
Compartimentos intracelulares e transporte 2008 - Portal IFSCanapaula/ffi710/2008/aula8.pdf ·...
41
Compartimentos intracelulares e transporte
Transcript of Compartimentos intracelulares e transporte 2008 - Portal IFSCanapaula/ffi710/2008/aula8.pdf ·...

Compartimentos intracelulares e transporte
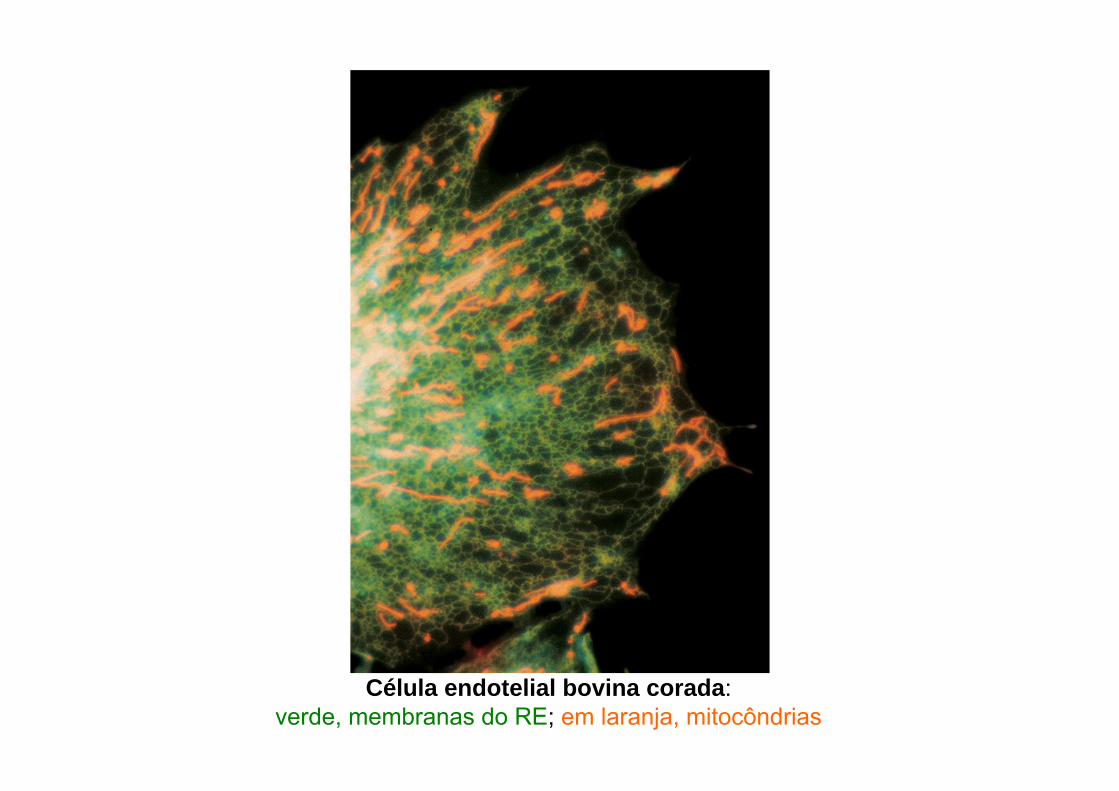
Célula endotelial bovina corada: verde, membranas do RE; em laranja, mitocôndrias

Compartimentos envoltos por membranas nas células eucarióticas
Síntese de ATP e fixação de CO2 pela fotossínteseCloroplastos
Sítio de fosforilação oxidativa.Mitocôndrias
Contém enzimas utilizadas em reações oxidativas que degradam lipídeos e destroem moléculas tóxicas.
Peroxissomos
Compartimento de distribuição de materiais endocitadosEndossomos
Pequenos sacos com enzimas digestivas. Degradam organelas esgotadas e materiais endocitados.
Lisossomos
Recebe e modifica proteínas e lipídeos vindos do RE e “despacha” para outros sítios na célula
Aparelho de Golgi
Sua membrana é contínua a membrana ext. nuclear. Sintetiza a maior parte de lipídeos e proteínas p/ distribuição
RE
Circundado pelo envelope nuclear com poros;Síntese de DNA e RNA
Núcleo

Visão geral da principal via de distribuição de proteínas em células eucarióticas

Mecanismos de importação pelas organelas
O destino de uma proteína sintetizada no citosol dependede sua seqüência de aminoácidos;
Sinal de distribuição
Se não há sinal: residem no citosol;Sinais diferentes podem direcionar p/:
núcleo, mitocôndrias, cloroplastos, peroxissomos ou RE.
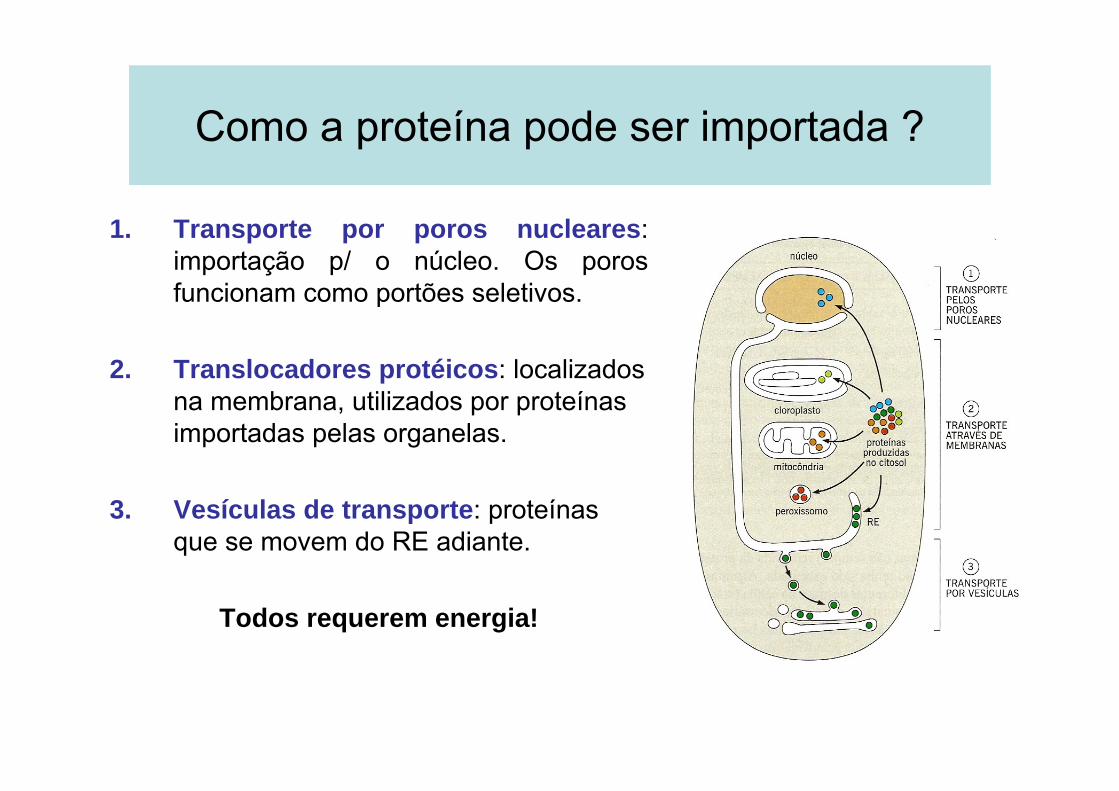
Como a proteína pode ser importada ?
1. Transporte por poros nucleares: importação p/ o núcleo. Os poros funcionam como portões seletivos.
2. Translocadores protéicos: localizados na membrana, utilizados por proteínas importadas pelas organelas.
3. Vesículas de transporte: proteínas que se movem do RE adiante.
Todos requerem energia!

