Absorción iónica celular - Genética16 · PDF fileseparación de...
15
FISIOLOGÍA VEGETAL: Tema 26.- Absorción iónica celular. Análisis cinético de la nutrición y compartimentación. Transporte Transcelular: Transporte pasivo, transporte facilitado, transporte activo. Factores que afectan al transporte iónico. Absorción y transporte de iones en la planta. Distribución y circulación de iones por las plantas. La endodermis. Pautas de distribución y circulación. Absorción foliar. 1 1 Absorción iónica celular A modo de introducción veamos el caso de los iones en la interfase suelo-raíz: Los aniones son ab sorbidos por las plantas de forma directa a partir de la disolución en el agua del suelo, mientras que los cationes están ad sorbidos por los puntos de carga negativa de las micelas de arcilla y la materia orgánica del suelo. Según la hipótesis del intercambio por contacto, los cationes pueden ser transferidos de los coloides arcillosos sin aparecer como iones libres en la disolución del suelo. Esto será el resultado de la oscilación de los iones adsorbidos en un reducido volumen de espacio. No todos los especialistas aceptan esta hipótesis. Otra posibilidad sería que los hidrogeniones provinieran del ácido carbónico originado en fase acuosa a partir de la respiración de la raíz. Los cationes liberados mediante este mecanismo difundirían a través de la disolución del suelo hasta la superficie radical, donde ocurriría la absorción. Es muy importante considerar el efecto de los potenciales eléctricos al estudiar la absorción de solutos: Los potenciales eléctricos pueden alterar profundamente el transporte iónico y provocar la absorción de iones contra gradiente de concentración. En las membranas se observan equilibrios dinámicos entre los potenciales eléctricos y los potenciales químicos. La mayoría de las membranas celulares vegetales aparecen cargadas eléctricamente (en los estudios con microelectrodos conectados a un voltímetro de alta impedancia –es necesaria una resistencia elevada en el circuito para evitar que el sistema membrana se descargue-). Los gradientes eléctricos están implicados en procesos de transporte iónico a través de la membrana y, a su vez, interaccionan con los flujos de agua que tienen lugar por ósmosis. Para un ión cualquiera el potencial eléctrico que puede originar este flujo pasivo se calcula mediante la ecuación de Nernst: Ce Ci zF RT E ln · =
Transcript of Absorción iónica celular - Genética16 · PDF fileseparación de...

FISIOLOGÍA VEGETAL: Tema 26.- Absorción iónica celular. Análisis cinético de
la nutrición y compartimentación. Transporte Transcelular: Transporte pasivo, transporte
facilitado, transporte activo. Factores que afectan al transporte iónico. Absorción y transporte de
iones en la planta. Distribución y circulación de iones por las plantas. La endodermis. Pautas de
distribución y circulación. Absorción foliar.
1
1
Absorción iónica celular A modo de introducción veamos el caso
de los iones en la interfase suelo-raíz: Los aniones son absorbidos por las
plantas de forma directa a partir de la disolución en el agua del suelo, mientras que los cationes están adsorbidos por los puntos de carga negativa de las micelas de arcilla y la materia orgánica del suelo. Según la hipótesis del intercambio por contacto, los cationes pueden ser transferidos de los coloides arcillosos sin aparecer como iones libres en la disolución del suelo. Esto será el resultado de la oscilación de los iones adsorbidos en un reducido volumen de espacio. No todos los especialistas aceptan esta hipótesis. Otra posibilidad sería que los hidrogeniones provinieran del ácido carbónico originado en fase acuosa a partir de la respiración de la raíz. Los cationes liberados mediante este mecanismo difundirían a través de la disolución del suelo hasta la superficie radical, donde ocurriría la absorción.
Es muy importante considerar el efecto de los potenciales eléctricos al estudiar la
absorción de solutos: Los potenciales eléctricos pueden alterar profundamente el transporte iónico y provocar la absorción de iones contra gradiente de concentración. En las membranas se observan equilibrios dinámicos entre los potenciales eléctricos y los potenciales químicos.
La mayoría de las membranas celulares vegetales aparecen cargadas eléctricamente
(en los estudios con microelectrodos conectados a un voltímetro de alta impedancia –es necesaria una resistencia elevada en el circuito para evitar que el sistema membrana se descargue-). Los gradientes eléctricos están implicados en procesos de transporte iónico a través de la membrana y, a su vez, interaccionan con los flujos de agua que tienen lugar por ósmosis.
Para un ión cualquiera el potencial eléctrico que puede originar este flujo pasivo se
calcula mediante la ecuación de Nernst:
Ce
Ci
zF
RTE ln·=

FISIOLOGÍA VEGETAL: Tema 26.- Absorción iónica celular. Análisis cinético de
la nutrición y compartimentación. Transporte Transcelular: Transporte pasivo, transporte
facilitado, transporte activo. Factores que afectan al transporte iónico. Absorción y transporte de
iones en la planta. Distribución y circulación de iones por las plantas. La endodermis. Pautas de
distribución y circulación. Absorción foliar.
2
2
• Donde E es el potencial eléctrico en mV
• z es la valencia con su signo
• F constante de Faraday: 96 485 C�mol-1
• R constante universal de los gases: 8,3143 J�mol-1� K-1
• Ce y Ci concentraciones externa e interna (mmol�L-1) Si el potencial eléctrico predicho para un determinado ión difiere
significativamente del valor observado mediante mediciones con electrodos, existen buenas razones para suponer que el ión está siendo transportado contra su gradiente electroquímico. Otro tipo de diferencia de potencial eléctrico que encontramos en los sistemas biológicos es la que aparece asociada con las cargas inmóviles o fijas de la fase sólida adyacente a una fase acuosa y que se denomina potencial Donnan. La región que contiene las partículas inmóviles se denomina fase de Donnan. Aparece este tipo de separación de fases en las paredes celulares que posee gran número de grupos carboxilo (—COO-) inmóviles asociados con pectina. También aparecen fases Donnan en el citoplasma, donde las cargas inmóviles se deben, fundamentalmente, a las proteínas.
