
Cría masiva de Diatraea saccharalis y su agente de control ... · Las estrategias que el hombre ha...
74
Cría masiva de Diatraea saccharalis y su agente de control biológico Cotesia flavipes en condiciones de laboratorio Tesis para optar al grado de MAGÍSTER EN CIENCIAS AGRARIAS (Orientación Producción Sostenible) Tesista: Lic. María del Pilar Medina Pereyra Director: Dr. Mariano Ordano Directora Asociada: Mg. Ing. Agr. Analía Ruth Salvatore Comisión de Supervisión: Ing. Agr. Adriana Jaime Dra. Marta Yasem de Romero Facultad de Agronomía y Zootecnia Universidad Nacional de Tucumán 2013
Transcript of Cría masiva de Diatraea saccharalis y su agente de control ... · Las estrategias que el hombre ha...

Cría masiva de Diatraea saccharalis y su agente
de control biológico Cotesia flavipes en
condiciones de laboratorio
Tesis para optar al grado de MAGÍSTER EN CIENCIAS AGRARIAS
(Orientación Producción Sostenible)
Tesista: Lic. María del Pilar Medina Pereyra
Director: Dr. Mariano Ordano
Directora Asociada: Mg. Ing. Agr. Analía Ruth Salvatore
Comisión de Supervisión:
Ing. Agr. Adriana Jaime
Dra. Marta Yasem de Romero
Facultad de Agronomía y Zootecnia
Universidad Nacional de Tucumán
2013

A mis padres,
Lucrecia y Victor Hugo,
por inculcarme el valor
del estudio y la superación.

“Mi pasado, Señor,
lo confío a tu misericordia,
mi presente a tu amor,
mi futuro a tu providencia”.
San Pío de Pietrelcina.

AGRADECIMIENTOS
Al Dr. Mariano Ordano, no sólo por dirigirme brindándome generosamente su
tiempo y conocimientos, sino también por alentarme constantemente a superar
obstáculos y limitaciones para encontrar los frutos en el trabajo realizado.
A la Mg. Ing. Agr. Analía Salvatore por la colaboración en la dirección de tesis
sumando un valioso aporte de conocimientos, experiencia y calidez humana.
A la Dra. Marta Yasem y a la Ing. Agr. Adriana Jaime, miembros de la comisión
de supervisión, por las sugerencias y cuidadosa revisión del manuscrito.
A la Fundación Miguel Lillo, al Ingenio y Refinería San Martín del Tabacal y a
la Dra. Carmen Reguilón, directora del Proyecto que dio marco para el desarrollo de
esta tesis, así como también al plantel técnico del mismo.
A los Sres. Marcos Pérez Visñuk, Luis Morales, Leopoldo Risso y José Luis
Moyano por la colaboración activa en los experimentos realizados.
Al Lic. Hugo Pablo Pereyra de la Fundación Miguel Lillo por la ilustración de la
portada.
A mi esposo Victor por su contención y apoyo en los momentos necesarios.

INDICE GENERAL
RESUMEN 1
SUMMARY 3
1. INTRODUCCIÓN GENERAL
1.1. Introducción 4
1.2. Sistema de estudio 5
1.2.1. Diatraea saccharalis: características 5
1.2.2. Daños e importancia económica 6
1.2.3. Cotesia flavipes: características 8
1.2.4. Antecedentes como agente de control biológico 9
1.3. Hipótesis general 9
1.4. Objetivos generales 9
1.5. Objetivos específicos 10
2. CICLO DE CRÍA MASIVA EN LABORATORIO 11
3. EFECTO DE LA DIETA
3.1. Introducción 16
3.2. Materiales y métodos 17
3.2.1. Composición de las dietas 17
3.2.2. Procedimiento en el laboratorio 18
3.2.3. Análisis de datos 20
3.3. Resultados 21
3.4. Discusión 27
4. EFECTO DE LA DENSIDAD Y PROPORCIÓN SEXUAL
SOBRE LA FECUNDIDAD DE DIATRAEA SACCHARALIS
EN CONDICIONES DE LABORATORIO
4.1. Introducción 29
4.2. Materiales y métodos 30
4.2.1. Procedimiento 30
4.2.2. Análisis de datos 34

4.3. Resultados 34
4.4. Discusión 36
5. EFECTO DEL NÚMERO DE PARASITOIDIZACIONES
5.1. Introducción 37
5.2. Materiales y métodos 38
5.2.1. Procedimiento 38
5.2.2. Análisis de datos 39
5.3. Resultados 40
5.3.1. Efecto de parasitoidismo simple o doble 40
5.3.2. Efecto del número de vasos utilizados para realizar
dos parasitoidizaciones
44
5.4. Discusión 46
6. EFECTO DE LA EDAD LARVAL DEL HUÉSPED
6.1. Introducción 48
6.2. Materiales y métodos 49
6.2.1. Procedimiento 49
6.2.2. Análisis de datos 50
6.3. Resultados 50
6.4. Discusión 55
7. CONCLUSIÓN GENERAL 56
8. BIBLIOGRAFÍA 59
9. ANEXOS
9.1. Anexo 1: Dieta artificial. Composición 67
9.2. Anexo 2: Preparación de la dieta 68

1
RESUMEN
Una herramienta del control biológico de plagas es la cría masiva de parasitoides
o enemigos naturales para la liberación en el campo. En este sentido la cría masiva en
laboratorio, tanto de Diatraea saccharalis, una plaga de gran interés económico, como
de su parasitoide Cotesia flavipes, ha dado lugar a estudios destinados a su ajuste y
mejoramiento, desde principios del siglo XX.
El presente trabajo aborda problemáticas de la cría masiva en laboratorio
tomando cuatro puntos esenciales dentro de una amplia gama de factores que se pueden
considerar en el proceso.
Para estudiar el efecto de la dieta artificial sobre la producción se llevaron a cabo
ensayos considerando once combinaciones de harinas y antibióticos incluidos en una
dieta base. Los resultados mostraron que dietas con harina de poroto y una dosis doble y
única de ampicilina como antibiótico se relacionan con una mayor supervivencia pupal
de D. saccharalis y una mayor cantidad y eficiencia en la producción de cocones de C.
flavipes.
El efecto de la abundancia y proporción sexual de D. saccharalis sobre la
fecundidad fue estimado mediante el conteo de adultos en jaulas de oviposición, en
relación al número de huevos para el total de posturas encontradas. Se observó que la
proporción sexual (número de hembras / número total) por jaula se relacionó
negativamente con el número de huevos predicho, mientras que el número de mariposas
por jaula mostró una relación no lineal con el número de huevos.
Para conocer el efecto del número de parasitoidizaciones sobre la producción de
cocones se realizaron experimentos considerando el efecto de una o dos
parasitoidizaciones de C. flavipes en larvas de D. saccharalis, realizadas en forma
manual y controlada. Los resultados sugieren el uso de parasitoidismo simple (una sola
oviposición).
Por último se estudió el efecto de la edad larval de D. saccharalis a la cual se
produce la parasitoidización, tomando como referencia cinco edades en días: 17, 18, 19,
20 y 21. Los resultados obtenidos mostraron que el éxito en la producción de cocones
fue más alto en larvas de 18 y 19 días, lo mismo que para el número de adultos y
hembras emergidas de C. flavipes. La proporción sexual de C. flavipes (H:M) fue
decreciente desde larvas de 17 a 21 días. Considerando un óptimo en larvas de D.
saccharalis de 17 a 19 días se encontró que larvas más jóvenes promueven una

2
proporción sexual sesgada a hembras, mientras que las de mayor edad favorecen la
producción de cocones. La combinación de resultados de éxito en producción de
cocones y proporción sexual sugiere el uso de larvas con 17 días.

3
SUMMARY
A tool of biological control of pests is the mass rearing of parasitoids and natural
enemies for release in the field. In this sense the mass rearing in the laboratory, both of
Diatraea saccharalis, a pest of great economical interest, and its parasitoid Cotesia
flavipes, has led to studies aimed at the adjustment and improvement, from the early
twentieth century.
This paper take up problems of mass rearing in the laboratory taking four
essential points in a wide range of factors that may be considered in the process.
To study the effect of artificial diet on production, trials were carried out
considering eleven combinations of flour and antibiotics included in a basal diet. The
results showed that bean meal diets and a double and single dose of antibiotic ampicillin
was associated with increased pupal survival of D. saccharalis and a greater number
and efficiency in production of C. flavipes cocoons.
The effect of abundance and sex ratio of D. saccharalis on fertility was
estimated by counting adults in oviposition cages in relation to the number of eggs for
total egg-layings found. It was observed that sex ratio (number of females / total of
butterflies) per cage was negatively related to the number of eggs predicted, while the
number of butterflies per cage showed a nonlinear relationship with the number of eggs.
To know the effect of number parasitoidism on the cocoons production
experiments were performed considering the effect of one or two ovipositions from C.
flavipes on D. saccharalis larvae, made by controlled form. Results suggest use of
parasitism simple (a single oviposition).
Finally was studied the effect of D. saccharalis larval age to which parasitoidism
occurs, with reference to five ages in days: 17, 18, 19, 20 and 21. Results showed that
success in cocoons production was highest in larvae of 18 and 19 days, as well as to the
number of adults and emerged females of C. flavipes. The C. flavipes sex ratio (H: M)
showed a decrease from larvae of 17 to 21 days. Considering an optimal larval D.
saccharalis 17 to 19 days it was found that youngest larvae promote a female biased sex
ratio, whereas oldest ones favored cocoon production. The combination of successful
results in cocoons production and sex ratio data suggest the use of 17 days larvae.

4
1. INTRODUCCIÓN GENERAL
1.1. Introducción
Las estrategias que el hombre ha desarrollado a lo largo de la historia frente a las
plagas que afectan los cultivos para su producción y consumo se vieron matizadas por
distintos tipos de control. Pero fue la generalización del uso de insecticidas,
especialmente desde la segunda mitad del siglo XX y atravesando una fase de control
químico indiscriminado, lo que planteó una creciente tendencia mundial a la reducción
y uso racional de los agroquímicos, como medida para disminuir los efectos nocivos
que pueden provocar tanto en el proceso productivo como en el medio ambiente y la
salud humana (Romero, 2004). De este modo, atravesando sucesivas etapas de control
químico, se fue gestando una nueva manera de concebir la fitoprotección,
desarrollándose el concepto de “Manejo Integrado de Plagas” (MIP). Este concepto fue
definido por la FAO en 1967 como “un sistema de manejo de plagas que, en el contexto
del medio ambiente asociado y las dinámicas de poblaciones de las especies plaga,
utiliza todas las técnicas y métodos adecuados de la manera más compatible posible y
mantiene las poblaciones plaga a niveles por debajo de aquellos que causan un daño
económico” (Dhaliwal et al., 2004). La Comunidad Económica Europea propone su
definición como “la aplicación racional de medidas biológicas, biotecnológicas,
químicas, de cultivo o de selección de vegetales que hacen que el uso de fitosanitarios
químicos se limite al mínimo necesario para mantener la población de la plaga en
niveles inferiores a los que producirían daños o pérdidas económicas inaceptables”
(Pérez Moreno, 2000).
Por su parte el control biológico es una práctica comprendida en el MIP y es
definida por la Organización Internacional de Lucha Biológica (OILB) como "la
utilización de organismos vivos, o de sus productos, para evitar o reducir las pérdidas o
daños causados por los organismos nocivos" (Guédez et al., 2008).
Se pueden distinguir tres tipos: el control biológico clásico comprende
frecuentemente la importación y colonización de parasitoides o predadores. Las
liberaciones se realizan con un pequeño número de insectos (inoculativas) como una
medida de control a largo plazo. Un segundo tipo es el control biológico natural referido
a la conservación de poblaciones de enemigos que ocurren naturalmente por medio de la
manipulación favorable de su ambiente cultural. El tercer tipo es el control biológico

5
aplicado, el cual se realiza por medio de liberaciones inundativas o aumentativas (Parra
et al., 2002). Es así que las liberaciones de agentes de control biológico (ACB) que se
realizan de esta manera en el campo requieren de una provisión abundante y constante
de estos organismos que en la mayoría de los casos se hace posible gracias a las crías
masivas de insectos en laboratorio. Este sistema ha sido adoptado en distintos países en
el contexto del conjunto de prácticas acordes a principios de desarrollo sustentable.
Enfocado en un caso particular de cría masiva de huésped-parasitoide, que ha
dado origen a distintos métodos de trabajo en diferentes países, en esta tesis se propone
investigar qué factores afectan el desempeño y la eficiencia de producción de Cotesia
flavipes Cameron (1891) a partir de su huésped Diatraea saccharalis Fabricius (1794)
en condiciones de laboratorio. Dado que la estabilidad de un sistema de control
biológico aumentativo basado en la cría masiva del ACB requiere que sea lo menos
costoso posible, es importante llevar a cabo un control de calidad y mejoramiento de la
eficiencia del proceso de cría masiva. Desde un enfoque biológico, este estudio
permitiría hacer más eficiente y rentable el proceso de producción del ACB C. flavipes,
de importancia para su uso como herramienta en el control biológico del gusano
barrenador de la caña de azúcar.
1.2. Sistema de estudio
1.2.1. Diatraea saccharalis: características
Diatraea saccharalis es un insecto lepidóptero que pertenece a la familia
Crambidae, se lo conoce comúnmente como el gusano perforador o barrenador de la
caña de azúcar, aunque éste no es el único cultivo que ataca. También causa daños en
cultivos de maíz, arroz, avena, sorgo y trigo. Es la especie más ampliamente distribuida
en América, prefiriendo zonas cálidas y húmedas, extendiéndose desde el sur de los
EE.UU. hasta el centro de Argentina (faja maicera). De las especies citadas para
Argentina, dos afectan a la caña de azúcar: D. saccharalis y D. dyari Box (1930)
(Kalnay, 1984).
Estos insectos, por ser holometábolos, cuentan con cuatro estados de desarrollo
en su ciclo de vida: huevo, larva, pupa y adulto. Los huevos de forma aplanada y
ovalada miden en promedio 1.15 mm de largo y 0.05 mm de ancho. El color crema

6
inicial de los embriones se torna más oscuro con la madurez (pasando por un color
amarillo hasta marrón) por efecto de la visualización de las cápsulas cefálicas de los
embriones en el interior. Las larvas son de tipo eruciforme, con tres pares de patas
torácicas, cuatro pares de pseudopatas abdominales y un par anal. Recién emergidas del
huevo tienen color amarillento, al crecer alcanzan 20 a 33 mm de largo y poseen dos
series de puntitos con pelos en el dorso. Las pupas son de tipo obtecta, de color marrón
y 16 a 20 mm de largo. Como característica poseen dos protuberancias cortas en la
cabeza. Esta polilla presenta dimorfismo sexual, siendo las hembras más grandes y con
un abdomen más voluminoso que el de los machos. La expansión alar es de 18 a 28 mm
en machos y 27 a 39 mm en hembras. Las alas anteriores son más pigmentadas en los
machos y poseen dos líneas oblicuas más destacadas que en las hembras, las alas
posteriores son blanquecinas (Layward, 1942; Willink, 1982; Pinto et al., 2006).
La duración de cada uno de los estados como del ciclo completo varía
principalmente de acuerdo a las condiciones climáticas. La cópula es nocturna, las
hembras oviponen luego de la misma durante tres o cuatro días depositando en las hojas
de plantas hospederas aproximadamente 300 huevos en grupos de 2 a 50 y de manera
imbricada. En estudios hechos en laboratorio, Linares (1987) señala un valor promedio
de 538,60 huevos por hembra a 26,06 ˚C. La duración de esta fase disminuye de seis a
cuatro días, con el aumento de temperaturas entre 26 a 30˚C. En el campo, una vez que
las larvas emergidas alcanzan el segundo estadio, perforan el tallo y abren una galería
donde se alimentan y crecen pasando generalmente por 4 estadios más. La etapa larval
dura de 25 a 30 días aproximadamente hasta alcanzar el estado de pupa. Luego de un
período de 5 a 9 días emerge el adulto que vive de 3 a 8 días (Willink, 1982; Capinera,
2001; Santillana et al., 2003).
1.2.2. Daños e importancia económica
Esta especie representa una plaga de gran importancia en el cultivo de la caña de
azúcar, debido a su amplia distribución y a la incidencia negativa en la productividad
del mismo (González, 1984). En las regiones cañeras de Argentina y en la provincia de
Tucumán, esta plaga adquiere una particular importancia si se toma en cuenta la
trascendencia económica y social que tiene este cultivo ya que la provincia es la
principal productora argentina de caña de azúcar, con el 66% de la producción nacional
(Pérez et al., 2007).

