
EFECTO DEL ESTRÉS SOBRE EL TEJIDO LINFOIDE...
33
1 EFECTO DEL ESTRS SOBRE EL TEJIDO LINFOIDE ASOCIADO A NARIZ EN EL RATN REGISTRO SIP: 20071310
Transcript of EFECTO DEL ESTRÉS SOBRE EL TEJIDO LINFOIDE...

1
EFECTO DEL ESTRÉS SOBRE EL TEJIDO
LINFOIDE ASOCIADO A NARIZ EN EL RATÓN
REGISTRO SIP: 20071310

2
RESUMEN El sistema inmunitario de las mucosas (MALT) comprende tejidos
asociados con la superficie de las mucosas respiratoria, digestiva y genitourinaria,
los cuales defienden el organismo de antígenos externos. El tejido linfoide
asociado a nariz (NALT) representa un compartimiento inductor de la respuesta
inmunitaria y se localiza en piso de la nariz de algunos roedores; mientras que la
lámina propia de la mucosa nasal representa al compartimiento efector. El estrés,
provoca un incremento de la secreción de hormonas glucocorticoides y
catecolaminas por activación del eje hipotálamo-hipófisis-adrenal. Estas
hormonas interactúan con receptores de las células inmunitarias ocasionando
cambios en su respuesta. El objetivo de este trabajo fue determinar los efectos del
estrés sobre la morfología del NALT, el número y distribución de sus células y la
secreción de IgA. Se utilizaron ratones Balb/c machos de 9 semanas de edad,
divididos en un grupo control y dos grupos problema, a los que se aplicó 3 horas
de estrés por inmovilización, durante 4 y 8 días. De cada animal, se obtuvo el
NALT y se realizaron tinciones con H-E para estudiar la morfología e
inmunohistoquímicas para cuantificar linfocitos B (IgM+, CD45+) y T CD3+ (CD4+
y CD8+) y macrófagos En otros grupos de animales se cuantificó la población de
linfocitos B y T CD3+, CD4+ y CD8+ por citometría de flujo.
En el análisis morfológico, no se observaron cambios en la arquitectura
del NALT, sin embargo durante el estrés de 4 días el volumen del órgano
disminuyó ligeramente, mientras que a los 8 días éste se incrementó. La cantidad
de macrófagos fueron semejantes en los tres grupos de estudio. La citometría de
flujo mostró que los linfocitos B y T CD8+ disminuyeron en los animales
estresados, sin embargo esta modificación no se observó en el análisis
morfológico. La citometría de flujo y el análisis morfológico mostraron una
disminución de los linfocitos T CD3+ y CD4+ en los animales estresados. Los
linfocitos intraepiteliales CD4+ y CD8+, disminuyeron significativamente con el
estrés de 4 días, incrementando su número con el estrés de 8 días. Se concluye
que el NALT es sensible al esquema de estrés utilizado, afectando tanto a las
células de la respuesta inmunitaria celular (linfocitos T) como a las de la respuesta
humoral (linfocitos B), posiblemente por la influencia de las hormonas secretadas
durante el estrés.

3
INTRODUCCIÓN SISTEMA INMUNITARIO DE LAS MUCOSAS
En las mucosas se encuentra la primera línea de defensa contra antígenos
externos, ya que un gran número de agentes patógenos las utilizan como vía de
entrada. El conjunto de tejidos linfoides organizados y difusos en las mucosas del
organismo, se denominan �tejidos linfoides asociados a las mucosas� (MALT) los
cuales a través de la inmunidad innata y adquirida mantienen una homeostasis
inmunitaria, a lo largo de la gran superficie interior de las vísceras, la cual
comprende las mucosas oral, nasal, respiratoria, urinaria y gastrointestinal.
(Tristram GP 2001)
Características particulares del sistema inmunitario de las mucosas
El sistema inmunitario de las mucosas, además de tener cierta independencia,
presenta características únicas que lo diferencian de la inmunidad sistémica, una
de ellas es el poseer células T con propiedades reguladoras o efectoras
especificas para la mucosa, entre las que se encuentran los LIEs CD4+ y CD8+
(TCR γδ / αβ). (Cheroutre H 2005, Okuda M y Pawankar R 1992)
Adicionalmente es un sistema inmunitario polarizado hacia las mucosas, el cual
permite a los linfocitos activados migrar de manera selectiva hacia los tejidos
linfoides difusos, situados por debajo del epitelio. (Csencsits KL y cols 1999)
ARQUITECTURA HISTOLÓGICA DEL NALT
El tejido linfoide asociado a nariz (NALT) forma parte del sistema
inmunitario de las mucosas (MALT). Es un órgano linfoide par que se desarrolla
en algunos roedores después de la primera semana de nacimiento (Mebius R
2003, Ying X y cols 2005, Harmsen A y cols 2002); tiene la forma de un cilindro de
aproximadamente 3 mm de longitud y descansa a ambos lados del tabique sobre
el piso de la cavidad nasal. (Asanuma y cols 1997, Heritage y cols 1998, Sminia T
y Kraal G 1999)
Debido a su localización anatómica, se considera un análogo del anillo de
Waldeyer humano (Boyaka NP y cols 2000) y funcionalmente se compara con la
placa de Peyer por presentar características inductoras similares. (Kiyono H y
Fukuyama S 2004)

4
La función atribuida al NALT, es proporcionar un sistema de defensa en las
vías aéreas superiores. (Csencsits KL y cols 1999, Boyaka PN y cols 2000). El
NALT es un compartimiento inductor y corresponde a la porción de células
linfoides organizadas.
En cuanto a su estructura, el NALT está compuesto principalmente por
linfocitos T (predominando los CD4+) y linfocitos B en cantidades similares. El
órgano se encuentra regionalizado de acuerdo al patrón de distribución de cada
tipo celular; los linfocitos B se localizan preferentemente en la porción central del
órgano, conformando la zona B o folicular, mientras que los linfocitos CD3+ se
distribuyen en la periferia de la zona B, conformando la zona T o parafolicular.
Otras poblaciones presentes son células dendríticas y macrófagos (Asanuma H y
cols, 1997).
El NALT, en una gran cantidad de trabajos, se ha reportado que juega un
papel muy importante en el sistema de defensa del tracto respiratorio alto en los
roedores, ya que se ha demostrado que puede provocar respuestas inmunitarias,
tanto locales como sistémicas, por lo que se ha considerado un modelo muy
efectivo para inducir inmunidad. (Tamura S y cols 1998, Zuercher AW y cols 2002,
Liang B y cols 2001, Balmelli C y cols 2002, Rojas-Hernández S y cols 2004).
ESTRÉS Y EL EJE HIPOTALAMO-HIPOFISIS-ADRENAL
Actualmente el estrés se define como un �estado de amenaza para la
homeostasis, durante el cual se activa una respuesta adaptativa compensatoria�.
(McEwen BS 1998). Cannon señala que el sistema nervioso simpático es esencial
para restaurar la homeostasis alterada por el estrés y para promover la
sobrevivencia del organismo, además mencionó la existencia de respuestas al
estrés, especificas para cada tipo de agente estresor. (Paca´k K y Palkovits M
2001).
De acuerdo a la duración, el estrés se clasifica en:
Estrés agudo: En el sentido de la lucha, se considera simple, intermitente y de
exposición por tiempo limitado.
Durante el estrés agudo se produce una respuesta adaptativa, la cual
provoca que los procesos fisiológicos inhiban o estimulen varios sistemas y/o sus
componentes, para movilizar las reservas energéticas hacia órganos prioritarios
como los sistemas muscular y nervioso.

5
Estrés crónico: (carga de tensiones acumulativas); es la exposición a agentes
estresantes en forma prolongada y continua.
El hipotálamo es un centro de integración que recibe y monitorea información
acerca del ambiente, y además coordina las respuestas a través del sistema
nervioso central, autónomo y endocrino. Las principal información que recibe es la
visual, olfativa, auditiva y sensitiva.
El sistema de estrés está constituido por el eje hipotálamo-hipófisis-adrenal
(HPA) y el sistema simpático sistémico, los cuales se encuentran controlados por
señales límbicas, circadianas, neurosensoriales y hormonales. Las situaciones de
estrés, tales como la hipoglicemia, procesos inflamatorios, fiebre, trauma, ejercicio
intenso, cirugía y problemas emocionales pueden activar este sistema. (Paca´k K
y Palkovits M 2001)
El estrés producido por estímulos fisiológicos, emocionales o cognitivos
negativos, incrementan en el hipotálamo la producción de CRH y en la hipófisis
anterior de ACTH, la cual a su vez controla la actividad de las zonas fasciculada y
reticular de la corteza suprarrenal. Específicamente en la zona fascicular, la ACTH
estimula la producción de cortisol y el incremento de esta hormona, provoca
hiperplasia de las células, lo que conlleva a hipertrofia de la glándula suprarrenal.
(Guyton AC 2003)
Además el estrés físico o mental puede ocasionar la estimulación de nervios
simpáticos que inervan la médula suprarrenal, lo que ocasiona la liberación de
catecolaminas a la circulación sanguínea.
La adrenalina y noradrenalina se encuentran muy relacionadas con las
reacciones de lucha o huida ante estímulos estresantes. La síntesis de adrenalina
se incrementa en situaciones de estrés físico, cólera y conductas de alto riesgo.
En cambio la noradrenalina se relaciona más con los estados de estrés psíquico
como ansiedad, miedo y angustia. (Guyton AC 2003)
Modelos animales de estrés.
Hans Selye (Paca´k K y Palkovits M 2001) en 1936, fue el primer
investigador que utilizó el modelo de estrés por inmovilización en ratas, las cuales
manifestaron varios signos que caracterizaron un síndrome ocasionado por el

