
Cactáceas - · PDF fileCACTÁCEAS Y SUCULENTAS MEXICANAS Tomo XLV año 45...
25
TOMO XLV No. 3 JULIO - SEPTIEMBRE 2000 ISSN 0526-717X Cactáceas Y SUCULENTAS MEXICANAS
Transcript of Cactáceas - · PDF fileCACTÁCEAS Y SUCULENTAS MEXICANAS Tomo XLV año 45...

TOMO XLV No. 3 JULIO - SEPTIEMBRE 2000 ISSN 0526-717X
C a c t á c e a s Y S U C U L E N T A S M E X I C A N A S

Figura 2. Planta juvenil temprana (cinco meses) de Fouquieria purpusii
Figura 3. Desarrollo de los juveniles tempranos de Fouquieria purpusii (cinco
meses). Las dos columnas de plantas a la izquierda corresponden al pretratamiento de asoleo, las dos columnas del centro al pretratamiento con imbibición en agua, y las dos columnas de la derecha al testigo.

CACTÁCEAS Y SUCULENTAS MEXICANAS
Tomo XLV año 45 No. 3
julio-septiembre 2000
Editor Fundador Jorge Meyrán
Consejo Editorial Anatomía y Morfología
Dra. Teresa Terrazas Colegio de Posgraduados Cultivo de Tejidos
Dr. Víctor Chávez Jardín Botánico IB-UNAM
Ecología Dr. Arturo Flores-Martínez
Escuela Nacional de Ciencias Biológicas, IPN Etnobotánica
Dr. Javier Caballero Nieto Jardín Botánico IB-UNAM
Evolución y Genética Dr. Luis Eguiarte
Instituto de Ecología, UNAM Fisiología
Dr. Oscar Briones Instituto de Ecología A. C.
Florística Dra. Raquel Galván
Escuela Nacional de Ciencias Biológicas, IPN Química
Dra. Kasuko Aoki UAM-Iztapalpa
Sistemas Reproductivos Dr. Francisco Molina F.
Instituto de Ecología Campus Hermosillo, UNAM Taxonomía y Sistemática
Dr. Fernando Chiang Instituto de Biología, UNAM.
Editores Biól. Jordan Golubov Figueroa
Dra. María del C. Mandujano Sánchez Instituto de Ecología, UNAM
Biól. Jerónimo Reyes S. Jardín Botánico IB-UNAM
Diseño Editorial Jorge Saldívar
Jardín Botánico IB-UNAM SOCIEDAD MEXICANA DE CACTOLOGÍA, A. C.
Presidenta Fundadora Dra. Helia Bravo Hollis
Presidente Biól. Jerónimo Reyes Santiago
Vicepresidente Christian Brachet
Secretario Gabriel García Tesorera
Roxana Mondragón Bibliotecario Raymundo García
Sociedades Filiales Querétaro, Hidalgo, Puebla, Oaxaca,
Jalisco, San Luis Potosí y Tamaulipas.
Fotografía de portada: Fouquieria purpusii
Foto: Jerónimo Reyes
Contenido
Ensayos de germinación en semillas de Fouqueria purpusii Brandegee. Luis M. Cervantes S. y Patricia Martínez..............................
52
Senecio praecox (Cav.) DC. var. praecox: toda una vida impresa en su arquitectura. Rubén Pérez Ishiwara y Miguel Franco Baqueiro...............................
62
Opuntia bradtiana en la zona calcárea del Bolsón de Mapimí, México. María C. Mandujano y J. Golubov......................................................
66
Euphorbia gumaroi Meyrán, una nueva especie del estado de Hidalgo, México. Jorge Meyrán García.........................................................................
68
Contents
Seed germination trials of Fouquieria purpusii Brandegee Luis M. Cervantes S. and Patricia Martínez........................................
52
Senecio praecox (Cav.): Life depicted in architecture Rubén Pérez Ishiwara and Miguel Franco Baqueiro ...........................
62
Opuntia bradtiana in the Calcarea of the Bolson de Mapimi, Mexico. María C. Mandujano and J. Golubov..................................................
66
Euphorbia gumaroi Meyrán, a new species of the state of Hidalgo, México. Jorge Meyrán García..........................................................................
68
www.cactus-mall.com/smc/index.html
Cactáceas y Suculentas Mexicanas es una revista trimestral de circulación internacional, publicada por la Sociedad Mexicana de Cactología, A. C. desde 1955, su finalidad es promover el estudio científico y despertar el interés en esta rama de la botánica.
El contenido de los artículos es responsabilidad exclusiva de los autores. Se autoriza su reproducción total o parcial siempre y cuando se cite la fuente.
The journal Cactáceas y Suculentas Mexicanas is a publication of the Mexican Society of Cactology since 1955.
Complete or partial copying of articles is permitted only if the reference is cited.
Correspondencia e información (contact information): Sociedad Mexicana de Cactología A. C., Instituto de Ecología, UNAM. Aptdo. Postal 70-275, 04510 México, D.F. México
Correo electrónico: [email protected]
La cuota de inscripción a la sociedad es de $175.00 para México y $25 USD para el extranjero. Pago de cuotas: Sociedad Mexicana de Cactología, A.C., Banamex Suc. 148 No. de cta. 631037-1. En el extranjero el pago se realiza por Orden de Pago. La orden de pago, recibo bancario y/o la documentación pertinente deberán ser enviados a la tesorera: Roxana Mondragón Aptdo. Postal 60-487, San Pedro de los Pinos, 03801, México, D.F.
Subscription rates: $25 USD. Payments by International Money Order to: Sociedad Mexicana de Cactología, A. C. c/o Roxana Mondragón Aptdo. Postal 60-487, San Pedro de los Pinos, 03801, México, D.F.

Cactáceas y Suculentas Mexicanas Tomo XLV Año. 45 No. 3 julio septiembre 2000 52
Introducción
La reserva de la biósfera del Valle Tehuacán-Cuicatlán, se caracteriza por tener un clima semiárido con precipitación media anual de 400 mm, así como la presencia de algunas especies suculentas que son exclusivas o endémicas para esta región. Una de estas especies, localizada en Oaxaca, en el Municipio de Tepelmeme, es
Fouquieria purpusii Brandegee la que es conocida en esta localidad con el nombre común de “jarrilla”. Esta planta es un arbusto o árbol pequeño de tallos suculentos, alargadamente cónicos, verdes y con ramas horizontales; flores tubulares blancas; semillas de color pardo rojizo, aladas, de 3 a 7 mm de largo por 2 a 3 mm de ancho (Ezcurra y Medina 1997).
Debido a la construcción de la supercarretera Cuacnopalan-Oaxaca en el año de 1994, el tramo que
1 Jardín Etnobotánico. Centro Cultural Santo Domingo, Oaxaca.
ENSAYOS DE GERMINACION EN SEMILLAS DE Fouquieria purpusii Brandegee. Luis M. Cervantes S. 1 y Patricia Martínez R.
Resumen En este trabajo se determinó la germinación de semillas y el desarrollo de plántulas de Fouquieria purpusii, especie considerada en peligro de extinción dentro de la norma oficial mexicana. Las semillas fueron obtenidas de plantas polinizadas mecánicamente y localizadas en la colección de plantas vivas del Jardín Etnobotánico del Centro Cultural Santo Domingo en Oaxaca. Dos pretratamientos de germinación fueron probados: imbibición en agua y asoleo. Los pretratamientos presentaron diferencias en la proporción de semillas germinadas, la imbibición de semillas en agua durante dos horas fue el más exitoso. En los cinco meses de observación en el desarrollo de las plantas germinadas no se registró mortalidad. En plántulas y juveniles tempranos se evaluaron algunos parámetros de desarrollo como la altura, grosor del tallo y hojas producidas; los individuos del pretratamiento de imbibición mostraron mayor desarrollo.
Abstract The loss of species habitat has contributed to the alteration of natural plant populations. In addition, illegal trade has increased species loss with serious consequences on population growth and genetic pool. A means of protecting natural populations from illegal extraction is by germinating the species in controlled environments to satisfy public demand. In this study, we determined the germination and growth rates of Fouquieria purpusii a species that is considered in danger of extinction. Seeds were collected from hand-pollinated plants and two germination pretreatments were tested: water imbibition and solar exposure. There were differences in the proportion of seeds that germinated, the seeds that had imbibed water for 2 hr were the most successful treatment. During the five months of growth that were followed, no mortality of seedlings was observed. The following parameters were measured on seedlings: plant height, as well as stem and leaf width. The individuals that had been pretreated with imbibition had the highest growth rates. Germination in controlled environments of F. purpusii is feasible and would help in decreasing the pressure on natural populations.

