
CAPÍTULO 14 recursos entre especies herbívoras, la...
23
CAPÍTULO 14 LA UTILIZACIÓN DE LOS PASTOS POR LOS GRANDES HERBÍVOROS: PRINCIPIOS BÁSICOS Y CASOS DE ESTUDIO R. García-González Introducción Tal como señalaba nuestro insigne ecólogo Ramón Margalef (1988), el pasto, al igual que el plancton, posee una alta tasa de renovación, lo que quiere decir, que la relación entre el valor de la producción y la biomasa es más alta que en otros ecosistemas (ver figura 11.2). Este excedente de producción es aprovechado por los herbívoros mediante el pastoreo, transformándolo en energía de mantenimien- to y en producciones, que en el caso de los animales domésticos y cinegéticos, interesan al hombre (carne, leche, lana, trofeos). Pero el pastoreo ocasiona a su vez efectos interactivos con el pasto, que pueden llegar a alcanzar bas- tante complejidad, tal como se expuso en el capítulo 3. Por esta razón, es difícil encontrar patrones universales que permitan modelizar la utilización del pasto por parte de los herbí- voros domésticos en pastoreo libre. En ocasio- nes es necesario recurrir a patrones generales de uso del hábitat observados en sus congéne- res salvajes. Sin embargo, intentaremos descri- bir en este capítulo ciertas tendencias o regu- laridades observables en el uso de los pastos, que en el caso de los pastos de montaña, adop- tan variaciones particulares. Dichas particula- ridades son debidas fundamentalmente al gra- diente climático que impone la altitud y a la heterogeneidad ambiental. Revisamos algunos de estos principios generales a través de cinco grandes temas interconectados entre sí: la optimización de la ingestión, la complementación espacial en el uso de los recursos pastables, la partición de recursos entre especies herbívoras, la selección de la dieta y la conducta de pastoreo. Sin em- bargo, previamente, es necesario realizar algu- nas consideraciones acerca de la jerarquiza- ción espacial del proceso selectivo (cambios de escala). Los cambios espaciales de escala En la interacción entre los herbívoros y sus recursos alimentarios, suelen reconocerse dis- tintos niveles o escalas espacio-temporales, de forma que la búsqueda y recolección del ali- mento es abordada como un proceso multi- escala. Las decisiones cambian notablemente cuando se pasa de un nivel a otro (propiedades emergentes), aunque conservando aspectos comunes que indican la existencia de conexio- nes e interdependencias entre los distintos niveles. Una misma decisión influye sobre varias escalas a la vez, pero de forma diferente. Cuando el herbívoro selecciona un tipo de forraje determinado, de forma simultánea está eligiendo aquel tipo de hábitat donde menor resulta ser el esfuerzo y coste energético de la búsqueda de dicho alimento. Por ello, muchas veces resulta difícil desvincular ambas escalas, aunque teóricamente sean claramente diferen- tes. Así por ejemplo, los cambios en la vegeta- ción (especialmente su diversidad), produci- dos por el pastoreo de ungulados, puede reper- cutir, en mayor o menor grado, de una escala a otra, dependiendo de la capacidad de persis- tencia (“resilience”) y de colonización de esa vegetación (Austrheim y Erikson, 2001). Dentro de la teórica gradación de esca- las posibles, la selección de los recursos natu- rales puede analizarse según distintos niveles espaciales (figura 14.1), que a su vez presentan un rango de desarrollo temporal diferente. Por un lado, una escala regional que sería una esca- la de poco detalle, en la cual existe una combi- nación de unidades paisajísticas. En el caso de cap.14 | 205
Transcript of CAPÍTULO 14 recursos entre especies herbívoras, la...

CAPÍTULO 14LLAA UUTTIILLIIZZAACCIIÓÓNN
DDEE LLOOSS PPAASSTTOOSS PPOORR
LLOOSS GGRRAANNDDEESS HHEERRBBÍÍVVOORROOSS::
PPRRIINNCCIIPPIIOOSS BBÁÁSSIICCOOSS
YY CCAASSOOSS DDEE EESSTTUUDDIIOO
R. García-González
IInnttrroodduucccciióónn
Tal como señalaba nuestro insigne ecólogo
Ramón Margalef (1988), el pasto, al igual que
el plancton, posee una alta tasa de renovación,
lo que quiere decir, que la relación entre el
valor de la producción y la biomasa es más
alta que en otros ecosistemas (ver figura 11.2).
Este excedente de producción es aprovechado
por los herbívoros mediante el pastoreo,
transformándolo en energía de mantenimien-
to y en producciones, que en el caso de los
animales domésticos y cinegéticos, interesan al
hombre (carne, leche, lana, trofeos). Pero el
pastoreo ocasiona a su vez efectos interactivos
con el pasto, que pueden llegar a alcanzar bas-
tante complejidad, tal como se expuso en el
capítulo 3. Por esta razón, es difícil encontrar
patrones universales que permitan modelizar
la utilización del pasto por parte de los herbí-
voros domésticos en pastoreo libre. En ocasio-
nes es necesario recurrir a patrones generales
de uso del hábitat observados en sus congéne-
res salvajes. Sin embargo, intentaremos descri-
bir en este capítulo ciertas tendencias o regu-
laridades observables en el uso de los pastos,
que en el caso de los pastos de montaña, adop-
tan variaciones particulares. Dichas particula-
ridades son debidas fundamentalmente al gra-
diente climático que impone la altitud y a la
heterogeneidad ambiental.
Revisamos algunos de estos principios
generales a través de cinco grandes temas
interconectados entre sí: la optimización de la
ingestión, la complementación espacial en el
uso de los recursos pastables, la partición de
recursos entre especies herbívoras, la selección
de la dieta y la conducta de pastoreo. Sin em-
bargo, previamente, es necesario realizar algu-
nas consideraciones acerca de la jerarquiza-
ción espacial del proceso selectivo (cambios
de escala).
LLooss ccaammbbiiooss eessppaacciiaalleess ddee eessccaallaa
En la interacción entre los herbívoros y sus
recursos alimentarios, suelen reconocerse dis-
tintos niveles o escalas espacio-temporales, de
forma que la búsqueda y recolección del ali-
mento es abordada como un proceso multi-
escala. Las decisiones cambian notablemente
cuando se pasa de un nivel a otro (propiedades
emergentes), aunque conservando aspectos
comunes que indican la existencia de conexio-
nes e interdependencias entre los distintos
niveles. Una misma decisión influye sobre
varias escalas a la vez, pero de forma diferente.
Cuando el herbívoro selecciona un tipo de
forraje determinado, de forma simultánea está
eligiendo aquel tipo de hábitat donde menor
resulta ser el esfuerzo y coste energético de la
búsqueda de dicho alimento. Por ello, muchas
veces resulta difícil desvincular ambas escalas,
aunque teóricamente sean claramente diferen-
tes. Así por ejemplo, los cambios en la vegeta-
ción (especialmente su diversidad), produci-
dos por el pastoreo de ungulados, puede reper-
cutir, en mayor o menor grado, de una escala a
otra, dependiendo de la capacidad de persis-
tencia (“resilience”) y de colonización de esa
vegetación (Austrheim y Erikson, 2001).
Dentro de la teórica gradación de esca-
las posibles, la selección de los recursos natu-
rales puede analizarse según distintos niveles
espaciales (figura 14.1), que a su vez presentan
un rango de desarrollo temporal diferente. Por
un lado, una escala regional que sería una esca-
la de poco detalle, en la cual existe una combi-
nación de unidades paisajísticas. En el caso de
cap.14 | 205

los animales domésticos, este nivel de selec-
ción (y a menudo el siguiente), son ejercidos
por el ganadero, a partir de las posibilidades y
limitaciones de las unidades administrativas y
de pastoreo. La escala paisaje incluiría unida-
des fisionómicas y topográficas de característi-
cas similares (laderas, pequeñas cuencas, etc.).
Correspondería aproximadamente a lo que en
la fauna silvestre se conoce como hábitat
(escala intermedia, referente al área de cam-
peo o dominio vital de una población). Le
seguiría el nivel de comunidad vegetal, donde
las unidades vegetales son consideradas de
forma separada, y el nivel de 'parcela'
(“patch”), dentro de una misma comunidad
vegetal (escala de mayor detalle, correspon-
diente a las “estaciones de alimentación (fee-
ding station)”). Se entiende por “estación de
alimentación” el área que alcanza la cabeza y
cuello del animal sin mover las patas delante-
ras. Por último, a una escala de mayor detalle,
tendríamos la selección a nivel de especies y de
partes de la planta. Cada una de estas escalas
está definida, tanto por la capacidad de per-
cepción de los recursos y del medio por parte
de los herbívoros y sus respuestas a los estí-
mulos ambientales, como por el tipo e intensi-
dad de control humano en los procesos en que
interviene.
En los sistemas pastorales pirenaicos,
sometidos a un importante control humano,
la influencia de este control sobre las decisio-
nes de los herbívoros presenta también rasgos
muy diferentes según sea la escala de análisis
considerada. Así, por ejemplo, cuando un
rebaño de ovejas es guiado por un pastor, las
decisiones de éste pueden determinar en gran
medida la selección de los recursos a escala
regional o paisajística, pero sin embargo, pue-
den no afectar en la selección a nivel comuni-
dad vegetal o de planta. En términos generales
podemos considerar que, en el uso del espacio
que realizan los animales domésticos en
pastoreo extensivo, la capacidad de selección a
escala regional es muy reducida o inexistente.
Sin embargo, dependiendo de las condiciones,
esta capacidad puede ser importante a escalas
menores.
LLaa ooppttiimmiizzaacciióónn ddee llaa ttaassaa
ddee iinnggeessttiióónn
Basándonos en la teoría de la dieta óptima,
por la cual los animales tratan de conseguir el
máximo de energía y nutrientes con el míni-
mo coste energético (Pianka, 1982), podría-
mos establecer que, uno de los principios bási-
cos del uso del pasto es que los herbívoros
siempre tratan de maximizar la tasa de inges-
tión de hierba (cantidad de biomasa por uni-
dad de tiempo) (Huston y Pinchak, 1991). A
ello, evolutivamente, se ha opuesto la tenden-
cia contraria por parte del pasto de “no dejarse
comer” (Malechek y Balph, 1987). Esta inter-
acción ha dado lugar a adaptaciones mutuas,
algunas de las cuales se han expuesto en
206 | cap.14
Fig.14.1. Jerarquía ecológica con la que se encuentran los grandesherbívoros cuando pastan. Las escalas están determinadas portasas y rendimientos de procesos ecológicos y de pastoreo. Los lími-tes entre unidades en cada escala están determinados por la con-ducta animal. (Adaptado de Senft et al., 1987)

