
INFORME ANUAL A INRENA - georgeolah.weebly.com · INTRODUCCIÓN El presente es el informe anual del...
58
“ECOLOGÍA REPRODUCTIVA Y USO DE COLPAS DE GUACAMAYOS EN MADRE DE DIOS” INFORME ANUAL A INRENA Autorización N o 044 S/C-2007-INRENA-IANP PhD. Donald J. Brightsmith Director de Investigaciones Research Assistant Professor - Texas A&M University [email protected] Blga. Gabriela Vigo Trauco, Jefe de Campo Noviembre 2008 Universidad Nacional Agraria La Molina [email protected] Zoólogo George Oláh, Jefe de Campo Diciembre 2008 a Enero 2009 Szent István University- Hungría [email protected] Blgo. Jesús Zumarán Rivera, Jefe de Campo Febrero a Marzo 2009 Universidad Nacional Agraria La Molina [email protected] M.V.Z Lizzie Ortiz Cam, Veterinaria de Campo Noviembre a Febrero 2009, Coordinadora 2009 Universidad Peruana Cayetano Heredia [email protected] LIMA, ABRIL 2009
Transcript of INFORME ANUAL A INRENA - georgeolah.weebly.com · INTRODUCCIÓN El presente es el informe anual del...

“ E C O L O G Í A R E P R O D U C T I V A Y U S O D E C O L P A S D E G U A C A M A Y O S E N
M A D R E D E D I O S ”
I N F O R ME A N UA L A I N REN A
Autorización No 044 S/C-2007-INRENA-IANP
PhD. Donald J. Brightsmith
Director de Investigaciones
Research Assistant Professor - Texas A&M University [email protected]
Blga. Gabriela Vigo Trauco, Jefe de Campo
Noviembre 2008
Universidad Nacional Agraria La Molina [email protected]
Zoólogo George Oláh, Jefe de Campo
Diciembre 2008 a Enero 2009
Szent István University- Hungría [email protected]
Blgo. Jesús Zumarán Rivera, Jefe de Campo
Febrero a Marzo 2009
Universidad Nacional Agraria La Molina [email protected]
M.V.Z Lizzie Ortiz Cam, Veterinaria de Campo
Noviembre a Febrero 2009, Coordinadora 2009
Universidad Peruana Cayetano Heredia [email protected]
L I M A , A B R I L 2 0 0 9

CONTENIDO
INTRODUCCIÓN 3
MARCO TEÓRICO 5
Área de estudio 6
Capítulo I: Éxito reproductivo 7
Capítulo II: Metodología de captura 12
Capítulo III: Guacamayos reintroducidos en TRC ”Los chicos” 18
Capítulo IV: Trabajo con profesionales y voluntarios 30
AGRADECIMIENTOS 32
BIBLIOGRAFÍA 33
ANEXOS 39

INTRODUCCIÓN
El presente es el informe anual del Proyecto Guacamayo, correspondiente a la autorización Nº
044 S/C-2007-INRENA-IANP, otorgada por el Instituto Nacional de Recursos Naturales con el fin
de realizar el trabajo de investigación “Ecología Reproductiva y uso de Colpas de Guacamayos en
Madre de Dios”. El trabajo de campo se lleva a cabo al interior de la Reserva Nacional
Tambopata, teniendo como base de operaciones al albergue turístico y centro de investigación
“Tambopata Research Center” (TRC).
En Noviembre del 2008 el Proyecto Guacamayo cumplió su noveno año de estudios de
psitácidos en TRC, bajo la dirección del Dr. Donald Brightsmith. Durante estos años el proyecto
ha logrado colectar datos sobre éxito reproductivo, crecimiento de pichones, uso de colpas,
concurrencia de psitácidos a la colpa, composición de grupos familiares, dieta, forrajeo,
disponibilidad de alimento, sobrevivencia de guacamayos criados a mano, abundancia de
psitácidos en el bosque, telemetría satelital, comportamiento de Ara macao en el nido, entre
otros estudios.
Como es conocido en el medio ornitológico, los psitácidos corresponden un grupo de aves que
debido a sus características intrínsecas (como la falta de comportamiento territorial,
vocalizaciones, movilización de largas distancias y su naturaleza de permanecer en el dosel) son
difíciles de estudiar, y son escasas las fuentes de información sobre estudios detallados de estas
especies.
Dentro de este contexto es que el Proyecto Guacamayo, a través de un trabajo de larga
trayectoria, busca continuar realizando estudios que brinden información que nos ayude a
entender aspectos ecológicos y de historia de vida de psitácidos; ya que consideramos a este
taxón de gran importancia principalmente debido a que incluye aves muy amenazadas y que son
consideradas un valioso recurso turístico.
Al igual que en años anteriores, a través de este documento se presentan los logros del Proyecto
Guacamayo obtenidos dentro del marco de la autorización 044 S/C-2007-INRENA-IANP.
Empezamos el reporte con los antecedentes. Brindando una visión general sobre el
conocimiento científico que se tiene sobre los psitácidos.

En el Capítulo I “Éxito reproductivo y descripción de nidos”, mostramos los resultados del
monitoreo continuo de nidos de Ara macao durante el año 2008 y 2009 (temporada
reproductiva 2009)
En el siguiente Capítulo II “Metodología de captura”, se describen las técnicas de captura que se
utilizó durante la temporada reproductiva 2009. En este capítulo se detalla el Sistema de
perchas y el uso de redes, así como los protocolos de cada método.
Dentro del Capítulo III “Guacamayos reintroducidos en TRC: Los Chicos” describe la situación
actual en la que se encuentran los guacamayos criados en el albergue TRC por seres humanos y
liberados entre los años 1992 y 1995. Su supervivencia hasta el momento, los avistamientos que
se han podido tener y el éxito reproductivo de los mismos.
Luego, el Capítulo IV “Trabajo con profesionales y voluntarios”, resume el esfuerzo humano de
asistentes “guacamayeros” que han ayudado a sacar adelante cada uno de los estudios
específicos que engloba el Proyecto Guacamayo.
Finalmente, se presentan los agradecimientos, la literatura citada y los anexos correspondientes
a cada capítulo desarrollado en el documento.

MARCO TEORICO
Los Psitácidos son notoriamente difíciles de estudiar en vida silvestre (Beissinger and Snyder
1992). Debido a su falta de comportamiento territorial, vocalizaciones, movilización de largas
distancias y su naturaleza de permanecer en el dosel, muchos estudios ornitológicos generales
no incluyen el muestreo adecuado de psitácidos (Casagrande and Beissinger 1997, Marsden
1999, Masello et al. 2006). Como resultado, estudios detallados de la comunidad de psitácidos
son raros (Roth 1984, Marsden and Fielding 1999, Marsden et al. 2000). Sin embargo,
información acerca de su historia natural es importante para el entendimiento y conservación
de ésta familia tan amenazada (Bennett and Owens 1997, Collar 1997, Snyder et al. 2000,
Masello and Quillfeldt 2002).
Las tierras bajas de la Amazonía oriental poseen algunas de las comunidades aviares más
diversas en el mundo (Gentry 1988), albergando a más de 20 especies de psitácidos
(guacamayos, loros y periquitos), (Terborgh et al. 1984, Terborgh et al. 1990, Montambault
2002, Brightsmith 2004). Las densidades de psitácidos en esta región pueden ser alcanzar los
cientos miles de loros congregados diariamente en las riveras de los ríos a comer arcilla
(Emmons 1984, Nycander et al. 1995, Burger and Gochfeld 2003, Brightsmith 2004).
Aparentemente, la arcilla consumida en éstas colpas representa una importante fuente de sodio
y protege a las aves de las toxinas presentes en la dieta (Emmons and Stark 1979, Gilardi et al.
1999, Brightsmith 2004, Brightsmith and Aramburú 2004).
Los psitácidos se alimentan predominantemente de semillas, frutos maduros e inmaduros, y
flores, suplementados ocasionalmente con cortezas y otros insumos (Forshaw 1989, Renton
2006). A diferencia de muchas otras aves, los psitácidos del Nuevo Mundo parecen no ser
capaces de modificar su dieta a predominantemente insectívora, razón por la que están
íntimamente ligados a los patrones de floración y producción de frutos.
Mientras que las fluctuaciones de clima en los trópicos es notablemente menor que en zonas
templadas, se conoce que la floración y producción de frutos varía estacionalmente en cada
lugar en el que han sido estudiadas (Frankie et al. 1974, Croat 1975, Lugo and Frangi 1993, van
Schaik et al. 1993, Zhang and Wang 1995, Adler and Kielpinski 2000). Pocas especies vegetales
florecen y producen frutos a lo largo de todo el año, significando para los psitácidos que
diferentes fuentes de alimento están disponibles en diferentes épocas del año. En adición, la
abundancia total de alimento fluctúa en respuesta a los patrones estacionales de precipitación
(van Schaik et al. 1993). Estas variaciones anuales en disponibilidad de alimento tienen
implicaciones importantes en los ciclos de vida anuales de psitácidos. Por ejemplo, la carencia
de alimento puede conllevar a algunos o todos los miembros de una especie a moverse a otras
áreas siguiendo los recursos alimenticios disponibles (Powell et al. 1999, Renton 2001). La
estación de anidamiento y el éxito reproductivo pueden también estar ligadas a los patrones de
floración y producción de frutos (Sanz and Rodríquez-Ferraro 2006).

ÁREA DE ESTUDIO
Todos los estudios fueron llevados a cabo en el albergue y centro de investigación Tambopata
Research Center (TRC) 13º 07′ S, 69º 36′ W, en la frontera entre la Reserva Nacional Tambopata
(275,000 ha) y el Parque Nacional Bahuaja-Sonene (537,000 ha) en el departamento de Madre
de Dios, al sureste del Perú.
El sitio se encuentra en el límite entre bosque húmedo tropical y bosque húmedo subtropical, a
250 m.s.n.m. y con una precipitación anual de 3,200 mm (Tosi 1960, Brightsmith 2004). La
estación seca se extiende de Abril a Octubre, con una precipitación mensual promedio de 90 a
250 mm (Brightsmith 2004).
El centro de investigación está localizado en un pequeño claro (<1 ha) rodeado por bosque
inundable maduro, bosque inundable sucesional, aguajal y restinga (Foster et al. 1994). Un largo
parche de bambú (Guadua sarcocarpa: Poaceae) cubre el área inmediatamente adyacente a la
colpa, el mismo que floreció y se secó entre los años 2001 y 2002 (Foster et al. 1994, Griscom
and Ashton 2003).
El área presenta poblaciones de guacamayos grandes (Ara ararauna, A. chloroptera y A. macao)
y predadores grandes (Harpia harpyja, Morphnus guianensis, Spizatus tyrannus, Spizatus
ornatus y Spizastur melanoleuca). La densidad de psitácidos en esta región puede alcanzar los
cientos, miles de loros congregados diariamente en las riveras de los ríos a comer arcilla
(Emmons 1984; Nycander, Blanco et al. 1995; Burger and Gochfeld 2003; Brightsmith 2004).
La colpa es un banco de arcilla de 500m de longitud y 25 a 30m de alto a lo largo del margen
occidental del Alto Tambopata aproximadamente a 1 km del centro de investigación. El
barranco está formado por la erosión causada por el Río Tambopata en la sedimentos aluviales
emergidos en la Era Terciaria (Räsänen and Salo 1990, Foster et al. 1994, Räsänen and Linna
1995). Los suelos de la colpa son ricos en arcilla con alta capacidad de intercambio catiónico y
altos niveles de sodio (Gilardi et al. 1999), DJB datos no publicados).

CAPÍTULO I:
ÉXITO REPRODUCTIVO
METODOLOGÍA
Los nidos son revisados periódicamente desde que los padres dan señales de uso del nido
(inicios del mes de Noviembre) hasta que vuelan los últimos pichones (fines de Abril). El
monitoreo consiste en escalar hasta los nidos mediante técnicas de ascensión en cuerda para
poder medir y pesar a los pichones en cada etapa de su desarrollo (Nycander et al. 1995).
Los nidos fueron inspeccionados por medio de técnicas de escalada de árboles utilizando sogas
(Munn 1991), con una frecuencia aproximadamente inter-diaria durante los 30 primeros días de
edad del individuo menor de la nidada y, posteriormente, dejando 2 días hasta que el nido se
encontró vació. Cada equipo de inspección de nidos fue conformado por dos personas: el
escalador que envía a los pichones hacia abajo, en donde una segunda persona lo recibe y toma
las medidas respectivas (Nycander, Blanco et al. 1995). Las medidas fueron colectadas por un
total de 24 equipos (3 equipos por estación reproductiva) conformados por asistentes
voluntarios del Proyecto Guacamayo de Tambopata durante las temporadas reproductivas de la
especie comprendidas entre los años 2000 y 2008.
Investigadores y guías trabajando en el área han registrado de manera casual nidos de otras
especies de psitácidos desde 1990. Sin embargo, sólo se dispone de cronología anual detallada
del anidamiento de A. macao y A. chloropterus, que muestran poca variación anual y gran
sincronización. Para la mayoría de las otras especies, hemos encontrado sólo unos pocos nidos
(Brightsmith 2005).
Lo que se presenta a continuación son los resultados obtenidos para la última temporada
reproductiva 2009.

