Efectos de la interacción entre células tecales y ...
162
Dirección: Dirección: Biblioteca Central Dr. Luis F. Leloir, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. Intendente Güiraldes 2160 - C1428EGA - Tel. (++54 +11) 4789-9293 Contacto: Contacto: [email protected] Tesis de Posgrado Efectos de la interacción entre Efectos de la interacción entre células tecales y granulosas sobre células tecales y granulosas sobre la esteroidogénesis en el folículo la esteroidogénesis en el folículo ovárico porcino ovárico porcino Lischinsky, Adriana Leonor 1983 Tesis presentada para obtener el grado de Doctor en Ciencias Químicas de la Universidad de Buenos Aires Este documento forma parte de la colección de tesis doctorales y de maestría de la Biblioteca Central Dr. Luis Federico Leloir, disponible en digital.bl.fcen.uba.ar. Su utilización debe ser acompañada por la cita bibliográfica con reconocimiento de la fuente. This document is part of the doctoral theses collection of the Central Library Dr. Luis Federico Leloir, available in digital.bl.fcen.uba.ar. It should be used accompanied by the corresponding citation acknowledging the source. Cita tipo APA: Lischinsky, Adriana Leonor. (1983). Efectos de la interacción entre células tecales y granulosas sobre la esteroidogénesis en el folículo ovárico porcino. Facultad de Ciencias Exactas y Naturales. Universidad de Buenos Aires. http://digital.bl.fcen.uba.ar/Download/Tesis/Tesis_1775_Lischinsky.pdf Cita tipo Chicago: Lischinsky, Adriana Leonor. "Efectos de la interacción entre células tecales y granulosas sobre la esteroidogénesis en el folículo ovárico porcino". Tesis de Doctor. Facultad de Ciencias Exactas y Naturales. Universidad de Buenos Aires. 1983. http://digital.bl.fcen.uba.ar/Download/Tesis/Tesis_1775_Lischinsky.pdf
Transcript of Efectos de la interacción entre células tecales y ...

Di r ecci ó n:Di r ecci ó n: Biblioteca Central Dr. Luis F. Leloir, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. Intendente Güiraldes 2160 - C1428EGA - Tel. (++54 +11) 4789-9293
Co nta cto :Co nta cto : [email protected]
Tesis de Posgrado
Efectos de la interacción entreEfectos de la interacción entrecélulas tecales y granulosas sobrecélulas tecales y granulosas sobrela esteroidogénesis en el folículola esteroidogénesis en el folículo
ovárico porcinoovárico porcino
Lischinsky, Adriana Leonor
1983
Tesis presentada para obtener el grado de Doctor en CienciasQuímicas de la Universidad de Buenos Aires
Este documento forma parte de la colección de tesis doctorales y de maestría de la BibliotecaCentral Dr. Luis Federico Leloir, disponible en digital.bl.fcen.uba.ar. Su utilización debe seracompañada por la cita bibliográfica con reconocimiento de la fuente.
This document is part of the doctoral theses collection of the Central Library Dr. Luis FedericoLeloir, available in digital.bl.fcen.uba.ar. It should be used accompanied by the correspondingcitation acknowledging the source.
Cita tipo APA:Lischinsky, Adriana Leonor. (1983). Efectos de la interacción entre células tecales y granulosassobre la esteroidogénesis en el folículo ovárico porcino. Facultad de Ciencias Exactas yNaturales. Universidad de Buenos Aires.http://digital.bl.fcen.uba.ar/Download/Tesis/Tesis_1775_Lischinsky.pdf
Cita tipo Chicago:Lischinsky, Adriana Leonor. "Efectos de la interacción entre células tecales y granulosas sobrela esteroidogénesis en el folículo ovárico porcino". Tesis de Doctor. Facultad de CienciasExactas y Naturales. Universidad de Buenos Aires. 1983.http://digital.bl.fcen.uba.ar/Download/Tesis/Tesis_1775_Lischinsky.pdf

UNIVERSIDAD DE BUENOS AIRES
FACULTAD DE CIENCIAS EXACTAS Y NATURALES
EFECTOS DE LA INTERACCION ENTRE CELULAS
TECALES Y GRANULOSAS SOBRE LA ESTEROIDOGENESIS
EN EL FOLICULO OVARICO PORCINO
AUTOR: Adriana Leonor Líschínsky
DIRECTOR: Dr. David Thomas Armstrong
Tesis presentada poro opfor oI TI'fqu de
DOCTOR EN CIENCIAS QUIMICAS
-I9831775qa.

AGRADECIMIENTOS
Quiero expresar mi gratitud al Dr. David T. Armstrong
por la oportunidad que me dio de trabaiar en su Laboratorio, y
por su conseío y apoyo durante el curso de estos estudios. Tam
bién deseo expresar el respeto y admiración que siento por él
como cientifico y como ser humano.
No tengo palabras para expresar mi gratitud al Dr.
Carlos P. Lantos por su apoyo incondicional en cada etapa de
mis estudios. Desde ¡975 en que me permitió trabaíar a su lado
por algunos meses hasta el presente ha sido la persona ala que
he recurrido siempre que se me presentara un problema, y de
quien siempre recibi especial atención.
Agradezco especialmente al Dr. Ronald Hobkirk por su
conseio asi como por permitirme usar las facilidades de su Labs
ratorio para la cromatografia en DEAE-Sephadex. Sin su ayuda,
todo el trabaío en coníugados esteroideos hubiera sido imposible
de realizar.

Estoy también agradecida a distintos miembros del MRC
Group in Reproductíve Biology de Canadó por su ayuda técnica
asï como por valiosïsímas discusiones.
Agradezco al Dr. Eduardo Charreau por su estímulo pa
ra que este trabaío fuera presentado.
Finalmente quiero agradecer a mis padres por ser como
son. Nada hubiera sído posible sin su aliento y apoyo íncondí
cional.

INDICE
pógina
AGRADECIMIENTOS.H... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
INDlCE....°...v.....,............ ....... ..........¡VABREVIATURASHHun“........ ....... .... v¡¡1.0 INTRODUCCION..... .. .. .t.y. . ..... .. . ..
l.l Estructura del foliculo ovórico y crecimientofolicular............“........c..u.... i1.2 Caminos para la esteroidogénesis en las góna
51.3 Biosintesis de esteroides en el foliculo ovóri
co 91.3.1 Unión de gonadotrofinas a células teca
les y granulosas......°................. . . . . .. 91.3.2 Capacidad esteroidogénica de células
granulosasl....°.b....°. 0I OfiOOI ICOOBÓOOICUQOUI1.3.3 Capacidad esteroidogénica de prepara
cionestecales..9..s......................... 131.3.4 Factores intraovóricos enla regulación
de la esteroidogénesis folicular . . . . . . . . .. ió1.3.4.1 Esteroides . . . . . . . . . . . . . . . . . . . . . . . .. ió1.3.4.2 Factores no esteroideos . . . . . . . . .. 19
1.3.5 Resumen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. 201.4 Formación y probable rol de los sulfatos de
esteroides en las glóndulas esteroidogénicas... 201.5 Propósitos de este trabaio . . . . . . . . . . . . . . . . . . . . .. 28
2.0 MATERIALES Y METODOS . . . . . . . . . . . . . . . . . . . . . . . . . . . .. 3l
2.] Animales . . yo. . . . . . . . . . . . a . . s . . . . . . . . . . . . . . . . . . . . . . 3]2.2 Aislamiento de células tecales y granulosas.... 3]2.3 Condiciones generales de cultivo................ 322.4 Determinación de esteroides . . . . . . . . . . . . . . . . . . 352.,5 Determinación de proteinas . . . . . . . . . . . . . . . . . . . . . .. 392.6 Extracción de esteroides de medio de cultivo.. 40

página
2‘7 Estudio del metabolismo de andrógenos . . . . . . . . . . . . .. 412.7.] Condiciones de cultivo . . . . . . . . . . . . . . . . . . . . . . .. 4]2.7.2 Preparación de las columnas de DEAE
Sephadex . . . . . . . . . er . . . . . . . . . . . . . . . . . . . . . . . . . .. 422.7.3 Cromatografía en DEAE-Sephadex A 25...... 432.7.4 Hidrólísis de sulfatos de esteroides . . . . . . . . .. 442.7.5 Cromatografíaen capa delgado.... .. . . 442.7.6 Cromatografía liquida de alta presión....... 452.7.7 Reducción con borohidruro de sodio . . . . . ..... 47297.8 Oxidación con el reactivo de Jones......... 47267.9 Determinación de la actividad de sulfo
transferasa....”...nunuunn................ 482.8 Efecto de testosterona sobre la utilización de
progesteronapor células granuiosas...“............. 492.9 Análisisde los resultados.............................. 50
3_o RESULTADOS...“...o...............n....°...9.............. 51
3.] Rol de las células granulosas enla sintesis deandrógenos....................°........9................ 5]3.1.1 Efecto de medio usado por células gra
nulosas sobre la sintesis tecal de andró
5]3.1.2 Efecto de medio usado extraido con car
bón-dextrón sobre Ia sintesis tecaI deandrógenos..,........7........‘.H......“........ 55
3.1.3 Concentración de esteroides en mediousadopor células granulosas................. 58
3.1.4 Efecto de C2¡-esteroides sobre la sintesistecaldeandrógenos....s. ó]3.2 Rol de las células granulosas en el metabolis
mo de andrógenos. . . . u. . a. 643.2.1 Capacidades relativas de los tipos ce
lulares foliculares para sintetizar andrógenos y estrógenos . . . . . . . . . . . . . . . . . . . . .. 64
3.2.2 Capacidades relativas de los tipos celulares foliculares para metaboiizarandrógenos.... . . . . . . . . . . . . . . . t,. . . . . . . . .. 67
3.2.3 Metabolismo de andrógenos por célu lasgranulosas.... 683.2.4 Actividad de sulfotransferasa en cé
Iulas granulosas . . . . . . . . . . . . . . . . . . . . ... 70

página
3.2.5 Estudios tendientes a la identificaciónde los metabolitos coniugados por lascélulas granulosas . . . . . . . . . . . . . . . . . . . . . . . . . .. 73
3.3 Efecto de la teca sobre la sintesis de progesterona por las células granulosas . . . . . . . . . . . . . . . .. 863.3.] Efecto de los andrógenos sobre la acu
mulación de progesterona por célulasgranulosas....... . . . . . . . .. . . . . . . .. . . . 86
3.3.2 Efecto de la testosterona sobre el metabolismo de progesterona . . . . . . . . . . . . . . .. 92
3.3.3 Efecto de 17 [5 -estradiol sobre la sintesis de progesterona. .. . .. . .. . . . 94
3.4 Mecanismo por el cual los andrógenos inhiben la sintesis de progesterona por célulasgranulosasen respuestaa FSH...... 963.4.] Relación temporal entre la acumula
ción de progesterona y l7I5-estradiolen cultivo en presencia de FSH y testosterona.... ................ 96
3.4.2 Efecto de un inhibidor de la aromatasa sobre la inhibición dela sintesisde progesterona por testosterona........ 99
3.4.3 Efecto de testosterona sobre la sintesis de progesterona estimulado por dibutiril-AMP ciclíco . . . . . . . . . . . . . . . . . . . . .. 102
3.4.4 Efecto de los andrógenos sobre la sintesis de pregnenolono por c'elulas granulosas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . lOó
4.0 DISCUSION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. 115
4.1 Rol delas células granulosas enla sintesisde andrógenos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. 115
4.2 Capacidades relativas de los tipos celularesfoliculares aislados y en recombinación para sintetizar andrógenos y estrógenos......... H9
4.3 Sintesis de Sulfatos de andrógenos . . . . . . . . . . . .. 1224.4 Efecto de la teca sobre la sintesis de proges
terona por las células granulosas . . . . . . . . . . . . .. 126
5.0 CONCLUSIONES . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. 138
6.0 BIBLIOGRAFIA . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. 144

ABREVIATURAS
A4A
4 a A
AMP cíclico
Androsterona
(Bu)2cAMP
C19-Esteroídes
C21-Esferoídes
DEAE-Sephadex
Dehídroepíandrosterona
(DHEA)
D.l.
Dïhídrotesfosferona (DHT)
Epíandrosferona
E.S.
Etíocolanolona
HEPES
HPLC
«4 ,[A -Androsfenedlona.
4-Acefoxí-androstenedíono.
3', 5'-Adenosína monofosfato cïclíco.,3 0*-Androsfan-3 Ci—o|-Ï7-ona.
N6, o2 -d¡bufírí| adenosína 3‘-5'
monofosfato cíclico.
Esteroides de 19 átomos de carbono.
Esteroides de 2] átomos de carbono.
DíetílaminoetíI-Sephadex.
5-Androsten-3 ¿ol-l7-ona.
Diámetro ínferno.
5 c<_Androstan—17¿'5-ol-3-ona.
5 Al-Androsfan-Ïi ,5 -oI-]7-ona.
Error standard.
5 á-Androstan-3 d-oI-I7-ona.
N-2-Hídroxíefilpíperazína-N'-2etanosulfóníco.
Cromatografía líquida de alta presión.
vii

M ¡Wedío usado por células gronuloscs
control.
MB ¡Medio usado por células granulosos
estímulodos con FSH.
MEIM Medio Esencial Mínimo.
RIA Rodíoínmunoensoyo.
T Testosterona.
vÍïí

1.0 INTRODUCCION
1.] Estructura del Foliculo Ovórico y Crecimiento Folicular.
La capacidad para la reproducción normal depende
del doble rol del ovario en proveer células germinales maduras
y hormonas esteroideas que regulan su crecimiento y preparan
al tracto reproductivo para el transporte de óvulos y la implan_
tación. Los componentes funcionales más importantes del ova
rio maduro son los foliculos y el cuerpo lüteo.
El foliculo ovóríco maduro es una estructura esférica
casi totalmente embebida en la matriz de teíido conectivo del
ovario, pero visible desde la superficie como una vesicula
translúcída (l). Su pared está compuesta por 3 capas:
l) La teca externa, compuesta de bandas concéntricas de fibro
blastos conectados entre si por tibrillas de colágeno embebi
das en mucopolisacórido, responsable de la elasticidad de la
pared folicular.
2) La teca interna, formada por una red de fibroblastos en que
están suspendidas células vacuoladas, secretoras de esteroi
des. Un plexo capilar denso y varios vasos linfáticos pro

veen la irrigación sanguinea y drenaíe linfático a las capas
externas de la pared folicular. Este aporte sanguineo es la
fuente de nutrientes necesarios para mantener a las células
granulosas y al ovocito, ya que no existen capilares luego
de la membrana basal que separa a la teca interna de las
células granulosas.
3) Las células granulosas, células columnares pseudoestratifi
cadas que encierran una cavidad llena de fluido donde se
encuentra el ovocito. Estas células son muy activas enla
biosintesis de proteinas (2, 3) y serian responsables dela
sintesis de moduladores de ciertas funciones toliculares a
demós de desempeñar un rol fundamental enla biosintesis
de esteroides.
Una de las caracteristicas notables del ovario es que
desde el nacimiento contiene una cantidad muy grande de cél_u
las germinales no proliferantes, en exceso al número que se n_e
cesitaró durante toda la vida reproductiva. En el humano, al
nacer, el ovario contiene 2.000.000 de ovocitos (4) de los cu_a
les un máximo de 400 ovularó durante todos los años reproduc
tivos activos. Cada dia un número de foliculos empieza a cre
cer; aunque las señales que hacen que un determinado folículo

empiece a crecer son desconocidas, una vez que el crecimien
to es iniciado, estos foliculos continuarón creciendo hasta que
ocurra atresia (degeneración) u ovulación (5, ó).
El crecimiento Folicular está caracterizado por la formación
dela capa granulosa, aparentemente mediado por influencias
inductivas locales del ovocito (7). Esto está seguido por la a
parícíón de la membrana basal, aparentemente segregada por
la capa de células granulosas, y lo organización de células e_s
tromales circundantes en una capa tecal externa a la membrana
basal El desarrollo folicular posterior involucra crecímie_n
ta del ovocito y proliferación de células tecales y granulosas.
Los cambios en estas células parecen ser interdependíentes y
secuenciales, parcialmente coordinados con, y dependientes
de, los cambios ciclicos en secreción ganadotrófica (8)..
Ovulación y luteínización son los estadios terminales de
diferenciación de foliculos grandes, preovulatarios. En respues
ta al pico gonadotrófico, uno o más foliculos, dependiendo de
la especie, presentan hipertrofia de células tecales y granulg
sas (9), disolución de la membrana basal, ruptura del contacto
célula-célula entre células granulosos (10), iniciación de me
iosís en el ovocito (ll). El folículo distendido se rompe, y el

óvulo es expulsado hacia la cavidad abdominal, tomado porlos
tubos uterinos y transportado al útero. Elfoliculo que se rompe
al ovular, se llena con sangre. Las células tecales y granulosas
proneranrópidamente, yla sangre coagulada esreemplazada
por célulaslüteas, ricas en lïpidos, esteroidogénicamente ac
tivas. Si ocurre implantación, el cuerpo lúteo persiste. Si no,
empieza a degeneransíendo reemplazado por el cuerpo albican
te.
Las hormonas esteroideas, secretadas por células teca
les y granulosas en respuesta a estimulación gonadotrófica, es
tón involucradas en varios aspectos de la diferenciación de los
componentes delfoliculo ovórico y son esenciales para el man
tenimiento de lastunciones gonadaleL También son componen
tes importantes de mecanismos de retroalimentación que regulan
la secreción gonadotrófica mediante acciones sobre el Hstema
hipotólamo-hipofisario (12a). Estudios mós recientes han demos
trado que los esteroides ovóricos también eiercen acciones sig
nificativas a nivel de las células esteroidogénicas influyendo
en el tipo y extensión de su respuesta a la estimulación gona
dotrófica (12 b). Debido a esto es importante entender el control
de su biosintesis.

1.2 Caminos para la Esteroidogénesis en las Gónadas.
Los caminos para la biosintesis de esteroides ganada
les estón bien establecidos; los caminos y mecanismos enzimóti_
cos han sido reseñados por Engel (13). Se esta de acuerdo en
que existen caminos esencialmente similares en todo teíido es
teroidogénico (gónadas, corteza adrenal y placenta) para la
formación de los precursores comunes a estos teiidos (incluyerl
do ácido mevalónico, escualeno, colesterol y pregnenolona).
Alguna divergencia en caminos ocurriria luego, en el camino
hacia los andrógenos, con pregnenolona siendo sometida 0174
hidroxilación directa y ruptura de cadena lateral (por C17-2o
líasa) (camino A5), antes de la conversión mediante el sistema
3 (5 -hidroxiesteroide deshidrogenasa- A, isomerasa, o siendo
convertida a progesterona por medio de este sistema enzimática
antes de experimentar 17 oí -hidroxilación y ruptura dela cade
na lateral (camino A”).
Androstenediona y testosterona son canvertidas a estro
na y 17‘",5-estradio|, respectivamente, por una serie de reac
ciones integradas que involucran l9-hidroxilación, escisión .del
metilo en carbono lO, y deshidrogenación del anillo A, referi
das colectivamente como aromatización' Estas reacciones son

catalizadas por una secuencia de enzimas referidas colectiva
mente como aromatasa. Los caminos generalmente aceptados pa
ra la biosintesis de estrógenos estón resumidas enla Figura l.
En lugar de ser secretados directamente, o convertidas
a estrógenos, androstenediona y testosterona pueden experimen
tar suertes metabólicas distintas dentro de testiculo y ovario.
Estas incluyen saturación de sus anillos A por la Álesteroíde
5d reductasa, dando 5 d-androstan-3,l7-diona y 17 A“-hi _
droxi - 54-androstan - 3 - ona (SX-dinidrotestosterona, DHT),
seguido por reducción estereospecifica de sus grupos 3 - ceto
por la 34-hidroxiesteroide deshidrogenasa, a androsterona y
54-androstan-3fl/, 17 5 —diol. Ya que las SOC-reductasas son
aparentemente irreversibles (14) y los componentes con anilla
A saturado no pueden experimentar aromatización (13), el me
tabolismo intragonadal de androstenediona y testosterona a lo
largo de este camino podria desempeñar un rol regulatorio en
la biosÏntesis de estrógenos, desviando a estos sustratos del ca_
mino de la aromatasa. Ademós, existe evidencia de que ciertos
andrógenos SKI-reducidos son potentes inhibidores de la aroma
tasa (15, ló), ofreciendo otro mecanismo potencial para la re
gulación de la formación de estrógenos.

FiguraÏ: ¿1: J: ó:
CaminosparaIabíosïntesisdeestradïolenlasgónadas.
enzimasderupturadecadenalateraldecolesterol.
A....,'\.
3 ‘-—'n¡drox¡estero¡dedeshxdrogenasa/“JS-¡somerasa. 1/:-.'—Ïrwídroxí|asa. IíOSÜ. 17í—hídroxíesteroide-deshídragenose.aromarasa.

nOrmmquOr
h É
vmeZmZOrOZb,
vaOmmquOZ.»
m
¿NR10111meZmZOrOZ>.NAIOIvaOOmmquOZb,.A
Mo
UmI_0xOm2>ZUwOm._.m_NOZ>Ill!>ZUmqumZmEOZ>IIIIIV3.302)
No
>ZUzOmHIuImZ|wAusG“¡EOrIl..|vqmqumquOZ)|||.|Vmm4z>U_O_.l._u\w

1.3 Biosintesis de Esteroides en el Foliculo Ovórico Porcino.
1.3.] Unión de Gonodotroiinos a Células Tecales y Granulo
SOS.
Las células granulosas poseen receptores para la hor
mona folículo-estimulante (FSH). Células derivadas de foliculos
pequeños (1-2 mm) unen mós FSH que células de foliculos media_
nos (3 —5 mm) y grandes (7 -12 mm) (17). Esta unión resulta
en mayor producción de AMP cíclico (18), y dibutiril AMP cicli
co puede simular los efectos esteroidogénicos de FSH en las cé
lulas granulosas (19).
Las células granulosas provenientes de foiiculos antra
les pequeños no poseen receptores para la hormona luteinízante
(LH); células granulosas derivadas de folicuios de tamaño medi-a
no unen cantidades significativas de gonadotrofina corióníca
humana (hCG; esta gonadotrofína se une alos receptores a LH),
mientras que células provenientes de toliculos grandes unen más
hCG (20, 2], 22).
No se han podido detectar receptores a FSH en células
tecales de las distintas especies estudiados. Sin embargo, el te
¡ido tecal une hCG; teiido proveniente de foliculos grandes
une mas hCG que el obtenido de foliculos pequeños del mismo

-10
o de otros ovarios (23).
1.3.2 Capacidad Esteroidogénica de Células Granulosas.
Los primeros estudios sobre biosintesis de esteroides
"in vitroII por células granulosas aisladas fueron realizados
en el ovario porcino, por Bíersing y Carstensen (24, 25). Estos
autores incubaron células granulosas provenientes de Foliculos
de tamaño mediano con distintos intermediarios esteroideos en
Ia cadena biosintética, radioactivos. Sus hallazgos más impor
tantes fueron:
1.- Las células granulosas contienen 3/5-hidroxiesteroide des
hidrogenasa y ZOX-hidroxiesteroide deshidrogenasa eficientes.
2.- Las células granulosas tienen baja actividad de ¡WK-hi
droxilasa y C¡7_20 Iiasa y, por Io tanto, son relativamen
te incapaces de sintetizar andrógenos.
3.- Las células granulosas poseen 175-hidroxiesteroide deshi
drogenasa (que convierte A4-androstenediona a testoste
rona) y aromatasa activas.
La mayor parte de estas actividades enzimóticas fueron
confirmadas independientemente mediante estudios histoquu'mi
cos (26).

