

Fisiología intrínseca del laberinto posterior1
41
Fisiología intrínseca del laberinto posterior Daniela Rodríguez R.
-
Upload
xikitita-yop -
Category
Documents
-
view
457 -
download
4
Transcript of Fisiología intrínseca del laberinto posterior1

Fisiología intrínseca del laberinto posterior
Daniela Rodríguez R.

Actividad refleja mediante la cual el ser humano puede mantener su postura corporal con
respecto a la gravedad y al medio inercial, sin caer,
tanto en reposo como en movimiento, y de esta forma puede percibir el
entorno teniendo la impresión de estar estable
y seguro.
Estado en que se encuentra un cuerpo sometido a fuerzas opuestas que se contrarrestan
exactamente
Equilibrio

Laberinto posterior o aparato vestibular
Propioceptiva
Visual Auditiva
Cerebelosa
Programa
motor responsable del EQUILIBRIO
Canal de
entrada de
información al SNC
• Que elaborado
con
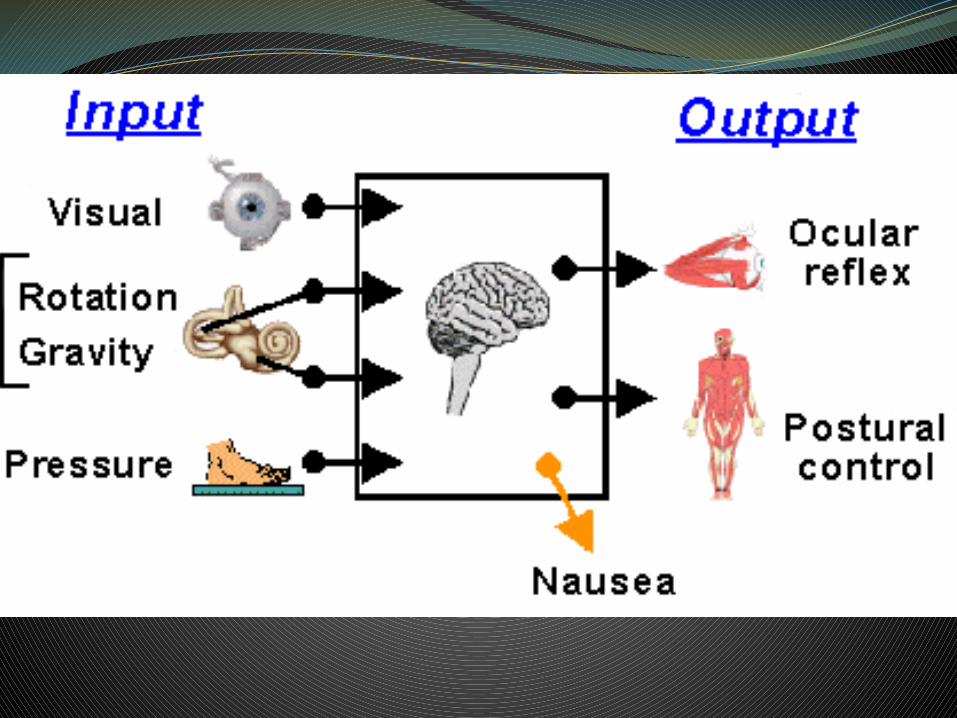


