
Detección, purificación y caracterización parcial de ... · interacción hospedero patógeno y a...
125
Detección, purificación y caracterización parcial de lectinas presentes en algas marinas colombianas Deisy Johana Hidalgo Roa Universidad Nacional de Colombia Facultad de Ciencias, Departamento Química Bogotá, Colombia 2017
Transcript of Detección, purificación y caracterización parcial de ... · interacción hospedero patógeno y a...

Detección, purificación y caracterización parcial de
lectinas presentes en algas marinas colombianas
Deisy Johana Hidalgo Roa
Universidad Nacional de Colombia
Facultad de Ciencias, Departamento Química
Bogotá, Colombia
2017

Detección, purificación y caracterización parcial de lectinas presentes en algas
marinas colombianas
Deisy Johana Hidalgo Roa
Tesis o trabajo de investigación presentado como requisito parcial para optar al título de:
Maestría en Ciencias Bioquímica
Directora
Dr Sc Nohora Angélica Vega Castro
Codirectora
Ph.D. Mónica Puyana Hegedus
Línea de Investigación:
Estudios de Lectinas de algas marinas del Caribe Colombiano
Grupo de Investigación:
Grupo de investigación en Proteínas (GRIP)
Universidad Nacional de Colombia
Facultad de Ciencias, Departamento Química
Bogotá, Colombia
2017

Dedicado a todas las personas que han creído en mí,
gracias a su voto de confianza hoy sé que he crecido un poco más.
“… No te rindas, por favor no cedas,
aunque el frio queme,
aunque el miedo muerda,
aunque el sol se esconda y se calle el viento,
aún hay fuego en tu alma,
aún hay vida en tus sueños… “
Mario Benedetti

Agradecimientos
A Dios por regalarme fuerza y voluntad para emprender cada día nuevos proyectos.
Agradecimientos especiales a mi directora de tesis Nohora Angélica Vega Castro por su
apoyo incondicional y por sus conocimientos, los cuales fueron primordiales para el
desarrollo y término exitoso de este trabajo. Al profesor Edgar Antonio Reyes Montaño un
maestro y ser humano excepcional. A la doctora Mónica Puyana Hegedus y el equipo de
trabajo que ayudo en la colección de las muestras y su posterior identificación. A mis
compañeros del grupo de investigación en proteínas por su colaboración en el proceso de
ejecución del proyecto.
También agradezco a Andrea Ruiz del grupo de Andrología de la Facultad de Veterinaria
de la Universidad Nacional de Colombia, por la ayuda en la obtención de las muestras de
sangre de animales para cada uno de los ensayos realizados. Al grupo de investigación
interacción hospedero patógeno y a el profesor Harold Duban Ardila Barrantes por su
colaboración.
A Colciencias por darme la beca pasantía “joven investigadora”, que me permitió realizar
la maestría en Ciencias bioquímica.

Resumen y Abstract
Resumen
El objetivo de este estudio fue realizar estudios enfocados en la búsqueda de lectinas de
algas marinas del caribe colombiano y estudiarlas bioquímicamente. Con este fin fue
evaluado un método para la obtención de extractos y su posterior determinación de
actividad aglutinante. Los resultados indicaron que lavados sucesivos de la harina de alga
con acetona al 97%, y diálisis de los extractos son dos pasos necesarios para obtener
mayores cantidades de proteína y además eliminar moléculas de naturaleza reductora,
polifenoles, terpenoides, y pigmentos de los extractos facilitando la cuantificación por el
método del ácido bicinconinico (BCA) y la evaluación de su actividad aglutinante usando
eritrocitos de oveja tripsinizados. Se determinó la presencia de lectinas en cinco extractos
de las siete especies evaluadas. Las especies Tricleocarpa cylindrica y Bryopsis ramulosa
aglutinaron eritrocitos de humano tipo O, mientras que especies como Caulerpa taxifolia,
Dictyota pinnatifida, Sargassum hystrix var buxifolium y B. ramulosa, eritrocitos de oveja
tratados enzimáticamente.
Se realizó la purificación y caracterización parcial de las lectinas de B. ramulosa
(Chlorophyta), tras una precipitación fraccionada del extracto salino con sulfato de amonio
entre el 0-50%s, cromatografía de intercambio iónico sobre DEAE-Sepharosa o Sephacel.
Los resultados mostraron la presencia de varias lectinas en el extracto. La fracción retenida
de intercambio fue purificada por cromatografía de afinidad sobre aMSB-Sepharosa 4B y
se obtuvo una fracción retenida con actividad aglutinante que mostró por PAGE-SDS una
banda en 18 kDa. En la fracción no retenida se encontró una lectina tetramerica con
subunidades de 15 kDa; mostró afinidad por L-fucosa, D- manosa y D-glucosa. Los
resultados in silico mostraron la lectina de 15 kDa podría tener un dominio de tipo F,
fucolectina, aunque no idéntico al encontrado en bryohealin mientras que para la lectina
de 18 kDa su especificidad hacia GalNAc, podría indicar que se trata de una proteína
diferente con respecto a las estudiadas en especies relacionadas a B. ramulosa.
Palabras clave: Lectina, Bryopsis ramulosa, Aglutinación, Algas marinas

Abstract
The objective of this study was to carry out studies focused on the research of seaweed
lectins from the Colombian Caribbean and to study them biochemically. To this end, a
method for obtaining extracts y their subsequent determination of agglutinate activity was
evaluated. The results indicated that successive washes of seaweed flour with 97%
acetone and dialysis of the extracts are two steps necessary to obtain higher amounts of
protein and eliminate molecules of reducing nature, polyphenols, terpenoids, y pigments
from the extracts facilitating the quantification by the bicinchoninic acid method (BCA) y
evaluation of its agglutinate activity using trypsinized sheep erythrocytes. The presence of
lectins in five extracts of the seven evalute species was determined. The species
Tricleocarpa cylindrica y Bryopsis ramulosa agglutinated human type O erythrocytes, while
species such as Caulerpa taxifolia, Dictyota pinnatifida, Sargassum hystrix var buxifolium
and B. ramulosa, sheep erythrocytes treated enzymatically.
Was carried out a Partial purification and characterization of B. ramulosa (Chlorophyta)
lectins, after the fractional precipitation of the saline extract with ammonium sulfate
between 0-50%s, ion exchange chromatography on DEAE-Sepharose or Sephacel. The
results showed the presence of several lectins in the extract. The retained fraction from
exchange was purified by affinity chromatography on aMSB-Sepharose 4B and was
obtained a retained fraction with agglutinate activity which showed by PAGE-SDS an 18
kDa by. In the non-retained fraction a tetrameric lectin with 15 kDa subunits was found;
Which showed affinity for L-fucose, D-mannose and D-glucose. The in silico results showed
the 15 kDa lectin could have a F-type domain, fucolectin, although not identical to that
found in bryohealin whereas for the 18 kDa lectin its specificity towards GalNAc might
indicate that it is a different protein with respect to other lectins studied in species related
to B. ramulosa.
Keywords: Lectin, Bryopsis ramulosa, agglutination, seaweed

Contenido
1. Marco Teórico ........................................................................................................... 3 1.1. Historia ............................................................................................................... 3 1.2. Lectinas .............................................................................................................. 4 1.3. Estrategias para purificar lectinas ..................................................................... 14 1.4. Algas ................................................................................................................ 15
1.4.1. Características citológicas de las algas marinas ............................................ 16 1.4.2. Lectinas en algas ........................................................................................... 17
2. Objetivos ................................................................................................................. 23 2.1. Objetivo general ............................................................................................... 23 2.2. Objetivos específicos ....................................................................................... 23
3. Capítulo 1. Preparación y evaluación de la actividad aglutinante en extractos de algas marinas del caribe colombiano. .................................................................... 24
3.1. Metodología. .................................................................................................... 24 3.1.1. Colección de muestras .................................................................................. 24 3.1.2. Obtención de los extractos y evaluación su actividad aglutinante .................. 24 3.1.3. Cuantificación de proteínas BCA ................................................................... 26 3.1.4. Ensayos de aglutinación ................................................................................ 26 3.1.5. Cuantificación de proteínas por el método de micro Kjeldahl. ........................ 26 3.1.6. Tratamiento estadístico de los datos .............................................................. 27
3.2. Resultados. ...................................................................................................... 27 3.2.1. Obtención de los extractos salinos ................................................................ 27 3.2.2. Cantidad de proteína en extractos de algas marinas ..................................... 28 3.2.3. Actividad aglutinante de extractos no dializados ............................................ 30 3.2.4. Actividad aglutinante con eritrocitos de oveja tratados enzimáticamente. ...... 31
3.3. Análisis ............................................................................................................. 33 3.3.1. Porcentajes de proteína soluble ..................................................................... 33 3.3.2. Ensayos de aglutinación ................................................................................ 37
3.4. Conclusiones .................................................................................................... 41
4. Capítulo 2. Purificación de la (s) lectina(s) presente(s) en Bryopsis ramulosa . 42 4.1. Metodología ..................................................................................................... 42
4.1.1. Colección de la muestra ................................................................................ 42 4.1.2. Obtención de la harina de B. ramulosa .......................................................... 42 4.1.3. Determinación porcentaje de humedad y contenido de proteína total de la harina de B. ramulosa .............................................................................................. 42 4.1.4. Obtención de los extractos proteicos ............................................................. 43 4.1.5. Cuantificación de proteínas ........................................................................... 43 4.1.6. Electroforesis unidimensional en geles de poliacrilamida. .............................. 43 4.1.7. Ensayos preliminares de purificación: precipitación fraccionada y cromatografía de intercambio iónico a pequeña escala. ........................................... 44 4.1.8. Ensayos de inhibición con carbohidratos ....................................................... 44 4.1.9. Purificación a gran escala de la (s) lectina(s) de B. ramulosa ........................ 45 4.1.10. Purificación por cromatografía de afinidad .............................................. 46
4.2. Resultados ....................................................................................................... 48 4.2.1. Ensayos preliminares de purificación: precipitación fraccionada y cromatografía de intercambio iónico a pequeña escala. ........................................... 48 4.2.2. Ensayos de inhibición con carbohidratos ....................................................... 51

4.2.3. Preparación y purificación de proteínas a gran escala del extracto de B. ramulosa. ................................................................................................................. 52 4.2.4. Purificación por cromatografía de afinidad ..................................................... 54
4.3. Análisis ............................................................................................................. 58 4.4. Conclusiones .................................................................................................... 63
5. Capítulo 3. Modelamiento molecular y docking de la lectina bryohealin, aislada del alga verde Bryopsis plumosa (clorophyta), frente a diferentes ligandos. .......... 64
5.1. Metodología ...................................................................................................... 64 5.1.1. Análisis de la secuencia y modelo tridimensional .......................................... 64 5.1.2. Docking molecular ......................................................................................... 65
5.2. Resultados y análisis ........................................................................................ 65 5.3. Conclusiones .................................................................................................... 76 5.4. Perspectivas ..................................................................................................... 76
6. Anexo 1. Tablas ...................................................................................................... 77
7. Anexo 2. Figuras .................................................................................................... 85
8. Bibliografía ............................................................................................................. 95

Contenido
Lista de figuras
Figura 1-1 Representación esquemática de la clasificación de lectinas dependiendo el
número de sitios de reconocimiento al carbohidrato.. ....................................................... 7
Figura 1-2 Clasificación de las lectinas en términos de plegamiento de dominio de unión
a carbohidrato. ................................................................................................................. 8
Figura 1-3 Comparación de la estructura terciaria de un monómero de proteína típica del
grupo Jacalina (Helianthus tuberosus) y de un dimero de Griffithsia sp. ......................... 20
Figura 3-1 Diagrama de flujo, procedimiento usado para elaboración y evaluación de los
extractos. ........................................................................................................................ 25
Figura 3-2 Eliminación de pigmentos con lavados sucesivos de acetona ...................... 27
Figura 3-3 Vista al microscopio de aglutinaciones obtenidas para diferentes extractos de
algas provenientes de harina tratada con acetona frente a eritrocitos de oveja. ............. 32
Figura 3-4 Eliminación secuencial de pigmentos para una muestra de harina de B.
ramulosa. ....................................................................................................................... 37
Figura 3-5 Región terminal del antígeno de Forssman. ................................................ 40
Figura 4-1 Esquema general del proceso de purificación de un extracto de B. ramulosa47
Figura 4-2 Perfil electroforético de extracto crudo de B. ramulosa. ................................ 48
Figura 4-3 Precipitación fraccionada con sulfato de amonio para un extracto de B.
ramulosa. ....................................................................................................................... 49
Figura 4-4 Cromatografía de intercambio iónico para B. ramulosa. ............................... 50
Figura 4-5 Cromatografía de intercambio iónico para el precipitado de sulfato de amonio
(0-50%) del extracto de B. ramulosa. ............................................................................. 53
Figura 4-6 Cromatografía de afinidad para fracción retenida DEAE-Sephacel. ............. 55
Figura 4-7 Cromatografía de afinidad para fracción no retenida DEAE-Sephacel .......... 56
Figura 5-1 Frecuencia de aminoácidos que componen bryohealin, determinado por
ProtParam. ..................................................................................................................... 66
Figura 5-2 Diagrama de Ramachandran para el modelo obtenido de bryohealin con I-
TASSER. ........................................................................................................................ 68
Figura 5-3 Alineamiento de plantillas usadas por I-TASSER para realizar la predicción de
la estructura terciaria de bryohealin, ............................................................................... 69
Figura 5-4 Predicción de estructura terciaria de bryohelian ........................................... 70
Figura 5-5 Interacciones atómicas entre bryohealin y diferentes ligandos. .................... 73

Lista de tablas
Tabla 1-1 Clasificación de las lectinas según los monosacáridos que reconoce ............. 11
Tabla 3-1 Coordenadas de colección Isla de Providencia y Santa Marta ........................ 24
Tabla 3-2 Porcentajes de proteína soluble cuantificados por el método de BCA y micro
Kjeldahl para los extractos provenientes de harina tratada con acetona y no tratada de
siete especies de algas marinas. .................................................................................... 28
Tabla 3-3 Ensayo preliminar de aglutinación en dos extractos ....................................... 30
Tabla 3-4 Actividad aglutinante en siete extractos de algas marinas previo a un proceso
de diálisis. ....................................................................................................................... 31
Tabla 3-5 Actividad aglutinante para los extractos de algas marinas usando eritrocitos de
oveja ............................................................................................................................... 32
Tabla 4-1 Condiciones de afinidad para cada uno de los ensayos de cromatografía ...... 46
Tabla 4-2 Extracto de B. ramulosa, pequeña escala. ...................................................... 48
Tabla 4-3 Precipitación fraccionada con (NH4)2SO4 en un extracto crudo de B. ramulosa.
....................................................................................................................................... 49
Tabla 4-4 Cromatografía de intercambio iónico sobre de DEAE Sepharosa .................. 50
Tabla 4-5 Ensayos de Inhibición de actividad aglutinante con carbohidratos para las
fracciones de intercambio iónico. .................................................................................... 51
Tabla 4-6 Proteína total de tres fracciones obtenidas al realizar precipitación fraccionada
con (NH4)2SO4 a un extracto crudo de B. ramulosa ......................................................... 52
Tabla 4-7 Contenido de proteína de las fracciones obtenidas por DEAE-Sephacel ........ 53
Tabla 4-11 Características moleculares de las lectinas aisladas de especies del género
Bryopsis . ........................................................................................................................ 62
Tabla 5-1Afinidad de cuatro ligandos a bryohealin ......................................................... 71
Tabla 5-2 Interacciones presentes entre los aminoácidos de bryohealin frente a
diferentes ligandos. ......................................................................................................... 72

Lista de símbolos y abreviaturas
p/ps peso/ peso seco
BCA Ácido Bicinconínico
BLAST Basic Local Alignment Search Tool
BSA Albúmina de Suero Bovino
Ca 2+ Iones de Calcio
Mn 2+ Iones Manganeso
DEAE Dietilaminoetilo
DTT Ditiotreitol
ExPASy Expert Protein Analysis System
Fuc Fucosa
Gal Galactosa
Glc Glucosa
GalNAc N-acetilgalactosamina
GlcNAc N-acetilglucosamina
Man Manosa
GRIP Grupo de Investigación en Proteínas
kDa Kilo Daltons
mA Miliamperios
NaCl Cloruro de Sodio
PBS Buffer Fosfato Salino
PDB Protein Data Bank
PM Peso Molecular
SDS-PAGE Electroforesis en gel de poliacrilamida con dodecilsulfato de sodio
aBSM asialo Mucina submaxilar bovina
Diá. Diámetro
ND No Determinado
ANOVA Análisis de la varianza
ConA Concanavalina A
LCA Lectina de Lens culinaris
RCA Lectina de Ricinus communis
ALL Lectina de Amaranthus leucocarpus
LTA Lectina de Lotus tetragonolobus
UEA Lectina de Ulex europeus
LPA Lectina de Limulus polyphemus
BTL Lectina de Bryothamnion triquetrum

HJA Lectina de Hypnea japónica
BTL Lectina de Bryothamnion seaforthii
HML Lectina de Hypnea cervicornis
HCA Lectina de Hypnea musciformis
ESA2 Lectina de Eucheuma serra
GRFT Lectina de Griffithsia sp.
CDR Dominio de reconocimiento
DS Desviación estándar
H Hemólisis
N.D. No determinado
L Lavado
NL No lavado
R Retentato
D Difusato
MWCO Molecular weight cut-off
HM Lectinas que reconocen manosa
DNA Ácido desoxirribonucleico
RNA Ácido ribonucleico
Col. Colección
AOAC Asociación de químicos Analiticos Oficiales

Introducción
Las lectinas son proteínas o glicoproteínas bioactivas ubicuas en la naturaleza de
respuesta no inmune que poseen dos o más dominios de unión a glicanos de la superficie
celular causando la aglutinación de las células sanguíneas, o precipitación de
glicoconjugados en suspensión (Singh et al., 2015). Esta definición describe
adecuadamente las propiedades y características de estas proteínas, de gran importancia
en la glicobiología moderna. Las lectinas pueden ser aisladas de casi todos los grupos de
organismos desde virus hasta seres humanos, aunque las lectinas más estudiadas han
sido en plantas superiores (Cheung et al., 2015). Su amplia distribución en los diferentes
grupos de organismos habla de su importancia primordial en procesos biológicos como las
interacciones célula-célula, hospedero-patógeno, planta-simbionte, entre otras muchas
(Sharon y Lis, 2003).
Las especies marinas constituyen alrededor de la mitad del total de la biodiversidad
mundial; por lo tanto, los océanos pueden suministrar nuevas moléculas activas con
diversas aplicaciones biológicas (Aneiros y Garateix, 2004), sin embargo a pesar de esta
gran biodiversidad, comparado con las lectinas aisladas de plantas superiores terrestres,
las lectinas de especies marinas se han estudiado a un ritmo mucho más lento (Chernikov
et al., 2013). Los estudios moleculares que se han realizado han mostrado que las lectinas
de algas marinas tienen alto contenido en aminoácidos ácidos, bajo peso molecular, alta
afinidad por glicoproteínas y no requieren de cationes divalentes para su actividad biológica
(Hori et al., 1990; Okamoto et al., 1990; Rogers y Hori, 1993; Teixeira et al., 2012), por otra
parte, se ha visto que poseen actividad anti-tumoral, anti-viral, anti-inflamatoria, anti-
micótica y anti-bacteriana, entre otras (Gonzaga do Nascimento-Neto et al., 2012; Swamy,
2011; Teixeira et al., 2012); por su bajo peso molecular se han convertido en moléculas
muy empleadas en glicopatología (Singh et al., 1999).
En el caribe colombiano existen alrededor de 575 especies de algas, de las cuales las
algas rojas (rhodophyta) son las más diversas, seguidas por las algas verdes (chlorophyta)
y en último lugar las algas pardas (phaeophyta) (Diaz-Pulido y Díaz-Ruíz, 2003) . Dada

2
esta gran biodiversidad de especies, el estudio de sus lectinas reviste gran importancia
motivo por el cual en este trabajo se hizo la detección de lectinas presentes en extractos
obtenidos a partir de algas del caribe colombiano y se realizó la purificación de lectinas
presentes en B. ramulosa. El presente estudio está enmarcado bajo la línea de
investigación “Estudios de Lectinas de algas marinas del Caribe Colombiano” del grupo de
investigación en proteínas GRIP de la Universidad Nacional de Colombia y financiado por
Colciencias (No 110148925106) y por la DIB dentro del proyecto “Evaluación preliminar de
organismos marinos (Pterois volitans, Bryopsis ramulosa y Conus geographus) como
fuentes potenciales de péptidos y /o lectinas con actividad biológica. Código QUIPU
201010026978

3
1. Marco Teórico
1.1. Historia
Las lectinas fueron conocidas como “hemaglutinas” o más comúnmente como
“fitohemaglutinas” por la aglutinación de eritrocitos siendo estudiadas inicialmente en
plantas (Sharon y Lis, 2003), sin embargo la presencia de estas moléculas se extendió
después a otros grupos de organismos desde bacterias y virus, hasta animales vertebrados
(Del Sol et al., 2006). El estudio de las lectinas fue iniciado por Hermman Stillmark en 1888
al describir el fenómeno de la hemoaglutinación en extractos de semillas de ricino (Ricinnus
comunis). Stillmark demostró que la toxicidad de las semillas se debía a la presencia de
un factor proteico aglutinante de eritrocitos llamado ricina (Del Sol et al., 2006).
Posteriormente H. Hellín, encontró una hemaglutinina tóxica "abrina", en los extractos de
Abrus precatorius; tanto la ricina como la abrina pronto estuvieron disponibles en el
mercado, lo que llevó a Paul Ehrlich, en el Instituto Real de Terapia Experimental
(Frankfurt) a emplearlas para detección de antígenos en estudios inmunológicos (Sharon
y Lis, 2004).
En 1919, James B. Sumner en la Universidad de Cornell (Ithaca, Nueva York) obtiene por
primera vez una hemaglutinina pura llamada concanavalina A (ConA). Sin embargo,
pasaron casi dos décadas antes de que Sumner y Howell (1936) informaran que la ConA
además de causar la aglutinación de eritrocitos y levaduras, también era capaz de
precipitar una solución de glucógeno. Fue de esta manera como se demostró que la
actividad aglutinante de ConA era inhibida por sacarosa, hecho que hablaba de la
especificidad de las lectinas por determinados azucares. Con mucha previsión, sugirieron
que la hemaglutinación inducida por ConA podría ser la consecuencia de una reacción de
la proteína vegetal con carbohidratos de la superficie de las células rojas (Sharon y Lis,
2003). Stillmark, también describió la especificidad de ricina por eritrocitos de diferentes
fuentes animales.
Esta idea fue corroborada en 1900 cuando Karl Lysteiner de la Universidad de Viena
describe los diferentes grupos sanguíneos en humanos y cuando Mäkelä (1957), estudió
extractos de semillas provenientes de 165 géneros y 743 especies de plantas de la familia
de las leguminosas; encontró actividad hemoglutinante en más de un tercio de las especies

4
que además exhibieron especificidad por un tipo sanguíneo. Los estudios mencionados
previamente, señalaron hacia la década de 1960, que estas proteínas disponibles en la
naturaleza podrían ser muy valiosas para el estudio de los carbohidratos simples y
complejos en solución y en las superficies celulares, así como para la caracterización de
células (Sharon y Lis, 2004). Los hallazgos sobre lectinas como la de germen de trigo
(WGA) (Aub et al., 1965), de Canavalia ensiformis (ConA) (Inbar y Sachs, 1969), y la de
soya (Sela et al., 1970), crearon una gran expectativa ya que aglutinaban células
transformadas, pero no tenían el mismo efecto sobre células parentales normales. Los
resultados proporcionaron pruebas convincentes que las células transformadas podrían
estar asociadas con un cambio en los azúcares de la superficie celular, una idea que sólo
unos pocos años antes había sido considerada completamente infundada, y que dio lugar
a la aplicación de las lectinas en la investigación del cáncer.
Para 1973, por lo menos veinte lectinas se habían caracterizado parcialmente, se conocía
la composición de aminoácidos de sólo unas pocas de ellas y solo se había resuelto la
estructura tridimensional de la ConA. Aun así, los datos disponibles en ese entonces
permitieron proponer que, "Las lectinas tienen muchas propiedades biológicas en común,
representan un grupo diversificado de proteínas con respecto al tamaño, la composición y
la estructura" (Sharon et al., 1990). Una predicción totalmente confirmada por cientos de
secuencias de lectinas y estructuras que incluso hoy en día aún no se han determinado.
En la actualidad se han usado otras formas de definir estas moléculas de gran interés, pero
hasta el momento las más aceptadas comparten que las lectinas son proteínas o
glicoproteínas de origen no inmune, que reconocen específicamente y reversiblemente
carbohidratos, con capacidad para aglutinar células y precipitar glicoconjugados
(Goldstein, 1980).
1.2. Lectinas
Actualmente, uno de los campos de más rápido crecimiento en bioquímica y biología
molecular es el estudio de la función de los glicanos y cómo pueden regular procesos
biológicos que van desde la coagulación (Shriver et al., 2004) hasta procesos de infección
viral y bacteriana actuando en diferentes tipos de células (Imberty et al., 2005). El patrón
de glicosilación de las proteínas es dependiente del tipo de célula en que se produce y
puede modificarse ante diversas situaciones biológicas o ante la presencia de

5
enfermedades; estas observaciones han llevado al desarrollo de la glicopatología y en este
contexto las lectinas han sido un instrumento muy útil (Clark y Mao, 2012). Las lectinas son
un grupo muy diverso de proteínas de origen animal o vegetal y se caracterizan por unir
específicamente de manera reversible monosacáridos u oligosacáridos (Lis y Sharon,
1998). Estas propiedades las han convertido en una herramienta fundamental en
glicobiología y su distribución, interacciones e importancia ha sido ampliamente
documentada (Damme et al., 1998; Sharon y Lis, 2003; Vasta y Ahmed, 2009). El estudio
de estas proteínas y su funcionalidad ha sido denominado “Lectinómica” y se espera que
los avances en su conocimiento permitan desarrollar aplicaciones biotecnológicas y
clínicas muy definidas (Akkouh et al., 2015; Gabius et al., 2002; Procópio et al., 2017; Vasta
y Ahmed, 2009). La lectinas se encuentran en toda clase de familias de organismos que
han sido examinados incluyendo bacterias y virus (Van Damme et al., 2008). El tejido y
distribución celular en la cual se encuentran es variable, y este podría ser afectado por
varios factores como el estado de desarrollo, edad y condiciones de patogenicidad del
organismo (Sharon y Lis, 2003). Esta gran ubicuidad se explica por el papel clave que
tienen en gran variedad de procesos celulares como el reconocimiento específico de
espermatozoide en el óvulo durante la fecundación, adhesión entre células y célula matriz
extracelular, control en la migración de linfocitos, inducción a la apoptosis, actividad
mitogénica y antiviral (Baruffi et al., 2017; Cui et al., 2017; Gabius, 1997; Mitchell et al.,
2017; Ponraj et al., 2017; Sharon y Lis, 2003; Singh et al., 2017; Varki y Lowe, 2009) estas
moléculas podrían jugar un papel importante en el sistema inmunológico humano para el
reconocimiento de microorganismos (Mayer et al., 2017; Wesener et al., 2017). En plantas
poseen un papel clave en los mecanismos de defensa contra fitopatógenos (Karimi et al.,
2012) o como mediadoras de la simbiosis con bacterias fijadoras de nitrógeno (Oldroyd y
Downie, 2008); también se ha visto que son importantes durante el proceso de
comunicación e infectividad microbiana (Neu y Kuhlicke, 2017; Sharon, 1987) y viral
(Greber y Bartenschlager, 2017; Liu et al., 2017).
En cuanto a su aplicación en campos como la genética, la biomedicina y la inmunología,
se ha aprovechado la propiedad que tienen para reconocer específicamente varios tipos
de glicoconjugados presentes en las superficies celulares y fluidos corporales (Hernández
et al., 1999; Procópio et al., 2017). Su actividad mitogénica ha sido bien aprovechada
permitiendo que se utilicen en estudios que tienen como base la proliferación de linfocitos
en cultivos y la producción de citoquinas (interferón e interleuquinas). La expresión de

6
receptores de citoquinas también se ha estudiado a partir de los sobrenadantes de cultivos
de linfocitos, que son estimulados con lectinas, y provienen de pacientes con
enfermedades de alto impacto social, como el síndrome de inmunodeficiencia adquirida,
la tuberculosis y la leishmaniasis, entre otras (Born et al., 1995; Cui et al., 2017; Ijichi et al.,
1996). Interacción entre virus, como por ejemplo entre el virus de inmunodeficiencia
humana y el virus de la hepatitis B, así como la susceptibilidad y resistencia a éstos
(Cisterna et al., 1995; Pinkston et al., 1995; Santos, 2017). La evaluación de la efectividad
de terapias antirretrovirales de acuerdo con la respuesta de los linfocitos a la estimulación
con lectinas antes y después de la terapia, como por ejemplo en terapias contra el VIH
(Casseb et al., 1994) y la detección de anormalidades cromosómicas (Tomassetti et al.,
1995) entre otros. Las lectinas forman parte de conjugados como lectina-lectina, lectina-
enzimas y lectina-anticuerpos, lo que ha permitido el desarrollo de técnicas
cromatográficas de afinidad para la purificación de glicoproteínas. Un ejemplo es la
purificación de IgG utilizando anti-IgG la cual es reconocida por la Con A-Sepharosa (Kong
et al., 1997), así como también la purificación de enzimas y de las propias lectinas
(Hernández Díaz et al., 1999). En algunas técnicas histoquímicas y de microscopía
electrónica se usan lectinas conjugadas a moléculas fluorescentes como marcadores para
estudiar la estructura y función de la membrana plasmática (Hartmut, 1988) y detectar
alteraciones en la expresión de moléculas presentes en la superficie celular (Kato et al.,
1995; Zschäbitz et al., 1995). Otra área importante en la cual se emplean las lectinas es la
detección de transformaciones malignas en células y actividad anticancerígena (Camby et
al., 1997; Kiss et al., 1997; Mohammed, 2016; Poiroux et al., 2017); además en muchas
investigaciones las lectinas han sido usadas para la búsqueda de biomarcadores que
sirvan para el diagnóstico y/o pronóstico de enfermedades (Plavina et al., 2007; Yang y
Hancock, 2004; Yang et al., 2005). Un ejemplo de ello es la detección del antígeno Tn
presente específicamente en células de carcinomas de seno y el cual es detectado con la
lectina de Vicia villosa isolectina B4 (Limpias et al., 2010). Con otras lectinas se han hecho
estudios enfocados en la búsqueda de biomarcadores para cáncer de próstata en muestras
de suero (Drake et al., 2006); mostrando su posible utilidad como marcadores tumorales o
histológicos. También son de gran utilidad para hacer la separación de glicoproteínas
(Schwarz et al., 1993), y en la caracterización de los grupos sanguíneos humanos o de
otras fuentes (Sharon y Lis, 2003).

7
Existen tres formas de clasificar las lectinas, una de ellas se basa en el número de sitios
de reconocimiento por el carbohidrato (Figura 1-1). Las merolectinas son proteínas
constituidas por un solo sitio de unión a carbohidratos y debido a esta característica estas
no poseen actividad aglutinante (Cruz et al., 2005), las hololectinas contienen dos o más
dominios del mismo tipo estructural y funcional; al menos dos de estos dominios presentan
actividad de unión a carbohidratos de modo que son capaces de generar aglutinación. En
el último grupo se encuentran las quimerolectinas, estas además de poseer un dominio de
unión a carbohidratos presentan otro(s) dominio(s) con actividad distinta de reconocimiento
a carbohidratos (actividad enzimática, inactivadora de ribosomas) y dependiendo de la
topología y sitios de unión a azucares pueden o no presentar hemoaglutinación (Damme
et al., 1998). Una segunda manera de clasificar las lectinas es de acuerdo a su
especificidad hacia el monosacárido que inhibe su actividad hemaglutinante (Tabla 1-1) y
hacia las estructuras oligosacarídicas que reconocen, clasificándolas en lectinas que
reconocen N-glicanos y O-glicanos (Makela, 1957).
Figura 1-1 Representación esquemática de la clasificación en lectinas dependiendo el número de sitios de reconocimiento al carbohidrato.
RIP; Ribosome, inactivate proteins. Adaptado de (Damme et al., 1998).
Por otra parte, el genoma y el análisis de transcriptoma proporcionan una amplia evidencia
de la ocurrencia en todas las plantas terrestres de numerosos genes con uno o más
dominio(s) bien definidos que permiten la clasificación de estas lectinas en 12 familias

8
(Figura 1-2) (Jiang et al., 2010; Van Damme et al., 2008), la presencia de residuos
conservados conlleva a un plegamiento determinado para cada uno de los grupos.
Figura 1-2 Clasificación de las lectinas en términos de plegamiento de dominio de unión a carbohidrato.
En forma muy breve se resumen a continuación los rasgos estructurales característicos de
cada uno de los plegamientos (Figura 10, Anexo se representan algunas de estas lectinas).
Dentro de esta clasificación, se destacan las aglutininas homologas de Agaricus bisporus;
en especies como Marchantia polymorpha, sus lectinas tienen dos dominios de
reconocimiento diferentes y con afinidad por el antígeno T(Galβ1,3GalNAc) y N‐
acetilglucosamina (Nakamura-Tsuruta et al., 2006). Cada uno de los monómeros consiste
de una estructura sándwich constituido por dos hojas β interconectadas por un motivo
hélice-loop-hélice (Carrizo et al., 2004). Las lectinas de tipo Amaranto fueron aisladas
inicialmente a partir de semillas de Amaranthus caudatus, y se caracterizan por ser
homodiméricas con un peso molecular de 33 kDa y con afinidad por el antígeno
T(Galβ1,3GalNAc) (Rinderle et al., 1989). Las subunidades de amarantina comprenden
dos dominios homólogos dispuestos en tándem de alrededor 150 aminoácidos. Cada
dominio tiene un plegamiento de β-trefoil formada por seis β-hebras antiparalelas cubiertas
Lectinas
Heveína
Amaranto
Jacalina
Leguminosas
Monoct. Man.
Quitinasa
Cianovirinas
LysM
Ricina B
Nictaba
Euonymus europaeus
Agaricus bisporus

9
por tres -hairpins en un barril corto (Transue et al., 1997). Las lectinas homólogas a
quitinasa, están representadas por una lectina aislada de Robinia pseudoacacia, la cual es
un homodimero compuesto por 337 residuos de aminoácidos. Aunque la lectina comparte
una alta identidad de secuencia con quitinasas de clase V, está desprovista de la actividad
enzimática, pero tiene una baja afinidad con N-glicanos tipo manosa. Aunque no se ha
cristalizado la proteína, los alineamientos realizados frente a otras proteínas indican que
esta podría adoptar el plegamiento tipo TIM-barril encontrado en la proteína chitotriosidasa
humana (Fusetti et al., 2002). La familia cyanovirina, fue reportada por primera vez con el
estudio de la lectina del alga azul Nostoc ellipsosporum. Esta es una proteína de bajo peso
molecular y con dos repeticiones internas en tándem cada una de 50 aminoácidos,
presenta cuatro residuos de Cys que forman dos puentes disulfuro, los cuales son
indispensables para su actividad (Akkouh et al., 2015).
Trabajos recientes en genómica revelan que la aglutinina de Euonymus europaeus (EEA)
presenta el plegamiento de un nuevo grupo de lectinas vegetales (Fouquaert et al., 2008).
En el caso la lectina de Galacthus nivales (GNA), cada monómero contiene dos sitios de
unión de carbohidratos y un tercer sitio se crea cuando se encuentra en forma tetrámerica,
dando como resultado un total de 12 sitios de unión de carbohidratos (Alen y Schols, 2012);
cada uno de los monómeros está conformado por cuatro hebras beta antiparalelas y unidos
por β harpin, y estas subunidades a su vez están organizadas en un plegamiento tipo B
prisma (Hester et al., 1995). Por otro lado existe un dominio característico para algunas
plantas denominado heveina, el cual está conformado por 43 aminoácidos y representado
por una lectina aislada de Hevea brasiliensis (Walujono et al. 1975), se ha descrito para
proteínas de este tipo entre 2 a 7 dominios de tipo heveina separados por secuencias de
5 a 10 residuos de aminoácidos (Van Damme et al., 2004). A nivel estructural este dominio
consiste de tres hojas beta antiparalelas (β1, β2, β3) asociadas por dos segmentos de alfa
hélice (α1, α2), el plegamiento se encuentra estabilizado por cuatro puentes disulfuro entre
ocho residuos de cisteina altamente conservados (Aboitiz et al., 2004). La familia Jacalina
se subdivide en un grupo con mayor afinidad a galactosa (en algunas especies de los
géneros Artocarpus, Maclura, y Morus) o afinidad a manosa (Castanea crenata, Parkia
platycephala, Brassica napus etc…), en general estas lectinas son tetraméricas,
glicosiladas, donde cada subunidad contiene una cadena pesada α y una cadena ligera β,
y está constituida por tres hojas β antiparalelas arregladas a manera de un prisma
triangular (Makela, 1957; Van Damme et al., 2007).

