
2.10.1.2 LEPIDÓPTEROS DIURNOS DIVERSIDAD BIOLOGICA · riqueza y diversidad biótica están...
15
571 2.10.1.2 LEPIDÓPTEROS DIURNOS DIVERSIDAD BIOLOGICA Se define como la variabilidad entre los organismos vivientes de todas las fuentes, incluyendo, entre otros, los organismos terrestres, marinos y de otros ecosistemas acuáticos, así como los complejos ecológicos de los que forman parte; esto incluye diversidad dentro de las especies, entre especies y de ecosistemas. El término comprende, por tanto, diferentes escalas biológicas, desde la variabilidad en el contenido genético de los individuos y las poblaciones, el conjunto de especies que integran grupos funcionales y comunidades completas, hasta el conjunto de comunidades de un paisaje o región. (Moreno 2001). Recientemente para los estudios de biodiversidad se ha propuesto tener en cuenta las relaciones con la latitud, clima, productividad biológica, heterogeneidad y complejidad de los hábitat’s, así como el efecto de los disturbios, el tamaño y la distancia de los fragmentos o islas (Ricklefs, R. E, 1993.) Los patrones de la riqueza y diversidad biótica están influenciados por los gradientes latitudinales, a menor latitud hacia la línea ecuatorial el número de especies aumenta, mientras que hacia los polos (mayor latitud) disminuye. El gradiente de altitud en los ecosistemas terrestres la diversidad de especies generalmente disminuye con la altura; en las cordilleras de Colombia este fenómeno ha sido documentado ampliamente (Cuatrecasa, 1934); El clima influye en la riqueza de especies; entre los factores climáticos, la precipitación quizás sea el factor que mejor se relaciona con los valores de diversidad. Colombia con menos del 1 % de la superficie continental del planeta, alberga en su territorio cerca del 14 % de las especies conocidas. Esta riqueza es el resultado de la ubicación del país como puente entre los continentes americanos, su historia geológica y la complejidad de su ecosistema. En términos biológicos ocupa el segundo lugar a nivel mundial, solo superado por Brasil, un país siete veces su área. La variabilidad de ecosistemas en el territorio colombiano comprende desde páramos y laderas andinos hasta selvas tropicales húmedas, llanuras y desiertos, esta variedad de ecosistemas intensifica la riqueza biológica, la cual se calcula en 10 % del total de especies del planeta (Vélez y Salazar, 1991). Esta diversidad se encuentra seriamente amenazada principalmente por la destrucción y fragmentación de sus bosques (Andrade, 1998). La enorme riqueza biológica de Colombia contrasta de manera evidente con la falta de recursos económicos para, al menos, conocer lo que tenemos y definir cuales áreas deben mantenerse al margen de los procesos de colonización (Fagua et al, 1999). Esta situación obliga a utilizar especies o grupos taxonómicos capaces de reflejar el estado de la biota, su diversidad, endemismo o grado de intervención, el cual es el principio fundamental de la utilización de bioindicadores (Kremen, 1994), especies o
Transcript of 2.10.1.2 LEPIDÓPTEROS DIURNOS DIVERSIDAD BIOLOGICA · riqueza y diversidad biótica están...

571
2.10.1.2 LEPIDÓPTEROS DIURNOS DIVERSIDAD BIOLOGICA Se define como la variabilidad entre los organismos vivientes de todas las fuentes, incluyendo, entre otros, los organismos terrestres, marinos y de otros ecosistemas acuáticos, así como los complejos ecológicos de los que forman parte; esto incluye diversidad dentro de las especies, entre especies y de ecosistemas. El término comprende, por tanto, diferentes escalas biológicas, desde la variabilidad en el contenido genético de los individuos y las poblaciones, el conjunto de especies que integran grupos funcionales y comunidades completas, hasta el conjunto de comunidades de un paisaje o región. (Moreno 2001). Recientemente para los estudios de biodiversidad se ha propuesto tener en cuenta las relaciones con la latitud, clima, productividad biológica, heterogeneidad y complejidad de los hábitat’s, así como el efecto de los disturbios, el tamaño y la distancia de los fragmentos o islas (Ricklefs, R. E, 1993.) Los patrones de la riqueza y diversidad biótica están influenciados por los gradientes latitudinales, a menor latitud hacia la línea ecuatorial el número de especies aumenta, mientras que hacia los polos (mayor