![Nucleo celular[1]](https://static.fdocuments.ec/doc/165x107/559e882a1a28ab5d778b45a5/nucleo-celular1.jpg)
Aspectos Del Nucleo Celular
13
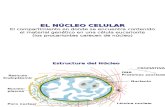
ASPECTOS GENERALES DE LA ANATOMIA Y FISIOLOGÍA DEL NÚCLEO. Organización estructural y fisiología del núcleo El núcleo es el órgano más notable en el citoplasma de casi todas las células animales y vegetales; está rodeado por una membrana, es esférico y mide unas 5 μm de diámetro. En la matriz del núcleo se encuentra la cromatina. La cromatina se organiza en cromosomas, son poco visibles, mientras que están bien delimitadas durante la división celular. El núcleo de las células eucariotas está rodeado por una membrana doble, y la comunicación con el citoplasma se presenta mediante poros nucleares. El nucléolo es una región donde se sintetizan partículas que contienen ARN y proteína que migran al citoplasma a través de los poros nucleares y luego se modifican para transformarse en ribosomas. La principal función del núcleo es controlar la síntesis de proteínas. En él tienen lugar procesos tan importantes como la replicación del DNA y la transcripción. La forma del núcleo es generalmente esférica o elíptica, aunque en algunas células es completamente irregular. Las células procariotas no poseen una membrana nuclear definida, pero si contienen ADN. El núcleo de una célula puede presentar cambios en su estructura que están con el ciclo celular, por ejemplo, la membrana nuclear desaparece y el nucléolo. El número de núcleos por célula es variable: es uno en la mayoría de las células; pueden ser dos, como en algunos hepatocitos, o muchos, como en los osteoblastos y las fibras musculares estriadas. Componentes del núcleo y función. Membrana nuclear http://www.javeriana.edu.co/Facultades/Ciencias/neurobioquimica/libros/celular/nucleo.html Es una envoltura nuclear formada por dos membranas concéntricas con poros nucleares, que limitan y separan al núcleo del citoplasma. A través de los poros se produce el transporte de moléculas entre el núcleo y el citoplasma. La membrana más externa se encuentra unida al retículo endoplásmico rugoso. Las membranas nucleares tienen la misma estructura de una membrana celular. La membrana interior proporciona soporte al núcleo. Está compuesta por proteínas llamadas lamininas, las cuales son proteínas relacionadas con el citoesqueleto celular.
-
Upload
jhancarlosflorezarteaga -
Category
Documents
-
view
62 -
download
0
description
en este documento encontrara todo lo que tenga que ver con el núcleo celular
Transcript of Aspectos Del Nucleo Celular
-
ASPECTOS GENERALES DE LA ANATOMIA Y FISIOLOGA DEL NCLEO.
Organizacin estructural y fisiologa del ncleo
El ncleo es el rgano ms notable en el citoplasma de casi todas las clulas animales y vegetales; est rodeado por una
membrana, es esfrico y mide unas 5 m de dimetro. En la matriz del ncleo se encuentra la cromatina. La cromatina
se organiza en cromosomas, son poco visibles, mientras que estn bien delimitadas durante la divisin celular. El ncleo
de las clulas eucariotas est rodeado por una membrana doble, y la comunicacin con el citoplasma se presenta
mediante poros nucleares. El nuclolo es una regin donde se sintetizan partculas que contienen ARN y protena que
migran al citoplasma a travs de los poros nucleares y luego se modifican para transformarse en ribosomas. La principal
funcin del ncleo es controlar la sntesis de protenas. En l tienen lugar procesos tan importantes como la replicacin
del DNA y la transcripcin.
La forma del ncleo es generalmente esfrica o elptica, aunque en algunas clulas es completamente irregular. Las
clulas procariotas no poseen una membrana nuclear definida, pero si contienen ADN. El ncleo de una clula puede
presentar cambios en su estructura que estn con el ciclo celular, por ejemplo, la membrana nuclear desaparece y el
nuclolo.
