
DESARROLLO Y APLICACIONES DE LAS CELULAS ...(fibroblastos STO inactivados en su mitosis o...
22
Wolf & Brem Biotecnología de la Reproducción www.reprobiotec.com 168 DESARROLLO Y APLICACIONES DE LAS CELULAS EMBRIONARIAS PRIMORDIALES E. Wolf & G. Brem Introducción A partir de la exitosa producción de líneas celulares pluripotentes de embriones de ratón a comienzos de la década del '80, las mismas se emplean intensamente como modelos para el estudio embrionario básico como así también como vectores para la transferencia de genes. Otros objetivos en la investigación de células primordiales embrionarias comprende su empleo como donantes de núcleos en experimentos de clonado como también la conservación ex situ de información genética. El presente capítulo presenta un resumen de los resultados en la investigación. Estos documentan el estado actual de la puesta a punto y empleo de las células embrionarias primordiales en animales de experimentación y de interés productivo. Definición y características Las células embrionarias primordiales son líneas celulares continuas de origen embrionario y totipotentes, a partir de las cuales se pueden desarrollar todos los órganos, incluyendo la hilera germinal. Las células pluripotentes de origen embrionario pueden obtenerse mediante diferentes métodos (ROBERTSON, 1987). Un camino posible lo constituye el aislamiento de células pluripotentes de teratocarcinomas espontáneos como también su producción a partir de localizaciones embrionarias ectópicas (DAMJANOV y col., 1987). Junto con esas alternativas existe actualmente la posibilidad de aislar células pluripotentes directamente de los embriones. Las células pluripotentes aisladas de teratocarcinomas se denominan células-EC, las obtenidas de embriones EK (BRANDLEY y col., 1984) o más comúnmente ES. Las células EC no pueden diferenciarse de las morfológicamente ES, en su comportamiento antigénico de superficie como tampoco en su diferenciación in vivo o in vitro (EVANS y KAUFMAN, 1981; MARTIN, 1981). No obstante las células totipotentes embrionarias cuentan con algunas ventajas frente a las del carcinoma embrionario: - Directa aislación a través de marcadores genéticos como determinadas isoenzimas (BRADLEY y col., 1984) o mutaciones (MARTIN y col., 1987) - La conservación de un número diploide de cromosomas en el cual se hayan establecido las líneas femenina (XX) y masculina (XY, NICHOLS y col., 1990) - Un porcentaje relativamente alto de quimeras después de la inyección de células ES en blastocistos con la posibilidad de formar quimeras de la hilera germinal (BRADLEY y col., 1984) - Permite la posibilidad de obtener células ES de embriones partenogenéticos que cuentan con una constitución genética homocigota diploide (ROBERTSON, 1987). Formación y cultivo La aislación de líneas totipotentes de embriones se basa en las siguientes condiciones (BRADLEY, 1990): - Las células deben estar presentes en el estadio del embrión - Las células deben ser protegidas de señales que desencadenen su diferenciación - Deben se capaces de reproducirse in vitro
Transcript of DESARROLLO Y APLICACIONES DE LAS CELULAS ...(fibroblastos STO inactivados en su mitosis o...

Wolf & Brem
Biotecnología de la Reproducción www.reprobiotec.com
168
DESARROLLO Y APLICACIONES DE LAS CELULAS EMBRIONARIAS PRIMORDIALES E. Wolf & G. Brem Introducción A partir de la exitosa producción de líneas celulares pluripotentes de embriones de ratón a comienzos de la década del '80, las mismas se emplean intensamente como modelos para el estudio embrionario básico como así también como vectores para la transferencia de genes. Otros objetivos en la investigación de células primordiales embrionarias comprende su empleo como donantes de núcleos en experimentos de clonado como también la conservación ex situ de información genética. El presente capítulo presenta un resumen de los resultados en la investigación. Estos documentan el estado actual de la puesta a punto y empleo de las células embrionarias primordiales en animales de experimentación y de interés productivo. Definición y características Las células embrionarias primordiales son líneas celulares continuas de origen embrionario y totipotentes, a partir de las cuales se pueden desarrollar todos los órganos, incluyendo la hilera germinal. Las células pluripotentes de origen embrionario pueden obtenerse mediante diferentes métodos (ROBERTSON, 1987). Un camino posible lo constituye el aislamiento de células pluripotentes de teratocarcinomas espontáneos como también su producción a partir de localizaciones embrionarias ectópicas (DAMJANOV y col., 1987). Junto con esas alternativas existe actualmente la posibilidad de aislar células pluripotentes directamente de los embriones. Las células pluripotentes aisladas de teratocarcinomas se denominan células-EC, las obtenidas de embriones EK (BRANDLEY y col., 1984) o más comúnmente ES. Las células EC no pueden diferenciarse de las morfológicamente ES, en su comportamiento antigénico de superficie como tampoco en su diferenciación in vivo o in vitro (EVANS y KAUFMAN, 1981; MARTIN, 1981). No obstante las células totipotentes embrionarias cuentan con algunas ventajas frente a las del carcinoma embrionario: - Directa aislación a través de marcadores genéticos como determinadas isoenzimas (BRADLEY y
col., 1984) o mutaciones (MARTIN y col., 1987) - La conservación de un número diploide de cromosomas en el cual se hayan establecido las líneas
femenina (XX) y masculina (XY, NICHOLS y col., 1990) - Un porcentaje relativamente alto de quimeras después de la inyección de células ES en blastocistos
con la posibilidad de formar quimeras de la hilera germinal (BRADLEY y col., 1984) - Permite la posibilidad de obtener células ES de embriones partenogenéticos que cuentan con una
constitución genética homocigota diploide (ROBERTSON, 1987). Formación y cultivo La aislación de líneas totipotentes de embriones se basa en las siguientes condiciones (BRADLEY, 1990): - Las células deben estar presentes en el estadio del embrión - Las células deben ser protegidas de señales que desencadenen su diferenciación - Deben se capaces de reproducirse in vitro

Células embrionarias primordiales
Biotecnología de la Reproducción www.reprobiotec.com
169
Dos grupos de investigación lograron por primera vez el aislamiento de líneas celulares totipotentes de ratón al comienzo de la década del 80. EVANS y col. (1981) cultivaron blastocistos de la cepa consanguínea 129, cuya implantación fue impedida a través de una ovariectomía del animal donante en el día 2,5 después del estro y un tratamiento simultáneo con progesterona (implante). Las células ES se obtuvieron los días 6-8 de comenzado el estro, fueron cultivadas durante 4 días, momento en el cual los blastocistos producidos eclosionan y las células trofoectodérmicas se diferencian en trofoblastos gigantes (grandes). La masa celular interna (MCI), estructura con forma oval-cilíndrica, ubicada en el centro del blastocisto fue aislada con un capilar de vidrio y tripsinada. Las células obtenidas fueron cultivadas en un medio DMDM (Dulbecco's modified Eagles medium) modificado, individualmente o en pequeños grupos sobre una línea continua de fibroblastos de ratón (STO) inactivados con mitomicina. Las colonias con una morfología típica ES (colonias claras, continuas, sin limites celulares visibles, foto 1) fueron recolectadas y colocadas sobre una capa de células nutrientes (feederlayer).
Foto 1: Célula embrionaria primordial totipotente (ES) MARTIN (1981) aisló MCIs de blastocistos expandidos a través de cirugía inmunológica (foto 2). A través de un tratamiento con Pronase (0,5%) en M2 (37o C, 3-10 min) se eliminó la zona pelúcida (foto 2a). Las MCIs fueron cultivadas en DMEM modificado con fibroblastos STO inactivados en su mitosis. El medio DMEM fue previamente condicionado con células de carcinoma de la línea PSA-1. Una información más exacta sobre el protocolo de trabajo se encuentra en el trabajo de ROBERTSON (1987). La aislación y cultivo de células ES se logró originalmente sólo con el empleo de células nutrientes (fibroblastos STO inactivados en su mitosis o fibroblastos embrionales primarios), cuyas funciones como la desintoxicación del medio en general, la fijación de las células y la inhibición de la diferenciación en particular fueron descriptas (ROBERTSON, 1987). La presencia de células nutrientes complica, sin embargo, la manipulación genética de las células ES como así también la caracterización de los factores de crecimiento y diferenciación. SMITH y HOOPER (1987) descubrieron un factor soluble en un medio condicionado con células BRL (buffalo Rat liver), que podía inhibir la diferenciación en forma reversible (differentiation inhibiting activity = DIA) aún cuando es cultivado sin células nutrientes. En forma pura DIA es una glicoproteína con un peso molecular de 43000 e idéntico al factor inhibidor de la leucemia (LIF = Leukeaemia inhibitory factor, SMITH y col., 1988; WILLIAMS y col., 1988). LIF está actualmente disponible biotecnológicamente (GEARING y col., 1989) y se ofrece en el mercado bajo la denominación ESGRO (Amrad, Australia).