7
La acción primaria del gusano perforador de la caña de azúcar es la formación de
galerías (Fig. 1), que trae como consecuencia una reducción del peso y número de tallos
por hectárea. Además las perforaciones actúan como puerta de entrada de hongos y
bacterias que son los responsables del deterioro de los tejidos que inciden
negativamente en la calidad, pureza y contenido de sacarosa del jugo de caña extraíble.
Figura 1. Daños causados por Diatraea saccharalis en caña de azúcar.
Cañas variedad LCP 85-384. Infestación natural forzada en campos experimentales de la Estación
Experimental Agroindustrial Obispo Colombres. Tucumán. Febrero de 2009.
Además existen otros daños que afectan la calidad vegetativa, como la aparición
de brotes laterales, atrofia de entrenudos y muerte de la yema apical, síntoma conocido
como “corazón muerto” (Risco, 1971; Willink et al., 1999).
Se han detectado focos de ocurrencia de esta plaga en nuestra provincia, los
cuales presentan un índice de infestación promedio de 5 a 8%, de acuerdo a datos
relevados durante las campañas 2003 al 2007 (Salvatore et al., 2008), llegando en
algunos casos a un 60%. En términos de producción azucarera la pérdida estimada es de
650 gramos de azúcar por tonelada de caña por punto porcentual de infestación
(Salvatore et al., 2009).
Aunque se conocen en otros países variedades de caña que presentan diferentes
grados de resistencia a D. saccharalis, por ejemplo las variedades CP70-321 y CP65-
357 en Louisiana, EE.UU. (Bessin et al., 1990), las variedades comerciales más
difundidas en Tucumán, tales como LCP 85-384; TUC 77-42; CP 65-357; RA 87-3 y L
75-33, presentan escasa o nula resistencia a la plaga y la susceptibilidad al ataque es
semejante entre ellas (Salvatore et al., 2008). A estos datos se puede agregar el hecho de

8
que el uso de productos químicos (insecticidas) sólo puede hacerse efectivo en los
primeros estadios larvales, es decir, antes de que la larva penetre los tallos (Schexnayder
et al., 2001), debido al hábito barrenador de esta larva que la protege de los controles
químicos.
1.2.3. Cotesia flavipes: características
Cotesia flavipes es un microhimenóptero, gregario, originario de la región Indo-
Australiana. Fue introducido en más de 40 países de los trópicos y subtrópicos en Asia,
África y América como parasitoide de larvas de barrenadores del género Diatraea,
Chilo y Sesamia (Polaszek y Walker, 1991).
Esta avispa pertenece a la subfamilia Microgastrinae dentro de una familia
considerada la segunda más numerosa del orden Hymenoptera, Braconidae. La gran
diversidad de esta familia incluye 15.000 especies a nivel mundial, de las cuales se
encuentran 318 aproximadamente en la Argentina. Todas ellas de hábito parasítico no
manifiestando preferencia por regiones tropicales o templadas o por ambientes húmedos
o áridos (Berta y Ovruski, 2008).
El cuerpo es de color negro y mide en promedio 4 mm de longitud. El largo de
las antenas filiformes, situadas por encima de una repisa en medio de los ojos
compuestos, permite diferenciar machos y hembras. Ambos poseen el mismo número de
segmentos antenales (18) siendo en las hembras visiblemente más cortos (Cueva et al.,
1980; Achterberg, 1990).
El ciclo de vida dura en promedio 20 días. Las hembras introducen sus huevos
en el hemocele de larvas preferentemente de tercero a sexto estadio, desarrollándose en
su interior, es decir, son endoparasitoides como las especies de la mayoría de las
subfamilias de Braconidae (81%) (CAÑAMIP, 2000). Además son koinobiontes porque
permiten el desarrollo del huésped por un tiempo luego de ser parasitoidizado. El
período de incubación es de tres a cuatro días, pudiéndose desarrollar más de un
parasitoide por huésped parasitoidizado, por una única hembra o por hembras de la
misma especie, condición conocida como parasitoidismo gregario (Anento y Selfa,
1997). La larva madura se forma en ocho a diez días y luego de pasar por 3 estadios
emerge a través de la cutícula del huésped para formar la pupa dentro de un capullo
(cocón) construido por la larva. Este período tiene una duración de dos a tres días, luego
de lo cual emergen los adultos que viven aproximadamente cinco días. C. flavipes tiene

9
mecanismo haplodiploide de determinación del sexo (o arrenotoquia), las hembras
ponen huevos fecundados (diploides) que dan lugar a hembras y huevos no fecundados
(haploides) que se desarrollan partenogenéticamente en machos (Berta y Ovruski,
2008).
1.2.4. Antecedentes como agente de control biológico
A nivel mundial la búsqueda de agentes de control biológico para su utilización
en D. saccharalis ha sido exhaustiva. Por los logros obtenidos, el microhimenóptero C.
flavipes, se ha considerado exitoso como controlador natural en relación a otros
parasitoides (Smith y Bellotti, 1996).
La primera introducción de C. flavipes en América ocurrió en Florida, en la
década del 60 (Gifford y Mann, 1967). Posteriormente, en otros países tales como
Trinidad y Tobago, Colombia, Costa Rica, México, Venezuela y Brasil fue introducida
y liberada en forma experimental en ensayos de control biológico aumentativo o
inundativo. Como ejemplo de la alta eficiencia que posee como controlador del gusano
perforador de la caña de azúcar, se pueden citar las liberaciones masivas iniciadas en
1977, en San Pablo, Brasil, que lograron disminuir el porcentaje de infestación de la
plaga de un 7% a un 2% (Botelho y Macedo, 2002).
1.3. HIPÓTESIS GENERAL
El incremento de la eficiencia de producción de los ACB en sistemas de cría
masiva en laboratorio depende del desempeño del huésped en condiciones particulares
de desarrollo y de las técnicas de manejo empleadas.
1.4. OBJETIVOS GENERALES
1- Evaluar la variación de parámetros biológicos en D. saccharalis y C. flavipes
en condiciones de laboratorio.
2- Determinar los factores que mejoran la producción de C. flavipes y
promueven un ciclo de producción más eficiente.

10
1.5. OBJETIVOS ESPECÍFICOS
1- Examinar el efecto del tipo de dieta artificial sobre la línea de cría de D.
saccharalis y la producción de cocones de C. flavipes.
2- Evaluar el desempeño de D. saccharalis en función de la proporción sexual
en jaulas de cría.
3- Examinar el efecto del número de parasitoidizaciones de C. flavipes en larvas
de D. saccharalis sobre la producción de cocones y la proporción sexual de parasitoides
emergidos.
4- Examinar el efecto de la edad larval a la cual son parasitoidizadas sobre la
producción de cocones y la proporción sexual de parasitoides emergidos.

11
2. CICLO DE CRÍA MASIVA EN LABORATORIO
Los experimentos se llevaron a cabo entre los años 2008 y 2009, en
dependencias del CIRPON (Centro de Investigaciones sobre Regulación de Poblaciones
de Organismos Nocivos), en donde se mantuvo un pie de cría de D. saccharalis y la
producción de cocones de C. flavipes, como parte del Proyecto “Control biológico de
Diatraea saccharalis con Cotesia flavipes en extensiones cañeras del NOA”, en
convenio entre la Fundación Miguel Lillo y el Ingenio y Refinería San Martín del
Tabacal (2007-2010). Dicho proyecto surgió con el objetivo de implementar el control
biológico del taladrador con un parasitoide. La cría utilizada en este estudio se inició a
partir de huevos de D. saccharalis y cocones de C. flavipes procedentes de la biofábrica
“Biocontrol” ubicada en Sertãozinho, São Paulo, Brasil.
Cotesia flavipes fue criado a partir de su huésped D. saccharalis (Fig. 2).
Figura 2. Ciclo de cría de Diatraea saccharalis y Cotesia flavipes en laboratorio.
Imágenes tomadas de la cría masiva mantenida en el CIRPON - Tucumán.
Huevos
Siembra en DIETA artificial
Parasitoidización de larvas de D. saccharalis con C. flavipes
Cocones de Cotesia flavipesCotesia flavipesCotesia flavipesCotesia flavipes
DiatraeaDiatraeaDiatraeaDiatraea saccharalissaccharalissaccharalissaccharalis
Adultos
Pupas
Larvas

12
La cría de D. sacharalis comenzó con la siembra de huevos, la cual se realizó
cortando con una tijera las posturas contenidas en las hojas de papel provenientes de los
“tubos jaula”. Las posturas se sumergieron en un baño de solución de sulfato cúprico
(7g/1000cm3 de agua) por tres minutos para su desinfección y luego fueron adheridas a
las paredes de los frascos de vidrio. Estos contenían una dieta artificial para alimentar
las futuras larvas (Fig. 3).
Figura 3. Siembra de huevos de D. saccharalis
Los frascos sembrados con aproximadamente 100 huevos cada uno fueron
llevados a cámara de incubación con una temperatura de 28 a 30ºC y humedad relativa
de 60 a 70% (Fig. 4).
Figura 4. Cámara de cría en el CIRPON Tucumán.
Cada cajón contiene frascos con larvas de D. saccharalis

13
Una parte de los lotes de siembra de huevos (se denomina lote al grupo de
frascos sembrados el mismo día), se utilizó para la parasitoidización con C. flavipes a
los 18-19 días.
La parte restante del lote se mantuvo en la cámara de cría hasta los 21 días y fue
destinada al mantenimiento del pie o línea de cría de D. saccharalis. Esto consistió en la
selección de las larvas de acuerdo a su tamaño y calidad, se colocaron seis en cada
cápsula de Petri conteniendo una dieta de mantenimiento. Estas cápsulas fueron
mantenidas en la cámara de cría hasta que las larvas alcanzaron el estado de pupa.
Luego las pupas se colocaron sobre bandejas de telgopor de 22 cm x 17 cm en el
interior de jaulones de estructura metálica de 60 cm de ancho x 90 cm de alto x 30 cm
de profundidad. Los jaulones fueron cubiertos por una malla plástica sujetada por velcro
y ubicados en una habitación con una temperatura de 25-26ºC y humedad relativa de
60-70% (Fig. 5).
Figura 5. Jaulones con pupas para emergencia de adultos de D. saccharalis.
Una vez emergidos los adultos fueron colectados de los jaulones y colocados en
tubos de PVC (“tubos jaula”) recubiertos en su interior por una hoja de papel en donde
las mariposas hembras colocaron sus huevos luego de la cópula (Fig. 6).

14
Figura 6. Jaulas individuales (“tubos jaula”) con adultos de D. saccharalis.
Las hojas se retiraron y reemplazaron diariamente por hojas nuevas para evitar la
superposición de posturas. Estas hojas se almacenaron en las mismas condiciones de
temperatura y humedad relativa hasta la maduración de los huevos.
La parasitoidización de larvas de D. saccharalis con C. flavipes se realizó de
forma manual. Esto consistió en la extracción de larvas de los frascos y su selección por
tamaño. Luego se tomó cada larva con los dedos o con ayuda de una pinza colocándola
de tal manera que la parte posterior quedara libre para facilitar que la avispa se posase e
introdujera sus huevos (Fig.7).
Figura 7. Vaso de adultos de C. flavipes utilizado para la parasitoidización manual
de larvas. Se observa la manipulación individual de larvas ofrecidas a los parasitoides.

15
Las larvas parasitoidizadas fueron colocadas en cápsulas de Petri con dieta
artificial y llevadas a cámara con una temperatura de 26-28ºC por 14 días, al cabo de los
cuales se extrajeron los cocones de C. flavipes. Una parte de estos se separó para el
armado de vasos que fueron utilizados en el ciclo siguiente. Estos vasos plásticos de 100
cm3 fueron preparados con 25 masas de cocones y almacenados a 26ºC por 48 horas
para permitir la emergencia y cópula de adultos de C. flavipes.

16
3. EFECTO DE LA DIETA
3.1. Introducción
Los requerimientos nutricionales de un organismo son parte fundamental para el
crecimiento, desarrollo y desempeño de las funciones vitales. Recrearlos en el
laboratorio empleando sustitutos a partir de la mezcla de distintos ingredientes ha
resultado en el desarrollo de dietas artificiales para la cría de insectos, adecuándolas
para obtener una cantidad necesaria y permanente de individuos con el objeto de
emplearlos en estudios biotecnológicos y bioecológicos (Luna et al., 2004). Los trabajos
llevados a cabo para comparar los efectos de distintos tipos de dietas sobre lepidópteros
incluyen observaciones sobre su ciclo de vida, tasas de supervivencia, fertilidad,
proporción de sexos, tamaño y peso de larvas y pupas (Acatitla Trejo et al., 2004;
Bavaresco et al., 2004; Meneguim et al., 1997; Murúa et al., 2003). Estas dietas, que
incluyen la variación de sus componentes, sean estos naturales o artificiales, han
demostrado tener influencia en los parámetros de interés del ciclo de vida (Lastra y
Gómez, 1988; Magrini et al., 1995; Boiça et al., 1997).
Los componentes comúnmente adicionados a los formulados comprenden
fuentes de nitrógeno (proteínas o aminoácidos), lípidos, carbohidratos y minerales.
También vitaminas B, como biotina y ácido fólico entre otras, que actúan como factores
de crecimiento y vitamina C (ácido ascórbico) como fagoestimulante y antioxidante.
Además se incorporan estabilizantes que modifican el pH (ácido acético, ácido
benzoico) y gelificantes como agar y caragenato que dan consistencia al alimento
(Cohen, 2004; Marín et al., 2006).
En condiciones de cría masiva, lo que se espera de una dieta artificial es que,
además de cubrir las necesidades nutricionales, sea capaz de atenuar los riesgos de
enfermedades y contaminación por patógenos, cuyos efectos podrían disminuir
drásticamente la población manejada en el laboratorio. En este sentido para prevenir la
contaminación microbiana se emplean antifúngicos (ácido sórbico, metil
parahidroxibenzoato) y antibióticos (Calderón, 1987; Estrada Hurtarte, 1992; Vargas et
al., 2001; Botelho y Macedo, 2002; Cano et al., 2006). De este modo, la optimización
de una dieta artificial contribuye al mantenimiento de la cría del huésped D. saccharalis
y a una producción eficiente del parasitoide C. flavipes.

17
Hipótesis Particular 1: Modificaciones en la dieta artificial por la variación de
sus componentes influyen en el desarrollo del huésped y en la producción del
parasitoide.
Objetivo Específico 1: Examinar el efecto del tipo de dieta de D. saccharalis
sobre la línea de cría de D. saccharalis y la producción de cocones de C. flavipes.
3.2. Materiales y métodos
3.2.1. Composición de las dietas
Para la formulación de las diferentes dietas artificiales se trabajó tomando como
base la dieta desarrollada por la Dra. Carmen Reguilón y colaboradores (comunicación
personal), cuyos componentes y preparación se describen en los Anexos 1 y 2
respectivamente del capítulo 9. Sobre esta composición general se introdujeron las
variantes. Se trabajó con dos tipos de harinas: de poroto blanco y de soja. En el caso de
la harina de poroto blanco se utilizaron dos tipos de molienda: fino y grueso, de acuerdo
a la disponibilidad de los proveedores comerciales.
La elección de los antibióticos se basó en la función bacteriostática de la
oxitetraciclina y la capacidad bactericida de la ampicilina y estreptomicina. Estos se
utilizaban en forma alternada entre períodos del año, de acuerdo a los niveles de
contaminación con patógenos observados durante la producción en el CIRPON.
Las diferentes formulaciones de las dietas artificiales fueron:
-Harina de poroto blanco molido grueso con:
oxitetraciclina y estreptomicina (A)
ampicilina y estreptomicina (B)
ampicilina y oxitetraciclina (C)
ampicilina, oxitetraciclina y estreptomicina (D)
ampicilina (E)
-Harina de soja texturizada con:
ampicilina y oxitetraciclina (F)
ampicilina, oxitetraciclina y estreptomicina (G)
18
-Harina de poroto grueso y fino con:
oxitetraciclina y estreptomicina (H)
ampicilina y estreptomicina (I)
-Poroto grueso y soja texturizada con:
ampicilina y oxitetraciclina (J)
ampicilina, oxitetraciclina y estreptomicina (K)
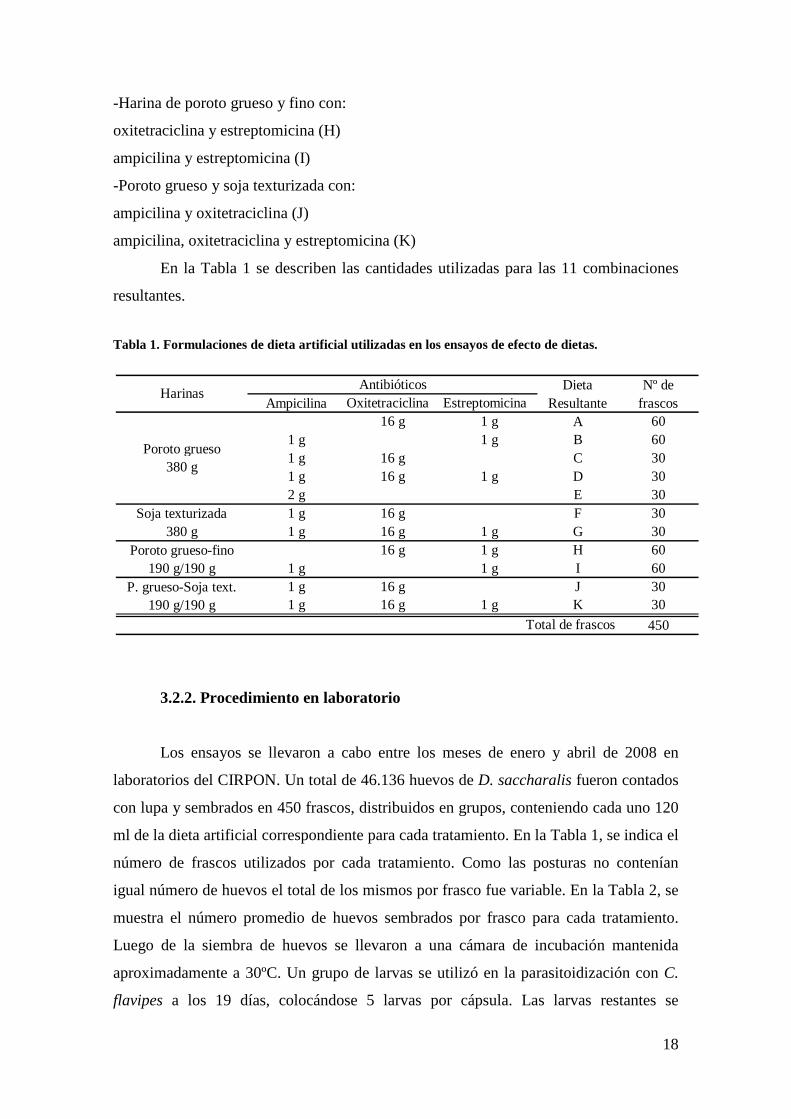
En la Tabla 1 se describen las cantidades utilizadas para las 11 combinaciones
resultantes.
Tabla 1. Formulaciones de dieta artificial utilizadas en los ensayos de efecto de dietas.
3.2.2. Procedimiento en laboratorio
Los ensayos se llevaron a cabo entre los meses de enero y abril de 2008 en
laboratorios del CIRPON. Un total de 46.136 huevos de D. saccharalis fueron contados
con lupa y sembrados en 450 frascos, distribuidos en grupos, conteniendo cada uno 120
ml de la dieta artificial correspondiente para cada tratamiento. En la Tabla 1, se indica el
número de frascos utilizados por cada tratamiento. Como las posturas no contenían
igual número de huevos el total de los mismos por frasco fue variable. En la Tabla 2, se
muestra el número promedio de huevos sembrados por frasco para cada tratamiento.
Luego de la siembra de huevos se llevaron a una cámara de incubación mantenida
aproximadamente a 30ºC. Un grupo de larvas se utilizó en la parasitoidización con C.
flavipes a los 19 días, colocándose 5 larvas por cápsula. Las larvas restantes se
Dieta Nº deAmpicilina Oxitetraciclina Estreptomicina Resultante frascos
16 g 1 g A 601 g 1 g B 601 g 16 g C 301 g 16 g 1 g D 302 g E 301 g 16 g F 301 g 16 g 1 g G 30
16 g 1 g H 601 g 1 g I 601 g 16 g J 301 g 16 g 1 g K 30
450
P. grueso-Soja text. 190 g/190 g
Total de frascos
HarinasAntibióticos
Poroto grueso 380 g
Soja texturizada 380 g
Poroto grueso-fino 190 g/190 g