latitud) disminuye. El gradiente de altitud en los ecosistemas terrestres la diversidad de especies generalmente disminuye con la altura; en las cordilleras de Colombia este fenómeno ha sido documentado ampliamente (Cuatrecasa, 1934); El clima influye en la riqueza de especies; entre los factores climáticos, la precipitación quizás sea el factor que mejor se relaciona con los valores de diversidad. Colombia con menos del 1 % de la superficie continental del planeta, alberga en su territorio cerca del 14 % de las especies conocidas. Esta riqueza es el resultado de la ubicación del país como puente entre los continentes americanos, su historia geológica y la complejidad de su ecosistema. En términos biológicos ocupa el segundo lugar a nivel mundial, solo superado por Brasil, un país siete veces su área. La variabilidad de ecosistemas en el territorio colombiano comprende desde páramos y laderas andinos hasta selvas tropicales húmedas, llanuras y desiertos, esta variedad de ecosistemas intensifica la riqueza biológica, la cual se calcula en 10 % del total de especies del planeta (Vélez y Salazar, 1991). Esta diversidad se encuentra seriamente amenazada principalmente por la destrucción y fragmentación de sus bosques (Andrade, 1998). La enorme riqueza biológica de Colombia contrasta de manera evidente con la falta de recursos económicos para, al menos, conocer lo que tenemos y definir cuales áreas deben mantenerse al margen de los procesos de colonización (Fagua et al, 1999). Esta situación obliga a utilizar especies o grupos taxonómicos capaces de reflejar el estado de la biota, su diversidad, endemismo o grado de intervención, el cual es el principio fundamental de la utilización de bioindicadores (Kremen, 1994), especies o

572
grupos taxonómicos que pueden reflejar el estado de una biota y su relación con otras áreas geográficas. Lo anterior implica que deben ser taxones o grupos de especies abundantes, estables y preferiblemente sedentarias dentro de un ecosistema, que sean fácilmente identificables, tengan una biología y taxonomía bien conocida. Su manipulación en campo y laboratorio debe ser sencilla, generalmente presentar ciclos de vida cortos y tener sensibilidad y fidelidad ecológica (Amat, 1993). Las mariposas cumplen en buena medida estos requerimientos y son ubicados como el mejor grupo bioindicador. Debido a que son organismos muy sensibles a los cambios de temperatura, humedad y radiación solar que se generan por disturbios en su hábitat, por lo cual el inventario de sus comunidades con medida de la diversidad y riqueza, constituye una herramienta validad para evaluar el estado de conservación y/o alteración del medio natural (Fagua et al, 1999). Además como polinizadores son un grupo muy importante para la reproducción de las plantas vasculares.
TAXONOMIA Las mariposas (Lepidóptero) son consideradas como el segundo orden de la clase Insecta más abundante y altamente diversificada en los ecosistemas terrestres; el término mariposas diurna se ha utilizado extensamente para designar aquellas especies que vuelan de día y se caracterizan por tener las antenas terminadas en una clava. Las llamadas polillas o mariposas nocturnas son las especies que vuelan en su mayoría de noche y se caracterizan por tener las antenas de forma muy variada. Se calcula que existen unas 255000 especies a nivel mundial de las cuales 20000 pertenecen a las mariposas diurnas (Constantino, 1997). Las mariposas diurnas se encuentran distribuidas en dos superfamilias, Hesperioidea y Papilionoidea. La primera comprende una sola familia Hesperiidae, con cinco subfamilias en el Neotropico (Pyrrhopyginae, Pyrginae, Heteropterinae, Hesperiinae y Magathyminae). La segunda contiene cinco familias Nymphalidae, Pieridae, Lycaenidae, Riodinidae y Papilionidae. De las cuales Nymphalidae presenta el mayor número de subfamilias en el neotrópico, 11 en total (Nymphalinae, Biblidinae, Ithomiinae, Danainae, Heliconinae, Morphinae, Charaxinae, Satyrinae, Limenitidinae, Lybitheinae y Apaturinae). En cuanto a Pieridae en Colombia, cuenta con tres subfamilias (Coliadinae, Pierinae y Dismorphiinae); al igual que Lycaenidae con (Lycaeninae, Theclinae y Polymmatinae); Riodinidae con Riodininae y Euselasiinae y Papilionidae con una sola subfamilia en el territorio colombiano (Papilioninae) (Lamas, 2004).

