FACULTAD DE CIENCIAS DE LA SALUD...
65
Mblgo. Luis Alberto Sánchez Angulo / Mblgo. José Luis Gutierrez Aponte Biología Celular y Molecular Universidad Los Ángeles de Chimbote - ULADECH i FACULTAD DE CIENCIAS DE LA SALUD DEPARTAMENTO ACADÉMICO DE QUÍMICA Y FARMACIA SECCIÓN DE CIENCIAS BIOLÓGICAS ASIGNATURA BIOLOGÍA CELULAR Y MOLECULAR CONTENIDO: • CAPITULO UNO : “LAS BASES BIOLÓGICAS Y QUÍMICAS DE LOS SERES VIVOS” o La biología y los seres vivos o Componentes químicos de la materia viviente • CAPITULO DOS : “ESTRUCTURA Y FISIOLOGÍA CELULAR” o Biología celular: Citología y teoría celular o Estructura celular: Membrana celular citoplasma y núcleo o Fisiología celular: Respiración celular • CAPITULO TRES : “EL FLUJO DE LA INFORMACIÓN GENÉTICA” o Reproducción celular: Ciclo celular, mitosis y meiosis o Flujo de la información genética: Replicación, transcripción y traducción o El código genético y la regulación genética • CAPITULO CUATRO : “GENÉTICA, BIOTECNOLOGÍA E INGENIERÍA GENÉTICA” o Genética: Conceptos básicos y genética mendeliana o Genética póstmendeliana. Citogenética general y humana o Biotecnología e ingeniería genética COMPILADORES: MG. BLGO. MBLGO. LUIS ALBERTO SÁNCHEZ ANGULO BLGO. MBLGO. JOSE LUIS GUTIERREZ APONTE
Transcript of FACULTAD DE CIENCIAS DE LA SALUD...

Mblgo. Luis Alberto Sánchez Angulo / Mblgo. José Luis Gutierrez Aponte Biología Celular y Molecular
Universidad Los Ángeles de Chimbote - ULADECH i
FACULTAD DE CIENCIAS DE LA SALUD DEPARTAMENTO ACADÉMICO DE QUÍMICA Y FARMACIA
SECCIÓN DE CIENCIAS BIOLÓGICAS
ASIGNATURA
BIOLOGÍA CELULAR Y MOLECULAR
CONTENIDO:
• CAPITULO UNO : “LAS BASES BIOLÓGICAS Y QUÍMICAS DE LOS SERES VIVOS” o La biología y los seres vivos o Componentes químicos de la materia viviente
• CAPITULO DOS : “ESTRUCTURA Y FISIOLOGÍA CELULAR” o Biología celular: Citología y teoría celular o Estructura celular: Membrana celular citoplasma y núcleo o Fisiología celular: Respiración celular
• CAPITULO TRES : “EL FLUJO DE LA INFORMACIÓN GENÉTICA” o Reproducción celular: Ciclo celular, mitosis y meiosis o Flujo de la información genética: Replicación, transcripción y traducción o El código genético y la regulación genética
• CAPITULO CUATRO : “GENÉTICA, BIOTECNOLOGÍA E INGENIERÍA GENÉTICA” o Genética: Conceptos básicos y genética mendeliana o Genética póstmendeliana. Citogenética general y humana o Biotecnología e ingeniería genética
COMPILADORES: MG. BLGO. MBLGO. LUIS ALBERTO SÁNCHEZ ANGULO
BLGO. MBLGO. JOSE LUIS GUTIERREZ APONTE

Mblgo. Luis Alberto Sánchez Angulo / Mblgo. José Luis Gutierrez Aponte Biología Celular y Molecular
Universidad Los Ángeles de Chimbote - ULADECH 1


Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 69
ESTRUCTURA Y FISIOLOGÍA CELULAR
LA CITOLOGÍA O BIOLOGÍA CELULAR
La citología o biología celular es la rama de la biología que estudia las células en lo que concierne a su estructura, sus funciones y su importancia en la complejidad de los seres vivos. Citología viene del griego κγτοs cavidad. Con la invención del microscopio óptico fue posible observar estructuras nunca antes vistas por el hombre, las células. Esas estructuras se estudiaron más detalladamente con el empleo de técnicas de citoquímica y con la ayuda fundamental del microscopio electrónico. La biología celular se centra en la comprensión del funcionamiento de los sistemas celulares, de cómo estas células se regulan y la comprensión del funcionamiento de sus estructuras. Una disciplina afín es la biología molecular.
La célula es la unidad esencial que tiene todo ser vivo. Es además la estructura funcional fundamental de la materia viva según niveles de organización biológica, capaz de vivir independientemente como entidad unicelular, o bien, formar parte de una organización mayor, como un organismo pluricelular. La célula presenta 2 modelos básicos: la procarionte y eucarionte. Su organización general comprende: membrana plasmática, citoplasma y ADN. RESEÑA HISTÓRICA Y APORTES
La primera referencia al concepto de célula data del siglo XVII cuando el inglés Robert Hooke utilizó este término (por su parecer a las habitaciones de los sacerdotes llamados Celdas) para referirse a los pequeños huecos poliédricos que constituían la estructura de ciertos tejidos vegetales como el corcho. No obstante hasta el siglo XIX no se desarrolla este concepto considerando su
estructura interior. Es en este siglo cuando se desarrolla la teoría celular, que reconoce la célula como la unidad básica de estructura y función de todos los seres vivos, idea que constituye desde entonces uno de lo pilares de la Biología moderna. Fue esta teoría la que desplazó en buena medida las investigaciones biológicas al terreno microscópico pues no son visibles a simple vista. La unidad de medida utilizada es el micrómetro (μm) existiendo células de entre 2 y 20 μm.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 70
La investigación microscópica pronto daría lugar al descubrimiento de la estructura celular interna incluyendo el núcleo, los cromosomas, el aparato de Golgi y otros orgánulos celulares así como la identificación de la relación existente entre la estructura y la función de los orgánulos celulares. Ya en siglo XX la introducción del microscopio electrónico reveló detalles de las ultraestructura celular y la aparición de la histoquímica y de la citoquímica. También se descubrió la base material de la herencia con los cromosomas y el ADN con la aparición de la citogenética.
Atendiendo a su organización celular, los seres vivos se clasificarán en acelulares (virus, viroides) y celulares, siendo estos a su vez clasificados en eucariotas y procariotas. CAMPOS DE ESTUDIO
Para alcanzar sus objetivos, los biólogos celulares se ven obligados a estudiar los componentes de la célula a nivel molecular (biología molecular). Componentes principales del estudio celular: membrana plasmática, citoesqueleto, núcleo celular, ribosomas, retículo endoplásmico, aparato de Golgi, mitocondrias, cloroplastos, lisosomas, peroxisomas, vacuolas, pared celular, tráfico intracelular de membranas.
TEORÍA CELULAR
La teoría celular es una parte fundamental de la Biología que explica la constitución de la materia viva a base de células y el papel que éstas juegan en la constitución de la vida. Robert Hooke había observado ya en el siglo XVII que el corcho y otras materias vegetales aparecen constituidas de células (literalmente, celdillas).
Dos científicos alemanes, Theodor Schwann, histólogo y fisiólogo, y Jakob Schleiden, botánico, se percataron de cierta comunidad fundamental en la estructura microscópica de animales y plantas, en particular la presencia de núcleos, que el botánico británico Robert Brown había descrito recientemente (1827).
Publicaron juntos la obra Investigaciones microscópicas sobre la concordancia de la estructura y el crecimiento de las plantas y los animales (Mikroskopische Untersuchungen über die Übereinstimmung in der Struktur und dem Wachstum der Tiere und Pflanzen, Berlin, 1939). Asentaron el primer principio de la teoría celular histórica:
“Todo en los seres vivos está formado por células o productos secretados por las células”.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 71
Otro alemán, el médico Rudolf Virchow, interesado en la especificidad celular de la patología (sólo algunas clases de células parecen implicadas en cada enfermedad) explicó lo que debemos considerar el segundo principio:
“Toda célula se ha originado a partir de otra célula, por división de ésta”.
Ahora estamos en condiciones de añadir que la división es por bipartición, porque a pesar de ciertas apariencias, la división es siempre, en el fondo, binaria. El principio lo popularizó Virchow en la forma de un aforismo creado por Francois-Vincent Raspail, «omnis cellula e cellula». Virchow terminó con las especulaciones que hacían descender la célula de un hipotético blastema. Su postulado, que implica la continuidad de las estirpes celulares, está en el origen de la observación por August Weismann de la existencia de una línea germinal, a través de la cual se establece en animales (incluido el hombre) la continuidad entre padres e hijos y, por lo tanto, del concepto moderno de herencia biológica.
La teoría celular fue debatida a lo largo del siglo XIX, pero fue Pasteur el que, con sus experimentos sobre la multiplicación de los microorganismos unicelulares, dio lugar a su aceptación rotunda y definitiva.
Se puede resumir el concepto moderno de teoría celular en los siguientes principios:
1. Todo en los seres vivos está formado por células o por sus productos de secreción. La célula es la unidad anatómica de la materia viva, y una célula puede ser suficiente para constituir un organismo.
2. Todas las células proceden de células preexistentes, por división de éstas (Omnis cellula e cellula).
3. Las funciones vitales de los organismos ocurren dentro de las células, o en su entorno inmediato, controladas por sustancias que ellas secretan. En una célula caben todas las funciones vitales, de manera que basta una célula para tener un ser vivo (que será un ser vivo unicelular). Así pues, la célula es la unidad fisiológica de la vida.
4. Cada célula contiene toda la información hereditaria necesaria para el control del desarrollo y el funcionamiento de un organismo de su especie y para la transmisión de la información a las siguientes generaciones celulares. Así que la célula también es la unidad genética.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 72
LA CÉLULA
La célula es la unidad esencial de todo ser vivo. Es además la estructura funcional fundamental de la materia viva según niveles de organización biológica, capaz de vivir independientemente como entidad unicelular, o bien, formar parte de una organización mayor, como un organismo pluricelular. La célula presenta 2 modelos básicos: la procarionte y eucarionte. Su organización general comprende: membrana plasmática, citoplasma y ADN (núcleo).
La teoría celular es la base sobre la que se sustenta gran parte de la biología. Si excluimos los virus, todos los seres vivos que forman los reinos biológicos están formados por células. El concepto de célula como unidad funcional de los organismos surgió en los años 1830 y 1880. Las investigaciones se vieron retrasadas por el poco avance de los microscopios ópticos.
Comparación entre la célula eucariota animal y la procariota. En la célula procariota, la cápsula no siempre se presenta.
CLASIFICACIÓN
Existen dos tipos básicos de células: procariotas y eucariotas.
Las células procariotas son estructuralmente simples. Conformaron a los primeros organismos del tipo unicelular. Éstos tenían un ADN cerrado circular, el cual se encontraba disperso en el
citoplasma ausente de núcleo. La célula no tenía organelos –a excepción de ribosomas- ni
estructuras especializadas. Como no poseen mitocondrias, los procariotas obtienen energía del medio mediante mesosomas o invaginaciones en la membrana. Sus mayores representantes son las bacterias.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 73
Las células eucariotas son más complejas que las procariotas. Surgieron de las células procariontes. Tienen mayor tamaño y su organización es más compleja, con presencia de organelos, lo que permite la especialización de funciones. El ADN está contenido en un núcleo permeable rodeado de membranas. A este grupo pertenecen protozoos, hongos, plantas y animales.
Diferencias entre una célula Eucariota y Procariota
ESTRUCTURA PROCESOS
EUCARIOTAS PROCARIOTAS
Nucleo Verdadero o definido Falso, primitivo o no definido
Membrana nuclear
Presente Ausente
ADN Combinado con proteínas (histonas) forman la cromatina
Desnudo y circular. Ubicado en la región nuclear o nucleoide.
Cromosomas Múltiples Único
División celular
Mitosis o Meiosis Fisión binaria
Mitocondria Presentes (con ribosomas 70S) Ausentes:
Los procesos bioquímicos equivalentes tienen lugar en la membrana citoplasmática.
Cloroplasto Presentes en células vegetales (con ribosomas 70S)
Ribosomas 80S (a 60S y 40S sus subunidades)
70S (a 50S y 30S sus subunidades)
Pared celular Presente: Presente, constituida por mureína

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 74
Vegetales (por celulosa)
Hongos (quitina, mananos y glucanos)
Artropodos (exoesqueleto:quitina)
o péptidoglicano.
Nucléolos Presentes Ausentes
Retículo endoplásmico
Presente Ausente
Órganos de locomoción
Cilios y flagelos que al corte transversal presentan una distribución característica de microtúbulos: 9 + 2
Flagelos sin estructura 9+2
Diferencias estructurales entre una célula animal (a) y una célula vegetal (b)

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 75
FORMA Y TAMAÑO
La forma de la célula es variada y relacionada a la función que realizan en los diferentes tejidos, algunas tienen formas típica, como las neuronas (células del tejido nervioso), son mas largas que anchas y otras, como las del parénquima (un tipo de célula de las plantas) y eritrocitos (glóbulos rojos de la sangre), son equidimensionales; otras, como los leucocitos, son de forma cambiante. Muchas células cuando se encuentran en medio líquido tienden a tomar la forma esférica y, cuando están agrupadas en grandes masas forma poliédrica.
El tamaño de la célula está en relación con su función. La mayor parte de las células eucariotas sólo son visibles con el microscopio estando su diámetro comprendido entre 10 y 100 micrones (salvo excepciones).
Por lo general el tamaño resulta constante para cada tipo celular e independiente del tamaño del organismo, es decir una célula del riñón de un caballo es del mismo orden que la de un ratón. La diferencia en el tamaño del órgano se debe al número de células y no al tamaño de las mismas.
Los huevos (o, por usar la palabra latina, ova) son muy grandes, a menudo son las células mas grandes que produce un organismo (no en todos los casos, algunos organismos ponen "su huevo en una sola canasta" mientras que otros ponen una plétora de pequeños huevos). El gran tamaño de muchos huevos es en realidad una excepción, hecho relacionado con el proceso de desarrollo que ocurre luego que el óvulo es fertilizado,
cuando el contenido (del ahora cigoto) es usado en una serie de rápidas divisiones celulares, que requieren una tremenda cantidad de energía obtenida de las reservas de la célula huevo. Mas tarde el organismo adquirirá su propia energía pero, en el principio tiene un "fondo energético acumulado”.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 76
ESTRUCTURA DE LA CÉLULA EUCARIOTA ANIMAL
ESTRUCTURA DE LA CÉLULA EUCARIOTA VEGETAL

