRH de maneradosis-dependiente,e incluso,
se ha sugeridoque in vivo, partede la retroalimentaciónnegativaque ejercenlas
hormonastiroideasestámediadapor el aumentode la sensibilidadde las células
tirotropasa la somatostatinahipotalámica(Spira, 1986).
En situacionesde hipotiroidismoinducidaspor la administraciónde PTU se
ha visto que seduplica la capacidadde los extractoshipotalámicospara inactivara
la SRIil, y se disminuye en el mismo grado el contenido hipotalámicode SRIH
(Berelowitz, 1980).
2. Neurotensina(NT).
La neurotensinaafecta a los niveles plasmáticosde ‘[Sil; algunos estudios
sugierenque una inyecciónintravenosade NT eleva la concentraciónplasmáticade
‘[SU en la rata, mientras que en otros una inyección de este factor en el tercer
ventrículo de la rata no cambia los nivelescirculantes de ‘[SU, o incluso los
disminuye(Vijayan, 1980).
3. Momoaminas
.
En la rata,la norepinefrinaestimulala secreciónde ‘[SU por la activaciónde
los receptoresa, mientrasque la dopaminainhibe su secreción(Spira, 1986).
23

Introducción
1.1.2.2.3.Hormonastiroideas.
La administraciónde hormonastiroideas,comoseha mencionado,inhibe los
niveles plasmáticos de ‘[SN basal y la estimulación de la liberación de la TSH
inducidapor ‘[Ril, como consecuenciade la reduccióndel númerode receptoresde
TRH en células que secretan hormona tiroestimulante.
1.1.3. Metabolismoy accionesde las hormonastiroideas.
1.1.3.1.Metabolismo.
El metabolismode las hormonastiroideasen tejidos extratiroideosinvolucra
a unaseriede reaccionesenzimáticascomplejascomoson: desyodación,conjugación
y/o desaminacióny descarboxilaciónoxidativa. No obstante, la principal ruta
metabólicaes ladesyodación,queestámediadaporuna familia de enzimaspresentes
en la glándula tiroidea y en tejidos extratiroideos (Leonard, 1990). Se han
identificadodos reaccionesgeneralesde desyodación:pérdidade un átomo de yodo
del anillo fenólico (externo), recibeel nombrede 5’-desyodación,y si el átomo de
yodo se libera del anillo tirosilo (interno) se denomina 5-desyodación.La 5’-
desyodaciónseconsideracomo unarutade activaciónporquegeneradirectamenteel
metabolitoactivo (‘[3) a partir de la “prohormona inactiva (‘[4) y, por el contrario,
la 5-desyodaciónes una ruta de inactivación porque da lugar a un metabolito
biológicamenteinerte (rT3). Se conocentres tipos de desyodasas:
1. Desvodasatipo 1: Estápresenteen tejidostiroideosy extratiroideos(riñón,
hígado, pulmón, músculo, piel) y poseeprincipalmenteactividad 5’-desyodasa,
aunqueen menormedidatambiéndesyodael anillo tirosilo (actividad5-D), genera
la mayorpartede la T~ liberadaa la circulación.
2. Desvodasatino II: Sóloposeeactividad5’-desyodasa,sehallapresenteen
el cerebro pituitaria y tejido adiposomarrón,y es la responsablede la mayoríade
la ‘[3 producidaa partir de la ‘[4 dentrode las células.
3. Desvodasatino III: Tiene sólo actividadsobreel anillo tirosilo y se expresa
en el córtexcerebraly piel de ratasadultasy en placentay otros tejidos fetales(St
Germain, 1994).
La importancia fisiológica de la desyodaciónsobre la economíade las
hormonastiroideasesgrande,y así, diversosestudiosenratasneonatalesy fetoshan
demostradoque, en situacionesde déficit de hormonastiroideas, se favorece la
conversiónde ‘[4 a ‘[3 en órganoscomo el cerebro lo que constituyeun mecanismo
24

Introducción
deprotecciónmuyeficaz,especialmenteduranteperiodoscríticosdel desarrollocomo
el perinatal, en el que el desarrollodel cerebrodependeen gran medida de las
hormonastiroideas(Ruiz de Olla, 1988; Aláez, 1992b; Porterfield, 1993).
1.1.3.2.Accionesbiológicasde lashormonastiroideas.
La primera acciónreconocidade las hormonastiroideas fue su efectoen la
generación de calor, que se refleja en un aumento del consumo de oxígeno y del
metabolismobasa);así, activanlos mecanismosoxidativosmitocondrialesen hígado,
riñón, músculosy corazón(vanHardeveld,1986).
Las honnonastiroideas intervienentambiénen la regulación de procesos
metabólicos como la síntesis y degradación de proteínas, tanto estructurales como
enzimáticas (a-glicerolfosfato deshidrogenasamitocondrial, enzima málico...),
incrementanel metabolismolipídico, afectandomásala degradaciónquea la síntesis,
intervienen en la regulación del metabolismo de carbohidratos (aumentan la
gluconeogénesisy la utilizaciónde la glucosa)y también,participanen la síntesisy
el mantenimientodel nivel adecuadode algunasvitaminas,y en su transformaciónen
cofactoresenzimáticos(Freake,1995; Venditti, 1997).
Además,entre los efectosque ejercencabedestacarel papelquejueganen
procesosde crecimiento, diferenciacióny desarrollo, y entre ellos cabe citar la
metamorfosisde los anfibios (Nikodem, 1990) o su actuacióndirectasobreel gen de
Gil (Yaffe, 1984).
La falta de hormonastiroideasduranteel desarrollode los mamíferoslleva
asociadamúltiples alteracionesen el crecimiento y maduraciónde muy diversos
tejidos. En etapastempranas,en el sistema nervioso central durante el periodo
crítico del desarrolloembrionarioy perinatal, la falta de hormonastiroideaspuede
producir dañosseverose irreversibles (Eayrs, 1968 ; Timiras, 1989), que en el
hombre conducena un retraso mental profundo (Dobbing, 1977; Morreale de
Escobar,1983; Legrand,1986).Esteperiodo, en el queel cerebroesmuy vulnerable
se denominacrítico, coincideconuna etapade mitosis activasy diferenciaciónde las
células nerviosas,y en el hombrese sitúaentreel final de la gestacióny los 2 años
de edad.
1.1.4. Interacciónentrehormonastiroidease insulina.
Las hormonastiroideasy la insulinaejercenaccionesmetabólicascontrarias,
no obstante,aún no se conocebien el efecto que ejercen la ‘[3 y la 1’4 sobre el
25

Introducción
metabolismo de la insulina. En trabajos anteriores, se ha mostrado que el
hipertiroidismo puedeaumentar(Beer, 1989; O’Meara, 1993), disminuir (Ikejiri,
1978; Escrivá, 1981>, o incluso, no alterar (Taylor, 1985> los niveles de insulina. El
hipotiroidismotambiénsemuestracomoun cuadrocontrovertido,aunqueaúnmenos
estudiado que el hipertiroidismo.
En el hombre, el hipotiroidismo usualmente se presenta con unas
concentracionesbasalesde insulina normalesy, raramentedisminuidas (Lenzen,
1984). Por otro lado, el hipotiroidismo experimentalpuedeir acompañadode una
variabilidad similar a la del hipertiroidismo en cuantoa los nivelesplasmáticosde
insulina. John y col. (1970) observaron en la rataquela induccióndel hipotiroidismo
mediantela tiroidectomíao la administraciónde ciertosgoitrógenos(C104, ReO¿,
MMI) disminuían los valores circulantes de insulina frente a los controles, mientras
que si se le administraba a las ratas fármacos antitiroideos como propiltiouracilo
(PTU) y SCN (isotiocianato), se producía un aumento en la insulinemia de estos
animales respecto a sus controles. Montes y col. (1977) indujeron en la rata un
hipotiroidismo experimental mediante tiroidectomia, e inicialmente observaron, a los
5, 10 y 15 días después de la operación, que los niveles circulantes de insulina no se
alteraban, mientras que si se prolongaba más el déficit de las hormonas tiroideas los
valores plasmáticos de esta hormona disminuían respecto a sus animales control. La
terapia de reemplazamiento con dosis fisiológicas de ‘[4, al igual que la
administración de Gil, normalizó los bajos niveles de insulina de estos animales
hipotiroideos(John, 1970; Montes, 1977).
A pesarde quela velocidadde secreciónde la insulinapuedeestaraumentada
(Lenzen,1984) o no alterada(O’Meara, 1993)en situacionesde excesoy déficit de
hormonastiroideas, pareceque en los pacienteshipertiroideos la respuestade la
insulinaala administraciónde glucosaintravenosau oral estámuy exagerada(Ahrén,
1981; O’Meara, 1993>, mientras que el hipotiroidismo produceun aumento,una
disminucióno no alteradicharespuesta.
La velocidad de desapariciónde la insulina aumentapor el exceso de
hormonastiroideas(Moghetti, 1994)y disminuyeporel déficit (Lenzen, 1984),con
lo que la duraciónde la acciónde la hormonase ve alteradapor el estadotiroideo.
Estas alteracionesen la secreciónde la insulina podrían hacer pensaren
cambiosa nivel de los islotespancreáticos,y así, se ha observadoque la hipotrofia
26

Introducción
de los islotes parece ser un efecto directo de las hormonas tiroideas sobre el páncreas
endocrino que resulta de unos cambios degenerativos tras situaciones de
hipertiroidismoprolongado(Lenzen 1976; Lenzen, 1978).En estesentidotambién,
Morovat y col. (1998) apuntanque los altos niveles plasmáticosde insulina
encontradosen cerdosjóvenestiroidectomizadospuedenser debidos,al menosen
parte, a un descensoen la sensibilidad de los tejidos a esta hormona, como
previamente se había sugerido en situaciones de hipertiroidismo (Abrén, 1981;
Mdller, 1988) e hipotiroidismo(Jayendra,1972; Katsilambros, 1972).
Como hemos visto, el estadotiroideo afecta a la secreción normal de la
insulina, así como a su metabolismo (Foss, 1990) y contenido pancreático
(Katsilambros,1972), lo queconducea unamodificaciónde la homeostasisglucídica
que podría llevar al desarrollo de una diabetes (más comúnmente en el
hipertiroidismoque enel hipotiroidismo).
1.2. FACTORESDE CRECIMIENTO SIMILARES A LA INSULINA (IGFs).
Los factores de crecimientosimilares a la insulina (“insulin-like growth
factors>IGF-I y II son polipéptidossimilaresestructuralmentea la proinsulina, de
hecho, aunqueson codificadospor genes individuales,se encuentranrelacionados
ancestralmentecon el gende la insulina y poseenefectosmetabólicossimilaresa esta
(ilumbel, 1990). Se sintetizanen numerosostejidos sobrelos que ejercenacciones
a nivel de los procesosde proliferacióny diferenciación, tanto locales (auto y
paracrinas)comoendocrinaspor la unión a receptoresespecíficosde membrana.
En la sangrey otros fluidos extracelulares,los IGFs forman complejoscon
unasproteínasespecíficas(‘insulin-like growth factor-bindingproteins’, IGFBPs),
que constituyenuna familia de péptidosrelacionadosestructuralmenteentre si.
1.2.1. Estructura,síntesisy transporte.
1.2.1.1.Estructura.Los IGFs son polipéptidos de cadena única de 70 y 67 aminoácidos (IGF-I y
JI, respectivamente>, con tres puentes disulfuro, y comparten entre sí una homología
del 62%, y con la proinsulina del 42% (Sara, 1990). En la estructura de estas
proteínas se han caracterizado varios dominios: A y B (similares a la insulina), C
(análogo al péptido de conexión de la proinsulina) y D (no está presente en la
insulina).
27

Introducción
La estructura primaria de los IGFs está altamente conservada entre especies.
El IGF-I de rata difiere del humano en tres aminoácidos, y el IGF-II tan solo muestra
cuatro sustituciones conservativas respecto al péptido humano (Daughaday, 1989). No
obstante, en el suero humano se han identificado variantes de estos péptidos de
distinta longitud, se han detectado formas truncadas y de alto peso molecular de
ambos IGFs, probablemente generadas por un procesamiento post-traduccional, dado
que no se han identificado ARNmque codifiquen para esas primeras formas.
Los genes que codifican para IGF-I y II en el hombre y en la rata comparten
numerosos elementos estructurales.
El gen que codifica para el IGF-I está compuestopor 6 exonesseparadospor
5 intrones de un tamaño de 1.9 a 5OKb. Los exones1 y 2 codificanel péptidoseñal
y una región 5’ no traducida; estos exones sufren un acoplamiento alternativo con el
exón 3 que codifica el resto del péptido señal y la mayoría del dominio B. El exón
4 codifica el resto del dominio B, C, A y D y los 16 primeros aminoácidos del
péptido E. El exón 5 codifica parte del péptido E, y el exón 6 el resto del péptido E
y una secuencia de la región 3’ no traducible (Lund, 1994). Este gen está regulado
por dos promotores, y como consecuencia de diferentes empalmes alternativos y de
quien lidera la transcripción, se generan dos tránscritos de IGF-I: ARNmde IGF-Ia
(carece de la secuencia del exón 5 que codifica para el péptido E) y ARNmde IGF-Ib
(contiene las 52 bases del exón 5 que codifican para el péptido E) (Hall, 1992).
El gen de IGF-II consta de 9 exones y excepto los exones 7, 8 y parte del 9,
que codifican para el péptido maduro, son exones no traducidos. Este gen está
controlado por cuatro promotores y, como consecuencia de la activación diferencial
de esos promotores y de los diferentes emplames alternativos se generan tres
tránscritos de IGF-JI.
1.2.1.2. Síntesis y transporte.
La expresión de los genes de los IGFs es muy ubicua, se ha detectado su
ARNm en diversostejidos como: hígado,pulmón, hipófisis, bazo, riñón, corazón,
músculo esquelético, ovario, útero, testiculo, cerebro, glándula mamaria y otros
(ilalí, 1992 y Lund, 1994), en medioscondicionadosde muchascélulas (ilumbel,
1990) y en la sangre. También se ha descrito la presencia de IGF-I y II o ambos en
otros líquidos corporales como orina, linfa, líquido cefalorraquideo, semen, saliva,
leche o líquido folicular. No obstante, el mayor productor de IGEs es el hígado, que
28

introducción
secreta más del 90% del IGF total circulante (Móller, 1991).
Los IGFs, a diferencia de la insulina, circulan por la sangre unidos,
prácticamente en su totalidad,aunasproteínastransportadorasespecíficas(IGFBPs)
formando dos complejos de unos 150 y 5OkDa,aproximadamente. Estas proteínas se
encuentran presentes también en el espacio extracelular y pueden controlar la
distribución tisular o celular de los IGFs, modificando su acción y su
biodisponibilidad; también pueden participar en la unión de los IGFs a su receptor.
1.2.2. Acciones biológicas de los IGFs.
1.2.2.1. Iii vitro.
Los efectos de los IGFs ¡it vitro pueden ser a corto plazo, como las acciones
metabólicas sobre las proteínas y carbohidratos, o a largo plazo, como las acciones
sobre el crecimiento y la diferenciación celular.
1. Efectos sobre la progresión del ciclo celular: El IGF-I funciona como un
factor de progresión del ciclo celular junto con los factores competentes que lo inician
y capacitan a las células para respondera los factoresdeprogresión.
Se ha visto que células quiescentes en G0 pueden ser inducidas a entrar en G1
por los llamados factores competentes (PDGFy FGF); sin embargo,el tratamiento
de estas células con dichos factores de competencia y otros de progresión como el
IGF-I, permite el avance hacia G1 y la continuación del ciclo celular, produciéndose
la síntesis de ADNy la proliferación celular (Jones, 1995).
2. Efectos sobre la vroliferación ce]u]ar: Los IGFs poseenuna actividad
mitogénica sobre una gran variedad de células (fibroblastos, condrocitos,
osteoblastos, células foliculares tiroideas, células neuronales, adipocitos, líneas
tumorales...) de ahí queparticipen en el crecimiento del embrión, feto y neonato, en
fenómenos de hipertrofia fisiológica o compensadora de ciertos órganos, en
reparación de heridas en nervio músculo y células endoteliales (ilunziker, 1994;
McCarthy, 1994).
3. Efectos sobre la muerte celular: Los IGFs poseen la capacidad de inhibir
la muerte celular por apotosis en ciertas células (hematopoyéticasy células de
carcinoma humano de pecho MCF-7); sin embargo, morfológicamente las células
mueren por necrosis más que por apoptosis, y en esos casos, los IGFs no han
demostrado aumentar la supervivencia celular de otros tumores (MDA-23 1, HEP-2
y KB).
29

Introducción
4. Efectos sobre la diferenciación celular: Los JGFs estimulan la
diferenciación de los mioblastos, osteoblastos, adipocitos y oligodendrocitos
(Hunziker, 1994; McCarthy, 1994); además,intervienenen la maduraciónde la
funcióntesticulary ovárica,inducenla eritropoyesis,estimulanel crecimientode las
neuritasy estánimplicadosen la sinaptogénesis.
5. Efectossobrela funcióncelular: Los IGFs regulanla secreciónhormonal
de un numerosotipo de células.
IGF-I y II estimulanla síntesisy secreciónhormonalde las células ováricas
y muestranefectossinérgicosen combinacióncon FSH y estrógeno,aumentanel
númerode receptoresde ACTH y en cultivos de célulassomatotropasde pituitaria,
inhiben la secreciónde Gil (Melmed, 1986). Tambiénposeenimportantesefectos
sobreel sistemainmune: potencianla función citotóxica de las células T (Clark,
1997).
1.2.2.2. lía vivo.
La administraciónde IGFs provocauna seriede efectosmetabólicosa corto
plazoy, crecimientoa largo plazo.
1. Efectosde la administraciónde IGFs en animales: En ratasneonatales,la
infusión de IGF-I provoca hipoglucemia por la estimulaciónde la captaciónde
glucosaen la periferia pero sin afectara su producciónhepática,y no altera la
concentraciónde los ácidos grasoslibres al contrario que la insulina. Recuperala
función renal y aumentalas proteínascorporalestotales y, a pesar de que no
incrementael pesode la carcasa,si aumentael del bazo, riñón, timo e intestino
(Gargosky,1994).
A nivel hormonal, la administraciónde IGF-I disminuye los nivelesde Gil,
pero aumenta el de catecolaminas (Glasscock, 1992).
Los efectos de las altas concentraciones de IGF-II no alteran el peso corporal
ni la longitud de los animales control, aunque la concentración sérica de IGF-I
disminuye (Glasscock, 1992>. Por el contrario, en las ratas hipofisectomizadas,
aumentasu pesoy longitud a la vez que desciendela urea en el suero(ilumbel,
1990).
2. Efectosde la administraciónde IGF-I enhumanos:Serecogenen la Tabla
1:
30

Introducción
Tabla 1.- Efectos de la administraciónde IGF-I iii vivoen humanos.
FUNCION
Metabolismode la glucosa
Metabolismolipídico
Anabolismoproteico
Punciónrenal
Hormonascontrarregulatorias
Crecimientolineal
Miscelánea
EFECTODEL IGF-I
captación de glucosa
producciónhepáticadeglucosa(humanos)
sensibilidadaparentea la insulina
Hipoglucemia(especialmentecon administracióniv)
cetonasséricas
4 ácidosgrasoslibres séricos(humanos)
4 triglicéridos
1 síntesisproteica
4 excreciónde nitrógeno
proteínascorporalestotales
pesocorporal/órgano,especialmentedetimo, riñón,bazo
e intestino
Mejorala cicatrización
1 velocidadde filtración glomerular
flujo plasmáticorenal
velocidad de recuperaciónde un fallo renal agudo
isquémico
4 GH
4 Glucagón
1 Catecolaminas
en hipopituarismoy sujetosinsensiblesa GH
Taquicardia,edemaperiférico, aumentode las parótidas
y pérdidade sensibilidad
(Jones,1995)
1.2.2.3.Accionesautocrinas,paracrinasy endocrinas.
Aunque el hígado es la principal fuente de RiUs, muchosórganosexpresan
estos factores (Hall, 1992; Lund, 1994). La ubicuidad de los IGFs sugiereunas
accionesautocrinas/paracrinasque puedensermuy relevantesparala diferenciación
y proliferación celular.
La importancia de estos efectos locales es más evidente en aquellos órganos
y tejidos con mayor expresión y concentración de IGFs como el hígado, riñón,
testiculo, células de la granulosa y sistema nervioso central, mientras que las acciones
endocrinas participarían en el crecimiento corporal global integrado (Daughaday,
31

Introducción
1989). No obstante, la importancia relativa de cada modode acciónvaríade un tejido
a otro y con la etapa de desarrollo.
En la etapafetal, seha sugeridoun papelpredominantede las accionesauto
y paracrinassobrelas endocrinas(Daughaday,1989).En el feto, los genesde IGF-I
y II se expresanen muchostejidos, pero sus niveles circulantesson bajos (Lund
1986). Porotro lado, en la etapapostnatal,existela posibilidadde una interacción
entrelas accionesauto/paracrinasy endocrinasde IGF-I, ya que tras el nacimiento
aumentanprogresivamentelos nivelesde IGF-I a la vezqueel númerode receptores
hepáticos de GH, mientras la expresión del gen de IGF-I se reduce (Rotwein, 1993).
1.3. PROTEINASTRANSPORTADORASDE IGFs (IGFRPs).
1.3.1. Estructura, síntesis y regulación.
1.3.1.1. Estructura.
Los IGFs circulan por el plasma unidos a una familia de proteínas
transportadorasrelacionadasestructuralmente,las IGFBPs. Se han donado y
secuenciadosiete polipéptidos (IGFBP-1 a 7), pero sólo se han estudiadoseis
(IGFBP-1 a 6), y 4 de ellos (IGFBP-1 a 4) seencuentranen el sueroen cantidades
importantes(Rechíer, 1998). Se trata de unas proteínas de un tamaño similar (201 a
289 aminoácidos)y de un pesomolecularentre21.5 y 31.4kDa.
La analogíamás destacadarespectoa la estructurade estos péptidoses la
conservaciónde 18 residuosde cisteinatantoen el extremocarboxicomoen el amino
terminal. Además, en el extremo C-terminal de todas las IGFBPs existe una secuencia
conservada arg-gly-asp que parece ser la secuencia mínima requerida en muchas
proteínas de matriz extracelular para unirse a receptores de membrana (Rosenfeld,
1990).
A pesar de estos aspectossimilares,hay variasdiferenciasentrelas secuencias
de aminoácidos de las IGFBPs, como el hecho de que las secuencias espaciadoras
existentes entre los residuos de las cisteinas no estánbienconservadas,la variabilidad
en la secuencia del péptido señal o la diferente glicosilación de las IGFBPs.
1.3.1.2. Síntesis.
Las IGFBPs son producidas por una gran variedad de tejidos y se encuentran
en varios fluidos biológicos.
En el suero, las IGFBPs circulan en forma de dos complejos, uno de 150-
32

Introducción
200kDa que contiene en cantidades equimolares la IGFBP-3 (GH dependiente) y una
proteína (subunidad ácido-lábil, ALS), y el complejo de 3OkDa que contieneotras
formas de IGFBPs (IGFBP-1 y 2).
Se han hallado también diversasformas de IGFBPsen numerososlfquidos
corporales: linfa, leche, liquido cefalorraquideo, líquido amniótico, plasma seminal,
líquido folicular, humor acuoso y vítreo, orina, así como en el medio condicionado
de numerosos cultivos celulares (Rosenfeld, 1990; Rajaram, 1997). En la leche
humana, por ejemplo, se han encontrado IGFBP-1, 2 y 3, mientras que en la de rata
también está presente la IGFBP-4; no obstante, en ambos casos, la proteína
predominante, como en el suero,es la IGFBP-3. En el fluido cerebroespinal,sin
embargo, la forma predominante es IGFBP-2, al igual queen la linfa, e IGFBP-1 es
la proteína mayoritaria en el líquido amniótico.
Las IGFBPs también se sintetizan en múltiples órganos y tejidos (Straus,
1991; Babajko, 1993; Gosteli-Peter, 1994); IGFBP-1 se expresaen hígado,placenta
o endometrio; IGFBP-2 abunda en hígado, riñón, intestino y pulmón, pero donde
mantiene unos niveles altos es en el cerebro (plexos coroideos). IGFBP-3, por su
parte, se expresa en hígado, placenta y riñón, mientras que el ARNmde IGFBP-4
se encuentra de manera abundante en hígado y los de IGFBP-5 y 6, en riñón y
pulmón, respectivamente.
1.3.1.3. Regulación.
1.3.1.3.1.Variaciones diurnas.
Los valores plasmáticos de IGFBP- 1 estánsujetosa variaciones diurnas que
se relacionan con los niveles de insulina; las cifras circulantes de esta proteína
transportadora son muy bajas por la tarde y a media noche, y muy altas por la
mañana; además, parece ser que este aumento coincide con un incremento en los
niveles de IGF-I (Holly, 1988).
IGFBP-2 y 3 son más estables y no muestran variaciones diurnas ni están
sujetas a cambios postprandiales.
1.3.1.3.2. Nutrición.
El efecto de la restricción proteico-calórica altera los niveles séricos y de
expresión de las IGFBPs, incluso en los fetos de madres ayunadas (Straus, 1991). En
las ratas neonatales, la malnutrición aumenta la expresión de IGFBP-1 y 2 en hígado,
aunque la realimentación reduce los niveles del ARNmde ambas proteínas a los
33

Introducción
valores control. La expresión hepática de IGFBP-3 disminuye ligeramentecon la
malnutrición, y la realimentación eleva sus niveles por encima de los control (Rivero,
1995; Goya, 1996).
1.3.1.3.3. Ejercicio.
En el hombre adulto, el ejercicio se acompaña de un aumento en los niveles
circulantesde IGFBP-1 y 3 y un descensoen los niveles de insulina e IGF-I
(Rajaram,1997).
1.3.1.3.4.Ontogenia.Durante el periodo fetal, IGFBP-1 y 2 son las proteínas transportadoras
predominantes en el hombre y la rata, se conservan altos niveles durante el periodo
neonatal, disminuyendo progresivamente a la vez que empieza apredominarIGFBP-3
que es la abundante en adulto (Babajko 1993).
En la pubertad, IGFBP-3 alcanza un pico, y disminuye durante el periodo
adulto. Por el contrario, IGFBP-1 y 2 muestran unos niveles altos en el nacimiento
y la senescencia y bajos durante la pubertad (Rosenfeld, 1990).
1.3.1.3.5.Hormonas.
El nivel de influencia de las hormonas sobre los valores de las IGFBPs
depende de: la magnitud de cambio en la producción de la correspondienteproteína
transportadoraen el/los tejido/s diana/sy de la contribuciónde ese/esostejido/s en
la secreciónde la proteínatransportadoray, además,las hormonaspuedenafectara
los nivelesséricosde unao másproteínastransportadoras.La insulina, porejemplo,
pareceser la principal reguladorade la producciónhepáticade IGFBP-1, mientras
que la GH parecealterar los nivelescirculantesde IGFBP-3 (aumentalos niveles
séricosde IGFBP-3e inducelaproducciónde ALS), al igual que los glucocorticoides
(disminuyen los niveles de IGFBP-1 y 2, pero aumentanlos de IGFBR3) (Luo,
1990). Las hormonastambiénregulanla expresiónde las proteínastransportadoras
por uno o múltiples mecanismos.
1.3.2. Tipos de IGFRPsy accionesbiológicas.
Los IGFs no sólo estánpresentesen la circulación,sino tambiénen el espacio
extracelular unidos a las IGFBPs (Baxter, 1993; Jones,1995; Rechíer, 1998). Las
IGFBPs ligan IGF-I e IGF-II, pero no insulina y así pueden controlar su distribución
tisular o celular, participar en su unión a sus receptores o regular su acción por la
degradación de estas IGFBPs por parte de unas proteasas (Fowlkes, 1997).
34

Introducción
1.3.2.1.Tipos de IGFBPs.
1. IGFBP-1: Es una proteína de 25kDa que se sintetiza de manera
predominanteenel hígado.Presentaunaforma desfosforilada,conbajaafinidadpor
IGF-I, que se cree que potencia la acción de IGF-I, y otra fosforilada,que inhibe las
acciones mitogénicas de este factor de crecimiento.Además,IGFBP-1 secomporta
como inhibidor de IGF-I cuando este factor se presenta en altas concentraciones, pero
actúa como estimulador si está presente en el mediode cultivojunto conbajosniveles
de plasma pobreen plaquetas(Jones,1995; Rechíer, 1998).
Las principales señales reguladoras de la IGFBP-1 son metabólicas; diversos
estudios han demostrado que la combinación insulina/glucosa es el mayor
determinante de sus niveles. Las concentracionescirculantesde IGFBP-1 están
elevadas en el ayuno, la malnutrición y la diabetes mellitus de ambos tipos (Cotterill,
1988; Suikkari, 1988; Yeoh, 1988; Straus, 1994; Rivero, 1995; Goya, 1996). Por el
contrario, la realimentación o la ingesta de carbohidratos y la infusión de insulina
durante el “clamp” de glucemia en pacientes de diabetes tipo II o el tratamiento de
las ratas diabéticas con insulina se acompaña de un rápido descenso de los niveles de
IGFBP-1 (Cotterill, 1988; Suikkari, 1988; Yeoh, 1988; Lewitt, 1994; Goya, 1996).
Este patrón de regulación es similar al de las hormonas contrarreguladorasde la
insulina, como el glucagón, lo que ha hecho que se sugiera que la IGFBP-1 podría
desempeñar un papel contrarregulador en el mantenimiento de la homeostasis
glucídica (Lewitt, 1991; Baxter, 1995). Además,en el suero humano, IGFBP-1
muestra, a diferencia del resto de las proteínastransportadorasbien caracterizadas,
un ritmo circadiano marcado que no depende del estatus secretor de Gil, sino que
está regulado por el metabolismo, probablemente por la secreciónde insulina(ilolly,
1988).
En la rata, se ha descrito una regulación similar en ciertos estados catabólicos
como la malnutrición y la diabetes; en ambas situaciones, los niveles de insulina y
Gil son bajos, pero se produce un incremento en los niveles hepáticos del ARNmde
IGFBP-1, lo que sugiere que, al menos durante la etapa neonatal en que la Gil no
ha establecido totalmente sus conexiones neuroendocrinas,es la insulinemia la
responsable del aumento en los tránscritos de IGFBP-1 (Rivero, 1995; Goya, 1996).
2. IGFBP-2: Esta proteína de bajo peso molecular (31-36kDa) ha sido menos
estudiada que la IGFBP-1. Su acción predominante es la inhibición de la acción de
35

Introducción
IGF-I e RIF-II, aunqueen ciertos tipos de células puedecomportarsecomo un
estimulador débil de la acción del IGF-I (Baxter, 1993; Jones, 1995; Rechler, 1998).
A pesar de que los niveles circulantes de IGFBP-2 son menos sensibles a los
cambios metabólicos agudos que IGFBP-1, se ha descrito una regulación por el
estadonutricional.En situacionesde malnutrición,en la rata,seproduceunaumento
tantoa nivel séricocomo de expresióndel ARNm hepáticode estaproteína(Straus,
1994; Rivero, 1995>; por el contrario, la realimentacióny el tratamientode los
animales diabéticos con insulina se acompaña de un descenso en las concentraciones
circulantes y los niveles de ARNmhepático de IGFBP-2 (Goya 1996), lo que ha
llevado a sugerir a ciertos autoresun importantepapel en la regulación de esta
proteína por parte de los nutrientes (Thissen, 1994).
3. IGFBP-3: Es la proteína transportadora predominante en el suero de rata
adulta y su peso molecular oscila entre 38 y 43kDa dependiendo del número de sitios
glicosilados. En la circulación, la IGFBP-3 se asocia con una proteína de 8OkDa
llamada ALS (“acid-labile subunit) y una molécula de IGF para formar un complejo
de l5OkDa. Este complejo consta de IGFBP-3 e IGF-I y II en una proporción
equimolecular, lo que sugiere que la mayor parte de esta proteína transportadora está
saturada en el suero (Rajaram, 1997).
La expresión tisular de IGFBP-3 es muy ubicua; los tejidos con mayor
abundancia de esta proteína transportadora son hígado, riñón, placenta, útero, ovario,
testículo, bazo, páncreas, estómago y corazón (Rajaram, 1997).
Existen evidencias que demuestran que IGFBP-3 puede potenciar e inhibir las
acciones de IGF-I iii vitro, indicando que esta proteína transportadora puede ser un
importante modulador de las acciones de IGF-I in vivo (Gargosky, 1994). La
presenciade IGFBP-3 en suero puede tener también un papel muy importante
aumentando la vida media de los IGFs, sin olvidar que esta proteína transportadora
está sujeta a una regulación por la acción de ciertas proteasas (Fowlkes, 1997).
El principal regulador hormonal de IGFBP-3, y por consiguiente del complejo
ternario, es la Gil. Se ha observado que los nivelescirculantesy de expresión
hepática de esta proteína transportadora son dependientesde GH, dado que ambos
valores se reducen tras la hipofisectomía de las ratas y aumentan despuésdel
tratamiento con Gil (Glasscock, 1991; Gargosky, 1994; Gosteli-Peter, 1994>. No
obstante, estudios más recientes han mostrado que el tratamiento con insulina de ratas
36

Introducción
deficientes en GH puede aumentar los niveles circulantes de esta proteína
transportadora(Butíer, 1996). Además,seha observadoque el ayunoprolongado,
la subnutrición y la diabetes provocada en la rata por la administración de STZ
causan una reducción de los valores séricos de IGFBP-3 (Rivero, 1995).
4. IGFBP-4: Es una proteína no glicosiladade 25kDay 32-36kDaque fue
aislada por primera vez en células de osteosarcoma, aunque está presente en otras
líneas celulares dondese han descritodistintas isoformas(Rajaram,1997). Se trata
de una proteína fundamentalmente inhibidora de las acciones de los IGFs, lo que
protege a las células de una sobrestimulación por parte de los factores de crecimento,
o permite una activación de rutas de señales transmembrana alternativas que son
inhibidas por la exposición a los IGFs (Rechíer, 1998).
5. IGFBP-5: Esta proteína se purificó primero a partir de suerode rataadulta,
y presenta la capacidad de unirse a la matriz extracelular. Su principal acción es
potenciarel efecto de los IGFS (Jones,1995).
6. IGFBP-6: Se expresaen múltiplestejidosy en suero,y su principal acción
es la inhibición de los efectos del IGE-Il.
1.3.2.2. Acciones biológicas.
Las IGFBPs parecen modular las acciones de los IGFs, dado que regulan su
disponibilidad en los tejidos diana. En este sentido, los niveles séricos de las
proteínastransportadoraspuedenjugar un importantepapel regulandolas acciones
endocrinas de los IGFs.
1. Modular las acciones de los IGFs: La actividad de los IGEs es neutralizada
como consecuencia de su unión a las IGFBPs, de ahí que se diga que estas proteínas
transportadorasposeenuna función puramenteinhibidora (Jones, 1995; Rechíer,
1998). No obstante la ausenciade las IGFBPsaumentaríael efecto de los IGFs y
desequilibraría la homeostasis glucídica, puesto que los IGFs no parecen estar
controlados por la glucemia. Sin embargo, se han descrito varios mecanismos capaces
de liberar a los IGEs de sus proteínastransportadoras,principalmentemedianteel
descenso de la afinidad de los factores de crecimiento por sus IGFBPs. Este
mecanismo incluye la unión de las IGFBPs a moléculas de la matriz extracelular
(Rajaram, 1997>, fosforilación de las proteínastransportadoras(Coverley, 1995) y
degradación proteolítica de las IGFBPs (Fowlkes, 1997).
2. Aumentar la semivida de los IGFs: Las IGFBPs pueden facilitar las
37

Introducción
acciones endocrinas de los IGEs por el aumento de su vida mediaen la circulación.
Esta estabilización de los IGFs dependefundamentalmentede la IGFBP-3, y aunque
otras proteínas transportadoras, como IGFBP-1 y 2 también participan sobre todo en
periodosinmaduros,desempeñanunpapelmenorporquepresentanunasvidasmedias
más cortas que la proteína transportadora de alto peso molecular (Rosenfeld 1990).
Así pues, la IGFBP-3 es la proteína que alterala relaciónde IGFs libres y ligados,
lo que indirectamente prolonga la semivida de estos factores de crecimiento o los
protege de su rápida degradación, y por tanto, a la vez, regula sus acciones
metabólicas.
3. Controlar el transporte de los IGFs desde el espacio vascular: Para obtener
la respuesta que estimula el crecimientoporpartede los IGFs, se necesitaque estos
factores séricos sean transportados desde los vasos al líquido extracelular, y para ello,
se requiere que las IGFBPs que llevan unidos a ellas IGFs, sean degradadas o
puedan cruzar la barrera endotelial.
La mayor parte de los IGFs circula unida al complejo de l5OkDa (ALS-
IGFBP-3-IGF) lo que hace que sea considerado como un reservorio de IGFs y cobre
gran importancia en el transporte de los factores de crecimiento fuera del
compartimento vascular (Rajaram, 1997). Así pues, para que esta salida se produzca,
y dado que el complejo no es capaz de atravesar el endotelio vascular, se necesita la
presencia de unas proteínas capaces de reducir la afinidad de la IGFBP-3 por los
IGFs (Fowlkes, 1997), o también se ha apuntado hacia una reducción en la afinidad
para la fonnación del complejo por la unión a esta proteína transportadora de alto
peso molecular de glucosaminoglucanos presentes en la superficie de las células
endoteliales (Jones, 1995).
El porcentaje de IGEs que no circulan unidos a la IGFBP-3 lo hacen unidos
a la fracción de 3OkDa que sí atraviesala barreracapilar, porello la abundancia de
estas proteínas en periodos inmaduros parece estar dirigida a facilitar el paso del
endotelio a los IGFs en momentos de desarrollo.
1.4. FACTORES ENDOCRINOS QUE REGULAN EL DESARROLLO:
HORMONAS TIROIDEAS E IGFs, SU INTERACCION.
Los niveles circulantes de los IGFs, como hemos venido viendo, están
influenciados por multitud de factores,entre los que se incluyen Gil, edad, nutrición,
38

Introducáón
sepsis, hormonas tiroideas y esteroides (Daughday, 1989; Sara, 1990), y están muy
implicados regulando el desarrollo en etapas inmaduras de los mamíferos. La
importanciade las hormonas tiroideas sobre el SNCes sobradamenteconociday ha
sido ya someramenteexpuesta.Por ello, el estudiode la interacciónde estosfactores
endocrinos con las hormonas tiroideas tiene una gran relevancia en estudios
perinatales.
1.4.1. Interacción entre las hormonas tiroideas y el axis IGFs/IGFJIPs.
En el hombre, el hipotiroidismo disminuye los niveles circulantes de IGF-I,
y tambiénsu actividadbiológica (Chernausek,1983; Cavaliere,1987; Mielí, 1993).
En la rata, este descenso del IGF-I aparece asociado a una disminución de la
expresión hepática del gen de este factor de crecimiento (Burstein, 1979; Gallo,
1991). Por el contrario, los pacienteshipertiroideosmuestranunosvalorescirculantes
de IGF-I altos o normales junto con una baja actividad biológica (Westermark, 1988;
Mielí, 1993). Sin embargo, estudios más recientes con ratas han demostrado que el
hipertiroidismono altera los nivelesséricostotalesde IGF-I, pero si disminuyesus
valores libres (Frystyk, 1995).
Entre el periodo adulto y el neonatalexistendiferenciasfundamentalesen
cuanto a la regulación del axis IGFs/IGFBPs. Se ha establecido que, en periodos
fetales, la secreción de IGFs en hígado es independiente de Gil, y en la etapa
neonatal de la rata, esta secreción parece regida también por un equilibrio
insulina/nutrientes de forma independiente de Gil (Rivero, 1995; Goya, 1996)
finalmente, en periodo adulto, aunque es indudable que la hormona de crecimiento
es el regulador principal (Straus, 1994; Thissen 1994), el equilibrio insulina/
nutrientestambiénparecejugarun papel(Goya,1996; Goya,en prensaenLife Sci.).
En este contexto, las acciones de las hormonas tiroideas sobre la secreción de los
IGFs estarán,sin duda en adulto, mediadaspor la Gil, ya que esconocidoque las
hormonas tiroideas son necesarias para la expresión normal de la Gil in vitro
(Shapiro, 1978; Geary, 1989; Ezzats, 1991; Ceda,1992) e in vivo (Hervás, 1975;
Samuels, 1989). Todo ello sugiere “a priori” que las hormonas tiroideas van a incidir
sobre la secreción de los IGFs de forma distinta en periodos inmaduros y en periodo
adulto segúnla secreciónde IGFs seamáso menosdependientede Gil (Glasscock,
1990; Glasscock, 1991; Kim, 1993). En cualquier caso, no se puede excluir un efecto
directo de las hormonas tiroideas sobre la secreción del IGF-I e independiente de Gil,
39

Introducción
puesto que la ‘[3 es capaz de incrementar la secreción de este factor de crecimiento
enhígadode rataperfundido(Ikeda, 1989); así pues,las accionesde las hormonas
tiroideas sobre la regulación del axis IGFs/IGFBPs presenta muchas lagunas de
conocimiento.
De igual forma se ha descrito que los desórdenes tiroideos son capaces de
inducircambiosen los nivelesde las IGFBPs.Se ha demostrado que, en el hombre,
el hipotiroidismo disminuye los nivelesde IGFBP-l y 3, y aumenta los de IGFBP-2
(Mielí, 1993; Mielí, 1994), mientras que la tirotoxicosis incrementalos niveles
circulantes de IGFBP-1 y 3 (Mielí, 1993).
Sehaestablecido que el efecto de las hormonas tiroideas sobre los IGFs y sus
proteínas transportadoras depende del estado de desarrollo en la rata (Nanto-Salonen,
1992).Duranteel periodoperinatal,las ratashipotiroideasmostraronun aumentoen
los niveles séricos y de su ARNmhepático para la IGFBP-2 que se normalizó por el
reemplazamientotemprano con hormona tiroidea (Nanto-Salonen,1991; Nanto-
Salonen,1992).
En las ratashipotiroideasadultas,los bajos nivelescirculantesde IGFBP-3 e
IGFBP-4 se asociaron a un descenso del ARNmde IGFBP-3 y a un aumento
paradójico del ARNmde IGFBP-4 (Nanto-Salonen, 1992). Este descenso en la
expresión hepática de IGFBP-3 de las ratas hipotiroideas se ha confirmado en otros
estudios en los que además se encontró un aumento en la abundancia del ARNmde
IGFBP-1 y 2 (Rodríguez-Arnao,1993a;Rodríguez-Arnao,1994).
El tratamiento de las ratas hipotiroideascon Gil normalizó los niveles de
IGFBP-3 sin afectara la expresiónde IGFBP-2e IGFBP-4 (Nanto-Salonen,1993),
lo que sugiere,de nuevo,que estosefectosde las hormonastiroideassobrelos genes
se producenpor mecanismosindependientesy que algunasvecessonmediadospor
la Gil.
Recientemente,se ha mostradoque las ratashipertiroideasincrementanlos
niveles séricos de IGFBP-1 e IGFBP-4 (Frystyk, 1995). Finalmente,no sepuede
excluir una acción directa de las hormonas tiroideas sobre las JGFBPs. Se ha visto
que la ‘[3 estimula, a nivel transcripcional, la formación de IGFBP- 1, 2 y 3 (Ceda,
1992; Schmid, 1992; Angervo, 1993a). Además, se ha demostrado en cultivos de
hepatocitos de ratas adultas que la T3 induce un efecto directo positivo sobre la
expresión de IGFBP-4 y, como consecuencia, sobre la actvidad de los IGFs (Demori,
40

Introducción
1997).
La importancia de las hormonas tiroideas durante el desarrollo es conocida,
y dada la implicación del axis IGFs/IGFBPs en la proliferación y diferenciación
celular,el estudiode la interacciónentreestashormonasy dichosfactoresposeeuna
importancia relevante en la regulación endocrina del desarrollo; sin embargo, como
se ha expuesto, dicha interrelación muestra numerosos aspectos desconocidos.
1.5. TRANSPORTADORESDEGLUCOSA(GLUTs).
La glucosa es la principal fuente de energíametabólicapara las células. El
carácter hidrofilico de esta moléculaimplica la necesidadde que existanproteínas
transportadoras, para que atraviese la bicapa lipídica, que constituyen los sistemas de
membrana. En el túbulo proximal del riñón y el lumen intestinal, la captación de
glucosa es activa porque se mueve en contra de gradiente, por lo que el proceso se
lleva a cabo mediante un cotransportador Na~/glucosa. En los restantes tejidos, el
transporte serealizapordifusión facilitada,y en esteprocesointervieneunafamila
de proteínas estructuralmente relacionadas llamadas GLUTs.
1.5.1. Estructura, función y expresión tisular.
En la actualidad, se han identificado seis isoformas de GLUT producto de
diferentes genes. Sus pautas de expresión son también distintas, tanto en lo que se
refiere al tejido en dondese encuentrancomo a su concentración.
Todos los transportadoresde glucosa tienen un tamaño similar (492-524
aminoácidos)y suestructuraaparecebastanteconservadaentrelas distinasisoformas
y entrediferentesespecies,si bien las regionesmásdivergentescorrespondena sus
extremosC y N (Gould, 1990>.Se trata de proteínasmuy hidrófilas; casi la mitad
de su secuenciaestá inmersa en la bicapalipídica, y allí forman 12 dominios
transmembranososconectadospor bucleshidrofílicos por fuera y por dentro de la
célula. Los extremosC y N soncitoplasmáticos.
- GLUT-1: Se expresaen mayor medida en los tejidos fetales (menos en
adultos), placenta y cerebro; en este último, facilita el paso de la glucosa a través de
la barrera hematoencefálica.También es la isoforma predominante en céiulas
cultivadas y en muchos tumores dado que prácticamente todos los mitógenos
estimulansu expresión(Hiraki, 1988). A pesarde queen algunode ellos lo estáen
cantidadmínima, el GLUT-1 se encuentracasi en todos los tejidos, por lo que se
41

Introducción
considera el responsable de la captaciónconstitutivade glucosa.
- GLU’[-2: La expresión de este transportador es más restringida: hígado
(media la captación y liberación por el hepatocito), membranas basolateralesdel
intestino delgado y riñón (participa en el transporte transepitelial de glucosa) y células
/3 (participa en la secreción de insulina inducida por glucosa).
- GLU’[-3: Su mayor expresión se produce en el tejido nervioso, y
concretamente en el cerebro, esto es, en células con un alto requerimiento de glucosa.
Se cree que este transportador es el que media el paso de esta hexosa a las neuronas
(Maher, 1993). Se encuentra también, pero en cantidades menores, en placenta,
hígado, riñón y corazón,pero no enmúsculoesquelético.La ubicuidad del GLUT-3
ha sugeridoque, junto con el GLUT-1, participeen el transporteconstitutivode la
glucosa.
- GLU’[-4: Su localización tisular es muy restringida, se encuentraen los
tejidosadipososblanco y marrón, músculo cardíaco, diafragma y músculo esquelético
(tejidos sensiblesa la insulina).
Se trata de un transportador insulino-sensible; el GLUT-4 se localiza en
vesículasintracelularesen situaciónbasal,pero la presenciade insulina provocasu
translocacióna la membranaplasmática(Barnard, 1992; Stephens,1995).
- GLUT-5: Es la isoforma más diferente en cuanto al grado de homología
(Gould, 1990) y presenta mayor afinidad por la fructosa que por la glucosa,por lo
que se cree que es el transportadorde la cetohexosade la dieta. Se expresa
mayoritariamenteen la membranaapical de los enterocitosdel yeyuno, aunque
tambiénsedetectaen los tejidos insulino-sensiblesmencionadosanteriormente,tejido
adiposoy cerebro.
- GLUT-6: No existe como proteínafuncional. Su pseudogenorigina un
tránscritohomólogoal del GLU’[-3, de expresiónubicua,peroconmúltiplescodones
stop y desplazamientosde la pautade lectura(Gould, 1993).
- GLUT-7: Este transportadorfacilita el paso de la glucosa-6-fosfato,
procedentede la glucogenolisisy la gluconeogénesis,desdeel citoplasmaal retículo
endoplásmatico.Selocalizaespecíficamenteen el retículodel hígadoy poseemuchas
concordanciascon la estructuradel GLUT-2.
1.5.2. Efecto del estadotiroideo sobrelos transportadoresde glucosa.
La expresión de los GLUTs, que se produce de manera específica en los
42

Introducción
tejidos, puede ser alterada por multitud de factores, y entre ellos se incluyen glucosa,
insulina, hormonas tiroideas,factoresde crecimientoy edad(Burant, 1991).
Las hormonas tiroideas aumentan la actividad metabólica tisular. Las ratas
tratadas con ‘[3 y ‘[4, en músculo y tejido adiposo, aumentan el metabolismo de la
glucosa, la velocidad de su captación y su transporte basal, debido al menos en parte,
al incremento del contenido de GLU’[-4, tanto en la membrana plasmática como en
fraccionesinternas,y tambiénse incrementala expresióndel transportador(Casia,
1990; Weinstein, 1991). No obstante,esteaumentode la velocidadde transporteen
el hipertiroidismo no sólo se asocia con un aumento en el número de transportadores
GLU’[-4, sino tambiéncon un aumentode su actividad funcional, al menosen los
adipocitos (Matthaei, 1995). Estos aspectosse han corroboradotambién in vitro:
célulasderivadasde hígado de rata ARLíS (Weinstein, 1990), en cardiomiocitos
(Gosteli-Peter,1996) y célulasClone 9, que sólo expresanGLUT-1 (Shetty, 1996).
El hipotiroidismo neonatal experimental,por el contrario, disminuye la
expresión de GLUT-4, así como de GLUT-1 tanto en el músculo esquelético como
en el adipocito (Weinstein, 1991; Matthaei, 1995). El hipotiroidismo, a pesardel
marcado descenso del contenido de GLU’[-1 y 4, no disminuye el transponede
glucosa estimulado por la insulina al menosen el adipocito(Matthaei, 1995), lo que
implicaría un aumento en la actividad funcional de los transportadores de glucosa.
En contra de las observacionesprevias en el músculo esquelético, el
hipotiroidismo aumenta la expresión y el contenido cardiaco de GLUT-1, y disminuye
el ARNmde GLUT-4, aunque mantiene los niveles en la membrana de esta proteína
transportadora. La administración de ‘[3 suprime el aumento de GLU’[-1, incrementa
la expresión de GLUT-4 y mantiene su contenido en los extractos membranosos
(Weinstein, 1992).
La expresión de los transportadores de glucosa se encuentra reguladapor la
edad. Durante la etapa fetal y al principio de la postnatal, el GLU’[-1 es la isoforma
predominantey se expresaen músculo,tejido adiposomarróny corazón(Santalucía,
1992; Postic, 1994). Después,en la vida postnatal,se produce una represióndel
GLU’[-1 en músculo y tejido adiposo marrón, que es concomitante con un aumento
en la expresión del GLUT-4 (Santalucía, 1992; Postic, 1994). Esta represión del
GLU’[- 1 y la inducción del GLUT-4 pueden estar mediadas por diferentes
mecanismos,entreellos, las hormonastiroideas. Se ha visto que el hipotiroidismo
43

Introducción
altera la transiciónnormal de GLUT-1 a GLU’[-4 desde los niveles fetales a los
neonatales en corazón y tejido adiposo marrón (Castelló,1994).Esteefectosepodría
producir de una manera directa puestoque seha descritola existenciadeunelemento
de regulación positiva en el gen de GLU’[-4 capaz de responder a las hormonas
tiroideas(‘[orrance, 1997b),resultadoquepodríaexplicarlos obtenidosin vivo, antes
mencionados.
El hígado juega un papel central en la regulación de la homeostasis glucídica
y tiene capacidad para almacenar glucógeno y para liberar glucosa a la circulacion.
El hígado es también el principal sitio gluconeogenético y el único tejido en que la
concentración intracelular de glucosa puede exceder a la de la circulación. La
isoformatransportadorade glucosapredominanteen el hígadoes el GLU’[-2, como
hemosvisto, y es la encargadade captary liberar la glucosa. La alteraciónde la
expresiónde estetransportadorde glucosamodificaríaengranmedidael metabolismo
de los carbohidratos,al igual que lo altera el estadotiroideo. El hipotiroidismo
experimentaldisminuyeel contenidoy la expresiónhepáticadel GLU’[-2, mientras
que el hipertiroidismolos incrementa(Weinstein, 1994).
Con respectoaotras influenciasendocrinassobrela expresiónde los GLU’[s,
la insulina parecejugar un papelcrucial. En el músculoesqueléticode la rata, la
hipoglucemiacrónica (subnutrición)aumentael contenidode GLUT- 1 sin alterarel
de GLU’[-4 (Rubio, 1996); sin embargo,el GLUT-4 también responde a los
estímulos insulinicos, y se ha visto que las ratas subnutridas presentanuna
translocaciónde estetransportadorpor la insulina en músculo más eficaz que los
animalescontrol (Escrivá,en evaluaciónenDiabetología).Encuantoal GLUT-2, la
insulina inhibe suexpresión(Postic, 1993a;Colomb, 1995)y la glucosala estimula
(Postic, 1993b; Rencurel,1996). No obstante,los cultivos de islotesde Langerhans
procedentesde ratas neonatalesy adultassubnutridasno alteransu contenido en
GLU’[-2, aunquesí aumentael de GLUT-1 (Martin, 1995).
Enestetrabajose intentaaclararsi la posibleregulaciónde los transportadores
de glucosapor las hormonastiroideas tendríamecanismossimilaresa los que rigen
la regulaciónde la secrecióndel axis IGFs/IGFBPspor dichashormonastiroideas.
1.6. PLANTEAMIENTO DEL ‘[EMA.
El programagenético rige el desarrollo de los mamíferos en periodo
44

Introducción
embrionario,pero después,factoresexternos,llamadosambientales,modulandicho
desarrollo de forma decisiva. Estos factores son estudiados en las llamadas
alteracionesadquiridasdel desarrollo,a las que nuestrogrupo de investigaciónha
dedicado su trabajo desde1970. Entre estos factores ambientaleslos nutrientes
ocupan un lugar preponderante no sólo por ser el sustrato energético para crecer,sino
porque modulan factores endocrinosgenéticamenteestablecidosque regulan el
crecimiento como son: la insulina, las hormonastiroideasy los lOEs (Aláez,1992b;
Escrivá, 1992; Rivero, 1995; Goya. 1996; Alvarez, 1997; Martín, 1997).
El trabajoquesepresentaenestatesisesun intentode estudiode las posibles
interaccionesque puedenexistir entrelas hormonastiroideas, insulina e lOEs enun
modelo de hipotiroidismo por tiroidectomiay en tres etapasde vida de la rata
(neonatal,destetaday adulta).
Los lOEs son polipéptidoscon accionesendocrinas,autocrinasy paracrinas
(Daughday, 1989), si bien su función pareceestar moduladapor sus proteínas
transportadoras(IGFBPs). Ambos, lOEs e IGFBPsson secretadospor el hígado,y
dicha secreciónestámoduladapor el estadonutricional y la 01-1 (Straus, 1994;
‘[hissen, 1994>. El axis IGFs/IGFBPsconstituyeun sistemaregulatoriocomplejoque
poseeun importantepapelenel crecimientoy la diferenciacióntisulares,y que está
implicadoennumerososprocesosduranteel crecimientoy tambiénduranteelperiodo
adulto.
En periodosinmadurosdeldesarrollo,seexpresanmayoritariamenteel lOE-II
y el complejo de IGFBPs de 3OkDa (IGFBP-1 y 2), pero despuéseste perfil es
reemplazadoen el animal adulto, mostrandoaltosniveles de IGF-I e IGFBP-3, y
reducidosde IGF-II, IGFBP-1 y 2. Seha descritoque estoscambiosseretrasanpor
la falta dehormonastiroideas(Gallo, 1991; Nanto-Salonen,1991), lo quesugiereque
la influenciade la ‘[3 y la ‘[4 sobreel complejoIGFs/IOFBPsesedad-dependiente
(Nanto-Salonen,1992).
Las hormonas tiroideas juegan un papel fundamental en el inicio y el
mantenimientodel crecimiento de los mamíferos en etapas inmaduras. Estas
hormonas,tambiénafectanacasi todoslos aspectosdel metabolismoy así, suacción
estimulalas rutascatabólicasy oxidativas porque modifican la síntesisde enzimas
clave del metabolismode lípidos y carbohidratoso porquealteran las accionesde
otras hormonasreguladoras(insulina, glucagón, catecolaminas)(Okajima, 1978;
45

Introducción
Venditti, 1997).Las hormonastiroideasy la Gil, a travésdelos IGFs, sonesenciales
para un crecimiento normal, no obstante,el contenido hipofisario y los niveles
circulantes de GHestán regulados por las hormonas tiroideasin vivo (Hervás, 1975;
Samuels,1989)e in vitro (De Fesi, 1984) y, además,tambiéncontrolanla expresión
del gen de Gil (Evans, 1982; Yaffe, 1984>. Por otra parte, se sabe que el axis
GI-I/IGFs actúacomoprincipal mecanismoparael crecimiento,ya que el efectorde
las accionesde la Gil sobreel desarrolloson los IGFs (Daughday,1989; Jones,
1995), por tanto, las accionesde la ‘[3 y la T4 sobreel sistema IGE se podrían
explicarporel efectodirecto de estashormonastiroideassobrela secreciónde Gil.
Sinembargo,hay indicios experimentalesque muestranqueno todaslas accionesde
las hormonastiroideassobrela secreciónde IGFs estánmediadaspor la Gil. La
perfusiónde honnonatiroideaenhígadoaumentala secreciónde IGFs (Ikeda, 1989),
y las dosis de Gil dadasa hipotiroideosno restauranlos bajos niveles de IGFs
(Nanto-Salonen,1993). Nuestro grupo,junto con otros autores,ha establecidoque
en periodos de inmadurez,el sistemaIGFs/IGFBPspareceestar reguladopor el
balanceinsulina/nutrientesmás que por la Gil (Rivero, 1995; Goya, 1996), ello ha
sido estudiadoen animalessubnutridosy diabéticos,y se ha establecidola existencia
de undescensoen los nivelescirculantesy de expresiónhepáticade los lOEs de estas
ratas(Rivero, 1995; Goya, 1996).
Las hormonastiroideasafectanglobalmenteal metabolismo,propiciandouna
compleja regulaciónen los niveles de insulina, no obstante,nunca había sido
estudiadala insulina como posible mediadorade las acciones de las hormonas
tiroideassobreel axis IGFs/IGFBPs.Se ha descritoque el hipotiroidismoproduce
un descenso en los nivelesde IOFs y que disminuyela biodisponibilidadde dichos
factores (Burstein, 1979; Mielí, 1993), mientrasque el hipertiroidismo produce un
aumentode los IGFs, aunquetambiéndisminuyesu biodisponibilidad(Mielí, 1993).
Dicha biodisponibilidaddepende,sin duda, de las variacionesen las IGFBPs,luego
las hormonas tiroideas parecen alterar el axis IGFs/IGFBPs;sin embargo,quedapor
aclarar si el mediador de sus acciones es siempre la GH, si se trata de una acción
específica de las hormonastiroideaso sonotras hormonaslas que mediansuacción.
La insulina parecetenerun papelcomo reguladoradel axis IGFs/IGFBPsen
etapasinmaduras,por ello, enestetrabajo, nospropusimosestudiar,en un mismo
modelo de hipotiroidismopor tiroidectomía,las variacionesa nivel circulantey de
46

Introducción
expresiónhepáticadel ARNm de IGFs e IGFBPsen tres poblacionesde animales:
neonatos(10-20 días), destetados(27-37 días) y adultos (77-87 días), y en él,
dosificamos además, la insulina, Gil y glucemia.El trabajose divide en tres partes:
1. Estudio de las alteracionesdel axis IGFs/IGFBPs en situacionesde
hipotiroidismo (tiroidectomía o tratamiento con MMI) desde la etapa neonatala la
adultay la posible implicaciónde la insulina como mediadorade ellas.
2. Estudiode la repercusiónsobreel axis IGFs/IGFBPs,a nivel circulantey
en la expresión hepática de su ARNm, de las mismas tres poblaciones
tiroidectomizadasdespuésde la rehabilitación con hormona tiroidea por dos vías:
inyección intraperitoneal de ‘[4 e implante de pellet subcutáneo para una absorción
continuada.
También se intenta, en una población de ratas diabéticas, la rehabilitaciónde
los IGFs y las IGFBPs por la administración de tiroxina, como estudio
complementario al anterior.
3. Estudio de las variaciones de los transportadores de glucosa en animales
neonatales hipotiroideos para establecer la posible existencia de correlación entre las
accionesde las hormonastiroideassobredichostransportadoresde glucosay sobre
el axis IGFs/IGFBPs.
47

MATERIAL YME TODOS

Materialy métodos
2. MATERIAL Y METODOS.2.1. ANIMALES.
A lo largo del trabajo que constituye la presente tesis, se han utilizado ratas
de la raza Wistar obtenidas del propio criaderodel Instituto de Bioquímica (Centro
mixto CSIC-UCM, ubicado en el departamento de Bioquímica y Biología Molecular
II de la Facultadde Farmacia).Criar nuestrospropios animalesha permitidoejercer
un estrictocontrol de sus condicionesnutricionalesy generales,tanto en las etapas
gestacionaly lactantecomo en la adulta; dicho control es imprescindibleen los
modelosexperimentalesque han sido objeto de este trabajo.
En el mencionadocriadero existe un acondicionadorde temperaturaque
mantieneestavariableambientalconstanteentre22 y 240C. Porotra parte,las ratas
estánsometidasaun ciclo de luz/oscuridadautomático,graciasaun reloj controlador
que ilumina la estanciaa las 7:00 h y la oscurecea las 19:00 h.
El inicio de la gestaciónfue establecidomediantela observación,por frotis,
de la presenciade espermatozoidesen la vaginade la rata(180-200g)al día siguiente
de su apareamientocon el macho(300-400g);dichacondición se confirmó a los 14
díasporpalpaciónabdominal.El díadel nacimiento(díaO de vida) todaslas camadas
se uniformaron,disponiéndose8 crías,con númeroequivalentede machosy hembras
encadauna. Las ratassedestetaronel día 22 de vida y se agruparonen jaulasque
contenían4 animales.
2.2. COMPOSICIONDE LA DIETA GENERAL
El alimento suministradofue el pienso SANDERMUS S-10, adquirido en
SANDERS; se presentaen forma de porcionesprensadascuya composición, en
gramos por ciento, es la siguiente:
Proteínas 18
Grasas 2.5
Almidón 35
Azúcarestotales 3.5
Cenizas 8
Fibra 5.5
Humedad 13
49

Materialy métodos
Calcio 1.2
Fosfato 0.65
NaCí 0.6
El resto de los ingredientesestáconstituidopor una mezclade mineralesy
vitaminas,cuya composiciónes la siguiente:
MINERALES (mg/Kg de pienso)
Cobalto 0.6
Yodo 1.5
Selenio 0.3
Cobre 15
Hierro 25
Zinc 50
Manganeso 80
VITAMINAS (porKg de pienso)
Vitamina A 10000 U.l.
Vitamina D3 2000 U.I
VitaminaE 15 mg
Vitamina K 1 mg
Vitamina Bí 1 mg
Vitamina B2 3.5 mg
Vitamina Bá 1 mg
Vitamina B12 15 gg
Acido fálico 0.3 mg
Biotina 15 ~g
Acido nicotínico 30 mg
Pantotenato cálcico 10 mg
Colina 250 mg
Considerandolas cifras convencionalesde equivalenciacalóricade los tres
principios inmediatos(4.1 Kcal/g paralos glúcidosy prótidos y 9.0 Kcal/g paralos
50

Materialy métodos
lípidos>, la mezcla descrita posee un valor de 2.54 Kcal/g.
2.3. HIPO’[IROIDISMO.
Se estudiaronde maneracomparativadospoblacionesde animales,controles
e hipotiroideas, compuestas por ratas de diferentes edades, que abarcarondesdela
etapa neonatal a la adulta. El estudio comprendió animales neonatales hasta 20 días
de vida, destetadas de 22 a 37 días, y adultas, hasta 87 días de edad.
Ratascontrol: Los animalescontrolprocedíande progenitoresalimentadosad
libitum en todo momento.Las controles-lactantesfueron amamantadaspor ratasque
dispusieronde piensolibremente,y lascontroles-adultasrecibierondirectamenteeste
tipo de dieta a partir del destete.
Estosanimalesno fueronsometidosa ningúntipo de tratamiento.No obstante,
a un grupo de estasratascontrol se les practicóuna operaciónsimuladapara que
pudierancompararsecon los animalestiroidectomizados,eliminando así en dicha
comparaciónposiblesconsecuenciasdebidasal stressde la intervención.
Ratashipotiroideas:El hipotiroidismoseprodujoportiroidectomíaquirúrgica,
aunquecon el fin de facilitar estaoperaciónsesuministróen el aguade bebida,y en
una proporción del 0.02% (p¡’v), 5-mercaptoimidazol(MMI) días antes de la
operaciónen las tres poblacionesestudiadas(neonatos,destetadasy adultas),lo que
provocó la hipertrofia de la glándulay posibilitó su eliminación. Se continuó con
dicho tratamientohastael momentodel sacrificio de los animalescon el fin de
mantenerunosnivelesbajos de hormonastiroideasy prevenirsu renovaciónpor ¡a
posibleexistenciade tejidos tiroideos residuales.
2.4. MODELOS DE HIPOTIROIDISMO. (Figura 1).
2.4.1. Animaleshipotiroideosneonatales.
En los estudiosrealizadosen periodoneonatal,el MMI se suministróen el
aguade bebida materna,con lo que los animaleslactanteslo recibieron por vía
indirecta a travésde la leche.
2.4.1.1.Ratasneonatalestiroidectomizadas.(Ti, Figura 1).
A estosanimales,a los 2 díasde vida, se les administróMMI en el aguade
bebida(0.02%,p/v), setiroidectomizarona los 5 díasde viday se sacrificarona los
5,10 y 15 díasdespuésde la operación(10, 15 y 20 díasde vida, respectivamente).
51

1’-e-)
toCM1-
CME’)
.0)
+
1—
~C4o
yy
0)Na-)
CM0)
CM1’-
lo(0
o
o
o10)
o)
•0)
1—
o-CMoo
[y
CMN
1—
Ni
CM
]Cf)
0)
No
)ESCM
14
)
(0
o
o
‘7
a-
3-o
y
+0
3N
.02
1~j~
j~
e4
..’tj~
~~
o,...
kk£
~&
~~
h~::
....~
~
.~
~tM
,.flva
,.
L~
e.
~j.s~
~~,~
e’e~
Oee
‘-~-
y
½t)
’~
uO
Ou
•eti
¿
~‘4
-.~
O~
co
oti
t::~
E~
.t~c.jt4
Co~
.S?
<~
~-~
~.2
2~~P~J’—
.~J
~O‘~~
;N.~
~e
%~
-ze
cx.e
k
en
N1—(1)E~0
-UEoo
0-
CM~
+0
.0)
‘4a—£9
-t
y
(u-Ua>tú(uoII)
CMCM
IP
o
¾—o
CM
.
07
E‘ooz
oa
ChCo6)
•1o
-Co
CM
oCMCM
CoC
o
U)
CoCo
-(1)
(oCo
3->
—a
~C
Mo
CMe-)
r-.CM
CMCM
U)
0-z
09
(O
—a
~u-)o
y
z
0)
yo[y
o

Material y métodos
El suministrode estebociógenose realizó en los animalestiroidectomizadospara
asegurarel hipotiroidismoen ellos, aunen casode la existenciade posiblesresiduos
deglánduladespuésde la operación.
Los animalestiroidectomizadosrecibierondesdeel díade suoperaciónlactato
de calcio al 1 %en el agua de bebida, junto con el MMI, para evitar la hipocalemia
debida al posible daño o extirpación que pudo sufrir la glándula paratiroides durante
la tiroidectomia.
2.4.1.2. Ratas neonatales tratadas con MMI. (Hipotiroideas-MMI, Figura
1).
En la población neonatal, se estableció también un segundo modelo de
hipotiroidismo que consistió tan solo en la administración de MMI al 0.05% (p/v>
desde el segundo día de vida hasta e! momento del sacrificio (10, 15 y 20 días de
vida). En este caso, no se sometió a los animales a la tiroidectomía,con lo que se
consiguió un hipotiroidismo que podría parecer más suave, aunque los niveles de las
hormonastiroideasfueronigualmentebajos respectoa las ratascontrol.
2.4.1.3.Ratasneonatalestiroidectomizadastratadascon STZ.
(T5 + S’[Z, Figura 1).
A un grupo de animalesneonatalestiroidectomizados,a los 10 días de la
operación(15 días de vida), se les bloqueó la secreciónde insulina mediantela
administraciónintraperitonealde una solución de estreptozotocina(STZ, Upjohn
Farmoquimica,SA, Madrid) en tampóncitrato0.OSM pH 4.5. La dosisadministrada
ffie de 7Omg/Kg de pesoy los animalessesacrificarona los 15 díasdespuésde la
tiroidectomía(20 díasde vida), esto es, a los 5 díasde la administraciónde S’[Z.
La comprobaciónde la falta de insulina en estapoblaciónse llevó a cabo
mediantela determinaciónde la glucemiae insulinemía.
2.4.2. Animaleshipotiroideosdestetados.
2.4.2.1.Ratasdestetadastiroidectomizadas.(‘[22, Figura 1).
En estapoblación de animales, la administraciónde MMI (0.02%,p/v) se
inició una semanaantesde] destete(15 díasde vida>. Posteriormente,a los 22 días
de vida, se tiroidectomizaron,y se sacrificarona los 5, 10 y 15 díasdespuésde la
operación(27, 32 y 37 días de vida, respectivamente).
53

Material y métodos
2.4.2.2.Ratasdestetadastiroidectomizadastratadascon insulina. (‘[22 +
1, Figura 1).
La glucemia de las ratas hipotiroideas destetadas, que estaba disminuida
respectoa las controles,sele normalizómediantelaadministraciónde insulinaLente
(mono componente de páncreasporcino y bovino) de Novo (amablemente
suministradapor Novo Nordisk PharmaSA, Madrid). Esta insulina esuna mezcla
formadaporun 30% de insulina de páncreasporcinoen estado“amorfo” y un70%
de insulina de páncreasbovino en forma cristalina.Su acciónes lenta, comenzando
a las 2.5 horas de su administración,alcanzandoun máximo a las 7-15 horas y
finalizandosu accióna las 24 horas.
Las dosisde insulina administradasy la pautaconcretade tratamiento,que se
describea continuación,se determinópor tanteo, aunquese consideróadecuadala
dosiscapazde mantenerla glucemiaenun rangonormal(80-l2Omg¡ml).
El tratamientose inició a los 10 díasde la tiroidectomía(32 díasde vida) y
consistióen la administracióndurante5 díasde 1 UI de insulina vía subcutáneapor
la mañana(9h). Seefectuaronmedidasdiariasde la glucemiaantesde administrarla
insulinaparacomprobarla evolucióndel tratamiento.Finalmente,los animalesfueron
sacrificadosa los 37 díasde vida (15 díasdespuésde la tiroidectomía).
2.4.3. Animaleshipotiroideosadultos.
2.4.3.1.Ratasadultastiroideetomizadas.(T72, Figura 1).
La pautaseguidacon estosanimalesfue la mismaqueen las ratasdestetadas,
peroenestecasose tomócomodíauno del experimentoaquelenel quelos animales
alcanzaronlos 120gde peso(65 díasde vida), iniciándoseentoncesla administración
del MMI (0.02%, p/v). Se sacrificaron a los 5, 10 y 15 días despuésde la
tiroidectomía(77, 82 y 87 díasde vida, respectivamente).
2.4.3.2.Ratasadultastiroidectoinizadastratadascon insulina. (‘[72 + 1,
Figura 1).
En estapoblación,comoen el casode las ratasdestetadastiroidectomizadas,
que también recibieron el tratamiento hormonal, la pauta de administraciónse
determinópor tanteoy teniendoencuentalos valoresde glucemia.
El tratamientose inició a los 10 días despuésde la tiroidectomía(82 díasde
vida). En estecaso, a los animalesse les administraron2 UI de insulina por vía
subcutáneadurante5 díasrepartidasendosdosis, unapor la mañana(9h) y otra por
54

Materialy métodos
la tarde (17h). También, como con los animalesdestetados,antesde procedera la
administración de insulina, se midió la glucemia. Al finalizar el tratamiento, los
animales fueron sacrificados (15 días después de la tiroidectomía, es decir a los 87
díasde vida>.
2.5. MODELOSDEHIPO’[IROIDISMO CON REHABILITACION. (Figura2).
2.5.1. Animaleshipotiroideostratadoscon tiroxina.
Una vez establecido el cuadro de hipotiroidismo en los animales neonatales,
destetadosy adultos, fueron tratadoscon ‘1’4 segúnlas pautasque se especificana
continuación.
En principio, la recuperacióncontiroxina serealizó mediantela inyecciónde
la hormona,pero con el fin de conseguirunaabsorciónmásgradual,y por tanto, una
rehabilitaciónmás fisiológica, se recurrióa la implantaciónde pelletsbajo la piel.
2.5.1.1.Rehabilitaciónmedianteinyecciónde ‘[4. (‘[5 + ‘[4 (R>, Figura2).
A los 10 días despuésde la operación,y con una población de animales
neonatalestiroidectomizados(‘[~), destetados(T22) o adultos(‘[72), seinició una pauta
de recuperaciónque consistíaen administrardurante5 días una dosis diaria, vía
intraperitoneal,de ‘[4 (1.Sgg/lOOgde pesopara las ratasneonatalesy 1 .75~íg/100g
de pesoparalos animalesdestetadosy adultos).
2.5.1.2. Rehabilitaciónmedianteimplantaciónde pellets.(T5 + T4 (RP),
Figura 2).
Otro grupodiferentede ratas neonatales,destetadasy adultasfue sometido a
un procesode rehabilitacióncon T4 por implantaciónde pellets(IRA, Sarasota,FL,
USA). Este dispositivo, que libera la hormona de forma continuada,se implantó
subcutáneamenterealizandouna pequeñaincisión en el lomo del animal. La dosis
administradaen esteprocesode rehabilitaciónfue la misma que enel caso anterior
(1 .Sgg/lOOgde pesoparala poblaciónneonataly 1 .75pg/lOOgdepesopara lasratas
destetadasy adultas,respectivamente),sin embargo,en el caso de las animales
destetadosy adultosel tratamientoserealizódurante5 ó 10 días.
2.6. MODELO DE DIABETES POR ESTREPTOZOTOCINA.(Figura 3).
El cuadro de diabetes mellitus se provocó mediante la inyección vía
intraperitoneal(ip) de una soluciónde STZ (Upjhon Farmoquimica,5. A., Madrid)
55

Población Días de vida Condición
0 2 5 15 20
N MMI(O•02p/v)N
T5
U------ 1~-i-14+T41 s
a----- - T5±14a
N
N
N
15 22 32 37
4
42
MMr<O.O2p/v) + T4
y — — — -
s
¡4
S
SS
0 65 72 82 87 92
N MMI(O•O2pIv) + T4 1
y — — -
a
a
a a
o(RP)
122±14 (R)
t= 14 (RF~)
o
14(R)14 (RP9
14 (RF~’0)
o
F¡g 2 Diseño experimentaldel estudioderehabilitacióncon T4 de ratas neonarales(N),destetadasCD) y adultas<‘A). Las abreviaru;asde las distintaspoblacionesson: 7’~+ T4 (7?), ratas neonatalestiroidectonilzadasy tratadascon 1.Sgg/lOOgdepesodeT4defonnadiaria vía IP; T~ + 714 (Rl’,), ratasneonata/esoperadasy tratadasconJ.Sgg/lOOgde pesode 714 <‘pc/id); 71,, + 71, (RL ratas destetadasoperadasytratadas con una inyecciónde 1. 75gg/lOOgde pesode T,,~ T,,±7% (Rl’), ¡-atasdestetadasoperadas, tratadascon 1. 7Spg/IOOgde pesode 714 (pellet) durante 5días;T,2 +71, (Rl’1<>), ratasdestetadasti¡-oidectomizadasy tratadascon 1. 7Sgg/]OOgdepesode T, (pc/leí) durante 10 días,~ T~, + 714 fl), ratas adultasoperadasa los72 díasy tratadas con una dosis ip de 1. ZSgg/lOOgde pesode 714 durante5 días;T~ + T, (Rl’), ratasadultastiroidectomizadasy ti-atadascon 1. 75ug/lOOgdepesode T
4 (pellet) durante 5 días,~Tr + 714 (RP
1~0, amínalesadultos tiro idectomizadosy tratadosdm-ante 10 días con 1 75pg/lOOgde pesodc 71, <‘pc/leQ; C animalescontrol; A, nacimiento;71, tiroidectomía;MMI, nwtimazoí;S, sacúficio.
(R>
D
A
¡4

uo,0~-o
Y5
~
00
oa
cdr5a)-D(>2censj-
U)
HoC’\JJCO
Lo
-
-Co
c~-Cfl
c,j
o.-z
‘~o
.c
O‘0
o‘o.—
oce---ce—
o-.-‘o
oo)1
-0
+0
Li
CA
+
o,
=1
=
E-E
Cf)o¡.4
0-2
1
o-(1)
-Cf>
‘u,
01
2
st’
r
o.e
jo8
-Co
Un
’—u
.R~
,ti~
u.
u~
Ntie
t-.~1’~~
OC
eou
e0
ti.e
uuC
O~
o-~
<-~
¼.
zze
uu
~~
O—
utiO
K
-CoCo
.2
z
H
o+
0o
(Ni
LO1<-
o,
21
Ch
E+
o,
I—E
00
o
z
CO21
-~e
CO
u~‘jo
-eO.
oO‘.5
-‘uu
eeOO‘.~
0
‘¿e
e‘joe
u~
-~.
j
ooee
—O
.OO
eO
tiy-.
u
t:e
e-.4
-‘.4O
-.—ti
~O
u~
ou
~-o
.4,
ee
u-e
.—e
~-,
o~
OO
e5-.
uO
.--.
o~O
eW
<O
.e
t~O
(-fl-=
ue
oe
=-~
-t
->O
u-~
-~
-~o<~,
o~
entE
z
—0o~0coo
oej
‘-n.r
r4)r(>0
oo
it-0oIt