RESULTADOS Y DISCUSIÓN
El Proyecto Guacamayo de Tambopata basa sus investigaciones con 3 tipos de nidos: (1) Nidos
artificiales de madera, (2) Nidos artificiales de PVC y (3) Nidos naturales. En total existen dentro
del área de estudio: 8 nidos de madera, 12 de PVC y 24 nidos naturales. Durante la temporada
reproductiva 2009 hubo 13 nidos activos de los guacamayos con 1 o 2 pichones. Respecto a los 3
tipos de nidos se tiene la siguiente distribución, indicando los nombres de los nidos y sus
números de pichones entre paréntesis (ver Gráfico 1.1):
- Nidos artificiales de madera: Hugo (1), Franz (1).
- Nidos artificiales de PVC: Invisible (1), PVC (1), Pukakuro (2), Odio (1), Huayranga (2).
- Nidos naturales: Vaginito (1), Intocable (2), Horizontal (1), Max (2), Ayahuasca alto (2), Arroz
(1).
Los guacamayos tuvieron 18 pichones en total hasta Febrero 2009. Había 2 pichones en 5 nidos
y el resto, 8 nidos, cada uno tuvo 1 pichón (ver Gráfico 1.2).
Distribución de nidos activos
Madera; 2; 15%
PVC; 5; 38%
Natural; 6; 47%
Madera PVC Natural
Gráfico 1.1: Distribución de nidos alrededor del área de
investigación del Tambopata Research Center (TRC)

a. Especies
La temporada 2009 dio como resultado 2 nidos de Guacamayo Cabezón (Ara chloroptera) y 11
nidos de Guacamayo Escarlata (Ara macao). Hasta Febrero 2009 los Guacamayos Escarlatas
tuvieron 14 pichones y 1 huevo, utilizando 12 nidos: 2 de madera, 4 de PVC y 6 naturales. En el
nido Pukakuro (nido PVC en trocha C) se reprodujeron 2 guacamayos criados a mano y tuvieron
2 pichones. En el nido Franz (nido madera en trocha C1) el macho de la pareja fue un guacamayo
criado a mano. En el nido Arroz (un nido natural en trocha B que descubrimos en esta
temporada) encontramos que uno de los padres del pichón es hijo de un guacamayo estudiado
en otra temporada, esto lo supimos hallando un anillo en una de sus miembros inferiores.
Esta temporada tuvimos gratas sorpresas y datos saltantes que nos brindan novedosa
información y a su vez, nos estimulan a continuar investigando: por primera vez se pudo
estudiar a dos parejas silvestres (y no criados a mano) de Guacamayo Cabezón empollando en
un nido artificial de PVC (con 1 pichón) y en un nido natural (con 2 pichones): “PVC” e
“Intocable” respectivamente.
Además, encontramos un árbol que muy posiblemente tenía un nido natural de Guacamayo de
frente castaña (Ara severa) en la trocha B, cerca del “Palm Swamp”.
También, por primera vez descubrimos un nido natural de Chirricles (Pionites leucogaster) en
trocha C2, al cual se pudo ascender con el uso de cuerdas y se pudo obtener datos fotográficos,
demostrando así que aquel nido contaba con cuatro pichones. Siendo este, el primer registro
que se obtiene de un nido de Chirricles.
0
1
2
3
4
5
6
7
8
nidos con 2 pichones nidos con 1 pichon
Numero de los pichones per nidos
Gráfico 1.2: Número de pichones por nido durante la
Temporada Reproductiva 2009 en TRC

b. Cámaras en nidos
Se instaló nueve cámaras en: dos nidos de madera, seis de PVC y por primera vez en un nido
natural en la isla de “Fish Pond”. Siete de estos nidos estuvieron activos durante toda la
temporada y se tuvo la oportunidad de observar a los guacamayos durante la incubación sin la
necesidad de perturbarlos, obteniendo con ello datos muy importantes del comportamiento de
los padres, el cuidado paternal de ellos con las crías al eclosionar y la interacción entre padres e
hijos y entre los hermanos. El principal objetivo de este subproyecto fue el de investigar el
porqué nunca sobreviven los terceros pichones y sólo raramente los segundos. Al realizar la
comparación con otros lugares y otros países, en TRC generalmente vuelan (abandonan el nido)
menos juveniles de guacamayos.
c. Distribución topográfica de los nidos en TRC
Catalogando los nidos según su distribución topográfica, conseguimos las siguientes áreas (Ver
Gráfico 1.3):
- Trocha A, “Bowl”: suroeste del albergue, detrás de la colpa; hay 1 nido artificial de madera y 7
nidos naturales; había 1 nido natural activo.
- Trocha B: oeste de TRC; hay 2 nidos de madera, 6 de PVC y 5 naturales; había 4 nidos activos (2
PVC, 2 naturales).
- Trocha C, C1, Albergue: estos nidos están la más cerca del albergue y entra el río y TRC; hay 3
nidos de madera, 5 de PVC y 7 naturales; había 8 nidos activos (2 madera, 3 PVC, 3 naturales).
- Isla del “Fishpond”: la isla en frente de la colpa; hay 1 nido de madera y 3 nidos naturales;
había 1 nido natural activo.
- Trocha C2, TRC: la área al norte y este del albergue; hay 2 nidos naturales y 1 de PVC; no había
nido activo.
En conclusión, los más nidos activos se encuentran alrededor del albergue y al oeste en trocha
B.

1
7
4
9
8
7
1
3
0
3
0
2
4
6
8
10
12
14
16
nu
me
ro d
e n
ido
s
Distribución topográfica de los nidos
Nidos pasivos 7 9 7 3 3
Nidos activos 1 4 8 1 0
Trocha A,
BowlTrocha B
Trocha C,
C1,
Albergue
Isla de
Fishpond
Trocha
C2, TRC
d. Leyenda reproductivo de nidos
Habían nidos que estaban activos (los guacamayos pusieron huevos) desde Noviembre o
Diciembre, pero luego fracasaron. En 7 casos encontramos huevos rotos en los nidos y en 3
casos huevos infértiles. En estos casos los guacamayos ya no regresaron a aquellos nidos,
excepto en el nido Franz (un nido de madera en la trocha C1) a donde posiblemente otra pareja
regreso y se reprodujeron.
Gráfico 1.3: Distribución Topográfica de nidos alrededor del área de
investigación del Tambopata Research Center (TRC)

CAPÍTULO II:
METODOLOGIA DE CAPTURA
SISTEMA DE PERCHAS
El Sistema de Perchas ha sido diseñado para ser utilizado para cualquier tipo de Psitácidos que
este presente en el momento de la Actividad de la Colpa. Sin embargo, no existe razón alguna
para pensar que este método es exclusivo para ellos, pues existe la posibilidad de que otras aves
puedan percharse en dicho sistema.
Además, se tuvo en consideración el tamaño del nylon a usar para los nudos; se utilizo nylon de
12 libras apuntando a capturar aves pequeñas, mientras que para las más grandes se uso nylon
de 20 libras (ya que es más difícil de romper). Esto se trabajo en base a horarios, considerando
que los Guacamayos grandes bajan, en mayor porcentaje, en un horario distinto a los
Guacamayos pequeños, loros y periquitos.
A. Metodología:
a) Preparación del Sistema de Perchas
Dentro del sistema de Perchas que se prepara para hacer las capturas, se hace uso de 3
partes: una fija, una semifija y una móvil.
La parte fija del sistema se denominará Tronco, esta es la que esta fijada al suelo y queda en
forma perenne (a menos que haya la necesidad de cambio por razones de antigüedad y
seguridad). Se usa un Tronco de aproximadamente unos 8 m de alto, este es “sembrado” frente
al Escondite (o Blind) que se usa para el trabajo. Con una excavadora manual se realiza un hueco
de 2 metros de profundidad donde se coloca el tronco. Posteriormente, este es rellenado con
piedras, arcilla y tierra; todo esto, para hacerlo más compacto y de mayor estabilidad. En la
parte superior y con la ayuda de clavos, se fija una base de madera con 2 argollas detrás de la
base; además, una argolla en el extremo superior del Tronco.
La parte móvil se denominara Rama, esta tiene un soporte que está fijado a la base colocada en
el Tronco. Dicho soporte está formado por dos pedazos de madera unidos por una bisagra, de
manera que la Rama pueda bajar sin moverse el soporte, evitando así su caída. La elevación de
la Rama se da mediante el uso de drizas; dos tiras de drizas son usadas para elevar el soporte y
la Rama hasta la parte superior del Tronco. Estas pasan por las argollas colocadas en el Tronco
de manera que se evite enredarlas. Además, en el extremo superior de cada Rama se perforan
dos huecos de manera que puedan ser colocadas las Perchas usando tornillos.

De la misma manera, una tercera driza es amarrada a la mitad de la Rama, esta pasa por la
argolla colocada en el extremo superior del Tronco y llega hasta el nivel del suelo, a través de las
cuales el equipo de captura puede levantarla y liberarla a voluntad.
La parte semifija se denominara Percha, esta se considera semifija, porque pueden ser ajustadas
o desajustadas de la Rama según se disponga, así como colocadas en diferente orden cada vez.
Se considera dos tipos de Perchas: las Perchas de Control y las Perchas con Nudos; las primeras
son perchas libres de nudos utilizadas con el objetivo de permitir que las aves se “acostumbren”
a percharse en ellas, mientras que las segundas perchas contienen nudos diseñados para su
propia captura.
Para el uso de las Perchas de Control, se debe escoger pedazos de ramas gruesas; de tal forma
que al taladrar la base de estas, se pueda insertar una base para tornillos sin romper la rama.
En cuanto a las Perchas con nudos, las ramas escogidas para este fin, deben ser taladradas a lo
largo, se debe dejar un centímetro de separación entre cada hueco; deberá hacerse dos hileras
de perforaciones paralelas (Lado Superior), pero en forma diagonal de manera que ambas
hileras tengan como salida final una sola hilera recta al otro lado (Lado Inferior). Sera el Lado
Superior el que contenga los Lazos de nylon y el Lado Inferior donde se coloca la Línea Base.
b) Preparación de los Nudos:
Para la preparación de la Línea Base:
1. Cortar un pedazo de hilo de nylon con un largo de una vez y media con respecto al largo de la Percha a usar.
2. Insertar un extremo del nylon por uno de los huecos realizados en la Percha en dirección a y amarrar con nudos simples. Realizar esto de manera que la longitud total del nylon quede en la parte donde los huecos mantienen una línea recta (parte “Inferior” de la Percha).
3. Realizar otro nudo a mitad de la Percha, manteniendo el nylon lo mas estirado posible. 4. Finalmente, anudar en el extremo final de la Percha. 5. Cortar el excedente de nylon.
Para la preparación de los nudos individuales:
El nudo a usar es denominado “Cuadrado Doble (Double Square Knot)”, el cual, será diseñado
para que se deslice y asegure al ser jalado con un mínimo de fuerza.
A continuación se describe los pasos a seguir para realizar dicho Nudo:
1. Cortar aproximadamente veinte centímetros de nylon del tamaño escogido. 2. Rodear el crochet de manera que un extremo del nylon tenga una longitud máxima de
ocho centímetros. 3. Usar el extremo pequeño del nylon cruzarlo por encima del extremo mas largo y pasarlo
tres veces entre el espacio dejado por los dos extremos de nylon (Ver Gráfico 2.1, Fig A).

4. Ajustar lo mejor posible. 5. Usando nuevamente el extremo pequeño del nylon, cruzarlo por encima del extremo
largo y pasarlo nuevamente tres veces por el espacio dejado por el lazo. 6. Una vez realizados los nudos, con el extremo pequeño del nylon, realizar un nudo
simple sobre si mismo, pegándolo lo máximo posible hacia los nudos realizados anteriormente (Ver Gráfico 2.1, Fig B).
7. Cortar el resto de nylon sobrante en el extremo usado para hacer los nudos.
Una vez realizado el Nudo Cuadrado Doble, quedará el otro extremo en forma de una “cola”
larga. Esta servirá para insertar los nudos en los huecos realizados en las “Perchas”; se tiene que
amarrar a la Línea Base (previamente colocada en la Percha) usando el mismo tipo de nudo
usado para explicado anteriormente (Nudo Cuadrado Doble) como se puede ver en la Figura C
(Ver Gráfico 2.1) Se debe tener cuidado de dejar aproximadamente un lazo de dos centímetros
de diámetro que será el lazo de trampa.
El nylon sobrante (en el extremo) debe ser cortado dejando el nudo y la Línea Base lo menos
expuesto posible.
Figure A
Figure B
Figure C
Gráfico 2.1 Preparación de Nudos individuales para perchas de captura
en TRC

Actividades en el Escondite (o Blind):
El equipo de Captura debe llegar aprox. 60 minutos antes del amanecer; esto asegura que la
presencia de los integrantes del grupo no perturbe la actividad de Colpeo de las aves.
Las actividades a seguir son las siguientes:
1. Bajar las “Ramas” y cambiar las “Perchas” de Control por las “Perchas” de trampa (con los nudos de Nylon).
2. Ingresar al Blind y preparar las bolsas de atrapadas e instrumentos necesarios para la medición de las posibles aves capturadas.
3. Esperar en el interior hasta el momento de la actividad. 4. Una vez comenzada la actividad, el líder del grupo determinara el momento preciso para
elevar las “Ramas”. Esto deberá ser cuando la actividad este en su mayor apogeo, de manera que las aves no se den cuenta de la elevación realizada.
5. Una vez arriba, mantener las Ramas en dicha posición amarrando la driza a la base del “Tronco”; mantenerse vigilando las Perchas.
6. Estar atento a las aves que se perchen, de manera que apenas se note que han “accionado” los nudos y han sido atrapadas, se libere la driza, dejándola correr y la Rama baje trayendo al Ave atrapada. La persona que haya “atrapado” un ave deberá informar al resto del equipo para poder obtener la ayuda correspondiente.
7. Una persona sale del escondite a coger al ave, mientras otra sale a cortar los nudos que han sido accionados para la atrapada.
8. Ingresar al Blind, colocar al Ave dentro de una bolsa de tela, cerrarla y dársela al grupo veterinario para proceder con el muestreo.
9. Una vez terminado el muestreo, se colocara al ave en un tronco colocado en el exterior del Blind (llamado Percha de Liberación).
10. El líder del grupo decide en que momento volver a subir la “Rama” o si fuera necesario bajar todas y terminar con el proceso de atrapadas(esto dependerá del tiempo de demora del muestreo o de cuantos individuos se tiene en espera).
11. Al finalizar el proceso de captura, se deberá esperar hasta que no haya actividad en la Colpa para poder retirarse del Blind. Bajar las “Ramas” y cambiar las “Perchas” con nudos por las de Control.
Una persona designada deberá tomar el tiempo de todo el proceso e informar acerca del tiempo
usado durante todo el proceso; el veterinario a cargo decidirá cuando es necesaria la liberación
de un ave antes de haber concluido con los muestreos necesarios.
Así mismo, en consideración de que existe la posibilidad de que caiga mas de un ave en un
mismo periodo de tiempo, un integrante del equipo deberá estar alerta para salir a apoyar al
Equipo de Captura; estos cogerán las aves mientras el apoyo corta los nudos.