El nmero de ncleos por clula es variable: es uno en la mayora de las clulas; pueden ser dos, como en algunos
hepatocitos, o muchos, como en los osteoblastos y las fibras musculares estriadas.
Componentes del ncleo y funcin.
Membrana nuclear
http://www.javeriana.edu.co/Facultades/Ciencias/neurobioquimica/libros/celular/nucleo.html
Es una envoltura nuclear formada por dos membranas concntricas con poros nucleares, que limitan y separan al ncleo
del citoplasma. A travs de los poros se produce el transporte de molculas entre el ncleo y el citoplasma. La
membrana ms externa se encuentra unida al retculo endoplsmico rugoso. Las membranas nucleares tienen la misma
estructura de una membrana celular. La membrana interior proporciona soporte al ncleo. Est compuesta por protenas
llamadas lamininas, las cuales son protenas relacionadas con el citoesqueleto celular.
-
Nucleosol
Tambin se conoce como jugo nuclear, nucleoplasma o carioplasma, el cual es el medio interno del ncleo donde se
encuentran suspendidos el resto de los componentes nucleares, como la cromatina y los nucleolos.
Nuclolo
Son masas densas y esfricas en las cuales se sintetiza el RNA ribosmico y en donde se producen las primeras fases
de ensamblaje de los ribosomas.
Poros nucleares
http://www.javeriana.edu.co/Facultades/Ciencias/neurobioquimica/libros/celular/nucleo.html
Son perforaciones de la membrana nuclear que permiten el intercambio de pquenos elementos entre el ncleo y el
citoplasma como por ejemplo iones, molculas polares y macromolculas como (protenas y RNA). Los mRNAs (RNA
mensajeros) que son sintetizados en el ncleo son transportados eficientemente al citoplasma donde permiten la sntesis
de protenas. Las molculas puede ser transportadas difusin pasiva (pequeas partculas de al menos de 20 kD) o por
transporte activo (protenas y mRNAs).
Cromatina
http://www.javeriana.edu.co/Facultades/Ciencias/neurobioquimica/libros/celular/nucleo.html
-
Constituida por DNA y protenas, aparece durante la interfase; es una estructura altamente condensada durante la
mitosis, pero cuando la clula entra en divisin la cromatina se organiza en estructuras individuales que son los
cromosomas. Durante la interfase la cromatina (heterocromatina) permanece muy condensada y es transcripcionalmente
inactiva mientras que la restante (eucromatina) se encuentra descondensada y se distribuye a travs de ncleo.
Transporte a travs de los poros nucleares Las protenas que forman parte del complejo del poro se denominan nucleoporinas. En las levaduras hay unas 30
protenas distintas en cada poro nuclear, mientras que en los metazoos pueden ser 40 o ms. Se cree que un poro de
una clula de mamferos contiene unas 400 nucleoporinas, ya que muchas de ellas se encuentran repetidas. Las
protenas que forman los poros nucleares se asocian para formar 8 bloques que configuran un octgono regular y se
organizan formando anillos: el anillo citoplasmtico orientado hacia el citoplasma, el anillo radial situado en el hueco que
deja la envuelta nuclear y es responsable de anclar el complejo del poro a las membranas de la envuelta nuclear y el
anillo nuclear que se encuentra en el nucleoplasma. Adems, desde cada bloque se proyectan fibrillas proteicas que van
hacia el citoplasma denominadas fibras citoplasmticas, y otras al interior del ncleo que reciben el nombre de fibras
nucleares. stas ltimas se conectan a otro conjunto de protenas que forman una estructura cerrada llamada jaula
nuclear o anillo distal.
