1. ANABOLISMO AUTÓTROFO - Blogs Maristas Segovia | Blogs ... · ANABOLISMO AUTÓTROFO REACCIONES...
20
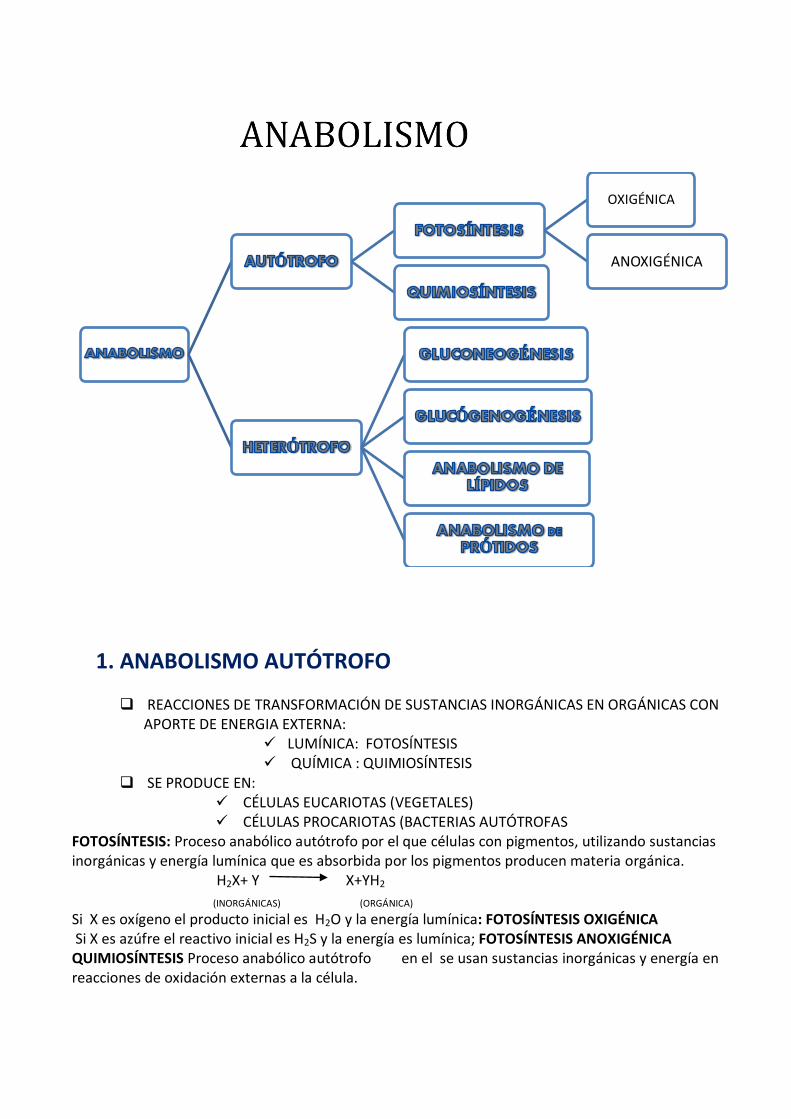
1. ANABOLISMO AUTÓTROFO REACCIONES DE TRANSFORMACIÓN DE SUSTANCIAS INORGÁNICAS EN ORGÁNICAS CON APORTE DE ENERGIA EXTERNA: LUMÍNICA: FOTOSÍNTESIS QUÍMICA : QUIMIOSÍNTESIS SE PRODUCE EN: CÉLULAS EUCARIOTAS (VEGETALES) CÉLULAS PROCARIOTAS (BACTERIAS AUTÓTROFAS FOTOSÍNTESIS: Proceso anabólico autótrofo por el que células con pigmentos, utilizando sustancias inorgánicas y energía lumínica que es absorbida por los pigmentos producen materia orgánica. H 2 X+ Y X+YH 2 (INORGÁNICAS) (ORGÁNICA) Si X es oxígeno el producto inicial es H 2 O y la energía lumínica: FOTOSÍNTESIS OXIGÉNICA Si X es azúfre el reactivo inicial es H 2 S y la energía es lumínica; FOTOSÍNTESIS ANOXIGÉNICA QUIMIOSÍNTESIS Proceso anabólico autótrofo en el se usan sustancias inorgánicas y energía en reacciones de oxidación externas a la célula. OXIGÉNICA ANOXIGÉNICA
-
Upload
phungtuyen -
Category
Documents
-
view
236 -
download
0
Transcript of 1. ANABOLISMO AUTÓTROFO - Blogs Maristas Segovia | Blogs ... · ANABOLISMO AUTÓTROFO REACCIONES...
1. ANABOLISMO AUTÓTROFO REACCIONES DE TRANSFORMACIÓN DE SUSTANCIAS INORGÁNICAS EN ORGÁNICAS CON
APORTE DE ENERGIA EXTERNA: LUMÍNICA: FOTOSÍNTESIS QUÍMICA : QUIMIOSÍNTESIS
SE PRODUCE EN: CÉLULAS EUCARIOTAS (VEGETALES) CÉLULAS PROCARIOTAS (BACTERIAS AUTÓTROFAS
FOTOSÍNTESIS: Proceso anabólico autótrofo por el que células con pigmentos, utilizando sustancias inorgánicas y energía lumínica que es absorbida por los pigmentos producen materia orgánica.
H2X+ Y X+YH2
(INORGÁNICAS) (ORGÁNICA) Si X es oxígeno el producto inicial es H2O y la energía lumínica: FOTOSÍNTESIS OXIGÉNICA Si X es azúfre el reactivo inicial es H2S y la energía es lumínica; FOTOSÍNTESIS ANOXIGÉNICA QUIMIOSÍNTESIS Proceso anabólico autótrofo en el se usan sustancias inorgánicas y energía en reacciones de oxidación externas a la célula.
OXIGÉNICA
ANOXIGÉNICA

1.1. FOTOSINTEIS OXIGÉNICA DEL CARBONO PROCESO DE SINTESIS DE MATERIA ORGÁNICA CARBONADA A PARTIR DE INORGÁNICA EN EL QUE SE LIBERA OXÍGENO
Mediante este proceso solo se producirán moléculas hidrocarbonadas puesto que como reactivos solo se usa agua y dióxido de carbono. Para producir moléculas como aa y nucleótidos, tendrán que incorporar nitratos y fosfatos. La ecuación química de la fotosíntesis es la inversa de la respiración, lo que nos indica la importancia de la fotosíntesis para mantener el equilibrio en los ecosistemas; ya que produce la materia orgánica y el oxígeno necesarios para que los seres vivos puedan realizar la respiración, aportando la materia y energía necesarias para los heterótrofos, y además, reciclan la materia inorgánica al transformarla de nuevo en materia orgánica, indispensable para cerrar así los ciclos biogeoquímicos. La aparición de la fotosíntesis en la Tierra permitió la acumulación de oxígeno en la atmósfera y la formación de la capa de ozono que nos protege de la radiación ultravioleta.