CAPTURA CON RED
Esta actividad fue desarrollada exclusivamente para atrapar Guacamayos Grandes, esto debido
a que el tamaño de los cocos de la Red son muy grandes para aprisionar a un ave pequeña;
además, el horario que se uso para este tipo de atrapada coincide con el horario de arribo de los
Guacamayos Grandes. La exclusión hacia las aves más pequeñas se da debido a que los objetivos
que se buscan alcanzar (colocación de collar satelital) son diferentes con respecto a los objetivos
en que se basa el Sistema anterior.
Para atrapar con redes, se debe considerar dos etapas de trabajo: Preparación del Sistema de
Red y la Ejecución del trabajo.
Preparación del Sistema:
1. Identificar el lugar donde se colocara la red. Este debe ser un lugar cercano a la zona de Colpeo; es importante asegurarse de que no este directamente sobre dicha zona para evitar ahuyentar a las aves. Además, debe tener arboles cercanos que servirán a modo de “Polea”.
2. Limpiar la zona escogida de pastos y malezas que superen el nivel de la rodilla y por una longitud aproximada de 15 metros. Esto es necesario para que al subir y bajar la red, esta no se enrede en dichas plantas.
3. Una vez escogidos los arboles que serán usados como “Poleas”, se debe hondearlos; con esto lograremos pasar drizas que nos servirán para subir y bajar el sistema de redes (“Drizas de Amarre”).
4. Cerca de los arboles “Poleas”, deberá acondicionarse un pequeño “escondite” para dos Atrapadores. Estos escondites deberán tener buena visibilidad hacia la red y estar camuflados por arbustos aledaños. En una zona un poco mas alejada, pero igual camuflada, pueden permanecer dos ayudantes mas (Atrapadores).
5. Cerca del escondite, deberá colocarse un palo a modo de “Poste” (podría ser también un tallo de un árbol o arbusto cercano) donde se pueda amarrar las Drizas de Amarre para asegurarlas.
6. Conseguir dos palos de aproximadamente unos 4 metros de altura (diámetro variable). Estos servirán como base para la red, y serán soporte para elevar la red.
7. Usar una driza de 12 m. para unir los extremos superiores de los Palos “Base” mediante nudos simples. Realizar lo mismo en la parte inferior, simulando así la forma de la red estirada. Esto será considerado como un “Control”(no siendo propiamente una Red).
8. Atar los cabos de las drizas hondeadas a la parte superior de los palos base. Utilizar nudos simples pero de gran resistencia, para que no vayan a caer con el peso. De la misma manera, atar una driza a la parte inferior de cada Palo base; estas drizas(“Drizas de Control”) servirán para estirar completamente la red una vez que esta sea elevada.
9. Dejar el Sistema puesto para que las Aves se acostumbren a su presencia.

Ejecución del Trabajo:
1. Tratar de llegar en el momento en el que la actividad es nula, usualmente después de que la actividad de la mañana ha terminado y recién empiezan a llegar los Guacamayos Grandes (entre las 7 – 7:30 a.m). Se puede consultar esto con los investigadores que están monitoreando la actividad en la Colpa dicho día.
2. Soltar la Driza de Amarre superior de uno de los Palos Base; sacar las Drizas de Control y colocar la red siguiendo el orden de los Lazos. A continuación estirar la Red a lo largo hasta llegar hasta el lado opuesto donde también se deberá soltar la Driza de Amarre superior y colocar la Red en el orden indicado.
3. Una vez hecho esto, deberá amarrarse nuevamente los extremos superiores de los Palos Base usando las Drizas de Amarre.
4. Estirar la Red lo mas que se pueda a lo largo y alto, revisando que el orden sea el adecuado y que no haya nada enredado en la red (pues esto podría hacer visible la Red y evitar que los Guacamayos se dirijan hacia ella).
5. Una vez terminado esto, los dos “atrapadores” deberán dirigirse a su respectivo escondite y esperar hasta que comience la actividad.
6. El líder es quien toma la decisión de subir la Red, basado en la actividad de la Colpa. Debido al silencio que debe ser mantenido, previamente se debe decidir como se hará la comunicación; es recomendable que el Líder suba la Red en el momento indicado y el otro Atrapador haga lo propio con la suya sin necesidad de intercambiar palabras.
7. Una vez alcanzada la altura máxima de la Red, se deberá asegurar las Drizas de Amarre al “Poste” que tienen cerca. Además deberán tensar las Drizas de Amarre de la parte inferior, de manera que la Red quede extendida en toda su longitud.
8. En el momento en que un Guacamayo caiga en la Red, ambos Atrapadores deberán liberar las drizas del “Poste” y bajar la Red de manera eficaz y segura.
9. Una vez abajo, asegurar las Drizas de Amarre de manera que la Red no caiga al suelo; el tercer y cuarto ayudante saldrán de su escondite, mientras uno coge al Guacamayo, el otro lo libera de la Red (ya sea removiendo la Red de su cuerpo, o en caso de estar muy enredado, cortar la Red).
10. Llevar al Guacamayo dentro del Blind para ser colocado en la bolsa de tela y ser muestreado.
11. Antes de ser liberado, se asegura de que no tenga restos de Red (en caso se haya cortado la Red).
12. Colocarlos en la Percha de Liberación y dejarlo ir. 13. El Líder es quien decide si se vuelve a subir la Red o no. Preferentemente, una vez
atrapado un Guacamayo, se deberá dar por terminado el día de Atrapadas. 14. Guardar la Red y volver a colocar el Sistema de Red Control hasta el siguiente día de
Atrapadas.
En caso de que cayera más de un Guacamayo en la Red, el quipo principal de captura saldrá a
apoyar; deberán coger a los Guacamayos que hayan caído mientras que uno pasara a cortar la
Red que los retiene. En este caso, es mejor cortar directamente la Red, pues el intentar soltarlos
de la Red haría que el segundo o eventualmente un tercer Guacamayo permanezca mucho
tiempo en la Red, ocasionándoles gran estrés.

CAPÍTULO III:
GUACAMAYOS REINTRODUCIDOS EN TRC: LOS “CHICOS”
INTRODUCCION
Un total de 31 guacamayos (20 Am, 5 Ara chloropterus y 6 Ara ararauna) fueron criados a mano
y liberados entre 1992 y 1995 en el bosque colindante al centro de investigaciones Tambopata
(TRC) en Madre de Dios. A estos individuos se les conoce con el apelativo de “Chicos”.
Debido a la técnica de crianza utilizada, estos guacamayos presentan un considerable grado de
improntaje hacia la figura humana (Brightsmith et al. 2005). Los individuos se caracterizan por
presentar poco temor y demasiado interés hacia las actividades humanas. Los guacamayos han
sido registrados perchados muy cerca de humanos y husmeando dentro de las instalaciones del
centro de investigación.
Desde el año 1999, se lleva un registro continuo de avistamientos de guacamayos criados a
mano y liberados con el fin de registrar su supervivencia y reproducción. Cada vez que un
guacamayo “Chico” es avistado ya sea en el centro de investigadores, en sus alrededores, en la
colpa, o en los monitoreos de nidos en el bosque colindante, se procede a colectar los siguientes
datos: (1) Especie, (2) Pata en la que presenta el anillo, (3) Código grabado en el anillo, (4)
Condiciones de plumaje, (5) Presencia de pareja, (6) Registro fotográfico y (7) Observaciones
particulares.
El presente reporte ha sido elaborado analizando los datos re organizados de los anos 2001 al
2007 y los nuevos datos colectados en el ano 2008.

RESULTADOS Y DISCUSION
SUPERVIVENCIA
De los 20 guacamayos Am liberados, 17 individuos sobrevivieron a su primer año de vida (85%)
(Brightsmith et al. 2005). De los sobrevivientes, 7 fueron no fueron vistos desde antes del 2001
por lo que se considera que no han logrado sobrevivir. Además, 2 individuos son considerados
“desaparecidos” puesto que no han sido vistos en los últimos 2 años. A su vez, otros 2 individuos
considerados “desparecidos” en el pasado son considerados “re aparecidos” puesto que fueron
vistos después de 3 y 6 anos de no haber sido avistados. Finalmente, se considera que 9
individuos (incluyendo los 2 “re aparecidos”) sobreviven hasta diciembre del 2008 (45%).
De los 5 individuos de Ara chloropterus liberados, 3 fueron vistos por última vez en el año 1995.
Uno de ellos es considerado desaparecido puesto que no ha sido visto desde fines del ano 2007.
Se tiene registros de 1 individuos avistados hasta diciembre del 2008 ( 20 %).
No se ha registrado ningún avistamiento de individuos de Ara ararauna desde el año 1994.
Gráfico 1: Sobrevivencia de guacamayos reintroducidos en TRC (“Chicos”)
Sobrevivencia de guacamayos reintroducidos en TRC
(2001-2008)
45
20
0
55
80
100
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Ara macao Ara chloropterus Ara ararauna
Especie
Sobrevivencia Mortalidad

Tabla 1: Detalle de guacamayos reintroducidos en TRC que sobreviven hasta Diciembre 2008
Nombre Sp # Foot 2001 2002 2003 2004 2005 2006 2007 2008 Total
visitas Ultima visita
Visto en
Nido Status
Mellizo I Ac 1 N N N N N N N N 0 Nunca Nunca No sobrevive
Mellizo II Ac N N N N N N N N 0 Nunca Nunca No sobrevive
Ascencio Ac 17 R Y Y Y Y Y Y Y Y 223 Dec 08 Mar 08 Sobrevive
Matías Ac 6 L Y Y Y Y Y Y Y Y 359 28-Dec-07 Nunca Desaparecido
Boquita Am 9 R Y N N N N N N Y 2 8-Jul-08 Nunca Reaparecido
Aborto Am 12 R N N Y N N N Y N 6 18-Nov-07 Nunca Reaparecido
Octagon Am 22 L Y Y N N N N N N 18 06-Ene-02 Nunca Desaparecido
Fernando Am 25 L Y Y N N N Y N N 48 27-Jun-06 Nunca Desaparecido
Mas Mas Am 1 R N N N N N N N N 0 Nunca Nunca No sobrevive
PVC Am 3 R N N N N N N N N 0 Nunca Nunca No sobrevive
Moro
Manzanilla Am 5 R N N N N N N N N 0 Nunca Nunca No sobrevive
Suenos Am 6 R N N N N N N N N 0 Nunca Nunca No sobrevive
Ted Am 23 L N N N N N N N N 0 Nunca Nunca No sobrevive
Chispa Am 51 R N N N N N N N N 0 Nunca Nunca No sobrevive
Tulio Am 10 R N Y Y Y Y Y Y Y 56 19-Oct-08 Nunca No sobrevive
Macario Am 4 R N Y Y Y Y Y Y Y 10 10-May-08 Mar 08 Sobrevive
Pukakuro Am 7* R Y Y Y Y Y Y Y Y 59 Dec 08 Ene 08 Sobrevive
Inglesita Am 16 L N N Y Y Y Y Y Y 78 28-Nov-08 Nunca Sobrevive
Avecita Am 14 L Y Y Y Y Y Y Y Y 360 7-Nov-08 Feb 08 Sobrevive
Tabasco Am 24 L Y Y Y Y Y Y Y Y 408 20-Nov-08 Feb 08 Sobrevive
Inocencio Am 8 R Y Y Y Y Y Y Y Y 466 Dec 08 Feb 08 Sobrevive
Chunchuy Am 11** R Y Y Y Y Y Y Y Y 513 Dec 08 Feb 08 Sobrevive

AVISTAMIENTOS
Durante el periodo comprendido entre los años 2001 y 2008 se registró 3215 veces la presencia
de un guacamayo “Chico” en las instalaciones del centro de investigación. Debido a que los
registros de estos avistamientos son tomados por asistentes voluntarios que permanecen por
periodos no mayores a 3 meses, muchas veces no fue posible reconocer individualmente al
“visitante”, por lo que únicamente se registro su especie (83 veces, 1.7%)
Gráfico 2: Número de avistamientos por año
En el periodo comprendido entre el 2001 y 2008 se registro en promedio 402 veces por año la
visita de un guacamayo “chico”, lo que significa que en promedio se vio a uno de ellos en el
centro de investigaciones al menos una vez al día (N=2922 días, Promedio=1.1 veces al día,
Promedio min= 0 avistamientos, Promedio máx. = 2 avistamientos). El 2004 el año se
registraron mas avistamientos (578 veces) y el año 2001 el año se registraron menos (217
veces). Es posible que el numero de registros de estos guacamayos este relacionado con la
presencia de asistentes en el centro de investigación. Algunas veces los guacamayos “chicos”
son vistos por personal del staff o personas ajenas al Proyecto Guacamayo y no son registrados
en las correspondientes hojas de datos. Es posible que el mayor numero de registros este
relacionado con los periodos de estadía largos de los asistentes (periodos mayores a 3 meses).
Al permanecer meses continuos en el centro de investigación, es posible aprender a diferenciar
a los individuos por características externas únicas (condición de plumaje, etc.) y no es tan
necesario leer el anillo de identificación. El menor numero de avistamientos registrado en el
2001 puede deberse al poco numero de asistentes con los que conto el proyecto en ese periodo;
debido a lo cual los asistentes permanecieron todo el día fuera del albergue, no siendo posible
registrar a los “chicos” que visitaron el centro de investigación durante el día.
217
436497
578
365 321
454
347
0
100
200
300
400
500
600
700
# d
e a
vis
tam
ien
tos
2001 2002 2003 2004 2005 2006 2007 2008
Ano
Avistamientos por ano de guacamayos reintroducidos en TRC
(2001-2008)