capítulos precedentes. Sin embargo, el herbí-
voro no puede consumir ilimitadamente aun-
que disponga de alimentos en abundancia. La
ingestión voluntaria de alimentos tiene un
límite, que se reconoce a través de la respuesta
funcional (figura 14.2).
Algunos de los factores que influyen en esa
limitación son la capacidad del retículo-
rumen del rumiante y la digestibilidad de la
hierba. El rumen es una cámara fermentativa
en donde se procesa parte de la fracción fibro-
sa de los vegetales. Antes de permitir la entra-
da de nuevo alimento, el rumen debe vaciarse
parcialmente, pasando parte de su contenido a
las cámaras digestivas siguientes (libro, cuajar)
y al intestino. El paso se hace a través del ori-
ficio retículo-omasal, pero éste actúa como
una válvula, que no permite el paso de conte-
nido ruminal si éste no tiene una textura
determinada (Beever et al., 2000). Cuando los
alimentos son muy fibrosos y lignificados
(baja digestibilidad), tardan más tiempo en
fragmentarse y en abandonar el rumen, con lo
cual la tasa de ingestión se reduce. De hecho,
existe una relación estrecha entre la tasa de
ingestión y la digestibilidad de la hierba, según
han puesto de manifiesto diversos autores
(figura 14.3).
A menudo la ingestión de alimentos por parte
de los grandes herbívoros se expresa como el
producto de tres factores:
I = TP · RB · TB
Siendo, I la ingestión diaria, TP el tiempo de
pastoreo (minutos dedicados a pastar al día),
RB el ritmo de bocados (número de bocados
por minuto) y TB el tamaño del bocado
expresado en gramos. El tamaño del bocado
(TB) está muy ligado a la estructura del pasto,
mientras que el tiempo de pastoreo (TP) y el
ritmo de bocados (RB) están más relaciona-
dos con el estado de hambre o saciedad del
animal. Se ha comprobado que el tamaño de
bocado depende de la altura de la hierba, pero
cuando ésta disminuye, la densidad del pasto
también es un factor importante (Coates y
Penning, 2000). Una vez establecido el tamaño
de mordisco, mediante el registro detallado de
la conducta ingestiva, puede determinarse el
número de mordiscos y el tiempo de inges-
tión, con lo cual puede estimarse el consumo
diario de hierba de los animales en pastoreo.
Los modelos de optimización de la tasa
de ingestión, desarrollados a escala de detalle
(altura del pasto), permiten también com-
prender y modelizar lo que es observable a
escala de paisaje. Por ejemplo, una práctica
conocida del pastoreo de ovejas en los puertos
de verano es el desplazamiento altitudinal en
cap.14 | 207
Fig. 14.2. Efecto de la altura del pasto sobre la tasa de ingestión enovejas Scottish Halfbred pastando en una pradera de Lolium peren-ne. Obsérvese que a partir de 6 cm de altura de la hierba la tasa deconsumo no se incrementa (según Penning, 1986).
Fig. 14.3. Relación entre el consumo de hierba y la digestibilidad dela materia orgánica (DOM) para varias comunidades pascícolas delas montañas de Cheviot (Reino Unido) (según Armstrong et al.,1986).

función del estado nutritivo de la hierba. En la
figura 14.4 se presenta un modelo idealizado de
utilización de la producción vegetal por parte
de las ovejas en los puertos de verano, en fun-
ción del tiempo y de la altitud. En altitudes
relativamente bajas (1600 m) la producción
primaria tiene dos máximos. El primero
corresponde al máximo primaveral típico, el
segundo, cuantitativamente menor, se produ-
ce por el rebrote de la vegetación tras las llu-
vias de final de verano y por el estímulo que
ejerce el pastoreo previo, sobre el crecimiento
vegetal. En años de sequía este segundo máxi-
mo puede faltar. A medida que se asciende en
altitud, estos dos picos de producción se hacen
más pequeños y más próximos en el tiempo.
Ello se debe a que la temperatura desciende
0,6º C como media por cada 100 m de ascenso
altitudinal y los máximos de producción se
producen más tarde cuanto más alto se ascien-
de. Cuando la altitud es relativamente elevada
(2400 m) las condiciones ambientales (fusión
tardía de la nieve, temperaturas bajas), no per-
miten más que un sólo pico de producción
herbácea estival. Así pues, la característica
diferencial de la montaña respecto al llano, es
que la producción herbácea se escalona a lo
largo del tiempo, produciéndose un desfase
fenológico cuando se progresa en altitud
(onda fenológica). En Junio se producen
máximos de floración y producción a 1600 m,
mientras que a 2400 m todavía persiste la
nieve (figura 14.5). En Agosto, cuando la hierba
se seca a 1600 m, a 2400 m se encuentra en su
momento óptimo vegetativo (de calidad y
producción). Se estima que este desfase es del
208 | cap.14
Fig. 14.4. Modelo idealizado de optimización del pastoreo estival en pastos supraforestales pirenaicos. Mediante el desplazamiento altitudinallos herbívoros consiguen acceder durante el mayor tiempo posible a los máximos de producción vegetal (según García-González et al., 1990).
Fig. 14.5. Onda fenológica en la Montaña de Sesa. En junio el pasto se encuentra verde y en su mejor calidad en las partes bajas, seco enaltitudes medias, y en las cumbres nevadas todavía no ha empezado a crecer. A mitad de agosto la situación se invertirá (cumbres verdes ypartes bajas secas). (Foto: M. Gartzia).