Fotosíntesis CO2 + H2O + energía lumi → materia orgánica + O2
Respiración materia orgánica + O2 → CO2 + H2O + energía (ATP) ESTE PROCESO SE RALIZA EN 2 FASES:
o LOCALIZACIÓN: EN LAS MEMBRANAS TILACOIDALES o PROCESOS:
o FOTOLISIS DEL AGUA(FORMACIÓN DE OXÍGENO) o FOTOFOSFORILACION (ATP)
o PRODUCE PODER REDUCTOR (NADP2H)
o LOCALIZACIÓN : ESTROMA o PROCESOS:
o REDUCCIÓN DEL DIÓXIDO DE CARBONO (forma mat. Orgánica) o UTILIZACIÓN DEL NADP2H Y EL ATP DE LA ETAPA LUMÍNICA

A. FASE LUMINOSA DE LA FOTOSÍNTESIS En la fotosíntesis intervienen cuatro componentes básicos:
• Fotosistema I (PSI) y Fotosistema II (PSII) • Cadena transportadora de electrones. • PARTÍCULAS F: Enzimas ATP- sintetasas.
El proceso se inicia en los fotosistemas, los fotones inciden sobre los fotosistemas y sus electrones pasan a la cadena trasportadora de electrones y los protones que son liberados son captados por el NADP y ayudan a la producción de energía, por las ATP- sintetasa. Todas estas moléculas se encuentran en la membrana de los tilacoides.
FOTOSISTEMAS Son agrupaciones de pigmentos fotosintéticos cuya función es captar la energía luminosa para utilizarla durante el transporte de electrones que servirá para sintetizar moléculas energéticas. Tipos de pigmentos que forman los fotosistemas.
► CLOROFILA a, b y c. ► CAROTENOIDES y xantofilas Cada pigmento incluye un cromóforo (grupo químico que absorbe a una determinada longitud de onda del espectro visible) Cada pigmento capta la luz de determinada longitud de onda permitiendo un amplio rango de captación energética. El sol emite muchas longitudes de onda pero el único rango que utilizan los organismos vivos es 380nm- 750nm ( fotosíntesis, visión, curvatura de tallos……..) Las clorofilas se ven de color verde porque reflejan la longitud de onda correspondiente al verde (650-700nm) Estructura de la clorofila: • Anillo de porfirina: absorbe la luz, los e- forman una nube a su alrededor. • Cadena hidrófoba de fitol mantiene la clorofila en la membrana tilacoidal.
Además de clorofilas en los fotosistemas nos encontramos , pigmentos accesorios como carotenos y xantofilas, pigmentos lipídicos que dan color rojo y amarillo respectivamente. Estos pigmentos se denominan accesorios y su función es transferir la energía de los fotones
a la clorofila. En algas aparecen otros pigmentos diferentes que se denominan ficobilinas; entre las que destacan: ficoeritrina (rojo) y ficocianina(azul) Cada fotosistema consta de: COMPLEJO ANTENA: formado por pigmentos accesorios (carotenos, xantofilas y varios tipos
de clorofilas(entre 250 y 400 moléculas de pigmentos) ) asociados a lípidos y proteínas de fijación a la membrana tilacoidal .
CENTRO DE REACCIÓN FOTOQUÍMICO: formado por una clorofila asociado a proteínas, un compuesto aceptor de electrones y un dador de electrones.

FUNCIONAMIENTO DEL FOTOSISTEMA: Los pigmentos accesorios captan un fotón de energía lumínica y la energía es transferida de una molécula a otra de pigmento por resonancia, canalizándola hasta el centro de reacción fotoquímico, en donde se produce la excitación de dos electrones.
Excitación (cambio en la distribución de e)
Transmiten energía a otro pigmento por resonancia
Cuando los pigmentos vuelven a su estado inicial se produce una liberación de energía química, calor y fluorescencia. Conceptos asociados a energía lumínica Foton de luz: cuanto de energía lumínica que posee la energía suficiente para hacer salta un electron desde un nivel energético inferior a un nivel energético superior. Radiaciones electromagnéticas emitidas por el sol:
Radiaciones ultravioleta (U.V.)= son las de menor longitud de onda, son las más energéticas. No son útiles para la vida. (producen mutaciones) Luz visible (380nm-750nm)= Funciones que dependen de ella son: fotosíntesis, visión, curvatura de tallos.
Luz infrarroja (+ de 750nm)= muy poco energética solo provocan alguna agitación térmica (vista de serpientes)
Forma de reacción de los pigmentos: Liberando energía en forma de luz y calor (fluorescencia)
Transfiriendo energía pero no un electrón a otro pigmento vecino, a esta forma de transmisión se le llama transmisión de energía por resonancia Perdiendo un electrón a un orbital de mayor energía, que será captado por un transportador cercano de electrones. Para que el pigmento retorne a su estado original tiene que retomar los electrones de un donador.
TIPOS DE FOTOSISTEMAS: FOTOSISTEMA I (P700) ; FOTOSISTEMA II (P680) Los fotosistemas de plantas se diferencian por la longitud de onda a la que absorben PSI (P700) y PSII(P680) , su localización y a quien ceden o de quien captan los electrones.
Fotosistema I (PS I): En membranas de tilacoides no apilados en contacto con el estroma. Centro Reacción: 2 clorofilas a P700 (máxima absorción a 700 nm de λ).Acepta los electrones de la plastocianina. Es el responsable de la reduccióndel NADP+, cede los electrones a un aceptor poco conocido (X)
Fotosistema II (PS II): En las membranas de los tilacoides de los grana Centro Reacción: 2 clorofilas a P680 (máxima absorción a 680 nm de λ). Acepta los electrones del agua, a través de un dador Z, está implicado en la liberación de oxígeno, cede los electrones a las quinonas.
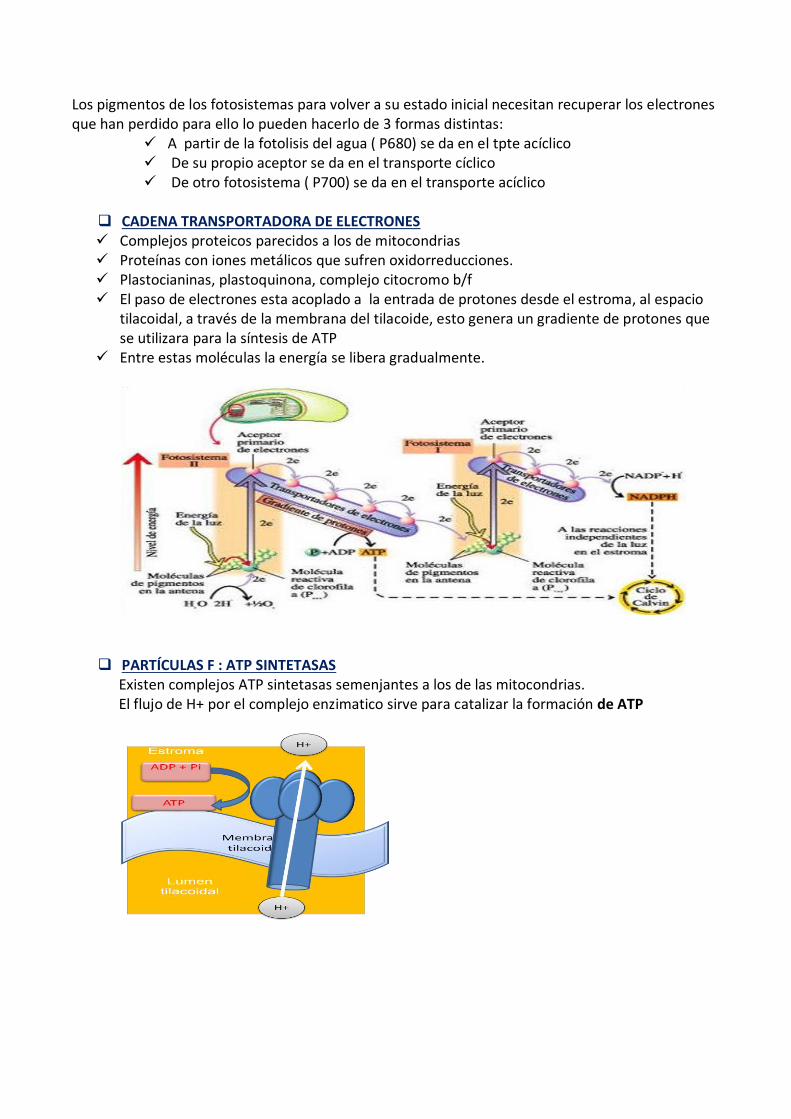
Los pigmentos de los fotosistemas para volver a su estado inicial necesitan recuperar los electrones que han perdido para ello lo pueden hacerlo de 3 formas distintas:
A partir de la fotolisis del agua ( P680) se da en el tpte acíclico De su propio aceptor se da en el transporte cíclico De otro fotosistema ( P700) se da en el transporte acíclico
CADENA TRANSPORTADORA DE ELECTRONES Complejos proteicos parecidos a los de mitocondrias Proteínas con iones metálicos que sufren oxidorreducciones. Plastocianinas, plastoquinona, complejo citocromo b/f El paso de electrones esta acoplado a la entrada de protones desde el estroma, al espacio
tilacoidal, a través de la membrana del tilacoide, esto genera un gradiente de protones que se utilizara para la síntesis de ATP
Entre estas moléculas la energía se libera gradualmente.