FRECUENCIA DE AVISTAMIENTOS
La frecuencia de visitas de los guacamayos “chico” varia a lo largo del ano. Existen meses (como
noviembre y diciembre) en los que los “chicos” visitan mas seguido el centro de investigación,
así como otros meses (febrero y marzo) en los que rara vez se ve a una de estas aves en la casa.
(Ver grafico 3)
Gráfico 3: Promedio de frecuencia de visitas por ano de guacamayos reintroducidos en TRC
Es posible que los “chicos “ visitaron mas seguido el centro de investigación en los meses en los
que la disponibilidad de alimento era baja. Este “baja disponibilidad” puede deberse a dos
razones. (1) El alimento es escaso en el bosque. Estudios de fenología del bosque colindante a
TRC muestran que el periodo comprendido entre abril y junio son los meses en los que pocas o
ninguna planta forrageada por guacamayos, presenta frutos o semillas.(Brightsmith, com.per) Es
posible que los “chicos” consideren el centro de investigación un sitio mas de “forrageo” y
regresen a la casa en busca de alimento. (2) Los guacamayos “chico” estuvieron defendiendo sus
nidos. Ocho de los nueve “chicos” sobrevivientes anidan en los alrededores del centro de
investigación. La temporada reproductiva se inicia a fines del mes de octubre. Durante este
periodo los guacamayos (silvestres y “chicos”) invierten gran parte del día defendiendo la
cavidad que desean utilizar para poner huevos y criar a sus pichones. Es posible que durante
estos meses, los “chicos” consideren que es mas probable o mas cerca conseguir alimento en el
centro de investigación que en el mismo bosque y por eso regresen a la casa en busca de
comida.
Promedio de frecuencia de visitas por ano de guacamayos
reintroducidos en TRC
2001-2008
0
10
20
30
40
50
60
70
Enero
Febre
ro
Mar
zoAbr
il
May
oJu
nio
Julio
Agosto
Setiem
bre
Octub
re
Nov
iem
bre
Diciem
bre
Mes
# v
isit
as
Guacamayos
defendiendo
nidos
Poco alimento en
el bosque

AVISTAMIENTOS POR ESPECIE
Debido al mayor numero de sobrevivientes de Guacamayos Escarlata (Ara macao) “chico” el numero de avistamientos de individuos de esta especie es considerablemente mayor. Sin embargo, el avistamiento de Guacamayos cabezones (Ara chloropterus) es resaltable debido a que solo dos invidividuos sobrevivieron hasta el 2007 y posiblemente solo uno quede vivo el 2008.
Gráfico 4. Avistamientos por especie entre los anos 2001 y 2008
Gráfico 5. Avistamiento mensual por especie entre los anos 2001 y 2008
Avistamientos mesnuales por especie de guacamayos reintroducidos
en TRC (2001-2008)
0%
20%
40%
60%
80%
100%
Enero
Febre
ro
Mar
zoAbr
il
May
o
Junio
Julio
Agosto
Setiem
bre
Octub
re
Nov
iem
bre
Diciem
bre
Ara chloropterus Ara macao
21%
78 %
0
20
40
60
80
%
Ara chloropterus Ara macao
Avistamientos por especie durante el periodo
comprendido entre el 2003 y 2008

AVISTAMIENTOS POR INDIVIDUO
Es importante resaltar que la identificacion de individuos “chico” en el caso de A.chloropterus
fue mucho mas sensilla que la correspondiente a los “chico” A.macao. El menor numero de
individuos de la primera especie mencionada, y el hecho de que cada uno de los sobrevivientes
llevara el anillo en patas diferentes (anillo de Mathias en pata izquierda y anillo de Ascencio en
pata derecha) hizo que los avistamientos de estos individuos fueran mas precisos. Para el caso
de los “chico” Ara macao la identificacion se hizo un tanto mas complicada, en especial para los
individuos que frecuentaban poco el centro de investigacion.
Grafico 6. Avistamientos por individuo de guacamayos reintroducidos en TRC (2001-2008)
Es interesante resaltar el caso del “chico” Avecita y el chico “Chuchuy”. Durante los anos 2001 y
2005 el nido de “Avecita” se ubico muy cerca del centro de investigacion. Este chico
selectivamente eligio nidos artificiales y cavidades colocados a 200 metros a la redonda. En el
ano 2005 el centro de investigacion se muda 500 metros dentro del bosque, ubicandose a
menos de 50 metros del nido de “Chuchuy”emparejada con el chico “Inocencio”.
Avistamiento de guacamayos reintroducidos "chicos" en TRC
(2001-2008)
513
466
408
360
359
223
78
59
56
48
18
10
6
2
0 100 200 300 400 500 600
Chunchuy
Inocencio
Tabasco
Av ecita
Matías
Ascencio
Inglesita
Pukakuro
Tulio
Fernando
Octagon
Macario
Aborto
Boquita
Ind
ivid
uo
s
# Visitas
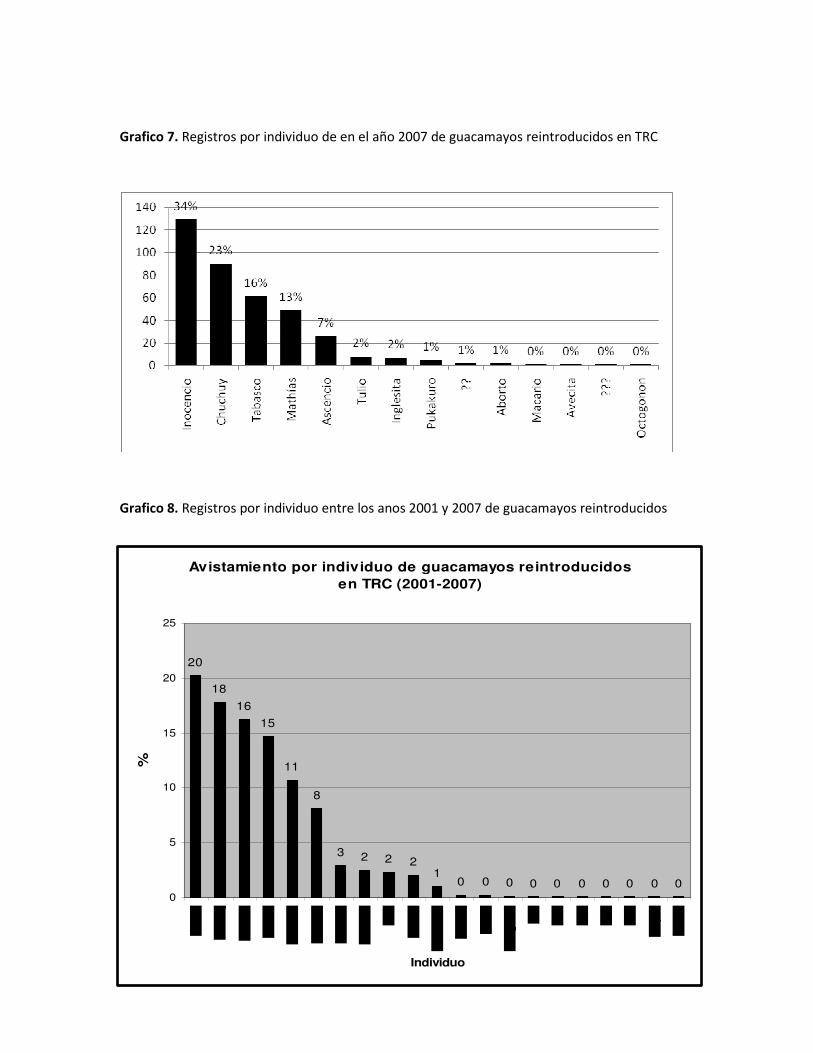
Grafico 7. Registros por individuo de en el año 2007 de guacamayos reintroducidos en TRC
Grafico 8. Registros por individuo entre los anos 2001 y 2007 de guacamayos reintroducidos
Avistamiento por individuo de guacamayos reintroducidos
en TRC (2001-2007)
20
18
16
15
11
8
3 2 2 21
0 0 0 0 0 0 0 0 0 00
5
10
15
20
25
Individuo
%

Comparando los dos gráficos anteriores, vemos que el individuo “Avecita” ya no encabeza la
lista para el 2007, y el número de avistamientos en este año se acerca al cero porcentual. En
contraste con los acumulados 2001-2006, “Avecita” ocupaba el mayor número de
avistamientos. Por el contrario, los avistamientos de “Chuchuy” y en especial de “Inocencio” se
incrementaron mucho.
Es posible que el hecho de que el nido de estos individuos se encuentre en las proximidades del
centro de investigación haya influenciado en esta variación en el numero de avistamientos.
Posiblemente, los guacamayos “chicos” consideren la casa en si como parte de su “territorio”.
Éxito Reproductivo de guacamayos reintroducidos en TRC
Desde el ano 2000 se viene incrementando las labores relacionadas con la estación reproductiva
de los guacamayos. El monitoreo de cavidades naturales y de nidos artificiales ha sido mas
intensivo así como la implementación de nuevas cavidades artificiales.
Se lleva un registro anual de que guacamayos “chico” fueron vistos anidando en una de las
cavidades que se monitorea. La identificación de estos individuos no es sencilla, puesto que el
anillo es apenas visible cuando están en vuelo. Sin embargo, el comportamiento de los
guacamayos “chico” cuando defienden sus nidos es muy diferente al de los guacamayos
silvestres. Un individuo silvestre es temeroso, da gritos de alarma y generalmente vuela del nido
cuando el trepador se aproxima al nido. Un individuo “chico” es aguerrido, permanece en el
nido , ofrece resistencia y pelea con el trepador de una manera muy parecida a la que lo haría
con otro guacamayo si es que este “invadiera su nido”.
El “chico” debe ser reconocido por medio de registros fotográficos (plumaje, señales
particulares, etc). Algunas veces se logro reconocer que el nido revisado estaba siendo usado
por un “chico”, pero no se pudo reconocer al individuo (3 individuos)

Tabla 2. Detalle del éxito reproductivo de guacamayos reintroducidos en TRC (2001-2008)
SPECIE # Foot Nombre
Huevos
puestos
Huevos que
eclosionaron
Pichones
que
volaron
Ara chloropterus 17 R Ascencio 23 12 10
Ara macao 4 R Macario 16 12 10
Ara macao 17 L NN 1 2 1 0
Ara macao 7 R Pukakuro 21 6 4
Ara macao 11 R Chuchuy 15 10 7
Ara macao 8 R Inocencio 6 4 1
Ara macao 14 L Avecita 18 8 3
Ara macao 16 L Inglesita 2 0 0
Ara macao 24 L Tabasco 15 12 8
Ara macao UNK-LR L NN 2 1 1 1
Ara macao UNK-RR R NN 3 2 2 2
TOTAL 121 70 50

Algunos guacamayo “chico” anidaron exitosamente pocas veces o solo una vez en los
alrededores del albergue, por lo que su éxito reproductivo parece ser muy alto (NN 2 y NN 3 con
100% de éxito de eclosión y éxito de vuelo) o muy bajo (Inglesita con 0 % de éxito de eclosión y
éxito de vuelo). Estos tres individuos solo fueron vistos anidando una vez entre el 2001 y 2008.
pero solo fueron vistos anidar una vez. Sin embargo, otros individuos (Avesita, Chuchuy,
Inocencio, Ascencio, Macario, Tabasco y Pukakuro) han sido monitoreados durante todo el
periodo entre el 2001 y 2008, siendo exitosos algunos anos y poco exitosos en otros.
Exito Reproductivo de guacamayos reintroducidos en TRC
2001-2008
83
83
67
70
25
38
0
67
0
100
100
0 20 40 60 80 100 120
Ascencio
M acario
Pukakuro
Chuchuy
Inocencio
Avecita
Inglesita
Tabasco
NN 1
NN 2
NN 3
Ind
ivid
uo
s
Porcentaje
% Eclosion % Vuelo

Guía de identificación de guacamayos reintroducidos en TRC
Con el objetivo de mejorar tanto la calidad como la cantidad del registro de avistamientos de
guacamayos “chico”, se elaboro una “Guía de identificación” por medio de la cual se brinda de
manera clara algunas pautas claves para la identificación de individuos específicos. Esta guía
esta escrita en ingles y muestra de manera didáctica como leer los anillos de identificación y
como reconocer a cada “chico” por medio de señales particulares en el rostro o el plumaje.

CAPÍTULO IV:
TRABAJO CON PROFESIONALES Y VOLUNTARIOS
TRABAJO CON VOLUNTARIOS EN EL PROYECTO
Durante el periodo de Noviembre 2007 a Noviembre del 2008, el Proyecto Guacamayo contó
con 17 voluntarios, tanto internacionales como peruanos. (Anexo 1).
El trabajo realizado por los asistentes voluntarios es de suma importancia para el Proyecto, ya
que sin él, no sería posible cumplir con nuestros objetivos, por lo que estamos realmente
agradecidos.
Diariamente se trabajan unas 10 horas en promedio, 6 días a la semana, comenzando las
actividades muy temprano en las mañanas, generalmente antes del amanecer; contando aves
en la orilla del río o en las colpas, censando loros, trepando nidos ubicados a 30 metros de
altura, ingresando datos en computadoras, tomando datos de temperatura y precipitación, sin
olvidar cumplir el rol de limpieza de la casa y área de trabajo. En sumatoria, un arduo trabajo,
para el que muchos voluntarios viajan a miles de kilómetros de sus países de origen.
El mejor resultado, es que muchos de ellos regresan. La experiencia vivida al apoyar en un
proyecto de investigación y conservación en el bosque tropical, hace que muchas veces estos
voluntarios repitan el viaje en la siguiente temporada, creando lazos de amistad y
compañerismo con colegas de países tan lejanos y diferentes.
El trabajo de los voluntarios forma una parte muy importante del Proyecto Guacamayo, y
esperamos seguir contando con personas interesadas en apoyar al proyecto.