6
estrés. Por otro lado cuando se compararon las regiones cerebrales activadas, por
diferentes situaciones estresantes como la inmovilización, la hipoglicemia, el frío,
la hemorragia, y el dolor, se observó que el modelo de estrés por inmovilización
es uno de los más efectivos para producir activación de una mayor cantidad de
regiones cerebrales. Además se encontró que cada agente estresante, utiliza vías
neuroendocrinas específicas.
HIPÓTESIS
Si el estrés provoca un incremento en la secreción de hormonas
glucocorticoides y catecolaminas y si estas tienen influencia sobre las células del
sistema inmunitario asociado a la mucosa nasal, entonces los ratones sometidos
a estrés, presentarán modificaciones en el tamaño del NALT, en el número y
distribución de las células inmunitarias
OBJETIVO GENERAL
Analizar los efectos del estrés, sobre la cantidad y distribución de las
células del sistema inmunitario en el tejido linfoide asociado a la nariz.
MATERIAL Y METODOS Animales
Se utilizaron ratones Balb/c machos con un peso entre 21-24 g
proporcionados por Harlan México, el manejo de los animales se realizo de
acuerdo a las normas de la Comisión de Bioética de la ESM. Los ratones se
dividieron en tres grupos de 7 animales cada uno; un grupo control, uno con
tratamiento de estrés de 4 días y otro con tratamiento de 8 días.
Reactivos necesarios para realizar las inmunohistoquimicas:
-Anticuerpos monoclonales de rata anti-ratón conjugados con biotina: anti
CD3+, anti CD4+, anti CD8+, anti CD45+ (B220+), anti-Mac-3+ (DB PharMingen
Technical Data Sheet).

7
-Anticuerpo policlonal de cabra anti-ratón conjugado con peroxidasa: anti-
IgM (Serotec)
-Diaminobencidina (Pierce)
-Estreptavidina conjugada con peroxido (Zymed Laboratories Inc).
Tratamiento de estrés.
Los animales fueron sometidos a un esquema de estrés por inmovilización
en contenedores cilíndricos apropiados al tamaño del animal (5 cm. de largo, 3 cm
de alto y 3.5 cm de diámetro) con ventilación adecuada. Antes y después del
periodo de estrés, los animales se mantuvieron en jaulas durante los días de
tratamiento, ingiriendo agua y comida a libre demanda. Los animales control se
mantuvieron en las mismas condiciones.
Los animales problema se sometieron diariamente a 3 horas de estrés,
bajo un horario establecido (8:00-11:00 AM). Para evitar el estado de adaptación
al estrés durante la inmovilización, los ratones se estimularon cada 30 minutos de
la manera siguiente: 1.-Agitación de los contenedores durante 10 segundos,
iniciando el ciclo a los 10 minutos. 2.-Rotación de los contenedores durante 10
segundos, iniciando el ciclo a los 20 minutos. 3.-Por ultimo, se sumergió la cola de
los animales en agua fría durante 10 segundos, iniciando el ciclo a los 30
minutos.
Día Agitación de los
contenedores
Rotación
de los contenedores
Inmersión de la cola en agua
fría
1 x x
2 x x x
3 x x
4 x x x
Sacrificar el grupo de ratones controles y estresados durante 4 días
5 x x
6 x x x

8
7 x x
8 x x x
Sacrificar el grupo de ratones estresados durante 8 días
Obtención y procesamiento del material biológico.
Posterior al tratamiento, los animales se anestesiaron profundamente con
vapores de éter, se sangraron por punción cardiaca directa y se sacrificaron por
decapitación, se removió la piel de la cabeza, se retiró el maxilar inferior y los
tejidos blandos, como lo describen (Asanuma y cols, 1997; Heritage y cols, 1998).
Con la ayuda de microscopio estereoscópico, se realizaron dos incisiones
verticales desde el vértice del paladar por la parte interna de los dientes incisivos,
siguiendo las porciones laterales hasta la base del paladar por la parte interna de
los dientes molares. Enseguida el colgajo fue sujetado por el vértice, con pinzas
de punta fina y se realizó tracción hacia la base del paladar, retirándolo por
completo.
El paladar extraído se colocó en contenedores de aluminio de 1 cm³, sobre
una base de medio de inclusión hidrosoluble (Tissue-tek), con el NALT en
posición ventral, éste fue cubierto con una segunda capa de tissue-tek y se
congeló en isopentano. Las muestras fueron almacenadas a �70° C, hasta su
corte en criostato.
Procesamiento
De las muestras del NALT congeladas se obtuvieron cortes de 7 µm de
espesor, se colocaron en series de portaobjetos previamente tratados con
gelatina la 1% y se fijaron en acetona durante 20 minutos.
Tinciones
En los cortes fijados en acetona se realizaron las tinciones
inmunohistoquímicas para identificar las siguientes poblaciones celulares: por
inmunohistoquímica directa, células plasmáticas productoras de IgM utilizando
anticuerpo policlonal de cabra anti-IgM de ratón conjugado con HRP (Serotec).

9
Utilizando el método de inmunohistoquímica indirecta se detectaron linfocitos T
CD3+, CD8+, CD4+, y por la expresión del antígeno Mac 3 en los macrófagos i
ICAM-1 (CD54) utilizando los respectivos anticuerpos monoclonales de ratón,
conjugados con biotina (Pharmingen). Posteriormente se aplicó estreptavidina
conjugada con peroxidasa (Jackson Immuno Reserch).
Análisis microscópico y cuantificación de las células.
Las células se contaron en áreas constantes de 12538 µm2, 25050 µm2 y
30013 µm2 para cada tipo celular: Para los linfocitos T CD3+, CD4+, CD8+ en las
zonas parafolicular y folicular 12538 µm2. Los linfocitos B IgM+ y CD45+ se
contabilizaron en la zona folicular, en una área de 25050 µm2, mientras que los
macrófagos F4/80+ se contabilizaron en la base del NALT, en una área 12538
µm2.
Para valorar la intensidad de reacción al marcador CD54+ (ICAM-1) en el
NALT, se ajustaron las propiedades de la cámara con los siguientes valores;
contraste 30%, Brillo 45%, saturación 63%; mientras que el microscopio se ajustó
a un objetivo de 20x, con una intensidad de luz y apertura del diafragma en 0. El
área utilizada fue 30013µm2, con un rango de intensidad media de color mínimo
de 10 y máximo de 150%.
Para la cuantificación de linfocitos intraepiteliales CD4+ y CD8+, se realizó
en la longitud promedio del epitelio respiratorio del NALT, la cual fue de 780 µm
lineales. Cada uno de los conteos se realizó por duplicado.
Obtención de linfocitos para citometría de flujo.
Después de remover el NALT, la obtención de linfocitos se realizó de la
siguiente forma: el paladar del ratón, conteniendo el NALT se colocó en una caja
de Petri con 10 ml de medio RPMI-1640, se realizó un raspado suave con una
espátula para separar el NALT del paladar, después se disgregó el NALT con el
émbolo de plástico de una jeringa y se recuperó la suspensión celular, la cual se
filtró en tela de organza de 10 × 10 cm. con una abertura de 1 mm y se colocó en
tubos cónicos de 15 ml. La suspensión celular se centrifugó a 1500 rpm/10 min. a
4° C, se desechó el sobrenadante y se resuspendió la pastilla en 1 ml de RPMI-

10
1640. Finalmente se ajustó a 1x106 células, para realizar las diferentes
inmunotinciones para citometría de flujo.
Inmunotinción de linfocitos para citometría de flujo
La tinción de los linfocitos se realizó con anticuerpos anti-ratón (Becton
Dickinson Technologies, Gaithersburg, MD) obtenidos de PharMingen (San Diego,
CA) con fluorocromos conjugados biotinilados. Para linfocitos (Lc) T CD3+
anticuerpos marcados con fluorocromos peridinin chlorophyll protein (PERCP), o
con fluorescein isothiocynate (FITC). Para controles de isotipo, anticuerpos anti-
IgG2a marcados con Phicoerithrin (PE) y Ab anti-IgG2b con (FITC), además se
utilizó el anti-CD4+ (FITC) y anti-CD8+ (PE) y para los Lc B el CD45 RO (B220)
con (PE). Los anticuerpos se diluyeron 1:100 con amortiguador de fosfatos con
albúmina sérica bovina p.H 7.4 (PBA) (10 mg/ml) y se colocaron 10 µl de cada
anticuerpo, por cada millón de células, incubando en oscuridad a 4°C por 30 min.
Al término de ese tiempo se lavaron con PBA y se desechó el sobrenadante, al
paquete celular se le agrego 400 µl de para-formaldehído al 1% y se guardo en
oscuridad a 4 º C hasta su lectura en el Citómetro de flujo FACSCAN (Becton
Dickinson, San Jose, CA), cada conteo se realizo en un mínimo de 10,000
eventos. En las muestras para lectura de CD4+ contra CD8+ y CD3+ contra B220,
se combinaron los anticuerpos marcados con diferentes fluorocromos, además de
utilizar un control. El análisis de resultados, se llevó a cabo mediante el software
(winMDI 2.8)
Análisis estadístico
Utilizando la prueba de ANOVA de una vía, se compararon los grupos
experimentales de cada protocolo con su respectivo testigo. Los datos se
analizaron mediante el software Sigma Stat y se graficaron mediante el software
Sigma Plot.

