
Acariosis del maíz en Navarra. Ecología de Tetranychus...
12
Bol. San. Veg. Plagas, 24: 609-620, 1998 Acariosis del maíz en Navarra. Ecología de Tetranychus turkestani Ugarov y Nikolski V. M. IRAOLA, M. L. MORAZA y R. BlURRUN Se ha estudiado el comportamiento ecológico de las poblaciones de Tetranychus turkestani Ugarov y Nikolski, tetraniquido fitófago que ataca al maíz en Navarra. Esta especie, que puede pasar el invierno en forma de hembra diapausica, comienza su acti- vidad sobre la vegetación espontánea del borde de las parcelas de maíz a partir del mes de Marzo (temperatura media 13 °C). Desde esta vegetación, se desplaza a las hojas in- feriores de las plantas de maíz del borde de la parcela a partir del mes de Junio, depen- diendo del clima y de la existencia o no de vegetación de ribazo. A partir de ese mo- mento, los ácaros siguen una doble dirección: hacia arriba, colonizando la planta y hacia los lados, colonizando las plantas vecinas. El estado fisiológico de la planta de maíz incide acusadamente en esta colonización. La semana previa y durante el estado de floración masculina, el crecimiento de T. tur- kestani se ve favorecido, produciéndose la colonización total de la planta por un lado, y de la parcela por otro. Las poblaciones de tetraníquidos se ven afectadas por: el estado fisiológico del maíz, el clima (temperaturas elevadas y humedad relativa baja les favo- recen) y los enemigos naturales (especialmente los ácaros fitoseidos) que controlan sus poblaciones. V. M. IRAOLA y M. L. MORAZA: Departamento de Zoología y Ecología. Universidad de Navarra. Apdo. 177. 31080 Pamplona R. BIURRUN: Instituto Técnico y de Gestión Agrícola S.A. Ctra del Sadar, Edificio El Sano. 31006 Pamplona Palabras clave: Maíz, acariosis, Tetranychus turkestani, dinámica de colonización, dinámica de poblaciones, Navarra INTRODUCCIÓN El cultivo del maíz en Navarra constituye casi la tercera parte de la superficie total de- dicada al regadío en Navarra, preferente- mente en la zona conocida como La Ribera (Valle del Ebro) (GOBIERNO DE NAVARRA, 1996). En la mayor parte de las parcelas de La Ribera Navarra el regadio se realiza por inundación (Fig. 1), siendo todavía escasas las parcelas que disponen de riego por as- persión. Los ácaros fitófagos no se han considera- do hasta recientemente como plagas impor- tantes del maíz, salvo en Estados Unidos donde los tetraníquidos (Acari: Tetranychi- dae) son desde hace tiempo un problema económico de primer grado en las grandes llanuras maiceras del Oeste, fundamen- talmente al haber desarrollado resistencias a los insecticidas organofosforados (LOGAN et al, 1983). En Europa, a partir de la década de los 80, se han agravado los ataques de estos ácaros al maíz. Por ejemplo, hasta 1983 en Francia, sólo se habían registrado ataques en dos departamentos, mientras que seis años más tarde el número se había ele- vado a 10 (sobre todo en el Sur), achacándo- se este aumento a la sequía que había favo- recido el desarrollo de los ácaros (BOUF- FARD, 1990). En España, los ataques ocurri- dos a partir de la década de los 80 en maiza-
Transcript of Acariosis del maíz en Navarra. Ecología de Tetranychus...

Bol. San. Veg. Plagas, 24: 609-620, 1998
Acariosis del maíz en Navarra. Ecología de Tetranychusturkestani Ugarov y Nikolski
V. M. IRAOLA, M. L. MORAZA y R. BlURRUN
Se ha estudiado el comportamiento ecológico de las poblaciones de Tetranychusturkestani Ugarov y Nikolski, tetraniquido fitófago que ataca al maíz en Navarra. Estaespecie, que puede pasar el invierno en forma de hembra diapausica, comienza su acti-vidad sobre la vegetación espontánea del borde de las parcelas de maíz a partir del mesde Marzo (temperatura media 13 °C). Desde esta vegetación, se desplaza a las hojas in-feriores de las plantas de maíz del borde de la parcela a partir del mes de Junio, depen-diendo del clima y de la existencia o no de vegetación de ribazo. A partir de ese mo-mento, los ácaros siguen una doble dirección: hacia arriba, colonizando la planta yhacia los lados, colonizando las plantas vecinas.
El estado fisiológico de la planta de maíz incide acusadamente en esta colonización.La semana previa y durante el estado de floración masculina, el crecimiento de T. tur-kestani se ve favorecido, produciéndose la colonización total de la planta por un lado, yde la parcela por otro. Las poblaciones de tetraníquidos se ven afectadas por: el estadofisiológico del maíz, el clima (temperaturas elevadas y humedad relativa baja les favo-recen) y los enemigos naturales (especialmente los ácaros fitoseidos) que controlan suspoblaciones.
V. M. IRAOLA y M. L. MORAZA: Departamento de Zoología y Ecología. Universidad deNavarra. Apdo. 177. 31080 PamplonaR. BIURRUN: Instituto Técnico y de Gestión Agrícola S.A. Ctra del Sadar, Edificio ElSano. 31006 Pamplona
Palabras clave: Maíz, acariosis, Tetranychus turkestani, dinámica de colonización,dinámica de poblaciones, Navarra
INTRODUCCIÓN
El cultivo del maíz en Navarra constituyecasi la tercera parte de la superficie total de-dicada al regadío en Navarra, preferente-mente en la zona conocida como La Ribera(Valle del Ebro) (GOBIERNO DE NAVARRA,1996). En la mayor parte de las parcelas deLa Ribera Navarra el regadio se realiza porinundación (Fig. 1), siendo todavía escasaslas parcelas que disponen de riego por as-persión.
Los ácaros fitófagos no se han considera-do hasta recientemente como plagas impor-tantes del maíz, salvo en Estados Unidosdonde los tetraníquidos (Acari: Tetranychi-
dae) son desde hace tiempo un problemaeconómico de primer grado en las grandesllanuras maiceras del Oeste, fundamen-talmente al haber desarrollado resistencias alos insecticidas organofosforados (LOGAN etal, 1983). En Europa, a partir de la décadade los 80, se han agravado los ataques deestos ácaros al maíz. Por ejemplo, hasta1983 en Francia, sólo se habían registradoataques en dos departamentos, mientras queseis años más tarde el número se había ele-vado a 10 (sobre todo en el Sur), achacándo-se este aumento a la sequía que había favo-recido el desarrollo de los ácaros (BOUF-FARD, 1990). En España, los ataques ocurri-dos a partir de la década de los 80 en maiza-

Fig. 1.-Parcela de maíz.
les de Extremadura, Andalucía, Aragón(ARIAS Y ALVARADO, 1983; ALVARADO et
al, 1984, 1986; ESTEBAN, 1990; LÓPEZ-BE-LLIDO, 1991) y Navarra (MACUÁ, 1990) pu-sieron de manifiesto la importancia de lostetraníquidos.