Qual o compartimento destino?
Seqüência sinal (15 a 60 aminoácidos)
Sinal de distribuição típico.Freqüentemente é removido após a síntese
As seqüências-sinal que especificam o mesmo destino podem variar.
Propriedades físicas mais importantes que a seqüência exata.

Destino: núcleo• Envelope nuclear: 2 membranas concêntricas.
O envelope nuclear é perfurado por POROS NUCLEARES
Formam canais para o trânsito de macromoléculas
Entram: proteínas nuclearesSaem: RNAm maduro (mRNPs= complexo mRNA+proteínas) e
subunidades ribossomais, tRNAs
Seqüência-sinal = 1 ou 2 seq. curtas contendo várias K ou RSinal de localização nuclear

Micrografia eletrônica (criofratuta) de uma ponta de raiz de cebola mostrando o núcleo e os poros
nucleares.
Fonte:http://www.bioweb.wku.edu
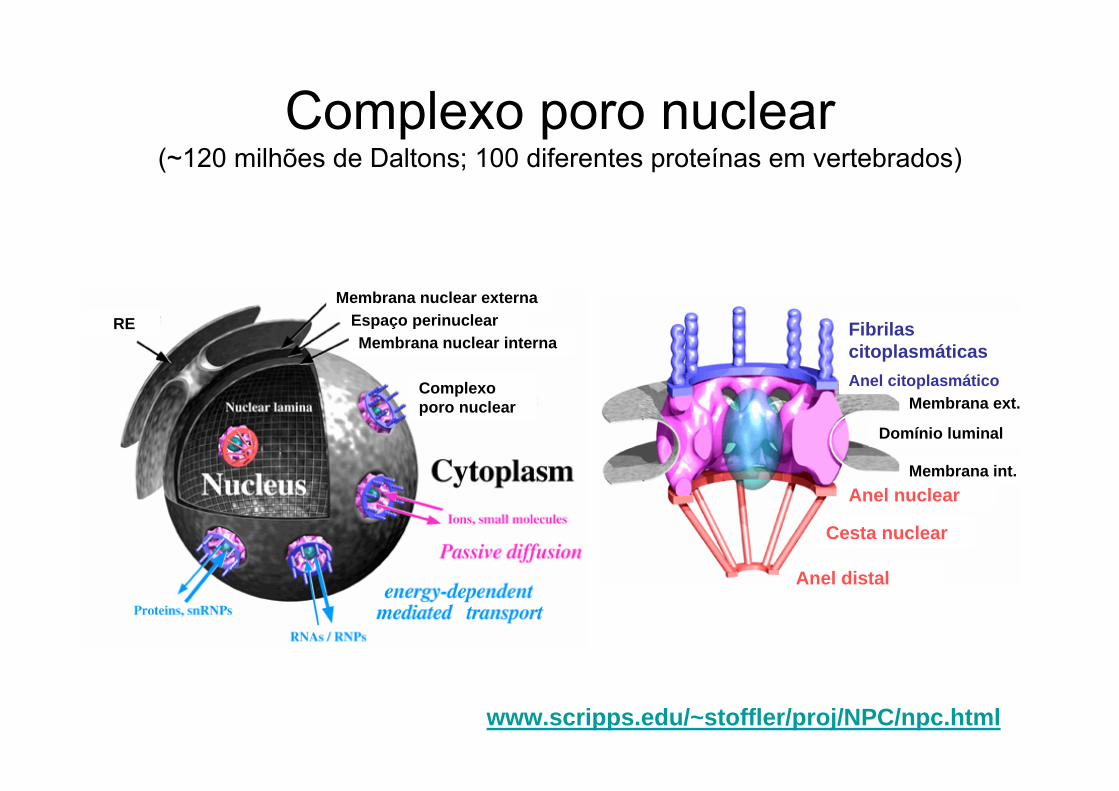
Complexo poro nuclear(~120 milhões de Daltons; 100 diferentes proteínas em vertebrados)
www.scripps.edu/~stoffler/proj/NPC/npc.html
Anel distal
Cesta nuclear
Fibrilas citoplasmáticas
Anel nuclear
Domínio luminal
Membrana int.
Membrana ext.Anel citoplasmático
Membrana nuclear externa
Membrana nuclear internaEspaço perinuclearRE
Complexo poro nuclear

Membrana nuclear externa
Membrana nuclear interna
Lâmina nuclearcesta
nuclear
Fibrilas citoplasmáticas
Fibrila nuclear

