
ELS LAGOMORFS (LAGOMORPHA; MAMMALIA) … · Experiments realitzats en captivitat mostren que es...
76
ELS LAGOMORFS (LAGOMORPHA; MAMMALIA) DEL PLEISTOCÈ INFERIOR D’ORCE (GRANADA) I ATAPUERCA (BURGOS) Roger Joan De Marfà Taillefer Treball de Recerca Tutelat de 2 on any Programa de Doctorat “Ciències de la Terra” (2004-2006). Paleontologia Departament d'Estratigrafia, Paleontologia i Geociències Marines Universitat de Barcelona. 4 de setembre del 2006
Transcript of ELS LAGOMORFS (LAGOMORPHA; MAMMALIA) … · Experiments realitzats en captivitat mostren que es...

ELS LAGOMORFS (LAGOMORPHA; MAMMALIA)
DEL PLEISTOCÈ INFERIOR D’ORCE (GRANADA)
I ATAPUERCA (BURGOS)
Roger Joan De Marfà Taillefer
Treball de Recerca Tutelat de 2on any Programa de Doctorat “Ciències de la Terra” (2004-2006). Paleontologia
Departament d'Estratigrafia, Paleontologia i Geociències Marines Universitat de Barcelona. 4 de setembre del 2006

2

3
ELS LAGOMORFS (LAGOMORPHA; MAMMALIA)
DEL PLEISTOCÈ INFERIOR D’ORCE (GRANADA)
I ATAPUERCA (BURGOS)

4
DEPARTAMENT D'ESTRATIGRAFIA, PALEONTOLOGIA I GEOCIÈ NCIES MARINES
UNIVERSITAT DE BARCELONA
ELS LAGOMORFS (LAGOMORPHA; MAMMALIA)
DEL PLEISTOCÈ INFERIOR D’ORCE (GRANADA)
I ATAPUERCA (BURGOS)
ROGER JOAN DE MARFÀ TAILLEFER
4 DE SETEMBRE DEL 2006
DIRECTORS
Dra. Rosa Domènech Arnal
Dr. Jordi Agustí Ballester
Dra. Gloria Cuenca Bescós

5
Sumari
1. Introducció......................................................................................................................06
1.1. Objectius............................................................................................................06
1.2. Problemàtica......................................................................................................07
1.3. Els lagomorfs del Plio-Pleistocè europeu: “State of the art”.............................12
2. Material i mètodes..........................................................................................................14
2.1. Els jaciments de Barranco León i Fuente Nueva 3............................................14
2.1.1. Localització geogràfica i geològica....................................................14
2.1.2. Estratigrafia i sedimentologia dels jaciments.....................................15
2.1.3. Paleoambient i datació........................................................................16
2.2. El jaciment de Trinchera Elefante.....................................................................18
2.2.1. Localització geogràfica i geològica....................................................18
2.2.2. Estratigrafia i sedimentologia dels jaciments.....................................21
2.2.3. Paleoambient i datació........................................................................22
2.3. Metodologia.......................................................................................................24
3. Biologia dels lagomorfs..................................................................................................27
3.1. Generalitats........................................................................................................27
3.2. Anatomia, etologia i hàbitat..............................................................................29
4. Sistemàtica......................................................................................................................36
- Prolagus sp.............................................................................................................36
- Oryctolagus cf. cuniculus.......................................................................................37
- Oryctolagus cf. lacosti...........................................................................................41
5. Conclusions.....................................................................................................................49
6. Agraïments......................................................................................................................50
7. Bibliografia......................................................................................................................51
Làmines i mesures..............................................................................................................59

6
1. Introducció.
1.1. Objectius.
En el desenvolupament d'aquest treball s'han tractat d'assolir tres objectius principals:
a) Descriure i identificar les associacions de lagomorfs del Pleistocè inferior presents en els
jaciments de Barranco León 5 i Fuente Nueva 3 (Orce, Granada), i Trinchera Elefante 9
(Atapuerca, Burgos).
Aquests tres jaciments són claus per entendre l’evolució dels humans i la seva dispersió per
Europa occidental. Certament, en ells s’han trobat diverses indústries lítiques, que són
proves irrefutables de presència humana. Per tant, és important conèixer amb quina fauna
van conviure aquests primers europeus, i més tenint en compte que els conills i les llebres
podrien haver constituït una font important d’aliment.
b) Comparar morfològicament i biomètrica aquestes associacions de lagomorfs.
L’interès d’aquestes comparacions resideix en el fet que els jaciments d’Orce i Atapuerca
s’ubiquen en àrees geogràfiques de caràcters climàtics diferents. D’aquesta manera, les
possibles dissimilituds observades en els fòssils podrien ser conseqüència directa de les
diferències climàtiques entre ambdues zones, l’una més meridional i mediterrània, i l’altra
més septentrional i continental.
c) Presentar una primera aproximació de l’edat relativa dels tres jaciments.
Tenint en compte l’evolució dels lagomorfs durant el Plio-Pleistocè i la seva distribució
paleobiogeogràfica, es proposa una datació relativa dels tres jaciments considerats, la qual
s'afegeix a la obtinguda per altres autors a través de l'estudi de rosegadors i insectívors.
Aquest estudi forma part d’un més complet sobre els lagomorfs del Plio-Pleistocè europeu,
el qual s’anirà desenvolupant en el marc de la Tesi Doctoral.

7
1.2. Problemàtica.
Els lagomorfs (conills, llebres i piques) són un grup de mamífers sempre abundants en els
jaciments continentals del Terciari i Quaternari, ja que tenen una taxa reproductiva molt
elevada i, a més, constitueixen una base alimentària important pels predadors, entre els
quals s'hi troba l'home (GUENNOUNI, 2001; CALLOU , 2003). No obstant això, es tracta d'un
grup relativament poc estudiat, sobretot a causa dels problemes que comporta l'elevada
variabilitat intra i interespecífica en la sistemàtica del grup. De fet, els lagomorfs han estat
sotmesos a nombroses revisions i fins i tot s'han requerit anàlisis bioquímiques per ajudar
en l'estudi de la seva evolució i la biogeografia històrica.
Part del problema radica en que el flux genètic entre poblacions de lagomorfs és acusat i
molt ràpid, com a conseqüència de la seva àmplia distribució geogràfica i alta velocitat de
dispersió. Aquest fet condueix a una marcada estabilitat morfològica del grup, la qual es
reflexa en una baixa diversificació i en el reconeixement de només 2 famílies ben
diferenciades des de la seva aparició a l'Eocè, Leporidae i Ochotonidae, amb uns 45-60
gèneres i unes 150 espècies (WOOD, 1957; DAWSON, 1967; LÓPEZ, 1989). Aquests taxons
s'han relacionat amb els Eurymiloidea del Paleocè asiàtic, tot i que el grau de parentesc
encara no ha estat suficientment establert (VAN VALEN, 1964). Tanmateix, recentment s'ha
identificat a Mongòlia un possible ancestre dels lagomorfs, Gomphos elkema, de 55 Ma
(ASHER, 2005).
Un dels problemes principals és que el gènere Lepus (llebres) està molt poc diferenciat, és
a dir, el procés d'especiació dintre del grup encara no ha finalitzat. Les espècies que s'han
distingit fins ara presenten una estructura parapàtrica (formen poblacions adjacents amb un
flux genètic entre elles) i constitueixen doncs una superespècie (LÓPEZ, 1989). Per aquest
motiu és complicat caracteritzar les associacions fòssils en relació a les espècies actuals.
De fet, les controvèrsies en la taxonomia de Lepus són molt acusades degut als caràcters
diagnòstics, ja que fins i tot en les llebres actuals la diferenciació interespecífica és petita si
es compara amb la variació individual i geogràfica. La causa podria raure en el fet que les
llebres actuals mostren una baixa variació adaptativa en l'alimentació, el comportament i la
locomoció i, conseqüentment, també en la morfologia de les estructures que serveixen per
realitzar aquestes funcions. Així, la diferenciació interespecifica no queda ben reflectida en
les diferències estadístiques corresponents a la mida, les proporcions i la coloració del
pelatge, les quals van lligades a la variabilitat geogràfica. De fet, l'elevada variabilitat

8
intraespecífica d'aquests caràcters provoca que les anàlisis quantitatives i qualitatives es
solapin entre les espècies, de manera que és possible que les llebres siguin en realitat
conspecífiques i que els caràcters diferenciadors consisteixin en mecanismes bàsicament
etològics (ANGERMANN, 1983). Experiments realitzats en captivitat mostren que es poden
obtenir híbrids entre espècies considerades vàlides, com Lepus europaeus i Lepus timidus.
Així doncs, sense barreres morfològiques ni kariològiques en les llebres, s'intueix que la
hibridació també existeix en condicions naturals (ANGERMANN, 1983).
Tota aquesta problemàtica en l'estudi de les llebres actuals comporta que la qüestió de
saber precisament quines espècies de llebre són presents en una mateixa àrea geogràfica és
complexa, doncs espècies considerades com a tal per un autor seran considerades com
subespècies per un altre (CORBET, 1983; FLUX , 1983, ANGERMANN, 1983; PALACIOS,
1983; SCHNEIDER et al., 1983; CALLOU , 1997, 2003). Per exemple, PALACIOS (1979, 1980)
utilitza mètodes bioquímics, morfològics i etològics per confirmar la descripció d'una nova
espècie, Lepus castroviejoi, a la Península Ibèrica, de manera que coexistiria amb les dues
espècies ja identificades en aquesta àrea geogràfica, Lepus europaeus i Lepus granatensis.
No obstant, per altres autors, aquestes diferències no són suficients com per separar fins a
tres espècies diferents en la Península Ibèrica i proposen l'existència d'una sola espècie de
llebre (SCHNEIDER & LEIPOLDT, 1983). El debat continua en l'actualitat.
Afortunadament, en els conills actuals la variabilitat que s'observa és menor, tot i que es
dóna que les diferències entre alguns gèneres monoespecífics, en particular Oryctolagus
(conill europeu) i Sylvilagus (conill americà), no són majors que entre algunes espècies
d'aquest últim gènere (CORBET, 1983).
D'altra banda, el conjunt d'estudis sobre els conills fòssils europeus ha conduït a alguns
autors a considerar l'existència de 2 subespècies, Oryctolagus cuniculus cuniculus i
Oryctolagus cuniculus huxleyi (o algirus). Tanmateix, per d'altres, els conills europeus del
Pleistocè Mitjà i Superior es poden dividir en 5 subespècies cronològiques: Oryctolagus
cuniculus lunellensis, O. c. grenalensis, O. c. baumensis, O. c. huxleyi i O. c. cuniculus.
Les recerques recents de conills actuals semblen portar de la subdivisió de les poblacions
en 2 subespècies a la distinció d'una sola població mostrant variacions clinals continues de
mida sense interrupció notable, o bé per una repartició d'aquestes poblacions en 2 grups
geogràficament separats diferenciats morfològicament i genètica (d'una banda, les formes
A, més petites, es trobarien al sud de la Península Ibèrica i les formes B, de mida més gran,

9
al nord de la Península Ibèrica i al sud de França) (BIJU-DUVAL , 1991; MONNEROT, 1994;
LOREILLE, 1997; CALLOU , 2003; GUENNOUINI, 2001). La diferència en la mida podria
radicar en les variacions climàtiques, sobretot en la taxa de precipitació (GUENNOUINI,
2001), o en la disponibilitat d'aliment (CALLOU , 2003).
Pel que fa a les dents, que són les peces més abundants en els jaciments, existeix una
elevada variabilitat morfològica dins de cada espècie, sobretot en el cas de les llebres
(ANGERMANN, 1966; PALACIOS, 1980). Les més usades per la sistemàtica són el P3 (tercer
premolar inferior) i el P2 (segon premolar superior), tot i que PETTER (1965) va donar
molta importància a la morfologia de l'incisiu superior en la seva revisió del gènere Lepus i
ho va usar per reunir en una sola espècie, L. capensis, totes les llebres europees i africanes.
El motiu d'aquesta variabilitat és que la mida i la morfologia de la superficie oclusal de les
dents dels lagomorfs està en funció del seu creixement continu (hipsodòncia), fet que
origina una important variació al llarg de la vida de l'animal (TOBIEN, 1978; LÓPEZ, 1989).
Així doncs, com que a la variació filogenètica se li afegeix una variació individual deguda
a la ontogènia i al desgast, l'ús dels paràmetres estadístics habituals (mitjana, variança,
coeficient de variació) és molt complex. Per l'estudi concret de les dents dels ochotònids,
LÓPEZ (1989) utilitza els coeficients alfa (alometria) i beta (constant de creixement inicial)
de les regressions logarítmiques de Y (longitud) sobre X (amplada); és a dir, els paràmetres
de les línies de regressió corba de cada població. La regressió caracteritza perfectament
cada població i depèn del seu desenvolupament ontogenètic, que varia notablement al llarg
de la filogènia del grup. Tanmateix, el comportament biomètric que s'observa en els P3 dels
lepòrids és diferent, ja que la relació existent entre les variables Y i X sembla constantment
isomètrica i les poblacions es fragmenten més que les dels ocotònids, agrupant-se en quatre
o més núvols de punts (LÓPEZ, 1989).
Un bon exemple de la variació de la morfologia dental en funció de l'ontogènia és el de
Hypolagus, el qual presenta un model tipus Paleolaginae (un reentrant posterointern i un
reentrant posteroextern) en estadis juvenils i un model Arqueolaginae (un únic reentrant
posteroextern que penetra fins la meitat de l'amplada del P3) en estadis més avançats
(KORMOS, 1934). En Trischizolagus del Pliocè del centre d'Europa també s'han observat
variacions importants i, segons alguns autors, la variació ontogenètica d'aquest gènere ha
conduït al model Leporinae (un sol reentrant posteroextern que travessa gairebé en la seva
totalitat el P3) característic de les llebres i conills actuals (AVERIANOV, 1995, 1997).

10
En incorporar el factor temporal, la sistemàtica dels lagomorfs encara es complica més,
ja que probablement existeixen diverses convergències evolutives. Així, per exemple, el
gènere Nekrolagus del Pliocè d'Amèrica del Nord presenta un patró i morfologia dental
similar a Sylvilagus (conill americà), Oyctolagus (conill europeu) i Lepus (llebre), pel que
es podria tractar del seu ancestre (HIBBARD, 1963). No obstant, Nekrolagus és molt similar
a Trischizolagus del Pliocè europeu i, per tant, aquest últim podria ser també l'ancestre
d'Oryctolagus i Lepus (LÓPEZ, 1989; AVERIANOV, 1995). Tanmateix, per altres autors, no
està clar que Lepus derivi de Nekrolagus i/o Trischizolagus, ja que els P3 de llebres amb un
model dental molt similar a aquests darrers no presenten l'istme que parteix el reentrant
posteroextern en el centre, sinó que està desplaçat lingualment (RADULESCO et al, 1967).
En relació a l'origen de Lepus, les dades moleculars indiquen clarament que les llebres
nord-americanes no constitueixen un clade monofilètic, sinó que molt probablement altres
llebres del vell món van envair secundàriament Amèrica del Nord a través de la connexió
continental de Bering (HALANYCH et al., 1999). Així doncs, l'origen de les llebres podria
ser bifilètic (GUREEV, 1964), tot i que altres autors aposten per un origen monofilètic,
vistes les variacions en la crenulació de les dents, la llargada del paladar, l'amplada de les
coanes i les sutures interparietals, caràcters que haurien aparegut independentment com a
adaptació a medis oberts (CORBET, 1983). Els taxons Sylvilagus i Oryctolagus es situarien
en línies evolutives paral·leles i tindrien un origen comú amb Lepus (BIJU-DUVAL , 1991;
HALANYCH et al., 1999; MATHEE et al., 2001, 2004) (fig.1).
Segons aquests mateixos estudis, el node de divergència entre els gèneres Oryctolagus,
Lepus i Sylvilagus podria situar-se fa uns 10 Ma a Amèrica del Nord o Àsia (ROBINSON et
al., 2002, 2005; MATHEE et al., 2004). És força probable que posteriorment Lepus s'hagués
dispersat amb la marcada reducció de massa forestal i la conseqüent obertura del paisatge,
esdeveniments que van coincidir amb la formació del casquet de gel a l'oest de l'Antàrtida
(ZACHOS, 2001). Aquest factor també va promoure l'establiment de ponts continentals que
van permetre la dispersió de la fauna entre Amèrica, Àsia, Europa i Àfrica.
No obstant això, per altres autors, la divergència entre aquests tres taxons, també per
criteris moleculars, es va produir més recentment, fa entre 6 i 8 Ma, i les dues formes de
conill europeu fa uns 2 Ma (BIJU-DUVAL et al., 1991).
La qüestió sobre l'origen d'Oryctolagus i la seva relació amb Lepus i Sylvilagus és un
motiu de debats intensos i importants en l'actualitat. S'ha suggerit que les formes del sud de

11
la Península Ibèrica haurien estat desplaçades en aquesta regió a l'època de les glaciacions i
les del sud de França haurien quedat refugiades allà i separades per les glaceres dels
Pirineus (BEAUCOURNU, 1980; BIJU-DUVAL et al., 1991).
Malauradament, el registre fòssil dels lepòrids és escàs i no ha ajudat fins ara a resoldre
amb eficiència tota la problemàtica relacionada amb l'origen i evolució dels lagomorfs
europeus actuals. A tot això cal afegir que en l'estudi dels lepòrids del Plio-Pleistocè hi
havia una dificultat afegida, i era la inexistència d'una descripció completa i detallada del
material original de O. lacosti de Perrier (França), espècie típica d'aquesta època i clau per
resoldre diversos interrogants sobre els lepòrids.
Finalment, una circumstància tafonòmica que cal tenir molt en compte durant el procés
d'excavació paleontològica, ja que pot alterar greument els resultats i la interpretació que
se’n faci, és que els conills construeixen caus en els que poden morir, fet que podria
suposar problemes relacionats amb la posició estratigràfica dels fòssils (Donard, 1982).
Figura 1. Filogènia dels gèneres de lagomorfs actuals basada en l'anàlisi molecular dels gens mitocondrials (ROBINSON & MATTHEE, 2005).

12
1.3. Els lagomorfs del Plio-Pleistocè europeu: Estat de la qüestió.
El Plio-Pleistocè europeu constitueix una època molt interessant de cara a l'estudi de la
família Leporidae (Ordre Lagomorpha), ja que és durant aquest interval temporal quan
augmenta notablement la presència de conills (Oryctolagus) i llebres (Lepus) en el registre
fòssil continental, en detriment dels representants de la família Ochotonidae (piques), que
fins aleshores eren els lagomorfs predominants.
Durant el Pliocè mitjà (MN15) apareixen els fòssils més antics de lepòrids del gènere
Oryctolagus, els quals van ser trobats a Layna (Soria, Espanya) i van ser descrits com una
nova espècie, O. laynensis (LÓPEZ, 1977, 1989). Pel que fa als ocotònids, a Europa
occidental només es troba ben representat el gènere Prolagus, el qual va aparèixer en el
Miocè inferior i va anar reduint la seva àrea de distribució geogràfica fins extingir-se a
l’Holocè (LÓPEZ, 1989, 2001). Aquest gènere és encara abundant durant el Pliocè i es troba
en diversos jaciments de la Península Ibèrica, com Gorafe 1 i 2 (Granada) (LÓPEZ, 1989 ),
Layna (Soria) (LÓPEZ, 1989 ) i Huéscar 3 (Granada) (MAZO et al., 1985).
En el Pliocè superior (MN16 i MN17) es citen restes d'Oryctolagus en els jaciments
francesos de Perrier (Auvergne) (POMEL, 1853; VIRET 1954), Montoussé (Midi-Pyrénnées)
(CHALINE et al., 2000), Saint Vallier (Drome) (VIRET, 1954; MARTÍN et al., 2004) i Senèze
(Haute-Loire) (SCHAUB, 1943), i en el jaciment italià de Valdarno (Toscana) (FORTELEONI,
1971; LÓPEZ, 1989). Aquestes restes s'han atribuït a Oryctolagus lacosti, una espècie amb
caràcters intermedis de conill i llebre, la qual presenta una forta variabilitat en els caràcters
diagnòstics més importants (paladar i morfologia del tercer premolar inferior) (LÓPEZ,
1989). La posició sistemàtica i filogenètica d'aquesta espècie és força problemàtica, de
manera que fins i tot ha sofert canvis a nivell de gènere per part de diversos autors, i alguns
d'ells n’allarguen la presència fins el Pleistocè mitjà (LÓPEZ, 1989). No obstant això, s'ha
de ser prudent en afirmar que es tracta d'una espècie diferent a la de Layna (Sòria), doncs
en les poblacions actuals de lepòrids existeixen importants diferències morfològiques i
biomètriques, sobretot en funció del clima i la disponibilitat d'aliment (GUENNOUNI, 2001;
CALLOU , 2003). D'altra banda, durant el Pliocè es troben a Europa dos altres gèneres de
lepòrids: Hypolagus i Trischizolagus. El primer ha aparegut en jaciments d'Europa central i
occidental, com Tegelen Clay (Holanda) (SCREUDER, 1936) i Montoussé (Midi-Pyrénnées,
França) (CHALINE et al., 2000), però no s'ha identificat mai fins ara a la Península Ibèrica.
El seu origen es situa en el Miocè mitjà d'Amèrica del Nord i es pensa que es va extingir

13
probablement per la competència amb Lepus (DAWSON, 1967; FOSTOWICZ, 2003). En
canvi, el segon només es troba durant el Pliocè a Europa central, en jaciments com
Malusteni i Beresti (Rumania) (RADULESCO & SAMSON, 1967), si bé durant el Miocè la
seva repartició era molt més estesa pel continent. Alguns autors el proposen com l'ancestre
d'Oryctolagus i/o Lepus (LÓPEZ, 1977, 1989; AVERIANOV, 1995, 1997).
Durant el Pleistocè inferior, els ocotònids d'Europa Occidental, representats pel gènere
Prolagus, queden relegats al litoral mediterrani, zona en la que també és força abundant
l'enigmàtic Oryctolagus lacosti (MEULEN, 1973; LÓPEZ, 1989; AGUSTÍ et al., en premsa).
El gènere Hypolagus sobreviu en determinades zones geogràfiques aïllades, com en el
Monte Pellegrino (Sardenya, Itàlia) (FLADERER & FIORE, 2003), però acaba desapareixent.
D'altra banda, és durant aquesta època que s'identifiquen els primers representants de
Lepus a diversos jaciments d'Europa Central, com Beremend 9, Nagyharsány i Villány
(Hungría) (KRETZOI, 1956) i Deutsch-Altenburg (Austria) (FLADERER, 1987), Itàlia (Monte
Peglia, Orvieto) (MEULEN, 1973) i Espanya (Huéscar 1, Granada) (MAZO et al., 1985).
Tanmateix, aquestes primeres llebres estan únicament documentades per restes dentals i,
observada la gran variabilitat existent en el material actual i fòssil, així com la probable
existència de convergències evolutives, és arriscat extreure'n conclusions fiables.
Finalment, en el Pleistocè mitjà, Prolagus s'extingeix pràcticament d'Europa i només
sobreviu fins l'Holocè a les illes de Còrcega i Sardenya (LÓPEZ, 1989). D'altra banda, al
jaciment de Valdemino (Savona, Itàlia) apareix O. burgi, una espècie que és considerada
per alguns autors com un possible descendent de O. lacosti (NOCCHI & SALA , 1997, 1998).
Pel que fa als lepòrids actuals d'Europa Occidental, les dades paleontològiques procedents
de diversos jaciments francesos i espanyols semblen indicar que el conill (O. cuniculus) i
les llebres (Lepus europaeus, Lepus timidus, Lepus granatensis) apareixen també durant el
Pleistocè mitjà (DONARD, 1982; LÓPEZ, 1989; GUENNOUINI, 2001; CALLOU , 2003).
En definitiva, les dades bibliogràfiques existents indiquen que en el Plio-Pleistocè es van
produir importants canvis en la evolució i la paleobiogeografía dels lagomorfs, fets
probablement associats als canvis ambiental que tingueren lloc durant aquesta època.

