
Comparación de modelos predictivos de ondulación geoidal ...
Upload
truongcongCategory
view
220download
0
ESTUDIO DE LA BIODIVERSIDAD DE
OLIGOQUETOS ACUÁTICOS SUBTERRÁNEOS
EN SISTEMAS KÁRSTICOS DEL PAÍS VASCO Y
ZONAS LIMÍTROFES - Fase I
Asociación Deportiva y Espeleológica Saguzaharrak, 2007

ESTUDIO DE LA BIODIVERSIDAD DE
OLIGOQUETOS ACUÁTICOS SUBTERRÁNEOS
EN SISTEMAS KÁRSTICOS DEL PAÍS VASCO Y
ZONAS LIMÍTROFES - Fase I
Autores:
AINARA ACHURRA
PILAR RODRIGUEZ
Memoria del proyecto presentada al Departamento de Medio Ambiente y
Ordenación del Territorio para subvenciones para proyectos de
investigación aplicada de la biodiversidad, en el marco de las previsiones
de la Estrategia Ambiental Vasca de Desarrollo Sostenible.

RESUMEN: El conocimiento de los oligoquetos acuáticos subterráneos
en la península Ibérica es aún muy escaso, pero los datos disponibles
sugieren que la región Cantábrica puede presentar un nivel importante de
biodiversidad y endemismo para la fauna acuática subterránea. Es por ello
que se han planteado como objetivos 1) contribuir al conocimiento de las
especies de oligoquetos acuáticos subterráneos en sistemas kársticos del
País Vasco y zonas adyacentes, 2) describir nuevos taxones y completar la
descripción de taxones interesantes, 3) identificar hotspots y analizar las
razones, 4) estudiar las relaciones filogenéticas entre taxones y 5) aportar
datos de biodiversidad animal que contribuyan a los planes de
conservación de los sistemas kársticos estudiados. Se han determinado 32
taxones de oligoquetos acuáticos subterráneos en el karst de Gorbeia,
tres los cuales posiblemente sean especies nuevas para la Ciencia, por lo
que se procederá a su descripción. Este grupo faunístico representa del 5
al 92 % de los invertebrados bentónicos en cuevas y surgencias de
Gorbeia, y los resultados revelan diferencias importantes en la
composición respecto al karst de Urdaibai. Entre los factores ambientales
estudiados se revela el papel influyente de la altitud o temperatura y la
conductividad en la composición de especies de oligoquetos. Finalmente,
se ha creado un ranking de conservación de cuevas y surgencias del karst
de Gorbeia, basado en la biodiversidad de oligoquetos acuáticos
subterráneos.

ÍNDICE:
1. INTRODUCCIÓN
1.1 ANTECEDENTES Y ESTADO ACTUAL DEL TEMA
1.2 INTERÉS GENERAL DEL PROYECTO
1.3 OBJETIVOS
2. METODOLOGÍA
2.1 SELECCIÓN DE LOS PUNTOS DE MUESTREO
2.2 REGISTRO DE LOS PUNTOS DE MUESTREO
2.3 MAPA DE LOS PUNTOS DE MUESTREO
2.4 CARACTERIZACIÓN DEL HÁBITAT
2.5 MUESTREO Y PROCESADO DE FAUNA ACUÁTICA SUBTERRÁNEA
2.6 ESTUDIO TAXONÓMICO
2.7 ANÁLISIS MOLECULAR Y ESTUDIO FILOGENÉTICO
2.8 ÍNDICES DE BIODIVERSIDAD Y MODELOS PREDICTIVOS
2.9 MEJORA DEL ÍNDICE DE VULNERABILIDAD
3. RESULTADOS
3.1 MUESTREO EN CAVIDADES Y SURGENCIAS KÁRSTICAS DEL PAÍS
VASCO Y ZONAS ADYACENTES Y PROCESADO DE LAS MUESTRAS
3.2 ESTUDIO TAXONÓMICO Y DESCRIPCIÓN DE NUEVAS ESPECIES
PARA LA CIENCIA
3.3 MEJORA DEL MODELO DE VULNERABILIDAD PARA ESPECIES
3.4 ESTUDIO FILOGENÉTICO Y EVOLUTIVO
3.5 IDENTIFICACIÓN DE ZONAS DE ALTA BIODIVERSIDAD O HOTSPOTS
Y ANÁLISIS DE RAZONES
3.6 CONTRIBUCIÓN A LOS PLANES DE CONSERVACIÓN DE LOS
SISTEMAS KÁRSTICOS ESTUDIADOS
4. DISCUSIÓN GLOBAL Y CONCLUSIONES
5. BIBLIOGRAFÍA
6. ANEXOS

1. INTRODUCCIÓN
1.1 ANTECEDENTES Y ESTADO ACTUAL DEL TEMA
El interés por el conocimiento de la fauna subterránea de oligoquetos acuáticos en
Europa se remonta principalmente a los estudios de Cernosvitov y Hrabe en varios países
del Centro de Europa en la primera mitad del siglo veinte. Más recientemente, a partir de
los años setenta diversos trabajos de Dózsa-Farkas y Dumnicka se han centrado
principalmente en la fauna de la familia Enchytraeidae, mientras que Giani, Juget,
Martinez-Ansemil, Sambugar y Rodriguez han estudiado distintas familias de oligoquetos
acuáticos en el Sur de Europa. Las contribuciones de estos últimos autores al conocimiento
de la fauna de oligoquetos acuáticos en hábitats subterráneos del sur de Europa, se recogen
en un trabajo publicado recientemente (Giani et al. 2001), donde se cuantifican hasta 58
especies de oligoquetos estigobiontes (habitantes específicos del medio acuático
subterráneo) del Sur de Europa. El proyecto europeo PASCALIS (2001-2004, Protocols
for the Assesment and Conservation of Aquatic Life In the Subsurface) ha impulsado el
estudio de la fauna acuática subterránea como base de planes de conservación para este
medio y se han establecido protocolos estándares de muestreo para estimar la
biodiversidad.
En la Península Ibérica, los primeros estudios se deben a Hrabe (1963) y Delay
(1973), pero éstos fueron trabajos aislados, aunque interesantes en cuanto a que contienen
la descripción de nuevas especies. La originalidad y alto grado de endemismo de los
taxones descritos durante los últimos veinte años a través de diversas contribuciones (14
nuevas especies: Rodríguez, 1988; Giani y Rodríguez, 1988,1984; Rodríguez y Giani,
1987, 1989, 1994) es relevante y demanda una mayor atención investigadora. El número
total de especies de oligoquetos acuáticos que habitan el medio subterráneo citadas en la
Península Ibérica alcanza 166 especies, de las cuales 47 se encuentran en la Reserva de la
Biosfera de Urdaibai (RBU). A escala del País Vasco y áreas limítrofes las especies

subterráneas representan un 44% del total de las especies acuáticas epigeas e hipogeas
conocidas en la actualidad (Achurra y Rodriguez, 2008, en prensa).
El grupo investigador ha estudiado la biodiversidad de oligoquetos acuáticos
subterráneos en la RBU y ha identificado el área kárstica como hotspot o punto caliente de
biodiversidad para este grupo taxonómico, como resultado de aplicar índices de
biodiversidad (riqueza de especies, rareza y vulnerabilidad) y criterios como número de
especis estigobiontes y endemicas (Achurra y Rodriguez, 2008, en prensa). Los resultados
obtenidos también contribuyen a la gestión para la conservación de las cuevas y surgencias
estudiadas.
Se han formulado distintas hipótesis para explicar el origen de los taxones animales
en el medio subterráneo, las cuales pueden ayudar a comprender las causas de la elevada
biodiversidad característica de este medio. En primer lugar, un número de especies
acuáticas habría colonizado el medio subterráneo desde el medio hiporréico, es decir, a
partir de las comunidades que habitan el medio intersticial de lechos fluviales. En segundo
lugar, dado que algunas especies acuáticas estigobíticas pertenecen a familias e incluso
géneros marinos, se estudia la hipótesis de una posible contribución a las comunidades
subterráneas de elementos marinos a través del medio intersticial litoral (marino y
anquihalino). La existencia de una paleogeografía regional dinámica, con numerosos
episodios de transgresiones y regresiones marinas en la región costera del Cantábrico en
épocas geológicas recientes, ofrece un escenario plausible de colonización del medio
subterráneo por esta vía, como sugirió Stock (1980) en su modelo de Regresión. Más
tarde, este modelo fue redescrito más detalladamente como “Two step model”
(Notenboom, 1991), incluyendo dos cambios biogeográficos y ecológicos: uno llamado
“transición vertical”, donde una especie pasa a ser estigobionte; y una segunda “transición
horizontal” donde la especie pasa del mar al agua dulce. Por último, otras especies habrían
colonizado activamente el medio subterráneo, usándolo como refugio durante condiciones
ambientales adversas (bajas o altas temperaturas) que se produjeron durante el Pleistoceno
(periodos glaciares e interglaciares) e incluso en épocas anteriores.

El grupo investigador ha puesto a punto la metodología de preparación de las
muestras (muestreo, identificación de especies, extracción de DNA y amplificación por
PCR), logrando con éxito la secuenciación de fragmentos de los genes 16S rRNA y COI
de algunas especies de oligoquetos. Estos métodos permitirán contrastar las hipótesis
filogenéticas de taxones que se encuentren durante el presente proyecto.

1.2 INTERÉS GENERAL DEL PROYECTO
Los sistemas kársticos de la costa Cantábrica se encuentran situados en el área que
engloba el hipotético cinturón de alta biodiversidad descrito para los taxones subterráneos
terrestres, entre los 42 y 46º N de latitud en el Sur de Europa, en áreas templadas de alta
productividad y alta densidad de cuevas (Culver et al. 2006). Se trata, por tanto, de una
zona potencialmente muy rica en especies subterráneas y que posiblemente tenga su
correspondencia con la menos conocida fauna acuática. Son imprescindibles estudios
intensivos en el área que evalúen la riqueza específica de la fauna acuática subterránea a
una escala regional, europea o incluso mundial. Esperamos identificar hotspost de
biodiversidad para este grupo faunístico en la zona del País Vasco y áreas limítrofes, así
como encontrar taxones nuevos para la Ciencia y contribuir en el estudio de la sistemática
de los oligoquetos.
La posición costera de algunas unidades kársticas permite abordar hipótesis acerca
de la historia evolutiva de las especies estigobiontes, que colonizaron el medio subterráneo
tanto desde el medio superficial como desde el marino. Un ejemplo de ello es la unidad
kárstica de la RBU, ya que su gran diversidad de oligoquetos aporta ejemplos para estudiar
las hipótesis evolutivas. Destaca la presencia del oligoqueto Aktedrilus argatxae, especie
endémica que pertenece a un género común en el litoral marino intersticial y que, sin
embargo, fue descubierta en el manantial de la cueva de Argatxa (Arteaga, Urdaibai). Este
descubrimiento, constituyó entonces la primera cita de este género en aguas dulces
subterráneas del mundo. Posteriormente se han localizado otras especies del mismo género
en aguas subterráneas de Italia, Omán y Australia. La cueva de Argatxa es también la
localidad tipo del oligoqueto Troglodrilus galarzai. La distribución fragmentada de esta
especie, presente únicamente en Bizkaia y la cuenca del Ródano (Francia), tiene un gran
interés ya que podría explicarse mediante la teoría refugio a partir de una distribución más
amplia en el sudoeste Europeo. Una nueva especie del género Lamadrilus hallada en
varias localidades de la RBU es también interesante desde el punto de vista evolutivo, ya
que los únicos representantes de este género que se conocen en la actualidad han sido