19
utilizaron como pie de cría a los 21 días, en número de 6 larvas por cápsula. El
procedimiento general fue el descripto para el ciclo de cría en el capítulo 2. En todos los
casos las cápsulas de Petri con larvas fueron identificadas para su seguimiento según el
tipo de dieta y destino (Fig. 8).
Figura 8. Identificación de cápsulas con larvas de D. saccharalis para su seguimiento.
Las variables evaluadas fueron definidas de la siguiente manera:
Para el pie de cría de D. saccharalis, los potenciales efectos de la dieta fueron
evaluados sobre la supervivencia de los estados huevo a larva (supervivencia larval) y
supervivencia de larva a pupa (supervivencia pupal).
Supervivencia larval: fue medida como número de larvas de D. saccharalis
aptas para parasitoidización dividido por el número de huevos sembrados por frasco.
Una larva apta fue considerada por su tamaño mediano a grande (20-25 mm de longitud
aproximadamente) y de comportamiento vivaz.
Supervivencia pupal: fue medida como número de pupas de D. saccharalis
sobre número de larvas aptas. En este caso se consideraron las mismas características
que para la parasitoidización a excepción del tamaño mayor y la menor vivacidad,
características de larvas próximas a empupar.
Para larvas destinadas a producción de C. flavipes, dichos efectos fueron
evaluados sobre la eficiencia de producción de masas de cocones o eficiencia de
cocones, considerando una unidad como masa de cocón producida por larva
parasitoidizada. Dado que sería deseable una combinación que incremente la eficiencia
de ambas líneas, se evaluó el efecto de la dieta sobre un estimador de eficiencia
combinando la supervivencia larval y pupal de D. saccharalis con la producción de

20
cocones. Esto se hizo multiplicando la supervivencia huevo-larva por la supervivencia
larva-pupa o la eficiencia de cocones. Esta idea se basó en los componentes
multiplicativos de éxito reproductivo (o fitness) a partir del artículo de Arnold y Wade
(1984). Definida como eficiencia global, se espera que esta variable sea máxima
significando que el proceso de siembra en una determinada combinación de dieta, fue
eficiente tanto para la línea de D. saccharalis como para la de C. flavipes en conjunto.
Por otro lado, la eficiencia global fue dividida por el costo de la dieta correspondiente,
constituyendo un índice de rentabilidad relativa . El costo fue calculado con base en
los precios de los insumos durante el período de estudio.
3.2.3. Análisis de datos
De acuerdo a cada modelo estadístico, se realizaron pruebas de normalidad de
Shapiro-Wilk. Cuando fue necesario, las variables de respuesta fueron transformadas
para alcanzar el supuesto de normalidad. El número de huevos por siembra fue
transformado como la raíz cuadrada de X, la supervivencia larval, la supervivencia
pupal, y la eficiencia de producción de cocones fueron transformadas como el arcoseno
de la raíz cuadrada de X. La eficiencia global y la eficiencia por unidad de costo no
fueron transformadas.
Dado que el número total de huevos por siembra puede afectar la supervivencia
larval por efectos de densidad, este fue considerado una covariable y además se estaría
controlando por un potencial efecto de la densidad inicial de larvas eclosionadas,
asumiendo que la variación en la proporción de eclosión fuera despreciable. Respecto
de las variables supervivencia pupal y eficiencia de producción de cocones, el efecto de
densidad fue fijo, ya que en la cría masiva se trabajó con 6 y 5 larvas por cápsula
respectivamente.
Se aplicó un modelo lineal general siendo el factor principal la composición de
la dieta, el número de huevos sembrados la covariable y las variables de respuesta la
supervivencia huevo-larva, la supervivencia larva-pupa, la eficiencia de producción de
cocones, la eficiencia global y la rentabilidad. Si bien se presume que estas variables
están relacionadas entre sí, a los fines de esta tesis se optó por aplicar modelos
univariados. Siempre que fue posible se evaluaron interacciones entre factores. Para
determinar diferencias significativas entre tipos de dietas, se aplicaron pruebas a
posteriori siempre que el factor principal fuera significativo. Para simplificar la

21
comparación entre los once tratamientos, en los resultados se referirá a los tipos de
dietas con los valores promedio más altos o más bajos. Todos los análisis fueron
llevados a cabo en STATISTICA 7.0 (StatSoft, Inc.; 2004).
3.3. Resultados
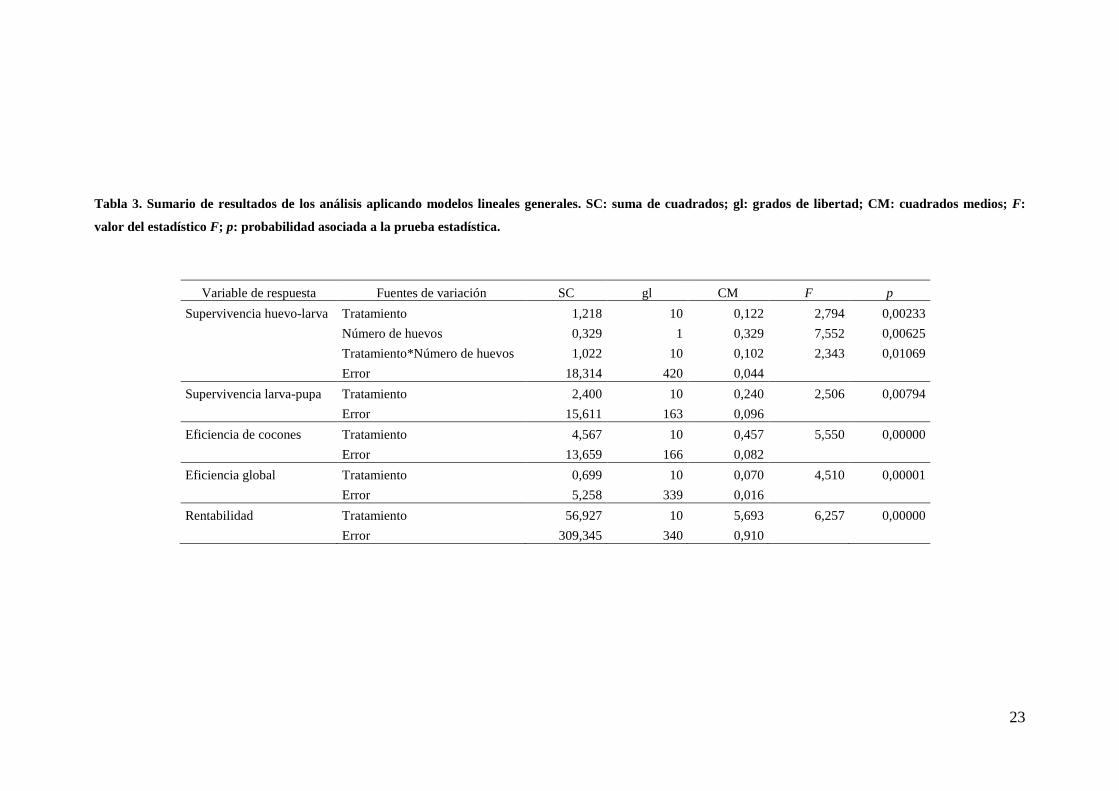
Un resumen de los valores promedio de cada una de las variables consideradas
por tratamiento se describe en la Tabla 2. La composición de la dieta afectó
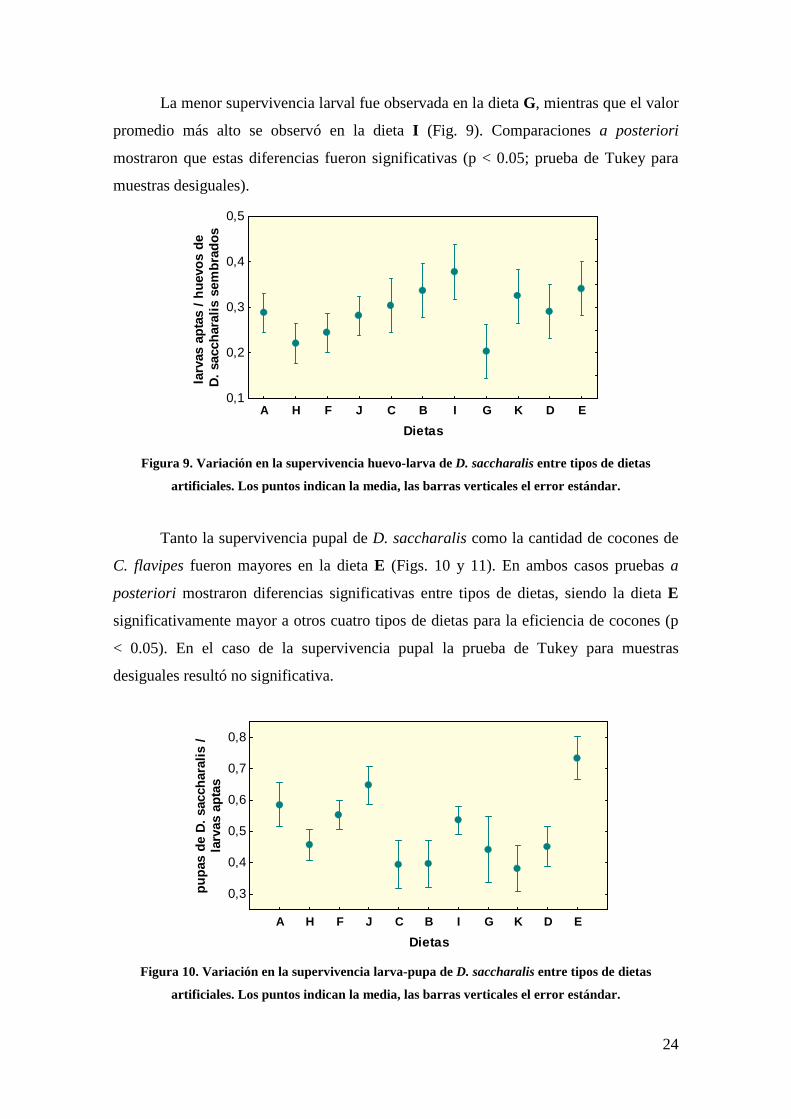
significativamente todas las variables de respuesta consideradas (Tabla 3). Esto es, al
menos un tipo de dieta difirió significativamente de las demás.
En el caso de la supervivencia larval, hubo un efecto significativo del número de
huevos sembrados y un efecto significativo de la interacción entre el número de huevos
sembrados y el tipo de dieta (Tabla 3). Esto indica que la covariación entre la
supervivencia larval y el número de huevos sembrados dependió del tipo de dieta.

22
Tabla 2. Tamaño de muestra (n), promedio (ŷ) y desvío estandar (σ) de cada una de las variables implicadas por tratamiento de dieta artificial.
Tratamientos A H F J C B I G K D E
Número de huevos n 60 58 59 58 30 30 30 30 30 30 30
ŷ 111,05 124,16 102,49 102,43 85,33 82,87 74,50 105,17 104,67 106,27 95,87
σ 27,97 52,79 23,00 30,49 28,48 24,34 23,61 15,86 15,97 14,76 15,49
Supervivencia huevo-larva n 58 57 59 58 30 30 30 30 30 30 30
ŷ 0,29 0,22 0,24 0,28 0,30 0,34 0,38 0,20 0,32 0,29 0,34
σ 0,15 0,15 0,14 0,16 0,19 0,15 0,24 0,14 0,13 0,15 0,21
Supervivencia larva-pupa n 15 28 32 18 8 14 13 8 15 13 10
ŷ 0,59 0,46 0,55 0,65 0,39 0,40 0,53 0,44 0,38 0,45 0,73
σ 0,27 0,26 0,26 0,26 0,22 0,28 0,16 0,30 0,28 0,23 0,22
Eficiencia de cocones n 29 16 19 20 14 15 9 13 13 15 14
ŷ 0,50 0,46 0,42 0,54 0,21 0,25 0,24 0,40 0,27 0,33 0,62
σ 0,22 0,19 0,28 0,26 0,13 0,25 0,16 0,19 0,26 0,24 0,18
Eficiencia global n 43 44 51 38 22 29 22 21 28 28 24
ŷ 0,17 0,13 0,15 0,19 0,10 0,12 0,20 0,12 0,12 0,13 0,29
σ 0,13 0,12 0,12 0,14 0,09 0,13 0,13 0,08 0,12 0,12 0,17
Rentabilidad n 44 44 51 38 22 29 22 21 28 28 24
ŷ 1,81 1,42 0,98 1,41 0,87 0,73 1,14 0,54 0,59 0,71 1,43
σ 1,42 1,30 0,76 1,02 0,76 0,78 0,78 0,35 0,61 0,64 0,84
23
Tabla 3. Sumario de resultados de los análisis aplicando modelos lineales generales. SC: suma de cuadrados; gl: grados de libertad; CM: cuadrados medios; F:
valor del estadístico F; p: probabilidad asociada a la prueba estadística.
Variable de respuesta Fuentes de variación SC gl CM F p
Supervivencia huevo-larva Tratamiento 1,218 10 0,122 2,794 0,00233
Número de huevos 0,329 1 0,329 7,552 0,00625
Tratamiento*Número de huevos 1,022 10 0,102 2,343 0,01069
Error 18,314 420 0,044
Supervivencia larva-pupa Tratamiento 2,400 10 0,240 2,506 0,00794
Error 15,611 163 0,096
Eficiencia de cocones Tratamiento 4,567 10 0,457 5,550 0,00000
Error 13,659 166 0,082
Eficiencia global Tratamiento 0,699 10 0,070 4,510 0,00001
Error 5,258 339 0,016
Rentabilidad Tratamiento 56,927 10 5,693 6,257 0,00000
Error 309,345 340 0,910
24
La menor supervivencia larval fue observada en la dieta G, mientras que el valor
promedio más alto se observó en la dieta I (Fig. 9). Comparaciones a posteriori
mostraron que estas diferencias fueron significativas (p < 0.05; prueba de Tukey para
muestras desiguales).
A H F J C B I G K D E
Dietas
0,1
0,2
0,3
0,4
0,5la
rvas
apt
as /
hue
vos
de
D.
sacc
har
alis
sem
brad
os
Figura 9. Variación en la supervivencia huevo-larva de D. saccharalis entre tipos de dietas
artificiales. Los puntos indican la media, las barras verticales el error estándar.
Tanto la supervivencia pupal de D. saccharalis como la cantidad de cocones de
C. flavipes fueron mayores en la dieta E (Figs. 10 y 11). En ambos casos pruebas a
posteriori mostraron diferencias significativas entre tipos de dietas, siendo la dieta E
significativamente mayor a otros cuatro tipos de dietas para la eficiencia de cocones (p
< 0.05). En el caso de la supervivencia pupal la prueba de Tukey para muestras
desiguales resultó no significativa.
A H F J C B I G K D E
Dietas
0,3
0,4
0,5
0,6
0,7
0,8
pupa
s de
D.
sacc
har
alis
/la
rvas
apt
as
Figura 10. Variación en la supervivencia larva-pupa de D. saccharalis entre tipos de dietas
artificiales. Los puntos indican la media, las barras verticales el error estándar.