Wolf & Brem
Biotecnología de la Reproducción www.reprobiotec.com
170
Foto 2: Aislación inmunológica de las MCIs de un blastocisto (ratón): a) eliminación de la zona pelúcida; b) Cultivo de los embriones (37oC, 20-30 min) en suero de conejo anti ratón, inactivado por medio de calor y lavado en M2; c) colocación de los embriones en gotas con complemento de cobayo (1:10) para destruir las células trofoectodérmicas; d) eliminación de las células lisadas con una pipeta. La flecha indica la MCI
Con el agregado en el medio de cultivo de LIF se logró la formación de líneas celulares ES diploides con o sin el empleo de células nutrientes (HENDYSIDE y col., 1989; NICHOLS y col., 1990). WELLS y col. (1991) pudieron establecer líneas celulares ES de embriones sometidos a un shock térmico (42o C durante 10 min) y a un tratamiento con Puromicina, con una eficiencia significativamente mayor que con embriones no tratados. Mientras que en la mayor parte de los ensayos para el desarrollo de células ES se emplearon embriones de la línea consanguínea 129 LEDERMAN y BURKI (1991) lograron establecer una quimera de la hilera germinal a partir de embriones de la línea C57BL/6. En ese trabajo, el aislamiento de las células ES se logró por medio de células con mitosis inactivada de la linea 5637 (carcinoma humano), con mayor frecuencia que con los fibroblastos embrionarios murinos. El trabajo de SMITH (1991) presenta una lista completa de los métodos disponibles y materiales necesarios para el cultivo de células ES. Diferenciación in vivo e in vitro de células embrionales totipotentes Cuando las células ES alcanzan una alta concentración en el cultivo tienden a diferenciarse espontáneamente, observándose en primer lugar células epiteloides como endodermo primitivo. Si las células ES son mantenidas sin células nutrientes, en cultivos de suspensión sobre sustratos no adhesivos se presenta un fenómeno de diferenciación aún más complejo (DOETSCHMAN y col., 1985). Como primer estadio se forman agregados de células ectodérmicas, rodeadas por una capa simple de células endodérmicas (= "Embryoid bodies" = cuerpos embrioides simples; foto 3a). Con un cultivo progresivo se forman estructuras quísticas que contienen también estructuras mesodérmicas (cuerpos embrioides complejos; foto 3b).

Células embrionarias primordiales
Biotecnología de la Reproducción www.reprobiotec.com
171
Foto 3: Cuerpos embrioides simples (A) y complejos (B)
Bajo las condiciones establecidas por DOETSCHMAN y col. (1985) se formaron "islas de sangre" y células miocárdicas en 30% de los cuerpos embrioides complejos. Si los cuerpos embrioides son colocados nuevamente sobre un substrato adhesivo se produce una amplia diferenciación en derivados de las tres capas dérmicas (por ejemplo, células nerviosas, musculares, pigmentarias, condrocitos, etc). Tabla 1: Producción de quimeras por medio de la inyección de células primordiales embrionarias * en
blastocistos (resultados no publicados)
Blastocistos NMRI C57Bl/6 Balb/c
Blastocistos inyectados y transferidos 225 520 56
Número de transferencias Blastocistos/transferencia
24 9.4
48 10.8
6 9.3
Preñeces 10 (42%)a 25 (52%)a 3 (50%)a
Nacimientos 33 (15%)b 87 (17%)b 6 (11%)b
Animales destetados 28 (85%)c 54 (62%)c 4 (66%)c
Número de quimeras de pelaje 9 (32%)d 24 (44%)d 4 (100%)d
Manifestación del quimerismo de pelaje < 20% bis 95 % bis 50%
Quimeras apareadas 9 22 4
Quimeras infértiles 0 7 1
Quimeras con más de 10 descendientes 9 11 3
Quimeras de la hilera germinal 0 6 1 *D3 y diferentes clones homólogos recombinados (gene disruption); Valores promedio de atransferencias, bembriones transferidos, canimales nacidos y danimales destetados El potencial de diferenciación in vivo de las células ES puede ser evaluado a través de un transplante sucutáneo en ratones singénicos. EVANS y KAUFMAN (1981) observaron en ese ensayo una alta

Wolf & Brem
Biotecnología de la Reproducción www.reprobiotec.com
172
incidencia de tumores de rápido crecimiento. En la evaluación histopatológica de los mismos se observó la participación de varios tejidos que fueron identificados como teratocarcinomas. Sin embargo el test definitivo de la totipotencia de las células embrionarias se lleva a cabo a través de la inyección de las células ES en blastocistos o mórulas de otra especie con el fin de producir quimeras (fig. 1).
Inyección de células ES en los blastocistos y transferencia de los embriones a receptoras seudográvidas
Nacimiento de las quimeras
Test para comprobar el quimerismo de la hilera germinal a través del cruzamiento de las quimeras con marcadores genéticos (marcadores de color de pelaje)
Nacimiento de la descendencia, proveniente de células ES
Fig 1: Evaluación in vivo de la pluripotencia de las células ES a través de la inyección en blastocistos Una amplia exposición de los materiales necesarios y de los diferentes pasos metodológicos se encuentran en el trabajo de BRADLEY (1987). Por esa razón nos referiremos a los puntos más importantes de acuerdo a nuestra experiencia. Las células ES se inyectan en blastocistos (d 3) recolectados de ratonas donantes superovuladas. Alternativamente se pueden utilizar mórulas (d 2). Estas deben ser tratadas, sin embargo, con citocalasina B en un medo libre de Ca++ y Mg++ para lograr un reversible aflojamiento del citoesqueleto. BRADLEY (1987) recomienda para la inyección una cámara enfriada (10o C), que de acuerdo a nuestra experiencia no es necesaria. De importancia es la calidad de las micropipetas de inyección y sujeción. Su construcción se presenta en las figuras 2 y 3. Los aparatos necesarios para ello en el capítulo X (fotos 5, 6 y 8).

Células embrionarias primordiales
Biotecnología de la Reproducción www.reprobiotec.com
173
Fig 2: Elaboración de las pipetas para inyección de lás células ES en blastocistos o mórulas. Los
capilares de borosilicato son estirados con un estirador de pipetas y cortados en diagonal en un diámetro de 18 a 22 m con un escalpelo sobre goma siliconada. Con cierta experiencia se obtiene el 50% de los capilares con la forma ideal (a). Pipetas muy largas (b), muy cortas (c) y con irregularidades (d) son inadecuadas para la inyección
Ambas pipetas se manipulan a través de micromanipuladores. El sistema de sostén es llenado con aire y controlado con la boca o una jeringa de 10 ml. El sistema de microinyección que contiene silicona líquida es accionado por medio de una jeringa Hamilton (fig. 4). La inyección se lleva a cabo como se indica en la fig. 8, siendo la mejor posición el límite entre 2 células trofoectodérmicas. Después de la inyección de 8-15 células ES en cada blastocisto ó 3-4 células ES en cada mórula, los embriones son cultivados 30 a 60 min y posteriormente transferidos en el útero de receptoras seudográvidas (d 2).
Fig 3: Elaboración de una pipeta de sostén. El capilar es estirado a mano hasta un diámetro de 80-120 m y apoyado sobre la esfera de vidrio de la microfragua (a). La esfera es calentada hasta que la pared del capilar y la esfera se funden ligera-mente (b). Se interrumpe a continuación la corriente eléctrica de forma tal que el filamento que calienta la esfera se contrae y el capilar se quiebra (c). A continuación se redondea y reduce el borde de la pipeta con ayuda de la esfera candente (d, e, f) a un diámetro final de 20 a 30 m. La eficacia de la producción de quimeras como así también el porcentaje de células ES en los tejidos somático y reproductivo dependen esencialmente de la calidad de las líneas celulares ES pero también del origen genético de los embriones receptores. De esta forma los blastocistos de la línea C57BI/6 se

Wolf & Brem
Biotecnología de la Reproducción www.reprobiotec.com
174
adecuan muy bien para la inyección de células ES de la línea 129 (SCHARTZ-BERG y col., 1989; resultados propios no publicados; foto 5, tabla 1). NAGY y col. (1990) pudieron producir quimeras a partir de células ES y embriones tetraploides. En las quimeras se pudo comprobar su completo origen de células ES. Los animales murieron, sin embargo, poco después del nacimiento por razones no establecidas. La colonización de la hilera germinal a través de células ES fue observada por primera vez por BRADLEY y col. (1984). Un factor de importancia en ese sentido es el cariotipo de las células ES (BRADLEY, 1990). Mientras las traslocaciones y trisomías sólo tienen un efecto reducido sobre el porcentaje de células ES en tejidos somáticos, la participación en los tejidos de la hilera germinal es muy improbable. Células embrionarias totipotentes de la hilera germinal A partir de las experiencias descriptas en el ratón, diferentes grupos de trabajo intentaron establecer líneas celulares totipotentes a partir de embriones bovinos, porcinos, caprinos y ovinos. Las primeras publicaciones sobre la aislación de líneas celulares contínuas de embriones bovinos y ovinos datan del año 1987.
Foto 4: Inyección de células primordiales embrionarias en un blastocisto, a) antes de la inyección; b) colocación de la pipeta en el límite entre dos células trofoectodérmicas; c) inyección de 8-10 células ES; d) el blastocisto se colapsa luego de retirar la pipeta
STRINGFELLOW y col. (1987) cultivaron un embrión bovino de 11 dias, obtenido en forma no quirúrgica, en medio Ham's F-20 (100 ml suplementado con 10 ml de suero fetal bovino, 11 mg Piruvato de Na, 0,5 mg insulina y 10 l EGF). Después de 13 días se estableció una monocapa (monolayer), que fue separada con 0,4% de Tripsina en medio PBS libre de Ca2+ y Mg2+ y nuevamente cultivada. Nueve días después 50% de las células mostraron confluencia. Esa línea celular pudo ser mantenida en el medio de cultivo hasta el pasaje 38 (1:2 respectivamente). Las células mono- y binucleares fueron consideradas morfológicamente similares a las trofoblásticas. Los mismos autores lograron, poco después, establecer una línea celular totipotente (BE12-6) bajo las mismas condiciones y con