-1]
Estudios subsiguientes sobre las capacidades esteroidg
génicas de las células granulosas consistieron enla determina_
ción de las esteroides producidos a partir de precursores endó
genos. En 1970, Channing (27) informó que las células granulci
sas producian progesterona en cultivo, siendo esta sintesis es
timulada por FSH y/o LH dependiendo del tamaño de ios folicu_
los de los que provenian.
La incapacidad de las células granulosas porcinas de
sintetizar cantidades apreciables de andrógenos a partir de pr_e
cursores endógenos ha sido confirmada por Tsang y col. (28) y
Evans y col. (29), y está de acuerdo con estudios en casi todas
las especies (para una descripción detallada ver 30).
Aunque las células granulosas parecen ser incapaces
de sintetizar cantidades significativas de andrógenos, pueden
convertir eficientemente andrógenos a estrógenos (28). La ca
pacidad de convertir testosterona a 17"6 —estradio| cambia du
rante e| desarrollo folicular. Anderson y col. (31) cultivaron
células granulosas derivadas de foliculos antrales porcinos pe
queños, medianos y grandes (obtenidos de material de matade
ro) con o sin andrógenos, Las células granulosas de todos los
tamaños de foliculos secretaron pequeñas cantidades de 17(7

-12
estradiol. La adición de testosterona o Lá-androstenediona re
sultó en un aumento de 8 a 420 veces en la secreción de estró
genos. Se observó la menor conversión con células granulosas
provenientes de foliculos pequeños y la mayor con células pro
venientes de foliculos medianos y grandes.
Adición de FSH iunto con testosterona no produio efe_c
to alguno sobre la secreción de 17 ¿e-estradiol por células gr_a
nulosas de foliculos pequeños y medianos, aunque aumentó la
secreción de estrógenos por células granulosas derivadas de f2
liculos grandes. Esto no puede ser explicado por cambios en los
receptores a FSH, que son menores en número en células granu
losas de Foliculos grandes. Los investigadores lo explicaron
considerando que tal vez el porcentaíe de receptores a FSH a
coplados a una respuesta que resulta en estimulación de la ac
tividad de aromatasa es mayor en células de foliculos grandes,
o que la aromatasa no es capaz de responder hasta que las cé
lulas granulosas se encuentren en un estado mós maduro. Usan
do un modelo distinto, la cerda prepúber tratada con PMSG
(gonadatrofina de suero de yegua preñada) para inducir el de
sarrollo folicular, Evans y col. (29) observaron menor conver
sión de andrógenos a estrógenos al madurar el foliculo, y nin
gún efecto de las gonadatrofinas sobre la aromatasa. La ausen

_¡3_
cia de un efecto de FSH sobre la aromatasa parece contrastar
con el rol de FSH enla estimulación dela aromatasa "in vivo“
e "in vitroll en células granulosas derivadas de roedores de la
boratorio (32, 33)e Sin embargo, no se han llevado a cabo es
tudios para determinar si en el cerdo, como enla rata, FSH i_n
duce actividad de aromatasa en células granulosas que no han
IIsido expuestas a FSH in vivo".
1.3.3 Capacidad Esteroidogénica de Preparaciones Tecales
Las preparaciones tecales sintetizan cantidades elevg
das de andrógenos (predominantemente A4-androstenediona),
siendo esta sintesis estimulado por LH en todos los estadios del
desarrollo (28, 29). La teca es deficiente en ¡7 ¡’5-hidroxiestí
roide deshidrogenasa y, por lo tanto, incapaz de convertir
grandes cantidades de A4—androstenediona a testosterona (25).
La capacidad del teiido tecal porcino de sintetizar
17 Ï-estradiol depende del estadiodel desarrollo folicular.
Stokiosowa y coli (34) separaron teca interna de foliculos
grandes (diámetro: 10-12 mm), ia dispersaron e incubaron co
mo monocapas; observaron que la producción de estrógenos por
'células tecales era mayor que la producción por células granu

-14
losas derivadas de los mismos foliculos.
Usando preparaciones tecales derivadas de foliculos
de tamaño mediano (4-ó mm), Tsang y col. (28) fueron incopg
ces de observar sintesis de 'l7¿"/-estradiol. Evans y col. (29)
observaron que la teca contribuye tanto como las células gra
nulosas ala sintesis folicular de estrógenos en los foliculos
más maduros, mientras que, durante los estadios mós tempranos
del desarrollo Folicular preovulatorio, la teca es incapaz de
convertir andrógenos a estrógenos.
Recientemente, Haney y Schomberg (35) han informa
do que, en toliculos porcinos de tamaño mediano (4-8 mm ), la
teca es responsable dela mayor parte de la producción folicu
lar total de estrógenosa Los autores llegaron a esta conclusión
porque la producción basal de estradiol fue mayor en cultivos
tecales que en los de células granulosas y porque no observa
ron un aumento significativo al co-cultivar ambos tipos celu
lares. Sin embargo, cuando compararon la capacidad de célu
las tecales y granulosas aisladas para aromatizar testosterona
exógena encontraron que, en este caso, la producción de es
tradiol por las células granulosas era mayor. Ya que en el to
liculo las células granulosas estan expuestas a niveles eleva
dos de andrógenos en el fluido folicular (36), esto implica

-15
que probablemente las células granulosas hacen una contribu
ción móssignificativa ala sintesis folicular de estradiol tam
bién en este estadio, como fue observado anteriormente (28).
La ausencia de efecto sobre la sintesis de estrógenos al C0"CL_J_l
tivar ambos tipos celulares podria deberse a que las células
granulosas metabolicen el andrógeno de origen tecal a otros
compuestos no aromatizablese
En conclusión, parece que la capacidad de la teca de
sintetizar estrógenos aumenta con el desarrollo tolicular. Estos
resultados están de acuerdo con estudios en toliculos ovinos
(37) pero en contraste con estudios en roedores de laboratorio
(38); en estos últimos la teca es incapaz de producir estróge
nos en todos los estadios del desarrollo folicular que han sido
examinados.
Los resultados obtenidos en distintas especies en ba
se a estudios "in vitro" con tipos celulares ovóricos aislados
han llevado al planteo dela hipótesis bicelular bigonadotróti
ca para Ia biosintesis folicular de estradiol: las células tec_a
les sintetizan andrógenos, baío la influencia de LH, que son
transportados a las células granulosas, donde son aromatizados
a estradiol, baío la influencia de FSH (30). Esta hipótesis se
ria vólida en roedores, en todos los estadios del desarrollo to

-16
licular pre0vulatorio y, en animales mós grandes, en los esta
dios más tempranos del desarrollo folicular preovulatorio.
1.3.4 Factores lntraovóricos en la Regulación de la Este
roidogénesis Folicular.
1.3.4.1 Esteroides.
Existe abundante evidencia de que los esteroides ovó
ricos podrian modular Ia bíosintesis ovóríca de otros esteroides
mediante acciones directas sobre las células ovóricas involucr_a
das.
En preparaciones tecales derivadas de foliculos Ovari_
cos porcinos de tamaño mediano, Tsang y col. (39) demostraron
que 17‘0 -estradiol ¡nhibia significativamente la producción
de testosterona y DHT de una manera dosis dependiente en pre
sencia de LH. Ya que estradiol no afectó la sintesis tecal de
AMPciclíco y progesterona, esto sugirió que el esteroide ac
tuarl'a inhibiendo el sistema l7m’ -hidroxilasa//C]7_20 líasa
o desviarïa alos C21-esteroides de este sistema estimulando
caminos alternativos de metabolismo. En la rata, Leung encon
tró que estradiol ínhibia la sintesis de andrógenos estimulado

_]7_
por LH tanto "in vivo" como "in vitro“ (40, 41). En esta espe
cie se demostró que estradiol estimularia el metabolismo a preg
nanos 5%—reducidos(42).
Se ha implicado a andrógenos y eHTógenos en el can
trol de la sfntesh de progesterona en células granulosas. Se op
servó (43) que los estrógenos sinergizaban con hCIG en la esti
mulación de la producción de progesterona por células granuLp
sas derivadas de foliculos de 5-8rnm. Schomberg y col.(44)
examinaron el efecto de estos esteroides en la producción de
progesterona por células granulosas derivadas de foliculos poL
cínosde 3-5 mm°DHTenimulósignHicaHvamentela acumula
ción de progesterona en cultivos de 2 dias mientras que testoí
terona no eíerció efecto alguno, y tanto dehídroepíandostero
na (DtlEA)como i7 Ü-estradiolinhibieron significativamente
la acumulación de progeflerona.Los auhnes concluyeron que
la ausencia de efecto estimulatoño de testosterona o DtlEA se
debia a aromatización de estos esteroides a estrógenos, ya que
las células granulosas eran capaces de aromatizar estos esteroí
des. Sin embargo, los autores no demostraron que esto estaba
ocurriendo. Luego informaron que también en células granulo
sas ahladas defolfculosde 8-10 mm, DHTculmulaba y 17? estradiol ínhibia la producción de progesterona (45).

-13
Otros autores (46, 47) observaron que testosterona y
1746 -estradiol inhibian la producción de progesterona en res
puesta a FSH por células granulosas derivadas de toliculos pe
queños. Dado que DHT, un andrógeno no aromatiza ble, no pro
duio efecto alguno, estos investigadores concluyeron que éste
era un efecto estrictamente estrogéníco y que testosterona pro_
bablemente actuaba luego de ser aromatizada a estradiol.
Cultivando por 2 dias células granulosas provenientes
de foliculos de tamaño mediano, Anderson y col. (3]) no obser
varon un efecto significativo de los andrógenos sobre la acum_u
loción de progesterona estimulada por FSH; en cambio, cuando
las células fueron mantenidas en cultivo por 4 dias, los andrá
genos actuaron sinergisticamente con FSH estimulando la pro
ducción de progesterona.
En conclusión, se ha encontrado que los andrógenos y
estrógenos podrian estimular o inhibir la producción de proge_s
terona por células granulosas "in vitro", dependiendo de la Ion
gitud delos cultivos, el tamaño del foliculo y/o condiciones
de cultivo.

-19
l.3.4.2 Factores no esteroideos.
Factores no esteroideos también han sido implicados
en el control de la producción de progesterona en células gra
nulosas. Ledwitz-Rigby y colo (48) observaron que fluido foli
cular derivado de foliculos porcinos pequeños y medianos, ex
traido con carbón-dextrón, inhibia la luteinización morfológ_i
ca y secreción de progesterona por cultivos de células granulg
sas de foliculos preovulatorios porcinos. El fluido folicular
también inhibió la acumulación de AMP ciclico basal y en re_s_
puesta a LH luego de un periodo de incubación de 24 a 48 ho
ras. En cambio, fluido folicular derivado de foliculos preovu
latorios no produio efecto alguno. Otros autores (49) hicieron
una observación semeiante al estudiar el efecto de fluido fol_i_
cular humano sobre la acumulación de AMP ciclico en ovario
de rata. Este inhibidor actuaria estimulando la actividad de
fosfodiesterasa, ya que adición de ¡sobutil-metil-xantina
(inhibidor de fosfodiesterasa) fue capaz de revertir la acción
del fluido sobre los niveles de AMP ciclica (50). También se
encontró un estimulador dela secreción de progesterona en
fluido folicular porcino (51). Este seria controlado por FSH.
Se ha observado la presencia en el ovario de factores

-20
que unen gonadotrofinas. Se observó (52)inhibición dela unión
de FSH a células granulosas por fluido folicular b0vino. Estu
dios en cuerpo lúteo de rata (53) y cerdo (54) sugieren Ia exi_s
tencia en este teiido de factores que unen LH. Se ha informado
que Ia adición de extractos de cuerpo lüteo a células granulo
sas porcinas inhibe la secreción de progesterona en presencia
o ausencia de LH exógeno (55).
1.3.5 Resumen.
Nuestro conocimiento actual de Ia biosïntesis de es
teroides por células tecales y granulosas del folículo ovórico
porcino esta resumido en la figura 2. Se ha omitido el efecto
de los andrógenos y estrógenos sobre Ia sintesis de progestero
na ya que Ia evidencia parece indicar que su efecto es apare_n
te sólo después de 2-4 dias en cultivo, cuando las células gra
nulosas ya han luteinizado (27).
104 Formación y Probable Rol de los Sulfatos de Esteroides
en las Glóndulas Esteroidogénícas.
Por mucho tiempo los sulfatos de esteroides han sido

Figura2:Roldecélulasfecalesygranulosasenlaesteroídogénesís enFolïculosporcinosdetamañomediano. Laslíneasenterosrepresentancaminosesferoídogénícosyacciones LaslíneasquebradasrepresentanHuioíntercelulordeesteroides.
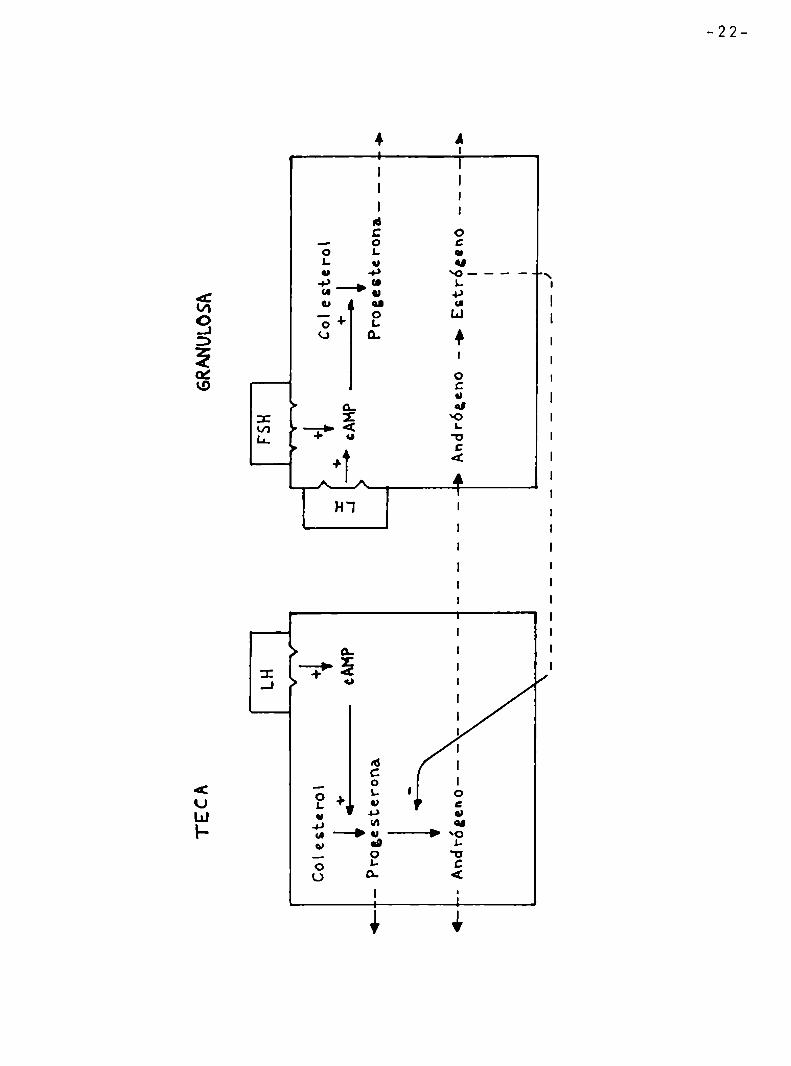
TECAGRANULOSA
LHIFSHl
ÏTY
Colesterol44 Colestero|
+I*+
i.————¿AMP.J—'cAHP-——>l
»—Progesterona
Progecterona___...>
_-Andrógeno——————————-———--—-45Andrógeno—->Butt-¿gano--4-I
I
-22

-23
considerados como meros productos metabólicos de hormonas e_s_
teroideas, sintetizados en el higado y excretados en la orina
o bilis (56). En i959, el sulfato de dehídroepiandrosterona fue
aislado de un tumor adrenal y se demostró que era segregado
por teíído hiperplástíco adrenal (57). Estudios posteriores de
mostraron que el sulfato de DHEA es uno de los principales es
teroides segregados por la glándula adrenal humana (58). Evi
dencia de secreción de este coníugado por una glándula este
roidogénica estimuló estudios de biosintesis y metabolismo de
conjugados esteroideos. La cantidad de estudios realizados en
el periodo 1960-1970 fue enorme. Sólo se mencionarán aquellos
trabaios relevantes para entender el probable rol de los sulfa
tos de esteroides en las glándulas esteroidogénicas, asi como
la mayor parte dela evidencia de SUbiosintesis y metabolismo
en el ovario.
Se ha observado la presencia de pequeñas cantidades
de sulfatasa mamifera en higado, testiculo, ovario y adrenal
(59). Su presencia en las glándulas esteroídogénicas ha suger_i
do un rol para los sulfatos de esteroides enla biosintesis de
esteroides asi como enla regulación de los efectos de los es
teroides en los teiidos efectores.

-24
Varios 5-en-3_Ï -hidroxiesteroídes coniugados con ¡o
nes sulfato como sulfato de colesterol, de pregnenolona, de
179€ —hidroxipregnenolona y de DHEAhan sido involucrados
en la biosintesis de hormonas esteroideas (60-63). Se ha demo_s
trado que la secuencia de conversiones de colesterol a DHEA,
via pregnenolona y i7qí-hidroxipregnenolona, tiene lugartag
to para los 5-en-3f3-hidroxiesteroideslibres (64-66) como pa
ra los correspondientes sulfatos de esteroides (60-63), en cuyo
caso el grupo sulfato permanece en la molécula a través de to
da la conversión. La participación y regulación de la actividad
de sulfatasa (67-70), asi como el rol de los sulfatos de estero_i_
des como precursores de hormonas activas (58, 60, 63) también
han sido estudiados extensamente. Esta participación ha sido
observada en glándula adrenal, testiculo y placenta. En esta
Última, existe una sulfatasa muy activa, y el sulfato de DHEA
es un precursor importante enla sintesis de estrógenos (7l, 72).
En la glóndula adrenal se observó que la sulfatasa es
la enzima limitante en la biosintesis de esteroides (70, 73).
En eflc glándula, ACTtlestimulaIa shnesh de corHcoides ac
tuando en una etapa previa a la formación de pregnenolona
(69).. Dominguez y col. (74) observaron que ACTH también est_i

-25
mula la actividad de sulfatasa por un mecanismo aparentemente
independiente de AMP ciclicos A partir de sus re5ultados espe
cularon que enla adrenal el mecanismo de acción de ACTH a
través de AMP ciclico ocurriria en condiciones de stress mien_
tras que la sulfatasa desempeñaria un rol en los mecanismos de
retroalimentación regulatorios asi como en aumentos pequeños
enla ACTHcirculante.
La importancia regulatoria de Suliotransferasas y Sul
fatasas ha sido estudiada extensamente en teiido testicular por
el grupo de Notation. Este grupo describió la formación de su_l
fatos de esteroides y su retención enla sangre periférica como
una función de almacenamiento (67). Esto proveeria a cada te
¡ido con una abundante cantidad de precursores monosulfatados
para regulación "in situ" de los niveles de esteroides. La uti
lización de estos monosulfatos pareceria estar baio influencia
esteroidea directa (68). Notation (75) presentó una reseña de
evidencias que apoyan la premisa de que, a pesar de la baia
actividad de la sulfatasa, la regulación de la hidrólisis de
los monosulfatos esteroideos es importante para la biosintesis
de andrógenos primariamente en el sitio efector. La sulfatasa,
independientemente del teiido del que proviene,parece ser una
unidad monomérica con un sitio de unión de sulfato y un sitio

-26
regulatorio de unión de esteroides. La asociación intima del
monómero o sus agregados con la membrana celular implica
transporte y metabolismo de hormonas esteroideas. El autor pre
senta un modelo en que los monosulfatos de esteroides (sustra
tos) contribuirian ala biosintesis de andrógenos por medio de
hidrólisis reguladas hormonalmente (modificadores o inhibidores
esteroideos alostéricos) en la membrana celular por sulfatasas
en el sitio de utilización dela hormona (producto esteroideo).
Esto ocurriria enla barrera sangre-órgano en los teiidos efec
tores a andrógenos y, en particular, como manera de regular el
nivel de andrógenos en los tÚbulos seminiieros.
Ha habido controversias con respecto ala capacidad
del ovario de formar sulfatos de esteroides. Estudios con homo
genatos de tejido ovórico humano demostraron sólo una pequeña
capacidad de sulfatación (76). Se ha observado formación de p_e
queñas cantidades de sulfatos de estrógenos en extractos de o
varios bovinos (77)o También se informó secreción de sulfato de
DHEA y de estrona (78, 79). Melville y Braunsberg (80) demos
traron formación de sulfatos de DHEAy de estradiol por sobre
nadante de teiido ovórico de sólo 7 de 12 ovarios humanos nor_
males, Sin embargo, otros autores (8]) no pudieron detectar se

-27
'creción o utilización de cantidades medibles de sulfatos de es
teroides por ovarios humanos normales en ambas fases del ciclo.
Estudios mas recientes demostraron formación de sulfatos de tes
tosterona, estrona y estradiol por teiido ovórico bovino y de
rata (82) y por ovarios humanos perfundídos "in vitro" (83-85).
La formación de Sulfatos de estrógenos era estimulado por gona_
dotrofinas. En todos los casos se observó sólo una pequeña con
versión a sulfatos, que los autores atribuyeron a baía actividad
de la sulfotransferasa.
También existe evidencia de utilización de sulfatos
esteroideos para la biosintesis de esteroides en el ovario.
Se encontró actividad de sulfatasa en ovario humano
(86). Se observó que el sulfato de DHEAes un precursor eficien_
te de andrógenos y estrógenos en ovarios humanos perfundidos
(87)o Luego de incubar ovarios de coneio con sulfato de DHEA,
Payne y Mason (88) aislaron A4-androstenediona y testosterona.
PatWordhCIny Lanthier (89) observaron que tanto el teiido fo
licular como el estroma de ovario humano podian utilizar sulfa
to de DHEA como precursor para la formación de andrógenos y
estrógenos.

-23
1.5 Propósitos de este Trabaio.
Los componentes del foliculo ovórico experimentan
cambios notables durante el crecimiento folicular. Aunque es
tos cambios dependen de la secreción ciclica de gonadotrofinos
hipofisarias, la concentración de gonadotrofinas a la que los
distintos foliculos estón expuestos no explica porqué en cada
ciclo sólo un número limitado de foliculos en crecimiento (uno
o mós dependiendo de la especie) ovula y Iuteiniza en respueí
ta al pico gonadotrófico, mientras que los demós degeneran.
Una explicación posible para esta respuesta diferencial de los
foliculos al mismo medio ambiente hormonal podria ser la con
centración intrafolicular de hormonas esteroideas.
Los estrógenos estimulan Ia proliferación de las célu
las granulosas y la sensibilidad del ovario a FSH (90, 9]),
mientras que los andrógenos, ademós de servir como sustratos
para la sintesis de estrógenos, eiercen un efecto opuesto sobre
el crecimiento folicular, promoviendo atresia (92). Por lo tan
to, el balance entre la acumulación de andrógenos y la forma
ción de estrógenos desempeñaria un rol significativo en deter
minar la suerte de un foliculoi Debido a esto, la sintesis y/o
el metabolismo de andrógenos deberian estar precisamente reguladas.