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 77
ORIGEN DE LAS CÉLULAS: Se cree que todos los organismos que viven sobre la Tierra, proceden de una única célula primitiva nacida hace varios miles de millones de años. Las similitudes entre todos los seres vivos parecen tan acusados que no se puede explicar de otra manera.
Las células vivas surgieron probablemente en la Tierra gracias a la agregación espontánea de moléculas, hace aproximadamente 3500 millones de años. Conociendo los organismos actuales y las moléculas que contienen, parece que debieron producirse por lo menos tres etapas antes de que surgiera la primera célula.
Debieron formarse polímeros de ARN capaces de dirigir su propia replicación a través de interacciones de apareamiento de bases complementarias.
Debieron desarrollarse mecanismos mediante los cuales una molécula de ARN pudiera dirigir la síntesis de una proteína.
Tuvo que ensamblarse una membrana lipídica para rodear a la mezcla autoreplicante de ARN y moléculas proteicas. En alguna fase posterior del proceso evolutivo, el ADN ocupó el lugar del ARN como material hereditario.
Hace unos 1.500 millones de años se produjo la transición desde células pequeñas con una estructura interna relativamente sencilla (células procariotas), hasta células más grandes, más complejas como las que componen los animales y las plantas (células eucariotas).
o Diferencias entre animales y vegetales
Vegetales
Animales
Poseen un pigmento verde, que constituye la clorofila indispensable para la fotosíntesis.
Como nutrientes utilizan el dióxido de carbono, agua con sales disueltas y energía solar para que por medio de la fotosíntesis puedan sintetizar compuestos orgánicos.
Por realizar la fotosíntesis tienen nutrición autótrofa, al elaborar sus propios alimentos.
Almacenan almidón.
En cuanto a la célula: o Membrana celular con pared celular
celulósica, que es de naturaleza rígida.
o Carecen de lisosomas (vegetales superiores).
o Carecen de centrosomas. o Las células de los tejidos se
comunican mediante aberturas finísimas denominadas
Carecen de clorofila.
Los animales y los hongos utilizan como alimento compuestos orgánicos portadores de energía química elaborados por las plantas.
Por los compuestos orgánicos ya preparados que utilizan, tienen nutrición heterótrofa.
Almacenan glucógeno.
En cuanto a la célula: o Sólo con membrana celular. o Poseen lisosomas para secreción de
enzimas digestivas. o Poseen centrosomas para la
reproducción de la célula. o Las células en los tejidos se
relacionan mediante barreras

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 78
plasmodesmos.
Los vegetales son seres fijos.
La irritabilidad (sensibilidad) es respondida con mucha lentitud y a través de simples movimientos de orientación (tropismos).
El crecimiento en longitud es ilimitado, teniendo lugar en el ápice y en los extremos de los órganos (yemas y raíces), durante la vida del organismo.
Los principales órganos de la planta son externos (raíz, tallo, hojas, etc.) y de una organización simple.
La conformación externa de los vegetales es muy ramificada.
intracelulares para la difusión, denominadas desmosomas.
Los animales tienen movimiento espontáneo al desplazarse en la búsqueda del alimento; a excepción de esponjas y corales.
Los animales responden con mayor rapidez, con respuestas más complicadas y visibles, por que la mayoría tienen sistema nervioso.
El crecimiento es limitado.
Los principales órganos son internos y protegidos dentro de cavidades. Estos órganos son de estructura compleja.
En los animales al ramificación de los órganos es interna, y es entre la masa orgánica del cuerpo.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 79
LA MEMBRANA CITOPLASMÁTICA
La membrana celular, citoplasmática o plasmática es una estructura laminar que envuelve el citoplasma de todas y cada una de las células, además de los orgánulos. Es una bicapa lipídica que sirve de "contenedor" para los contenidos de la célula, así como protección mecánica. Esta formada principalmente por lípidos y proteínas. Esta barrera presenta una permeabilidad selectiva, lo cual le permite "seleccionar" las moléculas que entran y salen de la célula. Tiene un grosor aproximado de 75 Å. Vista al microscopio electrónico presenta entre dos capas oscuras una central más clara.
En las células procariotas y en las de eucariotontes osmótrofos como plantas y hongos, se sitúa bajo otra capa, denominada pared celular.
Estructura de la membrana celular
Composición
La membrana plasmática está compuesta por proteínas, lípidos y glúcidos, cuyas masas guardan proporciones aproximadas de 50%, 40% y 10% respectivamente. Las moléculas más numerosas son las de lípidos, ya que se cree que por cada 50 lípidos hay una proteína. Sin embargo, las proteínas, debido a su mayor tamaño, representan aproximadamente el 50% de la masa de la membrana. Entre las proteínas, el 80% son intrínsecas, mientras que el 20% restantes son extrínsecas. De las proteínas se pueden encontrar las translocadoras o las enzimas asociadas a membrana, entre otras.
Los lípidos de la membrana son anfipáticos. Esto quiere decir que presentan un lado hidrófilo (que da la cara al agua) y un lado hidrofóbico (que no se junta con el agua). De entre los lípidos, los más importantes son los fosfolípidos y esfingolípidos, que se encuentran en todas las células; le siguen los glucolípidos, así como esteroides, como el colesterol. Estos últimos no existen o son escasos en las membranas plasmáticas de las células procariotas.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 80
Estructura
Su modelo estructural es conocido como mosaico fluido, El "mosaico fluido" es un término acuñado por S.J. Singer en 1971. Este consiste en una bicapa lipídica complementada con diversos tipos de proteínas. La estructura básica se mantiene unida mediante uniones no covalentes.
Esquema de una membrana citoplasmática según el modelo del mosaico fluido
Las proteínas de la membrana plasmática se pueden clasificar según cómo se dispongan en la bicapa lipídica.
• Proteínas integrales: Embebidas en la bicapa lipídica, atraviesan la membrana una o varias veces, asomando por una o las dos caras (proteínas transmembrana); o bien mediante enlaces covalentes con un lípido o a un glúcido de la membrana. Su aislamiento requiere la ruptura de la bicapa.
• Proteínas periféricas: A un lado u otro de la bicapa lipídica, pueden estar unidas débilmente por enlaces no covalentes. Fácilmente separables de la bicapa, sin provocar su ruptura.
Los glúcidos se hallan asociados mediante enlaces covalentes a lípidos, proteínas y generalmente forman parte de la matriz extracelular.
Otras sustancias pueden estar asociadas a esta estructura básica como diversos tipos de glúcidos que pueden unirse de forma covalente a lípidos (glucolípidos) o a proteínas (glucoproteínas). Las cadenas de estos glúcidos se disponen hacia el medio extracelular por la cara externa de la membrana y constituyen el glucocálix o matriz extracelular.
Esta estructura general -modelo unitario- se presenta también en las membranas de diversos orgánulos del interior de la célula: los del sistema de endomembranas, tales como retículo endoplasmático, aparato de Golgi y envoltura nuclear, y los de otros orgánulos, como las mitocondrias y los plastos, que proceden de endosimbiosis.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 81
Funciones
La función básica de la membrana plasmática reside en mantener el medio intracelular diferenciado del entorno. Esto es posible gracias a la naturaleza aislante en medio acuoso de la bicapa lipídica y a las funciones de transporte que desempeñan las proteínas. La combinación de transporte activo y transporte pasivo hacen de la membrana plasmática una barrera selectiva que permite a la célula diferenciarse del medio.
Los esteroides, como el colesterol, tienen un importante papel en la regulación de las propiedades, es decir que su rol es muy importante físico-químicas de las membranas regulando su resistencia y fluidez.
En el componente proteico reside la mayor parte de la funcionalidad de la membrana, las proteínas realizan funciones específicas y podemos clasificarlas según su función en:
• Estructurales: estas proteínas hacen de "eslabón clave" uniéndose al citoesqueleto y la matriz extracelular.
• Receptores de membrana: que se encargan de la recepción y transducción de señales químicas.
• Transportadoras a través de membrana: mantienen un gradiente electroquímico mediante el transporte de diversos iones.
Estas a su vez pueden ser:
• Proteínas transportadoras: Son enzimas con centros de reacción que sufren cambios conformacionales.
• Proteínas de canal: Dejan un canal hidrofílico por donde pasan los iones.
En el transporte transmembrana podemos hablar de:
• Transporte pasivo: Se produce sin consumo de energía y a favor de gradiente electroquímico.
• Transporte activo: Se produce con consumo de energía y en contra de gradiente electroquímico. El componente glucídico forma el glucocáliz, con funciones de cierta protección ante agresiones mecánicas y químicas, y la que parece más importante ya que permite diferenciar el exterior celular permitiendo un reconocimiento intercelular.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 82
EL CITOPLASMA
El citoplasma es la parte del protoplasma que en una célula eucariota se encuentra entre el núcleo celular y la membrana plasmática. Consiste en una emulsión coloidal muy fina de aspecto granuloso, el citosol o hialoplasma, y en una diversidad de organelos celulares que desempeñan diferentes funciones. Su función es mantener flotando los organelos celulares y al mismo tiempo ayuda al movimiento de los mismos. El citosol es la sede de muchos de los procesos metabólicos que se dan en las células.
Los cloroplastos (en las células vegetales) se encuentran en el citoesqueleto del citoplasma alrededor del citosol sublingual.
El citoplasma se divide en ocasiones en una región externa gelatinosa, cercana a la membrana, e implicada en el movimiento celular, que se denomina ectoplasma; y una parte interna más fluida que recibe el nombre de endoplasma y donde se encuentran la mayoría de los orgánulos. El citoplasma se encuentra en las células procariotas así como en las eucariotas y en él se encuentran varios nutrientes que lograron atravesar la membrana plasmática, llegando de esta forma a los orgánulos de la célula.
El citoplasma de las células eucariontas está subdividido por una red de membranas conocidas como retículo endoplasmático (liso y rugoso) que sirven como superficie de trabajo para muchas de sus actividades bioquímicas.
El retículo endoplasmático rugoso está presente en todas las células eucariontas y predomina en aquellas que fabrican grandes cantidades de proteínas para exportar. Es continuo con la membrana externa de la envoltura nuclear, que también tiene ribosomas adheridos.
CITOESQUELETO
El citoesqueleto es un entramado tridimensional de microtúbulos y microfilamentos que proveen el soporte interno para las células, anclan las estructuras internas de la misma e intervienen en los fenómenos de movimiento celular y en su división. Es una estructura dinámica que mantiene la forma de la célula, facilita la movilidad celular (usando estructuras como los cilios y los flagelos), y desempeña un importante papel tanto en el transporte intracelular (por ejemplo, los movimientos de vesículas y orgánulos) y en la división celular.
El citoesqueleto eucariota
Las células eucariotas tienen tres tipos de filamentos citoesqueléticos:
Microfilamentos (Actina y Miosina)
De unos 7 - 5 nm (nanómetros) de diámetro. Están formadas por una proteína globular llamada actina que puede presentarse de dos formas:
-Actina no polimerizada: la actina se encuentra asociada a la profilina que evita su polimerización. Representa la mitad de la actina de la célula y es utilizada para polimerizar microfilamentos cuando es necesario.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 83
-Actina polimerizada: es una doble hélice dextrógira de dos hebras de actina no polimerizada.
Esta actina se puede encontrar asociada a otras proteínas:
- Proteínas estructurales: que permiten la unión de los filamentos de actina. - Proteínas reguladoras: la más importante es la miosina que permite la contracción muscular al permitir que la actina se desplace sobre ella.
Las funciones de los microfilamentos de actina son la contracción muscular, la formación de pseudópodos, el mantenimiento de la morfología celular y, en la citocinesis de células animales, forma un anillo contráctil que divide la célula en dos.
Filamentos intermedios
Son filamentos de proteína fibrosa de unos 12 nm de diámetro, son los componentes del citoesqueleto más estables, dando soporte a los orgánulos (por sus fuertes enlaces), y heterogéneos. Las proteínas que conforman estos filamentos, la citoqueratina, vimentina, neurofilamentos, desmina y la proteína fibrilar acídica de la glía, dependen del tejido en el que se hallen. Su función principal es la organización de la estructura tridimensional interna de la célula (por ejemplo, forman parte de la envuelta nuclear y de los sarcómeros). También participan en algunas uniones intercelulares (desmosomas).
Las células eucariotas tienen tres tipos de filamentos citoesqueléticos:

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 84
Organización citológica:
Los microtúbulos se nuclean y organizan en los centros organizadores de microtúbulos (MTOCs), como pueden ser el centrosoma o los cuerpos basales de los cilios y flagelos. Estos MTOCs pueden poseer centríolos o no.
Además de colaborar en el citoesqueleto, los microtúbulos intervienen en el tránsito de vesículas (véase la dineína o la kinesina), en la formación del huso mitótico mediante el cual las células eucariotas segregan sus cromátidas durante la división celular, y en el movimiento de cilios y flagelos.
Farmacología:
Existen drogas que afectan a la estabilidad de los microtúbulos: El taxol, útil en los cánceres de ovario, a concentraciones bajas se une a los microtúbulos y los estabiliza, inhibiendo su acortamiento. La colchicina, o su derivado colcemida, se une a los dímeros de tubulina con alta afinidad, pero reversiblemente, lo que facilita que los dímeros envenenados se adhieran al extremo de un microtúbulo en crecimiento, impidiendo el agregado o pérdida de nuevas unidades. Así, el microtúbulo queda estabilizado. La colchicina se emplea ampliamente para sincronizar células, puesto que detiene la mitosis en metafase.
Microtúbulos
Los microtúbulos son estructuras tubulares de 25 nm de diámetro que se originan en los centros organizadores de microtúbulos y que se extienden a lo largo de todo el citoplasma. Se pueden polimerizar y despolimerizar según las necesidades de la célula. Se hallan en las células eucariotas y están formados por la polimerización de un dímero de dos proteínas globulares, la alfa y la beta tubulina. Cada microtúbulo está compuesto de tres protofilamentos formados por los dímeros de tubilina. Intervienen en diversos procesos celulares que involucran desplazamiento de vesículas de secreción, movimiento de orgánulos, transporte intracelular de sustancias, así como en la división celular (mitosis y meiosis). Además, constituyen la estructura interna de los cilios y los flagelos. Los microtúbulos son más flexibles pero más duros que la actina.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 85
ORGANELOS CELULARES
En biología celular, se denominan orgánulos llamados también organelas, organelos o mejor elementos celulares, a las diferentes estructuras suspendidas en el citoplasma de una célula eucariota, que tienen una forma y unas funciones especializadas bien definidas y diferenciadas. La célula procariota normalmente carece de orgánulos.
No todas las células eucariotas contienen todos los orgánulos al mismo tiempo, aparecen en determinadas células de acuerdo a sus funciones.
Principales organelos eucarioticos Orgánulo Función Estructura Organismos Notas
cloroplasto fotosíntesis posee doble-membrana
plantas, protistas
contiene algunos genes
retículo endoplasmático
síntesis y embalaje de proteínas y ciertos lípidos
puede asociarse con ribosomas en su membrana
eucariotes
aparato de Golgi transporte y embalaje de proteínas
sacos aplanados rodeado por membrana citoplasmática
la mayoría de eucariotes
en las plantas se conocen como dictiosomas
mitocondria producción de energía compartimiento de doble membrana
la mayoría de eucariotes
contiene algunos genes
vacuolas almacenamiento, transporte y homeostasis
sacos de membrana vesicular
plantas y hongos
núcleo mantenimiento de ADN y ARN, y expresión genética
rodeado por membrana doble
todos los eucariotes
contiene el genoma
Atendiendo a su génesis, los orgánulos se clasifican en dos grupos:
1. Orgánulos autogenéticos, desarrollados filogenética y ontogenéticamente de la complejización de estructuras previas.
2. Orgánulos de origen endosimbiótico, procedentes de la simbiosis con otros organismos.
Orgánelos autogenéticos
Las células eucariotas tienen un citoesqueleto complejo y dinámico. En esto se basa su capacidad para sostener estructuras membranosas complejas, así como para realizar desplazamientos internos y cambios de localización, orientación o forma de sus partes.
• Sistema de endomembranas. Es un conjunto de estructuras organulares basadas en vesículas o vacuolas como el retículo endoplasmático, liso o rugoso, los dictiosomas del aparato de Golgi o los lisosomas. La parte fundamental de la envoltura nuclear se interpreta como una vesícula del retículo endoplasmático y debe considerarse en este capítulo.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 86
• Estructuras especializadas del citoesqueleto. En este capítulo entran los centriolos y los relacionados corpúsculos basales, así como el axonema de los cilios y de los flagelos.
Orgánulos endosimbióticos
Son orgánulos incorporados a la célula eucariota inicialmente como bacterias endosimbiontes. Los orgánulos de origen endosimbiótico tienen su propio genoma, su propia maquinaria de síntesis proteica, incluidos ribosomas, y se multiplican por bipartición, de manera que si se extirpan experimentalmente de una célula no pueden volver a formarse.
• Mitocondrias.- Todos los eucariontes conocidos tienen mitocondrias, orgánulos derivados de ellas, como los hidrogenosomas, o al menos restos de genes mitocondriales incorporados al genoma nuclear.
• Plastos.- Hay dos clases de plastos, los primarios derivan de cianobacterias por endosimbiosis y los secundarios por endosimbiosis de células eucariotas ya dotadas de plasto. Éstos últimos son mucho más complejos. Los plastos se han designado muy a menudo con otros nombres en función de su pigmentación o del grupo en que se presenta. La denominación cloroplasto es usada habitualmente como nombre genérico para todos ellos.
Estructura de una célula eucariota
Las células eucariotas están formadas por diferentes orgánulos que desarrollan diversas funciones como son: Nucléolo. Núcleo celular. Ribosoma. Vesículas de secreción. Retículo endoplasmático rugoso. Aparato de Golgi. Citoesqueleto. Retículo endoplasmático liso. Mitocondria. Vacuola. Citoplasma. Lisosoma. Centríolo (Solo en la célula animal). Membrana citoplasmática. Cloroplasto (Solo en la célula vegetal y de las algas). Pared celular (Solo en la célula vegetal, de hongos y protistas).
Las células procariotas tienen el material genético disperso por el citoplasma y no en un núcleo diferenciado.
RETÍCULO ENDOPLASMÁTICO
El retículo endoplásmico, es una red de membranas interconectadas que forman cisternas, tubos aplanados y sáculos comunicados entre sí, que intervienen en funciones relacionadas con la síntesis protéica, metabolismo de lípidos y algunos esteroides, así como el transporte intracelular. Se encuentra en la célula animal y vegetal pero no en la célula procariota.
El retículo endoplasmatico rugoso se encuentra unido a la membrana nuclear externa mientras que el retículo endoplasmatico liso es una prolongación del retículo endoplasmatico rugoso.
• El retículo endoplasmático rugoso tiene esa apariencia debido a los numerosos ribosomas adheridos a su membrana mediante unas proteínas denominadas "riboforinas". Tiene unos sáculos más redondeados cuyo interior se conoce como "luz del reticulo" o "lumen" donde caen las proteínas sintetizadas en él. Está muy desarrollado en las células que por su función ceben realizar una activa labor de síntesis, como las células hepáticas o las células del páncreas.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 87
• El retículo endoplasmático liso no tiene ribosomas y participa en el metabolismo de lípidos.
Retículo endoplasmático rugoso
El retículo endoplasmático rugoso (RER), también llamado Retículo Endoplasmático Granular, Ergastoplasma o Retículo Endoplásmico Rugoso, es un orgánulo que se encarga de la síntesis y transporte de proteínas en general. Existen retículos sólo en las células eucariontes. En las células nerviosas es también conocido como Cuerpos de Nissl.
El término Rugoso se refiere a la apariencia de este orgánulo en las microfotografías electrónicas, la cual es resultado de la presencia de múltiples ribosomas en su superficie.
El RER está ubicado junto a la envoltura nuclear y se une a la misma de manera que puedan introducirse los ácidos ribonucleicos mensajeros que contienen la información para la síntesis de proteínas. Está constituido por una pila de membranas que en su pared exterior presentan adosados los ribosomas.
Funciones del Retículo Endoplasmático Rugoso
• Circulación de sustancias que no se liberan al citoplasma. • Síntesis y transporte de proteínas producidas por los ribosomas adosados a sus
membranas, pueden ser, proteínas de membrana, proteínas lisosomales o proteínas de secreción.
• Glicosilación de proteínas.
Las proteínas de secreción producidas, serán luego empaquetadas por el [aparato de Golgi] y serán liberadas al exterior de la célula para cumplir sus funciones (hormonales, enzimáticas, etc.). Las proteínas lisosomales también serán empaquetadas por el aparato de Golgi y

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 88
terminaran formando un lisosoma listo para cumplir sus funciones metabólicas intracelulares. Entre las enzimas producidas, se encuentran las lipasas, lasfosfatasas, las DNAasas, RNAsas y otras. Las proteínas de membrana pasarán a formar parte de la membrana plasmática o de la membrana de algún orgánulo.
El reticulo endoplasmático rugoso suele estar muy desarrollado en las células con alta actividad secretora de proteínas como son los plasmocitos, las células pancreáticas, etc.
Al evitar que las proteínas sean liberadas al hialoplasma, el retículo endoplasmático rugoso, consigue que estas no interfieran con el funcionamiento de la célula y sean liberadas solo cuando sean necesario, de otra manera, si por ejemplo quedaran libres en la célula proteínas enzimáticas que se encargan de la degradación de sustancias, las mismas destruirían componentes vitales de la célula.
Retículo endoplasmático liso
Conjunto de membranas que participan en el transporte celular y síntesis de triglicéridos, fosfolípidos y esteroides. También dispone de enzimas detoxificantes, que metabolizan el alcohol y otras sustancias químicas. En realidad los retículos endoplasmáticos lisos tienen diferentes variantes funcionales que sólo tienen en común su aspecto: los ribosomas están ausentes. Las cisternas del retículo endoplasmático liso son típicamente tubulares y forman un sistema de tuberías que se incurvan en el citoplasma.
Funciones del Retículo Endoplasmático liso
• En gónadas y corteza suprarrenal realizan la síntesis de hormonas esteroideas. • En el hígado detoxifican varios tipos de compuestos orgánicos como barbitúricos o etanol.
La detoxificación tiene lugar por una serie de enzimas oxigenasas entre las que se encuentra la citocromo P450 que dada su inespecificidad son capaces de detoxificar miles de compuestos hidrófobos transformándolos en hidrófilos, más fáciles de excretar.
• Liberación de glucosa a partir de Glucosa 6-fosfato via Glucosa 6-fosfatasa. • También secuestran los iones calcio y lo liberan regularmente en algunas células (retícula
sarcoplasmático).
Funciones del Retículo endoplasmático
• Síntesis de proteínas123: La lleva a cabo el retículo endoplasmático rugoso mediante los ribosomas. Estas proteínas serán transportadas al Aparato de Golgi mediante vesículas de transición donde dichas proteínas sufrirán un proceso de maduración para luego formar parte de los lisosomas o de vesículas secretoras.
• Metabolismo de lípidos: El retículo endoplasmático liso, al no tener ribosomas le es imposible sintetizar proteínas pero sí sintetiza lípidos de la membrana plasmática, colesterol y derivados de éste como los ácidos biliares o las esteroideas.
• Detoxificación: Es un proceso que se lleva a cabo principalmente en las células del hígado y que consiste en la inactivación de productos tóxicos como drogas, medicamentos o los propios productos del metabolismo celular, por ser liposolubles (hepatocitos)

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 89
• Glucoxilación: Son reacciones de transferencia de un oligosacárido a las proteínas sintetizadas. Se realiza en la membrana del retículo endoplasmático. De este modo, la proteína sintetizada se transforma en una proteína periférica externa del glucocálix.
APARATO DE GOLGI
• El Aparato de Golgi es un conjunto de dictiosomas (de 4 a 8 sáculos aplanados rodeados de membrana y apilados unos encima de otros). Funciona como una planta empaquetadora, modificando vesículas del retículo endoplasmático rugoso. El material nuevo de las membranas se forma en varias cisternas del Golgi. Se encuentra en el citoplasma de la célula. Dentro de las funciones que posee el aparato de Golgi se encuentran la glicosilación de proteínas, selección, destinación, glicosilación de lípidos y la síntesis de polisacáridos de la matriz extracelular.
• Debe su nombre a Camilo Golgi, Premio Nobel de Medicina en 1906 junto a Santiago Ramón y Cajal.
Se pueden diferenciar diferentes partes:
Cara externa, proximal o cis: Es la más próxima al retículo. De él recibe las vesículas de transición, que son sáculos con proteínas que han sido sintetizadas en la membrana del retículo endoplasmático rugoso (RER), introducidas dentro de sus cavidades y transportadas por el lúmen hasta la parte más externa del retículo. Estas vesículas de transición son el vehículo de dichas proteínas que serán transportadas a la cara externa del aparato de Golgi.
Cara interna, distante o trans: Es la que se encuentra más cerca de la membrana citoplasmática, de hecho sus membranas, ambas unitarias, tienen una composición similar. En el interior de los sáculos del dictiosoma la proteína puede sufrir una serie de modificaciones hasta su composición final. Cuando llega a la zona interna se transforma en una vesícula de secreción pudiéndose unir a otras formando un gránulo de secreción para salir mediante exocitosis al espacio extracelular.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 90
Funciones generales:
El aparato de Golgi también llamado complejo o cuerpo de Golgi, se encarga de la distribución y el envío de los productos químicos de la célula. Modifica proteínas y lípidos (grasas) que han sido sintetizados previamente tanto en el retículo endoplasmático rugoso como en el liso y los prepara para expulsarlos fuera de la célula.
• Modifica sustancias sintetizadas en el RER: En el aparato de Golgi se transforman las sustancias procedentes del RER. Estas transformaciones pueden ser agregaciones de restos de carbohidratos para conseguir la estructura definitiva o sufren la proteolisis lo que les confiere la forma activa.
• Ej: en el RER de las células acinosas del páncreas se sintetiza la proinsulina que debido a las transformaciones que sufre en el aparato de Golgi, tomará la forma o conformación definitiva de la insulina.
• Secreción celular: Las sustancias atraviesan todos los sáculos del aparato de Golgi y cuando llegan a la cara trans del dictiosoma, en forma de vesículas de secreción, será transportada a su destino fuera de la célula, atravesando la membrana citoplasmática por exocitosis.
• Producción de membrana citoplasmática: Los gránulos de secreción cuando se unen a la membrana en la exocitosis pasan a formar parte de esta.
• Participa en la síntesis de carbohidratos, como la celulosa.
• Forma los lisosomas primarios.
• Forma el acrosoma de los espermios.
EL LISOSOMA:
Los lisosomas son vesículas relativamente grandes formadas por el retículo endoplasmático rugoso y luego empaquetados por el complejo de Golgi que contienen enzimas hidrolíticas y proteolíticas que sirven para digerir los materiales de origen externo o interno que llegan a ellos.
El pH en el interior de los lisosomas es de 4,8 (bastante menor que el del citosol, que es neutro) debido a que las enzimas proteolíticas funcionan mejor con un pH ácido. La membrana del lisosoma estabiliza el pH bajo bombeando protones (H+) desde el citosol, y asimismo, protege al citosol y al resto de la célula de las enzimas degradantes que hay en el interior del lisosoma.
Las enzimas lisosomales son capaces de digerir bacterias y otras sustancias que entran en la célula por fagocitosis, u otros procesos de endocitosis.
Los lisosomas utilizan sus enzimas para reciclar las diferentes organelas de la célula, englobándolos, digiriéndoles y liberando sus componentes en el citosol. De esta forma los