La presencia de estos ácaros en las hojasde maíz se pone de manifiesto por la deco-loración que producen al alimentarse. Losácaros se alimentan succionando el jugo dela epidermis y de las células parenquimáti-cas, principalmente del envés de las hojas.Las células rotas y vacías, que ya no partici-pan en el metabolismo de la planta, formanpequeñas manchas marrones. Los principa-les daños que ocasionan son la destrucciónde hojas y un retraso, o una reducción, delperiodo de llenado de la mazorca, lo quehace disminuir la longitud de esta y el pesode los granos (FAUVEL et al, 1987; GOA-RANT, 1987; LÓPEZ-BELLIDO, 1991).
Frente a un problema de acariosis en elmaíz el tratamiento normal es la utilizaciónde productos acaricidas, existiendo trata-mientos curativos y preventivos. En los cu-rativos se recomienda el uso de dicofol mástetradifon, propargita o cihexaestan, mien-tras que en los preventivos se recomienda lautilización de hexitiazox, clofentezin o deazufre que tiene efecto frenante sobre los te-traníquidos (FAUVEL et ai, 1987; NAIBO,1989). Sin embargo, el tratamiento químicono está exento de problemas. El gran portede las plantas de maíz en los meses de Julioy Agosto (cuando es necesario el tratamien-to) obliga a que, en las parcelas que no dis-ponen de riego por aspersión, los tratamien-tos se realicen mediante avión o tractores«zancudos» con el consiguiente incrementoen los costes. Además, la frecuente ubica-ción de las explotaciones de maíz en Nava-rra en la vecindad de cultivos hortícolas deconsumo humano directo, hacen que no seadeseable, en muchos casos, la fumigacióndel maíz.
La introducción de enemigos naturales hasurgido como una herramienta alternativa ala utilización de los productos químicos. EnEstados Unidos y en Francia se han realiza-do ensayos de introducción artificial de po-blaciones de fitoseidos depredadores (Acari:Phytoseiidae) Phytoseiulus persimilis At-hias-Henriot y Amblyseius californicus (Mc-Gregor) (GiLSTRAP et al, 1977; PICKETT YGILSTRAP, 1986a, PICKETT et al, 1987; Go-ARANT, 1988). Otros campos de acción,dentro de la lucha integrada, son la posibili-dad de utilizar líneas de maíz resistentes alos ácaros (MANSOUR et al, 1993) y hongosentomopatógenos (Neozygites) que infectande forma natural a los tetraníquidos en maíz(DlCK Y BUSCHMAN., 1995).
En Navarra, al igual que en el resto de Es-paña, los graves ataques de ácaros de maíz apartir de 1987 originaron una preocupaciónsobre esta plaga. Esto motivó que se realiza-ra en 1989 un trabajo en el Departamento deZoología y Ecología de la Universidad deNavarra, financiado por el Gobierno de Na-varra, acerca de la especie responsable de la

plaga y de sus posibles enemigos naturales.A partir de este trabajo se realizó otro poste-rior (1991-93), conjunto entre el Departa-mento de Zoología y el Instituto Técnico yde Gestión Agrícola, incluido en el PlanSectorial de I+D del MAPA (n.° 9571) y fi-nanciado en parte por el Departamento deAgricultura, Ganadería y Montes del Go-bierno de Navarra. Los resultados que se ex-ponen a continuación son deudores de di-chos trabajos, habiéndose publicado con an-terioridad las especies de ácaros fitoseidosdepredadores presentes en maíz (IRAOLA etal, 1997).
MATERIAL Y MÉTODOS
Zona de estudio
Durante el año 1992 se realizaron unaserie de muéstreos en 43 parcelas de diferen-tes municipios del Sur de Navarra, donde serecogieron muestras de planta de maíz y/ode vegetación de ribazo. En el año 1993 fue-ron 22 las parcelas (de las que siete no se ha-bían muestreado el año anterior) y finalmen-te en 1994 se recogieron muestras de nueveparcelas no muestreadas anteriormente.
Las localidades estudiadas fueron (el nú-mero de parcelas por localidad figura entreparéntesis): Buñuel (7), Cabanillas (3), Ca-dreita (3), Caparroso (2), Carcastillo (1),Cáseda (2), Castejón (2), Cortes (4), Funes(8), Fustiñana (3), Larraga (2), Miranda deArga (1), Peralta (2), Ribaforada (2), Santa-cara (1), Valtierra (8) y Villafranca (8). Lasituación de estas localidades en Navarra seencuentra en la Figura 2.
Muestreo en planta de maíz
Para registrar los distintos estados fenoló-gicos del maíz se ha utilizado el código de-cimal empleado por GROOT et al. (1986)(cuadro 1).
En el año 1992, los muéstreos se realiza-ron en dos épocas. A comienzos de Julio y
Corles
Fig. 2.-Localidades donde se han efectuadolos muéstreos.
Cuadro 1 .-Códigos de los estados de desarrollodel maíz (según GROOT et al., 1986)
Estado fenológico
GerminaciónAparición y crecimiento de la plántula2 hojas totalmente desarrolladasAlargamiento del tallo. 4 hojas total-
mente desarrolladas6 hojas totalmente desarrolladas. Apa-
rición raíces de anclaje8 hojas totalmente desarrolladas10 hojas totalmente desarrolladas12 hojas totalmente desarrolladas13 hojas totalmente desarrolladas. Co-
mienzo aparición panícula masculina14 hojas totalmente desarrolladasFloración. Comienzo floración mascu-
lina50% de las flores femeninas emitiendo
filamentosFin de la floraciónGrano en estado acuoso inicial. Co-
mienzo secado filamentosEstado acuoso intermedioGrano en estado lechoso-pastosoGrano vitreoMadurez completa

en Septiembre, cuando la mayor parte de lasparcelas presentaban poblaciones elevadas,muestreándose cada parcela al menos unavez. En el año 1993 se concentraron los es-fuerzos en los meses de Julio y Agostomuestreándose dos veces o más, mientrasque en 1994 se recogieron muestras única-mente en el mes de Agosto.