El papel del SV se puede resumir en tres funciones necesarias para el mantenimiento del equilibrio:
• Formación de la sensación de orientación espacial.• Mantenimiento del equilibrio corporal reflejo en reposo y
movimiento. • Estabilización de la cabeza y de las imágenes retinianas.
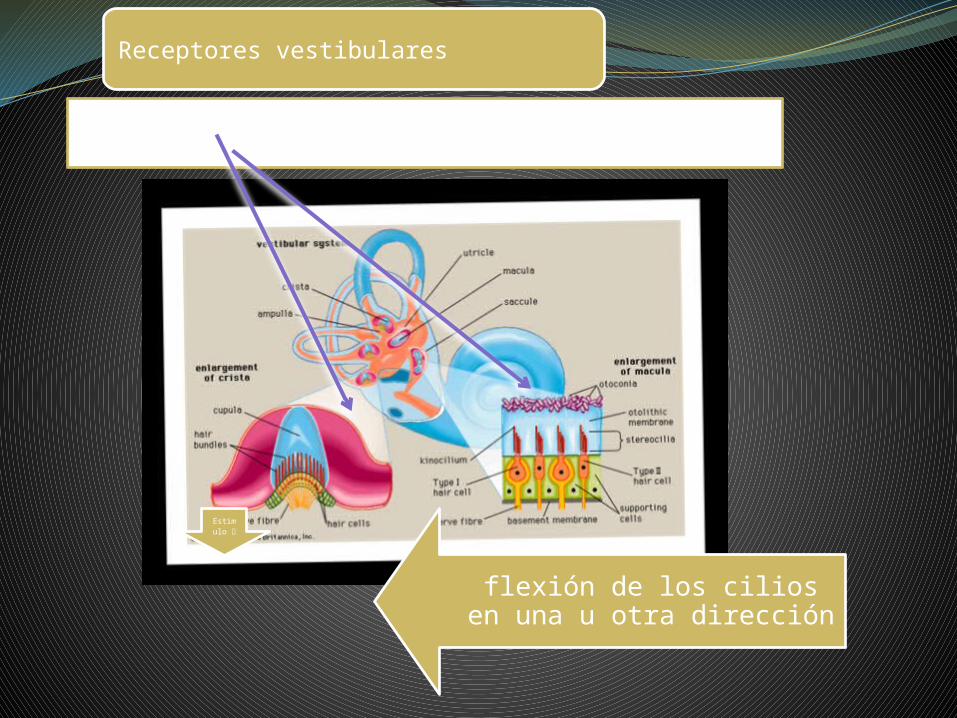
Receptores vestibulares
Estimulo
flexión de los cilios en una u otra dirección

Movimientos de la cabeza
Angulares (rotación de la cabeza) que estimularán a los
CS
Lineales (verticales y horizontales) que estimularán al
sistema otolítico o macular.

El sistema canalicular
Cada CS forma con su ampolla y el utrículo un anillo cerrado lleno de endolinfa.
Corriente endolinfática de inercia
Movimiento retrasado de la endolinfa con respecto ala
pared del CS cuando el anillo entra en rotación.
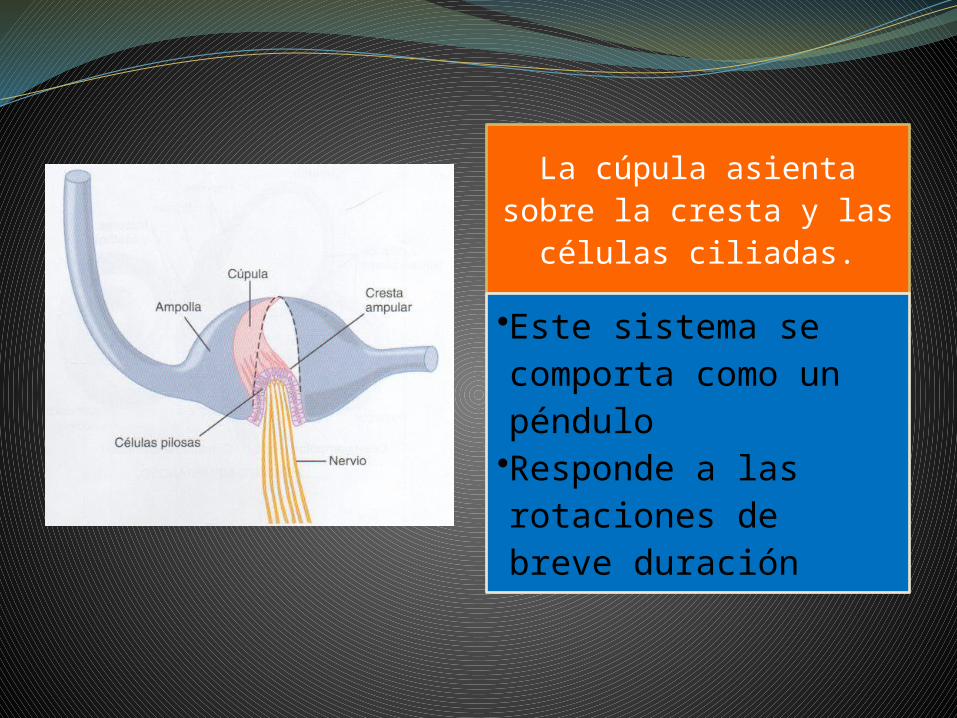
La cúpula asienta sobre la cresta y las
células ciliadas.•Este sistema se comporta como un péndulo•Responde a las rotaciones de breve duración

La cúpula solo puede desviarse hacia el
canal o hacia el utrículo
•La inclinación es proporcional a la intensidad del estímulo•Duración: 20-40s
Estos movimientos se denominan ampulípetos o utriculípetos y ampulífugos o utriculífugos
según su dirección.

