Análisis comparativo del sistema inmune de los anfibios ...
23
1 Análisis comparativo del sistema inmune de los anfibios, con especial atención a la respuesta inmune a la quitridiomicosis Maria Paula Rueda Mejía 1. Introducción Durante las últimas décadas se ha presentado un declive en las poblaciones de anfibios alrededor del mundo, donde alrededor del 40% de las especies presenta una disminución en sus poblaciones, llevando a muchas especies al riesgo de extinción (Stuart et al., 2004), entre las razones se ha propuesto la pérdida de hábitat y los cambios climáticos. Otro de los motivos importantes para este declive es la quitridiomicosis, una enfermedad infecciosa causada por el hongo Batrachochytrium dendrobatidis (Berger et al., 1998; Lips et al ., 2006) . Para muchas especies y poblaciones de anfibios la mortalidad a causa de este patógeno es muy alta, llegando a diezmar poblaciones o desaparecerlas en pocos años (Lips et al., 2006), en estudios donde se han realizado un conteos de individuos antes y después de la detección de Bd en el área se observa un declive de varios miles de individuos en especies susceptibles en una localidad a solo decenas en cuestión de 4-5 años (Vredenburg et al., 2010 ). Bd tiene un propágulo flagelado llamado zoospora (Longcore et al., 1999), este ataca las células epidérmicas de los anfibios, se interna en la piel queratinizada de las ranas, se enquista y allí se reproduce y produciendo más zoosporas. Estas son liberadas al medio y pueden reinfectar al mismo individuo o propagarse a individuos cercanos. Al enquistarse en la piel produce hiperqueratosis, pérdida de las funciones de la piel, como la osmoregulación y finalmente la muerte (Voyles et al ., 2009). El conocimiento de la cantidad de especies que presentan alta mortalidad y declives poblacionales causados por Bd ha llevado a entender la importancia de conocer mejor las dinámicas de esta enfermedad. Se han reportado especies de anfibios resistentes y susceptibles a la infección; algunas especies que no presentan signos de infección, otras presentan signos pero no síntomas de enfermedad, algunas presentan una enfermedad pero no se da un declive poblacional y otras presentan altas tasas de mortalidad (Alford & Richards, 1999; Parker et al., 2002; Kriger & Hero, 2006; Woodhams et al., 2007). Se han propuesto varias hipótesis para explicar este fenómeno, algunas relacionadas con los hábitos predominantemente terrestres o acuáticos de las diferentes especies, y otras con la virulencia de las diferentes variedades del patógeno (Berger et al. , 2005) y/o diferencias en la respuesta inmune del
Transcript of Análisis comparativo del sistema inmune de los anfibios ...

1
Análisis comparativo del sistema inmune de los anfibios, con especial atención a la
respuesta inmune a la quitridiomicosis
Maria Paula Rueda Mejía
1. Introducción
Durante las últimas décadas se ha presentado un declive en las poblaciones de anfibios alrededor del
mundo, donde alrededor del 40% de las especies presenta una disminución en sus poblaciones, llevando a
muchas especies al riesgo de extinción (Stuart et al., 2004), entre las razones se ha propuesto la pérdida
de hábitat y los cambios climáticos. Otro de los motivos importantes para este declive es la
quitridiomicosis, una enfermedad infecciosa causada por el hongo Batrachochytrium dendrobatidis
(Berger et al., 1998; Lips et al., 2006) .
Para muchas especies y poblaciones de anfibios la mortalidad a causa de este patógeno es muy alta,
llegando a diezmar poblaciones o desaparecerlas en pocos años (Lips et al., 2006), en estudios donde se
han realizado un conteos de individuos antes y después de la detección de Bd en el área se observa un
declive de varios miles de individuos en especies susceptibles en una localidad a solo decenas en cuestión
de 4-5 años (Vredenburg et al., 2010 ). Bd tiene un propágulo flagelado llamado zoospora (Longcore et
al., 1999), este ataca las células epidérmicas de los anfibios, se interna en la piel queratinizada de las
ranas, se enquista y allí se reproduce y produciendo más zoosporas. Estas son liberadas al medio y pueden
reinfectar al mismo individuo o propagarse a individuos cercanos. Al enquistarse en la piel produce
hiperqueratosis, pérdida de las funciones de la piel, como la osmoregulación y finalmente la muerte
(Voyles et al., 2009).
El conocimiento de la cantidad de especies que presentan alta mortalidad y declives poblacionales
causados por Bd ha llevado a entender la importancia de conocer mejor las dinámicas de esta enfermedad.
Se han reportado especies de anfibios resistentes y susceptibles a la infección; algunas especies que no
presentan signos de infección, otras presentan signos pero no síntomas de enfermedad, algunas presentan
una enfermedad pero no se da un declive poblacional y otras presentan altas tasas de mortalidad (Alford
& Richards, 1999; Parker et al., 2002; Kriger & Hero, 2006; Woodhams et al., 2007).
Se han propuesto varias hipótesis para explicar este fenómeno, algunas relacionadas con los hábitos
predominantemente terrestres o acuáticos de las diferentes especies, y otras con la virulencia de las
diferentes variedades del patógeno (Berger et al., 2005) y/o diferencias en la respuesta inmune del

2
hospedero. Es posible que todas estas interactúen para crear la variación en la susceptibilidad ante la
infección por Bd.
Para evaluar la importancia de la respuesta inmune del hospedero se han realizado estudios comparativos
de la respuesta inmune entre especies resistentes y susceptibles en estado de infección por Bd, con el
objetivo de encontrar los factores que afectan la sensibilidad a la infección. La mayoría de estos se
concentran en el aislamiento y comparación de la actividad de péptidos antimicrobianos frente al hongo ,
otros estudian de la misma forma la actividad de lisozimas y algunos en la respuesta mediada por
linfocitos, usando radiación para eliminar esta respuesta y comparando la eficacia de la defensa con y sin
actividad de linfocitos (Ramsey et al., 2010).
A partir el desarrollo de las nuevas tecnologías de secuenciación se han empezado a realizar estudios de
expresión genética diferencial frente a la infección por Bd, secuenciando el proteoma o transcriptoma de
los tejidos involucrados en la respuesta inmune. Tras obtener las secuencias de todas las proteínas o
transcritos presentes en la muestra en la mayoría de los casos se realiza una búsqueda en bases de datos de
estas secuencias, con el objetivo de identificar la secuencia por similaridad con otra ya identificada y
caracterizada en bases de datos. En la mayoría de los casos no hay otro análisis que se realice con las
secuencias que no fueron identificadas y estas quedan fuera del análisis.
Al realizar una búsqueda en bases de datos a partir de transcritos obtenidos en la piel de anuros se observa
que solo entre un 5% y 10% de los transcritos con homología, relacionados con el sistema inmune,
corresponden a proteínas caracterizadas en anfibios. Esto es claramente un efecto de la extensiva
investigación sobre el sistema inmune humano y los modelos animales usados en investigación médica, la
rata parda (Ratus novergicus) y el ratón doméstico (Mus musculus), las tres especies más representadas en
estas búsquedas en bases de datos. Esta situación plantea el problema de saber hasta que punto las
secuencias de aminoácidos en bases de datos representan los genes expresados por el sistema inmune de
la piel de una especie de anfibio estudiada.
Este documento revisa la información existente hasta el momento sobre el sistema inmune de los anfibios,
cuáles proteínas de que funciones del sistema están caracterizadas y secuenciadas y, de aquellas funciones
no estudiadas a profundidad en anfibios, las diferencias que existen dentro de los vertebrados y entre
mamíferos y anfibios en el sistema inmune. Esto para conocer si hay una posibilidad real de identificación
de las proteínas de una función inmune a partir de una muestra de piel de un anfibio así no estén
caracterizadas dentro de este grupo.