Cactáceas y Suculentas Mexicanas Tomo XLV Año. 45 No. 3 julio septiembre 2000 53
cruza la zona donde se localiza F. purpusii fue perturbado. Además, esta planta está considerada como una especie en peligro de extinción dentro de la norma oficial mexicana NOM-059-ECOL-1994; por lo que la situación actual de esta especie demanda realizar estudios sobre su biología, particularmente en sus aspectos reproductivos.
Una característica básica que debe determinarse en especies amena-zadas o en peligro de extinción, es la capacidad de germinación de sus semillas, lo que determina el poten-cial de propagación de la especie. También es importante precisar la latencia o tiempo de reposo que tienen las semillas sin perder su viabilidad. Acerca de la latencia, se pueden tener semillas con amplios periodos de reposo, a las que se les denomina semillas ortodoxas; o con periodos muy cortos de reposo, a las que se les llama semillas recalci-trantes (Vázquez-Yanes 1987).
Los objetivos planteados en este trabajo fueron: 1) Determinar el número de semillas germinadas y el tiempo transcurrido para iniciar la germinación, en semillas de F. purpusii obtenidas por fertiliza-ción artificial en plantas localizadas en la colección de plantas vivas (plantas ex-situ o cautivas) del Jardín Etnobotánico del Centro Cultural Santo Domingo. 2) Probar la respuesta de germinación para dos pretratamientos, uno con agua y otro con sol, ya que estas variables ambientales pueden jugar un papel en la germinación de esta especie en la región donde crecen
sus poblaciones silvestres y 3) Caracterizar la morfología general y el crecimiento de las plántulas obtenidas.
Método
Las flores producidas por 3 indivi-duos de F. purpusii, localizados en la colección de plantas vivas del Jardín Etnobotánico, fueron polini-zadas manualmente en el mes de febrero de 1999. Las semillas producidas se cosecharon durante abril del mismo año, almacenán-dose a temperatura ambiente en bolsas de papel. Las semillas obtenidas se separaron en dos grupos, uno destinado al banco de semillas y otro para experimenta-ción. Las semillas para experimen-tación fueron mezcladas homogé-neamente, lavándose con una solución de cloro al 10% durante cinco minutos; estas semillas a su vez se separaron en dos conjuntos. El primer conjunto fue distribuido en dos lotes testigo y cuatro lotes para la aplicación de pretrata-mientos de germinación por duplicado, cada lote tuvo treinta semillas. Los pretratamientos de germinación fueron los siguientes: 1) pretratamiento de imbibición, que consistió en mantener a las semillas sumergidas en agua durante dos horas, 2) pretrata-miento de asoleo, que consistió en la exposición de las semillas a sol directo durante ocho horas. Los lotes se sembraron en charolas germinadoras con sustrato esterili-zado en un horno de microondas por veinte minutos y preparado con una mezcla de arena fina, suelo

Cactáceas y Suculentas Mexicanas Tomo XLV Año. 45 No. 3 julio septiembre 2000 54
agrícola y suelo de monte, en la proporción 1 : 1 : 0.5 respectiva-mente. Las charolas germinadoras se colocaron en estantes para germinación, elaborados con madera y cubierta de plástico.
Para el pretratamiento que resultó más eficiente, se realizó una segunda repetición de germinación con el fin de determinar el momento de emergencia de la radícula. Para ello se utilizó el segundo conjunto de semillas para experimentación, distribuyéndolas en dos lotes testigo y dos lotes experimentales a los que se les aplicó el pretratamiento más exitoso, que fue el remojo en agua por dos horas; cada lote tuvo veinte semillas. Las semillas se sembraron sobre papel filtro en cajas de petri y fueron colocadas en los estantes de germinación.
Todos los lotes se regaron cada tercer día con agua purificada. Para impedir el crecimiento de hongos, el primer riego fue con una solución de Captan al 1%. Las semillas se observaron diariamente durante el proceso de germinación. Posterior-mente, del duplicado para cada lote experimental se tomaron al azar 25 plántulas para determinar sus características morfológicas y seguir su desarrollo hacia juveniles tempranos , tomándose l os siguientes parámetros: la altura y grosor del tallo, que fueron medidos con un vernier digital, y el conteo de hojas producidas; estas mediciones de crecimiento se realizaron en los siguientes intervalos: 25, 26, 31 y 38 días.
Con un termohidrógrafo digital se registraron la temperatura y humedad ambiental del interior del estante de germinación, tomándose diariamente en intervalos de 2 horas durante 10 horas en el día. Los resultados se analizaron con las siguientes pruebas estadísticas con α=0.05. El número de semillas germinadas se analizaron con una prueba de Χ ² (Zar 1974). Para la comparación de las tasas de germinación, se agruparon en una tabla de contingencia las semillas germinadas por pretratamientos y los lotes testigo, para distintas fechas, aplicándose una prueba de Χ ² de independencia (Levin y Rubin 1998). Los resultados de las medidas de crecimiento presentaron varianzas diferentes, por lo que se aplicó la prueba no paramétrica de Kruskal-Wallis (Levin y Rubin 1998; Zar 1974), comparando los resul-tados de las variables de crecimiento de los lotes agrupados por las fechas de registro de dichas variables.
Resultados
Para un periodo de cinco meses de observación y bajo las condiciones en que se llevaron a cabo los ensa-yos de germinación, se obtuvieron los siguientes resultados:
Germinación de semillas sembradas en sustrato
La proporción de germinación entre los tres lotes fue significativamente diferente (P<0.05). El mayor número de plántulas emergidas (salida de las plántulas del interior del sustrato hacia el nivel de la

Cactáceas y Suculentas Mexicanas Tomo XLV Año. 45 No. 3 julio septiembre 2000 55
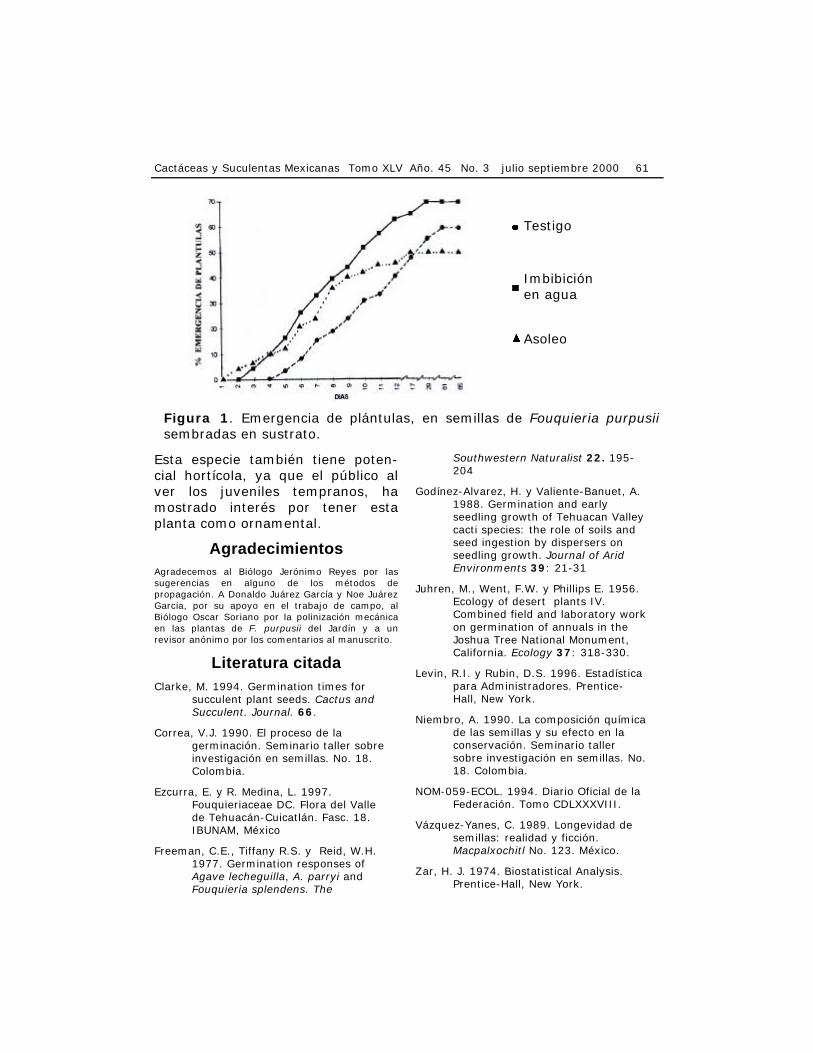
superficie del mismo), fue regis-trado en el pretratamiento de imbi-bición, seguido del lote testigo y por último el pretratamiento de asoleo. El tiempo de inicio de emergencia de las plántulas fue diferente en los tres casos: el pretratamiento de imbibición tardó tres días, el pretra-tamiento de asoleo dos días y el testigo cinco días. El número total de días para alcanzar la proporción final de plántulas emergidas también fue muy variable: el pretratamiento de asoleo presentó el lapso mas corto, pero con la más baja propor-ción de germinación; el pretrata-miento de imbibición ocupó la posi-ción intermedia y el testigo el rango de tiempo más amplio (Cuadro 1).
Las curvas de emergencia de plántulas, del testigo y del pretra-tamiento de imbibición (Figura 1), tienden a ser parecidas, mientras que la del pretratamiento de asoleo es ligeramente diferente. El resul-tado de la prueba de independencia de Χ ² aplicada al agrupamiento de fechas de germinación, no fue signi-ficativamente diferente en la tasa de germinación entre los tres lotes. Sin embargo, el pretratamiento de imbibición puede ser considerado como el más efectivo, al dar una mayor proporción de germinación en poco tiempo; siendo el testigo el más lento en responder y por último, el pretratamiento de asoleo alcanzó la menor proporción de germinación en un tiempo intermedio.
Germinación de semillas sembradas en cajas de petri
Considerando al pretratamiento de imbibición como el más efectivo,
éste se repitió con sus respectivos testigos, sembrando las semillas en cajas de petri para observar directa-mente la emergencia de la radícula y tomar el tiempo en que se da este proceso. En la siembra con sustrato no fue posible observar directa-mente la emergencia de la radícula. Los resultados obtenidos volvieron a indicar diferencias significativas entre el testigo y el pretratamiento, ya que este último siguió siendo el más efectivo, alcanzando el 100% de germinación en cinco días, mien-tras que en el testigo no germinaron todas las semillas y el máximo total de germinación fue alcanzado a los ocho días (Cuadro 2). En el pretra-tamiento de imbibición, la emergen-cia de la radícula fue más rápida ya que se dio dos días después de sembradas las semillas; en el testigo tardó dos días más.
Características morfológicas de las plántulas y juveniles
Los cotiledones de las plántulas en todos los lotes se desarrollaron durante los tres primeros días de germinación. Son de forma elíptica, suculentos y con venación aparente; alcanzan su mayor crecimiento hacia los treinta días, teniendo una longitud promedio de 10.5 ± 0.15 mm y un ancho de 5.6 mm (± 0.07). A partir de la primera semana de emergidas, las plántulas presentan un tallo que va engrosándose paulatinamente en su parte central. A los quince días, las raíces tienen una longitud promedio de 3.1 mm (±0.09), en las plántulas testigo y 8.6 mm (±0.28), en las plántulas de los pretrata-mientos; en las plantas juveniles de