Los conductos semicirculares se organizan en pares:
Los dos conductos horizontalesEl conducto superior y el conducto posterior
contralateralEl conducto posterior y el superior contralateral
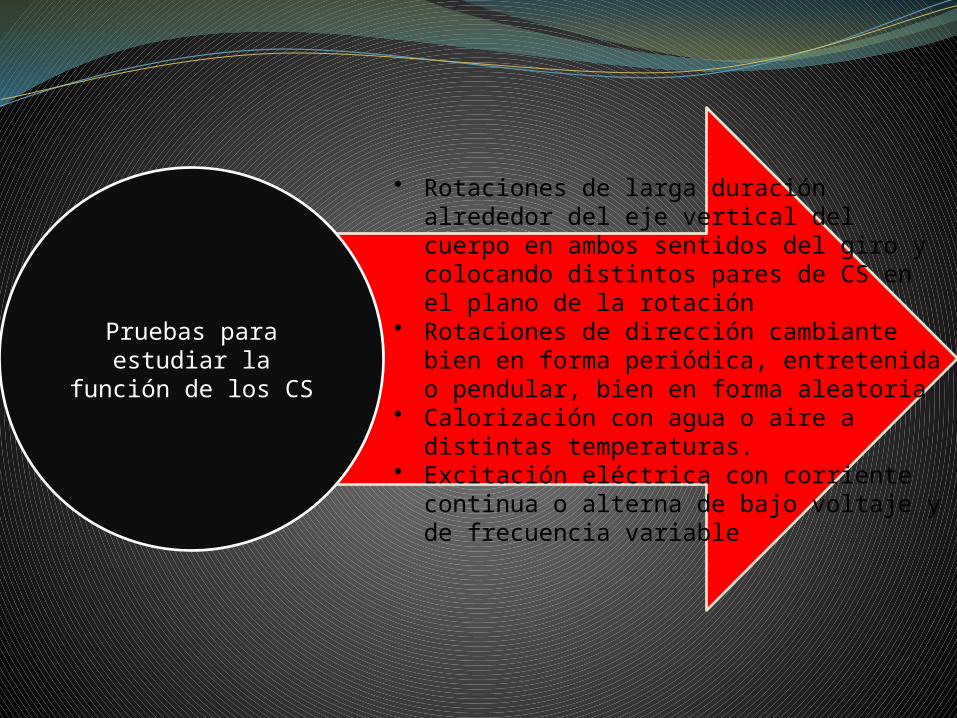
• Rotaciones de larga duración alrededor del eje vertical del cuerpo en ambos sentidos del giro y colocando distintos pares de CS en el plano de la rotación
• Rotaciones de dirección cambiante bien en forma periódica, entretenida o pendular, bien en forma aleatoria
• Calorización con agua o aire a distintas temperaturas.
• Excitación eléctrica con corriente continua o alterna de bajo voltaje y de frecuencia variable
Pruebas para estudiar la función de los CS
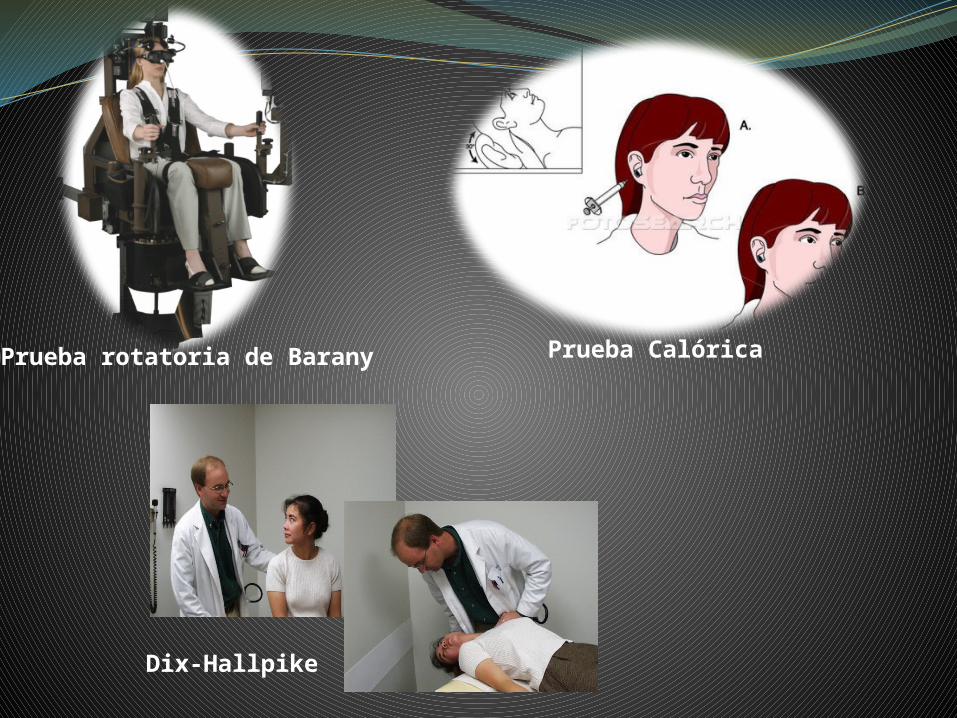
Prueba rotatoria de Barany Prueba Calórica
Dix-Hallpike