3
2. Sistema inmune innato
El sistema inmune innato es la primera línea de defensa contra organismos infecciosos, este tiene
mecanismos que reconocen patrones generales de estos organismos, no responden específicamente frente
a cada patógeno, y la mayoría de sus componentes están presentes, aunque algunos en conformación
inactiva, desde antes de que ocurra la infección (Goldsby, 2002).
Este sistema está compuesto por células fagocíticas que introducen material extracelular, que puede ser
patógenos o partes de ellos, y lo degradan y varias sustancias antimicrobianas.
2.1.Células fagocíticas y presentadoras de antígenos
La fagocitosis es el proceso de ingestión de material extracelular por parte de una célula, durante este
proceso la membrana de la célula fagocítica se extiende alrededor del material que será fagocitado, que
puede ser un patógeno o partes de el, lo engolfa y forma una vesícula que incluye el material, llamada
fagosoma (Goldsby, 2002). Estas células no solo se caracterizan por esta acción sino por la presentación
del antígeno fagocitado en la membrana, por medio de una molécula de membrana llamada MHC. Tras la
fagocitosis, dentro del fagosoma, la célula procesa el antígeno degradándolo, al unirse a un lisosoma que
contiene enzimas hidrolíticas. La mayoría de los productos de esta degradación del antígeno son
expulsados de la célula por exocitosis, pero algunos pueden formar un complejo con un MHC de clase II
y ser expuestos en la membrana para activar células del sistema inmune adaptativo (Goldsby, 2002). Las
células dendríticas y los macrófagos tienen esta función presentadora, mientras que los neutrófilos,
eosinófilos y basófilos solo realizan el proceso de fagocitosis.
Estas células están presentes en peces teleósteos y se han encontrado células con actividad fagocítica, así
como células con moléculas MHC en membrana y morfologías celulares similares a las observadas en
mamíferos de estas células en las especies estudiadas de todos los grupos de vertebrados.
Dentro de los anfibios, en Xenopus laevis y el género Rana se han identificado células presentadoras de
antígeno, que expresan en su membrana MHC de clase II, con morfología dendrítica, similares a CDs y a
las denominadas en mamíferos como células de Langerhans (Du Pasquier & Flajnik, 1990; Carrillo-Farga
et al., 1990). Se ha observado que los procesos de maduración y las células progenitoras son similares a
los que ocurren en mamíferos. Igualmente se han encontrado macrófagos en la piel de renacuajos de
Xenopus (Lehman, 1953)

4
2.2. MHC
El complejo mayor de histocompatibilidad (MHC, por su siglas en inglés) es una familia de proteínas de
membrana cuya función es presentar un antígeno procesado al interior de la célula que lo expresa
(Goldsby, 2002). Estas se dividen en dos grupos, los MHC de clase I, expresadas por casi todas las
células somáticas del organismo cuando están infectadas por un microorganismo, y los MHC de clase II,
expresadas por un grupo de células llamadas presentadoras de antígeno, que incluyen, pero no se limitan,
a células dendríticas (CDs), macrófagos y células o linfocitos B (Carvalho et al., 2009).
Ambos tipos de MHC están presentes en todos los vertebrados mandibulados (Flajnik & Kasahara, 2001).
Los genes que codifican para estas proteínas son similares y están conservados al comparar grupos, sin
embargo hay diferencias en la organización de estos. Se cree que hubo un rearreglo de los genes
involucrados en el procesamiento de antígeno y la expresión de MHC, bastante ligados en el genoma. En
mamíferos placentarios los genes encargados del procesamiento de antígenos de clase I al interior de las
células están físicamente entre los que codifican para el mismo proceso en clase II y para el MHC de
clase II como tal, mientras que el gen de MHC clase I se encuentra separado (Kaufman, 1999). En los
demás grupos de vertebrados, incluyendo anfibios, los genes de procesamiento de antígeno y expresión de
MHC de clase I están ligados entre si, al igual que los de clase II, se puede decir que el estado primitivo
de esta organización es el que presentan peces, anfibios y reptiles, y que en mamíferos se dio esta
translocación de genes de procesamiento clase I (Ohta et al., 2006).
En Xenopus el MHC está muy bien caracterizado (Flajnik & Du pasquier, 1990), este funciona igual al de
mamíferos, los genes que codifican para los distintos componentes de estas proteínas, clase I y clase II, en
otros vertebrados fueron buscados en el genoma de Xenopus y se encontraron 110 genes con alta
similaridad, así que se puede decir que el MHC y los genes relacionados con su producción y regulación
son conservados y funcionan en anfibios igual que en otros vertebrados (Ohta et al., 2006).
2.3.Respuesta innata molecular: Sistema complemento y lisozimas
El sistema complemento es una función del sistema inmune innato compuesta por más de 30 proteínas
séricas solubles con capacidad de unión a membrana celular, estas se encuentran en un estado inactivo,
distintos mecanismos activan el sistema, haciendo que estas proteínas tengan la capacidad para dañar las
membranas biológicas e intercalarse en ellas; destruyendo o facilitando la eliminación del patógeno por

5
medio de células fagocíticas (Goldsby, 2002). Además estas proteínas interactúan para dirigir la
activación de los demás procesos inmunes, tanto innatos como adaptativos, donde estos existen (Goldsby,
2002).
Las proteínas del sistema complemento pueden actuar como mediadores en la cascada de regulación del
sistema o ser parte de un complejo de ataque a las membranas de los microorganismos patógenos. Tanto
los mediadores como las proteínas del complejo están identificadas y su secuencia y función dentro de la
formación del complejo son conocidas.
El sistema complemento puede ser activado por la unión entre un antígeno y un anticuerpo, o
directamente por proteínas características de las membranas de los patógenos (Goldsby, 2002) y regula
otras funciones del sistema inmune como la inflamación y la acción de linfocitos B y T (Carroll, 2004).
Al comparar las proteínas de este sistema con las de otros organismos se ha encontrado que el sistema
complemento está presente en organismos con sistemas inmunes rudimentarios, en los que no hay
presencia de sistema inmune adaptativo. El gen codificante para la proteína C3, componente central del
sistema complemento, está presente en todos los deuterostomados (Miyazawa, 2001), dada la importancia
de esta proteína se la puede considerar un buen indicativo para la presencia de sistema complemento en
cualquier organismo en que se encuentre. Las proteínas del sistema complemento son entonces bastante
conservadas, así como la forma en la que interactúan.
Al realizar búsquedas de estos genes, caracterizados en mamíferos, en los genomas de pollo, Xenopus,
peces óseos y algunos invertebrados se ha encontrado que los vertebrados comparten un set de genes del
sistema complemento prácticamente igual, con alta homología entre grupos (Nonaka & Kimura, 2006).
En Xenopus varios de los componentes del sistema han sido identificados, aislados y caracterizados,
aunque no todos los componentes se han encontrado se asume que están presentes dado que es un proceso
altamente conservado dentro de los vertebrados (Mo et al., 1996).
Las lisozimas son una familia de proteínas solubles hidrolíticas presentes en secreciones mucosas y en las
lágrimas (Goldsby, 2002), estas tienen la capacidad de romper enlaces específicos en los peptidoglicanos
de la pared celular de las bacterias (Nonaka & Kimura, 2006). Las lisozimas están presentes en todos los
grupos de organismos, en animales se han identificado tres tipos de lisozimas, que difieren en secuencia
de aminoácidos y propiedades bioquímicas y enzimáticas: la tipo-c, cuyo modelo es la presente en claras

6
de huevo de pollo, la tipo-g, descrita en claras de huevo de ganso y la tipo-i, encontrada en insectos y
presente en varios grupos de invertebrados, no se encuentra en cordados (Callewaert & Michiels, 2010).
Dentro de los vertebrados la investigación se ha enfocado a la identificación y caracterización de
lisozimas de mamíferos y aves únicamente, la información disponible sobre otros grupos se limita a
comparaciones con estos.
El conocimiento de los patrones de expresión de lisozimas en anfibios es muy limitado, ya que la
información está basada en reconocimiento de proteínas identificadas previamente en otros grupos de
organismos, una lisozima tipo-c fue aislada en Bufo andewsi (Zhao et al., 2006), no se han identificado
lisozimas tipo-g en anfibios. En Rana pipiens y Bufo boreans se han encontrado indicios de presencia de
lisozimas en secreciones, para esto se han aislado proteínas que tienen alta homología con lisozimas
identificadas de otros organismos (Zhao et al., 2006), sin embargo no se han realizado estudios donde se
aíslen proteínas de secreciones mucosas en busca de actividad microbiana y potenciales lisozimas
específicas de anfibios. Teniendo en cuenta que hay una homología baja entre las clases de lisozimas que
se han encontrado en la naturaleza, y que dentro de cada clase hay una alta variabilidad, es posible que la
identificación por medio de búsqueda de homología con proteínas descritas en organismos de otros
grupos deje otras clases de lisozimas sin encontrar y se esté perdiendo potencial información específica
sobre el componente humoral de la respuesta inmune innata en las secreciones mucosas de los anfibios.
Tanto los componentes receptores de sistema complemento como las lisozimas tienen capacidad de
reconocimiento de patrones moleculares generales de los microorganismos patógenos.
2.4. Receptores tipo Toll (TLRs)
Los TLRs son una familia de proteínas de membrana que reconocen patrones moleculares generales
conservados en microorganismos, estos patrones pueden ser lipoproteínas, lipopolisacáridos, flagelina y
ácidos nucléicos (Miyake, 2007). Estos receptores se encuentran en células presentadoras de antígeno,
células dendríticas y macrófagos, y activan en estas una cascada de regulación que activa la producción
de varios mediadores de la respuesta inmune innata: citoquinas, quimioquinas, interferones, o que regulan
la respuesta adaptativa (Kawai & Akira, 2007).
Los TLRs son un mecanismo conservado en animales, estos cambian dentro de los vertebrados de
acuerdo con los ligandos de los microorganismos patógenos que los afectan. Por esto las regiones