encontrados en la región de Siberia. Por último, la presencia de otras especies, como por
ejemplo la del oligoqueto endémico Rhyacodrilus gernikensis, podría ser de gran utilidad
científica para explorar las relaciones evolutivas con la fauna intersticial hiporréica. En
este apartado, esperamos revelar nuevos datos sobre los escenarios evolutivos y su
antigüedad, utilizando datos moleculares y morfológicos.
El alto grado de especialización y la baja densidad en que se encuentran muchas
especies que habitan el medio subterráneo las hace especialmente vulnerables a la
contaminación del medio y a las alteraciones del hábitat. La contaminación de las aguas
puede producirse a partir de vertidos en dolinas o arroyos que pueden entrar en contacto
con el medio subterráneo directamente o aportar contaminantes por filtración o lixiviado.
Fertilizantes y pesticidas utilizados en tierras de labor situadas sobre el medio subterráneo
pueden ser también peligrosas fuentes de contaminación difusa, y eventualmente ocasionar
episodios catastróficos. Entre las alteraciones del hábitat, las más frecuentes son la
sedimentación no natural (por ejemplo por actividades forestales) y las modificaciones del
nivel freático. Los episodios de contaminación en algunas cuevas y manantiales de la RBU
probablemente hayan tenido consecuencias en las comunidades subterráneas. La Cueva de
San Pedro de Busturia es un ejemplo de ello. Durante los años 80 existía una población
estable de Trichodrilus diversisetosus que actualmente sospechamos que haya
desaparecido. Se trata de una especie estigobia, rara, muy vulnerable y endémica del norte
de la península y Francia. Nuevos estudios en esta localidad y en otras permitirán evaluar
el grado de afección de la fauna acuática subterránea a la contaminación del medio y
alteraciones del hábitat.
El estudio de la biodiversidad es la base para la gestión del medio acuático y su
efectiva conservación. Los sistemas karsticos constituyen una gran proporción del
territorio de la RBU y la protección efectiva de la fauna subterránea requiere no sólo un
control efectivo del acceso a las cavidades, sino también la conservación de todo el
sistema terrestre y acuático que alimenta a dichas cuevas, garantía de pervivencia de sus
poblaciones animales subterráneas. En este aspecto, es interesante mejorar el modelo de

vulnerabilidad para especies recientemente creado por parte del grupo investigador, ya que
nos aportará datos útiles para la conservación de las especies de oligoquetos acuáticos
subterráneos.
Por último, cabe destacar la importancia de integrar los estudios moleculares en la
conservación del medio, ya que ésta debería orientarse no solo desde el punto de vista de
conservación de especies y hábitats, sino también desde la idea de conservar el proceso
evolutivo del que depende toda la biodiversidad (Pullin, 2002). Debemos preguntarnos si
bajo la actual práctica de la conservación, las especies serán capaces de mantener o
modificar sus rangos de distribución como lo hicieran en el pasado. Para planificar una
estrategia de conservación para el futuro en un contexto evolutivo es imprescindible
obtener información sobre que sucedió en el pasado.

1.3 OBJETIVOS:
A continuación se resumen los objetivos generales del proyecto:
• Estudio de la biodiversidad de oligoquetos acuáticos subterráneos en sistemas
kársticos del País Vasco y zonas adyacentes
• Descripción de nuevos taxones
• Identificación de zonas de alta biodiversidad o hotspots y análisis de razones
• Estudio de las relaciones filogenéticas entre taxones utilizando datos moleculares y
morfológicos
• Contribución a los planes de conservación de los sistemas kársticos estudiados
Los objetivos concretos de esta primera fase del proyecto se centran principalmente en
el muestreo, el procesado de las muestras, identificación de especies de oligoquetos
acuáticos y la mejora del modelo de vulnerabilidad para especies subterráneas acuáticas:
Los objetivos generales de este estudio se enmarcan en un proyecto más amplio que
incluye la descripción de nuevos taxones y el estudio filogenético y evolutivo, los cuales
se espera completar en el año 2008 tras una segunda fase.

2. METODOLOGÍA
2.1. SELECCIÓN DE LOS PUNTOS DE MUESTREO
Con objeto de determinar la biodiversidad de oligoquetos acuáticos subterráneos en las
diferentes unidades hidrogeológicas, se han seleccionado puntos de muestreo en base a los
siguientes criterios:
• puntos muestreados con anterioridad
• puntos de fácil acceso
• puntos que puedan ser muestreados con técnicas de muestreo disponibles
Cada punto de muestreo se corresponde con una surgencia o una cueva con río endógeno.
a) Preselección de los puntos de muestreo
La selección de los puntos que pueden ser potencialmente muestreados (preselección)
se ha basado en la información disponible en:
• Mapas topográficos, hidrogeológicos, geográficos, etc…
• Inventarios de cuevas y surgencias
• Artículos científicos de fauna subterránea
• Conocimientos y experiencia del ADES
b) Visitas al campo
La prospección de los puntos preseleccionados asegura que dichos puntos puedan
ser muestreados con las técnicas de muestreo disponibles. Por ejemplo, algunas surgencias
no pueden ser muestreadas si han sido captadas para abastecimiento de agua.

2.2. REGISTRO DE LOS PUNTOS DE MUESTREO
Los puntos de muestreo se registran en una tabla, la cual contiene la información
mínima requerida para la localización espacial de dichos puntos. Este registro se adjunta
en el Anexo 1. A continuación se explica la información contenida en la tabla:
• Código: a cada punto de muestreo se le asigna un código, el cual es usado en el
muestreo, etiquetado de muestras de fauna, agua y sedimento, y análisis de datos. El
código consiste en 3 letras para la unidad hidrogeológica o karst y 2 números para el
punto de muestreo. Por ejemplo, el código “GOR01” se refiere al punto 01 en la
unidad hidrogeológica de Gorbeia.
U. H. de Santa Eufemia-Ereñozar: RBU
U. H. de Itxina: GOR
• Nombre: es el nombre del punto de muestreo, que preferiblemente coincidirá con el
que aparece en los mapas.
• Coordenada X: es la longitud en formato UTM, obtenida a partir de mapas
topográficos o medida en el campo.
• Coordenada Y: es la latitud en formato UTM, obtenida a partir de mapas
topográficos o medida en el campo.
• Altitud Z: es la elevación medida en metros sobre el nivel del mar.
• Hábitat: es una surgencia o un río endógeno en una cueva
• Tipo: especifica el grado de conocimiento de oligoquetos en el punto de muestreo,
distinguiendo entre:
1: punto suficientemente estudiado, no es necesario muestreo
2: punto estudiado con anterioridad que requiere muestreos adicionales
3: punto nuevo, es necesario muestreo completo

2. 3. MAPA DE LOS PUNTOS DE MUESTREO
Las figuras 1 y 2 muestran la distribución espacial de los puntos de muestreo en los
karst de la RBU y Gorbeia, respectivamente.
Figura 1. Mapa de la unidad hidrogeológica de Santa Eufemia-Ereñozar. Se
muestran en gris oscuro las subunidades, en gris claro la cuenca y rodeada con una
línea de puntos, el área de la Reserva de la Biosfera de Urdaibai. Los puntos de
muestreo se han marcado con un círculo negro y numerado.

Figura 2. Mapa de la unidad hidrogeológica de Gorbeia. Se muestran en gris oscuro
las subunidades. Los puntos de muestreo se han señalado con un círculo negro y
numerado.
2.4. CARACTERIZACIÓN DEL HÁBITAT
a) Descripción del punto de muestreo
Se han seleccionado un conjunto de variables que permitan describir el punto de
muestreo y den información sobre su hidrología, hidrogeología, geología y geomorfología.
Dichas variables se agrupan en el apartado “Caracterización del medio” de las hojas
creadas específicamente para los muestreos de surgencias y cuevas. Se adjuntan en el
Anexo 2. A continuación se explica la información contenida en dichas hojas:
Descripción de surgencias:
Zona: si la surgencia se localiza en el epikarst, la zona vadosa, la zona freática
Geología: es la unidad litológica principal
Morfología: referida al tipo de surgencia: reocrenon, limnocrenon helocrenon

Uso humano: abastecimiento, regardío, otros
Anchura (media): <1 m, 1-5 m, 5-25 m, 25-100 m , >100 m
Profundidad (media): < 0,1 m, 0,1-0,5 m, 0,5-1 m, 1-2 m, > 2 m
Pendiente (‰): <1 , 1-3, 3-7, >7
Velocidad de la corriente: turbulenta / rápida (≥ 0.3 m/s), estancada / lenta (< 0.3 m/s)
Corriente: perenne o intermitente
Descarga (estimación): 10 l/s o 100 l/s
Sustrato dominante: Bloques, cantos rodados, grava, arenas, limos y arcillas
Sustrato en %: Bloques, cantos rodados, grava, arenas, limos y arcillas
% de detritus
Condición del sustrato (%): Limpio, cubierto por hojarasca, materia orgánica, limo y
arcilla, otros
Estructura de las orillas: natural, bosque plantado (chopos, pinos, otros), canal de piedra,
canal de hormigón
Cobertura arbórea: % cobertura sobre el cauce
Sustrato muestreado: breve descripción
Vegetación acuática: algas, macrófitas, musgos, otros
Descripción de cuevas:
Zona: si la surgencia se localiza en el epikarst, la zona vadosa. la zona freática
Geología: es la unidad litológica principal
Longitud de la cueva
Profundidad de la cueva

Distancia del punto de muestreo al suelo, a la zona saturada y a la entrada de la cueva
Hábitat muestreado: infiltración lenta, sifón, lago, río endógeno, río exógeno
Velocidad de la corriente del río: turbulenta / rápida (≥ 0.3 m/s), estancada / lenta (<
0.3 m/s)
Corriente: perenne, intermitente
Descarga (estimación): 10 l/s o 100 l/s
Sustrato dominante: Bloques, cantos rodados, grava, arenas, limos y arcillas
Sustrato en %: Bloques, cantos rodados, grava, arenas, limos y arcillas
% de detritus
Condición del sustrato (%): Limpio, cubierto por hojarasca, materia orgánica, limo y
arcilla, otros
Sustrato muestreado: breve descripción
Así mismo, también se han tomado fotografías de los puntos de muestreo. Ver
Anexo 3 con las fotografías de los puntos del karst de Gorbeia.
b) Medida de parámetros físico-químicos

En cada ocasión de muestreo se ha medido en el campo las siguientes variables
físico-químicas: pH, temperatura (ºC), oxígeno disuelto (mg/l O2 y % de saturación), y
conductividad (µS/cm), en cada cuerpo de agua donde se toma una muestra de fauna.
Cada muestra se etiqueta con el código del punto de muestreo y la fecha.
En cada ocasión de muestreo y para cada cuerpo de agua donde se toma una muestra
de fauna, se ha recogido una muestra de agua, que se ha mantenido en frío hasta su análisis
químico, el cual deberá realizarse tan pronto como sea posible. El agua se filtró a través de
filtros de membrana 0.45 mm y se analizaron nitratos, fosfatos, calcio, magnesio y otros
iones (ICP: SGIker UPV/EHU).
c) Determinación de materia orgánica y granulometría del sedimento
Para cada punto de muestreo se ha recogido una muestra de sedimento y en el
laboratorio se realizó la determinación de la materia orgánica en el sedimento mediante el
cálculo del porcentaje de LOI (% Loss On Ignition) y el análisis de la granulometría en
porcentaje del peso seco de arenas gruesas (2.0 – 0.250 mm), arenas medias (250 – 125
µm), arenas finas (125 – 63 µm) y limos y arcillas (<63 µm), tras cribar el sedimento
previamente seco en la estufa por una columna de tamices.
2.5. Muestreo y procesado de fauna acuática subterránea
El muestreo de fauna acuática subterránea (oligoquetos y otros invertebrados) se ha
realizado con red Surber (tamaño de malla de 200 µm). Las muestras biológicas se lavaron
por un tamiz de 100 µm y se fijaron en el campo en formaldehído al 10%. Las muestras
para futuros estudios moleculares se recogieron en vivo y se transportaron al laboratorio
en hielo y en oscuridad. Cada muestra se etiqueta con el código del punto de muestreo y la
fecha.