En cada parcela se tomaron 10 hojas me-dias, cercanas a la mazorca, procedentes deplantas elegidas al azar de filas cercanas alborde de la misma. Las hojas se guardabanen bolsas de papel y estas en bolsas de plás-tico. Estas bolsas se introducían en una ne-vera portátil para su traslado al laboratorio.El método de extracción de los ácaros con-sistió en el lavado de las hojas en agua tem-plada con jabón y posterior filtrado a travésde tamices de luz de 0,4 y 0,2 mm. Losejemplares recogidos en la malla del tamizeran trasladados a placas de Petri para su es-tudio.
Para observar las dinámicas de los ácaros,se siguieron en detalle durante 1992, tres par-celas comerciales de tamaño medio (4.000-6.000 m2), con riego por inundación y sinningún tipo de tratamiento fitosanitario paraplagas. Estas parcelas estaban situadas enlos términos de Funes, Cadreita y Caparro-so. El periodo de muestreo se extendiódesde los meses de Julio a Septiembre conperiodicidad mensual. El tamaño de muestraconsistió en las dos hojas que rodean la ma-zorca de 20 plantas elegidas al azar de laparcela. En el caso de que no hubiese nacidola mazorca, se recogían las dos primerashojas verdes, contadas a partir de la base dela planta. Durante 1993 se siguieron, duran-te Julio y Agosto, trece parcelas con una pe-riodicidad semanal o quincenal. El tamañode muestra consistió en una hoja de posiciónmedia de 10 plantas elegidas al azar.
Para conocer el proceso de propagaciónde la infestación dentro de la parcela, en losaños 1992 y 1993 se eligió una parcela, si-tuada en la localidad de Funes, de una ex-tensión de 5.800 m2, dividida en 25 áreascuadradas iguales de 232 m2. En cada árease revisaron semanalmente, desde finales de
Junio hasta Septiembre, cuatro plantas alazar, anotando la existencia o no de ácaros,el estado fenológico del maíz y las hojas enlas que se encontraban los ácaros.
Con el fin de conocer la distribución delos ácaros en la hoja, se dividieron las hojasde 100 plantas en tercios y mediante una ob-servación directa, con ayuda de una lupa decampo, se procedió a anotar el número deácaros en cada tercio.
Muestreo en ribazo del maíz
Para conocer las especies de plantas de ri-bazo y adventicias en las que se refugian losácaros tetraníquidos, se han recogido mues-tras durante el invierno, primavera y veranode plantas del interior y de los bordes de laparcela. También se recogieron muestras derestos del cultivo de la campaña anterior. Enel año 1992 se muestreo, a finales del mesde Abril en 33 parcelas y a mediados deJunio en 15 parcelas, mientras que en 1993se recogieron plantas a lo largo del invierno,de Diciembre a Marzo, en 10 parcelas.
Los muéstreos del ribazo se realizaron re-cogiendo 10 plantas de la especie más abun-dante si era herbácea, o 40 hojas si era ar-bustiva o arbórea. En el caso de gramíneas yen los restos de la cosecha anterior se mues-treo una superficie de 40 x 40 cm. Las plan-tas se cortaron desde la base con unas tijeraso un cuchillo. El método de extracción utili-zado fue el embudo de Berlese-Tragardmanteniendo las muestras una semana.
Las especies de plantas de ribazo estudia-das y el número de muestras de cada unafueron: Achillea millefolium L. (4), Ama-ranthus retroflexus L. (2), Anthemis cothulaL. (1), Brachypodium ramosum R. Y S.(l),Capsella bursa-pastoris (L.)Medicus (1),Chelidonium majus L. (1), Chenopodiumalbum L. (1), Convolvulus arvensis L. (3),Diplotaxis erucoides (L.) DC (3), Elymusrepens (L.)Gould (1), Fraxinus excelsior L.(1), Galium sp. (1), Hedera helix L. (1),Lactuca serriola L. (1), Malva sylvestris L.(4), Prunus spinosa L. (1), Rubia peregrina

L. (1), Rubus sp. (4), Sinapis alba L. (2),Taraxacum officinale Webber (4), Trifoliumsp.(l), Ulmus minor Mill. (1), Veronica per-sica Poiret (1), diferentes especies de gramí-neas (45) y restos de la cosecha de maíz delaño anterior (23).
RESULTADOS
Especies de tetraníquidos presentesen plantas de maíz
La especie encontrada en todas las locali-dades muestreadas, a lo largo de todos los es-tados fenológicos del maíz, ha sido Tetrany-chus turkesíani Ugarov y Nikolski (Fig. 3).
Esta es la primera cita de T. turkestán! enel cultivo del maíz en España, ya que las es-pecies previamente citadas en este cultivoson T, cinnabarinus (Boisduval) o T. urticaeKoch (ALVAREZ, 1964; AL VARADO et al,1984, 1986; ESTEBAN, 1990; LÓPEZ-
BELLIDO 1991). ARIAS Y ALVARADO(1983) hablan de diversas especies de tetra-níquidos que atacan el maíz pero no llegan acitar las especies.
La ausencia de T. urticae en este estudiono quiere decir que no esté presente en maízen el Sur de Navarra, ya que se ha encontra-do en las mismas localidades muestreadas,aunque en otros cultivos y en menor propor-ción que T. turkestani. Ambas especies sehan encontrado cohabitando en maíz en elSur de Francia (NAIBO et al.t 1987; FAUVELet al., 1987; NAIBO, 1989), por lo que es po-sible que también coexistan en Navarra eneste cultivo, aunque la especie dominante enel sur sea T. turkestani. Un muestreo mássostenido, tanto geográfica como temporal-mente, podría demostrar esta suposición.
Entre los trabajos dedicados a la determi-nación de las especies de tetraníquidos enmaíz en otros países, destacan los realiza-dos en los Estados Unidos donde las espe-cies presentes, según numerosos artículos(EHLER, L. E., 1974; PICKETT y GILSTRAP,
1985, 1986b entre otros) son Oligonychuspratensis (Banks) y T. urticae. En África se
Fig. 3.-Aspecto de una hembra de Teíranychuiturkestani UGAROV Y NlKOLSKI.
señala a T. urticae como la especie princi-pal, siendo citadas otras como O. pratensisy O. gripus Baker y Pritchard (MEYER,1987). En la India se cita a O. indicus Hirst(GUPTA, 1991). En Europa se ha encontradoa T. urticae como plaga menor del maíz enGrecia (HATZINIKOLIS, 1969), Italia (comoprincipal ácaro fitófago) (RAGUSA Y PAO-LETTI, 1985) y en Francia (FAUVEL et al,1987; NAIBO et al, 1987, 1988) junto a T.turkestani.