7
intermembranal y citoplasmática del receptor son conservadas entre los grupos de vertebrados, pero los
sitios de unión extracelulares son variables (Zhu et al., 2013).
En mamíferos se han identificado seis tipos de TLRs en vertebrados, en la mayoría de las especies
estudiadas se ha encontrado por lo menos un representante de cada uno de los tipos, donde algunos de los
seis son más conservados y otros tienen gran cantidad de modificaciones en cada especie (Roach et al.,
2005).
El funcionamiento de los TLRs no ha sido estudiado en anfibios, pero se han realizado búsquedas de
genes que codifican para TRLs en humanos en el genoma de Xenopus tropicalis, alrededor de 20 tuvieron
un alta homología, incluyendo 9 péptidos con actividad antifúngica en la piel de mamíferos (Ishii et al.,
2007). Por lo demás no hay información suficiente para saber si los TLRs presentes en anfibios tienen
homología con las secuencias en bases de datos ya que es posible que los anfibios tengan TLRs
específicos con estructuras significativamente diferentes a los descritos hasta el momento en otros grupos.
2.5. Péptidos antimicrobianos (AMPs)
Los péptidos antimicrobianos son usualmente moléculas hidrofóbicas y catiónicas que, en un ambiente de
membrana biológica pueden formar hélices α y romper estas membranas de los microorganismos
patógenos (Yeaman & Yount, 2003).
Aparte de estas características generales no hay muchas similaridades en los AMPs presentes en la piel de
los anfibios. Al comparar especies cercanamente relacionadas hay semejanzas en el grupo de AMPs
presentes en la piel y otras mucosas, en general comparten familias de proteínas, pero no se ha encontrado
el mismo péptido en dos especies diferentes (Conlon et al., 2004). Esto implica que lo más probable es
que los AMPs de una muestra no sean detectados si se realiza una búsqueda basada en bases de datos, a
menos que los péptidos particulares de la especie estudiada estén aislados, caracterizados y secuenciados.

8
Tabla 1: Cantidad de péptidos antimicrobianos de la piel de anfibios descritos por especie.
Algunos de los AMP s de la piel se han aislado y caracterizado en los géneros Bombina, Xenopus, Rana,
Amolops, Hylarana, Phyllomedusa, Litoria, Limnonectes, Leptodactylus, Pseudis, Bufo e Hyla (Ver tabla
1) , hay una cobertura filogenética mayor en la caracterización de estos péptidos que de cualquier otra
función inmune en anfibios, la demás funciones solo se han estudiado en los géneros Xenopus, Bufo y
Rana. Al mismo tiempo esta es la función para cuyas proteínas están menos conservadas, lo cual hace
que, a pesar de la cobertura a nivel taxonómico, sea muy poco probable que los péptidos de la piel de un
anfibio sean identificado a partir de una búsqueda en bases de datos.

9
Tabla 1: Continuación
En los últimos años, con el desarrollo de la secuenciación de nueva generación se ha empezado a utilizar
la peptidómica para encontrar nuevos péptidos antimicrobianos (He et al., 2013), esto ha producido una
gran cantidad de datos en poco tiempo, y cada vez más péptidos nuevos se descubren con una sola
muestra de tejido.
Para enfrentar este problema al tener datos transcriptómicos se podría realizar una búsqueda
disminuyendo el porcentaje de identidad mínimo, o hacer un análisis diferente, buscando genes
candidatos.
3. Sistema inmune adaptativo
El sistema inmune de los vertebrados mandibulados es bastante complejo, tiene la capacidad de reconocer
patógenos y activar una respuesta específica para cada uno de ellos. Además este sistema genera una
memoria, que permite una eliminación de un patógeno específico más efectiva y rápida.
El sistema inmune adaptativo depende directamente de la capacidad de generar una gran diversidad de
receptores de antígenos, llamados inmunoglobulinas, expresados en la membrana celular de los linfocitos.
Cada uno de estas inmunoglobulinas es específica para un antígeno particular, cuando lo detecta, la célula

10
que lo expresa dispara la respuesta adaptativa donde más inmunoglobulinas específicas para el antígeno
se producen (Burnet, 1959), pero ya no como receptores de membrana sino en forma de secreción, en esta
conformación las inmunoglobulinas tienen el nombre de anticuerpos.
El mecanismo principal por medio del cuál se produce la gran diversidad de inmunoglobulinas es la
recombinación (Tonegawa, 1976), el mecanismo de recombinación que se ha descrito en mamíferos es
compartido y se ha encontrado en todos los grupos de vertebrados mandibulados. Un mecanismo de
recombinación diferente, que parte de genes distintos y es mediado por moléculas diferentes, se ha
encontrado en agnatos, pero no hay algo similar en los demás metazoos. Estos únicamente cuentan con un
sistema inmune innato. Se cree los sistemas de recombinación presentes en vertebrados mandibulados y
agnatos surgieron de forma independiente y son una convergencia evolutiva (Cooper & Alder, 2006).
3.1. Los anticuerpos
Las inmunoglobulinas, llamadas anticuerpos cuando son secretadas en su forma soluble, son moléculas
compuestas por dos cadenas pesadas y dos livianas. Las últimas tienen un sitio de unión al antígeno en el
extremo amino terminal, que es el que interactúa con la molécula del patógeno o sustancia para la cuál es
específico dicho anticuerpo.
Las inmunoglobulinas, tanto secretadas como en forma de receptor de membrana en linfocitos, son una
innovación de los vertebrados (Manning, 1979), presente en peces teleósteos, anfibios, reptiles, aves y
mamíferos.
El mecanismo por el cuál se produce el gran repertorio de inmunoglobulinas, la recombinación de genes
VSJ y VJ mediada por RAG, está presente en todos los vertebrados mandibulados (Litman et al., 1999;
Flajnik and Kasahara, 2001). Un mecanismo similar se encuentra en vertebrados no mandibulados, donde
se produce una gran diversidad de linfocitos, cada uno expresando un receptor con especificidad para un
antígeno diferente, pero los genes que codifican para estos receptores son diferentes, así como la
estructura del receptor (Pancer et al., 2004; Alder et al., 2005).
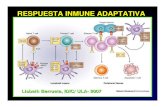
Hay varias clases de inmunoglobulinas llamadas isotipos (Ver Fig. 1), que difieren entre sí en la
estructura de la cadena pesada. El isotipo más común y con mayor concentración en suero es el IgM, su
acción es sistémica y se encuentra en suero de todos los vertebrados. En mamíferos y aves encontramos el
isotipo IgA, posteriormente se encontró en otros reptiles (Flajnik & Kasahara, 2010), de bajo peso