Materialy métodos
en tampón citrato 0.05M pR 4.5. Se estudiaronanimalescontrolesy diabéticosde
las poblacionesneonataly adulta.
La adminstraciónde la STZ (7Omg/Kg peso)serealizó a los 10 díasde vida
enel casode las ratasneonatales,o a los 72 días de vida, en las ratasadultas;tres
días más tarde se comprobóel estadode diabetesde dichos animalesmediantela
determinaciónde la glucemiay, posteriormente,de la insulinemia. Los animales
fueron sacrificadosa los 20 y 87 días de vida, respectivamente.
2.6.1. Tratamientode animalesdiabéticoscon T4. (D + ‘[4, Figura 3).
Con el fin de observarel efecto de las hormonastiroideas en los animales
diabéticosrespectoal axis IGFs¡IGFBPs,se administró‘[4 a unaspoblacionesde ratas
diabéticasneonatalesy adultas.
El tratamientohormonalde los animalesdiabéticosneonatalesy adultos,que
se realizómediantela implantaciónde pellets, se inició en el casode los primerosa
los 15 días de vida, y en el de los segundos, a los 82 días de vida, y las dosis
administradasfueronde 1 .5¡íg/lOOgdepesoy 1.75~g/100gdepeso,respectivamente.
La duracióndel tratamientofue de 5 días,con lo que los animalessesacrificarona
los 20 y 87 díasde vida, respectivamente.
2.7. ESTUDIO DE LOS TRANSPORTADORES DE GLUCOSA (GLU’[s) EN
RATASNEONATALESHIPOTIROIDEAS. (Figura 3).
El estudio de los GLUTs durante el desarrollo de la rata se llevó a cabo en
poblacionesneonataleshipotiroideasy controles,desdelos 2 a los 20 díasde edad.
La pautaque se siguió comenzócon la administraciónde MMI (0.02%,p/v)
enel aguade bebidade las rataspreñadasa partir del día 14 de gestación;las crías
fueron sacrificadasa los 2 y 4 díasde vida (hipotiroideas-MMI).
Por otro lado, los animalesneonatalesde 8 y 20 díasde edad(T5), que se
incluyeron en este estudio, fueron sometidosa la misma pauta que las ratas
neonatalestiroidectomizadas,esto es, se inició la administracióndel fármaco
bociógenoa los 2 días de vida (0.02%,p/v), se tiroidectomizarona los 5 días de
edady se sacrificarona los 3 y 15 díasdespuésde la operación(8 y 20 díasde vida,
respectivamente).
58

Material y métodos
2.8. RECOGIDA DE MUESTRAS.
Los animales neonatales y adultos empleados en el estudio fueron sacrificados
entre las 09:00 y las 11:00 horas, aunque para las ratas hipotiroideastratadascon ‘[4
seestablecióla hora del sacrificioen las 11:00 horas.
Las muestrasde sangre(mezclaarterialy venosa)se obtuvierondel tronco
del animal tras la decapitación y se recogieron en tuboseppendorfcon y sin heparina
para obtener suero y plasma, respectivamente. Se dejaroncoagularen hielo durante
30 minutos, y a continuación fueron centrifugados 10 minutos a 12.OOOrpm
(Microfuge 11, Beckman); el suero y el plasma se trasvasaron a otra serie de tubos
eppendorf que se conservaron a -800C hasta su posterior análisis.
Las muestras de tejidos (hígado, hipófisis, corazón, cuadriceps y
gastroenemius)se extrajeron inmediatamentedespués de la decapitación, se
sumergieronen N2 líquido y se conservarona -80
0C hasta el momento de su
valoración.
2.9. DETERMINACION RADIOINMUNOLOGICA DE IGF-1.
2.9.1. Fundamentodel radioinmunoanálisis.
El RIA es un caso particular del radioanálisiscompetitivo basadoen una
reacciónde competiciónentreun ligando dadoque actúacomo antígeno(hormona,
Ag) y otro antígeno marcado (hormona’, Ag’) por la unión a un receptor específico,
su anticuerpo (Ac). En esquema:
Ag’ + Ac > Ag’ - Ac.
+ Ag > Ag - Ac.
Esta reacción de competición tiene lugar durante la incubación de ambos
antígenos, marcado y no marcado (frío), con el anticuerpo, produciéndosela unión
del antígeno marcado al anticuerpo en proporción inversa a la concentración de
antígenofrío quelo desplazade la unión, ya que el antígenofrío presentaunamayor
afinidadporel anticuerporespectoal antígenomarcado,estoes,el antígenofrío tiene
mayor actividad biológica. Así pues, el grado de competición, y por tanto la
concentracióndel ligando a valorar, puede conocersepor los cambios en la
concentracióndel antígenomarcadolibre o del complejoantígeno - anticuerpo.A
continuación,se procedea la separaciónde la radiactividad libre del complejo
inmunológico para valorar la radiactividad ligada, y por comparacióncon la
59

Materialy métodos
radiactividad ligada en presenciade cantidadesconocidasy crecientesde antígeno
(curva patrón), puede conocerse su concentración en una disolución problema.
Este análisis presenta la sensibilidad de las técnicas isotópicas y posee la
especificidadpropia de los métodos inmunológicos, lo que pennite valorar un
componenteen una muestrade grancomplejidadcon un mínimo de interferencias.
Para obtener resultados fiables se debe tener en cuenta las siguientes
condiciones:
1. Que el anticuerpo obtenidoseaespecíficodel antígeno<sustanciaproblema)
y que sus diluciones sean adecuadas.
2. Que el antígeno se pueda marcar con un isótopo radiactivo de manera que
se consiga una alta actividad específica sin una disminución de su actividad biológica.
3. Que el método de separación de la radiactividad libre de la ligada en forma
del (complejo Ag-Ac) sea adecuado.
2.9.2. Filtración en gel.
Esta técnica permite la separación de las IGFBPs del suero y constituye el
paso previo para la determinación de la concentraciónde IGF-I medianteRIA y de
IGF-II por radiorreceptor (RRA). Además, este procedimientofue mostradocomo
el idóneo por nuestro grupo de investigación frente a la extracción en ácido fórmico-
acetona y a la de ácido-etanol con crioprecipitación, sobre todo para el caso de
muestras procedentes de ratas neonatales (Rivero, 1994).
2.9.2.1. Fundamento.
La cromatografía de permeabilidad o de tamiz molecular separa las moléculas
en función de su tamaño, forma y peso molecular mediante el paso de las muestras
a través de una matriz de pequeñas esferas de material inerte que está englobada en
una columna.
Esta técnica constituye el método de elección para la eliminación de
interferentes indeseables de distinto peso molecular respecto a los problemas por
diferentes razones:
a) es la cromatografía más sencilla,
b) no requiere muestras muy puras,
c) no se presentan fenómenos de adsorción, lo que la hace ideal para muestras
biológicas lábiles,
d) permiteseparacionesrápidassin necesidadde unosgradientesde elución,
60

Materia/y métodos
e) produce bandas estrechas en el cromatograma y, finalmente,
O permite correlacionar fácilmente el peso molecular con el volumen de
elución.
Entre los parámetrosa teneren cuenta en este tipo de cromatografíase
encuentra el coeficiente de disponibilidad(Ka) de una molécula, dado que representa
la fracción de la fase estacionaria accesible para la difusión de dicha molécula. Se
expresa como:
y-ye O
ve—ve
donde:
Ve (volumen de elución): volumen necesario para eluir una molécula
determinada.
V0 (volumen vacío): volumen de la fase móvil en el que se distribuyen las
moléculas de peso molecular suficientemente elevado como para no penetrar por el
tamiz molecular.
V, (volumen total): volumen de la columna (fase móvil y estacionaria) en el
que se distribuyen las moléculas de peso molecular muy bajo.
El K~ de una molécula es independiente de las dimensiones de la columna y
se relaciona de forma constante con su coeficiente de difusión.
2.9.2.2. Material y reactivos.
Columna. Se utilizaron columnas de vidrio borosilicado XK 16/100
(PharmaciaLKB Biotechnology,Uppsala,Suecia)de 100cm de altura y lámm de
diámetro.
Fase móvil. Está formada por ácido acético 1M y cloruro sódico 0.02M en
agua; se emplea para el equilibrado de la columna y como eluyente.
Fase estacionaria. Se utiliza Sephadex G-50 Fine (Pharmacia Fine Chemicals,
Uppsala, Suecia). Posee un rango de fraccionamiento de 1.600 a 30.OOODa, adecuado
por tanto, para la separación de IGFs (7.áOODa) de sus proteínas ligadoras
(aproximadamente 50. OOODa).
2.9.2.3. Calibración de la columna,
Para determinar con exactitud en qué fracciones eluyen los IGFs y sus
61

Materialy métodos
proteínasligadorasunas vez separadas,se filtré una alícuotade IGF marcado
radiactivamente con 21, se recogieron fracciones de unos 2m1 y, a continuación, se
procedió a su contaje en un contador PackardAuto-GammaCobra II. En las tres
columnas empleadas el IGF eluyó con un K,~ de 0.22.
El perfil de elución del péptido marcado mostró claramente dos picos de
radiactividad de IGF, el primerocorrespondíaa las IGFBPsy el segundoalos IGFs.
2.9.2.4.Filtración de los sueros.
Se diluyó la muestrade sueroenuna proporción1:1 enácidoacético2M, de
forma que la concentración final de ácido en la muestra fuera 1M, y para disociar los
complejos IGF/IGFBP se mantuvo la mezcla durante una hora a temperatura
ambiente. A continuación, la muestra se filtró por la columna a un flujo de 4m1/h y
se utilizó como eluyente ácido acético XMy clomro sódico O.02M. Finalmente, las
fracciones correspondientes a IGF se reunierony se liofilizaron. El liofilizado se
resuspendió en tampón de RIA y se repartió en alícuotas que se conservaron a -200C
hasta su análisis.
2.9.3. Marcaje radiactivo de IGF-I y II.
2.9.3.1. Fundamento.
Se persiguela obtenciónde péptidoscon suficienteradiactividadespecífica
pero sin una actividad biológica disminuida, de forma que la cuantificación de IGF
en la muestra posea una adecuada sensibilidad.
Para el marcaje radiactivo de IGFs se utilizó una modificación del método de
la cloramina‘[ (Hunter, 1962) que es ampliamente utilizado para marcar pequeñas
cantidades de proteína con una alta radiactividad específica que se empleará como
radioligando en el radioanálisis competitivo. La reacción que tiene lugar es una
oxidacióndel Na’251 en presenciade unaproteínaquecontieneresiduosde tirosina,
con la subsiguiente incorporacióndel radioyodoa dichosresiduos.La cloraminaT
es la sal sódica del ligando N-monocloro de la p-toluenosulfonamida y en disolución
acuosa se degrada lentamente formando ácido hipocloroso que es un agente oxidante.
Así, el Nal en presencia de cloramina T y a pH ligeramente básico (pH 7.5> es
oxidado, se generan iones yodinio (I~) que reaccionan con el anillo fenólico de la
cadena lateral de la tirosina y que se incorporan en orto respecto al hidroxilo.
El proceso se detiene por dilución de la mezcla de reacción en presencia de
una alta concentración de ESA (albúmina sérica bovina), ello altera las condiciones
62

Materialy métodos
de reaccióny haceque se incorporeel 1251 al BSA, que por ser una proteínade alto
pesomoleculares fácilmenteseparabledel IGE. Finalmente,se separala proteína
marcada del yodo radioactivo no incorporado,así como de la albúminamarcada
mediantefiltración engel.
2.9.3.2.Material y reactivos.
Hormona.SeutilizaronrhIGF-I y rhIGF-II (IGF-I y II humanorecombinante)
de Boehringer Mannheim (Leverkusen, Alemania) que vienen liofilizados. Se
reconstituyeron en ácido acético 0. 1M y se liofilizó una alícuota. Estos péptidos
tambiénseemplearonen la elaboraciónde la curvapatrónenel ensayode RIA y en
el de RRA.
Na’251. Se presenta a una concentración de lOOmCi/ml y es suministrado por
Amersham(AmershamIbérica SA., Madrid).
Columna. Se emplearon columnas de Sephadex G-50 Fine equilibradas antes
del marcaje con PBS 0.03M (tampón fosfato), BSA al 0.25% (p/v) y azida sódica
0.02% (plv) para el marcaje de IGF-I, o con ácido acético 1My USAal 0.25% (plv)
en el caso del marcajedel IGF-II.
2.9.3.3. Procedimiento.
La reacción se desarrolló en el mismo tubo donde se liofilizó el péptido que
se resuspendió en PBS 0.5M (tampón fosfato). A continuación,se añadióel yodo
radiactivo (Na125I) y la disolución de cloramina T (lmg/ml en PUS 0.O1M) recién
preparada y se dejaron transcurrir 30 segundos para que se produjera la yodación del
péptido.
La reacción se detuvo por la adición de USAal 5% (p/v) en PBS 0.O1M para
el marcaje de IGF-I, y con USAal 5% (p/v) en ácido acético LM en el marcaje de
IGF-II. A continuación,se transfirió la mezcla de reacción a la columna y se
recogieron fracciones de imí a un flujo de unos 30m1/h. Se contó una alícuota de
cadafracciónenun contadory (PackardAuto-GammaCobraII) paraconservarlas
fracciones interesantes que se mantuvieron congeladas a -20W y divididas en
alícuotas -
2.9.3.4. Cálculo del rendimiento y la actividad específica.
El rendimiento es el porcentaje del total de radiactividad presente en el tubo
de reacción que se incorpora al péptido, y se calcula como el cociente entre las cpms
del pico de IGF-I ó II y las cpms totales eluidas de la columna. (Osciló entre 47-78%
63

Materialy métodos
para el IGF-I y 45-81% para el IGE-Il).
La radiactividad incorporada al péptidose obtieneal multiplicar el rendimiento
y la radiactividad añadida al tubo de reacción corregida a la fecha del marcaje.
Finalmente,el cocientede la radiactividadincorporaday la masadel polipéptidonos
permite conocer la actividad específica en ¡íCi/¡xg. (Osciló entre9O-160gCi/~gpara
IGF-I y 9O-l74~tCiI~gpara el IGF-II).
2.9.3.5.Perfil de elución de radiactividad.
El perfil obtenido por el método de la cloramina ‘[ muestra tres picos de
radiactividad:el primerocorrespondeal USA que se yoda al detenerla reacción,el
segundo, es la hormona marcada y, por último, el tercero, se identifica con el yodo
libre que no ha reaccionado ni con la hormona ni con el BSA (Figura 4).
2.9.3.6. Repurificación del IGF marcado.
El péptidomarcadoconservasu actividadbiológicadurantemásde dosmeses.
Sin embargo,el ligamiento inespecíficoenel RIA aumentaal transcurrirel tiempo,
aunque este inconveniente se evita repurificando ciertas fracciones del marcaje; de
esta manera, se consigue separar la hormona marcada íntegra de la deteriorada por
la radiactividady del yodo radiactivodesprendidodel péptido.
La repurificaciónseefectúaen las mismascondicionesque el marcajey su
perfil de elución tambiénmuestratres picos: el primero probablementese debea
polímerosde IGF-I, el segundo,que es el mayor, se correspondecon la hormona
marcaday, finalmente,en tercer lugar, eluyeel yodo libre, desprendidodel péptido
marcado.
2.9.4. Desarrollo del RíA de IGF-I.
2.9.4.1.Material y reactivos.
Patrón. Se empleó IGF-I humano recombinante (rhIGF-I) de Boehringer
Mannheim que viene liofilizado. Se obtiene de E. coli y presenta una pureza superior
al 95%. También se utiliza estahormonaen los marcajes.
‘[ampón. Está compuesto por tampón fosfato 0.O1M pH 7.5, cloruro sódico
O.15M, azida sódica al 0.02% (p/v) y BSA al 0.5% (p¡v). Se utilizó para diluir el
patrón, las muestras filtradas, el radioligando, el primer anticuerpo y también para
reconstituirlas muestrasfiltradas despuésde su liofilización.
Radioligando.Se empleó 1251-IGF-I que sediluyó con tampónde RíA para
obtenerde 8.000 a 10.OOOcpm/tubo.
64
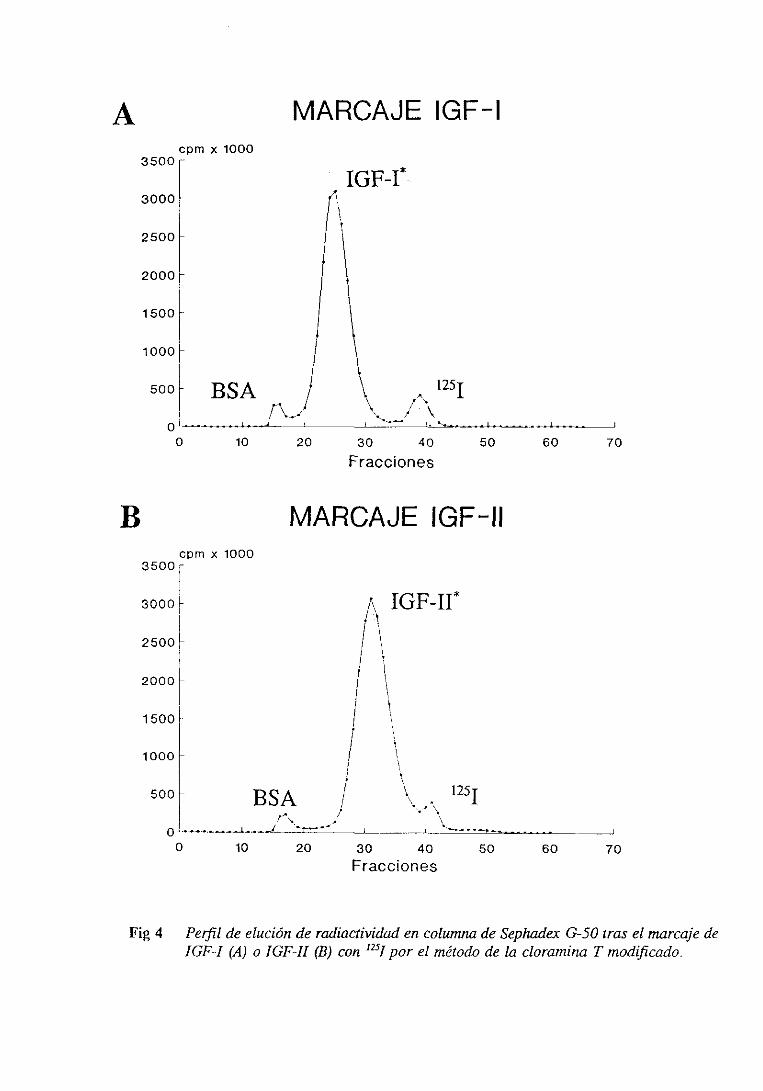
MARCAJE IGF-Icpm x 1000
o
epm x 1000
o
JGF~IJ*
1251
10 20 30 40 50 60 70
Fracc¡ones
Fig 4 Perfil de elución de radiactividaden columnade SephadexG-SO tras el marcajede
A3500
3000
2500
2000
1500
1000
500
o
RW-1 *
BSA 1251
B
10 20 30 40 50 60 70
Fracciones
MARCAJE IGF-II
3500
3000 -
2500 -
2000 -
1500-
1000 -
500 - PSAJA’
o
IGF-I (A) o IGF-II (B) con por el métodode la cloramnia 71 modificado.

Materialy métodos
Primeranticuerpo.Se utilizó un antisueropoliclonal (fracción globulínica)
anti-IGF-I obtenido en conejos inmunizados con hIGF-I y el fragmentoC-terminal
(residuos 5-57) de KabiGen AB (Estocolmo,Suecia)que sepresentaen forma de
liofilizado. La dilución empleada fue de 1:1.000.
Segundo anticuerpo. Se empleó una suspensión de complejos de anticuerpo
y microsefarosaobtenidosenovejay queprecipitaanticuerposde conejo(Pharmacia
DecantingSuspension3).
2.9.4.2.Preparaciónde las muestras.
Las muestrasde suerose analizarondespuésde ser sometidasa la filtración
en gel para separarlas proteínasligadorasde IGF. Se evaluarona unadilución 1:40.
2.9.4.3.Procedimiento.
La curvapatrónabarcaun rangode concentracionesde 100 a 02ng/mly se
analizópor duplicadoal igual que las muestras.
Parallevar a caboel análisis, se añadióel primer anticuerpoa los tubos de
las muestrasy a los patronesy, a continuación,el radioligando. Despuésde una
incubacióna 4W de 16-20h, se adicionó la suspensióndel segundoanticuerpo,se
volvieron a incubar durante 30 minutos ahora a temperatura ambiente y se les añadió
agua destilada (OSmí). Finalmente, los tubos secentrifugarondurante10 minutosa
1 .500g y 4”C, se descarté el sobrenadante y se procedió al contaje del precipitado
durante dos minutos en un contador de radiactividady (PackardAuto-GammaCobra
II).
2.9.4.4. Cálculos.
Para calcular la concentraciónde IGF-I en las muestrasse empleó un
programa en el que se construye la curva patrón según una representación
logarítmica,poniendoen abscisasel logaritmode la concentraciónde hormona,y en
ordenadas la función logarítmica de la proporción de ligamiento U/U0.
Esta proporción se calcula por la expresión:
E/Ej 8-
»
30—A’
donde, U es la radiactividad ligada de la muestrao patrón correspondiente,U0, la
media de la radiactividadde los tubosdondeel ligamientoesmáximo, y N, la media
66

Materialy métodos
de la radiactividad ligada a los tubos donde el ligamiento es inespecifico
La funciónlogarítmicaseexpresacomo:
logitR/B0=In B/B1—
Se calculó para cada muestralogit BIB0, se interpoló en la curvapatrón la
concentracióncorrespondientey se calcularonlas mediasde los duplicados.Por
último, el resultado se multiplica por el factor de dilución de cada caso.
2.10. DETERMINACIONRADIOINMUNOLOGICA DE 13 Y ‘[4.
La valoración de ‘[3 y ‘[4 en suero se realizó en el Centro de Investigaciones
Biomédicasdel CSIC, enel laboratoriode la Dra. GabrielaMorreale,mediantelos
RIAs específicosde alta sensibilidaddescritos por Weeke y Orskov (1975) y
modificadosparala ratapor Obregóny col. (1979).
Denuevo,damoslas graciasa la Dra GabrielaMorrealey a sus colaboradores
por la ayudaque nosprestaron.
2.10.1.Marcaje radiactivo cte ‘[3 y ‘[4.
El método se basa en una reacciónde oxidacióncon cloraminaT (Kjeld,
1975), dondeel sustrato(3’3 o ‘[2) se yoda con 1251 o 11311 a una yodotironinade un
grado superior de yodación. La reacciónse detienecon metabisulfito sódico y la
separaciónde las diferentesyodotironinas y el yoduro se lleva a cabo mediante
filtración en gel (SephadexG-25).
2.10.1.1.Material y reactivos.
Hormona. Se emplearon ‘[2 y ‘[3 reconstituidasenhidróxido sódico 0.05M,
de forma que se consiguierauna concentraciónfinal de íOpg/ml. Estos péptidos
tambiénseemplearonparaelaborarla curvapatrónde ¡os RIAs.
Na1251. Se presentacon una concentraciónde IOOmCí/ml y es suministrado
por Amersham(AmershamIbérica S.A., Madrid).
Columna. Se emplearon columnasde SephadexG-25 equilibradascon PUS
0.OSM pH 11.9.
2.10.1.2.Procedimiento.
FI procesose llevó a cabo a temperaturaambiente.A una alícuotade la
67

Materialy métodos
hormonase le añadióla solución de Na’251 y la de cloramina‘[ (4mg/ml en PUS
0.OSM pH 7.6). ‘[ranscurridos 20 segundosseparó la reacciónconuna disolución
de metabisulfito (2.Smg/ml en PBS 0.05M pH 7.6), se transfirió la mezcla de
reaccióna la columnay seeluyó con PBS 0.05M pH II» a un flujo de 40m1/h.
Se recogieronfraccionesde 2m1, y a cadauna deellasse les añadióuna gota
deácido clorhídrico 6N para queel pH final fuera neutro.A continuación,secontó
una alícuota de cadafracción en un contadorde radiactividad y (PackardAuto-
GammaCobra II). Los tubos de interés se protegieron con 2m1 de metanol y se
conservarona 4W.
2.10.1.3.Perfil de elución de radiactividad.
El perfil de elución tras el marcajeradiactivo de la hormonapresentótres
picos de radiactividad: el primero de ellos correspondióal yodo que no se ha
incorporadoal péptido,el segundo,a la T3 marcaday el tercero,a la 14 yodada.
2.10.2.Desarrollodel RíA dc ‘[3.
2.10.2.1.Material y reactivos.
Patrón.La hormonaempleadacomo standardfue la rT3 (T3 de rata)que se
resuspendióen hidróxido de sodio 0.05N para proporcionaruna concentraciónde
4~gIml.
‘[ampón. Seutilizó para la elaboraciónde la curva patrón y sirvió como base
parala preparaciónde otrostamponesdel análisis. Estáformadopor fosfatodisódico
0038M, fosfatosódico0.002M, mertiolato ft6mM y USA al 0.22% (tampónA).
En el ensayo,se empleótambiénestetampónsin albúmina(tampónC) y otro
quetienecomobaseal tampónA, peroqueademásincluye en su composiciónlOOmg
de cisteinay 600mgde ANSA (competidorde proteínas),esel tampón B.
Radioligando. Seutilizó 12511 quesediluyó en tampónU paraproporcionar
6.000-7.OOOcpm/tubo.
Primer anticuerpo. Se trata de un antisuero de conejo obtenido de Henning
Uerlin GMBH (Alemania). Se diluyó con tampón A para conseguir un título de
1:50.000.
Segundo anticuerpo. Para precipitar el complejo antígeno-anticuerpo, se
empleóunamezclaformadaporsuerode ternera,tampónC y polietilenglicolal 30%
diluido en tampónC en las proporciones:5:28:67.
68

Materialy métodos
2.10.2.2.Procedimiento.
La curva patrón abarcaun rango de concentracionesde O a Sng/ml y se
analizópor triplicado, al igual que las muestras.
En el análisis,paraigualarproteínas,se les añadióa los patrones,un volumen
desuerolibre de T3 (1O~l), y a las muestras,un volumende tampónA. Despuésse
adicionaron la hormona marcaday el primer anticuerpo. Tras una noche de
incubación,se les adicionóel segundoanticuerpo(1 <SmI), secentrifugóy seprocedió
al contajedel precipitado.
2.10.2.3.Cálculos.
Los niveles de T~ se conocieronpor la representacióndel logaritmo de la
concentraciónfrente a la relación BIT y la correccióndel valor por el factor de
dilución de las muestrasque fue 10.
*100cs
donde,C1 es la radiactividadde un tubo problema,Ch, la radiactividadde los blancos
y C~, la de los standardsde contaje.
2.10.3.Desarrollodel RIA dc ‘[4.
2.10.3.1.Material y reactivos.
Patrón. El standard empleado fue la rT4 (T4 de rata) que se disolvió en
hidróxido de sodio 0.OSN paraobtenerunaconcentraciónfinal de lOggIml.
Tampón. Está formado por fosfato sódico O.002M, fosfatodisódico0.038M,
mertiolato 0.6mM y USA al 0.05% (tampónA). Estetampón sin USA (tampónU)
se diluyó enunaproporción1:4 (tampónA: tampónU) y se empleóparaelaborarla
curva patróny diluir las muestras(tampónC). Así mismo,enel análisis se utilizó
otro tampónque se preparóa partir del tampónA con USA al 0.22%,y no 0.05%,
por la adición de lOOmg de cisteína/l y 600mg de ANSA!> (TampónD) y otro, a
partir del tampónC, al que sele añadieronlOOmg de cisteína/l (tampónE).
Radioligando. Se utilizó ‘251-T
4 diluida en tampón para conseguir 6.000-
7.Ooocpmltubo.
Primer anticuerpo. Se empleó una y-globulina de conejo a un título de
1:30000diluido en tampónC.
69

Material y métodos
Segundoanticuerpo.Consisteen unamezclade suerode ternera,tampónE
y polietilenglicol (PEG) al 30% diluido en tampónD (5:28:67).
2.10.3.2.Procedimiento.
La curva standardabarcaun rangode concentracionesde 0-12.Sng/mly, al
igual que los problemas,se analizópor triplicado. A los patronesseles añadióun
volumende standardy otro de suerolibre de ‘[4 (25~l), mientrasquelas muestrasse
diluyeron en tampón C en una proporción 1:33; a continuaciónse adicionaronel
radioligandoy el antisueroy se incubaronuna nocheenoscuridad.Al día siguiente,
tras la adición del segundoanticuerpo(1 SmI) y la centrifugaciónde los tubos, se
obtuvo un sedimento,cuya radiactividadse contó enun contador-y (PackardAuto-
GammaCobraII).
2.10.3.3.Cálculos.
La concentraciónde ‘1’4 se obtuvopor la interpolaciónen la rectaque resulta
al representarel logaritmode la concentraciónfrentea la relaciónU/T de los valores
obtenidostras el contaje.El factor de correcciónde los datosdebidoalas diluciones
fue 0.0667.
2.11. DETERMINACION RADIOINMIJNOLOGICA DE INSULINA.
La determinaciónde la insulina en muestrasde plasmase realizópor medio
de unoskits comercialesde la casaIncstar.
2.11.1.Material y reactivos.
Patrón.Estáconstituidopor insulinaporcinaliofilizada prediluidaentampón.
‘[ampón. Es un liofilizado compuestoporBSA, tampónboratoy que incluye
timerosalcomo conservante.
Radioligando.Se utilizó insulinaporcinamarcadacon 1251 y diluidaenBSA-
tampónboratoque ademáscontieneEDTA (ácidodianiinotetracético)y timerosal.
Antisuero.Se empleóun liofilizado de antisuerode insulinade cobayadiluido
enel tampóndel ensayo.
Complejo precipitante(GAGP-PPT).Estáformadopor un liofilizado quecontieneen su composiciónsuerode cobayanormal, precipitadocon antisuerode
cobayamachoy PEG. Al igual queen el casode los otros reactivos,se resuspendió
en aguadestiladaen el momentoen que se realizó el ensayo.
70

Material y métodos
2.11.2.Procedimento.
El primer día se preparó la curva standard, que abarca un rango de
concentracionesde 4 a lOOgU/ml (0.16-4ng/ml) y las muestras.A continuación,se
adicionóa todos los tubos primeroel antisuero(exceptoa los tubos que contenianla
radiactividadtotal -T- y a los del ligamiento inespecífico-NSB-) después,la insulina
marcada y se incubaron a 4W durante 16-20 horas. Se añadió el complejo
precipitantea todos los tubos exceptoa los T, semantuvieronunos15-20 minutosa
20-25W y transcurridoeste tiempo se centrifugarona 760g y 20-25W durante20
minutos.Porúltimo, secontóelprecipitadoen un contadory (PackardAuto-Gamma
CobraII) duranteun minuto.
2.11.3.Cálculos.
Al igual que en los otros RíAs descritos,en estecaso, se llegó a conocerla
concentración de insulina mediante la representacióndel logaritmo de la
concentración (abscisas) frente a la relación U/U0 (ordenadas) y la posterior
interpolaciónenestagráfica de los valoresde las muestras.
2.12. DETERMINACION RADIOINMUNOLOGICA DE HORMONA DE
CRECIMIENTO (GH).
2.12.1.Preparacionde las hipófisis parael RíA.
Para llevar a cabo el análisis de los niveles de GH en las hipófisis, sedebe
homogenizarpreviamenteel tejido. La homogenizaciónde las hipófisis se realizó
manualmentecon ayuda de un potter de vidrio en PUS 0.O1M pH 7.6 y el
homogenadoresultantede estaoperaciónse conservóa -20W. Las muestrasse
diluyeronen PUS 0.O1M pH 7.6 con USA al 1% en las proporcionesdeterminadas
por tanteoque ahorase detallanen funciónde la edady condicióndel animal:
- ratasneonatales:hipotiroideas,1:100y 1:250 y controles,1:250;
- ratasdestetadas:hipotiroideas,1:500 y 1:1.000y controles, 1:1.000;
- ratasadultas:hipotiroideas,1:5.000y 1:10.000y controles, 1:10.000.
2.12.2. Marcajeradiactivode GH.
El marcajeradiactivode la hormonade crecimientose llevó a cabosiguiendo
el método de la cloraminaT (Hunter, 1962). Como ya se explicó en el marcaje
radiactivode IGF-I, la reacciónquímicaque tienelugares la oxidacióndel Na’251 en
presenciade unaproteínaque contienerestostirosina, y es en estos residuosdonde
71

Materialy métodos
se incorpora el radioyodo. El procesose detienepor la adición a la mezcla de
metabisulfito,y a continuación,seseparala proteínamarcadadel yodoradiactivono
incorporadomediantefiltración engel (Sephadex0-50).
2.12.2.1.Material y reactivos.
Hormona:SeempleórGH (Gil de rata),queal igual quetodos los reactivos
(marcaje y RIA), son suministradospor el National Institute of Diabetes and
Digestiveand Kidney Diseases(NIDDK, California, USA). La rGH sereconstituyó
en NaHCO30.01N(lOOgg/ml>y serepartióen alicuotasde 25g1 que seconservaron
a -20W. Estepéptido tambiénse utilizó paraelaborarla curvapatrónen el MA.
Na125I: Se presentaconuna concentraciónde lOOmCi/ml y es suministrado
por Amersham(AmershamIbérica, SA, Madrid).
Columna:Seemplearoncolumnasde ScphadexG-50equilibradasconveronal
0.02M.
2. 12.2.2.Procedimiento.
El procesose llevó a cabo a temperaturaambiente.Así, a una alícuotade la
hormona,se le añadióla soluciónde Na’251 y la soluciónde cloraminaT (3 .Smglml
en PUS 0.05M) reciénpreparaday seagitó el tubo durante90 segundos,momento
enel que se produjo la yodacióndel péptido.La reacciónse detuvopor la adición
de una disolución de metabisulfito (1 .Ymg¡ml en PBS O.05M) que se preparó
extemporáneamente.La mezclade reacciónse transfirió a la columnapreviamente
equilibradacon veronal, tampónque seempleó paraeluir las fraccionesa un flujo
aproximadode 30m1/h. Se recogieronfraccionesde 0.5m1,sobreOlmí de BSA al
2% en PBS0.OIN, y de estasse contóunaalícuotaen un contadorde radiactividad
-y (PackardAuto-GammaCobraII). Las fraccionesde interésseconservarona -20W
en PBS-BSAal 2% (dilución 1:5>.
2.12.2.3.Cálculo del rendimientoy la actividadespecífica.
El cálculo del rendimiento y de la actividad específicase llevó a cabo
siguiendolo ya descritoparael marcajeradiactivode IGFs. El rendimientoobtenido
en los marcajesde Gil osciló entre un 85-95%, y la actividad específicade la
hormonase situó en torno a 75-S0~Ci/pg.
2.12.2.4.Perfil de elución de radiactividadtrasel marcajeradiactivode
OH.
El perfil de elución obtenidotrasel marcajeradiactivode la hormonapresenta
72

Materialy métodos
dospicos de radiactividad:el primero de ellos correspondea la hormonamarcada,
y el segundoal yodo libre que no ha llegadoa incorporarseal péptido.
2.12.2.5.Repurificaciónde la GH marcada.
La Gil marcadaconservasuactividadduranteunosdosmeses,sin embargo,
con el tiempo se produceun aumentodel ligamiento inespecíficoen el RIA. No
obstante,este inconvenientese superarepurificandolas fraccionesresultantesdel
marcaje.
La repurificación se realizóen las mismascondicionesen que se efectuóla
purificacióntras el marcaje,pero se empleóen este casouna columnade Sephadex
G-100y se recogieronfraccionesde mayorvolumen(2.5m1). Con estaoperaciónse
consiguió la separaciónde la hormona intacta (segundopico) de la deteriorada
(primerpico) y del yodo radiactivo(tercerpico) que liberael péptido conel pasodel
tiempo.
2.12.3.Desarrollodel RíA de GH.
2.12.3.1.Material y reactivos.
Patrón.La hormonautilizada como standardfue rGIvI que se presentóen
formade liofilizado y fue proporcionadaporel NIDDK, al igual que el primer y el
segundoanticuerpo.Seresuspendióen aguaparaproporcionarunaconcentraciónde
lOpg/pl.
Tampón.EstáconstituidoporPUS 0.O1M pH 76, EDTA O.025My USA al
1% (plv) y se empleó para diluir las muestras,la radiactividad,el primer y el
segundoanticuerpo.
Radioliganclo. Se utilizó ‘251-GH que se diluyó en el tampón del RIA para
proporcionarunas8.OOOcpm/tubo.
Primer anticuerpo. Se presentó como un liofilizado que sereconstituyó en
agua(dilución 1:50>. Se trata de un antisueroextraídode glándulapituitaria de rata
con aguay sulfato de amonio. Se diluyó en suerode mono al 1 % a un título de
1:500.000.
Segundo anticuerpo. Se utilizó una gamma-globulina antimono que fue
proporcionadapor el NIDDK por Antibodies, Inc. (California, USA). Precipitael
sueronormal de mono y estádisuelto en PUS.
2.12.3.2.Procedimiento.
El primerdíaseprepararonlas muestrasy la curva patrón,que se analizópor
-73

Materialy métodos
triplicado y abarcóun rango de concentracionesde 0.03 a l6ng/ml. Las muestrasse
analizaronporduplicado,y en el casode las hipófisis, a las diluciones reseñadas.
Los problemasy los patronesseincubaroncon los reactivos: se les añadió
la dilución del antisuero y el tampón, y se mantuvieron 6 horas a 37”C. A
continuación,se adicionóa los tubosel radioligandoy seincubaron18 horasa370C.
Al día siguiente,se añadióel segundoanticuerpoy se incubarondurante18 a 24
horasa 40C. Finalmente,el tercerdía, se lavaroncon 2m1 de PUS O.O1M pH 7.6,
se centrifugaron40 minutos a 760g, y se procedióal contajede radiactividaddel
precipitadodurante2 minutosen el contador-y (PackardAuto-GammaCobraII>.
2.12.3.3.Cálculos.
Como enel casode los otros RIAs, los valoresde Gil se llegarona conocer
por la representacióndel logaritmode la concentración(abscisas)frentea la relación
U/U0 (ordenadas) y la posterior corrección por el factor de dilución de las muestras;
en el caso de las hipófisis, los cálculosserefirieron a mg de tejido.
Destacamosque las determinacionesde GH fueron realizadas con la
colaboraciónde la Dra. CarmenAlvarez Escolá, miembro de nuestrogrupo de
investigación,y a quienreitero mi agradecimientoporsu ayuda.
2.13.DETERMINACION DELA CONCENTRACION SERICA DE IGF-II POR
ANALISIS DE RADIORECEPTOR (RRA).
2.13.1. Fundamento.
Al igual que el RIA, el RRA es un caso particular del
competitivo. La competiciónse estableceentreuna hormona(IGE—IL)
misma hormonasin marcarpor la unión a un receptorespecífico.
Los receptoresse obtuvierona partir de hígadode rata adulta
receptoresde IGF tipo II, y prácticamentecarecede receptorestipo 1.
2.13.2. Preparación de membrana plasmática particulada de hígado de
rata.
Se homogeneizaronlos hígadosde rataenunasoluciónde sacarosa0.25Mcon
PMSF 0.SmM en una proporción de 3.5mlIg de tejido. Se reunieron los
homogenadosy se filtraron a travésde una gasa. A continuación,se centrifugaron
durante10 minutos a 600g y 4W, se recogieron los sobrenadantesy de nuevo se
radioanálisis
marcaday la
que contiene
74

Materialy métodos
centrifugarona 17.OOOgdurante30 minutosa 40C. Los sobrenadantesqueresultaron
sereunierony seles adicionócloruro sódicoy sulfatode magnesiohastaalcanzaruna
concentraciónfinal de 0.1 y 1M, respectivamente.Despuésse llevaron a cabo dos
centrifugacionesa 40.OOOgdurante40 minutosy a 4W, y el sedimentoobtenidose
resuspendióy homogeneizóentampónTris 5OmM PH 7.7. Porúltimo, seseparóuna
alícuotade la preparaciónpara determinarla concentraciónde proteínassegúnel
métodode Uradford. Estapreparaciónse conservóa -80”C repartidaen alícuotas.
2.13.3.Desarrollodel RRA de IGF-II.
2.13.3.1.Materialy reactivos.
Patrón. Se empleó rhIGF-II (IGF-II humano recombinante) de Uoehringer
Mannheimque sepresentaen forma de liofilizado. Se obtiene de E. coli, presenta
una purezasuperioral 95% y seutilizó en los marcajes.
Tampón. Se empleó el tampón del RIA (PUS 0.O1M pH 7.5 con cloruro
sódico 0.15M, azidasádica0.02%y USA 0.5%). Se utilizó paradiluir el patróny
reconstituirlos liofilizados de las muestrasde suerosometidasa la filtración engel.
‘[ampón ‘[ris-HCl. Incluye en su composición Tris base 0.05M y USA al
0.5% (p/v) y su pH es 7.7. Seempleóparadiluir la preparaciónde membranasy el
radioligando,así como parael lavado de los tubos.
Radioligando.Se empleó‘251-IGF-II, marcadoporel métodode la cloramina
T (Hunter, 1962)y sediluyó en tampón‘[ris-HCí paraobtenerunas10.OOOcpm/tubo.
Membrana.Se utilizarondiluidasaunaconcentraciónde lmg proteína/mlen
tampónTris-HCI.
2.13.3.2.Procedimiento.
Este método se llevó a cabo en dos días. El primer día se prepararonlas
muestras,la curva patrón y se incubaron los reactivos, y en el segundodía, se
procedióa la separaciónde la radiactividadlibre y al contajede los tubos.
La curva patróncomprendeun rango de concentracionesde 100 a 0.2ng/ml
y, al igual que las muestras,se analizópor triplicado. A los tuboscon las muestras
y los patronesse les añadiósucesivamentela dilución del radioligandoy la de las
membranasy se incubaron20 horas a 40C. Al día siguiente, se centrifugaron10
minutosa 12.OOOrpmy 4W y seaspiróel sobrenadante.Se lavó el precipitadocon
tampón Tris-HCI frío (200~l), y de nuevo se centrifugaronlos tubos durante 5
minutosa 4W y l2.OOOrpm. Finalmente,secontó el precipitadoen un contadorde
75

Materialy métodos
radiactividad-y (PackardAuto-GammaCobraII).
2.13.3.3.Cálculos.
La concentraciónde IGE-lí en las muestrasse calculó por el procedimiento
usadoenel MA de IGF-I.
‘[ECNICAS ELECTROFORETICAS.
Bajo esteepígrafesereunenunaseriede técnicasbasadasen lamigraciónque
experimentanlas especiesiónicas en un solvente(en un medio líquido) por la acción
de un campoeléctrico.
v~g.E
donde, y1 es la velocidadlineal de migración, ji, la movilidadiónica y, finalmente,
E, el campoeléctricocontinuo.
Así pues, los diferentes iones de una muestrase desplazanbajo el efecto
continuodel campoeléctrico, haciael cátodoo el ánodosegúnel signo de su carga
y a una velocidadpropia de cadauno.
Parael análisisde las muestrasse empleansoportesinertes(gelesde agarosa,
poliacrilamida...)que posibilitan:
a) la utilización de pequeñascantidadesde muestras(del ordende jil) y su
clara visualización;
b) separaciónde los componentesde la muestracon gran resolución;
c) y, tiemposde análisiscortos.
2.14. ANALISIS DE LAS PROTEINAS LIGADORAS DE LGF: WES’[ERN
BLOT.
2.14.1.Fundamento.
En el ‘Western blot”, en primer lugar se lleva a cabouna electroforesisen
condicionesno reductoraspara no desnaturalizarlas proteínas(transportadorasde
glucosa o de IGEs), para separarlas distintas IGFBPs, y a continuación,dichas
proteínassetransfierenaunamembrana(de PVDF o nitrocelulosa)que se incubará
conanticuerposespecíficoscontradeterminadasIGFBPs(‘inmunoblot’) o con 1251
IGF-I ó II (“ligand blot”, Hossenlopp,1986).
2.14.2.Electroforesis.
Se realizó con el sistemaMini ProteanII de Bio Rad(Richmond,California).
76

Materialy métodos
2.14.2.1.Preparación del gel.
El gel de poliacrilamida tiene dos zonas diferentes: una breve con una
concentraciónde acrilamidadel 5% llamadagel acumulador(almea las muestras
aplicadas)y otra que contieneun 12% de acrilamidaque separalas proteínas(gel
separador)que correna sutravés.La composicióndel gel separadores la siguiente:
agua,‘[ris-HCI 0.375M pH 8.8, SDS 0.1% (p/v), acrilamida/bis40%, APS 0.05%
(p/v) y TEMED. El gel acumuladorincluye en su composiciónlos mismosreactivos
que el separador, sólo difieren en el porcentaje de poliacrilamida y en la
concentracióndel Tris-HCI empleado,0. 125M pH 6.8 en el gel acumuladorfrente
a 0.375M pH 8.8. enel gel separador.
2.14.2.2.Preparaciónde lasmuestras.
Las muestrasse prepararonmomentosantesde su aplicación. Se diluyeron
1:10 (Westernligand blot e “inmunoblot”) (Figura5) entampónLSB 2x (Laemmli
samplebufffer, Laemmli, 1970>queestáconstituidoporTris-ilCí 0.0625MpH 6.8,
glicerol 10% (y/y), SDS 2% (p/v) y como coloranteazul de bromofenol 0.005%
(p/v). A continuación,se calentaron5 minutos a 95W, y se dejó que tomaranla
temperaturaambienteantesde su aplicación.
Comomarcadorde pesomolecularseempleóun patrónde amplio rango(6.5-
200kDa)de BioRad.
Finalmente,seaplicaronal gel las muestrasy el patrón.
2. 14.2.3.Condicionesde electroforesis.
La electroforesisse efectuóen el senode unadisolución fonnadapor glicina
192mM, Tris base 2SmM, SDS 0.1% (p/v) en aguadestiladabajo una corriente
continuade 125 V que seprolongó hastaque el frente de tinción alcanzóel borde
inferior del gel.
2.14.3.Electrotransferencia.
La transferenciaelectroforéticase realizó en el sistema Mini-trans-blot de
BioRady se utilizó como soporteparala transferenciaunamembranade nitrocelulosa
de 0.2gm de tamañode poro.
Una vezconcluidala electroforesisseextrajeronlos gelesy se sumergieron
enel tampónde transferencia(Tris base2SmM, glicina 192mM y metanol20% (p/v)
en aguaa pH 8.3). A continuación,cadage] sepusoen contactocon la membrana
de nitrocelulosay se situaronentredos láminas de papelWhatmamm3MM y dos
77
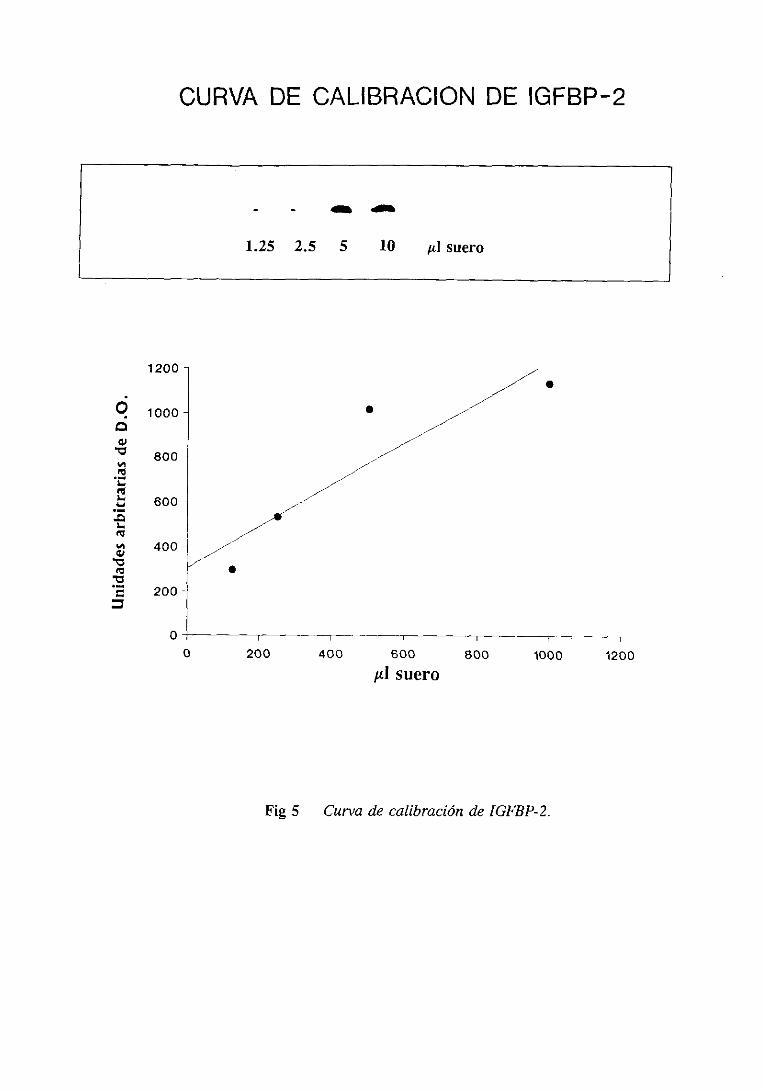
CURVA DE CALIBRACION DE IGFBP-2
- ea
1.25 2.5 5 10 ji1 suero
1200-
9
OcJ
y,
a-a-4-,
a-
y,
=
1000-
800 -
600 -
400 -
200 -
0-o
e
e
e
200 400 600
gI suero800 1000 1200
Fig 5 Curva de calibraciónde IGFBP-2.

Materialy métodos
almohadillas.Estemontaje se colocósobreun soporteparagelesque se introdujo en
la cubetade transferencia.El procesode transferenciase efectuóa 100 V durante2
horasenel tampónya señalado.
2.14.4.Western ligand blot.
2.14.4.1.Reactivos.
Los tratamientosposterioresde la membranase realizaronen un tampón
(“ligand blot buffer”, LUU) que estáformadopor ‘[ris-HCl 0.O1M pH 7.55, cloruro
sódico 0. 1SM, azidasódica0.1% (p/v) en agua. Estadisolución fue la basepara la
preparaciónde otras:
1. Solucionesparala preparaciónde la membrana:
a) Soluciónde Nonidet-P40 3% (y/y) en LBB.
b) Soluciónde USA al 1% (p/v) en LUB.
e) Solución de Tween-20al 0.1% (y/y) en LBB.
2. Solución para la incubación de la membrana. En ella se diluyó el
radioligandoy estuvoconstituidaporBSA al 1 ¼(p/v) y Tween-20al 0. 1 ¼(y/y> en
LBB.
3. Solucionesparael lavadode la membrana.
a) Solución de Tween-20al 0.1% en LBU.
b) LBB.
2. 14.4.2.Preparación de la membrana.
Antesde incubarla membranacon el radioligandosesometió a una seriede
lavadoscon los que se bloquearonsus sitios de unión libre por la exposicióna una
alta concentraciónde BSA. Así pues, la membranase lavó en primer lugar con
Nonidet-P40 a temperatuaambiente durante 30 minutos y ligera agitación, a
continuacióncon una solución de BSA durante2 horas, y finalmente, con una
disoluciónde ‘[ween-20 (10 minutos).
2. 14.4.3.Incubacióncon el radioligando.
Una vezfinalizadoslos lavados,la membranaseincubósin agitacióncon una
alícuotade unasoluciónde radioligandopreparadamomentosantes(1.5.1O~ cpm 1251
IGF-I ~ II> durante18-20horasa 4W.
2.14.4.4.Lavado y detecciónde las IGFBPs.
Finalizadala incubación,se procedióal lavadode la membranacon ligera
agitaciónparaeliminarel excesode radioligandono incorporado,primerocon LUB
79

Materialy métodos
y Tween-20(dosveces)y despuéscon LUB (3 veces)durante15 minutosa 4W. A
continuación,y una vez secala membrana,sepusoen contactocon una películay
entrepantallasintensificadorasa -800C, procediéndosedespuésa su revelado(Agfa
Curex 60).
La integraciónpor volumen de las bandasobtenidaspor el Densitómetro
Personalde Molecular Dynamics(Sunnyvale,California)se expresócomounidades
arbitrariasde la densidadóptica.
2.14.5.Western ¡nmunoblot.
2.14.5.1.Reactivos.
El posteriorprocesamientode la membranase llevó a caboen tampónTUS
(“‘[ris-buffered salme’)formadoporTris-HCI 0.05M PH 7.4 y cloruro sódico0.2M
en agua destilada.Esta disolución constituyó la base para la preparaciónde los
siguientestampones:
1. ‘[135-’[ween: TampónTUS con Tween-20al 0.75% (y/y).
2. Soluciónbloqueante:TUS-Tweenconlecheenpolvo desnatadaal5% (p/v).
Su preparaciónes extemporánea.
3. Soluciónde preincubación: TUS con 0.OSmg/ml de USA.
4. Anticuerpo primario: Se trata de un anticuerpopoliclonal obtenido en
conejos inmunizadoscontra bIGFBP-2 (de células de riñón). Se empleó al título
recomendadopor el proveedor(UBI, Inc., NY, USA), 1:2000.
5. Anticuerposecundario:Se obtiene en cabrasinmunizadascon anticuerpo
de conejo y es una Inmunoglobulina-G(U +L) conjugadacon fosfatasaalcalina
(BioRad).
6. Solucionesde revelado:
a) TampónTris-MgCI2constituidoporTris-ilCí 0. IM pH 9.5 y MgCI2
0.5mM en aguadestilada.
b) SoluciónNUT: 3Omg/ml enN,N-dimetilformamidaal 70% (y/y, en
aguadestilada).
c) SoluciónBCIP: l5mg/ml en N,N-dimetilformamida.
Las proporcionesutilizadas fueron: 100 ‘[ris-MgCI2: 1 solución NUT: 1
soluciónBCIP.
2.14.5.2.Preparaciónde la membrana.
Al igual que sucedeenel ‘ligand blot”, se debenbloquearlos sitios deunión
80

Material y métodos
libresde la membranaantesde incubaríaconel primeranticuerpo;esto se consiguió
exponiéndolaa una alta concentraciónde proteína,presenteen la lecheen polvo.
Todos los lavadose incubacionesse realizarona temperaturaambientey con
ligera agitación.Así, en primerlugar selavó la membranacon TBS-Tweendurante
5 minutos, para despuéssustituir estetampón por solución bloqueantedonde se
mantuvola membranadurante30 minutosy, finalmente,sehicierondos lavadosde
5 minutoscon ‘[US-’[ween.
2.14.5.3.Incubación con el primer anticuerpo.
Una vez finalizadoslos lavados,se incubó la membranadurante2 horascon
unaalícuotade soluciónde preincubacióndondeestabadiluido elanticuerpoprimario
al título deseadoy TUS-’[ween. Transcurridoel tiempo de incubación,se lavó la
membranauna vez en soluciónbloqueantey tres en TBS-Tween(5 minutos cada
lavado).
2.14.5.4.Incubación con el anticuerpo secundario.
Se incubóahorala membranacon unaalícuotadel segundoanticuerpoy TUS-
‘[ween duranteun tiempo que osciló entre 1 y 3 horas. A continuación,se lavó la
membranatres vecescon ‘[BS-’[ween y doscon TUS (5 minutos cadalavado) y se
secóentrepapelesde filtro.
2. 14.5.5.Detecciónde las IGFBPs.
El revelado se basa en una reacción química catalizadapor la fosfatasa
alcalinaconjugadaal anticuerposecundarioquegeneraun productoinsolubledecolor
púrpurasobrela membrana(Uers, 1985).
La membrana se sumergió en la solución de revelado durante el tiempo que
seconsideróconvenientesegúnel fondoy señaldeseados,y paradetenerla reacción
se mantuvoen un recipientecon aguadestiladadurante10 minutos.Finalmente,la
membranasedejó secarentrepapelesde filtro.
2.14.6. Western inmunoblot.
2.14.6.1.Reactivos.
Los tratamientosposterioresde la membranase realizaronen un tampón
(TUS) constituido porTris-HG lOmM pI-1 8 y cloruro sódico I5OmM. Estetampón
sirvió de baseparaotrasdisolucionesempleadasen el análisis:
1. Soluciónde bloqueo: TUS con lecheen polvo desnatadaal 5% y ‘[ween al
0.05%.
81

Material y métodos
2. Soluciónde bloqueo: Estáconstituidapor TUS y ‘[ween al 0.05%.
3. Anticuerposprimarios: Se trata de anticuerpospoliclonales(SantaCruz
Biotechnology,Inc., USA) obtenidosen carnerosinmunizadoscontrapéptidosque
correspondena los aminoácidos254-272del extremoC-terminaldel precursorde la
IGFBP-1 de ratóno a los aminoácidos288-305del mismoextremodel precursorde
la IGFBP-2 de ratón. Seemplearona un título de 1:100 diluidos enuna alícuotade
soluciónde bloqueo.
4. Anticuerpo secundario: Se trata de un complejo formado por una
Inmunoglobulina-G-peroxidasa(SantaCruz Uiotechnology)quesediluye en solución
de bloqueoa un titulo de 1:1.000.
En estos experimentos,se empleó como marcadorde peso molecularun
patrónpreteñidode amplio rango (4-25OkDa)de Novex.
2.14.6.2.Preparaciónde la membrana.
Parabloquearlos sitios de unión libre de la membrana,estafue tratadacon
una soluciónrica enproteínaslácteasdurante45 minutos,a temperaturaambientey
con ligera agitación.
2.14.6.3.Incubacióncon el anticuerpoprimario.
La incubaciónde la membranacon el correspondienteanticuerpose realizó
duranteuna nochea 40C y con ligera agitación.Despuéssesumergióla membrana
en una solucióncompuestapor ‘[US y Tweenal 0.05%y seefectuarontres lavados
de 5 minutos, a temperaturaambientey con agitación.
2. 14.6.4. Incubacióncon el anticuerposecundario.
La membranase puso en contactocon el segundoanticuerpoa temperatura
ambientedurante60 minutos, con ligera agitación. A continuación,serealizaron4
lavadosigualesa los anteriores,de 5 minutosde duración, a temperaturaambiente
y con agitación; los 3 primeros lavadosse efectuaroncon TUS y Tween al 0.05%,
y el último sólo con TUS.
2.14.6.5.Detecciónde las IGFBPs.
La visualización de las IGFUPs se logró mediante la técnica de
quimioluminiscencia.La peroxidasaligada al segundoanticuerpoen presenciadel
peróxido de hidrógenooxida al luminol; en este proceso se generauna molécula
intermedia emisorade luz que es capaz de impresionaruna película. La señal
obtenidaes proporcionala la cantidadde complejo imnunológicoquehay sobre la
82

Materialy métodos
membrana,esto es, a la cantidadde proteína(IGFBP-1, 2 o como veremosmás
adelante,GLU’[-1, 2 ó 4) previamentetransferidaa esta.
La membranasetratóconlos reactivosde unkit suministradoporAmersham
(ECL), y la intensidadde las bandasobtenidasse cuantificó en un Densitómetro
Personal(MolecularDynamics).
2.15. ANALISIS DE LOS TRANSPORTADORES DE GLUCOSA (GLUTs):
WES’[ERN INMUNOBLO’[.
El procedimientoseguidofue la técnicadel ‘Western blot”, cuyos detallesse
han descrito en el apartado anterior. En este caso, las condiciones para la
electroforesisy electrotransferencia,al igual que la composiciónde los geles de
poliacrilamida,fueronlas mismasqueallí sedescribieron.No obstante,cabedestacar
varios aspectosque fueron distintosenestecaso:
a) Las muestrasse pusieronen contactocon el LSB a temperaturaambiente
durante30 minutos,no se calentaron.
b) La electrotransferenciano se efectuósobreunamembranade nitrocelulosa
sino sobre una de difluoruro de polivinilideno (Immobilon, Millipore) que
previamentehabíasido sumergidaen metanol(30s)y, a continuación,enagua(60s>.
c) El marcadorde pesomolecularutilizado fue la fumarasa(48.500Da)por
su similitud con el pesomolecularde los GLUTs (Sigma).
2.15.1.Preparación de las muestras.
2.15.1.1.Reactivos.
La extracciónde las membranasse llevó a cabo en distintos tamponesen
funcióndel tejido que se tratara.
1. Tampónde homogeneizaciónde los tejidos muscularesy corazón: Está
constituidoporHEPES2SnM, sacarosa25OmM, EDTA 4mM, benzamidina2SmM,
PMSF 0.2mM, inhibidor de tripsina IU/ml y pepstatina lgM a pH 7.4. Esta
disolución tambiénse empleópara la resuspensiónde estostejidos.
2. Tampónde homogeneizacióndel hígado:EstáformadoporHEPES25mM,
sacarosa2SOmM, EDTA ImM, DTT 3mM, PMSF 0. ImM e inhibidor de tripsina
0.26U/ml a pH 7.4.
3. Tampónde resuspensióndel hígado:Se componede Tris SOmM, sacarosa
25OmM, cloruropotásico lOOmM y cloruro de magnesio5mM a pI-! 7.4.
83

Material y métodos
2.15.1.2.Extracción de las membranas de tejidos muscularesy corazon.
El procesoseguidopara obtenerla fracción membranosacruda (membranas
globales) fue descrito por Santalucíay col. (1992). Los tejidos (0.3-0.5g) se
homogeneizaronen 10 volúmenesde tampónde homogeneizacióny se centrifugaron
a 40C y 15.OOOg,durante20 minutosen el caso de los músculosesqueléticosy a
10.OOOg, 5 minutos cuando se trató de músculo cardíaco. Los sobrenadantes
obtenidosseenriquecieronenclorurode potasiohastaobteneruna concentraciónde
0.8M. Se incubaron 30 minutos a 40C con agitación y, a continuación, se
centrifugaron: el extracto cardíacoa 150.OOOg, durante2 horas, y los extractos
procedentesdel gastrocnemiusy el cuadricepsa 200.OOOg, 1.5 horas.El sedimento
obtenido se resuspendióen el tampón empleadopara la homogeneización,y tras
determinarla concentraciónproteica del extracto se almacenóa -800C hasta el
momentode suanálisis.
2.15.1.3.Extracciónde las membranashepáticas.
El método aplicadoparaextraer la fracciónmembranosafue una adaptación
del descritoporThorensy col. (1988).Separtióde unos0.2gde tejido, que unavez
homogeneizadosen 10 volúmenesde tampón, se centrifugarona 40C y 8.OOOg
durante10 minutos.El sobrenadantese volvió a centrifugardurante20 minutosa la
misma velocidad y el nuevo sobrenadantese centrifugó a 150.OOOg durante40
minutos. El sedimento obtenido se resuspendióen un tampón distinto al de
homogeneizacióny finalmenteseanalizósu concentraciónproteicay se almacenóa -
800C.
2.15.2.Westerninmunoblot. (Figura 6)
2.15.2.1.Reactivos.
1. Soluciónde bloqueo: PUS (NaCí 0.135M,KCl 2.7mM, KH2PO4 1.4mM,
Na2HPO48mM, PH 7.4) con leche en polvo desnatadaal 5%. Su preparaciónes
extemporánea.
2. Anticuerposprimarios: Se trata de anticuerpospoliclonales(Uiogénesis)
obtenidosen conejosinmunizadoscontra:
- un péptido que incluye los 12 aminoácidosdel extremo C-terminal del
transportador(anti-GLU’[-4);
- un péptidosintéticode secuenciabasadaen la previstadel GLU’[ delcerebro
de la rata(anti-GLUT-1);
84
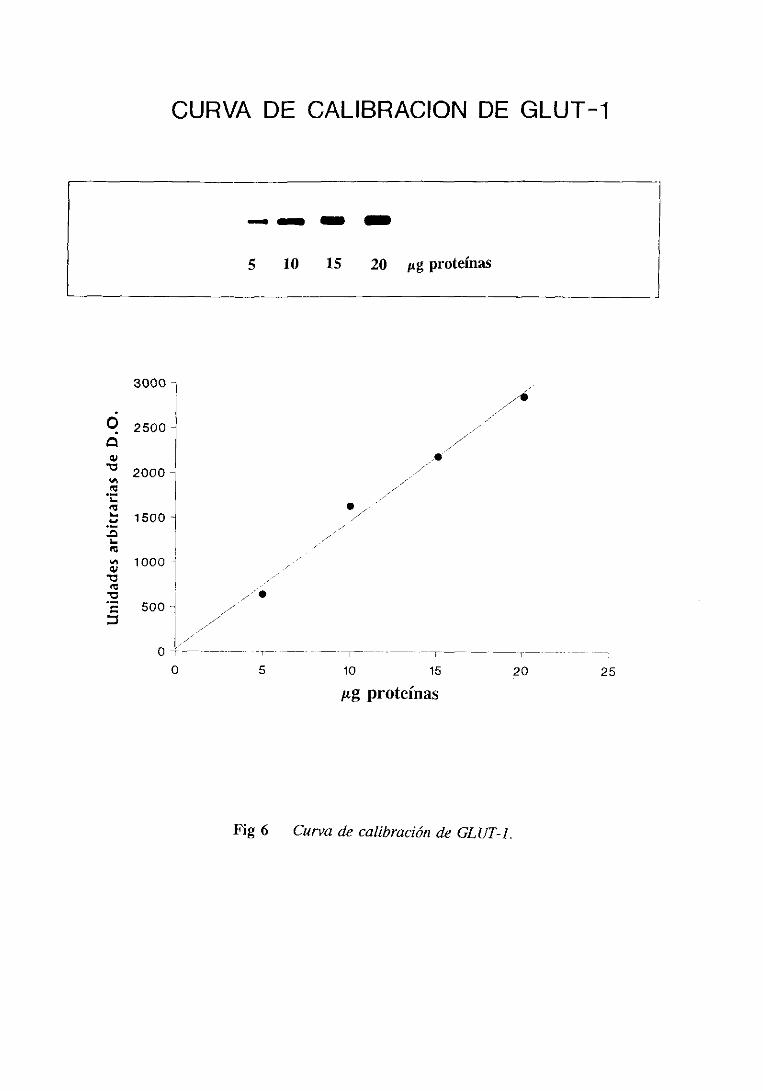
CURVA DE GALIBRACION DE GLUT-1
3000 -~
xx.
o
s7
e—,
.7
5 10 15 20 25
gg proteínas
oo4,
jA
a-a-4-
.0
y,4,
2500 -
2000 -
1500 —
1000 -
500-
Fig 6 Curva de calibración de GLUT-1.