Focos iniciales de infestación, plantasreservorio
T. turkestani puede pasar el invierno enforma de hembra adulta diapausica despuésde una serie de cambios que pueden afectartanto a la coloración del tegumento (pasandel verde oscuro a un naranja brillante),como a su fisiología y comportamiento (nose reproducen) (POPOV Y VEERMAN, 1996).Estas hembras se refugian debajo de corte-zas de árboles, de las plantas que rodean loscultivos o en las anfractuosidades del suelo(VEERMAN, 1985). Desde estos lugares dehibernación, una vez finalizado el invierno e

inducidos especialmente por el fotoperiodo,se desplazan a las plantas espontáneas en lasque comienzan a reproducirse. Otra posibili-dad es que pasen el invierno reproduciéndo-se como hembras normales. En este estudio,se han encontrado hembras diapausicas eninvierno (Diciembre y Febrero) en T. offici-nale, Rubus sp. y en Gramíneas, así comoen restos de maíz de la cosecha anterior,aunque siempre en número muy escaso. Nose han encontrado hembras no diapausicasdurante los meses de invierno.
En Navarra, el comienzo de la coloniza-ción de las especies vegetales del borde delas parcelas de maíz sucede en torno a me-diados del mes de Abril, con una temperatu-ra media en torno a los 13 °C. Esta mismatemperatura es la que FAUVEL et al. (1987)dan para el comienzo de la actividad porparte de las hembras diapausicas. Las espe-cies vegetales donde se han encontrado po-blaciones de Tetranychus y desde donde ini-cian la colonización del maíz han sido: A.retroflexus, C. bursa-pastoris, Chenopo-dium sp., C. arvensis., L. vulgare., M. syl-vestris, Rubus sp., S. alba., Trifolium sp., T.officinale y varias gramíneas como Digita-ria sanguinalis (L.) Scop., Echinocloa crus-galli (L.) Beauv., Setaria pumila (Poir.)Schultes y S. verticillata (L) Beauv.
Otras especies que han sido citadas comoreservorio de Tetranychus en el cultivo delmaíz son: Campsis radicans (L.) Seem, es-Bureau, Geranium sp., Lamium amplexicau-le L., Lonicera japonica Thumb., Polygo-num sp., Rubus spp., Trifolium pratense L.,Vicia sp. y diversas Umbelliferae, Gramine-ae y Malvaceae (BRANDENBURG Y KEN-NEDY, 1982; POPOV, 1983; MARGOLIES YKENNEDY, 1985; FAUVEL etal.,\9Sl).
tación espontánea a las plantas de MAÍZ(BRANDENBURG y KENNEDY, 1982; MAR-GOLIES Y KENNEDY, 1985; FAUVEL et al.,1987; NAIBO, 1989).
En 1992, las primeras colonias se detecta-ron a finales del mes de Junio (en la semana25) con el maíz en un estado de desarrolloentre 4 y 8 hojas (código decimal 2,00-3,00). En el año 1993 ocurrieron a mediadosdel mes de Julio (semana 28), en un estadode desarrollo entre 8 y 12 hojas (3,00-4,00).
Esta diferencia de tres semanas, en nues-tra opinión, fue debida a la climatología delaño 1993. En este año la temperatura mediaen las localidades estudiadas durante losmeses de Abril y Mayo fue inferior a la delaño 1992, lo que produjo un retraso en laevolución del maíz y en los incrementos delas poblaciones de tetraníquidos. Sin em-bargo, las temperaturas medias de las sema-nas en las que se detectaron los primerosataques fue de 17 °C en 1992 y de 20 °C en1993.
Los ataques más tempranos siempre ocu-rrieron en parcelas con abundante vegeta-ción de ribazo. En el caso de un ribazo esca-so, las colonizaciones se retrasaban a finalesdel mes de Julio.
Los datos obtenidos coinciden con losdescritos en Francia, donde las primeras co-lonizaciones suceden a partir de la segundaquincena de Junio (GOARANT, 1987). Lasfechas encontradas para T. urticae en laVega del Guadalquivir por ALVARADO et al.(1984, 1986) indican el comienzo de la co-lonización a principios de Mayo, aunque enel mismo estado de desarrollo del maíz queen nuestro caso (las diferencias climatológi-cas hacen que la plantación del maíz se anti-cipe en Andalucía).
Colonización de las plantas del maíz
Las primeras colonizaciones de las parce-las de maíz ocurrieron en las plantas delborde más cercanas a la vegetación espontá-nea. Numerosos estudios afirman que losácaros se desplazan andando, desde la vege-
Dinámica de la colonización de la parcela
La dispersión de los ácaros dentro de lasparcelas de maíz se ha descrito como en«mancha de aceite» (ALVARADO et al.,1986), es decir, una vez colonizadas lasplantas de maíz del borde del cultivo toman
una doble dirección: «vertical» colonizandola totalidad de la planta y «horizontal»,desde el borde y hacia el interior de la parce-la a medida que progresa la estación (BRAN-DENBURG y KENNEDY, 1982). El ácaro semueve buscando tanto un hospedador acep-table como un microclima idóneo para sudesarrollo, existiendo dos factores principa-les para que abandone el hospedador: el au-mento de densidad poblacional y el agota-miento de los recursos (MARGOLIES y KEN-NEDY, 1984, 1985).
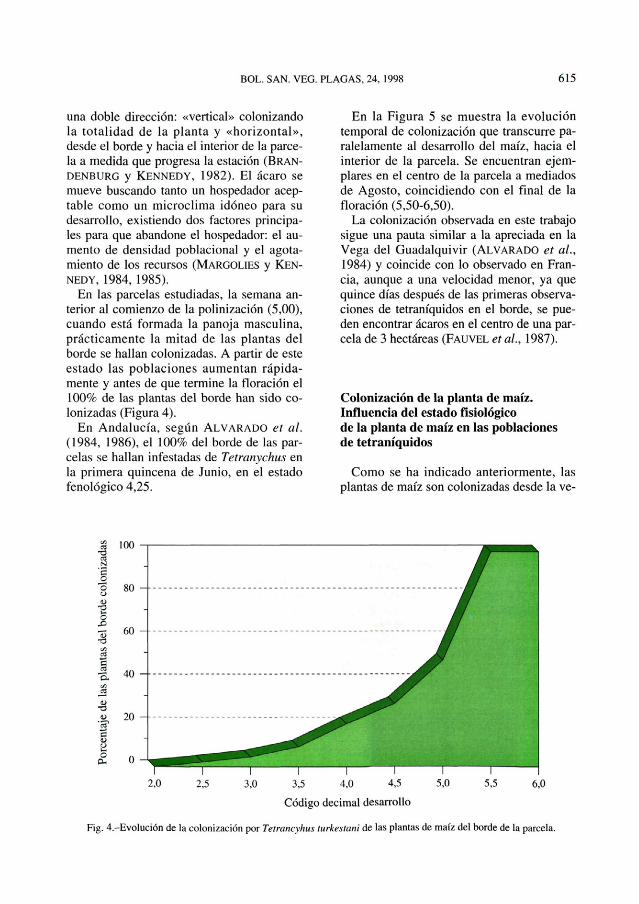
En las parcelas estudiadas, la semana an-terior al comienzo de la polinización (5,00),cuando está formada la panoja masculina,prácticamente la mitad de las plantas delborde se hallan colonizadas. A partir de esteestado las poblaciones aumentan rápida-mente y antes de que termine la floración el100% de las plantas del borde han sido co-lonizadas (Figura 4).