Células embrionarias primordiales
Biotecnología de la Reproducción www.reprobiotec.com
175
morfología comparable a la anterior que ha podido mantenerse en cultivo más de 200 pasajes y que actualmente es considerada inmortal (STRINGFELLOW y col., 1991).
Foto 5: Quimeras en la hilera germinal, producto de la inyección de células ES D3 (agutí) en blastocistos Balb/c (albino)
HANDYSIDE y col. (1987) trataron de establecer células ES de ovino. Los autores aislaron masas celulares internas (MCIs) de embriones de 6 y 8 días de la raza Welsh Mountain a través de inmunocirugía y las cultivaron intactas o desmenuza-das (3-5 min en 0,25% de Tripsina, 1 mM EDTA Na2, 1% suero de pollo en PBS libre de Ca2+ y Mg2+) con células fibroblásticas de la línea STO o de la piel de ovino (TGskin) inactivadas con mitomicina en 60% de medio CMß condicionado con BRL (microgotas de 20 l bajo parafina, a 37o C, 5% CO2/95% aire). Los autores emplearon en total 60 embriones (16 mórulas o blastocistos tempranos, 30 expandidos y 14 protruidos). Una evaluación morfológica de los embriones (cortes semifinos, microscopia electrónica) mostró diferencias entre las células de la MCI adyacentes al blastocele del resto de sus células. En las primeras los autores observaron estadios tempranos de diferenciación endodérmica en los blastocistos con zonas intactas, mientras que en los blastocistos protruidos fue posible reconocer un endodermo claramente diferenciado. Una aislación de la MCI a través de cirugía inmunológica fue más simple de llevar a cabo en blastocistos expandidos con zona intacta. Después de la adhesión de las células de la MCI (comprendiendo cerca de 43 células) se produce un aplanamiento de la colonia celular y no un cilindro oval como en el ratón. La división celular fue reducida en extremo comparada con el ratón. Algunos grupos de células, semejantes en un inicio a las células ES de ratón fueron cubiertas enteramente por células de apariencia endodérmica que formaron estructuras quísticas. Después del primer pasaje no se encontraron colonias semejantes a las células ES presentes. REXROAD (1990) cultivó embriones ovinos (d 7-9) y MCIs, obtenidas a través de cirugía inmunológica sobre fibroblastos STO o fibroblastos embrionarios ovinos. A partir de las MCIs se originaron con mayor eficacia colonias primarias que a partir de embriones intactos, independientemente de la edad de los embriones y del tipo de células nutrientes que se emplearon. PIEDRAHITA y col. (1988) llevaron a cabo ensayos orientados a aislar del cerdo células semejantes a las ES. Los autores cultivaron embriones de 7 a 8 dias (dia 0 = primer día del celo) en DMEM sobre fibroblastos STO inactivados por medio de rayos x. El medio fue suplementado con 10% SFB, 0,1 mmol/l 2-mercapto-etanol, 2mml/l L-glutamina y antibióticos. Colonias similares morfológicamente a las colonias ES fueron cambiadas de medio cada 7-10 días. De 118 MCIs 84 sobrevivieron las manipulaciones y 3 líneas pudieron ser mantenidas hasta el 10o pasaje. Una de las 3 líneas tenía morfología de las células ES, las otras 2 estaban compuestas de células similares a las ES y epiteliales. De 25 embriones cultivados se establecieron 8. Se pudieron establecer 2 líneas que sobrevivieron 10

Wolf & Brem
Biotecnología de la Reproducción www.reprobiotec.com
176
pasajes. Ambas líneas contenían células trofoblásticas como así también una pequeña parte de células similares a las ES. STROJEK y col. (1990) evaluaron el efecto de la edad de los embriones y diferentes medio de cultivo sobre la formación de células ES. Mientras que con embriones de 9 días no se obtuvieron resultados positivos se desarrollaron 14 colonias a partir de 69 embriones de 10 días de edad. Estas colonias provenían, según los autores, de la MCI. El cultivo de los embriones y su adherencia se lograron sobre fibroblastos de útero de cerdo y no sobre fibroblastos STO. Los autores sospecharon en una interacción química no definida aún como la razón de ese fenómeno. Los pasajes posteriores pudieron llevarse a cabo, sin embargo, sobre fibroblastos STO. El medio de cultivo más adecuado fue el DMEM suplementado con 10% de SFB, 10% de suero de cerdo, 0,2 mg/ml de insulina bovina y 0,1 mM 2-mercaptoetanol. En total se pudieron expandir 7 colonias mediante 3 a 5 pasajes y las líneas resultantes pudieron ser congeladas. EVANS y col. (1990) y NOTARIANNI y col (1991) emplearon blastocistos expandidos de la raza Large British White para producir células embrionarias totipotentes. Fueron cultivados los embriones intactos o las MCIs después de su aislación mecánica en medio DMEM modificado (+ 10% de suero de ternero + 5% de SFB + 10-4 mol/l 2-mercaptoetanol + antibióticos) sobre fibroblastos STO inactivados con mitomicina. Los autores observaron el crecimiento de los embriones y de las MCIs aisladas sobre las células nutrientes (fibroblastos STO), contrariamente a lo encontado por STROJEK y col. (1990). Al principio se observó el desarrollo de células epiteloides (grandes, aplanadas, translúcidas), mientras que después del primer pasaje (7-14 dias después del comienzo del cultivo) se observaron colonias de dos tipos. El primer grupo, formado por pequeñas colonias, se caracterizó por células grandes indiferenciadas de las cuales surgían células gigantes similares a las trofoblásticas. Las células del segundo grupo fueron caracterizadas de acuerdo a los criterios morfológicos (células con un núcleo translúcido claro, nucléolo visible y poco citoplasma) como células similares a las ES. Estas formaron grandes colonias. Como criterio bioquímico del estatus indiferenciado de esas células se demostró la falta del filamento intermediario Vimentin a través de una prueba de fluorescencia. Esas células pudieron ser mantenidas en cultivo cerca de un año, sin alteraciones morfológicas, con pasajes semanales. Los ensayos de diferenciación in vitro indicaron diferencias claras con respecto a la morfología de las cuerpos embrioides en relación con el ratón. Sin embargo las células se diferenciaron en las 3 capas embrionarias (epitelio, endotelio, células musculares y nerviosas) sobre un substrato adhesivo. La comprobación de la totipotencia in vivo de las células similares a las ES, a través de la producción de quimeras, no fue publicada hasta el momento. NOTARIANNI y col. (1990) lograron establecer, en la especie ovina, una línea similar a la ES empleando las mismas condiciones de cultivo. PIEDRAHITA y col. (1990a) estudiaron el efecto de diferentes células nutrientes sobre la eficiencia de la producción de líneas celulares embrionarias de cerdo. En ese experimento se emplearon fibroblastos STO, células BRL, combinación de esos cultivos (9:1 y 1:1), fibroblastos embrionarios primarios de cerdo (PEF) y ratón (MEF), PEF en combinación con células BRL, células epiteliales uterinas de cerdo y células epiteloides aisladas de embriones porcinos. Los fibroblastos STO fueron evaluados en combinación con medio condicionado con células BRL. La aislación de células epiteloides como también de líneas celulares similares a la ES se logró exclusivamente sobre células fibroblásticas STO con y sin medio condicionado con células BRL. Los autores no pudieron mejorar las condiciones de cultivo establecidas por EVANS y col. (1990). Mientras PIEDRAHITA y col. (1990a) confirmaban la aparición de diferentes tipos celulares después de la desagregación de MCIs proliferadas y adheridas, resultaron divergencias en el potencial de diferenciación de las líneas celulares de tipo epitelial y similares a ES. En las primeras se formaron estructuras en cultivos de suspensión que fueron consideradas análogas de los cuerpos embrioides de ratón. Las líneas celulares similares a la ES, por el contrario, no mostraron diferenciación in vitro alguna. En un estudio posterior PIEDRAHITA y col. (1990b) pudieron comprobar el status indiferenciado de las células similares a las ES dado que éstas no exprimieron Vimentina ni Citoqueratina 18. El intento de establecer paralelamente en el mismo trabajo líneas celulares ovinas fue menos exitoso, las células