10
Las lectinas con dominio leguminosa están constituidas por una hebra de siete laminas β
(que forman la cara interna del protómero) y seis hebras β curvas que forman la parte
frontal de la subunidad. Ambos conjuntos de hebras β se encuentran interconectadas por
giros o loops; el plegamiento se da en forma de β sándwich o jelly roll estabilizada por una
hoja β adicional de cinco hebras. Este tipo de lectinas se caracteriza por ser
metaloproteínas que incorporan en su estructura Ca2+ y Mn2+ (Edelman y Wang, 1978; Van
Damme et al., 2008).
El dominio LysM fue identificado por primera vez en proteínas de especies de legumbres,
y ha sido encontrado en plantas que interactúan con oligosacáridos de quitina
oligosacáridos o análogos estructurales con ácidos grasos. Aunque la estructura del
dominio aún no ha sido estudiada, se ha encontrado que hay proteínas relacionadas
presentes en bacterias que tienen una estructura β-α-α-β en la cual dos hélices se
empaquetan sobre el mismo lado de una hoja β antiparalela, se cree que posiblemente la
superficie de la proteína actúa como sitio de unión al carbohidrato (Bateman y Bycroft,
2000).
La familia Nictaba hace referencia a proteínas que poseen el dominio encontrado por
primera vez en la aglutinina de Nicotiana tabacum. Aunque las primeras lectinas
descubiertas se unían a quitooligosacáridos, las Nictaba también reconocen estructuras
tipo manosa y complejos N–glicosilados (Velasco Martín, 2016).
Finalmente, la familia de las lectinas tipo Ricina, tienen un dominio característico
encontrado por primera vez en Ricinus communis. La ricina es una proteína constituida por
una cadena A con actividad enzimática, unida mediante un puente disulfuro a una cadena-
B con actividad de lectina. Esta cadena-B está formada a su vez por dos dominios ricina,
responsables de la unión de la proteína a estructuras con galactosa (Velasco, 2016).

11
Tabla 1-1 Clasificación de las lectinas según los monosacáridos que reconoce
Monosacárido Lectina Abreviatura
α-D manosa, α-D glucosa Canavalia ensiformis
Lens culinaris
ConA
LCA
β galactosa
N acetil α-D galactosamina
Ricinus communis
Glycine max
Arachis hypogaea
Amaranthus leucocarpus
RCA
SBA
PNA
ALL
N-acetil-β-D-glucosamina Triticum vulgare WGA
α-D-fucosa Lotus tetragonolobus
Ulex europeus
LTA
UEA
α-N-acetilneuramínico Limulus polyphemus LPA
(Cruz et al., 2005)
Las lectinas de origen animal se caracterizan por la presencia de un CDR (dominio de
reconocimiento a carbohidratos) (Yerson et al., 2008; Lis y Sharon, 1998). Cada lectina
animal posee su propio CDR el cual tiene una secuencia motivo de 115 – 130 residuos y
cuatro cisteínas que se encuentran altamente conservadas y forman dos puentes disulfuro
(Yerson et al., 2008; Matsumoto et al., 2001). Inicialmente se consideró dividir las lectinas
de origen animal en dos grandes grupos: Las de tipo C (dependientes de Ca2+ y las lectinas
tipo S (tiol dependientes) (Drickamer, 1988). Tras un aumento en el número de secuencias,
la clasificación se modificó incluyendo muchos más grupos (Loh et al., 2017; Sanchez et
al., 2006). Su clasificación basada en la estructura del dominio CRD comprende:
Las lectinas tipo C las cuales requieren de Ca2+ para su actividad y tienen una secuencia
14 aminoácidos invariables y 18 residuos altamente conservados, presentan afinidad por
ligandos como la manosa, la fucosa, y glucosamina (Graham et al., 2012) se han
encontrado en macrófagos y algunas células endoteliales, y podrían estar involucradas en
eventos de reconocimiento hacia patógenos (Dambuza y Brown, 2015), endocitosis
(Cambi y Figdor, 2003), adhesión e inmuno-modulación (Gordon, 1990).
Las lectinas tipo P, poseen afinidad a manosa 6-fosfato y se ha visto que podrían estar
involucradas en procesos de endocitosis y unión intracelular de enzimas lisosomales
(Dahms y Hancock, 2002). En el grupo de lectinas de tipo S se encuentra el grupo de
galectinas, compuesto de homólogos estructurales del dominio de reconocimiento a
carbohidratos, que típicamente se unen a glicoconjugados que contienen β-galactosa.

12
Estas proteínas se expresan principalmente en células del sistema inmunológico y en las
células epiteliales. Las galectinas están implicadas en el reconocimiento de ligandos de
carbohidratos endógenos y también en la unión de glicanos en la superficie de los
microorganismos (Vasta, 2009). Las galectinas presentan un papel significativo en la
defensa del huésped contra las bacterias (Thurston et al., 2012).
Las lectinas tipo - I se expresan en como monocitos y macrófagos, y regulan la interacción
y diferenciación de células mieloides, un ejemplo son las sialo adhesinas (Varki y Angata,
2005).
Las lectinas tipo M, son proteínas con afinidad por manosa que están estrechamente
relacionadas con las -manosidasas residentes del retículo endoplasmático y Golgi; están
involucradas en modificaciones postraduccionales de proteínas N-glicosiladas. La región
CRD de estas lectinas corresponde a una estructura de tipo barril con hélices alfa y hojas
beta. Algunas de las lectinas de tipo M tienen extensiones C-terminales después de la
región CRD que no están relacionados con el plegamiento (Karaveg et al., 2005).
Las lectinas tipo L están presente en proteínas vegetales, fúngicas y animales, pero,
aunque están en vegetales y animales tienen secuencias divergentes y diferentes
propiedades moleculares. Las lectinas vegetales son secretadas y se encuentran en
tejidos especializados, mientras que las de origen animal son a menudo proteínas de
membrana o proteínas luminales y se encuentran en baja cantidad; estas diferencias
reflejan el hecho de que las lectinas vegetales y animales de tipo L pueden cumplir
funciones diferentes. Su estructura adquiere una forma de -sandwich con una hoja
cóncava de siete hebras β y una lámina convexa de cinco hebras β. La mayoría de los
CRD de tipo L requieren iones metálicos para la unión del ligando (Dodd y Drickamer,
2001).
Las lectinas de tipo quitinasa son proteínas de mamífero que pertenecen a la familia de
glicósidos hidrolasas 18. Se trata de una familia de proteínas solubles, intracelulares o
secretadas que tienen una estructura tipo barril de triosfato-fosfato (TIM), a veces con la
inserción de un pequeño dominio α/β (Sun et al., 2001). Las lectinas de tipo F son
fucolectinas y están filogenéticamente extendidas en diferentes grupos de organismos
como invertebrados marinos (artrópodos, equinodermos, moluscos y platelmintos) y en

13
vertebrados en algunos teleósteos, peces cartilaginosos, y anfibios (Vasta et al., 2008), se
ha reportado la presencia de este dominio en una lectina aislada del alga marina Bryopsis
plumosa (Kim et al., 2006); también se ha reportado en bacterias y han sido muy
estudiadas en Streptococcus mitis y Streptococcus neumoniae SP2159 (Bishnoi et al.,
2015). Por otra parte, una de las primeras estructuras tridimensionales descritas para una
hemaglutinina de Anguilla anguilla, sugiere que esta podría adoptar un plegamiento tipo -
barril (Bianchet et al., 2002a) con un bolsillo de unión cargado positivamente (Vasta et al.,
2008). Los plegamientos descritos recientemente corresponden a las lectinas tipo H o la
lectina del caracol romano Helix pomatia; este ha sido descrito en múltiples organismos
invertebrados y en la ameba Dictyostelium discoideum, y muestran afinidad por la GalNAc,
GlcNac y el antígeno T (Sanchez et al., 2006).
La información que existe sobre las lectinas de algas marinas a nivel bioquímico y
estructural es escasa y por tanto su clasificación funcional y filogenética aun no es clara.
Solo existe información estructural primaria completa para 14 lectinas aisladas de algas
marinas. Comparaciones filogenéticas de estas proteínas indican que las lectinas aisladas
de Bryothamnion triquetrum (BTL), Hypnea japonica (HJA) y B. seaforthii (BTL) pueden ser
consideradas como una primera familia de algas que no muestran similaridad con otro tipo
de organismos. La segunda familia de lectinas en algas está conformada por proteínas
aisladas de H. cervicornis (HML) y H. musciformis (Nagano et al., 2005).
La lectina de Griffithsia sp. (GRFT) es la única lectina de algas marinas para la cual se ha
establecido su estructura terciaria por cristalografía de rayos x (Ziółkowska, 2007), su
estructura primaria tiene 30% de identidad con proteínas tales como la Jacalina, miembros
de la familia de lectinas β-prisma en plantas (Mori et al., 2005; Ziółkowska et al., 2006).
Otras lectinas como las aisladas de Eucheuma serra, E. amakusaensis, E. cottonii
(Kawakubo et al., 1999) tienen pesos moleculares alrededor de 28 kDa, y poseen
similaridad con la secuencia de la isolectina 2 de E. serra (ESA2); pueden ser
consideradas como una familia diferente de lectinas presentes en algas rojas.
Las lectinas purificadas del género Ulva como U. pertusa y U. limnetica no presentan
similaridad con familias descritas de lectinas vegetales o animales (Mori et al., 2005; Wang
et al., 2004). En el caso de tres lectinas aisladas de B. plumosa especificas por GalNAc y
GlcNAc (Bryohealin, BPL3 y BPL4) y B. hypnoides, se ha encontrado que estas presentan
dominios característicos de organismos invertebrados (lectinas animales tipo F y tipo H)

14
(Han et al., 2012; Han et al., 2011; Kim et al., 2006). La lectina BPL2 aislada de B. plumosa
(específica por manosa) presenta una alta identidad con una lectina aislada de B. maxima
(Han et al., 2010). Otra lectina aislada de un alga verde, Boodlea coacta (BCA), tiene
secuencias similares a las encontradas en la familia de lectinas vegetales de Galanthus
nivalis (Sato et al., 2011). Esta información indica que no existe un plegamiento típico para
las lectinas de algas marinas.
1.3. Estrategias para purificar lectinas
Existen diversos métodos para detectar la actividad de una lectina; el método más común
consiste en determinar la actividad de aglutinación de los extractos salinos obtenidos a
partir del material biológico (Rüdiger, 1993), también hay métodos que incluyen la
precipitación de polisacáridos y /o glicoconjugados (Goldstein, 1972).
La hemaglutinación se lleva a cabo con la técnica de diluciones en placa y usualmente se
hace con eritrocitos de humano o de animales como conejo y oveja entre otros. Algunas
veces los eritrocitos son tratados con enzimas como tripsina, papaína, neuraminidasa y β-
glactosidasa entre otras, para exponer determinantes antigénicos y de esta forma observar
mejor la aglutinación. Con esta técnica se puede seguir el curso de la purificación de estas
proteínas y se puede determinar su grado de purificación. La interacción de la lectina y el
carbohidrato se dan por medio de interacciones débiles como puentes de hidrogeno,
interacciones electrostáticas e hidrofóbicas (Sharon, 1994; Sharon y Lis, 2003). Otros
métodos incluyen el ensayo de ELLSA donde se puede trabajar con lectinas biotiniladas
de especificidad definida y de esta forma realizar estudios actividad biológica con una
mayor sensibilidad (Pérez y Vega, 2007). Si se obtiene un resultado positivo es esencial
corroborar que la actividad es específicamente inhibida por un monosacárido,
oligosacáridos o glicoproteínas. Otros métodos de reciente aplicación para evaluar la
interacción de lectinas con carbohidratos incluyen los glicoarreglos con los que se puede
hacer un screening rápido de especificidad (Gabius et al., 1994), arreglos en sándwich de
anticuerpo-lectina los cuales permiten el estudio de biomarcadores (Haab y Yue, 2011) o
lectinas acopladas a columnas de afinidad (Drake et al., 2006).

15
El aislamiento de las lectinas comienza con la extracción empleando un buffer o una
solución salina a partir de los tejidos u órganos en los cuales está presente (Goldstein y
Poretz, 1986; Rüdiger, 1993). Cuando se trabaja con material vegetal, por lo general se
emplean solventes orgánicos para la remoción de pigmentos y lípidos (Rüdiger, 1993);
también en el buffer de extracción se pueden incluir inhibidores de proteasas para prevenir
la degradación de la lectina durante la extracción y purificación. De forma preliminar es
posible usar técnicas de precipitación fraccionada para obtener una fracción aislada de
proteína y eliminar otros constituyentes como polisacáridos en el caso de las plantas, con
este mismo fin es posible el uso de cromatografía de intercambio iónico. Una de las
técnicas más exitosas en las etapas finales de purificación es el uso de la cromatografía
de afinidad; se pueden preparar una amplia variedad de soportes según las propiedades
de reconocimiento de la lectina que se va a purificar (Alborés et al., 2016; Gardères et al.,
2016).
1.4. Algas
Las algas son un grupo diverso de organismos fotosintéticos que comúnmente se incluyen
dentro de las plantas, sin embargo no poseen las características principales de estas, tales
como las raíces, hojas y tejidos vasculares (Etcheverry, 1986). Este grupo presenta una
gran variedad de morfologías, desde formas unicelulares móviles o no, aparentemente
simples a formas pluricelulares, algunas parenquimatosas complejas; cada grupo de algas
tiene características estructurales y moleculares propias (Acleto y Zúñiga, 1998). Las algas
incluyen tanto organismos procariotas como eucariotas, agrupados en 12 phyla con
distribución cosmopolita, incluyendo ecosistemas marinos, salobres, agua dulce y
ambientes terrestres (Adey, 1998; Diaz-Pulido y Díaz-Ruíz, 2003).
Su papel biológico en la naturaleza es variado, algunas como las cyanophytas fijan el
nitrógeno atmosférico, a modo de fertilizante biológico participando en el ciclo del
nitrógeno, otras como productores primarios de materia orgánica, intervienen en la cadena
trófica de muchos organismos heterotróficos (Acleto y Zúñiga, 1998). Las algas bentónicas
juegan un papel clave en los ecosistemas marinos, por ejemplo contribuyendo a crear
hábitats propicios para otros organismos, y también participando en la construcción y
degradación de los arrecifes coralinos (Adey, 1998; Kelaher et al., 2003). En la industria,
las algas son de interés para la obtención de alimentos y polisacáridos valiosos como

16
alginatos, agar y carragenos (Acleto y Zúñiga, 1998; Etcheverry, 1986). La clasificación de
las algas en su conjunto o como grupos particulares, muestra un carácter cambiante y
dinámico a través del tiempo. Los pigmentos fotosintéticos son una característica de
clasificación más comúnmente empleada para diferenciar a los miembros de las distintas
divisiones de algas (Acleto y Zúñiga, 1998). Dentro de los grupos más representativos
encontramos las algas bentónicas, aquellas algas marinas macroscópicas que pertenecen
a los phyla Rhodophyta (algas rojas), Heterokontophyta (phaeophyceae o algas pardas) y
Chlorophyta (algas verdes) (Abbott y Hollenberg, 1992; Hoek et al., 1995).
A una escala global, existen alrededor de 4.100 especies descritas de algas rojas
(Womersley, 1994), 1.500 especies de algas pardas, y 8.000-9.000 especies de algas
verdes, de las cuales 10% son marinas (Clayton y King, 1990). En el caribe colombiano se
registra un total de 575 taxones (el archipiélago de San Andrés y Providencia es uno de
los tres sitios más biodiversos con 202 taxas) de algas marinas bentónicas de las regiones
oceánicas y costeras de las cuales las algas rojas (Rhodophyta) contiene la mayor
diversidad en número de especies, estando en segundo lugar las algas verdes
(Chlorophyta) y luego las algas pardas (Phaeophyceae) (Diaz-Pulido y Díaz-Ruíz, 2003).
La flora del caribe colombiano incluye casi la mitad de las especies conocidas en el
Atlántico Occidental Tropical y Subtropical (desde Carolina del Norte hasta Brasil) (Littler y
Littler, 2000) lo que sugiere que el caribe colombiano es altamente diverso especialmente
cuando se tiene en cuenta que constituye menos del 5% (1.600 Km) de la línea costera
comprendida desde Carolina del Norte hasta el sur del Brasil (Diaz-Pulido y Díaz-Ruíz,
2003).
1.4.1. Características citológicas de las algas marinas
Estas características conciernen a la estructura y naturaleza de la pared celular,
cromoplastos, pirenoides, pigmentos fotosintéticos, productos de reserva y otras
inclusiones citoplasmáticas (Etcheverry, 1986). La pared celular de los integrantes de algas
verdes, pardas y rojas es similar a la de plantas superiores. Esta es de naturaleza
celulósica y estructura microfibrilar con orientación variada; en la mayoría de las algas la
pared celular es pecto-celulósica, sin embargo, en algunos géneros como Halimeda,
Caulerpa, Penicillus, la celulosa es reemplazada por xilano y manano. En algas pardas y
rojas existen polisacáridos como alginatos, agar y carragenano, y deposiciones de

17
carbonato de calcio como en el caso de Halimeda y Corallina entre otras muchas (Lewin,
2013). Otro tipo de inclusión citoplasmática son los cromoplastos, estos llevan pigmentos
fotosintéticos característicos y están presentes en casi todos los grupos de algas; dentro
de sus componentes están las proteínas, lípidos, ribosomas, DNA y RNA en cantidades
pequeñas. A estos elementos se asocian los pirenoides que son cuerpos proteicos densos,
frecuentemente refractivos, de variada morfología y número según la especie (Acleto y
Zúñiga, 1998). En cuanto a los pigmentos fotosintéticos en las algas, estos se pueden
definir en tres grupos bioquímicos diferentes: clorofilas, carotenoides y biliproteínas (Lewin,
2013). Las clorofilas son pigmentos verdes solubles en alcohol y solventes orgánicos, y
está presente en todas las algas; la clorofila b se encuentra en Chlorophytas así como en
plantas superiores, la clorofila c es un pigmento de amplia distribución y se encuentra en
el grupo de las Phaeophytas, la clorofila d solo está presente en las Rhodophytas.
Los carotenoides (carotenos y xantofilas) son pigmentos amarillentos o anaranjados,
solubles en solventes orgánicos o en los lípidos. Los carotenos son hidrocarbonos lineales
no saturados mientras que las xantofilas son derivados oxigenados de los carotenos, el β-
caroteno solo está presente en las Chlorophytas y Rhodophytas mientras que las xantofilas
altamente diversas se encuentran en los diferentes grupos de algas. En último lugar están
las biliproteínas que son pigmentos proteicos complejos, conocidos antes como ficobilinas,
de color azul (ficocianina) o rojo (ficoeritrina) y se pueden encontrar en el grupo de las
Rhodophytas. La cantidad de pigmentos fotosintéticos es variable en cada grupo de algas
y proporciona a cada uno la coloración correspondiente, su proporción varía
considerablemente en relación con las condiciones ambientales (Etcheverry, 1986; Lewin,
2013; Montoya, 1984). En cuanto a los productos de reserva las algas poseen diferentes
moléculas que por lo general corresponden a productos de su metabolismo, las
Chlorophytas poseen almidón, las Rhodophytas almidón de florideas, compuestos
fenólicos y glicosidos como el floridósido. Algunas especies poseen compuestos de iodo y
bromo. Otras especies de algas pardas poseen paramilon y/o manitol (Etcheverry, 1986).
1.4.2. Lectinas en algas
La diversidad marina ha sido ampliamente explorada desde hace varias décadas en la
búsqueda de compuestos biológicamente activos y una amplia investigación sobre lectinas
se adelanta hoy en día. Las lectinas derivadas de organismos marinos son
estructuralmente diversas y también difieren de las identificadas a partir de organismos

18
terrestres. Estas proteínas tienen un tamaño relativamente pequeño, son estables debido
a la extensa formación de puentes disulfuro, y tienen una alta especificidad hacia
glicoconjugados en lugar de azúcares simples. En comparación con los organismos
terrestres, muchas de estas moléculas aún no han sido estudiadas (Cheung et al., 2015).
A pesar de los estudios recientes en estos organismos en lo que respecta a sus funciones
fisiológicas y las aplicaciones de sus lectinas, aún el conocimiento es muy escaso sin
embargo es un tema de gran interés. Estas proteínas se han encontrado en alrededor de
300 especies de origen marino que incluyen moluscos, ascidias, poliquetos, esponjas,
peces, equinodermos, artrópodos, y algas (Chernikov et al., 2013; Cheung et al., 2015).
Las algas marinas son reconocidas como una fuente rica en compuestos bioactivos que
podrían ser explotadas como ingredientes funcionales en el campo de la salud humana y
animal (Gupta y Abu-Ghannam, 2011). Son muy pocas las lectinas de algas marinas que
han sido aisladas y caracterizadas bioquímicamente, siendo la clase de las algas rojas
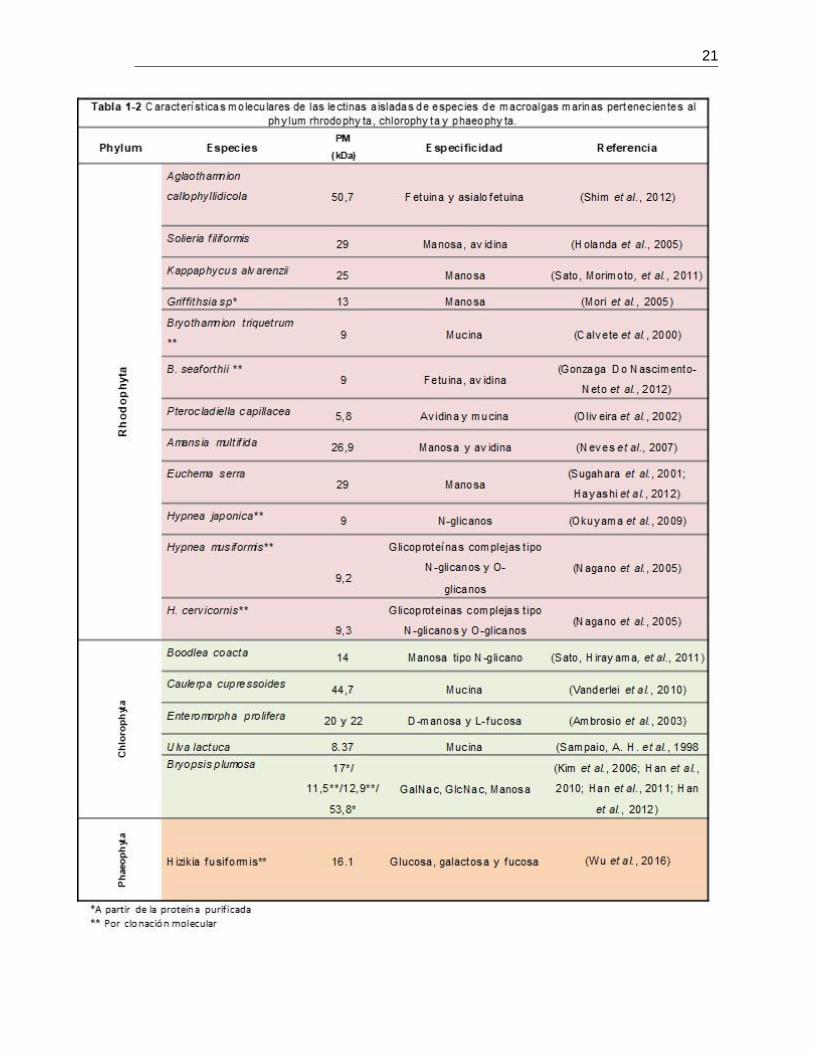
(Rhodophyta) la que presenta un mayor número de especies estudiadas (Tabla 1-2). Por
el contrario, en las algas verdes (Chlorophyta), a pesar de incluir 350 géneros y 2650
especies, el número de lectinas aisladas es bajo y restringido a unos cuantos géneros
(Nagano et al., 2005). En algas pardas (Phaeophyta), los estudios son muy escasos y se
deben principalmente a la cantidad de polifenoles presentes en los extractos crudos que
interfieren en la detección de la actividad aglutinante (Blunden et al., 1986).
Las publicaciones científicas que describen la existencia de lectinas en algas rojas, pardas
y verdes son pocas. Ello es debido a que a pesar de que las lectinas fueron descritas hace
más de un siglo, la actividad hemoaglutinante de extractos de algas marinas solo se
conoce desde hace 50 años (Boyd et al., 1966), en contraste con las lectinas vegetales
superiores, que han sido objeto de numerosos y detallados estudios bioquímicos y
estructurales (Loris, 2002). Los estudios han conducido a proponer que los eritrocitos de
oveja y conejo son un modelo ideal para la detección de lectinas en algas (Freitas et al.,
1997), adicional a ello resulta idóneo hacer un tratamiento enzimático de los eritrocitos, lo
cual permite la exposición de los residuos crípticos de la superficie de la membrana de los
eritrocitos (Sharon y Lis, 1972) y por tanto un aumento en el acceso para la interacción de
los glicanos y la lectina.

19
Estudios de actividad hemoaglutinante han sido reportados en extractos de especies,
como Bryopsis hypnoides (Niu et al., 2009), B. pennata (Ainouz y Sampaio, 1991), B.
plumosa (Han et al., 2010; Jung et al., 2010), Caulerpa cupressoides (Ainouz y Sampaio,
1991; Benevides et al., 2001; Freitas et al., 1997) C. setularioides (Chiles y Bird, 1989;
Fajarningsih et al., 2015), C. paspaloides (Fajarningsih et al., 2015), Dictyota dichotoma,
D. dentata (Chiles y Bird, 1989; Fajarningsih et al., 2015), Halimeda micronesica, H.
macroloba y Sargassum duplicatum, S. polycystum (Fajarningsih et al., 2015),
desafortunadamente solo se ha llegado a hacer una purificación y caracterización posterior
de estas moléculas de unas pocas especies.
La caracterización molecular de las lectinas de algas está empezyo, también su
clasificación estructural, funcional y muchos estudios filogenéticos. Los datos estructurales
disponibles de lectinas de algas verdes, B. plumosa (Kim et al., 2006), E. prolifera
(Ambrosio et al., 2003), U. pertusa (Wang et al., 2004), B. triquetrum (Calvete et al., 2000),
H. japonica (Hori et al., 2000) H. musciformis (Nagano et al., 2002), especies del género
Ptilota (Sampaio et al., 1998a; Sampaio et al., 1999), y Griffithsia sp. (Mori et al., 2005),
muestran la existencia de una gran variedad de familias de lectinas presentes en las
diferentes especies de las algas marinas estudiadas.
Por otra parte solo se han determinado las estructuras primarias de catorce lectinas, entre
ellas B. triquetrum (Calvete et al., 2000), H. japonica (Hori et al., 2000), H. cervicornis, H.
musciformis (Nagano et al., 2005), U. pertusa (Wang et al., 2004), Griffithsia sp. (Mori et
al., 2005), B. plumosa (Bryohealin) (Kim et al., 2006), BPL2 (Han et al., 2010), BPL3 (Han
et al., 2011), BPL4 (Han et al., 2012), B. hypnoides (Niu et al., 2009), E. serra (Hori et al.,
2007), Boodlea coacta (Sato et al., 2011) y en algas pardas el único estudio reportado
recientemente fue para Hizikia fusiformis (Wu et al., 2016). La estructura tridimensional
solo se ha determinado para la lectina perteneciente a Griffithsia sp. (Ziółkowska et al.,
2006; Ziółkowska et al., 2007a), esta proteína tienen un motivo -prisma en sus
subunidades que se encuentra en lectinas de tipo jacalina (Figura 1-3) (Shimizu y
Morikawa, 1996), sin embargo, a diferencia de otras lectinas en este homodímero la
interacción de sus subunidades es única (Ziółkowska et al., 2006).

20
Figura 1-3 Comparación de la estructura terciaria de un monómero de proteína típica del grupo Jacalina (Helianthus tuberosus) y de un dimero de Griffithsia sp.
A) Helianthus tuberosus obtenido de PDB (1C3K) B) Dímero Griffithsia sp en complejo
con manosa. Tomado de (Ziółkowska y Wlodawer, 2006).
La detección de lectinas en extractos de algas marinas fue reportada por (Boyd et al.,
1966) y desde entonces se denominan phycolectinas (Matsubara et al., 1996; Rogers et
al., 1977), presentando diferencias en sus características físicoquímicas con respecto a las
estudiadas en plantas. Estas proteínas se encuentran en forma monomérica y son de bajo
peso molecular, presentan un alto contenido de aminoácidos ácidos, con puntos
isoeléctricos (pI) entre 4-6, no requieren iones metálicos para sus actividades biológicas y
la mayoría de ellas muestran más especificidad por glicoproteínas que por monosacáridos
(Hori et al., 1990; Rogers y Hori, 1993; Shiomi et al., 1980). Estas pueden ser clasificadas
en tres categorías teniendo en cuenta las propiedades de interacción con las
glicoproteínas, las específicas por N-glicanos, las que reconocen residuos de manosa y
las que presentan ambos tipos de especificidad (Hori et al., 1990).
B A
21

22
Actualmente es poco lo que se conoce respecto a las aplicaciones biotecnológicas de las
lectinas de algas marinas, principalmente por los bajos rendimientos durante los procesos
de purificación. Sin embargo algunos estudios señalan que estas moléculas tienen un
beneficio adicional; ya que en su mayoría poseen pesos moleculares pequeños y además
la presencia de puentes disulfuro, las han considerado muy estables (Teixeira et al., 2012),
y podrían ser de utilidad para la detección de carbohidratos de la superficie celular y en la
orientación de fármacos (Nascimento et al., 2006; Pinto et al., 2009). En otros estudios se
ha encontrado que poseen actividad anti-cancerígena (Fukuda et al., 2006), anti-
nociceptiva y anti-inflamatoria (Bitencourt et al., 2008; Silva et al., 2010; Vyerlei et al.,
2010), anti-bacteriana (Holya et al., 2005; Liao et al., 2003; Teixeira et al., 2007),
cicatrizante (Gonzaga do Nascimento-Neto et al., 2012), anti-viral (Mori et al., 2005; Sato
et al., 2011; Sato et al., 2015), inductor de la migración de neutrófilos in vivo/in vitro
(Bitencourt et al., 2008; Neves et al., 2001), implicados en la liberación de óxido nítrico
(Lima et al., 2004), y tienen actividad mitogénica (Hori et al., 1987). Estas lectinas han
recibido mayor atención debido a su gran especificidad de interacción con oligosacáridos
(Okuyama et al., 2009). Los estudios que han sido mencionados solamente cubren una
porción insignificante del número total de especies de algas y por lo tanto es necesario el
avance en este tipo de estudios.

23
2. Objetivos
2.1. Objetivo general
Realizar estudios enfocados en la búsqueda de lectinas de algas marinas del caribe
colombiano y estudiarlas bioquímicamente.
2.2. Objetivos específicos
• Preparar extractos salinos a partir de diferentes especies de algas marinas
• Evaluar la actividad aglutinante de los extractos salinos obtenidos de algas marinas
• Purificar la lectina de una de las especies evaluadas y determinar sus características
moleculares.

24
3. Capítulo 1. Preparación y evaluación de la actividad aglutinante en extractos de algas marinas del caribe colombiano.
3.1. Metodología
3.1.1. Colección de muestras
Las áreas de recolección para este estudio fueron seleccionadas por la Dra Mónica Puyana
Hegedus, en la tabla 3-1 se presentan las coordenadas de la colección para los siete
especímenes.
Tabla 3-1. Coordenadas de colección Isla de Providencia y Santa Marta
Numero Col. Especie Sitio de colección
599056 Turbinaria turbinata Felipe’s Place. Costado W de la isla de
Providencia, a 3,25 Km de Aguadulce. 12-20 m de profundidad
599057 Dictyota pinnatifida
599058 Sargassum hystrix var buxifolium
599059 Bryopsis ramulosa Punta la Loma (11°07’31’’N, 74°14’00’’W) al occidente de la ciudad Santa Marta. Zona
intermareal
599049 Caulerpa taxifolia
599050 Halimeda opuntia
599051 Tricleocarpa cylindrica
El muestreo se realizó en el mes de mayo del 2015, y una porción de la muestra se usó
para su clasificación taxonómica (Littler y Littler, 2000) usando caracteres morfológicos. Se
dejó el correspondiente registro del voucher en el Instituto de Ciencias Naturales-Herbario
Nacional Colombiano (Col). El material biológico fue almacenado en agua de mar dentro de
bolsas plásticas, transportado y congelado a -200C hasta su uso.
3.1.2. Obtención de los extractos y evaluación su actividad aglutinante
Se siguió la metodología descrita por Nagano et al. (2005) para limpiar las diferentes
muestras. Estas fueron descongeladas en agua a temperatura ambiente, separadas de
epifitas y lavadas con abundante agua destilada, después fueron secadas sobre papel
absorbente durante 24 h a 18 ⁰C. Se maceraron 10 g de material de cada una de las
especies con nitrógeno líquido hasta la obtención de un homogenizado el cual
denominaremos “harina”. Posteriormente 5 g de harina se lavaron sucesivamente usando
un homogenizador manual de embolo con acetona fría al 97% hasta la obtención de un

25
polvo fino blanco; los 5 g restantes de la harina no fueron tratados con acetona fría (Figura
3-1). La harina tratada y no tratada con acetona fue pesada y resuspendida en una
proporción 1:10 en PBS 1X pH 7,2 (Na2HPO4 20 mM, NaH2PO4 20 mM, NaCl 150 mM) 5
mM tiourea, y se dejó en agitación constante durante 24 h a 40C. Los extractos fueron
centrifugados a 10000 rpm durante 5 min, y el sobrenadante fue separado del pellet y usado
para ensayos preliminares de actividad aglutinante y SDS-PAGE.
Figura 3-1 Diagrama de flujo, procedimiento usado para elaboración y evaluación de los extractos.
Una alícuota de 1 ml de cada extracto se dializó en una membrana de 3 kDa (MWCO),
realizando dos cambios con agua destilada (c/u 1 L) y uno contra NH4 HCO3 20 mM, el
producto de la diálisis se liofilizo durante 48 h y se resuspendió en 200 µl de NH4 HCO3 20
10 g material
biológico
Macerar
Nitrógeno liquido
Obtener harina homogénea
Pesar y Re suspender polvo seco proporción 1:10 en PBS 1X tiourea
Agitación constante x 24 h 4⁰C
Centrifugación 10 000 rpm x 5 min 4⁰C
Separar sobrenadante
(Extracto)
Ensayos de aglutinación
Cuantificación BCA
SDS-PAGE
Diálisis y liofilizado
1 ml extracto
Ensayos de aglutinación
Cuantificación BCA
SDS-PAGE
5 g de harina
Lavados sucesivos acetona fría al 97% a 4󠅵⁰C
(Homogenizador manual)
5 g harina
no lavados
Resuspender 200µl
NH4 HCO3 20 mM
Se repite el proceso
para todos los
extractos y se
cuantifica por micro
Kjeldahl

26
mM, se evaluó nuevamente concentración de proteína por el método de BCA, actividad
aglutinante y perfil electroforético por SDS-PAGE.
3.1.3. Cuantificación de proteínas BCA
Esta fue realizada por el método del ácido bicinconínico (BCA) descrito por Smith et al.
(1985) y en microplaca de 96 pozos según lo descrito por Almanza, (1999) usando como
patrón una solución de BSA (Albúmina Sérica Bovina) de 1,5 mg/ml. A partir de las
concentraciones obtenidas fue determinado el porcentaje de proteína soluble (peso /peso
seco) en los extractos.
3.1.4. Ensayos de aglutinación
Para determinar el modelo de evaluación de la actividad aglutinante se realizó un ensayo
preliminar usando los extractos de T. turbinata, y T. cilíndrica frente a suspensiones de
eritrocitos de oveja, vaca, caballo y eritrocitos humanos tipo O+ siguiendo el método
descrito por Navarro y Pérez (1978), de acuerdo con los resultados obtenidos fueron
seleccionadas las suspensiones al 2% de eritrocitos de oveja y humano tipo O para realizar
los subsecuentes ensayos de actividad biológica. La sangre de todos los organismos fue
suministrada por el grupo de Andrología de la Facultad de Veterinaria de la Universidad
Nacional de Colombia.
Para los ensayos de actividad aglutinante con eritrocitos tratados enzimáticamente las
células fueron tratadas con tripsina siguiendo la metodología descrita por Ainouz et al.
(1992) y con pronasa (Hori et al., 1988). La actividad aglutinante se reportó empleando una
escala cualitativa de (-) a +4 según el grado de aglutinación, donde (-) indica la no
aglutinación y +4 la máxima aglutinación. Los resultados fueron verificados al microscopio
mediante un frotis en placa, bajo el objetivo 10 y 40X. Las pruebas fueron realizadas por
duplicado, como blanco se usó el buffer de suspensión de la muestra.
3.1.5. Cuantificación de proteínas por el método de micro Kjeldahl.
Con el fin de corroborar los resultados de concentración de proteína obtenidos para los
extractos, se hicieron nuevos extractos (Figura 3-1). Se tomó 1 ml de cada una de las
muestras y se determinó por duplicado el contenido total de nitrógeno proteico por el método
de micro Kjeldahl siguiendo los métodos propuestos por la AOAC (1990).