Análisis cinético de la nutrición y compartimentación El movimiento de moléculas entre los diferentes compartimentos de los sistemas
vivos se conoce como transporte. El transporte en el ámbito celular condiciona todas las actividades de orden superior. Y, fundamentalmente, está controlado específicamente por las membranas plasmáticas.
Las fuerzas que dirigen el transporte biológico, como los gradientes de
concentración, los gradientes de potencial eléctrico y las presiones hidrostáticas se incluyen en el concepto global de potencial electroquímico. El transporte de solutos hacia potenciales electroquímicos más bajos (por ejemplo, por difusión) se denomina transporte pasivo. El movimiento de solutos contra gradiente de potencial electroquímico se denomina transporte activo y requiere gasto de energía.
El grado en que una membrana permite o restringe el movimiento de una sustancia
se denomina permeabilidad de la membrana. Las características de la membrana y las propiedades químicas del soluto son los dos factores fundamentales que determinan la permeabilidad de membrana. Cuando los aniones y cationes se mueven a diferentes velocidades a través de la membrana, el potencial eléctrico que aparece se denomina potencial de difusión.
Para cada ión, la relación entre la diferencia de voltaje de los dos lados de la
membrana y la distribución del ión en equilibrio se describe con la ecuación de Nernst, no obstante la suma de todos los potenciales de difusión de los diferentes iones se describe con la ecuación de Goldman:

FISIOLOGÍA VEGETAL: Tema 26.- Absorción iónica celular. Análisis cinético de
la nutrición y compartimentación. Transporte Transcelular: Transporte pasivo, transporte
facilitado, transporte activo. Factores que afectan al transporte iónico. Absorción y transporte de
iones en la planta. Distribución y circulación de iones por las plantas. La endodermis. Pautas de
distribución y circulación. Absorción foliar.
3
3
Tiene el aspecto de la ecuación de Nernst, pero con los sumatorios de todos los
iones (con sus cargas eléctricas) que entran y salen a través de la membrana. Todas estas condiciones de equilibrio pasivo son alteradas por los procesos de
transporte activo (bombas electrogénicas) que, además, varían el potencial de membrana desde el estado de reposo (o estado basal) a otro más elevado.
Las membranas tienen proteínas especializadas –canales, transportadoras y
bombas- que facilitan el transporte de solutos. Los canales son proteínas que forman poros de uno al otro lado de la membrana; a
través de estos poros los solutos difunden hacia potenciales químicos más bajos. Las proteínas transportadoras (carriers) enlazan un soluto en un lado de la
membrana y lo sueltan en el otro lado. En las plantas un grupo de ATPasas ligadas a bombas de protones suministran la
fuerza primaria para el transporte a través de la membrana. En el tonoplasto existen otros tipos de bombas electrogénicas.

FISIOLOGÍA VEGETAL: Tema 26.- Absorción iónica celular. Análisis cinético de
la nutrición y compartimentación. Transporte Transcelular: Transporte pasivo, transporte
facilitado, transporte activo. Factores que afectan al transporte iónico. Absorción y transporte de
iones en la planta. Distribución y circulación de iones por las plantas. La endodermis. Pautas de
distribución y circulación. Absorción foliar.
4
4
Cuando se comparan las permeabilidades de iones y moléculas en membranas bi-capa artificiales y en las biológicas aparecen semejanzas y diferencias: para moléculas no polares y moléculas polares pequeñas (agua, glicerol) los dos tipos de membrana tienen permeabilidades semejantes. Por el contrario, para iones y moléculas polares grandes (como los azúcares) las membranas biológicas presentan mucha mayor permeabilidad que las membranas artificiales. La diferencia radica en que las membranas biológicas contienen proteínas de membrana con función transportadora, que facilitan el paso de iones y de otras moléculas polares. Las moléculas que muestran permeabilidades semejantes con ambos tipos de membrana difunden, aparentemente, de forma directa a través de los lípidos de la membrana. Sin embargo, la mayoría de las sustancias que son importantes en la nutrición y en el metabolismo celular no pueden difundir directamente a través de los lípidos y deben ser transportados por proteínas de membrana.
Estas proteínas de transporte pueden agruparse en 3 grupos: proteínas canal,
proteínas transportadoras (carriers) y bombas. En general los Canales son proteínas transmembrana que funcionan como poros
selectivos. El tamaño del poro y la distribución de cargas en su interior determinan la especificidad del transporte. El movimiento de los iones a través del poro es muy rápido (del orden de 108 iones por segundo), pero los canales no permanecen abiertos indefinidamente, normalmente contienen compuertas (gates) que abren o cierran el poro en respuesta a señales. El transporte a través de canales es siempre pasivo y, debido a que la especificidad del transporte depende del tamaño del poro y su carga eléctrica, más que de una unión selectiva, el transporte por canales está normalmente limitado a los iones y al agua.
En el transporte mediado por una proteína transportadora (carrier) el soluto que se
transporta se une inicialmente a un sitio específico de la proteína. La unión origina un cambio conformacional en la proteína, que dirige la sustancia al otro lado de la membrana. El transporte se completa cuando el soluto se suelta del sitio específico. Debido al cambio conformacional de la proteína la velocidad de transporte en este caso es mucho más bajo que a través de los canales (100 a 1.000 iones por segundo, un millón de veces más lento que por un canal). La unión y liberación al sitio específico que ocurre en las proteínas transportadoras son similares a los que ocurren entre enzima y sustrato, de manera que se utilizan análisis cinéticos enzimáticos para describir los transportes debidos a proteínas transportadoras.