PARTÍCULAS F : ATP SINTETASAS Existen complejos ATP sintetasas semenjantes a los de las mitocondrias. El flujo de H+ por el complejo enzimatico sirve para catalizar la formación de ATP

A.1. FOTOFOSFORILACIÓN ACÍCLICA
Una vez que los fotones de la luz excitan los dos fotosistemas y producen la fotólisis del H2O:
Los 2 e- del H2O son recogidos por el PSII.
2 e- excitados salen del PSII y son captados por la FEOFITINA.
Los e- pasan a varias moléculas de PLASTOQUINONA y de ahí al CITOCROMO b6f Y PLASTOCIANINA.
Simultáneamente otros 2 e- han salido del PSI y han sido captados por la ACEPTOR.
Los 2 e- captados por la feofitina se desplazan hasta el PSI reemplazando los e- perdidos.
Los 2 e- captados por la ACEPTOR llegan hasta la FERREDOXINA AHÍ hasta hasta el NADP+ al cual llegan tambien los protones del agua y se forma NADP2H
El movimiento de los e- a través de los transportadores permite al citocromo b6f utilizar la energía que estos liberan para transportar H+ en contra de un gradiente electroquímico, desde el estroma hacia el espacio tilacoidal.
Este gradiente es aprovechado por la ATP-sintetasa para fosforilar ATP.(síntesis ATP)
A.2. FOTOFOSFORILACIÓN CÍCLICA
Si el cloroplasto tiene suficiente NADPH2 pero necesita más ATP, se produce el transporte cíclico de electrones, en el que sólo funciona el FSI, no se produce NADPH2 porque la ferredoxina en lugar de ceder los electrones al NADP se los cede al citocromo b6f que produce bombeo de H+ que servirá para producir ATP.
• Sólo interviene el fotosistema I.(P680) • No se reduce el NADP • No se rompe el H2O: no se libera O2. • Sí se sintetiza ATP. • El uso de una forma u otra de fosforilación se debe a:
Se activa cuando hay desequilibrio entre ATP y NADPH.
Si hay poco NADP2H: acíclico
Si hay mucho NADP2H y poco ATP: CÍCLICO
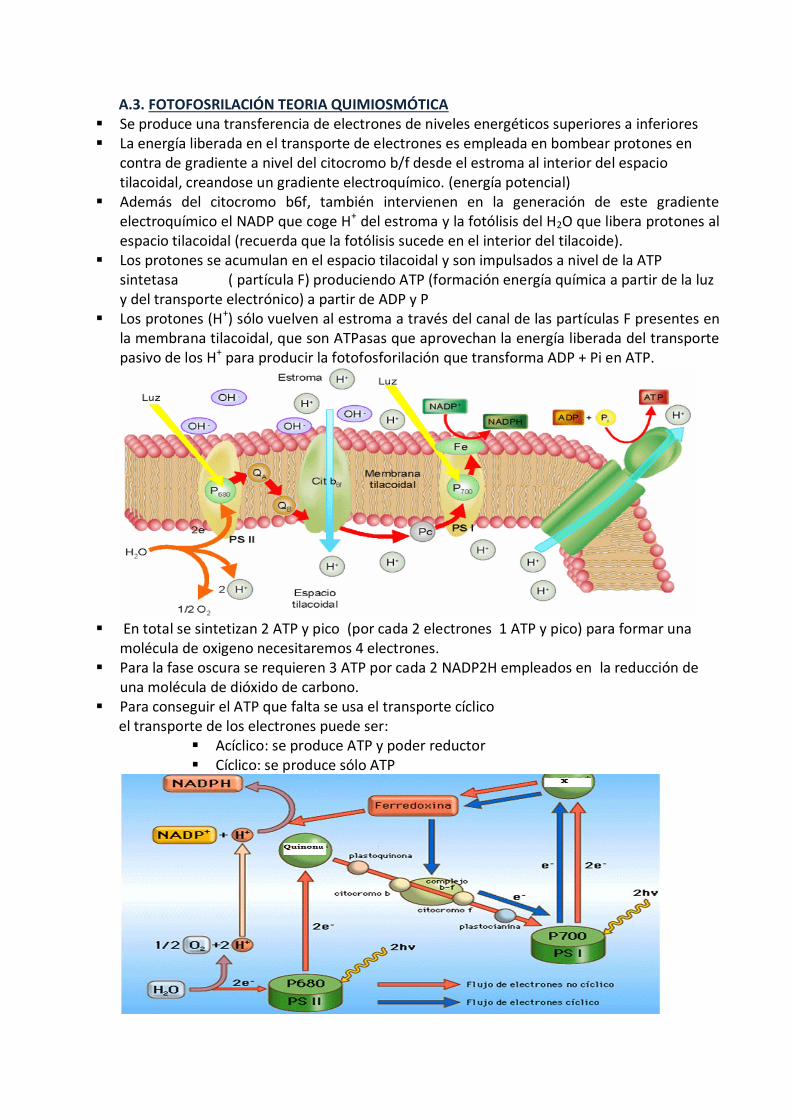
A.3. FOTOFOSRILACIÓN TEORIA QUIMIOSMÓTICA Se produce una transferencia de electrones de niveles energéticos superiores a inferiores La energía liberada en el transporte de electrones es empleada en bombear protones en
contra de gradiente a nivel del citocromo b/f desde el estroma al interior del espacio tilacoidal, creandose un gradiente electroquímico. (energía potencial)
Además del citocromo b6f, también intervienen en la generación de este gradiente electroquímico el NADP que coge H+ del estroma y la fotólisis del H2O que libera protones al espacio tilacoidal (recuerda que la fotólisis sucede en el interior del tilacoide).