-29
En toliculos de rata (93; y oveio '94, 37) se ha obser
vado que la eliminación de células granulosas de preparaciones
tecalesresulta en una disminución en la producción de andrógf
nos. Ya que las células granulosas son incapaces de sintetizar
cantidades apreciables de andrógenos, uno de los propósitos de
este trabaio es examinar el rol de las células granulosas enla
sintesis y/o el metabolismo de andrógenos. También se estudia
ró el efecto de estos andrógenos sobre la esteroidogénesis por
las células granulosas. Ambos estudios estarian comprendidos
dentro del obietivo general de ampliar nuestro conocimiento
sobre las interacciones existentes entre los tipos celulares to
liculares.
El modelo elegido para este trabaio ha sido el ovario
porcino, porque provee de un gran número de foliculos de torna
ño óptimo para realizarla separación de tipos celulares folicrilares.
El método usado ha sido el de cultivo de corto térmi
no "in vitro" de tipos celulares foliculares aislados y en co
cultivo, en presencia de distintas sustancias. La ventaio de e_s_
ta técnica es que permite la determinación del potencial este. I I l o a Irordogenico de cada compartimiento folicular y examinar Ia ma

-30
nera en que factores esteroideos y no esteroideos la regulan.
Sin embargo, esta técnica presenta variaslimitaciones. Aunque
las células granulosas son fácilmente obtenidas en forma pura
y homogénea, en elfoliculo, las células granulosas son avas
culares, mientras que, al ser aisladas e incubadas, estén ex
puestas a oxigeno; Elteiido tecal es heterogéneo y no fócilde
obtener en forma pura“ Otra limitación es que, al separar estos
tipos foliculares, se interrumpe la asociación estrecha existen
te entre estas células al formar parte del foliculo. Por lo tan
to, losresultados de este enfoque pueden indicar la capacidad
esteroidogénica de los distintos tipos celulares foliculares pe
ro no su verdadero rol en la esteroidogénesisfolicular. Sin em
bargo, éste es uno de los meíores métodos disponibles actual
mente para estudiar las interacciones que tienen lugar en el
foliculo ovóricou

-3]
2.0 MATERIALES Y METODOS.
2°] Animaleso
Se recolectaron ovarios de cerdas prepüberes inmedia
tamente después de su muerte en un matadero local; ovarios
con cualquier indicio de ovulación fueron descartados. Para
un solo experimento se obtuvieron ovarios de cerdas maduras
en estadios no determinados del ciclo estral. Los ovarios se
mantuvieron en Medio Esencial Minimo modificado (MEM, ver
mas adelante) suplementado con buffer HEPES (20 mM) a 4° C.
Foliculos de tamaño mediano (diametro = 4-6 mm) se aislaron
de los ovarios, y se limpiaron cuidadosamente de teiido inters
ticial circundante utilizando forceps finos baio un microscopio
de disección. Aquellos foliculos que presentaban señales visi
bles de atresia (células granulosas separadas de la pared folí
cular o dispersas en el antro) fueron descartados, y sólo se u
saron foliculos de aspecto sano.
2.2 Aislamiento de Células Tecales y Granulosas.
Los foliculos se cortaron en cuartos y las células gra

-32
nulosas se separaron de las tecales raspando suavemente el in
terior de los foliculos con una varilla de vidrio,cuyo extremo
habia sido estirado para dar un diametro de curvatura apropia_
do para el tamaño de los foliculos.
Los trozos de teca se combinaron y colocaron en so|u_
ción salina balanceada de Hanks, libre de magnesio y calcio,
suplementada con buffer HEPES (20 mM). Se agitaron suaveme_n
te sobre un agitador magnético por 5 minutos para liberar las
células granulosas remanentes, y asi obtener preparaciones te_
cales más limpias (95). Las células granulosas se reunieron y
recolectaron por centriiugación. Se lavaron dos veces resus
pendiendo en MEMmodificado y centrifugando, y se resuspen
dieron en MEM fresco.
2.3 Condiciones Generales deCultivo.
El medio de cultivo usado fue Medio Esencial Minimo
de Eagle (Flow Lab.), suplementado con glutamina (4 mM), a
minoácidos no esenciales (0,1 mM), fungizona (2,5 ug/ml ),
penicilina (100 unidades/ml) y estreptomicina (100 ug/ml) (al
que se refiere como MEMmodificado). Los cultivos se mantuvie

-33
ron por dhtintostiempos en un incubador humidificado a 36°C
baío una atmósfera de 5‘% C(32 en aire.
Alicuotas conteniendo un número de células granulo
sas correspondientes a un foliculo (alrededor de 5 x 105 célu
las viables; 0,20 mg de proteina) se cultivaron en suspensión
en lrnlde medhadeculeo entubosde cuHivo(tubosplóni
cosestérHes Falcon, 12 x 75 mnfl.
Cuatro trozos de teca (un foliculo equivalente de te
¡ido) se colocaron sobre reifllas de acero inoxidable en las cg
vídades de placas de plástico Falcon 3047 Nhfltiwell conteniej
do l ml de medio de cultivo, o 0,5 ml de medio fresco más 0,5
ml de medio usado por células granulosas (en el coso de deter
minar el efecto de las células granulosas sobre la sintesis tecal
de andrógenos)o en medio de culeo conteniendo célulosgra
nulosas (en el caso de co-cultivosL
Variassustancias, solas o en combinación, se agrega
ron ontesdel cuHivo,Estasfueronzlug/ml FSH(NlH-FSH-SIIL
l ug/ml LH (NlH-LH-B8L vaHos eHeroidesexógenos(Sterohï
ids) a distintas concentraciones, dibutiril AhAPciclico ((BU)2c
AhAP, Sigma). Los detalles de los diseños experimentales se de
rón en la sección de resultados.

-34
Al final del periodo de cultivo, los medios de cultivo
se guardoron a -20°C paro determinación posterior de esteroi
des.
Con el obieto de determinar el contenido tisular de
esteroides, alicuotas de etanol redestilado se agregaron a las
cavidades de las placas de cultivo que contenian células gra
nulosos adheridas, y las placas se agitaron para extraer los ei
teroides; el teiido tecal se colocó en l ml de etanol redestíl_a_
do y se ogitó por 2 minutos. Luego de centrifugar, alicuotas
del extracto etanólico se secaron y redisolvieron en buffer pci
ra determinación posterior de esteroides. El pellet se usó para
determinación de proteinas. En cada experimento, 2 o 3 mues
tras de teiido no cultivado se usaron para determinar el conte
nido inicial de esteroides; éste se sustraio del contenido en
teiido y medio al finalizar el cultivo para estimar la acumulg
ción de esteroides durante el tiempo de cultivo.
Los resultados se expresaron en términos de acumula
ción de esteroides en medio de cultivo y teíido por miligramo
de proteina o por foliculo equivalente, corregido por conten_i
do inicial tisular de esteroides. Análisis preliminar de los da_
tos expresados por miligramo de proteina dieron resultados simi

.lares a aquellos expresados por foliculo, y en algunos casos se
eligió esta última expresión para dar una comparación más sig
nificativa de la producción total de esteroides por cada tipo
celular.
2.4 Determinación de Esteroides.
Se determinaron esteroides en alicuotas del medio de
cultivo sin extraer, y en extractos etanólicos del teiido (eva
porados y resuspendidos en buffer) por radioinmunoensayo. Pa
ra cada esteroide se verificó la validez del ensayo directo del
medio de cultivo comparando los valores obtenidos por este mi
todo con los obtenidos luego de extracción de las mismas mue_s_
tras con éter.
El radioinmunoensayo de Aw-androstenediona (A4 A)
fue realizado según la técnica descripta por Leung y Armstrong
(42). El anticuerpo se produío en coneios usando ll °¿-hidroxi
androst-4-en-3, l7-diona-hemisuccinil-albümina sérica bovina.3 1 .El, 2, ó, 7 H_j-androstenedlona se obtuvo de New England
Nuclear (Boston, Massachusetts, USA). El titulo dando alrede
dor de 50 % de unión cuando l5.000 cpm de trazador eran usa
dos, fue de 1:2.000. El rango de la curva standard fue de 12,5

-36
I800 pg. El coeficiente de variación intraensayo fue de 3,3 °o
(n:3ó), El coeficiente de variación interensayo fue de 11,7%
(en 26 ensayos). El paralelismo se determinó ensayando andros_
tenediona en distintas alicuotas de extracto etanólico de ava
rios de rata. La regresión dela dosis en el tamaño de la alicu_o
ta se describió como Y = 1,886 Ï 0,058 x - 27,966, siendo el
coeficiente de regresión significativamente distinto de cero.
Las alicuotas deseadas de muestras se llevaron a 100
Í/wl con buffer fosfo-salino (PBS) pH 6,8 con lO % de gelati
na. Luego se agregaron lOO/ul de trazador (10.000-15.000 cpm)
y lOO/ul de anticuerpo atodos los tubos“ Luego de incubar por
8 - 20 horas a 4°C se agregó l ml de una suspensión de 25 %
carbón - 2,5 °/odextrón en PBS. Los tubos se mantuvieron a
4°C por l5 minutos y se centrifugaron 01.200 g por 15 minutos
a 4°C. Los sobrenadantes se decantaron en viales a los que se
agregó 5 ml de solución de centelleo. La radioactividad se de
terminó en un contador de centelleo liquido.
Los radioinmunoensayos para testosterona (T), proges
terona (P), 175 —estradiol (E2) y estrona se realizaron con an_
tisueros altamente especificos. Estos ensayos fueron validados
previamente en el laboratorio (4], 96). Las reacciones cruza
das de los anticuerpos empleados se muestran enla Tabla l.

TABLA l
-37
Reacciones cruzadas de algunos esteroides con los anticuerpos
utilizados.
Anticuerpo anti- A4-androstenediona
Esteroide °/oreacción cruzada
Anticuerpo anti-testosterona
A4-Androste nediona TOO,OO5 a<-Androstano-3, T7-díona 5, OOTestosterona 2, 00Otros Esteroides < 1,00
Esteroide %reacci6n cruzadc
Testosterona 100, 00DHT 53, 0054-Androstano-31, 30, 0017 {b-díol.5 (-Androstano-3É, T3,0017,6 -dío|
A 4 -Androste nediona < 0, 04
Anticuerpo anti- Progesterona Anticuerpo antí-Pregne nolona
Esteroide % reacción cruzada Esteroide °/oreacción cruzada
Progesterona ¡00, OO Pregnenolona TOO,005P-pregnano-3, 20-diona 9, 20 5- Pregnen-3fi, 25, 005“—pregnano-3,20-díona 8,60 204-diolSi-pregnano-3Óol-20-ona 2, 60 5-Pregnen-3P, 8,30Pregnenolona O,80 20,9-diolotros esteroides < 0, TO Progesterona 0,60
17"5-Estradío| < 0,20
Anticuerpo anti-1715-Estradiol
Esteroide % reacción cruzada
17/3-Estradio| TOO,00Estrona 3, 00Testosterona 4 0, 02Progesterona 4 0,01

-38
El porcentaie de reacción cruzada para cada uno de los estero_i
des probados fue calculado como la relación entre la masa de
esteroide necesaria para producir un desplazamiento del 50 %
en la curva, con respecto a la masa de esteroide en estudio ne
cesario para producir el mismo desplazamiento.
El anticuerpo contra testosterona da 50 °/ode reacción
cruzada con DHT, y las estimaciones de niveles de testostero
na pueden entonces incluir DHT.
El anticuerpo contra pregnenolona, preparado en con_e
¡os contra el coníugado de albúmina sérica bovina al 16°C
carboxi-etil tioéter de pregnenolona, fue donado por el Dr. G.
Niswender y el ensayo fue realizado como el descripto por Ing
ba y col. (97), excepto que se ensayaron alicuotas de medio
sin extraer. La validez del ensayo directo se verificó compa
rando valores obtenidos para cantidades conocidas de esteroi
de en medio de cultivo y en buffer fosfato; los valores estaban
significativamente correlacionados (r = 0,99 y y: 0,05 Ï 0,9] x)
Para determinar Ia contribución de otros metabolitos
presentes en el medio de cultivo a la pregnenolona medida en
este ensayo, pregnenolona fue aislada de algunas muestras por
cromatografía en columna. Alicuotas de las muestras se extra
¡eron tres veces con 3 volúmenes de éter dietilico. Se prepara

-39
ron columnas de LH-20 y se corrieron usando como solvente
heptano : benceno : metanol (85:10:5) (98). La recuperación. . , 3del esteroude se determlno agregando ( H) -pregnenolona an
tes de la extracción con éter. La fracción de pregnenolona se' o I O Íensayo por radnonnmunoensayo de allcuotas del extracto etereo.
En los ensayos de los distintos esteroides se incluye
ron alicuotas de las distintas sustancias presentes en los culti
vos para determinar su reacción cruzada con los anticuerpos u
tílizados.
2.5 Determinación de Proteinas.
Finalizados los cultivos, las células granulosas se in
cubaron en l ml de NaOH 0,lN por ló horas a temperatura ar:
bíente. El contenido de proteinas se determinó por el método
de Lowry y col. (99) usando albúmina sérica bovina (0-150/¿09
disueltos en agua) como standard.
A alicuotas de 0,3 ml se agregaron 3 ml de solución
4. 5 HZO 0,5 % en tartrato
de sodio y potasio l % + 49 ml de N02C03 2 % en NaOH 0,lN).
alcalina de cobre (l ml de Cu SO
Los tubos se agitaron y deíaron lO minutos a temperatura am

-40
bienteg Luego se agregaron 0,3 rnl de reactivo de Folin Cioca_l
teau 2N (Fisher Scientific Company) diluido 1:1 con agua. Se
agitó inmediatamente y luego de una hora a temperatura ambie_n
te, se determinó la absorbancía a óóO nm.
Las preparaciones tecales se hirvieron por 30 minutos
en l ml de NaOH lN. La determinación de proteinas se realizó
de la manera descripto para las células granulosas excepto que
se usó Na2CO3 2% en agua y la curva standard se preparó en
NaOH IN.
2.6 Extracción de Esteroides de Medio de Cultivo.
50 mg de carbón (Norít A, Fisher Scientific Company)
y 0,5 mg de dextrón (T 70, Pharmacia Fine Chemicals) se agre
garon a l ml de medio de cultivo. Luego de incubar por l5 mi
nutos a 4°C, el medio se centrifugó 01200 g por 15 minutos,
Este procedimiento se repitió 2 veces y el sobrenadante se fil
tró. La efectividad de este procedimiento para eliminar este
roides del medio se confirmó por radíoinmunoensayo de alicuo_
tas del medio extraido.

-4]
2.7 Estudio del Metabolismo de Andrógenos.
2.7.]. Condiciones de Cultivo.
Células granulosas, preparaciones tecales y co-culti_
vos se incubaron con 4,4 x 10-9 M (l, 2, ó, 7 3H) -andros
tenediona (New England Nuclear, H4 Ci/mmol) o 3,7 x 10-9M
l, 2, ó, 7, ló, l7 3H) -testosterona (New England Nuclear,
135 Ci/mmol) en MEM modificado por 24 hs a 36°C baío una
atmósfera de 5 % CO2 en aire. La cantidad de esteroide agre
gado correspondió en masa al contenido inicial de andrógeno
(A4A o T) del teíido tecal. Los blancos para cada experimento
consistieron de l ml de medio de cultivo conteniendo el sus
trato radioactivo, en ausencia de teíído.
Finalizados los cultivos, los esteroides se extraieron
del medio 3 veces con 3 volúmenes de éter dieti'líco. En algu
nos casos, los esteroides se extraíeron del teíído con etanol.
Esto no se realizó sistemáticamente ya que menos del 8 % de
la radíoactividad inicial se encontraba en el teiido al final
del cultivo.
La fase acuosa remanente luego de la extracción del
medio de cultivo con éter se trató con lO volúmenes de etanol

-42
redestilado para precipitar las proteinas. Luego de centrifugar,
el pellet se descartó; el etanol se eliminó del sobrenadante por
evaporación, y el residuo, disuelto en agua, se sometió a cro
matografia en DEAE-Sephadex.
2.7.2. Preparación de las columnas de DEAE-Sephadex.
Se mezcló Sephadex A 25 (tamaño de partícula: 40
120 micrones, Pharmacia Ltd., Montreal, Canadá ) con agua bi
destilada en una proporción de lO gramos por 150 ml. Esta mea
cla se agitó por lo menos ó veces, a intervalos; cada vez se dE
¡ó que se depositase por 30 minutos, se aspiró el sobrenadante
y se reemplazó con agua antes de volver a agitar.
Se utilizaron columnas de Sephadex (K 9/60) de dió
metro interno (D.l.) de 0,9 cm (Pharmacia Ltd.). En el extremo
inferior de las columnas se colocó un trozo de tubo de goma
que se cerró con una llave de salida, y la columna se llenó
hasta 4-5 cm del borde superior con agua. Se fue agregando
cuidadosamente la suspensión de gel. Cuando el extremo supe
rior estaba libre de A 25, el agua se eliminaba con pipeta
Pasteur y se reemplazaba con un volumen similar de la suspen

-43
sión0 Esto se repitió hasta que la mitad dela columna se llena
ra con gel sedimentado. Luego se abrió la llave y se permitió
que el liquido fluyera libremente, agregando continuamente
suspensión hasta obtener una altura de columna de 58 cm.
2.7.3 Cromatogratia en DEAE-Sephadex A 25°
Las cromatografias se realizaron a temperatura ambie_n
te según la técnica descripta por Hobkirk y Nilsen (100). Las
muestras, disueltas en 2 ml de agua, fueron aplicadas cuidadg
samente con pipeta Pasteur. El tubo que habia contenido Ia
muestra se lavó varias veces con 2-3 ml de agua, y esto tam
bién se agregó a la columna. La columna se conectó a un shte
ma de gradiente compuesto por dos frascos idénticos de ¡000
ml conectados entre si por un trozo de tubo de polietileno y u
no de ellos, el frasco mezclador, agitado magnéticamente. De
este último, un tubo llevaba el eluyente a la columna. El sis
tema de gradiente utilizado contenia 500 ml de agua en el tra-s
co mezclador y 500 ml de Na Cl 0,8 M en el envase donor.
Se deíó que la columna fluyera libremente baío 50 cm de solu
ción eluyente. Fracciones de eluido de iO ml se recolectaron

-44
y la radioactivídad se determinó en alicuotas por espectrome
tria de centelleo liquido.
2.7.4 Hidrólisis de Sulfatos de Esteroides.
Los coniugados fueron extraidos de solución satura
da de NaCl, 3 veces con 3 volúmenes de acetato de etilo. Se
agregó sulfato de sodio anhidro y, luego de filtrar, el acetato
de etilo fue evaporado. Al residuo se agregaron 0,25 ml de m_e
tanol y l ml de ácido acético glacial: acetato de etilo (l : 9)
y se incubó ló hs a 50°C (lOl). Luego de evaporar completa
mente, el residuo fue resuspendido en 2 ml de agua y extraido
3 veces con 3 volúmenes de éter dietilico.
2.7.5 Cromatografia en Capa Delgado.
Las muestras,disue|tas en el menor volumen de bence
no: metanol (1:1), fueron sembradas en placas de vidrio recu
biertas con gel de silica (GF 254, Brínkmann; 0,25 mm de es
pesor). Esteroides conocidos apropiados fueron sembrados en
bandas adyacentes. Las corridas fueron realizadas a 4°C en el

-45
sistema cloruro de metíleno : éter díetr'lico (5:2), como des
cripta por Armstrong y col. (102). Los esteroides standard con-o
cídos se localizaron por fluorescencia ultravioleta o por expcl
sición a vapores de iodo.
Al final de la corrida, bandas del centimetro fueron
raspadas de las placas y eluidas con etanol. La radioactividad
fue determinada en alr'cuotas del eluïdo con un contador de
centelleo liquido.
Todos los esteroides radioactivos utilizados en los di_s
tintos estudios descriptos en esta tesis fueron purificados me
diante esta técnica antes de su uso.
2.7.6 Cromatografía Líquida de Alta Presión.
Se utilizó un equipo adquirido de Waters Associates
(Canadá). El sistema de distribución de solvente consistió de
dos bombas modelo 6000 A. Las muestras fueron ínyectadas me
diante un inyector universal modelo UóK o un procesador de
muestra modelo 7108. Los detectores usados fueron un monitor
de longitud de onda ultravioleta variable modelo 450, un de
tector ultravioleta modelo 44] o un refractómetro modelo R401.

-46
La velocidad de fluio y Ia composición de solvente fueron con
troladas por un controlador de shtema modelo 770. Las aHMras
y áreas de los picos y los tiempos de retención se midieron por
un data module modelo 730. Las columnas usadas fueron C18U
Bondapak Uongítudz 30 cm; D.|; 3,9 mm);tamaño de paHicu
|a=iO/M), C18 RadialPak carhidge Uongitud= iO cm;D.h= 5
mni; tamaño de particula: lO/x), con un módulo de compresión
radial RCfW-ÏOOo microporasi|(longitud= 30 cm; D.L= 3,9 mn1L
(Zomofases móbíles se usaron mezclas binarias de sol
vente: agua-acetonitñlo, agua-metanol y cloroformo-isoocta_
no, degasHicadosbaío preüón reducida o porinmerflón en un
baño ultrasónico por 20 minutos. Las velocidades de fluio usa
das fueron de 1,5 ml/min para la columna C18 U Bondapak,
l mI/nfin para el C18 radial carhfldge y 3 ml/min para la cg
Iumna de microporasH.
Debido a la pequeña cantidad de masa presente en
las muestras a analizar, sus tiempos de retención se determina
ron evaluando la radioactividad en alicuotas de las fracccio
nes obtenidas.

-47
2.7.7 Reducción con Borohidruro de Sodio.
AI esteroide disuelto en 0,5 mi de isopropanol se a
gregaron 25 mg de borohidruro de sodio. Luego de incubar por
una hora a temperatura ambiente, Ia reacción se detuvo agre
gando 2 ml de metanol. Los productos de reacción se evapora
ron, resuspendieron en 2 ml de agua y extraíeron tres veces
con 3 volúmenes de éter dietilico.
2.7.8 Oxidación con el Reactivo de Jones.
EI reactivo se preparó disolviendo 26,72 gramos de
trióxido de cromio en 23 mI de ócido sulfúrico concentrado
llevado a 100 ml con agua.
AI esteroide se agregaron 2 mI de acetona y15 ¡url
del reactivo de Jones. La reacción se realizó 012-140C por
4 minutos con agitación constante. Se agregó i ml de metano]
para detener Ia reacción, y se evaporó. El residuo fue resus
pendido en agua y extraido con 3 volúmenes de éter dietilico.

-48
2.7.9 Determinación de la Actividad de Sulfotransterasa.
Se siguió la técnica descripto por Sturm y Hannapel
(82). Células granulosas o preparaciones tecales fueron homo
geneizadas en 5 volúmenes de buffer Tris HCI (0,]M; pH 7,4)
con 0,25 M de sucrosa. El homogenato se centrifugó 0100.0009
por 60 minutos.
La mezcla de incubación consistió en Tris HCl 0,] M
conteniendo 1,4 umol de ATP, 2,8 umol de Mg Cl2 y 7 umol
de K2504. La reacción se realizó a 37°C por 2 horas en pre
sencia de alicuotas del sobrenadante de 100.000 g y de distin
tos esteroides radioactivos como sustratos (New England Nuclear).
En algunos casos se agregó (355)-N02504 (New England Nuclear).
La reacción se detuvo sumergiendo los tubos en un baño a 4°C.
El esteroide libre se separó dela mezcla de reacción
por extracción con éter dietilico.
Los blancos consistieron en tubos conteniendo la mez
cla de incubación y el sustrato rodioactivo.
Como control se usó el sobrenadante de l00.000 g de
homogenatos de higado de rata.

-49
2.8 Efecto de Testosterona sobre la utilización de Progeste
rona por Células Granulosas.
Células granulosas se cultivaron por distintos tiemposl4
con lug/ ml de FSH (NlH-FSH-SII) y (4 - C) -progesterono
10-6M (New England Nuclear, 50 mCi/mmol) en presencia o
ausencia de testosterona (5 x 10-6M). Finalizado el cultivo,
los esteroides se extraíeron del medio de cultivo con éter die_
tilico. El extracto etéreo se evaporó baio nitrógeno seco y el
residuo se redisolvió en benceno: metanol (lzl). Esto se some
tió a cromatografia en capa delgada en el sistema descripto
anteriormente, agregóndose 25/44rgde progesterona a las mues_
tras como standard interno. Luego de la corrida, la banda co
rrespondiente a progesterona, ubicada por fluorescencia ultrE
violeta, se raspó de las placas y se eluyó con etanol. El elui_
do se evaporó y su radioactividad fue determinada. El sustrato
radioactivo utilizado se sometió al mismo procedimiento; la
cantidad de radioactividad recuperada en ambos casos fue
comparada, calculóndose de esta manera la desaparición de
progesterona.

-50
2.9 Anólisis de los Resultados.
La existencia de diferencias significativas enla acu
mulación de esteroides en respuesta alos distintos tratamien
tos se determinó mediante técnicas standard de análisis de va
rianza (103). Cuando la prueba de Bartlett reveló heterogenei
dad de varianza entre los valores no transformados, los anólisis
se realizaron sobre los valores transformados logaritmicamente.
Sin embargo, para uniformidad enla presentación, los resulta
dos se presentan usando valores no transformados. En el caso
de usarse porcentaies, los anólisis se realizaron sobre valores
transformados angularmente.
La prueba de rangos múltiples de Duncan se utilizó
para comparaciones especificas entre valores medios.
El efecto de medio usado por células granulosas y de
esteroides sobre la sintesis tecal de andrógenos se analizó me
diante la prueba no paramétrica de Friedman (¡04) debido a
considerable heterogeneidad de varianza de muchos valores.

-51
3.0 RESULTADOS
3.] Rol de las Células Granulosas enla Sintesis de Andróge
nos.
3.1.1 Efecto de Medio Usado por Células Granulosas sobre
la Sintesis Tecal de Andrógenos.
Células granulosas fueron cultivadas por 24 hs sin
hormonas exógenas (A) o con FSH Al final del período de
incubación los medios de cultivo se congelaron para ser usados
posteriormente en cultivos tecales.
Las preparaciones tecales fueran cultivadas en l ml
de medio fresco o 0,5 ml de medio fresco mós 0,5 ml de me
dio en que células granulosas habian sido cultivadas previame_n
te (medio usado A y B) en presencia o ausencia de LH. Al fina
lizar el periodo de cultivo se determinaron esteroides en medio
de cultivo y proteinas en teíido. Este procedimiento se repitió
en 7 experimentos distintos con ó replicados por tratamiento
en cada uno. Debido ala variabilidad observada en las canti
dades de testosterona y A4-androstenediona producidos por los

cultivos control en cada experimento, los resultados se expre
saron como porciento de los cultivos control. Los resultados
combinados de siete experimentos estén representados en las fi
guras 3 y 4. A pesar de las diferencias en valores absolutos o_b
tenidos en los distintos experimentos, los efectos de los disti_n
tos tratamientos fueron consistentes en cada experimento, y
las diferencias entre experimentos desaparecieron cuando los
resultados se expresaron como porciento de valores control.
LH significativamente estimuló de 2 a 4 veces la acu
mulación de testosterona y ¿34-androstenediona.
Células granulosas aisladas de los mismos foliculos si
cretaron cantidades muy pequeñas de ambos andrógenos durante
las 24 hs de cultivo (datos no representados).
En ausencia de LH, la cantidad total de andrógenos
(A4A y T) sintetizados dependió del medio presente en la in
cubación (P < 0,01). En cambio, el medio no afectó la sfntesís
de andrógenos estimulada por LH. La producción de andrógenos
por preparaciones tecales cultivadas en presencia de medio u
sado control (MA) no fue significativamente distinta delos
controles (P< 0,05)o Sin embargo, medio en que células gran_u
losas habian sido cultivadas en presencia de FSH (MB) causó

-53
50°" C] CONTROL
. a LHA soo
‘z‘ B
8 u 400t:
L’LJ 0U .m 300o _b; 3u zool- RV
|00>
1-!- 4- ..'
50% MA 50% MB
Figura 3: Efecto de medio usado por células granulosas y/o LH so
bre la producción tecal de testosterona (T) en 24 hs de cultivo. Los
valores representan el promedio Ï E.S. del porciento de cultivos
control.
Abreviaturas: C, control; MA,medio obtenido de células granulo
sas cultivadas en MEM; MB,medio obtenido de células granulosas
cultivadas en MEM con FSH (l ug/ml).
(Los valores en los cultivos control fueron 10,52 Ï 2,89; 18,32Ï
2,90; 4,74 t 0,73; 5,85 Ï1,35;12,óó t 5,28,- 3,92 Ï 0,56 ng/
mg proteina).

