DIVERSIDAD DE ESPECIES BRIÓFITAS: MUSGOS, …¡reas... · de acuerdo a las relaciones fi...
6
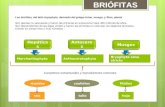
Musgos, hepáticas y antocerotes han sido considerados tradicionalmente clases de la división Bryophyta. Estos tres grupos (véase el cuadro 1), comparten las características de ser pequeñas plantas verdes, desprovistas de tejidos vas- culares que caracterizan a las plantas terrestres superiores (división Tracheophyta) y de mantener unidas, de manera permanente, las dos fases del ciclo de vida: la generación sexual, con los órganos productores de gametos (gametofi- to), y la generación asexual, productora de esporas (espo- rofito). Otra característica común a las briófitas es que la gene- ración dominante, verde y autótrofa, corresponde a la fase sexual que produce los gametos (gametofito), mientras que la planta asexual productora de esporas (esporofito) es una estructura que consiste de un pedicelo o seta que se ancla por su base al gametofito, dependiendo nutricionalmente de él, y que culmina en una cápsula que produce las esporas. Esto las diferencia de los helechos y plantas con semillas, donde la generación dominante e independiente es el es- porofito. HISTORIA DE LAS BRIÓFITAS EN CHILE El estudio de las briófitas chilenas comienza con la ex- pedición del Beagle, en la que el naturalista Charles Darwin recolectó material en el archipiélago de los Chonos y en el cabo Tres Montes (1839). El material recolectado por Darwin, depositado en herbarios europeos, fue estudiado por dife- rentes botánicos, entre otros, Taylor (1846), Dalton Hooker (1847) y Müller (1847, 1885). El naturalista francés, Claudio Gay, incluye en su obra “Historia Física y Política de Chile” un capítulo sobre mus- gos y hepáticas, realizado por C. Montagne (1850), donde se describen 117 especies de hepáticas y 179 de musgos para Chile. DIVERSIDAD DE ESPECIES BRIÓFITAS: MUSGOS, HEPÁTICAS Y ANTOCEROTES ELIZABETH BARRERA Y FELIPE OSORIO Cuadro 1. Diferencias entre hepáticas, musgos y antocerotes. Carácter Hepáticas Musgos Antocerotes Protonema Reducido Desarrollado Carecen Gametofito Taloso o folioso Folioso Taloso Rizoides Unicelulares Pluricelulares Unicelulares Disposición de filidios Dos hileras 3 o más hileras Células de filidios Isodiamétricas Generalmente alargadas Hidroides Ausentes Presentes* Ausentes Esporangio Sin estomas Con estomas* Con estomas* Dehiscencia de la cápsula Por fisuras longitudinales Sin peristoma Por opérculo* Con peristoma Por fisuras longitudinales Sin peristoma Columela Ausente Presente* Presente* Eláteres Presentes Ausente Pseudoeláteres * No en todos. Página derecha: Consideradas plantas exitosas, los musgos tienen facilidad para propagarse y obtener nutrientes de las lluvias y el rocío. Al evitar la erosión del suelo, cumplen un rol ecológico im- portante. Son además alimento de innumerables invertebrados y vertebrados. En las imágenes se observan Musgos (1 a 5), Antoce- rote (6) y Hepáticas (7 y 8). Fotos: Felipe Osorio. 340
Transcript of DIVERSIDAD DE ESPECIES BRIÓFITAS: MUSGOS, …¡reas... · de acuerdo a las relaciones fi...

Musgos, hepáticas y antocerotes han sido considerados tradicionalmente clases de la división Bryophyta. Estos tres grupos (véase el cuadro 1), comparten las características de ser pequeñas plantas verdes, desprovistas de tejidos vas-culares que caracterizan a las plantas terrestres superiores (división Tracheophyta) y de mantener unidas, de manera permanente, las dos fases del ciclo de vida: la generación sexual, con los órganos productores de gametos (gametofi -to), y la generación asexual, productora de esporas (espo-rofi to).
Otra característica común a las briófi tas es que la gene-ración dominante, verde y autótrofa, corresponde a la fase sexual que produce los gametos (gametofi to), mientras que la planta asexual productora de esporas (esporofi to) es una estructura que consiste de un pedicelo o seta que se ancla por su base al gametofi to, dependiendo nutricionalmente de él, y que culmina en una cápsula que produce las esporas. Esto las diferencia de los helechos y plantas con semillas, donde la generación dominante e independiente es el es-porofi to.