Los protones se acumulan en el espacio tilacoidal y son impulsados a nivel de la ATP sintetasa ( partícula F) produciendo ATP (formación energía química a partir de la luz y del transporte electrónico) a partir de ADP y P
Los protones (H+) sólo vuelven al estroma a través del canal de las partículas F presentes en la membrana tilacoidal, que son ATPasas que aprovechan la energía liberada del transporte pasivo de los H+ para producir la fotofosforilación que transforma ADP + Pi en ATP.
En total se sintetizan 2 ATP y pico (por cada 2 electrones 1 ATP y pico) para formar una
molécula de oxigeno necesitaremos 4 electrones. Para la fase oscura se requieren 3 ATP por cada 2 NADP2H empleados en la reducción de
una molécula de dióxido de carbono. Para conseguir el ATP que falta se usa el transporte cíclico
el transporte de los electrones puede ser: Acíclico: se produce ATP y poder reductor Cíclico: se produce sólo ATP

B. FASE OSCURA- CICLO DE CALVIN
Su finalidad es reducir compuestos químicos inorgánicos, principalmente el CO2 aunque también puede reducir sales minerales como NO3
-, SO42-... Para estas reducciones se usa el NADPH2
y el ATP formado en la fase luminosa y de este modo se fabrica materia orgánica. Sucede en el estroma de los cloroplastos. El ciclo de Calvin comienza con la ribulosa-1,5-
difosfato y acaba volviendo a regenerar dicha molécula. Para comenzar se fija el CO2 a una molécula de ribulosa-1,5-difosfato gracias a la enzima
ribulosa bifosfato carboxilasa- oxidasa más conocida como rubisco. La materia orgánica que se va formando en el ciclo de Calvin es siempre una molécula de 3
carbonos llamada gliceraldehído-3P (recuerda que el gliceraldehído es una aldotriosa , que aparece de intermediaria en la glucólisis), por lo que para regenerar la ribulosa-1,5-difosfato necesitamos fijar 3 CO2 (metemos 3 carbonos para sacar el gliceraldehído de 3 carbonos)
Por cada CO2 que se fija se gastan 3 ATP y 2 NADPH2, entonces por cada gliceraldehído-3P que sale del ciclo de Calvin, se han fijado 3 CO2 a 3 ribulosa-1,5-difosfato y se han gastado 9 ATP y 6 NADPH2 .
Al fijarse 3 CO2 a 3 ribulosa-1,5-difosfato debemos observar que hay 18 carbonos y al salir una molécula de 3 carbonos (gliceraldehído-3P), los 15 carbonos restantes vuelven a dar las 3 ribulosa-1,5-difosfato cerrando el ciclo. Los gliceraldehído-3P formados en el ciclo de Calvin se usan para formar otras moléculas orgánicas como glúcidos. Por ejemplo para formar una molécula de glucosa se deben fijar 6 CO2 para obtener 2 gliceraldehído-3P, es decir, 2 vueltas en el ciclo de Calvin (cada vez que se obtiene un gliceraldehido es una vuelta). Con los 2 gliceraldehído-3P (3 carbonos) formamos una glucosa (6 carbonos) y se han gastado 18 ATP y 12 NADPH2.
Hay que distinguir 3 etapas en el ciclo de Calvin:
1. Carboxilación: realizada por la enzima ribulosa difosfato carboxilasa 2. Reducción: el 3 fosfoglicerato se reduce a gliceraldehido 3 fosfato con consumo de ATP y
NAP2H 3. Regeneración: se regenera la ribulosa 1,5 difosfato mediante una serie de reacciones en las
que intervienen azúcares de 3 atomos de carbono y a partir del gliceraldehido 3 fosfato que no se libera.

Balance energético Para formar una molécula de glucosa (6C) se necesitan fijar 6 CO2 y gastar 18 ATP y 12 NADPH (formados previamente en la fase luminosa)
RuBisCO es la forma abreviada con que se designa a la ribulosa-1,5-bisfosfato carboxilasa-oxidasa. Cataliza dos procesos opuestos:
1. La fijación del CO2 (carboxilasa). 2. La fotorrespiración, en la que actúa como oxigenasa del mismo sustrato.
El que actúe con actividad oxidasa o carboxilasa depende de las concentraciones de oxígeno y de dióxido de carbono.
• Es un enzima compuesto de 8 ubunidades. • Sólo fija 3 moléculas de CO2 por segundo
(muy lenta comparada con otras enzimas). • Por esto es tan abundante en los cloroplastos
y puede alcanzar el 15% de sus proteínas, y de hecho, es la proteína más abundante en la biosfera.
Algunas de las cadenas peptídicas de esta enzima se catalizan a partir de ADN del cloroplasto (Con los ribosomas del cloroplasto)y otras a partir del ADN del núcleo ( con los ribosomas del citoplasma).
1.2. FOTOSÍNTESIS DEL NITRÓGENO Los organismos necesitan compuestos nitrogenados (aminoácidos, nucleótidos etc) Los vegetales captan nitrógeno en forma de nitratos (NO3-) del suelo, debiendo ser reducidos a NH3 para poder ser incorporado a las moléculas orgánicas como grupo amino Se utiliza para ello ATP y NADP2H producidos en las fase lumínica de la fotosíntesis.
Reducir el NO3
- (sal mineral absorbida por las raíces) a NO2-, mediante el enzima nitratorreductasa
y usando el NADPH2 obtenidos en la fase luminosa de la fotosíntesis. Reducir el NO2
- a NH3 (NH4+) mediante el enzima nitritorreductasa y usando el NADPH2 obtenidos
en la fase luminosa de la fotosíntesis.
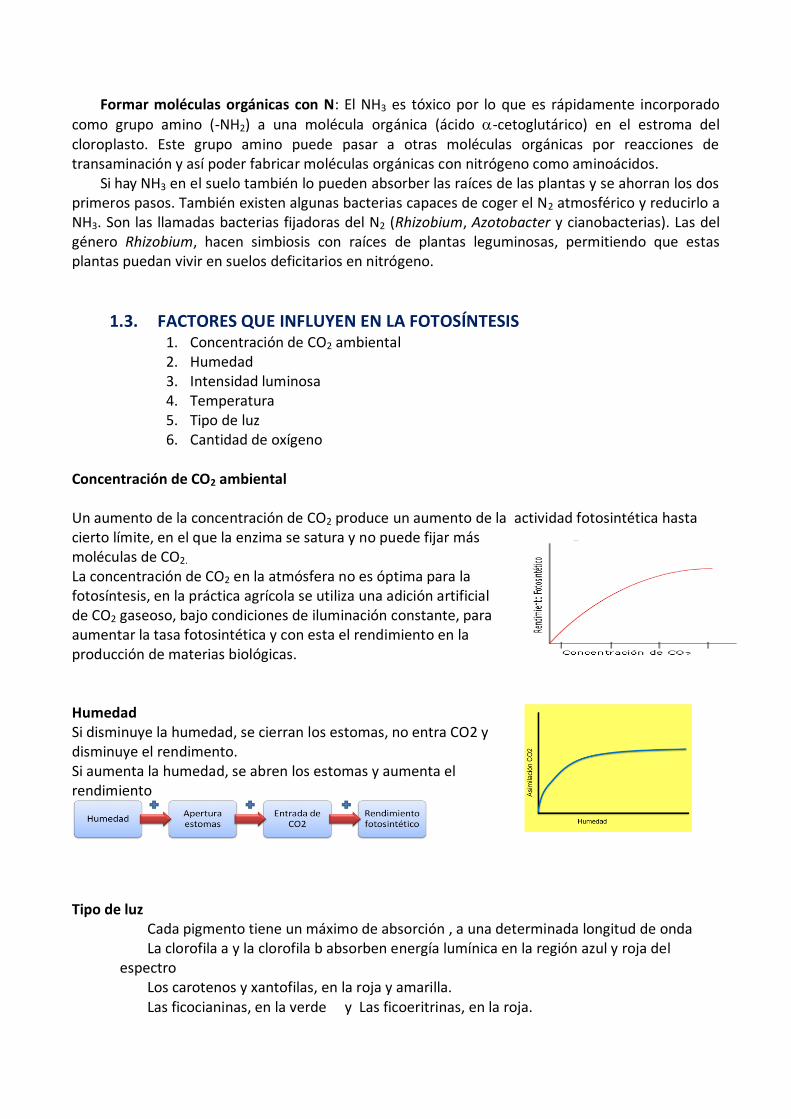
Formar moléculas orgánicas con N: El NH3 es tóxico por lo que es rápidamente incorporado
como grupo amino (-NH2) a una molécula orgánica (ácido -cetoglutárico) en el estroma del cloroplasto. Este grupo amino puede pasar a otras moléculas orgánicas por reacciones de transaminación y así poder fabricar moléculas orgánicas con nitrógeno como aminoácidos.
