
Ecofisiología de la tuna (Opuntia ficus-indica (L.) Mill.) · desde 1990, especialmente en Túnez...
16
Lozano, L. - Ecofisiología de la tuna (Opuntia ficus-indica (L.) Mill.) Disponible en www.horticulturaar.com.ar Avances en Horticultura - Review Ecofisiología de la tuna (Opuntia ficus-indica (L.) Mill.) L. Lozano Universidad Nacional de Salta, Cátedra de Horticultura. Av. Bolivia 5150 (4400) Salta. Recibido: 19/4/10 Aceptado: 13/9/11 Resumen Lozano, L. 2011. Ecofisiología de la tuna (Opuntia ficus-indica (L.) Mill.). Horticultura Argentina 30(72): 37-52. El uso hortícola de la tuna (Opuntia ficus-indica (L.) Mill.) no está difundido en Argentina, en general, ni en la provincia de Salta, en particular. Los cladodios jóvenes de esta Cactácea son ricos en Ca y fibra y, además de sus cualidades nutricionales, poseen propie- dades medicinales que ayudan a controlar la obesidad, la hiper- glucemia, el colesterol, etc. Pueden ser consumidos crudos en en- saladas, pero la mayoría de las personas los prefieren cocidos y combinados con otros alimentos como huevos, pimientos, cama- rones, etc. y hasta en salsas. En la presente revisión bibliográfica se consideran aspectos morfológicos y ecofisiológicos de las opun- tias, responsables de su adaptación ecológica y se analizan los fac- tores principales que afectan la toma de CO 2 y la acumulación re- sultante de la biomasa de las opuntias: el contenido de agua del suelo, la temperatura del aire, la luz y varios elementos del suelo. La gran plasticidad de las opuntias a las distintas condiciones de clima y suelo, otorgadas por sus características morfológicas y fi- siológicas, deberían incentivar su cultivo para la producción de jóvenes brotes destinados al consumo humano en zonas margina- les de la provincia de Salta, como los valles Calchaquíes y la Puna, introduciendo entre los pequeños productores las técnicas de su cultivo, de la cosecha y de la preparación de comidas de alto valor nutritivo. Palabras claves adicionales: opuntias, morfología, fisiología, cla- dodios jóvenes, adaptación. Abstract Lozano, L. 2011. Cactus pear ecophysiology. Horticultura Argen- tina 30(72): 37-52. The horticultural use of cactus pear (Opuntia ficus-indica (L.) Mill.) has not been spread in Argentina. The young cladodes are rich in calcium (Ca) and fibers; and besides their nutritional qua- lities they have medicinal properties that help to control obesity, hyperglycemia in blood, cholesterol, etc. They can also be consu- med raw in salads, but most people prefer them stewed and com- bined with other foods like eggs, peppers, shrimps, etc. and even in sauces. In this bibliographical review, morphologic and physio- logical aspects of the opuntias are considered in their ecological adjustment. The main factors that affect the uptake of carbon dio- xide (CO 2 ) and the accumulation of biomass are analyzed. They are: water content in the soil, air temperature, light and soil ele- ments. The great adjustment of the opuntias to different climatic and soil conditions, granted by their morphologic and physiologi- cal characteristics should stimulate their culture with commercial aims, especially at the Calchaqui valley and in the Puna, helping the farmers with the culture and harvest techniques, and food pre- parations with a high nutritional value. Additional keywords: Opuntia, morphology, ecophisiology, young cladodes, adaptation. Horticultura Argentina 30(72): May.-Ago. 2011 37 ISSN de la edición on line 1851-9342 1. Introducción La presente revisión bibliográfica tiene como ob- jetivo analizar los factores ambientales que inciden en la toma del CO 2 y en la acumulación de la biomasa de cladodios destinados a la producción de nopalitos cu- yo consumo como hortaliza está muy difundido en México. La provincia de Salta presenta características eda- foclimáticas adecuadas para el cultivo de las opuntias con fines hortícolas por lo que, desde el año 2006, la Cátedra de Horticultura de la Universidad Nacional de Salta trabaja en este tema. La provincia de Salta, ubicada en el Noroeste Ar- gentino, presenta dos zonas que se adecuarían al cul- tivo de la tuna para la producción de fruta, para la pro- ducción de brotes para la alimentación humana y para la producción de forraje, que corresponden a la zona de los Valles Calchaquíes y la Puna, que a continua- ción describimos brevemente. 1.1 Valles Calchaquíes Los Valles Calchaquíes, en la provincia de Salta, se ubican entre los 24° 30’ y 26° 30’ de latitud S y 66° 20’ de longitud O; su altitud varía entre los 1.680 m en Ca- fayate y 3.015 m en La Poma. Políticamente están in- tegrados por los departamentos La Poma, Cachi, Mo- linos, San Carlos y Cafayate (Paoli et al., 2002). La topografía de la zona de estudio es montañosa, sólo entre un 2 y 3 % de la superficie es utilizable con fines agrícolas. La precipitación anual varía entre los 95 mm en Sitio Argentino de Producción Animal 1 de 16
Transcript of Ecofisiología de la tuna (Opuntia ficus-indica (L.) Mill.) · desde 1990, especialmente en Túnez...

Lozano, L. - Ecofisiología de la tuna (Opuntia ficus-indica (L.) Mill.)
Disponible en www.horticulturaar.com.ar
Avances en Horticultura - Review
Ecofisiología de la tuna (Opuntia ficus-indica (L.) Mill.)
L. Lozano
Universidad Nacional de Salta, Cátedra de Horticultura. Av. Bolivia 5150 (4400) Salta.
Recibido: 19/4/10 Aceptado: 13/9/11
Resumen
Lozano, L. 2011. Ecofisiología de la tuna (Opuntia ficus-indica
(L.) Mill.). Horticultura Argentina 30(72): 37-52.
El uso hortícola de la tuna (Opuntia ficus-indica (L.) Mill.) no estádifundido en Argentina, en general, ni en la provincia de Salta, enparticular. Los cladodios jóvenes de esta Cactácea son ricos en Cay fibra y, además de sus cualidades nutricionales, poseen propie-dades medicinales que ayudan a controlar la obesidad, la hiper-glucemia, el colesterol, etc. Pueden ser consumidos crudos en en-saladas, pero la mayoría de las personas los prefieren cocidos ycombinados con otros alimentos como huevos, pimientos, cama-rones, etc. y hasta en salsas. En la presente revisión bibliográficase consideran aspectos morfológicos y ecofisiológicos de las opun-tias, responsables de su adaptación ecológica y se analizan los fac-
tores principales que afectan la toma de CO2 y la acumulación re-sultante de la biomasa de las opuntias: el contenido de agua delsuelo, la temperatura del aire, la luz y varios elementos del suelo.La gran plasticidad de las opuntias a las distintas condiciones declima y suelo, otorgadas por sus características morfológicas y fi-siológicas, deberían incentivar su cultivo para la producción dejóvenes brotes destinados al consumo humano en zonas margina-les de la provincia de Salta, como los valles Calchaquíes y la Puna,introduciendo entre los pequeños productores las técnicas de sucultivo, de la cosecha y de la preparación de comidas de alto valornutritivo.
Palabras claves adicionales: opuntias, morfología, fisiología, cla-dodios jóvenes, adaptación.
Abstract
Lozano, L. 2011. Cactus pear ecophysiology. Horticultura Argen-tina 30(72): 37-52.
The horticultural use of cactus pear (Opuntia ficus-indica (L.)Mill.) has not been spread in Argentina. The young cladodes arerich in calcium (Ca) and fibers; and besides their nutritional qua-lities they have medicinal properties that help to control obesity,hyperglycemia in blood, cholesterol, etc. They can also be consu-med raw in salads, but most people prefer them stewed and com-bined with other foods like eggs, peppers, shrimps, etc. and evenin sauces. In this bibliographical review, morphologic and physio-logical aspects of the opuntias are considered in their ecological
adjustment. The main factors that affect the uptake of carbon dio-xide (CO2) and the accumulation of biomass are analyzed. Theyare: water content in the soil, air temperature, light and soil ele-ments. The great adjustment of the opuntias to different climaticand soil conditions, granted by their morphologic and physiologi-cal characteristics should stimulate their culture with commercialaims, especially at the Calchaqui valley and in the Puna, helpingthe farmers with the culture and harvest techniques, and food pre-parations with a high nutritional value.
Additional keywords: Opuntia, morphology, ecophisiology,young cladodes, adaptation.
Horticultura Argentina 30(72): May.-Ago. 2011 37ISSN de la edición on line 1851-9342
1. Introducción
La presente revisión bibliográfica tiene como ob-jetivo analizar los factores ambientales que inciden enla toma del CO2 y en la acumulación de la biomasa decladodios destinados a la producción de nopalitos cu-yo consumo como hortaliza está muy difundido enMéxico.
La provincia de Salta presenta características eda-foclimáticas adecuadas para el cultivo de las opuntiascon fines hortícolas por lo que, desde el año 2006, laCátedra de Horticultura de la Universidad Nacional deSalta trabaja en este tema.
La provincia de Salta, ubicada en el Noroeste Ar-gentino, presenta dos zonas que se adecuarían al cul-tivo de la tuna para la producción de fruta, para la pro-
ducción de brotes para la alimentación humana y parala producción de forraje, que corresponden a la zonade los Valles Calchaquíes y la Puna, que a continua-ción describimos brevemente.
1.1 Valles CalchaquíesLos Valles Calchaquíes, en la provincia de Salta, se
ubican entre los 24° 30’ y 26° 30’ de latitud S y 66° 20’de longitud O; su altitud varía entre los 1.680 m en Ca-fayate y 3.015 m en La Poma. Políticamente están in-tegrados por los departamentos La Poma, Cachi, Mo-linos, San Carlos y Cafayate (Paoli et al., 2002).
La topografía de la zona de estudio es montañosa,sólo entre un 2 y 3 % de la superficie es utilizable confines agrícolas.
La precipitación anual varía entre los 95 mm en
Sitio Argentino de Producción Animal
1 de 16

Molinos, hasta los 200 mm en Cafayate; se producedurante los meses de verano (noviembre-marzo) perí-odo en que precipita el 80 a 85 % del total anual re-gistrado.
Las temperaturas máximas y mínimas absolutasmuestran que el clima de la región es templado paralas condiciones del valle, con notables variaciones tér-micas del aire en serranías y áreas elevadas (Paoli et
al., 2002).
1.2 PunaLa Puna constituye la prolongación austral del al-
tiplano peruano-boliviano en una faja de orientaciónnorte-sur. Limita al este con la Cordillera Oriental ylas Sierras Pampeanas (que la cierran también al sur)y al oeste con el límite político Argentina-Chile.
En la provincia de Salta, la Puna está integrada casien su totalidad por el departamento de Los Andes yparte de los departamentos de Iruya y Santa Victoriay tiene una superficie de 29.000 km2 (Paoli et al.,2002).
El clima de la Puna definido como árido, presentacaracterísticas de continentalidad debido a la presenciade la Cordillera Oriental, barrera natural que la limitalateralmente. La extrema sequedad del aire, la bajatemperatura, los pronunciados cambios de ésta entreel día y la noche, así como los fuertes vientos, hacenque la vida tanto animal como vegetal sea muy exigua.(Paoli et al., 2002).
La Puna salteña se ubica en la región “desértica”,donde las precipitaciones alcanzan sus más bajos re-gistros regionales (menores a 50 mm anuales), es laPuna de los salares que ocupa los fondos de los vallesy bolsones y también recibe el nombre de “desierto deescombros”.
En general, son muy escasos los días de lluvia enla Puna salteña y durante el invierno no se producenprecipitaciones en ninguna localidad. Aún en la épocade ocurrencia de lluvias, cuyo mes representativo esenero, el número de días con precipitaciones alcanzasólo a un día en la localidad de La Casualidad, ubicadaen la Puna desértica. Anualmente se contabiliza 6 díasde lluvias como valor medio anual (Paoli et al., 2002).
El régimen térmico depende, entre otros factores,de la latitud y principalmente de la altitud, que ejerceuna marcada influencia en la conformación regionalen el campo de las temperaturas locales.