Cactáceas y Suculentas Mexicanas Tomo XLV Año. 45 No. 3 julio septiembre 2000 56
cinco meses las raíces están profu-samente ramificadas, con una longitud promedio para todos los lotes, de 12.3 cm (±0.28). El primer par de hojas, opuestas y de forma elíptica linear surgen a los diez días de germinada la semilla, desarrollándose otros tres pares de hojas en las tres semanas subse-cuentes. Posterior a estos cuatro pares de hojas, las hojas siguientes van surgiendo sin definir pares, en este momento las plantas juveniles
tienen cinco meses, con una altura promedio de 19.5 cm (±0.46), para los lotes testigo y el pretratamiento de asoleo, y 22.6 cm (±0.10) para el pretratamiento de imbibición.
Los parámetros referidos como indicadores del crecimiento de las plántulas presentaron los siguientes resultados (Cuadro 3):
Número promedio de hojas por planta. (Cuadro 3): Se observó un notorio aumento del número de
Lote % Germinación a Tiempo Inicial (días) Tiempo Total b (días)
Testigo 57 5 29-45
Imbibición en agua 70 3 17-29
Asoleo 50 2 12-17
Lote % Germinación a Tiempo totalb (días)
Testigo 80 8
Imbibición en agua 100 5
Cuadro 1. Semillas de Fouquieria purpusii en sustrato. Proporción de semillas germinadas (germinación*), tiempo de emergencia de plántulas (tiempo inicial **) y tiempo máximo en la emergencia hacia la superficie del sustrato del total de plántulas registradas (tiempo total).
*Referida a todas las plántulas que salieron hacia el exterior del sustrato. **Tiempo que tardaron las primeras plántulas en salir del interior del sustrato hacía la superficie del
mismo. aSe obtuvieron diferencias significativas entre lotes P<0.05 (prueba de X2 de homogeneidad). bNo hubo diferencias significativas en las tasas de germinación entre lotes.
Cuadro 2. Semillas de F. purpusi sembradas en cajas de petri. Proporción de semillas germinadas (germinación*) y tiempo máximo de emergencia de la radícula en todas las semillas germinadas (tiempo total).
*Referida a las semillas con emergencia de radícula. aDiferencias significativas entre lotes, P<0.05 (prueba de X2). bDiferencias significativas en la tasa de germinación entre lotes, P<0.001 (prueba de X2 de
independencia).

Cactáceas y Suculentas Mexicanas Tomo XLV Año. 45 No. 3 julio septiembre 2000 57
hojas por individuo durante los cinco meses de observación (Fig. 2). El pretratamiento de imbi-bición tuvo el promedio más alto.
Altura promedio por planta (Cuadro 3). En general el aumento en altura no fue tan rápido en todos los lotes. Sin embargo, en el pretra-tamiento de imbibición las plantas alcanzaron una altura promedio mayor a la de todos los demás lotes (Figura 3).
Diámetro promedio del tallo por planta (Cuadro 3). El incremento del grosor del tallo es lento hasta los dos meses y medio, a partir de ese momento hay u n n o t a b l e a u m e n t o d e aproximadamente 200% en los individuos de todos los lotes.
Tamaño promedio de las hojas. Se definieron tres clases de hojas y en cada una de ellas se tiene la siguiente distribución: para la clase de hoja grande (41.1 mm), el testigo tiene cuatro individuos; el pretratamiento de imbibición, veinticinco y el pretratamiento de asoleo, nueve; en la clase de hojas intermedias (26.5 mm), el testigo tiene diecisiete individuos; el pretratamiento de imbibición, once y el pretratamiento de asoleo, catorce; en la clase de hojas chicas (17.9 mm), el testigo tiene veintiséis individuos, el pretratamiento de imbibición, seis y el pretratamiento de asoleo, ocho. Esta distribución en el tamaño de las hojas indica mayor desarrollo para los individuos p r o v e n i e n t e s d e s e m i l l a s remojadas durante dos horas.
Parámetros ambientales prome-dio en el interior del germina-dor. Durante las primeras once semanas de registro en la germinación y primeras fases de desarrollo de los juveniles, en los estantes germinadores se dio un incremento paulatino en la hume-dad atmosférica mínima promedio diaria, que inició con 30% (±3.8) en la última semana de mayo; hasta alcanzar 67.1% (±8.7) en la segunda semana de julio, volviendo a disminuir durante principios de agosto hasta 48% (±3.3). La tempe-ratura promedio máxima diaria tuvo un descenso de 40 ºC (±2.4) a finales de mayo, hasta 31.1 ºC (±2.4) a mediados de julio; volviendo a incrementar hasta 35.5 ºC, duran-te la primera semana de agosto.
Mortalidad
Del número inicial de plántulas germinadas en todos los lotes, durante los cinco meses de registro no se tuvo mortandad, por lo que la sobrevivencia hasta esa fecha fue del 100%.
Discusión
Los resultados de germinación, tanto en los lotes experimentales como en los testigos, indican que las semillas obtenidas, germinaron en aproximadamente 60%; el resto de las semillas que no germinaron pudieron haber continuado en un estado de latencia más prolongado o no haber sido maduras. Estos aspectos deberán determinarse con más precisión en estudios posteriores.
Cons iderando so lamente la respuesta de germinación de los

Cactáceas y Suculentas Mexicanas Tomo XLV Año. 45 No. 3 julio septiembre 2000 58
lotes testigo, que fue entre el 50% y 80%, se ve que aún sin pretrata-miento alguno las semillas tienen buen porcentaje de germinación; por lo que aunado a la pronta respuesta en su germinación, podrían considerarse tentativa-
mente dentro de la categoría de semillas de rápida germinación. Sin embargo, es necesario que a partir de un lote de semillas de la misma edad, se registre la respuesta de germinación para diferentes fechas de cultivo, tanto a nivel de
Días 24 41 72 112
X ± EE
(Rango)
X ± EE
(Rango)
X ± EE
(Rango)
X ± EE
(Rango)
Altura (mm)
Testigo 11.00 0.50
(9.99 - 12.10)
14.02 0.50
(13.00 - 15.04)
15.70 0.58
(4.50 - 16.90)
20.14 0.69
(18.71 - 21.57)
Imbibición 14.36 0.68*
(12.96 - 15.76)
16.81 0.78*
(15.19 - 18.43)
19.02 0.79*
(17.40 - 20.65)
23.91 1.03*
(21.79 - 26.03)
Asoleo 12.00 0.59
(10.78 – 13.21)
14.05 0.67
(12.67 – 15.43)
15.68 0.68
(14.28 – 17.08)
20.04 0.62
(18.77 – 21.32)
Grosor del tallo (mm)
Testigo 1.18 0.04
(1.10 – 1.25)
3.09 0.11
(2.86 – 3.33)
4.13 0.11
(3.90 – 4.36)
5.91 0.11
(5.68 – 6.14)
Imbibición 1.85 0.11
(1.62 – 2.08)
3.67 0.14
(3.37 – 3.96)
5.19 0.14
(4.90 – 5.48)
6.53 0.15
(6.23 – 6.84)
Asoleo 1.57 0.08
(1.40 – 1.74)
3.56 0.15
(3.26 – 3.87)
4.62 0.13
(4.34 – 4.90)
6.31 0.14
(6.02 – 6.61)
No. de hojas por planta
Testigo 0.64 0.19
(0.25 – 1.03)
3.8 0.24
(3.31 – 4.29)
5.8 0.18
(5.42 – 6.18)
10.44 0.46
(9.50 – 11.38)
Imbibición 1.20 0.23
(0.72 – 1.68)
5.20 0.19
(4.60 – 5.80)
7.76 0.41*
(6.92 – 8.60)
14.96 0.69*
(13.54 – 16.38)
Asoleo 0.96 0.20
(0.54 – 1.38)
5.20 0.19
(4.80 – 5.60)
7.00 0.22
(6.55 – 7.45)
13.92 0.49
(12.91 – 14.93)
Cuadro 3. Valores promedio de altura, grosor de tallo y número de hojas, en plántulas (n=25) de 24-41 días de edad, que posteriormente desarrollaron a juveniles tempranos (72-112 días de edad) de Fouquieria purpusii, germinadas ex situ.
* Diferencia significativa entre lotes P<0.05 (Prueba de Kruskal-Wallis)