27
3.1.6. Tratamiento estadístico de los datos
Para las pruebas estadísticas se usó el software STATGRAPHICS Centurion, se aplicó una
prueba de normalidad Shapiro-Wilk y las pruebas que evidenciaron normalidad se
analizaron con un ANOVA simple para observar diferencias significativas entre tratamientos
con un 95% de confianza. Para las cuantificaciones realizadas con BCA se realizaron tres
replicas experimentales, mientras que las determinaciones realizadas con el método de
Kjeldahl se realizaron por duplicado.
3.2. Resultados
3.2.1. Obtención de los extractos salinos
El método de extracción para elaborar los extractos usando PBS 1X-tiourea 5 mM (Van
Driessche et al., 1984) permitió obtener extractos con actividad aglutinante y fueron
desprovistos de pigmentación cuando la harina era tratada con acetona en comparación
con la harina que no fue tratada (Figura 3-2). Los extractos salinos fueron homogéneos, sin
precipitado, de viscosidad moderada y con el tiempo no se observó oxidación por la
presencia de la tioúrea en el PBS.
Figura 3-2 Eliminación de pigmentos con lavados sucesivos de acetona
La cantidad de lavados necesaria para limpiar 2,5 g de harina estuvo relacionada con el
grupo de algas con la cual se trabajó, por ejemplo especies de algas verdes tenían una
mayor cantidad de pigmentos requiriéndose entre 17 y 20 lavados con acetona, las algas

28
rojas requirieron entre 14 y 15 lavados mientras las algas pardas entre 7 y 10 lavados (Por
cada 2,5 g de alga fueron necesarios 3 ml de acetona en cada lavado)
3.2.2. Cantidad de proteína en extractos de algas marinas
Los porcentajes obtenidos de proteína soluble determinados por el método de BCA (Figura
1, anexo 2) para los extractos cuantificados antes de un proceso de diálisis, mostraron
valores que oscilaron entre un 4,27 ± 0,97 % p/ps (peso/peso seco) y un 10,22 ± 0,52 %
p/ps correspondientes a las especies C. taxifolia y T. turbinata. De las siete especies
evaluadas, no se obtuvieron datos para la especie H. opuntia indicando las más bajas
concentraciones y además para 10 de 14 extractos se obtuvieron porcentajes de proteína
soluble sobre el 5 %. Los resultados del ANOVA mostraron que existían diferencias
significativas (p < 0,05), entre los extractos de T. turbinata, D. pinnatifida, C. taxifolia y S.
hystrix var buxifolium provenientes de harina tratada y no tratada con acetona fría (Tabla 3-
2). En todos los casos se observó una tendencia en la que los extractos obtenidos a partir
de harina tratada con acetona tenían los mayores valores de proteína soluble.
Tabla 3-2 Porcentajes de proteína soluble cuantificados por el método de BCA y micro Kjeldahl para los extractos provenientes de harina tratada con acetona y no tratada de siete especies de algas marinas.
Extracto Antes de la diálisis BCA
Después de la diálisis BCA
Antes de la diálisis micro Kjeldahl**
Proteína soluble % ± DS
ANOVA Valor-p
Proteína soluble % ± DS
ANOVA Valor-p
Proteína soluble % ± DS
ANOVA Valor-p
T. turbinata (L) 10,22±0,52 0,020*
1,46 ± 0,03 0,0002*
0,57±0,113 0,6176
T. turbinata (NL) 7,35±0,25 0,54 ± 0,05 0,52±0,042
T. cylindrica (L) 9,39±0,21 0,552
0,39 ± 0,01 0,689
0,52±0,042 0,4226
T. cylindrica (NL) 8,65±1,46 0,38 ± 0,04 0,49±0,000
B. ramulosa (L) 6,76±0,41 0,161
1,33 ± 0,12 0,056
3,22±0,262 0,1663
B. ramulosa (NL) 5,80±0,47 0,86 ± 0,12 2,82±0,000
D. pinnatifida (L) 8,81±0,11 0,008*
0,55 ± 0,02 0,222
0,55±0,078 0,3165
D. pinnatifida (NL) 4,60±0,54 0,47 ± 0,06 0,47±0,035
C. taxifolia (L) 7,85±0,05 0,035*
0,50 ± 0,27 0,063
1,37±0,262 0,2938
C. taxifolia (NL) 4,27±0,97 1,32 ± 0,13 1,02±0,226
S. hystrix var buxifolium (L) 8,98±0,09 0,029*
0,10 ± 0,00 0,506
0,36±0,042 0,0543
S. hystrix var buxifolium (NL) 6,11±0,71 0,10 ± 0,00 0,60±0,071
H. opuntia (L) - -
- -
0,47±0,035 0,3165
H. opuntia (NL) - 0,24 ± 0,01 0,39±0,078
Los porcentajes obtenidos de proteína soluble estan calculados sobre peso seco total del material usado para cada uno de los extractos (Ver tabla 1, 2 y 3 anexo), *indica diferencias significativas entre harina
lavada con acetona (L) y no lavada (NL) con un p < 0,05, DS desviación estándar, - no se detectó. Nota:
Los extractos cuantificados por micro Kjeldahl corresponden a un pool diferente a los cuantificados por BCA. ** coeficiente usado Kjeldahl: 6,25. n: 3 (BCA) y n: 2 (Kjeldahl)

29
Por otro lado, los resultados arrojados por la cuantificación de BCA para los mismos
extractos después del proceso de diálisis, no superaron el 1,5 % lo cual difirió de lo
encontrado cuando los extractos no fueron dializados. Los resultados del ANOVA solo
mostraron diferencias significativas (p < 0,05) para los extractos de T. turbinata, también se
observó una tendencia donde los extractos provenientes de harina tratada con acetona
presentaron mayores porcentajes indicando que los lavados con acetona sobre la harina
no generaron perdida de proteína excepto para C. taxifolia y H. opuntia donde no se observó
esta tendencia.
La determinación realizada por el método de Kjeldahl con nuevos extractos provenientes
de la harina tratada y no tratada no mostró diferencias significativas. Además, cuando se
realizó una comparación con los resultados obtenidos para los extractos dializados y
cuantificados por BCA los valores fueron similares (no superaron el 3,22 %), por ejemplo,
para los extractos de T. turbinata (NL) y D. pinnatifida (L y NL) indicando que para cuantificar
por BCA es necesario realizar diálisis a los extractos. Solo se hallaron diferencias entre los
extractos de B. ramulosa (L y NL) y S. hystrix var buxifolium (L y NL) donde los porcentajes
de proteína determinados por Kjeldahl fueron mayores a los encontrados por BCA (tabla 4,
anexo 1), mientras en los extractos de T. turbinata (L) y C. taxifolia (NL) fueron mayores las
determinaciones hechas por BCA.
En el SDS-PAGE para los extractos que no fueron dializados, no se obtuvieron bandas a
pesar de que en cada uno de los carriles fueron sembrados 100 µg de proteína teniendo en
cuenta los datos de cuantificación (Figura 2, anexo 2). Con los extractos que fueron
dializados y nuevamente cuantificados, se observaron bandas para algunas especies a
pesar de tener 100 g de proteína total por carril (Figura 3, anexo 2); por ejemplo, para B.
ramulosa se observaron numerosas bandas de alto y bajo peso molecular, un perfil de
bandas similar al encontrado en C. taxifolia, especie perteneciente al grupo de algas verdes.
Para los extractos de algas pardas como T. turbinaria, y D. pinnatifida se observaron menos
bandas, las cuales se observaron alrededor de 95, 2 y 10 kDa, para las especies restantes
no se obtuvieron bandas. En general los perfiles electroforéticos donde se observaron
bandas definidas correspondieron a los extractos provenientes de harina lavada de
acetona.

30
3.2.3. Actividad aglutinante de extractos no dializados
Los resultados preliminares usando los extractos de dos especies (Tabla 3-3), mostraron
que la suspensión de eritrocitos de humano y los eritrocitos de oveja podrían ser usados
como modelo inicial para buscar actividad aglutinante en los siete extractos de algas. En el
caso de los eritrocitos de oveja aunque la actividad fue débil, se determinó seguir usándola
por los reportes de este tipo de eritrocitos como idóneos después de los eritrocitos de conejo
para detectar lectinas en extractos de algas (Hori et al., 1988).
Tabla 3-3 Ensayo preliminar de aglutinación en dos extractos
Especie Cantidad*
(µg)
Oveja
Máxima
aglutinación
Vaca
Máxima
aglutinación
Caballo
Máxima
aglutinación
Humano tipo
O Máxima
aglutinación
T. turbinata (L) 36,48 ˗+ ˗ ˗ ˗
T. cylindrica (L) 9,81 ˗ ˗ ˗ +3
T. turbinata (NL) 13,49 ˗ ˗ ˗ ˗
T. cylindrica (NL) 9,49 ˗ ˗ ˗ +3
(L) extractos provenientes de harina lavada con acetona, (NL) Extractos provenientes de harina no
lavada con acetona. Los colores representan los diferentes grupos de algas (pardas y rojas).
*Cuantificación por BCA para 25 µl de extracto usados en el ensayo (-+) aglutinación no
confirmada en diferentes campos
Cuando se hizo la evaluación de todas las especies, los extractos de T. cilíndrica aglutinaron
solamente eritrocitos tipo O+ con 9,49 µg de proteína total (Tabla 3-4). De forma similar,
extractos como los de C. taxifolia mostraron actividad aglutinante positiva frente a eritrocitos
tipo O+ y de oveja con 12,58 g de proteína. Vale la pena resaltar que, en este último caso,
se observó una mayor aglutinación (+3) con una cantidad de 32,92 µg de proteína total del
extracto obtenido de la harina sin ser tratada con acetona.

31
Los extractos de B. ramulosa y D. pinnatifida causaron hemólisis de los eritrocitos cuyos
extractos venían de la harina que no fue tratada con acetona, y su actividad aglutinante solo
pudo ser evidenciada en los extractos donde la harina recibió el tratamiento con acetona.
Con 33,23 µg de proteína total del extracto de B. ramulosa se observó una aglutinación
fuerte para eritrocitos de oveja (+4) y de humano tipo O+ (+3), mientras que para los
extractos de D. pinnatifida con 13,76 µg de proteína total se observa una aglutinación más
débil (+2) en eritrocitos de oveja y +1 eritrocitos de humano tipo O+. Para el extracto de S.
hystrix var buxifolium, se observó una actividad muy débil con eritrocitos de oveja, sin
embargo, estos resultados no pudieron ser verificados al microscopio tal como se observa
en la Figura 3-4.
3.2.4. Actividad aglutinante con eritrocitos de oveja tratados enzimáticamente.
Teniendo en cuenta que los extractos donde la harina recibió tratamiento de lavados con
acetona de siete especies, tres mostraron actividad aglutinante evidente usando eritrocitos
de oveja, mientras que con eritrocitos humanos solo la especie T. cylindrica mostró ser
especifica (Tabla 3-4), se decidió optimizar la detección de la actividad de lectina realizando
tratamiento enzimático a los eritrocitos de oveja siguiendo lo propuesto en otros estudios,
para aumentar la sensibilidad del ensayo (Ainouz y Sampaio, 1991; Ainouz et al., 1992;
Hori et al., 1988; Nagano et al., 2005).
Tabla 3-4 Actividad aglutinante en siete extractos de algas marinas previo a un proceso
de diálisis.
Especie
No tratada con acetona Tratada con acetona
Cantidad (µg) **
Oveja Máxima
aglutinación
Humano tipo O Máxima
aglutinación
Cantidad (µg) **
Oveja Máxima
aglutinación
Humano tipo O Máxima
aglutinación
T. turbinata 13,49 ˗ ˗ 36,48 ˗ ˗
T. cylindrica 9,49 ˗ +3 9,81 ˗ +3
B. ramulosa 21,40 H H 33,23 +4 +3
D. pinnatifida 11,84 H H 13,76 +2 +1
C. taxifolia 32,92 +3 +3 12,58 +2 +2
S. hystrix var buxifolium
2,45 ˗ ˗ 2,39 ˗ + ˗
H. opuntia 5,90 H H ND ˗ ˗
Blanco ˗ ˗ ˗ ˗
*lisis de eritrocitos, (-+) aglutinación no confirmada en diferentes campos (-) sin actividad, (ND) no se detectó proteína, (H) hemólisis. Los colores representan los diferentes grupos de algas (pardas, rojas y verdes). **Cuantificación por BCA para 25 µl de extracto usados en el ensayo

32
Tabla 3-5 Actividad aglutinante para los extractos de algas marinas usando eritrocitos de oveja
Especie Cantidad
(µg) *
Sin tratamiento enzimático
Eritrocitos tratados con tripsina
Eritrocitos tratados con pronasa
Máxima aglutinación
Título aglutinante
(µg)
Máxima aglutinación
Título aglutinante
(µg)
Máxima aglutinación
Título aglutinante
(µg)
T. turbinata 29,18 - - - - - -
T. cylindrica 7,85 - - - - - -
B. ramulosa 26,58 +4 6,65 +4 3,32 +4 3,32
D. pinnatifida 11,01 +2 5,50 +3 0,17 +3 1,38
C. taxifolia 10,07 +2 1,26 +3 0,63 +3 0,63
S. hystrix var buxifolium
1,91 +2 0,48 +3 0,06 +3 0,24
H. opuntia ND - - - - - -
Blanco - - - - - -
(-) sin actividad, (ND) no determinado. Los colores representan los diferentes grupos de algas (pardas, rojas y verdes). *Cuantificación por BCA para 20 µl de extracto usando en el ensayo
Figura 3-3 Vista al microscopio de aglutinaciones obtenidas para diferentes extractos de algas provenientes de harina tratada con acetona frente a eritrocitos de oveja.
A) B. ramulosa B) C. taxifolia C) D. pinnatifida, D) S. hystrix var buxifolium. Objetivo 10X
Los resultados mostraron que cuando los eritrocitos de oveja fueron tratados
enzimáticamente, la cantidad de proteína para observar aglutinación en todos los casos fue
menor con respecto a la cantidad requerida para observar aglutinación con eritrocitos no
tratados (Tabla 3-5). Para el extracto de B. ramulosa al usar eritrocitos tripsinizados o
tratados con pronasa, se requiere solo la mitad (3,32 µg) de proteína total con respecto a
la necesaria para aglutinar los eritrocitos sin tratamiento (6,65 µg). Se observó la misma
A B
C D

33
tendencia para el extracto de C. taxifolia y en el caso de extractos como los de D. pinnatifida
y S. hystrix var buxifolium se requieren muy bajas cantidades de proteína total para observar
actividad usando eritrocitos tripsinizados, 0,17 y 0,06 µg respectivamente. Algunas de las
aglutinaciones observadas al microscopio se muestran en la figura 3-4.
3.3. Análisis
3.3.1. Porcentajes de proteína soluble
Al observar el comportamiento de los datos de cuantificación de proteína soluble en los
extractos surgen dos grandes interrogantes; el primero ¿Por qué después de realizar un
proceso de diálisis, los porcentajes de proteína soluble son inferiores en todos los
extractos? y ¿Por qué se observa una tendencia de mayores porcentajes de proteína
soluble en los extractos que provienen de harina a la cual se le han realizado lavados
sucesivos con acetona?
Para responder estos dos interrogantes, debe considerarse que existen estudios
fitoquímicos donde se reporta que los extractos de algas marinas contienen no sólo
moléculas antioxidantes lábiles como el ascorbato, glutatión (Burritt et al., 2002; Indegaard
y Minsaas, 1991; Morgan et al., 1980), también moléculas más estables como los
carotenoides (Morgan et al., 1980; Okai et al., 1996; Yan et al., 1999), aminoácidos de tipo
micosporina (Nakayama et al., 1999; Yuan y Walsh, 2006), y además una variedad de
polifenoles como catequinas, y florotaninos (Nakamura et al., 1996; Yoshie et al., 2000).
También existe una correlación directa entre las actividades antioxidantes y el poder
reductor de ciertos extractos de plantas (Duh et al., 1999; Zhang et al., 2010). Las
propiedades reductoras se asocian generalmente con la presencia de un agente reductor,
el cual se ha demostrado ejerce acción antioxidante rompiendo la cadena de radicales libres
donando un átomo de hidrógeno (Gordon, 1990). La diferencia en los porcentajes de
proteína soluble en los extractos no dializados respecto a los dializados se puede
correlacionar con la presencia de este tipo de moléculas ya que su poder reductor puede
generar una sobreestimación en las concentraciones de proteína determinadas por el
método de BCA. Se presume que los iones Cu+2 podrían estar siendo reducidos a un estado
Cu+1, no solo por aminoácidos como la cisteína, cistina, triptófano y tirosina de las proteínas
(Brown et al., 1989; Wiechelman et al., 1988), también por estas moléculas antioxidantes
generando una sobreestimación de la cantidad de proteína presente en las muestras. Otras

34
moléculas como azucares reductores (Castellanos González et al., 2012), niacina, tiamina
(Lundin y Ericson), ácido ascórbico (Morgan et al., 1980), podrían llegar a estar involucradas
en este efecto, ya que han sido reportados en extractos acuosos y alcohólicos de diferentes
grupos de algas.
Los porcentajes superiores de proteína soluble en los extractos no dializados provenientes
de harina de alga lavada con acetona respecto a los provenientes de harina no lavada
pueden explicarse porque estas muestras estuvieron sometidas a un mayor proceso de
ruptura de pared celular (en el homogenizador manual), lo cual permitió la extracción de
una mayor cantidad de proteína y metabolitos que pueden interferir con el método de
cuantificación. Por lo tanto, los lavados sucesivos con acetona no estarían eliminando por
completo algunos metabolitos (por la cantidad y/o naturaleza química de las moléculas). Al
realizar el proceso de extracción de la harina, se incluyó en el buffer de extracción tiourea
para evitar la oxidación posterior de los polifenoles que pudieran permanecer en los
extractos (Van Driessche et al., 1984), los cuales son extraídos por maceración
mayoritariamente en algas pardas. Los polifenoles y sus productos de oxidación
(quinonas), se unen fuertemente a las proteínas enmascarando su actividad; además la
tiourea inhibe de la acción de polifenoloxidasas y por lo tanto evitando la oxidación los
extractos antes de los procesos cromatográficos (Loomis, 1974; Van Driessche et al., 1984).
Por otro lado compuestos de naturaleza reductora fueron eliminados efectivamente con un
proceso de diálisis tal como lo muestran los resultados (Tabla 3-2), y además las
electroforesis realizadas antes y después de este proceso, confirmaron que la cantidad de
proteína en los extractos no dializados fue sobreestimada pues a pesar de sembrar en cada
carril 100 µg de proteína y posterior tinción con plata no se observaron bandas (Figura 2,
anexo 2); mientras que con los extractos que fueron dializados y nuevamente cuantificados
se encontraron bandas de proteína ya que se aseguró una cantidad real de proteína por
carril (Figura 3, anexo 2). En otros estudios de extracción de proteínas a partir de algas
marinas no se incluyó la adición de agentes antioxidantes (Nagano et al., 2002; Wu et al.,
2016), ni el uso de acetona para obtener extractos activos y desprovistos de pigmentación
para los posteriores pasos de purificación. Extractos de plantas tratados con acetona,
permitieron eliminar polifenoles e inactivar las polifenoloxidasas (Araújo et al., 2015;
Sánchez‐Ferrer y García‐Carmona, 1992); otros reportes de obtención de extractos a partir
de semillas vegetales, mostraron que la adición de tioúrea reduce notablemente la

35
oxidación enzimática, manteniendo la actividad de las lectinas hasta en un 80% por ensayo
de ELLSA (Jiménez, 2000); lo anterior demuestra la utilidad del método que se empleó en
este trabajo. Otros estudios, como el de Nagano et al. (2002), permitieron eliminar
pigmentos como las ficobilinas del extracto de H. musciformis disminuyendo el pH a 1,0 con
HCl y posterior purificación por cromatografía de intercambio iónico sobre DEAE-Celulosa;
sin embargo, este tratamiento podría desnaturalizar las proteínas del extracto. Por otra
parte, aunque hay muy pocos estudios enfocados a la eliminación de pigmentos para
obtener extractos proteicos de algas marinas, algunos indican que la acetona permiten
eliminar una gran proporción de clorofilas y carotenoides (Torres et al., 2014) además de
algunos tipos de polifenoles, en especial taninos (Cox et al., 2010). Este paso es importante
para llegar a estudiar bioquímicamente las lectinas ya que los metabolitos endógenos
podrían interferir con los estudios de purificación (Storey y Beevers, 1978).
Otros estudios han determinado que hay un menor porcentaje de polifenoles en extractos
acuosos de algas con respecto a los hidroalcohólicos (agua-etanol); por ejemplo para Ulva
fasciata hay 3.72% de polifenoles en buffer salino pH 7,2 con respecto 7,44% que se
extraen en una mezcla de agua-alcohol (Valdés et al., 2008). Una situación diferente fue la
hallada para D. pinnatifida donde los mayores contenidos de polifenoles fueron hallados en
extractos acuosos (34.98%) respecto a extractos hidroalcohólicos (10,12%); por lo tanto, la
concentración y naturaleza de los polifenoles presentes en las dos especies es diferente.
De forma similar, un análisis fitoquímico realizado con extractos acuosos de Padina
gymnospora muestran la presencia de mayores cantidades de fenoles y taninos respecto
a las halladas en extractos hidroalcóholicos (Gonzalez et al., 2015). En este último caso
quizás sea uno de los motivos por los cuales en estudios donde se evalúa la cantidad de
proteína soluble en extractos crudos de algas marinas se opta por métodos como el de
Bradford (de Souza et al., 2009; Garateix et al., 2000; Valdés et al., 2008) y Kjeldahl (Frikha
et al., 2011) ya que con estos métodos la interferencia por agentes reductores no existe.
Nuestros resultados indicaron que el proceso de diálisis de los extractos provenientes de
harina tratada y no tratada, permitió eliminar una gran proporción de las moléculas
interferentes y determinar proteína soluble por el método de BCA, el cual tiene una
sensibilidad mayor a la reportada por otros métodos (Keller y Neville, 1986). Esta idea es
acertada y se observa que los porcentajes de proteína obtenidos por el método de Kjeldahl,
fueron cercanos a los valores arrojados por BCA (Tabla 3-2). Para la mayoría de los

36
extractos evaluados no se encontró una diferencia significativa al comparar los resultados
obtenidos por los dos métodos (Figura 3-3) excepto para los extractos de T. turbinata, y B.
ramulosa, donde esta diferencia puede ser atribuida a que en primera medida los
porcentajes de proteína determinados por Kjeldahl no fueron realizados sobre las mismas
muestras usadas para la cuantificación de BCA, aunque se mantuvieron las mismas
condiciones para prepararlos y reproducir el proceso de homogenización sobre la harina
tratada. También debe considerarse la presencia de nitrógeno no proteico proveniente de
nitratos, nitrógeno amoniacal, carbohidratos complejos además de aminoácidos libres
presentes en todos los extractos de algas pero que se podrían presentar diferencialmente
de una especie a otra (Lourenço et al., 2002). En un estudio realizado por Keller y Neville
en 1986, al determinar el contenido de proteína total en leche humana, encontraron que las
cantidades de proteína soluble halladas por métodos como Biuret, Lowry, y BCA eran
comparables a lo obtenido por Kjeldahl y aún más importante, no encontraron diferencias
estadísticamente significativas contra los resultados del BCA.
Un trabajo realizado en 2008 con algas colectadas del Archipiélago cubano, se determinó
para los extractos hidroalcohólicos de las especies D. pinnatifida, porcentajes de proteína
soluble (método de Bradford) del 11%, T. turbinata del 2,83%, S. fluitans del 6,64% y S.
natans del 8,44% (Valdés et al., 2008). Para otras especies de diferentes zonas del
archipiélago, al realizar la cuantificación de proteína por el método de Lowry se encontraron
porcentajes de proteína para S. fluitans del 31,00 %, y para H. opuntia de (3,97%); los
porcentajes hallados para S. natans difieren en los dos estudios, incluso entre zonas donde
se colecto la misma especie. Los resultados de este estudio también difieren de lo reportado
por estos autores para D. pinnatifida, T. turbinata y H. opuntia; esencialmente porque la
síntesis de proteínas depende de las condiciones ambientales a las que se encuentra
expuesta la especie; por ejemplo en respuesta al estrés oxidativo, por contaminación
orgánica o metálica, se da una mayor expresión de enzimas antioxidantes, compuestos
hidrosolubles y moléculas liposolubles antioxidantes (Contreras et al., 2005; Foyer et al.,
1997). Así cuando la producción de especies reactivas al oxigeno excede la capacidad
antioxidante, ocurre la oxidación de proteínas, lípidos, polisacáridos y ácidos nucleicos.
Debe considerarse además que el hecho que algunas especies requirieran una mayor
cantidad de lavados como las especies B. ramulosa, C taxifolia y H. opuntia, podría llegar
a tener un efecto en las concentraciones de proteína. Los altos porcentajes de proteína en
este estudio se explican debido al uso de soluciones hidroalcohólicas para la extracción de
proteínas por ejemplo los porcentajes de proteína para Ulva fasciata en extracto Etanol-

37
agua fue de 13,40 % mientras que en un extracto acuoso fue de 0,12 % (Valdés et al., 2008)
valor similar a los hallados en este estudio, considerando que las lectinas son proteínas
globulares y su extracción es eficiente en soluciones salinas.
Figura 3-4 Eliminación secuencial de pigmentos para una muestra de harina de B. ramulosa.
Los resultados obtenidos en este trabajo nos permiten sugerir que el método para la
extracción, cuantificación y evaluación de actividad aglutinante de múltiples extractos de
algas corresponde a la obtención de harina de alga, la cual debe ser sometida a lavados
sucesivos con acetona a bajas temperaturas (Figura 3-2 y 3-4), estos lavados deben
realizarse hasta que la harina se torne sin pigmentos. Una vez limpia y seca la muestra se
procede a obtener un extracto proteico usando PBS 1X- 5 mM tiourea; al final los extractos
deben ser dializados y cuantificados por el método de BCA para realizar las pruebas de
aglutinación.
3.3.2. Ensayos de aglutinación
Se encontró la actividad de lectina en cinco de siete especies estudiadas, entre ellas T.
cylindrica, B. ramulosa, D. pinnaifida, C. taxifolia y S. hystrix var buxifolium, y aunque para
T. turbinata existen reportes de aglutinación con eritrocitos de humano tipo O, A y B (Boyd
et al., 1966) y para H. opuntia con eritrocitos de conejo tratados enzimáticamente con
pronasa (Perez-Lorenzo et al., 1997), en el presente estudio no se encontró actividad en
estos extractos con eritrocitos de oveja ni humano tipo O.
En especímenes colectados en la región Antártica, se reporta para Halimeda sp. actividad
con eritrocitos de conejo tratados con tripsina usando 239,4 µg de proteína total (de Souza
et al., 2009), sin embargo vemos que la cantidad total de proteína en la cual se observó
aglutinación para las especies estudiadas es 40 veces menor. Aunque no está muy clara

38
cuál es la función que tienen las lectinas en este tipo de organismos, se han relacionado en
procesos reproductivos (Kim et al., 2006), defensa contra microorganismos y virus (Akkouh
et al., 2015; Balzarini et al., 2007; Cheung et al., 2015) y juegan un papel muy importante
en la simbiosis con otros organismos (Hori et al., 1988), lo cual explica que al igual que los
metabolitos secundarios, estas se expresen de forma diferencial en especies colectados en
épocas y localidades diferentes, por lo que esta puede ser una de las causas de los
resultados negativos obtenidos para las especies T. turbinata. y H. opuntia.
Para las demás especies evaluadas no existen reportes sobre actividad aglutinante, sin
embargo, se han llegado a estudiar, en algunos casos, representantes de los mismos
géneros. En algas verdes como C. racemosa no se llegó a detectar actividad, mientras que
con C. sertularioides se encontró un resultado positivo con eritrocitos de pollo (Ainouz et
al., 1992), en este mismo estudio para el alga parda S. vulgäre no se reportó actividad
positiva mientras que para S. filipendula se logró detectar aglutinación con eritrocitos de
oveja y conejo. En otras especies de algas pardas como D. dichotoma se reportó
aglutinación de eritrocitos de conejo y oveja, con cantidades de proteína de 0,92 µg y 0,4
µg respectivamente; extractos de Padina vickersiae y Spatoglossum schroederi aglutinaron
eritrocitos de conejo (Bird et al., 1993). Los trabajos realizados con algas pardas son menos
respecto a los desarrollados con algas rojas y verdes, lo cual se ha atribuido a la presencia
de una alta cantidad de polifenoles en este tipo de extractos que enmascaran la actividad
de las lectina (Loomis, 1974).
Los extractos de las algas verdes C. paspaloides y C. sertulariodes aglutinan eritrocitos de
conejo con 0,09 µg y de oveja con 2,9 y 31 µg de proteína total respectivamente. En el caso
de B. plumosa se obtuvo actividad con eritrocitos de oveja con 1,3 µg de proteína total (Bird
et al., 1993).
Eritrocitos de varias fuentes animales han sido empleados para la detección de lectinas en
extractos salinos debido al reconocimiento que hace la lectina por carbohidratos que se
encuentran en la superficie de las células (Eagle, 1930; Sumner y Howell, 1936), en el caso
de los extractos de algas marinas algunos estudios (Chiles y Bird, 1989; Hori et al., 1988)
muestran la utilidad del uso de suspensiones de eritrocitos de oveja y conejo cuando se
quiere hallar actividad aglutinante. También se hace degradación enzimática de proteínas
y/o carbohidratos de la superficie de la membrana de los eritrocitos para generar una mayor
accesibilidad para la lectina (Ainouz et al., 1992; de Souza et al., 2009; Hori et al., 1988;

39
Nagano et al., 2005; Nagano et al., 2002; Wu et al., 2016). Ainouz et al. (1992) encuentran
que la mayor cantidad de detecciones y en su mayoría los mejores títulos aglutinantes se
obtienen con eritrocitos de conejo tripsinizados; mientras que para Hori et al. (1998), las
mejores condiciones fueron determinadas usando eritrocitos de oveja tratados con pronasa.
Como no había un consenso sobre el tipo de tratamiento para obtener los mejores títulos
aglutinantes, en este trabajo se hicieron ensayos enfocados en determinar cuál de estos
tratamientos enzimáticos era el mejor. Algunos estudios han determinado que el mayor
componente que existe sobre la membrana de los eritrocitos de oveja corresponde al
antígeno de Forssman, seguido del componente Lacto-N neotetraosilceramida: Gal β (1,3)
GlcNAc β (1,3)Gal β (1,4) Glc β (1,1) ceramida (McKibbin, 1978; Momoi y Yamakawa, 1978).
El determinante de Forssman (Figura 3-6) es un glicolípido con la estructura terminal
GalNAcα(1,3)GalNAcβ(1,3)Galα(1,4)Galβ(1,4)Glc (Yamamoto et al., 2012) siendo
estructuralmente similar al antígeno A de eritrocitos humanos; también existen otros
componentes de la membrana de eritrocitos de oveja que aún no han sido bien
caracterizados, lo cual en cierta medida podría explicar porque el tratamiento con tripsina
sobre eritrocitos de oveja mejoró los títulos aglutinantes de especies como D. pinnatifida y
S. hystrix var buxifolium. La tripsina al cortar de manera selectiva sobre argininas (Arg) o
lisinas (Lys) de glicoproteínas expuestas en la superficie celular podría dejar algunos
residuos expuestos importantes para la interacción entre la lectina y ligando, mientras que
la pronasa los eliminaría en su totalidad. Los resultados de aglutinación indican que en el
caso de especies como D. pinnatifida, C. taxifolia, S. hystrix var buxifolium y B. ramulosa
podrían presentar lectinas que reconocen residuos de GalNAc; en el caso de B. ramulosa
y T. cylindrica, este reconocimiento también podría estar dado por residuos como la fucosa
o galactosa, presentes en el antígeno de eritrocitos humanos tipo O, llamado antígeno H:
Fuc α (1,2) Gal β (1,3/4) GlcNAc β (1,3) Gal (Hara et al., 2013).

40
Figura 3-5 Región terminal del antígeno de Forssman.
La presencia de polifenoles en las muestras podrían causar aglutinación de los eritrocitos y
de esta forma enmascarar la actividad de las lectinas de algas pardas (Rogers y Loveless,
1991), con la metodología realizada en este trabajo, se observó claramente la actividad de
los extractos de D. pinnatifida y S. hystrix var buxifolium debida a la presencia de lectinas.
En contraste con la actividad de aglutinación, se presentó lisis con los extractos que se
obtuvieron a partir de la harina no tratada con acetona, para las especies B. ramulosa, D.
pinnatifida y H. opuntia (Tabla 3-4); la presencia de saponinas (Gonzalez et al., 2015;
González et al., 2003) u otras moléculas de naturaleza terpenoide (Govindan et al., 1993)
son capaces de aumentar la permeabilidad de la membrana a iones pequeños, afectando
la estabilidad estructural de la membrana y desestabilizando el empaquetamiento de la
bicapa lipídica (Maguna et al., 2006), hecho por el cual a muchas de estas moléculas se les
atribuye actividad antibacteriana y en este caso causan la lisis de los eritrocitos (González
et al., 2003; Martínez et al., 2009). El tratamiento con acetona no generó perdida de la
actividad biológica de las lectinas de los extractos de las especies de algas evaluadas y es
un paso importante para encontrar la presencia de estas proteínas en las algas marinas.

41
3.4. Conclusiones
1. El tratamiento con acetona sobre la harina de alga antes de la elaboración de los
extractos disminuye la pigmentación entre otros contaminantes de las muestras.
Luego de este tratamiento en los extractos, no se observó una pérdida significativa
de proteína, siendo este un paso importante para la obtención de los extractos
proteicos.
2. Para realizar cuantificación de proteínas por el método del ácido bicinconinico
(BCA), se requiere hacer un paso adicional de diálisis de los extractos proteicos.
3. Los lavados con acetona no causan perdida de la actividad de las lectinas, al
contrario, con algunos extractos se pudo evidenciar la presencia de lectina y evito
la hemólisis de los eritrocitos.
4. Los extractos de T. cylindrica, B. ramulosa, D. pinnatifida y C. taxifolia reconocieron
eritrocitos tipo O de humano, mientras que B. ramulosa, D. pinnatifida, C. taxifolia y
S. hystrix var buxifolium eritrocitos de oveja siendo mejores las aglutinaciones con
este último tipo de células.
5. Para las especies T. turbinata y H. opuntia no se detectó actividad aglutinante debido
a la cantidad de proteína total empleada.
6. El tratamiento enzimático de los eritrocitos de oveja mejoro los títulos de aglutinación
y para algunos extractos como los de B. ramulosa y C. taxifolia no se encontró
ninguna diferencia mientras que con los extractos de las especies D. pinnatifida y S.
hystrix var buxifolium, los eritrocitos tripsinizados permitieron obtener mejores
títulos.
7. El extracto de Bryopsis ramulosa mostro buena actividad de aglutinación y una alta
cantidad de proteína, por lo tanto, es una buena candidata para iniciar los estudios
de purificación, caracterización de la estructura de sus lectinas.