25
A H F J C B I G K D E
Dietas
0,1
0,2
0,3
0,4
0,5
0,6
0,7
coco
nes
/ la
rvas
apt
as
Figura 11. Variación en la eficiencia de producción de cocones de C. flavipes por larva de D.
saccharalis parasitoidizada entre tipos de dietas artificiales. Los puntos indican la media, las barras
verticales el error estándar.
La mejor eficiencia global también fue resultado de un efecto del tipo de dieta E,
que fue significativamente superior a otros ocho tipos de dietas (p < 0.05; Fig. 12).
A H F J C B I G K D E
Dietas
0,08
0,12
0,16
0,20
0,24
0,28
0,32
efic
ienc
ia g
loba
l
Figura 12. Variación en la eficiencia global (supervivencia larva-pupa multiplicada por la eficiencia
de producción de cocones) entre tipos de dietas artificiales. Los puntos indican la media, las barras
verticales el error estándar.
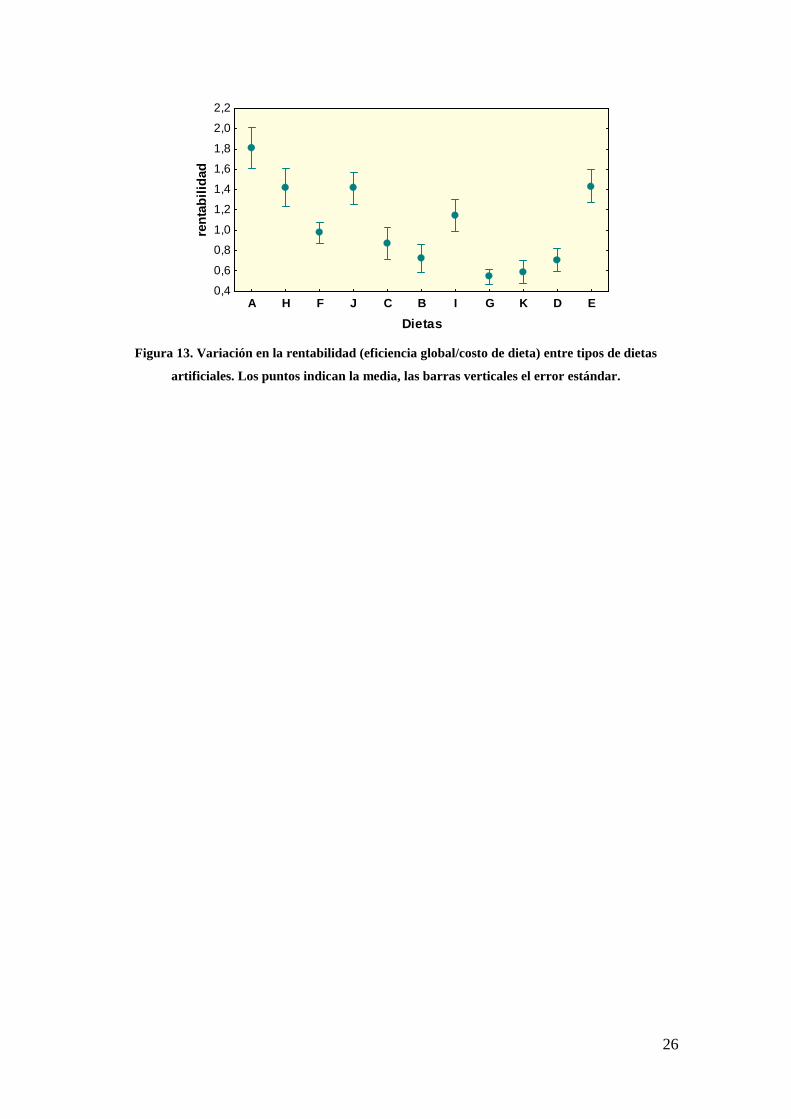
No ocurrió lo mismo con la rentabilidad, que evidenció que la dieta A fue más
rentable (basado en el costo de la dieta) respecto de otros seis tipos de dietas (p < 0.05;
Fig. 13).
26
A H F J C B I G K D E
Dietas
0,4
0,6
0,8
1,0
1,2
1,4
1,6
1,8
2,0
2,2
rent
abili
dad
Figura 13. Variación en la rentabilidad (eficiencia global/costo de dieta) entre tipos de dietas
artificiales. Los puntos indican la media, las barras verticales el error estándar.

27
3.4. Discusión
En este trabajo se evaluó el efecto de 11 combinaciones de harinas y antibióticos
sobre la supervivencia de D. saccharalis y la producción de cocones de C. flavipes en
condiciones de laboratorio.
Los resultados demostraron que las diferentes composiciones afectaron las
variables de estudio. Por una parte, respecto de la combinación de harinas utilizadas, los
mismos sugieren el empleo de harina de poroto como un componente más eficaz que la
harina de soja. Este punto sería contra-intuitivo si se considera que los productos de la
soja son ampliamente difundidos como componentes de la dieta en insectos, ya que de
ellos se destaca en la bibliografía un alto contenido proteico, vitamínico y mineral y un
perfil lipídico benéfico (Cohen, 2004).
En contraste con lo obtenido se encuentran las observaciones efectuadas por
Lastra y Gómez (1988), quienes compararon cuatro tipos de dietas con base en
zanahoria, soja, caseína y poroto sobre parámetros biológicos de D. saccharalis. Los
autores señalan como menos adecuada la dieta a base de poroto por proporcionar un
menor porcentaje de pupas y un menor número de larvas aptas para parasitoidización.
Una explicación a este hecho podría darse por un efecto en la variación de la textura
final de la dieta en las condiciones de los formulados utilizados. Teniendo en cuenta que
se parte de una textura muy fina en el caso de la harina de soja en comparación con el
poroto molido, ésta confiere una textura más compacta o menos aireada, que a su vez
puede incidir en la capacidad de captación de humedad. Un equilibrio en la textura de la
dieta con respecto a la humedad (dietas con exceso de humedad pueden ahogar larvas
recién emergidas y dietas secas dificultan la penetración de las mismas) es mencionado
por Murúa et al. (2003) y Luna et al. (2004) por jugar un papel importante en la
supervivencia de las larvas y en el aprovechamiento de los componentes nutritivos. Otra
posibilidad para explicar las diferencias observadas es la relación con el pH, el cual no
fue considerado como parte de los tratamientos en este estudio. Esta variable fue tomada
en cuenta en un experimento de Marín et al. (2006), quienes encontraron la mejor
relación rendimiento/costo en dietas a base de poroto para larvas del lepidóptero
Grapholita molesta Busck. Además señalaron que la dieta con óptima respuesta
biológica fue coincidente con un pH cercano al de la alimentación de las larvas en la
naturaleza.

28
Por otra parte, teniendo en cuenta los antibióticos incorporados, los mejores
resultados se obtuvieron con el empleo de ampicilina en diferentes dosis. Las
recomendaciones del uso de bactericidas o bacteriostáticos son variables dependiendo
del sistema de estudio y de los fines de la cría del insecto, incorporándose por lo general
un solo tipo de antibiótico. En trabajos de cría masiva de lepidópteros en laboratorio se
emplean bacteriostáticos que, al igual que la oxitetraciclina, pertenecen a la familia de
los tetraciclínicos. Es el caso de la tetraciclina en dosis de 5 g/1,6 L de agua para cría de
Spodoptera exigua Hubner y S. sunia Guenee (Estrada Hurtarte, 1992); clortetraciclina
0,3 g/0,85 L para Hypsipyla grandella Zeller (Vargas et al., 2001); aureomicina 0,39
g/L para D. saccharalis (Calderón, 1987). En biofábricas de Brasil se utilizan con éxito
para la cría masiva de D. saccharalis la terramicina 0,5 g/9,6 L, un equivalente a la
oxitetraciclina (Cano et al., 2006) o la estreptomicina 5 g/4,8 L (Botelho y Macedo,
2002), que al igual que la ampicilina posee actividad bactericida. Sin embargo no
existen registros del uso combinado de ambos o del empleo de ampicilina como único
antibiótico.

29
4. EFECTO DE LA DENSIDAD Y PROPORCIÓN SEXUAL SOBRE LA
FECUNDIDAD DE DIATRAEA SACCHARALIS EN CONDICIONES DE
LABORATORIO
4.1. Introducción
La obtención del parasitoide C. flavipes se realiza a partir de su huésped D.
saccharalis. Por este motivo, la disponibilidad de huevos de D. saccharalis es un punto
clave para la producción de larvas que serán parasitoidizadas con C. flavipes. Para
lograrlo sería deseable incrementar la eficiencia del sistema de producción. Promover el
éxito reproductivo de las hembras de D. saccharalis podría significar, para el sistema
productivo, un aumento en la calidad de las larvas, una disminución en el número de
adultos de D. saccharalis utilizados y/o un incremento en el número de huevos
disponibles.
Un factor relacionado a la fecundidad en las hembras de los insectos es el grado
de poliandria, esto es, su tendencia para copular varias veces con distintos machos. En
general la cópula múltiple eleva la fecundidad de la hembra por un incremento en la
cantidad neta de producto seminal recibido (Torres Vila et al, 2004; Torres Vila y
Jennions, 2005), además de tener un efecto benéfico sobre la oviposición y longevidad
de los adultos (Wedell et al., 2002). Si bien la poliandria es la estrategia reproductiva
más común para los lepidópteros existen algunas especies que, por el contrario tienden a
la monandria, es decir exhiben un solo evento de cópula. Entre estas últimas se
encuentra D. saccharalis (Pérez y Long, 1964; Walker, 1965; Linares, 1987).
Otro punto importante a tener en cuenta es la procedencia de los recursos que la
hembra utiliza para la producción de huevos. Estos pueden provenir principalmente de
reservas de la alimentación durante el período larval, o bien de la alimentación en la
fase adulta o de donaciones del macho durante la cópula (Boggs, 1986).
Milano et al. (2010) estudiaron el efecto de la alimentación de adultos sobre la
reproducción en seis especies de lepidópteros. Encontraron que las hembras de D.
saccharalis poseen una proporción alta de oocitos inmaduros al momento de emerger el
adulto. Este hecho hace necesaria la obtención de recursos posteriores para completar la
maduración de los oocitos. Sin embargo observaron que los adultos de esta especie no
se alimentan sugiriendo que las hembras utilizan los nutrientes adquiridos en la fase

30
larval para la reproducción y aquellos transferidos por los machos en la maduración de
nuevos oocitos.
Consecuentemente, cualquier variación en las condiciones de laboratorio que
favorezca la eficiencia sería ventajosa para el sistema de cría masiva de D. saccharalis y
C. flavipes. De aquí la importancia de evaluar el efecto de la proporción sexual y la
abundancia de adultos de D. saccharalis en tubos de oviposición sobre la fecundidad.
Hipótesis Particular 2: La densidad y proporción sexual de adultos de D.
saccharalis en jaulas de oviposición afecta la fecundidad.
Objetivo Específico 2: Evaluar el desempeño de D. saccharalis en función de la
abundancia y proporción sexual en jaulas de cría.
4.2. Materiales y métodos
4.2.1. Procedimiento
Los adultos de D. saccharalis emergieron en el interior de jaulones como se
describió en el capítulo 2. Diariamente fueron colectados y depositados en jaulas de
oviposición consistentes en tubos de PVC de 10 cm de diámetro y 18 cm de alto. Los
tubos fueron cubiertos en su interior por una hoja de papel de 80gr/m2 cuya función fue
oficiar de sustrato de oviposición en donde las hembras depositaron los huevos una vez
producida la cópula (Fig. 14). Los papeles con posturas fueron extraídos y marcados
para la posterior evaluación de la producción de huevos, que fue correlacionada con la
abundancia y proporción sexual de mariposas contenidas en las jaulas.

31
Figura 14. Jaulas individuales de oviposición.
La colecta de mariposas fue llevada a cabo por el operario con la ayuda de un
tubo de ensayo de 50 ml (Fig. 15), el cual fue llenado en sus 3/4 partes con mariposas y
vaciado tres veces consecutivas en las jaulas de oviposición. Es decir que los adultos no
se contaron individualmente ni se distinguieron por su sexo en el momento de armado
de las jaulas, esto respondió a una medida de practicidad para agilizar la tarea de quien
se encontraba a cargo de este sector, una medida típica en condiciones de cría masiva.
De otra manera requeriría mayor mano de obra, lo cual hace inviable en términos de
costo el trabajo en jaula de cría.

32
Figura 15. Recolección de adultos de D. saccharalis.
El trabajo se efectuó en condiciones de 26ºC y 60 % HR, tomando 25 jaulas
individuales, las que fueron etiquetadas para su diferenciación y seguimiento. En cada
jaula de cría se contó el número de mariposas distinguiéndose machos y hembras. Las
hojas con posturas retiradas llevaron la misma identificación de su jaula de origen. Sólo
se tomaron en cuenta las posturas del primer día posterior al armado de la jaula (jaulas
de 1 día). Esto permitió reducir el número de factores involucrados, asumiendo en base
a observaciones personales, que la fecundación del primer día sería un buen estimador
de la fecundación total.
Para estimar el número de huevos por hoja se utilizó un modelo de regresión
lineal en el que la variable de respuesta (número de huevos por hoja) se estimó a partir
de la relación entre el número de huevos contados por postura y el área de postura
correspondiente. Para esto se tomaron fotografías digitales de posturas individuales de
diferentes tamaños y se contaron los huevos que la conformaban utilizando como
herramienta el programa Image J (Rasband, 1997). Dado que las fotografías tenían
asociada una escala, fue posible estimar el área cubierta por cada postura en mm2. De
esta manera, el número total de huevos por hoja podría ser predicho por el área total
cubierta por huevos en la hoja de oviposición. Un análisis de regresión mostró que el
número de huevos puede ser estimado como: número de huevos = 3.93 (intercepto) +
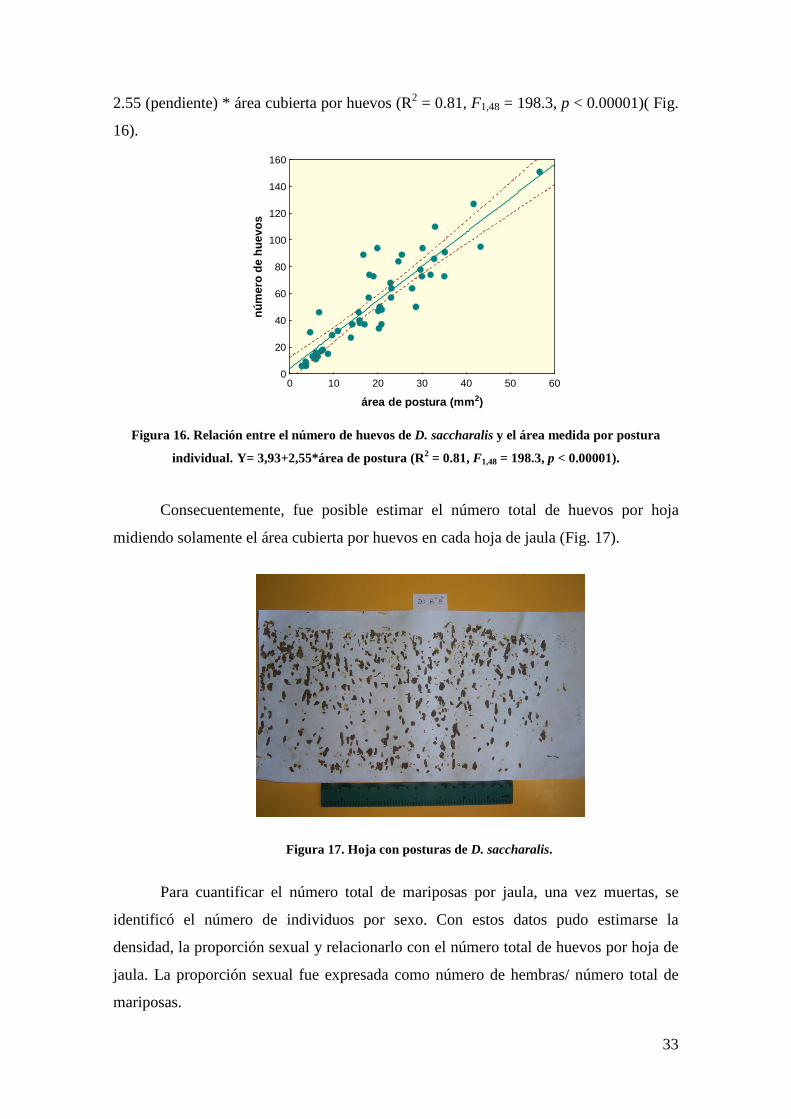
33
2.55 (pendiente) * área cubierta por huevos (R2 = 0.81, F1,48 = 198.3, p < 0.00001)( Fig.
16).
0 10 20 30 40 50 60
área de postura (mm 2)
0
20
40
60
80
100
120
140
160
núm
ero
de h
uevo
s
Figura 16. Relación entre el número de huevos de D. saccharalis y el área medida por postura
individual. Y= 3,93+2,55*área de postura (R2 = 0.81, F1,48 = 198.3, p < 0.00001).
Consecuentemente, fue posible estimar el número total de huevos por hoja
midiendo solamente el área cubierta por huevos en cada hoja de jaula (Fig. 17).
Figura 17. Hoja con posturas de D. saccharalis.
Para cuantificar el número total de mariposas por jaula, una vez muertas, se
identificó el número de individuos por sexo. Con estos datos pudo estimarse la
densidad, la proporción sexual y relacionarlo con el número total de huevos por hoja de
jaula. La proporción sexual fue expresada como número de hembras/ número total de
mariposas.

