
El Sistema Complemento (del Sistema Inmune) · Título original: El Sistema Complemento (del...
80
El Sistema Complemento (del Sistema Inmune)
Transcript of El Sistema Complemento (del Sistema Inmune) · Título original: El Sistema Complemento (del...

El Sistema Complemento(del Sistema Inmune)

Título original: El Sistema Complemento (del Sistema Inmune). Autor: Jorge Cabrera Muñoz.Especialidad: Técnico Superior de Laboratorio de Diagnóstico Clínico.Edita e imprime: FESITESS ANDALUCÍA
C/ Armengual de la Mota 37Ofi cina 1 29007 MálagaTeléfono/fax 952 61 54 61www.fesitessandalucía.es
Diseño y maquetación: Alfonso Cid IllescasEdición: Octubre 2011

Contenido
UNIDAD DIDÁCTICA I PRESENTACIÓN Y METODOLOGÍA DEL CURSO 5
1.1 Presentación, nornmas y procedimientos de trabajo 71.2 Sistema de cursos a distancia 71.3 Orientaciones para el estudio 81.4 Estructura del curso 9
UNIDAD DIDÁCTICA II INTRODUCCIÓN AL SISTEMA INMUNE 13
2.1 Introducción. 152.2 El sistema inmune. 152.3 Antígenos. 172.4 Células del sistema inmune. 192.5 Anticuerpos. 212.6 Respuesta inmune. 23
UNIDAD DIDÁCTICA III INTRODUCCIÓN AL SISTEMA COMPLEMENTO 25
3 .1 Introducción. 273.2 Historia del sistema complemento. 273.3 Importancia. 283.4 Funciones del complemento. 283.5 Componentes del complemento. 293.6 Métodos para la identi icación y medición de los componentes del complemento. 29
UNIDAD DIDÁCTICA IV VIAS DE ACTIVACIÓN DEL COMPLEMENTO 31
4.1 Vías de activación del complemento. 334.2 Causas, consecuencias y factores de la actividad del complemento. 40
UNIDAD DIDÁCTICA V ENFERMEDADES ASOCIADAS 47
5.1 Introducción. 495.2 De iciencias del complemento. 495.3 Efectos patológicos de un sistema del complemento normal. 50

UNIDAD DIDÁCTICA VI TÉCNICAS BASADAS EN LA REACCIÓN DE FIJACIÓN DEL COMPLEMENTO 53
6.1 Introducción. 556.2 Tipos de técnicas relacionadas con la reacción de ijación del complemento. 556.3 Muestras y reactivos necesarios. 576.4 Principales técnicas de ijación del complemento. 57
UNIDAD DIDÁCTICA VII TÉCNICAS BASADAS EN LA REACCIÓN DEL COMPLEMENTO PERO SIN FIJACIÓN 63
7.1 La turbidimetría. 657.2 La nefelometría. 657.3 Tipos de determinaciones. 66
UNIDAD DIDÁCTICA VIII CUESTIONARIO 69
8.1 Cuestionario 71

UNIDAD DIDÁCTICA IPRESENTACIÓN Y METODOLOGÍA DEL CURSO


7
El Sistema Complemento (del Sistema Inmune)
1.1 Presentación, nornmas y procedimientos de trabajo
Introducción
Antes de comenzar el Curso, es interesante conocer su estructura y el método que se ha de seguir. Este es el sen do de la presente introducción.
Si usted no conoce la técnica empleada en los Cursos a Distancia, le recomendamos que lea atentamente los epígrafes siguientes, los cuales le ayudarán a realizar el Curso en las mejores condiciones. En caso contrario, sólo ene que seguir los pasos que se indican en el siguiente índice:
Presentación
1. Sistema de Cursos a Distancia
En este apartado aprenderá una serie de aspectos generales sobre las técnicas de formación que se van a seguir para el estudio.
2. Orientaciones para el estudio.
Se dan una serie de recomendaciones generales para el estudio y las fases del proceso de aprendizaje propuesto por el equipo docente.
3. Estructura del Curso
Mostramos cómo es el Curso, las Unidades Temá cas de las que se compone, el sistema de evaluación y cómo enfrentarse al po test.
1.2 Sistema de cursos a distancia
1.2.1 Régimen de enseñanza
La metodología de Enseñanza a Distancia, por su estructura y concepción, ofrece un ámbito de aprendizaje donde pueden acceder, de forma fl exible en cuanto a ritmo individual de dedicación, estudio y aprendizaje, a los conocimientos que profesional y personalmente le interesen. Tiene la ventaja de estar diseñada para adaptarse a las disponibilidades de empo y/o situación geográfi ca de cada alumno. Además, es par cipa va y centrada en el desarrollo individual y orientado
a la solución de problemas clínicos.
La Formación a Distancia facilita el acceso a la enseñanza a todos los Técnicos Especialistas/Superiores Sanitarios.
1.2.2 Características del curso y del alumnado al que va dirigido
Todo Curso que pretenda ser efi caz, efec vo y efi ciente en alcanzar sus obje vos, debe adaptarse a los conocimientos previos de las personas que lo estudiarán (lo que saben y lo que aún no han aprendido). Por tanto, la difi cultad de los temas presentados se ajustará a sus intereses y capacidades.
Un buen Curso producirá resultados defi cientes si lo estudian personas muy diferentes de las inicialmente previstas.
Los Cursos se diseñan ajustándose a las caracterís cas del alumno al que se dirige.
1.2.3 Orientación de los tutores
Para cada Curso habrá, al menos, un tutor al que los alumnos podrán dirigir todas sus consultas y plantear las difi cultades.
Las tutorías están pensadas par endo de la base de que el aprendizaje que se realiza en esta formación es totalmente individual y personalizado.
El tutor responderá en un plazo mínimo las dudas planteadas a través de correo electrónico exclusivamente.

8
Técnico Superior Sanitario de Laboratorio de Diagnóstico Clínico
Diferenciamos para nuestros Cursos dos pos de tutores:
• Académicos. Serán aquellos que resuelvan las dudas del contenido del Curso, planteamientos sobre cues ones test y casos clínicos. El tutor resuelve las dudas que se plantean por correo electrónico.
• Orientadores y de apoyo metodológico. Su labor se centrará fundamentalmente en cues ones de carácter psicopedagógicas, ayudando al alumno en horarios, métodos de trabajo o cues ones más par culares que puedan alterar el desarrollo normal del Curso. El tutor resuelve las dudas que se
plantean por correo electrónico.
1.3 Orientaciones para el estudio
Los resultados que un estudiante ob ene no están exclusivamente en función de las ap tudes que posee y del interés que pone en prác ca, sino también de las técnicas de estudio que u liza. Aunque resulta di cil establecer unas normas que sean aplicables de forma general, es más conveniente que cada alumno se marque su propio método de trabajo, les recomendamos las siguientes que pueden ser de mayor aprovechamiento.
Por tanto, aún dando por supuestas la vocación y preparación de los alumnos y respetando su propia inicia va y forma de plantear el estudio, parece conveniente exponer algunos patrones con los que se podrá guiar más fácilmente el desarrollo académico, aunque va a depender de la situación par cular de cada alumno y de los conocimientos de la materia del Curso:
• Decidir una estrategia de trabajo, un calendario de estudio y mantenerlo con regularidad. Es recomendable tener al menos dos sesiones de trabajo por semana.
• Elegir el horario más favorable para cada alumno. Una sesión debe durar mínimo una hora y máximo tres. Menos de una hora es poco, debido al empo que se necesita de preparación, mientras que más de tres horas, incluidos los descansos, puede resultar demasiado y descendería el rendimiento.
• U lizar un si o tranquilo a horas silenciosas, con iluminación adecuada, espacio sufi ciente para extender apuntes, etc.
• Estudiar con atención, sin distraerse. Nada de radio, televisión o música de fondo. También es muy prác co subrayar los puntos más interesantes a modo de resumen o esquema.
a) Fase recep va.
• Observar en primer lugar el esquema general del Curso.
• Hacer una composición de lo que se cree más interesante o importante.
• Leer atentamente todos los conceptos desarrollados. No pasar de uno a otro sin haberlo entendido. Recordar que en los Cursos nunca se incluyen cues ones no ú les.
• Anotar las palabras o párrafos considerados más relevantes empleando un lápiz o rotulador transparente. No abusar de las anotaciones para que sean claras y signifi ca vas.
• Esquema zar en la medida de lo posible sin mirar el texto el contenido de la Unidad.
• Completar el esquema con el texto.
• Estudiar ajustándose al horario, pero sin imbuirse prisas o impacientarse. Deben aclararse las ideas y fi jarse los conceptos.
• Resumir los puntos considerados primordiales de cada tema.
• Marcar los conceptos sobre los que se tengan dudas tras leerlos detenidamente. No insis r de momento más sobre ellos.

9
El Sistema Complemento (del Sistema Inmune)
b) Fase refl exiva.
• Refl exionar sobre los conocimientos adquiridos y sobre las dudas que hayan podido surgir, una vez fi nalizado el estudio del texto. Pensar que siempre se puede acudir al tutor y a la bibliogra a recomendada y la u lizada en la elaboración del tema que puede ser de gran ayuda.
• Seguir paso a paso el desarrollo de los temas.
• Anotar los puntos que no se comprenden.
• Repasar los conceptos contenidos en el texto según va siguiendo la solución de los casos resueltos.
c) Fase crea va.
En esta fase se aplican los conocimientos adquiridos a la resolución de pruebas de autoevaluación y a los casos concretos de su vivencia profesional.
• Repasar despacio el enunciado y fi jarse en lo que se pide antes de empezar a solucionarla.
• Consultar la exposición de conceptos del texto que hagan referencia a cada cues ón de la prueba.
• Solucionar la prueba de cada Unidad Temá ca u lizando el propio cues onario del manual.
1.4 Estructura del curso
1.4.1 Contenidos del curso
• Guía del alumno.
• Temario del curso en PDF, con un cues onario po test.
• FORMULARIO, para devolver las respuestas al cues onario.
• ENCUESTA de sa sfacción del Curso.
1.4.2 Los cursos
Los cursos se presentan en un archivo PDF cuidadosamente diseñado en Unidades Didác cas.
1.4.3 Las Unidades Temáticas
Son unidades básicas de estos Cursos a distancia. Con enen diferentes pos de material educa vo dis nto:
• Texto propiamente dicho, dividido en temas.
• Cues onario po test.
• Bibliogra a u lizada y recomendada.
Los temas comienzan con un índice con las materias contenidas en ellos. Con núa con el texto propiamente dicho, donde se desarrollan las cues ones del programa. En la redacción del mismo se evita todo aquello que no sea de u lidad prác ca.
El apartado de preguntas test serán con los que se trabajen, y con los que posteriormente se rellenará el FORMULARIO de respuestas a remi r. Los ejercicios de po test se adjuntan al fi nal del temario.

10
Técnico Superior Sanitario de Laboratorio de Diagnóstico Clínico
Cuando están presentes los ejercicios de autoevaluación, la realización de éstos resulta muy ú l para el alumno, ya que:
• Tienen una función recapituladora, insis endo en los conceptos y términos básicos del tema.
• Hacen par cipar al alumno de una manera más ac va en el aprendizaje del tema.
• Sirven para que el alumno valore el estado de su aprendizaje, al comprobar posteriormente el resultado de las respuestas.
• Son garan a de que ha estudiado el tema, cuando el alumno los ha superado posi vamente. En caso contrario se recomienda que lo estudie de nuevo.
Dentro de las unidades hay dis ntos epígrafes, que son conjuntos homogéneos de conceptos que guardan relación entre sí. El tamaño y número de epígrafes dependerá de cada caso.
1.4.4. Sistema de evaluaciòn
Cada Curso con ene una serie de pruebas de evaluación a distancia que se encuentran al fi nal del temario. Deben ser realizadas por el alumno al fi nalizar el estudio del Curso, y enviada al tutor de la asignatura, con un plazo máximo de entrega para que pueda quedar incluido en la edición del Curso en la que se matriculó y siempre disponiendo de 15 días adicionales para su envío. Los tutores la corregirán y devolverán al alumno.
Si no se supera el cues onario con un mínimo del 80% correcto, se tendrá la posibilidad de recuperación.
La elaboración y posterior corrección de los test ha sido diseñada por el personal docente seleccionado para el Curso con la intención de acercar el contenido de las preguntas al temario asimilado.
Es IMPRESCINDIBLE haber rellenado el FORMULARIO y envío de las respuestas para recibir el cer fi cado o Diploma de ap tud del Curso.
1.4.5 Fechas
El plazo de entrega de las evaluaciones será de un mes y medio a par r de la recepción del material del curso, una vez pasado este plazo conllevará una serie de ges ones administra vas que el alumno tendrá que abonar.
La entrega de los cer fi cados del Curso estará en relación con la fecha de entrega de las evaluaciones y NUNCA antes de la fecha de fi nalización del Curso.
1.4.6 Aprendiendo a enfrentarse a preguntas tipo test
La primera u lidad que se deriva de la resolución de preguntas po test es aprender cómo enfrentarnos a las mismas y evitar esa sensación que algunos alumnos enen de “se me dan los exámenes po test”.
Cuando se trata de preguntas con respuesta po verdadero / falso, la resolución de las mismas está más dirigida y el planteamiento es más específi co.
Las preguntas po test con varias posibles respuestas hacen referencia a conocimientos muy concretos y exigen un método de estudio diferente al que muchas personas han empleado hasta ahora.
Básicamente todas las preguntas test enen una caracterís ca común: exigen iden fi car una opción que se diferencia de las otras por uno o más datos de los recogidos en el enunciado. Las dos palabras en cursiva son expresión de dos hechos fundamentales con respecto a las preguntas po test:
• Como se trata de iden fi car algo que va a encontrar escrito, no va a ser necesario memorizar conocimientos hasta el punto de reproducir con exac tud lo que uno estudia. Por lo tanto, no debe agobiarse cuando no consiga recordad de memoria una serie de datos que aprendió hace empo; seguro que muchos de ellos los recordará al leerlos formando parte del enunciado o las opciones de una pregunta de test.

11
El Sistema Complemento (del Sistema Inmune)
• El hecho de que haya que dis nguir una opción de otras se traduce en muchas ocasiones en que hay que estudiar diferencias o similitudes. Habitualmente se les pide recordar un dato que se diferencia de otros por ser el más frecuente, el más caracterís co, etc. Por lo tanto, este po de datos o situaciones son los que hay que estudiar.
Debe tenerse siempre en cuenta que las preguntas test hay que leerlas de forma completa y fi jándose en determinadas palabras que puedan resultar clave para la resolución de la pregunta.
La u lidad de las preguntas test es varia:
• Acostumbrarse a percibir errores de conceptos.
• Adaptarse a los exámenes de selección de personal.
• Ser capaces de aprender sobre la marcha nuevos conceptos que pueden ser planteados en estas preguntas, conceptos que se re enen con facilidad.
1.4.7 Envío
Una vez estudiado el material docente, se contestará la encuesta de sa sfacción, la cual nos ayudará para evaluar el Curso, corregir y mejorar posibles errores. Cuando haya cumplimentado la evaluación, envíe las respuestas a la dirección indicada.