Los poros nucleares contienen un canal acuoso de unos 90 nm de longitud y entre 45 y 50 nm de dimetro cuando est
en reposo, pero que se puede expandir cuando realiza transporte activo. El canal puede permitir el paso libre de
pequeas molculas (menores de 60 kDa) pero restringir el movimiento de otras de mayor tamao con potencial papel
fisiolgico. Aunque incluso algunas molculas menores de 20-30 kDa tales como las histonas, los ARNt o algunos
ARNm. stas molculas y otras ms grandes cruzan por transporte pasivo facilitado. Pero aunque sea pasivo facilitado
pueden viajar en contra de su gradiente, por tanto necesitan energa que es aportada por el gradiente de otras molculas
denominadas Ran-GTPasas.
http://webs.uvigo.es/mmegias/5-celulas/4-
Gradientes creados por las molculas
Ran entre el citoplasma y el
nucleoplasma. La energa se consume
en el nucleoplasma para crear Ran-
GTP a partir de Ran y fsforo
inorgnico, manteniendo la
concentracin de Ran-GTP elevada.
En el citoplasma las Ran-GTP son
rpidamente convertidas en Ran-GDP,
manteniendo la concentracin de
estas ltimas elevada. Mientras, la
hidrlisis del Ran-GDP a Ran,
mantiene la concentracin de Ran-
-
El transporte mediado por los poros nucleares est orquestado por las protenas Ran-GTPasas y por unas familias de
protenas denominadas importinas y exportinas. Las molculas Ran-GTPasas son trascendentales tanto para la
importacin como para la exportacin de molculas. Son las responsables de crear un gradiente que dirige el
transporte y crear este gradiente es la nica parte del transporte por parte de los poros nucleares que gasta energa.
Las molculas Ran pueden estar en tres estados: Ran-GTP, Ran-GDP y Ran. El paso de un estado a otro est
mediado por otras enzimas. En el nucleoplasma hay una mayor concentracin de Ran-GTP, mientras que en el
citoplasma abunda la Ran-GTP.
Las protenas que tienen que ser importadas al nucleoplasma necesitan poseer una secuencia de aminocidos o
pptido seal de entrada y las que tienen que ser exportadas un pptido seal de salida. Estas secuencias de
aminocidos sern reconocidas por las importinas o por las exportinas, respectivamente. Las protenas de los poros
nucleares no interaccionan directamente con las protenas transportadas sino son las importinas y las exportinas. Estos
complejos importina-protena o exportina-protena utilizan los gradientes de las molculas Ran-GTP o Ran-GDP para
cruzar los poros nucleares y llevar sus cargas al otro lado. Adems de protenas, las molculas de ARN deben tambin
atravesar los poros nucleares. El mecanismo que usan los distintos tipos de ARN para ser transportados difieren entre
s, pero todos estn mediados por un mecanismo de asociacin con protenas. El exporte de ARNt sigue un mecanismo
en el que es reconocido por una exportina denominada exportina-t, que tambin se une a una Ran-GTP. El mecanismo
de exportacin de los ARNr no se conoce muy bien. Los ARNm no utilizan, en su mayora el sistema de transporte
mediado por Ran-GTP sino el mediado est mediado por dos protenas que forman el complejo Tap/Nxt, el cual
interacciona con las nucleoporinas y posibilita el transporte con gasto de ATP. Una pequea cantidad de ARNm parece
usar las protenas Crm1, siendo en este caso un transporte dependiente del gradiente creado por las protenas Ran.
Cromatina clasificacin y funcin.
La cromatina es la sustancia fundamental del ncleo celular. Su constitucin qumica es simplemente filamentos de ADN
en distintos grados de condensacin. Estos filamentos forman ovillos. Existen tantos filamentos como cromosomas
presente la clula en el momento de la divisin celular. La cromatina se forma cuando los cromosomas se descondensan
tras la divisin celular o mitosis. Existen diversos tipos de cromatina segn el grado de condensacin del ADN. Este ADN
se enrolla alrededor de unas protenas especficas, las histonas, formando los nucleosomas (ocho protenas histnicas +
una fibra de ADN de 200 pares de bases). Cada nucleosoma se asocia a un tipo distinto de histona la H1 y se forma la
cromatina condensada.
La funcin de la cromatina es: proporcionar la informacin gentica necesaria para que los orgnulos celulares puedan
realizar la transcripcin y sntesis de protenas; tambin conservan y transmiten la informacin gentica contenida en el
ADN, duplicando el ADN en la reproduccin celular.