Si hay NH3 en el suelo también lo pueden absorber las raíces de las plantas y se ahorran los dos primeros pasos. También existen algunas bacterias capaces de coger el N2 atmosférico y reducirlo a NH3. Son las llamadas bacterias fijadoras del N2 (Rhizobium, Azotobacter y cianobacterias). Las del género Rhizobium, hacen simbiosis con raíces de plantas leguminosas, permitiendo que estas plantas puedan vivir en suelos deficitarios en nitrógeno.
1.3. FACTORES QUE INFLUYEN EN LA FOTOSÍNTESIS 1. Concentración de CO2 ambiental 2. Humedad 3. Intensidad luminosa 4. Temperatura 5. Tipo de luz 6. Cantidad de oxígeno
Concentración de CO2 ambiental Un aumento de la concentración de CO2 produce un aumento de la actividad fotosintética hasta cierto límite, en el que la enzima se satura y no puede fijar más moléculas de CO2. La concentración de CO2 en la atmósfera no es óptima para la fotosíntesis, en la práctica agrícola se utiliza una adición artificial de CO2 gaseoso, bajo condiciones de iluminación constante, para aumentar la tasa fotosintética y con esta el rendimiento en la producción de materias biológicas.
Humedad Si disminuye la humedad, se cierran los estomas, no entra CO2 y disminuye el rendimento. Si aumenta la humedad, se abren los estomas y aumenta el rendimiento
Tipo de luz Cada pigmento tiene un máximo de absorción , a una determinada longitud de onda La clorofila a y la clorofila b absorben energía lumínica en la región azul y roja del
espectro Los carotenos y xantofilas, en la roja y amarilla. Las ficocianinas, en la verde y Las ficoeritrinas, en la roja.
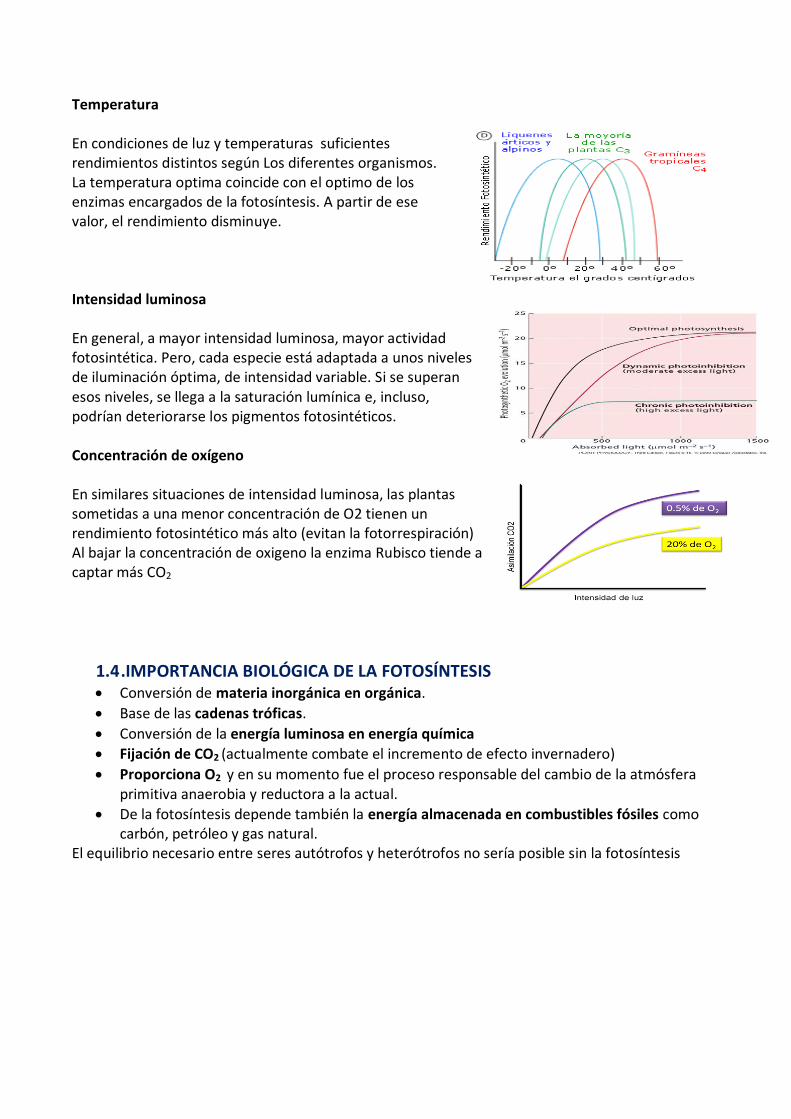
Temperatura
En condiciones de luz y temperaturas suficientes rendimientos distintos según Los diferentes organismos. La temperatura optima coincide con el optimo de los enzimas encargados de la fotosíntesis. A partir de ese valor, el rendimiento disminuye.
Intensidad luminosa
En general, a mayor intensidad luminosa, mayor actividad fotosintética. Pero, cada especie está adaptada a unos niveles de iluminación óptima, de intensidad variable. Si se superan esos niveles, se llega a la saturación lumínica e, incluso, podrían deteriorarse los pigmentos fotosintéticos. Concentración de oxígeno
En similares situaciones de intensidad luminosa, las plantas sometidas a una menor concentración de O2 tienen un rendimiento fotosintético más alto (evitan la fotorrespiración) Al bajar la concentración de oxigeno la enzima Rubisco tiende a captar más CO2
1.4 .IMPORTANCIA BIOLÓGICA DE LA FOTOSÍNTESIS Conversión de materia inorgánica en orgánica.
Base de las cadenas tróficas.