orden de un mes por cada 300-500 m de des-
nivel, para la misma especie vegetal. Ascen-
diendo progresivamente en altitud durante la
primera mitad del verano y descendiendo en
la segunda mitad, los herbívoros pueden
tener acceso al pasto de mejor calidad durante
todo el verano (figura 14.4; Chatelain y Troxler,
2005).
La altitud no es la única causa de varia-
ción fenológica, ya que la microtopografía
(contraste solana-umbría, concavidad-con-
vexidad) también la condiciona notablemente.
Todo ello produce un mosaico de vegetación
en diferentes estados fenológicos y producti-
vos, distribuidos heterogéneamente por el ter-
ritorio, tanto vertical como horizontalmente.
Una correcta estrategia de pastoreo, consiste
en aprovechar los máximos de producción
herbácea allí y cuando se producen. Por
eso los pastores adaptados a su ambiente,
mueven sus rebaños horizontal y vertical-
mente siguiendo la onda fenológica vegetal (o
los máximos de calidad y producción), tal
como han hecho los herbívoros salvajes desde
la prehistoria. Con ello se consigue optimizar
el aprovechamiento de los pastos de alta
montaña.
CCoommpplleemmeennttaarriieeddaadd eessppaacciiaall
eenn eell uussoo ddee llooss rreeccuurrssooss yy
ssuu sseelleecccciióónn aa eessccaallaa rreeggiioonnaall
((eell ssiisstteemmaa vvaallllee--ppuueerrttoo))
En un sistema interactivo, como es el del pasto
y los herbívoros que lo utilizan, los máximos
de producción de ambos elementos deberían
estar lo más ajustados posibles para que no se
produzcan desequilibrios, tanto espacial como
temporalmente. Así por ejemplo, en latitudes
templadas, la época de partos y cría de la
mayoría de los herbívoros salvajes sucede en
primavera, cuando los recursos vegetales son
más abundantes. Sin embargo, lo habitual es
que se produzcan desajustes entre las produc-
ciones vegetal y animal. A lo largo de un ciclo
anual, el periodo de crecimiento y reproduc-
ción vegetal suele ser breve y el periodo de
senescencia muy prolongado, especialmente
en climas fríos y de montaña. Sin embargo, los
herbívoros homeotermos deben satisfacer
unas necesidades energéticas mínimas a lo
largo de todo el año. Frecuentemente, el máxi-
mo productivo de la vegetación supera larga-
mente las necesidades del herbívoro en ese
momento, con lo cual se producen excedentes,
(que pueden ser recolectados y conservados
cap.14 | 209
Fig. 14.6. Evolución de las necesidades energéticas en Unidades Forrajeras/día (línea continua) y de las cantidades de energía ingerida(líneas a trazos) durante el ciclo de explotación de una vaca adulta (650 kg) de raza Charolais. Durante los meses del máximo crecimientode la hierba la oferta del pasto supera las necesidades (según Jarrige, 1990).
por el hombre). Durante el periodo de dia-
pausa de la vegetación su valor energético no
llega a cubrir las necesidades mínimas de los
herbívoros (figura 14.6) y el ganadero suministra
a sus animales alimentación suplementaria en
esa época de escasez.
En condiciones naturales los herbívoros
silvestres afrontan estos desfases productivos
de varias formas. Por ejemplo, ajustando su
densidad a la capacidad de carga del territorio,
o mediante adaptaciones morfo-fisiológicas
de su sistema digestivo, que les permiten apro-
vechar los alimentos ligno-celulósicos más efi-
cientemente. Una de las formas más habitua-
les de enfrentarse a estos ciclos temporales de
escasez-producción, es por medio de las
migraciones estacionales. Cuando los recursos
alimentarios empiezan a escasear en una zona,
los herbívoros se desplazan hacia otros territo-
rios en donde las condiciones ambientales
permiten que exista una producción vegetal
más elevada. A menudo estas migraciones son
cíclicas y entre los mismos territorios, y en
ellas juegan un papel muy importante el gre-
garismo, la capacidad de aprendizaje y la
transmisión cultural entre los animales. Son
bien conocidas las migraciones espectaculares,
como la de los ungulados del Serengeti o la de
los renos en los países árticos, pero este prin-
cipio de complementariedad de los recursos
vegetales, en el tiempo y en el espacio, también
es la base de la trashumancia de los animales
domésticos.
En la Península Ibérica, los sistemas
trashumantes más clásicos y conocidos suelen
establecerse entre el llano y la montaña; por
ejemplo, entre las dehesas extremeñas y la
Montaña leonesa, o entre los Pirineos y el
Valle del Ebro (ver capítulo 5). La trashumancia
es un buen ejemplo del principio de comple-
mentariedad mencionado anteriormente,
mediante el cual los rebaños son trasladados
espacial y temporalmente para aprovechar los
máximos de producción y calidad de la hierba
allí donde se producen. Este sistema de apro-
vechamiento tiene la misma base ecológica
que el que realizaban los herbívoros salvajes
(renos) hace milenios (capítulo 5) y que aun
persiste en la actualidad en latitudes árticas.
En la figura 4.8 se representa un esquema con
los desplazamientos estacionales de los reba-
ños estantes y trashumantes en los valles de
Broto, Vió, Añisclo, Escuaín y Pineta. La orga-
nización de estos movimientos ha dado lugar
210 | cap.14
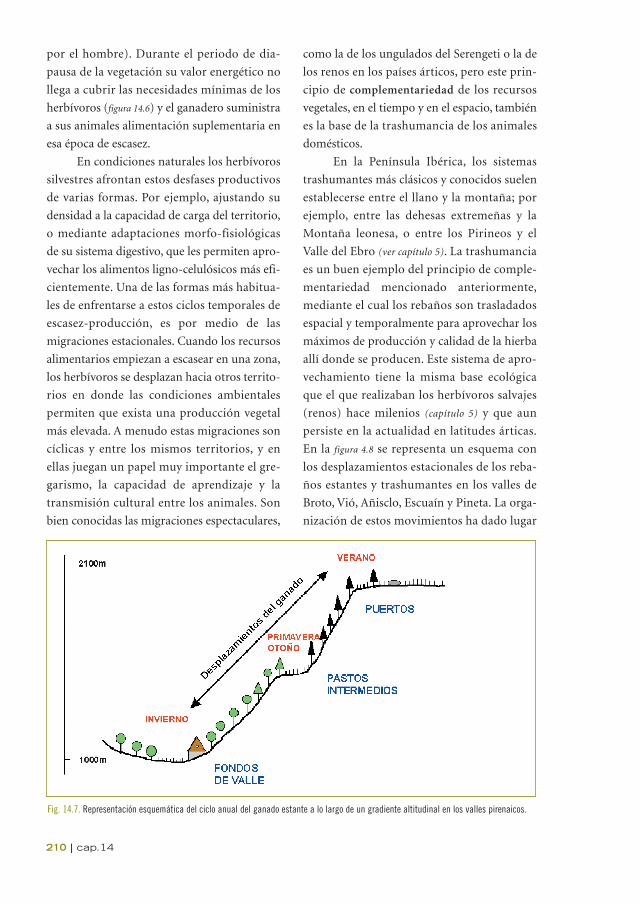
Fig. 14.7. Representación esquemática del ciclo anual del ganado estante a lo largo de un gradiente altitudinal en los valles pirenaicos.

a ordenanzas centenarias (Casa de Ganaderos
del Valle de Broto), que distribuyen las unida-
des de pastoreo (puertos), entre los diferentes
ganaderos, pueblos y vicos (conjunto de varios
pueblos), según un turno rotacional riguroso,
creando una cultura ganadera compleja y
adaptada a su ambiente (Aldezabal et al.,
1992).
Existen también una variedad de movi-
mientos migratorios menos aparentes,
espacial y temporalmente más cortos, y que
han recibido nombres particulares: trans-
terminancia, trashumancia inversa, etc. En la
figura 14.7 se representa sintéticamente un ciclo
anual de desplazamientos altitudinales para
rebaños estantes (no-migratorios) en un valle
típico pirenaico, también llamado sistema de
complementariedad valle-puerto.
En los Pirineos los pueblos suelen ubi-
carse en el fondo de los valles, en altitudes
que oscilan entre los 800 y 1300 m. En sus
proximidades acostumbran a encontrarse los
prados. En invierno los rebaños estantes per-
manecen en el establo y ocasionalmente salen
a pastar por sus alrededores si la ausencia de
nieve lo permite. Durante la primavera
ascienden en altitud y comienzan a pastar los
nuevos rebrotes de los pastos de zonas inter-
medias (panares, bajantes, puertos de tránsi-
to, etc), aproximadamente entre los 1300 y
1600 m de altitud. Estos pastos intermedios
suelen ser zonas llanas, en claros de bosque,
antiguamente aprovechadas para cereal, y en
los valles con suficiente rango altitudinal
puede encontrarse un barrio de bordas
(Chocarro et al., 1990). Durante el verano
ascienden a los puertos estivales, en donde el
pasto, ya libre de la nieve, ha iniciado su creci-
miento. En otoño, con los puertos agostados,
reemprenden el camino inverso, aprovechan-
do normalmente un segundo pico de produc-
ción que se produce en las zonas intermedias
(figura 14.4).
DDiissttrriibbuucciióónn eessppaacciiaall ddee llooss
hheerrbbíívvoorrooss eenn uunn ggrraaddiieennttee ddee llaaddeerraa
yy llaa sseelleecccciióónn aa eessccaallaa ddee ppaaiissaajjee
En la situación actual es frecuente que las uni-
dades de pastoreo estival pirenaicas (puertos)
sean pastadas simultáneamente por varias
especies de ungulados. Sin embargo, también
es frecuente que dichas especies no pasten
conjuntamente sino que cada una utilice áreas
diferentes. La tipificación de dichas áreas es la
resultante del manejo humano (desplaza-
mientos impuestos por los ganaderos) y las
características morfo-fisiológicas de los herbí-
voros, que les permiten utilizar mejor un tipo
u otro de superficies y pastos. La segregación
espacial de las diferentes especies de herbívo-
ros puede realizarse siguiendo un gradiente
vertical u horizontal.
En la figura 14.8 se presenta un caso real
de este sistema de aprovechamiento, en el gra-
diente altitudinal que ofrece un puerto de
verano del Pirineo occidental con un rango
altitudinal entre 1600 y 2600 m. El ganado
vacuno pastó generalmente entre altitudes de
1600 a 1800 m. Las ovejas pastaron entre 1800
y 2000 m en julio, primera quincena de agosto
y septiembre, mientras que durante la
segunda quincena de agosto fueron desplaza-
das entre 2200 y 2400 m de altitud. Los sarrios
(Rupicapra p. pyrenaica) se mantuvieron
altitudinalmente por encima de los domésti-
cos, sin embargo frecuentemente visitaban los
puntos de sal (“saleras”) del ganado (flechas).
Las áreas rayadas de los domésticos son más
geométricas que las de los sarrios indicando la
influencia del manejo humano. El rayado
horizontal indica el proceso de fusión de
nieve, el cual aumenta al avanzar la estación y
es retardado cuanto mayor es la altitud.
Paralelo a dicho proceso de fusión, se produce
la brotación del pasto tierno. Los herbívoros
salvajes, siguiendo la onda fenológica vegetal,
ascienden paulatinamente hasta las cumbres
cap.14 | 211