42
4. Capítulo 2. Purificación de la (s) lectina(s) presente(s) en Bryopsis ramulosa
4.1. Metodología
4.1.1. Colección de la muestra
La especie fue colectada en Punta la Loma, Santa Marta, Colombia tal como fue descrito
en la sección de métodos del capítulo 1 “Preparación y evaluación de actividad aglutinante
en extractos de algas marinas del caribe colombiano”.
4.1.2. Obtención de la harina de B. ramulosa
Para la limpieza de la muestra se siguió el método descrito por Nagano et al. (2005). El
material descongelado fue lavado con agua destilada y limpiado de epifitas. Posteriormente
se dejó secar a temperatura ambiente sobre papel absorbente por 24 h, transcurrido este
periodo de tiempo el material fue macerado en nitrógeno líquido hasta la obtención de una
harina homogénea la cual fue lavada sucesivamente con acetona fría al 97% en un
homogenizador manual. El polvo fino obtenido al final del proceso fue secado a temperatura
ambiente y pesado para la elaboración del extracto.
4.1.3. Determinación porcentaje de humedad y contenido de proteína total de la harina de B. ramulosa
Se siguió la metodología propuesta en la AOAC 1990 numeral 14004 para determinar el
porcentaje de humedad (Of, 1990). Se colocó 1 g de muestra en una cápsula de porcelana
previamente tarada, posteriormente la muestra fue introducida en una estufa de convección
a una temperatura de 85°C durante 72 h. Transcurrido este tiempo la capsula fue
depositada en un desecador con cloruro de calcio hasta que alcanzó la temperatura
ambiente; finalmente esta fue pesada. El porcentaje de humedad fue determinado por la
diferencia de los pesos, antes y después del calentamiento. La determinación fue realizada
por triplicado. Para la determinación de proteína total en la harina de alga, se tomaron dos
matraces de Kjeldahl para la digestión y a cada uno de ellos se adiciono 30 mg de la harina
(no tratada), a continuación, se adicionó 100 mg de mezcla catalizadora y se realizó la
digestión, destilación y titulación según el método establecido por el AOAC.

43
4.1.4. Obtención de los extractos proteicos
El polvo de alga seco y previamente lavado con acetona fría al 97%, fue pesado y
resuspendido en una proporción 1:10 en PBS 1X pH 7,2 (Na2 HPO4 20 mM, NaH2 PO4 20
mM, NaCl 150 mM) 5 mM tiourea, la solución fue dejada en agitación constante durante 24
h a 4oC, centrifugada a 10.000 rpm durante 5 min, el sobrenadante fue separado del pellet.
Para la preparación del extracto en pequeña escala solo fue realizada una extracción sobre
el pellet, y con el sobrenadante se realizaron las diferentes pruebas. En el caso de la
extracción a gran escala, se realizó tres veces la extracción sobre el mismo pellet con el fin
de obtener la mayor cantidad de proteína posible. Para cada una de las extracciones se
empleó la misma cantidad de buffer (1:10), al final del proceso los sobrenadantes de las
tres extracciones se unieron en un solo pool para obtener el extracto total.
4.1.5. Cuantificación de proteínas
Para cada una de las etapas de purificación, fue determinada la concentración por el
método de BCA (ácido bicinconínico) descrito por Smith et al. (1985) y según la metodología
de Almanza, (1999). La determinación se realizó empleando como patrón una solución de
BSA (Albúmina Sérica Bovina) y como blanco el buffer en el que se encontraba la muestra.
También se hicieron determinaciones por micro Kjeldahl siguiendo las especificaciones de
la AOAC 1990.
4.1.6. Electroforesis unidimensional en geles de poliacrilamida
En cada una de las etapas de purificación, fue realizada una electroforesis en condiciones
denaturantes según los métodos descritos por Laemmli (1970) en geles del 12,5% o 15%.
Las muestras fueron preparadas con (DTT) (40-100 mM) y desnaturalizadas por
calentamiento a 95°C durante 5 min. Los geles fueron fijados y teñidos con plata siguiendo
el protocolo empleado por el grupo de investigación (GRIP), y cuando se observó tinción
inversa para algunas bandas, se realizó tinción con azul de Coomassie G-250. Los geles
fueron desteñidos y estos fueron registrados con el foto-documentador compacto y
automatizado, usando el programa Quantity One o Image Lab de Bio-Rad.

44
4.1.7. Ensayos preliminares de purificación: precipitación fraccionada y cromatografía de intercambio iónico a pequeña escala.
Con 30 ml del extracto total se realizó una precipitación fraccionada según la metodología
descrita por Englard y Seifter (1990) usando sulfato de amonio entre el 0-50% y el 50-70%
de saturación; cada uno de los pellets obtenidos fueron resuspendidos en 3 ml de (NH4)
HCO3 20 mM, y fueron dializadas usando una membrana de 3 kDa contra agua desionizada,
realizando dos cambios (c/u 1 L) cada 8 h y uno final con (NH4) HCO3 20 mM.
Posteriormente se determinó la actividad aglutinante empleando una suspensión de
eritrocitos al 2% de oveja, concentración de proteína por BCA y se hizo SDS-PAGE en geles
al 12,5%.
2,9 ml del precipitado que mostró actividad aglutinante fue equilibrado en PBS 1X pH 7.2
(Na2HPO4 0,02 M, NaH2PO4 0,02 M, NaCl 150 mM) y fue sometido a una cromatografía de
intercambio iónico sobre DEAE-Sepharose (3ml de soporte en una columna de 1,5 cm de
diámetro X 9 cm de alto) usando un gradiente discontinuo de fuerza iónica. La fracción no
retenida se eluyó con un buffer PBS 1X pH 7.2 y la fracción retenida con el mismo buffer en
NaCl 500 mM. Las fracciones de mayor absorbancia fueron colectadas y luego dializadas
en una membrana de 3 MWCO contra agua desionizada, haciendo dos cambios (c/u 1 L)
cada 8 horas y uno final con (NH4) HCO3 20 mM. El producto de la diálisis fue liofilizado y
resuspendido en 3 ml de (NH4) HCO3 20mM. Luego se evaluó la actividad y título aglutinante
equilibrando la muestra con PBS 10X (Na2HPO4 0.2 M, NaH2PO4 0.2 M, NaCl 1500 mM) a
una concentración final de PBS 1X y usando una suspensión de eritrocitos de oveja
tripsinizados al 2%, también se determinó contenido de proteína por BCA, y se hizo SDS-
PAGE en geles al 12.5%.
4.1.8. Ensayos de Inhibición con carbohidratos
Para el ensayo de inhibición con carbohidratos fueron usadas 17 soluciones de diferentes
azúcares: Lactosa 0,3 M, D-Galactosa 0,3 M, D-Glucosa 0,3M, L-Fucosa 0,3 M, D-manosa
0,3 M, N-acetil α-D Glucosamina 0,3 M, N-acetil D Galactosamina 0,3M, P-Nitrofenil N-acetil
-D galactosamina 37 mM, P-Nitrofenil -D manopiranosido 37 mM, P-Nitrofenil N-acetil -
D galactopiranosido 37 mM, P-Nitrofenil α-D manopiranosido 37 mM, P-Nitrofenil N-acetil
α-D glucosamina 37 mM, P-Nitrofenil β-D glucosamina 37 mM, P-Nitrofenil β-D

45
galactopiranosa 37 mM, P-Nitrofenil β-D glucopiranosido 37 mM, Metil β-D glucopiranosido
37 mM y Metil α -D manosido 0,3 M (Vega, 2004).
En una placa de 96 pozos, 20 µl de la fracción retenida y no retenida se dejaron interactuar
durante 30 min con 20 µl de las soluciones de cada uno de los carbohidratos. Transcurrido
este periodo de tiempo fueron adicionados 20 µl de la suspensión de eritrocitos tripsinizados
de oveja al 2% y luego de 40 min se evaluaron los resultados de aglutinación. Cada uno de
los ensayos se realizó por duplicado. Como control fue usado buffer PBS 1X, la solución de
carbohidrato y la suspensión de eritrocitos. Los resultados fueron comparados con la
aglutinación obtenida para la fracción retenida y no retenida adquiridas del paso de
purificación por intercambio iónico (4.1.7).
Con los carbohidratos que mostraron una inhibición positiva de la aglutinación, se determinó
la concentración mínima inhibitoria del azúcar. Para ello a partir de 20µl de cada una de las
soluciones de los carbohidratos se realizaron diluciones seriadas con PBS 1X en una placa
de 96 pozos, en cada pozo se adicionaron 20 µl del extracto y se dejaron interactuar durante
30 min a temperatura ambiente. Transcurrido este tiempo se adicionó 20 µl de suspensión
de eritrocitos tripsinizados al 2% y 40 min después se tomaron los resultados. Como control
fue usado buffer PBS 1X, solución de carbohidrato y la suspensión de eritrocitos. Los
resultados fueron comparados con la aglutinación obtenida en la fracción retenida y no
retenida obtenidas de intercambio iónico (4.1.7).
4.1.9. Purificación a gran escala de la (s) lectina(s) de B. ramulosa
Siguiendo el método descrito en 4.1.2, fueron obtenidos 8,48 g de polvo fino y seco de B.
ramulosa sobre el cual se realizaron tres extracciones manteniendo una proporción 1:10
(p/v) con buffer PBS 1X (ver 4.1.4.). Se realizó cuantificación de proteína soluble por
duplicado con el método micro Kjeldahl. A continuación 244 ml de extracto crudo fueron
precipitados con (NH4)2SO4 entre el 0-50%s y 50-70%s en las mismas condiciones descritas
en 4.1.7. El exceso de sales fue eliminado por diálisis con membrana de 3 kDa realizando
dos cambios con agua y uno contra (NH4) HCO3 20 mM (c/u 1 L). A cada una de las
fracciones se les realizaron ensayos de aglutinación con una suspensión de eritrocitos de
oveja tripsinizados al 2%, concentración de proteína por micro Kjeldahl y SDS-PAGE en
geles del 12,5%. La fracción activa fue equilibrada y dividida en tres volúmenes de extracto
(10 ml, 20 ml, y 7,2 ml). Con cada una de estas fracciones se realizaron tres cromatografías

46
de intercambio iónico usando un soporte de DEAE Sephacel (Volumen de soporte 23 ml,
dimensiones: 2,10 cm diámetro. X 4,80 cm longitud) con flujo constante (3 ml/ 5 min). Se
obtuvieron dos fracciones, una no retenida y una retenida; cada una de las fracciones fueron
concentradas por ultrafiltración en membrana de 1kDa. Las fracciones fueron llevadas a un
volumen final de 40 ml; en el caso de la fracción retenida fue equilibrada en PBS 1X (NaCl
150 mM) por el mismo procedimiento. Tanto a los retentatos y a los difusatos se les
determinó concentración de proteína por BCA, actividad aglutinante con una suspensión de
eritrocitos de oveja tratados con tripsina al 2% y SDS-PAGE en geles al 12.5%.
4.1.10. Purificación por cromatografía de afinidad
Con las fracciones retenida y no retenida obtenidas en 4.1.9. se realizaron diferentes
ensayos de cromatografía de afinidad, las condiciones usadas en cada ensayo se describen
en la tabla 4-1. Cada uno de los soportes ensayados eran funcionales según pruebas
realizadas antes de su uso.
Tabla 4-1 Condiciones de Afinidad para cada uno de los ensayos de cromatografía Fracción Soporte Soluciones de elución
Retenida
Sepharosa 4B -GalNAc (1 cm Diá. X 2 cm Largo)
PBS 1X
Gly 50 mM HCl NaCl pH 2,5
Tris 50 mM HCl 0,1M, NaCl 0,5M pH 8,13
Tris 50 mM OH pH 11,4
Sepharosa 4B -aMSB (0,8cm Diá.X 1,5 cm Largo)
PBS 1X
Gly 50 mM HCl pH 3.0
Tris-HCl 50 mM HCl pH 6,8
Tris -HCl pH 1,5 M y pH 8,8
Sepharosa 4B- Galactosa (0,8 cm Diá. X 1,8 cm Largo)
PBS 1X
PBS1X D (+) Galactosa 0,2 M
No retenida
Sepharosa concanavalina A (1 cm Diá. X 1,5 cm largo)
NaCl 1%, 5 mM de Mg++ y Ca++
NaCl 1% Glucosa 0,2 M
Gly 50 mM HCl pH 3,0
Tris -HCl pH 1,5M pH 8,8
Sepharosa 4B GalNAc (1 cm Diá. X 2 cm Largo)
PBS 1X
Gly 50 mM HCl NaCl pH 2,5
Tris 50 mM HCl 0,1M, NaCl 0,5M pH 8,13
Tris 50 mM OH pH 11,4
Manosa-Agarosa (0,6 cm Diá. X 1 cm largo)
PBS1X
NaCl 1% D-manosa 0,2 M
Metil α D Manopiranosido 0,2 M
Gly 50 mM HCl NaCl pH 2,5
L Fucosa- Agarosa- (7 mm Diá. X 2,2 cm largo)
PBS 1X
Gly 50 mM pH 3,0
Tris 50 mM HCl pH 8,8
Agarosa-Glucosa (7 mm Diá. X 2,2 cm largo)
PBS 1X
PBS 1X Glucosa 0,2 M
Gly 50 mM pH 3,0
Tris 50 mM HCl pH 8,8
En cada una de las cromatografías de afinidad correspondientes a la fracción retenida fueron sembrados 0,99 mg de proteína total, mientras que en las cromatografías correspondientes a la fracción no retenida fueron sembrados en cada columna 1,11 mg de proteína total.
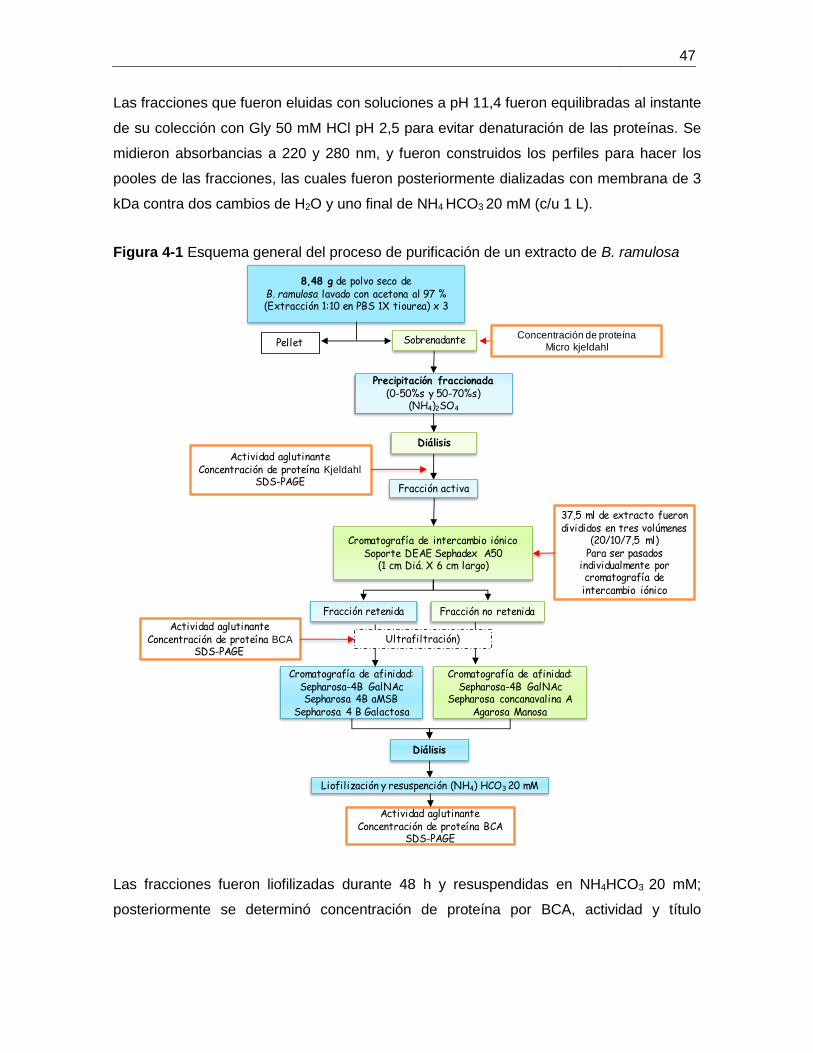
47
Las fracciones que fueron eluidas con soluciones a pH 11,4 fueron equilibradas al instante
de su colección con Gly 50 mM HCl pH 2,5 para evitar denaturación de las proteínas. Se
midieron absorbancias a 220 y 280 nm, y fueron construidos los perfiles para hacer los
pooles de las fracciones, las cuales fueron posteriormente dializadas con membrana de 3
kDa contra dos cambios de H2O y uno final de NH4 HCO3 20 mM (c/u 1 L).
Figura 4-1 Esquema general del proceso de purificación de un extracto de B. ramulosa
Las fracciones fueron liofilizadas durante 48 h y resuspendidas en NH4HCO3 20 mM;
posteriormente se determinó concentración de proteína por BCA, actividad y título
Cromatografía de afinidad:Sepharosa-4B GalNAc
Sepharosa concanavalina A Agarosa Manosa
8,48 g de polvo seco de B. ramulosa lavado con acetona al 97 %(Extracción 1:10 en PBS 1X tiourea) x 3
Pellet Sobrenadante
Precipitación fraccionada (0-50%s y 50-70%s)
(NH4)2SO4
Fracción activa
Fracción retenida
Cromatografía de intercambio iónicoSoporte DEAE Sephadex A50
(1 cm Diá. X 6 cm largo)
Cromatografía de afinidad:Sepharosa-4B GalNAc Sepharosa 4B aMSB
Sepharosa 4 B Galactosa
Fracción no retenida
Actividad aglutinanteConcentración de proteína BCA
SDS-PAGE
Actividad aglutinanteConcentración de proteína Kjeldahl
SDS-PAGE
Ultrafiltración)
Diálisis
Diálisis
Liofilización y resuspención (NH4) HCO3 20 mM
Concentración de proteínaMicro kjeldahl
37,5 ml de extracto fueron divididos en tres volúmenes
(20/10/7,5 ml)Para ser pasados
individualmente por cromatografía de intercambio iónico
Actividad aglutinanteConcentración de proteína BCA
SDS-PAGE

48
aglutinante con una suspensión de eritrocitos de oveja tripsinizados al 2%, y se realizó SDS-
PAGE en geles al 12.5%.
4.2. Resultados
4.2.1. Ensayos preliminares de purificación: Precipitación fraccionada y cromatografía de intercambio iónico a pequeña escala.
Los resultados indicaron que el extracto a pequeña escala presentó un porcentaje de
proteína soluble de 3,19 %, con una actividad aglutinante +4 (Tabla 4-2).
Por SDS-PAGE para el extracto crudo de B. ramulosa (Fig. 4-2) se observa la presencia de
numerosas bandas que se distribuyen en un amplio rango de pesos moleculares, el mayor
número de bandas y con una mayor intensidad se encuentran entre los 51-14 kDa tal como
lo muestra el análisis de densitometría (Figura 4-2) el cual se muestra la intensidad de las
bandas que tocan la línea amarilla.
Figura 4-2 SDS-PAGE. Perfil electroforético de extracto crudo de B. ramulosa.
Tabla 4-2 Extracto de B. ramulosa, pequeña escala. Polvo de alga
(g) Volumen del extracto
(ml) mg/ml* Cantidad
(mg) %
(p/ps) Máxima
Actividad**
3,02 30 3,22 96,6 3,19 +4 *Cuantificación por el método de Kjeldahl, este valor corresponde a la concentración reportada en la tabla 3 del anexo 1 para el extracto de B. ramulosa (L). **Actividad aglutinante suspensión de eritrocitos de oveja al 2% Nota: El extracto fue elaborado como se indica en 4.1.4.
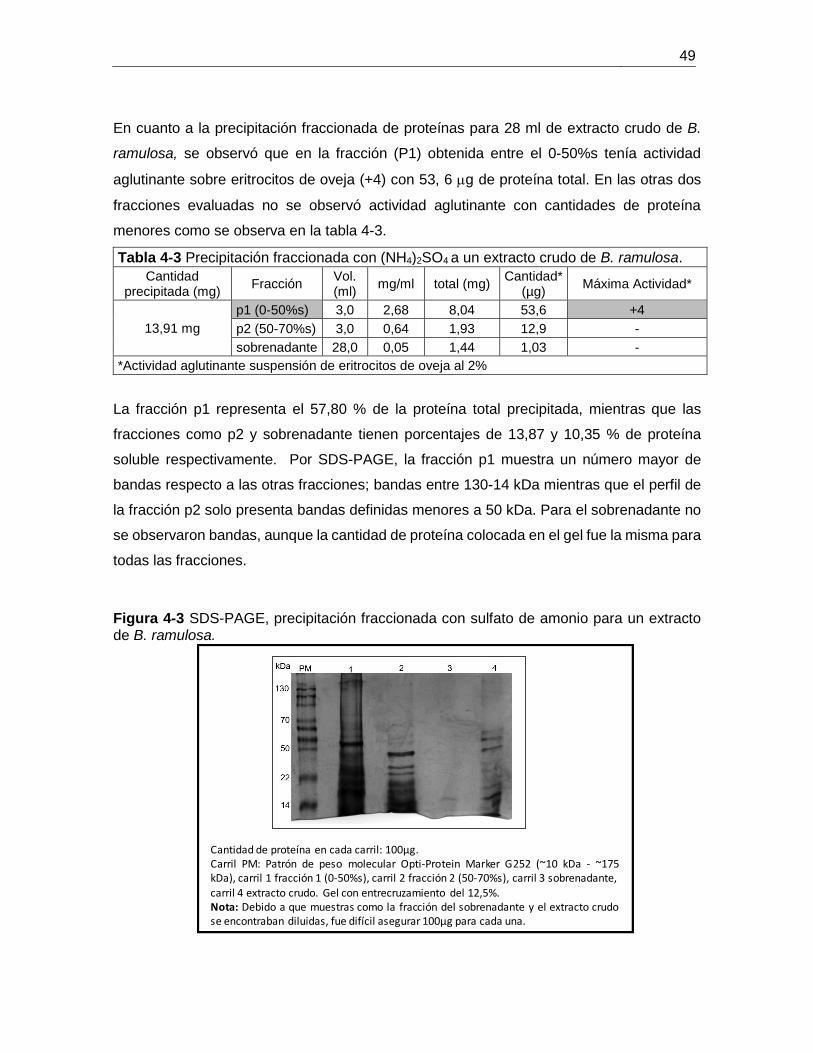
49
En cuanto a la precipitación fraccionada de proteínas para 28 ml de extracto crudo de B.
ramulosa, se observó que en la fracción (P1) obtenida entre el 0-50%s tenía actividad
aglutinante sobre eritrocitos de oveja (+4) con 53, 6 g de proteína total. En las otras dos
fracciones evaluadas no se observó actividad aglutinante con cantidades de proteína
menores como se observa en la tabla 4-3.
Tabla 4-3 Precipitación fraccionada con (NH4)2SO4 a un extracto crudo de B. ramulosa.
Cantidad precipitada (mg)
Fracción Vol. (ml)
mg/ml total (mg) Cantidad*
(µg) Máxima Actividad*
13,91 mg
p1 (0-50%s) 3,0 2,68 8,04 53,6 +4
p2 (50-70%s) 3,0 0,64 1,93 12,9 -
sobrenadante 28,0 0,05 1,44 1,03 -
*Actividad aglutinante suspensión de eritrocitos de oveja al 2%
La fracción p1 representa el 57,80 % de la proteína total precipitada, mientras que las
fracciones como p2 y sobrenadante tienen porcentajes de 13,87 y 10,35 % de proteína
soluble respectivamente. Por SDS-PAGE, la fracción p1 muestra un número mayor de
bandas respecto a las otras fracciones; bandas entre 130-14 kDa mientras que el perfil de
la fracción p2 solo presenta bandas definidas menores a 50 kDa. Para el sobrenadante no
se observaron bandas, aunque la cantidad de proteína colocada en el gel fue la misma para
todas las fracciones.
Figura 4-3 SDS-PAGE, precipitación fraccionada con sulfato de amonio para un extracto de B. ramulosa.
Cantidad de proteína en cada carril: 100µg.Carril PM: Patrón de peso molecular Opti-Protein Marker G252 (~10 kDa - ~175kDa), carril 1 fracción 1 (0-50%s), carril 2 fracción 2 (50-70%s), carril 3 sobrenadante,
carril 4 extracto crudo. Gel con entrecruzamiento del 12,5%.Nota: Debido a que muestras como la fracción del sobrenadante y el extracto crudose encontraban diluidas, fue difícil asegurar 100µg para cada una.
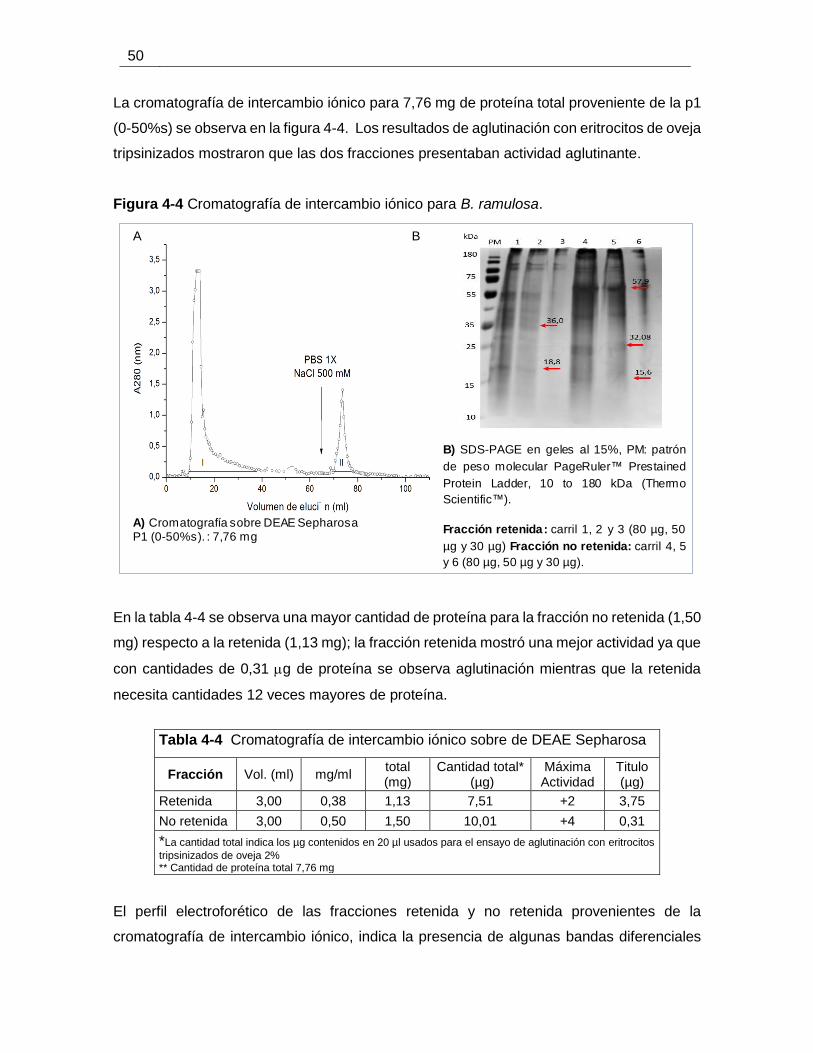
50
La cromatografía de intercambio iónico para 7,76 mg de proteína total proveniente de la p1
(0-50%s) se observa en la figura 4-4. Los resultados de aglutinación con eritrocitos de oveja
tripsinizados mostraron que las dos fracciones presentaban actividad aglutinante.
Figura 4-4 Cromatografía de intercambio iónico para B. ramulosa.
En la tabla 4-4 se observa una mayor cantidad de proteína para la fracción no retenida (1,50
mg) respecto a la retenida (1,13 mg); la fracción retenida mostró una mejor actividad ya que
con cantidades de 0,31 g de proteína se observa aglutinación mientras que la retenida
necesita cantidades 12 veces mayores de proteína.
El perfil electroforético de las fracciones retenida y no retenida provenientes de la
cromatografía de intercambio iónico, indica la presencia de algunas bandas diferenciales
Tabla 4-4 Cromatografía de intercambio iónico sobre de DEAE Sepharosa
Fracción Vol. (ml) mg/ml total (mg)
Cantidad total* (µg)
Máxima Actividad
Titulo (µg)
Retenida 3,00 0,38 1,13 7,51 +2 3,75
No retenida 3,00 0,50 1,50 10,01 +4 0,31
*La cantidad total indica los µg contenidos en 20 µl usados para el ensayo de aglutinación con eritrocitos
tripsinizados de oveja 2% ** Cantidad de proteína total 7,76 mg
A B
A) Cromatografía sobre DEAE SepharosaP1 (0-50%s). : 7,76 mg
B) SDS-PAGE en geles al 15%, PM: patrón
de peso molecular PageRuler™ Prestained
Protein Ladder, 10 to 180 kDa (Thermo
Scientific™).
Fracción retenida: carril 1, 2 y 3 (80 µg, 50
µg y 30 µg) Fracción no retenida: carril 4, 5
y 6 (80 µg, 50 µg y 30 µg).
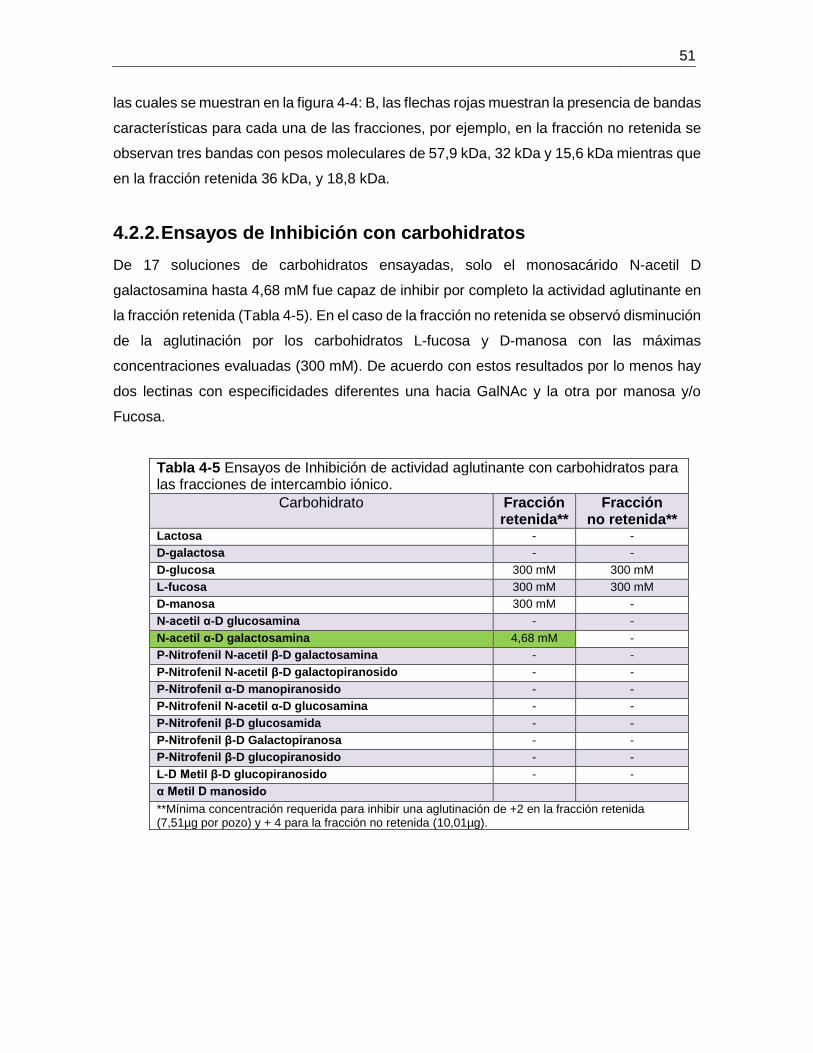
51
las cuales se muestran en la figura 4-4: B, las flechas rojas muestran la presencia de bandas
características para cada una de las fracciones, por ejemplo, en la fracción no retenida se
observan tres bandas con pesos moleculares de 57,9 kDa, 32 kDa y 15,6 kDa mientras que
en la fracción retenida 36 kDa, y 18,8 kDa.
4.2.2. Ensayos de Inhibición con carbohidratos
De 17 soluciones de carbohidratos ensayadas, solo el monosacárido N-acetil D
galactosamina hasta 4,68 mM fue capaz de inhibir por completo la actividad aglutinante en
la fracción retenida (Tabla 4-5). En el caso de la fracción no retenida se observó disminución
de la aglutinación por los carbohidratos L-fucosa y D-manosa con las máximas
concentraciones evaluadas (300 mM). De acuerdo con estos resultados por lo menos hay
dos lectinas con especificidades diferentes una hacia GalNAc y la otra por manosa y/o
Fucosa.
Tabla 4-5 Ensayos de Inhibición de actividad aglutinante con carbohidratos para las fracciones de intercambio iónico.
Carbohidrato Fracción retenida**
Fracción no retenida**
Lactosa - -
D-galactosa - -
D-glucosa 300 mM 300 mM
L-fucosa 300 mM 300 mM
D-manosa 300 mM -
N-acetil α-D glucosamina - -
N-acetil α-D galactosamina 4,68 mM -
P-Nitrofenil N-acetil β-D galactosamina - -
P-Nitrofenil N-acetil β-D galactopiranosido - -
P-Nitrofenil α-D manopiranosido - -
P-Nitrofenil N-acetil α-D glucosamina - -
P-Nitrofenil β-D glucosamida - -
P-Nitrofenil β-D Galactopiranosa - -
P-Nitrofenil β-D glucopiranosido - -
L-D Metil β-D glucopiranosido - -
α Metil D manosido
**Mínima concentración requerida para inhibir una aglutinación de +2 en la fracción retenida (7,51µg por pozo) y + 4 para la fracción no retenida (10,01µg).

52
4.2.3. Preparación y purificación de proteínas a gran escala del extracto de B. ramulosa.
Existen algunas pruebas para caracterizar el material con el cual se pretende trabajar, una
de ellas es el porcentaje de humedad el cual da una idea de la eficiencia que puede tener
un proceso de extracción. En el presente trabajo este porcentaje de humedad fue
determinado sobre la harina de alga la cual fue el punto de partida para realizar la
extracción. La caracterización realizada sobre la harina mostró que el contenido en
humedad en el material analizado fue de 54,05 ± 2,02 % (Tabla 5, anexo 1). Por otro lado,
el porcentaje de proteína soluble en el extracto fue de 4,46 % (p/ps) tal como se indica en
la tabla 6 del anexo 1. El ensayo de actividad aglutinante para el extracto mostró que con
una cantidad de 29,8 µg de proteína se obtenía una actividad aglutinante de +4, y titulo de
3,72 µg.
Tabla 4-6 Proteína total de tres fracciones obtenidas al realizar precipitación fraccionada con (NH4)2SO4 a un extracto crudo de B. ramulosa
Fracción Vol. (ml)
mg/ml * total (mg) Cantidad total
(µg) ** Máxima Actividad
Título µg
p1 (0-50%s) 37,5 2,30 86,4 46,1 +4 0,72
p2 (50-70%s) 27,0 1,91 51,5 38,1 +2 9,53
sobrenadante 179,5 1,22 218,6 24,3 - -
*Cuantificación realizada con el método de micro Kjeldahl
**La cantidad total indica los µg contenidos en 20 µl usados para el ensayo de aglutinación con eritrocitos
tripsinizados de oveja 2%. Concentración del extracto crudo 1,49 mg/ml, Cantidad de proteína precipitada: 363,56 mg.
En la fracción p1 (0-50%s) se obtuvo actividad aglutinante con 0,72 g, lo indica que la
mayoría de la lectina se concentró en esta fracción, mientras que en p2 (50-70%) la cantidad
de proteína que se requieren para aglutinar eritrocitos era mayor (9,53 g). Por otra parte,
en el sobrenadante quedo un porcentaje mayor (60%) de proteína sin precipitar, aunque
con 24 g no se observa actividad (Tabla 4-6). Para continuar el proceso de purificación se
siguió trabajando con la fracción p1(0-50%s) donde fue necesario dividir los 86,4 mg de
proteína total en tres volúmenes para ser sometidos a cromatografía sobre DEAE-Sephacel,
la cantidad sembrada en cada una de las cromatografías y los correspondientes perfiles
electroforéticos se muestran en la figura 4 del anexo 2. Estas graficas corroboraron el
resultado obtenido en los ensayos preliminares, en la figura 4-5: A se observa que la
fracción no retenida presenta las mayores absorbancias en contraste a la fracción retenida,
similar a lo observado en la figura 4-4. El proceso general muestra una cantidad de 40,95
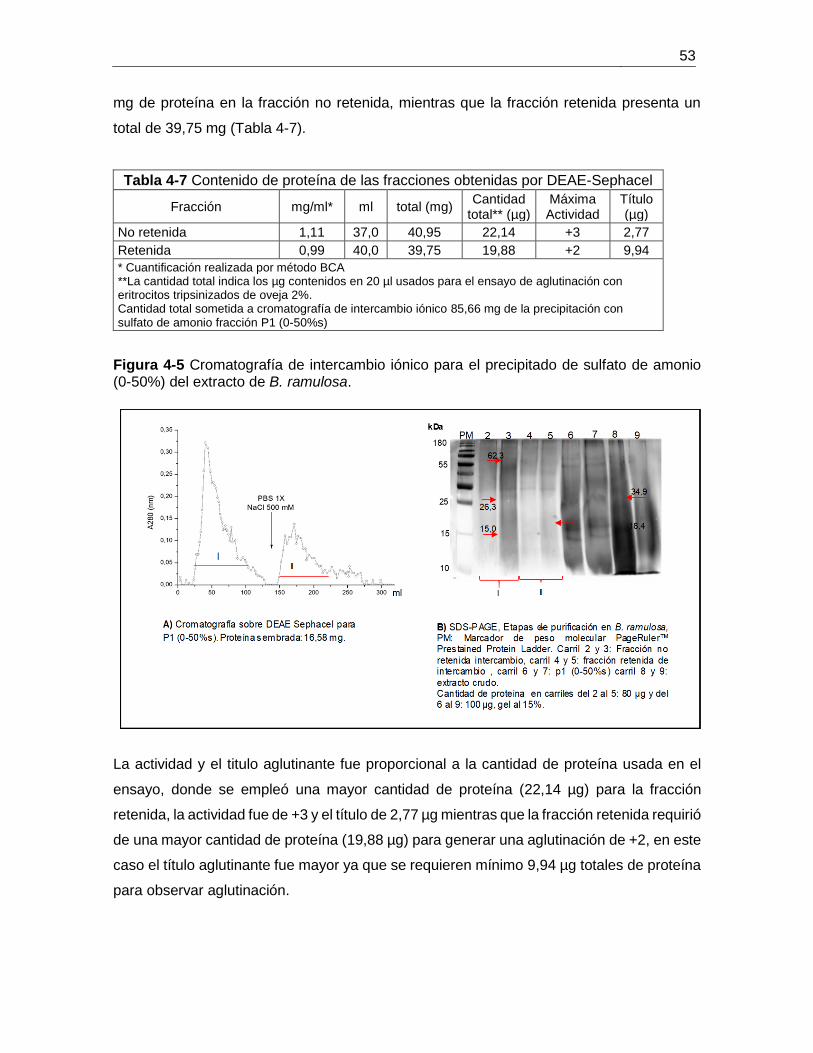
53
mg de proteína en la fracción no retenida, mientras que la fracción retenida presenta un
total de 39,75 mg (Tabla 4-7).
Tabla 4-7 Contenido de proteína de las fracciones obtenidas por DEAE-Sephacel
Fracción mg/ml* ml total (mg) Cantidad
total** (µg) Máxima Actividad
Título (µg)
No retenida 1,11 37,0 40,95 22,14 +3 2,77
Retenida 0,99 40,0 39,75 19,88 +2 9,94
* Cuantificación realizada por método BCA **La cantidad total indica los µg contenidos en 20 µl usados para el ensayo de aglutinación con eritrocitos tripsinizados de oveja 2%. Cantidad total sometida a cromatografía de intercambio iónico 85,66 mg de la precipitación con sulfato de amonio fracción P1 (0-50%s)
Figura 4-5 Cromatografía de intercambio iónico para el precipitado de sulfato de amonio (0-50%) del extracto de B. ramulosa.
La actividad y el titulo aglutinante fue proporcional a la cantidad de proteína usada en el
ensayo, donde se empleó una mayor cantidad de proteína (22,14 µg) para la fracción
retenida, la actividad fue de +3 y el título de 2,77 µg mientras que la fracción retenida requirió
de una mayor cantidad de proteína (19,88 µg) para generar una aglutinación de +2, en este
caso el título aglutinante fue mayor ya que se requieren mínimo 9,94 µg totales de proteína
para observar aglutinación.