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 91
orgánulos de la célula se están continuamente reponiendo. El proceso de digestión de los orgánulos se llama autofagia. Por ejemplo, las células hepáticas se reconstituyen por completo una vez cada dos semanas.
Las enzimas más importantes en el lisosoma:
• Lipasa, que digiere lípidos, • Glucosilasas, que digiere
carbohidratos (azúcares), • Proteasas, que digiere
proteínas, • Nucleasas, que digiere ácidos
nucleicos.
El lisosoma, sólo está presente en células animales.
Proceso de digestión celular:
La célula utiliza estos lisosomas para degradar biomoléculas complejas. Utiliza dos métodos: la endocitosis y la autofagia.
En la endocitosis los materiales son recogidos del exterior celular y englobados mediante endocitosis por la membrana plasmática que forma un fagosoma. El lisosoma se une al fagosoma formando un fagolisosoma y vierte su contenido en este, degradando las sustancias del fagosoma. Una vez hidrolizadas las moléculas utilizables pasan al interior de la célula para entrar
en rutas matabólicas y lo que no es necesario para la célula se desecha fuera de esta por exocitosis.
En la autofagia la célula digiere estructuras propias que no es necesario. El material queda englobado por vesículas que provienen del retículo endoplásmico y del aparato de Golgi formando un autofagosoma. Al unirse al lisosoma primario forma un autofagolisosoma y sigue el mismo proceso que en el anterior caso.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 92
LOS PEROXISOMAS:
Los peroxisomas son orgánulos citoplasmáticos muy comunes en forma de vesículas que contienen oxidasas y catalasas. Estas enzimas cumplen funciones de detoxificación celular. Como todos los orgánulos, los peroxisomas solo se encuentran en células eucariotas.
Estructura:
Los peroxisomas están envueltos por una membrana citoplasmática semipermeable. Se forman por gemación al desprenderse del retículo endoplasmático liso, aunque por sí mismos pueden abulatar cierta porción de su membrana produciendo nuevos peroxisomas sin derramar su contenido en el citoplasma. Dicha membrana protege la célula de los efectos dañinos del interior del peroxisoma. Las particulas de su interior suelen estar cristalizadas.
Función:
En los peroxisomas se degradan las purinas y otros compuestos. En las plantas son el asiento de una serie de reacciones conocidas como fotorrespiración. En los peroxisomas se produce agua oxigenada (H2O2, un producto tóxico del metabolismo celular) compuesto que es degradado rápidamente por una enzima oxidativa dentro del peroxisoma. Contienen una enzima llamada catalasa que participa en la degradación del peróxido de oxígeno a agua y oxígeno por intermedio de ciertas sustancias orgánicas (en la ecuación la variable R').
H2O2 + R'H2 R' + 2H2O
También tienen otras enzimas (catalasas) que utilizan este mismo peróxido para reacciones de oxidación, como por ejemplo, la oxidación de sustancias tóxicas como los fenoles, etanol, formaldehído, entre otros, las cuales van a ser posteriormente eliminadas. Tal es el mecanismo de detoxificación realizada por el hígado y los riñones, por ejemplo.
Debido a que los peroxisomas necesitan usar el peróxido, se han dotado de enzimas que la sintetizan a partir de oxigeno y removiendo hidrógeno de sustratos orgánicos específicos (en la ecuación la variable R).
RH2 + O2 R + H2O2
La beta oxidación también ocurre dentro de los peroxisomas, en la cual los ácidos grasos son comúnmente degradados a Acetil-CoA la cual es reciclada por la célula.
Ciertas deficiencias involucran a peroxisomas ineficaces, tales como la enfermedad de Zellweger.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 93
MITOCONDRIA
Las mitocondrias son los orgánelos que se encuentran en prácticamente todas las células eucariotas (también hay en células gaméticas), encargados de suministrar la mayor parte de la energía necesaria para la actividad celular; actúan por tanto, como centrales energéticas de la célula y sintetizan ATP por medio de la fosforilación oxidativa. La mitocondria presenta una membrana exterior permeable a iones, metabolitos y muchos polipéptidos. Eso es debido a que contiene proteínas que forman poros llamados Porinas o VDAC (canal aniónico dependiente de voltaje), que permiten el paso de moléculas de hasta 10000 Dalton y un diámetro aproximado de 20Å. La membrana mitocondrial interna presenta pliegues dirigidos hacia el interior llamados crestas, que contienen tres tipos de proteínas:
o Las proteínas que trasportan los electrones hasta el oxígeno molecular o Un complejo enzimático, la ATP-sintetasa que cataliza la síntesis de ATP (fosforilación
oxidativa). o Proteínas trasportadoras que permiten el paso de iones y moléculas a través de la
membrana interna.
Estructura y composición
Las mitocondrias se componen de dos bicapas lipídicas que separan tres espacios: el citosol, el espacio celular y la matriz de la mitocondria. La membrana externa es una bicapa lipídica lisa (no forma ondulaciones) con proteínas asocputasoras, lo que la hace poco permeable excepto a ATP, ADP, Ácido pirúvico, O2 y agua. Esta membrana forma ondulaciones, normalmente transversales al eje de la mitocondria, llamadas crestas mitocondriales. En la cara interna tienes proteínas ATP-sintetasas. Como consecuencia, el espacio intermembrana está compuesto de un líquido similar al hialoplasma. La matriz mitocondrial, sin embargo, contiene menos moléculas del citosol, aunque contiene iones, metabolitos a oxidar, ADN circular bicatenario muy parecido al de las bacterias, ribosomas tipo 70S similares a los de bacterias, llamados mitorribosomas, que realizan la síntesis de algunas proteínas mitocondriales y contiene ARN mitocondrial; es decir, tienen los orgánulos que tendría una célula procariota de vida libre. Al respecto de esto la científica estadounidense Lynn Margulis junto con otros científicos ha propuesto la teoría endosimbiótica.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 94
Según ésta, en un momento dado, el mitocondrio, una célula procariota capaz de obtener energía a partir del oxígeno, se fusionó en un momento de la evolución con lasos células eucariotas, proporcionándoles una fuente de energía de la que sacaron mucho partido, aprovechando el aumento de la concentración de oxígeno en la atmósfera terrestre. Morfología y función
La morfología de la mitocondria es difícil de describir puesto que son estructuras muy plásticas que se deforman, se dividen y fusionan. Normalmente se las representa en forma alargada. Su número depende de las necesidades energéticas de la célula. Al conjunto de las mitocondrias de la célula se le denomina condrioma celular Su principal función es la oxidación de metabolitos (glucólisis, ciclo de Krebs, beta-oxidación de ácidos grasos) y la obtención de ATP, que supone un porcentaje muy alto del ATP sintetizado por la célula. También sirve de almacén de sustancias como iones, agua y algunas partículas como restos de virus y proteínas. Orígenes
La científica estadounidense Lynn Margulis, junto con otros científicos, recuperó en torno a 1980, reformulándola como teoría endosimbiótica, una antigua hipótesis. Según esta versión actualizada, hace unos 1500 millones de años, una célula procariota capaz de obtener energía de los nutrientes orgánicos empleando el oxígeno molecular como oxidante, se fusionó en un momento de la evolución con otra célula procariota o eucariota primitiva al ser fagocitada sin ser inmediatamente digerida, un fenómeno frecuentemente observado. De esta manera se produjo una simbiosis permanente entre ambos tipos de seres: la procariota fagocitada proporcionaba energía, especialmente en forma de ATP y la célula hospedadora ofrecía un medio estable y rico en nutrientes a la otra. Este mutuo beneficio hizo que la célula invasora llegara a formar parte integral del organismo mayor, acabando por convertirse en parte de ella: la mitocondria. Esta hipótesis tiene entre sus fundamentos la evidencia de que las mitocondrias poseen su propio ADN y está recubierta por su propia membrana. A lo largo de la historia común la mayor parte de los genes mitocondriales han sido transferidos al núcleo, de tal manera que la mitocondria no es viable fuera de la célula huésped y ésta no suele serlo sin mitocondrias.
RIBOSOMA
Los ribosomas son orgánulos sin membrana, sólo visibles al microscopio electrónico debido a su reducido tamaño (29 nm en células procariotas y 32 nm en eucariotas). Están en todas las células vivas (excepto en los espermatozoides). Su función es ensamblar proteínas a partir de la información genética que le llega del ADN transcrita en forma de ARN mensajero (ARNm). La información genética está en el ADN. Esa información se transcribe en ARN. El ribosoma lee el ARN mensajero y ensambla la proteína con los aminoácidos suministrados por los ARN de transferencia, este proceso se denomina síntesis de proteínas. El ribosoma consta de dos partes, la subunidad mayor y una menor, estas salen del núcleo por separado. Por experimentación se puede decir que se mantienen unidas por cargas, ya que al

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 95
bajarse la concentración de Mg+2, las subunidades tienden a separarse. El ribosoma procariota tiene un coeficiente de sedimentación de 70s y está formado por dos subunidades (50s y 30s). El ribosoma eucariota tiene un coeficiente de sedimentación de 80s (formado por dos subunidades, una de 60s y otra de 40s). Este se puede encontrar unido al retículo endoplasmático rugoso (RER), que es la forma habitual en la célula eucariota, o encontrarlo en el citoplasma, donde recibe el nombre de polisoma o polirribosoma (forma habitual en la célula procariota). Este polisoma se encarga de sintetizar proteínas de localización celular, mientras que los ribosomas del RER se encargan de sintetizar proteínas de exportación, o sea que se irán de la célula hacia otro lugar donde se necesite.
Los ribosomas, son organelos celulares donde el ARNm es traducido. Está formado por dos subunidades, y cada una de ellas contiene ARNr y proteínas ribosomales.
VACUOLA
Una vacuola es una cavidad rodeada por una membrana que se encuentra en el citoplasma de las células, principalmente de las vegetales.
Se forman por fusión de las vesículas procedentes del retículo endoplasmático y del aparato de Golgi. En general, sirven para almacenar sustancias de desecho o de reserva (agua con varios azúcares, sales, proteínas y otros nutrientes disueltos en ella).
En las células vegetales, las vacuolas ocupan la mitad del volumen celular y en ocasiones pueden llegar hasta casi la totalidad. También, aumentan el tamaño de la célula por acumulación de agua.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 96
Están relacionadas con los lisosomas secundarios, ya que éstos engloban dos tipos de vacuolas, las heterofágicas o digestivas y las autofágicas. Contienen enzimas hidrolíticas y sustratos en proceso de digestión. En el primer tipo, los sustratos son de origen externo y son capturados por endocitosis; una vez producida la digestión, ciertos productos pueden ser reutilizados y los no digeribles (llamados cuerpos residuales) son vertidos al exterior por exocitosis. En el caso de las vacuolas autofágicas, lo que se digieren son constituyentes de la célula. Hay otro tipo de vacuolas, las pulsátiles o contráctiles, que aparecen en muchos protozoos, especialmente en los dulceacuícolas. Se llenan de sustancias de desecho que van eliminando de forma periódica y además bombean el exceso de agua al exterior.
VESÍCULA: En biología celular, una vesícula es un orgánulo que forma un compartimento pequeño y cerrado, separado del citoplasma por una bicapa lipídica igual que la membrana celular.
Las vesículas almacenan, transportan o digieren productos y residuos celulares. Son una herramienta fundamental de la célula para la organización del metabolismo. Muchas vesículas se crean en el aparato de Golgi, pero también en el retículo endoplasmático, o se forman a partir de partes de la membrana plasmática.
CILIO
Cilio o cilia (cilium, masculino; plural cilia) significa en latín "pestaña". Se llama cilio (y, especialmente en Latinoamérica, es frecuente hallar la denominación de cilia) a cada uno de los pequeños apéndices mótiles que cubren total o parcialmente la superficie de muchas células desnudas (sin pared). Los flagelos tienen una estructura esencialmente equivalente, aunque hay algunas diferencias morfológicas y muchas diferencias funcionales.
Los cilios tienen una forma cilíndrica, de diámetro uniforme en toda su longitud, con una terminación redondeada, semiesférica. Pueden ser descritos como una evaginación digitiforme (una prolongación en dedo de guante) de la membrana plasmática, con un contenido que es continuación del citoplasma.
Estos orgánulos están dotados de un armazón compleja, semejante a la de los flagelos, basada en microtúbulos y que se llama axonema. El axonema se continúa, en la base del cilio y por debajo de la membrana plasmática, con un corpúsculo basal, que tiene una estructura semejante pero más compleja.
Se mueven rítmicamente y de forma coordinada, cada uno con un movimiento semejante al del brazo de un nadador, retrocediendo en posición extendida, y en conjunto al de un trigal azotado por el viento (movimiento de batida coordinado). Mientras reciban la energía necesaria en forma de ATP los cilios siguen batiendo automáticamente. El efecto es un empuje neto, que da lugar a que la célula se desplace en su medio, como ocurre con ciertos protistas y animales muy pequeños; o que el líquido extracelular circundante sea impulsado, que es la función que cumplen los cilios en el epitelio de las vías respiratorias humanas.
La coordinación de los cilios entre sí, al fustigar el agua sobre la superficie de una célula, viene dada por la misma agua, movida por el cilio precedente. El que sigue en fila halla así una

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 97
dirección favorecida y se mueve por ella con un pequeño retraso, conducta llamada metacronismo.
El control de los cilios es fundamental en los protozoos ciliados que las emplean para cazar otros protozoos y alimentarse con ellos. Para seguirlos, alcanzarlos, poner su estoma o "boca celular" en posición, retrocediendo si es necesario, para luego comérselos, es menester controlar la natación. Este control se logra por medios eléctricos. Los valores del campo eléctrico en la membrana exterior del protozoo, de donde emergen los cilios o cilias, "dibujan" los movimientos de la presa cercana. Estos le son revelados por las presiones del agua, que interfieren con las oscilaciones u ondas eléctricas que realizan el control.
FLAGELO
Un flagelo es un apéndice con forma de látigo que usan muchos organismos unicelulares y unos pocos pluricelulares. Sin embargo, estos apéndices pueden también estar implicados en otros procesos. Este nombre cubre realmente tres estructuras diferentes encontradas en cada uno de los tres dominios.
Los flagelos bacterianos son los filamentos helicoidales que rotan como tornillos. Los flagelos de Archaea son superficialmente similares, pero son diferentes en muchos detalles y considerados no homólogos. Los flagelos de Eukarya - aquellos de células Protista animales y vegetales - son complejas proyecciones celulares que azotan hacia adelante y hacia atrás. En ocasiones, este último es llamado cilio o undulipodia para acentuar su distinción.
Esencialmente, la estructura del flagelo es igual a la del cilio, pero generalmente se complica con otras estructuras añadidas, resultando más grueso y más largo.
Los flagelos más estudiados son los de espermatozoides. En el espermatozoide de mamíferos, el flagelo (cola) está constituido por: un axonema (9 pares de microtúbulos periféricos y un par central) rodeado por las fibras externas densas 9 cilindros proteicos (uno por cada doblete) que intervienen en el movimiento del flagelo.
Por fuera de estas fibras, existen otras estructuras rodeando el complejo axonema-fibras: la vaina mitocondrial, si el corte es por la pieza intermedia, o la vaina fibrosa, si el corte se realiza en la pieza principal.
La vaina mitocondrial está constituida por mitocondrias dispuestas en hélice que proporcionan la energía necesaria para el movimiento del flagelo. La vaina fibrosa son pares de estructuras proteicas (cada una rodea la mitad de las fibras densas). Parece que intervienen en la protección del axonema y quizás también en el movimiento del flagelo.
Por fuera, de todo ello, se dispone la membrana plasmática. Los flagelos, que impulsan a los espermatozoides y a muchos protozoos, están diseñados para desplazar toda la célula a través de un fluido.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 98
NUCLEO CELULAR
El núcleo celular es la estructura más característica de las células eucariotas. Se rodea de una cubierta propia, llamada envoltura nuclear y contiene el material hereditario, que es la base del repertorio de instrucciones en que se basa el desarrollo y el funcionamiento de cada organismo, y cuya composición se basa en el ácido desoxirribonucleico (ADN).
Por la existencia del núcleo, en las células eucariotas se dan en espacios separados los procesos de replicación del genoma y transcripción del ARN, que ocurren dentro, y la biosíntesis de proteínas (traducción), que se produce fuera. Esta compartimentación es una de las condiciones de la complejidad del control funcional que distingue a los eucariontes de los procariontes.
El núcleo es una estructura dinámica, que en los organismos con mitosis abierta, se deshace durante el reparto cromosómico. Se llama núcleo interfásico al que se observa antes de la mitosis y después de ésta, ya duplicado; es decir, durante los momentos del ciclo celular que no corresponden a la mitosis. Cuando no se especifique otra cosa, las explicaciones siguientes se refieren al núcleo interfásico.
Además, el núcleo cuenta con una estructura que se tiñe con facilidad, el denominado nucléolo. Forma, tamaño y posición El núcleo es casi siempre una estructura esferoidal relativamente grande, cuando se la compara con los orgánulos citoplasmáticos comunes. En términos absolutos, puede medir menos desde 1 µm (en los llamados nanoeucariontes) hasta más de 20 µm. Su volumen guarda cierta proporcionalidad con el del citoplasma.
El núcleo tiende a ocupar una posición central, pero en las células adultas de las plantas se ve desplazado a la periferia por el importante volumen del vacuoma (conjunto de vacuolas). Número Lo típico es que cada célula eucariota contenga un núcleo, sin embargo son frecuentes e importantes las excepciones. En los hongos también es normal la condición dicariótica (dos núcleos).
En protistas es donde se observa mayor diversidad de casos, en éste como en otros temas básicos de la biología eucariótica. En los ciliados existen regularmente dos núcleos, el macronúcleo y el micronúcleo. Los eritrocitos (glóbulos rojos) maduros de casi todos los mamíferos carecen de núcleo.
Estructura El núcleo interfásico presenta al menos las siguientes partes diferenciadas:
Envoltura nuclear.- Se basa en una doble membrana (2 bicapas lipídicas) reforzada por el citoesqueleto. Está perforada por poros nucleares, a través de los cuales el interior del núcleo se comunica con el citosol. La envoltura presenta ribosomas adheridos

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 99
externamente y es la continuación del retículo endoplasmático rugoso. La envoltura nuclear se halla reforzada por dos armazones de filamentos intermedios, uno adosado a su superficie interna: la lámina nuclear. Y otro situado sobre la cara citosólica de la membrana externa.
Cromatina.- Es la forma que toma el material hereditario durante la interfase del ciclo celular. Consiste en ADN asociado a proteínas.
Nucleoplasma.- también llamado carioplasma o cariolinfa. Se trata del medio interno indiferenciado que llena el núcleo, semejante al citosol o hialoplasma, bañando a sus componentes.
Nucléolo(s).- Una o más estructuras esferoidales, relacionadas con la síntesis de las principales piezas de los ribosomas y con su ensamblaje parcial. Éste está conformado por ARN y proteínas básicas. Se distinguen dos porciones del nucléolo, la región granular, formada por gránulos de ARN, y la región fibrilar formada por filamentos de ARN. Una tercera región, muy difícil de observar es la denominada porción cromosómica del nucléolo, en ésta se encuentran filamentos de DNA.
Funciones
1. Dirige la actividad celular, ya que contiene el programa genético, que dirige el desarrollo y funcionamiento de la célula.
2. Es la sede de la replicación (duplicación del ADN) y la transcripción (síntesis de ARN), mientras que la traducción ocurre en el citoplasma. En las células procariotas todos esos procesos coinciden en el mismo compartimento celular.
ENVOLTURA NUCLEAR
También llamada carioteca, es la envuelta que rodea y delimita al núcleo propio de la célula eucariota. La envoltura nuclear está formada por dos membranas concéntricas, así que la expresión membrana nuclear, frecuentemente usada para referirse a ella, no puede considerarse apropiada.
Constitución