Cactáceas y Suculentas Mexicanas Tomo XLV Año. 45 No. 3 julio septiembre 2000 59
laboratorio como en el campo, para que de esta manera se puedan determinar con más detalle los lapsos de longevidad ecológica y de longevidad potencial que las semillas puedan tener (Vazquez-Yanes 1989).
El número total de días para alcan-zar la proporción final de germina-ción está dentro de rangos similares al de las especies de Fouquieria diguetii y F. splendens, así como en la categoría de los lapsos más cortos de germinación registrados dentro de un grupo de 233 especies de plantas suculentas (Clarke 1994).
Las semillas de F. purpusii no tienen testa dura que impida el paso de humedad y que ésta pudiera ser un factor que propiciara un estado de latencia prolongado. La rapidez de germinación, de dos a tres días, pudiera estar acoplada al ambiente donde crece esta especie, sitio de condiciones limitantes, principal-mente de humedad, ya que la región donde se desarrolla tiene poca precipitación pluvial marcada-mente estacional; por lo que las semillas podrían responder inmedia-tamente a la humedad del suelo aportada por las primeras lluvias. Juhren et. al. 1956 en un estudio de cinco años sobre la germinación de plantas anuales desérticas, mostra-ron que una especie no germina fuera de sus límites de tolerancia y que la germinación tiene una estre-cha correlación entre la cantidad y duración de la lluvia, lo que a su vez tiene que ver con la estación del año. De manera que una lluvia abundante de corto tiempo no
produce la mayor germinación de semillas, sino que esta se da más exitosamente con precipitaciones pluviales de intensidad moderada pero con mayor distribución a lo largo de la estación pluvial del año; asimismo, una cantidad adecuada de lluvia previa resultó ser impor-tante para estimular la germi-nación. En el caso de las semillas imbibidas por dos horas, podría considerarse que ese lapso de remojo hubiera sido como el equivalente de esa lluvia previa o de lluvias menos intensas pero constantes que puedan iniciar la germinación.
La respuesta de imbibición de las semillas en agua, fue similar a los resultados de los ensayos de pre-germinación aplicados a F. formosa, donde el pretratamiento de imbibi-ción presentó la mejor respuesta de germinación (Freeman et al. 1977). Es probable que el contacto con el agua durante el tiempo de imbibi-ción haya estimulado para dar una mayor proporción y rapidez de germinación, así como un mejor desarrollo de las plántulas, ya que cuando se aumenta el contenido de humedad que rodea a la semilla, hay un notable incremento en la actividad de enzimas hidrolíticas como resultado de la rehidratación, lo que da inicio al proceso de germinación (Niembro 1990; Correa 1990).
En el ensayo de remojo, el tiempo de imbibición de dos horas que fue probado, puede ser considerado como adecuado para las semillas de F. purpusii. De acuerdo a la experiencia de Correa (1990), el

Cactáceas y Suculentas Mexicanas Tomo XLV Año. 45 No. 3 julio septiembre 2000 60
remojo por poco tiempo estimula la germinación en algunas semillas, pero puede reducirla en otras espe-cies cuando es muy prolongado. Sin embargo, en otro tipo de plantas suculentas como en el caso de algu-nas especies de cactáceas ubicadas en el Valle de Tehuacán y que fueron pretratadas con diferentes lapsos de imbibición, no tuvieron diferencias significativas en su germinación con los lotes testigos (Godínez-Alvarez et al. 1998).
El pretratamiento de asoleo, tuvo la más baja proporción en germina-ción, es probable que la exposición prolongada de las semillas al sol haya causado un cierto deterioro de las mismas, logrando solo la sobre-vivencia de un 50%. En condiciones naturales es más frecuente que una semilla esté sujeta a temperaturas fluctuantes más bien que fijas, la variación puede ser diaria o esta-cional. La germinación ocurre normal-mente en un rango amplio de tempe-raturas siendo mejores las altas; sin embargo, a temperaturas extremas no hay germinación, ya que algunas semillas germinan mejor con temperaturas alternas (Correa 1988).
Es importante considerar los efectos de la fototemperatura establecida de acuerdo a las estación del año y que es característica para cada especie. Sin embargo, la combina-ción adecuada de humedad y tempe-ratura, puede establecer aquellas condiciones que propicien la germinación (Juhren et. al. 1956).
El desarrollo de las plántulas hacia juveniles tempranos (cinco meses
de edad) fue satisfactoria, además de que no hubo mortalidad en todos los lotes. Estos resultados indican que la especie tiene un alto potencial de sobrevivencia y sugiere las posibilidades de manejo que puede tener esta especie. Pero habrá que tener en cuenta que si se confirma la categoría como semilla recalcitrante o semirecalcitrante, serían más difíciles las posibilidades de su almacenamiento en un banco de semillas.
Los datos de los parámetros de crecimiento reafirman que el mayor desarrollo logrado hasta la última fecha de registro, fue en la mayoría de los individuos de los lotes del pretratamiento de imbibición, ya que estos fueron los más grandes, vigorosos y con mayor número de hojas.
Al tratarse de una especie que está dentro de la norma oficial, estos resultados son importantes, ya que dan ciertas bases para el diseño de posibles programas de propagación para la conservación de esta especie. Será necesario obtener semillas por polinización cruzada, para incrementar el acervo gené-tico, por lo que se debe colectar semillas en el campo para futuros programas de propagación de esta especie. Mediante este procedi-miento se cubren dos aspectos: disponer de material genético para la investigación básica o aplicada, y por otra parte, la posibilidad de regenerar, con germoplasma de su propia procedencia, zonas de poblaciones perturbadas de F. purpusii.
Cactáceas y Suculentas Mexicanas Tomo XLV Año. 45 No. 3 julio septiembre 2000 61
Esta especie también tiene poten-cial hortícola, ya que el público al ver los juveniles tempranos, ha mostrado interés por tener esta planta como ornamental.
Agradecimientos Agradecemos al Biólogo Jerónimo Reyes por las sugerencias en alguno de los métodos de propagación. A Donaldo Juárez García y Noe Juárez García, por su apoyo en el trabajo de campo, al Biólogo Oscar Soriano por la polinización mecánica en las plantas de F. purpusii del Jardín y a un revisor anónimo por los comentarios al manuscrito.
Literatura citada Clarke, M. 1994. Germination times for
succulent plant seeds. Cactus and Succulent. Journal. 66.
Correa, V.J. 1990. El proceso de la germinación. Seminario taller sobre investigación en semillas. No. 18. Colombia.
Ezcurra, E. y R. Medina, L. 1997. Fouquieriaceae DC. Flora del Valle de Tehuacán-Cuicatlán. Fasc. 18. IBUNAM, México
Freeman, C.E., Tiffany R.S. y Reid, W.H. 1977. Germination responses of Agave lecheguilla, A. parryi and Fouquieria splendens. The
Southwestern Naturalist 22. 195-204
Godínez-Alvarez, H. y Valiente-Banuet, A. 1988. Germination and early seedling growth of Tehuacan Valley cacti species: the role of soils and seed ingestion by dispersers on seedling growth. Journal of Arid Environments 39: 21-31
Juhren, M., Went, F.W. y Phillips E. 1956. Ecology of desert plants IV. Combined field and laboratory work on germination of annuals in the Joshua Tree National Monument, California. Ecology 37: 318-330.
Levin, R.I. y Rubin, D.S. 1996. Estadística para Administradores. Prentice-Hall, New York.
Niembro, A. 1990. La composición química de las semillas y su efecto en la conservación. Seminario taller sobre investigación en semillas. No. 18. Colombia.
NOM-059-ECOL. 1994. Diario Oficial de la Federación. Tomo CDLXXXVIII.
Vázquez-Yanes, C. 1989. Longevidad de semillas: realidad y ficción. Macpalxochitl No. 123. México.
Zar, H. J. 1974. Biostatistical Analysis. Prentice-Hall, New York.
Figura 1. Emergencia de plántulas, en semillas de Fouquieria purpusii sembradas en sustrato.
Testigo
Imbibición en agua
Asoleo