54
Aunque en el perfil electroforético no se obtuvo un perfil como el observado en el ensayo
preliminar, si se detectaron bandas características similares a las observadas en la figura
4-4: B. La fracción no retenida presenta tres bandas con pesos de 62,3 kDa, 26,3 kDa y 15
kDa, mientras que en la fracción retenida solo de observo una banda alrededor de 18,4
kDa; para las dos fracciones hay una banda de 34,9 kDa (Figura 4-5 B).
4.2.4. Purificación por cromatografía de afinidad
Los mejores resultados de inhibición con las soluciones de carbohidratos mostraron que la
fracción retenida obtenida de intercambio iónico era inhibida con una solución de GalNAc
4,8 mM; de acuerdo con lo anterior se realizaron los ensayos de purificación sobre un
soporte de Sepharosa 4B-GalNAc (Figura 5: A, anexo 2). Se obtuvieron fracciones
retenidas que eluyeron a diferentes condiciones de pH. Las únicas fracciones en la que se
pudo hacer la detección de la actividad aglutinante fue en las eluidas a pH=8.13, fracciones
IV y V, donde con 3,97 µg de proteína pura (Tabla 9, anexo) y un título aglutinante de 0,40
µg se evidenció la presencia de una lectina. Sin embargo, con este tipo de soporte no se
obtiene una buena cantidad de proteína aislada, por esa razón y teniendo en cuenta que
en algunos la purificación se hace a partir de glicoproteínas tipo mucina, se realizaron
ensayos sobre un soporte de aMSB – Sepharose 4B. La asialo mucina submaxilar bovina
(aMSB) es una glicoproteína de alto peso molecular que tiene un alto contenido de GalNAc
unido a serina o treonina (Vega y Pérez, 2006). Los resultados (Figura 4-6: A) nos
permitieron obtener fracciones retenidas, pero con una baja actividad para las fracciones IV
(eluida a pH 6,8), V y VI (eluidas a pH 11,4) (Tabla 4-8). Finalmente, se realizaron otros
ensayos con un soporte de afinidad sobre Galactosa–Sepharose 4B y no se obtuvo ninguna
fracción retenida (Figura 5: B, anexo 2). Por SDS-PAGE para las fracciones activas se
observaron dos bandas alrededor de 18,4 y 36,2 kDa (Figura 4-6: B). Para corroborar los
ensayos realizados se deben hacer columnas con una cantidad mayor de proteína para
realizar posteriores ensayos de caracterización. La cromatografía que presentó mejor
porcentaje de recuperación corresponde a la columna de GalNAc -Sepharosa 4B con un
14,68 % y el menor valor, 8,89 %, con la columna de galactosa-Sepharosa 4B (Tabla 9,
anexo 1).
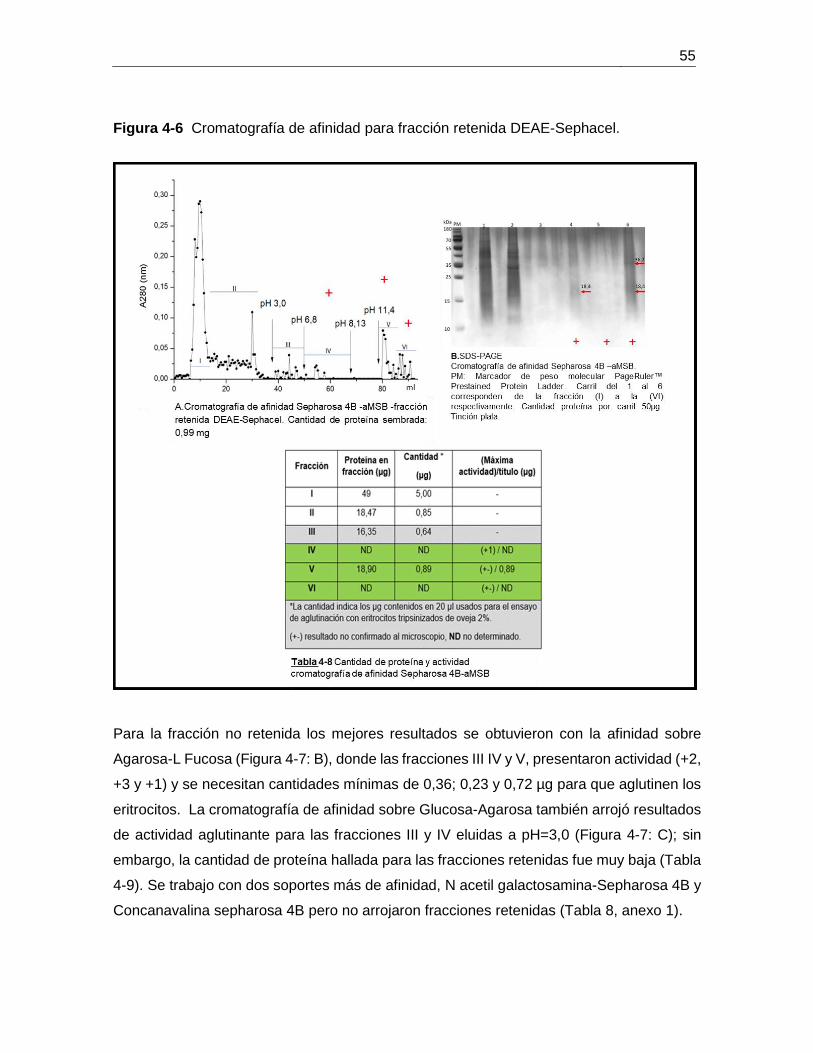
55
Figura 4-6 Cromatografía de afinidad para fracción retenida DEAE-Sephacel.
Para la fracción no retenida los mejores resultados se obtuvieron con la afinidad sobre
Agarosa-L Fucosa (Figura 4-7: B), donde las fracciones III IV y V, presentaron actividad (+2,
+3 y +1) y se necesitan cantidades mínimas de 0,36; 0,23 y 0,72 µg para que aglutinen los
eritrocitos. La cromatografía de afinidad sobre Glucosa-Agarosa también arrojó resultados
de actividad aglutinante para las fracciones III y IV eluidas a pH=3,0 (Figura 4-7: C); sin
embargo, la cantidad de proteína hallada para las fracciones retenidas fue muy baja (Tabla
4-9). Se trabajo con dos soportes más de afinidad, N acetil galactosamina-Sepharosa 4B y
Concanavalina sepharosa 4B pero no arrojaron fracciones retenidas (Tabla 8, anexo 1).
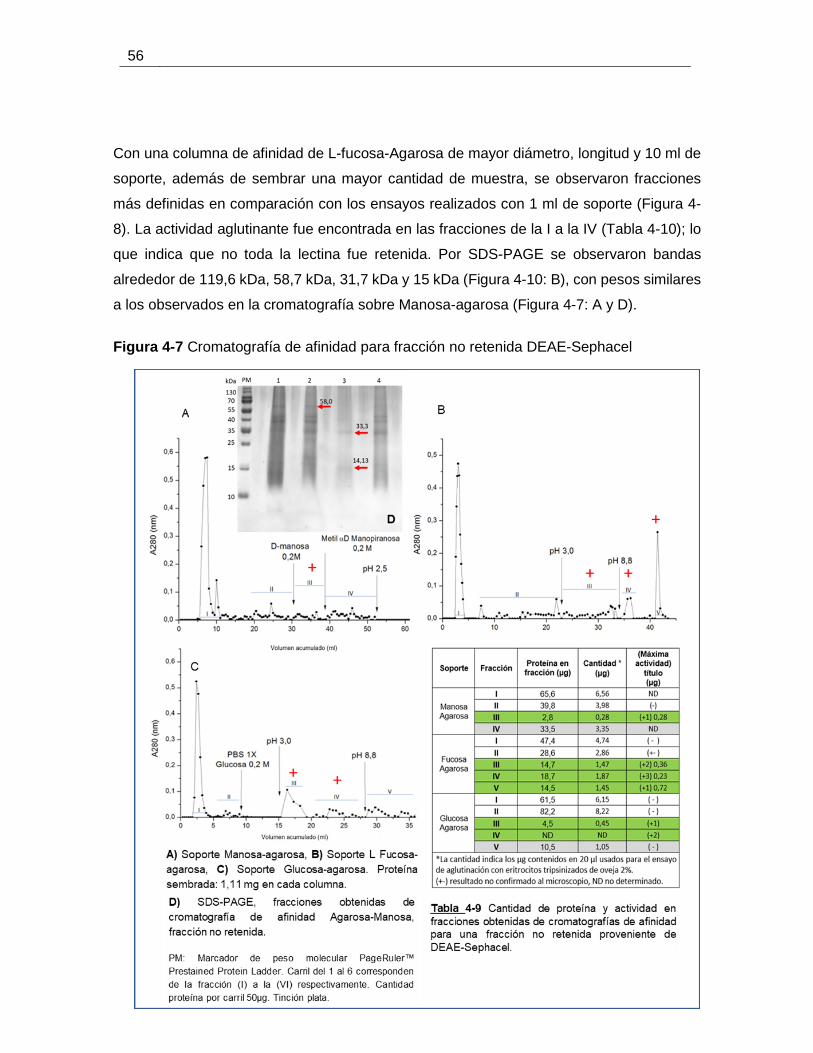
56
Con una columna de afinidad de L-fucosa-Agarosa de mayor diámetro, longitud y 10 ml de
soporte, además de sembrar una mayor cantidad de muestra, se observaron fracciones
más definidas en comparación con los ensayos realizados con 1 ml de soporte (Figura 4-
8). La actividad aglutinante fue encontrada en las fracciones de la I a la IV (Tabla 4-10); lo
que indica que no toda la lectina fue retenida. Por SDS-PAGE se observaron bandas
alrededor de 119,6 kDa, 58,7 kDa, 31,7 kDa y 15 kDa (Figura 4-10: B), con pesos similares
a los observados en la cromatografía sobre Manosa-agarosa (Figura 4-7: A y D).
Figura 4-7 Cromatografía de afinidad para fracción no retenida DEAE-Sephacel

57
Figura 4-8 Cromatografía de afinidad sobre soporte de Fucosa-Agarosa para fracción no retenida DEAE-Sephacel.
Los porcentajes de recuperación para las diferentes cromatografías de afinidad realizadas
sobre la fracción no retenida indican valores entre 11,16 y 14,31 %, siendo obtenido el
mayor porcentaje con la columna de glucosa y el menor con la columna de fucosa (Tabla
8, anexo 1).

58
4.3. Análisis
Los métodos empleados para purificar lectinas de algas marinas, incluyen métodos de
precipitación fraccionada (Chiles y Bird, 1989), y cromatografía de intercambio iónico y de
afinidad. La precipitación fraccionada con sulfato de amonio, es una buena estrategia en
las primeras etapas de purificación de las proteínas de interés, donde se tiene en cuenta la
solubilidad diferencial que tienen estas moléculas de acuerdo a su composición de
aminoácidos (Englard y Seifter, 1990).
En nuestro caso, la (s) lectina (s) presente (s) en el extracto de B. ramulosa fueron solubles
en PBS 1X y precipitaron con sulfato de amonio. Cuando se realizaron ensayos en pequeña
escala, la actividad aglutinante se encontró solamente en un rango de precipitación entre el
0-50%s, en esta fracción precipitó el 58 % de 13,91 mg de proteína total (Tabla 4-3). Sin
embargo, cuando se realizó el mismo procedimiento a gran escala (con 363,56 mg de
proteínas totales); se encontró que toda (s) la (s) lectina (s) no precipitaron en el mismo
rango de saturación y que entre el 50-70% había una actividad aglutinante residual (Tabla
4-6). Lo anterior se debe a las diferencias de concentración de los dos extractos (3,22 y
1.49 mg/ml respectivamente) ya que en el último caso por estar menos concentrado
requiere una mayor cantidad de sulfato de amonio para que precipite completamente, por
ejemplo, hasta el 60%.
Además, solamente precipito el 37,9 % de las proteínas solubles, quedando un 62,1 % de
proteínas en el sobrenadante. Lo anterior nos muestra que el extracto empleado para los
ensayos a gran escala estaba muy diluido y en ese sentido se necesita una mayor cantidad
de sales para que se dé la precipitación total de las proteínas o concentrar el extracto como
un paso previo al proceso de precipitación fraccionada. En otros trabajos realizados para
aislar lectinas de algas marinas, autores como Chiles y Bird (1989) realizaron una
precipitación fraccionada con sulfato de amonio para varios extractos entre el 0-20%s y 20-
75%s y encontraron que la actividad biológica se concentró en esta última fracción. Otros
autores optaron por realizar la precipitación en un solo rango de saturación entre el 0-70%s
con sulfato de amonio en un extracto salino de Hypnea musciformis (Nagano et al., 2002),
y entre el 0-80%s en extractos del alga roja Pterocladiella capillacea (Oliveira et al., 2002),

59
en estos últimos estudios no se realizó un fraccionamiento sino la precipitación total de
proteínas para separarlas de carbohidratos y pigmentos (Englard y Seifter, 1990).
En este estudio dependiendo de la concentración del extracto podríamos pensar que las
lectinas de B. ramulosa precipitan entre el 0-50%s con sulfato de amonio donde se observó
actividad aglutinante con 0,72 g de proteína total, mientras que en la fracción obtenida
entre el 50-70% la cantidad de proteína que se requiere para evidenciar aglutinación es 13
veces mayor. Por lo tanto, los pasos posteriores de purificación se hicieron con la fracción
de proteínas obtenidas en la precipitación con sulfato de amonio entre el 0-50%s. Esta
fracción fue sometida a cromatografía de intercambio aniónico sobre un soporte de DEAE-
Sepharosa para la extracción a pequeña escala y DEAE-Sephacel A-50 para la extracción
a gran escala (Figura 4-4, Figura 4-5), a pH 7.2 estos soportes se encuentran cargados
positivamente y solo interactúan con proteínas que tengan carga negativa, después estas
moléculas eluyen en la fracción retenida al competir por los sitios de interacción frente a la
solución PBS 1X NaCl 500 mM, de manera que siempre se espera obtener proteínas
diferentes en cada una de las fracciones, además estos soportes tienen una alta capacidad
de retención.
Por lo general las lectinas de algas marinas, tienen un alto porcentaje de aminoácidos
ácidos y por lo tanto puntos isoeléctricos menores a pH de 6,0 (Calvete et al., 2000), esta
situación conduce a que algunas lectinas se encuentren cargadas negativamente en PBS
1X (pH 7,2 -7,4), y de esta manera ellas pueden interactuar con el soporte. Sin embargo,
los resultados de aglutinación positiva obtenidos tanto en la fracción retenida como no
retenida indican la presencia de más de una lectina en el extracto de B. ramulosa con
propiedades moleculares diferentes. En especies cercanas taxonómicamente como B.
plumosa se han encontrado y purificado cuatro lectinas (Tabla 4-11), de medio y bajo peso
molecular.
Una de las características que se ha evidenciado en lectinas de algas, en comparación a
las de origen vegetal es su especificidad de unión a azucares complejos, lo cual se ha
aprovecha en algunos casos como estrategia para purificar especialmente lectinas de algas
verdes como algunas del género Codium (Fábregas et al., 1988; Rogers y Flangu, 1991;
Rogers et al., 1986) Ulva lactuca (Sampaio et al., 1998b) Caulerpa cupressoides (Benevides
et al., 2001), Enteromorpha prolifera (Ambrosio et al., 2003), Ulva pertusa (Wang et al.,

60
2004), B. plumosa (Han et al., 2010; Han et al., 2012; Han et al., 2011; Kim et al., 2006), B.
hypnoides (Niu et al., 2009), aunque existen algunos trabajos donde se han llegado a usar
en especies de algas rojas del género Ptilota (Sampaio et al., 1998a; Sampaio et al., 1999).
En el presente estudio los resultados de inhibición con carbohidratos mostraron que la
actividad aglutinante de la fracción retenida era inhibida por una solución GalNAc con una
concentración mínima 4.68 mM, por esa razón se realizaron dos cromatografías de afinidad
una con el monosacárido (GalNAC- Sepharosa 4B) y la segunda empleando un soporte con
una mucina desialilada (aMSB) (aMSB-Sepharosa 4B), esta glicoproteína tiene un alto
porcentaje de residuos del azúcar unidos a serina o treonina. En los dos casos se obtuvieron
fracciones retenidas con actividad, aunque la cantidad de proteína empleada para los
ensayos fue baja; con 0,89 µg de proteína pura se observó aglutinación (+) con eritrocitos
de oveja tripsinizados al 2% (Tabla 4-8). El perfil electroforético muestra en el carril 4 y 6
correspondientes a las fracciones con actividad de aglutinación IV y VI, la presencia de dos
bandas intensas con pesos moleculares de 18,4 y 36,2 kDa (Figura 4-6: B) las cuales
podrían corresponder al monómero y el dímero respectivamente. Esta consideración es
posible al tener en cuenta que para ninguna de las electroforesis se utilizó DTT pero si otras
condiciones denaturantes como la temperatura (95 ⁰C); lo anterior podría indicar que las
dos especies predominan en la proteína en equilibrio y que su disociación no requiere
condiciones especiales de calentamiento ni condiciones denaturantes.
En especies de algas rojas como P. serrata1 (Sampaio et al., 1999) se ha reportado una
lectina trimétrica con peso para su monómero de 18,39 kDa, esta mostró inhibición de su
actividad con galactosa y sus derivados junto con la glicoproteína aMSB. En P. filicina
también se reporta la presencia de un trímero (Sampaio et al., 1998a) con un peso para el
monómero de 19,32 kDa, también muestra afinidad por aMSB, y además una fuerte
inhibición de su aglutinación con P-nitrofenil-N-acetil-α D-galactosamina y P-nitrofenil-N-
acetil- D-galactosamina.
En B. plumosa, una especie relacionada taxonómicamente con B. ramulosa, se han
encontrado dos lectinas de bajo peso molecular de 11,5 y 12,9 kDa identificadas como
BPL3 y BPL4 (Tabla 4-11), estas son proteínas que reconocen N-acetil-D-galactosamina y
1 Estructura primaria y punto isoeléctrico no determinado.

61
N-acetil-D-glucosamina, con un dominio tipo H característico del caracol Helix pomatia (Han et
al., 2012). Las interacciones del dominio con el carbohidrato N acetil galactosamina están dadas
principalmente por la formación de puentes de hidrogeno donde el O4 del anillo de piranosa,
establece tres puentes de hidrogeno con Arg 63, Trp 83 y Asp 55, el residuo de Arg 63
también establece un puente de hidrogeno con O5 y O6 del anillo de GalNAc, el O3 forma
puentes de hidrogeno con el oxígeno de la cadena principal donde está la Gly 24 y además
hay formación de puentes a través de una molécula de agua a la cadena principal de la Arg
25, el oxígeno del grupo carbonil del grupo N acetil del azúcar, forma puentes de hidrogeno
con el nitrógeno de la cadena principal de Gly24. Además, His 84 establece una fuerte
contacto por una interacción de van der Waals con el grupo CH de C1 (Sanchez et al.,
2006).
Por otro lado, a pesar de que en la fracción no retenida proveniente de intercambio iónico
había una mayor concentración de proteína total y por tanto la actividad era mayor, los
resultados de inhibición con carbohidratos fueron inciertos para determinar el tipo de
cromatografía de afinidad. Por esa razón se ensayaron varios soportes de afinidad con
monosacáridos y se encontró afinidad sobre Fucosa-Agarosa, Glucosa-Agarosa y Manosa-
Agarosa los cuales permitieron aislar fracciones con actividad aglutinante. Las
electroforesis correspondientes a las cromatografías sobre manosa-agarosa y L fucosa-
agarosa mostraron la presencia de cuatro bandas alrededor de los 58,7 kDa, 31,7 kDa y 15
kDa (Figuras 4-7A y 4-8B) la especie dímerica. También la interacción esta mediada por
interacciones débiles ya que los pesos podrían corresponder con el monómero, dímero y
tetrámero; y en este caso predominaría ya que el ensayo se hizo solamente en condiciones
denaturantes y sin reductor.
En especies de algas verdes pertenecientes al orden bryopsidales como Codium fragile
ssp. atlaticum y C. fragile ssp. tomentosoides (Loveless y Rogers, 1985) se han purificado
lectinas tetraméricas, con monómeros de 15 kDa, y con afinidad por GalNAc; resultados
similares a los reportados Bird et al. (1993). Los resultados obtenidos de la afinidad sobre
concanavalina A (Tabla 8, anexo 1) corroboraron que estas lectinas no tienen una
glicosilación típica como la encontrada en lectinas vegetales las cuales tienen
oligosacáridos tipo N altos en manosa (Man9(GlcNAc)2) que son muy conservados en
plantas y animales; por otra parte, la gran mayoría de lectinas de algas marinas estudiadas
no están glicosiladas.

62
La lectina encontrada en la fracción no retenida de la cromatografía de intercambio iónico,
nos indica que su carga neta es positiva y por lo tanto hay un mayor porcentaje de
aminoácidos básicos en su estructura. Esta lectina no discrimina los epimeros de glucosa
y manosa, y además reconoce fucosa; esto indica que el OH en posición 6 no es importante
en el reconocimiento y que el grupo metilo podría estar implicado en interacciones de tipo
hidrofóbico con aminoácidos que se encuentran cercanos al sitio de reconocimiento del
carbohidrato. Una interacción similar ha sido descrita para lectina purificada de B. plumosa
(BPL2), un monómero de 17 kDa con un pI 7,3 y su actividad sobre eritrocitos de oveja es
inhibida con soluciones de D-manosa (1 mM), L fucosa (8mM) y D glucosa (125 mM) (Han
et al., 2010). Su purificación se realiza mediante dos pasos consecutivos de afinidad,
sobre N-acetil galactosamina-sepharosa para retirar lectinas con afinidad a este
carbohidrato y posteriormente la fracción no retenida es sometida a cromatografía de
afinidad sobre manosa-sepharosa.
Tabla 4-8 Características moleculares de las lectinas aisladas de especies del género Bryopsis sp.
Especie / Ref. Lectina
Especificidad Peso (kDa)
Subunidades pI
B. plumosa (Kim et al., 2006; Yoon et al., 2008)
Bryohealin N-acetil-D-glucosamina y N-acetil-D-galactosamina
53,8 Dímero 5,75
B. plumosa (Han et al., 2010)
BPL 2 D-manosa, α-metil-D-manosa y L-fucosa, D-glucosa
17 Monómero 7,3
B. plumosa (Han et al., 2011)
BPL 3 N-acetil-D-galactosamina y N-acetil-D-glucosamina
11.5 Monómero 7,0
B. plumosa (Han et al., 2012)
BPL 4 N-acetil-D-galactosamina y N-acetil-D-glucosamina
12.9 Monómero 6,0
B. hypnoides (Niu et al., 2009)
-
N-Acetil galactosamina, N-Acetil glucosamina y glicoproteína mucina submaxilar bovina
27 Monómero 5 - 6
B. máxima (Hori, 2007) patente.
BPL 17 Manosa 27,4 * - 5,75*
B. ramulosa (Este trabajo)
BRL-1 N-acetilgalactosamina 36.2** 18.4**
Dímero Monómero
N.D
B. ramulosa (Este trabajo)
BRL-2 Manosa, fucosa y glucosa 15.0** 31.7** 58.7**
Monómero Dímero
Tetrámero N.D
*Valores teóricos estimados. Obtenido con ProtParam http://web.expasy.org/cgi-bin/protparam/protparam **Por SDS PAGE en condiciones denaturantes ND: no determinado

63
BPL 2 no muestra homología en las bases de datos con otras proteínas, excepto con una
lectina de B. máxima que se encuentra patentada en European patent application (EPA no.
06714741.3) por Hori (2007) para la purificación de anticuerpos (Han et al., 2010) Un BLAST
realizado sobre la secuencia parcial de aminoácidos para la lectina de B. máxima, revela la
posible presencia de un dominio de tipo fucolectina (dominio FLD) (imagen 7 anexo 2), el
cual se encuentra ampliamente distribuido en la naturaleza, y también está presente en
lectinas aisladas de B. plumosa (Bryohealin) y B. Hypnoides (Tabla 4-11).
4.4. Conclusiones
1. De la especie B. ramulosa se aislaron dos lectinas con diferente especificidad,
una que reconoce residuos de N-acetilgalactosamina y otra, residuos de Manosa,
Glucosa y fucosa. Las dos proteínas presentaron diferentes características
moleculares que se evidenciaron del proceso de separación por cromatografía de
intercambio iónico y afinidad.
2. Por SDS-PAGE se encontró para la lectina especifica por N-acetilgalactosamina
(BRL-1), un peso molecular de 18,4 kDa para la forma monomérica y de 36,2 kDa
para la forma dímerica; las subunidades se asocian por interacciones débiles.
3. El monómero de la lectina especifica por manosa, glucosa y fucosa (BRL-2), tiene
un peso molecular de 15 kDa y se asocia en forma dimérica, trimérica y tetramérica
por enlaces no covalentes.

64
5. Capítulo 3. Modelamiento molecular y docking de la lectina bryohealin, aislada del alga verde Bryopsis plumosa (clorophyta), frente a diferentes ligandos.
Los resultados in vitro obtenidos en el capítulo 2 aportan información importante sobre
algunas características bioquímicas de las lectinas encontradas en B. ramulosa. Las
interacciones de BRL-1 frente a GalNac y de BRL-2 con manosa, glucosa y fucosa revelan
que estas proteínas podrían presentar dominio(s) tipo H o tipo F, tal como se ha encontrado
en lectinas aisladas en diferentes especies del género Bryopsis sp. (Tabla 4-11) de las
cuales no existe información sobre sus estructuras terciaria en bases de datos como PDB.
Por otro lado, sobre la lectina BPL-4 de B. plumosa se realizó el modelamiento de un plegado
típico de lectina tipo H (Han, 2012) usando la secuencia de aminoácidos. En el caso de bryohealin
una lectina dimérica con dominio tipo F aislada de esta misma especie no existe información que
revele como podría estar dado este plegamiento, ni las interacciones características que se
pueden presentar entre la proteína y diferentes carbohidratos. La obtención de esta información
permite hacer un acercamiento a la explicación desde el un punto de vista atómico, de los
resultados obtenidos en el capítulo 2 los cuales apuntan a que BRL-2 podría presentar un
dominio tipo F con afinidad principal por L-Fucosa. De acuerdo con este planteamiento se
propuso la siguiente metodología.
5.1. Metodología
5.1.1. Análisis de la secuencia y Modelo tridimensional
Se seleccionó una lectina presente en la especie B. plumosa DOI: 10.1111/j.1529-
8817.2006.00162.x. La secuencia fue descargada en formato FASTA de la base de datos
NCBI (https://www.ncbi.nlm.nih.gov/) (ACF05191.1). Posterior se realizó un análisis con el
servidor Sigma-P http://www.cbs.dtu.dk/services/SignalP-3.0/ para determinar la presencia
de péptido señal en la secuencia; luego el formato FASTA fue modificado de acuerdo con
el resultado. Una vez se tenía la secuencia se hizo un alineamiento con BLAST
https://blast.ncbi.nlm.nih.gov/Blast.cgi usando el algoritmo Blastp, matriz Blossum 62,
(penalización 11:1) y usando la herramienta ProtParam de Expasy

65
http://web.expasy.org/cgi-bin/protparam/protparam para determinar algunas características
moleculares de la lectina como composición de aminoácidos, peso molecular y punto
isoeléctrico. A partir de la secuencia de la proteína, con el servidor I -TASSER disponible
en http://zhanglab.ccmb.med.umich.edu/I-TASSER/, se obtuvo un modelo para la
estructura tridimensional de la lectina. Fue seleccionado el modelo con el mejor C-score y
el modelo óptimo según el diagrama de Ramachandra para lo cual se obtuvo con PBDsum
en http://www.ebi.ac.uk/.
5.1.2. Docking molecular
Para realizar el acoplamiento molecular se tuvieron en cuenta los resultados de posibles
ligandos y sitios de unión reportados por el meta-servidor COACH basado en la estructura
3D predicha por I-TASSER. Fue descargado el archivo PDB (Rep.) con las coordenadas
del sitio de unión al carbohidrato -L-Fucosa (C-Score:0.32). También fueron descargados
los ligandos de interés en formato PDB con la conformación ideal (GalNac, GlcNac, -D
glucosa, -D manosa, y GalNac-Ser). En Autodock-tools 1.5.6 los archivos fueron editados,
se eliminaron aguas y se adicionaron hidrógenos polares. Sobre la molécula que contenía
el ligando se definió la posición y tamaño del gridbox para realizar el docking (X center:
72.457; Y center: 4,175; Z center: 76,179 / Size x: 20; Size Y: 22; Size Z: 22). Para cada
uno de los ligandos se definieron los ángulos de torsión y se guardaron los respectivos
archivos en formato PDBQT, en el caso del receptor, este se definió como una estructura
rígida. Para la corrida del docking se usó el programa AutodockVina 1.1.2 (Trott y Olson,
2010), fue seleccionado para cada uno de los ligandos el modelo con la mejor energía de
interacción (valor más negativo). Para representar las interacciones entre los carbohidratos
y el ligando con una representación 2D y 3D se usó Discovery studio 2016.
5.2. Resultados y Análisis
Bryohealin es una lectina aislada de B. plumosa con un peso molecular de 53,8 kDa, de los
cuales en el NCBI se reporta una de sus subunidades con 278 residuos de aminoácidos de
los cuales Sigma P indica hasta la posición 23 la presencia de péptido señal (Figura 8,
anexo 2). Tras eliminar esta porción de la secuencia se obtuvo una proteína de 255
residuos, con peso molecular de 27,38 kDa y punto isoeléctrico teórico de 5.75 indicando
un alto contenido de aminoácidos ácidos. Según los datos aportados por ProtParam (Figura
5-1) se encontró que el 36,8% de los residuos eran no polares alifáticos, 9% aromáticos,
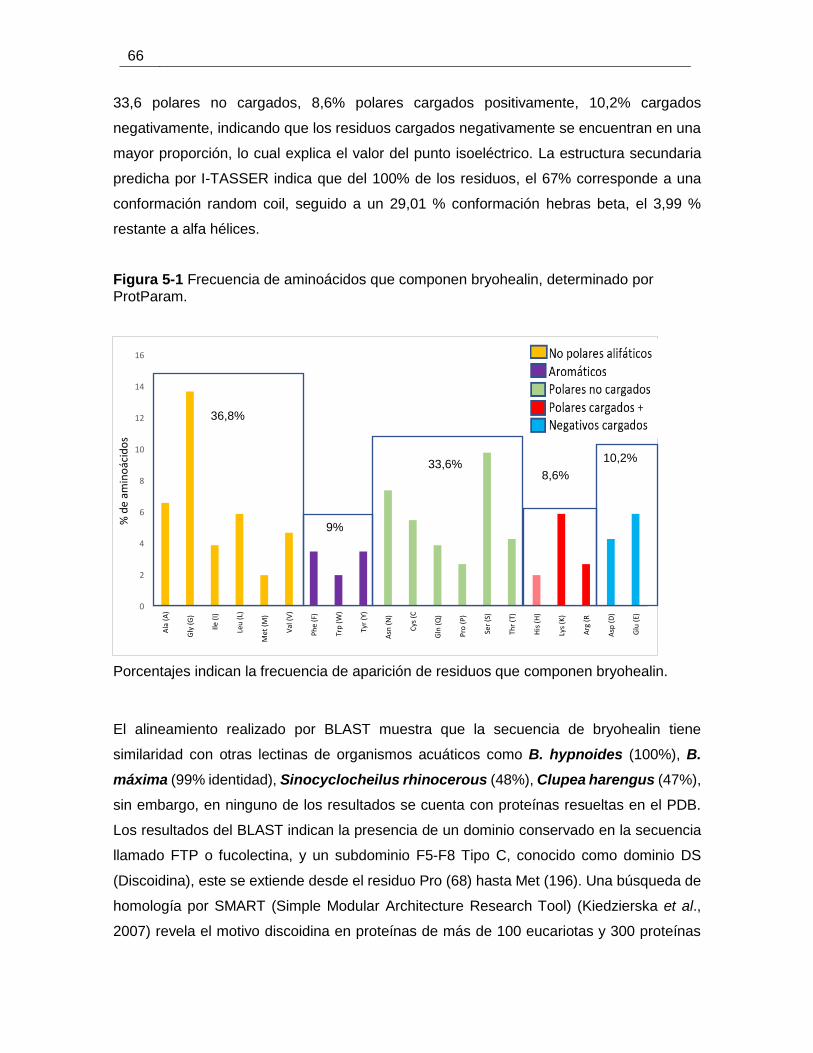
66
33,6 polares no cargados, 8,6% polares cargados positivamente, 10,2% cargados
negativamente, indicando que los residuos cargados negativamente se encuentran en una
mayor proporción, lo cual explica el valor del punto isoeléctrico. La estructura secundaria
predicha por I-TASSER indica que del 100% de los residuos, el 67% corresponde a una
conformación random coil, seguido a un 29,01 % conformación hebras beta, el 3,99 %
restante a alfa hélices.
Figura 5-1 Frecuencia de aminoácidos que componen bryohealin, determinado por ProtParam.
Porcentajes indican la frecuencia de aparición de residuos que componen bryohealin.
El alineamiento realizado por BLAST muestra que la secuencia de bryohealin tiene
similaridad con otras lectinas de organismos acuáticos como B. hypnoides (100%), B.
máxima (99% identidad), Sinocyclocheilus rhinocerous (48%), Clupea harengus (47%),
sin embargo, en ninguno de los resultados se cuenta con proteínas resueltas en el PDB.
Los resultados del BLAST indican la presencia de un dominio conservado en la secuencia
llamado FTP o fucolectina, y un subdominio F5-F8 Tipo C, conocido como dominio DS
(Discoidina), este se extiende desde el residuo Pro (68) hasta Met (196). Una búsqueda de
homología por SMART (Simple Modular Architecture Research Tool) (Kiedzierska et al.,
2007) revela el motivo discoidina en proteínas de más de 100 eucariotas y 300 proteínas
0
2
4
6
8
10
12
14
16
Ala
(A
)
Gly
(G
)
Ile (
I)
Leu
(L)
Met
(M
)
Val
(V
)
Ph
e (F
)
Trp
(W
)
Tyr
(Y)
Asn
(N
)
Cys
(C
Gln
(Q
)
Pro
(P
)
Ser
(S)
Thr
(T)
His
(H
)
Lys
(K)
Arg
(R
Asp
(D
)
Glu
(E)
% d
e am
ino
ácid
os
36,8%
9%
33,6% 8,6%
10,2%