La marcha anual de la temperatura media perteneceal denominado tipo “continental”, con el máximo enel mes de enero y el mínimo en julio. En las localida-des situadas a altitudes superiores a los 4.000 m.s.n.m.el máximo se desplaza al mes de febrero (Paoli et al.,2002).
2. Plantas CAM
La mayoría de las plantas abren sus estomas alamanecer y es cuando comienzan a tomar el CO2 dela atmósfera, el cual se incorpora a varios productosde la fotosíntesis que es un proceso que ocurre sola-mente en la luz. La apertura diurna de los estomas con-duce a una pérdida inevitable de agua desde dentro delas hojas y los tallos meristemáticos. En las opuntiasy otros cactus, el patrón de toma del dióxido de Car-bono (CO2) y pérdida de agua ocurren principalmenteen la noche, cuando la temperatura es más baja y lahumedad es más alta, lo que reduce la pérdida de agua.(Nobel, 1999).
La apertura de los estomas durante la noche signi-fica que la toma de CO2 ocurre en la oscuridad, lo cuales una extraña situación reconocida inicialmente aprincipios del siglo XIX. En 1813, Benjamín Haeynemordió una planta suculenta de Kalanchoe pinnata dela familia Crassulaceae durante varias veces al día, porla mañana las hojas tenían un sabor ácido que se re-ducía gradualmente durante el resto del día. Antes deeso, en 1804, Nicholas Théodore de Saussure habíadeducido que las opuntias toman su CO2 durante lanoche. Estas observaciones se relacionan con un pa-trón de intercambio de gases hoy conocido como elmetabolismo Ácido Crasuláceo (CAM). Las plantasCAM tienden a ser nativas de regiones áridas y semiá-ridas o de microhábitats que son secos periódicamenteincluyendo playas, sitios rocosos y lugares tropicalesdonde cactus epífitos crecen en árboles (Nobel, 1999).
Las plantas CAM comenzaron a ser usadas en Mé-xico por las antiguas civilizaciones mesoamericanas.Varias especies de Agave, Opuntia y Stenocereus for-maron parte de la dieta de tribus indígenas que habi-taron tierras áridas templadas, subtropicales y tropi-cales. Evidencias en excrementos fosilizados humanoshan mostrado que tanto ágaves como cactus fueronconsumidos por humanos desde hace al menos 9.000años (Nobel et al., 2002). Los jóvenes tallos eran co-cinados y mezclados con carnes de animales salvajes,como ciervos, armadillos, conejos y ratas y fueroncombinados con cebollas, tomates, chiles y semillasde zapallos, maníes y cacao. Los frutos eran consumi-dos frescos o secos al sol antes de ser almacenados pa-ra las épocas en que escaseaba la comida. Los frutosfueron también procesados para obtener diferentesproductos como miel, mermeladas y caramelos. Losconquistadores españoles, primero, y luego las pobla-ciones mestizas y criollas, adoptaron los tallos comocamuflaje protector (Nobel et al., 2002).
Las opuntias fueron introducidas en Europa desdeAmerica a principios de 1500, primero a España y más
Lozano, L. - Ecofisiología de la tuna (Opuntia ficus-indica (L.) Mill.)
Horticultura Argentina 30(72): May.-Ago. 201138 ISSN de la edición on line 1851-9342
Sitio Argentino de Producción Animal
2 de 16

tarde a otros países de climas templados de la regiónmediterránea. Los moros las llevaron desde España aÁfrica a fines de 1500. El cultivo de las opuntias al-canzó un desarrollo espectacular en la isla de Sicilia yallí comenzaron a cultivarse 100 años antes de las pri-meras plantaciones en México.
Jóvenes industriales suecos, de la industria del cue-ro, luego, introdujeron las opuntias desde Californiaal semiárido del nordeste brasileño en respuesta a lasevera sequía que afectó al ganado. El cultivo de lasopuntias proveyó forraje para el ganado, y así benefi-ció a la industria del cuero. Las opuntias han comen-zado a ser un importante cultivo para el nordeste bra-sileño, donde 400.000 ha han sido cultivadas para fo-rraje del ganado (Nobel et al., 2002).
Las opuntias son parte del paisaje natural y de lossistemas agrícolas de muchas regiones del mundo. Al-gunas especies de Opuntia son malezas naturalizadasen países como Australia y Sudáfrica, donde las con-diciones ambientales son particularmente favorablespara su crecimiento. Más de 600.000 ha han sido plan-tadas con opuntias en África del Norte para forrajedesde 1990, especialmente en Túnez y Argelia. Lamayor parte de las opuntias cultivadas en el mundo sedestinan a la obtención de cladodios usados como fo-rraje para vacas, ovejas y cabras (Nobel et al., 2002).
Los cladodios también son usados como hortalizaspara el consumo humano especialmente en México yen el sudoeste de Estados Unidos. Aunque solamente100.000 ha están involucradas, la producción de frutoses la principal finalidad del cultivo de las opuntias enla mayoría de los países. México e Italia lideran a másde 20 países que cultivan comercialmente Opuntia
ficus-indica para la producción de frutos (Nobel et al.,2002).
En Cuba se ha demostrado que el extracto de nopalpuede utilizarse a nivel local como coagulante primarioen la clarificación de aguas para consumo humano, pu-diendo remover más del 90 % de la turbidez y 80 %
Lozano, L. - Ecofisiología de la tuna (Opuntia ficus-indica (L.) Mill.)
Horticultura Argentina 30(72): May.-Ago. 2011 39ISSN de la edición on line 1851-9342
del color del agua, y que también es muy útil en la re-moción de coliformes, por lo que su aplicación en elcontrol de la contaminación del agua en áreas ruralespuede ser muy valiosa (Flores Hernández et al., 2006).
El género Opuntia en nuestro país está represen-tado por unos 20 taxones y en el Valle de Lerma, pro-vincia de Salta, hay siete especies, cuatro variedadesy dos formas (Kiesling et al., 2011).
O. ficus-indica (L.) Mill. f. ficus-indica es cultivadaen gran parte de la Argentina por sus frutos comesti-bles y también como forrajera de emergencia. En losúltimos años se han desarrollado cultivos más o menosextensos en Santiago del Estero, Catamarca, Córdoba,etc., con el objetivo de producir fruta tanto para serconsumida en el país como para exportar. En el Vallede Lerma se implantaron pequeños cultivos como fo-rrajera, y en menor medida como frutal para consumofamiliar (Kiesling et al., 2011).
A O. ficus-indica (L.) Mill. f. amyclaea (Ten.)Schelle se la encuentra frecuentemente en Catamarca,Chaco, Córdoba, Formosa, La Rioja, Salta y Santiagodel Estero, asilvestrada cerca de caminos y poblacio-nes. También se cultiva para la cría de la cochinilla dela grana y para aprovechamiento de sus frutos (Kies-ling et al., 2011).
Tanto O. ficus-indica (L.) Mill. f. ficus-indica comoa O. ficus-indica (L.) Mill. f. amyclaea (Ten.) Schellecuyos cladodios poseen pocas espinas y se adecuaríanpara el consumo humano, se presentan también en lazona de los valles Calchaquíes y en la Puna entre los 0a los 2.500 m.s.n.m. (Instituto de Botánica, 1997-2011).
3. Aspectos Morfológicos de las plantas CAM
Muchos de los rasgos morfológicos de las plantasCAM cultivadas, entre las que se encuentran las Cac-táceas y los Agaves, pueden estar emparentados a susrelaciones con el agua. Por ejemplo, los tallos son su-culentos, indicando que un gran volumen (V) de agua
Tabla 1. Aspectos morfológicos de las plantas CAM cultivadas. Fuente: Nobel et al., 2002.
EspecieVolumen/Área
del órganofotosintético (mm)
Grosorde la
cutícula (mm)
Frecuencia deestomas
(cantidad·mm-2)
Longitudde los poros
estomáticos (mm)
Grosor delclorénquima
(mm)
Profundidadpromedio
de la raíz (cm)
Agave mapisaja 12,4* 27* 29* 60* 1,4* 20-25
Agave salmiana 11,5* 29* 27* 71* 1,6* 20-25
Agave tequilana 3,3* 22* 70* 33* 1,5* 15-20
Opuntia ficus-indica 8,5* 13* 20 46* 3,0* 15-20*
Opuntia robusta 9,0 20 40 48 3,0 10-15*
Stenocereus queretaroensis 25,4* 15 36 36 2,0 20-30*
* Datos no publicados en agosto de 2001. Los datos son para plantas maduras y son de Nobel & Valenzuela (1987), Pimienta-Barrioset al. (1993, 2002a) y Jiménez López et al. (1995).
Sitio Argentino de Producción Animal
3 de 16

Lozano, L. - Ecofisiología de la tuna (Opuntia ficus-indica (L.) Mill.)
Horticultura Argentina 30(72): May.-Ago. 201140 ISSN de la edición on line 1851-9342
puede ser almacenado en sus órganos fotosintéticos enrelación al área superficial (A), a través de la cual lapérdida de agua por transpiración ocurre. Para las del-gadas hojas de las plantas C3 y C4, la relación V/A esgeneralmente 0,1 a 0,2 mm. La relación V/A, para losórganos fotosintéticos de seis plantas CAM cultivadasen México, oscilan desde 3,3 mm para A. tequilana a25,4 mm para S. queretaroensis (Tabla 1) (Nobel et
al., 2002).Estas especies CAM se caracterizan por tener cu-
tículas relativamente gruesas y por una capa cerosa so-bre la superficie vegetal que ayudan a prevenir la pér-dida de agua, oscilando desde 13 µm para Opuntia
ficus-indica a 29 µm para A. salmiana (Tabla 1) (No-bel et al., 2002). Estas cutículas son mucho más grue-sas que las típicas de las hojas de las plantas C3 y C4de 2 a 5 µm, y ayudan a cactus y agaves a tolerar ladesecación en ambientes de regiones áridas (menos de250 mm anuales de lluvias) y semiáridas (250 a 450mm anualmente). La frecuencia de los estomas es tam-bién relativamente baja para plantas CAM maduras,silvestres o cultivadas, oscilando desde 20 por mm2
para Opuntia ficus-indica a 70 por mm2 para A. tequi-
lana (Tabla 1) (Nobel et al., 2002), la cual es muchomás baja que las 150 a 300 por mm2 típicas para lashojas de plantas C3 y C4. Teniendo menor número deestomas por unidad de área en los órganos fotosintéti-cos, disminuye la tendencia para la pérdida de agua,pero el tamaño de los estomas está también involu-crado. En este sentido, la longitud de los poros esto-máticos oscila desde 33 µm a 71 µm (Tabla 1) y es si-milar a la de especies C3 y C4. El clorénquima (tejidoverde donde la fotosíntesis tiene lugar) es bastantegrueso para las plantas CAM cultivadas oscilando des-de 1,4 a 3,0 mm (Tabla 1) comparado con 0,2 a 0,4mm para hojas de especies C3 y C4. (Nobel et al.,2002).
Las raíces son bastante cortas y profundizan entre10 a 30 cm (Tabla 1). Esta característica favorece la
respuesta a las ligeras lluvias que caracterizan a las re-giones áridas y semiáridas. El crecimiento de las raíceses aparentemente durante la estación húmeda, cuandolas raíces finas desarrollan a partir de las permanentes.Las raíces finas de agaves y cactus están comúnmenteasociadas con micorrizas vesículo-arbusculares.Talasociación de las raíces con los hongos puede incre-mentar la toma neta de CO2 de carbono para O. ro-
busta y S. queretaroensis (Nobel et al., 2002).