Células embrionarias primordiales
Biotecnología de la Reproducción www.reprobiotec.com
177
pudieron mantenerse vivas sólo 3 pasajes como máximo. El estudio de MEINECKE-TILLMAN y MEINECKE (1991) con el objeto de establecer líneas celulares pluripotentes a partir de embriones ovinos, caprinos y porcinos comparó diferentes células nutritivas (fibroblastos de hígado, riñón y testículo fetales de bovino; células granulosas de porcino y bovino; células epiteliales de útero y oviducto de oveja, cabra y cerda como así también fibroblastos STO). Principalmente con el empleo de fibroblastos embrionarios de hígado de bovino los autores pudieron establecer líneas embrionarias totipotentes de las tres especies estudiadas e incluso mantenerlas por varios meses en cultivo. El trabajo no contiene, sin embargo, información más detallada de la caracterización de esas líneas celulares. STROJEK y col. (1991) emplearon embriones obtenidos el día 7 in vivo o producidos in vitro para la producción de células totipotentes. Los autores evaluaron el efecto de diferentes medios (DMEM y BME/Ham's F 10 (1:1), cada uno suplementado con 5 y 20% de SFB, 10% de suero de ternero neonato y 10-4 mol/l 2-mercaptoetanol), células nutrientes (fibroblastos embrionarios primarios de ratón y células primarias de la granulosa de vaca) y diferentes presiones de oxígeno (5 y 20%) sobre la protrusión, fijación y diferenciación de los blastocistos. Los mejores resultados de protrusión se obtuvieron sin células nutrientes en BME/Ham's F 10 con 5% de CO2 en atmósfera de aire a una temperatura de 39o C. Por otro lado el empleo de células nutrientes tuvo un efecto positivo sobre la fijación de los blastocistos protruidos. En ese sentido los autores diferencian 2 tipos de fijaciones: el tipo A caracterizado por un crecimiento radial de las células trofoectodérmicas (fig. 10), el tipo B mostró la fijación de la MCI con degeneración de las células trofoectodérmicas. Aunque bajo particulares condiciones de cultivo se produjo una fuerte proliferación de las células trofoectodérmicas, en ningún caso se observó una proliferación de la MCI. No se observaron diferencias entre los embriones producidos in vivo o in vitro. La falta de proliferación de las masas celulares internas de embriones bovinos fue publicada anteriormente por SCHELLANDER y col. (1989), quienes trabajaron bajo condiciones similares. En un ensayo posterior los autores observaron el efecto positivo de LIF sobre la proliferación de MCIs de blastocistos bovinos cuando éstos fueron cultivados sin células nutrientes en medio DMEM modificado. Sin embargo, el intento de establecer líneas celulares no tuvo éxito (HASSAN-HAUSER y col., 1990).
Foto 6: Blastocisto bovino después de 10 días de cultivo
(tipo de fijación A: células trofoectodérmicas de crecimiento radial, MCI en posición central)
STRELCHENKO y colaboradores lograron, por otra parte, aislar líneas celulares similares a las ES a partir de MCI de embriones bovinos (SRELCHENKO y col., 1991; SAITO y col., 1992). Los autores cultivaron 11 hemi-embriones, resultado de la división de 6 embriones de 7 días obtenidos por medios no quirúrgicos, en TCM 199 suplementado con 10% de suero de ternero neonato sobre fibroblastos embrionarios primarios de ratón, inactivados con mitomicina. Tres embriones se fijaron de acuerdo al tipo A, en los cuales no sólo se observó la proliferación de las células del trofoectodermo sino también un crecimiento de la MCI. Esta fue disociada con tripsina a los 7 días y colocada sobre células

Wolf & Brem
Biotecnología de la Reproducción www.reprobiotec.com
178
nutrientes frescas. Los autores emplearon, a partir de ese momento, medio MEM modificado con 10% SFB, 10-4 mol/l 2-mercaptoetanol como también 4,5 g/l de glucosa. Un 30% del volumen de ese medio fue condicionado sobre células LIF de la línea 5637 en proliferación. Después de 5 días las primeras colonias similares a las ES fueron visibles. A partir de ellas se pudieron producir 2 líneas que sobrevivieron 4 pasajes. Una serie tóxica de SFB eliminó las células producidas sin que se pudiese comprobar su totipotencia in vitro e in vivo. En la tabla 2 se presentan los diferentes medios de cultivo empleados exitosamente en la producción de líneas celulares similares a las ES a partir de embriones de animales de interés productivo. Es importante destacar, sin embargo, que los resultados alcanzados en la especie murina aún no han podido ser alcanzados por las especies domésticas. Como razón de esa diferencia se discute el diferente desarrollo después de la implantación entre el ratón y los ungulados. Mientras en el embrión de ratón ocurre una rápida proliferación de la MCI después de la implantación, el disco embrionario de las especies unguladas se mantiene inicialmente en inactividad mitótica (NOTARIANNI y col., 1990).

Células primordiales
Biotecnología de la Reproducción www.reprobiotec.com
179
Tabla 2: Ensayos exitosos para producir líneas celulares similares a la ES de animales domésticos
Autor EVANS Y COL. (1990)
NOTARIANI Y COL. (1990/1991)
PIEDRAHITA y col. (1990a,b) STROJEK Y COL.(1990) STRECHELKO y col. (1991)
Especie Porcina, ovina, bovina Porcina Porcina Bovina Material empleado Blastocistos y MCIs d 7-9 MCIs de embriones d 7-8 Embriones d 10 Embriones divididos d 7 Células nutrientes Fibroblastos STO Fibroblastos STO Fibroblastos de útero
porcino (FUP) Fibroblastos embrionarios murinos (FEM)
Medios de cultivo Medio base
DMEM DMEM DMEM TCM 199 (Para el cultivo inicial MEM-Alfa (Cultivos posteriores)
L-Gln [mmol/l] - 2 - -
2-ME [mmol/l] 0,1 0,1 0,1 0,1
Antibiótico Si Si Si S. i.
Otros agregados - Medio cond. BRL 0,2 g/l Insulina, Nucleótidos, aminoáci-dos no esenciales
4,5 g/l Glucosa Medio 5673-condionado
Temperatura [ C] S. i. S. i. S. i. S. i.
CO2 [%] S. i. S. i. S. i. S. i.
O2 [%] S. i. S. i. S. i. S. i.
S.i.: sin información; L-Gln= L=-Glutamina; 2-ME= 2-Mercaptoetanol; SF = suero de ternero; SFB = suero fetal bovino; SP = suero porcino

Wolf & Brem
Biotecnología de la Reproducción www.reprobiotec.com
180
Aplicaciones de las células embrionarias totipotentes Las células ES del ratón se emplean en la actualidad casi en forma de rutina como vectores en la transferencia de genes para la producción de modelos animales con modificaciones génicas determinadas. Dado que la integración del gen en las células de los mamíferos se produce al azar, la tasa de recombinación homóloga/integración al azar es de alrededor de 13-3 (FROHMAN y MARTIN, 1989; MELTON, 1990). Esa frecuencia de recombinaciones homólogas es muy baja para llevar a cabo la alteración del genoma a través de la microinyección del gen en el pronúcleo. Por esa razón se opta por el empleo de las células embrionarias totipotentes, a las que se les ha transfeccionado el gen deseado y se evalúa la integración en el lugar adecuado. En esos ensayos se emplearon, en la mayoría de los casos, líneas ES con cariotipo masculino (por ejemplo: B. E14TG2a, CCE, D3, CC1.2). Para lo cual existe una serie de razones (MANSUR, 1990): - Las líneas celulares masculinas ES parecen ser más estables en cultivo que las femeninas. - Solamente a partir de células ES masculinas pueden originarse espermatozoides funcionales. Dado
que la descendencia es más numerosa es más fácil detectar quimeras masculinas que femeninas, respecto a la presencia de células ES en la hilera germinal.
- Cuando células ES masculinas son inyectadas en un blastocisto femenino y las células totipotentes forman una parte importante del organismo se produce una conversión del sexo. Esto significa que se produce un macho. En ese caso la hilera germinal es formada en un 100% por células ES.
La electroporación es el método de elección para la transfección de células ES, dado que en condiciones experimentales óptimas se integra sólo una copia del vector de ADN (THOMAS y CAPECCHI, 1987). Los vectores deben estar acompañados de un gen marcador, dada la baja frecuencia de integraciones estables (10-5-10-2), a partir de la cual puedan ser seleccionadas las células. En la mayoría de los casos se emplea el gen bacteriano de la Neomicina fosfotransferasa (neo®). El medio de selección contiene G418. Como forma alternativa a la electroporación se empleó la microinyección de ADN (ZIMMER y GRUSS, 1989). Ese método permite producir un alto porcentaje de células transformadas estables (10-20%), sin embargo la ejecución es muy complicada. THOMAS y CAPECCHI (1987) desarrollaron 2 diferentes tipos de vectores, que emplearon para la disrupción del gen HPRT (fig. 5).
Fig 5: Vector para un transferencia génica con un objetivo determinado (según CAPECCHI, 1989)
A
reemplazo desecuencia
B
inserción de secuencia
hprt- G 4 1
1 2 3 4 5 6 7 9Neo
hprt
1 2 3 4 5 6 7 8 9
96
Neo
hprt
1 2 3 4 5 6 7 8 9
7
6 9
Neo
hprt- G 4 1
1 2 3 4 5 6 7 8 9Neo 6 7 8 9