11
molecular, principalmente en secreciones mucosas, se cree que precisamente el peso molecular es la razón
evolutiva para la existencia de este isotipo, ya que esto facilitaría el movimiento hacia las mucosas y fuera
del sistema vascular en vertebrados terrestres, que tienen mayor presión hidrostática y osmótica en las
venas y arterias (Manning & Turner, 1976).
En sangre en vertebrados en general se ha observado que al aumentar la cantidad de anticuerpos hay una
mejoría en la asimilación de productos extraños, estos desaparecen de la sangre más rápida y
eficientemente.
En peces teleósteos se han identificado tres isotipos de inmunoglobulinas: IgM, IgD e IgT/Z (Fletcher &
Grant, 1969), con gran prevalencia de IgM en plasma y piel, la función de IgD en peces es poco conocida
y no se ha identificado en piel, IgT, por otro lado parece tener una función importante en la respuesta
inmune en la mucosa intestinal, análogamente al IgA de mamíferos, siendo IgM encargado de inmunidad
de forma sistémica. IgT parece ser el isotipo de inmunoglobulina más antiguo encargado particularmente
de inmunidad en mucosas. El tejido linfoide asociado a la piel en peces es más similar al asociado al
intestino en mamíferos, por su condición de mucosa (Turner & Manning, 1974).
Solo un reptil no ave ha sido estudiado en busca de inmunoglobulinas, Anolis carolinenesis, en este se
encontraron tres isotipos IgM, IgD e IgY (Wei et al., 2009).
En Xenopus se han identificado cuatro isotipos: IgM, IgD, cuya función no está bien entendida, pero es
más similar, en cantidad de dominios, a el IgD de peces que al de mamíferos, IgX, presente en
secreciones mucosas (Hsu & Du Pasquier, 184; Amemiya et al., 1989) e IgY, análogo al IgG de
mamíferos, este se expresa en el vaso y solo como consecuencia de la respuesta mediada por linfocitos T.
En mamíferos se han identificado seis isotipos, 1)IgM, 2)IgD, cuya función, al igual que en peces, no está
bien entendida, pero es un isotipo antiguo cuya estructura ha variado entre grupos y se cree que también
lo ha hecho su función, a diferencia de IgM algunos grupos de vertebrados han perdido este tipo de
anticuerpo (Flajnik & Kasahara, 2010). 3)IgG, encargada de respuestas de memoria e IgE, presente en
funciones inflamatorias en epitelio. El origen de estos dos isotipos está en una proteína similar al IgY de
anfibios. 4)IgA, presente en mucus, y un IgG de cadena sencilla presente, hasta donde se sabe, solo en
camélidos.

12
3.2 Células linfoides
En todos los vertebrados mandibulados se han identificado dos tipos de linfocitos con funciones
claramente diferenciadas (Manning et al., 1979): Las células T,encargadas de inmunidad celular, y las
células B, encargadas de síntesis de anticuerpos.
Los progenitores celulares de los linfocitos, células madre hematopoyéticas, también se han identificado
en los diferentes grupos de vertebrados mandibulados y los genes que codifican para sus receptores,
receptor de células T (TCR, por sus siglas en inglés) y receptor de células B (BCR, por sus siglas en
inglés) (Cooper & Alder, 2006). En Xenopus el TCR se encuentra caracterizado (Haire et al., 2002).
Fig 1: Diferencias en el sistema inmune adaptativo entre las clases de vertebrados. Izquierda: Isotipos de
inmunoglobulinas representados con el número de dominios y clasificadas por función y similitud. Derecha:
Órganos linfáticos primarios y secundarios presentes en cada grupo.

13
Los BCR son inmunoglobulinas de membrana, por lo tanto tienen la estructura de dos cadenas livianas y
dos pesadas de los anticuerpos. En humanos hay dos tipos de cadena livianas, λ y κ, codificadas por dos
genes diferentes cada una, el κ en el cromosoma 2 y el λ en el cromosoma 22, en otros organismos se
identifican por la secuencia de aminoácidos de la proteína o de nucleótidos en el genoma. Estos dos tipos
de cadena se han encontrado en genomas de organismos modelo todos los grupos de vertebrados
(Criscitiello & Flajnik, 2007). En anuros se identificó otro tipo de cadena ligera, la σ, está presente en
peces y reptiles, se cree que los mamíferos y aves perdieron secundariamente este gen (Schwager et al.,
1991).
Las cadenas pesadas de los BCR generan una alta variabilidad con la capacidad de detectar
específicamente patrones proteicos de microorganismos patógenos para generar anticuerpos específicos y
una respuesta inmune adaptativa muy eficaz. Esta variabilidad se genera por medio del mecanismo de
recombinación mediado por RAG, este mecanismo funciona de la misma manera en todos los vertebrados
mandibulados, solo hay diferencias en la organización de los genes que codifican para las cadenas
pesadas de los BCR dentro del genoma entre los grupos de vertebrados, pero estos genes son muy
similares en secuencia al comparar entre taxa (Flajnik & Kasahara, 2010).
En cuanto a los receptores de células T (TCR) existen dos tipos con distinta función, que definen dos
tipos de linfocito T, según la clase de receptor que tengan. Un tipo, el más común, son los receptores αβ
que reconocen el complejo formado por un MHC y el antígeno que presenta en otra célula (Kozbor et al.,
1990). La estructura, de traslocón, posición y regulación de los genes que codifican para este tipo de
receptor son conservados desde peces óseos hasta mamíferos (Deng et al., 2013). En Xenopus se
comprobó que, al igual que en los demás grupos de vertebrados la respuesta de los linfocitos T con
receptor αβ depende de la detección de un MHC unido a un antígeno (Harding & Flajnik, 1993).
El otro tipo son los receptores γδ, son menos abundantes y se han encontrado principalmente en la
mucosa del sistema digestivo, también están presentes en otras mucosas, en muy bajas cantidades (Morita
et al., 1995). No está claro que tipo de moléculas son detectadas por estos receptores, pero se cree que son
capaces de reconocer antígenos solubles, de una forma similar a la de los anticuerpos, pero de una forma
menos específica, reconociendo patrones generales de los microorganismos, se diferencian del otro tipo
de TCR en que son independientes de MHC (Sciammas & Bluestone, 1999). Su organización de genes es
diferente a la que se presenta en los receptores αβ, pero la organización básica de los genes γδ es
conservada en gnatostomados (Flajnik & Kasahara, 2010).

14
3.3 Órganos linfoides
Tradicionalmente se conoce que en mamíferos las células linfoides son producidas en el timo, y la médula
ósea y, en aves, en la bursa de Fabricio (Cooper et al., 1965). De este conocimiento derivan los nombres
de los linfocitos T, por provenir del timo (thymus en inglés) y los linfocitos B, por provenir de la bursa de
Fabricio o médula ósea (bursa of Fabricius y bone marrow en inglés, respectivamente) (ver fig.1).
En la médula ósea de mamíferos también se producen las demás células hematopoyéticas, que circulan
por la sangre, eritrocitos, plaquetas y células NK entre otras.
La médula ósea ocurre irregularmente, y usualmente está ausente en peces y anfibios urodelos, mientras
que en anuros, reptiles, aves y mamíferos está presente de forma consistente (Manning, 1979). Se
desconoce donde se encuentran las poblaciones de células madre progenitoras de linfocitos en los
vertebrados que no tienen médula ósea. Sin embargo en Xenopus adultos la médula ósea no es la
productora de linfocitos B, esta produce neutrófilos, un tipo de célula fagocítica y presentadora de
antígeno (Hadji-Azimi et al., 1987).
Si bien las células progenitoras de los linfocitos son invariables a lo largo de vertebrados mandibulados ,
existen variaciones en los órganos donde se producen y maduran estas células.
El timo está presente en todos los vertebrados mandibulados, y en su epitelio se da la diferenciación de
linfocitos T, en Xenopus se demostró la función de este órgano por medio de una timectomía, que produjo
un daño general en las respuestas mediadas por linfocitos T, pero no en las mediadas por linfocitos B
(Tochinai & Katagari, 1975).
El bazo es en todos los grupos de vertebrados el principal centro de actividad linfoide, allí los antígenos
que se encuentran en sangre son filtrados y se transportan al sitio germinal del órgano, donde se sintetizan
anticuerpos IgM específicos para los antígenos filtrados. Además, en este órgano se producen moléculas
involucradas en la ayuda al proceso de fagocitosis. Las mismas funciones ocurren en este órgano en todos
los vertebrados mandibulados, las diferencias están en la complejidad de la compartimentalización y
arquitectura del órgano, ligeramente más desarrollada en mamíferos (Diener & Bryan, 1974). Este es un
órgano bastante conservado en estructura y función a lo largo de vertebrados mandibulados.