4. Respuesta del intercambio gaseoso al ambiente
El CO2 que entra por los estomas de las opuntiasdurante la noche se une a un compuesto de tres carbo-nos, el fosfoenolpiruvato (PEP) en una reacción cata-lizada por la enzima Fosfoenol Piruvato Carboxilasa(PEP carboxilasa); esto conduce a la formación de unácido de cuatro carbonos, oxaloacetato, que se con-vierte rápidamente en ácido málico que finalmente sealmacena en las vacuolas y es el responsable del saboramargo o ácido que se encuentra durante la noche. Du-rante el día los estomas de las plantas CAM tienden aestar cerrados, el málico se difunde fuera de las va-cuolas y se descarboxila liberando CO2 de carbono alcitosol de esa célula. El CO2 se fija en productos foto-sintéticos en los cloroplastos iluminados por la luz uti-lizando la enzima Ribulosa 1,5-bifosfatocarboxilasa/oxigenasa (Rubisco) de la misma manera que las plan-tas C3 (Nobel, 1999).
La toma del CO2 y la acumulación resultante de labiomasa de las opuntias depende de las condiciones am-bientales. Los cuatro principales factores son el conte-nido de agua del suelo, la temperatura del aire, la luz yvarios elementos del suelo (Nobel, 1999).
La toma neta del CO2 en plantas CAM consta decuatro fases. La primera fase describe la asimilaciónnocturna del CO2, involucrando la fijación inicial delCO2 por la Fosfoenol Piruvato Carboxilasa (PEP car-boxilasa); la fase 2 es el período de la asimilación del
Tabla 2. Temperatura media, flujo de fotones fotosintéticos total diario (PPF), potencial del agua del suelo y toma neta diaria de CO2
para plantas CAM cultivadas bajo condiciones controladas de laboratorio. Fuente: Nobel et al., 2002.
Especie
Temperaturadel aire
promedioDía/Noche
(°C/°C)
PPFdiario total
sobre órganofotosintético
(mol·m-2·día-1)
Potencialde aguadel suelo(MPa)
Toma netadiaria total
de CO2
(mol·m-2·día-1)
Períodos de tomaneta positiva
de CO2
Porcentajesde contribucióna la toma netadiaria de CO2
Día Noche Total Día Noche
A. salmiana 25/15 22 -0,2 481 5 12 17 3 97
A. tequilina 15/5 22 -0,1 298 6 12 18 30 70
O. ficus-indica 25/20 20 -0,1 550 3 12 15 10 90
S. queretaroensis 28/15 19 -0,2 317 6 12 18 14 86
Los datos son para temperaturas cercanas al óptimo, suelo húmedo y PPF indicado y son de Cui & Nobel (1994), Nobel & Pimienta-Barrios (1995) y Nobel et al. (1998).
Sitio Argentino de Producción Animal
4 de 16

Lozano, L. - Ecofisiología de la tuna (Opuntia ficus-indica (L.) Mill.)
Horticultura Argentina 30(72): May.-Ago. 2011 41ISSN de la edición on line 1851-9342
CO2 al alba y representa la transi-ción desde la PEP carboxilasa a lacarboxilación de la Ribulosa-1,4-bifosfato carboxilasa/oxidasa (Ru-bisco); el cierre de los estomas enla mitad del día y la descarboxila-ción de malato en la fase 3, luegogeneran una alta concentración in-terna de CO2 el cual es refijado porRubisco; al final de la tarde, los es-tomas pueden abrirse y comienza lafase 4, reflejando el paso de la car-boxilación de Rubisco a PEP carbo-xilasa. Todas las cuatro fases pue-den ocurrir para Agave mapisaja,A. salmiana, A. tequilana (Figura 1A, B y C), O. ficus-indica, O. ro-
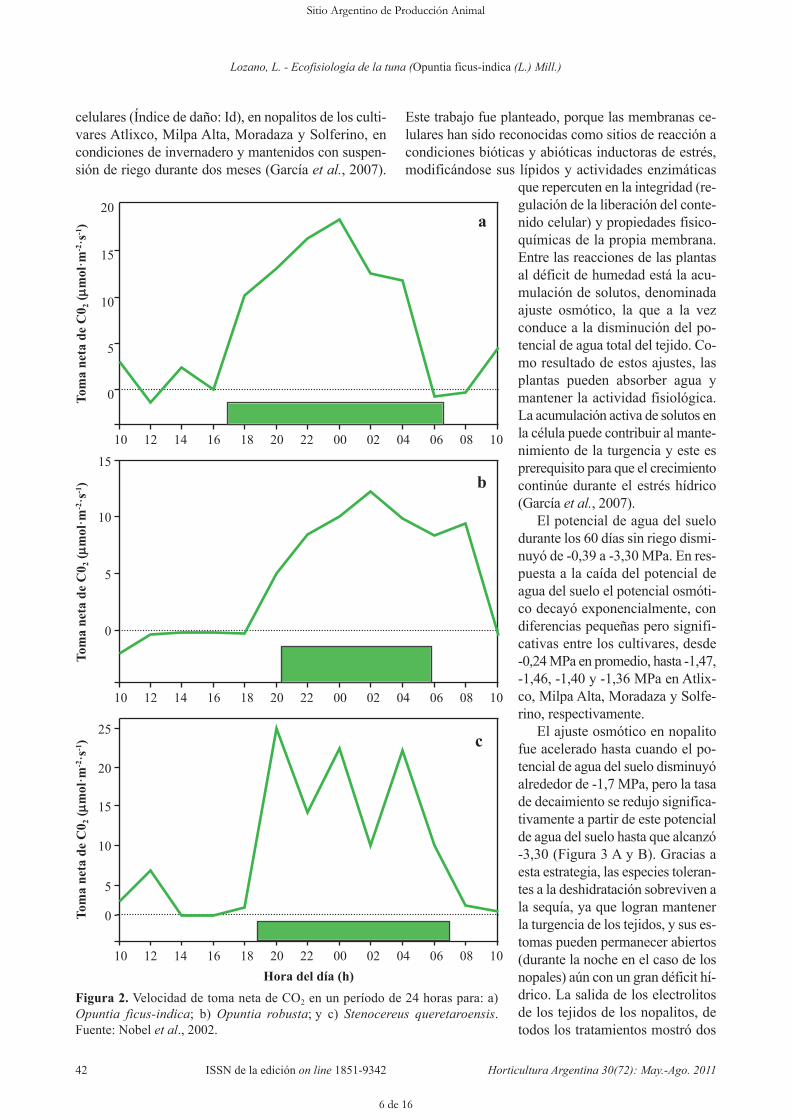
busta y S. queretaroensis (Figura 2A, B y C), aunque no bajo todas lascondiciones ambientales. Tanto pa-ra los estudios de campo y de labo-ratorio casi toda la toma neta deCO2 ocurre principalmente durantela fase 1 (en la noche), como es ca-racterístico de las plantas CAM(Tabla 2).
El hecho que las plantas CAMmuestren asimilación de CO2, tantode día como de noche, implica unaactivación sustancial tanto de laRubisco (durante el día) como de laFosfoenol Piruvato Carboxilasa (du-rante la noche), incrementando lademanda por el nitrógeno del suelopara la síntesis de más proteína(Nobel et al., 2002).
Las células del clorénquima delas plantas CAM contienen vacuo-las capaces de ocupar el 90 % omás del volumen celular, siendo ahídonde los ácidos orgánicos se acu-mulan y almacenan durante la no-che.
5. Agua del suelo
La toma de agua por una plantaocurre en condiciones húmedascuando la energía del agua o “po-
25
Tom
a ne
ta d
e C
0 2(m
mol
·m-2·s
-1)
20
15
10
5
0
10 12 14 16 18 20 22 00 02 04 06 08 10
30
Tom
a ne
ta d
e C
0 2(m
mol
·m-2·s
-1)
25
15
10
5
0
10 12 14 16 18 20 22 00 02 04 06 08 10
20
35
Tom
a ne
ta d
e C
0 2(m
mol
·m-2·s
-1)
30
20
10
5
0
10 12 14 16 18 20 22 00 02 04 06 08 10
Hora del día (h)
25
15
Figura 1. Velocidad de toma neta de CO2 en un período de 24 horas para: a) Agave
mapisaja; b) Agave salmiana; y c) Agave tequilana. Fuente: Nobel et al., 2002.
tencial hídrico del suelo” es mayor que la de las raíces.En las opuntias, cuyo sistema radicular es escaso, el po-tencial hídrico dentro de una raíz es generalmente esta-blecido por el tallo suculento cuyo potencial hídrico es
del orden de -0,3 a -0,6 MPa (Megapascal) (Nobel,1999).
Se evaluó el efecto del potencial hídrico del sueloen el potencial osmótico y alteración de las membranas
a
b
c
Sitio Argentino de Producción Animal
5 de 16
Lozano, L. - Ecofisiología de la tuna (Opuntia ficus-indica (L.) Mill.)
Horticultura Argentina 30(72): May.-Ago. 201142 ISSN de la edición on line 1851-9342
20
Tom
a ne
ta d
e C
0 2(m
mol
·m-2·s
-1)
10
5
0
10 12 14 16 18 20 22 00 02 04 06 08 10
15
Tom
a ne
ta d
e C
0 2(m
mol
·m-2·s
-1)
10
5
0
10 12 14 16 18 20 22 00 02 04 06 08 10
15
Tom
a ne
ta d
e C
0 2(m
mol
·m-2·s
-1)
20
10
5
10 12 14 16 18 20 22 00 02 04 06 08 10
Hora del día (h)
25
15
0
Figura 2. Velocidad de toma neta de CO2 en un período de 24 horas para: a)Opuntia ficus-indica; b) Opuntia robusta; y c) Stenocereus queretaroensis.Fuente: Nobel et al., 2002.
celulares (Índice de daño: Id), en nopalitos de los culti-vares Atlixco, Milpa Alta, Moradaza y Solferino, encondiciones de invernadero y mantenidos con suspen-sión de riego durante dos meses (García et al., 2007).
Este trabajo fue planteado, porque las membranas ce-lulares han sido reconocidas como sitios de reacción acondiciones bióticas y abióticas inductoras de estrés,modificándose sus lípidos y actividades enzimáticas
que repercuten en la integridad (re-gulación de la liberación del conte-nido celular) y propiedades fisico-químicas de la propia membrana.Entre las reacciones de las plantasal déficit de humedad está la acu-mulación de solutos, denominadaajuste osmótico, la que a la vezconduce a la disminución del po-tencial de agua total del tejido. Co-mo resultado de estos ajustes, lasplantas pueden absorber agua ymantener la actividad fisiológica.La acumulación activa de solutos enla célula puede contribuir al mante-nimiento de la turgencia y este esprerequisito para que el crecimientocontinúe durante el estrés hídrico(García et al., 2007).
El potencial de agua del suelodurante los 60 días sin riego dismi-nuyó de -0,39 a -3,30 MPa. En res-puesta a la caída del potencial deagua del suelo el potencial osmóti-co decayó exponencialmente, condiferencias pequeñas pero signifi-cativas entre los cultivares, desde-0,24 MPa en promedio, hasta -1,47,-1,46, -1,40 y -1,36 MPa en Atlix-co, Milpa Alta, Moradaza y Solfe-rino, respectivamente.
El ajuste osmótico en nopalitofue acelerado hasta cuando el po-tencial de agua del suelo disminuyóalrededor de -1,7 MPa, pero la tasade decaimiento se redujo significa-tivamente a partir de este potencialde agua del suelo hasta que alcanzó-3,30 (Figura 3 A y B). Gracias aesta estrategia, las especies toleran-tes a la deshidratación sobreviven ala sequía, ya que logran mantenerla turgencia de los tejidos, y sus es-tomas pueden permanecer abiertos(durante la noche en el caso de losnopales) aún con un gran déficit hí-drico. La salida de los electrolitosde los tejidos de los nopalitos, detodos los tratamientos mostró dos
a
b
c
Sitio Argentino de Producción Animal
6 de 16

Horticultura Argentina 30(72): May.-Ago. 2011 43ISSN de la edición on line 1851-9342
Lozano, L. - Ecofisiología de la tuna (Opuntia ficus-indica (L.) Mill.)
fases. La primera fue acelerada ybreve, y se presentó en los primeros10 minutos, y la segunda fase fuelenta y prolongada con duración en-tre 20 y 60 minutos, variable entrelas cultivares de opuntia y depen-dientes del potencial de agua delsuelo. La máxima liberación deelectrolitos entre los tratamientossin riego (hasta 79 mS·cm-1·g-1 detejido fresco) casi duplicó los valo-res generados por tejidos de los no-palitos sin restricción de humedaddel suelo. Lo anterior demostró quela disminución del potencial deagua del suelo, de -0,39 a -3,30MPa, en un período de 60 días,afectó parcialmente las membranascelulares de los tejidos de los nopa-litos, pero se mantuvo una propor-ción significativa de su integridadestructural y funcional, ya que eldaño máximo que podría alcanzarsees equivalente a un Id de 100 %(García et al., 2007).