La cromatina se clasifica en:
La eucromatina se tie dbilmente con distintas coloraciones y permanece dispersa (no condensada) durante la
interfase, momento donde ocurre la transcripcin del ARN. Los genes activos estn situados en la eucromatina.
La heterocromatina es la cromatina que se tie ms fuerte, es ms condensada y que se encuentra inactiva. Puede ser
de dos tipos: la de tipo constitutivo idntica para todas las clulas del organismo y que carece de informacin gentica,
incluye a los telmeros y centrmeros del cromosoma que no expresan su ADN. La heterocromatina facultativa diferente
-
en los distintos tipos celulares, contiene informacin sobre todos aquellos genes que no se expresan o que pueden
expresarse en algn momento. Incluye al ADN satlite y al corpsculo de Barr.
Organizacin estructural y funcin del nuclolo
El nuclolo es una regin del ncleo considerada como un orgnulo. La funcin principal del nuclolo es la produccin y
ensamblaje de los componentes ribosmicos. El nuclolo es aproximadamente esfrico y est rodeado por una capa de
cromatina condensada. El nuclolo, es la regin heterocromatica ms destacada del ncleo. No existe membrana que
separe el nuclolo del nucleoplasma.
La estructura del nuclolo como la de una esponja, con tabiques irregulares interconectados entre los cuales quedan
espacios vacos. Se distinguieron los siguientes componentes:
1. Parte amorfa. Corresponde a los espacios de escasa densidad a los electrones que forman cavidades
intercomunicadas en la parte densa. Contiene grnulos de DNA y equivale al nucleoloplasma.
2. Parte densa. Se corresponde con el nucleolonema. A su vez se distinguen en ella:
Parte granular. Formada por acumulaciones de grnulos de unos 25 nm de dimetro, que contienen
ribonucleoprotenas.
Parte fibrillar. Ms densa que la anterior, constituida por fibrillas de unos 8-10 nm, tambinformadas por
ribonucleoprotenas.
Centro fibrillar. Muy evidente en algunos nuclolos, donde puede haber varios centros fibrillares y de densidad inferior a
la de las partes granular y fibrillar. Consiste en finas fibrillas de 7-9 nm. Contiene DNA y algo de RNA. Debido al
contenido en DNA, se ha considerado que los centros fibrillares corresponden a organizadores nucleolares en fase activa
o de transcripcin. Algunos autores han discrepado de esta interpretacin debido a que los centros fibrillares no se
observan en muchos tipos celulares y no parecen indispensables para la correcta funcin del nuclolo, y han atribuido al
centro fibrillar otras funciones como la de almacn de reservas proteicas para la sntesis ribosmica. En muchos
nuclolos se observa tambin una masa fibrillar densa que contiene exclusivamente DNA. Es la heterocromatina
asociada al nuclolo y corresponde a la heterocromatina telomrica de los organizadores nucleolares.
La funcin principal del nuclolo es la biosntesis de ribosomas desde sus componentes de ADN para formar ARN
ribosomal."El nuclolo es una mquina productora de ribosomas cuyo destino final es el citosol" Est relacionado con la
sntesis de protenas.
Adems, investigaciones recientes, han descrito al nuclolo como el responsable del trfico de pequeos segmentos de
ARN. El nuclolo adems, interviene en la maduracin y el transporte del ARN hasta su destino final en la clula.
Aunque el nuclolo desaparezca en divisin, algunos estudios actuales aseguran que regula el ciclo celular. La estructura
granular de los nuclolos solo puede ser observada con microscopia electrnica.
Concepto y organizacin estructural del cromosoma Los cromosomas son estructuras en forma de bastn que aparecen en el momento de la reproduccin celular, en la
divisin del ncleo o citocinesis. Estn constituidos qumicamente por ADN ms histonas puesto que son simplemente
cromatina condensada. Su nmero es constante en todas las clulas de un individuo pero vara segn las especies. Un
-
cromosoma est formado por dos cromtidas (dos hebras de ADN idnticas) que permanecen unidas por un centrmero.