Complexo poro nuclear
Acima: microscopia-eletrônica de uma fina fatia do envelope nuclear (250 nm), não fixada e não corada revelando complexos poro-nucleares individuais.
Abaixo: mudanças estruturais reversíveis mediadas por cálcio visualizados por AFM. O anel distal (seta) pode agir como um diafragma, abrindo após a adição de quantidades milimolares de cálcio e fechando após a remoção de cálcio.
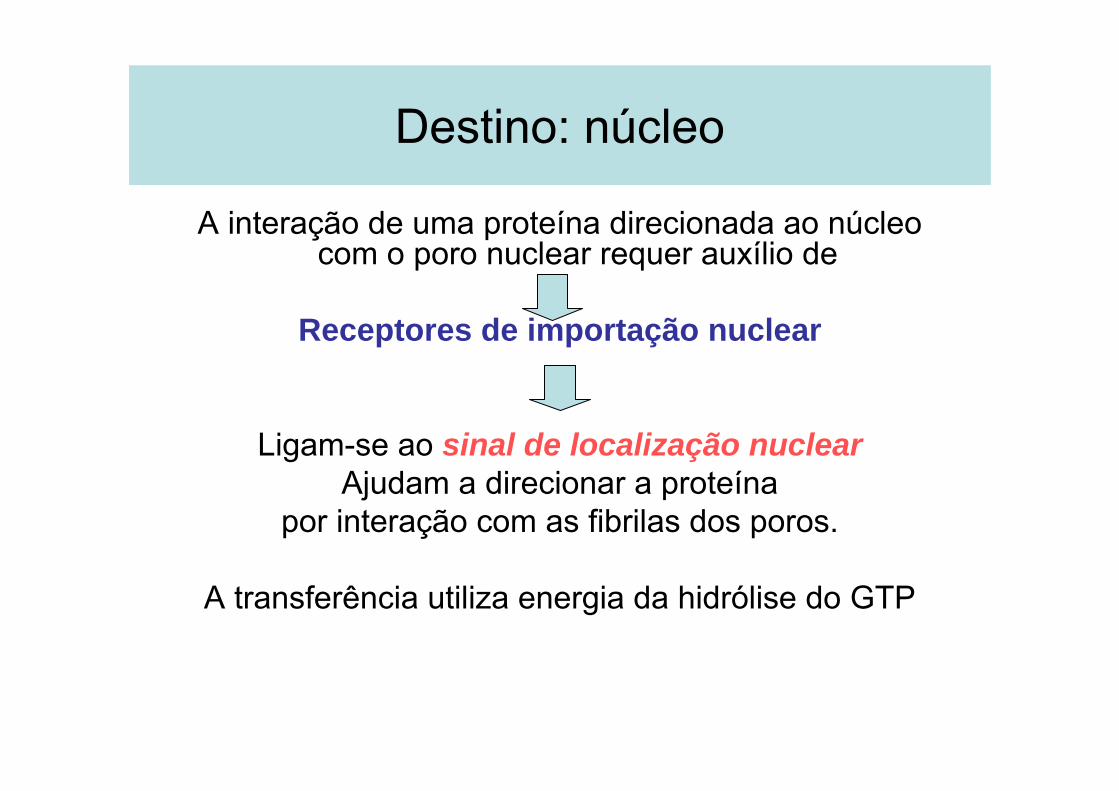
Destino: núcleo
A interação de uma proteína direcionada ao núcleo com o poro nuclear requer auxílio de
Receptores de importação nuclear
Ligam-se ao sinal de localização nuclearAjudam a direcionar a proteína
por interação com as fibrilas dos poros.
A transferência utiliza energia da hidrólise do GTP
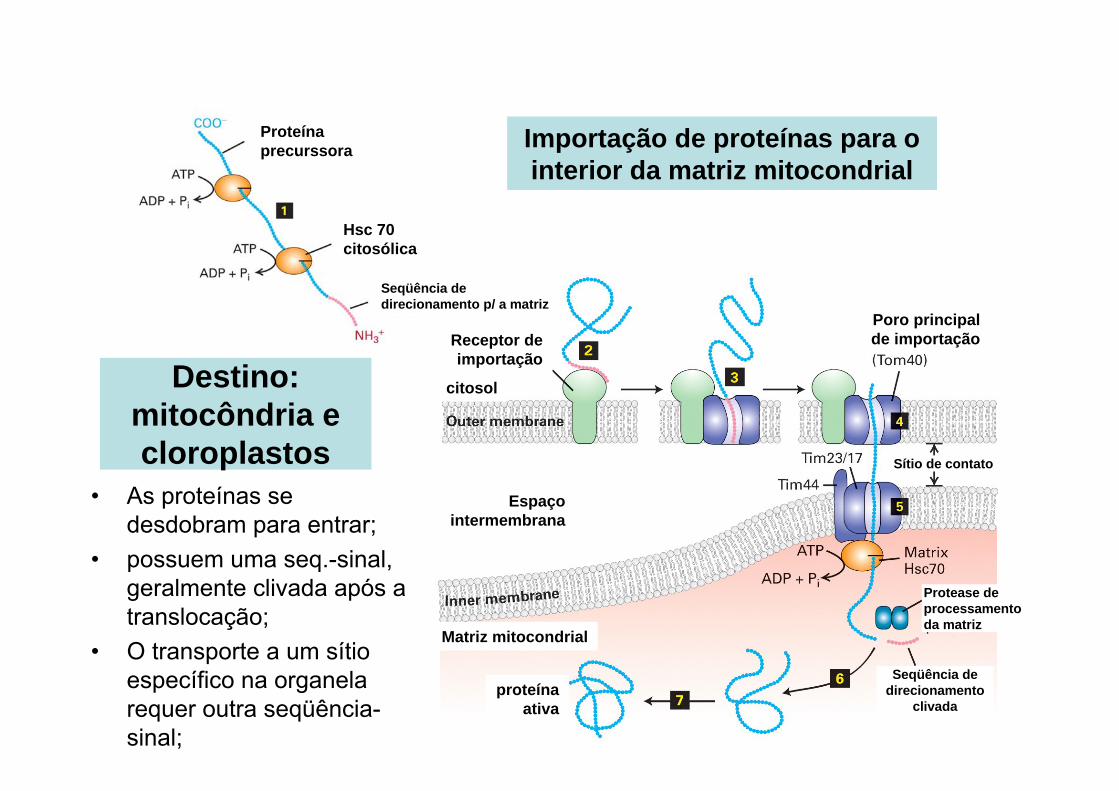