TRABAJO CON PROFESIONALES PERUANOS
Este último año, el Proyecto Guacamayo ha ampliado sus investigaciones, con el fin de mejorar
nuestro conocimiento de la historia natural y conservación de psitácidos en el Perú. Con esta
finalidad estamos contratando biólogos, veterinarios y otros profesionales jóvenes para
encabezar sus propios trabajos con la asesoría y financiamiento del Dr. Donald Brightsmith y
Texas A&M University (TAMU).
Bajo esta premisa se busca realizar estudios de diferentes temas relacionados con conservación
de los bosques tropicales.
El primero de estos estudios satélite: La Colpa de Palmeras de Sandoval: el rol de sodio, el uso
turístico y los efectos del incendio del 2005 (Autorización 004-C/C-2007-INRENA-IANP)
encabezado por la Bióloga Aimy Cáceres realizó actividades de campo entre los meses de marzo
y noviembre 2007 siendo la parte de análisis de datos y elaboración de informes preliminares
realizada entre diciembre 2007 y enero 2008. El documento científico correspondiente a esta
investigación está en preparación.
El estudio de Enfermedades Infecciosas en el Tráfico Legal e Ilegal de Aves Silvestres en Perú se
inició en el 2006 y es encabezado por Patricia Mendoza y Roberto Elías. Este estudio se
encuentra en su tercer año de muestreo. Los resultados de esta investigación buscan revelar
información para colaborar con las normas de sanidad en el comercio de aves.
El proyecto distribución Espacial de Colpas y Características Físicas de Colpas en Madre de Dios
(Autorización Nº011 S/C-2007-INRENA-IANP) fue encabezado por la Bióloga Gabriela Vigo Se
realizó las actividades de campo entre los meses de junio y noviembre 2007. El equipo
conformado por 3 investigadores recorrió las cuencas de 5 ríos de la región (Río Amigos, Río
Colorado, Río Tambopata, Río Piedras y Río Alto y Bajos Madre de Dios) en un total de 300 km
lineales recorridos recopilando información acerca de colpas.
El Proyecto Cámaras en Nidos, encabezado también por la Bióloga Gabriela Vigo. El estudio
busca descubrir el comportamiento entre padres y pichones y entre hermanos, para poder
determinar con certeza la causa de muerte de los segundos y terceros pichones.
El Proyecto Guacamayo de Tambopata sigue como la base de entrenamiento y el punto central
para esta red de investigadores e investigaciones. Muchos de estos investigadores, se iniciaron
como asistentes voluntarios y hoy son los directores de sus propias investigaciones.

Los profesionales peruanos que trabajan en estos nuevos sub proyectos están encargados de
formar sus equipos de trabajo, que muchas veces incluyen a jóvenes estudiantes de carreras
afines con interés en conservación e investigación. Comparten el derecho de autor de los datos
colectados, y cuentan con asesoría para publicar artículos en revistas internacionales, así como
presentar trabajos en Congresos y Conferencias.
El fin de emplear este modelo es asegurar que jóvenes investigadores peruanos aprendan los
pasos necesarios para llevar a cabo proyectos de investigación (incluyendo postular para
financiamiento, escribir propuestas, presupuestos, protocolos, y hojas de datos, diseñar
estudios factibles, colectar datos en campo, digitar y analizar datos, y escribir reportes y
publicaciones internacionales).
El director del Proyecto Guacamayo, Ph.D. Donald Brightsmith, tiene la confianza de que usando
este modelo se puede armar en el Perú un grupo de profesionales calificados en el estudio y
conservación de aves en general, y de los psitácidos en particular, animando a nuevos
estudiantes a participar en el proyecto, o desarrollando proyectos satélite.

AGRADECIMIENTOS
EL PROYECTO GUACAMAYO AGRADECE A TODOS LOS ASISTENTES QUE
BRINDARON SU APOYO A LO LARGO DE LA DURACIÓN DE LA AUTORIZACIÓN
044 S/C-2007-INRENA-IANP, ESPECIALMENTE A LOS AYUDANTES LISTADOS EN
EL ANEXO 1. AGRADECE TAMBIÉN AL SCHUBOT EXOTIC BIRD HEALTH CENTER
DE TEXAS A&M UNIVERSITY, EL EARTHWATCH INSTITUTE, WILDLIFE
PROTECTION FOUNDATION, RAINFOREST EXPEDITIONS, JANICE BOYD DE
AMIGOS DE LAS AVES USA PARROT CONSERVATION FUND QUIENES FINANCIAN
NUESTRO TRABAJO. SE AGRADECE AL GRUPO DE COLABORADORES DEL
TRABAJO DE TELEMETRÍA, ROBIN BJORK, DAVID WAUGH, GREG MATUZAK,
ROAN MCNAB, RONY GARCIA, FUNDACION LORO PARQUE, WILDLIFE
CONSERVATION SOCIETY, TELONICS Y NORTHSTAR SCIENCE AND TECHNOLOGY.
IGUALMENTE AGRADECEMOS AL PERSONAL DE INRENA LIMA Y PUERTO
MALDONADO POR SU APOYO EN EL PRESENTE ESTUDIO.

BIBLIOGRAFÍA
Abramson, J., B. L. .Spear, and J. B. Thomsen. 1995. The Large Macaws: Their Care, Breeding and
Conservation. Raintree Publications, Ft. Bragg, CA.
Adler, G. H., and K. A. Kielpinski. 2000. Reproductive phenology of a tropical canopy tree,
Spondias mombin. Biotropica 32:686-692.
Arms, K., P. Fenney, and R. C. Lederhouse. 1974. Sodium:stimulus for puddling behavior by tiger
swallowtail butterflies, Papilio glaucus. Science 185.
Beissinger, S. R., and N. F. R. Snyder. 1992. New World Parrots In Crisis. Smithsonian Institution
Press, Washington, D.C.
Bennett, P. M., and I. P. F. Owens. 1997. Variation in extinction risk among birds: chance or
evolutionary predisposition? Proceedings of the Royal Society of London B. 264:401-
408.
Renton, K. (2002). "Seasonal variation in occurrence of macaws along a rainforest
river." Journal of Field Ornithology 73(1): 15-19.
Birdlife International. 2006. Data zone. Accessed: 15 August 2006.
http://www.birdlife.org/datazone/species/index.html
Bjork, R. 2004. Delineating patterns and process in tropical lowlands: Mealy Parrot migration
dynamics as a guide for regional conservation planning. Ph.D. Oregon State University.
Brightsmith, D. J. 2004. Effects of weather on avian geophagy in Tambopata, Peru. Wilson
Bulletin 116:134 -145.
Brightsmith, D. J. 2005. Parrot nesting in southeastern Peru: seasonal patterns and keystone
trees. Wilson Bulletin 117:296-305.
Brightsmith, D. J., and R. Aramburú. 2004. Avian geophagy and soil characteristics in
southeastern Peru. Biotropica 36:534-543.
Brightsmith, D. J., J. Hilburn, A. Del Campo, J. Boyd, M. Frisius, R. Frisius, D. Janik, and F. Guillén.
2005. The use of hand-raised Psittacines for reintroduction: a case study of scarlet
macaws (Ara macao) in Peru and Costa Rica. Biological Conservation 121:465 - 472.
Brightsmith, D. J. 2006. The psittacine year: what drives annual cycles in Tambopata's parrots?
Pages 44-53. Proceedings of the VI International Parrot Convention, Puerto de la Cruz,
Tenerife, Spain.

Brightsmith, D. J., and J. Boyd. 2006. Testing satellite telemetry tags for psittacines in
Tambopata, Peru. Page 12. Schubot Exotic Avian Health Center, Texas A&M University,
College Station, Texas.
Burger, J., and M. Gochfeld. 2003. Parrot behavior at a Rio Manu (Peru) clay lick: temporal
patterns, associations, and antipredator responses. Acta Ethologica 6:23 - 34.
Casagrande, D. G., and S. R. Beissinger. 1997. Evaluation of four methods for estimating parrot
population size. Condor 99:445-457.
Clubb, K. J., D. Skidmore, R. M. Shubot, and L. S. Clubb. 1992. Growth Rates of Handfed
Psittacine Chicks.in R. M. Shubot, K. J. Clubb, and S. L. Clubb, editors. Psitacine
Aviculture - Perspective, tecniques and Research. Avicultural Breding and Research
Center, Florida.
Collar, N. J. 1997. Family Psittacidae. Pages 280-479 in J. d. Hoyo, A. Elliott, and J. Sargatal,
editors. Handbook of the Birds of the World. Lynx Edicions, Barcelona, Spain.
Croat, T. B. 1975. Phenological behavior on Barro Colorado Island. Biotropica 7:270-277.
Dunning, J. B. 1993. CRC handbook of avian body masses. CRC Press, London.
Emmons, L. H. 1984. Geographic variation in densities and diversities of non-flying mammals in
Amazonia. Biotropica 16:210-222.
Emmons, L. H., and N. M. Stark. 1979. Elemental composition of a natural mineral lick in
Amazonia. Biotropica 11:311-313.
Forshaw, J. M. 1989. Parrots of the world. Third edition. Landsdowne Editions, Melbourne,
Australia.
Foster, R. B., T. Parker, A, A. H. Gentry, L. H. Emmons, A. Chicchón, T. Schulenberg, L. Rodríguez,
G. Larnas, H. Ortega, J. Icochea, W. Wust, M. Romo, C. J. Alban, O. Phillips, C. Reynel, A.
Kratter, P. K. Donahue, and L. J. Barkley. 1994. The Tambopata-Candamo Reserved Zone
of southeastern Peru: a biological assessment. RAP working papers 6., Conservation
International, Washington, D.C.
Frankie, G. W., H. G. Baker, and P. A. Opler. 1974. Comparitive phenlogical studies of trees in
tropical wet and dry forests in the lowlands of Costa Rica. Journal of Ecology 62:881-
919.
Forshaw, J. M. (1989). Parrots of the World. Melbourne, Australia, Landsdowne Editions.
Gentry, A. H. 1988. Tree species richness of upper Amazonian forests. Proceedings of the
National Academy of Science. USA 85:156-159.
Gilardi, J. D., S. S. Duffey, C. A. Munn, and L. A. Tell. 1999. Biochemical functions of geophagy in
parrots: detoxification of dietary toxins and cytoprotective effects. Journal of Chemical
Ecology 25:897-922.

Griscom, B. W., and P. M. S. Ashton. 2003. Bamboo control of forest succession: Guadua
sarcocarpa in southeastern Peru. Forest Ecology and Management 175:445-454.
Holdo, R. M., J. P. Dudley, and L. R. McDowell. 2002. Geophagy in the African elephant in
relation to availability of dietary sodium. Jounal of Mammalogy 83:652-664.
Iñigo-Elias, E. E. (1996). Ecology and breeding biology of the Scarlet Macaw (Ara macao) in the
Usumacinta drainage basin of Mexico and Guatemala. Gainesville, FL, University of
Florida: 117.
Juniper, T. and M. Parr (1998). Parrots: a guide to parrots of the world. New Haven, Yale
University Press.
Karubian, J., J. Fabara, D. Yunes, J. P. Jorgenson, D. Romo, and T. B. Smith. 2005. Temporal and
spatial patterns of macaw abundance in the Ecuadorian Amazon. Condor 107:617-626.
Lack, D. 1968. Ecological adaptations for breeding in birds. Methuen, London.
Lugo, A. E., and J. L. Frangi. 1993. Fruit fall in the Luquillo Experimental Forest, Puerto Rico.
Biotropica 25:73-84.
Marsden, S. J. 1999. Estimation of parrot and hornbill densities using a point count distance
sampling method. Ibis 141:377-390.
Marsden, S. J., and A. Fielding. 1999. Habitat associations of parrots on the Wallacean islands of
Buru, Seram, and Sumba. Journal of Biogeography 26:439-446.
Marsden, S. J., M. Whiffin, L. Sadgrove, and P. G. Jr. 2000. Parrot populations and habitat use in
and around two Brazilian Atlantic forest reserves. Biological Conservation 96:209-217.
Masello, J. F., M. L. Pagnossin, C. Sommer, and P. Quillfeldt. 2006. Population size, provisioning
frequency, flock size and foraging range at the largest known colony of Psittaciformes:
the Burrowing Parrots of the north-eastern Patagonian coastal cliffs. Emu 106:69-79.
Masello, J. F., and P. Quillfeldt. 2002. Chick growth and breeding success of the burrowing
parrot. Condor 104:574-586.
Matuzak and Brightsmith. 2007. Roosting of Yellow-naped Parrots in Costa Rica: estimating the
size and recruitment of threatened populations. J. Field Ornithol. 78(2): 159-169.
Montambault, J. R. 2002. Informes de las evaluaciones biológicas de Pampas del Heath, Perú,
Alto Madidi, Bolivia, y Pando, Bolivia., Conservation International., Washington, D.C.
Munn, C. A. (1991). "Tropical canopy netting and shooting lines over tall trees." Journal of Field
Ornithology 62: 454 - 463.
Myers, M. C., and C. Vaughan. 2004. Movement and behavior of Scarlet Macaws (Ara macao)
during the post-fledging dependence period: implications for in situ versus ex situ
management. Biological Conservation 118:441-420.

Nettleship, D. N. 1972. Breeding success of the Common Puffin (Fratercula arctica L.) on
different habitats at Great Island, Newfoundland. Ecological monographys 42:239-268.
Nycander, E., D. H. Blanco, K. M. Holle, A. d. Campo, C. A. Munn, J. I. Moscoso, and D. G. Ricalde.
1995. Manu and Tambopata: nesting success and techniques for increasing
reproduction in wild macaws in southeastern Peru. Pages 423-443 in J. Abramson, B. L.
Spear, and J. B. Thomsen, editors. The large macaws: their care, breeding and
conservation. Raintree Publications, Ft. Bragg, CA.
O´Connor, R. J. (1984). The growth and development of birds. New York : Wiley.
Phillips, A., and L. S. Clubb. 2002. Psittacine Neonatal Development.in R. M. Shubot, K. J. Clubb,
and S. L. Clubb, editors. Psitacine Aviculture - Perspective, tecniques and Research.
Avicultural Breding and Research Center, Floris.
Powell, G., P. Wright, U. Aleman, C. Guindon, S. Palminteri, and R. Bjork. 1999. Research findings
and conservation recommendations for the Great Green Macaw (Ara ambigua) in Costa
Rica. Accessed: http://www.lapaverde.or.cr/lapa/lapa_resultados_en.htm
Powell, G. V. N., and R. Bjork. 1995. Implications of intratropical migration on reserve design: a
case study using Pharomachrus mocinno. Conservation Biology 9:354 - 362.
Powell, G. V. N., and R. Bjork. 2004. Habitat linkages and the conservation of tropical
biodiversity as indicated by seasonal migrations of Three-Wattled Bellbirds.
Conservation Biology 18:500-509.
Räsänen, M. W., and A. M. Linna. 1995. Late Miocene tidal deposits in the Amazonian foreland
basin. Science 269:386-390.
Räsänen, M. W., and J. S. Salo. 1990. Evolution of the western Amazon lowland relief: impact of
Andean foreland dynamics. Terra Nova 2:320-332.
Renton, K. 2001. Lilac-crowned parrot diet and food resource availability: resource tracking by a
parrot seed predator. Condor 103:62-69.
Renton, K. 2002. Seasonal variation in occurrence of macaws along a rainforest river. Journal of
Field Ornithology 73:15-19.
Renton, K. 2006. Diet of adult and nestling Scarlet Macaws in Southwest Belize, Central America.
Biotropica 38:280-283.
Rickels, R. (1973). "Patterns of growth in birds II. Growth rate and mode of
development." Ibis 115: 177 - 201.
Ricklefs, R. E. (1967). "A graphical methods of fitting equacion to growth curves". Ecology 48(6):
978 -983.
Ricklefs, R. E. (1968). "Patterns of growth in birds." The Ibis 110(4): 419 - 451.