573
• Hesperiidae. Son mariposas de tamaño muy variable, algunas son muy pequeñas y otras de mediano tamaño, su cuerpo es fuerte y robusto, cubierto de abundante vellosidad; sus tres pares de patas se encuentran bien desarrolladas y las antenas presentan la parte terminal curvada y delgada después del mazo (en forma de gancho (De la Maza 1987). Su coloración es muy poco llamativa predominando colores oscuros como el café y negro, sin embargo existen algunos con colores brillantes como el azul y verde metálico. Las principales plantas hospederas de las larvas pertenecen a las familias Fabaceae, Heliconiaceae, Maranthaceae y Poaceae, siendo el chusque (Chusquea scandens.), una fuente importante de alimento para muchas especies altoandinas. Algunas especies son plagas de importancia económica, como Urbanus proteus en cultivos de fríjol (García – Robledo et al, 2002) • Nymphalidae. Contiene unas 7250 especies haciéndola la familia mas diversa representando el 42% de las especies neotropicales. A nivel taxonómico el estudio de esta familia esta aún incompleto, la principal característica es que presentan el primer par de patas reducidas y cubiertas de vellosidades, y solo dos pares son aptas para la locomoción (Garcia – Robledo et al 2002). Se alimentan de variados tipos de plantas, como Crassuleaceas, Euforbiaceas, Urticaceas, Moraceas, y Ulmiaceas, entre otras. Los adultos prefieren zonas abiertas donde abundan flores y humedades (Siproeta, Biblis, Chlosyne, Marpesia, etc), otras prefieren zonas boscosas donde se alimentan de frutos en descomposición, secreciones de árboles y excrementos de anímales (De la maza, 1 987). • Pieridae. Los piéridos se reconocen por sus colores blancos, amarillos o naranjados, los cuales resultan de la incorporación de pigmentos en las escamas de las alas, tienen patas bien desarrolladas para caminar, uñas tarsales bífidas y celda discal cerrada en ambas alas. Los adultos presentan sexos similares en apariencia, pero algunos géneros como Dismorphia, Anteos y Phoebis, presentan caracteres sexuales secundarios bien notorios en los machos, sobre todo en las alas posteriores. Con frecuencia los machos forman grandes congregaciones en charcos y zonas lodosas a lo largo de los ríos (De la Maza, 1 987). Se han documentado migraciones en los géneros Phoebis y Ascia. Algunos géneros como Perrhybris y Dismorphia imitan bien especies de Heliconiinae e Ithomiinae con los que forman complejos miméticos. Las principales familias de plantas hospederas que utilizan en el Neotrópico son Brasicaceae, Capparidaceae, Fabaceae, Mimosaceae, Loranthaceae y Tropaeolaceae (De la Maza, 1 987). • Lycaenidae. Constituida por mariposas de tamaño pequeño conocidas localmente como mariposas listadas por las pequeñas colas que tienen la mayoría de los géneros en las alas posteriores. Es un grupo bastante grande y complejo de

574
mariposas que se encuentran actualmente en proceso de revisión taxonómica (De la Maza, 1 987). En los adultos el color predominante de la superficie superior de las alas es azul o cobrizo metálico, castaño oscuro o anaranjado, mientras que en la cara ventral presentan tonalidades más oscuras. En algunas especies el diseño de las alas posteriores imita una falsa cabeza con ojos simulados por ocelos negros dibujados en el margen anal de las alas posteriores y las colas que semejan antenas. Por lo general sus larvas son de color verdes o café, muchas de ellas han desarrollado una interesante simbiosis con hormigas conocida como “mirmecofilia” la cual esta medida por secreciones de glándulas epidérmicas especializadas, llamados órganos mircofilos. Las plantas hospederas de esta familia incluye Acantáceas, Annonaceae, Euphorbiaceae, Fabaceae, Orchidiaceae y hasta Cycadaceae como el caso de las mariposas pertenecientes al género Eumaeus que se alimentan de Zamia, el género de planta más primitiva que se conoce sobre la tierra (García –Robledo et al, 2002). • Riodinidae. Constituye una de las familias de colores y diseños más variados, son pequeñas y algunos de ellas diminutas. Presentan patrones de coloración llamativos en combinaciones de amarillo, rojo, naranja, blanco y negro, así como dibujos y manchas de colores metálicos en verde, azul, dorado y plateado. En los adultos las patas anteriores están reducidas en los machos, pero en las hembras son funcionales, aunque de menor tamaño que los otros dos pares, además se caracterizan porque se posan con las alas extendidas, ocultándose debajo del follaje en el sotobosque y en bordes de bosque por lo cual pasan desapercibidos (García –Robledo et al, 2002).Se conocen unas 1200 especies en la región Neotropical y unas 630 especies en Colombia, distribuidas en todos los pisos térmicos. Con altos niveles de endemismo, que alcanzan el 37 % en la costa Pacífica, 30 % en el Putumayo y el piedemonte de la cordillera Oriental y 27 % en el Magdalena Medio. Las plantas hospederas de la familia Riodinidae incluye una amplia gama de familias entre los que se destacan Anacardiáceas, Bromeliaceae, Cecropiaceae, Bombacaceae, Clusiaceae, Euphorbiaceae, Fabaceae, Hippocrataceae, Malpighiaceae, Loranthaceae, Melastomataceae, Myrtaceae, Orchidiaceae, Rubiaceae, Sterculiaceae, Ulmiaceae, e incluso musgos (García Robledo et al, 2002). Papilionidae. Son lepidópteros de tamaño mediano a grande. Generalmente de color blanco, amarillo o negro y de variadas formas en las que se pueden encontrar largos apéndices caudales. Presentan sus tres pares de patas bien desarrolladas, el tórax y el abdomen son de similar tamaño y la venación presenta la celda discal de tipo cerrado en ambas alas, con antenas cortas y recurvadas hacia arriba (De la

575
Maza, 1987). En Colombia, esta familia se encuentra representada por una sola subfamilia con 68 especies y 128 subespecies, distribuidas en ocho géneros (Battus, Parides, Mimoides, Protographium, Protesilaus, Heraclides, Papillo y Pterourus), con algunos géneros nuevos recientemente propuestos. Las plantas hospederas de esta familia incluyen las familias Aristolochiaceae, Anonaceae, Lauraceae, Magnoliaceae, Piperaceae, Rutaceae, Apiaceae y Cannelaceae (De la Maza 1987).