Receptor de importação
Espaço intermembrana
citosol
Matriz mitocondrial
Poro principal de importação
Proteína precurssora
Hsc 70 citosólica
Seqüência de direcionamento p/ a matriz
Seqüência de direcionamento
clivadaproteína
ativa
Sítio de contato
Importação de proteínas para o interior da matriz mitocondrial
Protease de processamento da matriz
• As proteínas se desdobram para entrar;
• possuem uma seq.-sinal, geralmente clivada após a translocação;
• O transporte a um sítio específico na organela requer outra seqüência-sinal;
Destino: mitocôndria e cloroplastos
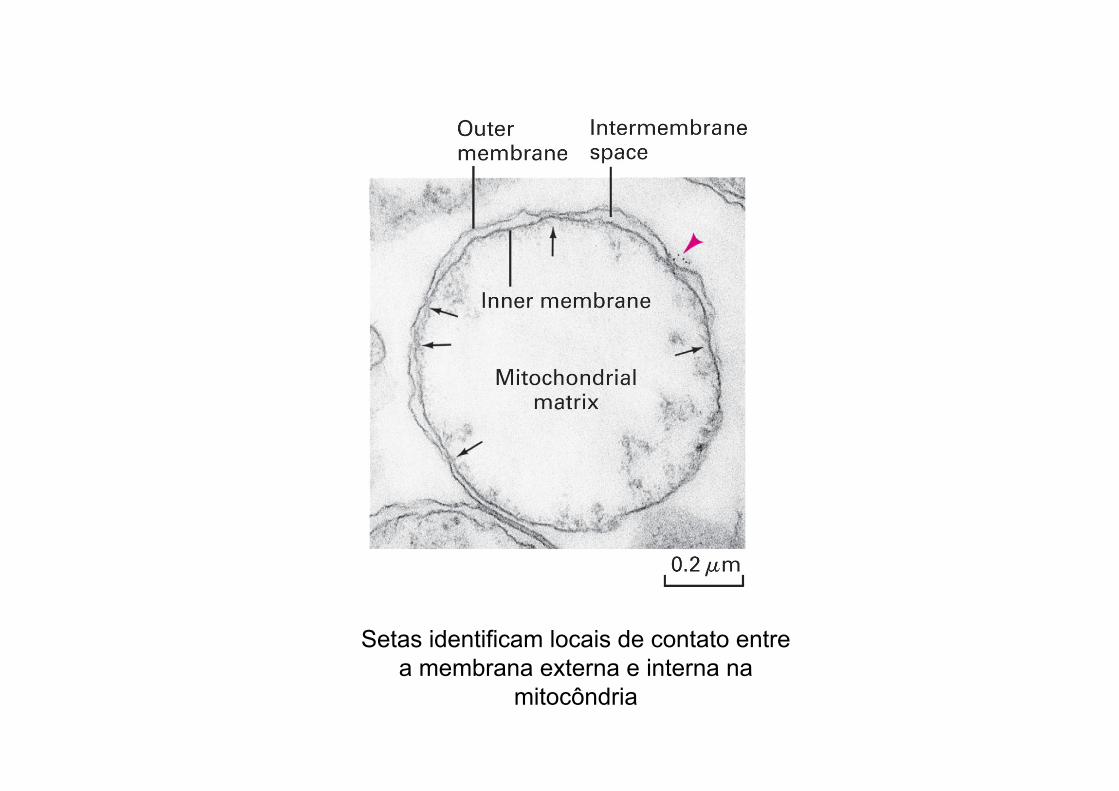
Setas identificam locais de contato entre a membrana externa e interna na
mitocôndria
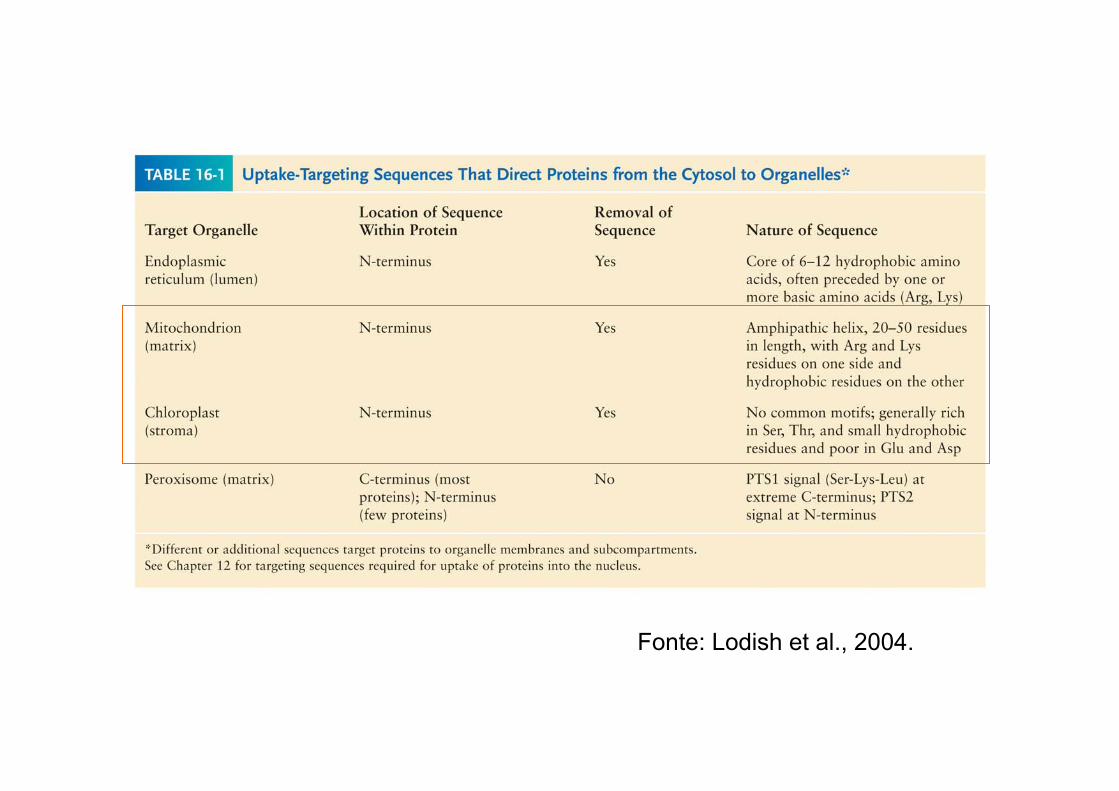
Fonte: Lodish et al., 2004.