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 100
La envoltura nuclear es una estructura compleja que se basa en una vesícula de retículo endoplasmático extendida alrededor del material hereditario nuclear (cromatina). Como tal vesícula, la envoltura aparece conformada por dos membranas: la membrana nuclear externa y la membrana nuclear interna. Por el lado de fuera queda el citoplasma y por el de dentro el contenido del núcleo. Por el lado del núcleo la membrana nuclear interna lleva adosada una estructura llamada lámina nuclear, la cual está formada por proteínas, como las llamadas laminas, a veces en forma de capa continua, a veces con la estructura de un panal. El hecho de que la envoltura sea una especialización del retículo endoplasmático se observa también en que suele aparecer recubierta de ribosomas (algo que es característico del retículo endoplasmático rugoso), los cuales fabrican precisamente proteínas que se incorporan a la composición de las membranas nucleares.
Funciones
La envoltura nuclear aparece atravesada de manera regular por perforaciones, los poros nucleares. Estos poros no son simples orificios, sino estructuras complejas acompañadas de una armazón de proteínas, que facilitan a la vez que regulan los intercambios entre el núcleo y el citoplasma. Se llama complejo del poro a cada una de esas puertas de comunicación. Por ahí salen las moléculas de ARNm producidas por la transcripción, que deben ser leídas por los ribosomas del citoplasma. Por ahí salen también los complejos de ARNr y proteínas a partir de los cuales se ensamblan en el citoplasma los ribosomas. Por los poros entran al núcleo las proteínas, fabricadas en el citoplasma por los ribosomas, que cumplen su papel dentro del núcleo. Dinámica: En las células con mitosis abierta, que son la mayoría, la envoltura nuclear desaparece al principio de la mitosis, para formarse de nuevo, ahora alrededor de dos núcleos hijos, al acabar aquélla.
El proceso depende de la alteración de las láminas, las proteínas de la lámina, por un complejo enzimático. Cuando el proceso de la mitosis termina, las láminas vuelven a su estado inicial, formándose primero dos láminas nucleares sobre las cuales, por extensión del retículo endoplasmático, terminan por formarse dos envolturas nucleares completas.
En las células con mitosis cerrada, una variante que se observa en muchos protistas, la envoltura nuclear no desaparece durante la mitosis, sino que se estira, estrángulándose, para terminar formando los dos núcleos hijos.
CROMATINA
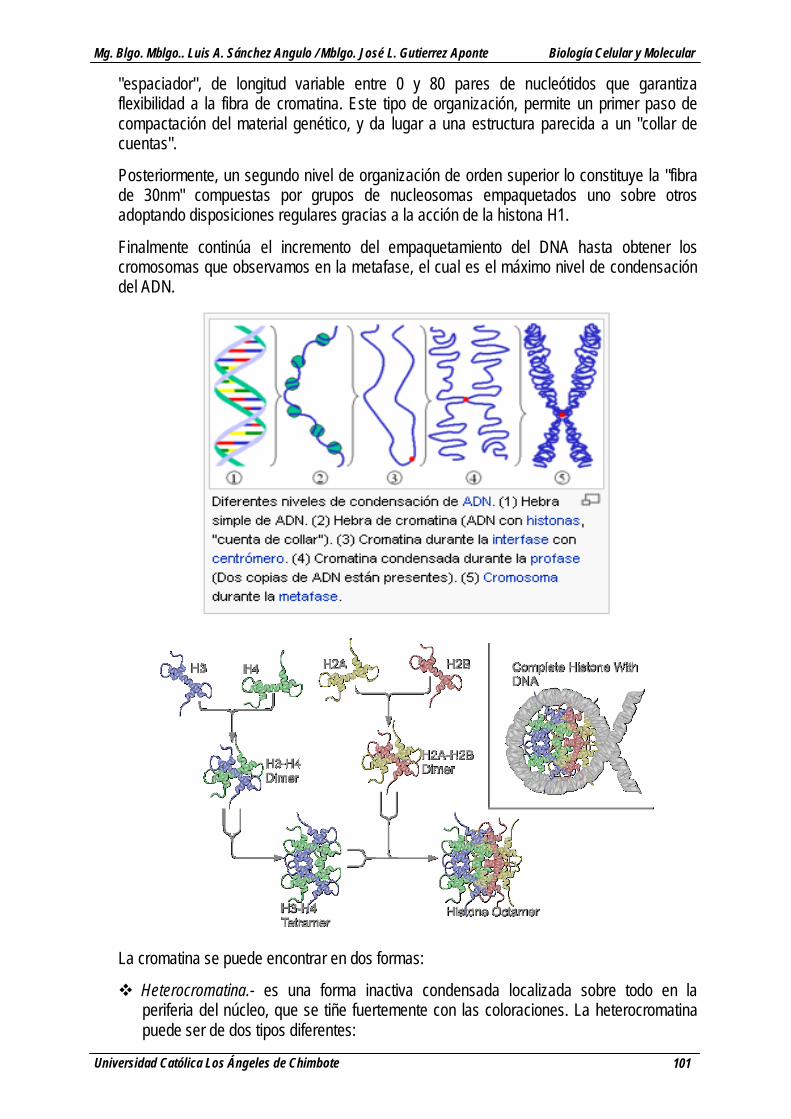
La cromatina es el conjunto de ADN, histonas y proteínas no histónicas que se encuentra en el núcleo de las células eucariotas y que constituye el cromosoma eucariótico.
Las unidades básicas de la cromatina son los nucleosomas. Éstos se encuentran formados por aproximadamente 146 pares de bases de longitud (el número depende del organismo), asociados a un complejo específico de 8 histonas nucleosómicas (octámero de histonas). Cada partícula tiene una forma de disco, con un diámetro de 11 nm y contiene dos copias de cada una de las 4 histonas H3, H4, H2A y H2B. Este octámero forma un núcleo proteico alrededor del que se enrolla la hélice de ADN (da aproximadamente 1.8 vueltas). Entre cada una de las asociaciones de ADN e histonas existe un ADN libre llamado ADN
Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 101
"espaciador", de longitud variable entre 0 y 80 pares de nucleótidos que garantiza flexibilidad a la fibra de cromatina. Este tipo de organización, permite un primer paso de compactación del material genético, y da lugar a una estructura parecida a un "collar de cuentas".
Posteriormente, un segundo nivel de organización de orden superior lo constituye la "fibra de 30nm" compuestas por grupos de nucleosomas empaquetados uno sobre otros adoptando disposiciones regulares gracias a la acción de la histona H1.
Finalmente continúa el incremento del empaquetamiento del DNA hasta obtener los cromosomas que observamos en la metafase, el cual es el máximo nivel de condensación del ADN.
La cromatina se puede encontrar en dos formas:
Heterocromatina.- es una forma inactiva condensada localizada sobre todo en la periferia del núcleo, que se tiñe fuertemente con las coloraciones. La heterocromatina puede ser de dos tipos diferentes:

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 102
o La constitutiva.- idéntica para todas las células del organismo y que carece de información genética, incluye a los telómeros y centrómeros del cromosoma que no expresan su ADN.
o La facultativa.- diferente en los distintos tipos celulares, contiene información sobre todos aquellos genes que no se expresan o que pueden expresarse en algún momento. Incluye al ADN satélite y al corpúsculo de Barr.
Eucromatina.- está diseminada por el resto del núcleo (menor condensación), se tiñe débilmente con la coloraciones (su mayor tinción ocurre en la mitosis y no es visible con el microscopio de luz. Representa la forma activa de la cromatina en la que se está transcribiendo el material genético de las moléculas de ADN a moléculas de ARNm, por lo que es aquí donde se encuentran la mayoría de los genes activos.
La distinción tajante entre eucromatina activa y heterocromatina inactiva es aceptable actualmente, pero con importantes reservas.
NUCLEOPLASMA
El nucleoplasma es el medio interno del núcleo celular, en el se encuentran las fibras de ADN, que asociadas con proteínas denominadas histonas forman hebras llamadas cromatinas y ARN conocidos como nucleolos.
NUCLÉOLO
El nucléolo es un suborgánulo del núcleo que tiene como principal función la síntesis de los ARNr. En la ultraestructura del nucléolo podemos distinguir:
o Centro fibrilar: es poco denso a los electrones. Se encuentra formado por fibrillas muy finas, complejos de preiniciación y factores de iniciación de la transcripción.
o Alrededor del centro fibrilar suele situarse un componente fibrilar más denso a los electrones, constituido por fibras más gruesas. Es la zona del ADN que se está transcribiendo activamente, formando árboles de navidad.
o Componente granular: formado por gránulos y ribonucleoproteínas (ANRprer).
o Intersticios: son zonas donde no se localiza ningún componente.
El gen que codifica los ARNr da lugar a un transcrito de 45 S, el ARN prerribosómico (ARNprer). En ese ARNprer pueden distinguirse dos regiones:
Las regiones no metiladas: se degradan mediante la introducción de proteínas prerribosómicas en el núcleo y asociación de éstas al ARNprer.
Las regiones metiladas: quedan libres como resultado de la lisis del ARNprer. Se forman los tres ARNr:
• El ARN 18 S junto con algunas proteínas da lugar a la subunidad pequeña inmadura, en forma de RNP. Ésta debe migrar al citoplasma por un poro. Esta salida ocurre muy rápidamente.
• Los ARN 28 S y 5’8 S se asocian a otras proteínas y a un ARN 5S sintetizado fuera del nucléolo. Los tres forman un gránulo que constituye la subunidad grande del ribosoma y que deberá igualmente atravesar la envoltura nuclear mediante un poro. Esta salida lleva hasta 30 minutos, ya que la estructura debe fragmentarse para atravesar el poro.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 103
Los genes que codifican este ARNprer 45 S se repiten en cinco pares de cromosomas: 13, 14, 15, 21 y 22. En los cromosomas acrocéntricos con constricción secundaria, es en ésta donde se asocia el gen. El ARN 5S se encuentra codificado en el brazo Q del cromosoma 1. En el nucléolo, además de formarse estas subunidades, se realizan otros procesos:
• Formación de proteínas telomerásicas y organización de la telomerasa. • En los nucleolos y en los cercanos cuerpos de Cajal, se maduran RNP que se
sintetizaron en el citoplasma y migraron al nucleolo para su maduración.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 104
LA RESPIRACION CELULAR Y FERMENTACIÓN
Virtualmente todas las células, eucariotas (de plantas, animales, hongos y protistas) poseen organelos complejos, llamados MITOCONDRIAS, que son el sitio de la RESPIRACIÓN CELULAR AEROBIA, proceso que incluye casi todas las reacciones que convierten la energía química de determinados alimentos en ATP. Constituyen uno de los ejemplos de integración morfofuncional más admirable, ya que proveen el andamiaje sobre el que asientan las innumerables moléculas que participan en las reacciones que transfieren la energía depositada en los alimentos a una molécula extraordinariamente versátil como es el ATP
CARACTERÍSTICAS GENERALES La respiración aeróbica necesita oxígeno y causa la liberación de átomos de carbono de las moléculas alimentarias, en la forma de dióxido de carbono (un producto de desecho). Las mitocondrias son más numerosas en células muy activas y que, por lo tanto, tienen altas necesidades de energía. Se han encontrado más de 1000 en una sola célula hepática. Las mitocondrias difieren en su tamaño, que va de 2 a 8 um. de longitud, son cilíndricas y experimentan cambios sutiles con rapidez en su tamaño y forma derivados de su actividad. Se encuentran ubicadas en las regiones de las células donde la demanda de energía es mayor, por lo que se desplazan de un lado a otro del citoplasma hacia las zonas necesitadas de energía. No obstante, en algunos tipos celulares, como los espermatozoides, las células musculares y las células grasas, las mitocondrias permanecen en lugares fijos. Por lo regular una mitocondria da origen a otra mediante crecimiento y ulterior división.
Cada MITOCONDRIA está limitada por una doble membrana, que forma dos compartimientos diferentes en el organelo; ESPACIO INTERMEMBRANOSO y la MATRIZ MITOCONDRIAL. El espacio intermembranoso es el compartimiento que se forma entre la MEMBRANA EXTERNA y la MEMBRANA INTERNA. La matriz mitocondrial, el compartimiento rodeado por la membrana interna, contiene enzimas que degradan moléculas alimentarias y convierten su energía en otras formas de energía química.
La MEMBRANA MITOCONDRIAL EXTERNA es lisa y permite el paso de muchas moléculas pequeñas presentes en el citosol, pero no de las macromoléculas. Esto debido a que en su bicapa lipídica presenta, entre otros componentes una proteína de transmembrana llamada “porina” que forman canales acuosos pequeños. En contraste, la MEMBRANA MITOCONDRIAL INTERNA se pliega repetidas veces y regula de manera estricta los tipos de molécula que pueden atravesarla; los pliegues, llamados CRESTAS MITOCONDRIALES, se extienden en el interior de la matriz