2.10.1.2.1 MATERIALES Y MÉTODOS
La metodología utilizada en este proyecto se encuentra dividida en dos partes, la primera esta constituida por una fase de campo, donde se llevo cavo la captura de los ejemplares y la toma de datos. La segunda fase consistió en actividades de laboratorio tales como el montaje, determinación de los ejemplares y análisis de datos. AREA DE ESTUDIO. Este estudio se llevó a cabo en la Cuenca Mayor del Río Totare, la cual tiene un área de 151474 Km. Localizada al norte del departamento del Tolima, flanco oriental de la cordillera Central (N 04º 48’ W 74º 49´). Forma parte de la región del alto Magdalena donde tienen influencia los municipios de Ibagué, Alvarado, Piedras, Venadillo, Murillo, Santa Isabel y Anzoátegui. Esta Cuenca se caracteriza por su diversidad de pisos altitudinales, por ser fuertemente quebrada, por presentar pendientes muy fuertes y prolongadas. (CORTOLIMA, 1998). MÉTODOS De Campo. Se establecieron 11 zonas de estudio comprendidas desde 270 a 3642 m cubriendo diversas zonas de vida (Tabla No 2.130). En cada zona, se establecieron transeptos de longitud no definida (tipo sendero) tratando de abarcar tres tipos de hábitat (pradera, borde de bosque y bosque). Adicionalmente se realizó un muestreo en las estaciones Palomar y La Estrella en el mes de Julio, estos datos fueron tenidos en cuenta en la composición general de la cuenca pero no en el procesamiento estadístico. • Temporalidad. Las zonas de muestreo fueron visitadas en los meses de
Febrero (época de sequía) y Mayo (época de lluvia) de 2007. Se establecieron 11 estaciones de muestreo cubiertas en su totalidad en 15 (15) días, con un esfuerzo de captura de 12 horas diarias/ persona. Las localidades el Humedal La Cascada y La Estrella solo fueron visitada una sola vez.

576
• Colecta de ejemplares. De acuerdo a la metodología propuesta por Fagua (2001) los ejemplares fueron colectados con red lepidopterologicas (red aérea) la cual posee un diámetro de 0.4 m. y una profundidad de 1.20 m; los ejemplares fueron sacrificados por presión digital al tórax y guardados en sobres de papel milano blanco para su posterior determinación. Por cada ejemplar capturado se anotó el número de captura, hábitat (bosque, borde, pradera), actividad y hora.
Tabla No 2.130 Localización de las estaciones de muestreo en la cuenca del Río Totare.
No. Altura Municipio Localidad Zona de vida Cordenadas
1 270 Piedras M. Rodríguez (bs-T) N 4º 35’ 58.7” W 74º 49’ 44.0”
2 276 Piedras Humedal (bs-T) N 4º 36´4´´ W 74º 50´58 ´´
3 310 Venadillo La Argelia (bs-T) N 4º 38’ 22.6” W 74º 53’ 32.7”
4 700 Venadillo Potrerito (bs-T) N 4º 41’ 49.1” W 74º 59’ 22.3”
5 750 Ibagué Chucuní (bs-T) N 4º28’5” W 75º4’25.8”
6 1800 Ibagué Ambalá parte alta (bh-P) N 4º28’5”
W 75º4’25.8”
7 2100 Santa Isabel Guaimaral (bh-P) N= 4º35’58.7” WO=74º49’44.0”
8 2100 Anzoátegui La Flor (bh-MB) N= 4º39’18.5” WO=75º7’3.3”
9 2642 Palomar La Casacada Páramo N 4º 39´00´´ W 75º14´32´´
10 2580 Santa Isabel La Estrella Páramo Nº4º 44´ 16´´ W 75º 14´ 22´´
11 3636 Palomar Palomar Páramo Nº 4º 39´4´´ W 75º 13´4´´
Fuente: Autores (2007). De Laboratorio Los ejemplares colectados fueron transportados hasta el Laboratorio de Investigación en Zoología de la Universidad del Tolima (LABINZO), donde se realizó su respectivo montaje y determinación taxonómica, contribuyendo de ésta manera a la ampliación de la Colección Zoológica de Lepidópteros diurnos (CZUT-Lp) de la Universidad del Tolima. • Montaje y determinación de organismo. Los ejemplares fueron montados y
etiquetados según las recomendaciones de De Vries (1987). Para la determinación se utilizaron claves e ilustraciones de revisiones taxonómicas (Andrade 1990; Andrade – C 1995; De Vries 1987; y Constantino 1997; De La Maza 1987; García-Robledo et al. 2002, García - P y Ospina - L 2004 y Peña 2007).