Materialy métodos
- un péptido sintético que incluye los 25 aminoácidosC-terminalesde la
secuenciadel GLUT-2 de rata y otro péptido que contiene los 13 aminoácidos
terminalesdel mismo tramo.
Se utilizaron a un titulo de 1:1.000parael GLU’[-2 y 4 y de 1:5.000 para
GLUT-1 diluidos en PUS con USA al 3% y azidasódicaal 0.02%.
3. Soluciónde lavado: PUS conTweenal 0.1%.
4. Anticuerpo secundario: Consiste en un complejo formado por
Inmunoglobulina-G-peroxidasa(Sigma)que se diluyó en PUS con leche desnatadaal
1% y’[ween al 0.05%.
2.15.2.2.Preparaciónde la membrana.
Despuésde la electrotransferenciase procedióal bloqueode los sitios de
ligadura inespecíficade la membranamedianteel tratamientocon una soluciónrica
enproteínaslácteasdurante2 horas,con ligera agitacióny a temperaturaambiente.
2.15.2.3.Incubacióncon el anticuerpoprimario.
Se incubó la membranaconel anticuerpocorrespondienteduranteuna noche
a 40C y conuna ligera agitación.A continuación,sesumergióla membranaen una
solución de PUS con Tween y se hicieron 4 lavadosde 10 minutos a 370C y con
agitación.
2. 15.2.4. Incubacióncon el anticuerposecundario.
Sepusoencontactoahorala membranaconel segundoanticuerpodurante60
minutos, a temperaturaambientey con ligera agitación.Transcurridoel tiempo de
incubaciónserealizaron4 lavadosigualesa los anteriores(10 minutos,37W y con
agitación).
2.15.2.5.Detecciónde los GLU’[s.
Paravisualizar los GLUTs se utilizó la técnicade quimioluminicencia,y la
cuantificacióndel volumende la manchase realizóen un densitómetro(Molecular
Dynamics).
Destacamosque la puesta a punto del método de determinaciónde los
transportadoresde glucosa fue realizadacon la colaboracióndel Dr. Fernando
EscriváPons,pertenecientea nuestrogrupode investigación,a quienagradecemos
de nuevo su ayudadesinteresada.
86

Materialy métodos
2.16. ANÁLISIS DE EXPRESION DE IGFs E IGFBPs: ENSAYO DEPROTECCION DE RIBONUCLEASAS.
2.16.1. Extracción del ARN total.
2.16.1.1.Preparación de las muestras.
El ARN total se preparóporhomogeneizaciónde los tejidosen tiocianatode
guanidinasegúnel métododescritoporChomczynskiy Sacchi(1987). Separtió así
de una pequeñacantidadde hígadoque se homogeneizóenuna disoluciónformada
porun 45% de soluciónD (tiocianatode guanidina4M, citrato sódico25miM pH 8
y lauril sarcosina0.5%),50% de fenol, 4% de acetatosódico2M y un 10% de CIA
(cloroformoy alcohol isoamílico en una proporción24:1). El ARN así obtenidose
purificó medianteuna segundaextracción,se resuspendióen EDTA 0.5mM y, por
último, seprocedióa su valoracion.
1. Valoracióndel ARN
.
a) Cuantitativa
.
El ARN total secuantificócomparandola absorbanciade la muestracon un
blanco(ED’[A 0.5mM y agua)a tres longitudesde ondadiferentes:260nm(máximo
de absorcióndel ADN y ARN), 2SOnm (máximo de absorciónde las proteínas)y
3 lOnm (máximo de absorciónde impurezas).Estaoperaciónnospermitió conocerla
concentraciónde la muestra (medida de la absorbanciaa 260nm), eliminar las
interferenciasdebidasa las impurezasy tambiénestimarla calidaddel ARN extraído
medianteel cocientede la absorbanciaa 260 y 280mn,de maneraque las muestras
cuya relación 260/280no oscilabaentre 1.7 y 2.2 fueron desechadas.
b) Cualitativa: Electroforesis
.
Las muestrasse sometierona una electroforesisengel de agarosa,lo que nos
permitió confirmar la integridaddel ARN y normalizarsu cantidaden las distintas
calles.
- Preparacióndel gel: Paraprepararel gel se hirvió la agarosadisueltaen la
solucióntamponadora(Mops 2OmM, acetatode sodio 5mM y ED’[A lmM) enuna
proporcióndel 1.1%. A continuación,se le adicionóformaldehído(15%) y seesperó
hastasu compactación.
- Preparaciónde las muestras:Las muestrasseprepararonmomentosantes
de su aplicación. Se tomóel volumennecesarioparaaplicar una cantidadde ARN
determinaday se igualaronvolúmenescon agua.A continuación,se le adicionóun
87

Material y métodos
volumende tampóndesnaturalizante(67% formamida,13% de formaldehídoy 20%
de Mops 0.2M, acetatode sodio 5OmM y EDTA lOmM) y un 4% de bromurode
etidio parateñir las bandasde ARN ribosomal 18 y 28 s. Finalmente,secalentaron
las muestrasdurante5 minutosa65Wy seles añadióun 10%deazuldebromofenol
(colorantede la electroforesis).
- Condicionesde la electroforesis:Paralaelectroforesisseempleóel sistema
SubcellElectrophoresisCelí de BioRad (Richmond,CA) y se empleócomo tampón
electrodo Mops (Mops 2OmM, acetato de sodio SmM, EDTA lmM). La
electroforesisseefectuó concorrientecontinuaa 50 V durante4 horas.
2.16.2.Ensayode protecciónde ribonucleasas.
2.16.2.1.Fundamento.
El análisissebasaen la hibridaciónen solucióndeuna ribosondaconel ARN
de unamuestra,la digestiónde los fragmentosno hibridadosy laposteriorseparación
de esos híbridos en geles de poliacrilamida. Para ello, se parte de un ADNc
plasmídicodonde se encuentrael inserto correspondientea los diferentesIGFs e
IGFBPs.Así, enpresenciade una ARN polimerasa,y a partir del promotordel ADN
plasmídico,se inicia la síntesisde una cadenade ARN marcadaradiactivamentecon32P-U’[P (ribosonda) que va a ser complementariaa la cadena de ADN. A
continuación,se adicionala ribosondaa la muestrade ARN dondehibridarácon los
tránscritosde ARN complementarios.Finalmente, los fragmentosde ARN no
hibridadossondegradadospor las ARNasasy los híbridos (ARN-ARN) seseparan
en gelesde poliacrilamida.
2.16.2.2.Preparaciónde lasmuestras.
Despuésde comprobarla integridaddel ARN y normalizadasucantidadpara
los distintos gruposexperimentales,se prepararonlas muestraspara su análisis.
La cantidadde ARN total hepáticoutilizado paralos diferentesestudiosfue
de 20jtg; se igualaron los volúmenesde las muestrascon aguaDEPC y sedejaron
precipitandoen 0,1 volúmenesde acetatosódico3M y 2.5 volúmenesde etanolpuro
a -20W hastael momentode su análisis.
2.16.2.3.Síntesisde la ribosonda.
2.16.2.3.1.Reactivos.
1. ARNasaA (50u/mg):enzimaextraídade páncreasporcinoy proporcionada
porBoehringerMannheim(Leverkusen,Alemania).
88

Materialy mÉtodos
2. ProteinasaK (20u/mg): se trata de una endopeptidasaobtenida del
Tritiriachum album (BoehringerMannheim).
3. BcoRI (lOu/gl): enzimade restricciónservidaporBoehringerMannheim.
4. Hind III (lOu/¡xl): enzimade restricción obtenidade Boebringer Mannheim.
5. SistemaribosondaGemini III Care: seempleóparagenerarlas sondasde
ARN (Promega, Madison, WI, USA) e incluía los componentesnecesariospara la
reacción de transcripción, como son: ‘[SC (tampón de transcripción), DT’[
(antioxidante),ARNsin (inhibidor de ARNasas),dN’[Ps (nucleótidosno marcados:
dA’[P, dC’[P, dGTP y dU’[P), ARN polimerasas(‘[7 y SP6).
6. ADNasa1 (lOu/gl): enzimaextraídade páncreasporcino, libre de ARNasas
que fue adquiridade UoehringerMannheim.
7. 32P-UTP(lOmCi/ml): nucleótidomarcado(Nuclear Ibérica, Madrid).
2.16.2.3.2.Obtencióndel plásmido.
1. Crecimientode las bacterias
.
Las bacteriasse sembraronenun medio constituidoporagary ampicilina, lo
que nospermitió el aislamientode las cepasde E. coli resistentesa esteantibiótico,
ya que sonestaslas que presentaronel plásmidoque incluía el ADNc de los IGFs y
las IGFBPs.
Una vez aisladas las bacterias, se cultivó una colonia en un caldo LU
(triptona, cloruro sódicoy extractode levadura)con ampicilina (SOgg/ml) a 37W
durante8 horas.
2. Aislamientodel plósmido
.
Paraextraerel ADNc plasmídicose rompió la paredbacterianamediantela
adición de una pequeñacantidadde lisozima, y la membranaplasmática,con ayuda
de la soluciónde lisis (hidróxido sódico2.4mM y SDS 0.12%)y se precipitaronlas
proteínasy el ADN cromosómicocon acetatopotásico 1.SM pH 4.5 y el ADN
plasmidicocon isopropanol.EsteADN plasmídicoasíobtenidose purificó mediante
la adición de ARNasaA (lOmg/ml) y proteinasaK (250gg/ml)que elimininaronel
ARN y las proteínas,respectivamente.Tras la adición de PMSF, se añadiófenol a
la mezcla y se reprecipitó el ADN plasmídicocon acetatosódico 0. I5mM y un
0.38% de etanol.
El ADN plasmídicose resuspendióen TE pH 8 (Tris base lOmM y EDTA
0.lmM) y se procedióa su valoración.
89

Material y métodos
3. Valoración del ADN
.
a) Cuantitativa
.
Al igual queenel casode la valoracióndel ARN, la concentracióndel ADN
secuantificópor la medida de las absorbanciasde la muestra(ADN y agua) frente
a un blanco(TE y agua> a las tres longitudesde ondaya mencionadas:260, 280 y
3lOnm. ‘[ambién, como en el casodel ARN ya mencionado,semostróuna especial
atenciónal cocienteresultantede la relación260/280.
b) Cualitativa: Electroforesis
.
Las muestrasdeADN plasmídicosesometieronaunaelectroforesisengel de
agarosapara confirmar su integridad.
- Preparacióndel gel: Sehirvió unaproporciónde agarosadel 1 ¼en solución
tampón(TAE pH 7.2, Tris base1.6M, acetatosódico0.8M, EDTA 4OmM) y cuando
la mezclaestuvotransparente,sedejó enfriarunosminutosy sevirtió enel soporte.
- Preparaciónde las muestras.Se tomóunapequeñacantidadde muestra(1-
1.5~¿l), se diluyó en el tampón de electroforesis(1:5) y se le adicionó un 10% de
colorante(azulde bromofenol).
En todoslos gelesseincluyó ademásun standard(LambdaDNA-Hind III) que
nos ayudóa conocerla concentraciónde cadabandapor su intensidady su peso
molecular.Estestandardfue tratadode la misma maneraque las muestras.
- Condicionesde la electroforesis: Parala electroforesisseempleóel sistema
Mini-Subcell ElectrophoresisCelí de BioRad (Richmond, CA) y se utilizó como
tampónde electrodoTAE con bromuro de etidio al 0.075%. La electroforesisse
realizócon corrientecontinuaa 70 y duranteuna hora.
2.16.2.3.3.Linearizacióndel plásmido.
Una vez comprobadala integridaddel ADNc se procedió a linearizar el
insertoen el que se encuentrael vectorqueexpresalos IGFs y las IGFUPs. Esto se
consiguióincubandouna cantidadde ADNc plasmidico(2OO~g-1mg)con la enzima
de restriccióny su tampónespecíficospara cadainserto(Tabla 2):
90

Material y métodos
Tabla 2.- Enzimasde restricción.
ADNe INSERTO ENZIMA DE RESTRICCION TAMPON
IGF-I pGEM-3 Hind III B
TOP-II pGEM-3 Hind III B
IOFBP-1 pGEM-3 Hiod III B
IGFBP-2 pOEM-4Z Hiud III B
IGFBP-3 pOEM-4Z EcoRí H
La incubacióncon la enzimase realizó a 37W durante8 horas y en las
proporciones:83 agua: 6 ADN: 10 buffer RE: 1 RE. A continuación,y una vez
terminada la digestión se procedió a purificar el plásmido mediante repetidas
extraccionescon fenol y porúltimo, por precipitacióncon 0.83% de etanol puroy
acetatosódico0. l2mM.
El ADNc plasmídicose resuspendióen TE y se valoró para comprobarsu
correctalinearización.En estecaso,el gel mostróuna únicabandacorrespondiente
al ADN línearizadoy e! standardLambda DNA-fUnd III (49O~g/ml, Promega)nos
permitió comprobarque su peso molecular correspondíaefectivamentea dicho
inserto.
2.16.2.3.4.Marcajede la ribosonda.
Estaetapaincluye dos partes:reacciónde transcripción(síntesisde la sonda
de ARN) y reacciónde la ADNasa (destruccióndel ADNc plasmídico).
1. Reacciónde transcripción
.
Las ribosondasse construyeronsegúnlas técnicasdescritasporLowe y col.
(1987). Segeneraronlas sondasmarcadaspor la incubacióndel plásmidolinearizado
(200-400ng/pl)con 32P-UTP(a) y con los demásnucleótidosfríos (dA’[P, dGTP y
dCTP) en presenciade la ARN polimerasa adecuada(Tabla 3) y de estamanera, se
construyeronribosondasantisentido(complementariascon el ADN).
91

Materialy métodos
Tabla 3.- ARN polimerasas.
RIBOSONDA ARN POLIMERASA TAMAÑO FRAGMENTO PROTEGIDO
IGF-I T7 IGF-Th 224bp y Ib 386bp
¡GE-II T7 500bp
IGFBP-I SP6 300 y lOObp
IGEBP-2 SP6 550bp
IGFBP-3 T7 343bp
2. Reacciónde la ARNasa
.
Se procedióa la eliminacióndel ADN mediantela digestióncon la ADNasa
1 y a continuación,a la purificacióndel ARN pormedio de extraccionescon fenol
y una primeraprecipitacióncon acetatosódico0.O6mM y 70% de etanol. Después
se volvió a precipitarcon acetatosódico0.O4mM y un 98% de etanoly seprocedió
al contajede unapequeñaalícuota(igl) en uncontadorde centelleo(1209Rackbeta
LKU Wallac).
2.16.2.3.5.Hibridación en solución.
2.16.2.3.5.1.Reactivos.
1. ARNasa T1 (l0O.OOOu/ml): enzima obtenida de Aspergillus oryzoe y
proporcionadaporUoehringerMannheim.
2. ARNt: seobtienea partir de la levadurade la cervezay ayudaa precipitar
el ARNm (UoehringerMannheirn).
2.16.2.3.5.2.Hibridación de la ribosonda.
El ARN total hepático(2Opg)obtenidode los diferentesgruposexperimentales
fue hibridadocon 500.OOOcpmde la ribosondamarcadadurante18 horasa 45W en
una solución que conteníaformamida al 75%, Tris SOmM, EDTA 4mM, cloruro
sódico 1.6M y SDS al 0.4%.
2.16.2.3.5.3.Digestión de la ARNasa.
La digestiónde los fragmentosde ARN no hibridados se realizó con 4Opg/ml
de ARNasaA y 2kg/ml de ARNasa‘[~ (Lowe, 1987; Hernández,1990)duranteuna
hora a 37”C. A continuación, se purificaron los fragmentosprotegidos, previa
eliminación de las ARNasascon ayuda de proteinasaK (0.2pg/ml), mediantela
extracción con fenol y cloroformo seguida de una precipitación con etanol.
Finalmente,los híbridosfueron sometidosa la electroforesis.
92

Material ymétodos
2.16.2.3.5.4.Electroforesis.
Los híbridosprotegidos(ARN-ARN) se sometierona electroforesisen gel
desnaturalizantey se empleóel sistemaSequi-genII SequencingCelí de BioRad.
1. Preparacióndel gel
.
La polimerización del gel seprodujo en el aparato, y se llevó a cabo por la
adición a una soluciónformadapor poliacrilamida8%, urea 8M y ‘[BE 5% (Tris
base 1M y ED’[A disódico 2OmM), un 0.037% de TEMED y APS 0.l3mM. El
espesordel gel fue de 0.2 mm.
2. Preparaciónde las muestras
.
Las muestrasfueron tratadasconun tampón que conteníaun colorante de
electroforesisconstituidopor xileno cianol al 1% y azul de bromofenolal 1 %. A
continuación,secalentaron3 minutosa 95Wy se enfriaronen hielo.
En todos los geles se incluyeron, ademásele las muestras,carriles para la
ribosondano tratadaconARNasas(controlnegativo)y parala ribosondadigeridacon
ARNasas(control positivo).
3. Condicionesde la electroforesis
.
Se efectuóen tampón de electrodoTBE (Tris base 1M, ácido bórico 1M,
ED’[A disódico2OmM) a 40 W hastaque el frentede tinción del colorantealcanzó
la parteinferior del gel.
2.16.2.3.5.5.Cuantificación densitométrica.
La autorradiografía serealizó a -800C poniendo en contacto el gel con una
película.Las bandas,querepresentanlos fragmentosprotegidos,secuantificaronpor
densitometría(Molecular Dynamics).
Destacamosy agradecemosde nuevo la colaboracióndel Dr. Luis Goya
Suárez,compañerode nuestrogrupo de investigación,por su ayudaen la puestaa
punto del análisisde protecciónde la RNasa.
2.17. OTRAS DETERMINACIONES ANALíTICAS.
2.17.1.Análisis de la glucemia.
2.17.1.1.Glucosaen sangre.
La determinaciónse realizó en un analizadorReflolux II M (Boebringer
Mannbeim); setratade un aparatoportátil frecuentementeutilizado por las personas
93

Materialy métodos
diabéticaspara el autocontrolde su glucemia. Es un fotómetro de refexión cuyo
funcionamientosebasaen la coloraciónde una tira de plásticoencuyoextremohay
una zona impregnadade reactivo. Dicha zona se divide en dos mitades que se
colorean con distinta intensidady son leídaspor un sistemade doblehaz, lo que
permite un amplio intervalo de medición (10-SOOmg/lOOml). El tiempo total
empleadoen la determinaciónfue de 2 minutos.
2.17.1.2.Glucosaen sangredesproteinizada.
La desproteinizaciónde la sangre(30¡íl) se efectuó conuna mezclaa partes
iguales(150M1) de sulfato de cinc e hidróxido de bario (mezclade Somogyi). A
continuación,se centrifugó durante5 minutos a 13.OOOrpmy se analizóla glucosa
porel métodode la glucosaoxidasa(GOD) en sOgl del sobrenadante.
Este análisis se basa en que la glucosa-oxidasacataliza la oxidaciónde la
glucosaa gluconato,reacciónque se acoplaa una oxidaciónde un cromógenoque
generaun compuestocoloreadopor la acciónde la peroxidasa(POD):
GOD
a) Glucosa + 02 + 1120 > Gluconato+ 11202
POD
b) 11202 + Cromógeno > Compuestocoloreado+ 1120
La medidade la absorbanciaa 420nm(máximodel compuestocoloreado)de
las muestrasse comparócon la ofrecida por una solución patrón de glucosa de
9. lmg/ml (UoehringerMannheim), lo que permitió el cálculo de la concentración
sanguíneade glucosasegúnla expresión:
AproblGma x 100= mg glucosa/loo ml sangreApatr6n
2.17.2.Determinación de las proteínas.
Las proteínasde los diferentesextractoscrudosde membranasedeterminaron
porel métodode Uradford(1976),utilizandounasoluciónde lmg/ml de ‘y- globulina
comostandard(Sigma).Se trata de un ensayocolorimétricobasadoen el cambiode
absorbanciaque experimentauna soluciónácidade CoomassieUrillant Ulue G-250
94

Materialy métodos
(465nm) cuando se pone en contacto con proteínas (595nm).
Se preparó una curva patrón a partir de la solución standard que abarcó desde
O-4Omg/ml. A continuación, a los tubos de la curva patrón y a los problemas se les
añadió imí de reactivo de coloración (BioRad Potein Assay) previamentediluido
(1:5) y filtrado. Por último, tras 5 minutos de incubación en la oscuridad, se procedió
a la lectura de los tubos en el espectrofotómetro a 595nm.
La concentración proteica de los extractos de membrana se conoció por la
interpolación de los valores de sus absorbancias sobre la recta de la curva patrón.
2.18. CÁLCULOSESTADISTICOS.
Los resultados se expresan como media + desviación standard (D.S.).
- > (~1 3~)2 n.s
.
n-1
El número de datos de cada población (u) se detalla en cada experimento
concreto.
En todos los casos, de manera previa al análisis estadístico pertinente, se
aplicó el test de la X2 para rechazar aquellos datos que se hallaban sensiblemente
alejados de la media y que no se incluían dentro del intervalo
x2z(1 x)~
donde X2 se determinó en las tablas correspondientes en función del número de datos
que quedaban fuera del intervalo definido.
2.18.1.Análisis de la varianza.
El grado de significación estadística de la diferencia entre las medias de dos
poblaciones se determinó mediante el análisis de la varianza (ANOVA> de una vía
(muestras independientes) o de dos vías (muestras apareadas). Este análisis consiste
en comparar las varianzas S~2 (variabilidad de los datos dentro de cada muestra
debida sólo al azar) y ~E2 (variabilidad entre las muestras debida al azar y, si las
95

Materialy métodos
mediasson distintas, a que las muestrasprocedende poblacionesdistintas) por el
procedimiento de la F de Snedecor(Snedecor,1956). Cuando el ANOVA es
significativo o se busca qué media son igualesy cuálesdistintas, se comparala
cantidadexperimental
texp= ‘321 1
$1, (— + —)n. n
1 .1
con una ta(f;k) con f= número de datos totales - número de grupos y k= número
de comparaciones a analizar. Esta cantidad tcx se determina en las tablas
correspondientes según el test a aplicar, que depende a su vez del objetivo y la
planificacióndel experimento.Una diferenciaseconsiderósignificativa a partir de
p<O.05.
2.18.2.Regresiónlineal y correlacion.
El análisis de regresión lineal se llevó a cabo por el método de los mínimos
cuadrados. Para verificar si la correlación obtenida era significativa se comparó el
valor experimental (texp) con una kv de n-2 grados de libertad:
texp= (n-2>r21 -r2
donder es el coeficientede correlacióny n el númerode parejasde datos.
Se consideróquehabíasignificacióna partir de p <0.05.
96

RESULTADOS

Resultados
3. RESULTADOS.
3.1. ‘[IROIDECTOMIA (‘[)Y TRATAMIENTO CON METIMAZOL (MMI) EN
PERIODO NEONATAL.
3.1.1. Evolución del pesocorporal.
Seestudiael pesocorporalde los animalesneonataleshipotiroideosde 10, 15
y 20 díasde vida, tanto de los tiroidectomizadosquirúrgicamente(5, 10 y 15 días
después de la operación) como de los tratados con MMI.
La Tabla 4 muestra el esperado incremento del peso de los animales en
relación a su edad, tanto en la población control como en las hipotiroideas
(tiroidectomizaday tratadacon MMI), aunque siempreel peso corporal estuvo
disminuido respectoa las ratas control en ambaspoblacioneshipotiroideas. No
obstante,cabedestacarqueestadisminuciónen el pesode los animaleshipotiroideos
fue más acusada en el caso de la población tiroidectomizada (a los 10 días de vida fue
de un 9.28% y, a los 20 días de vida, de un 40.19%) que en la tratada con MMI (a
los 10 días de vida fue de un 0.47% y, a los 20 días de vida, de un 17.45%) respecto
a los controles,sin embargo,en amboscasos,a los 10 díasde vida, la diferencia
encontrada respecto a la población control estaba ya disminuida.
3.1.2. Niveles séricosde ‘[~ y ‘[4 y plasmáticose hipotisariosde GH.
De un modo análogo a lo que sucedíacon el pesocorporal, las hormonas
tiroideas circulantes disminuyeron en ambos grupos de animales neonatales
hipotiroideosrespectoal control(Figura7) y ya, a los 10 díasde vida (5 díasdespués
de la operación),se encontróuna diferenciaentre los niveles de ‘[3 y ‘[4 de las
poblacioneshipotiroideas(tiroidectomizaday tratadacon MMI) respectoa la control.
La disminuciónen las cifras de ‘[3 a estaedad(10 díasde vida) fue de un 55.29%
paralas ratastratadascon MMI y de un 70.64%paralas tiroidectomizadasrespecto
a los animalescontrol, mientrasque en el casode la 14, el descensoen los valores
circulantesfue de un 92.77% y un 91.19% para aquellas ratas a las que se le
administró MMI y operadas, respectivamente, siempre respecto a los valores control
a los 10 días de vida; por consiguiente,los animalestratadoscon MMI también
mostraron unos niveles de hormonas tiroideas muy disminuidos respecto a los de la
poblacióncontrol,aunquelos nivelesde descensode T3 no fuerontanbajos como en
las tiroidectomizadas.
En los animales tiroidectomizados, de forma inesperada, los niveles
98
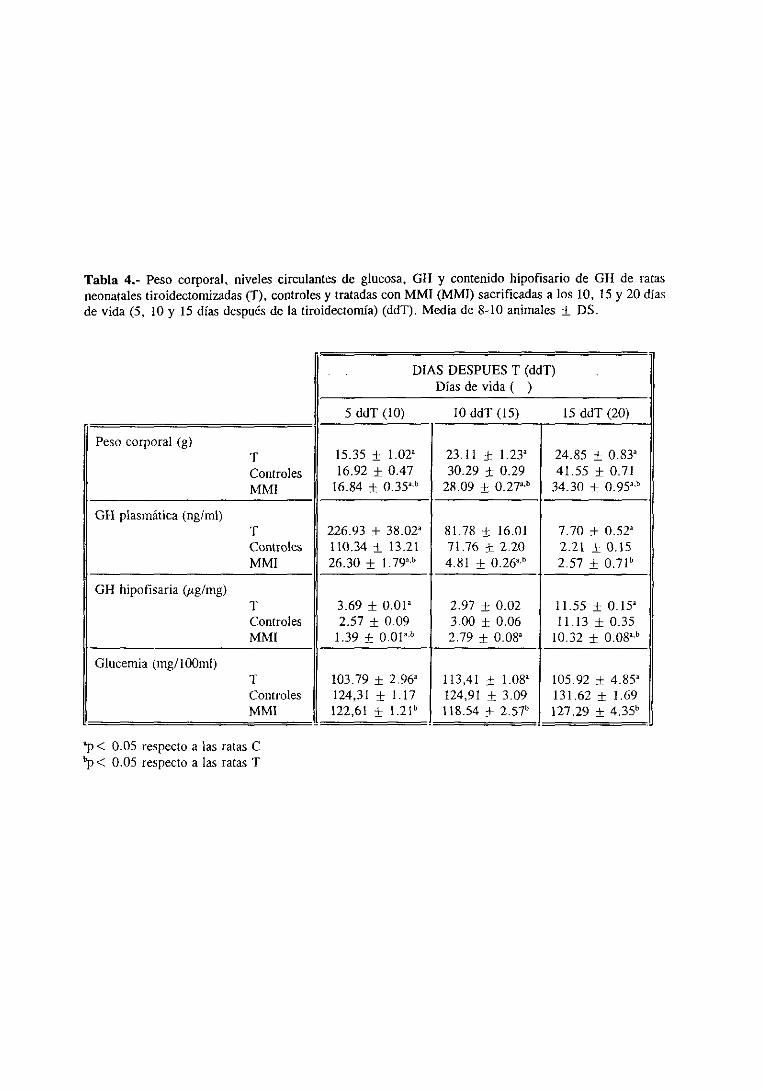
Tabla 4.- Pesocorporal, nivelescirculantesde glucosa,GH y contenido hipofisario de GH de ratasneonatales tiroidectomizadas (T), controlesy tratadascon MMI (MMI) sacrificadasa los 10, 15 y 20 díasde vida (5, 10 y 15 díasdespuésde la tiroidectomía)(ddT). Media de 8-10 animales±DS.
OlAS DESPUES1 (ddT)Díasde vida
Pesocorporal(g)TControlesMMI
15.35 + 1.02’16.92±0.47
16.84 ±035’b
2311 ±1.23’30.29 + 0.29
28.09 + 0.27a.b
24.85 ±0.83~41.55±0.71
34.30 + 0.95’’
GH plasmática(ng/mi)1Controles
MMI
226.93 ±38.02’110.34 + 13.21
26.30±1,79’,b
81.78 + 16.0171.76 + 220
4.81 + 0.26&b
7.70 ±0.52’211 ±0.15
2.57 ±07¡b
GH hípofisaria(pg/mg)1Controles
MMI
3.69 + 0.01’2.57 + 0.09
1.39 + 0•01a.b
2.97 + 0.023.00 + 0.06
2.79 + 0.08’
11.55 ±0.15’11.13 ±0.35
¡0.32 ±008’,b
Glucemia(mg/[OCmI)1ControlesMMI
103.79±2,96’124,31 + 1.17122,61 + 1.2V’
11341 + 1.08’12491 + 3.09118.54+ 257b
105.92±4.85’131.62±1,69127.29 ±4.39
5 ddT (10) 10 ddl (15) 15 ddT (20)
< 0.05 respectoa las ratasCbp < 0.05 respectoa las ratas1
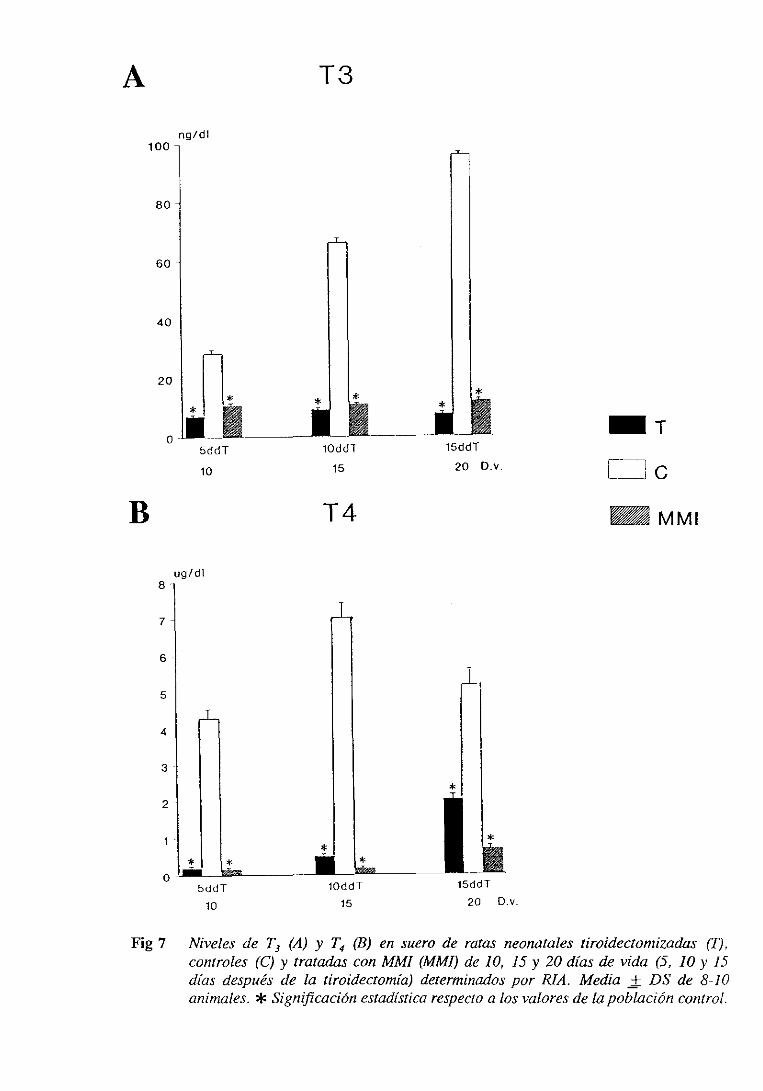
lOddT
15
l5ddT
20 Ox.mC
~ MMI
l5ddT
20 Dv.
Fig 7 Nivelesde T3 <A) y T4 (E) en suero de ratas neonatalestiroidectomizadas(T),controles(C) y tratadas con MMI (MMI) de 10, 15 y 20 díasde vida (5, 10 y ¡5días despuésde la tíroidectomía) determinadospor MA. Media + DS cte 8-JOanimales.* Significaciónestadísticarespectoa los valoresdela poblacióncontrol.
A T3
ng/di100
80
60
40
20
o
* *
5ddT
10
B 14
ug/dt8]
7—
6
5
4
3
2
o
*
*
*
5ddT lOddT10 15

Resultados
circulantesehipofisariosde GH aumentarona los 5 y 15 díasdespuésde laoperación
(10 y 20 díasde vida) respectoa los valorescontrol (Tabla4); sin embargo,a los 15
días de vida (10 días despuésde la tiroidectomía)no se encontrarondiferencias
significativas entre ambos grupos de animales, aunque se mantuvieronaltos,
probablementepor falta de población.Por el contrario, las ratastratadascon MMI
presentaronunosnivelesde Gil plasmáticainferioresa los de los animalescontrol
(‘[abla 4) en un 76.16%y un 93 .30% a los 10 y 15 díasde vida respectivamente,
aunquea los 20 días de vida, ambaspoblacionespresentaronuna concentraciónde
estahormonacirculantesimilar. Del mismomodo,a nivel hipofisario,estosneonatos
tratadoscon MMI presentaroncontenidosen Gil inferiores a los de la población
control.
En las tres etapasestudiadas,la poblacióncontrol mostró un incremento
progresivo asociado a laedaden los nivelesde T3 y el contenidode GR enpituitaria,
mientras que las cifras de Gil plasmática disminuyeron. Por otra parte, en los
animales hipotiroideos (tiroidectomizadosy tratados con MMI) se observóuna
alteraciónde la pautaseguidaduranteel desarrolloporestashormonas.
3.1.3. Glucemiae insulinemia.
Los valoresde glucemiade los animalestratadoscon MMI fueron similares
a los control,pero lascifras de glucosaen sangrede las ratasoperadasdisminuyeron
respecto a las de la población control, por lo que la glucemia de las ratas
tiroidectomizadastambiénfue inferior a la de aquellos animalesa los que se les
administró el fármacobociógeno(Tabla 4).
La insulina plasmática de la población hipotiroidea tratada con MMI
disminuyó respecto a los animales control en una proporciónque osciló desdeun
28.92% a los 10 díasde vida hastaun 37.55% a los 20 días de vida (Figura8). Sin
embargo,la insulinemiaaumentóen los animalesoperadosrespectoa los controles
(27.48% a los 10 días de vida, y 42.59% a los 20 días de vida), lo que podría
explicar el hecho de que los animales operadospresentaranniveles de glucosa
sanguíneainferioresa los de los animalescontrol.
3.1.4. Factores de crecimiento similares a la insulina (IGFs): niveles
séricosy expresión del ARNn’ hepático.
3.1.4.1.Niveles séricos.
Los nivelesde IGF-I en suerode rata control experimentaronun incremento
progresivoentre los 10 y 20 díasde vida, pasandode 178.05 a 255.SOng/ml(Figura
101
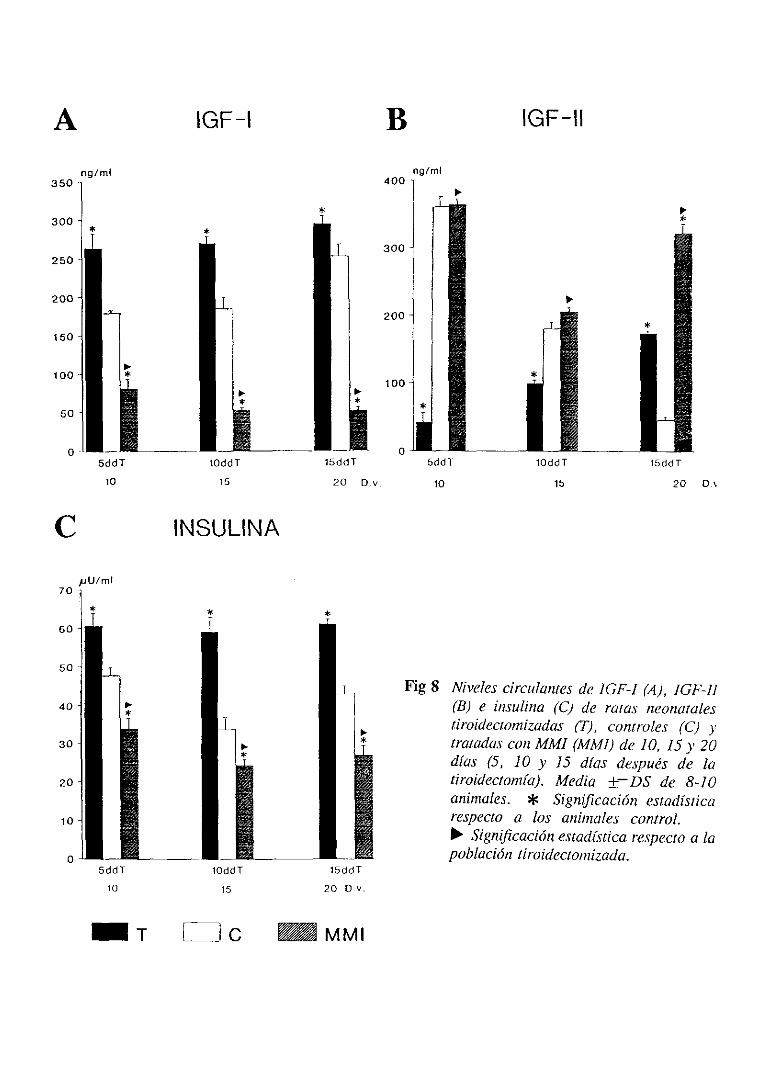
A IGF-I B ¡OF-II
ng/ml ng/ml350 400
300
250
200
200
150
1 00100
50
oISddT
20 0>
C INSULINA
,jU/mI70
60
50
Fig 8 Niveles circulantes de ¡CF-! (A), ¡CF-JI
40 (BJ e insulina (C) de ratas neonatalestiroidectomizadas (7), controles ((9 y
so tratadas con MMI (MMI) de 10, 15>’ 20días (5, 10 y 15 días despuésde latiroidectonila). Media + VS de 8-10
20animales. * Sign~caciónestadísticarespecto a los animales control.
lO ~ Significaciónestadistica respectoa la
población tiro idectonilzada.O
5ddT loddT lSddT
lO 15 20 DV.
-c MMI
300
o5ddT lOddT lSddT SddT 1OddT
lO 15 20 Dv. lo 15

Resultados
8). De manera análoga a lo ya descrito para la insulina, los animales
tiroidectomizadospresentaronnivelesdeIGF-I ensuerosignificativamentesuperiores
a los de sus controlesen todaslas edadesanalizadas.En las ratastiroidectomizadas,
los nivelesdel péptido fueronun 71.60%(10 díasde vida), un 128.49%(15 díasde
edad) y un 64.40%(20 díasde vida) superioresa los de sus controles.
Los niveles séricos de IGF-I de los animalestratadoscon MMI, por el
contrario, disminuyeronrespectoa los de los controles,de manerasimilar a lo que
ocurriócon la insulinemia.Estedescensodel IGF-I circulantesecifró enun 54.75%
a los 10 días de vida y alcanzóhastaun 65.12%a los 20 díasde vida.
En cualquiercaso, el hipotiroidismo causó, al igual que en el caso de la
insulina, un transtornoenel patrónde cambiosde los nivelesde IGF-I asociadoa la
edad.
En la Figura 8 serepresentan las concentracionesde IGF-II existentesen el
suero de las poblacionesde ratas neonataleshipotiroideas (tiroidectomizadasy
tratadascon MMI) y controles. En los animalescontrol, se apreciaun descenso
progresivo en los niveles de este péptido conforme aumenta la edad (359.92ng/ml a
45. 9Ong/ml).
En las ratas tiroidectomizadas, a los 10 y 15 días de vida (5 y 10 días después
de la operación), se encontró un descensoen los valorescirculantesrespectoa la
población control (88.34 y 44.93%, respectivamente), mientras que a los 20 días de
edad (15 días después de la tiroidectomía), la concentración sérica del péptido se
incrementó enormemente (278.04%) respecto a las ratascontrol. Por otro lado, los
animales neonatales que fueron tratados con MMIpresentaronunosnivelesde IGF-II
superiores a los controles desde los 20 días de edad.
3.1.4.2. Expresión del ARNmhepático.
La expresión hepática de IGF-I mostró un perfil similar al ya visto en suero.
Así, se observóun aumentoen el ARNmhepático de los neonatos tiroidectomizados
y un descenso en el de los tratados con MMI; dicha disminución en la expresión se
inició a los 15 días de vida, mientras que el aumento se aprecióya a los 10 díasde
vida (5 díasdespuésde la tiroidectomía)(Figura 9).
Al igual que se había mostrado a nivel circulante, la expresión del IGE-Il en
hígadodisminuyóal aumentarlaedaddel animal control (Figura9), sin embargo, las
ratas hipotiroideas (tiroidectomizadasy tratadascon MMI) mostraron un perfil
diferenteque semodificó dependiendodel tipo de hipotiroidismoconseguido.Así,
103
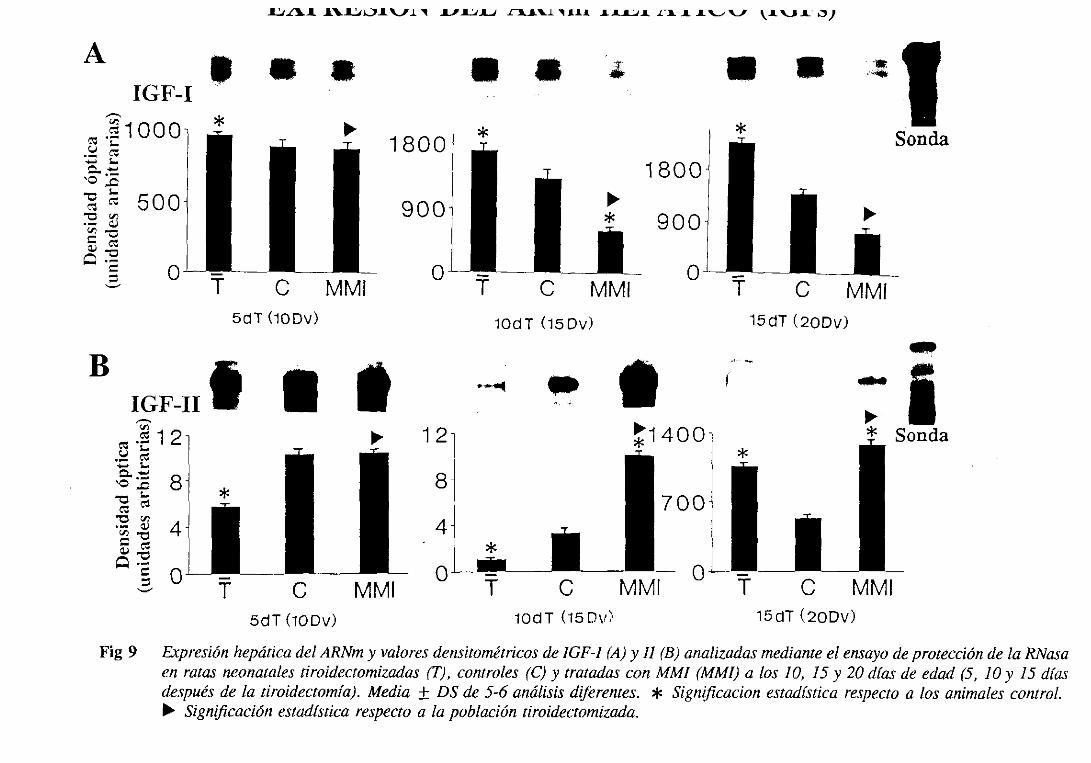
oo1-~0
U)
oo1*
*
1
oo
oo
oO)
4
1-
oo
oo
ca1~
oU)
oc
1-or
II—
o
22
o0
9
~0U)
1—
reou)
oo
y
casi-
OC
\J1~~
*
cesi-
u
“~‘n
E‘.5
22~
.o—
a>
-’o
o~
~t?~
1-:~
-~
.n‘~
II—
—o
-.
-,*
2-~
tt
2—o
oU).z.
—<
‘-~
1—~
o1~
t-..—
.~
E~~
22>o
o~
~.-
u-~
‘,.~1-.~‘5
i—•0
.~~—
U)
II—~
~
~“5t
~k
(sBp
lw!q
J13
Sa
pl3
piu
fl)
(sn~auJ}~qJEsapup!un)
E3!ldQpB
pisu3Q
reo(n
¡a
’23U
)
1~
eu1
e1~
1~
ooo
1o
ooLo
E3!1(I9P
UP
!StflN
iI

Resultados
en las poblacionesde animalestratadoscon MMI, los nivelesde ARNm del péptido
se vieron paulatinamenteaumentadosdesde los 15 días de vida respectoa la
población control y, por el contrario, en el caso de las ratas operadas
quirúrgicamente, el hipotiroidismo cursó con niveles de expresión de IGF-II
disminuidos(10 y 15 díasde vida) aunque,finalmente,aumentaron(20díasde vida).
De nuevo, seencontróun paralelismoentrelos nivelescirculantesy los de
ARNm de los péptidos, lo que sugiere la existencia de una regulacióna nivel
transcripcional en la secreción de los IGFs debido a la falta de hormonas tiroideas.
3.1.5. Estudiode regresiónlineal y correlación.
Se efectuóel análisisde regresióny correlaciónde diversosparámetrosde las
ratasen periodoneonatal(Figura 10). Paralos cálculosse reunieronen cadaanálisis
los datos de las tresedadesestudiadas.
Se encontróunafuertecorrelaciónpositiva entreJos niveles séricosde IGF-1
y los plasmáticosde insulinatanto en los animaleshipotiroideos (r= 0.83, p < 0.05
para las ratas tiroidectomizadasy r~ 0.89, p < 0.05 para las tratadascon MMI)
como en los controles(r= 0.96, p < 0.05). Sin embargo,cuandose representaron
los niveles circulantesde IGF-I frente a los de Gil, estaspoblacionesmostraronuna
correlación negativa (r= -0.58) o baja (r= 0.46 y 0.75) para los animales
tiroidectomizados,tratadosconel bociógenoy control, respectivamente.
3.1.6. Proteínasligadorasde IGFs (IGFBPs): niveles séricos y expresión
del ARNm hepático.
3.1.6.1. Niveles séricos.
3.1.6.1.1.Westernligand blot.
En todos los estadiosanalizadosse observóun patrónclaro constituidopor
tres gruposde bandasde un pesomolecularestimadoen45kDa, 3OkDay 24kDa,y
que se designancomo IGFBP-3, IGFBP-1 y 2 e IGFBP-4, respectivamente(Figura
11).
En las ratascontrol, la bandade 30kDadisminuyóconformelo hacia la edad
del animal, mientrasque la bandade 45kDaexperimentóun aumentode intensidad
paraleloal de la edad,lo cual no se apreciaconclaridaden la Figura 11 debido al
cambiode escalas.
Los valoresséricosde las proteínasde bajo pesomolecular(IGFBP-1 y 2)
aumentaronpor la falta de hormonas tiroideas desde los 10 días de vida, sin
embargo,dichoincrementofue másmarcadoen los animalestratadosconel fármaco
105
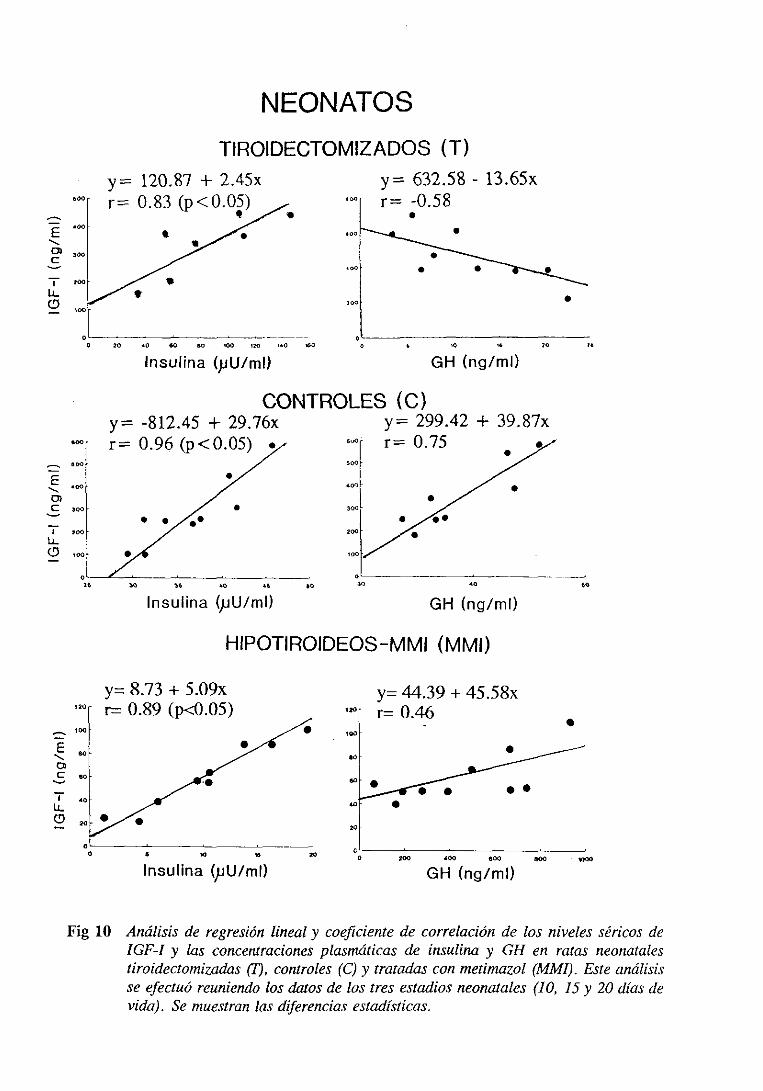
CONTROLES <C)y= -812.45 + 29.76x y 299.42r= 0.96 (p’zO.O5) . r~ 0.75
+ 39.87x
CH <ng/mi)
la’
y= 44.39 + 45.58xr= 0.46
loo-
SO
60
20
20
e
e e e cee
800 —0 200 400 600
CH (ng/mi)
Fig 10 Análisisde regresiónlineal y coeficientede correlación de los niveles séricosde¡GE-! y las concentracionesplasmáticasde insulina y Clii en ratas neonatalestiroidectomizadas(T), controles (C) y tratadascon metimazol(MMI). Esteanálisisse efectuóreuniendolos datos de los tres estadiosneonatales<10, 15y 20 díasdevida). Se muestranlas diferenciasestadísticas.
NEONATOS
TIROIDECTOMIZADOS (T)
y~ 120.87 + 2.45x•~~f r=~ 0.83 (p’C0.05)
e OOt
*
300
200’
y= 632.58 - 13.65xr= -0.58
e
E02
a
u-o
E0>
a
u-o
0 20 40 60 30 ‘00 20 40 60 0 1 0 6 20 E’
Insulina (pU/mi) CH (ng/mi)
00
e
e
300
200.
o31 30 36 ‘0 65 50 40
Insulina (pU/mi)
HIPOTIROIDEOS-MMI <MMI)
y= 8.73 + 5.09x“a’ r= 0.89 (p<O.05)loo
SoE02
a
LI-
o
ee
6 ~0
Insulina (pU/mi)
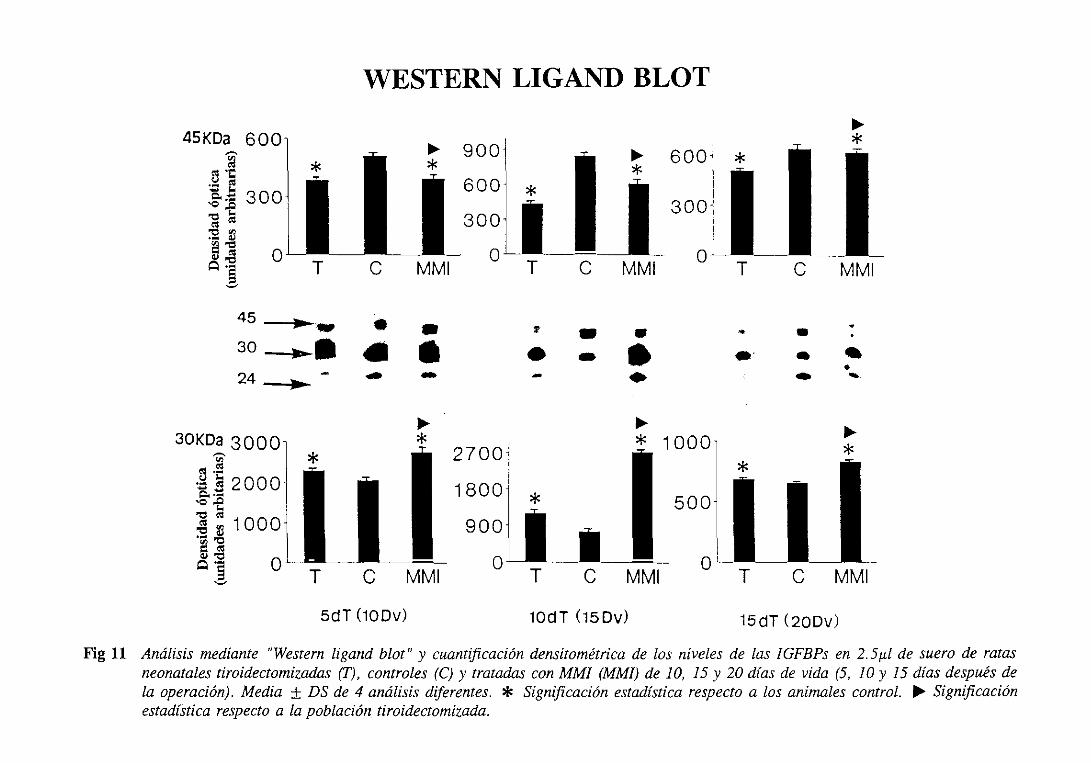
e•~
o1-
e,.
I~o
•8
oCo
00
00
00
0)C
OC
O
o
oo
Co
m(s4ffiJ;~qs~sapup¡un)
OEZU$dO
pEp!S
IM«
scLfl
3,
o~1
o
oo
o[1)
1~
o
2
—o
00
00
00
0N
caO
)C
M—
22se’
O1—
3,,Lo
01
‘4.C
iCN¿
—O
oo
oo
oo
oo
o~
flCM
1-
o
<o(sE
pE>~qiu
sap
up
¡un
)Osc
tpd
9pE
P;suaa
oCi
-~‘~
‘4~k.
‘~~
k~~
-sk.
Nu‘5
Oo~
CN~
1~—~—
‘‘5
ULO~
r
u
~‘~
~1
‘5~
’A>
—4
‘5e-.
OU)‘.4
si.~A
~0‘4
,
o1~
‘44u—
4.
>~
‘-4.
~
1—~
ULo
‘5c~j
-~
~‘5
~~E
~
—4
u~
bt
it
oo
oo
(0oz0z
Sr
ooCo
oo

Resultados
bociógeno,lo quepusode manifiestounadiferenciaentreestaspoblacionesde ratas
en cuantoa las cifras séricasdel complejode 3OkDa.
Los nivelesde IGFBP-3de las ratashípotiroideasexperimentaronun descenso
también desde los 10 días de edad,pero fue másacusadoenel casode los animales
neonatalestiroidectomizadosque en el de los tratadoscon metimazol, siempre
respectoa las ratascontrol; de nuevo, encontramosuna alteraciónen el patrónde
cambiode los nivelesde las IGFBPsen los animaleshipotiroideos.
3.1.6.1.2.Westerninmunoblot.
El análisismediante“Westernligandblot” mostróun aumentoen las proteínas
de bajo pesomolecular del suero (IGFBP-1 y 2), y para averiguar si dicho
incrementose debíaa ambasproteínaspor igual o al predominio de una de ellas
sobre la otra, se recurrió a otro tipo de análisis que nos permitiera conocerla
intensidadde la bandadebida a cada proteínapor separado;se trataron así las
membranascon anticuerposanti-IGFBP-1y anti-IGFBP-2específicospara rata.
El tratamientode las membranascon anti-IGFBP-1 nos reveló un aumento
significativo en los nivelesséricosde estaproteínade bajopesomolecularenaquellos
animalesque fuerontiroidectomizadosa 10, 15 y 20 díasde vida respectoa las ratas
control (Figura 12). Porel contrario, las poblacionesde ratashipotiroideasa las que
se les administró el fármaco bociógenomostraron un descensoen los valores
circulantesde estepéptido respectoa la población control en todos los estadios
analizados(10, 15 y 20 díasde edad).
Los nivelescirculantesde IGFBP-2se encontraronaumentadospor la falta de
hormonastiroideas(animalestratadosconMMI y tiroidectomizados)a partir de los
10 y 15 días de edad, respectivamente(Figura 12); no obstante,el incremento
observado fue mucho más acusado en el caso de las ratas tratadas con el fármaco
antitiroideorespectoal mostradoporaquellosanimalesque fueronoperadosen todas
las edadesestudiadas(10, 15 y 20 días de vida).
3.1.6.2.Expresióndel ARNm hepático.
Los nivelesde ARNm hepáticode la proteínade bajopesomolecularIGFBP-
1 seencontraronaumentadosen las ratastiroidectomizadasrespectoa las controles
en los tres estadiosanalizados(Figura 13), mientrasque dismuníanen los animales
hipotiroideos tratadoscon MMI tambiéndesdelos 10 días de vida, mostrandoun
paralelismocon los valorescirculantesde IGFBP-l por inmunoblot (Figura 12).
La expresiónde IGFBP-2 aumentótantoen los animalesneonatalestratados
los
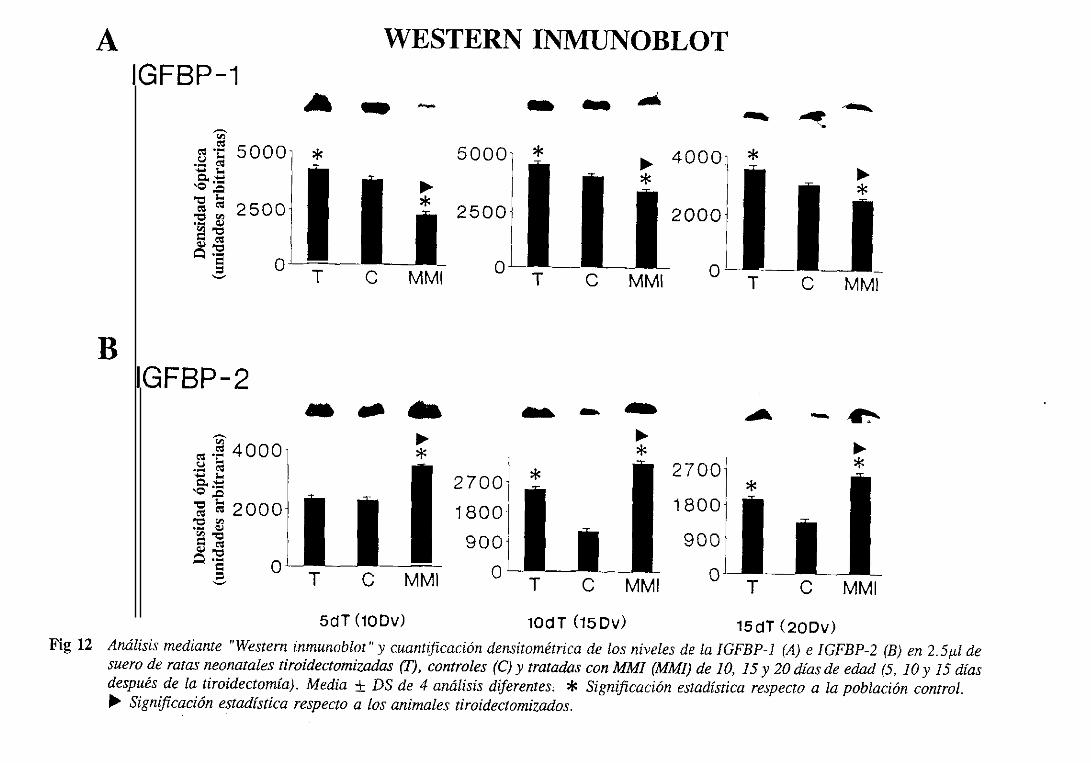
‘¡—
o
*oo
oo
ooCM
*oo
oo
OU
)
LOCM
tnqdopcpisuau
H
o
Io1—
a¡o
,o1—
9’
—o
eo
o-
caLI-
(5
oo
oo
ooCM
-~
‘n‘~
‘..~
‘4
‘—4
uC
N~
~
og
t$~~
E—
,-‘‘4
o“5‘5u
~
0-4
osi.0
‘~>
‘4‘.~
.~
1—~
~‘4
E
o
-S~
.
‘5
2
>oo—
4
..—c~
j-H~
1~—.0~
1—U)
.L..
..~-%
.
~>
~c‘5
E‘4”u,
~‘~
.~
‘~u
,’~A
o
(sB¡.was¡q.w
sap~p¡un)B
opdopE
pisua~
ti
e0
0O
00
oca
o)CM
1
o
41e1e4
o-
cau-(5
00
0
00
0
CM
oo
oo
OLO
U)
CM
(sB¡Ju.q~q.zusapup¡un)
it
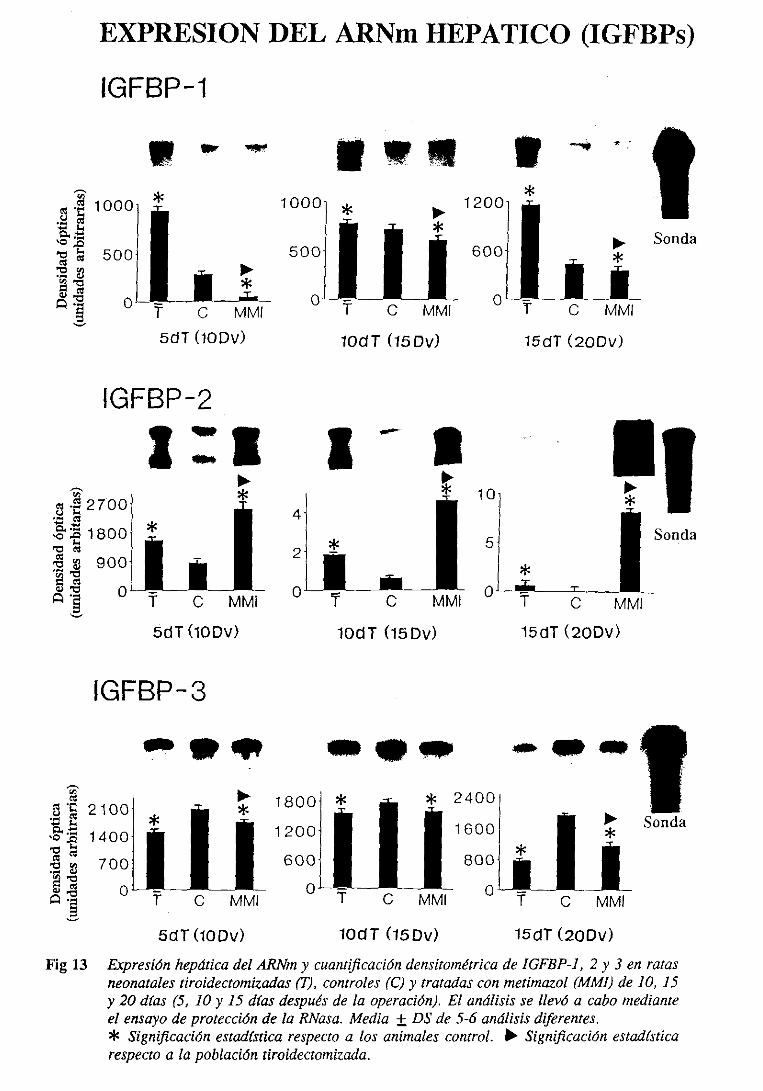
EXPRESION DEL ARNm HEPATICO (IGFBPs)
IGFBP-1
1000 *
500
1
w ~
1000
5dT (10Dv)
“e1200
6
*
-~ r~
1~ o MMI
l5dT (200v)
IGFBP-2
2700
18001¿I900
Y O MMI
5dT (100v) lOdT (150v) lbdT (200v)
IGFBP-3
en 000
1800 *
1200
rn600T O MMI
lOdT (wsDv)
sea
2400
160dMIM *800’LÍAI
T o MMI
l5dT (200v)
F¡g 13 Expresiónhepáticadel ARNmy cuantWcacióndensitométricade IGFBP-J, 2 y 3 en ratasneonatalestiroidectomizadas(2), controles(C) y tratadascon inetimazol(MMI) de JO, 15y 20 días (5, 10 y 15 díasdespuésde la operación).El análisisse llevó a cabo medianteel ensayode protecciónde la RNasa. Media + JiSde 5-6 análisisdiferentes.* Significaciónestadísticarespectoa los animalescontrol. > Significaciónestadísticarespectoa la poblacióntiro idectomizada.
u‘o ,~ Sonda
*
1 0 MMI
ladI (l5Dv)
¡‘o .0
A-
310
5
o
4
2
o
*
*
C MMI
Sonda
O MMI
2100
1400
700
o
.~ .~—S.S‘o
95dT (ioDv)
Sonda

Resultados
con MMI comoen los tiroidectomizadosrespectoa los controles(Figura 13); en la
poblaciónde ratasoperadasquirúrgicamente,el aumentoen los nivelesde ARNm de
IGFBP-2 se produjo a los 10 díasde vida (5 días despuésde la tiroidectomía),y
aunque,también se observóa los 10 días de edad, fue más pronunciadoen los
tratadoscon MMI, lo que está igualmentede acuerdocon los valores circulantes
encontradospor inmunoblot (Figura 12).
La IGFBP-3disminuyósuexpresiónhepáticaenambaspoblacionesneonatales
hipotiroideas(tiroidectomizadasy tratadascon MMI) respectoa la control desdelos
10 díasde vida, aunqueestadisminuciónparecióser másacusadaen el casode los
animalestiroidectomizadosy seobservóconmayorclaridada los 15 díasdespuésde
la operación(Figura 13).
A lavistade todosestosresultados,seobservala existenciade unparalelismo
entrelos nivelescirculantesde las IGFBPsy suexpresiónenhígado,lo que vuelve
a sugerir una posibleregulaciónde estasproteínasa nivel transcripcionalporparte
de las hormonastiroideas.
3.2. TRATAMIENTO CON ESTREPTOZOTOCINA (STZ) DE RATAS
NEONATALES TIROIDECTOMIZADAS.
Paraprofundizarenel papelde la insulinasobrela secreciónde los IGFs en
situacionesdehipotiroidismoneonatal,se realizóel siguienteexperimento.Las ratas
neonatalestiroidectomizadas,que presentabanaltos niveles de insulina, fueron
tratadascon STZ a los 15 díasde edad(10 díasde vida) parabloquearla liberación
de estahormonadespuésde la tiroidectomía.
3.2.1. Evolución del pesocorporal.
Las ratasneonatalesoperadastratadasconSTZ mantuvieronel pesode cuerpo
por debajo de los valores de los animalescontrol, si bien presentaronun ligero
aumentorespectoa los tiroidectomizados(Tabla5).
3.2.2. Nivelesséricosde T3 y T4 y plasmáticose hipofisariosde GH.
Los neonatos tiroidectomizados tratados con STZ el día 15 de edad
presentaronunos nivelescirculantesde hormonastiroideas inferiores a los de sus
controlesen un 86.00%para la ‘[3 y en un 94.95%para la ‘[4 (Tabla 5); de esta
manera,la concentraciónséricade las hormonastiroideasde estosanimalesresultó
ser también inferior a la de los controles como en las ratas neonatales
tiroidectomizadasno sometidasal tratamiento.
111
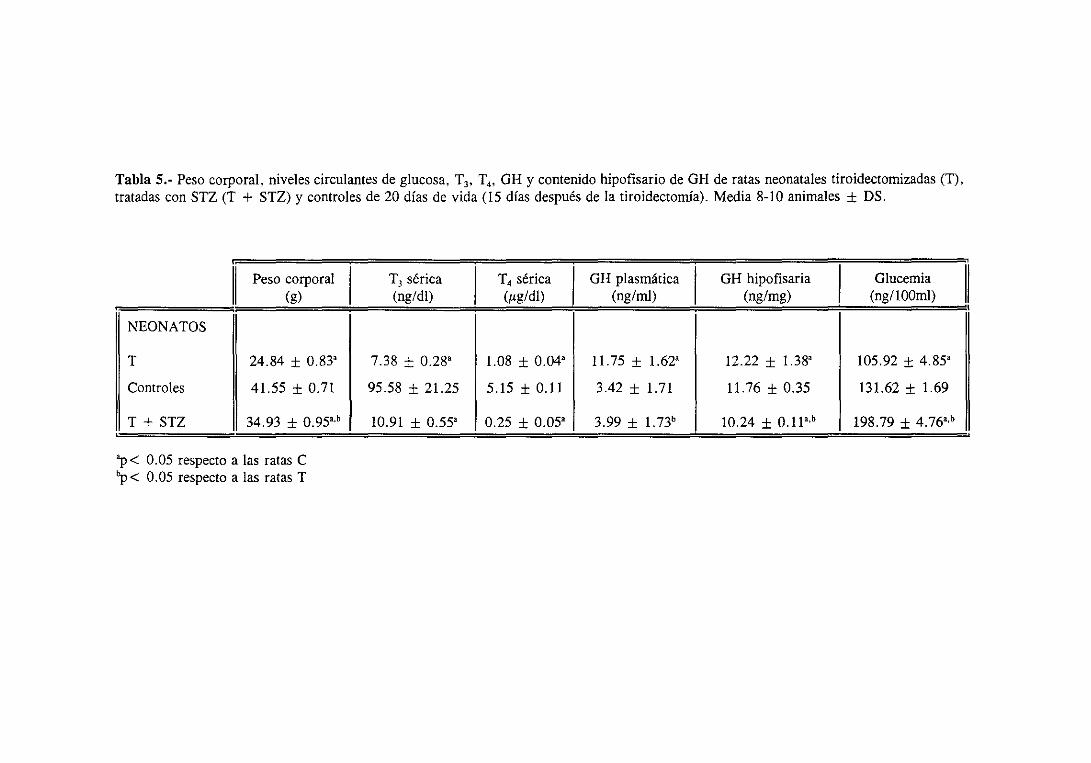
o—
.0
~fl~‘
E
00
‘0~0
4—
1~
-
+i
-H-H
cl
cl
~O
N
oe
n—
—Q
N
.0Ej
—e
n.
—
—t
cl
‘Orl
t~.
.rl
—.-
oN
.2
oo
.0en
.t~.:
r—
——
+1
~+
1
~N
QN
.~Q
N
—e
n~<;
—4
oA
-’—
—5-
.t‘4)—
.~
obfl
4.
0->
o‘—
o
00
0
+1
-H-4
-4
00
~flE
n
O—
Cl
,-~E
n8
‘ortto
En
cl
En.
EnN
-~8
o±í±
1+1
00—
(flEn
O~.~
oQN
—
1-Oco
n0OrAO
.>
.0
en
—
~0~.
En
.Q
N
00
8-4
-1-1
-4+
1
‘~E
n
En
en
..
QN
—4
rl 4
en
enoE—zoz
E’,
4)O5
-
OE-
U
NE-
en+E-
o”Onc-E
j
~S
+15
-—
‘0
j-44)
4-,
.-
cc~
20
‘00
0
ce
~O
)-u
O0
0
‘00
CE
-O
~
—‘0
‘ci
o>4
<~j
En
en-t
¿ci
~0o~oo”~
o-Ii
00
4)A
-’
ZN
CH
OOuN
oE~
~t00
di
O
ti>En
.Ofl
c~
c4
UE
-
c4c4
E~
5-
4/,
‘0
c~c4
00
OC
)0
4)
cn~
n4
)0)
1.4E
-
En
En
00
00
VV

Resultados
El bloqueode la secreciónde insulina provocóun descensoen los valores
plasmáticos de Gil superior al 65% respecto a la población de animales
tiroidectomizados,lo que igualó la concentraciónde Gil circulante de estas ratas
sometidasal tratamientoa los nivelesde los animalescontrol. Porotraparte,en estos
animalesinyectadoscon STZ, tambiéndisminuyó,incluso pordebajode los valores
control (19.57%), el contenido hipofisario de esta hormona, con lo que este
parámetrofue muy inferior en estas ratas respectoa los animales neonatales
tiroidectomizadosno sometidos al tratamiento (Tablas 4 y 5), mostrando un
paralelismoentreel descensode la insulinemiay dichosparámetroscomosucedíaen
los animalesdiabéticos.
3.2.3. Glucemiae insulinemia.
La administraciónde STZ causó,comoera de esperar,un gran aumentoen
los nivelescirculantesde glucosade los animalessometidosal tratamiento(Tabla5),y, de maneraparalela,un descensoen su insulinemiadel 70.44% respectoa los
valores control (Figura 14) debido al bloqueo de la secreciónde insulina por la
droga.
3.2.4. IGFs: niveles séricos y expresióndel ARNm hepático.
3.2.4.1.Niveles séricos.
Los nivelesde IGF-I, comosucediócon las ratasneonatalestiroidectomizadas
sin STZ, mostraronun perfil similar al ofrecidopor la insulina, estoes, los valores
circulantes del péptido de las ratas tratadas con STZ disminuyeron en un 76.32%respecto a la población tiroidectomizada y mostraron unos niveles similares a los de
los animalescontrol (Figura 14).
Laconcentraciónde IGE-Il ensueroaumentódespuésde la administraciónde
STZ respectoa la población control (Figura 14), e incluso, estos valores superaron
a los alcanzadospor los animales tiroidectomizados,cuyos nivelesaparecíanya muy
incrementadosrespectoa las ratascontrol.
3.2.4.2.Expresióndel ARNm hepático.
La expresióndel ARNm hepático de los tránseritos de IGF-I (Figura 14),que
estabaaumentadaen los neonatostiroidectomizados,disminuyó, de forma paralela
a lo que ocurríaen el suero,respectoa estapoblacióny a la control tras sometera
los animalesal tratamientocon STZ. Sin embargo,en estasratasneonatales,después
de la administraciónde STZ, los nivelesde ARNm de IGF-II aumentaronaún más
que en las tiroidectomizadasrespectoa la poblacióncontrol (Figura 14), igual que
113

Fig 14 A) NivelescirculantesdeJGF-J, insulina e IGF-¡I deratas neonatalesuiroidecromizadasrn. controles(C) y tiroidecromizadastratadas con 522 (1 + 572) de 20 días (15 días después de la operación).Media ±DS de 8-10 animales.B) Expresión hepática del ARNm y cuan¡~ficación densizométricadel IGF-I y II en ratastiroidectomizadas(7’), controles (C) y ¡iroidecromizadastratadascon STZ (T + 572)de 20días. Elanálisisseefectuómedianteel ensayode protección de ribonucleasas.Media + DSde 4 análisisdistintos.* Significación estadísticarespectoa los animales control. ~ Significaciónestadísticarespectoala poblaciónoperada.
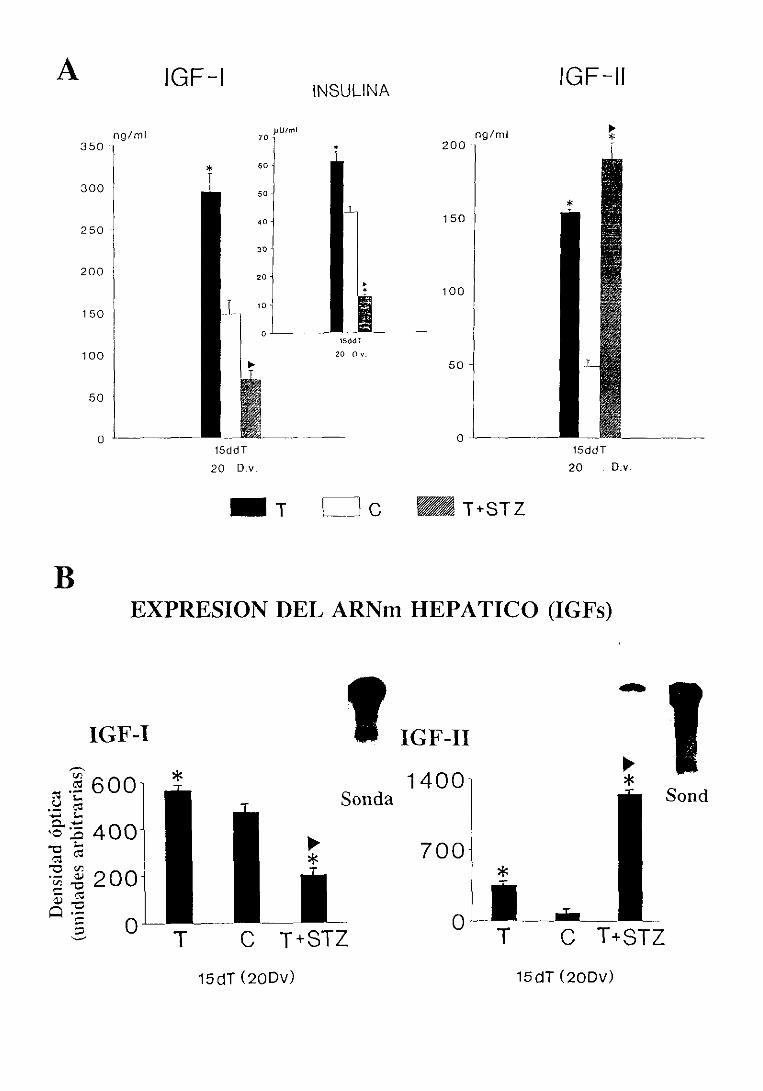
INSULINA
ng/ml200
50 H
100
50 -
o
u-
IGF—II
ph.
*
*
~1~~~
l5ddT
20 Dv,
c T+STZ
BEXPRESION DEL ARNm HEPÁTICO (IGFs)
Sonda
*~1~
a
IGF-II
1400
700-
oc T~STZ
*Sond
T O T+STZ
*
iGF-I
ng/rol PU,rnlza
A
350
300
250
200
150
100
50
o
20 0w
lSddT
20 0w
IGF-I
*
jCC
~-) :.-
-D
-o
~1”
600
400
200
0~T
l5dT (2aDv) l5dT (200v)