Riveros, J. C. (1999). Crecimiento y desarrollo postnatal del Pinguino del Humbolt Spheniscus
humboldti (Meyen, 1834). Biologia Lima, Universidad Nacional Agraria La Molina.
Biologo: 72.
Roth, P. 1984. Repartição do habitat entre psitacídeos simpátricos no sul da Amazônia. Acta
Amazonica 14:175 - 221.
Salcedo, J. C. R. (1999). Crecimiento y desarrollo postnatal del Pinguino del Humbolt Spheniscus
humboldti (Meyen,1834). Biología Lima, Universidad Nacional Agraria La Molina.
Biologo: 72.
Sanz, V., and A. Rodríquez-Ferraro. 2006. Reproductive parameters and productivity of the
Yellow-shouldered Parrot on Margarita Isalnd, Venezuela: a long-term study. Condor
108:178-192.
Smedley, S. R., and T. Eisner. 1996. Sodium: a male moth's gift to its offspring. Proceedings of
the National Academy of Science. USA 93:809-813.
Snyder, N. F. R., P. Mc Gowan, J. Gilardi, and A. Grajal. 2000. Parrots. Status survey and
conservation action plan 2000-2004. IUCN, Gland, Switzerland and Cambridge, UK.
Terborgh, J., J. W. Fitzpatrick, and L. H. Emmons. 1984. Annotated checklist of birds and
mammals species of Cocha Cashu Biological Station, Manu National Park, Peru. Fieldiana
(Zoology, New Series) 21:1 - 29.
Terborgh, J., S. K. Robinson, T. A. P. III, C. A. Munn, and N. Pierpont. 1990. Structure and
organization of an Amazonian forest bird community. Ecological Monographs 60:213-
238.
Tosi, J. A. 1960. Zonas de vida natural en el Perú. Memoria explicativa. sobre el mapa ecológico
del Perú. Instituto Interamericano de las Ciencias Agricolas de la Organización de los
Estados Americanos.
van Schaik, C. P., J. Terborgh, and S. J. Wright. 1993. The phenology of tropical forests: adaptive
significance and consequences for primary consumers. Annual Review of Ecology and
Systematics 24:353-377.
Wiley, A. S., and S. H. Katz. 1998. Geophagy in pregnancy: a test of a hypothesis. Current
Anthropology 39:532-545.
Wiedenfeld, D. A. (1994). "A new subspecies of Scarlet Macaw and its status and conservation".
Ornitologia Neotropical 5: 99-104.
Williams, M. (2001). Variacion en parametros de crecimiento en pollos de carne con diferentes
densidades energeticas en la dieta. Nutricion. Lima Universidad Nacional Agraria La
Molina.
Zhang, S. Y., and L. X. Wang. 1995. Comparison of three fruit census methods in French Guiana.
Journal of Tropical Ecology 11:281-294.

Anexo 1
Cronograma de Asistentes Nov 2007 – Nov 2008
Autorización N° 044 – S/C - 2007 – INRENA – IANP
Asistentes Nov Dic Ene Feb Mar Abr May Jun Jul Ago Set Oct Nov
Donald Brightsmith x x x x
Gabriela Vigo x x x x X
Jerico Solis x x x x
Richard Amable
Aimy Cáceres
Patricia Mendoza x X
Carolina Caillaux X x x x
Adrián Sánchez x x x x x
Renzoandre de la Peña x x
Elizabeth Shedd
Lizzie Ortiz
Alan Keneth Lee
Caterina Cosmópolis
Vilma Taipe
Jennette Fox
Kelly Jones
Jill Heatley
Erin Wilensky
Lucía Castillo
George Oláh
Jesús Zumarán
X
x
x
X
X
x
x
x
X
x
X
x
x
X
x
x
X
x
x
x
x
x
X
X
X
x
x
x
X
X
x
X
x
X
x
X
X
x
X
x
x
x
X
X
X
x
X
x
X
x
* Aimy Cáceres y Richard Amable no formaron parte de las actividades correspondientes al
periodo Noviembre 2007 – Noviembre 2008, por encontrarse trabajando en otros
proyectos.

Figure 1: Satellite
collars attached to Blue
and yellow Macaws in
southeastern Peru. The
Telonics unit is above
and the North Star unit
is below.
Anexo 2
SATELLITE TELEMETRY OF LARGE MACAWS IN
TAMBOPATA, PERU
Progress as of March 2009
Donald J. Brightsmith
Schubot Exotic Bird Health Center
Texas A&M University
Since 2003 a group of parrot researchers, conservation organizations, and
manufacturers have been working together to design and test satellite
collars for tracking large macaws (Appendix 2). To date, this consortium has
developed two prototypes, tested these prototypes on captive macaws,
tested a prototype for location accuracy in Peru (Brightsmith and Boyd
2006) and Guatemala (Garcia et al. 2007), attached two collars to birds in
Guatemala (Bjork and McNab 2007), and attached eight collars to birds in
Peru. In this report I will briefly describe the collars and the accuracy testing
in Peru, then discuss in depth the results from two collars attached to birds
in Peru in 2008.
DEVELOPMENT AND TESTING OF PROTOTYPES
To date, two different types of prototype satellite telemetry transmitters
have been designed and tested. The first was developed in 2005 by North
Star Science and Technology Inc., with the financial support of Amigos de las
Aves USA and the Loro Parque Foundation. The second was developed in
2007 by Telonics Inc. with the financial support of Amigos de las Aves USA.
The North Star unit is a 32 g adjustable collar (Fig.1). A dummy prototype
was tested for durability on captive Scarlet Macaws at Loro Parque, Spain.
The collar was not damaged by the birds, and despite the size and rigid
antenna, the birds were not hampered during the test period while moving

in and out of nest boxes and living in the cage environment (Matthias Reinschmidt pers. com.).
During September of 2006, a working prototype was tested at Tambopata Research Center to
determine its accuracy. The collar was programmed to transmit for 5 hours per day and hung in
emergent trees to simulate the favored perches of large macaws. Over the 15 day test, the
collar provided 0.87 good locations per day (with average errors of about 1.5 ± 1.5 km)
(Brightsmith and Boyd 2006). An additional 42 potentially useful locations with higher error
rates were also logged.
The Telonics unit is also an adjustable collar with a total
weight of 37.5 g (Fig 1). Two dummy prototypes
constructed by Telonics were tested for durability on
captive macaws at Texas A&M’s Schubot Aviary in July
2007. Over the course of a month, one collar was
completely destroyed by a Scarlet Macaw (A. macao) and
its mate a Red-and-green Macaw (A. chloropterus). On the
other collar, the antenna attachment was loosened by a
pair of Blue-and-yellow Macaws. The collars were returned
to Telonics to be hardened; however, the final unit sent to
Tambopata did not appear very different from the first
prototypes. This collar was placed on a Blue-and-yellow
Macaw on 25 January 2008, registered one unusable
location on 26 January and then ceased transmitting. In
September 2008 a redesigned dummy prototype was
tested on captive macaws at Texas A&M University and it
was determined to be sufficiently resistant. As a result, two
collars were placed on wild macaws at Tambopata
Research Center in early 2009.
COLLAR DEPLOYMENT AND PERFORMANCE 2008
In January 2008, my team at Tambopata Research Center captured three Blue-and-yellow
Macaws and attached a satellite collar to each. The birds were trapped using tall perches
covered with nylon nooses (Fig 2.). As mentioned above, the one collar manufactured by
Telonics Inc. was placed on a Blue-and-yellow Macaw on 25 January 2008, registered one
unusable location on 26 January and then ceased transmitting. Two collars manufactured by
North Star were deployed on 22 and 28 January and functioned in to November 2008. The
remainder of the discussion will focus exclusively on the results from these two North Star
collars.

Figure 2: Perch traps (long poles), trapping blind and close-up of nylon
nooses (below) used to capture Blue-and-yellow Macaws at Tambopata
Research Center, Peru.
A total of 245 useable locations were
generated in a total of 1222 broadcast
hours by the two transmitters combined
(Table A). Useable locations are defined
as those with location quality 3, 2, 1 or 0
and an estimated error of < 6 km. A total
of only eight highly precise locations
(class 3, < 1 km error) have been received
during the course of the study.
Table 1: Locations received from two satellite transmitters placed on Blue-and-yellow Macaws at
Tambopata Research Center in the lowlands of southeastern Peru. The broadcast schedule used is
presented in Table 3. Error locations are given in kilometers.
Location class
Month 3 2 1 0 A B Z Total
Broadcast
days
Jan 0 2 0 3 4 1 2 12 3
Feb 1 5 9 9 10 11 9 54 14
Mar 1 4 5 7 9 11 14 51 16
Apr 0 0 3 4 8 13 16 44 14
May 0 2 2 10 9 11 14 48 17
Jun 0 2 6 3 9 12 18 50 18
Jul 1 2 10 3 7 11 11 45 20
Aug 0 4 7 4 3 13 7 38 22
Sep 1 8 19 8 11 28 31 106 20
Oct 2 11 29 31 18 30 31 152 20
Nov 2 11 5 9 10 8 0 45 9
Total 8 51 95 91 98 149 153 645 173
Argos error >1.0 <1.0 <0.35 <0.15
No
estimate
No
estimate
Invalid
location
TRC error* 5.09 1.59 1.46 0.56 2.24 14.3 28.3
* TRC error locations from (Brightsmith and Boyd 2006).

Table 2: Numbers of useable locations (location class 3, 2, 1, and 0, estimated
error < 6 km) for each of two satellite transmitters attached to Blue-and-yellow
Macaws in the Tambopata region of southeastern Peru in January 2008.
Locations per hour refer to locations per hour the collar was broadcasting.
Tiny (#33) Libertad (#34)
Useable locations 107 138
Locations per day 1.33 ± 1.64 1.67 ± 1.67
Broadcast hours 639 538
Locations per hour 0.17 0.24
0.0
0.1
0.2
0.3
0.4
0.5
0.6
1 2 3 4 5 6 7 8 9 10 11 12
Good locations per broadcast hour
Month
Tiny Libertad
Figure 3: The number of good locations (<6 km average error) received per
broadcast hour for two satellite telemetry collars placed on Blue-and-yellow
Macaws captured at Tambopata Research Center in the lowlands of southeastern
Peru. Collars broadcasted 6 hours per day from January until mid September, after
which the collars broadcasted 8 hours per broadcast day.

Figure 5: Project staff Don Brightsmith, Gabriela Vigo and Lizzie Ortiz with bird #34,
Libertad on 28 January 2008 at Tambopata Research Center in southeastern Peru.
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
3-Jan-08 22-Feb-08 12-Apr-08 1-Jun-08 21-Jul-08 9-Sep-08 29-Oct-08 18-Dec-08
Battery voltage
Date
Voltage Tiny
Voltage Libertad
Figure 4: The voltage output of the batteries of two satellite telemetry collars
placed on Blue-and-yellow Macaws captured at Tambopata Research Center
in the lowlands of southeastern Peru. These are from a voltmeter inside the
collar. Collars broadcasted 6 hours per day from January until mid September,
after which the collars broadcasted 8 hours per broadcast day.