Células embrionarias primordiales
Biotecnología de la Reproducción www.reprobiotec.com
181
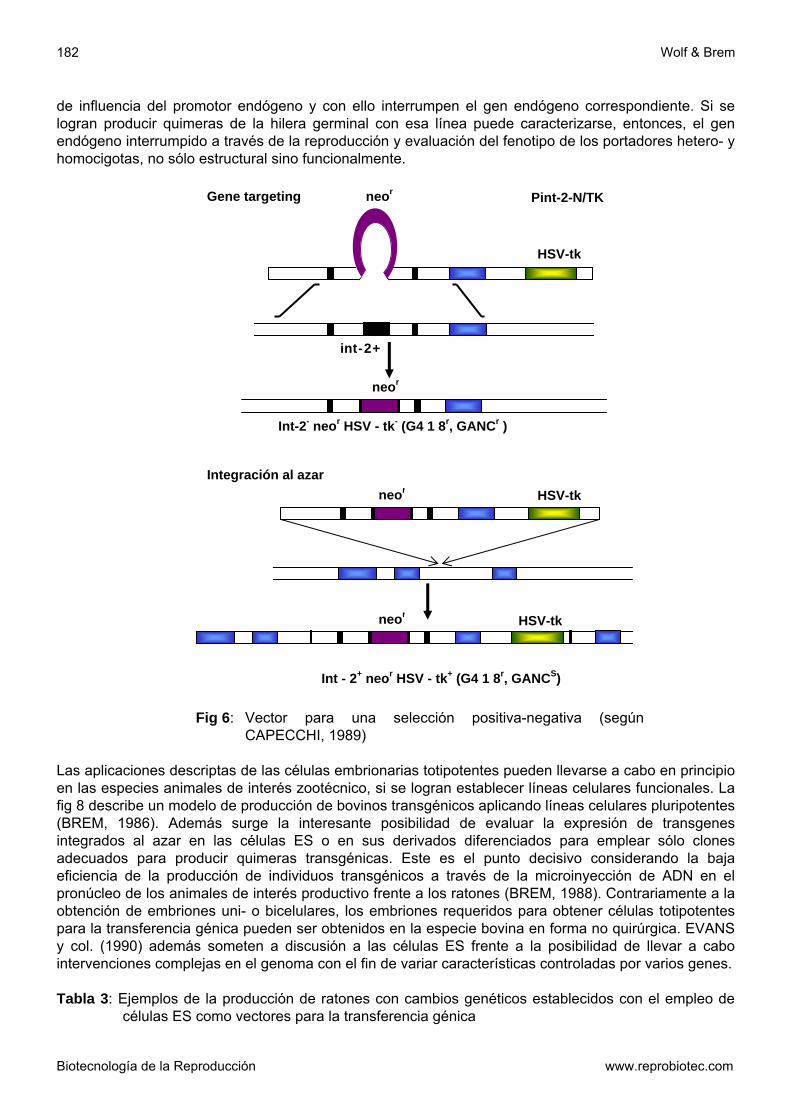
Los denominados genes de inserción tienen una fractura en la doble hélice en el lugar correspondiente a la secuencia homóloga del gen endógeno. Si se produce cross over en el momento de la yuxtaposición del gen endógeno, se integrará todo el vector en el gen endógeno y las secuencias de ambos se presentan en forma doble. Por otra parte existen vectores complementarios, co-lineales a la secuencia endógena, que contienen una determinada mutación y que reemplazan al gen endógeno a través de un cross-over doble. La frecuencia de clones homólogos recombinados puede ser aumentada en genes que ya son exprimidos en las células ES si se renuncia a las secuencias promotoras tanto en zona homóloga del vector como también del Gen neo®. Si se produce la recombinación las secuencias en el vector entran bajo el control del promotor del gen endógeno el gen neo® se exprimirá. De otra forma se logra la expresión con una integración al azar y gran parte de las células mueren durante la selección con G418 (MASOUR, 1990). Con genes que no se exprimen en las células ES se emplea otra estrategia conocida como selección positiva-negativa (SPN). Ese método propuesto por MASOUR y col. (1988) se basa en dos supuestos: - la integración al azar de un una estructura génica lineal ocurre al final de la estructura, mientras
la recombinación homóloga tiene lugar dentro de la zona homóloga a través de cross-over. - Estructuras génicas lineales que no contienen al final secuencias homólogas están en
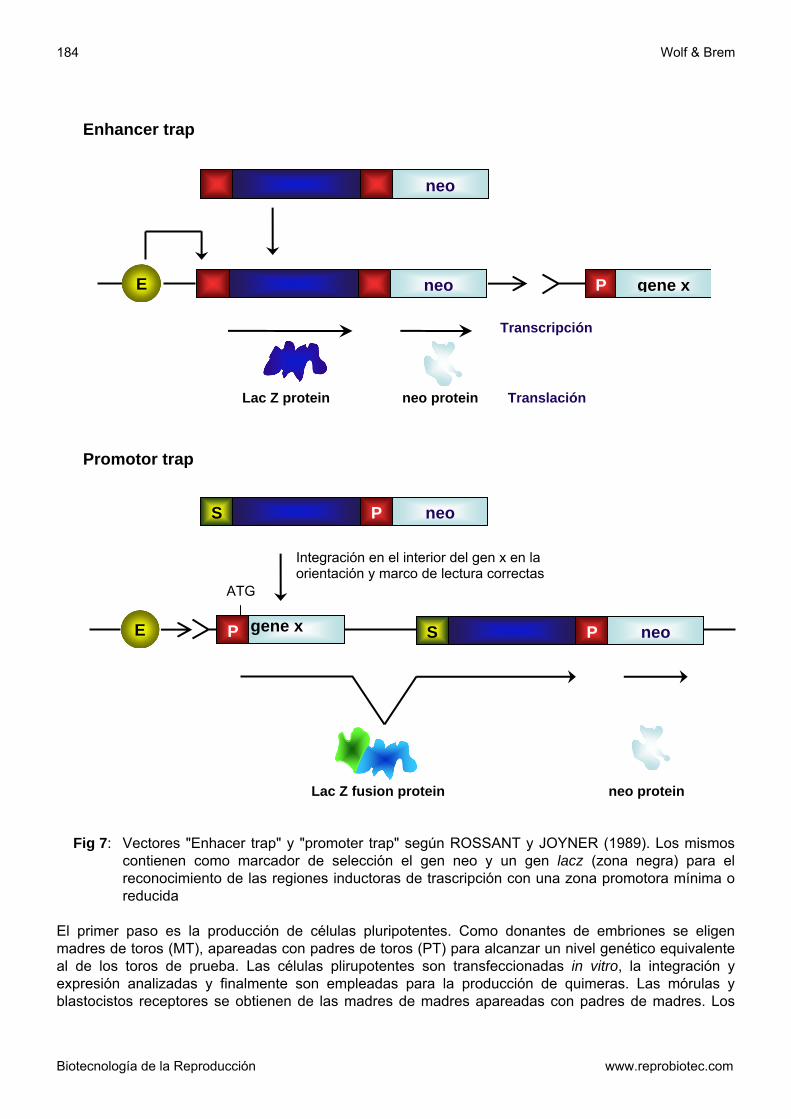
condiciones de recombinarse en forma homóloga eficientemente Un vector SPN contiene además del gen neo® como secuencia selectiva positiva dentro de la zona homóloga, un marcador contra el cual se puede seleccionar en forma negativa. Un ejemplo de ello es el gen de la Timidinquinasa (VHS-tq) del virus Herpes simplex que está ubicado distal a la región homóloga (fig. 6). Si se lleva a cabo la recombinación se pierde la zona que incluye el gen VHS-tq. En el caso de una integración al azar permanece esa región y las células que expresan el gen VHS-tq mueren mediante selección con Ganciclovir. Con ese método se pudo aumentar los clones recombinantes por el factor 10 hasta 12500 (MASOUR, 1990). Clones positivos se identifican con ayuda de la sonda de ADN (polimerase-chain reaction, PCR) como así también por medio del análisis Southern-Blot. Luego se reproducen para ser inyectados a blastocistos y producir quimeras transgénicas. Si las células embrionarias totipotentes en esos animales colonizan también la hilera germinal se produce descendencia heterocigota, después del apareamiento con animales no transgénicos. Esta puede ser empleada para producir animales y líneas transgénicas homocigotas. Cambios fenotípicos en portadores homocigotas de cambios genéticos inducidos permiten hacer deducciones sobre la función del gen en estudio y sus productos. Con ese método fueron inactivados y caracteriza-dos interesantes genes de importancia biológica y evolutiva en el ratón (tabla 3). Células ES fueron empleadas también para buscar importantes genes o secuencias regulatorias de ADN, hasta el momento desconocidos, para la diferenciación durante la ontogénesis (GOSSER y col., 1989; FIEDRICH y SORIANO, 1991). Con ese objetivo se transfeccionaron células ES con construcciones (estructuras) génicas, las cuales junto a un marcador selectivo positivo (en el mayor de los casos neo®) contienen el gen de la -galactosidasa bacteriana (lacZ) con una zona promotora mínima (enhacer trap) o bien sin promotor (promotor trap, fig 13). Los clones que integran esa estructura génica se emplean en la producción de quimeras. La expresión del gen lac Z puede hacerse visible por medio del cronogen X-gal, cuyo fraccionamiento provoca una coloración azul. De esta forma el gen puede ser determinado en diferentes estadios embrionarios. En el caso que se establezca un modelo de expresión interesante, es posible clonar las zonas responsables a partir del gen lac Z. En las estructuras del promotor Trap se produce la expresión del gen lac Z sólo si éstas se integran en la zona
Wolf & Brem
Biotecnología de la Reproducción www.reprobiotec.com
182
de influencia del promotor endógeno y con ello interrumpen el gen endógeno correspondiente. Si se logran producir quimeras de la hilera germinal con esa línea puede caracterizarse, entonces, el gen endógeno interrumpido a través de la reproducción y evaluación del fenotipo de los portadores hetero- y homocigotas, no sólo estructural sino funcionalmente.
Fig 6: Vector para una selección positiva-negativa (según
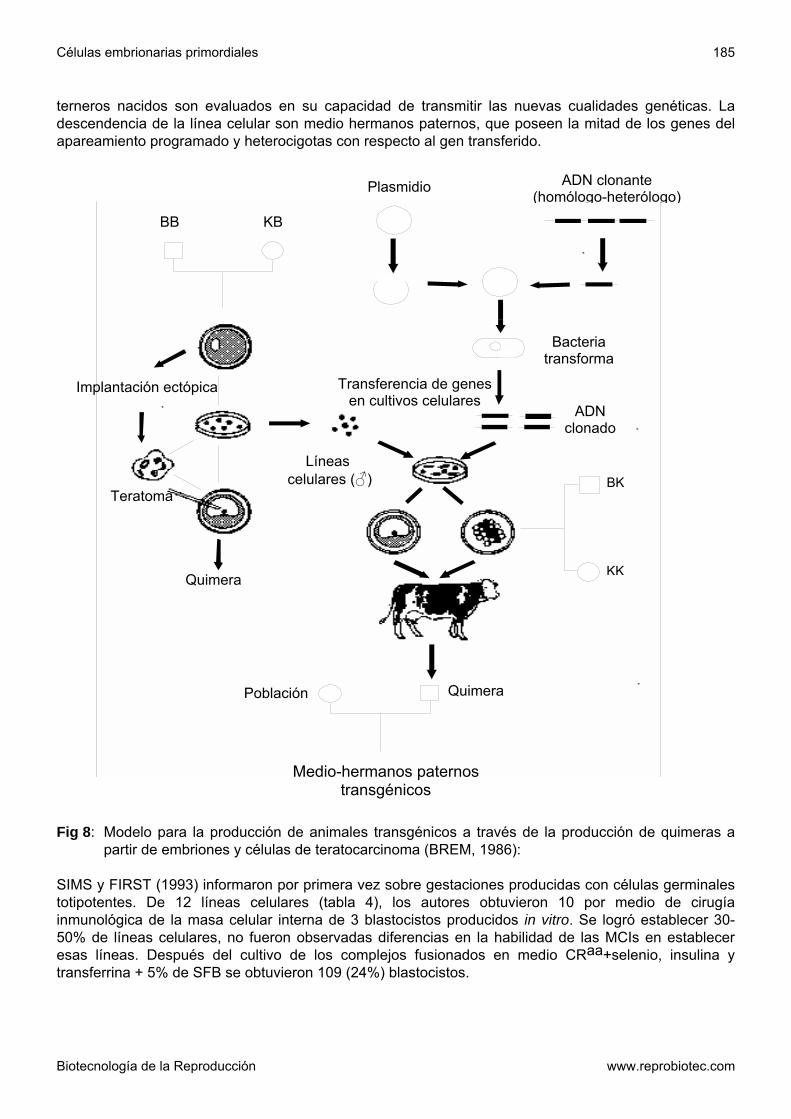
CAPECCHI, 1989) Las aplicaciones descriptas de las células embrionarias totipotentes pueden llevarse a cabo en principio en las especies animales de interés zootécnico, si se logran establecer líneas celulares funcionales. La fig 8 describe un modelo de producción de bovinos transgénicos aplicando líneas celulares pluripotentes (BREM, 1986). Además surge la interesante posibilidad de evaluar la expresión de transgenes integrados al azar en las células ES o en sus derivados diferenciados para emplear sólo clones adecuados para producir quimeras transgénicas. Este es el punto decisivo considerando la baja eficiencia de la producción de individuos transgénicos a través de la microinyección de ADN en el pronúcleo de los animales de interés productivo frente a los ratones (BREM, 1988). Contrariamente a la obtención de embriones uni- o bicelulares, los embriones requeridos para obtener células totipotentes para la transferencia génica pueden ser obtenidos en la especie bovina en forma no quirúrgica. EVANS y col. (1990) además someten a discusión a las células ES frente a la posibilidad de llevar a cabo intervenciones complejas en el genoma con el fin de variar características controladas por varios genes. Tabla 3: Ejemplos de la producción de ratones con cambios genéticos establecidos con el empleo de
células ES como vectores para la transferencia génica
Gene targeting
HSV-tk
Pint-2-N/TK
Integración al azar
Int - 2+ neor HSV - tk+ (G4 1 8r, GANCS)
neor
Int-2- neor HSV - tk- (G4 1 8r, GANCr )
int-2+
HSV-tk
HSV-tk
neor
neor
neor