14
2. Material i mètodes.
2.1. Els jaciments de Barranco León i Fuente Nueva 3 (Orce, Granada).
2.1.1. Localització geogràfica i geològica.
Els jaciments paleontològics de Barranco León i Fuente Nueva 3 es situen a Orce, una
petita localitat de la província de Granada ubicada en la conca neògena de Guadix-Baza
(fig.2). Aquesta conca té una alineació NE-SW, com tota la Serra Bètica, i presenta una
extensió d'uns 110 x 50 Km. Està limitada al nord per les Serres de Segura, Castril i la
Sagra; al est per les d'Orce-María, Estancias i Baza; al sud per la Serra Nevada y al oest per
la Serra Arana (ESPIGARES & ROS, 2003).
Figura 2. Esquema geològic general de la conca de Guadix-Baza (VERA, 1970).
A la conca es distingeixen dues cubetes o subconques, la de Guadix, localitzada en el
sector sud-ocidental, i la de Baza, en el sector nord-oriental. Ambdues estan separades
geogràficament i es caracteritzen per presentar successions litològiques diferents: a la
subconca de Guadix predominen sediments al·luvials, mentre que la de Baza està
fonamentalment constituïda per dipòsits lacustres (VERA, 1970, 1985; OMS, 1998; GARCÍA-
AGUILAR & MARTÍN, 2000).

15
Paleontològicament, cal destacar que la conca de Guadix - Baza presenta un excel·lent
registre estratigràfic continental del Pliocè i Pleistocè inferior i mitjà, amb més d’un
centenar de jaciments de vertebrats. Entre aquests destaquen els dos estudiats en aquest
treball i Venta Micena, també situat a Orce, en el qual es va exhumar un polèmic fragment
cranial que es va atribuir inicialment a Homo, tot i que podria pertànyer a un èquid, bòvid o
cèrvid (GIBERT et al., 1983; AGUSTÍ & MOYÀ, 1987; MOYÀ & AGUSTÍ, 1989; MOYÀ &
KÖLHER, 1997; GIBERT et al., 1999; CAMPILLO , 2002; MARTÍNEZ, 2002; GIBERT, 2004).
Pel que fa a la situació dels jaciments, Barranco León és un barranc orientat N-S que es
troba a uns 3 km del poble d’Orce, en direcció a María. Conegut des dels anys 80 com
jaciment paleontològic, va ser excavat l’any 1995 per l’equip de J. Gibert (GIBERT et al.,
1998), campanya durant la qual es va localitzar i definir un nivell arqueològic, identificat
com BL5 o capa D, en el qual es van documentar diverses peces lítiques associades a restes
d’hipopòtam. Tanmateix, un segon nivell arqueològic molt important és BL1 (capa C).
D'altra banda, el jaciment de Fuente Nueva 3 es troba a uns 5 km del poble d’Orce, també
en direcció María, prop de l’església en runes i de les antigues fonts que li donen nom. Va
ser descobert l’any 1992 com a conseqüència de la instal·lació d’una torreta d’electricitat, i
les primeres campanyes d’excavació es van organitzar als anys 1994 i 1995 sota la direcció
dels Drs. B. Martínez, llavors membre del Dipartimento di Scienze della Terra (Universitat
de Firenze, Itàlia) i Alain Turq, del Musée National de Préhistoire (Les Eyzies, França), en
codirecció amb el Dr. Josep Gibert, de l’Institut Paleontològic M. Crusafont de Sabadell
(TURQ et al., 1996; MARTÍNEZ et al., 1997). Durant aquestes primeres campanyes van
aparèixer diversos nivells arqueològics amb restes òssies i indústria lítica.
2.1.2. Estratigrafia i sedimentologia dels jaciments.
Les successions estratigràfiques dels jaciments de Barranco León i Fuente Nueva 3 (Orce)
pertanyen a la unitat litoestratigràfica anomenada Membre Calcari Superior de la Formació
Baza, constituïda fonamentalment pels dipòsits lacustres que ocupen la part oriental de la
conca (VERA, 1970).
Des del punt de vista paleogeogràfic, ambdues successions estratigràfiques pertenyen a una
gran fase d'expansió lacustre del Pleistocè inferior (GARCÍA-AGUILAR & MARTÍN, 2000).
Tanmateix, dins d'aquesta fase d'expansió es poden reconèixer fases menors de expansió-
retracció, així com diversos canvis de salinitat ben representats per diferents associacions
d'invertebrats, especialment ostràcodes (ANADÓN et al., 1994).

16
Els dipòsits de la zona d'excavació de Barranco León (BL) es van formar en un ambient
lacustre som, amb emersions locals i influències d'aports fluvials. En el jaciment es troben
dues importants capes arqueològiques: la unitat BL1 (Capa C), que correspon a un dipòsit
de calcarenites, i la unitat BL5 (Capa D), formada majoritàriament per materials detrítics
que contenen una rica associació de fòssils, així com indústria lítica (fig.3).
En el jaciment de Fuente Nueva 3 (FN3), els dipòsits sedimentaris de la zona d'excavació
es van originar en un ambient lacustre som i oligohalí, amb freqüents emersions que van
conduir a la formació de paleosòls. L'estratigrafia és més complexa que la de Barranco
León, ja que existeix un cert grau de deformació, però es poden diferenciar clarament una
calcària basal de color blanc, en tram amb gresos i lutites carbonatades que conté restes
fòssils i indústria lítica, un nivell margocalcari i un sostre amb calcàries compactes (fig.3)
(ANADÓN & JULIÀ , 2003; OMS et al., 1996).
2.1.3. Paleoambient i datació.
L'associació d'invertebrats que s'ha trobat a Barranco León 5 i Fuente Nueva 3, básicament
constituïda per ostràcodes i mol·luscs (Ilyocypris sp., Heterocypris sp., Candona sp.,
Pisidium sp., Melanoides tuberculata, Mercuria emiliana, Islamia minuta, Hydrobia acuta,
Planorbis planorbis, Trochoidea i Arionidae indeterminat, indicaria un medi aquàtic
oligohalí amb aigües netes i amb una certa quantitat de vegetació aquàtica (ANADÓN &
JULIÀ , 2003; DE MARFÀ, en premsa).
D'altra banda, l'associació de microvertebrats formada bàsicament per Leuciscus sp.,
Discoglossus sp, Pelobates cultripes, Bufo bufo, Rana cf. perezi, Chalcides sp., Anguidae
indet., Natrix maura, Erinaceus sp., Crocidura sp., Sorex sp., Galemys sp., Castillomys sp.,
Apodemus sp., Mimomys savini, Allophaiomys aff. lavocati, Allophaiomys sp., Oryctolagus
sp. i Prolagus cf. calpensis és un indicador que en el Pleistocè inferior de Barranco León i
Fuente Nueva 3 el clima era relativament més fresc i humit que l'actual en la regió, amb
presència de làmines d'aigua permanent envoltades per vegetació aquàtica, matolls i prats
(BLAIN , 2003; FURIÓ, 2003; DE MARFÀ, en premsa; AGUSTÍ et al., en premsa). L'associació
formada per una forma primitiva d'Allophaiomys sp., Mimomys savini i Castillomys sp.,
complementada amb la presència d'altres vertebrats i amb l'estudi magnetoestratigràfic de
la regió, indicaria una edat dels jaciments situada en el Pleistocè inferior, entre els nivells
de Venta Micena i Huéscar 1 (AGUSTÍ, 1986; AGUSTÍ & MADURELL, 2003; AGUSTÍ et al.,
2001; AGUSTÍ et al., en premsa; OMS et al., 2003).
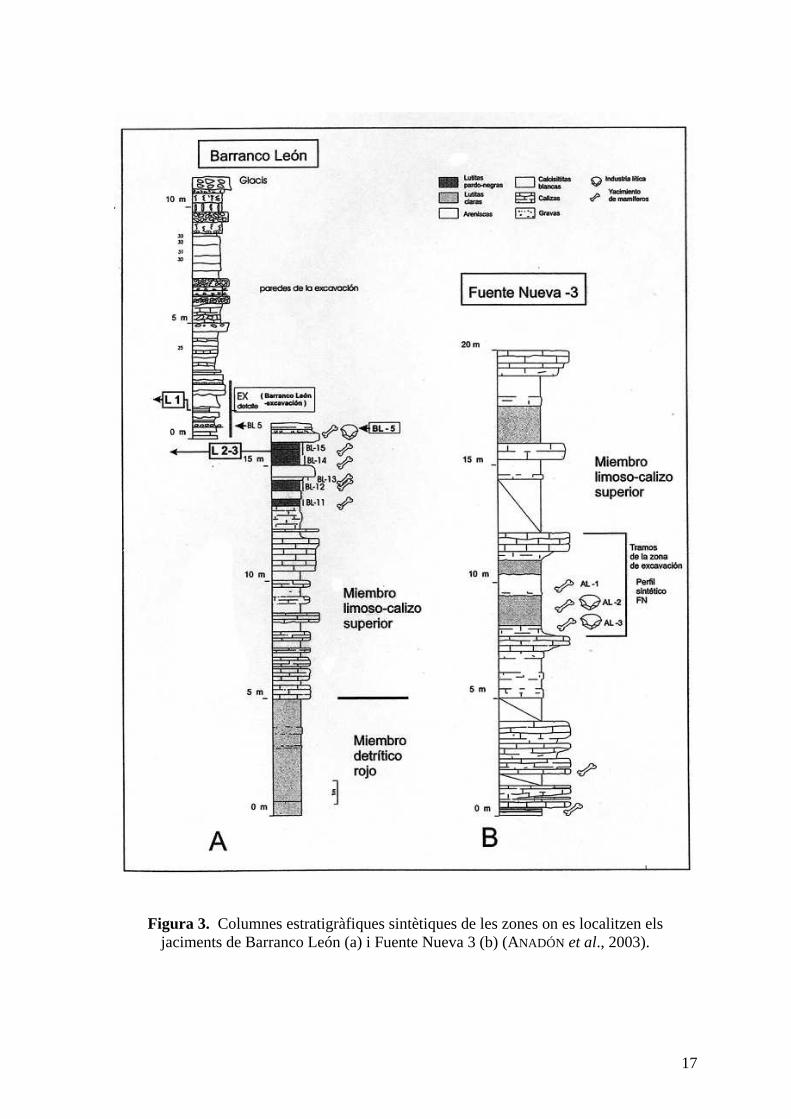
17
Figura 3. Columnes estratigràfiques sintètiques de les zones on es localitzen els jaciments de Barranco León (a) i Fuente Nueva 3 (b) (ANADÓN et al., 2003).

18
2.2. El jaciment de Trinchera Elefante (Atapuerca, Burgos).
2.2.1. Localització geogràfica i geològica.
El jaciment de Trinchera Elefante es troba a la Serra d'Atapuerca (Burgos), un petit conjunt
muntanyós situat entre les conques de l'Ebre i el Duero, i entre la Serralada Cantàbrica i el
Sistema Ibèric, en l'extrem oriental del corredor de la Bureba (fig.4). Aquest corredor ha
estat utilitzat al llarg de tota la seva existència com a pas principal fins l'interior de la
Península des d'Europa, ja que uneix la vessant mediterrània amb la vessant atlàntica. Així
doncs, per aquest corredor s'han produït continuats passos de fauna i flora, fet corroborat
per la gran riquesa de fòssils present en els jaciments d'Atapuerca (AGUIRRE, 1995;
CERVERA, 1998; CARBONELL et al., 1999; ARSUAGA et al., 2002; CUENCA et al., 2004).
Figura 4. Situació geogràfica de la Serra d'Atapuerca (CUENCA et al., 2004).

19
La Serra d'Atapuerca està constituïda per calcàries i gresos del Cretaci superior, les quals
estan recobertes de materials aportats pel riu Arlanzón, que ha format terrasses fluvials
durant l'època Quaternària. Els jaciments formen part d'un sistema kàrstic que va començar
a desenvolupar-se a finals del Terciari (Pliocè superior - Pleistocè inferior). Durant el
Pleistocè, les coves es van anar reomplint amb sediment, plantes, pol·len i restes d'animals,
producte de la denudació dels pendents circundants i del retrocés de les parets i sostres de
les coves, fins arribar a tapar completament les cavitats. Els productes de l'activitat
biològica dels depredadors, especialment les carcasses dels animals consumits, les
egagròpiles i els excrements són la principal font d'acumulació dels restes de vertebrats
fòssils que es troben en els jaciments kàrstics. Els materials de mida menor van entrar tant
per les entrades com per les nombroses fissures del sostre i les parets (PARÉS & GONZÁLEZ
1999; ARSUAGA et al., 2002; BENITO, 2004; CUENCA et al., 2004).
Algunes de les cavitats formen part encara d'un sistema tancat com les del complex Sima
de los Huesos - Cueva Mayor, i altres han estat exhumades per la construcció d'un
ferrocarril miner a finals del segle XIX, el que fa que se les conegui com els jaciments de
la Trinchera del Ferrocarril: Dolina, Zarpazos, Galería i Elefante (fig.5). Tots aquests
jaciments són excepcionals pel que fa a l'abundància, conservació i importància científica
dels fòssils que contenen, i la cronologia que abasten és molt àmplia, ja que corresponen a
reompliments dipositats durant el Pleistocè inferior-mitjà, és a dir, entre 1,5 Ma i 200 ka.
Ara bé, els jaciments més coneguts internacionalment són Trinchera Dolina, que conté les
restes d'Homo antecesor, l'homínid més antic d'Europa occidental (800 ka) (CARBONELL,
1995; BERMÚDEZ et al., 1997, ARSUAGA et al., 1999) i Sima de los Huesos, que presenta el
registre més complet del món d'Homo heidelbergensis (400 ka) (ARSUAGA et al., 1997).
Tanmateix, el material aquí estudiat procedeix del jaciment Trinchera Elefante (TE), que
conté una rica associació de fòssils de vertebrats i indústria lítica del mode 1 (olduvaià)
associada al Pleistocè inferior; així doncs, és una de les evidències més antigues de
activitat humana a Europa (ROSAS et al., 2001, 2004; CUENCA BESCÓS & ROFES, 2004).
Efectivament, a TE no s'han trobat fòssils humans, però l'activitat humana està reflectida
en el nivell TE9, ja que s'han trobat indústries lítiques (ROSAS et al., 2001, 2004; CUENCA
& ROFES, 2004). Així, tot sembla indicar que es va produir ben aviat una migració humana
cap a l'oest d'Europa probablement durant el període càlid Waalià, hipòtesi recolzada per la
troballa de lítica en altres dos localitats de la Península Ibèrica: BL5 i FN3 (Guadix-Baza).

20
La indústria lítica que s'ha trobat en aquests tres jaciments (TE9, BL5 i FN3) pertany
tecnològicament a l'Olduvaià o Mode 1. A més a TE9 s'han trobat restes de grans herbívors
amb marques de tall, fet que porta a considerar que els humans foren responsables d'una
part de les acumulacions.
Figura 5. Plànol amb la situació dels jaciments d'Atapuerca (CUENCA et al., 2004).
Així doncs, els jaciments paleontològics d'Atapuerca són jaciments kàrstics, en els que una
sèrie de coincidències geològiques i paleontològiques, climàtiques i històriques fan que
sigui el registre més complet de la història de la vida en medis continentals del Pleistocè
europeu. El ric contingut paleontològic de les coves de la Serra situen Atapuerca com un
important referent per la paleontologia i en general per la ciència espanyola. El complex
paleo-arqueològic està declarat Patrimoni de la Humanitat per la UNESCO (2000) i l'equip
investigador ha rebut el Premi Príncipe de Asturias de Investigación Científica (1997).

21
2.2.2. Estratigrafia i sedimentologia del jaciment.
El jaciment de Trinchera Elefante o Sima del Elefante (TE) es troba a uns 200 m al sud
dels jaciments de Trinchera Dolina i Galería. És també un jaciment de reompliment de
cova fossilitzada com els anteriors, en el que els materials van ser tallats per les obres del
ferrocarril i afloren en l'actualitat a ambdós costats de la trinxera. Té un gruix visible d'uns
11 m, tot i que les dades del sondeig realitzat al 2001 revelen que pot assolir els 19 m de
potència, de manera que seria el jaciment amb més gruix de tots els coneguts a la Serra
d'Atapuerca. Els materials de Trinchera Elefante tenen una edat compresa entre uns 1,5 Ma
i 200 ka (CUENCA & ROFES, 2004).
El reompliment de TE es divideix en 21 nivells estratigràfics (només afloren de E8 a E21)
formats durant 3 fases diferents (fig.6) (ROSAS et al., 2001):
Figura 6. Estratigrafia de Trinchera Elefante (ROSAS et al., 2001).

22
En la primera fase de reompliment la cova va omplir-se amb argiles i graves vermelles,
constituint la Unitat Roja Inferior, de 9 m de potència (amb blocs de calcària en alguns
trams i nivells amb copròlits de rat-penats), tots amb un alt contingut en restes fòssils
(nivells E8 - E14). En aquesta unitat es troba el nivell TE9 o “Toblerone”, objecte d'estudi
d'aquest treball.
En una segona fase, la cova sofreix un important procés de reactivació, amb una probable
erosió dels nivells inferiors, i hi ha un reompliment posterior format per una unitat blanca
superior (nivells 15 - 20, d'uns 9 m de gruix, formada fonamentalment per margues, llims i
nivells d'espeleotemes calcítics en el nivell 17) que cap a sostre augmenta en granulometría
i quantitat de clasts i presenta una coloració més vermellosa (E18 – E19).
Finalment, una tercera fase de reompliment correspon als dipòsits de gravetat que colmaten
i tanquen la entrada de la cova des d’un conducte vertical situat al sud de la secció. Aquest
reompliment final es produeix en el nivell 20 (ROSAS et al., 2001).
2.2.3. Paleoambient i datació.
En la campanya d'excavació del 1995 es va iniciar el mostreig sistemàtic de microfauna en
la Unitat Roja de TE en fer una excavació de prova de 1 m2 d'extensió, ampliada a 2 m2 en
el 1996, any en el que es va fer un sondeig de 3,5 m de fondària. En aquesta segona
campanya es van processar 16 mostres. En el 1999 es va iniciar l'excavació sistemàtica del
jaciment que continua en l'actualitat (ROSAS et al., 2001; CUENCA et al., 2004).
Les espècies de rosegadors determinades en els nivells E8 - E13 de la Unitat Roja Inferior
corresponen a una associació del Pleistocè inferior, tot i que els valors de presència /
absència i nombre d'individus varien d'un nivell a un altre, sent el més ric i divers el TE9.
La llista actualitzada de rosegadors és la següent: Allophaiomys chalinei, A. lavocati,
Iberomys aff. nutiensis-thenii-huescarensis, Ungaromys nanus, Pliomys episcopalis, cf.
Mimomys, Castillomys rivas, Apodemus sp., Glis sp., Eliomys cf. quercinus, Castor fiber i
Sciurus sp. (Cuenca, 2001; Cuenca et al., 2004). D'altra banda, els insectívors de la Unitat
Roja estan representats per diverses espècies agrupades en 3 famílies: Erinaceidae
(Erinaceus cf. europaeus); Talpidae (Talpa cf. europaea y Desmaninae indet.) y Soricidae
(Beremendia cf. minor:, Neomyinae ind., Sorex sp., Crocidura cf. russula, Crocidura sp.)
(CUENCA & ROFES, 2004; ROFES & CUENCA, en premsa).