UNIDAD DIDÁCTICA IIINTRODUCCIÓN AL SISTEMA INMUNE


15
El Sistema Complemento (del Sistema Inmune)
2.1 Introducción.Inmunología ciencia biológica encargada del estudio de todos los mecanismos fi siológicos de defensa de la
integridad biológica del organismo o lo que es lo mismo del estudio de inmunidad. Dicho mecanismo fi siológico consiste en la iden fi cación de lo extraño al organismo y su propia destrucción. Esta ciencia también se encarga del estudio de los factores inespecífi cos que ayudan a los anteriores a conseguir sus efectos fi nales que serán destrucción e iden fi cación de lo extraño al organismo.
Inmunidad conjunto de mecanismos fi siológicos de defensa que todos los seres vivos enen frente a agentes extraños, lo que le permite a estos reconocer sustancias extrañas a sus propios organismos y neutralizarlas o eliminarlas con o sin lesión para los tejidos. En el ser humano se adquiere al nacer y va madurando y consolidándose durante los primeros años de vida.
También podemos defi nirla como el estado de resistencia que presenta un organismo frete a una enfermedad infecciosa.
Inmunología clínica ciencia que se encarga del estudio del sistema inmune.
Sistema inmune conjunto de células y factores solubles desarrollados por los seres vivos para defenderse de todo lo que les es extraño, principalmente frente a infecciones por microorganismos patógenos tales como virus y bacterias etc.
2.2 El sistema inmune.
Los seres humanos al igual que el resto de los seres vivos disponemos de un mecanismo de defensa frente a microorganismos y sustancias extrañas al organismo (consideradas como patógenos), esto es lo que se conoce como Sistema Inmune. Cuando una sustancia extraña entra en contacto con el organismo, este el sistema inmune pone en marcha la respuesta inmune también conocida como el mecanismo mediante el cual el cuerpo se defi ende de patógenos, células tumorales ó sustancias toxicas.
2.2.1 El sistema inmune.
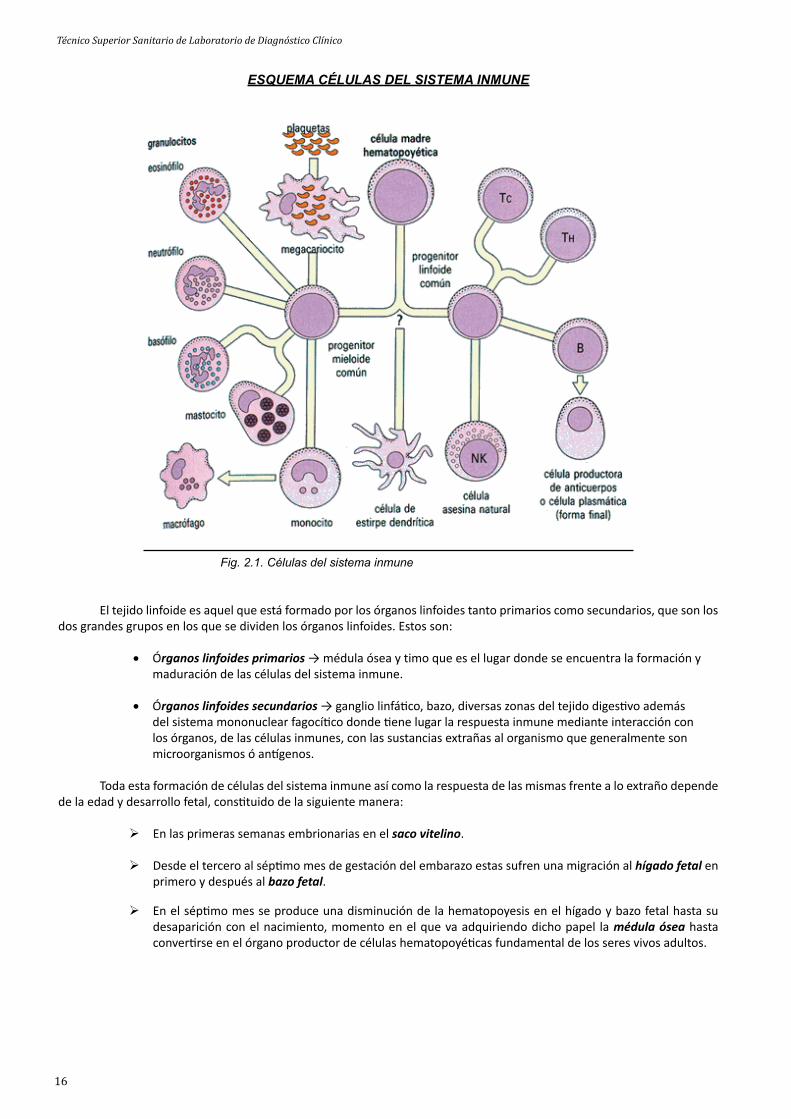
El sistema Inmune está formado por órganos, tejidos y células del propio sistema. En los órganos del sistema inmune es donde ene lugar la formación, maduración y diferenciación de las dis ntas células que lo componen, las cuales son conocidas como leucocitos, que pueden ser de dis ntos pos siendo los que se conocen como linfocitos los más abundantes y por tanto los de mayor importancia, que pueden ser a su vez de dis ntos pos. Por este mo vo a los órganos encargados de la formación de células se les conoce con el nombre de órganos linfoides. A parte de los leucocitos el sistema inmune consta de otro po de células llamadas plaquetas.
El proceso de formación de las células por parte de los órganos del sistema inmune más conocidos como órganos linfoides se denomina hematopoyesis. La hematopoyesis se defi ne como aquel proceso que consiste en la formación y diferenciación de células sanguíneas a par r de la célula madre pluripotencial, también conocida como stem cell. Estas células sanguíneas son las mismas que las del sistema inmune solo que el sistema inmune no presenta eritrocitos, células sanguíneas más abundantes en la sangre, encargadas de dar el color rojo caracterís co a la misma por encontrarse dentro de estas células una sustancia llamada hemoglobina encargada de dar ese color rojo caracterís co a la sangre.
Las células del sistema inmune se dividen en dos grandes grupos o líneas procedentes todas de la ya citada célula madre, dichas líneas son:
Serie mieloide → monocitos y macrófagos; neutrófi los y polimorfo nucleares; eosinófi los; básofi los y mástocitos (todos pertenecientes a los linfocitos); plaquetas, que son todas las células del sistema inmune restantes que no son linfocitos.
Serie linfoide → todos los pos de linfocitos T y B; linfocitos no T y no B donde tenemos los NK (natural Killer) y las ADCC (células efectoras de citotoxicidad o dependientes de an cuerpos).
16
Técnico Superior Sanitario de Laboratorio de Diagnóstico Clínico
ESQUEMA CÉLULAS DEL SISTEMA INMUNE
Fig. 2.1. Células del sistema inmune
El tejido linfoide es aquel que está formado por los órganos linfoides tanto primarios como secundarios, que son los dos grandes grupos en los que se dividen los órganos linfoides. Estos son:
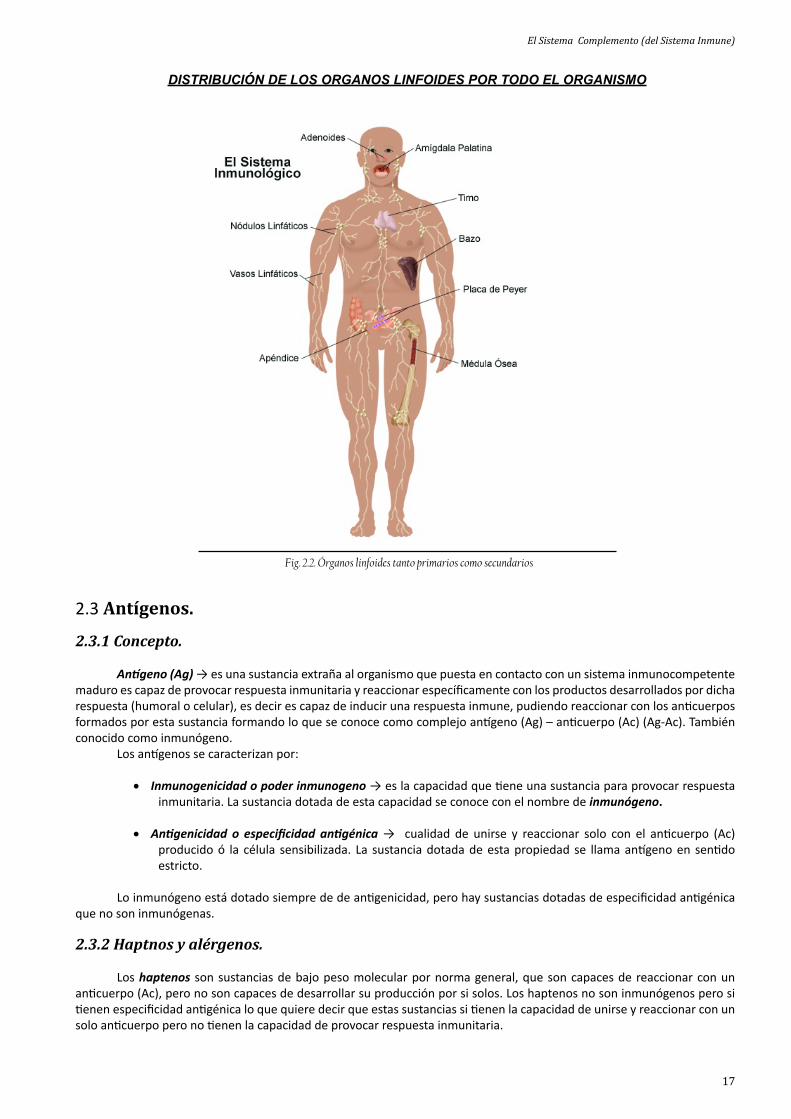
Órganos linfoides primarios → médula ósea y timo que es el lugar donde se encuentra la formación y maduración de las células del sistema inmune.
Órganos linfoides secundarios → ganglio linfá co, bazo, diversas zonas del tejido diges vo además del sistema mononuclear fagocí co donde ene lugar la respuesta inmune mediante interacción con los órganos, de las células inmunes, con las sustancias extrañas al organismo que generalmente son microorganismos ó an genos.
Toda esta formación de células del sistema inmune así como la respuesta de las mismas frente a lo extraño depende de la edad y desarrollo fetal, cons tuido de la siguiente manera:
En las primeras semanas embrionarias en el saco vitelino.
Desde el tercero al sép mo mes de gestación del embarazo estas sufren una migración al hígado fetal en primero y después al bazo fetal.
En el sép mo mes se produce una disminución de la hematopoyesis en el hígado y bazo fetal hasta su desaparición con el nacimiento, momento en el que va adquiriendo dicho papel la médula ósea hasta conver rse en el órgano productor de células hematopoyé cas fundamental de los seres vivos adultos.
17
El Sistema Complemento (del Sistema Inmune)
DISTRIBUCIÓN DE LOS ORGANOS LINFOIDES POR TODO EL ORGANISMO
Fig. 2.2. Órganos linfoides tanto primarios como secundarios
2.3 Antígenos.
2.3.1 Concepto.
An geno (Ag) → es una sustancia extraña al organismo que puesta en contacto con un sistema inmunocompetente maduro es capaz de provocar respuesta inmunitaria y reaccionar específi camente con los productos desarrollados por dicha respuesta (humoral o celular), es decir es capaz de inducir una respuesta inmune, pudiendo reaccionar con los an cuerpos formados por esta sustancia formando lo que se conoce como complejo an geno (Ag) – an cuerpo (Ac) (Ag-Ac). También conocido como inmunógeno.
Los an genos se caracterizan por:
Inmunogenicidad o poder inmunogeno → es la capacidad que ene una sustancia para provocar respuesta inmunitaria. La sustancia dotada de esta capacidad se conoce con el nombre de inmunógeno.
An genicidad o especifi cidad an génica → cualidad de unirse y reaccionar solo con el an cuerpo (Ac) producido ó la célula sensibilizada. La sustancia dotada de esta propiedad se llama an geno en sen do estricto.
Lo inmunógeno está dotado siempre de de an genicidad, pero hay sustancias dotadas de especifi cidad an génica que no son inmunógenas.
2.3.2 Haptnos y alérgenos.
Los haptenos son sustancias de bajo peso molecular por norma general, que son capaces de reaccionar con un an cuerpo (Ac), pero no son capaces de desarrollar su producción por si solos. Los haptenos no son inmunógenos pero si enen especifi cidad an génica lo que quiere decir que estas sustancias si enen la capacidad de unirse y reaccionar con un
solo an cuerpo pero no enen la capacidad de provocar respuesta inmunitaria.

18
Técnico Superior Sanitario de Laboratorio de Diagnóstico Clínico
Los alérgenos son ciertos an genos (sustancias extrañas al organismo) que inducen reacciones de hipersensibilidad, es decir reacciones alérgicas y se comportan como inmunógenos o como haptenos.
En esto podemos decir que los términos inmunógeno y an genos son sinónimos.
Existen muchas sustancias inmunógenas en la naturaleza que cons tuyen los an genos naturales. Lo mismo ocurre con algunas sustancias de origen sinté co que pueden comportarse como inmunógenos.
2.3.3 Epítopes y determinantes antigénicos.
La an genicidad de una sustancia no depende de toda la molécula del an geno, sino de una pequeña zona de ella llamada epítope, grupo determinante ó determinante an génico que es simplemente una conformación molecular de la superfi cie del an geno que es capaz de combinarse específi camente con una zona complementaria existente en la molécula del an cuerpo producido denominado zona combinante (formando lo que se conoce como llave-cerradura).
2.3.4 Inmunogenicidad.Las sustancias inmunógenas solo se comportan como tales si penetran en un organismo capaz de responder por
tener un sistema inmunitario maduro.
2.3.5 Antigenicidad
Los an cuerpos sólo reaccionan con el an geno que es mulo su formación o con an genos muy relacionados, parecidos.
Esta an genicidad deriva de la existencia de los determinantes an génicos o epitopes que podrán unirse a las zonas combinantes del an cuerpo que necesariamente deberá ser complementario de aquellos.
2.3.5.1 Factores determinantes de antigenicidad.
Número → es muy raro que una molécula an génica posea un solo grupo determinante, lo más frecuente es que su número sea múl ple y se producirán, tantos an cuerpos específi cos como grupos determinantes tenga el an geno de manera que a mayor número de epítopes mayor an genicidad.
Valencia an genica →es el número de moléculas igual al de an cuerpos con los que puede combinarse el an geno, lo que es igual al número de grupos determinantes idén cos (epítopes), aunque no siempre es así, pues la unión de un an cuerpo con su epítope correspondiente difi culta la unión del siguiente epítope. A este fenómeno se le llama difi cultad espacial ó inhibición estérica.
Existes epítopes para las dis ntas células del sistema inmune, siendo debido a su frecuencia los más importantes los de los linfocitos T y B.
2.3.6 Clasi icación de los antígenos.
Los an genos se encuentran ampliamente distribuidos por la naturaleza. Los podemos encontrar clasifi cados en dos grandes grupos fundamentales aunque existe otro tercer grupo.
2.3.6.1 Antígenos de células y tejidos. Se clasifi can atendiendo a dos criterios:
Según procedencia:
Xenoan genos → son an genos que proceden de dis nta especie animal.
Isoan genos → son también llamado aloan genos y son aquellos an genos que proceden de misma especie animal pero dis nta cons tución gené ca.
Autoan genos → moléculas propias del organismo reconocidas como extrañas por el propio sistema inmune.

19
El Sistema Complemento (del Sistema Inmune)
• Según especifi cidad:
An genos heterogené cos → sustancias an génicas presentes en organismos de especie no relacionadas entre sí, es decir sustancias dis ntas relacionadas entre sí por un hapteno común.
An genos específi cos de especie → an genos reconocidos en todos los individuos de una misma especie animal.
An genos específi cos de órganos o tejidos → dentro de un mismo organismo las proteínas que forman un órgano o tejido son dis ntas del resto de órganos o tejidos de dicho organismo (cual sea).
2.3.6.2 Antígenos bacteriano.
An genos cons tu vos → son aquellos an genos que forman parte de la propia estructura de la bacteria y son an genos capsulares, fl agelares y somá cos o de la pared.
An genos secretores → son los an genos que se encuentran en los productos de secreción de las bacterias por lo que pueden ser an genos de exotoxinas y an genos de endotoxinas.
2.3.6.3 Antígenos virales.
En él nos encontramos con las proteínas de la cáspside del virus, las subunidades de la envoltura cuando existe y los ácidos nucléicos asociados a nucleoproteínas también del virus.
2.4 Células del sistema inmune.
Como ya se comento las células del sistema inmune se dividen en 2 grupos que son la serie mieloide, compuestas por todas aquellas células del sistema inmune no pertenecientes a linfocitos y la serie linfoide, la más importante formada por como su nombre indica los linfocitos.
De esta serie la linfoide, diremos que es una serie formada por linfocitos, células del sistema inmune con papel fundamental en la respuesta inmune ya que permite el reconocimiento de an genos y su destrucción.
Se producen en medula ósea, llegando hasta los tejidos a través de la circulación sanguínea y la linfa.
Con morfología aproximada de entre 6-10 micras de diámetro, con núcleo de aspecto uniforme debido a la presencia de una densa croma na y citoplasma con escases de gránulos, lo que da un aspecto pálido a dichas células.
Existen tres pos de poblaciones diferentes:
1. Linfocitos T 2. Linfocitos B3. Linfocitos no T no B
Fig. 2.3. Clasifi cación de antígenos según estructura

20
Técnico Superior Sanitario de Laboratorio de Diagnóstico Clínico
Presenta los linfocitos una caracterís ca fundamental que los dis ngue del resto de células hematopoyé cas, ya que la maduración y diferenciación de los dis ntos pos de linfocitos no se produce de manera simultánea, sino en dos etapas, una primera en donde se producen los cambios celulares que dan lugar a la diferenciación de células T y B y una segunda etapa de diferenciación producida tras la es mulación an génica y que dará lugar a la formación de células plasmá cas a par r de linfocitos B o células T citotóxicas/ supresoras o auxiliares procedentes de los linfocitos T.
1. Linfocitos T
Los linfocitos T comienzan a formarse en la médula ósea, y terminan de madurar en el mo de ahí su nombre.
Para reconocerlos una de las formas más fi ables es mediante la recurrencia a los an genos de diferenciación leucocitaria, que son un conjunto de moléculas presentes en la membrana plasmá ca de dichas células, según estado de maduración de las mismas y que confi ere caracterís cas fundamentales bien defi nidas. Estas molécula se han estudiado en el empleo de an cuerpos monoclonales, las cuales reciben una nomenclatura internacional llamada siglas de Cluster of Diff erencia on CD (grupo de diferenciación con un numero). De estas moléculas las más importantes en superfi cie de linfocitos T son CD3, CD4, CD8, estando solo las CD3 presente en todos los linfocitos T. Las moléculas CD4 y CD8 no están presentes en todos los linfocitos T, lo que da lugar a la formación de dos subpoblaciones de este po de linfocitos tanto funcional como feno picamente diferentes.
Los linfocitos T que presentan moléculas CD4 son los conocidos como linfocitos T Helper (auxiliares o cooperadores), cuya función se encarga de inducir la interacción de células T y B con macrófagos (otro po células), también necesarias para inducir la proliferación de linfocitos B así como la diferenciación de células plasmá cas productoras de inmunoglobulinas (Ig). También produce sustancias de naturaleza proteica que permite la actuación sobre algunas células del sistema inmune, llamado linfoquinas y un interferón que con ene ac vidad an vírica.
Por otro lado los linfocitos T que presentan moléculas CD8, se conocen como linfocitos T supresores/citotóxicos, cumpliendo la función de frenado de respuesta inmune, además de lisar células diana e inac var células inductoras para suprimir la producción de inmunoglobulinas por los linfocitos B.
El complejo molecular proteico común en todos los linfocitos T, el CD3 ene un papel meramente estructural y funcional esencial, siendo responsable de la traducción de la señal al interaccionar el receptor TCR del linfocito T (otra molécula en superfi cie de dicho po de linfocitos, presente en todos los linfocitos T) para ac var asá a los linfocitos T, dando función de reconocimiento de an genos y de dirigir al an geno cuando este se encuentra asociado al complejo mayor de histocompa bilidad (MHC).
Existen otro po de moléculas receptoras presentes en los linfocitos T, pero que son de menor importancia que las anteriores.
2. Linfocitos B
Comienzan a formarse en el hígado fetal hacia las 8-9 semanas de gestación, para desaparecer y aparecer con posterioridad en médula ósea durante toda la vida.
En el transcurso de maduración se suceden una serie de etapas, que llevan a estas células a dar lugar a la producción de los dis ntos pos de inmunoglobulinas, cuando el linfocito B es capaz de agregar cualquier po de inmunoglobulina en la superfi cie de la membrana celular se dice que el linfocito B a alcanzado la madurez para poder contactar con los an genos.
Una vez ene lugar este proceso podemos encontrar dos pos de linfocitos:
Linfocitos B de memoria, que no se dividen.
Linfocitos B efectores, células efectoras capaces de sinte zar an cuerpos, por lo tanto ene capacidad de división y no presentan memoria.
3. Linfocitos no T no B
Células que no presentan los marcadores de los linfocitos T y B.Su estructura es granular y grande por presentar un gran citoplasma con gránulos electrodensos y núcleo con
croma na menos densa que el resto de linfocitos. La mayoría de ellos enen en su superfi cie receptores para el factor de complemento de las inmunoglobulinas G (IgG).