Las muestras biológicas fijadas se lavaron y se tiñeron con eosina alcohólica en el
laboratorio, lo cual facilita la separación de los oligoquetos de menor tamaño, así como la
del resto de invertebrados bajo microscopio estereoscópico. El material separado se
conserva en alcohol etílico de 70%. Las muestras recogidas en vivo se separaron bajo la
lupa binocular en el laboratorio y tras la identificación de algunas especies de oligoquetos
interesantes al microscopio óptico, se fijaron en alcohol de 96º, para posibilitar su estudio
molecular ulterior. Los invertebrados no oligoquetos encontrados en las muestras se
identificaron a nivel de familia con objeto de determinar la contribución de los oligoquetos
a la comunidad del bentos en abundancia relativa.
Para cada uno de los puntos de muestreo se ha recopilado la siguiente información
en la hoja de muestreo (Ver Anexo 2):
SITE
Nombre Cuenca hidrogeográfica Código Muestreador
Fecha Hora X Y Z
DATOS CLIMATICOS
Lluvia Sol Tª aire Periodo de aguas bajo /medio /
alto
TECNICA DE MUESTREO
Aparato tamaño de malla (um) área
tiempo réplicas (nº) Comentarios
DIBUJO DEL ÁREA DE MUESTREO

Fotografía 1. Red Surber para muestreo de fauna acuática subterránea en una de las
cuevas del karst de la RBU.
2.6. Estudio taxonómico:
La identificación de los ejemplares de oligoquetos se ha realizado con la ayuda del
microscopio óptico de contraste interferencial Nomarski, mayoritariamente mediante
observación de montajes in toto en glicerina. También se realizaron preparaciones
microscópicas de algunos ejemplares en polivinil-lactofenol y líquido de Hoyer como
medios de montaje, para la observación de estructuras de naturaleza quitinosa como quetas
y cutículas peniales. Cuando fue necesaria la observación de estructuras del aparato
reproductor, los ejemplares se tiñeron en hematoxilina de Erlich y se realizaron micro-
disecciones y montajes en bálsamo de Canadá después de ser deshidratados y aclarados en
creosota.

Para la identificación de los ejemplares se usaron principalmente las siguientes
guías:
• BRINKHURST, R.O. 1971. A guide for the Identification of British Aquatic
Oligochaeta. Freshwater Biological Association, Scientific Publication No. 22,
55pp.
• BRINKHURST, R.O. 1986. Guide to the Freshwater Aquatic Microdrile
Oligochaetes of North America. Department of Fisheries and Oceans, Ottawa,
259pp.
• BRINKHURST, R.O. y JAMIESON, B.G.M. 1971. The Aquatic Oligochaeta of the
World. Oliver & Boyd. Edinburgh, 860pp.
• NIELSEN Y CHRISTENSEN.1959. The Enchytraeidae, critical revision and
taxonomy of European species. Natura Jutl., 8-9: 1-160.
• SPERBER, C. 1948. A taxonomical study of the Naididae. Zool. Bidr., Upps., 28:
1-296.
• TIMM T. 1999. A guide to the Estonian Annelida, Naturalist´s Handbooks 1.
Estonian Academy Publishers, Tartu-Tallinn, 208pp.
Son imprescindibles, además, los artículos científicos en que se describen diversos
taxones subterráneos y endémicos de la Península Ibérica que se listan a continuación:
• GIANI, N. & RODRIGUEZ, P. 1988. Description de quelques espèces nouvelles de
Tubificidae (Oligochaeta) des grottes et sources karstiques de la Péninsule Ibérique.
Stygologia, 4 (2): 122-137.
• GIANI, N. & RODRIGUEZ, P. 1994. New species of the genus Trichodrilus
(Oligochaeta, Lumbriculidae). Zoologica Scripta 23(1): 33-41
• JUGET, J, M. DES CHATELLIERS & P. RODRIGUEZ. 2006. Troglodrilus
(Annelida, Oligochaeta, Tubificidae), a new genus from subterranean habitats in
southwestern Europe. Hidrobiología 564 (1): 7-17.

• RODRIGUEZ, P. 1988. Sur certaines espèces de Lumbriculidae (Annelida:
Oligochaeta) du nord de la péninsule ibérique. Annls Limnol. 24 (3): 203-211.
• RODRIGUEZ, P. & N. GIANI. 1984. Description de Varichaetadrilus bizkaiensis
n. sp., nouvelle espèce de Tubificidae (Oligochaeta) d'Espagne. Bol. Soc. Hist. Nat.
Toulouse,120:127-130
• RODRIGUEZ, P. & N. GIANI. 1987. Sur deux espèces de Lumbriculidae
(Oligochaeta) cavernicoles: Cookidrilus speluncaeus n. gen n. sp. et Trichodrilus
diversisetosus Rodriguez, 1986. Boletin de la Societé d’Histoire Naturelle de
Toulouse, 123: 45-59.
• RODRIGUEZ, P. & N. GIANI. 1989. New species of Phallodrilus (Oligochaeta,
Tubificidae) from caves of North of Spain and Southwest of France. Hydrobiologia,
180: 57: 63.
• RODRIGUEZ, P. & N. GIANI. 1994. A preliminary review of the taxonomic
characters of the genus Trichodrilus Claparède (Oligochaeta, Lumbriculidae).
Hydrobiologia, 278: 35-51.
2.7. Análisis molecular y estudio filogenético:
El ADN se extrajo de ejemplares enteros o de las colas de los ejemplares con el Kit
QIAamp DNA (Qiagen). Se amplificaron fragmentos de aproximadamente 500 pb del gen
16S rRNA por PCR (Polymerase Chain Reaction) con los cebadores universales de
Palumbi et al. (1991). También se amplificaron fragmentos de unos 685 pb del gen COI
por PCR con los cebadores de Folmer et al. (1994). La PCR se llevó a cabo en un
termociclador BioRad iCycler en condiciones adecuadas. Los productos se tiñeron con
bromuro de etidio y se migraron en un gel de agarosa al 1.5% para verificar los
amplificados. Éstos se secuenciaron con el Kit dRhodamine Terminator Cycle Sequencing
Ready Reaction Kit (Applied Biosystems) en un analizador ABI PRISM Modelo 3100.
Las secuencias obtenidas, junto a otras tomadas de la base de datos del GenBank, se
alinearon usando el programa CLUSTALX version 1.81 (Thompson et al., 1997) y luego

se adaptaron manualmente para reducir los desajustes. Las secuencias se analizaron en tres
grupos: 16S rRNA, COI y un grupo de ambas secuencias concatenadas (16S + COI). Las
secuencias obtenidas serán depositadas en la base de datos del GenBank con sus
correspondientes números de acceso, una vez realizada la publicación de los datos.
2.8. Índices de biodiversidad y modelos predictivos:
Para la identificación de áreas de gran valor de biodiversidad o hotspots se han
usado 4 índices: riqueza de especies (S), rareza (R), vulnerabilidad (V) y
complementariedad (C). La riqueza (S) se mide como el número total de especies para
cada localidad. La rareza (R) se refiere al rango de restricción de las especies, medido
como área de ocupación. La rareza de una especie i se mide como el inverso del número
de localidades donde está presente (1/ni). Para una localidad r, el índice de rareza es
( )∑=
=S
iinR
1/1 , donde ni el número de especies halladas en la localidad.
Para la mejora del modelo de vulnerabilidad de especies, se han introducido nuevos
criterios adecuados a la fauna acuática subterránea y en especial a los oligoquetos (ver
apartado siguiente). Posteriormente se aplicaron a los datos de biodiversidad disponibles,
calculando así un valor de vulnerabilidad para cada especie. Así mismo, se calculó un
índice de vulnerabilidad para cada localidad como ∑=
=S
iiVV
1 donde Vi es la puntuación de la
vulnerabilidad para la especie i.
El índice de complementariedad (C) se ha basado en la riqueza específica y se ha
calculado de la siguiente forma. Primero, se elige el punto con mayor riqueza; después, se
escoge el punto con mayor número de especies no representadas en el punto escogido
previamente; y se continúa de la misma forma hasta que todas las especies estén
representadas por lo menos una vez. Finalmente, el índice de complementariedad (C) para
cada punto se calcula como el porcentaje de nuevas especies incorporadas (n) del total de

la riqueza específica observada en el área de estudio (Sobs) (C = n * 100 / Sobs) (Baquero,
2001).
Se realizaron curvas de acumulación de especies y estimas de riqueza de especies
con el programa informático EstimateS (Colwell, 2004). Las curvas de acumulación de
especies se generaron utilizando el modelo de incidencia de curvas de acumulación de
especies propuesto por Colwell et al. (2004), que indica el número de especies esperado en
una colección de muestras aleatoria. Las estimas de riqueza de especies en los karst se han
realizado con las funciones de Chao SChao1, ICE (Incidence-based Coverage Estimator),
Bootstrap de Smith & van Belle (1984) y Jackknife de Burnham & Overton (1978)
(Gotelli & Colwell, 2001).
2.9. Mejora del índice de vulnerabilidad:
Hemos calculado Vi como la suma aritmética de las puntuaciones para 6 criterios
elegidos para determinar la vulnerabilidad de las especies de oligoquetos. Los criterios son
los siguientes: distribución geográfica a escala global, distribución geográfica a escala
local, endemismo, especificidad de hábitat, tamaño de la población local y riesgo de
pérdida de hábitat. La selección de estos criterios se ha basado en los aquellos propuestos
por Rabinowitz et al., (1986) y por Abellan et al., (2005b). Las puntuaciones para cada
criterio son de 0 a 3 y se explican a continuación:

Figura 3. Regiones biogeográficas aceptadas para la fauna acuática. Modificado de
Banarescu (Fig.9/1: 1992). I: Región Holártica. Ia: Subregión este de Norte América.
Ib: Subregión oeste de Norte América. Ic: Subregión central de México. Id:
Subregión Euro-Mediterránea. Ie: Subregión oeste de Mongolia. If: Subregión Artica
de Norte América. Ig: Subregión de Liberia. Ih: Subregión de Baikal. II: Región de
Sino-India. I-I: Área de transición del oeste de Asia. III: Región de Etiopia (Africa).
IV: Región de Madagascar. V: Región Neotropical (Sur América). I-V: Área de
transición de Centro América/Antillas. VI: Región de Australia. VII: Región de
Nueva Zelanda. VIII: Región Indo-Pacifico oeste.
• Distribución geográfica a escala global (GD): hemos definido 4 áreas basándonos
en las regiones para fauna acuática de Banarescu. La puntuación más alta (3) se ha
asignado a aquellas especies restringidas a la subregión Euro-Mediterránea (Id); a
las especies que habitan las subregiones Ie, Ig y/o Ih se les ha asignado una
puntuación de 2; a las especies que se encuentran en las regiones Holarctica (I) y
Sino-India (II) se las ha dado una puntuación de 1; y si la distribución de la especie
incluye por lo menos una de las regiones III, IV, V, VI, VII o VIII, se le ha dado
una puntuación de 0. La distribución de las especies se ha basado en la literatura.
• Distribución geográfica a escala local (LD): hemos dado la puntuación más alta (3)
a aquellas especies que habitan un solo punto de muestreo en el área de estudio; una

puntuación de 2 si habitan dos puntos; una puntuación de 1 si habitan 3 puntos; y 0
si habitan más de 3 puntos.
• Endemismo (E): solo se han puntuado las especies endémicas del área Euro-
Mediterránea. Las puntuaciones asignadas son: Bizkaia (3), norte de España y sur
de Francia (2); distribución extensa en el suroeste de Europa, definido por Giani et
al., (2001) (1); y distribución extensa en el área Euro-Mediterránea (0).
• Especificidad de hábitat (H): hemos puntuado las especies según la clasificación de
Juget & Dumnicka (1986); Giani et al., (2001) y nuestra propia experiencia. La
máxima puntuación (3) se ha asignado a las especies estigobiontes; 2 puntos a las
estigófilas; 1 punto a aquellas especies raras que han sido halladas tanto en el medio
acuático subterráneo como en el epigeo, pero todavía de dudosa preferencia
ecológica; y 0 puntos a las estigoxenas.
• Tamaño de la población local (P): hemos dado la puntuación más alta (3) a las
especies con las poblaciones locales más pequeñas, que tienen menos de 2
individuos por muestra (aproximadamente 20 individuos/m2) en ≥ 70% de los
casos y nunca más de 9 individuos por muestra (aproximadamente menos de 100
individuos/m2). Se han asignado 2 puntos a las especies de las que se hallaron
menos de 2 individuos por muestra en el 69-50% de los casos; 1 punto si en el 49-
30% de los casos; y el resto 0 puntos.
• Riesgo de Pérdida de Hábitat (RL): en un primer paso, hemos calculado una
puntuación de riesgo para cada punto de muestreo, basándonos en 3 criterios:
alteraciones humanas (turismo en cuevas o uso de surgencias para abastecimiento de
agua, basado en datos de campo), evidencia de contaminación agrícola (datos del
EVE, 1996) y proximidad a núcleo urbano o infraestructura importante (menos de
0.5 km). Se ha asignado la puntuación máxima (3) a un punto si cumple todos los
criterios; si solo 2 criterios, 2 puntos; y etc. En un segundo paso, se calcula para
cada especie la media de las puntuaciones de riesgo incluyendo todos los puntoss
donde dicha especie se ha hallado; y se asigna una puntuación de 0 a un valor medio
de 0; 1 punto si el valor medio es entre 0.1 y 1; 2 puntos si es entre 1.1 y 2; y 3
puntos si es entre 2.1 y 3.