577
63,16
19,14
3,035,90 6,70
2,07
0
10
20
30
40
50
60
70
NYMPHALIDAE PIERIDAE PAPILIONIDAE HESPERIIDAE RIODINIDAE LYCAENIDAE
FAMILIAS
% AR
De Análisis. Para el análisis de datos se calculó el porcentaje de abundancia relativa (AR %) para las familias, se determinó la riqueza específica (S) (Moreno, 2001) y la diversidad por localidad y época climática a través del índice de Shannon - Wiener (H´) (Magurran 1988) utilizando el programa Pastprogram versión 1.21 (2004). La similitud entre estaciones se obtuvo mediante coeficiente de comunidad de Bray - Curtis (Ludwig y Reynolds 1998) y las matrices resultantes del procedimiento anterior fueron agrupadas mediante UPGMA (Ligamento promedio por la media aritmética no ponderada), con el programa Pastprogram versión 1.21 (2004) obteniéndose los respectivos dendogramas de similitud.
2.10.1.2.2 RESULTADOS COMPOSICIÓN TAXONÓMICA GENERAL Con un esfuerzo de captura de 240 horas/persona se colectó un total de 665 ejemplares de mariposas diurnas representados en 6 familias, 110 géneros y 164 especies. La familia más abundante y con mayor número de especies fue Nymphalidae (63.16 %) (95 especies), seguida de Pieridae (19.14 %) (28 especies) (figura No 2.47), (Figura No 2.48 y 2.49), donde se encuentran las especies más abundantes Hamadryas feronia (2.71%), Pseudohaetera hypaesia (2.39 %), Heliconius melpómene (2. 23 %). Además se reportan dos nuevas especies para el departamento (Tatochila xantodice y Antirrhea geryon (Figura No 2.55) (Apéndice 2.10.1.2.1 Lepidópteros Diurnos). Figura No 2.47 Abundancia relativa para las familias de mariposas diurnas registradas en la Cuenca Mayor del Río Totare (Tolima) en el año 2007.
Fuente: Autores (2007).

578
0 500 1000 1500 2000 2500 3000 3500 4000
NYMPHALIDAE
PIERIDAE
PAPILIONIDAE
HESPERIIDAE
RIODINIDAE
LYCAENIDAE
Fam
ilias
Altura (m)
95
28
811
16
5
0
10
20
30
40
50
60
70
80
90
100
NYM PHALIDAE PIERIDAE PAPILIONIDAE HESPERIIDAE RIODINIDAE LYCAENIDAE
Familias
Nº
de esp
ecies
DISTRIBUCIÓN ALTITUDINAL DE LAS FAMILIAS En cuanto a la distribución altitudinal las familias Nymphalidae y Pieridae evidenciaron los mayores rangos de distribución (270 - 3642 m); las familias Papilionidae y Lycaenidae presentaron el menor rango de distribución (Figura No 2.48). Figura No 2.48 Número de especies para las familias registradas en la Cuenca Mayor del Río Totare (Tolima) en el año 2007.
Fuente: Autores (2007). Figura No 2.49 Distribución altitudinal de las familias de Lepidópteros diurnos encontradas en la Cuenca Mayor del Río Totare (Tolima) en el año 2007.
Fuente: Autores (2007).