El transporte efectuado por proteínas transportadoras (carriers) es primariamente
pasivo (aunque si está acoplado a otro proceso de transporte con gasto energético –acoplado a una bomba- puede ser activo de manera secundaria) y pueden transportar un rango amplio de sustancias. El transporte pasivo mediado por una proteína transportadora se suele denominar difusión facilitada, aunque se asemeje a la difusión, únicamente, porque la sustancia se transporta a favor de gradiente de potencial electroquímico (de modo pasivo, sin un gasto adicional de energía).
FISIOLOGÍA VEGETAL: Tema 26.- Absorción iónica celular. Análisis cinético de
la nutrición y compartimentación. Transporte Transcelular: Transporte pasivo, transporte
facilitado, transporte activo. Factores que afectan al transporte iónico. Absorción y transporte de
iones en la planta. Distribución y circulación de iones por las plantas. La endodermis. Pautas de
distribución y circulación. Absorción foliar.
5
5
Transporte Activo Primario, donde se acopla una fuente de energía metabólica (como la hidrólisis de ATP, una reacción de óxido-reducción en las cadenas de transporte de mitocondrias y cloroplastos, o la absorción de luz por una proteína transportadora –en halobacterias, la bacteriorodopsina-) con el movimiento contra
gradiente de una sustancia. Las proteínas de membrana que llevan a cabo el transporte activo primario se denominan bombas. La mayoría de las bombas transportan iones, como el H+ o Ca2+. Las bombas pueden ser clasificadas como electrogénicas o electroneutras. El transporte electrogénico se refiere al transporte de iones que implican un movimiento neto de carga a través de la membrana. Por el contrario, el transporte electroneutro, como indica su nombre, no origina movimiento neto de carga. Por ejemplo, la ATPasa Na+/K+que bombea 3 iones Na+ por cada 2 de K+ es una bomba electrogénica.
En las membranas plasmáticas de plantas, hongos y bacterias, así como en los
tonoplastos de plantas y otras endomembranas de plantas y animales, el protón (H+) es el principal ión que es bombeado a través de las membranas. La ATPasa de protones (ATPasa -H+) crea un gradiente de potencial electroquímico de protones a través de las membranas, mientras que la ATPasa-H+Vacuolar (V-ATPasa) y la Pirofosfatasa- H+ (PPasa-H+) bombean electrogénicamente protones al lumen de la vacuola y a las cisternas del aparato de Golgi.
En membranas plasmáticas vegetales, sólo el H+ y el Ca2+ parecen ser movidos por
bombas y la dirección del bombeo es hacia el exterior de la célula, no hacia el interior. Por tanto, se necesitan otros mecanismos para explicar el transporte activo de la mayoría de los nutrientes minerales.
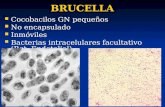
La forma más habitual es por acoplamiento de un transporte en contra gradiente,
con otro a favor de gradiente electroquímico. Este tipo de transporte acoplado mediado por proteínas transportadoras se denomina Transporte Activo Secundario y está movido indirectamente por bombas.
FISIOLOGÍA VEGETAL: Tema 26.- Absorción iónica celular. Análisis cinético de
la nutrición y compartimentación. Transporte Transcelular: Transporte pasivo, transporte
facilitado, transporte activo. Factores que afectan al transporte iónico. Absorción y transporte de
iones en la planta. Distribución y circulación de iones por las plantas. La endodermis. Pautas de