El cromosoma puede presentar constricciones primarias (centrmero) que origina los brazos del cromosoma y
secundarias que se producen en los brazos y originan satlites. Alrededor del centrmero existe una estructura proteica,
llamada cinetocoro, que organiza los microtbulos que facilitarn la separacin de las dos cromtidas en la divisin
celular.
Su esqueleto tiene dos partes, llamadas cromtidas, que estn unidas por un centrmero. Este ltimo es fundamental
para asegurar la correcta distribucin de los cromosomas duplicados en las clulas hijas durante las divisiones celulares.
En sus extremos estn los llamados telmeros, que se encargan de impedir que las terminaciones se enreden y adhieran
unos con otros. Adems, ayudan a que los cromosomas semejantes se emparejen y entrecrucen durante la meiosis.
En los humanos, cada clula contiene 46 cromosomas dispuestos en 23 pares. Las nicas excepciones son las clulas
sexuales (espermatozoide y vulo) que contienen 23 cromosomas, pero que al fecundarse crean una clula con una
dotacin completa de cromosomas, es decir, 46.
De los 23 cromosomas, los primeros 22 se denominan autosomas o autosmicos, y al par 23 -los cromosomas sexuales-
se les conoce como gonosomas o heterocromosomas (X e Y). Estos ltimos difieren del resto, ya que no siempre son
idnticos. La mujer posee dos cromosomas X idnticos y el hombre, un cromosoma X y un cromosomaY, que es ms
pequeo.
CICLO CELULAR
El ciclo celular es un conjunto ordenado de eventos que culmina con el crecimiento de la clula y la divisin en dos
clulas hijas. El ciclo celular es un proceso que se conforma de dos partes: la divisin nuclear (Cariocinesis) y la divisin
del citoplasma (Citocinesis). Ambos procesos pueden darse asociados, uno detrs del otro, o de forma independiente,
primero uno, y algn tiempo despus el otro. Para que pueda darse la divisin nuclear es necesario que se d
previamente otro proceso, que es la replicacin del ADN.
http://recursostic.educacion.es/ciencias/biosfera/web/alumn
o/2bachillerato/La_celula/contenidos15.htm
-
FASES DEL CICLO CELULAR
http://www.slideshare.net/Dagoberto48/mapa-conceptual-ciclo-celular
La cariocinesis es conocida como la mitosis y ocurre despus de completarse las tres fases preparatorias que
constituyen la interfase.
El primer proceso clave para que se de la divisin nuclear es la replicacin del ADN; esto ocurre antes de que inicie la
divisin, en un perodo del ciclo celular llamado Interfase. La interfase se divide en G1, S y G2. En fase G1 la clula
duplica su tamao y aumenta la cantidad de organelos, enzimas y otras molculas; en fase S se presenta la duplicacin
del DNA y protenas asociadas; existen ahora dos copias de la informacin gentica de la clula; y en la fase G2 las
estructuras necesarias para la divisin empiezan a conformarse, los cromosomas empiezan a condensarse.
-
http://www.curtisbiologia.com/b1957
MITOSIS
Clula madre en divisin Las dos cadenas de ADN idnticas se espiralizan y se convierten en CROMOSOMAS. En la
especie humana 2n = 46 cromosomas (formados por dos cadenas idnticas cada uno).
La mitosis es un proceso de divisin nuclear que sirve para repartir las cadenas de ADN de forma que todas las clulas
hijas que se originan tengan la MISMA INFORMACIN GENTICA que su madre y entre ellas.
La mitosis es continua, sin interrupciones (todo o nada), relativamente rpida, que para ser estudiada se suele dividir en
varias fases, que son: PROFASE, METAFASE, ANAFASE y TELOFASE.
PROFASE: Comienza con la conversin de la CROMATINA en CROMOSOMAS (1) por un proceso de espiralizacin de
las cadenas, seguiremos teniendo lo mismo, pero de forma diferente: las dos cadenas que son completamente idnticas
(ya que una se ha formado por replicacin de la otra) se espiralizan juntas originando las cromtidas del cromosoma. Se
duplican los centriolos (2). La membrana nuclear desaparece (3).