(ver apartado anterior). Los herbívoros do-
mésticos, especialmente ovejas y cabras, prosi-
guen poco después el mismo patrón de utili-
zación herbácea. El resultado final es que se
produce, durante el verano, una estratificación
vertical de los rumiantes a lo largo de las ver-
tientes, ordenada según su tamaño corporal.
Los ungulados más pesados en el fondo de
valle y laderas bajas, las ovejas en altitudes
medias y los más ligeros (sarrios) en las
cumbres y roquedos. Se trata de un esquema
general, basado en patrones ecológicos, pero
sujeto a numerosas excepciones y variantes
locales.
LLaa ccoommpplleemmeennttaarriieeddaadd yy llaa sseelleecccciióónn
aa eessccaallaa ddee ccoommuunniiddaadd vveeggeettaall
Siguiendo el proceso de jerarquización del
proceso selectivo indicado al principio de este
capítulo, veremos a continuación un ejemplo
de lo que puede ser la selección a escala de
comunidad vegetal en la misma unidad pasto-
ral del ejemplo anterior (Puerto de Aísa). Una
forma de visualizar la utilización que realizan
las diferentes especies de herbívoros de los dis-
tintos tipos de pastos, es superponiendo un
mapa de vegetación de la zona a la distribu-
ción espacial de las diferentes especies ani-
males (figura 15.5). El resultado se muestra en la
figura 14.9.
Las vacas utilizan principalmente los
pastos densos del tipo Bromion erecti y Nardion
strictae de producciones relativamente altas.
Las ovejas pastan principalmente la comuni-
dad de Nardion strictae, de entre la cual es
capaz de seleccionar las especies más nutri-
tivas (Agrostis capillaris, Trifolium alpinum) y
rechazar las menos digestibles (Nardus stricta).
También utiliza en gran medida los pastos del
tipo Primulion intricatae, pastos densos de
altura, de producción moderada pero de alto
valor nutritivo. Los sarrios son los que más
utilizan este último tipo de pastos pero tam-
bién frecuentan las comunidades de roquedos,
pastos pedregosos y pedrizas de entre las
cuales son capaces de seleccionar las especies
más nutritivas.
LLaa sseelleecccciióónn ddee llaa ddiieettaa
aa eessccaallaa ddee llaa ppllaannttaa
Los postulados de la dieta óptima (maxi-
mización de la entrada de energía neta con el
212 | cap.14
Fig. 14.8. Distribución altitudinal de varias especies de ungulados en el Puerto de Aísa (Pirineo occidental) durante el verano de 1986 (véaseexplicación detallada en el texto) (según García-González et al., 1990).

mínimo coste energético), fueron elaborados
pensando más bien en los carnívoros, para los
cuales, la energía adquirida se encuentra en
paquetes de calidad similar (presas) y el coste
de adquisición puede ser elevado (Malechek y
Balph, 1987). En el caso de los herbívoros, la
energía bruta disponible (vegetales) suele ser
elevada, pero su calidad (de la que depende su
transformación en energía neta) es muy varia-
ble. Por otra parte, el coste de adquisición
suele ser bajo (gastos de locomoción). Por esta
razón la alimentación de los herbívoros es un
compromiso entre la cantidad y la calidad de
los vegetales que deben consumir. Así pues, la
selección de la dieta por parte de los herbívo-
ros se convierte en un proceso de decisión en
el que intervienen múltiples factores, de los
cuales los principales se han representado en
la figura 14.10. El grosor de las flechas intenta
simbolizar la importancia de cada factor, aun-
que como se ha mencionado al principio del
capítulo, esta importancia puede variar según
la escala espacial y temporal en que se produce
la selección.
Factores dependientes del medio físico y de la vegetación disponibleEn parte inspirados por las predicciones de la
teoría de la dieta óptima, numerosos estudios
han puesto gran énfasis en la maximización de
la ingestión como estrategia de adquisición de
alimento por parte de los herbívoros. La maxi-
mización de la tasa de ingestión es directa-
mente proporcional a la cantidad de alimento
disponible, hasta un cierto punto (respuesta
funcional, figura 14.2). La estrategia de maximi-
zar el consumo conduciría a los animales a
seleccionar las especies más abundantes. Sin
embargo, como se dijo anteriormente, la
ingestión tiene un límite determinado por la
digestibilidad del alimento (figura 14.3). Por otra
parte, el organismo necesita que la dieta sea
equilibrada en cuanto a nutrientes (proteína y
minerales), para que no se produzcan trastor-
nos metabólicos. Ambas necesidades, digesti-
bilidad y nutrientes, conduciría a los herbívo-
ros a seleccionar por la calidad del alimento,
además de por la cantidad.
cap.14 | 213
Fig. 14.9. Porcentaje de superficie ocupada por las principales comunidades de pasto en el Puerto de Aísa y en las áreas utilizadas por cadaespecie de herbívoro. Las áreas sin vegetación y con cobertura menor al 1% no han sido tenidas en cuenta. Las majadas incluyen las alianzas fitosociológicas Rumicion pseudoalpini y Polygonion avicularis y las gleras comprenden principalmente las alianzas Iberidion spathulatae y Saxifragion praetermissae (según García-González et al., 1991).

Puede considerarse sin embargo, que el
principal factor de selección por parte de los
herbívoros es la disponibilidad de vegetales, lo
cual se refiere no sólo a su cantidad (biomasa)
sino a su asequibilidad (mayor o menor posi-
bilidad de obtenerlo). La disponibilidad de-
pende también de la estacionalidad, ya que los
vegetales siguen un ciclo vegetativo, con perio-
dos de crecimiento, reproducción y senes-
cencia (más marcados en los climas templa-
dos). Los principales factores que determinan
la disponibilidad vegetal son los factores abió-
ticos, especialmente suelo, agua y temperatura.
Estos factores actúan sobre el nivel de produc-
ción del pasto en la forma que se ha indicado
en capítulos precedentes (ver capítulo 11). Por
otra parte, estos factores físicos pueden actuar
indirectamente sobre el animal: las condicio-
nes climáticas adversas, además de modificar
la conducta de pastoreo del animal, puede
producir un incremento de su gasto metabóli-
co para compensar sus pérdidas de calor. Un
aumento de los requerimientos energéticos del
herbívoro puede modificar su conducta de
selección, buscando alimentos más energéti-
cos, o si no los hay (por ejemplo en invierno),
aumentando el tiempo de ingestión.
La calidad, como se ha dicho antes,
puede convertirse a veces en el principal factor
de selección, especialmente en periodos climá-
ticos desfavorables, en los cuales los vegetales
interrumpen o disminuyen su metabolismo.
En los periodos de senescencia la cantidad de
fibra vegetal aumenta considerablemente (ver
capítulo 12), especialmente la lignina, y la diges-
tibilidad se reduce proporcionalmente. Así, el
herbívoro puede encontrarse con gran abun-
dancia de vegetales pero no puede procesarlos
por su baja o nula digestibilidad. Se han
encontrado ungulados salvajes muertos de
inanición, pero con el rumen lleno de plantas
que no podían digerir. Los compuestos anti-
herbívoro pueden causar graves trastornos o
la muerte por intoxicación, pero en muchas
ocasiones actúan solamente reduciendo la
digestibilidad de la planta. Resinas, terpenos
y taninos son importantes depresores de la
digestibilidad y su acción es más dañina en
los periodos de escasez. Algunos autores han
sugerido incluso que la lignina no sólo se
214 | cap.14
Fig. 14.10. Esquema que representa los principales factores que actúan en el proceso de selección de la dieta por parte de los grandesherbívoros. El grosor de las flechas estaría relacionado con el grado de importancia de cada factor. Las líneas a trazos indican efectos
indirectos sobre la selección. En verde los factores dependientes de la planta; en rojo los dependientes del animal.

habría desarrollado evolutivamente en rela-
ción con la función de soporte de la planta,
sino también como compuesto defensivo con-
tra los herbívoros, dado su carácter indigesti-
ble. Por esa razón, ambos factores (calidad y
defensas antiherbívoro) actúan indirectamen-
te sobre la disponibilidad, lo cual se ha indi-
cado con una línea a trazos en la figura 14.10.
En la figura 14.11 se expone los resultados
de un estudio realizado en pastos estivales
pirenaicos, en el cual se trató de evaluar el nivel
de selección ejercido por los grandes herbívo-
ros sobre las principales especies que compo-
nen los pastos supraforestales (Gañán et al.,
2002). La disponibilidad (abundancia de es-
pecies) se determinó por medio de contactos
con una aguja (“point quadrat”) en transectos
lineales (ver métodos capítulo 8) y la utiliza-
ción por los herbívoros, por medio del registro
de las señales de mordiscos sobre las especies
contactadas en los mismos transectos lineales.
Dicho método no permite conocer la especie
animal responsable del consumo, por lo que
debe considerarse selección de alimento de
ovejas y vacas, conjuntamente. Como puede
observarse, existen dos patrones de selección:
un grupo de especies (que suelen ser las que
sobrepasan una abundancia media del 5%),
son seleccionadas proporcionalmente a su
abundancia (Festuca eskia, Nardus stricta),
mientras que otro grupo de especies (de abun-
dancias medias entre 2-7%), parece que son
seleccionadas independientemente de su abun-
dancia. Este segundo grupo probablemente es
seleccionado en función de su calidad (mayor
o menor contenido en fibra y proteína) o de la
presencia de compuestos antiherbívoro, aun-
que también puede existir una influencia de su
altura y posición en la estructura vertical del
pasto. Así pues, en la figura 14.11 puede observar-
se la influencia de los dos factores menciona-
dos (cantidad y calidad) en la selección de las
especies pascícolas por parte de los herbívoros
que pastan en los puertos estivales.
cap.14 | 215
Fig. 14.11. Relación entre la abundancia media de algunas especies características de los pastos pirenaicos y su selección por los herbívorosdomésticos. Las especies se indican con las tres primeras iniciales del nombre científico (género-especie). Las barras verticales indican loslímites de confianza al 95%. El índice de selección varía entre +1 (alta preferencia y -1 (rechazo total). El valor 0 indicaría indiferencia. Laselipses indican dos tipos de especies: las que son seleccionadas (positiva o negativamente) en función de su abundancia y las que sonindependientes de ella.