Variando el contenido de aguadel suelo, otro trabajo evaluó losefectos del número de cladodioshijos sobre el intercambio de gasesen especies cultivadas del generoOpuntia (Pimienta-Barrios et al.,2005). La ganancia diaria de CO2
por los cladodios madre fue redu-cida a medida que el número de cla-dodios hijos aumentó de ninguno(0) a ocho (8), especialmente durante la sequía. Esta fueacompañada por la disminución relativa del contenidode agua del cladodio madre, lo que sugiere movimien-tos de agua del cladodio madre a los cladodios hijos.La asimilación de CO2 fue más afectada en la fase 4de CAM (la toma neta de CO2 tarde en la tarde) porlos efectos combinados de los cladodios hijos (que du-rante los tempranos estadios de desarrollo presentanfotosíntesis C3 con apertura diurna de los estomas) yla sequía. La precipitación elevó el contenido de aguadel suelo disminuyendo los efectos de los cladodioshijos sobre la toma neta del CO2 por los cladodiosmadre: estos se rehidratan y la toma neta sustancial deCO2 se presenta en la fase 4. Los cladodios hijos acen-túan los efectos de la sequía sobre los cladodios madredebido a la competencia por el abastecimiento de aguay consecuentemente disminuyen la ganancia diaria deCO2 de los cladodios madre, principalmente por inhi-
Pot
enci
al o
smót
ico
(Mpa
) -0,3
-1,2
Potencial de agua del suelo (Mpa)
0
-0,9
-1,5
AtlixcoMilpa altaMoradazaSolferino
-3 -2 -1 0
Indi
ce d
e da
ño (
%)
10
Potencial de agua del suelo (Mpa)
30
20Atlixco
Milpa altaMoradazaSolferino
-3 -2 -1 00
Figura 3. a) Potencial osmótico en las membranas celulares de nopalito Opuntia
spp. dependientes del déficit de agua del suelo. b) Índice de daño en las mem-branas celulares de nopalito Opuntia spp. Fuente: García et al., 2007.
a
b
bición de la fase 4 CAM. (Pimienta-Barrios et al.,2005).
La sequía se define como la caída del potencial hí-drico del suelo por debajo del potencial hídrico de lasraíces y momento a partir del cual se detiene la absor-ción de agua desde el suelo; a partir de ese momentolas plantas dependen del agua almacenada en sus teji-dos, lo que conduce a una reducción gradual en laapertura de los estomas y, por tanto, en una reducciónen la toma diaria de CO2.
Durante la primera semana de sequía no se verifi-can cambios en la toma neta de CO2 en O. ficus-in-
dica, lo que indica que el suministro de agua es ade-cuado aún en suelos arenosos bien drenados. Despuésde tres semanas de sequía la toma diaria de CO2 co-mienza a ser aproximadamente la mitad de cuando setenía buena condición hídrica, y para las cinco sema-nas de sequía la toma de CO2 se reduce a aproxima-
Sitio Argentino de Producción Animal
7 de 16

Lozano, L. - Ecofisiología de la tuna (Opuntia ficus-indica (L.) Mill.)
Horticultura Argentina 30(72): May.-Ago. 201144 ISSN de la edición on line 1851-9342
damente el 10 % al de la condición húmeda (Nobel,1999).
A los efectos de investigar como influyen los cam-bios de la humedad del suelo en el desarrollo y com-posición de los nopalitos, se evaluó el efecto de ladisminución del potencial de agua del suelo (ΨA) du-rante 63 días (de -0,14 a -3,5 MPa) en el crecimientoy contenido de mucílago de los nopalitos de las culti-vares Copena, Moradaza y Solferino y una recolecciónsilvestre de P. streptacanta en condiciones de inver-nadero (O. Camacho et al., 2007).
La disminución de la humedad del suelo incre-mentó paulatina y significativamente la biomasa delos nopalitos de los cuatro tratamientos sin riego (5 a25 %), aumento que puede ser resultado de la acumu-lación neta de biomasa en respuesta a la sequía, efectoindirecto de la deshidratación de los tejidos o ambos.Sin embargo, disminuyó el espesor y la longitud delos nopalitos tanto de las cultivares como del tipo sil-vestre. Se incrementó (3 %) el contenido de mucílagoy hemicelulosa (8 a 16 % en el tejido seco) en Mora-daza, pero las pectinas y celulosa fluctuaron irregular-mente (O. Camacho et al., 2007).
Con humedad limitada en el suelo se incrementasignificativamente la proporción de biomasa de los no-palitos, ese incremento no es proporcional a la estabi-lidad del crecimiento, ni a los cambios en el contenidode polisacáridos estructurales. Lo anterior podría tenerrepercusión en la calidad de los nopalitos, pues se haseñalado que el contenido de mucílago afecta la acep-tabilidad de los consumidores y la proporción de pec-tina entre los nopales puede determinar su uso en laindustria farmacéutica, de alimentos, de cosmetología,etc. (O. Camacho et al., 2007).
Los hongos micorrizógenos arbusculares, desem-peñan un papel determinante en el establecimiento, re-laciones hídricas y crecimiento vegetal de Opuntia
streptacantha Lem., además de contribuir a mantenerel estado hídrico en condiciones de sequía. Por ello serecomienda micorrizar plántulas de esta especie enprogramas de restauración ecológica de zonas semiá-ridas deterioradas (González Monterrubio et al.,2005).
En Opuntia robusta, la inhibición de la coloniza-ción de hongos micorrizógenos arbusculares, usandoBenomyl no afectó la fotosíntesis, la toma de agua yla toma de P bajo condiciones húmedas y secas reve-lando que el desarrollo de estas especies CAM en unambiente altamente estresante podría ser regulado porotras estrategias fisiológicas y mecanismos tales comoel metabolismo CAM, además de la simbiosis conhongos arbusculares, particularmente bajo sequía pro-longada (Pimienta-Barrios et al., 2003).
6. Temperatura
La temperatura no solamente afecta los procesosmetabólicos y la absorción neta de CO2 sino que lastemperaturas extremas pueden inducir daños y aun lamuerte de las plantas. En este aspecto O. ficus-indica
es extremadamente tolerante a temperaturas altas delaire, pero no a temperaturas substancialmente abajodel punto de congelación (Nobel & De la Barrera,2003). Cuando las plantas son aclimatadas a tempera-turas altas del aire en los ciclos día/noche de 50/40 °C,las células del clorénquima no son seriamente dañadaspor la exposición de una hora a 60 °C, y la mayoríade las células sobrevive 1 hora a 65 °C. El daño portemperatura alta en O. ficus-indica es generalmenteobservado únicamente a nivel del suelo, donde lastemperaturas en los desiertos pueden alcanzar 70 °C,las plantas jóvenes o recién plantadas son especial-mente vulnerables al daño. En contraste, el daño ce-lular en el campo ocurre a temperaturas de conge-lación de 5 a 10 °C. El daño varía con la cultivar, conla rapidez con que se establece la congelación y conel contenido de agua en el tallo, debido a que un bajocontenido de agua conduce a una mejor tolerancia atemperaturas bajas del aire y del tallo (Nobel, 2003).
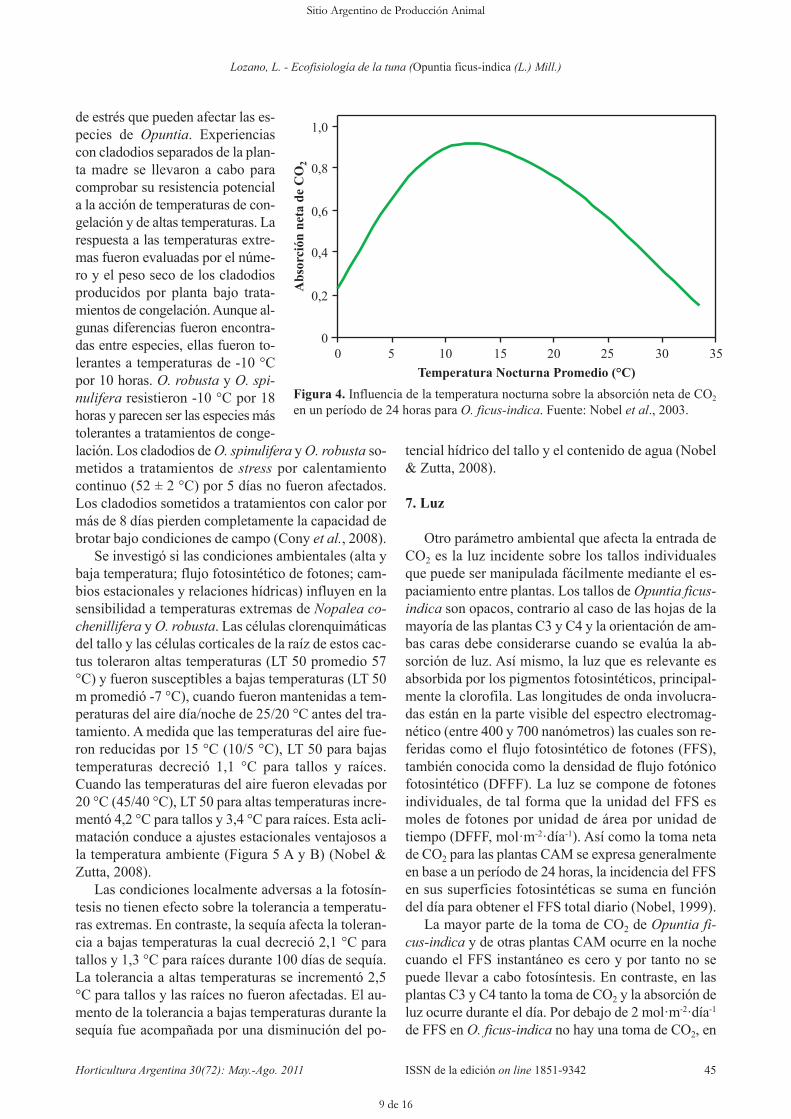
Debido a que la absorción de CO2 en las plantasCAM ocurre durante la noche, las temperaturas noc-turnas son mucho más importantes que las diurnaspara la absorción neta de CO2 de O. ficus-indica. Latemperatura nocturna óptima es relativamente baja, 15°C, y temperaturas de 5 a 20 °C conducen a un mínimodel 80 % de la entrada neta de CO2. Temperaturas tanbajas también se asocian con bajas tasas de transpira-ción. En O. ficus-indica conforme la temperatura noc-turna se eleva, los estomas tienden a cerrarse; porejemplo a 30 °C, los estomas solamente abren un 30 %de lo que se abren a 20 °C, lo que ayuda a reducir la en-trada neta de CO2 a la más alta temperatura (Figura 4).Excepto para temperaturas nocturnas sustancialmentedebajo de la de congelación o arriba de 30 °C, la tem-peratura no es generalmente un factor limitante impor-tante para la absorción de CO2 de O. ficus-indica, es-pecialmente durante las estaciones cuando la lluvia estádisponible (Nobel, 2003).
La tolerancia tanto a temperaturas de congela-miento como a altas temperaturas depende del órga-no de la Opuntia y de su edad, y puede ser mejoradapor un período de aclimatación que permite a las plan-tas ajustarse a los cambios de la temperatura del aire.La tolerancia a altas y bajas temperaturas aumentó conla edad del cladodio de hasta 10 años de edad (Nobel& De la Barrera, 2003).
Las temperaturas extremas están entre los factores
Sitio Argentino de Producción Animal
8 de 16
Lozano, L. - Ecofisiología de la tuna (Opuntia ficus-indica (L.) Mill.)