-54
5°°' [j CONTROL
É ra;A LHO - ¿ooH oo Ll; uÉ .u o woF ow8 3 moD 1'!
35¡:3 mo74
50%MA
Figura 4: Efecto de medio usado por células granulosas y/o LH
sobrela producción tecal de [34-androstenediona (¿X4A) en
24 hs de cultivo. Los valores representan el promedio Ï E.S.
del porciento de la cantidad de ¿34A en los cultivos controL
(Las cantidades producidas por los cultivos control fueron:
95,65 Ï 26,43; 99,67 Ï 3,65; 51,20 Ï 8,31; 58,68 Í 9,04;
198,06 t 41,29; 74,04 Í 15,90 ng/mg proteína).

-55
un incremento al doble enla acumulación de Á4-androstene
díona y al triple en la de testosterona. Este efecto no se debe
ala presencia de FSH en el medio ya que la misma cantidad de
FSH no produjo efecto alguno (P < 0,1) sobre la sintesis de ari
drógenos por las mismas preparaciones tecales (Tabla ll).
3.].2. Efecto de Medio Usado Extraido con Carbón-Dextrón
sobre la Sintesis Tecal de Andrógenos.
Para determinar sí el efecto del medio usado por cél_u
las granulosas se debí'a a un esteroide o a algún otro factor,
el medio se extraío con carbón-dextrón. El efecto de este me
dio sobre la sintesis tecal de andrógenos esta' representado en
la figura 5. MBestimuló significativamente la sintesis de an
drógenos (A4- androstenediona mas testosterona) (P < 0,01),
mientras que MBextraido con carbón-dextrón no afectó la sin_
tesis tecal de andrógenos (P < 0,5), Ya que la extracción de
medio fresco con carbón-dextrón no afectó la sintesis basal de
andrógenos (¡00 % del control), estos resultados sugieren que
la capacidad del medio usado de estimular la sintesis de andré
genos se debe a la presencia de alguna pequeña molécula.

TABLA ll
Efecto de FSH sobre lo sfntesh fecal de ondrógenos (ng/mg pro
fel'na).
4Ao To
Control 58,68 i 9,04 5,85 :1,35
FSH(1 ug/ml) ¿4,03 Í10,06 6,35 t1,51
a Promedio Ï E.S.de ó replicadoa

-57
300
ÉEADazooZu< cgg J
lE 35 IOOFzG
C 50% MB cVoe MB
Figura 5: Efecto de la extracción con carbón-dexfrán sobre la
capacidad del medio usado por células granulosas de estimular
_ . . ¿34 . ,Ia Sintesis fecal de androgenos( -androsfened¡ona mas fes
fosterona). Los valores representan el promedio Ï E.S. del por
ciento de cultivos control.
Abreviaturas: e MB, MBextraído con carbón-dextrón.

-58
Con el obieto de determinar si el efecto del medio u
sado sobre Ia sintesis de andrógenos se debia ala presencia de
esteroides, se midieron en éste las concentraciones de sustra
tos posibles para la sintesis de andrógenos.
3.1.3 Concentración de Esteroides en Medio Usado por Cé
lulas Granulosas.
Las concentraciones de A4-androstenediona, testostE
rona, pregnenolona y progesterona en los medios usados (MA
y MB) se muestran enla Tabla lll. Ambos medios contenian can
tidades muy pequeñas de los andrógenos. Las concentraciones
de los C21-esteroides fueron mayores en el medio usado B
(P4 0,001). El contenido de pregnenolona en ambos medios fue
mucho mayor que el de progesterona (P< 0,001).
Debido alos valores elevados de pregnenolona obterli
dos por radioinmunoensayo, se decidió determinar si otros me
tabolitos producidos por las células granulosas estarian inter
firiendo en el ensayo. Alicuotas de 10 muestras fueron extrai
das con éter dietilico y pregnenolona fue aislada por cromatg
grafía de los extractos en columnas de LH-20. La Tabla IV com

TABLA III
-59
Concentración de algunos esteroides en medio usado por células
granulosas (ng/0,5 mI).
Medio A4A 'T °
MA ND 0,09to,02
MB ND 0,15Ío,03
a Promedio Ï E.S. de 16 muestras.
Abreviaturas:
Pregnenolona Progesterona
41,04t2,21 3,77i 0,62
86,51Ï5,89 27,35Ï 3,81
MA: medio obtenido de células granulosas cultivadas en MEM.
MB: medio obtenido de células granulosas cultivadas en MEM
con Ï ug/ml FSH.
ND: no detecfable.

-60
TABLA IV
Concentración de pregnenolona (ng/ml, n=5) antes y después de
purificación del medio de cultivo por cromatografía en columnas
de LH-20.
Preparación dela Control FSHmuestra
Medio no extraÍd0(X) 46,96Ï 6,68 112,87Ï11,40
Extracto etéreo 53,48 Ïl4,25 Hó,7óÏ 37,14
cromatografíado en*
LH-20 (Y)
Y =41,36Ï0,45X r=0,9ló
* corregido por recuperación de esteroides de las columnas.

-6]
para la cantidad de pregnenolona ¡nmunoreactiva obtenida por
ensayo directo con la obtenida luego de extracción y cromato
grafia. Tanto en cultivos control como en aquellos tratados con
FSH, la estimación del contenido de pregnenolona obtenida lue
go de cromatografia fue similar a la obtenida por radioinmuno
ensayo directo del medio de cultivo.
3.1.4 Efecto de CZI-Esteroides sobre la Sintesis Tecal de
Andróge nos.
Se examinó la capacidad de pregnenolona y progeste
rona exógenas de incrementar la sintesis tecaI de andrógenos.
Preparaciones tecales fueron incubadas por 24 horas en presen
cia de 0-200 ng de pregnenolona o progesterona. Los resultados
combinados de dos experimentos distintos con 5 replicados por
dosis en cada uno estan representados en las Figuras ó y 7. Aun
que progesterona estimuló significativamente la producción de
A 4-androstenediona (P< 0,001) y de testosterona (P< 0,001),
los niveles presentes en el medio usado (Tabla lll) parecen ser
insuficientes para causar un incremento enla sintesis de andró
genos.

-62
aoo_
É‘0‘3 .S g;“1:, 00-150p- L 02“ avo: 1-!z\ EL“4:“ .3IC _‘27V JD4 p
¡.1
PROGESTERONA(nq/m0
Figura ó: Curva de dosis respuesta de cultivos fecales a proges
l'eronCI. Cada punto representa el promedio Ï E.S. de 9 incuba —
ciones.
( ——-—— , A4A;--A--, T)

-63
200"
<z AO f53 f.Lu o 5°"Z .uLJ 0C s.n
É a"2<5? mr< v . i a
o 1 a no ID al)
PREGNENOLONA(ng/nü
Figura 7: Curva de dosis respuesta de cultivos tecales a preg
nenolona. Cada punto representa el promedio Ï E.S. de 9 incu
bacíones.

-64
Pregnenolona, desde concentraciones tan bajas como
25 ng por cuHivo, eflímulósígniHcafivamenle (P< 0,00l)la
sintesis tecal de ¿34 - androstenediona. En presencia de preg
nenolona,la producción de testosterona fue variable. Sin embaj
go, dado que testosterona representa sólo lO 96 de la cantidad
de andrógeno total( A4Amás T) acumulada en los cultivoste
cales, los niveles presentes en NlBparecen ser Suficientes pa
ra provocar un incremento al doble en la sfiflesistotal de an
drógenos.
Aunque el término "andrógeno total" usado incluye
androstenediona, testosterona y DHT (ya que el RIA usado no
distingue entre estos dos Ülthnos compuestosL se reconoce que
otros andrógenos, como los 5‘(—androstanedioles y androsterg
na, también podrian ser producidos en cantidadessignificatiVOS.
3.2 Rol delas Células Granulosaseriel MetaboHsmo de
Andrógenos.
3.2.1. Capacidades Relativas de los Tipos Celulares Foli
culares para Sintetizar Andrógenos y Estrógenos.
Células tecales y granulosas derivadas de loliculos

-65
ovóricos de cerdas prepúberes se separaron y cultivaron, aislo_
dos o en recombinación, por 24 horas. Lo cantidad de esteroi
des sintetizado en cultivo por foliculo equivalente por los dis_
tintos tipos celulares estó representada enla Figura 8. Los cé
lulas granulosas produieron cantidades no detectables de todos
los esteroides determinados.
Las preparaciones tecales sintetizaron grandes canti
dades de andrógenos, predominantemente A4-androstenediona,
siendo la relación de A4-androstenediona a testosterona semi
¡ante a lO . La producción de 17(5 -estradiol por la teca fueescasa.
La acumulación de A4-androstenediona en los co-cul
tivos fue mucho menor (P< 0,001) que en los cultivos tecales.
La acumulación de testosterona no ditirió significativamente
(P > 0,05) entre co-cultivos y cultivos tecales. Si bien los co
cultivos produíeron 3 veces mós l7í/al-estrodiol que las prepa
raciones tecales, la cantidad total de andrógenos (A4A mós T)
y ¡7? -estradiol siguió siendo menor en los co-cultivos que
en los cultivos tecales. También se determinó estrona en algu
nas muestras pero ésto sólo representó alrededor del 15 °/ode
la producción total de estrógenos (estrona + E 2 ). Lo inclusión
de FSH (l ug/ml) en los co-cultivos no produío efecto alguno
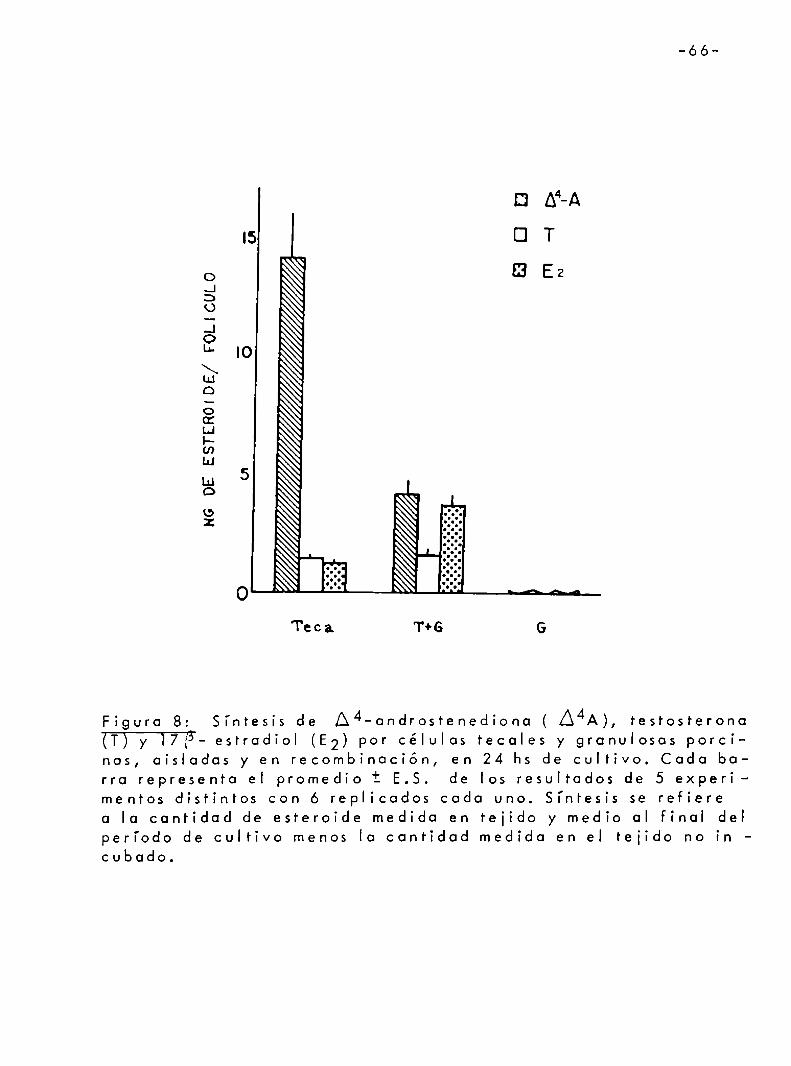
-óó
El A4-A
NGDEESTEROÍDE/FOLICULO
Teca T+G G
Figura 8: Sintesis de A4—androstenediona ( A4A), testosterona¡Ti y Í7¡3- estradíol (E2) por células tecales y granulosas porcinas, aisladas y en recombinoción, en 24 hs de cultivo. Cada barra representa el promedio Í E.S. de los resultados de 5 experimentos diantos con ó replicados cada uno. Sintesh se refierea la cantidad de esteroide medida en teiido y medio al final delperiodo de culeo menosla cantidad medida en elteiido noin cubado.

-67
sobre la acumulación de A4A y T (datos no mostrados).
3.2.2 Capacidades Relativas delos Tipos Celulares Folicu
lares para Metabolizar Andrógenos.
Dado que anteriormente se observó que adición de me
dio en que células granulosas habian sido cultivadas estimula
ba la sintesis tecal de andrógenos, la observación de un efec
to inhibitorio al co-cultivar células granulosas con preparaci_o
nes tecales sugirió que las células granulosas probablemente
metabolizaban el andrógeno fecal a otros productos. Para exa
minar esta posibilidad, los distintos tipos celulares foliculares
fueron cultivados en presencia de testosterona o A4-androste
nediona radioactivas, purificadas por cromatografia en capa
delgada. Al final del periodo de Cultivo, los esteroides se ex_
traíeron a éter dietilico para caracterización posterior, Ines
peradamente, la radioactividad adicionada alos cultivos no
pudo ser recuperada totalmente en los extractos etéreos en to_
dos los casos. Más aún, los porcentajes de radioactivídad ex
traibles al éter (esteroideslibres) dependieron del teiido pre
sente en las incubaciones (P < 0,001). Los porcentaies de radio

-68
actividad recuperada se muestran en la Tabla V. La mayor par
te de la radioactividad en los cultivos tecales pudo ser extraí
da con éter. Sin embargo, siempre que los cultivos contuvieran
células granulosas, mas del 50 % de la radioactividad inicial
permaneció en la fase acuosa, no habiendo diferencia alguna
entre los cultivos de células granulosas aisladas y los co-cult_í
vos. En vista de estos resultados se planteó la posibilidad de
que las células granulosas fueran capaces de formar coníugados
de esteroides, solubles en agua.
Al cultivar células granulosas con dehidroepiandroste
rona radioactiva, sólo ll % de la radioactividad permaneció
en la fase acuosa, mientras que al agregar androsterona o DHT
radioactivas, toda Ia radioactividad fue recuperada en el ex
tracto etéreo.
3.2.3 Metabolismo de Andrógenos por Células Granulosas.
Para obtener mayor evidencia de coniugación de este
roides y determinar qué tipo de coníugados se formaban, se es
” l . 3 A4 . ,tudio e metabolismo de ( H) - -androstenediona por celu
las granulosas. En 18 experimentos distintos, 60,4 Í 2,2 % de

-69
TABLA V
Porcenfoíes de rodíoacfívídod exfrou'ble ol éter luego de cultivo
con (3H)-ondrosfenedíono o (3H)-fesfosferono por 24 horas.
Típo Celular Cultivos con Cultivos con
(3H)-Androsfenedíono ° (3H)-Tesfosterono b
Teco 60,6Ï 4,5 90,9Ï 8,0
Granulosa 24,1i1,2 38,5 :1,4
Teco+Gronulosa 21,3Í 1,0 41,8Ï5,9
° Los valores representan el promedio Ï E.S. de lO replicados.
b Los valores representan el promedio Í E.S. de 5 replícodos .

-70
la radioactividad inicial permaneció en Ia fase acuosa luego
de extracción con éter dietilico. La tracción acuosa fue exa
minada por cromatografia en DEAE-Sephadex. La Figura 9 repr_e
senta el perfil de elución observado en un experimento repre
sentativo. 57,52 i 4,05 % (n = 18) de la radioactividad en la
fase acuosa eluyó en una posición similar del gradiente (Pico
ll, Figura 9). La Figura lO representa el perfil de elución ob
servado al cromatografiar en este sistema varios monosulfatos
de esteroides neutros y esteroides libres. Es evidente que el
principal metabolito de androstenediona no corresponde con
testosterona - i7-monosulfato o con androsterona-3 - monosulfa
to.
El material presente en las fracciones 4-5 (Pico l,
Figura 9) fue completamente extraido al éter. Luego de solvó
lisis, 98 % de la radioactividad en las fracciones 15-19 (Pico
ll, Figura 9) pudo ser extraido con éter, sugiriendo que 60 %
de la fracción acuosa probablemente consistia de C]9-esteroi_
des neutros coníugados a iones sulfato.
3.2.4 Actividad de Sulfotransferasa en Células Granulosas.
Se realizaron ensayos de actividad de sulfotransferasa
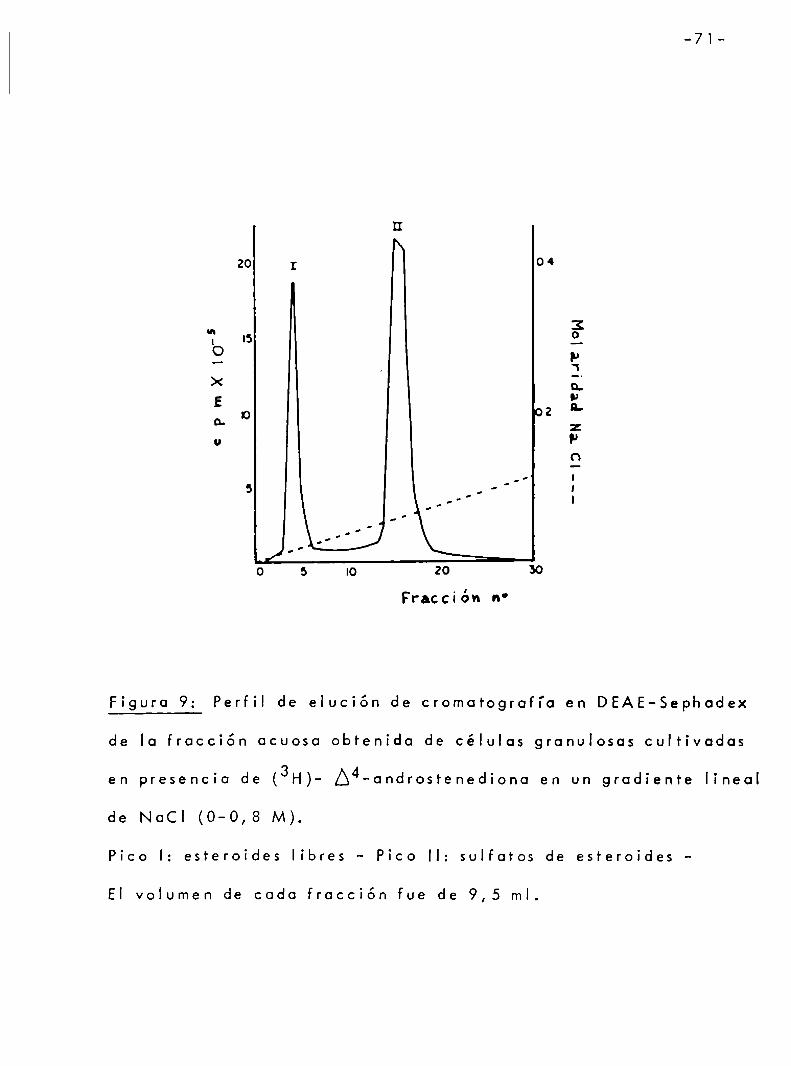
-7]
20 o 4
3'T Is 9.9 ¡vlx eE n.
o 2o. P zo w
ñ— I
5 tl
o :io
Fracción n'
Figura 9: Perfil de elución de cromatografía en DEAE-Sephadex
de la fracción acuosa obtenida de células granulosas cultivadas
en presencia de (3H)—A4-androstenediona en un gradiente lineal
de NaCl (O-O,8 M).
Pico l: esteroides libres - Pico ll: sulfatos de esteroides
El volumen de cada fracción fue de 9,5 ml.

-72
4
l -03'P 39 í>-<IO- .02E ,— io. 2 ¿,-— zU '_—
f — ' —o-I ”- z ' Q
a , ' la ' l
a , — lf 1 l l ll
IO 20 30Fracción h'
Figura 10: Perfil de elucíón de cromatografía en DEAE-Sephadex
en un gradiente lineal (0-0,8 M NaCI) de: pico 'l, DHEA; pico
2, sulfato de testosterona; píco 3, sulfato de DHEA;pico 4, sul
fato de androsferona.
El volumen de cada fracción fue de 10 ml.

-73
con el obieto de obtener evidencia independiente de formación
de sulfatos de andrógenos por células granulosas. La Figura ll
representa el perfil de elución en DEAE-Sephadex A25 del pro_
ducto de reacción obtenido al incubar sobrenadante de 100000 g
de células granulosas con (3H) -androsterona, en presencia de
(35 S) - Na2SO4, previa eliminación del sustrato en exceso por35 3extracción con éter dietilico. Se observa que S y H eluye
ron enla porción del gradiente correspondiente a androsterona
3-monosulfato, siendo la relación entre 355 y 3H constante
(parte superior de Figura ll). Estos resultados fueron repetidos
dos veces con el mismo sobrenadante de 100000 g. Sin embargo,
sobrenadantes obtenidos de distintos lotes de células granulosas
sólo coniugaron cantidades pequeñas de androsterona, aunque
algunos de ellos coniugaron testosterona. La inconsistencia de
estos resultados sugiere que la capacidad de conjugar estos ari
drógenos no es uniforme en todos los toliculos, o que las con
diciones óptimas para el ensayo enzimática en teíido ovórico
porcino difieren de aquellas para otros teiidos estudiados.
3.2.5 Estudios Tendientes a la Identificación de los Meta
bolitos Coníugados por las Células Granulosas.
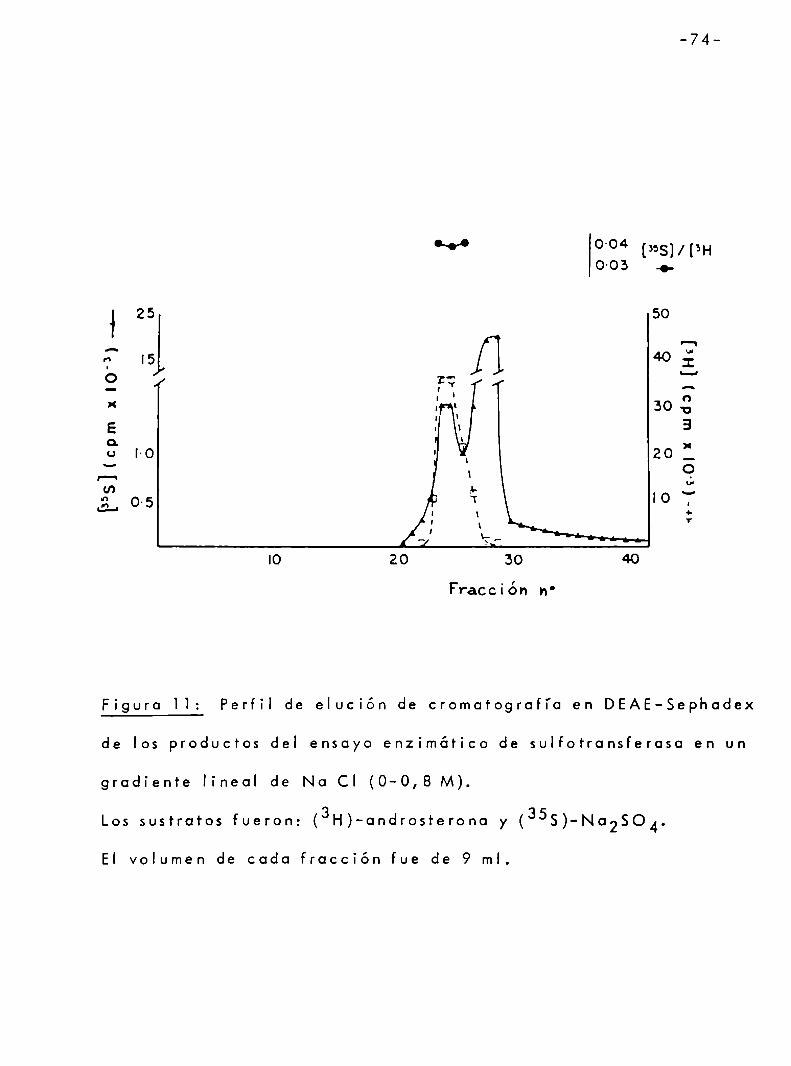
-74
0'03 .0.
+ 25 50A T7 IS 40 I° ÏÏ_. 303E 3o.u I-O 20 Zv O
H ¿a¿n 0-5 ¡o 7'
lO 40
Fracción n'
Figura H: Perfil de elucíón de cromatografía en DEAE-Sephadex
de los productos del ensayo enzímóríco de sulfotransferasa en un
gradiente lineal de Na CI (0-0,8 M).
Los sustratos fueron: (3H)-androsterona y (355)—F4025C)4.
EI volumen de cada fracción fue de 9 ml.