579
33
7
36
53 5144
28 30
6 4
93
11
70
128
88
100
55
69
1014
0
20
40
60
80
100
120
140
M. R
OD
RIG
UEZ
HU
MED
AL
ARG
ELIA
POTR
ERIT
O
CH
UC
UN
I
AMBA
LA
GU
AIM
ARAL
LA F
LOR
LA C
ASAC
ADA
PALO
MAR
Estaciones
Especies Individuos
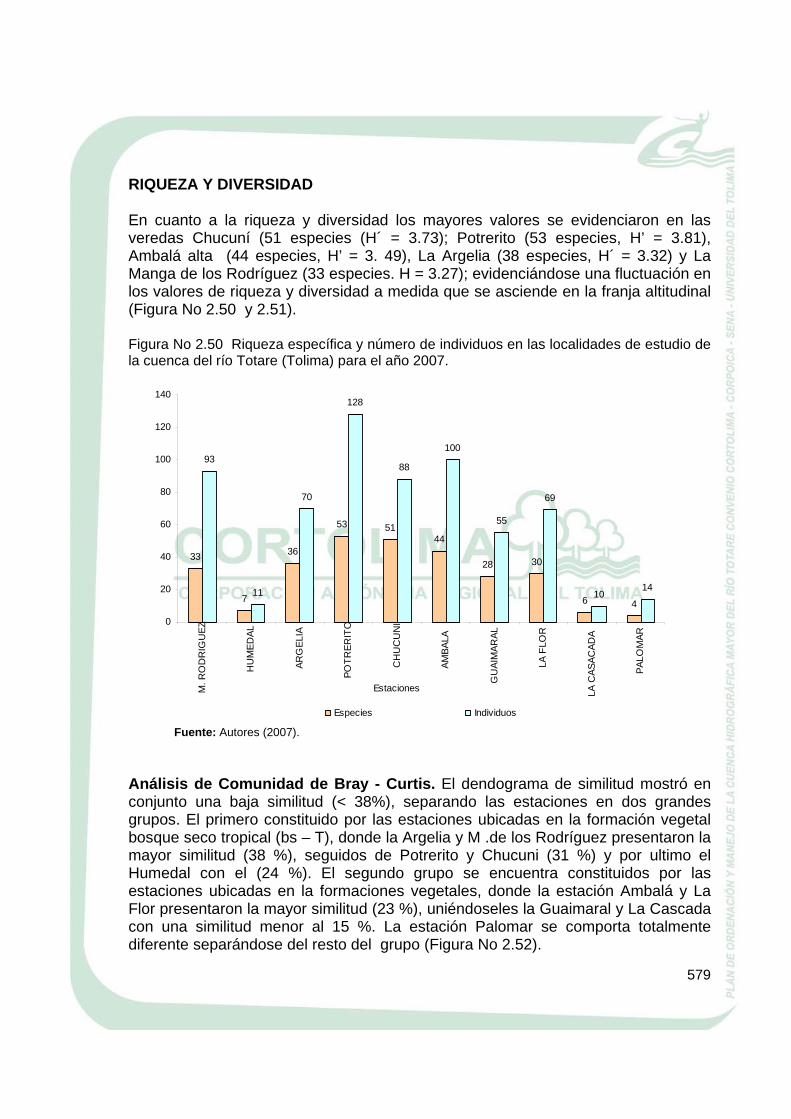
RIQUEZA Y DIVERSIDAD En cuanto a la riqueza y diversidad los mayores valores se evidenciaron en las veredas Chucuní (51 especies (H´ = 3.73); Potrerito (53 especies, H’ = 3.81), Ambalá alta (44 especies, H’ = 3. 49), La Argelia (38 especies, H´ = 3.32) y La Manga de los Rodríguez (33 especies. H = 3.27); evidenciándose una fluctuación en los valores de riqueza y diversidad a medida que se asciende en la franja altitudinal (Figura No 2.50 y 2.51). Figura No 2.50 Riqueza específica y número de individuos en las localidades de estudio de la cuenca del río Totare (Tolima) para el año 2007.
Fuente: Autores (2007). Análisis de Comunidad de Bray - Curtis. El dendograma de similitud mostró en conjunto una baja similitud (< 38%), separando las estaciones en dos grandes grupos. El primero constituido por las estaciones ubicadas en la formación vegetal bosque seco tropical (bs – T), donde la Argelia y M .de los Rodríguez presentaron la mayor similitud (38 %), seguidos de Potrerito y Chucuni (31 %) y por ultimo el Humedal con el (24 %). El segundo grupo se encuentra constituidos por las estaciones ubicadas en la formaciones vegetales, donde la estación Ambalá y La Flor presentaron la mayor similitud (23 %), uniéndoseles la Guaimaral y La Cascada con una similitud menor al 15 %. La estación Palomar se comporta totalmente diferente separándose del resto del grupo (Figura No 2.52).

580
3,274
1,894
3,324
3,702 3,7383,498
2,9893,133
1,609
1,055
0
0,5
1
1,5
2
2,5
3
3,5
4
M. R
OD
RIG
UEZ
HU
MED
AL
ARG
ELIA
PO
TRER
ITO
CH
UC
UN
I
AMBA
LA
GU
AIM
ARAL
LA F
LOR
LA C
ASAC
ADA
PALO
MAR
ESTACIONES
H´
12
34
56
78
910
11
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9 1
Similarity
PALOMAR
AMBALA
LA FLOR
GUAIMARAL
LA CASACA
ARGELIA
M. RODRIG
POTRERITO
CHUCUNI
HUMEDAL
Figura No 2.51 Índice de Shannon – Wiener (H’) para las localidades de estudio de la Cuenca Mayor del Río Totare (Tolima) para el años 2007.