Horticultura Argentina 30(72): May.-Ago. 2011 45ISSN de la edición on line 1851-9342
de estrés que pueden afectar las es-pecies de Opuntia. Experienciascon cladodios separados de la plan-ta madre se llevaron a cabo paracomprobar su resistencia potenciala la acción de temperaturas de con-gelación y de altas temperaturas. Larespuesta a las temperaturas extre-mas fueron evaluadas por el núme-ro y el peso seco de los cladodiosproducidos por planta bajo trata-mientos de congelación. Aunque al-gunas diferencias fueron encontra-das entre especies, ellas fueron to-lerantes a temperaturas de -10 °Cpor 10 horas. O. robusta y O. spi-
nulifera resistieron -10 °C por 18horas y parecen ser las especies mástolerantes a tratamientos de conge-lación. Los cladodios de O. spinulifera y O. robusta so-metidos a tratamientos de stress por calentamientocontinuo (52 ± 2 °C) por 5 días no fueron afectados.Los cladodios sometidos a tratamientos con calor pormás de 8 días pierden completamente la capacidad debrotar bajo condiciones de campo (Cony et al., 2008).
Se investigó si las condiciones ambientales (alta ybaja temperatura; flujo fotosintético de fotones; cam-bios estacionales y relaciones hídricas) influyen en lasensibilidad a temperaturas extremas de Nopalea co-
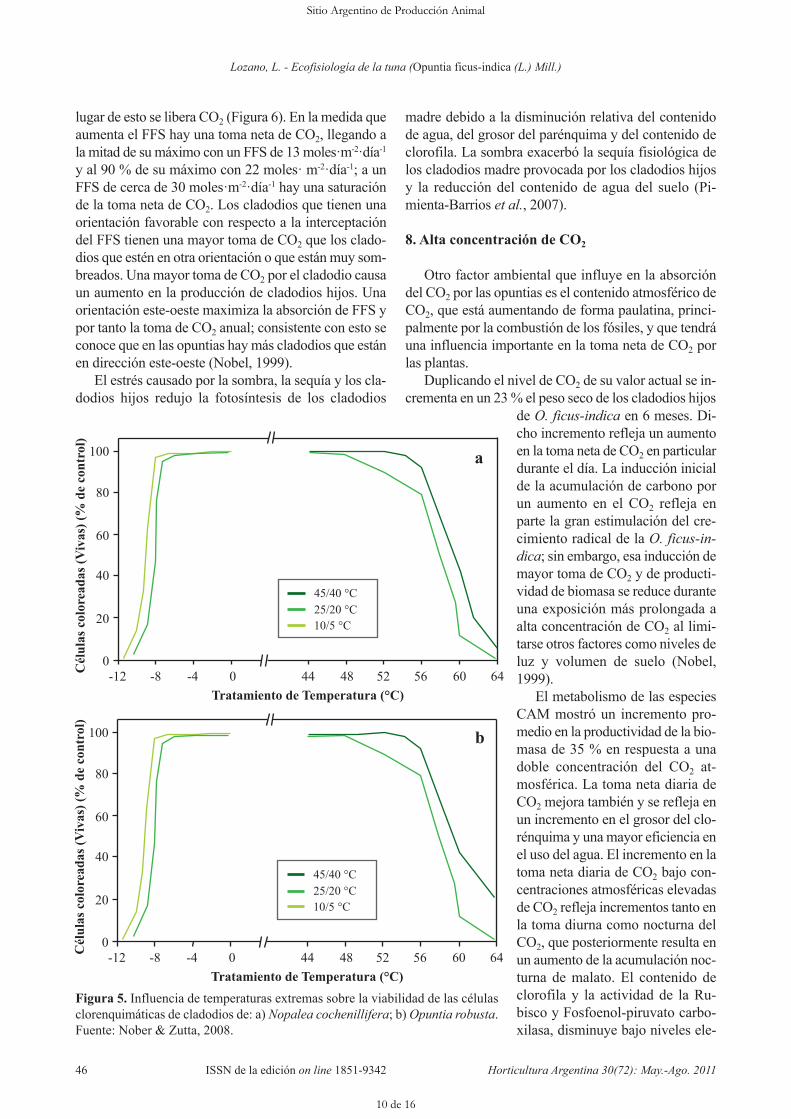
chenillifera y O. robusta. Las células clorenquimáticasdel tallo y las células corticales de la raíz de estos cac-tus toleraron altas temperaturas (LT 50 promedio 57°C) y fueron susceptibles a bajas temperaturas (LT 50m promedió -7 °C), cuando fueron mantenidas a tem-peraturas del aire día/noche de 25/20 °C antes del tra-tamiento. A medida que las temperaturas del aire fue-ron reducidas por 15 °C (10/5 °C), LT 50 para bajastemperaturas decreció 1,1 °C para tallos y raíces.Cuando las temperaturas del aire fueron elevadas por20 °C (45/40 °C), LT 50 para altas temperaturas incre-mentó 4,2 °C para tallos y 3,4 °C para raíces. Esta acli-matación conduce a ajustes estacionales ventajosos ala temperatura ambiente (Figura 5 A y B) (Nobel &Zutta, 2008).
Las condiciones localmente adversas a la fotosín-tesis no tienen efecto sobre la tolerancia a temperatu-ras extremas. En contraste, la sequía afecta la toleran-cia a bajas temperaturas la cual decreció 2,1 °C paratallos y 1,3 °C para raíces durante 100 días de sequía.La tolerancia a altas temperaturas se incrementó 2,5°C para tallos y las raíces no fueron afectadas. El au-mento de la tolerancia a bajas temperaturas durante lasequía fue acompañada por una disminución del po-
Abs
orci
ón n
eta
de C
O2
0,4
Temperatura Nocturna Promedio (°C)
1,0
0,6
5 15 25 350
0,8
0,2
0 10 20 30
Figura 4. Influencia de la temperatura nocturna sobre la absorción neta de CO2
en un período de 24 horas para O. ficus-indica. Fuente: Nobel et al., 2003.
tencial hídrico del tallo y el contenido de agua (Nobel& Zutta, 2008).
7. Luz
Otro parámetro ambiental que afecta la entrada deCO2 es la luz incidente sobre los tallos individualesque puede ser manipulada fácilmente mediante el es-paciamiento entre plantas. Los tallos de Opuntia ficus-
indica son opacos, contrario al caso de las hojas de lamayoría de las plantas C3 y C4 y la orientación de am-bas caras debe considerarse cuando se evalúa la ab-sorción de luz. Así mismo, la luz que es relevante esabsorbida por los pigmentos fotosintéticos, principal-mente la clorofila. Las longitudes de onda involucra-das están en la parte visible del espectro electromag-nético (entre 400 y 700 nanómetros) las cuales son re-feridas como el flujo fotosintético de fotones (FFS),también conocida como la densidad de flujo fotónicofotosintético (DFFF). La luz se compone de fotonesindividuales, de tal forma que la unidad del FFS esmoles de fotones por unidad de área por unidad detiempo (DFFF, mol·m-2·día-1). Así como la toma netade CO2 para las plantas CAM se expresa generalmenteen base a un período de 24 horas, la incidencia del FFSen sus superficies fotosintéticas se suma en funcióndel día para obtener el FFS total diario (Nobel, 1999).
La mayor parte de la toma de CO2 de Opuntia fi-
cus-indica y de otras plantas CAM ocurre en la nochecuando el FFS instantáneo es cero y por tanto no sepuede llevar a cabo fotosíntesis. En contraste, en lasplantas C3 y C4 tanto la toma de CO2 y la absorción deluz ocurre durante el día. Por debajo de 2 mol·m-2·día-1
de FFS en O. ficus-indica no hay una toma de CO2, en
Sitio Argentino de Producción Animal
9 de 16
Lozano, L. - Ecofisiología de la tuna (Opuntia ficus-indica (L.) Mill.)
Horticultura Argentina 30(72): May.-Ago. 201146 ISSN de la edición on line 1851-9342
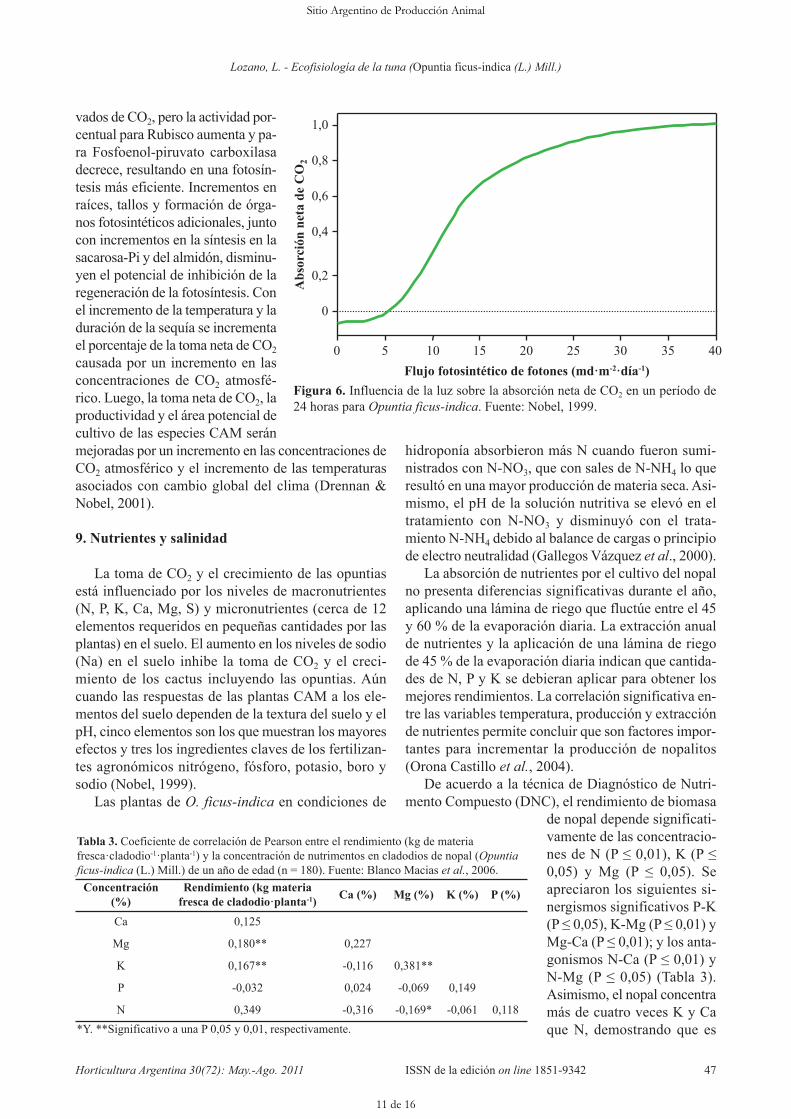
lugar de esto se libera CO2 (Figura 6). En la medida queaumenta el FFS hay una toma neta de CO2, llegando ala mitad de su máximo con un FFS de 13 moles·m-2·día-1
y al 90 % de su máximo con 22 moles· m-2·día-1; a unFFS de cerca de 30 moles·m-2·día-1 hay una saturaciónde la toma neta de CO2. Los cladodios que tienen unaorientación favorable con respecto a la interceptacióndel FFS tienen una mayor toma de CO2 que los clado-dios que estén en otra orientación o que están muy som-breados. Una mayor toma de CO2 por el cladodio causaun aumento en la producción de cladodios hijos. Unaorientación este-oeste maximiza la absorción de FFS ypor tanto la toma de CO2 anual; consistente con esto seconoce que en las opuntias hay más cladodios que estánen dirección este-oeste (Nobel, 1999).