21
El Sistema Complemento (del Sistema Inmune)
Son en su mayoría conocidas como células NK (Natural Killer o agresoras naturales) encargadas de la lisis (destrucción) de células tumorales, además de actuar en células atacadas por virus y Células ADCC (efectoras de citotoxicidad o dependientes de an cuerpos) que se encargan de la lisis de células a través de la unión de an cuerpos a células dianas.
2.5 Anticuerpos.
2.5.1 Conceptos
Los an cuerpos (Ac) son glucoproteínas que se forman en el organismo como respuesta al contacto con un an geno (Ag) y que reacciona específi camente contra él, también se puede defi nir como una sustancia producida por el organismo, de naturaleza proteica, inducida por otras sustancias con las que puede reaccionar de forma específi ca.
Debido al papel desempeñado en el sistema inmunitario y a que, en la electroforesis, migran junto a otras globulinas, también se les llama inmunoglobulinas (Ig), estas en la electroforesis enen la capacidad de separarse en la fracción gamma y por ello también son las inmunoglobulinas conocidas con el nombre de gammaglobulinas.
Las inmunoglobulinas no son más que globulinas plasmá cas que por su ac vidad de an cuerpos reciben el nombre este, por lo tanto los términos inmunoglobulinas (Igs) y an cuerpos (Ac) son sinónimos.
Las inmunoglobulinas son sinte zadas a par r de células plasmá cas que proceden de la diferenciación de los linfocitos B.
Las inmunoglobulinas cons tuyen un grupo heterogéneo de proteínas que cons tuyen el 10% de las proteínas del plasma.
Estos an cuerpos, las inmunoglobulinas se encuentran: en plasma, líquidos extravasculares, secreciones orgánicas y
también en la superfi cie de la membrana de los linfocitos B, incluso en el interior del citoplasma de dichas células.
En el ser humano existen varios pos de inmunoglobulinas que se hacen llamar de la siguiente forma: A, D, E, G y M las cuales se diferencian entre sí por su s propiedades sico-químicas, an génicas y funciones biológicas, estos serán los pos de inmunoglobulinas.
2.5.2 Características generales de las inmunoglobulinas (Igs).
Como ya se ha dicho anteriormente las inmunoglobulinas son glucoproteínas compuestas por proteínas en un 98% y el resto de hidratos de carbono, un 2%.
Cada molécula de inmunoglobulina está cons tuida por una unidad estructural básica o monómero, formada a su vez por cuatro cadenas polipep dicas unidas entre sí por puentes de disulfuro y otras uniones no covalentes.
Las cadenas polipep dicas formadas las podemos dividir y diferenciar en dos grupos que son:
Cadenas ligeras (L) → estas son las dos cadenas más pequeñas de las cuatro existentes por monómero, cuyo peso molecular aproximado es de 22000 μg las cuales están cons tuidas por la unión de 214 aminoácidos aproximadamente. Estos pueden ser de dos pos que se diferencian por la secuenciación de los mismos y son la cadena kappa (κ) y la cadena lambda (λ).
Estas cadenas son expresadas por genes que las codifi can los cuales se encuentran en el cromosoma 2 para el caso de la cadena kappa () y en el cromosoma 22 para el caso de la cadena lambda () en cada uno de los cuales se encuentran distintos diversos segmentos génicos que codifi can dichos genes.
En el ser humano se pueden dar una u otra cadena pero no ambas en una misma inmunoglobulina, de esta forma se encuentran 65% de k y 35% de de las inmunoglobulinas. Esto quiere decir que en las cadenas ligeras (L), las dos son k ó pero no una de un tipo y la otra de otro.
Cadenas pesadas (H) → estas son las dos cadenas más grandes de las cuatro existentes por monómero cuyo peso molecular es del orden de 50000-70000 μg y 450 aminoácidos, dicha secuenciación de aminoácidos de esta la cadena pesada (H) es lo que determina que la inmunoglobulinas sean de un po u otro así como las subclases dentro de cada po, esto es lo que hace que haiga tantas clases de
inmunoglobulinas (Igs) como clases de cadenas pesadas existentes.

22
Técnico Superior Sanitario de Laboratorio de Diagnóstico Clínico
Cada po de cadena pesada (H) existente se designa con una letra minúscula griega lo que corresponde a la letra mayúscula la na de la inmunoglobulina, estas cadenas pesadas son: cadenas alfa α (A), delta δ (D), exilón ε (E), gamma γ (G) y mu μ (M).
Además todos estos pos de cadenas pesadas H son expresados por una serie de genes que codifi can dichas cadenas, genes que se encuentran en el brazo corto del cromosoma 14 en el cuál se encuentran los dis ntos diversos segmentos génicos que codifi can dichos genes.
Tanto las cadenas ligeras (L) como las cadenas pesadas (H) al ser cadenas polipep dicas poseen dos extremos que son:
Extremo aminoterminal → NH3.
Extremo carboxiterminal → COOH.
También el monómero de las inmunoglobulinas posee puentes intercatenarios que unen dos cadenas polipep dicas y puentes intracatenarios que unen fragmentos polipep dicos de una cadena.
En toda cadena polipep dica tanto ligera como pesada existen dos zonas que son:
Zona constante → que corresponde con el extremo carboxi-terminal, conocido como CH y CL (constantes pesadas y ligeras).
Zona variable → que se corresponde con el extremo amino-terminal, conocido como VH y VL (variables pesadas y ligeras).
La parte constante de las dis ntas cadenas pesadas H de cada inmunoglobulina (Igs) es diferente según el po de inmunoglobulina, dado que de la parte constante de las cadenas pesadas H de las inmunoglobulinas es de lo que depende la existencia de una u otro po de inmunoglobulinas de los cinco existentes.
La parte variable de las inmunoglobulinas es la que une a las inmunoglobulinas con su an geno correspondiente.
Dentro de la zona o región variable de las inmunoglobulinas (Igs) se encuentran las regiones hipervariables, tanto de las cadenas ligeras L como de las cadenas pesadas H.
Estas las regiones variables constan de tres regiones en las que se concentra fundamentalmente la variabilidad y que se denominan como ya se ha mencionado anteriormente regiones hipervariables, estas se designan con las letras CDR de la siguiente forma: CDR1, CDR2, CDR3, además de estas regiones existen otras cuatro regiones con secuencias de aminoácidos rela vamente conservada denominada zona de entramado y que se designa con la letra FR de la siguiente forma: FR1, FR2, FR3, FR4.
Con ello podríamos decir que tanto la región hipervariable como la de entramado dentro de la región variable se colocan de la siguiente forma: FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4-CDR4.
En los CDR se encuentran las zonas de la región variable donde se determinan la forma del centro ac vo que permite el reconocimiento y la unión con el an geno, cada una de estas regiones que son tres y que ya han sido mencionadas anteriormente están compuestas por entre 17 y 20 aminoácidos de forma que pequeñas variaciones en la secuencia de estos implica la existencia de dis nto an cuerpo.
Estas cuatro cadenas se organizan entre sí cons tuyendo una estructura en forma de i griega llamada Región Bisagra que es la zona de unión de las dos partes que conforman el monómero, es decir la unión de una cadena ligera y una pesada con la otra cadena ligera y pesada, unidas quedando la estructura pica de las inmunoglobulinas que es en forma de Y.
Esta pica molécula con estructura en forma de Y mediante una enzima proteolí ca llamada papaína, hace que esta se rompa en tres partes:
Dos de ellos son idén cos y constan, cada uno, de una cadena ligera (llamada L) y la porción aminoterminal (N-terminal) de 1 cadena pesada (llamada H). Estos reciben el nombre de fragmento Fab (fragmento capaz de combinarse con el Ag).
El tercero está cons tuido por las porciones carboxiterminales (C-terminales) de las cadenas pesadas (aproximadamente la segunda mitad de las cadenas pesadas H) y se denomina fragmento Fc (fragmento que puede obtenerse en forma cristalina), esta es la fracción que no está dotada de capacidad de unión con el an geno.

23
El Sistema Complemento
La misma molécula, con misma estructura pero esta vez mediante otra enzima proteolí ca llamada pepsina, hace que esta molécula de inmunoglobulina (Igs) se rompa en los siguientes fragmentos:
Un fragmento F (ab)2, precipitante y que corresponde a dos fragmentos Fab unidos entre sí por conservación de puentes de disulfuro entre ambas cadenas.
Varios fragmentos polipep dicos correspondientes a la fragmentación del fragmento Fc.
Todo esto se observa en la siguiente fi gura a modo de resumen.
UNIDAD EXTRUCTURAL BÁSICA DE LA IGS O MONÓMERO
Fig. 2.4. Estructura básica de las inmunoglobulinas (monómero)
A parte de esta estructura básica las inmunoglobulinas presentan moléculas adicionales de glucoproteínas las cuales se conocen como cadena J y pieza de cola o secreción.
Estas moléculas adicionales junto a la estructura básica de las inmunoglobulinas, conocida como monómero y a la unión de varios monómeros, formaran los dis ntos pos de inmunoglobulinas conocidos, ya mencionados, así como los dis ntas clases de inmunoglobulinas dentro de cada po dado que no todas las inmunoglobulinas de cada po son estructuralmente iguales, lo que vendrían a ser las subclases dentro de cada po de inmunoglobulinas.
2.6 Respuesta inmune.
La respuesta inmune puede ser de 2 pos:
Respuesta humoral.
Respuesta celular.
2.6.1 Respuesta humoral
Depende de la interacción an geno-an cuerpo (Ag-Ac) protegiendo al organismo en la mayoría de los casos. Los an cuerpos enen capacidad de renacimiento de an geno siempre que tengan acceso a él en cualquier forma o circunstancia, es decir tanto solubles como en superfi cie sólida como pueden ser un microorganismo o una célula entre otras sustancias.
Los an cuerpos ac van sistemas muy efi caces de defensa frente a lo que le es extraño.

24
Técnico Superior Sanitario de Laboratorio de Diagnóstico Clínico
Aunque se llame humoral requiere células puesto que necesitan:
- Activación de linfocitos B (tipos de células del sistema inmune) para que estos se transformen en células plasmá cas ( po de células del sistema inmune) y puedan producir an cuerpos.
- La ac vación de linfocitos T que han de producir factores solubles que ac ven los linfocitos B.
- Atraer y dirigir células fagocí cas (células destructoras de sustancia de desechos) al lugar necesario para que estas puedan actuar.
Por lo tanto y en base a lo comentado, para que se lleve acabó la respuesta inmune humoral se necesitan tanto la ac vación de linfocitos B como la de los linfá cos T, dado que sin esta ac vación no se llevaría a cabo la respuesta inmune humoral.
2.6.2 Respuesta celular.
Se inicia tras la ac vación de los linfocitos T únicamente.
No intervienen ni an cuerpos ni linfocitos B sino únicamente linfocitos T.
Se desarrolla para hacer frente:
- A microorganismos en el interior de células fagocí cas que escapen a mecanismos lí cos tales como: bacterias intracelulares, hongos y determinados parásitos.
- Microorganismos en el interior de células sin capacidad para destruirlas y que u lizan la maquinaria de la propia célula para reproducirse (virus).
Dicha respuesta la celular da lugar a factores solubles que ac van células fagocí cas.

UNIDAD DIDÁCTICA IIIINTRODUCCIÓN AL SISTEMA COMPLEMENTO


27
El Sistema Complemento (del Sistema Inmune)
3 .1 Introducción.Podemos defi nir el complemento como un grupo aproximado de unas 30 proteínas que al igual que en mecanismos
de coagulación que se llevan a cabo por las plaquetas, forma un sistema enzimá co en cascada en el plasma, por lo que son consideradas como proteínas plasmá cas en su mayoría aunque una pequeña parte de ellas son proteínas de la membrana. En dicho grupo de proteínas se produce una respuesta rápida y ampliada frente a un es mulo an génico al igual que se ejerce una autorregulación para limitar el daño sular producido.
Muchas de estas proteínas se encuentran inac vas. La ac vación de dichas proteínas del complemento ocurre por varias vías, caminos que son:
La clásica o común
lec na
La alterna va
La lí ca o secuencia fi nal, o complejo de ataque a la membrana
La vía lí ca es la vía en donde convergen las otras tres vías en forma de secuencia común de fenómenos que conduce a la generación de un complejo molecular que causa la lisis celular de ahí que se conozca también como secuencia fi nal o complejo de ataque a la membrana.
De estas vías la común o clásica es aquella que se inicia por la unión de an cuerpos a una célula blanco; la reacciones de las inmunoglobulinas M (IgsM) y ciertas subclases de inmunoglobulinas (IgG) ac van esta vía.
Las otras vías tanto la de la lec na como la alterna va son independientes del an cuerpo. Estas vías se inician por la reacción de proteínas del complemento con moléculas superfi ciales de los microorganismos.
Además del papel fundamental en la lisis celular, el sistema complemento mediara la opsonización de bacterias, la ac vación de infl amación y la depuración de complejos inmunitarios. Podemos decir que esta son algunas de las principales funciones del complemento junto a la citolisis (destrucción de células).
Las interacciones de proteínas del complemento y fragmentación proteínica con receptores en las células del sistema inmunitario contralarán las reacciones inmunitarias innata y adquirida. Dichas proteínas se designan con la letra C seguidas de un número que es indica vo del lugar de la secuencia donde se ac va, a excepción del C4, que reacciona entre el C1 y C2.
Por otro lado En el proceso de ac vación del complemento se produce un desdoblamiento de proteínas en donde se le denomina “b” al componente de mayor tamaño y “a” al de menor.
De todos los componentes los que enen ac vidad enzimá ca son aquellos que llevan una línea sobre los números, el resto carecen de dicha ac vidad.
Todo ello puede dar al complemento capacidad para dañar al organismo huésped, mediante mecanismos reguladores pasivos y ac vos complejos. Con ello, consecuencias clínicas de defi ciencias hereditarias del complemento variaran desde un aumento de la suscep bilidad e infecciones al daño de tejidos causados por complejos inmunitarios.
3.2 Historia del sistema complemento.
El sistema complemento es el medidor humoral primario de las reacciones an geno-an cuerpo, principales reacción del sistema inmunológico frente a un agente extraño al organismo. Este sistema fue descubierto, a fi nales del siglo XIX, al comprobarse la capacidad bactericida del suero en fresco por Ehrlich, cien fi co que nació en Polonia y murió en Alemania cuya vida trascurrió entre mediados de siglo XIX y los primeros quince años del siglo XX, llegando a ser premio nobel de medicina en 1908, mediado por dos factores, an cuerpos específi cos frente a microorganismo o termoestable (resistentes) y otro termolábil (que se alteran con facilidad frente al calor).
Rodeny y R. Porter, (1916-1976) (bioquímico inglés) sobre todo, con posterioridad a Ehrlich, que gano el premio Nóbel en 1972, otorgado primordialmente por sus trabajos en inmunoglobulinas (Igs.). Aporto a las inmunoglobulinas adicionalmente en los úl mos 15 años bastante sobre para el esclarecimiento de los mecanismos de acción del complemento.

28
Técnico Superior Sanitario de Laboratorio de Diagnóstico Clínico
Así como, Jules Bordet (1870-1961) (medico inmunólogo y microbiólogo belga), que gano el premio Nobel de medicina en 1919 por sus trabajo en donde se mostraron como el complejo era bacterio co y como par cipaba en la reacciones hemolí cas.
Con esta inves gación se requiere recopilar información acerca del sistema del complemento y su importancia para el sistema inmunológico, ya que este cons tuye unos de los principales mecanismos efectores de la inmunidad, par endo de la base historia presentada.
El sistema complemento
El sistema complemento es el principal efector de la rama humoral del sistema inmunitario. La inves gación del complemento se inicio en la década de 1890, cuando Jules Bordet, ya mencionado, del ins tuto Pasteur en Paris en donde se demostró que el an suero de oveja introducido a la bacteria Vibrio Cholerae lisó (romper) la bacteria y el calentamiento del an suero destruyo su ac vidad bacteriolí ca. Como hecho sorprendente, se restableció la capacidad del suero calentado para lisar bacterias con la adición de suero fresco que no contenía an cuerpos dirigidos contra la bacteria por lo que fue incapaz de destruir la bacteria por sí mismo, con lo que Bordet llego a la conclusión de que la ac vidad bacteriolí ca requería de dos sustancias diferentes que eran, los an cuerpos an bacterianos específi cos que sobrevivían al proceso de calentamiento y el componente sensible al calor que causa la ac vidad lí ca. En base a estos resultados Bordet diseño una prueba bastante simple para la ac vidad lí ca, la lisis de glóbulos rojos recubiertos con an cuerpos que se detectan con facilidad, la denominada hemólisis.
Por otro lado Paul Ehrlich, en Berlín, llevo a cabo de manera independiente experimentos similares e ideo el terminó complemento y lo defi nió como la “ac vidad del suero sanguíneo que completa la acción del an cuerpo”. En los años que le siguieron, los inves gadores descubrieron que la acción del complemento era el resultado de la interacción de un grupo grande y complejo de proteínas.
3.3 Importancia.
El sistema complemento es un sistema de defensa y limpieza, cons tuido como ya se ha comentado por una serie de proteínas solubles, transportadas por la sangre. Puede ser ac vado por an genos (Ag) tras la reacción de estos con an cuerpos (Acs), mediante la unión Ac-Ag. Al hacerlo sus proteínas se solidifi can y adhieren a la superfi cie de las células, gérmenes o moléculas que deben ser destruidas liberando simultáneamente moléculas pequeñas que incrementa la fagocitosis y amplían los mecanismos de infl amación.
Los dis ntos mecanismos que se ponen en marcha con la ac vación del sistema complemento, complementan y refuerzan el mecanismo inmunológico de defensa. No obstante, su acción puede ser nociva cuando su ac vación ene lugar en forma extemporánea o cuando se prolonga innecesariamente, como el caso de las enfermedades autoinmunes.
3.4 Funciones del complemento. La inves gación sobre el complemento incluye en la actualidad más de 30 proteínas solubles unidas a células. Las
ac vidades biológicas de este sistema afectan a las inmunidades innata y adquirida y van mucho más allá de las observaciones originales de la lisis de bacterias y glóbulos rojos mediada por an cuerpos.
Las comparaciones estructurales de las proteínas que par cipan en las vías del complemento sitúan el origen de este sistema en microorganismos primi vos que poseen los sistemas inmunitarios inmunitarios innatos más rudimentarios.
En contraposición, el reconocimiento de que la interacción de receptores celulares con proteínas del complemento controla las ac vidades de las células, linfocitos B, proporciona a este sistema un papel en el sistema inmunitario adquirido y desarrollado en sumo grado. Como consecuencia existe un sistema entre la inmunidad innata y la adquirida que contribuye a cada una de forma diversa.
Después de dicha ac vidad inicial, interactuaran los diferentes componentes del complemento en una cascada muy regulada para llevar a cabo varias funciones básicas, algunas ya mencionadas, entre las que tenemos:
Lisis de células, bacterias y virus.
Opsonización, que promueve la fagocitosis de an genos par culados.

29
El Sistema Complemento (del Sistema Inmune)
Unión a receptores de complemento específi cos en células del sistema inmunitario, lo que desencadena funciones caracterís cas de dichas células, infl amación y secreción de moléculas inmunorreguladoras.
Depuración inmunitaria, para eliminar complejos inmunitarios de la circulación sanguínea y depositarlos en bazo e hígado.
3.5 Componentes del complemento.
El sistema complemento se compone de proteínas y glucoproteínas que se sinte zan sobre todo en los hepatocitos (células del hígado), aunque también producen en menor can dad, pero siendo esta también bastante signifi ca va en monocitos sanguíneos, macrófagos sulares y células epiteliales de los aparatos diges vo y genitourinario.
Estos componen en cuanto a peso cons tuyen el 5% de la fracción de la globulina sérica. Casi todos los componente circulan en suero del organismo como pro-enzimas formando funciones inac vas, o cimógenos (precursor inac vo de una enzima), hasta que ocurre la segmentación proteolí ca que eliminara un fragmento inhibidor y expondrá el si o ac vo, produciendo la ac vación de estas proenzimas, iniciando así la secuencia de reacción del complemento mediante la ac vación de una cascada enzimá ca.
Los componente del complemento de lo que ya se menciono algo antes, se designan con la letra C seguida de un numero hasta 9, es decir de C1-C9, con letras o números comunes como el factor D, o nombres comunes como el factor de restricción homologo.
Los fragmentos pep dicos que se formaran por ac vación de un componente se iniciaran, como ya se comento, con a letras pequeñas. En la mayoría de los casos, los fragmentos más pequeños que resultaran de la segmentación de un componente se designaran con la letra “a” y los más grandes con letra “b” quedando por ejemplo C3a, C3b,etc. De estos fragmentos los más grandes se unirán al blanco cerca del si o de ac vación y los más pequeños se difundirán desde el si o y podrán proporcionar reacciones infl amatorias localizadas por unión a receptores específi cos, Dichos fragmentos actuarán entre sí para formar complejos funcionales, de estos complejos los que tengan ac vidad enzimá ca se designaran con una línea sobre el numero o signo (ej.:.
3.6 Métodos para la identi icación y medición de los componentes del com-plemento.
Son necesarias ciertas precauciones para el manejo de las muestras de sangre en los estudios del complemento. Después de coagularse preferiblemente a temperatura ambiente, debe separarse el suero del coagulo de sangre lo más pronto pronto posible y congelarlo, a -70 ºC (nitrógeno liquido), para prevenir la perdida de ac vidad del complemento. Algunos estudios de laboratorio requieren la inac vación del complemento con el fi n de evitar hemólisis. Esto se logra normalmente por el calentamiento del suero a 56ºC durante 30 minutos, un procedimiento que destruye muchos componentes del complemento.
La ac vidad del complemento por lo general se mide determinando la dilución límite del suero (dilución sucesiva de una suspensión celular, hasta conseguir una alícuota que contenga una sola célula), que lisa los eritrocitos de carnero sensibilizados con an cuerpos an carnero de conejo (hemolisina). Las tulaciones del complemento de este po darán una medida global de la integridad de la vía clásica del complemento y del mecanismo de ataque a la membrana. Los valores se expresan como unidades 50% hemolí cas del complemento por ml (
Un sistema semejante de análisis se emplea para la medición de la ac vidad hemolí ca de los componentes individuales del complemento, en forma aislada o suero. En este po de sistema de ensayo todos los componentes, excepto el implicado, son proporcionados en exceso, imponiéndose ciertas condiciones rigurosas de reacción que son óp mas para componente. En tales condiciones, el número de unidades hemolí cas puede ser conver do, sobre una base ponderal, a la concentración absoluta del componente ac vo del complemento.
En los úl mos años se han ido desarrollando sistemas de valoración para detectar y cuan fi car caracterís cas específi cas del proceso de ac vación del complemento. Estos procesos incluyen análisis que cuan fi can los fenómenos de segmentación proteolí ca ligada que caracterizan a la ac vación de las vías clásica y alterna del complemento.
Existen otros métodos que no son cuan ta vos, pero que aportan evidencias de la ac vación del complemento. Una de estas técnicas incluye la medición del estado fi sicoquímico de los componentes individuales del complemento por técnica de inmunoelectroforesis, la cual mide dis ntas proteínas.