HISTORIA DE LAS BRIÓFITAS EN CHILE
El estudio de las briófi tas chilenas comienza con la ex-pedición del Beagle, en la que el naturalista Charles Darwin recolectó material en el archipiélago de los Chonos y en el cabo Tres Montes (1839). El material recolectado por Darwin, depositado en herbarios europeos, fue estudiado por dife-rentes botánicos, entre otros, Taylor (1846), Dalton Hooker (1847) y Müller (1847, 1885).
El naturalista francés, Claudio Gay, incluye en su obra “Historia Física y Política de Chile” un capítulo sobre mus-gos y hepáticas, realizado por C. Montagne (1850), donde se describen 117 especies de hepáticas y 179 de musgos para Chile.
DIVERSIDAD DE ESPECIES
BRIÓFITAS: MUSGOS, HEPÁTICAS Y ANTOCEROTESELIZABETH BARRERA Y FELIPE OSORIO
Cuadro 1. Diferencias entre hepáticas, musgos y antocerotes.
Carácter Hepáticas Musgos Antocerotes
Protonema Reducido Desarrollado Carecen
Gametofi to Taloso o folioso Folioso Taloso
Rizoides Unicelulares Pluricelulares Unicelulares
Disposición de fi lidios Dos hileras 3 o más hileras
Células de fi lidios Isodiamétricas Generalmente alargadas
Hidroides Ausentes Presentes* Ausentes
Esporangio Sin estomas Con estomas* Con estomas*
Dehiscencia de la cápsulaPor fi suras longitudinalesSin peristoma
Por opérculo*Con peristoma
Por fi suras longitudinalesSin peristoma
Columela Ausente Presente* Presente*
Eláteres Presentes Ausente Pseudoeláteres
* No en todos.
Página derecha: Consideradas plantas exitosas, los musgos tienen facilidad para propagarse y obtener nutrientes de las lluvias y el rocío. Al evitar la erosión del suelo, cumplen un rol ecológico im-portante. Son además alimento de innumerables invertebrados y vertebrados. En las imágenes se observan Musgos (1 a 5), Antoce-rote (6) y Hepáticas (7 y 8). Fotos: Felipe Osorio.
340

341
Capítulo II: Nuestra Diversidad Biológica
1. Ptychomnion cygnisetum (C. Müll.) Kinb. 2. Ancistrodes genufl exa (C. Müll.) Crosby.
3. Rigodium implexum Kunz., “lana del pobre”. 4. Hypopterygium arbuscula Brid, “paragüita del sapo”.
5. Dendroligotrichum dendroides Broth. ssp. dendroides, “musgo pinito”.
6. Megaceros sp.
7. Planaria sp. sobre Lepidozia chordulifera Taylor. 8. Cryptochila grandifl ora (Lindenb & Gottsche) Grolle.

342
Biodiversidad de Chile, Patrimonio y Desafíos
En el año 1866, Paul Gunther Lorentz registra los mus-gos colectados por el Dr. Krause en la costa valdiviana. Más tarde, en 1869, William Mitten describe nuevas especies de musgos chilenos en su obra Musci Austro-Americani.
Entre otros importantes botánicos, siempre extranjeros, que aportaron al conocimiento de la fl ora briofítica de Chile se puede citar a Brotherus (1924), Cardot y Brotherus (1923), Dusén (1903, 1905, 1906), Herzog (1938, 1954), Herzog y Hosseus (1938), Roivainen (1937). Thériot, entre los años 1915 y 1935, publicó numerosos trabajos sobre este grupo de vegetales.
SISTEMÁTICA Y TAXONOMÍA DEL GRUPO
Estudios modernos basados en caracteres molecula-res han demostrado que existen varios linajes de distintas edades dentro del grupo, ya diferenciados en el tiempo de colonización del ambiente terrestre, en el Paleozoico. Así, de acuerdo a las relaciones fi logenéticas entre los grupos de briófi tas propuesta por Boop y Capesius (1996), las gran-des hepáticas talosas complejas, la clase Marchantiopsida, precisamente aquellas que le confi eren el nombre al grupo, serían el linaje basal de las briófi tas y las más divergentes y antiguamente diferenciadas. Por otra parte, la clase Junger-manniopsida, el subconjunto de hepáticas talosas simples (subclase Metzgeriidae) y todas las hepáticas foliosas (sub-clase Jungermanniidae), conformarían el grupo hermano de las hepáticas talosas complejas, conjuntamente con los musgos y antocerotes.