El estrés causado por la sombra, la sequía y los cla-dodios hijos redujo la fotosíntesis de los cladodios
Cél
ulas
col
orea
das
(Viv
as)
(% d
e co
ntro
l)
40
Tratamiento de Temperatura (°C)
100
60
-8 44 52 640
80
20
-12 0 48 56-4 60
10/5 °C25/20 °C45/40 °C
Cél
ulas
col
orea
das
(Viv
as)
(% d
e co
ntro
l)
40
Tratamiento de Temperatura (°C)
100
60
-8 44 52 640
80
20
-12 0 48 56-4 60
10/5 °C25/20 °C45/40 °C
Figura 5. Influencia de temperaturas extremas sobre la viabilidad de las célulasclorenquimáticas de cladodios de: a) Nopalea cochenillifera; b) Opuntia robusta.Fuente: Nober & Zutta, 2008.
a
b
madre debido a la disminución relativa del contenidode agua, del grosor del parénquima y del contenido declorofila. La sombra exacerbó la sequía fisiológica delos cladodios madre provocada por los cladodios hijosy la reducción del contenido de agua del suelo (Pi-mienta-Barrios et al., 2007).
8. Alta concentración de CO2
Otro factor ambiental que influye en la absorcióndel CO2 por las opuntias es el contenido atmosférico deCO2, que está aumentando de forma paulatina, princi-palmente por la combustión de los fósiles, y que tendráuna influencia importante en la toma neta de CO2 porlas plantas.
Duplicando el nivel de CO2 de su valor actual se in-crementa en un 23 % el peso seco de los cladodios hijos
de O. ficus-indica en 6 meses. Di-cho incremento refleja un aumentoen la toma neta de CO2 en particulardurante el día. La inducción inicialde la acumulación de carbono porun aumento en el CO2 refleja enparte la gran estimulación del cre-cimiento radical de la O. ficus-in-
dica; sin embargo, esa inducción demayor toma de CO2 y de producti-vidad de biomasa se reduce duranteuna exposición más prolongada aalta concentración de CO2 al limi-tarse otros factores como niveles deluz y volumen de suelo (Nobel,1999).
El metabolismo de las especiesCAM mostró un incremento pro-medio en la productividad de la bio-masa de 35 % en respuesta a unadoble concentración del CO2 at-mosférica. La toma neta diaria deCO2 mejora también y se refleja enun incremento en el grosor del clo-rénquima y una mayor eficiencia enel uso del agua. El incremento en latoma neta diaria de CO2 bajo con-centraciones atmosféricas elevadasde CO2 refleja incrementos tanto enla toma diurna como nocturna delCO2, que posteriormente resulta enun aumento de la acumulación noc-turna de malato. El contenido declorofila y la actividad de la Ru-bisco y Fosfoenol-piruvato carbo-xilasa, disminuye bajo niveles ele-
Sitio Argentino de Producción Animal
10 de 16
Lozano, L. - Ecofisiología de la tuna (Opuntia ficus-indica (L.) Mill.)
Horticultura Argentina 30(72): May.-Ago. 2011 47ISSN de la edición on line 1851-9342
vados de CO2, pero la actividad por-centual para Rubisco aumenta y pa-ra Fosfoenol-piruvato carboxilasadecrece, resultando en una fotosín-tesis más eficiente. Incrementos enraíces, tallos y formación de órga-nos fotosintéticos adicionales, juntocon incrementos en la síntesis en lasacarosa-Pi y del almidón, disminu-yen el potencial de inhibición de laregeneración de la fotosíntesis. Conel incremento de la temperatura y laduración de la sequía se incrementael porcentaje de la toma neta de CO2
causada por un incremento en lasconcentraciones de CO2 atmosfé-rico. Luego, la toma neta de CO2, laproductividad y el área potencial decultivo de las especies CAM seránmejoradas por un incremento en las concentraciones deCO2 atmosférico y el incremento de las temperaturasasociados con cambio global del clima (Drennan &Nobel, 2001).
9. Nutrientes y salinidad
La toma de CO2 y el crecimiento de las opuntiasestá influenciado por los niveles de macronutrientes(N, P, K, Ca, Mg, S) y micronutrientes (cerca de 12elementos requeridos en pequeñas cantidades por lasplantas) en el suelo. El aumento en los niveles de sodio(Na) en el suelo inhibe la toma de CO2 y el creci-miento de los cactus incluyendo las opuntias. Aúncuando las respuestas de las plantas CAM a los ele-mentos del suelo dependen de la textura del suelo y elpH, cinco elementos son los que muestran los mayoresefectos y tres los ingredientes claves de los fertilizan-tes agronómicos nitrógeno, fósforo, potasio, boro ysodio (Nobel, 1999).
Las plantas de O. ficus-indica en condiciones de
Abs
orci
ón n
eta
de C
O2
0,4
Flujo fotosintético de fotones (md·m-2·día-1)
1,0
0,6
5 20 30 40
0
0,8
0,2
0 15 2510 35
Figura 6. Influencia de la luz sobre la absorción neta de CO2 en un período de24 horas para Opuntia ficus-indica. Fuente: Nobel, 1999.
hidroponía absorbieron más N cuando fueron sumi-nistrados con N-NO3, que con sales de N-NH4 lo queresultó en una mayor producción de materia seca. Asi-mismo, el pH de la solución nutritiva se elevó en eltratamiento con N-NO3 y disminuyó con el trata-miento N-NH4 debido al balance de cargas o principiode electro neutralidad (Gallegos Vázquez et al., 2000).
La absorción de nutrientes por el cultivo del nopalno presenta diferencias significativas durante el año,aplicando una lámina de riego que fluctúe entre el 45y 60 % de la evaporación diaria. La extracción anualde nutrientes y la aplicación de una lámina de riegode 45 % de la evaporación diaria indican que cantida-des de N, P y K se debieran aplicar para obtener losmejores rendimientos. La correlación significativa en-tre las variables temperatura, producción y extracciónde nutrientes permite concluir que son factores impor-tantes para incrementar la producción de nopalitos(Orona Castillo et al., 2004).
De acuerdo a la técnica de Diagnóstico de Nutri-mento Compuesto (DNC), el rendimiento de biomasa
Tabla 3. Coeficiente de correlación de Pearson entre el rendimiento (kg de materiafresca·cladodio-1·planta-1) y la concentración de nutrimentos en cladodios de nopal (Opuntia
ficus-indica (L.) Mill.) de un año de edad (n = 180). Fuente: Blanco Macias et al., 2006.
Concentración(%)
Rendimiento (kg materiafresca de cladodio·planta-1)
Ca (%) Mg (%) K (%) P (%)
Ca 0,125
Mg 0,180** 0,227
K 0,167** -0,116 0,381**
P -0,032 0,024 -0,069 0,149
N 0,349 -0,316 -0,169* -0,061 0,118
*Y. **Significativo a una P 0,05 y 0,01, respectivamente.
de nopal depende significati-vamente de las concentracio-nes de N (P ≤ 0,01), K (P ≤0,05) y Mg (P ≤ 0,05). Seapreciaron los siguientes si-nergismos significativos P-K(P ≤ 0,05), K-Mg (P ≤ 0,01) yMg-Ca (P ≤ 0,01); y los anta-gonismos N-Ca (P ≤ 0,01) yN-Mg (P ≤ 0,05) (Tabla 3).Asimismo, el nopal concentramás de cuatro veces K y Caque N, demostrando que es
Sitio Argentino de Producción Animal
11 de 16

Lozano, L. - Ecofisiología de la tuna (Opuntia ficus-indica (L.) Mill.)
Horticultura Argentina 30(72): May.-Ago. 201148 ISSN de la edición on line 1851-9342
una planta calciotrófica. Algo similar ocurre entre Mgy P, pues el primero se concentra cuatro veces más queel segundo (Blanco Macias et al., 2006).
Los coeficientes de correlación positivos sugierenque el rendimiento puede incrementar si se asocia conun aumento de concentración de esos elementos en loscladodios; es decir, es de esperar que los rendimientosaumenten al fertilizar con N, Mg y K.
En nopal, el Ca, Mg y K son los nutrimentos quese concentran en mayor proporción en los cladodiosde un año de edad, marcando la diferencia con res-pecto a otras plantas cultivadas, en las cuales el papelde macronutrientes, corresponden al N, P y K. (BlancoMacias et al., 2006).
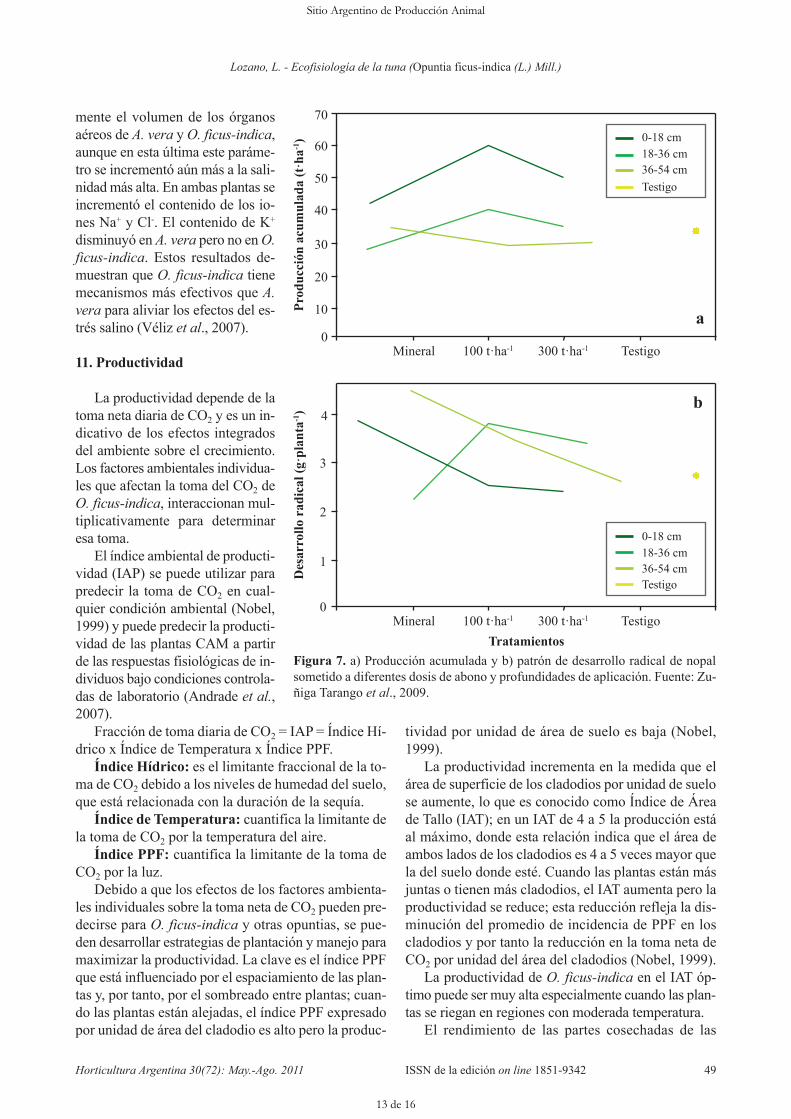
Según Zúñiga Tarango et al. (2009), –quienes es-tudiaron el efecto de las aplicaciones de estiércol bo-vino y fertilizante mineral a diferentes profundidadescon respecto al crecimiento radical, producción de ma-teria seca y concentración de nutrientes en nopalito–,la mayor abundancia relativa de raíces de las plantasde nopal fue en el primer estrato 0-18 cm con un 96%, seguido del estrato de 18 a 36 cm con 3 % (Figura7 A y B). El rendimiento de nopalitos, la producciónde materia seca y el numero de brotes fueron mayoresen las plantas sometidas al tratamiento de 100 t·ha-1
de estiércol aplicado en el estrato superior (0 a 18 cm)(Tabla 4). El contenido mineral de los cladodios mos-tró valores similares entre tratamientos y el testigo; sinembargo la concentración de fósforo y de microele-mentos fue mayor en las plantas sometidas a los trata-mientos de estiércol en el estrato de 0 a 18 cm. Lasaplicaciones de estiércol incrementaron el contenidomineral y de materia orgánica del suelo al final del ex-perimento (Zúñiga Tarango et al., 2009).
Tabla 4. Valores promedio del rendimiento de nopal sometido a diferentes dosis de abono y profundidades de aplicación. Fuente:Zúñiga Tarango et al., 2009.