11
RESULTADOS Linfocitos T CD3+
Las inmunotinciones para identificar a los linfocitos CD3+ mostraron, que
en los tres grupos de estudios, estos se localizan predominantemente en el área
parafolicular, con pocas células en el área folicular observándose además algunos
linfocitos CD3+ incluidos en el epitelio que recubre el NALT.
En la cuantificación de los linfocitos CD3+ un área de 12538 µm2 ubicados
en la zona parafolicular, se observó que los animales sometidos a estrés por 4 y
8 días tuvieron 249±7.5 y 223±10.6 células respectivamente, y el grupo control,
237±12.3.
Comparando estos tres grupos por ANOVA de una vía; se encontraron
diferencias significativas entre el grupo de control y el grupo de estrés de 8 días
(F2,15=9.525; P 0.002), este último también mostró diferencias significativas con el
grupo de estrés de 4 días (F2,15=9.525; P 0.035). Sin embargo entre el grupo
control y el grupo de estrés de 4 días no hubo diferencias significativas (P
0.0059). (Figura 1).
De la misma forma comparando el número de linfocitos CD3+ en el área
folicular, del grupo control, 25±2.9 con el número de células del grupo de 4 días,
18±1.5 y el de 8 días, 17±1.3. Se encontraron diferencias significativas del grupo
control con los grupos de estrés de 4 y 8 días (F2,15=23.902; P 0.001). Sin
embargo entre los dos grupos problema, no hubo diferencias significativas (P
0.766). (Figura 2). La disminución en la cantidad de estas células, ocurrió tanto
en el área parafolicular como en la folicular.

12
Linfocitos T CD4+
En la tinción inmunohistoquímica, no se alteró la distribución de los
linfocitos CD4+ en el NALT por el estrés. La mayoría de las células se
distribuyeron en el área T o parafolicular predominado hacia la base del NALT
mientras que el área folicular presentó pocas células
El número de linfocitos CD4+ en un área de 12538 µm2, fue el siguiente;
en el grupo testigo, el promedio de linfocitos CD4+ localizados en el área
parafolicular fue de 215±5.3; comparado con esta cifra, el número de células fue
menor en el grupo de animales sometidos a estrés durante 4 días, el cual mostró
un promedio de 173±12.3 células y aun menor en el grupo de 8 días, 159±7.6
células (Figura 3). Comparando el grupo control con los grupos de estrés de 4 y 8
días respectivamente, se encontraron diferencias estadísticas significativas
(F2,15=63.593; P 0.001 y P 0.001) así como entre los dos grupos problema
(F2,15=23.902; P 0.015).
En relación al área folicular, no hubo diferencias significativas entre los tres
grupos de estudio (F2,15=23.902; P 0.699). El grupo testigo mostró un promedio de
27± 1.5 células, mientras que los estresados durante 4 y 8 días, presentaron
4 DIAS
Num
ero
de c
élul
as C
D3+
en
área
par
afol
icul
ar
0
50
100
150
200
250
300
CONTROL 8 DIAS
P 0.035
P 0.002
4 DIAS
Num
ero
de c
élul
as C
D3+
en
área
folic
ular
0
5
10
15
20
25
30
35
CONTROL 8 DIAS
P <0.001
P <0.001
Figura 2. Cantidad de linfocitos CD3+ en área folicular. La gráfica compara el número de células de los dos grupos problema con el testigo. Con respecto al grupo control, el numero de linfocitos disminuyó con el estrés durante 4 y 8 días (F2,15=23.902; P 0.001).Entre los dos grupos problema no hubo diferencias significativas.
Figura 1. Linfocitos CD3+ del áreaparafolicular. La gráfica compara elnúmero de células entre los tresgrupos de estudio. Con respecto algrupo control, el número de linfocitosdisminuyó significativamente con elestrés de 8 días (F2,15=9.525; P0.002). También se encontrarondiferencias significativas entre losdos grupos problema