34
4.2.2. Análisis de datos
Para estimar los efectos de la densidad y proporción sexual se aplicó un análisis
de regresión múltiple con los componentes lineales y cuadráticos del número total de
mariposas por jaula y de la proporción sexual. Previamente estas variables fueron
estandarizadas como (promedio de X) – Xi / (desvío estándar de X). La variable de
respuesta fue el número de huevos estimado sin transformar. Este análisis fue llevado a
cabo en STATISTICA 7.0 (Statsoft, Inc., 2004).
4.3. Resultados
El análisis de regresión múltiple aplicado sobre el número de huevos estimado,
mostró que la cantidad de huevos que las mariposas oviponen en una jaula depende
linealmente de la proporción sexual y no linealmente del número de mariposas (Tabla
4).
Tabla 4. Sumario de resultados del análisis de regresión múltiple aplicado al número de huevos
predicho por hoja de jaula, en función del número de mariposas por jaula y la proporción sexual.
SC: suma de cuadrados; gl: grados de libertad; CM: cuadrados medios; F: valor del estadístico de
prueba; p: valor de probabilidad. Valores de SC y CM expresados en notación científica.
Variable SC gl CM F p
Número de mariposas 4.82E+06 1 4.8E+06 0.058 0.81229 Número de mariposas^2 3.49E+08 1 3.5E+08 4.193 0.05395 Proporción sexual 3.94E+08 1 3.9E+08 4.740 0.04163 Proporción sexual^2 2.37E+08 1 2.4E+08 2.854 0.10668 Error 1.66E+09 20 8.3E+07
(R2 del modelo completo = 0.38; F4,20 = 3.011; p < 0.05).
La proporción sexual por jaula se relacionó negativamente con el número de
huevos estimado (Fig. 18).
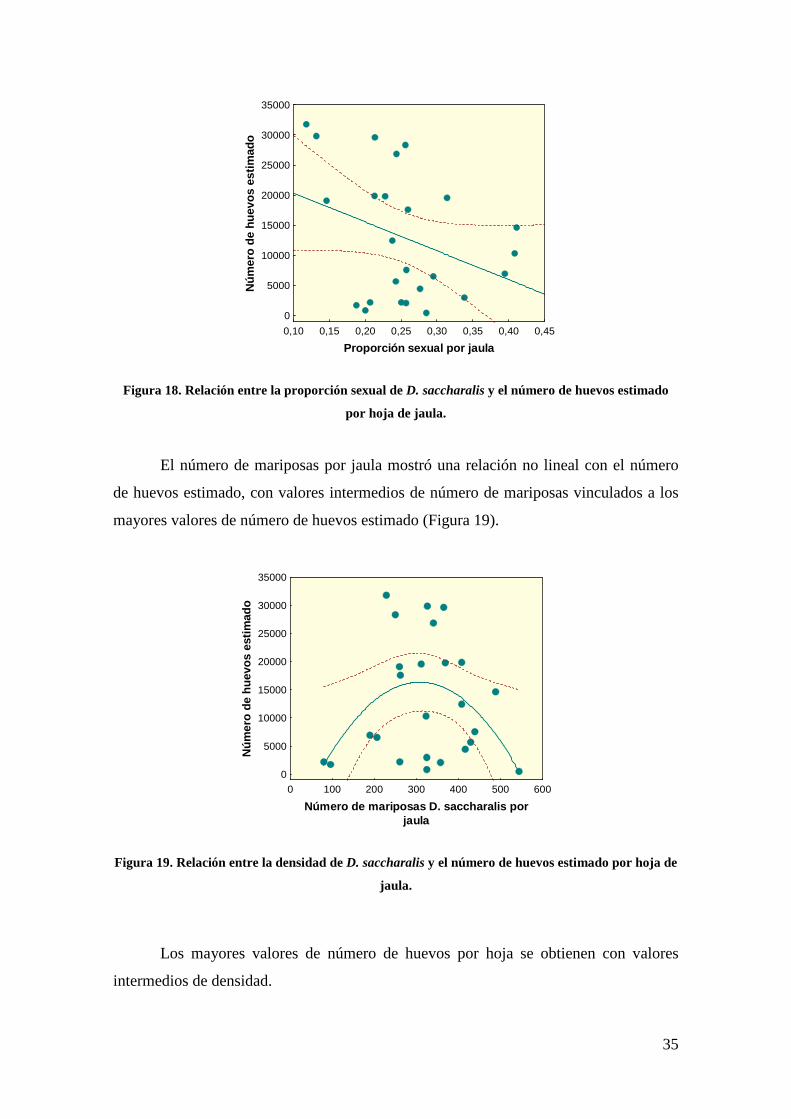
35
0,10 0,15 0,20 0,25 0,30 0,35 0,40 0,45
Proporción sexual por jaula
0
5000
10000
15000
20000
25000
30000
35000
Núm
ero
de h
uevo
s es
timad
o
Figura 18. Relación entre la proporción sexual de D. saccharalis y el número de huevos estimado
por hoja de jaula.
El número de mariposas por jaula mostró una relación no lineal con el número
de huevos estimado, con valores intermedios de número de mariposas vinculados a los
mayores valores de número de huevos estimado (Figura 19).
0 100 200 300 400 500 600
Número de mariposas D. saccharalis porjaula
0
5000
10000
15000
20000
25000
30000
35000
Núm
ero
de h
uevo
s es
timad
o
Figura 19. Relación entre la densidad de D. saccharalis y el número de huevos estimado por hoja de
jaula.
Los mayores valores de número de huevos por hoja se obtienen con valores
intermedios de densidad.

36
4.4. Discusión
Se evaluó el efecto de la abundancia y proporción sexual de adultos de D.
saccharalis sobre la fecundidad en jaulas individuales de oviposición. Los resultados
del experimento demostraron que existe una disminución en el número de huevos a
medida que aumenta la proporción sexual y que esta relación es lineal. De esta manera,
una proporción sexual sesgada a hembras (mayor número de hembras) disminuye la
cantidad de huevos obtenidos por hoja de oviposición. Esta relación puede explicarse
por el hecho que, bajo condiciones de laboratorio, la ocurrencia de reapareamiento en
las hembras es con frecuencia elevado debido a la alta densidad de población y la
proximidad forzada de ambos sexos. Una condición contraria ocurre en observaciones
realizadas en la naturaleza en donde hembras de D. saccharalis muestran una alta
incidencia de una cópula (Pérez y Long, 1964).
Una proporción sexual sesgada a machos aumenta la posibilidad de encuentros
de hembras con machos vírgenes favoreciendo la fecundidad. En este sentido Torres
Vila y Jennions (2005) encontraron que en 22 de 25 especies de lepidópteros estudiadas
la producción de hembras que copularon con machos vírgenes fue más alta que la de
aquellas que se aparearon con machos no vírgenes. Este efecto se debió al tamaño del
espermatóforo, machos vírgenes produjeron un espermatóforo más grande que los que
copularon previamente debido a que sufrieron una depleción de los precursores de esta
estructura luego de cada cópula.
Por otra parte se conoce que los machos no son un recurso limitado debido a que
su tasa reproductiva es mayor que la de las hembras resultando en una proporción
sexual sesgada a machos (Wedell y Cook, 2002). Dato concordante con lo observado
por Linares (1987) quien determinó una proporción de sexos para D. saccharalis de
1:0,87 machos/hembras respectivamente. Por esta razón existen valores intermedios en
relación a la abundancia de mariposas por jaula de oviposición y que estarían dados por
un efecto de hacinamiento y competencia de los machos.

37
5. EFECTO DEL NÚMERO DE PARASITOIDIZACIONES
5.1. Introducción
La parasitoidización de larvas de D. saccharalis con hembras de C. flavipes,
para la reproducción masiva del parasitoide, es un proceso que se realiza manualmente
en condiciones de laboratorio. En el sistema huésped – parasitoide están involucrados
factores que determinan la dinámica del proceso. Algunos de estos factores se
relacionan con el ambiente, la densidad de individuos huésped o recurso, o la capacidad
de búsqueda que posee el parasitoide. En particular este trabajo se enfoca en aquellos
factores que pueden resultar de interés en la ocurrencia del proceso en laboratorio, en
donde la parasitoidización se realiza en forma manual y por lo tanto, a diferencia de lo
que ocurre en el campo, se pueden controlar algunos parámetros tales como el número
de individuos que parasitoidizan una larva.
El superparasitoidismo puede tener consecuencias en el número de adultos del
parasitoide emergidos, la proporción sexual de los mismos, su rendimiento y longevidad
(Cueva et al., 1980; Scaglia et al., 2005; Pexton et al., 2008).
Así también el número de huevos que ovipone el parasitoide en cada huésped y
la capacidad de este último para sustentar la progenie pueden afectar la producción de
cocones en términos de calidad (Acosta y Galán, 1999).
Yamauchi et al. (1997) demostraron que si larvas de D. saccharalis reciben dos
oviposiciones sucesivas por una misma hembra, la producción de adultos de C. flavipes
aumenta respecto de aquellas larvas que recibieron una sola oviposición, sin modificar
la proporción sexual, sin embargo evidencian la disminución de otros parámetros de
interés para el desempeño como agente de control biológico tales como búsqueda de
huésped, dispersión y longevidad. La decisión de trabajar con 1 ó 2 parasitoidizaciones
en el contexto de una cría masiva representa para el operario una inversión de tiempo
que puede ser utilizado tanto para aumentar el número de larvas parasitoidizadas como
para obtener una mayor cantidad de parasitoides y en consecuencia, favorecer la
eficiencia del proceso. En este sentido la forma de parasitoidización y el conocimiento
del efecto que ésta produce es importante en una producción masiva.
Hipótesis Particular 3: El nivel de parasitoidismo ejerce un efecto en la
producción del parasitoide y en su proporción sexual.

38
Objetivo Específico 3: Examinar el efecto del número de parasitoidizaciones de
C. flavipes en larvas de D. saccharalis sobre la producción de cocones y la proporción
sexual de parasitoides emergidos.
5.2. Materiales y métodos
5.2.1. Procedimiento
Este trabajo se llevó a cabo en Junio de 2008 en las instalaciones del CIRPON.
Las larvas de D. saccharalis y adultos de C. flavipes procedían de la línea de cría
desarrollada en el CIRPON y originalmente de la biofábrica Biocontrol de Brasil. Todas
las larvas utilizadas provenían de un mismo lote de cría (dieta, día de siembra, y
condiciones de cámara iguales) y de 19 días de edad, contados desde la fecha de
eclosión. El trabajo se realizó con tres operarios, y cada uno de ellos realizó la
parasitoidización manual de 30 larvas con una sola oviposición y 50 larvas con dos
oviposiciones consecutivas, cada una. El modo de realizar la parasitoidización fue el
siguiente: se extrajeron las larvas de los frascos de vidrio, se las separó de la dieta y al
mismo tiempo se realizó una selección por tamaño. Luego se tomó una larva con los
dedos o con ayuda de una pinza colocándola de tal manera que la parte posterior
quedara libre para que la avispa fuera capaz de posarse e introducir sus huevos en la
misma (Fig.20).
Figura 20. Operarios realizando la parasitoidización manual de larvas
de D. saccharalis con C. flavipes.

39
Los dos vasos utilizados por cada operario fueron identificados como A y B.
Esta distinción obedeció a que, si bien la línea de cría fue la misma, las masas de
cocones contenidas en cada vaso provinieron de diferentes hembras. Se estima que esto
permitió considerar el potencial efecto de arrenotoquia sobre la variación en la
proporción sexual derivado de la endogamia. De existir este problema, la utilización de
un único vaso podría haberlo solapado. Se realizaron dos parasitoidizaciones con un
mismo vaso conteniendo adultos de C. flavipes, o con dos vasos (una parasitoidización
por vaso, de manera consecutiva). De esta manera, se pudo evaluar la variación entre
vasos que podría afectar la producción de cocones derivada del número de
parasitoidizaciones. Así el experimento quedó conformado como se indica en la Tabla
5.
Tabla 5. Diseño experimental para el número de parasitoidizaciones manuales de C. flavipes sobre
D. saccharalis en condiciones de cría masiva.
Número de parasitoidizaciones
Número de larvas
Vaso con Cotesia
Grupo de análisis
1 15 A 1 1 15 B 1 2 15 A 1 y 2 2 15 B 1 y 2 2 20 A y B 2
Las larvas luego de ser parasitoidizadas se colocaron individualmente en
cápsulas de Petri y se mantuvieron en cámara de cría a una temperatura de 26±1˚C por
14 días, al cabo de los cuales se revisaron registrándose: larvas de D. saccharalis
muertas, pupas, éxito de producción de cocones, adultos de C. flavipes emergidos y
proporción sexual de C. flavipes emergidos por larva de D. saccharalis parasitoidizada.
5.2.2. Análisis de datos
Se realizaron dos grupos de análisis. El primero comparó el efecto del número
de parasitoidizaciones utilizando como fuente de datos las derivadas de un único vaso
(A ó B). El segundo grupo de análisis se realizó comparando sólo las larvas
parasitoidizadas 2 veces, utilizando como fuente de datos aquellas derivadas de un
único vaso (A ó B) o de la mezcla de los dos vasos (A y B) (Tabla 5). Esta última

40
comparación se realizó para examinar si la variación entre 1 y 2 parasitoidizaciones
podría haber sido afectada por el hecho de utilizar un único vaso para realizar dos
parasitoidizaciones.
El éxito de producción de cocones fue considerado una variable de respuesta
binomial (0= ausencia de cocones; 1= presencia de cocones). Para examinar el efecto
del número de parasitoidizaciones sobre el éxito de producción de cocones se aplicó un
modelo lineal generalizado (Dobson, 1990) con función liga LOGIT y corrigiendo por
sobredispersión de los datos. Un modelo lineal generalizado es análogo a un modelo
lineal general; pero mientras que un modelo lineal general puede ser aplicado a datos
continuos y la distribución de los errores debe reflejar una distribución normal, en un
modelo lineal generalizado se pueden considerar efectos lineales y/o no lineales, tanto
para variables continuas como discretas. Para evaluar el efecto del número de
parasitoidizaciones sobre el número total de C. flavipes adultos emergidos por larva,
total de hembras emergidas por larva, y proporción sexual, se aplicó el mismo modelo,
pero utilizando una distribución normal y función liga LOG.
En el modelo para el primer grupo de datos se incorporaron los factores Operario
(3 niveles) y Número de parasitoidizaciones (2 niveles). En el modelo para el segundo
grupo de datos se incorporaron los factores Operario (3 niveles) y Número de Vasos (2
niveles): dos parasitoidizaciones provenientes de un solo vaso y dos provenientes de dos
vasos diferentes. En ambos grupos de análisis, el factor Operario es incorporado para
considerar dicha variación, ya que es conocido que en sistemas de cría masiva la
eficiencia de parasitoidización de distintos operarios varía significativamente.
5.3. Resultados
5.3.1. Efecto de parasitoidismo simple o doble
El éxito de producción de cocones fue significativamente mayor en larvas de D.
saccharalis parasitoidizadas una vez (media = 0.906 ± desvío estándar = 0.293, N =
149) que en larvas parasitoidizadas dos veces (0.840 ± 0.368, N = 131; Wald = 4.691, P
= 0.03; Fig. 21).
La tabla 6 muestra un sumario de resultados del análisis lineal generalizado
(univariado) para examinar el efecto del número de parasitoidizaciones (1 ó 2) con C.

41
flavipes sobre D. saccharalis en cuatro variables que describen la producción de C.
flavipes.
Tabla 6. Efecto de parasitoidización (1 o 2) con C. flavipes sobre D. saccharalis. Se indican valores
del estadístico Wald y de probabilidad (P). Para las cuatro variables de respuesta los grados de
libertad fueron: Número de Parasitoidizaciones = 1; Operario = 2; Interacción = 2.
Variable de respuesta Fuente de variación Wald P
Éxito de producción de cocones Número de Parasitoidizaciones 4,691 0,03032 Operario 8,827 0,01211
Interacción Número de Parasitoidizaciones * Operario 1,083 0,58182
Adultos emergidos Número de Parasitoidizaciones 0,821 0,36484 Operario 5,571 0,06170
Interacción Número de Parasitoidizaciones* Operario 0,675 0,71353
Hembras emergidas Número de Parasitoidizaciones 0,177 0,67422 Operario 6,352 0,04176
Interacción Número de Parasitoidizaciones * Operario 0,116 0,94355
Proporción sexual de adultos emergidos Número de Parasitoidizaciones 1,678 0,19523 Operario 1,731 0,42081
Interacción Número de Parasitoidizaciones * Operario 0,236 0,88869
1 2
Número de parasitoidizaciones
0,80
0,85
0,90
Éxi
to e
n pr
oduc
ción
de
coco
nes
Figura 21. Efecto del número de parasitoidizaciones de C. flavipes aplicadas manualmente a larvas
de D. saccharalis sobre el éxito de producción de cocones de C. flavipes.