Tabla 1. Criterios y puntuaciones para el cálculo de Vi.
Criterio
Puntuación
GD LD E H P RL
3 Subregión Euro-
Mediterránea
1 punto
de
muestreo
Bizkaia Estigobiontes Menos de 2
individuos por
muestra en ≥
70% de los
casos y nunca
más de 9
individuos por
muestra
2.1
- 3
2 Subregiones Ie,
Ig y/o Ih
2 puntos Norte de
España y Sur
de Francia
Estigofilas Menos de 2
individuos por
muestra en el
69-50% de los
casos
1.1
- 2
1 Regiones
Holarctica (I) y
Sino-India (II)
3 puntos Distribución
extensa en el
suroeste de
Europa
Especies raras que
han sido halladas
tanto en el medio
acuático subterráneo
como en el epigeo,
pero de dudosa
preferencia ecológica
49-30% de los
casos
0.1
- 1
0 Por lo menos
una de las
regiones III, IV,
V, VI, VII o VIII
Más de 3
puntos
Distribución
extensa en el
área Euro-
Mediterránea
Estigoxenas Otros 0

3. RESULTADOS
3.1. MUESTREO EN CAVIDADES Y SURGENCIAS KÁRSTICAS DEL
PAÍS VASCO Y ZONAS ADYACENTES Y PROCESADO DE LAS
MUESTRAS
Se han realizado muestreos de fauna acuática subterránea, agua y sedimento, así
como el procesado de dichas muestras, siguiendo los métodos descritos en el apartado
2, en 8 cavidades y surgencias del karst de Gorbeia y 3 de Urdaibai. Se han
completado estos datos con otros datos faunísticos previos de dichos karst. Así
mismo, se ha iniciado la preselección de puntos de muestreo en karst de Cantabria y
Asturias.
a) Resultados de fauna acuática subterránea:
Los oligoquetos representan del 5.4 al 91.7 % de los invertebrados en la
comunidad del bentos, compuesta también por cnidarios, nematodos, planarias,
gasterópodos, bivalvos, crustáceos y larvas de insectos (Figura 4). La densidad de
oligoquetos varía entre 311 y 767 individuos/m2 en cuevas y entre 244 y 5689
individuos/m2 en surgencias. La tabla 3 recoge los taxones identificados en el
presente estudio así como aquellos hallados en estudios precedentes (Giani &
Rodriguez, 1988; Giani et al., 2001).
Las comunidades de oligoquetos en el karst de Gorbeia están numéricamente
dominadas por los enquitreidos (familia Enchytraeidae), siendo la familia de
oligoquetos dominante en 6 de las 8 localidades estudiadas en otoño. Sin embargo,
los tubífícidos (familia Tubificidae) son la familia dominante en la surgencia GOR08,
y GOR04 siendo también importantes en términos de abundancia relativa en GOR06
y GOR07 con un representación superior al 30%. Los lumbricúlidos son claramente
más abundantes en cuevas que en surgencias y constituyen un bajo porcentaje del
total de oligoquetos en el karst de Gorbeia. Los lumbrícidos están presentes

únicamente en una localidad. No se han encontrado representantes de las familias
Haplotaxidae y Parvidrilidae (Ver Tabla 2).
Tabla 2. Abundancia relativa de las familias de oligoquetos acuáticos en las
localidades estudiadas en el karst de Gorbeia en otoño (se marcan en gris las
celdas con máxima abundancia relativas)
Tubificidae
(%)
Enchytraeidae
(%)
Lumbriculidae
(%)
Lumbricidae
(%)
Nº
individuos
por
muestra
GOR08 96.09 0 3.91 0 512
GOR01 5.00 62.50 32.50 0 40
GOR02 2.84 95.45 1.70 0 176
GOR05 0.83 99.17 0 0 121
GOR03 3.57 53.57 42.86 0 28
GOR04 63.64 27.27 9.09 0 22
GOR06 43.48 55.07 1.45 0 69
GOR07 31.88 53.62 13.04 1.45 69
Total 54.68 39.44 5.79 0.10 1037

Figura 4. Abundancia relativa de los oligoquetos en la comunidad de otoño del
karst de Gorbeia.
0% 20% 40% 60% 80% 100%
GOR08
GOR01
GOR02
GOR05
GOR03
GOR04
GOR06
GOR07
TOTAL
% fauna acompañante% oligoquetos

Tabla 3. Riqueza taxonómica de oligoquetos acuáticos subterráneos encontrados en cavidades y surgencias del karst de
Gorbeia. Datos correspondientes a muestreos de la campaña de 2007 y muestreos anteriores en los años 80.
LOCALIDADES
Riqueza específica
GOR01 GOR02 GOR03 GOR04 GOR05 GOR06 GOR07 GOR08 GOR09 GOR10 GOR11 GOR12
ENCHYTRAEIDAE 2 7 4 4 4 4 7 1 3 1 2
HAPLOTAXIDAE 1
LUMBRICULIDAE 1 1 3 1 1 1 1 1 1 1 1
TUBIFICIDAE 1 1 1 2 3 2 3 2 1
LUMBRICIDAE 1
Nº especies/localidad 4 9 8 7 8 7 12 3 4 3 2 3

Tabla 4. Riqueza específica encontrada en las cavidades de RBU10, RBU11 y
RBU06 del karst de la RBU correspondientes a muestreos de la campaña 2007 y
datos relativos a muestreos anteriores realizados en los años 80 y 2005.
Localidades RBU10 RBU11 RBU06
Campañas de muestreo 1985 2007 1985 2007 1984 2005 2007
Riqueza taxonómica 2 4 12 7 9 7 7
b) Características físico-químicas:
A continuación se muestran los resultados de los análisis físico-químicos
resumidos en forma de tabla para el karst de Gorbeia.
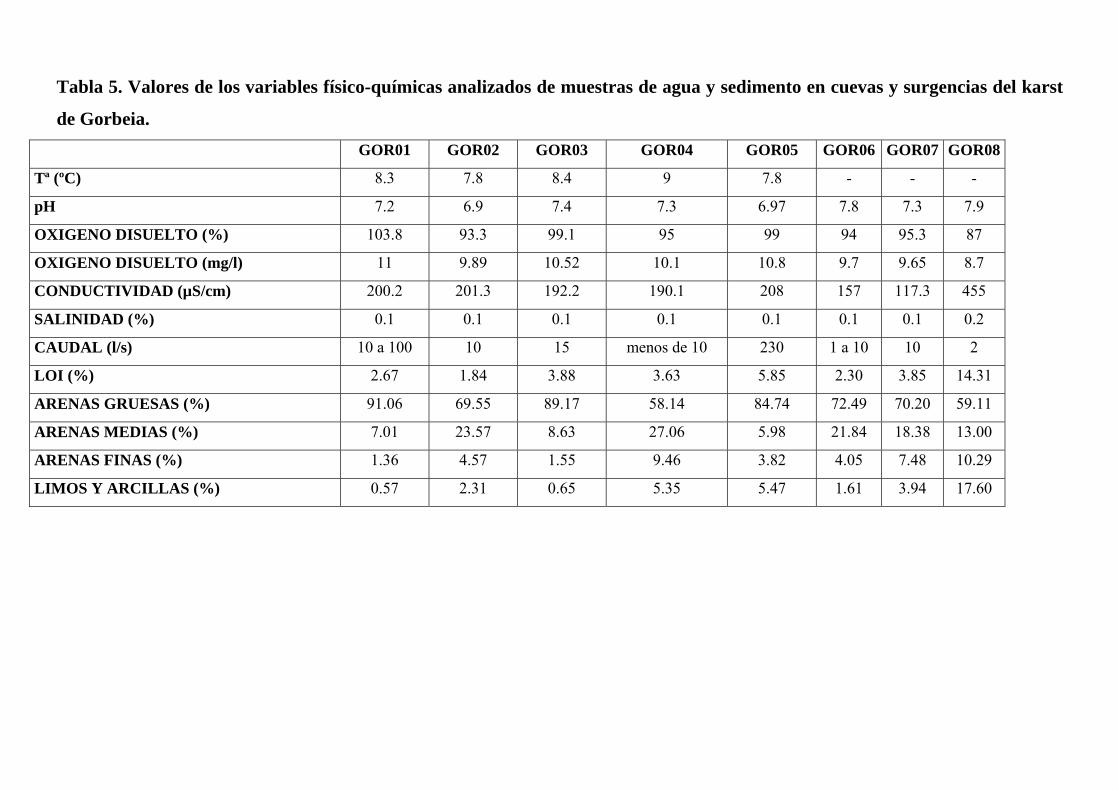
Tabla 5. Valores de los variables físico-químicas analizados de muestras de agua y sedimento en cuevas y surgencias del karst
de Gorbeia. GOR01 GOR02 GOR03 GOR04 GOR05 GOR06 GOR07 GOR08
Tª (ºC) 8.3 7.8 8.4 9 7.8 - - -
pH 7.2 6.9 7.4 7.3 6.97 7.8 7.3 7.9
OXIGENO DISUELTO (%) 103.8 93.3 99.1 95 99 94 95.3 87
OXIGENO DISUELTO (mg/l) 11 9.89 10.52 10.1 10.8 9.7 9.65 8.7
CONDUCTIVIDAD (µS/cm) 200.2 201.3 192.2 190.1 208 157 117.3 455
SALINIDAD (%) 0.1 0.1 0.1 0.1 0.1 0.1 0.1 0.2
CAUDAL (l/s) 10 a 100 10 15 menos de 10 230 1 a 10 10 2
LOI (%) 2.67 1.84 3.88 3.63 5.85 2.30 3.85 14.31
ARENAS GRUESAS (%) 91.06 69.55 89.17 58.14 84.74 72.49 70.20 59.11
ARENAS MEDIAS (%) 7.01 23.57 8.63 27.06 5.98 21.84 18.38 13.00
ARENAS FINAS (%) 1.36 4.57 1.55 9.46 3.82 4.05 7.48 10.29
LIMOS Y ARCILLAS (%) 0.57 2.31 0.65 5.35 5.47 1.61 3.94 17.60