23
L'evolució de les associacions de micromamífers sembla correspondre amb les 3 fases c-f-c
característiques del Waalià del nord d'Europa, un episodi en general càlid del Pleistocè
inferior (RZEBIK, 1995; ZAGWIN, 1998). El Waalià es troba dins el magnetocron Matuyama
i es caracteritza per una successió d'events càlid, fred, càlid (A, B, C) que tenen lloc entre
fa 1,5 i 1,2 Ma. Els estudis paleomagnètics confirmen la posició de la Unitat Roja dins del
cron Matuyama (ROSAS et al., 2001). Queda descartada una edat més antiga com Eburonià,
doncs aquesta representa un episodi fred respecte el Waalià i, per altra banda, la fauna de
rosegadors no sembla ser d'una edat superior als 1,5 Ma. A continuació del Waalià es troba
de nou un episodi fred, el Menapià, i a partir d'aquest el Bavelià (al voltant d'1 Ma), més
càlid i, finalment, el Cromerià, que engloba el límit Matuyama -Brunhes (780.000 anys).
La base del Bavelià conté el cron positiu Jaramillo dins del cron negatiu de Matuyama. No
sembla haver-hi canvis en la polaritat magnètica negativa de tota la Unitat Roja Inferior de
Trinchera Elefante, pel que es pot descartar que els nivells estudiats corresponguin al
Bavelià (ROSAS et al., 2001, CUENCA & ROFES 2004).
La fauna de rosegadors ens dóna una edat que podria anar des d'1 Ma, per compartir
espècies similars a les de jaciments europeus d'aquesta edat (Jaramillo), com Valerots,
Vallonet o Untermassfeld, fins 1,5 Ma, per les espècies presents en jaciments anteriors a
Jaramillo, com A. chalinei i Castillomys, taxó aquest últim propi del Pliocè espanyol que
s'extingeix durant el Pleistocè inferior (LAPLANA & CUENCA, 2000). També els insectívors
indiquen una edat antiga, ja que la majoria dels seus representants europeus són formes del
Plio-Pleistocè (CUENCA & ROFES, 2004). Aquesta unitat seria, com a mínim, tan antiga
com els altres dos jaciments considerats en aquest treball: Barranco León i Fuente Nueva
3, és a dir, d'una edat mínima de 1,2 Ma (MARTÍNEZ et al., 1997; AGUSTÍ et al., 2004).
Pel que fa al paleoambient, la base de Toblerone (9b) representa un moment amb un clima
càlid i relativament més sec que en els nivells superiors, fet interpretat a partir de la
abundància de crocidurins. En canvi, en el nivell superior (9a) es produeix un refredament i
augment de la humitat. Finalment, el clima es torna més càlid i menys humit en el nivell
TE10 (CUENCA & ROFES, 2004).

24
2.3. Metodologia.
Per realitzar aquest treball s'han realitzat els següents passos:
1) Treball de camp consistent en participar a la excavació paleontològica organitzada als
jaciments de Barranco León i Fuente Nueva 3 (Orce, Granada) durant la campanya d'estiu
del 2003. Aquests jaciments es treballen en àrees d'1 m2 i s'excaven seguint acuradament
les superfícies estratigràfiques. Els fòssils i la indústria lítica distingibles a simple vista es
cartografien mitjançant un sistema de coordenades tridimensional, anotant també les seves
dimensions i orientació relativa. D'altra banda, el sediment obtingut es col·loca dins d'uns
sacs etiquetats segons el jaciment, indicant l'àrea de procedència (identificada per una lletra
i un nombre) i el nivell estratigràfic mesurat en “UME” (Unitat de Mostreig Estratigràfic
basada en capes artificials d'entre 3 i 5 cm). També s'ha participat en el rentat i tamisat de
centenars de kilograms de sediment procedent dels dos jaciments, tasca que es realitza amb
aigua i amb tamisos d'una malla de 5, 2 i 0,5 mm. Els tamisos més petits són els més
importants per recuperar les restes de micromamífers (rosegadors, insectívors, quiròpters i
lagomorfs) donada la mida reduïda de les seves parts esquelètiques quan es troben aïllades.
Posteriorment el material resultant del procés es deixa assecar al sol i guarda novament en
sacs, separats per la granulometria (fotos 1 i 2).
No s'ha participat en la campanya paleontològica organitzada en el jaciment de Trinchera
Elefante (Atapuerca, Burgos), si bé la metodologia del treball de camp és molt similar.
2) Treball de laboratori consistent en triar els fòssils del material tamisat provinent de
Barranco León 5, Fuente Nueva 3 i Trinchera Elefante 9. Per seleccionar les restes fòssils
de lagomorfs s'han utilitzat unes pinces i una lupa binocular, i s'han separat les diferents
parts esquelètiques en capses diverses, cadascuna d'elles amb una etiqueta identificativa.
Les dents de lagomorfs s'han rentat amb l'ajuda d'un pinzell i s'han muntat sobre un suport
de plàstic amb una pasta adhesiva, de manera que la cara oclusal quedi horitzontal.
Aquesta part del treball s'ha realitzat al Museu Paleontològic Miquel Crusafont de Sabadell
i a la Facultat de Geologia de la Universitat de Saragossa, que es on es troben dipositades
les col·leccions de micromamífers dels jaciments d'Orce i Atapuerca respectivament.
Concretament, s'han obtingut i preparat pel seu estudi 113 peces dentals de Barranco León,
216 de Fuente Nueva 3 i 228 de Trinchera Elefante 9. També s'ha disposat de 2 calcanis, 7
astràgals, 3 fragments d'húmer, 1 fragment d'ulna i 2 fragments de maxil·lar de TE9.

25
Foto 1. Campanya paleontològica en el jaciment de Trinchera Elefante (2006).
Foto 2. Rentat i tamisat del sediment al riu Arlanzón (Atapuerca) (2006).

26
3) Treball de gabinet consistent en la identificació del material fòssil obtingut. Per dur a
terme aquesta tasca s'ha realitzat una exhaustiva recerca bibliogràfica i diverses consultes
de restes esquelètiques de conills i llebres actuals dipositats a les col·leccions del Museu de
Zoologia de Barcelona i al Museu Nacional de Ciències Naturals de Madrid. D'altra banda,
també s'ha consultat el material fòssil d'Oryctolagus lacosti del jaciment Pliocè de Perrier
(França), el qual es troba a la Universitat Claude Bernard de Lyon.
Simultàniament, s'han realitzat dibuixos de les peces dentals amb una cambra clara Wild i
diverses fotografies amb el microscopi electrònic de rastreig (MER) Hitachi S-2300 dels
Serveis Científico-Tècnics de la Universitat de Barcelona. També s'han mesurat totes les
restes fòssils amb l'ajuda d'un peu de rei digital i un binocular amb un sistema de medició
integrat i, posteriorment, s'ha procedit a la descripció del material. Pel que fa a les dents,
s'ha utilitzat la terminologia de STIRTON (1935) que anomena únicament els sinclinals, i
s'ha completat amb la de LÓPEZ MARTÍNEZ (1974, 1975), de manera que es simplifiquen al
màxim les descripcions. La terminologia i els paràmetres mesurats es mostren a la figura 7.
Finalment, s'han comparat els resultats obtinguts per cada jaciment amb la bibliografia,
s'han discutit els resultats i s'han extret les conclusions que aquí es presenten.
Figura 7. Terminologia i paràmetres mesurats sobre la superfície oclusal del tercer premolar inferior o P3 (a) i segon premolar superior o P2 (b). 1: paraflex; 2: anterocònid lingual; 3: anteroflèxid; 4: anterocònid labial; 5: protoflèxid; 6: protocònid; 7: hipoflèxid; 8: hipocònid; 9: mesoflex; 10: lagicon; 11: paraflex; 12: hipercon mesial; 13: hipoflex; X: amplada; Y: llargada.

27
3. Biologia dels lagomorfs.
3.1. Generalitats.
Els lagomorfs constitueixen un ordre de mamífers relativament restringit, format per 58
espècies distribuïdes per tot el món, ja sigui de forma natural o perquè han estat introduïdes
per l'home. Totes aquestes espècies es reparteixen en 11 gèneres agrupats en dues famílies:
la dels ocotònids (Ochotonidae), que inclou les piques, i la dels lepòrids (Leporidae), que
comprèn els conills i les llebres. Tanmateix, en les classificacions antigues els lagomorfs
s’atribuïen a l'ordre dels rosegadors, fins que van ser separats d'aquests per GIDLEY (1912).
Les diferències es troben en el nombre d'incisius superiors (dos parells en els lagomorfs i
un parell en els rosegadors), en l'estructura del paladar (més reduït en els lagomorfs), i en
altres caràcters postcranials. Ara bé, les similituds són tals que SIMPSON (1945) va agrupar
els lagomorfs i els rosegadors en el superordre Glires, classificació que encara és utilitzada
per alguns autors (DONARD, 1982; CUENCA, 1990).
Pel que fa a les piques, actualment hi ha 14 espècies repartides en un sol gènere, Ochotona,
habitant boreal de les estepes i altes muntanyes d'Amèrica del Nord i Àsia (DAWSON, 1974;
CUENCA, 1990), mentre que la família dels lepòrids es subdivideix en dos grups sense rang
taxonòmic: les llebres (amb un sol gènere, Lepus) i conills (amb 10 gèneres: Oryctolagus,
Sylvilagus, Pronolagus, Nesolagus, Poelagus, Pentalagus, Romerolagus, Bunolagus,
Brachylagus i Caprolagus). En total existeixen fins a 44 espècies de lepòrids àmpliament
repartides per tot el món (fig. 9; foto 3, 4 i 5).
A la Península Ibèrica hi ha citades 3 espècies de llebre, Lepus granatensis (al sud del riu
Ebre), Lepus castroviejoi (a la Serralada Cantàbrica) i Lepus europaeus (nord del riu Ebre)
(PALACIOS, 1983), així com la única espècie de conill europeu, Oryctolagus cuniculus, la
qual podria tenir el seu origen en la península. A la resta d'Europa occidental cal afegir la
presència d'una altra llebre, Lepus timidus, que viu en les regions muntanyoses i més fredes
(DONARD, 1982).
Tots els lagomorfs presenten característiques morfològiques i estructurals comunes, com
un pelatge generalment llarg i tou, unes extremitats totalment cobertes de pèl, unes orelles
llargues, uns ulls situats en una posició alta, la qual cosa els ofereix un camp de visió molt
ampli, i un nas tallat en nostrils que es poden obrir i tancar amb un replec del tegument.

28
El crani presenta una morfologia lleugerament arrodonida i les bulles timpàniques estan
molt ben desenvolupades. L'esquelet postcranial dels lagomorfs, com la majoria dels seus
caràcters, ha variat poc al llarg del temps, estabilitat que es veu reflectida en la baixa
diversificació que ha sofert l'ordre des de la seva aparició. Són animals mamífers terrestres
plantígrads, amb unes extremitats relativament curtes i desiguals, que mostren adaptacions
saltadores, corredores o cavadores (CUENCA, 1990).
Pel que fa a l'alimentació, els lagomorfs són herbívors, ja que s'alimenten de gramínies,
fulles, escorces, llavors i arrels, tot i que en certs casos s'ha constatat que algunes espècies
introdueixen insectes a la seva dieta. La dentició dels adults es caracteritza per l’existència
de dos parells d’incisius superiors i un parell d’inferiors, amb un llarg diastema que els
separa de la dentició jugal, la qual està formada per premolars i molars de creixement
continu (hipsodòncia). En els molars superiors, l'alçada de la corona és major en la cara
lingual que en la labial, motiu pel qual adquireixen una curvatura força important,
especialment accentuada pel desgast (TOBIEN, 1976). D'altra banda, els incisius es toquen
quan els molars estan en oclusió, fet que permet que les diferents accions de rosegar i
mastegar es puguin produir simultàniament. El moviment efectuat durant el procés de
masticació és bàsicament transversal, ja que les mandíbules són de menor amplada que el
maxil·lar, de manera que les sèries dentaries no coincideixen en posició oclusal. Finalment,
la fórmula dentaria dels conills i llebres és I2/1 C0/0 P3/2 M3/3, mentre que les piques
tenen un molar superior de menys a cada costat (fig.8) (LÓPEZ, 1989; CUENCA, 1990).
Figura 8. Dentició de Oryctolagus laynensis, del Pliocè de Soria. A: Sèrie superior dreta; B, C: Sèrie juvenil inferior dreta; D: Sèrie inferior esquerra (LÓPEZ, 1977).

29
Els lagomorfs constitueixen la base de l'alimentació d'un gran nombre de depredadors i, en
moltes espècies actuals, la depredació, les malalties i els factors climàtics són els agents
primaris d'unes taxes de mortalitat que poden afectar fins el 90% dels juvenils cada any.
No obstant això, aquesta elevada taxa de mortalitat és contrarestada per la gran capacitat
reproductora dels lagomorfs. Així, algunes espècies arriben a la maduresa sexual ben aviat
(el conill als 3 mesos d'edat) i el període de gestació sol molt curt: 40 dies per les espècies
del gènere Lepus i uns 30 dies per als altres representants de l'ordre (DAWSON, 1974).
Les restes fòssils indiscutibles més antigues de lagomorfs s'han identificat a l'Eocè superior
d'Àsia i Amèrica del Nord. Tanmateix, les restes asiàtiques són lleugerament més antigues
que les americanes, fet que sembla suggerir un origen asiàtic del grup. Ara bé, els lepòrids
típics es desenvoluparen a Amèrica del Nord durant l'Oligocè i el Miocè inferior, i no es
troben al vell continent fins a finals del Miocè. Efectivament, no habitaren Euràsia fins el
Pliocè, època en què van desenvolupar formes semblants a les llebres i conills actuals, que
van colonitzar ràpidament la majoria de continents. D'altra banda, les piques aparegueren a
Àsia a mitjans de l'Oligocè i es van dispersar per Amèrica del Nord i Europa durant el
Pliocè (DAWSON, 1974; LÓPEZ, 1989; CUENCA, 1990).
3.2. Anatomia, etologia i hàbitat.
Les piques són lagomorfs petits, amb orelles arrodonides i relativament grans, potes curtes
i cua curta i poc peluda. Mesuren fins 20 cm i assoleixen un pes que no passa dels 300 gr.
Habiten zones muntanyoses, estepàries i semidesèrtiques de l'oest d'Amèrica del Nord, l'est
d'Europa i àrees d'Àsia al nord de l'Himàlaia. Viuen habitualment entre 1.800 i 4.000 m
d'altitud en les latituds inferiors a 60º N, mentre que en latituds superiors baixen fins la
zona litoral (fig.9) (BONHOTE, 1904; HOWELL, 1924; MALATESTA, 1947; LESCA, 1975).
No obstant, aquestes dades no poden extrapolar-se directament al registre fòssil doncs, tot i
que la primera fase d'immigració del grup des d'Àsia es situa en un període de refredament
(Oligocè Mitjà), la seva persistència a Europa durant en Miocè va associada amb un clima
subtropical humit i un medi forestal estès per tot el continent. Però la fase de refredament
del Pliocè va empènyer els ocotònids, reduïts a un sol gènere, al litoral Mediterrani, de
manera que en el Pleistocè inferior apareixen confinats a la franja costanera de la Península
Ibèrica. Aquest fet s'atribueix bàsicament a les causes climàtiques, però és molt probable
que existeixin altres raons de tipus ecològic, com la competència, que hagin impedit als
ocotònids habitar localitats amb clima temperat més allunyades de la costa (LÓPEZ, 2001).

30
Figura 9. Distribució geogràfica dels lagomorfs actuals. Recentment, el conill europeu (Oryctolagus cuniculus) ha estat introduït per l'home a Europa Central, Austràlia, Nova
Zelanda i diverses illes del Mediterrani i Atlàntic (LÓPEZ, 1989).
Foto 3. Ochotona sp. Fotografia de Renee Franken.

31
Els conills i les llebres tenen el cos més allargat i les potes més desenvolupades i adaptades
a córrer per terrenys més o menys oberts; les seves orelles són llargues i la cua és curta i
estarrufada. Ocupen un ampli ventall d'hàbitats, ja que es troben des dels terrenys freds i
nevats de l'Àrtic fins zones semidesèrtiques, passant per les extensions herbàcies, d'Europa,
el bosc tropical d'Amèrica del Sud i els boscos muntanyosos de Sumatra (fig.9).
Foto 4. Oryctolagus cuniculus. Fotografia de Herminio Muñiz.
Foto 5. Lepus sp. Fotografia de J. Aragonés.

32
Ara bé, com ja s'ha comentat, la taxonomia dins de la família Leporidae encara té molts
punts per aclarir, tot i que la distinció entre Lepus i Oryctolagus està clarament recolzada
per nombrosos criteris genètics, etològics i morfològics. Així, el primer gènere posseeix 48
cromosomes, mentre que el segon en té 44, fet que impedeix l'existència d'híbrids entre els
dos gèneres (ROBINSON et al., 1980, 2005). D'altra banda, les llebres i els conills presenten
certs comportaments diferencials. Les cries de llebre neixen amb pèl i amb els ulls oberts,
de manera que al cap de pocs minuts ja poden començar a córrer. El part es realitza
directament sobre el terreny després d'uns 40 dies de gestació. En canvi, les cries de conill
neixen cegues i desvalgudes dins d'un niu preparat expressament per la mare, després d'un
període de gestació d'uns 31 dies. El seu elevat potencial reproductor provoca que puguin
assolir grans densitats poblacionals i els confereix una enorme capacitat colonitzadora.
Pel que fa al comportament i hàbitat, les llebres són animals més aviat solitaris que habiten
pràcticament tots els tipus de biòtops i, tot i que prefereixen terrenys plans i oberts, se'ls
pot trobar tant en zones boscoses de caducifolis i pinedes, com en prats alpins, fins una
altitud d'uns 3.700 m (VAN DER BRINK, 1967; DAWSON, 1974). Contràriament, els conills
són més sedentaris i viuen en societats jerarquitzades (LOMBARDI et al., 2003). Poden
ocupar biòtops diversos, tot i que per fixar la seva residència requereixen sòls sorrencs per
poder excavar els seus caus. Generalment prefereixen terrenys oberts, com garrigues, si bé
també poden habitar boscos amb clarianes, sense pujar gaire en altitud (900 metres com a
màxim) (DAWSON, 1974; LÓPEZ, 1989).
Des del punt de vista morfològic, les llebres són més grans i tenen les orelles més llargues
que el cap, mentre que els conills són més petits i arrodonits i tenen les orelles més curtes.
D'altra banda, les llebres tenen les potes més llargues, adaptades a la carrera, la qual cosa
els hi proporciona una gran velocitat de desplaçament, superant fins i tot els 50 km/h.
Les diferències osteològiques més importants, que constitueixen les claus de determinació
pels paleontòlegs, són les següents (fig.10a i 10b):
- La protuberància occipital és més ample en la llebre que en el conill (FAURE, 1992).
- L'interparietal només existeix en el conill (LESBRE, 1892).
- El marge caudal del parietal és còncau en el conill i convex en la llebre (FAURE, 1992).
- El procés zigomàtic de l'os frontal o apòfisi supra-orbitària és més ample i divergent en la
llebre que en el conill (FAURE, 1992).
- L'os frontal és més ample en la llebre que en el conill (FAURE, 1992).

33
- L'os nasal és llarg i estret en el conill, i ample i curt en la llebre (FAURE, 1992).
- L'obertura de les coanes és estreta en la regió esfenoidal en el conill, contràriament a la
llebre, en la qual és ample i no presenta cap estretament (FAURE, 1992).
- El procés palatí del maxil·lar és més estret transversalment i més llarg longitudinalment
en el conill que en la llebre (FAURE, 1992).
- El diastema de la mandíbula és més llarg i estret en la llebre (FAURE, 1992).
- El foramen mentonià és petit i allunyat de la sèrie dentaria inferior en la llebre, i més gran
i proper al P3 (tercer premolar inferior) en el conill (FAURE, 1992).
- El lòbul mesial de l'incisiu superior és més arrodonit en el conill (DONARD, 1982).
- En el P3, els anterocònids són de mida i morfologia similar en el conill, mentre que són
asimètrics en la llebre (DONARD, 1982). En les llebres l'anterocònid lingual sol ser més
petit que el protocònid i l'anteroflèxid és menys profund que en els conills (LÓPEZ, 1989).
- El procés hamatus de l'escàpula és llarg en el conill i curt en la llebre (LESBRE, 1892).
- El solc intertubercular de l'húmer presenta una amplada constant en la llebre mentre que
decreix distalment en el conill. A més a més, el tubercle medial de la tròclea distal és molt
més marcat en el conill, mentre que la tròclea és més gruixuda en la llebre.
- La cresta deltoidea de l'húmer s'atenua progressivament en la llebre, mentre que en el
conill s'interrrumpeix sobtadament (MARZIN, 1988).
- En vista medial, la diàfisi del radi és menys corbada en la llebre (MARZIN, 1988).
- La diàfisi de l'ulna, en la cara cranial, presenta un gruix constant en el conill mentre que
decreix progressivament en el sentit proximo-distal en la llebre. Cal afegir que en el conill
la cresta lateral del marge proximal de la tuberositat olecraniana és menor que la medial;
en canvi, en la llebre són de mida similar (MARZIN, 1988).
- En vista lateral, el marge de l'ala ilíaca del coxal forma un angle més arrodonit en la
llebre que en el conill (CALLOU , 1997).
- En vista cranial, el coll del fèmur és molt curt en el conill, mentre que en la llebre és més
allargat i el marge proximal és més rectilini (PELTIER, 1985). En la part distal, la tròclea del
fèmur del conill presenta dues crestes iguals; en la llebre, la cresta medial és més curta.
- Els dos tubercles que formen l'eminentia intercondylaris de la tíbia són més prominents
en la llebre. En vista caudal, el marge distal de la superfície articular descriu en el conill un
solc arrodonit regularment, mentre que en la llebre té un petit sobresortint (CALLOU , 1997).
- En vista proximal, la tròclea de l'astràgal és molt quadrada en la llebre i més rectangular
en el conill (CALLOU , 1997).