Cactáceas y Suculentas Mexicanas Tomo XLV Año. 45 No. 3 julio septiembre 2000 62
Distribución y hábitat
Senecio praecox (Compositae) es un arbusto suculento que se
Senecio praecox (Cav.) DC. var. praecox: toda una vida impresa en su arquitectura.
Rubén Pérez Ishiwara1 y Miguel Franco Baqueiro.
Resumen La arquitectura regular de Senecio praecox var. praecox permite reconstruir con detalle la historia de vida de la planta a partir del análisis de su patrón de crecimiento y ramificación. Los individuos adultos florecen y fructifican todos los años. La planta producto de una semilla posee un solo tallo, el cual alcanza la madurez sexual a los 10-15 años de edad. El crecimiento discreto anual del tallo puede ser fechado ya que los entrenudos de las hojas del final de la época de crecimiento son más cortos que los entrenudos del principio de la época de crecimiento, produciendo una cicatriz anular en el tallo. Cuando se alcanza la madurez sexual, el tallo produce una inflorescencia terminal. Esto hace que Senecio solo pueda reiniciar su crecimiento en la siguiente temporada a través de la liberación de uno o más de los meristemos laterales, localizados por debajo del meristemo terminal. Además, uno o varios años después de alcanzar la madurez sexual, S. praecox tiene la posibilidad de producir nuevos tallos a partir de la corona de la raíz. Estos tallos tienen el mismo comportamiento que el tallo original, excepto que alcanzan la madurez sexual a una edad más temprana. La presencia de una dicotomía en el tallo es evidencia de un evento reproductivo fechable. Por lo tanto, el crecimiento anual y la reproducción pueden ser cuantificados a lo largo de toda la vida de la planta. La arquitectura del Senecio contiene la historia demográfica de cada planta.
Abstract Senecio praecox (Compositae) is a succulent shrub distributed from Zacatecas state, in the North, to Oaxaca, in the South, including the states of Central Mexico. Senecio praecox var. praecox is found only in the Valley of Mexico. It grows on rocky substrate of volcanic origin, where soil is poorly developed and competition for light is low. Dominated by S. praecox, the vegetation corresponds to a Xerophytic Shrubland. The strikingly regular architecture of S. praecox var. praecox allows detailed reconstruction of the life history of each individual plant. Adult plants set flower and fruit every year. After germination, each plant consists of a single stem which reaches sexual maturity at 10-15 years of age. The discrete annual growth along the stem can be dated because, unlike the long internodes produced by leaves early in the summer season, the compacted internodes of the leaves produced late in the season leave a continuous scar around the stem. When reaching maturity, a single inflorescence is produced by the stem’s terminal meristem. After flowering, S. praecox can only continue its growth in the next summer season by liberating one or more lateral meristems situated below the terminal one. A few years after S. praecox reaches sexual maturity, it begins sprouting new stems from the root-crown. These stems have the same properties as the original one, except that sexual maturity is reached sooner. The combined processes of flowering-branching-cloning produce S. praecox’s characteristic candle-stick morphology. Branch dicotomy signals the occurrence of a past, datable reproductive event. Thus, annual growth and reproduction can be accurately quantified throughout the whole life of the plant. Senecio’s architecture hold the demographic history of each plant.
1 Instituto de Ecología, UNAM. Lab. de dinámica de poblaciones y evolución de historias de vida. Apdo. Postal 70-275, Ciudad Universitaria. México, D. F., 04510. México. Correo electrónico: [email protected]

Cactáceas y Suculentas Mexicanas Tomo XLV Año. 45 No. 3 julio septiembre 2000 63
distribuye principalmente en los Estados de Zacatecas, San Luis Potosí, Querétaro, Guanajuato, Hidalgo, Jalisco, Michoacán, Edo. de México, Distrito Federal, Puebla y Oaxaca.
La variedad S. praecox var. praecox se encuentra distribuida únicamente en el Valle de México (Xochimilco, Ajusco, Tlalpan y Ciudad Universita-ria). Se conoce localmente como “palo loco”, “palo bofo” y “candelero”, entre otros (Rzedowski y Rzedowski 1985). Por lo general habita en lugares rocosos de origen basáltico con suelos muy pobres, denominando al tipo de vegetación: “matorral xerófilo” (Rzedowski y Rzedowski 1979). Las poblaciones mas conservadas de S. praecox var. praecox se encuentran localizadas en el derrame de lava producto de la erupción del Xitle hace aproximadamente 2500 años (Carrillo 1995). Por la abundancia de esta especie la vegetación recibe t a m b i é n e l n o m b r e d e “Senecietum” (Rzedowski 1979). Se encuentra asociado con diversas especies de árboles entre las que destacan Schinus molle (pirúl), Buddleia cordata (tepozán) y, en las zonas de mayor altitud, con Quercus rugosa; también pueden citarse algunas herbáceas que son r e l a t i v amen t e abundan tes : Verbesina virgata, Wigandia caracasana, Dodonaea viscosa, y a últimas fechas ha aumentado la presencia de otra suculenta, Opuntia robusta.
Senecio praecox var. praecox florece y fructifica todos los años. Cada individuo adulto puede llegar
a producir 200 racimos. Cada racimo puede presentar en promedio 20 inflorescencias y cada inflorescencia presenta entre 15 y 20 flores hermafroditas en la zona del disco. Además presenta 5 flores liguladas (flores masculinas) alrededor del mismo (Rzedowski y Rzedowski 1979). La mayor parte de las semillas producidas no son viables. Observaciones personales han mostrado que en promedio solo se produce una semilla viable por inflorescencia. No se conoce con certeza algún efecto curativo de la planta, sin embargo, la infusión de hojas es utilizada para curar heridas y reumatismos (Rzedowski y Rzedowski 1979). Por otra parte, la disminución del hábitat ha provocado que las poblaciones de S. praecox var. praecox se vean severamente dañadas. El área original del “matorral xerófilo” ha sido reducida de 4000 ha. (el 50% del área original producida por el derrame del Xitle) a 124.5 ha. en la actualidad (Valiente-Banuet y De Luna García 1990). En conse-cuencia, el número de especies de plantas se ha reducido sustan-cialmente (de 350 especies a 226). A pesar de que el área de la comunidad ha sido reducida en un orden de magnitud, aún conserva el 50% de las especies originales (Alvarez et al 1989). La creación de la “Reserva del Pedregal de San Ángel” en el año de 1983 (125 ha.) surgió como una excelente alterna-tiva para conservar este gran recinto de riqueza florística en la Cd. de México.

Cactáceas y Suculentas Mexicanas Tomo XLV Año. 45 No. 3 julio septiembre 2000 64
Historia de vida y su dinámica poblacional
S. praecox var. praecox presenta atributos arquitectónicos que permiten reconstruir con detalle la historia de vida de la planta a partir del análisis de su patrón de rami f i cac ión (arqu i tec tura) . Durante mas de 12 años se ha venido registrando la dinámica poblacional del arbusto Senecio praecox en la Reserva del Pedregal de San Ángel, en Cd. Universitaria, al sur de la Cd. de México (entre 19O 15’ N. y 19O 17’ N. aprox.). S. praecox es un arbusto perenne que florece y fructifica en la primavera, germinando las semillas con la llegada de las lluvias en el verano. Durante varios años, las plántulas crecen y acumulan recursos en la raíz, antes de produ-cir un tallo aéreo no ramificado. Una vez producido, este tallo crece durante varios años (entre 10 y 15 años, obs. pers.) antes de alcanzar la madurez sexual (aprox. 30 cm. de altura). Alcanzada ésta, se produce una inflorescencia terminal (Fig. 2) que hace que Senecio praecox solo pueda reiniciar su crecimiento vegetativo a través de la liberación de uno o mas ( u sua lmen t e do s ) d e l o s meristemos laterales localizados por debajo del meristemo terminal. Además, uno o varios años después de haber alcanzado la madurez sexual, S. praecox produce otros tallos a partir de la corona de la raíz. Estos tallos tienen el mismo comportamiento del tallo original, excepto que crecen a una tasa mas acelerada y por ende alcanzan la
madurez sexual a una edad mas temprana (entre 3 y 6 años aprox., obs pers.). La producción de tallos a partir de la corona de la raíz, la liberación de mas de un meristemo lateral después de la floración y la repetición de este proceso de clonación-reproducción-ramificación durante varios años produce la forma de “candelabro” típica de esta especie, “Modelo arquitectural de Leewenberg” (Hallé, Oldeman y Tomlinson 1978; Fig. 1). Por lo anterior, la presencia de una dicotomía en la morfología de la rama es evidencia de haber produ-cido una inflorescencia. Por lo tanto, identificando el número de dicoto-mías a lo largo de todas las ramas, es posible conocer el número de eventos reproductivos acumulados por la planta a lo largo de toda su vida.
Otro atributo arquitectónico que destaca en S. praecox, es la posibi-lidad de conocer la edad de las ramas a partir del análisis de las mismas. Senecio praecox comienza a producir hojas una vez iniciada la temporada de lluvias (mayo-junio), manteniéndose hasta finales del verano (septiembre-octubre). Al final de la temporada de lluvias los entrenudos son tan cortos que las últimas hojas producidas en el año forman una especie de roseta y dejan una cicatriz casi continua alrededor del tallo, denotando así el final de la temporada de creci-miento para la planta. Ésta cicatriz, por lo general muy evidente a lo largo de toda la extensión de la rama, permite contabilizar el total de temporadas de crecimiento, y en