UNIDAD DIDÁCTICA IVVIAS DE ACTIVACIÓN DEL COMPLEMENTO


33
El Sistema Complemento (del Sistema Inmune)
4.1 Vías de activación del complemento.4.1.1 Introducción.
Existen dos vías de ac vación del sistema complemento que son:
Vía clásica → esta es la vía de ac vación donde interviene un complejo an geno-an cuerpo como ac vador de dicha vía que seguidamente será estudiada con más amplitud.
Vía de la lec na → esta es la vía, variante de la clásica que se ac va sin la necesidad de presencia de an cuerpos.
Vía alterna va → esta es la vía de ac vación en la que el propio microorganismo es el desencadenante del proceso.
De estas vías, podemos decir antes de profundizar más en ellas que la clásica enen un mecanismo más reciente en el desarrollo evolu vo, ya que esta es una respuesta inmune adapta va específi ca, la vía de la lec na es una variante de clásica (como ya se dijo) y la vía alterna va es menos reciente y forma parte de la inmunidad innata especifi ca de los organismos.
Todo esto convergerá en la formación de un componente, el C3, el cuál dará lugar en sucesivas reacciones a un complejo de ataque a la membrana para producir la histólisis (destrucción celular), también llamada vía lí ca.
4.1.2 Vía clásica.
La vía clásica puede ser ac vada por complejos an geno-an cuerpo o inmunoglobulinas agregadas.Las inmunoglobulinas humanas capaces de iniciar la vía clásica son:
Subclase IgG1, IgG2 e IgG3
Clase IgM
El resto de las inmunoglobulinas se quedaran inac vas.Entre las inmunoglobulinas mencionadas que se ac van en el caso de las IgG, de las tres subclases la IgG3 será la
más ac va seguida de la IgG1 e IgG2 (por ese orden). La ac vación de estas inmunoglobulinas se llevara a cabo a través de la combinación del primer componente del
complemento (C1) localizado en la región Fc. de las inmunoglobulinas G y M.La vía clásica puede tener ac vación no inmunológica mediante sustancias químicamente diversas, en donde
incluiremos el DNA, la proteína reac va C ciertas membranas celulares y enzimas semejantes a la tripsina. Esta ac vación ocurrirá por combinación directa de C1 con estas sustancias o en el caso de las enzimas como la fi brinoli ca plasmi ca, por ataque proteolí co directo sobre la molécula de C1.
Esta vía comprende los siguientes pasos de reacción de los componentes del complemento:
C1(primero)
C2 (segundo)
C4 (tercero)
C3 (substrato de C4)
Esta vía la clásica, puede dividirse en 2 unidades funcionales:
La primera perteneciente a la ac vación del complemento C1.
La segunda compuesta por dos enzimas relacionadas con el complejo del complemento:
C4b,2a
C4b,2a,3b

34
Técnico Superior Sanitario de Laboratorio de Diagnóstico Clínico
En esto diremos que la primera etapa de la vía clásica es la C1, etapa en la que se ac va el C1 después del ataque proteolí co en donde se construye la primera unidad funcional de la vía clásica del complemento.
C1
Este el C1 está formado por tres fragmentos de moléculas proteicas que son:
C1q
C1r
C1s
Ambas moléculas se mantendrán unidas por un enlace dependiente del calcio, que se encuentran en circulación formando una compacta macromolécula. El resto de subunidades solo podrán aparecer en situación patológica, por lo tanto la aparición de otras subunidades será indica vo de estado patológico del complemento.
Sin embargo, puede ser disociado y reabsorbido con facilidad por eliminación y reposición de los iones de calcio.De estas 3 moléculas del C1 tendremos:
C1q → Electroforé camente, por técnica de electroforesis que veremos más adelante en próximos capítulos, presenta un peso molecular de 400,000, siendo una de las proteínas mas ca onizadas del suero humano, que con ene grandes can dades de glicina, aminoácidos hidroxilazos hidroxiprolina e hidroxilisina y un contenido importante de carbohidratos cons tuidos en gran parte por residuos de glucosa y galactosa pegados en forma de disacáridos, al radical hidroxilo de la hidroxilisina. Además este es el fragmento del C1 que reconoce este complejo an geno-an cuerpo formado, dado que el complejo es capaz de combinarse con la porción Fc de las inmunoglobulinas siempre que estas sean de la clase IgM, IgG1 o IgG3 en los humanos debido a la incapacidad del C1 de unirse a otro po de inmunoglobulinas, siendo capaz de combinarse con aproximadamente 6 moléculas de IgG.
C1r → es una β-globulina cuyo peso molecular es 190,000 que actúa tras la fi jación de C1 a los ac vadores de C1q que se forman y que ene la capacidad de ac var enzimá camente a C1s, al obtener un C con ac vidad esterasa.
C1s → es una α-globulina con peso molecular de 87,000 que después de la ruptura de un solo enlace pep dico en esta molécula por C1r. adquiere ac vidad enzimá ca proteolí ca. Siendo la nueva enzima generada de po serinesterasa y por lo tanto inhibida por análogos del diisopropilfl uorofosfato, localizando el si o enzimá co en la cadena más pequeña de las dos producida por la ac vación proteolí ca de las moléculas. Dichas cadenas estarán ligadas entre sí mediante puentes disúlfuro. Con esta molécula enzimá ca se completa la fase C1 o fase inicial de la vía clásica del complemento en donde dejaran de ser necesarios los reac vos incluyendo el an geno, el an cuerpo, el C1q y C1r para el proceso de reacción del complemento. Este el C1s ac vara la fase siguiente de la vía clásica del complemento encargándose de romper a C4 en dos fragmentos, uno grande C4b y otro más pequeño C4a; donde las moléculas de C4a enen una pequeña capacidad de contracción del músculo liso y las C4b son las con nuadoras de la secuencia del complemento, uniéndose a superfi cies de membranas celulares más próximas.
Fig. 4.1. Primer componente de la vía clásica del sistema complemento

35
El Sistema Complemento (del Sistema Inmune)
C4 y C2
Seguidamente del C1 vendrán el C4 y C2 de los cuales el C4 es una -globulina y peso molecular de 206,000 y la C2 está formada por una –globulina con peso molecular de 117,000. La forma de este complejo bimolecular C2 y C4 ocurre después de que ambas moléculas hayan sido desdobladas por C1s o en el caso C4:
- C1 rompe un solo enlace pep dico localizado en la cadena más grande de las 3 cadenas polipep dicas de esta molécula.
- Ello conducirá a la formación de un si o lábil de fi jación en el fragmento mayor de C4, C4b, el cual lo capacita para unirse a los ac vadores por un periodo breve después de su generación.
- En los úl mos años se ha demostrado que la cadena C4 al igual que la C3 con ene un enlace roester interno del cual no se tenían antecedentes, formado entre un residuo glutámico y otro cisteinico.
- Eso puede permi r al grupo acilo del residuo glutámico formar un enlace covalente con un hidroxilo reac vo o un grupo amino sobre la superfi cie del ac vador.
- El C4a, pequeño pép do producido por C1 se encargara de romper a C4, produciendo ac vidad biológica.
Por otro lado la ruptura de C2 genera:
- Por C1 en C2, C2a, cuya composición química es desconocida pero permite fi jarse a C4b, requiriendo iones de magnesio para la formación del complejo C4b, 2a, cuyo peso molecular es de 280,000.
- El proceso C4b, 2a no será efi ciente.
Por ello la mayor parte de las moléculas C2 y C4 pierden sus si os lábiles de fi jación, antes de lograr la unión con la membrana o entre sí, difundiéndose como producto inac vo de reacción.
El C4b,2a que se formara, dará lugar a una enzima proteolí ca que asume el papel de con nuar una reacción progresiva del complemento, también llamado convertasa de C3, desdoblada y por ello ac va el siguiente componente reaccionante de la serie, C3. Residiendo el si o enzimá co en la fracción C2 del complejo.
C3
Tras formarse el sustrato C4b,2a en el fi nal de C2 y C4, formándose una molécula una molécula –globulina con peso molecular de 185,000 ocurre:
- La molécula del sustrato es desdoblada en un solo si o localizado cerca del aminoterminal del an cuerpo de la cadena mayor (pesada) de la molécula.
- Se forman como en los anteriores casos dos fragmentos C3a y C3b.
- El más pequeño de los dos fragmentos el C3a, es un pép do biológicamente potente.
- El fragmento mayor el C3b que genera un si o de enlace lábil, enlace que permite a esta molécula insertarse en las membranas celulares en si os cercanos, pero dis ntos, de aquellos u lizados por el an cuerpo y C4b,2a. La naturaleza química del si o lábil de fi jación se ha fi jado en los úl mos años.
La vía clásica del complemento consiste, por tanto en base a todo esto, en una serie de acciones reciprocas enzima-sustrato y proteína-proteína que conducen a la formación sucesiva de diversas enzimas del complemento. Esto provoca que las reacciones involucradas sean altamente especifi cas, sin hallarse otras moléculas que puedan sus tuir a los componentes requeridos del complemento.
En las reacciones enzimá cas que se dan en esta vía se producen cambios considerables en las moléculas C2, C3 y C4 en las etapas respec vas de reacción donde se produce un acumulo de productos libres de la reacción en plasma. Debido a la ac vidad biológica de algunos de estos productos resulta evidente que un es mulo rela vamente pequeño para la ac vación del complemento puede conducir a la generación considerable de estos productos biológicamente ac vos.

36
Técnico Superior Sanitario de Laboratorio de Diagnóstico Clínico
Fig. 4.2. Activación de la vía clásica del complemento
Fig. 4.3. Activación de los factores C3 y C5 de la vía clásica del sistema complemento
4.1.3 Vía de lectina.La vía de la lec na, es una variante de la vía clásica como ya se comento. Esta vía está formada por las lec nas que
son proteínas que reconocen blancos de carbohidratos específi cos y se unen a ellos.
La lec na que ac va el complemento se une a unos residuos de una sustancia llamada manosa, que algunos inves gadores designan como vía MB-lec na o vía lec na de unión a la mañana.
Al igual que la vía alterna va, que veremos a con nuación, la ac vación de la vía de la lec na no depende de an cuerpos. Sin embargo, el mecanismo es más similar al de la vía común, porque después de iniciarse prosigue a través de la acción de C4 y C2 para producir la convertasa C5.
Esta vía se ac va por la unión de la lec na que une manosa (MBL) a residuos de manosa en glucoproteínas o carbohidratos en superfi cie de microorganismos, incluidas ciertas cepas de Salmonella, Listeria y Neisseria, además del Crypto coccus neoformans y Cándida albicans. La manosa (MBL) es una proteína de fase aguda producida en respuestas infl amatorias. Teniendo un función similar en la vía del complemento a C1q, a cuya estructura se asemeja. Después de unirse MBL a la superfi cie de una célula o patógeno, lo hacen a MBL porretazas de serina relacionadas con MBL, MASP-1 y MASP-2. El complejo ac vo formado por esta vinculación causara segmentación y ac vación de C4 y C2.

37
El Sistema Complemento (del Sistema Inmune)
Por otro lado diremos que MASP-1 y MASP-2 presentan similitudes estructurales con C1r y C1s e imitan sus ac vidades. Esto signifi ca que la ac vación de los componentes C2-C4 para formar una convertasa de C5 sin necesidad de unión de an cuerpo específi co representa un mecanismo innato de defensa importante comparable a la vía alterna va (que veremos a con nuación), pero que u liza los elementos de la vía común, excepto las proteínas C5.
4.1.4 Vía alternativa
Fue descrita en su origen como el sistema de la properdina, un grupo de proteínas involucradas en la resistencia a la infección, sistema que era semejante al complemento, pero diferente a él. Se encontró que el sistema de la properdina intervenía en la destrucción de ciertas bacterias, en la neutralización de algunos virus y en la lisis de los eritrocitos de enfermos con hemoglobinuria paroxís ca nocturna. El sistema no parecía requerir de an cuerpos específi cos, varios de los factores involucrados en el sistema fueron iden fi cados y aislados en un estado parcialmente purifi cado.
Estos incluían:
Properdina
Factor A
Proteína de alto peso molecular semejante en ciertas propiedades a C4 que era destruida por el tratamiento del suero con hidracina.
Sustancia termolábil (factor B) semejante a C2 pero dis nto a él.
Las inves gaciones indican que la vía alterna va de reciente iden fi cación, es idén ca al sistema de la properdina iden fi cado anteriormente. Esta, la properdina fue aislada y tras ser aislada se comprobó que era una glucoproteína con peso molecular de 220,000.
La ac vación de la vía alterna va se realiza de manera diferente a la de la vía clásica. Un requerimiento inicial para su ac vación es la presencia de C3b, el cual es generado en forma con nua en pequeñas can dades en la circulación. Ac vación que se produce muy probablemente después de la ruptura inducida por agua del enlace oester C3, formando así C3*, el cual reaccionará con los factores B y D que también intervienen en esta vía para general un encima de fase liquida capaz de fragmentar a C3 en C3a y C3b.
Esta vía también puede ser ac vada por una proteína aislada del veneno de la cobra, dado que al parecer esta proteína representa el C3b de la cobra. Esta sustancia ha sido u lizada para disipar la ac vad del complemento in vivo con la fi nalidad de estudiar su función biológica. Esta proteína el factor veneno de cobra, forma un complejo fi rme con el factor B en presencia de los iones de magnesio. El factor B levemente alterado por la incorporación en el complejo, es suscep ble de ser desdoblado y ac vado por el factor D, así se generara una encima desdoblante de C3.
Con todo ello diremos que las proteínas del complemento ac vadoras de esta vía, la alterna va son:
Properdina
Factor A
Factor B
Factor D
Además está cons tuida dicha vía de ac vación por dos proteínas inhibidoras que son:
Factor H (b1H)
Factor (C3INA)
Al igual que la vía clásica dicha proteína de ac vación son β-globulinas de elevado peso molecular.
Las enzimas de esta vía se unen a sí mismas sobre la membrana de la célula diana, sin intervenir para nada el an cuerpo, para romper C3 en C3a y C3b, mediante proteínas que se encuentran en el veneno de la cobra.

38
Técnico Superior Sanitario de Laboratorio de Diagnóstico Clínico
Por otro lado los fragmentos factor B y factor H (inhibidor) compe rán entre sí para unirse al fragmento C3b, por parecer que el si o de unión es común. La prevalencia de uno y de otro depende de la naturaleza de la superfi cie donde se haya fi jado el C3b.
Existen en superfi cies, ac vadoras polisacáridos en su mayoría, que favorecen la unión del factor B al C3b con lo que se forma el complejo C3b,B.
Este complejo el C3b es escindido enzimá camente por la acción del factorD, de manera que es perdido un fragmento pequeño Ba y se forma el enzima C3b,Bb que es un C3 convertasa capaz de transformar C3 en C3b y con nuar con un proceso de retro alimentación.
Esta convertasa la C enen una vida media corta, de 5 minutos aproximadamente, pero que se puede alargar mediante la unión de properdina. De este modo el complejo C llegando a tener una vida media de 30 minutos, con lo que se potencia la acción enzimá ca en dicha vía.
Cuando el fragmento C3b está unido a la superfi cie no ac vadora, el factor H desplaza al factor B del lugar de unión y permite la actuación del factor I, dicho factor se encarga de romper las cadenas α del C3b perdiendo este entonces su ac vidad hemolí ca y su inmunoadherencia.
4.1.5 Secuencia inal, vía lítica o formación del complejo de ataque a la membrana (MAC).
La secuencia fi nal, que es la secuencia terminal de la ac vación del complemento incluye:
C5b
C6
C7
C8
C9
Todos ellos interactúan de manera secuencial para formar una estructura macromolecular denominada complejo de ataque a la membrana (CAM). Este complejo forma un conducto grande a través de la membrana de la célula blanco que permite que se difundan iones y moléculas pequeñas con libertad a través de la membrana.
C5 Y C6
El resultado fi nal obtenido de la ac vación de las vías común o clásica, de la lec na o alterna va es:
Producción de una convertasa C5 ac va. Esta enzima segmentara C5 que contendrá dos cadenas proteínicas, α y β.
Una vez formadas estas dos cadenas proteínicas se producirá la segmentación de α, tras la unión de C5 al componente C3b no enzimá co de la convertasa que segmentara el terminal amino de la cadena α.
Estas dos cadenas proteínicas producidas serán:
El fragmento pequeño C5a, que será difundido.
El fragmento grande C5b, que se unirá a la superfi cie de la célula blanco proporcionando un si o de unión para los componentes subsecuentes del complejo de ataque a la membrana.
De estos dos componentes en los que se fragmenta C5 el C5b será el que presente un extremo lábil que se inac vara en el transcurso de dos minutos a menos que se una a C6 y estabilice su ac vidad.