-75
Se realizaron estudios para determinar la identidad de
los esteroides coníugados al cultivar células granulosos en pre
sencia de A4-androstenediona. En la Figura 12 se muestra el
camino de metabolismo de andrógenos que supuestamente se si
gue en el ovario, en base a resultados obtenidos enla rata
(105, 106). Se llevaron a cabo estudios preliminares tendientes
a encontrar un sistema que permitiera una buena separación de
estos compuestos. El método elegido fue cromatografia liquida
de alta presión (HPLC). Se obtuvo una buena separación de es
tos compuestos usando una columna de microporasil (fase nor
mal) con elsistema cloroformozísooctano(6:4)(Figura 13). El
perfil de elución obtenido ol cromatografiar en este sistema al
producto de solvólisis ¡unto a otros esteroides carrier está re
presentado enla Figura l4. El pico radioactivo eluyó con un
tiempo de retención de l,2ó-l,40 relativo a A4-androstene
diona. La anchura del pico se debe probablemente a que el su_s
trato usado ( A4A) consiste de una población de isótopos. En
efecto, este sistema es capaz de resolver isótopos de un traza_
dor, como se eiemplifica enla Figura 15 para testosterona ra
dioactiva purificada. Alternativamente, el pico podria conte
ner varios compuestos, no resueltos por este sistema.
En la Tabla VI se da una lista de los esteroides cromo
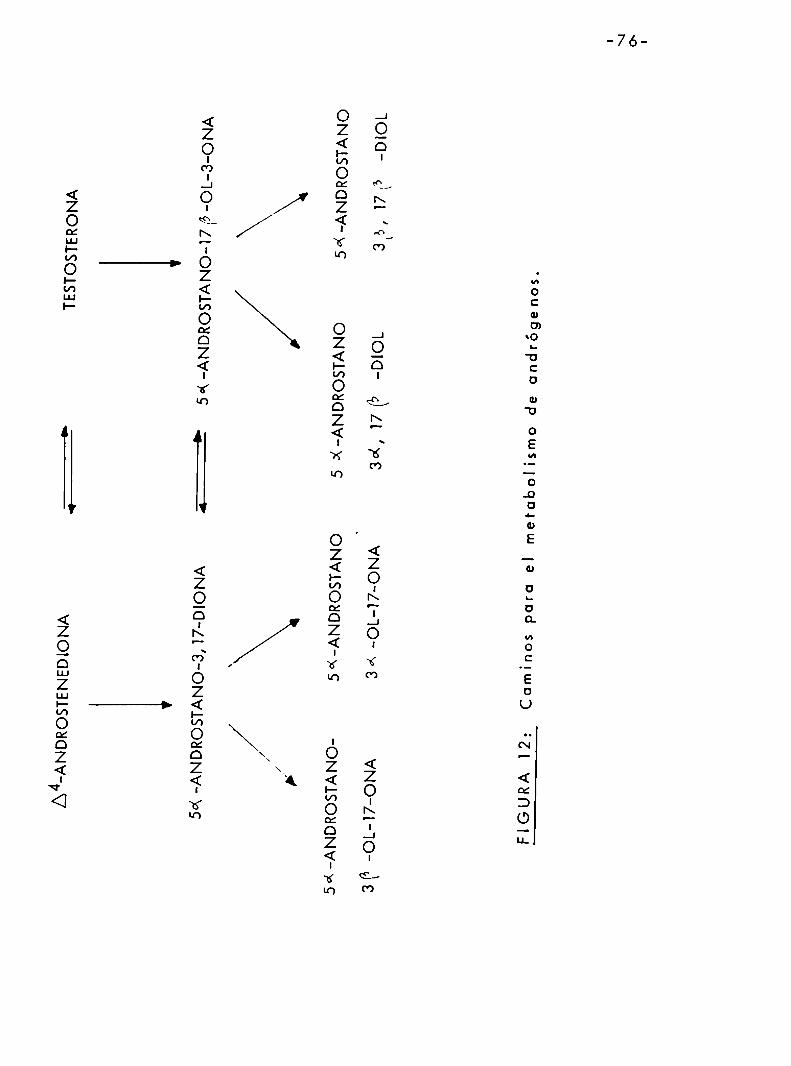
“VAL/2029:2862»,#202202»,
mi¡>ZUmOm._.>ZO¡u\_N¡D_OZ>
x/uen¡>ZOxOm._.>ZO¡
A“,uA¡>ZUxOMH>ZOLNm¡OrIuIOZ>
\/
mA¡>ZOmOmfipZOmx¡>Zox0m59ZO
mA¡>ZUmOm._,>ZO
un-OTG-oz>Ï-oT:-oz>mx.Gm-99un:Gm-99
ÏOCmb,dm"ñoamnounaaam.3m_.aco:u30amoaaqmmmaou.
-76

-77
AA
‘ DHT
&
A
¿
U 3P' dio' 3<-diol
o é ¡b ¡5 ' 26
Tiempo de Retención (minutos)
Figura 13: Peer de elucíón del cromatógrafo Iïquído de alfa
presión mostrando los Índices de refracción relativos para [x4
androstenedíono ( [X4A), díhidrotesfosferona (DPíT), androsfe_
rona (A), SOC-androsran-B [3 ,]7(a -dio| (3/5 -d¡o|) y 50( -an _
drostan-30<,17P —dío|(34-dío|L Loscueroídesfueronsepargdos en una columna de mícroporasH yel üsfema de solventes
cloroformozísoocfano ::ó:4; 3 mI/mín.

-78
y Peron!CHCIJ:nsooctano (6:4)
l 20 3-0 nal/min
"i" IUo W "x l: ¿ Epsondrostuono x|
;‘, OHT": Q Anarosterom m|| g I l
E '° 5': 2 :3 Icx :2 : 5': .". '
° 5 : í : 1 5';I I 1 ' ' |
: '. : : 1| .' x
i ‘. J E a J a
O 5
'T‘iernpode Retención (minutos)
Figura 14: Separación por HPLC de los productos de solvólïsís
de Ia fracción de monosulfafo neutro en una columna de micro
porasíl y el sistema de solventes cloroformo: ísoocfano z: ó :4,
3 mI/mín.

-79
l 1
7 3A 1°9 I x
x 5' ,5, CD
s 1: Io. :I .U l:
llll
L'x A5 ¡o
Tiempo de Retención (minutos)
Figura 15: Separación por HPLCde ¡sótopos de (3H)-testostero
na.
(l, 2, ó, 7, 16, 17 3 H)-Testosterona y testosterona no radio
activa fueron cromatografíados en una columna de microporasíl
con el sistema de solventes cloroformo: ísooctano :: ó : 4; 3 ml/
mín.

-80
TABLA VI
Tiempos de retención relativos de algunos esteroides de 19 ótomos
de carbono en HPLC (columna UPorosíI; CH Cl 3: ísooctono :: ó : 4,
3 mI/min).
Esteroíde Tiempo de Retención
Relativo a Androstenedíono = 1
Compuesto desconocido I,26-T,40
5<4-D¡hídrotestosterono (DtiT) 1,28
Epíondrosterono 1,45
Androsterona 1,67
Testosterona 2,68
5d-Androstono-3¡/¿,T7í/5 -díol 2,80Etiocolonolono 3,04
5 d -AndroMono-3c4-¡7fl -dio| 3,73
5 vc -Androst-Tó-en-30(-ol 0,59
ScÁ -Androstano-3,T7-díono 0,57
Sci -Androst-Tó-en-3-ono 0,55
Tiempo de retención de A4-Androstenedíono= 5 mÍnUÍOL

-3]
tografiados en este sistema con sus tiempos de retención en el
mismo. Aquellos esteroides con tiempos de retención semeian
tes al compuesto desconocido (DHT, epiandrosterona, androst_e
rona) fueron descartados por datos de recristalización. Al oxi_
dar este compuesto por el método de Jones, y someterlo a cro
matografia en este sistema, la rodioactividad eluyó con un
tiempo de retención semeíante a A4-androstenedíona. La ¡den
tídad del producto de reacción fue confirmada por recristaliz_a
ción con [14C]-A4-androstenediona a relación l4C/3H cons
tante.
La reducción con borohídruro de sodio dio lugar a dos
compuestos que se comportaron como dioles en este sistema.
Esto sugiere que el compuesto es un esteroide no saturado con
una función ceto y un hidroxilo.
Aunque el metabolito representó aproximadamente 40%
del producto solvolizado en los dos primeros lotes de células
examinadas, otros lotes produieron cantidades mucho menores
de este compuesto. Se decidió entonces emplear otros sistema
para determinar la identidad de los metabolitos producidos al
incubar células granulosas con A4-androstenediona.
En la Figura ló se ilustra el perfil de elución del pr_o
ducto solvolizado en una columna de fase reversa con el siste

-32
30 ILF
I'T 2°"2xEo.9 ID
1 jO lO 410
Tiempo de Retención (minutos)
Figura ló: Separación por HPLCde los productos de solvólhís de
Ia fracción de monosulfato neutro en una columna C18 Radial Pak
Cartridge y elsístema de solventes acetonítrHo
1 ml/mín.
:agua u 4 :ó,

-83
ma acetonitrilo:agua (4:6). Se observaron varios picos; el cor_n
puesto más abundante aparece con un tiempo de retención de
3,5-4,5 minutos (O,27-0,35 relativo a ¿4-androstenediona).
Contrariamente al metabolito anterior, la presencia de este
compuesto fue consistente en todas las incubaciones realizadas
(ló lotes de células obtenidos en distintos dias). Este producto
no eluye dela columna de microporasil con el sistema de sol
ventes usado debido a su alta polaridad.
Los tiempos de retención de varios esteroides en el
sistema de fase reversa empleado se encuentran en la Tabla Vll.
Es evidente que en este sistema sólo l9-hidroxi-androstenedio
na y l9-hídroxi-testosterona presentan tiempos de retención
milares al compuesto desconocido.
El mayor pico de la Figura ló (ll, tiempo de retención=
3,5-4,0 minutos) se cromatografió luego en una columna de fa
se reversa con el sistema metanolzagua (6:4), presentando un
tiempo de retención de 4 minutos. Baio las mismas condiciones,
l9-hidroxi-androstenediona y l9-hídroxi -testosterona eluyeron
a 3,90 y 3,75 minutos respectivamente.
Al ser cromatografiado en capa delgada, el metaboli
to radioactivo se comportó como un compuesto muy polar , con
un Rf semejante a l9-hidroxi-androstenediona standard. Sin em

-34
TABLA VII
Tiempos de retención relativos de algunos esteroides de 19 átomos
de carbono en HPLC (C18 RAD. CART. 10, 5 mm l.D.; ocetonítrilo:
agua :: 4: ó; 1 mI/mín).
Esteroíde Tiempo de Retención
Relativo o Androstenedíono= 1
Pico 1 de HPLC 0,19Pico 1| de HPLC 0,27-0,3519-Hídroxítestosterono 0,2219-Hídroxí- B4-Gndrostenedíono 0,264-Androsten-3,17';'-d101 0,67Testosterona 0,814-Androsten-3lb, 17‘1-dío1 0,825.a: -Androstono-3¿ ,17 5 —díol 0,854-Androsten-3«v._-Ï,17¡5 -dío| 0,864-Androsten-313—o|—17-ono 0,8650< —Androstano—3<x ,17¿’5—díol 1,08545 —Androstan—3|3—o|-17-ono 1,14Díhídrostestosterona (DHT) 1,2153/5-Androston-30<-o|-17-ono 1,27Androsterona 1,4950€,—Androstano-3,17-díono 1,515°! -Androst -1ó-en-3“/ -o| *5-“4-Androst -1ó-en-3 3-01 *
* No eluído luego de 20 minutos en este sistema.
Tiempo de retención de Ali-androstenedíona = 12,91 minutos.

bargo, recristalízación delcompuesto radioactívo con l9-hidroxi
androstenediona o l9-bidroxi-testosterona indicó que no ero nin_
guno de estos.
Por los resultados obtenidos en cromatografía en Sepha_
dex A25, en capa delgada y de alta presión, este compuesto pa
rece ser un hidroxi-esteroide, no saturado, no fenólico.
En este sistema, en algunas incubaciones se observó un
pico a un tiempo de retención de 1,2] respecto a A4-androste
nedíona, que correspondia al metabolito aislado en el sistema
de fase normal.
La identidad delos compuestos coniugados por las cé
lulas granulosas porcinas no pudo ser determinada. Los esteroi
des Sd-y 3M -reducidos, que son producidos por ovarios de o
tras especies (105, lOó) fueron descartados debido a sus tiem
pos de retención en los sistemas cromatogróticos usados (Tablas
Vl y Vll). El hecho de que ni androsterona ni DHT Fueron con
¡ugados por las células granulosas también sugiere que es imprí)
bable que los compuestos coníugados sean metabolitos 54-redg
cidos de A4-androstenediona. Asimismo, se descartó la posibi
lidad de esteroides 5 [5-reducidos. Es improbable que sean Aló
andrógenos, formados en grandes cantidades por teíido testicu
lar de cerdo (l07) debido a su compartamiento en HPLC de fase

-86
normal (Tabla VI) y reversa (Tabla Vll). Otros estudios usando
métodos fisicos en cantidades mayores de este coniugado serian
necesarios para elucidar sus estructuras completamente.
3.3 Efecto dela Teca sobre Ia Sintesis de Progesterona por
las Células Granulosas.
3.3.] Efecto de los Andrógenos sobre la Acumulación de
Progesterona por Células Granulosas.
Células granulosas fueron incubadas por 24 horas en
presencia de testosterona, A4-androstenediona, DHTo andros
terona en combinación con o sin l ug de FSH. Cada andrógeno
se incluyó a concentraciones de 0,5 uM y 5 uM. Finalizados
los cultivos, se determinó la cantidad de progesterona y 17,47
estradiol acumulada. Los experimentos se realizaron por lo me
nos 2 veces con distintos lotes de ovarios. En cada experimen
to, cada tratamiento se replicó 4 veces.
Experimentos preliminares indicaron que el contenido
de 17} -estradiol de las células luego del cultivo estaba por
debaío de los limites de detección del ensayo. Aunque el con

-87
tenido tisular de progesterona era mós significativo (aproxima
damente 25 %>de la acumulación totalL el contenido Hsular
estaba siempre altamente correlacionado con el contenido en
el medio de cultivo, independientemente de la presencia de
FSH o FSH y testosterona (controL r = 0,96;+ FSH, r = 0,99;
+ FSH + T, r = 0,99). Por lo tanto, los resultados se expresaron
en términos de acumulación de progesterona o l7 3 -estradiol
en el medio de cultivo por foliculo equivalente.
En ausencia de andrógenos, FSH conshtentemente esü
muló lO vecesla sintesü de progesterona. Ninguno de los an
drógenos afectó significativamerne la sintesis basal de proges
terona; por lo tanto estos resultados no se presentan. Los efec
tos de los distintos andrógenos sobre Ia acumulación de progeí
terona en respuesta a FSH se presentan en las Figuras 17-20;
en el caso de testosterona y Ót4-androstenediona, se incluyen
los datos para la acumulación de 17 5 -estradiol; dado que los
andrógenos no aromatizables, DHTy androsterona, no afecta
ron la acun1ulación de 17 5 -estradiol, estos datos no estón gng
ficados.
En la Figura 17 se observa que testosterona inhibió la
producción de progesterona en respuesta a FSH de una manera
dosis-dependiente (P< 0,001), mienhas quela acumulación

-88
u 75'aO
fi ESTRADÍOL'WB5';“ '3 507u u PROGESTERONAo \z 3o ‘4
5 E 25< c_J v
:2)< c0 0.5 5.0 O 0.5 5.0
MH TESTOSTÏRONA
Figura 17: Efecto de testosterona sobre la acumulación de pro
gesterona y17P-estradíol por células granulosas en cultivo en
presencia de l ug/ml de FSH (n: 14).
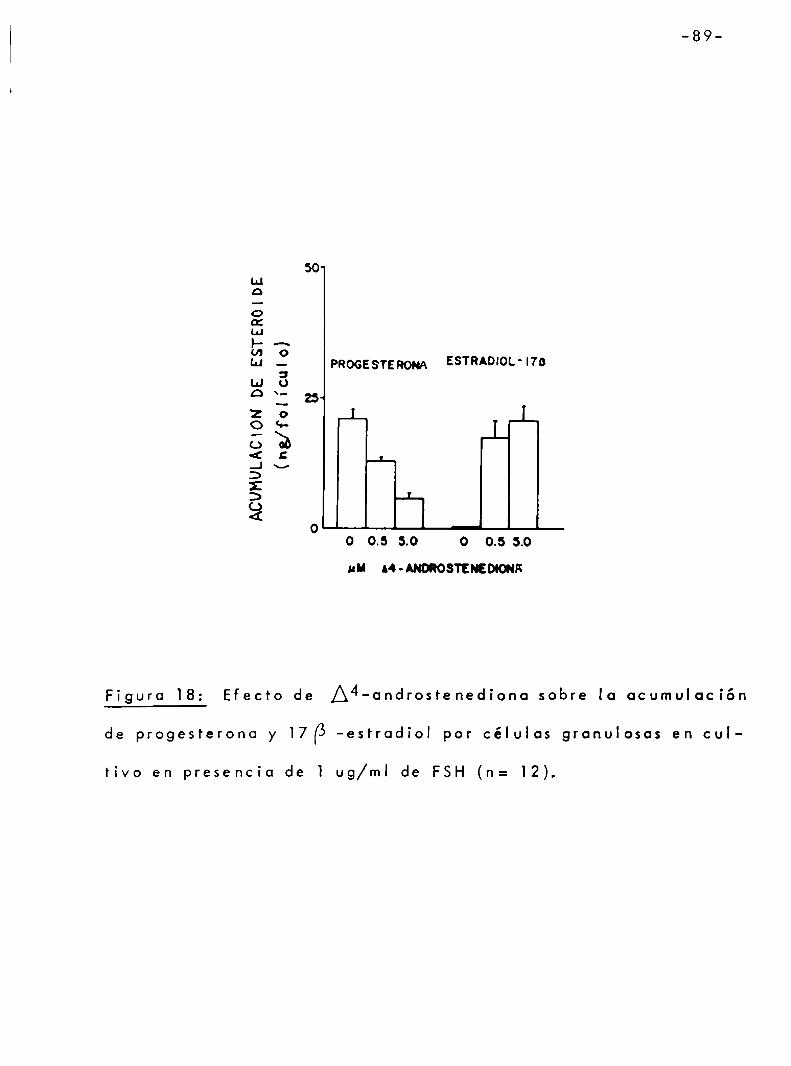
-89
uOo(zLAJ
i; 7;Lu .5. pRoGESTERONA ESTRÁDIOL'ITOLJ U
D 1-: a42 Oo ‘0u Ü< fi_J VD5:3<Í
O0 0.5 5.o o 0.5 5.0
un M-ANOROSTENEDMR
Figura 18: Efecto de A4—androstenedíona sobre la acumulación
de progesterona y 17(3 -estradíol por células granuloscls en cul
tivo en presencia de ï ug/ml de FSH (n: 12).

-90
25*
FL
PROGESTERONA(ng/folículo)
O 0.5 5.0
uM DHT
Figura 19: Efecto de díhïdrofesfosterona (DHT) sobre la acumu
lación de progesterona por células gronulosas en cultivo en pre
sencia de I ug/ml de FSH (n = 10).

-9]
50'
"3
_L IE\ÜÉ
V 25u‘z‘o5}_
:3Ü8O.
o 0 0.5 5.0
ANDROSTERONA
Figura 20: Efecto de androsterona (A) sobre la acumulación de
progesterona por células granulosas en cultivo en presencia de
lug/ml de FSH (n = 8).

-92
de 17? -estradiol aumentó al incluirse testosterona en los cu_l
tivos (P< 0,001). Similarmente, A4—androstenediona inhibió
Ia acumulación de progesterona en respuesta a FSH (P<0,001)
y aumentó la de 17:6-estradiol (P< 0,001) (Figura 18). DHT
(Figura 19) y androsterona (Figura 20) no produieron efecto al
guno sobre la acumulación de progesterona estimulado por FSH.
El efecto de testosterona sobre la sintesis de progest_e
rona en presencia de FSH en' células granulosas aisladas de o
varios de cerdas maduras se presenta enla Tabla Vlll; también
en este caso testosterona inhibió significativamente (P<0,05)
la sintesis de progesterona en respuesta a FSH.
3.3.2 Efecto de Testosterona sobre el Metabolismo de Proges
terona.
Con el obieto de determinar si testosterona causaba
una reducción en la acumulación de progesterona estimulado
por FSH mediante la inhibición de la sintesis de progesterona
ola estimulación de su metabolismo, células granulosas fueron
incubadas en presencia de (14C) -progesterona (luM) y FSH,
con o sin testosterona (SUM). Luego de un periodo de 24 horas

-93
TABLA VIH
Acunuflación de progesterona (ng/folículo equivalente) por célu
las granulosos obtenidas de foliculos de tamaño mediano de cer
dos sexualmente maduras en 24 hs de cultivo (n=4).
*
Progesferona
control 4,5 Ï 0,2 a
FSH 36,9Ï7,5bFSH+O,5uMT 20,9Ï4,7°
FSH + 5 UM T
Las diferencias entre a y b, a y c, y, b y c fueron significati
vas a P<.0,05 (Prueba de rangos mGIlees de Duncan).