Cactáceas y Suculentas Mexicanas Tomo XLV Año. 45 No. 3 julio septiembre 2000 65
consecuencia la edad de las ramas. En el Pedregal de San Ángel se han registrado ramas que presentan entre 35 y 40 años de edad, sin embargo, la planta en su conjunto es posible que viva varios cientos de años.
Estas características arquitectónicas, las cuales no se manifiestan de la misma forma en la mayor parte de las plantas, permiten reconstruir la histo-ria de crecimiento y reproducción de la planta. Para obtener el mismo tipo de información en algún otro tipo o grupo de plantas, sería necesario seguir a las poblaciones por varios años y/o décadas. Con S. praecox, un “simple” análisis de su arquitectura nos resume con detalle la historia de vida de cada individuo. Todo lo anterior es resultado de varios años de seguimiento, lo cual nos ha permitido interpretar la dinámica demográfica de la especie.
Literatura citada Álvarez, J., Carabias, J., Meave, J.,
Moreno-Casasola, P., Nava, D.,
Rodríguez, F., Tovar, C. y Valiente-Banuet, A. 1989. Proyecto para la creación de una Reserva en el Pedregal de San Ángel. Cuadernos de Ecología No. 1. Facultad de Ciencias, UNAM. México.
Carrillo, C. 1995. “El pedregal de San Ángel”, UNAM. México.
Hallé, F., Oldeman, R. y Tomlinson, P. 1978. Tropical trees and forests: an architectural analysis. Springer-Verlag, Berlín.
Rojo, A. 1994. Reserva Ecológica del Pedregal de San Ángel: ecología, historia natural y manejo. Ariel Rojo compilador. UNAM. México.
Rzedowski, J. y Rzedowski, G. 1979. Flora Fanerogámica del Valle de México, Vol. I. Ed. CECSA. México.
Rzedowski, J. y Rzedowski, G. 1985. Flora Fanerogámica del Valle de México, Vol. II. Ed. CECSA. México.
Valiente-Banuet, A. y De Luna García, E. 1990. Una lista florística actuali-zada para la Reserva del Pedregal de San Ángel, México, D.F. Acta Botánica Mexicana 9: 13-30.
Figura 1. Diagrama esquemático del modelo de crecimiento para Senecio praecox. Las dicotomías indican eventos reproductivos y las líneas entre
dicotomías indican los crecimientos anuales (edad de la rama).
Evento reproductivo
1997
1998
1999
2000

Cactáceas y Suculentas Mexicanas Tomo XLV Año. 45 No. 3 julio septiembre 2000 66
Introducción
Opuntia bradtiana es una especie endémica del desierto Chihuahuense con una distribución restringida y localmente abundante en Cuatro Ciénegas, Coahuila (Gold 1965, Bravo-Hollis 1978, Pinkava 1985, 1997). También se ha reportado para el estado de Durango y Chihua-hua sin precisar las localidades (Gold 1966, 1967). Esta especie fue descrita por primera vez por Coulter (1896) quien la clasificó como Cereus bradtianus. Britton et. Rose (1919-1923) mencionan que fue descrita en base a especímenes colectados por Anna B. Nickles en 1895, después se clasificó dentro de la subfamilia Opuntioideae (por la presencia de glóquidas) en un nuevo género como: Grusonia cereiformis, y fue clasificada como Opuntia por la presencia de hojas caducas, flores y frutos afines a los de otras especies de este género con el nombre de Opuntia bradtiana K. Brandegee, y
más tarde como Opuntia cereiformis Weber (Britton et Rose 1919-1923). Posteriormente, en la revisión taxo-nómica de la familia hecha por Britton et Rose (1919-1923), propu-sieron como nombre válido Grusonia bradtiana (Coulter) Britton et Rose. Sin embargo, Bravo-Hollis (1978) reconoció Grusonia a nivel de subgé-nero y retoma como nombre válido Opunt ia bradt iana (Coulter) K. Brandegee (Fig. 1).
Recientemente se ha registrado la posición geográfica de las localida-des de O. bradtiana en la región de Cuatro Ciénegas y rodeando la Sierra de La Paila, al norte y noro-este de Cuatro Ciénegas (C. Brachet y J. Reyes com. pers.), pero no se han reportado las localidades en las que ésta especie se localiza en el Bolsón de Mapimí (al oeste de las áreas tradicionalmente descritas como las localidades tipo de la especie).
Materiales y métodos
Se realizaron dos viajes a la zona calcárea en la Reserva de la Biosfera de Mapimí para ubicar a las
Opuntia bradtiana en la zona calcárea del Bolsón de Mapimí, México.
María C. Mandujano1 y J. Golubov2
Resumen Opuntia bradtiana es una cactácea endémica cuya localidad esta restringida al área oeste que rodea Cuatro Ciénagas, Coahuila. Se describen especies asociadas y frutos colectados. Para entender la baja producción de semillas de tales poblaciones es necesario llevar a cabo más investigaciones.
Abstract The endemic Opuntia bradtiana is locally restricted to the area surrrounding Cuatro Cinegeas, Coahuila. The locality of O. bradtiana in the Bolson de Mapimi (west of the type locality), the associated species and a description of collected fruits is described. More thorough research is needed to fully understand the low seed production of these populations.
1,2 Instituto de Ecología, UNAM. Apdo. Postal 70-275, Ciudad Universitaria. México, D. F., 04510. México. Correo electrónico: [email protected]

Cactáceas y Suculentas Mexicanas Tomo XLV Año. 45 No. 3 julio septiembre 2000 67
poblaciones de O. bradtiana que se habían reportado sin precisar la localidad (Cornet 1985). Los viajes a la calcárea se realizaron en mayo de 1999 y en marzo del 2000. En este último se localizó a la especie y se hicieron recorridos a pie en los dos cerros en los que se encontró (Cerro Cipriano y Cerro de enfrente). En ellos se estimó visualmente la densidad de plantas de O. bradtiana y se buscaron frutos y semillas. Además, se colectó al azar una muestra de 30 frutos, se registró la presencia de semillas y su número.
Resultados
Encontramos que O. bradtiana es relativamente más abundante en las zonas de bajada con pendientes de 2 a 4% que en la zona de pie de monte y ladera. En las planicies no se observó a la especie. En el cerro Cipriano (26° 42’ N, 103° 31’ W) la vegetación corresponde a matorral dominado por Larrea tridentata (DC.) Cov., y se observó Agave l e chug i l l a Tor r . , Euphorb ia antisyphilitica Zucc., Echinocactus horizonthalonius (Lem.) Doweld, Fouquieria splendens Engelm., Jatropha dioica Cerv., y Mammillaria spp. En el cerro de enfrente (26° 41’ N 103° 31’ W) la vegetación corres-ponde a matorral rosetófilo domi-nado por A. lechugilla y Hechtia glomerata Zucc. En las dos áreas se encontraron algunos individuos con frutos que en su mayoría estuvieron vacíos. De los 30 frutos colectados, 83% no presentaron semillas y se observó el parénquima del pericar-pelo muy desarrollado y en algunos casos todo el fruto presentaba una
coloración negra, posiblemente por la presencia de algún parásito. Los 5 frutos viables tuvieron en promedio 26 ± 4.6 semillas, siendo 13 el mínimo y 41 el máximo.
Discusión
Es necesario estudiar en detalle el estado de conservac ión de O. bradtiana en las localidades repor-tadas. Además, como esta especie presenta propagación clonal (Bravo-Hollis 1978, Hamilton 1970) se debe determinar si la alta proporción de frutos vacíos se debe a efectos de depresión por endogamia. Definitiva-mente es una especie muy locali-zada en las zonas calcáreas y de distribución restringida. Nuestras observaciones preliminares sugieren un bajo reclutamiento vía semilla, debido a que los frutos están vacíos o presentan pocas semillas (ver Hamilton 1970). Dada la escasa información y la importancia de esta especie, única en su forma de vida y endémica de algunas áreas del desierto Chihuahuense, hemos iniciado un estudio que abarca aspectos ecoló-gicos, demográficos y reproductivos de la misma.
Agradecimientos C. Montaña y A. Herrera nos guiaron y acompañaron en los recorridos. SEMPARNAP concedió el permiso de colecta (Oficio #3942). J. Reyes fungió como editor de esta nota. Agradecemos a Rubén Pérez Ishiwara y a un revisor anónimo sus sugerencias.
Literatura citada Bravo-Hollis, H. 1978. Las cactáceas de
México. Vol. I. Universidad Nacional Autónoma de México. D.F., México.
Britton, N. L. y Rose J. N. 1919-1923. The Cactaceae, 4 volúmenes. Carnegie Institute Washington Publication 248, Washington, D. C.