15
El riñón es un lugar donde ocurren las reacciones inmunes, un órgano linfoide secundario, por lo cuál
presenta acumulaciones de linfocitos, en el riñón de anuros y peces hay acumulaciones sencillas de
linfocitos localizadas cerca de los vasos sanguíneos y lugares donde las células pueden responder a la
presencia de un antígeno (Turner, 1973). En amniotas la actividad inmune en el riñón disminuye
considerablemente, aunque se siguen presentando acumulaciones de linfocitos, igualmente se observa que
hay desarrollo de otros órganos linfoides secundarios que no son activos inmunológicamente en anfibios
y peces.
Estos órganos linfáticos secundarios que encontramos en amniotas, altamente desarrollados en aves y
mamíferos, son llamados nódulos linfáticos y se encuentran distribuidos por el cuerpo del animal. Los
nodos linfáticos de aves y mamíferos se caracterizan por estar asociados con canales linfát icos que
transportan eficientemente las células. Los reptiles no aves tienen nódulos linfáticos repartidos, pero sin
estos canales (Manning, 1979). En los anuros donde se ha estudiado no hay presencia de nodos linfáticos
(Manning & Horton, 1982), en Xenopus y en la salamandra Pleurodeles waltlii se ha descrito la presencia
de estructuras similares a nodos linfáticos, sin embargo en ningún anuro se han observado acumulaciones
linfoides organizadas por fuera de los órganos principales ya descritos. En peces se observa presencia de
estructuras similares a nodos linfáticos a lo largo del tracto digestivo únicamente.
Es importante tener en cuenta que aquellos vertebrados que no poseen nódulos dos linfáticos verdaderos,
peces, anfibios y reptiles dependen más de otros órganos linfáticos secundarios, como el riñón y, sobre
todo, el bazo.
4. La piel como mucosa en anfibios
La piel de todos los vertebrados presenta una estructura similar compuesta de dos capas, la dermis y la
epidermis, con el mismo origen embrional a través de todos los taxa. En peces la epidermis actúa como
una mucosa, con las células epiteliales en directo contacto con el medio externo y no presenta
queratinización. En otros grupos vemos un desarrollo de estructuras protectoras, plumas, escamas o
pelaje, como adaptaciones al medio terrestre. En mamíferos la piel tiene adaptaciones como pelaje y
glándulas sudoríparas, por la adaptación al medio terrestre la piel ha perdido su función de mucosa.
La respuesta relacionada con células B y anticuerpos en peces es muy similar entre la piel y el intestino,
se podría sugerir un funcionamiento similar en las mucosas de la piel de los anfibios (Xu et al., 2013). IgT
está presente de forma similar que en el intestino, sobre todo en polímeros y está en mayor cantidad
respecto a IgM que en suero, pero IgM sigue siendo más abundante en esta mucosa, a diferencia del

16
intestino de mamíferos, donde el isotipo IgA es el más abundante. En peces el rol de IgT en la piel sería el
de ejercer control sobre la microbiota de la piel, el mismo patrón se observa con las células B (Salinas et
al., 2011) .
En anfibios la piel está queratinizada, pero, al igual que en los peces, esta cumple funciones de mucosa.
Las mucosas, además de actuar como barreras físicas, son sitios con respuesta inmune activa, tanto celular
celular como humoral, más compleja a nivel anatómico y molecular que la respuesta en el resto del
sistema (Brandtzaeg, 2009; Cerutti and Rescigno, 2008; Fagarasan, 2008; Macpherson et al., 2008). Es
importante tener esto en cuenta a la hora de estudiar la respuesta inmune de los anfibios frente a la
quitridiomicosis, ya que la repuesta de la piel será más parecida a la que se da en la mucosa gástrica que
en mamíferos.
Las inmunoglobulinas llevan a cabo su función entonces bajo unos principios que parecen conservados a
lo largo de todos los vertebrados. Y en general se observa que se requiere un tipo particular de
inmunoglobulina para regular la microbiota de las mucosas, diferente a la que trabaja a nivel sistémico.
5. Efecto de la quitridiomicosis en es sistema inmune
Los síntomas principales de la quitridiomicosis son hiperqueratosis, hiperplasia de la epidermis y
descamación excesiva de la piel (Parker et al., 2002). Usualmente no está asociada con lesiones severas o
inflamación considerable y si se ha encontrado que hay una disminución en la cantidad de neutrófilos y
eosinófilos en sangre (Woodhams et al., 2007).
En estudios de expresión genética se ha visto que en Silurana tropicalis 157 genes inmunes fueron
identificados en el hígado, al comparar individuos infectados con no infectados la respuesta frente a Bd
fué muy débil, presentándose una disminución en la transcripción de la mayoría de estos, algunos de los
que presentaron un aumento fueron identificados como genes supresores, por ejemplo genes reguladores
de apoptosis, y otros como genes que regulan la respuesta a estrés crónico (factores de crecimiento,
conexina y colágeno). Por otro lado, se observó un aumento en el gen que codifica para la queratina I
(Rosenblum et al., 2009).
Esto hace pensar que Bd tiene un efecto supresor sobre el sistema inmune, lo cuál puede ser una razón
para el avance rápido de la enfermedad en los individuos susceptibles. Se propone entonces que aquellos

17
anfibios resistentes a la infección no son los que tienen una respuesta inmune más efectiva sino los que
tienen un mecanismo que impide la inhibición del sistema por parte de Bd.
Habría que estudiar a fondo entonces las interacciones moleculares que ocurren entre productos
secretados por Bd y el sistema inmune de los anfibios enfocándose en la búsqueda de un posible
mecanismo de interferencia. Los estudios realizados hasta ahora comparando transcriptomas de
individuos infectados y no infectados se podrían revisar buscando cuáles funciones son suprimidas o
disminuidas y desde qué punto basal en las rutas metabólicas ocurre esta regulación negativa.
6. Conclusiones
La mayoría de las funciones del sistema inmune de los vertebrados están bastante conservadas, y están
descritas en muy pocos géneros de anfibios (Rana, Bufo y Xenopus), las respuestas celulares encontradas
en mamíferos son muy similares en todos los vertebrados, se han encontrado células con morfologías y
funciones iguales en todos los demás grupos. Los órganos linfoides varían en función específica y
organización, además no están presentes en todos los grupos de vertebrados, sin embargo las funciones de
estos órganos están cubiertas de algún modo en cada grupo y los productos de estas se mantienen
conservados.
La respuesta inmune humoral, mediada por compuestos extracelulares presenta una mayor variación. Por
un lado las inmunoglobulinas tienen una región que es la más variable de cualquier proteína en
vertebrados, y otra región, de mayor tamaño que es conservada dentro de los isotipos y hace posible una
identificación de anticuerpos si hay una secuencia del mismo isotipo, a pesar de esto sería recomendable
al analizar expresión de inmunoglobulinas en una muestra realizar varias búsquedas disminuyendo la
identidad mínima necesaria para homología y compararlas para no perder información, además de
asegurarse de que todos los isotipos identificados para el grupo trabajado estén caracterizados en alguna
de las bases de datos usadas.
Por otro lado los péptidos antimicrobianos (AMPs), a pesar de tener la mayor representación filogenética
de todas las funciones inmunes en anfibios, también es la más variable, los AMPs comparten unas pocas
características pero sus estructuras y secuencias de aminoácidos cambian de una especie a otra, para hacer
una búsqueda que incluya los AMPs presentes en la muestra sería recomendable, además de disminuir la
identidad mínima como con las inmunoglobulinas, realizar otros análisis con los transcritos sin homología
en bases de datos buscando genes candidatos para péptidos antimicrobianos, observando una predicción

18
de estructuras secundarias, especialmente con transcritos de menos de 200 pares de bases. También es
importante tener en cuenta tamaño de los péptidos antimicrobianos, que pueden tener menos de 20
aminoácidos, por lo que al ensamblar los transcritos hay que disminuir también la longitud mínima.
Los receptores tipo Toll están muy pobremente estudiados en anfibios, según estudios en otros grupos,
mamíferos y peces, de los 6 tipos identificados en mamíferos, algunos están presentes en todos los
vertebrados donde se ha buscado, incluyendo en el genoma de X. laevis, pero otros son excusivos a
mamíferos o no están presentes en todos los grupos. Los TLRs identificados en Xenopus solo están
identificados a partir de las secuencias de otros caracterizados en diferentes grupos, por esto no hay
certeza de que no exista un tipo de TLR con estructura particular en anfibios que no esté presente en bases
de datos. Esta sería otra función para la que valdría la pena realizar otro tipo de análisis aparte de la
búsqueda de homólogos, pero lo más importante es caracterizar mejor y directamente los receptores de
anfibios.
Es importante tener en cuenta al realizar las búsquedas las características de la piel de los anfibios, que
inmunológicamente actúa como una mucosa, así que es de esperar homología con productos del sistema
inmune característicos del tubo digestivo u otras mucosas en otros vertebrados terrestres.
Igualmente, dado que Bd puede tener un efecto supresor en el sistema inmune, es necesario realizar
análisis enfocados también en la interacción hospedero-patógeno, así como en una posible regulación
negativa por parte de Bd.
La transcriptomica es una herramienta útil, que nos permite obtener grandes cantidades de información
sobre el metabolismo de un organismo en un momento y bajo una circunstancias específicas, pero muchos
investigadores restringen su análisis a la búsqueda en bases de datos, sin saber si estas contienen
suficiente información para cubrir la función de interés en su organismo de estudio. Es muy importante
entonces conocer las limitaciones del método, entender la fisiología del organismo y saber cuál es la
cobertura pueden tener las secuencias de las bases de datos de la función o funciones de interés para el
organismo estudiado, así como hacer uso de otras herramientas informáticas que nos ayudan a plantear
genes y proteínas candidatos a partir de las secuencias no identificadas por homología en bases de datos.
7. Agradecimientos
Agradezco a Andrew Crawford por su apoyo y buenas recomendaciones para realizar este trabajo, y a
Pilar Rodríguez y Luisa Castellanos por su apoyo con bibliografía e ideas.