-94
de cultivo, el porcentaíe de recuperación de la radioactividad
inicial asociado con progesterona fue de 43,1Ï10,l %(n:3)
en presencia de testosterona, no significativamente distinto de
los controles sin testosterona (30,9 Í 5,2 %, n :3). Esto sugie
re que testosterona no estimula el metabolismo de progesterona,
y por lo tanto eíerceria su efecto sobre la acumulación de pro
gesterona inhibiendo su sintesis.
3.3.3 Efecto de 17;"?-Estradiol sobre la Sintesis de Proges
teronag
Dado que los andrógenos aromatizables podrian eier
cer su efecto ínhibitorio sobre la acumulación de progesterona
por via de su conversión 017/5-estradiol, fue importante deter_
minar si estradíol inhibia la sintesis de progesterona en este
sistema. Por lo tanto se incluyó estradíol en los cultivos a con_
centraciones de 0,5/uM y 5/uM. Como se observa enla Figura
2], la acumulación de progesterona en respuesta a FSH disminii
yó en presencia de 173 -estradiol (P4 0,001), confirmando que
estradíol es inhibitorio a estas concentraciones.
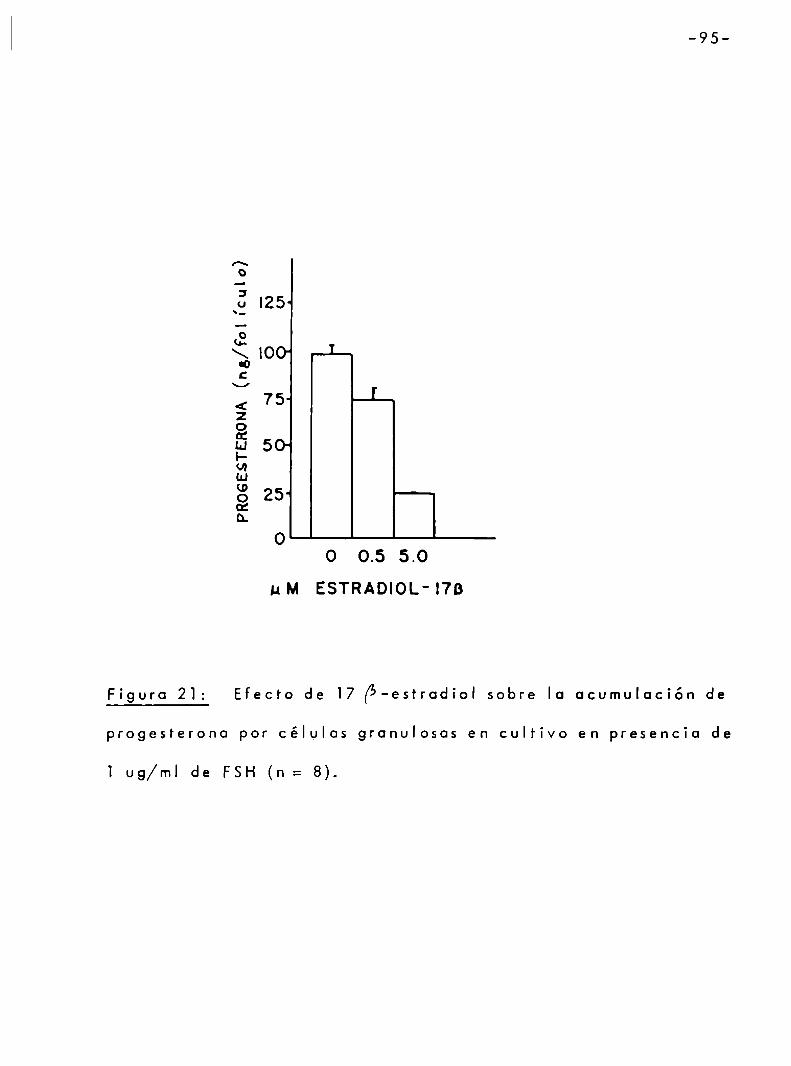
-95
l25<
¡00* r—LT
75
50
25*
PROGESTERONA(ng/Fai(culo)
0 0.5 5.0
HM ESTRADIOL'I'IB
Figura 21: Efecto de 17 fi-estradíolsobre Ia acumulación de
progesterona por células granulosas en cultivo en presencia de
I ug/ml de FSH (n= 8).

-96
3.4 Mecanismo por el cual los Andrógenos Inhiben la Sintesis
de Progesterona por Células Granulosas en Respuesta a
FSH.
3.4.]. Relación Temporal entre la Acumulación de Proges
terona y l7 .7-Estradiol en Cultivo en Presencia de
FSH y Testosterona.
Células granulosas fueron Cultivadas en presencia de
FSH solo o FSH y testosterona (SUM). Los cultivos se interrum
pieron alas 4, 8, ló o 24 horas. La acumulación de progester_o
na esta' representada en la Figura 22 y la de estradiol en la Fi_
gura 23. Los datos se analizaron por análisis de varianza en
dos direcciones. Los niveles de progesterona (P4 0,001) y de
17:5 -estradiol (P < 0,00l) aumentaron con tiempo en cultivo.
En presencia de testosterona, la acumulación de progesterona
fue inhibida al nivel de P <' 0,05 a todos los tiempos estudiados
(Prueba de rangos múltiples de Duncan) y, similarmente, la a
cumulación de ¡75-estradiol estuvo significativamente eleva
da (P < 0,05) a todos los tiempos estudiados. En ausencia de
testosterona, Ia acumulación de progesterona aumentó marcada
mente entre 4 y 8 horas de cultivo, pero, en presencia de tes
tosterona, no aumentó sustancialmente hasta después de 8 horas

-97
20q
A0Éu
M.
,3 FSH\0ÉV lo.< Jz z8 zlU Il... x{3 ,’FSH+TQ z8 _,—(L “'—
#_—*———_-_F’I I I 1—
o H 8 ¡6 2%
Tiempo en cultivo (horas)
Figura 22: Curva de tiempo para la acumulación de progesteroI O O Ína por celulas granulosas en cultivo. El medio contenla un fo
lículo equivalente de células, 1 ug/ml de FSH, y ningún este
roíde (línea entera) o testosterona 5 uN\(|ïneo quebrada)(n==4).

-98
g.Ao
3 /y l- FSH+'T' /8 //\ /o lS xV IJ q“ x0 /"' Í3 /a /IC; zu x| lQ ,L_-_J,
N ,1_ lz FSH/
A I Ï ' Í0 H 8 (6 aq
Tiempo en cuLtívo (horac)
Figura 23: Curva detïempo parala acumulación de 17/6-esna
díol por célula; granulosas en cultivo. Elrnedïo contenía un fo
lículo equivalente de células, 1 ug/ml de FSH, y ningún esteroí
de (línea entera) o testosterona 5 uN\(|ïnea quebrada) (n = 4).

-99
en cultivo. En presencia de testosterona, la acumulación de
17'?)—estradiol no se alteró entre las 4 y 8 horas de cultivo y
no aumentó notablemente hasta después de las 8 horas. Es decir
que la acumulación de progesterona fue inhibida por testostero_
na antes de un aumento sustancial en los niveles de estradiol;
3.4.2 Efecto de un lnhibidor dela Aromatasa sobre la
Inhibición dela Sintesis de Progesterona por Tes
tosterona.
Para obtener mayor evidencia de que el efecto inhib¡_
torio sobre la acumulación de progesterona era causado por tes
tosterona misma,sin previa conversión a 171%—estradiol, célu
las granulosas fueron cultivadas en presencia de FSH, con o
sin testosterona (0,5 u M y 5 uM) y con o sin l ug de 4-ace_
toxi-4-androsten-3,l7-diona( 4 a A). Este Último es un inhib_i
dor efectivo de la aromatasa (108). Los resultados estón repre
sentados en las Figuras 24 (acumulación de progesterona) y 25
(acumulación de 17 —estradiol). Anólisis de varianza en dos
direcciones demostró que testosterona promovia la acumulación
de estradiol (P< 0,001) y que 4 a A Ia inhibia efectivamente

-IOO
IZS
¡0° -4-ocfloxy-A “accion-A
PROGESTERONA
(ng/folículo)
u9,
' o 0.3 5.o o 0.5 5.o
uu TESTOSTERONA
Figura 24: Efecto de un inhibidor de la aromarasa sobre la acu
mulación de progesterona por células granuloscs en cultivo. El
medio contenía un foll'culo equivalente de células, 1 ug/ml de
FSH, 0 o i ug/ml de 4-0cetoxï-androstenediona, y las concen
traciones indicadas de testosterona. (n=9).

-]O]—
IOOï-4-ocoto¡y-A #4-ncotoly-A_J A
9. 3 75C) :1< Ua: ‘23 —— 504
w 5|Q ¡a 4N .C 25
00 0.5 5.0 0 0.5 5.0
HH TESTOSTERONA
Figura 25: Efecto de un inhibidor de la oromatosa sobre la o
cumulacíón de 17(5-estrodíol por células gronulosas en cultivo.
El medio contenía un folículo equivalente de células, 1 ug/ml
de FSH, 0 o] ug/ml de 4-ocetoxí-androstenedíona, y las con
centraciones indicadas de testosterona. (n- 9).

-102—
(P < 0,001); no habia interacción. Sin embargo, en presencia
de 4 a A, testosterona seguia inhibiendo Ia sintesis de proges
terona. Por lo tanto, 4 a A no pudo impedir la inhibición de
la sintesis de progesterona dependiente de testosterona.
3.4.3 Efecto de Testosterona sobre la Sintesis de Progeste
rona Estimulada por Dibutiril-AMP ciclico.
Los andrógenos podrian inhibir la producción de pro
gesterona en respuesta a FSH interfiriendo con la unión de
FSH a su receptor, interfíriendo con el acoplamiento del rece_p
tor ala adenilato ciclasa enla producción de AMPciclico,
estimulando la Fosfodiesterasa y/o inhibiendo las enzimas inv_o
lucradas en Ia biosintesis de progesterona. Para determinar si
los andrógenos actuaban antes o después de la producción de
AMPcíclico, células granulosas fueron cultivadas por 24 horas
en medio conteniendo dibutiril AMP ciclico timM) solo, testo_s
terona (5 UM) sola, o testosterona y dibutiril AMP cíclico, en
un experimento factorial 2x2. Como se observa enla Figura 26,
dibutiril AMPciclico estimuló significativamente (P < 0,001)
la sintesis de progesterona tanto en ausencia como en presencia

-103
ñ3 IOr'
3 r_J_'8s.\qc
5ï2gu¡.UIuÉa. C y
(Bu)2cAMP - 0 - .
T ‘ " t ‘
Figura 26: Efecto de cultivar células granulosas porcinas por
24 hs en medio solo (control), con testosterona (5 uM) y/o
(Bu)2 CAMP (1 mM) sobre Ia acumulación de progesterona (pro
medio Í E.S.; n :12).

—lO4
de testosterona, y testosterona eierció una inhibición signifi
cativa (P< 0,002) sobre la sintesis de progesterona. Una inte
raccíón altamente significativa (P<0,01) indicó que el efecto
de testosterona sobre la acumulación de progesterona era dife
rente dependiendo de la presencia o ausencia de dibutiril AMP
cíclico.
Células granulosas fueron cultivadas por 24 horas en
presencia de distintas cantidades de dibutiril AMPciclico y
testosterona. La acumulación de progesterona en el medio de
cultivo está representada en lo Figura 27. En ausencia de tes
tosterona, dibutiril AMP ciclico estimuló la producción de te_s_
tosterona (P< 0,001) de manera dosis-dependiente; una dosis
de 0,5 mMde dibutiril AMP ciclico fue suficiente para causar
una estimulación significativa (P< 0,05). La presencia de tes
tosterona inhibió significativamente (P<0,01) la sintesis de
progesterona estimulado por dibutiril AMP cicl ica, pero no a
fectó la sintesis basal de progesterona (P> 0,1). En presencia
de testosterona 0,5 uM, la menor concentración de dibutiril
AMPciclico necesaria para producir un aumento significativo
enla sintesis de progesterona fue de i mM, mientras que, en
presencia de testosterona 5 u M, se requirieron 2 mM de dibuLi
'ril AMP cíclico. La acumulación de progesterona en presencia

-105
A yo
— Control5U\..3É
C
V ¡1 OSuMT
U‘- tr_ _ - ' SUHT
É ' Ï
mM (Bu)z CAMP
Figura 27: Relación dosis-respuesta enla estimulación dela
acumulación de progesterona por díbutíril AMP cíclico en pre
sencía o ausencia de testosterona. Los puntos representan el
promedio Ï E.S. de 4 cultivos replicados.

-lOó
de l Ug/ml FSH fue de 18,5 Í 4,7 ng/cultivo, y fue disminui
da a 4,7 0,9 ng/cultivo en presencia de testosterona 5 uM.
Estos resultados sugieren que la inhibición es eiercida a un pa
so posterior a la formación de AMP ciclico.
3.4.4 Efecto de los Andrógenos sobre la Sintesis de Preg
nenolona por Células Granulosas.
Con el obieto de determinar la capacidad de las célu
las granulosas de sintetizar pregnenolona en cultivo, células
granulosas fueron cultivadas por distintos tiempos en presencia
o ausencia de FSH. Finalizado el periodo de cultivo, pregneng
lona fue determinada por radioinmunoensayo de alicuotas del
medio de cultivo y de los extractos etanólicos del teíido. La
cantidad de pregnenolona acumulada en el medio asi como el
contenido total de pregnenolona en medio y células al final de
los distintos periodos de incubación estan representados en la
Figura 28. Los cultivos control sintetizaron pregnenolona alo
largo de las 48 horas de cultivo, aunque la sintesis fue menor
luego delas primeras 24 horas. FSH estimuló significativamen
te (P< 0,001) tanto la acumulación en el medio de cultivo co
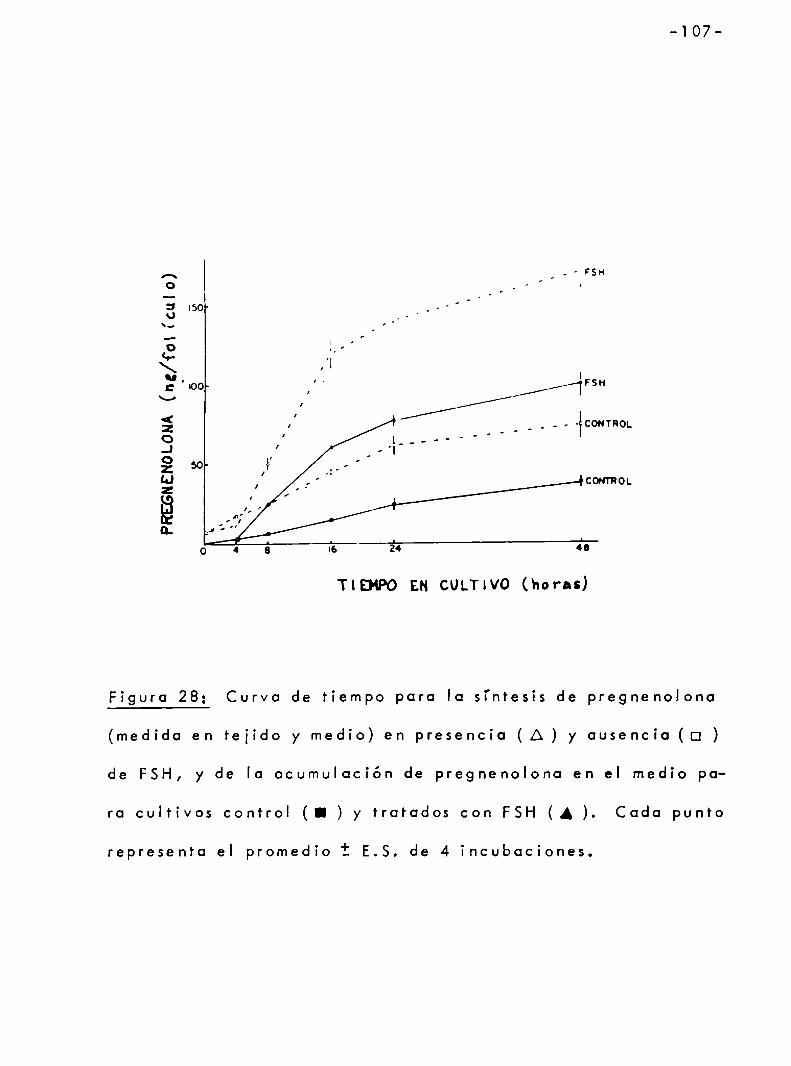
-lO7-—
—rsnA
Go .
Ao:3u
‘v
0c...hc 'noob /l/TFSHV /
I< I /z , ___---CONTROLo ’ 1__ _ - - ' 'á I ¡alZ 50" ,g .:"(i) l corme
I ,
1'“,a c'- Ic 4 á Iï ¿a ¿o
TlEHPO EN CULTIVO (horas)
Figura 28: Curva de tiempo para la sïnfesís de pregnenolona
(medida en feiído y medio) en presencia (A) y ausencia (c1)
de FSH, y de Io acumulación de pregnenolona en el medio pa
ra cultivos control (I ) y tratados con FSH (A Cada punto
representa el promedio Ï E.S. de 4 íncubacíones.

-108
mola cantidad total de esteroide sintetizado. Esta estimulación
fue evidente a partir de las primeras 4 horas de cultivo.
Como se observa enla Figura 28, la cantidad de preg
nenolona presente en el teiido fue alta tanto a tiempo cero c2
mo al final de los distintos periodos de incubación. Sin embar
go, eI perfil de acumulación en medio de cultivo no difirió
del de sintesis total (cantidad de esteroide presente en medio
y teiido al final del periodo de cultivo menos el contenido ti
sular inicial). Más aún, la cantidad total de pregnenolona sir_¡_
tetizada estaba altamente correlacionada con la cantidad pre
sente en el medio en los cultivos control (r: 0,970) y en los
tratados con FSH (r = 0,991). Dado que en presencia de testo_s
terona 5 uM (datos no mostrados), la cantidad de esteroide a
cumulada en el medio también estaba altamente correlacione
da con la cantidad total sintetizado ( r = 0,986), en experi
mentos subsiguientes sólo se determinó la acumulación en el
medio de cultivo.
Para determinar el efecto de los andrógenos aromati
zables sobre la sintesis de pregnenolona, células granulosas
fueron cultivadas por 24 horas en presencia de A4-androsten_e
diona o testosterona a concentraciones de 0,5uM y 5uM,

-109
con o sin FSH. Como se observa en la Figura 29, adición de
A4-androstenediona a células granulosas estimuló significat_i
vamente la sintesis basal de pregnenolona (P < 0,001). En pre
sencia de FSH, A4-androstenediona estimuló la sintesis de preg'
nenolona de uno manera dosis-dependiente (P < 0,05).
El efecto de concentraciones bajas (5 x lO-7M) y al
tas (5 x lO-óM) de testosterona sobre la producción de pregne
nolona por células granulosas en cultivo estó representado en
la Figura 30. La concentración alta causó un incremento del
40 % en la acumulación basal de pregnenolona (P< 0,01) y del
60 °/oenla acumulación de pregnenolona estimulado por FSH
(P < 0,05).
Para determinar si los andrógenos estimulaban la si'nte
sis de pregnenolona actuando antes o después de la formación
de AMPciclico, células granulosas fueron cultivadas por 24
horas en medio conteniendo dibutiril AMP ciclico (l mM) solo,
testosterona (5uM) sola o testosterona y dibutiril AMPciclico
en un experimento factorial 2 x 2. Como se observa enla Figura
3], dibutiril AMPciclico estimuló significativamente (P< 0,001)
la acumulación de pregnenolona en presencia o ausencia de tes
tosterona. Testosterona aumentó significativamente (P< 0,001)
la sintesis de pregnenolona. La ausencia de una interacción

-110
¡50
A a CONTROL
_°_ Ü FSH:1
O I\_
i D0cV<23o so»zuZoua:a fi
c 0..CONTROL 0-5 un A‘A 50M a‘A
Figura 29: Efecto de A4-cndrostenedíona ( A4A) y FSH sobre
la producción de pregnenolona durante un período de cultivo
de 24 hs. Los valores representan el promedio Ï E.S. de 8
cultivos.

-H]
3E3 CONTROL
CJ FSH
3.
PREGNENOLONA(ng/(ol{cuto)
son
o fi fi gCONTROL 05 UMT 50H T
Figura 30: Efecto de testosterona (T) y FSH sobre la acumula
ción de pregnenolono en 24 hs. Los valores representan el pro
medio Ï E.S. de 8 cuIt-ívos.

-112
po 6o
[7(BMZCAMP - o - 6
T - ' ‘ *
PREGNENOLONA(ng/folícul
Figura 3]: Efecto de testosterona (T , 5 x 10-6 M) y díbutí
ríl AMP cu'clíco (1 mM) sobre la producción de pregnenolona
en 24 hs. Los valores representan el promedio Ï E.S. de 12
cultivos.

-113
significativa indicó que este efecto era independiente de la
presencia de dibutiril AMPciclico.
Células granulosas fueron cultivadas por 24 horas en
presencia de distintas cantidades de dibutiril AMPciclico. La
acumulación de pregnenolona en el medio de cultivo está repr_e
sentada en la Figura 32. Dibutiril AMPciclico estimuló signi
ficativamente (P < 0,00l)la sintesis de pregnenolona desde la
menor concentración ensayada (0,5 mM). Testosterona causó un
incremento significativo (P< 0,001) en la sintesis de pregnen_o_
lona en respuesta alas distintas concentraciones de dibutiril
AMP ciclico. La mayor dosis fue efectiva a todos los niveles
de dibutiril AMP ciclico mientras que la menor sólo produío un
aumento significativo (P < 0,05) a una concentración de dibut_i
ril AMP ciclico de l mM. Estos resultados demuestran que los
andrógenos aromatizables aumentan la acumulación de pregne
nolona en cultivos control asi como en presencia de FSH o de
dibutiril AMPciclico. Dado que anteriormente se observó que
los andrógenos aromatizables inhibian la sintesis de progestero
na dependiente de FSH en los mismos Cultivos, la observación
de un efecto estimulatorío sobre la sintesis de pregnenolona su
giere que estos andrógenos actúan sobre la conversión de preg
nenolona a progesterona dependiente de FSH.
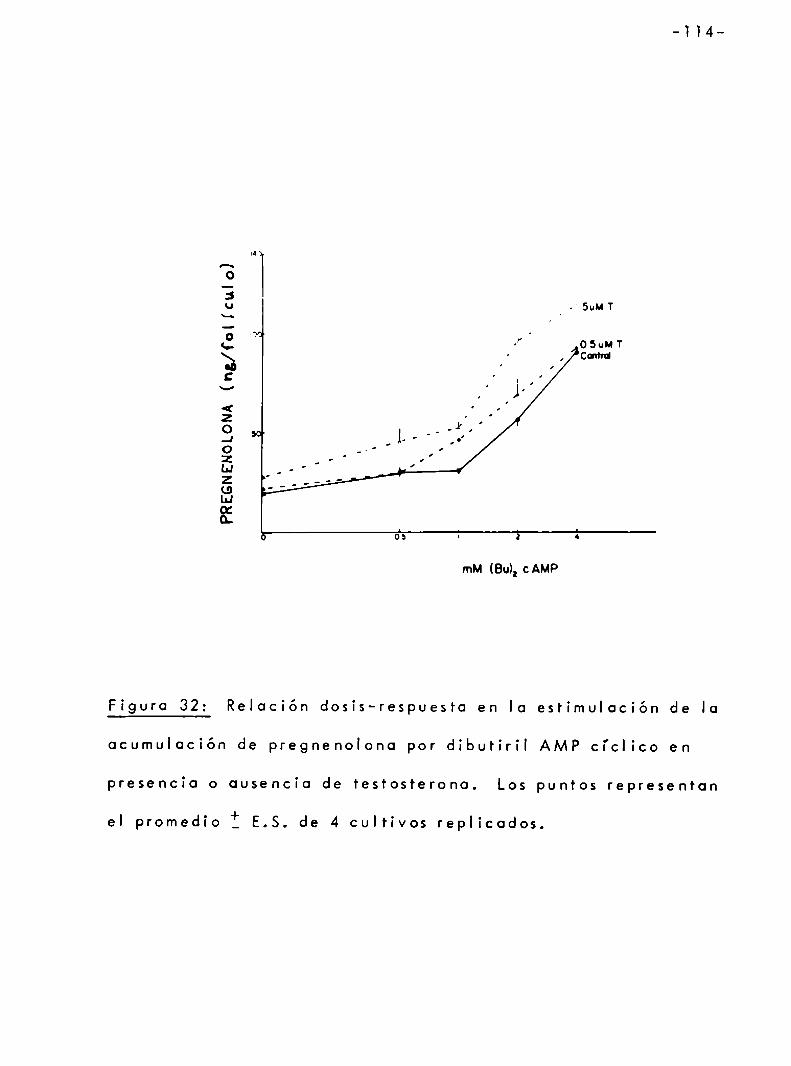
-114
PREGNENOLONA(ng/rolrculo)
rnM (Bu), CAMP
Figura 32: Relación dosis-respuesta enla estimulación de la
acumulación de pregnenolona por díbufíríl AMPcïclíco en
presencia o ausencia de testosterona. Los puntos representan
el promedio Í E.S. de 4 cultivos replicados.