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 105
mitocondrial y constituyen una superficie para las reacciones químicas que transforman la energía química de las moléculas alimenticias en energía química almacenada en el ATP. La membrana contiene la compleja serie de enzimas y otras proteínas necesarias para dichas, reacciones. Esta membrana presenta un alto grado de especialización. Los organismos autótrofos fijan la energía solar en forma de energía química contenida en los compuestos orgánicos, glucosa (carbohidrato), en particular. Esta energía, convenientemente liberada, será utilizada posteriormente por las partes de la planta que no tienen cloroplastos, como suele ser el caso de las raíces y tallos no verdes, o por toda la planta cuando falta la energía solar. Es también esta energía (presente en estas moléculas orgánicas: glucosa) la que permite la vida de los organismos heterótrofos. La respiración celular y las fermentaciones son las vías catabólicas más corrientes para la obtención de la energía contenida en las sustancias orgánicas. Ambas vías, no obstante, tienen una primera fase común: la glucólisis. La respiración celular es el conjunto de reacciones bioquímicas que ocurre en la mayoría de las
células, en las que el ácido pirúvico producido por la glucólisis se desdobla a anhídrido carbónico (CO2) y agua (H2O) y se producen 36 o 38 moléculas de ATP. En las células eucariotas la respiración se realiza en las mitocondrias y ocurre en tres etapas que son:
o Oxidación del piruvato. o Ciclo de los ácidos tricarboxílicos. o Cadena respiratoria y fosforilación
oxidativa del ADP a ATP. Los vegetales, mediante sus cloroplastos, fijan y almacenan la energía; sintetizando moléculas orgánicas (carbohidratos: glucosa). Posteriormente los animales heterótrofos; con la ayuda de sus mitocondrias transforman esta energía (almacenada en los carbohidratos: glucosa) en energía calórica y energía para el trabajo celular “ATP”.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 106
La respiración celular es una parte del metabolismo, concretamente del catabolismo, en el cual la energía contenida en distintas biomoléculas, como los glúcidos (carbohidratos), es liberada de manera controlada. Durante la respiración una parte de la energía libre desprendida en estas reacciones exotérmicas, es incorporada a la molécula de ATP, que puede ser a continuación utilizado en los procesos endotérmicos, como son los de mantenimiento y desarrollo del organismo (anabolismo).
La respiración celular podría dividirse en dos tipos, según el papel atribuido al oxígeno:
o RESPIRACIÓN AEROBIA: Hace uso del O2 como aceptor último de los electrones desprendidos de las sustancias orgánicas. Es la forma más extendida, propia de los organismos eucariontes (cuyas mitocondrias realizan dicha función) y algunos grupos de bacterias. Se llama aerobios a los organismos que, por este motivo, requieren O2.
o RESPIRACIÓN ANAEROBIA: No interviene el oxígeno, sino que se emplean otros aceptores finales de electrones, muy variados, generalmente minerales y, a menudo, subproductos del metabolismo de otros organismos. Un ejemplo de aceptor es el SO42- (anión sulfato), que en el proceso queda reducido a H2S:
La respiración anaerobia es propia de procariontes (p.ej.: bacterias) diversos, habitantes sobre todo de suelos y sedimentos, y algunos de estos procesos son importantes en los ciclos biogeoquímicos de los elementos. No debe confundirse la respiración anaerobia con la fermentación, que es una oxidación – reducción interna a la molécula procesada, en la que no se requiere ni O2 ni ningún otro aceptor de electrones.
La respiración anaerobia es un proceso biológico de oxidorreducción de azúcares y otros compuestos. La realizan exclusivamente algunos grupos de bacterias. En las bacterias anaerobias, la cadena de transporte de electrones es análoga a la de la respiración aerobia, ya que se compone de los mismos componentes (citocromos, quinonas, proteínas ferrosulfúricas, etc.). La única diferencia por tanto radica en el aceptor último de electrones.
Todos los posibles aceptores en la respiración anaerobia tienen un potencial de reducción menor que el O2, por lo que se genera menos energía en el proceso. LA RESPIRACIÓN CELULAR AERÓBICA
La respiración aerobia es un tipo de metabolismo energético el que seres vivos extraen energía de moléculas orgánicas, como la glucosa, por un proceso complejo en el que el carbono queda oxidado y en el que el oxígeno procedente del aire es el oxidante empleado. En otras variantes, muy raras, de la respiración el oxidante es distinto del oxígeno.
La respiración aerobia o normal, es el proceso

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 107
responsable de que la mayoría de los seres vivos, los llamados por ello aerobios, requieran oxígeno. La respiración aerobia es propia de los organismos eucariontes en general y de algunos tipos de bacterias.
El oxígeno que, como cualquier gas, atraviesa sin obstáculos las membranas biológicas, atraviesa primero la membrana plasmática y luego la mitocondrial, siendo en la matriz de la mitocondria donde se une a electrones y protones (que sumados constituyen átomos de hidrógeno) formando agua.
En esa oxidación final, que es compleja, y en procesos anteriores se obtiene la energía necesaria para la fosforilación del ATP.
En presencia de oxígeno, el ácido pirúvico obtenido durante la fase primera anaerobia o glucólisis, es oxidado para proporcionar energía (ATP), dióxido de carbono y agua. A esta serie de reacciones se le conoce con el nombre de respiración aerobia.
EL ADENOSIN TRIFOSFATO “ATP”
El trifosfato de adenosina (ATP) o adenosín trifosfato es una molécula que consta de una purina (adenina), un azúcar (ribosa), y tres grupos fosfato. Gran cantidad de energía para las funciones biológicas se almacena en los enlaces de alta energía que unen los grupos fosfato y se liberan cuando uno o dos de los fosfatos se separan de las moléculas de ATP. El compuesto resultante de la pérdida de un
fosfato se llama difosfato de adenosina, adenosín difosfato o ADP; si se pierden dos se llama monofosfato de adenosina, adenosín monofosfato o AMP, respectivamente.
ATP Y METABOLISMO
El acoplamiento entre las reacciones exergónicas (que liberan energía al medio) y endergónicas (que gastan energía del medio) en los seres vivos se realiza a través del ATP. Por eso se le conoce como moneda de intercambio energética celular. La mayoría de los organismos nos alimentamos de metabolitos complejos (proteínas, lípidos, glúcidos, etc.) que degradamos a lo largo del tracto intestinal. De modo que a las células llegan metabolitos complejos, pero no tan complejos como los ingeridos.
En la célula van a ser oxidados por una serie de reacciones químicas degradativas “catabolismo”. Como productos del catabolismo se obtienen metabolitos simples y energía. Ambos son los precursores para la síntesis de los componentes celulares. Todo el conjunto de reacciones de síntesis se llama “anabolismo”. En el catabolismo (oxidación) se produce una liberación de electrones que serán captados por unos transportadores de electrones como el NAD+ (que al aceptar electrones se reduce a NADH). Por otra parte, la energía liberada quedará retenida en su mayoría en el ATP. La síntesis (anabolismo) de los compuestos celulares se realizará con los metabolitos simples, utilizando la energía contenida en el ATP y los electrones contenidos en el NADH, ya que este es un proceso reductivo (toma electrones). El ATP es esa moneda de intercambio energético debido

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 108
a su estructura química. Cuando se hidroliza libera mucha energía que va a ser captada por las enzimas que catalizan las reacciones de biosíntesis.
¿Por qué tienen los enlaces del ATP tanta tendencia a hidrolizarse?
o Veamos la molécula de ATP y su hidrólisis a ADP + Pi: o Se puede representar así: A– P ~ P ~ P o Donde “~” son los enlaces anhídrido de ácido, que son de alta energía. En la hidrólisis
del ATP se está hidrolizando uno de esos enlaces anhídrido de ácido. Esto libera gran energía, concretamente 7'7 kcal/mol. Es decir: ΛG = -7,7 kcal/mol.
o Es una reacción muy exergónica. Su keq (constante de equilibrio) es 11. o Así se comprende que el ATP tiene tendencia a hidrolizarse de forma natural y liberar
energía.
Nicotinamida Adenín Dinucleótido “NAD”
La nicotinamida adenín dinucleótido (NAD) es una coenzima, de la vitamina B3 cuya función principal es la intercambio de hidrogeniones en la producción de energía de todas las células. El NAD forma el primer complejo en la captación de hidrógenos en la fosforilación oxidativa y aparece en múltiples reacciones del metabolismo. El NADH es la nicotinamida adenín dinucleótido reducida, siendo la forma activa. Cuando pierde el hidrógeno (deshidrogenación), cede energía. El NADP es la nicotinamida adenín dinucleótido fosfato, siendo la NADPH su forma reducida. Las formas reducidas del NAD se obtienen de la glucólisis y ciclo de Krebs principalmente.
Flavin Adenine Dinucleotido “FAD”
El grupo de la flavina interviene en muchas reacciones bioquímicas de transferencia de hidrógeno. Se encuentra a menudo unido a un grupo de adenosindifosfato ([ADP) en cuyo caso se suele conocer bajo su nombre inglés flavin adenine dinucleotide (FAD).
REACCIONES DE OXIDO – REDUCCIÓN
Las reacciones de reducción-oxidación (también conocido como reacción redox) son las reacciones de transferencia de electrones.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 109
Esta transferencia se produce entre un conjunto de especies químicas, uno oxidante y uno reductor (una forma reducida y una forma oxidada respectivamente). OXIDADO (FAD+) REDUCIDO (FADH2)
Oxidación – Reducción de la molécula de FAD
Para que exista una reacción redox, en el sistema debe haber una especie que ceda electrones y otra especie que las acepte:
1. El reductor es la especie química que tiende a ceder electrones de su estructura química al medio, quedando con carga mayor a la que tenía.
2. El oxidante es la especie que tiende a captar esos electrones, quedando con carga menor a la que tenía.
Cuando una especie química reductora cede electrones al medio se convierte en una especie oxidada, y la relación que guarda con su precursor queda establecida mediante lo que se llama un par redox. Análogamente, se dice que cuando una especie capta electrones del medio se convierte en una especie reducida, e igualmente forma un par redox con su precursor reducido. Las reacciones químicas de la respiración aeróbica de la glucosa pueden agruparse en cuatro etapas. En los eucariotas, la primera etapa (glucólisis) se realiza en el citosol, y el resto ocurren en el interior de las mitocondrias. La mayor parte de las bacterias también efectúan estos procesos, pero dado que sus células carecen de mitocondrias, todas las etapas se llevan a efecto en el citosol y en asociación con la membrana plasmática.
1. GLUCÓLISIS. Ocurre en el citosol, donde cada molécula de glucosa, con sus 6 átomos de carbono, se oxida parcialmente dando lugar a dos moléculas de piruvato (de 3 átomos de carbono). Se invierten dos ATP pero se generan cuatro, y 2 NADH.
2. FORMACIÓN DE ACETILCOENZIMA A. Cada molécula del piruvato entra en una mitocondria y se oxida para convertirse en una molécula de dos carbonos (acetato) que se combina con la coenzima A y forma acetilcoenzima A; se produce NADH2 y se libera C02 como producto de desecho.
3. CICLO DEL ÁCIDO CÍTRICO. El grupo acetato de la acetilCoA se combina con una molécula de cuatro carbonos (oxalacetato), y se forma una molécula de seis carbonos (citrato). En el transcurso del ciclo ésta se recicla a oxalacetato y se libera CO2 como producto de desecho. Se captura energía como ATP y los compuestos reducidos de alto contenido de energía NADH2 y FADH2. Todas estas reacciones ocurren en la matriz mitocondrial.
4. CADENA DE TRANSPORTE DE ELECTRONES Y QUIMIOSMÓSIS. Los electrones extraídos de la glucosa durante las etapas precedentes se transfieren de NADH2 y FADH2 a una cadena de compuestos aceptores de electrones. A medida que los

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 110
electrones pasan de un aceptor a otro, parte de su energía se emplea para bombear hidrogeniones (protones) a través de la membrana mitocondrial interna, con lo que se forma un gradiente de protones. En un proceso denominado quimiosmósis, la energía de este gradiente se usa para producir ATP.
Las cuatro fases de la respiración aeróbica. La primera fase, la glucólisis ocurre en el citosol. Su producto, el piruvato, entra en la mitocondria, donde continúa la respiración celular con formación de acetilcoenzima A, ciclo del ácido cítrico y la cadena transporte de electrones/quimiosmósis. La mayor parte del ATP se sintetiza por quimiosmosis.
La oxidación de la glucosa es una fuente principal de energía en la mayoría de las células. Cuando la glucosa se degrada en una serie de pequeños pasos por medio de enzimas, una proporción significativa de la energía contenida en la molécula vuelve a empaquetarse en los enlaces fosfato de las moléculas de ATP.
La PRIMERA FASE en la degradación de la glucosa es la glucólisis que se efectúa en el citosol de la célula. La SEGUNDA FASE es la respiración aeróbica, que requiere oxígeno y, en las células eucarióticas, tiene lugar en las mitocondrias. La respiración comprende la formación de la acetilcoenzima A, el ciclo del ácido cítrico (ciclo de Krebs) y el transporte terminal de electrones acoplado al proceso de fosforilación oxidativa. Todos estos procesos están íntimamente relacionados.
En condiciones anaeróbicas, el proceso de fermentación transforma al ácido pirúvico producido por la glucólisis o en etanol o en ácido láctico.
Es posible saber cómo y en qué cantidad la energía química, originalmente presente en la molécula de glucosa, se recupera en forma de ATP en el curso de la degradación de la molécula de glucosa. Así, es posible calcular el rendimiento energético global de la oxidación de la glucosa, que puede dar como resultado un máximo de 38 moléculas de ATP (aunque pueden ser también 36 ATP). La actividad de la glucólisis y la respiración están reguladas de acuerdo con las necesidades energéticas de la célula.
Hasta ahora nos hemos referido a la degradación de la molécula de glucosa, pero otras moléculas alimenticias, que incluyen a las grasas, los polisacáridos y las proteínas, pueden ser también degradadas a compuestos que pueden ingresar en las vías centrales -glucólisis y ciclo de Krebs- en diferentes pasos. La biosíntesis de compuestos orgánicos utiliza los compuestos precursores derivados de intermediarios en la secuencia respiratoria y es impulsada por la

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 111
energía derivada de esos procesos. Así, otras vías catabólicas y anabólicas están íntimamente interrelacionadas.
Esquema global de la oxidación de la glucosa
Hasta ahora se ha referido a la degradación de la molécula de glucosa, pero otras moléculas alimenticias, que incluyen a las grasas, los polisacáridos y las proteínas, pueden ser también degradadas a compuestos que pueden ingresar en las vías centrales -glucólisis y ciclo de Krebs- en diferentes pasos. La biosíntesis de compuestos orgánicos utiliza los compuestos precursores derivados de intermediarios en la secuencia respiratoria y es impulsada por la energía derivada de esos procesos. Así, otras vías catabólicas y anabólicas están íntimamente interrelacionadas.
Energía de carbohidratos, proteínas y grasas

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 112
En presencia de oxígeno, el ácido pirúvico entra en el ciclo de Krebs donde se sintetiza más ATP y se transfieren más electrones y protones a las coenzimas. Estas coenzimas aceptoras de electrones transfieren su carga a la cadena transportadora de electrones a lo largo de la cual, paso a paso, los electrones caen a niveles inferiores de energía. A medida que esto ocurre, se fabrica mucho más ATP. Al final de la cadena transportadora, los electrones se reúnen con los protones y se combinan con el oxígeno, formándose agua. En ausencia de oxígeno, el ácido pirúvico puede convertirse en ácido láctico o etanol. Este proceso, llamado fermentación, no produce ATP, pero regenera las moléculas de coenzima aceptoras de electrones, necesarias para que la glucólisis continúe.
EN LA GLUCÓLISIS LA GLUCOSA SE CONVIERTE EN DOS PIRUVATOS
La glucólisis, también denominada glicólisis o ruta de Embden-Meyerhof, es la secuencia metabólica en la que se oxida la glucosa. Consiste de nueve reacciones enzimáticas que producen dos moléculas de ácido pirúvico o piruvato y dos equivalentes reducidos de NADH, los que, al introducirse en la cadena respiratoria, producirán cuatro moléculas de ATP.
Cuando hay ausencia de oxígeno, la glucólisis es la única vía que produce ATP en los animales. Los organismos primitivos se originaron en un mundo cuya atmósfera carecía de 02 y, por esto, la glucólisis se considera como la vía metabólica más primitiva. Está presente en todas las formas de vida actuales. Es la primera parte del metabolismo energético y en las células eucariotas ocurre en el citoplasma (citosol).
La glucólisis (que literalmente significa “rotura de azúcar”) no necesita oxígeno y ocurre en condiciones aeróbicas o anaeróbicas. En la figura 04 se presenta un resumen sencillo de la glucólisis, en la cual una molécula de glucosa (compuesto de seis carbonos) se convierte en dos moléculas de piruvato (compuesto de tres carbonos). Se captura parte de la energía de la glucosa; hay una ganancia neta de dos moléculas de ATP y dos de NADH. Las reacciones de la glucólisis ocurren en el citosol, donde hay los componentes necesarios, como ADP, NAD+ y fosfatos inorgánicos, que flotan libres en el citosol.
La glucólisis es un proceso en el cual una molécula de glucosa de 6 carbonos se escinde en dos moléculas de 3 carbonos de ácido pirúvico. Este proceso da como resultado un rendimiento neto de dos moléculas de ATP (a partir de ADP y fosfato inorgánico) y dos moléculas de NADH (a partir de NAD+).
La glucólisis comienza con una molécula de glucosa. En este proceso, primero se invierte energía por transferencia de un grupo fosfato desde una molécula de ATP, una por cada paso, a la molécula de azúcar. La molécula de 6 carbonos luego se escinde y, de allí en adelante, la secuencia produce energía. En cierto momento se reduce una molécula de NAD+ a NADH y H+ almacenandose parte de la energía producida por la oxidación del gliceraldehído-3-fosfato. En los pasos finales las moléculas de ADP toman energía del sistema, fosforilándose a ATP.
Resumiendo: para iniciar la secuencia glucolítica es necesaria la energía de los enlaces fosfato de dos moléculas de ATP. Posteriormente se producen dos moléculas de NADH a partir de dos de NAD+ y cuatro de ATP a partir de cuatro de ADP:
Glucosa+2ATP+4ADP+2Pi+2NAD+=>2 Ácido pirúvico+ 2ADP + 4ATP + 2NADH2 + 2H+ + 2H2O
De esta forma, una molécula de glucosa se convierte en dos moléculas de ácido pirúvico. La ganancia neta, la energía recuperada, es dos moléculas de ATP y dos moléculas de NADH por molécula de glucosa. Las dos moléculas de ácido pirúvico contienen todavía una gran parte de la energía que se encontraba almacenada en la molécula de glucosa original. La serie de