Células embrionarias primordiales
Biotecnología de la Reproducción www.reprobiotec.com
183
Gen Linea
celular ES Donante de blastocistos
Quimera de la hilera germinal
Literatura
Hox-1.1
D3
C57Bl/6
ZIMMER & GRUSS, 1989
Hox-1.5 CC1.2 C57Bl/6 3 CHISAKA & CAPECCHI, 1991
Hox-1.6 D3 C57Bl/6 3 LUFKIN Y col., 1991
En-2 D3 CD1 - JOYNER Y col., 1989
C57Bl/6 -
ß2-micro- globulina
D3 C57Bl/6 6 ZIJLSTRA y col., 1989/90
N-myc D3 C57Bl/6 4 STANTON y col., 1990
HPRT E14TG2a C57Bl/6JLac x CBA/CaLac
19 HOOPER y col., 1987
HPRT CCE MF1 11 KUEHN y col., 1987
HPRT- E14TG2a C57Bl/6/Ola x CBA/Ca/Ola
1 THOMPSON y col., 1989
HPRT- ES98-12 C57Bl/6 2 KOLLER y col., 1989
Wnt-1 (int-1)
AB-1 C57Bl/6 19 McMAHON & BRADLEY, 1990
IGF II CCE.33 CD-1 - DeCHIARA y col., 1990/91
MF1 3
C57Bl/6 4
MHC Class II
D3 C57Bl/6 si COSGROVE y col., 1991
c-abl CCE CD-1 0 SCHWARTZBERG y col., 1989/91
MF1 0
C57Bl/6 6
c-abl CCE C57Bl/6 1 TYBULEWICZ y col., 1991
IL-2 E14 C57Bl/6 3 SCHORLE y col., 1991
c-src AB2.1 C57Bl/6 14 SORIANO y col., 1991
Wolf & Brem
Biotecnología de la Reproducción www.reprobiotec.com
184
��
Fig 7: Vectores "Enhacer trap" y "promoter trap" según ROSSANT y JOYNER (1989). Los mismos
contienen como marcador de selección el gen neo y un gen lacz (zona negra) para el reconocimiento de las regiones inductoras de trascripción con una zona promotora mínima o reducida
El primer paso es la producción de células pluripotentes. Como donantes de embriones se eligen madres de toros (MT), apareadas con padres de toros (PT) para alcanzar un nivel genético equivalente al de los toros de prueba. Las células plirupotentes son transfeccionadas in vitro, la integración y expresión analizadas y finalmente son empleadas para la producción de quimeras. Las mórulas y blastocistos receptores se obtienen de las madres de madres apareadas con padres de madres. Los
neo
P gene x
neoP
Integración en el interior del gen x en la orientación y marco de lectura correctas
S
neoE
Transcripción
Enhancer trap
Promotor trap
Lac Z fusion protein neo protein
Lac Z protein neo protein Translación
neoP SE P gene x
ATG
Células embrionarias primordiales
Biotecnología de la Reproducción www.reprobiotec.com
185
terneros nacidos son evaluados en su capacidad de transmitir las nuevas cualidades genéticas. La descendencia de la línea celular son medio hermanos paternos, que poseen la mitad de los genes del apareamiento programado y heterocigotas con respecto al gen transferido.
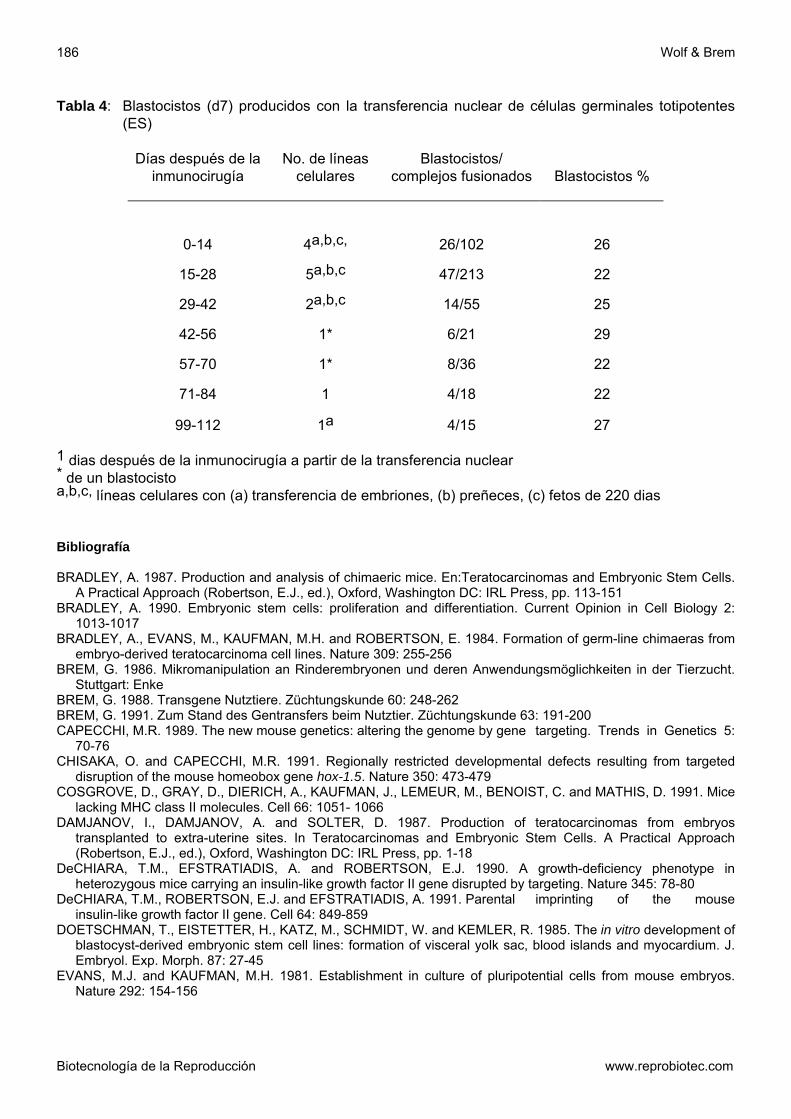
Fig 8: Modelo para la producción de animales transgénicos a través de la producción de quimeras a
partir de embriones y células de teratocarcinoma (BREM, 1986): SIMS y FIRST (1993) informaron por primera vez sobre gestaciones producidas con células germinales totipotentes. De 12 líneas celulares (tabla 4), los autores obtuvieron 10 por medio de cirugía inmunológica de la masa celular interna de 3 blastocistos producidos in vitro. Se logró establecer 30-50% de líneas celulares, no fueron observadas diferencias en la habilidad de las MCIs en establecer esas líneas. Después del cultivo de los complejos fusionados en medio CRaa+selenio, insulina y transferrina + 5% de SFB se obtuvieron 109 (24%) blastocistos.
BK
KK
ADN clonado
Bacteria transforma
Quimera
Teratoma
Plasmidio
BB KB
Transferencia de genes en cultivos celulares
Quimera
Medio-hermanos paternos transgénicos
ADN clonante (homólogo-heterólogo)
Implantación ectópica
Líneas celulares (♂)
Población
Wolf & Brem
Biotecnología de la Reproducción www.reprobiotec.com
186
Tabla 4: Blastocistos (d7) producidos con la transferencia nuclear de células germinales totipotentes (ES)
Días después de la
inmunocirugía No. de líneas
celulares Blastocistos/
complejos fusionados
Blastocistos %
0-14
4a,b,c,
26/102
26
15-28 5a,b,c 47/213 22
29-42 2a,b,c 14/55 25
42-56 1* 6/21 29
57-70 1* 8/36 22
71-84 1 4/18 22
99-112 1a 4/15 27 1 dias después de la inmunocirugía a partir de la transferencia nuclear * de un blastocisto a,b,c, líneas celulares con (a) transferencia de embriones, (b) preñeces, (c) fetos de 220 dias Bibliografía BRADLEY, A. 1987. Production and analysis of chimaeric mice. En:Teratocarcinomas and Embryonic Stem Cells.
A Practical Approach (Robertson, E.J., ed.), Oxford, Washington DC: IRL Press, pp. 113-151 BRADLEY, A. 1990. Embryonic stem cells: proliferation and differentiation. Current Opinion in Cell Biology 2:
1013-1017 BRADLEY, A., EVANS, M., KAUFMAN, M.H. and ROBERTSON, E. 1984. Formation of germ-line chimaeras from
embryo-derived teratocarcinoma cell lines. Nature 309: 255-256 BREM, G. 1986. Mikromanipulation an Rinderembryonen und deren Anwendungsmöglichkeiten in der Tierzucht.
Stuttgart: Enke BREM, G. 1988. Transgene Nutztiere. Züchtungskunde 60: 248-262 BREM, G. 1991. Zum Stand des Gentransfers beim Nutztier. Züchtungskunde 63: 191-200 CAPECCHI, M.R. 1989. The new mouse genetics: altering the genome by gene targeting. Trends in Genetics 5:
70-76 CHISAKA, O. and CAPECCHI, M.R. 1991. Regionally restricted developmental defects resulting from targeted
disruption of the mouse homeobox gene hox-1.5. Nature 350: 473-479 COSGROVE, D., GRAY, D., DIERICH, A., KAUFMAN, J., LEMEUR, M., BENOIST, C. and MATHIS, D. 1991. Mice
lacking MHC class II molecules. Cell 66: 1051- 1066 DAMJANOV, I., DAMJANOV, A. and SOLTER, D. 1987. Production of teratocarcinomas from embryos
transplanted to extra-uterine sites. In Teratocarcinomas and Embryonic Stem Cells. A Practical Approach (Robertson, E.J., ed.), Oxford, Washington DC: IRL Press, pp. 1-18
DeCHIARA, T.M., EFSTRATIADIS, A. and ROBERTSON, E.J. 1990. A growth-deficiency phenotype in heterozygous mice carrying an insulin-like growth factor II gene disrupted by targeting. Nature 345: 78-80
DeCHIARA, T.M., ROBERTSON, E.J. and EFSTRATIADIS, A. 1991. Parental imprinting of the mouse insulin-like growth factor II gene. Cell 64: 849-859
DOETSCHMAN, T., EISTETTER, H., KATZ, M., SCHMIDT, W. and KEMLER, R. 1985. The in vitro development of blastocyst-derived embryonic stem cell lines: formation of visceral yolk sac, blood islands and myocardium. J. Embryol. Exp. Morph. 87: 27-45
EVANS, M.J. and KAUFMAN, M.H. 1981. Establishment in culture of pluripotential cells from mouse embryos. Nature 292: 154-156