Tabla 6. Composición química de metales en agua, (mg/l) en localidades muestreadas en el karst de Gorbeia.
Mg l-1 Mg Al V Cr Mn Fe Co Ni Cu Zn As Se Ag Cd Sn Sb Hg Pb
GOR03 414.04 4.38 < 0.01 < 0.01 < 0.01 49.82 < 0.01 <0.01 0.18 37.90 0.33 <0.01 0.19 0.14 <0.01 <0.01 12.52 <0.01
GOR04 182.35 <0.01 <0.01 0.53 <0.01 19.00 <0.01 <0.01 <0.01 12.90 0.24 <0.01 0.18 0.10 <0.01 <0.01 3.60 <0.01
GOR01 192.05 0.60 <0.01 0.98 <0.01 28.11 <0.01 <0.01 <0.01 11.82 0.224 <0.01 0.18 0.10 <0.01 <0.01 5.14 <0.01
GOR08 1244.34 0.77 <0.01 2.04 0.32 78.93 <0.01 <0.01 <0.01 62.96 0.22 <0.01 0.18 0.13 <0.01 <0.01 1.72 <0.01
GOR07 408.91 6.21 <0.01 <0.01 <0.01 61.28 <0.01 <0.01 <0.01 70.60 0.27 0.03 0.18 0.13 <0.01 <0.01 1.55 <0.01
GOR06 267.88 2.36 <0.01 <0.01 <0.01 41.91 <0.01 <0.01 <0.01 28.96 0.23 0.06 0.18 0.10 <0.01 <0.01 1.54 <0.01
GOR02 988.03 3.38 <0.01 1.33 <0.01 62.40 <0.01 <0.01 <0.01 35.76 0.31 0.24 0.18 0.12 <0.01 <0.01 1.59 <0.01
GOR05 596.16 17.87 <0.01 1.22 <0.01 96.10 <0.01 <0.01 <0.01 38.01 0.37 0.28 0.18 0.11 <0.01 <0.01 1.49 <0.01

3.2. ESTUDIO TAXONÓMICO Y DESCRIPCIÓN DE NUEVAS ESPECIES
PARA LA CIENCIA
En las campañas de muestreo realizadas se han hallado tres taxones que
merecen mención especial ya que probablemente constituyen nuevas especies y
están en fase de estudio para la descripción de sus características morfológicas.
a) Stylodrilus sp.
Es destacable el hallazgo de una posible nueva especie para la Ciencia del
género Stylodrilus Claparede, 1862 en la localidades GOR01, GOR02, GOR03 Y
GOR12 del karst de Gorbeia. En la actualidad estamos describiendo la especie a
partir de 4 ejemplares maduros hallados en el muestreo de 2007 mediante técnicas de
microdisección y montaje. Existenn otros ejemplares de la misma especie en
muestreos anteriores., que permiten valorar positivamente la estabilidad de las
poblaciones.
La especie Stylodrilus sp. se diferencia de Stylodrilus parvus en varios
caracteres relacionados con su aparato reproductor, por lo cual se han estudiado las
características de esta especie en la Serie Tipo enviada por el Museo de Praga, que
nos servirá para establecer comparaciones.. Es por ello que tras un estudio de su
morfología más amplio, si se verifica como nueva especie, se procederá a su
descripción.
b) Troglodrilus galarzai (Giani y Rodriguez, 1988)
Se están estudiando características morfológicas de ejemplares de Troglodrilus
galarzai de localidades del karst de Gorbeia, con el objeto de determinar la
variabilidad de esta población con respecto a la del karst de la RBU y las poblaciones
francesas. Así, se ha determinado la variabilidad en caracteres morfológicos como

longitud y anchura de la cutícula penial, número y longitud de quetas, longitud y
anchura de la espermateca, del atrio y otros caracteres del aparato reproductor.
Estudios anteriores han realizado comparaciones con los ejemplares de la localidad
tipo y de poblaciones de la región del Ródano (Francia) ( Juget et al. 2006). El
análisis molecular de las poblaciones de Gorbeia y las del Ródano (enviadas por el
Dr. des Chatelliers) puede aportar una información muy valiosa acerca de las
distancias genética de ambas poblaciones, su historia evolutiva y relaciones
filogenéticas con otros taxones de tubifícidos.. Estudios previos (Achurra &
Rodriguez en prensa) han señalado la desaparición de esta especies de la localidad
tipo que se encuentra en la Reserva de la Biosfera de Urdaibai, por lo que esta
localidad del Gorbeia junto con otra localidad del karst de Sta. Eufemia, prospectada
en relación con este proyecto, son las dos localidades donde en la actualidad se puede
encontrar esta especie estigobia, en la Península Ibérica.
c) Protoberodrilus sp
Se han hallado dos ejemplares en GOR07 pertenecientes al tubifícido
Protoberodrilus sp, todavía sin confirmar la identidad de las especie, ya que son
necesarios más ejemplares para ampliar el estudio morfológico mediante estudio
histológico del aparato reproductor. Los ejemplares pertenecen al género
Protoberodrilus Giani y Martinez-Ansemil 1979 por la forma del atrio, presencia de
un único poroforo masculino, presencia de grandes quetas copuladoras en sacos
setígeros muy conspicuos en varios segmentos preatriales y postatriales. El género
Protuberodrilus es hasta la fecha monoespecífico y endémico de la región gallega,
donde ha sido hallado en tramos fluviales que no presentan características kársticas.
3.3 MEJORA DEL MODELO DE VULNERABILIDAD PARA ESPECIES
Hemos puntuado los 15 localidades de la RBU según el índice de vulnerabilidad
(V/S’) (Tabla 7). Los valores de V/S’ oscilan entre 2.33 (RBU04) y 9.00 (RBU10).

Cabe destacar que las localidades más vulnerables se corresponden con RBU01,
RBU02 y RBU10.
Tabla 7. Valores del índice de vulnerabilidad corregido (V/S’) para 15
localidades del karst de Santa Eufemia-Ereñozar (RBU).
Localida
d
V/S’
1 8.40
2 8.25
3 6.25
4 2.33
5 6.32
6 6.40
7 4.56
8 5.22
9 6.92
10 9.00
11 7.27
12 5.29
13 6.06
14 - *
15 6.50
* En este punto solo se hallaron ejemplares inmaduros y por tanto, no se pudo
calcular el índice corregido de vulnerabilidad.
Tabla 8. Valores de vulnerabilidad (Vi), distribución geográfica a escala global
(GD), distribución geográfica a escala local (LD), endemismo (E), especificidad
de hábitat (H), tamaño de la población local (P) y riesgo de pérdida de hábitat
(RL) para los taxones del karst de Santa Eufemia-Ereñozar (RBU). Taxones Vi GD LD E H P RL
ENCHYTRAEIDAE

Achaeta sp. 6 0 3 0 0 3 0
Cernosvitoviella atrata (Bretscher, 1903) 7 1 1 0 0 3 2
Cernosvitoviella cf. immota (Knöllner,
1935)
8 1 3 0 0 3 1
Cernosvitoviella palustris Healy, 1979 6 3 0 0 1 0 2
Cernosvitoviella sp. 6 1 3 0 0 2 0
Cernosvitoviella imm. - - - - - - -
Enchytraeus sp. 7 0 2 0 0 3 2
Fridericia bisetosa (Levinsen, 1884) 7 0 2 0 0 3 2
Fridericia imm. - - - - - - -
Marionina argentea (Michaelsen, 1889) 6 1 0 0 0 3 2
Marionina sp. 13 3 0 3 3 2 2
Marionina imm. - - - - - - -
Mesenchytraeus armatus (Levinsen,
1884)
7 1 3 0 0 3 0
HAPLOTAXIDAE
Haplotaxis sp. 6 0 2 0 0 3 1
Haplotaxis imm. - - - - - - -
LUMBRICULIDAE
Stylodrilus heringianus Claparède, 1862 4 1 3 0 0 0 0
Stylodrilus lemani (Grube, 1879) 3 2 0 0 0 0 1
Stylodrilus parvus (Hrabe and
Cernosvitov, 1927)
4 2 1 0 0 0 1
Trichodrilus campoyi Rodriguez, 1988 9 3 0 2 3 0 1
Trichodrilus diversisetosus Rodriguez &
Giani, 1986
14 3 2 2 3 2 2
Trichodrilus tenuis Hrabe, 1960 12 3 3 0 3 3 0
Trichodrilus strandi Hrabe, 1936 13 3 3 1 3 3 0
Trichodrilus sp. (moravicus group) 7 1 2 0 1 2 1
Lumbriculidae imm. - - - - - - -
TUBIFICIDAE
Phallodrilinae
Aktedrilus argatxae Giani and
Rodriguez, 1988
13 3 3 3 3 0 1
Gianius aquaedulcis (Hrabe, 1960) 10 1 3 0 3 3 0
Phallodrilinae indet. 7 0 3 0 0 3 1

Phallodrilinae imm. - - - - - -
Tubificinae
Lamadrilus sp. 11 3 0 3 3 0 2
Limnodrilus hoffmeisteri Claparède,
1862
7 0 2 0 0 3 2
Limnodrilus udekemianus Claparède,
1862
3 0 1 0 0 0 2
Limnodrilus imm. - - - - - - -
Tubifex ignotus (Stolc, 1886) 2 0 1 0 0 0 1
Potamothrix bavaricus (Oschmann,
1913)
6 0 3 0 0 3 0
Psammoryctides barbatus (Grube, 1861) 7 1 2 0 0 3 1
Spirosperma velutinus (Grube, 1879) 3 1 0 0 0 0 2
Troglodrilus galarzai (Giani and
Rodriguez, 1988)
14 3 2 2 3 2 2
Tubifex tubifex (Müller, 1774) 4 0 2 0 0 0 2
Varichaetadrilus bizkaiensis Rodriguez
& Giani 1984
9 3 2 3 1 0 0
Tubificinae imm. - - - - - - -
Rhyacodrilinae
Peristodrilus montanus (Hrabe, 1962) 7 3 2 1 1 0 0
Rhyacodrilus falciformis Bretscher,
1901
7 1 1 0 0 3 2
Rhyacodrilus gernikensis Giani and
Rodriguez, 1988
15 3 3 3 3 0 3
Rhyacodrilus okamikae Giani &
Rodriguez, 1988
14 3 2 3 3 2 1
Rhyacodrius sp. 7 0 3 0 1 3 0
Rhyacodrilinae imm. - - - - - - -
Naidinae
Amphichaeta sannio Kallstenius, 1892 5 1 3 0 0 0 1
Nais bretscheri Michaelsen, 1899 6 0 3 0 0 0 3
Nais communis/Nais variabilis Piguet,
1906
2 0 0 0 0 0 2
Nais elinguis Mülller, 1773 3 0 2 0 0 0 1
Pristina aequiseta Bourne, 1891 2 0 0 0 0 0 2

Pristina jenkinae (Stephenson, 1931) 3 0 0 0 0 0 3
Pristina longiseta Ehrenberg, 1828 6 0 2 0 0 2 2
Pristina menoni (Aiyer, 1906) 4 0 3 0 0 0 1
Pristina sima (Marcus, 1944) 9 0 2 0 2 3 2
Tubificidae imm. - - - - - - -
Tubificidae indet. - - - - - - -
LUMBRICIDAE
Eiseniella tetraedra (Savigny, 1826) 4 0 2 0 0 0 2
Lumbricidae imm. - - - - - - -
También los oligoquetos sido evaluados de acuerdo con sus puntuaciones de
vulnerabilidad (Vi) (Tabla x). Según los resultados de distribución geográfica local,
16 especies habitan una localidad exclusivamente. Por el contrario, Pristina jenkinae,
Spirosperma velutinus y Pristina aequiseta habitan 8, 7 y 6 localidades,
respectivamente, del total de 15 de ellas. Veinte taxones tienen una distribución
geográfica mundial, mientras que 13 están restringidos a la Euro-Mediterránea, y 5
son endémicos de Bizkaia. Once especies han sido clasificadas como estigobiontes y
solo una como estigófila. Dieciocho especies tienen un tamaño de población pequeño
en la unidad kárstica; sin embargo, la abundancia de Pristina jenkinae (hasta 2033
individuos/m2), Tubifex ignotus (hasta 1519 individuos/m2) y Nais communis (557
individuos/m2) es muy alta. P. jenkinae habita, en general, localidades con elevado
riesgo de pérdida de hábitat. El máximo valor teórico de vulnerabilidad (Vi) para una
especie en este estudio es 18, pero la máxima puntuación obtenida fue 15 y la mínima
2. Un enquitreido (Marionina sp.), 3 lumbriculidos (Trichodrilus diversisetosus, T.
tenuis y T. strandi) y 4 tubificidos (Aktedrilus argatxae, Troglodrilus galarzai,
Rhyacodrilus okamikae y R. gernikensis) son las especies más vulnerables.
3.4 ESTUDIO FILOGENÉTICO Y EVOLUTIVO
Hemos elegido la especie Troglodrilus galarzai para realizar el estudio
filogenético por su interés evolutivo. La especie fue hallada por primera vez en la
surgencia de Argatxa, karst de Urdabai (localidad tipo) y en la cueva de Goiketxe,
karst de Urdabai, en los años 1984-85. Posteriormente, se encontraron ejemplares de