Resultados
sucedíaa nivel circulante.
3.2.5. Estudio de regresión lineal y correlación.
La Figura 15 resume los resultados del análisis de regresión de algunos de los
parámetrosestudiadosenesteexperimentocon STZ que se acabade describir.Para
el análisis serecogieronlos datosobtenidosparalas tres poblaciones.
La correlaciónentre IGF-I e insulinemia fue alta y positiva (r= 0.97, p <
0.05),mientrasque el IGF-I secorrelacionóde formanegativacon los nivelesde Gil
en plasma(r= -0.83).
3.2.6. IGFBPs: nivelesséricosy expresión del ARNni hepático.
3.2.6.1. Niveles séricos.
3.2.6.1.1.Western ligand blot.
El hipotiroidismoprovocó, como ya seha mencionadoconanterioridad,un
aumentode las proteínastransportadorascirculantesde bajopesomolecular(IGFBP-I
y 2) respecto a la población control, y dicho incremento no fue corregido tras el
tratamientode las ratastiroidectomizadascon STZ (Figura 16).
Los niveles séricos de IGFBP-3, que disminuyeron en aquellos animales que
fueron operados,aumentaronsignificativamentedespuésde la administraciónde STZ
e incluso,alcanzaronunosvaloressimilaresa los de lapoblacióncontrol,comohabía
ocurrido con los niveles de GH (Figura 16).
3.2.6.1.2.Western inniunoblot.
El análisisefectuadocon el anticuerpoespecíficode rata anti-IGFBP-1 nos
mostróque los nivelesde estaproteínatransportadorase incrementaban,respectoa
los de las ratas control, tanto en animales que fueron tiroidectomizados como en los
que fueron sometidosal tratamiento(Figura 16).
La IGFBP-2presentóun perfil análogoal ofrecidopor IGFBP-1,estoes, sus
niveles séricos aumentaron en las ratas operadas y en las que se les administró STZ
cuandosecompararoncon los mostradospor los animalescontrol, y la SIZ no los
varió (Figura 16).
3.2.6.2.Expresión del ARNrn hepático.
La Figura 17 muestrala expresiónhepáticade IGFHP-1,2 y 3 en los animales
neonatalesantesy despuésde ser tratados con STZ. En estascondiciones,seobservó
un mantenimentoen los nivelesdel ARNm de IGFBP-1 en los animalesa los que se
les administró STZ respecto a los tiroidectomizados, lo que parece motivado por el
bloqueode la secreciónde insulina en estasratas, dado que esta proteínaligadora
115
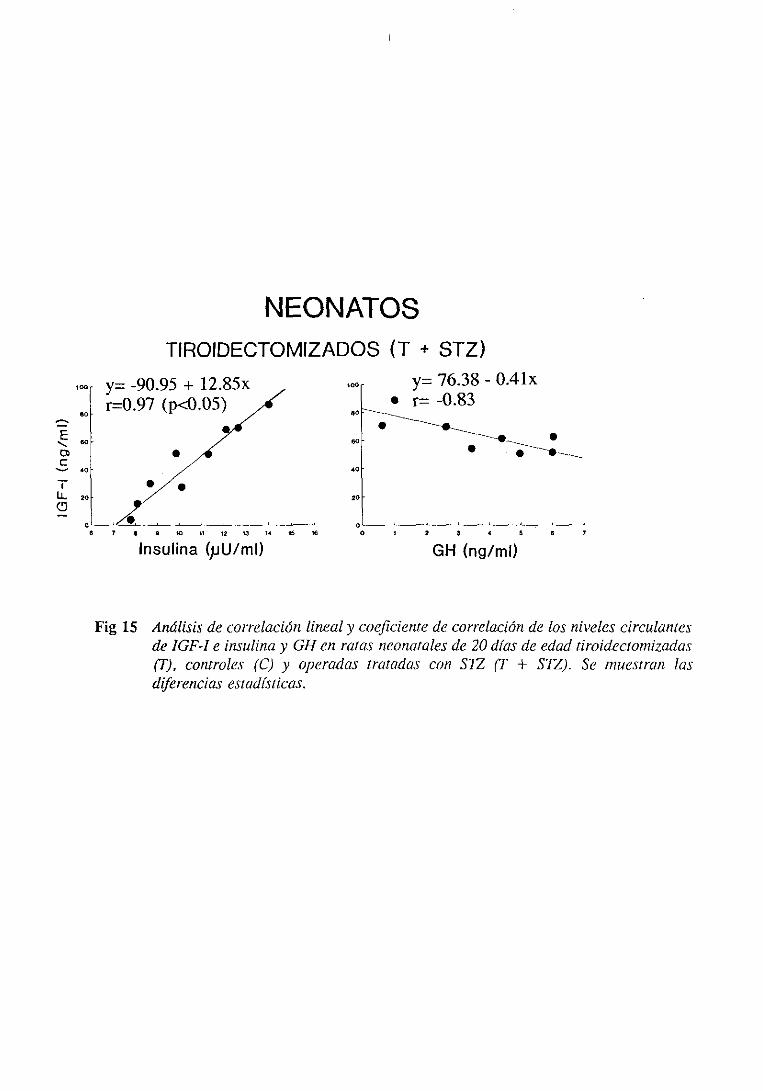
NEONATOSTIROIDECTOMIZADOS <1 + SIZ)
y= -90.95 + 12.85xr=0.97 (p<0.05)
e
e
e6 ~ 6 9 lO II Q II 14 16 Id
Insulina <pU/mi)
I00~
50
so
40
20
y= 76.38 - OAlxpr -0.83e
ee
0 1 2 2 4
GH <ng/mí>
Fig 15 Análisis de correlación lineal y coeficientede correlaciónde los nivelescirculantesde IGF-l e insulina y Gil en ratas neonata les de 20 días de edad tiro idectonzizadas(1,>, controles (C) y operadas tratadas con STZ (7’ + Si?). Se muestran lasc4ferenciasestadísticas.
loo
60
E
Li~CD
60
40
20
5 5 7
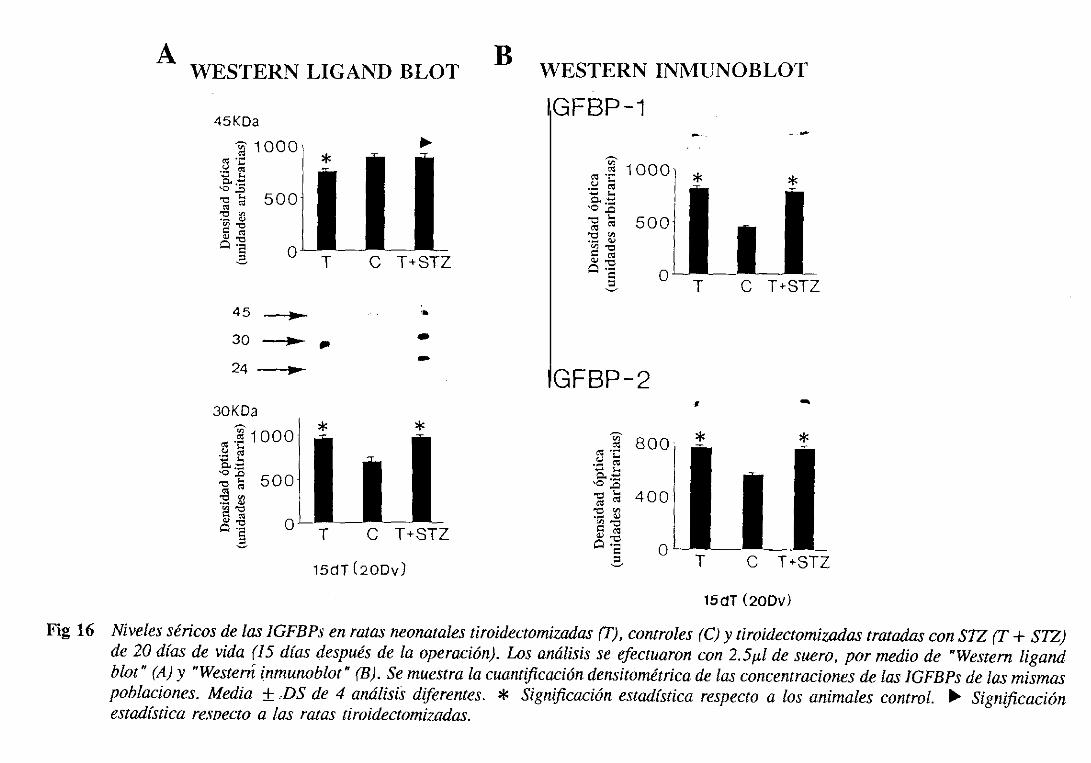
N1
HCID+1—
t~*
oo
oo
o1~~~
0It)
r
o-
A
(sr~iu.ii~q.insapEpliAn)
E3!$dOpP
plsuaU
oo
oo
oIt)
oEn
NCo+1—
E*
.*o
oo
ot.e’J
o-
LoLL0
.q•
~
o
r(su¡ar~~’qiusapupjun)
u~¡>dopEpISU9tJ
1
‘no
,~.~
Cl
sap
rp!u
n)
Eafldo
P~p!suaQ
C~
r
o~(srnIE
JpqJUsapup~ua)
OuaudpP~P!~I’~U
z~
u
t tcA
NE--ti)+E--
>oQ
CloLo—u
E--
—~
u.~
oE~
.~,~
.tj§~
éj~
E~
u
-~~
*u
Ni
1-C/D+I~ro1—n
1-
~~~
C~
~~
oo0zzCIDLL(5
oz0zCID
oo
oo
oo
OLI)
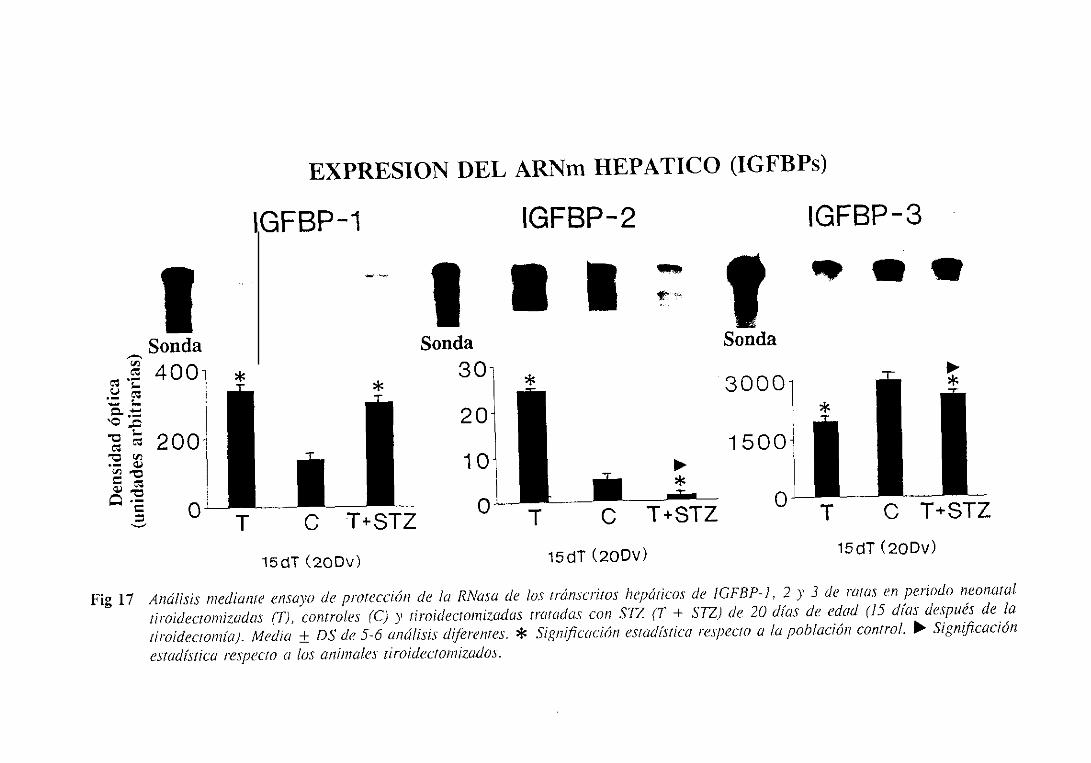
A*1
Co
¿-ucau
-o
t
I
ZL
L
zo
1~
~
a-
~O
2u
-(9
F
*1•
oo
o‘d
OO
~O
oCIDC
t)
N1-
Á*C
I)+1-o
*
OO
OO
—~
CO
C’JoUD
*1~
1~
*1-
oCID
OO
OO
Oc\J(suuxu~qJB
Sapuplun)
1—Co+H3
oo0
23
~‘o
E-
‘~‘
~
—u
U)
1~
1—rfl¾~
CN
(Z~
‘~
CMu
uu
~0
U)
~étz
r
1—
%.~
-~L
..‘~
u-~
1—Cou
—
+It
—‘
‘r—>1o
~-<9
o~
o—
~~->+1
~1—
~O~2
2’-2
~
1—-~
Nu-ju
u---
uu
0o
B3!)dQPEP!SU~G

Resultados
muestrauna dependencia,ya descrita,de estahormona:un descensoen la insulina
provocauna subidade los nivelesde IGFBP-1.
Porotro lado, la expresiónde IGFBP-2,que estabaaumentadaen los neonatos
tiroidectomizados,se vió disminuidaen estasratas neonatalesporel tratamientocon
STZ incluso hastaunos valoresinferioresa los control (Figura 17).
La expresióndel ARNm de IGFBP-3 sufrió un aumentosignificativo tras el
tratamientode las ratasoperadascon STZ respectoa la poblacióntiroidectomizada,
aunqueesteno fue tangrandecomo para equipararlosa los de los animalescontrol
(Figura 17).
El perfil de los tránscritosde las IGFBPs es similar al observadoa nivel
circulante: IGFBP-1 disminuyóe IGFBP-3 aumentó;sin embargo,IGFBP-2 ofreció
un comportamientodiferente en el caso de los animales tiroidectomizadosy
posteriormentetratadoscon STZ, y así, se encontró un aumento en los niveles
séricos, mientras que su expresión hepática disminuía.
3.2.7. Resumen de los resultados del estudio de la tiroidectomía y la
administración de MMI en ratas neonatales y de las ratas neonatales
tiroidectomizadas tratadas con STZ.
Los neonatostiroidectomizadospresentaronunosaltos nivelescirculantesde
insulina y Gil plasmática e hipofisaria, así como unos valores séricos y de expresión
hepática de IGF-I aumentados respecto a las ratas control. Por otro lado, las cifras
circulantes de IGF-II y los niveles de su ARNmen hígado aparecieron disminuidos
en estos animales tiroidectomizados, excepto a 20 días en que estos valores
aumentaron respecto a los animales control. Por el contrario, los neonatos tratados
con MMI presentaron una baja insulinemia junto con unos valores plasmáticos e
hipofisarios de GH, y unos niveles séricos y de ARNmhepático de IGF-I
disminuidos,mientras que los valores de IGF-II aumentaronrespectoa las ratas
control. En ambaspoblacioneshipotiroideas(tiroidectomizaday tratadacon MMI),
se encontró una correlación positiva por análisis de correlación lineal entre los niveles
circulantes de IGF-I e insulina.
Los valores séricos de IGFBP-3 disminuyeron en las dos poblaciones
hipotiroideas,al igual que la expresiónde su ARNm hepático;por el contrario,en
los neonatos tiroidectomizados, las IGFBPs de bajo peso molecular aumentaron sus
niveles en suero y de expresión en hígado, mientras que aquellas ratas que sólo
fueron tratadascon MMI mostraronun aumentoen los valores circulantesy de
119

Resultados
expresiónde los tránscritoshepáticosde la IGFBP-2, aunquela IGFBP-1 disminuía
a ambos niveles. Pareceque el aumentoen los animalestiroidectomizadosen el
complejo de 3OkDa fue debido a IGFBP-1 y 2, mientras que en el caso de los
animaleshipotiroideospor tratamientoconMMI, pareciódeberseal aumentosólo de
IGFBP-2.
Los animalestiroidectomizadosy tratadoscon STZ presentaronvaloresbajos
de insulinemia, Gil plasmáticae hipofisaria e IGF-I circulante, así como de su
ARNm hepático(Cuadro1). El IGF-II circulantey su ARNm aumentanal igual que
la glucemia;aumentanIGFBP-1 y 2, tanto circulante como ARNm hepáticoy la
IGFBP-3 circulanteno varia y disminuyesu ARNm hepático(Cuadro 1). Todos los
valoressecompararoncon los animalescontroleseutiroideos.
El resumende estosresultadosestárefejadoen el Cuadro 1.
Seestablecióunacorrelaciónpositivaentrelos nivelescirculantesde insulina
e IGF-I.
Cuadro1.- Resumende los resultadosde los estudiosde tiroidectomía(T),
administraciónde MMI (MMI) y tratamientoconSTZ (T + SIZ) en ratasneonatales
respectoa la poblaciónde animalescontrol.
T MMI T+STZGLUCEMIAINSULINEMIA t 4 ¿
GB CIRCULANTE 1 4OH HIPOFISARIA 1 ¿ ¿
IGF-I CIRCULANTE 1ARNm [GP-! t 4 4
IGF-Ií CIRCULANTE 1 1 1
ARNm IGF-I[ 1 1 1
IGFBP-1 CIRCULANTE 1 4
ARNm IGFBP-1 1 4 1
IGFBP-2 CIRCULANTE 1 1
ARNm IGFBP-2 1 1 4IGPBP-3CIRCULANTE 4 ¿
ARNm IGFBP-3 4 4 4
120

Resultados
3.3. TIROIDECTOMIA EN PERIODODE DESTETEY ADULTO.
En las ratasenperiodode destetey adulto, sellevó a caboun estudioanálogo
al efectuadoen la etapaneonatal.Se sacrificarona los 5, 10 y 15 díasdespuésde la
operación(27, 32 y 37 díasde vida para los animalesdestetadosy 77, 82 y 87 días
de vida, paralos adultos).
3.3.1. Evolución del pesocorporal.
En las Tablas6 y 7 se recogenlos resultadosdel pesocorporalde las ratas
destetadasy adultas.Seobservóen las dospoblacionesde animalestiroidectomizados
un descensoen los valoresde esteparámetro,que llegó a serde un 34.37%paralas
ratasdestetadasa tos 32 díasy de un 14.20%paralas adultasa los 87 días,respecto
a los animalescontrol.
Los tresestadiosdeedadesconsideradosparaestosanimalestiroidectomizados
y en ambaspoblaciones(destetaday adulta)presentabanel pesodisminuido respecto
a los controles.
3.3.2. Nivelesséricosde T3 y T4 y plasmáticose hipofisariosde GH.
Las ratas tiroidectomizadas,tanto destetadascomo adultas,mostraronunos
nivelescirculantesde T3 y ‘[4 disminuidosdespuésde la operación(Figura 18). Esta
disminuciónfue más acusadaa los 15 días despuésde la tiroideetomíaquea los 5
dias, y así, en el caso de los animalesdestetados,seprodujo una caída desdeun
86.70% (27 días de vida, 5 díasdespuésde la operación)a un 94.89%(37 díasde
edad,15 díasdespuésde la tiroidectomía)para ‘[3 y desdeun 90.81%a un 93.09%
para‘[4; paralas ratasadultas,dichodescensooscilóentreun91.49%(5 díasdespués
de la tiroidectomía,77 díasde edad)y un 92.24%(15 díasdespuésde la operación,
87 días de vida) para ‘[3, y desdeun 85.63% (77 días) a un 92.02%(87 días)para
‘[4.
Los nivelesplasmáticosde OH, de maneraparalelaa lo que sucedíacon el
pesocorporaly los valorescirculantesde T3 y T4, experimentaronun descensoen los
animalestiroidectomizadosdestetadosy adultos (Figura 18 y Tablas 6 y 7). Sin
embargo,estadisminuciónde OH en sangreseprodujoen las ratasdestetadasa los
15 díasdespuésde la tiroidectomía(37días), mientrasqueen las adultassemanifestó
ya a los 5 díasdespuésde la operación(77díasde vida).
El descensode OH en pituitaria a los 5 díasdespuésde la tiroidectomíafue
de un 35.00% en las ratasdestetadasy de un 53.00% en las adultas,aunqueesta
diferenciasuperóel65.00%en ambosgrupos15 díasdespuésde laoperación(Tablas
121
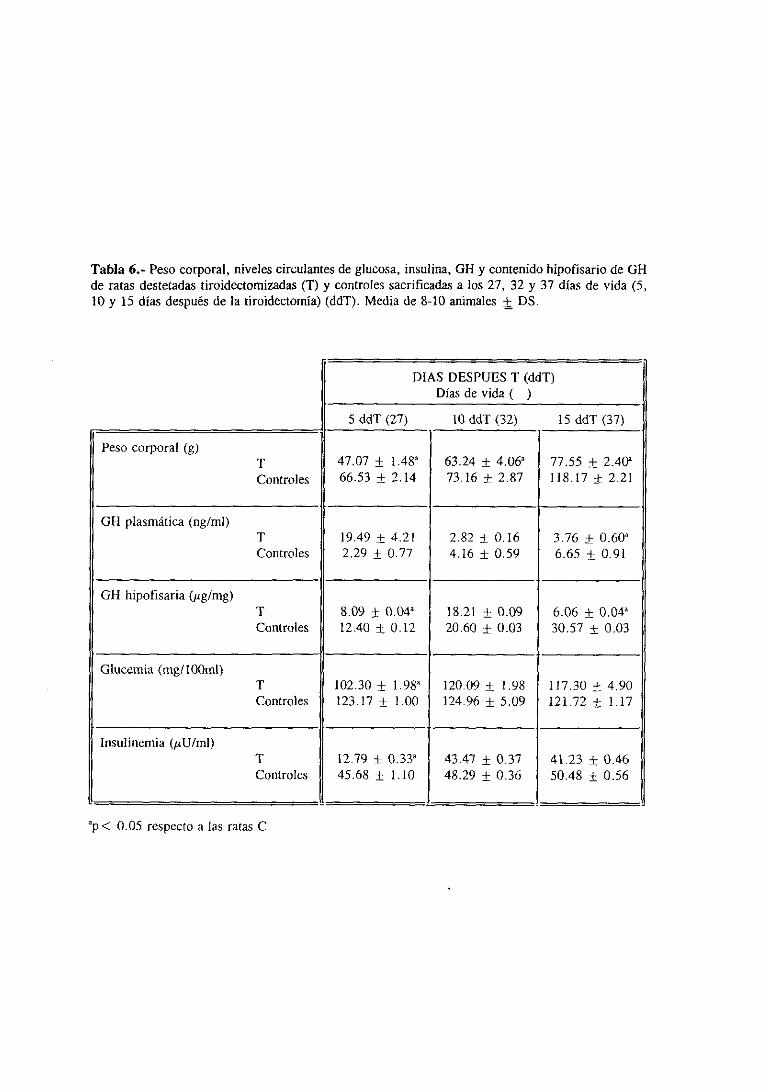
Tabla6.- Pesocorporal,nivelescirculantesde glucosa,insulina, OH y contenidohipofisariode OHde ratasdestetadastiroidectomizadas(T) y controlessacrificadasa los 27, 32 y 37 días de vida (5,10 y 15 díasdespuésde la tiroidectomía)(ddT). Media de 8-10 animales+ DS.
DíAS DESPUES1’ (ddT)Días de vida
Pesocorporal (g)TControles
4707 ±1.48’66.53 + 2.14
63.24 + 4.06’73.16±2.87
77.55 + 2.40’118.17 + 2.21
OH plasmática(ng/mí)TControles
19.49±4.212.29 + 0.77
282 + 0.164.16 + 0.59
376 + 0.60’6.65 + 0.91
OH hipofisaria(pg/mg)TControles
8.09 + 0.04’12.40 ±0.12
18.21 + 0.0920.60 ±0.03
6.06 + 0.04’30.57 + 0.03
Glucemia(mg/lOOml)TControles
102.30± 1.98’123.17 ±1.00
120.09 + 1.98124.96 + 5.09
117.30+ 4.90121.72 + 1.17
Insulinemia(1xU/ml>‘¡7Controles
12.79 ±0.33’45.68± 1.10
43.47 + 0.3748.29 ±0.36
41.23 + 0.4650.48 + 0.56
1 5 ddt (27) lO ddT <32) 15 ddT (37)
‘p < 0.05 respectoa las raíasC
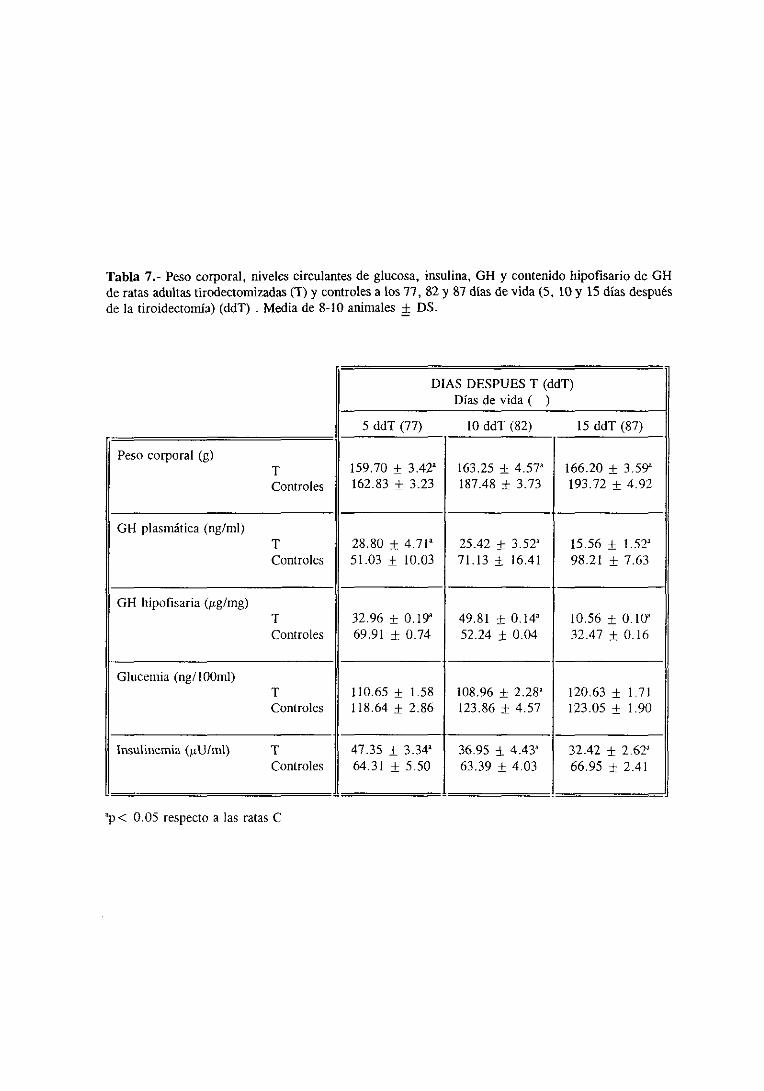
Tabla 7.- Pesocorporal,niveles circulantesde glucosa,insulina, OH y contenidohipofisario de OHde ratasadultastirodectomizadas(17)y controlesa los77, 82y 87 díasdevida (5, 10 y 15 díasdespuésde la tiroidectomía)(ddT) . Media de 8-10 animales+ OS.
DíAS DESPUESY (ddT)Días de vida
Pesocorporal (g)TControles
159.70 ±3.42’162.83 ±3.23
163.25 + 4.57’187.48±3.73
166.20 ±3.59’193.72±4.92
OH plasmática(ng/mí)1Controles
28.80 + 4.71’51.03 ±10.03
25.42 + 3.52’71.13 + 16.41
15.56 ±1.52~98.21 ±7.63
OH hipofisaria(pglmg)T
Controles
32.96 ±0.19’
69.91 + 0.74
49.81 + 0.14’
52.24 + 004
10.56±0.10’
32.47 + 0.16
Glucemia(ng!IGOmí)TControles
110.65 + 1.58118.64 ±2.86
108.96 + 2.28’123.86±4.57
120.63 + 1.71123.05+ 1.90
Insulinemia(pAl/mi) 17Controles
47.35±3.34’64.31 ±5.50
36.95 + 4.43’63.39 + 4.03
32.42 ±2.6T66.95 ±2.41
5 ddT (77) 10 ddT (82) 15 ddT (87)
‘p< 0.05 respectoa las ratasC
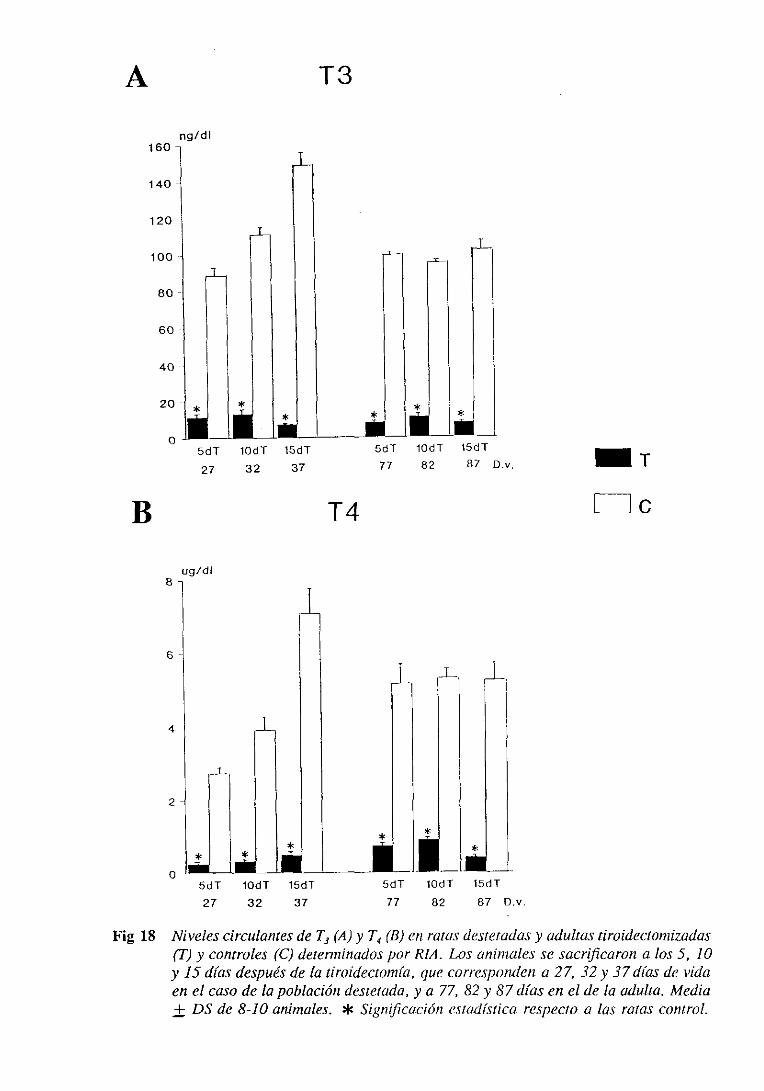
A
160
140
120
loo
80
60
40
20
o
13
ng/dl
B
8
e
4
2
o
T4 LIZo
Dv.
Fig 18 Nivelescirculantesde T3 (A) y 7% (13) en ratasdestetadasy adultastiro idectomizadasrn y controles(Q derenn¡nadospor RÍA. Los animalessesacrificaron a los 5, 10y 15 díasdespuésde la tiroidectomía,que correspondena 27, 32y 37díasde vidaenel casode la poblacióndestetada,y a 77, 82 y 87díasen el de la adulta. Media+ DSde 8-10animales. * Sign¿ficaciónestadísticarespectoa las ratas control.
5dT lOdT lSdT 5dT lOdT I5dT
27 32 37 77 82 87 Dv.
ug/dI
bdT lOdT l5dT 5dT lOdT l5dT
27 32 37 77 82 87

Resultados
6 y 7).
3.3.3. Glucemiae insulineinia.
Los nivelesde insulinadisminuyeronen los animalesdestetadosa los 5 días
despuésde la tiroidectomía, pero no a los 10 ni a los 15 días despuésde la
operación; las ratas adultas hipotiroideas, sin embargo, presentaronvalores
plasmáticosde estahormonainferioresa los de sus respectivoscontrolesdesdelos
5 díasdespuésde la operación(Tablas6 y 7).
La glucemiadescendiórespectoa las ratascontrol,en los animalesdestetados
a los 5 díasdespuésde la operacióny en los adultosa los 10 días, mientrasque en
los otros estadiosanalizados,los nivelescirculantesde glucosamostraronunascifras
similares a los de la poblacióncontrol (Tablas6 y 7).
3.3.4. IGFs: nivelesséricosy expresióndel ARNm hepático.
3.3.4.1.Nivelesséricos.
Los niveles de IGF-I en suero fueron más bajos en los animalesoperados
repectoa los controlestanto a los 10 díasdespuésde la tiroidectomía(40.46%a 32
días de vida y 82.55%a 72 días) comoa los 15 días (45.52%a 37 díasy 32.09%
a 87 días) (Figura 19). Sin embargo,sólo los animalesdestetadosmostraronuna
diferenciasignificativa en cuantoa los valores circulantesde IGF-J respectoa sus
controlesa los 5 días despuésde la operación,quizás por falta de población en
adultas.
El IGF-II sólo fue dosificadoen los animalesneonatales,ya que sus niveles
en las ratas destetadasy adultas son despreciables,se sitúan por debajo de la
sensibilidaddel método.
3.3.4.2.Expresióndel ARNm hepático.
La expresióndel IGF-I en hígadofue inferior para las dos poblacionesde
ratasdetetadasy adultastiroidectomizadasen los tres estadiosanalizadossiempre
respectoa los animalescontrol (Figura 19); este hecho, correlacionadocon los
nivelesséricosanteriormentemostrados,nos hacesugeriruna posibleregulacióna
nivel transcripcionalen la secreciónde los IGFs por la falta de hormonastiroideas.
3.3.5. Estudiode regresiónlineal y correlación.
La Figura 20 resumelos resultadosdel análisisde la regresiónde algunode
los parámetrosestudiadosen estosexperimentos.Paralos cálculos se reunieron¡os
datosobtenidosde los tres estadiosde las poblacionesdestetaday adulta.
Las ratasdestetadasy adultaspresentaronunaalta correlaciónentrelos niveles
125

Fig 19 Niveles séricos (A) y expresiónhepática del ARNm (fi) de ¡GF-J en ratas destetadasy adultastiroidectomizadas<W) y controles(C) a los 5, lOy 15 díasdespuésde la operación (27, 32 y 37 ó77, 82 y 87 días, respectivamente).
A) Media ±DSde 8 animales.B) Ensayodeprotecciónde la RNasay cuantf/¡cacióndensitométrica(5 análisisdiferentes).
* Sign¿/¡caciónestadísticarepectoa la población de animalescontrol.
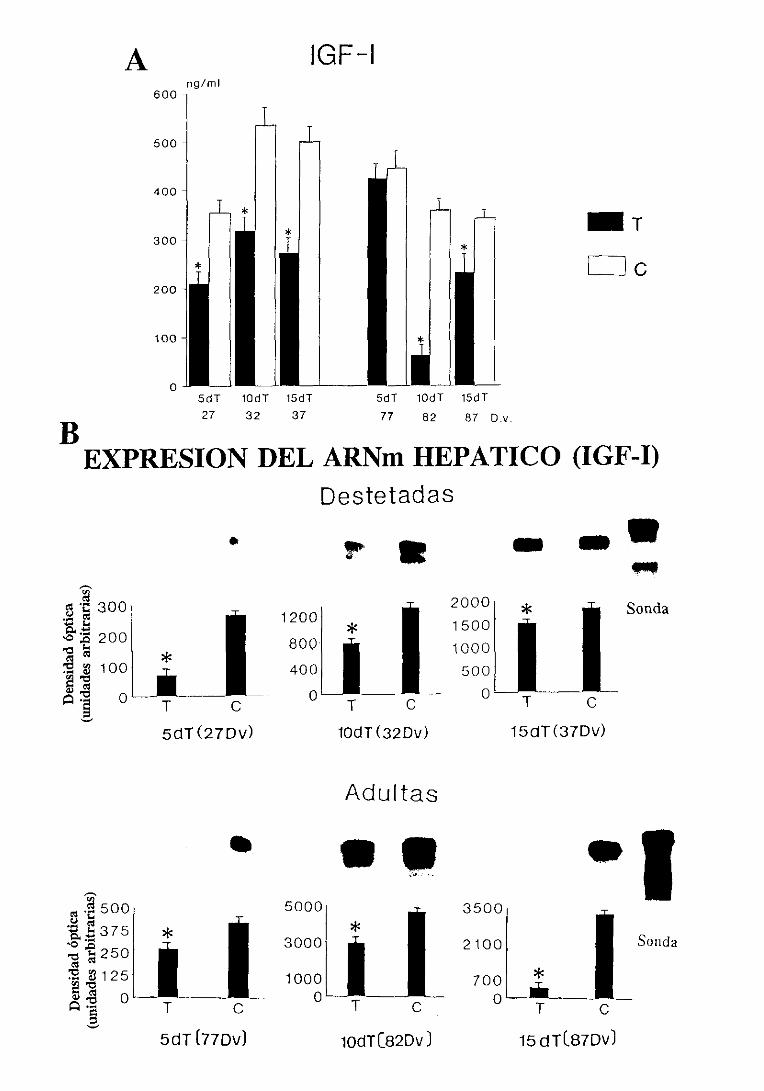
‘OF-’ng/ml
27 32 37 77 82 87 Dv.
EXPRESION DEL ARNm HEPATICO (IGF-I)
Destetadase ——e
u”
300
200
100
T O
12000
1500
1000
500
o
*jiT O
5dT (27Dv) lOdT(32Dv) l5dT(37DV)
Adultas
*Ka]1 0
5000 __
1* U300011 u1000 .1
o ¶ O
3500
2100
700
o
A600
500
400
300
200
loo
o
o
B5dT lOdT l5dT 5d7 lOdT l5dT
Senda
T O
500
~t -,375~0 ~
.~ ~250~ ~125
o
e
So¡ida
T o
5dT (770v] 1OdTC82DV 9 15 dTCBZDV)
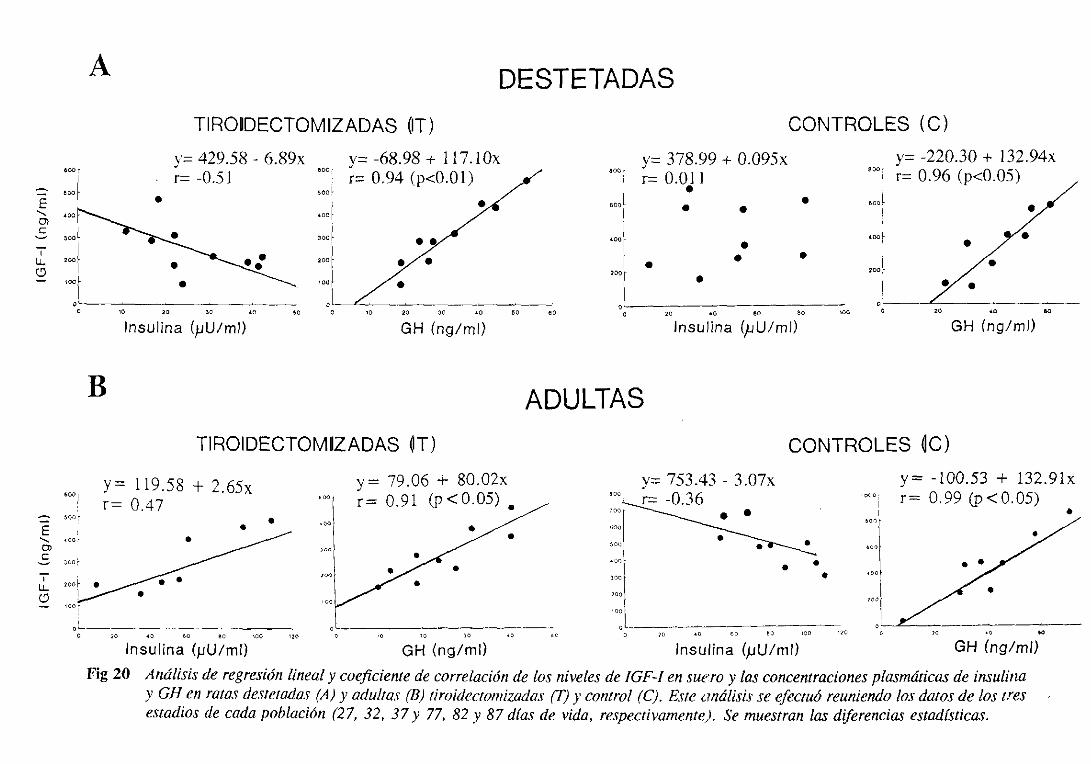
ci,-’
mr
—C
+o
CV
c’4c\
ci
oII—
‘t-
a
Éso&oo+&
—e
en
oII
1~
EO)
1(1J
ee
o
E
•e
e
CE
ee
a,E
e
oo
oo
oo
oo
o
o-4—ooo
-&‘ou
~
Ir0>
’J0
JI-‘¼
1-,-
e8
a0w-Jolx1—zoo
y-4
(n‘e
+0
yenC
cx
—4do
ji1-,
2
yen
enen
es
‘soyoII
CoCC-JDoCC
o-~O
>
JZ2
01
—
COci:o4N
l
o1—oLUoÓ
cl
oC
03E
eyCte.0
oy
+5
0~
ON
Ox
ho
IIti
;:—,
1-4
y‘so‘ocl
+00
Ir—o
II1-
282
3
2
E~
..%.
.~
o,‘—
EIt-ao
Ec~
~J
9E
t$~
~ti
-D~
~~
E
k.t~
‘02-9
2~
-.<
u
rN~
.
~e
t~
cc
3—
1
~en
~2
u“-a~
D-~
s...~
..—ti
ti
‘~
2~
‘C~
—.
~0
,
4
e
e38
oo—
ChLU-Jocr1—zoo
(1)CCOCCLU1—Cf>InO
1
-
COCCO4Nl
2o1-
ooO
e
ee
(IW/e
u)
1-191(íw
¡6u
)1
-Ao
l

Resultados
deOH enplasmay los de IGF-1 en suerotantoen los animalestiroidectomizados(r~
0.94, p< 0.01 y r= 0.91, pcZO.05, respectivamente)como en los controles(r=
0.96, Pc 0.05 y r= 0.99, p< 0.05) (Figura 20). Porel contrario, la correlación
encontradaentre la insulinemia y los valores circulantes de IGF-I en las ratas
destetadastiroidectomizadasy controlesfue muy bajae incluso, negativa(r= -0.51
y r~ 0.011, respectivamente),e igual ocurrióenel casode los animalesadultos(r~
0.47 y r= -0.36, respectivamente)(Figura 20).
3.3.6. IGFBPs: nivelesséricosy expresióndel ARNm hepático.
3.3.6.1. Nivelesséricos.
Las IGFBPs del suero se analizaron por ‘Western ligand blot”, y
posteriormentese hizo un estudiocomparativoempleando‘Westernligand blot” y
Westerninmunoblol’ paraidentificar IGFBP-2,dadoqueelantisueroanti-bIGFBP-2
fue el único de los ensayadosque dió reaccióncruzadacon las IGFBPs circulantes.
3.3.6.1.1.Westernligand blol.
Los niveles en suero de las proteínasligadorasde bajo peso molecular
(IGFBP-1 y 2) fueron inferiores en los animalestiroidectomizadosrespectoa sus
controlesen las dos poblacionesde ratas(destetadasy adultas)paratodaslas edades
analizadasa los 10 y 15 díasdespuésde la operación,y sólo a los 5 días despuésde
la tiroidectomíaparalos animalesadultos(Figuras21 y 22). Tambiénse encontróuna
disminuciónen la intensidadde las bandasde IGFBP-3 pata todos los animales
destetadosy adultos tiroidectomizadosrespectoa los controles (5, 10 y 15 días
despuésde la operación)(Figuras21 y 22).
3.3.6.1.2.Westernimnunoblot.
El análisis mediante Westerninmunoblot mostróque, tanto los animales
destetadoscomo los adultostiroidectomizados,y en todaslas etapasconsideradas,
presentabanun ligero aumentoen la intensidadde la bandacorrespondientea IGFBP-
2 respectoa la poblacióncontrol (Figuras21 y 22). Así pues,por lacomparaciónde
estos resultadoscon los obtenidosmediante“Western ligand blot, se podríadecir
que en estasratas(destetadasy adultas)seprodujoun incrementoen los nivelesde
IGFBP-2 circulante, y que este aumento fue menos marcado que el descenso
provocadoporel hipotiroidismosobrelos valoresséricosde IGFBP-1, puestoqueel
ligand blot, que muestra los niveles circulantes conjuntos correspondientesal
complejo constituido por las proteínasde bajo pesomolecular(IGFBP-1 y 2), nos
revelóunadisminuciónen la intensidadde estasbandasde 3OkDa(Figuras21 y 22).
128
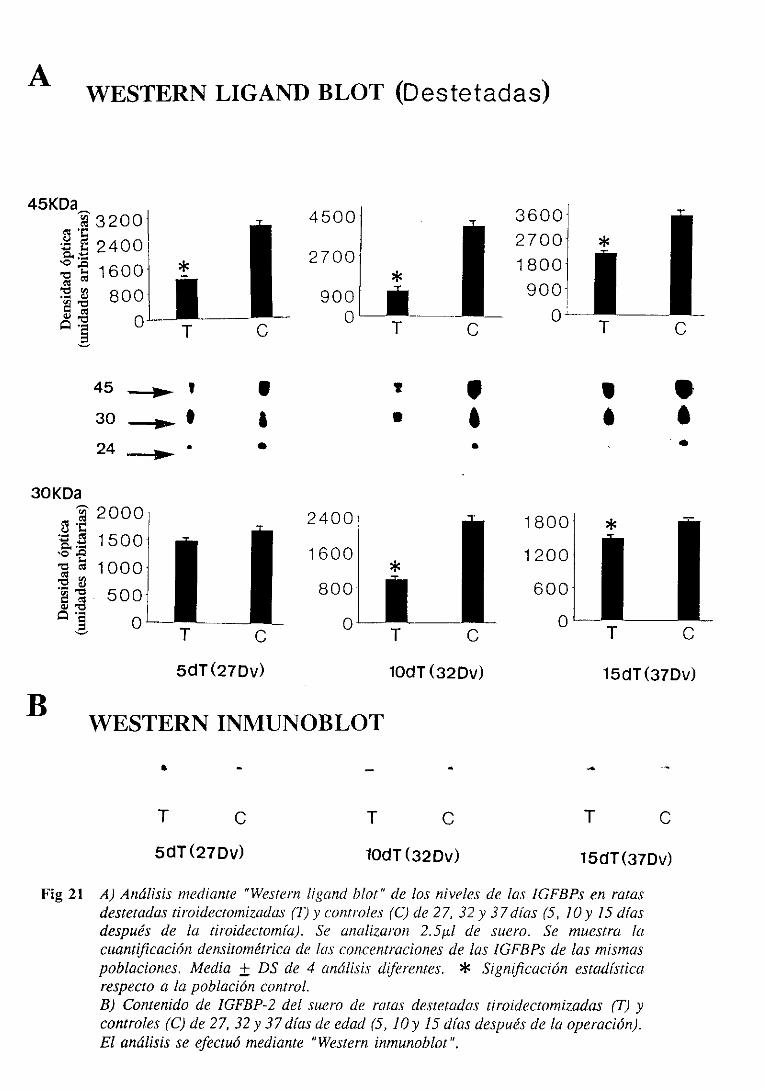
WESTERN LIGAND BLOT (Destetadas)
4u._T O
4500
2700
900o
T
36
27
189
00*
00 ¡00k00
T o
45 .—~h- Y
30 ~
24
.3
g3e4
e
*
T
1800
120 II60
T O
bdT (27Dv) lOdT (32Dv) 15 dT(37Dv)
WESTERN INMUNOB LOT
1 o T
5dT(27 Dv) lOdT (32 Dv) l5dT(37Dv)
Fig 21 A) Análisismediante“Western íígand bíot” de los nivelesde las JGFBPs en ratasdestetadastiroidectomizadas(T) y controles(C) de27, 32 y 37días(5, JOy ISdíasdespuésde la tiroidectomía~. Se analizaron 2.5i2 de suero. Se muestra lacuantWcacióndensitométricade las concentracionesde las IGFBPs de las mismaspoblaciones.Media + DS de 4 análisis diferentes. * Sign4ficaciónestadísticarespectoa la poblacióncontrol.B) Contenidode IGFBP-2 del suero de ratas destetadastiroidectomizadas(T3 ycontroles(C) de 27, 32 y 37 díasde edad(5, lOy 15 díasdespuésde la operación).El análisis se efectuómediante“Western inmunoblot”.
A
45KDa.~ .~
— 1-<c4:~‘0 .0
3 200
2400
1600
800
o
*
o
uae
3OKDa
tza‘0 .0
~
e4a
2000
1500
1000
500
o
2400
1600
800
o1 o
B
O
o T o
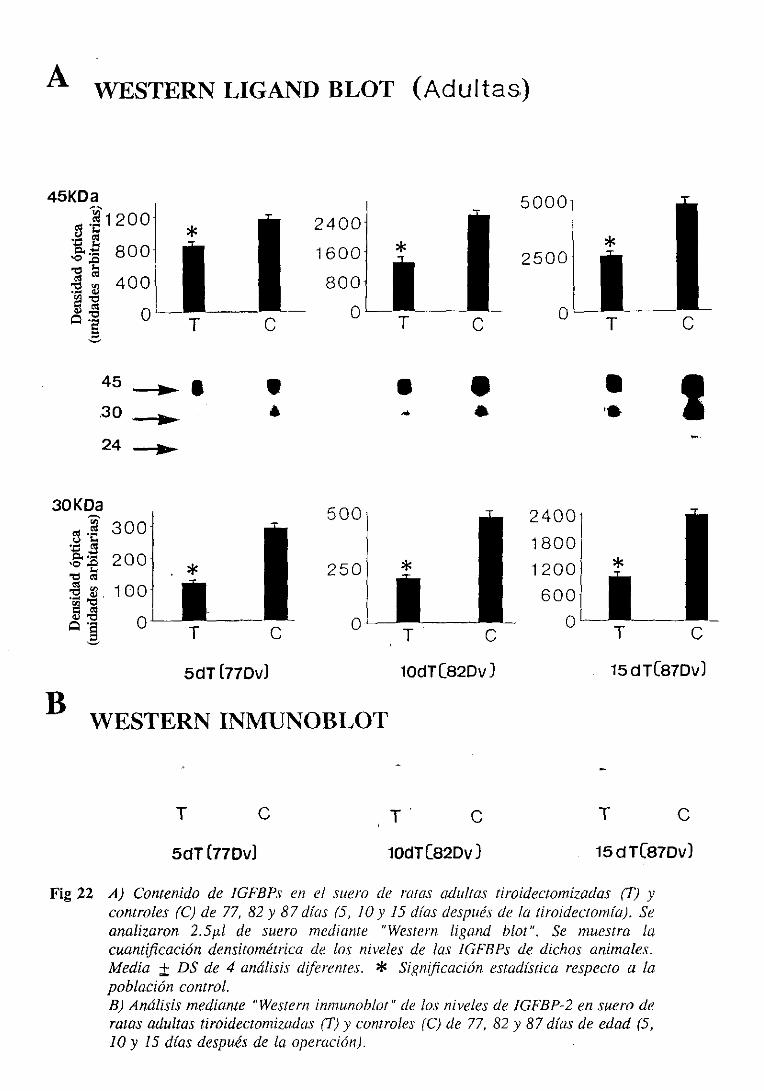
WESTERN LIGAND BLOT (Adultas)
4SKDa~1 200
.~ .~— Lt‘0 .0
o
45
30
800
400
—*1~.S4
2400
1600
800
o
ea
5000
2500
T O
a‘e
24
__¡o
500
250
oT O
WESTERN INMUNOBLOT
1 o
5dT (7lDv) lOdT CS2Dv :i 15 dTC87Ov)
Fig 22 A) Contenido de IGFBPs en el suero de raras adultas tiroidectomizadasaY ycontroles(Ql de 77, 82 y 87 días (5, 10 y 15 días despuésde la tiroidectonifa» Seanalizaron 2.5¡A de suero mediante “Western ligand blm”. Se muestra lacuat4ficacióndensirométricade los niveles de las ÍGFBPs de dichos animales.Media + DS de 4 análisis d¿ferenres. * Sign¿Iicacíónestadísticarespecto a lapoblacióncontrol.B,) Análisismediante Wesrerninmunobior” de los nivelesde IGFBP-2 en sueroderatasadultastiroidectomizadas(1) y controles(69 de 77, 82 y 87díasde edad (5,10 y 15 díasdespuésde la operación).
A
T O 1 o
3OKDa
‘0.05-
u300
200
100
o
*
T
24001800
1200600
o
B5dT(77DvJ lOdTE82DvJ
T o15 dTC87DV)
T o T o

Resultados
3.3.6.2.Expresióndel ARNm hepático.
Los niveles de ARNm hepático de IGFBP-1 disminuyeronen las ratas
destetadashipotiroideasa los 5, lay 15 díasdespuésde la tiroidectomía(Figura23).
Sinembargo,la poblaciónadultatiroidectomizadamostróun descensoen la expresión
de estaproteínade bajo pesomoleculara los 5 y 10 días despuésde la operación,
pero a los 15 díasdespuésde la tiroidectomíaaumentó,siemprerespectoa las ratas
control (Figura24). Así mismo,se observótambiéncomo a medidaque aumentaba
la edaddel animal control disminuíanlos nivelesde ARNm de estaproteína,dado
que estaIGFBP es constitutivade etapasinmaduras.
Al igual que sucedíacon los nivelesde ARNm de IGFBP-l en el hígadode
las ratascontrol, la expresiónhepáticade IGFBP-2 disminuíaconformeaumentaba
laedaddel animal,y estedescensose observómuy claramenteen la poblaciónadulta
(Figura24). Además,laexpresiónde IGFBP-2 se vió incrementadaa 5,10 y 15 días
despuésde la tiroidectomiaen las ratasdestetadasy a los 10 y 15 díasdespuésde la
operaciónen las adultas(Figuras23 y 24).
Finalmente,la expresiónhepáticade la proteínatransportadorade alto peso
molecular (IGFBP-3), característicade la edadadulta, disminuyóen los animales
destetadostiroidectomizadosa los 10 y 15 díasdespuésde la operación(Figura 23),
mientrasque en las ratasadultas,estedescensose observóya a los 5 díasdespués
de la intervenciónquirúrgica (Figura 24).
De nuevo,y al igual que ocurrió con los IGFs, encontramosla existenciade
un paralelismoentrelas fluctuacionesde los nivelesséricosde las IGFBPsy las del
ARNm hepáticode las proteínastransportadoras.
3.4. TRATAMIENTO CON INSULINA DE RATAS TIROIDECTOMIZADAS
DESTETADAS Y ADULTAS.
Paraprofundizarenel papelque desempeñala insulinaen la secreciónde los
IGFs, e intentar restaurarlos bajos valores circulantes de dicha hormona que
presentabanlos animalesdestetadosy adultosdespuésde tiroidectomizarlos,se les
inyectó insulina a los 10 días despuésde la operación (32 y 82 días de vida,
repectivamente).
3.4.1. Evolución del pesocorporal.
El pesocorporal,quedisminuyó en las ratastiroidectomizadas,aumentótras
el tratamientode estos animalescon insulinaenun 22.95%paralos destetadosy en
131
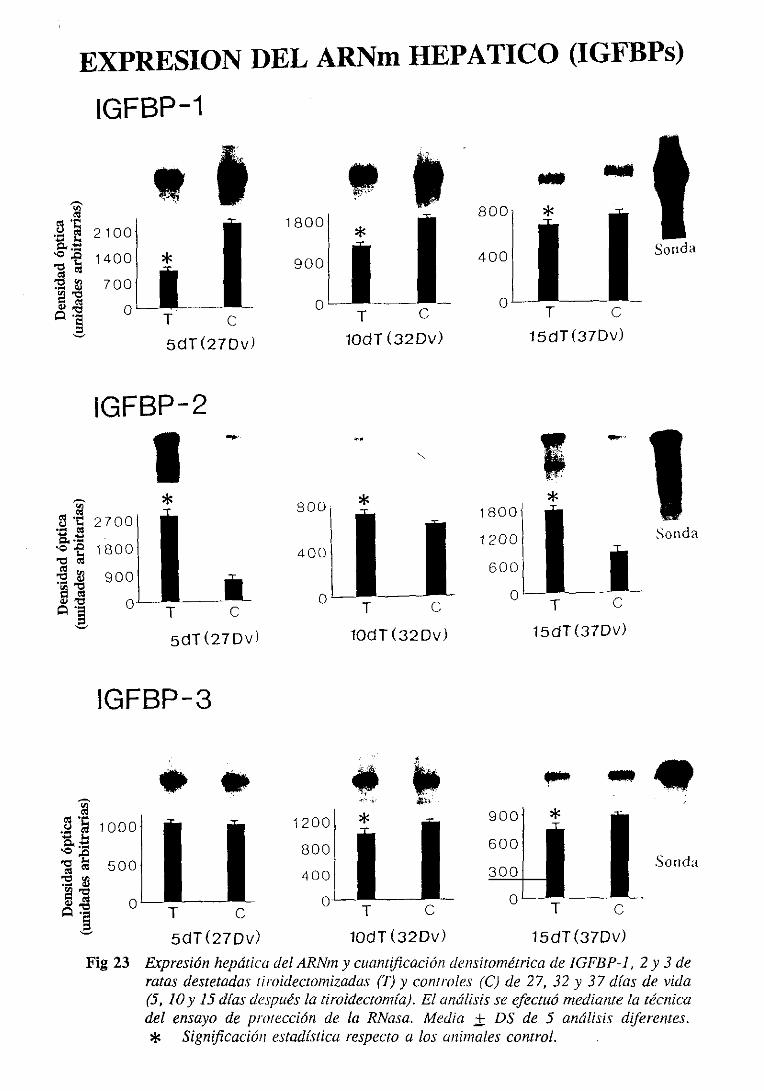
EXPRESION DEL ARNm HEPATICO (IGFBPs)
IGFBP-1
*2100
1400 * 1700
o T c5dT(27 Dv)
9~1’~¡
o1 0
lOdí (32 Dv>
a a
400800 *_¡0 0
15dT(37DV)
1
5dT (270 vi
uT C
lOdT(32 Dv)
*1800
1200
600 [j~1 C
I5dT(37DV)
IGFBP-3
*
T O
bdT (27 Dv)
*4
1200 *LT O
lOdT (32Dv)
u”
900 *
600
3003
T ol5dT(370V)
delARNmy cuantwcacióndensitométricade IGFBP-1, 2 y 3 deratas destetadastiroidecromizadas(1) y controles (6’,) de 27, 32 y 37 días de vida(5, 10y15 díasdespuésla tiroidecromía,).El análisisseefectuómediantela técnicadel ensayode prwección de la RNasa. Media + DS de 5 análisis diferentes.* Sign~caciónestadísticarespectoa los animalescontrol. -
.~ i—
‘0 .01~
Sonda
IGFBP- 2
u*
t4- —04~~‘o .05-
2700
1800
900
o T O
Soada
9i
.~ .~4- L~
‘0.0
1000
500
o
Fig 23 Expresiónhepática
Sorida
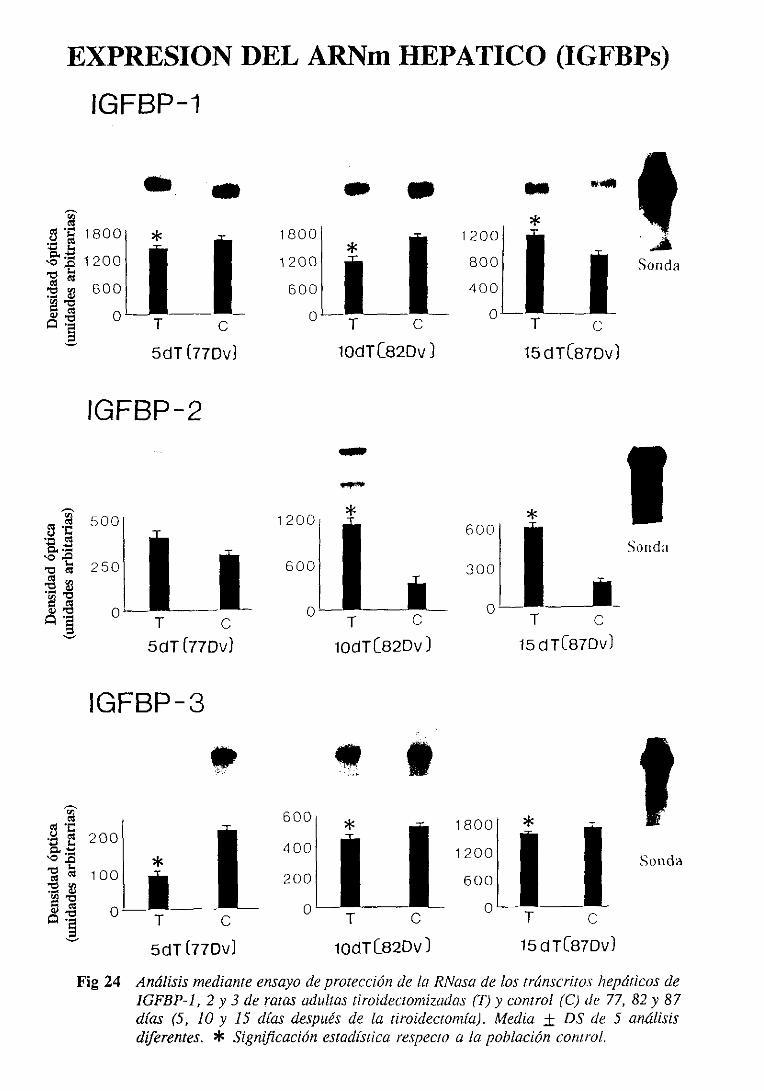
EXPRESION DEL ARNm HEPATICO (IGFBPs)
IGFBP-1
e e
1800
1200.3
600
T o
5dT (770v]
e e
800
12001 Li600
T O
1200
800
lOdTCB2Dv J
*
T o
15dTCB7Ov)
IGFBP-2e
500
250~IAT O
5dT (770v)
1200’
0— T
600
1~O
lOdTÉ82Dv J
¿300
o
*
15dTCS7DVJ
IGFBP-3
e
200
II-100o T O
5dT (770v]
e600
400
200 L1______O 1
1OdTC82DVJ
e1800
1200 ¡600
0 0
15 dTC87Ov)
O
Fig 24 Análisismedianteensayodeprotecciónde la RNasade los trónscriboshepáticosdeIGFBP-1, 2 y 3 de ratasadultas tiroidectomizadas(2’) y control (69 de 77, 82 y 87días <5, 10 y 15 días despuésde la tiroidecromía). Media + DS de 5 análisis
.~ .~4- 5-
‘0 .0
Cd
Sonda
— —o..,..‘0.0
‘4
~
Sonda
1 0
.~ .~
4- 3-Os.-.‘oE Senda
diferentes. * Sign¿ficaciónestadísticarespectoa la poblacióncontrol.

Resultados
un porcentajemenorparalos adultos(8.71%). No obstante,tanto a los 37 como a
los 87 días de vida (ratas destetadasy adultas, respectivamente)el pesode estas
poblacionestratadasfue inferior al de sus respectivoscontroles(Tabla 8).
3.4.2. Nivelesséricosde 123 y 124 y plasmáticose hipofisariosde GH.
Les valorescirculantesde las hormonastiroideasde las ratastratadascon
insulina permanecieronpor debajode los nivelesde la poblacióncontrol (Tabla 8).
La concentraciónséricade T> aumentórespectoa los animalestiroidectomizados,
tantoenel casode las ratasdestetadas(57.41%)comoenel de las adultas(27.55%),
sin embargo, los niveles de 124 no se modificaron respectoa los de la población
operadaen destetadasy bajaronen la adulta.
El tratamientocon insulinade las ratastiroidectomizadasprovocóun aumento
distinto enporcentajede los nivelesde OH circulantey que difirió segúnla edadde]
animal (Tabla 8): 25.43%paralas destetadasy 63.56%para las adultas,respectoa
las ratasoperadas.De un modo similar, el contenido de Gil en hipófisis se vió
incrementadoen un porcentaje superior al 35.00% para ambas poblaciones
hipotiroideasinyectadascon insulina respectoa las ratastiroidectomizadas,aunque,
como sucedió en el plasma, los niveles alcanzados continuaron siendo
significativamenteinferiores a los de los animalescontrol.
3.4.3. Glucemiae insulinemia.
La insulinemiade las ratastiroidectomizadassometidasal tratamientocon la
hormona,comoseesperaba,fue significativamentesuperior(61.26%en el casode
las destetadasy 48.73%enel de las adultas)a la de la poblacióncontrol(Figura25).
Sin embargo, en el momento del sacrificio, no se encontraron diferencias
significativasentre las glucemiasde estasratashipotiroideastratadascon insulina y
las de las ratascontrol, quizáporhaberserestauradoya la hipoglucemiaproducida
por la administraciónde la hormona(Tabla 8).
3.4.4. IGFs: nivelesséricosy expresióndel ARNni hepático.
3.4.4.1. Niveles séricos.
Los nivelesde IGF-I ensuerofueronmásbajosen los animalestratadoscon
insulina que en los controles,y semantuvieronigualesa los tiroidectomizados,a 37
y 87 díasde Vida (Figura25). Así, y a pesarde que la administraciónde la insulina
aumentósus propios niveles circulantes,no modificó los bajos valores de IGF-I
disminuidospor la tiroidectomía.
134
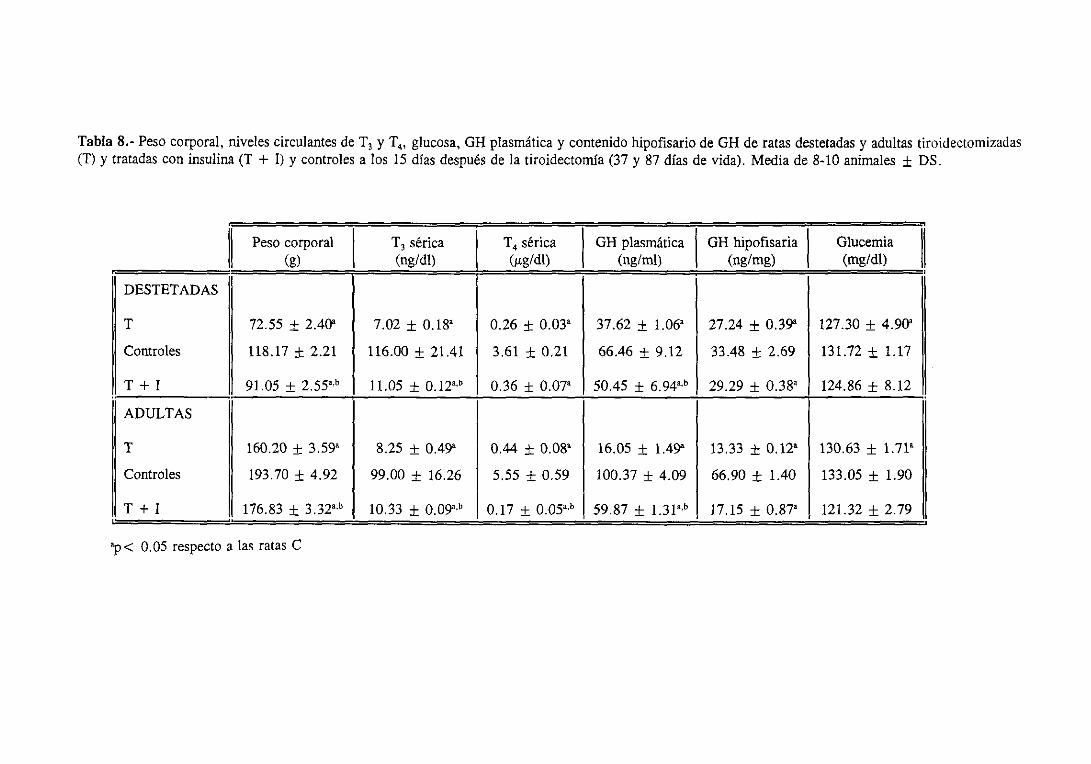
4.)uto~
(it
br-
c’Uo
st
——
+1-n
-n
2C
li—
—
~c
osr- 0
~-4
—el
-IAH
+4
Cflu-~
elVn8en—
——
ccE—
02zO
¿C
l8
-IH±I
+1
t00
osC
l~
Cl
r.<~
osC
lV
n
~o
~8
—8
en0
‘¿~—~
so—
—
clo
O
.0
C
0e
l4
Ot
os
4o
s
-H+
1±1
el
xt
en
~t
o
.0
~C
N~
.—
—-H
±1
2q
r-•
.00
0‘~
Oo
s—
—
-eVIÓfl
enOeM
0¿
0¿
-+4+~
-i-~‘0
‘so
ej‘~.
enO
rnO
.0
So
os
o.fl
¿0
oO+
4.~
<+
1
e-.
¿u
,~
‘oto
—a
~4
2
tel
.0
±1
-n-~-~
oe
lO
IC>
9~
¿9
4-
——
—
‘4os
O—
+1
-f~
./->O
en
C1
9o
co
sos
a.>
••
Cl
el1
-H
00
0tn
.
—I
os
Cl
.0¿Cl
en
en
’Ú±í-f4
~en
00
8~o
os
~o
——
00-<
-<E-
E-
00
‘o4>o—
4-.~
+o
E-U
E-
00
E-
VI4>o‘-4
oE
-U+E-
U“oclCd‘-4
VICdCdOUa.>VI
a.>14
oOy2<
cl-eclNEoo4>-eo
c~
—£0
‘e-’
clc
l
VIO
1-~
~4.>4
-~0
0CA4
>o
‘U-u
~‘e4>
>•d
4>o-e~‘4-4
or—
o17—
-de
n
00
oo
>,
4.>
cl:~
Cje
14‘c
lZ
CM
-U04‘ocl4
>
Uy,
;tfl
E-—
CV
I
ojO
CI’
—o
o>
~‘-4ci
—
Ce
o-O
oc
00
00
It,VI
Ve
d
cl.0
~~
-.
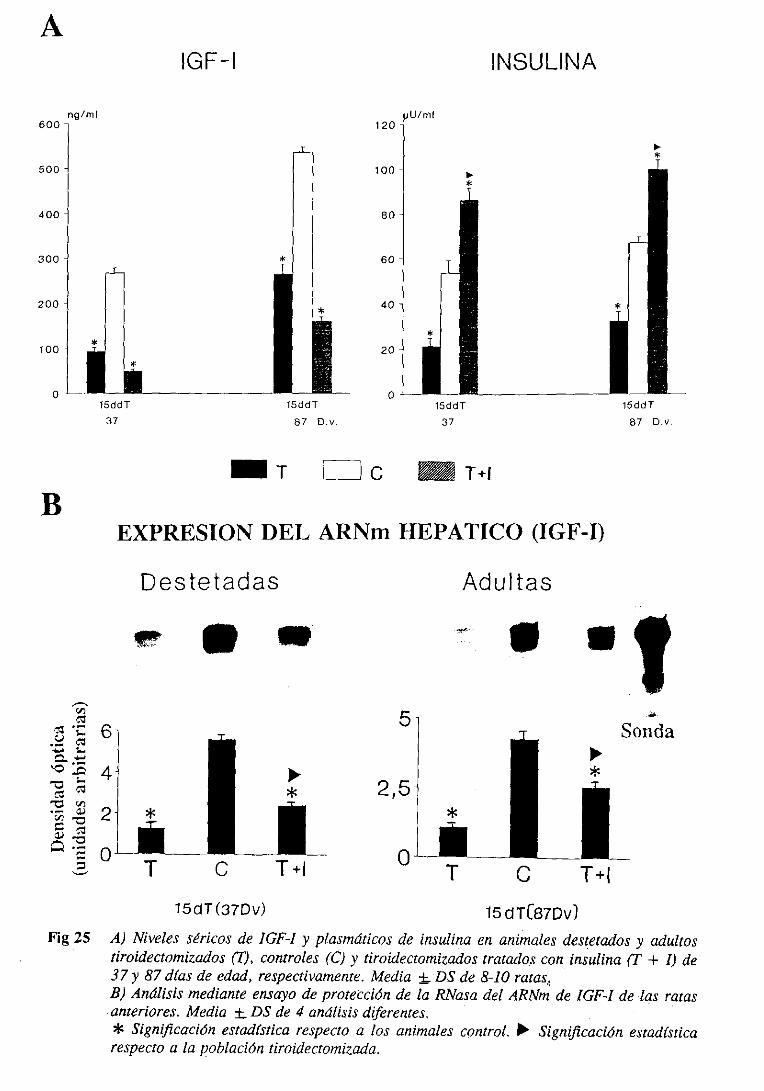
IGF-I
ng/mr
lSddT ISddT
37 87 Dv.
Lii
INSULINA
1 20 -100 -
80 -
60 -
40 -
20
0
U/ni
o PI
EXPRESION DEL ARNm HEPÁTICO (JGF-I)
Destetadas
*
T
Adultas
e
*
0 1+1
5
2,5
0~—
*
Tl5dT (370v)
u
Sonda
*
1+I
15 dTC87DvTiFag 25 A) Niveles séricosde JGF-J y plasmáticosde insulina en animalesdestetadosy adultos
tiroidectomizados(‘¡9, controles (6’) y tiroidectomizadostratadQscon insulina <1 + 1) de37y 87díasde edad> respectivamente.Media + DSde 8-JOrasas~.B) Análisismedianteensayode protecciónde la RNasadel ARNmde IGF-¡ de las ratas
• anteriores.Media + DSde 4 análisisdiferentes.* Significación estadísticarespectoa los animales control. > Significaciónestadísticarespectoa la población tiroidectomizada.
A
600
500
400
500
200
100
o
*
*
*
B
l5ddT lGddT
37 87 Dv.
.~ e~— %.
-~ r¡~.-4)~4D-o
6
4
2
o
~1~
o

Resultados
3.4.4.2.Expresióndel ARNm hepático.
La Figura 25 muestraque, tanto en las ratastiroidectomizadasdestetadas
comoen las adultas,laadministraciónde insulinaprovocóunaumentoen los niveles
del ARNm hepáticode IGF-I respectoa la poblaciónoperada,aunquedichosvalores
no llegarona alcanzarlos de las ratascontrol (sesituaronpor debajode ellos). De
nuevo, comoocurríaa nivel circulante,seobservócomo el tratamientocon insulina
no era capazde normalizarla bajaexpresiónde los tránscritoshepáticosde IGF-I
causadapor la tiroidectomía.
3.4.5. Estudiode regresiónlineal y correlación.
Parael análisisde correlaciónse recogierontodos los datoscorrespondientes
a las dospoblacionesde los dos estadiosy se muestranen la Figura 26
Al igual que enel caso de las ratasdestetadasy adultastiroidectomizadasy
controles, se volvió a encontraruna alta correlaciónentre los niveles de OH en
plasmay los de IGF-I en suero(r= 0.91 y r~ 0.98, p’Z 0.05 para las poblaciones
hipotiroideasdestetaday adulta inyectadascon insulina, repectivamente).Por el
contrario,la correlaciónhalladaparala insulinemiay los nivelesséricosde IGF-I fue
de nuevo,muy bajae incluso, negativaen los animalesdestetadosy adultostratados
con insulina (r= -0.038y r= 0.62, respectivamente)(Figura 26>.
3.4.6. IGFBPs: nivelesséricosy expresióndel ARNm hepático.
3.4.6.1.Niveles séricos.
3.4.6.1.1.Westernligand blot.
La tiroidectomíacausóun descensoen los niveles séricos de las proteínas
transportadorasde 30 (IGFBP-1 y 2> y 45kDa frente a la poblacióncontrol. No
obstante, la administraciónde insulina a estos animales operadosprodujo un
incrementoen los valorescirculantesdel complejoproteicode bajopesomolecular,
tanto en las ratasdestetadascomoen las adultas,aunqueen ningunade estasdos
poblacionesse llegarona alcanzarlos valorescontrol (Figura27).
Encuantoa la IGFBP-3,en los animalesmásjóvenes(destetados),la insulina
sí aumentósus niveleshastaalcanzarlos valorescontrol, aunqueen el caso de las
ratasadultas,el tratamientohormonalno alteró lascifras circulantesde estaproteína
transportadora(Figura 27), quizás porque la insulina tiene un mayor papel en la
regulacióndel sistemaIGFs/IGFBPsen etapasmás inmadurasdel desarrollodel
animal que en el estadioadulto, dondepareceserque la CH pasaa tenerunamayor
relevancia,o tambiénporquelos niveles de Gil despuésde la administraciónde la
137
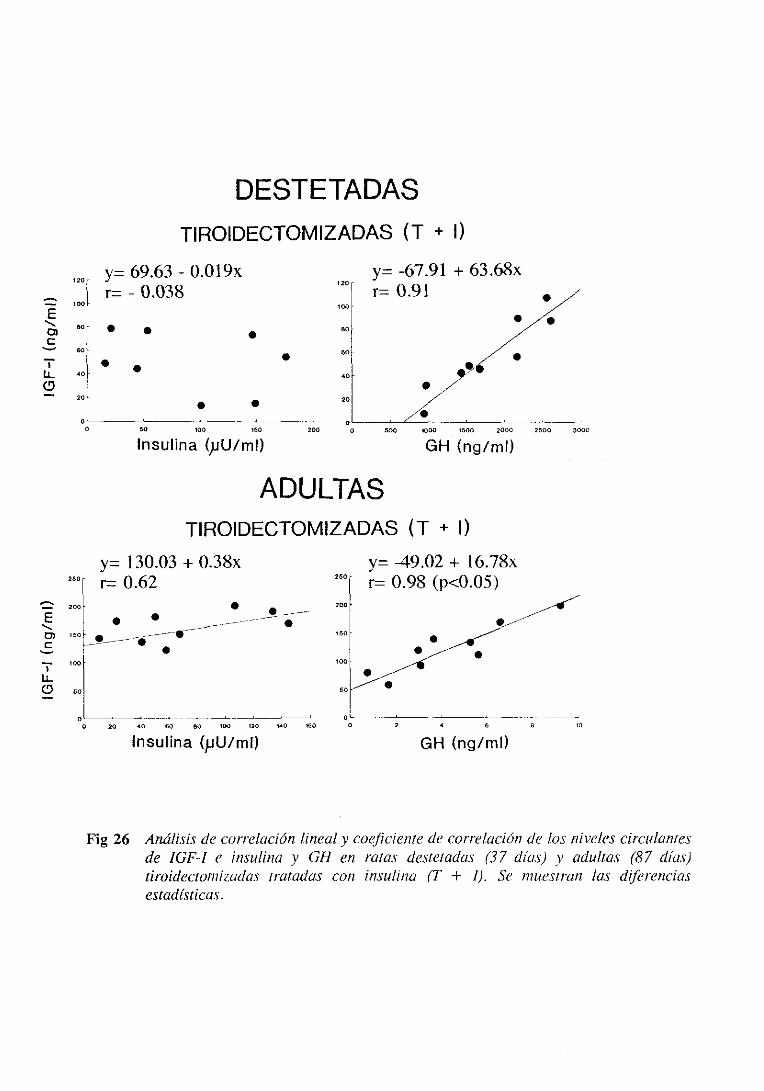
DESTETADAS
TIROIDECTOMIZADAS (T
y= 69.63 - 0.019x- 0.038
e e
20
y= -67.91 + 63.68xr= 0.91
100 -
50e
60ee
40
20e e
0—
o 50 lOO 150 200
Insulina (pU/mi)
e
e0 500 1000 1500 2000 2500 2000
GH <ng/mi)
ADULTASTIROIDECTOMIZADAS (T +
y= 130.03 + 0.38xr= 0.62
e
250 -
ee
200
50
eloo -
00
o •--
0 20 40 60 80 00 20 140 00
Insulina (pU/mi)
y= -49.02 + 16.78xr= 0.98 (p.cO.O5)
ee
ee
e
0 ---—~ •.——————— - -- -•
0 2 4 6 8 10
GH <ng/mi)
Fig 26 Análisisdecorrelaciónlineal y co«ficientede correlaciónde los nivelescirculantesde IGF—I e insulina y Gil en ratas destetadas~37días) y adultas (87 días)riroidectomizadastratadas con insulina (T + 1). Se muestran las diferencias
120 -
lOO
+ 1)
a
Yu-o
so
60
40
20
e
ee
e
e
250 -
200
lOO —•
Eo)c
1>~a
loo -
50
estadísticas.
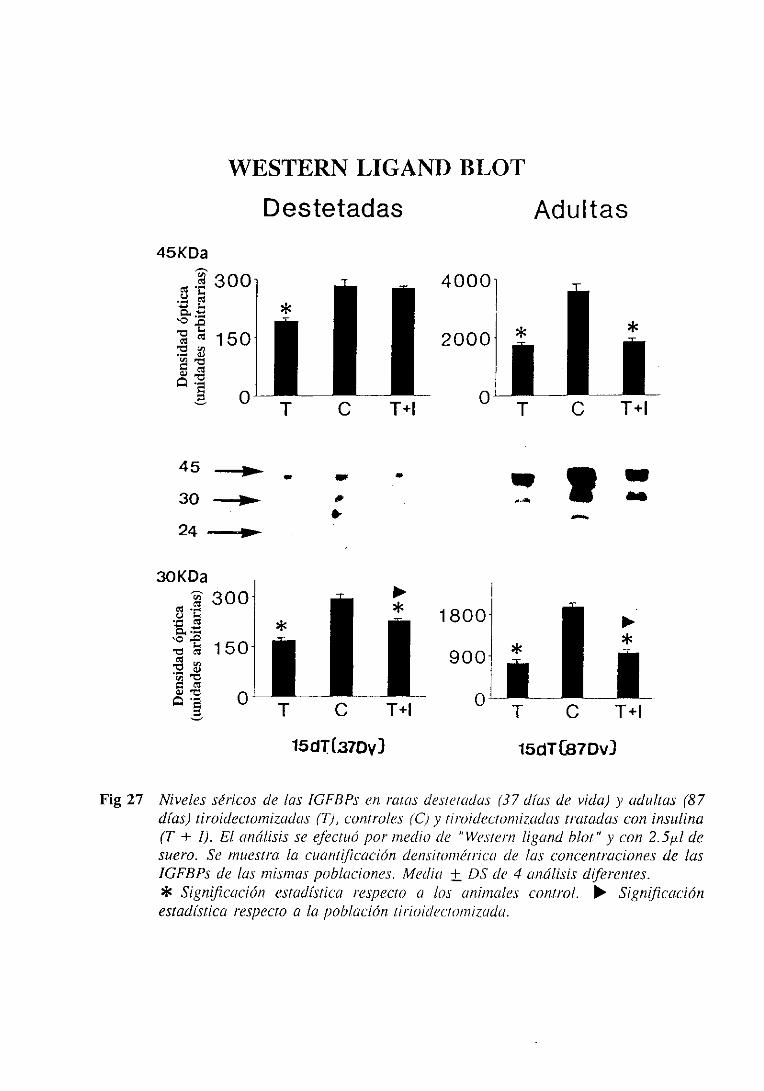
WESTERN LIGAND BLOT
Destetadas Adultas
T O Pi
45 —.*-.
30 —a..-
24 —~..-
3OKDa
~ t—Cd—4-.o.—‘o .04-.‘e CoCd
‘U
e
e’o,
a
300*
*150
011 0 T~I
‘u-4
1800
900~
0-
15d1(slDv)
a
iiiT O Pi
l5dTC8lDvJ
Fig 27 Nivelesséricosde las ZGFBPs en. ratas destetadas(37díasde vida) y adultas <‘87
días) tiroidectomizadas(T), controles (6’,) y timídectonilzadastratadascon insulina(T + 1). El análisis seeJéctuópor mediode “Western íigandbloC y con 2. S~~l desuero. Se muestra la cuant¿ficacióndensirométricade las concentracionesde lasIGFBPs de las núsinaspoblaciones.Media + DS de 4 análisis diferentes.* Signfficacíón estadística respecto a los animales control. ~ Sign¿ficaciónestadísticarespectoa la población.tírioídecro¿nizada.
45KDa
.~ •g4- 5-
300
150
0-
4000
2000
o1 0 Pi

Resultados
insulina fueron superioresen la poblacióndestetada.
3.4.6.2.Expresióndel ARNm hepático.
La IGFBP-1, como ya se ha mencionado,se encuentra reguladapor la
insulina, por ello, no fue extrañoel hechode que las ratassometidasal tratamiento
experimentaranunadisminuciónen la expresiónhepáticade estaIGFBP-1 respecto
a la poblacióncontrol, y que tal descensofuera incluso significativamentesuperior
al padecidopor los animalestiroidectomizados;estadisminuciónen los niveles de
ARNm de IGFBP-1 pareciósermásacusadaenel estadioadultoque enel destetado
(Figura 28).
Les nivelesde ARNm de IGFBP-3 aumentarony sehicieron comparablesa
los controles tras el tratamiento con insulina, tanto en el caso de las ratas
tiroidectomizadasdestetadascomo en las adultas, aunque en estas últimas, el
incrementofue mayory superóincluso los valorescontrol quizásporel aumentode
la GH (Figura 28).
En cuanto a la expresión hepática de IGFBP-2, en la población
tiroidectomizadadestetadaa la que se le administróla hormona,disminuyórespecto
a estosanimalesoperados,de manerasimilar a lo que sucedíaen el caso de los
neonatos tiroidectomizados tratados con STZ, mientras que en los animales
tiroidectomizadosadultostratadoscon insulina, los nivelesde ARNm de estaproteína
aumentaron(Figura28).
3.4.7. Resumende los resultadosdel estudiode tiroidectomíaen ratas
destetadasy adultasy de animalesdestetadosy adultosoperadostratadoscon
insulina.
La tiroidectomíacausóun descensoen la insulinemiay los nivelescirculantes
ehipofisariosdeGil de las ratasdestetadasy adultasrespectoa las control.El déficit
dehormonastambiénprodujo un descensoen los valoresséricosy de expresióndel
ARNm hepáticode IGF-I, IGFBP-1 y 3, mientrasque las cifras circulantesy las de
los tránscritosde IGFBP-2 aumentabanen estos animales destetadosy adultos
operadosrespectoa sus controles(Cuadro2).
La administraciónde insulinaa las ratasdestetadasy adultastiroidectomizadas
se acompañóde un incrementode su insulinemia,de los niveles circulantesde Gil
y de su contenidoen hipófisis respectoa los animalestiroidectomizados(Cuadro2).
No obstante,este tratamiento no modificó la expresión hepática ni los niveles
circulantesdel IGF-I respectoa las ratasoperadas.
140

flg 28 ExpresiónhepáticadelARNmdeIGFR¡‘-1, 2y 3 de ratasdestetadas(37días)y adultas(87días) ziroidectomizadas(79, controles CC) y tiroidectomizadastratadas con insulina (T +
1). El análisisseefectuómedianteel ensayodeprotecciónde la RNasa.Media + DSde4 análisis diferentes. * Sign~caciónestadística respecto a la población control.> Sign¿flcaciónestadísticarespectoa los animalestiroidectomizados.
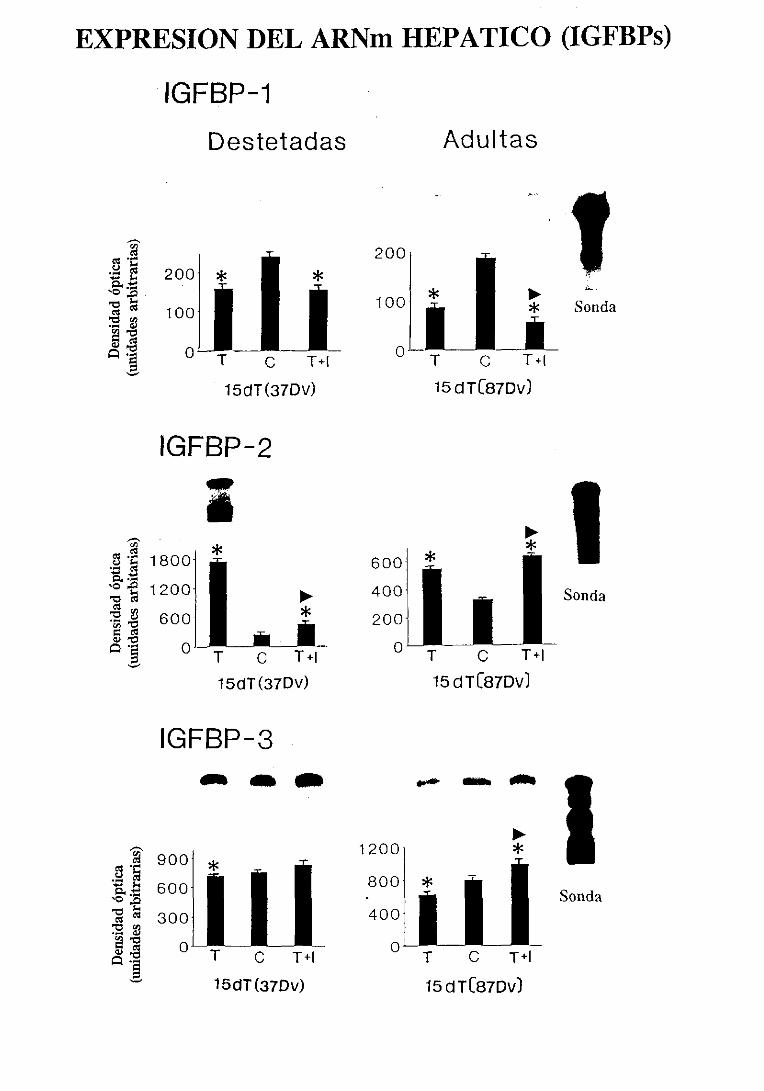
EXPRESION DEL ARNm HEPATICO (IGFBPs)
IGFBP-1
Destetadas
200~* Szloo]]]
C 141
l5dT(370v)
Adultas
200
100 >“ih*T 0 T~I
15 dTC87Dv)
IGFBP-2ea*
1800]é
1200]¡60dM *
1 0 fI1 5dT (37Dv)
600
400
200
o1 0 T~115 dTCB7DV)
IGFBP-3sea
T c 1+1
1200
800
400
o
•e 5
1 0 T~I
Cd
— 5-
Cd
4>p~
Sonda
t4- 4-a.—‘O .0
5-Cd
*
Sonda
i— I~
‘0 ~
Co
900
600
300
o
*
Sonda
l5dT(370v) 15dTC870v)

Resultados
En cuanto a las IGFBPs, la insulina aumentó los valores séricos de las
proteínasde bajo pesomoleculary tambiénIGFBP-3 respectoa las ratasoperadas,
aunquetan solo estaúltima alcanzólos valorescontrol en las ratasdestetadas.
Los tránscritosmuestranque la IGFBP-1 guardóun paralelismocon respecto
a valorescirculantes,mientrasque la IGFBP-2 disminuyeen destetadasy aumenta
en adultas.
La expresiónhepáticadel ARNm de IGFBP-3 pareceguardarun mayor
paralelismocon los valorescirculantes,aunqueseencuentraaumentadaen adultos
(Cuadro2).
Se establecióuna correlaciónpositiva entre los niveles circulantesde GH e
IGF-I.
Cuadro2.- Resumende los resultadosde los estudiosde tiroidectomía(T) y
tratamientocon insulina (T + 1) en ratasdestetadasy adultasrespectoa la población
control.
1 1+1
DESTETADAS ADULTAS
GLUCEMIA
INSULINEMIA 4 t 1
GH CIRCULANTE 4 4
CI-? HIPOFISARIA 4 4 4
IGF-I CIRCULANTE 4 4 4
ARNm IGF-I 4 4
IGFBP-1 y 2 CIRCULANTES 4 4 4
ARNm IGFEP-l 4 4 4
ARNm IGFBP-2 1 1 1
IGFBP—3 CIRCULANTE 4 4
ARNm IGFBP-3 4 1
3.5. TRATAMIENTO CON T4 DE RATAS TIROIDECTOMIZADAS
NEONATALES, DESTETADAS Y ADULTAS.
Con el objeto de continuar nuestro estudio sobre la influencia del
hipotiroidismoen el axis IGFs¡IGFBPs,se sometióa los animalestíroidectomizados
a una pautade rehabilitacióncon 14. Estaadministraciónse llevó a cabomediante
142

Resultados
la inyección de la hormonao por el implante de pellets para, de esta manera,
estableceruna comparaciónentre ambas vías de absorción,dado que con los
dispositivosde liberacióncontrolada(pellet)seconsigueunaformade administración
más fisiológica y continuada.El tratamientose inició a los 10 días despuésde la
tiroidectomía(15, 32 y 82 días de vida, para las ratas neonatales,destetadasy
adultas,respectivamente)y seprolongódurante5 días. Sin embargo,enel caso de
los animalesdestetadosy adultos, se estudiótambiénotra poblaciónen la que la
duraciónde la rehabilitaciónfue de 10 díasuna vez obtenidoslos resultadosde 5
días.
3.5.1. Tratamiento con T4 de las ratas neonatalestiroidectomizadas.
Comparaciónentredosisinyectadasy pellets.
3.5.1.1.Evolución del pesocorporal.
El tratamientode las ratastiroidectomizadascon T4 produjo un aumentode
supesocorporal respectoa la poblaciónoperada(Tabla 9), aunqueeste incremento
fue superiorcuandoseadministróal animal la hormonaen formadepellet (56.64%)
y no como inyección(31.28%);además,esto supusoque la ratatratadacon pellet
igualara su pesoal de la rata control frente a la rehabilitadacon la inyección
(21.52%).Se encontróuna diferenciaestadísticaentre los dos tipos de tratamiento.
3.5.1.2.Nivelesséricosde T3 y T4 y plasmáticose hipofisariosde GH.
Las hormonastiroideas séricas,como cabía esperar,disminuyeron en los
animalestiroidectomizados,aunqueel tratamientosustitutivo con tiroxina aumentó
los nivelescirculantesde T3 y T4 en estosanimalesoperados(Tabla 9). Así pues,
todos los gruposde ratastratadascon 114 mostraronun incrementoen los nivelesde
hormonastiroideas, que fue mayorcon la administraciónde pellets, pero que en
ningún casoresultó ser suficientepara alcanzarlos valorescontrol, y así, para las
ratastratadasconuna inyeccióndiaria de tiroxina, ladiferenciarespectoa lascontrol
se situó en un 84.78% para 113 y en un 73.74% para T4, mientrasque para las
recuperadasconpellet fue de un 66.30%parala 113 y de un 57.54%parala 114. Este
comportamientose debe a que las ratas neonatalespresentanuna elevaciónde
hormonastiroideasdurantela etapainmadura,y porello, enesteestadio,los niveles
de estas hormonasson superioresa los de los animalesadultos; así pues, en
experimentosfuturos intentaremosrestaurarlos nivelesde hormonastiroideasa los
valorescontrol mediante[a administraciónde dosis superioresde 114.
El tratamientode los animales tiroidectomizadoscon pequeñasdosis de
143
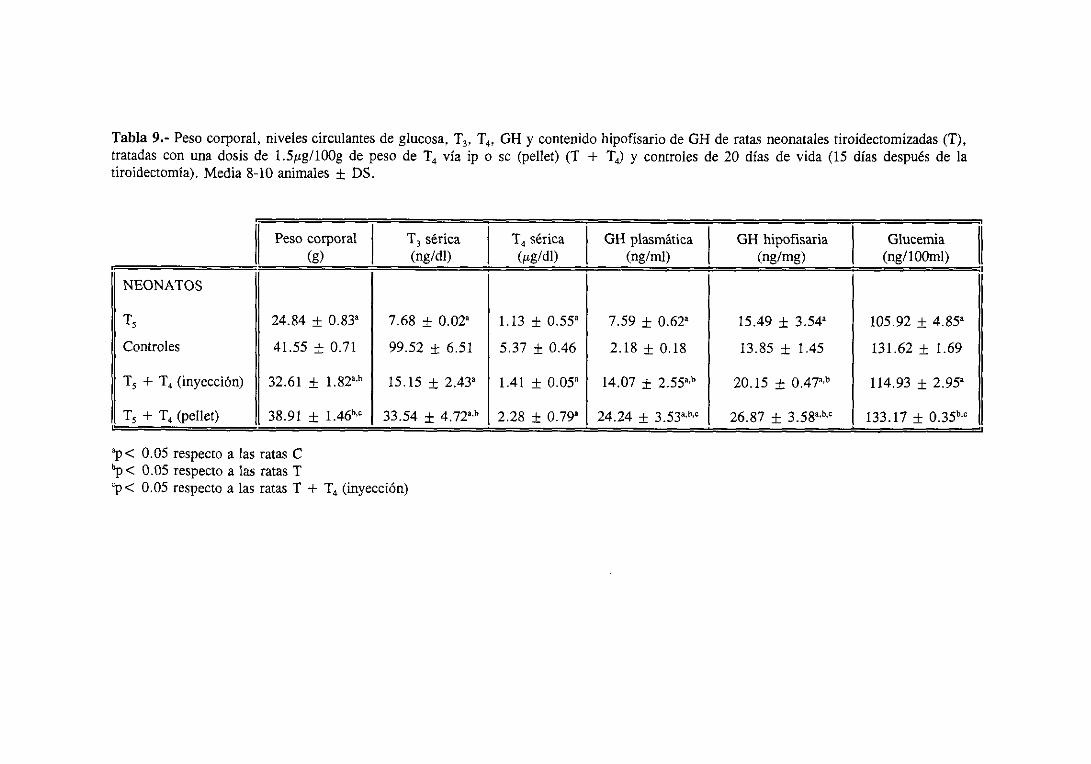
a—01)O
C
.e
n
+1
+1
-HA
H
rlC
i
2—
‘e—
ce
nZ
——
—e
n
cdCdcE
xO
.0-
.0~
tfl8
—‘e
oH
HH
-H-{j
O~
‘fl-t 0
R~
2r
~en
00
——
rl
Cde.>Cd
~~~01)
O
.0.0
iO<
8-
08
e
AH
+¡
AH
+1
00
—~
‘e—
~.
4—
rl
cdo,—‘-4
~0‘a.>V
ItO
E—
~8
u,-)
os
~‘Ú
jo
r—
80
00
+j
+1
+1
+1
en
r——
—~
~Úrl
——
Ci
Cdo,-.
¡
.08
—e
n
ei’n
-HC
i4
+1
rl~
AH
le0
0‘e
~
r-o’
t~e
n
EEt~
j
oen4>¡
.0l3
.0
00
t~-
Ci
‘C:
00
‘Ú—
—•
00
+¡
AH
AH
AH
,t‘~
>—
—•
00
‘e~
.O0
54
—C
i0
0.
Ci
Úe
ne
n
00oE-
zoz
VI
4>O‘.4
~O
E-U
‘0oO4>
E—+E:
4>cl
+
‘—4>
VIC
dV
Id
‘a.>cl
O‘O
0V
I
—‘O
o‘e
4-—
VI4>Cd
Cd
>0
4>
a.>t
e’oCd
Cd‘-40
~,
Ci
‘0‘UtO
CO
0
o0+
‘O•EE—
4>C04
.>o
—
E-
E—CdVIcE
-
4>
0
VIcl
4>00
•~tn
+I
~V
I4>
E4
>o
~~¡“UCO
00
VIo
~Cd
VI
A
u.o
Os
cd~E-
~-~
2‘0oo¡
4.>
E-:+
¡U
HF
-”
¡4
-4
~4
-.CO
COCd
‘-4u
-
00
00
00
4>
4>
0o.
cl
cl
VI
VIV
I4
>4
>4
>1-
•-u
-
00
00
00
vvv