34
Figura 10a. Elements cranials. Diferències entre el crani (A), mandíbula (B) i P3 (C) del conill europeu (Oryctolagus cuniculus) i la llebre (Lepus) (CALLOU , 1997).
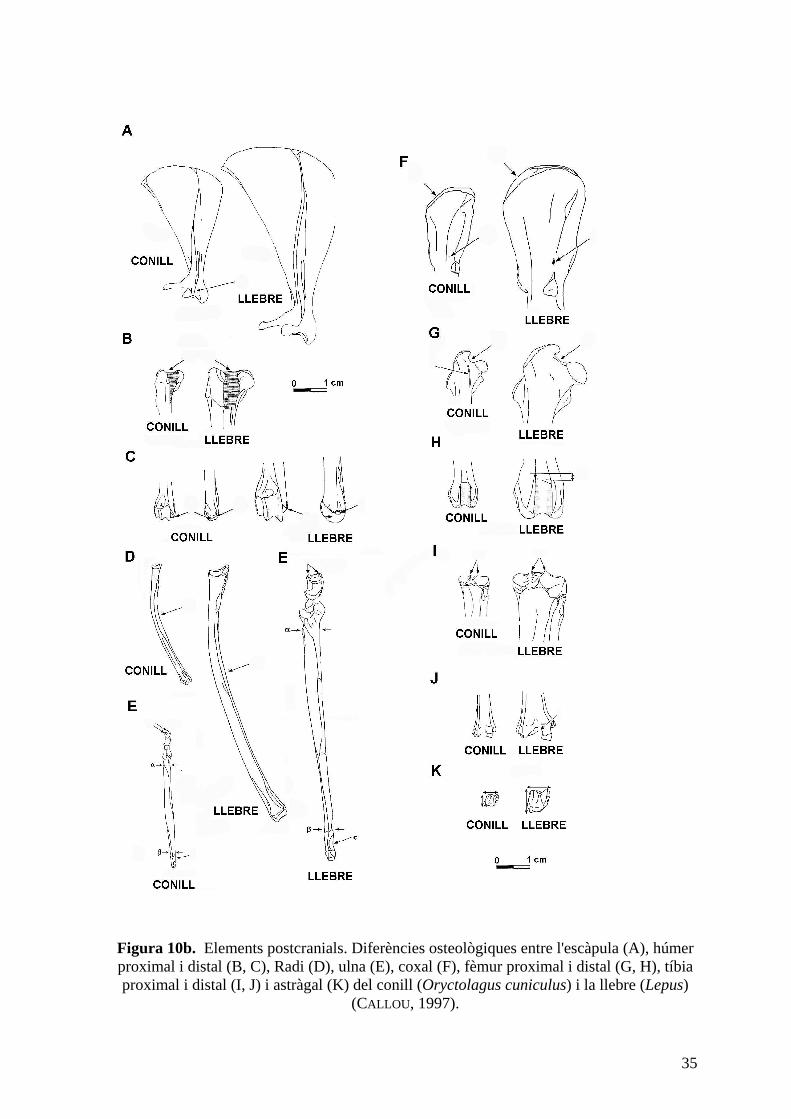
35
Figura 10b. Elements postcranials. Diferències osteològiques entre l'escàpula (A), húmer proximal i distal (B, C), Radi (D), ulna (E), coxal (F), fèmur proximal i distal (G, H), tíbia proximal i distal (I, J) i astràgal (K) del conill (Oryctolagus cuniculus) i la llebre (Lepus)
(CALLOU , 1997).

36
4. Sistemàtica.
Classe Mammalia LINNAEUS, 1758
Ordre Lagomorpha BRANDT, 1855
Família Ochotonidae THOMAS, 1897
Gènere Prolagus POMEL, 1853
Prolagus sp.
1975 Prolagus cf. calpensis MAJOR, 1905. López & Thaler, pp. 51, lam. 2.
1984 Prolagus cf. calpensis MAJOR, 1905. Gil & Sesé, pp. 88, fig. 2.
1987 Prolagus cf. calpensis MAJOR, 1905. Agustí et al., pp. 104, fig.7.
1989 Prolagus calpensis MAJOR, 1905. López, pp.147-148.
1989 Prolagus cf. calpensis MAJOR, 1905. López, pp.149-156, fig. 32, 34.
Material: Es disposa únicament d'un P4 de Barranco León 5 (làm. V, fig. m).
Descripció: Es tracta d'una peça de mida petita comparada amb els lepòrids procedents del
mateix jaciment, amb unes mesures màximes de 1,32 mm de longitud (Y) i 2,82 mm
d'amplada (X). Presenta un hipoflex relativament profund i dues fossetes desiguals, essent
la mesofosseta de menor mida i trobant-se rodejada per la parafosseta.
Repartició: Aquest gènere s’ha identificat en materials del Miocè inferior d’Alemanya,
França, Espanya i Portugal; Miocè mitjà d’Alemanya, França, Espanya i Turquia; Miocè
superior de França, Espanya, Portugal, Àustria, Grècia, Hongria i Turquia; Plio-Pleistocè
d’Alemanya, França, Hongria, Itàlia, Espanya, Marroc, Algèria i Tunis; Holocè de Còrsega
i Sardenya (LÓPEZ, 1989).
Discussió: El fet de disposar d'una sola peça dental i que, a més a més, aquesta no sigui
clau per la diagnosi, no permet atribuir-la a una espècie concreta. Tanmateix, és
relativament probable que es tracti de Prolagus calpensis, doncs aquest ocotònid apareix
associat a Oryctolagus en diversos jaciments espanyols del Pleistocè inferior, com Bagur 2
(Girona) (LÓPEZ & THALER, 1975) i Venta Micena (Granada) (AGUSTÍ et al., 1987).

37
Classe Mammalia LINNAEUS, 1758
Ordre Lagomorpha BRANDT, 1855
Família Leporidae GRAY, 1821
Gènere Oryctolagus LILLJEBORG, 1874
Oryctolagus cf. cuniculus (LINNAEUS, 1758)
1982 Oryctolagus cuniculus (LINNAEUS, 1758). Donard, pp. 19-25, lam. 39-42.
1989 Oryctolagus cuniculus (LINNAEUS, 1758). López, pp. 230-233, fig.60.
1997 Oryctolagus cuniculus (LINNAEUS, 1758). Callou, pp. 5-11, fig. 1-7.
2001 Oryctolagus cuniculus (LINNAEUS, 1758). Guennouni, pp. 28-34, fig. 12-15.
Material: Es disposa únicament de peces dentals, entre les quals hi ha 3 incisius, 8 P2, 15
P3, 46 molariformes superiors (P3-P4-M1-M2-M3) i 30 inferiors (P4-M1-M2-M3) procedents
del nivell D de Barranco León, i 14 incisius, 23 P2, 26 P3, 91 molariformes superiors y 28
inferiors dels nivells mitjà i superior de Fuente Nueva 3 (làm. V i VI). El nombre mínim
d'individus representats en la mostra que s'ha analitzat seria 9 a Barranco León 5 i 15 a
Fuente Nueva 3. En ambdós casos la peça majoritària és el P3 dret.
Descripció: Els incisius superiors (I1) presenten un lòbul mesial arrodonit, caràcter que
s'acostuma a associar al gènere Oryctolagus, tot i que en alguns exemplars és més aviat
punxegut. La morfologia del P2 és relativament variable, i la seva mida oscil·la entre 1,16-
1,44 mm de longitud i 2,42-2,99 mm d'amplada a Barranco León, i entre 1,19-1,54 mm de
longitud i 2,20-2,97 mm d'amplada a Fuente Nueva 3. El lagicon és de mida similar a
l'hipercon mesial i el paraflex penetra obliquament fins la meitat de la dent i pot presentar
alguna irregularitat en el seu marge anterior. El mesoflex forma un sinforme ben marcat i
l'hipoflex es troba netament definit i, tot i que en alguns exemplars és tan profund com el
mesoflex, acostuma a ser més curt. La resta de la dentició superior pot caracteritzar-se per
presentar un hipoflex arrissat que penetra una mica més que la meitat de l'amplada de la
dent; en general, el P3 i el P4 són els que desenvolupen més ondulacions en els marges
anterior i posterior de l'hipoflex. La dentició inferior només ofereix particularitats dignes
de menció a nivell del P3. Els anterocònids són de mida similar, tot i que el lingual pot ser
lleugerament més petit. L'anterocònid labial és allargat i força comprimit en els individus

38
joves mentre que en els adults presenta una morfologia més el·lipsoidal o bé rectangular.
L'anteroflèxid és recte i de profunditat variable i, en general, presenta marges convergents
cap a l'interior de la dent. El protoflèxid és ample i llis, formant un angle de 90º o superior,
i el protocònid és en general menys voluminós que l'anterocònid lingual, però de mida
similar a l'hipocònid. L'hipoflèxid és profund, travessant gairebé tota la dent, i el seu marge
anterior, a part de les dues inflexions centrals que corresponen a la antiga connexió entre el
trigònid i el talònid, pot ser llis o presentar algunes ondulacions. En canvi, el seu marge
posterior no acostuma a tenir irregularitats. D'altra banda, en la majoria dels exemplars no
existeix un paraflèxid ben marcat, sinó que es redueix a una suau inflexió. Finalment, la
mida del P3 oscil·la entre 1,78-3,1 mm de longitud i 1,82-2,8 mm d'amplada a Barranco
León i entre 2,02-3,1 mm de longitud i 1,99-3,2 mm d'amplada a Fuente Nueva 3.
Repartició: Aquesta espècie s'ha identificat en materials del Pleistocè mitjà de Cúllar de
Baza (Granada, España) (LÓPEZ, 1989), Lazaret (Alpes - Maritimes, França) (GUENNOUNI,
2001), Terra Amata (Alpes - Maritimes, França) (GUENNOUNI, 2001), Orgnac 3 (Ardèche,
França) (GUENNOUNI, 2001) i La Fage (Corrèze, França) (DONARD, 1982); Pleistocè
superior de diversos jaciments de la Península Ibèrica, França i Itàlia, com ara Zafarraya
(Màlaga, Espanya) (GUENNOUNI, 2001), Jaurens (Corrèze, França) (LÓPEZ, 1980), Grotte
de l'Hortus (Hérault, França) (DONARD, 1982) i Arma Manie (Liguria, Itàlia) (GUENNOUNI,
2001). En l'actualitat Oryctolagus cuniculus ha estat introduït a Europa central, Austràlia,
Nova Zelanda i diverses illes del Mediterrani i Atlàntic (LÓPEZ, 1989).
Discussió: El fet de no disposar de material cranial i postcranial dificulta la tasca d'atribuir
aquestes restes a una espècie concreta. Tanmateix, la morfologia de les dents trobades als
jaciments de Barranco León 5 i Fuente Nueva 3 és indistingible de la dels conills actuals
(làm. I), tot i que en l'actual estat de coneixements també es podria relacionar amb
Oryctolagus laynensis o Oryctolagus lacosti, l'enigmàtic lepòrid del Plio-Pleistocè. Ara bé,
com que es requereix una important revisió del material d'O. laynensis i O. lacosti, sembla
més adequat assignar els fòssils provisionalment a Oryctolagus cf. cuniculus. Certament,
O. laynensis i O. lacosti estan pobrament representats i no es descarta que en realitat es
puguin considerar sinònims. Cal afegir que l'holotip de Oryctolagus laynensis es troba en
parador desconegut i que l'espècie va ser descrita, sorprenentment, sense tenir en compte el
material original d'Oryctolagus lacosti de Perrier, el qual es figura per primera vegada en
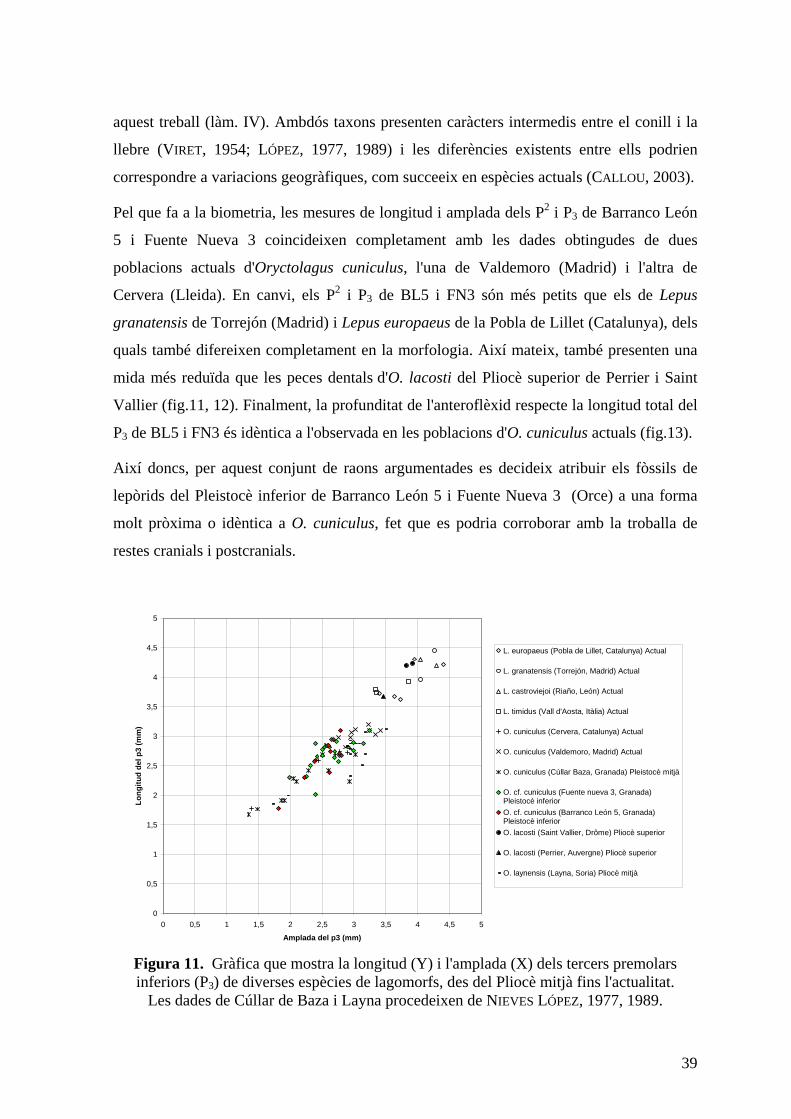
39
aquest treball (làm. IV). Ambdós taxons presenten caràcters intermedis entre el conill i la
llebre (VIRET, 1954; LÓPEZ, 1977, 1989) i les diferències existents entre ells podrien
correspondre a variacions geogràfiques, com succeeix en espècies actuals (CALLOU , 2003).
Pel que fa a la biometria, les mesures de longitud i amplada dels P2 i P3 de Barranco León
5 i Fuente Nueva 3 coincideixen completament amb les dades obtingudes de dues
poblacions actuals d'Oryctolagus cuniculus, l'una de Valdemoro (Madrid) i l'altra de
Cervera (Lleida). En canvi, els P2 i P3 de BL5 i FN3 són més petits que els de Lepus
granatensis de Torrejón (Madrid) i Lepus europaeus de la Pobla de Lillet (Catalunya), dels
quals també difereixen completament en la morfologia. Així mateix, també presenten una
mida més reduïda que les peces dentals d'O. lacosti del Pliocè superior de Perrier i Saint
Vallier (fig.11, 12). Finalment, la profunditat de l'anteroflèxid respecte la longitud total del
P3 de BL5 i FN3 és idèntica a l'observada en les poblacions d'O. cuniculus actuals (fig.13).
Així doncs, per aquest conjunt de raons argumentades es decideix atribuir els fòssils de
lepòrids del Pleistocè inferior de Barranco León 5 i Fuente Nueva 3 (Orce) a una forma
molt pròxima o idèntica a O. cuniculus, fet que es podria corroborar amb la troballa de
restes cranials i postcranials.
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
5
0 0,5 1 1,5 2 2,5 3 3,5 4 4,5 5
Amplada del p3 (mm)
Long
itud
del p
3 (m
m)
L. europaeus (Pobla de Lillet, Catalunya) Actual
L. granatensis (Torrejón, Madrid) Actual
L. castroviejoi (Riaño, León) Actual
L. timidus (Vall d'Aosta, Itàlia) Actual
O. cuniculus (Cervera, Catalunya) Actual
O. cuniculus (Valdemoro, Madrid) Actual
O. cuniculus (Cúllar Baza, Granada) Pleistocè mitjà
O. cf. cuniculus (Fuente nueva 3, Granada)Pleistocè inferior
O. cf. cuniculus (Barranco León 5, Granada)Pleistocè inferior
O. lacosti (Saint Vallier, Drôme) Pliocè superior
O. lacosti (Perrier, Auvergne) Pliocè superior
O. laynensis (Layna, Soria) Pliocè mitjà
Figura 11. Gràfica que mostra la longitud (Y) i l'amplada (X) dels tercers premolars inferiors (P3) de diverses espècies de lagomorfs, des del Pliocè mitjà fins l'actualitat.
Les dades de Cúllar de Baza i Layna procedeixen de NIEVES LÓPEZ, 1977, 1989.

40
0
0,5
1
1,5
2
2,5
3
0 0,5 1 1,5 2 2,5 3 3,5 4 4,5 5
Amplada del P2 (mm)
Long
itud
del P
2 (m
m)
L. europaeus (Pobla de Lillet, Catalunya) Actual
L. granatensis (Torrejón, Madrid) Actual
L. castroviejoi (Riaño, León) Actual
L. timidus (Vall d'Aosta, Itàlia) Actual
O. cuniculus (Cervera, Catalunya) Actual
O. cuniculus (Valdemoro, Madrid) Actual
O. cuniculus (Cúllar Baza, Granada) Pleistocè mitjà
O. cf. cuniculus (Fuente Nueva 3, Granada)Pleistocè inferior
O. cf. cuniculus (Barranco León 5, Granada)Pleistocè inferior
O. lacosti (Saint Vallier, Drôme) Pliocè superior
O. lacosti (Perrier, Auvergne) Pliocè superior
O. laynensis (Layna, Soria) Pliocè mitjà
Figura 12. Gràfica que mostra la longitud (Y) i l'amplada (X) del segons premolars superiors (P2) de diverses espècies de lagomorfs, des del Pliocè mitjà fins l'actualitat.
Les dades de Cúllar de Baza i Layna procedeixen de NIEVES LÓPEZ, 1977, 1989.
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
0 0,5 1 1,5 2 2,5 3 3,5 4 4,5 5
Longitud del p3 (mm)
Pro
fund
itat d
e l'a
nter
oflè
xid
(mm
)
L. europaeus (Pobla de Lillet, Catalunya) Actual
L. granatensis (Torrejón, Madrid) Actual
L. castroviejoi (Riaño, León) Actual
L. timidus (Vall d'Aosta, Itàlia) Actual
O. cuniculus (Cervera, Catalunya) Actual
O. cuniculus (Valdemoro, Madrid) Actual
O. cuniculus (Cúllar Baza, Granada) Pleistocè mitjà
O. cf. cuniculus (Fuente Nueva 3, Granada)Pleistocè inferior
O. cf. cuniculus (Barranco León 5, Granada)Pleistocè inferior
T. dumitrescuae (Malusteni, Rumania) Pliocèsuperior
O. lacosti (Perrier, Auvergne) Pliocè superior
O. laynensis (Layna, Soria) Pliocè mitjà
Figura 13. Gràfica que mostra la profunditat de l'anteroflèxid (Y) i la longitud (X) del tercer premolar inferior (P3) de diversos lepòrids, des del Pliocè mitjà fins l'actualitat.
Les dades de Cúllar de Baza i Layna procedeixen de NIEVES LÓPEZ, 1977, 1989.