13
respectivamente 26±1.4 y 27±2 células. (Figura 4). Los resultados muestran una
disminución en el número de linfocitos CD4+, principalmente en el área
parafolicular.
Linfocitos CD8+
En cuanto a los linfocitos CD8+; estos se situaron principalmente en el
área parafolicular con una mayor cantidad en la base del NALT y cercanos a la
Figura 4. Cantidad de Linfocitos CD4+ en área folicular. La gráfica compara el número de linfocitos CD4+ de los dos grupos problema con el testigo, no habiendo diferencias significativas entre los tres grupos de estudio.
4 DIAS
Num
ero
de c
elul
as C
D4+
en
área
par
afol
icul
ar
0
50
100
150
200
250
CONTROL 8 DIAS
P <0.001
P <0.001P 0.015
4 DIAS
Num
ero
de c
elul
as C
D4+
en
área
folic
ular
0
5
10
15
20
25
30
35
CONTROL 8 DIAS
Figura 3. Cantidad de linfocitos CD4+ en área parafolicular. La grafica compara el númerode células CD4+ entre los tres grupos de estudio. Con respecto al grupo control, el número delinfocitos disminuyó significativamente con el estrés durante 4 días y 8 días. (F2,15=63.593; P0.001 y P 0.001). Entre los grupos problema, también hubo diferencias significativas(F2,15=23.902; P 0.015).
14
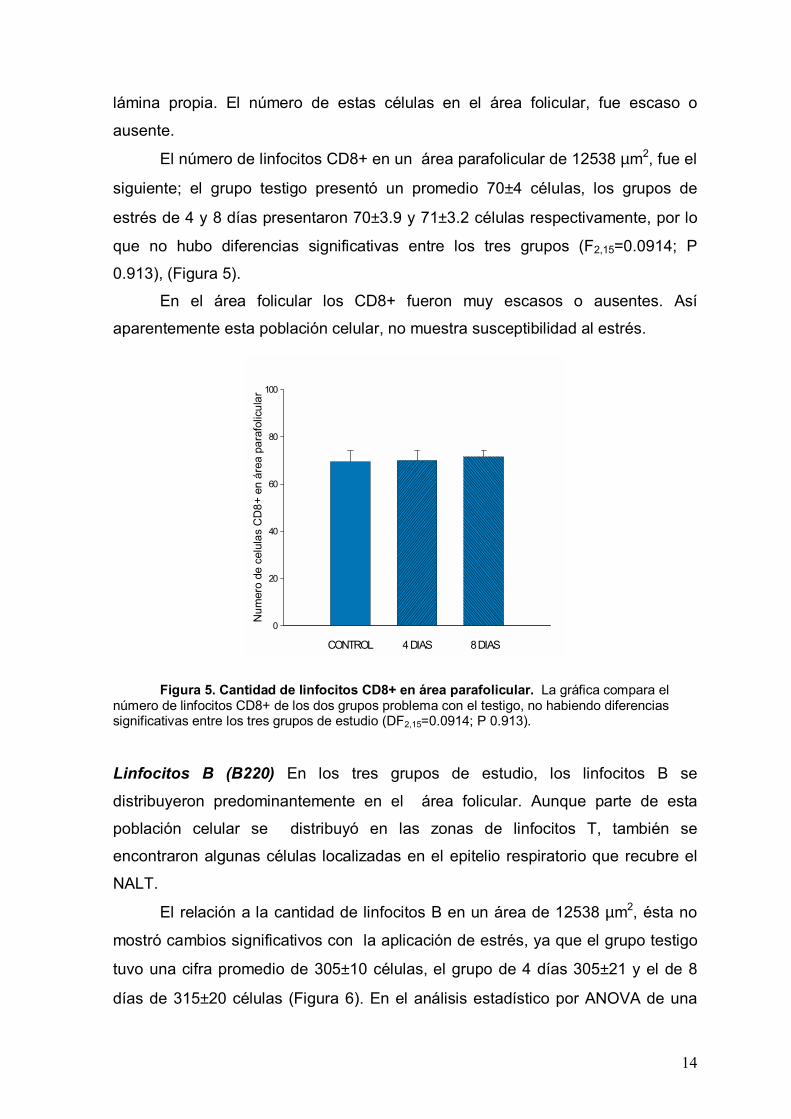
lámina propia. El número de estas células en el área folicular, fue escaso o
ausente.
El número de linfocitos CD8+ en un área parafolicular de 12538 µm2, fue el
siguiente; el grupo testigo presentó un promedio 70±4 células, los grupos de
estrés de 4 y 8 días presentaron 70±3.9 y 71±3.2 células respectivamente, por lo
que no hubo diferencias significativas entre los tres grupos (F2,15=0.0914; P
0.913), (Figura 5). En el área folicular los CD8+ fueron muy escasos o ausentes. Así
aparentemente esta población celular, no muestra susceptibilidad al estrés.
4 DIAS
Num
ero
de c
elul
as C
D8+
en
área
par
afol
icul
ar
0
20
40
60
80
100
CONTROL 8 DIAS Figura 5. Cantidad de linfocitos CD8+ en área parafolicular. La gráfica compara el
número de linfocitos CD8+ de los dos grupos problema con el testigo, no habiendo diferencias significativas entre los tres grupos de estudio (DF2,15=0.0914; P 0.913).
Linfocitos B (B220) En los tres grupos de estudio, los linfocitos B se
distribuyeron predominantemente en el área folicular. Aunque parte de esta
población celular se distribuyó en las zonas de linfocitos T, también se
encontraron algunas células localizadas en el epitelio respiratorio que recubre el
NALT.
El relación a la cantidad de linfocitos B en un área de 12538 µm2, ésta no
mostró cambios significativos con la aplicación de estrés, ya que el grupo testigo
tuvo una cifra promedio de 305±10 células, el grupo de 4 días 305±21 y el de 8
días de 315±20 células (Figura 6). En el análisis estadístico por ANOVA de una
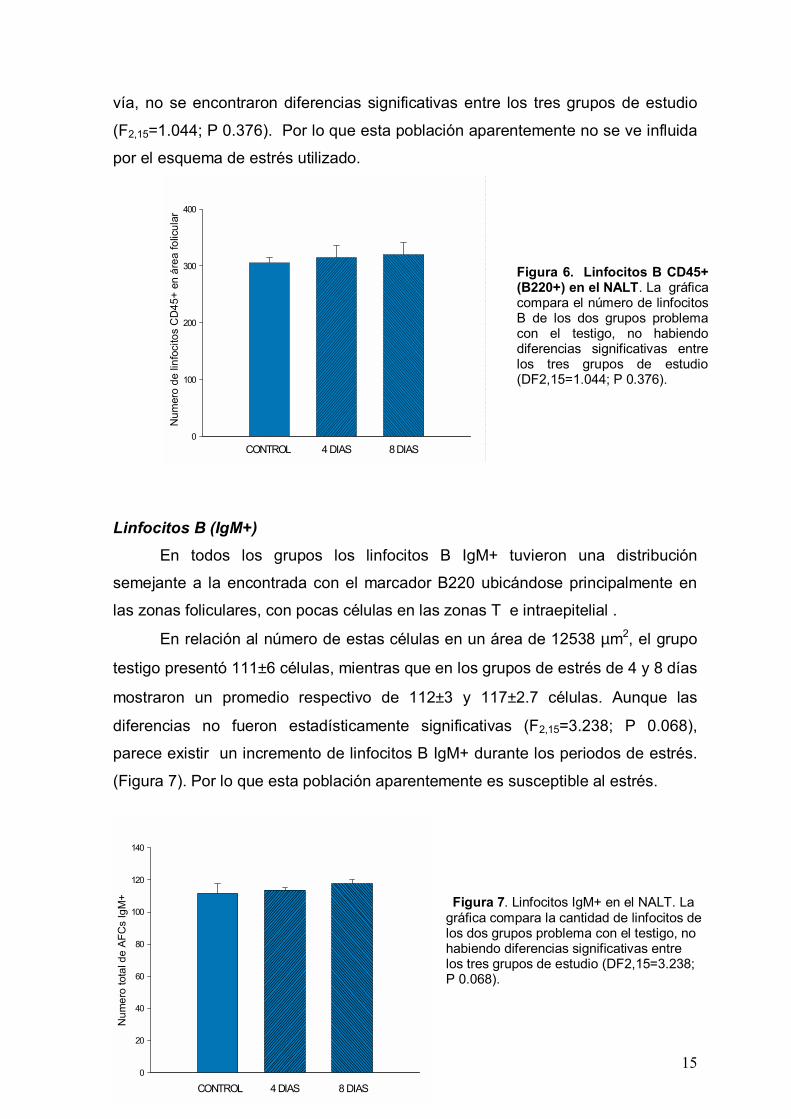
15
vía, no se encontraron diferencias significativas entre los tres grupos de estudio
(F2,15=1.044; P 0.376). Por lo que esta población aparentemente no se ve influida
por el esquema de estrés utilizado.
Linfocitos B (IgM+)
En todos los grupos los linfocitos B IgM+ tuvieron una distribución
semejante a la encontrada con el marcador B220 ubicándose principalmente en
las zonas foliculares, con pocas células en las zonas T e intraepitelial .
En relación al número de estas células en un área de 12538 µm2, el grupo
testigo presentó 111±6 células, mientras que en los grupos de estrés de 4 y 8 días
mostraron un promedio respectivo de 112±3 y 117±2.7 células. Aunque las
diferencias no fueron estadísticamente significativas (F2,15=3.238; P 0.068),
parece existir un incremento de linfocitos B IgM+ durante los periodos de estrés.
(Figura 7). Por lo que esta población aparentemente es susceptible al estrés.
Macrófagos
Ma
Macrófagos
Figura 6. Linfocitos B CD45+ (B220+) en el NALT. La gráfica compara el número de linfocitos B de los dos grupos problema con el testigo, no habiendo diferencias significativas entre los tres grupos de estudio(DF2,15=1.044; P 0.376).
4 DIAS
Num
ero
de li
nfoc
itos
CD
45+
en á
rea
folic
ular
0
100
200
300
400
8 DIASCONTROL
4 DIAS
Num
ero
tota
l de
AFC
s Ig
M+
0
20
40
60
80
100
120
140
CONTROL 8 DIAS
Figura 7. Linfocitos IgM+ en el NALT. La gráfica compara la cantidad de linfocitos de los dos grupos problema con el testigo, no habiendo diferencias significativas entre los tres grupos de estudio (DF2,15=3.238; P 0.068).
16
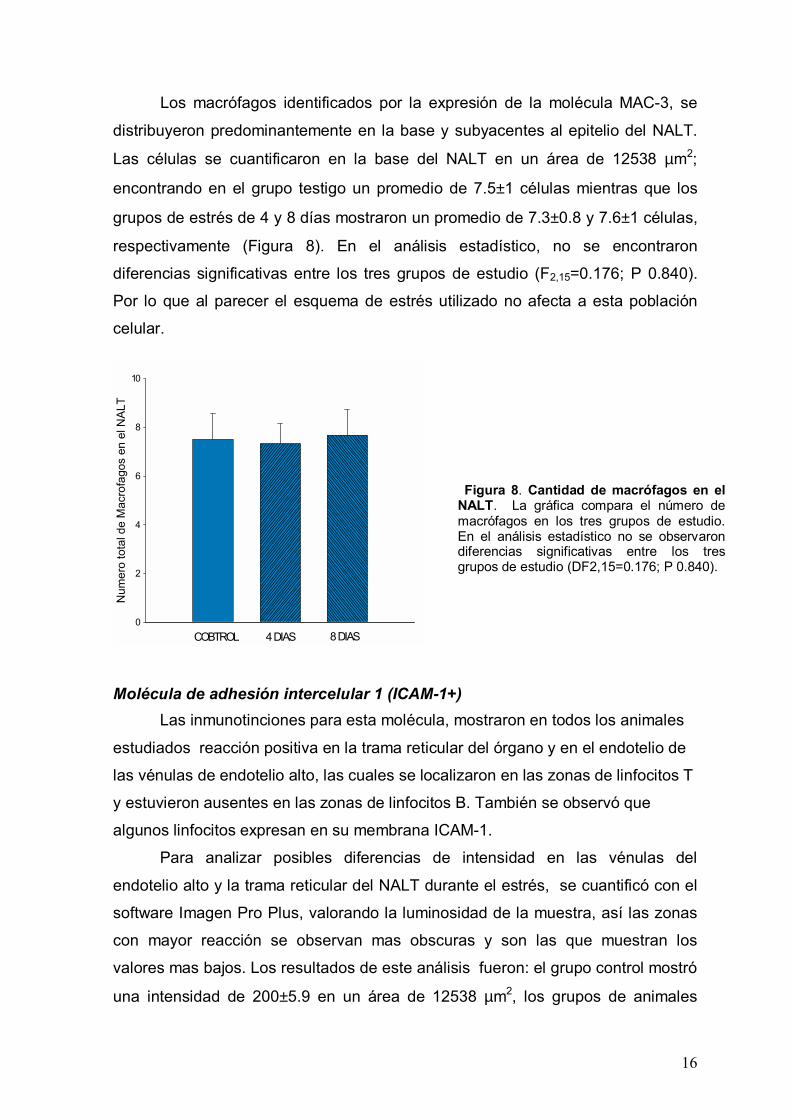
Los macrófagos identificados por la expresión de la molécula MAC-3, se
distribuyeron predominantemente en la base y subyacentes al epitelio del NALT.
Las células se cuantificaron en la base del NALT en un área de 12538 µm2;
encontrando en el grupo testigo un promedio de 7.5±1 células mientras que los
grupos de estrés de 4 y 8 días mostraron un promedio de 7.3±0.8 y 7.6±1 células,
respectivamente (Figura 8). En el análisis estadístico, no se encontraron
diferencias significativas entre los tres grupos de estudio (F2,15=0.176; P 0.840).
Por lo que al parecer el esquema de estrés utilizado no afecta a esta población
celular.
Molécula de adhesión intercelular 1 (ICAM-1+) Las inmunotinciones para esta molécula, mostraron en todos los animales
estudiados reacción positiva en la trama reticular del órgano y en el endotelio de
las vénulas de endotelio alto, las cuales se localizaron en las zonas de linfocitos T
y estuvieron ausentes en las zonas de linfocitos B. También se observó que
algunos linfocitos expresan en su membrana ICAM-1.
Para analizar posibles diferencias de intensidad en las vénulas del
endotelio alto y la trama reticular del NALT durante el estrés, se cuantificó con el
software Imagen Pro Plus, valorando la luminosidad de la muestra, así las zonas
con mayor reacción se observan mas obscuras y son las que muestran los
valores mas bajos. Los resultados de este análisis fueron: el grupo control mostró
una intensidad de 200±5.9 en un área de 12538 µm2, los grupos de animales
Num
ero
tota
l de
Mac
rofa
gos
en e
l NA
LT
0
2
4
6
8
10
COBTROL 4 DIAS 8 DIAS
Figura 8. Cantidad de macrófagos en el NALT. La gráfica compara el número de macrófagos en los tres grupos de estudio. En el análisis estadístico no se observaron diferencias significativas entre los tres grupos de estudio (DF2,15=0.176; P 0.840).