Resultados
tiroxina (1.5~gIlOOg de peso) incrementólos niveles plasmáticosde GH, que
aparecíanya aumentadosen las ratas operadasrespectoa la poblacióncontrol, en
mayormedida.La pautade recuperaciónconpellet superóenun 72.28%el aumento
registradopor la administraciónde la inyección(Tabla 9).
La administraciónde 114 aumentótambiénel contenidodeGH enhipófisis de
las ratastiroidectomizadas,incluso porencimade los valorescontrol enun30.08%
para la inyección y en un 73.47% para el pellet, con lo que se encontróuna
diferenciasignificativaentreambaspautasde recuperación(Tabla 9).
3.5.1.3.Glucemiae insulinemia.
Les nivelesde glucosacirculante aumentaronrespectoa los valoresde los
animalestiroidectomizadospor la administraciónde tiroxina cuandosellevó a cabo
el tratamientocon los pellets (Tabla 9), y mostraronuna glucemiacercanaa los
niveles control (133.17mg/dl), no así con la misma dosis de 114 inyectada
(1 14.93mg/dl).Estediferenteaumentoen la glucemiade los animalestratadoscon
una u otrapautaresultó serestadísticamentesignificativo, lo que de nuevo llama la
atenciónsobrela mayor eficaciadel pellet en la rehabilitaciónde estosanimales.
Las dosis de tiroxina (1.5¡xg/ 1 OOg de peso) administradas por vía
intraperitoneal(inyección) o subcutánea(pellet) no modificaron los altos niveles
circulantesde insulinaencontradosparala poblacióntiroidectomizada,con lo que las
diferenciasrespectoa los animalescontrol (Figura 29) se cifraron en un 114.20%
paralos animalesa los que se les inyectó la T4 y en un 170.96%paraaquellosa los
que seles administróen fonna de pellet.
3.5.1.4. IGFs: nivelesséricosy expresióndel ARNm hepático.
3.5.1.4.1.Niveles séricos.
El IGF-I, que aparecíaaumentadoen las ratasneonatalestiroidectomizadas,
disminuyócon la administraciónde 114, y así, ambaspautasofrecieronunosniveles
séricosdel péptido igualesa los valoresde los animalescontrol (Figura 29). Por
tanto, y contrariamentea lo que sucedíacon las cifras de insulinacirculante,ambos
tipos de tratamientoresultaronapropiadosparala rehabilitaciónde esteparámetro.
La recuperacióncon 14 de los aminaleshipotiroideoscausóun descensoen
sus aumentadosniveles de IGF-II, y los situó incluso por debajode los valores
control en un 65.08% y un 71.94% para la administraciónde 114 con inyección y
pellet, respectivamente(Figura 29).
145

Eig 29 4) Niveles circulantes de IGF-I, insulina e IGF-II en raías neonatatesde 20 días de vidauiroidectomizadas(7’), controles (C), tratadascon una dosisde 1.5¡ig/¡OOgdepesoadministradaenfonnade inyección (1?) y tratadascon la mismadosisde 7’4~ pero enforma depella (Rl>). Media ±DSde 8-10animales.II) Expresiónhepáticadel,4RNmy cuaní(JicacióndensizométricadelosIGF-¡ y ¡len ratasneonatalestiroidectomizadas(7).controles((2), tiroidectomizadasrecuperadasconuna inyecciónde 1.5~g/1OOgde peso de T4 (R) y con el pelleí de T4 (Rl>). El análisis se llevó a cabo medianteel ensayode
protecciónde la RNasaen animalesde 20 díasde vida (15 díasdespuésde la operación).Media ±DSde 4 análisis diferentes.* Signfficaciónestadísticarespectoa los animalescontrol. > Signficaciónestadísticarespectoalas ratas tiroidectomizadas.•Sign(ficaciónestadísticarespectoa la poblaciónde neonatostratadoscon la inyecciónde 1.Spg/lOOgdepesode L,.
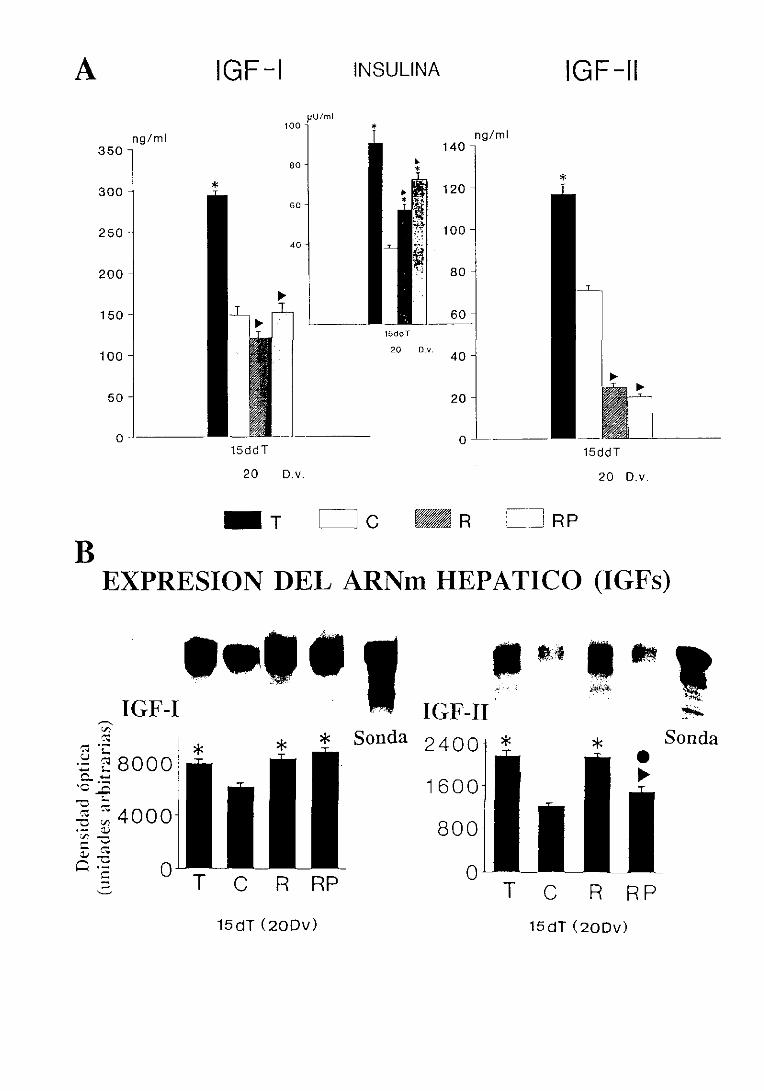
INSULINA ¡OF -II
ng/ml
*
20 Dv.
O _ IJRP
EXPRESION DEL ARNm HEPATICO (IGFs)
04
IGF-li
pnnn~.
‘O •~
-g ¿~ 4000
-ce 0--
*Sonda
TCRRP
2400-
1600-
800-
o-
* e
T o R PP
A ¡OF-!
ng/mlloo
360
300
260
200
160
loo
60
ol5ddT
20 Dv.
o
B
l5ddT
E
IGF-I
* * *-r
Sonda
l5dT (200v) 15d1 (200v)

Resultados
3.5.1.4.2. Expresióndel ARNin hepático.
Les nivelesde ARNm hepáticode IGF-I fueron, al igual queen los animales
tiroidectomizados,significativamentesuperiorestantoen los neonatosa los queseles
administróla T4 conuna inyecccióncomo los del pellet, respectoa los de animales
control (Figura29). Sin embargo,la expresiónhepáticade IGF-II disminuyótras el
tratamientohormonalconpellet (Figura 29), y no lo hizo con la inyección,siempre
respectoa la poblaciónde ratascontrol; seobservóasíunaclara diferenciaentreuna
y otrapautade reemplazamientoque, de nuevo,evidencióla mayoreficaciadel pellet
parala rehabilitaciónde los animalestiroidectomizados.
3.5.1.5.Estudiode regresiónlineal y correlación.
Seefectuóel análisisde regresióny correlaciónde los nivelescirculantesde
IGF-I frente a los de insulina y GH (Figura30).
Seencontróuna fuertecorrelaciónpositiva entrelos nivelesséricosde IGF-I
y los plasmáticosde insulinaen estosanimalestiroidectomizadostratadoscon 114, no
obstante,estacorrelaciónfue mayorcuandoel tratamientose llevó a cabomediante
la implantaciónde pellets (r=O.83) frente a la inyección (r=0.52). Sin embargo,
cuandose representaronlos valorescirculantesde RF-I frentea los de OH, estas
poblacionesmostraronunacorrelaciónbaja(r=O.50 y rzO.64paralas ratastratadas
con 114 vía intraperitonealy subcutánea,respectivamente).
3.5.1.6. IGFBPs: nivelesséricosy expresióndel ARNrn hepático.
3.5.1.6.1.Niveles séricos.
3.5.1.6.1.1.Westernligand blot.
Les animalesneonatalestiroidectomizadosexperimentaronun aumentoen los
niveles circulantes de las IGFBPs del complejo de 3OkDa, aunque los dos
tratamientosde recuperaciónprovocaronun descenso,muy similar, en sus valores
séricos(Figura 31) respectoa la poblaciónoperada,estohizo que ambosgruposde
animalestratadosalcanzaranunosnivelessimilaresa los de las ratascontrol.
La IGFBP-3 experimentóun aumento de sus niveles en suero tras la
rehabilitaciónde los neonatostiroidectomizadoscon T4 (Figura 31). Sin embargo,la
implantacióndel pellet, al igual que en otras ocasiones,consiguió restaurarlos
nivelesdel péptidode 45kDaa los valorescontrol, mientrasquela administraciónde
la inyecciónno consiguióun aumentoen los nivelesde estaproteínaligadorarespecto
a los animalestiroidectomizados.De nuevo, se observóuna mayor eficaciaen una
pautade tratamientosustitutivo respectoa la otra.
147
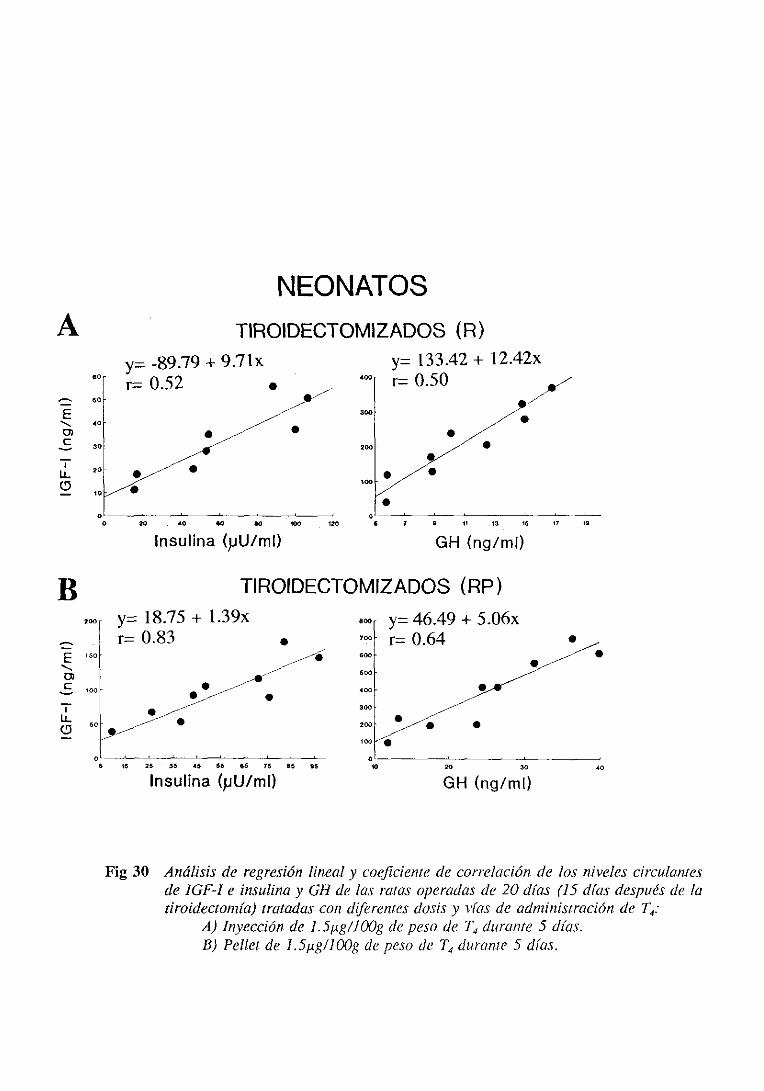
y= -89.79+ 9.71xr= 0.52
NEONATOSTIROIDECTOMIZADOS (R)
y= L33.42+ 12.42xrz 0.50
TIROIDECTOMIZADOS (RP)yr 18.75 + 1.39xr= 0.83 e
ee e
e e
5 15 25 25 45 65 65 75 85 95
Insulina (pU/mí>
Fig 30 Análisis de regresión lineal y coeficientede correlación de los niveles circulantesde IGF-I e insulinay CH de las ratas operadasde 20 días (15 días despuésde lariroidecíomla,.) tratadascon dWerentesdosisy vías de administraciónde 7%:
A) Inyecciónde ].5pg/lOOg de pesode T4 durante5 días.fi) Pcíletde l.Spg/lOOgde pesode 7% duranre 5 días.
A
Eo)c
LLo
60 ¡
40 ¡
20
20
lo
e
e e
B
20 40 40 40 lOO 120 6 7 9 II 53 15 17 15
Insulina (pU/mi) GH (ng/mi)
200
50
100 -50
Eo,
u-o
y= 46.49+ 5.06xr= 0.64
600
500,
400
000
200
ee
‘O 20 50 ~0
GH (ng/mi)
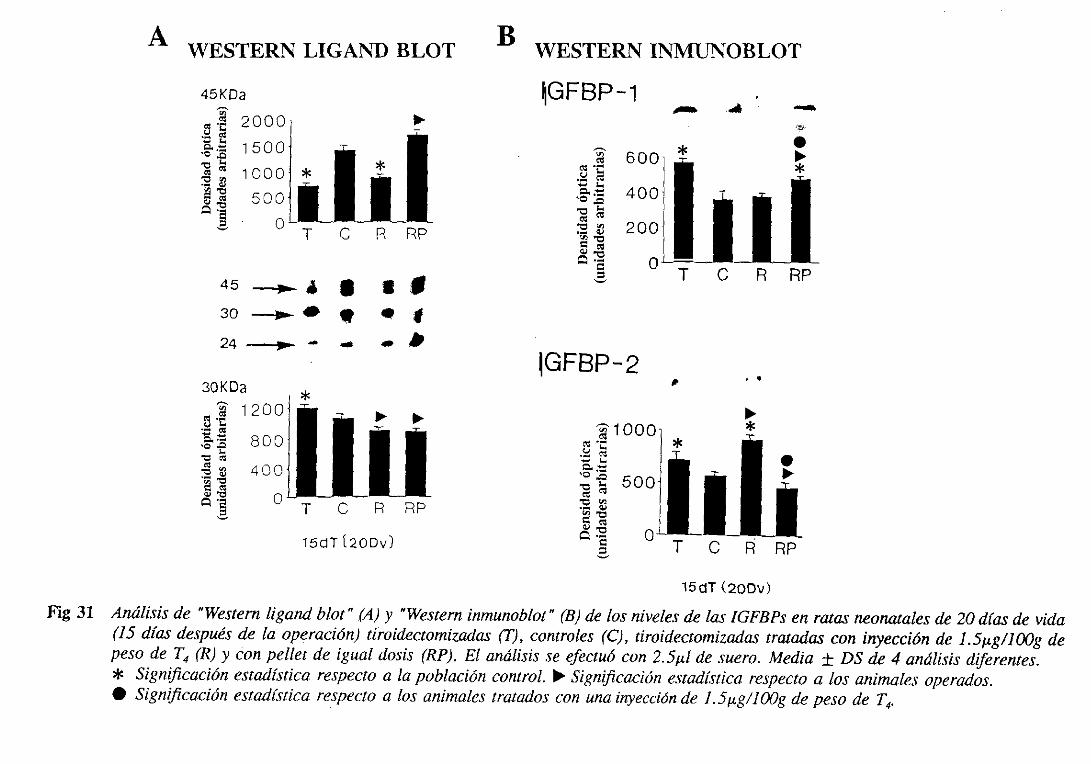
*1
o-
tro1-
oo
oo
oo
oC
O1
-c’J
(se
uE
s>
¡qa
esa
pe
pT
un
)133!ldO
pE
p¡s
ua
q
00
00
00
00
cg—
-
C\J
o-mLLo
o-ci
a:(em
t,
0
cl
s¿(su
uu
.zpq
susapupgnn)to~.
un
pd
pp
ep
~su
a~
j
1—•1
LOO
~-
aÁ
~o
-a:
•1a:
o
oo
oo
oo
to
(srqseipq.xesapepiun)~
3!1
d9
pupisuaq
oo
o-ir
OQq
o1
-u
00
00
00
0cg
ceti-
(suu
ns~
q~
esa
pe
pju
n)
ua
pd
9p
ep
wu
aq
‘t~~
‘-Nt
~—
a
;~~
,&~
~04
$U~2
-~~
~‘-a
~‘,~
~~
‘nt.5
t—
a.--~
<‘4
-a-a
U~
~—‘
~J.s
‘-~
-.-C’
—4
Q.~
~
-~~
-,k..-a
-~u
-~~
-~&
.k
-~1~
‘-4
—4
.4‘-4
E
~j.~
t‘,‘—
-~•-a—
4-~
*e
1111
~
o-
Loo
ozoz0zuj
A
*

Resultados
3.5.1.6.1.2. Western inmunoblot.
Los nivelesséricosde IGFBP-1, queaumentarondespuésde la tiroidectomía,
disminuyeronal tratar a estosanimalesoperadoscon una dosis de 1 .Sgg/lOOgde
pesode 114 (Figura 31). La administraciónintraperitonealde la hormonaigualó los
niveles de estaproteínade bajo pesomoleculara los valorescontrol, mientrasque
el tratamientocon pellet situó las cifras circulantesde IGFBP-1 en unos valores
intermedios y distintos a los mostrados por las poblacionestiroidectomizaday
control. Se encontróuna diferenciaentre los dos tipos de tratamiento,aunqueen
cualquiercaso, con ambaspautasde administraciónse encontróuna disminución
respectoa los nivelesalcanzadosdespuésde la tiroidectomia.
El tratamiento con T4 mediante la inyección intraperitonealprodujo un
aumentoaún mayorde los nivelesséricosde IGFBP-2 (Figura31). La implantación
de pellets,por el contrario, disminuyólos altosvalorescirculantesde estaproteína,
con lo que se alcanzaronunascifras similares a las mostradaspor los animales
control; de nuevo, se puso de manifiesto una diferenciaentre los dos tipos de
tratamiento.
3.5.1.6.2.Expresióndel ARNm hepático.
Les niveles de ARNm hepáticode IGFBP-1 disminuyeron al tratar a los
animales neonatalestiroidectomizadoscon T4 (Figura 32). La inyección de la
hormonaprovocó un gran descensoen la expresiónde estaproteínade bajo peso
molecular,y resultósermásacusadoque el producidopor la implantaciónde pellet,
que mostró unos niveles de ARNm similares a los de la población control; se
encontró así una diferencia estadísticamentesignificativa entre los niveles de
expresiónde IGFBP-1 de las ratastratadascon inyeccióny las control, y entrelas
dospautasde reemplazamiento.
La IGFBP-2 mostró una menorexpresiónhepáticadespuésde tratar a los
animalestiroidectomizadoscon el pellet (Figura 32). Por el contrario, la inyección
de 114 no sólo no provocóun descensoen los nivelesde ARNm de estaproteína,sino
que los aumentó,y los valores alcanzadosfueron muy superioresa los de los
neonatoscontrol e, incluso, a los de los tiroidectomizadosque los presentabanya
aumentados.El análisisestadísticomostró así una diferenciasignificativa entrelos
dos tipos de tratamiento.Este anómaloincrementoen la expresiónen hígadode la
IGFBP-2 asociado a la administración intraperitoneal de la T4 será estudiado
oportunamente.
150

ng 32 Análisis medianteensayodeprotecciónde la RNasade los tránscritoshepáticosde lGFBP-!, 2 y3 de razasenperiodo:
Neonatal(20días): ziroidectomizadas(2), controles ((2), tiroidectomizadastratadas con unainyecciónde 1..S~g/1OOgde peso(R,)ycon peZ/e:de igual dosis (Rl’,).Destetado(37 días)y adulzo(87días):tiroidectoniizadasrn. controles(C%tiroidectomizadascon una inyección de 1. 7Sgg/IOOgde peso de 7’., durante 5 días (R,), tiroidectomizadasrehabilitadasconpella de igualdosisduranteSdías(Rl>5), tiroidectomizadasrecuperadascon
pellet de la mismadosificacióndurante 10 días (RP¡a).*Signjficación esuidínicarespedoa la población control. > Sigt«ficaciónestadísticarespectoa los
animalestiroidectomizados.O Sign<ficación estadísúcarespectoa las ratas rehabilitadas con lainyecciónde 7’,.Semuestrael análisis densitomnñrieo.Media + DSde 4 análisis dferen¡es.
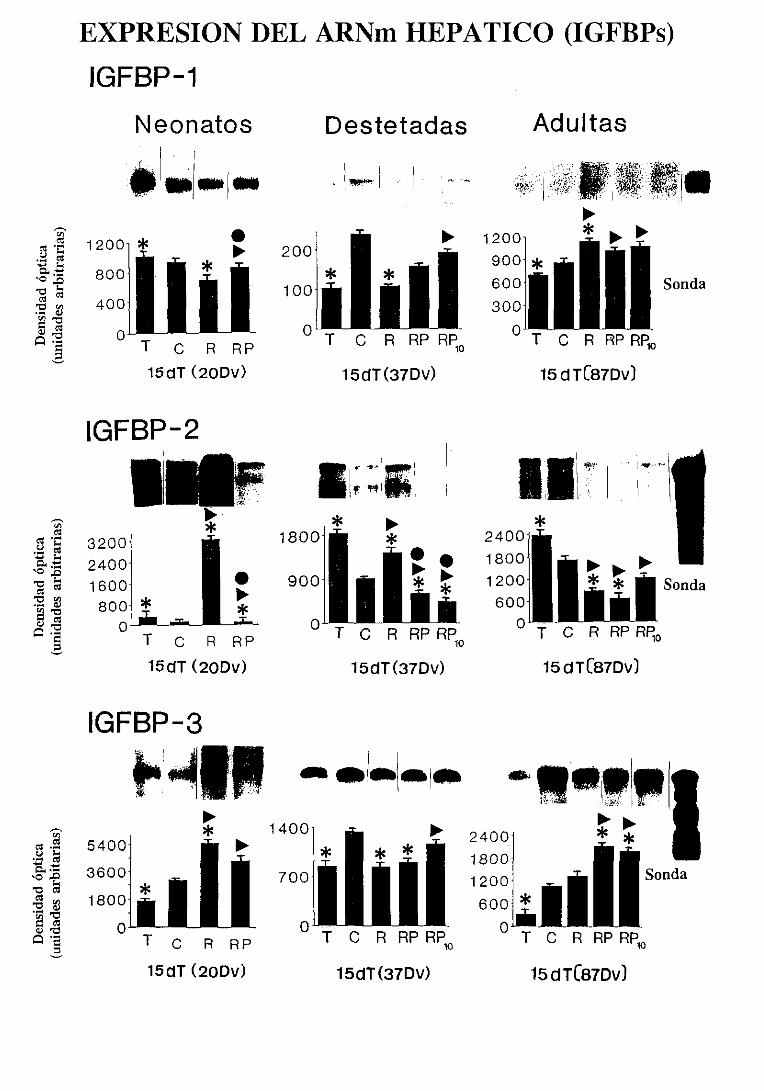
EXPRESION DEL ARNm HEPATICO (IGFBPs)
IGFBP-1
Neonatos Destetadas Adultas
k»
200
100
o
lbdT (2oDv)
T o R RPRF~Q
l5dT(37DV)
1200~
900j* .jMMM6001M mmm.20011111
T O R RPR9Q
15 dTC87Dv)
3200]
2400]
1600]800
oT
E1800
e*
RP
900
o
l5dT (200v) l5dT(370v)
2400
1800
1200600
O
*
Ir 1~~~
Sonda
11 C R RPR90
15 dTC87DW
IGFBP-3
H*
5400
3600iIII¡*1800 T o R RP
a e>ae~a
1400]
700hillT O R RPRP
lo
00CO
CO.~CO4- u-
u-CO
00~CO
1200
800
400
oT o R RP
5onda
IGFBP-2
00
CO
.~CO
‘0u-
‘~COCO
**1R nplo
PI.00CO
Cl.—u-4- 4-cl~
u-‘eCOCO
4> ‘e
**240
180120
60
Sonda
1 0 R FRPRP10
l5dT (2oDv) l5dT(37Dv) 15dTC87DV)

Resultados
La expresiónen hígadode la IGFBP-3 aumentódespuésde recuperara las
ratastiroidectomizadascon tiroxina, incluso superóa los valorescontrol cuandose
administró la hormonacomo inyección(Figura 32). A pesarde que no se encontró
ningunadiferenciaestadísticaentrelas dospautasde reemplazamiento,los nivelesde
ARNm de esta proteínade alto pesomolecularse aproximaronmás a los valores
controlcuandose llevó a caboel tratamientomediantela implantaciónde pellets.
A la vista de estosresultados,sepodríadecir que la administraciónde 114 a
ratasneonatalesrestauralas alteracionesproducidaspor la tiroidectomía.
3.5.1.7.Resumende los resultadosdel estudiodel tratamientocon T4 en
ratasneonatalestiroidectomizadas.
Los valores circulantes de IGF-I y II despuésde la administración de
1 .Sgg/lOOgde peso,tantoporvía intraperitonealcomo conpellet, desciendenal nivel
de los controlese igualmentela expresiónde suARN paraIGE-Il no paraIGF-I. Los
valores circulantesde insulina tambiéndisminuyen, quedandoaún por encimade
controles.Tambiéndesciendea nivel de controlesel complejode 3OkDa circulante;
por inmunoblotse muestraen los animalestratadosconpellet, que estadisminución
es mayoren la IGFI3P-2cosa que no semanifiestacon la inyecciónde 1 .5pg/íOOg
de peso.
La IGFBP-3 circulante no se altera con la inyección y remontaa valores
controlcon el pelletde 14. La expresiónde ARNm de las IGFI3Psde 3OkDamuestra,
igual que lo hizo el inmunoblot, que es mayorel descensoen IGFBP-2 despuésde
la implantacióndel pellet. Tambiénaumenta,alcanzandolos valorescontrol, el
ARNm de IGFBP-3; por tanto despuésde la implantacióndel pellet se muestrauna
recuperaciónparalelade los valorescirculantesy la expresiónhepáticadel ARNm
en las tres JGFBPsestudiadas,cosa que no sucedecon las mismasdosisde T4 por
inyección. Todo ello manifiestala no convenienciade estavía para rehabilitación
como ya se ve tambiénen los cambiosde los parámetrosde la Tabla 9 despuésde
la inyecciónde 114.
En los animalestratadosconpellet, la Gil plasmáticaehipofisariaaumentaron
respectoa las ratascontrolesy tiroidectomizadas,y la glucemiaalcanzalos valores
control, los mismos cambios se producenen los animalesinyectadoscon 14 pero
menosacusados.
Se establecióunacorrelaciónpositivaentre los nivelescirculantesde insulina
e IGF-I que esmayorcon pellet.
152