19
8. Bibliografía
Alder, M.N., Rogozin, I.B., Iyer, L.M., Glazko, G.V., Cooper, M.D., and Pancer, Z. (2005). Diversity and function of adaptive
immune receptors in a jawless vertebrate. Science 310, 1970–1973.
Alford, R, R., S.J. (1999). Global Amphibian Declines: A Problem in Applied Ecology. Annual Review of Ecology and
Systematics 30, 133–165.
Ali, M.F., Lips, K.R., Knoop, F.C., Fritzsch, B., Miller, C., and Conlon, J.M. (2002). Antimicrobial peptides and protease
inhibitors in the skin secretions of the rawfish frog, Rana areolata. Biochim. Biophys. Acta 1601, 55–63.
Amemiya, C.T., Haire, R.N., and Litman, G.W. (1989). Nucleotide sequence of a cDNA encoding a third distinct Xenopus
immunoglobulin heavy chain isotype. Nucleic Acids Res. 17, 5388.
Basir, Y.J., Knoop, F.C., Dulka, J., and Conlon, J.M. (2000). Multiple antimicrobial peptides and peptides related to bradykinin
and neuromedin N isolated from skin secretions of the pickerel frog, Rana palustris. Biochim. Biophys. Acta 1543, 95–105.
Batista, C.V., da Silva, L.R., Sebben, A., Scaloni, A., Ferrara, L., Paiva, G.R., Olamendi-Portugal, T., Possani, L.D., and Bloch,
C., Jr (1999). Antimicrobial peptides from the Brazilian frog Phyllomedusa distincta. Peptides 20, 679–686.
Berger, L., Speare, R., Daszak, P., Green, D.E., Cunningham, A.A., Goggin, C.L., Slocombe, R., Ragan, M.A., Hyatt, A.D.,
McDonald, K.R., et al. (1998). Chytridiomycosis causes amphibian mortality associated with population declines in the rain
forests of Australia and Central America. PNAS 95, 9031–9036.
Berger, L., Marantelli, G., Skerratt, L., and Speare, R. (2005). Virulence of the amphibian chytrid fungus Batrachochytrium
dendrobatidis varies with the strain. Diseases of Aquatic Organisms 68, 47–50.
Bevier, C.R., Sonnevend, A., Kolodziejek, J., Nowotny, N., Nielsen, P.F., and Conlon, J.M. (2004). Purification and
characterization of antimicrobial peptides from the skin secretions of the mink frog (Rana septentrionalis). Comp. Biochem.
Physiol. C Toxicol. Pharmacol. 139, 31–38.
Brand, G.D., Leite, J.R.S.A., de Sá Mandel, S.M., Mesquita, D.A., Silva, L.P., Prates, M.V., Barbosa, E.A., Vinecky, F., Mart ins,
G.R., Galasso, J.H., et al. (2006). Novel dermaseptins from Phyllomedusa hypochondrialis (Amphibia). Biochem. Biophys.
Res. Commun. 347, 739–746.
Brandtzaeg, P. (2009). Mucosal immunity: induction, dissemination, and effector functions. Scand. J. Immunol. 70, 505–515.
Callewaert, L., and Michiels, C.W. (2010). Lysozymes in the animal kingdom. J. Biosci. 35, 127–160.
Carrillo-Farga, J., Castell, A., Pérez, A., and Rondán, A. (1990). Langerhans-like cells in amphibian epidermis. J. Anat. 172, 39–
45.
Carroll, M.C. (2004). The complement system in regulation of adaptive immunity. Nature Immunology 5, 981–986.
Carvalho, L., Sun, J., Kane, C., Marshall, F., Krawczyk, C., and Pearce, E.J. (2009). Review series on helminths, immune
modulation and the hygiene hipothesis: mechanisms underlying helminth modulation of dendritic cell function.
Immunology 126, 28–34.
Carver, S., Bell, B.D., and Waldman, B. (2010). Does Chytridiomycosis Disrupt Amphibian Skin Function?
Cerutti, A., and Rescigno, M. (2008). The biology of intestinal immunoglobulin A responses. Immunity 28, 740–750.
Conlon, J.M., Sonnevend, A., Patel, M., Camasamudram, V., Nowotny, N., Zilahi, E., Iwamuro, S., Nielsen, P.F., and Pál, T.
(2003). A melittin-related peptide from the skin of the Japanese frog, Rana tagoi, with antimicrobial and cytolytic
properties. Biochem. Biophys. Res. Commun. 306, 496–500.
Conlon, J.M., Kolodziejek, J., and Nowotny, N. (2004). Antimicrobial peptides from ranid frogs: taxonomic and phylogenetic
markers and a potential source of new therapeutic agents. Biochim. Biophys. Acta 1696, 1–14.

20
Conlon, J.M., Abraham, B., Sonnevend, A., Jouenne, T., Cosette, P., Leprince, J., Vaudry, H., and Bevier, C.R. (2005).
Purification and characterization of antimicrobial peptides from the skin secretions of the carpenter frog Rana virgatipes
(Ranidae, Aquarana). Regul. Pept. 131, 38–45.
Conlon, J.M., Al-Ghaferi, N., Abraham, B., Jiansheng, H., Cosette, P., Leprince, J., Jouenne, T., and Vaudry, H. (2006).
Antimicrobial peptides from diverse families isolated from the skin of the Asian frog, Rana grahami. Peptides 27, 2111–
2117.
Cooper, M.D., and Alder, M.N. (2006). The evolution of adaptive immune systems. Cell 124, 815–822.
Cooper, M.D., Peterson, R.D., and Good, R.A. (1965). DELINEATION OF THE THYMIC AND BURSAL LYMPHOID
SYSTEMS IN THE CHICKEN. Nature 205, 143–146.
Criscitiello, M.F., and Flajnik, M.F. (2007). Four primordial immunoglobulin light chain isotypes, including lambda and kappa,
identified in the most primitive living jawed vertebrates. Eur. J. Immunol. 37, 2683–2694.
Deng, L., Luo, M., Velikovsky, A., and Mariuzza, R.A. (2013). Structural insights into the evolution of the adaptive immune
system. Annu Rev Biophys 42, 191–215.
Du Pasquier, L., and Flajnik, M.F. (1990). Expression of MHC class II antigens during Xenopus development. Dev. Immunol. 1,
85–95.
Fagarasan, S. (2008). Evolution, development, mechanism and function of IgA in the gut. Curr. Opin. Immunol. 20, 170–177.
Flajnik, M.F., and Kasahara, M. (2001a). Comparative genomics of the MHC: glimpses into the evolution of the adaptive
immune system. Immunity 15, 351–362.
Flajnik, M.F., and Kasahara, M. (2001b). Comparative genomics of the MHC: glimpses into the evolution of the adaptive
immune system. Immunity 15, 351–362.
Flajnik, M.F., and Kasahara, M. (2010). Origin and evolution of the adaptive immune system: genetic events and selective
pressures. Nat. Rev. Genet. 11, 47–59.
Flajnik, M.F., and Du Pasquier, L. (1990). The major histocompatibility complex of frogs. Immunol. Rev. 113, 47–63.
Fletcher, T.C., and Grant, P.T. (1969). Immunoglobulins in the serum and mucus of the plaice (Pleuronectes platessa). Biochem.
J. 115, 65P.
Gibson, B.W., Tang, D.Z., Mandrell, R., Kelly, M., and Spindel, E.R. (1991). Bombinin-like peptides with antimicrobial activity
from skin secretions of the Asian toad, Bombina orientalis. J. Biol. Chem. 266, 23103–23111.
Goraya, J., Wang, Y., Li, Z., O’Flaherty, M., Knoop, F.C., Platz, J.E., and Conlon, J.M. (2000). Peptides with antimicrobial
activity from four different families isolated from the skins of the North American frogs Rana luteiventris, Rana berlandieri
and Rana pipiens. European Journal of Biochemistry 267, 894–900.
Hadji-Azimi, I., Coosemans, V., and Canicatti, C. (1987). Atlas of adult Xenopus laevis laevis hematology. Dev. Comp.
Immunol. 11, 807–874.
Haire, R.N., Kitzan Haindfield, M.K., Turpen, J.B., and Litman, G.W. (2002). Structure and diversity of T-lymphocyte antigen
receptors alpha and gamma in Xenopus. Immunogenetics 54, 431–438.
Harding, F.A., Flajnik, M.F., and Cohen, N. (1993). MHC restriction of T-cell proliferative responses in Xenopus. Dev. Comp.
Immunol. 17, 425–437.
He, W., Feng, F., Huang, Y., Guo, H., Zhang, S., Li, Z., Liu, J., Wang, Y., and Yu, H. (2012). Host defense peptides in skin
secretions of Odorrana tiannanensis: Proof for other survival strategy of the frog than merely anti-microbial. Biochimie 94,
649–655.
Hou, F., Li, J., Pan, P., Xu, J., Liu, L., Liu, W., Song, B., Li, N., Wan, J., and Gao, H. (2011). Isolation and characterisation of a
new antimicrobial peptide from the skin of Xenopus laevis. Int. J. Antimicrob. Agents 38, 510–515.
Hsu, E., and Du Pasquier, L. (1984). Ontogeny of the immune system in Xenopus. Differentiation 28, 109–115.
saacson, T., Soto, A., Iwamuro, S., Knoop, F.C., and Conlon, J.M. (2002). Antimicrobial peptides with atypical structural
features from the skin of the Japanese brown frog Rana japonica. Peptides 23, 419–425.
Ishii, A., Kawasaki, M., Matsumoto, M., Tochinai, S., and Seya, T. (2007). Phylogenetic and expression analysis of amphibian
Xenopus Toll-like receptors. Immunogenetics 59, 281–293.