17
estresados durante 4 y 8 días presentaron intensidades de 210±9.8 y 209±8.4
respectivamente. (Figura 9). El análisis estadístico por no mostró diferencias
significativas entre los tres grupos de estudio (F2,15=1.676; P 0.220). Sin embargo
se puede decir que hay tendencia hacia una menor expresión de la molécula en
los animales sometidos a estrés.
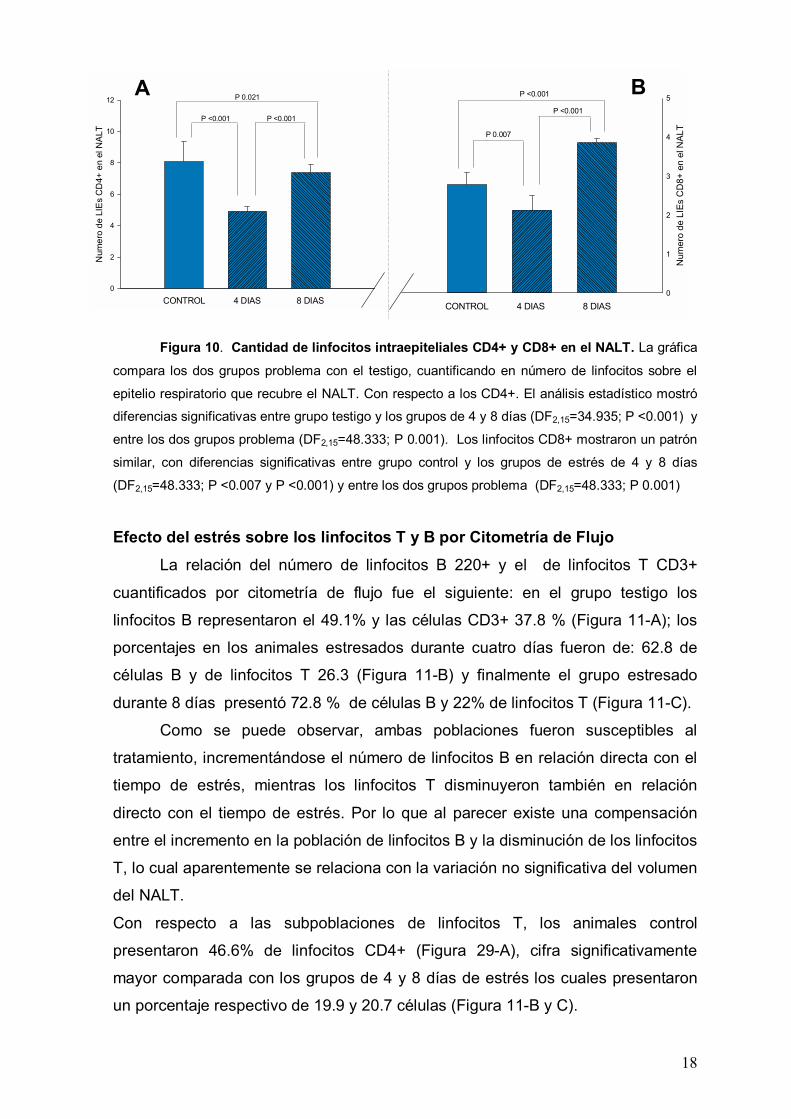
Linfocitos T CD4+ y CD8+ intraepiteliales.
Los linfocitos T intraepiteliales se localizaron preferentemente cerca de la
membrana basal del epitelio el cual presentó características normales en todos
los grupos de estudio. El número de linfocitos intraepiteliales se cuantificó en el
epitelio respiratorio de cortes transversales, a nivel de la porción media del NALT.
La longitud del epitelio en esta zona tuvo un promedio de 780.74±25.3 µm, la cual
se mantuvo constante en los tres grupos de estudio.
En el grupo testigo, el número de linfocitos CD4+ fue de 8.2±0.28 células,
en una constante de 780 micras lineales, mientras que los grupos de estrés de 4
y 8 días presentaron 4.9±0.6 y 7.2±1 células respectivamente (Figura 10-A). En
el análisis estadístico, hubo diferencias significativas entre el grupo testigo con
los grupo de estrés de 4 y 8 días respectivamente (F2,15=34.935; P <0.001 y P
0.021) y entre los dos grupos problema (F2,15=34.935; P <0.001).
Por lo que respecta a los linfocitos CD8+ su número fue menor que el de
los CD4+, los animales testigo presentaron un promedios de 2.7±0.3 células
mientras que los grupos de estrés de 4 y 8 días, tuvieron un promedio respectivo
de 2.2±0.25 y 3.7±0.19 células. (Figura10-B).
En el análisis estadístico, se encontraron diferencias significativas entre el
grupo testigo y los grupos de estrés de 4 y 8 días F2,15=48.333; P <0.007 y P
<0.001) y entre los dos grupos problema (F2,15=48.333; P 0.001).
El número de LIEs CD4+ y CD8+ fue muy pequeño, sin embargo fueron
susceptibles al estrés de 4 días, posiblemente disminuyendo su migración hacia el
epitelio.
18
4 DIAS
Num
ero
de L
IEs
CD
4+ e
n el
NA
LT
0
2
4
6
8
10
12
CONTROL 8 DIAS
P <0.001 P <0.001
P 0.021
Num
ero
de L
IEs
CD
8+ e
n el
NAL
T
0
1
2
3
4
5
CONTROL 8 DIAS4 DIAS
P 0.007
P <0.001
P <0.001
Figura 10. Cantidad de linfocitos intraepiteliales CD4+ y CD8+ en el NALT. La gráfica
compara los dos grupos problema con el testigo, cuantificando en número de linfocitos sobre el
epitelio respiratorio que recubre el NALT. Con respecto a los CD4+. El análisis estadístico mostró
diferencias significativas entre grupo testigo y los grupos de 4 y 8 días (DF2,15=34.935; P <0.001) y
entre los dos grupos problema (DF2,15=48.333; P 0.001). Los linfocitos CD8+ mostraron un patrón
similar, con diferencias significativas entre grupo control y los grupos de estrés de 4 y 8 días
(DF2,15=48.333; P <0.007 y P <0.001) y entre los dos grupos problema (DF2,15=48.333; P 0.001)
Efecto del estrés sobre los linfocitos T y B por Citometría de Flujo
La relación del número de linfocitos B 220+ y el de linfocitos T CD3+
cuantificados por citometría de flujo fue el siguiente: en el grupo testigo los
linfocitos B representaron el 49.1% y las células CD3+ 37.8 % (Figura 11-A); los
porcentajes en los animales estresados durante cuatro días fueron de: 62.8 de
células B y de linfocitos T 26.3 (Figura 11-B) y finalmente el grupo estresado
durante 8 días presentó 72.8 % de células B y 22% de linfocitos T (Figura 11-C).
Como se puede observar, ambas poblaciones fueron susceptibles al
tratamiento, incrementándose el número de linfocitos B en relación directa con el
tiempo de estrés, mientras los linfocitos T disminuyeron también en relación
directo con el tiempo de estrés. Por lo que al parecer existe una compensación
entre el incremento en la población de linfocitos B y la disminución de los linfocitos
T, lo cual aparentemente se relaciona con la variación no significativa del volumen
del NALT.
Con respecto a las subpoblaciones de linfocitos T, los animales control
presentaron 46.6% de linfocitos CD4+ (Figura 29-A), cifra significativamente
mayor comparada con los grupos de 4 y 8 días de estrés los cuales presentaron
un porcentaje respectivo de 19.9 y 20.7 células (Figura 11-B y C).
A B

19
En relación a los linfocitos CD8+, el grupo testigo presentó 8.7% de células,
el grupo estresado durante 4 días 6.9% y el de 8 días 5.7% de linfocitos.
Estos resultados muestran una mayor susceptibilidad de los linfocitos CD4+
al estrés, mientras que los CD8+, mostraron una mayor resistencia al estrés.
A B C
A B C
Figura 11. Cantidad de linfocitos B y T en el NALT por citometría de flujo. Se observa
que en relación al grupo control, los animales estresados presentaron un aumento de los linfocitos
B, por el contrario los linfocitos CD3+ mostraron una disminución.
DISCUSION
El estrés modificó la cantidad de linfocitos T del NALT
Los resultados de la cuantificación morfológica y por citometría de flujo,
mostró que los CD4+ son susceptibles a los efectos del estrés por inmovilización.
En los animales estresados el número de linfocitos T CD3+ disminuyó
significativamente tanto en el área parafolicular como la folicular; en cambio los
CD4+ solo disminuyeron de forma significativa en el área parafolicular, mientras
que el número de CD8+ tampoco mostró diferencias significativas.
Comparando los resultados del presente estudio con la literatura, el
predominio de células CD4+ encontrado concuerda con varios estudios que
reportan que la cantidad de linfocitos T CD4+ en el NALT murino, es
aproximadamente cuatro veces mayor que los linfocitos T CD8+ (Asanuma H y
cols 1997, Bienenstock J y McDermott MR 2005, Heritage PL y cols 1997,
Heritage PL y cols 1998)