39
El Sistema Complemento (del Sistema Inmune)
C7
Hasta este punto todas las reacciones del complemento se llevan a cabo en la superfi cie hidrófi la de membranas o en los complejos inmunitarios en la fase liquida. A medida que el complejo formado C5b6 se une a C7, este experimentará una transformación estructural hidrofi la-anfi fi la que expondrá regiones hidrófobas que servirán como si o de unión para fosfolipidos de membrana. Si la reacción ocurriera en la membrana de una célula blanco, los si os de unión hidrófobos permi rán que se inserte el complejo C5b67 dentro de la bicapa fosfolipida. Si por otro lado, la reacción tuviera lugar en un complejo inmunitario u otra superfi cie de ac vación no celular, entonces los si os de unión hidrófobos no podrán fi jar el complemento y este será liberado. Los complejos liberados, C5b67 podrán insertarse en la membrana de células cercanas y mediar la lisis. En condiciones normales, las proteínas reguladoras de la membrana impedirán que suceda lo anterior y se produzca la lisis; lo que ocurre es que en ciertas enfermedades el daño celular y sular puede ser efecto de la lisis.
C8 y C9
Los úl mos eslabones de la vía lí ca o de ataque la membrana o secuencia fi nal y de toda la vía del complemento son C8 y C9. En donde C8 al unirse a C5b67 y este a la membrana provocara un cambio de conformación en C8, de tal manera que este sufrirá una transformación hidrófi la-anfi fi lla que expondrá una región hidrófoba interactuante con la membrana plasmá ca.
La etapa fi nal del MAC es la encargada de la polimerización de la producción de C9, una molécula parecida a la perforina C5b678 que puede unirse y polimerizar de entre 10 a 17 moléculas de C9 por un solo complejo C5b678. Durante esta polimerización, las moléculas de C9 experimentaran una transformación hidrófi la-anfi fi la, de tal forma que también puede insertarse en la membrana. El MAC terminado, consiste en un complejo C5b678 rodeado por un complejo poli-C9. Debido a que los iones y moléculas pequeñas podrán difundirse con libertad a través del conducto central del MAC, las células no podrán conservar su estabilidad osmó ca entonces y se destruirán por la entrada de agua provocando la perdida de electrolitos.
Fig. 4.4. Vía lítica, secuencia fi nal del sistema complemento

40
Técnico Superior Sanitario de Laboratorio de Diagnóstico Clínico
Fig. 4.5. Esquema resumen del sistema complemento al completo
4.2 Causas, consecuencias y factores de la actividad del complemento.
4.2.1 Funciones de los factores del complemento.Estas funciones las realizan moléculas producidas durante el proceso de ac vación:
Factores producidos en la vía clásica
1. C4a → estos enen acción quimiotác ca, pero de muy baja potencia comparada con C3a y C5a que promueve la adherencia inmunológica, proceso por el cual células o plaquetas se adhieren a los an genos cubiertos con an cuerpos para facilitar la fagocitosis (destrucción).
2. C4b → es capaz de neutralizar virus, evitando su ac vación.
3. C2b → bajo la acción del alamín, el C2b genera moléculas con poderosa acción del po de las minas, llamadas C2 mínimas y que inducen la liberación de histamina. La C2 mínima incrementa la permeabilidad capilar en grado casi igual a la histamina pero sin sus otras acciones. Juega un papel importan en edemas angioneuro cos tanto congénitos como adquiridos.
Factores producidos en la vía alterna va
C3a y C5a
Son quimiotác cos, atraen leucocitos y son anafi lotoxides por excelencia, especialmente el C5a, que son cadenas formadas por 70 y tantos aminoácidos, los cuales al perder uno de ellos deja al descubierto ciertas sustancias que inducen a la contracción muscular lisa. Siendo C5a de10 a 20 veces más ac vo que C3a. El C5a como ya se dijo potente quimiotác co,

41
El Sistema Complemento (del Sistema Inmune)
ac va los neutrofi los, lo que provoca la acción de una sustancia, que induce a la granulación, produciendo contractura de musculatura lisa. Además ac va el sistema de coagulación al inducir la producción del ac vador de la tromboplas na, sustancia que interviene en la coagulación.
4.2.2 Receptores del complemento.
En los úl mos años se han intensifi cado los estudios sobre los receptores del complemento que poseen células del organismo y en especial las del sistema inmunitario. Se han conocido las funciones de varios de los receptores así como la patología que se origina por su carencia o defi ciencia gené ca.
Estos receptores reconocidos son:
1. Receptor C1q
2. Receptor CR1
3. Receptor CR2
4. Receptor CR3
5. Receptor CR4 (CD18/11c)
6. Receptor C5a
1. Receptor C1q
Este se une a las moléculas de C1q, manán, surfactam y coaglu ninas, ac vando su respuesta oxida va.
2. Receptor CR1
Este Reacciona con la fracciones C3b/C4b, que se conocen en la nueva nomenclatura de receptores de membrana de las células del sistema inmunitario como CD35, glucoproteína de membrana de 25 kD presente en eritrocitos, fagocitos, linfocitos B, células dentrí cas y tejidos como el glomérulo renal, codifi cados en genes del cromosoma 1, con cuatro variantes alo picas. Cada eritrocito presenta 500 moléculas de CR1. Sus funciones son promover la adhesión a células que lo poseen cuando se ac va el complemento, en eritrocitos C3b este receptor se encuentra como inac vo haciendo del glóbulo rojo un transportador de complementos inmunes hacia el sistema re culoendotelial.
3. Receptor CR2
Este receptor también es conocido como CD21, el cual reacciona con algunos componente del complemento que son los principales que son C3d, C3dg e iC3b, y se encuentra presente en linfocitos B y en células re culares de folículos de los ganglios linfá cos. Este receptor es igual que el de algunos virus como el Epstein-Barr.
4. Receptor CR3
Se encuentra integrado en algunas moléculas tales como CD11a, b, c y por la CD18, encontrándose en proteínas de la membrana nuclear, macrófagos y en linfocitos granulares. Reacción con el componente iC3b, el cual interviene en la adhesividad celular. Esto provoca desprendimiento tardío de cordón umbilical, suscep bilidad a infecciones, leucocitosis persistente y periodon s progresiva y otros síndromes como la defi ciencia de adhesión a los leucocitos.
5. Receptor CD4 (CD18/11c)
Presente en neutrófi los, monocitos, plaquetas y linfocitos B, reaccionando con las moléculas y ac vando la fagocitosis por macrófagos.
6. Receptor C5a
Es un receptor que capta las moléculas C5a en ac vación del complemento, con poderosa función quimiotác ca proteínas de la membrana celular que inducen a la liberación de las enzimas almacenadas en sus gránulos iniciando la producción e aniones.

42
Técnico Superior Sanitario de Laboratorio de Diagnóstico Clínico
4.2.3 Control de la actividad del sistema del complemento.
El sistema complemento es frenado en su acción por una serie de mecanismos enzimá cos, una vez logrados los niveles necesarios de ac vidad. Además del catabolismo (transformación de una biomolécula más compleja en otra más simple) normal del complemento, se han adquirido actividades enzimáticas, existiendo para ello varias proteínas reguladoras agrupadas en tres categorías:
Inhibidoras, que previenen la ac vación de varios de los factores del sistema.
Reguladoras, que desac van algunos de los factores
Protectoras, de células normales de los tejidos.
Inhibidoras, inac vadoras Las proteínas inhibidoras del complemento son:
1. Inac vador de C1 → enzima que inhibe la ac vidad enzimá ca del factor C1 e impide la función del factor Hageman ac vado (también conocido como factor XII de coagulación sanguínea). Cuando este está ausente por defi ciencia gené ca, se presenta una enfermedad llamada edema angioneuro co hereditario (rápida tumefacción de la piel, las mucosas y los tejidos submucosos) debido a una hiperproducción de moléculas con actividad vasodilatadora que incrementa la permeabilidad capilar.
2. Cofactor proteico de la membrana (MCP) o (CD46) → actúa sobre C4b y C3b.
ReguladorasLas proteínas reguladoras del complemento son:
1. Inac vador de anafi lotoxinas → es una carbosilpep dasa de 310 kD que desac va las moléculas C3a, C4a, C5a al removerle por la sustancia arginina terminal.
2. Acelerador del inac vador del C3b o Factor H → es una cadena de polipec dos de 150 kD, que acelera la disociación del complejo C3biBb.
3. Proteína ligadora del C4 o factor I → es una molécula heptamerica (formada por 8 monómeros), que al microscopio electrónico se ven en forma de araña y que acelera el catabolismo C4b2a y el iC3b (i porque depende de uno de los factores).
4. Inac vador del C5b → se conoce también como proteína-S o vitronec ca. Inac va el sistema impidiendo la unión del C5b a la membrana celular.
ProtectorasLas proteínas protectoras del complemento son:
1. Factor ac vador del catabolinos (DAF CD55) → es un factor inhibidor de lisis (MIRL), el cual se encarga de inhibir la asociación del C4b al C2 que se encuentra en membrana de los leucocitos y células endoteliales protegiéndolos de la acción del complemento, además de impedir que el complemento C5b7 se fi je a la membrana de las células.
2. Factor restrictor de homología (HRF) →provoca que la sustancia Clusterin, SP-40, 40 inhiba la formación del complejo de ataque a la membrana.
3. Factor inhibidor de lisis (MIRL) o CD59 →este se une al complejo C5b678 e impide que el factor 9 se pueda incrustar en la membrana. La molécula MIRL cubrirá el complejo C5b678 impidiendo que esta se una a las 6 moléculas de C9.
Todos estos factores protectores del complemento se anclan a la membrana por medio de una molécula de glucosilfosfa dilinositol (GPI), formando un conjunto que protege las células no blanco al inhibir la convertasa del C3. La carencia de GPI será la responsable de la hemoglobinuria paroxís ca nocturna (un po de anemia), en donde un clon de eritrocitos que carece del GPI experimenta la ac vación del complemento en su membrana y por consiguiente la lisis celular.

43
El Sistema Complemento (del Sistema Inmune)
4.2.4 Sistema complemento e in lamación.
Para controlar el sistema complemento en procesos infl amatorios se están evaluando compuestos endógenos como el inhalador C1(C-1NH), o receptores recombinantes solubles para factores del complemento, o an cuerpos, contra CD44 y CD59 para evitar la acción del complejo ataque a la membrana.
La infl amación se producirá mediante 3 procesos que son:
Movilización de leucocitos
Anafi laxis
Inmovilización de los leucocitos
1. Movilización leucocitaria → el fragmento C3e derivado del catabolismo del C3 será en parte responsable de la movilización de leucocitos de la medula ósea hacia el torrente circulatorio. El C3a y el C5a serán los encargados de la migración o salida de neutrofi los de los vasos hacia los tejidos.
2. Anafi laxis → aquí las fracciones C4a, C3a y C5a denominadas también anafi lotoxinas, por inducir a la liberación de histamina, lo que incrementa la producción de la sustancia eicosanodies, produciendo contractura de la musculatura lisa.
3. Inmovilización de los leucocitos → uno de los factores de la vía alterna va más concretamente el Ba se encarga de inmovilizar a los monocitos en el si o al cual fueron atraídos por el C5a.
Las funciones biológicas del sistema de complementos en cuanto a la infl amación se incluyen en dos categorías generales; la lisis celular por el complejo de ataque a la membrana (MAC), y los efectores biológicos de los fragmentos proteoroli cos del complemento.
Los factores derivados del complemento afectan a diversos fenómenos en infl amación aguda:
Fenómenos vasculares → los componente del complemento C3a, C4a y C5a producto de fragmentación del mismo, incrementa la permeabilidad vascular y producen vasodilatación mediante la liberación de la sustancia histamina, siendo esta una de las sustancias más toxicas del complemento. Por otro lado C5a también activa la vía conocida como lipocigenasa del metabolismo del acido araquidonico (AA) en los neutrofi los y monolitos, dando lugar a un incremento en la liberación de mediadores infl amataorios.
Adhesión, quimiotaxis y activación de los leucocitos → el C5a es un potente agente quimiotáctico para neutrofi los, monocitos, eosinofi los y basofi los. También se encarga de la adhesión de los leucocitos al endotelio mediante la activación de los propios leucocitos y el aumento de la intensidad de unión de las integrinas de superfi cie a su ligador endotelial.
Fagocitosis → C3b y C3bi, cuando se fi jan a la pared celular bacteriana, actúan como opsoninas favoreciendo la fagocitosis por parte de neutrofi los y macrófagos, que presentan receptores para C3b en su superfi cie.
Entre los componentes del complemento, el C3 y C5 son los mediadores infl amatorios más importantes. Su importancia radica en el hecho de que pueden ser ac vados por diversas enzimas proteo-lí cas presentes en el exudado infl amatorio. Entre ellas se encuentran la plasmina y las enzimas lisosomales liberadas por los neutrofi los. Por tanto, el efecto quimiotac co, del complemento y los efectos de ac vación del mismo que inducen los neutrofi los pueden instaurar un ciclo autoperpetuado de migración de neutrofi los.
4.2.5 Sistema complemento e infección.
El sistema complemento juega un papel muy importante en la defensa contra infecciones. El factor C3, produce el catabolismo del C3b, permite la salida de las proteínas de la membrana nuclear de la medula a los vasos. El C5a promueve la marginación y adhesión de estas células al endotelio vascular y su paso a los tejidos para legar al lugar de la infección.
Por otro lado el C3b adherido a bacterias es mula la fagocitosis. Una molécula de este factor reemplazara 100 de an cuerpos. El C5a ac va el metabolismo oxida vo de las proteínas membrana nuclear.

44
Técnico Superior Sanitario de Laboratorio de Diagnóstico Clínico
Ciertos factores como C3b, C3b1 y C5a es mulan la síntesis de varias citoquinas como la TNF y el GM-CSF que a su vez ac van la fagocitosis. Por este mecanismo indirecto, el complemento facilitara la inges ón de algunas bacterias como el Cryptococcus neoformans, que no es atacado directamente por el complemento.
Igualmente ac va por la misma vía a los potenciales membrana nuclear para la fagocitosis de plásmidos.
Por otro lado microorganismo como el gonococo y el meningococo pueden ser destruidos por el complemento en ausencia de proteínas de la membrana nuclear. Las cepas de algunas bacterias como el Staphilococcus albus y Epidermi dis ac van la vía alterna lo que hace que puedan ser fagocitadas sin necesidad de an cuerpos.
Algunos microorganismos usan el complemento en los procesos de patogenia. Así el virus de Epstein-Barr entrara en los linfocitos B por medio del Cr2. Por otro lado las microbacterias se fi jaran a las enzimas C2a en su membrana para cribar el C3.
También fi jaran igualmente el C3b generado, el cual se unirá al Cr3 en membrana de los macrófagos. Otras bacterias como el Schistosoma producen una molécula similar a la CD59 con la cual se inhibe la formación del complejo de ataque a la membrana.
4.2.6 Complemento y respuesta inmune humoral.
El complemento juega un importante papel en modular en la respuesta inmune humoral específi ca.
La carencia de C3 impide la producción de Inmunoglobulinas G, la unión de C3d al receptor CD21 de linfocitos B que se incrementa hasta en 1000 veces la respuesta de producción de an cuerpos.
Par cipa en la supervivencia de linfocitos B en los centros germinales, así como en el desarrollo de linfocitos de memoria a través del C3d y su ligando en los FdC. Igualmente infl uyen la tolerancia de los linfocitos B hacia an genos propios.
4.2.7 Consecuencias biológicas de la activación del complemento
El complemento sirve como medidor importante de la reacción humoral porque amplifi ca la respuesta y la convierte en un mecanismo de defensa efi caz para destruir microorganismos invasores. El complejo de ataque a la membrana media la lisis celular, en tanto que otros componentes del complemento o productos de segmentación par cipan en la reacción infl amatoria, la opsonización de an genos, la neutralización viral y la depuración de complejos inmunitarios.
Gran can dad de las ac vidades biológicas del sistema del complemento depende de la unión de fragmentos de complementos a receptores del mismo, expresado por diversas células. Además, algunos receptores del complemento enen una función importante en la regulación de la ac vidad del complemento por la unión de componentes del complemento, ac vo desde el punto de vista biológico, y su degradación en productos inac vos.
El complejo de ataque a la membrana formado por la ac vación del complemento puede lisar bacterias gram nega vas, parásitos, virus, eritrocitos y células nucleares. Puesto que las vías de ac vación alterna vas y de lec nas suelen ocurrir sin ninguna interacción inicial de an geno y an cuerpo, sirviendo estas vías de defensas inmunitarias innatas importantes contra microorganismos infecciosos. La necesidad de una reacción inicial de an geno y an cuerpo en la vía común completa estas defensas innatas inespecífi cas con un mecanismo de defensa más especifi co. En algunos casos, la necesidad de an cuerpo en el fenómeno ac vador pueden proporcionarla los llamados an cuerpos naturales, que se secretan contra componentes comunes de microbios.
Por lo general, el sistema del complemento se considera en términos del resultado fi nal de lisis celular, pero varios pép dos que se generan durante la formación del complejo de ataque a la membrana enen función decisiva en el desarrollo de una reacción infl amatoria efi caz. Los fragmentos más pequeños que resultan de la segmentación del complemento, C3a, C4a, C5a, llamadas anafi lotoxinas, se unen a receptores en células cebadas y basofi los sanguíneos e inducen a la desgranulación, con liberación de histamina y otros medidores con ac vidad farmacológica. Las anafi latoxinas inducen así mismo contracción del musculo liso y aumentan la permeabilidad vascular. Por lo tanto, la ac vación del sistema de complemento ene como resultado el ingreso de líquido que lleva an cuerpo y células fagocí cas al si o de entrada del an geno.
La importancia del sistema del complemento en la eliminación de complementos inmunitarios se observa en pacientes con la enfermedad auto-inmunitaria Lupus Eritematoso Sistémico.

45
El Sistema Complemento (del Sistema Inmune)
4 .2.8 Conclusión.
Tomando esta información como base se pudo determinar la importancia del sistema de complemento, ya que desempeña un papel importante en las enfermedades, como es el caso de enfermedades neurológicas infecciosas, autoinmunes y degenera vas, donde se encuentra involucrado en su fi siopatología como veremos a con nuación.
El sistema del complemento es un importante componente soluble del sistema inmunitario innato, compuesto por una serie de enzimas plasmá cas, proteínas reguladoras y proteínas que se ac van en forma de cascada, que dan lugar como resultado fi nal a la lisis celular.
Se ha podido determinar que en el proceso del sistema del complemento existen 4 vías importantes, que son:
La clásica → es mulada por el complejo inmunitario an geno-an cuerpo.
La de lec na → fi jadora de manosa es mulada por los microorganismos con grupos terminales de manosa.
La alterna va → que es es mulada por microorganismos o células tumorales.
La terminal, lí ca o de complejo de ataque a la membrana → es común para las otras 3 vías y origina como su propio nombre indica, complejo de ataque a la membrana que lisa a las células.
Por úl mo como ya comentemos antes, se ha podido determinar que el complemento ofrece protección contra la infección por bacterias piógenas, favorece la eliminación de complejos inmunes y de productos de la infl amación.