Figure 6: Bird #34, Libertad, being
given an electrolyte solution before
release. The Northstar satellite collar
is visible around its neck.
Libertad (transmitter #34) had a slightly higher average number of locations per
day (1.67 ± 1.67, N = 86 days) than Tiny (transmitter #33, 1.33±1.64, N = 88 days)
but this did not differ significantly (t-test: p = 0.17, Table 2).
The number of useable locations generated per broadcast hour declined from
deployment through August (Fig 3). However in mid September the number of
locations per broadcast hour increased dramatically. This switch corresponded
with the collars switching from a 6 hour transmission per broadcast day to an
eight hour broadcast day (see broadcast cycles below). The voltage readings
received from the transmitter batteries follows a similar trajectory, with initial
declines followed by an increase in mid September suggesting that there is a
causal relationship between the battery voltage and the number of good
locations.
BROADCAST CYCLES
The choice of the amount of time and the time of day the transmitter broadcasts and how long
it remains off between transmissions is a vitally important aspect of working with satellite
transmitters. These factors determine how long the transmitter will last, how many data points
are received per day, and the quality of those locations.
In this study we programmed the transmitters so that they would broadcast 6 or occasionally 11
hours per day once every 3 to 4 days (Table 3). Near the end of the deployment period, we
programmed the collars to transmit 8 hours per day once every four days. For all broadcast
days, the collar is programmed to start broadcasting at about 5:00 AM local time. This was
chosen because there was good satellite coverage predicted during the mornings over the study
area during the study period.
At the beginning of the study we programmed the collars to broadcast 6 hours per day once
every 4 days (6 hours on and 90 hours off, Table 3). This phase lasted for 136 days. At which
time the collars broadcast for 11 hours for one day then 6 hours per day once every 3 days for a
month before repeating the process. For the final cycle, during which the battery was predicted
to die, the collar was programmed to broadcast for 8 hours per day once every 3 days.
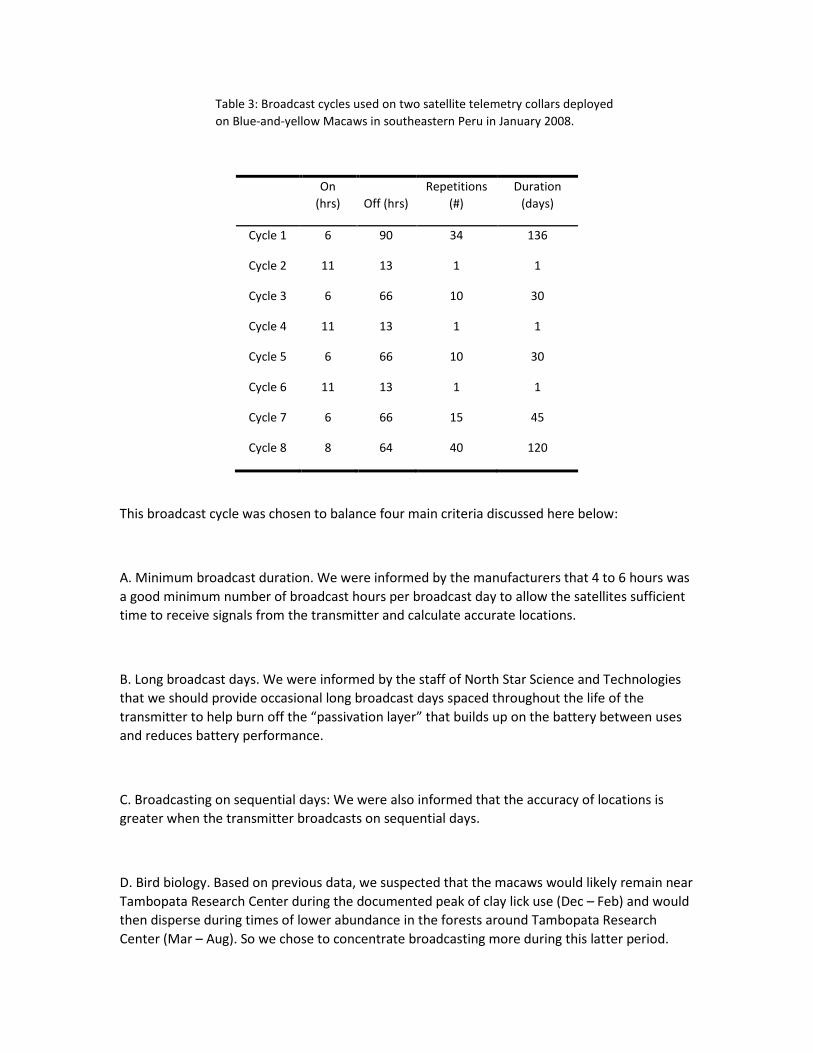
Table 3: Broadcast cycles used on two satellite telemetry collars deployed
on Blue-and-yellow Macaws in southeastern Peru in January 2008.
On
(hrs) Off (hrs)
Repetitions
(#)
Duration
(days)
Cycle 1 6 90 34 136
Cycle 2 11 13 1 1
Cycle 3 6 66 10 30
Cycle 4 11 13 1 1
Cycle 5 6 66 10 30
Cycle 6 11 13 1 1
Cycle 7 6 66 15 45
Cycle 8 8 64 40 120
This broadcast cycle was chosen to balance four main criteria discussed here below:
A. Minimum broadcast duration. We were informed by the manufacturers that 4 to 6 hours was
a good minimum number of broadcast hours per broadcast day to allow the satellites sufficient
time to receive signals from the transmitter and calculate accurate locations.
B. Long broadcast days. We were informed by the staff of North Star Science and Technologies
that we should provide occasional long broadcast days spaced throughout the life of the
transmitter to help burn off the “passivation layer” that builds up on the battery between uses
and reduces battery performance.
C. Broadcasting on sequential days: We were also informed that the accuracy of locations is
greater when the transmitter broadcasts on sequential days.
D. Bird biology. Based on previous data, we suspected that the macaws would likely remain near
Tambopata Research Center during the documented peak of clay lick use (Dec – Feb) and would
then disperse during times of lower abundance in the forests around Tambopata Research
Center (Mar – Aug). So we chose to concentrate broadcasting more during this latter period.

The balancing of these factors led us to choose the broadcast schedule presented in Table 3 and
led us to formulate the following hypotheses:
1. Days with longer broadcast duration would have more quality locations per broadcast hour than those with shorter broadcast durations
2. Broadcasting on sequential days should produce more high quality locations per broadcast hour on the second day of broadcasting compared to days without sequential broadcasting
3. Days following long broadcast days would have higher quality signals and stronger signal strength due to the burn off of the battery’s passivation layer
4. Birds would average further from TRC during the low season (Mar – Aug). This will be discussed in the section “Macaw movements from satellite data” (below).
1. Broadcast duration and number of quality locations: Each collar had three 11 hour broadcast
days during its life time. During these long broadcast days we have received an average of 2.17 ±
0.75 useable locations per day compared to 0.56 ± 0.86 on the 3 days immediately prior. This is
over five times more locations on the long broadcast days. Even when the number of locations is
corrected by the number of broadcast hours, the trend still holds, with 0.197 ± 0.07 usable
locations per broadcast hour on long broadcast days compared to 0.09 ± 0.14 usable locations
per broadcast hour on the three previous broadcast days. In both cases the number of locations
is statistically significantly1 greater during the long duration transmissions (t-test: N = 6, p <
0.04). However, the total number of good locations per broadcast hour was highest for 8 hour
transmissions (0.45 ± 0.24 good loc/broadcast hour, N = 41 days, t-test: p < 0.001).
2. Sequential transmission days and number of quality locations: On six occasions there have
been transmissions on sequential days. In all cases, long broadcast days were followed on the
next morning by a normal 6 hour transmission day. The day after the long transmission we
averaged 0.25 ± 0.22 useable locations per hour of transmission, compared to an average of
0.09 ± 0.067 locations on the three previous broadcast days. However, this trend is not
statistically significant (N = 6, P = 0.13) as only three times did the day following the long
transmission have more useable locations and the three previous days. With additional collars,
this trend may well prove to be statistically significant.
1 In this case the values from all six long transmission days are treated as independent. The author
acknowledges that this is a case of “pseudoreplication” and that more transmitters will be needed before a
more appropriate statistical analysis (ANOVA) can be used which takes in to account the repetitions within
each collar.

3. Improved performance following long transmissions: If battery passivation is reduced by long
transmission days, we predict that the broadcasts following these days would have stronger
signals and produce more usable locations. As predicted, satellites received stronger signals
from our transmitters on the days following long broadcast days (-123.7 ± 3.4 db, N = 6)
compared to the three previous broadcast days (-127.2 ± 3.4 db, N= 6, t-test: p = 0.04). However
this effect was apparently short lived, as the average signal strength received by the satellites on
the second, third and fourth broadcast days after the long broadcast day (-127.7 ± 1.4 db) did
not differ from the three broadcast days before the long broadcast day (-127.2 ± 3.4 db, N= 6, t-
test: p = 0.31).
As mentioned above, we received more usable locations on the day following the long broadcast
day than on the three previous broadcast days. However, this effect also appears to be short
lived as we did not average more locations on the second, third, and fourth broadcast days after
the long broadcast day (0.065 ± 0.042 loc/broadcast hour) when compared to the three
broadcast days immediately preceding the long broadcast day (0.04 ± 0.067 loc/broadcast hour,
N = 6, t-test: p = 0.41). The transmitter carries a battery volt meter, but it transmits battery
voltage readings intermittently so we are unable to directly test the effect of the long broadcast
day on battery voltage level.
Summary of broadcast cycle analyses
Given that our main biological interest in this study is to plot the seasonal movements of large
macaws, the ideal broadcast cycles would be one that produces one good location per
broadcast day and allows the transmitter to last as long as possible. The following discussion is
based on the definition of good locations as those with an estimated error of < 6km (location
classes 0, 1, 2, and 3). If the researcher were interested in locations with less than 2 km error it
would be a very different discussion.
Broadcasts of 6 hours per broadcast day produce on average 0.13 ± 0.14 good locations per
broadcast hour (N = 127 days). Long broadcast days (11 hours of continuous broadcast)
produced 0.20 ± 0.068 locations per hour (N = 6 days). However, the 8 hour broadcasts
produced 0.45± 0.24 locations per hour. This shows that the 8 hour broadcast periods likely
produce the most locations per broadcast. In addition, no useable locations were received
during 41% of the 6 hour broadcasts (N = 127), while only 5% of the 8 hour broadcasts gave no
useable locations.
Long transmissions apparently did improve battery performance, as the strength of the signal
received by the satellite on the subsequent transmission was greater, but this effect apparently
wore off rapidly as it was not evident from 4 – 10 days after the long transmission. However,

when the transmitter was broadcasting for one 8 hour period once every four days, the batter
performance was significantly improved.
Broadcasts on subsequent days may produce more locations than broadcasts once every three
days, but a larger sample size is needed to test this hypothesis effectively. This tendency, if real,
may be due to the improved battery performance mentioned above, the fact that the satellite
could better predict the transmitter’s location due to the locations calculated the day before, or
a combination of both.
Additional analyses are needed to determine the effects of the satellite locations during each
broadcast to tease out the effects of the weakening battery and the satellite availability in the
number of usable locations received.
COLLAR DURATION
The manufacturer Northstar conservatively estimated that the batteries in the satellite collars
should provide about 400 total hours of transmission. Both of the collars deployed in Peru
surpassed this value (639 hours for Tiny and 583 for Libertad). As a result, both collars lasted
from January to November, a period of over 41 weeks. This allowed us to track almost an entire
annual cycle for these birds.
SATELLITE COLLAR TESTS PLANNED FOR 2009
In February and March 2009 researchers at Tambopata Research Center hung two functioning
satellite telemetry collars in the canopy of a tree (nest tree Hormiguero). One collar was
manufactured by Telonics and one by Northstar. These over the coming months, these two
collars will rotate through a broadcast cycle of either 4, 6 or 8 hours per day once every 3, 4 or 5
days. This test will provide us with additional information on the most efficient broadcast
schedule. It will also allow us to compare the accuracy of the different classes of locations and
the different manufacturers.
TRAPPING AND COLLAR ATTACHMENT IN 2009
In January – February 2009 we attached an additional 5 collars to macaws by trapping them at
the clay lick at Tambopata Research Center. One Blue-and-yellow Macaw was trapped using the
perch traps covered by nylon nooses (see previous section for description of the trapping
methods). This bird was captured on 28 Dec in the early morning while trying to trap smaller

parrots. A Northstar collar was attached to the bird and it was released. The bird flew strongly
upon release. This collar broadcasted for only 18 days after release. Since that time no locations
have been received and we presume that the collar was destroyed by the bird. On 21 January,
we captured two birds. One, a Blue-and-yellow Macaw was captured in the perch traps. This
bird was affixed with a Telonics collar and released. The bird flew well upon release. This collar is
still broadcasting successfully as of March 2009. The second bird, a Scarlet Macaw, was captured
in a mist net hung approximately 20 m from a small section of the clay lick commonly used by
large macaws. While the bird was being removed from the net, it got wet in the water that was
under the net. The bird was affixed with a Northstar collar. Upon release the bird flew weakly
and went to the ground within about 30 m of the site where it was released. After resting briefly
it tried to fly again, flew weakly and fell in to the river. Two project assistants immediately dove
in to the river to rescue the bird. As they swam towards the bird, it was attacked and killed by a
caiman. As a result of this incident, we changed our capture procedures to ensure that birds
were not allowed to get wet during capture and that they were released further from the river
to ensure that they would not be able to fall in to the river during their first flights. On 2
February we captured an additional two birds, both Scarlet Macaws, in mist nets placed near
two different sections of the clay lick. One was fitted with a Northstar collar and one with a
Telonics collar. The bird fitted with the Telonics collar already had a band. It was a chick that had
fledged from the nest PVC in early 2005. Both birds flew well upon release and their collars are
transmitting well as of March 2009. In addition, both birds were photographed by a visitor
within 2 km of TRC on 14 and 16 Feb.
BLUE-AND-YELLOW MACAW NATURAL HISTORY
Counts at the clay lick have shown that most psittacine species have great seasonal fluctuations
in clay lick use, with use peaking during the breeding season. For Blue-and-yellow Macaws, lick
use peaks during breeding (Dec – Feb) and then is low from Mar through August. Point count
censuses show that the number of Blue-and-yellow Macaws in the forests around TRC are
lowest from May – September (Brightsmith 2006a, b). Our data also show that the food
abundance in the palm swamps and in the forest in general is lowest from Mar – August. This
suggests that some birds leave the area around TRC during the non-breeding season when
annual food supplies are at the lowest. Determining where these birds go is vital to our
understanding and conservation of this species and the clay licks in the region. The preliminary
results from the first two birds are presented here below.
MACAW MOVEMENTS FROM SATELLITE DATA
Using only the good locations from the satellite data, we estimated that the two macaws ranged
over areas of 1911 km2 (Tiny #33, N = 87 locations on 54 different days, Fig. 3) and 7341 km2
(Libertad #34, 117 locations on 66 days, Fig 4). The two birds used the landscape in different
ways. The area used by Libertad was nearly 4 times greater than the area used by Tiny.
However, there were fundamental similarities in the movements of both birds. Both birds

Figure 7: Blue-and-yellow Macaws perched near the clay lick at
Tambopata Research Center.
focused the majority of their activities in core areas of about 200 km2 within 25 km of the
capture site early in the season. These sites will be referred to as the core areas. Then as the
season progressed, both birds showed extensive “wanderings” which lasted for 2 to 6 weeks
during which they traveled > 50 km from their core areas. These periods of “wandering” usually
ended with the birds settling in a small area (< 20 km2) for about 2 weeks before moving to a
new site.
The long distance movements and settling in
distant areas occurred during the period of
March – June, a time with consistently low
food supply in the areas around TRC. The next
step in these analyses is to identify the
habitat features in the distant areas where
the birds are settling for these two week
periods. These sites may contain keystone
food resources which allow the birds to
survive the seasons of low food availability
near the capture site.
The majority of the locations (> 80%) for each
bird were within the Bahuaja Sonene National
Park – Tambopata National Reserve
protected area complex. However both left
the protected area complex at least once per year. Tiny crossed the border in to Bolivia where it
entered Madidi National Park. However, Libertad left the protected areas and even spent time
along the route of the proposed Interoceanic Highway.
The details of the movements of the two birds follow:
Tiny (#33): Over 85% of the locations for this bird were in an area of 324 km2 centered about 9
km southeast of where it was captured (capture location: 13° 08’ S, 69° 37’ W, Table 4, Fig. 3).
The bird used in this core area at least once in all months of the study except June. From 13 May
– 3 Jun, the bird ranged over an area of 727 km2 to the southeast of TRC (N = 7 locations over 4
days). From 3 Jun to 25 Jun (N = 5 locations on 4 days) the bird was in a small area (8.5 km2) 91
km to the southeast from TRC. This area was in the Madidi National Park in Bolivia. By on 7 July
the bird was 16 km north (still in Madidi, Bolivia) then returned the 90 km to the original core
area by 22 July where it remained until the collar stopped transmitting. In summary, the bird
was consistently in a small area southeast of the capture site for the majority of the study with
occasional visits to areas 7 to 15 km from the center of this area. However, over the period from
21 May – 22 July, the bird left the core area and ranged over 1,156 km2 (> 70% of the total area
recorded during this study).