T. galarzai en la cueva de Mairulegorreta, karst de Gorbeia, y en la cueva de Crotot y
la galería subterránea de Montgelas (Francia; ver Figura 5).
T. galarzai es una especie estigobia, es decir, limitada a las aguas subterráneas. En
el sur de Europa se cuentan actualmente 31 especies estigobias.
Durante el año 2007, hemos iniciado el estudio de esta especie, de la cual se han
recogido para los análisis de DNA, 3 ejemplares maduros y 11 inmaduros de
poblaciones francesas, x ejemplares de Gorbeia y x ejemplares de Urdaibai. La
desaparición de la población de T. galarzai en su localidad tipo (Argatxa), ha
dificultado la recolección de ejemplares de la población de Urdaibai, por lo que el
análisis de DNA ha sido pospuesto para el año 2008 (fase II del proyecto).
Se ha realizado un estudio morfológico de los ejemplares de T. galarzai hallados en
las localidades del karst de Gorbeia, que completa y reafirma la variación
intraespecífica geográfica de la especie estudiada por Juget et al., 2006.
Los resultados del análisis de DNA permitirán contrastar esta variabilidad. Por
otro lado, la distribución fragmentada de esta especie tiene un gran interés ya que
podría explicarse mediante la teoría refugio a partir de una distribución más amplia
en el sudoeste Europeo. Esperamos que los datos filogenéticos contrasten esta
hipótesis.

Figura 5. Mapa de la distribución actual de T. galarzai. 1 = karst de Urdaibai, 2
= karst de Gorbeia, 3 = Crotot, 4 = Montgelas.
3.5 IDENTIFICACIÓN DE ZONAS DE ALTA BIODIVERSIDAD O
HOTSPOTS Y ANÁLISIS DE RAZONES
Hemos comparado los dos karst estudiados, RBU y Gorbeia, en lo que se refiere a
variables físico-químicas, hidrológicas y geográficas, con el objetivo de buscar
diferencias que puedan ayudar a buscar razones que expliquen la existencia de un
hotspot de oligoquetos acuáticos subterráneos en un determinado karst.
Tabla 9. Valores medios de variables físico-químicas, hidrológicas y geográficas
de los karst de la RBU y Gorbeia.
GORBEIA* RBU Prueba t
VARIABLES media desvest media desvest p
Tª (ºC) 8.26 0.50 13.60 0.84 0.000
pH 7.27 0.30 7.07 0.22 0.167
Oxígeno disuelto (%) 97.07 3.75 89.09 9.82 -

Oxígeno disuelto (mg/l) 10.24 0.54 8.85 1.48 0.037
Conductividad (µs/cm) 180.87 32.54 509.37 74.08 0.000
Salinidad (%) 0.10 0.00 0.01 0.03 -
Altitud (m) 848.11 103.88 49.87 71.15 0.000
LOI (%) 3.43 1.33 3.70 1.24 0.739
Arenas gruesas (%) 76.48 12.12 80.20 10.22 0.475
Arenas medias (%) 16.07 8.71 11.05 5.22 0.268
Arenas finas (%) 4.61 2.96 5.45 3.99 0.708
Limos y arcillas (%) 2.84 2.09 3.30 2.65 0.754
*Se han excluido los valores de GOR08 para este cálculo.
No existen diferencias significativas (prueba de la t de Student, p>0.05) entre los
karst de Gorbeia y Urdaibai en composición granulométrica y porcentaje de materia
orgánica del sedimento, ni en el pH de las aguas. Por el contrario, ambos karst
difieren (p<0.01) en temperatura, oxígeno disuelto, altitud y conductividad medias.,
siendo las aguas de las localidades prospectadas en el macizo del Gorbeia más frías,
mejor oxigenadas, de mayor altitud y con menor contenido en sales, como cabía
esperar.
3.6 CONTRIBUCIÓN A LOS PLANES DE CONSERVACIÓN DE LOS
SISTEMAS KÁRSTICOS ESTUDIADOS
Se han puntuado las 12 localidades del karst de Gorbeia según los índices de
biodiversidad de riqueza de especies (S), rareza corregida (R/S’) y
complementariedad. Hemos corregido R por el número de taxones usado en el
cálculo del mismo (S’) para poder comparar los resultados de las distintas
localidades. Los valores de riqueza de especies (S) varían entre 2 (GOR11) y 12
(GOR07) y los valores de rareza (R/S’) varían entre 0.24 (GOR01) y 0.65 (GOR09).
Los resultados del índice de complementariedad (C) muestran que 8 de las 12
localidades comprenden todas las especies halladas hasta la fecha en el karst.

Tabla 10. Índices de biodiversidad para las localidades del karst de Gorbeia. S:
riqueza espefíca, R/S’: riqueza corregida, C: Complementariedad, E:
endemismo.
INDICES
LOCALIDADES
S R/S’ C E S
GOR01 4 0.24 0 0 0
GOR02 9 0.53 15.63 1 1
GOR03 8 0.41 3.13 1? 1?
GOR04 7 0.47 6.25 1 1
GOR05 8 0.64 21.88 0 0
GOR06 7 0.28 0 1 1
GOR07 12 0.57 34.38 1 / 2? 1 / 2?
GOR08 3 0.50 0 0 0
GOR09 4 0.65 9.38 0 0
GOR10 3 0.46 3.13 0 0
GOR11 2 0.29 0 0 0
GOR12 3 0.53 6.25 1? 1?
KARST 32 - - 1 / 3? 1 / 3?
Curva de estimación de especies y estimadores de riqueza específica
0
5
10
15
20
25
30
35
40
45
1 2 3 4 5 6 7 8 9 10 11 12
Nº de localidades
Nº d
e ta
xone
s Sobs (Mao Tau)Sobs 95% CI Lower BoundUniques MeanDuplicates MeanSobs 95% CI Upper Bound
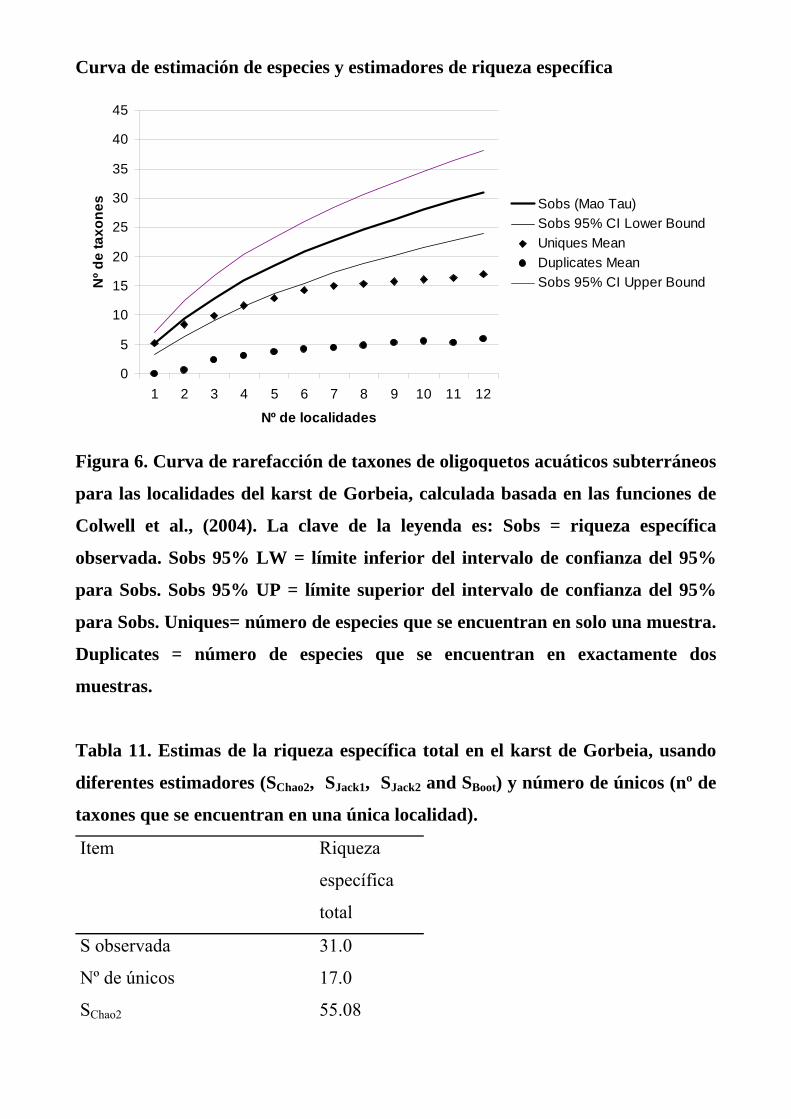
Figura 6. Curva de rarefacción de taxones de oligoquetos acuáticos subterráneos
para las localidades del karst de Gorbeia, calculada basada en las funciones de
Colwell et al., (2004). La clave de la leyenda es: Sobs = riqueza específica
observada. Sobs 95% LW = límite inferior del intervalo de confianza del 95%
para Sobs. Sobs 95% UP = límite superior del intervalo de confianza del 95%
para Sobs. Uniques= número de especies que se encuentran en solo una muestra.
Duplicates = número de especies que se encuentran en exactamente dos
muestras.
Tabla 11. Estimas de la riqueza específica total en el karst de Gorbeia, usando
diferentes estimadores (SChao2, SJack1, SJack2 and SBoot) y número de únicos (nº de
taxones que se encuentran en una única localidad).
Item Riqueza
específica
total
S observada 31.0
Nº de únicos 17.0
SChao2 55.08

SBoot 46.58
SJack1 56.2
SJack2 37.78
0
10
20
30
40
50
60
70
1 2 3 4 5 6 7 8 9 10 11 12
Nº de localidades
Nº d
e ta
xone
s
Sobs (Mao Tau)Jack 1 MeanJack 2 MeanBootstrap Mean
Figura 7. Curvas de estimación de especies y sus desviaciones estándar, basadas
en distintos estimadores: Jack1, Jack2 y Bootstrap.
La curva de rarefacción de especies del karst (Fig. 6 y 7) tiene una pendiente
relativamente pequeña; lo cual es debido al alto número de taxones únicos hallados.
Por otro lado, la curva no llega a formar una asíntota, lo que sugiere que existen
numerosos taxones en el karst aún por hallar. Esta hipótesis se refuerza con las
estimaciones de riqueza específica a partir de los modelos, los cuales sugieren que la
riqueza del karst es de 7 a 25 taxones mayor del valor observado (31 taxones), según
los modelos (Tabla 11).