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/genetica1/contenidos5.htm#cariocinesis
-
METAFASE: Es una fase breve en la que todos los cromosomas se encuentran situados en el ecuador (parte media) de
la clula, formando una figura muy caracterstica llamada PLACA ECUATORIAL (1). Tras colocarse aqu comienza la
siguiente fase.
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/genetica1/contenidos5.htm#cariocinesis
ANAFASE: Los centrmeros se dividen y las cromtidas se separan y se desplazan hacia los centriolos, al tiempo que
van desapareciendo las fibras del huso. En este momento ya se ha repartido el material hereditario (las cadenas de ADN)
de forma idntica en dos partes.
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/genetica1/contenidos5.htm#cariocinesis
TELOFASE: Es como una profase al revs. Los cromosomas se desespiralizan y se transforman en cromatina (2);
aparece la membrana nuclear (1), quedando una clula con dos ncleos. Aqu concluye la mitosis.
-
http://recursos.cnice.mec.es/biosfera/alumno/4ESO/genetica1/contenidos5.htm#cariocinesis
DIVISIN CITOPLASMTICA (CITOCINESIS)
No es una fase de la mitosis. Es la divisin del citoplasma en dos partes, con la reparticin aproximada de los orgnulos
celulares.
En las clulas animales se hace por estrangulacin, desde fuera hacia adentro, y en las vegetales se hace por
crecimiento de la pared celular desde adentro hacia afuera. El resultado final es que la clula madre se ha transformado
en dos clulas hijas idnticas genticamente.
MEIOSIS
La mitosis es la solucin al problema de la divisin
celular y la constancia en el nmero de cromosomas de
las clulas hijas. La meiosis resuelve el problema de la
presencia de dos padres (progenitores) en los
organismos sexuales y la constancia del nmero de
cromosomas entre generaciones.
La meiosis es la divisin celular que permite la
reproduccin sexual. Comprende dos divisiones
sucesivas: una primera divisin meitica, que es una
divisin reduccional, ya que de una clula madre
diploide (2n) se obtienen dos clulas hijas haploides (n);
y una segunda divisin meitica, que es una divisin
ecuacional, ya que las clulas hijas tienen el mismo
nmero de cromosomas que la clula madre (como la
divisin mittica). As, dos clulas n de la primera
divisin meitica se obtiene cuatro clulas n. Igual que
en la mitosis, antes de la primera divisin meitica hay
un perodo de interfase en el que se duplica el ADN. Sin
embargo, en la interfase de la segunda divisin meitica
no hay duplicacin del ADN.
http://www.curtisbiologia.com/b1957
-
Primera divisin meitica
- Profase I. Es la ms larga y compleja, puede durar hasta meses o aos segn las especies. Se subdivide en: leptoteno,
se forman los cromosomas, con dos cromtidas; zigoteno, cada cromosoma se une ntimamente con su homlogo;
paquiteno, los cromosomas homlogos permanece juntos formando un bivalente o ttrada; diploteno, se empiezan a
separar los cromosomas homlogos, observando los quiasmas; diacinesis, los cromosomas aumentan su condensacin,
distinguindose las dos cromtidas hermanas en el bivalente.
- Metafase I. La envoltura nuclear y los nucleolos han desaparecido y los bivalentes se disponen en la placa ecuatorial.
- Anafase I. Los dos cromosomas homlogos que forman el bivalente se separan, quedando cada cromosoma con sus
dos cromtidas en cada polo.
- Telofase I. Segn las especies, bien se desespiralizan los cromosomas y se forma la envoltura nuclear, o bien se inicia
directamente la segunda divisin meitica.
Segunda divisin meitica
Est precedida de una breve interfase, denominada intercinesis, en la que nunca hay duplicacin del ADN. Es parecida a
una divisin mittica, constituida por la profase II, la metafase II, la anafase II y la telofase II.