Sobre este fundamento, actualmente las briófi tas son cla-sifi cadas en tres divisiones (véase el cuadro 2) distintas de plantas (Crandall-Stotler y Stotler, 2000).
Cuadro 2. Clasifi cación según Crandall-Stotler y Stotler (2000).
División Clase Subclase
Marchantiophyta (=Hepatophyta) “hepáticas”
Marchantiopsida “hepáticas con talos complejos”
Jungermanniop-sida “hepáticas foliosas y talosas simples”
Metzgeriidae “hepáticas talosas simples”
Jungermanniidae “hepáticas folio-sas”
Anthocerotophyta (“antocerotes”)
Bryophyta (“mus-gos”)
A nivel mundial, los tres grupos de organismos mencio-nados anteriormente, suman más de 20.000 especies; por lo tanto, considerándolos en conjunto, representan a uno de los
grupos más diversos de plantas terrestres. Las hepáticas (Mar-chantiophyta) suman alrededor de 8.000 a 10.000 especies, distribuidas en 14 órdenes y 368 géneros (Crandall-Stotler y Stotler, 2000). Antocerotes (Anthocerotophyta) reúne alrede-dor de 400 especies en 10 géneros y los musgos (Bryophyta) agrupan alrededor de 13.000 especies, distribuidas en 700 géneros.
Estos grupos exhiben la mayor riqueza de especies y di-versidad de formas en los ecosistemas templado-lluviosos del sur de Sudamérica, debido a las condiciones favorables que allí existen, como el clima oceánico muy húmedo, la diversidad de hábitat y las temperaturas más o menos mode-radas por la infl uencia marina.
Si He, en 1988, entrega una lista de musgos para Chile donde cita 778 especies, 203 géneros y 63 familias.
Con relación a hepáticas y antocerotes, Hässel de Me-néndez (2003) y Hässel de Menéndez y Rubies (2004), con-feccionaron un “Catálogo de las Hepáticas y Antocerotes de Chile, Argentina y Uruguay”, entregando un número de 741 taxa para esta área; de este total, 549 taxa se encuentran re-presentados en Chile, los que principalmente corresponden a taxa endémicos del área de los bosques templado-lluviosos del sur del país.
Con el propósito de proporcionar una introducción al co-nocimiento de las hepáticas asociadas a estos ecosistemas, Villagrán y otros (2005) se concentraron durante los últimos tres años en las colecciones, determinaciones e ilustraciones de la fl ora de hepáticas y antocerotes del archipiélago de Chiloé (41°47’ – 43°30’S), considerado un destacado núcleo de biodiversidad de briófi tas en Chile (Villagrán et al. 2003), tanto por su riqueza fl orística como por su heterogeneidad vegetacional. Una de las causales de esta riqueza reside en el rol de refugio de fl ora subantártica que exhiben las islas durante las repetidas glaciaciones del Pleistoceno (Villagrán, 2001). La riqueza de especies de hepáticas y antocerotes re-conocidas a la fecha para Chiloé es de 79 géneros y 229 especies, números correspondientes a una buena proporción (85,9 por ciento de los géneros y 72,7 por ciento de las es-pecies) de la fl ora hepatológica de la Región de Los Lagos de Chile (Hässel de Menéndez y Rubies, 2004).
DIVERSIDAD Y DISTRIBUCIÓN
División Anthocerotophyta (antocerotes, hierbas con cuerno)
Se distribuyen de preferencia en hábitat húmedos y sombríos de las regiones subtropicales y templadas del glo-bo. El gametofi to es taloso, formado por láminas lobuladas que se disponen en rosetas cóncavas, de bordes sinuosos u ondulados, de 3 a 10 centímetros de diámetro. Se fi jan al sustrato por rizoides lisos y unicelulares. El talo es pluries-tratifi cado. Las especies pueden ser monoicas o dioicas, los órganos sexuales, anteridios y arquegonios, se encuentran hundidos en el talo. El esporofi to consta de un pie bulboso y una cápsula cilíndrica, generalmente larga y con aspecto de “cuerno”.