Tratamientos Número y fecha de corte / Rendimiento de nopalito (t·ha-1)
DosisProfundidad
(cm)1
06/08/982
02/09/983
08/10/984
19/11/985
25/01/996
13/02/997
22/03/998
06/05/99
100 t·ha-1 00-18 2,98a* 4,8ab 8,24a 5,75a 4,06a 5,10a 16,2a 13,5a
300 t·ha-1 00-18 1,98a 5,08a 6,81b 5,09a 2,6ab 4,32a 14,1ab 12,2ab
100 t·ha-1 18-36 2,36a 4,6abc 5,7bc 2,7bc 1,8bc 2,84b 11,1bc 10,6bc
Mineral 00-18 2,34a 3,3bcd 6,9ab 3,52b 1,1bc 1,34c 14,1ab 10,1cd
300 t·ha-1 18-36 1,98a 3,6bcd 4,4cde 1,8cd 0,83c 2,1bc 12,2bc 9,8cde
Mineral 18-36 2,02a 3,8abc 3,7cde 0,72de 0,82bc 1,10c 9,51c 7,48f
300 t·ha-1 36-54 2,34a 2,9cd 3,2de 0,49de 1,155c 1,15c 10,2c 9,9cde
Mineral 36-54 1,82a 3,1bcd 4,2cde 0,51de 1,9bc 1,7bc 12,0bc 8,2ef
100 t·ha-1 36-54 1,40a 3,7bcd 3,16e 0,283e 0,83bc 1,49c 10,1c 8,5def
Testigo 1,71a 1,99d 5,5cd 0,91de 0,97bc 1,54c 10,5c 8,3ef
*Medias con la misma letra en hilera, no difieren significativamente (Fisher LSD a P = 0,05).
El análisis del crecimiento radical en cuatro varie-dades de Opuntia ficus-indica (Liso forrajero, Villa-nueva, Jalpa y Copena V-I) se realizó a los efectos dehacer un uso más eficiente de agua y fertilizantes, paraindicar la manera adecuada de realizarse aplicacioneslocalizadas de estos insumos. Se determinó el tiempo(en días) en la emisión de raíces, la tasa de crecimientodiaria y las diferencias morfológicas (longitud, diáme-tro y peso seco) de sus raíces en las cuatro variedades.El tiempo en la emisión de raíces mostró diferenciasentre variedades, donde la variedad Liso forrajero fuela más precoz y Copena V-I la más tardía. Se encontróuna relación inversa en el tiempo de emisión de raícesy la tasa de crecimiento de raíces de nopal, dado queCopena V-I mostró la mayor tasa de crecimiento deraíces pero presentó un tiempo mayor en la emisiónde éstas, mientras que la variedad Liso forrajero mos-tró la tasa de crecimiento de raíces menor pero el me-nor tiempo en su emisión. La longitud, el diámetro yel peso seco de raíces mostraron diferencias significa-tivas entre variedades, donde la variedad Villanuevamostró los valores mayores de las tres variables (Váz-quez-Vázquez et al., 2007).
10. Salinidad
Tanto O. ficus-indica como Aloe vera son plantascultivables de alta productividad y rentabilidad paralas zonas costeras áridas y semiáridas. Sin embargo,estas plantas pueden verse afectadas por la salinidad:en el caso particular de O. ficus-indica, concentracio-nes bajas de NaCl redujeron el crecimiento y se res-tringió la acumulación de Na+ en los cladodios cuandose aumentó la salinidad. La salinidad afectó negativa-
Sitio Argentino de Producción Animal
12 de 16
Lozano, L. - Ecofisiología de la tuna (Opuntia ficus-indica (L.) Mill.)
Horticultura Argentina 30(72): May.-Ago. 2011 49ISSN de la edición on line 1851-9342
mente el volumen de los órganosaéreos de A. vera y O. ficus-indica,aunque en esta última este paráme-tro se incrementó aún más a la sali-nidad más alta. En ambas plantas seincrementó el contenido de los io-nes Na+ y Cl-. El contenido de K+
disminuyó en A. vera pero no en O.
ficus-indica. Estos resultados de-muestran que O. ficus-indica tienemecanismos más efectivos que A.
vera para aliviar los efectos del es-trés salino (Véliz et al., 2007).
11. Productividad
La productividad depende de latoma neta diaria de CO2 y es un in-dicativo de los efectos integradosdel ambiente sobre el crecimiento.Los factores ambientales individua-les que afectan la toma del CO2 deO. ficus-indica, interaccionan mul-tiplicativamente para determinaresa toma.
El índice ambiental de producti-vidad (IAP) se puede utilizar parapredecir la toma de CO2 en cual-quier condición ambiental (Nobel,1999) y puede predecir la producti-vidad de las plantas CAM a partirde las respuestas fisiológicas de in-dividuos bajo condiciones controla-das de laboratorio (Andrade et al.,2007).
Pro
ducc
ión
acum
ulad
a (t
·ha-1
)
40
70
50
Mineral 300 t·ha-1 Testigo
10
60
20
0100 t·ha-1
30
Testigo
36-54 cm18-36 cm0-18 cm
Des
arro
llo r
adic
al (
g·pl
anta
-1)
3
Tratamientos
Mineral 300 t·ha-1 Testigo
1
4
0100 t·ha-1
2
Testigo36-54 cm18-36 cm0-18 cm
Figura 7. a) Producción acumulada y b) patrón de desarrollo radical de nopalsometido a diferentes dosis de abono y profundidades de aplicación. Fuente: Zu-ñiga Tarango et al., 2009.
a
b
Fracción de toma diaria de CO2 = IAP = Índice Hí-drico x Índice de Temperatura x Índice PPF.
Índice Hídrico: es el limitante fraccional de la to-ma de CO2 debido a los niveles de humedad del suelo,que está relacionada con la duración de la sequía.
Índice de Temperatura: cuantifica la limitante dela toma de CO2 por la temperatura del aire.
Índice PPF: cuantifica la limitante de la toma deCO2 por la luz.
Debido a que los efectos de los factores ambienta-les individuales sobre la toma neta de CO2 pueden pre-decirse para O. ficus-indica y otras opuntias, se pue-den desarrollar estrategias de plantación y manejo paramaximizar la productividad. La clave es el índice PPFque está influenciado por el espaciamiento de las plan-tas y, por tanto, por el sombreado entre plantas; cuan-do las plantas están alejadas, el índice PPF expresadopor unidad de área del cladodio es alto pero la produc-
tividad por unidad de área de suelo es baja (Nobel,1999).
La productividad incrementa en la medida que elárea de superficie de los cladodios por unidad de suelose aumente, lo que es conocido como Índice de Áreade Tallo (IAT); en un IAT de 4 a 5 la producción estáal máximo, donde esta relación indica que el área deambos lados de los cladodios es 4 a 5 veces mayor quela del suelo donde esté. Cuando las plantas están másjuntas o tienen más cladodios, el IAT aumenta pero laproductividad se reduce; esta reducción refleja la dis-minución del promedio de incidencia de PPF en loscladodios y por tanto la reducción en la toma neta deCO2 por unidad del área del cladodios (Nobel, 1999).
La productividad de O. ficus-indica en el IAT óp-timo puede ser muy alta especialmente cuando las plan-tas se riegan en regiones con moderada temperatura.
El rendimiento de las partes cosechadas de las
Sitio Argentino de Producción Animal
13 de 16

opuntias (cladodios y frutos) depende del peso secoacumulado en los cladodios y a la vez depende de latoma neta de CO2 (Nobel, 1999).
Además de proporcionar información sobre la eco-fisiología de especies vegetales en el campo, el IAP(Índice Ambiental de Productividad) es un modelo nu-mérico que pueden utilizarse como herramientas depredicción con aplicaciones en agricultura y conser-vación. Por ejemplo, en Chile se ha utilizado el enfo-que del IAP para establecer plantaciones comercialesde nopal (Opuntia ficus-indica) altamente productivas(García de Cortázar et al., 1985; García de Cortázar& Nobel, 1992). También se han utilizado los IAP encombinación con datos climatológicos para determi-nar zonas adecuadas de cultivo de Agave tequilana enJalisco (Pimienta Barrios et al., 1999) y de Hylocereus
undatus y Opuntia ficus-indica en California, EstadosUnidos (Nobel et al., 2002). Recientemente se obtuvoun IAP para Mammillaria gaumeri, cactácea endémicay amenazada de la costa norte de Yucatán, con el finde identificar sitios adecuados para su reintroduccióny poder crear condiciones ambientales adecuadas parasu propagación en invernadero (Cervera, J.C. y An-drade, J.L., datos no publicados) (Andrade et al.,2007).
12. Discusión
Difundir el consumo de los jóvenes cladodios dela planta de tuna en zonas que presentan limitacionesclimáticas para el cultivo de otras hortalizas se funda-menta en varios aspectos: el valor nutritivo del broterico en fibra y Ca (Landero Esquivel & Cruz Saenz,2006), las adaptaciones que presentan las opuntias pa-ra sobrevivir a condiciones de escasez de agua (Garcíaet al., 2007; O. Camacho et al., 2007), su relativa to-lerancia a temperaturas por debajo de 0 °C (Nobel,2003; Nobel & De la Barrera, 2003) y la versatilidadde brote para la preparación de comidas dulces y sa-ladas con alto porcentaje de aceptabilidad (Quiroga &Yapura, 2008).
La producción intensiva de nopalitos en el Valle deLerma en camas de producción de reducida superficiees factible y se han obtenido rendimientos mayores a20.000 kg·ha-1 en períodos de producción de 7 meses(Lozano et al., 2009; Lozano et al., 2010), si bien lascondiciones climáticas de esta zona difieren de las delas zonas propuestas.
Tanto en la zona de los valles Calchaquies, comoen la Puna de la provincia de Salta, se observa la pre-sencia de Cactáceas y específicamente de opuntiashasta los 3.000 m.s.n.m. (Paoli et al., 2002; Trevisson& Demaio, 2006), cuyos cladodios se podrían utilizar
Lozano, L. - Ecofisiología de la tuna (Opuntia ficus-indica (L.) Mill.)
Horticultura Argentina 30(72): May.-Ago. 201150 ISSN de la edición on line 1851-9342
para iniciar la producción de nopalitos en camas deproducción intensiva, es decir, existe material de mul-tiplicación.
La zona de los valles Calchaquíes presenta una su-perficie de alrededor de 2.000 ha con suelos de pri-mera calidad para su uso agrícola, observándose prác-ticas no siempre adecuadas del uso del agua de rega-dío, pérdidas de humedad elevadas por evapotranspi-ración y altos valores de infiltración de agua en elsuelo (Valencia et al., 1970) factores negativos quepueden ser superados por las opuntias por su particularmorfología y fisiología.
En la Puna de Salta, la principal limitante está li-gada a las condiciones climáticas extremas y a la in-disponibilidad de agua superficial y/o subterránea encantidad y calidad. Pequeñas superficies para autocon-sumo (implantadas con papa, haba y maíz pueden sercultivadas en un ciclo agrícola limitado por un cortoperíodo libre de heladas que va desde mediados de no-viembre hasta fines de febrero (Paoli et al., 2002), pe-ríodo en el que la brotación de las pencas madres seproduciría favorecido por la benevolencia de las tem-peraturas. Se puede proponer para esta zona el uso demicrotúneles en las camas de multiplicación para mo-rigerar el efecto de las heladas sobre las pencas madrey lograr la brotación de éstas y la cosecha de jóvenescladodios (Luna Vázquez et al., 2001; Ramos Ríos &Quintana, 2004).
13. Conclusiones
La zona de los valles Calchaquíes y de la Puna dela provincia de Salta en Argentina presenta limitacio-nes climáticas relacionadas con la temperatura y la hu-medad (Paoli et al., 2002) que puede ser superada porlas opuntias allí presentes (Kiesling et al., 2011), porsus modificaciones morfológicas y fisiológicas. Algu-nas de ellas, como O. ficus-indica cuya existencia enesa zona ha sido comprobada por varios investigado-res puede ser cultivada para la producción de nopali-tos, por sus cladodios inermes (en su estado juvenil)para ser consumido como hortaliza de buen valor nu-tritivo (Landero Esquivel & Cruz Saenz, 2006).