42
El éxito de producción de cocones también varió significativamente
dependiendo del operario que lo realizara (Tabla 6). Sin embargo, esta variación no
modificó la variación debida al número de parasitoidizaciones, es decir, aunque la
variación entre operarios es evidente, el patrón de diferencias entre una y dos
parasitoidizaciones se mantiene; i.e., no hay interacción significativa (Tabla 6; Figura
22).
A B C
Operario
0,6
0,7
0,8
0,9
1,0
1,1
Éxi
to e
n pr
oduc
ción
de
coco
nes
1 parasitoidización 2 parasitoidizaciones
Figura 22. Efecto del operario sobre el éxito de producción de cocones de C. flavipes de acuerdo al
número de parasitoidizaciones de C. flavipes.
Respecto de las demás variables indicadoras de producción de Cotesia, no se
detectó efecto significativo alguno del número de parasitoidizaciones sobre el número
total de adultos de C. flavipes emergidos, el número de hembras emergidas, ni sobre la
proporción sexual (Tabla 6). En la Tabla 7 se indican los estadísticos descriptivos para
cada una de las variables. La única variación significativa detectada fue un efecto del
Operario sobre el número de hembras emergidas (Tablas 6 y 7).
Se encontró una pupa de D. saccharalis sobre 358 larvas parasitoidizadas y no
fue tenida en cuenta para el análisis. Es decir, la resistencia y/o fracaso de la
parasitoidización por C. flavipes es despreciable.
43
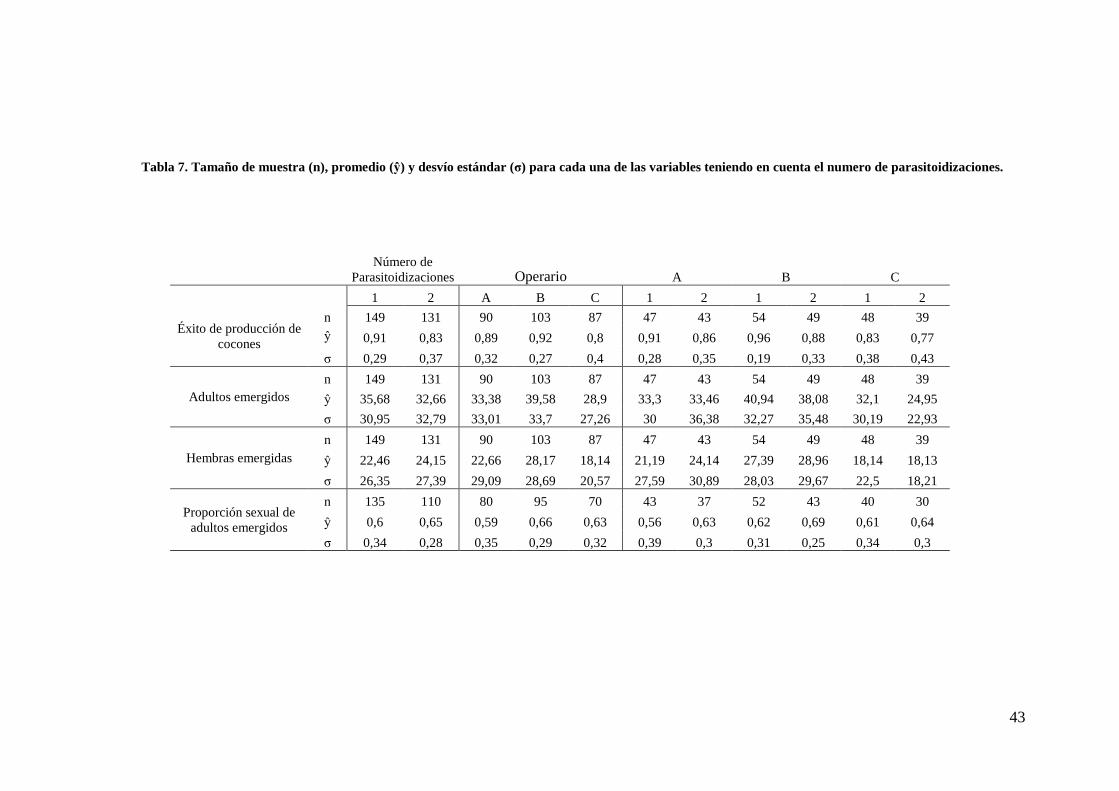
Tabla 7. Tamaño de muestra (n), promedio (ŷ) y desvío estándar (σ) para cada una de las variables teniendo en cuenta el numero de parasitoidizaciones.
Número de
Parasitoidizaciones Operario A B C
1 2 A B C 1 2 1 2 1 2
Éxito de producción de cocones
n 149 131 90 103 87 47 43 54 49 48 39 ŷ 0,91 0,83 0,89 0,92 0,8 0,91 0,86 0,96 0,88 0,83 0,77
σ 0,29 0,37 0,32 0,27 0,4 0,28 0,35 0,19 0,33 0,38 0,43
Adultos emergidos n 149 131 90 103 87 47 43 54 49 48 39
ŷ 35,68 32,66 33,38 39,58 28,9 33,3 33,46 40,94 38,08 32,1 24,95
σ 30,95 32,79 33,01 33,7 27,26 30 36,38 32,27 35,48 30,19 22,93
Hembras emergidas n 149 131 90 103 87 47 43 54 49 48 39
ŷ 22,46 24,15 22,66 28,17 18,14 21,19 24,14 27,39 28,96 18,14 18,13
σ 26,35 27,39 29,09 28,69 20,57 27,59 30,89 28,03 29,67 22,5 18,21
Proporción sexual de adultos emergidos
n 135 110 80 95 70 43 37 52 43 40 30
ŷ 0,6 0,65 0,59 0,66 0,63 0,56 0,63 0,62 0,69 0,61 0,64
σ 0,34 0,28 0,35 0,29 0,32 0,39 0,3 0,31 0,25 0,34 0,3

44
5.3.2. Efecto del número de vasos utilizados para realizar dos parasitoidizaciones
La variación en el éxito de producción de cocones no fue modificada
sustancialmente por la procedencia de las C. flavipes utilizadas en parasitoidización.
En la tabla 8 se indica un sumario de resultados del análisis lineal generalizado
(univariado) para examinar el efecto del número de vasos utilizados (1 solo ó 2
diferentes), teniendo en cuenta dos parasitoidizaciones con C. flavipes sobre D.
saccharalis en cuatro variables que describen la producción del parasitoide. Se indican
valores del estadístico Wald y de probabilidad (P). Además de un efecto del Operario
sobre el éxito de producción de cocones y el número de hembras de C. flavipes
emergidas, el único efecto significativo del vaso de parasitoidización fue detectado para
el número de adultos de avispas emergidos (Tabla 8).
Tabla 8. Sumario de resultados del análisis lineal generalizado (univariado) para examinar el efecto
del número de vasos utilizados. Para las cuatro variables de respuesta los grados de libertad
fueron: Vaso de Procedencia = 1; Operario = 2; Interacción = 2.
Variable de respuesta Fuente de variación Wald P Éxito de producción de cocones Número de Vasos 0.902 0.34231 Operario 8.648 0.01324 Interacción Número de Vasos * Operario 1.180 0.55428 Adultos emergidos Número de Vasos 5.508 0.01893 Operario 4.910 0.08587 Interacción Número de Vasos * Operario 1.253 0.53438 Hembras emergidas Número de Vasos 2.924 0.08726 Operario 10.066 0.00652 Interacción Número de Vasos * Operario 2.803 0.24618 Proporción sexual de adultos emergidos Número de Vasos 0.303 0.58174 Operario 2.054 0.35815 Interacción Número de Vasos * Operario 0.170 0.91868
El número de adultos emergidos fue aproximadamente el doble cuando los
operarios utilizaron un solo vaso para parasitoidizar una larva dos veces que cuando
utilizaron dos vasos para el mismo procedimiento (Tabla 9).
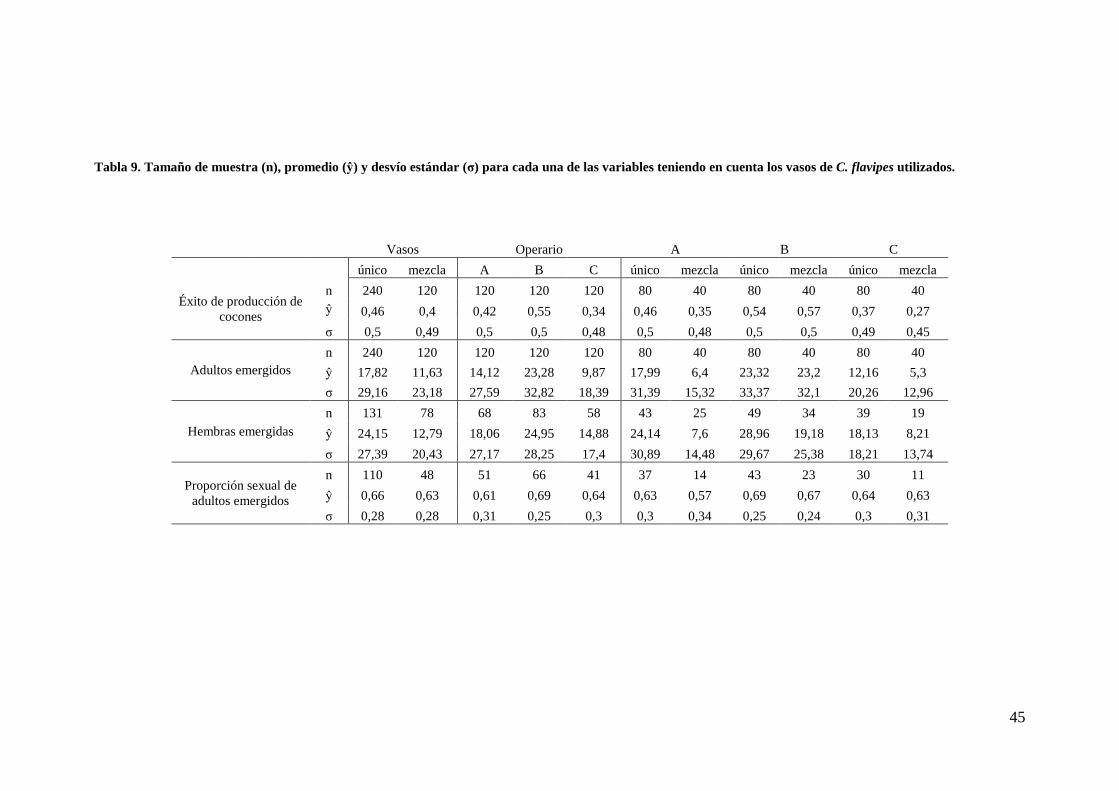
45
Tabla 9. Tamaño de muestra (n), promedio (ŷ) y desvío estándar (σ) para cada una de las variables teniendo en cuenta los vasos de C. flavipes utilizados.
Vasos Operario A B C
único mezcla A B C único mezcla único mezcla único mezcla
Éxito de producción de cocones
n 240 120 120 120 120 80 40 80 40 80 40 ŷ 0,46 0,4 0,42 0,55 0,34 0,46 0,35 0,54 0,57 0,37 0,27
σ 0,5 0,49 0,5 0,5 0,48 0,5 0,48 0,5 0,5 0,49 0,45
Adultos emergidos n 240 120 120 120 120 80 40 80 40 80 40
ŷ 17,82 11,63 14,12 23,28 9,87 17,99 6,4 23,32 23,2 12,16 5,3
σ 29,16 23,18 27,59 32,82 18,39 31,39 15,32 33,37 32,1 20,26 12,96
Hembras emergidas n 131 78 68 83 58 43 25 49 34 39 19
ŷ 24,15 12,79 18,06 24,95 14,88 24,14 7,6 28,96 19,18 18,13 8,21
σ 27,39 20,43 27,17 28,25 17,4 30,89 14,48 29,67 25,38 18,21 13,74
Proporción sexual de adultos emergidos
n 110 48 51 66 41 37 14 43 23 30 11
ŷ 0,66 0,63 0,61 0,69 0,64 0,63 0,57 0,69 0,67 0,64 0,63
σ 0,28 0,28 0,31 0,25 0,3 0,3 0,34 0,25 0,24 0,3 0,31

46
5.4. Discusión
En este trabajo se evaluó el número de parasitoidizaciones de C. flavipes en
larvas de D. saccharalis sobre la producción y proporción sexual de parasitoides. Los
resultados obtenidos muestran que la producción de cocones en condiciones de cría
masiva es más eficiente cuando los operarios realizan una única parasitoidización con
C. flavipes sobre larvas de D. saccharalis. Estos resultados coinciden con los
encontrados por Scaglia et al. (2005) quienes estudiaron la secuencia de oviposición
(primera a tercera) de C. flavipes sobre D. saccharalis destacando que hay un
incremento en la encapsulación de los parasitoides con las sucesivas oviposiciones.
También fue observado por Acosta y Galán (1999), el efecto de competencia larval
intrínseca de C. flavipes en larvas de D. saccharalis. Los distintos tratamientos
comprendían de 1 a 5 hembras de la avispa por larva huésped. Recién a partir de la
incidencia de 5 parasitoides por larva se manifestó un efecto resultando en la
disminución del número de cocones y adultos emergidos. Así también se produjo un
aumento en la mortalidad de larvas de D. saccharalis sin la obtención de masas de
cocones, sugiriendo una capacidad sobrepasada de la larva como recurso.
Por otra parte se deriva de este experimento que el nivel de parasitoidismo (1 o 2
oviposiciones) no incide en el número de adultos emergidos, así como tampoco en el
número de hembras ni sobre la proporción sexual.
Pexton et al. (2008), encontraron que el número de huevos depositados en
larvas de D. saccharalis parasitoidizadas dos veces fue aproximadamente el doble que
el correspondiente al parasitoidismo por una sola hembra de C. flavipes, pero su
descendencia disminuyó en número durante su desarrollo en el interior del huésped,
mientras que permaneció constante en larvas parasitoidizadas por una sola hembra. De
manera que el número de adultos resultante fue similar en ambos casos. La explicación
a este hecho es la existencia de un efecto de competencia, tanto por la cantidad de
alimento como por el espacio disponible, que ocurre en el interior de la larva, lo que
provoca la mortalidad de un cierto número de larvas del parasitoide durante su
desarrollo. Otro dato coincidente con lo observado por Pexton et al. (2008) es la
inexistencia de diferencias significativas en la proporción sexual resultante en ambos
tratamientos. A diferencia de lo estudiado por Cueva et al. (1980) quienes concluyen
que, cuando una misma hembra de C. flavipes ovipone 1 o 2 veces, la proporción sexual
(macho/hembra) aumenta de 1:2,18 a 1:1,2 respectivamente.

47
Es importante tener en cuenta si las oviposiciones sucesivas fueron producto de
una misma hembra o de diferentes hembras del parasitoide. Esto sería una explicación
posible al hecho que en el experimento de Yamauchi et al. (1996) se registraran un
número mayor de adultos de C. flavipes emergidos de larvas de D. saccharalis
parasitoidizadas dos veces por una misma hembra. En la progenie de diferentes hembras
coexistiendo en un mismo huésped se podría dar un efecto de competencia mayor al
existente en una progenie o descendencia de una misma hembra. Por lo que, al respecto
coinciden en el hecho que el número de larvas del parasitoide que no completaron su
desarrollo dentro del huésped fue mayor en el tratamiento con dos oviposiciones,
además de presentarse una disminución en la longevidad de los adultos emergidos.

48
6. EFECTO DE LA EDAD LARVAL DEL HUÉSPED
6.1. Introducción
Desde el momento de la oviposición los parasitoides inducen cambios en el
huésped para evadir la reacción del sistema de defensa y promover condiciones
adecuadas para el desarrollo de sus larvas en el interior del hemocele (Scaglia et al.,
2005; Hegazi y Khafagi, 2007). Estos cambios se ven reflejados en alteraciones de la
composición bioquímica de la hemolinfa y disminución de macronutrientes que se
correlacionan con los requerimientos nutricionales de los individuos inmaduros del
parasitoide (Salvador y Cônsoli, 2008).
Los parámetros biológicos del parasitoide varían por factores abióticos como la
temperatura o por la condición del huésped o calidad (Jiang et al., 2004).
Uno de los estimadores de calidad del mismo, es su tamaño. Esta característica
puede afectar la supervivencia de la descendencia del parasitoide, el tamaño y número
de individuos adultos que emergen (Campos Farinha y Chaud Netto, 2000; Chow y
Heinz, 2007; Hernández, 2010).
En la práctica en el laboratorio y bajo condiciones ambientales controladas las
larvas de D. saccharalis adquieren un tamaño que resulta adecuado para ser
parasitoidizadas por C. flavipes, en un rango comprendido entre 17 y 21 días. Sin
embargo, como podría ser esperado, existe variación tanto en el tamaño como en las
condiciones fisiológicas que podrían afectar los resultados del parasitoidismo.
La producción de cocones de C. flavipes puede ser incrementada aumentando la
calidad del huésped. Así también, dado que el mantenimiento de las larvas es muy
costoso, sería esperable que se optimicen las técnicas de cría masiva mediante una
relación de tamaño de larva/tiempo en cámara que a su vez garantice una producción
óptima de cocones.
Hipótesis Particular 4: La edad de la larva huésped ejerce un efecto en la
producción del parasitoide y en su proporción sexual.
Objetivo Específico 4: Examinar el efecto de la edad larval a la cual son
parasitoidizadas sobre la producción de cocones y la proporción sexual de parasitoides
emergidos.