UNIDAD DIDÁCTICA VENFERMEDADES ASOCIADAS


49
El Sistema Complemento (del Sistema Inmune)
5.1 Introducción.El sistema complemento puede intervenir en las enfermedades humanas de dos formas generales:
Forma primera →en esta primera forma, puede ocurrir una defi ciencia en cualquiera de los componentes proteicos que pueden causar patrones anormales de la ac vidad del complemento. La ausencia de un componente de la vía clásica o alterna va, así como la de la lec na (variante de la clásica) o ausencia de los pasos fi nales, vía lí ca o complejo de ataque a la membrana puede dar lugar a una ac vidad defi ciente del complemento. Si los que faltan son los reguladores, puede aparecer una ac vación excesiva del complemento en el momento o lugar equivocado, lo que puede provocar infl amación o lisis celular excesiva y defi ciencias de proteínas del complemento plasmá co por un consumo excesivo.
Forma segunda → en segundo lugar, en esta segunda forma, puede ocurrir que un sistema del complemento intacto, con funcionamiento normal, puede ser ac vado por un es mulo anómalo, como microorganismos persistentes, an cuerpos frente a an genos propios o depósitos de inmunocomplejos en los tejidos. En estas infecciones y enfermedades autoinmunitarias, los efectos lí cos e infl amatorios del complemento pueden contribuir signifi ca vamente a los cambios patológicos de la enfermedad.
En base a estas formas patológicas tendremos dos pos de patologías asociadas al sistema complemento:
Defi ciencias del complemento
Efectos patológicos de un sistema de complemento normal
5.2 De iciencias del complemento.
En los seres humanos se han descrito defi ciencias hereditarias y espontáneas de muchas proteínas del complemento. Entre las defi ciencias descritas tenemos:
1. Defi ciencias gené cas de los componentes de la vía clásica.
2. Defi ciencias de los componentes de la vía alterna va.
3. Defi ciencias de los componentes terminarles del complemento.
4. Defi ciencias de las proteínas reguladoras del complemento.
5. Defi ciencias de los componentes del complemento.
5.2.1 De iciencias genéticas de los componentes de la vía clásica
Se han descrito defi ciencias gené cos de algunos componentes de la vía clásica del complemento, tales como, C1q, C1r, C4, C2 y C3. De estas la de C2 es la defi ciencia observada con más frecuencia en los seres humanos.
Más del 50% de los pacientes con defi ciencia de C2 y C4 sufren una enfermedad similar al lupus eritematoso sistémico, una enfermedad autoinmunitaria. Se desconocen los mo vos de esta asociación, pero los defectos de la ac vación del complemento pueden hacer fracasar la eliminación de los inmunoclomplejos circulantes. Si los inmunocomplejos generados normalmente no se eliminan de la circulación se pueden depositar en las paredes de los vasos sanguíneos y de los tejidos, donde ac van a los leucocitos por las vías dependientes del receptor Fc produciendo una infl amación local.
Además las proteínas del complemento regularan las señales mediadas por los an genos recibidas por los linfocitos B; en su ausencia, los autoan genos pueden no inducir la tolerancia en los linfocitos B produciéndose autoinmunidad.
Aunque parezca sorprendente, la defi ciencia de C2 y C4 no suele asociarse a un aumento de la suscep bilidad a las infecciones, lo que indica que la vía alterna va y los mecanismos efectores mediados por el receptor Fc son sufi cientes para la defensa del huésped frente a la mayoría de los microorganismos.
La defi ciencia de C3 se asocia a infecciones bacterianas piógenas (protuberancia rojiza) graves que pueden ser incluso
mortales, lo que confi rma la función central de C3 y su importancia en la opsonización de la fagocitosis y la destrucción de estos organismos.

50
Técnico Superior Sanitario de Laboratorio de Diagnóstico Clínico
5.2.2 De iciencias de los componentes de la vía alternativa.
Defi ciencias de sustancias de la vía alterna va como la properdina y el factor D, provocan una mayor suscep bilidad a las infecciones por bacterias piógenas.
5.2.3 De iciencias de los componentes terminales del complemento.
En las defi ciencias de los componentes terminales del complemento, tales como C5, C6, C7, C8 y C9; curiosamente, el único problema clínico constante presente en los pacientes es una mayor propensión a las infecciones diseminadas por las bacterias del grupo Neisseria, tales como Neisseria meningi dis y Neisseria gonorrhoeae, lo que indica que la lisis bacteriana mediada por el complemento es especialmente importante para la defensa ante estos microorganismo.
5.2.4 De iciencias de las proteínas reguladoras del complemento.
Estas defi ciencias se asocian a una ac vación anómala del complemento y a varias alteraciones clínicas relacionadas.
Las defi ciencias del inhibidor C1 y el factor de desaceleración ya mencionados, en donde en los pacientes con defi ciencias del factor I, el factor C3 plasmá co se agota como consecuencia de la información no regulada de C3 convertasa de la fase líquida (mediante el mecanismo de ralen normal). La consecuencia clínica obtenida será el aumento de infecciones por bacterias piógenas. La defi ciencia del factor H es rara y se caracteriza por un exceso de ac vación de la vía alterna va, consumo de C3 y gromerulonefri s producida por la eliminación inadecuada de los inmunocomplejos y el depósito renal de productos del complemento. Los efecto de la falta de factor I y H serán similares a los de un autoan cuerpo denominado factor nefrí co C3 (), especifi co para la C3 convertasa de la vía alterna va (C3bBb).
CeNef se encargara por tanto de estabilizar a C3bBb y proteger al complejo de la disociación mediada por el factor H, lo que dará lugar a un consumo incontrolado de C3. Los pacientes con este an cuerpo a menudo presentan gromerulonefri s, posiblemente causada por una eliminación inadecuada de los inmunocomplejos circulantes.
5.2.5 De iciencias de los componentes del complemento.
Estas defi ciencias incluyen la ausencia del receptor del complemento po 3, CR3 y CR4, ambas debidas a mutaciones raras en el gen de la cadena beta (CD18), que es frecuente en las familias de moléculas CD11CD18 de las integrinas. La enfermedad congénita producida por este defecto gené co se llama defi ciencia de adhesión leucocitaria, enfermedad por defecto de la inmunidad innata que puede ser de po 1 y 2 (DAL-1, DAL-2) y que es autosómica recesiva rara, caracterizada por infecciones bacterianas y micó cas de repe ción y un deterioro de la cicatrización de las heridas. Además en estos pacientes, casi todas las funciones leucocí cas son anómalas. Por ello se dice que esta enfermedad es caracterizada como infecciones piógenas recurrentes, debidas a la adhesión incorrecta de los neutrófi los al endotelio en los tejidos de la zona de infección y un deterioro de la fagocitosis de bacterias dependiente de iC3b.
5.3 Efectos patológicos de un sistema del complemento normal.
Incluso cuando se regula correctamente y se ac va de forma apropiada, el sistema del complemento puede provocar un daño sular importante.
Existen algunas enfermedades asociadas a una defi ciencia gené ca de uno o varios componentes del sistema complemento que pueden provocar un daño sular importante. Algunos de los efectos patológicos asociados a la infecciones bacterianas pueden deberse a respuestas infl amatorias agudas a los microorganismos mediadas por el complemento.
En algunas situaciones, la ac vación del complemento se asociara a trombosis intravascular, produciendo lesiones isquémicas de los tejidos. Como ejemplo, los an cuerpos an endoteliales frente a los órganos trasplantados vascularizados y los inmunocomplejos, autoinmunitarias pueden unirse al endotelio vascular y ac var el complemento, produciendo así infl amación y generación de complejos de ataque a la membrana (CAM), lo que dañara la superfi cie endotelial y favorecerá la coagulación. También se ha comprobado que algunas proteínas fi nales del complemento pueden ac var la prontobinasa en la circulación, lo que indica trombosis independientemente de la lesión endotelial mediada por CAM.
Durante mucho empo se ha creído que el ejemplo más claro de trastornos mediados por el complemento son las enfermedades por inmunocomplejos, causantes de enfermedades producidas por an cuerpos, en donde la enfermedad se puede formar por an genos tanto propios como extraños, unidos al correspondiente an cuerpo, refl ejando la localización de los depósitos, pero sin depender de la procedencia celular del an geno, siendo esta sistémica con especifi cidad escasa o nula por tejidos u órganos concretos que aparecen en las enfermedades. Las vasculi s sistémicas y glomerulonefri s son

51
El Sistema Complemento (del Sistema Inmune)
debidas al depósito de complejos an geno an cuerpo en las paredes de los vasos y el glomérulo renal. El complemento ac vado por las inmunoglobulinas de estos inmunocomplejos iniciara respuestas infl amatorias agudas que destruirán las paredes de los vasos o los gromérulos y originaran trombosis, lesión isquémica de los tejidos y cicatrización.
Sin embargo, estudios realizados con ratones ha comprobado que estos carecen de proteínas del complemento C3 o C4 o receptores Fcγ, lo que señala que la ac vación de los leucocitos mediada por el receptor Fc puede provocar infl amación y daño sular como consecuencia del depósito de IgG incluso en ausencia de ac vación del complemento.


UNIDAD DIDÁCTICA VITÉCNICAS BASADAS EN LA REACCIÓN DE
FIJACIÓN DEL COMPLEMENTO


55
El Sistema Complemento (del Sistema Inmune)
6.1 Introducción.Estas técnicas las basadas en la reacción de fi jación del complemento están fundamentadas en la capacidad de unión
que enen los componentes de dicha reacción a los complejos an geno-an cuerpo, los cuales se encargan de evaluar la ac vidad de todos los componentes de este sistema el de complemento en general o la de algunos en par cular, en función del método empleado. También mediante estas técnicas se puede determinar los niveles de an genos o de an cuerpos en suero del paciente.
Existen otras técnicas que encargadas de la cuan fi cación de los componentes del complemento, como son el caso de las técnicas de nefelometría y turbidimetría que estudiaremos un poco más adelante en este mismo capítulo aunque estas no pertenezcan a la reacción de fi jación del complemento dado que pese a ello si son técnicas relacionadas con el sistema complemento, esto es debido a que no precisa la fi jación del sistema del complemento a un complejo an geno-an cuerpo. Estas son técnicas más sencillas que permiten más rapidez en el análisis dado que no necesitan ser fi jadas al sistema complemento, pero pese a ello son técnicas como ya se ha mencionado relacionadas con el sistema complemento aunque no con el de la reacción de fi jación del mismo.
Aunque las técnicas de fi jación del complemento han ido siendo desplazadas por pruebas más sensibles, como puede ser el caso del enzimoinmunoanálisis (EIA) que estudiaremos en capítulos posteriores, se siguen u lizando con frecuencia en el diagnos co microbiológico en determinadas infecciones producidas principalmente por los virus del grupo o po herpevirus, citomegalovirus y micoplasmas en donde se valorara la capacidad funcional del complemento.
Esta valoración en dichas técnicas las basadas en la reacción de fi jación del complemento se basan en la capacidad de unión de un componente del complemento formando en esto el complejo an geno-an cuerpo-complemento, dada la unión al complejo an geno-an cuerpo del complemento evitándose con ello la hemólisis al producirse la fi jación del complemento al complejo an geno-an cuerpo, que es lo mismo que dicho complemento produciría sobre un sistema de hema es A an hema es.
6.2 Tipos de técnicas relacionadas con la reacción de ijación del complemento.
6.2.1 Introducción.
En este po de técnicas el primer componente de la secuenciación del complemento se une a una molécula de an cuerpo por el segundo dominio constante de la cadena pesada. Esta unión sólo ocurre si el an cuerpo esta previamente fi jado a la molécula de an geno.
No todos los an cuerpos enen esta capacidad la de ac vación de la secuencia del complemento, pero si enen la capacidad de poder hacerlo como el caso de las inmunoglobulinas M que son las moléculas más efi cientes, debido a su estructura pentamérica.
Como norma general, en general todas las técnicas de este po u lizan una suspensión de hema es de carnero sobre los que actúa el sistema complemento, produciéndose una hemólisis con la liberación de hemoglobina. Esta hemoglobina se puede cuan fi car por espectrofotometría y es directamente profesional a la can dad de complemento de la muestra.
6.2.2 Técnica en una etapa.
Con estas técnicas basadas en la reacción de fi jación del complemento se puede cuan fi car el nivel total del sistema del complemento o cada uno de los componentes del mismo por separación.
Dicha técnica consiste en mezclar un volumen conocido de eritrocitos (normalmente de carnero), con un volumen conocido de an cuerpos an eritrocitarios de carnero, conocido también como hemolisina. Con ello una vez formados los complejos an geno-an cuerpo se les añade a estos el complemento.
Con todo esto por un lado si se quiere valorar la ac vidad total del complemento se les añaden diluciones diferentes del suero problema, pero si en vez de ser la ac vidad total es lo que se quiere valorar la ac vidad de uno solo de los componentes del complemento añadiremos un suero problema y a este un exceso de todos los componentes del complemento excepto el que interese estudiar, de modo que su concentración sea el factor limitante de la reacción.
En ambos casos se mide la hemoglobina liberada en la hemólisis de los eritrocitos por técnicas espectro fotométrica.

56
Técnico Superior Sanitario de Laboratorio de Diagnóstico Clínico
Fig. 6.1. Prueba de fi jación del complemento en una etapa
6.2.3 Técnica en dos etapas.
Esta técnica dentro de la fi jación del complemento se u liza para medir la concentración del an geno o an cuerpo presentes en un suero problema.
Con ello lo que acurre es que en una primera etapa se deja reaccionar el an geno con el an cuerpo correspondiente en presencia de can dad conocida de complemento.
De esta manera, si se da el caso de que exista la presencia del an geno o el an cuerpo buscado en el suero del paciente, se formarán complejos an geno-an cuerpo a los que quedaran fi jados el complemento.
Por otro lado en una segunda etapa se añadirá a la muestra una suspensión de eritrocitos de carnero sensibilizados con hemolisina.
La unión del complemento sobrante de la primera reacción daría lugar a una hemólisis, pero en este caso la hemoglobina liberada será inversamente proporcional a la can dad de an genos o an cuerpos presente en la muestra problema.
Fig. 6.2. Prueba fi jación del complemento en dos etapas

57
El Sistema Complemento (del Sistema Inmune)
6.3 Muestras y reactivos necesarios.
Dado que los an coagulantes pueden interferir en la acción del complemento, con lo que la muestra de elección corresponde a una muestra de suero.
Esto es importante dado que la muestra ha de estar sin homogeneizar, para evitar posibles errores en dicha determinación.
También hay que tener en cuenta los siguiente que es que si empleamos el método a dos etapas se debe inac var el componente del complemento presente en el suero problema mediante calentamiento a 56ºC durante 30 minutos.
Por otro lado si queremos medir la ac vidad del complemento en una muestra, esta se debe conservar adecuadamente, ya que el complemento se inac va con el almacenamiento. Estos grados son los siguientes: a 4ºC durante dos semanas o a -20ºC durante dos meses. Para periodos de empo más prolongados se empleara la congelación a -70ºC.
Los reac vos necesarios, como son los eritrocitos de carnero y la hemolisina deberán prepararse según métodos estandarizados y tulares antes de la prueba, con lo cual se convierte en una técnica complicada de realizar como análisis de ru na.
6.4 Principales técnicas de ijación del complemento.
Entre las principales técnicas de fi jación del complemento tenemos:
Serodiagnós co por reacción de fi jación del complemento (hemolisis en tubo).
Es mulación de la ac vidad del complemento mediante hemólisis en gel.
6.4.1 Serodiagnóstico por reacción de ijación del complemento mediante hemolisis en tubo.
Esta técnica se puede decir que es la única existente de forma manual o semiautomá ca dentro de la reacción de fi jación del complemento, la cual se da en dos etapas.
En la primera etapa lo que haremos será hace reaccionar los an genos correspondientes al agente estudiado con el suero problema en el que se sospecha la existencia de an cuerpos frente a esos an genos. En el caso de esta etapa de la técnica se producirá un complejo an geno-an cuerpo al cuál se le une el complemento añadido.
En la segunda etapa se añade el sistema hemolí co que no es más que una suspensión de eritrocitos de cordero sensibilizados con hemolisina que, si no encuentra complemento sobrante de primera etapa, precipita pero sin hemolizarse por lo tanto si no precipita quiere esto decir que ha encontrado el complemento sobrante.
Los materiales que u lizaremos serán tubos de hemólisis, pipetas y baño maría.
Los reac vos u lizados serán:
- An geno en suspensión.
- Suero control con an cuerpos frente al microorganismo buscado.
- Complemento de cobayo.
- Suero hemolí co de conejo (an hema es de cordero).
- Hema es de cordero al 50%.
- Tampón Veronal-Calcio-Magnesio (VCM) pH 7,2.
La muestra es suero, el cual es tratado mediante una técnica en la que debido a la presencia de complemento en el suero problema y suero control, es fundamental realizar una inac vación de ambos mediante calentamiento a 56ºC durante 30 min.
58
Técnico Superior Sanitario de Laboratorio de Diagnóstico Clínico
Una vez hemos realizado esto, procederemos a:
1. Titulación de suero hemolí co (SH)
2. Sensibilización de los hema es
3. Titulación del complemento
4. Titulación de suero problema
1. Titulación de suero hemolí co (SH)
Hacemos una primera dilución 1/100 con tampón VCM (Veronal-Calcio-Magnesio). De la siguiente manera:
a) Pipeteando 9,9 ml. de tampón y 0,1 ml. de SH, para obtener una disolución 1/100 de SH.
b) Pipeteando posteriormente 1 ml. de la dilución 1/100 del SH y añadiéndolo a un tubo con 9 ml. de tampón VCM, con lo que obtendremos una dilución 1/1000.
Tras obtener la dilución 1/100 (en ml.) preparamos 7 tubos con tampón VCM y SH
Tubos nº 1 2 3 4 5 6 7Diluciones fi nales 5500 6000 6500 7000 7500 8000 8500Tampón VCM-SH a
1/10004,5
1
5
1
5,5
1
6
1
6,5
1
7
1
7,5
1
Ahora prepararemos una suspensión de hema es de cordero al 2% en tampón VCM, con 4 ml de suspensión al 50% y 96 ml. de tampón.
Con ello haremos una dilución 1/25 del complemento también con tampón VCM. En donde realizaremos una nueva batería con 8 tubos (en ml.) de la siguiente manera:
Tubos nº 1 2 3 4 5 6 7 8Diluciones de SH 5500 6000 6500 7000 7500 8000 8500 Testigo
SHSH diluido
Tampón VCM
Complemento a 1/25
Suspensión de GR al 2%
0,25
0,40
0,2
0,25
0,25
0,40
0,2
0,25
0,25
0,40
0,2
0,25
0,25
0,40
0,2
0,25
0,25
0,40
0,2
0,25
0,25
0,40
0,2
0,25
0,25
0,40
0,2
0,25
0,25
0,60
-
0,25
Homogeneizamos todos los tubos y los ponemos al baño maría a 37ºC durante 1 h. Al cabo de este empo observaremos la hemólisis en los tubos y damos como tulo del SH el del úl mo tubo que presenta hemolisis total, y diremos que se corresponde con una unidad hemolí ca.
El tubo nº 8 que es el tes go de SH, no debe presentar hemolisis de lo contrario la prueba no sería válida.
2. Sensibilización de hema es
Prepararemos una disolución del SH obtenido anteriormente a 2 unidades hemolí cas, es decir si por ejemplo 1 unidad hemolí ca se corresponde a la dilución 1/8000, prepararemos una dilución a la mitad 1/4000 del SH.
Mezclamos entonces a partes iguales esta dilución del SH y la suspensión de hema es de cordero al 2% y dejamos la mezcla 15 minutos a 18-25ºC. Con ello obtendremos un sistema hemolí co que puede conservarse 24 horas a 2-8ºC.
59
El Sistema Complemento (del Sistema Inmune)
3. Titulación del complemento
Prepararemos la siguiente batería de tubos en ml.:
Nº de tubos 1 2 3 4 5 6 7 8 9Antígeno diluido
Tampón VCM
Complemento a 1/25
0,2
0,33
0,07
0,2
0,32
0,08
0,2
0,31
0,09
0,2
0,30
0,10
0,2
0,29
0,11
0,2
0,28
0,12
0,2
0,27
0,13
0,2
0,26
0,14
0,2
0,25
0,15Incubar 30 min. Al baño maría a 37ºCSistema hemolítico 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5Incubar 30 minutos al baño maría a 37ºC
En el primer tubo observaremos una hemolisis total lo que se corresponderá con una unidad del complemento. Para tular el suero problema necesitaremos 2 unidades de complemento en 0,2 ml. y calcularemos la dilución del complemento
según la siguiente fórmula, tomando como ejemplo el tubo 3:X= (0,2 ml.x 25)/(0,09 ml.x 2) = 27
En donde 0,2 ml. corresponderá al an geno diluido, 25 es debido a que el complemento es a 1/25, 0,09 serán los ml. del complemento al 1/25 y 2 por que se cogen 2 unidades, con lo cual la dilución del complemento será a 1/27 en tampón VCM.
4. Titulación del suero problema
Tras todo lo anterior diluiremos el suero problema y el suero control a ¼ con tampón VCM, y luego prepararemos una batería de tubos para cada suero problema en ml., de la siguiente manera:
Nº de los tubos 1 2 3 4 5 6 7 8 9 10Dilución obtenida
Testigo suero al 1/4
1/4 1/8 1/16 1/32 1/64 1/128 1/256 1/512 Testigo Ag
Tampón VCM
Suero a 1/4
Diluir
Antígeno al 3%
C´a 2 u
0,2
0,2
-
-
0,2
-
0,2
-
0,2
0,2
0,2
0,2
-
0,2
0,2
0,2
-
0,2
0,2
0,2
0,2
-
0,2
0,2
0,2
0,2
-
0,2
0,2
0,2
0,2
-
0,2
0,2
0,2
0,2
-
0,2
0,2
0,2
0,2
-
0,2
0,2
0,2
0,2
-
-
0,2
0,2
Homogenizamos cada una de las diluciones obtenidas a con nuación y dejamos durante 18 horas a 2-8ºC.
Luego añadimos 0,5 ml. de sistema hemolí co en cada tubo y lo llevamos al baño maría durante 30 minutos a 37ºC.
Lectura de resultados
Podemos observar tres resultados fi nales:
1. No hemólisis: fi jación total el complemento (4+)
2. Hemólisis parcial: fi jación parcial del complemento (3+ a 1+)
3. Hemólisis total: sin fi jación del complemento.