343
Capítulo II: Nuestra Diversidad Biológica
Muchas especies de este grupo presentan cavidades mucilaginosas que frecuentemente están ocupadas por co-lonias de cianófitos del género Nostoc.
División Marchantiophyta (hepáticas)El gametofito puede ser laminar (hepáticas talosas) o
estar formado por caulidios y filidios (hepáticas foliosas). Se unen al sustrato por medio de rizoides unicelulares. Las células tienen varios cloroplastos y en la mayoría de los casos tienen cuerpos oleiferos, que contienen aceites y esencias aromáticas. El esporofito, que en general es de vida efímera, consta de un pie, una seta y de una cápsula o esporangio.
Las hepáticas tienen un significativo interés biogeo-gráfico y evolutivo por su larga historia geológica, la que se remonta a los inicios de la colonización vegetal de la superficie terrestre, en la era paleozoica (hace 480 millo-nes de años), conjuntamente con el origen de las plantas vasculares. Estas plantas tienen dos clases de morfologías
básicas, ya sea ejes provistos de “hojitas” (hepáticas foliosas), o formas de mayor estatura, aplanadas y no diferenciadas en ejes y hojitas (hepáticas talosas).
División Bryophyta (musgos)En este grupo, el gametofito siempre es folioso y puede
medir desde unos pocos milímetros hasta 60 centímetros. Consta de un caulidio (“tallo”), erecto o rastrero que se fija al sustrato mediante rizoides pluricelulares, ramificados y con paredes transversales oblicuas, los filidios (“hojitas”) están dispuestos a su alrededor. Los esporofitos son diversificados y complejos y están formados por una seta y una cápsula o esporangio, la que, por lo menos en las primeras fases del desarrollo, está cubierta por la caliptra. La dehiscencia de la cápsula se produce mediante el opérculo; en algunos grupos no existe opérculo y las esporas salen a través de la rotura o descomposición de la pared de la cápsula. Las esporas, al germinar, originan un protonema pluricelular, filamentoso que, al desarrollarse, se transformará en gametofito.
Las turberas se caracterizan por tener depósitos esponjosos de turba, el crecimiento de árboles, arbustos y, dependiendo del tipo, un suelo cubierto por una alfombra gruesa de musgo. Los humedales son ecosistemas conformados por estratos subyacentes originados por acumulación de materia orgánica de origen vegetal. En la imagen se observa una turbera de Sphagnum spp., musgo conocido como Pon-Pón. Foto: Felipe Osorio.

344
Biodiversidad de Chile, Patrimonio y Desafíos
ECOLOGÍA DEL GRUPO
Son plantas exitosas, tienen facilidad para propagarse y obtener los nutrientes necesarios del agua de lluvia o del ro-cío. Son los primeros colonizadores de rocas y suelos desnu-dos pobres en nutrientes. Como acumulan materia orgánica, facilitan el asentamiento de plantas vasculares. Estas caracte-rísticas les permiten desarrollarse en lugares inhóspitos para otras plantas, como por ejemplo en la Antártica, donde se encuentran más de 150 especies de briófi tos, a diferencia de las plantas vasculares que sólo se hacen presentes con dos especies.
Cumplen importantes roles ecológicos puesto que evitan la erosión del suelo, fi jan en sus estructuras enormes canti-dades de C y N, ayudan a los bosques a mantener una sig-nifi cativa carga de humedad necesaria para la subsistencia de las especies, evitando así el estrés hídrico, y sirven de vivienda, cobijo y alimentación a innumerables invertebra-dos y vertebrados.
SINGULARIDADES
Los briófi tos, principalmente por ser muy sensibles a la contaminación, pueden tener un papel muy destacado como bioindicadores de la polución ambiental, al utilizarse para elaborar el índice de pureza atmosférica, el cual se basa en el número, frecuencia, recubrimiento y factor de resistencia de
las especies que viven en un determinado lugar. Este índice da una idea de los efectos de la polución en dicha área. Es fá-cilmente observable que los musgos no crecen en lugares cer-canos a áreas industriales, incluso en las grandes ciudades.