Fuente: Autores (2007). Figura No 2.52. Dendrograma de similitud de Bray – Curtis para las estaciones de muestreo en la Cuenca Mayor del Río Totare (Tolima) en el año 2007.
Fuente: Autores (2007).

581
16
0
23
43
37
28
2123
0 0
33
7
36
5351
44
2830
64
0
10
20
30
40
50
60
Estaciones
Riq
ueza
esp
ecific
a
Época de sequia Época de lluvia
Variación de la riqueza y diversidad en las épocas de sequía y lluvia. En cuanto a la estacionalidad la mayor riqueza y diversidad se registró en las localidades ubicadas por debajo de los 1700 m (zonas medias), siendo potrerito (700 m) la de mayor riqueza y diversidad en el período de sequía (febrero) y Chucuní (750 m) en el período de lluvias. (Ver figuras No 2.53 y 2.54). Figura No 2.53 Variación de la riqueza específica en las estaciones de muestreo de la Cuenca Mayor del Río Totare (Tolima) para el año 2007.
Fuente: Autores (2007).
Las estaciones bajas representadas por las estaciones La Argelia y manga de los Rodríguez también presentaron una alta riqueza y diversidad. No obstante estos patrones decrecen para las localidades que se encuentran por encima de los 2000 m, la Cascada y Palomar (Figura No 2.53 y 2.54).

582
2,57
0
2,98
3,58
3,43
3,07
2,78 2,
97
0 0
3,27
1,89
3,32
3,70 3,74
3,50
2,99 3,
13
1,61
1,06
0
0,5
1
1,5
2
2,5
3
3,5
4
M. R
OD
RIG
UEZ
HU
MED
AL
ARG
ELIA
POTR
ERIT
O
CH
UC
UN
I
AMBA
LA
GU
AIM
ARAL
LA F
LOR
LA C
ASAC
ADA
PALO
MAR
Estaciones
H´
Época de sequía Época de lluvia
Figura No 2.54 Variación de la diversidad (H’) en las estaciones de muestreo de la Cuenca Mayor del Río Totare (Tolima) para el año 2007.
Fuente: Autores (2007).
Figura No 2.55 Nuevos reportes para el departamento.
Fuente: Autores (2007).

583
Tatochial xantodice: especie encontrada en las montañas de los Andes colombinos, pon encima de los 3000 m, donde se le puede encontrar volando en áreas abiertas como potreros, a orillas de caminos y lagunas.
Antirrhea geryon: especie encontrada en áreas de con cierto grado de conservación, es una especie difícil de observar ya que es de hábitos crepusculares, observándose en horas de la tarde entre las 5:00 y 7:00 de la noche. 2.10.1.2.3 DISCUSIÓN
COMPOSICIÓN TAXONÓMICA GENERAL Al poseer el 5.46 % de las especies descritas para el país (3019) (Andrade – C & Amat 1996) y el 16.5 % de las descritas para la cordillera Central (1000), convierte a la Cuenca Mayor del Río Totare en un centro de riqueza y diversidad de mariposas diurnas para el departamento, ya que alberga a todas las familias descritas para Colombia (6) y dieciocho (18) de las 26 subfamilias descritas para el Neotrópico (Lamas 2004). La abundancia presentada por Nymphalidae se puede tribuir a que es la familia con el mayor numero de subfamilias en el Neotrópico y por ende de especies, dentro de estas se incluyen grupos generalistas y especialistas de baja especificidad de hábitat y de gran dispersión, confiriéndole a esta familia una gran ventaja en la explotación de los recursos disponibles frente a las demás familias, estos resultados concuerdan con los obtenidos por Andrade – C (1994) en dos zonas de bosque primario y secundario (Risaralda), Rodríguez et al (2000) en un relicto vegetal de Armenia, García-P & Ospina-L (2004) en la cuenca del río Coello, Peña (2007) en la

584
Cuenca del Río Prado. Las demás familias mostraron una menor abundancia probablemente debido a sus preferencias alimenticias ya que presentan especies en su mayoría nectarivoras. En cuanto a la abundancia presentada por las especies Hamadryas feronia, Heliconius melpómene, Chlosyne lacinia, Phoebis sennae, Dynamine posventa, Nyca falvilla silvestres, se debe a que son especies de baja especificidad de hábitat, con preferencias a zonas abiertas (potreros y bordes de caminos y de bosques); en cuanto a la abundancia presentada por Pseudohaetera hypaesia, se ha encontrado que es una especie común y abundante en bosque secundario con cierto grado de conservación, característica de los sitios donde fue registrada (Ambalá y La Flor). DISTRIBUCIÓN ALTITUDINAL DE LAS FAMILIAS El rango de distribución exhibido por Nymphalidae y Pieridae se debe posiblemente siguiendo el patrón de distribución de sus planta hospederas, ya que al ser insectos fitófagos su capacidad de adaptación va a estar determinada por la abundancia y distribución de