21
Jin, L.-L., Li, Q., Song, S.-S., Feng, K., Zhang, D.-B., Wang, Q.-Y., and Chen, Y.-H. (2009). Characterization of antimicrobial
peptides isolated from the skin of the Chinese frog, Rana dybowskii. Comp. Biochem. Physiol. B, Biochem. Mol. Biol.
154, 174–178.
Kaufman, J. (1999). Co-evolving genes in MHC haplotypes: the “rule” for nonmammalian vertebrates? Immunogenetics 50,
228–236.
Kawai, T., and Akira, S. (2007). Signaling to NF-kappaB by Toll-like receptors. Trends Mol Med 13, 460–469.
Kim, J.B., Halverson, T., Basir, Y.J., Dulka, J., Knoop, F.C., Abel, P.W., and Conlon, J.M. (2000). Purification and
characterization of antimicrobial and vasorelaxant peptides from skin extracts and skin secretions of the North American
pig frog Rana grylio. Regul. Pept. 90, 53–60.
Kindt, T.J., Goldsby, Osborne, B.A., and Kuby, J. (2002). Kuby immunology. (New York: W.H. Freeman).
Kriger, K.M., and Hero, J.-M. (2006). Survivorship in Wild Frogs Infected with Chytridiomycosis. EcoHealth 3, 171–177.
Kozbor, D., Cassatella, M.A., Lessin, S., Kagan, J., Finver, S., Faust, J. et al. (1990) Expression and function of cd - and ab -T
cell receptor heterodimers on human somatic T cell hybrids. J Immunol 14, 3677–3683
Kückelhaus, S.A.S., Leite, J.R.S.A., Muniz-Junqueira, M.I., Sampaio, R.N., Bloch, C., Jr, and Tosta, C.E. (2009). Antiplasmodial
and antileishmanial activities 50.of phylloseptin-1, an antimicrobial peptide from the skin secretion of Phyllomedusa azurea
(Amphibia). Exp. Parasitol. 123, 11–16.
Lai, R., Liu, H., Hui Lee, W., and Zhang, Y. (2002). An anionic antimicrobial peptide from toad Bombina maxima. Biochemical
and Biophysical Research Communications 295, 796–799.
Lee, W.-H., Zhang, J., Zhang, Y.-X., Jin, Y., Lai, R., and Zhang, Y. (2005). Maximin 9, a novel free thiol containing
antimicrobial peptide with antimycoplasma activity from frog Bombina maxima. FEBS Lett. 579, 4443–4448.
Lehman, H.E. (1953). OBSERVATIONS ON MACROPHAGE BEHAVIOR IN THE FIN OF XENOPUS LARVAE. Biol Bull
105, 490–495.
Li, J., Xu, X., Xu, C., Zhou, W., Zhang, K., Yu, H., Zhang, Y., Zheng, Y., Rees, H.H., Lai, R., et al. (2007). Anti-infection
peptidomics of amphibian skin. Mol. Cell Proteomics 6, 882–894.
Lips, K.R., Brem, F., Brenes, R., Reeve, J.D., Alford, R.A., Voyles, J., Carey, C., Livo, L., Pessier, A.P., and Collins, J.P. (2006).
Emerging infectious disease and the loss of biodiversity in a Neotropical amphibian community. PNAS 103, 3165–3170.
Litman, G.W., Rast, J.P., and Fugmann, S.D. (2010). The origins of vertebrate adaptive immunity. Nat Rev Immunol 10, 543–
553.
Liu, X., Liu, R., Wei, L., Yang, H., Zhang, K., Liu, J., and Lai, R. (2011). Two novel antimicrobial peptides from skin secretions
of the frog, Rana nigrovittata. J. Pept. Sci. 17, 68–72.
Longcore, J.E., Pessier, A.P., and Nichols, D.K. (1999). Batrachochytrium Dendrobatidis gen. et sp. nov., a Chytrid Pathogenic
to Amphibians. Mycologia 91, 219.
Lu, Y., Ma, Y., Wang, X., Liang, J., Zhang, C., Zhang, K., Lin, G., and Lai, R. (2008). The first antimicrobial peptide from sea
amphibian. Mol. Immunol. 45, 678–681.
Macpherson, A.J., McCoy, K.D., Johansen, F.-E., and Brandtzaeg, P. (2008). The immune geography of IgA induction and
function. Mucosal Immunol 1, 11–22.
Manning, M.J. and Horton, J.D. (1982) RES structure and function of the amphibia. In: N. Cohen and M. M. Siegel, eds., The
Reticuloendothelial System A Comprehensive Treatise 3. Phylogeny and Ontogeny. New York. Plenum Press
Manning, M.J. (1979). Evolution of the vertebrate immune system. J R Soc Med 72, 683–688.
Miyake, K. (2007). Innate immune sensing of pathogens and danger signals by cell surface Toll-like receptors. Semin. Immunol.
19, 3–10.
Miyazawa, S., Azumi, K., and Nonaka, M. (2001). Cloning and characterization of integrin alpha subunits from the solitary
ascidian, Halocynthia roretzi. J. Immunol. 166, 1710–1715.
Mo, R., Kato, Y., Nonaka, M., Nakayama, K., and Takahashi, M. (1996). Fourth component of Xenopus laevis complement:
cDNA cloning and linkage analysis of the frog MHC. Immunogenetics 43, 360–369.