Existen grupos de briófi tos que pueden retener de forma pasiva los contaminantes, de tal manera que al realizar aná-lisis químicos de la cubierta muscinal, se puede conocer la concentración de metales pesados como plomo, zinc, cad-mio y de isótopos radioactivos (Brugués, 1998).
La cubierta de musgos acumula fácilmente los granos de polen que precipitan del aire por acción de la gravedad o de la lluvia, durante largos períodos. Esto permite realizar estudios palinológicos, extrayendo de los musgos la “lluvia polínica” y conocer, cualitativa y cuantitativamente, los gra-nos de polen allí acumulados. De esta forma, los espectros polínicos obtenidos se pueden comparar con aquellos obte-nidos de sedimentos fósiles.
En jardinería los musgos se utilizan principalmente por su capacidad de retención de agua y son ocupados como sustratos para el desarrollo de otras plantas.
NECESIDADES DE INVESTIGACIÓN Y PROYECCIONES
Los estudios sobre musgos, hepáticas y antocerotes chile-nos por parte de botánicos nacionales han tenido un escaso desarrollo, situación que ya es preocupante, pero que reviste mayor gravedad cuando, como se sabe, especialistas extran-
Las briófi tas poseen características peculiares que les permiten sobrevivir en ambientes gélidos. Es el caso de este musgo de la especie Racomitrium lanuginosum (Hedw.) Brid, que puede encontrarse en lago Yulton, en la Región de Aisén. Foto: Víctor Ardiles.

345
Capítulo II: Nuestra Diversidad Biológica
jeros recolectan y estudian material chileno, muchas veces sin nuestro conocimiento y sin siquiera depositar ejempla-res duplicados en herbarios nacionales. Esta actitud frena el desarrollo de estudios por parte de jóvenes investigadores nacionales que comienzan a interesarse en estos grupos de plantas.
Bibliografía Boops M. & I. Capesius. 1996. “New Aspects of Bryophyte Taxo-
nomy Provided by a Molecular Approach”. Botanical Acta, 109: 368-372.
Brotherus, V.F. 1924. “Musci nonnulli Chilense a C. Skottsberg anno 1917 lectae”. Acta Horti Gothoburgensis, 1: 189-195.
Brotherus, V.F. 1924. “The musci of the Juan Fernández Islands”. In: Skottsberg, The Natural History of Juan Fernández and Easter Is-lands, 2: 409-448.
Brugués, M. 1998. “Briófi tos”. In: Izco, J. et al. (eds.), Botánica, McGraw-Hill, Interamericana, 781 pp.
Cardot, J. & V. F. Brotherus. 1923. “Les mousses, in botanische erge-binse der Schwedischen expedirtion nach Patagonien und dem Feurlande 1907-1909”. Bihang til Kongliga Svenska Vetenskaps. Akademien Handlingar, 63: 1-73.
Crandall-Stotler B. & R.E. Stotler. 2000. “Morphology and classifi ca-tion of the Marchantiophyta”. In: A.J. Shaw & B. Goffi net (eds.), Bryophyte Biology. Cambridge University Press.
Darwin, C. 1839. Journal of Researches into Geology and Natural History of the various countries visited by H.M.S. Beagle under the command of Captain Fitzroy, R.N. from 1832-1836. London, 615 pp.
Dusén, P. 1903. “Beitrage zur Bryologie der Magellanslander von Westpatagonien und Südchile I”. Arkiv för Botanik, K. Svenska Vetenskaps. Akademien Handlingar, 1(1): 441-466.
Dusén, P. 1903. “Patagonian and Fuegian mosses”. Part III. Reports of the Princeton University Expeditions to Patagonia. Botany, 8: 63-125.
Dusén, P. 1905. “Beitrage zur Bryologie der Magellanslander von Westpatagonien und Südchile II”. Arkiv för Botanik, K. Svenska Vetenskaps-Akademien, 4(1): 1-45.
Dusén, P. 1905. “Beitrage zur Bryologie der Magellanslander von Westpatagonien und Südchile III”. Arkiv för Botanik, K. Svenska Vetenskaps-Akademien, 4(13): 1-24.
Dusén, P. 1905. “Musci nonnulli novi e Fuegia et Patagonia reporta-ti”. Botaniska Notiser, 1905: 299-310.