49
6.2. Materiales y métodos
6.2.1. Procedimiento
El experimento se realizó entre los meses de Julio y Noviembre de 2008. La
parasitoidización se produjo utilizando larvas de D. saccharalis y adultos de C. flavipes
de la línea de cría del CIRPON, con 17, 18, 19, 20 y 21 días, todas parasitoidizadas una
sola vez. Cabe destacar que se tomó en cuenta la edad del huésped en días y no se
especificó estadio de desarrollo larval por considerarse, el primero, un dato más
relevante a los efectos de este estudio (minimizar el tiempo de cría reduce costos en
sistemas de producción masiva). Por cada día de parasitoidización cada operario (tres en
total), recibió 2 frascos escogidos al azar, de los que extrajeron las larvas para
parasitoidizar con C. flavipes, separando previamente larvas de buen tamaño (20 mm o
más aproximadamente) consideradas “buenas” y larvas de tamaño menor, consideradas
“malas”. Esta selección es la que normalmente se llevó a cabo en el sistema de cría
masiva desarrollado en el CIRPON.
De esta manera se contó el número de larvas “buenas” y “malas”, cuyos valores
se consignan en la Tabla 10, las que luego de ser parasitoidizadas manualmente, se
colocaron en cápsulas de Petri y fueron llevadas a cámara de cría con una temperatura
de 26±1˚C por un período de 14 días. De los cocones extraídos de cada cápsula se
registró número de adultos de C. flavipes emergidos, una vez muertos para facilitar el
conteo de los mismos, distinguiendo machos y hembras para obtener la proporción
sexual.
Tabla 10. Diseño experimental para las cinco edades larvales del hospedero D. saccharalis.
17 días 18 días 19 días 20 días 21 días
Operario Frasco
A Frasco
B Frasco
C Frasco
D Frasco
E Frasco
F Frasco
G Frasco
H Frasco
I Frasco
J
b m b m b m b m b m b m b m b m b m b m
A 70 58 54 43 142 36 94 43 68 63 80 42 56 77 54 30 72 30 80 22
B 77 51 82 - 83 42 118 26 133 33 20 15 110 57 54 24 84 23 102 25
C 130 35 91 46 58 80 85 52 53 87 119 75 36 63 64 41 112 30 78 29

50
6.2.2. Análisis de datos
Las variables de respuesta correspondientes a la evaluación del efecto de la edad
larval del huésped son de naturaleza no-normal. El éxito de producción de cocones se
consideró una variable de respuesta binomial, el número de hembras y adultos
emergidos una variable de conteo y la proporción sexual un índice. Por lo tanto, es
esperable que la distribución de los errores de los modelos asociados no cumplan con el
supuesto de normalidad. En consecuencia, para evaluar el efecto de la edad larval del
huésped sobre las variables mencionadas en C. flavipes, se utilizaron modelos lineales
generalizados, permitiendo de esta manera la consideración de la naturaleza del error.
La distribución asociada para el éxito de producción de cocones fue de tipo binomial,
mientras que para el número de hembras emergidas, el número de adultos emergidos y
la proporción sexual, se asoció un error normal. Los factores considerados en el modelo
fueron: 1) días de vida de la larva huésped, 2) operario que realizó las
parasitoidizaciones, 3) interacción entre días de vida y operario. Los resultados del
modelo lineal generalizado se expresan en una tabla de ANOVA, considerando el
estadístico Wald como el estadístico de referencia para los criterios de significancia.
6.3. Resultados
Hubo un efecto significativo de la edad larval de D. saccharalis en días sobre el
éxito en la producción de cocones (Wald = 103.117, P = 0.00), el número de adultos de
C. flavipes emergidos (Wald = 95.481, P = 0.00), el número de hembras emergidas
(Wald = 41.371, P = 0.00) y la proporción sexual (Wald = 37.893, P = 0.00). (Tabla
11).

51
Tabla 11. Efecto de la edad larval de D. saccharalis.
Variable de respuesta Fuente de variación Wald P
Éxito de producción de cocones Días de vida 103,117 0,00000
Operario 61,597 0,00000
Interacción Días de vida * Operario 52,665 0,00000
Adultos emergidos Días de vida 95,481 0,00000
Operario 20,147 0,00004
Interacción Días de vida * Operario 111,86 0,00000
Hembras emergidas Días de vida 41,371 0,00000
Operario 8,359 0,01530
Interacción Días de vida * Operario 70,561 0,00000
Proporción sexual de adultos emergidos Días de vida 37,893 0,00000
Operario 0,971 0,61534
Interacción Días de vida * Operario 43,040 0,00000
El éxito en la producción de cocones fue significativamente más alto en larvas
de 18 días (media = 0.72 ± desvío estándar = 0.45, N = 572) y 19 días (0.70 ± 0.46, N =
468) como se muestra en la Tabla 12 y Fig. 23.
Tabla 12. Tamaño de muestra (n), promedio (ŷ) y desvío estándar (σ) para cada una de las
variables teniendo en cuenta la edad larval en días de D. saccharalis.
Días de vida
17 18 19 20 21
Éxito de producción de cocones n 474 572 468 371 376 ŷ 0,53 0,72 0,7 0,39 0,6
σ 0,50 0,45 0,46 0,49 0,49
Adultos emergidos n 474 572 468 371 376
ŷ 19,53 31,87 33,59 10,99 11,62
σ 30,84 34,29 37,33 19,24 19,20
Hembras emergidas n 434 568 468 371 367
ŷ 13,92 16,94 17,24 5,93 5,51
σ 25,94 23,20 25,13 13,00 11,93
Proporción sexual de adultos emergidos n 265 407 328 152 214
ŷ 0,54 0,49 0,41 0,45 0,35
σ 0,32 0,31 0,31 0,32 0,31

52
17 18 19 20 21
Edad de larvas de D. saccharalis en días
0,3
0,4
0,5
0,6
0,7
0,8
Éxi
to e
n pr
oduc
ción
de
coco
nes
Figura 23. Éxito en la producción de cocones.
Lo mismo ocurre si consideramos el número de adultos emergidos: los mayores
valores se presentaron para larvas de 18 días (31.87 ± 34.29, N = 572) y larvas de 19
días (33.59 ± 37.33, N = 468). Para el número de hembras se mantiene la misma
tendencia: larvas de 18 días (16.94 ± 23.20, N = 568), larvas de 19 días (17.24 ± 25.13,
N = 468; Tabla 12. Figs.24 y 25).
17 18 19 20 21
Edad de larvas de D. saccharalis en días
5
10
15
20
25
30
35
40
Núm
ero
de a
dulto
s de
C
. fla
vip
es e
mer
gido
s
Figura 24. Número de adultos de C. flavipes emergidos.
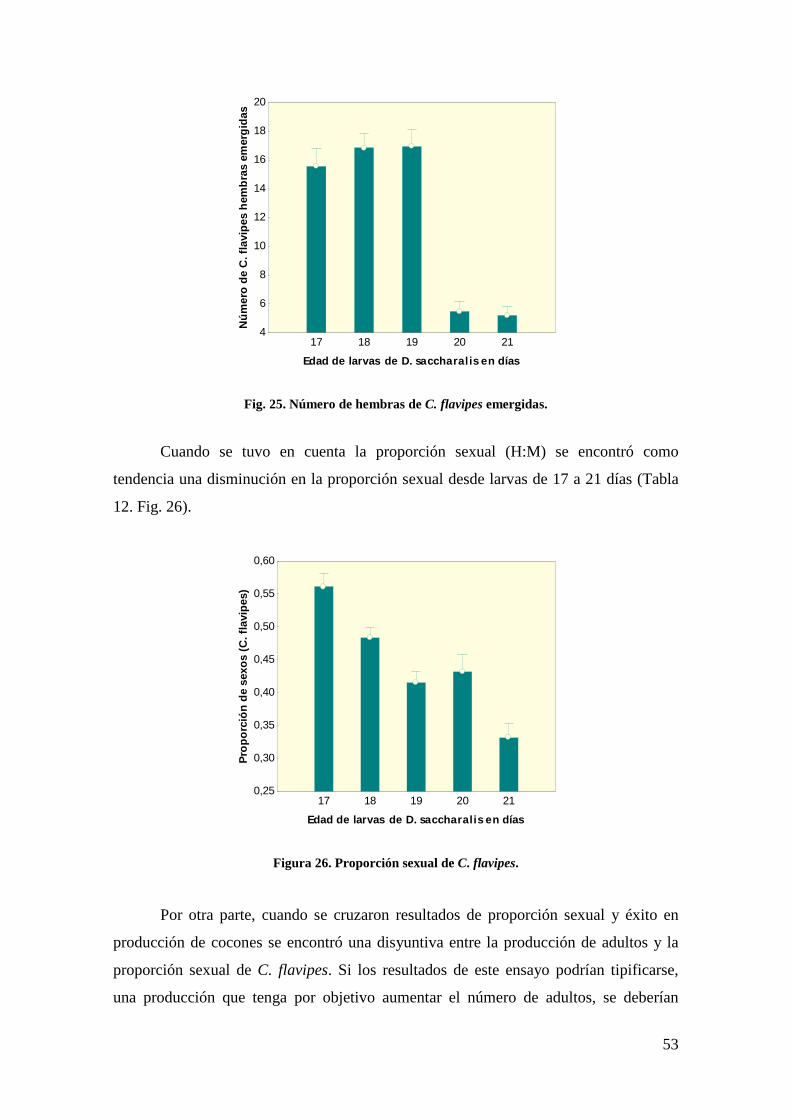
53
17 18 19 20 21
Edad de larvas de D. saccharalis en días
4
6
8
10
12
14
16
18
20
Núm
ero
de C
. fla
vip
es h
embr
as e
mer
gida
s
Fig. 25. Número de hembras de C. flavipes emergidas.
Cuando se tuvo en cuenta la proporción sexual (H:M) se encontró como
tendencia una disminución en la proporción sexual desde larvas de 17 a 21 días (Tabla
12. Fig. 26).
17 18 19 20 21
Edad de larvas de D. saccharalis en días
0,25
0,30
0,35
0,40
0,45
0,50
0,55
0,60
Pro
porc
ión
de s
exos
(C
. fla
vip
es)
Figura 26. Proporción sexual de C. flavipes.
Por otra parte, cuando se cruzaron resultados de proporción sexual y éxito en
producción de cocones se encontró una disyuntiva entre la producción de adultos y la
proporción sexual de C. flavipes. Si los resultados de este ensayo podrían tipificarse,
una producción que tenga por objetivo aumentar el número de adultos, se deberían

54
parasitoidizar larvas de 18 o 19 días. Sin embargo, si se tiene por objetivo sesgar la
producción a hembras (proporción sexual dominada por hembras), se deberían
parasitoidizar larvas de 17 días (Fig. 27).
17 18 19 20 21
Edad de larvas de D. saccharalis en días
0,25
0,30
0,35
0,40
0,45
0,50
0,55
0,60
0,65
0,70
0,75
Éxi
to e
n pr
oduc
ción
de
coco
nes
ypr
opor
ción
de
sexo
s
Figura 27. Disyuntiva entre el éxito en producción de cocones y la proporción sexual de C. flavipes.

55
6.4. Discusión
En este trabajo se evaluó el efecto de la edad larval del huésped sobre la
obtención de cocones de C. flavipes y la proporción sexual de los parasitoides. Se
encontró que tanto el éxito en la producción de cocones como el número de adultos
emergidos fue mayor para los grupos de larvas de D. saccharalis de 18 y 19 días.
Coincidente con lo encontrado en este experimento, es el trabajo de Hernández
(2010), quien observó una mayor emergencia de adultos de C. flavipes provenientes de
larvas D. saccharalis parasitoidizadas a los 18 días de edad que en larvas de 16 y 17
días. Campos Farinha y Chaud Netto (2000) proponen en su trabajo al considerar larvas
de tercero a quinto estadio, que un aumento registrado en el número de parasitoides
emergidos de C. flavipes se debería al aumento en el tamaño del huésped. Lo mismo
observaron Jiang et al. (2004), en el parasitoidismo de C. flavipes sobre Chilo partellus
Swinhoe, donde más parasitoides emergieron en larvas L4 que en L3. Además agregan
que la tasa de desarrollo de los parasitoides es más baja en larvas más pequeñas al
determinar que el tiempo de desarrollo de huevo a cocón fue mayor, dato que no fue
registrado en este ensayo.
Existen estudios que sugieren la dependencia del estadio larval con las
respuestas inmunes celular y humoral (Hegazi y Khafagy, 2007). Una disminución en la
producción de cocones y en el número de adultos observados para el grupo de larvas de
20 y 21 días podría deberse a los cambios internos en la larva relacionados a su
proximidad al estado de pupa, de tal manera que no hacen viable el desarrollo de los
parasitoides en su interior. Por otra parte la clara tendencia a la disminución en el
número de hembras a medida que aumenta la edad larval del huésped también tendría
que ver con este hecho y la capacidad que tendría la hembra de C. flavipes para inferir la
condición de sus huéspedes al momento de depositar los huevos. En relación a esta
situación, Chow y Heinz (2007) encontraron que hembras del parasitoide Diglyphus
isaea Walker (Hymenoptera: Eulophidae) produjeron una proporción sexual sesgada a
machos cuando se incrementó el tamaño del huésped.

56
7. CONCLUSIÓN GENERAL
Respecto de los cuatro puntos seleccionados como parte de los factores
inherentes a la cría masiva del parasitoide C. flavipes y su huésped D. saccharalis se
concluye que:
Composición de la dieta - La composición de la dieta suministrada a larvas de D. saccharalis tiene un
efecto sobre la producción de ambas líneas de cría, huésped y parasitoide.
- De los distintos tipos de dieta propuestos se obtienen los mejores resultados
con una combinación de harina de poroto y ampicilina como antibiótico, favoreciendo
la supervivencia de D. saccharalis y la producción de C. flavipes.
- De los resultados se puede inferir la posibilidad de adoptar un tipo de dieta de
acuerdo a las necesidades inmediatas en el estado de producción de la cría masiva. En el
caso de prevalecer un objetivo de levantar la línea de cría por sobre la necesidad de
obtención de los productos finales de la cría (cocones) se optaría por la dieta I, en donde
se obtuvo el valor promedio más alto para la supervivencia huevo-larva. Lo que sugiere
que en los primeros estadios larvales se puede reducir la dosis de ampicilina para
combinarla con otro antibiótico de efecto bactericida (estreptomicina). En caso de
optimizar los resultados para ambas líneas, es decir, pie de cría de D. saccharalis y
producción de cocones de C. flavipes la elección es a favor de la dieta E. La dieta A es
la opción si se toma en cuenta la rentabilidad, punto muy importante en el desarrollo de
la cría masiva con el objeto de su empleo en el control biológico. Por lo tanto, por
cuestiones prácticas la recomendación que se desprende de este estudio es realizar la
elección de la dieta teniendo en cuenta los costos y objetivos de la producción para
lograr ciclos de cría sustentables en el tiempo, y la capacidad logística del sistema de
cría masiva para producir varios tipos de dietas a la vez.
Cría de D. saccharalis - Existe una tendencia a la disminución de huevos de D. saccharalis con el
aumento de la proporción sexual expresada como número de hembras/número total de
adultos.

57
- La cantidad óptima de mariposas por jaulas individuales de oviposición es de
un promedio de 300 individuos.
- Un mayor número de machos aumenta la cantidad de huevos sugiriendo (como
se deriva de la figura 18), el armado de jaulas con una proporción sexual
hembras:machos de 1:3.
- Un futuro experimento podría realizarse controlando los niveles de densidad de
adultos, para estimar si existe además un potencial efecto de la interacción entre la
proporción sexual y el número de individuos por jaula de cría. Los resultados esperados
de un experimento de esta magnitud permitirían controlar y balancear la cantidad de
adultos de manera más eficiente para la obtención de huevos, permitiendo manejar
menos individuos con mejores resultados y con la consecuente disminución del tiempo
empleado en el proceso.
Número de parasitoidizaciones - Una parasitoidización única con C. flavipes sobre D. saccharalis realizada por
un operario es más eficiente para la producción de cocones del parasitoide, con respecto
a una parasitoidización doble.
- La parasitoidización manual doble o simple no tiene efecto sobre la proporción
sexual ni el número de adultos de C. flavipes emergidos.
- El adiestramiento de los técnicos en la selección de larvas que permitan el
normal crecimiento y desarrollo de los parasitoides hasta su emergencia es una medida
muy importante a tener en cuenta en una cría masiva para obtener resultados óptimos
que combinen eficiencia y calidad.
Edad de la larva para realizar la parasitoidización - La edad del huésped ejerce una influencia sobre la producción de parasitoides.
- El éxito en la producción de cocones así como un mayor número de adultos de
C. flavipes se obtienen a los 18 y 19 días de edad de larvas de D. saccharalis.
- La proporción sexual (hembras/machos) de individuos de C. flavipes
disminuye con el aumento de la edad larval del huésped D. saccharalis. Por lo tanto el
número de hembras de parasitoide emergidas es mayor en larvas más jóvenes.

58
- Para una línea de cría masiva se sugiere concentrar la parasitodización a los 18
días de edad larval del huésped, y a los 17 días cuando se requiera sesgar la proporción
sexual hacia las hembras.

59
8. BIBLIOGRAFÍA
1- Acatitla Trejo, C., N. Bautista Martínez, J. Vera Graziano, J. Romero Nápoles y H.
G. Calyecac Cortero. 2004. Ciclo biológico y tasas de supervivencia y reproducción de
Copitarsia incommoda Walker (Lepidoptera: Noctuidae) en cinco dietas artificiales.
Agrociencia 38: 355-363.
2- Achterberg, C. van. 1990. Illustrated key to the subfamilies of the Holarctic
Braconidae (Hymenoptera: Ichneumonoidea). Zoologische Mededelingen Leiden 64
(1): 1-20.
3- Acosta, S. y M. Galán. 1999. Efectos del superparasitismo de Cotesia flavipes Cam.
(Hymenoptera: Braconidae) sobre Diatraea saccharalis Fab. (Lepidoptera: Crambidae).
Revista de Protección Vegetal 14 (1): 55-58.
4- Anento, J. L. y J. Selfa. 1997. Himenópteros parásítica y control de plagas. En: Los
artrópodos y el hombre. Boletín de la Sociedad Entomológica Aragonesa 20: 151-160.
5- Arnold, S. J. y M. J. Wade. 1984. On the measurement of natural and sexual
selection: theory. International Journal of Organic Evolution 38 (4): 709-719.
6- Bavaresco, A., M. S. García, A. D. Grützmacher, R. Ringenberg y J. Foresti. 2004.
Adequação de uma dieta artificial para a criação de Spodoptera cosmioides (Walk.)
(Lepidoptera: Noctuidae) em laboratório. Ecology, behavior and bionomics. Neotropical
Entomology 33 (2): 155-161.
7- Berta, D. C. y S. M. Ovruski. 2008. Braconidae. En: Claps, L., Debandi, G., Roig-
Juñet, S. (Directores). Biodiversidad de artrópodos argentinos. Sociedad Entomológica
Argentina Ediciones. Vol. 2, 329-344.
8- Bessin, R. T., E. B. Moser y T. E. Reagan. 1990. Integration of management tactics
for Diatraea saccharalis (F.) (Lepidoptera: Pyralidae) populations in southern
Louisiana. Journal of Economic Entomology 83: 1563-1569.