41
Classe Mammalia LINNAEUS, 1758
Ordre Lagomorpha BRANDT, 1855
Família Leporidae GRAY, 1821
Gènere Oryctolagus LILLJEBORG, 1874
Oryctolagus cf. lacosti (POMMEL, 1853)
1853 Lepus lacosti POMMEL, 1853. Pommel, pp. 1-193.
1943 Oryctolagus cf. lacosti (POMMEL, 1853). Schaub, pp. 271-272.
1954 Oryctolagus lacosti (POMMEL, 1853). Viret, pp. 101-104, lam. 17.
1973 Lepus cf. terraerubrae KRETZOI, 1956. Meulen, pp. 26-28, fig. 8.
1976 Oryctolagus cf. lacosti (POMMEL, 1853). López, pp. 52-53, lam. 2.
1987 Oryctolagus cf. lacosti (POMMEL, 1853). Agustí et al., pp. 104, fig. 7.
1987 Lepus cf. terraerubrae KRETZOI, 1956. Fladerer, pp. 131-132, fig. 4.
1989 Oryctolagus lacosti (POMMEL, 1853). López, pp. 221-226, fig. 58.
1997 Oryctolagus burgi NOCCHI & SALA , 1997. Nocchi & Sala, pp. 19-38, fig. 1, 2.
2000 Oryctolagus cf. lacosti (POMMEL, 1853). Chaline et al., pp. 100-103, fig. 3.
2004 Oryctolagus lacosti (POMMEL, 1853). Martín & Mein, pp. 119-121, fig. 5.
Material: Es disposa de 188 dents (entre les quals hi ha 11 incisius superiors, 33 P2, 83
molariformes superiors, 4 incisius inferiors, 34 P3 i 23 molariformes inferiors), 2 calcanis
juvenils, 7 astràgals, 3 fragments distals d'húmer, 1 fragment d'ulna i 1 fragment de paladar
que inclou part del maxil·lar dret i del palatí. Tot aquest material prové de les capes A i B
del nivell 9 de Trinchera Elefante (TE9) així com del sondeig del 98 (làm. VII, VIII i IX).
El nombre mínim d'individus representats en la mostra que s'ha analitzat seria de 18.
Descripció: El fragment de paladar presenta una morfologia més similar a Oryctolagus
que a Lepus ja que el palatí és força llarg i s'intueix que les coanes són més aviat estretes.
Els incisius superiors tenen un lòbul mesial arrodonit, tot i que en alguns exemplars és
relativament més punxegut, i un solc en forma de V. Tanmateix, un dels incisius procedent
de la capa A és major que la resta i el seu solc anterior té forma de U. La morfologia del P2
és relativament variable, i la seva mida oscil·la entre 1,33-1,85 mm de longitud i 2,73-3,95
mm d'amplada. La majoria de P2 presenta una morfologia el·lipsoidal, tot i que hi ha alguns

42
exemplars més subovals i altres més triangulars. El lagicon sol ser de mida major o similar
a l'hipercon mesial, excepte en 2 exemplars, i el paraflex és el sinforme més profund,
penetrant fins la meitat de la longitud de la dent. En alguns exemplars en paraflex presenta
alguna irregularitat en el seu marge anterior. El mesoflex sol ser més aviat dèbil, en alguns
casos fins i tot gairebé absent, i l'hipoflex acostuma a ser més profund. Els molariformes
superiors no ofereixen tanta informació sistemàtica com els P2 i P3. No obstant això, és
interessant assenyalar que presenten un hipoflex que penetra fins unes 2/3 parts de la dent,
tot i que aquest és un caràcter que varia amb l'ontogènia. D'altra banda, la crenulació de
l'hipoflex augmenta del P3 al M2. Així, els P3 presenten entre 5 i 7 crenulacions en el marge
anterior i algunes irregularitats en el posterior, mentre que els M2 solen tenir uns marges
força més llisos. Alguns P3 posseeixen una mena de “ganxo” en la part lingual del marge
posterior de l'hipoflex. Finalment, cal destacar que alguns d'aquests molariformes superiors
presenten signes evidents de digestió. D'altra banda, la dentició inferior només ofereix
particularitats diagnostiques dignes de menció a nivell del P3. Aquesta peça dental mostra
una variabilitat extraordinària, podent-se classificar en 3 morfotips. El primer morfotip,
que és el més abundant, presenta una superfície oclusal relativament rectangular, amb uns
anterocònids lleugerament asimètrics. L'anterocònid labial és rectangular o bé el·lipsoidal,
mentre que el lingual acostuma a ser més petit i presenta una morfologia afilada, excepte
en un exemplar en què és molt arrodonit. L'anteroflèxid és recte i de profunditat variable i
els seus marges poden ser paral·lels o convergents cap a l'interior de la dent. El protoflèxid
és generalment llis, tot i que pot posseir una lleugera irregularitat en el marge anterior, i
forma un angle de 90º o inferior. El protocònid és similar o menor que l'anterocònid labial.
L'hipoflèxid és molt profund, travessant pràcticament tota la dent. El seu marge anterior, a
part de les dues inflexions centrals, pot ser llis o presentar algunes ondulacions. En canvi,
el seu marge posterior sol ser llis. No existeix paraflèxid ben marcat en cap dels exemplars,
sent el marge lingual arrodonit o sinuós. El segon morfotip que s'ha identificat correspon a
alguns P3 amb una morfologia molt arrodonida, amb uns anterocònids reduïts respecte el
trigònid i un anteroflèxid curt o bé inexistent. L'anterocònid labial és allargat i més petit
que el protocònid, mentre que el lingual és molt petit i afilat. El protoflèxid és d'un angle
variable i pot presentar diverses crenulacions. L'hipoflèxid és profund, llis o bé irregular, i
l'hipocònid és llarg i ample, major que el protocònid. Finalment, el tercer morfotip que s'ha
trobat a TE9 correspon a una minoria de P3 que presenten una superfície oclusal de perfil
subtriangular, amb un anteroflèxid de profunditat variable, un anterocònid labial major que

43
el lingual però similar al protocònid, i un hipoflèxid profund que en un exemplar està partit
en dos per un istme i en un altre s'intueix aquesta partició en un estat ontogenètic diferent.
La mida dels P3 de Trinchera Elefante 9 oscil·la entre 2,85-4,13 mm de longitud (Y) i
2,29-3,91 mm d'amplada (X).
Pel que fa a les restes poscranials, l'ulna presenta un olecranon relativament curt (12,8 mm)
i la cara dorsal o anterior de la diàfisi no és completament arrodonida, sinó que presenta un
acanalament poc marcat. Les crestes lateral i medial de la tuberositat olecraniana són de
mida similar. Els fragments distals d'húmer tenen una amplada que varia entre 9,5 i 10, 21
mm, amb una tròclea d'entre 4,70 i 4,90 mm. El tubercle medial de la tròclea és ample i la
cresta deltoidea s'atenua progressivament, és a dir, no forma un angle de 90º, sinó inferior.
Els dos calcanis que s'han recuperat pertanyen a individus juvenils, ja que l'os és porós i les
diàfisis no estan ben soldades, pel que el seu valor diagnòstic és restringit. La cara dorsal
és aplanada, el sustentaculum és arrodonit i la faceta medial és ampla i robusta. La seva
longitud és de 18,8 mm i 21,9 mm. Finalment, els astràgals, en vista proximal, presenten
una tròclea força rectangular, excepte en 2 exemplars, en els que és molt més quadrada.
Les seves dimensions oscil·len entre 12,0-14,28 mm de longitud, 5,65-6,91 mm d'amplada,
i 5,87-7,49 mm de gruix.
Repartició: La distribució geogràfica del problemàtic Oryctolagus lacosti i associacions
afins és típicament mediterrània occidental. Així, s'ha identificat en alguns jaciments Plio-
Pleistocens francesos, com ara Perrier (Auvergne) (POMEL, 1853), Senèze (Haute-Loire)
(SCHAUB, 1943), Saint Vallier (Drôme) (VIRET, 1954; MARTÍN & MEIN, 2004) i Montoussé
(Hautes-Pyrénées) (CHALINE et al, 2000) i espanyols. A Catalunya apareix presumiblement
en materials del Pleistocè inferior de l'Escala i Begur 2, i en el Pleistocè mitjà del Carmel
(LÓPEZ, 1977, 1989; LÓPEZ et al., 1976), mentre que al sud de la Península s'ha citat en el
Pleistocè inferior de la Unión (Murcia) (LÓPEZ, 1989) i Venta Micena (Granada) (AGUSTÍ
et al., 1987). Finalment, aquesta espècie també es localitzaria a Trinchera Dolina (Burgos)
(CUENCA et al., 1999). Si Oryctolagus laynensis (LÓPEZ, 1977, 1989), Lepus terraerubrae
(MEULEN, 1973) i Oryctolagus burgi (NOCCHI & SALA , 1997) s'acaben considerant
sinònims d'O. lacosti, caldrà afegir la seva presència en el jaciment del Pliocè mitjà de
Layna (Soria, Espanya), en el del Pleistocè inferior de Monte Peglia (Orvieto, Itàlia) i en el
del Pleistocè mitjà de Valdemino (Savona, Italia).

44
Discussió: La problemàtica principal en l'estudi dels lepòrids de Trinchera Elefante 9 rau
en l'elevadíssima variabilitat morfològica que mostren les seves restes, especialment les
dents que són més usades en el diagnòstic: el P2 i el P3. Moltes d'aquestes peces dentals
presenten un patró molt similar a Oryctolagus laynensis del Pliocè mitjà de Layna (LÓPEZ,
1977), Oryctolagus lacosti del Pliocè superior de Perrier (POMMEL, 1853) i Saint Vallier
(MARTÍN et al., 2004), i Oryctolagus burgi del Pleistocè mitjà de Valdemino (NOCCHI &
SALA , 1997), tot i que no es descarta que aquestes tres espècies siguin en realitat sinònims.
Tanmateix, en el material de TE9 hi ha algunes dents amb unes morfologies idèntiques a
Lepus terraerubrae del Pleistocè Inferior de Monte Peglia (MEULEN, 1973). No obstant,
s'ha de ser molt prudent en realitzar aquestes comparacions i en confirmar l'existència de
Lepus terraerubrae, ja que aquest taxó està exclusivament descrit amb unes poques restes
dentals aïllades, les quals podrien correspondre a una espècie amb una elevada variabilitat,
com la de TE9. De fet, cal més material cranial i postcranial per justificar la coexistència
d'Oryctolagus i Lepus durant el Plio-Pleistocè europeu.
D'altra banda, alguns P2 i P3 de TE9 recorden sorprenentment a les actuals Lepus timidus i
Lepus castroviejoi (làm. II i III), però també a Trischizolagus dumitrescuae del Pliocè
d'Europa central (fig.14) (AVERIANOV & TESAKOV, 1995, 1997). A més a més, aquest taxó
presenta una gran variabilitat morfològica, com succeeix en el material de TE9.
Figura 14. Variabilitat dental de Trischizolagus dumitrescuae del Pliocè d'Europa central (AVERIANOV, 1997).

45
Els paràmetres biomètrics de les dents també es situen entre Oryctolagus i Lepus, tant en la
mida com en la profunditat de l'anteroflèxid, i són força similars a T. dumitrescuae del
jaciment de Malusteni-Beresti (Rumania) (RADULESCO & SAMSON, 1967) (fig. 15, 16 i 17).
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
5
0 0,5 1 1,5 2 2,5 3 3,5 4 4,5 5
Amplada del p3 (mm)
Long
itud
del p
3 (m
m)
L. europaeus (Pobla de Lillet, Catalunya) Actual
L. granatensis (Torrejón, Madrid) Actual
L. castroviejoi (Riaño, León) Actual
L. timidus (Vall d'Aosta, Itàlia) Actual
O. cuniculus (Cervera, Catalunya) Actual
O. cuniculus (Valdemoro, Madrid) Actual
O. cuniculus (Cúllar Baza, Granada) Pleistocè mitjà
O. cf. cuniculus (Barranco León 5, Granada)Pleistocè inferior
O. cf. lacosti (Trinchera Elefante 9, Burgos)Pleistocè inferior
O. lacosti (Perrier, Auvergne) Pliocè superior
T. dumitrescuae (Malusteni, Rumania) Pliocèsuperior
O. laynensis (Layna, Soria) Pliocè mitjà
Figura 15. Gràfica que mostra la longitud (Y) i l'amplada (X) dels tercers premolars inferiors (P3) de diverses espècies de lagomorfs, des del Pliocè mitjà fins l'actualitat.
0
0,5
1
1,5
2
2,5
3
0 0,5 1 1,5 2 2,5 3 3,5 4 4,5 5
Amplada del P2 (mm)
Long
itud
del P
2 (m
m)
L. europaeus (Pobla de Lillet, Catalunya) Actual
L. granatensis (Torrejón, Madrid) Actual
L. castroviejoi (Riaño, León) Actual
L. timidus (Vall d'Aosta, Itàlia) Actual
O. cuniculus (Cervera, Catalunya) Actual
O. cuniculus (Valdemoro, Madrid) Actual
O. cuniculus (Cúllar Baza, Granada) Pleistocè mitjà
O. cf. cuniculus (Barranco León 5, Granada)Pleistocè inferior
O. cf. lacosti (Trinchera Elefante 9, Burgos)Pleistocè inferior
O. lacosti (Perrier, Auvergne) Pliocè superior
O. laynensis (Layna, Soria) Pliocè mitjà
Figura 16. Gràfica que mostra la longitud (Y) i l'amplada (X) del segons premolars superiors (P2) de diverses espècies de lagomorfs, des del Pliocè mitjà fins l'actualitat.
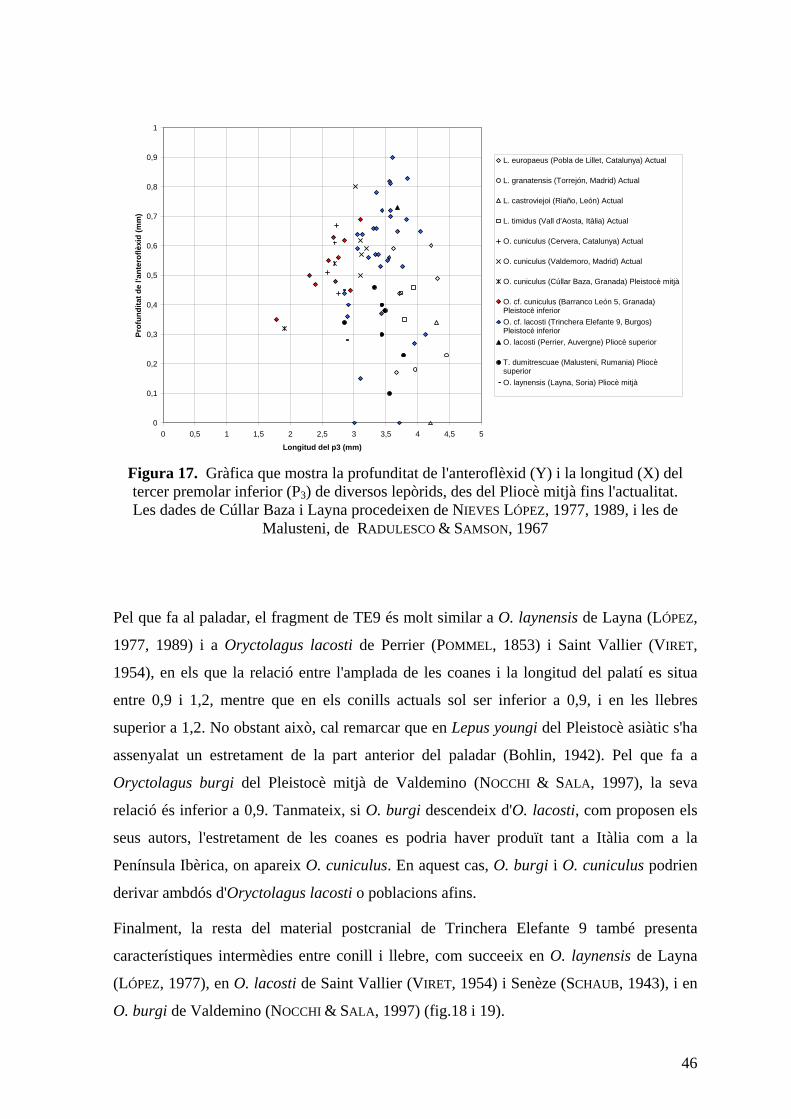
46
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1
0 0,5 1 1,5 2 2,5 3 3,5 4 4,5 5
Longitud del p3 (mm)
Pro
fund
itat d
e l'a
nter
oflè
xid
(mm
)
L. europaeus (Pobla de Lillet, Catalunya) Actual
L. granatensis (Torrejón, Madrid) Actual
L. castroviejoi (Riaño, León) Actual
L. timidus (Vall d'Aosta, Itàlia) Actual
O. cuniculus (Cervera, Catalunya) Actual
O. cuniculus (Valdemoro, Madrid) Actual
O. cuniculus (Cúllar Baza, Granada) Pleistocè mitjà
O. cf. cuniculus (Barranco León 5, Granada)Pleistocè inferior
O. cf. lacosti (Trinchera Elefante 9, Burgos)Pleistocè inferior
O. lacosti (Perrier, Auvergne) Pliocè superior
T. dumitrescuae (Malusteni, Rumania) Pliocèsuperior
O. laynensis (Layna, Soria) Pliocè mitjà
Figura 17. Gràfica que mostra la profunditat de l'anteroflèxid (Y) i la longitud (X) del tercer premolar inferior (P3) de diversos lepòrids, des del Pliocè mitjà fins l'actualitat. Les dades de Cúllar Baza i Layna procedeixen de NIEVES LÓPEZ, 1977, 1989, i les de
Malusteni, de RADULESCO & SAMSON, 1967
Pel que fa al paladar, el fragment de TE9 és molt similar a O. laynensis de Layna (LÓPEZ,
1977, 1989) i a Oryctolagus lacosti de Perrier (POMMEL, 1853) i Saint Vallier (VIRET,
1954), en els que la relació entre l'amplada de les coanes i la longitud del palatí es situa
entre 0,9 i 1,2, mentre que en els conills actuals sol ser inferior a 0,9, i en les llebres
superior a 1,2. No obstant això, cal remarcar que en Lepus youngi del Pleistocè asiàtic s'ha
assenyalat un estretament de la part anterior del paladar (Bohlin, 1942). Pel que fa a
Oryctolagus burgi del Pleistocè mitjà de Valdemino (NOCCHI & SALA , 1997), la seva
relació és inferior a 0,9. Tanmateix, si O. burgi descendeix d'O. lacosti, com proposen els
seus autors, l'estretament de les coanes es podria haver produït tant a Itàlia com a la
Península Ibèrica, on apareix O. cuniculus. En aquest cas, O. burgi i O. cuniculus podrien
derivar ambdós d'Oryctolagus lacosti o poblacions afins.
Finalment, la resta del material postcranial de Trinchera Elefante 9 també presenta
característiques intermèdies entre conill i llebre, com succeeix en O. laynensis de Layna
(LÓPEZ, 1977), en O. lacosti de Saint Vallier (VIRET, 1954) i Senèze (SCHAUB, 1943), i en
O. burgi de Valdemino (NOCCHI & SALA , 1997) (fig.18 i 19).

47
0
2
4
6
8
10
12
14
16
18
20
0 1 2 3 4 5 6 7 8 9 10
Amplada de l'astràgal (mm)
Long
itud
de l'
astr
àgal
(m
m)
L. europaeus (Pobla de Lillet, Catalunya) Actual
O. cuniculus (Cervera, Catalunya) Actual
O. cuniculus (Valdemoro, Madrid) Actual
O. cf. lacosti (Trinchera Elefante 9, Burgos)Pleistocè inferior
Figura 18. Gràfica que mostra la longitud (Y) i l'amplada (X) dels astràgals de diversos
lepòrids, des del Pliocè mitjà fins l'actualitat.
0
1
2
3
4
5
6
7
8
9
10
0 1 2 3 4 5 6 7 8 9 10
Amplada de l'astràgal (mm)
Gru
ix d
e l'a
strà
gal (
mm
) L. europaeus (Pobla de Lillet, Catalunya) Actual
O. cuniculus (Cervera, Catalunya) Actual
O. cuniculus (Valdemoro, Madrid) Actual
O. cf. lacosti (Trinchera Elefante 9, Burgos)Pleistocè inferior
Figura 19. Gràfica que mostra la longitud (Y) i l'amplada (X) dels astràgals de diversos
lepòrids, des del Pliocè mitjà fins l'actualitat.

48
En definitiva, la similitud dels P2 i P3 de Trinchera Elefante 9, així com del material de
Perrier, Saint Vallier i Montoussé 5, amb Trischizolagus, Lepus i Oryctolagus és acusada.
De fet, per AVERIANOV (1995, 1997), T. dumitrescuae podria ser l'ancestre de Lepus,
alhora que per NIEVES LÓPEZ (1977, 1989), Trischizolagus es situaria com el possible
avantpassat d'Oryctolagus. Així doncs, el lepòrid de Trinchera Elefante 9 podria ser un
descendent de Trischizolagus i, vistes la coexistència dels patrons dentals, es podria tractar
d’un ferm candidat a ocupar una posició evolutiva propera a la divergència entre Lepus i
Oryctolagus. En aquest sentit, Lepus hauria aparegut més recentment del que pressuposen
els estudis moleculars, i s’insinua un origen multiregional. A més a més, si es confirma que
el taxó del Plio-Pleistocè europeu és l'ancestre comú de Lepus i Oryctolagus, probablement
s'hauria de classificar com un gènere nou o bé com Lepus lacosti, tal i com va ser descrit
inicialment per POMMEL (1853) a partir del material de Perrier (França).

49
5. Conclusions.
- En aquest treball s’han estudiat per primera vegada les associacions de lagomorfs de
diferents nivells de les localitats de Barranco León i Fuente Nueva (conca de Guadix-Baza,
Granada) i de Trinchera Elefante (Serra d’Atapuerca, Burgos).
- En aquests jaciments s’han identificat els següents taxons de lagomorfs: Prolagus sp. i
Oryctolagus cf. cuniculus a Barranco León 5; Oryctolagus cf. cuniculus a Fuente Nueva 3,
i Oryctolagus cf. lacosti a Trinchera Elefante 9.
- La presència de Prolagus i Oryctolagus situa clarament els jaciments de Barranco León 5
i Fuente Nueva 3 en el Plio-Pleistocè. La similitud entre ambdues localitats indica que són
d’edat similar. D’altra banda, el lepòrid Oryctolagus cf. lacosti també ubica els materials
del jaciment de Trinchera Elefante 9 en el Plio-Pleistocè.
- Les associacions de lepòrids de Barranco León 5 i Fuente Nueva 3 (Orce) són diferents
morfològicament i biomètrica de les de Trinchera Elefante 9 (Atapuerca). Aquesta realitat
podria estar relacionada amb les diferències climàtiques d’ambdues regions geogràfiques,
la primera més mediterrània i l’altra més continental, hipòtesi a testar més endavant quan
es disposi de l’estudi d’un major nombre de jaciments i de material.
- Els lepòrids de Barranco León 5 i Fuente Nueva 3 presenten caràcters més moderns que
els de Trinchera Elefante 9, els quals mostren similituds amb formes del Pliocè, motiu pel
que, conseqüentment, aquest últim jaciment podria ser més antic. No obstant això, cal ser
prudent en comparar àrees geogràfiques diferents.
- La identificació d’Oryctolagus cf. cuniculus en els materials del Pleistocè inferior d’Orce
és la cita més antiga per aquesta espècie, el límit inferior de la qual fins ara era el Pleistocè
mitjà. La perllongació de la seva presència en el Pleistocè inferior coincidiria plenament
amb les prediccions de les anàlisis moleculars, que en situen l’origen al voltant dels 2 Ma.
- Les restes de lepòrids de Trinchera Elefante 9 constitueixen l’associació d’Oryctolagus
lacosti més completa d’Europa, i podrien representar la forma més propera a l’ancestre
comú d’Oryctolagus i Lepus trobada fins ara. A més a més, confirmen la relació evolutiva
entre Trischizolagus del Pliocè i els lepòrids actuals (Oryctolagus i Lepus).