Selección de categorías vegetales y partes de la plantaEn la Figura 14.12 se exponen los resultados de la
determinación de las dietas obtenidas por
diferentes herbívoros que pastan en el mismo
puerto de verano según un estudio realizado
en el Parque Nacional de Ordesa y Monte Per-
dido (Aldezabal, 2001). El régimen alimenta-
rio se obtuvo por medio del análisis microhis-
tológico de las heces de los animales, el cual se
basa en la determinación de fragmentos indi-
gestibles (principalmente cutículas y epider-
mis vegetales) al microscopio a partir de una
colección de referencia (Cuartas y García-
González, 1996). Las especies vegetales fueron
agrupadas por categorías vegetales: gra-
minoides, herbáceas y leñosas. Cada columna
corresponde a una especie de ungulado y es la
dieta promedio de los tres meses de verano. El
término graminoides comprende las familias
gramíneas, ciperáceas y juncáceas. El término
herbáceas comprende las herbáceas dicotile-
dóneas y monocotiledóneas no graminoides.
Como puede observarse, el mayor consumo
de graminoides correspondió a las yeguas,
seguido por las vacas, ovejas, cabras y sarrios.
Contrariamente, el consumo de herbáceas
siguió un orden inverso. Las cabras solas (en
rebaño monoespecífico) y los sarrios, consu-
mieron una buena proporción de leñosas,
mientras que en las demás especies y rebaños
fue de escasa importancia o inapreciable. Es
de destacar también las diferencias de las die-
tas de las cabras cuando pastan solas o junto
con las ovejas, las cuales se deben principal-
mente a que estas últimas pastaban una zona
más alta en la que las leñosas eran más escasas.
Excepto las ovejas, el resto de especies pasta-
ban libremente y en todos ellas la capacidad
de selección a escala de comunidad y especie
vegetal era alta.
Las graminoides y las leñosas contienen
normalmente mayor cantidad de fibra y son
menos digestibles que las herbáceas. En gene-
ral se observa una tendencia a consumir más
graminoides en las especies de tamaño grande
y más dicotiledóneas en las de tamaño más
pequeño. Las cabras hacen excepción a esta
tendencia y ello se debe probablemente, a que
las características de su sistema digestivo las
hacen comportarse más como herbívoros
comedores de alimentos “concentrados” (con
216 | cap.14
Fig. 14.12. Dietas promedio de los herbívoros que pastan en el Puerto de Góriz (P.N. Ordesa) durante el verano por categorías vegetales determinado mediante el análisis microhistológico de sus excrementos (adaptado de Aldezabal, 2001).
poca fibra), mientras que las ovejas, de tama-
ño algo inferior, se comportan más como
“pastadores”, mejor adaptados para el consu-
mo de alimentos fibrosos, tal como se verá en
el apartado siguiente.
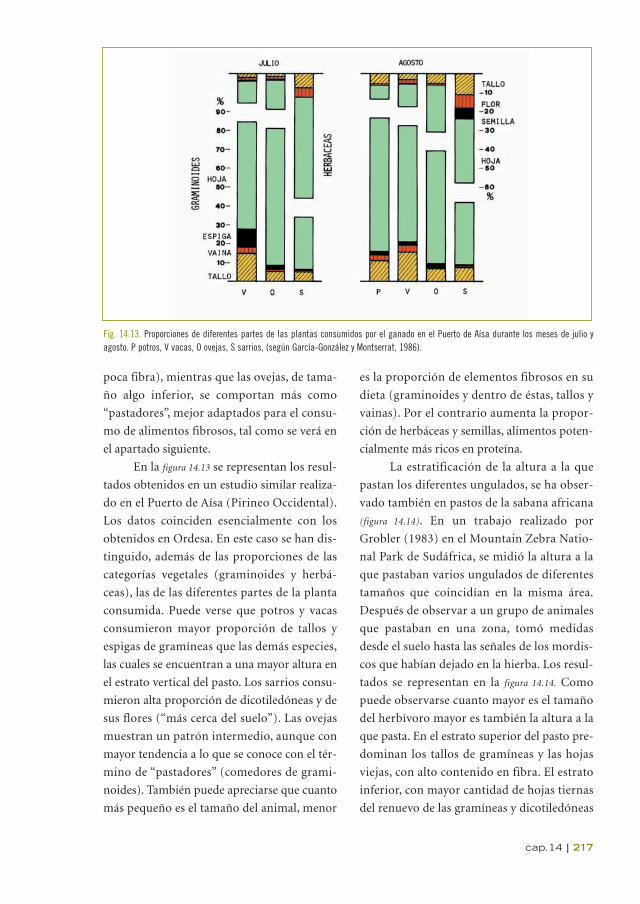
En la figura 14.13 se representan los resul-
tados obtenidos en un estudio similar realiza-
do en el Puerto de Aísa (Pirineo Occidental).
Los datos coinciden esencialmente con los
obtenidos en Ordesa. En este caso se han dis-
tinguido, además de las proporciones de las
categorías vegetales (graminoides y herbá-
ceas), las de las diferentes partes de la planta
consumida. Puede verse que potros y vacas
consumieron mayor proporción de tallos y
espigas de gramíneas que las demás especies,
las cuales se encuentran a una mayor altura en
el estrato vertical del pasto. Los sarrios consu-
mieron alta proporción de dicotiledóneas y de
sus flores (“más cerca del suelo”). Las ovejas
muestran un patrón intermedio, aunque con
mayor tendencia a lo que se conoce con el tér-
mino de “pastadores” (comedores de grami-
noides). También puede apreciarse que cuanto
más pequeño es el tamaño del animal, menor
es la proporción de elementos fibrosos en su
dieta (graminoides y dentro de éstas, tallos y
vainas). Por el contrario aumenta la propor-
ción de herbáceas y semillas, alimentos poten-
cialmente más ricos en proteína.
La estratificación de la altura a la que
pastan los diferentes ungulados, se ha obser-
vado también en pastos de la sabana africana
(figura 14.14). En un trabajo realizado por
Grobler (1983) en el Mountain Zebra Natio-
nal Park de Sudáfrica, se midió la altura a la
que pastaban varios ungulados de diferentes
tamaños que coincidían en la misma área.
Después de observar a un grupo de animales
que pastaban en una zona, tomó medidas
desde el suelo hasta las señales de los mordis-
cos que habían dejado en la hierba. Los resul-
tados se representan en la figura 14.14. Como
puede observarse cuanto mayor es el tamaño
del herbívoro mayor es también la altura a la
que pasta. En el estrato superior del pasto pre-
dominan los tallos de gramíneas y las hojas
viejas, con alto contenido en fibra. El estrato
inferior, con mayor cantidad de hojas tiernas
del renuevo de las gramíneas y dicotiledóneas
cap.14 | 217
Fig. 14.13. Proporciones de diferentes partes de las plantas consumidos por el ganado en el Puerto de Aísa durante los meses de julio y agosto. P potros, V vacas, O ovejas, S sarrios, (según García-González y Montserrat, 1986).

herbáceas, potencialmente tiene mayor conte-
nido en proteína y digestibilidad (ver también
figura 14.17).
Así pues, cuando se comparan los hábi-
tos alimentarios de diferentes ungulados que
pastan en el mismo área, se observa que pare-
ce existir una relación entre la altura a la que
pastan, la calidad de la dieta que ingieren, su
tamaño corporal, las características morfo-
fisiológicas de su sistema digestivo y su capaci-
dad de selección. Como se verá en el apartado
siguiente, cuanto menor es el tamaño del
animal, mayores son sus requerimientos
alimentarios (en términos relativos), adaptan-
do su dieta a dichas necesidades.
Factores de selección dependientes del animalUn factor de selección dependiente del animal
se refiere a la especie de herbívoro y a su abun-
dancia (figura 14.10). Hemos visto en apartados
precedentes como diferentes especies animales
obtienen dietas diferentes, aun pastando en la
misma área de pastoreo. Cuando considera-
mos a una única especie (pastoreo monoespe-
cífico), la selección alimentaria también puede
verse influida por el número de animales por
unidad de superficie (densidad o carga gana-
dera). A densidades altas la capacidad de dis-
criminación de los herbívoros se reduce y
suelen obtener dietas más homogéneas (de
menor diversidad). Por otra parte, a densida-
des altas la utilización y la presión que se
ejerce sobre el pasto son mayores, por lo que
la disponibilidad de hierba se reduce (inde-
pendiente de los cambios a medio plazo indu-
cidos en las comunidades pascícolas), afec-
tando de nuevo a la selectividad.
Las diferencias de selección entre espe-
cies de herbívoros se manifiestan principal-
mente a través de sus características morfo-
fisiológicas, entre las cuales, una de las más
importantes es el peso vivo. El peso vivo
determina las necesidades metabólicas del
animal, mayores en los de mayor tamaño.
Pero esta relación no es directamente propor-
cional sino que presenta alometría negativa,
siendo el coeficiente alométrico de 0,73
(aproximadamente la relación superficie/-
volumen de los cuerpos geométricos que
aumentan de tamaño). Todo ello significa que
los animales pequeños tienen necesidades
energéticas relativamente mayores que los
grandes, por lo cual necesitan un mayor nivel
de ingestión que los herbívoros de mayor
tamaño por unidad de peso. El peso metabó-
lico (Peso Vivo 0,73) homogeniza el peso de los
animales en función de su gasto metabólico y
sus necesidades energéticas. Para adquirir una
218 | cap.14
Fig. 14.14. Distribución de frecuencias de las alturas de pastoreoque utilizan cuatro ungulados en el Mountain Zebra National Parkde Sudáfrica (adaptado de Grobler, 1983; pesos de los unguladossegún Dorst y Dandelot, 1973)