4. DISCUSION GLOBAL Y CONCLUSIONES
La composición de familias en las comunidades de oligoquetos de otoño sigue
el mismo patrón general que la que se da en el conjunto de localidades estudiadas el
medio subterráneo de la región Sur de Europa (Giani et al., 2001), donde la familia
Enchytraeidae es la dominante en términos de abundancia relativa (50 % de los
individuos recogidos); las familias Tubificidae y Lumbriculidae representan casi
enteramente el resto de individuos de la comunidad. En general, los géneros más
abundantes en el Sur de Europa son los enquitreidos Enchytraeus, Cernosvitoviella y
Marionina y el lumbricúlido Trichodrilus. Este patrón es diferente en el karst de
Urdaibai, donde los tubifícidos dominan numéricamente la comunidad de oligoquetos
de otoño, mientras que los enquitreidos solo representan el 7% del total (Figura 8).
Podemos concluir que las comunidades de oligoquetos de otoño del karst de Gorbeia
y las del karst de Urdabai son diferentes en lo que se refiere a abundancia relativa de
las familias
0.00
20.00
40.00
60.00
80.00
100.00
% Lumbriculidae % Enchytraeidae % Lumbricidae % Tubificidae
GorbeaGorbea*Urdaibai
Figura 8. Abundancias relativas de las familias de oligoquetos en los karst de
Gorbeia y Urdaibai. Se ha representado también los datos para el karst de
Gorbeia excluyendo la localidad de Atxurdin, que aporta un porcentaje de
tubifícidos muy alto.
El karst de Urdaibai ha sido recientemente considerado como hotspot para los
oligoquetos acuáticos subterráneos (Achurra y Rodriguez, 2008). Si comparamos los

resultados de fauna del karst de Gorbeia con los de la RBU, observamos claramente
que la riqueza específica es menor en Gorbeia que en la RBU: 32 taxones frente a 47.
También el número de especies estigobiontes es muy inferior en el karst de Gorbeia:
1 especie frente a 11; lo mismo ocurre con el número de especies endémicas: 1
especie en Gorbeia frente a 5 en la RBU. Sin embargo el esfuerzo de muestreo
realizado en esta última zona es superior al realizado hasta la fecha en el Karst de
Gorbeia, por lo que deben realizarse nuevas campañas de muestreo para tener
conclusiones concluyentes. El. Karst de Gorbeia es un punto muy interesante para el
estudio de la biodiversidad de oligoquetos acuáticos ya que la presencia de la especie
estigobionte Troglodrilus galarzai, convierte al karst en una zona de estudio
indispensable ulteriores estudios evolutivos. Por otro lado, estamos actualmente
realizando el estudio morfológico de dos posibles nuevas especies halladas en la
surgencia GOR07, que de ser confirmadas, aportarían al karst un mayor valor de
endemismo y biodiversidad en general para este grupo faunístico, pudiendo variar la
clasificación de Vulnerabilidad de las cavidades estudiadas. Un mayor estudio en
estos dos karst y nuevos muestreos en otros karst adyacentes permitirían comprobar
si el alto valor de biodiversidad del karst de la RBU para los oligoquetos acuáticos
subterráneos es únicamente propio de dicho karst, constituyendo un hotspot de
biodiversidad o si, por el contrario, es una característica de toda la región Cantábrica,
como ha sido hipotetizado por Culver et al., 2006 para la fauna subterránea terrestre.
Un primer estudio de las posibles razones de la alta biodiversidad del karst de
la RBU señalan a las variables altitud, temperatura, cercanía al mar y conductividad
como posibles determinantes de su alta riqueza específica total y de especies
estigobias y endémicas, por comparación con el karst de Gorbeia. La altitud estaría
relacionada con las bajas temperaturas medias, y la conductividad con la composición
geológica. Una posible explicación relacionada las menor diversidad hallada en el
karst de Gorbeia podría estar relacionada con la colonización más reciente de las
cavidades de montaña trás el último periodo de glaciaciones. Esto explicaría que los
estigobiontes sean un componente más común de la fauna de oligoquetos acuáticos
subterráneos a baja altitud, en la RBU y que sin embargo, estén ausentes en áreas

elevadas de mayor altitud, en Gorbeia. Esta idea ha sido estudiada en el karst de Jura
Mountain, en Francia, para la fauna acuática subterránea en general. Sus resultados
muestran que las variables conductividad, elevación y distancia a los límites de la
última glaciación explican la distribución de especies estigobiontes (Castellarini et al,
2007). Es interesante continuar con este estudio y determinar también la influencia de
otras variables como puede ser el tamaño del karst o el número de cuevas, variables
hidrogeológicas, etc. Pueden tener sobre la composición taxonómica de las
comunidades.
Respecto a los nuevos muestreos en el karst de la RBU, cabe destacar la
confirmación de la persistencia de Rhyacodrilus. okamikae en la cueva de RBU11, ya
que se trata de una especie endémica del karst y estigobionte, únicamente hallada en
el mundo en dicha localidad. Por otro lado, es preocupante el no haber hallado la
especie R. gernikensis, endémica de la surgencia de RBU06 y estigobionte. Respecto
a Troglodrilus galarzai, es interesante el hallazgo de ejemplares en un nuevo punto
del karst de Santa Eufemia-Ereñozar, RBU10, ya que la población de la localidad tipo
(Argatxa, RBU) parece haber desaparecido. Son necesarios nuevos muestreos en la
surgencia de RBU06 y la cueva de RBU11 para constatar la ausencia de especies con
un valor importante como son R. gernikensis y T. galarzai.
Respecto a la conservación del medio acuático subterráneo del karst de
Gorbeia, proponemos un ranking de surgencias y cavidades basado en el índice de
complementariedad (C): GOR07 > GOR05 > GOR02 > GOR09 > GOR12 > GOR04
> GOR10 > GOR03 (de mayor a menor valor de conservación). Hemos seleccionado
este índice debido a que la protección de estas 8 localidades implicaría la
conservación de todas las especies de oligoquetos acuáticos subterráneos halladas en
el karst de Gorbeia hasta la fecha. Además la lista anterior incluye las localidades con
mayor valor de rareza (GOR09, GOR05 Y GOR07), así como la localidad con mayor
riqueza específica (GOR07). En una próxima fase en la que se incrementen los
muestreos, los resultados del índice de vulnerabilidad permitirán completar la lista de
surgencias y cavidades prioritarias a proteger.

Comparando ambos karst, respecto a su conservación, podemos discutir el
riesgo de pérdida de hábitat: según el EVE, no existen focos permanentes
contaminantes en Gorbeia, lo que conlleva que la calidad de las aguas subterráneas
sea buena. Las únicas incidencias señaladas son la de los núcleos de Urigoiti y
Ubidea, que están en los límites de la unidad y a cotas bajas y no afectan a los puntos
de muestreo. Sin embargo los depósitos de residuos procedentes de refugios en las
dolinas; y la actividad ganadera deben controlarse de forma estricta. Todas estas
actividades no suponen al parecer una alteración significativa de la calidad de las
aguas (EVE, 1996). Así, el promedio de riesgo de pérdida de hábitat en los puntos de
la RBU es 1.33, mientras que en Gorbeia es menor: 0.25, lo cual puede ser
consecuencia probable de la gestión que se lleva a cabo en el biotopo protegido de
Itxina y resto del parque de Gorbeia.

5. BIBLIOGRAFÍA CONSULTADA Abellán, P., D. Sánchez-Fernández, J. Velasco & A. Millán, 2005a. Conservation of freshwater biodiversity: a comparison of different area selection methods. Biodiversity and Conservation 14: 3457-3474. Abellán, P., D. Sánchez-Fernández, J. Velasco & A. Millán, 2005b. Assessing conservation priorities for insects: status of water beetles in southeast Spain. Biological Conservation 121: 79-90. Achurra, A. & P. Rodríguez, 2008. Biodiversity of grounwater oligochaetes from a karst unit in northern Iberian Peninsula: ranking subterranean sites for conservation management. Hydrobiologia: in press. ADES, 2007. Catálogo de Simas y Cuevas de Urdaibai. Servicio Central de Publicaciones del Gobierno Vasco, Vitoria-Gasteiz (In press). Banarescu, P., 1992. Zoogeography of Fresh Waters. Vol. 2. Distribution and dispersal of fresh water animals in North America and Eurasia. AULA-Verlag, Wiesbaden. Baquero, R. A., 2001. Riqueza de especies, rareza y endemicidad de vertebrados europeos: análisis e implicaciones para la conservación. Universidad Complutense de Madrid PhD Thesis. Bellés, X., 1987. Fauna cavernícola i intersticial de la Península ibérica i les illes Balears. Consell superior d’investigacions científiques-Editorial Moll, Mallorca. Benayas, J.M. & E. de la Montaña, 2003. Identifying areas of high-value vertebrate diversity for strengthening conservation. Biological Conservation 114: 357-370. Benayas, J.M., E. de la Montaña, J. Belliure & X. R. Eekhout, 2006. Identifying areas of high herpetofauna diversity that are threatened by planned infraestructura projects in Spain. Journal of Environmental Management 79: 279-289. Camacho, A.I., A.G. Valdecasas, J. Rodriguez, S. Cuezva, J. Lario & S. Sánchez-Moral., 2006. Habitat constraints in epikarstic waters of an Iberian Peninsula cave system. Annales de Limnologie - International Journal of Limnology 42: 127-140. Castellarini, F., F. Malard, MJ. Dole-Olivier & J. Gibert, 2007. Modelling the distribution of stygobionts in the Jura Mountains (eastern France). Implications for the protection of ground waters. Diversity and Distributions 13: 213-224. Cernosvitov, L., 1939. Études Biospéologiques 10 (1). Catalogue des Oligochètes hypogés. Bulletin du Musée Royal d'Histoire Naturelle de Belgique 15: 1-92.

Colwell, R.K., 2006. EstimateS: Statistical estimation of species richness and shared species from simples. Version 8.0. User´s Guide and application published at: http://purl.oclc.org/estimates. Colwell, R.K., C. X. Mao & J. Chang, 2004. Interpolating, extrapolating, and comparing incidence-based species accumulation curves. Ecology 85: 2717-2727. Crisp, M.D., S. Laffan, H.P. Linder & A. Monro, 2001. Endemism in the Australian flora. Journal of Biogeography 28: 183-198. Culver, D.C. & B. Sket, 2000. Hotspots of subterranean biodiversity in caves and wells. Journal of Cave and Karst Studies 62: 11-17. Culver, D. C., M. C. Christman, W.R. Elliot, H.H. Hobbs III & J.R. Reddell, 2003. The North American obligate cave fauna: regional patterns. Biodiversity and Conservation 12: 441-468. Culver, D. C., M. C. Christman, B. Sket & P. Trontelj, 2004. Sampling adequacy in an extreme environment: species richness patterns in Slovenian caves. Biodiversity and Conservation 13: 1209-1229. Culver, D.C., L. Deharveng, A. Bedos, J.J. Lewis, M. Madden, J. R. Reddell, B. Sket, P. Trontelf & D. White, 2006. The mid-latitude biodiversity ridge in terrestrial cave fauna. Ecography 29: 120-128. Danielopol D.L., R. Rouch & A. Baltanas, 2002. Taxonomic diversity of groundwater harpacticoida (Copepoda, Crustacea) in Southern France. Vie Milieu 52 : 1-15. Delay, B., 1973. Deux nouveaux Oligochètes Haplotaxidae troglobies d’Espagne: Haplotaxis navarrensis n. sp. et Haplotaxis cantabronensis n. sp. Anales de Spéléologie 28: 405-411. Dumnicka, E., 1990. Oligochaetes from subterranean waters of Italy and Greece. Mémoires de Biospéologie, Tome XVII: 163-168. Elliot, W.R., 2005. Protecting Caves and Cave Life. In Culver, D.C. & W.B. White (eds), Encyclopedia of Caves. Elservier, California, 458-467. EVE, 1996. Mapa hidrogeológico del País Vasco. Ente Vasco de la Energía, Bilbao. Ferreira, D., F. Malard, M-J. Dole-Olivier & J. Gibert, 2007. Obligate groundwater fauna of France: diversity patterns and conservation implications. Biodiversity and Conservation 16: 567-596. Gaston, K.J., 1994. Rarity. Chapman & Hall, London.