Existiendo material de propagación en las zonaspropuestas, la difusión de las técnicas de producción,precedidas por degustación de las preparaciones ela-boradas con nopalitos, pueden inducir a los pequeñosproductores de estas regiones para introducir el cultivode la planta de tuna con fines hortícolas.
La gran plasticidad de las opuntias a las distintascondiciones de clima y suelo, otorgadas por sus carac-terísticas morfológicas y fisiológicas, deberían incen-tivar su cultivo con fines comerciales y con diversos
Sitio Argentino de Producción Animal
14 de 16

propósitos: consumo humano y animal, fines medici-nales, cosméticos, industriales, etc. en las zonas mar-ginales de la provincia de Salta antes mencionadas.
6. Bibliografía
Andrade, J.L.; De la Barrera, E.; Reyes García, C.; Ri-calde, M.F.; Vargas-Soto, G. & Cervera, J.C.2007. El Metabolismo Ácido de las Crasuláceas:Diversidad, Fisiología Ambiental y Productivi-dad. Bol. Soc. Bot. Méx. 81: 37-50.
Blanco Macias, F.; Lara Herrrera, A.; Valdez Cepeda,R.D.; Cortés Bañuelos, J.O.; Luna Flores, M. &Salas Luevano, M.A. 2006. Interacciones nutri-mentales y normas de la técnica de NutrimentoCompuesto en Nopal (Opuntia ficus-indica L.Miller). Revista Chapingo Serie Horticultura.12:165-175.
Cony, M.A.; Guevara, J.C.; Trione, S. & Estevez, O.2008. Response to freezing and high temperatu-res of detached cladodes from Opuntia species.Journal of the Professional Association for Cac-tus Development p. 36-48.
Cui, M. & Nobel, P.S. 1994. Gas exchange and growthresponses to elevated CO2 and light levels in theCAM species Opuntia ficus-indica. Plant Celland Environment 17:935-944.
Drennan, P.M. & Nobel, P.S. 2001. Responses of CAMspecies to increasing atmospheric CO2 concen-trations. Plant Cell and Environment. 23:767-781.
Flores Hernández, A.; Acosta Rodriguez, G.F.; MurilloAmador, B.; Trejo Calzada, R. & Arreola Avila,J.G. 2006. Evaluación preliminar de la reservade nopal (Opuntia spp.) en la región Laguna -Chihuahua. Revista Chapingo 5:191-196.
Gallegos Vázquez, C.; Olivares Sáenz, E.; Vázquez Al-varado, R. & Zavala García, F. 2000. Absorciónde nitrato y amonio por plantas de nopal en hi-droponía. TERRA Latinoamericana. 18: 133-139.
García, M.; Peña-Valdivia, C.; Trejo, L.; Valle, S.; Co-rrales, J. & Sánchez-Urdaneta, A. 2007. Efectodel potencial hídrico del suelo en el potencial os-mótico y membranas celulares de nopalito(Opuntia spp.) Revista Facultad Agronomía(LUZ) 24:305-311.
García de Cortázar, V.; Acevedo E. & Nóbel, P.S. 1985.Modeling of PAR interception and productivityby Opuntia ficus-indica. Agricultural and ForestMeteorology 34:145-162.
García de Cortázar, V. & Nóbel, P.S. 1992. Biomass andfruit production for the prickly pear cactus,
Lozano, L. - Ecofisiología de la tuna (Opuntia ficus-indica (L.) Mill.)
Horticultura Argentina 30(72): May.-Ago. 2011 51ISSN de la edición on line 1851-9342
Opuntia ficus-indica. Journal of the AmericanSociety for Horticultural Sciencies 117: 558-562.
González Monterrubio, C.F.; Monroy-Ata, A.; García-Amador, E. & Orozco-Almanza, M. 2005. In-fluencia de hongos micorrizógenos arbusculares(HMA) en el desarrollo de plántulas de Opuntia
streptacantha Lem. sometidas a sequía, en con-diciones de invernadero. TIP Revista Especiali-zada en Ciencias Químico Biológicas. 8:5-10.
Instituto de Botánica Darwinion. 1997-2011. Flora delCono Sur. Catálogo de Plantas Vasculares.
Kiesling, R.; Saravia, M.; Oakley, L.; Muruaga, N.;Metzing, D. & Novara, L. 2011. Aportes Botá-nicos de Salta. Ser. Flora. Herbario MCNS Fa-cultad de Cs. Naturales. Universidad Nacionalde Salta.
Landero Esquivel, F. & Cruz Sáenz, E. 2006. Adapta-ción del Nopal (Opuntia ficus-indica L. (Mill.))para la producción de Nopal Verdura en la co-munidad Buena Vista del Sur, Dirimba, Cardozo.Tesis (Ing. Agr). Managua, Nicaragua. Univer-sidad Nacional Agraria, Facultad de Agronomía,Departamento de Producción Vegetal p. 1-9.
Lozano, L.; Tálamo, A.; López, N.; Ortín, P.; Visuara,M. & Toncovich, M. 2009. Efecto de la fertiliza-ción orgánica en la producción de nopalitos(Opuntia ficus-indica (L.) Mill.) en el valle deLerma, Salta. XXXII Congreso Argentino deHorticultura. Salta Argentina. p. 422 (resúmen).
Lozano, L.; Palavecino, R.; Tálamo, A. & Ortín, P.2010.The effect of the Organic fertilization ongreen cladodes (Opuntia ficus-indica (L.) Mill.)yield en Lerma´s valley, Salta, Argentina. SecondYear. The VII th International Congress on Cac-tus Pear & Cochineal. Agadir, Morocco. p.21(abstract).
Luna Vázquez, J. 2001. Producción Intensiva de Nopalde verdura. INIFAP./CIRNE A-200.
Nobel, P.S. & Pimienta-Barrios, E. 1995. Monthly stemelongation for Stenocereus queretaroensis: rela-tionships to environmental conditions, net CO2
uptake and seasonal variations in sugar content.Environmental and Experimental Botany 35: 17-24.
Nobel, P.S. 2003 Ecofisiología de Opuntia ficus-indica.El Nopal (Opuntia spp.) como Forraje. FAO Pro-ducción y protección vegetal. 169:17-24.
Nobel, P.S.; Pimienta-Barrios, E.; Zañudo-Hernández,J. & Ramirez-Hernández, B.C. 2002. Historicalaspects and net CO2 uptake for cultivated Cras-sulacean acid metabolism plants in Mexico. An-nals of Applied Biology 140:133-142.
Nobel, P.S. & De la Barrera, E. 2003. Tolerances and
Sitio Argentino de Producción Animal
15 de 16

Lozano, L. - Ecofisiología de la tuna (Opuntia ficus-indica (L.) Mill.)
acclimation to low and high temperatures for cla-dodes, fruits and roots of a widely cultivated cac-tus, Opuntia ficus-indica. New Phytologist 157:271-279.
Nobel, P.S.; Castañeda, M.; North, G.; Pimienta Ba-rrios, E. & Ruiz, A. 1998. Temperature influen-ces on leaf CO2 exchange, cell viability and cul-tivation range for Agave tequilana. Journal onArid Environment 39: 1-9.
Nobel, P.S. 1999. Biología Ambiental. Agroecologíacultivo y usos del nopal. FAO Producción y pro-tección vegetal 132: Falta página.
Nobel, P.S & Zutta, B.R. 2008. Temperature tolerancesfor stems and roots of two cultivated cacti, No-
palea cochenillifera and Opuntia robusta: Accli-mation, light, and drought. Journal of Arid En-vironments. 72:633-642.
O. Camacho, C.; Peña Valdivia, C.B. & Sanchez-Urda-neta, A.B. 2007. Efecto del potencial hídrico delsuelo en el crecimiento y contenido de polisacá-ridos estructurales de nopalito (Opuntia spp.) Re-vista. Facultad. Agronomía (LUZ) 24:254-259.
Orona Castillo, I.; Cueto Wong, J.A.; Murillo Amador,B.; Santamaría-César, J.; Flores Hernández, A.;Valdez Cepeda, R.; García Hernández, J. & Tro-yo Dieguez, E. 2004. Extracción nutrimental denopal verdura bajo condiciones de riego por go-teo. Journal of the Professional Association forCactus Development. 6: 90-96.
Paoli, H.; Bianchi, A.R.; Yañez, C.E.; Volante, J.N.; Fer-nández, D.R.; Mattalía, M.C. & Noé, Y.E. 2002.Recursos Hídricos de la Puna, valles y Bolsonesáridos del Noroeste Argentino. Convenio INTAEEA Salta-CIED.
Pimienta-Barrios, E.; Robles Murguía, C.; Ruiz Corral,J.A.; Nobel, P.S. & García Galindo, J. 1999. Re-giones Térmicas Optimas y Marginales para elCultivo de Agave tequilana en el Estado de Ja-lisco, Universidad de Guadalajara, Guadalajara.
Pimienta-Barrios, E.; Zañudo Hernández, J.; Rosas Es-pinoza, V.C.; Valenzuela Tapia, A. & Nobel, P.S.2005. Young daughter cladodes affect CO2 up-take by mother cladodes of Opuntia ficus-indica.Annals of Botany 95: 363-369.
Pimienta-Barrios, E; Castillo Cruz, I.; Zañudo Hernán-dez, J.; Méndez Morán, L. & Nobel, P.S. 2007.Effects of shade, drought and daugther cladodes
Horticultura Argentina 30(72): May.-Ago. 201152 ISSN de la edición on line 1851-9342
on the CO2 uptake by cladodes of Opuntia ficus-
indica. Annals of Applied Biology. 151:137-144.
Pimienta-Barrios, E.; Gonzalez del Castillo Aranda,M.E.; Muñoz Urias, A. & Nobel, P.S. 2003. Ef-fects of Benomyl and drought on the Mycorrhi-zal development and daily net CO2 uptake of awild Platyopuntia in a rocky semi-arid environ-ment. Annals of Botany 92: 239-245.
Quiroga, S.K. & Yapura, S.M.E. 2008. Estandariza-ción y elaboración de preparaciones con nopa-lito fresco (Opuntia ficus-indica (L.) Miller).Tesis (Licenciatura en Nutrición). Salta, Argen-tina. 118 p.
Ramos Rios, J. & V. Quintana, M. 2004. Manejo Ge-neral del Cultivo de Nopal. Manual del Partici-pante. Colegio de Posgraduados. México 81 p.
Trevisson, M & Demaio, P. 2006. Cactus de Córdoba yel centro de Argentina. 1° Edición. Bs.As, Argen-tina.
Valencia, R.; Lago, A.; Chafatinos, T.; Ibarguren, R.;Menegatti, R.; Ocaranza, A. 1970. Primera Parte-Estudio de Campo. Levantamiento de Suelos delos Valles Calchaquíes. Convenio Gobierno de laProvincia de Salta - Universidad Nacional deSalta.
Vázquez-Vázquez, C.; Zúñiga Tarango, R.; Orona Cas-tillo, I.; Murillo Amador, B.; Salazar Sosa, E.;Vázquez-Alvarado, R.; García Hernández, J.L.& Troyo Dieguez, E. 2007. Análisis del creci-miento radical en cuatro variedades de nopal(Opuntia ficus-indica (L.) Mill.) Journal of theProfessional Association for Cactus Develop-ment 11: 82-90.
Véliz, J.; Franco Salazar, V. & García, M. 2007. Efectode la salinidad en Aloe vera y Opuntia ficus-in-
dica: plantas CAM de interés comercial. RevistaFacultad Agronomía. (LUZ) 24:337-341.
Zúñiga Tarango, R.; Orona Castillo, I.; Vázquez-Váz-quez, C.; Murillo Amador, B.; Zalazar Sosa, E.;López Martínez, J.D.; García Hernández, J. &Rueda Puente, E. 2009. Desarrollo radical, ren-dimiento y concentración mineral en nopalOpuntia ficus-indica (L.) Mill. En diferentes tra-tamientos de fertilización. Journal of the Profes-sional Association for Cactus Development 11:53-68.
Sitio Argentino de Producción Animal
16 de 16


