En Andalucía, según ALVARADO et al.(1984, 1986), el 100% del borde de las par-celas se hallan infestadas de Tetranychus enla primera quincena de Junio, en el estadofenológico4,25.
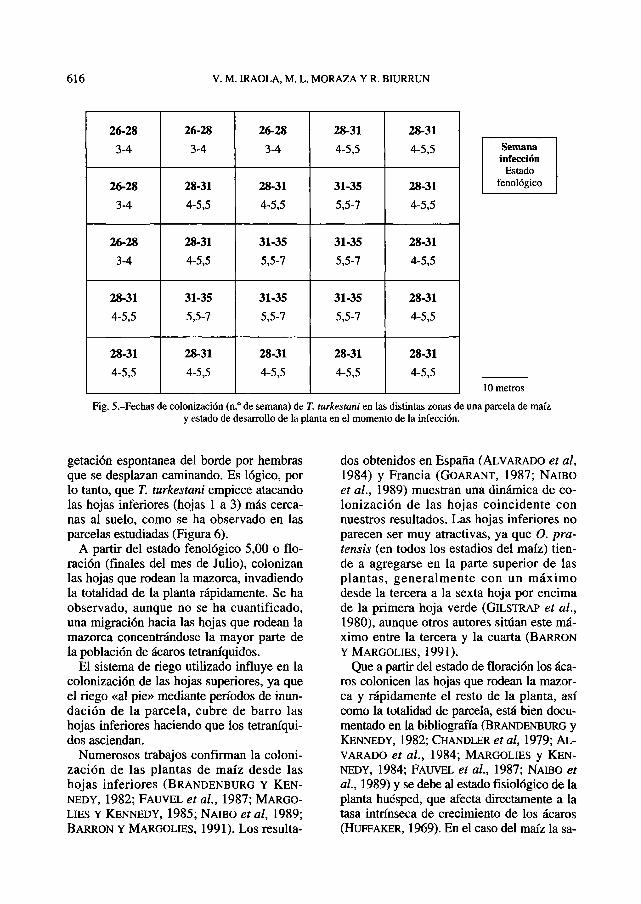
En la Figura 5 se muestra la evolucióntemporal de colonización que transcurre pa-ralelamente al desarrollo del maíz, hacia elinterior de la parcela. Se encuentran ejem-plares en el centro de la parcela a mediadosde Agosto, coincidiendo con el final de lafloración (5,50-6,50).
La colonización observada en este trabajosigue una pauta similar a la apreciada en laVega del Guadalquivir (ALVARADO et al.,1984) y coincide con lo observado en Fran-cia, aunque a una velocidad menor, ya quequince días después de las primeras observa-ciones de tetraníquidos en el borde, se pue-den encontrar ácaros en el centro de una par-cela de 3 hectáreas (FAUVEL et al, 1987).
Colonización de la planta de maíz.Influencia del estado fisiológicode la planta de maíz en las poblacionesde tetraníquidos
Como se ha indicado anteriormente, lasplantas de maíz son colonizadas desde la ve-
Fig. 4.-Evolución de la colonización por Tetrancyhus turkestani de las plantas de maíz del borde de la parcela.
getación espontanea del borde por hembrasque se desplazan caminando. Es lógico, porlo tanto, que T. turkestani empiece atacandolas hojas inferiores (hojas 1 a 3) más cerca-nas al suelo, como se ha observado en lasparcelas estudiadas (Figura 6).
A partir del estado fenológico 5,00 o flo-ración (finales del mes de Julio), colonizanlas hojas que rodean la mazorca, invadiendola totalidad de la planta rápidamente. Se haobservado, aunque no se ha cuantificado,una migración hacia las hojas que rodean lamazorca concentrándose la mayor parte dela población de ácaros tetraníquidos.
El sistema de riego utilizado influye en lacolonización de las hojas superiores, ya queel riego «al pie» mediante períodos de inun-dación de la parcela, cubre de barro lashojas inferiores haciendo que los tetraníqui-dos asciendan.
Numerosos trabajos confirman la coloni-zación de las plantas de maíz desde lashojas inferiores (BRANDENBURG Y KEN-NEDY, 1982; FAUVEL et al, 1987; MARGO-LIES Y KENNEDY, 1985; NAIBO et al, 1989;BARRON Y MARGOLIES, 1991). Los resulta-
dos obtenidos en España (ALVARADO et al,1984) y Francia (GOARANT, 1987; NAIBOet al., 1989) muestran una dinámica de co-lonización de las hojas coincidente connuestros resultados. Las hojas inferiores noparecen ser muy atractivas, ya que O. pra-tensis (en todos los estadios del maíz) tien-de a agregarse en la parte superior de lasplantas, generalmente con un máximodesde la tercera a la sexta hoja por encimade la primera hoja verde (GlLSTRAP et al.,1980), aunque otros autores sitúan este má-ximo entre la tercera y la cuarta (BARRONY MARGOLIES, 1991).
Que a partir del estado de floración los áca-ros colonicen las hojas que rodean la mazor-ca y rápidamente el resto de la planta, asícomo la totalidad de parcela, está bien docu-mentado en la bibliografía (BRANDENBURG yKENNEDY, 1982; CHANDLER et al, 1979; AL-VARADO et al, 1984; MARGOLIES y KEN-NEDY, 1984; FAUVEL et al, 1987; NAIBO etal, 1989) y se debe al estado fisiológico de laplanta huésped, que afecta directamente a latasa intrínseca de crecimiento de los ácaros(HUFFAKER, 1969). En el caso del maíz la sa-
Fig. 5.-Fechas de colonización (n.° de semana) de T. turkestani en las distintas zonas de una parcela de maízy estado de desarrollo de la planta en el momento de la infección.