distribución y circulación. Absorción foliar.
6
6
Cuando los protones son expulsados fuera de un compartimento celular por ATPasas electrogénicas, tanto en la membrana plasmática, como en la membrana de la vacuola (tonoplasto), se genera un potencial de membrana y de pH a expensas de la hidrólisis de ATP. Este gradiente de potencial electroquímico de protones, denominado fuerza protón-motriz (proton motive force, PMF) representa energía libre almacenada y se acopla con un transporte de muchas otras sustancias contra gradiente de potencial. Las proteínas de transporte pueden ser simportadoras o antiportadoras. El sodio es transportado fuera de la célula por una proteína antiportadora Na+-H+; pero el Cl-, NO3-
H2PO4-, sacarosa, aminoácidos y otras sustancias entran a la célula con proteínas de membrana simportadoras.
Hasta ahora se ha descrito el transporte en términos energéticos, sin embargo, el
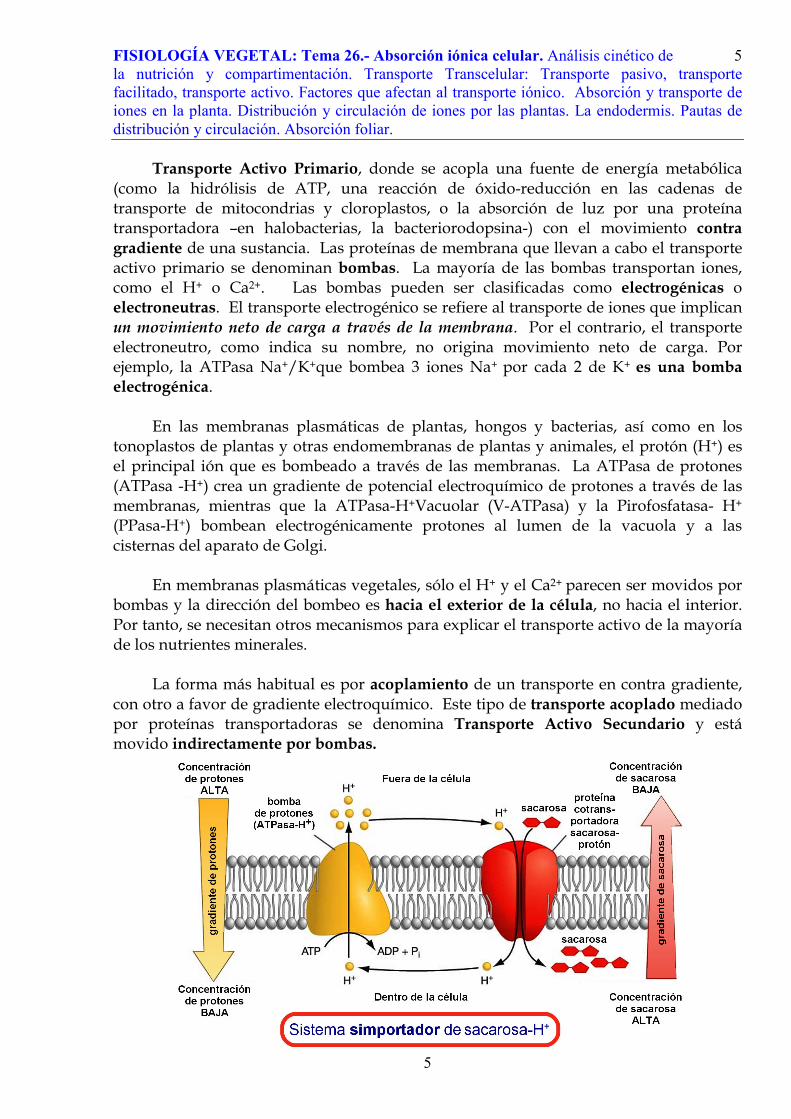
transporte celular puede estudiarse también mediante el análisis cinético de las proteínas transportadoras (carriers). En un transporte por difusión la entrada de un soluto es proporcional al gradiente de concentraciones a ambos lados de la membrana. Sin embargo, en el transporte mediado por proteínas transportadoras se tiende a alcanzar una velocidad máxima, (Vmax), que no puede ser superada, independientemente de la concentración de sustrato. La velocidad máxima se alcanza cuando los sitios de todas las proteínas transportadoras están ocupados y se vuelve limitante la cantidad de proteínas transportadoras, más que la concentración de sustrato. Las cinéticas de transporte son en este caso micaelianas (cinética de Michaelis-Menten) y muestran una Km (concentración de sustrato a la que se alcanza la mitad de Vmax) que refleja las propiedades (afinidad por el sustrato) del sitio de enlace de la proteína transportadora.
Usualmente, el transporte suele mostrar
propiedades tanto de movimiento mediado, como no mediado, por proteínas transportadoras. Por ejemplo, en la toma de sacarosa por protoplastos de cotiledones de soja, la toma aumenta rápidamente hasta saturarse alrededor de 10 mM de sacarosa. Por encima de 10 mM, la toma se vuelve lineal y no saturable. La inhibición de síntesis de ATP con venenos metabólicos bloquea el componente saturable, pero no el lineal. Esto se interpreta como que la captación de sacarosa externa a bajas concentraciones es un proceso de transporte activo secundario, que está mediado por proteínas simportadoras (sacarosa-H+). A concentraciones superiores, la sacarosa penetra en las células por difusión a favor de gradiente y éste no es un proceso sensible a los venenos metabólicos.
FISIOLOGÍA VEGETAL: Tema 26.- Absorción iónica celular. Análisis cinético de
la nutrición y compartimentación. Transporte Transcelular: Transporte pasivo, transporte
facilitado, transporte activo. Factores que afectan al transporte iónico. Absorción y transporte de
iones en la planta. Distribución y circulación de iones por las plantas. La endodermis. Pautas de
distribución y circulación. Absorción foliar.
7
7
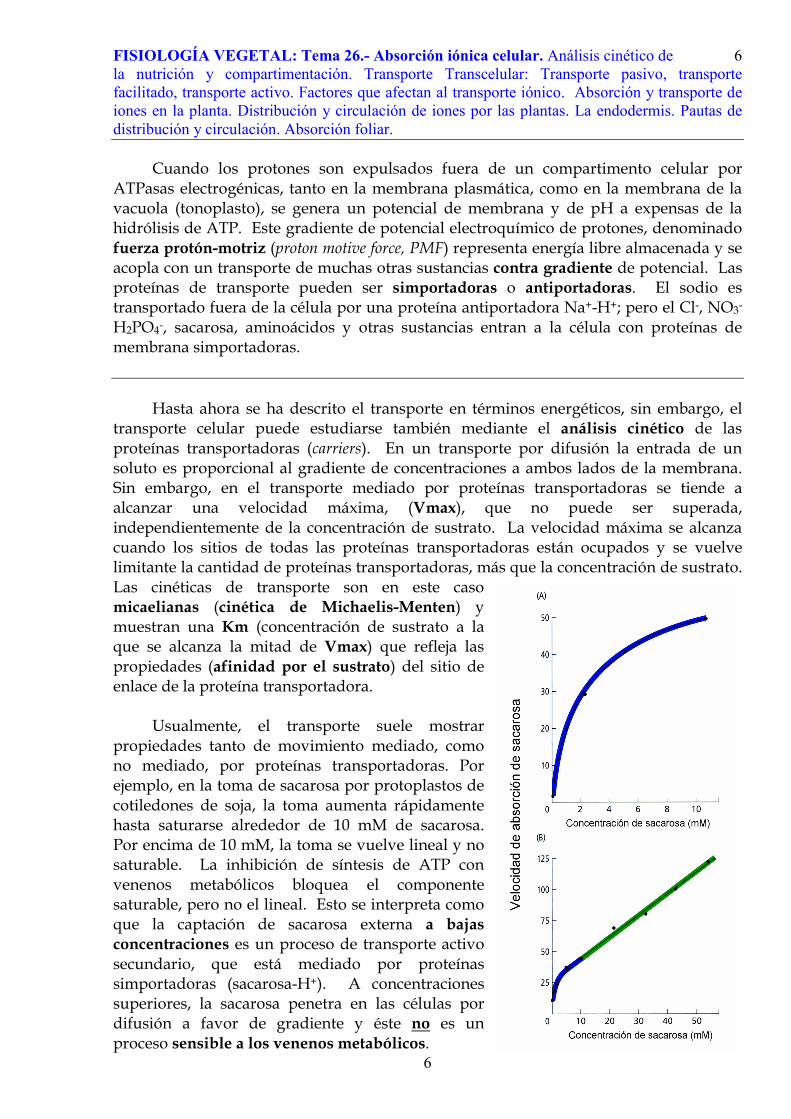
En el caso del potasio se observan curvas de saturación diferentes a bajas concentraciones (alta afinidad, se trataría de una proteína simportadora K+ - H+) y a altas concentraciones (baja afinidad, se trataría de canales específicos para el potasio).