Conversión de la energía luminosa en energía química
Fijación de CO2 (actualmente combate el incremento de efecto invernadero)
Proporciona O2 y en su momento fue el proceso responsable del cambio de la atmósfera primitiva anaerobia y reductora a la actual.
De la fotosíntesis depende también la energía almacenada en combustibles fósiles como carbón, petróleo y gas natural.
El equilibrio necesario entre seres autótrofos y heterótrofos no sería posible sin la fotosíntesis

1.5.MODIFICACIONES DEL CICLO DE CALVIN A. FOTORRESPIRACIÓN B. RUTA DE HATCH-SLACK DE LAS PLANTAS C4 C. PLANTAS CAM A. FOTORRESPIRACIÓN
Ocurre cuando escasea el CO2 y abunda el O2, esto se produce en situaciones de calor y déficit hídrico, ya que los estomas impiden que se libere el O2 y entre CO2 al estar cerrados para evitar la transpiración. En tal situación la rubisco en lugar de fijar CO2 a la ribulosa
bifosfato, le añade O2, actuando con forma oxidasa, se denomina fotorrespiración porque es un proceso que necesita la presencia deluz, y además se parece a la respiración porque consume oxígeno y libera CO2 . Ocurre en tres compartimentos celulars: cloroplasto, peroxisoma y mitocondria. En el cloroplasto se produce la oxidación de la ribulosa 1,5 difosfato produciendo ácidofosfoglicólico (2 Carbonos) y ácido 3 fosfoglicérico (3 Carbonos) En el peroxisoma el ácido fosfoglicólico anterior se transforma en glioxilato. En la mitocondria el glioxilato se descarboxila produciendo CO2. Por cada CO2 producido en la fotorrespiración se consumen 4 ATP y 5 NADP2H. La fotorrespiración es perjudicial para la célula, debido a que el fosfoglicérico se incorpora al ciclo de Calvin y se reduce el rendimiento a la mitad.
B. LAS PLANTAS C4
Las plantas C4 presentan una anatomía foliar peculiar, conocida como anatomía de tipo Kranz o en corona. En el corte transversal de estas hojas se observan dos tipos de células fotosintéticas: • unas grandes, que rodean a los haces conductores (a modo
de “corona”) formando una vaina • Otras que ocupan el mesófilo, menores y dispuestas por lo
general más o menos radialmente alrededor de la vaina. Vía C4
1. Es un proceso de fijación del CO2 atmosférico, captado en las células del mesófilo, pero en vez de ir al ciclo de Calvin, el CO2 reacciona con el PEP (Fosfoenolpiruvato).
2. El producto final entre el PEP y el CO2 es el ácido oxalacético, que luego se convierte en malato.
3. El malato es llevado a las células de la vaina, en donde es descarboxilado, produciendo el CO2 necesario para el ciclo de Calvin, además de ácido pirúvico.
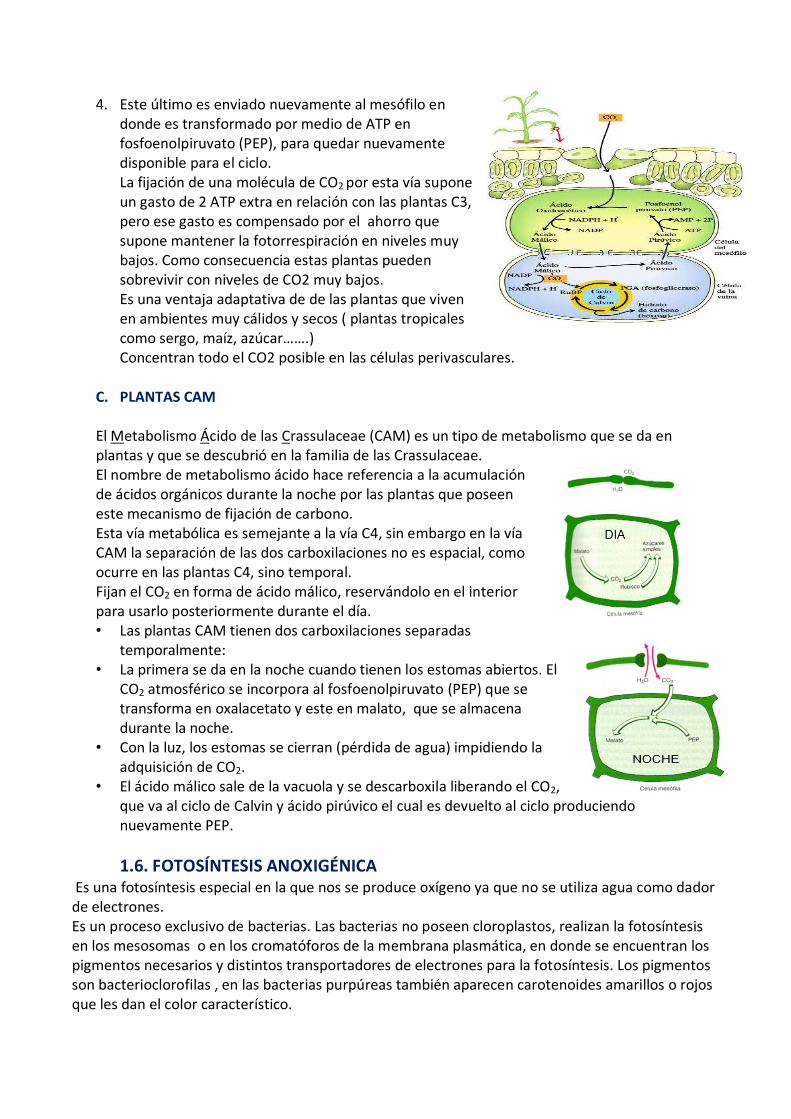
4. Este último es enviado nuevamente al mesófilo en donde es transformado por medio de ATP en fosfoenolpiruvato (PEP), para quedar nuevamente disponible para el ciclo. La fijación de una molécula de CO2 por esta vía supone un gasto de 2 ATP extra en relación con las plantas C3, pero ese gasto es compensado por el ahorro que supone mantener la fotorrespiración en niveles muy bajos. Como consecuencia estas plantas pueden sobrevivir con niveles de CO2 muy bajos. Es una ventaja adaptativa de de las plantas que viven en ambientes muy cálidos y secos ( plantas tropicales como sergo, maíz, azúcar…….) Concentran todo el CO2 posible en las células perivasculares.
C. PLANTAS CAM
El Metabolismo Ácido de las Crassulaceae (CAM) es un tipo de metabolismo que se da en plantas y que se descubrió en la familia de las Crassulaceae. El nombre de metabolismo ácido hace referencia a la acumulación de ácidos orgánicos durante la noche por las plantas que poseen este mecanismo de fijación de carbono. Esta vía metabólica es semejante a la vía C4, sin embargo en la vía CAM la separación de las dos carboxilaciones no es espacial, como ocurre en las plantas C4, sino temporal. Fijan el CO2 en forma de ácido málico, reservándolo en el interior para usarlo posteriormente durante el día. • Las plantas CAM tienen dos carboxilaciones separadas
temporalmente: • La primera se da en la noche cuando tienen los estomas abiertos. El
CO2 atmosférico se incorpora al fosfoenolpiruvato (PEP) que se transforma en oxalacetato y este en malato, que se almacena durante la noche.
• Con la luz, los estomas se cierran (pérdida de agua) impidiendo la adquisición de CO2.