60
Técnico Superior Sanitario de Laboratorio de Diagnóstico Clínico
En esto el tulo de an cuerpos se corresponderá con la inversa de la mayor dilución en la que se observa fi jación total del complemento (4+).
Interpretación clínica de los resultados obtenidos
Posi vo → este resultado se observa cómo no hemólisis, ya que el complemento se ha fi jado en la reacción Ag-Ac de la primera fase. Hay precipitación de hema es de cordero.
Nega vo → al no exis r an cuerpos en el suero problema, el complemento queda libre para hemolizar a los hema es sensibilizados. Hay una hemolisis total.
Tes go del an genos (Ag) → se produce hemolisis total ya que no hay suero con an cuerpos.
Tes go del suero → se produce hemólisis total ya que no hay an genos para reaccionar con los an cuerpos del suero.
Existen casos en que el suero ene poder an complementario, de manera que es capaz de fi jar el complemento en ausencia de an genos. Se producirá entonces en este tuvo una precipitación parcial de los hema es lo que signifi caría que no podemos interpretar los resultados de la técnica.
Para cada agente infeccioso habrá una interpretación del tulo de an cuerpos presentes en el suero del paciente.
Por ello debemos remi rnos a la que realice la casa suministradora del suero control.
6.4.2 Estimulación de la actividad del complemento mediante hemólisis en gel.
6.4.2.1 Introducción.
Esta es una variante, modifi cación de la técnica anterior, y una manera simple de es mular la ac vidad lí ca (ya comentada) global del sistema complemento, mediante la u lización de la lisis de eritrocitos que han sido incorporados en un gel de agarosa. Al agregar un suero que posee un complemento C ac vo a un hoyo del gel, que con ene eritrocitos sensibilizados con an cuerpos, el gel se difundirá radialmente y generara un halo circular de lisis fácilmente medido como podemos observar en la fi gura 6.3., expuesta al fi nal de la técnica.
El área y por tanto el diámetro del halo de lisis, guarda relación con la concentración y funcionalidad del complemento C de la muestra. Método determinado por algunos fabricantes como la variante de la técnica anterior de serodiagnós co, el del serodiagnós co, para indicar que evalúa el limite o borde donde la muestra pasa del 100% a 0% de lisis. Técnica más bien conocida como hemólisis en gel. En el mercado existen placas de agarosa/EA (eritrocitos de carnero con an cuerpos de conejo), disponibles comercialmente para esta determinación. Alterna vamente se puede preparar en el laboratorio de forma sencilla siguiendo los siguientes pasos que detallaremos a con nuación.
6.4.2.2 Procedimientos.
Para llevar a cabo esta técnica, prueba realizaremos los siguientes pasos en ese orden:
1. Comenzaremos realizando las etapas de preparación, ajuste y sensibilización de los eritrocitos de carnero con los an cuerpos de conejo (EA) son las mismas que en la técnica de serodiagnós co anterior, por lo tantos las etapas del procedimiento anteriores a la tulación de suero problema serán las mismas, siendo esta la única que cambie.
2. Tras realizar con la muestra todos estos pasos del procedimiento de la prueba anterior de serodiagnós co, para realizar esta técnica, prueba cogeremos y pondremos en un baño a 45ºC la mezcla de los de los eritrocitos de carnero con los an cuerpos de conejo (EA) con una sustancia conocida como agarosa de manera fundida, al 1% en amor guador de la sustancia de barbital, para que obtengamos un 2% de volumen de concentración fi nal, es decir pondremos 250 μl de eritrocitos de carnero con an cuerpos de conejo (EA) en 12,5 ml de agarosa. No deberemos sobrepasar la temperatura indicada, para evitar dañar los eritrocitos.
3. Homogeneizamos la suspensión cuidadosamente y vertemos de inmediato sobre una placa de plás co o vidrio, en este caso con capa de base, nivelada y dejamos gelifi car.

61
El Sistema Complemento (del Sistema Inmune)
4. Abrimos hoyos de 3 mm de diámetro en el gel (como podremos observar en la fi g. 6.3. del fi nal de la técnica, método), y cargamos cada hoyo con 10 μl de la muestra en cada uno, para lo cual u lizaremos micropipetas semiautomá cas.
5. Tras rellenar los hoyos taparemos la placa, que si es de plás co se tapara con tapa sellada, sino colocaremos una cámara húmeda (si es la placa abierta) e incubaremos durante 24 horas a 37ºC. Previamente antes de taparla incluiremos siempre en todas las condiciones una alícuota, muestra de suero control en cada lote de muestra, para asegurarnos que no obtenemos resultados erróneos.
6. Posteriormente leeremos el diámetro de los halos de la lisis, los cuales aparecerán como zonas translucidas en el opaco, cuyos valores normales serán establecidos por cada laboratorio según las variables u lizadas, la forma de la placa, el volumen, el grosor del gel, el diámetro de los hoyos, etc.
Tras la lectura del diámetro que puede variar de un laboratorio a otro por lo comentado, leeremos e interpretaremos los resultados, u lizando para ello los parámetros de la lectura de los resultados de la prueba anterior de serodiagnóstico (hemolisis en tubo), ya que como ya se comento ambas pruebas son similares y en ambas se miden los mismos parámetros.
Fig. 6.3. Determinación de la actividad lítica del complemento mediante hemólisis en gel
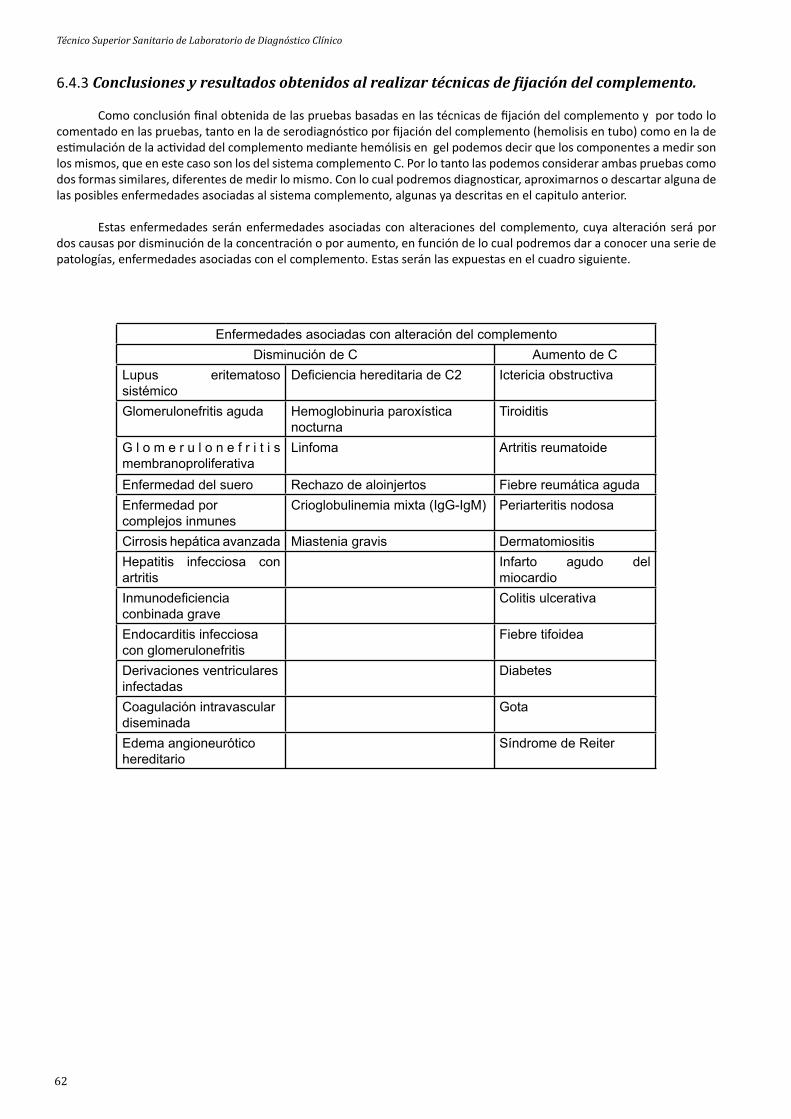
62
Técnico Superior Sanitario de Laboratorio de Diagnóstico Clínico
6.4.3 Conclusiones y resultados obtenidos al realizar técnicas de ijación del complemento.
Como conclusión fi nal obtenida de las pruebas basadas en las técnicas de fi jación del complemento y por todo lo comentado en las pruebas, tanto en la de serodiagnós co por fi jación del complemento (hemolisis en tubo) como en la de es mulación de la ac vidad del complemento mediante hemólisis en gel podemos decir que los componentes a medir son los mismos, que en este caso son los del sistema complemento C. Por lo tanto las podemos considerar ambas pruebas como dos formas similares, diferentes de medir lo mismo. Con lo cual podremos diagnos car, aproximarnos o descartar alguna de las posibles enfermedades asociadas al sistema complemento, algunas ya descritas en el capitulo anterior.
Estas enfermedades serán enfermedades asociadas con alteraciones del complemento, cuya alteración será por dos causas por disminución de la concentración o por aumento, en función de lo cual podremos dar a conocer una serie de patologías, enfermedades asociadas con el complemento. Estas serán las expuestas en el cuadro siguiente.
Enfermedades asociadas con alteración del complementoDisminución de C Aumento de C
Lupus eritematoso sistémico
Defi ciencia hereditaria de C2 Ictericia obstructiva
Glomerulonefritis aguda Hemoglobinuria paroxística nocturna
Tiroiditis
G l o m e r u l o n e f r i t i s membranoproliferativa
Linfoma Artritis reumatoide
Enfermedad del suero Rechazo de aloinjertos Fiebre reumática agudaEnfermedad por complejos inmunes
Crioglobulinemia mixta (IgG-IgM) Periarteritis nodosa
Cirrosis hepática avanzada Miastenia gravis DermatomiositisHepatitis infecciosa con artritis
Infarto agudo del miocardio
Inmunodefi ciencia conbinada grave
Colitis ulcerativa
Endocarditis infecciosa con glomerulonefritis
Fiebre tifoidea
Derivaciones ventriculares infectadas
Diabetes
Coagulación intravascular diseminada
Gota
Edema angioneurótico hereditario
Síndrome de Reiter

UNIDAD DIDÁCTICA VIITÉCNICAS BASADAS EN LA REACCIÓN DEL
COMPLEMENTO PERO SIN FIJACIÓN


65
El Sistema Complemento (del Sistema Inmune)
7.1 La turbidimetría.7.1.1 Características generales de la turbidimetría.
Cuando un rayo de luz colimado (aquel que atraviesa una suspensión homogénea de par culas) atraviesa una suspensión homogénea de par culas, una parte de esta dicha luz se absorbe, otra se refl eja, otra se tramite y otra úl ma es dispensada en todas las direcciones.
La turbidimetría consiste en medir la can dad de luz que se bloquea, es decir, can dad de luz no trasmi da por suspensión y por lo tanto no capaz de emerger de ésta en misma dirección que el rayo incidente.
Cuanto más luz bloqueada, es decir cuan fi cada en un espectrofotómetro (aparato u lizado para cuan fi car muestras) en forma de absorbancia, mayor será el numero de par culas presentes en suspensión y por lo tanto más alta la concentración de la sustancia problema que da lugar a las par culas y que está presente en la muestra.
Por un lado cuando se combinan un an geno con un an cuerpo en un medio líquido, se formarán agregados de inmunocomplejos que se quedarán en suspensión dado aspecto turbio a ese líquido.
Por otro lado cuando las par culas detectadas por estas técnicas las de turbidimetría sean inmunocomplejos, dichas técnicas recibirán el nombre de inmunoturbidemetría.
Fig. 7.1. Esquema general de un turbidimétro
7.2 La nefelometría.
7.2.1 Características generales de la nefelometría.
En el caso de la nefelometría lo que ocurre es que se mide la can dad de luz que se dispara por las par culas en suspensión, es decir se cuan fi ca la can dad de luz que es desviada por éstas, en determinado ángulo, con relación a dirección de rayo incidente.
El ángulo del rayo incidente u lizado, para evaluar la can dad de luz dispersada, se comprende entre los 30º y 90º C. Cuando las par culas en suspensión sean agregados de inmunocomplejos, dicha técnica será conocida como
inmunonefelometría.
Fig. 7.2. Esquema general de un nefelómetro
En estas técnicas la intensidad de luz que se dispersa es directamente proporcional a la can dad de agregados formados y por tanto a la concentración de an genos en la muestra que es u lizada para dicha técnica.