Leyes de la estimulación y de la respuesta del sistema vestibular
Ley de Flourens: las respuestas musculares o nistágmicas se producen en el mismo plano del conducto estimulado. La destrucción de un conducto provoca movimientos, desviaciones corporales y oculares en el plano de ese conducto.
Primera ley de Ewald: las respuestas musculares y nistágmicas tienen la misma dirección que la corriente endolinfática que las provoca. Según la segunda ley de Ewald: en el CSE las corrientes ampulípetas serían excitadoras y las ampulífugas inhibidoras. La excitación es más efectiva que la inhibición para cambiar la actividad laberíntica. Tercera ley de Ewald: en los CS verticales se invierte la segunda ley, es decir, es más activa la corriente ampulífuga que la ampulípeta.
Ley de Barany: el nistagmo prerrotatorio bate en el mismo sentido del giro que lo desencadena. El nistagmo posrotatorio bate en dirección opuesta al de la rotación precedente.

Sistema estatolítico
La mácula del utrículo queda en el plano horizontal de la superficie inferior del utrículo y determina la orientación de la cabeza cuando se encuentra
en posición vertical.La mácula del sáculo está situada verticalmente e informa la posición de la
cabeza cuando la persona está tumbada.

La gravedad se comporta como una aceleración lineal y debido a esto los
esteatolitos se dirigen por su peso
hacia abajo
Utrículo y sáculo
funcionan como una centrífuga
Esteatoconias mas densas y su masa es mayor que la
endolinfa
La membrana de esteatolitos se desplaza mas que la endolinfa y se desliza tangencialmente a la superficie del epitelio sensorial y flexiona los cilios en una u otra dirección


Posición de pie •Los esteatolitos de la mácula utricular descansan sobre el epitelio sensorial y no producen flexión alguna de los cilios.•En las máculas saculares sucede todo lo contrario.
Al ladear la cabeza sobre un hombro •La flexión de los cilios es máxima en el utrículo y mínima en el sáculo
Cuando el cuerpo realiza movimientos de traslación
•Membrana de esteatolitos se desplaza en determinadas zonas de las máculas, flexionando los cilios y excitando o inhibiendo las células sensoriales según su dirección

a•Contrarrotación ocular de Barany Toda inclinación de la cabeza en un sentido se acompaña de una desviación lenta de los ojos en sentido opuesto y en el mismo plano de la inclinación
b •Y Las desviaciones segmentarias de la musculatura esquelética (cuello, tronco, y miembros superiores)
Respuestas motoras de la activación del sistema esteatolitico
La activación del área vestibular de un lado condiciona el aumento del tono de los extensores y aductores en la mitad correspondiente
• En el lado opuesto se origina un predominio de los flexores y abductores

El funcionamiento del Aparato vestibular es un sistema de información muy sensible ya que no necesita sobrepasar un determinado umbral para entrar en actividad
Por eso carece de adaptación y fatiga fisiológicas
El umbral de activación del SNC que debe sobrepasarse para que aparezcan respuestas motoras visibles, registrables y cuantitativamente visibles.
Transformación del estímulo mecánico en excitación