Células embrionarias primordiales
Biotecnología de la Reproducción www.reprobiotec.com
187
EVANS, M.J., NOTARIANNI, E., LAURIE, S. and MOOR, R.M. 1990. Derivation and preliminary characterization of pluripotent cell lines from porcine and bovine blastocysts. Theriogenology 33: 125-128
FRIEDRICH, G. & SORIANO, P. 1991. Promotor traps in embryonic stem cells: a genetic screen to identify and mutate developmental genes in mice. Genes Devel. 5: 1513-1523
FROHMAN, M.A. and MARTIN, G.R. 1989. Cut, paste, and save: new approaches to altering specific genes in mice. Cell 56: 145-147
GEARING, D.P., NICOLA, N.A., METCALF, D., FOOTE, S., WILLSON, T.A., GOUGH, N.M. WILLIAMS, R.L. 1989. Production of leukemia inhibitory factor in Escherichia coli by a novel procedure andits use in maintaining embryonic stem cells in culture. Biotechnology 7: 1157-1161
GOSSLER, A., JOYNER, A.L., ROSSANT, J. and SKARNES, W.C. 1989. Mouse embryonic stem cells and reporter constructs to detect developmentally regulated genes. Science 244: 463-465
HANDYSIDE, A., HOOPER, M.L., KAUFMAN, M.H. and WILMUT, I. 1987. Towards the isolation of embryonal stem cell lines from the sheep. Roux's Arch. Dev. Biol. 196: 185-190
HANDYSIDE, A.H., O'NEILL, G.T., JONES, M. and HOOPER, M.L. 1989. Use of BRL-conditioned medium in combination with feeder layers to isolate a diploid embryonal stem cell line. Roux's Arch. Dev. Biol. 198: 48-55
HASSAN-HAUSER, C., SCHELLANDER, K., KORB, H., KNAUS, E., SCHLEGER, W. und MAYR, B. 1990. Langzeitkultivierung von Rinderembryonen im Hinblick auf die Erstellung totipotenter embryonaler Stammzellen. Reproduction in Domestic Animals 25: 22-32
HOOPER, M., HARDY, K., HANDYSIDE, A., HUNTER, S. and MONK, M. 1987. HPRT-deficient (Lesch-Nyhan) mouse embryos derived from germline colonization by cultured cells. Nature 326: 292-295
JOYNER, A.L., SKARNES, W.C. and ROSSANT, J. 1989. Production of a mutation in mouse En-2 gene by homologous recombination in embryonic stem cells. Nature 338: 153-156
KOLLER, B.H., HAGEMANN, L.J., DOETSCHMAN, T., HAGAMAN, J.R., HUANG, S., WILLIAMS, P.J., FIRST, N.L., MAEDA, N. and SMITHIES, O. 1989. Germ-line transmission of a planned alteration made in a hypoxanthine phosphoribosyltransferase gene by homologous recombination in embryonic stem cells. Proc. Natl. Acad. Sci. USA 86: 8927-8931
KUEHN, M.R., BRADLEY, A., ROBERTSON, E.J. and EVANS, M.J. 1987. A potential animal model for Lesch-Nyhan syndrome through introduction of HPRT mutations into mice. Nature 326: 295-298
LEDERMANN, B. and BÜRKI, K. 1991. Establishment of a germ-line competent C57BL/6 embryonic stem cell line. Exp. Cell Res. 197: 254-358.
LUFKIN, T., DIERICH, A., LEMEUR, M., MARK, M. and CHAMBON, P. 1991. Disruption of the Hox-1.6 homeobox gene results in defects in a region corresponding to its rostral domain of expression. Cell 66: 1105-1119
MANSOUR, S.L. 1990. Gene targeting in murine embryonic stem cells: introduction of specific alterations into the mammalian genome. GATA 7: 219-227
MANSOUR, S.L., THOMAS, K.R. and CAPECCHI, M.R. 1988. Disruption of the proto-oncogene int-2 in mouse embryo-derived stem cells: a general strategy for targeting mutations to non-selectable genes.Nature 336: 348-352
MARTIN, G.R. 1981. Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells. Proc. Natl. Acad. Sci. USA 78: 7634-7638
MARTIN, G.R., SILVER, L.M., FOX, H.S. and JOYNER, A.L. 1987. Establishment of embryonic stem cell lines from preimplantation mouse embryos homozygous for lethal mutations in the t-complex. Dev. Biol. 121: 20-28
McMAHON, A.P. and BRADLEY, A. 1990. The Wnt-1 (int-1) proto-oncogene is required for development of a large region of the mouse brain. Cell 62: 1073-1085
MEINECKE-TILLMANN, S. and MEINECKE, B. 1991. Experiments on the establishment in culture of pluripotential cell lines from sheep, goat and pig embryos. Proceedings of the 7th Scientific Meeting of the A. E. T. E., Cam-bridge 14.-15.09.1991, p. 178
MELTON, D.W. 1990. The use of gene targeting to develop animal models for human genetic diseases. Biochemical Society Transactions 18: 1035-1039
NAGY, A., GOCZA, E., DIAZ, E.M., PRIDEAUX, V.R., IVANYI, E., MARKKULA, M. and ROSSANT, J. 1990. Embryonic stem cells alone are able to support fetal development in the mouse. Development 110: 815-821
NICHOLS, J., EVANS, E.P. and SMITH, A.G. 1990. Establishment of germ-line- competent embryonic stem (ES) cells using differentiation inhibiting activity. Development 110: 1341-1348
NOTARIANNI, E., GALLI, C., LAURIE, S., MOOR, R.M. and EVANS, M.J. 1990. Derivation of pluripotent embryonic stem cell lines from porcine and ovine blastocysts. Proceedings of the 4th World Congress on Genetics applied to Livestock Production, pp. 58-64
NOTARIANNI, E., LAURIE, S., MOOR, R.M. and EVANS, M.J. 1991. Maintenance and differentiation in culture of pluripotential embryonic cell lines from pig blastocysts. J. Reprod. Fert., Suppl. 41: 51-56
PIEDRAHITA, J.A., ANDERSON, G.B., MARTIN, G.R., BONDURANT, R.H. and PASHEN, R.L. 1988. Isolation of embryonic stem cell-like colonies from porcine embryos. Theriogenology 29: 286