Libertad (#34): From 31 Jan – 3 March (N = 18 locations on 9 days), the bird was in a core area
of 190 km2 centered about 16 km NNW of the capture site (Table 5, Fig 4). From 3 March
through 19 March (N = 8 locations on 5 days), the bird ranged more widely, traveling 35 ± 14 km
between days and covering an area of 787 km2. During this time the bird left the protected area
complex and crossed the route of the Interoceanic Highway. From 19 March through 4 April (N =
5 locations on 4 days) it was about 12 km north of the core area in a small area (6.5 km) about
22 km north of the site of capture. From 12 to 16 April (N = 2 locations on 2 days) the bird
returned to the core area. From 16 April to 30 May (N = 9 locations on 6 days) the bird ranged
over an area of 4,853 km2 moving an average of 90 ± 40 km between days. During this time it
left the protected areas at least once and apparently spent one night roosting in the core area
before moving out again. From 30 May – 17 June (N = 8 locations on 4 days) the bird used an
area of 15 km2 about 77 km from its core area and 68 km to the SE of the capture area. From 26
June – 27 July the bird was in an area of 62 km2, about 19 km from its core area and 28 km NNW
of the capture site. This range is mostly outside the protected areas, and contains the route of
the Interoceanic Highway across its northern section and is bisected north to south by a route
used by miners to gain illegal access to the upper Malinowski River. Four of the locations are
within 1 km of roads where this bird may have been at risk from hunters. From 2 Aug to 2
September the bird settled in a small area (10 km2) just on the border of the reserve. Then from
8 September until the transmitter stopped functioning, the bird was back in the original home
range.
Table 4: Home ranges used by a Blue-and-yellow Macaw (Tiny, #33) captured and fitted with a
satellite telemetry collar at Tambopata Research Center in the lowlands of southeastern Peru.
The distance and direction from the trap site is given when the bird established in small
ranges, it is left blank for periods when the birds wandered widely throughout the landscape.
“Locations, Days” refers to the number of different days on which locations were received.
“Calendar days” gives the time period during which the bird used each area. For a graphical
representation of these data, see Figure 8.
Range From trap site Locations
Dates used
area
(km2)
Distance
(km) Direction Total Days
Calendar
days
26 Jan - 13 May, - 22
July – 14 Nov 324 9 120 75 45 222
13 May - 3 June 727 7 4 21
3 - 25 June 8.5 91 120 5 4 22
Total range 1911 87 54 292
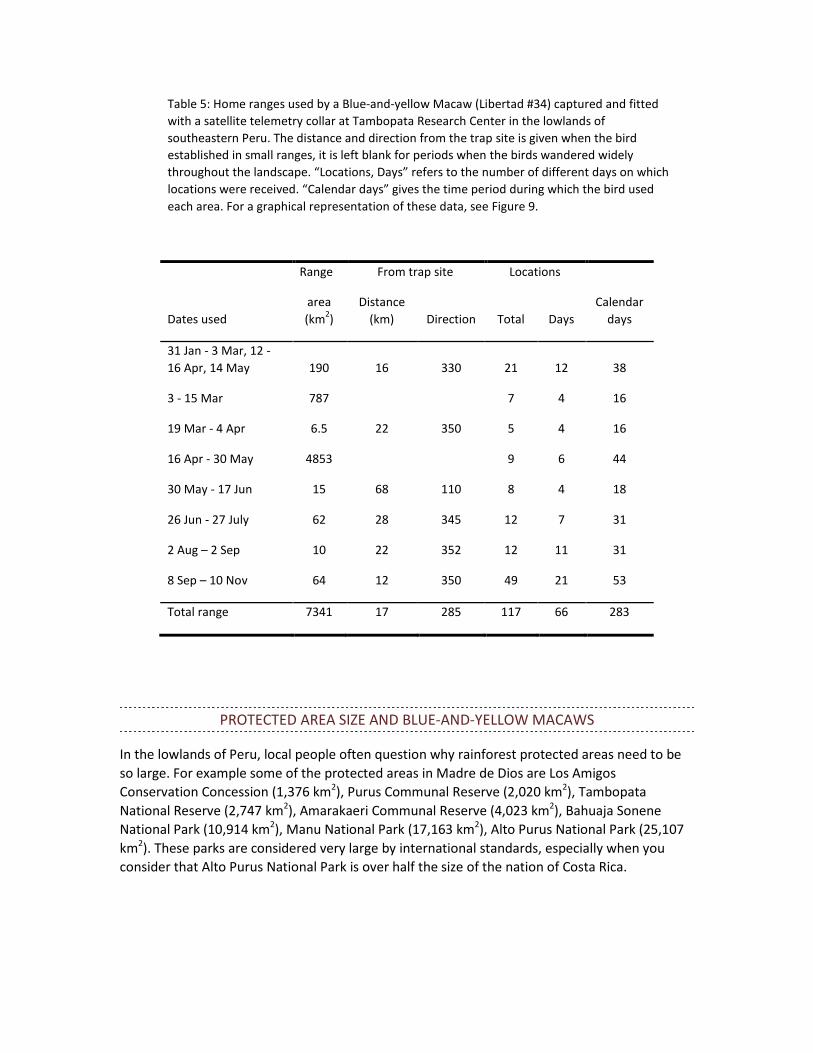
Table 5: Home ranges used by a Blue-and-yellow Macaw (Libertad #34) captured and fitted
with a satellite telemetry collar at Tambopata Research Center in the lowlands of
southeastern Peru. The distance and direction from the trap site is given when the bird
established in small ranges, it is left blank for periods when the birds wandered widely
throughout the landscape. “Locations, Days” refers to the number of different days on which
locations were received. “Calendar days” gives the time period during which the bird used
each area. For a graphical representation of these data, see Figure 9.
Range From trap site Locations
Dates used
area
(km2)
Distance
(km) Direction Total Days
Calendar
days
31 Jan - 3 Mar, 12 -
16 Apr, 14 May 190 16 330 21 12 38
3 - 15 Mar 787 7 4 16
19 Mar - 4 Apr 6.5 22 350 5 4 16
16 Apr - 30 May 4853 9 6 44
30 May - 17 Jun 15 68 110 8 4 18
26 Jun - 27 July 62 28 345 12 7 31
2 Aug – 2 Sep 10 22 352 12 11 31
8 Sep – 10 Nov 64 12 350 49 21 53
Total range 7341 17 285 117 66 283
PROTECTED AREA SIZE AND BLUE-AND-YELLOW MACAWS
In the lowlands of Peru, local people often question why rainforest protected areas need to be
so large. For example some of the protected areas in Madre de Dios are Los Amigos
Conservation Concession (1,376 km2), Purus Communal Reserve (2,020 km2), Tambopata
National Reserve (2,747 km2), Amarakaeri Communal Reserve (4,023 km2), Bahuaja Sonene
National Park (10,914 km2), Manu National Park (17,163 km2), Alto Purus National Park (25,107
km2). These parks are considered very large by international standards, especially when you
consider that Alto Purus National Park is over half the size of the nation of Costa Rica.

It is clear from our data from only two birds that Blue-and-yellow Macaws are moving with in a
single year over areas comparable to the sizes of the protected areas in Madre de Dios
Department. In a period of only six months, Libertad ranged over areas larger than four of the
protected areas mentioned above and used an area > 25% of the size of even the largest reserve
in the region. As a result, conservation of healthy populations of Blue-and-yellow Macaws likely
requires the birds be protected over areas of tens to hundreds of thousands of square
kilometers. However, before we can formally present this argument, we need additional data so
that we can have a clear and robust picture of the ranging patterns of these birds.
Figure 8: Home ranges used by a Blue-and-yellow Macaw (Tiny #33) captured and fitted with a satellite
telemetry collar at Tambopata Research Center in the lowlands of southeastern Peru. The point of
capture was the Colorado Clay lick (adjacent to TRC). Puerto Maldonado (~40,000 inhabitants) is the
capital of the department (Madre de Dios). The red line is the route of the Interoceanic Highway which
will connect Brazil to the coast of Peru and become a major international route for trucks. The locations in
the southeastern corner of the map are in Madidi National Park, Bolivia. For the approximate area of each
range, see Table 4.
25 km 16 April – 30 May 20 May – 17 Jun 3 - 19 Mar
26 Jun – 27 Jul 19 Mar – 4 Apr Puerto
Maldonado
Core 13 May – 3 June 3 – 25 June
25 km Puerto
Maldonado
25 km
13 May – 3 June
3 – 25 June
Core

Figure 9: Home ranges used by a Blue-and-yellow Macaw (Libertad #34) captured and fitted with a
satellite telemetry collar at Tambopata Research Center in the lowlands of southeastern Peru. The point
of capture was the Colorado Clay lick (marked as Colpa Colorado). For the approximate area of each
range, see Table 5. Puerto Maldonado (~40,000 inhabitants) is the capital of the department (Madre de
Dios). The red line is the route of the Interoceanic Highway which will connect Brazil to the coast of Peru
and become a major international shipping route. Deforestation around this road is expected to be
severe.
25 km 16 April – 30 May
26 Jun – 27 Jul
2 Aug – 2 Sep
30 May – 17 Jun
19 Mar – 4 Apr
3 – 19 Mar
Core

LITERATURE CITED
• BJORK, R., AND R. B. MCNAB. 2007. SUMMARY OF FIELDWORK: SATELLITE-
TAGGING SCARLET MACAWS IN THE MAYA BIOSPHERE RESERVE,
GUATEMALA, 25 JUNE - 5 JULY 2007. WILDLIFE CONSERVATION SOCIETY,
BOISE, ID.
• BRIGHTSMITH, D. J. 2006A. NATURAL HISTORY AND CONSERVATION OF
BLUE-AND-GOLD MACAWS IN PERU. PAGES 13-17 IN PROCEEDINGS OF THE
32ND ANNUAL CONVENTION. AMERICAN FEDERATION OF AVICULTURE,
DALLAS, TX, USA.
• BRIGHTSMITH, D. J. 2006B. THE PSITTACINE YEAR: WHAT DRIVES ANNUAL
CYCLES IN TAMBOPATA'S PARROTS?IN VI INTERNATIONAL PARROT
CONVENTION. LORO PARQUE FOUNDATION, LORO PARQUE, TENERIFE,
SPAIN.
• BRIGHTSMITH, D. J., AND J. BOYD. 2006. TESTING SATELLITE TELEMETRY
TAGS FOR PSITTACINES IN TAMBOPATA, PERU. UNPUBLISHED REPORT TO
THE LORO PARK FOUNDATION, NORTH STAR TECHNOLOGIES, AND AMIGOS
DE LAS AVES PSITTACINE CONSERVATION FUND, SCHUBOT EXOTIC AVIAN
HEALTH CENTER, TEXAS A&M UNIVERSITY, COLLEGE STATION, TEXAS.
• BROUWER, K., M. L. JONES, C. E. KING, AND H. SCHIFTER. 2000. LONGEVITY
RECORDS FOR PSITTACIFORMES IN CAPTIVITY. INTERNATIONAL ZOO
YEARBOOK 37:299-316.
• GARCIA, R., R. B. MCNAB, AND V. H. RAMOS. 2007. TESTING SATELLITE
TELEMETRY TAGS FOR PSITTACINES IN PETEN, GUATEMALA. UNPUBLISHED
REPORT, WILDLIFE CONSERVATION SOCIETY, FLORES, GUATEMALA.
• PEPPERBERG, I. M. 2006. ORDINALITY AND INFERENTIAL ABILITIES OF A
GREY PARROT (PSITTACUS ERITHACUS). JOURNAL OF COMPARATIVE
PSYCHOLOGY 120:205-216.

MEMBERS OF THE SATELLITE TELEMETRY DEVELOPMENT CONSORTIUM
SCIENTISTS
Robin Bjork: SalvaNATURA Programa de Ciencias para la Conservación, Colonia Flor
Blanca, 33 Av. Sur #640, San Salvador, El Salvador, [email protected]
Janice Boyd: Amigos de las Aves USA, 317 Thames Dr, Slidell LA 70458,
Donald J. Brightsmith, Schubot Exotic Bird Health Center, Department of Veterinary
Pathobiology, Texas A&M University, College Station, Texas 77843-4467,
Roan McNab: Wildlife Conservation Society-Guatemala Program, Avenida 15 de Marzo
No 3, Flores, Peten, Guatemala, [email protected]
Greg Matuzak: Amigos de las Aves USA, 10090 Skyline Drive, Grass Valley, CA 95945,
USA, [email protected]
David Waugh: Loro Parque Foundation, 38400 Puerto de la Cruz Tenerife, Canary
Islands, Spain, [email protected]
Rony Garcia: Wildlife Conservation Society-Guatemala Program, Avenida 15 de Marzo
No 3, Flores, Peten, Guatemala, [email protected]
Gabriela Vigo: Tambopata Macaw Project, 920 Av. Huaylas, Chorrillos, Lima, Peru,
Victor Hugo Ramos: Wildlife Conservation Society-CEMEC/CONAP, Avenida 15 de Marzo
No 3, Flores, Peten, Guatemala, [email protected]
Blake Henke: North Star Science and Technology LLC, Post Office Box 438, King George,
Virginia 22485, [email protected]
MANUFACTURERS
North Star Science and Technology, LLC
Telonics

FUNDERS
Amigos de las Aves, USA
Loro Parque Foundation
Wildlife Preservation Foundation
Schubot Exotic Bird Health Center at Texas A&M University
Rainforest Expeditions, S.A.C.
Wildlife Conservation Society – Guatemala


