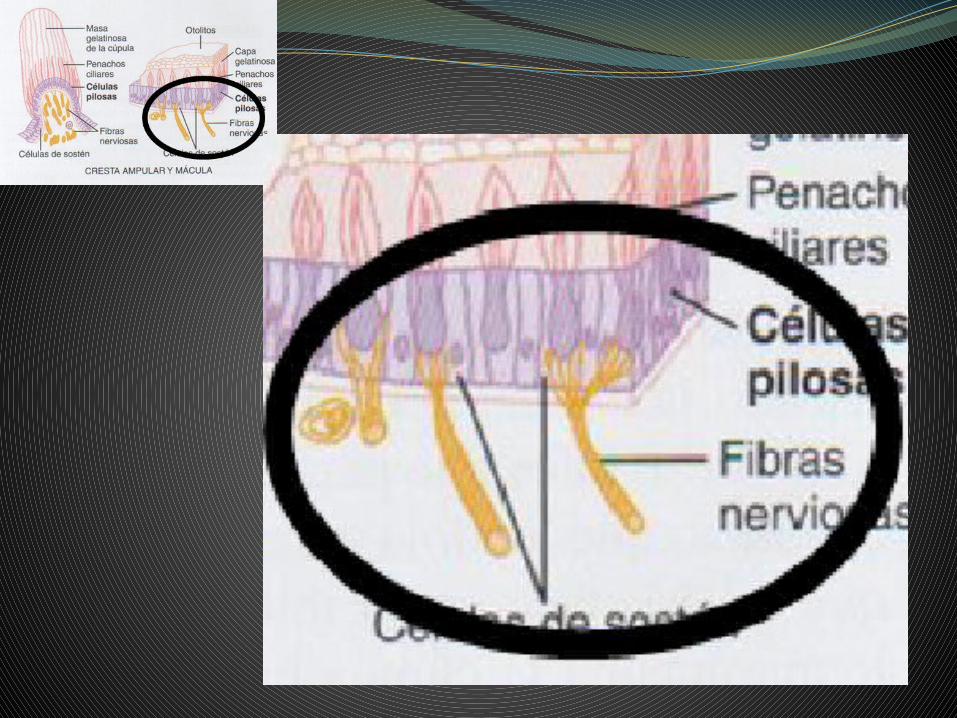
Estas conexiones sinápticas son diferentes para los dos tipos de células sensoriales de Wersall, cc tipo I y tipo II.
Las cc tipo I hacen sinapsis con una axón solamente, en las de tipo II un axón hace sinapsis con muchas cc. En este polo basal de las cc sensoriales nos encontramos también con fibras aferentes, llegadas de origen central, que tienen una función de control, modulando la sensibilidad dinámica de estas cc receptoras.
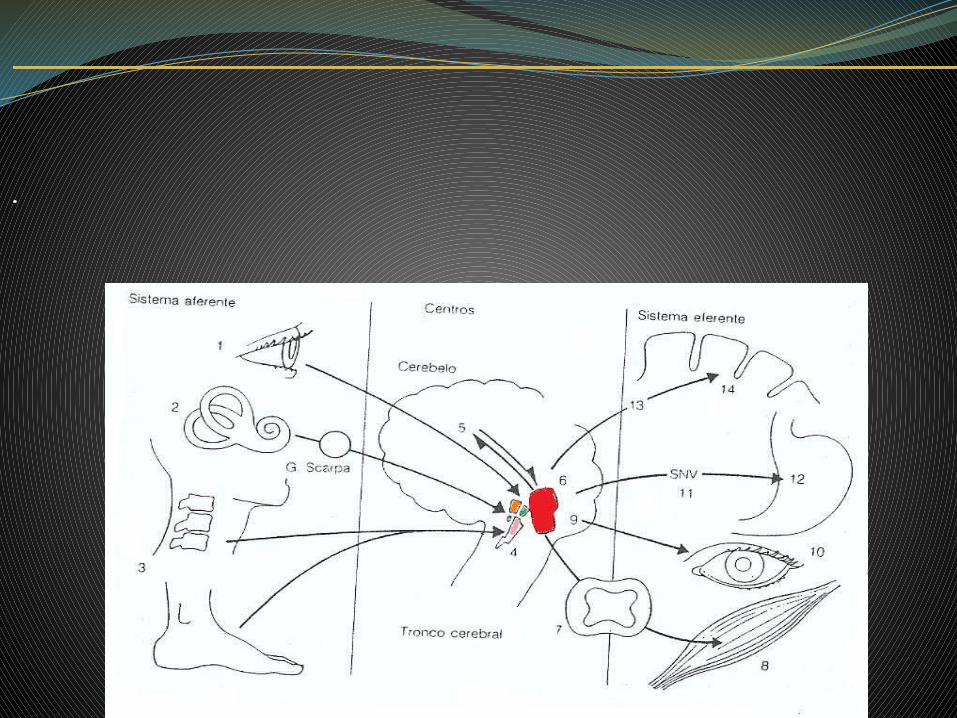
Las vías de conducción de los estímulos procedentes de los receptores del aparato vestibular llegan al ganglio vestibular(ganglio de Scarpa) en el conducto auditivo interno.