20
Se sabe que el timo posee 4 poblaciones de timocitos, definidos como
positivos simples CD4+ o CD8+, doble negativos o doble positivos (Smith L 1987)
y que algunas de estas células inmaduras, son exportadas desde el timo hacia
otros órganos linfoides secundarios como el bazo y nódulos linfoides (Antonica A
y cols 1996) por lo que es posible que el NALT también sea un órgano de llegada
de estas poblaciones celulares. Analizando en conjunto el numero de CD4+ y
CD8+, esto podría deberse a la presencia de linfocitos doble positivos (CD4+
CD8+) provenientes del timo.
La disminución de los linfocitos CD3+ causada por el estrés, coincide con lo
reportado por Steplewski y Vogel quienes aplicaron en ratas, un esquema de
estrés por inmovilización de 3 horas por 11 días observando que los animales
presentaron una disminución significativa de la cantidad de linfocitos T
supresores, en donde aparentemente el factor que influyó, fue la secreción de
glucocorticoides durante el periodo de estrés. (Steplewski Z y Vogel WH 1986).
Está reportado que pacientes asmáticos, a los que se les aplicaron
glucocorticoides sistémicos, presentaron 3 horas después una disminución en el
número de linfocitos circulantes CD3+CD4+, mientras que los CD3+CD8+ no
sufrieron modificaciones (Karagiannidis C y cols 2005). También el estrés no
controlado en ratas, provocó la disminución del número de linfocitos CD4+ y de la
proporción de CD4+/CD8+, así como un incremento de la población de linfocitos
CD8+ en el timo, bazo y sangre periférica (Nakata A y cols 1996).
Pero además, se encontró que sujetos sometidos a entrenamiento físico
moderado durante 4 semanas, presentaron aumento de los linfocitos CD4+
circulantes, así como un aumento en la excreción renal de cortisol y de la tasa
adrenalina/noradrenalina urinaria (Makras P y cols 2005) y que la adrenalina a
dosis fisiológicas, ocasiona disminución de la relación CD4+/CD8+ en sangre
periférica a los 60 minutos de aplicado el fármaco (Crary B y cols 1983). Por lo
que, tanto los glucocorticoides como las catecolaminas secretadas durante el
estrés pudieron ocasionar que las células linfoides fueran movilizadas desde el
NALT hacia la sangre, lo que explicaría el decremento celular local.
Como ya se explico, estos cambios en la movilización celular, a su vez
dependerían de modificaciones en la expresión de moléculas de adhesión y/o de

21
sus ligandos que permiten el alojamiento y la organización de los linfocitos en los
órganos linfoides, entre ellos el NALT. (Csencsits K y cols 1999, Xu B y cols 2003)
Además de la movilización, la apoptosis seria otra causa de la disminución
de CD3+ en el NALT, dado su incremento ocasionado por el aumento de
glucocorticoides durante el estrés. Se sabe que la señalización a través de los
receptores de estas hormonas, actúa como un potente inductor de apoptosis en
células linfoides (Ashwell JD y cols 2000, Cohen JJ 1992, Erlacher M y cols 2005)
y que por el contrario, la aplicación de antagonistas de sus receptores como (RU-
486), inhiben de manera dosis dependiente, el proceso en la placa de Peyer de
rata (Sudo N y cols 2001).
Al contrario de los linfocitos CD4+, La cantidad de CD8+ permaneció
prácticamente sin modificaciones en los tres grupos de estudio, lo que puede
indicar que esta población en el NALT es más resistente a los efectos del estrés o
sus hormonas, aunque un trabajo realizado en humanos menciona que el
incremento de glucocorticoides y catecolaminas después de un periodo de estrés
agudo, provoca leucocitosis moderada, la cual se debe en parte a un incremento
en la cantidad de linfocitos T CD8+ en sangre periférica. (Hennig y cols 2000,
2001), mientras que estudios en ratas, han mostrado que después de dos días de
confrontación social, los animales subdominantes, muestran un decremento tanto
en la población de CD4+ como de CD8+ en sangre periférica (Stefanski V y
Engler H 1998, Stefanski V 2001); efectos que fueron revertidos después de
realizar adrenalectomía (Engler H y cols 2004).
El estrés modificó la cantidad de linfocitos B del NALT
Los resultados obtenidos al contabilizar por área de tejido los linfocitos B
identificados por la expresión de IgM o la molécula B220, no mostraron
modificaciones significativas. Sin embargo, la cuantificación de la población
B220+ por citometría de flujo mostró un incremento en ambos grupos de animales
estresados. Aunque el conteo celular se realizó únicamente en áreas constantes
de 25050 µm2 a nivel de la porción media del NALT, es posible que ocurran
diferencias significativas si se contabiliza en el resto del órgano. Por otro lado, el
análisis por citometría, es más preciso.

22
Por lo que respecta a la proporción de linfocitos B220+/CD3+ la encontrada
en el presente trabajo fue de 1.3/1, ésta es ligeramente mayor a la reportada por
Asanuma y cols, pero esta diferencia puede explicarse por el número total de
células analizadas, ya que estos autores trabajaron con un total de 300,000
células (Asanuma H y cols, 1997), en cambio en el presente estudio, se trabajo
con un millón de células.
Con respecto al resultado por citometría, el incremento de linfocitos B
podría deberse a la modulación que ejerce el SNA-simpático por medio de los
receptores β-adrenérgicos. Está reportado que el estrés agudo moderado o la
aplicación de catecolaminas, ejercen un efecto estimulador en la proliferación
normal de linfocitos B en respuesta a mitógenos selectivos de linfocitos, debido al
incremento en el numero de receptores β-adrenérgicos en los linfocitos B, en
cambio el mismo tratamiento ejerció un efecto inhibidor en la proliferación normal
de linfocitos T. (Edgar VA y cols 2003). En contraparte, en otro trabajo realizado
en ratas, se observó que el estrés agudo, ocasionó disminución en el número de
linfocitos B circulantes. (Dhabhar FS y cols 1994). Estos resultados, pueden
significar alteración en los mecanismos de migración celular durante los periodos
de estrés debido a la aplicación de las hormonas relacionadas, lo que provocaría
disminución de linfocitos en sangre periférica por la migración de estas células
hacia los órganos linfoides entre ellos el NALT.
La mediación de los glucocorticoides en el efecto del estrés sobre los
linfocitos B, podría vincularse con los hallazgos de una investigación en ratones,
en la cual, una elevación de la corticosterona plasmática por medio de implantes
de liberación prolongada, provocaron al tercer día una disminución en el número
de células B220+ y células Ig+ en la medula ósea, lo cual correlacionó con una
disminución de 75% de las células B220+ en fase S después de 24 horas,
además de encontrarse gran cantidad de células en apoptosis (Garby Ba y cols
1993). En contraste con los resultados de este trabajo, es posible que los
procesos de apoptosis sean más acentuados con los corticosteroides exógenos,
los cuales se sabe que tienen efectos más potentes que los endógenos, sin
embargo los resultados anteriores, podrían tener una mayor relación con el estrés
crónico.

23
El estrés no modificó la población de macrófagos del NALT Cuantificados por área de tejido, el número total de macrófagos en el NALT
no presentó variaciones en los tres grupos de estudio, ya que el análisis de la
población de macrófagos se realizo únicamente sobre la porción media del NALT,
quedando la posibilidad de que en el resto del NALT, el número de células si se
haya alterado.
Al igual que los linfocitos, los macrófagos se redistribuyen entre los
compartimentos linfoides y la circulación periférica. A pesar de que está
reportado que el estrés disminuye la migración de los linfocitos y macrófagos
hacia los órganos linfoides (Dhabhar FS y McEwen BS 2001), la población de
macrófagos del NALT parece no ser afectada por el estrés por inmovilización. Esa
permanencia de los macrófagos en el NALT podría explicarse por el efecto del
MIF, citocina secretada por los linfocitos activados o por las células de porción
anterior de la glándula pituitaria y que actuaría como una hormona que
contrarresta los efectos de los glucocorticoides. Este factor, evita que los
macrófagos migren a un sitio de inflamación y/o se mantengan enclaustrados en
un sito particular, como los nódulos linfoides (Bucala R 1996, 1998, Calandra T y
Bucala R 1995, 1997). Otro trabajo que muestra que los glucocorticoides son
capaces de modular la expresión de agentes quimiotacticos vinculados con la
migración de los macrófagos Mizobe K y cols (1997), en donde después de
aplicación de peptona proteosa intraperitoneal en un modelo de estrés agudo en
ratones, se suprimió la migración de macrófagos y monocitos desde el bazo
hacia la cavidad peritoneal. Estos resultados se explicaron por la disminución en
la expresión de factores quimiotacticos como MCP-1/JE, los cuales fueron
revertidos con la aplicación de un antagonista del receptor de glucocorticoides
(RU-486) o después de la adrenalectomía.
Así es probable que también en el NALT, las hormonas del estrés modulen
la expresión de moléculas de adhesión en el NALT, impidiendo que los
macrófagos se movilicen.
Al igual que otras células, es probable que los macrófagos y los monocitos,
del NALT sean sensibles a apoptosis, sin embargo en caso de existir tal efecto, es