22
Morita, C.T., Beckman, E.M., Bukowski, J.F., Tanaka, Y., Band, H., Bloom, B.R., Golan, D.E., and Brenner, M.B. (1995). Direct
presentation of nonpeptide prenyl pyrophosphate antigens to human gamma delta T cells. Immunity 3, 495–507.
Nonaka, M., and Kimura, A. (2006). Genomic view of the evolution of the complement system. Immunogenetics 58, 701–713.
Ohta, Y., Goetz, W., Hossain, M.Z., Nonaka, M., and Flajnik, M.F. (2006). Ancestral organization of the MHC revealed in the
amphibian Xenopus. J. Immunol. 176, 3674–3685.
Pancer, Z., Amemiya, C.T., Ehrhardt, G.R.A., Ceitlin, J., Gartland, G.L., and Cooper, M.D. (2004). Somatic divers ification of
variable lymphocyte receptors in the agnathan sea lamprey. Nature 430, 174–180.
Parker, J.M., Mikaelian, I., Hahn, N., and Diggs, H.E. (2002). Clinical Diagnosis and Treatment of Epidermal Chytridiomycosis
in African Clawed Frogs (Xenopus tropicalis). Comparative Medicine 52, 265–268.
Prates, M.V., Sforça, M.L., Regis, W.C.B., Leite, J.R.S.A., Silva, L.P., Pertinhez, T.A., Araújo, A.L.T., Azevedo, R.B., Spisni,
A., and Bloch, C., Jr (2004). The NMR-derived solution structure of a new cationic antimicrobial peptide from the skin
secretion of the anuran Hyla punctata. J. Biol. Chem. 279, 13018–13026.
Ramos, R., Moreira, S., Rodrigues, A., Gama, M., and Domingues, L. (2013). Recombinant expression and purification of the
antimicrobial peptide magainin-2. Biotechnol. Prog. 29, 17–22.
Ramsey, J.P., Reinert, L.K., Harper, L.K., Woodhams, D.C., and Rollins-Smith, L.A. (2010). Immune Defenses against
Batrachochytrium dendrobatidis, a Fungus Linked to Global Amphibian Declines, in the South African Clawed Frog,
Xenopus laevis. Infect. Immun. 78, 3981–3992.
Rinaldi, A.C. (2002a). Antimicrobial peptides from amphibian skin: an expanding scenario: Commentary. Current Opinion in
Chemical Biology 6, 799–804.
Rinaldi, A.C. (2002b). Antimicrobial peptides from amphibian skin: an expanding scenario: Commentary. Current Opinion in
Chemical Biology 6, 799–804.
Roach, J.C., Glusman, G., Rowen, L., Kaur, A., Purcell, M.K., Smith, K.D., Hood, L.E., and Aderem, A. (2005). The evolution
of vertebrate Toll-like receptors. 78.Proc. Natl. Acad. Sci. U.S.A. 102, 9577–9582.
Rollins-Smith, L.A., Reinert, L.K., Miera, V., and Conlon, J.M. (2002a). Antimicrobial peptide defenses of the Tarahumara frog,
Rana tarahumarae. Biochem. Biophys. Res. Commun. 297, 361–367.
Rollins-Smith, L.A., Reinert, L.K., Miera, V., and Conlon, J.M. (2002b). Antimicrobial peptide defenses of the Tarahumara frog,
Rana tarahumarae. Biochem. Biophys. Res. Commun. 297, 361–367.
Rosenblum, E.B., Poorten, T.J., Settles, M., and Murdoch, G.K. (2012). Only skin deep: shared genetic response to the deadly
chytrid fungus in susceptible frog species. Molecular Ecology 21, 3110–3120.
Salinas, I., Zhang, Y.-A., and Sunyer, J.O. (2011). Mucosal immunoglobulins and B cells of teleost fish. Dev. Comp. Immunol.
35, 1346–1365.
Schwager, J., Bürckert, N., Courtet, M., and Du Pasquier, L. (1991). The ontogeny of diversification at the immunoglobulin
heavy chain locus in Xenopus. EMBO J. 10, 2461–2470.
Sciammas, R., and Bluestone, J.A. (1999). TCRgammadelta cells and viruses. Microbes Infect. 1, 203–212.
Simmaco, M., Mignogna, G., and Barra, D. (1998). Antimicrobial peptides from amphibian skin: What do they tell us? Peptide
Science 47, 435–450.
Sousa, J.C., Berto, R.F., Gois, E.A., Fontenele-Cardi, N.C., Honório, J.E.R., Jr, Konno, K., Richardson, M., Rocha, M.F.G.,
Camargo, A.A.C.M., Pimenta, D.C., et al. (2009). Leptoglycin: a new Glycine/Leucine-rich antimicrobial peptide isolated
from the skin secretion of the South American frog Leptodactylus pentadactylus (Leptodactylidae). Toxicon 54, 23–32.
Stuart, S.N., Chanson, J.S., Cox, N.A., Young, B.E., Rodrigues, A.S.L., Fischman, D.L., and Waller, R.W. (2004). Status and
Trends of Amphibian Declines and Extinctions Worldwide. Science 306, 1783–1786.
Tochinai, S., and Katagiri, C. (1975). COMPLETE ABROGATION OF IMMUNE RESPONSE TO SKIN ALLOGRAFTS AND
RABBIT ERYTHROCYTES IN THE EARLY THYMECTOMIZED XENOPUS. Development, Growth and
Differentiation 17, 383–394.
Tonegawa, S. (1976). Proceedings: Determination of the number of antibody structural genes by DNA-RNA hybridization.
Hoppe-Seyler’s Z. Physiol. Chem. 357, 617.

23
Turner, R.J., and Manning, M.J. (1974). Thymic dependence of amphibian antibody responses. Eur. J. Immunol. 4, 343–346.
Villa-Hernández, O., Hernández-Orihuela, L., del Carmen Rodríguez, M., Zamudio-Zuñiga, F., Castro-Franco, R., Pando, V., and
Batista, C.V.F. (2009). Novel antimicrobial peptides isolated from skin secretions of the Mexican frog Hyla eximia. Protein
Pept. Lett. 16, 1371–1378.
Voyles, J., Young, S., Berger, L., Campbell, C., Voyles, W.F., Dinudom, A., Cook, D., Webb, R., Alford, R.A., Skerratt, L.F., et
al. (2009). Pathogenesis of Chytridiomycosis, a Cause of Catastrophic Amphibian Declines. Science 326, 582–585.
Vredenburg, V.T., Knapp, R.A., Tunstall, T.S., and Briggs, C.J. (2010). Dynamics of an emerging disease drive large-scale
amphibian population extinctions. PNAS 107, 9689–9694.
Wang, G., Wang, Y., Ma, D., Liu, H., Li, J., Zhang, K., Yang, X., Lai, R., and Liu, J. (2013). Five novel antimicrobial peptides
from the Kuhl’s wart frog skin secretions, Limnonectes kuhlii. Mol. Biol. Rep. 40, 1097–1102.
Wang, M., Wang, Y., Wang, A., Song, Y., Ma, D., Yang, H., Ma, Y., and Lai, R. (2010). Five novel antimicrobial peptides from
skin secretions of the frog, Amolops loloensis. Comp. Biochem. Physiol. B, Biochem. Mol. Biol. 155, 72–76.
Wei, Z., Wu, Q., Ren, L., Hu, X., Guo, Y., Warr, G.W., Hammarström, L., Li, N., and Zhao, Y. (2009). Expression of IgM, IgD,
and IgY in a Reptile, Anolis carolinensis. J Immunol 183, 3858–3864.
Woodhams, D.C., Ardipradja, K., Alford, R.A., Marantelli, G., Reinert, L.K., and Rollins-Smith, L.A. (2007). Resistance to
chytridiomycosis varies among amphibian species and is correlated with skin peptide defenses. Animal Conservation 10,
409–417.
Wu, Y., Wang, L., Zhou, M., Ma, C., Chen, X., Bai, B., Chen, T., and Shaw, C. (2011). Limnonectins: a new class of
antimicrobial peptides from the skin secretion of the Fujian large-headed frog (Limnonectes fujianensis). Biochimie 93,
981–987.
Xu, B., Che, H., Kang, L., Zheng, S., Mu, S., and Wan, F. (2012). Molecular cloning and functional characterization of novel
antimicrobial peptides from the skin of brown frog, Rana zhenhaiensis. Zool. Sci. 29, 553–558.
Xu, Z., Parra, D., Gómez, D., Salinas, I., Zhang, Y.-A., von Gersdorff Jørgensen, L., Heinecke, R.D., Buchmann, K., Lapatra, S.,
and Sunyer, J.O. (2013). Teleost skin, an ancient mucosal surface that elicits gut-like immune responses. Proc. Natl. Acad.
Sci. U.S.A. 110, 13097–13102.
Yeaman, M.R., and Yount, N.Y. (2003). Mechanisms of antimicrobial peptide action and resistance. Pharmacol. Rev. 55, 27–55.
Zhao, Y., Jin, Y., Lee, W.-H., and Zhang, Y. (2006). Purification of a lysozyme from skin secretions of Bufo andrewsi. Comp.
Biochem. Physiol. C Toxicol. Pharmacol. 142, 46–52.
Zhou, J., McClean, S., Thompson, A., Zhang, Y., Shaw, C., Rao, P., and Bjourson, A.J. (2006). Purification and characterization
of novel antimicrobial peptides from the skin secretion of Hylarana guentheri. Peptides 27, 3077–3084.
Zhu, Z., Sun, Y., Wang, R., and Xu, T. (2013). Evolutionary analysis of TLR9 genes reveals the positive selection of extant
teleosts in Perciformes. Fish Shellfish Immunol. 35, 448–457.