Las que vienen de los núcleos dorsal, media y espinal se unen al fascículo longitudinal medial y se ponen en
relación con los núcleos óculomotores,
conduciendo así los impulsos de los
reflejos de nistagmo.
Los axones del núcleo medial y de los otros
núcleos van al cerebelo (nucleus fastigii) por el
fascículo vestíbulocerebeloso,
junto con fibras venidas directamente del nervio vestibular, reuniéndose así con todas las demás
regiones corporales
Las fibras secundarias que van desde los núcleos lateral y espinal a la
médula espinal, formando el fascículo
vestibulospinal, terminan en contacto con neuronas motoras y forman así el
arco reflejo para los reflejos posturales y de
enderezamiento de origen vestibular.

Modificaciones del equilibrio
Sensaciones vertiginosas
Movimientos oculares anómalos
Fisiopatología vestibular

Patogenia de las alteraciones del equilibrioMientras se conserve 2 de 3 elementos
aferentes, el ser humano es capaz de mantener correctamente el equilibrio
Será capaz de andar normal y
equilibradamente
•Gracias a la información visual y propioceptiva
1.

Será incapaz de andar normal y
equilibradamente
•Deberá ayudarse con la sensibilidad estereoceptiva
OSCURIDAD
2.

3.

4.
ÓRGANOS EFECTORES
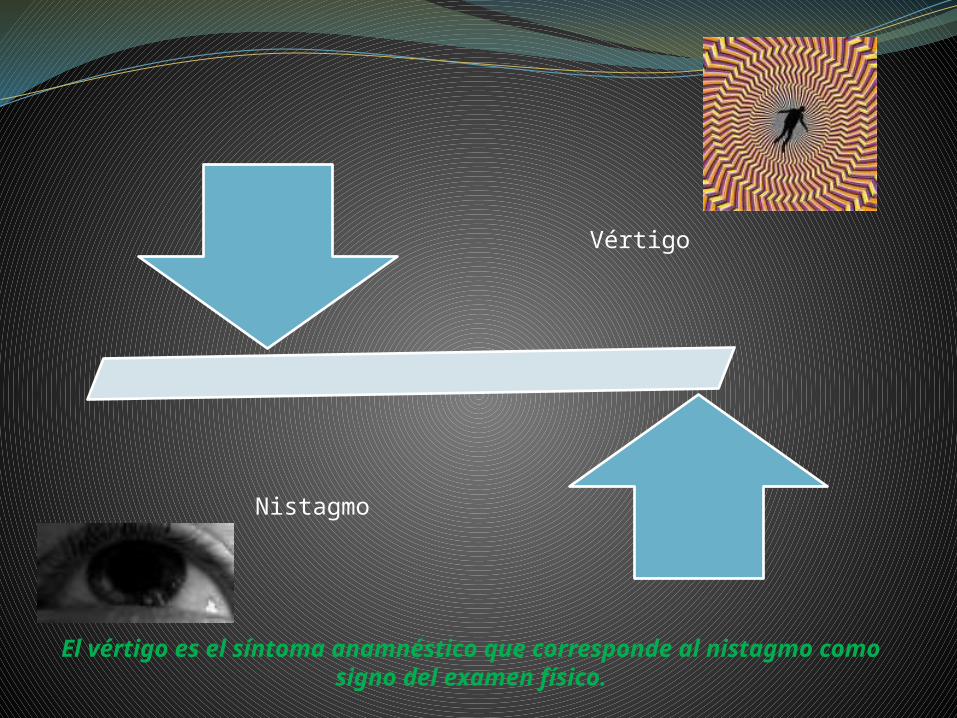
Vértigo
Nistagmo
El vértigo es el síntoma anamnéstico que corresponde al nistagmo como signo del examen físico.