sus plantas hospederas, la disponibilidad de hábitat y de recurso alimenticio, elementos fundamentales para el establecimiento de esto insectos. En cuanto a las demás familias su distribución se debe a que sus plantas hospedera se encuentran con más frecuencia y en mayor abundancia en zonas menores a los 2000 m. RIQUEZA Y DIVERSIDAD En general la riqueza y diversidad se vio favorecida en zonas bajas menores a los 1000 m, siendo el resultado de la heterogeneidad espacial que presentan estas localidades con gran intervención antrópica, constituidas por praderas, cultivos y pequeños relictos de bosques restringidos a las fuentes de agua, lo cual da como resultado una mayor heterogeneidad de hábitat, micro hábitat y de recurso alimenticio, condiciones favorables para el establecimiento de un mayor numero de especies. Además Connell (1978) propone que los valores elevados de diversidad pueden ser mantenidos por niveles intermedios de perturbación, los cuales varían en cuanto a frecuencia e intensidad para una zona; Emmel & Austin (1990) argumentan que los niveles de perturbación producidos por el hombre, entre otros factores, contribuyen de manera importante al mantenimiento de la diversidad de mariposas en diversas áreas. En cuanto a los picos de riqueza observados a los 1800 y 2100 m, puede ser el resultado del solapamiento de rangos altitudinales, lo cual va a generar una sobreposición de especies de zonas bajas y montanas, estos resultados también coinciden con los obtenidos por Fagua (1999) en la Cuenca del Río Guazunta

585
(Cundinamarca); García y Ospina en la Cuenca del Río Coello (Tolima); Peña (2007) en la Cuenca del Río Prado (Tolima). La disminución de la riqueza y diversidad por encima de los 2100 m, puede atribuirse a restricciones ambientales como temperaturas bajas, alta humedad, baja radiación solar, que aparecen a medida que se asciende altitudinalmente. Análisis de similitud de Bray - Curtis. Mediante el análisis de comunidad se encontró que la estaciones ubicadas por debajo de los 1000 m, evidenciaron una gran similitud, debido posiblemente a que comparten especies comunes y bien representadas en zonas bajas, posiblemente siendo el resultado de formación vegetal a la cual pertenecen estas localidades (bs – T) compartiendo especies con preferencias a zonas abiertas como Anartia amathea, Chlosyne lasinia, Dynamine posventa, Heliconius erato, H. melpomene. Thitorea harmonía y Phoebis sennae entre otras. No obstante la baja similitud presentada por las estaciones Ambla, La Flor, Guimaral y La Cascada, en relación a las zonas bajas es el resultado de encontrarse en otras zonas de vida (Bosque húmedo Premontano), generándose un cambio abrupto en la composición vegetal, haciendo que la comunidad de mariposas cambie compartiendo especies comunes como Leptophobia eleusis, Dannaus guilippus, Pseudohaetera hypaesia, Dismorphia medora, Pedaliodes manis, entre otras. Variación de la riqueza y la diversidad en los períodos de sequía y lluvia. Los altos valores de riqueza y diversidad observados entre los 310 – 2100 m (Argelia, Potrerito, Chucuni, Ambalá, Guaimaral y La Flor) en las dos épocas climáticas se debe a la gran heterogeneidad de hábitat y de recurso alimenticio (néctar, sales minerales y frutos en descomposición) que presentan estas localidades, constituidas por praderas enmalezadas, cultivos, relictos de bosque secundario restringido a las fuentes de agua y rastrojos. Además al estar la mayoría de estas localidades ubicadas en la zona cafetera, exhiben una gran diversidad de hábitat generando en conjunto una mayor variabilidad en el paisaje. McArthur y Wilson (1967), en su hipótesis de heterogeneidad de hábitat, asumen que la heterogeneidad estructural en los hábitat’s puede favorecer más nichos y diversas formas de explotar los recursos y de este modo, incrementar la diversidad de especies. No obstante los niveles de riqueza y diversidad mantenidos en la época de lluvia posiblemente se deban a que en los días de muestreos para esta época no se presentaron lluvias permitiendo de esta menear el vuelo de las especies, haciendo posible su captura. Ver Apéndice 2.11. Listado de especies encontradas en la Cuenca Mayor del Río Totare para el año 2007, con sus respectivas abundancias relativas.


