Nos puede sorprender que el transporte pasivo de cationes potasio por canales
muestre, también, como se puede observar en la gráfica, una saturación a concentraciones muy elevadas de potasio externo. Sin embargo, es un resultado lógico: el transporte por canales puede llegar a hacerse insuficiente y a “saturarse”, debido a que el número de canales en la membrana no es infinito, como no lo es su capacidad de transporte. En este caso de concentraciones externas de potasio experimentales –muy alejadas del rango de concentraciones fisiológicas normales- se llega a producir la saturación del transporte a través de los canales.
En algunos casos se han descritos transportadores en serie en la membrana
plasmática y en el tonoplasto. En estos casos es interesante destacar que se observan Km y Vmax bajas para el situado en el plasmalema y Km y Vmax altas en el sistema de transporte situado en el tonoplasto. Esto tiene sentido, porque el transportador del plasmalema trabaja a bajas concentraciones de soluto y requiere alta afinidad (Km baja), mientras que el del tonoplasto trabaja a concentraciones mayores y su velocidad de transporte es mayor, pero requiere menos afinidad (Km alta).
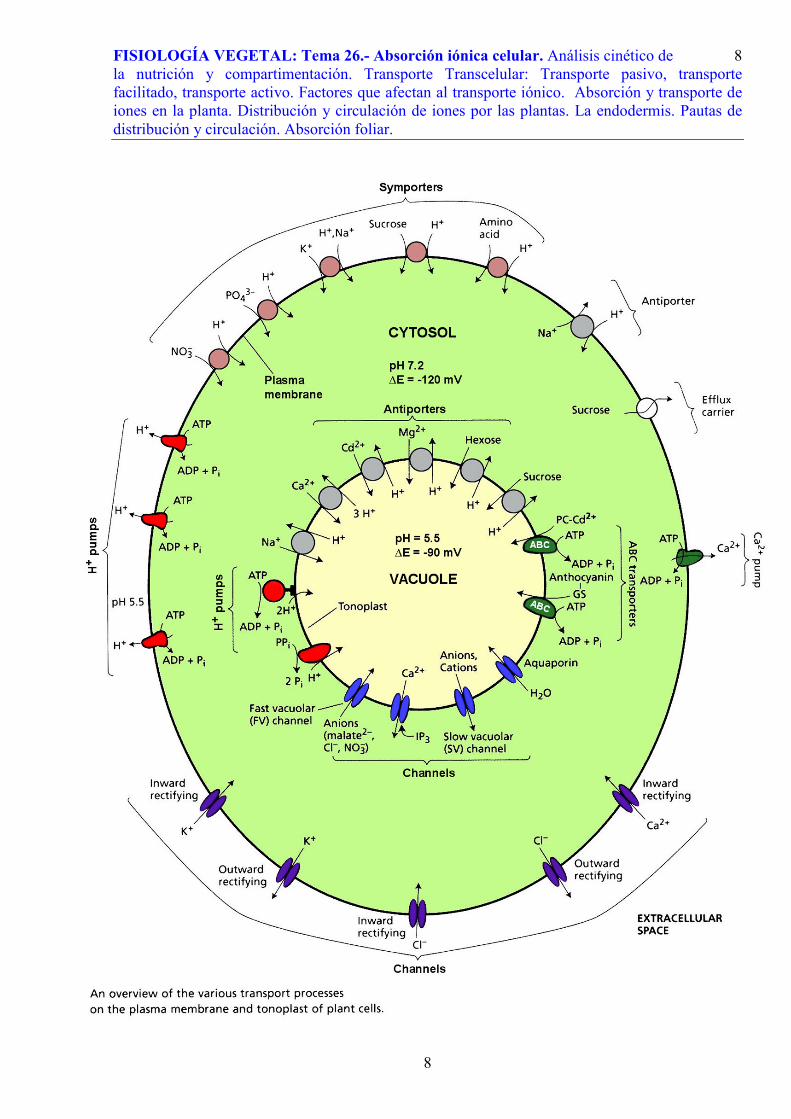
En la página siguiente se presenta un modelo de los tipos de transporte más
generalizados y estudiados en las células vegetales, entre el medio externo y los dos compartimentos celulares (el citoplasma y la vacuola).
FISIOLOGÍA VEGETAL: Tema 26.- Absorción iónica celular. Análisis cinético de
la nutrición y compartimentación. Transporte Transcelular: Transporte pasivo, transporte
facilitado, transporte activo. Factores que afectan al transporte iónico. Absorción y transporte de
iones en la planta. Distribución y circulación de iones por las plantas. La endodermis. Pautas de
distribución y circulación. Absorción foliar.
8
8

FISIOLOGÍA VEGETAL: Tema 26.- Absorción iónica celular. Análisis cinético de
la nutrición y compartimentación. Transporte Transcelular: Transporte pasivo, transporte
facilitado, transporte activo. Factores que afectan al transporte iónico. Absorción y transporte de
iones en la planta. Distribución y circulación de iones por las plantas. La endodermis. Pautas de
distribución y circulación. Absorción foliar.
9
9
Factores que afectan al transporte iónico Los procesos que más se ven afectados son los de transporte activo (tanto primario,
como secundario), fundamentalmente por su dependencia de la energía disponible por la célula. Es muy importante el efecto de la hipoxia en situaciones de encharcamiento de la raíz, por ejemplo. Los tejidos de la raíz están adaptados y suelen tolerar estas condiciones, en las que el nivel de oxígeno es bajo, durante periodos largos, pero pasan de un metabolismo aerobio (que permite producir y disponer de mucha energía) a otro fermentativo (anaerobio), donde la energía obtenida por las células y tejidos radicales suele ser muy inferior.