50
6. Agraïments.
Aquest treball ha sigut possible gràcies al suport del “Departament d'Universitats, Recerca
i Societat de la Informació de la Generalitat de Catalunya i del Fons Social Europeu”, a
través de la concessió d’una Beca de Formació d’Investigadors (2005-2009).
Ara bé, també desitjaria destacar l'ajuda que m'han ofert els meus tres directors, Dra. Rosa
Domènech (Departament d'Estratigrafia, Paleontologia i Geociències Marines, Universitat
de Barcelona), Dr. Jordi Agustí (Institut de Paleontologia Humana, Universitat Rovira i
Virgili) i Dra. Gloria Cuenca (Facultat de Ciències, Universitat de Saragossa).
Gràcies també als Drs. Jordi Martinell i Jordi Maria de Gibert (Departament d'Estratigrafia,
Paleontologia i Geociències Marines de la Universitat de Barcelona), Drs. Pierre Mein i
Abel Prieur (Universitat Claude Bernard de Lyon), Sra. Eulàlia Garcia (Museu de Zoologia
de la Ciutadella, Barcelona), Sra. Josefina Barreiro (Museu Nacional de Ciències Naturals
de Madrid) i a tota la meva família i amics.

51
7. Bibliografia.
AGUIRRE, E. (1995). Atapuerca (Burgos): su contribución a las ciencias del Cuaternario. Revista Española de Paleontología, 10 : 58-82.
AGUSTÍ, J. (1986). Continental Mammal Units of the Plio-Pleistocene from Spain. Memoria Società Geologica Italiana, 31 : 167-173.
AGUSTÍ, J., ARBIOL, S. & MARTÍN-SUÁREZ, E. (1987). Roedores y lagomorfos (Mammalia) del Pleistoceno inferior de Venta Micena (Guadix-Baza, Granada). Paleontologia i Evolució, Memòria Especial, 1 : 95-107.
AGUSTÍ, J. & MOYÀ, S. (1987). Sobre la identidad del fragmento craneal atribuido a Homo sp. en Venta Micena (Orce, Granada). Estudios Geológicos, 43 : 535-538.
AGUSTÍ, J., OMS, O. & PARES, J. M. (1999). Calibration of the Early - Middle Pleistocene transition in the continental beds of the Guadix - Baza Basin. Quaternary Science Review, 18 : 1409-1417.
AGUSTÍ, J., CABRERA, L., GARCÉS, M., KRIJGSMAN, W., OMS, O. & PARÉS, J. M. (2001). A calibrated scale for the Neogene of Western Europe. State of the art. Earth Science Reviews, 52 : 247-260.
AGUSTÍ, J. & MADURELL, J. (2003). Los arvicólidos del Pleistoceno Inferior de Barranco León y Fuente Nueva 3. Datos preliminares. En: El Pleistoceno Inferior de Barranco León y Fuente Nueva 3, Orce, Granada (Coord. Toro, I., Agustí, J. & Martínez, B.). Arqueología Monografías, 17 : 137-145. Junta de Andalucía, Granada.
AGUSTÍ, J., DE MARFÀ, R. & SANTOS, A. (En premsa). Roedores y lagomorfos del Pleistoceno Inferior de Barranco León 5 y Fuente Nueva 3. Arqueología Monografías. Junta de Andalucía, Granada.
ANADÓN, P., UTRILLA , R. & JULIÀ , R. (1994). Palaeoenvironmental reconstruction of a Pleistocene lacustrine sequence from faunal assemblages and ostracod shell geochemistry, Baza Basin. Palaeogeography Palaeoclimatology Palaeoecology, 111 : 191-205.
ANADÓN, P. & JULIÀ , R. (2003). Estratigrafía y estudio sedimentológico preliminar de diversos afloramientos en Barranco León y Fuente Nueva 3. En: El Pleistoceno Inferior de Barranco León y Fuente Nueva 3, Granada. (Coord. Toro, I., Agustí, J. & Martínez, B.). Arqueología Monografías, 17 : 47-72. Junta de Andalucía, Granada.
ANGERMAN, R. (1966). Beiträge zur Kenntnis der Gattung Lepus. I. Abgrenzung der Gattung Lepus. II. Der taxonomische Status von Lepus brachyurus und Lepus mandshuricus. Mitt. Zool. Mus. Berlin, 42 : 127-144, 321-335.
ANGERMANN, R. (1983). The taxonomy of old world Lepus. Acta Zool. Fen., 174 : 17-21.
ARSUAGA, J. L., MARTÍNEZ, I., GRACIA, A., CARRETERO, J. M., LORENZO, C., GARCÍA, N. &
ORTEGA, A. I. (1997). Sima de los Huesos (Sierra de Atapuerca, Spain). The site. Journal of Human Evolution, 33 : 109-127.
ARSUAGA, J. L., MARTÍNEZ, I., LORENZO, C., GRACIA, A., MUÑOZ, A., ALONSO, O. &
GALLEGO, J. (1999). The human cranial remains from Gran Dolina Lower Pleistocene Site (Atapuerca, Spain). Journal of Human Evolution, 37 : 431-457.

52
ARSUAGA, J. L., MARTÍNEZ, I., GARCÍA, N., CUENCA, G., PÉREZ, A., BERMÚDEZ, J. M., GRACIA, A., CARRETERO, J. M., FALGUÉRES, C. & CARBONELL, E. (2002). Los Yacimientos de la Sierra de Atapuerca. . En: F. M. Botín, ed. La memoria de la tierra: yacimientos que cambiaron la historia. Col. Historia y Documentos, Santander, Fund. M. Botín, 101-147.
ASHER, R., MENG, J., WIBLE, J., MCKENNA, M., ROUGIER, G., DASHZEVEG, D. & NOVACEK, M. (2005). Stem Lagomorpha and the antiquity of Glires. Science, 307 : 1091-1094.
AVERIANOV, A. O. (1995). Osteology and adaptations of the Early Pliocene rabbit Trischizolagus dumitrescuae (Lagomorpha, Leporidae). Jour. Vert. Paleo., 5 (2) : 375-386.
AVERIANOV., A. O. & TESAKOV, A. (1997). Evolutionary trends in Mio-Pliocene Leporinae, based on Trischizolagus. Paläont. Zeitschrift, 71 (1/2) : 145-153.
AVERIANOV, A. O. (1998). Homology of the cusps in the molars of the Lagomorpha (Mammalia) and certain general problems of homology in the morphological structures. Paleontological Journal, vol. 32, nº1 : 73-77.
BEAUCOURNU, J. C. (1980). Les ectoparasites du lapin de garenne, Oryctolagus cuniculus: apports à son histoire. Bull. Mens. off Nation. Chasse, nº Sp. Scien. Tech. : 23-36.
BENITO, A. (2004). Análisis geomorfológico y reconstrucción de paleopaisajes neógenos y cuaternarios en la Sierra de Atapuerca y en al Valle medio del río Arlanzón. Tesis doctoral. Universidad Complutense de Madrid.
BERMÚDEZ, J. M., ARSUAGA, J. L., CARBONELL, E., ROSAS, A., MARTÍNEZ, I. & MOSQUERA, M. (1997). A hominid from the Lower Pleistocene of Atapuerca, Spain: Possible ancestor to Neandertals and Modern Humans. Science, 276 : 1392-1395.
BIJU-DUVAL , C., ENNAFAA , H., DENNEBOUY, N., MONNEROT, M., MIGNOTTE, F., SORIGUER, R., GAAÏED , A. HILI , A. & MOUNOLOU, J. (1991). J. Molec. Evol., 33 : 92-102.
BLAIN , H. (2003). Los anfibios y reptiles de los yacimientos del Pleistoceno Inferior de Barranco León y Fuente Nueva 3. En: El Pleistoceno Inferior de Barranco León y Fuente Nueva 3, Orce, Granada (Coord. Toro, I., Agustí, J. & Martínez, B.). Arqueología Monografías, 17 : 159-172. Junta de Andalucía, Granada.
BOHLIN, B. (1942). A revision of the fósil Lagomorpha in the Palaeontological Museum, Upsala. Bull. Of the Geol. Instit. Of Upsala, Vol.XXX : 117-154. Upsala.
BONHOTE, J. L. (1904). On the Mouse-hares. Proc. Zool. Soc. London, vol. 2.
BRINK, VAN DER, F. H. (1967). Guide des mamifères sauvages de l'Europe Occidentale. Ed. Delachaux & Niestlé, Neuchatel.
CALLOU , C. (1997). Diagnose différentielle des principaux éléments squelettiques du Lapin (Oryctolagus) et du Lièvre (Lepus) en Europe Occidentale. Fiches d’ostéologie animale pour l’archéologie, Série B. Centre de Recherches Archéologiques, Valbonne.
CALLOU , C. (2003). De la garenne au clapier: étude archéozoologique du lapin en Europe Occidentale. 358 pp. Mémoire du Muséum d'Histoire Naturelle, nº189.
CAMPILLO , D. (2002). El cráneo infantil de Orce. El homínido más antiguo de Eurasia. Bellaterra Arqueología, Barcelona.
CARBONELL, E., BERMÚDEZ, J. M., ARSUAGA, J. L., DÍEZ, J., ROSAS, A., CUENCA, G., SALA , R., MOSQUERA, M. & RODRÍGUEZ, X. (1995). Lower Pleistocene Hominids and Artifacts from Atapuerca TD-6 (Spain). Science, 269 : 826-832.

53
CARBONELL, E., ESTEBAN, M., MARTÍN, A., MOSQUERA, M., RODRÍGUEZ, X. P., OLLÉ, A., SALA , R., VERGÈS, J. M., BERMÚDEZ, J. M. & ORTEGA, A. (1999). The Pleistocene site of Gran Dolina, Sierra de Atapuerca, Spain: a histoy of the archeological investigations. Journal of Human Evolution, 37 : 313-324.
CERVERA, J., ARSUAGA, J. L., BERMÚDEZ, J. M., CARBONELL, E. & TRUEBA, J. (1998). Atapuerca. Un millón de años de historia. 235 pp. Plot Ediciones, Madrid.
CHALINE , J., ERBAJEVA, M. & MONTUIRE, S. (2000). Upper Pliocene Leporids from Montoussé-5 (Hautes-Pyrénées, France). N. Jb. Geol. Pal. Mh., (2) : 93-106.
CORBET, G. B. (1983). A review of the classification in the family Leporidae. Acta Zool. Fennica, 174 : 11-15.
CUENCA, G. (1990). Glires (Roedores y Lagomorfos). En: Paleontología (Meléndez, B. Ed.) tomo 3, vol.1 : 269-312.
CUENCA, G., CANUDO, J. I. ARSUAGA, J. L., BAHAIN , J., BERMÚDEZ DE CASTRO, J. M., CARBONELL, E., CARRETERO, J., FALGUERES, C, GARCÍA, N., LAPLANA , C., LÓPEZ, N. & VAN DER MADE, J. (1999). 2nd Field Trip: The Sierra de Atapuerca sites (Pleistocene, Burgos, Spain). IV European Workshop on Vertebrate Paleontology, Albarracín, Spain, 6-1999. Programme and Abstracts Field Guide, 115-161.
CUENCA, G., CANUDO, J. I. & LAPLANA , C. (2001). La séquence des rongeurs (Mammalia) des sites du Pléistocène inferieur et moyen d'Atapuerca (Burgos, Espagne). L'anthropologie, 105 : 115-130.
CUENCA, G. & ROFES, J. (2004). Insectívoros (Mammalia), clima y paisaje de los niveles inferiores de Trinchera Elefante (Pleistoceno inferior, Atapuerca, Burgos). Miscelánea en homenaje a Emiliano Aguirre : 150-156.
CUENCA, G., MARTÍNEZ, I., ARSUAGA, J. L., BERMÚDEZ, J. M. & CARBONELL, E. (2004). Los yacimientos del Pleistoceno Inferior y Medio de Atapuerca. Guía de la excursión Pinilla – Atapuerca, Alcalá de Henares.
DAWSON, M. R. (1967). Lagomorph history and the stratigraphyc record. Essays in Paleontology & Stratigraphy, Raymond C. Moore commemorative volume : 287-316. University of Kansas.
DAWSON, M. (1974). Lagomorpha. Encyclopaedia Británica : 588-591. Ed. H. Benton.
DE MARFÀ, R. J. (En premsa). Microfauna del Pleistoceno Inferior de Barranco León y Fuente Nueva 3 (Orce, Granada, España): Estudio preliminar.
DONARD, E. (1982). Recherches sur les léporidés Quaternaires (Pléistocène Moyen et Supérieur, Holocène). 161 pp., 46 pl., 89 tab., 2 tomes. Thèse. Université de Bordeaux.
ESPIGARES, M. P. & ROS, S. (2003). Síntesis de los conocimientos sobre la geomorfología de la cuenca de Guadix-Baza. En: El Pleistoceno Inferior de Barranco León y Fuente Nueva 3, Orce, Granada (Coord. Toro, I., Agustí, J. & Martínez, B.). Arqueología Monografías, 17 : 33-45. Junta de Andalucía, Granada.
FAURE, A. (1972). Étude comparée du squelette céphalique du lapin domestique et du lièvre. Application à la diagnose différentielle. Thèse de Doctorat, 65 pp. Alfort.
FLADERER, F. A. (1987). Beitrag zur Entwicklung von Hypolagus und Lepus (Lagomorpha, Mammalia) im Pliopleistozän von Mitteleuropa. Aus den Sitzungsberichten der Österr. Akademie der Wissenschaften Mathem.-naturw. Kl., 196 : 123-138. Viena.

54
FLADERER, F. A. & FIORE, M. The Early Pleistocene insular hare Hypolagus peregrinus sp. Nov. From Northern Sicily. Palaeont. Italica, 89 : 37-63.
FLUX , J. E. C. (1983). Introduction to taxonomic problems in hares. Acta Zool. Fennica, 174 : 7-10.
FORTELEONI, G. (1971). The upper Valdarno lagomorph Lepus valdarnensis. Paleontolog. Italica, 55-68.
FOSTOWICZ-FRELIK, L. (2003) Species distribution and differentiation of Eurasian Hypolagus (Lagomorpha, Leporidae). In: Reumer, J. W. F. & Wessels, W. (eds), Distribution and migration of Tertiary mammals in Eurasia. A volume in honour of Hans de Bruijn,, Denisea, 10 : 197-216.
FURIÓ, M. (2003). Los insectívoros (Mammalia) del Pleistoceno Inferior de Fuente Nueva 3 y Barranco León. En: El Pleistoceno Inferior de Barranco León y Fuente Nueva 3, Orce, Granada (Coord. Toro, I., Agustí, J. & Martínez, B.). Arqueología Monografías, 17 : 147-157. Junta de Andalucía, Granada.
GARCÍA AGUILAR, J. & MARTÍN, J. (2000). Late neogene to recent continental history and evolution of the Guadix-Baza Basin. Rev.Soc. Geol.España, 13 (1) : 65-77.
GIBERT, J., AGUSTÍ, J. & MOYÀ, S. (1983). Presencia de Homo sp. en el yacimiento de Venta Micena. Paleontologia i Evolució, Mem. Especial. Sabadell.
GIBERT, J., IGLESIAS, A., MAESTRO, E. & GIBERT, L. (1998). Two Oldowan assemblages in the Plio-Pleistocene deposits of the Orce region, southeast Spain. Antiquity, 72 : 17-25.
GIBERT, J., CAMPILLO , D., EISENMANN, V., GARCÍA-OLIVARES, E., MALGOSA, D., WALKER, M., BORJA, C., SÁNCHEZ, F., RIBOT, F., GIBERT, L., ALBALADEJO, S., IGLESIAS, A. &
FERRÁNDEZ, C. (1999). Spanish late Pliocene and Early Pleistocene hominid, palaeolithic and faunal finds from Orce and Cueva Victoria (Spain). Human Evolution, vol.14, nº1-2.
GIBERT, J. (2004). El Hombre de Orce. 450 pp. Editorial Almuzara, Córdoba.
GIDLEY , J. W. (1912). The Lagomorphs, an independent order. Science, 36 : 285-286.
GUENNOUNI, K. (2001). Les lapins du Pléistocène moyen et supérieur de quelques sites préhistoriques de l’Europe méditerranéenne: Terra Amata, Orgnac 3, Baume Bonne, Grotte du Lazaret, Boquete de Zafarraya, Arma delle Manie. Étude paléontologique, taphonomique et archéozoologique. Thèse Doctorale. Institut de Paléontologie Humaine.
GIL , E. & SESÉ, C. (1984). Micromamiferos del nuevo yacimiento Villafranquiense de Casablanca I (Almenara, Castellón). Estudios Geológicos, 40 : 83-89.
GUREEV, A. A. (1964). Zaitzeobraznye (Lagomorpha). Fauna SSSR Mammalia, vol.3, nº10, 87 : 1-276.
HALANYCH , K., DEMBOSKI, J., JANSEN, B., KLEIN, D. & COOK, J. (1999). Cytochrome b phylogeny of North American hares and jackrabbits (Lepus, Lagomorpha) and the effects of saturation in outgroup taxa. Mol. Phylo. and Evolution, vol.11, nº2 : 213-221.
HIBBARD, C. W. (1963) The origin of the p3 pattern of Sylvilagus, Caprolagus, Oryctolagus and Lepus. Jour. Of Mammal., 44 (1) : 1-15.
HOWELL, A. H. (1924). Revision of American pika. N. A. Fauna, 47. US Dept. Agr.
KORMOS, T. H. (1934). Zur frage der Abstammung Eurasiatischer Hasen. Allat. Koziem., 31 : 65-78.

55
KRETZOI, M. (1956). Die altpleistozänen Wirbeltierfaunen des Villányer Gebirges. Geol. Hungar. Palaeont., 27 : 1-264.
LAPLANA , C. & CUENCA, G. (2000). Una nueva especie de Microtus (Allophaiomys) (Arvicolidae, Rodentia, Mammalia) en el Pleistoceno Inferior de la Sierra de Atapuerca (Burgos, España). Revista Española de Paleontología, 15 : 77-87.
LESBRE, F. (1892). Caractères ostéologiques différentiels des lapins es des lièvres. Bull. Soc. Anthrop. Lyon, 11 : 263-285.
LESCA, C., MOISAND, A. & PUGET, A. (1975). Characterization of mitochondrial DNA from the pika Ochotona rufescens. Zeits. für Natur., 37 (c) : 145-148.
LOMBARDI, L., FERNÁNDEZ, N., MORENO, S. & VILLAFUERTE, R. (2003). Habitat-Related differences in rabbit (Oryctolagus cuniculus) abundance, distribution and activity. Journ. Mammal., 84 (1) : 26-36.
LOPEZ MARTINEZ, N. (1974). Taux taxonomique d'évolution dans l'ordre des Lagomorphes. Bull. Soc. Géol. France, 16 (4) : 422-430.
LOPEZ MARTINEZ, N. (1974). Évolution de la lignée Piezodus-Prolagus (Lagomorpha) dans le Cénozoïque d'Europe Sud-Occidentale. Thèse. Université du Languedoc, Montpellier.
LOPEZ MARTINEZ, N. & THALER, L. (1975). Biogéographie, évolution et compléments à la systématique du groupe d’Ochotonidés Piezodus-Prolagus (Lagomorpha). Bull. Soc. Géol. France, 7 (17), 5 : 850-866.
LOPEZ MARTINEZ, N., MICHAUX , J. & VILLALTA , J.F. (1976). Rongeurs et Lagomorphes de Bagur-2 (Gérone, Espagne). Nouveau remplissage de fisure du début du Pleistocene moyen. Acta Geol. Hispanica, 11 (2) : 46-54.
LÓPEZ MARTÍNEZ, N. (1977). Revisión sistemática y bioestratigráfica de los lagomorfos (Mammalia) del Terciario y Cuaternario de España. Tesis doctoral, Univ. de Madrid.
LOPEZ MARTINEZ, N. (1980). Les lagomorphes du Pléistocène supérieur de Jaurens. Nouv. Arch. Mus. Hist. Nat. Lyon, 18 : 5-16.
LÓPEZ MARTÍNEZ, N. (1989). Revisión sistemática y bioestratigráfica de los lagomorfos (Mammalia) del Terciario y Cuaternario de España. Mem. del Museo Paleontológico de la Universidad de Zaragoza, nº3 (3).
LÓPEZ MARTÍNEZ, N. (2001). Paleobiogeographical history of Prolagus, an European ochotonid (Lagomorpha). Lynx, 32 : 215-231.
LOREILLE, O., MOUNOLOU, J. C. & MONNEROT, M. (1997). Histoire des lapins et ADN ancien. C. R. Soc. Biol., 191 : 537-544.
MALATESTA, A. (1951). Considerazioni sugli Ochotonidae fossili e viventi. Bull. Serv. Geol. Italia, vol. 71 (9) : 3-28.
MARTÍN-SUÁREZ, E. & MEIN, P. (2004). The Late Pliocene locality of Saint Vallier (Drôme, France). Micromammals. Geobios, 37 : 115-125.
MARTÍNEZ, B., TURQ, A., AGUSTÍ, J. & OMS, O. (1997). Fuente Nueva 3 (Granada, Spain) and the first human occupation of Europe. Journal of Human Evolution, 33 : 611-620.
MARTÍNEZ, B. (2002). The skull of Orce: parietal bones or frontal bones? Journal of Human Evolution, 42 : 265-270.