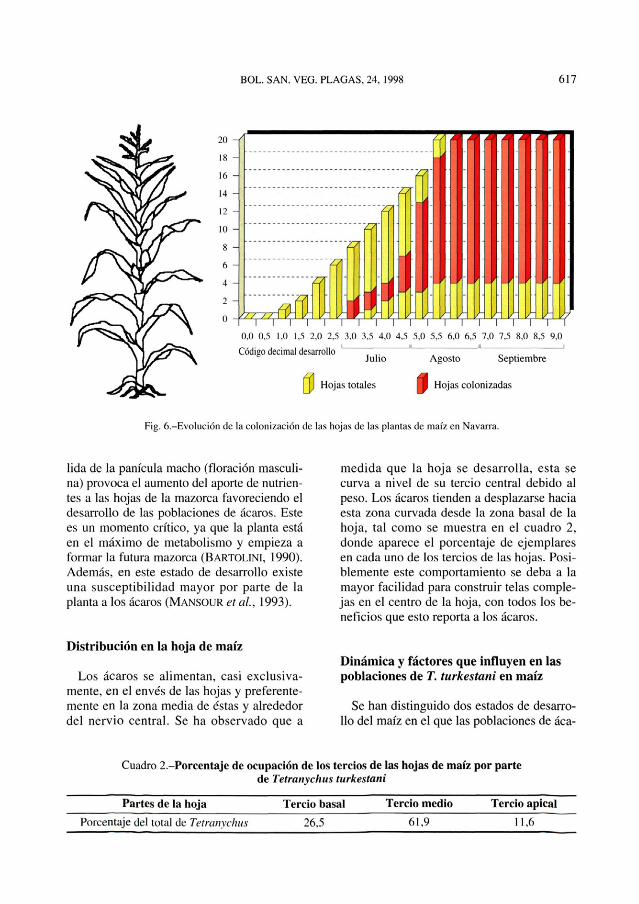
Fig. 6.—Evolución de la colonización de las hojas de las plantas de maíz en Navarra.
lida de la panícula macho (floración masculi-na) provoca el aumento del aporte de nutrien-tes a las hojas de la mazorca favoreciendo eldesarrollo de las poblaciones de ácaros. Estees un momento crítico, ya que la planta estáen el máximo de metabolismo y empieza aformar la futura mazorca (BARTOLINI, 1990).Además, en este estado de desarrollo existeuna susceptibilidad mayor por parte de laplanta a los ácaros (MANSOUR et ai, 1993).
Distribución en la hoja de maíz
Los ácaros se alimentan, casi exclusiva-mente, en el envés de las hojas y preferente-mente en la zona media de éstas y alrededordel nervio central. Se ha observado que a
medida que la hoja se desarrolla, esta securva a nivel de su tercio central debido alpeso. Los ácaros tienden a desplazarse haciaesta zona curvada desde la zona basal de lahoja, tal como se muestra en el cuadro 2,donde aparece el porcentaje de ejemplaresen cada uno de los tercios de las hojas. Posi-blemente este comportamiento se deba a lamayor facilidad para construir telas comple-jas en el centro de la hoja, con todos los be-neficios que esto reporta a los ácaros.
Dinámica y factores que influyen en laspoblaciones de T. turkestani en maíz
Se han distinguido dos estados de desarro-llo del maíz en el que las poblaciones de áca-
Cuadro 2.-Porcentaje de ocupación de los tercios de las hojas de maíz por partede Tetranychus turkestani
Partes de la hoja Tercio basal Tercio medio Tercio apical
Porcentaje del total de Tetranychus

ros tienden a aumentar. El primero es el pe-riodo de aparición de la mazorca que se hacomentado anteriormente. En Navarra, estemomento ocurre a comienzos del mes deAgosto. El segundo, cuando ya está formadala mazorca y los granos están en el estadoacuoso-pastoso (6,00-7,00), a comienzos deSeptiembre. Este incremento, también des-crito en la bibliografía (CHANDLER et al.,1979; GOARANT, 1988), puede ser muy va-riable y verse afectado por otros factores.
Los enemigos naturales, especialmente losácaros fitoseidos, si están presentes desde elcomienzo colonizando las parcelas de maízjunto a los tetraníquidos, reducen los aumen-tos de fitófagos en el mes de Agosto. El mo-mento crítico ocurre al comienzo de la flora-ción masculina, ya que si los enemigos natu-rales no están establecidos en esa fecha losfitófagos aumentan sus poblaciones especta-cularmente. La especie principal de fitoseidoen maíz, en Navarra, es A. californicus (IRA-OLA et al., 1997). Su abundancia, adaptaciónal medio y el hecho de que colonice el maízsincrónicamente con T. turkestani, asegurasu presencia en los momentos adecuados.Este fitoseido ha sido utilizado en Francia enensayos de suelta, observándose que produceuna disminución en las poblaciones de fitó-fagos, aunque su efecto nunca es de acarici-da de choque sino que funciona como unacaricida permanente, previniendo los au-mentos de las poblaciones de tetraníquidos(GOARANT, 1988).
El clima es un factor también importante.Las temperaturas altas y las bajas humeda-des favorecen a los tetraníquidos en general(WRENSCH, 1985) y a ! , turkestani en parti-cular (CAREY y BRADLEY, 1982). Histórica-mente, periodos prolongados de temperatu-ras altas y bajas precipitaciones han hechoaumentar los daños de los ácaros a los culti-vos. Además, el clima hace que el control delos fitoseidos sea más o menos efectivo.BERRY et al. (1991) con modelos de simula-ción de control de tetraníquidos por fitosei-dos en maíz, afirman que este control se pro-duce en condiciones de temperatura media yhumedad relativa alta. Los graves ataques de
tetraníquidos a los maizales de Navarra, a fi-nales de los años 80, coincidieron con un pe-riodo de prolongada sequía y altas tempera-turas en verano, lo que ocasionó que el con-trol por los enemigos naturales disminuyera.
Un factor a considerar es la existencia o node vegetación de ribazo en la lindes del culti-vo y las prácticas agronómicas que se realicensobre ella. La existencia de plantas espontáne-as en el borde de la parcela favorece la colo-nización en fechas tempranas, tanto de ácarosfitófagos como de fauna útil. La quema de lavegetación de los bordes de cultivo tiene undoble efecto; reduce la colonización de los te-traníquidos al mismo tiempo que limita la delos fitoseidos. Se ha observado que un retrasoen la entrada de los fitoseidos, respecto a la delos tetraníquidos, provoca que los fitófagosalcancen poblaciones elevadas.
Existe un factor, no planteado en este estu-dio, que son los tratamientos químicos. Losnuevos regadíos, con sistemas de riego quepermiten la utilización de aspersores, haránaumentar el número de hectáreas de maíztratadas, especialmente contra taladros Sesa-mia nonagrioides (Lefevbre) y Ostrinia nu-bilalis (HÜBNER). Estos tratamientos provo-can aumentos explosivos en las poblacionesde ácaros fitófagos, tanto por actuar sobre lafisiología de la planta como por eliminar losenemigos naturales (ALVARADO et al, 1986;SCHWEISSING, 1973; BYNUM y ARCHER,
1992; AYYAPPATH et al, 1996). La mayoríade los trabajos se refieren a O. pratensis y T.urticae y aunque no disponemos de trabajosrelacionados con el efecto de estos fitosani-tarios sobre T. turkestani, todo hace suponerque los efectos sean los mismos.