67
procariotas. Este dominio contiene un segmento de alrededor 40 residuos altamente
conservados los cuales se ubican hacia la región C terminal, siendo característica la
presencia de dos puentes disulfuros y un mayor número de triptófanos conservados,
además de detectar azucares modificados de la galactosa (Fuentes-Prior et al., 2002),
muchas de las proteínas que contienen el dominio DS están implicadas en eventos de
adhesión celular, migración o agregación, asociados principalmente con organogénesis
(vasculogénesis y angiogénesis) y otros procesos de desarrollo (Kiedzierska et al., 2007;
Vogel et al., 2006). En algunas especies de algas como B. plumosa (Han et al., 2010; Han
et al., 2011; Kim et al., 2001), Microdictyon umbilicatum (Kim et al., 2002), B. hypnoides (Niu
et al., 2009) se ha reportado que lectinas con este dominio se encuentran involucradas en
procesos reproductivos de tipo asexual. Cuando el talo del alga sufre un daño y el
protoplasma de la célula se extruye en agua de mar, los organelos celulares se agregan
rápidamente y se genera una nueva membrana plasmática alrededor de estos, después de
varias horas se da la formación de la pared celular originando varios individuos. Kim et al,
(2002) sugieren que este tipo de respuesta frente al daño del talo, puede ser interpretada
como un mecanismo de propagación y dispersión ya que la interrupción de una célula
monoparental resulta en la producción de cientos de nuevos organismos.
De los cinco modelos resueltos por I-TASSER con valores de C-score de -0.57, -2.53,-1.65,
-3.41 y -1.69 (Figura 9, anexo 2) fue seleccionado la primera opción con la mayor
significancia en la predicción (Roy et al., 2010; Yang et al., 2015; Zhang, 2008), el diagrama
de Ramachandran (Figura 5-2) favorece este resultado indicando que el modelo propuesto
es de buena calidad, ya que más del 90% de los residuos se encuentra en las regiones
favorecidas (conformación de ángulos phi y psi correcta), mientras tan solo un 0,6 % se
encontró dentro de regiones no permitidas. Este resultado concuerda con los porcentajes
de estructura secundaria predichos por I-TASSER, ya que en el diagrama Ramachandran
se observa una mayor nube de puntos en la zona Psi (45-180) y Phi(-180-(-45)
correspondiente a la conformación hojas beta, siendo los aminoácidos más frecuentes en
este tipo de estructura la Gly y Val (Smith et al., 1996); en este caso la Gly represento el
valor más alto (13,7%) frente a los demás residuos de la secuencia de la proteína según
ProtParam (Figura 5-1). Al analizar las 10 mejores platillas usadas por I-TASSER para
generar la predicción de estructura 3D de bryohealin (Figura 5-3), se observa que se usaron
lectinas provenientes principalmente de tres organismos: Morone saxatilis (plantilla con un
36% de identidad), Anguilla anguilla (30%), y Streptococcus pneumoniae (21%), con
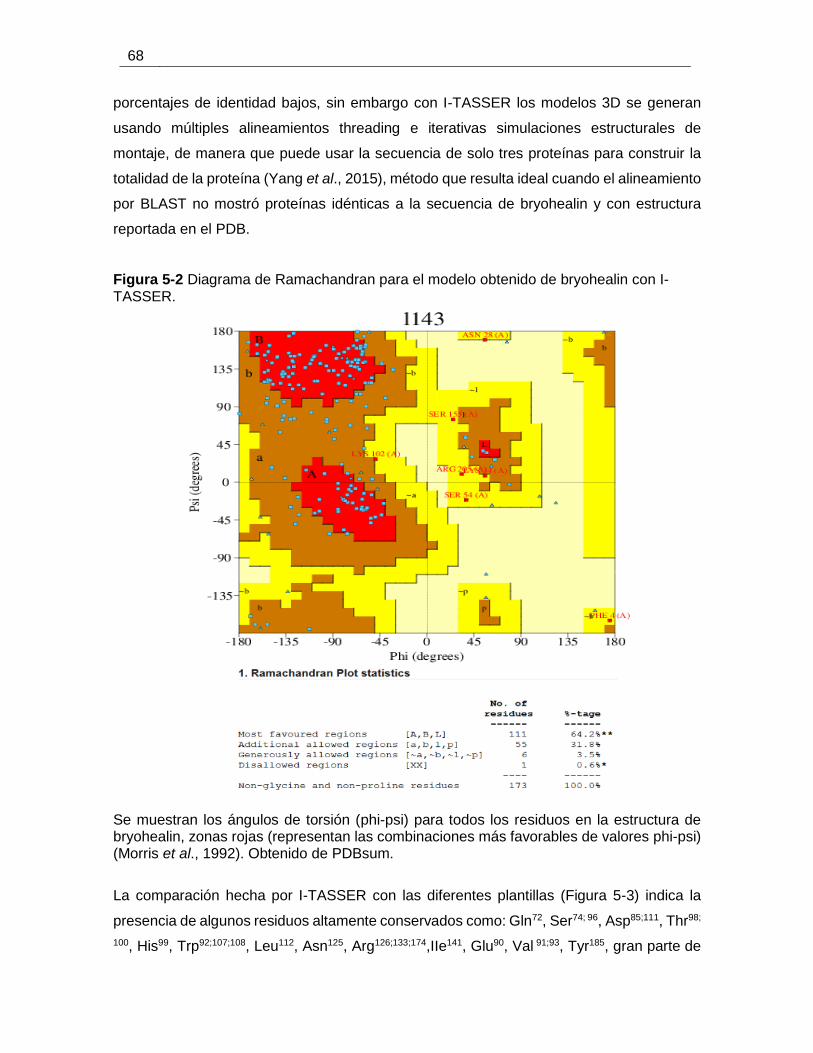
68
porcentajes de identidad bajos, sin embargo con I-TASSER los modelos 3D se generan
usando múltiples alineamientos threading e iterativas simulaciones estructurales de
montaje, de manera que puede usar la secuencia de solo tres proteínas para construir la
totalidad de la proteína (Yang et al., 2015), método que resulta ideal cuando el alineamiento
por BLAST no mostró proteínas idénticas a la secuencia de bryohealin y con estructura
reportada en el PDB.
Figura 5-2 Diagrama de Ramachandran para el modelo obtenido de bryohealin con I-TASSER.
Se muestran los ángulos de torsión (phi-psi) para todos los residuos en la estructura de bryohealin, zonas rojas (representan las combinaciones más favorables de valores phi-psi) (Morris et al., 1992). Obtenido de PDBsum.
La comparación hecha por I-TASSER con las diferentes plantillas (Figura 5-3) indica la
presencia de algunos residuos altamente conservados como: Gln72, Ser74; 96, Asp85;111, Thr98;
100, His99, Trp92;107;108, Leu112, Asn125, Arg126;133;174,IIe141, Glu90, Val 91;93, Tyr185, gran parte de
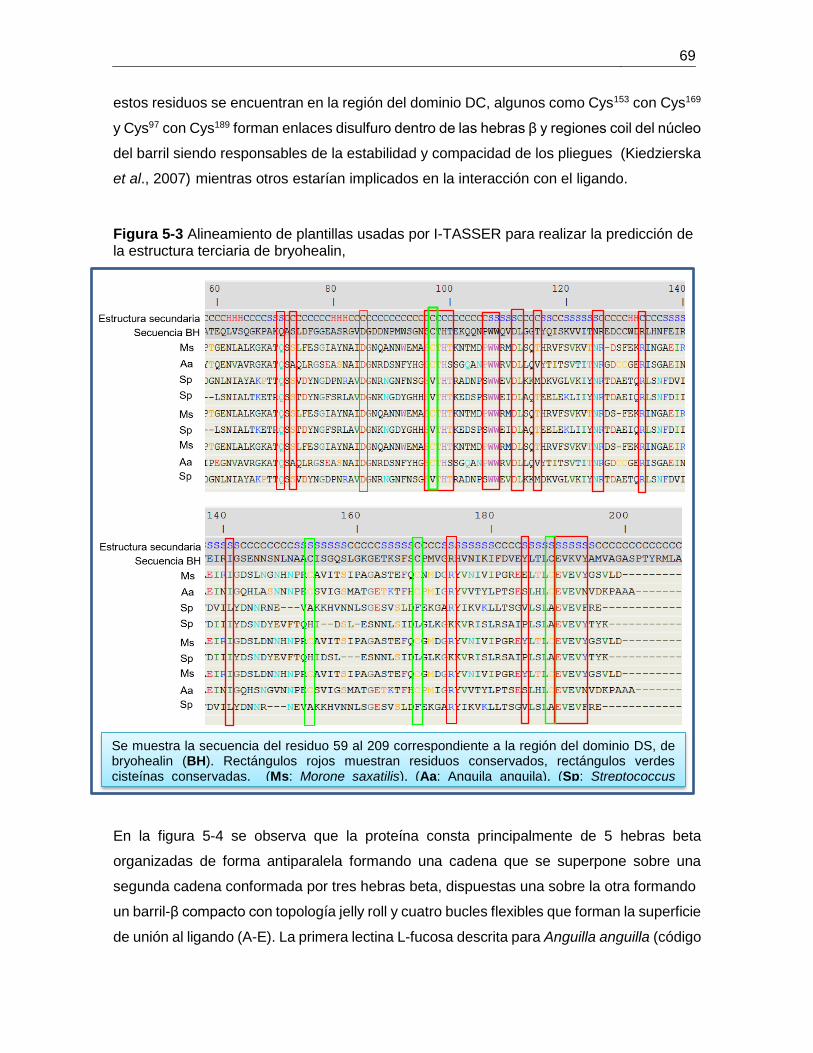
69
estos residuos se encuentran en la región del dominio DC, algunos como Cys153 con Cys169
y Cys97 con Cys189 forman enlaces disulfuro dentro de las hebras β y regiones coil del núcleo
del barril siendo responsables de la estabilidad y compacidad de los pliegues (Kiedzierska
et al., 2007) mientras otros estarían implicados en la interacción con el ligando.
Figura 5-3 Alineamiento de plantillas usadas por I-TASSER para realizar la predicción de la estructura terciaria de bryohealin,
En la figura 5-4 se observa que la proteína consta principalmente de 5 hebras beta
organizadas de forma antiparalela formando una cadena que se superpone sobre una
segunda cadena conformada por tres hebras beta, dispuestas una sobre la otra formando
un barril-β compacto con topología jelly roll y cuatro bucles flexibles que forman la superficie
de unión al ligando (A-E). La primera lectina L-fucosa descrita para Anguilla anguilla (código
Se muestra la secuencia del residuo 59 al 209 correspondiente a la región del dominio DS, de bryohealin (BH). Rectángulos rojos muestran residuos conservados, rectángulos verdes cisteínas conservadas. (Ms: Morone saxatilis), (Aa: Anguila anguila), (Sp: Streptococcus pneumoniae)

70
PDB 1K12), consiste del mismo barril β de ocho cadenas, donde el sitio de unión al ligando
es una bolsa de carga positiva superficial en un extremo del barril β, que está enmarcada
por cinco bucles, denominados las regiones determinantes de complementariedad (CDR1
a CDR5) (Bianchet et al., 2002a), características también observas para bryohealin (Figura
5-4: C).
Figura 5-4 Predicción de estructura terciaria de bryohelian
1 2
3 4 5
6
7
C-terminal
A B
C
D
E
C-terminal
1
2 3
4
5
6
7
A
B C
D
E
A) Estructura tridimensional de bryohealin resuelta con I-TASSER, se muestra sitio de unión al carbohidrato definido entre la región determinante (Bucles A-E) Obtenido en Discovery Studio 2016. Línea gris (random coil), cilindros rojos (hélices), flechas azules (hebras beta). B) Fucolectina proveniente de Anguila anguila Obtenido en Discovery Studio 2016. C) Sitio de unión al carbohidrato. Carga del bolsillo de unión de bryohealin frente al ligando N-acetil D glucosamina. Colores azules y violetas indican cargas positivas en la superficie del bolsillo de unión al ligando.
A B
C
8

71
Los resultados de docking molecular (Tabla 5-1) indicaron que la mejor energía de afinidad
fue obtenida con el antígeno Tn (GalNAC-Ser (Thr), seguido por GalNAc, GlcNAc, D-
manosa, D-glucosa y L-fucosa. Estos datos in silico concuerdan con resultados
experimentales de inhibición en la actividad aglutinante por diferentes carbohidratos en
bryohealin ya que los autores encuentran una inhibición de la aglutinación al usar N acetil
glucosamina y N acetil galactosamina, seguido por un bajo efecto de la D glucosa, L fucosa
y D manosa (Kim et al., 2006). La variabilidad de residuos críticos en el bolsillo de unión y
en los loops de interacción al ligando en las múltiples isoformas de lectinas tipo F, podrían
representar interacciones alternativas con un diversos tipos de carbohidratos (Bianchet et
al., 2002b).
Tabla 5-1Afinidad de cuatro ligandos a bryohealin
Energías de afinidad de cinco ligandos a la estructura de bryohealin, los valores más negativos indican mejores energías de unión.
Las interacciones obtenidas entre los ligandos y el sitio de unión al receptor (Figura 5-5)
muestran la importancia de ciertos residuos para el reconocimiento azúcar-proteína y que
son altamente conservados, tal es el caso de Arg126 uno de los H del grupo amino del
aminoácido interactúan con un oxigeno del anillo principal de la L fucosa y GalNAc. La
Arg133 fue importante para la interacción entre la L-fucosa, D-manosa y el antígeno Tn,
también interactuyo como donador de protón frente a un átomo de oxigeno del azúcar.
Residuos como el Trp92 fueron esenciales para la interacción de la proteína con GalNAc,
GlcNAc y el antígeno Tn, donde el H del grupo NH en la posición 1 del aminoácido forman
puentes de hidrogeno con GlcNAc y el Antigeno Tn (Figura 5-5: C, D, F). En el caso de
GalNAc fue el C2 del aminoácido Trp92 el que interactuó con el O6 del grupo OH del azúcar
(Figura 11:A anexo), para estos dos últimos casos se esperaba que la interacción fuera
similar ya que el antígeno Tn es una unidad de GalNAc unida a Ser, sin embargo la
presencia del aminoácido afecto el resultado del docking generando interacciones
Carbohidrato Afinidad Kcal/ml
Tn -4,4 GalNAc -4,2 GlcNAc -4,2 D manosa -4,0 D Glucosa -3,9 L fucosa -3,5

72
electrostáticas entre el extremo carboxílico de la Ser con Arg126, His99, Arg133 lo cual afecto
el valor final de energía libre (-4,4 Tabla 5-1), este resultado deja al GalNAc con el valor
más alto de interacción. Respecto a la interacción de Asp75 y Tyr185 no fueron importantes
contrario a lo observado para His99 donde un H del grupo NH en la posición 3 interactúa
con el O del grupo OH en el C6 del antigeno Tn, y al mismo tiempo Arg133 forma dos puentes
de hidrogeno adicionales con este mismo átomo del ligando, quedando al final una tría de
puentes de hidrogeno que estabilizan la molécula en el sitio de unión (Figura 11: B anexo)).
Residuos como la Asp75 frente a ligandos como la D-glucosa, GlcNAc y D- manosa, (Figura
5-5: B, C y E) actúa como aceptor de protones. A pesar de la sobre estimación en el valor
de energía libre encontrada con el antígeno Tn, los resultados indican que bryohealin podría
ser de utilidad en la detección del mismo en algunos tipos de carcinomas (Silva y Rangel,
2017), sin embargo aún no han sido realizados ensayos in vitro enfocados en la detección
del antígeno en líneas celulares o estudios histoquímicos.
Tabla 5-2 Interacciones presentes entre los aminoácidos de Bryohealin frente a diferentes ligandos.
Ligando Aminoácido de la proteína Átomos del ligando Distancia
L fucosa
TRP92 * FUC:C6 * 4,426
ARG126: NH2:H FUC: O5 2,278
ARG126: NH2:H FUC: O anillo 2,638
ARG133: NH2:H FUC: O3 2,997
HIS99:C2 FUC: O4 3,636
D-Glucosa HIS99: C2 GLC: O6 3,363
ASP75: COO-: O GLC: OH1: H 2,506
GalNAc
ARG126: NH2:H GALNAC: O6 2,783
TRP92: C2 GALNAC: O6 3,053
ASP128: COO-: O GALNAC: HO6 2,295
ARG133: NH2:H GALNAC: O amida 2,725
ARG133: NH2:H GALNAC: O amida 2,714
GlcNAc
CYS130:SH:H GLCNAC: HO6 2,974
TRP92: NH1: H GLCNAC: O amida 2,309
ASP75: COO-: O GLCNAC: HN2 2,432
D- Manosa
HIS99:C2 MAN: O5 3,635
ASP75: COO-: O MAN: HO1 2,479
ARG133: NH2:H MAN: O2 2,480
TYR185 MAN: HO2 2,701
Antígeno Tn
TRP92: NH1: H TN: O amida azúcar 2,075
HIS99:C2 TN: O anillo 3,287
HIS99:3NH:H TN: C6: OH: H 2,931

73
Aunque en el presente estudio el software usado (Discovery studio 2016) no permitió
detectar otras interacciones como aquellas de tipo electrostáticas, se sabe que en la
fucolectina típica (AAA) aislada de A. anguilla; interacciones importantes entre la L fucosa
y la proteína son los puentes de hidrogeno. También un trio de aminoácidos con cadenas
laterales de naturaleza básica permiten una interacción de van der Waals inusual entre un
puente disulfuro formado por cisteínas continuas (Cys82 y Cys83), y los carbonos del
esqueleto de azúcar C1, C2 y C6. Esta interacción faculta un libre acoplamiento en una
bolsa hidrófoba, apilándose contra los anillos aromáticos de dos residuos de His27 y Phe45
(Vasta et al., 2012). La ausencia de las cisteínas continuas (Cys82 y Cys83) en la estructura
de bryohealin explica la baja energía de unión reportada en el presente trabajo (Tabla 5-1)
frente a L fucosa.
Figura 5-5 Interacciones atómicas entre bryohealin y diferentes ligandos.
ARG133:NH2:H TN: C6: OH: H 2,608
ARG133:NH2:H TN: C6: OH: H 2,075
TYR185 TN: C6: OH: H 3,725
ASP75: COO-: O TN: O4 3,356 Las flechas indican la dirección en la que se da la donación del H+.
* Interacción hidrofóbica.
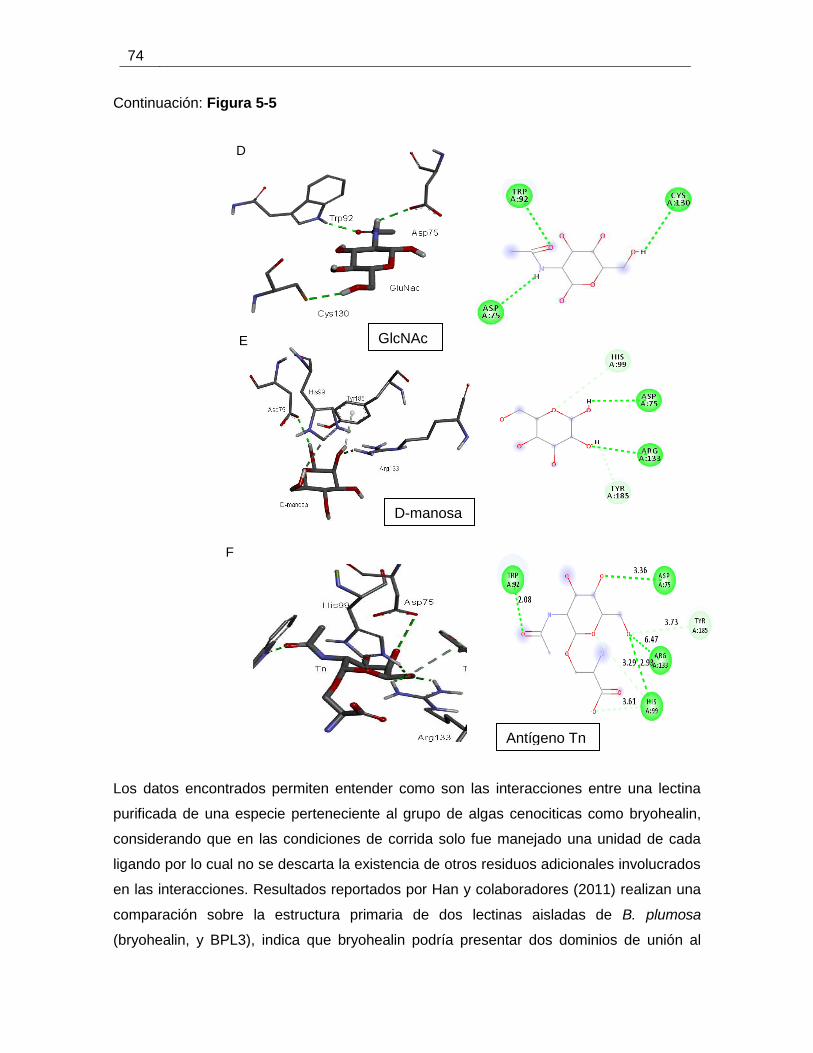
74
Continuación: Figura 5-5
Los datos encontrados permiten entender como son las interacciones entre una lectina
purificada de una especie perteneciente al grupo de algas cenociticas como bryohealin,
considerando que en las condiciones de corrida solo fue manejado una unidad de cada
ligando por lo cual no se descarta la existencia de otros residuos adicionales involucrados
en las interacciones. Resultados reportados por Han y colaboradores (2011) realizan una
comparación sobre la estructura primaria de dos lectinas aisladas de B. plumosa
(bryohealin, y BPL3), indica que bryohealin podría presentar dos dominios de unión al
D
E
F
GlcNAc
D-manosa
Antígeno Tn

75
carbohidrato, hipótesis que según los resultados obtenidos no es acertada nisiquiera al
tener en cuenta que el presente análisis solo fue realizado sobre una de las subunidades
del homodimero, esto último confirmado en datos experimentales (Kim et al., 2006). Han et
al. (2012) con la purificación de una nueva lectina denominada BPL4 y teniendo en cuenta
datos de ensayos de afinidad con GalNAc y GlcNAc establecieron que el dominio de unión
presente en bryohealin era diferente al encontrado en BPL4 y BPL3; las tres lectinas
aisladas de B plumosa muestran similitud con lectinas animales, en el caso de bryohealin
encontraron que esta podria pertenecer a lectinas de tipo-F mientras BPL3 y BPL4
pertenecen a lectinas tipo-H (aparecen en animales octocoral e invertebrados (Sanchez et
al., 2006) aunque este tipo de lectinas muestra afinidad por GalNAc y D-galactosa, mientras
BPL3 y BPL4 no muestran afinidad por este último azúcar lo cual puede ser atribuido a
pequeñas variaciones en residuos involucrados en las interacciones. La presencia de
diferentes tipos de lectinas especificas por el mismo carbohidrato podria representar una
estrategia para la agregación de los organelos celulares en un entorno de agua de mar
siempre cambiante (Han et al., 2012). Teniendo en cuenta que BPL3 y BPL4 no sean
consideradas como isoformas y que ademas se encontrara que BPL2 (Han et al., 2010) no
presenta homología con otras proteínas reportadas en las bases de datos y que tiene
afinidad por D-manosa, aunque en este último caso no se ha comprobado que esta lectina
contribuya a la formación de protoplasma.
El dominio FPT presente en bryohealin tambien ha sido hallado en eucariotas,
especificamente en proteínas de invertebrados como artrópodos, equinodermos, moluscos
platelmintos y en vertebrados como teleósteos con aletas de lóbulos, rayas, peces
cartilaginosos y anfibios. No se han encontrado en protozoos, nemátodos, ascidias, o
vertebrados superiores como reptiles, aves y mamíferos (Vasta et al., 2004), hongos y
plantas; en este ultimo caso bryohealin es una excepción ya que plantas superiores y algas
verdes estan relacionadas filogeneticamente (Lewis y McCourt, 2004). Este ultimo hecho
podria ser considerado como un claro ejemplo de evolucion paralela a traves de los limites
de las especies (Han et al., 2012).

76
5.3. Conclusiones
Se determino que la His99, Trp92, Arg 126;133 y Asp 75;128 son residuos indispensables para las
interacciones con los ligandos L fucosa, D glucosa, GalNAc, GlcNAc, D manosa y GlcNAc-
Serina.
Se encontró el dominio tipo F (fucolectinas) en bryohealin posee una mayor afinidad por
GalNAc-Ser, GalNAc y GlcNAc. Estos se evidencio por las energías determinadas en el
docking molecular.
5.4. Perspectivas
Hacer estudios enfocados en la determinación de la secuencia de aminoácidos de las dos
lectinas purificadas BRL-1 y BRL-2, y buscar si presentan homología con otras proteínas.
Esto permitirá hacer un primer acercamiento a sus estructuras terciarias, entender los
mecanismos de interacción a nivel atómico entre la proteína-carbohidrato y por lo tanto su
posible papel biológico en esta especie.

77
6. Anexo 1. Tablas
Tabla 1. Datos de cantidad y concentración de proteína por el método BCA, en extractos antes del proceso de diálisis.
Extracto
Peso alga (g)
Volumen Total (ml)
Concentración
�̅� (mg/ml)
Cantidad total (mg)
% de proteína
soluble en extracto
T. turbinata (L) 0,30 3,0 10,22 30,66 10,22
T. turbinata (NL) 0,65 6,5 7,35 47,77 7,35
T. cylindrica (L) 0,70 7,0 9,39 65,72 9,39
T. cylindrica (NL) 0,95 9,5 8,65 82,14 8,65
B. ramulosa (L) 0,50 5,0 6,76 33,79 6,76
B. ramulosa (NL) 0,80 8,0 5,80 46,43 5,80
D. pinnatifida (L) 0,40 4,0 8,81 35,22 8,81
D. pinnatifida (NL) 0,85 8,5 4,60 39,12 4,60
C. taxifolia (L) 0,67 6,7 7,85 52,56 7,85
C. taxifolia (NL) 0,90 9,0 4,27 38,40 4,27
S. hystrix var buxifolium (L) 0,70 7,0 8,98 62,88 8,98
S. hystrix var buxifolium (NL) 6,50 6,5 6,11 39,71 6,11
H. opuntia (L) 8,00 8,0 - - -
H. opuntia (NL) 7,00 7,0 - - -
Los porcentajes obtenidos de proteína soluble esta calculados sobre peso seco total del material usado para cada uno de los extractos. Harina lavada con acetona (L) y no lavada (NL). -: no se detectó proteína. n:3

78
Tabla 2. Datos de cantidad y concentración de proteína por el método BCA, en extractos después de un proceso de diálisis.
Extracto Peso alga (g)
Volumen Total (ml)
Concentración
�̅� (mg/ml)
Cantidad total (mg)
% de proteína soluble en extracto
T. turbinata (L) 0,30 3,0 1,46 4,38 1,46
T. turbinata (NL) 0,65 6,5 0,54 3,51 0,54
T. cylindrica (L) 0,70 7,0 0,39 2,75 0,39
T. cylindrica (NL) 0,95 9,5 0,38 3,61 0,38
B. ramulosa (L) 0,50 5,0 1,33 6,65 1,33
B. ramulosa (NL) 0,80 8,0 0,86 6,85 0,86
D. pinnatifida (L) 0,40 4,0 0,55 2,20 0,55
D. pinnatifida (NL) 0,85 8,5 0,47 4,03 0,47
C. taxifolia (L) 0,67 6,7 0,50 3,37 0,50
C. taxifolia (NL) 0,90 9,0 1,32 11,85 1,32
S. hystrix var buxifolium (L) 0,70 7,0 0,10 0,67 0,10
S. hystrix var buxifolium (NL) 0,65 6,5 0,10 0,64 0,10
H. opuntia (L) 0,80 8,0 - - -
H. opuntia (NL) 0,70 7,0 0,24 1,65 0,24
Los porcentajes obtenidos de proteína soluble esta calculados sobre peso seco total del material usado para cada uno de los extractos. Harina lavada con acetona (L) y no lavada (NL). -: no se detectó proteína. n:3
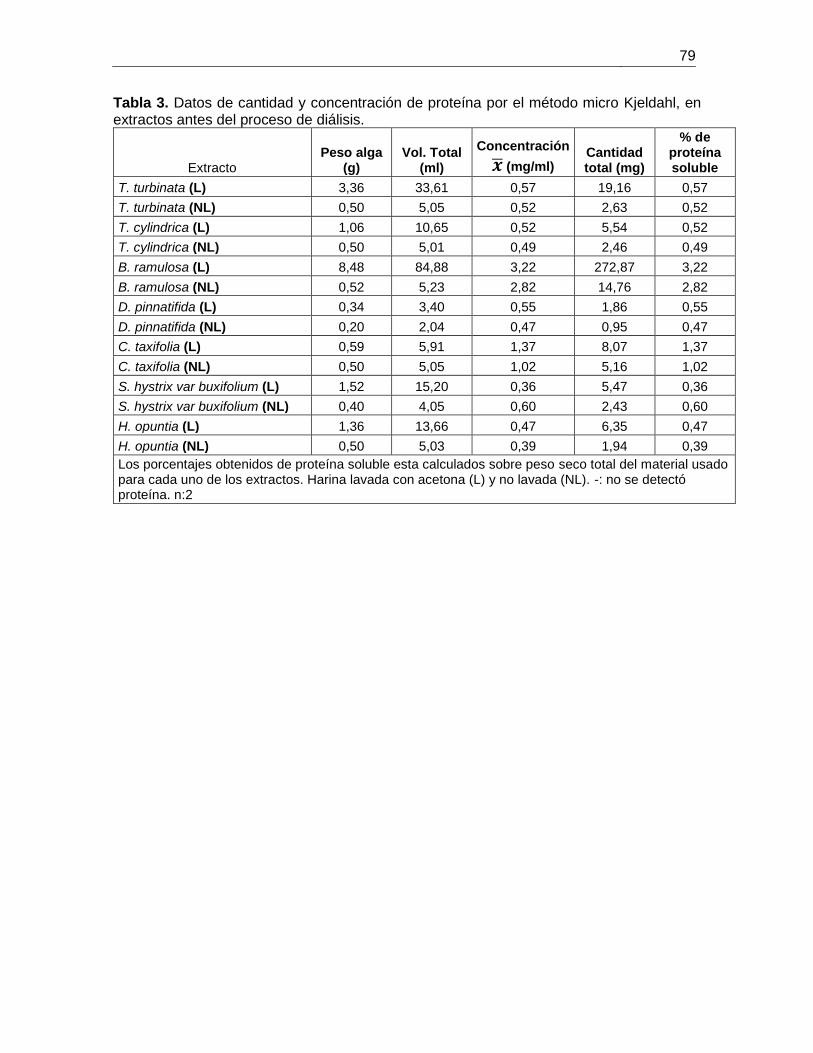
79
Tabla 3. Datos de cantidad y concentración de proteína por el método micro Kjeldahl, en extractos antes del proceso de diálisis.
Extracto Peso alga
(g) Vol. Total
(ml)
Concentración
�̅� (mg/ml) Cantidad total (mg)
% de proteína soluble
T. turbinata (L) 3,36 33,61 0,57 19,16 0,57
T. turbinata (NL) 0,50 5,05 0,52 2,63 0,52
T. cylindrica (L) 1,06 10,65 0,52 5,54 0,52
T. cylindrica (NL) 0,50 5,01 0,49 2,46 0,49
B. ramulosa (L) 8,48 84,88 3,22 272,87 3,22
B. ramulosa (NL) 0,52 5,23 2,82 14,76 2,82
D. pinnatifida (L) 0,34 3,40 0,55 1,86 0,55
D. pinnatifida (NL) 0,20 2,04 0,47 0,95 0,47
C. taxifolia (L) 0,59 5,91 1,37 8,07 1,37
C. taxifolia (NL) 0,50 5,05 1,02 5,16 1,02
S. hystrix var buxifolium (L) 1,52 15,20 0,36 5,47 0,36
S. hystrix var buxifolium (NL) 0,40 4,05 0,60 2,43 0,60
H. opuntia (L) 1,36 13,66 0,47 6,35 0,47
H. opuntia (NL) 0,50 5,03 0,39 1,94 0,39
Los porcentajes obtenidos de proteína soluble esta calculados sobre peso seco total del material usado para cada uno de los extractos. Harina lavada con acetona (L) y no lavada (NL). -: no se detectó proteína. n:2

80
Tabla 4. Comparación porcentajes de proteína soluble obtenidos para extractos por dos métodos de cuantificación BCA y Micro Kjeldahl.
Especie Método Concentración
�̅� (mg/ml) DS Valor P
T. turbinata (L) BCA 1,46 0,030
0,009* Micro Kjeldahl 0,57 0,113
T. turbinata (NL) BCA 0,55 0,066
0,690 Micro Kjeldahl 0,52 0,042
T. cylindrica (L) BCA 0,39 0,013
0,056 Micro Kjeldahl 0,52 0,042
T. cylindrica (NL) BCA 0,38 0,037
0,053 Micro Kjeldahl 0,49 0,000
B. ramulosa (L) BCA 1,33 0,119
0,011* Micro Kjeldahl 3,21 0,262
B. ramulosa (NL) BCA 0,86 0,116
0,002* Micro Kjeldahl 2,82 0,000
D. pinnatifida (L) BCA 0,55 0,016
0,932 Micro Kjeldahl 0,55 0,078
D. pinnatifida (NL) BCA 0,47 0,060
0,877 Micro Kjeldahl 0,46 0,035
C. taxifolia (L) BCA 0,50 0,275
0,0848 Micro Kjeldahl 1,36 0,262
C. taxifolia (NL) BCA 1,31 0,128
0,2479 Micro Kjeldahl 1,02 0,226
S. hystrix var buxifolium (L) BCA 0,09 0,000
0,0124* Micro Kjeldahl 0,36 0,042
S. hystrix var buxifolium (NL) BCA 0,09 0,000
0,0097* Micro Kjeldahl 0,60 0,071
H. opuntia (NL) BCA 0,23 0,012
0,1157 Micro Kjeldahl 0,38 0,078
H. opuntia (L) BCA 0,46 0,035 ND
Los porcentajes obtenidos de proteína soluble esta calculados sobre peso seco total del material usado para cada uno de los extractos *indica diferencias significativas entre valores obtenidos por dos métodos de cuantificación con un p < 0,05, DS desviación estándar, ND no determinado. Nota: Los extractos cuantificados por micro Kjeldahl corresponden a un pool diferente a los cuantificados por BCA. Harina lavada con acetona (L) y no lavada (NL).