Resultados
Cuadro3.- Resumende los resultadosde los estudiosde la administraciónde
T4 endosisde 1.5 ¡íg/lOOg de pesopor vía ip (11 +T4)~~ o vía sc (pellet) (T + T4)SC
en ratasneonatalesrespectoa la poblaciónde los animalescontrol (20 díasde vida).
(T + T4)1~ (T + T4),~
GLUCEMIA 4
INSULINEMIA t 1
GR CIRCULANTE 1 1
OH HIPOFISARIA 1 1
IGFt~I CIRCULANTE
ARNm IGF-I 1 1
LOE-II CIRCULANTE
ARNm LOE-II
IGFBP-1 CIRCULANTE
ARNm LGFBP-L
IGFBP-2CIRCULANTE
ARNm IGFI3P-2 1 4
IOPBP-3 CIRCULANTE
ARNm IGFBP-3 1
3.5.2. Tratamiento con 114 de las ratas destetadas y adultas
tiroidectomizadas.Comparaciónde dosisinyectadasy pellets.
Al igual que en los estudiosrealizadossobrela poblaciónneonatal,en las
ratas destetadasy adultastambiénsellevó a cabola rehabilitaciónde estosanimales
tiroidectomizadosmediante la administración ip (inyección) y sc (pellet) de la
tiroxina, para de nuevo estableceruna comparaciónentre las dos vías empleadas,
pero ahoraen estadiosmás madurosdel desarrollo.La recuperacióncon pellet se
realizódurante5 ó 10 días.
3.5.2.1. Evolución del pesocorporal.
Las ratastiroidectomizadaspresentaron,comoya seha visto, unpesocorporal
inferior al de las ratascontrol, sin embargo,la rehabilitaciónconT4 aumentóel peso
del animal operado. Este incremento fue significativo respecto a la población
tiroidectomizadacuandoel tratamientohormonalse realizó con el pellet, puestoque
la administracióndiaria de la inyecciónno modificó el pesode los animalestratados
con 114 respectoal de los operados,tanto en la etapadestetadacomo en la adulta
<Tabla 10). Cuandola recuperaciónse prolongódurante5 díasmás,el valor del peso
se igualó al de las ratas control, por tanto se ve un aumentoprogresivodel peso
153
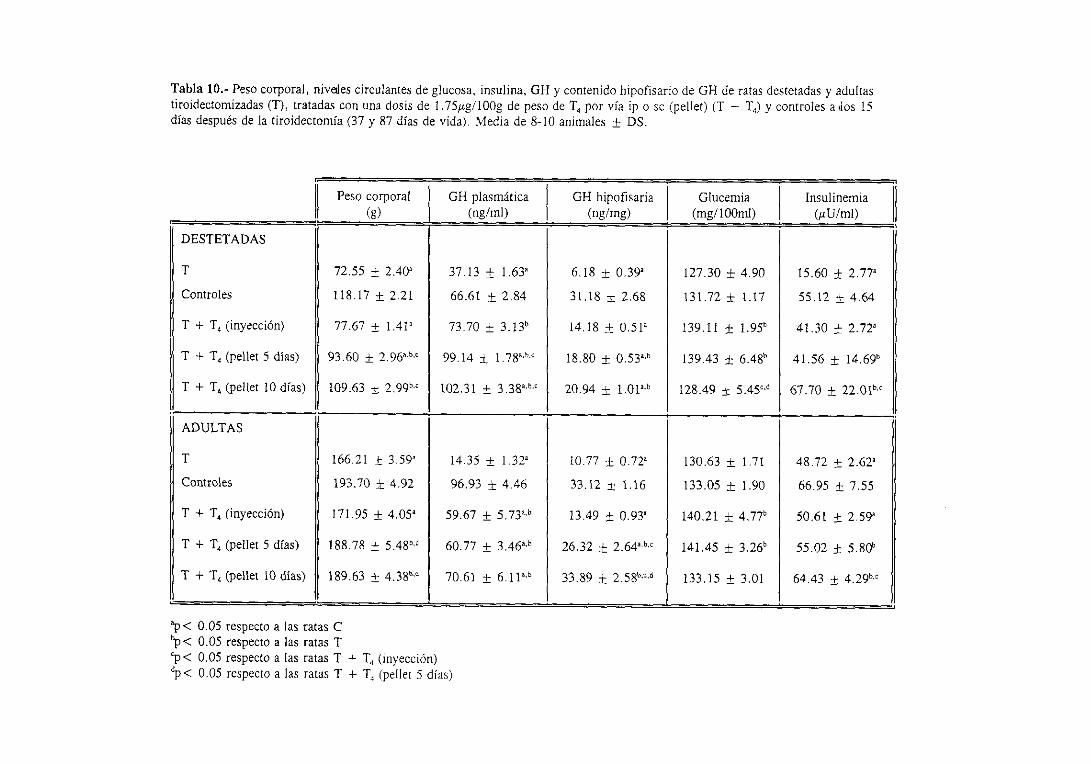
CO
~~-~b
k7‘0
~.-
.-
ci
yci
¡~1.n
~~o
c¡
41
+1
ci+1
¡4
1+
141
~+1
to~
.0
ci
‘o.-
‘0o
‘flU—
—,
‘o
Ci
Qn
e--
0’
~¿
QQQ
n
cio.Qnvn‘‘0
OU
-Q
n~
QN
—Q
N~
--,
O‘i
0=r-r-
-~sc
ci
OO
So
diOo
——
~
xt
-4+
1+
1+
t-fA
41
OC
i,-,
en
Co
C—y
±1
RA
CoQn
‘CO
441
..~ci
Co
~ir>y
Co
41
Qn
—
&~
22
22
Co
Co
OCO
--4
———
Co
Co
—OCCO
CO
QN
~C
oC
o.
Qn
o
o0
5.2
-Ci
I~+
~~
AA
‘0,—
—
~~~
QN
O~~Oci
-iA
00
Qn
+1
oo
ca
y—
-o
cQN
‘0C
o—
O-~
ci
¿C
o.-~
Co
QN
‘Ú~,-~
Ci
rn~CCi
QN
oc
Co
Co
COo‘CO
’
g0-ú
<~00
‘ooc~
—oc
Co—
C 4Co
‘0—
-c¿CorQn
-c~0
~Co
‘‘0
R~-Ñ
+1+
1-~
A~
~41
0.0
O
Co
‘~0
——
~U-~
Co
—.
Co
‘0U
-~‘
OQ
N
Qn
Co
0=
U-
‘06Qn
U-
U-
¿50
‘0¡
oU-
¡
--
o0
04-
ci
—C
o‘t
ÚÚ..,~
Cooc
y
o
Ci
-n-~
41
0-
+1
U-
Co
Qn
~0O
st
MA-H
—Oe-.
~r>
0=
41oce—
CCo‘0
di
oc
‘oci
—e
--.
U-
0-
Co
QN
—
‘0~
‘o0
’~
.4—
——
Co
E-
LLJ
CO
Ot
‘0o
-Q
n-4
ou~
di—
—di00
~d
i‘
OE
-HE
-~
++
+O
E-
(~)E
-E-
E-’
Co
E—-4
00
CO
e•o
‘0-Q
no
00
—di
‘~-‘
oa-
++
oE
-OE
-E
-
00
CO
Oo->diaE-”
+E-
COo
-OvnOo
sd
i—~
‘di--
a
++
UE
-E-E
-00
00
00
00
CO
CO
CO
CO
~CoC
OC
O0
00
00
00
00
00
COCO
CoCO
CO
Co
COCO
•0
00
0O
OO
Odi
di
di
di0
.a
o.
o.
0000
0000
didi
diO>
a-
a-
I~lo
Qn
Qn
Qn
Ir>0
00
00
00
0
vvvv
00
Qn
5o
4
00
0-O
OCo
Co
CO
—.
OO
CO1
.d
io
di0
•0>
,
CO
0z
di
di
—
o~.
loCOOo
oC
oa
.9-CO
o‘~
+1
-Ooo.~
eCE
—-—
od
iC
ot
o~
o—
o~
0t
adi
COd
i~0
.9-~
—C
odi
00
50
00
£0
di
Co
t~
o~
ni
di
dit
ni
t0
00
0C
Odi~
o-O
COn
iU
-O
CO
Oe
>N
oOC
o0
0O
’~d
iO
COdi;>.
COEO
—:
Co4>
Co1—
a-
o-0
O’—
~0
0.~
qC
O00
.0d
id
iCO
-Oa
00
E‘~
oO
a—
fl0
0
-o00
E--o
-u

Resultados
cuandoserehabilitanconpellet.
3.5.2.2.Nivelesséricosde 13 y 14 y plasmáticose hipofisariosde GR.
Los niveles de hormonastiroideasdisminuyeron despuésde la operación,
aunqueel tratamientosustitutivo con T4 produjo un incrementosignificativo en las
cifras circulantesde estashormonasen todos los casosestudiadosrespectoa los
animalesoperados(Figura 33). Este aumento,al igual que sucedíacon el peso
corporal,fue menorparaaquellasratasa las que se les inyectéla hormonarespecto
a las que se les implantéel pellet, tanto a los 5 comoa los 10 días de tratamiento.
El incrementoencontrado,que superóinclusolos valoresde los animalescontrol,en
las ratasdestetadastratadasconpelletdurante5 días, fue de un 13 1.05% para la T3
y de un 70.00% para la 14, mientras que para las adultasesteresultó ser más
modesto(5.20%parala T3 y 28.71%parala T4) e igualélos nivelesde las hormonas
tiroideas a los de la población control. Estos datos contrastaroncon los obtenidos
paraaquellosanimalesdestetadosy adultosinyectadoscon T4 diariamente,dadoque
el incrementofue menor, y en ningúncaso superaronlos valorescontrol (59.69%
para13 y 76.02%para114 en las ratasdestetadasy 72.42%paraT3 y 62.74%para
14 en las adultas);asípues,pareceque encuantoa la recuperaciónde las hormonas
tiroideasen ratasdestetadas,el tratamientoidóneoesel pellet durante10 días.
En los animalesadultos,el tratamientocon pelletsde T4 mantuvolos niveles
de hormonastiroideas igual a los controlestanto a los 5 como a los 10 díasdespués
del inicio de la administraciónhormonalporpellet (Figura 33). Sin embargo,y dado
que la dosis fue la misma para los animalesadultos y destetados,estos últimos
mostraronunosnivelesde hormonastiroideassuperioresa los de lapoblacióncontrol
a 5 días, probablementedebidoal menorpesode las ratasmás jóvenes.Por tanto,
cabedestacarque paralos animalesadultos,encuantoa las hormonastiroideaslas
dosis de 1 .75j¿g/lOOg de pesode 14 es la idóneatanto con 5 como con 10 días de
tratamiento.
La Gil circulanteaumentéal tratar con T4 a los animalestiroidectomizados
respectoa los operadosen ambaspoblaciones,superandolos valores control en
destetadasy no llegandoa ellos en adultos; igualmenteaumentóla Gil hipofisaria,
no llegandoa valorescontrol más que en las adultasde 10 díasde tratamientocon
pellet (Tabla10). Al igual quesucedíaparaelpesocorporaly las hormonastiroideas,
la rehabilitacióncon pellet durante10 días ofreció los niveles más cercanosa los
control.
155
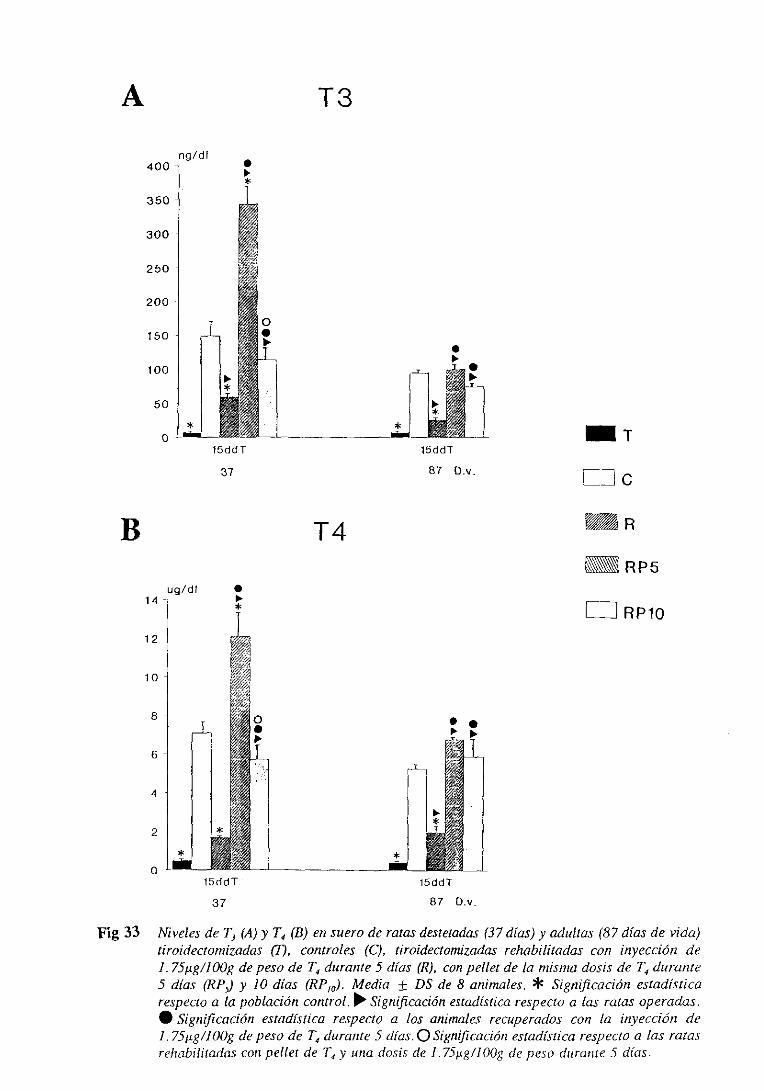
e
T3
ng/dr
T
ticrARW RP6
LII np-io
flg 33 Nivelesde T~ (A) y T4 (8) en suerode Tatasdestetadas(37días)y adultas (87díasde vida)¿‘iroidectomizadas(2), controles ((2), ¡‘iroidectomizados rehabilitadas con inyección de1. 7Spg/¡OOgdepesode T4 durante5 días <1?), conpellet de la mismadosisde 7’4 durante5 días (Rl>) y 10 días (Rl>,0). Media ±DS de 8 animales. * Signficación estadísticarespectoa la población control.~ Significaciónestadísticarespectoa las ratasoperadas.• Significación estadística respectoa los animales recuperadoscon la inyección de1. 7Sgg/lOOgdepesode 7% durante5 días,OSignificaciónestadísticarespectoa las ratasrehabífijadascon pelter de 7% y una dosisde l.75>g/lOOg de pesodurante 5 días.
T4
87 Dv.
lSddT lSddT
37 87 Dv.
A
400
350
300
250
200
150
loo
50
o
o
eefi’.
lSddT l5ddT
3~1
B
ug/drph.
*14-
12
10
8
6
4
2
o
* *

Resultados
La rehabilitaciónconpelleta lo largode 5 díaspareciósermásefectivaque
la inyección, que no modificó el contenidoen GH de estetejido respectoa las
poblacionestiroidectomizadasdestetaday adulta.A la vistade los resultadosse debe
destacarque la respuestaal tratamientodifirió segúnla edaddel animal; las ratas
destetadasrehabilitadascon pellet presentaronunos niveles de GH en plasma
superioresa los de los animalescontrol,perosucontenidohipofisariofue inferior al
de dichoscontroles,sin embargo,las ratasadultasa las que se les implantóel pellet
o se les inyectó la hormona,mostraronunos valoresde GH en plasmae hipófisis
inferiores o igualesrespectoa las ratascontrol; estasdiferenciaspuedenserdebidas
a la distinta madurezdel axis de la Gil, dadoque en las ratasdestetadaspuedenno
haberseestablecidolas conexionesneuroendocrinascompletamente.
3.5.2.3.Glucemiae insulinemia.
Los nivelesplasmáticosde insulina aumentaronal administrar114 a las ratas
tiroidectomizadas(Tabla 10) porambasvíasen destetadasy adultas,peroalcanzaron
valorescontrol sólo con el implantede pelletsa 5 o a 10 díasenambaspoblaciones
destetadasy adultas.A pesarde todo, y al igual queencasosanteriores,los valores
que fueron similares y más cercanosa los niveles control resultaronser los de
aquellasratastratadascon pellet y cuyo tratamientose prolongó durante10 días
(22.82y 3.76% paralas ratasdestetadasy adultas,respectivamente).
El tratamientocon tiroxina aumentólos nivelessanguíneosde glucosade los
animales tiroidectomizados (Tabla 10), y se observó de nuevo, un mayor
acercamientoa los valorescontrol en aquellasratastratadascon114 durante10 días.
3.5.2.4.IGFs: nivelesséricosy expresióndel ARNm hepático.
La recuperaciónde los animalestiroidectomizadoscon114 produjoun aumento
en los nivelescirculantesde IGF-I (Figura 34). Este incrementoen los valoresdel
péptidofue mayoral tratarcon pelletsdurante10 díasa ambaspoblacionesdestetadas
y adultas. El tratamientocon pellets durante5 días se mostró más efectivo en la
restauraciónde los nivelesde IGF-I que la inyecciónde 114 (Figura 34).
En los animalesdestetadostan solo el tratamientocon pelletdurante10 días
rehabilitó los nivelesde IGF-I a los controles,cosaque selogró a los 5 días en los
adultos.Sin embargo,el tratamientocon pellet ya a los 5 días rehabilitéa niveles
controlel ARNm hepáticode IGF-I en laspoblacionesdestetaday adulta, aunquelas
inyeccionesde igual dosis no tuvieronefecto (Figura 34).
Se observépuesun gran paralelismoentre los nivelesséricosde IGF-I y los
157
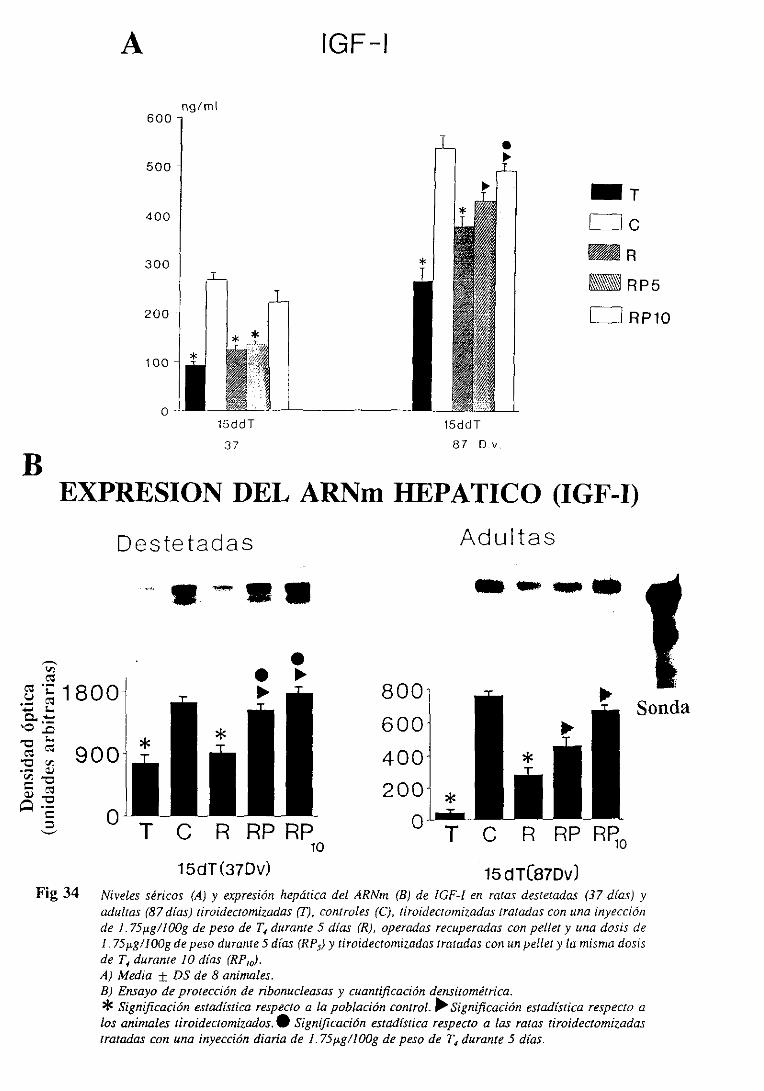
A IGF-l
600
500
400
300
200
100
o
ng/ml
LIllo
R Pb
LII RP1O
EXPRESION DEL ARNm HEPÁTICO (IGF-I)
37
Destetadas Adultas
u
Vr,CO
CO ~•~ ~180O— L.
‘o ~
COCCa>
Vi -~
CCOa>
=
900
—u.
e1~1~
ePI-
800600-400-200-
o RRPRP10
*
T
m — n S
NI-
*r
O RRPRI%
1 5dT (37Dv) 15 dTCS7Dv)Nivelesséricos <A) y expresiónhepáticadel ARNm (B) de [GE-! en ratas destetadas<37 días) yadultas(87días) tiroidecwmizadas(T), controles <ci» tiroidectomizadastratadasconuna inyecciónde 1. 7Spg/lOOgdepesode 7% durante5 días (R), operadasrecuperadascon pellety una dosisde1. 7SpgIJOOg de peso durantes días (Rl’5) y tiroidectomizadas tratadas conun pelter y fa mismadosisde T, durante10 días (Rl>,0).A) Media ±Di de 8 animales.B) Ensayodeprotecciónde ribonucleasasy cuantificacióndensitométrica.* Significaciónestadísticarespectoa la poblacióncontrol. ~ Significaciónestadísticarespectoalos animalesdroidectomizados.@ Significaciónestadísticarespectoa las ratas tiroidectomnizadastratadas con una inyeccióndiaria de 1. 75jsgIlOOg depesode 2% durante5 días.
e
*
Bl5ddT I5ddT
87 Dv.
*T
*~1~
Sonda
Fig 34

Resultados
de suexpresiónenhígado,lo que nos lleva unavezmása sugerirque las hormonas
tiroideas parecenactuar sobre el IGF-I a nivel transcripcional. Así mismo, se
encontróuna recuperaciónmás inmediatade la expresióndel ARNm hepáticode
IGF-I respectoa sus nivelesséricos,ya que tanto a los 5 como a los 10 díasdespués
del inicio del tratamientoconel pellet, las poblacionesdestetaday adulta igualaron
sus nivelesde ARNm a los valorescontrol.
3.5.2.5.Estudiode regresiónlineal y correlación.
Las Figuras35 y 36 resumenlos resultadosdel análisis de regresiónde los
nivelesséricosde IGF-I frentea los plasmáticosde insulina o GR.
Las ratas destetadasy adultaspresentaronuna alta correlación entre los
valores circulantesde Gil e IGF-I, tanto en las tratadascon inyecciónde 114 (r=
0.78, para las detetadasy r= 0.97, p’Z 0.05, para las adultas)como en aquellos
animalesa los que se les trató conel pellet durante5 días(r= 0.99, p<O.0S,para
los destetadosy r~ 0.97, p<O.OS,paralos adultos)como 10 días (r= 0.83 paralos
másjóvenes,y r= 0.99, p’c0.Ol, paralos adultos).Porel contrarío, la correlación
encontradaentrela insulinemiay los nivelesséricosde IGF-I en las ratasdestetadas
tiroidectomizadasy tratadascon114 fue muy baja,e inclusonegativa(r= 0.048, r= -
0.60 y r~ -0.85, para la administraciónde la hormonacomo inyección, pellet
durante5 díasy pellet durante10 días, respectivamente),e igual ocurrió en el caso
de los animalesadultos (r=r -0.62, r= -0.21 y r~ -0.28, para el tratamiento
intraperitonealde 114 y subcutáneode 5 y 10 días de duración, respectivamente)
(Figuras35 y 36).
3.5.2.6.IGFBPs: nivelesséricosy expresióndel ARNm hepático.
3.5.2.6.1.Niveles séricos.
3.5.2.6.1.1.Westernligand blot.
El hipotiroidismo,comoseha visto en la primeraparte,provocóun decenso
en los nivelescirculantesde las proteínasligadorasde alto y bajopesomolecular,sin
embargo, la recuperación de los animales tiroidectomizados mediante la
administraciónde T4 aumentélos valoresséricos de las IGFBPs de 30 y 45kDa
(Figura37>. Este incrementofue superiory supusouna rehabilitacióncompletade
los animalesdestetadosy adultos tratadoscon el pelletdurante10 días, encuantoa
los niveles de las IGFBPs de alto y bajo peso molecular. La administraciónde
tiroxina mediantela inyecciónde la hormonao por el implantedel pellet durante5
días no restauró los nivelesde la proteínade 45kDa (IGFBP-3) en la población
159
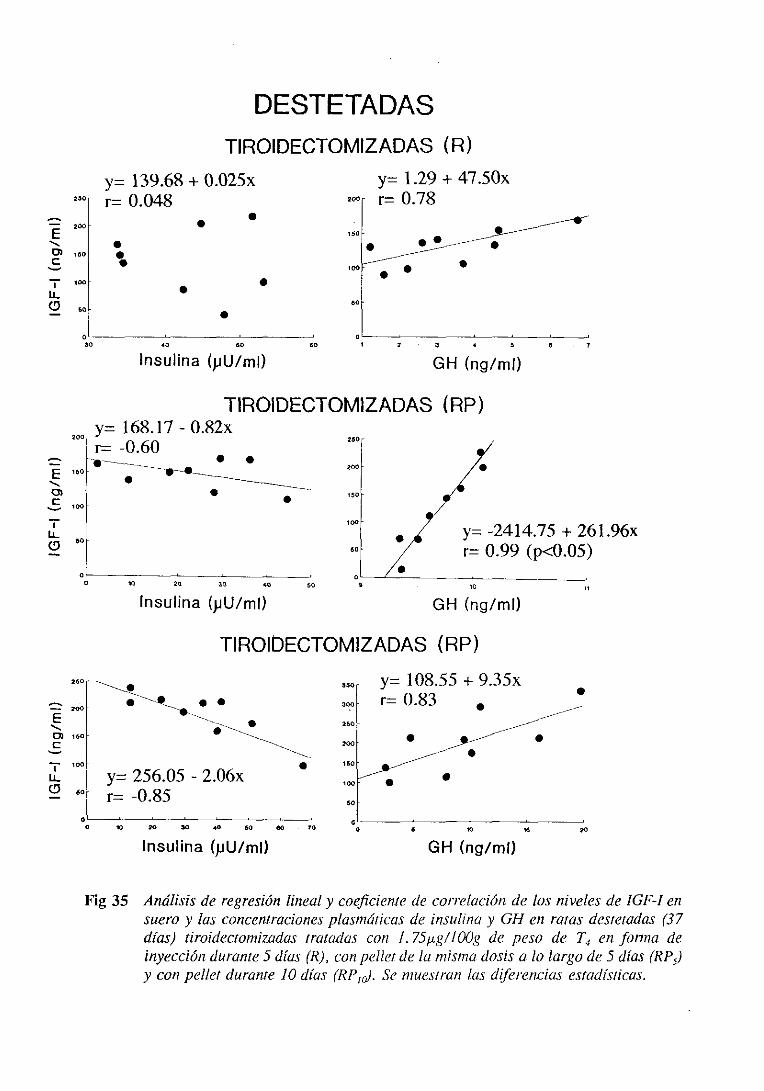
DESTETADASTIROIDECTOMIZADAS (R)
y= 139.68 + 0.025xr= 0.048
ee
e150
ee
eloo
e50
e
40 50
Insulina <pU/mi)
1.29r= 0.78
+ 47.50x
e
e e
e e e
9——e-
0~
60 1 2 0 4 5 6
OH (ng/mi)
TIROIDECTOMIZADAS (RP)
+ 261.96x
OH (ng/mi)
TIROIDECTOMIZADAS (RP)
e e
e
ey= 256.05 - 2.06xr= -0.85
S50.
>00
260
200
460
100
50o
O t 20 30 40 50 CC • 70
Insulina <pU/mi>
y= 108.55+ 9.35xr= 0.83
e e
-e
0 6 10
OH (ng/mi)
Fig 35 Análisis de regresión lineal y coeficientede co’-,-elaciónde los nivelesde IGF-I ensueroy las concentracionesplasmáticasde insulina y CH en ratas destetadast3~días) tiroidecronzizadastratadas con 1. 7Spg/]OOg de pesode T4 en forma deinyeccióndurante5 días (Rl, conpePe?de la misnza dosisa lo largo de5 días (Rl>)y conpellet durante JO días (RP1). Se muesa-anlas dlfe¡-enciasestadísticas.
250
E
c
ji-
CD
200
150
loo
50
o30
168.17 - 0.82xr= -0.60
e e—e
eII e
E
c
“-CD
‘0 20 50 40 $0 6 0
Insulina (pU/mi)
260
Eoc
E1~
CD
200
160
lOO:
60
16 20
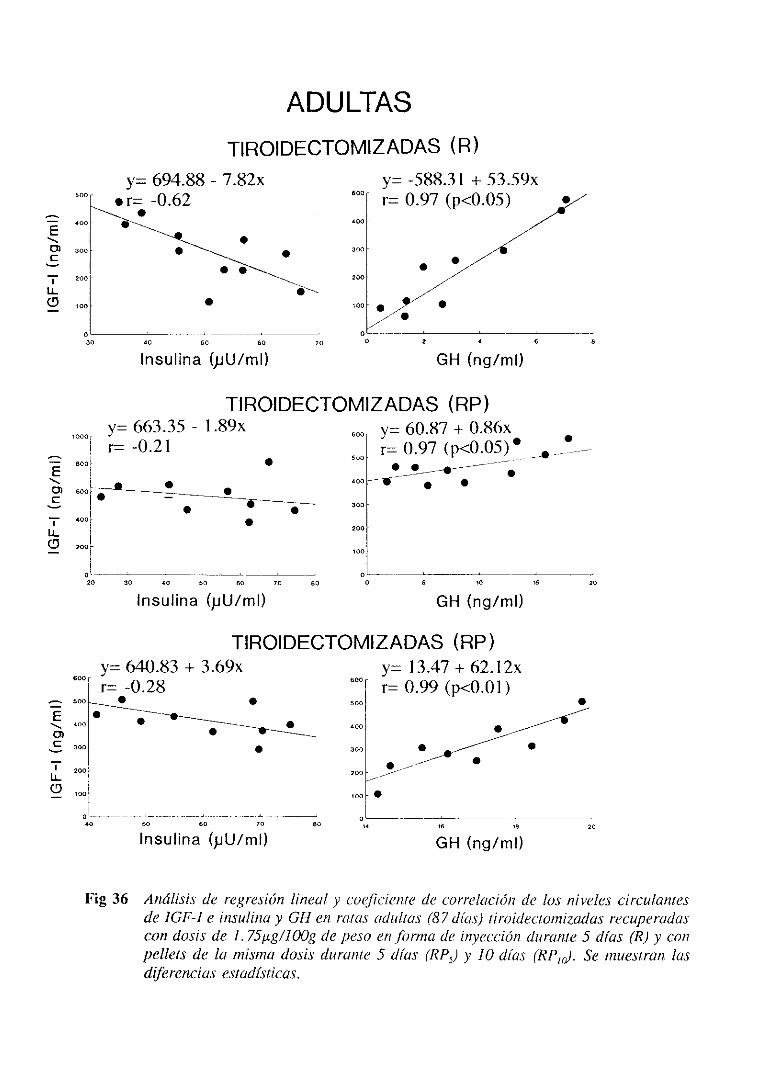
ADULTAS
TIROIDECTOMIZADAS <R)y= 694.88 - 7.82x
er= -062e
ee e
e.
e
60 60
insulina (pU/mi)
y= -588.31±53.59x
70 0
y= 663.35 - l.89xr= -0.21
TIROIDECTOMIZADAS (nP)600 y~ 60.87 + 0.86x
r~ 0.97 (p<O.O5)~e e ~ e
400~t ~ ee ee e ee
e—--e
300 -
200 -
loo
20 30 40 60 60
Insulina (pu/mi>70 60
0 ———- -~
0 5 lO 16 20
OH (ng/mi)
600 r y~z 640.83 + 3.69x
600 r= -0.28e e400 ee e——~~
e300
200 -
100
600
400 -
300 -
200 -
40 50 60 70 60
Insulina (pU/mi)
ee
e
e•
loo-
o$4 It 6 20
OH (ng/mi)
Fig 36 Análisisde regresiónlineal y c-ojicieníede cori-elación de los nivelescirculantesde ¡CF-! e insulina y CH en rotas ¿¡dulías (87días) tiro idee/onzizadasrecuperadascon dosisde 1. 7Syg/lOOgde pesoen fin-ma de inyeccióndurante5 días (R) y conpelleis de la mismadosis durante5 días (RP) y JO días (RP~. Se muestranlasd¿ferenciasestadísticas.
600
400
300
200
E5—
ItO
400
300
400
o30 40
4000
OH (ng/mi)
600
600
E‘5-o)
LI.
CD
400
200 -
o
TIROIDECTOMIZADAS (RP)
600- y= 13.47+62.12xr= 0.99 (p-cO.Ol)
E—5.o)
LI.CD
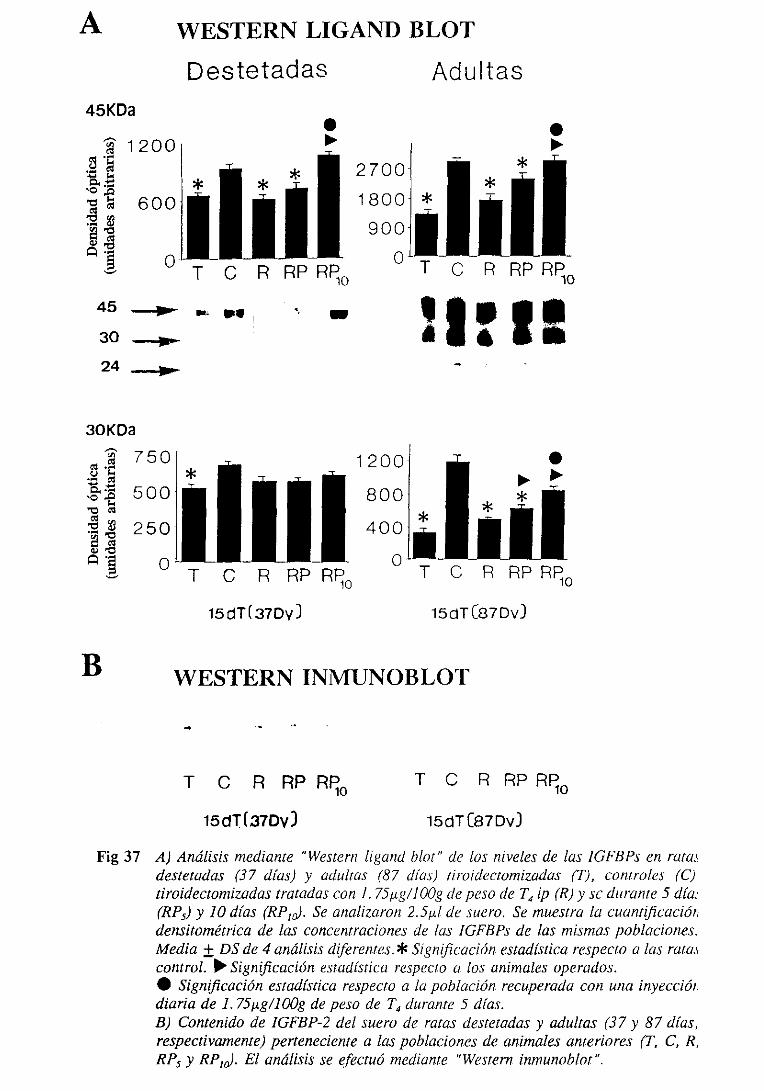
A WESTERN LIGANJ~ BLOT
Destetadas Adultas
45KDa e1200
600
0
—PI’— n. giu
e2700
1800 *
900 3RPR%
*
¡O RRP RP0
II? It24 ~
IIIO RRPRF%
15d1(37Dv)
1200
800
400
-r
*
OB-TC
15dTCB7DvJ
B WESTERN TNMUNOBLOT
T C RRPRF?O
l5dT(37Dy)
T O RRPRI~Q
15dTC87DvJ
Hg 37 A) Análisismediante“Wesíern ligand blw’ de los nivelesde las ¡CPBPs en ratasdestetadas<‘37 días) y adultas (87 días) íiroideuomizadas (7), conu-oíes ((2)tiroidectomizadastratadascon 1. 75yg/IOOgde pesode 7’4 ip (R9 y sedu¡ante5 día:(RP9y JO días (RPJJ. Se analizaron
2.Syíde suero. Se muestrala cuanrWcaciórdensitométricade las concentracionesde las WFBPsde las mismaspoblaciones.Media + DSde4 análisisdWerentes.* Sign~ficaciónestadísticarespectoa las rataácontrol. ~ Sign~caciónestadísticorespectoa los animalesoperados.• Sign¿tlcaciónestadísticarespectoa la población recuperadacon un.a inyecció¿diaria de 1, 7Sgg/lOOgde pesode 7’4 durante 5 días.B) Contenidode ICFBP-2 del suerode ratas destetadasy adultas (37y 87 días,respectivo,nente~pertenecientea las poblacionesde animalesanteriores (7’, (2, R,RP
5 y RP,9. El análisisse efectuómediante “Western inmunobíot”.
-~ .~
— ;~
‘o ,~
5.-
45
30
lCR
3OKDa
~ t~1~
‘o .~‘e
750*
500
250
01
e*
FR RPRF~0

Resultados
destetadani en la adulta. En estascondiciones,tampocoserecuperaronlos valores
del complejo de 3OkDa (IGFBP-1 y 2) en las ratas adultas, aunqueeste sí se
rehabilitócon los tres tratamientoshormonalesen la poblacióndestetada.
3.5.2.6.L2.Westerninmunoblot.
La administraciónde 114 a los animalesdestetadosy adultostiroidectomizados
provocóun decensoen los nivelesséricosde IGFBP-2(Figura37). Estadisminución
en la intensidadde las bandaspareció ser mayor en el tratamientode 5 días de
duraciónfrenteal de 10, que seaproximómás a los valorescontrol. Así pues,y por
comparaciónde estosresultadosconlos obtenidosmedianteel ‘Westernligand blot”,
sepodríasugerirque el descensoen la intensidadde las bandasde 3OkDaregistrado
mediante el “Western inmunoblot” se debió a la disminución de la IGFBP-2
circulante,puestoque el “Westernligand blot” mostróun aumentoen la cantidadde
complejode proteínasde bajo pesomolecular(IGFBP-1 y 2).
3.5.2.6.2.Expresióndel ARNm hepático.
La expresiónhepáticade IGFBP-1 disminuyóen las ratasdestetadasy adultas
tiroidectomízadasrespectoa suscontroles,sin embargo,laadministraciónde T4 a los
animalesoperadosprovocóuna restauraciónen los nivelesde su ARNm (Figura 38);
estarehabilitaciónse produjo ya con la inyecciónen las ratas adultasy no en las
destetadas.La rehabilitaciónfue igual a los controlescon 10 díasde tratamientoen
ambaspoblaciones.
Los niveles de IGFBP-2, que aparecieronaumentadosen los animales
tiroidectomizados,disminuyerontrasel tratamientocon 974 (Figura38). En las ratas
destetadas,el descensode la expresiónfue másacusadoal administrara los animales
los pellets, e incluso, los nivelesde los tránscritosde estaproteínafueron inferiores
a los mostradospor la poblacióncontrol. En los animalesadultos,sin embargo,y
quizás debido a los bajos niveles de IGFBP-2 que existen en ellos, los tres
tratamientosensayadosse mostraronefectivosparadisminuir la expresiónhepática
de la proteína, e incluso, como sucedíaen las ratas destetadas,estos valores
resultaronser inferiores a los mostradospor los controles.En ningunode los dos
gruposde animales(destetadosy adultos) se encontrarondiferenciassignificativas
entre las dospautasde rehabilitaciónllevadasa cabocon pelletsy sí respectoa los
de la inyección (destetadas).
En cuantoa la IGFBP-3, el tratamientocon pelletdurante10 días fue capaz
de restaurarlos niveles de ARNm de esta proteína en ratas tiroidectomizadasa
163

Fig 38 Análisis medianteensayodeprotecciónde la RNasade los tránscritoshepáticosde ICFBP-1, 2 y3 de ratas en periododesteuzdo(37días)y adulto (87días): ratas tiroidectornizadas (7), ccittroles
(Q.uiroideaomizadasy tratadascon una inyeccióndel. 7Sgg/lXgdepesode T4duranteSdías(RLtiroidectornizadasrehabilitadascon pellet de igual dosisdurante 5 días (Rl>), tiroidectomizadasrecuperadasconpetletde la mismadosfficacióndurante¡O días (RPJ4).* Significaciónestadísticarespectoa lapoblacióncontrol.NISignlficació,¿estadísticarespectoa losanimalestiroidecromizados.• Signqlcaciónestadísticarespectoa las ratas rehabilitadascon la inyecciónde T4 (1. 75p.g/lOOgdepeso).Semuestra el análisis densitométrico.Media + DS de 4 análisis dWerentes.
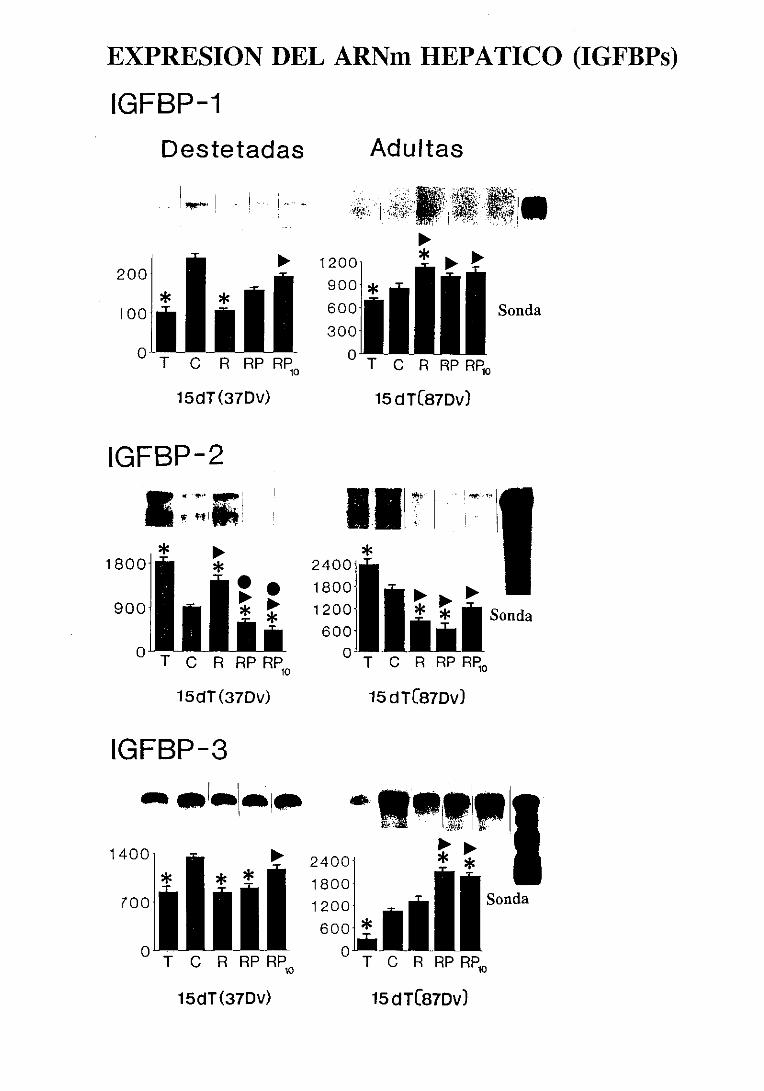
EXPRESION DEL ARNm IBIEPATICO (IGFBPs)
IGFBP-1
Destetadas
200¡ijilloo
T O R RPRPlo
1 5dT <37Dv)
IGFBP-2
1800
900
o
3;;,;
Adultas
PI.120090:tS600300
T O R RPRP10
Senda
15 dTC87Dv)
¡ 7t - y240180120
60
1 0 R RPRP10
1 5dT (37Dv)
Senda
15 dTC87Dv)
IGFBP-3
e e1ea~e
1400
Mli700 T O R RPRP1Q
e
2400
1800
1200
600
en’Senda
1 0 R RPRP10
*
1 0 R RPRI90
*
l5dT(37Dv) 15dTC87Dv)

Resultados
valores control (destetadas>o incluso superiores(adultas) (Figura 38>. En los
animalesadultos, los tres tratamientosensayadosaumentaronla expresiónhepática
de la IGFBP-3, y aunquelos pellets lo hicieron incluso por encimade los valores
control, la inyección mostróunos nivelessimilares a la poblacióncontrol. En las
ratasdestetadas,sin embargo,sólo la pautallevada a cabo mediantela implantación
del pellet durante 10 días, fue capaz de aumentarla expresiónhepáticade esta
proteínade alto peso molecularhastavalores control, y a pesar de ello, no se
encontrarondiferenciasestadísticasentreningunode los tres distintostratamientos.
3.5.2.7.Resumende los resultadosdel estudiodel tratamientocon 114 en
ratasdestetadasy adultastiroidectomizadas.
El IGF-I circulanteen animalesdestetadosserestauraigual a controlescon
pelletdurante10 díasy en adultaspor igual vía de administracióntantoa 5 como 10
días. La expresióndel ARNm hepáticoes rehabilitadaigual a controlescon peflet
tantoa 5 como 10 díasen ambaspoblaciones.El pesocorporal y la Gil plasmática
e hipofisariamuestranuna mejor restauración,a pesarde que no alcanzanvalores
controlmás que enel contenidoen pituitaria de adultasa los 10 díasde tratamiento
conpellet, mientrasla insulinaesrestauradaigual a controlesconpelleta 5 ó 10 días
enambaspoblaciones.
La glucemiaalcanzanivelescontrolescon cualquier tratamientode 114. En
estas condiciones, las bandas proteicas de bajo peso molecular circulantes se
restaurarontan solo conpellet 10 días en destetadasy adultasy la IGFBP-3 mejor
con 10 días de pellet en adultas. La expresiónde su ARN muestra que dicha
restauraciónpodríaserdebidaa IGFBP-1, ya que la expresiónde IGFBP-2disminuía
con 114 por cualquier tratamientotanto en destetadascomo adultas. El ARN de
IGFBP-3serehabilitóconpelletdurante10 díassubiendoporencimade los controles
en adultas.
165

Resultados
Cuadro4.- Resumende los resultadosde los estudiosde la administraciónde
114 en dosisde l.75¡íg/lOOgdepesoporvía ip (T ±974)duranteSdíaso vía sc a lo
largo de 5 ((T + 114)5) o 10 días ((97 + Tj~) en ratasdestetadas(37 días)y adultas
(87 días) respectoa la poblacióncontrol.
1 + 14
DESTETADAS
4INSULINEMIA
GLUCEMIA
GH CIRCULANTE -
GIL HIPOFISARIA ¿
IGF-I CIRCULANTE
ARNmIGF-I
IGFBP-1 y 2 C[RCULANTES=
ARNm IGFHP-1 4
ARNm IGFBP-2 1
IGFBP-3 CIRCULANTE ¿
ARNm IGFEP-3 ¿
ADULTAS
4
4
4
4
t
4
(1 +
DESTETADAS ADULTAS
1 4
4 4
4 —
(T + T~
DESTETADAS ADULTAS
t
4
4
4
1 1
NEONATAL Y ADULTO YREHABILITACION3.6-DIABETESENPERIODO
CON t.El modelo de diabetespor STZ se provocó por inyección
estasustanciaa los 10 díasde vida (neonatos)o a los 72 díasde
animalesfueron divididos en dosgruposy sacrificadosa los 20 y
el primer grupo fue sometidodurante5 díasa un tratamientocon
de LS y l.75jig/lOOg de pesoparalas ratasneonatalesy adultas,
y el segundoactué de control.
3.6.1. Evolución del pesocorporal.
Las ratasdiabéticaspresentaronun pesoinferior al de sus controlestantoen
la etapaneonatal(25.45%)como en la adulta (31.56%)(Tabla II).
En la etapaneenatal,el tratamientocon 114 aumentóel pesocorporalde estos
animalesdiabéticosa los que se les implantó el pellet (12.13%)frente a los no
tratados,aunque persistió la diferencia respectoa la población control. Por el
contrario, las ratas adultasno modificaron su peso tras la administraciónde T4
respectoa los animalescontrol.
intraperitonealde
vida (adultos). Los
87 días de edad:
unasdosisde T4
respectivamente,
166
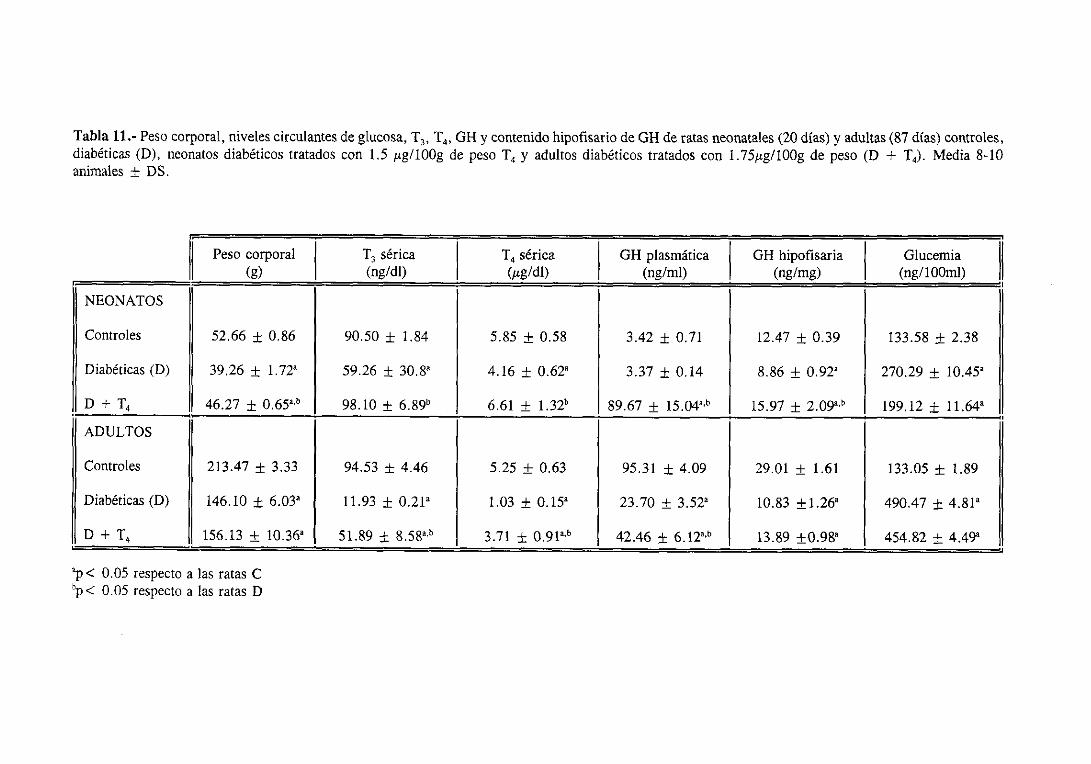
ca—
0$
00
flt
en
t‘0
o—
e-
-+
41
i+
40
0~
C1
OS
t-1cA
<Nj
—e
n.
.O
Os
—r—
os
0~
~-.
&N
00
00
s1~
—4
4
-U+
4-H
~r—
No
~.
o~
-4
en
~fl
05
60
—
‘4Cd00
A~
N
cE
5.—
o
05
.4en
Nó
~‘j9
oti
+1+
4‘o
r—
00
05ti
.—
00
~0
00
-ti
O~
——
O+
1+
1+
1•-
en
O’
000
00
os
Oe
nti
——
cdoo
——
~~
.—
o8
o+4
+4—
tir—
+1~
r—en
en‘9
tiO
-‘e:
—en
~+1
+1
—eng
‘005
en‘08
8+1
+1~
‘eci~
—9
‘0—
en
ca
zt
‘e‘4)
5.-.cnC
O
~ti
‘0~
en8
8—
~
+4
+4te
~—
~-t
60
~‘0
!
‘-4~0
“3—
00
000
05
-0
0oe
n‘o
-l-I+
r+
¡
O‘o
o~
0-
oO~
t~
,,—00
e4
o±1
-uen~
044
ocOS
——Ql
-~o
~o
‘o
—0
+4
+4
‘o~
~r—
ci
‘e
en
~O
en
o«
be
A~
¿o
-u+
4—-u
2e
n
—4
en
~——Ir,
00ozoz
e0000
cd4>
-~
4-‘4
)
o-—
oQ
fi+
00oE
-‘-4Q
e00
00cd.~
3-4‘4>
~ouQ
+
0,0
4)
—~1o
00
3-44
-.cd
e.—
cd
00‘aflQ
4)
00
4>
‘eco
00
e—
lo
004)
bO
~-.
60cd
t—eO
—4
)o
~003
-40
00
0
‘e.~
t.0
cd
cd
te00
0O
tO
o‘e
o)
CE—
O
-to
‘~1O
-ca
o60
cica4>
0‘e
o00
00g
oe
‘e
(34
.-.4
--
—0
0
00Ci
—4-4
4>‘4>.0
cte
a~
±4
•0
0—
Cd00
—(3
0
0000
Cd
Cd
cd
cd
4-
4-
0000
Cd
C~
00
oc.,
4)4
)
0000
4)4
)s~
4-
06
00
00
0
VV
ftP-

Resultados
3.6.2. Nivelesséricosde 113 y 114 y plasmáticose hipot’isar¡osde GH.
La diabetesprovocóun descensoen los niveles circulantesde las hormonas
tiroideas (Tabla 11). Esta disminuciónfue más acusadapara los animalesadultos
(85.57% para T3 y 82.16% para 114) que para los neonatos(34.52% para 113 y
28.89%para114) respectoa los controles.
La administración de T4 provocó el esperadoaumento en los niveles
circulantesde las hormonas tiroideas (Tabla 11). En el caso de los animales
diabéticosneonatalestratadoscon l.5p¿g/lOOgpesode tiroxina, el incrementofue de
un 65.54%para T~ y un 58.65%para14 respectoa las ratasdiabéticasno tratadas,
lo quesituó las concentracionesséricasde las hormonasenunosvaloresigualesa los
de las ratascontrol. Al igual quesucedíaconlas ratasneonatalesque recibíanT4, los
animalesadultosdiabéticossometidosal tratamientohormonalexperimentaronun
aumentoen los valorescirculantesde las hormonastiroideas (262.84%y 27132%
paraT3 y 14, respectivamente)frentea las ratasdiabéticas,perono sealcanzaronlos
valoresde la poblacióncontrol.
Los nivelesplasmáticosde OH disminuyeroncon el estadode diabetesen los
animalesadultos (75.13%)respectoa los control (Tabla 11). Sin embargo,las ratas
neonatalesdiabéticasexperimentaronun ligero descenso(no significativo) en los
valorescirculantesde estahormonarespectoa los de la poblacióncontrol.
El contenidohipofisariode OH, al igual quelos nivelescirculantes,disminuyó
en mayor medidaen los animalesadultosdiabéticos(62.67%)que en los neonatos
(28.95%)respectoa las ratascontrol (Tabla 11).
El tratamientode los animalesdiabéticoscon T4 provocó un aumentoen la
concentracióncirculantede OH y en su contenidohipofisario(Tabla 11). Los niveles
plasmáticosde OH en las ratas neonatalestratadascon la hormonasufrieron un
aumentoespectacular,e incluso sobrepasaronlos valoresde la poblacióncontrol,
mientrasqueen los animalesadultosdiabéticos,a los que tambiénse les adminstró
la tiroxina, esteincrementofue másdiscretoy situó los nivelesde estasratasen unas
cifras intermediasa las de las poblacionescontrol y diabética.A nivel hipofisario,
el aumentoprovocadoen el contenidode OH por la administraciónde 114 en las dos
poblacionesde animalesno redujo la diferenciaexistenterespectoa los animales
control, aunqueeste incrementoresultó ser ligeramentesuperioren el caso de las
ratasadultas(28.25%)respectoa las neonatales(28.07%).
168

Resultados
3.6.3. Glucemiae insulinemia.
La glucemiade las ratasdiabéticasfue de 2 a 4 veces superiora la de los
animales control (Tabla 11). No obstante, el tratamiento con 114 disminuyó
significativamentelos nivelessanguíneosde glucosade los neonatosdiabéticosque
recibieronunadosisde tiroxina de 1 .Sgg/lOOgde peso,aunqueno se alcanzaronlos
vaioresde la poblacióncontrol.
Los niveles de insulina en plasmade los animalesdiabéticosneonatalesy
adultos,comoera de esperar,fueronmuy inferioresa los de sus controles(55.63 y
84.81%, respectivamente)(Figura 39>. Estasdiferenciasrespectoa las poblaciones
control se mantuvierona pesardel tratamientohormonalcon T4 de estosanimales
diabéticosneonatalesy adultos(Figura 39).
3.6.4. IGFs: nivelesséricosy expresióndel ARNm hepático.
3.6.4.1. Nivelesséricos.
La concentracióncirculante de IGF-I disminuyóen los animalesdiabéticos
neonatalesy adultos respectoa los control en una proporcióncercanaal 50% en
ambaspoblaciones(Figura 39).
Las ratas neonatalesdiabéticas que recibieron la tiroxina aumentaron
significativamentelos nivelesséricosde IGF-I respectoa los animalesdiabéticos,de
modo que la concentracióndel péptido fue igual a la de la poblacióncontrol. En
cuantoa los animalesadultos,la implantacióndel pellet a los animalesdiabéticosno
pareciótenerningúnefectosobrelos nivelesséricosde IGF-I.
Los valores de IGF-ll en el suero de las ratas diabéticas neonatales
disminuyeron frente a los de la poblacióncontrol (43.57%) (Figura 39), y el
tratamientocon 1 .5¡tg/lOOgde pesode T4, quetaneficazsemostróen la restauración
de los nivelesde IGF-I, no tuvo ningúnefectoen la recuperaciónde la concentración
circulantede IGF-Il.
3.6.4.2.Expresióndel ARNm hepático.
Los valoresde ARNm de IGF-I en hígadode las ratasneonata]esdiabéticas
disminuyeronrespectoa la poblacióncontrol,al igual que sucedíaa nivel circulante
(Figura39).La administraciónde 114 a estosanimalesdiabéticosaumentólaexpresión
del péptido (Figura 39), pero no sealcanzaronlos nivelescontrol.
En la etapaadulta, como sucedíaen la neonatal,los nivelesde ARNm de
IGF-I disminuyeronen las ratasdiabéticas,pero en estecaso,el tratamientollevado
a cabocon T4 no tuvo ningúnefecto sobrelaexpresiónhepáticadel péptido (Figura
169
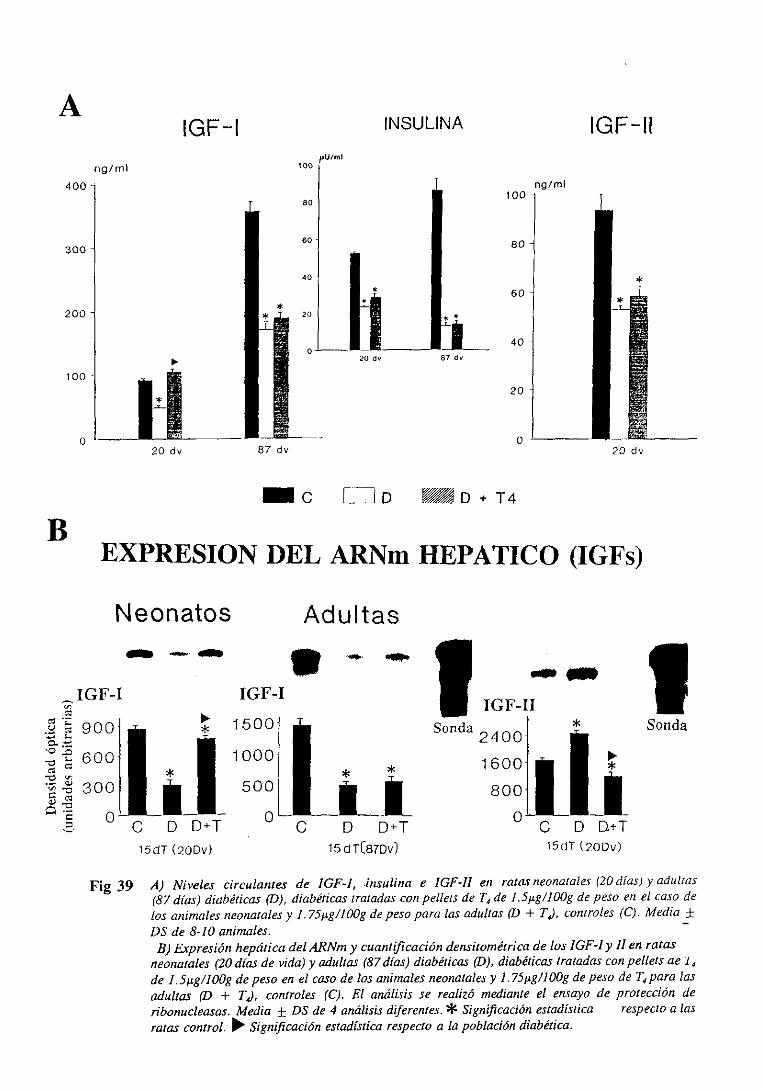
AIGF-I
ng/ml
400
300
200
1 00
o
INSULINA IGF-II
ng/ml~180
60
L 40 -
20
o
o Y”40+T4
BEXPRESION DEL ARNm HEPÁTICO (IGFs)
Neonatos Adultasa — a
IGF-I
*9001.
300
D D~T15dT (200v)
IGF-I
1500
1000 U500 1* *AL
O D 0~T15 d TtiSYOv)
e—
IGF-Ii
Sonda24001 *
1600*LII800 O D B±T
l5dT (200v)
Fig 39 A) Niveles circulantes de lOE-!, insulina e ¡OF-II en ratosneonatales(20 días)y adultas(87 días) diabéticas<71), diabéticastratadascon pelleis de 7’~ de 1 .5~g/lOOgde pesoen el caso delos animalesneonatalesy 1. 7sjsg/lOOgdepesopara lasadultas(D + T4). controles(C). Media +
VS de 8-10 animales.E) Expresiónhepáticadel ARNniy cuantificacióndensitométricadelos¡GE-!y ¡len ratas
neonatales(20 díasde vida)y Multas (87días) diabéticas(D). diabéticastratadasconpelleis ae 14del.5~¿g/1OOgdepesoene!casode losanimalesneonatalesy 1.7Spg/1003 depesode T4para lasadultas <~ + T4, controles (C). El análisis se realizó medianteel ensayode protección deribonucleasas.Media ±DS de 4 análisis diferentes.* Significaciónestadística respectoa lasratas control. ~ Significaciónestadísticarespectoa la poblacióndiabética.
,U~mIloo
*
20 dv
2Qdv S7dv 20 dv
CCcd
Cdcd
*4 1-‘0.0-e 3--
cd-e a-.>
CC
Sonda

Resultados
39).Los nivelesde ARNm hepáticode IGF-II de las ratasneonatalesaumentaron
enunasituaciónde diabetesrespectoa la poblacióncontrol(Figura39). No obstante,
la administraciónde 114 produjo un descensoen la expresiónde dicho péptido, e
incluso, fueron inferiores a los mostradospor la poblacióncontrol.
3.6.5. IGFBPs: nivelesséricosy expresióndel ARNm hepático.
3.6.5.1.Nivelesséricos.
3.6.5.1.1.Westernligand blot.
Enla etapaneonatal,la diabetesprodujoun descensoen los nivelescirculantes
de las proteínasligadorasde IGFs, tanto en el complejo de 3OkDa (IGFBP-1 y 2>
como en el de 45kDa (IGFBP-3), de los animales(Figura 40>. Sin embargo,el
tratamientohormonalllevadoa cabocon14 no modificó los nivelesséricosde dichas
proteínas,con lo quelos valorescirculantesde estospéptidossemantuvieroniguales
a los de la poblaciónneonataldiabética(Figura40).
Los animalesdiabéticosadultos,por su parte, tambiénmostraronun descenso
en los nivelescirculantesde los doscomplejos(30 y 45kDa),y tampocoen estecaso
el tratamientoefectuadocon 114 aumentólos niveles circulantesde estasproteínas
respectoa la poblacióndiabética(Figura40).
36.5.1.2.Westerninmunoblot.
La poblaciónde ratasneonatalesdiabéticasmostróun aumentoen los niveles
circulantesde IGFBP-1 respectoa los animalescontrol (Figura40), mientrasquelos
valoresde IGFBP-2 no sevieron afectados.
La administraciónde T4 a las ratasneonatalesdiabéticasprovocóun descenso
en los niveles séricosde IGFBP-l respectoa la población de animalesdiabéticos
(Figura 40)- Estadisminuciónigualó los valoresde las poblacionestratadascon la
hormonaa los de la control. Los nivelescirculantesde IGFBP-2 no se modificaron
por la administraciónde T4 a los animalesdiabéticos(Figura 40).
3.6.5.2.Expresióndel ARNm hepático.
De maneraanálogaa lo que sucedíaa nivel circulante, la diabetesprovocóun
granaumentoen la expresiónhepáticade la IGFBP-1 en lasratasneonatalesy adultas
respectoa sus poblacionesde animalescontrol (Figura41).
El tratamientocon T4 de los animalesneonatalesdiabéticosdisminuyó los
niveles de ARNm hepático de IOFBP-1 (Figura 41) y, aunque los valores
descendieronrespectoa los de las ratasdiabéticas,no sellegarona alcanzarlas cifras
171
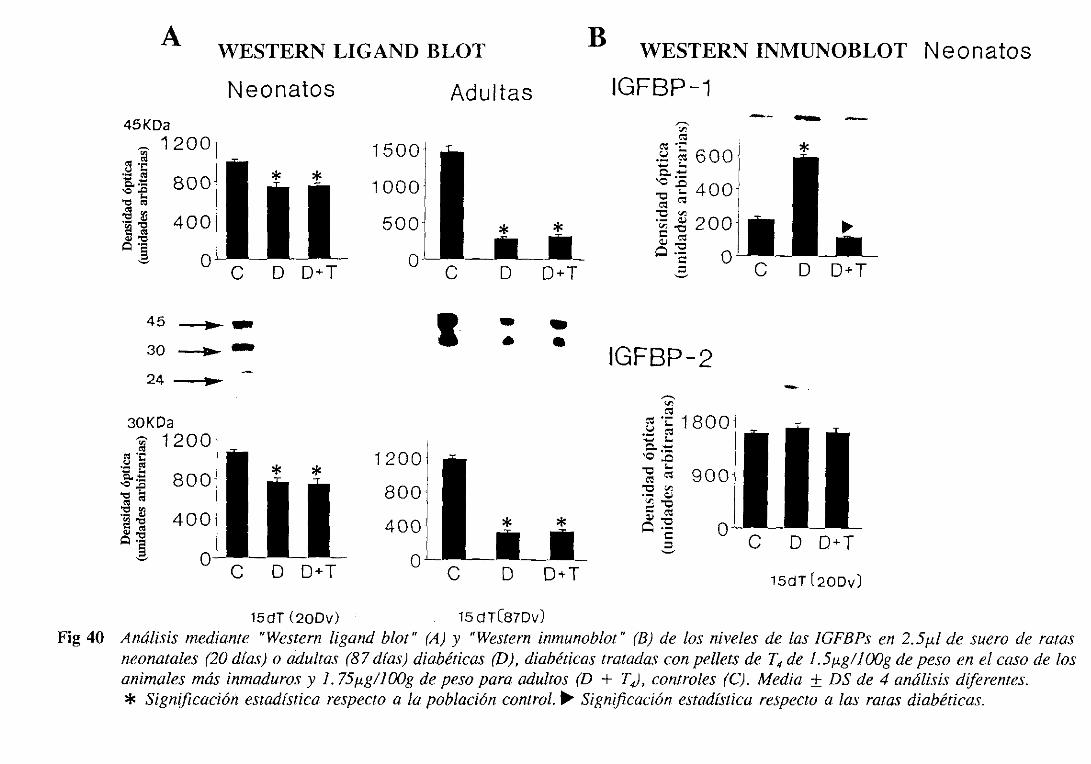
+o
00
00
00
0co
t
O
1—
—o
+
9o
(su~Je.i~iqJ¡~sa
pu
piu
n)
U3fldOp
Ep
!SU
OQ
LLO
~u7
oo
oo
Co
O)
o0
4
a.(sn
~n
tuíq
ausa
pu
plu
n)
cnB
31)dQP
EpIs~~aU
LL
O
9.
o
“3-,
3-,~~.>
-~(.3
-~‘JI
2‘5
...3-,
oO~
~a1—
o~
—4
‘-.4--4
3-’
~•:::
~
E‘~.>
~
~C
j~.>
o—
.Á~~
s%
~>
Dj~.
;L~
3-
.~
+o
oO
k~
.,.—
~3
-,
oo~
t‘~
E
o
~.29
t~
~
Y-*
+‘o
*Uo
BE
o
00
00
00
0LS)
OU
)r
r
oo
o0
00
ejca
~r
o 1—+Oo
mm,ff‘o
o•t
~.ci
c’q
00
00
00
0ej
CO•Ú
r
oo
co
00
ejca
t-
o(s
4xu
s$
~q
nsap¡~p~un)
«~inqdo
pup¡suaq
Coo-4-J
ceaoo,z
11
oozzzCC
Co(~14
-’
D-Do0zCID
Coo.4-’
ceaoo,z
oS¿(stu
~¿
~q
nsapepjun)
‘o~e
~p
dp
PeP
!suaU
o.I~1.

Fig 41 ExpresiónhepáticadelARNmy cuantfficacióndensitométricade IGFBP-1, 2y 3 enratasneonatales(20días)y adultas(87días) diabéticas<D), diabéticastratadascon1.Sgg/lOOgdepesoen el casode los animalesneonatalesy 1. 7Sgg/lOOgdepesopara los adultos <V + T4), controles (C). Media + DS de 4 análisis distintos.* Sigtgficaciónestadísticarespectoa las ratas control. > Sign~caciónestadísticarespectoa la población diabética.
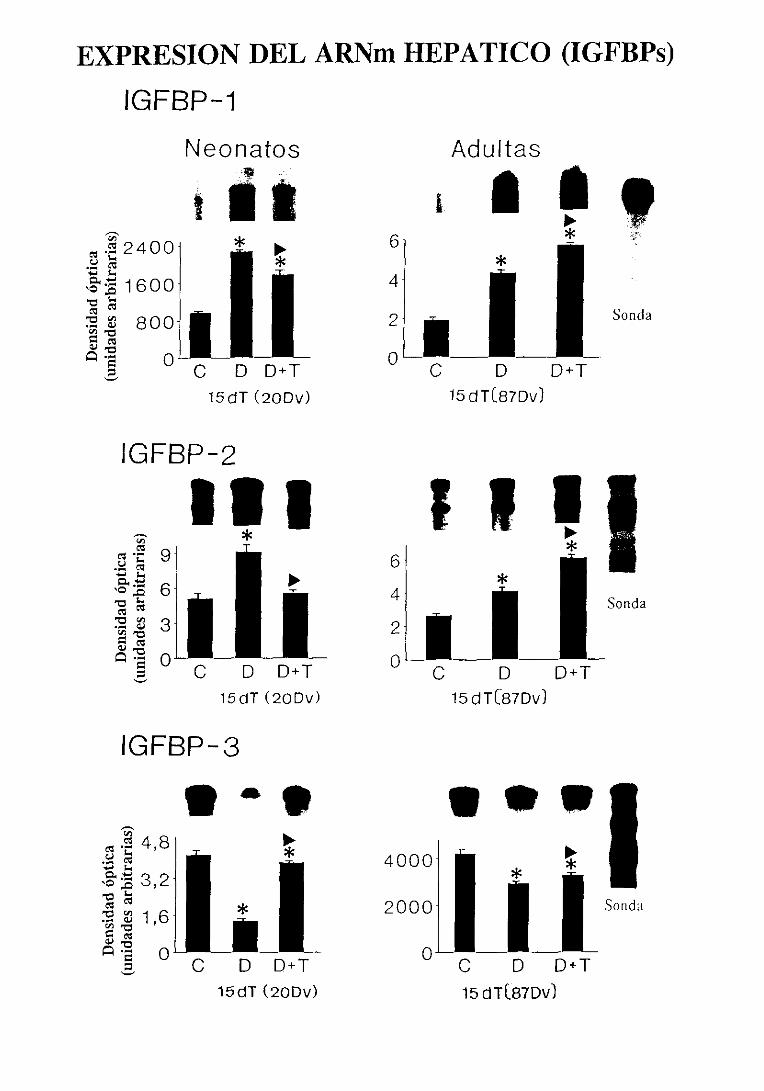
EXPRESION DEL ARNm HEPÁTICO (IGFBPs)
IGFBP-1
Neonatos
1ca
~ .~ 2400— %~
‘0 ~
‘e ca0, ‘e
4> ‘e o5
1600
800
3 aAdultas
6
4
2
oD D~T
l5dT (200v)
¡
O D D~T
64
2
lbdT (2oDv)
E
15 d1 E 87 Dv)
1
D D~T15dTC87Dv)
IGFBP
a.
*
ilO D D~T
e,ph-
4000 *
2000 ¡_II~~O D D~T
ph-*
*
O DD-’-T
Sonda
o
IGFBP-2
u*C.1~
c~-~ c~— ~-
‘0
‘e 0,.— 4)
CC ‘e
4> ‘e
6-
3-0-
ph-
*
Sonda
O
—3
CC
‘0.0‘e 1-<c~c~‘e 4)
4,8
3,2
1,6
o
Sond~i
lbdT (200v) 15 dTE87DV)

Resultados
de la poblacióncontrol.
En la etapa adulta, el tratamientocon 114 aumentóaún más los niveles de
ARNm de IGFBP-1 respecto a los de las ratas diabéticas (Figura 41), y la
administraciónde tiroxina por tanto no restaurósus niveles a los de los controles.
La expresiónhepáticade IGFBP-2aumentóen las ratasdiabéticasneonatales
y adultasrespectoa las control (Figura 41). Sin embargo,en la etapaneonatal,el
tratamientohormonalde estosanimalesdiabéticosdisminuyó los nivelesde ARNm
de IGFBP-2 hastaalcanzarunos valores similares a los de la poblacióncontrol
(Figura 41), y por el contrario, los animalesdiabéticosadultos tratadoscon la
hormonamostraronun aumentosignificativo de la expresiónde IGFBP-2 respectoa
la poblacióndiabética(Figura 41).
En cuanto a los niveles de ARNm hepático de IGFBP-3, disminuyeron
significativamenteenuna situaciónde diabetestanto en la etapaneonatalcomo en la
adulta (Figura 41). No obstante, la administraciónde 14 a las ratas neonatales
diabéticasaumentóla expresiónhepáticade la IOFBP-3,aproximandolos nivelesde
ARNm a los control (Figura41). Porúltimo, en la etapaadulta, laadministraciónde
114 a los animalesdiabéticosaumentóla expresiónhepáticade IGFBP-3 y situó sus
nivelesenvaloresintermediosy distintosa los mostradospor las poblacionescontrol
y diabética(Figura 41).
36.6. Resumende los resultadosdel estudio de la diabetesen ratas
neonatalesy adultasdiabéticasy tratadascon T4.
La administraciónde STZ a las ratas provocó un bloqueoen la secreciónde
insulina, por lo que los niveles plasmáticosde estahormonafueron muy bajos, y el
tratamientocon 14 no mejoró la situación.En los animalesdiabéticos,las dosis de
T4 rehabilitaronlos nivelesde IGF-I circulantey tambiénsu expresiónde ARNm
hepáticosin incrementarla insulina,cosa queno hicieronen adulto. El IGF-II no se
restauróa nivel circulante pero el ARNm, que lo tienen aumentadolos animales
neonatalesdiabéticos,disminuyódespuésdel tratamiento.Las IGFBPscirculantes,
que estándisminuidas en ambos animalesdiabéticosneonatalesy adultos, no se
rehabilitaroncon T4. En neonatossí que descendióla IGFBP-1 que tenía niveles
mayorescon respectoa controles.
Las dosis de 114 en neonatosdisminuyeron la expresión de IGFBP-1 y 2
aumentadasen los animalesneonatalesy adultosdiabéticos,pero no lo hicieron en
la población adulta. La expresióndel ARNm en neonatosy adultos se rehabilta
174

Resultados
despuésde las dosisen neonatosigual a controlesy no en adultas.Seconcluyela no
regulacióna nivel transcripcionaldel complejo de 3OkDa (IGFBP-1 y 2> en los
animalesdiabéticosneonatalesy adultos.
Cuadro5.- Resumende los resultadosde los estudiosde la administraciónde
en dosisde 1.5 y 1 .75$g/lOOgde pesopor vía sc (D + 114) en ratasneonatales
y adultasdiabéticas(20 y 87 díasde vida) respectoa la poblacióncontrol.
NEONATOS ADULTAS
D D±~ O D+T,
GLUCEMIA 1 1 t 1
INSULINEMIA 4 ¿ ¿
OH CIRCULANTE ¶ 4 4
OH HIPOFISARIA — t 4 4
IOF-l CIRCULANTE —
ARNm lORd 4 4 4 4
¡OF-II CIRCULANTE 4 4
ARNm IGF-II ¶ 4
IGPBP-1 Y 2 CIRCULANTES 4 4 4
IGFEP-I CIRCULANTE 1
ARNm IGFEP-1 1 1 1 1
IOFBP—2 CIRCULANTE —
ARNm IOFBP-2 1 — 1 1
IGFBP-3CIRCULANTE 4 4 4
ARNm IGfl3P-3 4 4 4 4
3.7. HIPOTIROIDISMO, IGFs, IGFBPs Y TRANSPORTADORES DE
GLUCOSA (GLUTs) EN PERIODONEONATAL.
En el hipotiroidismocomo se ha mostradosealterael axis IGFs/IGFBPsy en
dichasalteracionesparecetenerun papel importantela insulina. Por otra parte,está
descritoque el pasode GLUT-1, transportadorde glucosaabundanteen periodos
inmaduros,a GLUT-4, transportadorde glucosaen etapaadulta,pareceestarregido
por las hormonastiroideas (Castelló, 1994). Con el fin de ver si ambos procesos,
regulación del axis IOFs/IGFBPs y de GLUTs por dichas hormonas, están
relacionados,se compararonestosestudios.Se diseñóun protocolode trabajo enel
que se analizaronlas posiblesalteracionesen los niveles de los IGFs, IGFBPs y
GLUTs de los animaleshipotiroideos(tratadoscon MMI y tiroidectomizados)a lo
175

Resultados
largo del periodoperinatal(2, 4, 8 y 20 días de vida).
3.7.1. Evolución del pesocorporal.
El peso corporal de los animalestiroidectomizadosfue inferior al de los
controlesa los 4, 8 y 20 días de vida, y a esasedadesla variaciónde dicho peso
encontradaosciló frente a las ratascontrol entreun 13.28%, 21.75% y un 40.19%,
repectivamente(Tabla 12).
Finalmente,en ambaspoblaciones(hipotiroideay control), se observóun
aumentoprogresivodel pesocorporal con la edada lo largo de la etapaestudiada.
3.7.2. Niveles séricosde 113 y 114 y plasmáticose hipofisariosde GH.
Los niveles circulantesde 973 y 14 disminuyeron en todos los animales
hipotiroideos(tratadoscon MMI y tiroidectomizados)(Tabla 12). No obstante,las
ratasneonatalesde 2 y 4 días de vida, que fueron tratadascon MMI (0.02%,p/v)
desdeel día 14 de gestación,presentaronunos valores circulantes de hormonas
tiroideasligeramenteinferioresa los de las ratascontrol a 4 días(38.47%).Porotro
lado, los animalesque fueron operadospresentaronunosvalores séricosmuy bajos
de 113 (81.95% y 93.58% a los 8 y 20 díasde vida, respectivamente)y 114(84.21%
y 78.96%a los 8 y 20 díasde edad)frentea los mostradospor la poblacióncontrol.
Los nivelescirculantesde Gil disminuyeronen los animaleshipotiroideosde
2 (58.27%) y 4 días de vida (51.89%) respectoa la población control, pero
aumentaronde un modo significativo en las ratastiroidectomizadas,tantoa los 8 días
de edad(3 díasdespuésde la tiroidectomía,84.17%)comoa los 20 díasde vida (15
días despuésde la operación,248.42%),siemprerespectoa los neonatoscontrol
(Tabla 12) comosehabíaencontradoen la primerapartede estetrabajo.
El contenidode Gil en pituitaria de las ratashipotiroideasse alteró en el
mismo sentido, respectoa la poblacióncontrol, a como lo hicieron los niveles
plasmáticosde estahormona(Tabla 12): disminuyóa los 2 y 4 días de vida en un
60.53 y un 85.11%,respectivamente,y se incrementóa los 8(10.84%)y 20 díasde
edad(3.78%).
3.7.3. Glucemiae insulinemia.
Los animalestiroidectomizadospresentaronun descensosignificativo en los
nivelessanguíneosde glucosa(8 y 20 díasde vida) (Tabla 12).
Los valores plasmáticosde insulina no se modificaron en los animales
neonatalestratadoscon MMI de 2 días de vida, aunquea los 4 días de edad se
observóunadisminuciónen las concentracionesde estahormona(Figura42). Porel
176
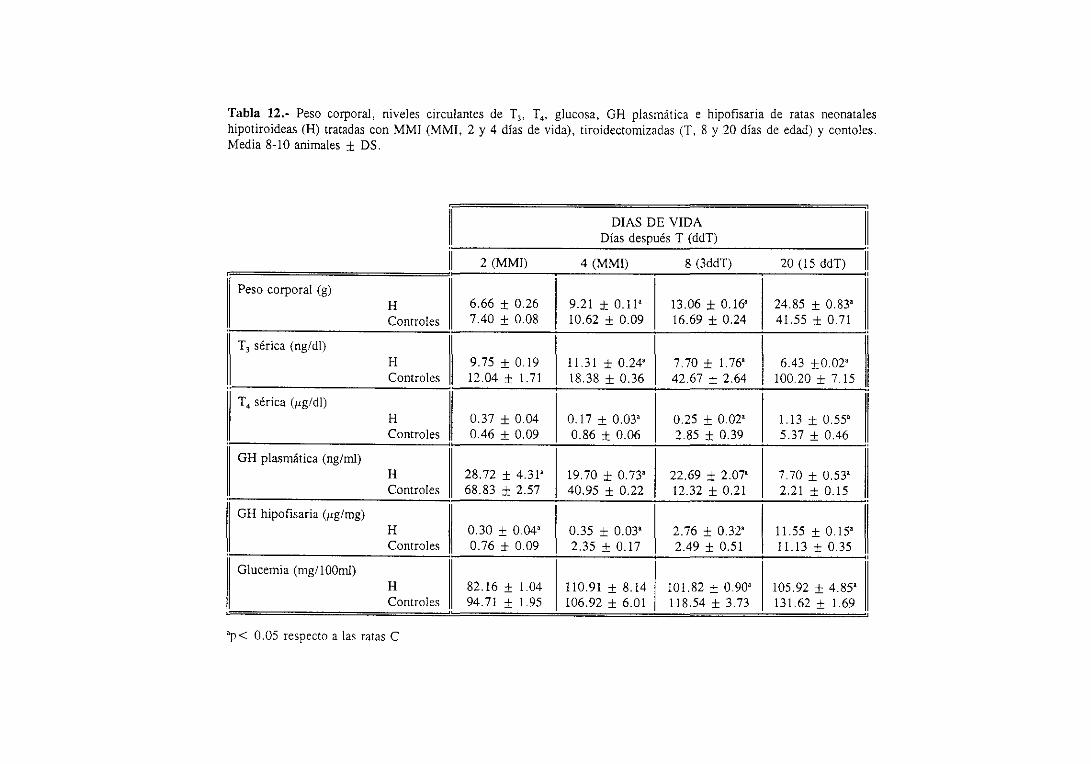
p‘0<
-e
~0~
e-o4
)<
-e—
CCo
o
-eVNo(-4
-u-uCo
00
oo
00
+131
VNVN
00
VN
04
-t
VN0
4O
ro
±1
+1o0
4‘0
‘00
VN
’0
VN‘t
00
+1+
1C
o—
Co
—VN
~0VN
VN00
11+1
0~
e—~
y- 0
4
~VN
—C
o
¿0
±1
+1
~tfl
VN-
00
~
4—31
+10
40
4Q
N~
VN—
oC
o
—~
t1~
~
00No
’~e
nV
—~
443
11e—
~.~
i-—,~
~t~rl
O
ÑÑO
31
31
ON
~
~
(flVN
±1
~ccl
EC
O
0—
-Q
N0
-oo3
13
104
—V
0
QN
—
~rV0
4C
od
é
31
3100
coC
o,..~
00
rn~
0-ó
031
31
e--
~.¿
o
Z..o
iÑ
N~03
13
1O
VN
y—Q
N~
o...‘~
t
Co
’~03
1+
1VN
VN
Co
<2
d~
00
V
CN
QN
0V0
04
00
Cdo&o<.3o4)
ca
‘00
00
10
00
+1+
1‘0
0V
tV
r—
CCa)
ooo
‘000
eCdoa-‘a->
E-
QN
0~
+1+
1o-0
4QN
CC4)
ooou
•0oca)-
Cdcaa—‘O
)
‘tQ
N
00
00
+1+
1
Co
00
z
4)
o4-.
ou
00
eCda)
‘CdECCCd
o
—e--
Co
VN
31310
4en
t’-0
0
00
00
04
V
CC
4)
ooou
00E00CdCdL0Co0
-
o
oS
do
1-1+1C
VC
o~
dO
4)
oa-
ou
-tVN
oQ
N
+1+
1‘0—
1’-
CMÚ
OO
QNUNa)Ooo
oo00
4)
e->
5
~C
C—
a)Cd
—~
0cd00
00
(30
eCd~C
d
4)4
)-eC
dQN
~Cd
VE
CO
o~
o--e0
0a
,
Cd
E-
-4va
‘CdCd
~o
¿2
ci
-e,—
’—
Cd
00
-u-ua)
-c‘t
CCa)0
4eCd(-4
Eeo
--a
->e-O
~U
NOCd
CC<a-
.5.n•~
O0
Cda-’
uCdCdcdCd
o‘-aa->0-
a)
VNooy

Resultados
contrario, las ratasque fueron operadasmostraronunosniveles superioresa los de
suscontrolesen un 64.86%y un 42.59%a los 8 y 20 díasde vida, respectivamente
(Figura42) igual a lo halladoen la primerapartede estetrabajo.
En ambas poblaciones(hipotiroideasy control), se observó un aumento
progresivoen los nivelesde insulina circulante,conunpico habitualy claro a los 4
días de vida en las ratas control que no aparece tan claro en las ratas
tiroidectomizadas.
3.7.4. IGFs: nivelesséricosy expresióndel ARNni hepático.
3.7.4.1.Nivelesséricos.
Los niveles de IGF-I en suerode las ratascontrol (Figura 42) mostraronun
incrementoprogresivo (de 54.84ng/ml a 148.O9ng/ml) de maneraanáloga a lo ya
descritopara la insulina.
Los animalestratadosconMMI presentaronunosvaloresde IGF-I inferiores
a los de sus controles a los 4 días de edad (47.28%). Por el contrario, las ratas
tiroidectomizadasmostraronun aumentoen los nivelescirculantesdel péptido que
llegó a ser de un 98.77% a los 20 días de vida y que resultó ser paralelo al
incrementomostrado por la insulinemia en todos los casos,exactamentecomo
sucedióen la primerapartedel trabajo.
La concentraciónde IGF-II en el suerode las ratascontrol experimentéun
descensoconformeaumentabala edaddel animal (Figura 42). Las ratasneonatales
de 2 y 4 díasdevida, que fuerontratadascon MMI, asícomolas de 20 díasdeedad,
quefuerontiroidectomizadas,mostraronun aumentoenlos nivelescirculantesdeeste
péptido frentea la poblacióncontrol(69.97,57.33y 241.00%,respectivamente).Sin
embargo,a los 8 díasde vida se igualaronlos valoresséricosde IGF-II de las ratas
operadasy controles.
3.7.4.2.Expresióndel ARNm hepático.
Los animaleshipotiroideosmostrarona 2 y 4 díasde vida (tratadosconMMI)
un descensoen los nivelesdel ARNm hepáticode IGF—I (Figura42). Sin embargo,
a los 8 y 20 díasde edad,en los neonatostiroidectomizados,se observóun aumento
en la expresióndel péptido enhígado,siemprerespectoa la poblacióncontrol.
Los nivelesde ARNm hepáticode IGF-II (Figura42)aumentaronenlas ratas
a las que se les administró el fármaco bociógeno(2 y 4 díasde vida), así como en
aquellasde 20 días de edadque fueron tiroidectomizadas,mientrasque a los 8 días
de vida (3 díasdespuésde la operación) la expresióndel MF-II fue menor en la
178
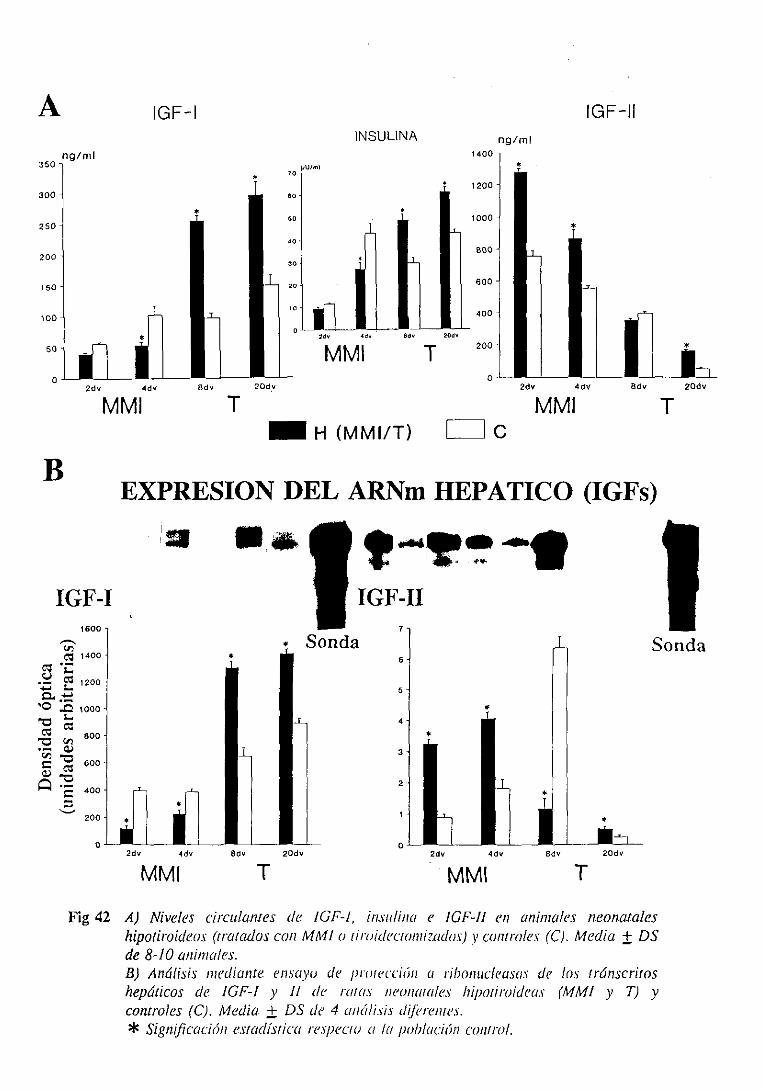
IGF-IAINSULINA
40
30
le
1400
H (MMI/T)
1200
1000
600
600
400
200
IGF -IIng/ml
LIZo
EXPRESION DEL ARNm HEPÁTICO (IGFs)
u U.
1
Sonda6
5
4
a
2
o
9.a¿~ea
IGF-II
Soda
circulantes de IGF-I, ¡¡isulina e ¡UF-II en anima/es neonatales(ti-citados &>¡¡ MMI o t¡/uidecloiniziaclos) y controles (C). Media + DS
Fig 42 A) Niveleshipoti’-oideosde 8-JO animales.fi) Análisis mediante ensayo de p¡uíec-c-¡án a ribonucíeosc¡s de los H-ánscritoshepáticos de ¡GP—! y 1/ dc irnos necn,auíles h¡poÑvideas ~MMl y 7) ycontroles (Q. Media + DS de 4 análisis ¿/ifr rentes.* Sign¿ficoc-ión <‘stúdíst¡&i /Y’5/WCIC) CI 1(1 ¡>oblctc¡óii c-ontrol.
¡ml‘u,.,”350
300
250
200
1 50
100
50
o
70
eo
so
20
2dv 4dv 8d-~ 2Odv
MMI T
Ido 44o 04’ 204’
MMI T
B
*
2dv 4dv Bdv 2Odv
MMI
IGF-I1 600
ca~ 1400
-~ E~ 1200
o’.-.‘o •~ ~
800
ca “e 000
E 400
200
o2dv 4dv Bdv 20dV
M MI2dv 4dv Sdv 20dv
MMI T

Resultados
poblaciónde animalestiroidectomízadosrespectoa la control.
Se destacaasí la existencia de un gran paralelismo entre las variaciones
encontradasparaestospéptidos(IGF-I y II) a nivel séricoy de expresión,lo que nos
hacesuponerla existenciade unaregulaciónpor partede las hormonastiroideasa
nivel transeripelonal.Además,se observóuna gran similitud de resultadosen estas
poblacionescon respectoa las igual tratadasen la primeraparte(apartado3. 1.), con
lo cual ratificamostotalmentenuestrosprimerosresultados.
3.7.5. Estudiode regresiónlineal y correlación.
El análisis de regresióny correlaciónentreIGF-I e insulina y Gil se efectuó
reuniendoen cadaanálisis los datosde las diferentesedadesestudiadas.
La correlaciónfue muy buenay positiva entreIGF-I e insulina, tanto para
poblaciónhipotiroidea(r~ 0.89, pC 0.001) como para la control (r= 0.84, p<
0.01) (Figura43). Sin embargo,estaspoblacionesmostraronunacorrelaciónnegativa
(rr= -0.12, p< 0.05 y r~ -0.93, p<O.Ol paralas ratas hipotiroideasy controles,
respectivamente)cuandose representaronlos nivelesde IGF-I frente a los de GH
(Figura 43), igual que lo obtenidoen la primeraparte.
3.7.6. IGFBPs: nivelesséricosy expresióndel ARNm hepático.
3.7.6.1.Nivelesséricos.
3.7.6.1.1.Westernligand blot.
Los animalesque fueron sometidosal tratamientoconel fármacobociógeno
presentaronunapautadiferente,respectoa las fluctuacionesobservadasen los niveles
circulantesde las IGFBPs, frentea aquellasratasque setiroidectomizaron.
Los nivelesséricosde las proteínasde 3OkDa de los neonatoshipotiroideos
disminuyerona los 2 y 4 días de edad(poblacionestratadasconMMI desdeel día
14 de gestación), mientras que aumentarona 8 y 20 días de vida (animales
tiroidectomizados)frentea las ratascontrol (Figura44).
La IGFBP-3 disminuyóen los animaleshipotiroideos(tratadoscon MMI y
tiroidectomizados)en todaslas edadesconsideradasenel estudio(Figura 44).
La poblacióncontrol, comoya se ha señaladoen otrasocasiones,mostrólos
cambiosde las proteínastransportadorasasociadasal desarrollo;estoes, los niveles
de IGFBP-1 y 2 disminuyeron al aumentarla edaddel animal, mientras que la
proteínade alto pesomolecularsiguióelpatróncontrario(aumentóconformelo hacía
la edadde la rata).
180
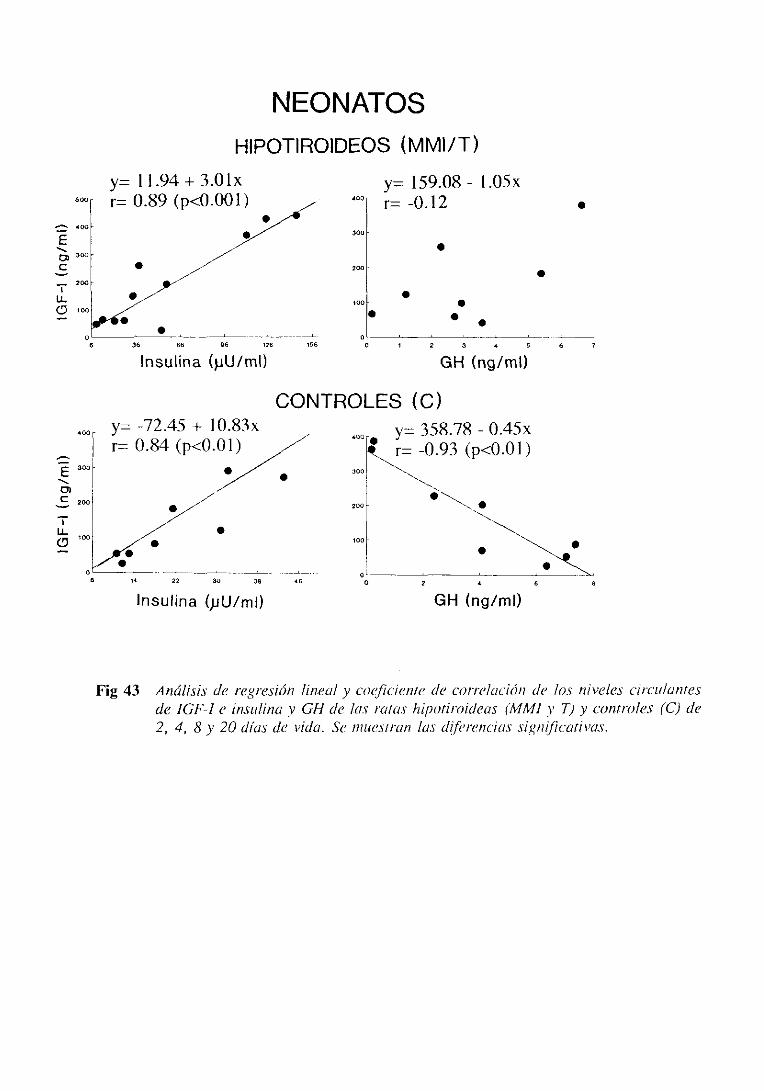
NEONATOSHIPOTIROIDEOS (MMI/T)
y= 1L94±3.Olx~ r= 089 (p<OOOI)400
200
1001
0~
y= 159.08- l.05x400 -
r= -012 e
$00 -
$00
loo -
o
e
ee
e ee
e0 1 2 3 4 5 6 7
Insulina <pU/mi) OH <ng/mt)
CONTROLES (O)
0.84 (p<O.0l)-72.45 + 1083x
e
ee
eee
14 22 30 38 46
$00
e 358.78 - 0.45x-0.93 (p<O.OI)
e$00 -
loo -
00 2
insulina (pU/mi) OH (ng/mi)
Fig 43 Análisis de regí-esión lineal y co«/¡cieme de corí-elacion de los niveles ch-tu/untesde fUE! e insulina y CH de las ‘-atas hiporíroideas (MMI y 7) y cono-o/es (C) de
eE
300
c
U.o
e
36 66 96 126 150
400
300 -
200 -
Eo)c
LI.
a loo -
oe
e
ee
• 6 8
2, 4, 8 y 20 días de vida. Se muestran. las diférencia.s sígn ¿¡¡cari vas.
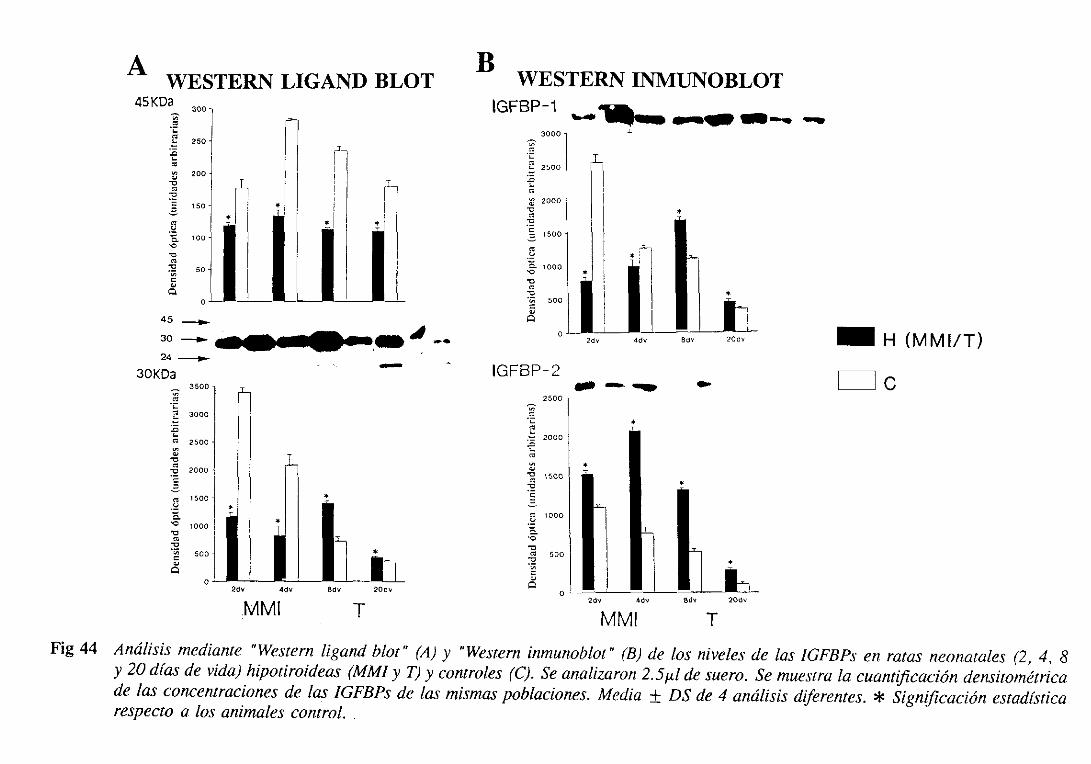
loII
a
__
_a)
__
__
__
__
__
__
_u
3I~
Nt
o
($ClÁ
lcilL(j113
SO
pZpiU
Il)~
!1dQp
op
!sUa
U
o_
__
__
__
_
(no
02
00
OO
a~o
l
<~(sr{in
ui¡q
an
S3pU
’p~Ufl)
O3~)dQ
PEP!SuaU
sc
ot
1~
e”o-
u-CD
e1_
__
__
__
__
__
_1
~—
1ff~o
nr,
o
(s6~[JUJ1~qJoS
dpUp!un)
~3!16I9pEp~su$Q
1
1-A
1a
-Sl
.u‘.4
%
e--,‘.4
e--,
1-.-~J
~-)
‘t~~J
ttt
~‘a,
ca,
1—~
0-~4
‘a~
01
o
---:2--~
o-‘—
1—~
01-~
—‘-.4
~—
.t
Oe
’Ca
-=‘.4
-~c.
‘~N
~o-
1£oE
¡
oo
oo
oo
oo
oo
OO
Oro
-mu
-o
oo
oo
oo
oo
alo
,1O
oo
oo
oo
oo
oo
o5
a,0
01
clCa
Ca-
—
I<,
(st~brJ1LC¡.Iesapup!ufl)
~~
!ld9PCP!SUaU
co
flscoci
‘1~

Resultados
3.7.6A1. Westerninmunoblot.
Los nivelescirculantesde IGFBP-1 se comportaronde modo muy distinto
segúnel protocoloa que fue sometidoel animal. Las poblacionesa las que se les
administróMMI (2 y 4 díasde vida) presentaronunascifrasséricasde estaproteína
disminuidas,mientrasque la tiroidectomía(8 y 20 díasde edad)produjoun aumento
en sus niveles,siemprerespectoa los neonatoscontrol (Figura44), siguiendoigual
pautaque la insulina.
El tratamientode las membranascon anti-IGFBP-2 nos revelóun aumento
significativo en los nivelesséricosde estaproteínade bajo pesomolecularen todos
los animaleshipotiroideos(tratadosconMMI y tiroidectomizados)respectoa las ratas
control (Figura44).
3.7.6.2. Expresiónhepática.
Los nivelesde ARNm hepáticode IGFBP-1 se encontrarondisminuidosenlos
neonatoshipotiroideosque fueron tratadoscon MMI respectoa los controlesy, por
el contrario, los animalestiroidectomizadosmostraronun aumentode la expresión
de IGFBP-1, tanto a los 8 como a los 20 días de vida (Figura 45), al igual que
ocurría con los nivelescirculantes.
Los animaleshipotiroideosmostraronunosnivelesde ARNm de IGFBP-2en
hígado (Figura 45) superioresa los de sus controlesen todaslas edadesestudiadas,
exceptoa los 2 díasde vida, en que no se modificaroncon respectoa controles.
La expresiónde la proteínade 45kDa (IGFBP-3)enhígadodisminuyóen las
ratashipotiroideasrespectoa las controlesen todaslas edadesconsideradas,excepto
a los 4 díasde vida (Figura45).
Estosresultadosnos muestran,por tanto, la existenciade un paralelismoentre
los valoresséricosy laexpresióndel ARNm hepáticode las proteínastransportadoras
encontradasparaestosanimales.Además,seencuentratambiénunperfil análogoen
cuantoa las alteracionesen los nivelesde las IGFBPsentrelos animalesde 10, 15
y 20 díasde vida tratadosconMMI (ver apartado3.1.)y los de 2 y 4 díasdeedad.
Sedebedestacartansolo que las ratasde 2 y 4 díasde vida presentaronunosniveles
en el complejoproteicode 3OkDa (Westernligand blot) disminuidosfrentea sus
controles,mientrasque los animalesde 10, 15 y 20 díasde vida, que fuerontratados
también con MMI, mostraron unos valores de IGFBP-1 y 2 aumentados;esta
diferenciaquizásse debaa un descensomás acusadoen los valores individualesde
IGFBP-1, a 2 y 4 días de edad, respectoa 10, 15 y 20 días de vida o a que 2 y 4
183

Fig 45 ExpresiónhepáticadelARNmy cuan4ficacióndensitométricade IGFBP-1, 2 y 3 enneonatoshipotiroideos(MMI y T) y controles(C) de 2, 4, Sy 20 días. El análisisseefectuómedianteel ensayo de protección de ribonucleasas.Media + DS de 4análisis deferentes.* Sign~ficacián estadística respecto a la poblacióncontrol.
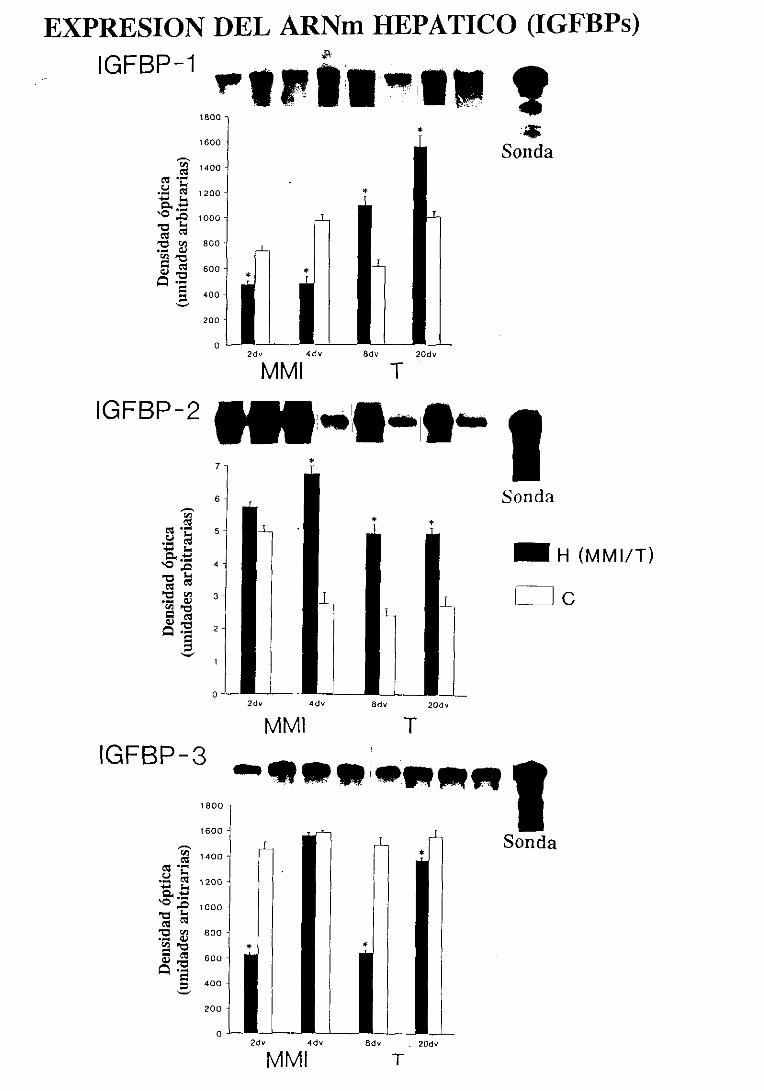
EXPRESION DEL ARNm HEPÁTICO (IGFBPs)IGFBP-1 wg7fl
0800
1600
cn
— ¡-1
‘0 .,~‘e 1~e~c~‘e cn.— ca
r4~
ca
1400
1200
1000
800
600
400
200
o
IGFBP-2
6
-—— ~
‘e 1~
ca
2
MMI
e
Sonda
H (MMI/T>
LIZo
IGFBP-3
1800
1600
Ci,
— 1..
‘0 .0‘e ¡-c~c
1‘e Ci,-— cau>ca ‘e
1400
1 200
1000
800
600
400
200
o
a 99~
sgSonda
2dv 4.iv Sdv 2Odv
MMI T
a
2dv 4dv Sd-s 2Odv
T
Sonda
2dv 4dv Bdv 2Odv
MMI 1

Resultados
díasson edadesmuy inmaduras.
3.7.7. Nivelesde GLUTs en membranacruda.
3.7.7.1. Corazón.
El corazónes un tipo especial de músculo esqueléticoque, como se ha
mencionado,cuentacon dos tipos de transportadoresde glucosa,el GLUT-1 y el
GLUT-4.El GLUT-1, que disminuyeal aumentarla edaddel animal en la ratacontrol
(Figura46), incrementósus nivelessignificativamenteen las membranasplasmáticas
de estetejido por la falta de hormonastiroideasa los 4 y 8 díasde edadrespectoa
los animalescontrol.A los 2 y 20 díasde vidano seencontródiferenciaentrelas dos
poblacionesconsideradas(hipotiroideasy controles)-
En los animalescontroly, tambiénen los hipotiroideos,el GLUT-4 aumentó
con la edad de la rata (Figura 46). Los niveles de este transportadorde glucosa
disminuyerontantoen los animalestratadoscon MMI como en los tiroidectomizados
respectoa los controles,exceptoa los 2 días de vida en que sus valores fueron
igualesa los de la población control.
A la vista de los resultados,se podría sugerir que la falta de hormonas
tiroideasprovocaun retrasoo una alteraciónenel patrónde cambiode los GLUTs,
se produceunadisrupciónen el incrementode los niveles de GLUT-4 (propio del
animal adulto).
3.7.7.2.Hígado.El GLUT-1, en el hígado,aumentóen los animaleshipotiroideosrespectoa
los controlesa los 4, 8 y 20 díasde vida, al igual queen el corazón.No se encontró
diferenciaestadisticamentesignificativa en los nivelesde estetranportadorentrelas
dos poblaciones(hipotiroideay control) a los 2 díasde edad(Figura46).
El GLUT-2, que es el transportadorde glucosamayoritarioen el hígado,
aumentóal hacerlo la edaddel animalen las ratascontrol (Figura 46). En las ratas
hipotiroideas,los nivelesde GLUT-2 aumentarona los 2 y 8 díasde vida, pero a los
20 díasde edad,momentoen que los animalesllevan un largo tiempo sufriendolas
alteracionesmetabólicasdebidasa los bajosnivelesde hormonastiroideas,mostraron
un descensosignificativo que, quizá, no seobservóantespor la necesidadde una
situación de hipotiroidismo prolongado para que se produzca una adaptación
(reducciónde los niveles de GLUT-2) en este órgano clave para el metabolismo
glucídico. De nuevo, pareceque las situacionesde hipotiroidismo provocanuna
185
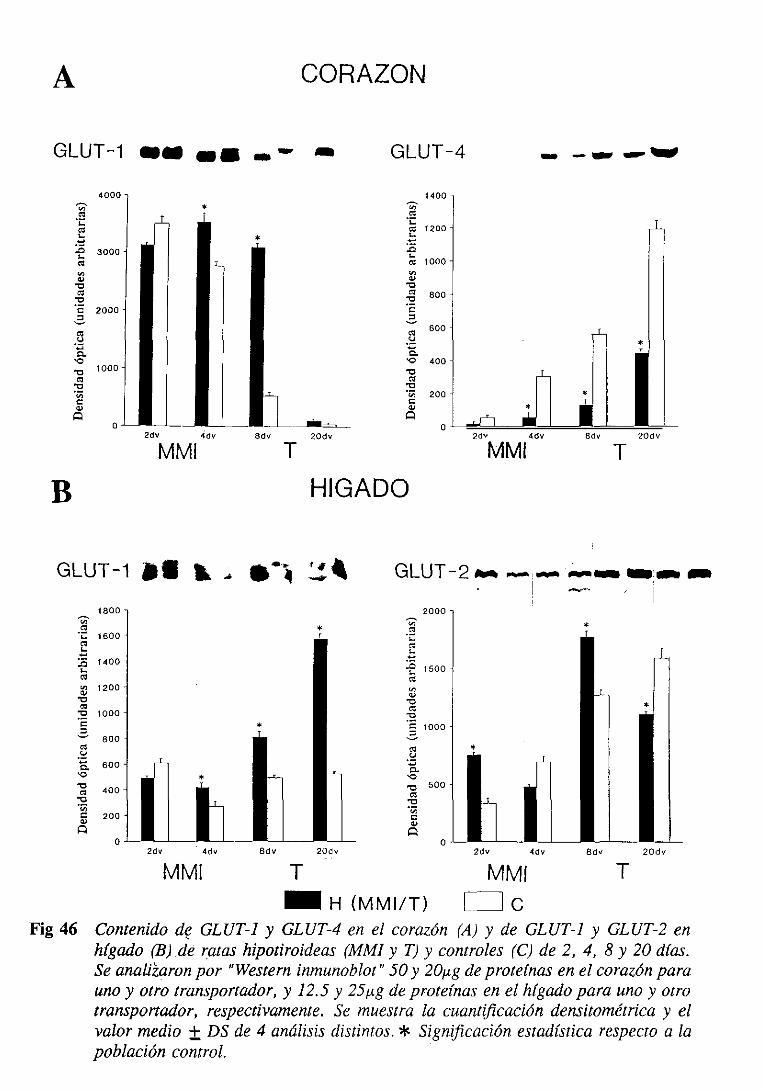
CORAZONA
GLUT—1 eS eS a — GLUT-4 — - a —
*
vaCd0-cd1-
1-Cd(a4>
Cd
a)
COCa
o-‘o
Cd
UN
4>
1400 -
1200
1000-
800-
600 -
400 -
200
0-3d, 2Odv
T
U
~J2t2dv 4dv 8dy
MMI 1
HIGADO
GLUT-1 58 1
UNCd0-
CO1-
-O‘aCdEn4>
-a)CO
•0a)
CdCa
o-‘o-o
Cd
joo4>
1800
1600
1400
1200
1000
800
600
400
200
o2dv 4dv
MMI
-a tl GLUT-2
UNCd‘a
e--
‘aCdva4>
Cd-v
CdCa
o-‘o
Cd
UNa)4>
2000
1600
1000
500
O
1
— —
MMI
— ea —
TH(MMI/T) Lic
F¡g 46 Contenidode GLUT 1 y GLUT-4 en el corazón (A) y de GLUT-1 y GLUT-2 enhígado (B) de ratas hipotiroideas (MMI y T) y controles(C) de 2, 4, 8 y 20 días.Seanalñaronpor “Western inmunoblot” SOy2Optg deproteínasen el corazónparaunoy otro transportador,y liS y 2Sggdeproteínasen el hígadopara uno y otrotransportador, respectivamente.Se muestrala cuantfficacióndensitométricay elvalor medio + DSde 4 análisis distintos.* Sign~caciónestadísticarespectoa lapoblacióncontrol.
Cd‘aCd1-
‘aCdCo4>Cd-co
o-‘O
-vCO
UNo4>
4000 -
3000
2000 -
1000-
0
*
2dv 4dv
MMI
B
Bdv 2Odv 2dv 4dv 8dÚ 2Odv

Resultados
disrupciónen el patrónde incrementode estetransportador,al igual quesucedíacon
el GLUT-4 en el corazón.
3.7.7.3. Músculosesqueléticos:cuadricepsy gastrocnemius.
En las ratastratadascon MMI, a los 2 díasde vida, seencontróun aumento
en los niveles de GLUT-1 respectoa la poblacióncontrol; este incremento fue
estadisticamentesignificativo enel cuadriceps,pero no en el gastrocnemius(Figura
47).Sin embargo,a los 4 díasde edad,el descensoen los valoresdeGLUT-1 de los
animales hipotiroideos fue mayor en el gastrocnemiusque en el cuadriceps.
Finalmente,el contenidoenGLUT-1, tanto en las membranasde cuadricepscomo
en las de gastrocnemius,aumentóa los 3 y 15 díasdespuésde la tiroidectomía(8 y
20 díasde vida).
El GLUT-4disminuyóen ambosmúsculosa los 4 díasde vida en los animales
hipotiroideosque fueron tratadosconMMI (Figura 47). Porel contrario, a los 3 y
15 díasdespuésde la operación(8 y 20 díasde edad) se encontróun aumentoen el
contenidodeestetransportadoren la fracciónmembranosade ambostejidos para la
población de ratasoperadas.
No se encontródiferenciaentre los niveles de GLUT-4 de los dos músculos
esqueléticosentrelas ratashipotiroideasy control a los 2 díasde vida.
3.7.8. Resumende los resultadosdel estudiodel hipotiroidismoenratas
neonatales.
Los resultadosmostradospor estos animalesneonatalesfueron igualesa los
ofrecidospor las ratasde 10, 15 y 20 días tratadascon MMI (2 y 4 días en este
estudio)y tiroidectomizadas(8 y 20 días) (apartado3.2.7.).
187
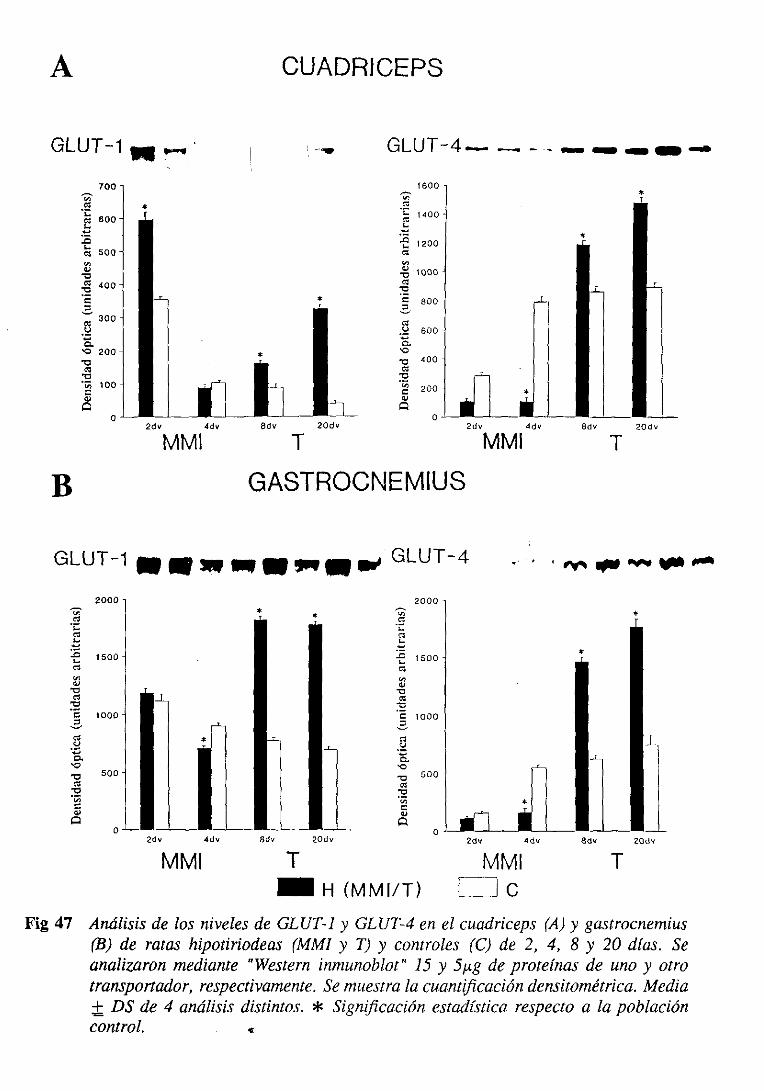
CUADRICEPS
----e GLUT—4--- — -
1600 1jo~- 1400Cd‘a
-ff 1200Cdjo -4>
~1000 -
Cd
~ 800
Cd•~ 600-0.‘o~ 400
CO“o~ 2004> ¡
0-2dv 4dv
MMI
GASTROGNEMIUS
GLUT-1 “ GLUT-4
H (MMI/T)
e,1-Cl‘a
.0‘aCdCo4>CO-oO
ce4
>
0.‘o
“oCd
‘oe4>
2000 -
1500
1000 -
500 -
0- 14LMMI
LicFig 47 Análisis de los nivelesde GLUT-l y GLUT-4en el cuadriceps (A) y gaserocnemius
(B) de ratas hipotiriodeas (MMI y T) y controles (C) de 2, 4, 8 y 20 días. Seanalizaronmediante “Western inmunoblot»15 y Spg deproteínasde uno y otrotransportador, respectivamente.Semuestrala cuantificacióndensitométrica.Media+ DS de 4 análisis distintos. * Sign¿ñcaciónestadísticarespectoa la poblacióncontrol.
A
*
700<tICd1..c~ 600‘a
‘aCd 500UN4>
Cd 400
COcd
300CO‘a>
0.‘o 200
CO
jo4>
4
*
*
*
100
o
MMI2dv 4dv Bdv 2Odv
T
B
8i1V 20dv
T
UNCl‘ael‘a-
a-clUN4>Cl
COcd
ClCa4.40.‘o
Cl-ojoCO4>
2000
1500
1000
500
o
4
*
2dv 4dv Sdv 2Odv
MMI Tatt ZOd<
T
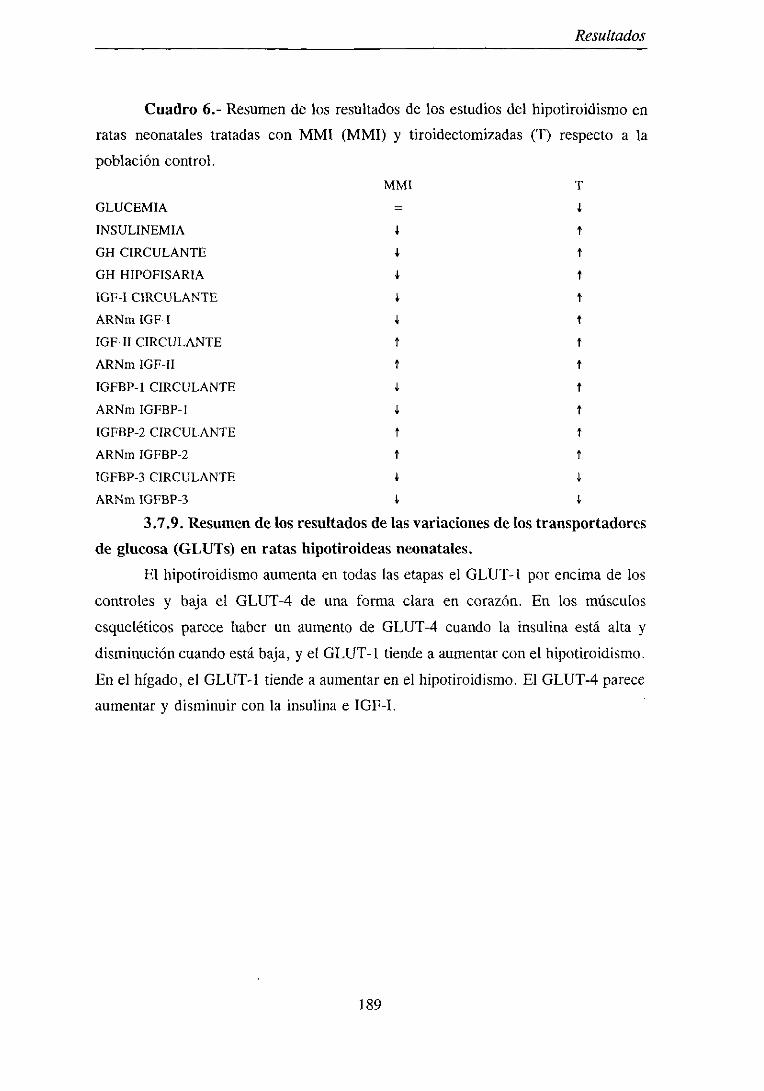
Resultados
Cuadro6.- Resumende los resultadosde los estudiosdel hipotiroidismoen
ratas neonatalestratadascon MMI (MMI) y tiroidectomizadas(T) respectoa la
poblacióncontrol-
MMI T
GLUCEMIA ¿
INSULINEMIA 4 t
Gil CIRCULANTE 4 1
Gil HIPOFISARIA 4 1
IGF-I CIRCULANTE
ARNm 1GW! 4 1
IGF-fl CIRCULANTE t 1
ARNm IGE-Il 1 1
IGFBP-l CIRCULANTE 1
ARNm IGFBP-1 4 1
IGPBP-2CIRCULANTE 1 1
ARNm IGFBP-2 1 1
IGFBP-3 CIRCULANTE 4 4
ARNm IGFBP-3 4 ¿
3.7.9. Resumende los resultadosde lasvariacionesdelos transportadores
de glucosa(GLUTs) en ratashipotiroideasneonatales.
El hipotiroidismoaumentaen todas las etapasel GLUT- 1 por encimade los
controles y baja el GLUT-4 de una forma clara en corazón. En los músculos
esqueléticosparecehaber un aumentode GLUT-4 cuando la insulina está alta y
disminucióncuandoestábaja, y el GLUT-I tiendeaaumentarconel hipotiroidismo.
En el hígado,el GLUT-1 tiendea aumentaren el hipotiroidismo.El GLUT-4 parece
aumentary disminuir con la insulina e IGF-I.
189

DISCUSION

Discusión
4. DISCUSION.4.1. ESTUDIO DE LAS ALTERACIONES DEL AXIS IGFs/IGFRPs EN
SITUACIONESDEHIPOTIROIDISMODESDELA ETAPA NEONATAL A LA
ADULTA.
En la actualidad,hay un númerocrecientede datos que sugierenque las
hormonastiroideasestáníntimamenteinvolucradasenla regulacióndel axisGH/IGF
a varios niveles. A pesarde que las hormonastiroideasejercenaccionessobre la
pituitaria, bien alterandoel númerode células somatotropas(Kitauchi, 1998), bien
influyendosobrela expresióndel gende GH (Evans, 1982),todos los efectosde las
hormonastiroideassobreel sistemaIGFs/IGFBPsno parecenestarmediadospor la
GH (Gaspard,1978; Ikeda,1989; Nanto-Salonen,1993).Enrealidad,la hormonade
crecimiento,y no la insulina, ha sido el objetivo de la mayoríade los estudios,dado
queseconsiderabacomoel posiblemediadorde los efectosde las hormonastiroideas
sobreel sistemaIGF/IGFBP(Burstein, 1979; Nanto-Salonen,1993; Mielí, 1994), a
pesarde que la T3 y la T4 afectana la mayoría de los aspectosdel metabolismo
modificandomuchashormonascomo la insulinay las catecolaminas(Azam, 1990),
y aunquetambiénla secreciónde los IGFs estáreguladapor el estadonutricional y
metabólico (Straus, 1994; Thissen, 1994). En este estudio, las ratas se
tiroidectomizaronparaconseguiruna reducciónmáximade los nivelesde hormonas
tiroideasy, además,paraprevenirlos distintosefectossobrela insulinemiamotivados
por las drogas bociógenas (John, 1970; John, 1974). Los resultados que se
desprendende este estudio, obtenidos pocos días despuésde la tiroidectomía,
muestran los efectos a corto plazo de unos niveles de hormonas tiroideas