Por lo tanto, una gestión adecuada del cul-tivo puede evitar problemas de acariosis.Salvo condiciones climáticas extremas (pro-longadas sequías, por ejemplo), los enemi-gos naturales van a mantener las poblacio-nes de ácaros fitófagos a unos niveles bajos.Facilitar la colonización de los depredadores(A. californicus, en este caso) al mismotiempo que los fitófagos, así como evitar lostratamientos químicos injustificados, puedegarantizar el control de los tetraníquidos.

AGRADECIMIENTOS
Al Dr. Fauvel del I.N.R.A. por su ayudaen la búsqueda bibliográfica.
A todos los propietarios de las fincas agrí-colas donde se realizaron los muéstreos.
A los componentes del departamento deZoología y Ecología de la Universidad deNavarra que participaron en los muéstreos.
ABSTRACT
IRAOLA, V. M.; MORAZA, M. L. y BIURRUN, R., 1998: Spider mites on corn in Na-varra. Ecology of Tetranychus turkestani UGAROV Y NlKOLSKI. Bol. San. Veg. Plagas,24(3): 609-620.
Ecological populations behavior of Tetranychus turkestani UGAROV Y NIKOLSKI,phytophagous spider mite, infesting corn in Navarra has been studied. This species,which can spend winter time as diapausic female, starts its activity on spontaneous ve-getation of the corn plots borders at the beginning of March (mean temperature 13 °C).From here on, and beginning with June, mites move to corn basal leaves of the plotsborder depending of weather and existence of slope vegetation. Beginning with thismoment, mites follow a double direction: upward colonizing corn plants and towardevery direction colonizing neighboring corn plants. Corn plants physiological state fallsthis colonization. During the masculine flowering state and previous week to this time,T. turkestani population increase is favored and total plant and total plot colonizationoccur. Spider mite populations are affected by: corn physiological state, clime (hightemperatures and relative low humidity favour the mite) and natural enemies (predace-ous phytoseiid mites over all) which control spider mite populations.
Key words: Corn, spider mites, Tetranychus turksetani, colonization dynamic, po-pulation dynamic, Navarra.
REFERENCIAS
ALVARADO, M.; ARANDA, E.; ALAMEDA, A. y DURAN,J. M., 1984: La araña roja del maíz en la vega delGuadalquivir. IV Simposium Nacional de Agroquí-micos. Sevilla, 18 pp.
ALVARADO, M.; DURAN, J. M.; ALAMEDA, A.; CABE-ZAS, J.; BARBAS, L.; ARANDA, E. y DE LA ROSA, A.,1986: Ácaros-taladros en el maíz de la vega del Gua-dalquivir. (Sevilla-Córdoba). IV Jornadas Técnicasdel Maíz- Lérida, 18 pp.
ÁLVAREZ, R.; BESNIER, F.; GÓMEZ, J. A.; GRANDE, J.A.; QUEIPO, J.; CHICO, S.; URQUIJO, A. y VADELL,M., 1964: Diez temas sobre el maíz. Servicio de Ex-tensión Agraria. Ministerio de Agricultura. Madrid,144 pp.
ARIAS, A. y ALVARADO, M., 1983: El cultivo del maízy su situación fitosanitaria en España. Evaluación depérdidas por «taladros». Ministerio de Agricultura,Pesca y Alimentación, Madrid, 25 pp.
AYYAPPATH, R.; WlTKOWSKI, J. F. y HlGLEY, L. G.,1996: Population changes of spider mites (Acari: Te-tranychidae) following insecticide applications incorn. Environ. Entomoi, 25(5): 933-937.
BARRON, J. A.; MARGOLIES, D. C , 1991: Within-plantdispersal of Banks grass mite (Acañ.Tetranychidae)on corn. J. Kansas Entomoi. Soc, 64(2): 209-215.
BARTOLINI, R., 1991: El maíz. Mundi-Prensa, 270 pp.BERRY, J. S.; HOLTZER, T. O. y NORMAN, J. M., 1991:
Experiments using a simulation model of the Banks
grass mite (Acari: Tetranychidae) and the predatorymite Neoseiulus fallacis (Acañ:Phytoseiidae) in acorn microenvironment. Environ. Entomoi., 20(4):1074-1078.
BOUFFARD, M. G., 1990: Acarien, quand tu nos tiens.Phytoma, 416: 14-16.
BRANDENBURG, R. L. y KENNEDY, G. G., 1982: Inter-crop relationships and spider mite dispersal in acorn-peanut agro-ecosystem. Entomoi. exp. appl.,32: 269-276.
BYNUM, E. D. y ARCHER, T. L., 1992: Banks grassmite (Acaú:Tetranychidae) response to selected in-secticides used for control of Southwestern cornborer. J. Agrie. Entomoi., 9(3): 189-198.
CAREY, J. R., BRADLEY, J. W., 1982: Developmentalrates, vital schedules, sex ratios, and life tables forTetranychus urticae, T. turkestani and T. pacificus(Acariña: Tetranychidae) on cotton. Acarologia, 23:333-345.
CHANDLER, L. D.; ARCHER, T. L.; WARD, C. R. yLYLE, W. M., 1979: Influences of irrigation practi-ces on spider mite densities on field corn. Environ.Entomoi., 8: 196-201.
DICK, G. L. y BUSCHMAN, L. L., 1995: Seasonal ocu-rrence of a fungal pathogen, Neozygites adjarica(Entomophtorales: Neozygitaceae), infecting BanksGrass Mites, Oligonychus pratensis, and twospottedspider mite, Tetranychus urticae (Acaú:Tetranychi-

dae), in field corn. J. Kansas Entomol. Soc, 68(4):425-436.
EHLER, L. E., 1974: A review of the spider-mite pro-blem on grain sorghum and corn in West Texas.Texas Agrie. Exp. Station Bulletin, 11149: 15 pp.
ESTEBAN DURAN, J., 1990: Enfermedades parasitariasy plagas del maíz. En: Vademécum del Maíz. Carlosde Liñan y Vicente (ed), Madrid: 254 pp.
FAUVEL, G.; NAIBO, B.; LE GOFF, CH. y TESSIER, P.,1987: Les acariens tétranyques (in mais). Phytoma;388: 23-27.