Wolf & Brem
Biotecnología de la Reproducción www.reprobiotec.com
188
PIEDRAHITA, J.A., ANDERSON, G.B. and BONDURANT, R.H. 1990a. Influence of feeder layer type on the efficiency of isolation of porcine embryo derived stem cell lines. Theriogenology 34: 865-877
PIEDRAHITA, J.A., ANDERSON, G.B. and BONDURANT, R.H. 1990b. On the isolation of embryonic stem cells: comparative behavior of murine, porcine and ovine embryos. Theriogenology 34: 879-901
REXROAD, C.E., Jr. 1990. Production of cell colonies from ovine blastocysts. Theriogenology 33: 305 ROBERTSON, E.J. 1987. Embryo-derived stem cell lines. In Teratocarcinomas and embryonic stem cells. A
Practical Approach (Robertson, E.J., ed.), Oxford, Washington DC: IRL Press, pp. 71-112 ROSSANT, J. and JOYNER, A.L. 1989. Towards a molecular-genetic analysis of mammalian development. Trends
in Genetics 5: 277-283 SAITO, S., STRELCHENKO, N. and NIEMANN, H. 1992. Bovine embryonic stem cell-like cell lines cultured over
several passages. Roux's Arch. Dev. Biol. 201: 134-141. SCHELLANDER, K., HASSAN-HAUSER, C., FÜHRER, F., KORB, H. and SCHLEGER, W. 1989. Culture of bovine
embryos for stem cell production. Theriogenology 31: 254 SCHORLE, H., HOLTSCHKE, T., HÜNIG, T., SCHIMPL, A. and HORAK, I. 1991. Development and function of T
cells in mice rendered interleukin-2 deficient by gene targeting. Nature 352: 621-624 SCHWARTZBERG, P.L., GOFF, S.P. and ROBERTSON, E.J. 1989. Germ-line transmission of a c-abl mutation
produced by targeted gene disruption in ES cells. Science 246: 799-803 SCHWARTZBERG, P.L., STALL, A.M., HARDIN, J.D., BOWDISH, K.S., HUMARAN, T., BOAST, S., HARBISON,
M.L., ROBERTSON, E.J. and GOFF, S.P. 1991. Mice homozygous for the ablm1 mutation show poor viability and depletion of selected B and T cell populations. Cell 65: 1165-1175
SIMS, M.M and N.L. FIRST. 1993. Production of fetuses from totipotent cultured bovine inner cell mass cells. Theriogenology, 39: 313.
SMITH, A.G. 1991. Culture and differentiation of embryonic stem cells. J. Tiss. Cult. Meth. 13: 89-94 SMITH, A.G., HEATH, J.K., DONALDSON, D.D., WONG, G.G., MOREAU, J., STAHL, M. & ROGERS, D. 1988.
Inhibition of pluripotential embryonic stem cell differentiation by purified polypeptides. Nature 336: 688-690 SMITH, A.G. and HOOPER, M.L. 1987. Buffalo rat liver cells produce a diffusible activity which inhibits the
differentiation of murine embryonal carcinoma and embryonic stem cells. Dev. Biol. 121: 1-9 SORIANO, P., MONTGOMERY, C., GESKE, R. and BRADLEY, A. 1991. Targeted disruption of the c-src
proto-oncogene leads to osteopetrosis in mice. Cell 64: 693-702 STANTON, B.R., REID, S.W. and PARADA, L.F. 1990. Germ line transmission of an inactive N-myc allele
generated by homologous recombination in mouse embryonic stem cells. Mol. Cell. Biol. 10: 6755-6758 STRELCHENKO, N., SAITO, S. and NIEMANN, H. 1991. Towards the establishment of bovine embryonic stem
cells. Theriogenology 35: 274 STRINGFELLOW, D.A., GRAY, B.W., TOIVIO-KINNUCAN, M., GALIK, P., RIDDELL, K.P., BROCK, K.V. &
KEMPPAINEN, R.J. 1991. A continuous cell line established from a preimplantation bovine embryo. Theriogenology 35: 275
STRINGFELLOW, D.A., RHODES, P.J., THOMSON, M.S., LAUERMAN, L.H. and BIRD, R.C. 1987. A cell line established from a preimplantation bovine embryo. Theriogenology 27: 279
STROJEK, R.M., REED, M.A., HOOVER, J.L. and WAGNER, T.E. 1990. A method for cultivating morphologically undifferentiated embryonic stem cells from porcine blastocysts. Theriogenology 33: 901-913
STROJEK-BAUNACK, R., BÜRKLE, K., BURICH, K., HENSE, S., REINTJES, C. und HAHN, J. 1991. Kokultivierung von Rinderblastozysten unter verschiedenen Bedingungen in vitro. Fertilität 7: 77-84
THOMAS, K.R. and CAPECCHI, M.R. 1987. Site-directed mutagenesis by gene targeting in mouse embryo-derived stem cells. Cell 51: 503-512
THOMPSON, S., CLARKE, A.R., POW, A.M., HOOPER, M.L. and MELTON, D.W. 1989. Germ line transmission and expression of a corrected HPRT gene produced by gene targeting in embryonic stem cells. Cell 56: 313-321
TYBULEWICZ, V.L.J., CRAWFORD, C.E., JACKSON, P.K., BRONSON, R.T. and MULLIGAN, R.C. 1991. Neonatal lethality and lymphopenia in mice with a homozygous disruption of the c-abl proto-oncogene. Cell 65: 1153-1163
WELLS, D.N., WILMUT, I., McWHIR, J. and HOOPER, M.L. 1991. Is an epigenetic modification required for the isolation of murine embryonic stem cells into culture? Proceedings of the 7th Scientific Meeting of the A. E. T. E., Cambridge 14.-15.09.1991, p. 218
WILLIAMS, R.L., HILTON, D.J., PEASE, S., WILLSON, T.A., STEWART, C.L., GEARING, D.P., WAGNER, E.F., METCALF, D., NICOLA, N.A. and GOUGH, N.M. 1988. Myeloid leukaemia inhibitory factor maintains the developmental potential of embryonic stem cells. Nature 336: 684-687
ZIJLSTRA, M., BIX, M., SIMISTER, N.E., LORING, J.M., RAULET, D.H. and JAENISCH, R. 1990. 2-microglobulin deficient mice lack CD4-8+ cytolytic T cells. Nature 344: 742-746

Células embrionarias primordiales
Biotecnología de la Reproducción www.reprobiotec.com
189
ZIJLSTRA, M., LI, E., SAJJADI, F., SUBRAMANI, S. and JAENISCH, R. 1989. Germ-line transmission of a disrupted 2-microglobulin gene produced by homologous recombination in embryonic stem cells. Nature 342: 435-438
ZIMMER, A. and GRUSS, P. 1989. Production of chimaeric mice containing embryonic stem (ES) cells carrying a homeobox Hox 1.1 allele mutated by homologous recombination. Nature 338: 150-153


