Destino: Retículo Endoplasmático (RE)
• Porta de entrada p/ : Golgi, endossomos, lisossomos e secreção;
• Recebe 2 tipos de proteínas do citosol:
– Hidrossolúveis, que são completamente translocadas pela membrana do RE e liberadas no lúmen secreção.
– Transmembrânicas, que são parcialmente translocadas pela membrana do RE e tornam-se embebidas nela.
As proteínas entram enquanto são sintetizadas...

A seqüência-sinal é guiada
1. Por uma partícula reconhecedora de sinal(PRS ou SRP)presente no citosol, liga-se àseqüência-sinal de RE exposta
2. Pelo receptor de SRP, embebido na membrana do RE.
Há sítios moleculares de posicionamento conectando os ribossomos que estão sintetizando proteínas aos canais de translocaçãodisponíveis.
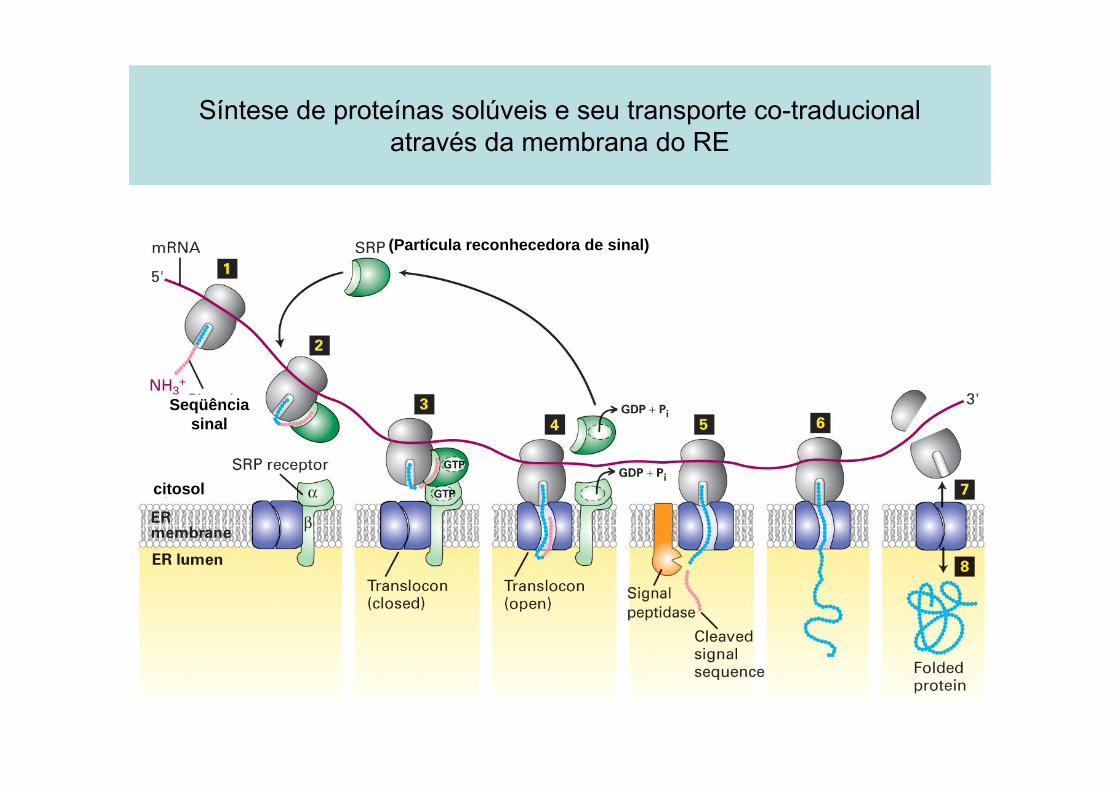
Síntese de proteínas solúveis e seu transporte co-traducionalatravés da membrana do RE
(Partícula reconhecedora de sinal)
Seqüência sinal
citosol

Síntese de proteínas transmembranasna bicamada lipídica do RE
Nem todas as proteínas que entram no RE são liberadas no lúmen...Algumas permanecem na membrana do RE (ou outras organelas)
unipasso multipasso
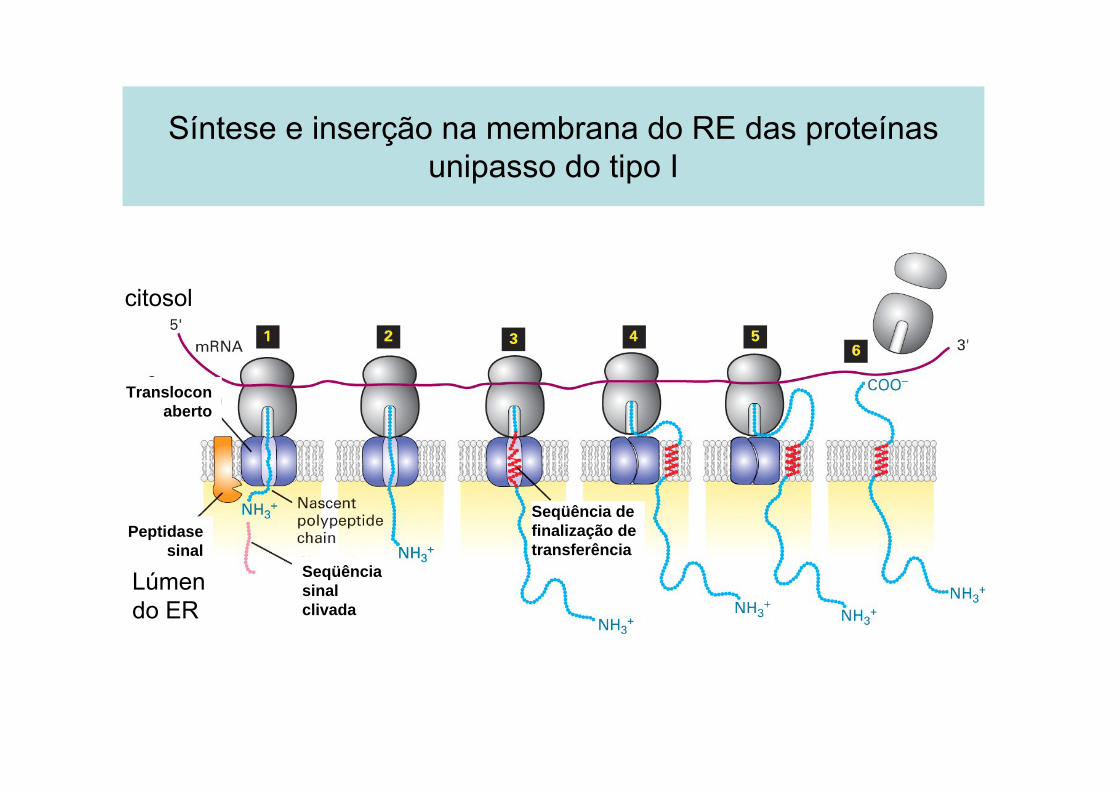
Síntese e inserção na membrana do RE das proteínas unipasso do tipo I
citosol
Lúmen do ER
Transloconaberto
Peptidase sinal
Seqüência sinal clivada
Seqüência de finalização de transferência

Síntese e inserção na membrana do RE das proteínas unipasso do tipo II
citosol
Sequênciasinal-âncorainterna
Lúmen do ER

Arranjo das sequências topogênicas nas proteínas demembrana unipasso e multipasso inseridas na membrana do RE