56
MARTÍNEZ, B., ESPIGARES, M. P. & ROS, S. (2003). Estudio preliminar de las asociaciones de grandes mamíferos de Fuente Nueva 3 y Barranco León 5 (Orce, Granada). En: El Pleistoceno Inferior de Barranco León y Fuente Nueva 3, Orce, Granada (Coord. Toro, I., Agustí, J. & Martínez, B.). Arqueología Monografías, 17 : 115-136. Junta de Andalucía, Granada.
MARZIN, I. (1988). Diagnose différentielle des os longs du membre thoracique chez quelques espèces de petits mammifères européens. Thèse de Doctorat, 124 pp. Nantes.
MATTHEE, C., JANSEN, B., BELL, D. & ROBINSON, T. J. (2004). A molecular supermatrix of the rabbits and hares (Leporidae) allows for the identification of five intercontinental exchanges during the Miocene. Syst. Biol., 53 (3) : 433-447.
MAZO, A., SESÉ, C., RUIZ BUSTOS, A. & PEÑA, J. (1985). Geología y paleontología de los yacimientos Plio-pleistocenos de Huescar (Depresión de Guadix-Baza, Granada). Estudios Geol., 41 : 467-493.
MEULEN, A. J. VAN DER. (1973). Middle Pleistocene smaller Mammals from the Monte Peglia (Orvieto, Italy) with special reference to the phylogeny of Microtus (Rodentia). Quaternaria, 17.
MONNEROT, M., CASANE, D., HARDY, C. & VIGNE, J. (1994). Evolution of Oryctolagus: relationships within the Lagomorpha and access to populations with ancient DNA. Pol. Ecol. Studies, 20 (3-4) : 543-551.
MOYÀ, S. & AGUSTÍ, J. (1989). Una reinterpretación del fragmento craneal de Orce: Equus stenonis. En: Los restos humanos de Orce y Cueva Victoria (Gibert, Campillo, García Olivares Eds.). Institut de Paleontologia “Dr. M. Crusafont”. Sabadell.
MOYÀ, S. & KÖLHER, M. (1997). The Orce skull: anatomy of a mistake. Journal Human Evolution, 33 : 91-97.
NOCCHI, G. & SALA , B. (1997). Oryctolagus burgi n. sp. (Mammalia, Lagomorpha) from the Middle Pleistocene levels of Grotta Valdemino (Borgio Verezzi, Savona, north-west Italy). Paleontologia i evolució, t.30-31 : 19-38.
OMS, O., DINARÈS, J. & PARÉS, J. M. (1996). Resultados paleomagnéticos iniciales de la sección Plio-Pleistocena de Fuente Nueva (Cuenca de Guadix-Baza, Cordilleras Béticas). Revista de la Sociedad Geológica de España, 9 : 89-95.
OMS, O. (1998). Magnetoestratigrafia i litoestratigrafia de la conca de Guadix-Baza i altres punts del Neògen continental de les Serralades Bètiques. Ph D Thesis. Departament de Geologia, Universitat Autònoma de Barcelona.
OMS, O., PARÉS, J. & AGUSTÍ, J. (2003). Datación magnetoestratigráfica de los yacimientos de Fuente Nueva 3 y Barranco León 5 (Orce, Granada). En: El Pleistoceno Inferior de Barranco León y Fuente Nueva 3, Orce, Granada (Coord. Toro, I., Agustí, J. & Martínez, B.). Arqueología Monografías, 17 : 105-114. Junta de Andalucía, Granada.
PALACIOS, F. (1979). Análisis cromosómico, carga de DNA y electroforesis de las liebres españolas. Doñana Acta Vert., 6 : 203-215.
PALACIOS, F. & LÓPEZ, N. (1980). Morfología dentaria de las liebres europeas (Lagomorpha, Leporidae). Doñana, Acta Vertebrata, 7 (1) : 61-81.
PALACIOS, F. (1983). On the taxonomic status of the genus Lepus in Spain. Acta Zool. Fennica, 174 : 27-30.

57
PARÉS, J. M. & PÉREZ, A. (1999). Magnetochronology and stratigraphy at Gran Dolina section, Atapuerca (Burgos, Spain). Journal of Human Evolution, 37 : 325-342.
PELTIER, P. (1985). Étude ostéologique comparée du membre pelvien du lapin et du lièvre. Application à la diagnose différentielle. Thèse de Doctorat, 100 pp. Alfort.
PETTER, F. (1965). Eléments d'une révision des lièvres africains du sous-genre Lepus. Mammalia, 23 (1) : 41-67.
POMEL, A. (1853). Catalogue méthodique et descriptif des vertébrés fossiles découverts dans le bassin hydrographique supérieur de la Loire et de l’Allier. J. B. Baillière, 1-193.
RADULESCO, C. & SAMSON, P. (1967). Contributions à la connaissance du complèxe faunique de Mâlusteni-Beresti (Pleistocène inférieur), Roumanie. N. Jb. Geol. Pal. Mh., 9 : 544-563. Stuttgart.
ROBINSON, T. J. (1980). Comparative chromosome studies in the family Leporidae (Order Lagomorpha, Mammalia). Cytogenet. Cell. Gen., 28 : 64-70.
ROBISON, T. J., YANG, F. & HARRISON, W. R. (2002). Chromosome painting refines the history of genome evolution in hares and rabbits (Order Lagomorpha). Cytogen. Gen. Res, 96 : 223-227.
ROBINSON, T. J. & MATTHEE, C. A. (2005). Phylogeny and evolutionary origins of the Leporidae: a review of cytogenetics, molecular analyses and a supermatrix analysis. Mam. Rev, vol.35, nº3-4 : 231-247.
ROFES, J. & CUENCA, G. (En premsa). First evidence of the soricidae (mammalia) Asoriculus gibberodon in the Pleistocene of North Iberia. Riv. Italiana Paleo. Estrat.
ROSAS, A., PÉREZ, A., CARBONELL, E., VAN DER MADE, J., SÁNCHEZ, A., LAPLANA , C., CUENCA, G., PARÉS, J. M. & HUGUET, R. (2001). Le gisement pléistocène de la Sima del Elefante (Sierra de Atapuerca, Espagne). L'Anthropologie, 105 (2) : 301-312.
ROSAS, A., HUGUET, R., PÉREZ, A., CARBONELL, E., VALLVERDÚ , J., VAN DER MADE, J., ALLUÉ, E., GARCÍA, N., MARTÍNEZ, R., RODRÍGUEZ, J., SALA , R., SALADIE , P., SIMÓN, G., MARTÍNEZ, C., BASTIR, M., SÁNCHEZ, A. & PARÉS, J. M. (2004). Initial approach to the site formation and Paleoecology of the “Sima del Elefante”: a Pleistocene karst locality at Atapuerca Hill; En: Baquedano E. & Rubio Jara S. (eds.) - Miscelánea en homenaje a Emiliano Aguirre. Volumen I: Geología, Zona Arqueológica 4. Museo Arqueológico Regional: 134-155. Alcalá de Henares.
RZEBIK-KOWALSKA B. (1995). Climate and history of European shrews (family Soricidae). Acta Zoologica Cracoviensia, 38 : 95-107.
SCHAUB, S. (1943). Die oberpliocaene Säugertierfauna von Senèze (Haute-Loire) und ihre verbreitungsgeschichtliche Stellung. Eclog. Geol. Helv., 36 (2) : 270-289.
SCHNEIDER, E. & LEIPOLDT, M. (1983). DNA relationship within the genus Lepus in SW Europe. Acta Zool. Fennica, 174 : 31-33.
SCHREUDER, A. (1936). Hypolagus from the Tegelen Clay with a note on the recent Nesolagus. Arch. Néerl. Zool., 2.
SESÉ, C. & SEVILLA , P. (1996). Los micromamíferos del Cuaternario peninsular español: cronoestratigrafía e implicaciones bioestratigráficas. Revista Española de Paleontología, special issue 1, 278-287.

58
SIMPSON, G. G. (1945). The principles of classification and a classification of Mammals. Bull. Americ. Mus. Nat. Hist., 85.
SOUTHERN, H. N. (1940). The ecology and population dynamics of the wild rabbit Oryctolagus cuniculus. App. Biol., 27 : 509-526.
STIRTON, R. A. (1935). A review of the Tertiary beavers. Bull. Dept. Geol. Scienc. Univ. California, 23 (13) : 116 pp.
SYCH, L. (1965). Fossil Leporidae from the Pliocene and Pleistocene of Poland. Acta Zool. Cracov., 10 (1) : 2-62. Cracovia.
TOBIEN, H. (1976). Brachyodonty and Hypsodonty in the Evolution of European Tertiary Lagomorphs (Mammalia). Evolutionary Biology : 177- 191.
TORO, I., MARTÍNEZ, B., TORO, M. & FAJARDO, B. (2003). La excavación arqueológica. En: El Pleistoceno Inferior de Barranco León y Fuente Nueva 3, Orce, Granada (Coord. Toro, I., Agustí, J. & Martínez, B.). Arqueología Monografías, 17 : 15-31. Junta de Andalucía, Granada.
TORO, I., DE LUMLEY , HENRY, BARSKY, D., CELIBERTI, V., CAUCHE, D., MONCEL, M., FAJARDO, B. & TORO, M. (2003). Las indústrias líticas de Barranco León y Fuente Nueva 3. Resultados preliminares. In: El Pleistoceno inferior de Barranco León y Fuente Nueva 3, Orce, Granada (Coord. Toro, I., Agustí, J. & Martínez, B. Arqueologia Monografias, 17. Junta de Andalucía.
TURQ,A., MARTÍNEZ,B., PALMQVIST ,P., ARRIBAS,A., AGUSTÍ,J. & VIDAL ,J. (1996). Le Plio-Pleistocene de la région d’Orce, province de Grenade, Espagne: Bilán et perspectives de recherche. Paleo, 8 : 161-204.
VAN VALEN, L. A (1964). A possible origin for rabbits. Evolution, 18 (3) : 484-491.
VERA, J. A. (1970). Estudio estratigráfico de la Depresión de Guadix - Baza. Boletín Geológico Minero, 84 : 429-462.
VERA, J. A., FERNÁNDEZ, J., LÓPEZ, A. & RODRÍGUEZ, J. (1985). Geología y estratigrafía de los materiales Plio-Pleistocenos del sector Orce-Venta Micena (Granada). Paleontología i Evolució, 18 : 3-11.
VIGO, M. (2002). Guia dels mamífers terrestres de Catalunya. 240 pp. Ed Pòrtic, Barcelona.
VIRET, J. (1954). Le loess à bancs durcis de Saint-Vallier (Drôme) et sa faune de Mammifères Villafranchiens. Nouv. Archives Mus. Hist. Nat., 4. Lyon.
WOOD, A. E. (1957). What, if anything, is a Rabbit? Evolution, 11 (4) : 417-425.
ZACHOS, J., PAGANI , M., SLOAN, S., THOMAS, E. & BILLUPS, K. (2001). Trends, Rhythms and Aberrations in Global Climate 65 Ma to present. Science, vol.292 : 686-693.
ZAGWIJN W. H. (1998). Borders and boundaries: a century of stratigraphical research in the Tegelen-Reuver area of Limburg (The Netherlands). The Dawn of the Quaternary. En: Kolfschoten T. & Gibbard P.L. (eds). Proceedings of the SEQS. EuroMam Symposium (1996) 60: 19-34.

59
LÀMINES

60
Làmina I. Morfologia del P2 (1e, 1f, 1g, 2d) i P3 (1a, 1b, 1c, 1d, 2a, 2b, 2c) de conills actuals.1: Oryctolagus cuniculus (Cervera). Museu de Zoologia de Barcelona; 2: Oryctolagus cuniculus (Valdemoro). Museu Nacional de Ciències Naturals de Madrid.

61
Làmina II. Morfologia del P2 (1f, 1g, 1h, 2c), P3 (1i, 1j, 2d) i P3 (1a, 1b, 1c, 1d, 2a, 2b) de
llebres actuals. 1: Lepus europaeus (Pobla de Lillet). Museu de Zoologia de Barcelona; 2: Lepus granatensis (Torrejón). Museu Nacional de Ciències Naturals de Madrid.

62
Làmina III. Morfologia del P2 (1c, 2c, 2d, 3b, 3c) i P3 (1a, 1b, 2a, 2b, 3a) de llebres actuals. 1: Lepus castroviejoi (Riaño); 2: Lepus europaeus (Alemanya); 3: Lepus timidus
(Vall d'Aosta, Itàlia). Museu Nacional de Ciències Naturals de Madrid.
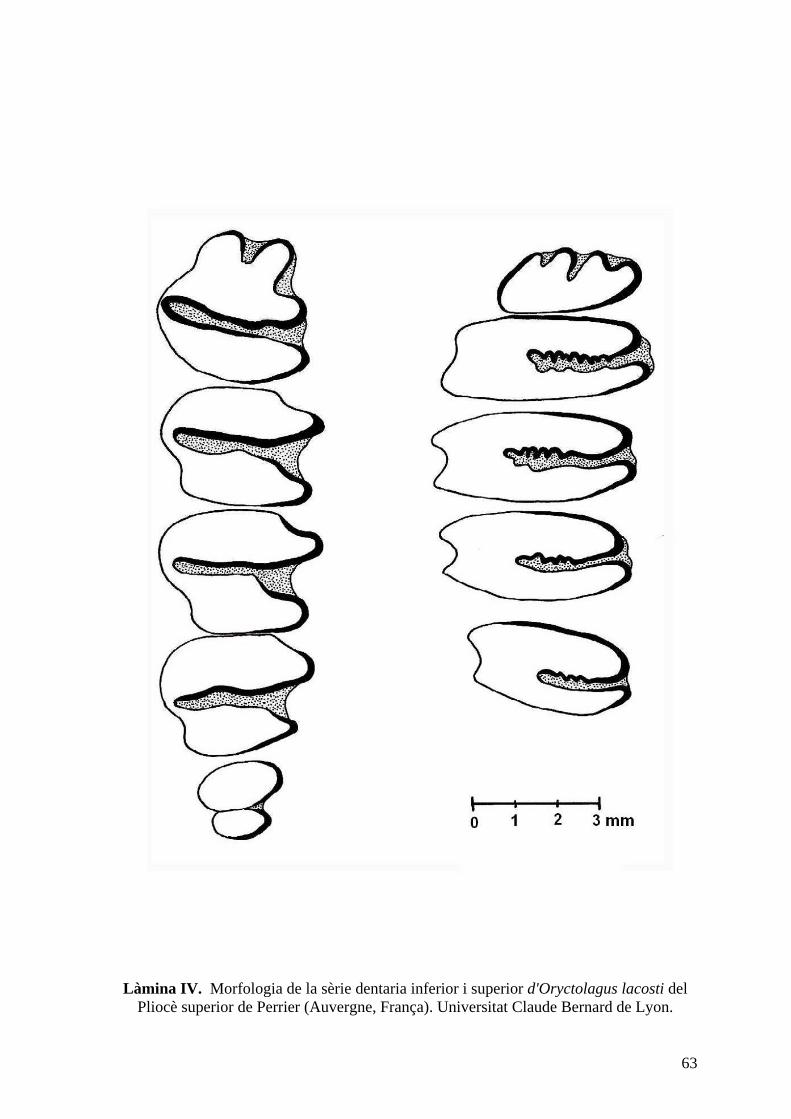
63
Làmina IV. Morfologia de la sèrie dentaria inferior i superior d'Oryctolagus lacosti del Pliocè superior de Perrier (Auvergne, França). Universitat Claude Bernard de Lyon.

64
Làmina V. Lagomorfs de Barranco León 5. Morfologia del P2 (a, b, c d), P3 (i), M2 (k) i P3 (e, f, g, h) d'Oryctolagus cf. cuniculus i del P4 (m) de Prolagus sp.
Col·lecció Institut de Paleontologia de Sabadell.
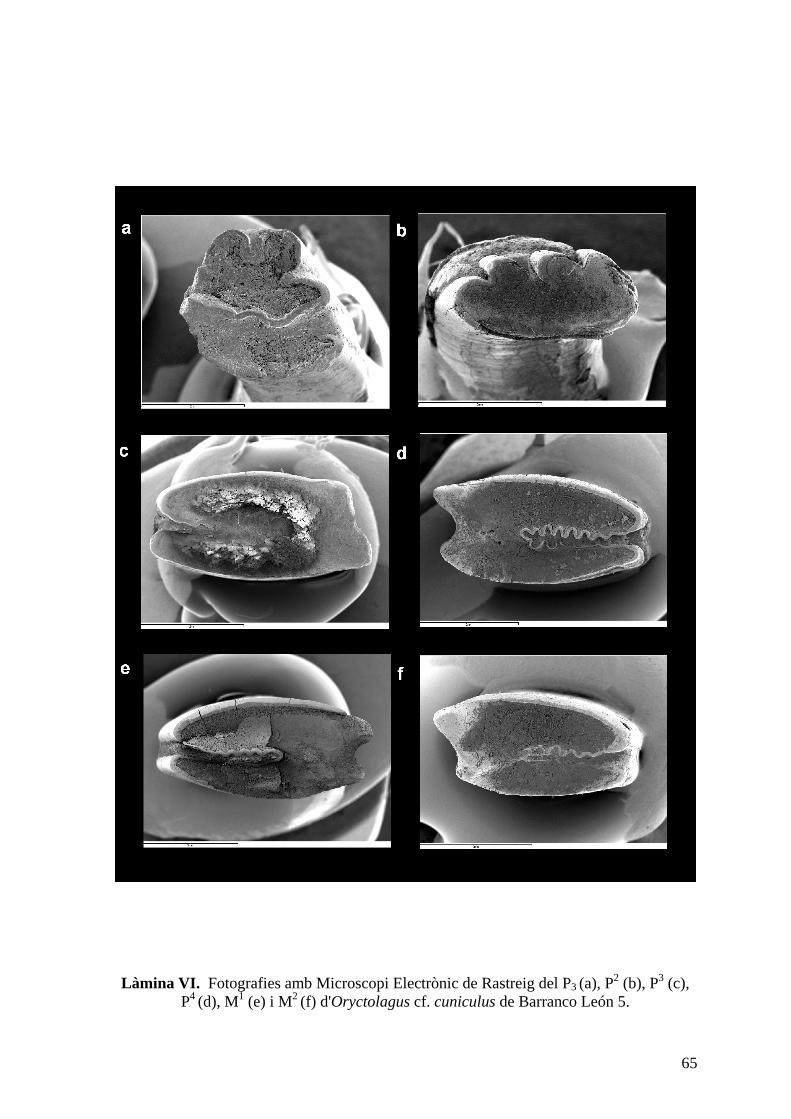
65
Làmina VI. Fotografies amb Microscopi Electrònic de Rastreig del P3 (a), P2 (b), P3 (c), P4 (d), M1 (e) i M2 (f) d'Oryctolagus cf. cuniculus de Barranco León 5.
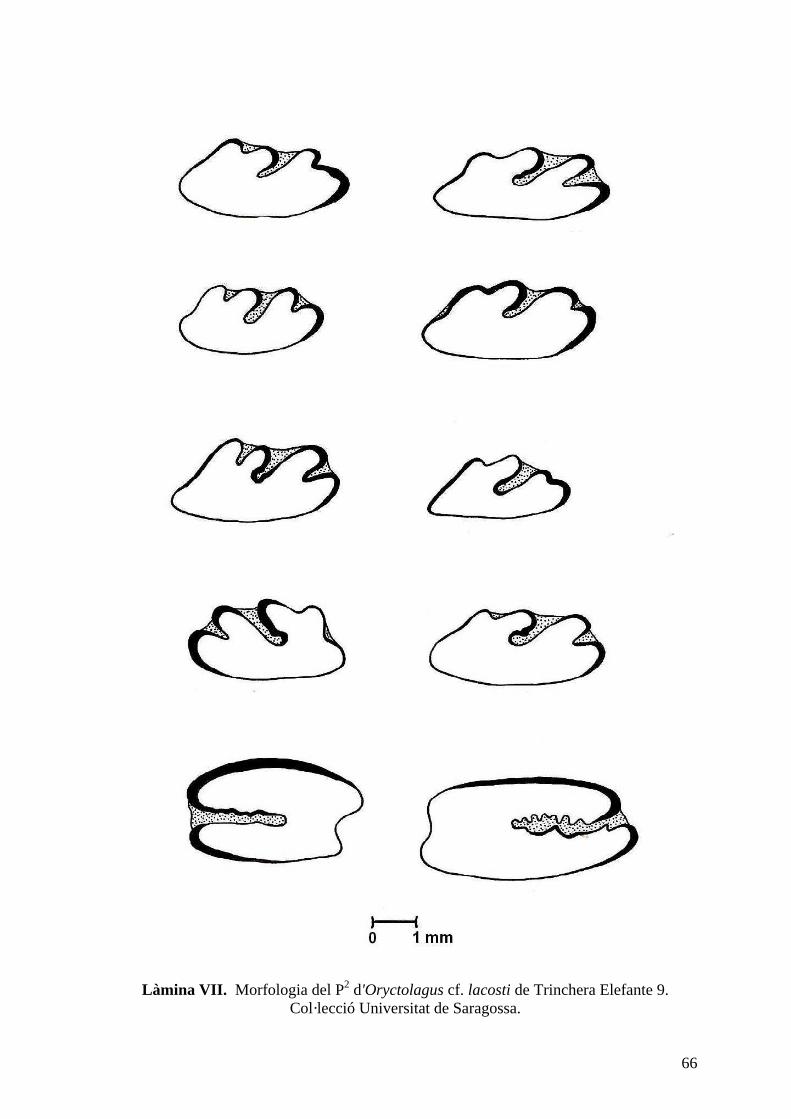
66
Làmina VII. Morfologia del P2 d'Oryctolagus cf. lacosti de Trinchera Elefante 9.
Col·lecció Universitat de Saragossa.
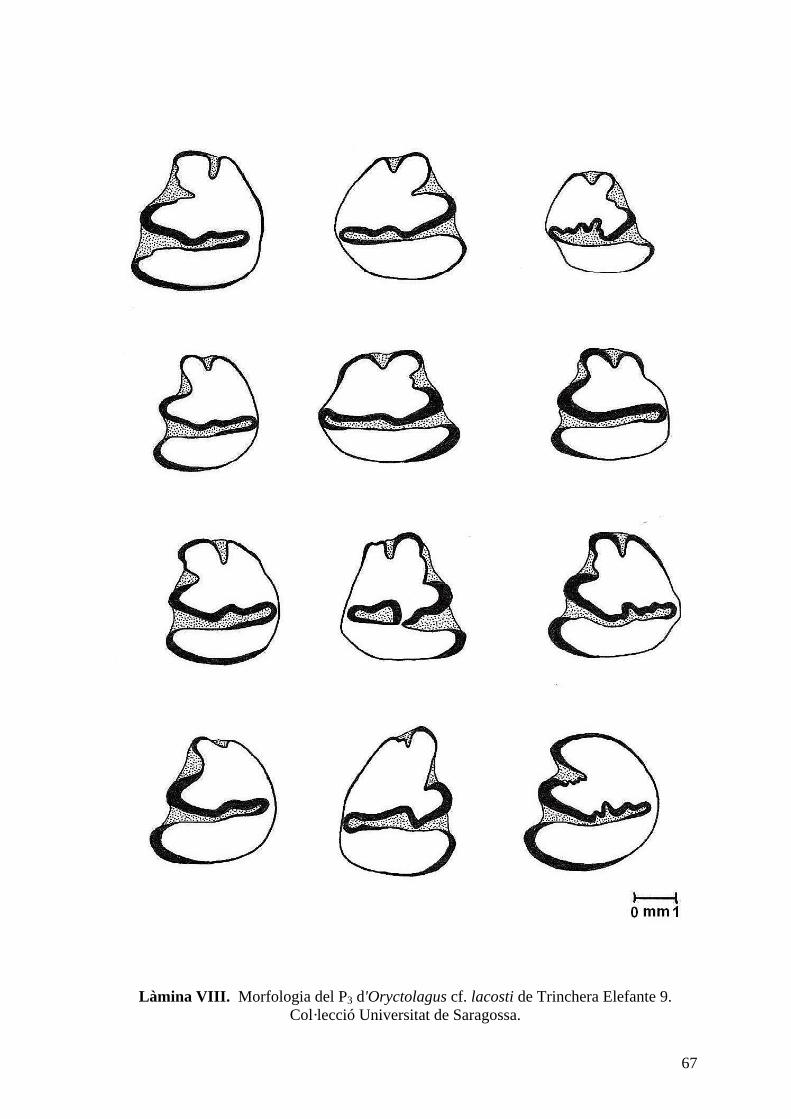
67
Làmina VIII. Morfologia del P3 d'Oryctolagus cf. lacosti de Trinchera Elefante 9. Col·lecció Universitat de Saragossa.
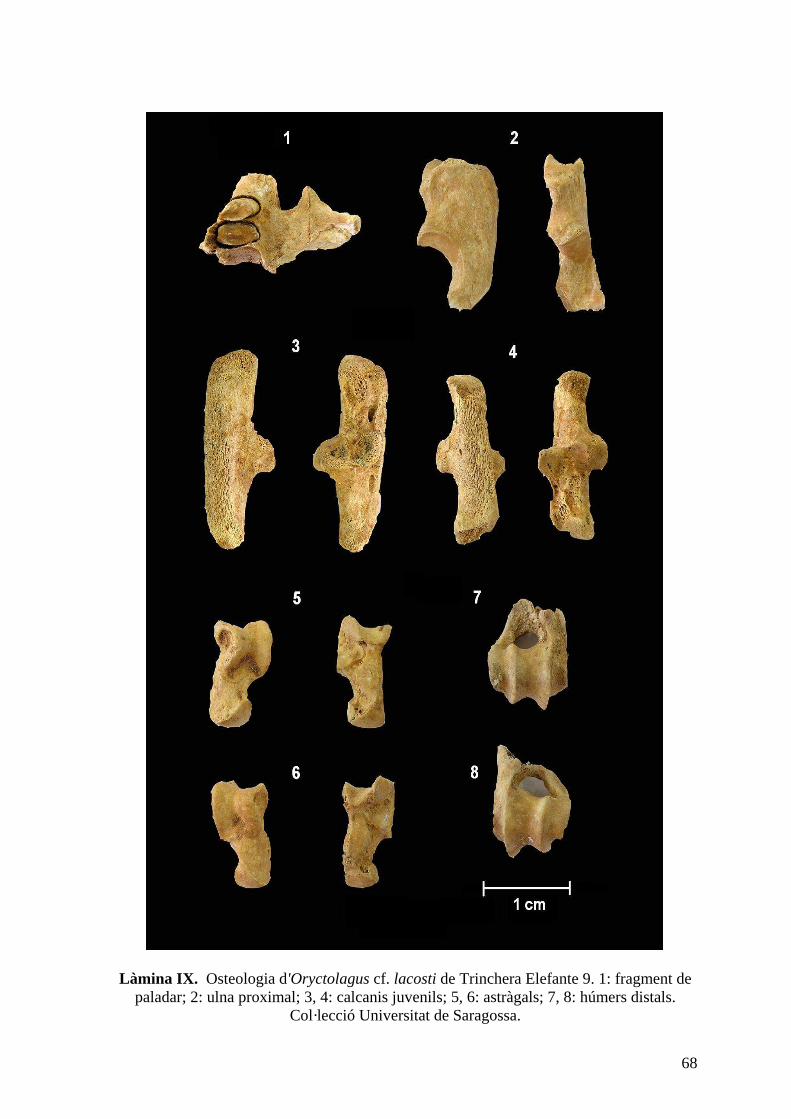
68
Làmina IX. Osteologia d'Oryctolagus cf. lacosti de Trinchera Elefante 9. 1: fragment de
paladar; 2: ulna proximal; 3, 4: calcanis juvenils; 5, 6: astràgals; 7, 8: húmers distals. Col·lecció Universitat de Saragossa.