GiLSTRAP, F. E.; FRIESE, D. D.; SUMMY, K. R. yARMSTRONG, A. A., 1977: Persistence of Ambly-seius californicus, Phytoseiulus persimilis and Typh-lodromus occidentalis released in field populationsof Banks grass mite in West Texas. Southwest. Ento-mol. Suppl, 2: 159-163.
GILSTRAP, F. E.; SUMMY, K. R.; CHANDLER, L. D.;ARCHER, T. L. y WARD, C. R., 1980: Within plantdistribution of Banks grass mite on corn in WestTexas. Environ. Entomol, 9: 546-548.
GOARANT, G., 1987: Les acariens du mais. DocumentService Protection des Vegétaux, Languedoc-Rousi-llon, 34 pp.
GOARANT, G., 1988: Les acariens du mais. DocumentService Protection des Vegétaux, Languedoc-Rousi-llon, 10 pp.
GOBIERNO DE NAVARRA, 1996: Superficies y produc-ciones. Coyuntura agrara, 119: 15-25.
GROOT, J.; KROPFF, M.; VOSSEN, F.; SPITTERS, C. yRABBINGE, R., 1986: A decimal code for the deve-lopmental stages of maize and its relation to acumu-lated heat units. Neth. J. Agrie. Sci., 34: 67-73.
GUPTA, S. K., 1991: The mites of agricultural impor-tance in India with remarks on their economic status.En: «Modern Acarology», Eds Dusbábek andBukva. Prague, 1: 509-522.
HATZINIKOLIS, E., 1969: Preliminary notes on tetrany-choid and eriophyid mites infesting cultivated plantsin Greece. Proceedings. 2nd International. Congressof Acarology, Nottingham 1967 (ed AkademiaiKiado), 161-167.
HUFFAKER, C. B.; VAN DE VRIE, M. y MCMURTRY,J. A., 1969: The ecology of tetranychid mites andtheir natural control. Ann. Review Entomol., 14:125-174.
IRAOLA, V. M.; MORAZA, M. L.; BlURRUN, R. y FE-RRAGUT, F., 1997: Fitoseidos (Acari: Phytoseiidae)en maíz y en vegetación en ribazo en Navarra. Den-sidades y composición de especies. Bol. San. Veg.Plagas, 23: 209-220.
LOGAN, J. A.; CONGDON, B. D. y ALLDREDGE, J. K.,1983: Ecology and control of spider mites on corn inNortheastern Colorado. Col. St. Univ. exp. St., 585S.41 pp.
LÓPEZ BELLIDO, L., 1991: Cultivos herbáceos. Vol I.Cereales. Mundi-Prensa, Madrid: 514 pp
MACUÁ, J. L., 1990: Maíz grano. Navarra Agraria, 90:7-23.
MANSOUR, F.; BAR-ZUR, A. y ABO-MOCH, F., 1993:Resistance of maize inbred lines to the carmine spi-der mite Tetranychus cinnabarinus (Acari: Tetrany-chidae): evaluation of antibiosis of selected lines atdifferent growth stages. Maydica, 38: 309-311.
MARGOLIES, D. C. y KENNEDY, G. C , 1984: Popula-tion response of the twospotted spider mite, Tetrany-chus urticae, to host phenology in corn and peanut.Entomol. exp. appl., 36: 193-196.
MARGOLIES, D. C. y KENNEDY, G. C , 1985: Move-ment of the twospotted spider mite, Tetranychus ur-ticae, among hosts in a corn-peanut agroecosystem.Entomol. exp. appl., 37: 55-61.
MEYER, M. K. P. S., 1987: African Tetranychidae(AcarkProstigmata), with reference to the world ge-nera. Entomol. Mem. Dep. Agrie. South Africa, Pre-toria, 69: 175 pp.
NAIBO, B., 1989: Protection du mais contre les aca-riens Tetranychus urticae et T. turkestani. Colloquesur les acariens des cultures. Montpellier., 2,1/1:429-435.
NAIBO, B.; LE GOFF, C. y ALGANS, J. L., 1987: Nuisi-bilite des acariens du genre Tetranychus sur mais.Contribution à la mise au point d'une stratégie dulutte. Conference Internationale sur les ravageurs enagriculture, Paris: 255-262.
NAIBO, B.; ALGANS, J. L. y LE GOFF, C , 1988: Straté-gie de lutte contre les acariens du mais. La Defensedes Vegétaux, 249-250: 21-24.
PICKETT, C. H. y GILSTRAP, F. E., 1985: Dynamics ofspider mite species (Acariña: Tetranychidae) com-position infesting corn. /. Kansas Entomol. Soc, 58(3): 503-508.
PICKETT, C. H. y GILSTRAP, F. E., 1986a ¡Inoculativereleases of Phytoseiids (Acari) for the biologicalcontrol of spider mites (Acari: Tetranychidae) incorn. Environ. Entomol., 15: 790-794.
PICKETT, C. H. y GILSTRAP, F. E., 1986b: Dispersionpatterns and sampling of spider mites (Acari:Te-tranychidae) infesting corn in the Texas High Plains.Environ. Entomol., 15: 335-341.
PICKETT, C. H.; GILSTRAP, F. E.; MORRISON, R. K. yBOUSE, L. F., 1987: Release of predatory mites byaircraft for the biological control of spider mites in-festing corn. J. Econ. Entomol., 80: 906-910.
POPOV, S. YA., 1983: Weed hosts of the Turkestanispider mite. Zashchita Rastenii (Moscow), 3: 48.
POPOV, S. YA. y VEERMAN, A., 1996: Behavioural res-ponse and winter survival of mated and unmateddiapausing females of the Tetranychus atlanticus-ur-ticae complex (Acari: Tetranychidae). Exp. Appl.Acaroi, 20: 167-175.
RAGUSA, S. y PAOLETTI, M. G., 1985: Phytoseiid mites(Parasitiformes, Phytoseiidae) of corn and soybeanagroecosystems in the low-laying plain of Véneto(N-E Italy). Redia, LXVIII: 69-89.
SCHWEISSING, F. C , 1973: Approaches to the manage-ment of the Banks grass mite on corn. Col. St. Univ.Exp. St. Progress Report, 41: 3 pp
VEERMAN, A., 1985: Diapause. En: World Crop Pests.Their biology, natural enemies and control. W. Helle& M.W. Sabelis (eds.), 1A: 279-310.
WRENSCH, D. L., 1985: Reproductive parameters. En:World Crop Pests. Their biology, natural enemiesand control. W. Helle & M.W. Sabelis (eds.), 1A:165-168.
(Recepción: 20 abril 1998)(Aceptación: 16 junio 1998)


