Giani, N. & E. Martinez-Ansemil, 1981. Observaciones acerca de algunos Tubificidae (Oligochaeta) de la Península Ibérica, con la descripción de Phallodrilus riparius n. s. Annales de Limnologie 17: 201-209. Giani, N. & P. Rodriguez, 1988. Description de quelques espèces nouvelles de Tubificidae (Oligochaeta) de grotes et de sources karstiques de la péninsule ibérique. Stygologia 4: 121-137. Giani, N. & P. Rodriguez, 1994. New species of the genus Trichodrilus (Oligochaeta, Lumbriculidae). Zoologica Scripta 23: 33-41. Giani, N., B. Sambugar, P. Rodriguez & E. Martinez-Ansemil, 2001. Oligochaetes in southern European groundwater: new records and an overview. Hydrobiologia 463: 65-74. Gibert, J. & L. Deharveng, 2002. Subterranean ecosystems: a truncated functional biodiversity. Bioscience 52: 473-481. Gotelli, N. & R.K. Colwell, 2001. Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecology Letters 4: 379-391. Hrabe, S., 1963. On Rhyacodrilus lindbergi n. sp., a new cavernicolous species of the family Tubificidae (Oligochaeta) fron Portugal. Boletim da Sociedade Portuguesa de Ciências Naturais 10: 52-56. Juberthie, C., 2005. Conservation of subterranean habitats and species. In Wilkens H, D.C. Culver & W.F. Humphreys (eds), Caves and Other Subterranean Ecosystems. Ecosystems of theWorld 30. Elsevier, Amsterdam: 691-700. Juget, J. & E. Dumnicka, 1986. Oligochaeta (including Aphanoneura) des eaux souterraines continentals. In Botosaneanu, L. (ed.), Stygofauna Mundi. E. J. Brill/Dr W. Backhuys, Leiden: 234-244. Kerr, J. T., 1997. Species Richness, Endemism, and the Choice of Areas for Conservation. Conservation Biology 11: 1094-1100. Notenboom, J., 1986. The species of the genus Pseudoniphargus Chevreux, 1901 (Amphipoda) from northern Spain. Bijdragen tot de Dierkunde 56: 75-122. Rabinowitz, D., S. Cairns & T. Dillon, 1986. Seven forms of rarity and their frequency in the flora of the British Isles. In Soulé, M. (ed), Conservation Biology. The Science of Scarcity and Diversity. Sinauer, Mass: 182-204. Reid, W.R., 1998. Biodiversity hotspots. Tree 13: 275-280.

Rodriguez, P., 1986a. Nuevos resultados acerca de la fauna de oligoquetos acuáticos del País Vasco y cuenca alta del Ebro. 1. Haplotaxidae, Naididae y Tubificidae. Munibe Ciencias Naturales 38: 75-80. Rodriguez, P., 1986b. Nuevos resultados acerca de la fauna de oligoquetos acuáticos del País Vasco y cuenca alta del Ebro. 2. Enchytraeidae. Munibe Ciencias Naturales 38: 81-87. Rodriguez, P. & J.C. Armas, 1983. Contribution à la connaissance de la faune d’Oligochètes aquatiques du pays basque et zones limitrophes. Annales de Limnologie 19: 93-100. Rodriguez, P. & N. Giani, 1984. Description de Varichaetadrilus bizkaiensis n. sp.; nouvelle espèce de Tubificidae (Oligochaeta) d’Espagne. Bulletin de la Société d’Histoire Naturelle de Toulouse 120: 127-130. Rodriguez, P. & N. Giani, 1986. Description de trios espèces nouvelles d’Oligochètes aquatiques du Pays Basque (Espagne). Hydrobiologia 139: 269-276. Rodriguez, P. & N. Giani, 1987. Sur deux espèces de Lumbriculidae (Oligochaeta) cavernicoles: Cookidrilus speluncaeus n. gen., n. sp. et Trichodrilus diversisetosus Rodriguez, 1986. Bulletin de la Société d’Histoire Naturelle de Toulouse 123: 45-49. Rodriguez, P. & N. Giani, 1989. New species of Phallodrilus (Oligochaeta, Tubificidae) from caves of northern Spain and southwestern France. Hydrobiologia 180: 57-63. Schneider, K. & D.C. Culver, 2004. Estimating subterranean species richness using intensive sampling and rarefaction curves in a high density cave region in West Virginia. Journal of Cave and Karst Studies 66: 39-45. Sket, B.,1999a. High biodiversity in hypogean waters and its endangerment. The situation in Slovenia, the Dinaric karst, and Europe. Crustaceana 72: 767-779. Sket, B.,1999b. The nature of biodiversity in hypogean waters and how it is endangered. Biodiversity and Conservation 8: 1319-1338.

6. ANEXOS ANEXO 1: REGISTRO DE LOS PUNTOS DE MUESTREO CÓDIGO NOMBRE X
(UTM) Y (UTM)
Z (m)
HÁBITAT TIPO
GOR01 UBEGI 1 0516296 4765996 960 Surgencia 3 GOR02 UBEGI 2 0516270 4766001 973 Surgencia 3 GOR03 LAPURZULO 0515327 4765332 890 Cueva 2 GOR04 LAPUR ABAJO 0515247 4765134 850 Surgencia 3 GOR05 ALDABIDE 0514409 4769875 702 Surgencia 2 GOR06 ARTZEGI 0520052 4762908 807 Surgencia/cueva 2 GOR07 ZUBIALDE 0520044 4762955 818 Surgencia 3 GOR08 ATXURDIN 0521578 4764247 693 Surgencia 3 GOR09 MAIRULEGORRETA Cueva 2 GOR10 AITZITURRI 0514852 4767605 940 2 GOR11 AZOLETA 2 GOR12 ITXINGOTE 1 GOR13 LEIXAKORTA 0520905 4765100 740 Surgencia 2 GOR14 GORBEIA 0520830 4761830 695 Cueva 2 RBU01 AXPE 524325 4803500 25 Cueva 1 RBU02 APRAIZ 524225 4801350 22 Surgencia/Cueva 1 RBU03 MUNIENE 0525790 4799363 30 Surgencia 1 RBU04 LAIDA 525650 4805450 6 Surgencia 1 RBU05 ARGATXA 527790 4800925 2 Surgencia 1 RBU06 REKALDE 528750 4800575 25 Surgencia 2 RBU07 OXIÑA 529825 4801300 38 Surgencia 1 RBU08 BOLLAR 531250 4800075 220 Surgencia 1 RBU09 BOLUNTA 532280 4797490 115 Cueva 1 RBU10 OKAMIKA 0536852 4797504 -5 Cueva 2 RBU11 GOIKOETXE 0537014 4797642 40 Cueva 2 RBU12 AULESTIA 0536494 4794430 66 Surgencia 1 RBU13 LAMINAK Cueva 1 RBU14 ONDARO Cueva 1 RBU15 OMAERREKA Surgencia 1 RBU16 ATXURRA 0541022 4797184 59 Cueva 3

ANEXO 2: CARACTERIZACIÓN DEL HÁBITAT
HOJA DE CAMPO - SPRINGS SITE
Nombre Cuenca hidrogeográfica Código Muestreador
Fecha Hora X Y Z
DATOS CLIMATICOS
Lluvia Sol Tª aire Periodo de aguas bajo /medio / alto
TECNICA DE MUESTREO
Aparato tamaño de malla (um) área tiempo réplicas (nº) Comentarios
CARACTERISTICAS DEL MEDIO
Zona: epikarst vadosa freatica
Geología:
Morfología: rheocrene limnocrene helocrene
Uso humano: beber regar otros
Anchura (media): <1 m 1-5 m 5-25 m 25-100 m >100 m
Profundidad (media): < 0,1 m 0,1-0,5 m 0,5-1 m 1-2 m > 2 m
Pendiente (‰): <1 1-3 3-7 >7
Velocidad de la corriente: turbulenta / rápida (≥ 0.3 m/s) estancada / lenta (< 0.3 m/s)
Corriente: perenne intermitente
gravity spring artesian spring
Descarga (estimación): 10 l/s 100 l/s
Naturaleza del sustrato:
Dominante: Bloques cantos rodados grava arenas limos y arcillas
%: Bloques cantos rodados grava arenas limos y arcillas
% Detritus Fase 1 Fase 2 Fase 3
Condición del sustrato (%): Limpio…… cubierto por hojarasca…… materia orgánica…… limo y arcilla…… otros……
Estructura de las orillas Natural Bosque plantado (chopos, pinos, otros)
Canal de piedra Canal de hormigón
Izda
Drcha
Cobertura arbórea (% cobertura sobre el cauce): …………………
Sustrato muestreado: ………………………………………………………………………………………………………………
………………………………………………………………………………………………………………………………………
Vegetación acuática: ausente(0) rara (1) moderada (2) abundante (3)
Algas Macrofitas Musgos Otros Centro del cauce

Orillas Usos del suelo en las inmediaciones del área de muestreo Agrícola Forestal Pastos Urbano Industrial Contaminación (indicar presencia / ausencia y características: olor, color, turbidez, espumas, aceites, etc): ……………….………………………………………………………………………………………………. Medidas físico-químicas
Tª (ºC) pH Oxígeno disuelto (%) Oxígeno disuelto (mg/l)
Conductividad (uS/cm)
Salinidad (‰)
Dibujo del área de muestreo:
CARACTERISTICAS DEL SUSTRATO Componentes inorgánicos: Fondo de roca fija, Bloques (>256 mm ø), Cantos rodados (64-256 mm ø), Grava (2-64 mm ø), Arena (0.06-2 mm ø), Limos (0.004-0.06 mm ø), Arcillas (< 0.004 mm ø). Componentes orgánicos: Detritus Fase 1 (Madera, palos y material grueso de origen vegetal sin descomponer), Detritus Fase 2 (Restos vegetales variados, en diferente grado de descomposición, verde a pardo), Detritus Fase 3 (Materia orgánica completamente descompuesta, color negro y grano fino).

HOJA DE CAMPO - CAVES SITE
Nombre Cuenca hidrogeográfica Código Muestreador
Fecha Hora X Y Z
DATOS CLIMATICOS
Lluvia Sol Tª aire Periodo de aguas bajo /medio / alto
TECNICA DE MUESTREO
Aparato tamaño de malla (um) área tiempo réplicas (nº) Comentarios
CARACTERISTICAS DEL MEDIO
Zona: epikarst vadosa freatica
Geología:
Longitud de la cueva: Profundidad de la cueva:
Distancia del punto de muestreo a: suelo zona saturada la entrada de la cueva
Hábitat muestreado: infiltración lenta sifón lago río endógeno río exógeno
Velocidad de la corriente del río: turbulenta / rápida (≥ 0.3 m/s) estancada / lenta (< 0.3 m/s)
Corriente: perenne intermitente Descarga (estimación): 10 l/s 100 l/s
Naturaleza del sustrato:
Dominante: Bloques cantos rodados grava arenas limos y arcillas
%: Bloques cantos rodados grava arenas limos y arcillas
% Detritus Fase 1 Fase 2 Fase 3
Condición del sustrato (%): Limpio…… cubierto por hojarasca…… materia orgánica…… limo y arcilla…… otros……
Sustrato muestreado: ……………………………………………………………………………………………………………… Usos del suelo en las inmediaciones del área de muestreo Agrícola Forestal Pastos Urbano Industrial Contaminación (indicar presencia / ausencia y características: olor, color, turbidez, espumas, aceites, etc): ……………….………………………………………………………………………………………………. Medidas físico-químicas
Tª (ºC) pH Oxígeno disuelto (%) Oxígeno disuelto (mg/l)
Conductividad (uS/cm)
Salinidad (‰)
Dibujo del área de muestreo:
CARACTERISTICAS DEL SUSTRATO Componentes inorgánicos: Fondo de roca fija, Bloques (>256 mm ø), Cantos rodados (64-256 mm ø), Grava (2-64 mm ø), Arena (0.06-2 mm ø), Limos (0.004-0.06 mm ø), Arcillas (< 0.004 mm ø). Componentes orgánicos: Detritus Fase 1 (Madera, palos y material grueso de origen vegetal sin descomponer), Detritus Fase 2 (Restos vegetales variados, en diferente grado de descomposición, verde a pardo), Detritus Fase 3 (Materia orgánica completamente descompuesta, color negro y grano fino).

ANEXO 3: FOTOS DE LOS PUNTOS DE MUESTREO GORBEIA
GOR 01
GOR 02

GOR 03
GOR 04

GOR 05
GOR 06

GOR 07
GOR 08



