Obviamente, cualquier otro proceso que reduzca la actividad de las ATP-asas de
membrana, como los venenos metabólicos u otros factores que limiten la obtención de energía en mitocondrias o cloroplastos, condiciona los procesos de transporte activo. Por ello, son muy relevantes todas las variables ambientales adversas (como baja iluminación, temperaturas muy bajas o altas y el estado hídrico de la planta).
Hay que recordar que las proteínas transportadoras tienen un coeficiente cercano a
2 en su dependencia de la temperatura (VT+10/VT ≈ 2; dentro, por supuesto, del rango fisiológico) porque las ATP-asas son enzimas y las proteínas transportadoras realizan procesos de transporte semejantes a los procesos enzimáticos.
Los procesos de sequía afectan tanto a nivel de transporte transcelular (a “corta
distancia”) por sus efectos sobre la energía disponible y su efecto negativo en la configuración de las membranas, como a los de “larga distancia” (a través de los conductos de xilema y floema). En los procesos de sequía los estomas se cierran y la corriente transpiratoria prácticamente se anula, con lo que se paraliza todo el sistema circulatorio del vegetal: afectando tanto al xilema como al floema.
Absorción y transporte de iones en la planta Los nutrientes minerales absorbidos
por la raíz se llevan hasta la parte aérea por la corriente transpiratoria a través del xilema.
En las raíces el proceso de captación es
altamente específico y bien regulado. Así mismo, lo es el movimiento posterior de iones desde la superficie de la raíz por el córtex y al interior del xilema. El transporte a través de la raíz obedece a las mismas leyes biofísicas que gobiernan el transporte

FISIOLOGÍA VEGETAL: Tema 26.- Absorción iónica celular. Análisis cinético de
la nutrición y compartimentación. Transporte Transcelular: Transporte pasivo, transporte
facilitado, transporte activo. Factores que afectan al transporte iónico. Absorción y transporte de
iones en la planta. Distribución y circulación de iones por las plantas. La endodermis. Pautas de
distribución y circulación. Absorción foliar.
10
10
transcelular. Sin embargo, como hemos visto en el tema 24, al hablar de la absorción de agua por la raíz, la anatomía de la raíz impone algunas restricciones en el camino del movimiento de iones desde la superficie de la raíz a los elementos conductores del xilema.
Desde la superficie de la raíz a los elementos conductores del xilema Como ya sabemos, el espacio intercelular y las paredes celulares ofrecen poca
resistencia al movimiento de agua y solutos. Por eso, se denomina espacio libre o apoplasto. Se puede determinar el volumen apoplástico de un fragmento de planta comparando la toma de agua marcada con tritio (3H) y la de manitol marcado con carbono 14 (14C). El manitol es un azúcar-alcohol que no atraviesa las membranas celulares, pero se distribuye uniformemente por el espacio libre. Por otra parte, el agua tritiada penetra las membranas celulares y también se distribuye plenamente por el espacio libre. Las medidas de este tipo demuestran que del 5 al 20% del volumen de un tejido está ocupado por las paredes celulares y zonas intercelulares (espacio libre).
Del mismo modo que las paredes celulares forman un espacio continuo, también los
hacen los citoplasmas de las células próximas, conectados por plasmodesmos: lo que constituye el simplasto. Las vacuolas no forman parte del simplasto: la membrana vacuolar, el tonoplasto, supone una limitación al movimiento de agua y solutos (en cualquier caso la vacuola es un compartimento en paralelo al flujo simplástico).
Los plasmodesmos
son conexiones citoplasmáticas de 20 a 60 nm de diámetro. Cada plasmodesmo está envuelto en membrana plasmática y contiene un túbulo estrecho (el desmotúbulo) que es una continuación del retículo endoplásmico. En los tejidos en los que existe un transporte intercelular intenso se observan también muchos plasmodesmos (hasta 15
por µm2 de superficie celular). Las células secretoras especializadas, como los nectarios florales y las glándulas secretoras

FISIOLOGÍA VEGETAL: Tema 26.- Absorción iónica celular. Análisis cinético de
la nutrición y compartimentación. Transporte Transcelular: Transporte pasivo, transporte
facilitado, transporte activo. Factores que afectan al transporte iónico. Absorción y transporte de
iones en la planta. Distribución y circulación de iones por las plantas. La endodermis. Pautas de
distribución y circulación. Absorción foliar.
11
11
de sal, tienen un gran número de plasmodesmos y, así mismo, las células en el ápice de la raíz, donde tiene lugar la mayor parte de la absorción de nutrientes.
Mediante experimentos en los que se inyectan colorantes o se miden resistencias
eléctricas en células con muchos plasmodesmos, los investigadores han demostrado que los iones y los solutos pequeños pueden moverse de célula a célula por estos poros. El agua también se mueve por los plasmodesmos. Sin embargo, las moléculas grandes, como las proteínas, no se mueven fácilmente, ya que cada plasmodesmo está ocluido parcialmente por el desmotúbulo y sus proteínas asociadas. En general se considera que para el agua y los iones los plasmodesmos actúan como conexiones que forman un simplasto continuo.
La absorción de iones por las raíces es más elevado en la zona pilífera que en la
zona apical o en la zona de elongación. Las células de la zona pilífera han completado su crecimiento primario, pero no así el secundario. Los pelos radicales son simplemente extensiones de células epidérmicas especiales que aumentan de forma enorme el área de absorción de la raíz.