4.0 DISCUSION
4.1 Rol delas Células Granulosas enla Sintesis de Andróge
nos.
La mayor parte de la evidencia acumulada en base a
estudios "in vitro" con tipos celulares ovóricos aislados sugie
re que, al menos en los estadios mas tempranos del desarrollo
folicular preovulatorio, las células tecales y granulosas actúan
sinergisticamente enla biosintesis de estrógenos. La naturaleza
de este sinergismo parece ser el de un pasaíe de sustratos : las
células tecales producen andrógenos que son luego transporta
dos alas células granulosas, donde son aromatizados a estrógi
nos (38).
Esta bien establecido que las células tecales son res
ponsables dela sintesis folicular de andrógenos y que LH es la
gonadotrofina responsable de su control (109-114). Los resulta_
dos de la primer parte de esta investigación indican que, en
ausencia de LH, las células granulosas proveerian el sustrato
(pregnenolona), para la sintesis tecal de andrógenos; esto pro
porciona evidencia de interacción entre ambos tipos celulares

-lló
en la sintesis basal de andrógenos.
Medio en que células granulosas habian sido cultiva
das previamente en presencia de FSH estimuló la sintesis tecal
basal de andrógenos ( A4-androstenediona y testosterona).
Estudios en diversas especies han indicado la presen
cia en el fluido folicular de factores no esteroideos (probable
mente péptidos) que regularfan la maduración del ovocito, la
luteinización de las células granulosas y la unión de FSH a
células granulosas, asr' como la secreción pituitaria de FSH
(reseñado recientemente, 115). Dado que las células granulo
sas parecen ser las células folículares responsables dela sinte
sis de por lo menos dos de estos factores (lló, H7), pareció
razonable suponer que el efecto de las células granulosas so
bre la sintesis tecal de andrógenos es ejercido mediante la producción de un factor similar.
Al ser extraido con carbón-dextrón, el medio usado
por células granulosas perdió su capacidad de estimular la sin
tesis de andrógenos, indicando que alguna pequeña molécula
presente en dicho medio seria la responsable de esta estimula
ción. El hecho de que cultivos tratados con 50 °/ode medio u
sado extraido con carbón-dextrón produieran cantidades signi_
ficativas de andrógenos (similares a los cultivos control) indi

—ii7
c6 que la ausencia de efecto del medio extraido no se debia a
cantidades limitantes de algún factor necesario para la super
vivencia delas células tecales en cultivo. Aunque la extrac
ción con carbón-dextrón podria eliminar otras moléculas pequ_e
ñas, se examinó la posibilidad de que las células granulosas
actuaran proveyendo un precursor esteroideo.
Se sabe que las células granulosas porcinas producen
grandes cantidades de progesterona, siendo esta sintesis esti
mulada por FSH (y LH) (27). La sintesis de pregnenolona por
cultivos de células granulosas porcinas no ha sido determinada
anteriormente. Los resultados de este estudio indican que Ia
sintesis de pregnenolona inmunoreactiva es incluso más eleva
da que la de progesterona. También se observó que el fluido
folicular porcino contiene niveles mas elevados de pregnenol_o
na que de progesterona (datos no publicados). Aunque Ia iden
tidad del material semeíante a pregnenolona no fue confirma
da quimicamente, Ia relación observada entre los niveles de
pregnenolona inmunoreactiva medida en estos experimentos en
el medio usado por células granulosas, y la capacidad del me
dio de estimular Ia sintesis tecal de andrógenos, respalda nue_s
tra interpretación de su identidad.
Si bien progesterona exógena fue capaz de incremen

-118
tar la sintesis tecal de andrógenos, los niveles de este esteroi
de presentes en el medio usado por Cultivos de células granulo
sas parecieron ser baios para causar el aumento observado. Los
niveles de pregnenolona presentes en MB (medio usado por cé
lulas granulosas cultivadas en presencia de FSH) podrian cau
sar este aumento. Sin embargo, las cantidades relativas de am
bos andrógenos fueron distintas; medio usado por cultivos de
células granulosas estimuló la sintesis de testosterona al do
bl I d A4 d d' I ' Ie y a e -an rostene ¡ona a trip e, en tanto que una
cantidad equivalente de pregnenolona exógena causó un aumen
to de 2 veces en la producción de A4-androstenediona y un au
mento menor enla de testosterona.
Estos resultados, que demuestran utilización de preg
nenolona exógena para lasintesh de andrógenos, estén de a
cuerdo con informes previos sobre estudios "in vivo" e "In vr
tro”. Young Lai y Short (118) inyectaron pregnenolona radioac
tiva dentro de un foliculo ovórico de una yegua y encontraron
. ., . . . N4 .¡ncorporacuon dela radioactiwdad en z. -androstenediona en
sangre de vena ovórica. Lacroix y col. (H9) observaron con
ut . - .versnon de pregnenolona radioactlva a -androstenediona por
preparaciones tecales bovinas en cultivo. Fortune (i20) demos
tró interacciones entre células tecales y granulosas bovinas

-ll9
C -esteroides, en presencia de LH.enla sintesis de Cl9-y 2]Nuestras observaciones indican que, en el ovario porcino, las
células granulosas podrian proveer el sustrato C21-esteroideo
en ausencia de estimulación por LH. De este modo las células
granulosas podrian controlar la sintesis tecal de andrógenos
cuando LH no está disponible. Ya que las células granulosas d_e
rivadas de foliculos medianos de cerdo son capaces de aromat_i_
zar andrógenos a estrógenos (29, 31), el hecho de que estimu
len la sintesis tecal de andrógenos plantea la interesante pos_i_
bilidad de un rol para las células granulosas en controlar la
sintesis del sustrato que usan para la aromatización.
4.2 Capacidades Relativas de los Tipos Celulares Foliculares
Aislados y en Recombinación para Sintetizar Andrógenos
y Estrógenos.
Las preparaciones tecales produíeron grandes cantida_
des de andrógenos, predominantemente androstenediona. Las
preparaciones de pared folicular (foliculo en que se hizo una
incisión para eliminar el fluido folicular, deíando intactas la
capa tecal y la membrana granulosa) produíeron cantidades más

-120—
elevadas de testosterona que las células tecales (observaciones
no publicadas). Esto está de acuerdo con evidencia en cerdo
(25), hamster (121))! oveia (37) de que las células granulosas
poseen una actividad de l7fi-hidroxiesteroide deshidrogenasa
más elevada que las tecales. Las preparaciones tecales produ
¡eron cantidades pequeñas de estrógenos; adición de células
granulosas causó un incremento al triple enla producción de
estrógenos. Estos resultados son similares a aquellos obtenidos
por Evans y col. (29) en la cerda prepüber. Aunque estos auto
res no han incubado células tecales y granulosas en co-cultivo,
sus resultados para la producción de estrógenos por células gr_a
nulosas aisladas, en presencia de testosterona exógena, indi
can que en su sistema también ocurriria sinergismo.
Recientemente, Honey y Scbomberg (35) informaron
que en foliculos porcinos de 4-5 mm la teca es responsable de
la mayor parte de la producción total de estrógenos, no obser
vando un aumento significativo en su sintesis al agregar célu
las granulosas alos cultivos. Podria ser que foliculos deriva
dos de animales sexualmente maduros se comportaran de distin
ta manera, aunque también es posible que las diferencias en
resultados se deban a técnicas de separación de tipos celulares
o a medios de cultivo empleados, Sin embargo, aunque dichos

-i2l
autores no observaron un aumento en la sintesis de estrodiol al
co-cultivar ambos tipos celulares, sus resultados demuestran
mayor conversión de andrógenos exógenos por células granulo
sas que por preparaciones tecales.
En conclusión, los resultados obtenidos del cultivo de
tipos celulares folículores respaldan la teoria bícelular bigong
dotrófica para la biosintesis de estradiol en foliculos porcinos
de tamaño mediano. En las condiciones usadas, fuimos incapa
ces de observar un efecto estimulatorio de FSH sobre la aroma
tización de andrógenos a estrógenos, confirmando los resulta
dos obtenidos por Evans y col. (29). Sin embargo, no se ha ex_a
minado la posibilidad de que FSH pueda inducir actividad de ci
romatasa en foliculos menos maduros.
Además, los resultados obtenidos del co-cultivo de t_i
pos celulares foliculores en este estudio sugieren un rol para
las células granulosas en el metabolismo de andrógenos. AI co
cultivar células tecoles y granulosas se observó una disminució
drástica enla acumulación de andrógenos (A4-androstenediona
más testosterona), que no estaba correlacionada con un aumen
to enla sintesis de l7fe>-estradiol. Ya que las células granulo
sas son capaces de aumentar la sintesis tecal de andrógenos pr_c_>
veyendo sustrato esteroideo, un efecto inhibitorio de las célu

-i22
las granulosas sobre la sintesis de andrógenos pareció improba
ble. En cambio, se examinó la capacidad de las células granul a llosas porcunas de metabollzar androgenos.
4.3 Sintesis de Sulfatos de Andrógenos.
Los resultados de la presente investigación proporcio
nan evidencia para la sintesis de sulfatos de andrógenos por
células granulosas porcinas. Existen varios factores que respa_l_
dan esta evidencia.
(1)Al cultivar células granulosas en presencia de All-andres
tenediona o testosterona, se obtienen derivados de estos ej
teroides,solub|es en agua.
(2) Luego de precipitar las proteinas, este compuesto se com
porta en columnas de DEAE-Sephadex A 25 como un monosu_l_
tato de esteroide neutro (Figuras 9 y lO; 122). Los sulfatos
de estrógenos son retenidos más tiempo en estas columnas
(123, 124).
(3) Luego de solvólisís se obtiene un compuesto soluble en éter.
Esto respalda la evidencia de la presencia de un sulfato de

-l23—
C19-esteroide, debido a que los glucurónidos de esteroides
no son hidrolizados baio las condiciones usadas (lOl).
La formación de sulfatos de C19-esteroides en el ovario ha
sido descripto anteriormente. Se observó formación de Sulfato
de dehidroepiandrosterona por homogenatos de ovarios humanos
(76, 80). Sturm y Hannapel (82) observaron formación de sulfg
to de testosterona por homogenatos de foliculos y cuerpos lü
teos de rata y bovinos. Experimentos con ovarios humanos de
la fase folicular, perfundidos, (85) mostraron conversión de
sólo l % del sustrato radioactivo a sulfatos esteroideos, sien
do un tercio de estos,sulfatos de estrógenos. En este estudio
se observó una mayor conversión (alrededor del 40 °/o) a mono
sulfatos de andrógenos. La baía conversión observada por Sturm
y col. (85) podria deberse a la presencia de grandes cantida
des, no fisiológicas, de sustrato, aunque también es probable
que se deban a diferencias entre especies. Más aün, nuestras
observaciones sugieren que los células granulosas son el lugar
en que ocurre la formación folicular de sulfatos de andrógenos.
Se ha especulado que, en los teiidos endócrinos, los
sulfatos de esteroides desempeñan una variedad de funciones,
como productos de desactivación, como precursores de hormo
nas activas, asi como estando involucrados en los mecanismos

424
regulatorios dentro de estos tejidos manteniendo niveles de es
teroides libres por interacción con la enzima limitante, la
sulfataso (74, 75). Se podria especular que, en el ovario, la
Formación de sulfatos podria controlar la cantidad intrafolicu
lar de andrógeno libre, evitando la atresia del foliculo. EstosI Í oandrogenos podrian tal vez ser usados, posteriormente, para
la sintesis de estrógenos, ya que aparentemente no son esteroi
des 5°( - o 5P - reducidos. Esto no requeriría acción previa
de la sulfatasa ya que existe evidencia de aromatización direc
ta de sulfatos de esteroides neutros (87).
El hecho de que 20 °/ode la radioactividad inicial so
Iuble en agua se comportara consistentemente como esteroides
libres en columnas de DEAE-Sephadex, luego de precipitar lasC o I n I Iproteinas (Figura 9), sugiere que estos estaban inICIalmente
unidos a proteinas. En efecto, en fluido folicular de diversas
especies, se han encontrado proteinas que unen esteroides
(124). Estas proteinas podrian originarse en células granulosas,I O O Í I Íya que estas celulas son CCl’lVGSenla blosmteSis de proteinas
(3, 4).
Separación cromatogrófica del producto solvolizado
de los distintos lotes de células granulosas confirmó que los
esteroides coniugados no son ninguno de los productos conoci

-l25
dos del metabolismo de andrógenos en el ovario. Si bien el me
tabolito muy polar (aislado en las columnas de fase reversa) a_
pareció en todas las incubaciones, la presencia del metabolito
menos polar (aislado en las columnas de fase normal) no fue
consistente. Podria ser que este Último no fuera estable baio
las condiciones de hidrólisis usadas. Los procedimientos solvo
liticos pueden ser quimicos o enzimáticos. Los procedimientos
quimicos (altas temperaturas, condiciones ácidos) podrian alt_e
rar la estructura del esteroide, introduciendo artificios (125).
Aunque los procedimientos enzimáticos no presentan este pro
blema, a nuestro conocimiento, no existen preparaciones de
sulfatasa pura disponibles. Alternativamente, podria ser que
las células granulosas metabolizaran el andrógeno a varios pro
ductos, variando las cantidades relativas de cada uno de estos
con el medio ambiente hormonal al que hubieran sido expuestas
en el foliculo. Aunque en todos los experimentos se usaron fo
liculos de aproximadamente el mismo tamaño, podria ser que
estos hubieran sido expuestos a distintas concentraciones de ggnadotrofinas
Los resultados inconsistentes obtenidos en los ensayos
enzimáticos de sulfotransferasa también podrian deberse a una
capacidad diferencial de las células de usar los distintos sus

-l26—
tratos. Las sulfotransferasas son enzimas muy especificas(126)
y es posible que ninguno de los esteroides empleados como sus
tratos sea el Sustrato especifico para la enzima presente en cÉ
lulas granulosas porcinos. Ademós, no pudo realizarse un estu
dio completo (usando distintos sustratos) de la actividad de la
enzima debido a que la cantidad de material obtenido en cada
experimento no fue suficiente para llevar a cabo más que dos
ensayos enzimáticos. Sin embargo, cuando (355)-Na2SO4 fue
incluido en la mezcla de reacción, se pudo obtener 2 veces
(usando el mismo lote) incorporación en el coniugado, con
(3H) -androsterona como sustrato, obteniendo evidencia inde
pendiente dela capacidad de las células granulosas de coníu
gar andrógenos a iones sulfato.
4ü4 Efecto de la Teca sobre la Sintesis de Progesterona por
las Células Granulosas.
La hormona folículo-estimulante (FSH) se une a recep
tores especificos en células granulosas derivadas de foliculos
porcinos de tamaño mediano (17) estimulando la producción de
AMP ciclico (l27) y la sintesis de progesterona (27). Los resul

-127
tados de este estudio demuestran que los andrógenos aromatiza
bles, androstenediona y testosterona, inhiben esta acumulación
de progesterona en respuesta a FSH.
En ratas, se ha establecido que tanto los andrógenos
aromatízables como los no aromatízables aumentan la sintesis
de progesterona por células granulosas aisladas,estimuladas por
FSH (128-130). La potencia de testosterona y DHTha sido relg
cionada con su velocidad de metabolismo dentro de las células,
y la potencia de otros andrógenos con su grado de conversión
a testosterona y DHT (131). Por otra parte, se ha implicado a
los andrógenos en la inhibición del desarrollo ovóríco en ratas
inmaduras hipofisectomizadas tratadas con dietílestilbestrol
(132). De acuerdo con esto se ha dicho que, en ratas, los an
drógenos estimulan la sintesis de progesterona y son "atretogé
nicos" (133).
En cultivos de células granulosas humanas, testosterg
na inhibió la biosïntesis de progesterona cuando las células
provenïan de foliculos grandes (133) pero no de foliculos pe
queños (134).
En cerdos, Thanki y Channing (47) observaron que te_s
tosterona inhibia la producción de progesterona estimulada por
FSH, mientras que Schomberg y col. (44, 45) informaron que

—128
los andrógenos estimulaban la sintesis basal de progesterona
en 48 horas de cultivo, siendo esta estimulación significativa
en el caso de DHT pero no en el de testosterona. La discrepan
cia aparente entre los resultados de SchOmberg y los obtenidos
ción de la sintesis de progesterona observada en este estudio
sólo fue evidente cuando FSH estaba presente durante el culti, , 4vo; no se observo efecto alguno de los androgenos (A -andros
tenediona, testosterona, androsterona y DHT)sobre la sintesis
basal de progesterona en 24 horas de cultivo. Nos pareció ade
cuado incluir FSH en los cultivos ya que las células granulo
sas están expuestas a FSH "in vivo" Cultivando células gran!
losas en presencia de FSH, Anderson y col. (3]) observaron
que testosterona no eiercia un efecto significativo sobre la
sintesis de progesterona después de 2 dias en cultivo, aunque,
en cultivos de 4 dias testosterona actuaba sinergisticamente
con FSH enla estimulación de la sintesis de progesterona. Da_
do que luego de 2 dias en cultivo, las células granulosas por
cinas presentan caracteristicas morfológicas de células lutei
nizadas (27),parecerïa que testosterona es capaz de estimular
la sintesis de progesterona por células luteinizadas. En este
estudio no se prolongaron los cultivos por más de 24 horas pa

-l29
ra poder obtener una aproximación de lo que ocurre cuando las
células forman parte del foliculo.
Las diferencias observadas también podrian deberse a
los medios de cultivo y al tipo de animales usados como fuente
de células granulosas. En este estudio se usaron células prove
nientes de foliculos ovóricos de cerdas prepúberes, que son r3
lativamente inmaduras aunque capaces de desarrollar respues
tas esteroidogénicas normales (29). Los otros autores (3], 44,
45) también usaron foliculos relativamente inmaduros, aunque
aislados de ovarios de cerdas sexualmente maduras. Usando cé
lulas granulosas obtenidas de cerdas maduras observamos que la
estimulación de la sintesis de progesterona por FSH era similar
a la obtenida con foliculos comparables de cerdas prepüberes,
y que testosterona inhibia esta respuesta en forma análoga (Ta
bla VIII). Por lo tanto, las diferencias en resultados no pueden
ser explicadas por diferencias en la fuente de células granulo
sas.
Es improbable que la inhibición observada en el pre
sente estudio se deba a un efecto tóxico de los andrógenos so
bre las células granulosas ya que células cultivadas por 24 ho
ras en presencia de testosterona y FSH fueron capaces de produ
cir niveles elevados de progesterona durante un periodo subsi

—i30
guiente de 24 horas en ausencia de hormona exógena (Tabla IX).
Los datos presentados sugieren que la acumulación di_s
minuida de progesterona en presencia de testosterona se debe
a inhibición de la sintesis de progesterona y no a estimulación
de su metabolismo. Los andrógenos podrian inhibir la sintesis
de progesterona interfiriendo con la unión de FSH a su recep
tor, interfiriendo con el acoplamiento del receptor a la adeni_
lato ciclasa, estimulando la fosfodiesterasa y/o inhibiendo las
enzimas involucradas en la biosintesis de progesterona. Para de
terminar si la inhibición ocurria a un paso anterior o posterior
a la formación de AlúP ciclico, células granulosas fueron inc!
badas con dibutiril AMP ciclico en presencia o ausencia de d'_s
tintas concentraciones de testosterona. Se observó que testost_e
rona también ¡nhibïa la acumulación de progesterona en respue_s
ta a dibutiril AMP cfclíco. Mós aún, la extensión de la inhibl
ción fue la misma: una disminución de 70 % enla sintesis de
progesterona en respuesta a dibutiril AMP ciclico (4 mM) y de
75 % en la sintesis en respuesta a FSH (l ug/ml).
Esto sugirió que los andrógenos actuaban a una etapa
posterior a la formación de AMP ciclico. Por lo tanto, los an
drógenos inhibirian alguna enzima en el camino biosintético y/o
estimularian la actividad de fostodiesterasa.

-i3i
TABLA IX
Acumulación de progesterona (ng/foliculo equivalente) por célu
las granulosas mantenidas en cultivo por 48 hs (n = 8). Las célu
las fueron cultivadas por 24 hs en presencia o ausencia de FSH
(i ug/ml) y testosterOna (5 UM); a las 24 hs el medio fue reemplg
zado totalmente por medio fresco sin hormonas exógenas.
Hormonas Presentes Durante
las Primeras 24 hs de Cultivo 0-24 hs 24-48 hs
2,5 t 0,2 I,ót0,1
FSH+T 6,3 t 0,5 2,9to,3

-i32
Estudios preliminares con un inhibidor de fosfodieste
rasa (3-isobutil-metil-xantina, IBMX)demostraron que, aunque
IBMX(0,] mM) aumentaba la sintesis basal de progesterona por
células granulosas en cultivo, adición de IBMXa células cul
tivadas en presencia de FSH o dibutiril AMP ciclico resultaba
en una acumulación disminuida de progesterona, aunque la a
cumulación de pregnenolona en los mismos cultivos no era afef
tada.
Esto está de acuerdo con observaciones de que niveles
altos de AMPciclico inhiben notablemente ala 3i’5-hidroxies
teroide deshidrogenasa-Asisomerasa en teíido adrenal (135) y
ovórico (136) de, rata.
Células granulosas produíeron cantidades elevadas de
pregnenolona, siendo esa sintesis estimulada por FSH (de 2 a
3 veces). Los andrógenos (¿4A o T) aumentaron aún más la sirL
tesis de pregnenolona en respuesta a FSH. Dibutiril AMP cicli
co estimuló la sintesis de pregnenolona de una manera dosis de
pendiente, siendo esta sintesis también aumentada por testost_e
rona. Ya que baio las mismas condiciones los andrógenos inhi
bian la sintesis de progesterona, se concluyó que los andróge
nos aromatizables afectan la conversión de pregnenolona a pre
gesterona.

—]33
La conversión de pregnenolona a progesterona es cata
Iizada por un sistema deA5-3ífl5-hídroxiesteroide deshidrogena
sa / AS- 3 - cetoesteroide isomerasa, que es activo en células
granulosas porcinas (25); Recientemente, Dorrington y Armstrong
(137) informaron que FSH estimula marcadamente la actividad
de este sistema enzimático en células granulosas de rata. Evi
dencia indirecta confirma que también en células granulosas
de cerdo FSH actúa a este nivel,ya que FSH produio un porcen
taje mós alto de estimulación de sintesis de progesterona que
de pregnenolona, aunque la cantidad absoluta de pregnenolo
na producida por encima de los valores control fue mayor.
Por lo tanto, parece que la sintesis disminuida de progestero
na en respuesta a FSH en presencial de andrógenos se debe a u
na inhibición dela AS- 3P -hidroxiesteroide deshidrogenasa
isomerasa. La inhibición de esta enzima por esteroides ha sido
informada anteriormente. En Ia placenta humana, Wiener y
Allen (138) observaron inhibición dela 135-3? -hidroxiesteroi
de deshidrogenasa por 20°C-hidroxí-4-pregnen-3-ona, mientras
que Townsley (139) observó que los ¿34-3-cetoesteroides eran
potentes inhibidores dela enzima. La inhibición por A4-andros_
tenediona y testosterona era de tipo no competitivo, y sólo po
dia ser interrumpida por metabolismo del inhibidor y/o su eli

-l34
minación de los alrededores de la enzima. Aunque no se han
realizado ensayos enzímóticos, la observación de una disminu
ción enla acumulación de progesterona y un aumento enla de. 4 . . .pregnenolona sugieren que los A -3-cetoestero¡des Inhlben a
la A5-3P-hidroxiesteroide deshidrogenasa -L_\5-ísomerasa en
células granulosas de folI'CUlo ovórico porcino como enla pla
centa.
. A4 .Las concentracuones de -androstenedlona o de tes
tosterona necesarias para producir inhibición de la sintesis de
progesterona o aumento dela acumulación de pregnenolona pa
receri'an ser elevadas. Los niveles de andrógenos en el fluido
Folicular parecen ser menores (36, 140 , Lischinsky y Armstrong,
datos no publicados). Sin embargo, las preparaciones tecales
porcinas sintetizan grandes cantidades de andrógenos. Por lo. 4 .tanto, las concentracnones de A -androstenedlona y testostero
na en los alrededores delas células granulosas adyacentes a la
capa tecal podrian ser mucho mós elevadas. Además, dado que
los efectos inhibitorios de estos esteroides parecen ser acumu
lotivos (139), la extensión de la inhibición refleiaria la con
centración total local de inhibidores.
Nuestros resultados indican que los andrógenos afec
tan principalmente la actividad dela enzima estimulado por

-l35
FSH, eíerciendo un efecto menor sobre su actividad basal. Da
do que el aumento en la acumulación de pregnenolona excedióo I 0' I Íen masa a la disminucron en los niveles de progesterona, podria
ser que los andrógenos también inhiban el metabolismo de preg
nenolona a lo largo de otros caminos. Alternativamente, los an
drógenos podrian estimular la sintesis de esteroides de 21 óto
mos de carbono en un paso anterior a la formación de pregneno
lona.
No se sabe silos andrógenos producen esta inhibición
por si mismos o luego de ser metabolizados a otro producto. YaÍ ' o 4que solo los androgenos aromatizables testosterona y A -andros
tenediona, pero no DHTy androsterona, fueron capaces de pro
vocar esta inhibición, fue necesario determinar si la reducción
en la acumulación de progesterona se debia ala conversión de
los andrógenos a estrógenos. 1703 -Estradiol inhibió la acumulal —
0' Icron de progesterona en este srstema, de acuerdo con resultados
obtenidos por otros investigadores en células granulosas porci
nas (45, 47) y bovinas (¡41). Sin embargo, nuestros resultados
indican que testosterona inhibe la producción de progesterona
en respuesta a FSHsin previa aromatización a 17P-estradiol.
La inclusión de testosterona en el medio de cultivo inhibió
notablemente la acumulación de progesterona en 8 horas de

-l3ó
cultivo, mientras que los niveles de estradiol no aumentaron
sustancialmente hasta después de 8 horas. Aunque el contenido
de esteroides en el medio no es necesariamente un fiel refleio
de los niveles intracelulares, e incluso niveles relativamente
baíos de l7p-estradiol podrian causar efectos fisiológicos,
parecería que testosterona eierce un efecto inhibitorio sobre
la producción de progesterona en ausencia de un aumento im
portante en la sintesis de estradiol. Si bien esta evidencia no
es concluyente, también se demostró que una reducción sustan_
cial en los niveles de estradiol provocada por la presencia de
un inhibidor de Ia aromatasa era incapaz de prevenir la inhib
ción de la acumulación de progesterona por testosterona. En
realidad, el inhibidor de la aromatasa por si mismo causó una
disminución en lasintesh de progesterona; elinecanismo de er
ta acción es desconocido, pero 4-acetoxi-androstenediona es
un andrógeno y podria eiercer efectos inhibitorios semeiantes
a testosterona y A4-androstenediona.
El hecho de que ni DtlT ni androsterona inhiban la sin
tesh de progesterona también indica que la reducción en la po
sición 5 no es probablemente un paso obligatorio enla acción
de testosterona y Af-androstenediona.
Las células granulosas de rata contienen receptores

-i37
a andrógenos que pueden unir tanto DHT como testosterona
(142). Por lo tanto, testosterona podria actuar luego de unir
se a su receptor, sin conversión previa a otros intermediarios
activos. Alternativamen'te, la observación de que los esteroides
que inhiben la sintesis de progesterona en respuesta a FSHson
los mismos que pueden ser.con¡ugados por las células granulo
sas, plantea la interesante posibilidad de un rol regulatorio
de los conjugados de andrógenc en las funciones foliculares.
La modulación de la sintesis de C2¡-esteroides por
andrógenos es otro eiemplo de interacción entre células gran_u
losas y tecales en la sintesis folicular de esteroides en el avg
rio porcino. Es interesante que, en foliculos porcinos de tama_
ño mediano, las células granulosas pueden controlar la conce_n
tración intrafolicular de andrógenos proporcionando una parte
del sustrato (pregnenolona) para la sintesis tecal de andróge
nos y coníugando los andrógenos a iones sulfato. Estas intera_c
ciones demuestran la importancia de los mecanismos intraovór_i
cos en la regulación de la función folicular.