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 113
reacciones que constituyen la glucólisis se lleva a cabo virtualmente en todas las células vivas, desde las células procarióticas hasta las células eucarióticas de nuestros propios cuerpos.
Glucólisis o ruta de Embden-Meyerhof: transformación de la glucosa en dos moléculas de piruvato (ácido pirúvico)

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 114
Panorama general de la glucólisis. La fase de inversión de energía de la glucólisis (columna izquierda) lleva al desdoblamiento del azúcar; ATP y NADH se producen durante la fase de captura de energía (columna derecha). Durante la glucólisis cada molécula de glucosa se convierte en dos piruvato, con rendimiento neto de dos moléculas de ATP y dos de NADH.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 115
Glucólisis
EL PIRUVATO SE CONVIERTE EN ACETILCOENZIMA A
En los eucariotas, las moléculas de piruvato formadas en la glucólisis entran en las mitocondrias, donde se convierten en acetilcoenzima A (acetilCoA). Estas reacciones ocurren en el citosol de los procariotas aerobios. En esta serie de reacciones, el piruvato experimenta un proceso denominado descarboxilación oxidativa. En primer término, un grupo carboxilo se disocia en la forma de dióxido de carbono, que se difunde hacia el exterior de la célula. Luego, se oxida el fragmento residual de dos carbonos, y el NAD+ acepta los electrones que se disociaron durante la oxidación. Por último, el fragmento oxidado de dos carbonos, un grupo acetilo, se une a la coenzima A, de lo que resulta acetilcoenzima A.
La coenzima A transfiere grupos derivados de ácidos orgánicos. En este caso se trata de un grupo acetilo, relacionado con el ácido acético. La coenzima A se sintetiza en la célula a partir de una de las vitaminas del complejo B, el ácido pantoténico. La fijación del grupo acetilo a la coenzima A es catalizada por un complejo multienzimático, que incluye varias copias de cada una de tres enzimas distintas. La reacción global para la síntesis de acetilcoenzima A es como sigue: 2 Piruvatos + 2 NAD+ + 2 CoA 2 AcetilCoA + 2NADH2 + 2C02

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 116
Nótese que la molécula original de glucosa se ha oxidado parcialmente en dos grupos acetilo y dos moléculas de C02. Los átomos de hidrógeno disociados redujeron el NAD+ a NADH.
En este punto de la respiración aeróbica, se han formado cuatro moléculas de NADH por el catabolismo de una sola de glucosa; a saber: dos durante la glucólisis y otras tantas al formarse la acetilCoA a partir del piruvato. Formación de acetilcoenzima A.- El piruvato, una molécula de tres carbonos que es el producto terminal de la glucólisis, ingresa a la mitocondria y experimenta descarboxilación oxidativa. En primer término, el grupo carboxilo se disocia en la forma de dióxido de carbono. Luego, se oxida el fragmento residual de dos carbonos, y sus electrones se transfieren al NAD+. Por último, el grupo de dos carbonos oxidado, que es un grupo acetilo, se une a la coenzima A. Esta tiene un átomo de azufre, que forma un enlace muy inestable (indicado mediante una línea ondulada) con el grupo acetilo. EL CICLO DEL ÁCIDO CÍTRICO OXIDA LA ACETILCOENZIMA A
Panorama general del ciclo del ácido cítrico. En el ciclo entran dos grupos acetilo por cada glucosa. Cada grupo acetilo, de dos carbonos, se combina con oxalacetato, de cuatro carbonos, para formar

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 117
citrato, de seis carbonos. Las dos moléculas de CO2 se regeneran para formar oxalacetato, y en el proceso se captura energía en la forma de un ATP, tres NADH y un FADH2 por grupo acetilo (o dos ATP, seis NADH y dos FADH2 por glucosa).
El ciclo del ácido cítrico, también llamado ciclo de los ácidos tricarboxílicos (ciclo TCA) o ciclo de Krebs (en honor de Sir Hans Krebs), ocurre en las mitocondrias. En la figura se presenta un resumen sencillo de este ciclo, y sus ocho pasos se ilustran en la figura 10. Cada reacción es catalizada por una enzima específica.
Ciclo del ácido cítrico. La vía catabólica principal de la acetil-CoA en los organis mos aerobios. La acetil-CoA, el producto del catabolismo de los carbohidratos, las proteínas y los lípidos, ingresa al ciclo junto con H20 y se oxida a C02 con liberación de equivalentes reductores (2H).

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 118
La primera reacción del ciclo ocurre cuando la acetilCoA transfiere su grupo acetilo de dos carbonos al oxalacetato, compuesto de cuatro carbonos, con lo que se forma citrato, que posee seis carbonos. El ciclo del ácido cítrico (ciclo de Krebs). Oxalacetato + AcetilCoA Citrato + CoA Compuesto Compuesto Compuesto de cuatro carbono de dos carbonos de seis carbonos
Luego, el citrato experimenta una sucesión de transformaciones químicas, en que pierde primero un grupo carboxilo y luego otro, en la forma de C02.
La mayor parte de la energía que queda disponible con los pasos oxidativos de este ciclo se transfiere en la forma de electrones de alto contenido de energía al NAD+, lo que forma NADH2. Por cada grupo acetilo que entra el ciclo del ácido cítrico se producen tres moléculas de NADH2. También se transfieren electrones al aceptor de electrones FAD, con lo que se produce FADH2.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 119
En el curso del ácido cítrico, se disocian dos moléculas de C02 y el equivalente a ocho átomos de hidrógeno (ocho protones y ocho electrones), con la formación de tres moléculas de NADH2 y una de FADH2. Se generan más hidrógeno del que entró en el ciclo con la molécula de acetilCoenzima A. Estos átomos de hidrógeno provienen de moléculas de agua que se agregan durante las reacciones del ciclo. El C02, producido corresponde a los dos átomos de carbono del grupo acetilo que entró en el ciclo ATC. Al final se ha regenerado en oxalacetato (de cuatro carbonos), y puede comenzar un nuevo ciclo.
Debido a que se producen dos moléculas de acetilCoA por cada una de glucosa, se requieren dos ciclos por molécula de glucosa. Al término de dos vueltas del ciclo la glucosa original ha perdido todos sus átomos de carbono y podría considerarse como totalmente consumida. Para resumir, el ciclo del ácido cítrico genera 4 C02, 6 NADH2, 2 FADH2 y 2 ATP por molécula de glucosa.
Al final del ciclo del ácido cítrico, la glucosa se ha catabolizado por completo. Solamente cuatro moléculas de ATP se ha formado por fosforilación al nivel del sustrato: dos durante la glucólisis y dos en el ciclo del ácido cítrico. Esta vez, la mayor parte de la energía de la glucosa se encuentra en la forma de electrones de alta energía en NADH2 y FADH2.Su energía servirá para impulsar la síntesis de más trifosfato de adenosina (ATP) mediante la cadena de transporte de electrones y la quimiosmósis.
El ciclo de Krebs resumido El ciclo de Krebs contituye la segunda etapa del catabolismo de carbohidratos. La glucolisis rompe la glucosa (6 carbonos) generando dos moléculas de piruvato (3 carbonos). En eucariotas el piruvato se desplaza al interior de la mitocondria (gracias a un transportador específico de membrana interna). En la matriz mitocondrial produce acetil-CoA que entra en el ciclo de Krebs.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 120
En el catabolismo de proteínas, los enlaces peptídicos de las proteínas son degradados por acción de enzimas proteasas en el tracto digestivo liberando sus constituyentes aminoacídicos. Estos aminoácidos penetran en las células, donde pueden ser empleados para la síntesis de proteínas o ser degradados para producir energía en el ciclo de Krebs. Para su entrada al ciclo deben eliminarse sus grupos amino (terminales y laterales) por acción de enzimas aminotransferasas y desaminasas, principalmente.
Leyenda de colores: enzimas, coenzimas, nombres de sustratos, iones metálicos, moléculas inorgánicas, inhibición, estimulación
En el catabolismo de grasas, los triglicéridos son hidrolizados liberando ácidos grasos y glicerol. En el hígado el glicerol puede ser convertido en glucosa vía dihidroxiacetona fosfato y gliceraldehído-3-fosfato, por la gluconeogénesis (ruta anabólica). En muy diversos tejidos, especialmente en músculo cardiaco, los ácidos grasos son degradados en la matriz mitocondrial mediante sucesivos ciclos de beta oxidación que liberan unidades de acetil-CoA, que pueden incorporarse al ciclo de Krebs. En ocasiones, el ciclo de Krebs puede rendir propionil-CoA (3 carbonos), que puede emplearse para la síntesis de glucosa en la gluconeogénesis hepática.
El ciclo de Krebs siempre es seguido por la fosforilación oxidativa. Este proceso extrae la energía en forma de electrones de alto potencial de las moléculas de NADH y FADH2, regenerando NAD+ y FAD, gracias a lo cual el ciclo de Krebs puede continuar. Los electrones son transferidos a moléculas de O2, rindiendo H2O. Pero esta transferencia se realiza a través de una cadena transportadora de electrones capaz de aprovechar la energía potencial de los electrones para bombear protones al espacio intermembrana de la mitocondria. Esto genera un gradiente electroquímico de H+, que es utilizado para la síntesis de ATP mediante la enzima

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 121
ATP sintetasa De este modo el ciclo de Krebs no utiliza directamente O2, pero lo requiere al estar acoplado a la fosforilación oxidativa. Por cada molécula de glucosa la energía obtenida mediante el metabolismo oxidativo, es decir, glucolisis seguida del ciclo de Krebs, equivale a unas 38 o 36 moléculas de ATP. LA CADENA DE TRANSPORTE DE ELECTRONES ESTA ACOPLADA A LA INTESIS DE ATP
A continuación se considera el destino de todos los electrones disociados de una molécula de glucosa durante la glucólisis, la formación de acetilCoA y el ciclo del ácido cítrico. Estos electrones se transfieren a los aceptores primarios de hidrógeno NAD+ y FAD, con la formación de NADH2 y FADH2.
Tales compuestos reducidos entran en este punto de la cadena de transporte de electrones, donde los electrones de alto contenido de energía de sus átomos de hidrógeno se transfieren de una molécula aceptora a otra.
Representación detallada del transporte de electrones y la quimiosmósis. La cadena de trasnporte de electrones en la membrana interna de la mitocondria incluye tres bombas de protones que se localizan en tres de los cuatro complejos de transporte de electrones. La energía liberada durante dicho transporte se utiliza para llevar protones (H+) de la matriz mitocondrial al espacio intermembranoso, donde se acumula una alta concentración de protones. Se impide que estos se difundan de nuevo hacia la matriz, excepto en conductos especiales en la cintasa del ATP en la membrana interna. El flujo de protones a través de la cintasa genera ATP.
La cadena de transporte de electrones es una serie de portadores de electrones incluida en la membrana interna de las mitocondrias en los eucariotas, y en la membrana plasmática de los procariotas aerobios. Como NADH y FADH2, cada portador puede existir en una forma oxidada o una reducida. Los electrones pasan por la cadena de transporte en una serie de reacciones redox: cada molécula aceptora se reduce cuando acepta electrones y se oxida cuando los cede. Los que entran en el sistema de transporte de electrones tienen contenido de energía relativamente alto, y pierden parte de ella en cada paso en la cadena de portadores.
La tercera etapa de la respiración celular es la cadena de transporte de electrones. Tal como hemos visto, la cadena obtiene electrones del transportador de hidrógeno NADH+ + H+, la forma reducida del NAD+. Otro transportador de hidrógeno relacionado, denominado FAD+ (flavín

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 122
adenina dinucleótido) también transporta algunos electrones desde el ciclo de Krebs hasta la cadena transportadora de electrones. La forma reducida del FAD+ es el FADH2.
De esta forma, la glucólisis y el ciclo de Krebs son etapas liberadoras de energía que extraen electrones de las moléculas de los metabolitos mientras los degradan hasta formar compuestos más sencillos como el CO2 o el agua.
De la glucólisis, el ciclo de Krebs, la etapa previa al ciclo de Krebs, la beta oxidación de ácidos grasos y la oxidación de aminoáidos se obtienen moléculas de NADH+ + H+ y FADH2 llamadas moléculas de poder reductor.
Cada molécula de NADH+ + H+ cede sus dos electrones a un complejo llamado NADH dehidrogenasa compuesto por flavín mononucleótido (FMN) que se encuentra oxidado. Se reduce con los dos electrones cedidos por el poder reductor que ahora está oxidado y que vuelve a las rutas catabólicas. Estos dos electrones pasan por una serie de compuestos que se van oxidando y reduciendo (cediendo electrones para oxidarse y aceptándolos para reducirse) hasta llegar a un aceptor final que es el oxígeno. El FADH2 se incorpora a la cadena más tarde, lo que explica su menor rendimiento energético.
Esto implica que cada complejo tiene mayor potencial redox que el sigiente, pero menos que el anterior, hasta llegar al aceptor final, el átomo de oxígeno, que se transforma en ión óxido y se une a dos protones para formar agua. Del complejo NADH deshidrogenasa y de los cuatro citocromos se expulsan protones H+, que servirán para formar ATP gracias a la fosforilación oxidativa.
Sin el oxígeno como aceptor final la cadena no podría ceder los electrones al último aceptor, asÍ que los complejos no podrían volver a su estado oxidado reiniciando la cadena: el proceso es aerobio. Las rutas catabólicas y la cadena de transporte electrónico dependen la una de las otras, ya que sin las rutas catabólicas no existiría poder reductor que comenzara la cadena y sin cadena respiratoria el poder reductor no se podría volver a oxidar. Todo este proceso se produce en la membrana interna de la mitocondria. Ya fue mencionado que la fosforilación oxidativa es la transferencia de electrones de los equivalentes reducidos NADH, NADPH, FADH, obtenidos en la glucólisis y en el ciclo de Krebs hasta el oxígeno molecular, acoplado con la síntesis de ATP. Este proceso metabólico está