Algumas proteínas são presas à membrana por uma âncora fosfolipídica
Glicosilfosfatidilinositol
(GPI)
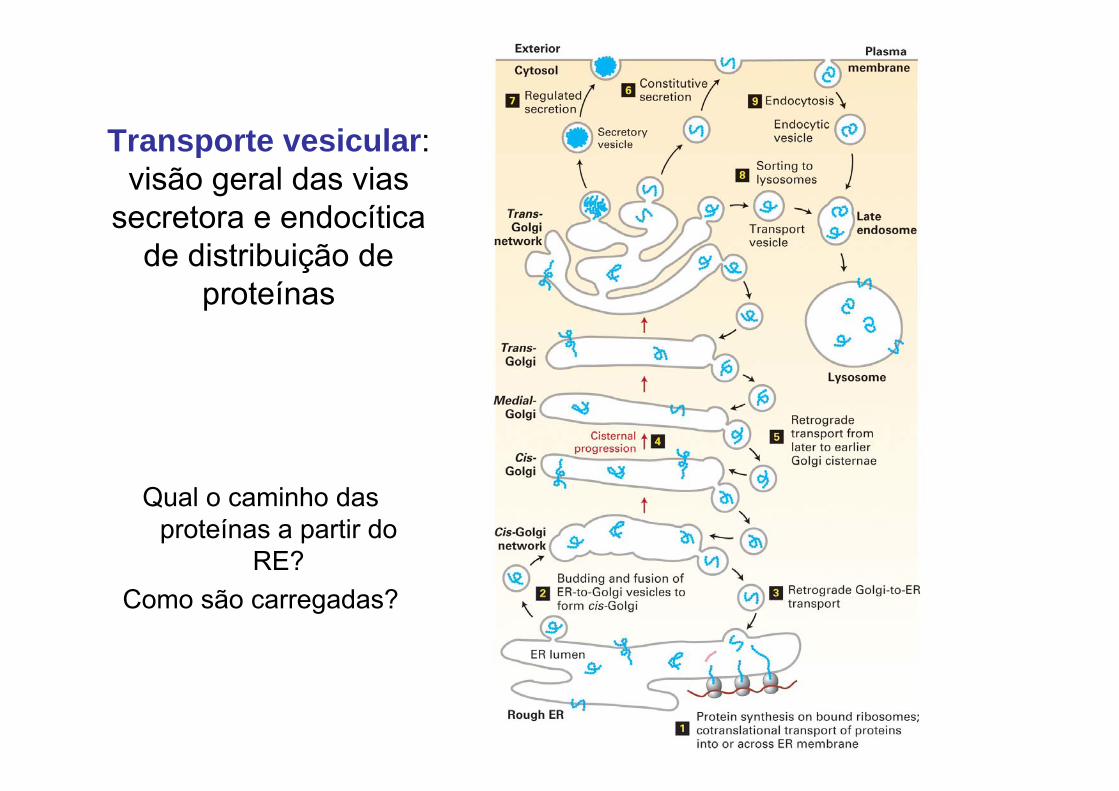
Transporte vesicular: visão geral das vias
secretora e endocíticade distribuição de
proteínas
Qual o caminho das proteínas a partir do
RE? Como são carregadas?
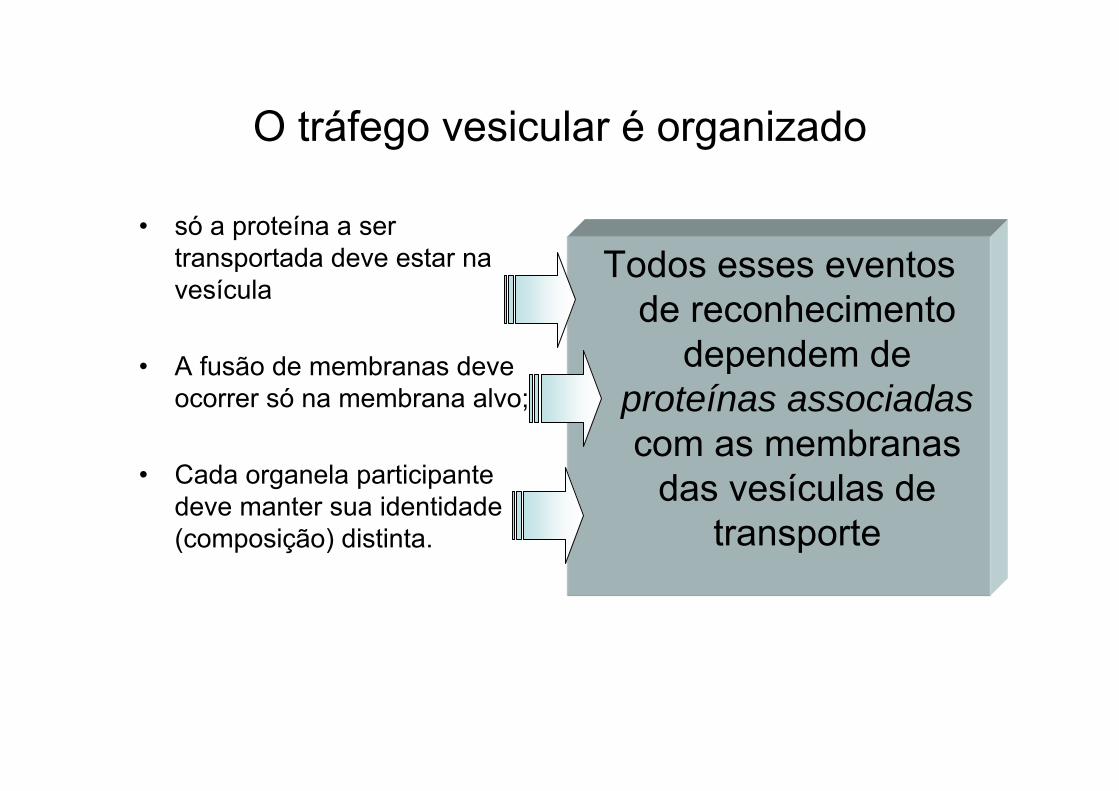
O tráfego vesicular é organizado
• só a proteína a ser transportada deve estar na vesícula
• A fusão de membranas deve ocorrer só na membrana alvo;
• Cada organela participante deve manter sua identidade (composição) distinta.
Todos esses eventos de reconhecimento
dependem de proteínas associadascom as membranas
das vesículas de transporte