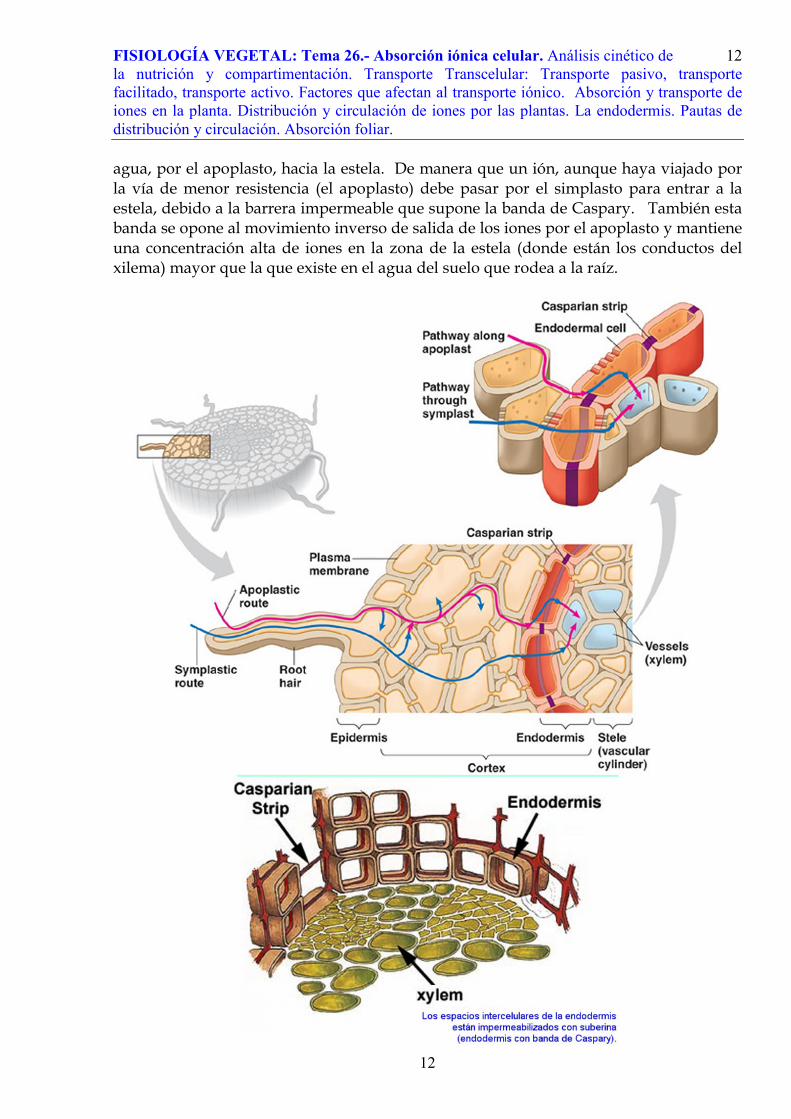
El apoplasto forma una fase continua desde la superficie de la raíz al córtex. En el
límite entre el cilindro vascular (o estela) y el córtex existe una capa de células especializadas: la endodermis. Aquí la capa de células suberizadas (banda de Caspary, descrita por el botánico alemán Johann Xaver Robert Caspary) bloquea la entrada de
FISIOLOGÍA VEGETAL: Tema 26.- Absorción iónica celular. Análisis cinético de
la nutrición y compartimentación. Transporte Transcelular: Transporte pasivo, transporte
facilitado, transporte activo. Factores que afectan al transporte iónico. Absorción y transporte de
iones en la planta. Distribución y circulación de iones por las plantas. La endodermis. Pautas de
distribución y circulación. Absorción foliar.
12
12
agua, por el apoplasto, hacia la estela. De manera que un ión, aunque haya viajado por la vía de menor resistencia (el apoplasto) debe pasar por el simplasto para entrar a la estela, debido a la barrera impermeable que supone la banda de Caspary. También esta banda se opone al movimiento inverso de salida de los iones por el apoplasto y mantiene una concentración alta de iones en la zona de la estela (donde están los conductos del xilema) mayor que la que existe en el agua del suelo que rodea a la raíz.
FISIOLOGÍA VEGETAL: Tema 26.- Absorción iónica celular. Análisis cinético de
la nutrición y compartimentación. Transporte Transcelular: Transporte pasivo, transporte
facilitado, transporte activo. Factores que afectan al transporte iónico. Absorción y transporte de
iones en la planta. Distribución y circulación de iones por las plantas. La endodermis. Pautas de
distribución y circulación. Absorción foliar.
13
13
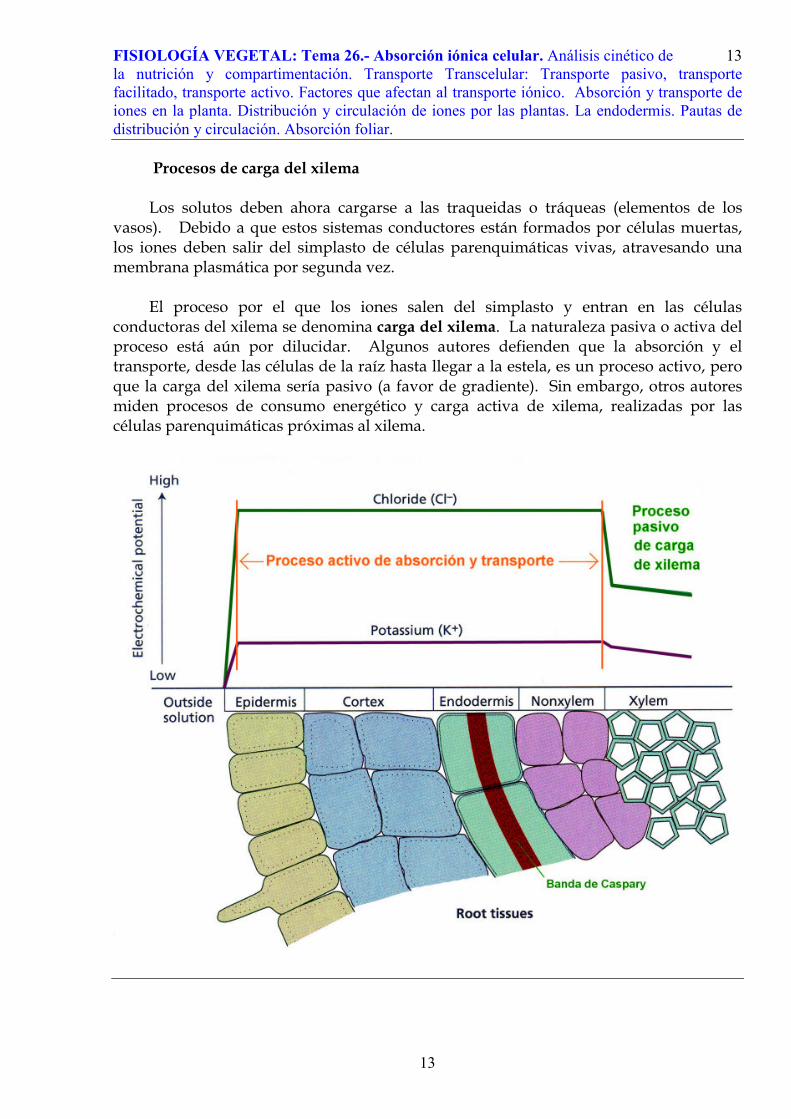
Procesos de carga del xilema Los solutos deben ahora cargarse a las traqueidas o tráqueas (elementos de los
vasos). Debido a que estos sistemas conductores están formados por células muertas, los iones deben salir del simplasto de células parenquimáticas vivas, atravesando una membrana plasmática por segunda vez.