69
MESURES

70
BARRANCO LEÓN 5
Peça Dental Zona-Capa Número
Longitud (Y) mm
Amplada (X) mm Y/X
Profunditat paraflex
P2 izq N56 UME2 LBL1' N-32 1,41 2,99 0,47 P2 izq J53 Capa D LBL5 J-6 1,16 2,42 0,48 0,65 P2 der I55 UME4 LBL5 I-3 1,35 2,98 0,45 0,77 P2 der M52 UME7 LBL5 M-2 1,32 2,77 0,48 0,61 P2 der M53 UME7 LBL5 M-16 1,27 2,59 0,49 0,53 P2 der M53 UME7 LBL5 M-17 1,4 2,56 0,54 0,63 P2 der M55 UME7 LBL5 M-22 1,44 2,99 0,48 0,72 P2 izq M55 UME7 LBL5 M-23 1,34 2,96 0,45 0,65
Peça Dental Zona-Capa Número
Longitud (Y) mm
Amplada (X) mm Y/X
Profunditat anteroflèxid
p3 esq N56 UME2 LBL1' N-29 2,94 2,68 1,1 0,45 p3 dret N56 UME2 LBL1' N-30 2,6 2,4 1,08 0,55 p3 esq N56 UME2 LBL1' N-31 2,85 2,6 1,1 0,62 p3 dret L60 CA LBL2 L-1 2,3 2,22 1,04 0,5 p3 esq L51 UME5 LBL2 L-4 2,71 2,77 0,98 0,48 p3 esq L51 UME5 LBL2 L-5 3,1 2,79 1,11 0,69 p3 esq L51 UME10 LBL2 L-7 0,56 p3 dret K60 A LBL2 K-11 1,78 1,82 0,98 0,35 p3 dret K61 C LBL2 K-15 0,52 p3 dret M53 UME7 LBL2 M-15 2,39 2,61 0,92 0,47 p3 dret Capa D LBL2 D-6 2,68 2,8 0,96 0,63 p3 esq Capa D LBL2 D-7 2,75 2,63 1,04 0,56 p3 dret N54 UME7 LBL2 N-49 2,58 2,38 1,08 p3 dret N54 UME9 LBL2 N-54 p3 dret O56 UME3 LBL2 O-1 FUENTE NUEVA 3
Peça Dental Zona-Capa Número
Longitud (Y) mm
Amplada (X) mm Y/X
Profunditat paraflex
P2 dret M86 UME4 LFN7 M-6 1,45 2,84 0,51 P2 dret N86 UME1 LFN7 N-2 1,47 2,49 0,59 0,72 P2 dret Q93 UME1 LFN7 Q-2 1,42 2,91 0,49 0,72 P2 dret M86 UME4 LFN7 M-23 1,52 2,6 0,58 0,76 P2 esq N86 UME4 LFN7 N-24 1,35 2,9 0,46 0,66 P2 dret Q86 UME1 LFN7 Q-9 1,34 2,83 0,47 0,68 P2 esq Q86 UME1 LFN7 Q-10 1,54 0,81 P2 dret N86 UME2 LFN7 N-37 1,43 2,66 0,54 0,71 P2 esq N86 UME2 LFN7 N-38 1,39 2,84 0,49 0,75 P2 esq N86 UME2 LFN7 N-42 1,43 2,96 0,48 0,75 P2 esq N87 UME3 LFN7 N-43 1,19 2,73 0,43 0,64 P2 dret O86 UME3 LFN7 O-17 P2 esq O86 UME3 LFN7 O-18 0,77 P2 dret O86 UME3 LFN7 O-20 1,42 2,95 0,48 0,66 P2 esq N87 UME3 LFN8 N-56 1,44 2,65 0,54 0,81
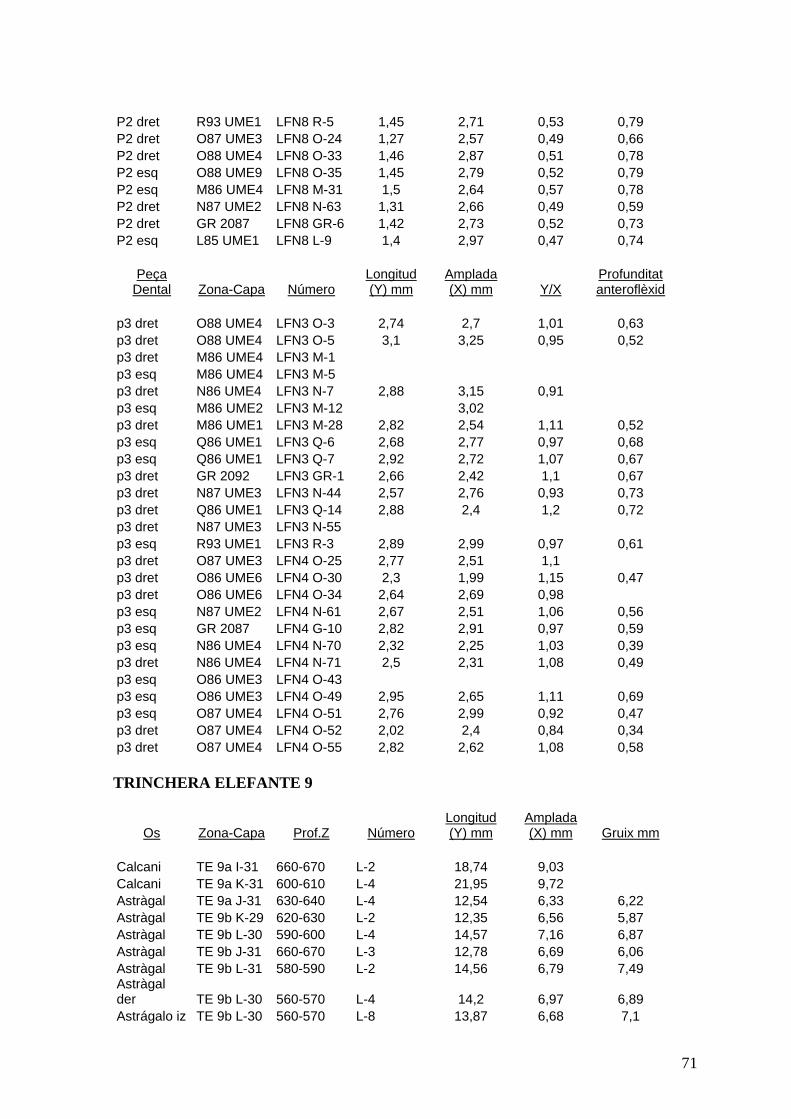
71
P2 dret R93 UME1 LFN8 R-5 1,45 2,71 0,53 0,79 P2 dret O87 UME3 LFN8 O-24 1,27 2,57 0,49 0,66 P2 dret O88 UME4 LFN8 O-33 1,46 2,87 0,51 0,78 P2 esq O88 UME9 LFN8 O-35 1,45 2,79 0,52 0,79 P2 esq M86 UME4 LFN8 M-31 1,5 2,64 0,57 0,78 P2 dret N87 UME2 LFN8 N-63 1,31 2,66 0,49 0,59 P2 dret GR 2087 LFN8 GR-6 1,42 2,73 0,52 0,73 P2 esq L85 UME1 LFN8 L-9 1,4 2,97 0,47 0,74
Peça Dental Zona-Capa Número
Longitud (Y) mm
Amplada (X) mm Y/X
Profunditat anteroflèxid
p3 dret O88 UME4 LFN3 O-3 2,74 2,7 1,01 0,63 p3 dret O88 UME4 LFN3 O-5 3,1 3,25 0,95 0,52 p3 dret M86 UME4 LFN3 M-1 p3 esq M86 UME4 LFN3 M-5 p3 dret N86 UME4 LFN3 N-7 2,88 3,15 0,91 p3 esq M86 UME2 LFN3 M-12 3,02 p3 dret M86 UME1 LFN3 M-28 2,82 2,54 1,11 0,52 p3 esq Q86 UME1 LFN3 Q-6 2,68 2,77 0,97 0,68 p3 esq Q86 UME1 LFN3 Q-7 2,92 2,72 1,07 0,67 p3 dret GR 2092 LFN3 GR-1 2,66 2,42 1,1 0,67 p3 dret N87 UME3 LFN3 N-44 2,57 2,76 0,93 0,73 p3 dret Q86 UME1 LFN3 Q-14 2,88 2,4 1,2 0,72 p3 dret N87 UME3 LFN3 N-55 p3 esq R93 UME1 LFN3 R-3 2,89 2,99 0,97 0,61 p3 dret O87 UME3 LFN4 O-25 2,77 2,51 1,1 p3 dret O86 UME6 LFN4 O-30 2,3 1,99 1,15 0,47 p3 dret O86 UME6 LFN4 O-34 2,64 2,69 0,98 p3 esq N87 UME2 LFN4 N-61 2,67 2,51 1,06 0,56 p3 esq GR 2087 LFN4 G-10 2,82 2,91 0,97 0,59 p3 esq N86 UME4 LFN4 N-70 2,32 2,25 1,03 0,39 p3 dret N86 UME4 LFN4 N-71 2,5 2,31 1,08 0,49 p3 esq O86 UME3 LFN4 O-43 p3 esq O86 UME3 LFN4 O-49 2,95 2,65 1,11 0,69 p3 esq O87 UME4 LFN4 O-51 2,76 2,99 0,92 0,47 p3 dret O87 UME4 LFN4 O-52 2,02 2,4 0,84 0,34 p3 dret O87 UME4 LFN4 O-55 2,82 2,62 1,08 0,58 TRINCHERA ELEFANTE 9
Os Zona-Capa Prof.Z Número Longitud (Y) mm
Amplada (X) mm Gruix mm
Calcani TE 9a I-31 660-670 L-2 18,74 9,03 Calcani TE 9a K-31 600-610 L-4 21,95 9,72 Astràgal TE 9a J-31 630-640 L-4 12,54 6,33 6,22 Astràgal TE 9b K-29 620-630 L-2 12,35 6,56 5,87 Astràgal TE 9b L-30 590-600 L-4 14,57 7,16 6,87 Astràgal TE 9b J-31 660-670 L-3 12,78 6,69 6,06 Astràgal TE 9b L-31 580-590 L-2 14,56 6,79 7,49 Astràgal der TE 9b L-30 560-570 L-4 14,2 6,97 6,89 Astrágalo iz TE 9b L-30 560-570 L-8 13,87 6,68 7,1

72
Os Zona-Capa Prof.Z Número Amplada (X) mm
Amplada Tróclea
Húmer distal Te9b K30 580-590 L-1 10,1 4,9 Húmer distal Te9b L31 570-580 L-1 10,2 4,8 Húmer distal Te9b L31 570-580 L-8 9,5 4,7
Os Zona-Capa Prof.Z Número Alçada
Olecranon Amplada
(mm) Ulna Prox Te9b L-31 580-590 L-1 12,8 11
Peça Dental Zona-Capa Prof. Z Número
Longitud (Y) mm
Amplada (X) mm
Profunditat paraflex
P2 der TE 9a I-31 660-670 L-1 1,4 3,12 0,75 P2 der TE9a M31 540-560 L-3 1,68 3,88 0,82 P2 TE9a L30 550-560 L-4 1,82 3,36 1,08 P2 der TE 9b I-30 670-680 L-15 1,69 3,71 0,8 P2 der TE9b M-29 530-540 L-2 1,68 3,81 0,84 P2 TE9b J30 650-660 L-8 1,82 3,58 0,81 P2 Te9b L-30 600-610 L-4 1,68 3,63 0,9 P2 der TE9b M-30 520-530 L-3 1,76 3,85 0,78 P2 Te9b K-30 610-620 L-1 1,51 3,42 0,67 P2 Te9b J-30 640-650 L-4 1,76 3,72 0,93 P2 Te9b L-31 580-590 L-8 1,79 3,95 0,82 P2 Te9b L-30 560-570 L-14 1,84 3,62 1,07 P2 Te9b J-30 640-650 L-1 1,6 3,73 0,93 P2 Te9b J-30 640-650 L-2 1,33 2,73 0,49 P2 Te9b J-30 650-660 L-10 1,52 3,13 0,82 P2 Te9b J-30 650-660 L-12 1,85 3,72 0,83 P2 Te9b J-30 650-660 L-13 1,76 3,77 1,02 P2 Te9b J-30 650-660 L-14 1,68 3,25 0,88 P2 izq ATA 98 Sondeo L-13 1,4 3,02 0,73 P2 izq ATA 98 Sondeo L-14 1,33 2,91 0,66 P2 izq ATA 98 Sondeo L-15 1,76 3,34 0,85 P2 der ATA 98 Sondeo L-16 1,31 3,02 0,55 P2 der ATA 98 Sondeo L-17 1,71 3,51 0,86 P2 izq ATA 98 Sondeo L-18 1,08 2,29 0,65 P2 der ATA 98 Sondeo L-19 1,36 3,08 0,82 P2 der ATA 98 Sondeo L-20 1,67 3,87 0,99 P2 der ATA 98 Sondeo L-21 1,57 3,48 0,73 P2 izq ATA 98 Sondeo L-22 1,97 3,57 1,05 P2 der ATA 98 Sondeo L-23 1,7 3,81 0,93 P2 izq ATA 98 Sondeo L-24 1,75 3,65 0,86 P2 der ATA 98 Sondeo L-25 1,77 3,83 0,85 P2 izq ATA 98 Sondeo L-26 1,85 3,67 1,02 P2 der ATA 98 Sondeo L-27 1,8 3,8 0,78 P2 der ATA 98 Sondeo L-28 1,82 3,66 1,04

73
Peça Dental Zona-Capa
Prof.Z - Núm
Longitud (Y) mm
Amplada (X) mm Y/X
Profunditat anteroflèxid
p3 izq TE 9b I-30 670-680 -16 3,84 3,62 1,06 0,83 p3 izq TE 9b I-30 670-680 -17 3,42 3,29 1,04 0,53 p3 izq TE 9b I-30 670-680 -18 3,23 2,98 1,08 0,56 p3 izq TE9b M-29 530-540 L-3 3,83 3,6 1,06 0,69 p3 izq TE 9b J-31 160-170 L-1 3,01 2,29 1,31 0 p3 der TE9b L-30 590-600 L-3 3,31 0,66 p3 der Te9b L-30 600-610 L-3 3,34 3,34 1 0,57 p3 Te9b K21 590-600 L-1 3,69 3,27 1,13 0,65 p3 izq Te9b J-31 660-670 L-4 3,35 3,12 1,07 0,78 p3 der Te9b J-31 660-670 L-5 3,53 3,75 0,94 0,55 p3 izq Te9b L-30 560-570 L-1 3,38 3,23 1,05 0,57 p3 izq Te9b L-30 560-570 L-2 3,57 3,69 0,97 0,81 p3 der Te9b J-30 650-660 L-9 2,85 3,01 0,95 0,44 p3 der Te9b J-30 650-660 -11 3,13 3,76 0,83 0,64 p3 der TE9a L-30 420-430 L-7 4,04 3,91 1,03 0,65 p3 izq TE9a L30 550-560 L-6 3,44 3,48 0,99 0,72 p3 Te9a L30 550-560 L-1 3,35 3,02 1,11 0,66 p3 izq Te9a L30 550-560 L-2 3,05 3,1 0,98 0,64 p3 izq Te9a L30 550-560 L-3 3,48 3,6 0,97 0,38 p3 der Te9a L30 550-560 L-4 2,91 2,75 1,06 0,4 d3 der ATA 98 Sondeo L-1 1,92 1,44 1,33 d3 izq ATA 98 Sondeo L-2 1,96 1,56 1,26 p3 der ATA 98 Sondeo L-3 3,61 3,87 0,93 0,9
p3 izq ATA 98 TE9 Sondeo L-5 3,56 3,26 1,09 0,82
p3 izq ATA 98 Sondeo L-6 3,06 3,14 0,97 0,59 p3 der ATA 98 Sondeo L-7 2,9 3,46 0,84 0,36 p3 izq ATA 98 Sondeo L-8 3,77 3,56 1,06 0,53 p3 izq ATA 98 Sondeo L-9 3,1 2,71 1,14 0,15 p3 der ATA 98 Sondeo -11 3,55 3,83 0,93 0,56 p3 izq ATA 98 Sondeo -12 3,57 3,34 1,07 0,72 p3 der TE9a J-31 630-640 L-3 3,58 3,33 1,07 0,7 p3 der ATA 98 Sondeo L-4 4,13 3,24 1,27 0,3 p3 TE9a L-31 540-550 L-1 3,43 3,27 1,05 0,37 p3 der Te9a L31 550-560 L-0 3,95 3,35 1,18 0,27 p3 izq ATA 98 Sondeo -10 3,71 3,56 1,04 0

74

75

76


