extremadamentereducidossobre diferentesparámetros,analizadosdesde la etapa
neonatala la adulta,y señalanel hechode quelos efectosde la funcióntiroideasobre
el sistema IGFs/IGFBPs dependende la etapa de desarrollo del animal, como
previamentehabíanmostradootros modelosexperimentales(Gallo, 1991; Nanto-
Salonen,1992).
4.1.1. Nivelesde IGFs en sueroy expresiónde su ARNm en hígadoen
animalesneonataleshipotiroideos.
El pesocorporal disminuyó en las tres poblacioneshipotiroideas(neonatal,
destetaday adulta)y los valoresde hormonastiroideasfueronmuy bajosa los 5 días
despuésde la tiroidectomía,de acuerdocon estudiosprevios realizadosporCoiro y
191

Discusión
col. (1979)y Walker y Dussault (1980).Las ratasdestetadas,y másclaramentelas
adultas,mostraronun descensodelpesocorporal acompañadode unadisminuciónde
las hormonastiroideas y Gil circulantes,así como del contenidohipofisario de
hormonade crecimiento,y porel contrario,enlas ratasneonatalestiroidectomizadas,
el descensodel peso corporal no se acompañóde una reducción en los niveles
plasmáticosde GH o séricosde IGF-I; por tanto, estosresultadosapoyaronel papel
crucialde las hormonastiroideasen la regulacióndelcrecimientodurantelosestadios
inmadurosdel desarrollo,como previamentese habíadescritoen ratasneonatales
hipofisectomizadas(Glasscock,1992), y en contrastecon la eficacia limitada de la
T4 para favorecerel crecimientoen ratasadultas hipofisectomizadas(Glasscock,
1981). En las ratas destetadas,se encontró un descensoen los niveles de OH
circulanteehipofisariaa los 15 díasdespuésde la tiroidectomíade acuerdoconotros
autoresque previamentehabíanmostradoun descensoa los 10 díasdespuésde la
operación(Coiro, 1979); sin embargo,en las ratasadultas,se encontróunareducción
en los valores hipofisariosy plasmáticosde Gil ya a los 5 días despuésde la
tiroidectomía. Una posible explicación para esta diferenciaes el hecho de que el
mecanismoregulatoriohipotalámicoparala liberaciónde Gil de la pituitaria se está
desarrollando durante el periodo neonatal en la rata (Kitauchi, 1998) y
probablemente,la falta de sensibilidada las hormonastiroideasen la hipófisis para
la síntesisde GH maduraduranteel periodoneonataly estáya plenamenteestablecida
en la etapaadulta. Glydon y col. (1957)demostraronque el plexo primario de la
eminenciamediano se forma en la rata hastael quinto día de vida postnatal,y que
la concentraciónde somatostatinaaumentaduranteel periodoneonatalhastaalcanzar
un pico a los 28 díasde edad.Esta inmadurezdel sistemaendocrinopodríaexplicar
el hecho de que los neonatosde 10 días (5 días despuésde la tiroidectomía)
mostraranun aumentoinesperado,quese mantienehastalos 20 días (15 díasdespués
de la operación),en los niveles de Gil plasmáticae hipofisariay de insulina, en
contrade lo que seobservóen los adultos.Paraconfirmarlasdiferenciasencontradas
entreneonatosy adultos,fue necesariollevar a caboun estudiode ratasneonatales
tratadascon MMI, que se discutirá másadelante.
Se encontróun paralelismoentre los niveles plasmáticosde GH e insulinaen
todas las poblaciones, ambos parámetrosaumentaronen las ratas neonatales
tiroidectomizadas,mientrasquedisminuyeronen los animalesdestetadosa los 15 días
192

Discusión
despuésde la tiroidectomía,y en los adultos,entodoslos estadios.Asípues,insulina
y GH modificaron los valores en la misma dirección en todos los grupos
hipotiroideos,lo que estáde acuerdoconel hechode que bajosnivelesde hormonas
tiroideasen animalesadultosdisminuyenlos valoresplasmáticosde GH e insulina
(Montes, 1977; Bedo, 1991), y además,la diabetes,enadultos,secaracterizaporun
efecto inhibitorio sobrela secreciónde Gil, mientrasqueel tratamientocon insulina
normalizalos nivelesplasmáticosehipofisariosde GR (González,1985; Robinson,
1987). Además,el paralelismoen el cambiode los nivelescirculantesde insulina y
GH, así como las concentracionesde GR hipofisaria observadasen este estudio,
apoyaronla validezde las variacionesplasmáticasde Gil obtenidasenunasolatoma
de muestra,dadoquees bien sabidoque la secreciónde Gil es episódicay sigue un
ritmo circadiano (Tannebaum,1976). No obstante,el mecanismopor el que las
hormonastiroideascontribuyena la regulaciónde la homeostasisde la glucosay la
insulinaescomplejo(Lenzen,1984).El hipotiroidismoexperimentalseacompañade
niveles de insulinabasal normales,aumentadoso disminuidos(Lenzen, 1984),y las
variacionesse puedenexplicarpordiferentesmecanismosde adaptaciónmetabólica
a la falta de hormonastiroideas,depediendodel estadiode desarrollo,dado que la
somatostatinaaumentaduranteel periodo neonatal (Walker, 1977) e inhibe la
secreciónde insulina (Alberti, 1973). Lenzeny col. (1976)encontraronun aumento
en la secreciónde insulina de los islotes pancreáticosen las ratas despuésde la
tíroidectomía,comomostraronnuestrosresultadosparalos animalesneonatales,pero
otros autoreshandescritoun descensode la sensibilidada la insulinaen situaciones
dehipotiroidismo(Morovat, 1998).Con el fin de evaluarsi el incrementode la GH
y la insulina en las ratas neonatalespodría ser explicado simplementepor la
inmadurezdel animal, o sí estáinvolucradala ausenciade glándula tiroideaenesta
adaptación, se estudió una población de ratas neonatales tratadas con MMI
(hipotiroideas-MMI)en las mismasetapasque los animalestiroidectomizados.
Las ratashipotiroideastratadascon MMI mostraronunosbajos niveles de
hormonas tiroideas, junto con unos niveles plasmáticose hipofisarios de OH
disminuidos, y una insulinemia también inferior a la de las ratas control. Estos
resultadossugierenque,enetapasinmadurasde desarrollodel sistemahipotalámico,
el desequilibrioprovocadopor la falta de glándula tiroidea aumentalos niveles
plasmáticosde OH e insulina, y que los valoresaumentadosde insulina y OH, que
193

Discusión
habíamosencontradoen los animalesneonatalestiroidectomizados,no sedebena la
inmadurezdel animal,puestoque en los animalesneonatalestratadoscon MMI la
insulina desciende. La estimulación /3-adrenérgica aumenta la concentración
plasmática de insulina en los organismos eutiroideos (Okajima, 1978), y el
tratamientode las ratasconglucocorticoidesaumentala insulinemiabasal(Aránguez,
1986); no obstante,el estudio de las consecuenciasde las adaptacionesal stress
quirúrgico bajo una situación de ausenciade hormonastiroideas será objeto de
investigacionesfuturas. Además,el posibleefectode los bociógenosperse sobrela
insulina plasmática(John, 1970;John, 1974) podríajugarun papelen la reducción
de los nivelescirculantesde estahormonaencontradaen las ratastratadascon MMI.
En cualquiercaso,el estudiocomparativode dos poblacionesneonatalesen
las que se provocó el hipotiroidismo por distintos medios, permitió encontrar
diferenciasclarasentre ambosgrupos: los neonatostiroidectomizadosmostraronun
aumentoen los niveles séricos y de expresióndel ARNm hepáticode IGF-I en
presenciade unos niveles plasmáticosde insulina altos y, por el contrario, se
encontróuna reducciónen los tres parámetrosanteriorespara el caso de las ratas
hipotiroideastratadascon MMI. Aunqueen los animaleshipotiroideostratadoscon
MMI ya se habíadescritouna reduccióndel IGF-I (Gallo, 1991; Nanto-Salonen,
1991; Nanto-Salonen,1993),nuncase habíandeterminadolos nivelesde insulina, ni
se había calculadola correlaciónexistenteentre esta hormonay el IGF-I en estas
condiciones-
La correlaciónalta y positiva encontradaentre los niveles circulantesde
insulina e IGF-I en la etapaneonatalse confirmó porel análisis de regresiónlineal,
dondeademás,semostróuna correlaciónpobreentre los valoresplasmáticosde GH
y los séricosde IGF-I (Figura 10). Se lialló unacorrelaciónpositiva igualmentealta
entre insulina e IGF-I en las ratas hipotiroideastratadascon MMI, lo que apoyó
resultadosprevios de nuestro grupo y otros autoresque parecíanindicar que el
balanceinsuhina/nutrientes,más que la Gil, regulabala secrecióndel IGF-I en
estadiosinmadurosde] desarrollo(Johnson,1989; Bóni-Schnetzler,1991; Rivero,
1995; Goya, 1996).
Por otro lado, tanto la tiroidectomíacomo el tratamientocon MMI alteró el
perfil normalde descensoprogresivodel IGF-II asociadoa la edadmostradopor las
ratascontrol(Donovan,1989).Lasdiferenciasencontradasen la regulaciónde IGF-II
194

Discusión
entrelos dosgruposde animalesneonataleshipotiroideosse puedenexplicarpor el
efecto reguladorde la glucemiasobreel IGF-II que habíasido sugeridoenperiodos
inmaduros(Rivero, 1995; Goya, en evaluaciónen J. Biol. Chem.),dadoque los
nivelesdeglucosaen sangreestabandisminuidosen las ratastiroidectomizadas,pero
no en las tratadascon MMI.
4.1.2. IGFs enanimalesdetetadosy adultostiroidectomizados.
En esteestudio, las ratashipotiroideasdestetadasy adultasmostraronunos
valoresséricosy deexpresióndel ARNm hepáticode IGF-I disminuidosen todoslos
estadios,juntocon un descensode los nivelesde GH e insulinade acuerdocon lo que
sehabíadescritopreviamente(Rodríguez-Arnao,1993b).
Los análisis de regresión lineal en estos estadios, destetadoy adulto,
mostraronuna correlaciónalta y positiva entrelos niveles plasmáticosde Gil y los
séricosde IGF-I, aunquedichacorrelaciónfue pobrepara la insulina. En cualquier
caso,el descensode los nivelesde la GIl hipofisariay plasmáticay de la insulinemia
tras la tiroidectomíafue más rápidoen la etapaadulta que en la destetada,lo que
sugiereque las ratasdestetadasconstituyenunaetapaintermediaenel desarrollodel
sistema reguladorentre la inmadurez,periodo neonatal,y el estadoadulto. Todos
estos resultadosllevan a la conclusiónde que los efectosde las hormonastiroideas
sobre la secreciónde los IGFs dependende la edad, como se había sugerido
previamentepara las IGFBPs (Nanto-Salonen,1992). Estadependenciade la edad
parece estar causadano sólo por la inmadurez del sistema endocrinopara la
adaptación,sino tambiénporun mecanismoregulatoriodistintoentreambosestadios.
Pareceque la mediadorade los efectosde las hormonastiroideassobrela secreción
de IGFs en etapasinmadurases la insulina, pasandoluego a ser la GEl en etapa
adulta. Sin embargo,para confirmar nuestrahipótesisy excluir el posible papel
reguladorde la Gil, que cambiabaen igual sentidoque la insulina, se bloqueóla
secreción de la insulina en las ratas neonatalestiroidectomizadasmediante la
administraciónde STZ, y las ratasdestetadasy adultasoperadasse trataron con
insulina.
4.1,3. IGFs en animales tiroidectomizadosneonatalestratados con
estreptozotocina(STZ) y en destetadosy adultostratadoscon insulina.
Los animales neonatales tiroidectomizadostratados con STZ, como se
esperaba,presentaronunos niveles bajos de insulina 5 días despuésde iniciar el
195

Discusión
tratamiento(20 días), pero las cifras de Gil en plasmase mantuvieronen unos
valoressimilaresa las ratascontrol, y su contenidohipofisariocomenzóa disminuir
como sucedíaen las situacionesde diabetesen la rata (Montes, 1977; Giustina,
1994). En estascondiciones(hormonastiroideasdisminuidas,nivelesplasmáticosde
Gil elevadosy reducidosde insulina),seencontróun descensoen los valoresde IGF-
1, queya había sido descrito en nuestrolaboratorioen trabajosrealizadoscon ratas
diabéticas(Rivero, 1995; Goya, 1996), y se mostró la existenciade unacorrelación
positivaentreinsulinaeIGF-I (r= 0.98, p < 0.05>. Encuaquiercaso,el tratamiento
conSIZ causóun aumentoen los nivelesséricosy en la expresióndel ARN de IGF-
II en los neonatostiroidectomizados,lo que sugiere, como previamentese ha
mencionadopara los neonatostratadosconMMI, el papel reguladorde la glucemia
sobreel IGF-II. Dehecho,Goyay col. (en evaluaciónen J. Biol. Chem.),denuestro
propio grupo, han demostradomedianteestudiosin vitro con hepatocitosfetalesde
rata que la glucosaejerceun doble efecto sobrela expresióndel gen de estefactor
de crecimiento, dado que induce su transcripcióne incrementala estabilidaddel
ARNm, y además,el hipotiroidismo pareceretrasarla aparicióndel cambiode la
expresiónde las formas fetalesde los JGFs a las adultas(Gallo, 1991).
Por otro lado, la administraciónde insulina aumentóel pesocorporalde las
ratasdestetadasy adultastiroidectomizadasrespectoa los animalesoperados,dado
que la hiperinsulinemiaseacompañade un aumentode las necesidadesenergéticas
(Melian, 1997). Sin embargo,no altera los niveles de Gil, lo que sugiere la
necesidadde un tratamientode mayor duración o de dosis superioresde dicha
hormonaparaaumentarlos nivelescirculantesde GIl (Ortiz-Caro, 1984).
4.1.4. Consideracionesfinalesdel estudiode las variacionesde los IGFs.
Los datospresentadossugierenque la insulina, y no la Gil, medialos efectos
de las hormonastiroideas sobre la secrecióndel IGF-I en la etapa neonatal del
desarrollo;encualquiercaso,dadoqueen el estadoadultoel sistemaneuroendocrino
ha maduradoy se han establecidolas conexiones,la Gil media los efectostiroideos
sobre la secrecióndel IGF-I. La regulación del sistema hormonatíroidea/IGF es
compleja,pero se clarificaráen la segundapartede estetrabajopor el estudiode las
IGFBPs en estas poblaciones y por los experimentos de rehabilitación de
hipotiroidismomediantela administraciónde T4.
Pensamosque el papel de la insulina, que ha sido olvidado, deberíaser
196

Discusión
consideradoen los estudiosde las relacionesentrela función tiroideae IGF-I en las
etapasinmadurasdel desarrollo. El papelpreciso de la insulina en el organismo
adulto, cuandola regulacióndel IGF-I por la GH estátotalmenteestablecida,será
objeto de investigacionesfuturas, ya que el equilibrio insulina/nutrientesparece
interarde forma sutil con laGil en adulto sobrela regulacióndel axis IGFs/IGFBPs
(Goya,enprensaenLife Sci.).
4.1.5. IGFBPs: nivelesséricosy expresióndel ARNmhepáticoenanimales
hipotiroideosneonatales.
En el suero, las IGFBPsjueganun importantepapel en la regulaciónde las
accionesde los IGFs en situacionesfisiológicasy patológicas(Rajaram, 1997), así
algunosestudiossobrela regulaciónde estasproteínastransportadorashanmostrado
que,endichassituaciones,algunashormonaspuedeninfluir sobrelos nivelesséricos
de una o varias IGFBPs (Jones, 1995; Rajaram, 1997). Además, se sabe que la
regulacióndel axis IGFs/IGFBPsdependedel estadonutricional (Clemmons, 1991;
Thissen, 1994), con lo que la influenciade las hormonastiroideas, que afectana
todos los aspectosdel metabolismo,sobredicho axis serácompleja.
Las hormonastiroideas, al igual que los IGEs (IGF-1 y II), jueganun papel
fundamental en el inicio y el mantenimientodel crecimiento somático en los
mamíferos,y las IGFBPsmodulanla actividadbiológicade los IGFs (Rechíer,1993;
Jones, 1995; Rajaram, 1997). Anteriormente,y en las tres poblacionesde ratas
(neonatal,destetaday adulta), se han estudiadolas variacionesde los JGFs y su
expresióndel ARN hepático, y se ha establecidoel papel de la insulina como
mediadorade los efectosde las hormonastiroideassobrela secreciónde los IGFs en
etapasinmaduras,pasandoa dependeren el adulto dichos efectosde la Gil. Las
variacionesséricasde las IGFBPs se han estudiadoen situacionesde alteración
nutritiva (Straus, 1994; Thissen, 1994), y también se han descrito variaciones
diurnas, sobre todo para IGFBP-l (Baxter, 1987), lo que estárelacionadocon la
ingesta,dado que el estadometabólico de un individuo se refleja en el nivel de
insulina, el cual,como se ha descrito,regulala secreciónde IGFBP-1 (Lewitt, 1994;
Baxter, 1995).
Las variacionesde la glucosa,insulina, GH, T3, T4 e IGF-I circulantes,así
como el contenidode Gil en hipófisis y el pesocorporal, respectoa los animales
neonatales,destetadosy adultoseutiroideos,puedenverse en las Tablas4, 5 y 6,
197

Discusión
respectivamente.Lasratasneonatalespresentarondossituacionesdiferentesencuanto
a los valoresde insulina, Gil e IGF-I circulante,puestoque estos nivelesfueron
superioresa los de los animalescontrol enla poblacióntiroidectomizada,e inferiores
en la tratadacon MMI, tanto a 10 y 15 comoa 20 días de vida (5, 10 y 15 días
despuésde la tiroidectomíaen la primerapoblación).Sin embargo,el pesocorporal
y las hormonastiroideasséricasdisminuyeronen ambosgruposde animalesrespecto
a los controles. Estas diferentesvariacionesencontradaspara las dos poblaciones
neonataleshipotiroideas,en cuanto a los nivelesplasmáticosde insulina y Gil,
sugierenque enetapasinmadurasdel sistemahipotalámico,el desequilibriocausado
por la extripacióndel tiroides aumentalas concentracionescirculantesde insulina y
GIl, quizás por una estimulación adrenérgica (Okajima, 1978) o por los
glucocorticoides(Aránguez, 1986); tambiénalerta sobre el posible efecto de los
bociógenosper se sobrela insulinemia(John, 1970; John, 1974),que podríanjugar
un papelenel descensode los nivelesde la insulinacirculanteen la poblacióntratada
con MMI. Sin embargo,ello permitió estudiar, como se hizo con los IGFs, las
variacionesen la secreciónde las IGFBPsen dos poblacionesde ratashipotiroideas
que presentaronun cuadrometabólicodiferente.
4.1.5.1. IGFBPsen los animalesneonataleshipotiroideosy tratadoscon
STZ.
En estas condiciones, las poblacioneshipotiroideas (tratada con MMI y
tiroidectomizada)presentaron,en el suero, unos niveles altos para el complejode
3OkDa (IGFBP-1 y 2) y bajos, para el de 45kDa (IGFBP-3>. Sin embargo,este
aumentofue mayor en los animalestratadoscon el fármacobociógenoque en los
tiroidectomizados(Figura 11), y tanto el análisis de los niveles circulantespor
“Western inmunoblot como el de los tránscritoshepáticos,parecieronmostrarque
dicho incrementoen los valoresdel complejode 3OkDa fue debidoa la IGFBP-2 en
las ratastratadascon MMI, de acuerdocon lo observadopor otros autores(Nanto-
Salonen,1991; Nanto-Salonen,1992;Nanto-Salonen,1993;Rodríguez-Arnao,1994).
Sin embargo, nuestros resultados encontrados para las ratas neonatales
tiroidectomizadasparecieronmostrarqueel aumentode IGFBP-2 no era debidoa la
deprivaciónde las hormonas tiroideas circulantesdirectamente,como se había
pensado,ya que en ambaspoblacionesneonataleslos nivelesde T3 y 3’4 estánmuy
bajoscon respectoa la control (Tabla4) y no aumentatantoIGFBP-2en la población
198

Discusión
tiroidectomizadacomoen la quefue tratadacon el bociógeno;quizás,en lapoblación
hipotiroideatratadacon MMI el aumentode IGFBP-2 esdebidoa una situaciónde
diabetes (baja insulina y glucosa igual que controles, Tabla 4), ya que se ha
establecidoque la IGFBP-2 aumentadurantelos estadosde diabetessuave (Ooi,
1990; Bereket, 1995; Strasser-Vogel,1995), y tambiénse ha visto que IGFI3P-2 se
regulademanerainversapor la insulina y queel tratamientocon insulina normaliza
sus niveles(Lewitt, 1994>.
Se ha establecidoque la insulina está involucrada en la regulación de los
nivelesséricosde JGFBP-1(Holly, 1988; Lewitt, 1994), y que los valoresde esta
proteínatransportadoravarianporla ingestade alimentos(Yeoh, 1988),descendiendo
despuésde la comidapor el efectodirecto de la insulina. Sin embargo,en las ratas
tiroidectomizadas,que presentaronaltos niveles de insulina circulante, IGFBP-1
aumentó,no descendió,tanto a nivel circulante como de su ARN enhígado,frente
a los animalescontrol, y en la poblaciónhipotiroideatratadacon MMI, que mostró
una baja insulinemia, IGFBP-1 disminuyó, no aumentó,tanto en el suerocomoen
suexpresiónhepática;ello se podríaexplicarporqueenlos niños, seha visto quelas
alteracionesen los valorescirculantesde IGFBP-1 no estánligadasa cambiosen la
secreciónde insulina (Counts, 1992; Smith, 1995), sino que estaríanreguladaspor
su estadometabólicoglobal (Cotterill, 1988).No obstante,la dependenciainversade
la insulina sobre la secreciónde IGFBP-1, ya descrita en pacientesdiabéticos
(Suikkari, 1988; Quin, 1994; Muñoz, 1996) y en estudiosrealizadoscon animales
(Lewitt, 1994), si se manifestóen la poblaciónneonataltiroidectomizadadespuésde
recibir STZ. Además, el hechode que los nivelescirculantesy la expresiónen
hígadode estaproteínavariarandel mismo modo resultóindicativo de que el efecto
de la insulinasobrelaproducciónde IGFBP-1 estuvoreguladoa nivel transcripcional
en el hígado(Pao, 1992).
Los altos valores circulantes de IGFBP-2 de estos animales neonatales
tiroidectomizadostratados con STZ (Rajaram, 1997) se acompañaronde una
expresiónhepáticadisminuida,del mismomodo enquehabíasido mostradoen ratas
neonatalesdiabéticas(Rivero, 1995).Estadiscrepanciaentrelos nivelesséricosy de
ARNm de estaproteínaligadoraevidencióun hecho que ya habíasido descritopor
otros autores,y esque la regulaciónde la expresióndel gen de IGFBP-2 serealiza
a nivel post-transcripcional(Ooi, 1992a).
199

Discusión
Los nivelescirculantesy la expresióndel ARNm hepáticode IGFBP-3,menos
abundante en etapas inmaduras, disminuyeron en los animales hipotiroideos
(tiroidectomizadosy MMI) respectoa los controles,al igual que sucedíaen las ratas
subnutridasy diabéticas(Philipps, 1989;Donovan,1991;Rivero, 1995;Goya, 1996).
Estasvariacionesde IGFBP-3 fueronanálogasen ambaspoblacioneshipotiroideas,
tiroidectomizaday tratadacon MMI, quepresentaronnivelesde insulina, Gil e IGF-I
aumentadosy disminuidos,respectivamente,frentea las ratascontrol; ello pareció
mostraruna independenciade la IGFBP-3 respectoa Gil e IGF-I en estasetapas
¡maduras,y de forma contrariaa lo establecidoen adultos (Rechíer,1993; Jones,
1995), lo que sugeriríauna posibleacción directade la deprivaciónde hormonas
tiroideas.
Por otro lado, los niveles de IGFBP-3 séricos disminuyeron en las ratas
neonatalestiroidectomizadastratadascon STZrespectoa los animalescontrol,aunque
su expresiónen hígado, que era inferior a la de la población control, aumentó
ligeramenterespectoa los valoresmostradospor las ratas tiroidectomizadas.Se ha
postuladoque el déficit de insulina aumentala actividad proteasade la IGFBP-3
(Rechíer, 1998), y ello explicaría la disminución encontradaen los animales
neonatalestratadoscon STZ.
4.1.5.2. Consideracionesfinales del estudio de las variacionesde las
IGFBPs.
Creemos pues que estos resultados, en dos poblaciones neonatales
hipotiroideas,ambasconnivelesmuy reducidosde hormonastiroideas y diferentes
cuadros metabólicos(niveles circulantesdistintos de Gil, insulina y glucosa),
muestranque las variacionesdel complejode 3OkDa (IGFBP-1 y 2), que esel más
abundanteenelperiodoneonatal(Donovan,1989; Rivero, 1995),parecenregidasno
poruna accióndirectade la falta de hormonastiroideas,sino por la repercusiónque
ello tiene en el estadometabólicodel animal; por tanto, el aumentodel complejode
3OkDa respectoa los valorescontrol parecedebidoa IGFBP-1 y 2 en los animales
tiroidectomizadosy sólo a IGFBP-2 en los tratadoscon MMI. Todo ello además,
ponede relieve la importanciade la insulina en la regulacióndel complejode 3OkDa
en los animalesneonatales,que ya fue puestade manifiestopara la regulaciónde
IGF-I. Por otra parte, en ambas poblaciones hipotiroideas neonatales
(tiroidectomizadasy tratadas con MMI) existe un paralelismoentre los niveles
200

Discusión
circulantesobtenidosparaIGFBP-1 y 2 y su expresióndel ARN hepático(Figuras
11, 12 y 13), sugiriendounaregulacióntranscripcional(Rodríguez-Arnao,1994). La
regulacióndel complejode 3OkDa, enestasratas,concuerdaademáscon resultados
realizadosen tejidos fetales y adultos, que muestranque IGFBP-2 aumentaen
situacionesde diabetesligera, mientras que IGFBP-1 sólo se incrementaanteuna
insulopeniasevera,enla diabetes(Ooi, 1990),y sugiereunpapelbiológicoespecífico
para IGFBP-1 y 2.
Se destacael hechode que la variaciónde las IGFBPsenanimalesneonatales
hipotiroideoscoincidecon las alteracionesencontradasen neonatossubnutridos,y las
de los animalestratadoscon STZ en este trabajo coinciden con las de las ratas
diabéticas(Philipps, 1989; Donovan, 1991; Rivero, 1995; Goya, 1996), lo cual
resaltala importanciadel estadometabólicodel animal en sus regulaciones.
4.1.6. IGFBPs en animales destetadosy adultos tiroidectomizadosy
tratadoscon insulina.
Las Tablas 5 y 6 muestranque las tatashipotiroideasdestetadasy adultas
presentaban,respectoa las controles,unosnivelescirculantesde insulina,Gil e IGF-
1 disminuidos, junto con variaciones mínimas en la glucemia y muy bajas
concentracionesde hormonastiroideas.Así pues, los animalesdestetadosy adultos
tiroidectomizadosmostraronuncuadrometabólicosimilaral quepresentaronlas ratas
neonataleshipotiroideas tratadas con MMI; en estas condiciones, los niveles
circulantes del complejo de 3OkDa y la IGFBP-3 estándisminuidos, pero el
tratamientocon insulinade estosanimaleshipotiroideosrestituyó los valoresséricos
de IGFBP-3 sólo a 37 días de vida, y aumentóligeramenteel complejode 3OkDa,
aunque no se llegaron a alcanzar los niveles control en ninguna de las dos
poblaciones,destetaday adulta. La expresióndel ARN hepáticode IGFBP-1 y 3
presentóunasvariacionesparalelasa las mostradasporlos nivelescirculantes,lo que
sugeríala existenciadeunaregulacióntranscripcional(Ooi, 1990; Villafuerte, 1996;
Villafuerte, 1997).Sin embargo,el ARN hepáticode IGFBP-2aumentótanto en los
animales destetadoscomo los adultos, y los ensayos hechos por “Western
inmunoblot” pusieronde manifiestotambiénun incrementodedichaproteína,como
sucediócon los neonatoshipotiroideostratadoscon MMI, quepresentabanun cuadro
metabólicomuy similar (Tabla 4); ello pareció mostrarde nuevo que, en dichas
condicionesde diabetesligera (Ooi, 1990), la IGFBP-2tambiénaumentaen adultos.
201

Discusión
4.1.6.1. Consideracionesfinales del estudio de las variacionesde las
IGFBPsen animaleshipotiroideos.
Estosresultadosmuestranqueexisteunaregulacióndiferentey específicapara
IGFBP-1, 2 y 3 en las ratashipotiroideaspara cualquieredadque se considere,y
apoyan la conclusión (Nanto-Salonen,1992) de que las consecuenciasde la
deprivaciónde hormonastiroideassobrela secreciónde las IGFBPsdependede la
edad.En las ratasneonatales,las variacionesdel complejode 3OkDa,sin embargo,
parecenseruna consecuenciade las alteracionesdel cuadrometabólicoprovocadas
por el déficit de hormonastiroideas,y no una acción específicadirecta de estas.
Tambiénestosresultadossugierenquela IGFBP-2, entodos los periodosestudiados,
parece tener un papel homeostáticoen cuadros que cursan con alteraciones
metabólicas,como se habíasugeridopara la rataadultaduranteestadoscatabólicos
(Orlowski, 1990).Así mismo,se ratificó, por los resultadosexpuestos,el importante
papel de las hormonas tiroideas como reguladorasdel crecimiento en etapas
inimaduras,no sólo por sus accionesespecíficasconocidassobreel sistemanervioso
central, sino por sus accionesmetabólicas,sobretodo a través de las variacionesde
los niveles de insulina que se produceny que parecenregir alteracionesen el
complejode 3OkDa que modulala biodisponibilidadde los IGEsen estasetapas.De
nuevo, comose describióen los animalessubnutridosy diabéticos(Rivero, 1995;
Goya, 1996), todo ello parece producirse en estas etapasinmadurasde forma
independientede Gil, como se ha sugerido tambiénen estudios con animales
hipofisectomizados(Glasscock,1991).Estosresultados,junto con los obtenidospara
los IGFs, alertan además sobre la importancia de la vía de producción del
hipotiroidismopara los estudiosde la influenciade las hormonastiroideassobre la
regulacióndel eje IGFs/IGFBPs.
4.2. ESTUDIO DE LAS ALTERACIONES DEL AXIS IGFs/IGFBPs EN
SITUACIONES DE IIIPOTIROIDISMO (TIROLDECTOMIA) DESDE LA
ETAPA NEONATAL A LA ADULTA DESPUESDE LA REHABILITACION
CON T4.
El retrasodel crecimientoes una de las primerasrespuestasfisiológicasdel
hipotiroidismo, por ello todos los animalestiroidectomizadosmostraronun menor
pesocorporal respectoa sus controles(Aláez, 1992a). No obstante,el tratamiento
202

Discusión
con T4 normalizócompletamenteel crecimientomedianteel implantede pellet, de
acuerdo con lo que se ha descrito ampliamenteen la bibliografía, que la
administraciónde GR o T4 restaurael crecimientode las ratastiroidectomizadas
(Nanto-Salonen,1992; Nanto-Salonen,1993).
La administraciónde una dosis de 1.5p¿g/IOOgde pesode T4 a estas ratas
tiroidectomizadasno igualó los nivelesde las hormonastiroideascirculantesen estos
animalesa los de la poblacióncontrol. Las ratasneonatalestienenunosnivelesen
suero más elevados de hormonas tiroideas de forma fisiológica, por ello,
probablemente,ladosisde T4 utilizada(1.Spg/íOOgde peso)resultóinsuficientepara
la rehabilitaciónde los nivelesde las hormonastiroideas.No obstante,estacantidad
de tiroxina sí resultó adecuadapara incrementar,incluso por encimade los valores
control, los niveles plasmáticose hipofisariosde Gil, lo que evidenció la gran
sensibilidad de la pituitaria de estos animalesneonatalesa la regulaciónpor las
hormonastiroideas(Kim, 1993).
La tiroidectomíacausó un incrementoen la insulinemia de los animales
neonatales,perola administraciónde T4 no disminuyólos nivelesplasmáticosdeesta
hormonaalos valorescontrol, quizásporqueladosisde tiroxinaadministradaresultó
insuficiente,ya que se habíadescritocon anterioridadque el tratamientocon T3 de
ratashipotiroideasdisminuyelos nivelesde insulina circulante(Azam, 1990).
Las hormonastiroideas, como se ha mencionadoen repetidas ocasiones,
poseenuna gran importanciaen el crecimiento, y además,los niveles de Gil son
dependientesdel estado tiroideo, de ahí que los animalesdestetadosy adultos
operadospresentaranunmenorpesocorporaly nivelesdeGIl circulantee hipofisaria
disminuidosrespectoa las ratascontrol.No obstante,la administraciónde T4 produjo
una normalizaciónde los valoresde hormonastiroideas,junto conuna recuperación
del peso (Aláez, 1992a) de los animalesdestetadosy adultos que pasó por un
aumentoen el contenidoy los nivelescirculantesde Gil (Nanto-Salonen,1991).
Glasscock y col. (1991) han descrito una dependenciahormonal similar del
crecimientoen ratashipofisectomizadas,conunacorreccióncompletadel crecimiento
por el tratamientocombinadode T4 y Gil, y una correcciónsólo parcial con la
administraciónde hormonade crecimiento.Además,se ha visto queel hipotiroidismo
disminuyela secreciónpituitaria de Gil y sus nivelesde ARNm, pero senormalizan
tras el tratamientocon T, (iones, 1990), y tambiénse ha demostradoque la T3
203

Discusión
aumentala transcripcióndel gen de Gil (Martial, 1977).
Por otro lado, Hervás y col. (1975) demostraronque los efectosde las
inyeccionesde T4 sobreel contenidohipofisariode Gil de las ratastiroidectomizadas
serelacionabacuantitativamentecon la dosisde tiroxina empleada,dichosefectosse
observabanmáspronto con dosisaltasde 14, y segúnel tiempo transcurridodesde
la administraciónde la hormona;demostrarontambiénque la concentraciónde GH
enpituitaria aumentabaantesquesusvalorescirculantes.Asípues,no resultaextraño
que el tratamientollevado a cabo durante10 días, en ratasdestetadasy adultas,
mostraranivelesmáscercanosa los presentadospor los animalescontrol, y lo mismo
sucediócon el contenidohipofisariode Gil en ratasadultasrehabilitadas.De nuevo,
sedebedestacarel hecho de que las ratasdestetadasmostraranun comportamiento
intermedioentre la etapaneonataly la adulta, puestoque al igual que las primeras,
presentaron,tras el tratamientocon T4~ unos nivelesde GH circulantessuperioresa
los de sus respectivoscontroles. Además, la normalizaciónde los niveles de las
hormonastiroideasen los animalesadultosse acompañóde un aumentogradualen
los valores plasmáticosde GR e insulina, mientras que en el caso de las ratas
destetadas,el tratamientocontiroxina, quecausóun fuerte incrementoen los niveles
séricosde ‘[3 y T4, produjo unaumentorápido y bruscoen las cifras circulantesde
Gil e insulina. Se encontróun paralelismoentre los niveles plasmáticosde GH e
insulina ya descrito con anterioridad(Montes, 1977; Bedo, 1991) en todas las
poblacionesestudiadas.
Se debedestacarel hecho de que el tratamientohormonal llevado a cabo
mediantepellets, que constituye una pauta de administraciónmás continuaday
fisiológica que la inyección, ofrezca una normalizaciónde los valorespara estos
parámetrosendocrinosbasales,dato que se deberátener en cuentapara futuros
estudios. Así pues, esta restitución de los parámetrosendocrinosdespuésde la
rehabilitacióncon T4 de los animalesdestetadosy adultostiroidectomizados(Tabla
9) estuvodeacuerdocon lo esperadosegúnlos resultadosanteriormenteestablecidos
en la literaturacientífica.
4.2.1. IGFs e IGFBPsen neonatos.
El tratamientocon 14 disminuyó los niveles séricos de IGF-I y II en los
neonatostiroidectomizados,aunque la expresiónhepáticadel ARNm permaneció
elevada para ambos factores respectoa las ratas control, como sucedíaen la
204

Discusión
poblacióntiroidectomizadaantesdel tratamiento.Sólo se igualó la cantidadde los
tránscritosde IGF-II a los animalescontrol cuandolas ratasfueron tratadascon el
pellet. Este hechopareció sugerir una vez más que, en la poblaciónneonatal,la
alteraciónen los nivelesde IGF-I asociadaal hipotiroidismo no era secundariaa la
secrecióndeGil, ya que las dosisde T4 dadasrestaurabanporencimade los valores
control los nivelesde Gil. Sin embargo,los nivelesde insulina, aunquedisminuyen
conrespectoa los animalesno tratados,siguenaltosrespectoa los control; en dichas
circunstancias,los valorescirculantesde IGF-I se rehabilitanigual a los controles,
pero no la expresiónde IGF-I. Dicha rehabilitaciónparcial de IGF-I apuntaa una
accióndirectade las hormonastiroideasa travésde la GH y al margende la insulina,
que sigue alta en los animalesneonatales.
El descensode IGF-II despuésde recibir tiroxina parecemostrarla capacidad
de la hormonade acelerarel procesode maduraciónde IGFs, ya queel IGF-II esel
IGF abundanteenperiodofetal, y ha sido descritopor otros autores,que el pasoa
valorescirculantesaltosde IGF-I (IGF del periodoadulto),parecereguladopor las
hormonastiroideas (Gallo, 1991; Nanto-Salonen,1993). Tambiénsemuestraen los
animalestiroidectomizadosneonatalesque la elevaciónde laglucemiajuegaun papel
en la restauracióndel IGF-II, como sehabíadescrito(Rivero, 1995).
En cualquiercaso, la disminuciónde los nivelesde insulinaen los neonatos
tiroidectomizadosrehabilitados,respectoa los animalesoperados,parecejugar un
papelen la restauraciónde los valoresséricosde IGF-I; consecuentemente,las ratas
neonatalessometidasal tratamientocon 14 mostraronunacorrelaciónalta y positiva
entrelos nivelescirculantesde insulina e IGF-I, aunquedicha correlaciónfue mayor
cuandola administraciónde la T4 se realizópormedio de pelletque por inyección.
Porotro lado, el coeficientede correlaciónfue pobreparala Gil, pero no tanbajo
como se encontrabaen los animalestiroidectomizadossin rehabilitar,mostrandoque
en este caso, los valores altos de Gil circulante parecenjugar un papel en la
rehabilitación.De nuevose repitió el hechode quela correlaciónfue mejor al tratar
a los animalescon el pellet frenteal otro tratamiento.Estoshechosapoyanuna vez
más que, a pesar de que el IGF-I parecereguladopor la insulina en periodos
inmadurosdel desarrollo(Bóni-Schnetzler,1991; Rivero, 1995; Goya, 1996),la GH
juegatambiénun papelendeterminadascircunstancias.
La rehabilitacióncon T4 causó un descensoen las proteínasligadorasde
205

Discusión
3OkDa (IGFBP-1 y 2) y un aumentoen IGFBP-3 en neonatos,mientrasaumentaron
endestetadasy adultasambasproteínas,esdecir, seproduce,en las trespoblaciones,
unarehabilitacióna nivel circulantede estasIGFBPsdespuésdel tratmientocon 14.
La implantacióndel pellet de 14 a animalesneonatalestiroidectomizadosse
acompañódeunadisminuciónen los valoresséricosy de expresióndel ARNm de las
dos proteínastransportadorasdel complejo de 3OkDa; las variacionesfueron más
pronunciadasen la transcripciónque a nivel circulante.
La IGFBP-1 se encuentraregulada por la insulina (Baxter, 1995), sin
embargo,y quizásdebido a que los valores plasmáticosde esta hormona no se
restaurarontotalmentea los nivelescontrol, la administraciónde T4 no fue capazde
normalizarcompletamentela secrecióny producciónde estaproteínatransportadora;
de hecho, se ha descritoin vitro que la adición de la T3 no afectaa la expresiónde
IGFBP-1 y 2, pero sí a la de IGFBP-4(Demori, 1997).La normalizaciónde IGFBP-
2 muestraclaramentela idoneidaddel tratamientoconpelletparasu recuperación.
La concentraciónséricay de ARNm en hígadode IGFBP-3 aumentócon el
tratamientocon T4, si bienesteincrementofue mayorenel casode la administración
porpelletquedandola inyección. Estaproteínade alto pesomolecularestáregulada
fundamentalmentepor la Gil, por ello otros autores, al administrarhormona de
crecimientoa las ratashipotiroideasigualaronsu nivel a los valorescontrol (Nanto-
Salonen,1993).
Porotraparte,cabedestacarel hechode que a pesarde que la rehabilitación
de los IGEs y sus IGFBPs no fue muy buena en los animales neonatales
tiroidectomizados,sobretodo paralos IGFs, ello fue, sin duda, debidoa la bajadosis
de 14 empleadaen el experimento.
En todos los casos,se encontróun paralelismoentre las alteracionesde los
nivelescirculantesy de los tránscritoshepáticosde estasIGFBPs,lo que sugeríala
existenciadeunaregulacióna nivel transcripcional(Nanto-Salonen,1992; Latimer,
1993).
4.2.2. IGFs e IGFBPs en destetadosy adultos.
La tiroidectomíase asociaconunosbajosniveles circulantesy de expresión
del ARN hepáticode IGF-I (Rodríguez-Arnao,1993b),pero la administraciónde T4
los aumenta(Chernausek,1983; Nanto-Salonen,1991),tal como sucedeen nuestros
resultadospara animalesdestetadosy adultos. Se ha demostradotambiénque el
206

Discusión
estadotiroideo puedealterar la expresióndel IGF-I en otros tejidos ademásdel
hígado: la 13 aumentalos efectosde IGF-I encultivos de osteoblastos(Schniid, 1992;
Varga, 1994)y decélulasde Sertoli (Palmero,1990).No obstante,el tratamientocon
14 mostró unos resultadosdispares:la administraciónde la hormonamediantela
inyecciónrehabilitósólo parcialmentelos nivelesde expresióny séricosdeestefactor
de crecimiento(van Buul-Offers, 1994), aunqueel pellet sí los normalizó, lo que
pareceindicar que la eficaciade la pautahormonaldependeno sólo de la dosiso la
duracióndel tratamiento,sino tambiénde la forma de administracióno de la vía
(Latimer, 1993). El paralelismoencontradoentre los niveles circulantesy los del
ARN hepáticodelIGF-I enanimalesdestetadosy adultostiroidectomizadosy tratados
con 14 sugiereel hechode que la regulaciónde lashormonastiroideassobreel IGF-I
parecehacersea nivel transcripcional(Wolf, 1989), siendo,en la etapaadulta, la
GIl, la hormonaque actuacomomediadoraprincipal de las hormonastiroideas,y
que escapazde alterarla síntesisy/o estabilidadde los tránscritosdel ARN de IGF-I
en el hígadode rata (Roberts, 1986). Consecuentemente,los análisis de regresión
lineal en estosestadios,destetadoy adulto,mostraronuna correlaciónalta y positiva
entre los nivelesplasmáticosde Gil y los séricosde IGF-I, y dichacorrelaciónfue
pobreparala insulina. En cualquiercaso,en las ratasadultas,el coeficientefue más
alto cuandola administraciónse realizócon pellet frentea la inyección, y aumentó
máscuantomayorfue la duracióndel tratamiento,lo que pareceindicar unavezmás
a la GIl en la etapaadultacomomediadorade la acciónde las hormonastiroideas
sobrela secreciónde los IGFs (Kitauchi, 1998).En cuantoa las ratasdestetadas,el
coeficiente de correlación mejoró con el tratamiento por pellet respecto a la
inyección.
La administraciónde T4 a las ratasdestetadasy adultasoperadascausóun
aumentoen los nivelesséricosde las dos proteínasdel complejode 3OkDa y de la
expresiónhepáticade IGFBP-1. La expresióndel gende estaproteínatransportadora
está regulada por la insulina (Ooi, 1990), y la insulina aumentó despuésdel
tratamiento;quizás, además,estoscambiosen los nivelescirculantesde IGFBP-l,
probablemente,estaríanrelacionadoscon accionesdirectasde la 14, al menos en
parte,puestoque se ha visto que la T3 escapazde aumentarel ARNm y la secreción
de estaproteína in vivo y en cultivos de células de hepatomahumano(Rodríguez-
Arnao, 1993b;Mielí, 1994).No obstante,estoscambios,en nuestroestudio,parecen
207

Discusión
mediadospor el aumentoprogresivode insulina (Rivero,1995; Goya, en prensaen
Life Sci.).
La altaexpresióndel ARNm de IGFBP-2senormalizóporel tratamientocon
14 tambiénenel suero,de acuerdoconNanto-Saloneny col. (1992).Porotro lado,
Nanto-Saloneny col. (1993) mostraron que la administraciónde Gil a ratas
hipotiroideasno afectabaa la expresióndel gende IGFBP-2, con lo queparecíaque
la regulaciónde estaproteínainvolucrabaotros mecanismos,y entreellos estarían
las hormonastiroideas(Nanto-Salonen,1993); unaevidenciade estamodulaciónde
la IGFBP-2 por las hormonastiroideasse ha mostradoen cultivos de osteoblastos,
dondeaumentabala produccióny secreciónde esta proteínatransportadorapor la
administraciónde 13 (Schmid, 1992).
Las concentracionesséricay de ARNm hepáticode IGFBP-3,quedisminuían
despuésde la tiroidectomía,aumentarondespuésdel tratamientocon T4, al igual que
habíanestablecidootrosautores(Nanto-Salonen,1993).De nuevo,parecióestarclaro
que esteefectoera debido a un aumentoprogresivode la secreciónde GIl. Se ha
descritoque la administraciónde CH a ratashipotiroideasnormalizabalos nivelesde
IGFBP-3 (Nanto-Salonen,1993), lo que estableceque la expresiónde estaproteína
ligadoraestáreguladapor los nivelesde CH.
El hecho de que los niveles circulantesy de ARNm en hígado de estas
IGFBPsvariende formaparalela,sugirió la existenciade una regulacióna nivel de
la transcripciónporpartede la 14, cosaque previamentehabíasido mostradaen los
animalesdestetadosy adultospor otros autores,y tambiénen humanos(Latimer,
1993; Nanto-Salonen,1992; ; Angervo, 1993b; Mielí, 1994).
Finalmente,cabedestacarel hechode queel efectode la 14 sobrelos niveles
de las IGFBPs parece ser específico del órgano (Latimer, 1993). En nuestros
resultados,la restauraciónen la expresiónde estasproteínasvarió dependiendode la
vía deadministraciónempleaday de la duracióndel tratamiento,ofreciendomejores
resultadosel tratamientomáslargoy cuandola liberaciónde la tiroxina administrada
fue continuada.
4.3. ESTUDIODE LA REHABILITACION CON 14 EN SITUACIONES
DE DIABETES EN PERIODONEONATAL Y ADULTO.
La diabetescausó un descensodel peso corporal (Maiter, 1989) que se
208

Discusión
acompañóde unadisminuciónen los nivelescirculantesde insulina, 13, T4~ Gil, y
de esta última hormona también a nivel hipofisario. Se ha descrito que esta
enfermedadalterael axis hipotálamo-hipófisis-tiroides;se produceunareducciónen
los nivelesdel TRH hipotalámicoy plasmático(González,1980; Aláez, 1992a)y de
la TSH hipofisaria y circulante (Aláez, 1992a; Rodríguez, 1992), disminuye la
velocidadde secreciónde ‘[Sil, la respuestade la TSH a la TRI-I (Pastor, 1983;
Aláez, 1992a) a pesar de que el metabolismoperiférico de la ‘[Sil es normal, y
tienenlugarnumerososcambiosestructuralesen la glándulatiroidea(Tontis, 1990).
Además,desciendela producciónde hormomastiroideas (Tontis, 1990; Rodríguez,
1992)y la desyodaciónde T4 a 13 enlos tejidosperiféricos(Ortiz-Caro,1984; Aláez,
enprensaenMetabolism;Calvo, 1997)y, comoconsecuenciade ello, seproduceuna
disminuciónen las concentraciónde ambasyodotironinasen la mayoríade los tejidos
(Aláez, 1992a; Ortiz-Caro, 1984). lodos estos cambiosen el estadotiroideo se
considerancomo una respuestaadaptativacausadapor la limitadadisponibilidadde
energíaa nivel celular.
En la rata, la diabetesinducidapor STZ se acompañade un descensoen los
nivelesplasmáticosy el contenidohipofisario de Gil (Rodríguez,1992; Giustina,
1994;Flyvbjerg, 1995).Estedefectopodríadeberseaun aumentode la somatostatina
hipotalámica,quemotivaríauna disminuciónen la velocidadde transcripcióndelgen
de GH, aunqueesto sólo no explicaría totalmenteel descensoen los niveles de
ARNm y de Gil en pituitaria observadoen la diabetes(Martial, 1977; Rodríguez,
1992). Se podría pensar, por tanto, que la administraciónde T4 a estas ratas
diabéticasnormalizaría los niveles de la hormona de crecimiento circulantese
hipofisarios,así como suexpresiónen esta glándula,pero estono esasí, dadoque
Bedo y col. (1991)describieronque para conseguiruna regulaciónnormal del gen
de la GIl por las hormonas tiroideas es necesariala insulina. Este hecho se
compruebaen los animalesdiabéticosadultosde nuestrotrabajo, en los cuales,las
dosis de T4 no aumentanla insulina y tampoco la GH a los valorescontrol; sin
embargo,no sucedelo mismo en los neonatosdiabéticosde nuestroexperimiento,en
los que el aumentode Gil despuésde la administraciónde T4 se produce sin un
incrementoen la insulina.
4.3.1. IGFs e IGFBPs: nivelesséricosy expresióndel ARNm hepático.
La diabetesse asociacon unos bajos niveles circulantesy de expresióndel
209

Discusión
ARN hepáticode IGF-I (Ishii, 1994; Rivero, 1995). Sin embargo,estaenfermedad
altera laexpresiónde estefactorde crecimientotambiénenotros tejidoscomoriñón,
músculoesqueléticoo glándulasadrenales(Luo, 1991; Bornfeldt, 1992; Ishii, 1994).
No seconocetotalmentela causaporla que disminuyela expresióndel IGF-I
en la diabetes,aunqueparecedependerde la interacciónde variashormonas,entre
las que seincluyen la insulina y la Gil (Miller, 1981). En estaenfermedad,sesabe
que se produceuna resistenciaa la GR enel hígadode los animales(Maiter, 1989),
pero tambiénlos efectosdirectoso indirectosde la insulina parecensernecesarios
para mantenerlos niveles normalesdel ARNm de IGF—I (Bornfeldt, 1992).
A pesarpues,de que el cuadrometabólicoestotalmentedistinto enunarata
diabética y en una hipotiroidea,nosotros quisimos estudiar hasta qué punto la
rehabilitacióncon hormonatiroideadelaxis IGFs/IGFBPsguardabao no paralelismo
conlo quehabíaocurridoenla rehabilitaciónde los animaleshipotiroideos.El hecho
de que se encuentreun paralelismoentre los nivelescirculantesy los del ARNm
hepáticode IGF-I sugiereque, en la ratadiabéticala expresióndel gen de IGF-I se
regulaa nivel transcripcional,lo que estuvode acuerdocon lo descritopor Adamo
y col. (1991)y Pao y col. (1992).
Las ratasneonatalesdiabéticasque recibieronel tratamientohormonalcon T4
aumentaronlos niveles séricos y de expresiónen hígado de IGF-I, aunque la
administraciónde T4 a los animalesadultosno pareciótenerningúnefectosobrelos
valoresséricos,de acuerdocon Ikeda y col. (1990),y de expresiónenhígadode este
factor de crecimiento.Sin embargo,la administraciónde insulina a niños diabéticos
elevalos valoresde IGF-I antesde incrementarlos nivelesde receptoresde GH y sin
aumentarlas cifras de hormonade crecimiento,lo que sugeriría,en la diabetes,un
efectoestimulatoriodirecto de la insulina sobrela producciónhepáticade IGF-I para
las edadestempranasdel desarrollo(Bereket,1995).Sin embargo,ennuestrotrabajo,
obtenemosunarestauraciónde IGF-I en los animalesneonatalestratadoscon tiroxina
sin un aumentode insulina, lo cualpruebaque, en estascircunstancias,el aumento
de IGF-I seha producidopor una accióndirecta de T4 a través,probablemente,de]
aumentoproducidoen la Gil. En los animalesadultos, cuya secreciónde IGF-I y
expresiónde su ARNm hepáticohemosdemostradoque dependede los nivelesde
Gil, es natural que las dosisdadasa nuestrosanimalesdiabéticos,que no aumentan
la Gil, no restaurentampocolos valoresde IGF-I.
210

Discusión
El modelo de diabetespor STZ mostró unos nivelescirculantesde IGF-II
disminuidosy una expresiónen hígadoaumentadarespectoa las ratascontrol. Sin
embargo,existenmuy pocasreferenciasbibliográficas, y con resultadosdispares,
acercade la regulacióndel IGF-II en situacionescomo la diabetes;así, los distintos
autoresmostraronunos niveles de IGF-II elevados(ilalí, 1989), no alterados o
disminuidos(Merimée,1984; Rivero, 1995).La administraciónde 14 a los animales
diabéticosno alteró los bajosnivelesde IGF-II mostradospor las ratasno tratadas,
y bajó su ARNm, de acuerdocon lo observadopor Nanto-Saloneny col. (1993).
La diabetesdisminuye las proteínastransportadorasde 30 y 45kDa, o sea
IGFBP-I y 2 e IGFBP-3.El estudiorealizadomediante“Westerninmunoblot’mostró
que,enel periodoneonatal,la diabetesaumentalos nivelescirculantesde IGFBP-1,
y de IGFBP-2 de acuerdocon Ooi y col. (1990)y Untermany col. (1990), e igual
ocurre en el hombre(Quin, 1994; Bereket, 1995). Esteaumentoen la expresiónde
IGFBP-1 se debe a la regulacióninversadescritaentre la insulina sobre IGFBP-1
(Baxter, 1995).
Las alteracionesenlos valoresséricosdel complejode 3OkDaseacompañaron
de unasmodificacionesa nivel de la expresiónen hígadoen sentidoinverso, esdecir,
seprodujo un aumentoen los valoresde ARNm de IGFBP-1 y 2 tanto en las ratas
neonatalescomoen las adultas(Ooi, 1990; Rivero, 1995), lo que sugiereque no se
están regulandoa nivel transcripcional.Sin embargo,convendría,en un futuro,
revisar dicha aseveraciónrepitiendo el Western ligand blot’ con una población
diabéticamás amplia.Ooi y col. (1991)demostraronque el aumentoen los niveles
del ARNm de IGFBP-1 se debíaa un incrementoenla velocidadde transcripcióndel
gen de estaproteína,y así mismo,en estadosdiabéticos,se ha descritola magnitud
y velocidaddel descensoen la transcripciónde IGFBP-1 despuésdel tratamientocon
insulina (Ooi, 1992b; Pao, 1992).
A pesarde que los nivelesde ARNm de IGFBP-2 tambiénaumentabanen el
hígadode los animalesdiabéticos,no sehaencontradoun aumentoen la transcripción
del gende estaproteína,porlo quese ha sugeridounaregulaciónpost-transcripcional
(Tseng,1992).No obstante,cabedestacarque la expresiónde IGFBP-2estáregulada
de manera específica y diferente en cada órgano en condiciones metabólicas
anormalescomo la diabetes(Chen, 1994).
Los niveles de IGFBP-3 fueron inferiores a los controles en las ratas
211

Discusión
diabéticasneonatales,adultas (Rivero, 1995; Rodgers, 1995) y en el hombre
(Brismar, 1994; Bereket, 1995; Hanaire-Broutin, 1996). Estos bajos valores
circulantesse asociarona una disminuciónen la abundanciadel ARNm de esta
proteína(Rivero, 1995), todo lo cual estáde acuerdocon nuestrosresultados,y el
tratamientocon 14 no pareceaumentarni los valorescirculantesde las IGFBPsni la
expresiónhepáticade su ARNm a los controles,aunquesu aumentocon respectoa
los animalesdiabéticosesevidenteen nuestrosresultados.Villafuerte y col. (1996)
han demostradoen cultivos de hepatocitosque la insulina estimulala transcripción
del gen de IGFBP-3, y que aumentala liberaciónde la proteína,pero ello no lo
hemosencontradoen nuestrotrabajo. Así pues, la IGFBP-3 duranteel periodo
adulto, en la diabetes,disminuyetanto en suerocomo la expresiónde suARNm, y
por tanto, parecereguladaa nivel transcripcional.Además, recientementese ha
demostradoqueenla diabetesseproduceun aumentoen actividadproteolíticade esta
proteína(Fowlkes, 1997; Rechíer,1998), lo queexplicaríatambiénlos bajosniveles
de IGFBP-3 encontradosen los animalesdiabéticos.
Hay que remarcarpues,que los resultadosobtenidoscon animalesdiabéticos
neonatalesy adultos,muestranunadisminuciónde IGF-I y de IGF-II en sueroenel
periodo neonatal, y de IGF-I en periodo adulto, con reducción en ambos de su
expresiónhepática,y que ello seacompañade una granreducciónde insulina y de
Gil circulanteen adulto,y enneonato,tansolo de insulina, aunque,en ambos,bajan
las hormonastiroideas. Las dosis de 14 restauranla Gil circulante y pituitaria en
neonatospero no la insulina, y rehabilitanel IGF-I en suero,así como la expresión
de su ARNm, cosaque no sucedeen adulto, ya que la Gil no aumenta,y por tanto,
no se restauranlos IGFs. Ello ratifica el papelde la GH como mediadorade las
accionesde las hormonastiroideassobrela secreciónde IGF-I enperiodoadulto, y
muestraque,enperiodoneonatal,en circunstanciasen lascualesla insulinaestábaja,
comoen los neonatosdiabéticostratadoscon T4, las hormomastiroideasconsiguen
rehabilitar los niveles de IGF-I a través de un aumentode Gil (Mathews, 1986;
Bichell, 1992).Estosdatos,junto con los obtenidosen ratasneonataleshipotiroideas,
parecenmostrarque en etapaneonatal,las hormonastiroideasejercensus acciones
sobre la secreciónde IGF-I en primer lugar, a través de la insulina y sólo, en
segundotérmino, cuandola insulinaestábloqueada,diabetesconSTZ, la mediadora
pasaa serla Gil, sin embargo,es la Gil siemprela mediadoraen periodoadulto.
212

Discusión
Estos resultadossobre animales diabéticosratifican pues los obtenidosen ratas
hipotiroideastratadascon 14, ya que los animaleshipotiroideosneonatalestratados
contiroxina tambiénrehabilitanparcialmenteel IGF-I conunaumentograndede GR.
Las IGFBPs, tanto las de 3OkDa como la IGFBP-3, parecenregularsesegún
unestadometabólicomuy complejoen ladiabetesquesedeberíaestudiara más largo
plazo, por lo cual sepuedeconcluir pocode esteestudio; tan solo se ve claramente,
una vez más, que en situaciónde francadiabetes,la IGFBP-1 seregula de forma
inversaa los nivelesde insulina.
4.4. ESTUDIO DE LAS VARIACIONES DE LOS TRANSPORTADORESDE
GLUCOSA (GLUTs) EN ANIMALES NEONATALES HIPOTIROIDEOS.
POSIBLE CORRELACION ENTRE LAS ACCIONES DE LAS HORMONAS
TIROIDEAS, GLUTs Y EL AXIS IGFs/IGFBPs.
Este estudio pretendeestablecerla posible existencia de una conexión,
mediadapor las hormonastiroideas,entre el patrón ontogénicode cambio de los
GLUTs y los IGEs. Se ha descrito que la transición del GLUT-1 (forma
predominanteen etapasinmaduras)al GLUT-4 (transportadortípico de la edad
adulta),en músculoy tejido adiposo,tiene lugarel día4 de vida (Santalucía,1992),
y quelas hormonastiroideasparecenretrasarestapautaontogénica(Castelló,1994).
Por otro lado, tambiénse sabeque el cambioenel predominiode la forma fetal de
los IGFs (IGF-II) a la adulta(IGF-I) se produceen la segundasemanade desarrollo
y que está influenciado por las hormonastiroideas (Gallo, 1991; Nanto-Salonen,
1991; Nanto-Salonen, 1992); así pues, se decidió estudiar un modelo de
hipotiroidismoa 2 y 4 díasde vida (momentodel cambiode los GLUTs) y a 8 y 20
días deedad(transiciónde los IGFs) paraevaluaren él las posiblesalteracionesen
los cambiosde estasproteínas(GLUTs e IGFs> y el papelquelas hormonastiroideas
desempeñabanendichamodificación de estaspautas.
El pesocorporaly los nivelescirculantesde hormonastiroideasdisminuyeron
enlosanimalesneonataleshipotiroideos,exceptoa los 2 díasde vida (Nanto-Salonen,
1991),aunqueel descensode estosparámetrosfue másacusadoenaquellosa los que
se les practicó la tiroidectomía,poblaciónde 8 y 20 días, respectoa aquellosque
recibieronel fármaco bociógeno,población de 2 y 4 días. Estos resultadosya se
habíanobtenido en aquellas poblacionesde animalesneonatales,10-20 días, que
213

Discusión
fueron sometidasa la operaciónquirúrgica frentea las que recibieronel MMI en la
primerapartede estetrabajo;así pues,enesteexperimentovamosapoderhablarde
dos grupos de animales hipotiroideos neonatales que van a presentar un
comportamientoparalelo al de las dos poblacionesantes citadas: tratadascon el
fármacoantitiroideo(MMI) y sacrificadasa 2 y 4 díasy tiroidectomizadas,población
sacrificadaa 8 y 20 días. El hechode que las ratasde 2 días no presentenun menor
pesocorporalpuedeestarmotivado por los cambioshormonalesdel momentodel
nacimientoy tambiénporquenecesitenun periodode latenciaentreel comienzode
la administracióndel MMI y la manifestaciónde sus efectos(Flórez, 1992). No
obstante, todos los neonatos hipotiroideos, incluidos los de 2 días de vida,
presentaronuna reducciónen los nivelesplasmáticosy el contenidohipofisariode
Gil, lo cual apoyael papel crucial de las hormonastiroideasen la regulaciónde la
síntesis de Gil (Seo, 1981). Se encontróun descensoen la insulinemia de los
animalesde 4 días, y un aumentoen las ratasneonatalessacrificadasa 8 y 20 días;
todo ello concuerdatotalmentecon lo que obtuvimosen la primerapartedel trabajo,
y ratifica nuestrosresultadosanteriores.
4.4.1. IGFs e IGFBPs: nivelesséricosy expresióndel ARNni hepático.
Se puso de manifiesto la existencia de un paralelismoentre los niveles
circulantesde insulina e IGF-I, de modo que los neonatostratadoscon MMI
presentaronunos valores inferioresa los de sus controles,mientrasque los animales
tiroidectomizadosmostraronunascifras de ambospéptidossuperioresa los de sus
controles,de modo análogoa lo que se habíadescritoparalas poblacionesneonatales
(apartado4.1).
Los animalesde 2 días de vida, que recibieronMMI, mostraronun pesoy
unosnivelescirculantesde hormonastiroideas,insulina eJGF-Isimilaresa los de las
ratascontrol y una GH disminuida, lo que parecesugeriruna vez más que, en las
etapasinmadurasdel desarrolloes la insulina, y no la Gil, la hormonaqueregulala
síntesisy secreciónde los IGFs (Rivero, 1995; Goya, 1996),tal como sesugiereen
la primerapartedel trabajo.
Por otro lado, tanto la tiroidectomíacomo el tratamientocon MMI, aunque
guardanel perfil normalde descensoprogresivode IGF-II asociadoa la edad,sus
valoresestuvieronporencimadelos controles(Latimer, 1993;Nanto-Salonen,1993).
La diferenciaentre los dos gruposde ratashipotiroideas,en cuantoa nivelesde IGF-
214

Discusión
II, se puedeexplicar por el efecto reguladorde la glucemiasobre el IGF-II en
periodosinmaduros(Rivero, 1995; Goya, 1998),dado quelos nivelesde glucosaen
sangredisminuyenen los animalestiroidectomizados,aunqueno en los tratadoscon
el fármacoantitiroideo.
De nuevo se encontró una correlación alta y positiva entre los niveles
circulantesde insulina e IGF-I, que se confirmó medianteel análisis de regresión
lineal, que además,ratificó la existenciade una pobrecorrelaciónentrelos valores
plasmáticosde Gil e IGF-I en estas poblaciones. Así pues, estos resultados
corroboraronla importancia de la insulina, y no la Gil, en la regulaciónde la
secreciónde IGF-I en estadios inmadurosde desarrollo (Bóni-Schnetzler,1991;
Rivero, 1995; Goya, 1996).
En la rata,existeuna secuenciade apariciónde las IGFBPs(Donovan,1989;
Rivero, 1995); enel periodoadulto, la mayorpartede los IGFs estápresenteen la
circulacióncomoun complejoproteicode l5OkDa (ALS) que prolongala semivida
en el plasmade los IGFs y proporcionauna reservade dichos factorespara los
tejidos. Sinembargo,enetapasinmaduras,el complejopredominante(5OkDa), ya no
estáformadopor IGFBP-3 sino por IGFBP-1 y 2, que no sondependientesde la Gil
comola IGFBP-3(Rivero, 1995;Goya, 1996). Estasproteínas,IGFBP-1 y 2, pueden
ejerceraccionesmetabólicasindependientesdelos IGFs (Lewitt, 1991;Baxter, 1995;
Rajaram,1997).
El hipotiroidismo, como hemos visto a lo largo de estetrabajo, altera los
niveles de las IGFBPs, y por tanto, la actividadbiológica de los IGFs. También,
como en otras ocasiones,tenemosdos grupos de animalesneonatales:las ratas
hipotiroideasde 2 y 4 días de vida, que muestranun comportamientosimilar al de
los neonatostratadosconMMI (descensode los nivelesde IGFBP-1 y 3, y aumento
de los de IGFBP-2), y los animalestiroidectomizadossacrificadosa 8 y 20 días de
edad (desciendeIGFBP-3 y aumentanIGFBP-1 y 2); al igual que sucedíaen la
primera parte del trabajo, y ello se produce tambiéncon un cuadro endocrino
diferente: uno, que cursa con insulopeniay normoglucemia(animalestratadoscon
MMI y sacrificadosa 2 y 4 días),y otro, con altosnivelesde insulinae hipoglucemia
(ratasde 8 y 20 días tiroidectomizadas).
Sin embargo,los animalesde 2 y 4 díastratadoscon MMI no mostrabanlos
valorescirculantesde 3OkDa aumentados,sino disminuidos;ello esporqueel cuadro
215

Discusión
metabólico, niveles de insulina y OH, no coincide totalmentecon los animales
neonatalesMMI de 10 días de la primera partedel trabajo. Sin embargo,si se
encuentra,por “Western inmunoblot’, los valoresde IGFBP-I disminuidosy los de
IGFBP-2 aumentados.Los animalessacrificadosa 8 y 20 días tiroidectomizadosse