• El ácido málico sale de la vacuola y se descarboxila liberando el CO2, que va al ciclo de Calvin y ácido pirúvico el cual es devuelto al ciclo produciendo nuevamente PEP.
1.6. FOTOSÍNTESIS ANOXIGÉNICA Es una fotosíntesis especial en la que nos se produce oxígeno ya que no se utiliza agua como dador de electrones. Es un proceso exclusivo de bacterias. Las bacterias no poseen cloroplastos, realizan la fotosíntesis en los mesosomas o en los cromatóforos de la membrana plasmática, en donde se encuentran los pigmentos necesarios y distintos transportadores de electrones para la fotosíntesis. Los pigmentos son bacterioclorofilas , en las bacterias purpúreas también aparecen carotenoides amarillos o rojos que les dan el color característico.

Solo tienen un fotosistema . El transporte de electrones es cíclico para la producción de ATP en las ATP sintetasas. El poder reductor cuando es necesario se obtiene por un transporte acíclico y es en forma de NADH, no de NADPH Sulfobacterias o bacterias verdes del azufre; usan como dador de electrones SH2 o H2. El S resultante no lo acumulan en su interior sino que lo expulsan al exterior. Bacterias purpúreas; usan como dador de electrones SH2 y el S resultante lo acumulan en su interior.
Los dos tipos de bacterias son Fotolitotrofos.
1.7. QUIMIOSÍNTESIS • Fuente de carbono CO2 ambiental • Nutrición autótrofa no fotosintética • La energía procede de reacciones de oxidación de sustancias inorgánicas • Exergónicas o exotérmicas • Exclusiva de bacterias
La diferencia entre la fotosíntesis y la quimiosíntesis está en la forma de obtener la energía, En la fotosíntesis la energía necesaria para reducir el CO2 durante el ciclo de Calvin procede de la luz, mientras que en la quimiosíntesis procede de reacciones redox exergónicas o exotérmicas, en las que se oxidan compuestos inorgánicos. La síntesis de materia orgánica se realiza, en los dos casos, a través del ciclo de Calvin. La quimiosíntesis es un tipo de nutrición autótrofa. Consiste en la obtención de materia orgánica a partir de inorgánica, utilizando como fuente de energía la liberada en reacciones químicas redox exergónicas o exotérmicas. La quimiosíntesis se divide en dos fases, equivalentes a las fases lumínica y oscura de la fotosíntesis:
• Obtención de energía. En la quimiosíntesis la energía se obtiene de reacciones químicas inorgánicas y exergónicas o exotérmicas, en las que se produce una oxidación que desprende energía en forma de ATP y coenzimas reducidas (NADH).
• Producción de materia orgánica. El ATP y el NADH obtenidos en la fase anterior se utilizan para la síntesis de materia orgánica por medio del ciclo de Calvin.
¿Qué características definen a los organismos quimiosintéticos? Los organismos quimiosintéticos presentan una serie de características comunes: Son procariotas autótrofas. Viven de una fuente inorgánica: agua, sales, O2, CO2 y compuestos inorgánicos de cuya
oxidación obtienen energía.

Obtienen la energía de una reacción química específica. Solamente crecen con compuestos específicos de origen inorgánico, o producidos por la actividad de otros organismos (descomposición, excreción).
Son aerobios. Utilizan el oxígeno como último aceptor de electrones. Sintetizan materia orgánica por medio del ciclo de Calvin. CLASIFICACIÓN BACTERIAS QUIMIOSINTÉTICAS
Las bacterias quimiosintéticas se agrupan, según el tipo de compuesto químico del que extraen la energía, en los siguientes grupos:
¿Qué función realizan los organismos quimiosintéticos en la naturaleza? Importancia biológica Los organismos quimiosintéticos desarrollan una función fundamental en la naturaleza, puesto que participan como elementos clave de los ciclos biogeoquímicos. Un ejemplo del papel de una bacteria quimiosintética en un ciclo biogeoquímico es la Nitrosomonas, que participa en el ciclo del nitrógeno: Transforma el amoniaco en nitritos (El amoniaco del suelo procede de los restos de seres vivos y de la fijación del N2 atmosférico que realizan algunas bacterias) en una reacción llamada nitrosación. Produce la transformación del amoniaco en nitratos, proceso llamado nitrificación. Nitrobacter realiza la transformación de nitritos en nitratos nitratación.
2. ANABOLISMO HETERÓTROFO GLUCONEOGÉNESIS GLUCOGENOSÍNTESIS AMILOSÍNTESIS BIOSISTESIS DE LÍPIDOS BIOSINTESIS DE PROTEÍNAS BIOSINTESIS ÁCIDOS NUCLÉICOS
El anabolismo heterótrofo consiste en la formación de materia orgánica más compleja a partir de materia orgánica más sencilla. Se da tanto en organismos autótrofos como en heterótrofos. Existen diferencias entre organismos autótrofos y heterótrofos a la hora de obtener los monómeros. Los heterótrofos los tomas del medio mientras que los autótrofos los pueden obtener a partir del ciclo de Calvin.
Tanto autótrofos como heterótrofos pueden obtener los monómeros de sus reservas.

2.1 GLUCONEOGÉNESIS
Consiste en la formación de glucosa a partir e pirúvico por inversión de la mayor parte de la s reacciones glucolíticas.
El pirúvico se obtiene a partir de: Glucosa ( por glucolisis) Glicerol ( por hidrólisis de triglicéridos) Ácido láctico Aminoácidos Nunca se obtiene a partir de de ácidos grasos.
Reacciones no reversibles de la glucolisis: 1. De pirúvico a fosfoenolpirúvico: No hay ningún enzima que pueda hacer este paso
directamente; por tanto, hay que hacer la vuelta siguiente: el ácido pirúvico entra en la mitocondria, ya que en este orgánulo está el enzima capaz de transformarlo en ácido oxalacético. Esta molécula no puede atravesar la membrana interna de la mitocondria, por eso se tiene que transformar en ácido málico, que sale al citosol, donde se transforma en ácido oxalacético, y éste finalmente se transforma en ácido fosfoenolpirúvico. (piruvato carboxilasa)
2. Transformación de fructosa-1,6-difostato en fructosa-6-fosfato: La realiza un enzima diferente al que actúa en la glucólisis que provoca la hidrólisis de un grupo fosfato sin haber síntesis de ATP. (difosfofructosa fosfatasa)
3. Paso de glucosa-6-fosfato a glucosa: El enzima que cataliza el paso inverso en la glucólisis actúa sólo en un sentido, por tanto, es necesario otro enzima, que separa el grupo fosfato sin que tampoco haya síntesis de ATP. (Glucosa 6fosfato fosfatasa)
En el caso de organismos heterótrofos este proceso ocurre principalmente en el hígado. En plantas la glucosa puede venir igual que en heterótrofos, pero además puede proceder del ciclo de Calvín o de la degradación de ácidos grasos.