81
Tabla 5. Datos de la determinación de porcentaje (%) de humedad en tres muestras de B. ramulosa.
Tabla 6. Extractos a gran escala de B. ramulosa
Harina (g)
Extracción (ml)
Volumen total
pool (ml)
Concentración* (mg/ml) ± DS
Cantidad total (mg)
% (p/ps)
Cantidad µg
Máxima Actividad
Título µg
8,49
84,87
254,62 1,49 ± 0,00 379,38 4,46 29,8 +4 3,72 84,87
84,87
*Determinada por método de micro Kjeldahl. n:3
Tabla 7. Contenido de proteína soluble en fracciones obtenidas de tres cromatografías de intercambio iónico.
Cantidad sembrada
(mg)
Cantidad total
sembrada (mg)
Fracción mg/ml ml Cantidad total (mg)
Cantidad total* µg
Máxima Actividad
Título µg
23,03
85,66
No retenida R 1,11 37,0 40,95 22,14 3 2,77
46,06 No retenida D 0,06 ND ND 1,29 -
16,58 Retenida R 0,99 40,0 39,75 19,88 2 9,94
Retenida D 0,00 ND ND 0,05 -
R: retentato, D: difusato
Crisol Peso
crisol (g) Peso alga Inicial (g)
Peso Crisol + Alga (g)
Peso post-tratamiento
Crisol + Alga (g)
Peso post-tratamiento
Alga (g) % humedad �̅� %
DS
1 25,2967 1,0014 26,2981 25,7726 0,4759 52,6165329
54,059251
2,024127
2 20,2109 1,0003 21,2112 20,6476 0,4367 56,3730971
3 18,5682 1,0006 19,5688 19,0372 0,469 53,1881231
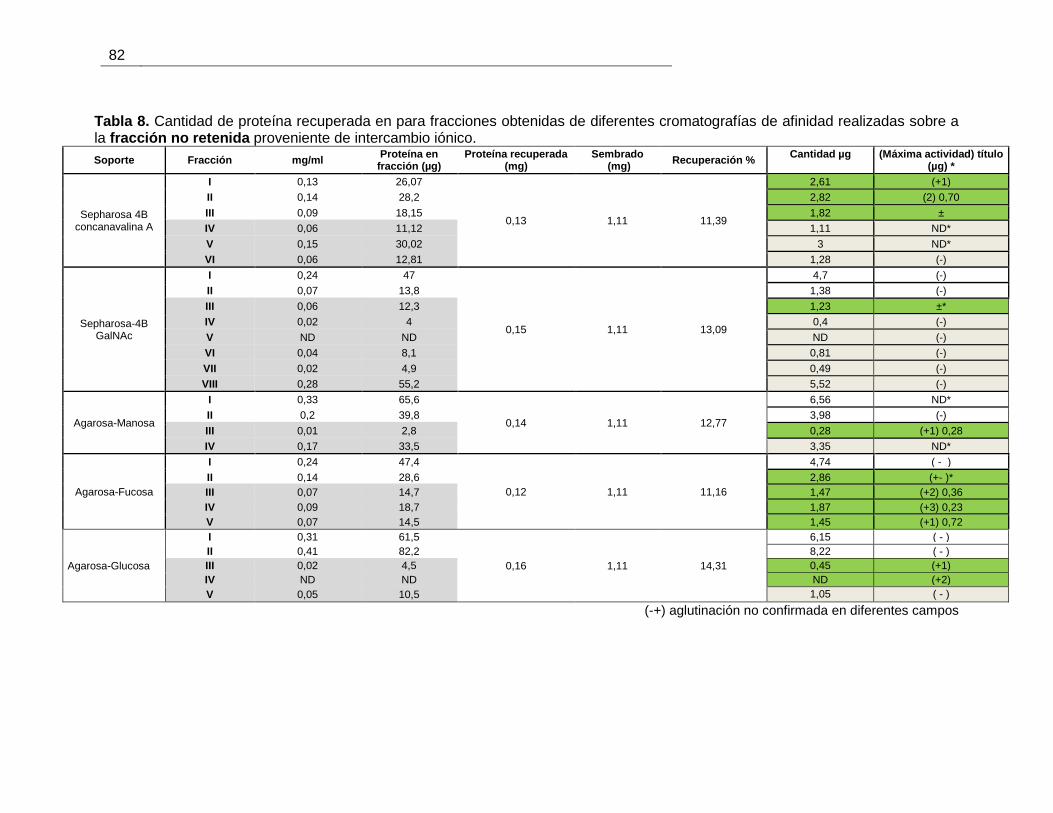
82
Tabla 8. Cantidad de proteína recuperada en para fracciones obtenidas de diferentes cromatografías de afinidad realizadas sobre a la fracción no retenida proveniente de intercambio iónico.
Soporte Fracción mg/ml Proteína en
fracción (µg) Proteína recuperada
(mg) Sembrado
(mg) Recuperación %
Cantidad µg (Máxima actividad) título (µg) *
Sepharosa 4B concanavalina A
I 0,13 26,07
0,13 1,11 11,39
2,61 (+1)
II 0,14 28,2 2,82 (2) 0,70
III 0,09 18,15 1,82 ±
IV 0,06 11,12 1,11 ND*
V 0,15 30,02 3 ND*
VI 0,06 12,81 1,28 (-)
Sepharosa-4B GalNAc
I 0,24 47
0,15 1,11 13,09
4,7 (-)
II 0,07 13,8 1,38 (-)
III 0,06 12,3 1,23 ±*
IV 0,02 4 0,4 (-)
V ND ND ND (-)
VI 0,04 8,1 0,81 (-)
VII 0,02 4,9 0,49 (-)
VIII 0,28 55,2 5,52 (-)
Agarosa-Manosa
I 0,33 65,6
0,14 1,11 12,77
6,56 ND*
II 0,2 39,8 3,98 (-)
III 0,01 2,8 0,28 (+1) 0,28
IV 0,17 33,5 3,35 ND*
Agarosa-Fucosa
I 0,24 47,4
0,12 1,11 11,16
4,74 ( - )
II 0,14 28,6 2,86 (+- )*
III 0,07 14,7 1,47 (+2) 0,36
IV 0,09 18,7 1,87 (+3) 0,23
V 0,07 14,5 1,45 (+1) 0,72
Agarosa-Glucosa
I 0,31 61,5
0,16 1,11 14,31
6,15 ( - )
II 0,41 82,2 8,22 ( - )
III 0,02 4,5 0,45 (+1)
IV ND ND ND (+2)
V 0,05 10,5 1,05 ( - )
(-+) aglutinación no confirmada en diferentes campos

83
Tabla 9. Cantidad de proteína y actividad aglutinante en fracciones obtenidas de diferentes cromatografías de afinidad realizadas sobre tres soportes para la fracción retenida DEAE-Sephacel.
*Concentración de proteína determinada por BCA, ND no determinado, (-+) aglutinación no confirmada en diferentes campos. ** Cantidad de proteína en 20 µl usados para el ensayo de aglutinación usando suspensión de eritrocitos tripsinizados al 2%
soporte Fracción (mg/ml) * Proteína en
fracción (µg)
Proteína recuperada (mg)
Sembrado (mg)
Recuperación (%)
Cantidad ** (µg)
(Máxima actividad) /título (µg)
Sepharosa-4B GalNAc
I 0,24 47
0,15 0,99 14,68
4,7 -
II 0,07 13,82 1,38 -
III 0,06 12,33 1,23 -
IV 0,02 3,97 0,4 (+2) / 0,40
V ND ND ND (+1) / ND
VI 0,04 8,15 0,81 -
VII 0,02 4,87 0,49 -
VIII 0,28 55,21 5,52 -
Sepharosa-4B aMSB
I 0,25 49
0,10 0,99 10,37
5,00 -
II 0,09 18,47 0,85 -
III 0,08 16,35 0,64 -
IV ND ND ND (+1) / ND
V 0,09 18,90 0,89 (+-) / 0,89
VI ND ND ND (+-) / ND
Sepharosa -4B Galactosa
I 0,33 66
0,09 0,99 8,89
6,6 (+3) / 1,65
II 0,11 22 2,2 ±

Tabla 10. Cantidad de proteína y actividad aglutinante en fracciones obtenidas de cromatografía de afinidad sobre soporte de L-fucosa. fracción no retenida DEAE-Sephacel.
Fracción (mg/ml) * Proteína en fracción (µg)
Proteína recuperada (mg)
Sembrado (mg)
Recuperación (%)
Cantidad * (µg)
Máxima actividad) título
(µg)
I 1,91 952,76
1,11 11,10 10,01
38,11 (+3) 0,07
II 0,06 29,50 1,18 (+1) 0,15
III 0,10 48,68 1,95 (+1) 0,12
IV 0,13 79,86 3,19 (+2) 0,05

85
7. Anexo 2. Figuras
Figura 1. Curva de calibración BSA para cuantificación de proteína por el método de BCA.
Concentración de la solución de BSA: 1,5 mg/ml
y = 0,0455x + 0,0097R² = 0,9963
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1
1,1
1,2
0 1,5 3 4,5 6 7,5 9 10,5 12 13,5 15 16,5 18 19,5 21 22,5 24 25,5
Ab
sorb
anci
a (n
m)
(µg) de proteína

86
Figura 2. SDS-PAGE, comparación de perfiles electroforéticos no dializados de extractos
de algas marinas provenientes de harina tratada y no tratada con acetona. Proteína por
carril: 100 µg.
A) Carril 1: Patrón de peso molecular Opti Protein Marker G252. Carril 2: T. turbinaria (L), carril 3: T. cylindrica (L), carril 4: B. ramulosa (L), carril 5: D. pinnatifida (L). B) carril 2: T. turbinaria (NL), carril 3: T. cylindrica (NL), carril 4: B. ramulosa (NL), carril 5: D. pinnatifida (NL). C) carril 2: C. taxifolia (L), carril 3: S. hystrix var buxifolium (L), carril 4 y 5: H. opuntia (L). D) carril 2: C. taxifolia (NL), carril 3: S. hystrix var buxifolium (NL), carril 4 y 5: H. opuntia (NL). Tinción azul de comassie y plata. Marcador de peso molecular Opti Protein Marker G252. Geles del 12,5%
Figura 3. SDS-PAGE, comparación de perfiles electroforéticos dializados de extractos de
algas marinas provenientes de harina tratada y no tratada con acetona. Proteína por carril:
100 µg
A) carril 1: patrón de peso molecular Opti Protein Marker G252. Carril 2 y 3: T. turbinaria (L), carril
4: B. ramulosa (L), carril 5: D. pinnatifida (L), carril 6: C. taxifolia (L), carril 7: B. ramulosa (NL). B)
carril 2: T. cylindrica (L), carril 3: S. hystrix var buxifolium (L), carril 4: H. opuntia (L), carril 5: T.
turbinaria (NL), carril 6: T. cylindrica (L), carril 7: H. opuntia (NL). C) carril 2: T. turbinaria (NL), carril
3: D. pinnatifida (NL), carril 4: C. taxifolia (NL), carril 5: S. hystrix var buxifolium (NL), carril 6: H.
opuntia (NL). Tinción azul de comassie y plata. Geles a 12,5 %.
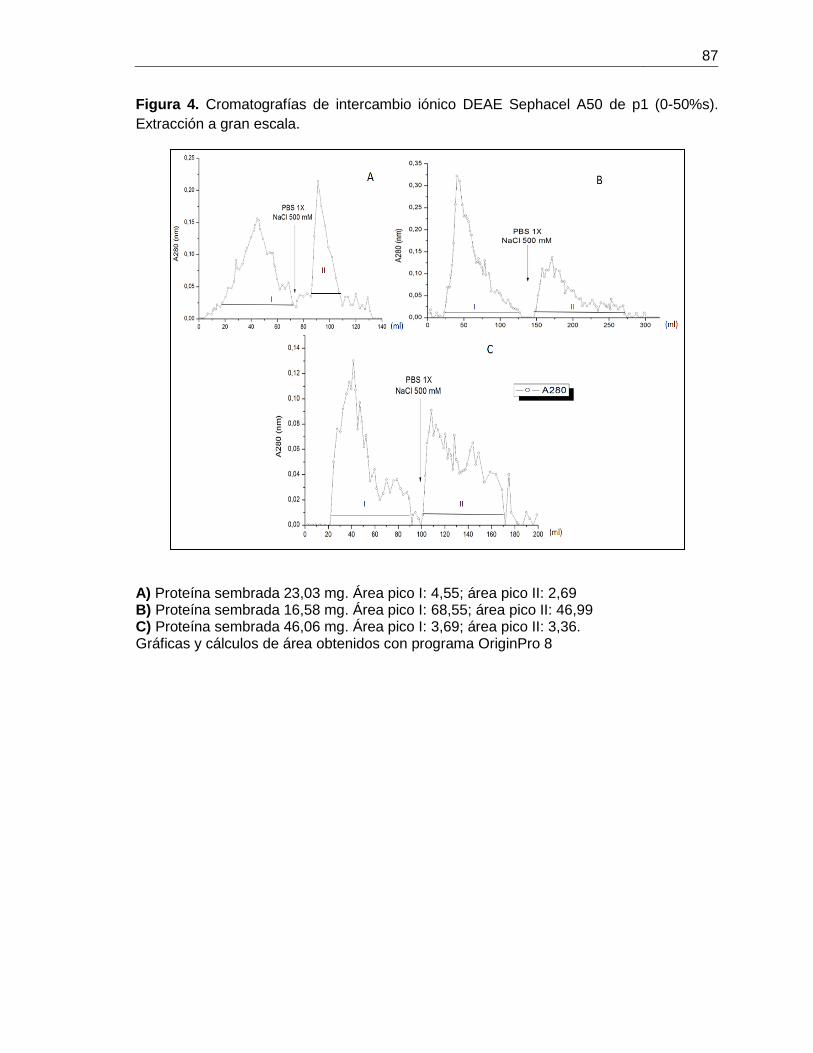
87
Figura 4. Cromatografías de intercambio iónico DEAE Sephacel A50 de p1 (0-50%s).
Extracción a gran escala.
A) Proteína sembrada 23,03 mg. Área pico I: 4,55; área pico II: 2,69 B) Proteína sembrada 16,58 mg. Área pico I: 68,55; área pico II: 46,99 C) Proteína sembrada 46,06 mg. Área pico I: 3,69; área pico II: 3,36. Gráficas y cálculos de área obtenidos con programa OriginPro 8

88
Figura 5. Cromatografías de afinidad - fracción retenida DEAE-Sephacel.
A) Afinidad sobre Sepharosa-4B GalNAc. B) Afinidad sobre Sepharosa 4B Galactosa.
Cantidad de proteína sembrada 0,99 mg.

89
Figura 6. Perfiles para cromatografías de afinidad: fracción no retenida
Perfiles de elución obtenidos para cinco cromatografías de afinidad donde fue sembrada la fracción no retenida proveniente de cromatografía de intercambio iónico. Eje de las ordenadas muestra absorbancia 280 (A280), eje de las abscisas volumen de elución (ml), números romanos indican fracciones colectadas. A) Cromatografía de afinidad soporte sepharosa concanavalina A, B) Soporte sepharosa-4B GalNAc, C) Soporte Manosa-Agarosa, D) Soporte L Fucosa-Agarosa, E) Soporte Glucosa.
B
C D
E

90
Figura 7. BLAST realizado para secuencia de B. maxima. Dominio tipo fucolectina.

91
Figura 8. Análisis péptido señal para la secuencia reportada en NCBI, en bryohealin.
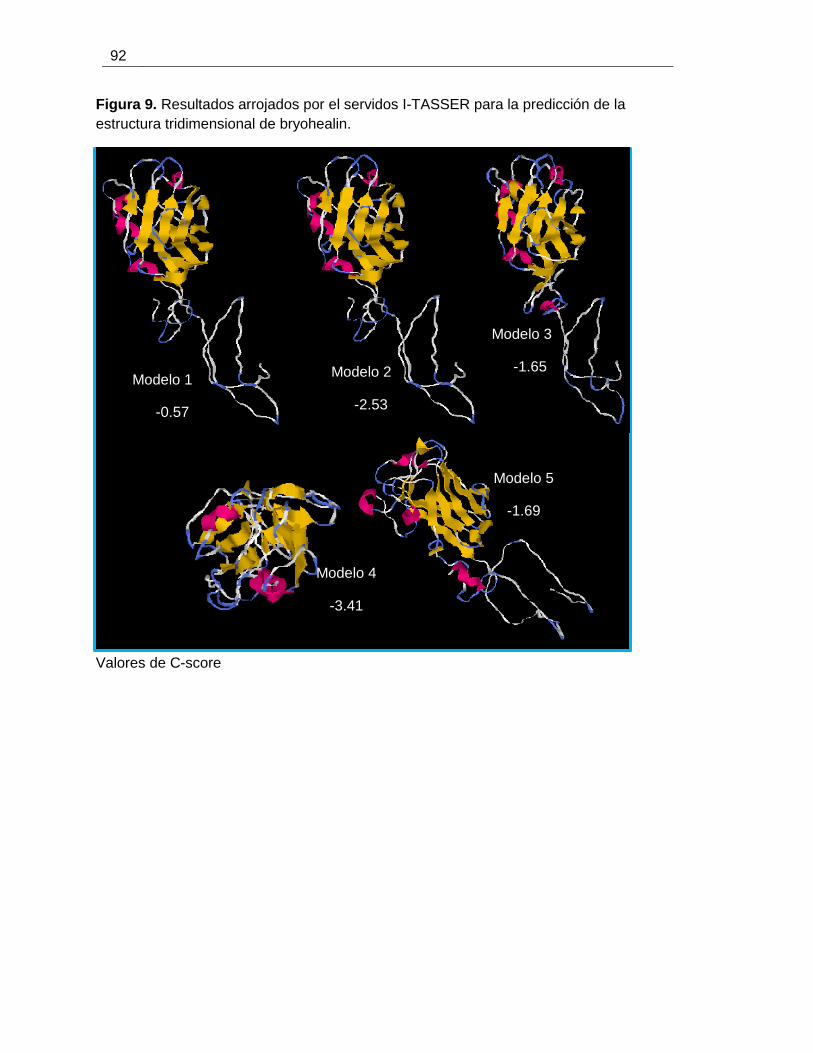
92
Figura 9. Resultados arrojados por el servidos I-TASSER para la predicción de la
estructura tridimensional de bryohealin.
Valores de C-score
Modelo 1
-0.57
Modelo 2
-2.53
Modelo 3
-1.65
Modelo 4
-3.41
Modelo 5
-1.69
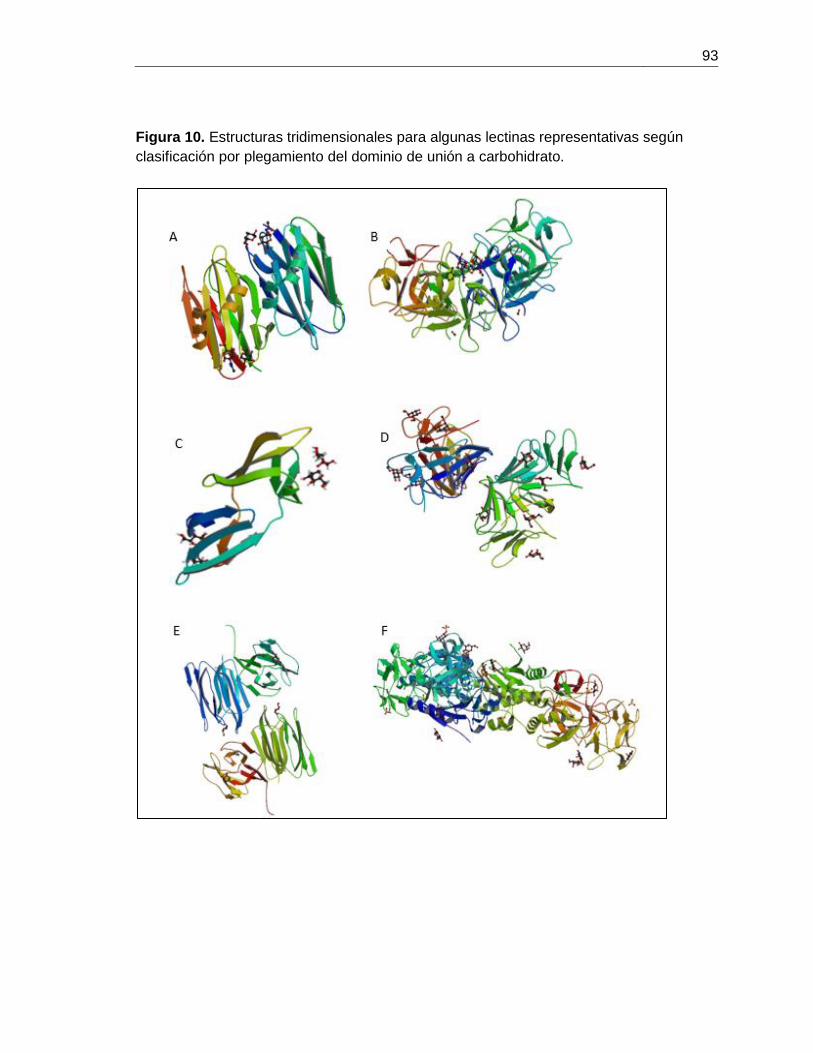
93
Figura 10. Estructuras tridimensionales para algunas lectinas representativas según
clasificación por plegamiento del dominio de unión a carbohidrato.

94
Figura 11. Interacción de aminoácidos importantes en el sitio de reconocimiento de
bryohealin frente a GalNAc-Ser.
Trp92 NH:H
O grupo amida: C2
Tyr 185 Anillo
O:C6
Arg133 NH2:H
A
B

95
8. Bibliografía
Abbott, IA, y Hollenberg, GJ (1992): Marine algae of California. Stanford University Press. Aboitiz, N, Vila‐Perelló, M, Groves, P, Asensio, JL, Yreu, D, Cañada, FJ, y Jiménez‐
Barbero, J (2004): NMR y Modeling Studies of Protein–Carbohydrate Interactions: Synthesis, Three‐Dimensional Structure, y Recognition Properties of a Minimum Hevein Domain with Binding Affinity for Chitooligosaccharides. ChemBioChem 5, 1245-1255.
Acleto, OC, y Zúñiga, R (1998): Introducción a las algas. Editorial Escuela Nueva. Adey, WH (1998): Review—coral reefs: algal structured y mediated ecosystems in
shallow, turbulent, alkaline waters. Journal of Phycology 34, 393-406. Ainouz, IL, y Sampaio, AH (1991): Screening of Brazilian marine algae for
hemagglutinins. Botanica Marina 34, 211-214. Ainouz, IL, Sampaio, AH, Benevides, NMB, Freitas, ALP, Costa, FHF, Carvalho, MR, y
Joventino, FP (1992): Agglutination of enzyme treated erythrocytes by Brazilian marine algal extracts. Bot Mar 35, 475-479.
Akkouh, O, Ng, TB, Singh, SS, Yin, C, Dan, X, Chan, YS, Pan, W, y Cheung, RCF (2015):
Lectins with anti-HIV activity: a review. Molecules 20, 648-668. Alborés, S, Moros, M, Cerdeiras, MP, de la Fuente, JM, Grazu, V, y Fraguas, LF (2016):
A Lectin Purified from Blood Red Bracket Mushroom, Pycnoporus sanguineus (Agaricomycetidae), Mycelium Displayed Affinity Toward Bovine Transferrin. International journal of medicinal mushrooms 18.
Alen, M, y Schols, D (2012): Broad antiviral activity of carbohydrate-binding agents
against dengue virus infection: Carbohydrates-Comprehensive Studies on Glycobiology y Glycotechnology. InTech.
Ambrosio, AL, Sanz, L, Sánchez, EI, Wolfenstein-Todel, C, y Calvete, JJ (2003): Isolation
of two novel mannan-y L-fucose-binding lectins from the green alga Enteromorpha prolifera: biochemical characterization of EPL-2. Archives of biochemistry y biophysics 415, 245-250.
Yerson, K, Evers, D, y Rice, KG (2008): Structure y function of mammalian carbohydrate-lectin interactions: Glycoscience. Springer, pp. 2445-2482.
Aneiros, A, y Garateix, A (2004): Bioactive peptides from marine sources:
pharmacological properties y isolation procedures. Journal of Chromatography B 803, 41-53.
Araújo Milanez de Sena Yrade, R, Sucupira Maciel, MI, Pinheiro Santos, AM, y de
Almeida Melo, E (2015): Optimization of the extraction process of polyphenols

96
from cashew apple agro-industrial residues. Ciência e Tecnologia de Alimentos 35.
Aub, JC, Sanford, BH, y Wang, L-H (1965): Reactions of normal y leukemic cell surfaces
to a wheat germ agglutinin. Proceedings of the National Academy of Sciences 54, 400-402.
Balzarini, J, Van Herrewege, Y, Vermeire, K, Vanham, G, y Schols, D (2007):
Carbohydrate-binding agents efficiently prevent dendritic cell-specific intercellular adhesion molecule-3-grabbing nonintegrin (DC-SIGN)-directed HIV-1 transmission to T lymphocytes. Molecular pharmacology 71, 3-11.
Baruffi, MD, Morani, EdSC, Roncoletta, M, del Cistia Yrade, C, y Rodrigues, LC (2017):
Methods for increasing the embryo implantation rate in mammals. Google Patents. Bateman, A, y Bycroft, M (2000): The structure of a LysM domain from E. coli membrane-
bound lytic murein transglycosylase D (MltD). Journal of molecular biology 299, 1113-1119.
Benevides, NMB, Holya, ML, Melo, FR, Pereira, MG, Monteiro, ACO, y Freitas, ALP
(2001): Purification y partial characterization of the lectin from the marine green alga Caulerpa cupressoides (Vahl) C. Agardh. Botanica Marina 44, 17-22.
Bianchet, MA, Odom, EW, Vasta, GR, y Amzel, LM (2002a): A novel fucose recognition
fold involved in innate immunity. Nature Structural & Molecular Biology 9, 628-634. Bianchet, MA, Odom, EW, Vasta, GR, y Amzel, LM (2002b): A novel fucose recognition
fold involved in innate immunity. Nature Structural & Molecular Biology 9, 628. Bird, KT, Chiles, TC, Longley, RE, Kendrick, AF, y Kinkema, MD (1993): Agglutinins from
marine macroalgae of the southeastern United States. Journal of applied phycology 5, 213.
Bishnoi, R, Khatri, I, Subramanian, S, y Ramya, TNC (2015): Prevalence of the F-type
lectin domain. Glycobiology 25, 888-901. Bitencourt, FdS, Figueiredo, JG, Mota, MRL, Bezerra, CCR, Silvestre, PP, Vale, MR,
Nascimento, KS, Sampaio, AH, Nagano, CS, y Saker-Sampaio, S (2008): Antinociceptive y anti-inflammatory effects of a mucin-binding agglutinin isolated from the red marine alga Hypnea cervicornis. Naunyn-Schmiedeberg's archives of pharmacology 377, 139-148.
Blunden, G, Rogers, DJ, Loveless, RW, y Patel, AV (1986): Haemagglutinins in marine algae-lectins or phenols. Lectins 5, 139-145.
Born, J, Uthgenannt, D, Dodt, C, Nünninghoff, D, Ringvolt, E, Wagner, T, y Fehm, H-L
(1995): Cytokine production y lymphocyte subpopulations in aged humans. An assessment during nocturnal sleep. Mechanisms of ageing y development 84, 113-126.
Boyd, WC, Almodóvar, LR, y Boyd, LG (1966): Agglutinins in marine algae for human
erythrocytes. Transfusion 6, 82-83.

97
Brown, RE, Jarvis, KL, y Hyly, KJ (1989): Protein measurement using bicinchoninic acid:
elimination of interfering substances. Analytical biochemistry 180, 136-139. Burritt, DJ, Larkindale, J, y Hurd, CL (2002): Antioxidant metabolism in the intertidal red
seaweed Stictosiphonia arbuscula following desiccation. Planta 215, 829-838. Calvete, JJ, Costa, FHF, Saker-Sampaio, S, Murciano, MPM, Nagano, CS, Cavada, BS,
Grangeiro, TB, Ramos, MV, Bloch Jr, C, y Silveira, SB (2000): The amino acid sequence of the agglutinin isolated from the red marine alga Bryothamnion triquetrum defines a novel lectin structure. Cellular y Molecular Life Sciences CMLS 57, 343-350.
Cambi, A, y Figdor, CG (2003): Dual function of C-type lectin-like receptors in the immune
system. Current opinion in cell biology 15, 539-546. Camby, I, Salmon, I, De Decker, R, Pasteels, J-L, Brotchi, J, Danguy, A, y Kiss, R (1997):
Lectin histochemistry of astrocytic tumors y in vitro characterization of lectin-induced modifications on the proliferation of the SW1088, U373 y U87 human astrocytic cell lines. Journal of neuro-oncology 34, 111-122.
Carrizo, ME, Capaldi, S, Perduca, M, Irazoqui, FJ, Nores, GA, y Monaco, HL (2004): The
antineoplastic lectin of the common edible mushroom (Agaricus bisporus) has two binding sites, each specific for a different configuration at a single epimeric hydroxyl. Journal of Biological Chemistry.
Casseb, JS, Benard, G, Saito, R, Brigido, LF, Tanaami, D, Joaquim, ES, y Duarte, AJ
(1994): The value of the lymphocyte proliferation test with phytohemagglutinin in the immune evaluation of Brazilian HIV-infected patients. Journal of investigational allergology & clinical immunology 5, 347-349.
Castellanos González, ME, Moreira González, AR, y León Pérez, AR (2012):
Caracterización fitoquímica de las macroalgas marinas Gracilaria caudata J. Agardh, Ulva lactuca L. y Ulva flexuosa subsp. flexuosa Wulfen de la bahía de Cienfuegos, Cuba. Algas, Boletín de la Sociedad Española de Ficología 46, 4-8.
Chernikov, OV, Molchanova, VI, Chikalovets, IV, Kondrashina, AS, Li, W, y Lukyanov, PA
(2013): Lectins of marine hydrobionts. Biochemistry (Moscow) 78, 760-770. Cheung, RCF, Wong, JH, Pan, W, Chan, YS, Yin, C, Dan, X, y Ng, TB (2015): Marine
lectins y their medicinal applications. Applied microbiology y biotechnology 99, 3755-3773.
Chiles, TC, y Bird, KT (1989): A comparative study of animal erythrocyte agglutinins from
marine algae. Comparative Biochemistry y Physiology Part B: Comparative Biochemistry 94, 107-111.
Cisterna, R, Campelo, C, Gorrino, T, Malave, C, Sarria, L, Lardelli, P, de Aranguiz, AF, y
Madariaga, L (1995): Association between HIV y other DNA viruses in vitro. European Journal of Clinical Microbiology y Infectious Diseases 14, 591-596.

98
Clark, D, y Mao, L (2012): Cancer biomarker discovery: lectin-based strategies targeting glycoproteins. Disease markers 33, 1-10.
Clayton, MN, y King, RJ (1990): Biology of marine plants. Longman Cheshire. Contreras, L, Moenne, A, y Correa, JA (2005): Antioxidant responses in Scytosiphon
lomentaria (phaeophyceae) inhabiting copper‐enriched coastal environments. Journal of Phycology 41, 1184-1195.
Cox, S, Abu-Ghannam, N, y Gupta, S (2010): An assessment of the antioxidant y
antimicrobial activity of six species of edible Irish seaweeds. Cruz, PH, Campos, EP, Martínez, LM, Ortiz, B, y Martínez, G (2005): Las lectinas
vegetales como modelo de estudio de las interacciones proteína-carbohidrato. Revista de Educación Bioquímica 24, 21-27.
Cui, B, Li, L, Zeng, Q, Lin, F, Yin, L, Liao, L, Huang, M, y Wang, J (2017): A novel lectin
from Artocarpus lingnanensis induces proliferation y Th1/Th2 cytokine secretion through CD45 signaling pathway in human T lymphocytes. Journal of natural medicines 71, 409-421.
Dahms, N. M., & Hancock, M. K. (2002). P-type lectins. Biochimica et Biophysica Acta
(BBA)-General Subjects, 1572(2), 317-340. Dambuza, IM, y Brown, GD (2015): C-type lectins in immunity: recent developments.
Current opinion in immunology 32, 21-27. Damme, EJMV, Peumans, WJ, Barre, A, y Rougé, P (1998): Plant lectins: a composite of
several distinct families of structurally y evolutionary related proteins with diverse biological roles. Critical Reviews in Plant Sciences 17, 575-692.
de Souza, BWS, Teixeira, DIA, Yrade, FK, Melo, MRS, Muñoz, AM, y Freitas, ALP
(2009): A survey of antarctic algae for agglutinins. Oecologia Australis 11, 122-130.
del Sol, FG, Nagano, CS, Cavada, BS, Sampaio, AH, Sanz, L, y Calvete, JJ (2006):
Lectinas. Investigación y Ciencia, 59. Diaz-Pulido, G, y Díaz-Ruíz, M (2003): Diversity of benthic marine algae of the Colombian
Atlantic. Biota Colombiana 4, 203-246. Dodd, RB, y Drickamer, K (2001): Lectin-like proteins in model organisms: implications for
evolution of carbohydrate-binding activity. Glycobiology 11, 71R-79R. Drake, RR, Schwegler, EE, Malik, G, Diaz, J, Block, T, Mehta, A, y Semmes, OJ (2006):
Lectin capture strategies combined with mass spectrometry for the discovery of serum glycoprotein biomarkers. Molecular & Cellular Proteomics 5, 1957-1967.
Drickamer, K (1988): Two distinct classes of carbohydrate-recognition domains in animal
lectins. J. biol. Chem 263, 9557-9560.

99
Duh, P-D, Du, P-C, y Yen, G-C (1999): Action of methanolic extract of mung bean hulls as inhibitors of lipid peroxidation y non-lipid oxidative damage. Food y Chemical Toxicology 37, 1055-1061.
Eagle, H (1930): Specific Agglutination y Precipitation I. The Mechanism of the Reactions.
The Journal of Immunology 18, 393-417. Edelman, GM, y Wang, JL (1978): Binding y functional properties of concanavalin A y its
derivatives. III. Interactions with indoleacetic acid y other hydrophobic ligys. Journal of Biological Chemistry 253, 3016-3022.
Englard, S, y Seifter, S (1990): [22] Precipitation techniques. Methods in enzymology 182,
285-300. Etcheverry, H (1986): Algas marinas bentónicas de Chile. Fajarningsih, ND, Yamin, DF, Yunita, I, Fahriza, A, Praseptiangga, D, Sarnianto, P, y
Chasanah, E (2015): Penapisan Senyawa Hemagglutinin dari Makroalga Asal Pantai Binuangeun, Banten, Indonesia. Jurnal Pascapanen dan Bioteknologi Kelautan dan Perikanan 10, 19-26.
Foyer, CH, Lopez‐Delgado, H, Dat, JF, y Scott, IM (1997): Hydrogen peroxide‐y
glutathione‐associated mechanisms of acclimatory stress tolerance y signalling. Physiologia Plantarum 100, 241-254.
Freitas, ALP, Teixeira, DIA, Costa, FHF, Farias, WRL, Lobato, ASC, Sampaio, AH, y
Benevides, NMB (1997): A new survey of Brazilian marine algae for agglutinins. Journal of applied phycology 9, 495-501.
Frikha, F, Kammoun, M, Hammami, N, McHirgui, RA, Belbahri, L, Gargouri, Y, Miled, N, y
Ben-Rebah, F (2011): Chemical composition y some biological activities of marine algae collected in Tunisia Composición química y algunas actividades biológicas de algas marinas recolectadas en Túnez. Ciencias Marinas 37, 113-124.
Fuentes-Prior, P, Fujikawa, K, y Pratt, KP (2002): New insights into binding interfaces of
coagulation factors V y VIII y their homologues-lessons from high resolution crystal structures. Current Protein y Peptide Science 3, 313-339.
Fukuda, Y, Sugahara, T, Ueno, M, Fukuta, Y, Ochi, Y, Akiyama, K, Miyazaki, T, Masuda,
S, Kawakubo, A, y Kato, K (2006): The anti-tumor effect of Euchema serra agglutinin on colon cancer cells in vitro y in vivo. Anti-cancer drugs 17, 943-947.
Fábregas, J, Muñoz, A, Llovo, J, y Carracedo, A (1988): Purification y partial
characterization of tomentine. An N-acetylglucosamine-specific lectin from the green alga Codium tomentosum (Huds.) Stackh. Journal of experimental marine biology y ecology 124, 21-30.
Gabius, H-J, Yré, S, Danguy, A, Kayser, K, y Gabius, S (1994): [4] Detection y
quantification of carbohydrate-binding sites on cell surfaces y in tissue sections by neoglycoproteins. Methods in enzymology 242, 37-46.

100
Gabius, H-J, Yré, S, Kaltner, H, y Siebert, H-C (2002): The sugar code: functional lectinomics. Biochimica et Biophysica Acta (BBA)-General Subjects 1572, 165-177.
Gabius, HJ (1997): Animal lectins. European Journal of Biochemistry 243, 543-576. Garateix, A, García, T, Buznego, MT, Ruenes, K, Valdés, O, Laguna, A, Hernández, Y,
Guzmán, F, Palmero, A, y Pérez-Saad, H (2000): Efectos neurofarmacológicos de extractos de algas marinas.
Gardères, J, Domart-Coulon, I, Marie, A, Hamer, B, Batel, R, Müller, WEG, y Bourguet-
Kondracki, M-L (2016): Purification y partial characterization of a lectin protein complex, the clathrilectin, from the calcareous sponge Clathrina clathrus. Comparative Biochemistry y Physiology Part B: Biochemistry y Molecular Biology 200, 17-27.
Goldstein, II, y Poretz, RD (1986): Isolation, physicochemical characterization, y
carbohydrate—binding specificity of Iectins. The Lectins. Properties, Functions, y Applications in Biology y Medicine, 233-247.
Goldstein, IJ (1972): Use of concanavalin A for structural studies. Methods Carbohydr
Chem 6, 106-119. Goldstein, IJ (1980): What should be called a lectin? Nature 285, 66. Gonzaga do Nascimento-Neto, L, Carneiro, RF, Da Silva, SR, Da Silva, BR, Arruda, FVS,
Carneiro, VA, do Nascimento, KS, Saker-Sampaio, S, da Silva, VA, y Porto, ALF (2012): Characterization of isoforms of the lectin isolated from the red algae Bryothamnion seaforthii y its pro-healing effect. Marine drugs 10, 1936-1954.
Gonzalez Giro, Z, Fuentes Reyes, M, Batista Corbal, P, Campos Castro, A, y Vera Perez,
Y (2015): Preliminary phytochemical analysis of two extracts of Padina gymnospora algae. BIOTECNIA 17, 26-29.
González, MEC, Pérez, ÁRL, y González, ÁM (2003): CARACTERIZACIÓN QUÍMICA DE
LA AGARÓFITA Gracilaria blodgettii HARVEY EN LA BAHÍA DE CIENFUEGOS, CUBA. Rev. Invest. Mar 24, 185-192.
Gordon, MH (1990): The mechanism of antioxidant action in vitro: Food antioxidants.
Springer, pp. 1-18. Govindan, M, McClean, AE, y Brown, KA (1993): Chemical constituents of the Caribbean
marine alga Bryopsis plumosa. Microchemical journal 47, 193-196. Graham, LM, Gupta, V, Schafer, G, Reid, DM, Kimberg, M, Dennehy, KM, Hornsell, WG,
Guler, R, Campanero-Rhodes, MA, y Palma, AS (2012): The C-type lectin receptor CLECSF8 (CLEC4D) is expressed by myeloid cells y triggers cellular activation through Syk kinase. Journal of Biological Chemistry 287, 25964-25974.
Greber, UF, y Bartenschlager, R (2017): An expyed view of viruses. FEMS microbiology
reviews 41, 1-4.