Dusén, P. 1906. “Beitrage zur Bryologie der Magellanslander von Westpatagonien und Südchile IV”. Arkiv för Botanik, K. Svenska Vetenskaps-Akademien, 6(8): 1-40.
Hässel de Menéndez, G. 2003. “Hepáticas y Anthocerotas del Sur de Sudamérica”. Boletín de la Sociedad Argentina de Botánica, 38: 5-6 (Suplemento).
Hässel de Menéndez, G. & M.F. Rubies. 2004. Catalogue of Mar-chantiophyta and Anthocerotophyta of Chile, Argentina and Uruguay. (En edición).
He, S. 1998. “A Checklist of the mooses of Chile”. J. Hattori Bot. Lab. 85: 103-189.
Herzog, T. y C. Hosseus. 1938. “Contribución a la Flora Briófi ta del sur de Chile”. Archivos de la Escuela de Ciencias Médicas de Córdoba, 7: 3-95.
Herzog, T. 1954. “Zur Bryophytenfl ora Chile”. Revue Bryologique et Lichenologique, 23: 27-99.
Hooker, J.D. 1847. Botany of Fuegia, the Falklands, Kerguelen’s land etc. The cryptogamic Botany of the Antarctic Voyage of H.M. Discovery Ships Erebus and Terror, 2: 219-574.
Lorentz, P.G. 1866. “Musci frondosi in Chile prope Valdiviam et pro-pe Corral lecti per Dr. Krause”. Botanische Zeitung (Berlin), 24: 185-189.
Mitten, W. 1869. “Musci austro-americani enumeratio omnium aus-tro-americanorum auctori hucusque congnitorum. Journal of the Linnean Botany Society”, Botany, 12: 1-659.
Montagne, C. 1850. “Musgos y Hepáticas”. In Gay. C. Historia Física y Política de Chile, tomo VII: 5-327.
Müller, C. 1847. “Des muscis nonnullis novis vel minus cognitis exo-ticis”. Botanische Zeitung (Berlin), 5: 801-806, 825-830.
Müller, C. 1885. “Bryologia fuegiana”. Flora, Jena, 68 (21-23): 391-429.
Roivainen, H. 1937. “Bryological investigations in Tierra del Fuego”. 1. Sphagnaceae-Dicranaceae. Annales Botanici Soc. Zoologicae Botanicae Fennicae ‘Vanamo’, 9: I-X: 1-58.
Taylor, 1846. “New Hepaticae”. London Journal of Botany, 5: 258-284, 365-417.
Theriot, I. 1915. “Sur quelques mousses recueillies au Chili par le Prof. Carlos Porter”. Rev. Chil. Hist. Nat. 19: 30-37.
Theriot, I. 1917. “Contribution a la fl ore bryologique du Chili”. Rev. Chil. Hist. Nat. 21: 6-37, 1918. 22: 79-94, 1921. 25: 289-312, 1923. 27: 9-15, 1924. 28: 129-139, 1925. 29: 287-292, 1926. 30: 341-361, 1927. 31: 30-37, 1928. 32: 252-255, 1929. 33: 515-517, 1930. 34: 258-262, 1934. 38: 83-85, 1935. 39: 16-21.
Villagrán, C. 2001. “Un modelo de la historia de la vegetación de la Cordillera de la Costa de Chile central-sur: la hipótesis glacial de Darwin”. Revista Chilena de Historia Natural, 74: 793-803.
Villagrán, C. y E. Barrera. 2002. Los Musgos del Archipiélago de Chi-loé. CONAF, Master Print Ltda. Puerto Montt, 24 pp.
Villagrán, C., E. Barrera y C. Medina. 2002. Las Hepáticas del Ar-chipiélago de Chiloé. CONAF, Master Print Ltda. Puerto Montt, 26 pp.
Villagrán, C., E. Barrera, J. Cuvertino y N. García. 2003. “Musgos de la Isla Grande de Chiloé, X Región, Chile: Lista de especies y rasgos fi togeográfi cos”. Boletín del Museo Nacional de Historia Natural, Chile, 52: 17-44.
Villagrán, C., G., Hässel y E. Barrera. 2005. Hepáticas y Antocerotes del Archipiélago de Chiloé. Una introducción a la fl ora briofítica de los ecosistemas templado-lluviosos del sur de Chile. Corp. de Amigos del Museo Nacional de Historia Natural, 160 pp.