dieta más energética los herbívoros pequeños
seleccionan (si pueden) alimentos muy diges-
tibles (concentrados), con alto contenido en
proteína y bajo en fibra. Contrariamente, los
herbívoros grandes pueden sobrevivir con
dietas menos concentradas, o con alimentos
más fibrosos como las gramíneas y por eso se
les llama “pastadores” (del inglés “grazers”).
Hofmann (1989) propuso una clasifica-
ción de los rumiantes según su tendencia a
comer más o menos alimentos fibrosos (pas-
tadores), o alimentos de alta digestibilidad
(comedores de concentrados, también llama-
dos ramoneadores), o situados en una posi-
ción intermedia entre ambos extremos (tipos
intermedios) (figura 14.15). El autor mencionado
afirma que las especies correspondientes a un
mismo tipo digestivo poseen además deter-
minadas adaptaciones morfo-fisiológicas que
les capacitan para un mayor o menor aprove-
chamiento de la fibra vegetal. Así, los tipos
pastadores poseen un volumen ruminal y un
tiempo de tránsito del alimento proporcional-
mente mayor que los comedores de concen-
trados, lo que les capacitaría para un mejor
procesamiento de la fibra. Además, la selec-
ción de vegetales menos fibrosos supone un
menor intervalo entre dos tomas, ya que el
rumen sencillo de las especies comedoras de
concentrados, dispone de menos estructuras
para diferir el avance de los alimentos que el
de especies pastadoras.
Otros autores como Gordon y Illius
(1994) conceden más importancia al peso vivo
del herbívoro como factor de selección de
dieta, ya que aseguran que es dudoso que exis-
tan diferencias específicas en la capacidad de
digestión de la fibra vegetal. Incluso existiendo
dichas adaptaciones, que sean lo suficiente-
mente fuertes como para hacer preferir, por
ejemplo, a los pastadores, alimentos pobres
teniendo a su disposición alimentos ricos.
cap.14 | 219
Fig. 14.15. Clasificación de las especies de rumiantes europeos por su forma de alimentarse según Hofmann (1989). Cuanto más seaproxima una especie a la derecha de la figura, mayor es su capacidad para digerir fibra en el rumen.

Según estos autores, las diferencias en cuanto
a necesidades energéticas impuestas por el
peso corporal, serían las responsables de que
algunos rumiantes puedan sobrevivir mejor
con dietas fibrosas con respecto a los peque-
ños. Sin embargo, existen excepciones nota-
bles, por ejemplo, herbívoros grandes que se
comportan como ramoneadotes (jirafa, alce) y
herbívoros pequeños que se comportan como
pastadores (oveja). Dichas excepciones encon-
trarían mejor acomodo con la propuesta de
los tipos digestivos de Hofmann.
Otra de las adaptaciones morfológicas
relacionadas con la selección de la dieta, es la
de la anchura de la arcada de incisivos de la
mandíbula o el índice anchura/profundidad
mandibular (figura 14.16). El pastoreo de los
rumiantes de boca ancha (tipo “pala”) como
las vacas, se asemeja más al de una guadaña,
mientras que los de índice de arcada pequeño
tienen un aparato bucal semejante a una
“pinza” que les permite seleccionar mejor el
alimento. Los herbívoros de “boca ancha”
encuentran sus condiciones óptimas de pasto-
reo en pastos de gramíneas altas, que frecuen-
temente tienen más contenido en fibra que los
pastos cortos. Este tipo de herbívoros suelen
ser de tamaño grande y corresponder al tipo
digestivo de los “pastadores”, y como su siste-
ma bucal les permite conseguir mayor canti-
dad de forraje por bocado, se benefician de
una mayor ingesta en términos absolutos,
aunque más fibrosa. Por el contrario los
rumiantes con sistema bucal tipo “pinza” sue-
len ser ramoneadores (comedores de concen-
trados), de tamaño pequeño, por tanto con
mayores requerimientos metabólicos relativos
(Janis y Ehrhart, 1988). Su sistema bucal les
permite seleccionar aquellas especies o partes
de la planta más nutritivas (por ej. entresacar
los folíolos de Trifolium alpinum entre las
macollas densas de Nardus stricta o de Festuca
eskia). Parecen preferir céspedes cortos con
alta proporción de dicotiledóneas.
Por último, se han descrito también
como factores importantes de la selección de
la dieta lo que podríamos llamar característi-
cas relacionadas con la organización social
(figura 14.10), de los cuales podríamos conside-
rar dos aspectos: los estrictamente relaciona-
dos con la organización social (gregarismo,
preferencia de hábitat) y los relacionados
con el comportamiento (conducta de apren-
dizaje). Entre los primeros se ha mencionado
la tendencia de los animales comedores de
concentrados a preferir el bosque o los hábi-
tats densos y a vivir en grupos pequeños. Con-
trariamente los pastadores preferirían es-
pacios abiertos (praderas, sabanas) y vivirían
en grupos numerosos (Geist, 1974). En cuanto
al comportamiento, se ha comprobado que los
rumiantes poseen una alta capacidad para
recordar y seleccionar posteriormente los
mejores pastos. Se han descrito incluso meca-
nismos fisiológicos por medio de los cuales se
produciría este reforzamiento de la conducta
(Provenza, 1995). Esta experiencia es transmi-
tida por los adultos a las crías que pastan
junto a ellos, incrementando su capacidad de
selección. En los animales domésticos suele ser
importante la existencia de los animales
“guía”, que son aquellos que actúan como líde-
res conduciendo al resto del grupo o rebaño.
El conocimiento que tienen estos animales de
las mejores manchas de pasto o de los puntos
220 | cap.14
Fig. 14.16 Tipos de arcada labial en los Rumiantes. A la izquierda,tipo “pala” (buey), a la derecha tipo “pinza” (ciervo ratón,Tragulus) (según Boué, 1970).

de agua, permite al resto del grupo mejorar
su eficiencia alimentaria. Se trata de un aspec-
to poco estudiado por los investigadores de
la conducta, pero hay autores que sugieren
que la selección y mejora de este tipo de
animales, e incluso su equipamiento con
GPS para su seguimiento por teledetección
(Bailey et al., 2004), serían interesantes líneas
futuras de trabajo para el progreso de la
ganadería extensiva.
AApprrooxxiimmaacciióónn aa uunn mmooddeelloo ddeessccrriippttiivvoo
ddee uuttiilliizzaacciióónn hheerrbbíívvoorraa
iinntteeggrraannddoo ddiiffeerreenntteess eessccaallaass::
bbaasseess ddee llaa rreeppaarrttiicciióónn
ddee rreeccuurrssooss yy eell ppaassttoorreeoo mmiixxttoo
Como se ha mencionado, es difícil encontrar
nexos de unión entre las diferentes escalas de
integración de la interacción pasto-herbívoro,
de tal forma que permitan establecer modelos
generales de aplicación en todas ellas. En el
cap.14 | 221
Figura 14.17. Gradientes de variación de la fibra vegetal, su distribución espacial y su utilización por parte de los herbívoros a escala deladera (A) y a escala pasto (B). Las flechas indican el incremento de la magnitud correspondiente. El color rojo y verde indica mayor o menorproporción de fibra, respectivamente.

caso que nos ocupa (pastoreo extensivo en
pastos pirenaicos), podemos, sin embargo,
establecer ciertas tendencias o “regularidades”
en el uso de los recursos vegetales, que pueden
ser de aplicación a diferentes escalas. Dichas
tendencias, que se esquematizan en la figura
14.17, están basadas en la disposición de los
pastos en montaña, su estructura vertical y las
especialidades tróficas de las diferentes espe-
cies de herbívoros. Como tales tendencias
generales, tienen un carácter meramente apro-
ximado y descriptivo, estando sujetas a nume-
rosas excepciones motivadas por la gran hete-
rogeneidad física y estructural de los pastos en
montaña.
La distribución espacial de los grandes
herbívoros a escala de ladera, mostrada en la
figura 14.17A, corresponde a situaciones obser-
vadas en diferentes casos de estudio ya
expuestos en apartados anteriores (figura 14.8).
Esta distribución altitudinal no parece que se
produzca al azar y coincide esencialmente con
el modelo descrito en los años 70 para los
ungulados africanos en la sabana con pen-
dientes (modelo de la “catena” de Bell, 1971).
Según dicho modelo, la causa de esta estratifi-
cación se basa en las diferentes adaptaciones
que poseen los grandes herbívoros para utili-
zar diferencialmente el pasto. Los de tamaño
grande poseen un sistema digestivo (figura 14.15)
y un aparato bucal (figura 14.16) más capacitado
para utilizar la fibra (celulosa, hemicelulosa),
por eso pastan en las zonas bajas en donde la
relación fibra/proteína es mayor. En los puer-
tos pirenaicos, en las partes bajas de la ladera
abundan los pastos del tipo Bromion y
Nardion, cuya producción es relativamente alta
(capítulo 11) y los herbívoros de mayor tamaño
pueden pastar más eficientemente. Los herbí-
voros de tamaño pequeño poseen una relación
superficie/volumen más alta, por tanto un
gasto metabólico relativamente mayor y mayo-
res necesidades de un alimento rico en
proteína y energía. Por ello les conviene pastar
en las partes altas del puerto, en donde en
general el pasto es más nutritivo (mayor rela-
ción herbáceas/graminoides y menor relación
fibra/proteína). Además de características
digestivas, en la alta montaña hay que añadir
las adaptaciones de locomoción particulares,
que impiden a los grandes animales caminar
por roquedos y pedrizas, tal como lo hacen los
sarrios, las cabras e incluso las ovejas.
La disposición de las partes de la planta y
de su consumo (figura 14.13), en el espacio refe-
rido a la estructura vertical del pasto, nos daría
un esquema parecido al de la figura 14.17B. Dicho
esquema permite apreciar ciertas coincidencias
entre el gradiente vertical de la distribución de
la fibra en las plantas y su utilización por los
herbívoros. Las partes altas de las gramíneas
(espigas, tallos, extremos apicales de las hojas
viejas) suelen contener mayor concentración
de fibra que las hojas basales, por lo general
más jóvenes y ricas en proteína (figura 12.1).
También las dicotiledóneas, -especialmente los
hemicriptófitos con rosetas basales abundantes
en pastos muy utilizados-, poseen menor can-
tidad de fibra que las gramíneas. Ello da como
resultado una estratificación en el uso del
pasto, pastando los animales grandes (equinos
y vacunos) en alturas del pasto intermedias*,
superiores a los pequeños (ovino) (Illius y
Gordon, 1987; Osoro 1990). Todo ello condu-
ciría a la obtención de dietas más fibrosas por
parte de los herbívoros grandes y de dietas más
“concentradas” (con más contenidos celulares)
en los pequeños. Cuando la estructura del
pasto es como la representada en la figura 14.17B,
lo cual sucede en pastos con presión de
pastoreo relativamente alta o en determinadas
222 | cap.14
* En realidad lo que ocurriría es que el rango de altura al
que pastan las vacas es mayor que el de las ovejas y por
tanto acceden no sólo a las partes fibrosas altas de tallos y
espigas, sino a muchas hojas secas que se acumulan en la
parte basal. A cambio obtienen una mayor ingesta de bio-
masa que suele ser mayor en las partes bajas del pasto.