60
9- Boggs, C. L. 1986. Reproductive strategies of female butterflies: variation in and
constraints on fecundity. Ecological Entomology 11: 7-15.
10- Boiça Jr., A. L., F. M. Lara y M. P. Bellodi. 1997. Influência de variedades de cana
de açúcar, incorporadas em dieta artificial, no desenvolvimento de Diatraea saccharalis
(Fabr.) e no seu parasitismo por Cotesia flavipes (Cam.). Anais da Sociedade
Entomológica do Brasil 26 (3): 537-542.
11- Botelho, P. S. y N. Macedo. 2002. Cotesia flavipes para o controle de Diatraea
saccharalis. En: Parra, J. R., P. S. M. Botelho, B. S. Correa Ferreira y J. M. S. Bento.
Controle biológico no Brasil. Parasitóides e predadores. Editorial Manole, São Paulo.
Cap. 25, 409-426.
12- Calderón, C. P. 1987. Cuatro dietas artificiales para la crianza masiva de Diatraea
saccharalis F. Revista Peruana de Entomología 29: 51-54.
13- Campos Farinha, A. E. de C. y J. Chaud Netto. 2000. Biologia reprodutiva de
Cotesia flavipes (Cameron) (Hymenoptera: Braconidae) V. Avaliação do número de
posturas, prole e razão sexual do parasitoide em relação ao tamaño do hospedeiro
Diatraea saccharalis Fabricius (Lepidoptera: Pyralidae). Arquivos do Instituto
Biologico, São Paulo 67 (2): 249-252.
14- Cano, M. A., E. M. dos Santos y A. de S. Pinto. 2006. Produção de Cotesia flavipes
para o controle da broca da cana. En: Controle de pragas da cana de açúcar. Boletim
Técnico Biocontrol 1. Cap. 3, 21-24.
15- CAÑAMIP. 2000. Manejo integrado de barrenadores en caña de azúcar.
CENGICAÑA, Guatemala. 26 pp.
16- Capinera, J. L. 2001. Sugarcane borer, Diatraea saccharalis (Fabricius) (Insecta :
Lepidoptera: Pyralidae). Document of Featured creatures. Entomology and Nematology
Departemnt, Florida Cooperative Extension Service. Institute of Food and Agricultural
Sciences, University of Florida. 5pp.

61
17- Chow, A. y K. M. Heinz. 2007. Use of host-size-dependent sex ratio theory in
parasitoid mass rearing. En: Hopper, K. R., K. M. Kester y K. A. Hoelmer.XIV
International Entomophagous Insects Workshop. Journal of Insect Science 7 (16), 25
pp.
18- Cohen, A. C. 2004. Insect diets. Science and technology. CRC Press. 324 pp.
19- Cueva, M., G. Ayquipa y V. Mescua. 1980. Estudios sobre Apanteles flavipes
(Cameron), introducido para controlar Diatraea saccharalis (F.) en el Perú. Revista
Peruana de Entomología 23 (1): 73-76.
20- Dhaliwal, G. S., O. Koul y R. Arora. 2004. Integrated pest management: retrospect
and prospect. En: Integrated pest management. Potential, constraints and challenges.
Cap. 1, 1-20.
21- Dobson, A. J. 1990. An introduction to generalized linear models. New York:
Chapman & Hall. 174 pp.
22- Estrada Hurtarte, R. E. 1992. Producción masiva de Spodoptera sunia y S. exigua.
CEIBA. Revista Técnica y Científica de la Escuela Agrícola Panamericana de
Zamorano, Honduras 33 (1): 161-175.
23- Gifford, J. R. y G. A. Mann. 1967. Biology, rearing and a trial release of Apanteles
flavipes in the Florida Everglades to control sugarcane borer. Journal of Economic
Entomology 60 (1): 40-47.
24- González, V. 1984. Perspectivas en la investigación sobre control del taladrador
(Diatraea spp.) y la candelilla (Aeneolamia spp.) en caña de azúcar. II Seminario sobre
los problemas de la candelilla y el taladrador en caña de azúcar y pastos. Barquisimeto,
Venezuela. 375 pp.
25- Guédez, C., C. Castillo, L. Cañizales y R. Olivar. 2008. Control biológico: una
herramienta para el desarrollo sustentable y sostenible. Academia. Trujillo, Venezuela.
7 (13): 50-74.

62
26- Hegazi, E. y W. Khafagy. 2007. Are the earlier instars better defended
immunologically tan later instars? A case study of immune dynamics in the Microplitis
rufiventris – Spodoptera littoralis system. En: Hopper, K. R., K. M. Kester y K. A.
Hoelmer.XIV International Entomophagous Insects Workshop. Journal of Insect
Science 7 (16), 25 pp.
27- Hernández, D. 2010. Estudio de algunos aspectos biológicos de Cotesia flavipes
Cameron (Hymenoptera: Braconidae) parasitoide de Diatraea saccharalis Fabricius
(Lepidoptera: Crambidae). Entomotrópica 25 (2): 69-81.
28- Jiang, N., M. Sétamou, A. J. Ngi Song y C. O. Omwega. 2004. Performance of
Cotesia flavipes (Hymenoptera: Braconidae) in parasitizing Chilo partellus
(Lepidoptera: Crambidae) as affected by temperatura and host stage. Biological Control
31: 155-164.
29- Kalnay, R. A. 1984. Diatraea spp. Lepidópteros minadores de la caña de azúcar.
Serie Didáctica. Ciencias Agrarias. Facultad de Ciencias. Agrarias. Universidad
Nacional de Jujuy. Publicación Nº1. 23 pp.
30- Lastra, L. A. y L. A. Gómez. 1988. Efecto de algunas dietas artificiales sobre la
biología del D. saccharalis F. (Lepidoptera: Pyralidae) y sus parásitos. Revista
Colombiana de Entomología 14 (2): 9-14.
31- Layward, K. J. 1942. El gusano chupador de la caña de azúcar (Diatraea
saccharalis F.). Revista Industrial Agrícola de Tucumán 32 (10-12): 315-325.
32- Linares, B. A. 1987. Influencia de la temperatura en el desarrollo de Diatraea
saccharalis Fabricius. Caña de Azúcar. Revista del Instituto Nacional de
Investigaciones Agropecuarias, Venezuela 5 (2): 43-66.
33- Luna, L. H., S. Vega Trompeta, A. Núñez y O. Carrazana. 2004. Elaboración de
uma dieta para la cria artificial de Utetheisa ornatrix venusta (Dalm.) (Lepidoptera:
Arctiidae). Revista Del Centro Agrícola, Cuba 31 (1-2): 9-12.

63
34- Magrini, E. A., J. R. P. Parra, M. L. Haddad y P. S. M. Botelho. 1995. Comparaçao
de dietas artificiais e tipos de milho, para criaçao de Anagasta kuehniella (Zeller, 1879)
(Lepidoptera: Pyralidae). Scientia Agricola, Piracicaba 52 (1): 60-64.
35- Marín, M. S., C. C. Sáez, A. E. Caballero y M. J. Quercetti. 2006. Grapholita
molesta. Evaluación de dietas larvarias artificiales. Revista de la Facultad de Ciencias
Agrarias de la Universidad Nacional de Cuyo 38 (2): 1-7.
36- Meneguim, A. M., J. R. P. Parra y M. L. Haddad. 1997. Comparação de dietas
artificiais, contendo diferentes fontes de ácidos graxos, para criação de Elasmopalpus
lignosellus (Zeller) (Lepidoptera: Pyralidae). Anais da Sociedade Entomológica do
Brasil 26 (1): 35-43.
37- Milano, P., E. B. Filho, J. R. P. Parra, M. L. Oda y F. L. Cônsoli. 2010. Efeito da
alimentação da fase adulta na reprodução e longevidade de espécies de Noctuidae,
Crambidae, Tortricidae e Elachistidae. Ecology, Behavior and Bionomics. Neotropical
Entomology 39 (2): 172-180.
38- Murúa, M. G., E. G. Virla y V. Defagó. 2003. Evaluación de cuatro dietas
artificiales para la cría de Spodoptera frugiperda (Lep.: Noctuidae) destinada a
mantener poblaciones experimentales de himenópteros parasitoides. Boletín de Sanidad
Vegetal. Plagas 29: 43-51.
39- Parra, J. R. P., P. S. M. Botelho, B. S. Correa Ferreira y J. M. S. Bento. 2002.
Controle biológico: terminologia. En: Parra, J. R., P. S. M. Botelho, B. S. Correa
Ferreira y J. M. S. Bento. Controle biológico no Brasil. Parasitóides e predadores.
Editorial Manole, São Paulo. Cap. 1, 1-16.
40- Pérez, D., C. Fandos, J. Scandaliaris, L. Mazzone, F. Soria y P. Scandaliaris. 2007.
Estado actual y evolución de la productividad del cultivo de caña de azúcar en Tucumán
y el noroeste argentino en el período 1990-2007. Publicación Especial Nº34. Estación
Experimental Agroindustrial Obispo Colombres. Tucumán. 22 pp.

64
41- Pérez, R. y W. H. Long. 1964. Sex attractant and mating behavior in the sugarcane
borer. Journal of Economic Entomology 57 (5): 688-691.
42- Pérez Moreno, I. 2000. Fundamentos teóricos del manejo integrado de plagas.
Boletín de la Sociedad Entomológica Aragonesa 27: 127-133.
43- Pexton, J. J., J. G. de Boer, G. E. Heimpel, L. E. M. Vet, J. B. Whitfield y P. J.
Ode. 2008. Competition and brood reduction: testing alternative models of clutch-size
evolution in parasitoids. Behavioral Ecology 403-409.
44- Pinto, A de S., M. A. V. Cano y E. M. dos Santos. 2006. A broca da cana, Diatraea
saccharalis. En: Controle de pragas da cana de açúcar. Boletim Técnico Biocontrol 1.
Cap. 2, 15-20.
45- Polaszek, A. y A. K. Walker. 1991. The Cotesia flavipes species-complex:
parasitoids of cereal stemborers in the tropics. Redia 74: 335-341.
46- Rasband, W. 1997. Image J. National Institutes of Health, USA.
http://rsb.info.nih.gov/ij/
47- Risco, S. H. 1971. Control biológico de los insectos de la caña de azúcar en el Perú.
Primer Congreso Latinoamericano de Entomología. Cusco. Perú. Boletín de la Sociedad
Entomológica del Perú 6 (2): 69-75.
48- Romero R, F. 2004. Manejo integrado de plagas: Las bases, los conceptos, su
mercantilización. Primera edición. Universidad Autónoma Chapingo, México. 103pp.
49- Salvador, G. y F. L. Cônsoli. 2008. Changes in the hemolymph and fat body
metabolites of Diatraea saccharalis (Fabricius) (Lepidoptera: Crambidae) parasitized
by Cotesia flavipes (Cameron) (Hymenoptera: Braconidae). Biological Control 45: 103-
110.
50- Salvatore, A. R., G. López, E. Willink, M. Ahmed y L. Varela. 2008. Ataque del
“gusano perforador de la caña de azúcar” Diatraea saccharalis en Tucumán: análisis

65
prezafra de las campañas 2003 a 2007. Revista Avance Agroindustrial. Estación
Experimental Agroindustrial Obispo Colombres, Tucumán 29 (1): 12-13.
51- Salvatore, A. R., G. López y E. Willink. 2009. Plagas en el cultivo de caña de
azúcar. En: Romero, E., Digonzelli, P. A. y Scandaliaris, J. (Editores). Manual del
cañero. Estación Experimental Agroindustrial Obispo Colombres. Cap. 11, 117-122.
52- Santillana, E. de L., H. Medrano, L. J. Galán Wong, K. Arévalo Niño, L. H.
Morales. 2003. Desarrollo y evaluación de formulados de B. thuringiensis contra
Diatraea saccharalis. Ciencia UANL. Universidad Autónoma de Nuevo León.
Monterrey, México. 6 (4): 484-490.
53- Scaglia, M., J. Chaud Netto, M. R. Brochetto Braga, S. A. Ceregato, N. Gobbi y A.
Rodrigues. 2005. Oviposition sequence and offspring of mated and virgin females of
Cotesia flavipes (Hymenoptera: Braconidae) parasitizing Diatraea saccharalis larvae
(Lepidoptera: Crambidae). Journal of Venomous Animals and Toxins including
Tropical Diseases 11 (3): 283-298.
54- Schexnayder, H. P. Jr., T. E. Reagan y D. R. Ring. 2001. Sampling for the
sugarcane borer (Lepidoptera: Crambidae) on sugarcane in Louisiana. Journal of
Economic Entomology 94 (3): 766-771.
55- Smith, L. y A. C. Bellotti. 1996. Proyectos acertados de biocontrol con énfasis
sobre el geotrópico. Proceedings Cornell Communications Conference on Biological
Control. Cornell University Press. 12 pp.
56- StatSoft, Inc. 2004. STATISTICA (data analysis software system), version 7.
www.statsoft.com.
57- Torres Vila, L. M., M. C. Rodríguez Molina y M. D. Jennions. 2004. Polyandry
and fecundity in the Lepidoptera: can methodological and conceptual approaches bias
outcomes? Behavioral Ecology and Sociobiology 55: 315-324.

66
58- Torres Vila, L. M. y M. D. Jennions. 2005. Male mating history and female
fecundity in the Lepidoptera: do male virgins make better partners? Behavioral Ecology
and Sociobiology 57: 318-326.
59- Vargas, C., P. J. Shannon, R. Taveras, F. Soto y L. Hilje. 2001. Un nuevo método
para la cría masiva de Hypsipyla grandella. Hoja Técnica N°39. Manejo Integrado de
Plagas, Costa Rica 62: 1-4.
60- Walker, D. W. 1965. Bionomics of the sugarcane borer Diatraea saccharalis (Fab.).
Adescription of the mating behavior. Proceedings of the Entomological Society of
Washington. 67 (2): 80-83.
61- Wedell, N., C. Wiklund y P. A. Cook. 2002. Monoandry and polyandry as
alternative lifestyles in a butterfly. Behavioral Ecology 13 (4): 450-455.
62- Willink, E. 1982. Especies de Diatraea en cañaverales del noroeste argentino.
Estación Experimental Agroindustrial Obispo Colombres. Tucumán. Boletín Nº140. 38
pp.
63- Willink, E., H. Salas, G. Erimbaue y G. Olaiz. 1999. El gusano perforador de la
caña de azúcar, Diatraea saccharalis. Nuevos focos en Tucumán. Revista Avance
Agroindustrial. Estación Experimental Agroindustrial Obispo Colombres, Tucumán 13-
15.
64- Yamauchi, M. N., N. Gobbi, J. Chaud Netto y A. E. Campos Farinha. 1996.
Relationship between number of ovipositions of Cotesia flavipes (Cam.) and number of
descendants emerged from its host Diatraea saccharalis (Fabr.). Anais da Sociedade
Entomológica do Brasil 26(1): 87-91.

67
9. ANEXOS
9.1. Anexo 1: Dieta artificial. Composición
La composición de la dieta artificial básica utilizada en la cría masiva en
laboratorio del CIRPON fue la siguiente:
Harina de poroto (poroto molido)………………380 g.
Harina de maíz (polenta)………………………..420 g.
Harina de soja…………………………………...380 g.
Levadura………………………………………...352 g.
Germen de trigo…………………………………240 g.
Formol…………………………………………..12 ml
Complejo Vitamínico (vía seca): 2 dosis/litro de agua
Biotina 20 mg
Niacinamida 1000 mg
Pantotenato de calcio 1000 mg
Riboflavina 500 mg
Clorhidrato de tiamina 250mg 250 mg
Clorhidrato de piridoxina 250 mg
Ácido fólico 100 mg
Cloruro de colina………………………………...6 g
Vitamina B12……………………………………120 ml
Induvet ………………………………………….3 cdtas
Metil parahidroxibenzoato (Nipagín)……………95 g
Ácido sórbico……………………………………18 g
Ácido ascórbico…………………………………35,7 g
Caragenato (gelificante)…………………………95 g
Agua…………………………………………….6 litros
Antibióticos:
Oxitetraciclina…………………………………..16 g
Estreptomicina…………………………………..1 g

68
9.2. Anexo 2: Preparación de la dieta artificial
La dieta se preparó primero mezclando por separado ingredientes secos y
líquidos. El agua caliente a una temperatura de aproximadamente 60ºC fue utilizada
para cocinar las harinas, luego se agregaron los componentes líquidos. Al mismo tiempo
se calentó agua por separado para disolver el caragenato con ayuda de una batidora.
Estas dos preparaciones se juntaron y licuaron para obtener una consistencia cremosa.
Esta dieta líquida se virtió en frascos de vidrio de 200 ml de capacidad que fueron
previamente esterilizados en autoclave a 120ºC y 1 atm de presión durante 50 minutos.
Figura 28. Frascos de vidrio con dieta artificial para larvas de D. saccharalis
Una vez llenado cada frasco con 120 ml de dieta aproximadamente se taparon
con tapas ahuecadas cubiertas con algodón o malla de alambre con papel secante (de
acuerdo a la disponibilidad de material) y previamente esterilizadas en estufa a 200ºC
durante 30 minutos. Harina de poroto y germen de trigo también fueron esterilizados en
autoclave de la misma manera que los frascos de vidrio. Una vez que los frascos
conteniendo dieta se enfriaron a temperatura ambiente, la misma se solidificó, quedando
listos para la siembra de huevos de D. saccharalis.


