101
Gupta, S, y Abu-Ghannam, N (2011): Bioactive potential y possible health effects of edible brown seaweeds. Trends in Food Science & Technology 22, 315-326.
Haab, BB, y Yue, T (2011): High-Throughput Studies of Protein Glycoforms Using
Antibody–Lectin Sywich Arrays. Protein Microarrays: Methods y Protocols, 223-236.
Han, JW, Jung, MG, Kim, MJ, Yoon, KS, Lee, KP, y Kim, GH (2010): Purification y
characterization of a D‐mannose specific lectin from the green marine alga, Bryopsis plumosa. Phycological research 58, 143-150.
Han, JW, Yoon, KS, Jung, MG, Chah, K-H, y Kim, GH (2012): Molecular characterization
of a lectin, BPL-4, from the marine green alga Bryopsis plumosa (Chlorophyta). Algae 27, 55-62.
Han, JW, Yoon, KS, Klochkova, TA, Hwang, M-S, y Kim, GH (2011): Purification y
characterization of a lectin, BPL-3, from the marine green alga Bryopsis plumosa. Journal of Applied Phycology 23, 745-753.
Hara, A, Imamura, A, Yo, H, Ishida, H, y Kiso, M (2013): A new chemical approach to
human ABO histo-blood group type 2 antigens. Molecules 19, 414-437. Hartmut, F (1988): Advances in lectin research. Advances in lectin research. Hernández Díaz, P, Martín González, O, Rodríguez de Pablos Vélez, Y, y Ganem Báez,
FA (1999): Aplicaciones de las lectinas. Revista Cubana de Hematología, Inmunología y Hemoterapia 15, 91-95.
Hester, G, Kaku, H, Goldstein, IJ, y Wright, CS (1995): Structure of mannose-specific
snowdrop (Galanthus nivalis) lectin is representative of a new plant lectin family. Nature Structural & Molecular Biology 2, 472-479.
Hoek, C, Mann, D, y Jahns, HM (1995): Algae: an introduction to phycology. Cambridge
university press. Holya, ML, Melo, VMM, Silva, L, Amorim, RCN, Pereira, MG, y Benevides, NMB (2005):
Differential activity of a lectin from Solieria filiformis against human pathogenic bacteria. Brazilian journal of medical y biological research 38, 1769-1773.
Hori, K, Matsubara, K, y Miyazawa, K (2000): Primary structures of two hemagglutinins
from the marine red alga, Hypnea japonica. Biochimica et Biophysica Acta (BBA)-General Subjects 1474, 226-236.
Hori, K, Matsuda, H, Miyazawa, K, y Ito, K (1987): A mitogenic agglutinin from the red alga Carpopeltis flabellata. Phytochemistry 26, 1335-1338.
Hori, K, Miyazawa, K, y Ito, K (1990): Some common properties of lectins from marine
algae. Hydrobiologia 204, 561-566. Hori, K, Oiwa, C, Miyazawa, K, y Ito, K (1988): Evidence for wide distribution of
agglutinins in marine algae. Botanica marina 31, 133-138.

102
Hori, K, Sato, Y, Ito, K, Fujiwara, Y, Iwamoto, Y, Makino, H, y Kawakubo, A (2007): Strict specificity for high-mannose type N-glycans y primary structure of a red alga Eucheuma serra lectin. Glycobiology 17, 479-491.
Ijichi, S, Yamano, Y, Osame, M, y Hall, WW (1996): A kinetic comparative study on
lymphocyte responses to superantigen y phytohemagglutinin: reciprocal presentation of superantigen on the surface of activated lymphocytes. Cellular immunology 173, 312-316.
Imberty, A, Mitchell, EP, y Wimmerová, M (2005): Structural basis of high-affinity glycan
recognition by bacterial y fungal lectins. Current opinion in structural biology 15, 525-534.
Inbar, M, y Sachs, L (1969): Interaction of the carbohydrate-binding protein concanavalin
A with normal y transformed cells. Proceedings of the National Academy of sciences 63, 1418-1425.
Indegaard, M, y Minsaas, J (1991): Seaweed resources in Europe: uses y potential. John
Wiley & Sons Ltd.: Chichester, UK, pp. 21-64. Jiang, S-Y, Ma, Z, y Ramachandran, S (2010): Evolutionary history y stress regulation of
the lectin superfamily in higher plants. BMC evolutionary biology 10, 79. Jiménez, S (2000): Obtención y caracterización parcial de las subunidades de la lectina
Tn específica de Salvia bogotensis: Trabajo de grado. Universidad Nacional de Colombia.
Jung, MG, Lee, KP, Choi, H-G, Kang, S-H, Klochkova, TA, Han, JW, y Kim, GH (2010):
Characterization of carbohydrate combining sites of Bryohealin, an algal lectin from Bryopsis plumosa. Journal of applied phycology 22, 793-802.
Karaveg, K, Siriwardena, A, Tempel, W, Liu, Z-J, Glushka, J, Wang, B-C, y Moremen, KW
(2005): Mechanism of class 1 (glycosylhydrolase family 47) α-mannosidases involved in N-glycan processing y endoplasmic reticulum quality control. Journal of Biological Chemistry 280, 16197-16207.
Kato, H, Kogure, K, Araki, T, y Itoyama, Y (1995): Graded expression of
immunomolecules on activated microglia in the hippocampus following ischemia in a rat model of ischemic tolerance. Brain research 694, 85-93.
Kawakubo, A, Makino, H, Ohnishi, J-i, Hirohara, H, y Hori, K (1999): Occurrence of highly
yielded lectins homologous within the genus Eucheuma. Journal of applied phycology 11, 149-156.
Kelaher, BP, Underwood, AJ, y Chapman, MG (2003): Experimental transplantations of
coralline algal turf to demonstrate causes of differences in macrofauna at different tidal heights. Journal of Experimental Marine Biology y Ecology 282, 23-41.
Keller, RP, y Neville, MC (1986): Determination of total protein in human milk: comparison
of methods. Clinical chemistry 32, 120-123.

103
Kiedzierska, A, Smietana, K, Czepczynska, H, y Otlewski, J (2007): Structural similarities y functional diversity of eukaryotic discoidin-like domains. Biochimica et Biophysica Acta (BBA)-Proteins y Proteomics 1774, 1069-1078.
Kim, GH, Klochkova, TA, Yoon, KS, Song, YS, y Lee, KP (2006): Purification y
characterization of a lectin, bryohealin, involved in the protoplast formation of a marine green alga Bryopsis plumosa (CHLOROPHYTA) 1. Journal of phycology 42, 86-95.
Kim, GH, Klotchkova, TA, y Kang, Y-M (2001): Life without a cell membrane: regeneration
of protoplasts from disintegrated cells of the marine green alga Bryopsis plumosa. Journal of Cell Science 114, 2009-2014.
Kim, GH, Klotchkova, TA, y West, JA (2002): From protoplasm to swarmer: regeneration
of protoplasts from disintegrated cells of the multicellular marine green alga Microdictyon umbilicatum (Chlorophyta). Journal of Phycology 38, 174-183.
Kiss, R, Camby, I, Duckworth, C, De Decker, R, Salmon, I, Pasteels, JL, Danguy, A, y
Yeaton, P (1997): In vitro influence of Phaseolus vulgaris, Griffonia simplicifolia, concanavalin A, wheat germ, y peanut agglutinins on HCT-15, LoVo, y SW837 human colorectal cancer cell growth. Gut 40, 253-261.
Kong, Z-L, Chiang, L-C, Fang, F, Shinohara, K, y Pan, P (1997): Immune bioactivity in
shellfish toward serum-free cultured human cell lines. Bioscience, biotechnology, y biochemistry 61, 24-28.
Lewin, RA (2013): Physiology y biochemistry of algae. Lewis, LA, y McCourt, RM (2004): Green algae y the origin of ly plants. American journal
of botany 91, 1535-1556. Liao, WR, Lin, JY, Shieh, WY, Jeng, WL, y Huang, R (2003): Antibiotic activity of lectins
from marine algae against marine vibrios. Journal of Industrial Microbiology y Biotechnology 30, 433-439.
Lima, RF, Criddle, DN, Souza, EP, Sampaio, AH, Nascimento, KS, Cavada, BS, y
Assreuy, AMS (2004): Red marine alga Bryothamnion triquetrum lectin induces endothelium‐dependent relaxation of the rat aorta via release of nitric oxide. Journal of pharmacy y pharmacology 56, 1415-1421.
Limpias, C, Pérez, G, Acosta, J, Vega, N, y Ricaurte, O (2010): Using Vicia villosa lectin (B4 isolectin) for detecting Tn antigen in epithelial tumours. Revista de la Facultad de Medicina 58, 293-305.
Lis, H, y Sharon, N (1998): Lectins: carbohydrate-specific proteins that mediate cellular
recognition. Chemical reviews 98, 637-674. Littler, DS, y Littler, MM (2000): Caribbean reef plants. Offshore Graphics. Liu, K, Qian, Y, Jung, Y-S, Zhou, B, Cao, R, Shen, T, Shao, D, Wei, J, Ma, Z, y Chen, P
(2017): mosGCTL-7, a C-type lectin protein, mediates Japanese encephalitis virus infection in mosquitoes. Journal of Virology 91, e01348-16.

104
Loh, SH, Park, J-Y, Cho, EH, Nah, S-Y, y Kang, Y-S (2017): Animal lectins: potential
receptors for ginseng polysaccharides. Journal of ginseng research 41, 1-9. Loomis, WD (1974): [54] Overcoming problems of phenolics y quinones in the isolation of
plant enzymes y organelles. Methods in enzymology 31, 528-544. Loris, R (2002): Principles of structures of animal y plant lectins. Biochimica et Biophysica
Acta (BBA)-General Subjects 1572, 198-208. Lourenço, SO, Barbarino, E, De‐Paula, JC, Pereira, LOdS, y Marquez, UML (2002):
Amino acid composition, protein content y calculation of nitrogen‐to‐protein conversion factors for 19 tropical seaweeds. Phycological Research 50, 233-241.
Loveless, RW, y Rogers, DJ (1985): Biochemical-studies on the lectins from subspecies
of Codium-fragile, pp. 188-188. Lundin, H, y Ericson, LE On the occurrence of vitamins in marine algae, pp. 39-43. Maguna, FP, Romero, AM, Garro, OA, y Okulik, NB (2006): Actividad Antimicrobiana de
un grupo de Terpenoides. Comunicaciones Científicas y Tecnológicas en Internet. Argentina: Facultad de Agroindustrias.
Makela, O (1975): Studies in hemagglutinins of leguminosae seeds. 35:1-133. Martínez, A, Arias, LA, Rueda, JL, Diaz, MC, y Bula-Meyer, G (2009): Estudio de la
actividad antimicrobiana de los extractos alcohólicos de algunas macroalgas del Caribe Colombiano. Vitae 9.
Matsubara, K, Sumi, H, y Hori, K (1996): Platelet aggregation is inhibited by phycolectins.
experientia 52, 540-543. Matsumoto, J, Nakamoto, C, Fujiwara, S, Yubisui, T, y Kawamura, K (2001): A novel C-
type lectin regulating cell growth, cell adhesion y cell differentiation of the multipotent epithelium in budding tunicates. Development 128, 3339-3347.
Mayer, S, Raulf, M-K, y Lepenies, B (2017): C-type lectins: their network y roles in
pathogen recognition y immunity. Histochemistry y cell biology, 1-15. McKibbin, JM (1978): Fucolipids. Journal of lipid research 19, 131-147. Mitchell, CA, Ramessar, K, y O'Keefe, BR (2017): Antiviral lectins: Selective inhibitors of
viral entry. Antiviral research. Mohammed, S (2016): Novel fusion proteins based on recombinant lectins for delivery of
a cytotoxic peptide specifically to cancer cells. PhD thesis, University of Salford. Momoi, M, y Yamakawa, T (1978): Glucosamine-containing sphingoglycolipids from
sheep erythrocytes. The Journal of Biochemistry 84, 317-325. Montoya, H (1984): Algas de la Laguna de Villa (Lima): Cyanophyta, Chlorophyta y
Chrysophyta (Xanthophyceae). Boletín de Lima 31, 75-89.

105
Morgan, KC, Wright, JLC, y Simpson, FJ (1980): Review of chemical constituents of the
red algaPalmaria palmata (Dulse). Economic Botany 34, 27-50. Mori, T, O'Keefe, BR, Sowder, RC, Bringans, S, Gardella, R, Berg, S, Cochran, P, Turpin,
JA, Buckheit, RW, y McMahon, JB (2005): Isolation y characterization of griffithsin, a novel HIV-inactivating protein, from the red alga Griffithsia sp. Journal of Biological Chemistry 280, 9345-9353.
Morris, AL, MacArthur, MW, Hutchinson, EG, y Thornton, JM (1992): Stereochemical
quality of protein structure coordinates. Proteins: Structure, Function, y Bioinformatics 12, 345-364.
Moulaei, T, Shenoy, SR, Giomarelli, B, Thomas, C, McMahon, JB, Dauter, Z, O'Keefe,
BR, y Wlodawer, A (2010): Monomerization of viral entry inhibitor griffithsin elucidates the relationship between multivalent binding to carbohydrates y anti-HIV activity. Structure 18, 1104-1115.
Nagano, CS, Debray, H, Nascimento, KS, Pinto, V, Cavada, BS, Saker‐Sampaio, S,
Farias, WRL, Sampaio, AH, y Calvete, JJ (2005): HCA y HML isolated from the red marine algae Hypnea cervicornis y Hypnea musciformis define a novel lectin family. Protein science 14, 2167-2176.
Nagano, CS, Moreno, F, Bloch Jr, C, Prates, MV, Calvete, JJ, Saker-Sampaio, S, Farias,
WRL, Tavares, TD, Nascimento, KS, y Grangeiro, TB (2002): Purification y characterization of a new lectin from the red marine alga Hypnea musciformis. Protein y peptide letters 9, 159-165.
Nakamura, T, Nagayama, K, Uchida, K, y Tanaka, R (1996): Antioxidant Activity of
Phlorotannins Isolated from the Brown Alga Eisenia bicyclis. Fisheries science 62, 923-926.
Nakamura-Tsuruta, S, Kominami, J, Kuno, A, y Hirabayashi, J (2006): Evidence that
Agaricus bisporus agglutinin (ABA) has dual sugar-binding specificity. Biochemical y biophysical research communications 347, 215-220.
Nakayama, R, Tamura, Y, Kikuzaki, H, y Nakatani, N (1999): Antioxidant effect of the
constituents of Susabinori (Porphyra yezoensis). Journal of the American Oil Chemists' Society 76, 649-653.
Nascimento, KS, Nagano, CS, Nunes, EV, Rodrigues, RF, Goersch, GV, Cavada, BS, Calvete, JJ, Saker-Sampaio, S, Farias, WR, y Sampaio, AH (2006): Isolation y characterization of a new agglutinin from the red marine alga Hypnea cervicornis J. Agardh. Biochemistry y cell biology 84, 49-54.
Neu, TR, y Kuhlicke, U (2017): Fluorescence Lectin Bar-Coding of Glycoconjugates in the
Extracellular Matrix of Biofilm y Bioaggregate Forming Microorganisms. Microorganisms 5, 5.
Neves, SA, Dias-Baruffi, M, Freitas, ALP, y Roque-Barreira, MC (2001): Neutrophil
migration induced in vivo y in vitro by marine algal lectins. Inflammation Research 50, 486-490.

106
Niu, J, Wang, G, Lü, F, Zhou, B, y Peng, G (2009): Characterization of a new lectin
involved in the protoplast regeneration of Bryopsis hypnoides. Chinese Journal of Oceanology y Limnology 27, 502-512.
Of, A (1990): cial Methods of Analysis. Association of Official Analytical Chemists,
Arlington. Okai, Y, Higashi-Okai, K, Yano, Y, y Otani, S (1996): Identification of antimutagenic
substances in an extract of edible red alga, Porphyra tenera (Asadusa-nori). Cancer letters 100, 235-240.
Okamoto, R, Hori, K, Miyazawa, K, y Ito, K (1990): Isolation y characterization of a new
hemagglutinin from the red algaGracilaria bursa-pastoris. Experientia 46, 975-977. Okuyama, S, Nakamura-Tsuruta, S, Tateno, H, Hirabayashi, J, Matsubara, K, y Hori, K
(2009): Strict binding specificity of small-sized lectins from the Red Alga Hypnea japonica for core (α1-6) fucosylated N-glycans. Bioscience, biotechnology, y biochemistry 73, 912-920.
Oldroyd, GED, y Downie, JA (2008): Coordinating nodule morphogenesis with rhizobial
infection in legumes. Annu. Rev. Plant Biol. 59, 519-546. Oliveira, SRM, Nascimento, AE, Lima, ME, Leite, Y, y Benevides, NM (2002): Purification
y characterisation of a lectin from the red marine alga Pterocladiella capillacea (SG Gmel.) Santel. & Hommers. Rev Brasil Bot 25, 397-403.
Perez-Lorenzo, S, Levy-Benshimol, A, y Gomez-Acevedo, S (1997): Presence of lectins,
tannins y protease inhibitors in venezuelan marine algae. Acta cientifica venezolana 49, 144-151.
Pinkston, P, Pelletier, N, Arena, C, Schock, J, Garly, R, y Rose, RM (1995): Quantitative
culture of HIV-1 from bronchoalveolar lavage cells. American journal of respiratory y critical care medicine 152, 254-259.
Pinto, V, Debray, H, Dus, D, Teixeira, EH, De Oliveira, TM, Carneiro, VA, Teixeira, AH,
Nagano, CS, Nascimento, KS, y Sampaio, AH (2009): Lectins from the red marine algal species Bryothamnion seaforthii y Bryothamnion triquetrum as tools to differentiate human colon carcinoma cells. Advances in pharmacological sciences 2009.
Plavina, T, Wakshull, E, Hancock, WS, y Hincapie, M (2007): Combination of abundant
protein depletion y multi-lectin affinity chromatography (M-LAC) for plasma protein biomarker discovery. Journal of proteome research 6, 662-671.
Poiroux, G, Barre, A, van Damme, EJM, Benoist, H, y Rougé, P (2017): Plant Lectins
Targeting O-Glycans at the Cell Surface as Tools for Cancer Diagnosis, Prognosis y Therapy. International Journal of Molecular Sciences 18, 1232.

107
Ponraj, T, Paulpyi, M, Vivek, R, Vimala, K, y Kannan, S (2017): Protein regulation y Apoptotic induction in human breast carcinoma cells (MCF-7) through lectin from G. beauts. International journal of biological macromolecules 95, 1235-1245.
Procópio, TF, Moura, MC, Albuquerque, LP, Gomes, FS, Santos, NDL, Coelho, L,
Pontual, EV, Paiva, PMG, y Napoleão, TH (2017): Antibacterial lectins: action mechanisms, defensive roles y biotechnological potential. Antibacterials: Synthesis, Properties y Biological Activities, Nova Science Publishers Inc., New York, 69-89.
Rinderle, SJ, Goldstein, IJ, Matta, KL, y Ratcliffe, RM (1989): Isolation y characterization
of amaranthin, a lectin present in the seeds of Amaranthus caudatus, that recognizes the T-(or cryptic T)-antigen. Journal of Biological Chemistry 264, 16123-16131.
Rogers, DJ, Blunden, G, y Evans, PR (1977): Ptilota plumosa, a new source of a blood-
group B specific lectin. Medical laboratory sciences 34, 193. Rogers, DJ, y Flangu, H (1991): Lectins from Codium species. Brish Phycol 26, 95-96. Rogers, DJ, y Hori, K (1993): Marine algal lectins: new developments. Hydrobiologia 260,
589-593. Rogers, DJ, Loveless, RW, y Balding, P (1986): Isolation y characterization of the lectins
from sub-species of Codium fragile. Lectins 5, 155-160. Roy, A, Kucukural, A, y Zhang, Y (2010): I-TASSER: a unified platform for automated
protein structure y function prediction. Nature protocols 5, 725-738. Rüdiger, H (1993): Isolation of plant lectins: Lectins y glycobiology. Springer, pp. 31-46. Sampaio, AH, Rogers, DJ, y Barwell, CJ (1998a): A galactose-specific lectin from the red
marine alga Ptilota filicina. Phytochemistry 48, 765-769. Sampaio, AH, Rogers, DJ, y Barwell, CJ (1998b): Isolation y characterization of the lectin
from the green marine alga Ulva lactuca L. Botanica Marina 41, 427-434. Sampaio, AH, Rogers, DJ, Barwell, CJ, Saker-Sampaio, S, Costa, FHF, y Ramos, MV
(1999): A new isolation procedure y further characterisation of the lectin from the red marine alga Ptilota serrata. Journal of Applied Phycology 10, 539-546.
Sanchez, J-F, Lescar, J, Chazalet, V, Audfray, A, Gagnon, J, Alvarez, R, Breton, C,
Imberty, A, y Mitchell, EP (2006): Biochemical y structural analysis of helix pomatia agglutinin a hexameric lectin with a novel fold. Journal of Biological Chemistry 281, 20171-20180.
Santos, ACTGd (2017): Modulation of HIV-1-host cell pathways: from virus entry to
latency. Tesis doctoral. Universidad de Lisboa. Sato, Y, Hirayama, M, Morimoto, K, Yamamoto, N, Okuyama, S, y Hori, K (2011): High
mannose-binding lectin with preference for the cluster of α1–2-mannose from the green alga Boodlea coacta is a potent entry inhibitor of HIV-1 y influenza viruses. Journal of Biological Chemistry 286, 19446-19458.

108
Sato, Y, Morimoto, K, Kubo, T, Sakaguchi, T, Nishizono, A, Hirayama, M, y Hori, K
(2015): Entry Inhibition of Influenza Viruses with High Mannose Binding Lectin ESA-2 from the Red Alga Eucheuma serra through the Recognition of Viral Hemagglutinin. Marine drugs 13, 3454-3465.
Schwarz, FP, Puri, KD, Bhat, RG, y Surolia, A (1993): Thermodynamics of
monosaccharide binding to concanavalin A, pea (Pisum sativum) lectin, y lentil (Lens culinaris) lectin. Journal of Biological Chemistry 268, 7668-7677.
Sela, B-A, Lis, H, Sharon, N, y Sachs, L (1970): Different locations of carbohydrate-
containing sites in the surface membrane of normal y transformed mammalian cells. The Journal of membrane biology 3, 267-279.
Sharon, N (1987): Bacterial lectins, cell-cell recognition y infectious disease. FEBS letters
217, 145-157. Sharon, N (1994): Lectin-carbohydrate interactions at the atomic-level, pp. 17-CARB. Sharon, N, y Lis, H (2003): Lectins. Springer Science & Business Media. Sharon, N, y Lis, H (2004): History of lectins: from hemagglutinins to biological recognition
molecules. Glycobiology 14, 53R-62R. Sharon, N, Lis, H, y Lotan, R On the structural diversity of lectins, pp. 693-709. Shimizu, T, y Morikawa, K (1996): The β-prism: a new folding motif. Trends in biochemical
sciences 21, 3-6. Shiomi, K, Yamanaka, H, y Kikuchi, T (1980): Biochemical properties of hemagglutinins in
the red alga, Serraticardia maxima. Bulletin of the Japanese Society of Scientific Fisheries 46: 1369-1373
Shriver, Z, Raguram, S, y Sasisekharan, R (2004): Glycomics: a pathway to a class of
new y improved therapeutics. Nature Reviews Drug Discovery 3, 863-873. Silva, LMCM, Lima, V, Holya, ML, Pinheiro, PG, Rodrigues, JAG, Lima, MEP, y
Benevides, NMB (2010): Antinociceptive y anti-inflammatory activities of lectin from marine red alga Pterocladiella capillacea. Biological y Pharmaceutical Bulletin 33, 830-835.
Silva, MLS, y Rangel, MGH (2017): A Vicia villosa agglutinin biosensor for cancer-
associated Tn antigen. Sensors y Actuators B: Chemical. Singh, RS, Thakur, SR, y Bansal, P (2015): Algal lectins as promising biomolecules for
biomedical research. Critical reviews in microbiology 41, 77-88. Singh, RS, Tiwary, AK, y Kennedy, JF (1999): Lectins: Sources, activities, y applications.
Critical reviews in biotechnology 19, 145-178.

109
Singh, RS, Walia, AK, Khattar, JS, Singh, DP, y Kennedy, JF (2017): Cyanobacterial lectins characteristics y their role as antiviral agents. International Journal of Biological Macromolecules 102, 475-496.
Smith, LJ, Fiebig, KM, Schwalbe, H, y Dobson, CM (1996): The concept of a ryom coil:
Residual structure in peptides y denatured proteins. Folding y Design 1, R95-R106.
Storey, RD, y Beevers, L (1978): A simple procedure for rapid separation of enzymes y
lectin from other cellular metabolites. Plant Science Letters 11, 251-259. Sumner, JB, y Howell, SF (1936): Identification of hemagglutinin of jack bean with
concanavalin A. Journal of Bacteriology 32, 227. Sun, Y-J, Chang, N-CA, Hung, S-I, Chang, AC, Chou, C-C, y Hsiao, C-D (2001): The
crystal structure of a novel mammalian lectin, Ym1, suggests a saccharide binding site. Journal of Biological Chemistry 276, 17507-17514.
Swamy, M (2011): Marine algal sources for treating bacterial diseases. Advances in food
y nutrition research 64, 71-84. Sánchez‐Ferrer, A, y García‐Carmona, F (1992): A simple procedure for purifying
polyphenol oxidase for a practical biochemistry course. Biochemistry y Molecular Biology Education 20, 178-179.
Teixeira, EH, Napimoga, MH, Carneiro, VA, De Oliveira, TM, Nascimento, KS, Nagano,
CS, Souza, JB, Havt, A, Pinto, VPT, y Gonçalves, RB (2007): In vitro inhibition of oral streptococci binding to the acquired pellicle by algal lectins. Journal of applied microbiology 103, 1001-1006.
Teixeira, EH, Sampaio, AH, Cavada, BS, da Silva, BR, Nagano, CS, Arruda, FVS, do
Nascimento, KS, y Carneiro, VA (2012): Biological applications of plants y algae lectins: An overview. INTECH Open Access Publisher.
Thurston, TLM, Wyel, MP, von Muhlinen, N, Foeglein, Á, y Ryow, F (2012): Galectin-8
targets damaged vesicles for autophagy to defend cells against bacterial invasion. Nature 482, 414.
Tomassetti, P, Cometa, G, Del Vecchio, E, Baserga, M, Faccioli, P, Bosoni, D, Paolucci,
G, y Barbara, L (1995): Chromosomal instability in multiple endocrine neoplasia type 1 cytogenetic evaluation with DEB test. Cancer genetics y cytogenetics 79, 123-126.
Torres, PB, Chow, F, Furlan, CM, Myelli, F, Mercadante, A, y Santos, DYACd (2014):
Styardization of a protocol to extract y analyze chlorophyll a y carotenoids in Gracilaria tenuistipitata Var. Liui. Zhang y Xia (Rhodophyta). Brazilian Journal of Oceanography 62, 57-63.
Transue, TR, Smith, AK, Mo, H, Goldstein, IJ, y Saper, MA (1997): Structure of benzyl T-
antigen disaccharide bound to Amaranthus caudatus agglutinin. Nature structural biology 4, 779-783.

110
Trott, O, y Olson, AJ (2010): AutoDock Vina: improving the speed y accuracy of docking
with a new scoring function, efficient optimization, y multithreading. Journal of computational chemistry 31, 455-461.
Valdés, O, Hernádez, Y, Fernández, MD, Hernández, I, Rodríguez, M, Cano, M, Laguna,
A, Díaz, C, y Cabrera, B (2008): Actividad antioxidante de algas y plantas marinas de la plataforma insular cubana.
Van Damme, EJM, Barre, A, Rouge, P, y Peumans, WJ (2004): Potato lectin: an updated
model of a unique chimeric plant protein. The Plant Journal 37, 34-45. Van Damme, EJM, Lannoo, N, y Peumans, WJ (2008): Plant lectins. Advances in
botanical research 48, 107-209. Van Damme, EJM, Rougé, P, y Peumans, WJ (2007): Carbohydrate-protein interactions:
plant lectins Comprehensive Glycoscience—From Chemistry to Systems Biology, 563-599 New York, NY. USA Elsevier.
Van Driessche, E, Beeckmans, S, Dejaegere, R, y Kanarek, L (1984): Thiourea: the
antioxidant of choice for the purification of proteins from phenol-rich plant tissues. Analytical biochemistry 141, 184-188.
Vyerlei, ESO, Patoilo, KKNR, Lima, NA, Lima, APS, Rodrigues, JAG, Silva, LMCM, Lima,
MEP, Lima, V, y Benevides, NMB (2010): Antinociceptive y anti-inflammatory activities of lectin from the marine green alga Caulerpa cupressoides. International immunopharmacology 10, 1113-1118.
Varki, A, y Angata, T (2005): Siglecs—the major subfamily of I-type lectins. Glycobiology
16, 1R-27R. Varki, A, y Lowe, JB (2009): Biological roles of glycans. Vasta, GR (2009): Roles of galectins in infection. Nature reviews. Microbiology 7, 424. Vasta, GR, y Ahmed, H (2009): Animal lectins: A functional view. CRC Press. Vasta, GR, Ahmed, H, Bianchet, MA, Fernández‐Robledo, JA, y Amzel, LM (2012):
Diversity in recognition of glycans by F‐type lectins y galectins: molecular, structural, y biophysical aspects. Annals of the New York Academy of Sciences 1253.
Vasta, GR, Ahmed, H, y Odom, EW (2004): Structural y functional diversity of lectin
repertoires in invertebrates, protochordates y ectothermic vertebrates. Current opinion in structural biology 14, 617-630.
Vasta, GR, Odom, EW, Bianchet, MA, Amzel, LM, Saito, K, y Ahmed, H (2008): F-type
lectins: A new family of recognition factors. Animal Lectins: A Functional View; Vasta, GR, Ahmed, H., Eds, 465-474.

111
Vega, N (2004): Caracterización bioquímica, funcional y biológica de la lectina de Salvia bogotensis y evaluación de su aplicación para la detección del antígeno Tn: Tesis de doctorado. Universidad Nacional de Colombia.
Vega, N, y Pérez, G (2006): Isolation y characterisation of a Salvia bogotensis seed lectin
specific for the Tn antigen. Phytochemistry 67, 347-355. Velasco Martín, F (2016): Lectinas vegetales: utilización en histoquímica y terapia
antitumoral. Tesis doctoral. Universidad de Lisboa. Vogel, WF, Abdulhussein, R, y Ford, CE (2006): Sensing extracellular matrix: an update
on discoidin domain receptor function. Cellular signalling 18, 1108-1116. Walujono, K, Scholma, RA, Beintema, JJ, Anton, M, y Hahn, AM (1975): Amino acid
sequence of Hevein [of latex]. Agricultural Information Bank for Asia, South-East Asian Regional Center for Graduate Study and Research in Agriculture.
Wang, S, Zhong, F-D, Zhang, Y-J, Wu, Z-J, Lin, Q-Y, y Xie, L-H (2004): Molecular
characterization of a new lectin from the marine alga Ulva pertusa. Acta biochimica et biophysica Sinica 36, 111-117.
Wesener, DA, Dugan, A, y Kiessling, LL (2017): Recognition of microbial glycans by
soluble human lectins. Current Opinion in Structural Biology 44, 168-178. Wiechelman, KJ, Braun, RD, y Fitzpatrick, JD (1988): Investigation of the bicinchoninic
acid protein assay: identification of the groups responsible for color formation. Analytical biochemistry 175, 231-237.
Womersley, HBS (1994): The Marine Benthic Flora of Southern Australia. Rhodophyta.
Part IIIA. Flora of Australia Supplementary Series Number 1. Australia Biological Resources Study, Canberra.
Wu, M, Tong, C, Wu, Y, Liu, S, y Li, W (2016): A novel thyroglobulin-binding lectin from
the brown alga Hizikia fusiformis y its antioxidant activities. Food Chemistry 201:7-13.
Yamamoto, M, Cid, E, y Yamamoto, F (2012): Molecular genetic basis of the human
Forssman glycolipid antigen negativity. Scientific reports 2, 975. Yan, X, Chuda, Y, Suzuki, M, y Nagata, T (1999): Fucoxanthin as the major antioxidant in
Hijikia fusiformis, a common edible seaweed. Bioscience, biotechnology, y biochemistry 63, 605-607.
Yang, J, Yan, R, Roy, A, Xu, D, Poisson, J, y Zhang, Y (2015): The I-TASSER Suite:
protein structure y function prediction. Nature methods 12, 7-8. Yang, Z, y Hancock, WS (2004): Approach to the comprehensive analysis of
glycoproteins isolated from human serum using a multi-lectin affinity column. Journal of chromatography A 1053, 79-88.

112
Yang, Z, Hancock, WS, Chew, TR, y Bonilla, L (2005): A study of glycoproteins in human serum y plasma reference styards (HUPO) using multilectin affinity chromatography coupled with RPLC‐MS/MS. Proteomics 5, 3353-3366.
Yoon, KS, Lee, KP, Klochkova, TA, y Kim, GH (2008): Molecular characterization of the
lectin, bryohealin, involved in protoplast regeneration of the marine alga Bryopsis plumosa (Chlorophyta). Journal of phycology 44, 103-112.
Yoshie, Y, Wang, W, Petillo, D, y Suzuki, T (2000): Distribution of catechins in Japanese
seaweeds. Fisheries Science 66, 998-1000. Yuan, YV, y Walsh, NA (2006): Antioxidant y antiproliferative activities of extracts from a
variety of edible seaweeds. Food y chemical toxicology 44, 1144-1150. Zhang, Y (2008): I-TASSER server for protein 3D structure prediction. BMC bioinformatics
9, 40. Zhang, Z, Wang, F, Wang, X, Liu, X, Hou, Y, y Zhang, Q (2010): Extraction of the
polysaccharides from five algae y their potential antioxidant activity in vitro. Carbohydrate Polymers 82, 118-121.
Ziółkowska, NE, O'Keefe, BR, Mori, T, Zhu, C, Giomarelli, B, Vojdani, F, Palmer, KE,
McMahon, JB, y Wlodawer, A (2006): Domain-swapped structure of the potent antiviral protein griffithsin y its mode of carbohydrate binding. Structure 14, 1127-1135.
Ziółkowska, NE, Shenoy, SR, O'Keefe, BR, McMahon, JB, Palmer, KE, Dwek, RA,
Wormald, MR, y Wlodawer, A (2007a): Crystallographic, thermodynamic, y molecular modeling studies of the mode of binding of oligosaccharides to the potent antiviral protein griffithsin. PROTEINS: Structure, Function, y Bioinformatics 67, 661-670.
Ziółkowska, NE, Shenoy, SR, O'Keefe, BR, y Wlodawer, A (2007b): Crystallographic
studies of the complexes of antiviral protein griffithsin with glucose y N‐acetylglucosamine. Protein science 16, 1485-1489.
Ziółkowska, NE, y Wlodawer, A (2006): Structural studies of algal lectins with anti-HIV
activity. Acta Biochim Pol 53, 617-626. Zschäbitz, A, Weiser, H, Stofft, E, Krahn, V, Gabius, HJ, Khaw, A, y Biesalski, HK (1995):
Characterization of glycoconjugate expression during development of Meckel's cartilage in the rat. Anatomy y embryology 191, 47-59.

113


