comunidades (Fanlo et al., 1991; Remón,
2004), se daría una coincidencia entre el
gradiente vertical de fibra en la fitomasa aérea,
la altura media a la que pastan los diferentes
herbívoros, su peso vivo y su supuesta
eficiencia para digerir la fibra (pastadores o
comedores de concentrados).
En relación con estos patrones de utili-
zación, varios autores (Montserrat, 1964;
McNaughton, 1976; Gordon, 1988) han men-
cionado también la manifestación de un fenó-
meno conocido como facilitación, por medio
del cual la acción de unos herbívoros mejora
las condiciones del pasto para que lo utilicen
otras especies. El pastoreo de los grandes, ade-
más de eliminar el estrato más alto del pasto y
permitir el acceso a las partes más bajas a los
pequeños ungulados, permite la entrada de
luz que estimula el crecimiento y la genera-
ción de nuevos brotes, más tiernos y menos
fibrosos. La acción de los herbívoros pastado-
res (vacunos y équidos) eliminando las partes
altas y fibrosas del pasto, permitiendo el acce-
so a las partes bajas del pasto y estimulando el
crecimiento de renuevos ricos en proteína y
nutrientes, favorecería el pastoreo posterior de
los ungulados más pequeños, más selectivos y
con mayores requerimientos metabólicos.
Así pues, podría existir una sucesión temporal
de herbívoros en el pastoreo de determinadas
zonas, con la cual los últimos se beneficiarían
de la acción mejorante de los primeros
(modelo de la “catena” de Bell, 1971).
RRiittmmooss ddiiaarriiooss ddee aaccttiivviiddaadd yy uussoo
ddeell ttiieemmppoo ppoorr ppaarrttee ddee llooss hheerrbbíívvoorrooss
Al igual que existe un patrón diferencial en
el uso del espacio y de las comunidades
vegetales por parte de las diferentes especies
de herbívoros, existe también un uso diferen-
cial del tiempo, tanto a lo largo del día como
en el conjunto del periodo de pastoreo. En las
figuras 14.18 y 14.19 se expone un ejemplo de
dichos patrones del uso del tiempo.
Durante la estancia del ganado en los
puertos de verano del Parque Nacional de
Ordesa y Monte Perdido, se llevaron a cabo
registros de los distintos tipos de conducta
(pastoreo, desplazamiento, reposo y otras) en
las cinco especies de herbívoros presentes
(Aldezabal et al., 1992). La vegetación consiste
en pastos supraforestales compuestos por un
mosaico de comunidades entre 1900 y 2400 m
de altitud. A intervalos de media hora se
registró la conducta de 20 animales de cada
especie, junto con otras variables ambientales,
durante los tres meses de verano (julio, agosto
y septiembre) tres días a la semana. El porcen-
taje de animales desarrollando un tipo
concreto de actividad, agrupados en intervalos
de dos horas, nos da una representación del
ritmo diario de actividades y sus variaciones
para el total del verano (figura 14.18).
Puede apreciarse dos patrones diferentes
en los ritmos de actividad: a) algunas especies
(vacas, cabras y sarrios) presentan máximos
de pastoreo por la mañana temprano y a últi-
ma hora de la tarde, con un periodo de reposo
al mediodía (patrón bimodal); b) otras espe-
cies presentan un reparto de actividades más
uniforme a lo largo del día, con dos máximos
de pastoreo suavizados (ovejas) o con varios
máximos (yeguas) (patrón multimodal). El
patrón natural para los ungulados durante la
estación cálida (verano) suele ser el modelo
bimodal (Arnod y Dudzinski, 1978). Las des-
viaciones de este patrón mostradas por yeguas
y ovejas, podrían deberse a razones diferentes
en cada especie. Los équidos, al poseer diges-
tión cecal, no presentan la misma eficiencia
digestiva que los rumiantes (Janis, 1976). El
tránsito del alimento por el tracto digestivo es
más rápido en los primeros, lo que les obliga a
pastar durante más tiempo de forma más
continua, sin intercalar grandes periodos de
cap.14 | 223

reposo. Las desviaciones del patrón bimodal
mostrada por las ovejas se debe al manejo de
los pastores, los cuales impusieron un ritmo
de pastoreo más constante al rebaño a lo largo
del día, en este ejemplo concreto. Por otra
parte, el periodo de pastoreo se inicia más
tarde ya que el rebaño es retenido en la maja-
da hasta que la hierba se desprende del rocío
de la noche. De esta forma los pastores preten-
den evitar el contagio de parásitos y los tras-
tornos digestivos (García-González et al.,
1990). Las cabras en pastoreo monoespecífico
y libre, mostraron un marcado patrón bimo-
dal de pastoreo, con mucho tiempo dedicado
al reposo, mientras que las cabras que acom-
pañan al rebaño de ovejas mostraron unos
patrones intermedios entre éstas y las cabras
que pastan solas. La conducta de estás ultimas
podría deberse a que consiguen una dieta más
energética por ramoneo o bien a cierta activi-
dad nocturna que no fue registrada.
La suma de todas las observaciones de
conducta a lo largo del verano, nos da la
distribución temporal de actividades para
cada especie. En la figura 14.19 se representa el
porcentaje del tiempo total dedicado a cada
tipo de conducta durante las horas de luz.
Puede observarse que el pastoreo es la acti-
vidad en la que se invierte más tiempo (más
del 50% en todos los casos). Sin embargo
existen diferencias significativas entre espe-
cies. Las yeguas son las que invierten más
tiempo en pastoreo, probablemente por causa
de las limitaciones digestivas mencionadas
224 | cap.14
Fig. 14.18. Distribución de los ritmos diarios de actividad para diferentes especies de herbívoros en los puertos estivales del Parque Nacionalde Ordesa durante el verano de 1991. El eje vertical muestra el porcentaje de animales ocupados en una actividad determinada. El eje horizontal muestra las horas del día (hora solar). Verde: pastoreo; amarillo: desplazamiento; azul claro: reposo echado; azul oscuro:reposo de pié; negro: otras actividades (según Aldezabal et al., 1992).

anteriormente. Entre los rumiantes, el tiempo
de pastoreo disminuye inversamente al
tamaño corporal, lo cual parece concordar
con las desiguales necesidades metabólicas
impuestas por la relación superficie/volumen:
a menor tamaño, mayor requerimiento en
energía por unidad de peso. Cabe destacar
también la importancia del reposo en las vacas
(40% del tiempo total) y contrariamente, la
alta proporción dedicada al desplazamiento
por parte de las ovejas, lo cual es un reflejo de
la influencia del manejo en la distribución del
tiempo: las vacas pastan libres y las ovejas
guiadas por pastores. Una gran parte del tiem-
po de reposo de las vacas fue dedicado, en este
caso concreto, a las “esperas de turno” en los
abrevaderos, lo cual pone de manifiesto la
influencia de la disponibilidad de agua en la
conducta total de pastoreo en esta especie. En
todos los casos estudiados las ovejas invierten
más del 30% de su tiempo diurno en el
desplazamiento, lo cual comporta un gasto
energético apreciable. El correcto manejo que
ejerce el pastor sobre su rebaño puede hacer
disminuir este gasto en beneficio de la pro-
ducción de los animales.
AA mmooddoo ddee ccoonncclluussiióónn
La utilización de la hierba por parte de los
herbívoros es un sistema interactivo complejo
(capítulo 3), pero es posible encontrar ciertas
regularidades o patrones de uso. Estos patro-
nes nos conducirían al establecimiento de
ciertos principios: unos basados en los prin-
cipios de la física y relacionados con la trans-
ferencia de energía (necesidades metabólicas),
y otros relacionados con el nivel de organiza-
ción y con la complejidad del sistema (bio-
diversidad). Entre éstos últimos estarían el uso
diferencial de los recursos vegetales, basado en
las características genéticas, fisiológicas y
conductuales de las especies. Este uso dife-
rencial permite la complementariedad entre
cap.14 | 225
Fig. 14.19. Distribución del tiempo total dedicado a varias actividades durante el periodo diurno por los herbívoros estivantes en el ParqueNacional de Ordesa durante el verano de 1991.