Brotamento de vesículas
Vesículas que brotam das membranas têm uma capa protéica
VESÍCULAS REVESTIDAS
Depois de formada, a vesícula perde seu revestimento, permitindo sua interação com a membrana destino
Funções da capa: - dar forma à membrana no brotamento
- ajudar na captura de moléculas p/ o transporteEx. vesículas com capas de clatrina; COPIs.
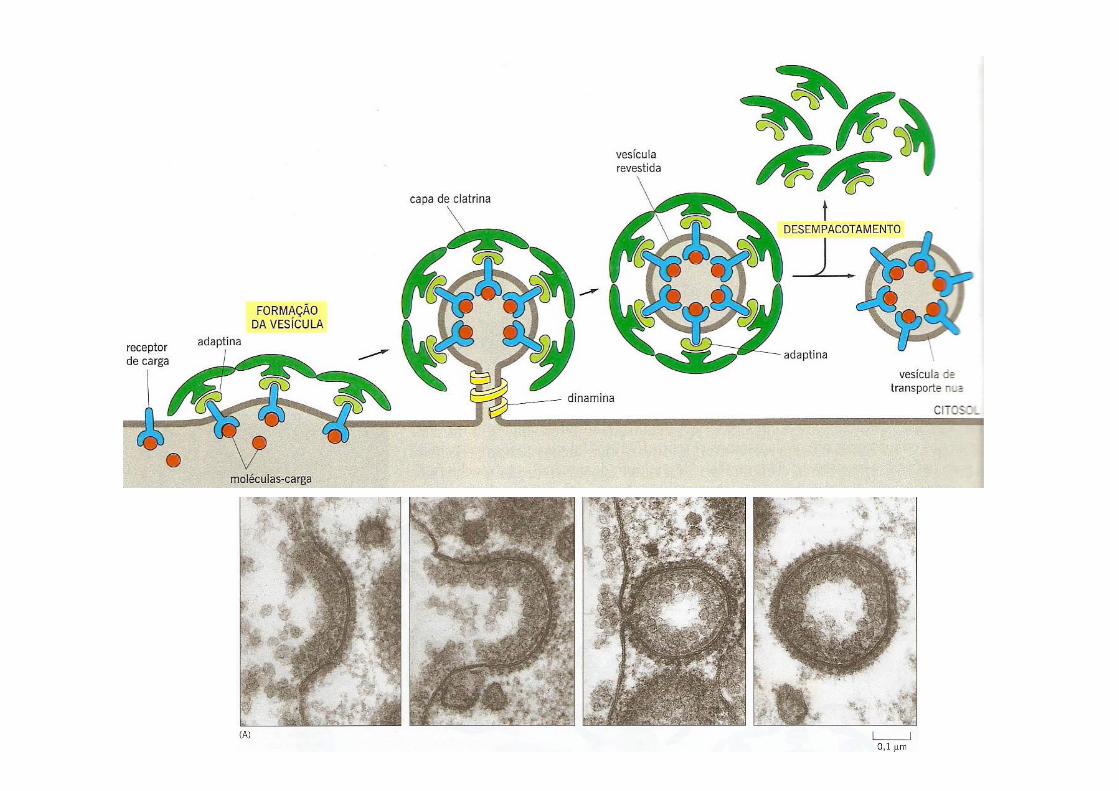
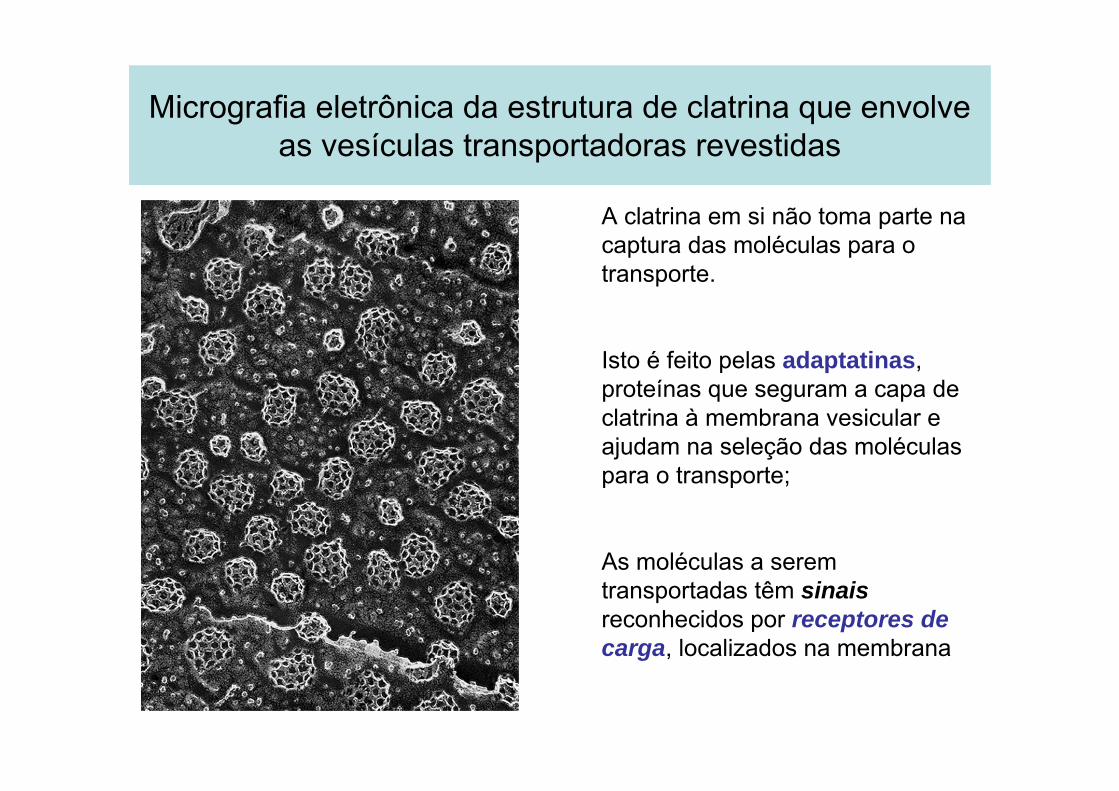
Micrografia eletrônica da estrutura de clatrina que envolve as vesículas transportadoras revestidas
A clatrina em si não toma parte na captura das moléculas para o transporte.
Isto é feito pelas adaptatinas, proteínas que seguram a capa de clatrina à membrana vesicular e ajudam na seleção das moléculas para o transporte;
As moléculas a serem transportadas têm sinais reconhecidos por receptores de carga, localizados na membrana

vesícula
dinamina

Como a vesícula encontra seu destino?
• A vesícula é transportada ativamente ao longo do citoesqueleto;
• Atingida a organela alvo a vesícula tem que reconhecê-la e ancorar nela;
• A especificidade de ancoramento depende das SNAREs
Proteínas transmembrana que certificam a seleção e auxiliam no ancoramento.

Modelo de fusão mediado por SNAREs. v-SNARE em verde, t-SNARE em rosa. Adaptado de Cell 92 (6).
O modelo postula que a fusão se dá pela interação das SNAREs forçando as membranas a um contato próximo.
As SNAREs formam um extenso coiled coil ancorado por regiões transmembranas de v- e t-SNAREs. A interação deve induzir um estresse
local na membrana, resultando na fusão.

Figura adaptada de: Littleton, J. T. et al. Synaptotagmin mutants reveal essential functionsfor the C2B domain in Ca2+-triggered fusion and recycling of synaptic vesicles in vivo. J. Neurosci. 21, 1421–1433 (2001).

Rotas secretoras


Modificações nas proteínas e controle de qualidade no RE
As proteínas que passam pelo RE sofrem 4 modificações principais antes de alcançarem seu
destino final:
1. Adição e processamento de carboidratos no RE e Golgi (S e T, O-ligados e N, N-ligados);
2. Formação de pontes dissulfeto no RE3. Dobramento apropriado e montagem de subunidades
no RE;4. Clivagens proteolíticas específicas no RE, Golgi e
vesículas secretoras.