REGULACIN DEL CICLO CELULAR
La regulacin del ciclo celular esta mediada por unas protenas, las cuales velan porque cada paso se haya realizado
correctamente y en el caso de presentarse una falla, detienen el ciclo hasta que el dao sea reparado o de lo contrario se
induce a la apoptosis. Las principales protenas que regulan el ciclo celular son:
KdC (kinase dependiente de ciclinas, agrega fosfato a una protena), junto con ciclinas son las mayores llaves de control
para el ciclo celular, causando que la clula se mueva de G1 a S o G2 a M.
FPM (Factor Promotor de la Maduracin) incluye la KdC y ciclinas que desencadenan la progresin del ciclo celular.
p53 Es una protena que funciona bloqueando el ciclo celular si el ADN est daado. Si el dao es severo esta protena
puede causar apoptosis (muerte celular).
1. Los niveles de p53 estn incrementados en clulas daadas. Esto otorga tiempo para reparar el ADN por bloqueo del
ciclo celular.
2. Una mutacin de la p53 es la mutacin ms frecuente que conduce al cncer. Un caso extremo de esto es el
sndrome de Li Fraumeni dnde un defecto gentico en la p53 conduce a una alta frecuencia de cncer en los individuos
afectados.
p27 Es una protena que se une a ciclinas y KdC bloqueando la entrada en fase S.
-
http://www.curtisbiologia.com/b1957
APOPTOSIS
En la formacin de un individuo, la muerte celular o apoptosis es tan importante como la divisin celular. La mayora de
las clulas fabrican las protenas que forman parte de una maquinaria para su propia destruccin. Esta maquinaria letal
est compuesta por enzimas capaces de degradar protenas (proteasas) cuya activacin produce, directa o
indirectamente, cambios celulares caractersticos. Las clulas que entran en apoptosis se encogen y se separan de sus
vecinas; luego las membranas celulares se ondulan y se forman burbujas en su superficie; la cromatina se condensa y
los cromosomas se fragmentan; finalmente, las clulas se dividen en numerosas vesculas, los cuerpos apoptsicos, que
sern engullidas por clulas vecinas.
Las enzimas involucradas en el proceso de apoptosis permanecen normalmente inactivas en las clulas, respondiendo a
mecanismos de control estrictos. Los mecanismos de control son los responsables de activar la maquinaria letal en
momentos particulares de la vida de la clula, respondiendo a seales externas o internas. Cualquier alteracin en estos
mecanismos de control puede tener consecuencias nefastas para el organismo, creando estados patolgicos producidos
tanto por la prdida de clulas normales como por la sobrevida de clulas que deberan entrar en apoptosis.
Cuando una clula muere por dao o envenenamiento, proceso denominado necrosis, normalmente se hincha y explota,
derramando su contenido en el entorno. Como consecuencia, se produce una inflamacin que recluta leucocitos, y que
puede lesionar el tejido normal que la circunda. La apoptosis, a diferencia de la necrosis, es un tipo de muerte activa, que
requiere gasto de energa por parte de la clula y es un proceso ordenado en el que no se desarrolla un proceso
inflamatorio.
GAMETOGNESIS La formacin de gametos por meiosis se llama gametognesis y es diferente en hombres y en mujeres.
-
En los machos, la gametognesis tiene como resultado la formacin de clulas espermticas y se llama espermatognesis. La produccin de espermatozoides ocurre en los testculos, rganos reproductores masculinos. En la hembra, los vulos se forman en los ovarios que son los rganos reproductores. La formacin de gametos en las hembras se llama ovognesis. Tanto la ovognesis como la espermatognesis tienen como resultado la formacin de gametos monoploides. Sin embargo, hay diferencias entre ambos tipos de gametognesis, la espermatognesis forma cuatro espermatozoides del mismo tamao y la ovognesis forma un vulo grande al igual que tres cuerpos polares que se desintegran y solo el vulo es un gameto funcional.
http://ww2.educarchile.cl/UserFiles/P0001/Image/PSU/Contenidos2011/Modulos_Biologia_2011/MOD