Referencias bibliográficasAldezabal, A. 2001.- El sistema de pastoreo del Parque
Nacional de Ordesa y Monte Perdido (PirineoCentral, Aragón). Interacción entre la vegetaciónsupraforestal y los grandes herbívoros. 317 pp.Public. Consejo Protección Naturaleza Aragón nº28, Zaragoza.
Aldezabal, A., Bas, J., Fillat, F., García-González, R.,Garin, I., Gómez, D. y Sanz, J.L. 1992.- Utilizaciónganadera de los pastos supraforestales en elParque Nacional de Ordesa y Monte Perdido.Informe inédito. CSIC-ICONA, Jaca-Huesca.
Armstrong, R.H., Common, T.G. y Smith, H.K. 1986.- Thevoluntary intake and in vivo digestibility of herbageharvested from indigenous hill plant communities.Grass and Forage Science 41: 53-60.
Arnold, G.W. y Dudzinski, M.L. 1978.- Ethology of free-ranging domestic animals. 198 pp. Elsevier,Amsterdam.
Austrheim, G. y Erikson, O. 2001.- Plant species diver-sity and grazing in the Scandinavian mountains -patterns and processes at different spatial sca-les. Ecography 24: 683-695.
Bailey, D.W., Keil, M.R. y Rittenhouse, L.R. 2004.-Research observations: daily movement patternsof hill climbing and bottom dwelling cows.Journal of Range Management 57: 20-28.
Beever, D.E., Offer, N. y Gill, M. 2000.- The feedingvalue of grass and grass products. In: A.Hopkins (ed.), Grass: Its production and utiliza-tion, pp. 140-195. British Grassland Society &Blackwell Sci., Oxford.
Bell, R.H.V. 1971.- A grazing ecosystem in theSerengeti. Scientific American 225: 86-93.
Boué, C. 1970.- Morphologie fonctionnelle des dentslabiales chez les Ruminants. Mammalia 34:696-711.
Chatelain, C. y Troxler, J. 2005.- Gardiennage perma-nent des moutons à haute altitude. Analyse decinq alpages et recommandation de gestion.Revue suisse Agric 37: 151-160.
Chocarro, C., Fanlo, R., Fillat, F. y Marin, P. 1990.-Historical evolution of natural resource use in theCentral Pyrenees of Spain. Mountain Researchand Development 10: 257-265.
Coates, D.B. y Penning, P. 2000.- Measuring AnimalPerformance. In: L. 't Mannetje and R.M. Jones(eds.), Field and Laboratory Methods forGrassland and Animal Production Research, pp.353-402. CABI Publishing, Oxon.
Cuartas, P. y García-González, R. 1996.- Review of avai-lable techniques for determining the diet of largeherbivores from their faeces. Oecologia Montana5: 47-50.
Dorst, J. y Dandelot, P. 1973.- Guía de Campo de losMamíferos Salvajes de Africa. 302 pp. Ed.Omega, Barcelona.
Fanlo, R., Chocarro, C. y Fillat, F. 1991.- Distribución dela biomasa aérea, en el primer corte, de unprado de siega pirenaico. XXXI ReuniónCientífica de la S.E.E.P. pp. 257-261. Consejeríade Agricultura, Ganadería y Pesca de la Regiónde Murcia, Murcia.
Gañán, N., Hernández, Y., Aldezabal, A., Gómez,D. y García-González, R. 2002.- Plant selec-tion by large herbivores in supraforestalPyrenean pastures. REU Technical Series,FAO 66: 86-88.
especies y la repartición de los recursos trófi-
cos y espaciales, permitiendo la disminución
de la competencia entre ellas. Por esta razón
muchos autores proponen que los sistemas de
pastoreo extensivo sean mixtos o multi-espe-
cíficos, ya que con ellos teóricamente se pro-
duciría un mejor aprovechamiento de los
recursos pascícolas.
El esquema de la figura 14.17 podría ser
generalizable, con más o menos variaciones, al
resto de la cadena pirenaica, ya que se basa en
patrones generales relativos a la complementa-
riedad de los recursos, la optimización de la
ingestión, las especialidades tróficas de las dife-
rentes especies de ungulados y sus adaptacio-
nes específicas para desplazarse por pendientes
pronunciadas. Sin embargo deben entenderse
tan sólo como tendencias generales, sujetas a
numerosas excepciones. En primer lugar por-
que las montañas no son figuras geométricas
perfectas, sino que su relieve es irregular, pro-
duciéndose una heterogeneidad estructural y
ambiental en ocasiones muy compleja. En
segundo lugar, porque el manejo humano
impone unas restricciones espaciales en cuanto
a las unidades pastorales y distribución de los
rebaños en ellas, produciéndose a menudo un
solapamiento espacial y temporal entre las
diferentes especies. Sin embargo, estos patrones
y principios generales están basados en adapta-
ciones naturales de los animales y en ciclos
vegetativos y ecológicos de carácter general, y
su conocimiento puede ser útil para mejorar la
gestión del pastoreo en régimen extensivo.
226 | cap.14

García-González, R., Gómez, D. y Remón, J.L. 1991.-Application of vegetation maps to the study ofgrazing utilization: a case in the WesternPyrenees. Phytocoenology 3: 251-256.
García-González, R., Hidalgo, R. y Montserrat, C.1990.- Patterns of time and space use by lives-tock in the Pyrenean summer ranges: a casestudy in the Aragon valley. Mountain Researchand Development 10: 241-255.
García-González, R. y Montserrat, P. 1986.-Determinación de la dieta de ungulados estivan-tes en pastos supraforestales del PirineoOccidental. Actas XXVI Reunión Científica de laS.E.E.P. pp. 119-134. Consejería de Agriculturay Pesca., Oviedo.
Geist, V. 1974.- On the relationship of social evolutionand ecology in ungulates. Amererican Zoologist14: 205-220.
Gordon, I.J. 1988.- Facilitation of red deer grazing bycattle and its impact on red deer perfomance.Journal of Applied Ecology 25: 1-10.
Gordon, I.J. y Illius, A.W. 1994.- The functional signifi-cance of the browser-grazer dichotomy inAfrican ruminants. Oecologia 98: 167-175.
Grobler, J.H. 1983.- Feeding habits of the Cape moun-tain zebra Equus zebra zebra Linn. 1758.Koedoe 26: 159-168.
Hofmann, R.R. 1989.- Evolutionary steps of ecophysio-logical adaptation and diversification of rumi-nants: a comparative view of their digestivesystem. Oecologia 78: 443-457.
Huston, J.E. y Pinchak, W.E. 1991.- Range AnimalNutrition. In: R.K. Heitschmidt and J.W. Stuth(eds.), Grazing Management. An EcologicalPerspective, pp. 27-63. Timber Press, Inc.,Portland.
Illius, A.W. y Gordon, I.J. 1987.- The allometry of foodintake in grazing ruminants. J. Anim. Ecol. 56:989-999.
Janis, C. 1976.- The evolutionary strategy of theEquidae and the origins of rumen and cecaldigestion. Evolution 30: 757-774.
Janis, C.M. y Ehrhart, D.E. 1988.- Correlation of relativemuzzle width and relative incisor width with
dietary preference in ungulates. ZoologicalJournal of the Linnean Society 92: 267-284.
Jarrige, R. 1990.- Gastos, necesidades y capacidad deingestión. In: INRA (ed.), Alimentación de bovi-nos, ovinos y caprinos, pp. 19-28. EdicionesMundi-Prensa, Madrid.
Malechek, J.C. y Balph, D.F. 1987.- Diet selection bygrazing and browsing livestock. In: J.B. Hackerand J.H. Ternouth (eds.), Nutritive quality offorage, pp. 121-132. Academic Press, Sidney.
Margalef, R. 1988.- Evolución de los macrófitos y sucoevolución con los herbívoros. Monografías delInstituto Pirenaico de Ecología, Jaca (Homenajea Pedro Montserrat) 4: 637-642.
McNaughton, S.J. 1976.- Serengeti migratory wildebe-est: facilitation of energy flow by grazing. Science191: 92-94.
Montserrat, P. 1964.- Ecología del Pasto. (Ecología delos agrobiosistemas pastorales). P. Cent. pir.Biol. exp. 1: 1-68.
Osoro, K. 1990.- Recientes avances y futuro de lainvestigación en el manejo de los sistemas pas-torales en zonas húmedas. XXX ReuniónCientífica de la S.E.E.P. pp. 309-360. SanSebastian, 4-8 Junio.
Penning, P.D. 1986.- Some effects of sward conditionon grazing behaviour and intake in sheep. In: O.Gudmundsson (ed.), Grazing Research atNorthern Latitudes, pp. 219-226. Plenum Press,NATO Series, New York & London.
Pianka, E.R. 1982.- Ecología Evolutiva. 365 pp. Ed.Omega, Barcelona.
Provenza, F.D. 1995.- Postingestive Feedback as anElementary Determinant of Food Preference andIntake in Ruminants. Journal of RangeManagement 48: 2-17.
Remón, J.L. 2004.- Estructura y producción de pastosen el Alto Aragón occidental (Aísa y Borau,Huesca). 236 pp. Public. Consejo ProtecciónNaturaleza Aragón nº 47, Zaragoza.
Senft, R.L., Coughenour, M.B., Bailey, D.W.,Rittenhouse, L.R., Sala, O.E. y Swift, D.M.1987.- Large Herbivore Foraging and EcologicalHierarchies. BioScience 37: 789-799.
cap.14 | 227


