Líquidos laberínticos
•Líquido de tipo exocelular con una constitución parecida al del LCR
Perilinfa
•Líquido de tipo intracelular
Endolinfa

Perilinfa
Los intercambios entre LCR y perilinfa se
realizarían a través del acueducto del caracol.
•Hay una transmisión de los aumentos de presión del LCR y perilinfa.•Maniobra de Queckensted (la compresión de las venas del cuello produce aumento de la presión del líquido cefalorraquídeo en el individuo normal, aumento que desaparece al cesar la compresión; pero si por cualquier causa hay bloqueo del conducto espinal, la compresión de las venas no produce ningún efecto)
Existe una diferencia en el nivel de proteínas entre LCR y perilinfa.
TEORIA 1.

TEORIA 2.
La perilinfa
•se deriva de la endolinfa

TEORIA 3.
La perilinfa se origina en los vasos que recubren las paredes de las cavidades
perilinfáticas.
•Mecanismo de producción: trasudación que da lugar a un ultrafiltrado pobre en proteinas.•Capilares perilinfaticos constituyen una barrera hematoperilinfática

ENDOLINFACONSTITUCIÓN •Líquido rico en potasio y pobre en sodio y albúmina.
•Llena todas las cavidades que forman el laberinto membranoso
ORIGEN •La secreción se localiza en ciertos segmentos del epitelio laberíntico (estria vasculaar)•En las paredes del utrículo y el sáculo existen formaciones idénticas a la estría vascular (planum semilunatum)

Reabsorción de la endolinfa
Tiene lugar en el saco endolinfático y / o a lo largo de todo el conducto coclear
La destruccion del saco endolinfático no provoca alteración de la endolinfa

Circulación de la endolinfa
Endolinfa secretada en la estría vascular
Sáculo (a través del conducto
reuniens
Circulación longitudinal (Guild)

Circulación radial (Naftalin y Harrison)
Perilinfa Memb. De Reissner
Endolinfa
Estría vascular
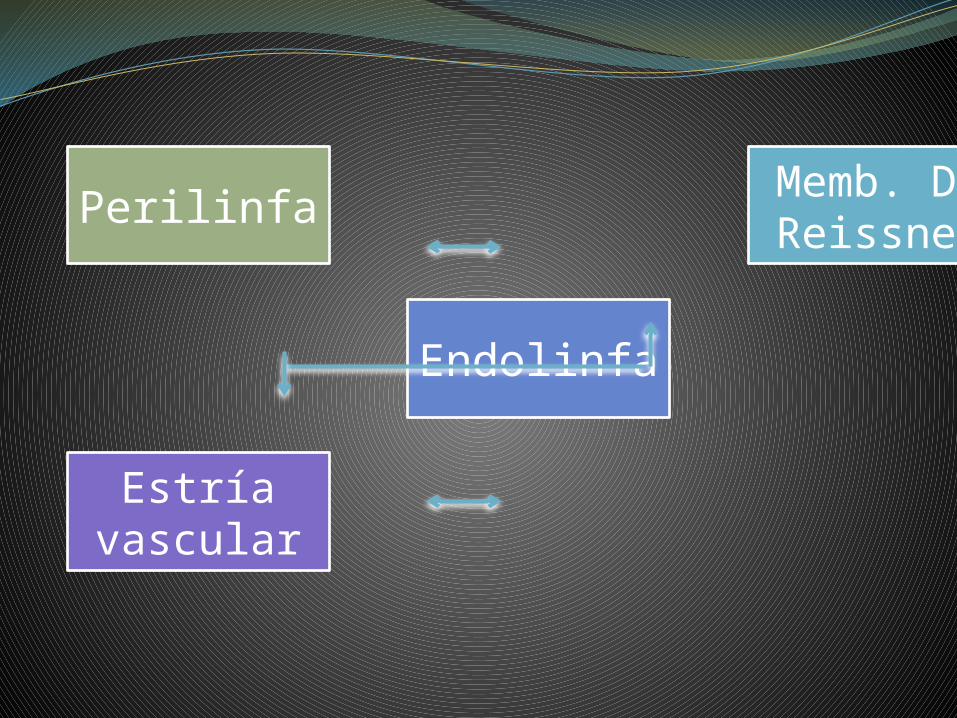
Perilinfa Memb. De Reissner
Endolinfa
Estría vascular


