Cactáceas y Suculentas Mexicanas Tomo XLV Año. 45 No. 3 julio septiembre 2000 68
Euphorbia antisyphylitica Meyrán, sp. nov.
P l a n t a s u c u l e n t a , caulesdecumbentes, ondulati, usque ad 60 cm longis, 3-5 mm diametro;
nodi quibusque 2-3.5 cm; rami preasertim in base et nonnumquam in superioribus partibus, virides; folia opposita, oblanceolata vel orvicularia, apex subacutum, obtusum, base angosta vel petiolata; flores in apice vel in primo
Cornet, A. 1985. Las cactáceas de la Reserva de la Biosfera de Mapimí. Instituto de Ecología, A. C. México, D.F., México.
Coulter, J. M. 1896. Preliminary revision of the North American species of Echinocactus, Cereus y Opuntia. Contr. U. S. Nat. Herb. 3:355-462.
Gold, D. B. 1965. Las Cactáceas del Estado de Coahuila. Cactáceas y Suculentas Mexicanas 10:102-104.
Gold, D. B. 1966. Las Cactáceas del Estado de Chihuahua. Cactáceas y Suculentas Mexicanas 11:65-67.
Gold, D. B. 1967. Las Cactáceas del Estado de Durango. Cactáceas y Suculentas Mexicanas 12:17-20.
Hamilton, M. W. 1970. Seedling development of Opuntia bradtiana (Cactaceae). American Journal of Botany 57: 599-603.
Pinkava, D. 1985. Vegetation and flora of the Bolson of Cuatro Cienegas region, Coahuila, Mexico: IV. Summary, endemism and corrected catalogue. Journal of the Arizona-Nevada Academy of Science. 19: 23-47.
Pinkava, D. 1997. Cactaceae: Opuntia Mill. Cholla, Prickly-pear, Nopal. En Henrickson, J. y M. C. Johnston (editores). A Flora of the Chihuahuan desert region. Draft version 1.2, privatley published by Hendrickson, Los Angeles, 1 vols.
Euphorbia gumaroi Meyrán, una nueva especie del estado de Hidalgo, México.
Jorge Meyrán García1
Resumen En este estudio describo una nueva especie de Euphorbia localizada al norte de Tepatepec, en el estado de Hidalgo, México, asociada a matorral xerófilo. Describo la especie y las diferencias más grandes con la especie más cercana del género E. antisyphilitica. Dados los caracteres morfológicos, encontrados, considero a esta planta como nueva. Las especies del género Euphorbia han crecido en los últimos años y se encuentran 11 especies y dos variedades. Esta especie fue nombrada en honor al Sr. Gumaro, descubridor de la localidad. El holotipo fue depositado en el Herbario Nacional MEXU.
Abstract This paper describes a new species Euphorbia gumaroi found in the northern portion of Tepatepec, Hidalgo, Mexico, associated to desert shrubland. We describe the species as well as the main differences between this species and E. antisyphilitica. The specimen warranted a species status due to the profound differences with its closest relative E. antisyphilitica. The number of species of the genus Euphorbia has gradually increased in Mexico to 11 species and two varieties, and is currently being expanded. The holotype of the specimen was deposited in MEXU.
1 Editor Fundador de la Revista Cactáceas y Suculentas Mexicanas.
Cactáceas y Suculentas Mexicanas Tomo XLV Año. 45 No. 3 julio septiembre 2000 69
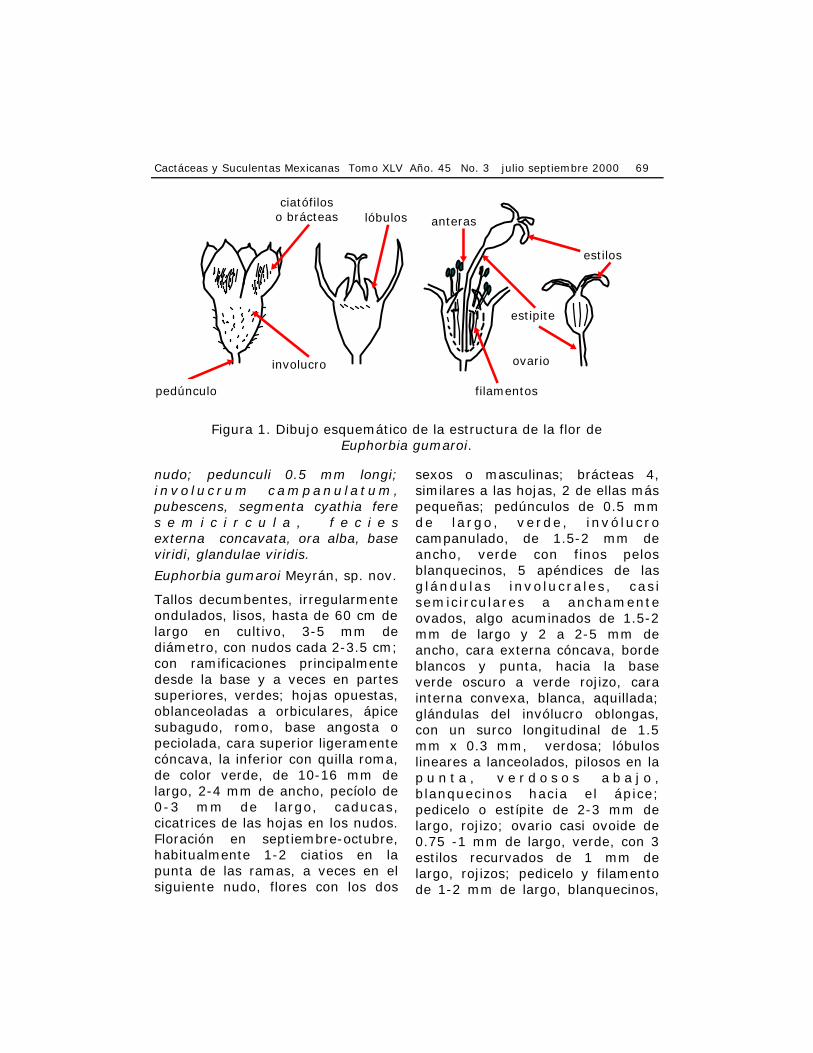
nudo; pedunculi 0.5 mm longi; i n vo l u c r um campanu l a t um, pubescens, segmenta cyathia fere s e m i c i r c u l a , f e c i e s externa concavata, ora alba, base viridi, glandulae viridis.
Euphorbia gumaroi Meyrán, sp. nov.
Tallos decumbentes, irregularmente ondulados, lisos, hasta de 60 cm de largo en cultivo, 3-5 mm de diámetro, con nudos cada 2-3.5 cm; con ramificaciones principalmente desde la base y a veces en partes superiores, verdes; hojas opuestas, oblanceoladas a orbiculares, ápice subagudo, romo, base angosta o peciolada, cara superior ligeramente cóncava, la inferior con quilla roma, de color verde, de 10-16 mm de largo, 2-4 mm de ancho, pecíolo de 0-3 mm de largo, caducas, cicatrices de las hojas en los nudos. Floración en septiembre-octubre, habitualmente 1-2 ciatios en la punta de las ramas, a veces en el siguiente nudo, flores con los dos
sexos o masculinas; brácteas 4, similares a las hojas, 2 de ellas más pequeñas; pedúnculos de 0.5 mm de largo, verde, invólucro campanulado, de 1.5-2 mm de ancho, verde con finos pelos blanquecinos, 5 apéndices de las glándulas involucrales, casi semicirculares a anchamente ovados, algo acuminados de 1.5-2 mm de largo y 2 a 2-5 mm de ancho, cara externa cóncava, borde blancos y punta, hacia la base verde oscuro a verde rojizo, cara interna convexa, blanca, aquillada; glándulas del invólucro oblongas, con un surco longitudinal de 1.5 mm x 0.3 mm, verdosa; lóbulos lineares a lanceolados, pilosos en la p u n t a , v e r d o s o s a b a j o , blanquecinos hacia el ápice; pedicelo o estípite de 2-3 mm de largo, rojizo; ovario casi ovoide de 0.75 -1 mm de largo, verde, con 3 estilos recurvados de 1 mm de largo, rojizos; pedicelo y filamento de 1-2 mm de largo, blanquecinos,
filamentos
ciatófilos o brácteas
involucro
estilos
lóbulos anteras
estipite
ovario
pedúnculo
Figura 1. Dibujo esquemático de la estructura de la flor de Euphorbia gumaroi.