24
probable que sean más resistentes a los efectos de los glucocorticoides, ya que
no hubo diferencias significativas. (Ottonello L y cols 2005).
El estrés por modificó el número de linfocitos intraepiteliales (LIE) en el
NALT
En este trabajo, la cantidad de linfocitos intraepiteliales únicamente se analizó
en el epitelio que recubre el NALT, a nivel de su porción media. Aunque el número
de LIEs fue muy pequeño, tanto los LIEs CD4+ como los CD8+ disminuyeron
significativamente con el estrés de 4 días y se incrementaron con el estrés de 8.
En la literatura no se encontraron reportes acerca de los linfocitos
intraepiteliales en el NALT, y solo se han realizado estudios en otras zonas de la
mucosa respiratoria como los cornetes, las amígdalas y adenoides en humanos
(Boyaka PN y cols 2000, Okuda M y Pawankar R 1992, 1993) o bien a nivel del
epitelio intestinal en ratones (Guy-Grand D y cols 2003)
Estas células son importantes, porque se encuentran incluidas y dispersas
entre el epitelio del tracto respiratorio, formando parte de la primera línea de
defensa, por su localización estratégica en un sitio de mayor exposición a
antígenos y siendo las primeras células inmunitarias en encontrar patógenos
invasores, (Cheroutre H 2005).
En relación al escaso número de LIE encontrado en el presente estudio,
Sminia T y Kraal G, refieren que los linfocitos son abundantes a nivel del epitelio
intestinal y se encuentran escasos en el tracto respiratorio. (Sminia T y Kraal G
1999). Con respecto a la relación de células intraepiteliales CD4+/ CD8+, se
encontró ser de 2:1, lo cual no concuerda con otros estudios que señalan a los
CD8+ como predominantes en la mucosa respiratoria (Beagley Kw y Husband AJ
1998, Graeme CF y cols 1993, Jahnsen FL y cols 1998, Okuda M y Pawankar R
1992, Pawankar R y Okuda M 1993), en cambio los resultados del presente
trabajo, coinciden con el reporte de Sminia T y Kraal G, en donde la proporción
LIEs CD4+ fue mayor que la de CD8+. Sin embargo, es posible que el número de
estas células sea mayor y pueda variar la proporción CD4+/CD8+, si se
contabilizan en el resto del epitelio que recubre el NALT.
El resultado de disminución de los LIEs con el estrés durante 4 días,
tendría el mismo mecanismo que para otras células, con disminución del tráfico
por efecto de las hormonas relacionadas con el estrés sobre las moléculas de

25
adhesión y sus ligandos, como se ha encontrado en intestino (Erle DJ y Pabst R
2000), o también por el aumento de la apoptosis, por las mismas hormonas
(Murosaki S y cols 1997), ya que la adrenalectomía o el tratamiento de los ratones
con antagonistas de glucocorticoides, incrementó significativamente la sobrevida
de los linfocitos. (Brunner T y cols 2001).
Finalmente, el ligero incremento de células CD4+ y CD8+ en los grupos de
estrés de 8 días, puede deberse a la adaptación de los animales a la
manipulación.
El estrés no modificó la expresión de la molécula ICAM-1 en el NALT
Se sabe que la molécula ICAM-1, se expresa continuamente a nivel del endotelio
vascular alto de los tejidos linfoides secundarios y en células endoteliales
cercanas a los procesos inflamatorios, en donde su principal finalidad es la de
dirigir la diapédesis de los leucocitos (Carlson SL 2001, Muller WA 2003, Shaw
SK y cols 2004).
La expresión de ICAM-1 (CD54), se analizó para determinar su posible relación
con el tráfico de linfocitos desde otros compartimientos hacia el NALT. A pesar
de no encontrar reportes de la expresión de la molécula sobre este órgano, esta
molécula se identifico en las vénulas de endotelio alto y a nivel de la trama
reticular del NALT; el grado de expresión de ICAM-1 sobre estas estructuras, fue
prácticamente igual en los tres grupos de estudio.
Su expresión en el NALT, puede indicar su participación en la
redistribución de las células linfoides, al igual que MAdCAM-1 y PNAd se sugiere
permiten el tráfico, la organización y el alojamiento de los linfocitos en varios
tejidos linfoides secundarios (Csencsits KL 1999). Los resultados indican que
esta molécula no se afecta por el modelo de estrés utilizado y por tanto las
variaciones en la cantidad de linfocitos, puedan deberse a cambios en la
expresión de integrinas leucocitarias (ligandos de ICAM-1), como ha sido
reportado en humanos, en donde el estrés psicológico provocó disminución en la
expresión de antígeno 1 asociado a función de linfocitos (LFA-1), el principal
ligando de ICAM-1, mientras que esta ultima no presentó cambios significativos
(Mills PJ y Dimsdale JE 1996).

26
CONCLUSIONES
1. Se comprueba la hipótesis de influencia del estrés sobre el tejido linfoide
del NALT.
2. El esquema de estrés utilizado, no provocó cambios significativos en el
tamaño y forma del NALT.
3. Las poblaciones celulares que disminuyeron con estrés por inmovilización
de 4 días, fueron los linfocitos T CD3+, CD4+ y CD8+, mismas que
mostraron un incremento con el estrés de 8 días, probablemente debido a
una adaptación.
4. Se sugiere que los efectos del estrés sobre las poblaciones celulares del
NALT, se ejercen a través de la influencia de las hormonas, catecolaminas
y glucocorticoides, por mecanismos como apoptosis o a su redistribución
de las células inmunitarias debido a modificaciones en la expresión de
moléculas de adhesión y/o apoptosis.
IMPACTO Y APORTACIONES DEL PROYECTO.
Ya que la mucosa nasal es una vía de entrada y primer sitio de contacto para
múltiples antígenos tanto aéreos como bucales y siendo el NALT es una
estructura linfoide organizada que se encuentra situada estratégicamente en la
porción del tracto respiratorio alto, que tiene la capacidad de iniciar respuestas
inmunitarias tanto locales como sistémicas las cuales han sido poco estudiadas.
El estrés y su relación con las alteraciones de la respuesta inmunitaria son
conocidas a nivel sistémico; pero es poco lo que se conoce de su relación con el
sistema inmunitario de las mucosas y menos aún a nivel del tejido linfoide de la
nariz (NALT).
Por lo que en nuestro conocimiento, este es el primer trabajo en donde se
investiga los efectos del estrés sobre la estructura del NALT.

27
Nuestros resultados mostraron que el estrés afecta la distribución de las células
tanto de la respuesta inmune celular y humoral así como en la secreción de IgA
principal la inmunoglobulina de las mucosas.
Por lo tanto nuestro trabajo es un antecedente importante para la compresión
de el porqué el estrés hace a los sujetos mas susceptibles a las infecciones
respiratorias.

28
BIBLIOGRAFIA
Asanuma H, Hodson AT, Iwasaki T, Sato Y, Inaba Y, Aizawa C, Kurata T, Tamura S. Isolation and characterization of mouse nasal-associated lymphoid tissue. J Immunol methods. 1997; 202: 123-131. Ashwell JD, Lu FW, Vacchio MS. Glucocorticoids in T cell development and function. Annu Rev Immunol. 2000; 18: 309-345. Balmelli C, Demotz S, Acha-Orbea H, De Grandi P, Nardelli-Haefliger D. Trachea, lung, and thacheobronchial lymph nodes are the major sites where antigen-presenting cells are detected after nasal vaccination of mice with human papillomavirus type 16 virus-like particles. Immunol Rev. 2002; 76: 12596-12602. Beagley KW, Husband AK. Intraepithelial lymphocytes: origins, distribution, and function. Crit Rev Immunol. 1998; 18: 237-254. Bienenstock J, McDermott MR. Bronchus and nasal-associated lymphoid tissues. Immunol Rev. 2005; 206: 22-31. Boyaka PN, Wright PF, Marinaro M, Kiyono H, Johnson JE, Gonzales RA, Ikizler MR, Werkhaven JA, Jackson RJ, Fujihashi K, Di Fabio S, Staats HF, and McGhee JR. Human nasopharyngeal-associated limphoreticular tissues, functional analysis of subepithelial and intraepithelial B and T cells from adenoids and tonsils. Am J Pathol. 2000; 157: 2023-2035. Brunner T, Arnold D, Wasem C, Herren S, Frutschi C. Regulation of cell death and survival in intestinal intraepithelial lymphocytes. Cell Death Differ. 2001; 8: 706-714.
Bucala R. MIF re-discovered: pituitary hormone and glucocorticoid-induced regulator of cytokine producction. Cytokine growth factor rev. 1996; 7:19-24. Bucala R. Neuroimmunomodulation by macrophage migration inhibitory factor (MIF). Ann N Y Acad Sci. 1998; 1: 78-82. Calandra T, Bucala R. Macrophage migration inhibitory factor: a counter-regulator of glucocorticoid action and critical mediator of septic shock. J Inflamm. 1995; 47: 39-51. Calandra T, Bucala R. Macrophage migration inhibitory factor (MIF): a glucocorticoid counter-regulator within the immune system. Crit Rev Immunol. 1997;17: 77-88. Carlson SL. Neural influences on cell adhesion mollecules and lymphocyte trafficking. Phychoneuroimmunology. 2001; 1: 231-237. Cheroutre H. IELs: enforcing law and order in the court of the intestinal epithelium. Immunol Rev. 2005; 206: 114-131.