Destino : Golgi

RE ou Golgi
O que determina se a proteína fica no RE ousegue a rota de exocitose?
• Só proteínas enoveladas saem do REaquelas não ou parcialmente enoveladas são retidasProteínas mal enoveladas são direcionadas para o citosol para degradação proteassômica.
• Proteínas que devem funcionar no RE: têm sinal de retenção no RE, reconhecido por uma proteína receptora ligada à membrana do RE.

Rota endocítica• Captura fluidos, moléculas grandes e pequenas.• O material a ser ingerido é progressivamente capturado
numa vesícula de membrana plasmática (endocítica)• Pode ser mediada por receptor ou não;• O pH ácido dos endossomos pode levar a dissociação
do receptor da carga:→ a carga é entregue aos lisossomos eo receptor é reciclado.→ se houver a dissociação a carga seguirá
outro destino.
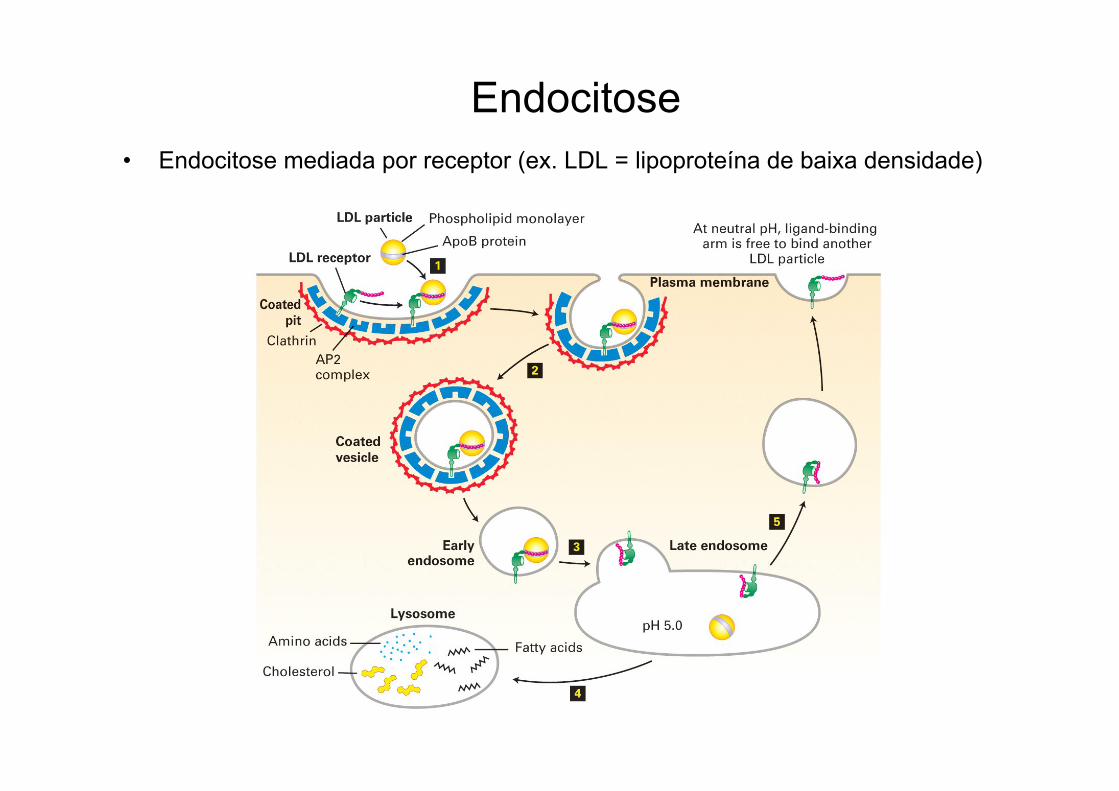
Endocitose• Endocitose mediada por receptor (ex. LDL = lipoproteína de baixa densidade)
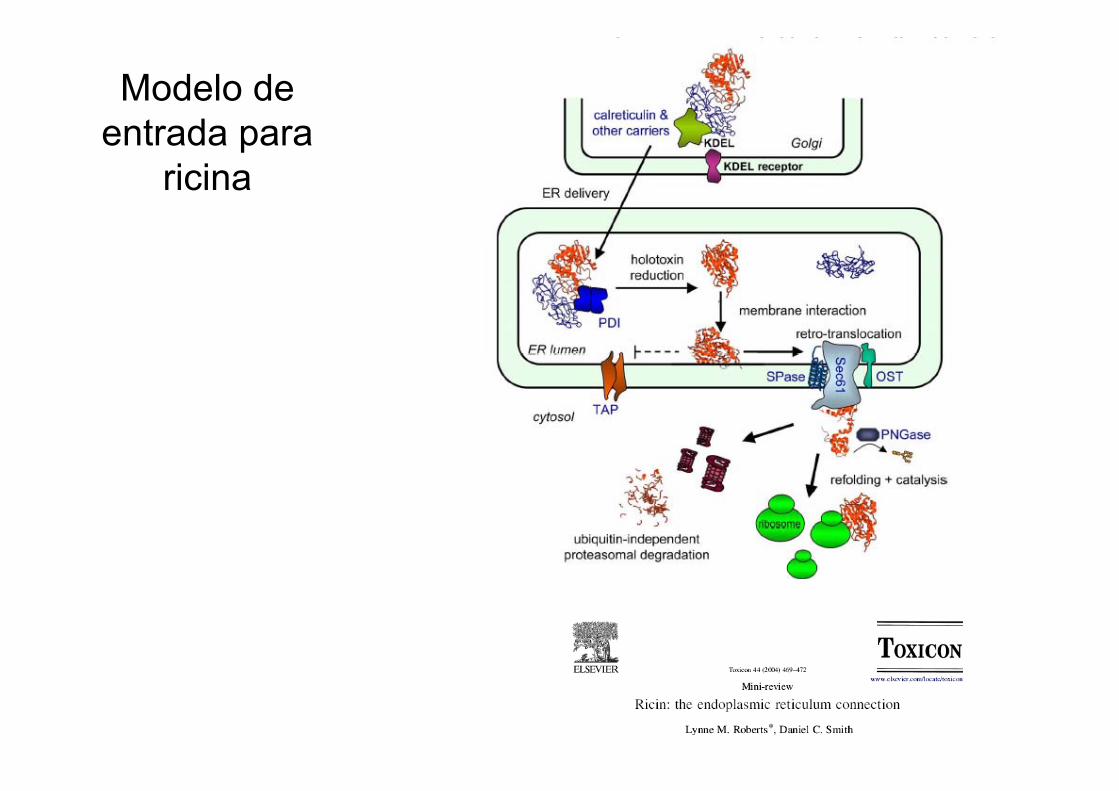
Modelo de entrada para
ricina


