comportan como los tiroidectomizadosde la primera parte en cuanto a las
concentracionescirculantey de ARNm de IGFBP-1 y 2, motivadopor la regulación
que ejerció la insulinasobreesasproteínastransportadoras(Baxter, 1995); esdecir,
encontramosresultadossimilares a los de la primera partedel trabajoen ambos
gruposde animalesneonatalestiroidectomizadosen cuantoa IGFBPs.
Las IGFBPsde bajo pesomolecular(IGFBP-1 y 2) disminuyeronconforme
aumentabala edaddel animal en los controles.Los nivelesde IGFBP-3 en sueroy
la expresiónde su ARNm en hígadodescendieronen el hipotiroidismo, lo cual es
debido a los bajos nivelesde Gil registrados(ilervás, 1975).
4.4.2.GLUTs:nivelesenmembranacrudadecorazón,hígado,cuadriceps
y gastrocnemius.
La regulaciónde los transportadoresde glucosapor las hormonastiroideasu
otros factoresesdiferenteen función del tejido de que setrate, e incluso, depende
del transportadorde glucosa considerado(Postic, 1993b). En el corazón,en los
animalescontrol,nuestrosresultadosmuestranclaramenteel descensoenel contenido
de GLUT-1 y el incrementode GLUT-4 asociadoa la edad (Santalucia,1992). El
hipotiroidismocausóun descensoen el contenidode GLUT-4 y aumentóla cantidad
de GLUT-1; otros autoreshan mostradotambiénestasmismas variacionesen la
expresiónde estos transportadoresdurantela etapaperinatal(Castelló, 1994). Se ha
descritoun aumentodosis-dependientede la expresiónde GLUT-4 porel tratamiento
con 13 en cardiomiocitosprocedentesde ratas adultas(Gosteli-Peter,1996). Por
tanto, parececlaro que el déficit de hormonastiroideasconducea un desajustedel
descensode GLUT-1 que ocurre en la etapaperinatal, a la vez que disminuye el
aumentoen la cantidadde GLUT-4 típico del periodoadulto; seproduceun retraso
en el cambiodel patrónontogénicode expresiónde los transportadoresde glucosa.
El hígadojuega un papel fundamentalen la regulaciónde la homeostasis
glucídica, y en él, el GLUT-2 está mayoritariamentesituado en la membrana
plasmática(Stephens,1995) y el transportede glucosava a dependerdirectamente
de la cantidadde GLUT-2.
216

Discusión
Nuestrosresultadosmuestranclaramentetresaspectos:1)el GLUT-2hepático
seva incrementandoa lo largo del desarrollode acuerdocon Rubio y col. (1997)y
Posticy col. (1994)en los controles;2) el hipotiroidismoafectaal contenidohepático
de este transportadorde glucosa de acuerdocon Weinstein y col. (1994); 3) el
GLUT-1 aumentaen hígadoen los animaleshipotiroideosrespectoa los valores
control. Luegoparece,que en estosanimaleshipotiroideos,tanto en corazóncomo
en hígado, se produceun retardo en la aparición de ambos transportadores.Sin
embargo,esdifícil e imposiblecon los datosactualespoderrelacionarlos nivelesde
IGF-I alterados en dichos animales y las variaciones encontradas en los
transportadoresde glucosa,realmenteparecenprocesossin correlaciónentreellos.
En los músculosesqueléticos(cuadricepsy gastrocnemius),al igual queenel
corazón y todos los tejidos insulino~sensibles,se expresandos isoformas de
transportador:GLUT-4 y GLUT-1, siendoel primeroel másabundante(80-90%)y
responsablede másdel 85% de la captaciónde glucosaestimuladapor la insulina.
Del mismomodoqueenel corazón,pudimosobservaren los animalescontrolcomo
durantela etapaneonatal,el transportadorpredominanteera el GLUT-1, pero fue
sustituidoporel GLUT-4 conformeavanzabala edaddel animal (Santalucía,1992).
La expresiónde los transportadoresdel músculoesqueléticoestácontrolada
por varios factores. Se habíadescrito que la administraciónde T3 aumentabala
expresióndel GLUT-4 en el músculoesqueléticode las ratasadultas(CasIa, 1990;
Weinstein, 1991; Torrance, 1997a), e inducía la diferenciaciónde esascélulas in
viti-o (Shimokawa,1997)y, se habíademostradoque las hormonastiroideasposeían
una acción directa sobrela expresiónde estetransportador,puestoque el gen del
GLUT-4 poseeunelementode respuestaa estashormonas(Ezaki, 1997; Shimokawa,
1997). No obstante,tambiénla insulina parecíatenerun importantepapel en este
tejido y sobreestetransportador,al igual que enel corazón(Fischer,1997>; bajo el
efecto insulínico,unapequeñaproporciónde GLUT-1 intracelularse relocalizabaen
la membranaplasmáticaen la fibra muscular(Napoli, 1995), aunque,el fenómeno
tiene muchamás importanciaparael GLUT-4, dadoque la insulinapuedeaumentar
suproporciónen la superficie celularde 15 a 20 veces,en tantoque el GLUT-1 sólo
incrementade 3 a 5 veces(Camps,1992; Gould, 1993). Además,estudiosin viti-o
(miocitosL6) mostraronque el GLUT- 1, frenteal GLUT-4, era menossensiblea la
regulaciónpor glucosaque por la insulina (Koivisto, 1991).
217

Discusión
La regulaciónde los transportadoresde glucosaparece, por tanto, que se
realizacon la participaciónde las hormonastiroideasy de la insulina. Asípues,el
hechode que encontremosun incrementoen la cantidadde GLUT-1 y GLUT-4 en
las ratastiroidectomizadas,que presentaronaltosniveles de insulinemia, podríaser
explicadopor el fuerteefectoestimulantede estahormonasobreel transportador.A
los 4 díasde vida se produjoun descensoenel contenidode los dos transportadores,
puestoque disminuyeron los nivelesde las dos hormonasestimulantes,hormonas
tiroidease insulina (Shetty, 1996). Por último, a los 2 días de edad, cuandolos
animaleserannormoinsulinémicosy eutiroideos,sólo seprodujo un aumentoen el
contenidode GLUT-1 en el cuadriceps(se encontróuna tendenciaanálogaen el
gastroenemius)que podría ser debido a un retraso en el cambio del patrón
ontogénico.
Se podría pensar que el aumento en los animales tiroidectomizadosy
sacrificadosa 8 y 20 díasde vida del GLUT-4y GLUT-1, y el descensodel GLUT-4
a 2 y 4 días,podríaestarpropiciadoporel aumentoy descensodel IGF-I que, como
él, va asociadoa unavariaciónparalelade la insulina,perosin duda,esestahormona
lacausantede los cambiosen los transportadores.Por tanto, los datosestudiadosde
GLUT-1 y GLUT-4 en los animaleshipotiroideosparecenmostrarque no existeuna
relación directa entre las variacionesde GLUTs y de IGFs en los animales
tiroidectomizados,sino que ambasvariacionesenperiodoneonatal,dependende los
nivelesde insulinaalteradosa su vezpor Ja falta de hormonastiroideas.
218

CONCLUSIONES

Car~mes
5. CONCLUSIONES.
Animaleshinotiroideosneonatales,destetadosy adultos
.
Neonatales
1. La comparaciónde resultadosde los parámetrosestudiadosmuestraun cuadro
metabólico distinto entre las dos poblacionesneonataleshipotiroideasestudiadas,
según se produzcala deficienciade hormonastiroideaspor tiroidectomía,o por
tratamiento con MMI. Gil e insulina están altas en los animales neonatales
tiroidectomizados,y disminuidas en los tratados con MMI, la glucemia está
disminuidaen los tiroidectomizadose igual a los controlesen los MMI. Con dicho
cuadrometabólico, los animalestiroidectomizadosaumentansu IGF-I circulante,
mientrasdesciendeen los tratadoscon MMI. La expresiónde su ARNm hepático
guarda un paralelismo con sus niveles en suero, sugiriendo una regulación
transcripcional-
2. El IGF-II circulanteparecereguladopor la glucemiade formapositiva; disminuye
en los animalestiroidectomizadosy no se alteraen los hipotiroideos-MMI.El estudio
de la expresiónde suARNm sugiereuna regulacióntranscripcional.
3. El análisis de correlaciónlineal de ambaspoblacionesneonatales,entrevalores
plasmáticosde insulina e IGF-I, muestraunacorrelaciónpositiva concoeficientede
correlaciónmuy alto, r=0.91, mientras la correlaciónes muy baja para valores
plasmáticosde Gil.
4. El estudiode las proteínasligadorasde IGFs muestraque el aumentoencontrado
del complejo de 3OkDa (IGFBP-1 y 2) en los animaleshipotiroideosneonataleses
más grandeen los hipotiroideos-MMI que en los tiroidectomizados,y tanto los
resultadosde “Western immunoblot’, como la expresiónde su ARNm hepático,
parecenmostrarque ello se debea un aumentode IGFBP-2, propiciado, sin duda,
porel cuadrometabólicocaracterísticode los animalesMMI. La IGFBP-1 no parece
regularseen los animalesneonatalesde forma inversa a la insulina comoocurre en
adultos. La IGFBP-3 disminuyeen ambaspoblacioneshipotiroideasneonatalesy,
como la IGFBP-2, parecesugerirseuna regulacióntranscripcional.
220

5. Por último, los estudios realizadosbloqueandola secreciónde insulina con
estreptozotocina(SIZ), en los animalesneonatalestiroidectomizados,ratifican la
correlaciónpositiva encontradaentrevalorescirculantesde insulina y de IGF-I, así
comomuestranqueel IGF-II aumentacon la hiperglucemia,y enambos,se sugiere
una regulacióntranscripcional.En cuantoa las proteínasligadoras,la IGFBP-3 no
varíaen plasma,y prácticamentetampocola expresiónde su ARNm. En estecaso
de clara diabetes,la IGFBP-1 aumentaal disminuir la insulina, y la IGFBP-2vuelve
a mostraruna sutil regulaciónsegúnel cuadrometabólicodel animal.
Destetadosy adultos
6. Estosanimalestiroidectomizadosdestetadosy adultosmuestranunadisminución
de IGF-I debidaa la falta de hormonastiroideas,y cuyacausapareceser la hormona
de crecimientodisminuida. La regulaciónparecehacersea nivel transcripcional.
7. El análisisde correlaciónlineal muestraunacorrelaciónalta entrenivelesde Gil
y de IGF-1, y bajacon la insulina.
8. En los animalestiroidectomizados,destetadosy adultos,disminuyeel complejode
3OkDa (IGFBP-l y -2), y tambiénlaproteínaligadoraIGFBP-3. Las dosisde insulina
administradasa los hipotiroideosadultosno alteran los nivelesde IGEs ni de las
proteínasligadoras.
9. Se concluyeque el mediadorde las accionesde las hormonastiroideassobreel
axis IGFs/IGFBPsen periodoneonatalpareceser la insulina, pasandoa ser la Gil
en periodoadulto.
Rehabilitación
.
LO. Los experimentosde rehabilitacióncon tiroxina, en periodoneonatalo enperiodo
adulto, muestranque la vía idóneade rehabilitaciónesel pellet,y no la inyecciónde
tiroxina.
Li. Los animalesneonatalestratadoscon 1.SggIlOOgde pesode 14 durante5 días
no consiguenrehabilitarsus valorescirculantesde IGF-I ni los de IGF-II. Tampoco
consiguenrebajara los valorescontrol su insulinaalta ni su GH plasmática,asícomo
221

wh&ones
la glucemiadisminuida;todo ello porquetampocorehabilitan,con la dosisutilizada,
los nivelesde las hormonastiroideas.
12. Las proteínastransportadorasde 3OkDa y de 45kDa se rehabilitanen la etapa
neonatalcon las dosisde T4 empleada.
13. Los animalesdestetadosy adultostiroidectomizadostratadoscon 1 .75~g/10Ogde
pesode tiroxina durante5 días rehabilitantodoslos parámetrosdelaxis IGFs/IGFBPs
estudiados.
14. El análisisde correlaciónlineal entreinsulinay nivelesde IGF-I enneonatos,así
como entrenivelesde GH e IGF-I en adultos,muestrauna correlaciónpositiva alta
en estosanimalesrehabilitados.Dicha correlaciónes mayoren los animalestratados
conpelletsen todaslas poblacionesy másclara en animalesadultos.
Diabetes
.
15. Los animalesneonatalesdiabéticosporSTZ tratadoscon tiroxina rehabilitanlos
valorescirculantesde IGF-I y la expresiónde su ARNm hepáticode IGF-I y II, así
como susproteínasligadoras,con aumentode GH y no de insulina. Sin embargo,los
animalesdiabéticosadultostratadoscon T4 no rehabilitan,con la dosisadministrada,
los parámetrosde IGFs, ya que no aumentantampoco la Gil ni las hormonas
tiroideashastalos nivelescontrol.
Transportadoresde 21uc0sa
.
16. Las alteracionesencontradasendiversostejidosde los transportadoresde glucosa
en animalesneonataleshipotiroideos, muestranque no pareceexistir correlación
directa entre las variacionesde IGFs y de GLUTs, sino que ambas alteraciones
parecendependientesde los nivelesde insulina, a su vez alteradospor el déficit de
hormonastiroideas-
En resumen,esteestudio realizadoen animalestiroidectomizados,desdela
etapaneonatala la adulta de la rata, muestraque la influencia de las hormonas
tirodeassobreel axis IGFs/IGFBPsvaría segúnla edaddel animal queseconsidere,
siendo el mediadorde dichaacción la insulina en periodosneonatales,y pasandoa
222

Conch&ones
ser la GH la hormonamediadoraenperiodoadulto. El estudiode las variacionesde
las proteínastransportadorasenestosanimalestiroidectomizadosponede manifiesto
la importanciadel estadometabólicoensuregulación,sobretodo, paralas proteínas
ligadorasde bajopesomolecularabundantesen etapaneonatal.La IGFBP-3 parece
mostrar una regulaciónmediadapor la hormonade crecimiento, sobre todo en
periodo adulto. Los estudios con animales hipotiroideos, neonatalesy adultos,
rehabilitadoscontiroxinaratificaronlos resultadosanteriores,no obstante,en periodo
neonatalestosestudios,junto con los realizadosdandotiroxina a animalesdiabéticos
neonatales,muestranque las hormonastiroideasen situacioneslímites de falta de
insulina tambiénactuanrehabilitandolos nivelesde IGF-I a travésde su acciónsobre
la hormonade crecimiento.Estos datos,y los obtenidossobrela regulaciónde los
transportadoresde glucosa,ponende relieve que las hormonastiroideassonfactores
endocrinosdecisivosen la regulacióndel desarrollo, no sólo por sus conocidas
accionesespecificassobre el Sistema Nervioso Central, sino porque producen
desequilibriosenvertientemetabólica,alterandootras hormonascomo la insulina y
laGil, quea su vezprovocanvariacionesen la secreciónde factoresde crecimiento,
IGFs, totalmenteimplicadoscomo reguladoresde] crecimientoen los mamíferos.
223

BIBLIOGRAFíA

Bibliografla
6. BIBLIOGRAFIA.
- AdamoML, Ben-HurH, RobertsCT and LeRoithD. Regulationof start site
usage in the leader exons of the rat insulin-like growth factor-I gene by
development,fasting and diabetes.Mol. Endocrinol. 5: 1677-1686<1991).
- AhrénB andLundquist1. Insulin secretoryresponseto different secretagogues
in hyper- and hypothyroidmice. Acta Endocrinol. 97: 508-513<1981).
- Aláez, C. Tesisdoctoral.Actividad 5’-desyodasay regulacióndel axis tiroideo
en ratasubnutrida.E. Farmacia. U. Complutense.Madrid <1992a).
- Aláez C, Calvo R, Alvarez C, Goya L, Martín MA, Obregón MJ and
Pascual-LeoneAM. Influenceof thyroid hormonedeiodinationon theregulation
of the thyroid axis in diabretic rats. Metabolism (enprensa).
- Aláez C, Calvo R, ObregónMJ and Pascual-LeoneAM. Thyroid hormones
and 5’-deiodinase activity in neonatal undernourishedrats. Endocrinology
130:773-779<1992/,).
- Alberti KGMM, ChristensenSE, IversonJ, Seyer-HansenK, Cliristensen
NJ, Prange-HansenAA, LunkbaekK and Orskow H. Inhibition of insulin
secretionby somatostatin.Lancet2:1299-1301(1973).
- AlvarezC, MartínMA, GoyaL, Bertin E, PorthaB andPascual-LeoneAM.
Contrastedimpact of maternal rat food-restriction on the fetal endocrine
pancreas.Endocrinology138: 2267-2273(1997).
- Angervo M, Leinonen P, Koistinen R, Julkunen M, SeppalaNl. Tri-
iodothyronineand cyclohexamideenhanceinsulin-like growth factor binding-
protein-l geneexpressioninhumanhepatomacelis.J. Mol. Endocrinol. 10: 7-13
(1993a).
225

Bibliografla
- AngervoM, ToivonenJ, LeinonenP, Valimaki M andSepliláM. Thyroxine
withdrawalis accompaniedby decreasedcirculatinglevelsof insulin-like growth
factorbindingprotein-1in thyroidectomizedpatients.J. Clin. Endocrino!. Metab.
76: 1199-1201 <1993b).
- Aránguez1, GoyaL and Pascual-LeoneAM. Changesin blood glucose,liver
glycogen,ketonebodiesandplasmainsulin in suckling ratstreatedwith a single
high cortisol doseoneday afterbirth. Acta Endocrinol. (Copenlz.)113: 598-603
<1986).
- Azam M, GuptaRL and BaquerNZ. Effect of hyperthyroidismon rat blood
celí insulin receptorsand catecholamines:relationship with cellular ageing.
Biochem.mt. 21: 135-144<1990).
- Babajko5, Hardouin5, SegoviaB, GroyerA andBinoux M. Expressionof
insulin-like growth binding protein-1 and -2 genesthroughthe perinatalperiod
in the rat. Endocrinology 132: 2586-2592(1993).
- Barnard RJ and YoungrenJF. Regulationof glucose transport in skeletal
muscle.FASER1. 6: 3238-3244<1992).
- Baxter RC. Circulating binding proteinsfor the insulin-like growth factors.
TrendsEndocrinol. Metab. 4: 91-96 <1993).
- Baxter RC. Insulin-like growth factors binding proteins as glucoregulators.
Metabolism44 Suppl4:12-17(1995).
- BaxterRC and Cowell CT. Diurnal rhythm of growth hormoneindependent
binding protein for insulin-like growth factors in human plasma. J. Clin.
Endocrino!. Metab. 65: 432-440(1987).
- Bedo G, SantistebanP, John 1 and Aranda A. Expressionof the growth
hormonegeneand the pituitary-specifictranscriptionfactor GHF-1 in diabetic
226

Bibliograjía
rat. Mo!. Endocrino!. 5: 1 730-1739 (1991).
- Beer SF, ParrJH, TempleC and HalesCN. ‘[he effect of thyroid diseaseon
proinsulinand C-peptidelevels. C!in. Endocrino!. 30: 379-383 (1989).
- BeltchezPE, Gredley G, Bird D and Himsworth RL. Regulationof ‘[SH
secretionby negativefeedbackof 13 on the hypothalamus.J. Endocrino!. 76:
439-448 (1978).
- BereketA, Lang CH, BlethenSL, GelatoMC, FanJ, FrostRA andWilson
lA. Effect of insulin on the insulin-like growth factor systemin children with
new-onsetinsulin-dependentdiabetesmellitus. J. Clin. Endocrino!. Metab. 80:
1312-1317(1995).
- BerelowitzM, MaedaK, Harris 5 and FrohmanLA. The effect of alteration
in thepituitary-thyroidaxis on hypothalamiccontentand in vitro releaseof SRIF-
like immunoreactivity.Endocrinology 107: 24-29 (1980).
- BersG andGarfin D. Proteinand nucleic acid blotting and immunochemical
detection.Riotechnique3: 275 <1985).
- Bichelí DP, Kikuchi K, Rotwein P. Growth hormonerapidly activatesIGF-I
genetranscriptionin vivo. Mo!. Endocrino!. 6:1899-1908(1992).
- Bóni-SchnetzlerM, Schmid C, Mejer PJ and FroeschER. Insulin regulates
insulin-like growthfactor-I mRNA in rat hepatocytes.Am.J. Physiol. 260:E846-
E851 (1991).
- Bornfeldt KE, SkottnerA and Arnqv¡st HJ. In vivo regulationof messenger
RNA encodinginsulin-like growth factor-I (IGF-I) and its receptorby diabetes,
insulin and IGF-I in rat muscle.J. Endocrino!. 135: 203-211 (1992).
- BradfordMM. A rapidandsensitivemethodfor thequantificationof microgram
227

Bibliograjía
quantities of protein, utilizing the principle of protein dye binding. Anal.
Biochem. 72: 248-254 (1976).
- Brismar K, Fernqvist-ForbesE, WahrenJ aud Hall K. Effect of insulin on
thehepaticproductionof insulin-likegrowthfactorbindingprotein-1(IGFBP-1),
IGFBP-3, and IGF-I in insulin-dependentdiabetes..1. C!in. Endocrino!. Metab.
79: 872-878 (1994).
- Burant CF, Siv¡tz WI, FukumotoH, Kayano T, Nagamatsu5, Seino 5,
Pessin SE and Belí GI. Mammalian glucose transporters: structure and
molecularregulation.RecentProg. Horm. Res. 47? 349-388<1991).
- Burstein PJ, Draznin B, Jolison CJ and Schalcb DS. The effect of
hypothyroidism on growth, serum growth hormone, the growth hormone-
dependentsomatomedin,insulin-like growthfactor, andits carrierproteinin rats.
Endocrinology104: 1107-1111(1979).
- Butíer AA, GallaherBW, Ambler GR,CluckmanPD andBreier BH. Insulin-
like growth factor-I and IGF binding protein-3 in plasmaof GH deficient rats.
J. Endocrino!. 150: 67-76 <1996).
- Calvo R, Morrealede EscobarG, Escobardel Rey F and ObregónMJ.
Maternal nonthyroidalillnessand fetal thyroid hormonestatus,asstudied in the
streptozotocin-induceddiabetesmellitus rat model. Endocrinology 138. 1159-
1169 (1997).
- CampsM, Castelló A, Muñoz P, Monfar M, Testar X, Palacín M and
Zorzano A. Bffect of diabetesand fasting on GLUT-4 (muscle/fat)glucose-
transporterexpressionin insulin-sensitivetissues. Biochem. J. 282: 765-772
(1992).
- CasiaA, Rovira A, Wells JA and Dohm GL. Increasedglucosetransporter
(GLUT-4) proteinexpressioninhyperthyroidism.Biochem.Biophys.Res.Comm.
228

Bib!iograJla
171: 182-188(1990).
- CastellóA, Rodríguez-ManzanequeJC, CampsM, Pérez-CastilloA, Testar
X, PalacínM, SantosA andZorzanoA. Perinatalhypothyroidismimpairs the
normal transition of GLUT4 and GLUT1 glucose transportersfrom fetal to
neonatallevelsin heartandbrownadiposetissue.1. Rio!. Chem.269:5905-5912
<1994).
- CavaliereII, Knobel M, Medeiros-NetoG. Effect of thyroid hormonetherapy
on plasmainsulin-like growth factor 1 levels in nonnal subjects,hypothyroid
patientsand endemiccretins.Horm. Res. 25: 132-139 <1987).
- CedaGP, FielderPJ, DonovanSM, RosenfeldRG, Hoffman AR. Regulation
of insulin-like growth factor binding proteinsexpressionby thyroid hormonein
rat Gil3 pituitary tumorcelís.Endocrinology130: 1483-1489<1992).
- Clark R. Thesomatogenichormonesandinsulin-likegrowthfactor-Istimulations
of lymphopoiesisand immune function. Endocr. Rey. 18: 157-179 <1997).
- ClemmonsDR and UnderwoodLE. Nutritional regulationof IGF-I and IGE-
binding proteins.Ann. Rey. Nutr. 11: 393-412<1991).
- Coiro y, BravermanLE, ChristiansonD, FangSL and GoodmanM. Effect
of hypothyroidismand thyroxine replacementon growth hormone in the rat.
Endocrino!ogy105: 641-646(1979).
- ColombV, LeturqueA, GuihotG, LoizeauM, Lavie 5, Colomer5, Ricour
C andGirard J. Role of nutrient delivery affectsinsulin sensitivity and liver
glucose transporter expression in rat. Am. J. Physio!. 269: E827-E833
<1995).
- Cotterill AM, Cowell CT, BaxterRC, McNeill D and Silinik M. Regulation
of thegrowthhotmone-independentgrowth factor-bindingproteinin children.J.
229

Ribliograjía
C!in. Endocrino!. Metab. 67: 882-887 (1988).
- CountsDR, GwirtsmanH, CarlssonLM, LesemM and Cutíer GB Jr. The
effect of anorexianervosaand re-feedingon growth hormone-bindingprotein,
the insulin-like growth factors (IGFs) and the IGF binding proteins.J. Clin.
Endocrino!. Metab. 75: 762-767(1992).
- CoverleyJA andBaxter RC. Regulationof insulin-like growth factor (IGF)
binding protein-3 phosphorylationby IGF-I. Endocrino!ogy 136: 5778-5781
<1995).
- ChenY andArnqvist HJ. Differential regulationof insulin-like growthfactor
binding protein-2 and -4 mRNA in muscle tissues and liver by diabetes or
fasting. J. Endocrino!. 143: 235-242(1994).
- ChernausekSD, Underwood LE, Utiger RD and van Wyk JJ. Growth
hormonesecretionandplasmasomatomedin-Cin primary hypothyroidism.Clin.
Endocrino!. 19: 337-344(1983).
- ChomczynskiP and SacchiN. Single step methodof RNA isolation by acid
guanidiniumthiocyanate-phenol-chloroformextraction.Anal. Biochem.162.- 156-
159 (1987).
- DaughadayWH and RotweinP. Insulin-like growth factors 1 and II. Peptide,
messengerribonucleicacidandgenestructures,serum,andtissueconcentrations.
Endocr. Rey. 10: 68-91 <1989).
- Demori 1, Bottazi C, Vocí A, Gallo G, Scbarf JG and FugassaE. Tri-
iodothyronineincreasesinsulin-like growth factor binding protein-4expression
in rat hepatocytes.J. Endocrino!. 154: 155-165 (1997).
- Dobbing J. The latter growth of the brain and its vulnerability. Pediatrics 53:
2-6 (1977).
230

Bibliografla
- Donovan SM, Atilano LC, H¡ntz RL, Wilson DM and Rosenfeid RG.
Differential regulationof te insulin-like growthfactors(IGF-I and -II) and IGF
bindingproteinsduring malnutritionin theneonatalrat. Endocrino!ogy129: 149-
157 <1991).
- DonovanSM, Oh Y, PhamH and RosenfeidRG. Ontogenyof te insulin-like
growthfactor-bindingproteinsin therats. Endocrino!ogy125:2621-2627<1989).
- EayrsJI. Developmentalrelationshipsbetweenbrainand thyroid. En: Michae!
RP <ed) Endocrino!ogyandhumanbehaviour,London:Oxford UniversityPress,
239-255<1968).
- Escrivá F, Agote M and Pascual-LeoneAM. Effects of undernutritionon
muscleglucosetransporterproteinin therat. Diabetologia (en evaluación).
- EscriváF and Pascual-LeoneAM. Decreaseof blood glucose, liver glycogen,
and insulin and changes in glucose tolerance in suckling rats treated
neonatallywith high L-thyroxine doses.Endocrinology108: 336-340(1981).
- EscriváF, RodríguezC, CachoC, Alvarez C, PorthaB andPascual-Leone
AM. Glucoseutilization and insulin actionin adult rats submittedto prolonged
food restriction.Am. J. Fhysiol. 263: EI-E7 (1992).
- EvansRM, BirnbergNC, RosenfeldMG. Glucocorticoidandthyroid hormones
transcriptionally regulategrowth hormonegeneexpression.Proc. Natí. Acad.
Sci. USA 79: 7659-7663<1982).
- Ezaki O. Regulatory elements in the insulin-responsiveglucose transporter
(GLUT4) gene.Riochem.Biophys. Res. Commun.241: 1-6 <1997).
- Ezzats 5, Laks D, OsterJ and Melemd 5. Growth hormomeregulation in
primary fetal and neonatal rat pituitary celí cultures: the role of thyroid
hormones.Endocrino!ogy128: 937-943(1991).
231

Rib!iograjla
- DeFesi CR, FeIsEc, SurksMI. ‘[riiodothyronine stimulatesgrowthof cultured
GC celísby actionearly in theG1 period: evidencefor mediationby thenuclear
13 receptor.Endocrinology116: 2062-2069<1985).
- FiseherY, ThomasJ, SevillaL, MuñozP, BeckerC, HolmanG, Kozka IJ,
PalacínM, Testar X, Kammermeier H and Zorzano A. Insulin-induced
recruitmentof glucosetransporter4 (GLUT4) and GLUT1 in isolatedrat cardiac
myocytes.1. Rio!. Chem.272: 7085-7092<1997).
- Flórez J and Amado JA. Hormonastiroideas y fármacosantitiroideos. En.-
Farmaco!ogíahumana.Ed: Masson-Sa!vatMedicina 2a<ed): 825-834 (1992).
- FlyvbjergA, LandauD, DomeneII, HernándezL, GronbaekH and LeRoith
D. Ihe role of growth hormone, insulin-like growth factors (IGFs) and IGF-
binding proteins in experimentaldiabetickidney disease.Metabolism44 Supp!
4: 67-71 <1995).
- FossMC, PaccolaGMGF, SaadMJA, PimentaWP, PiccinatoCEand Iazagi
N. Peripheral glucose metabolism in human hyperthyroidism. J. Clin.
Endocrino!. Metab. 70.- 1167-1172(1990).
- Fowlkes JL. Insulinlike growth factor-binding protein proteolysis. Trends
Endocrino!. Metab. 8: 299-306<1997)
- FreakeHC, OppenheimerJH. ‘[hermogenesisandthyroid function.Ann. Rey.
Nutr. 15: 263-291 (1995).
- Frystyk J, Gronbaek 1-1, SkjaerbaekC and Flyvbjerg A. Effect of
hyperthyroidismon circulatinglevelsof freeand total IGF-I and IGFBPsin rats.
Am. J. Physiol. 269: E840-E845(1995).
- Gallo G, deMarchisM, Voci A andFugassaE. Expressionof hepaticmRNAs
for insulin-like growth factors-I and -II during the developmentof hypothyroid
232

Bib!iografta
rats. J. Endocrino!. 131: 367-372 <1991).
- GargoskySE, TapanainenP and RosenfeldRG. Administration of growth
hormone (GH), but not insulin-like growth factor-I (IGF-I), by continuous
infusion can induce the formation of the 150-kilodaltonIGF-binding protein-3
complex in Gil-deficient rats. Endocrino!ogy134: 2267-2276<1994).
- Gaspard 1, Wondergem R, Hamamdzic M, KI¡tgaard HM. Serum
somatomedin stimulation in thyroxine-treated hypophysectomized rats.
Endocrinology120: 606-611 <1978).
- Geary ES, Lim M, Ceda GP, Ro 5, Rosenfeld RG, Hoffman AR.
Triiodothyronine regualtesinsulin-like growth factor-I binding to cultured rat
pituitary celís. J. Neuroendocrino!.1: 179-184(1989).
- GershengornMC. Thyrotropin releasing hormone stimulation on pituitary
hormonesecretion.Ann. Rey. Physio!. 48: 515-526(1986).
- GiustinaA andWehrenbergWB. Growthhormoneneuroregulationin diabetes
mellitus. TrendsEndocrino!. Metab. 5: 73-78 <1994).
- GlasscockGF, GelberSE, LamsonG, McGee-TekulaR and RosenfeldRG.
Pituitary control of growth in the neonatal rat: effects of neonatal
hypophysectomyon somaticandorgangrowth, seruminsulin-like growthfactors
(IGF)-I and -II levels, and expressionof IGF binding proteins.Endocrinology
127: 1792-1803<1990).
- GlasscockGF, Gin KKL, Kim JD, Hintz RL and RosenfeldRG. Ontogeny
of pituitary regulationof growth in thedevelopingrat: comparisonof effectsof
hypophysectomyand hormonereplacementon somaticandorgangrowth,serum
insulin-like growth factor-I (IGF-I) and IGE-Il levels, and IGF-binding protein
levels in theneonatalandjuvenile rat. Endocrino!ogy128: 1036-1047(1991).
233

Ribliografi’a
- GlasscockGF, Hein AN, Miller JA, Hintz RL audRosenfeldRG. Effectsof
continuous infusion of insulin-like growth factor-I and II, alone and in
combination with thyroxine or growth hormone, on the neonatal
hypophysectomizedrat. Endocrinology130. 203-210(1992).
- GlasscockGF and Nicolí CS. Hormonal control of growth in the infant rat.
Endocrinology109: 176-184 (1981)
- Glydon R andSr J. ‘[he developmentof the bloodsupply of thepituitary in the
albino rat, with specialreferenceto theportalvessels.J. Anat. <London) 91: 237
<1957).
- González C and John 1. Effect of streptozotocindiabetes and insulin
replacementon growth hormonein rats. J. Endocrinol. Invest. 8: 7-11 <1985).
- GonzálezC, Montoya E andJohn1. Effect of streptozotocindiabeteson the
hypothalamic-pituitary-thyroidaxis in the rat. Endocrino!ogy 107: 2099-2103
<1980).
- Gosteli-PeterMA, Schmid C and ZapfJ. Triiodothyronineincreasesglucose
transporter isotype 4 mRNA expression,glucose transport, and glycogen
synthesis in adult rat cardiomyocytesin long-termculture. Biochein. Riophys.
Res. Comm. 221: 521-524<1996).
- Gosteli-PeterMA, Winterhalter KH, Schmid C, FroeschER and Zapf J.
Expressionandregulationof insulin-like growthfactor-I (IGF-I) andIGF-binding
proteinmessengerribonucleicacid levels in tissuesof hypophysectomizedrats
infusedwith IGF-I andgrowthhormone.Endocrino!ogy135: 2558-2567<1994).
- Gould GW andBeil GI. Facilitativeglucosetransporters:an expandingfamily.
TIRS 15: 18-23 <1990).
- GonidGW andHolmanGD.Theglucosetransporterfamily: structure,function
234

Bibliografla
and tissuespecific expression.Riochem.J. 295: 329-341 (1993).
- GoyaL, dela PuenteA, Ramos5, Martín MA, EscriváF andPascual-Leone
AM. Regulationof IGF-I and -II by glucosein primary cultures of fetal rat
hepatocytes.J. Rio!. Chem. <en eva!uación).
- GoyaL, Rivero F, Martín MA, Alvarez C, Ramos5, de la PuenteA and
Pascual-leoneAM. Liver mRNA expressionof IGF-I and IGFBPs in adult
undernourisheddiabeticrats. Lije Sci. (en prensa).
- GoyaL, Rivero F, Martín MA, ArahuetesR, HernándezER andPascual-
LeoneAM. Effects of refeeding of undernourishedand insulin treatmentof
diabetic neonatalrats on IGF and IGFBP. Am. J. Physiol. 271.- E223-E231
<1996).
- de GreefWJ, RondeelJMM, vanHaasterenGAC, Klootwijk W andVisser
TJ. Regulationof hypothalamicTRITI productionand releasein therat. AMA 19.-
77-79 <1992).
- Guyton AC. Hormonastiroideasmetabólicas.En: Tratadode Fisiologíamédica,
ed: Interamericana-McGraw Hill 7Ú (ed): 888-899<1986).
- Hall K, JohanssonBL, Povoa G, Thalme B. Serum levels of insulin-like
growth factor (IGF) 1, II and IGF-bindingprotein in diabeticadolescentstretaed
with continuoussubcutaneousinsulin infusion. J. mt. Med. 225: 273-278
(1989>.
- Hall U, KajimotoY, Bichelí D, Kim S-W,JamesPL, CountsD, Nixon LI,
Tobin G andRotweinP. Functionalanalysisof the rat insulin-like growthfactor
1 geneand identificationof an IGF-I genepromoter. DNA and Celí Rio!. 11:
301-313 (1992).
- Hanaire-BroutinII, Sallerin-CauteB, PoncetMF, TauberM, Bastide R,
235

Bibliograjía
ChaleJJ, RosenfeidR, TaubertJP. Effect of intraperitonealinsulin delivery
on growth hormonebinding protein, insulin-like growth factor-I (IGF-I), and
IGF-binding protein-3 in IDDM. Diabeto!ogia39: 1498-1504<1996).
- HernándezER, RobertsCT, Hurwitz A, Le Roith D andAdash¡EV. Rat
ovarian insulin-like growth factor II geneexpressionis theca-interstitial-cell-
exclusive: hormonalregulationand receptordistribution. Endocrinology 127:
3249-3251<1990).
- llervásF, MorrealedeEscobarG, Escobardel ReyF. Rapid effectsof single
small dosesof L-thyroxineandtriiodo-L-thyronineongrowthhormoneasstudied
in the rat by radioimmunoassay.Endocrinology97: 91-101 <1975).
- Hiraki Y, Rosen OM and B¡rnbaum MJ. Growth factors rapidly induce
expressionof the glucosetransportergene.J. Rio!. Chem. 263: 13655-13662
<1988).
- Holly JMP, BiddlecombeRA, DungerDR, EdgeJA, Amiel SA, Howell R,
Chard 1, ReesLH and WassJAH. Circadianvariation of GH-independent
IGF-binding protein in diabetesmellitus and its relationshipto insulin. A new
role for insulin?. Clin. Endocrino!. 29.- 667-675(1988).
- HossenloppP, Seurin D, Segovia-QuinsonB, Hardouin 5 and Binoux NI.
Analysis of serum insulin-like growth factor binding proteins using Western
blotting: useof themethod for tritiation of the binding proteinsand competitive
binding studies.Anal. Riochem.154: 138-143<1986).
- HumbelRE. Insulin-like growthfactors1 andII. Eur. J. Riochem.190:445-462
(1990).
- Hunter WM and GreenwoodFC. Preparationof iodine-131 labelledhuman
growth hormoneof high specific activity. Nature 194: 495-496<1962).
236

Ribliograjia
- HunzikerER, WagnerJ andZapfJ. Differentialeffectsof insulin-likegrowth
factor-I and growth hormone on developmentalstagesof rat growth plate
chondrocytesiii vivo. J. C!in. Invest. 93: 1078-1086<1994).
- IkedaT, FujiyamaK, TakeuchiT. Effect of thyroid hormoneon somatomedin
C releasefrom perfusedrat liver. Experientia45: 170-171 <1989).
- Ikeda1, FujiyamaK, HoshinoT, Takeuchi1, MashibaH andTominagaM.
Possiblerole of thyroid hormonein decreasedsomatomedin-Clevelsin diabetic
and starvedrats.Ann. Nutr. Metab. 34: 8-12 <1990).
- Ikejiri K, VamadaT and Ogura H, Matsumoto and Kobe. Age-related
glucoseintolerancein hyperthyroidpatients.Diabetes27: 543-549(1978).
- Ishii DN, GuertinDM and WhalenLR. Reducedinsulin-like growth factor-I
mRNA content in liver, adrenal glands and spinal cord of diabetic rats.
Diabetologia 37- 1073-1081 <1994).
- JayendraHS and Cerchio GM. Hypoinsulinemiaof hypothyroidism. Arch.
Intern. Med. 132: 657-661 <1973).
- John 1 and Montes A. ‘[he different effectsof thyroidectomy, KCIO4 and
propilthiouracilon insulin secretionand glucoseuptakein the rat. Endocrino!ogy
94: 1502-1507<1974).
- John1, MorrealeG and Escobardel rey F. Differential effectsin the rat of
thyroidectomy, propylthiouracil and other goitrogens on plasma insulin and
thyroid weight. Endocrino!ogy87- 99-110 (1970).
- JohnsonIR, Blossey BK, DenkoCW and han J. Expressionof insulin-like
growth factor-I in cultured rat hepatocytes:effects of insulin and growth
hormone.Mo!. Endocrino!. 3: 580-587 <1989).
237

Rib!iografta
- JonesJI and ClemxnonsDR. Insulin-like growth factors and their binding
proteins:bologicalactions.Endocr. Rey. 16: 3-34 (1995).
- JonesPM, Burrin JM, GhateiMA, O’HalloranDJ, Legon5 andBloom SR.
‘[he influenceof thyroid hormonestatuson thehypothalamo-hypophysealgrowth
hormoneaxis. Endocrino!ogy126: 1374-1379(1990).
- KatsilambrosN, Ziegler R, SchatzII, Hinz M, Maier y and Pfeiffer EF.
Intravenousglucosetoleranceandinsulinsecretionin the ratafterthyroídectomy.
Horm. Metab. Res. 4: 377-379 <1972).
- Kim JD, Nanto-SalonenK, SzczepankiewiczJL, Rosenfeld RG aud
GlasscockGF. Evidencefor pituitary regulationof somaticgrowth, insulin-like
growth factors-I and -II, and the binding protein in the fetal rat. Pediatric Res
32: 144-151 <1993).
- Kitauchi 5, Yamanouchi H, Hirano N, Toné 5 and Sh¡ino M. Effect of
neonatalthyroidectomyon growth hormonesecretionin the rat. J. Endocrino!.
157.- 245-250<1998).
- Kjeld JM, Kuku SE, DiamantL, FraserIR, Joplin GE andMashiterK.
Productionand storageof 125J thyroxineand 1251 triiodothyronineof high specific
activity. Clin. Chim. Acta 61: 381-389(1975).
- Koivisto U-M, Martínez-ValdezH, Bilan PJ, BurdettE, Ramíal1 and Klip
A. Differential regulationof theGLUT-1 andGLUT-4 glucosetransportsystems
by glucoseand insulin in L6 musclecelís in culture. J. Rio!. Chem. 266: 2615-
2621 <1991).
- Laemmli UK. Cleavageof structural proteinsduring the assemblyof the head
of bacteriophage14. Nature 227: 680-685(1970).
- Latiiner AM, HausmanGJ, McCuskerRH andBuonomoFC. Theeffectsof
238

Rib!iograjla
thyroxineon serumand tissueconcentrationsof IGFs (IGF-I and -II) and 1GW
binding proteinsin the fetal pig. Endocrino!ogy 133: 1312-1319 (1993).
- LegrandJ. Morphogeneticactionof thyroidhormones.TrendsNeurosci.2.- 234-
236 <1979).
- Legrand J. Thyroid hormone effects on growth and development. En:
HennemannG <eds) Thyroid hormonemetabolism, New York: Marce! Dekker,
503-534 (1986).
- Lenzen5 andRailey C. ‘[hyroid hormones,gonadaland adrenocorticalsteroids
and the functionof the islets of Langerhans.Endocr. Rey.5: 411-434<1984).
- Lenzen5, JoostHG andHasselblattA. The inhibition of insulin secretionfrom
the perfusedrat pancreasafter thyroxine treatment.Diabetologia 12: 495-500
<1976).
- Lenzen5 andKlóppel G. Insulinsecretionandthemorphologicalandmetabolic
characteristicsof pancreaticislets of hyperthyroid ob/ob mice. Endocrino!ogy
103: 1546-1555 (1978).
- LeonardJL. Identification and structureanalysisof iodothyroninedeiodinases.
En: GreerMa <ed) Thethyroid g!and, New York: RayenPress, 285-305<1990).
- Lewitt MS and RaxterRC. IGFBP-1: a role in glucosecounterregulation?Mo!.
and Ce!!. Endocrino!. 79: C147-C152 <1991).
- Lewitt MS, SaundersH, PhyvalJL andBaxterRC. Regulationof insulin-like
growth factor-bindingprotein-1 in rat semm.Diabetes43: 232-239 <1994).
- Lowe WL, RobertsCT, Lasky SR andLe Roith D. Differential expressionof
alternative5’-unstranslatedregions in mRNAs encodingrat insulin-like growth
factor 1. Proc. Nat. Acad. Sci. USA 84: 8946-8950(1987).
239

Bibliografía
- Lund PK. Insulin-lilcegrowthfactor1: molecularbiologyandrelevanceto tissue-
specific expressionand action. RecentProg. Horm. Res. 49: 125-148<1994).
- Lund PK, Moats-StaatsBM, HynesMA, SimmonsJG, JansenM, D’Ercole
AJ andvanWyk JJ.Somatomedin-C/Insulin-like growthfactor-I and insulin-
like growth factor-II mRNAs in rat fetal and adult tissues.J. Rio!. Chem. 261:
14539-14544 (1986).
- Luo J andMurphy U. Differential expressionof insulin-like growth factor-I
and insulin-like growth binding protein-1 in the diabetie rats. Mo!. Ce!!.
Riochem.103.- 41-50 (1991).
- Uno J, ReidRE and Murphy U. Dexamethasoneincreaseshepaticinsulin-like
growth factor binding protein-1 (IGEBP-1) mRNA in serum IGFBP-1
concentrationsin the rat. Endocrino!ogy127- 1456-1462<1990).
- Magner JA. Thyroid-stimulating hormone: biosynthesis, celí biology and
bioactivity. Endocrin. Rey. 11: 354-385 (1990).
- Maher F, Vannuci SJ, Simpson lA. Glucose transporterproteins in brain.
FASER.1 8k- 1003-1011(/994).
- Maiter D, Fliejen T, UnderwoodLE, MaesM, GerardG, DavenportML
and KetelslegersJM. Dietary proteinrestrictiondecreasesinsulin-like growth
factor1 independentof insulin andliver growthhormonebinding. Endocrino!ogy
124: 2604-2611(1989).
- Martial JA, RaxterJD, GoodmanHM and SeeburgPH. Regulationof growth
hormonemessengerRNA by thyroid and glucocorticoidhormones.Proc. Natí.
Acad. Sci. USA 74: 1816-1820 <1977).
- Martín MA. Tesis doctoral. Secreciónde insulina en animalessubnutridos:
estudioin vivo e in viti-o. F. Biología. U. Complutense.Madrid <1995)
240

Bibliografía
- Martín MA, Alvarez C, GoyaU, PorthaB and Pascual-LeoneAM. Insulin
secretionin adultrats that hadexperienceddifferentunderfeedingpatternsduring
their development.Am. J. Physio!. 272: E634-E640 <1997).
- MathewsLS, NorstedtG and Palmiter RD. Regulationof insulin-like growth
factor-I gene expressionby growth hormone.Proc. Natí. Acad. Sci. USA 83:
9343-9347<1986).
- Matthaei5, Trost B, HamannA, KauschC, BeneckeII, GretenH, Hóppner
W andKlein HH. Effectof in vivo thyroid hormonestatuson insulinsignalling
and GLU’[1 and GLUT4 glucose transport systems in rat adipocytes. J.
Endocrino!. 144. 347-357 (1995).
- Mc Carthy1, Casinghino5, CentrellaM and CanalisE. Complexpatternof
insulin-like growth factor binding proteinexpressionin primary rat osteoblast
enriched cultures: regulation by prostaglandinE2, growth hormone, aud the
insulin-like growth factors.J. Ce!!. Physiol. ¡60: 163-175 <1994).
- Melian E, VelascoII, Barrios R and Sánchez-FrancoF. Basal and growth
hormone-inducedhepaticmessengerribonucleicacid expressionof insulin-like
growth factor-I (IGF-I) and IGF-binding protein-3 is independent of
hyperinsulinemiaandincreasedenergystautsin thegeneticallyobeseZuckerrat.
Endocrinology 138: 1066-1071<1997).
- Melmed5 andYaniasitaS. Insulin-like growthfactor-l action on hyperthyroid
rat pituitary celís: supressionof T3-induced growth hormone secretion anó
messengerribonucícieacid levels. Endocrinology 118: 1483-1490(1986).
- MeriméeTJ, GardnerDF, ZapfJ, FroeschFR. Effectof glycemiccontrol on
serum insulin-like growth factors in diabetesmellitus. Diabetes 33: 790-793
<1984).
- Miel! JP, Taylor AM, Zini M, MaheshwariHG, RossRJM and Valcavi R.
241

Bibliografía
Effects of hypothyroidismand hyperthyroidismon insulin-like growth factors
(IGFs) and growth honnone-and IGF-binding proteins. J. Clin. Endocrino!.
Metab. 76: 950-955 (1993).
- Mielí JP, Zini M, Quin JD, JonesJ, Portioli 1 andValcavi R. Reversible
effectsof cessationand recommencementof thyroxinetreatmenton isulin-like
growth factors (IGFs) and IGF-binding proteins in patients with total
thyroidectomy.J. Clin. Endocrino!. Metab. 79: 1507-1512(1994).
- Miller LL, Schalch 5 and Draznin B. Role of the liver in regulating
somatomedinactivity: effects of streptozotocindiabetesand starvationon the
synthesisand releaseof insulin-like growth factor and its carrier proteinsby
isolatedperfusedrat liver. Endocrinology108: 1265-1271(1981).
- Moghetti P, CastelloR, Tosi F, Zentí MG, MagnaniC, Bolner A, Perobelli
L andMuggeoM. Glucosecounterregulatoryresponseto acute hypoglycemia
in hyperthyroidhumansubjects.J. Clin. Endocrino!.Metab. 78: 169-173 <1994).
- Móller C, ArnerP, Sonnenfeld’[ andNorstedtG. Quantitativecomparisonof
insulin-like growth factor mRNA levels in humanand rat tissuesanalysedby a
solutionhybridizationassay.1 Mo!. Endocrino!. 7: 213-222<¡991).
- MontesA, HervásF andJohn 1. Effectsof thyroidectomyand thyroxineon
plasmagrowth and insulin leveles in rats. Horm. Res. 8: 148-158<19~~)-
- Morley J. Neuroendocrinecontrolof thyrotropinsecretion.Endocr. Rey. 2: 396-
436 <1981).
- MorovatA andDauncetMJ. Effectsof thyroid stautson IGF-I, Gil and insulin
are modified by food intake. Eur. J. Endocrino!. 138: 95-103 (1998).
- Morreale de Escobar G, Ruiz-Marcos A, Escobar del Rey F. Thyroid
hormone and the developing brain. En: Dussault JI!, Walker P (eds)
242

Bibliografía
Congenital hypotlzyroidism, New York : Marce! Decker, 85-126 <1983).
- Milíler MJ, Acheson KJ, Jequier E and Burger AG. Effect of thyroid
hormoneon oxidative and nonoxidativeglucosemetabolismin humans.Am. J.
Physiol. 255: E146-E152 (1988).
- Muñoz MT, Barrios Y, Pozo J and Argente J. Insulin-like growth factor 1, its
binding proteins 1 and 3, and growth hormone-bindingprotein in children and
adolescentswith insulin-dependentdiabetes mellitus: clinical implications.
Pediatr Res. 39: 992-998 (1996).
- Nanto-Salonen K, Glasscock GF and Rosenfeld RG. The effects of thyroid
hormoneon insulin-like growth factor (IGF) and IGF-binding protein (IGFBP)
expressionin the neonatal rat: prolonged high expressionof IGFBP-2 in
methimazole-inducedcongenitalhypothyroidism.Endocrinology129:2563-2570
<1991).
- Nanto-Salonen K, Muller HL, Hoffman AR, Vn TH and Rosenfeld RG.
Mechanismof thyroid hormoneactionon theinsulin-like growth factor system:
alí thyroid hormoneeffectsare not growth hormonemediated.Endocrino!ogy
132: 781-788<1993).
- Nanto-SalonenK and RosenfeidRG. Insulin-like growth factor binding protein
expressionin the hypothyroidrat is agedependent.Endocrinology131: 1489-
1496 <1992).
- Napoli R, Hirshman MF and Horton ES. Mechanism and time course of
impairedskeletalmuscleglucosetransportactivity in streptozotocindiabeticrats.
J. Clin. Invest. 96: 42 7-437 (1995).
- Nikodem VM, Petty KJ, Mitsuhashi T, DesvergneB. Structureandmechanism
of action of thyroid hormonereceptors.En: Greer MA <ed) The thyroid gland,
New York: RayenPress, 307-321 <1990).
243

Bibliografía
- Obregón MJ, Pascual A, Morreale de Escobar G, Escobar del Rey F.
Pituitary and plasma thyrotropin, thyroxine, and triiodothyronine after
hyperthyroidism.Endocrino!ogy 104: 1467-1473 <1979).
- Okajima F and Vi M. Adrenergic modulation of insulin secretion in vivo
dependenton thyroid states.Am. J. Physio!. 234: E106-EJIJ (1978).
- O’Meara NM, BlackmanJD, Sturis 41 and PolonskyKS. Alterations in the
kinetics of C-peptide and insulin secretion in hyperthyroidism. J. Clin.
Endocrinol. Metab. 76. 79-84 <1993).
- Ooi GT, Orlowsk¡ CC, Brown AL, Becker RE, Unterman TG and Rechíer
MM. Different tissue distribution and hormonal regulation of messengerRIJAs
encodingrat insulin-likegrowthfactorbindingprotein-1and -2. Mo!. Endocrinol.
4: 321-328(1990).
- Ooi GT, TsengLYH, RechíerMM. Post-transcriptionalregulationof insulin-
like growth factor binding protein-2 mRNA in diabetic rat liver. Riochern.
Biophys. Res. Commun. 189: 1031-1 03 7 (1992a).
- Ooi GT, íseng LYH, Iran MQ and Rechíer MM. Insulin rapidly decreases
IGFBP-1 gene transcription in streptozotocin-diabetic rats. Mo!. Endocrino!. 6:
2219-2228(1992b).
- Orlowski CC, Brown AL, Ooi GT, Yang IWH, Tseng LYH and Rechíer
MM. Tissue, developmentaland metabolic regulation of messengerribonucleic
acidencodingarat insulin-like growthfactorbindingprotein.Endocrino!ogy126:
644-652 (1990).
- Ortiz-Caro J, Obregón MJ, Pascual A and John 1. DecreasedT4 to T3
conversionin tissuesof streptozotocin-diabeticrats.Acta Endocrino1. 106: 86-91
<1984).
244

Bibliografía
- Palmero 5, Prati M, Barreca A, Minuto F, Giordano G and FugassaE.
Thyroid hormonestimulatestheproductionof insulin-likegrowthfactor-J(IGF-I)
by inmaturerat Sertoli celís.Mo!. Ce!. Endocrinol. 68: 61-65 (1990).
- Pao C-I, Farmer PK, Begovic 5, Goldstein 5, Wu G and Philips LS.
Expressionof hepaticinsulin-like growthfactor-I and insulin-like growth factor
binding protein-1 gene is transcriptionally regulatedin streptozotocin-diabetic
rats.Mo!. Endocrino!. 6: 969-977(1992).
- Pastor RM and John1. Peripheralmetabolismand secretionrateof thyrotropin
in streptozotocin-diabeticrats. Endocrinology 112: 1454-1459 (1983).
- Philipps A, Drakenberg K, PerssonB, Sjbgren B, EklÉif AC, Hall K and
Sara V. The effectsof altered nutritional status upon insulin-like growth factors
and their binding proteinsin neonatalrats. Pediatr. Res. 26: 128-134 <1989).
- Portertield JP and Hendrich CE. The role of thyroid hormones in prenatal and
neonatalneurologicaldevelopment-currentperspectives.Endocr. Rey. 14:94-106
(1993).
- Postic C, Burcelin R, Rencurel F, Pegorier J-P, Loizeau M, Girard J aud
Leturque A. Evidence for a transient inhibitory effect of insulin on GLUT2
expressionin the liver: studiesin vivo and in viti-o. Riochem.J. 293: 119-124
(1993a).
- PostieC, Leturque A, Printz RL, Maulard P, Loizeau M, Granner DK and
G¡rard J. Developmentand regulationof glucosetransporterand hexokinase
expressionin rat. Am. J. Physiol. 266: E548-E559(1994).
- Postic C, Leturque A, Rencurel F, Printz RL, Forest C, Granner DK aud
Girard J. Ihe effectsof hyperinsulinemiaand hyperglycemiaon GLUT4 aud
hexokinaseII mRNA and protein in rat skeletal muscle and adipose tissue.
Diabetes42: 922-929(1993b).
245

Bibliografía
- Quin JD, Fisher BM, MacCuish AC, BeastalíGH, SmaIl M, Holly JMP and
Cotterill AM. Insulin-like growth factor binding protein-1 responseto acute
insulin inducedhypoglycaemiain type 1 diabetes.Clin. Endocrino!. 41: 225-229
<1994).
- Rajaram 5, Baylink DJ and Mohan 5. Insulin-like growth factor-binding
proteinsin serumand otherbiological fluids: regulationand functions.Endocr.
Rey. 18: 801-831 <1997).
- ReclerMM. Insulin-like growthfactorbinding-proteins.VitaminsandHormone
47- 1-114 <1993).
- Rechíer MA and Clemmons DR. Regulatory actionsof insulin-like growth
factor bindingproteins. TrendsEndocrino!.Metal’. 9:176-183 (1998).
- RencurelF, WaeberG, AntoineB, RocchiccioliF, MaulardP, Girard Jand
Leturque A. Requirementof glucose metabolism for regulation of glucose
transportertype2 (GLUT2) geneexpressionin liver. Biochem. J. 314: 903-909
<1996).
- Rivero F, Goya L, Aláez C and Pascual-LeoneAM. Effects of undernutrition
anddiabeteson semmliver mRNA expressionofIGFs andtheirbindingproteins
during rat development.1 Endocr ¡45: 427-440 <1995).
- Rivero F, Goya L and Pascual-LeoneAM. Comparison of extraction
methodsfor insulin-like growth factor-bindingproteinsprior to measurementof
insulin-like growth factor-I in undernourishedneonataland adult rat serum. J.
Endocrino!. 140: 257-263 <1994).
- Roberts CT Jr, Brown AL, Graham DE, Seelig5, Berry5, GahbayKH anO
Rechíer MM. Growth hormone regulates the abundanceof insulin-like growth
factor-IRNA in adult rat liver. .1? Rio!. Chem. 261: 10025-10028 (1986).
246

Bibliografía
- Robinson ICAF, Clark RO and Carlsson LMS. Insulin, IGF-I and growth in
diabetierats. Nature326: 549 (1987).
- RodgersBD, Bautista RM and Nicol! CS. Regulation of insulin-like growth
factor binding proteinsin rat with insulin-dependentdiabetesmellitus. P. S. E.
R. M. 210: 234-241 (1995).
- Rodríguez M, Rodríguez F and John 1. Effect of restricted feeding, fasting
and diabeteson the retationshipbetweenthyroid hormonereceptoroccupancy,
growth hormone induction, and inhibition of thyrotropin release in
thyroidectomizedrats. Endocrinology 131: 1612-1 618 <1992).
- Rodríguez-ArnaoJ, Mielí 41, ThomasM, McGregor AM, RosRJM. Increased
hepatic IGFBP-2 geneexpressionin hypothyroid rats. Endocrinology137<Supp!)
P80 <1993a).
- Rodríguez-ArnaoJ, Miell J, Thomas M, MeGregor AM, Ros RJM. Changes
in hepaticinsulin-like growth factor-bindingproteins-1,-2 and -3 mRNA levels
in rats with alteredthyroid status.J. Endocrinol. 140: 251-255 <1994).
- Rodríguez-Arnao J, Miell J and Ross RJM. Influence of thyroid hormoneson
the GI-1-IGF-I axis. Trends Endocrino!. Metab. 4:169-173(1993b)
- Rondeel JJM, de Oree!’ WJ, Klootwijk W, Visser 141. Effects of
hypothyroidismon hypothatamicreleaseofthyrotropin-releasinghormonein rats.
Endocrinology 130: 651-656 (1992).
- Rondeel JJM, de Oree!’ WJ, van der Schoot P, Karels B, Klootwijk W,
VisserTi. Effect of thyroid status and paraventricular area lesions on the release
of thyrotropin-relesinghormone and catecholaminesinto hipophysealportal
blood. Endocrinology123: 523-527 <1988).
- Rondeel JJM, de Greef WJ and Visser TJ. Effect of tliyroid status on release
247

Bibliografla
of hypothalamicthyrotropin-releasinghormone.Horm. Metab. Res. 23 <Suppl):
1-4 <1990).
- Rosenfeid RO, Lamson O, Pham H, Oh Y, Conover C, de Leon DD,
Donovan SM, Ocrant 1 and Giudice L. Insulinlike growth factor-binding
proteins.RecentProg. Horm. Res. 46: 99-163 <1990).
- Rotwein P, Bicheil DP aud Kikuchi K. Multifactorial regulation of IGF-I gene
expression.Mal. Reprod. Dcv. 35: 358-364 <1993).
- Rubio E. Memoria de Licenciatura. Acción insulínica en ratas adultas
realimentadastras una etapade subnutrición. E. Farmacia. U. Complutense.
Madrid (1997)
- Rubio E, AgoteM, Escrivá F and Pascual-LeoneAM. GLUT-4 and GLU’[-1
contentand in vivo insulin responsein musclefrom undernourishedand refed
rats. Ars Pharmaceutica 37: 957-969 <1996).
- Ruiz de Olla C, Obregón MJ, Escobar del Rey F, Morreale de Escobar G.
Developmentalchangesin rat brain 5’-deiodinaseand thyroid hormonesduring
the fetal period: the effects of fetal hypothyroidism and maternal thyroid
hormone.Pediatr. Res. 24: 588-594(1988).
- SamucísMH, Wierman ME, Wang C, Ridway FC. The effect of altered
thyroid statuson pituitary hormonemessengerribonucleicacid concentrationin
the rat. Endocrinology 124: 2277-2282 <1989).
- Santalucía’[, Camps M, Castelló A, Muñoz P, Nuel A, Testar X, Palacín M
and ZorzanoA. Developmentalregulationof GLUT-1 (erythroid/HepG2) and
GLUT-4 (muscle/fat)glucosetransporterexpressionin rat heart,skeletalmuscle,
and brown adiposetissue.Endocrinology130: 837-846(1992).
- Sara VR and Rail K. Insulin-like growth factors and their binding proteins.
248

Bibliografía
Physiol. Rey. 70: 591-614 <1990).
- Schmid C, Scblapfer 1, Puto E, Waldvogel JS, Zapf 3 and Froesch ER.
Triiodothyronine (T3) stimulates insulin-like growth factor (IGF)-1 and IGF-
binding protein (IGFBP-2) production by rat osteoblasts in vitro. Acta
Endocrino!. 126: 467-473 (1992).
- Seo H, Wunderlich C, Vassart O and Refeto!’!’ 5. Growth hormone to thyroid
hormonein the neonatalrat. J. Clin. Invest. 67: 569-574 <1981).
- Shapiro LE, SamucísHM, Valle BM. Thyroid and glucocorticoids hormones
synergisticallycontrolgrowthhormonemRNA in culturedGUI celís.Proc. Natí.
Acad. Sc!. USA 75: 45-49 <1978).
- Shetty M, Kuruvilla AK, Ismail-Beigi F and Loeb JN. Stimulation of glucose
transport in Clone 9 celís by insulin and thyroid hormone: role of GLUT-1
activation.Riochim. Biophys. Acta 1314: 140-146(1996).
- Shimokawa T, Kato M, Shioduka K, Irie 3 and Ezaki O. Effect of
triiodothyronine on muscle celí differentiation and blood glucose level in
hyperglycemicKKmice. Riochem.Biophys.Res.Commun.235: 790-793<1997).
- Smith WJ, Underwood LE, Clemmons DR. Effects of caloric or protein
restriction on insulin-like growth factor-I (IGF-I) and IGF-binding proteins in
children and adults. J. Clin. Endocrino!. Metab. 80: 443-449 (1995).
- Snedecor GW and Cochran O. Statistical Methods, edn 5. Ames: Iowa State
Universizy Press.
- Spira O and Gordon A. Thyroid hormone feedback effects on thyroid-
stimulatinghormone.En:HennemannO (eds)Thyroidhormonemetabolism,New
York: Marce! Dekkermc, 535-578<1986).
249

Bibliografía
- StephensJM and Pilch PF. The metabolic regulation and vesicular transpon of
GLUT-4, themajorinsulin-responsiveglucosetransporter.Endocr. Rey. 16.- 529-
546 <1995).
- StGermain DL. Iodothyroninedeiodinases.TrendsEndocrino!.Metal’. 5:36-42
<1994)~
- Strasser-VogelB, Blum WF, PastR, KesslerU, Hoeflich A, Meiler B, Kiess
W. Insulin-like growth factor (IGF)-I and -II and IGF-binding proteins-1, -2 and
-3 in childrenand adolescentswith diabetesmellitus: correlationwith metabolic
controlandheightattainment.J. Clin. Endocrino!. Metal’. 80:1207-1213 (1995).
- Straus DS. Nutritional regulation of hormones and growth factors that control
mammaliangrowth.FASERJ. 8: 6-12 <1994).
- Straus DS, Ooi GT, Orlowski CC and Bechier MM. Expressionof the genes
for insulin-like growthfactor-I (IGF-I>, IGF-fl, andIGF-bindingproteins-1and -
2 in fetal rat under conditions of intrauterine growth retardationcausedby
maternalfasting.Endocrinology128: 518-525<1991).
- Suikkari AM, Koivisto VA, RutanenEM, Yki-J~irvinen H, Karonen S-L and
Seppiilii M. Insulin regulates the serum levelsof low molecular weight insulin-
like growth factor-bindingprotein. J. Clin. Endocrino!. Metal’. 66: 266-272
<1988).
- Tannenbaum GS, Martin JB and Coite E. Ultradian growth hormone rhythm
in the rat: effectsof feeding,hyperglycemiaand insulin inducedhypoglucemia.
Endocrinology 99.- 720-72 7 <1976).
- Taylor R, McCulloh Aj, Zeulen 5, Gray P, Clark F and Alberi KGMM.
Insulin secretion, adipocyte insulin binding and insulin sensitivity in
thyrotoxicosis.Acta Endocrinol. 109: 96-103 (1985).
250

Bibliografía
- ThissenJP, KetelslegersJM and Underwood LE. Nutritional regulation of the
IGFs. Endocr. Rey. 15: 81-101 <1994)
- Thorens B, Sarkar HK, Kaback HR and Lodish HF. Cloning and functional
expressionin bacteriaof a novel glucosetransporterpresentin liver, intestine,
kidney and j3-pancreaticislet celís. CeIl 55: 281-290 <1988).
- Timiras PS, NzekweEU. Thyroid hormones and nervoussystemdevelopment.
Biology of theneonate55: 376-385 (1989).
- Tontis DK, Bestetti GE, Reymond MJ, Lemarchand-Béraud 1 and Rossi
GL. The ageat onsetof diabetesinfluencesfunctional and structuralchangesin
thepituitary-thyroid axis of streptozotocin-diabeticmale rats. VirchowsArchiv.
B Cel!. Patho!. 59: 205-213 <1990).
- ‘[orrance CJ, Devente JE, Jones JP and Dohm OL. Effects of thyroid
hormoneon GLUT4 glucosetransportergene expressionand NIDDM in rats.
Endocrino!ogy138: 1204-1214<1997a).
- Torrance CJ, Usala SJ, PessinJE and Dohm L. Characterizationof a low
affinity thyroid hormone receptor binding site within the rat GLUT4 gene
promoter.Endocrinology 138: 1215-1223 <1 997b) -
- TsengLY-H, Ooi 01, Brown AL, StrausDS andRechíerMM. Transcription
of the insulin-like growth factor bindingprotein-2 genein increasedin neonatal
and fastedadult rat liver. Mo!. Endocrino!. 6:1195-1201 (1992).
- UntermanTG, PatelK, Kumar MahathreV, RajamohanG, OehlerDl and
Becker RE. Regulationof low molecular weight insulin-like growth factor
bindingproteinsin experimentaldiabetesmellitus.Endocrino!ogy 126:2614-2624
(1990).
- Van Buul-Offers SC, Bloemen RJ, Reijnen-Greinigt MG, van Leiden HA,
251

Bibliografía
HoogerbruggeCM and van den Brande JL. Insulin-like growth factor-I and -
II and their binding proteins during growth hormone and thyroxine therapy. J.
Endocrino!. 143: 191-198 <1994).
- Van Hardeveid C. Effects of thyroid hormone on oxygen cosumption, heat
production,and energy economy.En: HennemannG (eds) Thyroid hormone
metabolism,New York: Marce! Dekkermc, 579-608(1986).
- Varga F, Rumpler M and Klaushofer K. Thyroid hormones increaseinsulin-
like growth factor mRNA levels in the clonal osteoblasticcelí line MC3T3-E1.
FEBSLet.345:67-70<1994).
- Venditti P, Balestrieri M, di Meo 5 and de Leo’[. Effect of thyroid state on
lipid peroxidation,antioxidantdefences,and susceptibilityto oxidative stressin
rat tissues.1. Endocrino!. 155: 151-15 7 (1997).
- Vijayan E aud McCann SM. Effects of substanceP and neurotensin on growth
hormone and thyrotropin release in vivo and in viti-o. Lije Sci. 26: 321-327
(1980).
- Villafuerte B, Zhang W-N and Phillips LS. Insulin and insulin-like growth
factor-I regulatehepaticinsulin-like growthfactor bindingprotein-3by different
mechanism.Mo!. Endocrino!. 10: 622-630 <1996).
- Villafuerte BC, Zhao W, Herington AC, Saffery R and Phillips 5.
Identificationof an insulin-responsiveelementin the rat insulin-like growth Ñr
binding protein-3gene. 1? Rio!. Chan. 272: 5024-5030 <1997).
- Yaffe BM and SamuelsHH. Hormonal regulation of the Gil gene. J. Rio!.
Chem. 259: 6284-6291 (1984).
- Yeoh SI and Baxter RC. Metabolie regulation of the growth hormone
independentinsulin-like growth factor binding protein in humanplasma.Acta
252

Bibliografía
Encocrinol. 119: 465-473 <1988).
- Walker P and DussaultJH. Hypothalamic somatostatinand pituitary and serum
growth hormoneconcentrationsduring postnataldevelopmentin rats exposed
chronically to propilthiouracil or a 10w iodine diet. J. Dey. Physiol. 2: 111-117
<1980).
- Walker P, DussualtJH, Alvarado-Urbina G and Dupont A. The development
of the hypothalamo-pituitaryaxis in the neonatalrat hypothalamicsomatostatin
andpituitary and serumGH concentrations.Endorinology 101: 782-787 <1977).
- Weeke J and Orskov II. Ultrasensitive radioimmunoassay for direct
determinationof free triiodothyronineconcentrationin serum. Scand. J. Clin.
Lal’. Invest. 35: 237-244(1975).
- Weinste¡n SP, O’Boyle E, Fisher M and Haber RS. Regulation of GLUT2
glucose transporter expressionin liver by thyroid hormone: evidence for
hormonalregulationof thehepaticglucosetransportsystem.Endocrinology 135:
649-654 (1994).
- Weinstein SP and Haber RS. Diflerential regulation of glucose transporter
isoformby thyroid hormonein rat heart.Biochem.Biophys.Acta1136: 302-308
<1992).
- Weinstein SP, Watts J, GravesPN and Haber RS. Stimulation of glucose
transportby thyroid hormonein ARLíS celís: increasedabundanceof glucose
transporterproteinand messengerribonucleicacid. Endocrinology 126: 1421-
1429 <1990).
- Weinste¡n SP, Watts J and Haber RS. Thyroid hormoneincreasesmuscle/fat
glucosetransportergeneexpressionin rat skeletalmuscle.Endocrino!ogy129:
455-464(1991).
253

Bibliografía
Westermark K, Alin J, Skottner A aud Karlsson A. Growth factors and the
thyroid: effectsof treatmentfor hyper- and hypothyroidismon serumIGF-I and
urinary epidermalgrowthfactor concentrations.Acta Endocrino!. 118: 415-421
<1988).
Wolf M, Ingbar SH and Moses AC. Thyroid hormone and growth hormone
interactto regulateinsulin-like growth factor-I messengerribonucleicacid and
circulating levels in the rat. Endocrinology 125: 2905-2914 <1989).
254