66
Técnico Superior Sanitario de Laboratorio de Diagnóstico Clínico
7.3 Tipos de determinaciones.
7.3.1 Tipos de determinaciones tanto turbidimétricas como nefelométricas.
7.3.1.1 Determinaciones a punto inal.
En las determinaciones a punto fi nal tanto para las determinaciones turbidimétricas como nefelométricas, tras el procesamiento de la muestra se dejará transcurrir un empo sufi ciente para que se formen todos los agregados.
Tras ello, se medirá la can dad de luz bloqueada o dispersada por la muestra y se restara a la obtenida en un blanco preparado solo a base de reac vos (estos serán los u lizado para la muestra cuál sea pero sin la misma).
Finalmente, se determina la concentración del an geno cuál sea que estuviese presente en la muestra, introduciendo el valor calculado con anterioridad de la can dad de luz, en una curva de calibrado, en la que se trazara con los datos que se consigan a par r de varias diluciones un patrón del an geno cuya concentración se conozca previamente.
7.3.1.2 Determinaciones cinéticas.
En las determinaciones ciné cas lo que se pretende y se hace es calcular la velocidad con la que se forman los agregados, en la muestra.
Esta velocidad está directamente relacionada con la can dad de an geno presente en la muestra.
En este po de determinaciones no se necesita por tanto ni blanco reac vos ni tan siquiera un patrón.
Como conclusión a ambos pos de técnicas tanto ciné ca como de punto fi nal son técnicas que pueden ser u lizadas para hacer otras mediciones a parte de las inmunológicas en suero tales como las determinaciones lipídicas, proteicas, de glucosa, enzimá cas etc. todas ellas en suero aunque algunas pueden también ser determinadas de la misma forma en orina además de otras tales como la determinación de hemoglobina en sangre total, entre otras determinaciones realizadas por estas técnicas.
7.3.1.3 Factores para mejorar estas técnicas.
Los principales factores que se encargan de mejorar estas técnicas son:
A menor longitud de ondas (λ) del rayo incidente, mayor será la intensidad de la luz dispersada lo que aumentara la sensibilidad de la técnica.
Esta se incrementa aún más si se emplea, como rayo de luz incidente, una luz fl uorescente o láser. En dicho caso el aparato de medida no necesita ni colimador ni selector de longitud de ondas.
Los demás componentes de la muestra no deberán absorber luz, de forma apreciable, a la longitud de ondas trabajada. Dicho problema no exis rá si la determinación es realizada de forma ciné ca.
El an cuerpo deberá poseer alta afi nidad y especifi cidad por el an geno, y todos los reac vos deberán de tener alto nivel de transparencia. Por eso se dice que en estas técnicas los reac vos han de ser de grado nefelométrico.
Normalmente se prefi eren muestras diluidas para trabajar, para evitar que se formen con rapidez grandes agregados, que den lugar a dispersión de la luz de forma demasiado irregular.
Los agregados se cons tuirán de forma más veloz y serán más estables, si se añade a los reac vos un polímero hidrosoluble de polie lengricol (PEG).
No se deberá transcurrir demasiado empo entre el procesamiento de la muestra y la medición, para evitar la sedimentación de los agregados. La velocidad de sedimentación de estos podrá ser disminuida incorporando goma arábica o gela na a los reac vos.
Las determinaciones ciné cas requerirán una agitación constante de la muestra previamente procesada, para que se mantenga homogénea la suspensión de agregado que se forme en la misma.
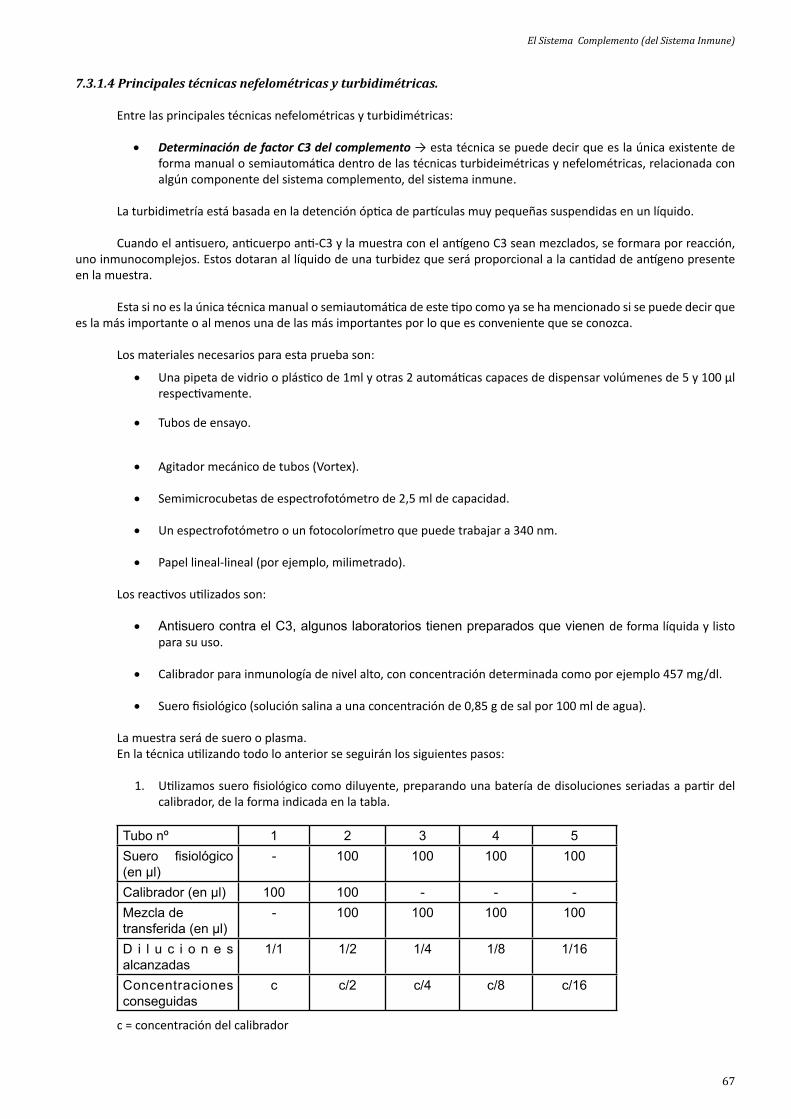
67
El Sistema Complemento (del Sistema Inmune)
7.3.1.4 Principales técnicas nefelométricas y turbidimétricas.
Entre las principales técnicas nefelométricas y turbidimétricas:
Determinación de factor C3 del complemento → esta técnica se puede decir que es la única existente de forma manual o semiautomá ca dentro de las técnicas turbideimétricas y nefelométricas, relacionada con algún componente del sistema complemento, del sistema inmune.
La turbidimetría está basada en la detención óp ca de par culas muy pequeñas suspendidas en un líquido.
Cuando el an suero, an cuerpo an -C3 y la muestra con el an geno C3 sean mezclados, se formara por reacción, uno inmunocomplejos. Estos dotaran al líquido de una turbidez que será proporcional a la can dad de an geno presente en la muestra.
Esta si no es la única técnica manual o semiautomá ca de este po como ya se ha mencionado si se puede decir que es la más importante o al menos una de las más importantes por lo que es conveniente que se conozca.
Los materiales necesarios para esta prueba son:
Una pipeta de vidrio o plás co de 1ml y otras 2 automá cas capaces de dispensar volúmenes de 5 y 100 μl respec vamente.
Tubos de ensayo.
Agitador mecánico de tubos (Vortex).
Semimicrocubetas de espectrofotómetro de 2,5 ml de capacidad.
Un espectrofotómetro o un fotocolorímetro que puede trabajar a 340 nm.
Papel lineal-lineal (por ejemplo, milimetrado).
Los reac vos u lizados son:
Antisuero contra el C3, algunos laboratorios tienen preparados que vienen de forma líquida y listo para su uso.
Calibrador para inmunología de nivel alto, con concentración determinada como por ejemplo 457 mg/dl.
Suero fi siológico (solución salina a una concentración de 0,85 g de sal por 100 ml de agua).
La muestra será de suero o plasma. En la técnica u lizando todo lo anterior se seguirán los siguientes pasos:
1. U lizamos suero fi siológico como diluyente, preparando una batería de disoluciones seriadas a par r del calibrador, de la forma indicada en la tabla.
Tubo nº 1 2 3 4 5Suero fi siológico (en μl)
- 100 100 100 100
Calibrador (en μl) 100 100 - - -Mezcla de transferida (en μl)
- 100 100 100 100
D i l u c i o n e s alcanzadas
1/1 1/2 1/4 1/8 1/16
Concentraciones conseguidas
c c/2 c/4 c/8 c/16
c = concentración del calibrador

68
Técnico Superior Sanitario de Laboratorio de Diagnóstico Clínico
2. Mezclar bien el an suero.
3. Depositar 1 ml de an suero en el interior de 6 tubos de ensayo.
4. Transferir 5 μl de cada una de las soluciones de la batería previamente preparada 5 de los tubos de ensayo dispuestos anteriormente.
5. Transferir 5 μl de la muestra al 6º tubo restante.
6. Mezclar el contenido de cada tubo, mediante agitador mecánico.
7. Incubar la mezcla a temperatura comprendida entre 25 y 37ºC, durante al menos 10 minutos.
8. Ajustar el espectrofotómetro a 340 nm.
9. Medir, con el espectrofotómetro, la absorbancia (A) de la luz, en un blanco preparado solamente a base de an suero, para restarlas de las absorbancias obtenidas posteriormente (lectura frente al blanco).
10. Medir y anotar las absorbancias logradas con las mezclas del calibrador y con la de la muestra.
Lectura de los resultados
Se trazara una curva de calibración o de referencia, en papel milimetrado. Para ello, se situara en una grafi ca con eje de ordenadas y abscisas, en donde en el eje de ordenadas se pondrán los valores de A obtenidos con las 5 mezclas preparadas a par r de la batería de diluciones del calibrador y en abscisas los valores de concentración que les correspondan.
Posteriormente, se señalaran en la grafi ca los puntos que confl uirán cada valor de A con su concentración correspondiente y se dibujara la recta que mejor unirá los 5 puntos.
Finalmente, se interpola el valor A logrado con la mezcla preparada a par r de la nuestra en la resta anteriormente trazada.
Interpretación clínica de los resultados obtenidos
La concentración sérica de C3 y C4 aumentan en los procesos infl amatorios, por lo que ambos serán considerados como reactantes de la fase aguda.
Por el contrario, las concentraciones séricas de uno o/y de otro disminuyen en procesos congénitos o adquiridos. En el siguiente cuadro expondremos las principales enfermedades en las que desciende algunas de estas concentraciones, o de ambas:
C3 C4Enfermedades autoinmunes con consumo de complemento D DEnfermedades por complejo inmune D DLupus eritematoso sistémico D DPérdida selectiva de proteínas D DGlomerulonefritis aguda D DGlomerulonefritis membrano- proliferativa D NEnfermedad hepatocelular D NHepatitis crónica activa D DEndocarditis Bacteriana D DVasculitis diseminada D DCoagulación intravascular D N
D = concentración disminuida. N = concentración normal
La observación del nivel de descenso de estos componentes puede u lizarse para evaluar el grado de ac vidad de esa enfermedad. Para detectar o descartar alguna de estas posibles enfermedades.

UNIDAD DIDÁCTICA VIIICUESTIONARIO


71
El Sistema Complemento (del Sistema Inmune)
8.1 Cuestionario
1. ¿Cuál es la función principal desempeña el sistema inmune?
a. Regulación de la temperatura corporal del organismo.b. Defensa frente a microorganismos y sustancias extrañas al organismo, conocidos como patógenos.c. Permi r la entrada y actuación en el organismo a sustancia externas al mismo.
2. Las células del sistema inmune se dividen en :
a. Serie linfoide y plasmá ca.b. Serie mieloide y plasmá ca.c. Serie linfoide y mieloide.
3. Los órganos linfoides primarios están formados por:
a. Médula ósea.b. Timo.c. Ambas son correctas.
4. ¿Qué entendemos por antígenos?
a. Sustancia extraña al organismo sin reacción con el sistema inmune.b. Sustancia extraña al organismo que puesta en contacto con un sistema inmunocompetente maduro es
capaz de provocar respuesta inmunitaria.c. Sustancia extraña al organismo capaz de producir an cuerpos.
5. ¿Qué es un epitope o determinante an génico?
a. Es una pequeña zona de la molécula del an geno que es capaz de combinarse con una parte de molécula del an cuerpo formando la estructura conocida como llave-cerradura.
b. Es la zona de la molécula del an geno que no es capaz de combinarse con la molécula del an cuerpo.c. Es el an geno que no es capaz de combinarse con el an cuerpo.
6. Los linfocitos son células del sistema inmune cuya función principal es:
a. Reconocimiento del an geno.b. Destrucción del an geno.c. Reconocimiento y destrucción del an geno.
7. En los linfocitos B ene lugar:
a. La producción de los dis ntos pos de inmunoglobulinas.b. La producción de un solo po de inmunoglobulinas.c. No es capaz de producir inmunoglobulinas.
8. Cuáles son los pos de inmunoglobulinas existentes:
a. A, D y M.b. A, D, E, G y M.c. A y D.

72
Técnico Superior Sanitario de Laboratorio de Diagnóstico Clínico
9. ¿Qué dos zonas presentan tanto las cadenas ligeras L como las pesadas H de las inmunoglobulinas y como se les conoce a sus extremos?
a. Una zona variable y dos zonas constantes para las cadenas pesadas H y una zona variable y otra constante para las cadenas ligeras L, en donde encontraremos en la zona constante un extremo carboxiterminal COOH y en la zona variable un extremo aminoterminal de ambas cadenas.
b. Dos zona variable y dos zonas constantes para las cadenas pesadas H y una zona variable y otra constante para las cadenas ligeras L, en donde encontraremos en la zona constante un extremo carboxiterminal COOH y en la zona variable un extremo aminoterminal de ambas cadenas.
c. Dos zona variable y dos zonas constantes para las cadenas pesadas H y una zona variable y dos constante para las cadenas ligeras L, en donde encontraremos en la zona constante un extremo carboxiterminal COH y en la zona variable un extremo aminoterminal de ambas cadenas.
10. Además de las cadenas polipep dicas, de la estructura básica de las inmunoglobulinas, esta presentan las siguientes moléculas adicionales:
a. Cadena J.b. Pieza de cola o secreción.c. Ambas.
11. Podemos defi nir el complemento como:
a. Grupo aproximado de 20 proteínas cuyo mecanismo se asemeja al de la coagulación de las plaquetas, formando un sistema enzimá co en cascada en el plasma, dando a conocer estas proteínas como proteínas plasmá cas aunque exista una pequeña porción que se encuentren en la membrana.
b. Grupo aproximado de 30 proteínas cuyo mecanismo se asemeja al de la coagulación de las plaquetas, formando un sistema enzimá co en cascada en la membrana celular, dando a conocer estas proteínas como proteínas de membrana aunque exista una pequeña porción que se encuentren en el plasma.
c. Grupo aproximado de 12 proteínas cuyo mecanismo se asemeja al de la coagulación de las plaquetas, formando un sistema enzimá co en cascada en el plasma, dando a conocer estas proteínas como proteínas plasmá cas aunque exista una pequeña porción que se encuentren en la membrana.
12. Muchas proteínas se encuentran inac vas en el complemento. La ac vación es llevada a cabo por una serie de vías que son:
a. Vía clásica ó común y alterna va.b. Vía clásica ó común, de la lec na, alterna va y lí ca, secuencia fi nal o complejo de ataque a la membrana.c. Ninguna de las dos son correctas.
13. En que radica la importancia del sistema complemento
a. En la defensa y limpieza del organismo provocando un incremento de la fagocitosis y ampliando los mecanismos de infl amación.
b. Nutrición de células del sistema inmune.c. Aumentar la producción de células del sistema inmune y con ello la energía y vitalidad del organismo.
14. Principales funciones del complemento:
a. Evitar la entra en el organismo de bacterias y virus, almacenamiento de proteínas del complemento.b. Lisis de células, bacterias y virus, opsonización, unión a células del sistema inmunitario de receptores
específi cos del complemento, lo que provoca en dichas células infl amación y secreción de moléculas inmunoreguladoras, depuración inmunitaria.
c. Lisis de células, bacterias y virus, opsonización, depuración inmunitaria y almacén de menoría inmunológica.

73
El Sistema Complemento (del Sistema Inmune)
15. ¿Qué precauciones debemos tomar si manejamos una muestra para estudio del complemento?
a. Una vez coagulada la sangre a temperatura ambiente, deberemos separar el suero del coagulo lo antes posible y congelarlo a -70ºC en nitrógeno liquido para evitar la pérdida de ac vación del complemento.
b. Una vez coagulada la sangre a temperatura ambiente, deberemos separar el suero del coagulo lo antes posible y congelarlo a -20ºC liquido para evitar la pérdida de ac vación del complemento.
c. Mantendremos la sangre sin coagular, para lo que obtendremos una muestra sanguínea en un tubo con an coagulante po EDTA o similar y lo congelaremos en nitrógeno liquido a -70ºC hasta su análisis, salvo que el análisis sea de inmediato, evitando con ello la perdida de ac vación del complemento.
16. La vía clásica de ac vación del complemento puede ser ac vada por el complejo an geno-an cuerpo ó inmunoglobulinas (Ig) agregas, de estas inmunoglobulinas, ¿Cuáles son capaces de iniciar la vía clásica?
a. La IgM y la IgG.b. La subclase IgG1, IgG2 e IgG3.c. La subclase IgG1, IgG2 y IgG3 así como la IgM.
17. Tras la ac vación de C1 de la vía clásica del complemento, vendrán C4 y C2 ambas ac vadas por C1, siendo estas moléculas de –globulinas, tanto C4 como C2, cuyo peso molecular es:
a. Mayor en C4 que en C2.b. Menor en C4 que en C2.c. Mismo peso molecular en C4 que en C2.
18. En su origen la vía alterna va se presentaba como un grupo de proteínas involucradas en:
a. Resistencia a infecciones.b. Lisis celular.c. División celular.
19. La úl ma etapa, o vía de ac vación del complemento se conoce como:
a. Vía alterna va.b. Secuencia fi nal, vía lí ca o formación del complejo de ataque a la membrana (MAC).c. ni a ni b son correctas.
20. Los receptores del complemento son sustancias que poseen las células del organismo y en especial las del sistema inmunitario, existen varios de los cuales el receptor CR3 provoca:
a. Desprendimiento tardío del cordón umbilical, suscep bilidad a infecciones, leucocitos persisten y periodon s progresiva y otros síndromes como la defi ciencia de adhesión a los leucocitos.
b. Protege a los leucocitos de posibles ataques derivados de infecciones.c. Provoca un desprendimiento excesivamente temprano del cordón umbilical.
21. Los inhibidores de la ac vación del complemento son:
a. El inac vador C3 y el cofactor proteico de la membrana (MCP) o (CD46).b. El inac vador C7 y el cofactor proteico de la membrana (MCP) o (CD28).C. El inac vador C1 y el cofactor proteico de la membrana (MCP) o (CD46).

74
Técnico Superior Sanitario de Laboratorio de Diagnóstico Clínico
22. Entre las conclusiones fi nales de la ac vación del complemento podemos decir:
a. Que desempeña un papel importante en enfermedades neurológicas infecciosas, autoinmune y degenera vas, donde se encuentra involucrado en su fi siopatología, además de protección contra infecciones por bacterias piógenas, lo que favorece la eliminación de complejos inmunes y de productos de la infl amación.
b. Que desempeña un papel importante en protección contra infecciones por bacterias piógenas, lo que favorece la eliminación de complejos inmunes y de productos de la infl amación, evitando la aparición de la enfermedad.
c. Que no desempeña un papel importante en protección contra infecciones por bacterias piógenas, lo que no favorece la eliminación de complejos inmunes y de productos de la infl amación, por lo que no evita la aparición de enfermedad, sino que provoca simplemente la lisis de microorganismo, virus y bacterias cuando estos han realizado su ciclo provocando la enfermedad y están muertos.
23. De que dos formas puede intervenir el sistema complemento en las enfermedades humanas:
a. De una primera forma produciendo defi ciencia en cualquiera de los componentes proteicos que pueden causar patrones anormales en la ac vidad del complemento y de una segunda forma mediante un sistema complemento inac vo con funcionamiento normal que puede ser ac vado por un es mulo anómalo.
b. De una primera forma produciendo defi ciencia en los componentes de la vía clásica del complemento que pueden producir patrones anormales en la ac vidad del complemento y de una segunda forma mediante un sistema complemento inac vo con funcionamiento normal que puede ser ac vado por un es mulo anómalo.
c. De una primera forma produciendo defi ciencia en los componentes de la vía clásica del complemento que pueden producir patrones anormales en la ac vidad del complemento y de una segunda forma mediante un sistema complemento inac vo con funcionamiento anormal ac vado por un es mulo anómalo.
24. Las técnicas basadas en la reacción de fi jación del complemento se fundamentan en:
a. La capacidad de unión que enen los componentes de dicha reacción a los complejos an geno-an cuerpo, los cuales se encargan de evaluar la ac vidad de todos los componentes del complemento en general o alguno en par cular, en función del método empleado.
b. La capacidad de evaluar la ac vidad de todos los componentes del complemento en general o alguno en par cular, en función del método empleado, tanto en la unión a los complejos an geno-an cuerpo como por separado fuera de la unión a estos complejos.
c. La capacidad de evaluar la ac vidad de todos los componentes del complemento en general o alguno en par cular, en función del método empleado, tanto en la unión a los complejos an geno-an cuerpo como por separado fuera de la unión a estos complejos.
25. Si queremos conservar una muestra de suero congelada a -20ºC para poder medir la ac vidad del complemento en dicha muestra, durante, ¿cuánto empo como máximo deberemos mantenerla congelada para poder medir dicha ac vidad?
a. 1 mes.b. 2 meses.c. 3 meses.
26. Las principales técnicas de fi jación de fi jación del complemento son:
a. Serodiagnós co por reacción de fi jación del complemento mediante hemolisis en tubo y es mulación de la ac vidad del complemento mediante hemolisis en gel.
b. Ninguna de las dos es correcta.c. Solo el serodiagnós co por reacción de fi jación del complemento mediante hemolisis en tubo es correcta.
27. La principal prueba diagnós ca basada en la reacción de fi jación del complemento es el serodiagnós co por reacción de fi jación del complemento. Esta se fundamenta en la técnica en una y en dos etapas de dicha reacción de fi jación. Tras llevar a cabo estas dos técnicas la de una etapa y dos etapas, ¿cuantos pasos deberemos realizar antes de llevar a cabo la lectura de los resultados?
a. 2b. 3c. 4

75
El Sistema Complemento (del Sistema Inmune)
28. ¿Cuáles son los dos grupos de técnicas principales basadas en la reacción del complemento pero sin fi jación?
a. Nefelometría y turbidimetría.b. Solo una de las dos de a es correcta.c. Ninguna de las dos de a son correcta.
29. ¿En qué consiste la turbidimetría?
a. Es capaz de medir la can dad de luz que se bloquea.b. Es capaz de medir la can dad de luz que se refl eja.c. Es capaz de medir la can dad de luz que se traspasa.
30. ¿Cuáles son los principales componentes del complemento medidos mediante técnicas nefelométricas y turbidimétricas?
a. C1 y C2.b. C1 y C4.c. C3 y C4.