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 123
formado por un conjunto de enzimas complejas que catalizan varias reacciones de óxido-reducción, donde el oxígeno es el aceptor final de electrones y donde se forma finalmente agua.
La fosforilación oxidativa es un proceso bioquímico que ocurre en las células. Es el proceso metabólico final (catabolismo) de la respiración celular: la glicólisis y el ciclo del ácido cítrico. De una molécula de glucosa se obtienen 38 moléculas de ATP mediante la fosforilación oxidativa.
Dentro de las células, la fosforilación oxidativa se produce en las membranas biológicas. En procariotas es la membrana plasmática y en eucariotas es la membrana interna de las dos que forman la membrana mitocondrial. El NADH y FADH2, moléculas donadores de electrones que "fueron cargadas" durante el ciclo del ácido cítrico, se utilizan en un mecanismo intrincado (que implica a numerosas enzimas como la NADH-Q reductasa, la citocromo “c” oxidasa y la citocromo reductasa), gracias a la bomba H+ que moviliza los protones contra un gradiante de membrana.
Un gran complejo proteico llamado ATP-sintetasa situado en la membrana, permite a los protones pasar a través en ambas direcciones; genera el ATP cuando el protón se mueve a favor de gradiente, y consume una molécula de ATP para bombear un protón en contra de gradiente. Debido a que los protones se han bombeado al espacio intermembranoso de la mitocondria en contra de gradiente, ahora pueden fluir nuevamente dentro de la matriz mitocondrial y mediante la vía ATP-sintetasa, se genera ATP en el proceso.
Cada molécula de NADH contribuye suficientemente a generar la fuerza motriz de un protón que produzca 2.5 moléculas de ATP. Cada molécula de FADH2 produce 1.5 moléculas de ATP. Todas juntas, las 10 moléculas de NADH y las 2 FADH2 contribuyen a través de la oxidación de la glucosa (glucólisis, conversión de piruvato en acetil-CoA y ciclo de Krebs) a formar 34 de las 30 moléculas totales de ATP transportadoras de energía. Hay que decir que estos valores de moléculas de ATP son máximos. En realidad cada molécula de NADH contribuye a formar entre 2 y 3 moléculas de ATP, mientras que cada FADH2 contribuye a un máximo de 2 moléculas de ATP. LA RESPIRACIÓN AERÓBICA DE UNA MOLECULA DE GLUCOSA GENERA CUANDO MUCHO 36 A 38 MOLECULAS DE ATP
Se analiza a continuación en que puntos de la respiración aerobia se captura energía biológicamente útil además de calcular el rendimiento total de energía que resulta de la oxidación completa de la glucosa.
1. En la glucólisis, la glucosa se activa con la adición de fosfatos procedentes de dos moléculas de ATP y se convierte por último en 2 piruvatos + 2NADH2 + 4 ATP, con la generación de dos moléculas de ATP.
2. Las dos moléculas de piruvato se metabolizan en 2 acetilCoA + 2 C02 + 2 NADH. 3. En el ciclo del ácido cítrico, las dos moléculas de acetilCoA se transforman en 4
CO2 + 6 + NADH2 + 2 FADH2 + 2 ATP.
La oxidación del NADH2 en la cadena de transporte de electrones genera hasta tres moléculas de ATP por cada una de NADH2, de modo que las 10 NADH2 pueden producir hasta 30 ATP. Sin embargo, las dos moléculas de NADH2 provenientes de la glucólisis originan cada una dos o tres de ATP. Esto se debe a que determinados tipos de células eucariotas deben invertir energía para

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 124
desplazar el NADH2 resultante de la glucólisis a través de la membrana mitocondrial.
Las células procariotas carecen de mitocondrias, de modo que no necesitan transferir moléculas de NADH2. Por este motivo, las bacterias son capaces de generar tres ATP por cada NADH, aún los producidos en la glucólisis.
Así el número máximo de moléculas de ATP formadas con la energía del NADH es de 28 a 30. La oxidación de cada molécula de FADH2 genera dos de ATP, de manera que las dos moléculas de FADH2 producidas en el ciclo del ácido cítrico dan origen a cuatro de ATP.
4. Si se suman todas las moléculas de ATP producidas (dos en la glucólisis, dos en el ciclo del ácido cítrico y 32 a 34 en el transporte de electrones y la quimiosmósis), se puede apreciar que el metabolismo aerobio completo de una molécula de glucosa produce como máximo 36 a 38 de ATP. Nótese que la mayor parte del ATP se genera por fosforilación oxidativa, en la que participan la cadena de transporte de electrones y la quimiosmósis. Sólo cuatro ATP se forman por fosforilación a nivel de sustrato en la glucólisis y el ciclo del ácido cítrico.
LANZADERAS A TRAVES DE LA MEMBRANA MITOCONDRIAL
La membrana mitocondrial interna es impermeable al NADH2, que es una molécula grande. Por tanto, el NADH2 producido por el citosol durante la glucólisis no puede difundirse al interior de las mitocondrias para transferir sus electrones a la cadena de transporte de electrones. A diferencia de ATP y ADP, el NADH2 no tiene una proteína portadora que lo lleve de un lado a otro de la membrana. En vez de ello, han aparecido por evolución varios sistemas que transfieren a las mitocondrias los electrones del NADH2, más no las moléculas del NAD.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 125
En las células de hígado, riñón y corazón, un sistema “LANZADERA” especial se encarga de transferir los electrones del NADH2 a través de la membrana mitocondrial interna a una molécula de NAD+ en la matriz. Estos electrones son transferidos a la cadena de transporte de electrones de la membrana mitocondrial interna, con producción de tres moléculas de ATP por cada par de electrones.
En las células de músculo esquelético, encéfalo y otras opera un tipo distinto de lanzadera. En éste se requiere más energía que en el de hígado, riñón y corazón, de manera que los electrones están en un nivel de energía más bajo cuando entran en la cadena de transporte de electrones. Los acepta la coenzima Q (ubiquinona), en vez del NAD+, de modo que se genera un máximo de dos moléculas de ATP por cada par de electrones. Por eso el número de moléculas de ATP que produce la respiración aerobia de una molécula de glucosa en las células del músculo esquelético es de 36, no de 38.
NOTA IMPORTANTE: Si bien la manera correcta de escribir la forma reducida del NAD+ es NADH + H+, por sencillez dicha forma se expresa simplememte como NADH2 en la presente separata.
LA FERMENTACIÓN
La fermentación es un proceso catabólico de oxidación incompleto, siendo el producto final un compuesto orgánico. Estos productos finales son los que caracterizan los diversos tipos de fermentaciones.
La fermentación típica es llevada a cabo por las levaduras. También algunos metazoos y plantas menores son capaces de producirla.
El proceso de fermentación anaeróbica se produce en ausencia de oxígeno como aceptor final de los electrones del NADH producido en la glucólisis (que funciona como proceso anaerobio). La necesidad de un aceptor final, para los electrones procedentes del NADH, distinto del oxígeno hace que se emplee un compuesto orgánico que se reducirá para poder reoxidar el NADH. El compuesto orgánico que se reduce (acetaldehído, piruvato, etc.) es un derivado del
sustrato que se ha oxidado anteriormente. En los seres vivos, la fermentación es un proceso anaeróbico y en él no interviene la cadena respiratoria. Son propias de los microorganismos, como las bacterias y levaduras. También se produce la fermentación en el tejido muscular de los animales, cuando el aporte de oxígeno a las células musculares no es suficiente para el metabolismo y la contracción muscular.
Reacción enzimática que produce ácido láctico anaeróbicamente a partir de ácido pirúvico en las células musculares.
Desde el punto de vista energético, las fermentaciones son muy poco rentables si se comparan con la respiración, ya que a partir de una molécula de glucosa, sólo se obtienen 2 moléculas de ATP, mientras que en la respiración se producen 38 moléculas de ATP a partir de una molécula de glucosa. Esto se debe a la oxidación del NADH2, que en lugar de penetrar en la cadena respiratoria, cede sus electrones a compuestos orgánicos con poco poder oxidante.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote 126
En la industria la fermentación puede ser oxidativa, es decir, en presencia de oxígeno, pero es una oxidación aeróbica incompleta, como la producción de ácido acético a partir de etanol.
Las fermentaciones pueden ser: naturales, cuando las condiciones ambientales permiten la interacción de los microorganismos y los sustratos orgánicos susceptibles; o artificiales, cuando el hombre propicia condiciones y el contacto referido. Pasos por los cuales el ácido pirúvico, formado en la glucólisis, se convierte anaeróbicamente en etanol. Usos:
El beneficio primario de la fermentación es la conversión, ej. convertir el mosto en vino, cebada en cerveza y carbohidratos en dióxido de carbono para hacer pan. De acuerdo con Steinkraus (1995), la fermentación de los alimentos sirve a 5 propósitos generales:
o Enriquecimiento de la dieta a través del desarrollo de una diversidad de sabores, aromas y texturas en los substratos de los alimentos.
o Preservación de cantidades substanciales de alimentos a través de ácido lácteo, alcohólico, ácido acético y fermentaciones alcalinas.
o Enriquecimiento de substratos alimenticios con proteína, amino ácidos, ácidos grasos esenciales y vitaminas.
o Detoxificación durante el proceso de fermentación alimenticia. o Una disminución de los tiempos de cocinado y de los requerimientos de combustible.
La fermentación tiene algunos usos exclusivos para los alimentos. Puede producir nutrientes importantes o eliminar antinutrientes. Los alimentos pueden preservarse por fermentación, la fermentación hace uso de energía de los alimentos y puede crear condiciones inadecuadas para organismos indeseables. Por ejemplo, avinagrando el ácido producido por la bacteria dominante, inhibe el crecimiento de todos los otros microorganismos. Dependiendo del tipo de fermentación, algunos productos (ej. alcohol de fusel) pueden ser dañinos para la salud. En alquimia, la fermentación es a menudo lo mismo que putrefacción, queriendo decir, permitir el pudrimiento o la descomposición natural de la sustancia.

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote
FACULTAD DE CIENCIAS DE LA SALUD DEPARTAMENTO ACADÉMICO DE QUÍMICA Y FARMACIA
SECCIÓN DE CIENCIAS BIOLÓGICAS
ASIGNATURA
BIOLOGÍA CELULAR Y MOLECULAR
Contenido: • CAPITULO DOS : “ESTRUCTURA Y FISIOLOGÍA CELULAR”
o Biología celular: Citología y teoría celular o Estructura celular: Membrana celular citoplasma y núcleo
Fisiología celular: Respiración celular

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote
CUESTIONARIO DE AUTOEVALUACIÓN
CAPÍTULO DOS
01. En que parte de la célula se lleva a cabo el proceso de GLUCÓLISIS: .......................................................
02. En el ciclo de los ÁCIDOS TRICARBOXILICOS, los NADH2 que se producen, en cadena respiratoria cuantos ATP generan: ...................................................
03. Es un organelo de la célula que se encuentra generalmente en las regiones donde se necesita alta concentración de energía: ....................................................
04. Son un grupo de organelos intracitoplasmáticos cuya función principal es la digestión del material intracelular o provenientes del exterior: .............................................
05. En el proceso de GLUCÓLISIS cuántos ATP se producen en forma total, incluyendo los NADH2 que van a cadena respiratoria: ...........................................
06. Cuantos ATP se producen en el CICLO DE KREBS, cuando se metaboliza media molécula de glucosa (un ácido pirúvico): .........................................
07. En que parte de la célula se lleva a cabo el CICLO DE LOS ÁCIDOS TRICARBOXILICOS: .................................
08. En CADENA RESPIRATORIA cuantos ATP se producen en total (sin tener en cuenta el gasto de energía) cuando se metaboliza por completo una molécula de glucosa: ..........................................
09. Son organelos presentes en todos los tipos celulares, de forma ovoide y limitados por una sola membrana. Contienen enzimas oxidativas y una de las más comunes es la catalasa: ...........................
10. Son estructuras citosólicas que se encargan de la síntesis proteica: ......................................................
11. Son estructuras que se encuentran generalmente en los extremos de los cromosomas: A. Cromátidas B. Cromatina C. Telómeros D.Histonas E. N.A
12. Ocupa alrededor del 10% del volumen total de la célula y en él se halla confinado el DNA, y está rodeado por una doble membrana que es atravesada por poros: A. Nucleolo B. Ribosoma C. Núcleo D. Lisosoma E. Peroxisoma
13. Es un elemento celular que se caracteriza por ser un armazón, mantiene la forma celular y también es responsable del movimiento intracelular: A. Mitocondrias B. Microtúbulos C. Peroxisomas D. Cloroplastos E. Enzimas RESPUESTAS:
1. Citosol
2. 18
3. Mitocondria
4. Lisosomas
5. 10
6. 1
7. Mitocondria
8. 40

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote
9. Peroxisomas
10. Ribosomas
11. Telómeros
12. Núcleo
13. Microtúbulos

Mg. Blgo. Mblgo.. Luis A. Sánchez Angulo / Mblgo. José L. Gutierrez Aponte Biología Celular y Molecular
Universidad Católica Los Ángeles de Chimbote
REFERENCIAS BIBLIOGRAFICAS Y WEBGRAFÍAS
o Alberts, B.; Bray, D.; Lewis, J.; Raff, M. ; Roberts, K. y J. Watson. 1983. Biología
Molecular de la Célula. 2da edición. ediciones Omega, S.A. Barcelona – España. o Asociación Educativa ADUNI. 2004. Biología Una perspectiva Evolutiva. 2da. Edición.
Lumbreras Editores, Lima – Perú.
o Curtis, H. y S. Barnes. 2002. Biología. 6ta. Edición. Editorial Panamericana. Buenos Aires – Argentina.
o De Robertis, E. y E. De Robertis 1997. Biología Celular y Molecular. 12ava edición.
Editorial El Ateneo. Buenos Aires – Argentina. o Gardner, E. Principios de Genética. 5ta Edición. Editorial Limusa S.A. de C.V. México
D.F. 1985.
o Jimeno, A. Biología. Editorial Santillana S.A. Madrid – España. 1983.
o Karp, G. 2003. Biología Celular y Molecular. McGraw – Hill Interamericana. México D. F.
o Solomon, E.; Berg. L. y D. Martin. 2001. Biología. 5ta. Edición. McGraw-Hill Interamericana.
o Sánchez, L. y J. Gutierrez. Manual de Prácticas de Biología Celular y Molecular. 2da
Edic. ULADECH. Chimbote – Perú. 2006.
o Villee, C. Biología. 7ma edición. Nueva Editorial Interamericana, S.A., México D.F. 1995.
o Aula virtual de Biología: Universidad de Murcia. Departamento de Biología
http://www.um.es/molecula/indice.htm
o Facultad de Ciencias Biológicas. Introducción a la Biología. http://www.uc.cl/.../bio100/html/portadaMIval2.6.1.html
o Biología Bachillerato. http://www.web.educastur.princast.es/.../INDICE.htm
o Proyecto biósfera. Ministerio de Educación y Ciencia (Biología y Geología). España.
http://www.recursos.cnice.mec.es/.../recursos_galeria2.htm