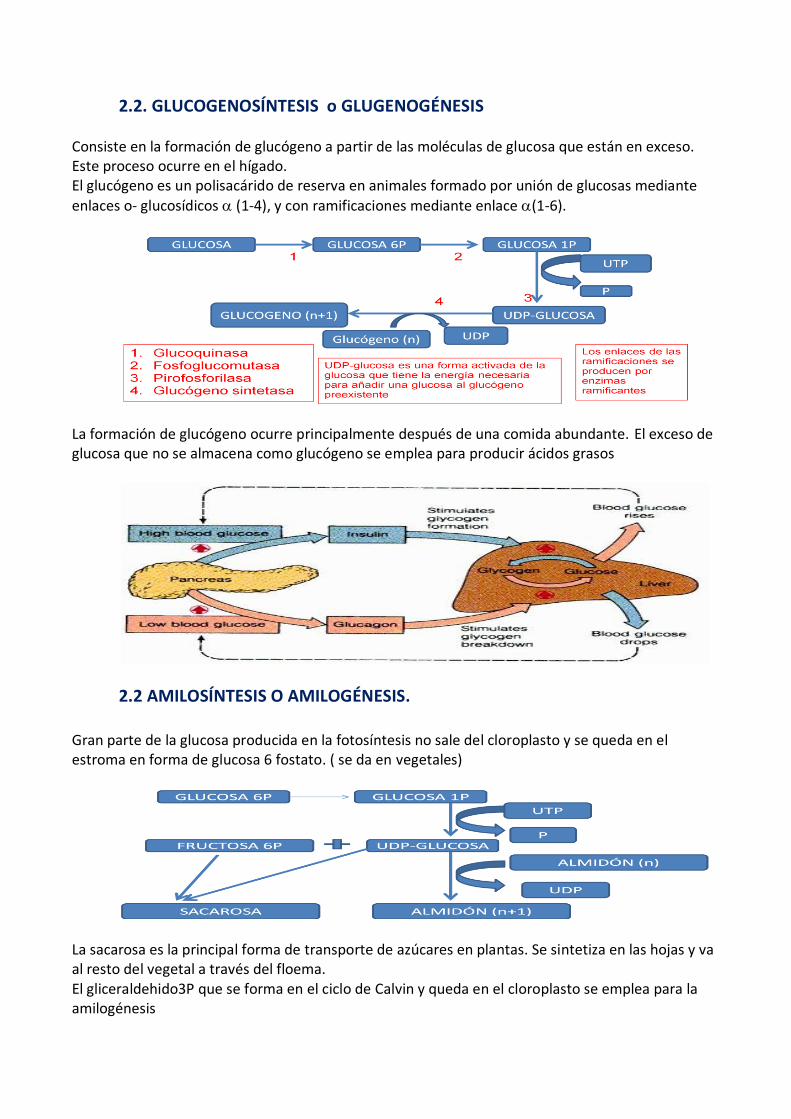
2.2. GLUCOGENOSÍNTESIS o GLUGENOGÉNESIS
Consiste en la formación de glucógeno a partir de las moléculas de glucosa que están en exceso. Este proceso ocurre en el hígado. El glucógeno es un polisacárido de reserva en animales formado por unión de glucosas mediante
enlaces o- glucosídicos (1-4), y con ramificaciones mediante enlace (1-6).
La formación de glucógeno ocurre principalmente después de una comida abundante. El exceso de glucosa que no se almacena como glucógeno se emplea para producir ácidos grasos
2.2 AMILOSÍNTESIS O AMILOGÉNESIS.
Gran parte de la glucosa producida en la fotosíntesis no sale del cloroplasto y se queda en el estroma en forma de glucosa 6 fostato. ( se da en vegetales)
La sacarosa es la principal forma de transporte de azúcares en plantas. Se sintetiza en las hojas y va al resto del vegetal a través del floema. El gliceraldehido3P que se forma en el ciclo de Calvin y queda en el cloroplasto se emplea para la amilogénesis

2.3. BIOSINTESIS DE LÍPIDOS
Sólo veremos la biosíntesis de acilglicéridos. Se necesita entonces:
A) glicerol B) ácidos grasos
Ambos compuestos se unirán después por esterificación y de esa forma se producirán los acilglicéridos.
A) Obtención del glicerol
Se obtiene por reducción del gliceraldehido3P o dihidroxiacetonaP ( intermediarios de la glucolisis) formando glicerol3P ( reducción a cargo de NAD). También puede venir de la degradación o saponificación del triglicérido.
B) Biosíntesis ácidos grasos
Se realiza por una vía inversa a la oxidación, pero que se localiza en el citoplasma no en la mitocondria. Esto se produce principalmente en células hepáticas y células del tejido adiposo (adipocitos) El acetilCoA obtenido en la mitocondria (degradación de azúcares, aa,….) sale al citoplasma. (recordamos que la membrana interna mitocondrial no es permeable al oxalacetato) Para esta salida el aceltilcoA se une al oxalacetato, se libera CoASH y se forma citrato, el cual sale al citoplasma y se une al CoASH citoplásmico y forma acetilcoA y oxalacetato (citoplasma)

El acetilCoA ya en el citoplasma forma malonilcoA por carboxilación. A continuación al malonilcoA se une otro acetilCoA y se libera CO2( dercarboxilación) y CoASH, formándose un compuesto de 4 carbonos unidos al CoA, a continuación se producen dos reducciones a cargo de NADP que proviene de la ruta del fosfoglucanato y finalmente se forma un acetilCoA de 4 átomos de carbono. Si quisiéramos seguir alongando la cadena a este ácido graso de 4 átomos de carbono se uniría otro malonilcoA que añadiría otro par de atomos de crbono y así sucesivamente. De esta forma conseguimos ácidos grasos de un número par de átomos de carbono, sobre ellos se producirán transformaciones para conseguir ácidos grasos insaturados y de número impar de átomos de carbono. Algunos ácidos grasos no podrán ser sintetizados y habrá que ingerirlos en la dieta (ácidos grasos esenciales)
Faltan reacciones El triglicérido finalmente se formara por la esterificación de los ácidos grasosactivados en forma de acetilcoA con el glicerol3P
2.3 ANABOLISMO DE PROTEÍNAS
La síntesis de aminoácidos es compleja porque cada aminoácido sigue una ruta metabólica distinta, además incluso un mismo aminoácido puede ser sintetizado de formas diferentes en las distintas especies CELULARES. No todas las células sobre todo las heterótrofas pueden sintetizar todos los aminoácidos, algunos tiene que ser ingeridos en la dieta (aa esenciales). Aunque cada uno tiene una ruta distinta la biosíntesis tiene en común: Se realiza en el citosol mediante reacciones de transaminación, inversas a las que tienen lugar en la degradación de los mismos, en las que el grupo amino del ácido glutámico es transferido a diversos esqueletos carbonados presentes en la célula, los cuales proceden del ciclo de Krebs o de la glucolisis. El ensamblaje de los aminoácidos para formar proteínas se lleva a cabo en los ribosomas siguiendo las instrucciones cifradas en la secuencia de nucleótidos del DNA. Las plantas pueden sintetizar todos los aa; los animales sólo algunos, el resto los incorpora con la dieta.

2.4 ANBOLISMO DE ÁCIDOS NUCLEÍCOS Aunque los nucleótidos, o sus componentes moleculares, que proceden de la hidrólisis de unos ácidos nucleicos generalmente se reciclan para sintetizar otros, a veces puede ser necesario sintetizarlos "ex novo". La ribosa y la desoxirribosa se obtienen en la ruta de las pentosas. El ácido fosfórico es un componente habitual de las células. Las bases nitrogenadas se sintetizan mediante complejas secuencias de reacciones que parten de los esqueletos de diversos aminoácidos.