4)
5)
tempranos del desarrollo folicular preovulatorio,
-i38
5.0 CONCLUSHDNES
Los resultados de esta investigación indican que:
Las células granulosas porcinas sintetizan grandes cantida
des de pregnenolona, siendo esta sintesh estimulado por
FSH.
hAediousado por células granulosas cultivadas en presencia
de FSH estimula la sintesh tecal de andrógenos.
Pregnenolona y progesterona pueden estimularla sintesh
tecal de andrógenos.
Las células granulosas pueden coníugar los andrógenos a
¡ones sulfato.
Los andrógenos aromatizables inhiben la sintesis de progeí
terona en respuesta a FSH en 24 horas de cultivo, actuando
probablemente aifivel dela converüón de pregnenolona a
progesterona.
ovario porcino, en los estadiosDe este modo, en el
las células
granulosas podrian regular la concentración intrafolicular de
andrógenom
l) Estimulando la producción tecal de andrógenos; 2) coníugan
do el andrógeno en exceso.

-i39
Se ha implicado alos andrógenos intraovóricos enla
regulación de la maduración folicular, Louvet y col. (132) ob
servaron que la administración de hCG a ratos hembras inma
duras hipofisectomizadas, tratadas con estrógenos, conduce a
atresia folicular, que puede ser revertida por antagonistas de
la acción de los andrógenos (143). De este modo, la capacidad
de las células responsables de la aromatización, las células
granulosas, de controlar la cantidad de andrógeno libre dentro
del foliculo proveeria un medio para seleccionar los foliculos
destinados a ovular o a sufrir atresia.
El hecho de que los andrógenos ( A4-androstenediona
y testosterona) inhiban la conversión de pregnenolona a proge_s
terona en respuesta a FSH sugiere la existencia de un mecanis
mo de retroalimentación positiva del andrógeno sobre su propia
sintesis. El andrógeno podria luego ser aromatizado a estrógeno,
que puede inhibir la subsiguiente sintesis tecal de andrógenos
(39). Alternativamente, se puede especular que, cuando los ni
veles de andrógenos estón en exceso a la cantidad que puede
ser aromatizado, o cuando la concentración de estrógenos es
alta, las células granulosas podrian coníugar el andrógeno. OE
servaciones recientes por HiIIíer y col. (144) sugieren que esto

—l40
último podria ocurrir; los niveles de andrógenos aromatizables
libres en el fluido folicular de foliculos humanos que contenian
células granulosas con baia actividad de aromatasa fueron simi
lares a aquellos de foliculos preavulatorios con células con al
ta actividad de aromatasa.
El hecho de que los andrógenos,producidos por la teca,
inhiban la sintesis de progesterona, sugiere que los andrógenos
desempeñan un rol en el control intrafolicular dela esteroido
génesis en esta especie. Este rol podria involucrar la preven
ción de luteinización prematura de las células granulosas, par
ticularmente antes de la inducción de la actividad de aromata
sa y aumento concomitante en los niveles de l7¿5—estradiol.
También es posible que los andrógenos aromatizables sean atre
togénicos.
Como resultado de este y otros estudios, el modelo de
biosintesis de esteroides en foliculos medianos de cerdo mostra
do en la Figura 2 puede ser puesto al dia como se representa
en la Figura 33.
Las células tecales sintetizan andrógenos, predomi
nantemente A4-androstenediona. Esta sintesis es estimulada
por LH. Aunque las células tecales no poseen receptores para
FSH, las células granulosas unen FSH y responden a esta hor

Figura33:¡Modeloparalaesferoídogénesísenfolïculosporcinosde tamañomediano. Laslíneasenterasrepresentancaminosbíosinféfícosyaccionesinfra ovórícasdehormonas:+estimulación;-¡nhíbícíón. LasIïneasquebradasrepresentanfluioíntercelulardeesteroides.

TECA
FL<—-CAMP
Colesterol
i #
Pregnenolona+-——
1Proqesferona+———
4 lAndrógeno--————
—+—Andrógeno
GRANULOSA
FSH 4+cAMPL»
Colesterol
¿ &
—————Pregneno|ona
1
-————Progesïerona.
/Es’rrogeno-— \Andrógeno
Sulf0to

-i43—
mona sintetizando grandes cantidades de pregnenolona y pro
gesterona. Estos esteroides difunden a Ia capa tecal donde sir
ven como sustratos para la sintesis de andrógenos en ausencia
de LH.
4A -Androstenediona puede ser convertida a testoste_
rona en la teca. Sin embargo, la teca no es eficiente en esta
conversión. Asi, la mayor parte de A4-androstenediona difun
de a la capa granulosa donde es convertida a testosterona y
luego a 17(17-estradiol, o coníugada a iones sulfato.
Además del pasaie de sustratos de un tipo celular al
otro, los esteroides sintetizados en un tipo celular pueden m2
dular la producción de esteroides en el otro. Los estrógenos,
producidos en las células granulosas, pueden inhibir la sinte
sis de andrógenos estimulada por LH. Los andrógenos, deriva
dos de la teca, pueden inhibir la conversión de pregnenolona
a progesterona en células granulosas estimuladas por FSH.

12a.
ó.0 BIBLIOGRAFIA
(i973)._ En : Handbook of Physiology; SectionE.B.
vol.
Lipner, H.7: Endocrinology. Editores: Greep, R.O. y Astwood,American Physíological Society, Washington, D.C.,ll, parte I, póg. 409.
Baker, T.G., Beaumont, H.M. yFranchi, L.L. (i969). J.Cell Sci. 4: 655.
Pool, W.R. y Lipner, H. (1969). Endocrinology 84 : 71].
Baker, T.G. (i963). Proc. R. Soc. London. Ser B. i582417.
Peters, H. (i969). Acta Endocrinol. (Kbh) 62: 98.
Peters, H., Byskov, A.G. y Grinsted, J. (i978). Clinics.Endocr. Metab. 7 : 469.
Eshkol, A. y Lunenfeld, B. (1972). En: "Gonadotropins".Editores: Savena, E.B., Beling, C.G. y Gandy, H.M. JohnWiley and Sons, |nc., póg. 335.
Richards, J.S. y Mídgley, R. (i976). Biol. Reprod. 14 : 82.
Bassett, D.L. (i943). Arn. J. Anat. 73: 25].
Richards, J.S. (i975). Endocrinology 97: ¡174.
Lindner, H.R., Tsafrírí, A., Lieberman, M.E., Zor, U.Koch, Y., Baumínger, S. y Barnea, A. (i974). Rec. Progr.Horm. Res. 30:79.
Kalra, P.J., Krulich, L., Quiiada, M., Kalra, S.P.,Fawcett, C.P. y Mc Cann, S.M. (i971). En: "HormonalSteroids". EditOres: James, V.H.T. y Martini, L. ExcerptaMedica, Amsterdam, póg. 708.

20.
21.
22.
23.
24.
25.
b
445
.-Leung, P.C.K. y Armstrong, D.T. (1980). Ann. Rev. Phys.42 : 7].
Engel, L.L. (1973). En: Handbook of Physiology, Section7: Endocrinology. Editores: Greep, RCO. y Astwood, E.B.American Physíologícol Society, Washington, D.C., vol.II, porte I, pág. 467.
Mc Guíre, J.S., Hollis, V.W. Jr. y Tomkíns, G.M. (1960)J. Biol. Chem. 235 : 3112.
Schwarzel, W.C., Kruggel, W.G. y Brodíe, H.J. (1973).Endocrinology 92 : 866.
Híllíer, S.G., van der Boogoord, A.M.J., Reichert, L.E.Jr. y von Hall, E.V. (1980). J. Endocr. 84: 409.
Nokono, R., Akahorí, T., Kofoyama, K. y Toio, S. (¡977).J. Reprod. Fert. 51 : 23.
KolenO, J. y Chonníng, C.P. (1972). Endocrinology 90:1543.
Chonning, C.P. y Seymour, J.F. (1970). Endocrinology87 z 165.
Channing, C.P. y Kommerman, S. (1973). Endocrinology92 : 531.
Schomberg, D.W. y Tyrey, L° (1972). Biol. Reprod. 7: 127.
Stouffer, R.L., Tyrey, L. y SchOmberg, D.W. (1976)°Endocrinology 99: 516.
Chonning, C.P. y Kommermon, S. (1974). Biol. Reprod.10: 179.
Biersíng, L. y Carstensen, H. (1964). Bíochím. Bíophys.Acta 86: 639.
Biersíng, L. y Corstensen, H. (1967). J. Reprod. Ferf.14: 101.

26.
27.
28.
29.
30.
3].
32.
33.
34.
35.
36.
37.
38.
39.
-i4ó
Biersing, L. (1967). Acto Endocrinologico Supp|.125:1.
Chonning, C.P. (1970). Endocrinology 87: 156.
(1978).Fed.Tsang, B.K., Moon, Y.S. y Armstrong, D.T.Proc. 37 : 723.
Evans, G., Dobias, M., King, G.J. y Armstrong, D.T.(198]). Biol. Reprod. 25 : 673.
Armstrong, D.T. y Dorrington, J.H. (i977). En :"RegulotoryMechanisms Affecting Gonadal Hormonal Action". Editores:Thomas, J.A. y Singhal, R.L. University Pork Press, Boltimore, vol. 3, pág. 217. _
Anderson, L.D., Schaerf, F.W. y Chonning, C.P. (i979).Adv. Exp. Med. Biol. 112 : 187.
Dorrington, J.H., Moon, Y.S. y Armstrong, D.T. (i975).Endocrinology 97: 1328.
Erickson, G.F. y Hsueh, A.J.W. (i978). Endocrinology102 : i275.
Stoklosowa, 5., Bahr, J. y Gregoraszczuk, E. (i978).Biol. Reprod. 19 : 712.
Honey, A.F. y Schomberg, D.W.(1981). Endocrinology109 : 97].
Eiler, H. y Nolbondov, A.V. (1977). Endocrinology 100:33].
Armstrong, D.T., Weiss, T.J., Selstom, G. y Seomark, R.F. (¡981). J. Reprod. Fert. Suppl. 30: 143.
Leung, P.C.K. y Armstrong, D.T. (i980). Ann. Rev. Phys.42: 7].
Tsong, B.K., Leung, P.C.K. y Armstrong, D.T. (i979).Molec. Cell. Endocr. 14: 13].

40.
4].
42.
43.
44.
45.
46.
47.
48.
49.
50.
5].
52.
53.
-147—
Leung, P.C.K., Goff, A.K.,D.T. (1978). Biol. Reprod, 19:
Kennedy, T.G. y Armstrong,641.
Leung, P.C.K. y Armstrong, D.T. (1979). Endocrinology104:]411.
Leung, P.C.K. y Armstrong, D.T. (¡979) Biol. Reprod. 2]:1035.
Goldenberg, R.L., Brídson, W.E. y Koh'ef, P.O. (1972).Bíochem. Bíophys. Res. Commun. 48: 101.
Schomberg, D.W., Stouffer, R.L. y Tyrey L. (1976).Bíochem. Biophys. Res, Commun. 68:77.
Honey, A.F. y Schomberg, D.W. (1978) Biol. Reprod. 19:242.
Thonkí, K.H. y Chonníng, C.P. (1976). Endocr. Res. Commun.3: 319.
Thankí, K.H. y Chonníng, C.P. (1978). Endocrinology103 : 74.
Ledwírz-Rigby, F., Rígby, B.W., Gay, V.L., Sfefson, M.,Young, J. y Channíng, C.P. (1977). J. Endocr. 74: 175.
Druker, B. y Lunenfeld, B. (1978). J.78 : ÏÓÏ.
Kraíem, Z.,Endocrinol.
324.Ledwifz-Rígby, F. (1980). Bíol. Reprod. 23:
Ledwífz-Rígby, F. y Rígby, B.W. (1977). J. Cell. Biol.75 : 178 a.
Darga, N.C. y Reicherr, L.E. Jr. (1978). Biol. Reprod.19: 235.
Yong, K.P., D.N. (1979).Endocrinology104:
N.A. y Ward,552.
Samoan,

54.
55.
56.
57.
58.
59.
óO.
ól.
62.
63.
64.
65.
óó.
ó7.
68.
69.
-l48
Sakai, C., Channing, C.P. y Engel, B. (¡977). Proc. Soc.Exp. Biol. Med. 155 : 373.
Kumari, L., Tucker, s. y Channing, C.P. (1979). Biol.Reprod. 21 1043.
Roy, A.B. (l960). Adv. in Enzymology 22 : 205.
Baulieu, E. (1960). Compt. Rend. 251 142].
Baulieu, E., Corpéchot, C., Dray, F., Emiliozzí, P.,Lébeau, M.C., Jarvis, P.M. y Robel, P. (1965). Rec.Progr. Horm. Res. 2] :411.
Bursfein, S. y Dorfman, R.J. (1963). J. Biol. Chem. 238:1656.
H.|., Van de Wiele, R.L. (1963).: 648.
Calvin, y Lieberman, S.Biochemisl’r)l 2
:259.Calvin, H.l. y Lieberman, S. (1964). Biochemisfry 3
Lébeau, M.C., Albergo, A. (1964).Biochem. Biophys. Res. Commun. 17:
y Baulieu, E.E.570.
Roberts, K.D., Bandi, L., Calvin, H.|., Drucker, W.D. yLieberman, S. (1964). J. Ame Chem. Soc. 86 : 958.
Weliky, l. y Engel, L.L. (1962). J. Biol. Chem. 23712089.
Ryan, K.J. y Smith, O.W. (1965)a Rec. Progr.Horm. Res.2] : 367.
Dominguez, O.V., Loza, A.C., Rangel, C.L. y Zárate,T.A. (1970). Archs. lnvo Med. (Mex) l : 187.
Notarion, A.D. y Ungar, F. (1968). Can. J. Biochem. 46:1185.
Notafion, A.D‘ y Ungar, F. (1972). Endocrinology 912852.
Fry, J.M. y Koritz, S.B. (1972). Endocrinology 91 : 852

70.
71.
72.
73.
74.
75.
76.
77.
78.
79.
80.
81.
82.
83.
84.
-149
Domínguez, O.V., Loza, A.C.,A.S. (1974). J. St. Biochem. 5:
Morón, L.Z. y Valencia,867.
J.C. y Timberlake, C.E. (1962). J. Clin.Merab. 22: ll48.
Warren,Endocrinol.
Warren, J.C. y Timberlake, C.E. (1964). Obstet. Gynecol.23 : 689.
Chem. 245:Korítz, S.B. y Kumar, A.M. (1970). J. Biol.152.
Domínguez, O.V., Valencia, S.A. y Loza, A.C. (1975).J. St. Bíochem. ó : 301.
311.Nofatíon, A.D. (1975). J. Sferoíd Biochem. ó:
Wallace, E. y Sílberman, W. (¡964). J. Biol. Chem. 239:2809.
Holcenberg, J.S. y Rosen, S.W. (1965). Archives Biochem.Biophys. llO : 551.
Flíckínger, G.L., Murarvec, T. y Touchsfone, J.C. (1965)Fed. Proc. 24: 162.
Aakvaag, A. y Fyllíng, P. (1968). Acta Endocrinologíca57: 447.
Melvílle, E. y Braunsberg, H. (1973). Bíochem. Soc.Trans. | : 757.
Kallíala, K., Laatikaínen, T., Luukkaínen, T. y Víhko,R. (1970). J. Clin. Endocr, Metab. 30: 533.
Srurm, G. y Hannappel, E. (1978). J. Endocr. 79: 29].
Sl’urm, G., Stahler, E. y Daume, E. (1974). J. SferoídBíochem. 5: 313.
Storm, (3., Sfahler, E. y Daume, E. (1975). Acta Endocrinologíco. Suppl. 193 : 104.

85.
86.
87.
88.
89.
90.
9].
92.
93.
94.
95.
96.
97.
98.
99.
100.
-]50
Grosch, B., Daume, E., Stohler, E.1401.
Sturm l C3-1R. (1980). J. Steroíd Bíochem. 13:
Sandberg, E.C. y Jenkins, R.C. (1966). Bíochím. Bíophys.Acta H3: 190.
Oertel, G.W., Treíber, L. (1967). Specialía15 : 97.
y Rindt, W.
Payne, A.H. y Mason, M. (1969). Steroíds 13 z 213.
Patwardhon, U. y Lonthíer, A. (1977). J. Endocr. 75:445.
Goldenberg, R.L., Voítukoítís, J.L. y Ross, G.T. (1972).Endocrinology 90: 1492.
Payne, R.W. y Helboum, A.A. (1955). Endocrinology 57:193.
Hillíer, S.G. y Ross, G.T., (1979). Biol. Reprod. 20 :261.
Fortune, J.E. y Armstrong, D.T. (1978). Endocrinology102 : 227.
Moor, R.M. (1977). J. Endocr. 73: 143.
Tsang, B.K., Henderson, K.M. y Armstrong, D.T. (1979).Am. J. Obstet. Gynecol. 133 : 256.
Walton, E.A. y Armstrong. D.T. (1981). Biol. Reprod. 25:272.
lnabo, T., Wiest, W.G. y Níswender, G.D. (1979).Steroíds 34: 663.
Corr, B.R., Míkhaíl, G.Clin. Endocrinol. Metob. 33
y Flickínger, G.L.(1971). J.:358.
Lowry, O.H., Rosebrough, N.J., Forr, A.L. y Randall, R.J. (1951). J. Biol. Chem. 193 : 265.
Hobkírk, R.337.
y Nilsen, M. (Ï970).Anolytícal Bíochem. 37:
y Buchholz,

IO].
102.
103.
104.
105.
iOó.
107.
108.
109.
110.
Hi.112.
H3.
H4.
115.
Loriaux, D.L., Ruder, H.J. y Lípsett, M.B. (197]).Steroid518z4ó3.
Armstrong, D.T., Kraemer, M.A. y Híxon, J.E. (1975).Biol. Reprod. 12 : 599.
Steel, R.G.D. y Torrie, J.H. (i960). Principles andProcedures of Statistics. Mc Graw-Hill, New York.
Siegel, S. (¡956). Nonparametric Statistics. Mc GrawHiII, New York.
Suzuki, K., Kawakura, K. y Tamaoki, B. (1978).Endocrinology102:i595.
Armstrong, D.T. (1979). Biol. Reprod. 21 1025.
Ahmad, N. y Gower, D.B. (1968). J. Endocrinol. 108:233.
Brodíe, A.M.H., Wu, J.T., Marsh, D.A. y Brodíe. M.J.(i978). Biol. Reprod. 18 : 365.
Endocrinol. 45: 297.Channíng, C.P. (i969). J.
Makris, A. y Ryan, K.J. (i975). Endocrinology 96 : 694.
Makris, A. y Ryan, K.J. (i980). Steroids 35: 53.
Erickson, G.F. y Ryan, K.J. (i976). Endocrinology 99:452.
Fortune, J.E. y Armstrong, D.T. (i977). Endocrinology100:134i.
Tsang, B.K., Moon, Y.S.,, Simpson, C.W. y Armstrong,D.T. (1979). J. Clin. Endocrinol. Metab. 48: 153.
Channing, C.P., Schaerf, F.W., Anderson, L.D. yTsatriri, A. (1980). En : Reproductíve Physíology lll international Review of Physíology, vol. 3 Editor: Greep,R.C’. University Park Press, Baltimore, pág. 117.

Hó.
H7.
118.
119.
120.
121.
122.
123.
124.
125.
126.
127.
128.
129.
—152
Erickson, G.F. y Hsueh, A.J.W. (i978). Endocrinology103: i960.
TsaFriri, A. y Channing, C.P. (i975). Endocrinology 96:922.
Younglai, E.V. y Short, R.V. (i970). J. Endocr. 47 :321.
Lacroix, E., Eechaute, W. y Leusen, I. (i974). Steroids23: 337.
Fortune, J.E. (i981). Biol. Reprod. 24, Suppl n° i: 39A.
Makris, A. y Ryan, K.J. (i977). Steroíds 29:65.
Hobkirk, R. y Davidson, S. (i971). J. Chromat. 54 : 431.
Hobkírk, R., Freeman, D.J., Harvey, P.R.C., Nilsen, M.y Jennings, B. (i977). Can. J. Biochem. 55: 390.
"The Vertebrate Ovary".New York, póg. 215.
K.P. (i978). En:R.E. Plenum Press,
Mc Natty,Editor: Jones,
Bradlow, H.L. (i970). En: "Chemical and BiologicalAspects of Steroíd Coníugatíon". Editores: Bernstein, S.y Solomon, S. Springer-Verlag, New York-HeidelbergBerlin, póg. 131.
Jakoby, W.B., Sekura, R.D., Lyon, E.S., Marcus, C.J.y Wang, J.L. (i980). En: "Enzymatic Basis of Detoxication"Editor: Jakoby, W.B. Academic Press, New York, vol 2,póg. 199.
y Channing, C.P. (197]). Bíochím. Biophys.601.
Kolena, J.Acta 252:
(i976). EndocrinologyArmstrong, D.T. y Dorrington, J.H.99: i411.
Nimrod, A. y Líndner, H. (i976). Molec. Cell. Endocr.5 : 315.

.130.
13].
132.
133.
¡34.
135.
136.
137.
138.
139.
140.
141.
142.
143.
¡44.
-153—
Lucky, A.W., Schreíber, J.R., HílI'íer, S.G., Schulman,J.D. y Ross, G.T. (1977). Endocrinology1002128.
Nimrod, A., Rosenfeld, R.L. y Otto, P. (1980). J.SteroídBiochem. 13 z 1015.
Louvet, J.P., Harman, S.M. y Ross, G.T. (1975).Endocrinology 97: 1179.
Anderson, L.D. y Betto, S.K. (1978).375.
Chonníng, C.P.,Clínícs ïn Obst. Gyn. 5:
Batta, S.K., Coulston Wentz, A. y Channíng, C.P. (1980).J. Clin. Endocrinol. Metob. 50: 274.
Korítz, S.B., Yun, J. y Ferguson, J.J. (1968).Endocríno|ogy82 : 620.
Sulímovicí, S. y Boyd, G.5. (1969). European J. Bíochem.7 : 549.
Dorríngton, J.H. y Armstrong, D.T. (¡979). Rec. Progr.Horm. Res. 35: 30].
Wiener, M. y Allen, S.H.G. (1967). Steroíds 9 : 567.
Townsley, J.D. (1975). Acto Endocrinologico 79: 740.
Aínsworth, L., Tsong, B.K., Downey, B.R., Marcus, G.Y. y Armstrong, D.T. (1980). Bíol. Reprod. 23 : 621.
Fortune, J.E. y Hansel, W. (1979). Endocrinology104:1834.
Schreíber, J.R., Reid, R. y Ross, G.T. (I97ó).Endocr¡nology98: 1206.
Louvet, J.P., Harman, S.M., Schreïber, J.R. y Ross, G.T.(1975). Endocrinology 97: 3óó.
Híllíer, S.G., Reichert, L.E. Jr. y von Hall, E.V.(1981)J. Clin. Endocrinol. Metob. 52 : 847.