Cactáceas y Suculentas Mexicanas Tomo XLV Año. 45 No. 3 julio septiembre 2000 70
con dos anteras amari l las, bractéolas lineares, pilosas de 1 mm largo, blanquecinas (Fig. 1).
Localidad tipo: Cerca de Tepatepec, Hidalgo, colectada por el Sr. Gumaro Manzo, en un matorral xerófilo y cultivada en la Ciudad de México (Meyrán 5745). Holotipo: MEXU.
El número de especies mexicanas suculentas del género Euphorbia ha ido creciendo lentamente, Miguel Cházaro, durante una conferencia dada en una sesión de la Sociedad Mexicana de Cactología (22 de mayo de 1997), señaló 10 especies y 2 variedades.
Entre las especies de Euphorbia mexicanas, la más relacionada con E u p h o r b i a g u m a r o i e s E. antisyphilitica. Las principales diferencias entre las dos especies se
presentan en el cuadro 1.
Esta especie (Fig. 2) es nombrada en honor al Sr. Gumaro Manzo, entusiasta cactófilo, miembro de la Sociedad Mexicana de Cactología, filial Hidalgo, quien encontró por primera vez esta interesante planta.
Agradecimientos El autor agradece al M. en C. Jaime Jiménez por su gentileza en la revisión y correcciones pertinentes a este trabajo, a Omar González Sórzano la traducción de la diagnosis al latín, al Biól. Jordan Golubov por su ayuda en la traducción a inglés, al Biól. Jorge Saldívar por ayuda en la elaboración del dibujo.
Referencias
Jacobsen H. 1960. A Handbook of Succulent Plants. Blandford Press, London.
Radford , A. E., W.C.Dickison, J.E. Massey & C.R. Bell. 1974.Vascular Plant Systematics. Harper and Row, New York.
E. antisyphilica E. gumaroi
Hábito erecto decumbente
Tallos rectos irregularmente ondulados
cicatrices de las hojas en espiral. sin nudos
cicatrices de las hojas en los nudos nudos cada 2-3.5 cm.
Ciatios surgen a 10 o más cm del ápice de las ramas
nacen en el ápice o en el primer nudo de las ramas
Brácteas involucrales o Ciatófilos.
forma redondeada semicirculares a anchamente ovadas
borde superior dentado entero
color blancos o rosados hacia arriba y base rojo oscuro
blancos base verde oscuro a verde rojizo
Lóbulos del involucro con pelos en la cara externa, rosados
sin pelos en la cara externa, sólo en la punta, verdosos
Estilos bífidos erecto o algo extendido simples recurvados
Cuadro 1. Principales diferencias entre Euphorbia antisyphilica y E. gumaroi

Fig. 2. (Meyrán) Euphorbia gumaroi planta en floración del estado de Hidalgo, México.
Foto: Jorge Meyrán.
Fig. 2. (Pérez y Franco) Senecio praecox en la Re-serva del Pedregal de San Angel, México.
Foto: Rubén Pérez.

Opuntia bradtiana (Coulter) K. Brandegee, (Cactaceae). Opuntia bradtiana pertenece a la tribu Opuntieae, género Opuntia y subgénero Cylindropuntia, que se caracteriza por presentar tallos cilíndricos en sección transversal (Hamilton 1970). Es una especie endémica del desierto Chihuahuense, tiene una distribución restringida en zonas calcáreas en el bolsón de Mapimí y en Cuatro Ciénegas, y es mas abundante en las zonas de bajada. O. bradtiana mide ca. de 1 m de altura con múltiples ramificaciones, presenta tallos rastreros, espinosos, de color verde claro, de 4 a 7 cm de diámetro, con 8 a 10 costillas longitudinales, poco profundas y tuberculadas. Las areolas nacen en los extremos de los tubérculos, los cuales forman costillas (coalescen) tempranamente en el estado de plántula. De cada areola nace una hoja pequeña y decídua, y se considera que tiene glóquidas solamente en las areolas florales. Las flores crecen en los ápices de las ramas y son amarillas, abren de mayo a junio. Los frutos son de 28 mm por 13 mm, espinosos, acostillados y no presentan absición. Algunos frutos son estériles y otros son la fuente de alimento de animales. Los frutos viables presentan en promedio 26 semillas. Las semillas son de 2-3 mm de diámetro, redondas y aplanadas; la testa externa tiene drusas que ocasionan que sean difíciles de germinar. O. bradtiana se propaga a partir de tallos caídos que enraizan. Las poblaciones de O. bradtiana han sido dañadas por la extracción de candelilla (Euphorbia antisyphylitica), una especie económicamente importante por su cera. O. bradtiana es una especie que no está considerada en el CITES, sin embargo, es una especie endémica, de forma de vida única, con distribución restringida y cuyas poblaciones han sido dañadas, por lo cual debería ser protegida.
Dolores Rosas Barrera. Instituto de Ecología UNAM.
Inflorescencia de Opuntia bradtiana en Cuatro Ciénagas Coah., Méx.

CACTACEAS Y SUCULENTAS MEXICANAS es una revista trimestral de circulación internacional que publica la Sociedad Mexicana de Cactología A. C. Esta revista esta disponible para toda contribución original científica o de divulgación sobre las Cactáceas y otras plantas suculentas.
Texto
Presentarlo en hojas blancas tamaño carta a doble espacio (incluyendo cuadros), con márgenes de 2.5 cm, numeradas consecutivamente, sin errores tipográficos, usando la fuente Times New Roman de 12 puntos. Las contribuciones pueden ser en español o en inglés. Los nombres científicos se anotarán con cursivas citando el género sin abreviar y el autor de la especie la primera vez que se mencione en el cuerpo del texto, en las subsecuentes podrá abreviarse. Los encabezados de las secciones deberán estar en negritas y centrados. El texto deberá incluir los siguientes puntos: Título, Autor(es) nombre y apellido (sin negritas) e indicar con superíndices numerados la referencia a la institución de adscripción. El nombre y dirección de la institución del autor(es) debe incluirse como nota al pie de página, incluyendo el teléfono, fax y correo electrónico del autor de correspondencia. Resumen: en español, máximo de 300 palabras. Abstract: en inglés debe proporcionar información detallada del trabajo, hasta 450 palabras que mencione el objetivo, la especie y sitio de estudio, breve metodología, resultados y conclusión. NOTA.- Si la contribución es en inglés, el Resumen en español deberá ser el más extenso. Palabras clave (seis). Introducción, Material y métodos, Resultados, Discusión, Agradecimientos (en forma breve), Literatura citada, Cuadros, Pie de figuras, y Figuras. Se deberá usar el sistema internacional de medición (SI) con las siguientes abreviaciones: min (minutos), h (horas), d (días), mm (milímetro), cm (centímetro), m (metro (s)), km (kilometro (s)), ml (mililitro), l (litro); para los símbolos estadísticos EE (error estándar), g.l. (grados de libertad), N (tamaño de muestra), CV (coeficiente de variación) y poner en cursivas los estimadores (p. ej. r2, prueba de t, F, P). Enviar 3 copias del texto de buena calidad y una copia en disco de 3.5” para PC, en formato Word, 6.0 ó posterior, formato, ASCII o RTF.
Cuadros
Cada cuadro debe empezar en una hoja nueva e ir numerado consecutivamente conforme se hace referencia en el texto. La leyenda del cuadro debe contener información suficiente para entenderse sin ayuda del texto. La primera letra del título debe de ir en mayúsculas al igual que la primera letra de cada entrada en cada columna o renglón.
Leyendas de figuras
Las leyendas van a doble espacio. Escriba las
leyendas en forma de párrafo y separadas de las figuras.
Ilustraciones
Los mapas, diagramas y otras ilustraciones se presentarán en hojas separadas, numeradas y en tinta negra (líneas con un mínimo de 2 puntos). Anote el nombre del primer autor y el número de la figura al reverso de cada una. Las fotografías pueden enviarse en papel o transparencia de buen contraste, etiquetadas y numeradas, los pies de figura se anexarán en la hoja de leyendas.
Literatura Citada
La literatura citada en el texto debe seguir el siguiente formato: un autor Buxbaum (1958) ó (Buxbaum 1958), 2 autores Cota y Wallace (1996) ó (Cota y Wallace 1996), tres ó más autores Chase et al. (1985) ó (Chase et al. 1985). Referencias múltiples deben de ir en orden cronológico (Buxbaum 1958; Chase et al. 1985). La literatura citada deberá estar en orden alfabético según el siguiente formato:
Bravo-Hollis, H. y Sánchez-Mejorada, H. 1991. Las Cactáceas de México. Vol. 3, UNAM, México. 643 pp.
Buxbaum, F. 1958. The phylogenetic division of the subfamily Cereoideae, Cactaceae. Madroño 14: 177-206.
Cota, J. H. y Wallace, R. S. 1996. La citología y la sistemática molecular en la familia Cactaceae. Cactáceas y Suculentas Mexicanas 41: 27-46.
Milligan, B. 1998.Total DNA isolation. Página 29-64. En A. R. Hoelzel (ed.). Molecular genetic analysis of populations. IRL Press, second edition. Oxford, England.
Las 3 copias del manuscrito deben enviarse al editor, anexando fotografías e ilustraciones a la siguiente dirección: Dra. María C. Mandujano. Instituto de Ecología, Apartado Postal 70-275, Ciudad Universitaria, UNAM. México, D. F. 04510, México. Correo electrónico: [email protected]
El autor de correspondencia debe conservar una copia para cualquier aclaración. Los manuscritos serán revisados por académicos especializados en el área de investigación, designados por el comité editorial.
La Sociedad Mexicana de Cactología proporciona sobretiros al autor(es) solo en formato electrónico (pdf). El Comité Editorial se reserva el derecho de rechazar cualquier contribución o solicitar al autor(es) modificaciones a su trabajo, así como hacer cambios menores en el texto sin consultar al autor(es).
NORMAS EDITORIALES PARA LOS AUTORES


