29
Cohen JJ. Glucocorticoid-induced apoptosis in the thymus. Semin Immunol. 1992; 4: 363-369 Crary B, Hauser SL, Borysenko M, Kutz I, Hoban C, Ault KA, Wqeiner HL, Benson H. Epinephrine-induced changes in the distribution of lymphocyte subsets in peripheral blood of humans. J Immunol. 1983; 131: 1178-1181 Csencsits KL, Jutila MA, Pascual DW. Nasal associated lymphoid tissue: phenotypic and functional evidence for the primary role of peripheral node addresin in naive lymphocyte adhesion to high endothelial venules in a mucosal site. J Immunol. 1999; 163: 1382-1389. Dhabhar FS, Miller AH, Stein M, Mcewen BS, Spencer RL. Diurnal and acute stress-induced changes in the distribution of peripheral blood leukocytes subpoblations. Brain Behav Immun. 1994; 8: 66-79. Dhabhar FS y McEwen BS. Bidirectional effects of stress and Glucocorticoid Hormones on Immune function: Possible explanations for Paradoxical observations. Psychoneuroimmunology. 2001; 1: 301-330. Edgar VA, Silverman DM, Cremaschi GA, Zieher LM, Genaro AM. Altered lymphocyte catecholamine reactivity in mice subjected to chronic mild stress. Biochem Pharmacol. 2003; 65: 15-23. Engler H, Dawils L, Hoves S, Kurth S, Stevenson JR, Schauenstein K, Stefanski V. Effects of social stress on blood leukocyte distribution: the role of alpha- and beta-adrenergic mechanisms. J Neuroimmunol. 2004; 156: 153-162. Erlacher M, Knoflach M, Stec IE, Bock G, Wick G, Wiegers GJ. TCR signaling inhibits glucocorticoid-induced apoptosis in murine thymocytes depending on the stage of development. Eur J Immunol. 2005; 35: 3287-3296. Erle DJ, Pabst R. Intraepitelial lymphocytes in the lung; A Neglected lymphocyte population. Am. J. Respir. Cell Mol biol. 2000; 22: 398�400. Garby Ba, King LE, Telford WG, Morford LA, Fraker PJ. Chronic elevation of plasma corticosterone causes reductions in the number of cycling cells of the B lineage in murine bone marrow and induces apoptosis. Immunology. 1993; 80: 587-592. Graeme-Cook F, Bhan AK, Harris NL. Immunohistochemical characterization of intraepithelial and subepithelial mononuclear cells of the upper airways. Am J Pathol. 1993; 143: 1416-1422. Guy-Grand D, Azogui O, Celli S, Darche S, Nussenzweid MC, Kourilsky P, Vassalli P. Extrathymic T Cell Lymphopoiesis : Ontogeny and Contribution to Gut Intraepithelial Lymphocytes in Athymic and Euthymic Mice. J Exp Med. 2003; 197: 333-341

30
Guyton AC, May J. Tratado de fisiología Medica. 10ª ed. México. McGraw-Hill-Interamericana; 2001. Harmsen A, Kusser K, Hartson L, Tighe T, Sunshine MJ, Sedwick JD, Choi Y, Littman DR, Randall TD. Cutting edge: Organogenesis of nasal-associated lympoid tissue (NALT) occurs independently of limphotoxin-α (LTα) and retinoic acid receptor-related orphan receptor-γ, but the organization of NALT is LTα dependent. J Immunol. 2002; 168: 986-990.
Hennig J, Netter P, Voigt KH. Mechanisms of changes in lymphocyte numbers after psychological stress. Z Rheumatol. 2000; 59: 43-48 Hennig J, Netter P, Voigt KH. Cortisol mediates redistribution of CD8+ but not of CD56+ cells after the psychological stress of public speaking. Psychoneuroendocrinology. 2001; 26: 673-687. Heritage PL, Underdown BJ, Arsenault AL, Snider DP,. McDermott MR. Comparison of murine nasal-associated lymphoid tissue and Peyer�s patches. Am J Respir Crit Care Med. 1997; 156: 1256-1262. Heritage PL, Brook MA, Underdown BJ, McDermott. Intranasal immunization with polymer-grafted microparticles activates the nasal-associated lymphoid tissue and draining lymph nodes. Immunology. 1998; 93: 249-56. Jahnsen FL, Farstad IN, Aanesen JP, Brandtzaeg P. Phenotypic Distribution of T Cells in Human Nasal Mucosa Differs from That in the Gut. Am J Respir Cell Mol Biol. 1998; 18: 392-401 Karagiannidis C, Ruckert B, Hense G, Willer G, Menz G, Blazer K, Schmidt-Weber CB. Distinct leucocyte redistribution after glucocorticoid treatment among difficult-to-treat asthmatic patients. Scand J Immunol. 2005; 61: 187-196. Kiyono H, Fukuyama S. NALT-versus peyer�s-patch-mediated mucosal immunity. Nat Rev Immunol. 2004; 4: 699-710. Liang B, Hyland L, Hou S. Nasal-associated lymphoid tissue is a site of long-term virus-specific antibody production following respiratory virus infecction of mice. J virol. 2001; 75: 5416-5420. Makras P, Koukoulis GN, Bourikas G, Papatheodorou G, Bedevis K, Menounos P, Pappas D, Kartalis G. Effect of 4 weeks of basic military training on peripheral blood leucocytes and urinary excretion of catecholamines and cortisol. J Sports Sci. 2005; 23: 825-834. McEwen BS. Protective and damaging effects of stress mediators. New England Journal Med 1998: 338; 171�179.

31
Mebius RE. Organogenesis of lymphoid tissues. Nat rev Immunol. 2003: 3; 292-303. Mills PJ, Dimsdale JE. The effects of acute psychologic stress on cellular adhesion molecules. J Psychosom Res. 1996; 41: 49-53. Mizobe K, Kishihara K, Ezz-Din El-Naggar R, Madkour GA, Kubo C, Nomoto K. Retraint stress-induced elevation of endogenous glucocorticoid supresses migration of granulocytes and macrophages to an inflammatory locus. J Neuroimmunol. 1997; 73: 81-89. Muller WA. Leukocyte-endothelial-cell interactions in leukocyte transmigration and the inflammatory response. Trends Immunol. 2003; 24: 327-334. Musosaki S, Inagaki-Ohara K, Kusaka H, Ikeda H, Yoshikai Y. Apoptosis of intestinal intraepithelial lymphocytes induced by exogenous and endogenous glucocorticoids. Microbiol Immunol. 1997; 41: 139-148.
Nakata A, Arari S, Tanigawa T, Sukurai S, Yokoyama M, Effects of uncontrollable and controllable electric shocks on T lymphocyte subpopulations in the peripheral blood, spleen, and thymus of rats. Neuroimmunomodulation. 1996; 3: 336-341. Okuda M, Pawankar R. Flow cytometric analysis of intraepithelial lymphocytes in the human nasal mucosa. Allergy. 1992; 47: 255-259. Ottonelo L, Bertolotto M, Montecucco F, Dapino P, Dallegri F. Dexametasone-induced apoptosis of human monocytes exposed to immune complexes. Intervention of CD95 and XIAP-dependent pathways. Int J Immunopathol Pharmacol. 2005; 18: 403-415. Paca´k K. and Palkovits M. Stressor specificity of central neuroendocrine responses: implications for stress-related disorders. Endocr Rev. 2001: 4; 502�548 Pawankar R, Okuda M. A comparative study of the characteristics of intraepithelial and lámina propria lymphocytes of the human nasal mucosa. Allergy.1993; 48: 99-105. Rojas-Hernández S, Rodríguez-Monroy MA, López-Revilla R, Resendiz-Albor A. A, Moreno-Fierros L. Intranasal coadministration of the cry1Ac protoxin with amoebal lysates increases protection against naegleria fowleri meningoencephalitis. Infec Immun. 2004; 72: 4368-4375. Shaw SK, Ma S, Kim MB, Rao RM, Hartman CH. U, Froio RM, Yang L, Jones T, Liu Y, Nusrat A, Parkos CH. A, Luscinskas FW. Coordinated Redistribution of Leukocyte LFA-1 and Endothelial Cell ICAM-1 Accompany

32
Sminia T, Kraal G. Nassal-associated lymphoid tissue. Mucosal immunology. 1999; chapter 21: 357-364. Smith L. CD4+ murine T cells develop from CD8+ precursors in vivo. Nature. 1987; 326: 798-800. Steplewski Z, Vogel WH. Total leukocytes, T cell subpopulation and natural killer (NK) cell activity in rats exposed to restraint stress. Life Sci. 1986; 38: 2419-2427 Stefanski V, Engler H. Effects of acute and chronic social stress on blood cellular immunity in rats. Physion Behav. 1998; 64: 733-741. Sudo N, Oyama N, Yu XN, Kubo C. Restaint stress-induced elevation of endogenous glucocorticoids decreases Peyer´s patch cell numbers via mechanisms that are either dependent or independent on apoptotic cell death. Neuroimmunomodulation. 2001; 9: 333-339 Tamura S, Iwasaki T, Thompson AH, Asanuma H, Chen Z, Susuki Y, Aisawa C, Kurata T. Antibody-forming cells in the nasal-associated lymphoid tissue during primary influenza virus infection. J Gen Virol. 1998; 79: 291-299.

33


