El proceso por el que los iones salen del simplasto y entran en las células
conductoras del xilema se denomina carga del xilema. La naturaleza pasiva o activa del proceso está aún por dilucidar. Algunos autores defienden que la absorción y el transporte, desde las células de la raíz hasta llegar a la estela, es un proceso activo, pero que la carga del xilema sería pasivo (a favor de gradiente). Sin embargo, otros autores miden procesos de consumo energético y carga activa de xilema, realizadas por las células parenquimáticas próximas al xilema.

FISIOLOGÍA VEGETAL: Tema 26.- Absorción iónica celular. Análisis cinético de
la nutrición y compartimentación. Transporte Transcelular: Transporte pasivo, transporte
facilitado, transporte activo. Factores que afectan al transporte iónico. Absorción y transporte de
iones en la planta. Distribución y circulación de iones por las plantas. La endodermis. Pautas de
distribución y circulación. Absorción foliar.
14
14
Absorción foliar La vida vegetal comenzó en los mares. En las plantas primitivas como las algas la
absorción se hace por todo el talo (los rizoides cumplen, esencialmente, una función de anclaje, pero no son órganos específicos de absorción, a diferencia de las raíces en las plantas superiores). Esta capacidad generalizada de absorción persiste aún en la parte aérea de muchas plantas superiores, siempre que no se trate de zonas muy suberizadas o con cutículas céreas muy gruesas.
Se tiene noticia que un suministro foliar de nutrientes comenzó a ser utilizado en Europa y en Brasil en el siglo XIX: se usaron sales de hierro, como también de nitrógeno, fósforo y potasio. A finales del mismo siglo, el agrónomo F.W. Dafert hizo ensayos con plantas de café, aplicando, nitrógeno, fósforo y potasio, a las hojas.
La absorción foliar se realiza en tres pasos, después de disponer los nutrientes en disolución sobre las hojas:
• (1) penetran la cutícula y las paredes epidérmicas por difusión (se necesitan agentes surfactantes –aquí actúan como humectantes- para favorecer este paso).
• (2) son absorbidas por el plasmalema a través de plasmodesmos especiales orientados hacia la superficie: ectodesmos (ectodesmata) y entran al citoplasma.
• (3) pasan a la vía simplástica y pueden distribuirse por el floema.
Nota: el empleo de surfactantes ≈ humectantes (wetting agents) es necesario cuando se aplica un fertilizante o un producto fitosanitario, disueltos en agua, en nebulización sobre la superficie de la parte aérea de las plantas (que está recubierta por una capa cérea impermeable, hidrófoba). Los surfactantes son sustancias de tipo detergente (son moléculas anfifílicas, con una parte polar y otra apolar): disminuyen la tensión superficial del agua y hacen posible que ésta se adhiera (y que las sustancias disueltas puedan luego penetrar) a una superficie hidrófoba, inicialmente repelente a sustancias polares, como el agua.

FISIOLOGÍA VEGETAL: Tema 26.- Absorción iónica celular. Análisis cinético de
la nutrición y compartimentación. Transporte Transcelular: Transporte pasivo, transporte
facilitado, transporte activo. Factores que afectan al transporte iónico. Absorción y transporte de
iones en la planta. Distribución y circulación de iones por las plantas. La endodermis. Pautas de
distribución y circulación. Absorción foliar.
15
15
El interés de la fertilización foliar está relacionado con los siguientes aspectos:
• puede ser empleada con carácter complementario al abonado del suelo. • es la manera más rápida de corregir deficiencias, particularmente de
microelementos, como hierro, zinc, cobre, boro, manganeso y algunos macroelementos, como magnesio o calcio.
• a veces es la única forma factible de aportar microelementos metálicos a las plantas en suelos alcalinos o cálcicos, donde estos elementos se hallan inmovilizados.
La toma de nutrientes por las hojas tiene su contrapartida en la pérdida por lavado o lixiviación foliar, que se favorece en condiciones ambientales de clima muy húmedo persistente (por lluvia, nieve, rocío o niebla). En este proceso la planta pierde nutrientes minerales y sustancias orgánicas como azúcares o aminoácidos.
También se pueden excretar inhibidores fenólicos y compuestos que originan, como
veremos al estudiar el Metabolismo Secundario, efectos alelopáticos (sustancias excretadas inhibidoras o dañinas para otras especies de plantas vecinas: es la “guerra química Planta-Planta”).
Muchas plantas adaptadas a los bosques tropicales lluviosos tienen cutículas céreas
muy gruesas e impermeables para disminuir las pérdidas por lixiviación.